
Title: Evolution
Author: James A. S. Watson
Release date: July 14, 2016 [eBook #52571]
Most recently updated: October 23, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/52571
Credits: Produced by MWS, Adrian Mastronardi, Christopher Wright
and the Online Distributed Proofreading Team at
http://www.pgdp.net (This file was produced from images
generously made available by The Internet Archive)
Transcriber's Note:
The image for "The Evolution of the Horse", surrounded by dashed blue lines, is a thumbnail for a larger format image. Click the thumbnail to load the larger image.
"THROUGH THE EYE"
SERIES
EVOLUTION
"THROUGH THE EYE" SERIES
THE FIRST TWO VOLUMES
EVOLUTION. By J. A. S. Watson, B.Sc.
THE CIVILIZATION OF THE ANCIENT EGYPTIANS.
By A. Bothwell Gosse.
Other Volumes in Preparation
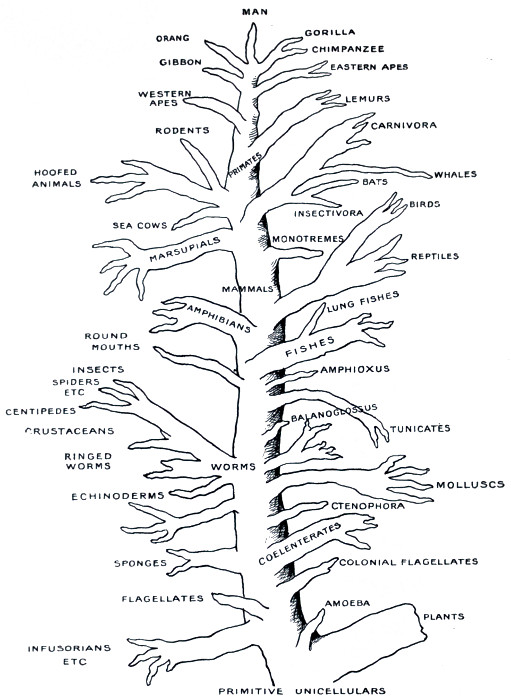
Fig. 1.—Tree illustrating the probable course of animal evolution.
BY J. A. S. WATSON, B.Sc.

THROUGH THE · EYE
Published by T. C. & E. C. JACK, Ltd.
35 & 36 PATERNOSTER ROW, LONDON, E.C.
AND AT EDINBURGH
Printed in Great Britain
| PAGE | ||
|---|---|---|
| CHAPTER I | The Evidence for Evolution | 1 |
| CHAPTER II | Unicellular and Multicellular Animals | 23 |
| CHAPTER III | The Worms and some of their Posterity | 58 |
| CHAPTER IV | The Early Vertebrates and the Fishes | 79 |
| CHAPTER V | The Conquest of the Land | 104 |
| CHAPTER VI | The Mammals and Man | 123 |
For many of the illustrations the Publishers are indebted to the Deutsche Verlags-Anstalt (from Günther's Vom Urtier zum Menschen) and Herr Wilhelm Engelmann (from Haeckel's Anthropogenie). Acknowledgment is also made to Messrs. Watts & Co., London, the Publishers of the English Edition of Haeckel's Anthropogenie, entitled The Evolution of Man.
EVOLUTION
THE EVIDENCE FOR EVOLUTION
The idea of Evolution is an old one. It is older than the Darwinian hypothesis; it is older than Lamarck, who published his particular theory in 1809, the year that Darwin was born; it is older than Buffon or Kant. In a fairly definite form it is as old as Aristotle. The Evolution idea has thus itself evolved, and is the product of many centuries of thought. Yet it was only the last generation that began to give the idea serious consideration, and it is perhaps only the present that has granted it any general measure of acceptance; and it was Darwin who wrought this change, who raised the conception of Evolution from the status of a vague speculative idea to that of a well-grounded theory, which appeals to the majority of educated minds as satisfactory and reasonable.
We do not here propose to sketch the development of the idea, either before or after Darwin; but only, in the first place, to state the grounds on which the belief in Evolution is based, and, in the second, to trace roughly the lines along which animal Evolution has proceeded. In the first few pages of this book, then, we shall endeavour to bring forward some of the evidence on which the modern Evolution theory rests.
Evolution
| First Appearance of Types. | Dominant Types. | |||||
|---|---|---|---|---|---|---|
| Modern | ... | Man | } | Post-tertiary, 1/2 per cent. |
||
| Diluvium | Man | ... | ||||
| Pliocene | ... | } | Mammals | } | Tertiary or Cænozoic, 2-1/2 per cent. |
|
| Miocene | Monkeys | |||||
| Oligocene | ... | |||||
| Eocene | Lemurs | |||||
| Cretaceous | Higher mammals | } | Reptiles | } | Secondary or Mesozoic, 11 per cent. |
|
| Jurassic | { | Birds Marsupials |
||||
| Triassic | Monotremes | |||||
| Permian | Reptiles | Amphibians | } | Primary or Palæozoic, 32 per cent. |
||
| Carboniferous | Amphibians | } | Fishes | |||
| Devonian | Lung fishes | |||||
| Silurian | Lower fishes | ... | ||||
| Cambrian | ... | ... | } | Archäen, 54 per cent. |
||
| Laurentian | ... | ... | ||||
Fig. 2.—Table showing the chronological succession of the stratified rocks, the subdivision of geological time, the approximate position of the earliest fossils of each of the main types of vertebrates, and the period of domination of each group.
As our first witness, we may call the rocks which constitute
the outer portion of the earth, and ask them to tell us what they
remember of the history of life upon the planet. We cannot hope
[2]
[3]
[4]
[5]for the whole truth from them, for their memory is imperfect;
and yet they can tell us a great number of important facts.
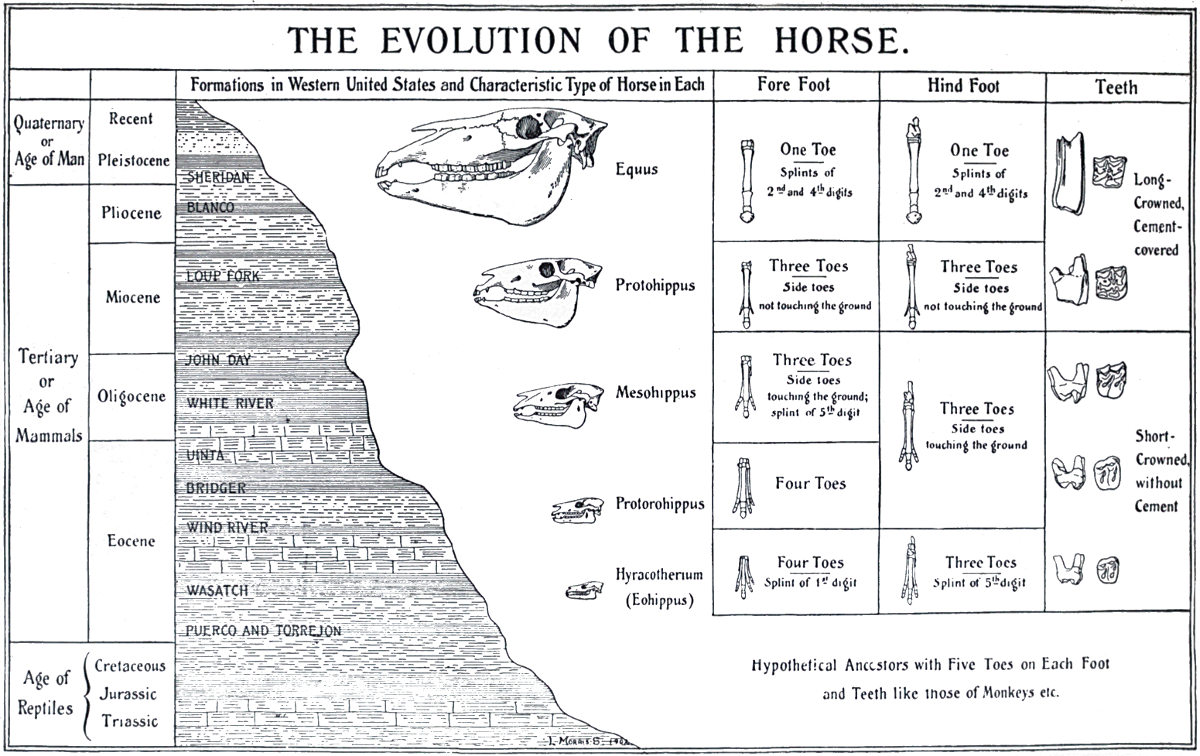
THE EVOLUTION OF THE HORSE
Fig. 3.
From The Guide to the American Museum of Natural History.
From the time when the world was sufficiently cooled for
water to condense on its surface, a continual process of unbuilding
and rebuilding of rocks has gone on. Wind and water, heat and
cold have laid their hands to the work, making sand and dust
and gravel out of solid stone, and these products of their labours
have been carried off to other places, laid down, and cemented
together into new rocks. We do not know the exact age of any
particular rock that has been made in this way, nor how long the
process has been going on. At a rough guess it may be three or
four hundreds of millions of years. The chronological succession
of the different rock formations is, however, known, and their
relative ages may be judged with considerable accuracy. Here
and there, as time went on, the body of a plant or an animal
was deposited in the sand or mud or chalk, and has remained
in the resulting rocks, in the form of a fossil, through all the
ages. If, then, we study the occurrence of fossils in this succession
of deposits, we ought to get some indications as to
the inhabitants of the globe at various stages of its history.
And if we do so, we meet unmistakable evidence that the
lower and simpler types, both of animals and of plants,
were in existence before the higher. Fig. 2 shows the facts
with regard to the vertebrates, the great upper class of the
animal kingdom. The first appearance of vertebrate fossils is
in the Upper Silurian rocks, that is to say, somewhere after
the middle of geological time. The fossils represent the
lowest group of fishes. In the next great formation, the
Devonian, fossils of two higher groups of fishes are to be found.
The first land vertebrates, the amphibians, are doubtfully represented
in the upper or newer layers of the same formation, and
definitely so in the next, the Carboniferous. Towards the end of
the Carboniferous or early in the Permian epoch, the first reptiles
appear, and in the following period, or after about three-fourths
of geological time had passed, the earliest fossils of mammals
occur. The significance of this sequence will become plainer when
[6]
[7]
[8]the differences and likenesses of these various groups are explained.
Each of these great groups in turn formed the dominant animal
population of the globe, and each in turn was superseded, although
not entirely, by the next. The mammal group itself appears
to be on the wane, overcome in the struggle for dominance by
its own latest and most remarkable member, man himself.

Fig. 4.
From The Feathered World.
Centre—Rock Doves.
1. Carrier.
2. Pouter.
3. Almond Tumbler.
4. Trumpeter.
5. Barb.
6. Fantail.
7. Jacobin.
8. Capuchin.
9. Dragoon.
10. Modena.
11. Scandaroon.
12. Turbit.
13. English Owl.
14. Nun.
15. Mottle Tumbler.
16. Saddle Tumbler.
17. English Beard.
18. Baldhead.
19. Runt.
20. Magpie.
21. Show Homer.
22. Archangel.
23. Oriental Roller.
24. Norwich Cropper.
25. Cumulet.
26. Tippler.
27. African Owl.
28. Working Homer.
29. Mane.
30. Domino.
31. Oriental Turbit.
32. Blondinette.
33. Satinette.
34. Shortfaced Antwerp.
35. Priest.
36. Fairy.
37. Frillback.
38. Swallow.
39. Suabian.
40. Fire Spot.
The broad facts in the history of living things upon the earth are, then, in accordance with the theory of Evolution. The chain of types is indeed a broken one, the gaps being many, and some of them wide. But this is readily to be understood from the comparative scarcity of fossils, and the imperfection of the geological record.
In certain particular instances, however, very complete series of fossil forms have been discovered, connecting, by small gradations, modern animals with greatly different extinct types. One of the most complete of such series has been discovered for the horse. The changes that have occurred in the evolution of this animal have been mainly in three directions—increase in size, reduction in the number of toes from the original five to the final one, and deepening of the crowns of the teeth, so as to render them capable of longer wear. From the Eohippus of early tertiary times, an animal of about the size of a fox terrier, with five toes behind, and four with the vestige of the fifth in front, there is a complete connecting series reaching up to the modern horse, with its single remaining toe and the vestiges of two others. A few of the main links in this chain are illustrated in Fig. 3. It is impossible to regard such a series without having the idea of Evolution strongly suggested to the mind.
In the second place, there is evidence for Evolution in the
fact that marked changes can and do occur in the characters of
living races of organisms. There is ample evidence, for example,
that all our modern breeds of pigeons are descended from the wild
rock-dove. How markedly some of these differ from their wild
ancestor, and among themselves, may be seen from Fig. 4.
The size of some is twice as great as that of others. The bill in
some is greatly increased in length, is almost ludicrously reduced
[9]
[10]
[11]in others. Colour, feathering, build, even the instincts and the
voice, vary enormously as between different varieties. In short,
there is hardly any obvious character that has not, in one or other
of the breeds, undergone great modification. As Darwin remarked,
any naturalist coming upon such a group of forms in nature
would have no hesitation in placing them in different species or
genera, or even perhaps in different families. Even granting
that the conditions of domestication are peculiar, we must admit
that if such large changes can occur in a few centuries, it is possible
that man has evolved from the lowest of living organisms during a
period some hundreds of thousands of times as long.

O. LATA O. LAMARCKIANA O. NANELLA.
Fig. 5.
Mutation in Oenothera lamarckiana. The parent species (in the middle) with two of the 'sports' from it.
From De Vries, The Evolution Theory. By permission of The Open Court Publishing Co.
But marked changes of type occur not only under conditions of domestication; nor is it necessary to infer the occurrence of any such changes without actual direct evidence. The formation of new types occurs in nature, and has taken place under the very eyes of scientific observers. Perhaps the most striking case that can be quoted is that of Lamarck's Evening Primrose, which, under the observation of Prof. De Vries in Amsterdam, produced some half-dozen of 'sports' which seem well entitled to rank as new species. Fig. 5 shows the parent plant and two of the new types that were produced by it. One is a dwarf in habit, the other is characterised by the greatly increased breadth of its foliage. Others showed different peculiarities. One might quote many other instances of violent changes of type—of the appearance of six-fingered children, whose peculiarity was afterwards inherited; of web-footed pigeons, and of new varieties of fruits, flowers, and vegetables. The causes of such 'sports' or mutations are unknown, but their moderately frequent occurrence is abundantly demonstrated. Such facts show, at all events, that the old conception of species as permanently fixed, unchanging types, can no longer reasonably be held.
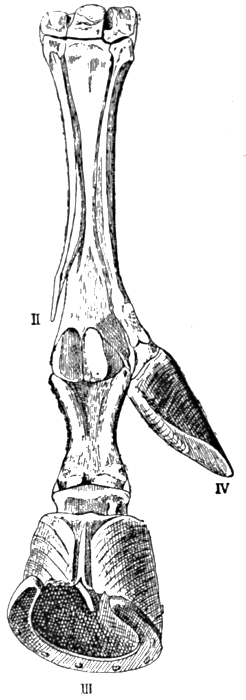
Fig. 6.—Horse's Foot, with well-developed Side Digit.
From Bateson's Materials for the Study of Variation (Macmillan).

Fig. 7.—Persistent Coccyx in Man.

