Title: Journal of Entomology and Zoology, Vol. 11, No. 4, December 1919
Author: Various
Release date: October 5, 2011 [eBook #37632]
Language: English
Other information and formats: www.gutenberg.org/ebooks/37632
Credits: Produced by Larry B. Harrison, Diane Monico, and the Online
Distributed Proofreading Team at http://www.pgdp.net
PUBLISHED QUARTERLY BY
POMONA COLLEGE DEPARTMENT of ZOOLOGY
CLAREMONT, CALIFORNIA, U. S. A.
| Page | |
| Notes on the Behavior of the Social Wasp Polistes—Horace Gunthorp | 63 |
| Biology of the North American Crane-Flies. V. The Genus Dicranoptycha—Charles P. Alexander | 67 |
| The Central Nervous System of Nucula and Malletia—W. A. Hilton | 75 |
Entered Claremont, Cal., Post-Office Oct. 1, 1910, as second-class matter, under Act of Congress of March 3, 1879
EDITED BY POMONA COLLEGE, DEPARTMENT OF ZOOLOGY
Subscription $1.00 to domestic, $1.25 to foreign countries.
This journal is especially offered in exchange for zoological and entomological journals, proceedings, transactions, reports of societies, museums, laboratories and expeditions.
The pages of the journal are especially open to western entomologists and zoologists. Notes and papers relating to western and Californian forms and conditions are particularly desired, but short morphological, systematic or economic studies from any locality will be considered for publication.
Manuscripts submitted should be typewritten on one side of paper about 8 by 11 inches. Foot notes, tables, explanations of figures, etc., should be written on separate sheets. Foot notes and figures should be numbered consecutively throughout. The desired position of foot notes and figures should be clearly indicated in the manuscript.
Figures should be drawn so that they may be reproduced as line cuts so far as possible. An unusually large number of half tones must be paid for in part by the author. Other more expensive illustrations will be furnished at cost. Figures for cuts should be made to conform to the size of the page when reduced, that is, 5 by 7-1/2 inches or less. The lettering should be by means of printed numbers and letters pasted on the drawings, in most cases.
Authors of articles longer than a thousand words will receive fifty reprints of their publications free of cost. If more than this are desired, the order should be given with the return of the proof sheets. Extra copies and special covers or special paper will be furnished at cost. Authors of short contributions will receive a few extra copies of the number containing their articles.
Manuscripts should be sent by express or registered mail.
Address all communications to
The Journal of Entomology and Zoology
William A. Hilton, Editor
Claremont, California, U. S. A.
HORACE GUNTHORP
Washburn College, Topeka, Kans.
One day last September the writer picked up a nest of the common social wasp, Polistes, which had been detached from its support, and placed it upon his desk. A short time later he was attracted by a scratching sound, and discovered that one of the wasps was just beginning to cut the cap from its cell preparatory to emerging. During the next few days a series of observations were made and notes taken covering the behavior of the wasps which emerged from their cells during that period. Miss Enteman[A] has made a careful study of the instincts of the social wasps, and while the observations recorded in the present paper are largely corroborative of her work, some interesting details are here added.
The cutting of the cap of the cell occupied some time, and extended around four-fifths of its circumference, the remaining one-fifth being gnawed and partially chewed through so that it was flexible enough to act as a hinge for the cap. After the cap was sufficiently cut away, the wasp started to slowly work itself out, pushing up the top of the cell like a trap door as progress was made. A good deal of effort was required to get the body out until the front legs were freed. Then the wasp had more purchase and progress was somewhat faster until the second pair of legs came out. After this slight effort seemed to be necessary for the completion of the operation.
For the next thirty minutes careful observations were made of the movements of this wasp in order to ascertain its first reactions. It is evident that they would be somewhat modified from what they are here recorded if the colony had contained the queen and other workers, as this specimen had the run of the entire nest, and none of its movements were effected by those of other individuals. It is equally evident that all stimuli came from within, or from contact with the nest, and not from suggestions received from other individuals or from contact with them. The following is the record made at one minute intervals, beginning with the time the specimen left its cell:
8:06. Specimen emerged from its cell.
8:07. Cleaned its front legs in its mouth and its antennæ with its front legs.
8:08. Moved around some. Rubbed its wings with its hind legs and spread them out twice.
8:09. Cleaned antennæ and front legs.
8:10. Swung abdomen back and forth, and brushed its wings. Moved around the nest rapidly and waved the antennæ, but all movements were jerky.
8:11. Explored nest, occasionally rubbing abdomen with legs.
8:12. Explored nest.
8:13. Explored nest. Movements unsteady. Cleaned antennæ and front legs.
8:14. Explored nest, in the course of which it went over the edge on to the back side, but immediately returned to the under side. Cleaned the front legs and antennæ, and then the hind legs.
8:15. Spread out the wings. Cleaned the antennæ.
8:16. Cleaned abdomen.
8:17. Crawled on top or back side of nest again and stayed there. Cleaned wings and abdomen.
8:18. Explored top. Cleaned front legs and antennæ.
8:19. Stood still. Occasional movement of head, antennæ or abdomen.
8:20. Same as 8:19.
8:21. Began to explore again, becoming quite lively. Antennæ constantly waving.
8:22. Same as 8:21, but extended its travels to the under (cell) side of the nest.
8:23. Left the nest entirely and began to walk around the surface of the desk.
8:24. Started to climb a bottle that was some six inches from the nest. Antennæ still waving.
8:25. On the neck of the bottle, two inches above the surface of the desk. Cleaned front legs and antennæ.
8:26. Quiet except that it spread its wings once.
8:27. Still on neck of bottle. Moved its head and antennæ back and forth.
8:28. Slight change in position. Antennæ were still waving. Rubbed its wings, spread them, and then rubbed them again.
8:29. Rubbed its hind legs together vigorously.
8:30. Spread wings once, then rubbed them and the abdomen with the hind legs. Rubbed the hind legs together, and finally rubbed the right wings vigorously.
