
Title: Botany: The Science of Plant Life
Author: Norman Taylor
Release date: June 14, 2015 [eBook #49211]
Most recently updated: October 24, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/49211
Credits: Produced by Juliet Sutherland, Chuck Greif and the Online
Distributed Proofreading Team at http://www.pgdp.net
Contents. (In certain versions of this etext, in certain browsers,
clicking on this symbol (etext transcriber's note) |

The prickly pear (Opuntia chlorotica santarita) of the
desert in the American Southwest.
(This painting was kindly loaned by Dr. David Griffiths of the United
States Department of Agriculture and reprinted here through the courtesy
of the Journal of the International Garden Club, where it first
appeared.)
Courtesy Journal of the International Garden Club
Popular Science Library
EDITOR-IN-CHIEF
GARRETT P. SERVISS
AUTHORS
WILLIAM J. MILLER HIPPOLYTE GRUENER A. RUSSELL BOND
D. W. HERING LOOMIS HAVEMEYER ERNEST G. MARTIN
ARTHUR SELWYN-BROWN ROBERT CHENAULT GIVLER
ERNEST INGERSOLL WILFRED MASON BARTON
WILLIAM B. SCOTT ERNEST J. STREUBEL
NORMAN TAYLOR DAVID TODD
CHARLES FITZHUGH TALMAN
ROBIN BEACH
ARRANGED IN SIXTEEN VOLUMES
WITH A HISTORY OF SCIENCE, GLOSSARIES
AND A GENERAL INDEX
ILLUSTRATED
![]()
VOLUME THIRTEEN
P. F. COLLIER & SON COMPANY
NEW YORK
Copyright 1922
By P. F. Collier & Son Company
MANUFACTURED IN U. S. A.
The Science of Plant Life
BY
NORMAN TAYLOR
Curator, Brooklyn Botanic Garden
![]()
P. F. COLLIER & SON COMPANY
NEW YORK
THIS book is for those who want some general knowledge of the plant world, without necessarily caring for the technical details upon which such knowledge is based. If it leaves the reader with an impulse to follow the subject further than has been possible here, it will have more than fulfilled its mission.
Throughout the book, it has often been convenient to refer to plants or their behavior in terms implying reasoning faculties. Of course, plants are never reasoning things, reasonable as many of their actions appear to be, and to ascribe such qualities to them is to saddle them with attributes perfectly foreign to the plant world. But the description of them in the terms of our everyday speech, the translation of plant behavior into the current conceptions of mankind, does so fix these in our minds that they cease to be among those interesting things that nearly everyone forgets. I have followed this method deliberately, understanding perfectly the objections to it, but believing, with the late C. E. Bessey, that in popular books “it is an admirable way of looking at some botanical things.”
All of the half-tone illustrations, except two, are from the photographic collections of the Brooklyn Botanic Garden, and I am under the greatest obligation to the director of that institution for permission to publish them here. The illustration of the living and fossil algæ has been taken from Prof. Henry Fairfield Osborn’s “Origin and Evolution of Life,” with his kind permission. The illustration of desert vegetation is from a photograph by the late E. L. Morris, and kindly loaned from the collection of the Brooklyn Museum. All the line cuts in the text are from drawings made specially for this book by my wife, Bertha Fanning Taylor.
While grateful and particular acknowledgments can be made for the illustrations, it is difficult or impossible to properly express my indebtedness to all those who, through their books and pamphlets, have indirectly aided in the making of this book. It would involve the mention of most of the better known writers of the books found in the larger botanical libraries. It is a pleasure to acknowledge help from Dr. M. A. Howe of the New York Botanical Garden on the literature of fossil and hot-spring algæ, and from Dr. Orland E. White of the Brooklyn Botanic Garden for helpful criticism of the section dealing with “How Plants Change Their Characters.”
Norman Taylor.
Brooklyn Botanic Garden
20th October, 1920
| CHAPTER | PAGE | |
| Introductory—Plants and Our Daily Needs | 9 | |
| I. | What Plants Are | 13 |
| II. | Plant Behavior | 76 |
| III. | How Plants Produce Their Young | 116 |
| IV. | The Families of Flowering Plants and Their Relationship | 168 |
| V. | Uses of Plants to Man | 208 |
| VI. | Garden Plants | 267 |
| VII. | History of the Plant Kingdom | 298 |
| VIII. | Distribution of Plants | 337 |
PERHAPS few of us realize that without plants all our modern civilization would be swept away and that upon plants has been built all that we have so far accomplished and everything that we may yet become. The overthrow of any king or republic, the wiping out of all money and finance or any of the manifold evidences of our modern world could not for a moment be compared to what would happen to us with the sudden destruction of plant life from the earth.
Food and drink, the very houses we live in and heat, medicines and drugs, books and pictures, musical instruments and tires for automobiles, all these and hundreds of our daily needs depend upon the fact that plants of many different kinds grow upon the earth and in sufficient amounts to be of importance. It is easy to say in opposition to this that we get much food from animals, that we can drink water, and that neither of these comes from plants. But water would soon be lost to us if forests did not conserve it, and upon pasturage most of our food animals depend for their life. The discovery of a single tree in the mountains of northern South America made possible for white man the malarial regions unfit for him before the advent of quinine. Just before Shakespeare’s time sugar and tea and coffee became regular articles of commerce. Not until the discovery of America was tobacco, Indian corn, chocolate, the pineapple or the potato known to man. Upon the spruce forests in the north depends much of our paper supply, from cotton we get clothes and explosives, from hemp and sisal ropes, from a single kind of Brazilian tree most of our rubber, and from rice a food that sustains nearly half the world.
While it is thus plain enough that life depends upon plants now present upon the earth it may not be so obvious that from certain ancient forests has come the greatest source of artificial heat in the world. Coal is nothing but the partial decomposition of vast forests, living ages before man was first found upon the earth, subsequently buried, and under the earth’s pressure forming soft coal, or where the pressure was severe enough hard coal. When it is remembered that a dead, partly decayed tree is only a fraction of its living size and that coal is found in many parts of the world in tremendous quantities we get a partial glimpse of what our debt is to a great forest that lived in luxuriance no one knows how many millions of years ago, reached its climax, and upon whose embedded remains we depend for heat.
Later on in this book will be given in greater detail some of the plants useful to man and just how we have used them. Hardly any part of the study of the plant kingdom has so much of interest as that dealing with our utilization of the things that grow about us. From the earliest struggles of our half-savage ancestors to grow definite crops rescued from the wild down to our modern nut butter made from the partly fermented meat of the cocoanut and shipped half round the world before it is refined, man has constantly striven to use for his advantage the plants most likely to prove valuable. Countries and empires have been built upon such facts. Even to-day rubber from the Straits Settlements and palm oil from Africa are deciding the economic life of those countries.
But man’s use of plants, in fact his absolute reliance upon them, is not the only reason for attempting to find out more about them, what they are, where they came from, how they live and produce their young. A knowledge of even a small part of such a science opens up a rich field of inquiry involving a concept of plant life of greater interest than mere bread and butter. For those with an eye to see and knowledge to interpret, a landscape with its trees or flowers or marshes may contain a host of hidden secrets of dramatic import. Unfolded before one may be found a spectacle of struggle and strife, quiet tragedies of the forest, the inexorable pressure of plants upon their neighbors, the woods upon the prairies or an apparently forlorn hope of some plant living in a hot desert or upon some icy mountain peak. And while these rather obvious things are happening how much more is hidden of the adjustments that leaves or flowers or roots or other organs of the plant are constantly making to the conditions about them. Upon the perfection of such adjustments to light, heat, or water, for instance, depends their very existence. Mistakes are fatal, the forces of nature seem peculiarly relentless, and it is literally a case where many are called but few chosen. Of the untold millions of seeds produced each year few ever grow, yet out of this enormous wastage springs all that makes the earth not only habitable but the beautiful panorama of vegetation to which we are so accustomed that it is nearly taken for granted.
The study of botany attempts to answer some of the questions raised above and many others. Subsequent parts of the book will deal with what plants are, the behavior of them, with the life histories of some of the better known ones, with the grouping of plants in families and their relationships one to another, with their uses to man, with the history of their development from the earliest times, and finally with their distribution over the earth. The latter will be discussed last because it is the most important of all the phases of plant life. How plants are distributed, whether as forest or prairie or thickets or what not, depends upon the response of individual plants and their organs to the conditions about them. The type of vegetation in different parts of the world has been dictated by the success of the survivors in meeting existing conditions and of having met them in the past. Upon this fact rests our civilization to-day. Upon this fact there has been reared a study the cultural, esthetic, and practical value of which may well outweigh any other.
While the study of botany is necessarily a technical one with a language all its own, its terms, though generally unfamiliar, are unexcelled for their purpose. They will be avoided here as much as a clear understanding of the subject will permit. The few that must be used will be explained where they first occur and it is assumed that the reader will understand their subsequent use.
WHAT we commonly call plants, such as corn or buttercups or an oak tree, are so familiar that a definition of what plants are may seem needless. It would be quite so if these generally recognized examples comprised all the plant kingdom. Actually what are ordinarily thought of as plants make up only a fraction of the great plant world. The fact that our familiar roadside and garden plants produce blossoms followed by a fruit and seed, such as peas and beans and all the ordinary flora of any region, separates them at once from that other large group of plants that do not. Common examples of the latter class are the green scum on the ponds, moss, seaweed, the rust on wheat, yeast, disease-causing bacteria, the smallest of all known plants, and many others. Most of these organisms are so small that they can be distinguished only under the higher powers of the microscope. Some of them in their habits and growth are like the minute animals described in the volume of this series devoted to that subject. In fact there are organisms about which scientists are still in doubt as to their animal or vegetable character.
One or two characteristics common to most plants, however, separate them from animals and these are their method of getting food and their practically immovable mode of life. Animals, however simple, do eat and digest their food, plants take various mineral substances from the earth or air or water in the form of inorganic matter such as oxygen, carbon, nitrogen, and all the food materials found in the soil, and transform them, with the aid of sunshine, into the structure that characterizes each particular form. Plants, then, may be defined as any living organism that, with minor exceptions, has the power to assimilate inorganic substances and transform them into organic matter. Nothing else in all the realm of nature has this power. It is a possession beyond all price, this ability to take from the soil and air and water simple chemical substances and under the magic of sunlight transform them into the wonderful plant life of the world. But this faculty has its handicaps, for it is usually, though not always, associated with inability to move from place to place, which, in some measure, even the simplest animals can accomplish.
It will be readily understood that this definition of plants includes many other things that are commonly attributed to the plant kingdom. For our purpose the discussion of these relatively unfamiliar forms of plant life will be left until later. A rough and ready distinction between those plants that everyone recognizes as such and those others like yeast and microbes, which are plants to most of us only by virtue of the definition given above, is that the first group produce flowers and seeds and the second do not. It should not be forgotten, and it will be shown later, that this is not a true distinction, but for practical purposes of dividing the plant kingdom it suffices.
The fact of outstanding importance to everyone who really looks at most plants is that part of them are above ground and part below. This simple observation carries with it the recognition of a fundamental difference of plant structure, namely roots and stem. Most plants bear obvious leaves, and at some time in their life flowers, inevitably followed by fruits and seeds. The ideally perfect plant would consist, then, of root, stem, leaves, flowers, fruit, and seed. These are subject to many changes of form, sometimes they are put to strange uses, and occasionally one or two may be lacking, as the stem is apparently from many violets, and leaves from some cactus and from the Tjemoro tree of Malaya. In fact, so varied are the different forms of these common attributes of most plants, so important are these differences of structure that no right understanding of plant life can be had without examining each in some detail.
The obvious purpose of the roots of plants is to serve as an anchorage or holdfast. Their other and equally important function is to secure food for the plant, a process that will be described in the part devoted to Plant Behavior. Certain plants bear no roots and attach themselves to the roots of other plants in which case they literally steal their food, as does the mistletoe and some others.
Roots are of various kinds, depending upon the soil in which they grow and upon the kind of plant to which they are attached. In the case of annuals, which live only one year, as does the purslane, and biennials, which live only two years, as does the fringed gentian, the roots are mostly fibrous (Figure 1) and apt to be only slightly under the surface. In perennials, which live many years, such as the dandelion, the root is deeper and forms what is known as a taproot (Figure 2). In shrubs and trees they are harder, woodier, and often penetrate to great depths.

If we examine the roots of a tree, we find a large part of them are woody, often as thick as the smaller branches, and it is only toward their extremities that they branch out into the multitude of rootlets

that creep through the earth seeking food for the tree. Some, as in the spruce or hemlock, do not go very deep but spread great distances through the soil in search of food, others, like the hickory, go nearly straight down. The interesting feature of these tree roots is that the part nearest the trunk is all but dead, and acts mostly as an anchor, while the fibrous rootlets or even finer subdivisions known as root hairs at the extremities are the food gatherers. At the very end of all rootlets and of roots is a rootcap (Figure 3), harder than the rest of the threadlike rootlet. This rootcap is, if not quite dead, at least useless as a food gatherer, but serves as a tiny pioneer wedge which forces its way among stones or other obstructions, so allowing the living root hairs just behind to gather the food to which it leads the way. In certain of the rocky islands of the Bahamas wild fig trees may be seen, growing on the bare rocks, their roots sprawling in every direction in search of a crevice through which the rootcap can force its way. Such roots may extend thirty or forty feet from the trunk of the tree over the bare rock in search of a favorable crack where they plunge to the cool depths and secure the food and water necessary for life.

Roots are not always of this common type. Sometimes, particularly in certain biennials, they are swelled to form great thickened portions, often weighing many pounds. The sweet potato is a familiar example, and a related plant, one of our morning-glories, has an enormous perennial root, known to weigh as much as fifty pounds. This swelling of the roots of plants is a quite common characteristic of certain kinds and has great commercial significance. Carrots, turnips, rutabagas, beets, and parsnips are familiar examples. The purpose of such roots is to store food for the plant, and this thrifty habit of some roots has been turned by the gardeners to our advantage.
It is a common sight to see parts of a sidewalk heaved up by a tree root and their force in this respect is tremendous. One of our common ferns has been known to raise a weight of over 500 pounds, and even to break through a concrete walk. Such is the force exerted by the roots of plants that we can truly think of roots as pushing through the earth almost regardless of obstacles, binding the soil together and not only serving the needs of the plants, but actually holding the soil on steep slopes. Where fire or ruthless lumbering has stopped this natural process the washing away of the soil and exposure of the bare rocks leaves desolation behind it.
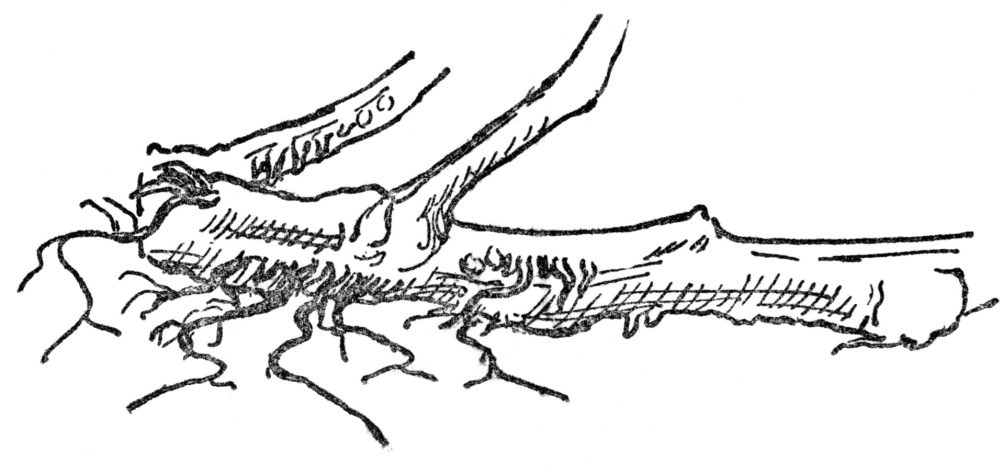
While most roots live under the surface many grow in the air, and a few grow from stems that are injured. The tomato vine often produces roots at the joints or where it has been injured. Such roots, known as adventitious roots (Figures 4 and 5), are fairly common in many plants, the common garden practice of making cuttings, which take root under favorable conditions, being based upon this fact.

But some plants produce roots in the air, as in poison ivy and the trumpet creeper, without injury or the gardener’s skill, and are known as aërial roots. They are some of the most peculiar and fantastic of nature’s devices for allowing plants to grow in apparently unfavorable places. In many orchids, some relatives of the pineapple, and a few other air-inhabiting plants, the roots live wholly in the air, the plants being fastened to a tree or even to a telegraph wire. Such plants live on the air and water vapor, and are mostly inhabitants of moist tropical regions.
Quite the most extraordinary of aërial roots are those produced in certain kinds of fig trees. Starting perhaps a hundred feet in the air and no thicker than a lead pencil they appear first as slender vine-like streamers blown hither and yon by the wind. Eventually they reach the ground and penetrate it, grow often to a great size and even form trunklike connections with the tree top. The banyan tree of India is the best known case of this peculiar habit. One which started from a single trunk, subsequently sending out great numbers of aërial roots, has now spread to such a size that it is over 2,000 feet in circumference, has 3,000 trunks, and once sheltered 7,000 soldiers.
A variation of this habit is the case of a giant fig tree of the West Indies in which a bird may deposit the seed of another tree. The seedling soon develops, sending out long, at first threadlike, aërial roots which are wrapped around the tree trunk. As the roots increase in size and further encircle the trunk they ultimately reach the ground, where they are frequently a foot in diameter. Then the true nature of the process becomes evident. For these apparently innocent aërial roots, as they reach the ground, have so completely inclosed the old trunk and their pressure is so great that they literally strangle the tree from which they started. It is slower but just as deadly as the strangulation of an animal by a boa constrictor, for these encircling roots cut off by strangulation the ascent of the sap, thus killing the tree. Fate sometimes overtakes them, however, as it is a common sight to see the strangler meet the same end. Some of nature’s most ruthless battles are fought out in this way, very silently, but very effectively.

Nothing dies harder than generally accepted delusions, particularly those regarding plant lore, and of all such incorrect impressions the one that a potato is a root, is the hardiest and most difficult to kill. Yet, the “eyes” of a potato give it away if one stops for a moment to reflect that the eyes are only buds and buds grow only on stems. That is one of the chief uses of stems—to support in the air the leaves and flowers that come from its buds, and no matter if the stem, as in the potato and many other plants, be ever so deeply buried their true stem nature cannot be mistaken. Sometimes these underground stems are not thickened but lengthened out, in which case, notably in common garden iris, they are called rootstocks. Again, these buried stems may be swollen, as in the potato, when they are known as tubers. Onions and the jack-in-the-pulpit bear still other kinds of underground stems, and there are many more, but they cannot be mistaken for roots, for it will be seen from Figures 6-9 that on their under sides they bear roots themselves. Besides this they bear buds or shoots, which no true root ever does.

Stems above ground, which is the most usual form for them, are of many kinds, all serving the purpose of support to the leaves and flowers, and as a means of carrying sap from the roots or underground stems to the upper part of the plant, and also to carry certain foods to the roots from the leaves, of which more anon. In the case of herbs, like goldenrod or daisy, the stem may be apparently all pith on the inside, with only a thin outer coating of harder substance, not unlike bark, but usually green. If we examine the cut-off trunk of a tree, a quite

different structure is apparent. Any lumberman can point out at once “heartwood” and “sapwood” (Figure 78), and his distinctions are just as good as those of the scientist, for he says in these two words as plainly as can be said that heartwood is the oldest and sapwood the youngest. The sapwood is nearer the bark and is honeycombed with passages which serve to carry the sap from the roots to the tree top, while just under the bark is the bright, green, living layer, known as cambium, which is renewed each year. The phloem is the carrier for the food made in the leaves to the roots. It is the successive layers of cambium, year after year, that gives to tree trunks their annual rings. The age of almost all trees can be reckoned exactly by counting these, one representing a year’s growth, and the tree’s rate of growth estimated from the closeness of the rings. Fires or droughts, perhaps long forgotten, here find a lasting record in rings so close together as to be all but invisible. The part nearest the center of the trunk is the heartwood, usually quite lifeless, yet in its maturity furnishing us with lumber. It may be and often is completely decayed, without injuring the flow of sap or the life of the tree for many years.

These two streams of sap, one going up and the other returning to the roots, each in its proper channel, are interspersed with air chambers that extend from the center of the tree out toward the bark, where they end in inconspicuous dots called lenticels. It is as though nature had provided an air-cooling device for the constant activity of these diverse currents. These lenticels are prominent on the bark of cherry, but whether obvious or not they are found in nearly all woody stems and insure a constant supply of fresh air to the busy interior.
In palms, sugar cane, corn, bamboo, and many other plants there is not any distinction between heartwood and sapwood (Figure 82), and in place of bark there is nothing but an outer rind, harder than the interior tissue. Such stems do not usually rot first at the center, have no cambium, and have no annual rings. This method of growth and structure is associated nearly always with definite leaf and flower forms peculiar to it and differing from most other plants. So fundamental are these characteristics, so uniform their occurrence and so clear are the distinctions between them and other plants that botanists have divided all flowering plants into those belonging to this group or to some others. More will be said of this in the chapter on the Families of Plants and Their Relationship.
The stems of some plants, such as the Big Trees of California, for instance, are among the oldest and most permanent of living things. “General Sherman,” one of the biggest in that most famous grove, was nearly three thousand five hundred years old when Columbus discovered America; it has lived through all the great periods of modern history, and to-day it is over 270 feet high and 35 feet in diameter. No living thing is so large or has lived so long. In Australia are great forests of blue gum trees even taller than our Californian Big Trees, but not so old nor so thick.
In the Pacific, off the coast of Oregon and British Columbia, a seaweed is commonly found with stalks over 500 feet long, and in India the rattan palm climbs over the tree tops for great distances, a single stem not much thicker than a broomstick measuring over 700 feet long. The search by leaves for light and air results in the stems of some plants performing almost incredible feats. Whether it is one of the Big Trees with a great massive trunk, or the rattan palm with its sinuous winding through the topmost heights of the tropical forests of India, the result is always upward to a “place in the sun.” This struggle for sunlight has taken many forms in different plants, the ordinary vines like morning-glory or grapevine, for instance, where the climbing stem is of great advantage. Some vines always twine to the left, as the hopvine, others to the right, as in the morning-glory, all seeking support from something else, each adopting its own most useful way of getting its leaves in the most advantageous position to catch the life-giving sunshine. If we could look down on any forest from an aeroplane, the striking efforts of nearly all plants, whether herbs, shrubs, vines, or trees, to get the utmost sunshine for their leaves would be evident at once. No apparently impossible twisting or bending of tree trunks or reaching out of stems of vines but is to be found in the inexorable struggle of stems to fulfill their task of giving the plant its chance to reach “a place in the sun.” Sometimes mere climbing or twining does not seem sure enough—it seems as though winds or the elements might break loose the vine from its support and thereby kill its chances. In certain vines this contingency appears to have been foreseen, and as if to clinch their opportunity of growing onward they are provided with special helps. Slender green tendrils, delicate prolongations of the stem, begin, almost insidiously, to catch hold of the nearest support and by a couple of turns about it and subsequent strengthening of their tissues make a permanent holdfast. The grapevine is a case in point. And as if this were not enough, certain other plants, such as the Boston ivy, have small disks which attach themselves to bare walls or tree trunks. This is to make assurance doubly sure, and it is this that makes the Boston ivy so useful to the gardeners for covering walls.
Some stems accomplish their purpose not by holding fast to a support in the air, but by creeping along the ground, as in the running blackberry, and often in the Virginia creeper. The purpose is the same, and, as if to confirm it, a few otherwise quite prostrate vines have their tips turned upward to the light, notably in the case of the creeping speedwell.
In certain plants the stem may assume curious forms due to special conditions under which they live and to which adjustment is necessary for the plant’s existence. In deserts, for instance, the cactus produces practically no leaves (Figure 10) and the green stem performs not only the function of leaves but acts as a storage for water. Where water is scarce this is of tremendous advantage, a single cactus having been known to store up 125 gallons. A similar habit of the cactuslike spurges in South Africa gives as weird an atmosphere to parts of their landscapes as we find in Arizona. It is as if the stem of such plants, being unable to push its leaves (it has none) up into the light, takes over some of the functions of leaves and makes up the deficiency by adopting other methods

to secure the plant’s survival. Other stems, looking and acting like leaves, reveal their true nature by producing buds, and the curious feature of the common butcher’s-broom (Figure 11), often colored scarlet for Christmas decorations, bearing flowers from the middle of what is apparently a leaf, but is actually a modified stem, is explained by this ability of stems to modify their habits to suit conditions. The butcher’s-broom is an inhabitant of dry regions along the Mediterranean, where a reduction or

absence of leaf surface is a decided advantage. In many partly desert or dry regions this production of leaflike stems or branches is common, an excellent garden example being asparagus, which came originally from Europe and the feathery growth of which is all stem. In Tasmania a kind of yew tree produces no leaves, all the foliage being modified stem, which is true of many kinds of spurge in the West Indies, where an almost impenetrable scrub is largely made up of a shrub which is apparently covered with leaves, all actually part of the branches and stems.

While stems, such as the Big Trees or the giant cactus, may be among the largest of nature’s creations they may be also the smallest, as the duckweed that floats on ponds is the tiniest of all flowering plants and its flat expanded surface is wholly stem. Figure 12 on this page better illustrates this strange modification of a stem than words could do.
From what has been read it will be seen that stems are not “just stems”—they are among nature’s most ingenious devices to secure the survival of the plant. Whether buried in the ground, and producing, almost by stealth, buds that develop into mature plants, or thrusting leaves to the utmost limits of their reach, or climbing by an intricately varied mechanism, or changing their character to suit desert conditions, or floating on the water—it matters not. Each modification of form or use secures to the individual plant its chances to survive; and in most cases its only chance, as anyone may see by the sudden death which follows a series of changes which prevents a stem from performing its proper tasks.
As the palm reader is supposed to be able to tell your history and future from veins in your hand, and as the veins in the wing of a butterfly tell their story to an entomologist, so the veins of a leaf are more significant than almost any other characteristic of a plant. Most leaves have their veins, or skeleton, with a single midrib and many branches off it on each side, which themselves break up into a fine network of veins. Such leaves are netveined (Figures 13-24). Others, such as corn and grass, have the veins running side by side from one end of the leaf to the other, sometimes with small branches off them, but instead of the veins forming a network they are parallel, and such are called parallel-veined leaves (Figure 38). In the chapter on Plant Families and Their Relationship more will be said as to the amazing regularity with which netveined leaves are associated with certain kinds of flowers and parallel-veined with other kinds, how these distinctions have been recognized since hundreds of years before Christ, long before their true import was understood. There are variations from both these

Banyan Tree (Ficus bengalensis). A fig tree of India, whose adventitious roots make frequent connection between the tree top and the ground. Starting as thin, whiplike streamers these roots ultimately form new trunks. (Courtesy Brooklyn Botanic Garden.)

Laceleaf (Aponogeton fenestralis). A submerged aquatic plant, with permanently skeletonized leaves, and an inhabitant of forest pools in Madagascar. (After Engler & Prantl. Courtesy of Brooklyn Botanic Garden.)

Root Hairs with Fine Soil Particles Attached. Note soil, water, and air spaces. Much magnified. (After U. S. Department of Agriculture. Courtesy of Brooklyn Botanic Garden.)

Fig. 13. A linear leaf with an acute tip. Fig. 14. Lanceolate leaf with an acuminate tip. Fig. 15. Oblanceolate leaf broadest above the middle. Fig. 16. Ovate, broadest below the middle. Fig. 17. Spatulate, broadest above the middle and with an elongated base. Fig. 18. Elliptical. Fig. 19. Obovate in which the general shape is ovate, but broadest toward the tip. Fig. 20. Oblong. Fig. 21. Orbicular or nearly round. Fig. 22. Deltoid or somewhat triangular, an ovate leaf with a broad base. Fig. 23. Kidney-shaped or reniform with heart-shaped base. Fig. 24. Peltate leaf of common garden nasturtium; note circular blade with leafstalk attached to the center.
types, but in nearly every case, once the difference is noted—and scarcely any other character of a plant is so much worth notice—they cannot be mistaken.
During the winter nearly all leaves are folded in various ways in a bud for protection from the elements. Nature shows herself in some of her wisest moods in the selection of methods to accomplish this. In some buds, notably those of the horse-chestnut, the bud is coated with a sticky substance to protect the tender young leaves inside. In others there is a hard outer coat, as in the hickory, impregnable to the most driving sleet, others again have the leaf rolled so tightly and pointed so sharply at the end, as in the beech, that water cannot cling to the bud nor soak in, until the warmth of spring gives the signal for the annual miracle of the bursting out of foliage. Leaf buds are sometimes hard to find on certain plants, as they are formed at the base of a leafstalk and covered by it during the growing season. It is only as the leaf falls in the autumn that the hollow base of its stalk is seen to have hidden during the summer the young bud for the following season. The plane tree or sycamore is a good example of a plant where no leaf buds can be found until the falling of the leaves in autumn.
The forms of leaves are infinite in their variety, and the reasons for some of their peculiarities in this respect are not yet understood. The average netveined leaf is obviously composed of a blade (Figure 25), and at the base a stalk known as a petiole. Sometimes at the base of the petiole—which is lacking in many leaves—there are two tiny leaflike appendages, called stipules, which are of no apparent use to the plant, and, as if in recognition of this fact, they often fall off long before autumn. In some plants, however, stipules are permanent, while in certain others they are never found, as, for instance, in the horse-chestnut tree.

Fig. 25. Simple leaf with blade, leafstalk (petiole), and two stipules at the base. Margins of the leafblade serrate or saw-toothed. Fig. 26. Leaf with a sagittate base, or shaped like an arrowhead, the lobes pointing downward, and with entire margins. Fig. 27. Retuse or emarginate tip, somewhat indented. Fig. 28. With the base auriculate or with rounded basal lobes. Fig. 29. Hastate, like an arrowhead but the lobes pointing outward. Fig. 30. With cuneate base (wedge-shaped). Fig. 31. Cuspidate tip with a usually hard and stiff point. Fig. 32. Perfoliate, the leaf bases joined and the stem passing through them. Fig. 33. Truncate, the top flattened. Fig. 34. Pinnately lobed, with deep indentations cut toward the midrib. Fig. 35. Palmately lobed, out toward the top of the leafstalk.
The outline of leaves is as varied as nature itself. Some of the common kinds are shown in drawings (Figures 13-24), which tell more of the story than pages of description could do. Their margins, too, their tips, their bases (Figures 25-35), all parts of them, in fact, are so variable and yet in each kind of plant so uniform, that in the description of the plants of any region the botanist has used these characteristics of leaves as one method of identifying the particular plant in hand.
The terms used to designate these different kinds of leaf margins or forms of blade are precise, nearly universally used, but need to be studied only by those who, because of special fondness for the subject, are likely to need them in using books which are beyond the scope of the present one. If, for instance, the reader is interested in finding out what his native roadside plants are, he would need a book describing them, and there are many for different parts of the country. In such books he would find these terms, which say so much in a single word (there are other sets of terms for flowers, fruits, and seeds) totally unfamiliar and quite likely to disgust him at the start. A little study may open up to him that most interesting and easily accessible of recreations, a first-hand familiarity with the wild flowers of one’s own neighborhood.
All leaves are not as simple as the figures show them to be. In many the midrib or principal vein is much elongated and there are small leaflets, sometimes even scores of them, all fastened to a common stalk. Such are called compound leaves (Figures 36-37), which may be found in ash, hickory, rosebushes, blackberries, peas, beans, and thousands of other plants.

Fig. 36. Palmately compound leaf, the five leaflets all arising from the tip of the common leafstalk. Fig. 37. Pinnately compound leaf, the leaflets arising from the sides of the common leafstalk. Fig 38. A parallel-veined leaf. All the other leaves figured are netted-veined.
While leaves are literally factories in which one of the most wonderful things in the world is produced, it is so much a part of what plants do or their behavior that the story of it will be given in the chapter on Plant Behavior. Sunlight is absolutely necessary for the process, and to reach this sunlight leaves are attached to their stems in a variety of ways. Some are always opposite each other, as in the common privet, lilac, or honeysuckle; others always alternate, as in the mustard or the rose. There are many variations of these simple arrangements, but in every case the process results in giving each leaf the utmost exposure to the light without which the plant must wither and die. So vital is this exposure to light that in some plants parts of the leaves produce tendrils, as in the case of peas, in order that some near-by support may be used. In one African relative of our lily, this change of leaf form has been so great that its long slender leaf tip is wonderfully adapted to reaching up and catching by its curved tip some support to lift it from the gloom of the tropical forest floor.
Looking down from above on any small plant or bush, or from the sky on a forest, about all that can be seen are the thousands of leaves, all so arranged that it is as though some celestial photographer asked every one of them to so place themselves that they would all be “in the picture.” The competition between leaves on the same plant and between leaves on rival plants is infinitely keener than the friendly pushing of a crowd to get in a picture, and it lasts forever. Furthermore, failure to get in means certain death. So intricate is the method of leaf arrangement, so marvelous the adjustments that all plants must make to insure ample light, that it is not inaptly called leaf mosaic. As we shall see in the chapter on Plant Distribution, particularly in forests, certain variations or partial failures of the process have far-reaching results.
If leaves did not perform this most important function to perfection, all animals, including man as well, would perish, and it would almost seem that their obligation to us and the plant world might stop there as long as their success in reaching the light is so overwhelming. But there are no union hours of labor, no regulation as to the kind of work leaves may perform, and some actually reach out for new tasks to do, and do them. In one, our common pitcher plant, the leaf, as is implied by the name, is formed into a slender hollow pitcher, wide at the mouth, but narrow at the base. Inside the pitcher are slender downward-pointing hairs so arranged that an insect may crawl in, but never out. The lower down the luckless insect gets the more certain is its death, and, to clinch matters, there is a tiny pool at the bottom where it is not only drowned, but, due to the composition of the mixture in the pool, digested. Only a very few plants can do this; only a minute fraction of the world’s vegetation can digest animal matter. Some experiments on the pitcher plant, which grows in bogs, show that it will digest bits of beefsteak dropped into the liquid at the base of the pitcher.
In the East Indies and in Africa there is a pitcher plant—in fact, scores of varieties of them—which grows up on the branches of trees. In this case the pitcher may be as long as some of our American kinds, often twelve to eighteen inches, and many of them are attached to a slender leafstalk two to three feet long, by which they hang suspended. Insects, literally by the thousands, are caught in these gaudy traps, for many of the pitchers are beautifully colored, and near the opening they secrete a sweetish liquid that lures their prey. They are, in fact, such curious and handsome plants that they are commonly grown in greenhouse collections.
Nature sometimes finds still other ways of using strange and curious-shaped leaves, and in our American bogs is a group of plants, also insect digesters, still more unusual than the pitcher plants. In bright sunny places in open bogs one may often find small reddish, glistening plants, called sundews, usually only a few inches tall, covered with sticky hairs. In fact, the glistening is due to the secretion of the sticky substance, a tiny drop of which may be found at the end of each hair. Flying insects are caught in these leaves, and, as a fly on fly paper, the greater the struggle the more involved does the insect become among the sticky threads. Once caught by such a plant, escape is practically impossible.
Lying in ambush for chance insects, as these sundews and pitcher plants do, may seem nearly the limit of what is to be expected of leaves. Merely to be always on the job, with a plentiful supply of insect digester, might seem to be all that could well be expected from what, after all, are only modified leaves. But nature’s devices are infinite, and there are still other ways to accomplish the apparently impossible. In a small section of the southeastern States there grows a plant that not only lies passively in wait for insects, but actually captures them. This flycatcher, known as Venus’s flytrap, has two valves to the leaf blade, supported on a stout broad stalk so arranged that their fringed surfaces face each other. If an insect—and many do—alights between these valves, they close together rapidly and the prisoner is hopelessly caught by the interlocking marginal bristles that fringe each valve. In this case there are glands on the face of the valves, against which the live insect is tightly pressed, and which secrete a digestive fluid. When nothing remains the valves slowly open and are ready for the next victim. They may be made to close by slight irritation with a lead pencil, and it is the impact of the insect that releases one of the most curious examples of movement in leaves known to us. There are a few other plants in different parts of the world that by still other modifications of their leaves catch and digest insects, but none of them are to be considered as “insect eaters,” or other names implying that they have definite designs on the life of passing insects. The process is sufficiently remarkable, the success of the operation so sure, that there is nothing gained by attributing to such plants, as many have done in the past, malignant characters that are possibly confined only to man. The whole wonderful process is more reasonably explained by realizing that all these insectivorous plants are so by virtue of necessity, that many of them are bog plants, which are often hard put to it to get suitable food, and that the extraordinary change of shape and function is but one more contribution of leaves to the economy of nature.
In dry or desert regions, where the conservation of moisture is essential to plant growth, water storage by leaves is nearly as great an aid to the plant as we have seen it to be in the stems of cactus, South African spurges, etc. Our common century plant, whose leaves are, in some kinds, a hundred times thicker than in ordinary foliage leaves, is a good example of leaves adapted to water storage. In our southwestern deserts hundreds of species of plants can exist only by virtue of the fact that their leaves are so changed in their form or structure that they serve as reservoirs for water storage. This may be accomplished by thickening, or it is more often contrived by a thick coating of hairs. The surfaces of thousands of different kinds of leaves are clothed with hairs either on the upper or lower side, or sometimes on both sides. In many cases they are quite obviously protection from too rapid drying out of the leaf. In others, as in the nettle, the hairs secrete a stinging substance which seems to insure the plant against grazing animals.
Leaves, then, are for something more than to provide the beautiful foliage which is their most spectacular accomplishment. So varied is this in its beauty, from plain green leaves to the wonderful coloring found in begonias, coleus, and many other garden plants, that the sheer beauty of the panorama of foliage is likely to blind us to the more important uses of leaves. First of all must we consider them the factories, in which night and day are produced the food of all plants and most animals. Then in certain cases we have seen that, by every ingenious device known to nature, they perform other special work, such as helping the plant to climb where that is necessary, catching or even capturing insects and digesting them when that peculiar service is demanded of them, and, finally, serving as storage reservoirs in regions where water is scarce. Probably no part of the plant works so unceasingly each season at its varied tasks. In the autumn, dropping to the forest floor, its decomposition furnishes still other food for the plant, and, to crown all, this busy life and by no means unprofitable death leaves behind it, as a promise for the continuance of the work, a snugly protected leaf bud which will repeat the process the next season.
While the plant’s and, consequently, our debt to the leaf is seen to be tremendous, it cannot be ignored that, if plants produced nothing but leaves, the end of all plant life would come with the death from old age or disease of the present generation of plants. Except for those kinds that reproduce themselves by division or extension of their rootstocks, which bear buds, there would be no provision for increase. As only a comparatively small number of plants can reproduce by this method, it is obvious that something more must be provided to secure new generations of plants. Flowers, and the fruits and seeds which inevitably follow them, do this. All plants, with some exceptions to be noted later, produce flowers at some time in their life. In the case of the century plant, only once, after which they die. But except for ferns, mushrooms, seaweed, yeast, bacteria, and some other forms of so-called flowerless plants, a flower or blossom is to be found at some stage in the life of all plants.
If we examine the leaves of a goldenrod, we find that they are large below and diminish in size toward the top. Just below and among the flower clusters they are so much reduced in size and often changed in color that they cease to be ordinary foliage leaves, and are known as bracts. The occurrence of bracts is nearly universal in flowering plants, and they form not only an apparently transitional stage between leaves and flowers, but an actual one.
In a complete and perfect flower there are, at the bottom of it, a row of green leaflike sheaths which

Fig. 39. A perfect and complete flower. A, petals, all of them forming the corolla; B, sepals, all of them forming the calyx; C, the stamen, composed of (C) the filament, and (C1) the anther, which produces the pollen; D, the pistil, consisting of the swollen base (D) the ovary, a slender shank (D1) the style, and the swollen or branched tip (D2) the stigma. (H. D. House, “Wild Flowers of New York.”) Fig. 40. Typical flower of the pea family. Two petals unite to form the keel (below), two more unite to form the wings (center), the remaining and larger petal forms the standard. In most plants of this family the stamens and pistils are concealed within the keel. Fig. 41. Two-lipped inequilateral flower, common in such plants as Salvia, Snapdragon, etc. Note the united calyx and corolla. Fig. 42. Gamopetalous or united and regular corolla of the Fringed Gentian. Figs. 43, 44, and 45, flowers of the Compositæ or daisy family. Many small flowers grouped in heads and usually surrounded by one or more series of bracts. Fig. 43. Flowers all tubular, the small one at the left being an individual flower. Common examples are Boneset and the common garden Ageratum. Fig. 44. Flowers both tubular and with rays, the tubular in the center and the rays on the margin. Below is an individual tubular flower on the right, and on the left an individual ray flower. Note that its five united divisions correspond to the five petals in other plants. Common examples are the daisy, sunflower, black-eyed Susan, etc. Fig. 45. Flowers all ray flowers, an individual one at the right. The Compositæ with only ray flowers usually have a milky juice and have often been grouped in a separate family, the Cichoriaceæ. Common examples are dandelion, chicory, and lettuce.
surround and often half inclose the brightly colored petals within. This outer covering of flowers is called calyx (Figure 39 B), the individual parts of it, where they are separated, sepals. Their chief use is to protect the interior petals while they are inclosed in the bud. The calyx may or may not have bracts just underneath it, as it does very conspicuously in the case of the flowering dogwood, whose white “flowers” are really only brightly colored bracts. The transition between bracts and calyx is not difficult to see in many plants, and where it is impossible the evidence from their internal structure confirms what our eye might be inclined to doubt.
Just inside the calyx is what most people call the “flower,” which is really composed of more highly colored sepals, but which we call petals (Figure 39A). Where these are joined together the collection, which forms tubular flowers like the lily of the valley, is called a corolla. It is, of course, the petals or corollas of flowering plants that give our landscapes their greatest beauty, their most gorgeous coloring. While this from one point of view amply justifies a prodigal nature in strewing the earth with beautiful flowers, the true value of the color to the plant is in quite other directions, which will be explained a little later.
Toward the base of the corolla, or sometimes on the petals or sepals, may be found a series of slender appendages, usually threadlike or a little thicker, crowned at the top by a distinctly large knob. The individual appendage is known as a stamen (Figure 39 C), its threadlike portion a filament (Figure 39 C), and the knoblike top an anther (Figure 39 C1).
Directly in the middle of the flower there is still another organ, usually swollen at the base, slender in the shank, and either thickened or branched at the tip. This central part of nearly all flowers is called collectively a pistil (Figure 39 D), its swollen base an ovary (Figure 39 D), the slender shank the style (Figure 39 D1), and the thickened or branched tip a stigma (Figure 39 D2). A perfect and complete flower, then, is composed as follows:
| Calyx or sepals, |
+ | Corolla or petals, |
+ | Stamens consisting of filaments and anthers, |
+ | Pistil consisting of ovary, style, and stigma. |
The stamen is the male organ of reproduction and the pistil the female. The actual process of fertilization, pregnancy, the forming of the fruit and later the seed, and the latter’s birth of a new plant, comprise one of the most fascinating of those provisions of nature which secure the perpetuation of the plant world. In the life history of even the commonest weed along the roadside there is this constant renewal of life by sexual reproduction, just as in animals and in man. In the chapter on “How Plants Produce Their Young” will be found some account of this supreme function of flowers, after which, as if their usefulness were over, they wither and perish.
Not all flowers are perfect or complete. Some lack petals, as the buckwheat, where the colored calyx replaces petals. Others have neither calyx nor corolla, as in the sycamore or plane tree. Most plants, however, have both calyx and corolla. In some very few plants certain of the flowers have no stamens, when they are said to be pistillate or female flowers, and certain others have no pistils, when they are called staminate or male flowers. In other words, the sexes are in different flowers in the same cluster or plant, as is true of the walnut and hickories, when they are said to be monœcious. In still others the sexes are on entirely different plants, in which case they are diœcious, as in practically all willows. In the latter case there are pistillate or female plants and staminate or male plants.
While it is a commonplace that peas do not look like daisies, nor a carnation like a rose, this simple observation does not begin to tell us of the wonderfully different flower shapes and colors that are to be found along any roadside. The perfect and complete flower that we have been studying is quite regular, composed as often as not of four or five petals, as many sepals, with five or ten stamens and perhaps a single pistil. Yet there is literally no limit to the variations from this scheme, and some of these must be understood here in order that the life-histories and behavior of plants discussed in later chapters may tell their full story.
The figures on page 44 show a regular flower, with five separate petals and sepals (Fig. 39). Such flowers are said to be polypetalous, i.e., separate petals. Sometimes three of the petals are larger, two smaller, in which case the flower is lopsided or, as it is said, inequilateral. Again all the petals are united to form a regular and equilateral tube, as in lily of the valley, when they are gamopetalous, i.e., united petals (Figure 42). As we shall see in the chapter on Plant Families, this is a distinction between two great groups of plants, as important in their classification as negro and white man are in classifying humans.
In peas, beans, the locust tree, and related plants the petals are much changed to form an irregular flower, with a keellike or prow-shaped part made from the uniting of two petals. Two more unite to form the wings, and the remaining and larger petal forms the standard. Figure 40 and the explanation under it illustrate this unusual form of flowers.
Our common garden salvia shows still another type of flower, which is tubular and irregular (Figure 41). There is an arching, hoodlike structure at the top overhanging a lower lip. This kind of irregularity is common in thousands of different sorts of plants and, usually, it is a device to insure fertilization of the flower by insect visitors. So necessary are these for pregnancy in many plants, that an orchid, once discovered in Madagascar with a tube eighteen inches deep, puzzled the botanists, who were unable to understand how the plant produced seed in the absence of any known insect with a tongue as long as that. Darwin said at once that such an insect would one day be discovered on that island. Years after, Baron von Humboldt, a German naturalist, found the insect and explained the mystery.
Perhaps there is no feature of plant life that shows such an amazing amount of variation as the forms of flowers, and while only a few of the simplest deviations from the normal have been discussed here, it must not be forgotten that this infinite variety is a reflection of the ingeniousness of nature in securing a plentiful supply of seed. Form, color, the secretion of sweetish nectar, the night or day blooming of different kinds of flowers, every device that will make fertilization certain, by the flower itself, by insects, or even by the wind, is used in such prodigal fashion, that we come to see the importance of it to all plants only by a realization of the complexity of it and the provisions against its failure.
One apparently most lavish method of securing fertilization is the arrangement of flowers in clusters. While many flowers are quite solitary, the great mass of individual plants produce a few or dozens, or even hundreds of flowers—in fact, certain relatives of the common carrot may produce over a thousand flowers in a single cluster. The form and plan of arrangement of these clusters follows a rather definite scheme, and here, as in the case of leaves and parts of individual flowers, the figures tell the story better than words. In the common dandelion and daisy, and their thousands of relatives, the “flower” (Figures 43-45), as commonly understood, is really composed of scores or even hundreds of true flowers in each head. In the case of the daisy the yellow center, if picked apart, is seen to be really made up of scores of tiny tubular flowers, each just as truly a flower as a single rose. The rays, or what are incorrectly called “petals,” which fringe the golden center with white, if carefully separated and examined closely, will be found to be also a complete flower, the true petals of which are all joined to make the strap-shaped ray. If one looks sharply, the united edges of these petals may be seen by the ridges or channels that represent their joined edges. Because plants of this sort produce two sets of flowers in each head, one conspicuous by its brightly colored rays and with another tubular set in the center which makes doubly certain the fertilization and seed supply, they are considered the most highly developed of all plants. It is not a close aristocracy, nor an exclusive one, for over eleven thousand different kinds of plants, scattered all over the world, have their flowers arranged in this fashion or some slight modification of it. They possess, above all others, the certainty that there will be no slip in their fertilization, pregnancy, and subsequent birth of a new generation. Because this is the great object of all flowers, and these daisylike plants have brought it to such perfection, they are most surely to be classed as the highest type upon the earth to-day.

Fig. 46. A spike, the individual flowers attached directly to the common stalk. Fig. 47. A raceme, a spikelike cluster where individual flowers are stalked. Fig. 48. An umbel, the individual flower stalks all arising from one point. Fig. 49. Individual flower stalks of different lengths but the cluster usually flat-topped (corymb). Fig. 50. A flower cluster in which the end of the stem is terminated by a flower from the base of which side branchlets similarly tipped with flowers arise (cyme).
While highly irregular flowers are common in nature, conspicuous examples being the orchids in any florist’s window, or the milkweeds along the roadside, they can nearly always be seen to have various changes in the shape of their petals, or sepals, or stamens, or pistils, which are adaptations to their mode of life, but which always result in fertilization. Some plants, true monstrosities of nature, are not only far from having the usual arrangement of flower parts, but they even produce increased numbers of one part at the expense of others.
Double buttercups, and hundreds of our most beautiful garden blossoms, have been rescued by cultivation or the arts of the gardeners. Some roses seem to be practically all petals, but for every increase of petals there must be a decrease of some other part of the flower, and more often than enough it is the stamens and pistils that lose out in this transformation. Just as there is a decrease almost to the vanishing point in the birthrate when people become too effete and cultivated, so in plants there seems to be a point beyond which they cannot be pushed without suffering partial or often complete inability to produce young. The more highly they have been developed, oftentimes the greater their beauty, the less able are they to see to it that the chief function of flowers is accomplished. Such garden plants are increased by root division, cuttings and other arts of the gardener. Naturally true double flowers are almost unknown in wild plants, and the habit seems to have been brought about by too easy a time of it, too little struggle, too much food, or by any other of those things that produce effete but beautiful things, charming in their way, but of no significance in the sturdy struggle for existence that all wild plants must meet or perish. Another curious modification of a flower bud is cauliflower. Here the bud has been so developed, its calyx, sepals, etc., so transformed that the large, cabbagelike head, produced at the apex of the main stem of the plant, has by so much lost all semblance of a flower that it is actually a vegetable.