Fig. 8.—Persistent Gill Slits in Man.
Not all of the abnormalities which thus suddenly appear, we know not how or wherefore, are new. Many recall characters in lower or older groups, and may reasonably be interpreted as 'reversions.' Thus the horse's leg shown in Fig. 6 bears a well-developed side toe, in place of the small vestige that is normally[12] present. Horses with this peculiarity have occurred with some frequency, probably before, and certainly since, the most famous of their kind, which Julius Cæsar rode. It seems reasonable to regard this peculiarity as a return to the old ancestral condition illustrated before, in which the side toes were well developed. The same applies to the instance of a persistent tail and persistent gill slits in man (Figs. 7 and 8), and to many[13] other instances that might be quoted. One must indeed deal carefully with such cases, for it is always difficult to say what changes are new departures, and what are returns to ancestral types. There is danger of arguing in a circle—of supposing the ancestry from the abnormality, and of terming the latter a reversion because it suggests the supposed ancestry. Nevertheless, when variations occur, suggesting characters which are believed, on other grounds, to be ancestral, they must tend to strengthen the other evidence as to the evolution of the type in question.
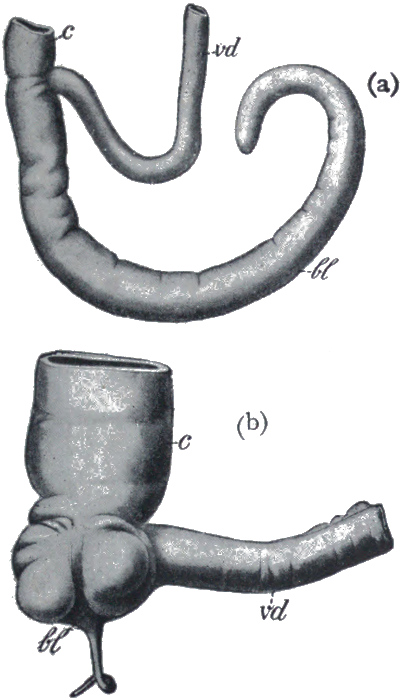
Fig. 9.
(a) The blind-gut of a kangaroo (bl), and (b) the corresponding reduced structure, the vermiform appendix in man (bl)
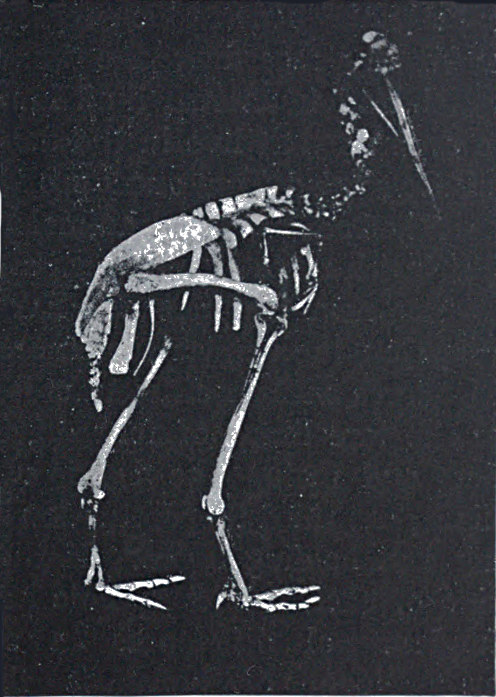
Fig. 10.—Skeleton of Cassowary, showing reduced wing-bones (a piece of black paper is placed under them).
From Dendy's Outlines of Evolutionary Biology (Constable).
Another and a very strong evidence of Evolution is to be found in what are termed vestigeal structures, two of which are illustrated in Figs. 9 and 10. They are, for the most part, obviously useless, and their occurrence has never been satisfactorily explained except by supposing them to be remnants of organs[14] that were functional in the past history of their possessors' race. The appendix of man, for instance, is not only useless, but is frequently a source of danger. But its presence is readily explained by supposing that it represents the blind-gut, which is large and functional in many of the lower animals. Again, how should we account for the presence of small functionless wing-bones in the cassowary, unless by supposing that its ancestors were accustomed to fly like ordinary birds? How should we explain the bones which represent the hind limbs of the whale, unless by regarding the whale as descended from an animal which had functional hind limbs, or the representatives of eyes in animals that live in the dark, unless by supposing that these are descended from ancestors which saw? It has been well said that the bodies of many animals are veritable antiquarian museums, filled with relics of their own ancestors.
The next argument for Evolution to which we would refer is
based on the similar structure and origin of organs or members
that have entirely different uses. In Fig. 11 are figured the bones
of the fore limbs of four different mammals, a whale, a bat, a
dog, and man. The first is used for swimming, the second for
flight, the third for locomotion on land, and the fourth as a grasping
and holding organ. If these organs had been specially designed,
each for its specific purpose, we should expect to find
fundamental differences in structure. Actually the general
arrangement of bones is the same in each case. A fact
like this points strongly to a common origin of the four types
mentioned, and to a general primitive arrangement of the bones
of the limb. This primary type, it seems natural to suppose, has
been modified for various special purposes in many different
directions, the general features remaining recognisable. Many
other cases of homology, or similarity of structure and origin,
in organs whose function is dissimilar, might be quoted. Thus
the poison gland of the poison snakes is not an organ which has
been specially developed, but is a modified portion of one of the
salivary glands. The hoof of the horse and the finger nail of man
can evidently be satisfactorily explained as modifications of a[15]
[16]
[17]
general type of terminal claw, and the scales of the scaly ant-eater
and the quills of the porcupine are only modified hairs.
The significance of facts like these, when carefully considered, is
very great.
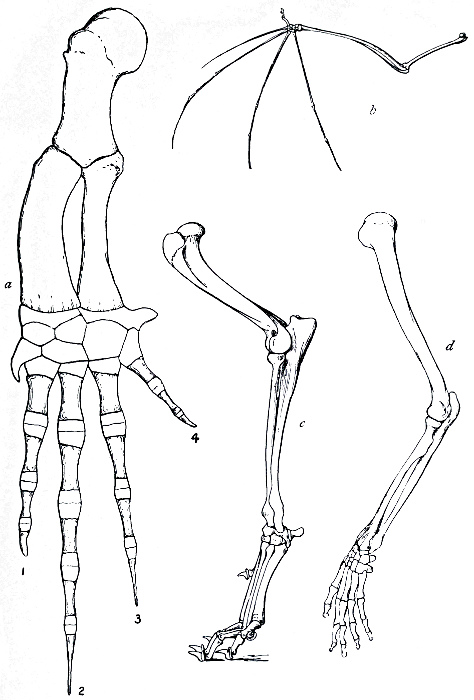
Fig. 11.—The bones of the fore limbs of (a) whale, (b) bat, (c) dog, and (d) man, showing essential similarity in arrangement.
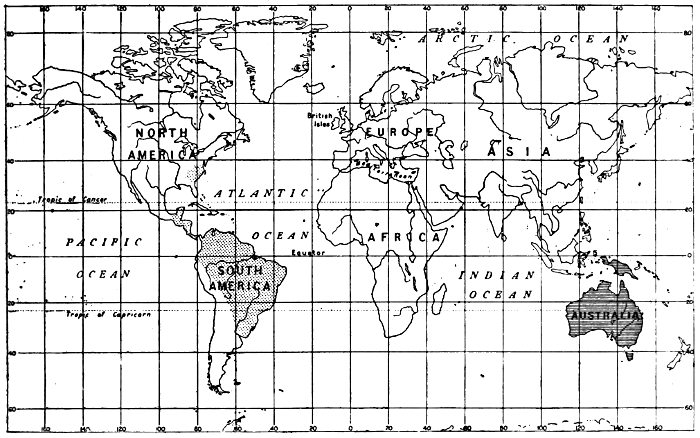
Fig. 12.—Distribution of Marsupials or pouch-bearing animals.
Australia, New Guinea, etc. 36 Genera. 144 Species.
America 3 Genera. 28 Species.
The study of the geographical distribution of animals has brought forth a great mass of facts which, considered by themselves, seem chaotic and meaningless, but which, in the light of Evolution, are full of significance. Observe, for example, the distribution of the Marsupials or pouch-bearing animals, shown on the accompanying map (Fig. 12). Australia is full of them, while they are relatively meagrely represented in a few other parts of the world. At the same time the greater and higher group of mammals was represented in Australia, at the time of its discovery, only by the bushman and his dog and a few species of mice. It is not as if the Australian environment were specially well adapted for marsupials, or specially ill-adapted for higher[18] mammals; for the sheep has proved itself splendidly adapted for the conditions, and the rabbit most inconveniently so. Why, then, this curious state of affairs? It is an undoubted fact that the marsupials are both lower in their position in the animal kingdom, and older, than the main group to which all our European mammals belong. Now it is believed that Australia was once connected by land with the Asiatic Continent, and that it was finally separated from it before the higher mammals were in existence. The great step of further progress occurred elsewhere than in Australia, and the mammals of the latter continent were left in their obsolete condition, preserved through lack of competition of that higher type which elsewhere became dominant.
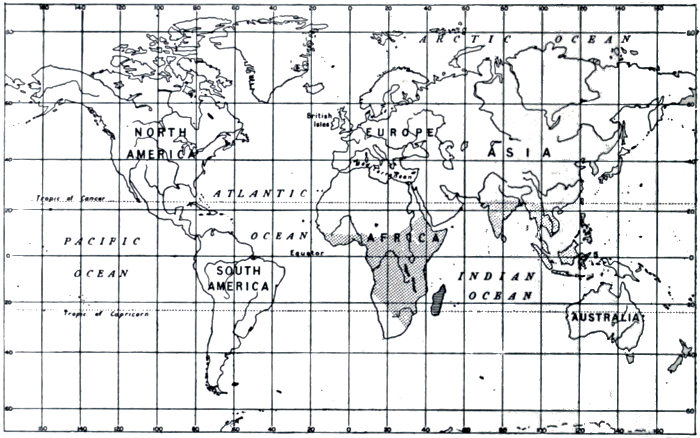
Fig. 13.—Distribution of Lemurs.
Madagascar 12 Genera. 36 Species.
Africa, India, Malay 5 Genera. 12 Species.
Madagascar offers a similar case. It abounds with forest vegetation and seems to offer a highly suitable environment for the monkey tribe. Yet there are no apes on the island. Their place is occupied by the Lemur tribe, which, there is every reason[19] to believe, is the older group of the two, and that from which the apes have sprung. It is supposed, then, that Madagascar was separated from Africa before the ape had evolved. The lemurs thenceforward were free from the competition of their more highly developed relatives, and have branched out into a great variety of types, while still remaining on a relatively low plane of intelligence and specialisation. The distribution of the Lemurs is shown in Fig. 13.
In Dr. Alfred Russel Wallace's book on Island Life there are set forth a great number of interesting facts on the subject of the animal population of islands, and many striking interpretations of these facts in the light of the Evolution theory. Coral islands, and those caused by volcanic eruptions, are peopled with inhabitants which have accidentally come thither by flight, or have been brought, for example, on floating timber by ocean currents. On the other hand, islands which represent separated fragments of continents have usually a fauna of the same general type as that of the continent of which they have formed a part. But the actual species are frequently different, and if the separation is of more ancient date, the differences are still more marked. The fact of this divergence of an isolated animal population from that from which it has originated is sufficiently striking, and would remain an inexplicable problem, were we without an Evolution theory. According to the Evolution hypothesis, however, the restricted and somewhat special environment favours a modification of the original types with which the island was provided, and a satisfactory explanation is offered.
Finally, we may mention the evidence that has been gathered
from the study of embryology and development. It has been
stated, in a metaphor which is perhaps more clever than it is
exact, that every animal climbs up its own ancestral tree; and
while it would be absurd to say, for instance, that a mammalian
embryo resembles successively a fish, an amphibian, and a reptile,
still many of the broad facts in the evolution of a race seem to
be repeated, in a more or less blurred and indistinct fashion, in
the development of the individual. Thus, for example, gill-slits
[20]
[21]
[22]and a tail are possessed in common by the embryos of all higher
animals, only afterwards to disappear in those types in which
the adult animal is without these structures. The heart of the
mammal or bird is at first simple, then two chambered like that
of a fish, then three chambered like an amphibian's, and finally
four chambered. Some of the main phases in the development
of the rabbit and of man are shown in Figs. 14 and 15 respectively.
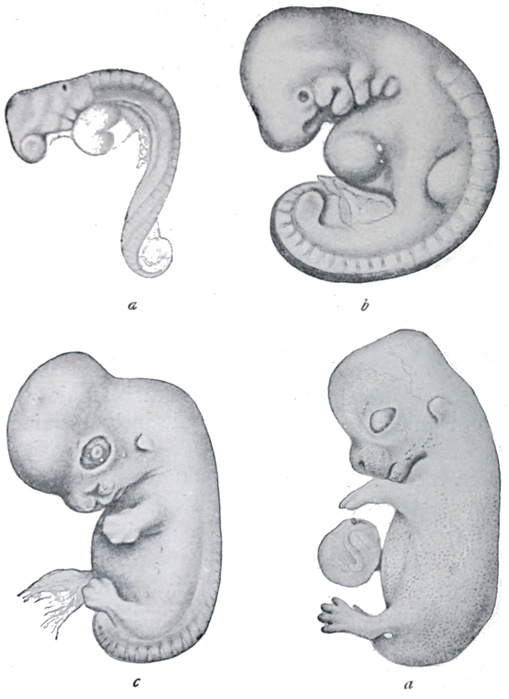
Fig. 14.—Stages in development of embryo of rabbit.
a, 10 days; b, 11 days; c, 15 days; d, 17 days old.
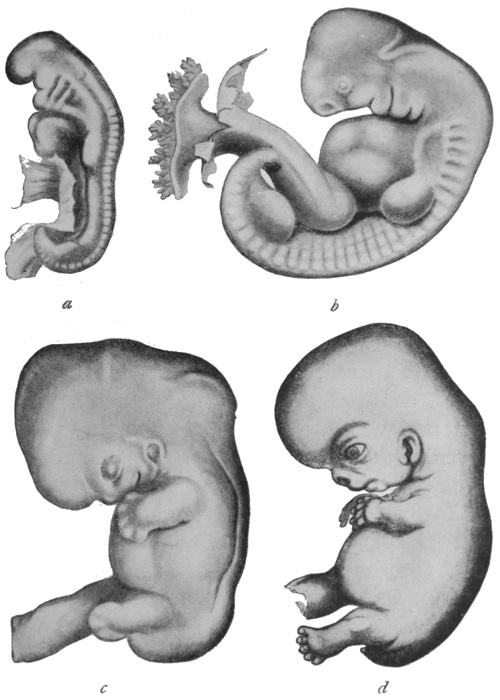
Fig. 15.—Stages in development of human embryo.
a, 18-21 days; b, 27-30 days; c, 35 days; d, 52-54 days old.
The young flat-fish is like an ordinary member of the fish tribe, with an eye on either side of its head, and its body built on the ordinary symmetrical lines. It is only later, when it begins habitually to be upon one side on the sea bottom, that the eye from the under side wanders round to the opposite aspect beside its fellow, and the upper side becomes pigmented, while the lower remains white.
In similar fashion a primitive form of kidney is, as it were, sketched in, in the development of the higher animals, only to be erased at a later stage and replaced by a better form. The human child has a complete body covering of hair, which disappears soon after birth. In these and many more instances, one cannot avoid the impression that the organism has not been specially designed for what it finally comes to be. It cannot forget, and must needs repeat, or so it seems, some considerable part of the history of its race.
Manifestly, then, all this evidence, gleaned from many different sources, points to a common origin of living things, and to the gradual evolution of the higher from the lower types. It may also be said that there is no scientific evidence against such a view.
UNICELLULAR AND MULTICELLULAR ANIMALS
We must now turn to the main project of this book, which is to attempt to trace out the lines along which animal Evolution has proceeded, with special reference to that particular line which leads up to man. Indeed, we shall have to stick somewhat closely to this one main highway, and can but barely pause to glance along the numerous branch roads, interesting though the travelling there might be.
It is perhaps necessary to say, at the outset, that the history of the Evolution of man cannot be written as a plain, matter-of-fact tale. Many portions of this history are tolerably well understood, but there are other periods, in some of which notable steps of progress were made, of which no record has ever been discovered. We must therefore expect occasionally to be reduced to speculation, and here and there to meet with controversy and with opposing theories.
It is not proposed here to enter into any full discussion as to the origin of life. It may shortly be said that in the existing state of knowledge, no very definite theory is possible. We know that life is associated with a jelly-like or semi-fluid substance called protoplasm, which consists of a very complex mixture of albuminoids. These albuminoids are continually undergoing changes and interactions of a complex kind, the sum total of which constitutes life. Many of these reactions have been reproduced, or imitated, artificially, and have been shown to be purely chemical or physical. The chemical nature of the albuminoids is indeed so complex that some considerable time must yet elapse before it can be completely investigated; and until such time it is obvious[24] that we cannot hope for any very definite conceptions as to the nature of life. Broadly, however, the majority of physiologists regard life as a highly intricate series of purely physical and chemical processes, and if such a view be accepted, there is no insuperable objection to a general theory of the origin of living from non-living matter. By this it is not intended to imply that the manufacture of living matter is an immediate possibility; for even according to such a theory as we have indicated, it would be supposed that living substance came into being by a very slow process of Evolution, which it is hardly conceivable could ever be repeated in the laboratory. Knowing, as we do, that there was a time when no life existed upon the earth, and believing, as there is good reason to believe, that there is no fundamental distinction between living processes and ordinary chemical and physical reactions, we may logically regard life itself as a product of a natural process of Evolution.
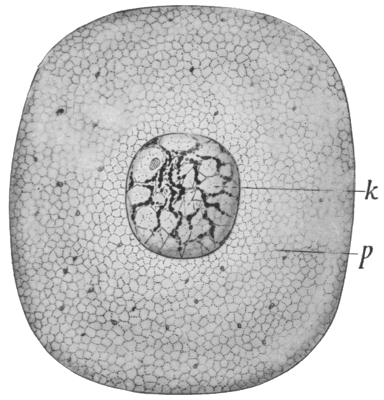
Fig. 16.—A typical cell (greatly magnified).
(k) Nucleus; (p) cell protoplasm.
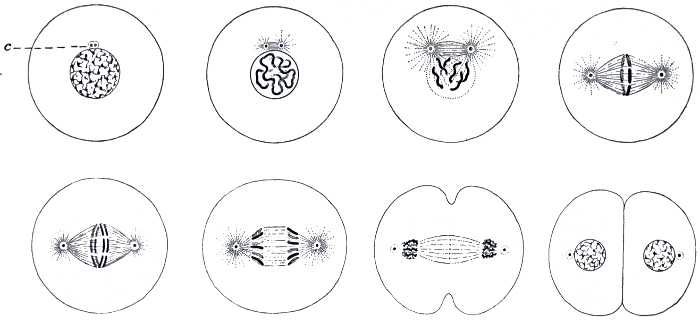
Fig. 17.—The process of cell division.
c, The centrosome, the body which divides first, and which controls the division of the nucleus.
To begin at the beginning of our tale, we may ask ourselves what are the lowest, simplest, living things that are known. The question does not admit of any very definite answer. For as we look around among a number of the most simple forms, we find ourselves handicapped in our attempt to judge between them, by a lack of knowledge of their nature. We come upon organisms so small that they appear, even under the most powerful microscope, only as the tiniest specks; whose size is to be measured in hundredths of thousandths of an inch. We even find good evidence that living things exist which we are unable, in any manner whatsoever, to see. Among the smallest known forms, and also among some of the larger, we find organisms that we can only describe as practically structureless, that appear as specks of almost homogeneous protoplasm; but it seems reasonable to suppose that this appearance is due rather to our imperfect observation than to an actual absence of differentiation.
It is certain, however, that the lowest of the great groups is that of the one-celled organisms. As all the higher types are built up of large numbers of cells, essentially similar to those which constitute the unicellular forms, it is important that we should know something of the nature of this organic unit. A typical cell is illustrated in Fig. 16. It consists of a mass of protoplasm, with a distinctly differentiated portion called the nucleus. The function of the nucleus is that of directing and controlling the activities of the cell; if it is removed, the remaining portion of the cell soon dies; while, on the other hand, a small portion of the cell, if it contains the nucleus, may frequently live, and build up new protoplasm to replace what was lost. Cells are formed only from previously existing cells, by a process of division, which is usually simply one of halving. This process is begun in the nucleus; it undergoes a complex rearrangement of its parts, the object of which appears to be to insure an absolute equality in the halves, and finally divides in two. The bulk of the protoplasm then separates into two portions, a portion remaining round each of the nuclei. The process of cell division is illustrated in Fig. 17.
Now it is a somewhat remarkable fact that we do not know whether or not all the humbler forms of life possess a nucleus. It was formerly believed that a considerable number of one-celled organisms were devoid of the body in question, but in most of such it has been shown that nuclear matter is present, though it may be distributed, in small portions, throughout the cell. If organisms do exist which consist of a cell without a nucleus, we must regard them as the simplest of living things. In any case, the formation of a nucleus, a process by which a kind of central government was formed, was probably one of the great early steps of Evolution.
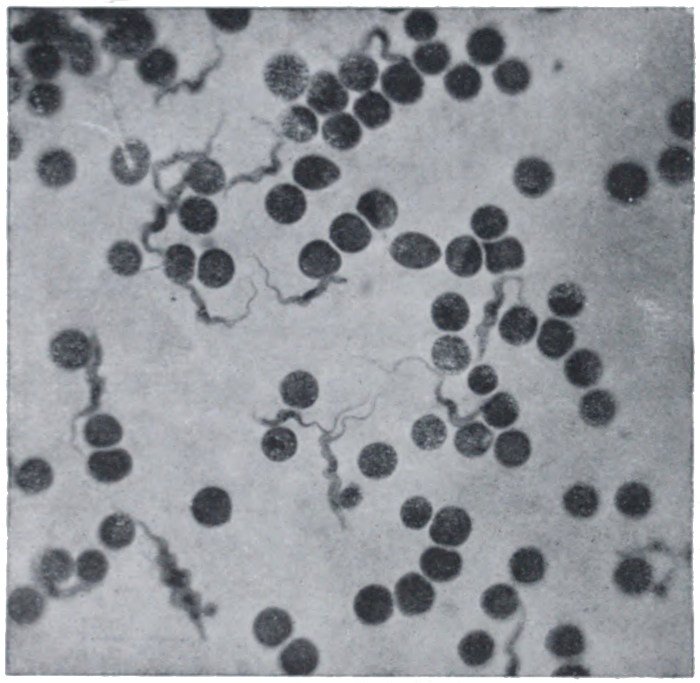
Fig. 18.—Organism of sleeping sickness in blood. The round bodies are red blood corpuscles.
Photo: F. Martin Duncan.
The life-history of an ordinary one-celled organism may be briefly summed up. It absorbs nourishment and energy, adds to its substance until it reaches a certain fairly definite size, and then divides in two, the halves separating, and going each its own way. In the world of one-celled organisms there is no 'death from natural causes.' The individual is potentially immortal, except in so far as we may regard the individual life as ceasing when division takes place. Death occurs only, as we say, accidentally—for example, from starvation or from the attacks of enemies. A number of simple unicellular organisms are shown in Figs. 18, 19, and 20.
The reader will have observed that we have referred to the group under consideration in general terms, and without endeavouring to classify its members as plants or animals. And indeed it is impossible to carry this great distinction down to the lowest group of the organic world. This stands below the first great forking of the tree of life; its members remain in what has[29] been described as a condition of 'chronic indecision,' neither clearly vegetable nor definitely animal. But very soon, in the march of progress, the forking of the roads was reached, and whosoever was bent on journeying farther had perforce to make the choice. We must here briefly consider what this choice was, and wherein the fundamental distinction between a plant and an animal consists; for, strange as the statement may seem, the basis of this distinction is by no means generally appreciated.

