Title: Physiology and histology of the Cubomedusæ
Author: E. W. Berger
Contributor: Franklin Story Conant
Release date: March 3, 2017 [eBook #54276]
Most recently updated: October 23, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/54276
Credits: Produced by Donald Cummings, Bryan Ness and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive/American Libraries.)
Transcriber’s Note: The images contained within black borders are clickable for a larger version, if you are using a browser/device that supports this functionality.
Memoirs from the Biological Laboratory
OF THE
JOHNS HOPKINS UNIVERSITY
IV, 4
WILLIAM K. BROOKS, EDITOR
INCLUDING
Dr. F. S. Conant’s Notes on the Physiology
A DISSERTATION PRESENTED TO THE BOARD OF UNIVERSITY STUDIES OF THE JOHNS
HOPKINS UNIVERSITY FOR THE DEGREE OF DOCTOR OF PHILOSOPHY
BY
E. W. BERGER
BALTIMORE
The Johns Hopkins Press
1900
PRINTED BY
The Lord Baltimore Press
THE FRIEDENWALD COMPANY
BALTIMORE, MD., U.S.A.
This Memoir is a continuation of the work upon the Cubomedusæ which was begun by the late Dr. Franklin Story Conant, and it contains his notes of physiological experiments, as well as new results which have been obtained by Dr. E. W. Berger from the study of material which had been collected by Dr. Conant, who had hoped to make it the object of further study.
In order that this work may be made public as a continuation of Dr. Conant’s researches, his sister, Grace Wilbur Conant, has, with the coöperation of other members of his family, made an adequate and generous provision for its publication.
For this gift, which is at once a contribution to science and a memorial of an able and promising investigator, lately student and fellow in this institution, the Johns Hopkins University returns its grateful acknowledgments.
DANIEL C. GILMAN, President.
W. K. BROOKS, Professor of Zoölogy.
| PAGE | |
| INTRODUCTION. | |
| History | 1 |
| Epitome of Anatomy | 2 |
| PHYSIOLOGICAL. | |
| Charybdea. | |
| Light and Darkness | 5 |
| Concretions | 8 |
| Sensory Clubs | 9 |
| Velarium and Frenula | 11 |
| Pedalia, Interradial Ganglia, Tentacles | 12 |
| Stomach, Suspensoria, Proboscis, Subumbrella | 13 |
| Margin, Radial Ganglia, Nerve | 15 |
| Stimulation | 17 |
| Activity of Charybdea | 17 |
| Temperature | 17 |
| Food and Feeding | 18 |
| Occurrence of Charybdea | 18 |
| Aurelia and Polyclonia (Cassiopœa) | 19 |
| Summary | 22 |
| DR. CONANT’S NOTES. | |
| Charybdea. | |
| Light and Darkness | 24 |
| Sensory Clubs | 26 |
| Nerve | 29 |
| Side, Subumbrella | 30 |
| Pedalia, Velarium, Ganglia | 31 |
| Tentacles | 32 |
| Proboscis, Stomach, Phacelli | 33 |
| Temperature | 33 |
| Food and Feeding | 33 |
| Occurrence of Charybdea | 33 |
| Activity of Charybdea | 34 |
| Aurelia and Polyclonia | 35 |
| Cassiopœa | 39 |
| [vi]Aurelia | 39 |
| HISTOLOGICAL. | |
| Method | 40 |
| Anatomy | 41 |
| Distal Complex Eye— | |
| General | 41 |
| Cornea | 42 |
| The Lens | 42 |
| The Capsule | 44 |
| The Retina | 45 |
| (a) The Prism Cells | 46 |
| (b) The Pyramid Cells | 48 |
| (c) The Long Pigment Cells | 50 |
| (d) Subretinal Nerve Tissue | 53 |
| (e) Discussion of Literature | 53 |
| (f) Function of the Retinal Cells, Patten’s Theory, and further Literature | 56 |
| The Proximal Complex Eye | 60 |
| The Simple Eyes | 61 |
| Lithocyst and Concretion | 63 |
| The Epithelium of the Clubs | 64 |
| Network and Multipolar Ganglion Cells | 67 |
| The Nerve Tissue | 67 |
| The Supporting Lamella | 68 |
| Epithelium of Ampulla and Floating Cells | 68 |
| The Endothelium of the Peduncle | 73 |
| The Tentacles and Pedalia— | |
| The Ectoderm | 74 |
| (a) Thread Cells | 74 |
| (b) Muscle Fibers | 74 |
| (c) Ganglion Cell | 75 |
| The Endoderm | 75 |
| Summary | 77 |
| LITERATURE | 78 |
| REFERENCE LETTERS | 80 |
| DESCRIPTION OF FIGURES | 81 |
This paper may be regarded as a continuation of the Cubomedusan studies pursued by Dr. F. S. Conant while in Jamaica, in 1896 and 1897, with the Johns Hopkins Marine Laboratory. His systematic and anatomical results have since been published as his Dissertation (“The Cubomedusæ”) by this University. Conant described this paper as Part I, hoping soon to add a second part on the physiology and the embryology, for which he had some notes and material at hand. Returning, however, to Jamaica with the laboratory, in 1897, he continued his physiological experiments, and preserved material for histological purposes. Upon the untimely death of Conant, his material and notes were placed in my hands by Professor Brooks, to whom I here take the opportunity of expressing my appreciation and sincere thanks for the honor thus conferred and for the many favors received.
In this paper I shall note at some length Conant’s physiological results and append his notes. I shall also add my results on the histology of the eyes and the sensory clubs in general, with some few facts on the histology of the tentacles. The embryology will be reserved for a future paper.
