
With the kind regards of Franklin Story Conant.
THE HELIOTYPE PRINTING CO. BOSTON
Title: The Cubomedusæ
Author: Franklin Story Conant
Release date: February 26, 2017 [eBook #54241]
Most recently updated: October 23, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/54241
Credits: Produced by Donald Cummings, Bryan Ness and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive/American Libraries.)
Transcriber’s Note: The images contained within black borders are clickable for a larger version, if you are using a browser/device that supports this functionality.
Memoirs from the Biological Laboratory
OF THE
JOHNS HOPKINS UNIVERSITY
IV, 1
WILLIAM K. BROOKS, EDITOR
A DISSERTATION PRESENTED FOR THE DEGREE OF DOCTOR OF PHILOSOPHY,
IN THE JOHNS HOPKINS UNIVERSITY, 1897
BY
FRANKLIN STORY CONANT
A MEMORIAL VOLUME
BALTIMORE
The Johns Hopkins Press
1898
PRINTED BY
The Lord Baltimore Press
THE FRIEDENWALD COMPANY
BALTIMORE, MD., U. S. A.
With the kind regards of Franklin Story Conant.
THE HELIOTYPE PRINTING CO. BOSTON
This Treatise is printed after the author’s death, as a Memorial by his friends, fellow-students and instructors, with the aid of the Johns Hopkins University. It consists of his Dissertation, reprinted from the copy which was accepted by this University at his examination for the degree of Doctor of Philosophy in June, 1897.
As he had made many notes on the embryology of the Cubomedusæ, and had hoped to complete and publish them together with an account of physiological experiments with these medusæ, he had described the Dissertation on the title-page as Part I, Systematic and Anatomical, and he went to Jamaica immediately after his examination to continue his studies and to procure new material, and he there lost his life.
Franklin Story Conant was born in Boston on September 21, 1870, and he died in Boston on September 13, 1897, a few days after his arrival from Jamaica, where he had contracted yellow fever through self-sacrificing devotion to others.
He was educated in the public schools of New England; at the University of South Carolina; at Williams College, where he received the degree of Bachelor of Arts in 1893; and in the Johns Hopkins University, where he received the degree of Doctor of Philosophy in 1897, and where he was appointed a Fellow in 1896 and Adam T. Bruce Fellow in 1897.
Most of his instructors have told us that they quickly discovered that Conant was a young man of unusual intelligence and energy and uprightness, and as his education progressed he secured the esteem and the affectionate interest of all who had him in charge, so that they continued to watch his career with increasing pride and satisfaction.
He entered the Johns Hopkins University in the spring of 1894, and at once joined the party of students in zoology who were working, under my direction, in the marine laboratory of the University at Beaufort, North Carolina; and from that time until his death he devoted himself continually, without interruption, to his chosen subject—spending his winters in the laboratory in Baltimore, and devoting his summers to out-of-door studies at Beaufort and at Wood’s Holl, and in Jamaica.
It is as a student and not as an investigator that we most remember Conant, for most of his time was given to reading and study on subjects of general educational value; although he had begun, before his death, to make original contributions to science and to demonstrate his ability to think and work on independent lines.
His study of the Chaetognaths was undertaken only for the purpose of verifying the account of their anatomy and development in the text books, but it soon showed the presence at Beaufort of several undescribed species. Without interrupting his more general studies, he employed his odd moments for three years in their systematic analysis, and at last published two papers, “Description of Two New Chaetognaths,” and “Notes on the Chaetognaths,” which show notable power of close and[x] accurate observation and of exact description; and, while short, are valuable contributions to our knowledge of this widely distributed but difficult group.
As he appreciated the value to one who has devoted himself to zoology of thorough acquaintance with physiological problems and the means for solving them, he wished, after he had completed his general course in physiology, to attempt original research in this field; and, at the suggestion of Professor Howell, he, in company with H. L. Clark, his fellow student, undertook and successfully completed an investigation of which Professor Howell gives the following account:
In connection with Mr. H. L. Clark, Mr. Conant undertook to investigate the character of the nervous control of the heart beat in decapod crustaceans. They selected the common edible crab, Callinectes hastatus, and made a series of most careful experiments and dissections which resulted in proving the existence of one inhibitory nerve and two accelerator nerves passing to the heart on each side from the thoracic ganglion. They not only demonstrated the physiological reaction of these nerves, but traced out successfully their anatomical course from the ganglion to the pericardial plexus. It seemed hardly probable from an a priori standpoint that in an animal like the crab there should be any necessity for an elaborate nervous mechanism to regulate the beat of the heart, but their experiments placed the matter beyond any doubt, and have since served to call attention to this animal as a promising organism for the study of some of the fundamental problems in the physiology of the heart. As compared with previous work upon the same subject it may be said that their experiments are the most definite and successful that have yet been made.
His chief completed work, the Dissertation on The Cubomedusæ, is here printed; and through it the reader who did not know Conant must decide whether he was well fitted, by training and by natural endowments, for advancing knowledge. I myself felt confident that the career on which he had entered would be full of usefulness and honor. I was delighted when he was appointed to the Adam T. Bruce Fellowship, for I had discovered that he was rapidly becoming an inspiring influence among his fellow students in the laboratory, and I had hoped that we might have him among us for many years, and that we might enjoy and profit by the riper fruits of his more mature labors.
Immediately after his examination for the degree of Doctor of Philosophy in June, 1897, he set out for Jamaica to continue his studies at the laboratory which this University had established for the summer at Port Antonio, and he there worked for nearly three months on the development, and on the physiology of the sense-organs, of the Cubomedusæ.
His notes and specimens are so complete that I hope it will be possible to complete in Baltimore, at an early day, the work which he had expected to carry on this year.
After the sudden and alarming death of the director of the expedition, Dr. J. E. Humphrey, Conant took the burden of responsibility upon himself, and while he fully appreciated his own great danger, he devoted himself calmly and methodically to the service of others who, in their afflictions, needed his help, and he fell in the path of duty, where he had always walked, leaving behind him a clear and simple account of all the business of the laboratory and of his scientific work, and of his own affairs, complete to the day before his death.
Immediately after the opening of the University in October his friends and companions and instructors assembled to express the sorrow with which they had heard the sad news of his death, and to record their love and esteem for the generous, warm-hearted friend who in all the relations of life had proved himself so worthy of their affectionate remembrance. At this meeting those who had worked at his side in our laboratories recalled his steadfast earnestness in the pursuit of knowledge, and the encouragement they had found in his bright example; while those who had been his instructors spoke of him as one who had bettered their instruction and enriched all that he undertook by sound and valuable observations and reflections. While all united in mourning the untimely loss of one who had shown such rich promise of a life full of usefulness and honor and distinction, it was pointed out with pride that his end was worthy of one who had devoted it to the fearless pursuit of truth, and to generous self-sacrifice and noble devotion to others; and it was resolved, “That we prize the lesson of the noble life and death of Franklin Story Conant.”
1. Description of Two New Chaetognaths. Johns Hopkins University Circulars, No. 119, June, 1895.
2. Notes on the Chaetognaths. Johns Hopkins University Circulars, No. 126, June, 1896.
3. The Inhibitory and Accelerator Nerves to the Crab’s Heart (an abstract), by F. S. Conant and H. L. Clark. Johns Hopkins University Circulars, No. 126, June, 1896.
4. On the Accelerator and Inhibitory Nerves to the Crab’s Heart, by F. S. Conant and H. L. Clark. The Journal of Experimental Medicine, Vol. I, No. 2, 1896.
5. Notes on the Cubomedusæ (an abstract). Johns Hopkins University Circulars, No. 132, November, 1897.
6. The Cubomedusæ. (This was accepted in June, 1897, as his thesis for the degree of Doctor of Philosophy, and it is here printed.)
| PAGE | |
| INTRODUCTION | 1 |
| PART I: SYSTEMATIC | 3 |
| Family I: Charybdeidæ | 3 |
| Charybdea Xaymacana | 4 |
| “ II: Chirodropidæ | 4 |
| “ III: Tripedalidæ | 5 |
| Tripedalia cystophora | 5 |
| PART II: GENERAL DESCRIPTION OF THE ANATOMY OF THE CUBOMEDUSÆ | 7 |
| A. Charybdea Xaymacana | 7 |
| a. Environment and Habit of Life | 7 |
| b. External Anatomy | 8 |
| 2. Form of Bell | 8 |
| 3. Pedalia | 8 |
| 4. Sensory Clubs | 9 |
| 5. The Bell Cavity and its Structures | 10 |
| (a) Proboscis | 11 |
| (b) Suspensoria, or Mesogonia | 11 |
| (c) Interradial funnels, or funnel cavities | 11 |
| (d) Velarium | 12 |
| (e) Frenula | 12 |
| (f) Musculature | 12 |
| (g) Nerve ring | 13 |
| c. Internal Anatomy | 13 |
| 6. Stomach | 13 |
| 7. Phacelli | 14 |
| 8. Peripheral Part of the Gastro-Vascular System | 14 |
| (a) Stomach Pockets (Valves and Mesogonial Pockets) | 14 |
| (b) Marginal Pockets | 17 |
| (c) Canals of the Sensory Clubs and Tentacles | 17 |
| 9. Reproductive Organs | 19 |
| [xvi]10. Floating and Wandering Cells | 20 |
| B. Tripedalia Cystophora | 22 |
| a. Habitat | 22 |
| b. External Anatomy | 23 |
| c. Internal Anatomy | 24 |
| PART III: DESCRIPTION OF SPECIAL PARTS OF THE ANATOMY | 27 |
| A. Vascular Lamellæ | 27 |
| B. Nervous System | 37 |
| LITERATURE | 57 |
| TABLE OF REFERENCE LETTERS | 58 |
| DESCRIPTION OF FIGURES | 60 |
Jelly-fish offer to the lover of natural history an inexhaustible store of beauty and attractiveness. One who has studied them finds within him a ready echo to Haeckel’s statement that when first he visited the seacoast and was introduced to the enchanted world of marine life, none of the forms that he then saw alive for the first time exercised so powerful an attraction upon him as the Medusæ. The writer counts it a rare stroke of fortune that he was led to the study of a portion of the group by the discovery of two new species of Cubomedusæ in Kingston Harbor, Jamaica, W. I., while he was with the Johns Hopkins Marine Laboratory in June of 1896.
The Cubomedusæ are of more than passing interest among jelly-fish, both because of their comparative rarity and because of the high degree of development attained by their nervous system. One fact alone suffices to attract at once the attention of the student of comparative morphology—that here among the lowly-organized Cœlenterates we find an animal with eyes composed of a cellular lens contained in a pigmented retinal cup, in its essentials analogous to the vertebrate structure. Perhaps this and other facts about the Cubomedusæ would be more generally known, had they not been to a certain extent hidden away in Claus’s paper on Charybdea marsupialis (’78), which, while a record of careful and accurate work, is in many respects written and illustrated so obscurely that it is very doubtful whether one could arrive at a clear understanding of its meaning who was not pretty well acquainted with Charybdea beforehand.
Before Claus’s paper was received at this laboratory, H. V. Wilson went over essentially the same ground upon a species of Chiropsalmus taken at Beaufort, N. C. When the article on Charybdea marsupialis appeared, however, the results were so similar that Wilson did not complete for publication the careful notes and drawings he had made.
Haeckel’s treatment of the Cubomedusæ in his “System” (’79) in the Challenger Report (’81) is much more lucid than Claus’s; but the extended scope of his work and the imperfect preservation of his material prevented a detailed investigation, and for a more complete and readily intelligible[2] account of the structure of the Cubomedusæ a larger number of figures is desirable.
In the foregoing facts lies whatever excuse is necessary for repeating in the present paper much that has already seen print in one form or another.
It seems advisable first of all to establish the systematic position of the two newly found species, Charybdea Xaymacana and Tripedalia cystophora. Haeckel’s classification, as given in his “System der Medusen,” is an excellent one and will be followed in this case. One of the new species, however, will not classify under either of Haeckel’s two families, so that for it a new family has been formed and named the Tripedalidæ. In showing the systematic position of the two new forms, an outline of Haeckel’s classification will be given, so far as it concerns our species, together with the additions that have been made necessary.
Characteristics: Acraspeda with four perradial sensory clubs which contain an auditory club with endodermal otolith sac and one or several eyes. Four interradial tentacles or groups of tentacles. Stomach with four wide perradial rectangular pockets, which are separated by four long and narrow interradial septa, or cathammal plates. Gonads in four pairs, leaf-shaped, attached along one edge to the four interradial septa. They belong to the subumbrella, and are developed from the endoderm of the stomach pockets, so that they project freely into the spaces of the pockets.
Cubomedusæ with four simple interradial tentacles; without marginal lobes in the velarium, but with eight marginal pockets; without pocket arms in the four stomach pockets.
Charybdeidæ with four simple interradial tentacles with pedalia; with velarium suspended, with velar canals and four perradial frenula. Stomach flat and low, without broad suspensoria. Four horizontal groups of gastric filaments, simple or double, tuft or brush-shaped, limited to the interradial corners of the stomach.
Bell a four-sided pyramid with the corners more rounded than angular, yet not so rounded as to make the umbrella bell-shaped. The sides of the pyramid parallel in the lower two-thirds of the bell, in the upper third curving inward to form the truncation; near the top a slight horizontal constriction. Stomach flat and shallow. Proboscis with four oral lobes, hanging down in bell cavity a distance of between one-third and one-half the height of bell; very sensitive and contractile, so that it can be inverted into the stomach. The four phacelli epaulette-shaped, springing from a single stalk. Distance of the sensory clubs from the bell margin one-seventh or one-eighth the height of bell. Velarium in breadth about one-seventh the diameter of the bell at its margin. Four velar canals in each quadrant; each canal forked at the ends, at times with more than two branches. Pedalia flat, scalpel-shaped, between one-third and one-half as long as the height of bell. The four tentacles, when extended, at least eight times longer than the bell. Sexes separate. Height of bell, 18-23 mm.; breadth, about 15 mm. (individuals with mature reproductive elements); without pigment. Found at Port Henderson, Kingston Harbor, Jamaica.
As may be seen from the above, C. Xaymacana differs only a little from the C. marsupialis of the Mediterranean. Claus mentions in the latter a more or less well defined asymmetry of the bell, which he connects with a supposed occasional attachment by the proboscis to algæ. In C. Xaymacana I never noticed but that the bell was perfectly symmetrical. C. Xaymacana is about two-thirds the size given by Claus for his examples of C. marsupialis, which were not then sexually mature. It has 16 velar canals instead of 24 (32), as given by Haeckel, or 24 as figured by Claus. Difference in size and in number of velar canals are essentially the characteristics upon which Haeckel founded his Challenger species, C. Murrayana.
Cubomedusæ with four interradial groups of tentacles; with sixteen marginal pockets in the marginal lobes of the velarium, and with eight pocket arms, belonging to the exumbrella, in the four stomach pockets.
This family is represented in American waters by a species of Chiropsalmus, identified by H. V. Wilson as C. quadrumanus, found at Beaufort, North Carolina.
Cubomedusæ with four interradial groups of tentacles, each group having three tentacles carried by three distinct pedalia; without marginal lobes in the velarium; with sixteen marginal pockets; without pocket arms in the stomach pockets.
For the present the characteristics of family and genus must necessarily be for the most part the same. The genus is distinguished by having twelve tentacles in four interradial groups of three each; velarium suspended by four perradial frenula; canals in the velarium; stomach projecting somewhat convexly into the bell cavity, with relatively well-developed suspensoria; four horizontal groups of gastric filaments, each group brush-shaped, limited to the interradial corners of the stomach.
Shape of bell almost exactly that of a cube with rounded edges; the roof but little arched. The horizontal constriction commonly seen near the top of the bell in the Cubomedusæ not present. Proboscis with four oral lobes; hanging down in the bell cavity generally more than half the depth of the cavity and at times even to the bell margin. In the gelatine of the proboscis an irregular number (15-21) of sensory organs resembling otocysts, from the presence of which comes the specific name. Phacelli brush-shaped, composed of from seven to thirteen filaments springing from a single stalk in each quadrant, or rarely from two separate stalks in one of the quadrants. Distance of the sensory clubs from the bell margin about one-fifth or one-fourth of the height of bell. Breadth of velarium about one-sixth the diameter of bell at margin; with six velar canals in each quadrant; the canals simple, unforked. Pedalia flattened, shaped like a slender knife blade, about half as long as the height of the bell. Tentacles at greatest extension observed two and a half times the length of pedalia. Sexes separate. Height of bell in largest specimens (reproductive elements mature) eight or nine mm. Breadth same as height or even greater. Color a light yellowish brown, due in large part to eggs or embryos in the stomach pockets. The reproductive organs especially prominent by reason of their similar color. Found in Kingston Harbor, Jamaica.
It will be seen from the above that Tripedalia possesses two of the[6] characteristics of the Charybdeidæ and two of the Chirodropidæ. The family was named from the prominent feature of the arrangement of the tentacles, in groups of three with separate pedalia. The small size of T. cystophora is worthy of note in connection with the fact that of the twenty species of Cubomedusæ given by Haeckel in his “System” only two are smaller than 20 mm. in height, and those are the two representatives of Haeckel’s genus Procharagma, the prototype form of the Cubomedusæ, without pedalia and without velarium. While Tripedalia has both pedalia and velarium, it may be perhaps that its small size, taken in connection with characteristics just about midway between the Charybdeidæ and the Chirodropidæ, indicate that it is not a recently acquired form of the Cubomedusæ.
1. The Cubomedusæ are generally believed to be inhabitants of deep water which come to the surface only occasionally. Both of the Jamaica species, however, were found at the surface of shallow water near the shore, and only under these circumstances. Whether these were their natural conditions, or whether the two forms were driven by some chance from the deep ocean into the Harbor and there found their surroundings secondarily congenial, so to speak, can be a matter of conjecture only. C. Xaymacana was taken regularly a few yards off-shore from a strip of sandy beach not ten minutes row from the laboratory at Port Henderson. It was seen only in the morning before the sea-breeze came in to roughen the water and to turn the region of its placid feeding-ground into a dangerous lee-shore. Some of the specimens taken contained in the stomach small fish so disproportionately large in comparison with the stomach that they lay coiled up, head overlapping tail. The name Charybdea, then, from the Greek χαρύβδις (a gulf, rapacious), seems to be no misnomer. It is worth mentioning that the digestive juices left the nervous system of the fish intact, so that from the stomach of a Charybdea could be obtained beautiful dissections, or rather macerations, of the brain, cord, and lateral nerves of a small fish.