Fig. 19.—The bacillus of bubonic plague (× 1000).
Photo: F. Martin Duncan.
The typical plant lives by absorbing carbon dioxide gas, water, and mineral salts from the surrounding media. These substances, by means of energy which it gathers from the rays of the sun, the plant builds up into organic substances, to be used in the maintenance of life, and for growth and reproduction. This process of chemical construction occurs only in the green, exposed parts of the plant, and indeed can occur only in the presence of chlorophyll, the green colouring matter of the leaves.
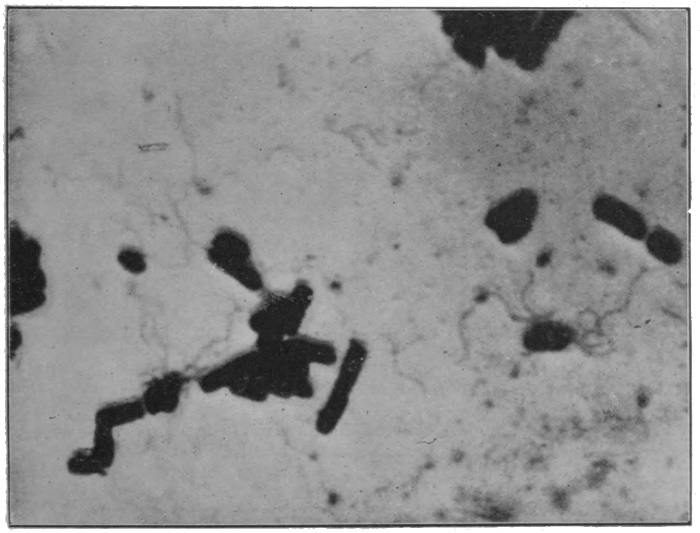
Fig. 20.—The bacillus of typhoid (× 2500 diameters).
Photo: F. Martin Duncan.
The animal, on the other hand, lives by appropriating, either directly or indirectly, what the plant has produced. All flesh is indeed grass, in a different sense from that originally intended by the statement. It is this essential difference which lies at the[30] root of all the plain and obvious distinctions between animals and plants. The plant has neither the necessity to go forth in search of its food materials, which nature brings to it, nor has it to spare of its painfully collected energy for the labour of locomotion. Hence it remains stationary. The animal must of necessity go to seek its more elaborate fare, therefore it moves. Moreover, to be successful in its search, the animal obviously requires a nervous system to direct and control its movements, which system, except in the simplest and crudest forms, is absent from the plant. In the main, then, the plant builds up and saves, the animal breaks down and spends. The plant is the producer, the animal the consumer.
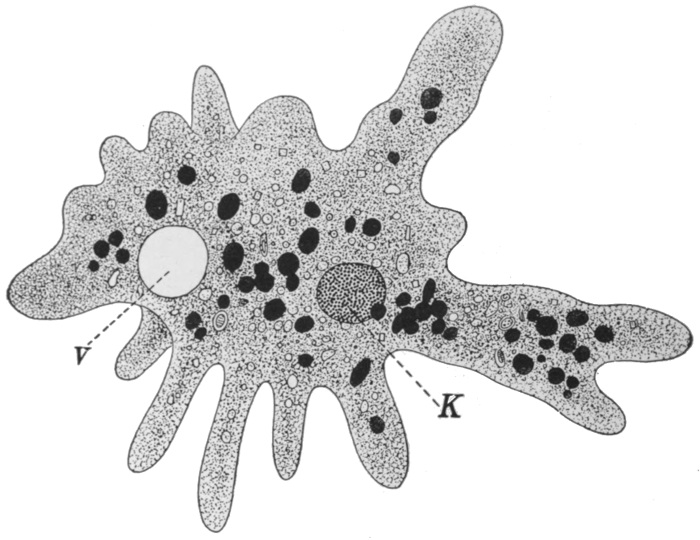
Fig. 21.—Amœba.
K, Nucleus; V, contractile vacuole.
Turning now to those of the lower organisms that are somewhat more definitely animal in nature, we may describe the common Amœba. Microscopic in size, this creature consists of a speck of semi-liquid protoplasm, which is irregular and ever-changing in[31] shape. It is continually pushing out finger-like projections from various parts of its surface, feeling, in a dim, vague way, for its food. It moves, if but slowly, by withdrawing its substance in one direction and pouring it forth in another. It indulges in such fare as bacteria or particles of dead organic matter and feeds by the simple method of surrounding the food particle with its protoplasm, and gradually digesting and absorbing whatever it contains of nutriment. Undigested portions are simply left behind as the creature moves on. The waste products are drained into a simple cavity in the protoplasm called the contractile vacuole, which empties itself periodically to the outside. The Amœba reproduces by the ordinary process of simple fission, illustrated, with the creature in its ordinary condition, in Figs. 21 and 22.
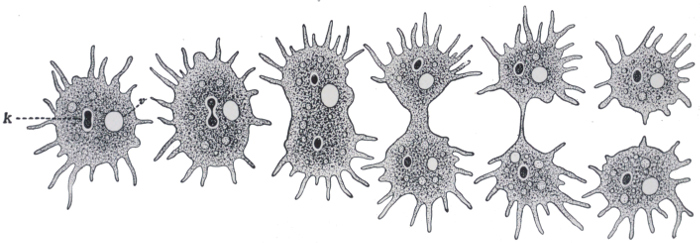
Fig. 22—Stages in division of Amœba.
K, nucleus.
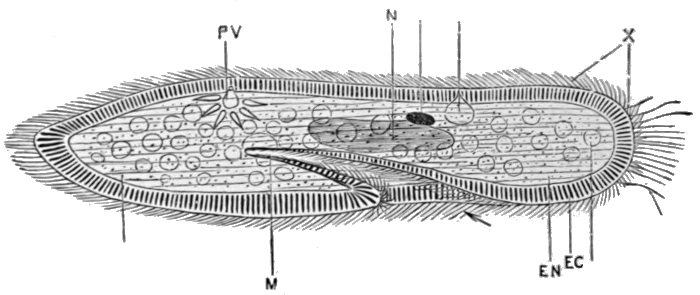
Fig. 23.—Paramœcium.
EC, Denser outer layer; EN, inner protoplasm; N, nucleus; PV, contractile vacuole; M, mouth; X, cilia.
From Marshall and Hurst's Practical Zoology (Smith, Elder & Co.).
Somewhat higher than the Amœba, and apparently along the main line of progress, stands the group which includes the slipper[32] animalcule, Paramœcium, shown in Fig. 23. This creature, barely visible to the naked eye, is found in pools of water, or, for example, in drops of rain or dew on plants, and it can generally be obtained in great numbers by soaking a little hay in water for a day or two. It has, as may be seen from the illustration, an elongated shape, with a depression, the mouth, about the middle of one side. The progress made good from the stage of the Amœba has been largely in the direction of a more efficient method of locomotion. Instead of crawling, with painful slowness, the Paramœcium swims freely and rapidly by means of the numerous whip-like projections or cilia which cover it, and with which it lashes the water. An advance is also to be recognised in the fact that the organism is surrounded by a dense outer wall; and that its shape is consequently fixed. Hence also the Paramœcium cannot take in food at any part of its surface, as the Amœba can, but only through the special depression already mentioned. Excretion is carried on in the same manner as in the Amœba. The Paramœcium is a water animal, yet it can resist drying, and remain alive in the absence of water, for a long period. This it accomplishes by becoming encysted, that is, by contracting into a ball and surrounding itself with a resistant shell, from which it can emerge when suitable conditions for active life return. It is worth passing notice that there exist a number of forms occupying a position intermediate between the two types which we have[33] described, and indicating that the second has, in all probability, been derived from the first. One of these is shown on Fig. 24.
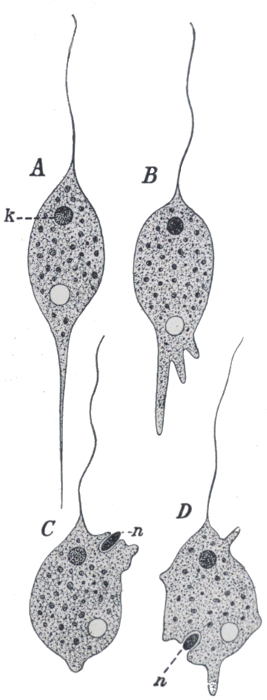
Fig. 24.—Cercomonas, a form intermediate between the crawling Amœba type and the free-swimming Paramœcium type.
There is another interesting fact in connection with Paramœcium. Under natural conditions, division and redivision continue in the ordinary way for a large and indefinite number of generations. But very occasionally, a process known as conjugation occurs. Two individuals lay themselves side by side, and partially unite; they exchange portions of their nuclear substance, and finally separate again, simple division afterwards proceeding as before. Conjugation, although distinctly different from the ordinary process of sexual reproduction, appears to serve the same purpose. Until quite lately its meaning, and that of the process of sexual reproduction in general, seemed to bid fair to remain a perpetual puzzle to biologists. But at last we seem to be approaching the solution. The characters of a species are determined, it is tolerably certain, by the constitution of the cell nucleus, and accordingly as this varies from one individual to another, so the characters of the individuals will vary. Now, if simple division were to continue indefinitely, successive generations would be produced on the same plan, and the racial characters would in the main remain constant. But conditions of life vary from time to time and from place to place, and the particular type which succeeds best under one set of circumstances may be ill adapted for another. It is therefore an advantage to a race to be capable of variation. And the process[34] of sexual reproduction, by continually bringing about a mixture of the nuclear substance, ensures the regular production of a variety of types. Of these various combinations of characters the few that are suited to the prevailing conditions will, for the time being, constitute the dominant types. When conditions change, fresh types will be available to replace them. The process of conjugation is illustrated in Fig. 25.
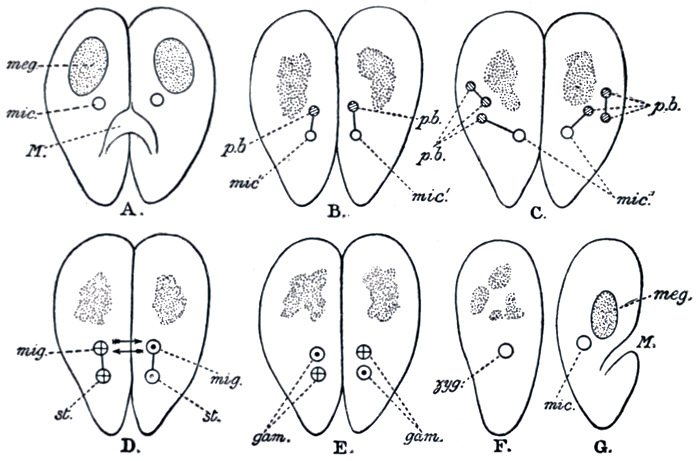
Fig. 25.—Stages in conjugation of Paramœcium.
meg., The meganucleus; mic., the micronucleus, which divides, and half of which is exchanged; p.b., Polar bodies, which the micronucleus throws off, and which disappear.
From Dendy's Outlines of Evolutionary Biology (Constable).
There are many groups of one-celled animals other than those typified in the Amœba and the Paramœcium, but they do not appear to have any significance so far as the descent of the higher animals is concerned, and they therefore do not immediately concern us.
We have already mentioned that water is the life medium of the slipper animalcule. It was destined to remain the natural element, both of animals and of plants, throughout many subsequent stages of progress. The reason of this is not far to seek. Active protoplasm consists to the extent of about three-fourths[35] of water, and a plentiful supply of this is one of the essentials for the continuance of active life. Therefore, before the conquest of the dry land could be accomplished, devices had to be evolved both for maintaining and for conserving the water supply—roots in the plant; in the animal, some method of locomotion by land or air, so that water could be frequently reached; protection against evaporation, in the form of a skin, in both; and numerous other special devices. Add to this the fact that locomotion on land presents much greater difficulties than that in water, and it will hardly occasion surprise that vast ages were yet to be required before the Evolution process could produce a land animal.
A striking analogy may be drawn between animal Evolution, from this point onwards, and social Evolution. In the latter case we begin with men, brought by a slow process of Evolution to a high state of individual perfection, living in a state of savage individualism. Each thinks and acts for himself, provides his own food, raiment, and dwelling; constitutes his own standing army and police. From this condition of affairs there has gradually been developed the modern social arrangement, by which each individual helps to carry out some distinctly special part of work for the community—be it wheat-growing, cloth-weaving, bricklaying, or the arresting of burglars—and trusts to the community for his requirements in all other directions. These requirements themselves have so multiplied during the course of social Evolution that innumerable forms of activity have sprung up between those occupations which provide the original necessities of life. The essence of the whole process has been co-operation and the division of labour.
In the story of animal Evolution we have reached a point where a highly perfected individual cell has been produced, which carries out for itself, and for itself alone, all the activities of life. From now onwards, co-operation and specialisation are the watchwords of progress. There is a clubbing together, first of a few cells, then of hundreds, and finally of millions upon millions, to form the bodies corporate which we recognise as individual higher animals. Division and distribution, subdivision and further[36] distribution of the life activities proceed at the same time, until we reach the condition prevailing in the higher animals, where the degree of specialisation almost passes conception. In such there is, to begin with, a vast frontier army of skin cells, occupied in securing peace, as far as possible, for the industries that go on within. There are directors and controllers of these industries—the brain cells—with a myriad of workers under their guidance, and a great and complex telegraph system between. The workers themselves are of all descriptions—common labourers like the cells of the muscles; transport workers like those of the circulatory system; skilled factory hands like those of the glands; even scavengers in the shape of the sweat gland and kidney cells. Nay, there is even a numerous police force, of white blood corpuscles, which patrol everywhere, arresting intruders and disposing of them by the summary method of swallowing them whole.
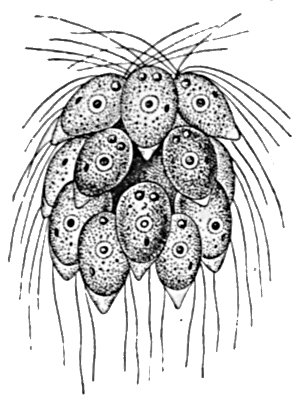
Fig 26.—Spondylomorum, a small colony of flagellates.

Fig. 27.—Magosphæra, a colonial flagellate.
Our information regarding the early history of this co-operative movement is fragmentary and incomplete, for only an odd species or so seems to survive of the group which we regard as the earliest of multicellular animals. In certain forms which are still essentially unicellular, such as the Spondylomorum shown in Fig. 26, there is a tendency to form smaller or larger cell colonies. When the individual cell divides, the two daughter cells do not separate, but remain somewhat loosely attached to each other, and the process of division without separation continues until a considerable group is produced. From this colony occasional individuals break away and proceed to form new colonies. From such[37] a type it is a comparatively easy step to the Magosphæra described by Haeckel and illustrated in Fig. 27. This consists of a simple ball of ciliated cells, which reproduces by the occasional breaking away of an individual member, which divides and redivides until a new sphere is produced. Unfortunately this animal has only once been discovered, and many hold that it has not been sufficiently investigated. No other of the same type is known.
If we turn to the plant kingdom, however, we find a comparatively common organism of somewhat similar form. This is the Volvox, a plant which consists of some thousands of cells, and reaches a size of about a pin-head. It has the form of a hollow sphere, the wall of which is one cell thick, and the cavity of which contains only water. The cells bear whip-like cilia on their outer surface, by whose means the organism is able to move, swimming by a rotary motion round a definite axis. The individual cells are separated by layers of a gelatinous substance, through which, however, pass connecting strands of protoplasm. The cells, of course, contain the green colouring matter common to plants in general. Distributed among the ordinary cells occur a few that are distinguished by their larger size, and by the fact that they lack cilia. These are the special reproductive members of the colony. When the Volvox reaches maturity, these cells begin to divide, and form new growths which take the form of hollow sacs, which project into the cavity of the parent sphere. Later they separate from the wall of the parent, and begin to move about, in the internal cavity, by means of the cilia which they have developed. Finally the parent breaks up and dies, and the progeny are set free to commence life for themselves.
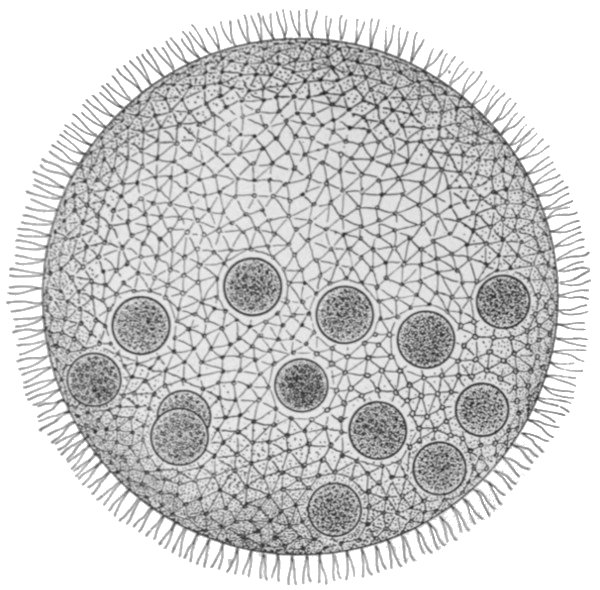
Fig. 28.—Volvox. A female, showing egg cells.
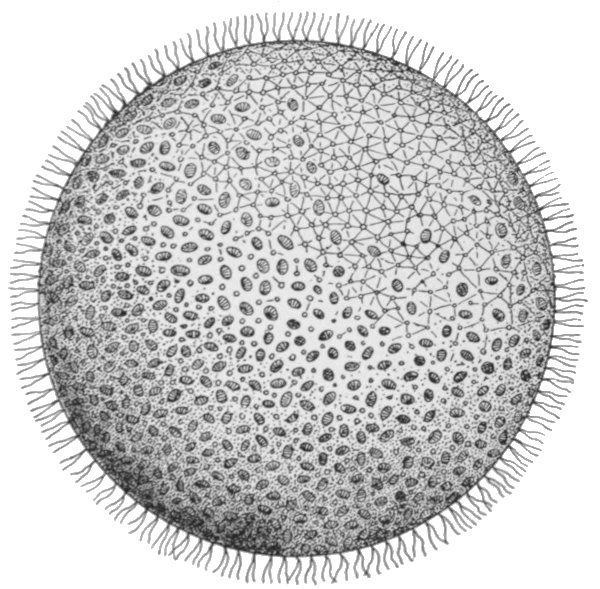
Fig. 29.—Volvox. Male, showing packets of sperm cells.
The fundamental importance of this type is that we have already a division of the life activities. The majority of the cells are concerned in the nutrition of the individual as a whole. These ultimately perish. A minority, however, are fed and protected by them, and these in return secure the perpetuation of the race. This division into a mortal 'body' portion and an immortal reproductive portion is the first and most important division of the life activities, whether in the animal or in the[38] plant kingdoms. The body cells, modified in various directions for their special purposes, could not, and do not, reproduce complete new individuals. Therefore a generalised type of cell is maintained for the express purpose of the propagation of the race. It is to be observed, now, that the process of reproduction in Volvox is not always such as we have described. Sometimes the reproductive cells are of two kinds. The one type divides into a great number of small ciliated cells, which escape separately and directly to the outside of the sphere, and swim away. These free-swimming individuals do not form new colonies, but seek out the reproductive cells of the other type, which latter still form part of the organism which has produced them. One of the free-swimming cells enters each of those of the other kind, and the nuclei of the two merge into one. The cell so produced, after a longer or shorter rest period, commences to divide and redivide[39] in the manner already described, forming a new colony. The process that we have described is that of sexual reproduction, and its essential features are the same as in Volvox throughout the whole animal kingdom. The small free-swimming cells are the male reproductive bodies or sperms, the others are the female or egg cells. The union of the two produces the fertilised egg, and the process of union is termed fertilisation. In Volvox, the male and female elements are sometimes produced by the same individual, at other times by different ones. Separation of the sexes is no necessary accompaniment of the process of sexual reproduction, and indeed it is only in the higher groups of animals that separate sexes are the rule. The various conditions in Volvox are illustrated in Figs. 28, 29, and 30.
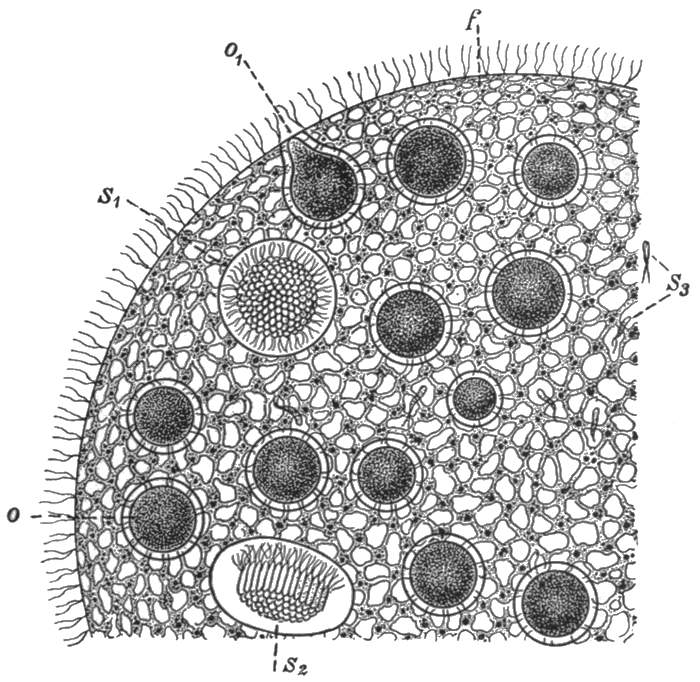
Fig. 30.—Volvox. Portion of a hermaphrodite individual, showing egg cells (O, O1), and sperms (S1 S2 S3).
The next great groups of animals are, on the one hand, that of the sponges, and, on the other, that which includes the sea-[40]anemones, jelly-fishes, corals, etc. At first sight their structure seems vastly different to that of the Volvox, from some form similar to which they have probably been derived. The evidence obtained from the study of their individual development, however, strongly suggests a process by which we suppose that they evolved from Volvox-like ancestors. We shall therefore briefly describe the earlier stages of the development of a coral. The sexually produced individual starts life as a single cell, the fertilised egg. This divides and redivides until a hollow ball of cells is produced, which cells, like those of the Volvox, bear cilia. Although simply spherical in shape, the creature moves by rotating round a definite axis, like a planet. Moreover, nutriment is absorbed not by any or every part of the surface, but only by a small area round the lower pole. Now as development proceeds, the cells at this pole divide more rapidly than the rest, with the natural result that the ball begins to get out of shape. The distended portion, however, develops to the inside, so that one part of the sphere is, as it were, pushed into the other. When this process has been completed, the original internal cavity is almost entirely eliminated, and a form is produced which resembles a double-walled flask or vase. Such a form may be taken as the fundamental architectural type of the groups that we are now to consider. The meaning of this further step of Evolution is again specialisation. The inner layer of cells takes on the functions of digestion and absorption of food, there having been evolved, in fact, the simplest possible form of mouth and stomach. Such other functions as those of locomotion, protection, and support are exercised by the outer layer. This process is illustrated in Fig. 31.
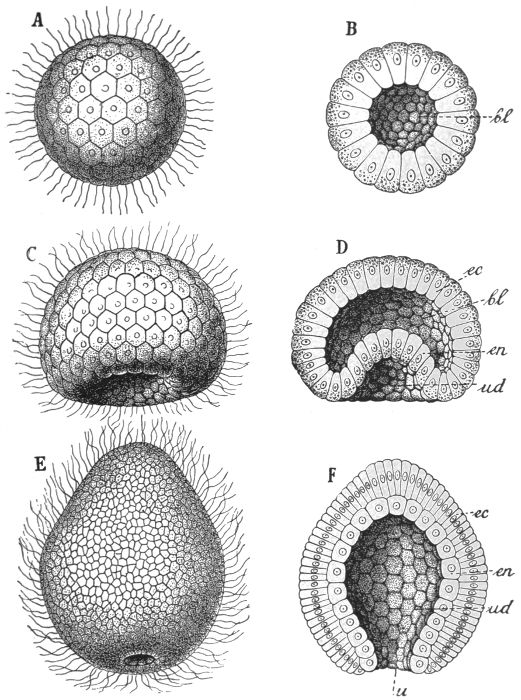
Fig. 31.—Process of gastrulation in a coral.
A, B, Blastula, or simple hollow ball; C, D, intermediate condition; E, F, gastrula, or double-walled flask condition.
But there is no known type of animal which, in its adult form,
shows quite the simple structure that we have described. Perhaps
the nearest approach is to be found in the lower sponges, in
which two modifications of the original plan have already been
introduced. In the first place, the creature is sedentary, being
fixed, in an inverted position, to some solid basis. It has, so to
speak, ceased to be a hunter, and is become a fisher. Secondly,[41]
[42]
[43]
its wall is pierced in many places, so as to permit of a freer circulation,
through the digestive cavity, of the water which contains
the food material. The water passes in through these
numerous perforations, and out through the main central opening
or 'mouth.' The sponges do not appear to represent a stage
in the main line of Evolution, but lead us almost immediately into
a cul-de-sac. We therefore cannot pause to describe fully the
many peculiar and interesting developments which occurred in
the group. An ordinary 'sponge,' by the way, bears the same
relation to the creature which produces it as does a 'coral' to
the coral animal. It represents, that is to
say, the skeletons of a large colony of individuals.
The structure of a sponge is shown
in Fig. 32.
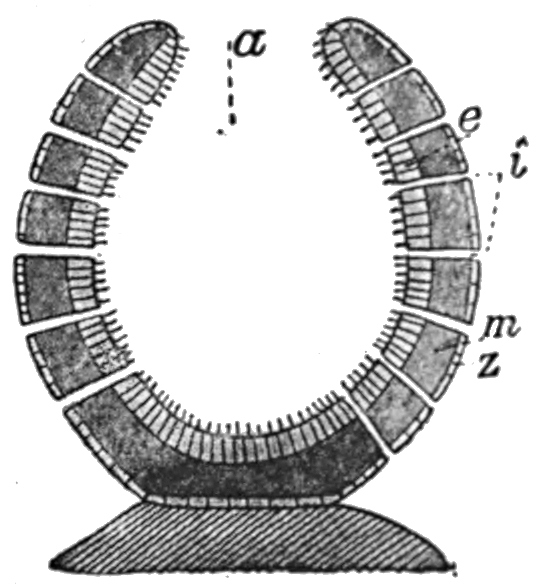
Fig. 32.—Diagrammatic section of lower sponge.
e, inner cell layer.
m, middle jelly-like layer.
z, outer cell layer.
a, digestive cavity.
i, perforations in the wall.
The other great group of primitive multicellular animals is that of the Cœlenterata, and as an example of the most primitive of these we may take the common freshwater Hydra. The Hydra reaches a length of nearly half an inch, and is to be found attached to water-weed and the like in streams. It consists of a hollow tube-shaped body which is fixed by the so-called 'foot.' Two layers of cells form the wall of this tube, these being separated by a thin membrane of gelatinous material. At the upper end is the mouth, which leads immediately into the internal cavity or stomach. The mouth is surrounded by a ring of from six to eight tentacles, which are outgrowths of both cell layers. The cells of the inner layer are large, and bear cilia that protrude into the internal cavity. Their functions are those of digestion and absorption. Part of the protoplasm of the outer cells is modified into a fibrous, contractile substance, which represents the beginnings of muscle tissue. The outer layer also forms a protective skin-like covering. In the outer layer also occur a large number of stinging cells, each of which has a complex mechanism for injecting a fluid poison into any creature which should happen[44] to come in contact with them. These 'nettle cells' occur in much greater numbers in the tentacles than elsewhere, and here they are brought into play against the animals, such as minute Crustaceans, which form the Hydra's prey. Coming in contact with the tentacles, such creatures are caught, paralysed by means of the stinging cells, and are gradually transferred into the mouth by a slow contraction of the tentacles. The Hydra reproduces, for the most part, by a simple process of budding. Small lateral outgrowths are formed, which gradually develop mouth and tentacles of their own. Ultimately these separate and are carried off by the water, later to settle down and become attached to some fixed object. Sometimes, however, sexual reproduction occurs. The reproductive cells are produced, male and female on the same individual, among the ordinary cells of the outer layer. These are set free, fertilisation occurs in the water, and the egg develops in the same manner as that of the coral. The Hydra is able, by means of the fibrous protoplasm of its outer cells, to show well-marked movements. It can bend its body in this direction or that, can contract its whole body into a small oval mass, and is even able, by performing a number of slow somersaults, to change its position. The structure and the methods of reproduction in Hydra will be readily understood from the illustrations of the creature on Figs. 33 and 34.
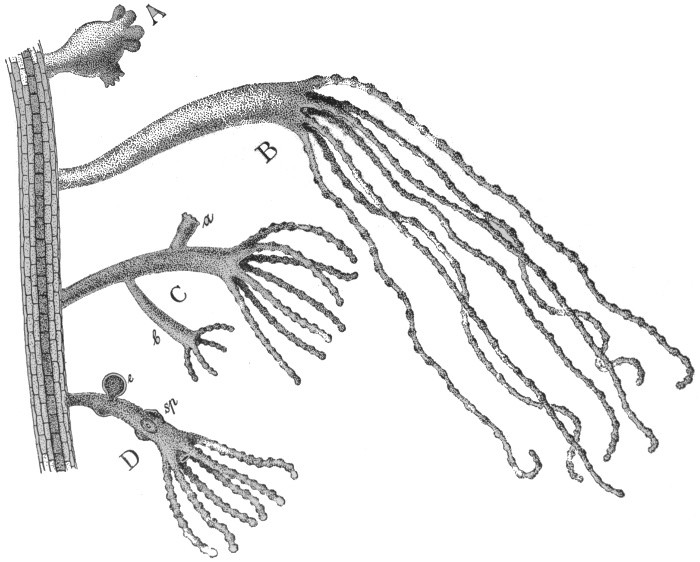
Fig. 33.—Specimens of Hydra on green water-weed.
A, Contracted; B, extended; C, specimen with vegetative buds; D, specimen with sex cells; sp, sperm cells; e, egg.
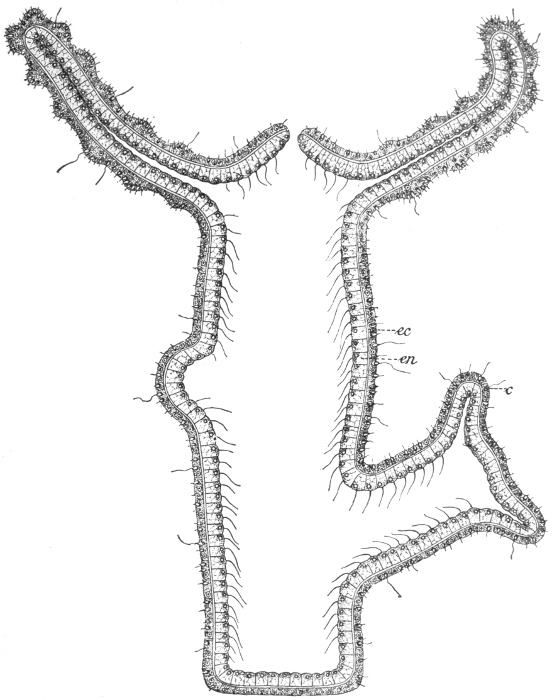
Fig. 34.—Diagrammatic section of Hydra.
en, Inner cell layer; ec, outer cell layer; c, nettle cell.
If now we make a brief general survey of the group to which
the Hydra belongs, we find in it two somewhat strikingly different
types. On the one hand are sedentary forms that resemble, in a
general way, the Hydra; that consist of a tube-shaped body, with
the mouth, surrounded by a ring of tentacles, at the upper end.
The sea-anemones and corals are examples of this type, in which,
however, the structure shows various complexities as compared
with that of the Hydra, which complexities we cannot here pause
to describe. On the other hand is the well-known Medusa form,
of which the common jelly-fish is a typical example. This
creature, as is well known, is mushroom shaped, with tentacles
round the edge. The mouth is in the middle of the lower aspect,
at the end of a short 'stalk.' This type is very different in
[45]
[46]
[47]
[48]
[49]general appearance from the Hydra or sea-anemone, yet the one
form may be somewhat easily derived from the other; we have
only to imagine that a Hydra is turned upside down, that it is
squashed, vertically, until the internal cavity is greatly reduced,
and the circumference, especially in the region of the tentacles,
greatly increased, and we should have something resembling a
Medusa. That the two types are actually closely related is
shown by the fact that there is in the life-history of one group
of Cœlenterates a regular alternation between the one and the
other.
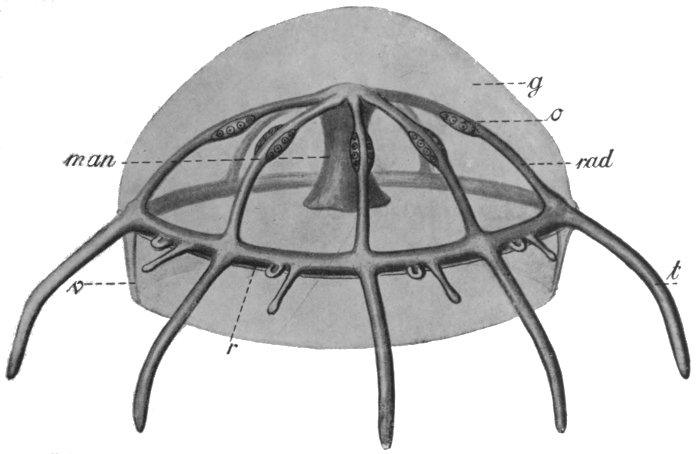
Fig. 35.—Diagram of Medusa.
rad, Radial canals, with reproductive bodies, o; r, ring canal; t, tentacle canal.
If the above general conception of the structure of the Medusa be borne in mind, its details will be easily understood. The internal cavity, instead of being simple, has become complicated, through the obliteration of certain parts of it, where the upper and lower walls come in contact. What is left is a comparatively small cavity immediately above the mouth, a number of symmetrically arranged canals radiating out from this, and a ring canal connecting the ends of these with each other. Another[50] special characteristic is that there is a great mass of gelatinous substance between the outer and inner cell layers. The reproductive cells, as in the sea-anemones, are produced by the inner cell layer, and escape by the mouth.
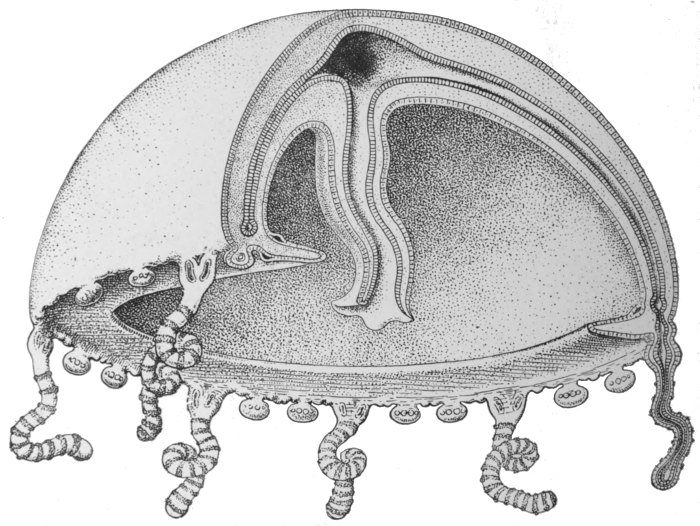
Fig. 36.—Diagrammatic section of Medusa.