The forms used in the physiological experiments were Charybdea Xaymacana, one of the two species (see Literature V, a and b) first found and described by Conant; Aurelia aurita; Polyclonia and Cassiopœa. The greater number of Conant’s notes are on Charybdea, and were left by him just as taken at the time of experimenting. Many of these notes are highly interesting and in the main fit in well with Romanes’[I] and Eimer’s[IV] results.
Dr. Conant’s work on Charybdea, in 1897, was wholly done at Port Antonio, Jamaica. At first Conant had only varying success in obtaining Charybdea, scouring the harbor and neighboring water at all hours, only to obtain but few specimens. It was on the forenoon of August 7th, while we were dredging at the head of East Harbor with a steam launch, that many Charybdeæ were brought up in the dredge. This gave Conant a clue to their whereabouts and to the means of obtaining them, and from that time on he was able to[2] obtain them in abundance. His first physiological experiments were begun on August 4th and continued thereafter at intervals of several days until his departure from Jamaica on September 6th.
Dr. Conant usually performed his experiments during the second half of the forenoon, after the animals had stood for a few hours in the laboratory.
The building that was rented at Port Antonio for a laboratory had, in the basement, a photographer’s dark-room, which was of great service to Conant in his experiments.
The experiments on Aurelia, in 1897, were also performed at Port Antonio, between August 6th and 9th. The experiments on Cassiopœa were probably made at Port Antonio, where specimens were occasionally obtained.
The notes on Aurelia and Polyclonia, in 1896, were taken at Port Henderson, between May 12th and June 27th.
In his notes Conant speaks of Polyclonia and Cassiopœa. It is at present undetermined whether he really had both forms or whether he uses the two names for the same form. It seems likely that in 1896 he thought the form to be Polyclonia, while for some reason, in 1897, he supposed it to be Cassiopœa. I have examined several specimens of these medusæ brought from Port Antonio and find that they all have twelve marginal bodies and twenty-four radial canals, according to which (V, Haeckel’s System), they should be Polyclonia. Conant, however, speaks of removing sixteen marginal bodies, which seems to indicate that he had Cassiopœa. A careful classification of this form of medusæ found about Jamaica seems to be a desideratum. I suppose, however, that for our purpose in this paper it will make little difference which name is used, the two forms being so similar in form and structure. I have, therefore, decided to retain both the names used by Conant.
For the complete anatomy of Charybdea the reader is referred to Dr. Conant’s dissertation, “The Cubomedusæ” (8b), or the Johns Hopkins University Circulars (8a), both published by the Johns Hopkins Press. But, for the convenience of those who may be less familiar with Cubomedusan anatomy, the following brief summary of the anatomy of Charybdea is given:
The Cubomedusæ, as the name implies, approximate cubes, with their tentacles (four in Charybdea) arranged at the four corners of the lower face of the cube. These tentacles are said to lie in the[3] interradii. Half way between any two points of attachment of the pedalia (the basal portions of the tentacles) and a little above the margin of the bell (cube), in a niche, hang the sensory clubs, one on each side, four in all. Each sensory club hangs in a niche of the exumbrella and is attached by a small peduncle whose axial canal is in connection with one of the four stomach-pockets and in the club proper forms an ampulla-like enlargement.
Each club is said to lie in a perradius, and, like the tentacles, belongs to the subumbrella. This is shown by the course of the vascular lamellæ, bands of cells that, stretching through the jelly from the endoderm to the ectoderm all around the margin, form the line of division between sub- and exumbrella.
Each club has six eyes. Two of these on the middle line of the club facing inwards are called the proximal and distal complex eyes, to distinguish them from the four simple eyes that are disposed laterally, two on each side of the line of the two complex eyes. All of these eyes look inwards into the bell cavity through a thin transparent membrane of the subumbrella. Besides the eyes and the ampulla already mentioned, a concretion fills the lowermost part of the club, and a group of large cells, having a network-like structure and called network cells by Conant, fill the uppermost part of the club between the proximal complex eye and the attachment of the club to its peduncle (Plate II, Fig. 13). What is evidently nerve tissue, fibers and ganglion cells, fills the rest of the club, with two groups of large ganglion cells disposed laterally from the network cells. A sensory (flagellate) epithelium covers the club.
Most Cubomedusæ, among them Charybdea, have a velarium (comparable to the velum of the Hydromedusæ), a membrane of tissue that extends inwards at right angles all around the margin. This velarium, like a velum, has a central opening through which the water is expelled from the bell-cavity when the animal pulsates. In the perradii and in the angle between the velarium and the body wall, are the frenula, which give support to the velarium much like brackets support a shelf, except that here the brackets are above the shelf instead of below.
In the upper part of the bell is the stomach, with the phacelli in its interradii, and continued ventrally into the manubrium, or the proboscis. The cavity of the stomach is continued in the perradii through the four gastric ostia into the four stomach pockets, which[4] occupy the sides of the bell and extend to the margin. Immediately below the gastric ostia, and in the bell cavity, are the suspensoria, one in each perradius. These support the floor of the stomach much as the frenula support the velarium, except that the suspensoria are placed under the shelf (to continue Conant’s figure) and not above it as are the frenula.
A nerve ring, underneath the epithelium of the subumbrella, passes from near the origin of each pedalium at the margin to the origin of the peduncles of the sensory clubs, a little above the margin, giving off a branch to each club. Eight ganglia are found in the course of this nerve. The four pedal ganglia lie near the bases of the pedalia, and are hence interradial; the four radial ganglia lie near the bases of the peduncles of the clubs, and are perradial. A small nerve, radial nerve, can be traced a short distance upwards from each radial ganglion. Underlying the epithelium of the frenula and the suspensoria are ganglion cells and nerve fibers in larger numbers than elsewhere (excepting the ganglia mentioned) in the subumbrella. Otherwise, ganglion cells and nerve fibers underlie the epithelium of the subumbrella, including the inner surface of the velarium, as also do muscle fibers, except in the perradii and in the region of the nerve, where the latter become interrupted.