In size C. Xaymacana agrees very well with the average of the genus. The four single tentacles characteristic of the genus are very contractile, varying from two or three to six or seven inches in length, and probably if measurements could be taken while the animal was swimming freely about, the length would be found to be greater still. Charybdea is a strong and active swimmer, and presents a very beautiful appearance in its movements through the water, the quick, vigorous pulsations contrasting sharply with the sluggish contractions seen in most Scyphomedusæ. With its tentacles streaming gracefully behind, an actively swimming Charybdea presents a fanciful resemblance to a[8] comet or meteor. When an attempt is made to capture one, it will often escape by going down into deeper water—as indeed do other jelly-fish. Escape from observation is all the more easy by reason of the entire absence of pigment excepting for the small amount in the sensory clubs. The yellowish or brownish color usually stated as common in the Cubomedusæ is nowhere present in C. Xaymacana.
2. Form of Bell. C. Xaymacana shows the typical division of the external surface into four almost vertical perradial areas (Figs. 1-3, p), separated by four stoutly arched interradial ribs or bands (Figs. 1-3, i). These ribs thus play the part of corners to the Cubomedusan pyramid. They are formed by the thickenings of the jelly of the exumbrella, and serve to give the necessary strength to the four interradial corners, each of which bears one of the four tentacles at its base. Each rib is further divided into two longitudinal strips by a vertical furrow lying exactly in the interradius (Fig. 2, ifr). The surface of the exumbrella is thus marked by twelve longitudinal furrows, as seen in the same figure (2). Of these, four are the interradial furrows just mentioned; the other eight are the adradial (afr) furrows, which set off the four perradial surfaces of the pyramid from the four interradial ribs or bands of the corners, each of which is again subdivided, as mentioned above, by the shallower interradial furrows. Each interradial furrow ends above the base of the corresponding pedalium, at about the level of the sensory club; each adradial furrow diverges toward the perradius in the lower third of its course, and thus with its companion furrow narrows down the perradial surface of the pyramid in the lower part of the bell to an area of not much greater width than the niches in which the sensory clubs lie. The projecting interradial corners are of course correspondingly enlarged in the lower part of the bell, and in this way the contours of the surface are changed from those figured in the view of the bell from above (Fig. 2) to those of Fig. 3, which represents a view of the bell margin from below.
3. Pedalia. From the base of the interradial corner bands spring the four pedalia (Fig. 1, pe), gelatinous appendages of the margin having much the same shape as the blade of a scalpel. These in turn bear on their distal ends, as direct continuations, the long, contractile, simple tentacles. The relatively stiff pedalia have the same relation to the flexible tentacles that a driver’s whip-stock has to the long lash. In the living animal the pedalia are found attached to the margin at an angle[9] of about 45° with the longitudinal axis of the bell. In the preserved specimens they are bent in toward the axis by the contraction of the strong muscles at their base, in which position they are figured by Claus for C. marsupialis (’78, Taf. I., Figs. 1 and 2).
The pedalia are in reality processes belonging to the subumbrella, as will be shown in the section treating of the vascular lamella. They are composed chiefly of gelatine covered with thin surface epithelium and carrying within the gelatine the basal portion of the tentacle canals. They have received various names at the hands of the writers. Gegenbaur called them “Randblätter.” Claus gave them the name of “Schirmlappen,” and incorrectly homologized them with the marginal lobes of other Acraspeda. Claus’s error was corrected by Haeckel, who termed them “Pedalia” or “Gallertsockel,” and homologized them with the pedalia of the Peromedusæ. Besides furnishing a base of support for the tentacles they may perhaps also serve as steering apparatus, a function for which their thin blade-like form would be admirably adapted.
Internal to the base of each pedalium, between it and the velarium, is found a funnel-shaped depression of the ectodermal surface. This is shown in Fig. 5 (ft) in longitudinal section, and in cross-section in Fig. 16. In the latter figure the epithelium of the outer wall of the funnel (mt) is shown much thickened, the result of a stout development of muscle fibres. These are the muscles that in the preserved specimens cause the inward contraction of the pedalia referred to above.
4. Sensory Clubs (marginal bodies, rhopalia). In spite of their position above the bell margin, the four sensory clubs, representing as they do transformations of the four perradial tentacles, are properly classed with the pedalia and interradial tentacles as appendages of the margin. They lie protected in somewhat heart-shaped excavations or niches in the perradial areas of the exumbrella. Each sensory niche is partially roofed over by a covering scale, a hood-like projection from the exumbrella. Below the covering scale the water has free access to the niche and to the sensory club within it. The sensory club consists of a hollow stock directly homologous with tentacle and canal, and a terminal, knob-like swelling, the sensory portion proper. The latter contains on its inner surface—the surface turned towards the bell cavity—two complicated unpaired eyes with lens, retina, and pigment, lying one above the other in the median line; and at the sides of these, two pairs of small, simple, pigmented, bilaterally symmetrical eye spots. At the end of the club, that is, on its lowermost point, lies a sac that contains a[10] concretion and is usually considered auditory. The canal of the stalk is directly continuous with the gastro-vascular system. In the swollen knob of the sensory club it forms an ampulla-like terminal expansion.
As was pointed out by Claus, the bottom of the sensory niche—by bottom is meant the vertical wall that separates the space of the niche from the bell cavity—is formed from the subumbrella only. This arrangement of parts, apparently impossible for a structure so far removed from the bell margin as the sensory niche, will be explained more fully under the special topic of the vascular lamellæ, or cathammal plates. It is sufficient at this point to refer to Fig. 44, which shows the shield-shaped area mapped out by a vascular lamella that connects the endoderm of the stomach pocket with the ectoderm of the bottom of the niche. By this the exumbrella is completely cut off from any part in the formation of the bottom of the niche. Cross and vertical sections through the niche (Figs. 39 and 37) help to a better understanding of these relations. Since the base of the stalk of the sensory niche lies within the ring of vascular lamella, the whole organ as well as the bottom of the niche belongs to the subumbrella, and so in spite of its position some distance upwards from the bell margin the sensory club is very properly called a “marginal body” (Randkörper).
The epithelium of the sensory niche consists entirely of the flattened ectodermal surface layer common to the whole exumbrella. No differentiation suggestive of nervous function in addition to that of the sensory clubs can be discovered, although it would be quite natural to expect to find something of the sort, as intimated by Claus (’78, p. 27).
It is worth while to mention again the fact that the eyes are directed inwardly toward the cavity of the bell. The larger and lower of the two median eyes looks into the bell cavity horizontally; the smaller upper eye is turned upward toward the region of the proboscis. This is in the normal pendant position of the sensory club. The stalk, however, is very flexible, and a range of other positions of the sense organs is possible, although nothing was observed to suggest that such positions were within the control of the animal. The eyes evidently have as their chief function to receive impressions of what is going on inside the bell, not outside. Perhaps the strongly biconvex, almost spherical lenses of the median eyes also point to a focus on near and small objects.
5. The Bell Cavity and its Structures. In general, the bell cavity repeats the external form of the bell, being almost cubical. In cross-section it appears very nearly square with the angles in the interradii as[11] seen in the series of drawings that figure sections of the whole jelly-fish at different levels (Figs. 6-16). Above, the bell cavity is roofed over by the stomach; below, it is open freely to the water, the opening being narrowed somewhat by the diaphragm-like velarium (Fig. 3, v); the four flat perradial sides are bounded by the walls of the four broad stomach pockets, to be described when we come to the internal anatomy.
(a) The Proboscis. From the stomach there hangs down into the bell cavity the proboscis or manubrium, which consists of a short funnel-shaped stalk bearing on its distal end the four mouth lobes or lips. The latter are somewhat broadly V-shaped processes lying in the perradii with the convexity directed outwards, and with the concavity on the inside forming the beginnings of four perradial furrows that are continued upwards to the stomach. The four furrows are shown in the stalk of the proboscis in Fig. 11, which represents a section taken a little above the level of the mouth lobes. The same cross-shaped section of the stalk shows the four perradial prominences or ridges overlying the furrows, which are the direct continuations of the four projecting mouth lobes.
(b) The Suspensoria or Mesogonia. The stomach (leaving out of consideration the proboscis) hangs down into the bell cavity as a slightly sagging saucer-shaped roof (Figs. 4 and 5). In the four perradii it is attached to the lateral walls of the subumbrella by four slenderly developed mesentery-like structures, the suspensoria or mesogonia. These are simple ridges of gelatine, covered of course with the epithelium of the bell cavity, which serve to keep the stomach in position much in the way that a shelf is supported by brackets (Fig. 4, su). The suspensorium accordingly has two parts, curved so as to lie at right angles with each other: a vertical portion lying along the wall of the subumbrella, and a horizontal which passes over from the vertical on to the basal wall of the stomach. In Fig. 10 the suspensorium in each quadrant is shown cut across just below the angle between the two parts, so that the two appear in the section as projections on the wall of the stomach and on the wall of the subumbrella.
(c) The Interradial Funnels or Funnel Cavities. It will be seen at once that the four suspensoria serve as partitions to divide the upper portion of the bell cavity, the part that lies between the stomach and the lateral walls of the subumbrella, into four compartments. These compartments extend upwards in the four interradii like inverted funnels, whence their name. In the series of cross-sections they can be traced[12] upwards with constantly diminishing area from the level of the suspensoria, Fig. 10 (if), to Fig. 6, which is taken very near the top of the bell. Homologous structures exist in all the Scyphomedusæ, and in some of the Lucernaridæ they are continued up even into the stalk of the attached jelly-fish.
(d) The Velarium. Charybdea, like most of the Cubomedusæ, possesses a velum-like structure around the opening of the bell cavity (Fig. 3, v). The velarium is a thin muscular diaphragm, resembling the true velum in position and essential structures, but differing from the velum in its origin, and in the possession of diverticula from the gastro-vascular system, the velar canals. Of these there are in C. Xaymacana very regularly sixteen, four in each quadrant. Their outline is seen in Fig. 3 to be forked with small irregular accessory processes. As for its origin, the velarium of the Cubomedusæ is commonly accounted to have arisen by fusion of marginal lobes, as in the case of the velarium of the Discomedusæ. Pending decisive ontological evidence, the slight notches in the four perradii seen in Fig. 3 may perhaps be taken as slight indications of a primitive unfused condition, but the question will be brought up again when the vascular lamellæ are discussed.
(e) The Frenula. Just as the stomach is attached to the walls of the subumbrella in the four perradii by the suspensoria, so in the lower part of the bell cavity the velarium is attached to the wall of the subumbrella in the perradii by four structures similar to the suspensoria, the frenula velarii. The frenula, like the suspensoria, resemble the brackets of a shelf, with the difference that in the case of the frenula the bracket is above the shelf, their purpose being evidently to keep the velarium stiff against the outflow of water produced by the pulsations of the bell. According to the greater need of strength in this case, we find the frenula stouter, more buttress-like than the suspensoria. The gelatinous ridge that gives them the necessary firmness is thickened so as to be triangular in section, as shown in Fig. 16 (frn).
(f) Musculature. As is general in medusæ, the muscular system, so far as known, is restricted to the subumbrella. It has a very simple arrangement, consisting of a continuous sheet of circular (i. e. horizontal) striated fibres, which is interrupted only in the four perradii by the radially directed muscle fibres of the suspensoria and the frenula. In each quadrant, between the muscle of the suspensorium above and that of the frenulum below, in an area just internal to the sensory niche, there lies a space free from muscle. This interruption of the muscle[13] layer is shown in Fig. 39. Under the head of musculature belonging to the subumbrella must be included also the radial, or longitudinal muscles at the bases of the pedalia, which were mentioned before (Fig. 16, mt). The mouth lobes and proboscis also are highly contractile and muscular.
(g) Nerve Ring. It is in the possession of a clearly defined nerve ring that the Cubomedusæ differ from all other Scyphomedusæ whose nervous system has been carefully studied. The nerve ring shows very plainly on the surface of the subumbrella as a well-defined clear streak. Its course is zig-zag or festoon-like. In the interradii, at the basis of the tentacles, it lies not far from the bell margin. In the perradii it rises to the level of the sensory clubs. This very striking arrangement is understood at once when it is remembered that the sensory clubs represent the four perradial primary tentacles, and were originally situated on the margin. When all the rest of the margin grew down and away from the four sensory clubs, fusing below them to form the present intact edge of the bell, the four portions of the nerve ring that lay in the perradii were left at the level of the sensory clubs, and the originally straight nerve ring was thus bent into a bow in each quadrant. The finer structure of the nerve will be treated of in the special part to be devoted to the nervous system.
6. Stomach. The shape of the stomach is approximately that of a biconvex lens, as seen in Fig. 4, which represents a Charybdea cut in halves longitudinally in the perradius. The lumen of the proboscis (the buccal stomach according to Haeckel’s terminology) communicates directly by a funnel-shaped enlargement with the stomach proper, or central stomach of Haeckel. The term basal stomach is carried over by Haeckel from the Stauromedusæ, where it has considerable significance, to the Cubomedusæ, and applied to the upper part of the central stomach. In the stalkless Cubomedusæ, however, it has no significance so far as actual structure goes, and our knowledge of the development of the Cubomedusæ is as yet too simple for us to say that the upper part of the main stomach represents what remains of the basal stomach of an earlier pedunculated stage.
The epithelium of the roof of the stomach is not specially differentiated and apparently has little or no part in digestion. The epithelium of the floor, on the other hand, is composed chiefly of very high and thickly crowded columnar cells which are usually described as coarsely granular,[14] but under high powers appear to be filled with vacuoles surrounded by a network of cell substance. Thickly interspersed among these columnar cells are goblet cells filled with mucus. The floor is thrown into numerous wrinkles by ridges in the supporting gelatine resulting in increase of digestive surface. The four perradial grooves of the proboscis are continued in the perradii along the floor of the stomach as four fairly deep furrows, which lead directly to the gastric ostia and stomach pockets—structures to be described presently. These furrows are lined with crowded columnar cells, smaller and denser than the other cells of the digestive epithelium, containing no granules and but little beside the relatively large, compact, deeply staining nuclei. The furrows probably represent special ciliated courses.
7. Phacelli. Lying in the four interradial corners of the stomach are the four phacelli or tufts of gastral filaments to the number of thirty or thirty-five in each tuft. The filaments are attached to a single stalk, like the fringe of an epaulette or the hairs of a coarse brush. The stalk bearing the filaments is an outgrowth of the lower wall of the stomach just at the point where it fuses with the upper. The phacelli are therefore structures of the subumbrella, proof of which will be found under the special topic of the vascular lamellæ. The stalk, an indication of which appears in sph. Fig. 6 (the section being a little below the axis of the stalk, which lies horizontally), consists of a firm core of gelatine covered with the high columnar epithelium of the floor of the stomach. The filaments themselves are slender processes repeating the structure of the stalk and having a central axis of gelatine for support covered with glandular epithelium, which in the case of the filaments bears numerous nettle cells. These processes are extremely contractile, and in the living animal show a continuous, slow, squirming movement like a mass of worms. The section just referred to (Fig. 6) shows diagrammatically three of these filaments (fph) cut across in each quadrant.
8. Peripheral Part of the Gastro-vascular System. The proboscis and stomach proper comprise the central part of the gastro-vascular system. In direct communication with the central is a peripheral part composed of pouches or pockets lying in the vertical sides of the cube-shaped bell, just as the central stomach lies in its roof. The peripheral part may be subdivided for convenience of description into the stomach pockets, the marginal pockets, and the canals of the tentacles and sensory clubs.
(a) Stomach Pockets. These are four broad, thin pouches lying between the exumbrella and the subumbrella in the four perradii (e. g.[15] Fig. 9, sp) and separated from one another in the interradii merely by four thin vertical strips of vascular lamella (ivl) or fusion between the two endodermal surfaces of a primitively single undivided peripheral cavity. The structure is exactly that which we should have if in a Hydromedusa, for example Liriope (Trachomedusæ), the four radial canals broadened out and the intervening cathammal plates correspondingly narrowed, until the relations in size were just reversed, and instead of four narrow radial canals separated from one another by four broad cathammal plates, we had four broad radial canals or pouches separated by four narrow cathammal plates.
The stomach pockets communicate at their top with the central stomach by means of four moderately large openings, the gastric ostia. These are seen in a side view of the whole animal as triangular spaces (Fig. 1, g. o.) near the top of the broad perradial sides. In Figures 7 and 8 they are seen in cross-sections, in Fig. 4 in vertical section.
The communication between the stomach and each stomach pocket is guarded by a valve that can cut the one entirely off from the other. The valve is simply the flexible lower margin of the gastric ostium, a thin vertical fold of the floor of the stomach, semilunar in shape, just at the point where it is passing over into the stomach pocket. A longitudinal section, such as is shown in Fig. 4, gives the best idea of the form and position of the valve that can be obtained from any simple section. Internal to the valve is seen a depression of the stomach wall, almost worthy to be called a pocket. The valve itself lies as a wall across the end of this depression, obstructing a free course to the stomach pocket. It will be seen at once that any pressure of fluids in the stomach against this vertical wall, or valve, would serve only to press it against the inner surface of the exumbrella, and thus effectually close the entrance into the stomach pocket. Such a closure would both keep the juices of the stomach from entering the pockets and the embryos in the pockets from entering the stomach before the proper time.