Fig. 37.—Group of Cœlenterates—Medusæ, Sea-anemones, and Corals.
Something remains to be said regarding the specialisation of
tissues in this group. We have already mentioned the stinging
cells, and the beginnings of muscular tissue, in Hydra. The
former are a constant feature of the Cœlenterates, while the latter
reaches a very considerable development in the higher forms, as
may be judged from the surprising rapidity with which the Medusa
can swim, or from the strength with which the sea-anemone
can retract its tentacles and draw itself together. Important,
further, is the nerve tissue. This consists of cells whose business
[51]
[52]
[53]it is to receive and transmit stimuli. They have long fibrous
projections connecting them with each other, so that there is a
network of communication throughout the whole animal. In
the Medusa, where co-ordinated movements of various portions
is necessary, there is a concentration of nerve cells into a double
ring near the edge.
Here also there are
special organs, probably
of sight and of
the sense of balance;
but as these cannot be
regarded as the forerunners
of the analogous
organs in higher
animals, we need not
pause to describe them.
The anatomy of the
Cœlenterates will be
better understood if
the reader will study
the diagrams in Figs.
35 and 36, while some
idea of the beauty and
variety met with in the
group may be obtained
from Fig. 37.
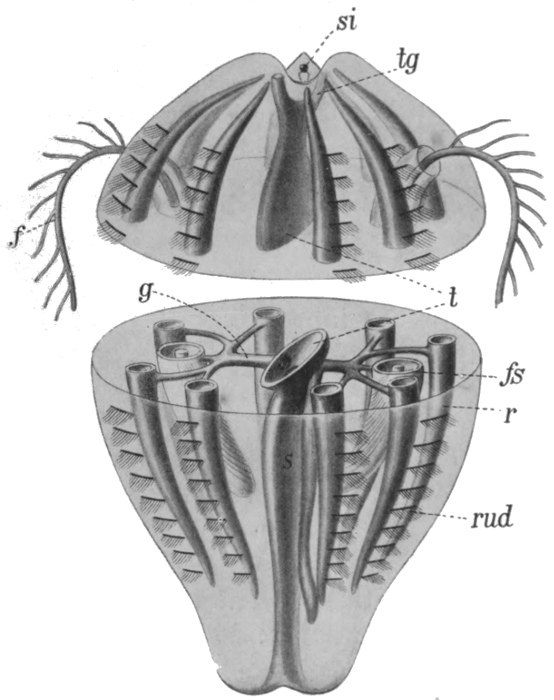
Fig. 38.—Diagram of Ctenophore.
f, Tentacle; fs, tentacle sac; t, central cavity; tg, upper canal; rud, plate bearing cilia; g, radial canal; r, longitudinal canal; si, sense organ.
There is another group of jellyfish-like marine animals which have been given the name of Ctenophora. By some they are regarded as a divergent sub-class of the Cœlenterates, by others as a distinct main group; in any case they appear to be important from our point of view. The structure of a typical member is shown in Fig. 38, and a few other forms are illustrated in Fig. 39. Our typical example is pear-shaped, with the mouth at the lower pole. The internal cavity is complex, but is on a different plan[54] from that of the Medusa. There is a central cavity communicating with the outside not only by the mouth but also by two canals opening near the upper pole. There are two radial canals, each of which divides into four, the branches of which lead at right angles into other canals, running from pole to pole and blind at both ends. There are two tentacles, as shown, which can be withdrawn into special sacs. At the opposite end from the mouth are sense organs, seemingly of smell and balance respectively. On the outer surface, above each of the longitudinal canals, is a row of small plates bearing cilia. It is by the movement of these cilia, like a multitude of minute oars, that the animal swims—a method of locomotion which does not occur in the true Cœlenterates. An additional feature is the formation, at an early stage of development, of a definite third layer of cells between the outer and the inner. This layer ultimately forms the greater part of the jelly-like mass of the body.
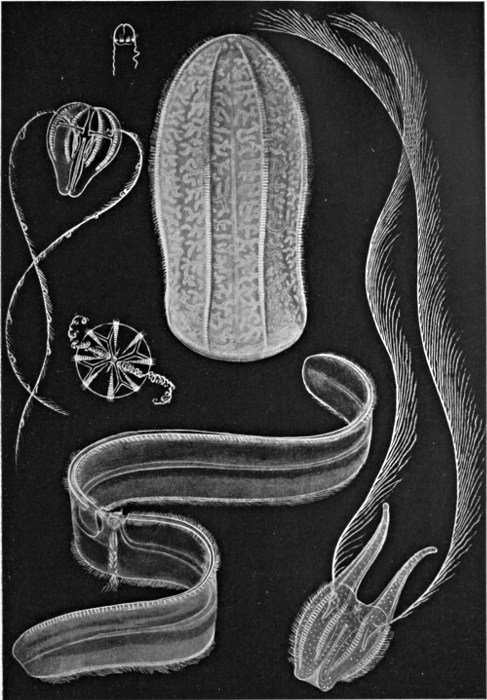
Fig. 39.—Group of Ctenophora.
Regarding the interrelationships of the various types that
we have described, and their respective importance with reference
to the descent of man, opinions are somewhat divided. Some
believe the Ctenophora to have been derived from the Medusa
form, but the more probable view seems to be that they have
evolved separately from some earlier and more primitive type
than any existing Cœlenterate, and that their ancestors have all
been free-swimming and ciliated. Now the Ctenophora are considered,
on good grounds, to be somewhat nearly akin to the
lowest worms, and thus to stand fairly close to the main line of
Evolution. If this view be correct, the whole group of existing
Cœlenterates forms a side branch of the Evolution tree. This
fact, however, does not take away the importance of the group
in relation to the theory of the descent of the higher animals,
for the Cœlenterates have certainly retained many of the characters
which were possessed by the direct ancestors of man, such, for
instance, as the simple digestive cavity, the primitive type of body,
consisting of two cell layers, the diffuse and elementary nervous
system, and the radial arrangement of parts. Moreover, the
course of Evolution in the group, leading from the Hydra to the
[55]
[56]
[57]sea-anemone and the Medusa, has probably been in many respects
parallel to that which started from some primitive extinct form,
and led up to the Ctenophora. Therefore the study of the group
has thrown much light on the earlier history of the animal world.
Regarding the age of the group, it may be mentioned that fossil
corals, etc., are found, along with Crustaceans and Molluscs, in
the earliest known fossil-bearing beds, belonging to the Cambrian
age.
THE WORMS AND SOME OF THEIR POSTERITY
The somewhat miscellaneous collection of animals that have been thrown together and termed worms is of the greatest importance for our theory of descent. Indeed, it seems probable that all of the four great groups which we have yet to mention have descended directly from worm ancestors. This, at all events, is the view of Haeckel, although it must be admitted that many other theories have been proposed. Nor can it be taken as a matter for surprise that agreement concerning this part of our history should be hard to reach, for the difficulties which are met in it are many and perplexing.
The worms comprise many greatly divergent groups, and the difference between the lowest and the highest of these has been produced by many important steps in Evolution. Of these groups but few immediately concern us; the first and lowest of those which do, is that of the Turbellarians, a section of the Platodes or flat-worms. The Turbellarians are small or microscopic tongue-shaped organisms, of which the majority of species live on the sea-floor, others however being found in fresh water. The surface of the body is covered uniformly with cilia, which serve, in the smaller forms, as organs of propulsion, while in the larger they appear to have the function of maintaining a flow of fresh water over the surface, and thus of assisting respiration. In some respects there has been little advance from the condition of the Cœlenterate. The digestive cavity is a simple or more or less divided sac, communicating with the exterior only by means of the mouth. Unlike the condition of affairs in the Cœlenterates and Ctenophora, however, the sex glands do not discharge the[59] reproductive bodies into the digestive cavity, but directly to the exterior by means of a special opening. Each individual has a pair of male and a pair of female reproductive glands, but the eggs are not self-fertilised; nor is the fertilisation of the eggs trusted to chance and the sea-water, as in the lower groups. Instead there is a definite exchange of sperms between two individuals, and the eggs are fertilised before they leave the body. They are also frequently supplied with a store of nutritive material by a pair of special yolk glands. A distinct step of progress can thus be recognised in the arrangements for reproduction. Between the outer skin and the inner digestive layer is developed a considerable mass of cells, forming muscular and connective tissue, etc. It will readily be understood that the development of such thick tissue masses occasions two distinct new difficulties in the animal economy; for where cells are in direct contact neither with the digestive layer nor with the exterior, their nutrition and the removal of their waste products can no longer be efficiently carried on without special devices. Hence on the one hand a circulatory system, for the transport of food materials, and on the other an excretory system, become necessary. The first of these new departures was not destined to be made until the next stage of progress; the Turbellarians seem to have temporarily got over the difficulty, like the Ctenophora, by developing a complex and ramifying digestive cavity. An excretory system, however, makes its appearance here. Indeed, the beginnings of such a system can be seen in the Ctenophora, in which there are small excretory organs opening into the digestive cavity. The corresponding organs in the worms, as in all subsequent types, open directly to the outside. In the Turbellarians these organs, which are termed nephridia, are two in number, and consist of long tubes which branch and ramify throughout the body, the small branches terminating in special excreting cells, and the whole constituting a complete and thorough drainage system. The nervous system consists of one or two small masses of nerve cells termed ganglia in the front region, with a somewhat complex network of nerves connecting them with various parts of the[60] body. There are frequently two pairs of sense organs, probably rudimentary eyes and ears respectively. The main features of the digestive, reproductive, excretory, and nervous system are shown in the figures in Fig. 40.
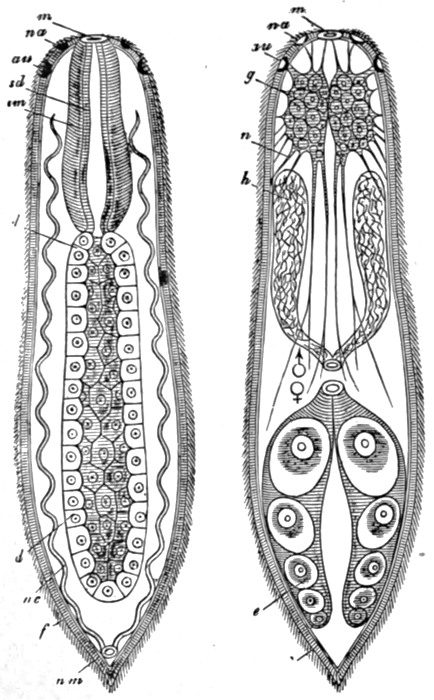
Fig. 40.—A simple Turbellarian—Rhabdocœlum (diagrammatic).
m, Mouth; d, digestive cavity; nc, nephridia; au, eyes; na, sense organs; g, brain; n, nerves; h, male, and e, female, reproductive glands.
The Turbellarians probably arose from Ctenophora or from some nearly related form, a view that receives support from the occurrence of several apparently intermediate types. The differences may, in fact, be partly accounted for as adaptations to meet the change of habitat from that of the upper waters to that of the sea floor. A spherical, or pear, or bell shape is suitable enough for a swimming animal, but would be impossible for one that was to crawl. The first change, then, we may imagine, was a flattening, which produced a disc-shaped animal, with the mouth in the centre of the lower aspect and the sense organs in the middle of the upper. Secondly, a definite mode of progression, by which one part of the body continually went first, would be an advantage, as permitting of a better co-ordination of movements, and an elongation of the body in the line of movement would have the effect of diminishing resistance and of making progression easier. Finally the sense organs, like the scouts of an army, would be best in front, and would migrate thither, and the mouth, in order to get the full benefit of the food which the sense organs[61] sought out, would gradually shift to a position beside them. These adaptions, it is obvious, have produced a complete change in the architecture of the animal. Our sea-anemone, or Medusa, or Ctenophore is radially symmetrical. That is to say, its parts are arranged like the spokes of a wheel, and it may be divided into two equal halves by each of several planes passing through the main axis. It has an upper and a lower surface, but no head and tail ends. The lowest of the worms now can be divided into two halves only in one direction, that which separates the right and left sides. They are, in scientific language, bilaterally symmetrical. The change to this type of architecture was a very important step of Evolution, particularly in relation to locomotion. Bilateral symmetry was destined to remain a constant feature of three of the four great groups that evolved from the worms. The star-fishes reverted to the earlier condition.
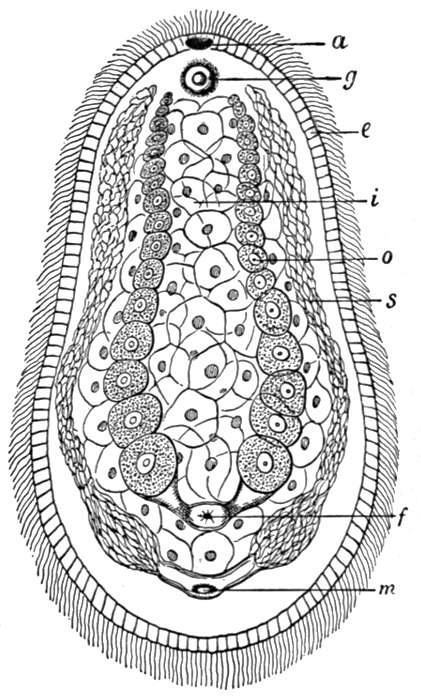
Fig. 41.—A primitive flat-worm—Aphanostomum (× 50).
a, Mouth; g, sense organ; i, internal digestive tissue; s, male, and o, female, reproductive glands; with m and f, external openings.
The next class of worms with which we have to deal is that of the Rotifera. In their general structure, and in their excretory and sensory-nervous systems, the Rotifers do not differ essentially from the Turbellarians. They do differ, however, in that the digestive cavity has a second opening to the exterior, at the end opposite to the mouth. The advantage of this arrangement, which was retained in the subsequent stages of Evolution, is obvious, for it renders possible a much more regular and thorough digestive process. Instead of the food passing in, and[62] the undigested remains passing out, by the same opening, and instead of the contents of the digestive cavity being a general mixture of food material in all stages of digestion, there is now a regular stream of food passing through the cavity in one direction, and being digested as it goes. A near relative of the Rotifers is shown in Fig. 42.
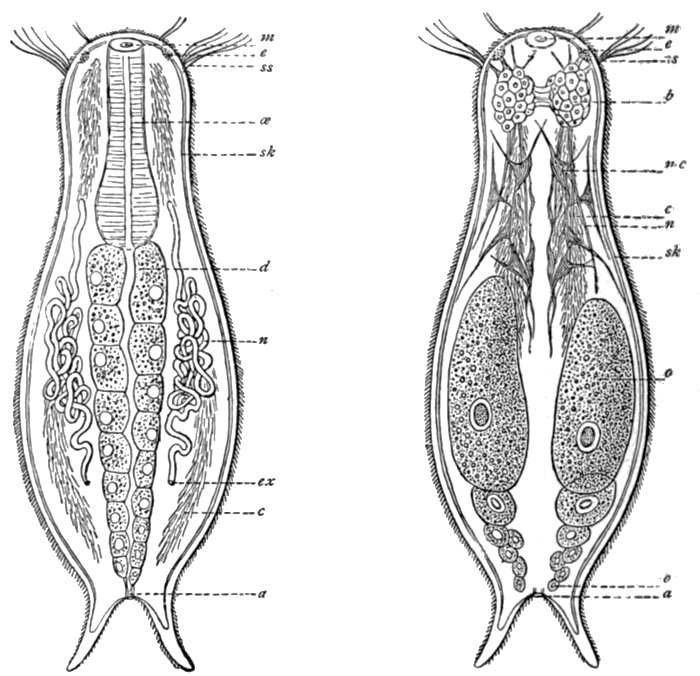
Fig. 42.—Chænonotus, a lower worm.
m, Mouth; e, eye; ss, sensory hairs; œ, œsophagus; sk, skin; d, digestive canal; n, nephridia; ex, excretory opening; c, cilia; a, anus; b, brain; mc, muscle cells; n, nerves; o, ovary.
Thirdly, we must briefly allude to the Nemertines. These are a group of flattened thread-like worms of very variable size, found both in fresh and salt water. The most notable advance in this group is to be seen in the occurrence of a special circulatory system. It has already been indicated that the gastric cavity of the lower forms has the double function of digestion and of the[63] transport of nutritive substances to the various parts of the body. In the Nemertines the second of these functions is carried out by the blood system, which consists of two or three vessels that run parallel throughout the length of the body and anastomose at either end. There is no indication of any enlarged or specially contractile portion of any of these, no indication, that is to say, of a heart. The blood conveys not only nutritive substances, but also, as in the higher animals, oxygen. Some Nemertines have indeed red blood, containing true hæmoglobin, which is well known as the oxygen-carrying material in the vertebrates. A typical Nemertine is shown in Fig. 44, and a diagram showing some features of the anatomy in Fig. 45. It will be seen that the nervous system is of the same type as in the worms already described. There are two pairs of sense organs, one pair being eyes, and the other probably having the function of gauging the chemi[64]cal nature of the water. The Nemertines possess a peculiar organ in a snout or proboscis, which they can protrude or withdraw into a special sac. The snout is armed with a sharp sting, and forms an effective weapon whether against the creature's enemies or its prey.
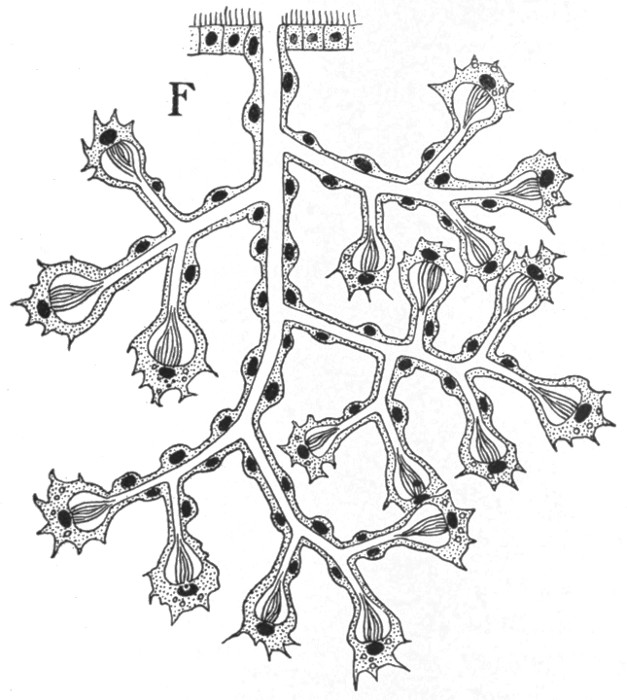
Fig. 43.—Nephridium of a Turbellarian.
About this stage of Evolution, the exact point being somewhat difficult to fix, there appears the body cavity. This, which is altogether distinct from the digestive cavity, is a familiar feature of the anatomy of the higher animals. In it are suspended the heart and lungs and the whole of the digestive organs and glands. The question of the origin of the body cavity and the blood system is a very difficult one, and a thorough theoretical discussion would take us too far.
Before proceeding to the question of the origin of the vertebrates, we may pause briefly to consider the other groups to which the worms appear to have given rise. First of these we may take the Echinoderms, which include the well-known star-fishes and sea-urchins, and the very beautiful feather stars. As already indicated, it is believed that the radial symmetry, which is so characteristic of this group, is not a primitive feature, but that, in fact, the Echinoderms are descended from bilaterally symmetrical ancestors. One reason for this view is that the larval or immature form is always markedly bilaterally symmetrical. In an ordinary star-fish, which we may take as typical of the group, the mouth is in the middle of the lower aspect, and the excretory opening of the digestive cavity in the upper side just opposite. There is no blood system, or excretory organs, and no concentration of nerve cells into any form of brain. Eyes, however, are present, and sensitiveness to light may be easily demonstrated. The most remarkable feature of the group is the water-vascular system, consisting of a series of radial canals, one in each ray, which join a circular one situated in the central portion of the body. The system of canals communicates with the exterior by means of a sieve-like plate on the upper surface, and it is kept full of water by the continual pumping action of cilia on the walls of the tube which leads down from the sieve plate.