Fig. 51. The outer leaflike tubular or hooded spathe surrounds in our common Jack-in-the-Pulpit a clublike spadix, upon which are crowded the tiny flowers.
No feature of a landscape gives us more pleasure than its flowers, over which poets have sung and artists have painted their most charming pictures, even a musician has composed a very beautiful piano piece, “To a Water Lily.” But their true place in the scheme of nature has a deeper significance: the wonderful color and symmetry of their parts, the plan of their arrangement, their transformation into curious forms, like the Madagascar orchid, and hundreds of others—all these point to their supreme function, an act of self-sacrifice comparable only to the fall of a leaf when its task is done. Petals, too, wither and die when the fertilized ovary, already a mother, begins the slow process of maturing its young and the end of the flowering stage is reached. Such a climax is this in certain plants that the whole plant dies, as we have already noted in the case of the century plant. The toddy or wine palm of India, often sixty or seventy years old and more than a hundred feet tall, flowers only once, and, as if in recognition of the fact that it has done that for which it grew, slowly dies as the seed ripens. More humble annuals, like buckwheat, and hundreds of others, live only one brief growing season, produce flowers and seeds, and then die, leaving behind them the only means of perpetuating their kind. The dormant seed carries over the winter the life they were themselves unable to maintain, as perennials and woody plants do in their buds.
The number of different kinds of fruits that one can buy even in the greatest markets in the world is so small, compared to all fruits that are annually produced by plants, that they might almost be likened to an ear of corn as against a Missouri cornfield. If, as we have seen, all flowering plants must produce fruits, then what we commonly call such can be only a fraction of what actually makes up nature’s annual harvest. It follows that fruits often occur in unfamiliar disguises and, as we shall see presently, some of the things we have been calling fruits may be so only partly, if at all.
Disregarding what we call fruits and looking at it from the plant’s point of view, a fruit is anything in which, or upon which, a seed is developed or ripened quite without regard as to whether it is edible by man or not. As the ovary is the female organ of reproduction and contains the yet undeveloped seed, it follows also that fruits are practically always a development of some part or modification of the ovary or the upper end of the flower stalk upon which it rests and from which it is often scarcely separable.
Familiar enough is the distinction between dry fruits, such as a pea pod and fleshy ones like oranges, and this quality of being fleshy or dry is practically universal. Among fleshy fruits a few well-known types may be mentioned, such as the orange, tomato, grape, gooseberry, and cranberry, all true berries. There are, of course, thousands of less familiar examples of berries, but, whether with a hard rind as in the orange or not, they are a direct development, or often a mere swelling of the ovary, with sometimes the adhering calyx, and contain the seed. In apples and pears, known as pomes, the fleshy part is a development of part calyx and part the receptacle upon which the ovary is supported while still in the flower. The ovary in these fruits is the parchmentlike interior which contains the seed. Plums and cherries, which have a single stone, instead of numerous seeds buried in the flesh, are known as drupes. These familiar examples are matched by thousands of others of which we hear nothing, all drupes and all formed directly from the ripened ovary and without much change, except the increase of size, juiciness and large development of the tiny immature seed, now transformed into a stone. In the watermelon, pumpkin, and related plants, is still another kind of fleshy fruit, called a pepo. All of this, including the hard rind, is transformed ovary and calyx completely incorporated, and forming in the pumpkin perhaps the largest fleshy fruit known. In a considerable number of plants there is not a single ovary, but several, or in some cases many. These occasionally all develop into what is called an aggregate fruit, of which examples are the blackberry, mulberry, magnolia, and many others.
While it would be logical to think that these fleshy fruits were designed to make delicious food for man, that, in the light of what we have seen to be the real function of the flower, is an assumption which, while flattering, is far from the truth. It is much more certain that fleshy fruits help plants in the dispersal of their seeds and that this fleshy, juicy character is just one more device of nature to see to it that not only do plants produce seeds, but that the seeds are carried and so spread the plant over considerable areas. Birds and animals eat such fruits in enormous quantities and, in fact, bird migrations are thought to be not so much response to winter cold as to the fact that fruits are scarce then. When it is remembered that some birds make tremendous flights, often over 10,000 miles in a few days, their capacity to spread seeds through their droppings may be imagined. In the chapter on plant distribution some truly remarkable cases of such seed dispersal will be given.
The chance of having seed carried great distances, because it is embedded in a fleshy, often brightly colored fruit, would seem to put plants having dry fruits at a disadvantage. Birds and animals cannot be expected to look after the dispersal of those fruits that are neither tempting to the sight nor to the taste. And it must be confessed that quite other qualities in dry fruits insure their dispersal. Some are so nutritious, like the acorn, that thrifty squirrels store them over the winter, as they do many other seeds which are harvested from dry fruits. Various grains are often so stored by man, and rice, wheat, buckwheat, and other cereals are common cases. In nearly all grains the seed fills so completely the fruit that cereals are very generally, but mistakenly, called seeds. A grain of wheat or corn is just as complete a fruit as a watermelon. Only its outer coat and inner seed are so closely welded together as not to be usually recognized as a fruit, with the seed inside.
One of the commonest types of dry fruit is the capsule (Figure 53), well named, as it is almost an exact counterpart of the capsule of the druggist, in that it is in many cases composed of a lower part and an upper, usually merely a domed lid. Others again, instead of splitting around the sides, split from top to bottom. Still others, as peas and beans, known as legumes (Figure 57), are pods that not only split lengthwise, but have no central partition, as do many other fruits of the same general type. When the seed is ripe nearly all pods and legumes finally split open, and the seed or seeds tumble out. A few, as in the violet and touch-me-not or jewelweed, apparently realizing that merely to spill out ripe seeds at the proper time will not spread the species very far, open their fruits with a sudden explosion and literally shoot their seeds considerable distances. The artillery plant, commonly grown in greenhouses, a delicate feathery herb from tropical America, opens its flowers with a report like a toy popgun and shoots its small pollen grains for several feet, but not its seeds as stated by some.
But many fruits do not open at all and seem to be at the greatest disadvantage in the effort to insure

Fig. 52. The strawberry. The fleshy part consists of the modified upper end of the flower stalk or receptacle, while the true fruits are the dry achenes on or embedded in the surface and popularly called the seeds. Fig. 53. A three-celled capsule splitting lengthwise as in the common Iris. Fig. 54. Fruit of the cocklebur, the hooked prickles of which are admirably adapted for clinging to the fur of animals. Fig. 55. Pods of a plant of the Mustard family, which split down both edges, unlike the true peas, which split down only one edge. Fig. 56. Two types of achenes of the daisy family tipped with plumed bristles, greatly aiding their carriage by the wind. Fig. 57. Common garden pea—a typical legume. Note that it splits only on one side. Fig. 58. The samara or two-winged fruit of the maple. Fig. 59. The samara or single-winged fruit of the ash. Fig. 60. The dry two-pronged and bristly fruit of the unicorn plant (Martynia), admirably adapted for dispersal by animals.
dispersal of their seeds. Greater food value to birds and animals overcomes this in some kinds, and another help is that some fruits of this sort are covered with hooked prickles or barbs (Figures 54 and 60). The common weedy burdock, the barbed fruits of which may often be found sticking to the fur of animals in great quantities, is a case in point. There are whole groups of plants that rely on this method for seed dispersal, notably the avens, tick-seed, tick trefoil, and many shrubs in the tropical regions.
Where the fruits are neither barbed nor very good to eat, and so apparently doomed to be more or less permanent stay-at-homes, nature has provided some of them with the proper equipment for flight through the air. Winged fruits like the maple are to be seen on any windy day during their season scurrying before the breeze, and consequently spreading their kind over considerable distances. In the maple there are two wings, joined at the base where the seeds are embedded in the wings, and the fruit is known as a samara (Figure 58), or key fruit, from a slight resemblance to an old-fashioned key. Ash trees bear fruits that are a slight modification of this type and may be carried considerable distances by the wind (Figure 59).
In the dandelion, daisy, and nearly all its thousands of relatives, this faculty of setting sail in the air has been carried to the greatest perfection, just as we saw its flowers were. In this family of plants, the largest in the world, the fruit is mostly tipped or surrounded by a small collection of very fine bristles. The fruit, known as an achene (Figure 56), is so light that with the added buoyancy of this tiny collection of down it can be transported great distances. Some have been known to fly hundreds of miles in severe storms, and, as we shall see in the chapter on plant distribution, these tiny plant balloons have played a conspicuous part in spreading their kind over the face of the earth. Cat-tails also, together with many other plants, have this faculty and make up by its possession for the lack of fleshy or otherwise desirable fruits that might be carried. All achenes are not winged, those which dot the surface of the strawberry being imbedded in the luscious flesh, which is not really fruit at all. Only the achenes on the strawberry are true fruits, the fleshy part being merely a development of the upper part of the flower stalk and not of the ovary (Figure 52).
Fruits, then, cannot be restricted to the common understanding of them. They are transformations of the ovary, in which or upon which seeds are nursed, and upon which most plants depend for the dispersal of their seeds. We shall see later on how fruits have fulfilled their destiny, how some are fit for their true function only when they have been eaten by birds, and when some digestive juice has released them from the impotence they would suffer without being eaten, how a whole forest has been changed in the West by the busy activity of squirrels upon the fruits and seeds of a single kind of fir tree; how the fruit of the coconut palm has been spread throughout the tropical world because it can float in the sea securely protected from injury from salt by the impervious coverings of its fruits.
As the final stage in the development of all plants is their seed, with the dropping of which they bid good-by to their fellows, it is not perhaps remarkable that in the seed of all flowering plants is the germ for the new generation. To seeds which may be as small as the mustard, so often mentioned in the Bible, or as large as the coco de mer, or double coconut, from the Seychelles Islands, often fifty pounds or over, is intrusted by cunning nature the one final and most important act in the whole kingdom of the plant world. Nearly all plants would die off forever if seeds did not have in them the germ of life, apparently quite dead, but actually only dormant. This living germ may persist for years, sometimes even a hundred years, and yet with the proper conditions it never fails to sprout.
Seeds have inside them a tiny plantlet folded and ready to grow when the seed splits to release it. Also, in the seed is stored up food to sustain the new plant until such time as its own roots begin to act. This young plantlet is known as the embryo, and to this all actions of the seed are subservient.
As the seed splits, and the young plant develops its first leaves and rootlets, there is shown one of the most remarkably uniform tendencies in plant life. In all plants with net-veined leaves the young plantlet starts life with two leaves, or cotyledons, as these first leaves are called, and this whole group of plants are thus known as dicotyledons. In plants with parallel-veined leaves the young plantlets start out with a single cotyledon and are therefore called monocotyledons. In only the pines, spruce, and a few other evergreen trees the seedling plants have several cotyledons and are known as polycotyledons. All the flowering plants in the world belong to one of these groups, so that merely to see the germinating seed tells the story at once. The linking of parallel-veined leaves and a single seed leaf, and net-veined leaves with two seed leaves, is also associated with very definite arrangement of their flower parts, their method of growth and other characters. Something has already been said of this in the discussion of stems and leaves, and more will be found in the chapter on plant families. No more beautiful example of the plan or scheme of nature is to be found than these characteristics of all plants, and in seeds we find the first hint as to which army the plant will join, under which banner it will fight, and under what generalship it will develop. Nothing tells us so much as these first seed leaves, pushing their way up through the soil and revealing, as they burst above ground, to what place in nature their destiny will consign them.
Flowering plants, which make up the bulk of the vegetation of the earth, have been discussed in some detail, not only because they furnish us with all the things that make life possible, but also because they show perhaps better than anything else the division of labor, all striving for one end. Roots, the food gatherers. Stems, the framework for the foliage and its means of reaching the light, or as a storage house for reserve food. Leaves of many kinds, all factories working night and day to make the necessary food. Flowers of every hue and shape to lure insects, or by other means secure union of male and female. Fruits to ripen the result of this mating of the sexes. And, finally, the seed carrying with it the yet unborn life. Each part occasionally losing itself in order that the end may be accomplished, many of them changing their form or even their function where that is of advantage, all in their separate ways doing their task, the end of which they cannot see, and the fruits of which they will never enjoy. Nowhere is it so true as in plants that to save oneself there must be the capacity to give oneself. Untold millions of leaves fall, or trees crash down, or seeds are developed, each fulfilling their destiny which is to insure the perpetuation of their kind. As we shall see later on, there are many mistakes, many apparently futile attempts, thousands are wiped out that one may be saved, and in the past multitudes have gone out forever. Yet the result of it all is the plant world as we know it to-day, each kind struggling to increase its sphere of influence, or to cover more of the earth’s area. The combat between different kinds is inexorable, yet the capacity for sacrifice on the part of different organs, in order that a certain individual kind may win, is literally beyond belief.
In the light of what has been said about flowers it may well be questioned how anything can be a plant and still have no flower. The fact is that flowers as we commonly understand them are unknown in the plants about to be discussed, but that what corresponds to a flower, and performs the function of a flower all plants must and do have. In the case of most flowering plants the possession of flowers is one of the beauties of nature in its most resplendent mood, while in the so-called flowerless plants the functions of flowers are performed by tiny microscopic organs, even the existence of which has been only recently discovered. Because flowering plants produce their sexual organs in such a gorgeous setting, for all the world to see their matings they have been called phanerogams, which means literally visible marriage, while the flowerless plants which perform similar functions in more secret ways are called cryptogams, meaning hidden marriage.
These cryptogams or flowerless plants occur in far greater numbers in the world than flowering plants, but their size in most cases is very much less. Many individuals are so small, as in the case of bacteria, that a single one can only be seen after it has been magnified many hundreds of times by the microscope. Of the cryptogams some of the largest, certainly the most beautiful, and probably the best known are
In nearly all woods one may find delicate feathery plants with graceful, usually much divided leaves that nearly always start up from the ground like a slowly opening, but somewhat fuzzy coil. (Figure 62.) Ferns, at least most of those that grow in America, uncoil their leaves in this way, almost without exception. The accompanying figure shows the procedure, and in addition to this character one may hunt in vain for flowers.
While they bear no flowers we already know that nature could not leave them with no means of reproduction without abandoning them to a childless old age and the consequent extinction of the race of ferns. So far from the truth is this that ferns make up a goodly proportion of the world’s vegetation, and there are many hundreds of different kinds known. The lack of flowers, of course, explains why ferns do not bear seeds which are matured in a fruit or ripened ovary.
On the back of the leaves of most ferns, along or near the edges of the finer subdivisions, one may

Fig. 61. A general view. Fig. 62. Its uncoiling spring condition. Fig. 63. The back of one of the smaller divisions of the leaf showing the collection of spore cases (sori). These are sometimes borne on special leaves, but in most of our American kinds on the backs of ordinary foliage leaves.
find, at the proper season, collections or rows of tiny, usually brownish dots. These contain often thousands of microscopic objects known generally as spores, and from this fact the dots are called spore-cases, or more technically sori. (Figure 63.) The process by which new plants are formed is a

Aerial Roots of Fig Trees Hanging Over the Edge of a Cave in the Rain Forest, San Lorenzo, Santo Domingo. (Photo by the author. Courtesy of Brooklyn Botanic Garden.)

Venus’s Flytrap, an Insectivorous Plant of the Southeastern United States. The fringed valves of its leaves close together when an insect alights between them. (Courtesy of Brooklyn Botanic Garden.)
somewhat complicated one, but the spores in these brown dots are the agency which makes reproduction possible, and the actual mechanism of it, one of the most interesting achievements in plant life, will be described in the chapter on “How Plants Produce Their Young.” Sometimes the spores are not borne on the backs of ordinary foliage leaves but on special leaves that bear, very often, nothing else.
Ferns are much like ordinary flowering plants; except for their lack of flowers, they have all the root, stem, and leaf characters of their more showy neighbors. While most of them have compound leaves, even sometimes twice or thrice compounded, a few have simple, narrow leaves without teeth, and one kind in tropical America has threadlike leaves. In many tropical rain forests, so called from their dripping wet condition, ferns form large trees, and these tree ferns are among the most graceful and feathery of all plants. There are, too, a few climbing kinds—one, called the climbing fern, is a native of the eastern United States. Then there is the walking fern, that seems to upset the statement that plants do not move as animals do. It sends out delicate runners that, rooting at the tips, form new plants, often several feet from the parent plant.
The characteristic of having, even in the simplest form, stems, leaves, and roots, with all that this implies in their internal structure, marks them off at once from all other flowerless plants. In ferns there is always some internal equipment for carrying food from one part of the plant, the roots, to another, and this ability is possessed by virtue of ducts or vessels through the stem and leaves. This system, found in all flowering plants and ferns, but nowhere else in the plant world, is called the vascular system, or literally, a vessel system. We shall see how important was the acquirement of this system of vessels, when we get to the chapter on the History of the Plant Kingdom. Its appearance upon the earth marks as important a stage in the development of plants as the dawn of a definite backbone did upon animal life.
Ferns, then, are vascular cryptogams because they do have conducting vessels in their stems, and they produce their young by a process of hidden marriage which will be described later. All other cryptogams or flowerless plants are without this system of vessels and are called therefore non-vascular cryptogams. Numerically they are tremendously important; upon them depend many manufacturing processes like bread making, brewing, and all arts using fermentation. But they are hardly recognized as plants by the general reader, and because of their size and the necessity of studying them with a microscope in order to understand their structure they will be treated here only briefly.
The remaining flowerless plants, having no duct system in their make-up, are, as we know, called non-vascular cryptogams. This is a general term for a very large group of plants, some quite obvious and well known like a mushroom, for instance; others so small or of such uncertain structure that they are not even well known by experts. This great mass of plant life, more numerous than all the other kinds of plants combined, contains many different forms, some of which are of gigantic size. A single plant of a certain Pacific Coast seaweed regularly exceeds in length the height of the tallest

Fig. 64. A moss plant. Fig. 65. A mushroom, a common type of the fungi, which include also puffballs, molds, and many disease-causing microscopic organisms. Fig. 66. A common seaweed, a representative of the algæ, which include the green scum on the top of ponds, and the kelp from which fertilizer is now being made. Fig. 67. A lichen, a common cryptogamous plant on logs and rocks. Our native kinds are usually grayish-green in color.
known trees. And yet other inhabitants of the water, certain kinds that float freely, are microscopic in size. The latter occur in such enormous numbers that their tiny decomposed skeletons after dropping to the bottom of the sea form the diatomaceous earth, so much used in polishing machinery. The commercial product now comes from deposits of these skeletons laid down in past ages, which, due to changes in the land and water surfaces of the earth, are now found in Virginia, Nevada, California, and in Bohemia. All these must have been in the bed of waters long since gone, which teemed with these microscopic organisms. To-day there are over ten thousand different kinds known, yet so small are they that their dimensions are measured in thousandths of an inch!
Somewhat lower in the scale of life—and by this we mean simpler in structure—than the ferns are the mosses. (Figure 64.) There are thousands of different kinds, but everyone is familiar with the collective growth of the commoner sorts which makes the velvety mossy carpet in our woods. The individual plants are small, but in many kinds sufficiently large to be seen without a microscope. Most important of all, practically every one of them has the ordinary green color of the better known plants, and as we shall see in the section devoted to “Leaves as Factories for the Making of Food,” that stamps them at once as plants, if other things did not.
Mosses are almost infinite in their habits, some growing on the dry rocks or trunks of trees, many growing in moist woods, some in the water, and immense quantities of certain kinds in bogs. The peculiar bog mosses, known as sphagnum, play an important part in forming peat and perhaps coal. While mosses are otherwise not of much commercial importance, they are among nature’s most beautiful ground covers, carpeting many a nook and dell with a soft, velvety, almost cushionlike growth.
Although they are rather small, they appear to have a somewhat definite stem and tiny leaflike appendages of it, without, however, having the vascular system found in all ferns. Mosses might almost be considered miniature ferns, of which they are perhaps only simple ancestors. Their vegetative or green parts vary much in shape, size, and the arrangement of the tiny leaflike appendages, and while most of them are a beautiful bright green, nearly all the bog or sphagnum mosses are rather ashy gray in color. In most of the typical mosses there arises from among the vegetative growth of them a slender stalk, at the top of which is a small capsulelike organ. This contains the spores, and it is upon this long slender stalk and its spore-filled capsule, really marvelous in its internal structure and mechanism for the discharge of the spores, that mosses depend for their reproduction. As in the case of the ferns this process will be considered later, along with that of some other plants. This whole story of how plants produce their young, perhaps the most fascinating of any part of the study of plant life, is so fundamentally a part of their history and shows nature in her most maternal moods, that a special chapter will be devoted to it. There we shall see, as a whole, how these vastly different acts of fertilization and reproduction are, in different groups of plants, all responses to that insistent command for life, more life, in a never-ending stream.
The chief characters to remember about mosses are that they are very simple, but practically always green plants that have some differentiation into stem and leaf; that, while they have no vascular system, their structure and particularly the mode of reproduction suggests that they are not very distant from the ferns, and quite likely simple ancestors of them. These characters are of more importance than appears on the surface, as we shall presently see, for they mark mosses off from many other nonvascular flowerless plants which have quite different structure and altogether different mode of life.
If you will turn to the chapter on Plant Behavior and read particularly the sections on “Leaves as Factories for the Manufacture of Food” and “Borrowing from the Living and Robbing from the Dead,” you will see in the food habits of the plants there noted the great difference that exists between plants, like mosses and ferns, that have green coloring matter in them, and those we are about to mention that never do. The lack of this green coloring substance tells us at once that plants of this sort live only on the dead remains of other plants. In the case of these nonvascular flowerless plants there are certain modes of growth that, in some forms at least, are always associated with this scavenger-like food habit.
The common mushroom (Figure 65) is the best known of that large group of plants, called generally fungi, which produce no green coloring matter, have no leaves attached to a stem, and always live on decayed vegetable, or sometimes inhabit living animals, even man himself. The mushroom with its brownish stalk and buttonlike dome is familiar enough, but there are literally thousands of different kinds, a common sort forming “brackets” on the trunks of trees. While perhaps everyone would recognize these as plants, peculiar as they are in their often weird shapes and unusual as they nearly always are in their color, there are many minute kinds of fungi that scarcely anyone would even think of as a plant, and yet for better or worse they are incomparably the most powerful plants in the world. For upon these microscopic fungi man depends for many things. It is certain kinds of them that make the manufacture of cheese possible. They turn milk sour (pasteurizing milk is merely stopping their work), give to yeast its power of “raising” bread, all brewing depends upon them, every process of fermenting the juice of fruits for wine making or for whatever else, the decay of wood—all these processes and scores of others, whether for the good or evil of mankind, depend upon the work of these plants, any one of which is so small that a single individual must be magnified hundreds of times to detect it. Many of them are the “germs”—better called bacteria—that cause diseases like tuberculosis, cholera, typhoid, anthrax, and diphtheria. All surgeons wage incessant warfare against a host of them that attack wounds and form pus. They live in our intestines and have much to do with digestion, and unhappily with indigestion, so that we may be said to carry about with us a whole flora of them! Nearly all the diseases of plants, like the blight of potato and the rust on wheat, are caused by them. Some other kinds live in the soil, and many flowering plants depend absolutely for getting their food upon the work of these fungi. Unfortunately their minute size and consequently obscure mode of life demand technical skill and the use of the microscope to detect them, so we must leave them here, always keeping in mind that these smallest of all plants are charged with a power for good or evil; so far as man’s life is concerned, greater perhaps than all other plants.
While most fungi, particularly those familiar ones like mushrooms and puffballs, are inhabitants of the land, the remaining group of nonvascular flowerless plants are nearly all water plants. Most of the better known ones live in the sea, and as the wrack or tangle washed up on the shore we recognize them as seaweed. The algæ (Figure 66), which is a general name for such plants—and they live in the sea, in fresh water, and even on dry land—are, so far as structure is concerned, the simplest of all plants.
Those that are fastened to rocks are often beautifully colored, much branched, and many kinds bear small bladders that act as buoys. These coast seaweeds are generally of different colors, those nearest the surface being generally greenish, the deeper water kinds reddish or brown. None of these seaweeds are found at great depths, because the really deep parts of the ocean are almost, if not quite, dark. Seaweeds, and in fact all the algæ, have green coloring matter in them, even where this is masked by reds and browns, as is the case in some particularly showy kinds. As you will find in the section on “Leaves as Factories for the Making of Food,” no plant with green coloring matter can live in the dark. That is why seaweeds are not found in the great deeps of the sea, some of which are several miles below the shore line along the coasts, and are so cold and dark that neither plants nor animals can grow in them.
Those seaweeds that grow along the coast, and are uncovered by the retreating tides, are well known by everyone, but by far the greater number of algæ float without anchorage of any kind. One kind that has been torn from its anchorage occurs in such enormous quantities that off the coast of America it has formed literally a floating island composed entirely of dense mats of a species of seaweed. This place, known as the “Sargasso Sea” from the name of the seaweed forming it, was the terror of old mariners and Columbus’s ship was fouled in it for two weeks. The area occupied by the weed is several hundred miles long and wide, and while old sea yarns about ships being caught in it and never escaping are gross exaggerations, it is certainly one of the most curious of plant growths, due entirely to a nonvascular cryptogam.
Of those kinds that are never anchored the number is legion, and in addition to those forming the diatomaceous earth, already mentioned, there are many more. They form almost the only food of hosts of creatures of the sea, but because of their floating freely in the water, the consequent difficulty of collecting them, and their unusually minute size, little is likely to be known of them, except by the experts.
Other algæ are always found in fresh water and form the scum found on stagnant pools. Individuals of any of these are so minute that, while under the microscope they are of the greatest beauty, their structure must remain for most of us a sealed book.
We have now traced, in only the briefest fashion, the outlines of what plants are, reversing the order of nature in beginning with those most complex but best known, the flowering plants. As we shall see later, these are the climax of prodigal nature and are to be considered the end rather than the beginning of plant life on the earth. Then, and still more briefly, have we stopped to see those less known plants that produce no flowers, such as the ferns, mosses, fungi, and finally the seaweeds or algæ. These are all to be considered as the ancestors of flowering plants, the ferns the nearest to them and the algæ probably the most distant relatives. The development of plants from the minutest alga up to our most gorgeous flowering plant, is an infinitely slow and painful process. With many mistakes, with its pathway strewn with the wreckage of forlorn hopes and false starts, it is incomparably the most dramatic story in the plant world. Some of its details will be told in the chapter on the “History of the Plant Kingdom.”
Nor can we leave the discussion of what plants are without some mention of the thing that really makes up their structure, whether it be a microscopic bacterial organism or the Big Tree of California. For the unit of all animal and plant life is the cell. In its simplest form it is merely a minute sac with a definite wall and inside the wall is a substance known as protoplasm, literally protos, first, and plasma, thing formed. It is protoplasm that forms the living tissue of all plants and animals; it is life itself. No one has ever succeeded in making any, notwithstanding that many learned men have tried for years. Its inclosure in the cell wall, its power of self-division and consequent multiplication of the units, make up those first things about which most of us can never know much, but the end of which we recognize in the beauty of plant life all about us. For only under the highest powers of the microscope may cells be seen and studied. Just as bankers reckon mills as a definite unit of a cent, and yet none of them has ever seen a mill, so we must think of cells as the definite unit of all living things, although most of us will never see a cell. But, unlike the mill, cells may be seen by those equipped to see them, and this study, the development and grouping of them to form all the varied objects that inhabit the plant world, is known as histology. It is literally the internal history of plants and animals, and lies quite outside the scope of this book. What we must never forget is that whatever knowledge we have gained, either from the foregoing account of what plants are, or from our observation of them, is, after all, only a partial notion of them, as unsatisfactory as our estimate of what people really are, from merely looking at the outside of the houses in which they live. The outer form we may know and admire, the inner substance must ever remain for most of us a secret treasure house the value of which is certain, but the key to which we do not possess.
PRACTICALLY all that has been said in the first chapter relates to what plants are, their organs, or what we may call the architecture or plan of their framework. But what they do with this elaborate structure is as important as what we do with a house that may contain every modern improvement but is never a home until these things have been put to use. One of the chief concerns of any architect is to see to it that the house has as much sunshine by day and as attractive illumination by night as possible. Nature, that greatest of all architects, also sees to it that plants get the utmost necessary sunlight, but for a much more important reason than the mere attractiveness of sunshine, be that ever so beautiful. For light, the life-giver of all green things, is so absolutely essential to plants that experiments to grow them in the dark have always failed, and many gardeners now use electric light in greenhouses in order to prolong the short daylight of winter. It is the lack of light that makes celery blanch.
Plants grown in the house inevitably turn toward the windows, even plants growing against a wall turn their leaves away from it—nowhere can one find living green things that do not find the light as surely and persistently as men try to get their food or their mates. Many examples of this could be given and must have been noticed by everyone.
Sometimes seeds germinate under a barn floor for instance, and the puny pale little plantling reaches out slender stems, all of which turn, as a compass turns to the north, to perhaps a crack of light in one corner of the building. We have already seen how the search for light will carry the slender rattan palms of India hundreds of feet to the topmost leaves of the forest. Individual plants, and, as we shall see later, whole forests make desperate efforts to get to the light. We know already, that the struggle for light is just as bitter as the struggle for food by roots. And finally if, as we have many times proved by experiments, plants die when grown in a dark room, what is it that light does for plants and how is a process carried on that everything leads us to think is of the greatest possible importance? Quite obviously it is not the mere beauty of sunshine dancing upon the landscape, as entrancing a picture as that may be any summer afternoon, with the play of sunshine and shadow on the tracery of foliage. That green color of the foliage, the almost universal green of so much of the earth’s vegetation, restful to tired eyes, providing us with the most pleasant shade, has wrapped up within it the secret of just what sunshine does for plants. For under the magic of light acting upon this greenery one of the most important industries in the world, the manufacture of food, is constantly going on.
It must be clear enough from the start that to call a leaf a factory for the making of food forces us to decide at once whether this is a mere way of speaking, or whether, incredible as it may seem, anything as thin as a leaf can really produce food. As we eat lettuce, and millions of cattle graze every day, leaves as food producers win handily on that score. But to understand how food is produced in such a tiny factory demands that we walk about in it for a bit, study the inside of it and especially its many small chambers within which is not only the machinery, but some of the finished product stored up for later use.
Unlike modern factories there are many entrances, from any one of which we can begin our tour of inspection. On the under side of nearly all leaves and on the upper side of some there are scores or even hundreds of small pores called stoma, so small that only with a microscope can they be seen. These entrances through the factory wall, are carefully guarded by a pair of watchmen whose business it is to see neither too much dry air gets in nor too much of the product of the factory gets out. They see to it, also, that waste products are thrown out at the proper time. These watchmen, or guard cells, as they are called, are constantly on the job, work almost automatically, but their chief function is connected with the proper ventilation of the place, and will be discussed later under “How Plants Breathe.”
Once past the entrance it is obvious that we are in one of the strangest of all factories, for none of the rooms are truly square or oblong and their irregularity as to outline would drive your average foreman into profanity. Yet they are certainly divided into distinct classes, at least as to size and as to what the rooms contain. Some are apparently filled with nothing but air and have direct connection through the stoma with the outdoors. These are called intercellular spaces. Others, and these are most important, are filled mostly with the green coloring matter that gives the leaf its color. This substance is known as chlorophyll, its individual units as chloroplasts, or literally, chlorophyll bodies. Quite independently of these chlorophyll cells or rooms, or the intercellular spaces which correspond to halls, there are some large and many small tubes. These are the veins of the leaf and their finer branches and by their direct connection through the stem to the roots, serve as the ducts through which some of the raw materials are brought into the factory.
This green coloring matter or chlorophyll is perhaps the most important substance in nature. Without it all except a very few plants would die, and even in those beautifully colored leaves like coleus or caladium chlorophyll is always found, but in these colored leaves it is merely obscured by other coloring substances. It is in the chlorophyll that the ability resides to take the inorganic substances through the roots or from the air, and by the aid of sunlight transform them into organic substances like starch and sugar. Nothing else in all nature can do it; without this faculty, which the commonest green leaf possesses, the earth would prove uninhabitable within a single year. Just what chlorophyll is chemically is not yet thoroughly known, but the thing of chief interest is that it is hardly ever found in parts of the leaf not exposed directly to the sunlight, and that during the autumnal coloring and before the fall of the leaf chlorophyll is carried to other parts of the plant, and quite possibly stored for use the following season.
While the composition of chlorophyll is not surely known, iron is certainly one of its constituents, as plants deprived of iron lose their green color. It also is known to contain oxygen, carbon, hydrogen, and nitrogen, but merely to catalog what we know about its make-up does not tell us that it is a living green substance and that sunshine sets it in motion. Just exactly how light acts on chlorophyll no one really knows; we merely know that it does so act and that the result is one of the marvelous secret processes of nature, perhaps like the secret of life itself forever hidden from man. In our tiny factory, then, we have raw products coming from the roots and through the stoma from the air; machinery of the most efficient type, for chlorophyll works night and day, and constantly renews itself while producing the finished products; energy from the sun; and finally the complete manufactured products which are foods in the shape of starch and sugar. During the growing season there is no banking of the fires, no stoppage of this most important of all industries, no strikes or lockouts. Each part of the whole works smoothly and with the nicest precision—in fact so perfectly does this process keep on going, so complete is the orderliness of the place, and so regular are the completed products turned out, that no modern factory manager or workman but can learn something from a rather close study of this smallest but most efficient factory in the world.
Some of the raw products are delivered to the leaf from the roots where they have been absorbed by another process that will be considered a little later. These consist of water and the inorganic substances dissolved in it, popularly called sap. Carbon and oxygen come mostly from the air, sometimes separately, more often in the form of a combination called carbon dioxide which is one of the chief constituents of the gas thrown off by man as he breathes out. Now these inorganic substances, contained in the sap or derived from the air, are literally mixed by the chlorophyll and form, always with the aid of sunlight, substances known as carbohydrates, the commonest example of which is sugar. Some form of sugar is one of the earliest results of this process, but sugar is quite easily dissolved in the sap which has contributed to its manufacture, and the excess sugar is thus removed. Otherwise it would clog the machinery and prevent the production of fresh supplies. This first step in the manufacturing process has not inaptly been called photosynthesis, the meaning of which photos, light, and synthesis, combining by means of, suggests in a word the necessity of light and the combination of the inorganic substances mentioned above. Of course this process of photosynthesis is not as simple as the brief account of it suggests, for it is actually a complicated chemical process only part of which is yet understood. It is fairly certain that it goes step by step; it is quite certain that the beginning is inorganic and the end organic compounds like sugar. Something is known also of the wear and tear on the chlorophyll, its waste products, and how it keeps itself not only fit but provides for its own constant renewal. One of the excess or by-products in this initial manufacture of sugar is oxygen. This is either used in other ways by the plant, or more generally it is thrown off through the stoma into the outer air. Oxygen, as one of the necessary constituents of the air that man breathes in, is thus thrown off, while, as we have seen, carbon dioxide, a poisonous gas which we breathe out, is a necessity for this manufacturing process in all green plants. Hardly any trick of nature so completely fulfills the wants of animal and plant life as this mutual exchange of by-products—in the case of animals it is the waste of respiration, in plants it is the wastage of sugar making and some other changes that go on in the plant just after this stage.
The amount of sugar made, carbon dioxide taken in, and oxygen given off by this process suggests that while leaves may be very tiny factories they are among the most efficient in the world. Assuming an area of leaf surface equal to about a square yard the amount of sugar made would be about one-third of an ounce in a day or nearly three pounds in a single growing season. Carbon dioxide withdrawn from the air would average from the same area of leaf surface about two gallons a day or over three hundred gallons for the season. As an equal amount of oxygen is given off by the leaf, it becomes clear that as all of this interchange must go through the stoma the functioning of these and their guardians must be nearly one hundred per cent perfect. As we shall see a little later, they perform still other duties with even greater perfection. When we stop to reflect what an absurdly minute fraction one square yard of leaf surface is to the total leaf surface in the world, we come to some realization of the gigantic proportions of this process of manufacturing sugar and exchange of gases mutually useful to animals and plants. While in the United States most of the leaves fall in the autumn, the great bulk of the vegetation of the world holds the greater part of its leaves all the year, notably in the vast evergreen forests in the north, and of course practically all tropical vegetation. Chlorophyll in such places works continually and what the total of sugar production may be no man can even guess.
Sugar, although the first step in the process, is not the final one, and the leaf has still other tasks to complete. Some of the sugar is used up in the process of renewing the chlorophyll, some of it is moved to other parts of the plant where in sugar cane it forms the world’s chief sugar supply; but the remainder is transformed into starch, a substance that is not dissolved by the water of the sap, and is therefore capable of permanent storage either in the leaf itself or in other parts of the plant, notably in the tubers of the potato, the solid part of which is nearly all starch. The conversion of sugar to starch, which is really a means of contriving to properly store the product of the factory, is done by certain ferments known as enzymes. Just what enzymes are or even how they work is not well known, but apparently they have the faculty of converting certain substances like sugar, and in the process they neither use up nor materially change their own composition. It is certain that the conversion of sugar to starch is an elaborate chemical process, but it is accomplished by these enzymes, the very existence of which has only recently been discovered. Enzymes not only do this, but they convert starch which is insoluble into a kind that may be dissolved and thus carried to different parts of the plant. Upon this power depends the storage of starch in roots, tubers, seeds, or wherever else it is found in the plant, and it is of course upon this power man depends for the food supply of the world. Wheat or corn, potatoes, rice, all the foods that are rich in starch produce none in that part of the plant harvested by man. All of it has come by the process which is only sketched in its briefest outlines in the foregoing paragraphs. All of it must come from that green coloring matter of nearly all plants which, while mostly confined to leaves, is not always so. And wherever chlorophyll is found this process goes on even in the simplest plants. Because it is so overwhelmingly a characteristic of leaves and, as we have seen, leaves are the one organ of the plant upon which man pins his only hope of future food supply, the leaves of all plants may be truly likened to a factory the work of which is never ending, the product of which the leaf will never use, but the result of which has far-reaching consequences to us all.
Now that we understand the importance of light to all except a very few plants, and its very close relationship to the green coloring matter of all leaves, many things about the arrangement and position of leaves, and indeed of the whole plant, may be understood, which, without this knowledge, seems the result of mere caprice or chance. It would seem as though the habit of plants growing toward the light, and against the pull of gravity, a character almost universal, no matter from what mountain declivity or rocky cliff it may spring, might be the result of the “pull” exerted by light on the green coloring matter in the leaves. While light does aid in plants having a generally erect habit it is not the cause of it, as we have many times proved by experiments. As a seed sprouts and the roots go down into the earth, the shoot, before it has broken through the surface and while still in the dark, always grows upward. This property of growing in two opposite directions at the same time, the roots always with gravity and the shoot nearly always against it, is known as geotropism. In the case of vines or other trailing plants there is the same tendency exhibited, even though the plant is not erect. We must think of geotropism as a growth habit of all plants, not caused by light, for it has been shown to act in the dark, but of the greatest advantage to all plants in their initial start toward the light. If this were not the case, it may be imagined into what chaos the vegetable world would be thrown. We are so accustomed to roots going down and shoots going up that we are not apt to think of it as the result of two antagonistic growth habits, the true cause of which is not understood, the result of which is common knowledge. Geotropism is one of those mysteries with which the book of nature is crowded, and merely to describe it and realize its force is by no means to arrive at its true inwardness.
But, quite independently of this peculiar growth habit, the stems and often whole plants do show response to light and many times the response, in its effects, cannot be distinguished from geotropism. Perhaps the most homely illustration of this is the common house geranium which, no matter how often it is turned, always grows toward the window, and if not turned at all becomes hopelessly lopsided, with the leaves all bending sharply toward the light. Trees growing on a cliffside, while always growing upward, nearly always may be seen bending away from the cliff where light is scarce and toward the unobstructed light. The position of hundreds of twigs and branches on any tree have been dictated by their exposure to light, and the habit of practically all trees in the forest of being clear of branches for many feet from the ground is another illustration of the profound effect of light. In the latter case the taller the trees the farther from the ground are the first branches, and in the big trees of California the first branches are frequently over a hundred feet from the ground. In their young stages all these trees were furnished with branches, the leaves of which in their day performed their appointed tasks. But in the strife and hurry of the crowns of the forest to overreach their neighbors these lower branches, from the bottom upward, gradually die off. So inexorable is the plant’s demand for light, that these lower branches, in spite of being nearest the source of their food from the roots, are doomed to be killed. Nature plays no favorites and these lower branches, once the pride and support of the young tree, are ruthlessly dropped off when they can no longer play the game. This wholesale slaughter of lower branches in a forest, more complete than any pruning by man could ever be, gives us, if the story of the factory leaf has not already done so, some conception of the part played by light in the plant world.
The shade of certain trees is so much denser than others that they have been planted for this purpose, notably the horse-chestnut and Norway maple. Foresters have long recognized this difference in trees and it would be strange if nature had not taken advantage of it also. If certain trees can still maintain themselves in the forest without producing a dense crowd of leaves, such as the silver maple for instance, they would have a decided advantage over a tree like the sugar maple which casts a much denser shade. A walk through any forest will show scores of examples of trees that live and produce seeds by virtue of the fact, not that they demand all available light, as their more vigorous neighbors do, but that by a compromise, by an almost diabolical cunning, their light demands, and of course their leaf exposure, have been cut down to a point where the tree can grow in a place impossible for trees that lack this ability. It is, of course, not a trick which any individual tree can perform at will. Rather is it a characteristic found in all individuals of certain kinds, where the comparative disadvantage of making less food and having less leaf exposure is more than overcome by the enormous advantage of being able to fight their way into a forest that would otherwise be impossible for them. We shall see, in the chapter on Plant Distribution, how this peculiar response to light has had effects of considerable significance upon forests, particularly after forest fires, lumbering, or other disturbance of the natural conditions. Trees in the forest, and the shrubs and herbs under them are not the quiet stately things about which the poets are so fond of singing. They are places, on the contrary, of intense warfare, and perhaps some of the greatest casualties occur in the battle for light.
Leaves, as being the most directly involved in the matter of utmost exposure to light, show the greatest amount of response to it, by their shape sometimes, by their position nearly always, and very often by the character of their leafstalks. In many herbs the first young leaves are relatively short-stalked, while as the plant grows upward the lower leaves are progressively longer stalked, which is a direct response to the fact that the upper leaves take their full share of light, leaving little or nothing for the lower ones. To avoid complete shading their leafstalks are often many times the length of their more fortunately placed neighbors above them. In those plants like the garden primrose or common weedy plantain, which bear all their leaves in a close cluster or rosette at the level of the ground, we see an almost fiendish cleverness in their earlier and later habits of growth. When the leaves first start, as they nearly always do among grasslike vegetation in which these plants usually have to fight for a chance of life, the leaves grow straight up, so that they may get above the level of the surrounding grass. Once there, and the precious light an assured fact, they gradually flatten out their leaves to form a rosette, of course cutting off the light from the grass about them and killing it just as certainly as though it were pulled up by the roots. Hundreds of different kinds of plants do this, apparently with the utmost cruelty to their inoffensive neighbors, with whom they start upon nearly equal terms in the race for life. If they began at once to spread out their rosette while it was still in its small spring state, the upward pointing grasses would smother it, and as if in anticipation of this the leaves grow up with the grass, only to flatten out when the proper time comes for them to show their true colors.
Light not only affects leaves in their habits of growth but it actually causes movements in some leaves which are as regular as clockwork. The best known cases are those in the pea family and wood sorrels, all of which bear compound leaves. During the day these leaflets are spread out in the ordinary way and catch the light, but at sundown, as though this were a quite useless exertion for the night, they fold up and the leaf “goes to sleep.” On cloudy days they partly fold up, as if in recognition of the fact that for their business of getting light it is an off day; but also if the sun comes out they hurriedly expand their leaflets. It is not yet certain whether these apparently intelligent movements of leaves in relation to light are of any real advantage to the plant as a whole or not. They are surely one of the most interesting things to watch and may be seen in locust trees and wood sorrel any night.
Just as we can have too much of a good thing, it is possible for plants to have too much direct sunlight. In open spaces, where the struggle for life centers not about the fight for light but over other matters, we find leaves actually protecting themselves against too much exposure, and by a variety of ingenious ways. The texture of the upper or lower side, the kind of hair growing on their surface, and the number and size of their pores, are the most usual ways of leaves arming themselves against an oversupply of the one thing that their neighbors in the cool forest fight to the death to obtain. There seems to be a fatality against which plants, like ourselves, are nearly helpless. Their attempts to overcome it, again like our own struggles against an apparently overmastering fate, develop those characteristics that insure survival to the fittest, death to the puny or unaccommodating.
We could hardly leave the subject of light and plants’ relation to it without mentioning, perhaps, the most remarkable case of adaptation to peculiar light conditions. All those aquatic plants that grow beneath the surface of the water need and get much less light than ordinary land plants. But from the island of Madagascar comes the lace leaf or water yam, which grows in quiet pools that are mostly in the depths of the tropical forest. Add to the dense shade cast upon the gloomy surface of such ponds the amount of light naturally lost in its passage through the water, and we get some notion of the singularly secluded home of this aquatic plant. What, now, is nature’s response to these peculiar conditions? How do the leaves of this well-shaded inhabitant of quiet pools behave? Their leaves are about a foot long and three or four inches wide, quite unnecessarily large for a submersed aquatic, but they consist wholly of veins. There is no “meat” to the leaf, none of that soft, green tissue so familiar in ordinary leaves. The conditions under which it is doomed to live almost seem as if it recognized the futility of having a broad expanse of the usually constituted leaf blade to expose to a light which is not there. It is significant that this skeletonized condition is permanent, the leaf functions much as ordinary aquatic leaves do, but its network of quite naked veins almost seems a mute protest against its fate. The delicate, lacelike “foliage” of this aquatic adds a touch of beauty to one of the most curious plants in the world.
If we could stretch an apparently impervious membrane, like the inner white skin just inside an eggshell, or a piece of parchment, and so form a wall through the middle of a glass box, and then pour into one of the compartments pure water and in the other a mixture of water and molasses, a very curious result would follow within a comparatively short period. We should find that presently there would be a gentle filtering of the water through the membrane toward the molasses water, and similar gentle current in the other direction. In other words, fluids of different density, if separated by a membrane, tend to equalize each other. This equalization may not be very rapid, and at first it will be more speedy from the less dense to the more dense, but eventually it will make the different fluids of a common density. This purely mechanical property of the equalization of fluids separated by a membrane is known as osmosis, and it is upon the possession of the equipment necessary for this that roots depend for getting food and water from the soil.
In our discussion of roots in Chapter I, we found that they end in very fine subdivisions, which are themselves split up into practically invisible root hairs. These root hairs are the only way that roots can absorb the food and water in the soil, and they are able to do this because they are provided with a membrane which permits osmosis to act between the solution inside the root hair and the water in the soil. The solution in the root hair is mostly a sugary liquid, some of that surplus sugar made in the leaves, and it is denser than the soil water, so there is apparently nothing to prevent an equalization of the liquids on different sides of the membrane. If this actually happened, as it would in the case of the simple experiment noted above, then roots would exchange a fairly rich sugary liquid for a much more watery one, and we should find that plants did not get their food from the soil, but really have it drained away from them by osmosis. But nature has a cunning device for stopping such robbery, which is prevented by the membranes of root hairs being only permeable to the extent of letting water in, not permeable enough to allow sugar to escape. As we have seen, osmosis is a purely mechanical process which, if left to operate without interference, would not aid but injure the plant. Surely, nothing with which plants are provided is so important to them as this delicate membrane of the root hairs which, while allowing osmosis to act in a one-sided fashion, preserves to the plant the sugary liquid that alone makes the absorption of soil water possible.
As root hairs are very much alive and work constantly, they must be provided with air, without which no living thing can exist. And here, again, it seems as though nature, with almost uncanny foresight, had deliberately planned for this requirement of roots. And, in this case, not by interfering with a physical process by an adjustment of plant structure, but by the arrangement of soil particles and the way in which water is found in all soils. Soil particles, even in the most compact clay, do not fill all the space occupied by the soil as a whole. There are tiny air spaces all through the soil, which insures a constant supply of fresh air. That is one reason why gardens are cultivated, to see to it that plenty of fresh air is allowed to permeate the soil. Around the finest soil particles there is always an almost incredibly thin film of water, which is renewed as soon as it is lost by its absorption by the root hairs or by evaporation. This renewal of the water film is itself a mechanical process, called capillarity, best illustrated by putting a few drops of water on a plate and placing on them a lump of sugar. The water will spread all through the lump of sugar in a few seconds and the capillarity that forces it up through the lump is the same as sees to it that the tiny film of water surrounding the finest soil particles is constantly renewed from the lower levels of the soil.
Little do we dream, as we walk over the commonest weed, that buried at its roots are these delicate arrangements for securing food and water. Osmosis allowed to act so that the “exchange” of liquids is all to the advantage of the plant, capillarity providing a constant water supply, and the very piling together of the soil so contrived that the life-giving air filters all through it—does it not seem as if all this were, if not a deliberate plan, certainly a more perfect one than mere man could have devised?
If you will turn back for a moment to the beginning of the description of how plants get their food, you will find that in osmosis the weaker liquid tends to permeate the denser one more rapidly than the denser one does the weaker. As we have just seen, the sugary liquid in the root hairs is denser than the soil water outside, and, furthermore, none of it is allowed to escape. This comparatively greedy process of taking everything and giving nothing results in a constant flow of soil water into the root hairs. When the flow of liquids in osmosis is not at once equalized, a gentle pressure is brought to bear to make them so. This is what is called osmotic pressure, and it is this pressure that forces the absorbed liquid through the roots and part way up the trunk of even the tallest trees. While we have just said it is a gentle pressure, that is true only in the case where the osmosis has free play, and the pressure is stopped with the perfect mixing of the two liquids. But what if they can never mix? What may not the accumulated osmotic pressure amount to in such a one-sided process as goes on in root hairs with everything coming in and nothing going out. Cut-off stems, with a pressure gauge attached to them, indicate that in some plants the pressure is from 60 up to 170 pounds!
Another result of this pressure is that it keeps leaves and the fleshy stems of plants in their ordinary position. The actual solid part of nearly all leaves is scarcely 5 per cent of their bulk and all the rest is water. The constant pressure of this water from the roots is sufficient to keep leaves comparatively stiff and rigid, how stiff is quickly realized if the pressure stops and the leaf wilts or withers. Sometimes this osmotic pressure, particularly during rainy weather, becomes so great as to cause injury to the plant, the splitting of tomatoes and occasionally of plums, being due to it. This osmotic pressure, together with the extra pull given by the leaves, is sufficient to account for the rise of water to the tops of the tallest trees. The tallest trees in the world are certain kinds of blue gum in Australia which frequently reach a height exceeding 300 feet. What the combined osmotic pressure and leaf pull must be to carry such a heavy thing as water to such a great height is easier to imagine than to calculate.
The root hairs, then, by the process already described, absorb the water from the soil, but plants can no more live on water alone than we can. As we have seen, the membrane in the root hairs cannot allow the passage of even the tiniest particle of solid matter. In fact the root hair itself is so small that it can only be seen through the microscope, and of course the membrane is smaller still. Plant foods, then, can never be solids, but must always be such materials as can be dissolved in water. The chief of these are chemical substances, such as lime, potash, nitrogen, magnesium, phosphorus, sulphur, and iron. Hydrogen is also necessary, but as this makes up half the composition of water, there is a permanent supply of that provided by the soil water. These things make up the great part of plant foods taken in through the roots, and it is from these that leaves, by a process you already understand in its essential details, manufacture sugar and starch.
But neither starch nor sugar, important as they are to the plant, and absolutely necessary as they are to us, are the only things made by plants. Leaves may well be called factories, but plants are themselves the most wonderful chemical laboratories, beside which any built by man are as play-things. For plants, by processes too complicated to be explained here, work over their accumulation of starch and sugar, recombine some of their constituents, and store up in various parts of the plant the results, which are often such food ingredients as protein. This is the really essential food substance in wheat, as it is in eggs and meat. No chemist has ever succeeded in making a single scrap of it, yet it is such an everyday occurrence in practically all plants that it, with starch and sugar, forms the great food supply of the world. Not protein alone, but all the amazing plant products like the oils from the olive and the resin from pine, rubber, the drugs of plant origin, even tobacco—all these and hundreds of others are made by plants from those few simple foods absorbed through the roots, literally pumped up to the leaves and there, under the magic of sunlight, combined and recombined, worked over and changed utterly in their make-up. Nothing could be more perfect than the marshaling of forces and contrivances to secure the result; let there be even the least bungling, and for us the world would cease to be worth fighting for.
Nor does the work of plants stop here. If it did, they would be not unlike a commission merchant who had gathered from the four corners of the earth a supply of eggs only to find he could not or more likely would not sell them all at once, and yet had failed to provide himself with proper storage. Plants, too, have times in their life when adequate storage is necessary for them. So true is this that unless there is food enough stored in seeds to give a start to seedlings before their own roots begin to work, they would die almost at once. In seeds and in many nearly dormant parts of plants these foods are stored away for future use. The tubers of potatoes and all our root crops, like beet and parsnip, are common examples of this. Even the manufacture of wood in the trunks of trees is a storage appliance on the part of the plant, for wood is just as much one of the food products of a plant as wheat or rice.
With such a beautifully perfected mechanism for getting food it might seem as though all plants would be satisfied to lead that life of independence for which they are so splendidly equipped. Some of them, however, are like men in one respect: there seems to be no end to the chase after getting something for nothing. Those that stand on their own roots, get their food honestly, and take nothing for which they do not make prodigal returns, make up the great bulk of the vegetation of the earth. Their independence has dubbed them with the title

Indian Pipe. (Monotropa uniflora). A saprophytic plant inhabiting rich woods in eastern North America. (Courtesy of Brooklyn Botanic Garden.)

The Partridge Berry (Mitchella repens), a trailing vine of northern forests. (Courtesy of Brooklyn Botanic Garden.)

Rafflesia. One of the Strangest Products of the Rain Forest. It consists only of a giant flower, the largest in the world, which is attached directly to the roots or stems of relatives of the grape, upon which it is parasitic. (After Kerner and Oliver. Courtesy of Brooklyn Botanic Garden.)
autophytes, literally solitary or self-providing plants, and this thrifty mode of life is called autophytic. But a few kinds of plants, actually many millions of individuals, have more devious ways of getting their food and provide strong contrast to their sturdier associates.
These baser modes of life appear to have been rather insidiously developed, as though there had been some hesitation at even the smallest departure from the normal. Of course we must not forget that plants, while living things, are never reasoning ones, and that good and evil and all other qualities that are ascribed to plants are perfectly foreign to them. Throughout this book, and in many others, the habits of plants are spoken of as base, for instance, or good. What is actually the fact is that nature works in truly marvelous ways, and to our reasoning faculties these adjustments seem clothed with attributes they do not really possess. But the description of them in the terms of our everyday speech, the translation of their behavior into the current conceptions of mankind, does so fix them in our minds that they cease to be “just plants,” and we no longer put their habits in the category of those interesting things that nearly everyone forgets.
One of the first signs of departure from the usual methods of getting food is the association of certain minute organisms at the roots upon which plants, otherwise autophytic, depend for aid in securing nourishment. This characteristic is fairly common, notably in all the plants of the pea family, such as peas, beans, locust trees, vetch, clover, and hundreds of others. If the roots of any of these be examined, it will be seen that attached to the smaller divisions of them are small tubercles from the size of a pinhead to a pea, depending on the kind. These tubercles or galls are caused by and infested with bacteria, the smallest of all plants. The bacteria have the extraordinary power of changing nitrogen into nitrates, which is the only form in which nitrogen can be absorbed by roots. Not only do they accomplish this, but excess nitrogen is stored in the roots by the same agency. It is this fact that has resulted in the planting of vetch and kindred plants for soil enrichment, as each year there is a residue of nitrogen left in their roots and by so much they add plant food to the soil. For hundreds of years farmers have done this, but only quite recently have we known why they did so. The occurrence of bacteria or microbes at the roots of plants is much more common than was formerly supposed to be the case, and many other plants than those of the pea family depend, at least in part, upon them in getting food from the soil. While not wholly autophytic, such plants do make some return for what they gain, as some of them at least pay dividends in extra nitrogen, and all of them provide opportunity for the bacteria to live. The latter play an important part in populating the soil, which is not the comparatively sterile thing it appears to be. Actually it is infested with organisms that play a mighty, if rather inconspicuous, part in the work of preparing the soil for plant growth. These organisms are so minute and the chemical nature of their work is so complicated that merely to mention their existence must suffice here. This close association of certain roots and bacteria, which, as we have seen, is of mutual advantage, is known as symbiosis. It is really only a kind of exchange, not unlike the storybook community that helped out by taking in each other’s washing. Unlike that community the association between the two works to the actual advantage of both, but the process is undeniably a step away from those wholly autophytic plants which live free and independent of such aid.
A much more gruesome habit of certain plants is their reliance for food only upon the dead. In the Indian pipe, some kinds of shinleaf, and in many other plants their roots and root hairs are changed or often nearly lacking, and we find them growing only on the dead bodies of other plants. One peculiarly repulsive characteristic of such plants is that they secrete at their roots a substance that hastens the decay of the dead, and, as if this were not rapid enough, there are associated with them certain kinds of minute fungus organisms that also speed up decomposition. Plants with this charming mode of life are known as saprophytes, literally sapros, rotten, and phytes, plants. “Rotten plants” they may be in their mode of life, but the pearly white stems and flowers of the Indian pipe have a certain ghostly charm, an almost statuesque beauty among the normal greenery of the gloomy dark woods in which they always grow. It is not without significance that Indian pipe bears no leaves, has none or almost none of the life-giving green coloring matter which we have seen to be the almost priceless possession of plants which lead a different, and perhaps a better life. The great bulk of saprophytes bear no leaves, and some only partially wedded to the habit appear to be midway between bearing normal green leaves and bearing none, or much reduced ones that are quite unlike the busy factories we know normal green leaves to be. Plants with this method of getting their food, must of course grow in places where dead and decaying vegetation is plentiful, and often as such soil is turned up there may be noted a peculiar dank odor, suggestive not only of its origin, but of the fact that these “rotten plants” make their home in it. Some of our most beautiful orchids grow in this fashion, but even there, in spite of flowers that for beauty of form are without rivals, the plants have no green coloring matter in their leaves, which are often reduced or even wanting altogether.
It might almost seem as if demoralization, so far as food habits are concerned, had reached its lowest point in these plants that literally rob the dead, but there are still lower depths to which certain plants have been reduced. This consists of robbing the living, and such plants are called parasites, a word perfectly familiar in other connections. Parasitic plants have no roots, but attach themselves to the roots of other plants, somewhat generously called hosts, from which they derive their food. The best known case is the common Christmas mistletoe, and the dodder (Figure 68), but there are hundreds of others. Nothing in all the realm of plant life so perfectly fits the action to the word as plants of this type, flourishing when the host flourishes, dying when it dies. Producing flowers and seeds, and often, by an irony of fate, perfectly green leaves, they are nevertheless the most debased of all plants in their mode of life.
These successive steps in the degradation of food habits, are not always the clean-cut things they might be inferred to be from the foregoing. There are many intermediate stages; it may even prove to be the case that some plants are wholly autophytic at certain stages of their life, and slip partially into more devious practices at other stages. The whole affair is not yet thoroughly understood and may well be the result of competition, as it is quite conceivable that if the getting of food in normal ways became difficult or impossible plants may have had to resort to other methods.