8:31. Moved around some, occasionally stopping to rub the right wings.
8:32. Explored the neck of the bottle.
8:33. Same as 8:32. Cleaned antennæ.
8:34. Same as 8:33.
8:35. Stood still but continued to clean antennæ and front legs.
8:36. Climbed up and explored the cork of the bottle.
8:37-8:40. Stood still on the cork, occasionally moving its jaws.
At 8:40 the nest was placed against the cork and the wasp immediately crawled onto it, but seemed restless. As the nest has a faint, but distinct, odor of honey, it was probably attracted to it through the sense of smell.
The next morning the specimen was nowhere in sight, but forty-eight hours later it fell out of a loose-leaf binder that had been lying on the desk. It seemed to be as active as when seen two days before. Some time during the second night after the appearance of the first specimen, that is, when it was some thirty hours old, a second individual emerged. This one was discovered on a pile of books two feet from the nest where it had evidently crawled soon after emerging.
As soon as the first specimen was rediscovered, that is, when it was sixty hours old, the second wasp then being thirty hours old, the two were placed on the nest, and this in turn was placed on a book. They both started on tours of observation, and every time they came in contact with each other they made sudden starts and jumps to avoid an evidently startling new object, meanwhile violently waving their antennæ and often cleaning these organs after such contact. Dr. Enteman says, "All wasps possess the instinct of fear. This ... is readily overcome by the frequent appearance of the awe-inspiring object." This is true, because they were evidently on familiar terms with each other in half an hour, and paid very little attention to the frequent meetings which before had apparently distressed them. They wandered freely over their nest and the top surface of the book on which it was placed, but did not attempt to climb off the latter.
At 12 o'clock, four hours later, a third wasp had appeared, and none of the specimens seemed to be disturbed by the presence of the others. When the nest was first picked up, one cell containing a well formed pupa was uncapped. This specimen was then alive, but it may have been dead at the time of this observation. In either case, it had been dragged out of its cell, decapitated, and the front legs torn off. No trace of the head was found, but the body and legs were on the book about one inch from the nest. Whether this act was connected with the hunger of the wasps themselves or[Pg 65] with the first development of the instinct of feeding the larvæ in the nest, which Miss Enteman says begins without imitation, is not clear.
At 2 p. m. (two hours later) the colony was placed out of doors, still on the book. Two of the wasps soon left the latter, and settled near it, keeping very quiet for half an hour. The third kept climbing over and around the nest. At 2:30 one of the two wasps returned to the nest.
At 3 p. m. two of the specimens were on the ground near the porch. They made only short flights, resembling jumps with the wings assisting, this being true even when they were disturbed. The third wasp was beside the colony, chewing on the decapitated pupa, probably getting some nourishment from it in the process.
During the afternoon the nest was disturbed, and at 6 p. m. all three specimens had gone from the porch. One was found wandering aimlessly on a canna leaf near by. It did not seem to be able to fly well. The other two had disappeared entirely.
The nest was saved and several days later a fourth wasp appeared. It was a very lively specimen, and spent the first few hours actively exploring the nest. It seemed of a very nervous disposition, being more easily disturbed than any of the others had been. Every time the nest was picked up, it would start for the fingers or forceps holding it. At one time it was observed with its whole body in a cell, head downward, evidently examining the interior. After staying close to the nest for a day, it began to fly around the floor of the room, paying no more attention to its former home. Even when it was placed on or near it, it would almost immediately crawl or fly away. Its flying was erratic, and seemed to lack power, but it got along much better than any of the other three had done.
From the above observations it would appear that the movements of the wasp recorded at one minute intervals after emergence from its cell were probably reactions due to the discomfort of the drying and hardening of the tissues. At first the wasps apparently had very little, if any, home instinct. The only things to indicate that they had any were the facts that the first specimen so readily left the cork on which it was sitting and went back to its nest when the latter was held near it, and the fourth wasp stayed on or near the nest for the first twelve hours. But all the specimens observed left the nest the first night and showed no intention or disposition to return. The presence of a second wasp seemed to bring the home instinct into existence more forcibly, as the first and second wasps stayed with the nest for six or seven hours when they were returned to it together, while the fourth one repeatedly left the empty nest almost at once when it was returned to it. But this instinct was seemingly not very strong, as they soon wandered away when placed out of doors. They seemed to have no idea as to how to carry on the work of the colony, but wandered aimlessly over it. Perhaps this was due to the fact that they were too young, as Miss Enteman says the development of the nursing instinct is usually manifested "any time after the first half day of imaginal life," but was observed in some neuters as young as four hours, while in others it was delayed for two weeks.
While the above observations are admittedly too few from which to draw definite conclusions, they seem to warrant the following assumptions, the first three of which are quoted from Miss Enteman, and hence are simply corroborative of her work:
1. "All wasps possess the instinct of fear. This is especially strong the first few days after emergence, but is readily overcome by the frequent appearance of the awe-inspiring object.[Pg 66]
2. "In a sense, the wasp remembers. This is indicated by the manner in which it accustoms itself to the sight of strange objects, and by its behavior when a change is made in its nest or surroundings.
3. "It shows considerable individual variability, both as to time and manner of its response to stimuli."
4. After emergence, the first reactions are associated simply with the discomfort of the hardening of the tissues.
5. It has marked curiosity, as shown by its repeated inspection of its nest and other familiar objects.
6. The "home instinct" seems to be slight when the wasp is alone, but becomes stronger when two or more are on the same nest.
7. The olfactory sense is closely associated with the early instincts of the wasp.
[A] Minnie Marie Enteman. "Some Observations on the Behavior of the Social Wasps." Pop. Sci. Mo., 61: 339-351, 1902.