Fig. 44.—A Nemertine—Tetrastemma.
Actual length about 1-1/2 inch.
Fig. 45.—Diagram of Nemertine—Nemertopsis.
cg and sg, Sense organ; a, eyes; gh, brain; bl, blood vessels; n, nerve cord; d, alimentary canal; ex, nephridia; r, snout, withdrawn.
The ordinary star-fish is carnivorous, and lives largely on ordinary mussels, which it bridges over with its arms, and opens by a steady and long-continued pulling, the soft parts being then sucked up by the partially protruded stomach. A few types of Echinoderms are shown in Figs. 46, 47, 48.

Fig. 46.—Star-fishes.

Fig. 47.—Feather Star.
Photo: Harold Bastin.
The group of the Mollusca includes such common forms as cuttle-fishes, whelks, slugs, snails, mussels, and oysters. These, it will be observed, comprise marine, freshwater, and land forms. The molluscs, like the next two groups with which we have to deal, have made a conquest of the land, though in the present instance it cannot be regarded as very complete. The anatomy of the group shows much variation, and only a few of the leading features can be alluded to. The digestive system is highly developed. The mouth is provided with a jaw or jaws, and with a tongue-like ribbon, which is covered with rows of teeth, like a file, and by whose action the food is torn and disintegrated. A gullet leads from the mouth to a stomach,[68] which is followed by an intestine. Salivary glands and a large hepatic gland or liver are present. Respiration occurs partly through the skin, but special organs also exist for this function, gills in the water forms, and a lung cavity in those which breathe air. There is a well-developed blood system, and generally a heart; the blood is pumped direct from the heart to the general body tissues, and returns to it by way of the kidneys or nephridia, which purify it of waste materials, and the respiratory organs, where it is freed of carbon dioxide and supplied with oxygen. The nervous system varies greatly, but a pair of cerebral ganglia—a brain—is usually present. There is a particularly keen sense of smell, and taste and hearing may also easily be shown to exist. Some forms are blind, from which condition there is a regular series of stages of development of the eye, up to forms in which it becomes a highly perfected organ, with cornea, iris, lens, and retina. The close similarity between this and the ordinary vertebrate eye, which must have evolved quite separately, is one of the strangest coincidences of Evolution. Thus in many ways the molluscs are to be regarded as highly specialised types. But in two important directions, in intelligence and in their arrangements for locomotion, they stand as a group on a low plane of development. Figs. 49, 50, and 51 illustrate some of the forms met with in the group. The origin of the molluscs, as well as that of the Echinoderms, is wrapped in obscurity. That each group is derived from some form of worm is probable, yet some zoologists hold even such a general statement as this to be lacking in support.

Fig. 48.—Sea Urchin.
1, With spines broken off; 2, with spines on.

Fig. 49.—Molluscs—Univalves.

Fig. 50.—Molluscs—Bivalves.

Fig. 51.—Molluscs—Cuttle-fish, with eggs.
Our third great group is that of the Arthropods (literally
'jointed footed'), including the Crustaceans (crabs, lobsters,
shrimps, etc.), spiders and mites, centipedes and insects. The
Arthropods are sometimes classed together with their ancestors,
the ringed worms (such as the common earth-worm), as Articulata,
a name which refers to a very obvious feature, the repetition of
similar segments in a regular series from front to rear. This
is perhaps most apparent in the ringed worms and centipedes,
but it is to be seen in all members of the group. This same
[69]
[70]
[71]tendency to reduplication of parts in a regular series may be
observed in the vertebrates, as we shall see. Slight indications
of it are also to be found in the Nemertines. Numerous theories
have been proposed which derive the vertebrates from some of
the Articulata—from the ringed worms or the Crustaceans, and
even from the air-breathing members; and at first sight such
theories seem attractive, for in some of their more obvious
characters there is a certain resemblance between the two groups.
But there are also many and fundamental differences, and few
zoologists have accepted any hypothesis of this type. We may
briefly allude to some of these differences.

Fig. 52.
1, Marine swimming ringed worm; 2, giant centipede; 3, Peripatus.
Photo: Martin Duncan, Berridge, and Bastin.
In the Arthropods, where the body consists of hard and soft parts, the 'skeleton' is an external one, and encloses the soft parts. Respiration occurs by means of the skin or of gills, or, in air-breathing forms, by 'trachea,' which are small branching tubes opening on the sides of the body. But in no case has the mouth or the digestive tract any connection with the respiratory system, a condition of affairs very different from that obtaining in the vertebrates. The nervous system consists of a brain, situated above the gullet, a nerve ring round the latter, and a double nerve cord running along the body, below the digestive canal. This is obviously the opposite position to that occupied[72] by the main nerve cord in the vertebrates, an important point of difference.
The Arthropods are an extraordinarily successful group. A multitude of forms of Crustaceans populate the waters, and they are excelled in numbers and variety only by the insects upon land. While the individual size appears to be somewhat strictly limited, probably by the nature of the respiratory and blood systems, many types show exceedingly high development in various directions—in intelligence, in social and parental instincts, etc. The insects are of course to be regarded as the highest Articulata, and have, like the highest vertebrates, the mammals and birds, almost completely forsaken the water for the dry land and the air. An interesting member of the Articulata, from the standpoint of the Evolution theory, is the Peripatus, shown with a ringed worm on Fig. 52 (3). It gains its interest for us from the fact that, while classed as an Arthropod, it stands very nearly half-way between the ringed worms (Fig. 52 (1)) and the true Arthropoda, and thus forms a solitary link between the two types. In Fig. 52 (2) and Figs. 53 to 58 are shown a number of types of Arthropods.

Fig. 53.—Arthropods—The Lobster.

Fig. 54.—Arthropods—Scorpion.
Photo: Leonard Bastin.

Fig. 55.—Arthropods (insects)—Stag-horn beetle.
Photo: Harold Bastin.

Fig. 56.—Arthropods (insects)—Dragon-fly.
Photo: Harold Bastin.

Fig. 57.—Arthropods (insects)—Mantis.
Photo: Harold Bastin.

Fig. 58.—Arthropods (insects)—Swallow-tail butterfly and larva.
Photos: Harold Bastin.
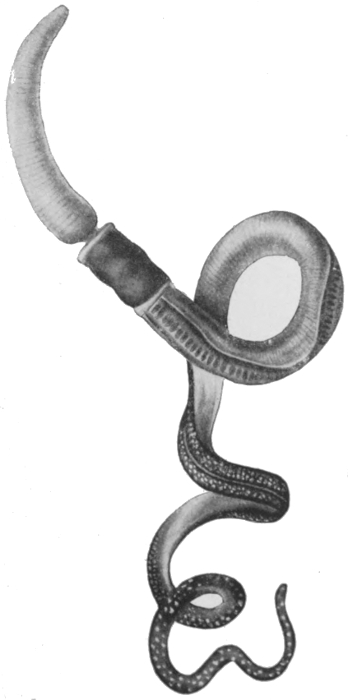
Fig. 59.—Balanoglossus.
We must now go back and take up the main thread of our
story. The next stage that falls to be described is that of a
highly interesting group of worms known as Enteropneusta, a
name signifying 'gut breathers.' This group contains a very
small number of worm forms, which are to be found burrowing
in the sand of the sea floor. A typical example is the Balanoglossus,
a worm of some four inches in length, whose general
appearance is illustrated in Fig. 59. The creature has, as will
be observed, a large muscular snout or proboscis, behind which
follows a small portion called the collar, and behind this again
the long body. The most noteworthy feature of the group, as
the name implies, is the respiratory system. The mouth, which
is situated in the region of the 'collar,' leads into a gullet, which
is partially divided into an upper and a lower canal by means of
two inwardly projecting longitudinal folds, one on either side.
Only the lower of these canals is used as a food passage; the
[73]
[74]
[75]
[76]
[77]upper communicates with the outside by means of a large number
of transverse slits on its sides. Water is continually being taken
in by the mouth and passed along this upper canal, to reach the
outside by way of the gill slits, and in doing so it passes over the
gills, where the blood is circulating in fine capillary vessels.
Here the blood is supplied with
oxygen from the water, and is at
the same time relieved of carbonic
oxide. This, it will be observed,
is the same method of respiration
as that of the fishes. Behind the
last gill slit the digestive canal
becomes a simple tube, with two
digestive glands or liver sacs.
There are two main blood vessels,
the larger running along above the
digestive canal, and the smaller
below it, the two being connected
by means of numerous branches.
There is a swelling of the dorsal
vessel—a heart—at its forward extremity,
in the base of the proboscis.
The nervous system is
peculiar; it consists of two nerve
cords, the smaller below the gut
and the larger above it—the latter
therefore occupying a position similar
to that of the spinal cord in
the vertebrates. Thus in two
respects, in its respiratory and nervous systems, Balanoglossus
must be regarded as a highly extraordinary member of the
worm group, and in both its peculiarities it shows an approach
to the vertebrate. There can be little doubt as to the position
of this group as an important connecting link between the ordinary
worms and the vertebrates. Fig. 60 illustrates the main features
of the anatomy.
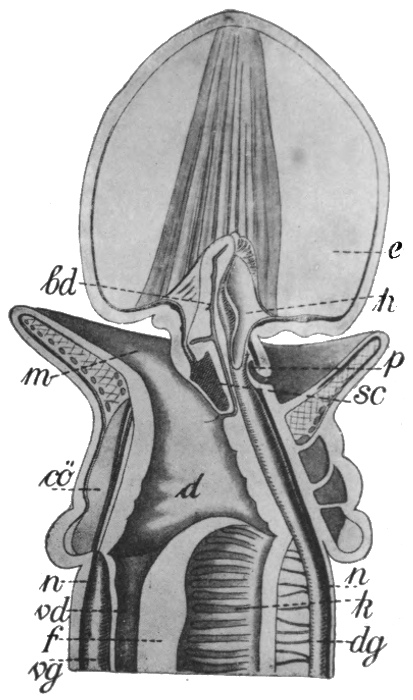
Fig. 60.—Section of front end of Balanoglossus.
e, Snout; m, mouth; h, heart; cö, body cavity; d, alimentary canal; n, n, nerve cords; vg, ventral blood vessel; dg, dorsal blood vessel; f, fold dividing the alimentary canal; vd, food canal; k, gill slits.
As to the origin of the Enteropneusta, opinions are somewhat divided. Their blood system and their development would seem to suggest a descent from the ringed worms. On the other hand, their possession of a snout, and their very slight indication of division into segments, would seem to separate them from the group mentioned and to connect them rather with the Nemertines. The latter view is perhaps the more probable.
THE EARLY VERTEBRATES AND THE FISHES
The lowest of the vertebrates—if indeed it can be called a vertebrate at all, seeing that it has no vertebræ—is the lancelet, Amphioxus. The common species of this animal (there are some eight in all) occurs in the sea off our own coasts, and is usually to be found half buried in the sand or mud of the sea floor. It is some two inches in length and has the shape of a laterally flattened cigar, and one of its very obvious features is the arrangement of the muscles in regular layers from front to back, in the same manner as those of a fish.
To describe some of its features in detail, the alimentary canal bears a somewhat striking similarity to that of Balanoglossus. There is a round, simple mouth, unprovided with jaws, and surrounded by a number of projecting bristles. This leads into a large pharynx, through the walls of which, on either side, pass a large number of gill slits. The pharynx is not divided into an upper and a lower canal, but there is a shallow groove along the bottom which serves the same purpose as the food canal in Balanoglossus. The remaining, digestive, part of the gut is practically a simple tube, with a blind sac attached, representing the liver. The gill slits do not open directly to the exterior, but into the so-called peribranchial chamber, formed by the junction below the body of two flap-like outgrowths, one from the upper part of either side. This chamber opens to the outside by a single pore.
Above the gut lies a straight, cylindrical rod of cartilage, pointed at either end. This is the highly important structure known as the Notochord, which is present in all the vertebrates, although in the higher forms it is replaced during development[80] by the vertebræ, the bony segments of the backbone. Above the notochord again lies the main nerve cord, a position which it retains throughout the whole vertebrate group. The nerve cord is simple in structure, with only a very slight swelling at the front end, representing the brain. There are two main blood vessels, an upper and a lower, which expand and contract alternately throughout their whole length, and thus maintain the circulation. The blood passes forward in the ventral vein, is pumped through the fine vessels of the gills, and collected into the upper artery. From this it is distributed throughout the body by branch vessels, to be re-collected into the ventral vein. If the reader will refer to the illustrations in Figs. 61, 62, and 63, the relationships of these parts will be more easily understood. There is a single small eye-spot, a single organ of smell, but no hearing organs. It seems probable that this extremely ill-developed condition of the sensory system is due in some measure to degeneration, and is not a primitive characteristic. There are numerous pairs of simple nephridia, which open into the peribranchial chamber, and bear a close resemblance to those of the worms.
The lancelet forms a most important link between the lower and the higher animals. It is in all probability derived from some form similar to Balanoglossus, and it certainly leads up to the round-mouths, which form the next step in the ladder.