Some one has said that one day without water would make men liars, in two days they become thieves, and after the third or fourth day they would kill to get water. In the Army Records at Washington is a report of one of our expeditions, which in chasing Indians got lost in a desert, and in which the soldiers fought among themselves for even the most repulsive liquids. It hardly needs these gruesome examples, however, to confirm what everyone who has ever been mildly thirsty knows, that water is an essential for all animals, and that to be without it is to suffer torture. Air of the proper kind is just as important, and because its absence or impurity causes more sudden agony and a quicker death, the need of it is that much more acute. Plants rely even more upon these two essentials of life, and in getting them they behave in ways just as ruthless as do men who are suddenly deprived of either of them.
As we have already seen in “How Plants Get Their Food and Water from the Soil,” the water is the carrier of the food elements from the soil, but water as such does much more for the plant than act as a carrier. Osmotic pressure, a never-ending pump, keeps sending up a steady stream of water to the limits of its power. In everything except trees it seems fairly certain that this pressure is sufficient to drive water into the remotest leaves. It finally reaches these tiny rooms in the leaf about which we read in the account of Leaves as Factories. And just here a very curious thing happens. Each room is, as we have seen, a very busy place, crowded with all the necessary equipment to make sugar, and yet there is still room for water which is just as necessary as the other fittings; in fact so necessary is it that the whole interior of the room is bathed in water. This irrigation system works so well that the walls of the room literally bulge with the pressure of the water in them. If they did not—a condition known as turgor—the plant would at once wilt, and if no new supply came it would wither and die.
But water cannot stay in this condition of pressure and stagnation for even a brief period. That would be as if a leaf were like a toy balloon which, after inflation, had the entrance pinched and so remained inflated. And while we have all along spoken of factories for making sugar, and pressure pumps for forcing up food and water, it must never be forgotten that this marvelously adjusted mechanism is a living thing. Constantly growing, even producing their own means of falling in the autumn, leaves must be thought of as living machines, just as we are still more highly developed machines. In other words the accumulated water in the cells of the leaf must be removed, after it has served its use, and replaced by fresh supplies. The removal is carried on by its evaporation into the halls, or, in the more precise terms of our account of leaves as factories, into the intercellular spaces. It will be recalled that these are connected with the outside air through the pores or stoma. When the air outside is hot and dry it might easily suck out by evaporation all the water vapor in these intercellular spaces and wilting follow at once. This would actually happen if the guard cells, already mentioned, were not constantly on the job. They control the size of the opening just as certainly as a steam valve does, and maintain, with a few exceptions, just the proper amount of water loss not only to maintain turgor, but to see to it that transpiration, as this process is called, goes on rapidly enough to insure fresh supplies of water being sent to the leaf. The opening and closing of the stoma by the guard cells is a nicely balanced operation dependent upon root pressure, turgor, and atmospheric conditions. Guard cells have, because of this, been much studied in spite of the fact of their microscopic size. We now know that they allow greater openings during the night and reduce them during the day. When we reflect that the constant removal of water in the leaf, both as such, and as the only carrier of food supplies from the roots, depends in such large measure upon the functioning of these guard cells, then we come to some realization of their importance to the plant.
They do not always work unaided, for in many places the transpiration, even with their best efforts, would exceed the rise of water in the plant and death must follow if such a condition exists for long. This may be the case in certain bog plants, where, even with their roots in the water, they actually are in danger of drying out because the composition of bog water makes it partially unfit for most plants. And, again, in very open dry or windy places, such as deserts or the mountain tops above timber line, the actual supply of water may be insufficient. Many plants growing in such places have their leaves, particularly the under surfaces of them, clothed with various kinds of hairs. These may be quite velvety or cottony, but in any event, either by their texture or their color, they tend to reduce transpiration. An extreme case is a desert plant from Arizona where the whole leaf surface is covered with an ashy gray velvety coating, which, of course, absorbs less heat than a normal green leaf, and in addition there are much fewer pores through which transpiration could be carried on. In ever so many leaves nature has provided them with a thick coating of hairs in early spring, which they lose later in the summer. Shrubs and herbs, especially those that start earlier than the trees under which they grow, very often may be found with a dense woolly or silky covering in early spring. As the shade becomes denser and the need of the protection less, the wool or silk is shed, sometimes completely. Some of the most conspicuous cases of this are certain kinds of our common shadbush, which in April are covered with a beautiful grayish-white silky coat, but by August are practically the ordinary green color of other leaves. The hairy covering of leaves is well worth observation, as it may hide not a few facts about transpiration and, in some leaves, has had much to do with their preservation from grazing animals. Some, like the common mullein, are never touched, and may be found standing like sentinels in fields otherwise cropped short.
In many leaves there is conflict between those forces that result in the leaf getting the utmost possible exposure to light and those that prevent too rapid transpiration. On the one hand there is the absolute necessity for light, on the other the ever-present danger that the response of leaves to this necessity will result in a transpiration rate too rapid to be held in check by the guard cells. The compromise between these two forces, each pulling in opposite directions, gives to some leaves a series of movements that are among the most interesting things in nature. One of the most marked examples is the common wild lettuce, a weedy plant of our roadsides introduced from Europe. In bright sunlight the leaves are turned so that the edge of the blade faces upward, and the surface is thus protected from the direct rays of the sun, but during cloudy weather or in the shade the leaves turn into the ordinary position of most foliage leaves. It is difficult to avoid the inference that photosynthesis, which, as we have seen is an absolute necessity to the leaf, is in the wild lettuce retarded by transpiration, to avoid the too rapid rate of which the leaf is turned on edge. In this plant the leaf base, as though to be ready for whatever change transpiration or photosynthesis may demand, is so attached to the stem that such changes are made with the least possible delay or wrenching. In one of the many kinds of blue gum trees of Australia all the leaves turn one way in the light, and another in shade or on cloudy days. Ever so many plants have partial movements of their leaves, a good many of which are in response to these opposing demands, one pulling the leaf into the greatest possible light, the other holding it away from that condition. There are other movements of leaves, of parts of the flower, or even of the whole plant that are not so certainly the result of the conflict between light requirements and the necessity of conserving water supply. They will be considered presently.
While most plants are well provided with methods of losing water, so well provided in fact that in very hot or very long dry periods it is a common sight to see many plants literally panting for more water, there are some apparently more cautious individuals, who reverse this process. All throughout tropical America hundreds of relatives of the pineapple have their leaves so formed and arranged that they catch and hold considerable quantities of water. In one kind, called Hohenbergia, the long leaves are joined together toward their base into a water-tight funnel, which will hold a quart or two of water over a period of drought. In Africa the extraordinary traveler’s-tree, a giant herb growing twenty to thirty feet tall, has the overlapping leaf bases so arranged that they hold many gallons of water. And we have already seen how the giant cactus of our own Southwest will hold 125 gallons. The most remarkable case is the Ibervillea from the deserts of Arizona. In riding over this country one may find objects that look not unlike a burned pudding, about two feet in diameter and nearly as high. From the center comes a delicate stalk with the finest feathery foliage and tiny flowers. Of roots there appear to be almost none, and these curious objects, which are very hard and woody, might almost be taken for stones. But they are actually plants not distantly related to squash and pumpkin, and one of them collected years ago and brought into a museum behaved in quite the most thrifty fashion of any plant yet discovered. It was carefully cleaned and put in a museum case and locked up as a curiosity for the wondering public to gaze at. But suddenly, almost miraculously, it sent out its delicate growth which grew its appointed time and then withered. Imagine the astonishment of the curators of this museum to find it doing the same thing the next year, and the next. Finally after putting forth its shoot for five years it actually died and is now a peaceful museum specimen. No other such case of water storage is known, but thousands of plants have this remarkable ability to a less degree, all in response to conditions that would mean destruction to plants not so providently equipped.
This conservation of water on such a great scale offers striking contrast to the truly prodigal habits of certain plants that actually drip water, so charged are they with this precious liquid, and so little stress do their conditions of life put them under in this respect. Where water is plentiful and turgor maintained almost to the bursting point, evaporation in a moist or chilly atmosphere does not suck out water vapor fast enough. Sometimes, around the edges of the leaves of the common garden nasturtium, drops of water may be found, literally forced out as drops, rather than transpired as water vapor. This happens to a considerable number of plants, during the night when transpiration is laggard, and such drops are usually mistaken for dew. The latter is actually the condensation of moisture in the air upon the leaves of plants which cool down more rapidly than the air, and seldom due to the forcing out of drops of water from leaves, although in rare cases it may be. In tropical forests, where the humidity is very heavy and water supply from the roots copious, certain leaves leak water so fast and are so constructed that this excess is prevented from accumulating on the leaf. The pipal tree of India has long drip tips to its leaves that conduct the excess water from the blade to the end of the slender tip where it drips off. The advantage of these dripping points is obvious, for in regions so humid that water is forced out of the leaf, the coating of the leaf with this extra moisture would by that much retard transpiration. Dripping points, which in less exaggerated forms than in the pipal tree are common in many parts of the world, are thus of decided advantage.
Whether it be desirous to retain water or to lose it by gradual evaporation, or expel an excess of it, each species of plant has developed the apparatus to best preserve its individual life. While only the barest outline of these adjustments to the water requirements of plants has been given here, the details form an almost dramatic picture of struggle of the different kinds of plants for survival. The extremes are the desert plants on the one hand and those of the rain forests in the tropics on the other. The chapter on Plant Distribution will show how important these water requirements of plants have been in determining what grows on the earth to-day.
With carbon dioxide going in, oxygen, water vapor and, as we have seen, even liquid water coming out of the stoma of leaves, it might be surmised that these busy little pores and their guard cells had done work enough for the plant. And yet there is still one more act to play and the stoma have much to do with it. For this process of photosynthesis and the closely related one of supplying food and water to the leaf cannot go on without respiration, which is quite another thing. In plants respiration or breathing has no more to do with digestion than it does in man. Digestion in man is not unlike photosynthesis in plants, except that plants make food in the process while men destroy it. But plants must breathe just as we do, and, as we need oxygen to renew our vital processes, so do they. While respiration is a necessary part of plant activity it is not such an important part as photosynthesis, for which it is often mistaken. The thing to fix in our minds is that photosynthesis makes food, uses the sun’s energy and releases oxygen in the process, while respiration uses oxygen and might almost be likened to the oil of a machine—necessary but producing nothing.
In walking through the quiet cathedrallike stillness of a deep forest or over the fields and moors, perhaps our chief thought is how restful the scene is, and what a contrast the quiet, patient plants make to the darting insects or flitting birds that our walk disturbs. We found at the beginning of this book that ability to get about is one of the main differences between animals and plants. Like so many first thoughts, this is, however, only a half truth, for while most plants, seemingly by a kind of fatality, are anchored forever to the place of their birth, many of them do move certain parts of themselves and that quite regularly. While some of these movements have already been hinted at as a possible response to transpiration or too intense light, there are others where the advantage to the plant, if any, has yet to be demonstrated. These other movements, perhaps because their cause has never been discovered, seem the more mysterious as they certainly are more weird and interesting than almost any other of the curious things that plants do.
Perhaps the most difficult thing in the world is to keep an active growing child perfectly still for more than a few moments at a time. There seems to be some impelling force that makes young growing things in a constant state of restlessness, and it is perhaps not so extraordinary, after all, that practically all young plants are restless in the sense that they are never quite still. And, like many grown-up people who do not know what repose in their waking moments really means, there are a goodly number of plants that are restless until the day they die.
Charles Darwin, perhaps the greatest man that the last century produced, wrote a book in two volumes on these restless plants, and proved by a series of experiments illustrated by charts which the plants themselves drew for him, that there were perhaps no plants that do not move at least some part of themselves during the early stages of their career. While he never could explain the cause of these movements he left in that book an imperishable record of the amount and direction of these mysterious movements, which are almost to be likened to the growing pains of young children.
The tips or growing shoots of many plants will point in one direction in the early morning, a different way at noon and still a different one by nightfall. Hundreds of totally unrelated plants seem to have this habit of moving their tips through a definite cycle during each day and this restlessness does not appear to be of the slightest use to them. It cannot be response to the moving of the sun through the sky, for often the movement may be away from the direct sunshine, and sometimes the motion goes on in the dark, as experiments have proved.
It is hard to see the movement of the whole upper part of a plant, although it is well known that they do move in many cases. But in the tendrils the movement is often easy to observe and even to induce. Some of these slender aids to climbing plants, if they happen to be swinging freely in the air, do actually make slow circular movements, that even if they were designed for the purpose could not more perfectly accomplish their obvious intent, which is to catch the nearest favorable support. These circular movements are to the left in the hop, honeysuckle and many other plants, to the right in the climbing beans, morning-glory and some others. When the tendril reaches a support it almost immediately turns about it, in the same direction as its free movements through the air have been. It is thus this apparently aimless swinging of tendrils through space that determines whether the vine is going to twine to the right or left. The speed with which a tendril will take its first turns about a support is so comparatively rapid that, once the support is caught there is scarcely a chance of the vine being torn away by the wind or other agency as would surely happen if tendril movements were the leisurely things that some folks think they are. In the case of one Passion-flower vine, which are gorgeous climbers mostly from the tropics, the tendril made a complete turn in two minutes after it first touched a possible support. And there is a quite noticeable movement in thirty seconds if the tip of the tendril be ever so lightly touched. Teasing tendrils to see how much or how fast they will coil has resulted in some extraordinary cases of the “comeback” of some of them. Darwin irritated a tendril for a few moments and induced a partial coiling which straightened out when the object causing it was withdrawn. To see how long the plant would stand this sort of thing and still not be literally tired of coiling he succeeded in making the plant partially coil, and by withdrawing the incentive uncoil again, over twenty times in fifty-four hours. An impulse to coil of such persistence as this naturally results in vines forming the impenetrable thickets they do in many forests. It emphasizes how restless are the growing points of these climbers, and serves as a striking illustration of those gradual movements of many other plants that seem to have some relation to growth, but in a way not yet understood. For while it is an obvious advantage for the vine to swing its tendrils through the air this advantage has not yet been proved the cause of the swinging. In fact if all possible supports are removed the tendril will often coil anyway, a perfectly futile proceeding, that looks almost like disgust.
This general restlessness, which by the imaginative has been thought of as a mild protest by plants at their otherwise fixed position, is not so spectacular as that of certain other plants, notably the poplars. A flattened instead of a round leafstalk makes the leaves of these trees flutter in the lightest air and in a gale the tree is a mass of animated foliage. No use has ever been found for this curious habit and it is not certain that it is of the least advantage to the tree. If anything, the constant movement may have the decided disadvantage of increasing transpiration.
In our common wood sorrel the leaflets on cloudy days or during the night regularly “go to sleep.” That is, they are folded at such times, rather than spread out in the ordinary way. These sleep movements may have something to do with transpiration, but whether or not this is true they are very regular and in certain plants the habit is remarkably and rather mysteriously uniform. Why, for instance, do the leaflets of these wood sorrels, the beans, lupine, locust tree and licorice plant, always fold downward while the clovers, vetch, peas, and bird’s-foot trefoil are always folded upward? Such movements and their direction are among the unsolved problems of botany, and merely to know of them or observe them leads us nowhere as to their true inwardness.
But quite apart from these merely restless plants, and there are thousands of different kinds which are known to move slightly, at least during their young stages, are a few more decidedly active ones that are seemingly irritable. At least they show peculiar movements if touched, and at night. One of the best known is the sensitive plant from tropical America. Its twice compound leaf is composed of many tiny leaflets which upon the slightest touch close up and apparently wither on their stalk at once. In five seconds after the leaf is touched it will appear like a wilted wreck. If the jar is sharp enough the whole plant will droop, and the response to a sudden jar is almost electrically quick in its action. And yet all this sudden wilting, actually caused by a quick loss of turgor, is slowly repaired and the plant carries on quite normally again until another shock renews its irritable response. This plant does the same thing gradually during the night, except that the leaflets recover their normal position only with the rise of the sun.
From India comes the most remarkable of all plants so far as movements are concerned. For in the telegraph plant the movements are so regular and long continued that irritability might almost be said to be continuous. The plant is a low shrub or herb with compound leaves, and the terminal leaflet, which is much larger than its neighbors on either side of their common stalk, performs a motion that describes with its tip an irregular oval or ellipse. But the movement is not steady; it goes by a series of slight but perfectly distinct jerks. It takes about two minutes for the leaf to complete its cycle, and it is this jerky movement that has given the plant its name. During the night its leaflets stop this apparently quite useless performance, the cause of which is quite unknown. It is often grown in greenhouse collections where its strange movements may be seen on any sunny day.
Many other cases of the restlessness or irritability of plants could be given, and nothing has been said here of the curious movements of some insectivorous plants as they have already been mentioned. The very considerable movements of certain flower and fruit organs will also be considered elsewhere.
It cannot have escaped the thoughtful reader that all of this chapter on plant behavior has dealt with those functions of plants in which roots, stems, or leaves play the chief part. These purely vegetative actions of plants, what might almost be called their bread and butter activities, would never lead to perpetuating their kind. For while all of these functions are necessary, except certain apparently wayward movements which still remain unexplained, they are in a sense only the preparation for an infinitely more important act, the reproduction of their kind. What the poetic have called the love of the flowers, or in more prosaic but perhaps more truthful words the fertilization, pregnancy, and birth of the new race, will be considered in a separate chapter. No other act of the plant world is so interesting as the mechanism of reproduction, the almost endless devices for securing it, and the ingenuity of nature in seeing to it that there are no flukes.
THERE is perhaps no device of nature that more perfectly accomplishes its purpose than the one with which all living things are endowed—the instinct for the renewal of life. In man the dawn of the mating instinct has ever been the theme of poets, and some of its manifestations are the despair of ascetics. Through it some of the noblest of man’s emotions have arisen, and because of its perversion our daily newspapers chronicle the basest and most sordid tragedies.
But whether noble or ignoble, this instinct for mating is, in its simplest terms, only a provision of nature that all life contains within itself the means of renewing life. Without this, life, so far as we know it, would end with the present generation. Perhaps our understanding of this decree of an all-wise nature to increase and multiply will be heightened by looking at it not only from its familiar manifestations in man, but more broadly. Seen from this broader viewpoint, it is the inherent legacy of all living things from the dawn of life on the earth down to the present. Even the simplest one-celled organisms have the faculty of increasing. In all plants, both the flowerless ones and those producing flowers, the process is carried to a perfection almost unbelievable in its intricacy and in provisions against its failure. From the matings of flowers much may be gleaned; even man himself can learn from them the capacity for sacrifice, the sinking of individual aims and pleasures in the greater scheme of conforming to that necessity for renewal of the race upon which all progress must be based.
The equipment which different flowers have developed for this purpose, their almost uncanny devices to make certain that only the distant and foreign male can ever impregnate the female, the enormous wastage of both unfertilized females and males that will never become fathers, and the overwhelming effectiveness of it all, in spite of this prodigality—these manifestations of the production of young in the plant world will take up the rest of this chapter. All the first part will tell of this process in flowering plants, while the second shows how flowerless plants accomplish the same end in more secret ways. Finally, in a brief third part, we shall see how, without mating of the sexes, nature has still one other way to see to it that there is a constant supply of young.
We have already made clear that all plants are divided upon the basis of whether they bear flowers and their mating goes on before the world, or whether they bear none and the process is accomplished in more secret ways. Because flowers are so much better known, and it is simpler to see how the act is consummated in them than in the cryptogamous plants, we shall first consider the phanerogams or flowering plants, and in the second section of this chapter the cryptogams or flowerless plants.
In the first chapter, under the section devoted to flowers, we found that the stamens are the male and the pistils the female organs of reproduction. As the period for mating draws near there is developed in the anther, which is the enlarged tip of the stamen, a fine, usually yellow, powder known as pollen. This matures in the anther, and when ripe is discharged from tiny pores.
Pollen is made up of individual pollen grains, which are very often stuck together so that we see only the mass, not the individual pollen grain. Sometimes the pollen is not sticky, as in the case of pine trees or in the ragweed—a fertile cause of hay fever. In these, and hundreds of other plants, the wind will blow great clouds of pollen through the air. When we stop to consider that a single, or at most a very few pollen grains are all that are necessary—in fact, are all that can be of real service—the enormous wastage of the male fertilizing substance, in order that mating be secured, gives us some idea of how prodigal is nature in this supreme function.
The pistil, or female organ of reproduction, is more cautious in the expenditure of its resources. As we have seen, it is composed of a swollen base, the ovary, a slender shank, the style, and a swollen or branched tip, the stigma. In some plants the ovary is divided into several compartments or cells, each with one or more ovules, which are only immature or unfertilized seeds, often very tiny, but usually quite easily seen if the ovary is cut open. It is the entrance of the pollen grain into this ovule that consummates the act of fertilization. As the ovule is carefully secreted within the ovary of the flower, and as the male fertilizing stuff or pollen is found only on the anther, it is obvious that some method of bringing the two together must be provided for.
In some plants this is accomplished by the anthers being just above the stigma, and when the pollen is ripe and the ovule ready, the stigma is found to be covered with a sticky substance. As the falling pollen grains touch the stigma, they are caught in this sticky substance just as surely as flies are caught once they touch a fly paper. But just here one of the most wonderful processes of nature begins. The pollen grain begins, slowly at first, to grow, and in the act it penetrates the outer coat of the stigma with a minute pollen tube. This slender threadlike tube, carrying with it the male germ, grows straight down through the stigma, into the narrowed style, and through this to the ovule. Once the pollen is caught on the stigma, nothing is so sure of fulfillment as that this male fertilizing stuff will ultimately reach the ovule. For the hitherto virgin ovule this impregnation starts a new phase in life. It means the beginning of the end, but in the process fruit and seed will be developed, and the young bride, already a mother, has triumphantly accomplished that for which she exists.
If fertilization of all flowers were as simple as this, there would be no need of what follows, but actually in surprisingly few plants are the stamens and pistils so arranged, the ripening of the pollen and readiness of the ovule for impregnation so timed that the act can be accomplished in such direct fashion. For it is quite obvious that in flowers in which the whole drama of mating goes on within the petals, without the interference or help of any outside agency, the result will be a crop of young who know no other characters than those of the parents, and have nothing to look forward to but a closely inbreeding progeny, very little, if at all different from themselves. In other words, such plants are pure bred, they lack the usually obvious virility that comes from crossing the male of one plant with the female of another. There are so many devices to prevent self-fertilization in flowers, so marvelous are the contrivances to see to it that only cross-fertilization can be effective, and, finally, the experience of breeders that strength and virility often or usually result from impregnating the ovules of one plant with the pollen of another, that we are forced to the conclusion that absolute purity in the sexual relations of flowers is rare indeed. It occurs, without peradventure of a doubt, only in those flowers whose petals never open and where fertilization is consummated, if not in private, at any rate without external help. In many violets the showy violet blossoms are often nearly infertile, while down near the ground are inconspicuous flowers which never open, but within which fertilization is so successful that the crop of seeds is far more plentiful than in the more showy ones that most people think are the only flowers ever borne by violets. These flowers that never open, or at any rate open so slightly that their sexual processes are modestly completed without intrusion, are known as cleistogamous flowers (Figure 69). They have been found in a few plants, but overwhelmingly the greater number of flowers not only do, but must, rely on some outside agency to insure fertilization.
Certain structural features of flowers have been so developed that fertilization of the ovary by the pollen of the same flower is impossible. The commonest case is in those flowers where the stamens are shorter than the pistils, as they always are in the common snowdrop, hyacinth, the sassafras tree, and in hundreds of others. There can be no consummation of the reproductive process in such flowers without some outside aid. More futile still without this aid are those flowers where the stamens are well above the pistils, but the time of maturing in both differs by a few days or even hours. Nothing could be more helpless than the pistil under these circumstances, for if its instinct for maternity were ever so strong, it would be doomed to barren sterility by the premature development of the males. Sometimes, too, the female is prematurely ripe for impregnation, and the stamens lag behind a day or two. Her time passes and with it her only chance of fertilization—by her own haste she has rendered impotent the now useless pollen which appears doomed to fall aimlessly upon the unreceptive stigma.

But, perhaps, the most hopeless of all is the well-known partridge berry, whose red berries are common in the woods during August and September. This seems as though it fought off any chance of securing a mate by a flower structure and behavior that would certainly so result if some way out of the difficulty were not at hand. The partridge berry bears two kinds of flowers that outwardly look much alike, but whose sexual organs differ in this way: in some flowers the stamens are all shorter than the pistil, and in others the pistil is much outtopped by the stamens. The extraordinary feature of it is not so much this structural difference, however, but the fact that pollen from the short-stamened flower is useful only to its neighboring short-styled relative, while the pollen from this long-stamened but short-styled neighbor is nearly useless where it is found and really useful only on the long-styled plant. By this device, but again only with outside aid, this plant does not prevent maternity, but increases its chances of being fruitful, for, as we have already seen, cross-fertilization appears to be the rule rather than the exception, and the partridge berry not only needs it, but can exist only when its offspring are the result of such crosses.
In all those plants that bear the different sexes in different flowers on the same plant, as in the hickory, or even on different plants, as in the willow, there must, of course, be some method arranged for cross-fertilization or they would promptly die out. So general is this cross-fertilization, so much a part of the economy of nature does it appear to be, that we can only think that there must be in the production of this vast horde of the cross-fertilized some advantage. Besides securing greater virility, which almost certainly results from this promiscuity, greater variability is promoted. If virility is the result, the price paid for it is tremendous, for the hindrances to self-fertilization are so many and so effective that most flowers would inevitably die as perfectly pure but ineffectual virgins if that fatality were not prevented. How they are saved from such a sterile fate, how they finally secure a mate by devices that outshine the most bewitching tricks of the daughters of Eve, is one of the most fascinating stories in all the history of the plant world.
For, of course, flowers do secure a mate, and they are aided in this enterprise by the most formidable array of helpers, one might almost call them conspirators. The chief of these are insects, thousands of different kinds of which are constant flower visitors. Some of the smaller birds, and even snails, also help flowers to meet their mates. The wind, too, bears pollen through the air to some expectant bride-to-be. And, finally, in the water, by a series of acts the like of which no one could improve for cunning, the cross-fertilization of certain aquatic plants is consummated. It would take a book larger than the present one to give even the briefest account of how these different aids to maternity do their work and how the flower responds to this help. As that is quite out of the question, only some of the best-known examples of cross-fertilization will be given, and these will be grouped according to what agency the flower is indebted for its chance of doing that for which it is created.
On any summer day, especially when the sun is shining brightly, we may see bees and butterflies flitting from flower to flower, busy as the proverbial bee. We already know enough about nature’s ways of doing things to be certain that these, and hundreds of other kinds of insects, do not come for nothing, and that the flower must have something to offer. Bees, especially, are thrifty creatures whose business demands exacting and prolonged toil. They would not waste five seconds upon idle flower calling if the blossoms did not yield a rich store. And thousands of flowers do yield the sweetest and richest kinds of stores of nectar or honey, which is, in fact, by the help of insects who alone can extract it, our sole source of honey. Many flowers which produce no nectar do have such plentiful stocks of pollen that the bees come for that alone. In the peony, for instance, over three million pollen grains are produced in each flower, only a minute fraction of which can ever fertilize an ovule. All the rest would be wasted were not pollen in itself a particularly nutritious food for young bees, and consequently much sought after by the careful bee mothers. They are the only insects that feed their young on pollen, or beebread, as it is called by the beekeepers, so that the enormous overproduction of this male fertilizing agent, from the point of view of the flower, is a decided attraction, one might almost call it a trap, to insure constant visitations from bees. For they are perhaps the most useful of all insects in the great game of securing cross-fertilization, as we shall presently see. Many other kinds of adult insects eat pollen directly and so add to the number of insect visitors.
No one has ever been able to explain the beautiful coloring of flowers, except that it serves as an attraction for insects and small birds. Like the honey or nectar, it seems to play no real part in the home economy of the flower, to be of not the least use otherwise. While honey and the gorgeous colors of flowers are a delight to man, that would be no sufficient reason for the ability to produce them. Both of these attributes of flowers, as attractions for insect visitors, are, however, so absolutely essential to cross-fertilization that we must think of them as having grown up out of that demand. As we shall see a little later, even the structure of some insects has been modified so that they can reach the nectar or pollen only by automatically doing for the flower what it cannot do itself.
While color of flowers seems as though it were attraction enough, it is very likely that their fragrance or perfume is still more seductive in its power of luring insect visitors and repelling useless ones. Poets have called this perfume the soul of the flower, and in its almost intangible beauty it might well be so called were it not for the fact that it appears to be of not the slightest use, except as a lure. In all the equipment of seduction there is none like this fragrance of flowers for attracting insects.
Flowers, then, have things to offer to insects which the latter need. Nectar and pollen are the chief, and where these merely bread and butter objects are not enough, or sometimes in addition to them, the flower is dressed out in gorgeous colors or perfumed with a fragrance beyond the dreams of the fairest bride. What insects do to complete the fertilization of such a legion of beauties makes up the romance of the flowers. Perhaps not even in man himself is this creation of new life so surrounded with beautiful ideas. Also, as in man, it sometimes is bound up with an almost fiendish cruelty and cunning. Some of these visitors and what they do, but unfortunately only a very few, can be mentioned here. They must serve as types or examples of what is going on all about us on any summer day.
The common blue columbine, much grown in gardens for its beautiful blossoms, always has the flowers hanging upside down, a habit that admirably serves to keep its pollen from rain. The opening and closing of many flowers in cloudy weather, or at night, may be for the same reason. Everyone knows the five blue spurs into which the petals of columbine are produced. At the very end of each spur, which is always curved, the flower secretes a considerable quantity of honey. This, one of the greatest attractions to bees, leads inevitably to a visit from one. The bee, in order to reach the honey, hangs on to the inverted flowers, clutching the base of the spur with its foreleg, and further securing itself by the mid or hind legs, which grasp the slender column into which, in the columbine, the stamens and pistils are crowded. In its anxiety to reach the honey the bee pokes its head as far into the spur as possible, but it gets in only a fraction of the full length of the tube. To reach the honey it extends its sucking apparatus, which is a complicated mechanism for this purpose on the head of nearly all insects, and which will hereafter be called by its true name of proboscis. It happens that bees can easily bend the proboscis downward or toward their own body, but only with considerable difficulty can they bend it in the opposite way. And yet the honey in the curved tip of the columbine can only be reached by curving the proboscis to fit the tube, and in this process the bee’s body for nearly half its length is forced to touch the anthers. While these are close to the stigma, they produce pollen only on their outside surface, where it is, of course, scarcely likely to reach the stigma, but must be brushed off by the contortions of the bee’s body in reaching the honey. The hairy body of the bee, coated with pollen, goes next to perhaps an older flower of the columbine. Heedless of any change in the flower the bee goes straight for the honey in one of the spurs, again catches hold of the only available support in the center of the flower. But this time, instead of brushing pollen off the exposed anthers, it brushes it off its own body to the stigmas, which, at a slightly later stage than in the one just described, are branched and perfectly adapted for collecting the pollen with which the bepowdered bee can hardly avoid dusting them. Cross-fertilization is of course assured, but it seems a precarious business at best, as the number of bees with a proboscis long enough to do the work is limited. The columbine, by a kind of uncanny foresight, is so constructed that bees or other insects that try to reach the honey and are not provided with a sufficiently long proboscis, nevertheless in further attempts upon other flowers, inevitably cross-fertilize them without reaping their reward. One or two kinds of bees, as though in retaliation for this subterfuge of the columbine, make short work of the honey by biting a hole in the spur and forthwith sucking out the honey without so much as touching anther, pollen, or stigma. The reply of the columbine to this ravaging of its chief attraction is that finally, as a last resort, and by a new movement of its reproductive organs, it is self-fertilized. Here the shape of the flower, the original position of the pistil and anthers, the exposure of pollen only in such a direction that, while a chance of cross-fertilization still exists, it can hardly ever fertilize its own stigmas, all point to cross-fertilization as the plant’s greatest requirement. And yet failing this, it falls back on self-fertilization rather than endure barren sterility.

While the columbine by its spurs and other interior structure succeeds nearly always in holding a bee long enough to insure its being dusted with pollen, the common barberry bush of Europe (Figure 70), also much planted in American gardens for ornament, actually drives bees away by sharp blows of its stamens, so that self-fertilization shall not result from the visit. In this shrub the petals partly cover the stamens unless the latter are disturbed, and, in fact, the curved tip of the petals forms a kind of socket into which each of the six stamens are fitted. The position of these is such that any insect

Plants of the Palm Family, Palmaceæ, in Ceylon. They are the talipot palm (Corypha umbraculifera), the fiber from the wood of which is used in India for paper making. (After Reinhardt. Courtesy of Brooklyn Botanic Garden.)

A Giant Herb of the Banana Family, Musaceæ. It is the travelers’ tree (Ravenala Madagascariensis), the sheathing leafstalks of which hold considerable quantities of water—hence the name. Grown throughout the tropical world. (Courtesy of Brooklyn Botanic Garden.)
can go straight to the honey glands which are at the base of the flower, without touching the anthers. But their filaments are broadened out at the base, so much so that their edges touch. The honey glands are so placed that the insect must touch the broad bases of at least two filaments, between which, in fact, it must force its proboscis in order to reach the honey. The moment any particular pair of filaments are irritated by the bee, two pollen-dusted stamens fly out from their position among the petals and the anthers strike the bee with a sharp blow. Many observations prove that almost never does the bee go on with his honey sucking after this rude interruption, which has resulted in at least its head being dusted with pollen. The low, sticky stigma is so placed that it is one of the first things the bee’s head strikes as it reaches the center of the flower. Because of the position of the stamens, while they are undisturbed, it is impossible that pollen from them could have been brushed off at the bee’s entrance of the flower. And by an almost miraculous adjustment of the power of the blow by the irritated stamens, this drives off the intruder only after he has brushed his pollen-laden head over the stigma. His head at this stage is, of course, covered only with foreign pollen gathered elsewhere, but just as soon as the bee tries to get what he came for, sometimes even before he gets his reward, out fly the pair of stamens, thoroughly dusting the bee, and seeing to it that the blow is just sufficient to drive off the pollen-laden insect. No device to secure cross-fertilization could be more effective. If the blow of the stamen were only ever such a slight fraction less than it is, the bee would only stop a moment and then go on honey sucking; which, because of the release of the pollen by even the gentlest blow, would result in self-fertilization by the aid of the bee, rather than cross-fertilization. Very few, if any, bees will stand this gentle reminder to go, however, and it is a little curious that such intelligent creatures as they are supposed to be, should not realize that it is all the clever but quite harmless trick of an apparently still more intelligent flower to secure fertilization from any pollen but its own.
To attract insects and then repel them seems a little like using them as some flirts notoriously use men, only to throw them over when they are no longer interesting. In the large-flowered magnolia tree from the southeastern United States, insects, however, fare somewhat better than this. In this magnolia, which has flowers several inches long, self-fertilization is impossible as the stigmas are ready to receive a mate several days before the laggard stamens are provided with the wherewithal. Without some insect or other outside help there would be only a childless old age for this particular tree. The flower opens rather early in the season, while the nights are still cool, and as a protection from the cold, rose beetles habitually fly into them. They find a pleasant shelter under the three inner petals which arch over the honey-coated stigma, and form a snug little chamber so much warmer than the outer air that its heat is appreciable to the touch. The rose beetles, once they are inside this warm shelter, cannot get out and are often held for a few days. Then, as the stigma passes its period and the stamens are furnished with pollen, the chamber opens by the gradual withering of the petals. But the insects, in their efforts to get out, have raised a perfect dust storm of pollen with which they are naturally covered. Just as soon as they are released they are free to seek another warm shelter where the process is repeated. Thus they always enter the flower with foreign pollen, use it up impregnating the waiting ovule, and are held until the flower’s own pollen gives the signal for their release properly dusted for a renewal of the work. The premature timing of the stigma, the tardiness of the anther in producing pollen, the generation of heat and secretion of honey which frequently covers the whole stigma—there could scarcely be a better equipment for securing cross-fertilization. And without it the magnolia would be simply sterile.
There are some other flowers that hold visiting insects in a trap until cross-fertilization has been completed, and all of them by no means furnish their visitors such a snug little heated chamber as the magnolia. One vine from the eastern United States, known as the Dutchman’s-pipe, or sometimes as the pipevine (Figure 71), is singularly ruthless in this respect. Its flowers are of such evil odor that only carrion-loving insects, such as certain kinds of flies and gnats, ever visit them. The flower is of very peculiar structure, being formed of a hollow tube bent from its stalk first downward, and then upward. The upper part ends at the opening which is provided with a three-lobed lip or doorway. Through this the insects crawl, and they finally reach the bottom of the curved part of the flower. Behind and above them is the entrance through which they have just come. And above them, in the other curved part of the flower, is the stigma. As in the magnolia, this matures several days before the pollen from its surrounding stamens is ripe, so that self-fertilization is never possible. What is now the

plight of the insect at the bottom of the upward-pointing tubes, one leading to the organs of reproduction, the other to the exit? By an almost diabolical cunning the inside of this flower is so smooth that no insect can crawl up its slippery sides. It takes some time for the prisoner to find this out, and, in the meantime, it has explored every nook and corner of the flower by flying. In the course of this exploration it reaches and covers the stigma with pollen, for as we shall see presently, it always comes into the flower pollen-laden. Evidently becoming panicky about getting out, the insect then flies with very considerable force in every direction. Toward the true exit it naturally flies the most, and by a refinement of cruelty this is the lighter end of the flower, and therefore the obvious mode of escape for it. But the three lips and the entrance they cover are not flat across the flower, as the cover of a lunch box would normally be, but turned at such an angle as the lunch box would be if set on end. The insect, in flying toward the light, invariably hits the smooth surface just inside the three lips and falls to the bottom of the flower because there is nothing rough enough for it to cling to. Throughout most of the day, and sometimes for several days, the insect will keep up this ceaseless struggle to escape, flying first up one tube and then the other. These frantic efforts would end in exhaustion if kept up too long, and before that happens, but after the impregnated ovules have no further use for the flies, the anthers give forth plentiful supplies of pollen. Of course, the insect can scarcely avoid becoming covered with this, and then, but only then, the flower begins to wither and up its now wrinkled sides the pollen-laden prisoner can at last crawl to liberty.
There appears nothing very romantic about the cross-fertilization of the Dutchman’s pipe, in fact, the whole affair seems but a sordid and, it must be confessed, a very efficient trick to get what the flower needs from the insect, rewarding it by many hours of apparently hopeless captivity. But most flowers do have something that insects want, and none so well fulfill the expectations of butterflies as the meadow pink. This is a graceful little perennial native in the fields in central Europe, but often grown in American flower gardens. It has beautiful pink flowers, with a long tubular calyx, at the bottom of which are rich honey glands, accessible only to the long proboscis of different kinds of butterflies. No other insects, and many try, are able to get the honey from this plant. It begins flowering by opening its five beautifully fringed petals, all of which are marked with lines from their edge toward the center. These obvious “pathfinders” are common on many other flowers and all seem to be there to act as a guide for alighting insects, and, as it were, steer them to the center of the flower. As a butterfly alights he finds five protruding anthers covered with pollen. These are of no use to him, but nevertheless his head is covered by their pollen. In fact, many other visitors, who can never reach the honey, find the bright color and good stocks of pollen sufficient attraction for them, and after a pollen feast fly off to other flowers. If the butterfly comes at this state of the flower’s life, it pokes its long proboscis down into the tube of the calyx, but finds the passage almost blocked by another set of five stamens not yet ready to discharge pollen, and, as though ashamed of the fact, quite hidden from view. Also, the proboscis is very nearly stopped by the style, which, if its stigmas were ready for mating, would then and there become impregnated. But they are rolled together into a tight spiral, and their pollen-receptive faces tightly pressed together in addition to the whole structure being twisted, so that the very probably pollen-laden proboscis of the butterfly finally gets to the honey without leaving one grain of pollen where it could possibly self-fertilize the flower. Honey-laden, it now flies away, and so far as this particular flower is concerned, simply nothing has happened but to coat several insects with pollen. A little later the anthers of the first set of stamens cease work and drop off. Then the five stamens hitherto hidden in the calyx burst out and furnish a second crop of pollen, but the stigmas are still safely coiled away from the possible danger of self-impregnation. During this second stage the flower may repeatedly be visited, but until this second crop of anthers become useless there is not the slightest risk of the stigmas becoming self-fertilized. For the hole through which the second set of stamens has come is so small that still only the proboscis of the butterfly can penetrate it. When finally the second crop of anthers also fall off, then the style slowly uncoils and thrusts its now receptive stigmas above the calyx and in plain view of passing visitors. These come, pollen-laden of course, from other flowers, and cross-fertilization is assured. Here the flower furnishes two crops of pollen, plentiful supplies of honey, and asks only that in gathering these the butterflies see to it that its stigma be covered only with foreign pollen. Keeping carefully out of the way while they are about their business and there is danger of self-fertilization, they come out boldly once that danger is past. In this meadow pink self-fertilization is simply impossible; everything in the production of its young it owes to the butterflies, to which it surely makes adequate returns.
While such plants as the meadow pink and thousands of others have lost, if they ever possessed, the power of self-fertilization, and rely absolutely on insect visitors for their perpetuation, there are many hundreds of kinds that apparently hope for cross-fertilization, but, in default of it, due to their inability to absolutely compel it, they finally accept self-fertilization as a last resort. Darwin once said that “Nature abhors perpetual self-fertilization,” and the frequent visits of insects and their rôle in preventing it, together with the flowers’ adaptations to such visits, support the contention, which has never been seriously questioned. But some flowers appear to have left the back door open, as it were, so that failing cross-fertilization, they may still rely upon self-fertilization. A geranium from the Pyrenees, a relative of our common woods geranium, is of this type. A day or two before its stigmas are ripe it produces first one set of five anthers, all pollen-coated, and then another. If these have not been brushed clean, as often or usually happens, then the stigmas ripen and, of course, are impregnated by their pollen. If they are brushed clean of pollen, then the stigmas must rely on foreign pollen, of which it is assured a supply from the visits of pollen-laden insects from other flowers, which are still attracted by the flowers’ color and honey. Perpetuation is assured, in any case, but the preference is still for cross-fertilization.
A more remarkable case of leaving one final chance for self-fertilization is the gas plant. It exhales such a strong and peculiar odor that only certain kinds of insects will visit it. In fact, the odor is so strong and is so heavily charged that a lighted match held near it has been known to slightly ignite—hence the plant’s name. The flower bears a low, squat stigma, profusely covered with honey, which is perfectly accessible to any insect visitor. It has ten stamens, which at first are quite out of the way of insects, two being folded back in each of the five yellow petals. First one stamen begins moving gently from the shelter of its petal, and the anther, pollen-coated, hovers over the stigma, which would inevitably lead to self-fertilization if the stigma were only ready. It never is, and, as though realizing this, the stamen gently moves back out of the way, still, in most cases, retaining some of its pollen. Then another tries and, again, as though realizing the futility of impregnating the unreceptive stigma, it also moves back. So it goes with all the other eight stamens, each of which moves gently out over the stigma, and gently back again, all of them failing to fertilize the laggard stigma. But there has been during all this time a constant procession of insects coming for the honey, and never for a moment have the stamens been off guard, so that each visitor goes off with at least some pollen clinging to it. Finally the stigma, after all the faithful ten are folded back among the petals, comes into its period, and is cross-fertilized by the insects which come from other flowers laden with pollen, as we now know. But if, by one of those accidents of nature, such as bad weather or what not, no insect ever does come, what then is to be the fate of the stigma? Has it staked all only to lose out in the end? It would almost seem that to ignore the steady attentions of the willing ten might make barren sterility a fitting punishment. But, in spite of what has happened, the stamens come to the rescue, if all insects fail, and this time, in a body, they rise up and shed upon the stigma from the half-withered anthers their few remaining pollen grains. There could be no finer example of having an anchor to windward.
One of the largest families of flowering plants is the pea family (Figure 72), with over five thousand members, practically all of which rely on bees for cross-fertilization. In some kinds, where bees occasionally fail them, the flowers wither without self-fertilization and, of course, no seed are then produced. In such a large family of plants there are

naturally many different adaptations for securing cross-fertilization—some of them of such extreme complexity that they could hardly be included here. All the family have the characteristic pealike flowers familiar enough in the sweet pea, which have already been described and figured on page 44. In all of these stamens and pistils are hidden inside the keel, at least in the early stages of the flower. In some, such as clovers, for instance, the organs emerge from the keel, and after fertilization by insects re-enter their retreat. There are scores of different plans for securing the desired object, but the common alfalfa, with a few other related plants, has the most startling. The flower begins life with its stamens and pistils concealed within the keel, which is apparently impregnable. When a bee alights on the flower and begins work he is welcomed by a small but violent explosion. When the dust of this clears away, and it is actually dusty with pollen, the dazed bee is seen to have fertilized with almost instantaneous rapidity the stigma of the exploding flower, which springs violently out of the burst keel. This is so arranged that, as it flies out of its trap, due to the explosion touched off by the bee, it strikes the under side of the bee’s body a distinct blow, brushing off in the process the pollen nearly always found there. This pollen has come from other plants. But, as in most plants of the pea family, the stamens closely surround the stigma, but are shorter than it. When the explosion occurs, the stigma, because it is slightly longer than the stamens, is the first thing to strike the bee’s body. Already impregnated, it is then indifferent to the cloud of pollen from the stamens of its own flower, which only a fraction of a second later also strikes the bee’s body. The first flower that a bee visits cannot, of course, be cross-fertilized notwithstanding the explosion which results, no matter from what angle the bee attempts to insert its proboscis. But in this case, as the stigma is unfertilized by foreign pollen, its own performs that service. In the vast majority of cases cross-fertilization is assured by certainly the most novel of processes, and, in the rare event of the bee not being covered with foreign pollen, self-fertilization is still possible.
In many of the plants already noted cross-fertilization is accomplished by virtue of the fact that the stamens mature before the stigma. But in the common strawberry the reverse is true. As the insect stands on the white petals, it must, in order to reach the honey at the base of the flower, put its head down so that if pollen were available there could only result self-fertilization. But while the stigma is ready, the pollen at this stage never is, and the insect which comes usually laden with pollen from other flowers cannot avoid impregnating the stigma with this foreign pollen. Later the stamens mature and, as insect visits continue so long as honey is to be found, they become dusted with pollen which is used for the fertilization of other plants. Scores of other plants also produce ripened stigmas before the pollen matures, and they must rely on insects to cross-fertilize them.
The cross-fertilization of the strawberry is such a comparatively simple process—seems in fact almost inevitable—that we are lost in wonder at the almost mathematical complexity of the act in the common purple loosestrife, which has been introduced into American gardens from Europe, and sometimes runs wild. In this plant there is a long terminal spike of showy, purplish-pink flowers, the color of which is sufficient to attract many insects from even a fairly swift flight. The petals are streaked with “pathfinders” toward the center of the flower. This consists of a tubular calyx; at the bottom of this is the honey, which secures the insect’s further interest once the color has attracted it. But it finds a condition of the reproductive organs almost without parallel. In some plants the style is hidden down in the calyx tube, while one set of stamens just peep out of the end of the tube, and a second set are still further and quite obviously protruded. In a neighboring plant the style will be found outside the tube, one set of stamens hidden in it, and the other set outtopping everything, except the petals. In still a third type of the loosestrife, the style exceeds everything but the petals, one set of stamens just emerge from the tube and the lowest set are hidden. It would appear as if the loosestrife could scarcely escape self-fertilization, except possibly in that form where the stigma outtops all the stamens, and this would result always if the pollen were indiscriminately useful from all three lengths of stamens. But it never is, only that from the short-stamened plants will fertilize the short-styled one, the mid-stamened ones the mid-styled counterpart, and the long-stamened ones the long-styled flower. The pollen grains of the different-lengthed stamens are even of different size and color. If this were not so, it is a simple mathematical problem that, with three different sets of style lengths and six sets of stamens, two in each flower, eighteen different crosses might be possible. As it is, only six crosses are ever possible and these only by the aid of insects, for it must be remembered that stamens and pistils of the same length are never found in the same flower. By an adjustment of the size of the body of the different insect visitors it works out so that, while all three body sizes frequently visit each flower, only that particular size of insect suited to the carrying of pollen from stamens of one definite length to a style of similar length actually accomplishes the cross-fertilization of that particular flower. And this without interfering with the visit of another different-sized insect which will accomplish the work for its particular set of stamens and pistils. So marvelous is the adjustment of style length and stamen length to each particular body size of the visiting insects, so perfect is the arrangement of the organs in each flower, that each contributes to the fertilization of its neighbors, never to its own, nor does it interfere with the process in other lengthened styles. Cross-fertilization is thus almost always accomplished in the six different combinations possible in this truly remarkable case of adaptation between insects and flowers.
From the almost mathematical complications of the cross-fertilization of the loosestrife it seems a far cry indeed to that of the Italian honeysuckle. This often runs wild over fences, but is unlike the more widely known Japanese honeysuckle, in that its stem passes through the different pairs of opposite, bluish-green leaves, which are joined together at the base. The Italian honeysuckle falls back on the more simple seductions of odor and honey for securing its really important insect visitors. It has such a long tube that only certain night-or evening-flying moths or butterflies can reach the honey. There is, even during the day, such a plentiful supply of this that it frequently fills half the tube, but even then it is quite out of reach of bees which never succeed in getting any. Toward evening, particularly on quiet, still evenings, the flower begins to send off in much increased quantity a heavy rich-scented odor almost overpowering in its sweetness. The butterflies and moths of the dusk having a long proboscis, succumb to this really enchanting lure, which, with the large store of honey, insures quantities of eager suitors. The stigma, while ripening simultaneously with the anthers, protrudes beyond them, so that the butterflies touch pollen only after touching the stigma, which of course is impregnated with pollen from an earlier visit of the insect to a different flower. So assiduous are the butterflies, that on a still night there will be not one grain of pollen left on the much-brushed anthers. If the night is cold or windy much pollen remains and cross-fertilization is left to pollen-eating insects such as mother bees or flies. While these can never get the honey they often do accomplish cross-fertilization and, sometimes by misadventure, self-pollination. It is only those moths and butterflies which, forcing their long proboscis down the honey-laden tube, must accomplish cross-fertilization. But failing this, the plant is more or less at the mercy of hosts of insect triflers, mere pollen eaters, who may or may not insure cross-fertilization, but in any case provide for self-fertilization.
Another use which certain plants make of honey, besides acting as a lure of insects, is found in the common lilac. The flower in this has considerable honey at the bottom of the tube, which can only be reached by insects with a proboscis sufficiently long to reach it. The lilac has only two stamens inserted near the top of the tube, the passage to which they very nearly obstruct. The stigma is hidden in the tube, and it matures simultaneously with the ripening of the pollen. As the insect inserts its proboscis between the stamens no pollen clings to it due to the character of the pollen grains. But as the proboscis is withdrawn from the tube its lower end is covered with honey to which pollen sticks. If a needle is inserted between the stamens and pushed only far enough to be still clear of the honey, no pollen will be found on it when withdrawn, but if pushed all the way down, its honey-coated point will catch considerable pollen. In the lilac, if insect visitors do not accomplish the work of cross-fertilization, the flower is self-fertilized ultimately by the protruding of the stigma far enough out of the tube to catch some of the remaining pollen grains.
It is perhaps useless to multiply instances of flowers which by various devices secure the cooperation of insects in getting pollen from a foreign source. To recapitulate some of those devices it is necessary only to recall what some flowers have done to force cross-fertilization. The heated chamber of the magnolia, the cruelty of the prison cell in Dutchman’s-pipe, the blow from the stamen of the barberry, the faithful rotation of the ten stamens of the gas plant, the explosive flower in some members of the pea family, the lure of honey and seductive odor of the Italian honeysuckle, the mathematical complexity of the loosestrife—these and hundreds of others all point to the necessity of cross-fertilization and a means to produce it almost beyond belief. There is the best of evidence that not only flowers but insects themselves have been modified in this great work, and that for every flower needing cross-fertilization some agency has been developed to secure it. Insects, beyond all other animal life, do this work, but it is accomplished by humming birds often, and in one plant even by a snail.
Two of the very largest plant families, not so far mentioned in this account, depend almost absolutely upon insects. In the daisy family, with over eleven thousand members, the large heads of flowers, often containing scores of individual flowers, are constantly brushed, over and over again, by the pollen-coated bodies of insect visitors. And in all or nearly all orchids (Figures 73-75), comprising over five thousand kinds, the same process is accomplished. In these plants, in fact, the act is, if possible, more

complicated than in any so far noted. Darwin’s book, “On the Various Contrivances by Which British and Foreign Orchids Are Fertilized by Insects,” reads like a fairy tale. Yet it is the result of years of patient observation by incomparably the greatest naturalist of recent times. To it the reader must go for the details of a drama of absorbing interest, but too long to sketch even briefly here. Perhaps one illustration may be mentioned of how far the principle of cross-fertilization has been carried, and to the deadly effects of its failure in at least one case. In a certain orchid from Brazil, known as butterfly orchid, the pollen is nearly always carried out of the flower by an insect visitor, but, if by mischance it is not, and falls on the stigma, not only does it fail to fertilize the ovules, but it kills the pistil forthwith. There may be a few other cases of such drastic results of self-fertilization, but in any case, and disregarding these apparently suicidal fanatics, cross-fertilization is so very nearly universal that nature must find it of enormous advantage. Only in this way is it possible to explain the intricate adjustments of insects and flowers, which work together in such wonderful harmony that cross-fertilization hardly ever fails in those flowers where it appears to be necessary.
It must have struck many thoughtful readers to ask a rather obvious question at this point. Why, if untold millions of insects are constantly flitting from flower to flower, does not the pollen get mixed, as it is quite certain that they will not fly from a

certain kind of geranium to another similar one for instance, but perhaps to a rose? The answer to this is simple enough, but its implications are limitless. Only pollen of a certain species or variety is useful to the stigmas of that variety. To practically all others the stigma is simply unreceptive, except in those closely related plants that may all have a common parentage. When crosses between such closely related plants do occur the result is known as a hybrid, which will be considered elsewhere. To this extent, then, flowers are peculiarly exclusive in their matings and promiscuity occurs in the vast number of cases only in plants of the same species. We do not yet understand the impotence of pollen of one species upon another; all that we do know, which has many times been proved by experiments, is that it fails to act. If it did act, no one could picture the chaos into which the vegetative world would be thrown.
While, as we have seen, thousands of plants rely upon insects for producing their young, still other thousands put everything to the hazard of the wind. Pollen is so light that it can easily be blown very great distances, and while the wastage is enormous, the process works so well that the greater part of the vegetation of the earth is thus fertilized. This is true not as to the number of different kinds of plants, for in that respect insect fertilization is more important than that accomplished by the wind. But in the number of individual plants concerned the wind is incomparably the greatest fertilizing agency that is known to us. This for the reason that all grasses and sedges, most catkin-bearing trees such as oak, hickory, birch, practically all pine trees and their relatives rely wholly on the wind for fertilization.
For reasons that will be enlarged upon in another chapter, all of these great groups of plants must be considered as of simple structure, some, like the pines, relics of a remote past when no flowering plants, as we know them to-day, existed on the earth. In any event the reliance upon the wind is certainly hazardous, and while it of course insures nearly universal cross-fertilization, it may well result in scanty fertilization or, in exceptional cases, complete failure of it. Quite obvious also is the amount and direction of the wind in the process, for in very open and windy places grasslike vegetation, or at least a predominance of species fertilized by wind, is likely to be found, rather than those plants that rely upon insects, that, unable to stand the full force of the wind, seek more sheltered places. While such a thing is not the cause of prairies, or the predominantly grasslike vegetation along sand dunes, or the exclusive spruce forests of the bleak and windy north country, it unquestionably aids in maintaining the often exclusive nature of such pure associations of plants. Over thousands of square miles on our own great plains or on the steppes of Russia, both subject to violent winds, the great bulk of the vegetation is wind fertilized. It could hardly be expected that pollen, once in the grip of such a wayward and shifting thing as the wind, should not be wasted in great quantities. This is particularly true of pine trees, which at pollen time may often be seen giving off golden clouds of dust, of which perhaps 95 per cent is wasted.
Those submerged aquatic plants upon which neither the winds nor honey-seeking insects can work the magic of cross-fertilization, seem to be about the poorest equipped for perpetuating their kind through impregnation of their tiny flowers. And yet, for at least two of them, which will be described presently, the process is accomplished by an adaptation of their mode of life to their watery environment that seems incredible. These two have been selected as illustrating two peculiar adaptations in the weight of pollen or pollen-holding flowers that is common to some other submerged aquatic plants. In one the male flower, or pollen from it, with the very nicest adjustment of function to environment in all the realm of the plant world, is just of the right specific gravity to float to the surface with dramatic suddenness and perfectly timed effectiveness. In the other the pollen is just enough heavier than the water to float betwixt the surface and the bottom, so that at the proper moment it is where it can fulfill its destiny.
The common eelgrass or tapegrass is a submerged aquatic which roots in the mud and has long grasslike leaves which may often be seen waving gently in the current of many quiet streams in this country and in Europe. Down near the base and in among its swaying verdure, it bears tiny flowers which have no petals, and in which, as if recognizing the futility of display in such a secluded watery home, even its calyx is reduced to small scales. Some of these minute flowers are females, others again all males, and as they appear in their early stages it looks as though never the twain could meet. And the hopelessness of their ever meeting is increased as the maturing female begins slowly to uncoil the fine stalk upon which it grows. Steadily but surely the loose spirals of the stalk of this ever more mature female flower uncoils, until, when quite ready for the pollen, it is at last upon the surface. The male flowers, in the meanwhile, are down near the bottom with their small freight of pollen ready to perform their function, but firmly anchored to a stalk absurdly inadequate to reach the surface where alone they can be of service. A great Belgian, Maurice Maeterlinck, who studied this plant with more sympathetic vision than any botanist has yet been able to equal, wrote in one of his essays on “The Intelligence of Flowers” the solution of this little drama of apparent hopelessness. No other words can ever convey the meaning of what happens to the eelgrass quite so well as his. “Is there any more cruel inadvertence or ordeal in nature? Picture the tragedy of that longing, the inaccessible so nearly attained, the transparent fatality, the impossible with not a visible obstacle! It would be insoluble, like our own tragedy upon this earth, were it not that an unexpected element is mingled with it. Did the males foresee the disillusion to which they would be subjected? One thing is certain: that they have locked up in their hearts a bubble of air, even as we lock up in our souls a thought of desperate deliverance. It is as though they hesitated for a moment; then, with a magnificent effort, the finest, the most supernatural, that I know of in all the pageantry of the insects and the flowers, in order to rise to happiness, they deliberately break the bond that attaches them to life. They tear themselves from their peduncle, and, with an incomparable flight * * * dart up and break the surface of the water. Wounded to death, but radiant and free, they float for a moment beside their heedless brides and the union is accomplished, whereupon the victims drift away to perish, while the wife, already a mother, closes her calyx, in which lives their last breath, rolls up her spiral, and descends to the depths, there to ripen the fruit of the heroic kiss.”
In the eelgrass it is the specific gravity of the male flower, or, the secreted air bubble, which makes the flower lighter than the water, and actually causes the flight from the depths to the surface. Because of this, fertilization can only take place on the surface, although the flowers and fruits otherwise mature under water. But in sea wrack, in Naias, and in ditch grass, all submerged aquatics, the flowers are even fertilized under the water. Pollen in such plants is much modified, and instead of being in the ordinary form of pollen grains, it is, at the time of ripening, lengthened out into tubular, hairlike structures. These delicate prolongations of the male fertilizing stuff are carried by the currents of the water, just as a thread would be, but with the difference that the pollen threads are so beautifully weighted to fit their watery environment that they float, suspended, in the depths of the water at or near the level of the female flowers. The pollen is set free at maturity, just as it is in the eelgrass, but to meet the female, which never rises, it must float with the current of the stream. There must, as in the wind-carried pollen, be a tremendous wastage, yet sufficient quantities of it do fertilize the females, particularly in the ditch grass, which fruits very freely.
Whether it be any of the various contrivances for insect fertilization, or by the winds, or, as in the eelgrass, by the water, the climax of the flower’s life is always reached in this act. For all annuals the plants, also, begin to die down then, a process that is completed with the production of seed, which is, of course, the object of all those varied modes of fertilization. Perhaps no answer to the question of why plants do not always self-fertilize themselves is so eloquent as the hundreds of ways they have adopted to avoid doing so, a few of which we already know. Many volumes have been written on this subject, but all of them, intricate as the methods they describe nearly always are, merely confirm what we have already seen—that rather than submit to self-fertilization, plants will adopt almost undreamed-of expedients. Sometimes, as in the eelgrass and in the visits of nocturnal insects to those night-blooming flowers that carry on their matings in the glamour of moonlight or in the dusk of eventide, the drama, in the eyes of imaginative writers, is one of singular beauty and charm. And, on the other hand, we have seen the well-nigh heartless cruelty of the Dutchman’s-pipe in keeping as prisoners its absolutely necessary insect deliverers. Even this is outranked for matchless ruthlessness by a wild arum, a relative of our jack-in-the-pulpit, from the East Indies. It produces a club-shaped inflorescence composed of tiny flowers that need cross-fertilization, but so offensive is the odor of the flower that no insects will tolerate it. A snail, a voracious eater of foliage, is attracted to the flower partly by the fine fleshy leaves, but mostly by a juice secreted at the apex of the flower column. To this the snail crawls, and fertilizes the tiny flowers over which it drags its body. When this is accomplished it speeds on hungrily to the juice just above it and eagerly devours the poison. Death follows almost immediately. The secretion of this murderous liquid to lure the only creature that will visit such an offensively malodorous plant, which, without it, would very likely be itself destroyed by the foliage-eating snails, is a gruesome contrast to that happy flitting of butterflies which completes the fertilization of most flowers in equally effective but more pleasing fashion.
Once impregnation of the ovule has been consummated, it begins a slow process of change, involving sometimes the modification of the ovary, or of the calyx, and very often of the swollen apex of the flower stalk upon which these organs are borne, known technically as the receptacle. We have seen, in the first chapter, what greatly different types of fruits are developed from different ovaries, and they of course produce seeds in varying size and amount. In the coco de mer, a palm from the Seychelles, the seed often weighs forty or fifty pounds, while in some orchids a single capsule will contain over a million almost microscopic seeds. Some of the devices of fruits and of seeds to secure the utmost spreading of the species over the earth will be considered in another chapter. All the devious methods of plants in producing their young become significant, so far as the earth’s vegetation is concerned, only when we find out what this enormous progeny has done with their opportunity. The chapter on the Distribution of Plants will tell us how well that opportunity has been used.
As we stated in the first chapter cryptogams, while they produce no flowers, must bear organs that perform the functions of flowers in the reproduction of new individuals. Because, generally speaking, the process is more hidden in its manifestations, and nearly always requires the aid of the microscope to detect it, it is not so well known as the reproductive processes of flowering plants by those who have not the opportunity to manipulate such instruments. The act, however, is just as interesting, and, as we shall presently see, it may well be considered the ancestor of those more showy methods of producing young, which have been all too inadequately treated in the preceding pages. While the parts having to do with reproduction in flowerless plants are microscopic in size, it is possible to understand the broad outlines of what goes on and perhaps the life history of such plants is as well illustrated in ferns as in anything else.
In the discussion of ferns in the first chapter we found that on the back of some of their leaves, or occasionally on special leaves devoted to the purpose, were many small brownish or dark spots, arranged in rather definite fashion, and known as sori. (Figure 63.) Each sorus contains many minute bodies known as spores, not unlike very miniature seeds in general appearance, but quite unlike them in behavior and mode of life. No better idea of their size can be gleaned than to record the fact that in each sorus there may be about one hundred small, often short-stalked spore cases, known as sporangia, and that in each sporangium well over forty, and sometimes over sixty, spores will be crowded. A healthy specimen of many of our common ferns will bear about ten or a dozen leaves, each of which is divided into many divisions, and among these divisions of the leaf there may be at least fifty that bear from fifteen to twenty sori. It can be easily figured from this that a healthy plant of this fern may and usually does produce over forty-five million spores, each of which contains within it the opportunity of developing into a new plant. There is thus a prodigality in producing the means of renewal of life among ferns that far outstrips the production of seeds in even the most prolific of flowering plants.
When the spores in the sporangium are mature and therefore ready for the next stage in their life history several things must happen. With somewhere about six thousand of them crowded together under each sorus, more room to develop is obviously the first consideration. This is provided for by the fact that when the spores are ripe the sporangia have the ability to throw them considerable distances; then of course the wind can carry them much farther. To be of any use they must fall upon damp ground, for some degree of moisture is absolutely necessary for what is about to happen to them. In nature countless millions never do fall in a favorable location, or, if they did, such an enormous production of fern spores would soon make the world exclusively a fern garden. The comparatively minute fraction of them that ever do find congenial surroundings, once they are expelled from the spore case, then begin a process that is not unlike the germination of a seed. For the spore must take in water from the soil, which by osmotic pressure finally bursts it open. From the burst spore a minute tube, known as the protonema, or literally first thread, begins to develop. It is, of course, of microscopic size, and yet near its base there is a branch tube formed, differing from it in structure and ultimately forming rhizoids, which are rootlike hairs. Both the protonema and the rhizoids begin growing, the first forming, usually flat on the ground, an often heart-shaped body having the characteristic green coloring matter of all plants. The rhizoids multiply and look not unlike roots. This young, still microscopic plant, grows apace, and may soon be distinguished with the naked eye. It looks not unlike a heart-shaped mass of greenish tissue quite flat on the ground, and is called a thallus.
Up to this point, then, we may trace the story of any fern which has thrown off its cloud of spores and from which develops this tiny thallus, looking not in the least like a fern nor as though it could ever be modified into one. Because this thallus is,