The depression of the floor of the stomach just internal to the valve may possibly be a structure of some morphological significance. In one series of sections it was found that in two of the quadrants the depression was deeper than that represented in Fig. 4, and extended perceptibly into the outer or vertical portion of the suspensorium. Fig. 32 is a diagram giving a vertical reconstruction in the perradius of the cross-sections in which this deepened depression was noticed. Fig. 31 is a drawing (the outline by camera lucida) of one of the cross-sections, through the lowermost[16] point of the depression. The figure gives the wall of the stomach lined with high columnar epithelium (ens), and the wall of the stomach pockets, with the suspensorium (su) connecting them. The section is taken just above the broad angle that lies between the two parts of the suspensorium, that is, in a plane parallel to the arrow a-b in Fig. 32, but a little lower down. At the points to which the reference letter x (Fig. 31) refers are seen the first indications of the division into two parts, i. e. of the apex of the angle. The next section or two lower down show the relation seen in Fig. 10 (su). There can be no doubt in this case that the depression or pocket lies in the outer vertical limb of the suspensorium. It is the position that gives it at least the appearance of some morphological significance. In two genera of Lucernaridæ named and described by Clark (’78), Halicyathus and Craterolophus, the mesogonia or suspensoria in all four perradii contain broad pockets. These mesogonial pockets in the Lucernaridæ have given rise to considerable misunderstanding owing to the fact that in some forms the reproductive organs bulge out from the stomach pockets in which they structurally lie, and come to take up a secondary position in the walls of the mesogonial pockets. The sections of Charybdea above referred to indicate that among the Cubomedusæ we may have the same structure reduced to its lowest terms, and may be a feather’s weight in favor of the view that the Cubomedusæ are descendants of an attached Lucernaria-like form.
Two more diagrams, Figs. 33 and 34, are added in order to give a more complete understanding of a gastric ostium and its neighboring structures, the mesogonial pocket and the valve. Fig. 33 is a view of the gastric ostium and valve from the stomach side, and represents the appearance that would be given by a thick section through the arrow x-y in Fig. 32, in a plane at right angles to the paper. The heavy lines outlining the gastric ostium (enr and enfl) represent the place where the plane of the section has cut across the epithelium of the roof of the stomach above the ostium and the epithelium of the floor of the pocket-like depression internal to the valve. The continuation of the two heavy lines in either side of the ostium represents the region where the roof and floor of the stomach meet; i. e., the edge of the lens-shaped stomach. The semilunar outline of the valve (vg) is shown by a light line just above the epithelium of the depression. As is seen by the reference arrow in Fig. 32, the valve lies a little external to the immediate plane of the section, and hence it is that its inner surface is seen in Fig. 33 and not a section of it. The vertical part of the suspensorium (su) is seen in section below the epithelium[17] of the depression. The reference numbers 1, 2, 3 and 4 denote the same points in Figs. 32 and 33. Fig. 32 referred to Fig. 33 would lie in a plane at right angles to the paper through the reference arrow x-v of the latter figure.
Fig. 34 represents a horizontal section through the gastric ostium at the level of the arrow a-b in Fig. 32, or arrow c-d in Fig. 33. The reference numbers 5, 6 and 7, 8 denote similar points in the two figures 33 and 34. Fig. 32 as referred to Fig. 34 is through the arrow e-f; Fig. 33 is through the arrow c-d. In the series of cross-sections, Fig. 9 is taken at a level a little below that of Fig. 34, and passes through the basal part of the valve (vg).
(b) Marginal Pockets. The part of the peripheral portion of the gastro-vascular system in each quadrant which is called the stomach pocket extends downwards as far as the sensory niche. Here by the coming together of the walls of the exumbrella and subumbrella the space between them is obliterated (Fig. 15) in the immediate perradius. From the sensory niche downward to the margin each stomach pocket is thus divided into two smaller pouches, the marginal pockets (mp). In each side of the Cubomedusan cube there are, then, in Charybdea two marginal pockets; or in all eight, a characteristic of the family Charybdeidæ. The marginal pockets as the name implies extend downwards to the bell margin, and are continued into the velarium as the velar canals. Of these (Fig. 3) there are two from each marginal pocket, or sixteen in all. The constancy in their number is one of the characteristics that distinguish C. Xaymacana from the very closely related C. marsupialis of the Mediterranean. (Compare Fig. 3 with the similar one by Claus for C. marsupialis, ’78, Taf. I., Fig. 6.) The forked shape, while to be sure the common form in C. marsupialis, is an almost invariable characteristic in C. Xaymacana. It may be mentioned again that the presence of these canals is one of the chief features that distinguish the velarium of the Scyphomedusæ from the velum of the Hydromedusæ.
(c) Canals of the Sensory Clubs and Tentacles. The four interradial definitive tentacles and the four perradial transformed tentacles, the sensory clubs, are hollow, and their canals communicate directly with the peripheral part of the gastro-vascular system. The canal of the sensory club in each quadrant leads directly out from the stomach by a somewhat funnel-shaped opening formed by the approximation of the two walls of the stomach pocket. The relation of the canal of the sensory club to the stomach pocket is seen at a glance in Fig. 37. It is given by means of cross-sections in Figs. 12-14. Figure 12 shows the inner[18] walls of the stomach pocket approaching the outer at two points, leaving between them a concavity freely open to the rest of the stomach pocket above and at the sides. Fig. 13, a little lower down, shows the two walls fused together at two points, making the interspaces a definite canal communicating with the stomach pocket above only. This canal lies directly over the sensory niche, and in the next figure (No. 14) the canal is seen to have passed through the roof of the sensory niche and to have entered the base of the stalk of the sensory club. In the enlarged end of the club, the part which bears the sensory structure, the canal widens into a terminal ampulla-like sac.
The endoderm lining the canal of the sensory club is specially differentiated. In the stalk it is more columnar than the epithelium of the stomach pockets, and is made up of cells containing a brightly staining nucleus with very little trace of cytoplasm. The cell bodies appear as if filled with a clear, non-staining fluid. Perhaps these cells give the stalk elasticity to act in connection with the thin layer of longitudinal muscle-fibres that are found just external to the supporting lamella. The epithelium of the terminal enlargement of the canal is composed of very high narrow cells, many of which show two nuclei of equal size and staining quality lying side by side.
In continuation of the specialized epithelium of the perradial furrows in the floor of the stomach the inner wall of the stomach pocket shows a strip of similar densely crowded columnar cells leading from the gastric ostium downwards to the canal of the sensory club. As in the other case, the strip probably represents a specially ciliated tract, and perhaps in it we see the reason why the canal of the sensory club is almost always found to contain either spermatozoa which are shed by the male reproductive organs directly into the stomach pocket, or else floating cells of the kind to be described in the next section.
The canals of the interradial tentacles arise from the peripheral gastro-vascular system much lower down than those of the sensory clubs, since these tentacles have preserved their primary positions with reference to the bell margin. Figure 16 represents a section taken at the level of the base of the pedalia which gives the connection of the tentacle canals with the gastro-vascular system. At the level below the sensory niche the four broad stomach pockets have been divided, as we have seen, into the right marginal pockets (mp). The figure shows that in the interradial corners the longitudinal septa (ivl, in the preceding figures), or lines of fusion between the two walls of the peripheral gastro-vascular[19] space, which divide the primitively simple space into the four stomach pockets, have come to an end, leaving a connecting canal (cc) in each corner as all that remains of the primitive uninterrupted communication between all parts of the peripheral system. It is from these four connecting canals that the tentacle canals take their origin. From this point of origin each tentacle canal passes downwards, surrounded by the gelatine of the pedalium, into the tentacle proper.
The connecting canals are of morphological importance in that they are supposed, with much reason, to represent in the Cubomedusæ the circular canal of the Hydromedusæ.
9. Reproductive Organs. The sexes are separate in Charybdea. In both sexes the reproductive organs consist of four pairs of long leaf-like bodies, each leaf attached along one edge to the wall of the subumbrella in an interradius (see Fig. 1, r), and hanging free in the stomach pockets. From this position in the stomach pockets it is evident that the reproductive organs are endodermal. The lines of attachment of each pair is just internal to the longitudinal vascular lamella that fuses the outer and inner walls of the stomach pockets together in the interradius (ivl), and the reproductive organs are therefore structures belonging to the subumbrella. It is interesting to note how careful examination of the medusan organization takes away from the importance of the outer cup, the exumbrella, and adds to that of the inner, the subumbrella. We have seen that the phacelli and the sensory clubs, from whose position it would be supposed that they belonged to the exumbrella, are organs of the subumbrella, and that there is no muscle-tissue in the exumbrella; we find now that the reproductive organs belong to the subumbrella, and it will be shown later that the tentacles, like the sensory clubs, are structures of the subumbrella also. To the exumbrella are left only the functions of support and covering.
The mature reproductive organs extend very nearly throughout the entire vertical length of the bell, and are therefore found in the series of cross-sections in all but the uppermost and lowermost (Figs. 7-15 r). The organs consist of germ cells within, covered by an epithelium of columnar cells that shows here and there nettle cells. The ova are found with different amounts of yolk, according to age, surrounding a large nucleus almost devoid of chromatin and an intensely staining nucleolus. In young ova there appears very plainly in every case at least one small deeply staining body inside the nucleus, which very much resembles the nucleolus. These are probably so-called yolk nuclei, and while I have not[20] made a special study of the ovogenesis, I infer that the constant presence of at least one, points to an origin of the ovum from a syncytium (of at any rate two cells), similar to that which has been recently shown by Doflein (’96) to occur in the formation of eggs in Tubularia. In the nearly mature ovary each ovum is surrounded by a layer of gelatine, which comes from the gelatinous sheet that enters the leaf-like ovary for its support along its line of attachment just internally to the interradial septum. It seems as if the ova, arising in the epithelium on the surface, pushed their way into the gelatine inside and there completed their development entirely surrounded by a slight investment of gelatine, which grows thinner around each ovum as it increases in size. In males the testes always show a similar division into compartments by gelatinous meshes, the compartments thus mapped out being filled with the small brightly staining spermatocytes. Ova and spermatozoa when mature are set free in the stomach pockets.
10. Floating and Wandering Cells. In the stomach pockets, the canals of the sensory clubs, and even in the stomach itself, are found in varying numbers freely floating cells having the appearance of young ova. They vary in size, the smallest being of the size and having the general aspect of the small ovocytes found in the ovary. The largest (Fig. 70) have exactly the same structure as the young ovarian eggs before they have begun to accumulate yolk. The granular deeply staining cytoplasm, the clear non-staining nucleus with its bright nucleolus and the nucleolus-like yolk nucleus, all show beyond doubt that these freely floating cells originate in the ovary.
In some of my preparations these cells are found not only floating free, but wandering through the tissues. Fig. 70 shows two such wandering cells fixed just as they were making their way either through the digestive epithelium into the gelatine of the floor of the stomach, or from the gelatine into the epithelium. The former seems the more probable, though why they should want to get into the gelatine is not very easy to conceive.
Perhaps there is some connection between this and the appearance that the young ovarian eggs have of pushing their way from the epithelium into the gelatine of the ovary. And of course it is not impossible that the whole phenomenon is abnormal, due to rupture of the ovaries which sets free young ova to exhibit their amœboid tendencies under new conditions. Against such an explanation, on the other hand, might be urged the fact that what seem to be the small floating cells are found[21] occasionally in males as well as females, and that in the females a series can be traced with a good degree of certainty between the small floating cells like those found in the walls, and the larger ones which have all the characteristics of young ova.
However that may be, this amœboid action of cells having the structure of ova brings to mind the remarkable form of asexual reproduction described by Metschnikoff for Cunina proboscidea, under the name of “Sporogonie.” Unfortunately Metschnikoff’s original paper was not accessible to me, so that I was unable to obtain more particulars on the subject than those given in Korschelt and Heider’s text-book (p. 33). The reproductive organs of both males and females of Cunina proboscidea are said to produce, besides the usual distinctively sexual elements, neutral amœboid germ cells, which wander into the endoderm of the stomach and circular canal, and also penetrate into the gelatine of the subumbrella. These amœboid cells divide parthenogenetically. One of the two cells of the first cleavage continues to divide and eventually forms an embryo of Cunina; the other remains amœboid and serves for movement, attachment and nourishment of the embryo.
Charybdea, however, has shown no sign of any such reproductive process on the part of its floating and wandering cells. The only indication that I get as to their use points to a possible nutritive function. The enlarged terminal portion of the canal of the sensory club almost invariably contains a number of the small-sized floating cells. These have a vacuolated, half disintegrated appearance, with the nucleus always compact and brightly staining. Now, examination of the high columnar cells that line the enlargement of the canal shows the presence in the cells of bodies of exactly the same appearance as those in the lumen. In one case a floating cell was found just at the end of an epithelial cell, to all appearance half ingested. The identity of the bodies inside the cells and those in the lumen is shown very clearly in some sections of material fixed in formalin, which preserves nuclei, cell walls and general outlines well enough, but does not retain the cytoplasm, and hence is useless for most purposes of histology. In the endodermal cells of the terminal enlargement thus preserved are found all the more distinctly the bright, compact, degenerated nuclei of the ingested cells, while in the lumen are seen other bright, compact nuclei with the poorly preserved remains of cell substances around them. In addition to the evidence from the appearance of the floating cells themselves and their ingestion by the endodermal cells, a little collateral evidence may perhaps be brought in[22] from the Tripedalia about to be described. From the ovaries in this form are detached masses of cells (Fig. 71) which float free in the stomach pockets among the developing embryos, and to judge from the vacuolation that appears, are used up in their favor. These cell masses are described more fully in the part on Tripedalia.
The species upon which the new family was founded was obtained in great abundance in one locality in Kingston Harbor in the summer of 1896. The environment was even more unlike that in which Cubomedusæ have been found heretofore than in the case of Charybdea Xaymacana. On the west side of the Harbor there is a part more or less cut off from the main body of water, and so from the ocean, by a peninsula. This sheltered bay is dotted with small mangrove islands which toward the head of the bay become so numerous as virtually to convert it into a mangrove swamp. The water is shallow and discolored with organic matter, showing that the tide does not exercise much influence here, and the bottom is for the most part a black mud, deep enough to make wading very uncomfortable but not impossible near shore. The islands rise but slightly above the level of the waters, and the thick vegetation that covers them, for the most part mangroves, grows out into the water on all sides, forming a fringe of overhanging boughs. It was here in the shelter of the boughs, among the roots and half-submerged stems of the mangroves, that the small Cubomedusa was found to thrive. It could be obtained in great abundance almost any day, and of all sizes from the largest adults with stomach pockets filled with eggs or embryos down to small specimens only about two millimeters in diameter. In but one other place was Tripedalia found, and that was a similar region of half landlocked water skirted with mangroves, situated near Port Royal, across the harbor from the locality just mentioned. It would be hard to find places in which the conditions of life were more strikingly different from those of the pure deep sea in which the Cubomedusæ have been generally found before. The slight brownish yellow pigment made the small medusæ a little difficult to see in the discolored water, but like the pellucid Charybdea in the clear water of the harbor, their active movements gave away their presence. The swimming was very vigorous and was effected by quick, strong pulsations (as many as 120 per minute were counted), very different from the slow, rhythmic contractions of the[23] Discomedusan Cassiopea which was found in the same region over by Port Royal. Whether or not the animal made intentional efforts to escape capture could not be decided satisfactorily, but certain it was that they did escape often enough by swimming quickly below the surface of the semi-opaque water.
Tripedalia endured captivity much more hardily than the Charybdea, and would live in aquaria happily enough for a number of days—no attempt was made to see how long. Specimens with their stomach pockets filled with ripe spermatozoa, or with young at any stage from egg to planula, were taken in plenty from the latter part of June to the latter part of July. In each female the young were all at the same stage. The embryos were thrown out in the aquaria as free-swimming planulæ, which settled down on the bottom and sides of the glass in a day or two, and quickly developed into small hydras with mouth and typically with four tentacles (and four tænioles, W. K. B.), though three and five were by no means uncommon. In this condition they lived for three weeks without essential change, and they were still giving no promise of further development when the laboratory broke up and the jars had to be emptied.
The structure of the Cubomedusæ seems to be that of a type well established, and accordingly offers no very wide range of diversity among the different genera. The Charybdea that has just been described is a very typical form and will serve well as a standard with which to compare our species of Tripedalia. The resemblances are so close that a detailed account of the anatomy of the second form would involve much needless repetition. It is hardly necessary to do more than merely point out in what points Tripedalia resembles Charybdea and in what points it differs.
The form of the bell is less pyramidal than in Charybdea. Some measurements even gave the breadth greater than the height. The external surface is divided, as typical for the Cubomedusæ, into the four perradial sides and the four convex interradial ridges, and the furrows that separate these areas are with one small exception exactly the same as those of Charybdea, as may be seen by comparing the series of sections of Tripedalia (Figs. 21-30) with those of Charybdea (Figs. 6-15). The exception is almost too slight to mention. The adradial furrow in each octant which sets off the corner rib from the perradial surface in[24] the lower part of the bell is not directly continuous, as in Charybdea, with the corresponding furrow in the upper part of the bell—that is, the afr´ of Figs. 24-27 is not continuous with the afr of Figs. 22 and 23, as is seen by both being shown in Fig. 24. The upper furrow (afr) is continued only a short distance, however, below the starting point of the lower (afr´).
The pedalia conform entirely to the description given those of Charybdea, except that there are three attached to the bell margin in each interradius instead of one, and that the blade of each pedalium is much narrower.
The sensory clubs also show exactly the same relation to the bell and exactly the same structure.
In the bell cavity the proboscis has a longer and better defined stalk than that of Charybdea, and has the further and more important difference of possessing special sensory organs, to the number of fifteen or twenty. The suspensoria are much more developed than in Charybdea, so that the interradial funnels lying between are more marked. In a corresponding way the frenula are larger and stouter (Figs. 28, 29, frn). The musculature shows no new features and differs only in being comparatively more strongly developed and having a more pronounced striation. The nerve ring follows the same looped course from the margin in each interradius up to the level of the sensory clubs in the perradius.
The stomach offers no peculiarities, and the phacelli also agree with those of Charybdea except in having a smaller number of filaments in each tuft. The stomach pockets are not guarded by such well-developed valves as those described for Charybdea, though the valvular nature of the lips of the gastric ostia is indicated and the valvular functions undoubtedly performed. The gastric ostia are smaller (cf. Figs. 7 and 22), and this makes highly developed valves less necessary. No trace of anything corresponding to mesogonial pockets was noticed.
In the matter of the marginal pockets, however, we find that the agreement with Charybdea is no longer continued. The regions that correspond to the eight marginal pockets of Charybdea are formed, as in that genus, by the coming together of the exumbrella and subumbrella at the sensory niche (Figs. 25-28), but each of these regions is subdivided, as it is not in Charybdea, into two marginal pockets, a larger (mp, Figs. 28-29) and a smaller (mp´). In this way sixteen marginal pockets are[25] formed as in the Chirodropidæ. Furthermore, as happens in the latter family but does not in the Charybdeidæ, the marginal pockets extend into the velarium. From each of the larger marginal pockets are given off two velar canals, while each of the smaller gives rise to but one short one (Fig. 18). Fig. 30 represents one of the last sections of a Tripedalia cut transversely, in which nothing but the pedalia and the velarium appear, and in it are shown the velar canals (vc), which come from the larger marginal pockets. The velarium appears in four segments because it is drawn upwards in the four perradii by the frenula (see Fig. 20). That the canals from the smaller pockets do not appear in the section is due to their shortness and to the fact that they are pulled upwards above the level of the sections by the frenula, together with that portion of the velarium.