Fig. 61.—The Lancelet—Amphioxus.
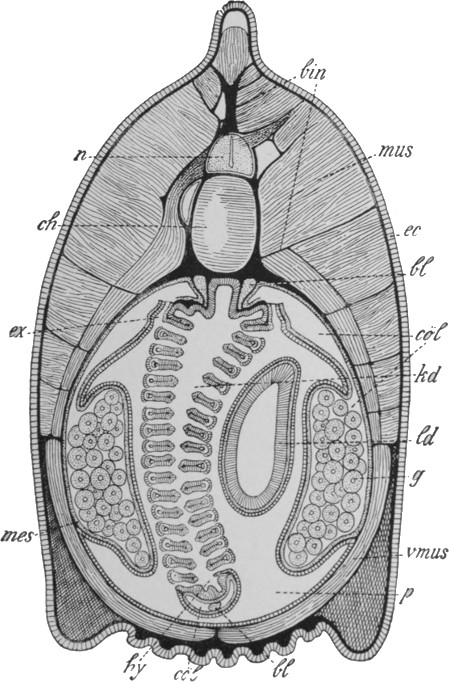
Fig. 62.—Diagrammatic cross-section of Amphioxus.
n, Nerve cord; ch, notochord; mus, muscle tissue; ec, skin; bl, blood vessel; cöl, body cavity; kd, pharynx; ld, liver sac; g, reproductive gland; p, peribranchial chamber.
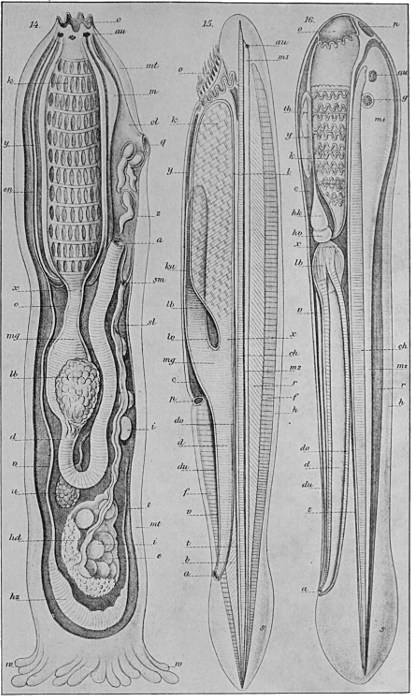
Fig. 63.—Diagrams of Tunicate (on the left), Amphioxus (centre), and young Lamprey (right).
o, Mouth; au, eye; c, peribranchial chamber; ch, notochord; d, alimentary canal; g, ear; hz, heart; k, gills; lb, liver; m, nerve cord; m, brain; mg, stomach; mt, mantle; z, tunicate embryos.
Before describing these latter, however, we must briefly allude
to the highly remarkable group of the tunicates or sea squirts,
one of which is shown in Fig. 64. They are sedentary creatures
found attached to rocks or weeds on the sea floor, and in appearance
they remind one rather of misshapen potatoes than of higher
animals. They are in fact regarded by the fishermen who bring
them to the surface as plants, and they were for long looked upon
by zoologists as akin to the molluscs. The only definite external
features of the tunicates are two apertures at the upper end, one in
the centre and one somewhat on one side. The absence of any other
definite external characters is due to the fact that the creature is
enclosed in a mantle of cellulose. The central opening is the mouth,
[81]
[82]
[83]
[84]
[85]which leads into a large pharynx, the walls of which are perforated
by numerous gill slits. This is
surrounded by the mantle cavity,
which connects with the outer
water by means of the second pore.
The gut is continued into a simple
stomach and intestine, the latter
bending back upon itself and opening
into the bottom of the mantle cavity,
as shown in the diagram in Fig. 63.
In the adult animal there is no trace of
the notochord, and only a remnant of
the nerve cord; and there are either no
special sense organs or only traces of
these. On the other hand, the tunicates
possess a centralised heart. They
are hermaphrodite, and, very curiously,
a number of forms multiply like corals,
by a simple process of budding.
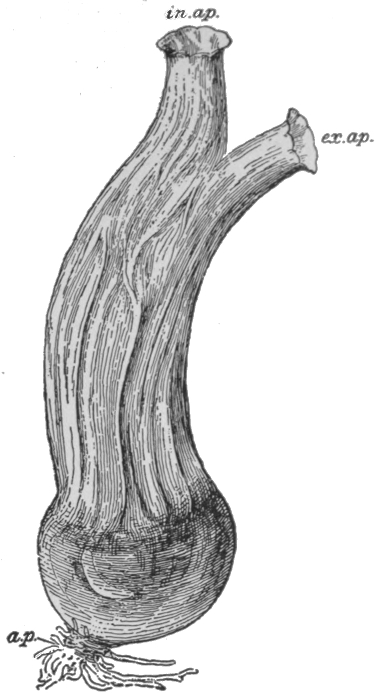
Fig. 64.—A Tunicate.
in.ap., Opening leading to mouth; ex.ap., opening of peribranchial chamber.
From Dendy's Outlines of Evolutionary Biology (Constable).
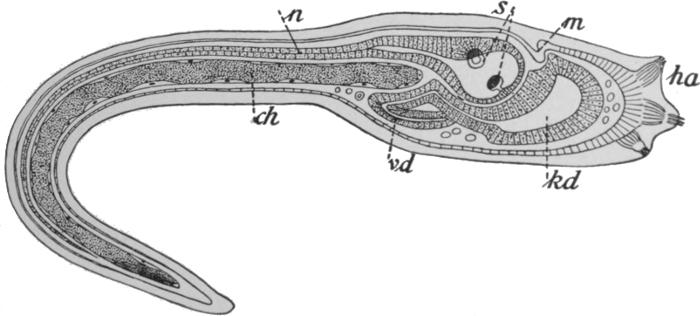
Fig. 65.—Larva of Tunicate.
n, Nerve cord; s, sense organs; m, mouth; kd, pharynx; vd, alimentary canal; ch, notochord.
Now the remarkable fact has been made out that the young tunicate (see Fig. 65) bears a most striking resemblance to the immature lancelet. It is a free-swimming, tadpole-like creature, and possesses a[86] notochord and nerve cord and in general the same characters that we described for Amphioxus. It is only later that the creature settles down and assumes its final degenerate sedentary form. There can be no doubt that the tunicates have been derived from some lancelet-like form, but the course of their evolution has been unique. The type is the lost brother of the vertebrate family, who has chosen a distinctly downward path in life; yet who has come to no miserable end, but lives on, more or less successfully, in his lower social sphere.
The round-mouths, including the lampreys and the hag-fish, stand midway between the lancelet and the fishes, and therefore constitute for us an important group.
The lamprey is a fairly generally known eel-like creature, of which there is a smaller fresh-water, and a larger salt-water species, the latter reaching a length of about a yard. It is found attached to, and feeding on, the dead bodies of fish, and less frequently on living specimens. The hag is much more definitely parasitic in its habits, and often occurs in the body cavity of living fish. These forms were for long regarded as fishes, and are sometimes even yet included in that group, but all their characteristics point to a very much lower position in the animal world than such a classification would indicate. One of the most striking external differences is that the round-mouths have nothing to represent the two pairs of fins which occur uniformly in the fishes, and which are, in a true sense, the forebears of our own arms and legs. Even more important than this is the absence of jaws. The mouth in this group is a simple round opening, whose edges are armed with pointed teeth, the latter, however, bearing no real resemblance to the teeth of the higher animals. By means of these teeth, and a pointed, tongue-like organ, and by suction, the round-mouths are able to bore into the tissues of the animals on which they prey. The absence of jaws and of extremities is, of course, a feature which they share with the lancelet. Turning to the internal structure, we may first observe that there is still no vertebral column, but only a simple, rod-like notochord, similar in its[87] shape, and in its relations to other parts, to that of Amphioxus. There is, however, an additional development of cartilage in the region of the head, forming, in particular, a sheath-like covering for the brain and also a kind of basket-work support for the pharynx and gills. The original tube-like form of the dorsal nerve cord is easily recognisable, but it is distinctly distended at its front end into a brain, which shows a division into a series of three distinct portions, called respectively the fore, mid, and hind brain. This division, it is interesting to observe, is the first process in the development of the brain in all the higher animals. There is a pair of well-developed hearing organs, and in the lamprey a pair of similarly well-developed eyes. In the hag fish the latter are greatly reduced, a condition which is explained by the creature's mode of life. The nostril is unpaired, a condition which is probably primitive. Respiration is carried out by means of gills, which are situated in a series of six to eight pouches, each of which opens into the gullet and again directly to the outside, the external openings being an obvious feature of the animal. There is a very distinct, simple heart, which pumps the blood to the gills, from whence it is collected and distributed throughout the body. The digestive canal is a simple tube, provided, however, with a liver and a pancreas, the two most important digestive glands in the higher animals. The sexes are separate, but traces of a previous hermaphrodite condition seem to persist. Henceforward in the vertebrate group the sexes are always separate. A character of the sex organs which is to be regarded as primitive is that they are unconnected with the excretory system, whereas in the higher vertebrates the two systems are always strangely interconnected. As in all the higher types, there is but one pair of male or female reproductive glands. Finally, the round-mouths differ markedly from the lancelet in the structure of the skin. In the latter animal the skin is composed of a single layer of cells. In the former it consists of an epidermis, some three or four layers thick, and an underlying cuticle or 'true skin'; in other words, the skin has the same general structure as that of the higher types. The[88] lamprey and the hag are illustrated in Figs. 66 and 67, and some of the main anatomical features of the group are shown in Figs. 63, 68, and 69.

Fig. 66.—The Lamprey—Petromyzon.

Fig. 67.—The Hag-fish—Myxine.
There remain, even after the most thorough investigation of Balanoglossus, the lancelet, and the round-mouths, some questions with regard to the origin of the vertebrates that are still unanswered. It must, however, be regarded as an extremely fortunate circumstance that representatives have come down to us of the three ancient groups to which these three types respectively belong. This is the more fortunate in that the groups in question[89] are known only from their few living members, a circumstance which is of course easily accounted for by the absence of any hard parts capable of being preserved as fossils. From this point onwards there is a skeleton, and we are consequently enabled to draw valuable information from fossils. Partly in consequence of this, the Evolution chain from this point onwards is much more complete than the portion that we have dealt with thus far.
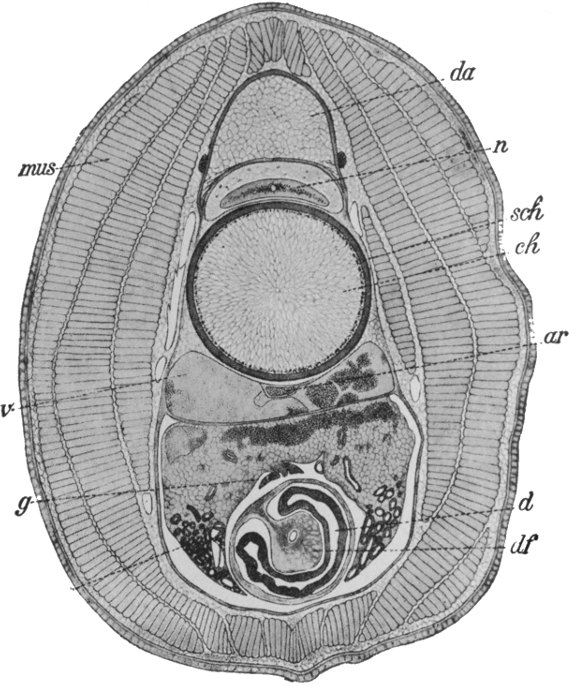
Fig. 68.—Diagrammatic cross-section of lamprey larva.
n, Nerve cord; ch, notochord; ar, artery; v, vein; g, reproductive body; d, alimentary canal.
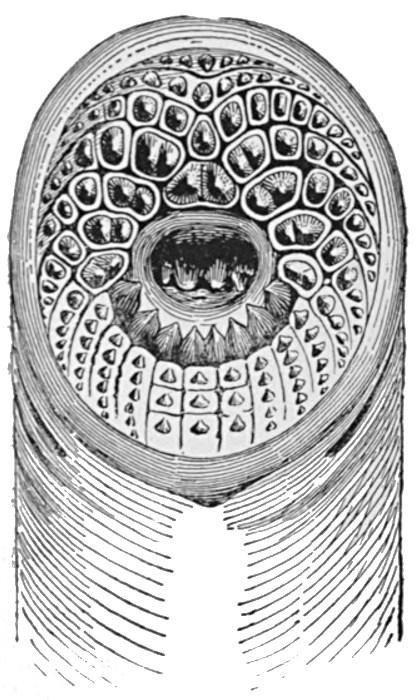
Fig. 69.—Mouth of lamprey.
We have already observed that the true fishes, to which we
must now direct our attention, differ from the round-mouths in
several important characters. They possess two pairs of extremities,
the pectoral and pelvic fins; they have a pair of nostrils;
there is also a well-developed skull, which includes a series of
cartilaginous or bony arches situated in the wall of the gut and[90]
between the successive gill clefts. These branchial arches bear a
certain resemblance to the basket-work arrangement of cartilage
in the round-mouths, but for various reasons are not regarded as
having been derived from the latter. It is from the first pair of
these arches that the jaws are formed, organs which make their
first appearance in the lower fishes. The skeleton shows great
development in other directions. The notochord is present in
its primitive condition during the earlier stages of development,
but it becomes surrounded, and in many cases largely suppressed,
by the portions of the vertebræ. Each
vertebra consists of an upper and a lower
portion, the upper forming an arch round
the nerve cord and the lower bearing
lateral processes or ribs. In the higher
forms the two portions become united
round the notochord, and the resulting
vertebra may encroach inwards until it
becomes solid, the notochord then remaining
only as a series of small pieces of
cartilage between the successive units of
the vertebral column. There is also, of
course, a skeleton in connection with the
limbs, but this does not yet correspond in
detail to that of the other classes of vertebrates.
The brain is much more highly
developed than in the round-mouths; in
many forms, particularly, there is a considerable development of the
cerebral hemispheres, a portion of the fore brain, and the seat of the
higher intelligence. The eyes and ears show the same main features
as in the higher groups. The ear has three semicircular canals, the
same number as in man, as against two in the lamprey and one in
the hag. Fishes are possessed of a peculiar 'sixth sense,' the
organs for which are situated in two lines running along the sides
of the body, the latter forming a familiar feature of a cod or
whiting. The nature of this sense is not definitely known, but it
appears to be of the nature of a very refined appreciation of wave
[91]
[92]
[93]motions in the water. It is probably by means of these 'lateral
line' sense organs, for instance, that fishes are so easily able to
avoid obstacles when swimming in the dark.
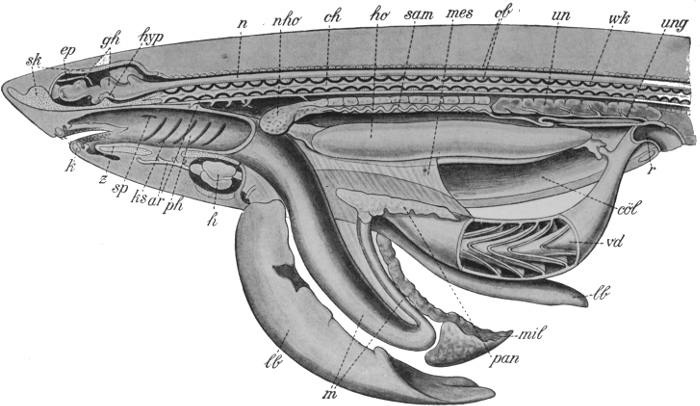
Fig. 70.—Diagrammatic dissection of dog-fish (Scyllium).
sk, Skull; gh, brain; n, nerve cord; ch, notochord; ho, reproductive gland; un, kidney; wk, vertebræ; k, jaw; z, tongue; ks, gill slits; ar, arteries; ph, pharynx; h, heart; lb, liver; m, stomach; mil, spleen; pan, pancreas; vd, intestine, with spiral fold; cöl, body cavity; r, rectum.
The heart has one auricle and one ventricle, except in a single group which we shall afterwards mention. The heart is situated immediately behind the gills, to which the blood is pumped directly by the ventricle. From the gills, the blood is collected and distributed throughout the body, is re-collected and returned to the auricle. The circulatory system is provided with a set of blood-glands, essentially similar to those in man himself. There is a spleen, a thymus and a thyroid gland, and a pair of suprarenal bodies. The several functions of these glands form an extremely difficult chapter of physiology, but, broadly speaking, they are concerned in the formation of the white blood corpuscles, the removal of worn-out red corpuscles, and in certain obscure but important chemical changes in the composition of the blood. The blood itself consists of a fluid plasma in which float white and red blood corpuscles, the latter being flat and oval, and containing the same oxygen-carrying substance, hæmoglobin, as is found in mammals.
The alimentary canal is simple. The mouth cavity is succeeded by the pharynx, the walls of which are perforated by the gill clefts. Next follow the gullet, the stomach, and the intestine, the division into the three portions being apparent often only after close examination. There are generally gastric glands, of simple form, a large liver, and almost always a pancreas. The kidneys and the reproductive organs open to the exterior by a common duct. A further characteristic feature of the fishes is their external covering of scales. True teeth, comparable to those of the higher vertebrates, appear first in this group. Some of the main features that we have mentioned are illustrated in Fig. 70.
Careful study of the fishes makes it evident that they have very much in common with the higher groups of vertebrates. It is not too much to say, with Haeckel, that there is far more difference between Amphioxus and the fishes than between the fishes and man.
There are four main divisions of the fish group. The first, that of the Elasmobranchs, comprises the sharks and dog-fishes, the skates and the rays. The second group, the Ganoids, includes the sturgeon and a few less well-known forms. The third, the so-called bony or food fishes, includes the vast majority of ordinary species, such as the salmon and trout, the cod, herring, eel, and all our ordinary freshwater species. The fourth, the 'lung fishes,' consists of three very remarkable species, which we shall later describe in detail. The mutual relationships of these groups is well understood, and it is possible to make fairly definite statements regarding their evolution.
The Elasmobranchs are at once the most primitive and, so far as is known, the oldest of the four. From these evolved the lower Ganoids, which then divided into two main branches, the first of which led up to the higher Ganoids and through them, at a comparatively late date, to the bony fishes. The second led to the lung fishes and, either through them or along a somewhat parallel line, to the amphibians and the land vertebrates generally. It is with the second line, therefore, that we shall be mainly concerned.
The Elasmobranchs are characterised by the fact that the gill slits open individually to the exterior, there being no gill cover, such as is found in the other groups. Their scales are simple, tooth-like projections, and in fact there is no essential difference between them and the teeth. The skull is more primitive than in the other groups, but a discussion of its details would necessarily be very involved. The living members of the group show a fairly high stage of development of the vertebræ—considerably higher, in fact, than that found in the lung fishes—but some extinct members showed a very primitive condition with regard to this point. In the fossil skeleton shown in Fig. 71, for instance, it is apparent that the notochord was present as a simple continuous rod. The skeleton in question is from the Permian and belongs to what is regarded as the most primitive type of fish known. Two specimens of Elasmobranchs are shown in Figs. 72 and 73, and the teeth of a shark in Fig. 74.
Fig. 71.—Fossil skeleton of Pleuracanthus Decheri, a primitive Elasmobranch.

Photo: Thiele.
Fig. 72.—Spotted dog-fish.

Photo: Thiele.
Fig. 73.—Blue Shark.
Fig. 74.—Teeth of Shark.
In the Ganoids and bony fishes there is a gill cover, and in all but a few Ganoids there is some formation of true bone, whereas[98] in the Elasmobranchs the skeleton is wholly cartilaginous. One of the most striking anatomical features of these groups, and one which distinguishes them from the Elasmobranchs, is the presence of a swim bladder, a large sac-like outgrowth from the upper part of the gut. The function of the swim bladder is that of regulating the specific gravity of the fish, which becomes greater or less according as air is expelled or taken in. The Ganoids and bony fishes are illustrated in Figs. 75 to 79.

Photo: Underwood.
Fig. 75.—Ganoid fishes—The Sturgeon.
The lung fishes or Dipnoi present a curious mixture of primitive and of highly advanced characters. In their persistent notochord and their inconsiderable formation of bone, they are much more primitive than the food fishes. On the other hand, an extremely important departure is seen in the adaption of the swim bladder as a respiratory organ. In one of the three existing species this organ is single, in the others it is double. The wall of the swim bladder is thick, and contains considerable muscle tissue. Its inner surface is covered with a complex system of pits and blind sacs, the walls of which contain numerous capillary vessels.

Fig. 76.—Ganoid fishes—Polypterus bihir.

Fig. 77.—Ganoid fishes—American bony pike, Lepidosteus.

Photo: Thiele.
Fig. 78.—Bony- or food-fishes—The Salmon.
Fig. 79.—Bony- or food-fishes—The Common Eel.
Photo: Underwood.

Fig. 80.—Lung fishes—Australian lung fish, Ceratodus.

Fig. 81.—Lung fishes—Protopterus (Africa).

Fig. 82.—Lung fishes—Lepidosiren (South America).
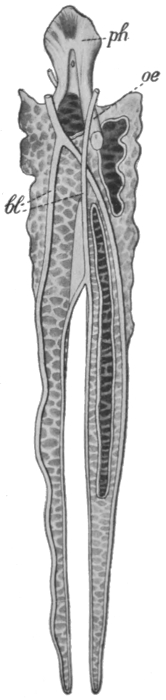
Fig. 83.—Swim bladder (lung) of Protopterus.
There are three living species of lung fishes, one of which
is found in Australia, another in Tropical Africa, and a third in
the tributaries of the Amazon. All live under conditions which
make ordinary respiration by gills difficult. The Australian
species inhabits rivers which become reduced, in the dry season,
to stagnant pools of foul water, in which ordinary fish frequently
fail to survive. Under such circumstances the creature comes
periodically to the surface to breathe. The other two species
live in rivers which actually dry up in summer, and the fishes bury
[99]
[100]
[101]
[102]
[103]themselves in the mud, and remain in a torpid condition, breathing
air by their lungs, until the rainy season comes round, perhaps
four or even six months later. Correlated with the special
method of respiration is a special type of blood system, whereby
part of the blood is pumped direct to the lungs, and returns
direct to the heart. There are two auricles, to receive the
blood from the lungs and the general circulation respectively,
but only one ventricle, in which the two streams become
mixed. Figs. 80 to 82 illustrate the three existing Dipnoi, and
the structure of the lung is shown in Fig. 83.
It is obvious, from the distribution of the lung fishes, and also from geological evidence, that the group was once very plentifully represented, and has only been preserved from total extinction by peculiar circumstances.
Regarding the position of the group, some zoologists regard them as the direct ancestors of the Amphibians. Others believe that the group had a common origin with the bony fishes and the Amphibia in some early form of Ganoid. In any case, the Dipnoi possess an extraordinary interest as showing the beginnings of adaption to a life out of the water.
THE CONQUEST OF THE LAND
The Amphibia are the oldest and the lowest group of vertebrates
that are able to lead an active existence on land, and the characters
which distinguish them most definitely from the fishes are all
to be interpreted as adaptions to the new mode of life. One of
the most obvious external differences between the two groups
is in the structure of the extremities, the fish having fins, while
the amphibian has limbs constructed on the same general lines
as our own arms and legs. The fish's fin is to be regarded as an
extremity with a very great number of fingers or toes. It has
the function of a paddle, and is obviously useless whether for
supporting or propelling the body on land. The first obvious
necessity for a land existence is some mechanism by which the
limb can be alternately pushed forward and, being fixed to some
solid object, drawn upon, so as to pull the body after it. A
different arrangement of bones and muscles, so as to give a much
more complex lever system than that of a fin, and some kind
of clawing arrangement at the end, were thus necessary. The
similarity in the limbs of all the land vertebrates is very striking,
as is indicated by the comparison of human and a frog's limbs
on Fig. 84. In each case there is a single bone in the upper
arm or thigh, which is attached to a bony girdle in the trunk.
There are two elements in the forearm and in the lower leg respectively,
below which, in either case, is a group of small bones
constituting a complex joint at the wrist or ankle. Then follows
the set of five bones in the foot or hand, to each of which is
attached a jointed finger or toe. We have no reason to believe
that this particular arrangement, and the particular number of
[105]
[106]
[107]digits, was arrived at except by accident. Once arrived at,
however, the arrangement was adhered to with considerable
strictness. For although the number of digits is in some groups—in
the birds especially—reduced, the primary design is almost
always readily recognisable. The second function of the limb,
that of supporting the body, was developed very slowly. In the
amphibians and reptiles, and even in the lower mammals, the
legs are comparatively weak and sprawling, and the creature
crawls on the belly.
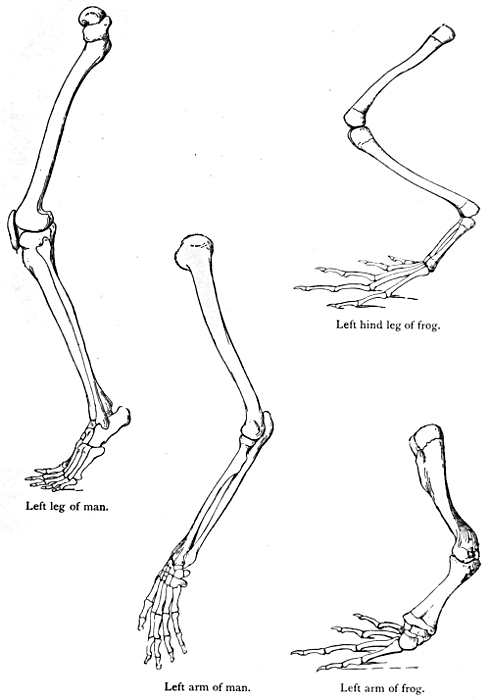
Left hind leg of frog.
Left leg of man.
Left arm of man.
Left arm of frog.
Fig. 84.
Fig. 85.—Development of the Frog.
The second great change which required to be made was of course in the method of breathing. An ordinary fish, when taken out of the water, dies of suffocation, because its gills become[108] inefficient for respiration as soon as they become dry. An entirely new type of organ had therefore to be evolved, and this occurred on the same lines as in the Dipnoi, by the development of a pair of sacs from the upper part of the digestive canal, in which the blood is made to circulate, and which are kept filled with air taken direct from the atmosphere. It is of course very well known that an ordinary amphibian is not a lung breather throughout its whole life. The metamorphosis of a gill-breathing tadpole into a lung-breathing frog, illustrated in Fig. 85, is a phenomenon with which everyone is familiar. And this condition, in which a change in the mode of life is made by each individual in the course of its development, is the typical one. But the modern amphibians include types ranging from completely water to perfect land forms. Some, like the Austrian Olm (Fig. 86), are gill breathers throughout their whole life. One which is normally of this type, the Axolotyl, illustrated in Fig. 87, can be made to acquire lungs and assume a land mode of life. Others, which normally make the metamorphosis, can be prevented from doing so by being confined to the water, and complete their life-histories in the condition of gill breathers. In still other forms (e.g. the Cœcilians, Fig. 89) the change is made before the young creature leaves the egg, and the independent life is commenced in the condition of a land animal.
Correlated with the development of the lungs is a change in the structure of the nostrils, from the condition of blind sacs, as they occur in the fishes, to that of air passages, communicating with the upper part of the alimentary canal, and thence with the lungs.