(A1) archegonia, (A2) antheridia, and (A3) the rhizoids. B: Prothallus, showing the young plant with its first leaf (B1), its own roots (B3) and the rhizoids of the prothallus (B2). Drawing and legend for it slightly altered from Kraemer.
in the truest sense, merely a preparation for the process that will produce another fern, it is always known as a prothallus. The prothallus is thus the first stage in the reproduction of ferns, a very simple stage, with only the faintest indication that the thallus might be considered the vegetative and its rhizoids perhaps the rootlike counterparts of foliage and roots of mature ferns. As we shall see presently, even this differentiation has not the significance that such a structure in flowering plants would indicate. There is not, as yet, the faintest indication of sexes that need to mate in order to produce their young. The spore has so far only produced a tiny flat body of green tissues with a few rootlike threads, so unlike the fern from which it started that its true significance, or even the fact that it had ought to do with ferns was not known until about the middle of the last century.
This green cushiony prothallus keeps on growing, its heart-shaped mass becoming divided into an obviously left and right hand side and the rhizoids multiplying in number. They are always borne on the lower side next the ground, or next whatever the prothallus may be growing on. Near the notch of the heart-shaped prothallus are developed a few flask-shaped bodies which contain within them an egg cell or single ovum, the female reproductive body. By a series of changes this egg cell becomes embedded in a mucilaginous material. This flask-shaped body with the female egg cell inside is known as the archegonium. From among the rhizoids there may, at about the same time, be found developing small globular organs that have in them a number of tiny cells, each of which has attached many minute threadlike tails. The globular organs, with their minute, tailed cells are known as antheridia, and comprise the male reproductive equipment. Just as in flowering plants, neither the archegonia (female) nor the antheridia (male) can produce offspring without mating and the method by which this marriage is accomplished differs tremendously both in practice and in its significations from that in phanerogams. In the first place, the male and female reproductive cells are separated by a considerable distance, they are both inclosed in structurally different casings, and the whole operation is so microscopic that insects can be of no service. Nor can the wind do for them what we have seen that it does for the pollen of pines and grasses.
Of the aids to fertilization there remains then only the water, which plays such an important part in the mating of the eelgrass and ditch grass among flowering plants. But in these ferns a very different drama is about to be enacted. The male cells, as we have seen, are provided with slender tails, which are movable. They move, in fact, to such good purpose that the male cell can actually swim in the water. Of course its minute size demands only the merest drop of water, in which it will take the only excursion of its brief life. For just as soon as it is mature, a heavy dew or the tiniest particle of water will set free the little male messengers. The water too has not been without effect on the female cell. More remarkable still, this mucilaginous matter contains in it a substance that acts as a lure to the swimming male cells. In any event they do swim directly to the entrance of the female cell’s abode, through it and to her, when the union is effected. At once there is thrown across the entrance a membrane that excludes all other males, and the fertilization is complete. From this union of the male and female cells a true young fern begins to develop. First a young leaf and roots, finally a stem and in the end, of course, a full-grown fern producing spores, ready to renew the whole process.
Some ferns do not follow all the steps exactly as we have outlined, for all of them have not the structure of the typical one whose life history has been sketched above. In the adder’s-tongue fern, for instance there is a stalklike prolongation from the base of the only leaf the plant bears, on which all the spores are borne. In certain others, as in the ostrich fern, the spores are borne on leaflike growths that serve only this function. Most ferns, however, bear spores on otherwise unmodified foliage leaves and the great bulk of them on the under side of such leaves.
There are several things about the life history of a fern that differ fundamentally from any flowering plant and perhaps the chief is what is known as the alternation of generations. A spore, for instance, can never produce a fern as a seed will always produce a flowering plant. In this respect they are like many insects that always have two or sometimes three different stages in their life history. Only by the complicated method of first a spore then the prothallus, from which archegonia and antheridia are produced, followed by the free swimming male cells fertilizing the female, can a fern reproduce itself. As we shall see in the chapter on the History of the Plant Kingdom, this alternation of generations, the absolute necessity of water in which to carry on the fertilization, and above all the ability of the male cells for free swimming in the water, are all landmarks in the development of plant life. In its simplest form fertilization in flowerless plants is characterized by one or all these processes, as it is in the ferns, while in the flowering plants, the act is accomplished by processes, discussed previously, which, in the development of the plant kingdom, mark a period only comparable, in the history of man, to such tremendous achievements as the acquirement of speech or the ability to make a fire.
Ever since the war, the peat-forming mosses, known as sphagnum, have become more widely known to the general public than any of the ten or twelve thousand mosses known to grow on the earth. Its power of absorption, greater than linen bandages, made it extensively used to pad surgical dressings. Hundreds of thousands of these sphagnum

A Small Cloud of Wind-Blown Pollen of the Japanese Red Pine (Pinus densiflora). (Photo by C. Stuart Gager. Courtesy of Brooklyn Botanic Garden.)

A Coconut Grove in the Philippine Islands. The people of tropical regions have more uses for this plant than there are days of the year. Its fruits will float in the sea for months without injury and it is thought to have been distributed all over the tropical world by ocean currents. Its true wild home is not certainly known, but is probably tropical America. See chapter V for an account of the tree. (Courtesy of Brooklyn Botanic Garden.)
dressings were made, and the collection of sphagnum from the bogs in which it nearly always grows was the task of many who could render no other service.
The reproduction of sphagnum is not unlike that of ferns already described. There is the same necessity of a film of water in which the free swimming male can fertilize the female. But some other things about their reproduction of young differ from ferns.
In the first place sphagnum is a nonvascular cryptogam, in that its leaves have no veins or ducts in them and its minute stem is also without those conducting passages that characterize all ferns, and the flowering plants, which are considered the most highly developed of all plant life. (See Chapter I for a discussion of this point, in the section devoted to “Flowerless Plants.”)
In this moss, also, there are small branches, some of which bear only the tiny leaves, but some bear leaves and the reproductive organs. The female or archegonia are much like those in the ferns, and the antheridia or male are also, as in the ferns, minute globular organs in which are the male cells. The branches bearing males are greenish, yellow, or even reddish, quite unlike the ashy gray foliage leaves which give to sphagnum its characteristic ashy gray color. Unlike the ferns, the male cells of sphagnum have only two tails, but they nevertheless swim, tail first, to the female, when the time for fertilization comes. The female branches are found mostly toward the upper end of the plant and bear the archegonia at their extremities.
From what we know of the reproductive stages in the ferns it is now obvious enough that in sphagnum moss, as we ordinarily see it, we have, because it bears antheridia and archegonia, a quite different condition from the ordinary spore-bearing leaves of ferns. For as yet spores have not been developed on the moss. The mating of male and female cells, directly on the plant, proves that in this “plant,” at least, our ordinary notion of this moss is mostly confined to a stage in its life history comparable in ferns to the production of archegonia and antheridia on the fern prothallus. From this mating of the male and female cells there results, as in the ferns, the production of a spore-bearing structure. This consists of a spore case, matured for the most part in the chamber occupied by the fertilized female cell, but ultimately its cap is carried upward. Later on the spore case ruptures, releasing the spores. As in the ferns, these germinate, forming a short green protonema followed by a prothallus. From this a short leafy branch develops, which completes the life cycle, as this is the young moss plant.
In other words, sphagnum, as we ordinarily see it, produces, on the plant, male and female cells which unite to form a spore case with spores in it. These are shed, develop into a protonema which is followed by the prothallus and from this the young moss plant develops. In ferns the conspicuous well-known stage is the spore-bearing one, in sphagnum it is the production of male and female cells directly on what appears to be the mature plant.
There are many other kinds of mosses than sphagnum, and their life histories differ in slight degrees from it. But they all agree in this, that the greenish, feathery little moss plant is a stage in its life history bearing male and female cells, the mating of which produces a spore-bearing contrivance. In most of the familiar green mosses this is a capsulelike body on a short stalk, usually well elevated above the green mass of plants. From this the spores are shed and develop into a protonema or “first thread” just as in ferns. Unlike them, and unlike sphagnum, the green mosses produce no thallus, and the young leaves of the moss are developed directly from this protonema.
The common mushroom that we eat is easily enough divided into a thick stalk, known as a stipe, and a broad hood called a pileus. The under side of the pileus is seen to be composed of thin plaits set closely together and radiating from the center toward the edge. These are known as gills. From among the gills the spores are shed when they are mature, usually foretold by the changing of the color of the gills from whitish to purplish and even to brown or blackish. The spores are then shed and ready for the next stage. From what we already know about ferns and mosses, it is clear that from these spores a mushroom cannot develop without the production of male and female cells and all the rest of that process of hidden marriage that characterizes all flowerless plants. But in most mushrooms no one has ever seen, nor have the most carefully conducted experiments ever demonstrated the germination of the spore. So far as we know at the present, many mushrooms may or may not produce their young through the germination of their spores in their native fields and meadows and the subsequent production of male and female reproductive organs. But if their spores do produce such organs, which all our knowledge of spores makes probable, it is, in a truer sense than in most cryptogams, a case of hidden marriage. The process of producing their young is thus a secret one that scientists have not yet been able to disclose. Of course it is a common practice of mushroom growers to purchase spawn from seedsmen which under favorable conditions will produce many young mushroom plants. This, however, is the production of young without mating of the sexes, a fairly common characteristic of many other plants which will be considered presently.
As we saw in the section devoted to Flowerless Plants in Chapter I, there are many other kinds of fungi than the familiar edible mushroom and their close relatives, the often deadly poisonous toadstools. The reproductive processes in these other fungi are fairly well understood, but they can hardly be included here. In the mold on bread, the yeast used in baking, the rust of wheat and the diseases of other plants and of animals, the individual organism is so minute that it can only be detected under the microscope. Their reproductive processes are, of course, on such a minute scale that they could be followed with profit only by those equipped to study them. They have been described in many botanical textbooks, and those interested in them should consult such books.
In recapitulating the reproductive processes in cryptogamous plants the thing that distinguishes them from all flowering plants is that they bear, in some stage of their life history, a spore. From this, in the great bulk of them, a mature plant never develops. Only by the production from the spore of some contrivance for bearing male and female cells, which may, as in some seaweeds, even be on different plants, can a mating of these be accomplished, and from this union will develop the mature plant. There are many modifications of this plan, but in nearly all of them the presence of water, for the free swimming of the male cell to its mate, is essential. Just as in flowering plants and in all the larger animals, however, the reproduction of young in cryptogams is a sexual process depending on the union of male and female. While in phanerogams that process may well be spoken of as visible marriage, with all the pageantry of insects and beautifully colored flowers, in cryptogams the process is not only a hidden marriage, its ways are sometimes so secret that, even in the common mushroom, the actual mating is conjectured rather than demonstrated.
It is so generally true in all plants that a union of male and female is necessary for the production of young, and, as we have seen in most of them, the process is so uniformly successful that still another mode of producing them seems almost unnecessary. Yet in a surprisingly large number of plants new individuals, both of flowering and flowerless plants, are regularly produced without such a union and where sexuality has nothing to do with the increase.
In the life plant—a thick-leaved shrub from Mexico commonly grown in greenhouses—the leaves are wavy margined. From their edges, especially when injured, many tiny new plants will often start to grow. Even if the leaf is cut up into fairly small pieces many of these will develop young plants, and in various forms of the common rex begonia the leaves are usually cut into small pieces by gardeners for the production of young plants which always sprout from such pieces. It is useless to multiply such cases, as everyone knows of the production of young plants from the ends of strawberry runners, the cutting up of potatoes, the universal garden practice of making cuttings, and the sprouting of willows, all of which are effective by virtue of this faculty of plants to produce young quite without the intervention of different sexes. Not so well known are the cases of a liverwort, a small relative of the mosses, which, if chopped into fine pieces, each will develop into a new plant. We have already spoken of the spawn of mushrooms; and even on sphagnum moss, in addition to its sexual reproduction, it produces sterile branches that will root and, after separation from the old plant, form a new one.
Wherever this tendency is found, whether it be in a microscopic seaweed, some of which know no other means of reproduction, or in the showy begonia, it depends for its success upon a property of the ultimate unit of its structure, the cell. Sometimes, as in bacteria or the most minute seaweeds and in some other kinds, the whole plant consists of a single microscopic cell, when it is said to be a unicellular plant. All others, in which the grouping or modifications of the cell makes more complex structures, such as trees or shrubs and all the plants that grow, both flowering and flowerless, are called multicellular plants. Whether they be of one or many cells, these have the faculty of dividing, and by this division making two where one existed before the division. This division of cells is what happens in the normal growth of plants and it is this division, in more unusual ways, that results in the production of new plants without mating of the sexes. As cells are themselves microscopic, of course their division is equally so, and cannot be described in detail here. It has been many times described and pictured both in books on plants and animals, as it is the ultimate unit of the structure of both.
Plant life, then, seems to be better provided with means to renew itself than most animals, for, as we have seen, it has several methods to rely on. These may be divided into sexual, which includes both that in flowering plants with their visible mating and in flowerless plants with invisible mating, and asexual, literally without sex. In the latter are all those unicellular plants that reproduce themselves by simple division of the cell, and also those flowering plants that either naturally, as in life plant, or by the gardener’s art of making cuttings, produce new plants quite without the intervention of the sexes. Whether it be sexual or asexual, nature has more than fulfilled its obligation to the plant world in providing it opportunities for self-renewal. No matter what apparently unfavorable condition arises and often in spite of an almost unbelievable wastage of potential life stuff, the renewal goes on, or else there is the total disappearance of the species. So strong is this tendency to provide for renewal of their kind that many plants, if injured or cut by a mower, will almost in their last gasp hurriedly flower and set seeds, and we have already seen that the little liverwort, even if cut to pieces, also obeys that nearly universal law of nature: “Be fruitful and multiply.”
THERE are perhaps over 150,000 different kinds of flowering plants known in the world to-day, but the flowerless ones are fewer than these in numbers. No one really knows how many thousands of the cryptogams there may be in the world, for all of them have not yet been described, and there are doubtless thousands of which we merely suspect the existence. Flowering plants are so much better known, and have for 2,000 years been the subject of scientific writings, that their relationships and obvious groupings into families are fairly definite and often easily recognizable.
In our ordinary discussions or gossip of neighbors or relatives, the absolutely necessary starting point is to know their name. Their acquirement of this by christening, or by the adoption of it through the usage of parents, settles for life what they will be called. Plants are also christened, and that ceremony is one of the most important events in our subsequent discussion of them.
As we always have at least two names, one to show that we are a Smith, for instance, and another to fix us as John Smith, so all plants have two names, sometimes three.
And because plants come from all over the world and are studied and loved by people of many different languages, it became necessary very early in the descriptive writings about them to hit upon some device that should insure the name of a particular plant being the same all over the world, whether used by a student in the Imperial University of China, or by a garden enthusiast in Connecticut. At the time when this need for christening plants with names that would pass current throughout the world was getting to be a crying necessity, the language of all learned men was Latin, so it was natural that they should give Latin names to plants. That practice has continued to the present day, and there are even now some botanists that cling to the old custom of describing the newly christened plant in Latin. In the olden days this was always done, so that much of our knowledge of plants has come down to us from early books written wholly in Latin. The unfamiliarity of Latin to most of us, and the terrifyingly difficult spelling of some plant names, has resulted in many people saying: “God made the flowers, but the devil gave them names.” Nevertheless, these Latin names are the only ones we can use without endless confusion, just as we bear the names assigned to us by our parents, and no others.
If we were to go out into the country and pick up a wild rose which seemed to be different from any other rose, it would be necessary, in order to talk about it subsequently, to give it a name. After carefully searching through all the books about roses and finding out that it really is a new kind of rose rather than merely being new to us, we should then be ready to christen our new find. As we have said, all plants bear two names. One of these is a general one, like Smith, for instance, and the other more specific, like John. These general names of plants, and they are always their first names, are, because they fix the plant as belonging to a particular group, known as generic names. The generic name of violets, for instance, is Viola; of buttercups, Ranunculus; of wheat, Triticum; of corn, Zea; and of roses, Rosa. Our new rose then bears, without any act of ours, the generic name Rosa, which was applied to roses many years ago, and must therefore be used for all subsequently discovered roses. This generic name of Rosa, like all other generic names, tells us that roses are a well-recognized group of plants, all more like one another than like blackberries, for instance, and because of this they are said to all belong to the same genus. A genus (plural, genera) is a group of different plants, all more like one another than like anything else. To go back to our new Rosa, we must now apply its second and more specific name. If it were a white rose, and had never before been described, we should almost certainly use for its second name something signifying its color and assign alba as the obvious Latin equivalent of white. The second name is always called the specific name, because it shows us that from all other roses our new Rosa alba differs in being white. It is of the genus Rosa, but it is also and forever after a recognized member of Rosa, to which a specific name has been applied—in other words, Rosa alba is a species of rose. Species are thus plants more like one another than they are like any other member of the genus to which they belong. Rosa alba is a species quite unlike Rosa lucida, or Rosa carolina, or all the other scores of roses already known or described. At the time of christening Rosa alba, we should not only enter its name in a book, as ours would be in a parish register, but do much more than that. We should so carefully describe it and, preferably, illustrate it with a picture that no one coming after could ever mistake Rosa alba for any other rose. It can be readily seen that the christening of new plants is very nearly as serious an affair as christening babies, and furthermore, it is only to be attempted by experts. Because this has not always been done, many plants have been christened two or three times. Of course, these subsequent christenings do not seriously matter, for plants, like ourselves, should have only one specific name, the first applied to them. But their subsequent christenings by the careless and ignorant have enormously increased the difficulty of talking or writing about plants. These spurious names are common throughout the literature of botany and are known as synonyms.
The thing to remember about plants, so far as our need for classifying them is concerned, is that they belong to different species which might almost be considered the unit or simplest recognizable category into which they may be sorted. For convenience, we sort species into genera which may well be considered the next highest category in which plants are grouped. The grouping of genera into tribes, of tribes into families, and of families into still larger categories, has nothing to do with their names, but everything to do with our understanding of how they are related to one another, and what these different categories mean in the great collection of plants all about us. In other words, it reduces to a definite system an apparently hopelessly mixed-up mass of plants that, without some contrivance of the sort, would simply be a lot of totally unrelated specimens of plant life. Actually they are grouped in fairly definite categories, some of which are easily recognizable, and all of which fit into that great scheme of nature where everything may seem chaotic, but to the observant it is really a very pattern of order. What it all means and how plants have been grouped into families will be explained, now that we understand how they have acquired their generic and specific names.
A scientist once visiting in Bulgaria noticed that the peasants in that country frequently lived over a hundred years and, in trying to find out the reason, he discovered that they drank large quantities of sour milk. This is alive with a definite kind of bacterium that is of great benefit to the digestive apparatus, and therefore helps in the prolongation of life. In Bulgaria, in other words, a certain food habit of the people has resulted in a definite prolongation of life and fixes that population as of somewhat different characteristics from people not addicted to sour milk. In Japan a whole race lives largely on fish and rice, and while this is not the cause of their yellow skin, it is almost surely the cause of their generally small stature. Many of the English are tall, light-haired, and blue-eyed people, fond of outdoor life and sports, and among the most highly developed of the peoples of the earth. The climate of that island, their generally large consumption of meat and the outdoor life of so many of them, have resulted in quite definite characteristics that make the typical Englishman an easily distinguishable type.
In studying man we are able not only to divide him into such broad divisions as white, black, and yellow races, but due to their particular country or mode of life there are scores of racial subdivisions of these larger categories that everyone recognizes. Such differences are often based on stature, shape of head, mental characters and many others, but those still finer shades of difference between, for instance, a Connecticut Yankee and a plantation owner in the South, are, while noted by everyone, very difficult to accurately describe.
In attempting to find such major differences in plants, some structural character that would set off one large group of plants from every other group, the botanist has a harder task than the person studying man. For all those differences of language and mentality that make up such a large part of our common knowledge of the different peoples of the earth are characters that are foreign to plants. We are thus thrown back on structure as the chief way in which plants differ, and because their reproductive organs are their most important ones, and therefore least likely to vary, it is upon certain characters of these organs that all flowering plants have been divided.
In the chapter on “How Plants Produce Their Young,” we found that most flowering plants have their ovules in an ovary which, after fertilization, develop into fruit and seed. But some plants, while they have ovules, only bear them naked or between scales, never inclosed in an ovary. This is true in all pines, spruces, hemlocks, and all the host of their generally evergreen relatives. Such trees bear cones, between the scales of which are perfectly naked ovules that develop into seeds (Figure 77) that have never been hidden in an ovary, as have the vast majority of the seeds of other plants (Figure 53). These naked-seeded plants are known as gymnosperms or literally gymnos, naked, and sperma, seed, and comprise all the cone-bearing trees in the world, the larger part of which are always evergreen. In some past ages such trees made up the bulk of vegetation of the earth, but at present they are much reduced in numbers. Familiar examples of these Coniferæ, or cone-bearing trees, are larch, spruce, fir, pine, hemlock, juniper, and yew.

Most of these are evergreen, which does not, of course, mean that they bear the same leaves always, but that only a few drop off at a time and are so constantly renewed that the tree is actually ever green.
All other flowering plants always bear their ovules in an ovary and, because of this fact, are called angiosperms, literally angeion, a vessel, and sperma, seed. These inclosed seeded plants comprise the great bulk of the vegetation of the earth to-day. So far as the temperate zone is concerned, nearly all of them drop their leaves in the fall, and the trees belonging to the angiosperms are thus said to be deciduous trees.
No better idea of the present size and importance of these two groups of plants can be gained than to state the fact that perhaps not over 500 different kinds of gymnosperms, all of which are trees and shrubs, are known. All the rest of the flowering plants in the world, comprising over 150,000 different kinds of herbs, shrubs, and trees of infinite variety, are angiosperms and therefore bear ovules in an ovary, followed by seeds in or on some sort of a fruit. It would almost seem as though the simplest way to dispose of this great mass of plants would be to sort them into trees, shrubs, and herbs. For all of them belong to one of these types of plant growth, and the ancient students of plants, just before the time of Christ, actually divided all flowering plants into these three classes. This, of course, threw the coniferous trees in with all other kinds and, as we have already seen, they differ from all other kinds in the important character of having naked ovules.
Here, again, in order to get some system out of apparent chaos, we must fall back on some fundamental character. And, again, it is the product of the reproductive process in all this host of angiosperms which furnishes the clue. In the seeds of many of them the young embryo has folded up within it two seed leaves, while in all the rest only one. As we saw in Chapter I, these seeds germinate either with a single seed leaf, like corn (Figure 85), or with two seed leaves, like beans (Figure 81). Every one of these angiosperms belongs to one of these classes or the other, and perhaps more extraordinary still is the fact that in those with one seed leaf there are associated certain leaf and flower characters, while those with two seed leaves are always very different.
In the monocotyledons, or plants with a single seed leaf, the leaves are practically always parallel veined (Figure 83), like corn and grass, and lilies and palms, and hundreds of others. Also, they nearly always have the parts of their flowers in threes (Figure 84). That is, they have three sepals, petals, stamens, and often pistils, or multiples of three. The common trillium or wake-robin, for instance, has three sepals, three petals, six stamens, and three styles. With a few exceptions, and nature seems to delight in producing a few such, all monocotyledons have this parallel-veined leaf character and flower parts in threes or multiples of three.
Plants which send up two seed leaves (Figure 81), on the other hand, bear practically always netted-veined leaves (Figure 79), and the parts of their flowers are nearly always in fours or fives or multiples of these numbers (Figure 80). The well-known wild geranium has five sepals, five petals, ten stamens, and a five-lobed or five-celled ovary. There is some individual variation from this plan, sometimes one organ and sometimes another having more or less than the regular number. But so overwhelmingly true are these distinctions that dicotyledons, or plants with two seed leaves, and monocotyledons,

Dicotyledonous and Monocotyledonous growth habits contrasted. Figs. 78-81. The trunk of a dicotyledonous tree showing division of the wood into heartwood, sapwood, and cambium, which the removal of a piece of outer bark exposes. Note the net-veined leaf (79), the seedling with two seed leaves (81), and with the parts of the flower in 5’s (80). Figs. 82-85. Monocotyledonous plant. Note the lack of zones of wood, cambium and corky bark. Such plants have parallel-veined leaves (83), parts of their flowers in 3’s or 6’s (84), and germinate with a single seed leaf (85).
or plants with a single seed leaf, have been for hundreds of years the two great classes into which all angiospermous flowering plants are divided.
Our general view of all the flowering plants may be summarized then as follows:
1. Gymnosperms, or naked seeded plants, include all cone-bearing plants, mostly evergreen and always trees or shrubs. The pine is a familiar example.
2. Angiosperms, or inclosed seeded plants, include all other flowering plants of whatever kind. Divided into: (a) Monocotyledons. Sprouting with one seed leaf, and leaves practically always parallel-veined. Parts of the flower in threes or multiples of three. Familiar examples are corn, grass, sugar-cane, palms, cannas, and lily of the valley (Figures 82-85). (b) Dicotyledons. Sprouting with two seed leaves, and the leaves practically always netted-veined. Parts of the flower in fours or fives or multiples of these numbers. Includes all the remaining flowering plants and is a larger group than the monocotyledons and the cone-bearing plants combined (Figures 78-81).
No matter from what part of the world a totally unfamiliar plant may come, it is always possible to decide into which one of these groups it belongs. That in itself tells us a good deal about its ancestors and its future, “places” it, in fact, in one of those major groups into which all plants are divided. No other characters that plants possess are so important in determining their true position in the scale of plant life as those we have briefly outlined. But merely to sort plants into these large groups does not tell us all we need to know about them. For all plants not only belong to monocotyledons, or dicotyledons, or gymnosperms, but also to smaller divisions of these groups. Just as white men are divided into Englishmen, Frenchmen, etc., so there is the greatest necessity of dividing our large plant groups into smaller and more precise categories.
Some of the chief subdivisions of these large groups have been decided upon the fact that a considerable number of plants in them have some character in common, not found in the remaining plants of the group. Among the monocotyledons, for instance, there is a large class of plants that have tiny flowers between dry, chaffy scales, bear no true petals or sepals, all wind pollinated and are all commonly, though incorrectly, called grasses. These include, strictly speaking, two groups; one, the true grasses in which the stem is mostly hollow and the fruit a grain, while the other, with solid stems and bearing achenes for fruits, are the sedges. The grasses form one family and the sedges another, but while they differ in the characters just mentioned they agree in having flowers of the same general type. Families of plants are thus groups of genera, placed together in the scheme of classification, because they are more like one another than like any other such group. Among the grasses, for instance, are corn, wheat, rice, bamboo, orchard grass, Kentucky blue grass, sugar cane, and hundreds of others, all belonging to different genera, but all those genera grouped into a single family because of their generally similar flowers. Just as the Kentucky blue grass has a generic name (Poa) and a specific one (pratensis), the families of plants must also bear names, usually derived from the generic name of one of the chief genera in it. Because Poa is a large and important genus of the grasses, the family is named after it, with the addition of ceæ. Poaceæ is thus the family name of all the grasses. Among the sedges one of the commonest genera is Cyperus, including many species of the galingale or earth almond. From this genus the sedge family has been named Cyperaceæ (Figure 87). So the rose family is the Rosaceæ, the violet family is Violaceæ, and so on through all the three hundred or more families which contain all the flowering plants so far discovered. Going back for a moment to the Poaceæ and Cyperaceæ, the fact that these two large families are different from each other, but have some characters in common, fixes them as both belonging to one order. Orders are thus groups of one or more plant families, all differing one from another, but obviously related and having some characters in common. The order containing the grasses and sedges is named for one of the families in it with the ending ales. Thus Poales include Poaceæ and Cyperaceæ. Rosales include Rosaceæ and several families.
In other words, individual plants are grouped in species, species into genera, genera into families, and families into orders. These orders are themselves grouped into still larger divisions; there are, for instance, twelve orders comprising all the monocotyledons, and about twenty orders comprising all the dicotyledons. Once we have decided that any plant is a monocotyledon or a dicotyledon, our next step should be as to which order it belongs, then its family, its genus, and finally its species. Needless to say, such studies are necessarily of a technical nature, and while the details of them lie outside the scope of this book, the general plan or scheme of flowering plant classification is as we have outlined it above.
This scheme of plant classification has been developed not only for our convenience in sorting plants into definite categories, but more important still to show, if possible, the relationships, and particularly the development from the simplest to the most complex types of plant life. Thus the monocotyledons begin with the cat-tails, which have mere bristles for calyx and corolla, and lead by infinite gradations to the showy and highly complex orchids, which are considered the climax of the monocotyledonous families. While no general account of the plant families can be attempted here, some of the more interesting in both the monocotyledonous and dicotyledonous groups will be briefly discussed.
Of the simple plants of this group the Grass Family, or Poaceæ (Figure 86), is the most important, for in it are all our turf grasses, the bamboo and sugar cane, besides scores of others. Over 4,500 species are known, and they inhabit every region of the globe. The steppes of Russia and our Great Plains are predominately grassy; in the wonderful bamboo forests in the tropics are also woody representatives of this family. Certain kinds in the tropics grow as vines, with great hooked spines at the joints, so that nearly every kind of growth-form is to be found in the Poaceæ. All agree in having very small flowers, arranged in tiny spikelets, which are themselves grouped in various ways, although the inflorescence is usually some form of spike, or raceme or panicle. The individual flowers are between chaffy scales, of which several make up each spike. Always the lowest two scales are empty, and the flowers begin in the third from the bottom, or

Fig. 86. Blue-joint grass, a common grass of North America and a member of the Poaceæ. Fig. 87. Wool-grass, a tall swamp sedge popularly but incorrectly spoken of as grass. It is a member of the Cyperaceæ or Sedge family, which have usually triangular solid stems, whereas grasses have hollow round stems.
sometimes even above that. The flower is so simple that there is neither calyx nor corolla, only three stamens and one to three styles. The fruit is a grain and the Poaceæ, therefore, are the chief source of cereals. Wheat, rice, corn, oats, barley, millet, and rye, all come from grasses, and all, except corn, are natives of the Old World. They were grown for countless ages before the discovery of America, when Europeans first saw corn used by the Indians. As they are wind-pollinated, the flowers of grasses produce no honey nor colored petals, and the vast majority of them have no odor. Most of them reproduce, not only by seeds, but by rootstocks, and many of them grow so closely together that they form turf. In nearly all of them the stem is hollow, and in the largest of them, the bamboo, these hollow stems are used as water and sewer pipes, especially in India. An exception to the hollow stem is the sugar cane, from whose solid stem the juice is pressed out, that is the chief source of sugar; and our common Indian corn.

A plant of the lily family (Liliaceæ). Note the tendency to net-veined leaves in a monocotyledonous plant. Such instances are common in nature and net-veined leaves are found in certain species of smilax and most of the plants of the Arum family, containing the jack-in-the-pulpit, both monocotyledons.
Much more highly developed than the grasses is the lily family or Liliaceæ (Figure 88), but comprising less than 1,500 species in about 125 genera. They are nearly always herbs, but the Spanish bayonet forms a woody trunk, while the dragon tree of the Canary Islands is an extraordinary plant for a lily relative, one giant specimen of this being 80 feet tall and over 45 feet in circumference.[1] The flowers in the Liliaceæ are nearly always perfect, that is, stamens and pistils are found in the same flower. Its perianth segments are nearly always six, sometimes distinguishable as petals and sepals, but more often, as in the tulip, all colored similarly. The fruits are practically always a capsule that splits lengthwise. Perhaps the different plants in the Liliaceæ, as well as any others, illustrate the fact that plants of any particular family need not look like one another in order to be included in the same family. Nothing could be farther from resemblance than the bulb-bearing onion, the tulip, the Easter lily, the Spanish bayonet, and the dragon tree. Yet they and hundreds of other plants belong to the Liliaceæ. It cannot be overemphasized that it is flower and fruit characters that determine inclusion in any plant family, and similarity of leaves or habit may or may not accompany such characters. Among other well-known plants in the family, which is found throughout the world, are the crocus, the day lily, the dogtooth violet, hyacinth, and colchicum and aloes used in medicine. Many of them produce bulbs, such as onion, tulip, and lily and some of these contain valuable foods and drugs. The great majority of them are insect fertilized and are therefore wonderfully colored, and some furnish rich stores of honey.
But the most highly developed and interesting of all the monocotyledonous plants are the orchids (Figures 89-92). This family, Orchidaceæ,

Fig. 89. Rose pogonia. A native bog orchid with purplish-pink flowers.
Fig. 90. Yellow-fringed orchid. A bog and meadow orchid of the eastern
United States.
comprises over 6,000 species and many varieties, the overwhelming proportion of which live in the tropics. Perhaps 90 per cent of them are epiphytes, or air plants, which are perched high up on the branches or bark of trees, and take all their food and water from the air. All the native orchids of temperate North America grow in the ground, however, and their food habits are unique. They depend for food upon a microscopic organism found inside the roots of all orchids, and which helps them to take in the food from the soil. So many of these orchids are partial saprophytes, and without the associated organism they could not grow. Almost uniformly the

Fig. 91. Whorled pogonia. A woodland orchid with the leaves and flowers whorled at the apex of the stem. Fig. 92. Arethusa. The most beautiful of our bog orchids, with a fringed lip and pinkish-purple flowers which bloom about Decoration Day. Note the highly irregular flowers in this and Figs. 89-91.
Orchidaceæ have only a very few sheathing leaves, entirely without marginal teeth, and some kinds are practically leafless. The flowers, among the most gorgeous in the world, are always irregular in the sense that there is no obvious series of sepals and petals. Both these are so much transformed as to be nearly unrecognizable as such, but in some orchids there appear to be three sepals. More often of the three inner segments of the flower two are somewhat alike, while the third is quite unlike them and is known as the lip; it is among the most variable of any parts of the orchid flower. As adapted to insect visitors, the flowers of orchids are the most wonderfully developed of all plants. Because of their beauty and strange shapes, orchids have been much sought after by collectors, and explorations of tropical, fever-ridden forests have not infrequently ended in death to orchid hunters. New and rare species of them are constantly being gathered by these collectors. One expedition to New Guinea found over 1,000 kinds never before known, and in the last few decades over 4,000 new orchids have been discovered. For these plants orchid fanciers pay large sums, and a single plant of a rare one sold in London at auction for over $500. The chances of collecting such species made expeditions to the tropics frequent during the latter half of the last century.
These three families, Poaceæ, Liliaceæ, and Orchidaceæ, are perhaps the most important of all the monocotyledons, although commercially the palms, or Palmaceæ, are extensively used. It is impossible to describe or even mention all the monocotyledonous families, but a list of the more important is added. The families are arranged in the order that seems to reflect the development from simpler ones to the most complex, and is the sequence of such families used by nearly all botanists in describing the plant families of the world:
Typhaceæ—The cat-tails. Tall, reedlike swamp plants found throughout the world. One genus and about ten species.
Pandanaceæ—The screw pines. Shrubs or trees with stout, woody trunks and mostly prickly margined, long sword-shaped leaves. Confined to the Old World tropics.
Poaceæ—The grasses. Noted above.
Cyperaceæ—The sedges. Grasslike plants with solid, often triangular stems. Very often inhabitants of wet places. Throughout the world, Crex rugs are made from a species of Carex, the largest genus in the family. About 75 genera and 3,200 species.
Palmaceæ—The palms. All trees or shrubs, or sometimes climbing vines. Includes the coconut and palm-oil trees, two palms of tremendous economic importance. Inhabitants of tropical and warm regions, and only very few found in the United States. Over 130 genera and 1,200 species.
Araceæ—The arums, of which the jack-in-the-pulpit is our best-known native representative. They are nearly always herbs, often of giant size, and the great majority are found in the tropical regions. Flowers very minute, crowded together on a central column (the spadix), and this often surrounded or having at its base a leaflike appendage (the spathe). Calamus root and the skunk cabbage are also native representatives. About 105 genera and over 900 species.
Liliaceæ—The lily and related plants, noted above.
Smilaceæ—Smilax. Mostly prickly vines; our native kinds often called cat briers. Sarsaparilla comes from at least four species of Smilax. Three genera and about 300 species, mostly natives of tropical, but a few of temperate regions.
Amaryllidaceæ—The amaryllis family, noted chiefly for the sisal fiber that comes from a species of Agave, which is one of the many different kinds of century plant. The family has usually capsular fruits and black seeds, and the narcissus and amaryllis of our gardens are well-known members. About 70 genera and 800 species from tropical and warm countries; a few in temperate regions, mostly herbs.
Iridaceæ—The iris, the source of orris root, and containing some our most beautiful garden plants, the blue-eyed grass of fields, and over 50 genera and 100 species are found in this family. Nearly throughout the world, and nearly all herbs.
Musaceæ—The banana and traveler’s-tree. Giant herbs, in the banana having the largest leaves known, frequently twelve feet long and two wide. Natives of tropical and warm regions, and 4 genera and 75 species are known. Flowers often very irregular, and in Strelitzia gorgeous.
Orchidaceæ—The orchids, already noted.
While there are many thousands of plants contained in these monocotyledonous families and in the others not mentioned here, they make up only about one-third of the total number of different kinds of plants known in the world. But in grasses and sedges, in the rushes and a few other families, the number of individuals is greater than in probably any other plant family.
All the great bulk of the flowering plants not included in the monocotyledons or the gymnosperms belong to about two hundred plant families that are included in the dicotyledons. In all of them the seed sends up two seed leaves, there are generally netted-veined leaves and the parts of the flower are in fours or fives or multiples of these numbers. In such a large aggregation of plant families there are three well-marked divisions, namely, those that bear no petals or sepals, those that do bear them and where they are separated to form individual sepals or petals, and those where the petals are united to form some sort of a tubular or at least connected corolla. These divisions are perhaps best shown thus:
(a) Apetalæ—Including families where the petals are never present, and in some there is even no calyx. Examples: walnut, hickory, willow, and oaks.
(b) Polypetalæ—Petals present but separate, not forming a tubular or connected corolla. Examples: buttercup, rose, pea, apple, geranium.
(c) Gamopetalæ—Petals united and forming some sort of a tubular or connected corolla. Examples: garden primrose, gentian, salvia, mint, snapdragon, and the daisy family.
Any attempt to describe the families contained in these three divisions of the dicotyledons would take all the rest of this book and crowd out other things about plant life that must not be omitted. All that can be done here is to outline briefly a few families in each division so that we shall have fixed in our minds what the general principles of plant classification are and how these are illustrated by well-known plants. There are many books that deal with this subject in great detail and to them the student should go for further elaboration of the subject. It is one of the most interesting phases of the study of botany, but it demands a longer and more intensive study than can be included here.
(a) Apetalæ—These include families of plants that are the simplest in structure of all the dicotyledons. In all of them there are no petals and in some both petals and sepals are lacking, leaving only essential organs. Taking first those families that have neither petals nor sepals we find that most of them bear their flowers in catkins, a flower cluster familiar enough in the pussy willow. Some of these families are the following:
Juglandaceæ—Trees with compound leaves, flowers in catkins and fruits inclosed in a thick husk. Examples: walnut and hickory. Six genera and over 30 species.
Salicaceæ—Shrubs or trees with simple leaves, flowers in catkins and capsular many-seeded fruits, but no nuts. Containing only willows and poplars. Two genera and over 200 species. (Figure 93.)
Myricaceæ—Shrubs or trees with simple usually fragrant leaves, flowers in catkins and fruits one-seeded. The bayberries. Two genera and about 35 species. (Figure 94.)

These and some other families of close relationship are the least developed, in their flower structure, of any of the dicotyledons. All of them bear only the essential organs of reproduction in their very simple flowers. In the walnuts and hickories the different sexes are in different flowers on the same plant, in the willows they are even on different plants, and in Myricaceæ they are often found both ways. All of these flowers are wind-pollinated, so that they bear no honey, usually have no odor, and of course their need for showy petals for attracting insects is nil, although some pollen-eating insects visit them.

Somewhat higher in the scale of plant life are those families that, while still lacking petals, do have sepals. Their flowers are for this reason much better protected against rain or other inclement conditions, which means that they are so much the more likely to reproduce their kind. At least two of these slightly more developed families cling to the habit of bearing some of their flowers in catkins, however. Other families are also found in other parts of the world, but in North America this group is represented by:


Betulaceæ—The birch, alder, hazelnut, and hornbeam. Both staminate and pistillate flowers in catkins. Fruit a small one-seeded nut or a winged samara in the birch. All wind-pollinated. Six genera and about seventy-five species, nearly all from north temperate zone.

Fagaceæ—The oaks, beech, and chestnut. All trees or shrubs with at least their staminate flowers in catkins. Fruit inclosed in a bur (chestnut and beech) or borne in a cup (the acorns of oaks). At least five genera and about 375 species, widely distributed.
The habit of bearing some or all their flowers in catkins which flower usually before the leaves appear, and of having such flowers wholly at the mercy of precarious winds, is, if not lost, at least much less frequent in the remaining families of the apetalæ. All the others, while still without petals, do have sepals and some of these are colored so that insect visitors are likely. There are too many of these families to be enumerated here, but two of the chief are:
Ulmaceæ—The elms and hackberry. Trees or shrubs with minute greenish or yellowish flowers crowded in small clusters or in spikes. Fruit a dry nut or one-seeded and winged; or in the hackberry a drupe, one of the first evidences of even a slightly fleshy fruit in the dicotyledons. About 13 genera and 140 species, widely distributed.
Polygonaceæ—The buckwheat, knotweeds, common dock, and many other genera. Sepals often colored white or pink so the flowers are sometimes at least insect-pollinated. Flowers small and crowded in various clusters, often in a spike. Fruit an achene, a dry fruit familiar enough in the buckwheat. About 40 genera and over 800 species, mostly herbs or vines here, but often trees in the tropics.
From here on plant families leave, with some exceptions, the greenish or otherwise inconspicuous flower color, and somewhere about here they begin to rely more upon insect fertilization for the perpetuation of their kind. None of those so far mentioned have any petals to their flowers, but in the pink family or Caryophyllaceæ we find the first evidences on any considerable scale of the presence of sepals and petals, the latter usually beautifully colored. Familiar representatives of this family are the pink, carnation, chickweed, corn cockle, and the stichwort. There are over 50 genera and 1,000 species, nearly all in temperate regions.
The apetalous families appear to show a development from catkin-bearing trees with the sexes separated, and with neither petals nor sepals, through the Polygonaceæ, with often colored sepals, and the beginnings of insect fertilization. In Caryophyllaceæ, the most highly developed of them all, there are, besides the sepals, often or usually petals, and the reliance on insect fertilization is nearly complete. There are many transitional stages which cannot be included here, but they show step by step the development of the apetalous families from perfectly naked reproductive organs to the next larger group, the polypetalæ, where the process of increasingly complex flower development will now be sketched.
(b) Polypetalæ—In this large group of plant families the petals are free and quite separate, but as if they had not yet lost all the characters of the apetalæ, some families show incompletely the general characteristics of their more stable neighbors. There are, for instance, no petals in many species of the buttercup family, none in the sweet-gum tree nor in the maples, and a few others. But in spite of occasional exceptions this large group of polypetalous families do usually bear separate petals and sepals, and are among the most important of all the plant families. As they number over a hundred and contain thousands of species, all that can be done here is to mention a few typical or important ones. Just as in the apetalæ the families in this large group appear to show definite stages in development from simpler to more complex forms. But the steps are harder to trace and what appears simple characters in some plants are very complex in others.
While all the families in this group have separate petals some of them show a tendency to have united sepals, a character of perhaps some advantage and certainly very common among the still more developed gamopetalæ. Some of the families that have separate sepals agree in having the stamens inserted below the ovary. Of these the following three families may serve as types.
Nymphæaceæ—The water lilies. Aquatic plants with usually large showy flowers in which the calyx, corolla, and stamens often merge one into the other so that it is sometimes difficult to know where one series ends and the other begins. Five genera and 45 species throughout the world.
Ranunculaceæ—Buttercup family. Includes buttercups, clematis, columbine, meadow rue, golden seal, marsh-marigold, hepatica, and scores of other native plants. All herbs, except a few semiwoody vines like clematis. Sepals always present, and where no petals are found, as in marsh-marigold, colored like them. Highly irregular flowers are not uncommon, as in columbine and monkshood. The fruits are berrylike in some genera and in others dry capsules. Thirty-five genera and over 1,000 species throughout the world, but most abundant in temperate regions.
Lauraceæ—Laurel or sassafras family. Includes besides them the guava and cinnamon and camphor trees, all tropical, and the native spice-bush. All trees and shrubs with small, yellow, or greenish-yellow flowers and usually aromatic juice. Fruit a one-seeded drupe or a berry. About 40 genera and over 1,000 species, nearly all tropical, but a few in the United States.
At this point, in the sequence of plant families, there are two or three families that bear quite different fruits than any heretofore noted, and in one of them, at any rate, the four petals are in the form of a cross. So uniformly is this true that the family was for years known as the Cruciferæ, but is better known as Brassicaceæ, from Brassica, the generic name of the mustard. This large mustard family mostly has fruits known as a silique or silicle, which are pods that split into two valves; and yellow or white, rarely pinkish flowers. The juice is always somewhat acrid, familiar through the pleasant pungent taste of water-cress, but none of the family is poisonous. There are over 200 genera and nearly 2,000 species of wide distribution, and common representatives include the cress, mustard, horse-radish, garden stock, sweet alyssum, cabbage, cauliflower, brussels sprouts, radish, and turnip.
Between the Brassicaceæ and the following families there are many others that cannot be mentioned here. Somewhat farther along in the sequence are a group of families, large and important, and all having their stamens inserted around or even above the ovary, and in which the sepals are partly or wholly united. They include some of our most beautiful flowers and useful fruits. Of the many closely related families that agree in these characters the two most important are:
Rosaceæ—Rose family. In the broad sense including, besides the rose, the strawberry, blackberry, apple, pear, peach, plum, besides many herbs with wholly dry fruits. There are always five petals, five lobes to the partly united calyx, but numerous stamens. They may be herbs, shrubs, or trees, with simple or compound leaves, but these are nearly always alternately arranged. There are over 100 genera and nearly 2,000 species. Because of the size of the Rosaceæ and differences in fruit, the apple and its relatives are often included in a separate family, the Malaceæ (Figure 96), and the peaches and plums in Amygdalaceæ. The general structure of the flower is sufficiently uniform, however, for them all to have been included in Rosaceæ (Figure 97).

A prickly shrub related to the apple, which, with the plums, cherries, pears, strawberry, blackberry and hundreds of other plants are all grouped in the Rosaceæ or rose family.
Papilionaceæ—Pea family. A large family having characteristic pealike flowers, a description of which has already been given in Chapter I. They all bear legumes, a pod that splits down one side, not both, as in Brassicaceæ. Besides the pea, the bean, vetch, alfalfa, lentil, locust tree, and dozens of valuable timber trees in the tropics belong here. Flowers all showy and absolutely dependent upon insects for fertilization. Seeds highly nutritious in many genera, and the roots of nearly all have bacteria associated with them. (See chapter on Food Habits of Plants.) Three hundred and twenty-five genera and over 5,000 species of wide distribution, but most frequent in the tropics.

There are many other smaller families in different parts of the world which hover, as it were, about these two giant plant families that make definite landmarks in the scheme of plant classification. The character of having partly united sepals and numerous stamens inserted around, or even above the ovary, give to all the flowers of the Rosaceæ and related plants a general family resemblance that is very striking. The pea family, and its relatives, also have flower and fruit characters of remarkable uniformity, considering the tremendous number of species.
From this point on to the end of the polypetalæ there are scores of plant families, all agreeing in having a compound ovary, that is, one that is more than one-celled, and in also having quite distinct and separate sepals. Their agreement in these characters, however, ends all other evidences of relationship, and it is beyond the scope of this book to go into the details of each, or even a few of them. A list with some brief notes on those most important must suffice here.
Geraniaceæ—Geranium family. Includes also the common garden as well as the wild geranium. Fruit splitting into five parts. Leaves always divided or even dissected; 12 genera and 470 species, all herbs.
Anacardiaceæ—Includes the tree of heaven, sumac, poison ivy, and in the tropics, the mahogany, all trees, shrubs, or vines; 60 genera and 500 species, mostly in the tropics.
Malvaceæ—Mallow family, including, besides the marshmallow, the rose of Sharon, and cotton. They all have the stamens united into a column or tube which surrounds the style. About 40 genera and 900 species of herbs, shrubs, or trees of wide distribution.
Cactaceæ—Cactus family. Nearly all desert plants, with no leaves or practically none, and greenish stems that function as leaves and also store water. Of the greatest variety of form and always bearing numerous petals and fleshy fruits, of which the prickly pear is familiar enough. Of 40 genera and over 1,000 species, all, but a handful, come from North and South America.
Umbelliferæ (often called Ammiaceæ)—The climax of the polypetalous families, and nearly always bearing flowers in umbels. There are usually many flowers, sometimes several hundred in each cluster. Familiar examples are parsley, celery, parsnip, Queen Anne’s lace, and many others. The seeds often contain an aromatic oil, as in caraway, and some are violently poisonous, as the water hemlock. About 250 genera and over 2,000 species, all herbs, widely distributed, but most common in temperate regions.
We have seen from the foregoing the probable development of dicotyledonous plants from those simplest ones, where, as in the pines, there is merely a naked ovule between scales, through the catkin-bearing trees, without petals or sepals, and all wind-pollinated, to families where just an inconspicuous and, subsequently, a colored calyx is found, and after this the dawn of those plants that have complete and perfect flowers. Among the latter all those so far noted have separate petals, but after the Ammiaceæ, or carrot family, there appears a new character, setting off practically all other dicotyledonous plants from those already treated. This new character—and hints of it are found before it reaches the perfection found in the subsequently described families—is that of the petals being united to form some sort of a connected or, more often, a tubular corolla. The petals are represented merely by the lobes of the corolla, mostly four or five, and in many families of this group, known as the gamopetalæ, literally, united petals, this tubular corolla is irregular and often beautifully formed. In salvia, for instance, there is a hoodlike upper part overhanging the lower tubular part. Other familiar examples of these irregular corollas are the garden snapdragon, Oswego tea, skullcap, pentstemon, and many others.
(c) Gamopetalæ—The earlier families among those generally having united petals seem not yet quite sure of their new character, for a few of them hark back to the condition of having, in some genera, quite separate petals. One of the first families in this series, the Ericaceæ (Figure 98), or heath family, has several genera in which this is true, notably in the Labrador tea and the sand myrtle among native plants, and some foreign relatives. The Ericaceæ are almost exclusively shrubs or trees, but some of our native sorts, such as trailing arbutus and wintergreen, are practically herblike, although they are, strictly speaking, woody plants. The family is remarkable for containing beautiful flowered garden plants, such as the hundreds of species of South African heaths, the heather, the azaleas and rhododendrons, and our beautiful native Rhodora, about which Emerson wrote one of his most beautiful poems. The flowers in the heath family are often perfectly regular and bell-shaped, but sometimes irregular, as in azaleas and several other genera. Nearly all the family rely on microscopic organisms to get their food, and some close relatives, like the Indian pipe, are saprophytes. There are over 70 genera and 1,200 species widely distributed. Central Asia is the home of most rhododendrons and azaleas, scores of species being found in the upper reaches of the Himalayas.
The Ericaceæ are typical of many families in the first group of the gamopetalæ, in that all of them, with a few exceptions like the cranberry, have a superior ovary. That is, the petals and sepals arise from the base or below the ovary, and consequently the mature fruit in such plants is never crowned with the remains of the withered calyx, as blueberries always are and all other gamopetalæ that have an inferior ovary. The character of having an inferior or superior ovary separates the gamopetalæ into two large groups of families, the heath family and many others, with superior ovary, and a few but numerically very important families that always have an inferior ovary.