Light and Darkness—Experiments 1-9, 10, 33, 34.—As already stated in the Introduction, a part of Conant’s experiments were performed in a photographer’s dark-room, with the animals in a deep glass jar. In the dark a fair proportion of the animals became nearly quiescent on the bottom, but upon lighting a lamp many started up immediately, while others took a longer time to come to the surface and swim. These experiments were tried a number of times and on different occasions with very similar results. Some medusæ, however, tried immediately after being brought in, seemed not to react so well upon being placed in the dark-room, nor would they become quiescent. This, probably, was due to the fact that the animals had not yet recovered from the effects of being caught and placed in new surroundings. (Experiments 1, 2, 3.)
Other experiments (4-8, 33, 34) were tried by carrying the jar with the animals from the weaker light of a room into the more intense light of outdoors or into direct sunlight. The usual result was an inhibition of pulsation and a settling to the bottom, while the medusæ immediately became active again upon returning with them to the room. These results were so marked that no doubts can be entertained as to their cause, though some exceptions occurred in which animals placed in the sun continued to swim on the surface or soon recovered pulsation. In some experiments, too, no animals responded to the inhibitory stimulus of the brighter light or all very soon recovered. (See, however, Temperature.)
Reducing the light by placing a coat over the jar produced the same effect in some experiments (8, 9, 10) as did reducing the light in other ways, while removing the coat produced the same effect as exposure to brighter light. In these instances it appears to be the transition from weaker to stronger light that inhibits pulsation, rather than the actual intensity of the light; and vice versa. It must be noted, too, that when left for some time in any one place[6] the animals changed, some coming to the surface and others going to the bottom.
These experiments show beyond doubt that Charybdea is sensitive to light, and that it is moderate light that stimulates the animals to activity, while darkness and strong light inhibit activity. While the individual exceptions, as Conant himself suggests, are well explained on the supposition of individual diversity, yet it appears that other conditions, such as the time of day, temperature, etc., may have been responsible for some of the exceptional experiments in which no animals responded as expected.
While light of any intensity seems to have stimulated Romanes’[I] Sarsia and Tiaropsis (Hydromedusæ) to activity, we note that it is moderate light that stimulates Charybdea. This fact is evidently correlated with the circumstance that Charybdea usually lives upon or near the bottom.
It may further be added in regard to Romanes’ Tiaropsis polydiademata, that when it was suddenly exposed to light it went into a spasm preceded by a long latent period during which there was a “summation of stimulating influence” in the ganglia. Sarsiæ would congregate toward the source of light and in general were more active in light than in the dark, while sudden darkness often inhibited a swimming bout. Romanes proves for Sarsia that the marginal bodies are the seat of luminous stimulation and that it is the light rays and not heat rays that stimulate. He also remarks that he has obtained similar results on the covered-eyed (Scyphomedusæ) medusæ, namely, that they respond to luminous stimulation.
It may here be of interest to note a few observations made by myself at Wood’s Holl, Mass., on a beautiful Olindiad, which is abundant in the Eelpond at the above place. I found that in a room, in the ordinary light of evening, the animals swam actively; but the moment the electric light was turned on they stopped swimming and settled to the bottom or attached themselves to a branch of some weed or stem suspended in the water. This was the result in every trial. It is found, further, to be little active during the brighter parts of the day, when one must dip quite deep with a net in order to obtain it. A similar observation is also made by Murbach[II], who further states that this medusa may be deceived into laying its eggs by placing it in the dark.
One cannot help but remark how analogous is the behavior of medusæ, in respect to light and darkness, to the behavior of many of the higher animals,—and medusæ are among the most lowly organized of the animal creation.
Were one to conclude from the behavior of Charybdea in light and darkness in the laboratory, that it remained on or near the bottom in the daytime but became more active near or at the surface evenings, nights and early mornings, one would probably not be far from the truth. Dr. Conant, while towing near the bottom with a weighted net, in water four to five feet (1.2-1.5 m.) deep not far from shore and deeper farther out, found Charybdea in abundance mornings and afternoons, but very few in the evening. In the evening some few were usually taken in the surface tow. (See Introduction, Occurrence and Activity.)
Again, who knows but that Charybdea is active during the day, on the bottom where it was dredged (the light there would only be moderate), and quiet at night. This supposition would seem to be true, at least, for those forms of Cubomedusæ that live in deep water. We can hardly suppose that they should regularly rise to the surface from great depths and become active. This much we do know that bright light inhibits Charybdea’s activities, while it probably would not be active in perfect darkness.
I do not know just what interpretation to put upon Conant’s finding Charybdea at Port Henderson at the surface during the early part of the forenoon, before the sea-breeze roughened the water (“Cubomedusæ” p. 7). This fact hardly fits in with my conclusions above. Perhaps Charybdea’s habits vary with its habitat.
Finally, while I find no experimental evidence in Conant’s notes about what parts of Charybdea are sensitive to light, yet it would seem preposterous, from histological evidence and from Romanes’ results on Sarsia, to doubt that the eyes of the marginal bodies are the seat of this stimulation.
Dr. Conant further experimented by cutting off certain organs and parts from the Cubomedusan bell. These excisions consisted chiefly in cutting out the concretions of the sensory clubs, cutting off the whole club, eliminating a part or whole of the margin and the velarium, cutting the bell into sectors, excising the stomach and parts connected with it, and other parts.
Concretions—Experiments 10, 11.—The four concretions were removed from each of four animals. Two of these (Experiments 10, and another (X), not appended, to save space) seemed to be little if at all affected by the operation. One of the two (10) swam actively, at first up and down more changeably than those intact, but later mostly near the surface. The other one also swam actively and showed nothing to indicate weakened sense-perception. The other two (11) did not stand the operation well, as Conant remarks, and immediately went to the bottom, where they remained, one swimming, while eight hours later one was still in good condition.