Fig. 86.—The Olm—Proteus anguincus.

Fig. 87.—Axolotyl. The ordinary gill-breathing form, and the artificially developed land form.
Fig. 88.—Amphibians—The Fire Salamander.
The living forms of the Amphibia differ considerably from the
types which constituted the group in those long-distant ages when
it was in the heyday of its prosperity. The latter forms were
characterised especially by a system of armour-plating over the
head, which frequently extended under the breast and even covered
the greater part of the lower surface, and which appears to have
formed a protection against the multitude of sharks which
populated the waters in which the amphibians partly lived. The
[109]
[110]
[111]armour-plated type has
long ago become extinct,
but it is in it, rather than
in the modern forms of
Amphibia, that we must
look for the direct ancestor
of man. A fossil of this
earlier group is shown on
Fig. 90.
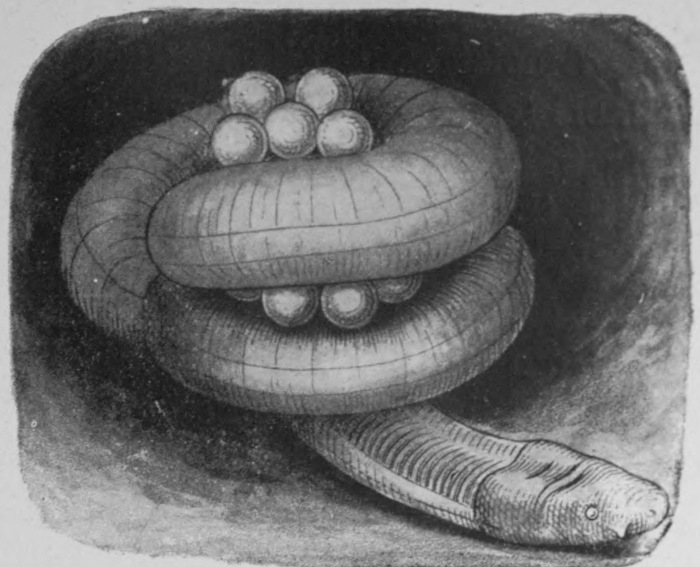
Fig. 89.—The Ceylon Cœcilian, Ichthyophis glutinosa, with eggs.
Fig. 90.—Fossil amphibian, Brachiosaurus, from the Permian.
Our next group in the order of Evolution is that of the reptiles, the main differences between which and the Amphibia are of the nature of more complete adaptions to a life on land. The reptiles have in fact completed the conquest of the land which was undertaken by the previous group, and many of them, living as they do in dry and hot deserts, are as independent of the water as any form of animal life. Thus whereas the amphibian has a thin skin, which is kept moist by the secretions of numerous skin glands, the reptile has a body-covering of scales, which form an effective protection against a too rapid loss of moisture. Evidently with the same object, the reptile egg is enclosed in a hard and resistant shell. Correlated with the change in the skin is a much more perfect development of the lungs, for[112] while the amphibian breathes to a considerable extent through its thin moist skin, this method of assisting respiration is not available to the reptile.
Again connected with the improvement of the respiratory process, there is a partial development of a septum dividing the ventricle or pumping chamber of the heart. The value of this division of course lies in the fact that the purified and oxygenated blood from the lungs is prevented from mixing with the venous blood from the body. The course of the blood is from the body to the right auricle, thence to the right ventricle, and thence to the lungs. The pure blood from the lungs returns to the left auricle, passes thence to the ventricle on the same side to be pumped to the general circulation. The disadvantage of a single ventricle, such as occurs in the Amphibia, and the advantage of the regular double circulation, such as that in man, are sufficiently obvious. The division of the ventricle into two chambers is less complete in the lizards and snakes, very nearly perfect in the crocodiles.
The reptiles, like the Amphibia, are 'cold blooded,' by which is meant, not that their blood is necessarily cold, but that its temperature varies with that of the surroundings, while that of the blood of the mammals and birds is practically constant.
A very important feature of the reptiles, which they possess in common with the mammals and the birds, is that the embryo produces two membranous outgrowths called respectively the amnion and the allantois, which completely envelop it, and which have important functions in connection with nutrition, respiration, and excretion during the period when the young creature is enclosed in the egg. It is, of course, not until we reach the higher mammals that these membranes assume their greatest importance.

Fig. 91.—Imaginative European landscape in the Cretaceous period, with reconstructions of typical reptiles.
Plesiosaurus (swimming, up to 50 ft. long).
Three Ichthyosaurians.
Pterodactyls (flying).
Iguanodon.
Megalosaurus (40 ft. long).
Rhamphorhynchus.
For a considerable time in the world's history the reptiles
were the dominant vertebrate class, and in the chalk period
especially they were represented by a great variety of forms,
and by a number of species of colossal stature, one at least of which
was over a hundred feet long. In those times the reptiles were
by no means all condemned to crawl on their bellies, for they
[113]
[114]
[115]included a large number of marine forms, comparable to the
porpoises and whales among the mammals, and flying forms
whose aspect must have resembled, and been equally terrifying
with, that of our mythical dragons. A few reconstructions of
these are shown in Fig. 91.

Photo: Berridge.
Fig. 92.—Reptiles—A Chameleon.

Photo: Berridge.
Fig. 93.—Reptiles—Indian Python.

Fig. 94.—Reptiles—Turtle and Tortoises.

Fig. 95.—Reptiles—Alligator.
All living reptiles, with a single exception, belong to the four
comparatively modern types of the lizards, snakes, tortoises, and
crocodiles, of which examples are illustrated in Figs. 92 to 95. None
of these are closely related to the mammals or birds. For the
common ancestor of all these types we must go back to some
primitive reptile form. Fortunately such a type is represented[116]
at the present day in a single peculiar species found in New
Zealand, which bears the Maori name of the Tuatara. It was
formerly found commonly on the mainland, but is now confined
[117]
[118]
[119]to a few small islands in the Bay of Plenty, North Island, where
it enjoys Government protection. It is, as the illustration in
Fig. 96 shows, a lizard-like creature, and reaches a length of
about two feet. It lives in burrows near the shore, and feeds on
small animals that are left behind by the tide. The Sphenodon,
as zoologists have named it, has apparently been preserved
owing to the absence of competition by the mammals, and
by adopting the rather curious mode of life just described.
In all its features, but especially in the primitive condition
of its vertebræ, it is very much lower than any other living
reptile, and it connects the higher groups with the Amphibia.
Many closely related fossil species are known, one of which is
shown in Fig. 97.
Fig. 96.—The Tuatara, Sphenodon punctatus.
Fig. 97.—Homeosaurus pulchellus. A fossil early reptile from the Jurassic.
To the lay mind the distinctions between the Amphibia and the reptiles are not very obvious, and indeed in the older classifications the former group was not separated from the latter. The differences between a reptile and a bird, on the other hand, are very striking. It might therefore be regarded as a matter for surprise that zoologists now make the greater distinction between the Amphibia and the reptiles, grouping the former in one great class with the fishes, the latter in a second great section with the birds. But in fact there are many fundamental points of agreement between reptiles and birds, and it is impossible to doubt that the latter have sprung from a reptilian stock. Indeed, a most interesting connecting link is known, in the fossil Archiopteryx shown in Fig. 98, of which only two specimens have been found, and which is the only creature of its type of which we have any record. In all its skeletal features, the Archiopteryx is reptilian, and it would undoubtedly have been classed as a new type of reptile but for the obvious and unmistakable traces of feathers. From what particular class of reptiles the birds have sprung is not known.
The birds have assumed the position of almost unquestioned masters of the air, but like other great groups they show possibilities of evolution in other directions wherever opportunity offers, and types like the kiwi and the penguin shown in Figs. 100[120] and 101 have forsaken their native element—the one for the land, the other for the water.
The birds agree with the mammals in the development of a four-chambered heart, in their warm blood, in their external covering for the skin, and in the development of arrangements and instincts for the parental care of the young. Their line of Evolution has thus been to some extent parallel to that of the mammals. On the other hand, they differ obviously in the structure and function of their fore limbs, in the absence of a diaphragm, and in their special methods for the care of the young, and there can be no doubt that the two groups have had quite different origins in the reptile class.
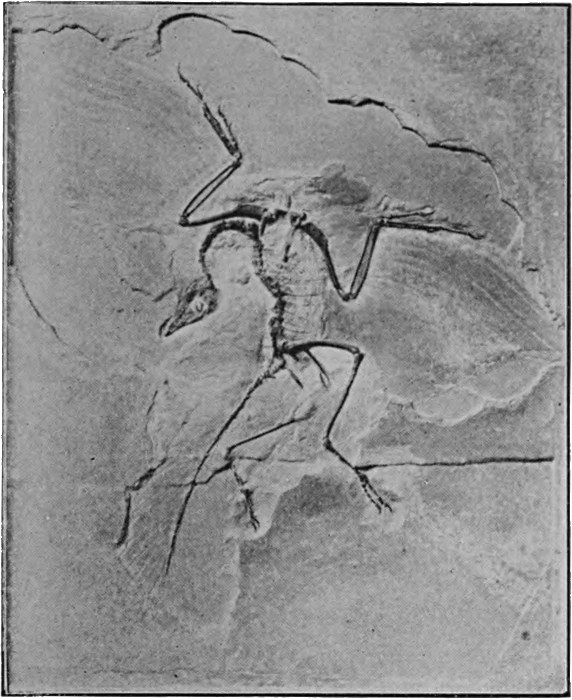
Fig. 98.—Archiopteryx, a fossil lizard-like bird.
Photo: Underwood.

Fig. 99.—Eagle, with prey.

Fig. 100.—The Kiwi, Apteryx. A practically wingless running bird.
Photo: W. P. Dando.

Fig. 101.—The King Penguin. A highly specialised diving bird.
Photo: W. P. Dando.
THE MAMMALS AND MAN
The subject of our discussion is now narrowed down to the group of the mammals. The mammals are characterised by two very obvious features: a body-covering of hair, and a set of special glands in the female which secrete milk for the nourishment of the young. These are constant characters, and neither is ever found in any other group. As to how the hair originated, nothing definite is known; but (while there are certain difficulties in regard to the theory) it is on the whole reasonable to suppose that the mammalian hair arose, as the bird's feather undoubtedly did, as a modification of the reptile's scale. The mammary gland appears to represent a modification of other skin glands, either of sweat glands or more probably of the oil glands which exist in connection with the hairs.
Another important character, already mentioned at the end of the last chapter, is the diaphragm, a muscular partition separating the body cavity into a thoracic and an abdominal portion. The diaphragm has important functions in connection with the mechanism of breathing. By means of it the thoracic cavity can be increased or diminished in size, and air thus drawn into or expelled from the lungs. It is interesting to observe that a similar partition, with the same function, occurs in the Crocodiles, but this has different relations to the abdominal organs, and has evidently evolved quite independently.
Very characteristic of the mammals are, further, their teeth, for whereas the teeth of the reptiles are indefinite in number, and generally very numerous, those of the mammal are relatively few, and each species has a definite normal number. Moreover,[124] the teeth of the reptile are all of a kind, and they may be replaced almost any number of times during the animal's life, whereas those of the mammal show differentiation according to their respective functions, and are only once changed, that is, when the milk teeth are replaced by the permanent set. The teeth of the mammal are of four kinds: incisors, or chisel-like cutting teeth, canines, which are especially well developed in carnivorous species, and which are used in the tearing up of flesh, etc., and two groups of grinders—premolars, which are replaced during the animal's life, and molars, which occur only as permanent teeth.
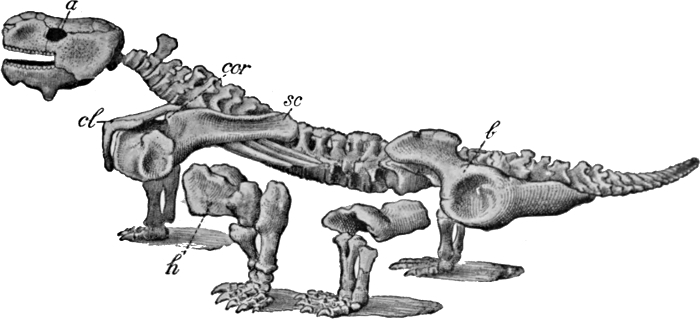
Fig. 102.—Pareiosaurus Baini. Fossil skeleton from the Permian of South Africa.
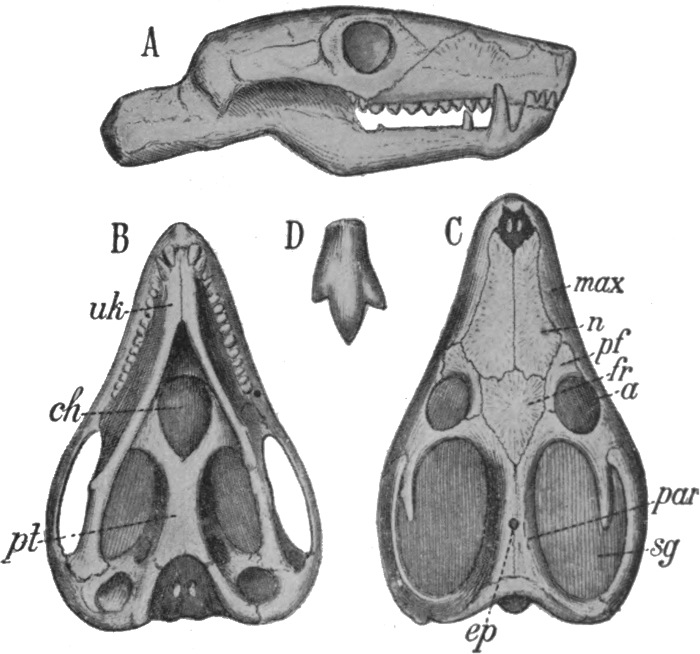
Fig. 103.—Skull of Galesaurus, from the Permian of South Africa.
A, From side; B, from below; C, from above; D, back tooth.
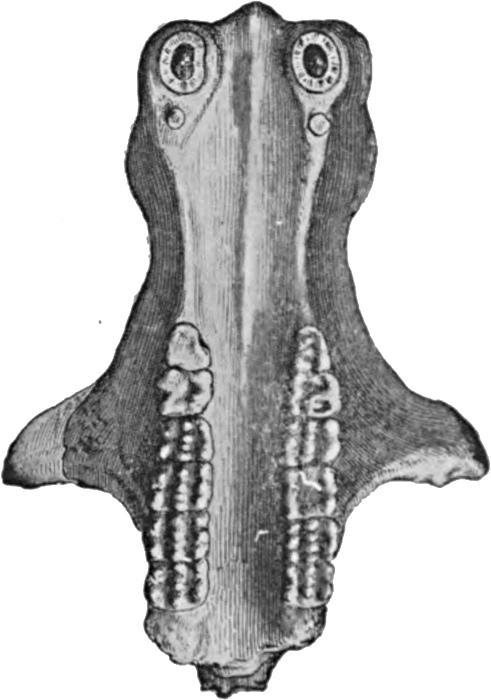
Fig. 104.—Skull of Tritylodon from below.
The mammals in all probability arose from a particular group of reptiles which flourished in the Permian and Triassic periods, and which disappeared very shortly afterwards. These pass under the name of the Theromorpha, and a typical specimen, Pareiosaurus, is illustrated in Fig. 102. The skull of another, illustrated in Fig. 103, shows distinct indications of a mammal-like differentiation of the teeth. The Tritylodon, whose skull is shown in Fig. 104, is an intermediate type between this group and the mammals, some zoologists regarding it, as it were, as the last reptile, others as the first of the mammals.
As to how the special mammalian features arose, or what special conditions called them into existence, we are of course without definite knowledge, for neither hair nor mammary glands are recognisable in fossils. But it seems likely that the warm blood and the hairy covering evolved in correlation with one another, and as adaptions to meet a gradual cooling of the climate. It is certain at all events that the present-day mammals (and also the birds) are far better adapted for a cold environment than the reptiles, which very easily get frozen to death; and it is also known that ice periods occurred in South Africa, where many fossil Theromorpha and the Tritylodon are found, at the time when these creatures existed; both of which facts support the theory indicated.
At the present time the mammals are the highest and on the
whole the most successful of the vertebrate groups. They include
the largest and strongest, the swiftest-footed and the most
[125]
[126]
[127]intelligent of the animal kind. They show refinements of the
senses of sight, hearing, and smell such as are met with nowhere
else. They range from the Equator to the coldest regions of the
earth in which any food is to be found; they people alike the
forest and the plain, and have their representatives both in the
air and in the sea.

Fig. 105.—The Australian Duck-mole or Duck-billed Platypus.
The most lowly of the mammals are the Monotremes, which include the well-known Australian duck-mole or duck-billed platypus, and two species of spiny ant-eaters, one of which is found in Australia, the other in New Guinea. The two types are shown in Figs. 105 to 107. The best-known and most striking fact concerning these is that, like the birds and reptiles, and unlike all other mammals, they lay eggs. Beyond this feature they show many affinities with the reptiles, in their skeleton for example, and particularly in their reproductive organs. Another interesting fact is that the blood temperature, of the ant-eater at least, is low, and varies considerably. It has been found to[128] range between 80 and 93 degrees Fahrenheit, whereas all other mammals, so far as is known, have a blood temperature between 97 and 105, and moreover one that is remarkably constant for each species during normal health. The brain development and the intelligence are also much lower than in other mammals, though distinctly superior to those met with among the reptiles.[129] The method of rearing the young in the case of the ant-eater (that of the duck-mole not being yet fully known) is that the single egg is placed, as soon as it is laid, in the pouch under the belly of the female. Here it hatches in a very short time, and here the young animal remains for the first two or three months of its life, being nourished by milk produced by the mammary glands, which open into the pouch. It is certainly owing to the absence of the competition of the higher mammals in the regions where they are found, that these two creatures, so interesting from the standpoint of Evolution, have been preserved to us.

Fig. 106.—New Guinea Spiny Ant-eater.

Fig. 107.—Australian Spiny Ant-eater.
The next group, the Marsupials, is the lowest in which we get the true mammalian characteristic of the bearing of living young. For while occasional members of other groups, of the reptiles especially, produce live young, the actual state of affairs is fundamentally different. In these latter the egg is merely retained in the genital duct until the young creature emerges. It is merely hatched inside the body of the mother instead of outside. But in the Marsupials the developing young receive nourishment from the mother, during prenatal life, other than what is contained in the yolk. This nourishment is obtained in the form of a secretion from the wall of the uterus, there being as yet (with a single partial exception) no real connection between mother and young.
The peculiar method of nourishing and protecting the young Marsupial after birth is of course well known. The young are born in a very immature and helpless condition, and are placed by the mother in her pouch. The mouth becomes permanently attached to the nipple of the dam, and the young creature remains thus for a considerable time, the milk being pumped into it by the mammary gland rather than sucked in by the efforts of the creature itself.
The Marsupials are further characterised by the possession of an extra pair of bones in the pelvis, which function as supports for the pouch; by the peculiar and primitive arrangement of the reproductive organs; by their still poorly developed brain;[130] and by their generally large number of teeth, which reach a total of fifty or fifty-two in some species, whereas forty-four is the ordinary maximum number in the main mammal group. This last is to be regarded as a character derived from the reptiles.

Fig. 108.—Kangaroo, with young.

Fig. 109.—Newborn young of Kangaroo.
It is very interesting to observe how, in Australia, where
the Marsupials have been free from the competition of other
mammals, they have evolved along many of the same general
lines as the higher group. We have indeed no marsupial whales,
bats, or seals, but there is a mole, very similar in its appearance
and habits to our European species, and a carnivorous type which
closely resembles a wolf or jackal; again we have bandicoots,
occupying the same place in nature as our rabbits and other
rodents; tree-dwelling, squirrel-like forms; and kangaroos,
which compare in their mode of life, if not in their appearance,
with the cattle, deer, and antelopes of other countries. The Marsupials
are illustrated in Figs. 108 to 113. This group, so far as
can be judged from fossils, is considerably older than that of
[131]
[132]
[133]the higher mammals, and everything points to some marsupial
type as a connecting link between the egg-laying Monotremes
and the placental mammals.
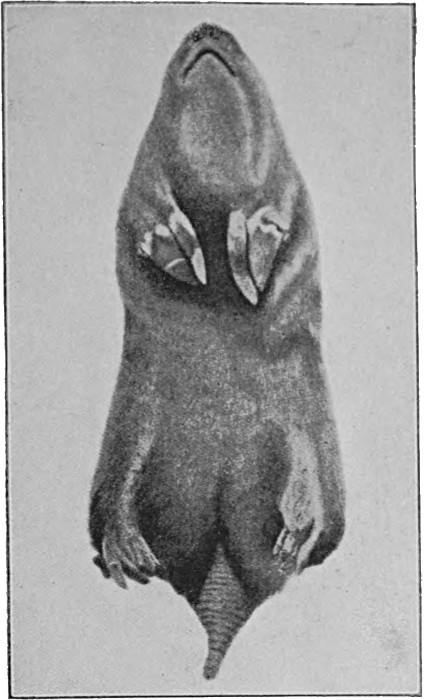
Fig. 110.—The Marsupial Mole.
Fig. 111.—Tasmanian Wolf, a carnivor-like marsupial.

Fig. 112.—Marsupials—Long-nosed Bandicoot.

Fig. 113.—Squirrel-like marsupial (Phascologale penicillata).
As has previously been indicated, the most important characteristic of the third great group is a modification of the membranes of the embryo to form a connection between it and the wall of the uterus. The allantois develops as a highly vascular membrane, the small blood vessels of which are brought into very close contact with those in the wall of the uterus. So that, while the blood of the mother does not actually mix with that of the child, the two fluids are separated only by thin membranes, through which nutritive substances easily pass. The broad advantage of this is, of course, that the young animal passes the earlier stages of its life inside the mother's body, where it is exposed to a minimum of risk, is efficiently nourished, and from which it is not sent forth into the world until it is tolerably well able to look after itself.