Taking first the families that all have a superior ovary, we must, for lack of space, exclude most of them from here. A few of the most important, or typical, after the Ericaceæ, are:
Primulaceæ—Primrose family. All herbs in which the stamens are as many as the lobes of the corolla and inserted on them. The flowers are quite regular. They all have some form of a capsule for fruit, which in most generally split lengthwise. Familiar examples include the garden primrose (not the wild evening primrose), yellow loosestrife, the star flower, pimpernel, shooting star, and the beautiful cyclamens. A few members of the family are slightly luminous in the dark, apparently an attraction to night-flying insects. About 28 genera and over 400 species, mostly from the northern hemisphere, a few in temperate South America and South Africa.
Gentianaceæ—Gentian family. Over 700 species in 70 genera, all bitter herbs, with opposite leaves, quite without teeth and beautiful, sometimes fringed, always regular flowers. In this and related families the stamens are of the same number as the lobes of the corolla, and always alternate with them. Gentian and sea or marsh pinks are our best-known native representatives, while some related plants are medicinal.
There are many other families in this part of the scheme of plant classification that have minor differences among themselves, but agree pretty generally in the number and position of their stamens, their superior ovary, and, on the whole, in the regular flower. Irregular and regular flowers may be recognized at once by cutting them lengthwise through the middle. In regular flowers there would be as much on one side as on the other of the dividing line, and in irregular ones quite obviously more on one side than the other. The character of all the genera in a family having irregular flowers begins to occur here with greater and sometimes exclusive frequency. In the mint family, or Lamiaceæ, nearly all of its 160 genera and over 3,000 species have two-lipped or irregular flowers. The garden salvia well illustrates the type.
The Lamiaceæ or Labiatæ, as they are often called from the two-lipped corolla, are herbs locally, but in the tropics often shrubs or trees. Almost universally they have four-sided stems and opposite leaves without stipules. The flowers may be solitary, much more often they are crowded into various kinds of clusters. The four stamens are borne on the corolla tube, and nearly always there are two long and two short ones. The family is universally fertilized by insects, and some of the flowers are wonderfully adapted for this end. Common examples, besides the salvia, are mint, thyme, skullcap, hyssop, bugle, blue curls, catnip, hedge-nettle, coleus, and Oswego tea. Most of the genera contain heavy odorous oils in their foliage, from which oil of mint, pennyroyal, lavender, rosemary, marjoram, savory and balm are the best known. These volatile oils give to members of the family their characteristic and often very beautiful odors.
There are many other families of plants, some with irregular and others with regular flowers, that appear to group themselves around the Lamiaceæ, all of which agree, in spite of individual differences, in having a superior ovary. The remaining families of the gamopetalæ, however, always have an inferior ovary, usually obvious by the insertion of petals above the ovary, and in the fruit often conspicuous by the remains of the withered calyx still clinging to the top of the fruit. Only two of the scores of families, with inferior ovary and gamopetalous corollas, will be mentioned here, both of which are important.
Rubiaceæ—Madder family. Common examples are the creeping bedstraw, the sweet woodruff, partridge berry, button bush, and bluets or quaker-ladies. All, except one of these, are herbs, but in the tropics the Rubiaceæ are nearly all shrubs or trees. Among those are the coffee, quinine, and ipecac. All the family have opposite leaves (a few verticillate) and stipules, regular flowers, with stamens as many as the corolla lobes, and alternate with them. The fruits are a drupe, berry, or capsule. Over 340 genera and 6,000 species are known almost throughout the earth.
Compositæ or Carduaceæ—The daisy family and the largest and most complex of all the plant families. As the culmination of the scheme of plant classification, they show the greatest perfection in the arrangements for cross-fertilization. For a description of their flower structure, see Chapter I, page 44, and Figures 43-45. Some of the Compositæ have no ray flowers, others are all ray flowers, but the great bulk of them bear both tubular and ray flowers in a single head. This may be single, or more commonly it is arranged in various kinds of clusters. Each head is surrounded at its base by one or more series of usually tightly overlapping bracts, incorrectly called a calyx by the unobservant. The Compositæ include over 900 genera and 11,000 species from all parts of the world. Most of them in America are herbs. Daisy, dandelion, dahlia, chrysanthemum, sunflower, boneset, chicory, lettuce, and scores of other examples could be cited, all herbs. In the tropics the Compositæ are more often trees and shrubs. The family contains many economic plants such as arnica, chamomile, artichokes, inula, and many others.
This all too brief account of the grouping of plants in families, and the sequence of these from the comparatively simple naked-seeded pines, through monocotyledons, the apetalous dicotyledons, followed by the polypetalous dicotyledons, and culminating in the Compositæ among the gamopetalous families, gives us merely a hint of what are the characters upon which plants are divided. While the details are necessarily omitted, the gradual development from naked-seeded plants, wholly at the mercy of the elements, up to those which are marvelously provided with contrivances to insure cross-fertilization, has been traced. Perhaps no other phase of botanical study offers such a rich opportunity as this, for upon the solution of some of the problems of plant classification depends the answer to many questions about the history of the earth and man’s ability to live on it. Certain of these plant families have lived on the earth hundreds of thousands of years before man first came. Others have apparently arisen comparatively recently. Many botanists believe that all the monocotyledons should be placed after the dicotyledons, as the latter may be a more ancient type than the former. How these different plant structures, some very ancient and others more recent, help to show us some of the history of the earth, will be treated, among many other evidences of a plant nature, in the chapter on “The History of the Plant Kingdom.”
FOR perhaps the largest number of readers the chief value of plants is what they furnish in the way of food, clothing, fuel, and so forth, and from this standpoint alone the study of them is more than worth while. It is unnecessary here to enumerate all the thousand and one things that we get from plants, and no attempt will be made to do so in the following pages. But certain plants like wheat, corn, cotton, jute, rubber producers, and tobacco have so shaped the life of the people, so absolutely dictated the development of whole regions of the earth’s surface that their stories are part of the history of mankind. What our cotton fields of the South, the wheat and corn fields of the Middle West, the jute in India, and the coconut palm and sugar cane in the tropics have done to dictate the economic destiny of those regions is common knowledge. Hundreds of less important plants throughout the world contribute their quota to the huge debt that man owes to the plant world. Probably no other feature of plant life offers such attractions as the study of man’s uses of plants, which is known as Economic Botany, and for which our Government maintains a large staff of experts. Some of the publications of this bureau are textbooks of the greatest value to those who grow or import plants or their products. What that amounts to in the aggregate no one can readily estimate. It certainly exceeds all other commerce combined.
Those early ancestors of ours that roamed over northern and central Europe between the periods of ice invasion, which at times made all that country uninhabitable, tell us by the relics of them found in caves that agriculture was then unknown. Living mostly by the chase and on a few wild fruits picked from the forest these half-wild and savage people wandered wherever game was plentiful and the continental glacier would permit. But there came a day when one of these races began the cultivation of some of the wild plants about them and with that day dawned the real beginning of man’s use of plants. And with that day also these simplest of our ancestors stopped their wanderings in large part and became farmers, albeit very crude ones, as their primitive stone implements show. They did not give up the chase, but their collection into more or less permanent camps or villages began with their cultivation of plants. Just when this happened no one can say, but most estimates of the time since the last ice age indicate that it could not have been much less than forty thousand years ago. And considerably before this, and long before the use of metals by man, we find these stone implements of agriculture and the probable beginnings of that great reliance upon plant life which the modern world has carried to such tremendous lengths. Unfortunately we do not know what plants these “Men of the Old Stone Age” grew in their primitive gardens, and it is thousands of years after this, and after man’s discovery of the use of metals, that we know definitely what plants he grew and how he used them. Unquestionably some of the early uses of plants, such as dyes for the face or for “rock pictures” are very ancient and are found long before any sign of agriculture, but as food in the sense of being produced food rather than that gathered from the wild, there are only the faintest traces until, in the remains of the lake dwellers in Austria, a single grain of wheat was discovered. Their metal instruments showed them to have been familiar not only with this, but with other plants, and it is well to remember that these people lived far longer ago than our most ancient historical records such as the Egyptians or Chinese. Both the latter, so far as our oldest records of them show, were an agricultural people who had enormously developed man’s uses of plants as compared with the men of the stone or bronze ages, whose agriculture must perhaps forever be a secret of the past.
The discovery of the grain of wheat in the remains of the lake dwellers tells us some things about men’s travels even in those early days, for wheat is not a wild plant there and must have come to central Europe from a great distance. Researches upon the home and antiquity of wheat are not very definite, but its occurrence as a wild plant somewhere in Mesopotamia or the vicinity appears to be indicated. The Chinese grew it 2700 B.C. and the earliest Egyptians spoke of its origin with them as due to mythical personages such as Isis, Ceres, or Triptolemus. From its ancient and perhaps rather restricted home it has gone throughout the temperate parts of the earth and now forms perhaps the most important source of food. Although many different kinds of wheat are raised in different parts of the world most of them have been derived from one wild ancestor, Triticum sativum. Forms known as hard and soft wheat and dozens of others suited to different regions or market conditions have been developed by plant breeders. As the most important of all the cereals it has been much studied, and its cultivation in America is on such a tremendous scale that we furnish a large part of the world’s supply. Russia, Argentina, and the southern part of Australia also raise large quantities. The plant is a grass and the “seed” is really a grain or fruit in which the outer husk tightly incloses the true seed.
It were perhaps well to note here that popular stories about the germination of grains of wheat taken from Egyptian mummies are not true. Wheat and even corn are sometimes given to travelers, and it is taken from these ancient Egyptian tombs. But it was not put there by the early Egyptians, as the presence of corn proves only too well. For this cereal is an American plant unknown before Columbus and 1492. Arabs and others have recently inserted various seeds in these mummies, some of which undoubtedly have germinated—hence the fable. The early Egyptians did put seeds in their mummy cases, but none have ever germinated.
The grass family furnishes this second most important cereal to all Americans and Europeans, although among inhabitants of tropical regions rice is perhaps more important than either wheat or corn. With the discovery of America the early travelers found the North American Indians, the Mexicans, and the Peruvians all growing corn and using it on a considerable scale. It must have been grown for hundreds of years before that time, as its wide distribution and many varieties testified even at that date. Its true home nor its actual wild ancestor has never been certainly determined, but a wild plant very closely related to our modern corn is found in the northern part of South America, and either there or in Central America is apparently the ancient home of corn. So much had corn entered into the life of the early Mexicans that the first Europeans to visit that country found the Mexicans making elaborate religious offerings to their corn goddess. And, as in Egypt, the tombs of the Incas of Peru contain seeds of the cereal most prized, which in the case of corn consists of several varieties. While their civilization is not as old as certain Old World races, the cultivation of corn must date back to the very beginnings of the Christian era. It is now spread throughout the world in warm regions, and as early as 1597 it was grown in China, a fact that led to the erroneous notion that China was its true home. Perhaps no fact is more conclusive as to its American origin than that corn belongs to a genus Zea, which contains only the single species mays, with perhaps one or two varieties, and that until the discovery of America Zea mays or Indian corn was unknown either as a wild or cultivated plant. Such an important cereal, if it actually were wild in the Old World, would have spread thousands of years ago as wheat did, and Columbus and his adventurous successors would not have brought from the New World a food that has since become second only to wheat.
Field corn of several different sorts, pop corn, and sweet corn were all developed by the Indians from the ancient stock, but comparatively recently the juice of the stem has been used for making corn sirup. The use of the leaves for cattle feed is known to all farmers, and from its solid stems it is now likely that some fiber good for paper making will be extracted.
Both wheat and corn are grasses that are cultivated in ordinary farm soils, but rice is derived from a grass that is nearly always grown for part of its life in water. It is taller than wheat, but not so tall as corn, and its wild home is in the tropical parts of southeastern Asia. It is still grown there in greatest quantity, and in the Philippines, while only a small part of the world’s supply comes from the New World. There are perhaps more people that rely upon rice for food than upon wheat and corn combined. It still is the principal article of diet of the inhabitants of China, Japan, India, and dozens of smaller Old World regions, while its use as a vegetable in tropical America is practically universal. A considerable part of the starch manufactured in Europe still comes from rice, and in India the intoxicating beverage arrack is made from it. The Japanese saké, a sweetish intoxicating liquor, is also made from rice. Notwithstanding its wide use it is not as nutritious as wheat or corn, being much lower in proteins than either of them.
More than 2800 years before Christ the Chinese cultivated rice, for at that time one of the emperors instituted a ceremony in which the grain plays the chief part. It has been grown on land useful to almost no other crops as it is usually subject to inundations. Some varieties, however, have been developed which will grow on uplands and these are grown even on terraced land both in China and the Philippines. It needs a heavy rainfall, however, and grows best in lands that are flooded. It is occasional dry seasons that produce the famines of India when the crop fails. The botanical name of rice is Oryza sativa, and it is known now as a wild plant in India and tropical Australia. Its introduction into Europe must have been long after wheat, for rice is not mentioned in the Bible, and was unknown in Italy before 1468, when it was first grown near Pisa. Rice paper, which some people think is made from this grain, comes from the pith of Aralia papyrifera, a tree of the rain forests of Formosa, related to our temperate region Hercules’-club.
In the chapter on what plants do with the material they take from the air and soil we found that sugar was one of the first fruits of that process. In at least two plants the overproduction of sugar is on such a great scale that our chief supplies of this substance now come from these two plants—the sugar cane, which is a tall grass, and the sugar beet. Hundreds of other plants produce surplus sugar, but for commercial purposes these two, and the sap of the sugar maple (Acer saccharum), are our chief sources of supply.
Cane sugar is an Old World grass known as Saccharum officinarum, frequently growing twelve feet high, and with a solid woody stem, quite unlike our ordinary grasses. It looks not unlike corn on a stout stem, and it is the stem which is cut and from which the sweet juice is pressed out between great rollers. The pressed-out juice goes through various processes in the course of which first molasses, then brown sugar, and finally white granulated sugar are produced.
Our consumption of sugar is now on such a scale that we scarcely realize that before the days of Shakespeare it was very scarce and expensive. Even as recently as 1840 it regularly sold in England for forty-eight shillings per hundred pounds, wholesale. At that time the total consumption in the world was only slightly over a million tons, while to-day it is over fifteen times that amount. The plant is native in tropical Asia, but just where is not known, nor are wild plants found in any quantity. It has been much modified by long cultivation, and has been reproduced by root-stocks for so long a period that it is rare for the plant to bear flower and seed. It has been known in India since before the Christian era, and was taken from there to China about 200 B. C. Neither the Greeks nor Romans knew much about it, nor do the Hebrew writings mention it. Somewhere in the Middle Ages the Arabs brought it into Egypt, Sicily, and Spain. Not until the discovery of the New World was it cultivated on any considerable scale, when the climate of Santo Domingo and Cuba and the African slaves imported to those islands afforded conditions that resulted in Cuba at least being one of the world’s chief sugar-producing countries. Sugar cane is now grown all over the earth in regions with a hot, moist climate, India and neighboring countries producing over half the world’s supply. Practically all the sugar produced in India is used there, however, so that the American tropics furnish to Europe and America about one-third of the world’s total consumption of cane sugar.
In 1840 under fifty thousand tons of beet sugar were produced, while in 1900 more sugar from this plant was made than from sugar cane. Considerably more than half this beet sugar was grown in three countries, Russia, Austria, and Germany, which explains what the great war has done to the sugar market. The plant from which beet sugar is derived is botanically the same as the common garden beet, Beta vulgaris, which is wild on sandy beaches along the Mediterranean and Caspian seas, and perhaps in India. Much cultivation has made this slender-rooted plant into the large-rooted vegetable we now have and its sugar content was much increased by Vilmorin, a French horticulturist. Many garden varieties are known, and some of these are grown in the United States, where beet sugar is produced, although in 1910 less than half a million tons were made here as against over four million tons in Europe. While the beet as a vegetable was known perhaps a century or two before the time of Christ, it was not until 1760 that its sugar content was understood, and it was nearly eighty years later before beet sugar became commercially important. Its cultivation in England on any considerable scale did not begin before the beginning of the present century.
Among the largest herbs in the world are the ordinary banana plants, now cultivated throughout the tropical regions, but originally native in the Malay Archipelago. From there it spread into India, and the early Greeks, Latins, and Arabs considered it a remarkable fruit of some Indian tree. It is actually a giant herb with a tremendous fleshy stem, formed mostly of the tightly clasping leaf bases, the blade of which is frequently ten to twelve feet long. In nature the blade splits into many segments due to tearing by the wind, a process that the plant not only tolerates but aids. The leaf has a thinner texture between its principal lateral veins, and along these weaker parts the leaf tears so that normal plants are usually almost in ribbons. The leaf expanse, without this relief, is so great that tropical storms would doubtless destroy the plant.
Many wild species of the banana are still found in tropical Old World countries, the genus Musa to which the banana belongs having over sixty-five species. There are at least three well-marked types of banana used to-day, two of them, our common yellow one and a smaller red sort being fruits of almost universal use. The remaining type is usually larger than the kinds sent to northern markets, is picked and used while still green and is always cooked before using, usually boiled as a vegetable. In this form it is known as the plantain, and is a good substitute for the potato in regions where the latter cannot be grown. Plantains are used on a large scale in all tropical countries, much more so than the yellow and red bananas which are familiar enough in northern markets. These are too sweet to be used as a staple diet, and the plantain is practically the only such diet which millions of the poorer people in the tropics ever get. There is almost no native hut but has its plantain field.
The flowers of the banana plants, all of which appear to be derived from the single species Musa sapientum or possibly also from Musa paradisiaca, are borne in a large terminal cluster which ultimately develops into the “hand” of bananas familiar in the fruit shops. The plant then dies down and a new one develops from a shoot at the base of the old stem. For countless ages this has been the only method of reproduction, and usually the banana produces no seeds. The plant is easily grown in greenhouses, one in the conservatory of the Brooklyn Botanic Garden producing 214 pounds in a single cluster consisting of 300 bananas.
Sir Walter Raleigh is usually credited with the introduction of the potato into Europe, although it appears as though the Spaniards were the first to bring the plant from America. It was brought to Ireland in 1585 or 1586 and from its wide use there became known as the Irish potato. Its native home is in southern South America, and although Columbus did not mention it after his first and second voyages, subsequent Spanish adventurers found natives on the mainland making extensive use of it. There are now several wild relatives of it in South America, but their tubers are not so large as those of Solanum tuberosum from which all the different varieties of potato have been derived. The plant is too well known to need description here, but its edible tuber, actually a stem organ, is often wrongly called a root. Figure 8 shows the tubers and true roots of the plant.
The sweet potato, which in early writings was often confused with Solanum tuberosum, is a very different plant. Its edible portion is the root of a vine very like our common morning-glory or convolvulus, and its Latin name is Ipomœa batatas. The specific name is taken from a native American word, which due to early confusion was corrupted into potato, and applied to the “Irish” potato. No one certainly knows where the sweet potato is native, but probably in tropical America. It belongs to a section of the genus Ipomœa, all the other species of which are American, and before Columbus and his followers its cultivation was unknown in the Old World. It was very soon carried by the Portuguese to Japan and other parts of the Old World, and for a time it was thought to be native there. America, however, is in all probability its ancient home, although no really wild plant has ever been found there or anywhere else. Its cultivation from very early times in America is indicated, and Columbus upon his return from the New World presented sweet potatoes to Queen Isabella.
It has been said many times that there are more uses for this plant than there are days in a year. Wood, thatch, rope, matting, an intoxicating beverage, and scores of other things are derived from different parts of this palm, but it is as a food and beverage that its chief value lies. The coconut palm is a tall tree with a dense crown of feathery but stout leaves and inhabits all parts of the tropics. It is found apparently wild along sandy shores, but its ancient home, while still unknown, is probably America. Each year the tree bears from ten to twenty fruits which are at first covered with a green and very tough fibrous husk, inside which is the seed, the coconut of commerce. In the early stages of the fruit the white meat is preceded in large part by a delicious milky liquid much used by the natives, but only rarely found in any quantity in the coconuts shipped to our markets. The meat is highly nutritious and is used on a great scale as food by millions of tropical peoples. Within the last few years a method of taking out the meat of the coconut and shipping it in a state of arrested fermentation to the north has been discovered. This product, known as copra, is produced in enormous quantities, both in the Old and the New World, particularly in India and the Philippines. From this copra a palm oil is refined, which is the chief source of the nut butter now so widely sold. Some idea of the extent of the cultivation of coconuts may be gleaned from the fact that in India and the Philippines the trees are counted by the hundreds of millions. The oil from the nuts is also largely used in cookery, in making candles, for burning in lamps, and in making certain kinds of perfume. The tree belongs to the Palmaceæ, a monocotyledonous family of plants of great commercial importance. It is known as Cocos nucifera, and the genus has over a hundred species, all of tropical American origin. Whether Cocos nucifera is American or not is still a disputed point. From the fact that it will float in sea water without injury to the seed it has been supposed that it was carried great distances by currents. It is found both wild and cultivated throughout the tropical world, and its use appears to have been known to the Asiatics probably four thousand years ago. The curious fact remains that it is the only palm that, in its wild state, is known both in the Old and New World, all others being peculiar to one hemisphere or the other. Perhaps its capacity for floating in the sea without injury may explain what is otherwise still a good deal of a mystery.
There are many other foods derived from plants, besides all the fruits and vegetables too numerous to be noted in detail here. One fact of significance seems to stand out from a study of the uses of plants by man. There are three distinct regions from which the great bulk of our food and many other useful plants have apparently come. One is the area of which Indo-China is approximately the center, and which is the ancestral home of rice, the banana, tea, sugar cane, and many other valuable plants. Somewhere in this southeastern corner of Asia there must have been a highly developed agriculture which rescued these plants from the wild, and from which they have spread throughout the world. The second region, somewhere near Mesopotamia, appears to be the cradle of wheat and a few other useful plants. And the third region is the western part of America from southern Mexico to northern Chile, where corn, tobacco, the pineapple, sweet potato, potato, the red pepper, and the tomato were all discovered with the discovery of this continent.
Alphonse de Candolle, from whose studies much of our information on the origin of cultivated plants is derived, once prepared a list of our common vegetables showing their ancient homes, their wild ancestors, and the length of time during which they have been in cultivation. With some recent additions and corrections by Dr. Orland E. White of the Brooklyn Botanic Garden, the list is printed below:
The letters indicate the probable length of cultivation.
(a) A species cultivated for more than 4,000 years.
(b) A species cultivated for more than 2,000 years.
(c) A species cultivated for less than 2,000 years.
(d) A species cultivated very anciently in America.
(e) A species cultivated in America before 1492 without giving evidence of great antiquity of culture.
(f) A species or subspecies of very recent domestication.
The following list of the common fruits also gives their native country, period of cultivation, and some additional notes about them. Those marked with a star were found in the markets of New York City by Dr. White, who also revised this list. The letters for the dates are the same as in the list of vegetables:

A Banana Plantation in Fruit. The banana is now grown throughout the tropical world, but native in tropical southeastern Asia. (Courtesy of Brooklyn Botanic Garden.)

Rice Terraces in China. In many regions where the forests have been destroyed and all the soil washed into the valleys, agriculture has to be carried on under conditions of great difficulty. Soil is brought up these slopes and held there by the artificially made terraces. (Photo by Bailey Willis. Courtesy of Brooklyn Botanic Garden.)
The operation of the Eighteenth Amendment to our Constitution will stop the manufacture in this country of the chief beverages that were made here from plants. All wines and brandies were from the juice of the grape, whiskey from rye and some other cereals, and beer from hops and barley. Our three remaining beverages of practically universal use are none of them produced in the United States, with the exception of a little tea grown in a more or less experimental way.
It is related in an old legend that a priest going from India into China in 519 A.D. who desired to watch and pray fell asleep instead. In a fit of anger or remorse he cut off his eyelids which were changed into the tea shrub, the leaves of which are said to prevent sleep. Unfortunately for the story tea was known in China more than three thousand years before the date of that legend, and it is very doubtful if it was ever brought from India to China. The wild home of the tea is apparently in the mountainous regions between China and India, but the plant will not stand the frost, so that its cultivation is now mostly in parts of China, Japan, India, Ceylon, Java, and some in Brazil.
The plant is mostly a shrub or occasionally a small tree, with white fragrant flowers and evergreen oval-pointed leaves. All the different kinds of tea are derived from the single species Camellia Thea, the differences in color and flavor being due to processes of culture or curing of the leaves.

While the use of tea has been known to the Chinese for over four thousand years, its introduction into Europe dates from the days of the Dutch East India Company, who brought some to Holland about 1600, and by the English East India Company, who sent some from China to England in 1669. It fetched at that time 60 shillings per pound for the common black kinds and as much as £5 to £10 per pound for the finer kinds. It was almost fifty years, or about 1715, before the price fell to 15 shillings per pound. From that time until the present there has been a tremendous increase in its use, although then as now the great bulk of the world’s tea is used by the Mongolians and Anglo-Saxons. Just before the war over 700 million pounds comprised the annual crop of tea. As its use became general the English put a tax upon its importation into Great Britain or its colonies, with results here that we all know.
The cultivation of tea is restricted to those regions where there is a large and frequent rainfall as well as a high temperature. It will not grow in marshy places such as rice prefers, but needs light, well-drained soils. The plant is propagated only from seeds which are sown in nurseries, and the young plants set out in the tea fields about four and a half feet apart each way. In two years they are bushes from four to six feet tall when they are cut back to a foot high. The increased vigor of the bush from this severe cutting back results in a dense bush, from which leaves are plucked from the third year in small quantity. Not until after the sixth or seventh year is there a normal yield, which in an average year would be from four to five ounces of finished tea. A poor yield of leaves would average about 400 pounds of tea per acre, good yields going as high as a thousand pounds or even more than that. The tea fields must be kept free of weeds, a tremendous task in a moist tropical region, which demands cultivation about nine times a year. The expense of properly setting out and maintaining a tea plantation is therefore considerable.
The plucking of tea leaves is a fine art beginning with the starting of new growth and continuing every few days until growth stops. In certain regions growth is practically continuous and plucking also, but in most regions the plant has an obvious resting period, when it is pruned back. A properly cared for plant may last as long as forty or fifty years. In a modern tea plantation the only part of the process of tea making that involves handling is the plucking of the leaves, largely done by women and children. The leaves are then spread on racks and allowed to partly wither, after which they are put between rollers so as to crush the tissue, thereby allowing the more rapid escape of water. After rolling, all black teas are again spread out when oxidation of their juices changes their color, but green teas omit this second spreading out and sometimes even the first. The oxidation of black teas is produced by an enzyme in the juice, in green tea this process is stopped by subjecting the leaf at once to steam. This kills the enzyme but preserves the green color of the leaf. In the black tea the enzyme is allowed to work for two or three hours when the leaves are again slightly rolled to seal in the juices and the leaves are then subjected to a current of air progressively warmer until it reaches a temperature well above boiling point. Once the temperature reaches about 240 degrees the process goes on only for about twenty minutes when the leaves are perfectly dry and crisp. The different sized leaves, buds, twigs, etc., are then sorted by mechanical sifters and the finished tea is ready for packing. Experts declare that there is no difference between broken and unbroken leaves, and if there is any the flavor is probably better from broken leaves. From the upper three leaves and their bud the finest teas are made, but from adjoining plantations, even from the same plants at different seasons or different pluckings, vastly different teas are often produced. In different regions the process varies slightly in its details, and different soils and culture undoubtedly affect the flavor of tea, just as they do other crops. Some of these local conditions are of great value, and the skillful handling of the leaves is as much of a fine art as it is a science. Unlike wines, tea is best when fresh and much of the romance of the sea centers around the China clippers which made remarkably swift passages between China and England around the Cape of Good Hope. With the opening of the Suez Canal competition for increased speed became still more keen, but steam vessels took the romance out of the trade. Much of the tea used in the United States comes from Japan and does not go through London, which for over two hundred years was the tea market of the world.
The thing for which we drink tea is an alkaloid in its leaves that is pleasant to the taste and refreshing to the senses. It is released in boiling water in a very few minutes, but if tea is allowed to stay in water longer than this, tannic acid is also released. This is a substance found in the bark of certain trees and is used in tanning leather. As 10 per cent of the leaf of tea consists of this substance it may readily be seen how easily improper methods of making tea will render it not a refreshing and delightful beverage but an actual poison to the digestive tract.
Like tea and chocolate, coffee also comes from a plant that can only be grown in the tropics. Its original home was in or near Arabia and its botanical name is Coffea arabica. There are, however, other species of the genus that produce coffee, but Coffea arabica is still its chief source. The plant is

now grown throughout the tropical world, but it does not thrive so well along the coast as it does at elevations of a thousand feet or so, where, in America at least, the best coffee is produced. The plant is a shrub or small tree, usually not over 12 to 15 feet tall, with opposite leaves and small tubular flowers, followed by a bright red berry, which contains the coffee “beans.” The flowers and berries are in small clusters in the axils of the leaves. Its use among the natives appears to date from time immemorial, but the Crusaders did not know it, nor was it introduced into Europe much before 1670. Its annual consumption is now well over two billion pounds, nearly half of which is used in this country. We use over ten pounds a year for each man, woman and child in the country, or nearly ten times the per capita consumption in England. Brazil produces over half the world’s total supply and consequently controls the coffee markets of the world. The plant was first brought into South America by the Dutch, who in 1718 brought it to Surinam. From there it spread quickly into the West Indies and Central America.
The coffee berries are collected once a year and spread out to dry, after which the two seeds are taken out. This is the simple method of all the smaller plantation owners, but a modern Brazilian coffee plantation follows a very different procedure. The berries are put in tanks of water, or even conveyed by water flues from the fields, and allowed to sink, which all mature berries will do. They are then subjected to a pulping machine which after another water bath frees the beans from the pulp. The former are still covered by a parchmentlike skin which, after drying of the beans, is removed by rolling machines. The coffee is then ready for export, but not for use until it is roasted. This is a delicate operation not understood except by experts, and should not be done until just before the coffee is ready to be used.
The average yield per plant is not over two pounds of finished coffee a year, but larger yields from specially rich soils are known. The plant is rather wide-spreading and not over five or six hundred specimens to the acre can be grown.
The so-called Mocha coffee is obtained in Arabia, where Turkish and Egyptian traders buy the crop on the plants and superintend its picking and preparation, which is by the dry method. Not much of this ever reaches the American markets, and the total amount of coffee now produced in Arabia, its ancestral home, is negligible.
Unlike tea and coffee, chocolate is a native of the New World and was noticed by nearly all the first explorers. It grows wild in the hot, steaming forests of the Orinoco and Amazon river basins, although it was known in Mexico and Yucatan as a cultivated plant from very early times. By far the largest supply still comes from tropical America, although it is grown in the Dutch East Indies, Ceylon, and West Africa. It must have been cultivated for many centuries before the discovery of America, as scores of varieties are known, all derived from one species. This is Theobroma cacao, and both the generic and specific names are interesting. Theobroma is Greek for god food, so highly did all natives regard the plant, and cacao is the Spanish adaptation for the original Mexican name of the tree. Throughout Spanish or Portuguese-speaking tropical America the tree is always spoken of as cacao.
The chocolate tree is scarcely over twenty-five feet tall, has large glossy leaves and bears rather small flowers directly on the branches or trunk. This unusual mode of flowering, common in rain forests, results in the large sculptured pods appearing as if artificially attached to the plant. Each pod, which may be 6 to 9 inches long, contains about fifty seeds—the chocolate bean of commerce.
When the beans first come out of the pod they are covered with a slimy mucilaginous substance and are very bitter. To remove this the beans are fermented or “sweated,” usually by burying in the earth or piled in special houses for the purpose. After several days, sometimes as long as two weeks, the beans lose the mucilage, most of their bitter flavor and often change their color. After this they are dried and are ready for roasting, which drives off still more of their bitter flavor. Chocolate is made from the ground-up beans containing nearly all the oil, which is the chief constituent, while cocoa is the same as chocolate with a large part of the oil removed. As in tea and coffee, there is an alkaloid in chocolate for which, with its fat, the beverage is mostly used.
Very little chocolate is now collected from wild plants, and cacao plantations are important projects in tropical agriculture. Because of its many varieties, some nearly worthless, the business was rather speculative until a few good sorts were perpetuated. Much valuable work on this plant and the isolation of many good varieties has been done by the Department of Agriculture in Jamaica, British West Indies. Cacao plantations are usually in moist, low regions near the coast, preferably protected from strong winds, to which the plant objects. The trees are set about ten to fifteen feet apart each way and begin bearing after the fifth or sixth year. The young plants are always shaded, often by bananas, which are cut off as the trees mature. The tree will not thrive unless the temperature is about 80 degrees or more and there should be preferably 75 inches of rainfall a year, about twice that at New York. Chocolate-growing regions are apt to be unhealthy for whites, and native labor is practically always used. The business is very profitable, but still somewhat speculative.
Not only do plants furnish us with food and drink, but most of our clothing is made from plant products. There is annually produced twice as much cotton as wool, while linen is made from the fibers in the stem of the flax plant, which is also the source of linseed oil. Fibers occur in many different parts of plants, but most often in the stem, or in the bark of the stem. Some occur in the wood itself, as for instance that in spruce wood, from which news paper is made. Others are found in the attachments of the seed, such as cotton. Some are very coarse, such as that of Carex stricta, a swamp sedge from which Crex rugs are woven. Others like that of the leaves of the pineapple are as fine as silk, and in the Philippine Islands where much pineapple fiber is produced, some of the most beautiful undergarments and women’s wear are made from it. Again, others, such as Manila hemp, furnish us with cordage of great strength.
It has been stated, perhaps a little rashly, that the value of the cotton crop in our Southern States exceeds all other agricultural products of the country. Whether this be true or no matters not, as cotton production and manufacture is certainly one of the most important industries of the world. Our own New England mills and those in Lancashire total an enormous volume of manufactured cotton goods, and what the stoppage of the cotton crop means to these industrial centers was shown even so far back as the Civil War, when the “cotton riots” in Lancashire were noised all over the world. Cotton is the most important of all fiber plants.
There are several different kinds of commercially important cottons, and perhaps dozens of others, all derived from the genus Gossypium, a relative of our common garden mallows belonging to the Malvaceæ. By far the most valuable is Sea Island cotton, derived from Gossypium barbadense, which is probably a native of the West Indies, although really wild plants are yet to be discovered. It is the kind, of which scores of varieties are known in cultivation, that is grown mostly along our southeastern coastal States. Next in value, but cultivated in greater quantity because larger areas are suited to it, is Gossypium hirsutum. The fiber is a little shorter, but the total amount of cotton derived from this species probably exceeds that from all other kinds. It is the cotton grown mostly in upland Georgia, Louisiana, and Texas, and the wild home of this species is supposed to be America, although it, too, has never been found in the wild state. The third cotton plant is Gossypium herbaceum, a native of India, and the origin of many varieties now grown in that country. It has a shorter fiber and is worth about one-third the price of Sea Island cotton. From Abyssinia and neighboring regions comes the fourth important cotton plant, Gossypium arboreum, differing from the others in being a small tree. All the others are shrubby, while G. herbaceum is merely a woody herb. These different plants have been tried in the countries suited to cotton raising, but, generally speaking, the chief crop from each is produced in the country nearest the supposed wild home of it.
In all of them the fiber is really an appendage of the seeds, and each pod as it splits open is found to be packed full of a white cottony mass of these fibers with the seeds attached. These white masses of cotton, or bolls, have to be picked by hand, as no really successful machine has ever been found for this purpose. Women and children do a large part of the picking, and the wastage due to careless picking is tremendous. The whole value of the cotton crop depends upon an invention by Eli Whitney, an American, of a machine to separate the cotton from its seed. This “ginning” machine is now much perfected and, in America at least, is the chief method of separation of fiber and seed. In India and for certain other varieties a different type of machine, known as the Macarthy gin, is employed. The latter is used in America also for some of the long-fiber Sea Island cottons. With the baling of the cotton the work of the grower is over and the product is ready for the manufacturers. The resulting seed, after ginning, once little valued, is now an important plant product, cottonseed oil, cattle feeds, soap, cottolene, fuel oil, and fertilizers being derived from it. Its value in the United States now totals millions of dollars annually.
In growing cotton in America seeds are sown in April, and the beautiful yellow flowers with a red center bloom about June or July, followed in August by the pod. This splits open and is ready for picking by September and October. The plants are grown in rows four feet apart and are set one foot apart in the row. Clean cultivation is absolutely necessary, and in first-class plantations all weeds are kept out. The plant needs a rich deep soil.
There are a variety of plants which furnish products known as hemp, but commercially only three are of much importance, the plant universally known under that name, the Manila hemp, and sisal. All of them are used chiefly for cordage.
The hemp of the ancients is a tall annual related to our nettles, with rough leaves, and a native of Asia. For centuries an intoxicating drink was made from the herbage of this plant, and this with the narcotic hashish, which is made from a resin exuded by the stems, obscured the fact that Cannabis sativa is a very valuable cordage plant. The coarse fibers are found in the stem, and these are cut and retted, the retting or rotting process separating the fibers from the waste portions of the stem. The fibers are so long and coarse that only cordage, ropes, and a rough cloth are made from them, but enormous quantities are raised for this purpose, especially in Europe. As hashish is now a forbidden product in many countries, due to its dangerous narcotic effects, the hemp plant is more cultivated for fiber than for the narcotic. But in the olden days hashish had a tremendous vogue in the Orient and was known at the time of the Trojan wars, about 1500 B.C. Fiber from the plant was almost unknown to the Hebrews, and it was not until the beginning of the thirteenth century that it came into general use. It is now probably as important as sisal, but not as Manila hemp, the most valuable of all cordage plants. The hemp is diœcious and the female plants are taller and mature later than the male. Two cuttings are therefore necessary in each field.
The Manila hemp is derived from a banana (Musa textilis), that is a native of tropical Asia and is much grown in the Philippines. While the fruits of this plant are of very little or no value, the fiber from the long leafstalk is the best cordage material known. Also the finer fibers near the center of the stalk are made up into fabrics, which are rarely seen here, but are said to be almost silky in texture. As a cordage plant, however, Musa textilis is now easily the most important, and from a commercial point of view the Philippine Islands is the only region to produce it in quantity. It has been tried with not much success in India and the West Indies. Methods of extracting the fibers are still very primitive, as it is nearly all scratched out by natives with saw-toothed knives made for the purpose. After the fleshy part of the leafstalk has been separated from the fiber this is merely put out on racks to dry. The finished product has so much value for large cords and ropes that the fiber makes up about half the total exports of the Philippine Islands. Its great strength may be judged from the fact that a rope made from it, only about one inch thick, will stand a strain of over four thousand pounds. No other fiber is anywhere near this in strength and yet of sufficient length to be of use as cordage. There are still thousands of acres suitable for its culture in the Philippines, but the extraction of the fiber awaits some inventive genius who will make a machine for that purpose. Many have tried, but so far the primitive scratching out by natives is the only method in use and it is admitted that it wastes nearly one-third of the fiber. The so-called Manila or brown paper is often made from old and worn-out ropes of Manila hemp, but, as in the case of cordage itself, adulteration with cheaper fibers is common.
From Yucatan, the Bahamas, and some other regions of tropical America comes the most valuable American cordage plant, known as sisal. The fiber is extracted from the thick coarse leaves of a century plant, known as Agave sisalina or Agave rigida, which looks not unlike the century plant so common in cultivation. The plant belongs to the Amaryllis family and is native in tropical America. Thousands of acres are planted to sisal in Yucatan and a machine for scratching out the fiber is in general use. The plant produces each year a crown of eight or ten leaves from three to five feet in height, each tipped with a stout prickle. Unlike the common century plant of our greenhouses there are no marginal prickles on the leaves of the sisal. After extraction the fiber is stretched out on racks to dry and is then ready for manufacture into rope.
During the late war the Germans were reported to be sending flour and sugar to their armies pressed into large bricks for the want of bags to ship them in the ordinary way. Gunny sacks, or jute bags, as they are more often called, are made literally by the hundreds of millions, as practically all sugar, coffee, grains and feeds, and fertilizers are shipped in them. Jute is a tall herb, a native of the Old World tropics, but suitable for cultivation in many other tropical regions. Practically all the world’s supply now comes from India, probably because of the cheapness of labor rather than any peculiar virtue of the soil or climate of that country. The plant has been experimentally grown in Cuba with entire success, but labor conditions made cheap production of the fiber impossible.
The jute plant, known as Corchorus capsularis or C. olitorius, grows approximately six to nine feet tall and is an annual, often branching only near the top. They are not very distantly related to our common linden tree. At the proper maturity the whole plant is harvested and the stems are tied into bundles ready for the retting process. Of all fiber processes this is the most difficult, largely because no machine or chemical has yet been found to

The fiber is mostly derived from Corchorus capsularis and from Corchorus olitorius.
extract the fiber of jute, or flax, and this is accomplished by placing the stems in water, which rots out the fleshy part of the stem, leaving the fiber. Some notion of the difficulty of this task in such plants as jute is gained by realizing that over twelve million bales of finished fiber are produced each year, and that the retting may take from two days to a month. The retting process is aided by certain organisms of decay in the water, by the temperature, and by some other factors not yet understood. The process is allowed to go on only long enough to separate flesh from fiber, which makes frequent inspection of the bundles in the filthy water an absolute necessity. At the proper time the natives are able to split off the bark, which contains the fiber, from the stem, and while standing up to the waist in the water, he picks or dashes off with water the remaining impurities. The fiber is then dried on racks and subsequently, under enormous pressure, packed in bales of four hundred pounds each. An average crop would be about two and one-half bales from an acre of jute, so that in India there must be considerably over five million acres devoted to the cultivation of the plant. While for many years this tremendous output of fiber was sent to England for manufacture, power looms were set up in India about the middle of the last century. There are now over three-quarters of a million spindles there, and some jute is sent to the United States for manufacture here.
Next to cotton jute is probably the most important fiber plant in the world. For hundreds of thousands of people in India and in England it is the only source of livelihood. To the inventor who can eliminate or reduce the costly retting process of jute, or of flax, which goes through essentially the same operation, there is waiting a golden future, for it is largely the cheapness of labor and willingness of its natives to stand in the retting pools that has made India the jute region of the world.
Lack of space forbids mention of the many other fiber plants, some of which, like flax, are of large importance. Their fibers are used in a variety of ways and are found in different parts of the plant. A few of these, together with the names of the plants and the regions where they are native, are as follows:
| Native | Product | Name of Plant |
| Bowstring hemp. Sansevieria, several species. | Bowstrings and cordage. | Tropical Africa and Asia. |
| Coconut palm. Cocos nucifera. | Coir. | Tropical America (?) |
| Flax. Linum usitatissimum. | Linen. | Europe and Asia. |
| Kapok. Eriodendron anfractuosum. | Kapok, for stuffing. | India. |
| New Zealand flax. Phormium tenax. | Cordage. | New Zealand. |
| Paper mulberry. Broussonetia papyrifera. | Paper pulp in Japan. | Japan. |
| Pita. Bromelia Pinguin. | Pita fiber, fabrics. | Tropical America. |
| Queensland hemp. Sida rhombifolia. | Jute substitute. | Tropical regions. |
| Raffia. Raphia ruffa. | Cloth and for tying. | Madagascar. |
| Ramie. Boehmeria nivea. | Ramie cloth. | Tropical Asia. |
| Rattan cane. Calamus rotang. | Rattan, cordage, and coarse cloth. | India. |
| Rush. Juncus effusus.[2] | Matting in Japan. | North temperate regions. |
| Sedge. Carex stricta. | Rugs and mattings. | Northern North America. |
| Spruce. Picea rubens, canadensis, etc. | Paper pulp. | Northern North America. |
| Willows. Salix, many species. | Basketry. | Temperate regions mostly. |
No mention can be made here of the hundreds of fiber plants used by the natives of various parts of the world, some of them probably having great commercial possibilities. The extraction of these fibers by machinery or chemically will open up a large commerce in such plant products, the value of which is now unsuspected or ignored. While the value of cotton, jute, and Manila hemp is reckoned in the hundreds of millions, some of these native fibers are found in plants whose wild supply is almost inexhaustible, and some of which are quite as capable of cultivation as the better known fiber plants. Few fields of inquiry offer greater possibilities to the economic botanist than fibers.
Along the north coast of Haiti, particularly near Cap Haiti and Puerto Plata, there are scattered a few plantations devoted to rubber growing; and it is not without interest that Columbus on his first voyage landed at about this precise spot in December, 1492, and found the natives playing a game of ball made of rubber. He wrote: “The balls were of the gum of a tree, and although large, were lighter and bounced better than the wind balls of Castile.” This is apparently the first notice of the use of rubber, a substance now of world-wide importance, and derived from many other plants than the tree mentioned by Columbus. This is Castilla elastica, a native of certain islands of the West Indies and the adjacent mainland, and a relation of our common mulberry.
For over three centuries rubber, or caoutchouc as it was often called, was only of very casual use before the process of vulcanizing was discovered by Charles Goodyear, an American, in 1839. This combination of rubber with sulphur transformed a material much subject to heat and cold, and of almost no manufacturing value, into one from which hundreds of articles of daily use are now manufactured. Previous to this it had been used mostly, and in fact almost exclusively, as a waterproofing material for cloth, a process much developed by the firm of Charles Macintosh & Co., who appear to have taken out the first patent for a waterproofing process in 1791, in England. The rubber tree found by Columbus is still grown in considerable quantities and is a valuable source of rubber, but it has been greatly overshadowed by a Brazilian tree which now produces over two-thirds of all the rubber in the world.

This Brazilian tree, a native of the rich rain forests of the Amazon, is Hevea brasiliensis, and a relative of our common spurges of the roadsides and of the beautiful crotons of the florist, all belonging to the family Euphorbiaceæ. The first important notice of this rubber appears to be by the astronomer C. M. de la Condamine, who was on an astronomical trip to the Amazon in 1735. He described Para rubber, as it has since been called, and by 1827 the export of this gum had grown to 31 tons a year. In 1910 Brazil exported over 38,000 tons, nearly all of which was collected from wild trees. After the discovery of vulcanization the demand for all kinds of rubber increased by leaps and bounds and it became obvious that the wild trees, although tapped regularly, would not supply all of the necessary amount. For years the Brazilian Government protested the export of seeds or other means of growing the plant out of the Amazon, but in 1876 H. A. Wickam chartered a steamer and loaded her with 70,000 seeds of para rubber trees and some crude rubber, and had the ship passed by the Brazilian port authorities as loaded with “botanical specimens.” He safely transported the cargo to the Kew Gardens, London, where only about 4 per cent of the seeds ever germinated. From there the young plants were sent to India, where now, and in the Straits Settlements and the adjacent islands, there are huge plantations of para rubber. From the wildest speculation in rubber shares on the London Stock Exchange, which followed the successful introduction of the plant into British possessions, the industry has now settled down to be one of the most profitable in plant products of the East.
The rubber of both Hevea and Castilla is produced from the milky juice or sap of the trees and is actually a wound response. As the trees are tapped the latex, as the milky juice is called, runs out of the wound and upon reaching the air coagulates. This material is removed and a new wound made, a process which is repeated for several years. There is still work to be done upon the problem of how often plantation trees should be tapped to get the greatest flow of latex without injuring the tree, but in many plantations it is done every day or every other day in the season, some rubber planters allowing a resting period during leaf fall, others again tapping almost continually. The actual wound is made by removing just enough bark to induce a flow of latex, but not until the wound is completely healed, a process taking from four to six years, can that particular part of the bark be cut again. With almost daily tappings the problem of finding fresh pieces of bark into which the cut may safely be made has been developed into a fine science. In the wild trees of Brazil it is still done by natives, probably rather wastefully. Rubber plantations in the tropical regions of Asia now total over a million acres, as compared to only slightly over 200,000 acres in the American tropics and from all other non-Asiatic sources. Of this probably 100,000 acres are in Africa. At the present time nearly half the world’s supply of rubber comes from these plantations, the balance still coming from Brazil.
There are two other rubber-producing plants, neither of which are as important as Hevea and Castilla. One is the common rubber plant so much grown as a house plant and said to be much cherished for that purpose in Brooklyn. It is a kind of fig tree, known as Ficus elastica, a native of India and the source of India rubber. It was used for years, before the days of vulcanization, mostly for lead-pencil erasers. Thousands of acres of it in India were recently destroyed to make room for Hevea, although it still produces a respectable amount of rubber, inferior, however, to Hevea and Castilla. In Mexico a new source of rubber is the guayule rubber plant, a small shrub native of the drier parts of the Mexican uplands. It does not produce latex, as practically all other rubber plants do, but particles of rubber are found directly in its tissues, mostly in the bark. While of some importance, this shrub, known as Parthenium argentatum, a member of the Compositæ or daisy family, is not likely to become a dangerous rival of either Para or Castilla rubber. Wild sources of guayule rubber are already reported as diminishing rapidly, so that its permanent success will depend upon cultivation. At least a score of other plants are known to produce rubber of a kind, but none of them have yet been much developed. From most plants with a milky juice, such as dogbane or spurge, rubber of some sort can usually be recovered. The production of this substance is directly due to a response of the plant to wounds, and there are still great fields of research necessary on this phase of plant activity, upon which a great industry has already been built.
Nearly all the drugs and medicines of importance are of vegetable origin, and from the days of Theophrastus the study of plants as possible medicines has been one of the chief phases of botanical research. In the early days all that was known about plants was learned by men interested in medicine, and some of their quaint old books are interesting relics of a bygone day. At present, pharmacognosy, or the science of medicinal plant products, is a highly developed specialty taught in medical and pharmacy schools. And yet the greatest medical college in this country has recently issued instructions to its staff of doctors and nurses to pay particular attention to “old wives’ remedies,” most of which consist of decoctions of leaves and other parts of plants. They have done this because all the knowledge of the scientists regarding medicinal plants has its origin in the habit of simple people turning to their local plants for a cure. The accumulation of the ages, aided and guided by the scientist, has resulted in the wonderful things that can now be done to the human body through the different drugs, nine-tenths of which are of plant origin.
There is almost no part of a plant that, in some species at least, has not been found to contain the various acids, alkaloids, oils, essences, and so forth, which make up the chief medicinal or, as the pharmacists call it, the active principle of plants. In certain of them the most violent poisons are produced, such as the poison hemlock which killed Socrates, and the deadly nightshade. And again the unripe pod of one plant produces a milky juice so dangerous that traffic in it is forbidden in all civilized countries, and yet later the seeds from that matured pod are sold by the thousands of pounds to be harmlessly sprinkled on cakes and buns by the confectioners. Earlier in this book it was said that plants are chemical laboratories, and nowhere has this alchemy been carried to such a pitch of perfection as in the hundreds of drugs produced by different plants. Reference to special books on that subject should be made by those interested, as only a few of our most important drug plants can be mentioned here.
The Spanish viceroy of Peru, whose wife, the Countess del Chinchón, was dangerously ill of a fever in that country about 1638, succeeded through the aid of some Jesuit priests in curing the malady, with a medicine which these priests had gotten from the natives. It was a decoction from the bark of a tree, and its fame soon spread throughout the world as Peruvian bark. Even in those days malaria was the curse of the white man in tropical regions, and since then it is hardly too much to say that the discovery of this drug has made possible for white colonists the retention of thousands of square miles of tropical country that without it would in all probability be unfit for occupancy. To the natives, too, quinine has been one of the greatest blessings, and in some of the remote regions of the tropics the writer has found quinine more useful than dollars in getting help from fever-ridden natives, too poor and too remote from civilization to get the drug.
For more than a hundred and fifty years after the discovery of Peruvian bark it was always taken as a liquid, usually mixed with port, and an extremely noxious and bitter drink it made. The fine white powder which we now use in tasteless pellets or pills has made the drug even more useful than before.
The trees from which the bark is used all belong to the genus Cinchona, named for the first distinguished patient to benefit by it, and belong to the Rubiaceæ or madder family. There are at least four or five different species. For many years Peru and near-by states were the only source of the bark, and the English became convinced that the great trade in the drug would exterminate the tree, so in 1880 they introduced the plant into India. These cinchona plantations now provide most of the world’s demand, and some idea of what that means may be gleaned from the fact that in Ceylon alone over fifteen million pounds of the drug are produced annually. In India itself the plantations are largely government owned and quinine is sold in the post offices at a very low rate. In cinchona plantations strips of the bark are removed, and after a proper period of healing, the process is repeated. It takes about eight years before it is safe to begin cutting the bark.
Most doctors use aconite for various purposes and it is mentioned here chiefly as illustrating a common characteristic of many medicinal plants. The whole plant is deadly poisonous, and in the root there appears to be a concentration of this poisonous substance, which makes the plant one of the most dangerous known. All of the drug aconite is derived from Aconitum Napellus, a monkshood, belonging to the buttercup family. It is a perennial herb with beautiful spikes of purple-blue flowers, not unlike a larkspur, and is a native of temperate regions of the Old World. A related Indian species has been used for probably thousands of years by the natives. They poison their arrows with it and so deadly is the drug that a tiger pricked by such an arrow will die within a few minutes. It is conceded to be the most powerful poison in India. Numerous accidents have resulted in Europe from careless collectors of the roots of horseradish, who sometimes get aconite roots mixed with that condiment, usually with fatal results.
So many other plants are violently poisonous, and yet yield the most valuable drugs, that the greatest care has to be used in their collection and preparation. The habit of many children of eating wayside berries should be discouraged, as some of our most innocent-looking roadside plants are actually deadly if their fruits or foliage are eaten. Fortunately only a very few plants are poisonous to the touch, notably poison ivy and poison sumac, and some of their relatives.
Almost no plant product has caused more misery and relieved more pain than the juice of the unripe pod of the common garden poppy, Papaver somniferum. From it opium is extracted, the chief constituent of which is morphine. If tea can be said to have precipitated our war of independence, opium was indirectly the cause of the opening of China to the western world. The degrading effects of opium had become so notorious that the Chinese in 1839 destroyed large stocks of it, mostly the property of British merchants, and prohibited further importations. In the subsequent negotiations which ended in war, China was opened up to trade. No civilized country now openly permits the sale of opium, although there is still a good deal of it used in practically all parts of the world. The effects of lassitude, subsequent ecstasy, and stupefaction are due to an alkaloid, the continued use of which forms a drug habit of serious consequences. Parts of China, Turkey, Persia, and Siam are said to be still large users of opium which is chewed, or more often smoked. Until comparatively recently fifteen out of every twenty men in some of these countries were regular users of the drug. The legitimate use of morphine by physicians has done more than almost anything else, with the possible exception of cocaine, to relieve suffering, and there is consequently a considerable trade in the drug.
The home of this poppy is unknown, as it has been cultivated from the earliest days. The ancient Greeks and Romans knew it and the Egyptians grew it for opium. It is still grown in India and China, where, notwithstanding vigorous governmental measures, there is a large opium consumption. After maturity the pods of the poppy, from whose milky juice in its earlier stages the drug is obtained, produce many seeds. From these an oil is pressed which is widely used as a cooking oil in the East, and is perfectly harmless. The seeds are also used as bird seed and by confectioners.