(Tipulidæ, Diptera)
V. The Genus Dicranoptycha Osten Sacken
BY CHARLES P. ALEXANDER, Ph.D. (Cornell)
Larva. Form very elongate, terete; integument smooth, glassy, transparent; abdominal segments two to eight with a basal transverse band or area of microscopic chitinized points on the ventral surface; segment eight with a similar band on the dorsum. Spiracular disk surrounded by four lobes, the lateral pair more slender than the blunt ventral pair; dorsal lobe very low or lacking; spiracles small, widely separated; a triangular brown mark on the disk between the spiracles; anal gills a fleshy protuberant ring surrounding the anus. Head-capsule compact, massive, the præfrons large with a few marginal punctures; externo-lateral plates very broad. Labrum large, flattened, pale; antennæ two-segmented, the apical segment almost as long as the basal segment, narrowed to the blunt tip; mandibles with a blunt dorsal and two blunt ventral teeth; maxillæ generalized in structure; hypopharynx a rounded cushion; mentum deeply split behind but not completely divided, with three principle teeth and a small lateral tooth on either side.
Pupa. Cephalic crest low, depressed, setiferous; labrum tumid; labial lobes oval, contiguous; antennal sheaths ending opposite the base of the wing. Pronotal breathing-horns microscopic, represented only by tiny triangular tubercles; mesonotum unarmed; wing-sheaths ending opposite the middle of the third abdominal segment; leg-sheaths ending opposite the base of the fifth abdominal segment, the tarsi terminating on a level, or nearly so. Abdominal tergites and sternites each with four transverse rows of microscopic setæ; lateral spiracles on segments two to seven.
The genus Dicranoptycha was erected by Osten Sacken in 1860 (Proc. Acad. Nat. Sci. Phila. for 1859, p. 217). The genus includes a small group of crane-flies with a Holarctic distribution, there being about six species in North America and two, or possibly three, in Europe. As I have indicated elsewhere, D. signaticollis v.d.W. of Java is undoubtedly a species of Libnotes. Of the American species, D. germana O.S. is characteristic of the Canadian life-zone of northeastern America. D. sobrina O.S. is widely distributed in the United States and southern Canada, usually occurring in the Transitional and Upper Austral life-zones. So far as known at present it is the only species of the genus occurring on the Pacific slope. The remaining American species (nigripes O.S., winnemana Alex., tigrina Alex. and minima Alex.) are Austral in distribution, occurring in the southeastern and south central United States. A more detailed account of the distribution of the species is given in another paper by the writer which may be consulted (Proc. Acad. Nat. Sci. Phila. for 1916, pp. 496, 497). All of the known species are generally similar to one another in appearance and are separated by relatively slight differences of size, color and structure.
Nothing has ever been written concerning the immature stages of this peculiar[Pg 68] group of crane-flies. The species described hereinafter were reared at Lawrence, Kansas, and the general conditions under which they occur may be briefly discussed:
North Hollow, on the Campus of the University of Kansas, is a typical dry Austral woodland traversed by a small stream that is entirely dry during the months of midsummer drought. The soil consists of a rich black humus that is soft and mellow except during the period of greatest dryness, being overlain by a varying depth of vegetable debris and leaf-mold. It is in this relatively dry soil that the larvæ of Dicranoptycha occur. The forest cover consists of Carolina poplar, Populus deltoides Marsh; black walnut, Juglans nigra L., white elm, Ulmus americana L.; Kentucky coffee-tree, Gymnocladus dioica (L.) Koch; honey locust, Gleditsia triacanthos L.; red bud, Cercis canadensis L.; yellow wood, Cladrastis lutea (Mx.f.) Koch; tree-of-heaven, Ailanthus glandulosa Desf., etc. The principle shrubs are the goose-berry, Ribes gracile Mx.; poison ivy, Rhus Toxicodendron L.; wahoo, Evonymus atropurpureus Jacq.; bladder-nut, Staphylea trifolia L.; coral-berry, Symphoricarpos orbiculatus Moench.; blackberried elder, Sambucus canadensis L., etc. The herbage is made up of tall grasses, composites and, in the spring, the all-dominant cleavers, Galium. In addition to the above, great tangles of lianas (Smilax, Vitis, Ampelopsis, etc.) are found.
In situations such as the above these Austral species of Dicranoptycha spend their entire lives. The first larvæ of D. winnemana were found here on March 20, 1918, by the writer and his wife. At this time they were well grown (length 16 mm.; diameter 0.9 mm.). They occurred just beneath the cover of fallen leaves and other debris in the upper layers of soil. Here they were associated with pupæ of Tipula angustipennis Lw., larvæ of Sciara (Mycetophilidæ); Psilocephala hæmorrhoidalis Macq. (Therevidæ), numerous beetle larvæ, centipedes, etc. By their elongate form and glabrous shiny skin they are very characteristic and easily recognized. The glassy appearance of the body suggests the shiny shells of a small coiled molluscan whose dead fragments occurred in some numbers in the same situations. These larvæ were placed in rearing and the first adults appeared in the breeding-cages on May 6, and from that time on continued to appear in large numbers. It was over a month later that the first individuals were taken in the field. The pupal duration could not be determined closer than ten days, and this may be the usual length of time required for this stage. The first larvæ of D. minima were found on July 2, 1918, in similar situations in North Hollow. At this time they were only about one-half grown. On July 11 much larger larvæ of this species were secured and placed in rearing, emerging as adults on July 21. The larvæ, like these of D. winnemana, live just beneath the layer of leaf-mold in the upper zone of black soil. They are usually quite sluggish in their motions but at other times are quite active. The larvæ are herbivores and feed on the rich organic earth in their haunts. When ready to pupate, they encase themselves in earthen cells (10 mm. × 3.5 mm.), firm in texture, rather thick-walled but without silk. There is a small opening at either end. The length of the cavity is but little greater than the pupa itself. In this cavity the pupa rests and matures. As in other insects, the teneral pupæ are very pale yellow but gradually darken in color until, at emergence, they are of a dark brownish-black. When newly transformed the teneral flies rest on the ground and on the leaves of low plants nearby.