Several attempts with stronger light by removing the coat from the jar made no difference in the behavior of 10; it continued to swim as heretofore. Upon a final trial, however, with removing the coat, it went to the bottom, thus showing a possible reaction to light; but when next seen it was keeping to the bottom.
That the concretions should function as organs of light sensation, as the first of the above animals might seem to indicate, I believe is out of the question.[a] The fact, too, that this same animal (10), together with another (X), swam actively, immediately changing their course upon coming to the surface, in reality behaving quite as normal animals, hardly permits us to conclude from the behavior of the other two (11) that the concretions function directly as organs of equilibrium or space relations. May these concretions not function simply as weights for keeping the sensory clubs with their eyes properly suspended? Since these concretions lie at the lowermost part of the clubs and in closed sacs and unsupported by cilia, it would seem that the above suggestion as to their being weights is not improbable. Direct observation (Experiment 20) by Conant shows, furthermore, that the clubs always hang with a tendency for the concretions to be lowermost, regardless of the position of the animal.
Again, while they may function as weights, as just explained, the fact that the epithelium of the clubs is flagellated (a flagellum, continued as a nerve fiber, to each cell—see Histology), the supposition lies near that these flagella are the ones influenced by the concretions as the clubs bear against one side of the sensory niche or the other.[9] A somewhat similar view seems to be held by other observers and is noted by Lang in his text-book (“The outer epithelium of the auditory body carries the auditory hairs”). It seems, then, that in functioning as weights for suspending the clubs, they may also serve at the same time for making the pressure of the club against the niche greater than if they were absent, and thus in part serve in equilibrium. On this supposition we should expect, furthermore, that after the removal of the concretions the animal would be little, if at all, affected, since the clubs themselves, without the concretions, would still be of sufficient weight to be influenced by gravity and thus to bear against the walls of the sensory niche. It must be noted, however, that Conant’s experiments upon equilibration in Charybdea are negative. Also, that Charybdea has any auditory sense is negatived by two attempts of Conant’s with a violin—one attempt with the violin near the animals, and another with it in contact with the dish. (From an unpublished note.) Hence, some other word such as sensory or equilibrating should perhaps be substituted for “auditory” in the above quotation.
Removing the concretions from Aurelia gave negative results very similar to those on Charybdea. (Experiment 42.)
Sensory Clubs—Experiments 12-19, 20, 24.—The entire sensory clubs were removed from a number of animals. A paralysis of pulsation followed by a rapid recovery was the usual result. In some instances, however, there was no paralysis, while in others no recovery followed paralysis. This is true in a general way whether one club only or all were removed. While no permanent paralysis followed the removal of one or two clubs, yet permanent paralysis did occur after the removal of a third club, as, of course, also after the removal of a fourth. It is evident, too, that as the removal of the clubs progressed recovery seemed to be weaker after each cutting, except in one case when pulsation seemed to be quickened after the removal of a second club. The pulsations after recovery seemed to be not so strong and regular, often quite feeble, and in one instance in groups. Pieces of tissue with a club attached and pulsating regularly, ceased pulsating after removal of the club, in one instance, however, still giving occasional contractions.
These results are quite the same as those of Romanes[I] on Aurelia, Cyanæa, etc., and of Eimer[IV] on Aurelia, Rhizostoma,[10] Cotylorhyza, etc.[b] In these forms Romanes sometimes obtained complete paralysis after the removal of the sensory clubs only, as also after the removal of the whole margin, though this was not marked in Aurelia. In Cyanæa and other forms motor centers seemed to be more abundant than in Aurelia, so that paralysis was oftener followed by recovery. He concludes that while the principal motor centers reside in the lithocysts, other centers doubtless exist that may function vicariously, but that the centers of the margin are more definitely limited to the marginal bodies in the Scyphomedusæ than in the Hydromedusæ, in which the whole margin seems to be replete with centers. He feels positive, furthermore, that no motor centers exist in Aurelia’s margin outside of the marginal bodies (lithocysts). Eimer’s results are essentially the same as Romanes’, so that for a more detailed comparison of the two, Romanes’ works should be consulted.
Romanes’ conclusion for the Hydromedusæ is that the motor centers are not so definitely localized in the marginal bodies, but in the margin generally, the excision of the marginal bodies alone producing only partial paralysis, as would also the removal of the margin from between the marginal bodies, but not so marked. For the Hydromedusæ he concludes, then, that all the centers of spontaneity are definitely localized in the margin, but not limited to the marginal bodies. To this he mentions one exception, namely, Staurophora laciniata, in which another center is found near the margin and two others in two opposite arms of the proboscis.
I made the remark in an abstract (VI) on Conant’s notes that Romanes did not obtain recovery of pulsation after removal of all the lithocysts in Aurelia. As noted above, he did obtain recovery, so that Conant’s results on Charybdea and also Aurelia (see Polyclonia and Aurelia) are quite in agreement with Romanes.
The paralysis following the removal of the clubs in Charybdea is evidently, primarily, the result of a loss of a part of its nervous mechanism (motor centers), and, secondarily, of nervous shock, and points to the existence of a definite nervous mechanism in the clubs. The histological evidence is here, as usual, corroborative of the physiological.
Another interesting phenomenon observed after the removal of[11] one or all of the clubs was the strange behavior of the proboscis. This would reach from side to side, expanding and contracting its lips as if trying to grasp something. This behavior is very similar to that of the proboscis of Tiaropsis indicans when Romanes stimulated any part of its subumbrella, or of Limnocodium sorbii, a little fresh-water medusa, when he stimulated its margin or the region of the radial canals. (Ib., p. 242.)