The smaller velar canals, a pair in each perradius, seem to have in the males some function in connection with the storing of matured spermatozoa. In specimens with ripe testes they are very often found crowded to distension with spermatozoa, while the other velar canals may or may not contain them, and generally do not. The epithelium lining them is, like that of the others, composed of columnar cells higher on the wall turned toward the bell cavity than on that turned towards the exterior, but otherwise not specially differentiated. I searched in vain for any trace of opening by which the spermatozoa might gain the exterior. Fig. 29 shows another point which may be mentioned in passing, namely, that the canal of each of the three tentacles opens into the peripheral gastro-vascular system independently. The central tentacle of each group is the homologue of the single tentacle of Charybdea, and is formed in Tripedalia before the two lateral tentacles appear. Its communication with the peripheral pocket system is higher up than the openings of the lateral tentacles, so that in the section drawn the latter are just beginning to be indicated (ct´).
It remains only to speak of the reproductive organs of Tripedalia. The sexes are separate in this form also, and ovaries and testes have the same structure as is found in other Cubomedusæ. The development of floating masses of cells in the females, however, is a feature which, so far as I know, has not been observed before. These masses, of which a small one is represented in section by Fig. 71, are apparently developed along with the eggs, and repeat the structure of the ovary to all intents the same as if they were various-sized fragments of it broken loose. They consist mostly of high, columnar epithelial cells surrounding a few[26] central cells and showing here and there a nettle cell just as the reproductive organ does. The epithelial cells differ from those of the ovary in containing one or more large vacuoles, and this vacuolation increases as the embryos, among which the masses float, develop. The idea naturally suggests itself, therefore, that they serve for nourishing and perhaps for protecting the embryos while they are developing in the stomach pockets of the mother individual.
In Medusæ it is a common thing to find that in certain definite places of the gastro-vascular system two endodermal surfaces that were primarily separated by a space have come together and fused into a single lamella or plate. Such a structure is called indifferently a cathammal plate, an endodermal lamella, or a vascular lamella. In the adult animal the vascular lamellæ are by virtue of their very nature formations “with a past.” They are scaffolding left in the completed structure, giving us clues as to the way in which that structure was brought about; and in the Cubomedusæ, whose development is as yet unknown, they therefore afford an unusually interesting subject for special consideration.
The vascular lamellæ that are found in Charybdea and Tripedalia may for convenience be described as forming two systems, the internal and the marginal. The former comprises the endodermal fusions that separate the stomach from the stomach pockets (except for the spaces of communication left free, the gastric ostia) and those that separate the stomach pockets from one another. The marginal system consists of the lamella that connects endoderm of the gastro-vascular system with ectoderm of the surface in a ring all around the bell margin, and with it also the vascular lamella of the sensory niche, which has already been referred to in the general description of Charybdea. The lamellæ of the internal system have been described by previous writers, and especially by Claus in his paper on Charybdea, but they are still in need of comprehensive and clear treatment. The lamellæ of the margin and of the sensory niche have also been described by Claus, but not thoroughly or with entire accuracy, nor did he recognize the vascular lamellæ of the sensory niche as originally a part of the lamellæ of the margin. This last was first determined by H. V. Wilson upon specimens of Chiropsalmus quadrumanus obtained at Beaufort, North Carolina. Professor Wilson’s unpublished notes on Chiropsalmus were very kindly placed in my[28] hands, and so far as the vascular lamellæ are concerned my own work is only a confirmation and amplification of his, since Charybdea and Tripedalia in this respect agree with Chiropsalmus.
The vascular lamellæ of the internal system are the most prominent and morphologically the most important. They comprise the four vertical strips of fusion that separate the four stomach pockets in the interradii (ivl in the figures of the series of cross-sections of Charybdea and Tripedalia, Nos. 6-15 and 21-29), and four curved horizontal cross-pieces at the top of these which separate the stomach from the stomach pockets, and would make the separation complete did they not leave in each perradius a free space between their ends, which makes possible the gastric ostia.
The arrangement of this internal system of vascular lamellæ is simple. What they amount to is a certain definite number of linear adhesions between the two walls of an originally undivided gastro-vascular space, by which that space is divided up into a central stomach and a peripheral portion, and the peripheral portion thus further divided into the four stomach pockets. Perhaps the idea may be conveyed by likening the whole medusa to a couple of bowls fitting closely one within another and plastered together at the margins. The exumbrella then would correspond to the outer bowl, the subumbrella to the smaller inner bowl, and the original undivided gastro-vascular space to the space between the two. If now the walls of the space be cemented together in four horizontal curved lines just in the plane where the bottoms are bending round to become the sides of the bowls, leaving four interspaces between the ends of the lines, we should have the original space divided into a central horizontal somewhat lens-shaped region between the bottoms of the two bowls that would correspond to the central stomach, and a peripheral vertical portion between the sides of the bowls that would correspond to the peripheral gastro-vascular system; central and peripheral portions would communicate by the four interspaces between the lines of fusion, which would correspond to the four gastric ostia. If, further, the vertical peripheral portion be subdivided by four more lines of fusion running vertically at equal distances apart, each connecting above with the middle point of the corresponding horizontal line of fusion, we should have the simple peripheral portion divided into four parts, corresponding to the stomach pockets, by four vertical lines of fusion, corresponding to the four interradial vascular lamellæ, the ivl of the figures.
These mutual relations of stomach, stomach pockets and lamellæ will perhaps be made clearer if a comparison is drawn between them and the similar structures of a Hydromedusa. Liriope, one of the Trachomedusæ, is a good form to take for such a comparison, since by reason of its direct development from the egg it is free from the complications of hydroid medusæ. The young medusa has at first a simple, undivided gastro-vascular cavity which later is divided up into the central stomach and the typical radial to circular canals of the Hydromedusæ by means of fusions between the two endodermal surfaces. Diagrams a, b and c of Fig. 35 represent very schematically this process of division into stomach and canals. In a we have a projection upon a plane surface of the primary, undivided gastro-vascular cavity, as seen from above; b shows the first four points of fusion in the interradii; c represents those four points expanded by growth in all directions into broad cathammal plates in such a way as to leave the stomach in the centre, the radial canals in the perradii, and the circular canal in the periphery as all that remains open of the primary simple cavity. These broad plates of vascular lamella, separating the narrow radial canals, persist in the adult Liriope to tell the tale of the formation of the definitive gastro-vascular system. It seems to me that we are justified by analogy in drawing a similar conclusion for the Cubomedusæ. In d of Fig. 35 is represented a projection of a Cubomedusa, in which the homology of the stomach pockets with the radial canals of the Hydromedusa, and of the narrow strips of fusion with the broad cathammal plates, is shown at a glance. To make the comparison more perfect we have only to remember that in the Cubomedusæ there exists below each interradial vascular lamella a connecting canal (Figs. 16, 29 and 35 d, cc) uniting the two separate adjacent pockets. This, as has been pointed out by other writers, is the representative of the circular canal of the Hydromedusæ. Practically the only difference between the structure of the gastro-vascular system of the Cubomedusæ and that of a form such as Liriope, is that in the latter the fused areas have broadened out at the expense of the radial canals, while in the Cubomedusæ on the contrary they have become long and narrow.
One is strongly tempted by the foregoing comparison to speculate a little as to whether the reproductive organs of the Cubomedusæ, which lie in the stomach pockets and are generally supposed to be endodermal, may not bear some closer relation to those of the Trachomedusæ, which lie “in the course of” the radial canals (Lang’s Text-book) and by common[30] consent are ectodermal. And while we are being led by facts such as those just mentioned above to wonder just a little whether after all the position of the Cubomedusæ among the Acraspeda is so firmly assured—doubting some, yet in the frame of mind of one who “fears a doubt as wrong”—the velarium suggests itself as another point in question. Haeckel does not hesitate to state emphatically that the velarium of the Cubomedusæ and the velum of the Craspedote medusæ are only analogous, but the reasons that he gives (sie sind unabhängig von einander entstanden, und ihre Structur ist zwar ähnlich, aber keineswegs identisch; namentlich das Verhalten zum Nervenring ist wesentlich verschieden: System, p. 426) somehow do not produce so much impression upon one as the very velum-like appearance of the velarium itself. The origin from the fusion of marginal lobes is not as yet a matter of observation, and the relation to the nerve ring is not essentially different from that of the velum to the lower (i. e. inner) nerve ring in the Craspedotæ. The four frenula and the diverticula from the gastro-vascular system seem to be the chief differences in structure after all, and these Haeckel evidently did not think worth mentioning. This speculation, as to the possible relation of the Cubomedusæ to such forms of the veiled medusæ as Liriope, though it may be very tempting, is scarcely fruitful enough to repay much effort on the part of either reader or writer. The whole subject must remain uncertain until the facts of the development of the Cubomedusæ are known.
If the structure of the vascular lamellæ of the internal system has been made clear, the appearances of the vertical and horizontal components in the figures will be understood without much further explanation. The four vertical strips in the interradii (ivl) have been already referred to in the figures of the cross-sections of both Charybdea and Tripedalia. In the longitudinal sections of the two jelly-fish through the interradii, the vertical lamellæ are cut throughout their entire length from stomach to connecting canals (Figs. 5-20, ivl). The horizontal cross-pieces at the tops of the vertical lamellæ also appear in several of the figures. Fig. 36 represents the appearance that would be given by a longitudinal section taken through any portion of the upper part of the bell except in the interradii, or in the perradii, through the gastric ostia. The horizontal vascular lamella (hvl) is shown connecting the endoderm of the stomach (ens) with that of the stomach pocket (enp). In a longitudinal section directly through an interradius (Fig. 5 or 20) the horizontal lamella is cut just at the point where it joins the vertical, so that[31] the two are not differentiated. In a section through the region of a perradius (Fig. 4 or 19) the horizontal lamella is of course not cut, since the section passes through the gastric ostium, whose existence is conditional upon fusion not having taken place between the endodermal surfaces.
The first figure in each of the series of cross-sections (Figs. 6 and 21) also shows the horizontal vascular lamella, cut across slantingly twice in each quadrant as it passes between the gelatine of the ex- and of the subumbrella to connect the epithelium of the stomach with that of the stomach pocket. The fact that more of the lamella does not appear in such a cross-section only shows that its course is not perfectly horizontal.
The region in which the same lamella lies is indicated in the surface view of the top of the bell of Charybdea (Fig. 2) by the bent line hvl in each quadrant. The figure manifests the appropriateness of Claus’s name for the horizontal lamella—“bogenförmige Verwachsungs-Streifen.” Haeckel calls the same structures “Pylorus-Klappen,” and in his account of Charybdea Murrayana in the Challenger Report, speaking of the three divisions of the stomach (buccal, central and basal) which he traces upwards from the stalked forms of Scyphomedusæ, he says: “The central stomach in this Charybdea, as in most Charybdea, is joined to the basal stomach, as the pyloric stricture between the two is not developed and only faintly indicated by the slightly projecting pyloric valves.” Again, in speaking of the valves of the gastric ostia, he says: “These four perradial ‘pouch valves’ alternate with the interradial pyloric valves.” It is difficult to understand, however, how the “bogenförmige Verwachsungs-Streifen” of Claus, which are undoubtedly the same structures as those which I have called the horizontal lamellæ, and are only strips of endodermal fusion, can be “projecting pyloric valves,” or indeed can properly be spoken of as valves at all. Possibly Haeckel was not quite able to understand Claus’s description, and in his desire to find something in the stomach of Charybdea which would serve to set off a central from a basal part, such as is found in the Lucernaridæ, hit upon Claus’s “Verwachsungs-Streifen.” I have elsewhere given it as my opinion that in such of the Cubomedusæ as I have studied there is no structure in evidence that would properly serve to mark a limit between a basal and a central portion of the stomach.
We have next to describe the marginal system. The vascular lamellæ mentioned above in every case connected endoderm of one cavity with endoderm of another; those of the margin have the noteworthy difference that they run from endoderms of some part of the[32] gastro-vascular system to ectoderm of the surface. The outermost cells of the endodermal lamellæ make direct connection with the ectodermal cells, without the usual intervention of a layer of gelatine.
The marginal lamella of Charybdea lies, as the name implies, just on the bell margin where the edge is curving round into the velarium. All around the whole circumference of the bell it is found (in Charybdea) at this same horizontal bend, except in the eight principal radii, where the tentacles and the sensory clubs have brought about modifications. In any place except these a vertical section through the margin will show the marginal lamella connecting the endoderm of the marginal pocket with the ectoderms of the surface, as represented by vlm in Fig. 38, which is a vertical section through the sensory niche a little to one side of the perradial axis.
In the interradii the marginal lamella undergoes modifications due to the fact that the bases of the pedalia are situated a little upwards from the exact margin, and that the lamella follows the outline of the bases. Fig. 1 shows one of the interradial corners of the bell margin looked at directly from the surface, so that the curved outline of the junction of the base of the pedalium with the exumbrella is seen. The trace made by the lamella where it meets the surface ectoderm follows this outline. The lamella is also shown in the vertical section through the interradius (Fig. 5 or 20, vlm), where it is seen running from the connecting canals (cc), which joins the two adjacent marginal pockets, upwards and outwards to meet the surface ectoderm. Its course from canal to surface is not in a direct line, but curved with the concavity upwards. Hence, in cross-sections at certain levels through the interradial corner it is met more than once and gives rise to appearances that seem at first sight too complicated for it to be just the same structure as the simple marginal lamella described above. That it is the same, and that the complication is only due to the insertion of the pedalia above the margin, can be determined by following through a series of cross-sections, the essential ones of which, as I hope, are given in Figs. 40-43. The levels of these are shown on Fig. 5 by the letters w, x, y and z, respectively. Fig. 40 shows the lamella cut but once, just below its highest part. The section is above the level of the connecting canal and hence still shows the vertical interradial lamella ivl. Fig. 41, at the next lower level (x), shows the same portion of the lamella intersected a little nearer the interior, while the junction with the endoderm of the connecting canal is shown still further inside. Fig. 42 is at level y, just through the[33] bend of the loop, so that in part of its course the lamella is cut almost horizontally, i. e. in its own plane. Fig. 43 finally shows the lamella as it appears below the level of the connecting canal, cut twice, each portion joining endoderm of marginal pocket with ectoderm of surface. It thus bears exactly the same relations that it had when we first met it in Fig. 38 (vlm), except that here in Fig. 43 one finds that a cross-section cuts it at right angles instead of a vertical as in Fig. 38, as a result of its being pushed upwards from its former position on the margin by the insertion of the pedalium above the margin.
The vascular lamella of the sensory niche has already been alluded to as part of the marginal system, and brief reference has been made to it in the section on the sensory clubs. Like the rest of the marginal lamella, it connects endoderm with ectoderm. The line that its fusion with the ectoderm traces on the surface frames in a shield-shaped area at the bottom of the sensory niche, which is seen in the drawing of the outlines of the niche, Fig. 44 (vls). This lamella was observed by Claus, and was figured by him both in surface view and in cross-section through the niche. Apparently, however, he omitted vertical sections through the niche, so that he supposed that the outline traced by the lamella was not continuous above, i. e. over the stalk of the sensory club (’78, Fig. 41; text, p. 28). That the outline is closed above, though masked in surface view by the roof of the sensory niche, is seen at once in vertical sections, such as Figs. 37 and 38, one of which is directly through the perradius, the other a little to one side. Both show the vascular lamella of the sensory niche (vls) intersected twice, above and below the sensory club, and completely cutting off the exumbrella from any share in the bottom (or inner wall) of the sensory niche. Fig. 39, which is a cross-section through the upper part of the niche, and is essentially like the similar figure of Claus, shows in like manner that the bottom of the sensory niche belongs to the subumbrella. H. V. Wilson was the first to point out, in his unpublished notes, that the lamella of the niche is complete all round.
In the adult structure of Charybdea and Tripedalia the lamella of the niche is connected with that of the margin by a vertical strip of endodermal fusion that does not come to the surface like the rest of the marginal system, but remains just internal to the gelatine of the exumbrella, connecting the two adjacent marginal pockets. In the cross-sections of Charybdea it is seen in Fig. 16 (vlc); in those of Tripedalia it is seen in Figs. 28 and 29. In vertical section it is found in Figs. 4, 19[34] and 37. In Fig. 44, which represents the bell margin and velarium of Tripedalia arranged as if the velarium were vertical and pendant from the margin (instead of suspended by the frenulum so as to be at right angles to the vertical plane), the connecting lamella is shown as a dotted line (vlc)—dotted because it does not come to the surface—joining the lamella of the niche with that of the margin (vlm).
The same figure (No. 44) shows a characteristic difference between the marginal lamella of Tripedalia and that of Charybdea. While in Charybdea, as Claus points out, the marginal lamella keeps at one level, just a little above the bell margin, all the way round (except where disturbed by the special modifications of the tentacles and the sensory clubs), and never descends into the velarium, in Tripedalia on the other hand it describes a sinuous course, following the outlines of the marginal pockets, as is indicated in the figure by the light parallel line vlm. The course as it would be seen in a surface view is obscured just at each side of the interradius by the overhanging of the bases of the two lateral pedalia. This is why the lamella is not indicated at these points in the diagram. The course is seen to lie almost wholly on the velarium, that is, in the figure below the line which represents the bell margin proper, the line at which the angle comes when the velarium is in its normal position, horizontal to the vertical side of the bell.
In this sinuous course of the marginal lamella we have another point of resemblance between Tripedalia and the Chirodropidæ. H. V. Wilson worked it out in his sections of Chiropsalmus, and the reconstruction which I have given in the figure under discussion is in all essentials similar to his for Chiropsalmus. The differences lie only in the fact that Chiropsalmus has more velar canals, and that the chief marginal pocket in each quadrant is not forked peripherally, as is that of Tripedalia (mp), but presents its distal margin parallel to the edge of the velarium. The two smaller marginal pockets in the perradii (mp´) are on identically the same plan in both.