Fig. 114.—Oeba Armadillo.
Photo: Berridge.

Fig. 115.—Sea-cows.
We shall now ask the reader to conceive a primitive placental[134]
mammal type, and to consider briefly the relationships of the
various groups that have sprung from it. The group whose
evolution has been perhaps least progressive is that of the Edentates,
including the sloths, armadillos, and American ant-eaters
(Fig. 114). Then, on the one hand, we have the herbivorous
types, including the remarkable sea-cows (Fig. 115), the rodents
(Fig. 116), and the hoofed animals, leading up from such comparatively
primitive forms as the coneys (Fig. 117) to such
highly specialised types as the giraffe (Fig. 118). The other series
of groups are believed to be more nearly related to each other
than to the aforementioned, and the nearest approach to their
[135]
[136]
[137]common ancestor, among modern mammals, is probably to be
found in the Insectivora (Figs. 119 and 120). Differing sharply
from the Insectivora in their possession of wings, but otherwise
closely similar, are the bats (Fig. 121). From this same
insectivor-like type have probably evolved the whales (Fig. 122),
the seals, etc. (Fig. 123), the carnivora (Fig. 124), and finally
the primates, leading up through the lemurs to the monkeys
and man. Thus merely indicating the many and devious roads
of mammalian Evolution, we turn again to the one particular
line that we set out to follow.

Fig. 116.—Porcupine.

Fig. 117.—Coney (Hyrax).

Fig. 118.—Giraffe.
Photo by Sport General.

Fig. 119.—Centetes ecaudatus, a primitive insectivor.

Fig. 120.—Hedgehog.
Photo: Berridge.
The Insectivora are by many of their features recognisable as low types of placental mammals. They have a primitive type of skull, and frequently show a rather marked similarity between the several classes of teeth. The brain is relatively ill-developed, reminding one rather strongly of the Marsupials,[138] and the hemispheres show little if any tendency to develop those wrinkles and fissures which always accompany the higher type of intelligence. Probably the most primitive member of the whole group is the Centetes from Madagascar, shown in Fig. 119. The hedgehog and the shrews are its best-known examples. An interesting point regarding the hedgehog is that, like few other mammals, it has persisted almost unchanged from early tertiary times. It is thus to be regarded as a very antique type, not only in its main features, but in its details.

Fig. 121.—Vampires.

Fig. 122.—Porpoises.

Fig. 123.—Sea Lions.

Fig. 124.—Indian Leopard.
Photo: Underwood.
The Lemurs are an interesting group, standing, as they do, midway between the primitive placental forms and the monkeys. Their special home, as already mentioned, is Madagascar, to which some thirty-six of the fifty known species are confined, but they occur also in Africa and in South-Eastern Asia. They are arboreal and mostly nocturnal in habits, and their food consists partly of fruit, etc., partly of insects. They were formerly much more widely distributed, and many fossils have been unearthed, for example, in North America. They show certain characters of a distinctly primitive kind, such, for instance, as their habit of hibernation. Their typical number of teeth is thirty-six, the same as in the lower monkeys, but fossil forms are known which possessed the full number of forty-four.
In their general build they show marked adaptation to their
arboreal life, and approach, some more and some less, the appearance
of the monkeys. The fore-limbs are considerably modified
from the condition in which they occur in ordinary mammals, in
which they are placed vertically under the body. They are
placed in a more lateral position, so that they can be moved
through greater angles, and extended over the head. In common
language, they are ceasing to be legs, and are becoming arms. As
in the monkeys, the thumb and the great toe are opposed to the
other digits so as to render the hand and foot more efficient as
grasping organs. Hence the Lemurs may be included with the
apes as 'Quadrumana,' or four-handed animals. The fingers
and toes either bear claws, as in the lower animals, or flattened
nails like those of the higher apes and man, many species possessing
[139]
[140]
[141]the two types of structures on different digits. The face is fox-like,
and lacks the human expression that is seen in the monkeys.
The brain shows considerable variation, being in some species
far more primitive, in others rather more highly developed, than
in the lower monkeys.

Fig. 125.—Ring-tailed Lemur.

Fig. 126.—The Slow Lemur or Loris.

Fig. 127.—The Tarsier (Lemur).
The history of the group has been very completely made out from fossils, and it is possible to work back to forms which, apart from their known subsequent evolution, could not be definitely separated from the ancestors of other mammal groups. The Lemurs are illustrated in Figs. 125 to 127.
The monkeys, to which we now turn, are divided into two[142] groups, the American or Western and the Eastern or Old-World types, each of which is definitely confined to the regions indicated by the names. The Western apes (Figs. 128 and 129) are by much the lower group of the two, and in fact they lead back to fossil forms which cannot be definitely distinguished from the Lemurs. They are a side branch of the monkey stem, but are in all probability the nearest living representatives of the first of our ape ancestors. They are characterised by a wide septum between the nostrils, which makes the latter open in an outward direction, a feature which enables them to be distinguished at a glance from the other group. They have further a prehensile tail, which is of much use in climbing, and generally thirty-six teeth, or four more than the Old-World group. The lowest of them are the well-known Marmosets, which have claws on all the digits except the great toe, which last has a nail. These are further characterised by the presence of four nipples, all other monkeys having only two. Correlated with this feature, the Marmosets normally bear two or three young at a birth, whereas in all other apes one is the usual number.

Fig. 128.—A Marmoset (Hapale jacchus).
The Eastern monkeys have thirty-two teeth, the same number
as in man; the septum between the nostrils is narrow, so that
these open downwards and forwards; the tail is never prehensile
and is frequently absent. They include such well-known forms as
the baboons, the Gibraltar ape, the sacred Hanuman of India,
the Diana monkey (Fig. 130), and the comical-looking Nasalis
[143]
[144]
[145](Fig. 131). These represent a further step towards the final climax
of the primate group.

Fig. 129.—Squirrel Monkey (Chrysothrix sciurca).

Fig. 130.—The Diana Monkey (Cercopithecus Diana).
Fig. 131.—The Proboscis Monkey (Nasalis larvatus).

Fig. 132.—Gibbon (Hylobates leuciscus).

Fig. 133.—Orang-Utan.

Fig. 134.—Young Chimpanzee.

Fig. 135.—The Gorilla.
A special class has to be made for the four genera of Old-World apes illustrated in Figs. 132 to 136. These are the Gorilla and Chimpanzee, which are African in distribution, and the Gibbon and Orang, which are Asiatic. They are termed the Anthropoid or man-like apes, and there can be no question that they are the nearest living relatives of the human species. This is seen in their general build, which is man-like in a high degree; by many similarities, even in the minutest details, in their skeleton and muscular system; and by the fact that their brain, while still greatly inferior to that of man, is by as much superior to that[146] of any other animal. The voice is frequently well modulated, and the expression of the emotions, whether by it or by the countenance, is very man-like. The position of the group was summed up by Huxley, after a most thorough investigation of their anatomy, in these words: "Thus, whatever system of organs be studied, the comparison of their modification in the ape series leads to one and the same result—that the structural differences which separate man from the Gorilla and Chimpanzee are not so great as those which separate the Gorilla from the lower apes." An interesting physiological proof of the close relationship between the anthropoids and man has more lately been discovered. It is found that the blood serum of any animal destroys the blood corpuscles of any other when these are mixed with it, except those of closely related species. Now the human blood serum is destructive of the corpuscles of all the lower animals, so far as is known, except those of the anthropoids.
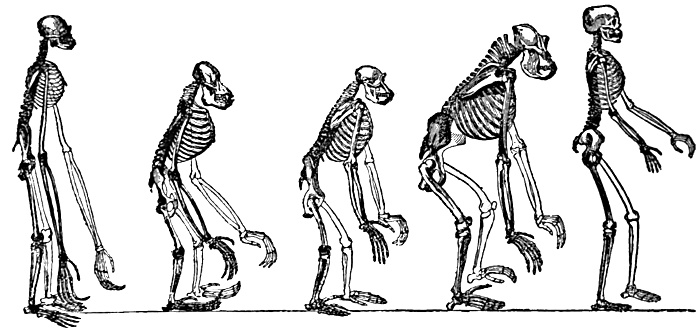
Fig. 136.—Skeletons (left to right) of Gibbon, Orang-Utan, Chimpanzee, Gorilla, and Man.
From Huxley's Man's Place in Nature.
As regards the inter-relationships of the four species, it is certain that the Gibbon is the lowest, and the nearest to the common ancestor of the other three and of man. It has indeed the man-like characteristic of walking in the erect position; but it has thirteen or fourteen pairs of ribs, as against the normal[147] twelve in man; it has long arms, like the lower Old-World apes, and large canine teeth. It is further the smallest of the group, and in habits the most like the lower monkeys.
The Gorilla and Chimpanzee are closely related. The Gorilla is not only the largest ape of the four, but in shape and build the most man-like, which is accounted for by the fact that it is less strictly arboreal than the others, and confines itself largely to the ground. Its skull is superficially much less human in appearance than that of the Chimpanzee, due to the strongly developed crests, which serve as attachments for the powerful muscles of the lower jaw. The teeth of the Chimpanzee are more uniform in size, and the skull smoother. There is a marked difference in temperament between the two species, the Gorilla being fierce and gloomy and quite untamable, while the Chimpanzee is of a pleasant and lively disposition, and can, as is well known, be trained to wear clothes, eat with a fork and knife, etc.
The Orang-Utan is found inhabiting forest ground in Borneo and Sumatra, and living largely in trees, in which it builds nests as temporary sleeping-places. It is a clumsy-looking animal, supporting itself on the knuckles of its hands when travelling along the ground, and moving but slowly. It has twelve pairs of ribs, the same number as in man, and one fewer than in the Gorilla and Chimpanzee.
Practically all the important anatomical distinctions between man and the anthropoids are reducible to two causes: the change from the arboreal existence to a life on the ground and in the open country, and the great development of the intelligence. When the change in the mode of life occurred, it is obvious that an erect carriage would possess an advantage over the stooping gait of the ape, in which it is neither definitely a biped nor a quadruped. The erect position necessitated a better-developed heel, stronger calf and hip muscles, and a more parallel position and stronger development of the great toe. It also brought about a shortening of the fore-arms and a widening of the pelvis. Much has been made of the difference in the foot, between the condition of a grasping organ, with an opposable great toe, and that seen in man.[148] But the change is just what we should devise in order to make the foot stronger for propelling the body on the ground. Moreover, the difference is more obvious than fundamental, and it is well known that in armless persons, who develop the possibilities of their feet, the latter members can still be turned into wonderfully efficient 'hands.'
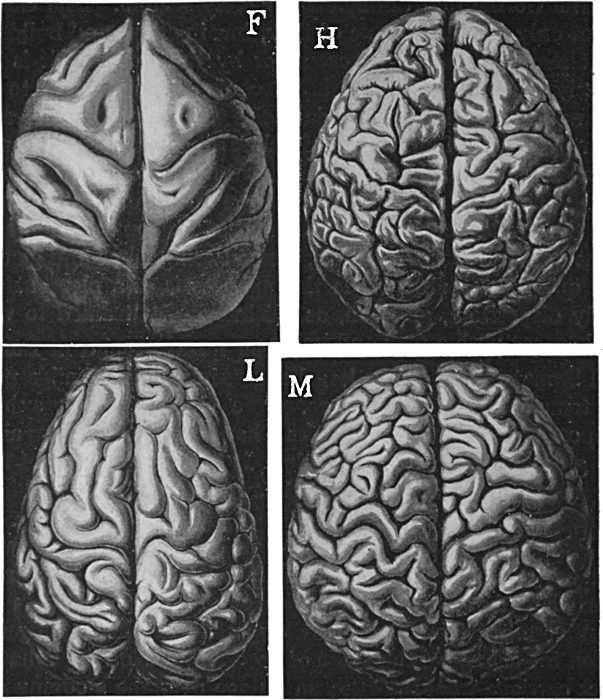
Fig. 137.—Brains of (F) Cercopithecus (Eastern Monkey); (H) Australian Bushman; (L) Chimpanzee; (M) European.
In brain development the anthropoids are, as already mentioned, greatly inferior to man; in the Gorilla, the largest-[149]brained type, the cranial capacity never, so far as is known, exceeds 600 cubic centimetres, whereas the average in man is about 1500, and the smallest known about 930. Apart from the difference in size, however, there is a surprisingly close similarity between the anthropoid and the human brains, which may be followed even in the particular arrangement of the fissures. That the gap between the two types in respect of the character in question is by no means extraordinarily wide may be seen from Fig. 137, in which a series of brains are depicted. The size of the hemispheres and the wrinkling of their surface are the characters from which we judge the brain development, and it is obvious that in these respects the step from the lower monkeys to the Chimpanzee is greater than that between the Chimpanzee and the lowest human type. In the teeth also, as is shown on Fig. 138, there is a gradual change from the lemur to the human type.
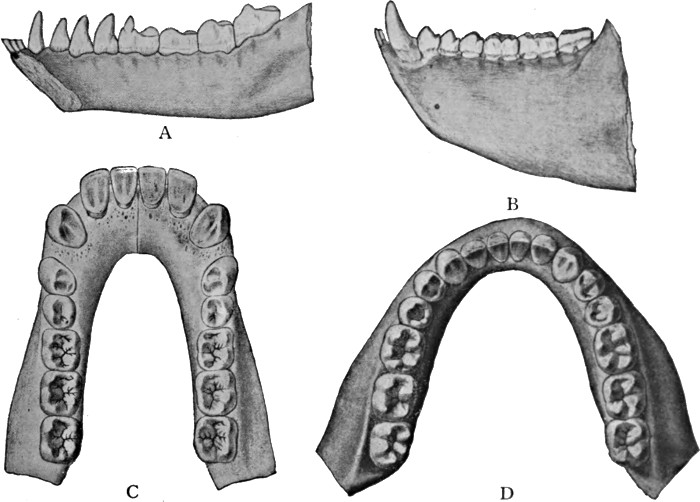
Fig. 138.
A, Lower jaw of Pelycodus, a primitive extinct lemur, with eleven teeth on either side; B, Lower jaw of red howling monkey (a Western ape), with nine teeth on either side; C, Lower jaw of chimpanzee, with eight teeth on either side; D, Lower jaw of man.
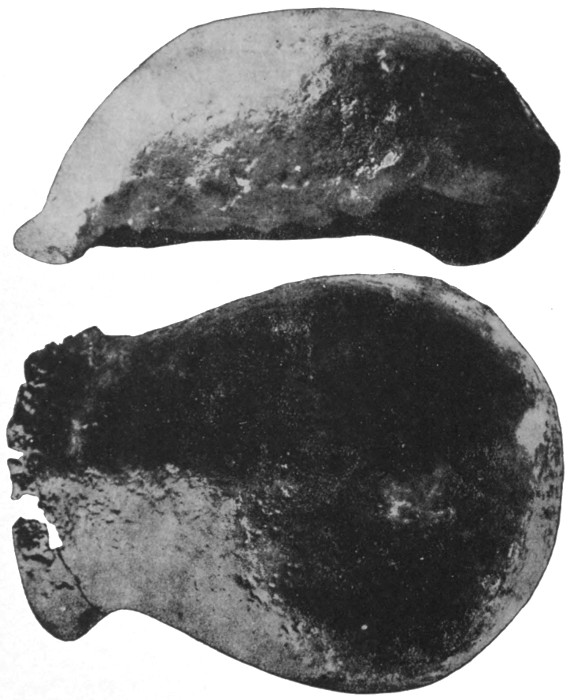
Fig. 139.—The Pithecanthropus skull from the side and from above.
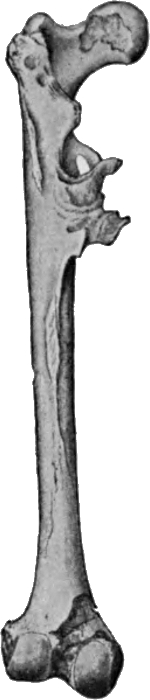
Fig. 140.—The Pithecanthropus femur, from behind. The bone shows an exostosis, evidently caused by an injury.
Thus there can be no reasonable doubt that man has evolved from an ancestor which, if it existed to-day, we should without hesitation class as an anthropoid ape. Could any doubt have remained, it would have been set aside by the discovery, in 1891, of a being occupying a position about midway between the highest apes and the most primitive known man. This is the Pithecanthropus, whose remains were discovered in Java in a volcanic deposit of somewhat doubtful age, but probably belonging to an era when a primitive type of man was already in existence. The remains were indeed somewhat scanty, consisting of the roof of the skull, a thigh bone, and a fragment of the lower jaw, the former two of which are illustrated in Figs. 139 and 140. From these[151] fragments of the skeleton numerous deductions have been drawn, of greater or less probability. It may be said with practical certainty, however, that this ape-man was of the size of a smallish man, and that he was accustomed to walk and stand in the characteristically human erect attitude. The cranial capacity has been calculated at from 850 to 1000 cubic centimetres, or considerably greater than the highest existing apes, and about equal to that of the lowest known human specimens. His statue, as his discoverer conceived him, is illustrated in Fig. 141.

Fig. 141.—Imaginative statue of Pithecanthropus erectus.
A restoration of the skull is shown in Fig. 142, and it is apparent that in respect of the shape of the roof, at least, the Pithecanthropus stands just about midway between the Chimpanzee and the most primitive living man. The gradual approach to the human type, as we move upwards in the primate scale, is very striking. The Pithecanthropus is by some regarded as the result of an abortive attempt at 'man making,' by others as a true transition form. We cannot in any case be very far from the truth if we hang up his picture among the portraits of our ancestors, for the transitional form would necessarily be closely similar to him in its main features.
The next ancestor of whom we have any knowledge is definitely a human being. This is the primitive man who inhabited Europe in earlier Diluvial times, particularly in the interval between the first and the second great ice ages. To him, from the place of his first discovery, the name of the Neandertal man has been applied, and he is classed by scientists as belonging to a different species from modern man, the latter being named Homo sapiens, while he is given the less flattering name of Homo primigenius. He was characterised, as may readily be seen from the skull illustrated in Fig. 143, by a very low and receding forehead,[152] with heavy ridges of bone over the eyes. In shape of head he stands midway between the Javan ape-man and the Australian native, the lowest type existing at the present day. Further ape-like peculiarities are the prominence of the lower part of the face, the very large and massive lower jaw, and the receding chin, shown in Fig. 144. From the skulls which have been found it is possible to form a good idea of the man's appearance, which idea has been expressed by a German sculptor in the bust illustrated in Fig. 145.
One may be permitted to hazard a guess at the cause of the process "running to brain," which is the main feature of the last phases of man's Evolution. The most probable theory seems to be that man came into existence owing to the disappearance of forest over an area inhabited by some high anthropoid ape. Ill-adapted as this ape would certainly be for a life on the plains, it was saved from extinction only by its high intelligence. And as cunning and reason would now, in the new environment, be the most important assets, the process of natural selection made for progress chiefly in respect of these characters.
The faculty of articulate speech, which we must regard as an accidental result of the great brain development, has given the human species that great advantage which it possesses over all other animals of being able to accumulate knowledge and experience from generation to generation. It is this mass of experience, which is not inherent in man's nature, but has to be impressed afresh on each successive generation, which accounts for man's unique position in the animal world. But it is no part of the scheme of this book to deal with the evolution of language or invention or culture, and we must conclude.
Mankind are divisible into many types and races, some of
which, like the Australian aborigines and the Veddas of Ceylon,
are relatively primitive, others like the Germanic races being
undoubtedly high in the series. None of the differences are
sufficient, however, to make it necessary to regard mankind except
as members of a single species. The lower races have from time
to time disappeared before the higher, and the process continues
[153]
[154]
[155]
[156]
[157]at the present time. However much we may regret it, this process
has undoubtedly been a great factor in the progress of the species
as a whole.
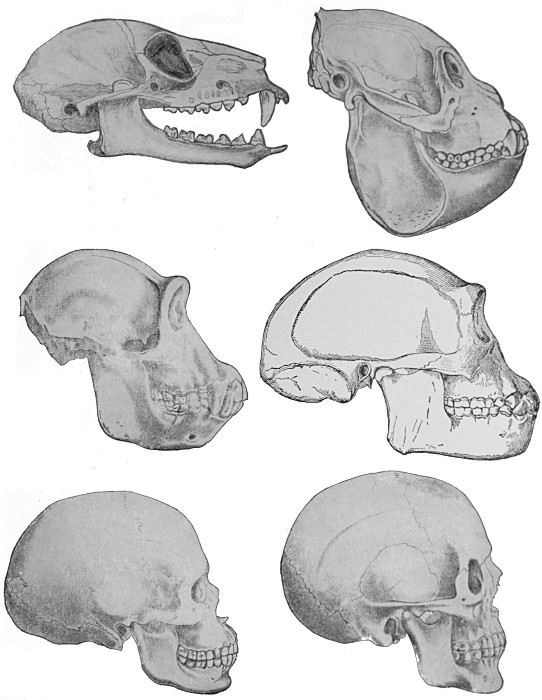
Fig. 142.
Skulls of
Lemur.
Chimpanzee.
Australian Bushman.
Skulls of
Howling monkey (Mycetes).
Pithecanthropus (restored).
European man.
Fig. 143.—Skull of Primitive Man, from Le Moustier.

Fig. 144.—The Heidelberg lower jaw (Homo primigenius).
Fig. 145.—Bust of Homo primigenius, by Hyatt Meyer.
Fig. 146.—Australian Bushman.
Within the cultivated races, however, man has practically ceased to evolve, at least in so far as concerns the main lines on which his Evolution has thus far proceeded. For in creating the artificial conditions of civilisation, he refuses any longer to be governed by the stern law of nature, which decides that the fit shall live and multiply, and the unfit surely perish. There is in fact evidence that conditions of civilisation are making for retrogression rather than for progress, a state of affairs that is worthy of the most serious consideration. The only rational and scientific remedy that has been offered for this state of affairs is the institution of some moderate system of artificially guiding man's further Evolution.
PRINTED IN GREAT BRITAIN.
Minor punctuation and printer errors repaired.
Every effort has been made to replicate this text as faithfully as possible, including obsolete and variant spellings and other inconsistencies.