I may add that I observed a very similar movement of the proboscis of the Olindiad, before mentioned. When I pulled off pieces of its gonads by means of quick jerks, with a small forceps, it would continually reach toward the injured part of its subumbrella. This medusa is generally quite active with its proboscis and can occasionally be seen to reach with it.
Romanes states in one place that the proboscis is not affected by the excision of the margin. This is evidently not the case in Charybdea, in which excision of the sensory clubs (which really belong to the margin—see “Cubomedusæ”) decidedly stimulated the proboscis to active movements. This, furthermore, points to the marginal bodies as being organs of considerable importance in giving information in the life of Charybdea. In Romanes’ Sarsia and other medusæ, however, the proboscis did respond to the stimulation of the tentacles and the marginal bodies, as also would the bell respond to a stimulation of the proboscis (manubrium), thus showing a reflex nervous connection between these regions of the bell, similar to that described for Charybdea.
Velarium and Frenula—Experiments 18, 29, 30, 41c.—“The power of originating contractions” to use Conant’s own words, “evidently resides in the velarium or in ganglion cells of the frenula, just as it does in the proboscis and the floor of the stomach.” Isolated pieces of the velarium contracted by themselves as did the whole velarium when all other tissue had been removed. An isolated velarium with the margin and the pedalia attached gave irregular contractions. When the pedalia with the interradial ganglia were removed it still contracted; and when all the other tissue was cut off contractions continued.
Cutting the velarium caused the pedalia to be strongly contracted inwards so that the tentacles were brought inside the bell. Cutting away the velarium did not interfere with the pulsations of the bell, but progress was much retarded.
Cutting the frenula caused the pedalia to contract but seemed not to affect the ability to swim. Comparing the velarium of the Cubomedusæ with the velum of the Hydromedusæ, I recall no observations similar to the ones here noted, though it seems that the two may have quite similar functions. It seems somewhat probable that the velum, and also the velarium, may function in obtaining food,—and this besides their function in swimming. Their probable function in swimming, as is well known, is evidently to narrow the mouth of the bell and thus to cause the water to be forced out in a smaller but more rapid stream, giving the animal a steady and more prolonged movement through the water at every contraction of the bell. In regard to taking food, I observed that a small crustacean, in the process of being swallowed by an Olindiad, seemed to be held by the velum being firmly contracted about it while the proboscis was working itself over the crustacean. It would seem, furthermore, that my supposition is supported for Charybdea by the fact that the pedalia and tentacles were contracted so as to be brought inside the bell when the velarium was cut. The stimulus of cutting the velarium may be comparable to a stimulus from some object touching it, and thus cause the pedalia and tentacles to come reflexly to aid in capturing or holding the object, a fish, crustacean, or such, to be captured.
Pedalia, Interradial Ganglia, Tentacles—Experiments 15, 23, 27-31, 41b.—When the pedalia were removed, the power of the animal to guide itself was completely gone. When one pedalium was cut the others contracted, while stroking the outer edge of the pedalia, touching the sensory clubs, or sharply pricking the subumbrella, often produced the same result. (See also Nerve.) The upper part of the subumbrella seemed not so sensitive and more seldom produced the reflex of the pedalia, while the base of the stomach did not give it at all. Stroking the outer edge of the pedalia of Tripedalia cystophora, the second of the two species of Cubomedusæ described by Conant, also caused the pedalia to be contracted inwards. I may note here that the muscle fibers under the ectoderm of the pedalia are specially well developed at and near the inner and outer edges, both in Charybdea and Tripedalia. On the flattened sides of the pedalia the muscle fibers are fewer.
When the pedalia were cut off far enough up to remove the interradial ganglia, coördination was not affected and the animal[13] could pulsate well enough but with little progress. (See above under Velarium and Frenula.)
An isolated tentacle is capable of squirming contractions, and when stimulated at either end, it would contract wholly or in part only.
The pedalia, then, it would seem, serve also as a steering apparatus, for which they are admirably fitted, considering their blade-like thinness.
Considering, now, the reflexes noted under this head and the preceding one, we find that there is an intimate nervous connection between the velarium and frenula, subumbrella, sensory clubs, nerve, and a single pedalium, on the one hand, and the pedalia on the other hand. This is born out fully, furthermore, by the histological evidence—(See Introduction and “Cubomedusæ”). Considering the subumbral plexus of ganglion cells and fibers, including the velarium and the frenula, which is in connection with the nerve ring and this again with the sensory clubs and the interradial ganglia at the bases of the pedalia, we have a basis for these reflexes. While Conant failed to demonstrate nerves (“Cubomedusæ”) from the interradial ganglia to the pedalia, yet, that a nervous connection exists between the pedalia and the bell is well shown by his physiological experiments. I have, furthermore, demonstrated ganglion cells under the ectoderm of the tentacles (see Histology).
Romanes obtained quite similar results in the Hydromedusæ. He found that when a tentacle of Sarsia was slightly stimulated, it alone would contract, but when it was more strongly stimulated the other tentacles also would respond as also the manubrium. I find no evidence in Conant’s notes of any such response of the manubrium of Charybdea, except when the clubs were cut off.
The reflex obtained on stimulating the subumbrella of Charybdea, when the pedalia would contract, is somewhat different from that obtained by Romanes, who found that the most sensitive part of the subumbrella in producing a reflex of the margin was at the junction of the manubrium to the bell and that the subumbrella below this point did not give the reflex.
Stomach, Suspensoria, Proboscis, Subumbrella—Experiments 12, 18, 19, 24-26, 29, 31.—The proboscis and the stomach with the phacelli when cut out, contracted with or without the lips removed. The isolated lips also contracted (twitched).