Tripedalia, having three tentacles joining the umbrella in each interradius, shows a disturbance of the course of the marginal lamella in these regions by just so much the more complicated than in Charybdea. The plan, however, is exactly the same. The lamella is pushed upwards from the margin by each of the bases of the three pedalia just as is done by the base of the single pedalium of Charybdea. Fig. 29 shows the lamella in the same relation to the canal of the central tentacle (ct) that it has in the similar sections of Charybdea (Figs. 16 and 43); and in[35] addition the first appearances (as the series is traced downwards) of the arches of the lamella over the two lateral tentacles (ct´), which are inserted a little lower down than the middle one of the group. As concerns these lateral tentacles, the relations of the vascular lamella at this level are the same as that in the level of Fig. 40 for Charybdea.
It has been stated more than once already that the vascular lamella of the sensory niche is a part of the lamella that runs round the margin, and so far the only evidence given has been the strip of endodermal fusion running from the marginal lamella to that of the niche. This strip, however, as has been described, does not come to the surface and consequently seems at first sight to be a different structure from the lamella of the margin. That it is not, however, I found very prettily shown in a series of sections of one of my youngest Tripedalia. In this the lamella of the niche as it was traced in successive sections downwards, was found not to form a closed ring at the bottom of the niche, but each side was continued directly and separately downwards to the margin, where it passed into the corresponding part of the marginal lamella. A reconstruction of the condition, similar to that of Fig. 44, is given in Fig. 45, and I think explains itself at a glance. Evidently the vascular lamellæ that connect the lamella of the sensory niche with that of the margin at first come to the surface, like the rest of the marginal system, but as the animal grows older come to lie within the gelatine. In this way the condition found in cross-sections just through the margin of my very small Tripedalia, and represented in Fig. 46, becomes that of the adult seen in the corresponding portion of Fig. 29. It is as complete a demonstration as could be required that the lamella of the sensory niche is at first only a loop of the marginal lamella, a conclusion that had been already reached by H. V. Wilson on theoretical considerations, based upon the facts of the adult structure as he found them in Chiropsalmus.
As Wilson pointed out in his notes, these facts have a close bearing upon the question of the origin of the velarium. Sixteen marginal pockets are found in both Chiropsalmus and Tripedalia, and all of them extend into the velarium. It is not unnatural to suppose that these belong to sixteen marginal lobes, and that these lobes have fused together to form the velarium. In the Chirodropus figured by Haeckel (Taf. XXVI) in his “System” gelatinous lobe-like thickenings are shown in the velarium, corresponding to the sixteen marginal pockets. In Tripedalia no special gelatinous thickenings are found, but the arrangement of the marginal pockets is the same as that of the Chirodropidæ, and[36] perhaps I ought, when treating of the systematic relations of Tripedalia (p. 5, Fam. III), to have recognized the analogy to the extent of saying that marginal lobes may not be completely absent from the velarium of Tripedalia. At any rate the gelatinous lobes in the case of Chirodropus on the one hand, and on the other hand the sinuous outline of the margin still mapped out by the lamella in Chirodropus, Chiropsalmus and Tripedalia, are certainly very suggestive of an ancestral Cubomedusa in which there was no velarium, but sixteen free marginal lobes instead. Two more indications favor slightly the same view. In both Charybdea and Tripedalia a small notch is seen in the edge of the velarium in the perradius (Fig. 44). Its constancy suggests that it may not be a chance or meaningless feature. The second point is the small size of the two marginal pockets adjoining the perradius. These are in the position of the ephyra lobes of the Discomedusæ, which always lie on either side of each sensory club, and which do not keep pace with the other marginal lobes in development. In the Rhizostome jelly-fish especially they are found much smaller than the other lobes, as will be seen by a glance at such figures as Haeckel’s for Lychnorhiza (System, Taf. XXXIV Fig. 2), or for Archirhiza (Taf. XXXVI, Fig. 5), or Hesse’s figure of the margin of Rhizostoma Cuvieri (’95, Taf. XXII, Fig. 22). The resemblance between such margins and that of Tripedalia (Fig. 44), with its simple, unbranched velar canals, is very suggestive. On the other hand it must be remembered that in considering the vascular lamellæ of the internal system we found the indication pointing rather more to Hydromedusan affinities than to any other. Charybdea throws no light on the question, since it has no marginal lobes on the velarium and the marginal pockets end strictly at the margin, so that the only diverticula of the gastro-vascular system in the velarium are the velar canals.
Before leaving the subject of marginal lobes and pockets I must answer a possible objection that may occur to some careful reader. It may seem that I am wrong in holding that there are two marginal pockets in each octant instead of three, that just as there is one velar canal from each of the smaller perradial pockets (mp´, Fig. 44), so each prong of the forked larger pocket (mp), since it is continued into a velar canal, ought to be called a marginal pocket likewise, the whole number of marginal pockets then being twenty-four instead of sixteen. Such a revision of the terminology would not be without some reason in its favor, and perhaps a study of more forms would show it to be correct. But for the present, at any rate, it seemed to me best to abide by the analogy of[37] Chiropsalmus, in which the peripheral edge of the larger marginal pocket in each octant is not bow-shaped, but runs parallel to the edge of the velarium. A revision of the terminology of the marginal pockets such as implied in the suggestion above would also give rise to complications when applied to Charybdea, since the latter has no marginal pockets in the velarium.
As to the functions of the vascular lamellæ, there is too little known to say much. It is rather improbable that structures retained so definitely should be mere scaffolding left over from a previous stage of usefulness. Claus has found in Chrysaora that the lamellæ form a kind of capillary network in communication with the gastro-vascular system, and he with others supports the view that they perform an accessory function in the nutrition of the tissues they penetrate. Upon this point I have no observations of my own to add.
The marginal vascular lamella is regarded by Claus as perhaps the vestige of a circular canal around the bell margin. On this subject, too, I have nothing to add. A lamella of endoderm that connects directly with the ectoderm of the surface along its whole course is a structure whose meaning I am wholly unable to understand or even to guess at. A similar lamella is described by Hesse (’95, p. 430) as occurring in the ephyra lobes of his Rhizostoma, and he mentions Eimer as the first to discover this structure, probably meaning the first to discover it in the Discomedusæ. Whether the lamella is found all around the margin is not stated. Hesse refers it to the ephyra, and remarks that the investigation of it in the ephyra would undoubtedly give interesting results.
I will close this part upon the vascular lamellæ with a very pertinent suggestion made by Professor Brooks to the effect that the usual way of speaking of the sensory clubs as having moved up from the margin is looking at the matter in the wrong way. The level of the sensory clubs undoubtedly represents the original margin, which elsewhere has grown down and away from its former level, leaving the sensory clubs like floatage stranded at high-tide mark. Only in this way can the lamella of the sensory niche have any meaning.
The nervous system of the Cubomedusæ is the most highly developed that is found in any of the jelly-fishes. If the position of the group among the Acraspeda is established, it alone is ample to prove that the Hertwigs had not sufficient evidence when they stated in their monograph[38] on the nervous system of the Medusæ (’78) that the Acraspeda show a much lower nervous organization than the Craspedota.
The system naturally groups itself under three heads, the nerve ring, the sensory clubs, and the motor plexus of fibres and ganglia that underlies the epithelium of the subumbrella. The general relations of the nerve ring and of the sensory clubs have been given before in the description of Charybdea Xaymacana, so that we may pass at once to the consideration of the finer details of the nervous tissues.
In the structure of the nerve ring I have found myself unable to come to the same results as those given by Claus, who so far as I know is the only one that has studied the nerve with special reference to its histology. Our difference amounts to this, that he finds two distinct types of cells in the epithelium of the nerve, sensory and supporting, which would make it a receiving as well as transmitting organ, while I have not been able to demonstrate satisfactorily the sensory cells, and, therefore, so far as my own observation is concerned, I am disposed to attribute to the nerve simply the function of conducting impulses. I do not know just how much weight to assign to my inability to find evidence in my sections of the sensory type of cells. Eimer (mentioned by Hesse, ’95, p. 420), the Hertwigs (’78) and Claus (’78) have independently discovered the two types in one medusa or another, and the Hertwigs, at least, have demonstrated them by macerated preparations. So far as Charybdea is concerned, however, Claus had only preserved material and had to rely upon sections, as have I, since the material which I had preserved with especial reference to maceration did not turn out well. The results that we get from sections vary enough for me to believe that Claus interpreted his sections very much by analogy with other forms—as indeed, is suggested by his own words (’78, p. 22): “Da es mir nicht geglückt ist die durch die längere Conservirung in Weingeist fest vereinigten Elemente zu isoliren, habe ich das muthmassliche Verhältniss beider Elemente nach Analogie der mir für die Acalephen bekannt gewordenen Verhältnisse, welche O. und R. Hertwig so schön auch am Nervenring der Carmarina zur Darstellung gebracht haben, zu ergänzen versucht.” There can be no doubt of our having the same structures to deal with, for C. Xaymacana is so much like C. marsupialis as to be perhaps more worthy of being called a variety of the latter than a distinct species.
The structure of the nerve as I conceive it is given in Figs. 47 and 48. The former represents a cross-section, and shows, as others have pointed out, that the layer of circular muscle fibres (cm) is interrupted by[39] the nerve. It is evident that the tissues which elsewhere on the subumbrella were differentiated into muscle epithelium and muscle fibre have here become nerve epithelium and nerve fibre, a point that has not been remarked upon before, so far as I remember, and that may be of interest in connection with the neuro-muscular theory. The epithelium of the nerve (scn) is seen to be made up of cells whose inner ends narrow down into a kind of stalk or process that runs to the gelatine of the supporting lamella (gs) and there joins a little cone of the gelatine that juts out to meet it. The cells are smaller in general than those that overlie the muscle layer, especially on the two lateral margins of the nerve, where they are more crowded together and overarch the nerve-fibres. The fibres are seen in cross-section between the processes of the cells. They apparently must lie imbedded in some clear, watery fluid that does not show in the preserved material. The processes of the epithelial cells give the fibres the appearance of lying in alveoli, or being divided into strands, and one of these strands (ax) is always discernible among the others by reason of its more numerous or finer or more compactly massed fibres. This is the “axis” of Claus. Here and there in its course appear ganglion cells having their long axis in the longitudinal direction of the nerve. Elsewhere, in the nerve as well, and usually nearer to the surface, are found other ganglion cells, mostly bipolar, some multipolar, which are readily distinguishable from those of the axis by the fact that their long axis lies across the nerve. One of these cells is shown in the figure (gc). Here and there in the epithelium alongside the nerve are found mucous cells (mc), distinguished by their clear contents and by the small exhausted-appearing nucleus at the base with a few threads of protoplasm.
In Fig. 48 I have tried to represent the structure of the nerve by means of a series of five different views such as would be given by focusing at five successive levels. In the first (1) we have the epithelium of the nerve (scn in Fig. 47) in surface view, the cells appearing polygonal in outline, with here and there a mucous cell. In (2) we find a very slight layer of ganglion cells and fibres having a transverse direction (gc and fp in Fig. 47). These are continuous with the plexus of fibres and ganglion cells which lie above the muscle layer all over the subumbrella, and which represent the motor part of the nervous system. This connection with the nerve shows how co-ordination is effected. At the same level are found fibres of the axis also having a longitudinal direction. In (3) is seen the main body of fibres, divided in the osmic preparation[40] from which the drawing was made into irregular wavy strands which are in all probability largely the result of preservation, but are in part also due to the separation by processes of the epithelial cells, as was seen in Fig. 47. The axis is seen with one of its longitudinally directed bipolar ganglion cells; and at the sides the fibres of the circular muscle of the subumbrella. These show a slanting direction to the nerve, due to the fact that the nerve, as mentioned before, has a sinuous course from the margin in interradius to the level of sensory club in perradius. At the next focus (4) we come to the gelatine of the subumbrella (gs in Fig. 47), and below this (5) to the larger polygonal outlines of the endodermal cells of the stomach pocket (enp, Fig. 47), which like the ectoderm show mucous cells at irregular intervals.
A comparison, now, with Claus’s figures (’78, Taf. II, Figs. 19-21) will show that, except for the rather unimportant matter of the mucous cells, which he finds regularly and thickly disposed on each side of the nerve (’78, Fig. 21), our only essential difference lies in the matter of sensory cells in the epithelium. His figures show a multitude of spindle-shaped sensory cells whose central ends are continued in processes that bend around into the mass of fibres of the nerve. In his Fig. 20 a relatively small number of nuclei, just one-third as many, are seen attached nearer to the surface, which represent the supporting cells. The plan of structure (as shown in his Fig. 20) is an alternation of (1) supporting cells offering a broad peripheral end to the surface and having the central end continued as a supporting fibre to the gelatinous lamella, and (2) spindle-shaped sensory cells with nuclei at a lower level, which send their peripheral process up between the supporting cells to the surface, while the central process becomes continuous with the nerve fibres, often branching into two processes. In my sections I have not been able to see either a regular alternation of nuclei at different levels, or central processes which unmistakably bend round into the nerve fibres. In every case in which I could trace the central process of a cell clearly it ran to the supporting lamella, and this whether the nucleus of the cell lay near the surface of the nerve or deeper down, as in the somewhat spindle-shaped cell seen on the left of the centre of the nerve in Fig. 47. Of course in many cases the central process could not be traced in a section, and this leaves room for the supposition that such were always the sensory cells. From my inability to demonstrate sensory cells in the nerves of Charybdea, I by no means wish to deny their existence; for that remains to be proved, or disproved, by macerations. At any rate,[41] they cannot be so numerous as has been supposed. The position of the nuclei shows that.
The epithelium of the nerve is said by Claus to be ciliated. It has been suggested by Schewiakoff that probably in such cases the sensory cells bear one long cilium, while the supporting cells have many smaller cilia. Unfortunately, I made no observations upon the ciliation of the nervous structures of the living animal, and the traces of cilia that are shown in preparations of preserved material are a poor basis to speculate much on. Claus considers the sensory cells of the epithelium of the nerve a special seat of tactile sensation.
The way in which the nerve reaches the sensory clubs is interesting. Under the topic of the vascular lamellæ it was explained that the sensory clubs and the bottom of the sensory niche from which they spring are parts of the subumbrella. Fig. 37 reminds at a glance better than any other one drawing how the bottom or inner wall of the niche is completely cut off from the exumbrella by vascular lamellæ above and below the stalk of the club. From this figure, now, it will readily be understood that the nerve in order to pass to the base of the stalk has simply to traverse the gelatine of the subumbrella. This fact, which seems surprising enough at first sight in view of the position of the clubs on the external surface of the umbrella, was correctly pointed out and explained by Claus, but one or two figures will serve perhaps to give a clearer idea of it.
Fig. 49 is a diagram of the nervous structures in the region of the sensory niche, as they would be seen on the surface of the subumbrella turned toward the bell cavity. The outline of the sensory niche as it is seen through the tissue of the animal is represented by the line osn. The sensory club (scl), and its stalk with a conical basal portion are given by the lightly dotted outline and are also imagined as seen through the animal. The nerve (n), being on the surface of the subumbrella, is shown as a heavy line describing an arch over the outline of the niche. In the middle point of the arch is a slight thickening of the nervous tissue (rg) which shows in section a large increase in the number of ganglion cells, and is the radial ganglion of Claus. The same is seen, exaggerated in size, in Fig. 12. From it there extends upward a slender strand of nervous tissue (rn), the radial nerve of Claus. In Charybdea this can be traced but a very short distance. In Tripedalia it is much more distinct and traceable for a longer distance, and I might say in passing that this and the sensory organs in the proboscis are the only differences[42] I have noted between the nervous systems of Tripedalia and Charybdea.
Nerve ring, radial ganglion and radial nerve all lie on the bell cavity surface of the subumbrella. The way, now, in which the nerve ring reaches the base of the stalk is simply by sending two roots through the gelatine of the subumbrella to the conical base of the stalk. These roots are seen in the diagram at rns. After passing through the gelatine the roots come together on the inner side of the base—that is, the side turned toward the bell cavity—and then pass downwards (nst) on the inner side of the stalk of the club to the mass of nervous tissue at its end.
This passage of nervous tissue through the gelatine in order to reach the sensory club is a little hard to grasp at the first, and I have tried to render it more intelligible by a couple of drawings of sections. Fig. 50 is a transverse section through the upper part of the region of the sensory niche, not quite horizontal (i. e. parallel with the bell margin), but slanting so as to lie on the plane of the reference arrow x-y in Fig. 49. The plane passes just through the top of the niche, and in two areas has cut through the roof with its epithelium of ectoderm (ece, ecs) so that the space of the sensory niche (sn) appears. The vascular lamella of the sensory niche (vls) is shown, as in Figs. 13 and 14, running on each side from the endoderm that lines the canal of the sensory club (enc) to the endoderm of the adjacent stomach pocket (enp). By it the gelatine of the exumbrella is separated from that of the subumbrella, and one sees that it is only through the latter that the nerve has to pass in order to reach the base of the sensory club. It is also seen that one part of the roof of the niche which is cut through lies outside of the ring of lamella and is therefore lined with ectoderm of the exumbrella (ece) while the other lies within the ring and is lined with ectoderm of the subumbrella (ecs). Owing to the slanting direction of the cut only the root on one side is cut through. The other is indicated, however, on the right side of the drawing. In this method of passage of nerve fibres, together with the accompanying ganglion cells, directly through the gelatine to the stalk of the sensory club my work is only confirmation and explanation of Claus.
Fig. 51 is a vertical section through the base of the stalk in the plane of the reference arrow w-z in Fig. 49, and therefore passing through one of the roots of the nerve of the stalk. Here again the region is seen to be cut off from the exumbrella by the vascular lamella of the sensory niche (vls), and the nerve is seen passing through the gelatine[43] of the subumbrella from the surface of the bell cavity (sc) to the base of the stalk hanging in the sensory niche (sn). One of the ganglion cells (gc) that accompany the nerve is seen to have two nuclei, a not infrequent occurrence which has been pointed out by others.