The adult flies of D. germana usually occur in the immediate neighborhood of running or stagnant water and may be swept from the rank vegetation in such places. The flies rest on the upper surface of the leaves of tall herbs and low shrubs. In[Pg 69] eastern Kansas, the flies of D. winnemana, D. tigrina and D. minima often occur together. In June, D. winnemana appears on the wing and is found associated with Tipula morrisoni Alex., T. mingwe Alex., etc.; in July, D. minima appears, together with Tipula flavibasis Alex., T. unimaculata Lw., etc.; still later in July D. tigrina emerges and all three species fly together during August and into September when they fly with Tipula ultima Alex., T. unifasciata Lw., etc. It is curious that no other species of Limnobiinæ occur in the thamnophytic association frequented by Dicranoptycha. All three species of this genus as discussed above have habits that are generally similar to one another. They are usually found resting quietly on the upper surface of the leaves but fly readily and on slight disturbance. Pairs in copulation are often found resting, the bodies directed away from one another and the wings folded over the abdomen. While thus united they fly readily, sometimes the female taking the initiative, sometimes the rather smaller male. The eggs are deposited in the soft earth in these situations.
In the Monographs (1869) Osten Sacken included the genus Dicranoptycha in his tribe (section) Limnobina anomala, or, as it subsequently became known, the Rhamphidini, and still later the Antochini. A recent survey of the immature stages of several Antochine genera has shown that the tribe is merely an artificial grouping based on superficial resemblance of the adult flies. This heterogeneous assemblage includes representatives of at least three other tribes, Dicranoptycha, together with Antocha, Elliptera, Rhamphidia, etc., showing an undeniable affinity with the Limnobiini, whereas Teucholabis, Elephantomyia, etc., show an equally clear relationship with the Eriopterini. Moreover a close phylogenetic relationship with the lowermost subtribes of the Hexatomini (Ularia, Epiphragmaria, etc.), is easily apparent.
Dicranoptycha shows the closest affinities with Antocha and Rhamphidia. The larvæ of these three genera, each of which typifies a division, show the following common characters:
Abdominal segments with basal transverse creeping welts or areas of microscopic points. The massive compact head-capsule with the præfrontal sclerite large, distinct, the externo-lateral plates large, mussel-shaped and very thin. The mentum is not completely divided medially. The maxillæ are large and of primitive structure, the cardines and stipites distinct, the two distal lobes large, subequal in size, covered with hairs and bearing sensory organs. Mandibles with one or more dorsal and two or more ventral teeth in addition to the apical point.
The differences between these allied divisions are best indicated by a key.
1. Spiracular disk with only the two long ventral lobes remaining; spiracles lacking or vestigial; abdominal segments with both dorsal and ventral welts; strictly aquatic.Antocharia.
Spiracular disk surrounded by four or five short lobes; spiracles large and functional; abdominal segments with ventral welts only (except the dorsum of segment eight); terrestrial or semiaquatic.
2. Body moderately elongated and covered with a long dark pubescence; spiracular disk squarely truncated, surrounded by five subequal stout lobes; mentum with five subequal teeth, the lateral one of either side not conspicuously reduced.Rhamphidaria.
Body very long and slender, glabrous; spiracular disk obliquely truncated, surrounded by four slender naked lobes; mentum with three subequal primary teeth and a much reduced lateral tooth on either side.Dicranoptycharia.
1. Pronotal breathing-horns branched; aquatic.Antocharia.
Pronotal breathing-horns not branched; semiaquatic or terrestrial.
2. Pronotal breathing-horns distinct, elongate-cylindrical.Rhamphidaria.
Pronotal breathing-horns apparently lacking, microscopic.Dicranoptycharia.
A Key to the Species of Dicranoptycha
1. Spiracular disk with the dark markings less extensive; the mark of the lateral lobes not contiguous with the spiracle or the triangular area on the disk; dorsal marking indistinct or lacking.D. winnemana Alex.
Spiracular disk with the dark markings more extensive; the mark of the lateral lobes suffusing the ventral inner margin of the spiracle and usually closely approximated or nearly contiguous with the triangular area on the disk; dorsal marking black, transversely rectangular.D. minima Alex.
Description of the Species.
1916 Dicranoptycha winnemana Alexander; Proc. Acad. Nat. Sci. Phila., pp. 500, 501; Pl. 25, fig. 12.
Larva.—Length, 20-22 mm.
Diameter, 0.9-1.1 mm.
Coloration varying from white to almost black depending on the nature and amount of the food eaten which shows clearly through the transparent integument. The fat-bodies likewise show through and give a white color to the larva especially after death.
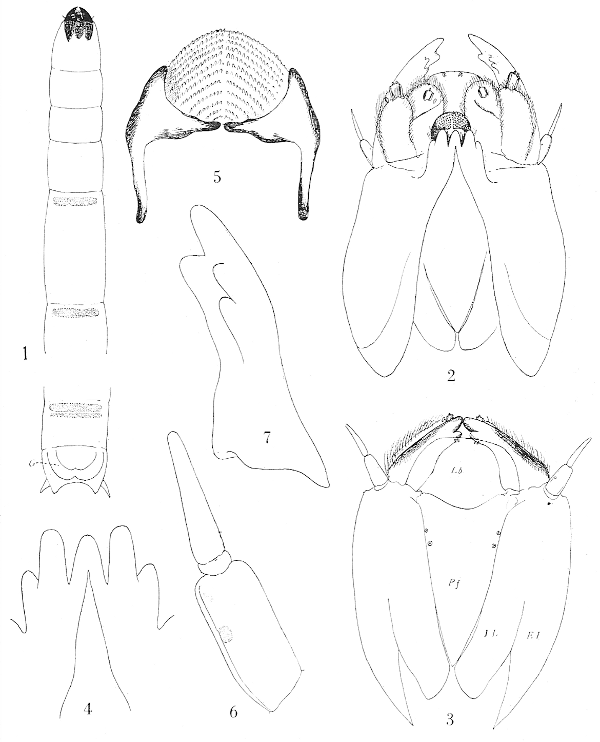
Form very elongate (fig. 1), body terete; integument very glabrous, transparent and glassy. Prothoracic segment a little longer than the mesothorax which, in turn, slightly exceeds the metathorax. The intermediate abdominal segments are elongated. The basal ring of sternites two to eight bears a transverse band or area of microscopic chitinized spicules, the one on the eighth segment split lengthwise by a capillary line. A similar band occurs in the same position on the dorsum of the eighth segment but the pleural region is devoid of such a band.