In the northern part of South America the Peruvians and some of their neighbors were discovered by early explorers to be chewing the leaves of a native shrub, apparently with much profit and no evil after effects. It served them much as the betel nut does to the natives of India and other regions of the tropical East. With scanty or no food this apparently harmless intoxicant will carry both men and women over periods of severe fatigue. The shrub bearing these leaves is not over four or five feet tall and has bright green foliage and small white flowers.
Quite different in its effects has been the drug which has been extracted from this plant, known as Erythroxylon Coca. Far from being a beneficial and harmless stimulant, cocaine is now one of the drug evils of our time. Its use, outside that prescribed by physicians, is forbidden practically everywhere, but its consumption in this country, aside from its great and legitimate use as a relief from pain, is still very large.
The number of drug plants is legion, so large in fact, that volumes have been filled with descriptions and notes about them. A few of the most important, omitting those already mentioned, are listed below:
| Drug | Derived from the | Native |
| Betel nut. | Seed of Areca Catechu. | India. |
| Calamus. | Rootstock of Acorus Calamus. | Eastern U. S. and in Asia. |
| Sarsaparilla. | Roots of various species of Smilax. | Tropical America. |
| Saffron. | Stigmas of Crocus sativus. | Europe. |
| Arrowroot. | Rootstock of Maranta arundinacea. | Tropical America. |
| Cubeb. | Unripe fruit of Piper Cubeba. | Old World tropics. |
| Creosote. | Wood of Fagus americana and F. sylvatica. | North America and Europe. |
| Hydrastis. | Rootstock of Hydrastis canadensis. | Eastern N. America. |
| Star Anise. | Fruit of Illicium anisatum. | Southern China. |
| Camphor. | All parts of Cinnamomum Camphora. | China and Japan. |
| Witch-hazel. | Leaves and bark of Hamamelis virginiana. | Eastern N. America. |
| Licorice. | Underground parts of Glycyrrhiza glabra. | Europe. |
| Cascara Sagrada. | Bark of Rhamnus Purshianus. | Western United States. |
| Ginseng. | Rootstock of Panax quinquefolium. | Eastern N. America and Asia. |
| Wintergreen. | Leaves of Gaultheria procumbens. | Eastern N. America. |
| Nux vomica. | Seeds of Strychnos Nux vomica. | India. |
| Digitalis. | Leaves of Digitalis purpurea. | Europe. |
| Ipecac. | Root of Uragoga Ipecacuanha. | Brazil. |
| Castor oil. | Seeds of Ricinus communis. | Africa or India. |
Not until 1492 was the use of tobacco known to the Europeans, when Columbus found the natives of Cuba and Santo Domingo both chewing and smoking it. Subsequent Spanish explorers of the mainland found its use almost universal both in North and South America. It had apparently been used there for countless ages, as smoking it formed part of the most solemn ceremonial rites both of the natives’ religion and their political gatherings. Brought to England in 1586 by Ralph Lane and Sir Francis Drake, the smoking of tobacco spread with the great speed that such a comfortable habit might be expected to exhibit. Notwithstanding violent opposition by certain priests and physicians and other more intolerant opponents of the weed, its use increased throughout the world. To-day, in spite of our modern anti-tobacco fanatics, over two billion pounds are produced annually, and in the United States there is a per capita consumption of over five pounds per year, greater than any country in the world, save Belgium.
All of the many different forms in which tobacco is used are derived from the leaves of Nicotiana tabacum, or perhaps one or two other species of the genus Nicotiana, which belongs to the Solanaceæ or potato family. There are many other species, all natives of the New World, but the actual home of the tobacco plant is in some doubt. As in so many cultivated plants, which have been grown for countless ages, wild specimens are practically unknown. The plant seeds freely and consequently frequently escapes from cultivation, so that in many parts of America apparently wild plants are to be found that

Temperate Forest on Gardiner’s Island, Long Island, N. Y. Note the open vista through the trees, and lack of undergrowth, due to the forest canopy, and contrast with the profusion of the under vegetation in the rain forest (Courtesy of Brooklyn Botanic Garden.)

Rain Forest. Root-climbing lianas on a tree stem in the south Mexican rain forest (State of Chiapas). Below: Sarcinanthus utilis, with bipartite leaves. Farther up: Araceæ. Highest of all: epiphytic shrubs are visible near leaves of Araceæ. Around the stem, the cord-like aerial roots of Araceæ on the branches of the tree. (A photograph by G. Karsten.) (After Schimper. Courtesy of Brooklyn Botanic Garden.)
trace their origin to cultivated plants. The antiquity of its culture may be gauged by the fact that in the most ancient Aztec tombs elaborately carved tobacco pipes have been found.
The plant is grown as a field crop in rows from one and one-half to three feet apart and set about fifteen inches apart in the row. From the time the seed is sown until the harvesting of the leaves is usually three or four months, during which the plants demand the best of culture. In the United States thousands of acres of tobacco are now grown under cheesecloth shades, an expensive process which is more than compensated for by the improved flavor.
Once the tobacco is cut there begin chemical changes in the leaves that are of great importance to its subsequent flavor and use. These are aided or induced first by a process of curing, which is accomplished by suspending the wilted leaves in the sun, a process that has been practically abandoned for curing by artificial heat. The leaves are hung in a building where slow fires bring the temperature up to 150 degrees F., which is maintained for a few days. The cured tobacco is then gathered into small bundles which are stacked or packed so closely that fermentation begins, often generating a temperature of 150 degrees F. The bundles are then reshifted and the process allowed to start again, which may be done several times, depending upon the quality of the leaf, flavor desired, and commercial requirements. Enzymes and bacteria play a large part in the fermentation process and inoculation of poor grades of tobacco with the organisms of finer grades has been tried. After fermentation has been stopped practically all tobaccos are aged for at least two years, some for longer periods.
In Cuba, where its use was first noticed, the finest tobacco in the world is still produced, notably in the province of Pinar del Rio. It is still something of a mystery as to what peculiar combination of soil, climate, or handling the unquestionable superiority of the Cuban leaf is due. For one thing it is grown in the open, without shade, and is never cured by artificial heat. Nor is the very excellent cigarette tobacco of Turkey ever artificially cured. But attempts to imitate the conditions under which these finest grades of tobacco are produced outside Cuba and Turkey have never been really successful, so that those countries have practical monopolies on the production of the finest cigars and cigarettes. The weed is cultivated nearly throughout the world, even Canada producing considerable quantities, but the best kinds and greatest production is in warmer regions. It is second only to the sugar crop in Cuba, and the United States produces over one-third the world’s total supply. Immense quantities are grown, however, in India and Sumatra, and in the Philippines.
Perhaps the most unusual and localized conditions of climate and subsequent handling are found in the production of perique tobacco. All the world’s supply is grown on a ridge at Grand Point, in the parish of St. James, Louisiana. The leaves are subjected to great pressure and the expressed juice, after oxidation, is reabsorbed by the leaves after the pressure is removed. The peculiar flavor is apparently due to this and to the damp climate. Perique is now used throughout the world as an ingredient of the better kinds of pipe tobacco.
The diseases and breeding of different strains of tobacco are commercial factors of tremendous importance to the industry. With a yearly value of well over two billions of dollars, the crop is one of the most important plant products, outside of foods. The capital invested in America alone is over five hundred million dollars.
As we have seen many of our most valuable food plants are natives of and are now cultivated in temperate regions, but “sugar and spice and all things nice” mostly come from the tropics. What we usually know as spices such as nutmeg, vanilla, ginger, mace, cloves, allspice, and cinnamon are practically all confined to the tropical regions in or near the East Indies, only vanilla being of American origin. The trade in these spices has been for hundreds of years a practical monopoly in the hands of Dutch and British traders, and for hundreds of years before that the caravans from the Far East came laden with precious freight from the then mysterious country beyond the Mediterranean.
The long pods of two climbing orchids native in Central America and the West Indies furnished for many years our only supplies of this flavoring extract. But in 1891 a process of making vanilline chemically from sugar was perfected so that the vanilla trade is not what it was years ago. Vanilla planters, however, have been able to keep up the price of the plant product because of its unquestionable superiority over the manufactured article. But the latter has enormously increased the general use of vanilla, while the total plant output scarcely exceeds four hundred tons a year. Nearly all this is grown in the Old World tropics, as tropical America, where the plant is common enough as a forest orchid, has not greatly developed its culture.
In both species, known as Vanilla pompona and Vanilla planifolia, the orchid has flat leaves and a fleshy climbing stem that hugs tree trunks or other supports, always in the dense shade of the tropical forests. It needs a hot moist climate, but if there be too much rain as the pods are ripening they drop off, so that only certain localities are suited to its cultivation. Various islands are apparently better suited to the plant than the mainland, Tahiti producing alone nearly half the world’s supply. The species most cultivated is Vanilla planifolia, which came originally from southern Mexico, where considerable plantations are still maintained. The pods are about as thick as a thumb or finger and from five to seven inches long, and yellow when ripe. The ripening process takes several months and when completed the pod is still without the delicious fragrance for which it is famous. Curing by dipping in boiling water or by fermentation, a very delicate process requiring long experience, brings out the flavor. In some regions the pods are plunged into ashes and left there until they begin to shrivel when they are cleaned off, rubbed with olive oil, and tied at their lower end to prevent splitting. Still another process demanding that the pods be plunged in rum is followed, but only in limited degree, owing to the expense. In all of them the result is the same—that of inducing chemical changes in the pod which are responsible for its subsequent flavor.

A native of southeastern tropical Asia. The fruit, somewhat enlarged here, consists of an inner part, the nutmeg. Around this is a “splendid crimson network” which is removed by hand and forms the mace of commerce.
A small tree of the tropical regions of eastern Asia, known as Myristica fragrans, or perhaps better as Myristica moschata, is the source of both nutmeg and mace which come from different parts of the same plant. The genus contains over one hundred species, belongs to the Myristicaceæ, and is scattered all over the Malayan region. Almost none of its relatives, however, have the fragrance of the nutmeg and none is used as a spice. Both nutmeg and mace have been known in Europe only from about 1195 A.D., when in a poem about the entry of the Emperor Henry VI into Rome, the streets were described as being perfumed by the burning of nutmegs and other fragrant plants. It was not until the rise of the Dutch, who burned large stores of it at Amsterdam in 1790 in order to keep up a falling price, that nutmegs came into general use. The trees are now chiefly cultivated in the Dutch East Indies, a small fraction of the supply coming from the West Indies, which is alleged to be an inferior product.
The trees produce male and female flowers, usually on different plants, but sometimes on the same one, yellow in color and aromatic. From the females are developed the fruit which is a drupe about two inches long with a thick fleshy husk which splits upon ripening. The seed inside is the nutmeg, but from its base is an outgrowth which covers the nut with a “splendid crimson network.” This covering or network is removed by hand and forms the mace of commerce.
In the family Myrtaceæ, which contains hundreds of plants from all over the world, mostly all shrubs and trees of tropical regions, however, there is a large genus, Eugenia. From the unopened flower buds of Eugenia caryophyllata, a small tree native only on a few islands in the Moluccas, the widely used spice known as cloves is derived. It appears to have been known to the Chinese at least two hundred years before Christ, and was regularly imported into Europe from the eighth century by caravans. Not until the Dutch began to import it by ship did it become cheap enough to have general use, but in 1609 a Dutch vessel reached England with over a hundred thousand pounds on board.
While the tree is of very local distribution, it has been introduced on a considerable scale into Penang, Zanzibar, and even to the West Indies. Trees are set out thirty feet apart each way, and in from four to eight years, depending on the locality, they begin to flower. After the full bearing stage is reached, a tree will produce from five to seven pounds of dried cloves, an average crop being about 375 pounds to an acre. The flowers are produced in small clusters not over an inch and a half long, so that hand picking is the only method of collection. As the buds become blood red they are usually in a fit state for picking, after which they are either sun dried or, more rarely, by artificial heat. Nothing further is done to them before shipment. Zanzibar and Pemba now produce more cloves than nearly all the rest of the world put together. Oil of cloves, largely used in perfumery, is pressed out of the dried cloves.

One of the commonest trees in the lowland parts of Ceylon is Cinnamomum zeylanicum, a tree of the family Lauraceæ, which also contains our native sassafras. From the bark of this tree is derived cinnamon, and from a related Japanese tree, Cinnamomum Camphora, camphor is taken. Practically all the Lauraceæ are aromatic shrubs or trees, most of them tropical. Ceylon was occupied by the Portuguese in 1536 for the cinnamon then growing on it, which they forced the native king to supply them. Later the Dutch completely controlled the cinnamon, often burning it in Holland to keep up the price. The British, who took Ceylon in 1796, made a government monopoly of cinnamon, but subsequently turned the plantations over to private interests. The tree is now grown on an extensive scale, not only in Ceylon, but in Java and India. Ceylon still controls the cinnamon market, however.

While the wild cinnamon trees reach considerable heights, the cultivated plants are cut so regularly that they almost always throw up a lot of young shoots from the roots, and it is the bark of these that furnishes the spice. When the bark is fit for peeling, the natives cut off the shoots, and strip the bark from them by hand, but with a specially constructed knife. After removal the bark is kept moist and in a day or two the outer skin is scraped off and the bark stretched over a stick, to form the familiar pipes or quills of cinnamon. These are graded, cut to uniform length, and after drying are ready for shipment. All of this is as yet hand work.
Other spice plants and condiments are of wide use, but can scarcely be mentioned in detail here. A few of the more important are the following:
| Name | Derived from the | Native |
| Allspice or pimento | Unripe fruits of Pimenta officinalis Pimenta officinalis | West Indies. |
| Cassia bark | Bark of Cinnamomum Cassia | Eastern Asia. |
| Black pepper | Fruit of Piper nigrum | Tropical Asia. |
| Cardamoms | Fruit of Elettaria Cardamomum | Malabar. |
| Capsicum or Cayenne pepper | Fruits of species of Capsicum | Tropical Asia. |
| Coriander | Fruits of Coriandrum sativum | Europe. |
| Cumin | Fruits of Cuminum Cyminum | Mediterranean region. |
| Dill | Fruits of Peucedanum graveolens | Europe and northern Africa. |
| Ginger | Rootstock of Zingiber officinalis | (?) |
| Turmeric | Rootstock of Curcuma longa | Tropical Asia. |
| Mustard | Seed of Brassica nigra and alba | Old World. |
| Thyme | Foliage of Thymus vulgaris | Southern Europe. |
| Caraway | Fruit of Carum Carui | Europe. |
| Caper | Seeds of Capparis spinosa | Southern Europe and Asia. |
This brief review of what the plant world provides us with in the shape of foods, beverages, fibers, drugs, rubber, spices, and tobacco, does not begin to tell us what man’s debt to plants really is. Thousands of plants, used by natives all over the world, may well provide future generations with unsuspected sources of plant products. No mention has been made of timbers nor all the forest products, except paper, which in the aggregate total an enormous sum. Perhaps no better idea of the tremendous value of plants, of the absorbing interest their utilization has always had for man, can be gained than to refer the reader to incomparably the best book on the subject, so far as tropical plants are concerned. Sir George Watt, in his “Dictionary of the Economic Products of India,” a book of several volumes, most of which deals with plants, has left an imperishable record of man’s struggles to tame the wild plants of the forest to his needs.
A few more economic plants not yet noted are listed below, and with this our account of plants as they are used by man must close:
| Plant Product | Derived from the | Native of |
| Absinthe | Foliage of Artemisia absinthium | Europe. |
| Brazil nut | Seeds of Bertholletia excelsa | Tropical S. America. |
| Camomile | Flowers of Anthemis nobilis | Europe. |
| Cassava | Roots of species of Manihot | Tropical America. |
| Water chestnuts | Fruits of Trapa natans | Southern Europe. |
| Cohune nut | Fruits of Attalea Cohune | Central America. |
| Cork | Bark of Quercus Suber | Southern Europe and northern Africa. |
| Gamboge resin | Stems of species of Garcinia | Southeastern Asia. |
| Gum arabic | Exudation of species of Acacia | Tropical Asia and Africa. |
| Hops | Female flowers of Humulus Lupulus | Eastern Europe. |
| Indigo | Whole plant of Indigofera tinctoria | India. |
| Mushroom | Whole plant of Agaricus campestris | Temperate regions. |
| Teak | Wood of Tectona grandis | Southeastern Asia. |
| Japanese varnish or lacquer | Sap of Rhus vernicifera | China. |
NOT only does the plant world furnish us with all the multitudinous products that we have already noticed, but it makes possible the beautifying of our homes and parks. For with plant materials, anyone with the knowledge and taste necessary for work of this kind may paint living landscape pictures that grow in beauty as their individual units reach maturity.
It lies outside the scope of this book to tell you the principles of design upon which such landscape pictures must be based to be really effective—that is the function of the landscape architect. But every one of us knows when a house looks and is bare of vegetation about it, and consequently has the earmarks of being merely a house, but not a home. A walk through any suburb of a large city or through most of our American villages would convince the lover of gardening that we are still miles behind England and many other countries in the love and appreciation of that kind of beauty in our home surroundings which plant life alone can furnish. How unnecessary this is anyone can see by visiting certain distinguished exceptions to the general indifference to plant life about our homes. Such suburbs as Brookline near Boston, Garden City and Morristown near New York, Guilford at Baltimore, Germantown near Philadelphia and many places on the Pacific Coast show what can be done to transform an otherwise indifferent landscape into a beauty spot. While these are on the whole the homes of the wealthy, money is not what has made them, for thousands of cottage gardens in England are just as beautiful and have been made by people who live a busy industrial life, but whose desire for beautiful surroundings makes them spend their brief leisure in tending their flowers.
While large garden schemes demand somewhat expert advice as to their planning and arrangement, it is perfectly simple for anyone to begin planting his own home grounds if he has in his mind’s eye the ultimate picture which he wishes his house and garden to become. But the habits of plants, their growth requirements, their stature, and particularly their colors are so various that, with the best will in the world, a garden enthusiast without some knowledge of these things will get a wholly disappointing result. Certain plants will grow in some sections of the country, but fail in others; some flower in the south, but will not do so in the north, and a few set seeds in certain places, but never do in less favored regions. In the different sections of this chapter a few good garden plants will be noted according to the regions to which they are suited, but it must not be forgotten that some are suited also to other regions than the one in which they are listed.
Those who have read the earlier chapters already know the difference between annuals, biennials, and perennials which comprise all the herbaceous plants upon which we depend mostly for cut flowers and in large measure for giving color to the garden. The woody plants are the ones upon which most garden pictures depend for their permanent value—trees, shrubs and vines of infinite shape and foliage character. In the case of trees, there are two major classes, those that drop their leaves in fall and are therefore deciduous, and of value chiefly during the growing season; and the evergreens, which retain their foliage all the year and make winter landscapes of great beauty. The garden enthusiast will very soon learn that evergreen plantings, while in many ways the most beautiful, are much the most expensive and are never suited to regions near big cities, for they will not stand smoke and other fumes as many deciduous trees will do. Nor will they stand violent winds, small rainfall, and great summer heat such as characterize the central parts of the country. Their best development is therefore found east of the Mississippi and west of the Rockies, and generally speaking, their use in the garden should be confined to this region.
So much of what makes landscapes permanently beautiful depends upon trees that first place must always be given to them in any scheme of planting. The location and ultimate spread of these trees will infallibly make or mar any garden picture so that great care should be used in selecting and planting them. The actual planting details such as preparation of the soil and all the after care of plants cannot be dealt with here, but many nursery catalogues give accurate directions and there are hosts of books on the practice of gardening which give the necessary information. In listing the different trees, symbols will be put before the names, indicating in which region they are likely to grow best, as follows:
* Suited to the region east of the Mississippi and north of the frostless region of the Gulf States, but not all hardy in the northern part of United States and adjacent Canada.
** Suited to the same general region, but most at home in the northern part of the area.
Those that have no symbol before the name are understood to be, generally speaking, hardy throughout the country, with, of course, exceptions such as the desert and alkali regions of the country.
| * | White Pine, Pinus Strobus. |
| * | Austrian Pine, Pinus Austriaca. |
| * | Scotch Pine, Pinus sylvestris. |
| * | Pitch Pine, Pinus rigida. |
| ** | Red Pine, Pinus resinosa. |
| * | Umbrella Pine, Sciadopitys verticillata. |
| * | White Fir, Abies concolor. |
| ** | Fraser’s Fir, Abies Fraseri. |
| * | Nordman’s Fir, Abies Nordmanniana. |
| * | Norway Spruce, Picea excelsa. |
| ** | White Spruce, Picea alba. |
| ** | Red Spruce, Picea rubens. |
| * | Koster’s Blue Spruce, Picea pungens glauca. |
| * | Engelmann’s Spruce, Picea Engelmannii. |
| * | Juniper. Different species of the genus Juniperus, mostly low growing and suitable for ground covers. |
| * | Japanese Cypress, Retinospora obtusa. There are many garden varieties. |
| * | Southern Cypress, Taxodium distichum. Not hardy in the northern part of the area. Best in wet places. |
| * | Lawson’s Cypress, Chamæcyparis Lawsoniana. |
| * | English Yew, Taxus baccata. |
| ** | American Yew, Taxus canadensis. |
| * | Hemlock, Tsuga canadensis. |
Of these the Austrian pine, hemlock, and the firs have the densest foliage and should be used for such effects. Almost nothing will grow under the evergreen trees, so close is their foliage. Lack of light and the acid leached out of their bark by rains, stop the growth of nearly all herbs underneath them.
Planted mostly for their foliage masses, but a few bear showy flowers and such will be noted. The same symbols apply.
| * | American Beech, Fagus ferruginea. |
| * | European Beech, Fagus sylvatica. |
| * | White Oak, Quercus alba. |
| * | Red Oak, Quercus rubra, the most rapid grower of all the oaks. |
| * | Scarlet Oak, Quercus coccinea. |
Horsechestnut, Aesculus Hippocastanum. | |
| * | Norway Maple, Acer platanoides. |
| * | Red Maple, Acer rubrum. Prefers moist places. |
Sugar Maple, Acer saccharum. | |
| * | Silver Maple, Acer saccharinum. Fine tree with interesting branching, but brittle. |
| * | Tulip Tree, Liriodendron tulipifera. Showy orange-green flowers. |
| * | American Plane Tree, Platanus occidentalis. A native tree, but not so satisfactory as |
| * | Oriental Plane Tree, Platanus orientalis. |
| * | Sweet Gum, Liquidambar styraciflua. |
White-leaved Poplar, Populus alba. | |
Balsam Poplar, Populus balsamifera. | |
| * | Flowering Plum, Prunus Pissardi and many other varieties and species of the genus Prunus such as Japanese Flowering Cherries and Plums. |
| * | American Elm, Ulmus americana. |
| * | English Elm, Ulmus campestris. |
| * | Maidenhair Tree, Ginkgo biloba. Not hardy in the northern part of the area. |
Black Locust, Robinia Pseudacacia. Showy pink flowers. | |
European White Birch, Betula alba. | |
Black Alder, Alnus glutinosa. | |
Ash, Fraxinus americana. | |
| * | Dogwood, Cornus florida. Showy white bloom. Tree will not grow well unless in partial shade. |
| * | Cornelian Cherry, Cornus Mas. Covered with yellow flowers before the leaves come out in early spring. |
| * | European Hornbeam, Carpinus Betulus. |
Weeping Willow, Salix Babylonica. | |
Box Elder, Acer Negundo. | |
| * | Magnolias. Different trees and some shrubs of the genus Magnolia all with showy flowers. Rather tender and cannot be grown without considerable care, especially when young. |
Honey Locust, Gleditsia triacanthos. | |
Catalpa, Catalpa speciosa. Showy flowers. | |
| * | Paulownia imperialis. Showy flowers, but not hardy in northern part of the area. |
There are many other deciduous and evergreen trees that might be listed and which will be found in the nursery catalogues of dealers in different parts of the country. Some of these require special conditions of soil and climate and should not be planted unless these conditions are understood. In the frostless region of the country many plants can be grown that are of tropical or near-tropical origin, but no list of them will be included here. Some of them are hardy as far north as Washington, D. C., and are worth trying by anyone living in this region, as they give us effects not possible with the trees noted.
While trees make the major feature of any garden, shrubs are chiefly used to fill in between them, or in small gardens the only woody plants that can be used are often shrubs. Within the last two or three years the Government has prohibited the importation of plants from abroad, upon the ground that various insect pests and fungous diseases were likely to be carried into the country upon such plants. For this reason American gardeners will have to propagate their own plants and we shall have to use more native plants than European and Asiatic species, which made up the bulk of our gardening material in the past.
There are excellent reasons for using native shrubs upon quite other grounds than the difficulty or danger of importing foreign ones. Native plants fit into the natural landscape better than introduced sorts, and very often the garden enthusiast can go out into the country and dig out small specimens instead of buying them.
In the list of native American shrubs given below, there are directions of where to use them, their heights, their flower color and other information about them that will help the amateur gardener to select his shrubs for definite effects. All of the shrubs listed can be gown in most parts of the country, and from the list nearly every wish of the garden planner may be gratified. This list is a practical one and has been used by landscape architects and others. It was written by the author for “The Garden Magazine” whose publishers, Doubleday, Page & Company, have kindly allowed its use here.
It will be noted that under each month group the names are arranged in botanical sequence so that allied plants are brought together. All the ninety-four species are offered for sale in American nurseries. Those in the column “Remarks and Notes” as well as about twenty others not included, must be collected in the wild.
A word now as to cultivation and care. Most of the shrubs, except those so noted, can either be planted in the spring or fall, as this is a matter that should be determined by the planter’s convenience. In digging the holes make them twice as wide and deep as the size of the roots apparently demand. Note carefully the column “Preferred Habitat,” so that the shrubs may find congenial surroundings. Pack the soil well around the roots, water thoroughly, and frequently if the weather is dry and windy. The first winter or two a heavy mulch of leaves, or leaves and manure mixed, to be dug in the following spring, will well repay the expense and trouble.
It will be noted that some of the shrubs are marked with an asterisk (*). These all belong to the heath family and require special treatment. A soil composed of rotten sods and leafmold, about half and half, is most essential for the successful cultivation of these plants. They require peculiar acid soil conditions well approximated by the above mixture, and a mulch, preferably of red-oak leaves, or the leaves of the mountain laurel if available. Never disturb the roots of these plants by digging in the mulch, which is better left on indefinitely. Soils with much lime in them must also be avoided when growing these heath-family plants.
It is often somewhat difficult in arranging a shrubbery planting to group the plants according to the color of their flowers. For the greater ease in using the larger table, and so that one can arrive at the relative frequency of the various colors desirable for use in the scheme, the following table is appended. The numbers refer, of course, to those in the table below. The figures given in parentheses are the total of plants in each division.
By color of flowers. Yellow-green (10): 1, 8, 9, 10, 30, 31, 36, 44, 49, 86. Brown-green (10): 2, 3, 4, 11, 12, 13, 14, 15, 27, 39. Yellow (5): 5, 25, 41, 84, 92. Pink-purple (4): 7, 74, 75, 83. White (35): 6, 17, 18, 19, 20, 21, 22, 24, 28, 29, 32, 40, 42, 43, 45, 46, 47, 51, 55, 57, 58, 59, 69, 70, 71, 72, 76, 78, 79, 80, 81, 82, 88, 89, 90. Green-white (11): 16, 23, 33, 50, 52, 63, 64, 65, 66, 87, 94. Pinkish-white (10): 26, 37, 38, 54, 56, 61, 67, 73, 77, 93. Pink (5): 34, 60, 62, 85, 91. Lilac (1): 35. Violet-purple (2): 48, 68. Orange-red (1): 53.
It often happens, too, that we have some definite spot, such as a small stream or swamp, a dry hill-side, or a shaded wood, that we wish to beautify. Therefore:
By preferred habitat of shrubs. Moist places (19): 1, 2, 4, 9, 15, 17, 30, 36, 42, 50, 75, 78, 82, 83, 84, 85, 87, 88, 89. Indifferent (32): 3, 5, 6, 7, 12, 14, 24, 29, 32, 35, 37, 38, 41, 43, 45, 47, 48, 51, 52, 59, 60, 61, 62, 68, 70, 71, 72, 79, 80, 81, 90, 91. Shaded woods (13): 8, 16, 23, 25, 26, 27, 31, 63, 65, 67, 69, 74, 92. Dry places (19): 10, 13, 18, 20, 21, 22, 34, 39, 46, 49, 53, 54, 56, 58, 64, 77, 86, 93, 94. Swamps (6): 11, 40, 55, 66, 73, 76. Thickets (1): 19. Cool woods (4): 28, 33, 44, 57.
In planning a screen for an unsightly fence or building, or to cover up some small landscape importunity, it is often essential to know, en masse, the heights of shrubs for such purposes. The following table gives the dimensions of the shrubs, normal individuals averaging about midway of the extreme heights given.
By height of shrubs. One to four feet (13): 2, 6, 21, 27, 34, 56, 62, 75, 77, 83, 84, 91, 93. Two to five (24): 10, 11, 13, 16, 22, 25, 31, 36, 37, 42, 44, 53, 57, 60, 61, 65, 67, 69, 73, 81, 82, 85, 90, 94. Three to nine (29): 3, 4, 8, 12, 14, 15, 17, 20, 26, 28, 32, 33, 35, 38, 39, 41, 43, 45, 48, 54, 58, 59, 70, 72, 74, 78, 80, 86, 88. Six to fifteen (21): 1, 5, 7, 18, 19, 29, 40, 46, 47, 49, 50, 52, 55, 64, 68, 71, 76, 79, 87, 89, 92. Ten to eighteen (7): 9, 23, 24, 30, 51, 63, 66.
PLANTING TABLE FOR OUR NATIVE SHRUBS[3]
| Common and Latin Names | Height (feet) | Color of Flowers | Preferred Habitat | Remarks and Notes | |
| Flowering in MARCH-APRIL | |||||
| 1. | Pussy willow (Salix discolor) | 7-12 | Yellow-green | Moist places | Flowers before the leaves come out. S. cordata, a larger bush, with broad leaves is worth cultivating. Not in the catalogues. |
| 2. | Dwarf willow (Salix tristis) | 1-4 | Brown-green | Moist places | Useful in masses. Can be made to grow in all sorts of places. One of the very earliest flowering shrubs. |
| 3. | Hazelnut (Corylus americana) | 3-6 | Brownish-green | Indifferent[4] | Nuts edible much gathered by the squirrels, The catkins out before the leaves. European hazelnut is a better plant. |
| 4. | Alder (Alnus rugosa) | 5-9 | Brownish-green | Moist places | Will grow in other situations. The fruits, not very strong, will stay on all winter. Useful in masses along brooks. |
| 5. | Spice bush (Benzoin odoriferum) | 6-15 | Yellow | Indifferent | Flowers much before the leaves, very fragrant, Near N. Y. usually not over 10 feet, larger southward. |
| 6. | Red chokeberry (Aronia arbutifolia) | 2-4 | White | Indifferent | Common from N. Y. southward. A. atropurpurea, with black fruit is worth while. A. arbutifolia has red fruit. |
| 7. | Red bud (Cercis canadensis) | 4-15 | Pink-purple | Indifferent | Magnificent masses of color before the leaves appear. Sometimes almost a tree. Rare as a wild plant, but easily cultivated. |
| 8. | Fragrant sumac (Rhus canadensis aromatica) | 3-8 | Yellowish green | Rocky woods | Will grow in unlikely places and an excellent shrub for wild effect. Flowers half hidden by compound leaves. |
| APRIL-MAY | |||||
| 9. | Shiny willow (Salix lucida) | 10-18 | Yellow-green | Low places | Will grow almost anywhere. S. myrtilloides a shrub 3½ feet, not in the trade, is handsome with yellow catkins. |
| 10. | Prairie willow (Salix humilis) | 3-6 | Yellow-green | Dry places | Will grow almost anywhere. Flowers out much before the leaves. Useful only in mass effects. |
| 11. | Sweet gale (Myrica Gale) | 3-6 | Incons-picuous | Swamps and bogs | Ash colored fruits effective all winter. Will grow in many other situations besides the preferred one. |
| 12. | Bayberry (Myrica carolinensis) | 3-8 | Not showy | Indifferent | Grows equally well in sand loam, or swampy places. Leaves shining green, long persistent. Fruits whitish; all winter. |
| 13. | Sweet fern (Comptonia asplenifolia) | 3-5 | Golden-brown | Dry hillsides | Golden catkins very showy before the leaves. Whole plant very fragrant. Can be grown almost anywhere. |
| 14. | Beaked hazelnut (Corylus rostrata) | 3-6 | Brown-yellow | Indifferent | Along streams it makes effective screens and borders. The long beak quite distinct from No. 3. Occasionally 8 feet. |
| 15. | Hoary elder (Alnus incana) | 4-9 | Greenish-brown | Moist places | Leaves pale green beneath. With Nos. 3, 4, 14 and 89, it can be used effectively along shores of streams and ponds. |
| 16. | American black currant (Ribes americana) | 3-5 | Green-white | Shaded woods | R. lacustre and R. rubrum, the latter with reddish purple flowers are very fine. Neither in the trade. |
| 17. | Juneberry (Amelanchier Botryapium) | 5-12 | White | Moist places | As individual plants very shapely, but rather ungainly in close formation. A. spicata (1-4 ft.) good, but not in the trade. |
| 18. | English hawthorn (Cratægus Oxyacantha) | 5-15 | White | Dry hillsides | The May. Much cultivated and now run wild. The American C. rotundifolia common on L. I. and N. J., but not for sale. |
| 19. | Scarlet thorn (Cratægus coccinea) | 5-15 | White | Thickets | The closely related C. Mollis, with scarlet fruits is effective in autumn. Not in the catalogues. |
| 20. | Dwarf thorn (Cratægus uniflora) | 2-8 | White | Dry sandy place | Quite indifferent as to locality when cultivated. C. macracantha with long spines is often 10 to 15 feet. Not in the trade. |
| 21. | Beach plum (Prunus maritima) | 1-4 | White | Sandy places | Fruit makes excellent jelly. Very successful near the sea. P. cuneata better grown near moist rocks. |
| 22. | Sand cherry (Prunus pumila) | 3-6 | White | Dry places | Splendid in masses or small hillocks. Will grow in almost pure sand. P. Gravessii not in the trade. |
| 23. | Prickly ash (Xanthoxylum americium) | 6-18 | Green | Shaded places | Will also grow in ordinary garden soil. The large compound leaves give splendid foliage effects. |
| 24. | Bladder nut (Staphylea trifolia) | 6-20 | White | Indifferent | Usually about 10 feet in our latitude. The showy pods stay on most of the winter. Flowers not showy. |
| 25. | Leatherwood (Dirca palustris) | 2-5 | Yellowish | Shaded places | In masses under trees or along shaded walks it is most welcome. Useful in a shaded rockery. |
| 26. | Pinkster flower (Azalea nudiflora) | 2-7 | Pinkish-white | Shaded wood[5] | A blaze of color when planted in masses with other Azaleas. Can also be grown successfully in the open. |
| 27. | Deerberry (Vaccinium stamineum) | 1-4 | Purple-green | Dry woods[5] | Flowers not showy, but purple fruits are attractive. Best not disturbed or transplanted after setting out. |
| 28. | Red-berried elder (Sambucus pubens) | 3-10 | White | Cool woods | Easily grown in the garden but most successfully under trees or along the north side of the house. |
| 29. | Black haw (Viburnum prunifolium) | 5-18 | White | Indifferent | After becoming a small tree. A magnificent snowy shrub in the spring. Fruits black. |
| MAY | |||||
| 30. | Silky willow (Salix sericea) | 6-15 | Yellow-green | Moist places | With the other willows useful for filling in low moist places. Leaves ashy beneath. Catkins showy. |
| 31. | Wild gooseberry (Ribes Cynosbati) | 3-5 | Greenish-yellow | Rocky woods | Better grown in the shade and in rich soil. The bristly fruits are odd persistent features of this shrub. |
| 32. | Black chokeberry (Aronia nigra) | 3-8 | White | Indifferent | Shiny black fruit stays on until December or January. Somewhat scraggy, except in masses. |
| 33. | Mountain holly (Ilex monticola) | 4-10 | Greenish-white | Cool shade | Flowers not showy but the red fruits showy all the autumn. Do not attempt to grow in hot, dry places. |
| 34. | Blue huckleberry (Vaccinium vaccillans) | 1-4 | Pink | Dry soil[5] | The profusion of tiny bell-like flowers appearing with the leaves makes this attractive. Fruits purple-black. |
| 35. | Common lilac (Syringa vulgaris) | 4-10 | Lilac | Ordinary garden soil | Cultivated everywhere and sometimes escaped from gardens. There are scores of attractive hybrids and forms. |
| 36. | American fly honeysuckle (Lonicera ciliata) | 2-4 | Greenish-yellow | Moist woods | Easily grown in ordinary garden soil, but prefers shade. Best planted along shaded walks. |
| 37. | Fly honeysuckle (Lonicera Xylosteum) | 2-6 | Pinkish-white | Indifferent | Sometimes an escape from cultivation. The scarlet berries are showy in the early fall. |
| 38. | Tartarian bush honeysuckle (Lonicera Tatarica) | 3-8 | Pinkish-white | Indifferent | L. oblongifolia with purplish-yellow flowers in May and June is attractive. Not in the catalogue. |
| MAY-JUNE | |||||
| 39. | Chinquapin (Castanea pumila) | 5-8 | Brownish-green | Dry soil | Apt to be affected with the chestnut blight. The long catkins and fruit are interesting but not showy. |
| 40. | Magnolia (Magnolia glauca) | 4-10 | White | Swamps and bogs | Can also be grown very well on dry ground and in any garden soil. Fruits rose red. |
| 41. | Common barberry (Berberis vulgaris) | 3-8 | Yellow | Common garden soil | Often an escape from cultivation. The well-known scarlet berries showy in autumn. |
| 42. | Virginian willow (Itea virginica) | 2-4 | White | Moist places | When massed either alone or with Clethra alnifolia it makes attractive patches of white. |
| 43. | Syringa (Philadelphus coronarius) | 4-10 | Cream-white | Indifferent | Many horticultural forms of this are in the trade. All are useful. Fruits brownish. |
| 44. | Fetid currant (Ribes prostratum) | 3-6 | Greenish-yellow | Cool moist places | Not easily grown as it grows naturally on the cool mountain slopes. Fruits red. |
| 45. | Opulaster (Spiræa opulifolia) | 3-9 | White | Indifferent | Splendid masses of flowers, as it is a profuse bloomer. Often from 3-6 feet wide and very bushy. |
| 46. | Cockspur thorn (Cratægus Crusgalli) | 6-14 | White | Dry soil | One of the most commonly cultivated of our native shrubs. Very thorny and a good hedge plant. |
| 47. | Pear haw (Cratægus tomentosa) | 4-12 | White | Indifferent | The dull red fruits cling on most of the winter. A profusely flowering shrub. |
| 48. | Bastard indigo (Amorpha fruticosa) | 4-10 | Violet-purple | Rich soil | A gorgeous flowering shrub, which in masses is unrivaled. Repays good cultivation and care. |
| 49. | Staghorn sumac (Rhus typhina) | 6-15 | Green | Dry places | Autumnal coloring magnificent. On a low hill very effective in large masses. |
| 50. | American holly (Ilex opaca) | 6-15 | Greenish-white | Moist woods | Best transplanted in the spring, when all the evergreen leaves should be knocked or clipped off. |
| 51. | Buckthorn (Rhamnus cathartica) | 8-16 | White | Indifferent | This and No. 52 both European shrubs that have run wild in this country. Neither is showy in flower. |
| 52. | Alder buckthorn (Rhamnus Frangula) | 4-11 | Greenish-white | Indifferent | Its natural home is in swamps and bogs, but generations of garden culture has made it at home. |
| 53. | Flame azalea (Azalea calendulacea) | 2-7 | Orange-yellow | Dry woods[5] | The showiest of all our native shrubs. Not very common in the wild state. |
| 54. | Mountain laurel (Kalmia latifolia) | 4-10 | Red | Dry woods[5] | In masses under the shade of trees a wonderfully effective shrub. Prefers rich soil. |
| 55. | Swamp huckleberry (Vaccinium corymbosum) | 6-15 | Pinkish-white | Swamps and wet woods[5] | Flowers not very showy, but the fruits are the finest of the tribe. Will not tolerate dry places. |
| 56. | Low blueberry (Vaccinicum Pennsylvanicum) | 1-4 | White | Dry or sandy soil[5] | Often grows in almost pure sand in the pine-barrens. Neither flower nor fruit showy. |
| 57. | Hobble bush (Viburnum alnifolium) | 3-6 | Pinkish-white | Cool, moist shade | The outer circle of flowers in each cluster very much larger than the inner. Does not like hot places. |
| 58. | Dockmackie (Viburnum acerifolium) | 3-8 | White | Dry woods | Looks like a small maple tree. Useful as it will grow almost anywhere. Fruits black. |
| JUNE-JULY | |||||
| 59. | Hydrangea (Hydrangea arborescens) | 4-9 | White | Indifferent | Thoroughly hardy and often easier grown than the more showy exotic species. |
| 60. | Meadow rose (Rosa virginiana blanda) | 2-4 | Pink | Indifferent | Along paths and roadsides it scrambles everywhere with apparent cultural indifference |
| 61. | Sweetbrier (Rosa rubiginosa) | 3-6 | Pinkish-white | Indifferent | Well repays good treatment when it often becomes a bushy shrub 4 to 5 feet in diameter. |
| 62. | Pasture rose (Rosa humilis) | 1-3 | Pink | Indifferent | The beautiful large petals very evanescent. It can be best grown in a moist place. Showy. |
| 63. | Water ash (Ptelea trifoliata) | 6-18 | Greenish-white | Shade | Flowers inconspicuous but the compound leaves make it a good foliage plant. Wood very brittle. |
| 64. | Black sumac (Rhus Copallina) | 5-15 | Greenish-white | Dry places | The large compound leaves a beautiful scarlet in the autumn. Profuse bloomer and fruits persistent. |
| 65. | Inkberry (Ilex glabra) | 3-6 | Greenish-white | Moist woods | The more rare I. mucronata of swamps is interesting botanically but must be collected from the wild. |
| 66. | Winterberry (Ilex verticillata) | 6-18 | Greenish-white | Swamps | Splendid scarlet fruits cling on in large clusters most of the winter. Often easily grown in the garden. |
| 67. | Strawberry bush (Euonymus americanus) | 3-7 | Greenish-pink | Low woods | Flowers small and inconspicuous but followed by red fruits that last until December. A slender plant. |
| 68. | Burning bush (Euonymus atropurpureus) | 8-15 | Purple | Indifferent | European shrub much cultivated and now widely established as a wild plant. Fruits red and showy. |
| 69. | New Jersey tea (Ceanothus americanus) | 2-6 | White | Shade | Effective as massed plantings. The leaves the source of tea in Revolutionary times. A profuse bloomer. |
| 70. | Kinnikinik (Cornus Amomum) | 3-10 | White | Indifferent | Purple twigs effective in winter. The reddish-twigged C. asperifolia effective but not on sale. |
| 71. | Red osier dogwood (miscalled kinnikinik) (Cornus stolonifera) | 3-12 | White | Indifferent | Twigs reddish-purple; and fine in masses for its winter color harmonies. Easily grown from cuttings. |
| 72. | Cornel (Cornus alternifolia) | 3-10 | White | Indifferent | The bright green twigs which keep their color all winter make it attractive grouped with Nos. 70 and 71. |
| 73. | Swamp honeysuckle (Azalea viscosa) | 3-6 | Pink and white | Swamps[5] | Rather shy of dry places but easily replaced in such places by the A. canescens, which must be collected. |
| 74. | Rhododendron (Rhododendron maximum) | 4-18 | Rose-white-purple | Woods[5] | Old plants, almost treelike, should never be disturbed. Be careful to nip all fruits as soon as they appear. |
| 75. | Sheep laurel (Kalmia angustifolia) | 1-3 | Purple-crimson | Low, moist places[5] | The rare K. glauca, not in the trade, is very much worth while. Neither is happy in open dry places. |
| 76. | Leucothoe (Leucothoe racemosa) | 5-12 | Cream-white | Swamps[5] | The glossy practically evergreen leaves make an effective winter showing. Can be grouped with Nos. 74, 77 and 78. |
| 77. | Staggerbush (Pieris Mariana) | 1-4 | Pinkish-white | Sandy soil[5] | Isolated plants are apt to be sprawling, but when massed the delicate flowers make attractive patches of color. |
| 78. | Privet andromeda (Xolisma ligustrina) | 4-9 | White | Moist places[5] | Leaves partially evergreen, and dark glossy green in color. A profuse bloomer with persistent fruits. |
| 79. | Elderberry (Sambucus canadensis) | 5-15 | White | Indifferent | In large clusters most effective as a screen. Will grow very well along a stream or pond. Fruits “mussy.” |
| 80. | Cranberry bush (Viburnum Opulus) | 3-12 | White | Indifferent | Profuse masses of flowers and large clusters of scarlet berries make it most useful all the year. |
| 81. | Withe rod (Viburnum nudum) | 2-8 | White | Indifferent | The C. pubescens of rocky woods equally good but not offered for sale. Fruits blue-black. |
| 82. | Appalachian tea (Viburnum cassinoides) | 2-8 | White | Moist places | V. Lentago with black fruits useful, but must be collected. Leaves of V. cassinoides glossy and dark green. |
| JULY-AUGUST | |||||
| 83. | Hard hack (Spiræa tomentosa) | 1-4 | Pink-purple | Low ground | The ashy underside of the leaves, contrasted with the pinkish-purple flowers is a novel combination. |
| 84. | Shrubby cinquefoil (Potentilla fruticosa) | 2-4 | Yellow | Moist places | One of the yellow-flowered shrubs that are used. Sometimes winter-kills near New York. |
| 85. | Swamp rose (Rosa Carolina) | 4-7 | Rose-colored | Moist places | Can also be successfully grown in ordinary garden soil, well manured. Flowers soon withering in open sunlight. |
| 86. | Smooth sumac (Rhus glabra) | 3-12 | Greenish | Dry places | Grouped with Nos. 49 and 64, it gives a wild touch to the landscape. Autumn color gorgeous. |
| 87. | Hercules’s club (Aralia spinosa) | 6-15 | White | Low ground | Large compound leaves 3 to 4 feet long, make this the foliage plant par excellence. Flowers inconspicuous. |
| 88. | Sweet pepper bush (Clethra alnifolia) | 3-8 | Cream-white | Low ground | Fragrant flowers followed by persistent fruits, a sturdy habit and bushy outline make this a favorite. |
| 89. | Buttonbush (Cephalanthus occidentalis) | 5-15 | Cream-white | Moist places | Best not attempted much away from water, and in such situations often becoming almost treelike. Flowers fragrant. |
| 90. | Snowberry (Symphoricarpos racemosus) | 2-6 | White | Indifferent | The flowers are not showy but the conspicuous white berries stay on all winter, thus valuable for winter effect. |
| 91. | Coralberry (Symphoricarpos vulgaris) | 1-4 | Pink | Indifferent | Much like the preceding but the red fruits are not so persistent. Forms a wide-spreading bush. |
| SEPT.-DEC. | |||||
| 92. | Witch hazel (Hamamelis virginiana) | 5-15 | Yellow | Moist shade | Flowers later than any other native shrub, often after all the leaves have fallen off, and the first frost arrives. |
| 93. | Heather (Calluna vulgaris) | 1-2 | Pinkish-white | Sandy places | Rather shy in its few American localities. Near the coast from Massachusetts to southern New Jersey it should do well. |
| 94. | Groundsel tree (Baccharis halimifolia) | 2-5 | Whitish-green | Dry soil | Best transplanted in the spring as its late flowering makes autumnal activity too great for easy transplanting then. |
Rhododendrons and azaleas, hundreds of varieties of which are known and admired by all garden lovers, are mostly derived from Asiatic species, and under the new law have become rare and expensive in this country. A few American nurserymen are able to propagate them so that we can still get plants of these showiest of all shrubs. They should not be used in regions where there is scant rainfall, very hot summers, high winds or extreme winters. Their use is practically confined to the region east of the Alleghenies.
The foregoing lists of shrubs and trees give sufficient information so that all garden enthusiasts can at least make the broad outlines of a garden picture. It must never be forgotten that these woody plants are the only really permanent things in the scheme and should therefore be placed with more care and thought than the herbs, which can be moved at will. In selecting herbs, two chief divisions of them should be kept in mind; annuals which are planted for quick effects and which die down at the end of the year, and perennials which live year after year and produce flowers usually after the second year. It is from perennials that the great bulk of our fine garden plants are derived. Planted in beds or better yet among shrubbery or on shrub-backed borders they are the most beautiful and most satisfactory of all herbaceous plants.
Most of them may be started from seed in August of any year and grown along through the balance of that growing season, after which they should be covered up with straw or manure. The following season at least three-quarters of them will flower and by their third season all of them will do so.
A study of the following list will show any garden amateur how he may group his perennials in borders or beds and how he can get different color effects at different seasons and in plants of different heights. This admirable list of perennial plants was prepared by Charles Downing Lay, a landscape architect, and published by “Landscape Architecture.” The editors of that publication and Mr. Lay have kindly consented to having the list reprinted here.
| Color | HARDY GARDEN PERENNIALS | Season | Height | |||||||||||||||||||||
| Pink | White | Yellow | Orange | Red | Blue | Lavender | Purple | April | May | June | July | August | September | October | 6 inches | 1 foot | 2 feet | 3 feet | 4 feet | 5 feet | 6 feet | |||
| —— | Achillea Ptarmica fl.-pl., The Pearl | —— | —— | —— | ||||||||||||||||||||
| —— | Millefolium, Cerise Queen | —— | —— | — | — | |||||||||||||||||||
| — | — | Aconitum autumnale. Monkshood | — | — | — | — | ||||||||||||||||||
| —— | —— | —— | —— | Althæa rosea. Hollyhock | —— | —— | ||||||||||||||||||
| —— | Alyssum saxatile compactum. Golden Tuft | —— | ||||||||||||||||||||||
| —— | Anchusa italica var. Dropmore. Alkanet | — | — | — | —— | — | ||||||||||||||||||
| —— | sempervirens. Alkanet | — | — | — | — | |||||||||||||||||||
| —— | Anemone japonica. Japanese Windflower | — | —— | —— | — | —— | ||||||||||||||||||
| —— | japonica, Queen Charlotte. Japanese Windflower | — | —— | —— | — | —— | ||||||||||||||||||
| —— | Anthemis tinctoria. Marguerite | — | — | — | — | |||||||||||||||||||
| —— | —— | Aquilegia Helenæ. Columbine | — | —— | — | — | —— | |||||||||||||||||
| —— | —— | canadensis. Columbine | — | —— | — | —— | — | |||||||||||||||||
| —— | chrysantha. Columbine | — | —— | — | — | — | ||||||||||||||||||
| —— | —— | —— | formosa hybrida. Columbine | — | —— | — | —— | — | ||||||||||||||||
| —— | haylodgensis. Columbine | — | —— | — | — | —— | ||||||||||||||||||
| —— | vulgaris nivea grandiflora. Columbine | — | —— | — | —— | — | ||||||||||||||||||
| —— | Arabis alpina. Alpine Rock Cress | — | — | —— | — | |||||||||||||||||||
| —— | Armeria plantaginea. Sea Pink | — | —— | —— | ||||||||||||||||||||
| —— | Artemisia lactiflora. White Mugwort | — | — | — | —— | — | ||||||||||||||||||
| —— | Asclepias tuberosa. Butterfly Weed | — | — | — | —— | |||||||||||||||||||
| — | — | Aster alpinus. Alpine Aster | — | — | —— | |||||||||||||||||||
| —— | — | novæ-angliæ. New England Aster | — | — | — | —— | — | |||||||||||||||||
| — | — | tataricus. Late-blooming Aster | — | —— | — | — | ||||||||||||||||||
| —— | Thomas S. Ware | — | — | — | — | |||||||||||||||||||
| —— | Baptisia australis. False Indigo | — | — | — | —— | —— | ||||||||||||||||||
| —— | —— | Bellis perennis. English Daisy | — | — | —— | |||||||||||||||||||
| —— | Boltonia asteroides. False Chamomile | — | —— | —— | — | — | —— | —— | —— | —— | ||||||||||||||
| — | — | latisquama nana. Dwarf False Chamomile | — | —— | —— | — | — | |||||||||||||||||
| —— | —— | Campanula carpatica. Carpathian Harebell | — | —— | — | —— | ||||||||||||||||||
| —— | —— | —— | Medium. Canterbury Bells (Biennial) | — | — | — | —— | — | ||||||||||||||||
| —— | —— | persicifolia grandiflora. Peach Bells | — | — | — | — | ||||||||||||||||||
| —— | —— | pyramidalis. Chimney Bellflower | —— | — | — | — | ||||||||||||||||||
| —— | rapunculoides | —— | — | — | —— | — | ||||||||||||||||||
| —— | Centaurea montana. Mountain Bluet | — | —— | —— | — | — | —— | |||||||||||||||||
| —— | Cerastium tomentosum. Snow-in-Summer | — | —— | —— | ||||||||||||||||||||
| —— | —— | —— | —— | —— | Chrysanthemum, Hardy Pompons | — | — | — | —— | — | ||||||||||||||
| —— | maximum | — | —— | —— | —— | — | ||||||||||||||||||
| —— | Clematis heracleæfolia (C. tubulosa) | — | — | — | — | |||||||||||||||||||
| —— | recta. Herbaceous Clematis | — | — | — | — | |||||||||||||||||||
| —— | integrifolia. Herbaceous Clematis | — | —— | —— | ||||||||||||||||||||
| —— | Convallaria majalis. Lily of the Valley | — | —— | —— | ||||||||||||||||||||
| —— | Coreopsis lanceolata grandiflora. Tickseed | — | —— | —— | — | — | — | |||||||||||||||||
| — | — | Delphinium belladonna. Larkspur | —— | —— | — | — | — | |||||||||||||||||
| —— | formosum. Larkspur | —— | —— | — | — | — | ||||||||||||||||||
| —— | —— | Delphinium grandiflorum chinense., Larkspur | — | —— | —— | |||||||||||||||||||
| —— | — | — | Kelway’s Hybrids | — | — | —— | —— | — | — | |||||||||||||||
| —— | Dianthus barbatus. Sweet William | — | — | — | — | |||||||||||||||||||
| —— | barbatus, Newport Pink | — | — | — | — | |||||||||||||||||||
| —— | barbatus, Scarlet Beauty | — | —— | — | — | — | ||||||||||||||||||
| —— | plumarius. June Pink, Clove Pink | —— | — | — | ||||||||||||||||||||
| —— | plumarius, Mrs. Sinkins | —— | — | —— | — | |||||||||||||||||||
| —— | plumarius, White Reserve | —— | — | —— | — | |||||||||||||||||||
| —— | Dicentra eximia | — | —— | —— | — | — | — | |||||||||||||||||
| —— | spectabilis. Bleeding-Heart | —— | — | — | — | |||||||||||||||||||
| —— | —— | Dictamnus Fraxinella. Gas Plant | — | —— | —— | |||||||||||||||||||
| —— | —— | Digitalis purpurea. Foxglove | — | —— | — | — | —— | — | ||||||||||||||||
| —— | sibirica. Siberian Foxglove | — | —— | — | — | — | ||||||||||||||||||
| —— | Doronicum plantagineum excelsum. Leopard’s Bane | —— | — | — | — | |||||||||||||||||||
| —— | Echinops humilis. Globe Thistle | —— | — | — | —— | — | ||||||||||||||||||
| —— | Erigeron macranthus. Flea Bane | — | —— | —— | — | |||||||||||||||||||
| —— | Eryngium amethystinum. Sea Holly | — | —— | — | — | |||||||||||||||||||
| —— | Eupatorium cœlestinum. Mist-Flower | — | —— | — | — | |||||||||||||||||||
| — | — | Filipendula hexapetala. Meadow Sweet | — | — | — | — | ||||||||||||||||||
| —— | Funkia Fortunei. Day Lily | —— | — | — | — | |||||||||||||||||||
| —— | lancifolia. Day Lily | — | — | — | — | |||||||||||||||||||
| —— | —— | ovate. Day Lily | — | — | — | — | ||||||||||||||||||
| —— | subcordata grandiflora. Day Lily | —— | —— | — | — | |||||||||||||||||||
| variegata. Day Lily | — | —— | — | — | — | |||||||||||||||||||
| — | — | Gaillardia grandiflora. Blanket Flower | — | —— | —— | — | — | |||||||||||||||||
| ——— | Gypsophila paniculata. Baby’s Breath | ———— | —— | —— | ||||||||||||||||||||
| ——— | paniculata fl.-pl. Double-flowering Baby’s Breath | —— | —— | —— | —— | |||||||||||||||||||
| —— | repens. Creeping Baby’s Breath | —— | ——— | ———— | ||||||||||||||||||||
| ——— | —— | Helenium autumnale. Sneezeweed | —— | ——— | —— | —— | ||||||||||||||||||
| ——— | Helianthus moltis. Woolly Sunflower | ——— | — | — | —— | |||||||||||||||||||
| ——— | orgyalis. Tall Sunflower | —— | ——— | —— | ——— | — | ||||||||||||||||||
| ——— | rigidus, Miss Mellish | ——— | — | —— | ——— | ——— | ||||||||||||||||||
| ——— | Maximilianii. Late-flowering Sunflower | —— | ———— | — | —— | |||||||||||||||||||
| ——— | Hemerocallis Dumortieri. Dwarf Day Lily | — | —— | —— | —— | |||||||||||||||||||
| ——— | flava. Yellow Day Lily | —— | — | —— | —— | |||||||||||||||||||
| ——— | Florham. Giant Day Lily | —— | — | —— | —— | |||||||||||||||||||
| ——— | fulva. Tawny Day Lily | ——— | — | —— | —— | |||||||||||||||||||
| ——— | fulva var. Kwanso. Double Orange Lily | ——— | —— | —— | —— | |||||||||||||||||||
| ——— | fulva, Gold Dust | —— | — | ——— | ||||||||||||||||||||
| ——— | Thunbergii. Japanese Day Lily | —— | — | — | — | |||||||||||||||||||
| —— | Heuchera brizoides. Hybrid Coral Bells | —— | — | —— | —— | |||||||||||||||||||
| —— | sanguinea. Coral Bells | —— | — | —— | —— | |||||||||||||||||||
| —— | ——— | Hibiscus militaris. Marsh Mallow | ——— | ——— | — | ——— | —— | |||||||||||||||||
| —— | ——— | Moscheutos, Mixed. Marsh Mallow | ——— | ——— | —— | ——— | —— | |||||||||||||||||
| —— | Moscheutos, Giant Red. Marsh Mallow | ——— | — | —— | ——— | —— | ||||||||||||||||||
| ——— | Iberis sempervirens. Evergreen Candytuft | —— | — | —— | — | |||||||||||||||||||
| ———— | Iris cristata. Dwarf Blue Iris | — | — | ———— | ||||||||||||||||||||
| —— | germanica. German Iris | —— | —— | ——— | ——— | |||||||||||||||||||
| —— | ——— | —— | ———— | —— | Kaempferi. Japanese Iris | —— | — | —— | — | |||||||||||||||
| ——— | ——— | —— | Kniphofia Hybrids. Red-Hot-Poker Plant | —— | ——— | ——— | —— | |||||||||||||||||
| —— | —— | Lespedeza. Japanese Bush Clover | — | — | — | — | ||||||||||||||||||
| —— | Liatris pycnostachya. Blazing Star | — | —— | — | —— | — | ||||||||||||||||||
| —— | epicata. Gay Feather | — | —— | — | —— | — | ||||||||||||||||||
| —— | Lilium candidum. Madonna Lily | —— | —— | |||||||||||||||||||||
| — | — | tigrinum splendens. Tiger Lily | —— | — | —— | |||||||||||||||||||
| —— | —— | —— | Lupinus polyphyllus. Lupine | — | — | —— | ||||||||||||||||||
| —— | Lychnis Flos-cuculi plenissima semperflorens. Campion | — | —— | —— | ||||||||||||||||||||
| —— | chalcedonica. Campion | —— | — | —— | ||||||||||||||||||||
| —— | —— | Haageana. Campion | —— | — | —— | |||||||||||||||||||
| —— | — | viscaria. Campion | — | — | — | — | ||||||||||||||||||
| —— | Lycoris squamigera. Fragrant Spider Lily | —— | — | —— | ||||||||||||||||||||
| —— | Megasea cordifolia. Saxifrage | — | —— | — | — | |||||||||||||||||||
| —— | Mentha piperita. Peppermint | — | — | |||||||||||||||||||||
| —— | Monarda didyma. Oswego Tea | —— | — | —— | ||||||||||||||||||||
| —— | Myosotis palustris semperflorens. Everblooming Forget-me-not | —— | — | —— | ||||||||||||||||||||
| —— | Œnothera fruticosa, Youngli. Evening Primrose | — | —— | — | — | — | ||||||||||||||||||
| —— | Opuntia vulgaris. Prickly Pear | —— | —— | |||||||||||||||||||||
| —— | Pæonia, Couronne d’Or | — | — | —— | ||||||||||||||||||||
| —— | Delachei | — | — | —— | — | |||||||||||||||||||
| —— | delicatissima | — | — | —— | — | |||||||||||||||||||
| —— | — | Duchesse de Nemours | — | — | —— | — | ||||||||||||||||||
| —— | Felix Crousse | — | — | —— | — | |||||||||||||||||||
| —— | festiva maxima | — | — | —— | — | |||||||||||||||||||
| —— | Mme. Crousse | — | — | —— | — | |||||||||||||||||||
| —— | Queen Victoria | — | — | — | — | |||||||||||||||||||
| —— | Pæonia, Richardsen’s Dorchester | —— | — | — | ||||||||||||||||||||
| —— | rubra superba | —— | —— | — | ||||||||||||||||||||
| —— | —— | sinensis, Mixed | — | — | —— | — | ||||||||||||||||||
| —— | Triomphe de l’Exposition de Lille | — | — | —— | — | |||||||||||||||||||
| —— | —— | —— | Papaver nudicaule. Iceland Poppy | — | —— | —— | — | — | ||||||||||||||||
| —— | orientale. Oriental Poppy | —— | — | — | ||||||||||||||||||||
| —— | Paradisea Lillestrum. St. Bruno’s Lily | — | — | — | — | |||||||||||||||||||
| —— | Penstemon barbatus Terreyi. Scarlet Beardtongue | — | — | —— | ||||||||||||||||||||
| —— | Polygonatum giganteum. Solomon’s Seal | —— | —— | |||||||||||||||||||||
| —— | Phlox paniculata, Baron Van Dedem | — | —— | —— | — | —— | ||||||||||||||||||
| —— | paniculata, B. Comte | — | —— | — | —— | |||||||||||||||||||
| — | — | paniculata, Beranger | — | —— | — | —— | ||||||||||||||||||
| —— | paniculata, F. G. von Laseburg | — | —— | —— | —— | |||||||||||||||||||
| —— | paniculata, Independence | —— | —— | —— | — | —— | ||||||||||||||||||
| —— | paniculata, Rheinlander | —— | —— | — | — | — | ||||||||||||||||||
| —— | paniculata, Von Hochberg | — | —— | —— | —— | |||||||||||||||||||
| —— | suffruticosa, Miss Lingard | — | —— | —— | —— | — | — | |||||||||||||||||
| —— | subulata alba, Moss Pink | — | —— | —— | ||||||||||||||||||||
| —— | subulata lilacina | — | —— | —— | ||||||||||||||||||||
| —— | subulata rosea | — | —— | —— | ||||||||||||||||||||
| — | — | Physalis Franchetii. Chinese Lantern Plant | — | —— | — | —— | ||||||||||||||||||
| — | —— | Physostegia virginica. False Dragonhead | — | —— | — | — | ||||||||||||||||||
| —— | virginica alba. False Dragonhead | — | —— | — | — | |||||||||||||||||||
| —— | —— | Platycodon grandiflora. Balloon-Flower | — | —— | —— | |||||||||||||||||||
| —— | —— | —— | —— | Primula veris. English Primrose | — | —— | —— | — | ||||||||||||||||
| —— | —— | —— | Pyrethrum roseum | — | —— | —— | ||||||||||||||||||
| —— | Ranunculus acris fl.-pl. Double-flowered Buttercup | — | — | — | — | |||||||||||||||||||
| —— | Rudbeckia laciniata. Golden Glow | —— | — | —— | ||||||||||||||||||||
| —— | purpurea. Giant Purple Coneflower | — | — | —— | ||||||||||||||||||||
| —— | Sagina subulata. Pearlwort | — | — | —— | ||||||||||||||||||||
| — | — | Salvia azurea grandiflora. Meadow Sage | —— | — | —— | |||||||||||||||||||
| —— | — | pratensis. Meadow sage | —— | — | —— | |||||||||||||||||||
| —— | Sedum acre. Stonecrop | — | — | —— | ||||||||||||||||||||
| —— | spectabile. Stonecrop | —— | — | — | — | |||||||||||||||||||
| —— | Silphium perfoliatum. Cup plant | —— | — | —— | ||||||||||||||||||||
| —— | Spiræa (Astilbe) Arendsii, Ceres | — | — | —— | ||||||||||||||||||||
| —— | Statice latifolia. Great Sea Lavender | — | —— | —— | ||||||||||||||||||||
| —— | Stokesia cyanea. Cornflower | —— | —— | — | — | — | ||||||||||||||||||
| —— | cyanea alba. White Cornflower | —— | —— | — | — | — | ||||||||||||||||||
| —— | Tradescantia virginica. Spiderwort | — | —— | —— | ||||||||||||||||||||
| —— | virginica alba. Spiderwort | — | —— | —— | ||||||||||||||||||||
| —— | Trollius europæus. Globeflower | — | —— | — | — | — | ||||||||||||||||||
| — | — | Valeriana officinalis. Garden Heliotrope | —— | —— | —— | —— | ||||||||||||||||||
| —— | Veronica longifolia subsessilis. Blue-Jay Flower | —— | —— | — | — | — | ||||||||||||||||||
| —— | amethystina. Speedwell | —— | — | —— | ||||||||||||||||||||
| —— | longifolia. Japanese Speedwell | —— | —— | —— | ||||||||||||||||||||
| —— | Viola cornuta alba. Tufted Pansy | —— | —— | —— | — | —— | ||||||||||||||||||
| —— | cornuta, G. Wermig. Tufted Pansy | — | —— | —— | — | —— | ||||||||||||||||||
| —— | Violet, Double Russian | —— | — | —— | ||||||||||||||||||||
| —— | Yucca filamentosa. Spanish Bayonet | —— | — | — | —— | |||||||||||||||||||
For those who prefer growing flowers merely to pick, the quickest way of getting them is to plant summer-blooming annuals. A list of thirty popular annuals, all of them easily grown, is given below.
BLOOMING IN JULY
| Lavatera | Lavatera trimestris |
| Clarkia | Clarkia elegans |
| Large-flowered Godetia | Œnothera Whitneyi |
| Early Cosmos | Cosmos bipinnatus |
| Sweet Alyssum | Alyssum maritimum |
| Marigold | Tagetes patula |
| Nicotiana | Nicotiana alata |
| Sander’s Nicotiana | Nicotiana Sanderæ |
| Arctotis | Arctotis grandis |
| Stock, gillyflower | Matthiola incana var. annua |
| Annual larkspur | Delphinium Ajacis |
| Bedding Lobelia | Lobelia Erinus |
| Wishbone flower | Torenia Fournieri |
| Phacelia | Phacelia congesta |
| African marigold | Tagetes erecta |
| California poppy | Eschscholtzia californica |
| Giant tulip | Hunnemannia fumariæfolia |
| Annual Gaillardia | Gaillardia pulchella |
| Scarlet sage | Salvia splendens |
| Youth-and-old-age | Zinnia elegans |
| Rose moss | Portulaca grandiflora |
| Balsam | Impatiens balsamina |
| Painted tongue | Salpiglossis sinuata |
BLOOMING IN AUGUST
| Gilia | Gilia capitata |
| Three-colored chrysanthemum | Chrysanthemum carinatum |
| Mourning bride | Scabiosa atropurpurea |
| China asters | Callistephus chinensis |
| Everlasting | Helichrysum bracteatum |
| Didiscus | Trachymene cœrulea |
THE quotation from the second sentence of the first chapter of Genesis tells us more in eight words than could very well be said in as many chapters. Not only have we biblical authority for this early absence of life on the earth, but all the accumulated knowledge of the ages points in the same direction. We have already seen that plants, because they can take inorganic substances from the earth and air and transform them into organic food, must in all probability have come on the earth before animals which, directly or indirectly, all rely upon plants for their food. Even those animals that eat only flesh devour other animals which depend upon plants for food. It may safely be repeated, then, that upon plants all animal life depends, and that, in the dim beginnings of things on the earth, it must have been some form of plants that were the first living things. Extremely simple unicellular animals, however, are known to date from early times.
In the volume devoted to that subject in this series, you will find that at the very earliest stages of what we know as our globe there was a segregation of land and water somewhat different from our great oceans and continents to-day in extent and area, but differing mostly in this—that much of the water was fresh and very nearly boiling hot. We have still the remnants of those great reservoirs of hot water, as our hot springs and shooting geysers only too well prove. And if all plants were as quickly killed by hot or boiling water as the common garden geranium, we should not expect any plant life to have developed upon the earth until all those great bodies of water had cooled. To have waited for that would have been to delay the appearance of plants for no one knows how many millions of years, and there is some fairly good evidence that long before normal conditions of heat and cold were established there already flourished certain kinds of plants. What those plants were is something of a speculation, and indeed exactly what they were no one knows. But in our present hot springs grow certain plants, microscopic in size, but quite obviously related to the algæ or to certain bacterialike organisms. They live with apparent comfort and reproduce themselves freely in water so hot that no other form of life will maintain itself. While there is no proof that these present plant inhabitants of hot springs, common in the West, are descended from infinitely ancient progenitors, it is a fair assumption that some organism capable of growing in warm or hot water was the first living thing to appear in a world otherwise “without form and void.”
This great question of how plants came on the earth, and particularly how from these apparently simplest organisms our whole wonderful vegetation has arisen, has always been one of the most interesting things in the history of the world. There are many different ways of studying this, and in the very earliest stages of plant development we are forced to reason, not so much by actual records or buried skeletons of the plants that probably existed then, for only a very few have ever been found, but by our knowledge of the physical and chemical requirements of unicellular plants, and those slightly more developed, and of their individual life histories. It is, for instance, certain that the first plants must have been aquatic, as no real land plants are known for hundreds of thousands of years after the earth was quite capable of maintaining plant life. The absolute necessity of water to complete fertilization in nearly all cryptogams also makes it fairly certain that water plants, and these of the simplest nonflowering type, were the first living things to be found on the earth. And it is more than a fair inference that these were inhabitants of warm or hot water. Subsequently, as the water cooled, they may well have been not unlike the green scum found on ponds to-day.
Of course, the actual origin of life itself is still as much of a riddle as it was when the ancient philosophers began to speculate about it years before the Christian era. Protoplasm, the unit or basis of all life, while its composition and growth requirements are fairly well known, has never been made in any laboratory. Nor have scientists ever been able to decide what the combination of physical and chemical forces must have been to originate it. But that from a perfectly sterile, probably steaming hot globe, there did finally develop some form of life, and that this must have been aquatic plant rather than animal life, seems not only certain, but the only hypothesis upon which all subsequent development of life must have been based. It is not necessary to ascribe the origin of life to providential inspiration nor to the meddling of strange and outlandish deities, as all savage tribes did and some more civilized peoples still do. There can, however, be no escaping the fact that life is more than the combination of physical and chemical conditions which sustain it, and that its origin has never, and may never be “explained” by merely describing the conditions which unquestionably favored its appearance. In other words, the origin of plant life throws us back upon things believed but incapable of proof, and is none the less wonderful because we cannot yet understand the probable progression of forces and materials to which it owes its origin.
Assuming, then, and we must all start with this proposition, that aquatic plants certainly, and warm or hot water plants probably, were the first living things upon the earth, what are the next steps in the history of the plant kingdom? The answer to that question involves a few simple facts in geology and, particularly, in the making of fossils, which must be understood before we can see those steps or their significance. The geological changes which have resulted in the present condition of the earth’s surface are described in the volume devoted to that subject, and will not be repeated here. But some mention must be made of the formation of fossil plants, particularly as it is upon the evidence of these that the story of the development of plant life is literally written in the rocks.
If a leaf or twig drops into shallow water with a clay or mud bottom, it ultimately sinks, and if then a film of clay or silt is brought down by freshets or what not, it will bury the leaf or twig, of course, filling in every slight depression. If then the buried object were raised so that it dried out and could be split open, we should find a perfect impression of the veins and other outward characters of the leaf etched in the clay. This is often such a perfect process that every detail of the leaf is left in the mud impression, and only the opportunity for this impression to become hardened into rock is needed for us to have a fossil. For these are merely the final hardened rock stages of a process that began as we have indicated, and the thousands of fossils that have been dug out of the earth prove how common the conditions for their formation must have been in certain periods of the world’s history.
But of the untold millions of fossils that have been made most have been destroyed, for the geologists tell us that the earth’s crust has been subjected to much upheaval. Mountain chains thrown up, inland seas formed, great river systems carved out, and tremendous periods of vulcanism or fire action have made the earth’s crust, at different periods, a mighty restless place. And these changes, so slow that often millions of years have elapsed before they were completed, have sometimes been favorable to the making of fossils and sometimes to their destruction. Darwin once wrote about the fossil record that he saw it “as a history of the world imperfectly kept, and written in a changing dialect; of this history we possess the last volume alone relating only to two or three countries. Of this volume, only here and there a short chapter has been preserved; and of each page, only here and there a few lines. Each word of the slowly changing language, more or less different in the successive chapters, may represent the forms of life, which are entombed in our consecutive formations, and which falsely appear to have been abruptly introduced.” And yet it is upon the evidence of this fossil record that most of our knowledge of the history of the plant kingdom is based.
The difficulty of getting any true picture of the beginnings of plant life is great, for those earliest stages of development were unquestionably water-inhabiting plants, whose tissue is mostly too soft and too easily decayed to make fossil impressions of them likely to be preserved. Yet fossil algæ have been found in rock strata so old that no fern or flowering plant had yet made its appearance. It is not too much to picture the world then as peopled only by cryptogamous plants of simple structure, living in the water, and land plants which to-day make up the bulk of our vegetation as not yet developed. Furthermore, there is in these earliest stages no trace of plants with any kind of a vascular system, such as all ferns and flowering plants possess. No stretch of our imagination can readily picture the earth as it in all probability was in that period, with no trees or vines or flowers, the land wholly bare of vegetation, and in the water, along its sterile shores, only unicellular or slightly more developed, wholly nonflowering plants. The conditions supporting such plant life existed for many millions of years, and some geologists have claimed that this period of time exceeded all the subsequent ones combined, so that algæ and some other unicellular plant types are the oldest in the world; and they still exist in enormous numbers.
Much later than this, fossil algæ of comparatively complex structure have been found, showing by their frequency and more highly developed characters a more advanced stage in the development of the plant kingdom. So common were these various types of what we now call seaweeds, although most of them apparently lived in fresh water, and so widespread was their occurrence, that this Pre-Cambrian period has often been called the reign of algæ. As yet no other plants had been developed and none of these ancient types had invaded the land, which for millions of years more must have been entirely without vegetation.
After the reign of algæ and other cryptogamous water plants, our knowledge of which is so unsatisfactory because of the incompleteness of the fossil record, there appeared the first evidences of plants that were able to live with “one foot on the land and the other in the water,” so to speak. How many transitional stages there may have been, and what relation any of these may bear to existing plants, is not known, or is, at any rate, so little understood that it is a disputed point. But somewhere about this period there did appear plants capable of living at least part of their life on the land, and possessing in their vascular system a structure of enormous advantage over their predecessors. It is pure speculation as to what this first land plant was derived from, or from what particular group of water-inhabiting plants it took some of its characteristics. Its appearance, in any case, was a dramatic event of the first importance. Not the least interesting feature of it is that the very plants of which we have indisputable evidences of being the first land plants have come down through the ages to the present day. For it is practically impossible to separate our modern representative from its ancient ancestor, despite all the tremendous changes that have been going on both in the history of the earth’s crust and in the vicissitudes of the vegetation in meeting those changes.
There is the best of evidence that these first land plants were of the club moss family, which are relatives of the ferns. One of them, representing our present Lycopodium Selago (Figure 107) so closely as to be practically indistinguishable, is a common type, as revealed to us in the fossil record. The present plant inhabits rich, moist, and mostly evergreen woods in the northern part of the globe, and is common in the Adirondack and White Mountains.