The same figure shows that the axis (ax) of the nerve has penetrated the gelatine with the other fibres. Here at the base of the stalk it takes a horizontal course and becomes directly continuous with the similar structure of the other root, as Wilson, I believe, first pointed out. This part of the nervous tract which runs horizontally along the base of the stalk between the two roots (Fig. 49, rns) has been considered by Claus the representative in Charybdea of the upper nerve ring of the Craspedota, which therefore exists in Charybdea in four separate portions. Seeing, however, that the region in which it is found belongs to the subumbrella, the homology seems very doubtful. Moreover, the fact that the axis of the nerve ring runs through this outer portion, instead of remaining on the inner surface of the subumbrella and passing to the radial ganglion, rather indicates that the outer portion is part of the original course of the nerve ring, while the portion that remains on the inner surface is perhaps a later formation.
A very interesting feature of the nervous system occurs in the same region in the form of a tract of fibres underlying the endoderm, and separated from the other fibres by the gelatine of the supporting lamella. It is seen in vertical section in Fig. 52 (enf), which is a section through the base of the stalk in just about its median plane, and, therefore, to one side of the arrow w-z in Fig. 49 and the corresponding drawing, Fig. 51. In cross-section it is represented also in Fig. 50 (enf). It varies in size and prominence very much in different specimens. Fig. 52 is a camera drawing of it in the case that showed it most developed. Ganglion cells are found in it, but comparatively infrequently. In some cases the tract itself can hardly be found with certainty. Hesse has described in a Rhizostome a much more highly developed tract in a corresponding position on the base of the marginal body. Fibres from the “outer sensory pit” pass through the gelatine to the sub-endodermal tract, which is described as surrounding the epithelium of the canal of the marginal body like a collar and is most thickly developed on the under surface of the canal, at the place that just corresponds with the point where, and where only, I find the tract in Charybdea. Hesse thinks that fibres then pass from this region to the nervous epithelium of the “inner sensory pit” lying underneath the base of the marginal body, which[44] contains a rich supply of ganglion cells and is considered by him to be the centre of the nervous system of the medusa. A close comparison cannot be drawn with Charybdea in this matter, however, since Charybdea has nothing to correspond with the “outer” and “inner” sensory pits. Moreover, the endodermal tract is not found encircling the canal of the sensory club, nor could I trace fibres passing from it through the supporting lamella into the fibres of the nerves.
Claus has figured (’78, Taf. V, Fig. 45, Fb) a small bundle of fibres in the stock of the sensory club lying between the endoderm cells of the canal and the supporting lamella. The same bundle is found in both Charybdea and Tripedalia and can be traced in cross-sections up the stalk to a point which must correspond with that at which the endodermal tract is seen in Fig. 52. Downwards it can be traced only as far as the entrance of the stalk into the knob of the club where it invariably becomes lost to view. According to Hesse (’95, p. 427) Schäfer found under the endoderm cells of the whole stalk of the marginal body a fibrous layer like that under the endoderm cells which he refers to slender processes from the cells of the crystalline sac. Although Hesse, as we have seen, finds the layer more limited in extent than Schäfer gives it, and does not trace it to the same source, the observation of Schäfer seems to me worthy of mention here, inasmuch as the trend of the fibrous bundle under the endoderm cells of the stalk in Charybdea and Tripedalia suggests quite strongly that the fibres come from the crystalline sac, as Schäfer thought to be the case in his medusa.
Besides the radial ganglion situated in the course of the nerve ring at its four perradial points there are four other similar ganglia on the subumbrella. These lie in the interradii, at the four lowermost points of the nerve’s course, and undoubtedly send off nerves into the pedalia at whose bases they are situated. F. Müller (’59), whose work was not accessible to me, is quoted by Claus as recording two ganglia opposite the base of each pedalium which gave off a great number of nerves partly into the velarium, partly into the tentacles. Claus observed nothing of the kind in Charybdea and states that even the interradial ganglia do not exist.
That they do, however, is shown without doubt in sections of both C. Xaymacana and Tripedalia, but nerves to the velarium or to the tentacles I was unable to find.
On the two sides of each frenulum and of each suspensorium are found subepithelial ganglion cells in greater numbers than elsewhere on[45] the subumbrella, and I am inclined to ascribe to them also the importance of special ganglia controlling the musculature of the frenula and suspensoria. Certainly such ganglia would not be out of place.
It has been mentioned that the greater prominence of the radial nerve and the possession of special sensory organs in the proboscis were the only points of difference I had noted between the nervous systems of Charybdea and Tripedalia. These sensory organs remain to be described. They are simple ciliated cysts containing a concretionary mass, and are situated in the gelatine of the proboscis, irregularly disposed of at any level, from the lips to the beginning of the stomach, and in any radius. In one series of the adult animal fifteen were counted, of which seven were situated about interradially, four perradially, two adradially and two subradially. In another, twenty-one were counted, twelve in the perradii and nine situated between the sub-and perradii. The one shown in Fig. 24 is in the perradial position, often seen. In the sections of the very young Tripedalia in which the vascular lamella had not reached the adult condition the sensory organs of the proboscis were not found, although the sensory clubs showed practically no difference from the adult. Their structure is very simple—merely a round or oval sac lined with ciliated cells which bear up and keep in constant motion an irregular coarsely granular concretion. Fig. 53 is a sketch made in Jamaica from the living specimen. Sections were somewhat disappointing in that they added but little. Fig. 55 was drawn to show that now and then a mucous cell (mc) is found among the other cells of the sensory epithelium. An irregular-shaped mass (rc) was always found inside the cysts as the organic remains of the concretion. It gave no trace of cellular structure and offered no evidence whether the concretion was the product of one or few or of all the cells of the cyst. The latter would be unique among the medusæ. Even if the otocyst is the result of the activity of only one or a few cells, it is, so far as I know, the only case known for the jelly-fish of a free, unsuspended concretion.
As to whether the cysts are of ectodermal or endodermal origin could not be determined, but there was some evidence in favor of the latter. Fig. 56 is a drawing of one seen in optical section in a whole mount of part of a proboscis, and shows a definite connection with the endoderm of the proboscis. This was the only case when such connection was satisfactorily established, but in sections it was not uncommon to find what seemed to be the remains of the broken stalk, as in Fig. 54 (rs?). No connection could be traced between the cysts and any other[46] part of the nervous system. As to function, the idea that they serve to give perception of space relations suggests itself as readily as any other hypothesis.
We come now to the consideration of the terminal knob of the clubs, the sensory portion proper. A complete and detailed account of the complex structure of these organs would fill many pages and involve much useless repetition. Claus (’78) has described them with accuracy, but not in great detail, and since then Schewiakoff (’89) has given a careful general description and has supplemented Claus’s work by observations upon the finer structure made with the aid of more recent technique. It seems in place for me, therefore, to give in the briefest possible way a general idea of their structure, and to pass then at once to the points in which my work has led me to different conclusions from those of Claus and Schewiakoff. In brief, then, the knob of the sensory club consists of a thick, complex mass of nerve fibres, more or less imbedded in which lie the special sensory organs, surrounding the ampulla-like terminal enlargement of the canal. The surface between the special organs is covered with less specialized sensory epithelium. The sensory organs are seven in number. Of these, four are simple invaginations of the surface epithelium arranged in two pairs symmetrically to the median line in the proximal end of the knob (the end where the stalk enters) and having pigment developed in the cells so invaginated, while the space of the invagination is filled with a gelatinous refracting secretion. These are considered simple eyes. Two more of the organs are complex eyes situated on the median line of the inner surface of the knob, the upper one smaller than the lower, but having almost exactly the same structure. Each has a cellular lens over which extends a superficial, corneal layer of cells; below the lens a refractive “vitreous body”; and below this a retina with pigmented cells. The seventh organ is the crystalline sac, which lies almost at the end of the knob opposite to the stalk and contains a large concretion. In view of the fact that the sensory clubs in toto have been abundantly figured by Claus and Schewiakoff, it is my intention to give but one simple figure of the general relations, and I justify that one in that it was made from the fresh material. Fig. 57 is a camera sketch of the outlines given by a sensory club seen in optical section from the side. The smaller upper and the larger lower complex eyes which are situated on the mid-line, are seen in profile, while the two small simple eyes give the outlines that they would in a surface view of their side of the knob. Of course it is understood[47] that two similar ones would appear on the other side, since the four simple eyes are symmetrically paired on either side of the mid-line. The sketch seems to show at least this much, that even in the living state the lens of the larger eye projects out beyond the other contours of the surface, so that the marked convexity ascribed to it in descriptions is not to be attributed to the preservation.
It is in reference to the structure of the retina and vitreous body of the complex eyes that I have found myself unable to come to the same conclusions as Claus and Schewiakoff. Since the work of the latter goes much further into the detail of the subject than does Claus’s paper, it will be sufficient for me to compare my results simply with those of Schewiakoff.
The latter finds that the retina is composed of two kinds of cells, corresponding to the supporting and sensory cells referred to in the description of the nerve ring. These he figures (’89, Taf. II, Figs. 12 and 13) as alternating regularly. The two kinds of cells differ as follows:
(1) Shape. The supporting cells like those referred to before, are cone-shaped, having a proximal fibrous process that runs into the underlying stratum of nerve fibres, and on the surface of the retina a broad distal pigmented termination. The sensory cells are spindle-shaped, the proximal processes becoming continuous with fibres of the underlying nervous mass, while the distal process runs up to the surface of the retina (the part toward the lens) in between the ends of the supporting cell. The two kinds of cells are accordingly designated as pigment and visual.
(2) Position of nucleus. This comes in as a corollary of the shape. The nuclei of the visual cells lie in the enlarged central part of the spindle-shape, and, therefore, at a lower level than the nuclei of the alternating pigment cells.
(3) Processes in the vitreous body. The distal processes of the spindle-shaped visual cells are continued through the vitreous body to the cells of the lens as rod-like visual fibres which lie in canals in the (supposedly) homogeneous vitreous body. The pigment cells on the other hand have no fibres passing from them through the vitreous body, but in the latter are situated cone-shaped masses of pigment whose bases rest upon the broad ends of the pigment cells without, however, being a part of the cell.
(4) Pigment. The distal ends of the pigment cells in the retina are strongly pigmented, as the name implies. The processes of the visual[48] cells, which alternate with these, are pigmented likewise, but the pigment is not so abundant and lies in the periphery of the cell body, leaving free a highly refracting central axis.
If the relation of these cells to each other has been made sufficiently clear, it will be understood that, in accordance with Schewiakoff’s scheme of the structure, sections that cut the retinal cells transversely give very different appearances at different levels. A section through the very tops of the retinal cells, that is, the last section of the retina before striking the vitreous body, would show large polygonal areas of heavy pigment (the ends of the pigment cells), in between which would lie the much smaller, less pigmented, highly refracting ends of the visual cells (’89, Taf. II, Fig. 19). A section lower down in the retina, that is, more toward the centre of the club, would strike the low-lying enlarged central portion of the visual cells with their contained nuclei, and the smaller, proximal ends of the pigment cells. It would, therefore, give the reverse appearance from the preceding section, namely, that of large unpigmented (or but slightly pigmented) areas (the swollen bodies and nuclei of the spindle-shaped cells), and in between them smaller pigmented areas, the ends of the proximally tapering pigment cells (’89, Taf. II, Fig. 20). A section on the other side of the one first described, that is, one of the first through the vitreous body, would show pigment areas of the same size as the large ends of the pigment cells (the cone-shaped streaks of pigment in the vitreous body which according to Schewiakoff are associated with the pigment cell), and in between them the cross-sections of the rod-like processes from the visual cells, lying in canals in the clear homogeneous ground-substance of the vitreous body (’89, Taf. II, Fig. 18).
Let me give a resumé of Schewiakoff’s conception of the structure of the retina.
a. There is an alternation of pigment and visual cells, the nuclei of the spindle-shaped visual cells lying at a lower level than those of the cone-shaped pigment cells.
b. From the visual cells extend rod-like processes into the vitreous body, lying in canals in the latter.
c. In the vitreous body a cone-shaped streak of pigment overlies each pigment cell of the retina, which is not a part of that cell.
d. Apart from these pigment streaks and the rod-like processes of the visual cells the vitreous body is structureless, probably a secretion of the pigment cells.
My own work, now, has led me to a different conception, so that my conclusions on the same points would be as follows:
a. There is not good evidence of an alternation of cone-shaped pigment cells and spindle-shaped visual cells, with the nuclei of the latter at a lower level than those of the former.
b. From some of the retinal cells otherwise not distinguished, there extend rod-like processes into the vitreous body, such as described by Schewiakoff.
c. The cone-shaped streaks of pigment in the vitreous body belong to the underlying pigment cells, in fact are direct continuations of them, and at their distal ends they are prolonged into fibrous processes lying in canals of the vitreous body exactly like the visual fibres of Schewiakoff.
d. The vitreous body is not a homogeneous secretion, but is composed of prisms of refracting substance, each with a denser central fibre.
Let us go over these four points in detail.
(a) As to the first, the question whether there is an alternation of pigment and visual cells, I am not prepared as yet to make a positive statement, since my not seeing both kinds as they are described has little evidential value against the fact that Claus and Schewiakoff both claim to have seen them. Perhaps proof could be obtained one way or the other by maceration of fresh or of specially prepared material, which none of us had. My evidence for not confirming alternation rests wholly upon sections. Fig. 58 represents a radial section through part of the larger eye of Charybdea, made from an osmic preparation which in this case showed two advantages over the material fixed in corrosive-acetic (usually by all odds the best), namely, that the vitreous body (vb) was not shrunken away from the retinal cells, as almost invariably happens, and that the retinal cells were contracted apart from one another in some places in such a way as to be almost equal to a macerated preparation. Now, in the figure it is seen that there is an apparent alternation of two kinds of cells, more regular than I usually find, but the ones that are undoubtedly the pigment cells of Schewiakoff are the ones that show the fibrous processes like his visual cells, and the pigment streaks in the vitreous body are seen to be integral parts of the cells, not cone-shaped masses lying in the vitreous body, merely associated with the pigment cells. If these are the pigment cells of Schewiakoff, the shorter cells in between must be his visual cells, yet they can by no means be said to conform to a spindle-shaped type, nor are their nuclei always at a lower level than (that is, internal to) those of the pigment cells. If the long cells with the fibres are, on the other hand, considered[50] the visual cells of Schewiakoff, then again we find nonconformity to a spindle-shaped type, and nuclei not always at a lower level. The matter of alternation of nuclei at different levels seems to me any way too slight a distinction upon which to base a difference in function. It is a necessary mechanical consequence of the crowding together of many cells on one surface. And in many cases in perfectly radial sections through the retina I find the nuclei fewer in number and arranged in very nearly a single level. The retina of the smaller eye represented in Fig. 69 shows this. In sections further along in the same series the nuclei are found at different levels, due without doubt to the slanting cut.
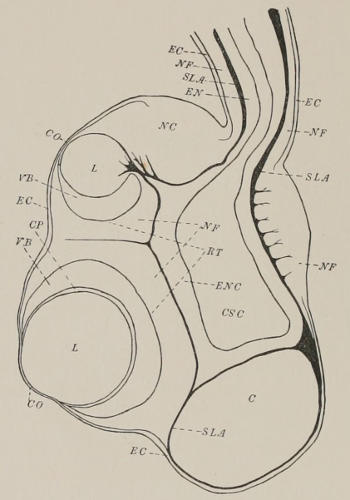
[Dr. Conant did not complete Fig. 72, and the accompanying outline of Fig. 7 of Schewiakoff’s memoir (Beiträge zur Kenntnis des Acalephenauges, Morph. Jahrb., Bd. XV, H. 1) has been substituted.—Editor.]
Explanation of Letters in Text Figure.—C—concretion cavity; CO—cornea; CP—capsule of lens; CSC—cavity of sensory club; EC—ectoderm; EN—endoderm; ENC—endoderm of sensory club; L—lens; NC—network cells; NF—nerve fibres; RT—retina; SLA—supporting lamella; VF—vitreous body.
Fig. 72 is a horizontal section through the large eye, and shows that here, too, when the sections pass through the eye just radially, the[51] nuclei are not found at different levels sufficiently definite to suggest two kinds of cells.
In the inner corner of the retina in the same figure (69) are seen cells without pigment which show nuclei undoubtedly at different levels. These cells in this position are a regular feature in the retina of the smaller eye. Schewiakoff considers them purely visual, because of the lack of pigment. In so doing it seems to me he forgets his own standard for discriminating between pigment and visual cells. The pigment cells of the retina, according to him, are the same thing as the cone-shaped supporting cells found elsewhere in the nervous epithelium, and are, therefore, distinguished from the visual cells primarily by shape and by position of nucleus, secondarily by the greater development of pigment. When on the ground of pigmentation alone he calls the cells in the corner of the retina visual, he judges them by only the second test, and in so doing virtually admits, as it seems to me, that shape of cell and position of nucleus are matters of no great moment. His own standards place him in a dilemma. If on the other hand he judges by the lack of pigment, the cells are visual; if by shape of cell and position of nucleus, they are both visual and pigment cells without the pigment or supporting cells. What use there would be for simple unpigmented cells in one limited region of the retina is hard to see, so he naturally takes the other horn of the dilemma and calls them visual because they have little or no pigment.
The distinction, then, between pigment and visual cells is brought down to one of pigmentation only. Schewiakoff’s test for this is that in the visual cells “Das Pigment durchsetzt aber nicht das ganze Protoplasma des centralen Zellenabschnittes, sondern ist auf seine Oberfläche beschrankt (Fig. 19, sz), so dass der innere, axiale, stark lichtbrechende Theil vollkommen frei von demselben ist.” (’89, p. 37.) That is, in a section through the ends of the retinal cells each pigment cell will appear as a uniformly pigmented area, while each visual cell will appear as a light, strongly refracting spot with a ring of pigment around its periphery. This is the arrangement given in his Fig. 19.
An arrangement so definite ought to be easily made out in sections, yet I have not been able to find it so. My sections show considerable difference in the amount of pigmentation even in material preserved with the same killing agent. If the retina is heavily pigmented the ends of the cells have the appearance shown in Fig. 62, which represents a portion of a cross-section. The ends are seen as clearly defined[52] polygonal areas differing among themselves in size, but not showing two types of size, or two kinds of pigmentation, the one uniform, the other a ring of pigment around a highly refracting central portion. If the retina is but slightly pigmented—and some were so light as to make depigmentation unnecessary—a difference is seen in the pigment, as shown in Fig. 63, but in no case were areas found that showed a highly refracting centre surrounded by a ring of pigment. (The unexplained structures in Fig. 63 will be referred to a little later.)