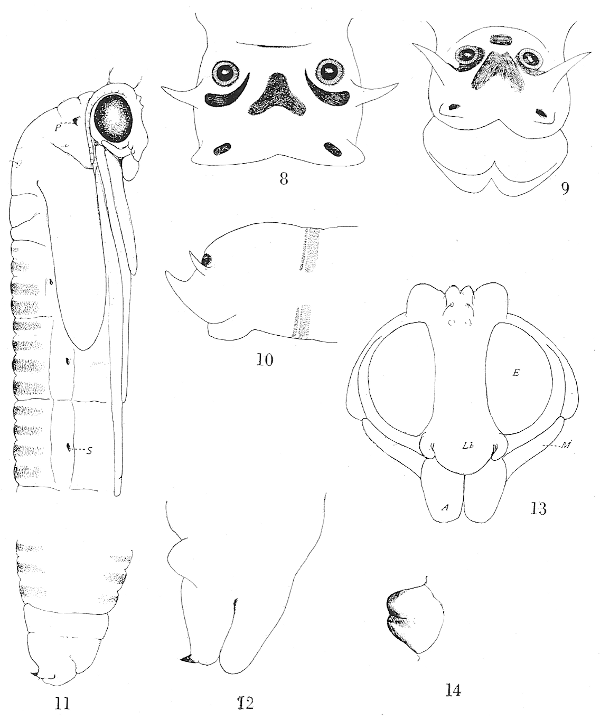
Spiracular disk (fig. 8) moderate in size, obliquely truncated, surrounded by four lobes, a pair of small, slender, lateral lobes and short, broader ventral lobes. The usual dorso-median lobe is lacking but its position is indicated by a gently rounded convexity. The inner face of the lateral lobe bears a narrow semi-lunate black mark with the concavity toward the spiracle, the proximal end acutely pointed. The ventral lobes bear a similar but smaller subrectangular black mark. A pale and usually indistinct dusky mark occupies the inner face of the dorsal lobe. On the disk between, and slightly below the level of, the spiracles is a large brown triangular or V-shaped mark. The spiracles are small, separated from one another by a distance equal to about 2.5 to 3 times the diameter of one; the center-piece of the spiracle is black, the[Pg 71] ring yellow surrounded by an outer dusky margin. Anal gills fleshy and protuberant as a blunt ring surrounding the anus (fig. 10).
Head-capsule (fig. 2) of the compact, massive type of the Limnobiini; præfrontal sclerite (fig. 3) large and distinct; the sclerite broad with the sides subparallel to about midlength, thence tapering gradually to the tip which is entire; there are two or three punctures at the margin before midlength. Interno-lateral plates narrow, a little longer than the præfrons; externo-lateral plates very broad, thin and flattened with the posterior margin very obtuse and the inner ventral portions continuous with the mental plate. Labrum (fig. 3) very broad and extensive, flattened, pale in color, the anterior margin with about two sense-organs. Mentum (fig. 4) deeply split behind but not completely divided, the anterior margin with three primary teeth that are subequal in size or the middle one a little smaller; a much reduced lateral tooth on either side. Præmentum smaller than the hypopharynx, in outline roughly oval or semicircular with the two labial palpi surrounded by hairs at the base. Hypopharynx (fig. 5) consisting of two chitinized arms that are contiguous but not fused medially, the concavity between them filled with a rounded cushion that is covered with tubercles arranged in more or less distinct oblique parallel rows. Antennæ (fig. 6) two-segmented, the basal segment cylindrical with an auditory plate on the face at beyond midlength; apical segment long and slender, in length but slightly less than the basal segment, tapering gradually to the bluntly rounded apex. Mandibles (fig. 7) simple with the teeth blunt; apical point longer than the lateral teeth; dorsal tooth single, broad, very flattened and obtusely pointed; ventral teeth two, a little smaller than the dorsal tooth. Maxillæ (fig. 2) of a generalized structure, the cardines distinct and feebly chitinized; distal lobes of the organ consisting of a subequal inner and outer lobe; the outer lobe with an abundance of long, delicate hairs and bearing a few sensory papillæ including one larger palpiform organ.
Pupa.—Length, 9.1-12.8 mm.
Width, d.-s., 1.6-1.8 mm.
Depth, d.-v., 1.6-1.9 mm.
Thoracic dorsum shiny light brown; in very old pupæ the color is much darker, but still retains a much brighter color than the leg and wing-sheaths; abdomen pale becoming darker in age, especially on the pleura.
Cephalic crest (fig. 13) low and depressed, inconspicuous, lying between the antennal bases which extend beyond it; there are four small setigerous lobes, the larger pair of which are posterior in position. Front between the eyes broad, subparallel. Two blunt tubercles on either side of the forehead. Eyes large, with coarse ommatidia. Labrum semicircular in outline, tumid. Labial lobes large, oval, contiguous with one another, at the tip of the labrum. Maxillary palpi moderately long and slender, nearly straight, gradually narrowed to the tip which ends opposite the knee-joint of the fore legs. Antennæ with the basal segments separated only by the cephalic crest, the sheaths ending about opposite or a little before the lateral angle of the thorax.
Pronotal breathing-horns (fig. 14) very small, almost microscopic; when viewed from the dorsal aspect appearing as tiny triangular tubercles. Mesonotum moderately convex, unarmed, the V-shaped suture distinct; a few setæ on the mesonotum, including one near the end of each scutal lobe. Wing-sheaths rather short, but narrow, ending about opposite midlength of the third abdominal segment. Leg-sheaths ending opposite[Pg 72] the base of the fifth abdominal segment, the tips of the tarsi ending about on a common level or those of the fore legs a trifle longer.