A club moss which has come down through the ages almost unchanged from the days when coal was being formed. Grows to-day in the north temperate zone, particularly in mountains.
Nor was the earth peopled wholly by this ancestor of Lycopodium Selago, for we find at this time, or just after it, a great development of plants of this type. Some of these were giant, treelike club-mosses that have been so well preserved as fossils that even their internal structure and spore-bearing characters are well known. Many other strange relatives of our modern club mosses flourished in those days, some of which have wholly disappeared, as have all the treelike forms. These highly organized club mosses, quite unlike any modern representative of the family, appear to have been crowded off the earth by other and subsequent types of vegetation, while Lycopodium Selago, and about thirty related species, have persisted to the present day; not precisely in all cases as they were in this dawn of a land flora, but in many cases with modern structure and reproductive processes so close to the ancestral types as to be nearly identical.
Perhaps nothing gives one a better impression of the tremendous time that must have elapsed before the appearance of these ancient club mosses than the very slight modifications from their ancient condition which their structure at the present time exhibits. While nothing is certainly known of their origin, when they first appeared they were plants with a well-developed vascular system, having stems and leaves quite unlike any of their predecessors’, and a reproductive process almost precisely like their modern descendants. In other words, if they have changed so slightly in all the millions of years since our rock-written records of them first occur, what an infinitely greater period must have elapsed down the dim vista of the ages before their appearance. Of this period, with the exception of fossil algæ, we know practically nothing, and, worst of all, the actual transition from a wholly water-inhabiting flora to these certainly land-inhabiting club mosses may never be known. For, added to the difficulty of water plants being preserved as fossils, already mentioned, is the fact that as they are the oldest, they are found in the deepest strata and, consequently, the hardest to find; and due to changes in the earth’s crust, these ancient fossil-bearing strata have often been much disturbed.
The conclusion appears to be indicated that the origin of a land flora came about with the appearance of these ancient club mosses, which are not mosses in our present-day interpretation of those plants, and that at about the same period many other plants also were found, the whole vegetation resembling nothing that exists at the present time, but many of the different kinds of this ancient flora showed unmistakable evidence of being the progenitors of many plants that exist to-day. What these were, and particularly what they accomplished, both in the history of the plant kingdom and in making the world habitable for man who did not come for millions of years after they were preparing the way, will be considered in
The carboniferous time, or the period when the earth was covered with huge forests of strange shrubs and trees, most of which were unlike their modern successors, apparently had a climate so nearly uniform and seasonless that fossil remains of these plants have been found throughout the world. Even in the Arctic the rock strata show the flourishing of forests that must have needed a climate very different from the frigid condition there to-day, and furnishing indisputable evidence of a warm, most probably frostless, climate practically throughout the world.

A modern horsetail or scouring rush, common in the north temperate zone. Ancestors of these formed huge forests at the time that coal was being formed.
The giant club mosses have already been mentioned, with their persistence to the present day in much reduced number, and vastly reduced sizes. No one can picture the grandeur of those ancient forests, peopled with queer animals long since extinct and with dragon flies known to have a wing-spread of two feet or more. But with the club mosses were giant horsetails, which in somewhat changed form have also come down to our times, but in much reduced stature and frequently are familiar enough as weedy plants along railway embankments, and sometimes in more natural environments. Most of our modern representatives of the genus Equisetum (Figure 108), or horsetails, are low herbs, but one South American kind still retains the ancient habit of growing to considerable heights, as specimens up to twenty feet high are known. Related to the ancient treelike horsetails were queer vines with slender twining stems, which, judging from their fossil remains, must have been very common. Both the ancestors of our club mosses and the horsetails must have occupied vast swampy areas, as their stem structure indicates a fondness for water, to which, as we have already seen, their still more ancient ancestors were always confined.
Vigorously competing with these plants for occupancy of those great swamps were vast quantities of plants that have been called cycadlike ferns from their likeness to ferns on the one hand and plants like the so-called sago palm on the other. The sago palm, or Cycas revoluta, is a modern representative of these ancient forms, and retains the remarkable characteristic of having its male fertilizing cell capable of movement as we know to be the case in nearly all cryptogamous plants. Yet Cycas, with its related genera, which are found in nearly all the warmer parts of the earth to-day, are true flowering plants which bear cones. We see, therefore, in these old cycadlike ferns one of the first, almost experimental, evidences of the seed habit, and consequently the breaking away from the spore habit which overwhelmingly characterized the reproductive processes of its ancient associates.
The inhabitants of higher parts of that dim, mysterious world, of which we know only that part revealed in the fossil record, were largely ancestors of our modern cone-bearing evergreens. They are known as Cordaitales and have long since disappeared. Forming forests of huge size and making long, slender trunks with a crown of leaves at the top not unlike some modern conifers to which they are, of course, related, these progenitors of our pines and spruces must have been striking objects of that strange landscape. Rooted stumps of these ancient trees have been uncovered, and their narrow leaves, often three feet long, are common as fossils. By some our present conifers and the Cordaitales are both thought to be descendants from a still more ancient group, of which the existence is only conjectured.
We can, perhaps, best summarize our sketch of the plant life existing at the close of this period by saying that all the forms show unmistakable evidence of being crytogamous so far as their reproductive processes are concerned, or else, as in the progenitors of our conifers and cycads, the beginnings of a definite seed habit are indicated. Most of the lowland representatives of this flora were cryptogamous in their characters and ancestry, while some of them, and nearly all the flora of drier sites, appear to have shown the beginnings of flower production. Some of these flowers, which are always cones, are unmistakable as such, and pollen in tremendous quantities has been found among their buried remains. These cones belong to trees that are actual gymnosperms or obvious ancestors of them, for no herbs are known as yet. Nor are angiospermous flowering plants known from this period, nor have any ever been found in strata millions of years younger than the fossil-bearing strata of this age of the ancestors of our modern ferns, conifers, or cycads.
Nor must we picture the development of these different plant inhabitants of that time as passing from one to another in orderly sequence, for that would give us the impression of a regular progression from simple to complex, which may or may not be the truth. There appears to be such a sequence, and the internal structure of the remains of many of these ancient types of plant life have greatly aided our ability to understand their relationships. But with the possibility of various reproductive processes or other structures appearing in quite unrelated forms, and with the comparative paucity of the fossil record in mind, no one can say for certain what are the true lines of descent. The necessity for water in the reproductive act of nearly all the crytogams, the origin of the vascular structure, and the consequent ability to live upon the land, and finally the production of a conelike flower structure with pollen, and all that that implies, are all found during this period.
To the vegetation inhabiting the swamps during this period, man owes a debt perhaps as great as to our modern food plants, for it is upon this, and some later plant remains, that we rely for coal. This period has been well called the Carboniferous, for its chief claim to attention, outside the realms of botanical research, was the deposition of those great collections of plant remains, which, as coal, contain as high as 90 per cent of carbon and furnish the fuel of the world. This is scarcely the place or time to go into the composition of different kinds of coal, but some mention of the conditions under which these ancient swampy forests were transformed into that valuable substance may well conclude the account of a vegetation period the history of which has in large part been found written in the very strata from which coal itself is derived.
In the lowest and wettest parts of those forests there occurred, just as there may occur to-day, a large accumulation of fallen trees and other vegetable refuse. In the ordinary way these would simply rot, due to the work of insects and the fungi of decay, and in a few score years there would be nothing to show. “Dust to dust” would be, and is, the history of so many living things that it is only some machinery for arresting this process which will give us very different results. In the case of coal formation the original impetus appears to be certain microscopic organisms, probably saprophytic fungi, fossil remains of which have been identified, which work upon the fallen mass of vegetation and start its decay, but which can only do so while their prey is still within the influence of the air. The initial stages of decay must, therefore, have been going on while the water was low enough for these organisms to work. But in many parts of that ancient landscape the water level was a fluctuating quantity, due to local conditions or to changes in the earth’s crust. So that many times partially decomposed vegetation masses would become submerged, stopping the work of these organisms of decay that demand air, but providing the only conditions under which certain others could complete the transformation. These bacterial organisms that will work only when deprived of air continue the process, but in a different way. For one thing, the lack of air delays decomposition or almost stops it, as witness the resistance of logs under water, some of which are known to be hundreds of years old. And forest stumps off the coast of Cape May, in New Jersey, are in nearly as perfect a state as when first submerged, over 40,000 years ago. In the production of coal these anærobic (living without air) bacteria release oxygen and hydrogen from the partly decayed mass, leaving as a residue a substance known as peat, which is largely carbon. The transformation of peat into coal depends upon requisite pressure of the strata that may be laid down on top of the peat bed, and probably upon chemical changes that go on after such covering strata have been laid down.
The fact that coal is sometimes found only in thin veins, with layers of shale and other material between, tells us that its origin must often have been a precarious affair, where alternate emergence and submergence would permit first the vegetation to develop and then its transformation to peat, followed by the deposition of fine sands or silt covering the bed. Several such cycles occurred, sometimes separated by untold ages of time, or again by much briefer periods. Certain mines, however, contain over 200 feet of solid coal. The length of time necessary for such a vast accumulation, or how many generations of these ancient plants went into their making, is beyond calculation. With the mining of coal running into the hundreds of millions of tons yearly, we get some idea of how great were those Carboniferous forests, and how extensive they were is proved from the widely separated localities in which coal mines are found.
The Carboniferous age of fern, cycad, and conifer ancestors was by no means a quiet, orderly period, as from geological evidence it appears to have been much subject to alternate emergence and submergence of great tracts of land. Compared with what followed, it actually was a period of comparative quietness, however, and it must, in at least most parts of the world, have permitted the slow development of certain of its plant groups to a state of perfection never reached since. This is particularly true of the ancient relatives of our club mosses and horsetails.
Perhaps one of the most obvious questions to ask about these plants is how long ago they lived, and upon the answer to such a question depend many others. What, for instance, is the position of the Carboniferous as compared to what preceded it and came after? How old is the earth and when did life first appear on it? The evidence upon which such questions are answered comes from the estimates of physicists as to the age of the earth; from students of fossil animals and plants; from astronomers, from geologists and other students. A compromise of these different estimates, and one that has consequently been widely accepted, gives the age of the earth, dating from the time of its having a definite crust with land and water masses, as somewhere near a hundred million years. Such figures are beyond our comprehension and consequently mean almost nothing, but the proportion in time of the different stages of the development of plants may be stated with greater certainty. Taking the total age of the earth as 100 per cent, the period when there is no record of life of any sort may be set down as about 45 per cent of the total, the reign of algæ and development of land plants about 8 per cent, the carboniferous or coal-forming plants about 28 per cent, which leaves only 19 per cent from that distant time to the present. And many things happened in this comparatively brief fifth of the plant world’s history, among them the origin of some plants that have come straight down to us, without discoverable change.

Found in most fossil strata and in a practically unchanged condition from the upper part of the Carboniferous to the most recent fossil records. Now unknown as a wild tree and preserved for us through its cultivation in ancient temple gardens in eastern Asia.
We could hardly leave the Carboniferous time without at least brief mention of the ginkgo tree (Figure 109), or, as some call it, the maidenhair tree. From the upper strata of the Carboniferous it is common, as it is in practically all subsequent fossil accumulations down to the most recent. And yet the tree has never been found wild, although its frequency in temple gardens in China and Japan, always as a cultivated tree, suggests that its disappearance as a wild plant must have come since the priests began preserving it, which can be only a matter of a few thousand years at most. In other words, we have just missed seeing in the ginkgo what has so many times happened to these very ancient types of vegetation, namely, their final extinction. This must have occurred within historic times, and, judging by its frequent use as a temple tree in eastern Asia, that region was its last outpost after its long journey from the dim past. Thousands of other ancient plants have completely disappeared, and one cycad from New South Wales is at this moment putting up a losing fight against modern competitors, but in the ginkgo tree the actual twilight and extinction of its wild existence has missed observations by modern plant geographers by only a brief period. It is almost as though we had waited all our life to see some great event and then missed it by a few moments. Fortunately the tree is now common in cultivation, and not the least interesting feature of it is the fact that its male fertilizing cell retains its power of movement, which dates back to its early associates. Among modern flowering plants only the ginkgo and the relatives of the sago palm or cycads retain this relic of an overwhelmingly cryptogamous ancestry.
The end of the Carboniferous or coal-forming ages was marked by great changes in the earth’s surface, some of them cataclysmic in their effects. What they were in detail is described in the volume on geology and need not be repeated here. What happened to the development of the plant kingdom after this will be considered in the next section of this chapter.
The vegetation at the ending of Carboniferous times was much affected by the great changes in the earth’s surface which happened then. The thrusting up of great mountain chains, the slow encroachment of continental glaciers, and the other phenomena characterizing that period could not but be reflected in the plant population. For one thing the giant club mosses and horsetails were much reduced in extent and finally disappeared, leaving only the immediate ancestors of our present-day forms. Cordaitales gave place to trees not unlike some of the modern yew trees. True ferns as well as the cycadlike ferns with seeds appear to have lived side by side with true cycads, which subsequently supplanted their obvious cycadlike fern ancestors. There was an obvious dwindling of ancient Carboniferous forms, some of which, however, persisted in considerable numbers. Many other plants existed then, some of which died out there, and some of which still survive in descendants, particularly among our conifers and ferns. But there happened toward the upper end of this period an event in the history of the plant kingdom so dramatic, of such far-reaching results, that its appearance might be likened to the overthrow of the Czar in Russian history or to the downfall of the Kaiser in Germany. For with it dawned a new era for the plant world, the effects of which we see all about us to-day.
Somewhere in the rock strata of this period we find the first angiosperm, or plant that matures its seed in a closed ovary, and with the origin of that habit there began such a development of plants of this type that its impetus has not yet been lost. It is impossible to tell at this distance from the origin of that first angiosperm from what it developed, nor how many ages it may have existed before the accident of its preservation as a fossil revealed its presence. It is certainly not without significance that it bore conelike fruits, such as all its associates and predecessors among flowering plants had done, but its possession of large, showy petals is the first evidence of a flower characteristic that was destined to make our present vegetation the lovely thing it is. This exceedingly interesting plant was a Magnolia (Figure 110), or so like our present plants of that genus as to be their obvious ancestor. Somewhere here, too, must have arisen the insect fertilization of flowers which we have seen to be such an important part of flower economy at the present time. Most of this ancient magnolia’s associates must have relied on wind pollination for seed production, as many modern plants still do, but the origin of insect fertilization appears to have come with the appearance of the first really petaliferous flower.

With this Magnolia were found other flowering plants which soon increased tremendously in numbers of individuals and differences of structure, such as our sassafras, the tulip tree, the poplar tree, and some others. All of these are trees or shrubs and we do not yet know whether herbs grew in the strange surroundings of that ancient forest or not. Their soft tissue may have prevented their preservation as fossils, but, at any rate, no herb has left its rock-written record from as early in this period as the trees and shrubs. All of these ancient trees have been recorded only in the northern hemisphere and it may be true that this part of the earth was the cradle of all those hosts of the flowering plants that now number over 150,000 species.
There must have been a mighty struggle for occupancy of the really desirable plant sites soon after the rise of these immediate ancestors of our modern plants. For there is every evidence of the progressive dwindling of those still more ancient holdovers from the Carboniferous, and the steady encroachment of the newly arisen and obviously vigorous young race. As we get higher up in the strata, or, in other words, nearer to the present, there are literally thousands of these immediate ancestors of our modern flora, and it is not very long before herbs, particularly grasses and sedges, begin to be common, together with other monocotyledonous plants such as palms. One not unlike our coconut palm has been found in some of these strata in France.
While this period records the origin of hundreds, and there are probably thousands of unrecorded species which are very near our modern descendants of them, it was also a period when the earth’s crust was in an almost constant state of restlessness. Ice periods, huge inland seas, great volcanic upheavals, and the thrusting up of mountain chains such as the Alps, Himalayas, and some others, were only a few of the disturbances to the orderly procession of this wholly new type of vegetation that doomed the older kinds and subsequently conquered the world. The spread of this new element in the plant kingdom was greatly helped and sometimes greatly hindered by land connections between continents, now separated by the oceans. The giant redwoods, now isolated in a few localities on our Pacific Coast, were found then nearly throughout the world. Because of these changes of land areas and some others of even greater influence on plant growth, such as climate, there was a constant shuffling of floral and, of course, animal elements, so that by the end of this period the new type of flora had spread throughout the world, but with here and there very local occurrence of certain genera and families, some of which have persisted to the present day. As we shall see in the last chapter of this book, certain whole families are confined to restricted areas; the cactus and pineapple family, and the genus Helianthus, or sunflowers, for instance, are, with one or two trifling exceptions, wholly American. And we have already seen how many food and other useful plants were first found here by the Spaniards—chocolate, tobacco, corn, the potato, and others.
It would fill the rest of this book to enumerate the plants that flourished toward the end of this period, and, in fact, it might almost be said that the flora of those days was not very different from our own, only it was distributed in different ways and mixed in very different proportions. With the disappearance or partial dwindling of more ancient groups, the rise of the plants that immediately preceded our own ushered in a new era in the history of the plant kingdom.

Fossil and Living Algæ Compared. C. A living algal pool colony near the Great Fountain Geyser, Yellowstone Park. (After Walcott.) B. Fossil calcareous algæ. Cryptozoön proliferum Hall, from the Cryptozoön ledge in Lester Park, near Saratoga Springs, N. Y. These algæ, which are among the oldest plants of the earth, grew in cabbage-shaped heads on the bottom of the ancient Cambrian sea and deposited lime in their tissue. The ledge has been planed down by the action of a great glacier which cut the plants across, showing their concentric interior structure. (Photographed by H. P. Cushing. Pictures and explanations of them from “The Origin and Evolution of Life,” by Professor Henry Fairfield Osborn, who kindly permitted their reproduction here.) (Courtesy of Brooklyn Botanic Garden.)

Probable Landscape in the Carboniferous Age. About the Time Coal Was in the Making. (After Patonie) 1. Tree fern 2. Giant ancestors of our horsetails. 3 and 4. Ancestors of our club mosses. 5. Cordaites, a primitive type, or perhaps even the ancestor of our modern evergreens. At this time no herbs and no plants with petals, were known, nor for ages after this period. (Courtesy of Brooklyn Botanic Garden.)
At the end of this period an event of commanding interest occurred, because it happened only some 40,000 years ago. With it came the encroachment from the north and south poles of the last of the great continental glaciers. There had been many before, stretching over a past period of time, but as the last of these great ice invasions it is the most interesting to us. It crowded all these temperate and even subtropical plants that then grew up in the far north toward the equator, and scraped clear of vegetation every part of the earth which it covered. In the volume on geology you will find an account of the extent and thickness of this great ice sheet, which ultimately receded to its present home. As it went back the plants crowded forward to occupy the freshly released land, the far northern or glacial first, followed by waves of other kinds. Some of the glacial or northern plants were left on the tops of the highest mountains, where to-day they persist in complete isolation, nearly all their friends and associates of that greatest of all winters having left them for points farther north. Many students of plant geography think that wave of plant life creeping northward to occupy the region uncovered by the retreating ice is still going on, and recent studies appear to show in at least one isolated mountain in the Adirondacks that the survivals of the ice age which have been isolated on its rocky peak ever since are in considerable danger of being crowded out by invaders from the lowlands.
Not all the geological changes which have remodeled the earth’s surface have been mentioned in this brief history of those plants that preceded our own, nor have anything like all the plants occurring in the different strata been even hinted at. But the thing which has been stressed and for us to fix in our minds is that all our present vegetation literally has its roots deep down in the earth. Some, as Lycopodium Selago, go back no one knows how many millions of years; others, like the flowering herbs, are much more recent. We come to understand how recent we are and what a comparatively brief flash in the pan all our modern development both of plants and in man has been since the last glacial period only by looking for a moment at what has happened in the past. In the account of the Carboniferous plants we found that there remained after that period only about 19 per cent of the earth’s age in which all the changes since then could have come about. If, as may well be possible, this period has been about 19,000,000 years, then the mere 40,000 years since the last Ice Age seems a brief period indeed. As some one has written, to contrast all man’s historic period, back to the days of the most ancient Chinese manuscript, with that long journey from the dim past which the plant world has slowly accomplished, is to realize that we are “as the flashing of a meteor through the sea of night.”
Fossil plants then, and this delving into the dead past of the plant world, reveals to us as nothing else can how much the modern plant kingdom is literally built upon a mighty race of ancestors. Some perished as did the Cordaitales, but left descendants who themselves gave rise to other groups that survive to-day. To look over a list of the fossil plant genera of the different strata is to visualize a drama the like of which no one living will ever see replayed, the results of which are recorded all over the world with its changing panorama of vegetation.
There are living to-day somewhere about 150,000 species of flowering plants; half a hundred conifers or gymnosperms; about 3,500 ferns; 500 club mosses; over 70,000 bacteria, fungi, and lichens, and probably over 20,000 species of algæ. The estimates of the last three divisions are more or less uncertain, as many species are still being discovered.
From what has just been read regarding the plants of earlier periods it is at once clear how completely the flowering plants have conquered all their ancient forbears, and what a pitifully small remnant of once proud and ancient forests are now represented by our club mosses and horsetails. That process of crowding, of the dominance of one kind to the exclusion or even extinction of others is still going on, and, as we shall see in the last chapter, often on a great scale.
If the period just after plants were first known on the earth may be called the Reign of Algæ, and subsequent periods were typified by still other kinds of plants, then our present period is par excellence the reign of flowering plants. In numbers of individuals they are still far outclassed by such cryptogams as the fungi, bacteria, algæ, etc., but their dominating influence in the plant world is unquestioned.
While all our present vegetation must have been derived from preexisting types, all of it is not necessarily directly descended from species which from their fossil records we know to have existed in geological periods older than our own. While the fossil record of the times immediately preceding the last ice age is a much more complete one than for many other periods, it fails to account directly for the great bulk of our varied flora of to-day. While ferns in great variety, gymnosperms, and hundreds of flowering plants are known quite definitely, they total only a minute fraction of these groups to-day. Even granting the always imperfect nature of the fossil record, and we have seen what an accident the formation of a plant fossil may be, and it is common knowledge how few comparatively have ever been recovered—even granting all this, there still remains a large part of our present flora of which the origin probably dates from comparatively recent times. So overwhelmingly true is this that of the Compositæ, or daisy, family, now numbering over 11,000 species, scarcely a handful of fossil species have been found. And in all collections of fossils the woody plants far outnumber the herbs, perhaps because of the greater probability of their being thus preserved rather than to any actual scarcity of herbs in the upper strata. And yet herbs to-day outnumber woody species over two to one. While it is true, then, that our present flora must have been derived from preexisting races, it is also true that much of it is apparently derived from plants that do not date very far back into the past. A few main types of flowering plants unquestionably are to be linked with fossil genera, but these types have now branched out into a wealth of detail that may not have existed and is certainly not recorded in the fossil record.
Some of these types stand out with remarkable clearness, notably magnolia, willows, poplars, walnuts, birches, oaks, figs, sassafras and its relatives, the rosales, the pea family, the spurges, maples, grapes, linden, myrtle, ginseng, and some others. All these, and in not very different aspect from their modern representatives, have been found in the fossils of the different and usually more recent strata before the last Ice Age. But the total fossil record of even these well-known genera is only a fraction of their modern development, and we are constantly confronted with the apparent dilemma of accounting for a present wealth of forms based upon an obvious poverty of ancestry. While the whole race of flowering plants is certainly a new one, as such things are reckoned fossilwise, there has been a fecundity in the origin of new species among these lusty upstarts that is simply amazing. How that, in part at least, has been accomplished will be considered in the final section of this chapter. Not only among these present-day plants, but all through the story of the development of the plant kingdom, we have been reading and writing of the changes of form and structure, some of which have been of far-reaching consequences. It is clear enough that if new types of vegetation and different races of plants have come into being and so modified the complexion of the plant kingdom, those changes must have first arisen in individuals which had within them some capacity for change, and furthermore the ability to use the change to their advantage. While, as we have seen, the losses have been tremendous, no one, with even this brief history of their development in mind, can doubt that there has been progress toward our present perfection of plant life.
It was with something very different in mind than the changing of plant characters that Cardinal Newman once said: “To live is to change, and to be perfect is to change often.” And yet nothing better expresses the facts of plants’ ability to change and the results of it than this reply of a great churchman to critics who could not or would not understand the truth of his now famous reply.
It is perhaps best to begin any discussion of the changes in plants by remembering a few simple facts regarding changes in ourselves. “Like father like son” is something more than an old saw which we repeat for centuries without stopping to think whether it is true or only half true. As with so many speeches of the sort, this is just precisely a half truth, for while sons are more apt to be like their fathers than other men, we all have within us the capacity, whether expressed or not, to change very considerably. In other words, all living things may be said to be a reflection or, perhaps better, the result of two divergent tendencies, one of which tends to make like produce like, and the other to produce something different.
Upon the ability of like to produce like rests the continuity of those plant groups, well exemplified by Lycopodium Selago and the ginkgo, which, through all the changing panorama of the history of the plant world, have steadily produced individuals so close to the ancestral type as to be essentially indistinguishable from it. It is upon the possession of this ability that all the different races of plants depend for the unchanged perpetuation of their kind. And, as we shall presently see, it is also upon this very ability that the new forms that do arise, rely for holding fast to their differences.
While it is true, then, that like tends to produce like, it is also and perhaps even more true that they do not precisely do so. In fact, they never do absolutely, and it is the degree of divergence from the type that different plants or animals exhibit, which is the measure of their ability to vary, or “produce something different.” Upon this capacity to vary, from whatever cause, rest all the changes which have occurred in the plant world, and, as we have seen in previous chapters, that has been by no means an insignificant affair. We know, in fact, that while one plant of Lycopodium Selago, than which scarcely any other now living has had greater opportunity to become fixed in its characters, is much like another, no two of them are actually identical. Nor are any two plants of the same species ever precisely alike, any more than two children, even of the same parents, are. The tendency for like to produce like is matched then, or sometimes exceeded, by an almost equally strong tendency to vary.
Heredity on the one hand and variation on the other are the two forces upon which the origin of new species or kinds of plants is based. Both of these work in rather definite ways, some of which are fairly well understood, but many of which are still among the things that scientists are striving to clear up. As the capacity to inherit characters from one generation to another reflects itself in the generally stable conditions which the plant world exhibits, while the capacity to vary is the only source of new forms, it is quite naturally the variations of plants from one generation to another which have been most studied. And the study of variation in plants is not the simple thing we might assume it to be, having in mind only the well-known fact that no two organisms are exactly alike. Wherein do they differ? Are their perhaps temporary or even quite casual differences passed on to their progeny? These and many other questions about variation make it at once the most complicated and often one of the most fruitful subjects of plant research. It is clear enough that with the bewildering variety of different plants in nature it is next to impossible not only to record accurately the amount of variation or its probability of being handed on, but least of all to arrive at any clue as to the origin of that variation. Because of this, and still more because practically no one has ever seen the actual origin of a new species in nature, for we only see the finished product, practically all our knowledge of the laws of variation has been derived from studying cultivated plants. The ease of controlling them and of recording thousands of observations of their characteristics has made the work of the plant breeder, and others who study variation in the vegetable world, much more of an exact science to-day than the mass of often interesting but usually unrelated data that crowd the pages of older botanical literature.
One of the main facts about variation is that it is itself a very variable thing, and the nature of those “fluctuating variations” which are so common in nature well illustrate the point. Within what we know as a species there are many individuals that vary one way or the other from a fairly central, we might almost call them normal, mass of individuals which are typical of that species. Nearly all these forms on the fringe of the species due to the environmental changes and not to changes of hereditary constitution will, if left to themselves, tend through their progeny to become more like the central mass as time goes on, while their position, or some other equally nontypical edge of the fringe, will be taken up by other variants from the average conditions. The amount of this fluctuating variability among plants is beyond calculation, and its action has often been likened to the swinging of a pendulum, which of course spends twice the time passing through the center of its arc, that it does on the limits of it. This very nearly expresses the proportion of fluctuating variants to the mass of typical individuals in many species of plants. In many others, often peculiarly unstable species, the number of individuals at the fringe is very large indeed. Sometimes there may be one or more that do hold their characteristics, in which case we know that they are not true environmental variations but have actually a different constitution. These will be considered presently under another and different sort of variability. But, speaking generally, these fluctuating or environmental variants are merely forms of the species, and, other things being equal, they will not actually originate new species.
It should be emphasized here, and before we go further in our discussion of variation, that species and varieties are after all largely creations of the mind of man rather than the reflection of actuality in nature. When we speak of a “species” it is merely a term which through usage by botanists becomes the symbol of a group of plants more like one another than like anything else. It is obvious that it is therefore necessarily an inaccurate designation of the actual conditions found among plants, which might almost be considered as all belonging to one great group of which, for our convenience in referring to them, we mark off units (families, genera, species, etc.), much as units are marked off on a rule. Species, then, and varieties of plants, notwithstanding the utmost refinement of method used in designating and describing them, and this is historically the most ancient and the most widely developed phase of botanical science, cannot reflect the true conditions, and for a number of reasons. The chief one is that species differ in usually several characters one from another and in large genera there is often a bewildering recombination of characters of the genus in the species belonging to it. Species and varieties are concepts of convenience, nay of absolute necessity, in talking or writing about plants, but hardly expressions of exact truth.
With this in mind we can appreciate the position of those plant breeders who insist that the basis of differences in all plants are the simplest, so-called, factor expressions, which can be isolated and studied with some approach to exactitude in experimental cultures. A factor may be defined as the hereditary determiner or base, which, either singly or in conjunction with other factors, is expressed as a character, such as tallness in peas, or brown eyes in human beings. Such studies have built up a body of information about variation in plants that show it to be of several kinds and with different chances of being passed on from one generation to another.
It was noted in the paragraph before the last that fluctuating variants were sometimes so far off the usual that they might almost be considered distinct forms or varieties. Many such changes appear to be the result of different conditions of the local environment, due to changed conditions rather than to any internal difference in the constitution of the plant itself. A familiar illustration of environmental variation may be seen in lima beans. In any considerable number of plants one often finds smaller and larger pods, either sparsely or well filled with beans. If the beans from the small-podded, few-seeded variants are planted they will produce, apparently quite indiscriminately, large and small podded progeny, just as there will result a mixed progeny if only beans from the well-filled and large-podded kind are sown. In other words the plant fluctuates about a general average which typifies the usual or mass characteristics of the species. One should not, however, regard all variations of the character in lima beans, or any other plant as environmental or fluctuating ones, for some of them may be due to differences in hereditary constitution. And these could only be determined by breeding tests.
Environmental variations are as frequent as the ever-changing conditions of plant growth may determine, and it is common knowledge that such diversity of the environment and the variants resulting from it are extremely frequent.
A much more fruitful source of new forms of plant life results from natural cross-fertilization, which, as we saw in an earlier chapter, is the nearly universal condition in the plant world. If species and varieties can be distinguished only by factor differences, as the plant breeders no doubt correctly insist, it becomes obvious enough that we have in cross-fertilization to consider not alone the factor differences of the pistillate or female, but also of the staminate or male contribution to the union, and how these are reflected in the progeny. Our knowledge of this has practically all been based on work done on cultivated plants under control conditions and it shows some interesting developments which occur from crossing.
If garden peas with, let us say, reddish-purple flowers are crossed with white-flowered ones, the progeny will not be a mixture of these colors but all reddish-purple. If all danger of subsequent cross-fertilization is excluded this first generation of reddish-purple progeny will themselves produce reddish-purple and white progeny in the ratio of three to one. But the extraordinary part of it is that in the third generation all the white and about one-third of the reddish-purple plants will breed true to color. The balance of the reddish-purple plants, which comprise about two-thirds of the second generation, will, if their seeds are germinated, produce colored as against white-flowered progeny in the three-to-one ratio. In other words, these artificial crosses, made by the plant breeder, and this splitting up of hybrids which has been many times verified, are seen to be very fertile causes of the origin of new forms of plant life, if only the factor and character differences in the ancestry be sufficiently complex. With no two plants precisely alike, with cross-fertilization so nearly universal, and with all characters, not a single character or factor expression, as in control conditions likely to be affected by the cross, it may be seen how fruitful a source of new forms this natural crossing may be. It is, in fact, not surprising that plants vary, but that the force of heredity will hold them into such recognizable categories that the red maple, or white ash, or blue cohosh are, with thousands of other species, after all fairly definite designations without which talking and writing about plants would be all but impossible. Some of our most beautiful garden plants have arisen either as the result of natural crossing, or crosses deliberately made by the plant breeder. The scores of forms of the common garden lilac have mostly come about by such crosses, although many other garden plants have arisen by still another kind of variability.
The effects of crossing which have been so briefly noted were not understood, as indeed the cause of them is still unknown, before 1865, when Gregor Mendel, an Augustinian monk, published the results of his work on peas, which furnished the basis for all subsequent work on this kind of variability. His work was neglected until 1900, when what is now known as Mendel’s law, involving the Mendelian ratio already noted, was rediscovered by three independent workers. It is now practically universally accepted as the way in which natural or induced hybrids transmit their characters.
There remains still another type of variability which has been noticed from very early days, and received the name sport, because quite suddenly, from a crop of otherwise similar specimens, one or a few plants showed marked and permanently transmitted differences from the average condition. Such sudden offshoots, which occur rather frequently in many plants, are known as mutants, the process as mutation. Hugo de Vries, a Dutch botanist of world-wide fame, was the chief modern figure who drew attention to mutants, and explained how they differ from fluctuating variants in that while these tend to revert to the average or mass conditions the mutant, once it appeared, held true to type. A well-known example of mutation is the cabbage, brussels sprouts, cauliflower, and kohl-rabi, all of which are sports or mutants from a weedy seaside plant of the mustard family, native in Europe. Since their appearance, hundreds of years ago, they have held their essential characters. If they had been environmental variants they would in all probability have reverted to their weedy ancestor. Hundreds of sports or mutants have been recognized and isolated, so that many of our most valuable garden plants have arisen through this ability of plants to vary in often sudden and rather startling degree. The gardener and horticulturist, from long observation and a keen sight for valuable novelties, have always known that sports are fruitful sources of new forms of plants, but De Vries first scientifically studied them and worked out the principles by which they apparently react. The cause of them is still unknown.
While the cause of mutants has not yet been revealed, we have already seen that the two remaining kinds of variability are due to changes in the environment, or to crossing. Charles Darwin when he published his “Origin of Species,” than which no other book has so completely revolutionized modern thought, did not state the cause of those variations of which he was our greatest observer. He did state the now universally accepted law of the “Survival of the Fittest” which explains how, once these variations make their appearance, the inexorable conflict of nature would automatically weed out the unfit. We have seen all through the course of this history of the plant kingdom how whole types of vegetation have been overthrown to give way to other types better fitted to survive. That process is going on with just as inexorable results to-day as it has down through the ages. While Darwin never claimed that such a purely selective process could initiate new species, many of his partisans who waged battle for him during the first years of the tremendous opposition his views encountered, did so claim, and probably wrongly. The actual cause of the origin of new species, except those demonstrated to result from new combinations of already known characters, through crossing, cannot be explained through the natural selection of plants or animals which exhibit favorable variations. We see their effects; it is obvious enough, that those of value tend for the survival of plants having such variations, and it was natural enough that older students of the problem should mistake these effects for the cause of them. The process of selective elimination constantly going on does tend to fix certain favorable variations and untold millions of plants have had their day in the past due to their possession of such, and the killing off of their less fortunately provided associates. We speak of this great march from the simplest organism up to our most complex plants and animals as their evolution, but we must never forget that it has gone step by step, by one or the other methods by which we have seen that plants vary, or perchance by some undetected method, and that while the results of it are for all to see, the causes of that infinitely slow and quite often wayward variation are not understood. Upon such a conception our modern plant life is seen to be a development of plants that have gone before, that all existing life is derived from preexisting, and not from providential interposition or special creations. All through the long marches of plant evolution there appears to be a definite and final goal toward which it tends, but we do not know the direction, least of all the object, of that goal. In fact, there may be many goals, just as there are the diversity of ambitions among human beings. In tracing the present ascendancy of our flowering plants from their links with the past perhaps we shall find no better statement of their present condition or destiny than to repeat Cardinal Newman’s reply: “To live is to change, and to be perfect is to change often.”
WE have seen in the previous chapters how many and how varied are the activities of the plant world and in this final one we shall get a glimpse of what these activities have produced. All the delicate mechanism of food getting and the manufacture of starch; the fertilization of flowers by insects, the wind or water; the response to changing light and to climate—these and scores of other activities of plants have resulted in the present vegetation of the world being what it is. Here we see the final reflection or register of not one but all the kaleidoscopic evidences of plant response and activities and history working in harmony, or, as we shall see presently, sometimes in violent conflict, and leaving as the result the wonderfully varied vegetation that now covers the earth. If we could read aright the story of which the vegetation of any particular country is the silent narrator, it would tell us not only what happened in the past but what is likely to happen in the future.
Plants, by what amounts to a kind of fatality, are rooted to the spot where they grow so that, unlike animals, their rapid distribution appears to be almost impossible, and yet the tremendous distances that some species have traveled seem like a pretty successful protest against the fact of the anchorage of individuals to the point of their origin. It is more than a successful protest, for it amounts in many species to an active campaign for dominance, to the exclusion or extinction of less aggressive neighbors, so that in any field or meadow or forest there are silent struggles constantly going on. Some of these are so inexorable in their results that they change not only the frequency of occurrence of the individuals involved, but sometimes the whole type of vegetation.
The competition to occupy just as much of the favorable plant sites as possible has been much aided by many species possessing means for the dispersal of their seeds or fruits that are ingenious in the extreme. Some of these are written plainly enough in the structure of the seed and its wonderful adaptability for the peculiar conditions to which it will be subjected. Before considering some of these structures we may profitably see how some plants look after the dispersal of their seeds within their own limited sphere of action.
In hundreds of plants the ripened pods, instead of being erect as their flowers have been, are pointed downward about the time the seeds are ready to be released, and their harvest is sown, sometimes by deliberate movements, in the immediate vicinity. No great areas are captured by such plants, except by the slow process of successive generations extending their range a few inches or at most a few feet a year. The great bulk of all seeds never do grow into new plants, but in those that only shed their seeds close to the parent plant the opportunity to reach new sites is by that much restricted. The chance of the species getting very far afield except by slow invasion of the neighboring region is limited. A few of such plants show remarkable ingenuity in reaching the utmost distance possible, perhaps the most effective cases being those that shoot their seeds by explosive bursting of their pods. Nearly all the violets do this, often shooting seeds several feet from the parent plant. Many plants of the pea family have pods that are twisted, which upon splitting release the previously pinched seeds so suddenly that they are shot considerable distances. In the common witch-hazel (Figure 111), the seed is shot through the air often as much as thirty feet.