Figures 59-62 are a series of four successive sections drawn with the camera lucida for comparison with Schewiakoff’s Figs. 20 and 19, and to show that the presence of two types of cells plainly marked within the retina by the position of the nuclei at different levels is at least not clearly demonstrated. Only the nuclei are drawn, since the cell bodies are not easily distinguished from the surrounding fibres. The eye is the same as that from which Fig. 72 was made. Fig. 59 shows a relatively small number of nuclei of slightly larger size than usual. These I take for two reasons to be nuclei of the ganglion cells that are found in the fibres at the base of the retinal cells (Figs. 58, gc, 69 and 72). They are the first nuclei struck in tracing sections toward the retina, and in the series from which Fig. 58 was taken similar nuclei appeared in both transverse and radial cuts through the retina stained brightly and clearly with hæmatoxylin, whereas the nuclei of the retinal cells proper were stained a diffuse brownish-yellow from pigment that had evidently gone into solution. Fig. 60 shows the closely aggregated, smaller nuclei of the retinal cells surrounded by the nuclei of the outlying ganglion cells. Schewiakoff’s corresponding drawing (’89, Fig. 20) shows at this level a definite alternation of the bodies and nuclei of unpigmented visual cells, with the smaller, pigmented, proximal processes of the pigment cells. In the next section (Fig. 61) the pigmented ends of a few of the cells have been struck, and the following section (Fig. 62) shows that, in this heavily pigmented specimen at least, there is no good evidence within the retina itself of two kinds of cells, so that it is apparent that at any rate we cannot accept Schewiakoff’s conception of the structure.
(b) Yet the fibres that Schewiakoff observed and associated with special visual cells occur beyond question. Fig. 64 is a drawing of the first cut through the vitreous body of Charybdea, and in among the sections of the pigment streaks are seen sections of processes lying within clear spaces exactly as Schewiakoff figures his visual fibres (’89, Taf. II, Fig. 18). That the fibres occur is indisputable, but as to the cells[53] to which they belong I can say nothing except that from such evidence as I have given in the preceding paragraph I conclude that they come from pigmented retinal cells of not very different type within the retina from the others, if different at all.
(c) On the third point, that the pigment streaks in the vitreous body belong to underlying cells and are continued distally into fibrous processes like the visual fibres of Schewiakoff, the evidence is decisive. Fig. 58 has already shown it, and if this were not enough, a case of unusual stoutness of the fibres drawn in Fig. 67 is conclusive. The preparation from which the section is taken was one preserved with corrosive-acetic, and I have drawn the outlines with the camera in order to avoid exaggeration of the fibres as far as possible, and also to show the shrinkage of the vitreous body (vb). It is the shrinkage of the vitreous body that makes it so difficult to determine the exact relation of structures seen in the vitreous body to the retina. The fibrous processes run through the vitreous body to the “capsule” of the lens (cp) (see also Fig. 72), a layer of homogeneous substance much resembling that of the vitreous body, which is classed as a part of the vitreous body, but usually in the shrinking adheres to the lens. The capsule is therefore regarded by Schewiakoff as a secretion of the lens cells. Some fibres were found by him to have the appearance of branching upon reaching the surface of the capsule, others of passing through it and of seemingly ending among the cells of the lens. The same appearances were given in my sections. It is altogether impossible in the distal portion of the vitreous body to distinguish between the fibres of Schewiakoff and those that come from the long pigment cells. (Figs. 64-66 represent the appearance of the vitreous body at successive levels, and are from the same series of sections as Figs. 59-62 and 72.) In Fig. 64 the sections of the processes that Schewiakoff calls visual are easily distinguished from the sections of the long pigment cells. In Fig. 65, which is two or three sections nearer the lens, the pigment cells are shown by their cross-sections to be tapering down, and in Fig. 66, nearer still to the lens, the two kinds of processes are no longer to be distinguished from each other. In a few cases I have found pigment in a fibre which but for this would be called one of the visual fibres of Schewiakoff. Such considerations as these, the similar appearance in cross-section, the finding of pigment in a few cases, and the inability to trace to any readily distinguished special type of retinal cell, make me wonder whether the visual fibres of Schewiakoff are anything more than the distal processes of pigment[54] cells, into which the pigment granules happened not to be produced at the moment of fixation.
Fig. 63, however, where the retina was only slightly pigmented, rather speaks against this view, for the number of darkly pigmented areas seen here (which are shown beyond question by radial sections to belong to the long pigment cells) is not great enough to account for the number of both pigment areas and visual fibres of Schewiakoff seen in such a section as Fig. 64. This would throw the visual fibres of Schewiakoff back upon some of the slightly pigmented cells of Fig. 63, otherwise not distinguished. I think the question cannot be settled without the maceration of fresh material, and experiments upon eyes killed in the light and in the dark.
In such cases as that of Fig. 63 it would seem conclusively shown that the long pigment cells must belong to a different type from the short, but as I have already said I can find no regularity in either their shape or in the position of their nuclei. And on the other hand Fig. 58 shows that the reverse relation may obtain and the long cells be less deeply pigmented on the edge of the retina than their shorter neighbors, so that it looks as if all the short cells had to do was to project half their pigment out into the vitreous body in order to become exactly like the long ones. This they could do if, as is possibly the case, they are prolonged into “visual fibres” of Schewiakoff that have escaped observation and so do not appear in the drawing.
Fig. 58 shows one more thing that is worthy of remark in passing. In the preparation in which the vitreous body (at this point at any rate) was not shrunken away from the retina, the fibre from each long pigment cell does not lie in a clearly defined space or “canal,” such as is usually described as a constant structure of the vitreous body. Very likely these canals are formed only by shrinkage around the fibres, and the irregular shape of the spaces around the three fibres in Fig. 67 rather bears out the same supposition.
As to the structure of the vitreous body, apart from the fibres and pigment streaks already mentioned, I find it to be made up of prisms extending from retina to capsule of lens, each containing a central axis or fibre. Fig. 64 shows that the space around the pigment areas and “visual fibres,” instead of being homogeneous, is wholly filled with the polygonal cross-sections of these prisms. In Charybdea they are generally more difficult to perceive than in my best material of Tripedalia which was killed in acetic acid. In this the polygonal areas stood apart[55] from each other more plainly. Curiously enough I have been unable to demonstrate in Tripedalia the “visual fibres” of Schewiakoff. Here and there were found spaces that at first sight reminded of them (Fig. 69, sh), but they contained no central fibre, and were probably due to shrinkage. The polygonal areas themselves, however, often contained a clear spot in the centre, at one side of which would be found the cross-section of the fibre, as is shown in many cases in Fig. 68. The clear spot is here undoubtedly due to shrinkage of the gelatinous substance of the prism.
I think that these prisms and fibres are the direct continuations of retinal cells. In a section such as that drawn in Fig. 63, which takes just the very tops of the cells of a slightly pigmented retina, in the centre of the section just grazing the space that lies between the retina and the shrunken vitreous body, most of the cells toward the middle (where especially the extreme tips are taken) show in their centres a dot exactly corresponding to the dots in the polygonal areas of the vitreous body. In the exact middle of the section, where only the cell walls appear, slightly indicated, a dot is seen in each case. The size and shape of the ends of the cells correspond with those of the polygonal areas in the vitreous body, and I do not doubt that the latter are continuations of the former. The vitreous body, then, instead of being homogeneous, is composed of the clear highly refracting outer ends of retinal cells. The assumption lies near that these are the true visual rods, but of course it is assumption only.
To give a brief review, the points in which my conclusions differ from those of Schewiakoff are as follows: I find (1) that the long pigment streaks are parts of retinal cells continued into processes like his visual rods; (2) that the vitreous body is composed of prisms with central fibres proceeding from retinal cells; (3) that I am unable to get satisfactory evidence of two types of cell distinguishable within the retina, and at any rate find considerable evidence against the two types he distinguishes.
These results are not wholly satisfactory, for they leave us with three kinds of fibrous processes in the vitreous body which for the present we are unable to trace to three, or even two distinguishable types of cell in the retina. It would be more pleasing if we could confirm Schewiakoff’s simple conception of the structure, with its one set of visual rods in the vitreous body referable to a clearly marked type of sensory cells in the retina, but I think the evidence that has been brought up justifies the conclusion that in some respects he saw too much, in[56] other respects too little. This is not to be wondered at, since his material, to judge from a single statement, consisted of but twelve marginal bodies, and, moreover, the work on Charybdea forms but one portion of a paper that is excellent for the clearness of its descriptions and illustrations.
Before leaving the subject I must mention that Wilson suggested from his observations on Chiropsalmus that the vitreous body had a prismatic structure, but he was probably mistaken when he thought he found evidence of nuclei in it. Claus says that the retina is composed of pigment and rod cells alternating, and Wilson agrees with him, but under a sketch of a sense cell from the nerve he makes the express statement “not very well preserved.” It seems very probable, therefore, that he followed Claus’s interpretation rather than independent observations, and Claus interpreted his results very much by analogy of what had been found in other forms.
The smaller complex eye which is represented in Fig. 69 agrees in structure very closely with the larger. The chief differences are that sections do not show pigment extending into the vitreous body, that there is no “capsule” to the lens, and that the lens seems to be supported by a kind of stalk formed by a thickening of gelatine of the supporting lamella (sl). The gelatinous thickening lies between the lens and an outgrowth of endodermal cells (en) from the canal of the club. This outgrowth is a constant feature, figured by Claus and Schewiakoff for Charybdea, and by Wilson for Chiropsalmus, and found in Tripedalia also. The regularity of its appearance in all three genera leads one to suspect that it may have some significance not yet understood.
Just above the smaller eye there lies a mass of cells of peculiar structure (Fig. 69, nc). They are of a rounded polygonal contour, with a comparatively small circular nucleus in the centre, and are found in this region only. In and amongst them bundles of fibrous tissue are found in the sections, which pass from the surface cells to the supporting lamella. Claus describes the contents of these cells as coarsely granular protoplasm and says they cannot be taken for ganglion cells. He is inclined to believe that they play the part of a special supporting tissue. Schewiakoff, on the other hand, is convinced that they are ganglion cells, and finds processes passing out from them (’89, Taf. II, Fig. 22). I find, however, that the cell contours are perfectly regular and clearly without processes, and it is incomprehensible to me how, if his material was at all well preserved, he could for a moment have taken them for the same[57] thing as the big multipolar ganglion cells with large nucleus and nucleolus which lie in about the same region and were correctly described and figured by Claus but are not specially mentioned by Schewiakoff. I cannot agree with Claus, however, that their contents are composed of coarsely granular protoplasm. That which appears such by low magnification shows itself under high powers to be a beautiful network with thickenings at the nodes of the meshes, which is brought out very plainly by a cytoplasmic stain such as Lyons blue. Around the nucleus is seen a more or less well-defined clear zone. What the function of the cell is remains as unknown to me as to Claus and Schewiakoff.
There is left one more point in reference to the nervous system upon which I wish to say a word. Claus and Schewiakoff both describe the wall of the crystalline sac as structureless, formed by the bare supporting lamella. The credit is due to H. V. Wilson of finding in Chiropsalmus that it has a special lining of epithelial cells, which he figures as a continuous, flattened layer. In both Charybdea and Tripedalia I find traces of the same in nuclei here and there, but whether they are the remains of a once continuous layer or not the sections do not show satisfactorily.
This ends the account of what it seemed worth while to say at present upon the nervous system. In concluding, the writer wishes to express his thanks for the help afforded by Dr. Wilson’s notes, in particular on the subject of the vascular lamellæ, and desires to make especial acknowledgment of his indebtedness to Professor Brooks, whose suggestions, based upon many years of experience with the Medusæ, have been most welcome and helpful, and whose evidences of unfailing kindliness, both in Jamaica at the time the material was obtained and in Baltimore when it was being studied in the laboratory, take a most honored part in the pleasant memories associated with the work.
Clarke, H. J. ’78. Lucernariæ and their Allies. Washington: Smithsonian Institution.
Claus, C. ’78. Ueber Charybdea marsupialis. Arb. aus d. Zool. Inst. d. Univ. Wien, Band II, Heft 2.
Doflein, F. ’96. Die Eibildung bei Tubularia. Zeitsch. f. wiss. Zool., Bd. LXII, Heft 1.
Haeckel, E. ’79. Das System der Medusen. Jena.—’81. Challenger Report on the Deep-sea Medusæ. Vol. IV.
Hertwig, O. and R. ’78. Das Nervensystem und die Sinnesorgane der Medusen. Leipzig.
Hesse, R. ’95. Ueber das Nervensystem und die Sinnesorgane von Rhizostoma Cuvieri. Zeitschr. f. wiss. Zool., Bd. LX, Heft 3.
Müller, F. ’59. Zwei neue Quallen von St. Catherina (Brasilien). Abhandlungen der naturf. Gesellschaft zu Halle.
Schewiakoff, W. ’89. Beiträge zur Kenntniss des Acalephenauges. Morph. Jahrb., Bd. XV, Heft 1.
Wilson, H. V. Unpublished notes.
| afr | = | adradial furrow. |
| afr´ | = | furrow in Tripedalia that separates perradial from interrad. regions in lower half of bell. (In Charybdea the same furrow is directly continuous with afr.) |
| ax | = | axis of nerve. |
| c | = | concretion. |
| cc | = | canal underneath ivl, connecting the two adjacent marginal pockets. |
| ccl | = | circular canal. |
| ci | = | cilia. |
| cm | = | circular muscle. |
| co | = | cornea. |
| cp | = | capsule of lens. |
| cs | = | covering scale of niche. |
| csc | = | canal of sensory club. |
| ct | = | canal of tentacle. |
| ct´ | = | beginning of canals of lateral tentacles in Tripedalia. |
| ec | = | ectoderm. |
| ece | = | ectoderm of exumbrella. |
| ecs | = | ectoderm of subumbrella. |
| ed | = | distal paired eye. |
| el | = | larger unpaired eye. |
| en | = | endoderm. |
| enc | = | endoderm of sensory club. |
| enf | = | tract of nerve fibres underlying endoderm. |
| enfl | = | endoderm of floor of stomach. |
| enp | = | endoderm of stomach pockets. |
| enr | = | endoderm of roof of stomach. |
| ens | = | endoderm of stomach. |
| ep | = | proximal paired eye. |
| es | = | smaller unpaired eye. |
| fc | = | funnel leading into canal of sensory clubs. |
| fp | = | fibre from subepithelial plexus of subumbrella. |
| fph | = | filaments of phacellus. |
| frn | = | frenulum. |
| ft | = | funnel-shaped depression in ectoderm axial to base of tentacle. |
| [59]g | = | gelatine. |
| gc | = | ganglion cell. |
| ge | = | gelatine of exumbrella. |
| go | = | gastric ostium. |
| gs | = | gelatine of subumbrella. |
| hvl | = | horizontal vascular lamella. |
| i | = | interradius. |
| if | = | interradial funnel of bell cavity. |
| ifr | = | interradial furrow. |
| ivl | = | interradial vascular lamella. |
| l | = | lens. |
| lv | = | lip of valve. |
| m | = | bell margin. |
| mc | = | mucous cell. |
| mep | = | mesogonial pocket. |
| mo | = | mouth. |
| mp | = | marginal pocket. |
| mp´ | = | smaller marginal pockets, in Tripedalia. |
| mst | = | muscle of stock of sensory club. |
| mt | = | muscle at base of tentacle. |
| n | = | nerve. |
| nc | = | network cells, in sensory club. |
| nf | = | nerve fibres. |
| nm | = | nematocyst. |
| nst | = | nerve of stalk of sensory club. |
| osn | = | outline of sensory niche. |
| p | = | perradius. |
| pe | = | pedalium. |
| ph | = | phacellus. |
| pr | = | proboscis. |
| r | = | reproductive organ. |
| rc | = | remains of concretion. |
| rcl | = | radial canal. |
| rg | = | radial ganglion. |
| rm | = | radial muscle. |
| rn | = | radial nerve. |
| rns | = | root of nerve of sensory club. |
| rs? | = | remains of stalk (?) of sensory organ. |
| rt | = | retina. |
| s | = | stomach. |
| sc | = | bell cavity. |
| scl | = | sensory club. |
| scn | = | supporting cell of nerve. |
| se | = | sensory epithelium. |
| sh | = | shrinkage space. |
| sl | = | stalk of lens. |
| sla | = | supporting lamella. |
| sn | = | sensory niche. |
| so | = | sensory organ in proboscis of Tripedalia. |
| sp | = | stomach pocket. |
| sph | = | stalk of phacellus. |
| ss | = | stalk of sensory organ, in proboscis. |
| [60]st | = | stalk of sensory club. |
| su | = | suspensorium. |
| sub | = | subumbrella. |
| tl | = | lateral tentacle. |
| tm | = | median tentacle. |
| v | = | velarium. |
| va | = | vacuole. |
| vb | = | vitreous body. |
| vc | = | velar canals. |
| ve | = | edge of velarium. |
| vfs | = | visual fibres, according to Schewiakoff. |
| vg | = | valve of gastric ostium. |
| vl | = | vascular lamella. |
| vlc | = | vascular lamella connecting vls with vlm. |
| vlm | = | vascular lamella of margin. |
| vls | = | vascular lamella of sensory niche. |
| vlst | = | vascular lamella of sensory niche at base of stalk. |
| wc | = | wandering cells. |
| w-x-y-z | = | successive levels of Figs. 40-43 on Fig. 5. |
Fig. 1. Charybdea Xaymacana, from one of the four interradial sides.
Fig. 2. The same from above.
Fig. 3. The same from below, the four tentacles cut off.
Fig. 4. The same cut in halves vertically (or radially) through a perradius.
Fig. 5. The same out in halves vertically (or radially) through an interradius.
Figs. 6-16. Diagrams of horizontal (or transverse) sections through C. Xaymacana at successive levels.
Fig. 17. Tripedalia cystophora, from one of the four interradial sides.
Fig. 18. The same from below.
Fig. 19. The same cut in halves vertically through a perradius.
Fig. 20. The same cut in halves vertically through interradius.
Figs. 21-30. Diagrams of horizontal sections through T. cystophora at successive levels.
(The following are of Charybdea, except when specially stated otherwise.)
Fig. 31. Horizontal section through the suspensorium.