Abdominal segments (fig. 11) subdivided into four annuli that bear transverse bands of microscopic setæ; these bands increase in width from the basal to the apical. Spiracles on the pleural region of segments two to seven, lying opposite the third annulus and close to the ventral margin of the pleura. No spiracles are discernible on the dorsum of the eighth segment. Male cauda (fig. 11) with the ventral lobes very blunt, rounded; the dorsal lobes very small, terminating in a sharp spine that is directed dorsad and bears a weak seta near its base. Female cauda (fig. 12) with the ventral lobes a little longer than the dorsal lobes; the latter at the outer angle of the apex with a short stout spine that is directed dorsad as in the male.
Nepionotype (type larva), Lawrence, Kansas, April 2, 1918.
Neanotype (type pupa), with the type larva, May 6, 1918.
Paratypes, larvæ and pupæ, about fifty from the type locality, March 20 to May 20, 1918.
1919 Dicranoptycha minima Alexander; Ent. News, Vol. 30.
The larva is very similar to that of D. winnemana as described above, but is slightly smaller. The spiracular disk (fig. 9) has the dark markings much more extensive. The mark of the lateral lobes is contiguous with the spiracles and is also closely approximated to the large triangular brown mark on the disk. There is a large transverse rectangular mark occupying the inner face of the dorsal lobe. The marking of the ventral lobe is about as in D. winnemana.
Nepionotype, Lawrence, Kansas, July 11, 1918.
Neanotype, Lawrence, Kansas, July 21, 1918.
Paratypes, a few larvæ from the type-locality.
A—Labial Lobes; E—Eye; EL—Externo-lateral Plate; G—Anal Gills; IL—Interno-lateral Plate; Lb—Labrum; M—Maxillary Palpus; P—Pronotal Breathing-horn; Pf—Præfrons; S—Spiracle.
WILLIAM A. HILTON
These bivalve forms are grouped among the simplest of the molloscs. It is especially from the condition in Nucula as described by Pelseneer '91, that the conception of the most anterior ganglion being composed of four ganglia, has its chief support. Drew '01, who has also studied Nucula, believes that the lobes of the ganglion in Nucula are superficial and that the four connectives coming from the ganglion may be interpreted in another way. That is, that one pair of nerves may represent an otocystic branch partly fused with the connective. This view seemed reasonable to him as Stempel '99 in Solenyma found the otocystic nerves arose directly from the cerebral ganglion.
The two species of this group used for study were collected at Laguna Beach. Nucula castrensis Hinds, occurs abundantly at low tide under rocks. It is rather small for dissection, but very good complete series were obtained and stained in hematoxylin. Malletia faba Dall, was much less abundant. Specimens were obtained from holdfasts or from dredging. Although this was a larger species, gross dissection was not very easily carried out on any of the specimens, but good series were made.
The ganglia of Nucula are easily studied in section. The cerebral mass seems composed of one main mass, partly divided into four subdivisions, the two central most completely fused, and the lateral quite distinct in places. The central portion might represent the cerebral ganglia and the lateral, the pleural if we take that interpretation. The pedal ganglion is made of right and left parts quite completely fused except at the margins. The pedal mass is the smallest of the three chief ganglionic areas. The visceral ganglia are quite widely separated and a little larger than the pedal mass.
The ganglia of Malletia are in general plan similar to those of Nucula, the greatest differences being in the cerebral mass. The cerebro-pleural mass seems almost one. In most sections it is very compact and a little more complicated in structure than the ganglion of Nucula. However there are two small ventral ganglionic branches or small ganglia attached to the ventral side of the cerebral mass. These small ganglia may represent the visceral. Farther back in a cross section series as the cerebral mass disappears two other small branches take origin and run parallel to the nerves from the ganglionic cords. These two branches on each side seem to run together before the pedal ganglia are reached. Neither of these pairs of nerves seems connected with an otocyst.
At the cephalic end of the cerebro-pleural ganglion the large ganglionic cords are in evidence. A little distance from the cephalic end on the dorsal side there are quite large groups of cells down from the surface and surrounded by nerve fibers. The course of the fibers here is quite complex. On the ventral lateral sides of the ganglia are paired light areas of fibers which may be traced into the fibers of the ganglionic cords.
The pedal ganglion is small and much as in Nucula. The visceral ganglia are larger and widely separated.
In both Nucula and Malletia young specimens were used for study. In Nucula[Pg 76] there was more the appearance of four ganglia in the cerebro-pleural mass, and the ganglia seem less complex than in Malletia. This last species has more separate pleural ganglia, if the ganglionic cords can be so regarded.
In neither of the species studied were all parts of the connectives easy to follow, so it was impossible to test the suggestions of Drew, but in both species there is some indication of two lateral lobes of the cerebral mass, and in Nucula there is good evidence of two central ganglia as well as the smaller lateral ones. The lateral ganglia of the cerebral mass are most clearly separated in Malletia. In Nucula the lateral ganglia are larger in proportion and the distribution of the gray and white matter is more irregular.
Drew, G. A.1901
The life history of Nucula delphinodonta. Quart, jour. sc. vol. 44, pt. 3.
Pelseneer, P.1891
Contribution á l'étude des Lamellibranchs. Arch. d. biol. xi.
Stempell1899
Zur Anatomie von Solrmya togata. Zool. Jahrb. Bd. xiii.