But with even the greatest ingenuity and the most explosive bursting of pods, most plants could never capture much new ground, and their very existence as a species is often contingent on their ability to spread, if these various methods by which plants shed their seeds were not aided by outside help. Some of these have had conspicuous and almost startling results.
The seeds or fruits of those plants that are used for animal food are often carried considerable distances, while the thrifty squirrels’ burying of acorns is everyday knowledge to those who have seen them busily engaged in the making of winter stores, or the planting of new trees, often many rods from the parent tree. In years of plentiful seed production squirrels have been known to plant great quantities of seeds of the Douglas fir, thereby hastening the establishment of one of the greatest evergreen forests in the west.
While some seeds are destroyed by passing through the digestive tract of animals many are not harmed in the least. From over two hundred and fifty different kinds of plant seeds fed to a variety of birds over 80 per cent germinated perfectly after passing through their digestive tracts. And perhaps the most remarkable case is the seed of a pondweed said to be incapable of germination until it has passed through a bird. This plant grows in fresh water ponds in great quantities and is much eaten by wild ducks. From the stomach of one bird over three hundred such seeds were recovered. In the Eastern States a common feature of our farms is the red cedar or juniper scattered along fence rows, nearly all of which are due to birds roosting on fence rails and dropping the seeds after passage through their digestive tract.
We get some idea of the part birds play in plant dispersal when we realize the enormous number of them that make their flights twice a year, often over great distances. Wild ducks, in untold millions, travel from the far north to the tropics, each carrying their freight of seeds, sometimes as food and often mechanically clinging to their feet. The writer once saw at Gardiner’s Island and Montauk, Long Island, hundreds of thousands of tree swallows which feed on the fruits of the bayberry (Myrica carolinensis). So dense was the flock that they covered nearly every inch of the bayberry patches, and after eating no one can calculate how many million seeds they started off toward their winter home. Stopping as they do each day on their long flight southward, is it any wonder that the bayberry is one of the commonest bushes along the Atlantic coast?
With flights of birds stretching from the Arctic to the Antarctic, sometimes a single species making such a flight during its migration period, and hundreds of species making shorter flights twice a season, it is easily seen how birds can carry seeds for long distances. That they do so carry them is common knowledge and in eleven wild ducks examined by H. B. Guppy, he found nearly 300 seeds of bur reed, forty-one of pondweed, 270 sedges and 222 seeds that he could not identify. Nearly all these seeds germinated when sown, some sprouting more quickly than if they had not passed through the bird’s stomach. Some few seeds which would usually be dormant for one, two or even three years, have their germination unquestionably hastened by passage through birds, who may be looked on in some cases at least as “flying germinators.”
Some sea-flying birds have been captured over five hundred miles from land and seeds of a buttercup and of the sea blite recovered from their stomachs. One student of the flora of Spitzbergen has stated that nearly all the plants of that cold region have come from the northern part of the Scandinavian peninsula through their carriage by birds. One of the commonest plants of that region is the crowberry, which is a favorite food of birds. The plant is found throughout the Arctic regions and on high mountain tops to the south of it, such as the Alps, our own Adirondacks and White Mountains, and many others. There are some islands in the Pacific known to have received a few of the plants now growing on them by birds carrying the seeds from other regions, often thousands of miles away. Several species of birds are known to make sustained oversea flights from Labrador to South America, and one from Alaska to Hawaii. In such flights stopping is impossible, so that the carrying of fresh seeds and spores is always likely and probably more quickly accomplished than by any other means of transport. In New Zealand a northern European bird was once found with the seeds of two species of marsh arrow grass in its stomach, both of which germinated. The plants are native in the cooler parts of the north temperate zone.
Not far from New York, at Montauk Point, Long Island, there was found a few years ago some plants of the cloudberry, a kind of blackberry with amber-colored fruits, which is otherwise unknown in that region but is common enough in the Arctic and on mountain tops northward. That point of land, extending out to sea, is a favorite stopping place for migrating birds and to them was undoubtedly due the introduction of the plant so far south of its true home. Scores of similar cases could be cited which confirm the observations of naturalists all over the world that birds are among the greatest aids to plants in securing wide dispersal. Of course the factors of favorable or unfavorable conditions, once the seed is deposited in the new home, operate to keep plants from the tropics from settling permanently in colder regions and vice versa, but we cannot escape the conclusion that the sky is filled during certain seasons with millions of seed carriers that have, in many cases, populated their stopping places with foreign plants.
Other animals than birds carry fruits and seeds, besides those that eat them. Some fruits like the Martynia (Figure 60), are so arranged that no animal with fur can avoid catching some if they come in contact. The prickly fruits of cocklebur resulted in one species of that weed from the steppes of Russia being carried all over southern Europe in a comparatively few years. Tickseeds, Bidens, and all the hosts of plants that have prickles, burs, spines or what not attached to their fruits, have, by the possession of such devices, a better chance for dispersal of their fruits than those not so provided. Others, again, have various coatings of mucilage which stick to animals and thus help plants to overcome their chief drawback to dispersal—their anchorage at the place of birth.
The great trade winds, the violent hurricanes and monsoons of Asia, are all active and constant aids to plant dispersal. The fruits of many plants are provided with various devices to insure buoyancy in the air, such as maple, ash, and most seeds of pines and their relatives, birches, poplars, and willows, all of which may be carried distances of a few hundred feet in ordinary winds. In the buttonball tree, the milkweeds, fireweed, and many other plants there are feathery attachments or plumes that insure their seeds or fruits being carried very long distances indeed in regular or violent winds. But in the daisy family or Compositæ we find nearly all the eleven thousand species provided with a plumelike attachment of their light fruits, familiar enough in the dandelion, which may explain their being more widely distributed than any other plants. Some of them, however, are carried by animals, as they produce, in certain genera, barbed or hooked achenes.
The whole seed of some plants is so small that it can be lifted bodily by the wind, for instance Rhododendrons and many other heaths, one of the hanging pitcher plants of tropical Asia, many orchid seeds, and of hundreds of other plants. Many of these are less than one ten-thousandth of a gram in weight and, with dust and the spores of nearly all cryptogams, may be transported thousands of miles. When it is recalled that the dust from a volcanic eruption at Krakatoa in the Pacific was picked up on London window sills, and that some seeds and nearly all spores are as light as most dust, the wind as a plant dispersal agent becomes significant. In the western part of the United States over 800 million tons of dust are carried over 1,400 inches in a single year, according to estimates by J. W. Evans in his article on “The Wearing Down of Rocks.” Some effects of wind dispersal of seeds and spores furnish interesting data to the plant geographer.
In the Bahamas the natives speak of “hurricane grass” as a plant that was unknown on Great Bahama Island before August 13, 1890, when there was a great hurricane. Soon after this the sedge, which had been blown over from another island, began to be common on Great Bahama, where it is now thoroughly established. On Krakatoa near Java a violent eruption in 1883 completely destroyed the vegetation, covering the island with volcanic material. In thirteen years over sixty species of plants had arrived on the island, of which about twenty had been blown there as seeds or spores. Birds had carried about 7 per cent and the remainder had come by other means. No plants with sticky burs or other devices for catching in the coats of animals were found.
It is among the Compositæ or daisy family that the wind is seen to work most effectively as distributing agents of its light-plumed fruits. In the Falkland Islands, St. Helena, and Prince’s Island, all from 300 to 1,500 miles from the nearest land, species of Senecio or groundsel have been found that may well have been wind-driven onto these remote islands from the mainland. In fact the whole genus Senecio, consisting of over two thousand species, and of world-wide distribution, has in all probability been spread largely by its wind-borne achenes. They are not edible, nor are they suited to carriage by the ocean currents, but some of them are known to stick in the plumage of birds.
But it is hardly necessary to cite these far-flung examples of the wind’s action in distributing seeds, for there are many interesting cases much closer home. The tumbleweeds, such as the false indigo and certain grasses, are familiar sights scudding before the wind over prairies in the West and open places in the East. By a kind of foresight the winged seeds of the pines are so weighted, due to lack of symmetry, that instead of sailing quickly to the ground, they tumble and flutter about, thus prolonging the time of flight. And in the linden the curious winglike attachment from which the stalk of the fruit arises is admirably fitted to slide over the snow and ice upon which, through their often tardy falling, they are deposited. Scores of such adaptations of structure to function are known, where there seems direct response to conditions and there is the temptation to say that such adaptations are caused by the wind or other agency. Nothing could be farther from the truth, as plants do not produce winged seeds or luscious fruits to insure seed dispersal, but the seeds of plants having such devices, from whatever cause, are naturally favored in the ceaseless struggle to occupy new land, which is quite another thing.
The scattering of seeds along streams is too common a process to need more than a mere mention here, for it is to be seen along any fresh water stream at harvest time. But seeds that may be carried by ocean currents have a much greater influence on the distribution of plants, such as the coconut palm, already mentioned among the food plants, and now common throughout the tropical world.
The number of seeds that will float in sea water and still keep the power of germinating is not very great. Most seeds sink at once; many will float for months, but are useless when they reach land; but those that will both float and grow afterward have worked some curious changes in the floras of different islands. Sometimes the great ocean currents, like the Gulf Stream, appear particularly futile in the fruits they carry such long distances, as the pods of a tropical vine from the West Indies are not infrequently found on the coast of Norway and even of Nova Zembla, of course uselessly. But many ocean currents, particularly in the Pacific, have carried fruits and seeds thousands of miles, they have even carried so-called floating islands of vegetation bodily.
Perhaps the most remarkable case is that of Entada scandens, a tropical vine of the pea family, bearing large pods, sometimes several feet long. The plant is not typically a seaside plant, and there is evidence that fruits matured in the shade of its usual forest home will not float. Those that grow nearer the coast and more in the open develop, through partial drying, a small air chamber inside upon which the seed depends for buoyancy. Its original home is apparently somewhere in Central America, from the west coast of which it spread over the Pacific to the shores of the Indian Ocean. Over the Atlantic it has reached the shores of tropical Africa, and in fact wherever ocean currents cast up their refuse on lonely beaches parts of the giant pod or individual seeds of Entada scandens are found. Partly fossilized remains of them have been taken from peat bogs along the coast of Norway, of course dating since glacial times, but showing by their presence there how long this water-borne seed of the pea family has been attempting to populate the earth. Of course all that do not reach tropic shores are lost, but nearly throughout the Pacific Islands, with some exceptions, the vine is now established. The extraordinary feature of it all is that scarcely half the seeds of the plant will float at all, nearly all inland forms sinking at once in the fresh water rivers into which they may chance to fall. Only those that grow near the sea, in mangrove swamps and the like, or at any rate near brackish water, will float. These, however, apparently float indefinitely without loss of germinating power. There is abundant evidence that many plants of oceanic islands have similar characteristics so far as their inland and seaside forms are concerned.
While Entada scandens has spread in spite of unfavorable adaptation for seed floating, there are some plants whose seeds always float, and in spite of the sea water retain their power of germination. H. B. Guppy, from whom much of the above data are taken and who has experimented for years on the buoyancy and germinating power of sea-borne seeds, reckons about two hundred species that may have spread by ocean currents. Many of these are nearly world-wide in their distribution within their climatic requirements, and most are confined to the tropics.
It is perhaps a natural enough question why such elaborate and effective methods of seed dispersal are necessary and why plants, once they grow in any particular locality are not satisfied to stay there. The answer to this is that individual and racial competition is so great that without means of dispersal, which may be looked on as equipment for seeking a more favorable site, species would often be crowded out.
There is no better place to see this than at the edge of a forest and grassland. The presence of the forest tells us at once that what may be described as the forest type of climate must have existed in the past to have produced the woods. In clearing off parts of this forest the openings will usually be grassland at first, but never in the end if nature is allowed to work out the solution. Along the edge of such a forest will be found a host of pioneers pushing out among the grasses, making ready, by conditions of shade, protection from drying winds and other influences, for the seedlings of the forest trees that creep slowly but resistlessly out to capture areas that by right of previous occupancy belong to them. In practically all parts of the world, with a few local exceptions, wherever the forest was the original type there is this ceaseless struggle to reclaim the open places, often or usually peopled by grass. It may be set down as almost a rule that if the open places produce herbs with broader leaves than grasses, the forest will capture the area many years sooner than if grass alone is the temporary tenant. Grass by its exclusive growth, its complete monopolization, so far as low seedlings are concerned, of much light and nearly all available surface water, is singularly well able to take care of itself once it is thoroughly established. But plotted and marked areas of this contact between forest and artificial clearings in it, show that in the end the forest will win, often at the rate of five hundred feet in a hundred years, sometimes much quicker than this.
If, on the other hand, what may be called the prairie type of climate has resulted in the formation of grassland, which has happened in our own West, in the steppes of Russia and less extensively in many other places in the world, forests can hardly ever get a foothold. Where, as in river bottoms, they sometimes flourish, the line between forest and grassland is sharp and apparently an impassable barrier for trees.
This invasion of immediately adjoining territory is going on constantly not only by different types of vegetation but by the units of it. The frequency of different plants in different years, their final ascendancy or extinction, all point to the struggle for expansion which in a score of ways the plant world is constantly waging. In many cases we are not yet able to see the struggle, but only its results, while in some places the bitterness of it may be gauged by the dead and dying that strew these silent battle fields.
The dominance of certain species of plants, such as grasses on a prairie, the fir and spruce in the coniferous forests of our North, the blue-gum trees in parts of Australia and the giant dipterocarp forests in the Philippines, are all based on the ability of individuals to spread from their point of origin. All species of plants must have one day been of very local distribution and confined to the region where they were born, but from their often very modest beginnings some of them at any rate have gone to the ends of the earth. The common bracken fern is found in nearly every country in the world from far northward through the tropics to the antipodes, and yet no one knows where its original home might have been. Somewhere up in the northern Andes it is supposed that the first ancestor of the huge daisy family had its origin. From there its many descendants, now ramified into hundreds of genera and over eleven thousand species, have spread to the very limits of plant growth. And the daisy family is one of the most recent of all the families of plants.
The distribution of plants is of many kinds depending on local conditions of climate and soil, on individual and racial competition, on methods of fertilization or other means of propagation or seed dispersal, and particularly upon the distribution of the plant or its ancestors in past ages. While it is often difficult or impossible to determine upon which of these factors, or upon what combination of them the distribution of any particular species is based, certain facts of plant dispersal appear to be indicated by a study of existing floras. A few instances must suffice here to illustrate the principles by which many plants are scattered over the earth, or else restricted to localized regions, and which, without knowledge of the factors involved, seem merely the wayward caprice of nature.
In the flora of eastern North America there are many genera that appear to be endemic there (found nowhere else), but are actually duplicated in eastern Asia, if not as to species, at any rate by plants so closely allied as to be of obviously common origin. These plants are unknown in Europe, or on our own western coast. The skunk cabbage, sassafras, twinleaf, May apple, Canada moonseed, spice-bush, ginseng, sour gum, trailing arbutus, fringe tree, lopseed and many others, all fairly common in eastern North America, are unknown between this and eastern Asia where, if not the identical species, which often happens, closely related forms are duplicated. The explanation of such discontinuous present distribution appears to be that at some time in the distant past there was a land connection between Asia and the western coast of America, the remnants of which form the Aleutian Islands, and over which there was a constant migration of plants and animals. With subsequently changing climatic conditions on our own western coast, due to warm ocean currents, most of those Asiatic migrants, or it may have been a migration in the opposite direction, were crowded out by later types which now dominate the Pacific coast. There is small chance that these plants were spread by birds, as an east-west bird migration is hardly likely. Nor is there any record that the seed of these plants, even where they might float, would survive ocean transport.
Sometimes the ancestors of now widely separated species or genera once covered all or nearly all the intervening area, and again sometimes only a minute fraction of the ancient distribution is left at the present time. Our own Big Trees of California were once known to grow in England, Iceland, all through central Europe and eastern Asia, Australia, New Zealand, southern Chile, and from Texas to Alaska. In the face of such widespread occurrence in the past their present distribution over an area of a few square miles is merely a pitiful relic of ancient grandeur. Scores of cases are known where, instead of a single outpost as in the Big Trees, there are only a few widely scattered survivors from a probably much more continuous distribution in the past. In northeastern North America there grows along pond sides and fresh-water beaches the shore-weed, a relative of our common weedy plantain. The only other species, and a close relative, comes from the southern tip of South America.
Many other species now existing are to be viewed only as relics of a bygone, often much more continuous and more widespread dispersal. In the persimmons, of which over a hundred different kinds are now known, the original distribution covered all North America, all Europe and Asia, except the Scandinavian Peninsula, all of Africa, northern Australia and South America. To-day the genus is restricted mostly to tropical Asia, southern Africa,

Some of the Commoner Plants of the Desert in the Southwest. The fanlike branches at the left are the ocotillo (Fouquieria), the two short tree cacti and choya cactus (Opuntia) and the leafless tree in the central background the palo verde (Parkinsonia). (Photo by the late Edward L. Morris, released for publication here by the Brooklyn Museum.)

Grassland and Tree Vegetation. All over the world there is a contest between grassland and tree vegetation for dominance wherever both occur. In this thorn veld in Natal the struggle is particularly keen. (After Bews. Courtesy of Brooklyn Botanic Garden.)
northern South America, and in North America it has dwindled to a handful of species confined to the region south of the Great Lakes and generally east of the Mississippi. In Mexico and south of it the genus is better represented. But with the persimmon, as in so many other types, our existing species are the remnants of preexisting ones, without a knowledge of which their present dispersal would be impossible to explain.
It is easy to reason from the foregoing that widely separated but related species of plants are all either very ancient, or directly descended from ancient ancestors, and that other things being equal a widely dispersed species is older than one with a restricted distribution. It is most certainly true that relics are unquestionably very ancient, and do actually represent the last outposts of a preexisting condition. But many isolated plants are relatively very new, so far as their immediate origin is concerned, as witness the hosts of young species of the daisy family, some of which have spread scarcely at all from their obvious point of origin. In judging of the distribution of even the commonest tree or shrub of our woodlands there are these links with the past, as well as response to present conditions, to be weighed if we are to understand the story aright.
Once a plant reaches a new and for it a strange country it is remarkable how quickly it will often capture the new territory. In the United States over six hundred of our commonest weeds have come from Europe and Asia. The daisy, dandelion, wild carrot, many hawkweeds, dozens of wild mustards, and many others are among the somewhat undesirable immigrants that now reach over a great part of the country—all brought over by the early settlers of America. From their home in subtropical Asia the lemon, lime, and orange have invaded every part of the tropical world. Once, in the most remote part of the Sierra Maestra Mountains in eastern Cuba, where only by the most arduous cutting could a passage through the dense tropical forest be forced, the writer found within a few square rods an orange and a tree from tropical India. Such cases could be multiplied, and all over the world we see this endless struggle of plants to conquer new territory, often at the expense of existing vegetation. On Long Island, New York, the introduced locust tree, brought from the southeastern United States about a hundred years ago, has completely routed the native trees in many places along the north shore of the island. And in Hawaii, some seeds of a screw pine, washed up among refuse along the beach or brought in by early aborigines, have made this Malayan plant a common tree thousands of miles from its home.
The geographic distribution of species of plants may be, as we have seen, the result of the geological changes of the past, of bird migrations, of more or less fortuitously water-borne seeds, or more usually of the slow spreading by invasion of those species apparently not so well supplied with external helps to dispersal. But no matter where plants grow, nor how they got there, they must fit the particular environment in which they find themselves or perish. This home economy of plants, or how they meet the environment and each other, is called Plant Ecology, a phase of botany now much studied, for it tells us more directly than most other plant research what the actual response to various factors of the environment may be. Just as plant distribution is the reflection of many, usually widely operating forces, so ecology narrows down to individual plants or groups of them the impact of the immediately surrounding conditions upon vegetation.
The basis for all study of the response of plants to the conditions under which they grow must rest upon the response of their different organs to those factors, just as our general movements are dictated by sufficient food or air or water to keep ordinary bodily functions going in the ordinary way. But the study of such plant response has shown that certain kinds of environmental conditions have resulted in quite similar response nearly throughout the world. Often totally unrelated plants assume characteristically similar growth forms where the conditions in widely separated areas are climatically or otherwise similar. In our own Southwest we have the dominant cactus vegetation, matched in parts of South Africa by giant cactuslike spurges. In Mexico we find the wealth of century plants, which are confined to the New World, matched in the Old World dry regions by the aloe, a group of succulent plants nearly as well suited to such areas. The species of plants characterizing peculiar regions may well be the result of geographic distribution that rests on more widely operating factors such as we saw in the previous section of this chapter, but the type of plants growing in a particular place hardly ever fails to be dictated by the local condition. With this in mind, a vast amount of time has been spent in studying the various factors of the environment, such as climate, soils, altitude, light, etc. And an equally valuable study has been the response of individual plants or their organs to such conditions. From this great body of information, obviously impossible to include here, we all recognize certain well-marked societies or groupings of plants which, wherever they occur, exhibit similarity of general response to the different conditions responsible for their occurrence. Once these typical plant societies or groupings are understood we can recognize them wherever they may occur, and we shall see that they are as widespread as are plants themselves.
Just as societies or races of men have often obscure beginnings, reach a climax, and afterward die, so these plant societies may be considered as exhibiting a similar progression. What these plant societies are, at least the more important of them, will be considered in the next section of this chapter. It should never be forgotten that the species of plants making up the dominant plant societies in different parts of the world are dictated by quite other conditions than those that result in the dominance of the society itself. Perhaps as good an illustration as any is the aristocratic type of mankind, recognizable throughout the world by the possession of finer qualities than the common run, but differing in individuals as much as the best type of Americans, the British peerage, and the samurai of Japan differ one from the other.
Not the least interesting feature of these plant societies is that we must view them as associations of plants, often of widely differing origin due to the vicissitudes of plant distribution, but all taking their part in the society to which they belong and often, as individuals, losing their life that the society may live. Upon such a conception a wood or prairie, or river bank, or salt marsh or alpine garden upon a mountain summit are, with many other plant societies, places of intense conflict. More cruel than any human society, these plant communities exist under conditions where only the individually strong survive, and only those societies are destined to reach their climax which can take advantage of every aid, quite without regard for severe losses or even death to the individual members of it. It is as if we poured into a crucible molten metals from many different sources, and after the incredible and relentless forces of manufacture had worked their magic upon them there resulted a product, purified and cleared of all dross. So the inexorable and relentless processes of nature work over the materials found in these plant communities, the results of which are the dominant types of vegetation in the world to-day. With this understanding of the part they play in plant distribution we may now consider a few of the most widely recognized plant societies and see how they have affected the vegetation, sometimes even the history of the regions in which they are found.
No one who has ever seen both our temperate forests and those in the tropics can fail to be impressed with the difference between them. Not only for the different plants in them, but for their wholly different aspect, tropical and temperate forests stand far apart as an expression of the forest covering the earth. Not all of us realize, however, that the heat of the tropics is not the deciding factor in the luxuriance of those dim jungles, and that a rainfall far above anything occurring in the United States is even more important. Upon the distribution of rainfall depends the occurrence not only of the two forest types that will be mentioned here, but of most of the other chief plant societies.
A small section along the lower side of the Gulf of Mexico, the northeastern edges of Cuba and Santo Domingo, nearly all of the region drained by the Orinoco and Amazon Rivers; in the Congo, Zanzibar, and Madagascar in Africa; all of southeastern Asia, including the East Indies and part of the north coast of Australia—these comprise the regions of the tropical rain forest. All of them have, besides continuous heat, a nearly continuous or in some places a periodic rainfall, averaging over, and usually much over, eighty-five inches a year, as compared with about half that near New York. There are, of course, other places in the world where these rain forests, so called from their abundant moisture and some of the effects of it, are found. But in the regions mentioned they are at once the most wonderful and to most white men the most awesome manifestations of the plant world.
Such forests seem, and actually are, pulsating with life, as instruments stationed in them have many times proved. With some kinds of bamboo growing over two feet a day, and a eucalyptus tree in Java forty-five feet in three years—and these are not isolated cases—the tremendous annual increase in the amount of vegetation can be glimpsed. Of course not all the plants in them grow at any such rate, but the great heat and abundant moisture does make tropical rain forests irresistible in their power. Plantation owners, and railways that have been run through such forests, wage constant warfare against the recapture by the teeming forest of man’s intrusion of it. The writer once saw in Santo Domingo a railway cut through such a jungle and abandoned only two or three years before. Not a trace of the roadbed could be found, ties, rails, and switches all covered with a dense vegetation, and overhead the canopy of the forest had closed over the opening and was already sending down hundreds of adventitious roots that would complete the obliteration of man’s handiwork. Everywhere there is the evidence of vegetable life run riot, ever crowding and pushing to close up openings made by the crashing down of old trees or the artificial clearings of man. Those living on the edges of such forests speak and think of them as dim, mysterious places where strange creatures and the ever-present fevers join forces with the vegetation to keep out humankind. That they are places of actual danger everyone knows who recalls that Stanley’s trip through equatorial Africa cost one hundred and seventy lives, many of which were sacrificed to disease and strangely enough nine were lost through starvation. While the tropics supply much of the food used there, these jungles produce almost none of it and because of the scarcity of edible fruits, the extraordinary difficulty of getting about and collecting what does grow, starvation faces anyone who goes into them without adequate supplies.
In the Amazon grows the largest water lily in the world, Victoria regia, with giant leaves upon which a moderate-sized man may stand in safety. It produces a flower over a foot in diameter, and it is surrounded by a forest the like of which it is difficult to describe. H. H. Rusby, who spent two years in this region hunting for medicinal plants, has described the country a few hundred miles below where Victoria was discovered. He writes: “Passing down the river Madeira to the lower Amazon, we come into a region of such grandeur in its vegetation that it is difficult of comprehension, even by one who is an eyewitness. Everything is in such proportion that one is apt in its size to miss the gigantic. Many of the trees of this region are undoubtedly many centuries old and appear to be good for many centuries more. Most of them have enormous buttresses at the base, and these buttresses often begin as high above the ground as are the tops of ordinary forest trees in our land. All are bound together with an impenetrable mass of tough vines. Running through these swamps are the most beautiful little bayous or canals. Nothing can exceed in interest and delight a day’s canoeing among these narrow waterways, although there is great danger that the inexperienced boatman will hopelessly lose his way. In the rainy season this river rises sixty feet or more above its low water mark and the boatman travels among the tree tops which a few weeks before were high above his head.”
The abundant water supply in the rain forests results in an atmosphere saturated with water vapor and in some of them it is a common sight in the morning to see the forest rising out of an unbroken blanket of mist. As this dries up under the heat of the day, or if there occurs one of the torrential downpours to which such regions are frequently subjected, there rises from the forest in plainly visible waves a vast quantity of water vapor. It is this that has so often made them be described as steaming forests. The water requirements of the plants are more than supplied, nay, there is such a surfeit of available water in all these forests, that there are numberless devices to get rid of the excess. Dripping points to the leaves, already described in an earlier chapter, are common. But in addition many plants have wonderfully colored leaves such as Begonias, some relatives of our jack-in-the-pulpit belonging to the Arum family, many orchids, and other plants. The colored leaves in the predominantly dark green and gloomy rain forest, because of their greater absorption of light and consequently higher transpiration rate, are of decided advantage.
While there is thus very little or in fact almost no struggle for water in the rain forest, the struggle for light is intense. In the deepest and most luxuriant of them the gloom of the forest floor is notorious and it was by no means a figure of speech for Stanley to describe his trip through equatorial Africa as “Through the Dark Continent.” So dark are most rain forests, and, as we have seen in a previous chapter, so inexorable are the plants’ demands for light, that the various devices to insure it are perhaps the one great difference between these forests and those of temperate regions. One effect of the struggle for light is the enormous production of vines often running hundreds of feet through the tree tops. In India the Calamus, or rattan palms, with stems no thicker than a walking-stick, will completely interlace the foliage of the canopy. Thousands of slender whiplike roots and stems of such plants descend from the topmost heights of the forest canopy, where the plants to which they are attached make such an inextricable tangle among the tree tops that orchid collectors have been known to travel considerable distances over the matted vegetation, with, it must be confessed, considerable danger. These vines or lianes as they are called, are however, often as thick as a man’s body and armed with great hooked prickles, an obvious aid in catching some support to reach that essential light for which all plants in such places are ever striving.
Besides the bewildering tangle caused by these lianes, the rain forest is further impeded by hosts of epiphytes or plants that are mechanically attached to tree trunks, branches, or anything else that will raise them to the light. Of all the plants of such regions the epiphytes are the most light-demanding. They must not be mistaken for parasites, as they have roots of their own through which they absorb nourishment, mostly as water vapor, but also as liquid water held in the bark and refuse in which they grow. Thousands of orchids are epiphytes, also ferns, and, only in the American tropics, thousands of different relatives of the pineapple. Many of the latter are among the most gorgeously colored of all plants, their superb foliage being much sought after and the specimens largely grown in our greenhouses. In most rain forests every available inch of space is covered by these epiphytes, so that no bark, scarcely any branches, are to be seen but those clothed in this motley array of plants that use the support to get the utmost possible light. Many of these epiphytes have rosettes of leaves arranged for holding water, and after a sharp thunder shower followed by fresh wind the writer has seen the ground strewn with thousands of relatives of the pineapple which, with the added supply of water, were unable to stand the strain and were consequently wrenched from their lofty perch. So enormous is the combined weight of these epiphytes, together with the lianes, that many trees crash down under the strain long before their time. Perhaps no sight of the rain forest so convinces one of the struggle for light as to see one of these forest monarchs come crashing down loaded with thousands of plants that have been using it for support, and to escape which it has pushed its canopy to the utmost limits of its growth. Such contests are common in a forest of which only the barest outlines can be conveyed to those who have never seen it. To those who have had that good fortune any description palls beside the wonderful actuality.
It is scarcely to be wondered at that these steaming rain forests with their gloom, and, as they were once described, “all hung about with fever trees,” should be dreaded by many, and the subject of fabulous tales to the credulous. The almost incredible difficulty of getting through them, not to mention the savage animals that inhabit many of them, have not lessened the tendency to exaggerate about these great forests. But the truth about them is so far beyond belief, the strange plants that intrepid explorers have brought out of them so almost incredible, that it only excited a temporary wonderment when the largest flower in the world was discovered in such a forest in the Malayan Archipelago.
Sir Stanford Raffles and Dr. Arnold, while exploring in Sumatra during the year 1818, discovered what was called “the greatest prodigy of the vegetable world,” and no flower since found equals its size. The plant, without stem or leaves, consists wholly of one gigantic flower about nine feet in circumference, and was subsequently called Rafflesia Arnoldii. It aroused a sensation in England which was not abated by knowledge of the fact that the flower is a parasite on the stems of certain tropical plants related to the grape. That such a huge flower should be the product of a parasitic mode of life is one more illustration of how this and related irregularities occur in widely separated families of plants, and under varying conditions. Relatives of it have since been found in India, some parasitic on roots, others, as in Rafflesia Arnoldii, on the stems of vines. The sticky seeds are in all probability carried from place to place on the hoofs of elephants, to which they have been known to cling. Only if they are deposited on a bruised or otherwise exposed tissue of their future host can they grow. These curious plants have been actually cultivated in the greatest tropical botanical garden in the world, at Buitenzorg, Java.
The original collectors of Rafflesia Arnoldii could scarcely credit their senses when they saw for the first time this extraordinary plant, whose whole life is spent in producing this great flower and fruit. As one of them says: “Had I been alone, and had there been no witnesses, I think I should have been fearful of mentioning the dimensions of this flower, so much does it exceed every flower I have ever seen or heard of.” The odor of the flower is repulsive, and, with its great size and curious mode of growth in the dark rain forest, it is surely one of the strangest productions of the vegetable world.
But as the utmost development of the plant world, and producing the greatest profusion and richness of plant life, these rain forests are, beyond the sporadic occurrence of such wonders as Rafflesia and some others, places of extraordinary interest. With every inch of space occupied by plants, the very epiphytes often having on their leaves still smaller plants, we see here what nature will produce when the maximum conditions for plant growth are so nearly perfect. Theodore Roosevelt in his book, “Through the Brazilian Wilderness,” gives a vivid picture of the rain forest there, and it may well end our account of those most interesting of all plant societies:
“In one grove the fig trees were killing the palms, just as in Africa they kill the sandalwood trees. In the gloom of this grove there were no flowers, no bushes; the air was heavy; the ground was brown with moldering leaves. Almost every palm was serving as a prop for a fig tree. The fig trees were in every stage of growth. The youngest ones merely ran up the palms as vines. In the next state the vine had thickened and was sending out shoots, wrapping the palm stem in a deadly hold.
“Some of the shoots were thrown round the stem like the tentacles of an immense cuttlefish. Others looked like claws that were hooked into every crevice, and round every projection. In the stage beyond this the palm had been killed, and its dead carcass appeared between the big, winding vine trunks; and later the palm had disappeared and the vines had united in a great fig tree. Water stood in black pools at the foot of the murdered trees, and of the trees that had murdered them. There was something sinister and evil in the dark stillness of the grove; it seemed as if sentient beings had writhed themselves round and were strangling other sentient beings.”
Many other forests in the tropics, where the rainfall is less, or less regularly distributed, are not unlike our own, having rather regular periods of leaf-fall that come with the dry season rather than with the autumn. The trees are of course never the same as ours, but the general aspect is not very different from that of temperate forests.
The transition from the tropical rain forest to our own woodlands is one of the most interesting, as it is certainly the most gradual in nature. Lack of space prevents our stopping to note those strategic points along this pathway from a hot, steaming forest to the cool shade of our open woods, where traveling, in at least a virgin forest, may be done easily on horseback. As we come northward, and if we could travel continuously through the forest, we should lose first the epiphytes, then most of the lianes, and finally all the condition of vegetation crowding into every inch of space suitable for it. While trees in our virgin forests are as thick as they can be, the forest floor is open and on it grow only a few herbs that will stand lack of sunshine.
But the really great difference is the long, unfavorable season in temperate regions where the forest must drop all its leaves, after, in at least our own Eastern States, the most gorgeous foliage coloring of any forest in the world. The winter months when the woody vegetation is practically fully exposed to the elements, are particularly severe in their effects. Leaf-fall, which is such a common sight as to arouse scarcely any interest, is the only device by which the great bulk of our forest trees survive, and only in the southern part of the region are there found such woody plants as the mountain laurel, rhododendron, American holly, and a few others which are evergreen but not cone-bearing ones, and are the only reminders of the truly evergreen forests of the tropics. The winter winds farther north and in the central treeless part of the United States prove too much for many kinds of trees, for instance, all the oaks and sassafras, none of which go very far north.
There is sufficient rainfall to produce forests much farther north than they are found, but lower temperatures prevent trees from growing just as too little water stops their growth altogether. Toward the northern limits, or upon high mountains, the upper limits of the forest, we get the best idea of how persistent woody vegetation is in the general forest area of the eastern United States. Stunted, wind-swept and weather-beaten trees are often found only a couple of feet high and over sixty years old. Sometimes they will be flattened out on the ground or on bare rock, making great patches of bushy growth quite unlike their lofty relations in the lowlands. The growth rate for such plants is so slow that their annual rings are all but obscured. With such persistence in the production of these elfin forests, high up on mountains under the most unfavorable conditions, it is little wonder that below this are trackless woods, and that the northeastern United States has one of the finest developments of the temperate or summer forest in the world.
Nor are all our woods of this general type made up of the same species, for everyone knows about the endless spruce and fir forests of the north, exclusively evergreen, and in the summer nearly always moist. This spruce belt stretches practically across the continent, where, in the West, other and our most gigantic evergreens, replace the eastern spruce and fir. A little farther south is the region of the white pine now nearly unknown as a virgin forest type, as its great value led to early and ruthless cutting. The white pine region is generally the area from New England southward along the Alleghenies and westward to Illinois. But the most characteristic of the temperate forest types is our summer forest, so called from its general lack of evergreens and its beautiful green foliage of summer and its bare branches in winter. Beech and birch and maple, in different proportions according to local conditions, predominate in such woods. These hard-wooded trees, with many others that are scattered through them, have been among the most valuable of all the natural plant products of our country and their destruction has been upon such a scale that only in a few places may the virgin forest be seen at the present time. Where it does occur we find the forest floor often with nothing growing on it except a mass of spring flowers which are half matured before the leaves of the forest canopy close out nearly all the light and much water and put them to rest until another year. The great preponderance of spring-flowering herbs in Eastern North America is due to their early warming up before the foliage of the trees cuts off their light. And in some virgin forests of this sort, particularly where there is a large mixture of oak, the writer has seen hundreds of square rods without undergrowth or herbaceous vegetation of any kind. Such places, very rare indeed at the present time due to senseless and wicked cutting, are rather dark, perfectly open to view for hundreds of feet ahead, and dotted only with the huge trunks of the trees that characterize this climax type of the temperate or summer forest.
The absence of direct sunlight and interception of much rain under the forest canopy has other effects besides stopping the growth of herbs and shrubs which are common enough along the edges, or where openings are made by the fall of old trees. It prevents the germination and growth of nearly all the seeds falling from such trees, and in a really virgin forest of this sort, almost no seedlings will be found. Upon cleared or open land thousands of saplings will cover much of the ground, but nearly all these will die off due to crowding, and leave as the climax only enough trees to close over the forest canopy.
Forests may be found in all stages of succession from those just beginning the process to those final forest monarchs which, having won out in the race, are, until one of them falls, often slow to perpetuate the type. For, as often as not, a new growth will spring up once a very large tree falls, and a very different kind of growth from the climax forest. At once a host of species, that one might almost say had been waiting for the tragedy of the monarch’s fall, will rush in and convert the opening into a nondescript brush patch, out of which will rise another tree that means business. As it grows to maturity, it kills off these smaller triflers one by one, until, when the canopy is finally closed, all of them will have disappeared, or, as often happens, retreated to other parts of the forest, where they wait for another chance. This succession of different kinds of growth in a temperate forest is so well known that in England they have for centuries practiced it, for commercial or pleasure purposes. In their oak-hazel copses they cut the trees enough to partly open the canopy, which permits a dense growth of hazel bushes and other plants. Every twelve or fifteen years the latter are cut down for various purposes, and will gradually spring up again to renew the dense growth. The spacing of the trees is sufficient for them to branch freely and yet not close the canopy enough to kill off the hazel. The trees are cut off a few at a time, not oftener than one or two hundred years in any one spot. By this procedure the owner gets a regular crop of hazel once in twelve or fifteen years, occasional big trees, and on many places a cover for pheasants. Under the hazel there is a regular progression of herbs, very plentiful just after the bushes are cut, and decreasing almost to nothing when the end of the growth period of the bushes is near.
The English oak-hazel copse, now much less grown than formerly, and the general lack of undergrowth in our own virgin forests, are both responses of the forest to light and other factors that are related to an open or closed canopy. As we stated a page or two back, it is not impossible, it is even frequent in some parts of the country, for a forest to produce, by its own growth, conditions inimical to its perpetuation. Where the casual falling of a forest giant is the only opportunity which that forest offers to perpetuate its type, it may well be said to be a climax forest, incapable of further development. But in some such woods a curious provision of nature insures an invasion of the gloomy forests by trees less light-demanding than the dominant ones. And often these trees that can get along with less light will capture considerable parts of forests that light-demanding trees could never conquer.
If the soil in which these temperate forests happen to grow is sandy or otherwise poor in plant food, the broad-leaved trees that make our woods such a delight in summer are replaced almost universally by pines. Along the sandy stretches of the coastal plain from Long Island, New York, to the Gulf, there are immense tracts of these pine forests, different species often being locally dominant, such as the pitch pine in the pine barrens of New Jersey and the long-leaf pine farther south. Almost throughout the world there is this monopolization of the poorer and drier forest sites by pines, which maintain the forest plant society in regions where the broad-leaved rapidly transpiring trees could not grow.
No account of forests, however brief, can omit some mention of the greatest agency for their protection in North America, the United States Forest Service. With corporation and individual cutting and attendant fire hazard upon a scale almost beyond belief in its ruthless disregard of our chief natural plant product, the Government soon found that Federal ownership or control of forests was the only policy that would maintain even a partially adequate timber supply. National forests, set aside either for pleasure or profit, now total more than the area of France or than all the New England and most of the Middle Atlantic States. These huge tracts, in every part of the country where forests are found, are well managed, properly planted, and most important of all, constantly guarded against fire. Forest fires not only destroyed over $25,000,000 worth of timber annually, but leaving devastation behind them, depleted the water supply in many parts of the country. Nothing but forests will hold the rainfall, to release it slowly through a thousand rivulets and springs that are the source of countless rivers. With the forest cut or burned off these streams are dry most of the summer and raging torrents for a few weeks in the spring, washing out all the priceless accumulation of the ages which the forest has conserved for its own and our benefit. While the reservation of these great national forests has worked individual hardship, experience for many years back in India and Germany shows Federal ownership or control the only wise policy.
Forest covering, whether temperate or tropical, depends for its occurrence all over the world upon an adequate rainfall. As we have seen in the tropics, this may be so great that coupled with the heat it produces a wealth of vegetation beyond the powers of description. Where it is less and the country cooler, the forests are of a different type, but even there the forest covering is, without interference, practically complete. Where, as in parts of Chile, southeastern Australia, and of Japan, there is a heavy rainfall but cool climate, there is a so-called temperate rain forest. Such forests are cold, drab, wet woods of peculiar aspect and extreme interest. For in them grow trees sometimes related to our own, but, due to the special conditions, producing a forest landscape quite unlike anything in America. It would seem as if we might almost plot the distribution of forests in our own country with a weather map showing rainfall, and such is actually the case. When the rainfall becomes less than will maintain forest growth it stops, often very abruptly. Generally speaking, the region west of the Mississippi, and some just east of it, westward to the mountains, is entirely devoid of forest, except in the river valleys. The forests give place to an entirely different type of vegetation—the prairie or grassland.
In nearly every region in the world there is an absence of forests and a replacement of them by grasslands, where the rainfall is less than about twenty-five inches a year, and where the winter winds, often far below the freezing point, are hostile to trees. The distribution of the rain mostly through the growing season also makes a condition peculiarly unfavorable for trees during the winter. Someone has said that the nations have fought since the days of the Romans for the belt of grassland which these climatic conditions have produced all round the world, and it is certainly true that these naturally grass-covered areas have produced the cereals of the world, all of which, except rice, grow to best advantage in such regions.
With of course different species of grass and quite different associated herbs, these grasslands are now found in our own prairies, the steppes of Russia, the plains of Hungary, the pampas of southern South America, the grasslands of Australia, the veld in Natal and in many other but mostly less extensive developments. Some of the grassland regions are warm, but without more rainfall than characterizes such areas the greater heat does not produce a forest. Usually, but not always, these grasslands are not found near the coast, where, as in America, the rainfall is double that of the plains and produces the forests that clothe the Atlantic and Pacific sections of the country. From the east westward there is a gradual decrease in the rainfall, until from about the Mississippi to the mountains it falls below the point where trees can compete with the prairie.
Another characteristic of prairies that once they have started tends to keep out trees is their almost annual firing. Tree seedlings cannot survive this, and we know that the Indians fired huge tracts of prairie every year, not to mention fires started by lightning which may set fire to grasslands and actually does set fire to forests every year.
The prairies in the United States—perhaps the most extensive in the world—are characterized chiefly by several grasses, buffalo grass (Buchloë dactyloides), gama grass (Bouletoua oligostachya), and several prairie grasses, such as Sporobolus asperifolius, Koeleria cristata, and some others. Among these, depending on the soil, are hundreds of prairie flowers which, during different parts of a single season, give quite different aspects to the region. Both the grasses and their associated herbs are well protected against too violent transpiration which their exposure to nearly continuous sunshine, high summer heat, and very considerable winds makes particularly active. In many places where the country is rolling, the lower and moister sites, besides developing more luxuriant growth of prairie plants, permits low shrubs, and in river bottoms even trees to flourish. But climatic conditions of small rainfall, high winds, and bitter winters make anything like a forest development out of the question.
In some regions, both in America and central Europe, a rainfall that is high enough to permit trees to exist and low enough to favor at the same time a grassland formation has resulted in the parklike landscape that creates the most beautiful scenic features of the regions where it occurs. In such places there are irregular patches of forest and grassland, and the struggle for supremacy as between the different types depends, not upon the general climatic conditions to which they both respond, but to local conditions of available water supply and soil conditions and often upon fires. Naturally such regions are places of intense strife for dominance, and in them some remarkable collections of plants have been found.
One of the most interesting of these struggles between grassland and woody vegetation in a region climatically able to produce both, is in Natal. Large sections of that country are grasslands or veld, as the people there call it. Scattered through the veld are various species of acacia trees, locally called thorn, with feathery compound leaves. These do not shut out enough light to prevent the development of grasses directly under their shade, yet the annual firing of the veld prevents, except accidentally, the production of the acacias. But the seeds of this tree, whether from long usage to this burning or not, are actually hastened in their germination by the firing, and it is a common practice in that country to roast or partly boil the seeds of the tree to hasten germination. The presence of the climatically favorable environment for both trees and grassland results in the latter being the dominant type of vegetation over large tracts of the country, largely because fires destroy tree seedlings, and yet the tree seeds, by a quite extraordinary fitness for their peculiar environment, offer a measure of insurance against the total destruction of woody vegetation by the grassland.
The pampas of the Argentine have been vividly described by P. G. Lorentz, who, in writing of the drier parts of it, says: “Viewed from a distance, these grasses seem to form a close grassy covering, and the pampa presents the appearance of extensive grassy tracts whose coloring varies with the seasons: coal black in the spring, when the old grass has been burned; bright green, the color of the mature grass; finally—at flowering time—when the silvery white spikes overtop the grass, over wide tracts it seems like a rolling, waving sea of liquid silver.
“After the Gramineæ (grasses), the family of plants that is represented in the pampas by the greatest number of individuals is that of Compositæ (daisy family); usually twiggy undershrubs with inconspicuous flowers, a bright yellow Solidago (golden-rod) alone gleams out from among the others.”
Here, as in the other grasslands of the world, if a local water supply above the general requirements of the grass exists, there is always a small element of woody plants, low, thick-leaved shrubs usually, but where water is more plentiful, trees, as in the region in Natal, already mentioned, and in many other parts of the world, notably parts of Australia, China, Brazil, and many sections of the western part of the United States.
We see in grasslands a plant response to rainfall and other climatic conditions which, with a little more rainfall, or in locally wet places, always produces woody growth, either as tongues of woods in river bottoms or the parklike landscape, already mentioned. If, however, the rainfall is too low to produce even the grasses and their associated herbs, an entirely different type of vegetation usurps these still more dry regions, resulting in some of those strange plants of the deserts, among which water storage is practically universal, as is the ability to live for long periods without rainfall.
Of absolute deserts there are none in North America, for no part of it is so dry that plants of some sort do not grow, and in fact in hardly any part of the world are there regions of any considerable extent where plant life of any kind is lacking. There is a small section of northern Chile and adjacent regions on the western slopes of the Andes where nothing grows, and the traveler is met with a cheerless landscape of bare ground and sandy or stony soil. There is no record of it ever having rained in such places, and if there be only a single rain consisting of a fraction of an inch a year, a few plants, usually scraggly low herbs with thick leaves or else quick-flowering annuals will be found.
But there are hundreds of thousands of square miles of the earth’s surface where the rainfall is sufficient to produce plant life, but where it hardly exceeds five or at most six inches in a year, and in some regions is less than two inches. From several different stations in the desert area of our own Southwest the average annual rainfall is about five and a half inches. Such regions produce a desert type of vegetation and are popularly, if not properly, called deserts. The largest is of course the Sahara in Africa, but there are huge tracts in Arabia, China, Thibet, Australia, South Africa, and in some other countries where the conditions for plant life are so unfavorable that only in the better sites are plants found at all. In all deserts there are very large areas entirely without plants, due to shifting sand or other local conditions, which, added to the generally unfavorable climate, make plant life impossible.
With rainfall so low and in most of the regions the temperature so high, the vegetation must be insured against too rapid transpiration. Perhaps the best illustration of how high transpiration both in plants and in man may be in such regions is gained by a statement of D. T. MacDougal, who writes of man traveling in the desert that “the amount (of water) thrown off the skin is correspondingly great, and if the loss is not made good, thirst ensues and ten hours’ lack of water may thicken the tongue so that speech is impossible.”
Under such conditions it is not surprising that desert plants are among the most curious and weird members of the plant world. Every device both to retard transpiration and to store up water to last over a completely rainless period, may be found. In America, to which all the hundreds of different species of cactus are practically confined, we find giant forms, often covered with spines and prickles which prevent their destruction by cattle, and many others that hug the dry sandy soil with curious tortuous branches. None of them have leaves such as plants of the forest or grassland possess, for desert plants cannot afford the luxury of foliage that, because of too rapid water loss, would destroy their chance to survive. Cacti do produce tiny leaves at the ends of their joints, but as if recognizing the inhospitable world into which they are born, practically all of them drop off, so that for the great bulk of the life of most cacti only the bare branches are evident. In most kinds these branches are green, assume the functions of leaves, such as transpiration and the manufacture of food by photosynthesis.
While in cacti and in the giant cactuslike spurges of South Africa the ability to store water is tremendously developed—our giant cereus or saguaro often holding 125 gallons—most desert plants rely upon retarding transpiration for their existence. Leafless shrubs and trees whose often spiny branches are green and perform, on a much-reduced scale, the function of leaves, are among the most common characteristics of desert plants. Some, as Parkinsonia microphylla or paloverde, have tiny leaves which they put out during the spring showers, but quickly lose them as it gets hotter and drier toward midsummer. Many of the plants that do produce leaves regularly have the surface of them so shiny as to appear varnished, or so thickly coated with hairs as to simulate cotton or wool, both of which reduce transpiration. There are many plants, some of which do not even live in a desert but in a locally dry habitat, that also have the utmost development of structure to prevent transpiration. One of the most extraordinary is the vegetable sheep in New Zealand. An inhabitant of dry rocky places, its water supply, although rainfall is fairly abundant, is precarious due to drainage and the failure of the rock to prevent run-off. The different species of Raoulia, of which R. eximia is one of the best known, are admirably adapted to exist under such conditions. L. Cockayne, an authority on the flora of that island, writes of these strange plants: “Perhaps the most striking denizens of rocks are the various kinds of vegetable sheep (species of Raoulia), which form hard cushions, mostly white, but occasionally green—and of enormous size. The raoulia cushions are all constructed on the same plan. Above, the stems branch again and again, and toward their extremities are covered with small woolly leaves, packed as tightly as possible. Finally stems, leaves, and all are pressed into a dense hard convex mass, making, in the case of Raoulia eximia, an excellent and appropriate seat for a tired botanist. Within the plant is a peat made of rotting leaves and branches, which holds water like a sponge, and into which the final branchlets send roots. Thus the plant lives in great measure on its own decay, and the woody main root serves chiefly as an anchor. The vegetable sheep are not inaptly named, for at a distance a shepherd might be misled.” The genus Raoulia belongs to the daisy family, and furnishes another illustration of the remarkable diversity of this largest and probably most recent of the families of flowering plants, which appears to have originated in the Andes and now covers the world.
In Damaraland, Southwest Africa, the most remarkable desert plant was discovered years ago, growing in a sandy and stony plain where the rainfall for fourteen years has not averaged more than two and a quarter inches annually. There are sea fogs, however, upon the condensation of which upon sheets of glass the discoverers of Welwitschia mirabilis relied for some of their scanty water supply. The plant, whose woody stem is deep buried in the ground with only the top appearing above the surface, looks not unlike “the burnt crust of a loaf of bread.” To this only two large leaves are attached. These are many feet long and split into several sections which undulate over the ground very like the tentacles of an octopus. With such strange products of the desert scattered over the plains it is little wonder that Welwitschia caused a sensation only equaled by the discovery of Rafflesia in the rain forest of Sumatra.
While deserts seem to be the most unfavorable places in which plants can exist, and their very existence in many deserts is often a precarious affair, it should be kept in mind that the soil of such places is often by no means sterile. As we found in the section of this book on “How Plants Get Their Food,” water is absolutely necessary for the absorption of food through the root hairs. Where, as in an oasis in the desert, water is locally plentiful a luxuriant vegetation springs up, and one of the most fertile parts of our Southwest was transformed from a desert by irrigation. Then, too, in many deserts there is a pronounced rainy season during which there is a marvelous development of showy flowering herbs that die down as rain ceases or becomes too slight, to wait for another opportunity to make the desert blossom into often gorgeous coloring.
Rain forests, temperate forests, grassland, and deserts—all are immense developments of plant societies depending upon climatic differences for their occurrence. There are some other plant communities which also depend for their development on still other differences of climate. Two such are the vegetation of mountain tops in the tropics, and that strange tundra vegetation near the poles which lives all its life on the ice, only the roots and soil in which it grows thawing out during the brief summer. Temperature rather than rainfall is the cause of these and some other plant societies of more local occurrence.
But what of such well-known plant societies as bogs, in which peat is formed, or the plants growing along the sea beaches all over the world? These, and scores of other plant communities play their part in the distribution of plants, but nearly all of them depend not upon climate, but upon usually purely local conditions of soil. Sandy, actually nearly sterile soil, the acidity of cranberry bogs, the alkaline regions in our own West, the salt lakes and inland seas, regions below sea level, the serpentine outcrops, all the hundred and one differences which local conditions exhibit—all these have a very direct bearing upon plant distribution. It is impossible here to go into the details of the different sorts of plant societies which inhabit such specialized places, nor into the truly wonderful adaptations of certain species to peculiar conditions. But in looking at the vegetation of regions through which one travels it must never be forgotten that its general type, such as forest, or grassland, or desert, or what not, is the result of usually widely operating climatic forces, while many, often quite extensive, plant societies in the region are the result of the local environment. There is often an active struggle as to the dominance of the type dictated by the climate of the place, and the local conditions of soil that tend to nullify general response to it. On Long Island, New York, for instance, there are areas which climatically should produce dense woods of the summer forest type so general all through the Northeastern States. Actually the water-worn sands and gravels that covered the south side of the island in glacial times, are so poor in plant food, that many square miles of this region are now covered only by low scrub oaks and other plants suited to poor soils.
A final word of caution is necessary to those who see in the foregoing brief account of some of the chief causes of plant distribution an answer to questions that many of us ask about why plants or vegetation are of such and such a kind in a particular locality. It has been convenient—nay, it has been necessary—to consider these various factors one by one, but the distribution of almost no individual, and certainly of no widely spread plant community, is the result of any one of these factors operating singly.
The geological history of the region, the links with the past of the species composing the vegetation, the climate, the cooperation of various outside agencies in seed dispersal, the conflict of different species, and of different vegetation types, these and scores of other factors, operating to-day, or having operated in the past—it is all these that are reflected in the plant covering of the earth. The variety and beauty of that covering are too well understood to need further mention here. The extraordinary efforts that the plant world makes to keep all but a minute fraction of the earth clothed with some sort of vegetation we have seen in the pages just turned. No other phase of the study of plant life is so replete with interest as plant distribution. Rightly understood, it is a study, “the cultural, esthetic and practical value of which may well outweigh any other.”
We have now traced, all too briefly and with the many omissions that such a general account as this makes necessary, the broad outlines of plant life. From the architecture of their outer characteristics, which takes up the first chapter, we have gone step by step into the story of what goes on within the plant, and how it reproduces its kind. These actions or behavior of plants have resulted in many things of great practical importance as well as being of absorbing interest in themselves. What some of these results have been, we see reflected in the uses of plants to man, in the history of their development, and, most of all, in the way they are distributed over the earth to-day.
If plants are still “just plants” to most readers, this book has been written in vain. Those who have gleaned from its pages some conception of what a fundamental thing plant life is, will doubtless want more information than could be included here.
If any considerable proportion of the readers of this volume feel that they have already outgrown it, and that they have many questions about the plant world for which they will have to go to more specialized works for the answer, then this book has more than fulfilled its mission.
FOOTNOTES:
[1] After living probably as long as the Big Trees of California, the most famous dragon tree in the world was destroyed by a great storm. It has been replaced by seedlings.
[2] A process recently discovered in England for extracting the fiber of this rush by a chemical bath has greatly increased the fiber possibilities of this common rush. Heretofore it has been used only for coarse weaving of rugs and mattings in Japan. By the new process a fine fiber capable of spinning is extracted that may eventually compete with jute.
[3] Copyright, 1912. Doubleday, Page & Company.
[4] The term “indifferent” in this connection is used to signify that the plant will adapt itself to average conditions.
[5] Plants marked thus belong to the heath family and require special conditions as indicated in text.