Pieces of the sides connected only with the stomach and suspensoria, or with the margin (Experiment 47 (?)) twitched spontaneously, but seldom did so when these were removed. In one instance the whole side was cut out so as to exclude the radial ganglion but still connected with a portion of the suspensorium. This pulsated, or contracted, but on being halved transversely, the lower half ceased to contract while the upper half connected with the suspensorium, continued to contract.
Cutting off the whole stomach end of the animal excited to very rapid pulsations of the remaining part, with the stream of water stronger out the aboral end than past the velarium.
Conant says, “It seems I get no good evidence of the subumbrella without connection with special nerve centers being able to contract by itself.” The piece in which he did get contractions he suspects may have been intimately associated with some part of the frenula or the suspensoria. In Polyclonia no such doubt exists, for small pieces of subumbrella were seen to contract. A small piece of subumbrella of Charybdea with a sensory club attached could contract by itself.
From the above it would seem that a center capable of inciting to contractions resided in the suspensoria as well as in the sensory clubs, and this may be one of the centers that becomes potent upon the removal of the clubs. This is further supported by Conant’s observation (Introduction and “Cubomedusæ”) that an extra large number of ganglion cells is found under the epithelium of the suspensoria. A somewhat similarly located center of spontaneity described by Romanes for Staurophora laciniata (Hydromedusa) has already been noted.
As to the rapid pulsations of the bell after cutting out the stomach end, this also is similar to Romanes’ results on Aurelia and other Scyphomedusæ, when he cut off parts of the manubrium or an aboral ring out of the bell. In these instances, however, Romanes soon obtained a slackening of the rhythm following the temporary acceleration. The temporary acceleration he attributes to the stimulus of cutting, and the slackening to a lack of some afferent stimulus from the removed tissue. Conant obtained the same results on Polyclonia by removing the oral arms (see Polyclonia) but says nothing about a slackening of the rhythm in Charybdea. I believe the increased rhythm in Charybdea was in part due to the decreased[15] amount of labor necessary to force the water out of two openings instead of one, namely, past the velarium. Just how much this observation bears upon Romanes’ theory of rhythmic contraction, that the rhythm is due to an alternate exhaustion and recovery of the contractile tissue, as opposed to the ganglionic theory of rhythm of physiologists, one does not wish to speculate much. Yet, I feel that the observation rather supports this theory. The tissue having to do less work, would become less exhausted at each contraction and require less time for recovery and hence have a more rapid rhythm.
I here sum up Romanes’ theory in a few words. The ganglia liberate a constant and comparatively weak stimulus, one perhaps about minimal. This stimulus sets off the contractile tissue; but as the tissue contracts and becomes exhausted the constant stimulus becomes, in relation to it, sub-minimal, and it does not contract again until it has recovered and the stimulus is again strong enough to set it off. The ganglionic theory of rhythmic contraction supposes that the ganglia liberate stimuli to the contractile tissue at successive intervals. Romanes had this theory suggested to him by the rhythmic contractions he succeeded in obtaining by subjecting deganglionated bells to a continuous but weak faradic stimulus, or by placing them into weakly acidulated water, or into 5 per cent. glycerine. Romanes claims that his theory better explains muscular tonus and the contraction of involuntary muscle. He does not, however, hold this theory to the exclusion of the ganglionic theory, since only too often does he speak in terms of the latter. He further brings in his support the fact that the frog’s tongue, in which no ganglia have been demonstrated, can be made to contract rhythmically when subjected to a weak and continuous stimulus. He also calls attention to the rhythmic contractions seen in the Protozoa, the snail’s heart, etc. Finally, physiologists are much inclined to explain the rhythmic contraction of the heart and other involuntary muscles, in part, at least, as due to a property of the contractile tissue.
Margin, Radial Ganglia, Nerve—Experiments 18, 21-23, 30.—Complete removal of the margin did not stop pulsation; but the removal of the radial ganglia stopped it permanently. While this experiment seems to have been tried only once, yet, taking into consideration the results of other operations, it would seem that the principal centers of spontaneity reside in these ganglia. (It should[16] here be remembered that the interradial ganglia were probably removed at the removing of the margin.)
Cutting the nerve in the eight adradii caused the pedalia to bend inwards at right angles to their normal position but did not in the least affect the coördination of the sides. When, however, the sides were cut in the eight adradii to the base of the stomach, coördination for the main part ceased, and each side pulsated in its own rhythm.
I have said that the principal centers of spontaneity reside in the radial ganglia. Upon further thought this hardly seems warranted. No doubt, among the principal motor centers must be placed the ganglionic masses of the clubs, and the radial ganglia, together with the homologous interradial ganglia, represent centers of equal value. I speak of these two sets of ganglia as homologous, since strictly speaking, they both belong to the margin, and the clubs at whose bases they lie probably represent modified tentacles. Conant’s experiments leave us in the dark as to the function of these ganglia. Next in order, it would seem, are the ganglion cells in the suspensoria, as is suggested by the contractions of an isolated side with a portion of a suspensorium attached. (See previous head.) While we have seen that the frenula and the velarium can contract by themselves, yet, I find no evidence that these can impart their contractions to any adjacent tissue.
Conant’s results on cutting the nerve eight times and then continuing the cuts to the base of the stomach are quite the same as Romanes and Eimer obtained upon Aurelia. Romanes, however, concludes that in his Sarsia, Tiaropsis, etc., coördination was broken when only short incisions were made in the margin. Charybdea appears, then, to agree with Aurelia rather than with the Hydromedusæ. Yet, since Romanes at first obtained similar results to those of Charybdea on Sarsia, but on further experimenting concluded that coördination had really been destroyed at the first cutting, we cannot speak with certainty that coördination had not been destroyed in Charybdea before the cuts had been continued to the base of the stomach. I say not with certainty, because the injury to the bell being slight, coördination may have been maintained on the principle of a simultaneously (simultaneous for the octants) alternate exhaustion and recovery of the contractile tissue on the principle of Romanes’ theory.