(Contribution from the Zoological Laboratory of Pomona College)
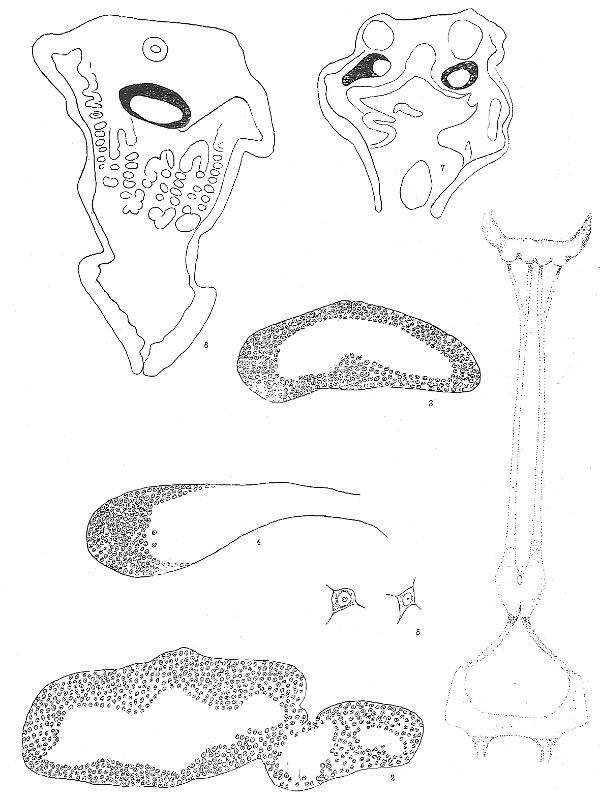
Fig. 1. Diagram of the ganglia of Nucula castrensis, reconstructed from serial sections. The probable position of the connectives is shown and the proportionate distances between ganglia are given. The upper ganglion is the cerebro-pleural with large nerves leading off from the ganglion which is itself lobed into four chief lobes. The pedal ganglion is next. In section the pedal ganglion at one place seems to be made up of four parts which may correspond to four connectives from the cerebro-pleural although only one pair of connectives was clearly determined. The visceral ganglion is connected with the pedal below. ×70.
Fig. 2. Cross section of cerebro-pleural ganglion. On the right side one of the lateral ganglia is shown. The one of the other side does not show because the section is not straight across. The dorsal side is up. ×300.
Fig. 3. Section of the pedal mass of Nucula, through the center. The dorsal side is up. ×300.
Fig. 4. Left side of the visceral mass of Nucula. Dorsal side up. ×300.
Fig. 5. Nerve cells from the central nervous system of Nucula. ×450.
Fig. 6. Section through the body of Nucula showing the position of the cerebro-pleural ganglion cut through the center. Dorsal side up. The cellular portion of the ganglion is black. ×70.
Fig. 7. Section through the body of Nucula at the level of the visceral nerves which are shown on either side of the section. The area of nerve cells is shown in black. ×70.
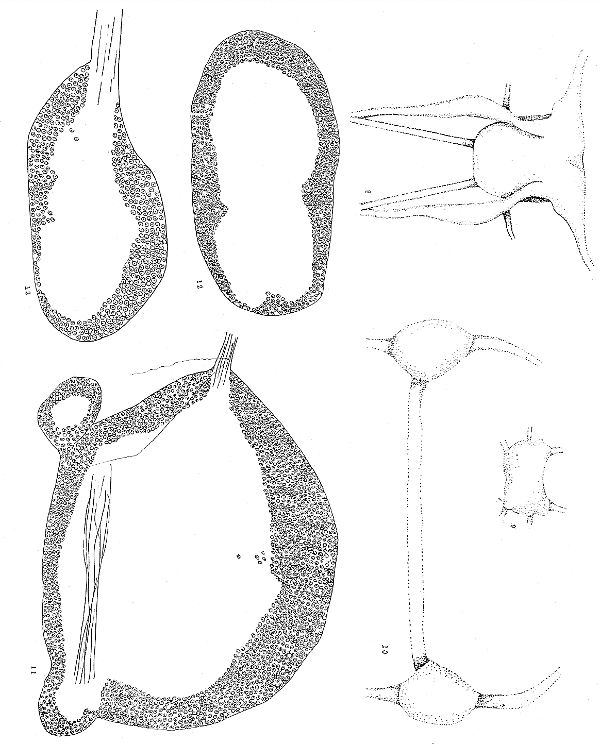
Fig. 8. Reconstruction from serial sections of the cerebro-pleural mass nerves and connectives of Malletia faba. The drawing is a ventral view, the cephalic side is at the top. ×70.
Fig. 9. Reconstruction of pedal ganglion of Malletia from the ventral side. Cephalic side at the top. ×70.
Fig. 10. Reconstruction of visceral ganglia of Malletia. ×70.
Fig. 11. Section through cerebro-pleural mass of Malletia. The dorsal side is up. On the ventral side to the left and right are the beginnings of the lateral lobes or ganglionic cords which may represent the pleural ganglia. In this species the cerebral ganglia are not separated into right and left halves as in Nucula. ×300.
Fig. 12. Section through the central part of the pedal mass of Malletia. The dorsal side is up. ×300.
Fig. 13. Section through one visceral ganglion of Malletia. The dorsal side is up. ×300.
Edited by
WALTER E. COLLINGE, M. Sc., F. L. S., F. E. S.
The Gatty Marine Laboratory
The University, St. Andrews, Scotland
The subject matter is strictly confined to original zoological research—systematic and anatomical. Fully illustrated by lithographic plates and text figures.
Each volume will consist of 4 parts, price $5.
All subscriptions should be forwarded to
Messrs. Dulau & Co., Ltd.
37 Soho Square, London, W., England
GRIFFITH
Incubators

A simple, well constructed bacteriological incubator
GAGE—The Microscope
An Introduction to Microscopic Methods and to Histology
By SIMON H. GAGE.
Twelfth Edition. Entirely rewritten, and with many new illustrations.
Price, postpaid, $3.00.
This work aims to give help to everyone who uses the microscope, whether he is a beginner or an advanced worker.
COMSTOCK—A Manual for the Study of Insects
By JOHN HENRY COMSTOCK, Professor of Entomology in Cornell University,
and ANNA BOTSFORD COMSTOCK, member of the Society of
American Wood-Engravers. 8vo. cloth, IX. +701 pages, 797 figures in the
text, and six full page plates. Nearly all of the figures were engraved
especially for this work. Postpaid $4.07; net $3.75.