Fig. 32. Diagram of a gastric ostium seen from the stomach side.
Fig. 33. Diagram of a vertical section through a gastric ostium.
Fig. 34. Diagram of a horizontal section through a gastric ostium.
Fig. 35. Diagram to illustrate the formation of the central and peripheral gastro-vascular systems of a Hydromedusa (a, b, and c) and a Cubomedusa (d).
Fig. 36. Vertical section through the upper part of the bell, adradial, to show horizontal vascular lamella.
Fig. 37. Vertical section through the perradius, to show vascular lamella of the niche of the margin.
Fig. 38. Vertical section a little to one side of the last, to show same structure.
Fig. 39. Horizontal section through the upper part of the sensory niche, to show vascular lamella of the niche.
Figs. 40-43. Horizontal sections through the base of a pedalium at successive levels, w-x-y-z, Fig. 5, to show marginal lamella.
Fig. 44. Diagram to show relations of sensory niche, of bell margin and velarium in adult Tripedalia. The velarium represented as pendant.
Fig. 45. To show the same structure in a young Tripedalia.
Fig. 46. Horizontal section through the last just at the margin, to compare with Fig. 29.
Fig. 47. Cross-section through the nerve ring.
Fig. 48. The structure of the nerve as seen by focusing at successive levels.
Fig. 49. Diagram to show the relation of the nerve ring to the sensory club.
Fig. 50. Horizontal section through the upper part of the sensory niche, to show passage of nerve root through gelatine of subumbrella to stalk of sensory club.
Fig. 51. Vertical section through base of stalk of sensory club, to show same passage.
Fig. 52. Similar section to last, but nearer to perradius, to show sub-endodermal tract of nerve fibres.
Fig. 53. Sensory organ in proboscis of Tripedalia, as seen from surface in living animal.
Figs. 54 and 55. Sections of same sensory organ.
Fig. 56. Vertical section through one side of proboscis, to show sensory organ attached to endoderm. (Tripedalia.)
Fig. 57. Diagram of the outlines of sensory club seen from the side, by camera lucida.
Fig. 58. Part of retina of larger complex eye cut radially.
Figs. 59-62. Four sections in direct sequence through retinal cells transversely, larger eye.
Fig. 63. Transverse section through the tips of cells of a slightly pigmented retina, larger eye.
Figs. 64-66. Three transverse sections through vitreous body at different levels. All from same series, but not in direct sequence; larger eye.
Fig. 67. Radial section through retina, to show fibres from the long pigment cells; larger eye.
Fig. 68. Transverse section through vitreous body of Tripedalia near retina.
Fig. 69. Vertical section through smaller complex eye.
Fig. 70. Wandering cells, Charybdea.
Fig. 71. Floating mass, from stomach pocket of Tripedalia.
Fig. 72. Horizontal section through larger complex eye. (See text figure, p. 50.)
CUBOMEDUSÆ. PLATE I.
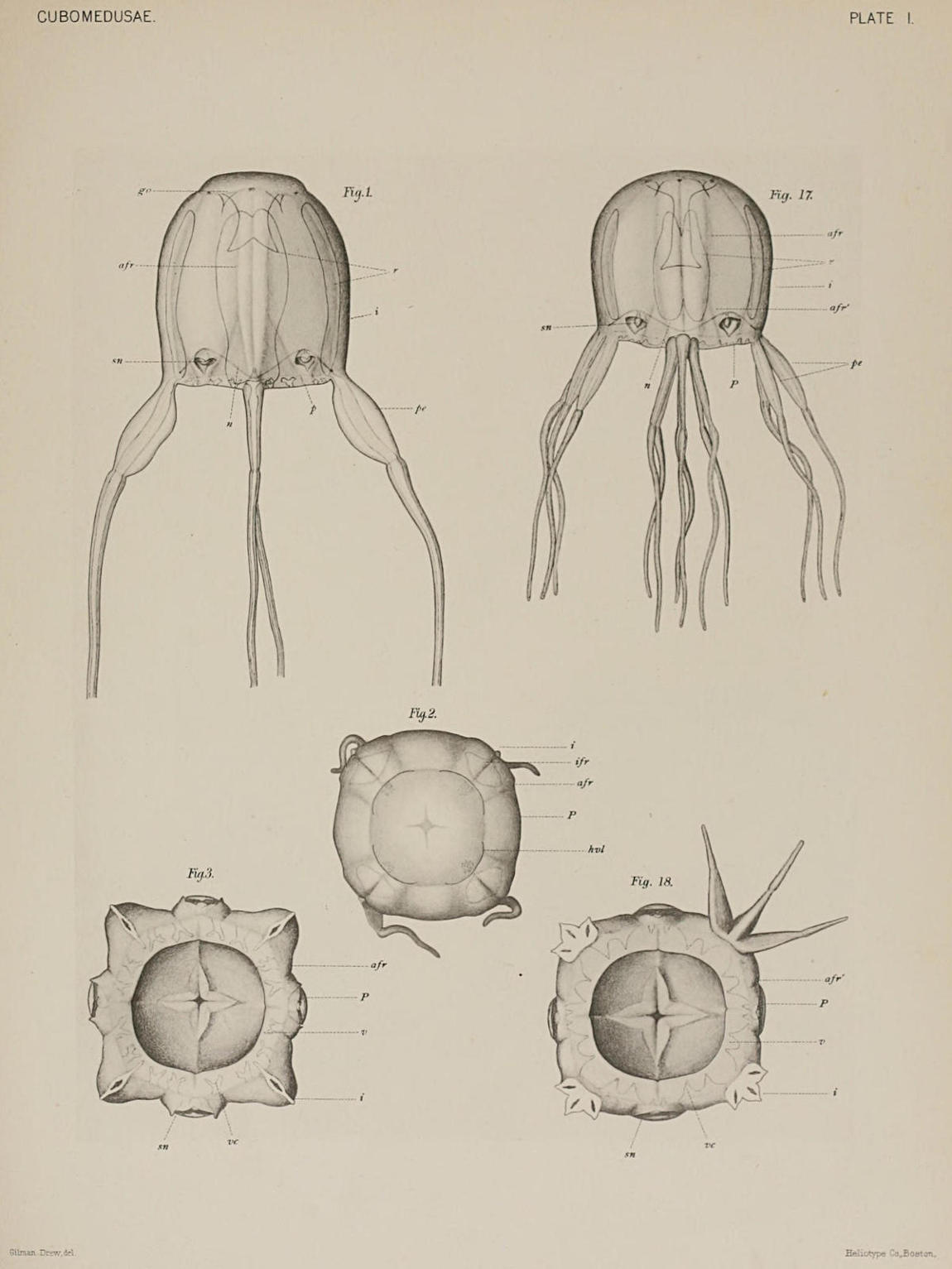
Gilman Drew, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE II.

Conant & Crew, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE III.
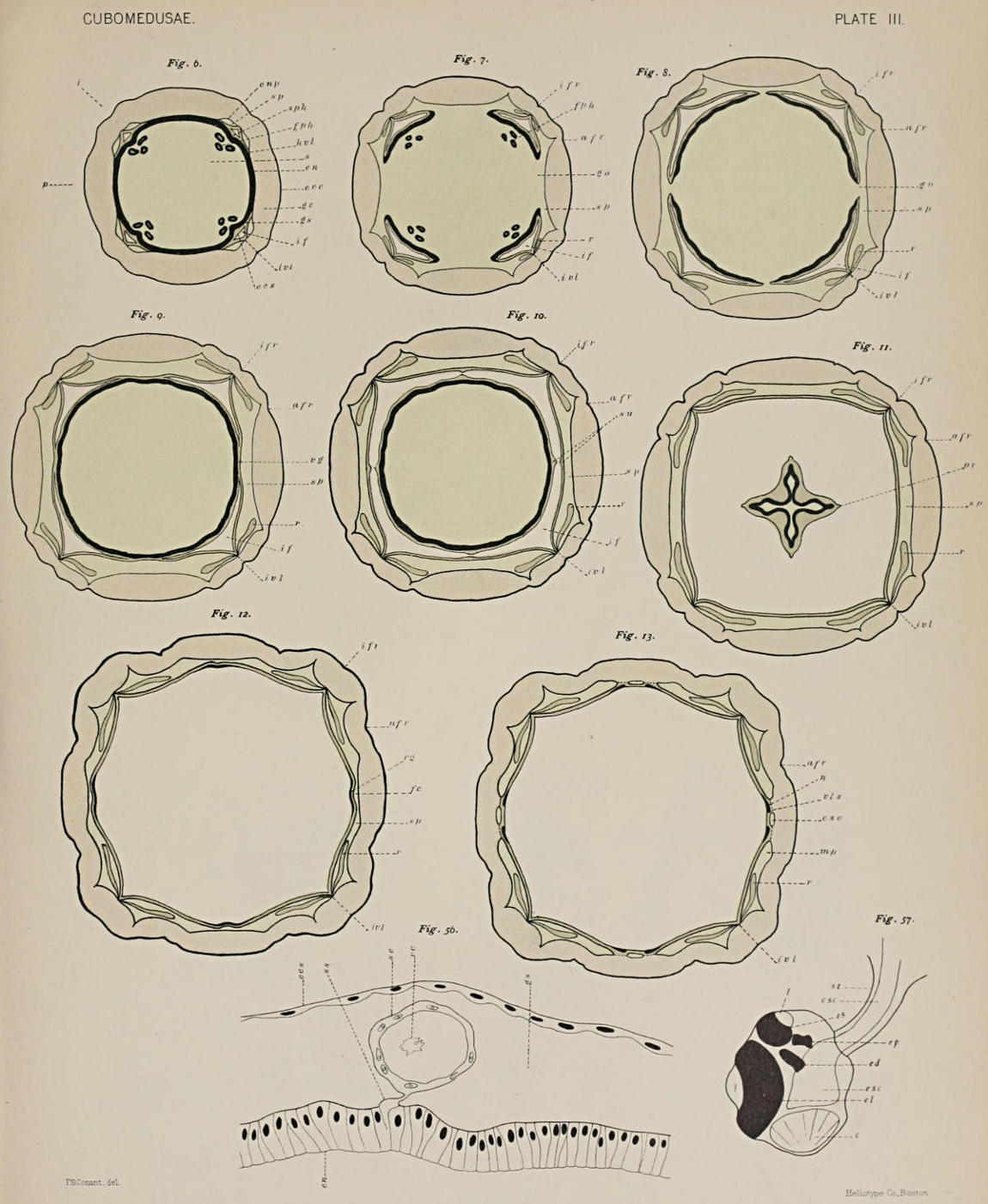
F.S. Conant, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE IV.
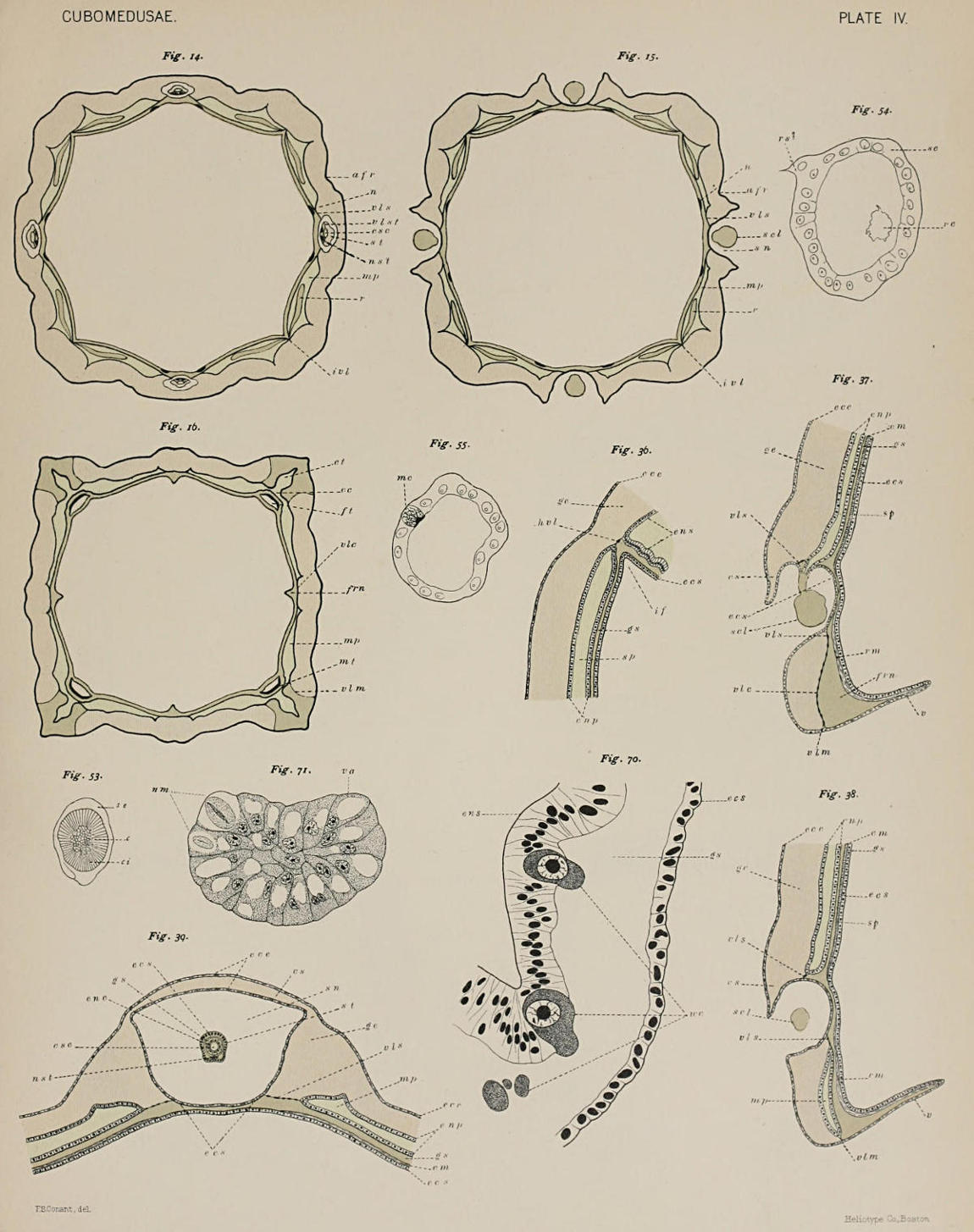
F.S. Conant, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE V.

F.S. Conant, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE VI.
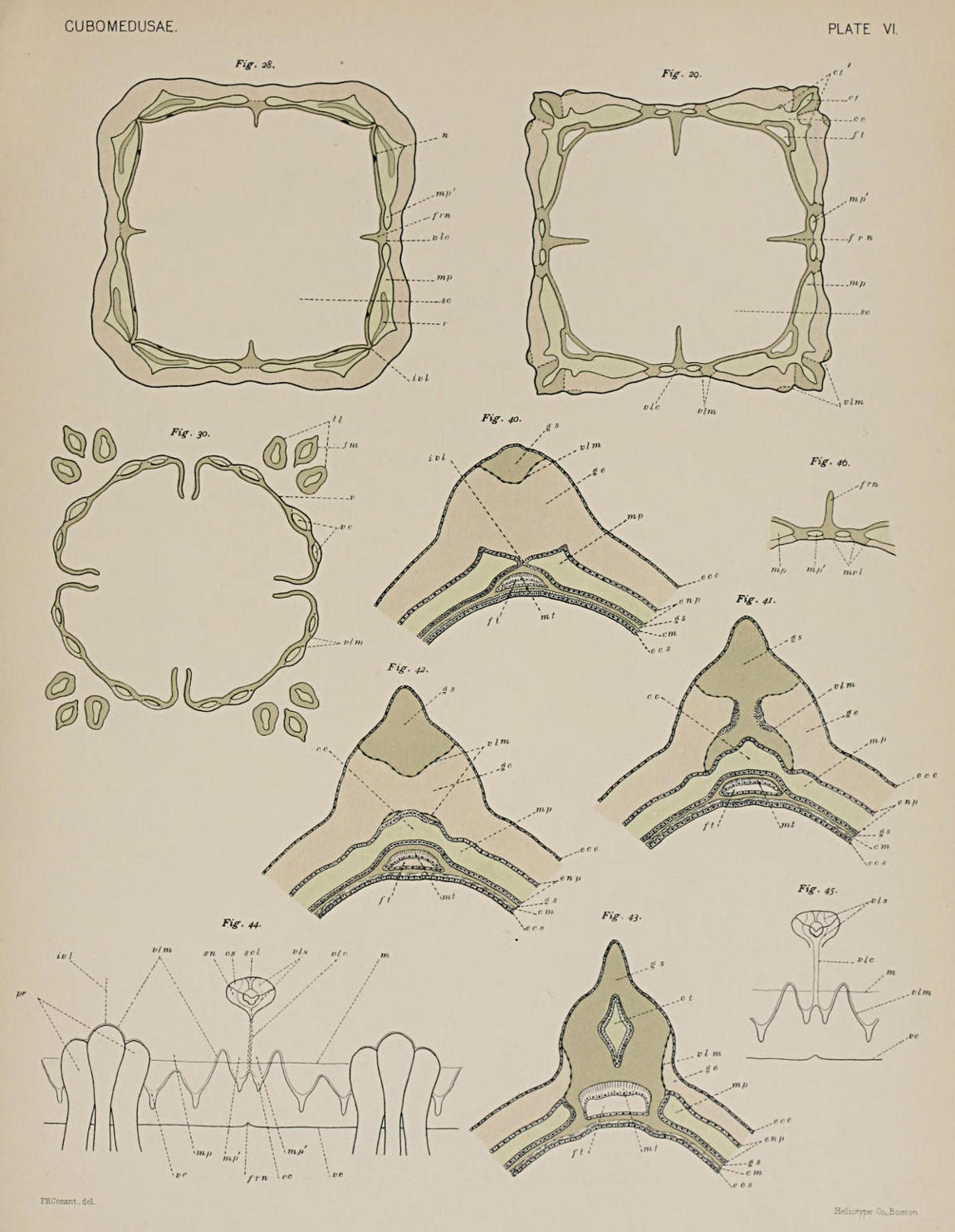
F.S. Conant, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE VII.

F.S. Conant, del. Heliotype Co., Boston.
CUBOMEDUSÆ. PLATE VIII.

F.S. Conant, del. Heliotype Co., Boston.