Stimulation.—Romanes found when he stimulated a deganglionated bell of a Hydromedusa, that it responded by a single contraction, while that of a Scyphomedusa responded with several quite rhythmic contractions. Charybdea in this respect agrees with the Scyphomedusæ. Romanes’ results were also verified on Aurelia. (Experiments 12c, 15, 50, 51.)
Activity of Charybdea.—In speaking of the activity of Charybdea, I cannot do better than refer the reader to the notes. (Experiment 41.) Conant remarks in his dissertation what an active swimmer Charybdea is, and this is further borne out by his later observations.
Temperature.—Ice in the water seemed to have no effect, except when held against an animal, when a slowing of pulsation followed in a few instances. On some pulsating actively in the sun the temperature of the water was found to be 92° F. (Experiments 33-35.)
Conant does not tell us how cold the water became when he placed ice in it, but judging from his results, it seems that he might have obtained a decided slowing of pulsation if the water in which the medusæ swam had been permitted to approach anywhere near the freezing point, say 35-40° F. Romanes obtained decided slowing of pulsation, and even complete inhibition, on a bell of Aurelia, as also a lengthening of the latent period on some strips cut from a bell of Aurelia, by lowering the temperature of the water. Replacing Aurelia in warmer water had the effect of immediate recovery and increased rhythm. In Aurelia, raising the temperature increased the rhythm but diminished it when the temperature of the water became 70-80° F. After a slowing of pulsation due to such a rise of temperature, it would not quicken again when the animal was placed in water of its normal temperature. Romanes explains this by supposing that the tissue of the medusa had been permanently injured by the abnormally high temperature. It would be interesting to observe how the tropical Aurelia behaved under such treatment, seeing that Charybdea pulsated actively and without apparent injury in water at 92° F. Limnocodium, noted by Romanes, and probably a tropical species, lived happily in water at 85° F. in the lily house of the Royal Botanical Society. The temperature of the water could be raised to 100° F. before it proved fatal to this medusa. Such facts point to a decided difference in the constitution of the protoplasm of tropical and[18] temperate medusæ. Romanes’ Sarsia became frantic when placed in milk-warm water.
While writing the above, I was led to wonder whether the temperature of the water may not have been the stimulating influence in those experiments on light (previously noted) in which the medusæ continued to swim actively in the sunlight.
Food and Feeding.—See Experiment 36.
I again make note of a few observations made by myself on the Olindiad. A crustacean became entangled in the tentacles of a medusa; apparently this wished to retain it, for the proboscis reached in the direction of the crustacean, which, however, got away. I then placed, by means of a needle, another small crustacean against one of the tentacles. This was seized but not retained, for the animal pulsated and it was washed away by the water. Twice I saw a good-sized crustacean in the proboscis. In one instance the velum appeared to hold the part of the crustacean not yet in the proboscis. I noticed another with a crustacean wholly in the proboscis, which was much lengthened out, the upper part of the crustacean being in the stomach. The next morning the crustacean was wholly in the stomach and the proboscis normal. At 5.30 P. M. the crustacean was ejected, nothing but the shell and some rubbish remaining.
These medusæ seem to pay no attention to being touched by one of their kind, except to give a pulsation or two.
The proboscis appears very “intelligent” in its actions.[c] First, some of the tentacles can be seen to contract and to bend inwards, then the side next the tentacles contracts and the proboscis is seen to reach in that direction. I could not see, however, what the irritant was.
Occurrence of Charybdea—Experiments 37-40.—Dr. Conant’s remarks (“Cubomedusæ”) on the occurrence of Charybdea at the surface of quite shallow water and near the shore (which is quite at variance with former observations, that the Cubomedusæ are essentially deep-sea forms) are further borne out by his observations at Port Antonio. As already noted in the Introduction, Charybdea was here found in abundance in quite shallow water and near shore, but on the[19] bottom instead of at the surface as at Port Henderson. It is possible that the animals had been active near the surface earlier in the morning and that some unknown conditions determined their settling to the bottom earlier in the former place than in the latter.
Conant’s conjecture, “whether these were their natural conditions, or whether the two forms,” Charybdea and Tripedalia, “were driven by some chance from the deep ocean into the harbor and there found their surroundings secondarily congenial, so to speak,” seems to be borne out in favor of the former supposition (for Charybdea at least),—that these are their natural conditions and that Charybdea Xaymacana is essentially a shore form.
Experiments 42-53.
Many of the observations on these forms relate to the rate of pulsation. In an Aurelia, following the removal of a lithocyst, there was a pause followed by pulsations. In about two minutes rhythmic pulsations were renewed. Four minutes after the operation there were nineteen pulsations to the half minute, while twenty minutes after there were only nine, and these in groups of six and three. The normal rate of pulsation was twenty-five to the half minute.
Polyclonia behaved much in the same manner as Aurelia. Upon the removal of lithocyst pulsations continued, but in groups with short pauses. The normal rate of pulsation was twenty-seven to the half minute, while three minutes after the operation it was seventeen, and eleven minutes after, fifteen to the half minute. The tissue connected with a removed lithocyst gave contractions. Placing a Polyclonia in fresh sea-water more than doubled the rate of pulsation, which, however, soon fell to the normal rate, and lower in one instance. In small individuals the rhythm is decidedly more rapid than in those of larger size. The few observations on this point would seem to show that it is in inverse proportion to the squares of the diameters of the bells.
The removal of a single oral arm or of the whole eight, in Polyclonia, had much the same effect as the removal of a lithocyst: there was a decided slowing of the rate of pulsation, while the immediate effect of cutting was an acceleration or a return to near the normal rate. About a day later this same animal had quite[20] regained its normal rate of pulsation and continued to live over two weeks. A long latent period followed the cutting of an arm, before the stimulation of cutting manifested itself.