This hand-book is designed to meet the needs of teachers in the public
schools and of students in high schools and colleges.
NEEDHAM—General Biology
A book of outlines and practical studies for the general student
By JAMES G. NEEDHAM, Professor of Limnology and General Biology in Cornell University. Cloth 8vo. XIV. +542 pages; 288 figures, mostly original. Postpaid $2.00.
This book is expressly designed to help the general student obtain a comprehensive grasp of the principles of biology.
NEEDHAM & LLOYD—The Life of Inland Waters
A Text-Book of Freshwater Biology
By JAMES G. NEEDHAM, Ph.D., Professor of Limnology and General Biology in Cornell University, and JOHN T. LLOYD, A.B., Assistant in Limnology in Cornell University.
This book is a broad presentation of the field of fresh-water biology, primarily in its scientific aspects, also in its relation to commercial, civic, aesthetic and public health interests of man. There is no book in the English language covering the field. Copiously illustrated with photographs from Nature. 438 pages; 244 figures. Price $3.00.
GAGE—Optic Projection
By SIMON HENRY GAGE, Professor Emeritus of Histology and Embryology in Cornell University, and Henry Phelps Gage, Ph. D.
This work of over 700 pages and with over 400 figures is of especial interest to workers in all fields of Biology in that it deals especially with the use of the Projection Microscope for demonstrations and for drawing. It also gives the fundamental principles of all the forms of projection. A 16-page circular will be sent on request. Postpaid, $3.00.
RILEY—Handbook of Medical Entomology
By WM. A. RILEY, Ph. D., Professor of Insect Morphology and Parasitology in Cornell University and O. A. JOHANNSEN, Ph. D., Professor of Biology in Cornell University.
A concise account of poisonous, and disease-carrying insects and their allies, including descriptions and illustrations of the principal species, with keys for their determination, and method of control. Bound Library Buckram, medium 8vo. Nearly 375 pages. Price $2.00 net.
Send for Descriptive Circular
The Comstock Publishing Company
Cornell Heights, Ithaca, N. Y.
for Lepidoptera and other insects
 Odontolabis wollastoni, Java
Odontolabis wollastoni, Java
We are able at last to offer our customers an all glass mount that we believe comes nearer to meeting the requirements of museums and private collectors than any previously put on the market.
No cut can properly show this mount, which must be seen to be appreciated. Write for Circular 192, which describes fully what we are offering.
When in need of Entomological supplies remember that we have the best insect pins on the market and that our hand made Schmitt insect boxes with three ply tops and bottoms are unsurpassed. Catalogue No. 33 free on application.
Prompt shipments can be made from our large stock of United States and exotic insects of all orders. We also prepare 150 different metamorphoses of beneficial and injurious insects. Pupae now on hand are listed in circular No. 198. Other circulars and price lists free upon request.
Ward's Natural Science Establishment
84-102 College Ave. Rochester, N. Y.
A forty-eight page illustrated magazine, published monthly except August and
September, devoted to the study of INSECT LIFE. It contains a list
of the titles of the current Entomological Literature, and also
articles by the leading Entomologists in the United
States and Canada. Valuable information
for the beginner, the economic
entomologist and the
systematist.
To new subscribers, $1.90; Renewals, $2.00; payable in advance.
Single copies 25 cents. Address
ENTOMOLOGICAL NEWS
1900 Race Street, Philadelphia, Pa.
1135 Newberne Avenue
RALEIGH, N. C., U. S. A.
Twenty-one years' experience
Price List on Application
A Quarterly Devoted to Medical Zoology
This journal will be a medium for the prompt publication of briefer papers and research notes on animal parasites. Emphasis laid on the morphology, life history and biology of zooparasites and the relations of animals to disease.
Subscription, $2.00 a Volume
Managing Editor, HENRY B. WARD;
Univ. Illinois, Urbana, Illinois
Do Business by Mail
It's profitable, with accurate lists of prospects. Our catalogue contains vital information on Mail Advertising. Also prices and quantity on 6,000 national mailing lists, 99% guaranteed. Such as:
| War Material Mfrs. | Wealthy Men | Fly Paper Mfrs. |
| Cheese Box Mfrs. | Ice Mfrs. | Foundries |
| Shoe Retailers | Doctors | Farmers |
| Tin Can Mfrs. | Axle Grease Mfrs. | Fish Hook Mfrs. |
| Druggists | Railroad Employees | Feather Duster Mfrs. |
| Auto Owners | Contractors | Hotels |
Write for this valuable reference book.
Ross-Gould, 1027H Olive Street, St. Louis.
Located in one of the most healthful and beautiful parts of the west coast. The mountains reach an elevation of ten thousand feet within a few miles of the college and these with the nearby ocean afford many special advantages for the study of things not in books. Special advantages are afforded by the fact that the college limits its attendance, the freshman class being restricted to two hundred applicants. The success of the college is particularly indicated by the large proportion of the graduates who proceed to advanced work in the large universities. In addition, well-manned departments of music and art afford exceptional advantages.
For further information, address
Secretary of Pomona College
Claremont, California
Page 64: Changed * * * to ... (preferred form for ellipsis).
Originally: This * * * is readily overcome by the frequent
Page 64: Changed "placd" to "placed".
Originally: surface of the book on which it was placd,
Page 68: Changed "X" to "×".
Originally: cells (10 mm. X 3.5 mm.)
Page 70: Changed "chitinizd" to "chitinized".
Originally: Changed area of microscopic chitinizd spicules,
Page 71: Changed "Lengh" to "Length".
Originally: Pupa.—Lengh, 9.1-12.8 mm.
Page 75: Retained "molloscs", as a possible spelling variant for
"molluscs". However, it may be a typo.
Originally: forms are grouped among the simplest of the molloscs.
Pages 75, 76: Retained "Stempel" and "Stempell" spelling variations.
Page 76: Changed "once" to "one".
Originally: the pedal ganglion at once place seems to be made up
Page 76: Changed all instances of "X" to "×" to indicate magnification.