An Aurelia, with all its lithocysts removed, still gave spontaneous and coördinated contractions after allowing time for recovery from the operation. This was the result in one instance, while in several others only a few contractions were observed. Removal of the sixteen marginal bodies (lithocysts) in a Cassiopœa produced paralysis for a time but recovery soon followed. A Polyclonia with its entire margin removed was paralyzed but had so far recovered in a day as to be able, at intervals, to give spontaneous pulsations.
The removed margin of a Polyclonia pulsated vigorously. This margin was then split so as to make a ring within a ring but connected at one point by a small bridge of tissue. The waves of contraction, which always originated on the ring with the lithocysts, passed the bridge to the inner ring quite as Romanes experienced. The outer ring was next split so as to separate the exumbral portion from the subumbral, when it was found that the contractions always originated from the latter. Seven days after its removal, this same margin was still alive and pulsating vigorously, and broken-off pieces of the subumbral portion were pulsating by themselves. Fifteen of the ganglia were removed. It was then found that while most of the pulsations originated at the remaining ganglion, now and then contractions originated in other parts where no ganglion remained. Two days later this margin was still alive with contractions originating as often from other parts as from the ganglion. A similar observation was made on a margin of Cassiopœa.
A Polyclonia with the eight lithocysts of one side removed, to compare with a normal one, gave no evidence of affected coördination.
An oral lobe from an Aurelia could give contractions some minutes after removal.
In another Aurelia a circular cut was made about the base of the oral lobes through the epithelium of the subumbrella. The animal could pulsate well enough but coördination seemed a little affected, while in another one with a like cut but semicircular, no effect was noticed.
These results on the removal of the lithocysts (and margin in Polyclonia) in Aurelia, Polyclonia and Cassiopœa agree quite with those on Charybdea and, of course, also with Romanes’ and Eimer’s[21] results as to paralysis and recovery following the removal of the lithocysts, or margin, in Aurelia, Cyanea, etc. I recall no similar observations, however, on removing a single lithocyst, and the question of an explanation for the slowing of the rhythm thus brought about arises. Romanes gives as an explanation for the slowing of the rhythm (Aurelia, Cyanea, etc.) following the temporary acceleration upon removing the manubrium or a portion from the center of the bell, as due to a lack of an afferent stimulating influence upon the ganglia from the excised tissue. May a similar explanation not serve to explain the slowing following the removal of a single lithocyst, above noted? The removed lithocyst could no longer give its efferent stimulus to the remaining ganglia nor to the tissue, so that the former would have a weaker stimulating influence, in consequence of which the latter (the contractile tissue) would be deprived of a part of the original stimulus of the remaining ganglia as also of that of the removed ganglion. The whole would thus result in giving to the contractile tissue a weaker stimulus, which, again, would require longer and greater recovery on the part of the tissue in order to be set off by the stimulus at hand. This explanation is given on the basis of Romanes’ theory of rhythmic contraction previously explained.
Of course, it may be suggested that the musculature had lost tonus, due to the lack of influence of the removed ganglion (lithocyst), in consequence of which there was a lowering of irritability on the part of the contractile tissue. This would require a greater summation of stimulating influence (Ganglionic theory of contraction) on the part of the remaining ganglia to set it off. Again, the loss of irritability on the part of the contractile tissue may have been due to a lack of nutritive influence from the removed ganglion.
Romanes’ explanation, that the slowing of the rhythm following the removal of the manubrium and central parts of the bell in Aurelia and Cyanea is due to a lack of an afferent stimulus on the ganglia from the removed tissue, likewise explains the similar results obtained by Conant by removing the oral arms from Polyclonia.
The fact that a margin of Cassiopœa and also of Polyclonia, connected with but one ganglion, often originated contractions in other parts as well as from the ganglion, seems to show that motor centers resided in the margin outside of the ganglia. This would be somewhat at variance with Romanes’ conclusion, that no[22] such centers existed in the Scyphomedusæ. Conant does not state whether the Polyclonia margin in question was kept in fresh sea-water or whether the water was not changed during the seven days. If the latter is the case, then some poisonous compounds may have been formed that acted as a stimulus much as weakly acidulated water served Romanes in producing rhythmic contractions in deganglionated bells.
Again, while it is true that no ganglia are known to exist in the margins of the Scyphomedusæ outside of the ganglia in the marginal bodies, yet, ganglion cells and nerve fibers are found in the subumbral part of the margin as well as in the rest of the umbrella. And as I know no reason why scattered ganglion cells may not function as ganglia, it is possible that the contractions in question were spontaneous.
Finally, is it possible that the remaining ganglion originated the contractions in different parts of the margin, thus acting at a distance from the points at which contractions originated? Romanes gives an instance in which he believed to have evidence that this was the case. Upon a final consideration I am inclined to this latter explanation.
Summing up for Charybdea, we have seen that it is very sensitive to light, strong light as also darkness inhibiting pulsations, while moderate light stimulates it to activity. Also, a sudden change from weaker to stronger light, or vice versa, may inhibit or stimulate to activity respectively. This behavior of Charybdea seems to be correlated with its habit of life on the bottom. We have no reason to doubt but that the eyes of the sensory clubs are the seat of light sensation.
The experiments on equilibration are negative, giving us no certain light on the function of the concretions, though it appears that they may serve, in part at least, for keeping the sensory clubs properly suspended. Their function in giving the animal sensations of space relations is not, however, excluded.
Excision of the sensory clubs demonstrates that they are the seat of important ganglionic centers, the removal of which results in temporary paralysis and weakness. That they also are the seat of organs (eyes, network-cells, concretions) that are of importance in[23] giving information in the life of Charybdea, is evident from the reaching motion of the proboscis after the removal of the sensory clubs. Other centers of spontaneity in their order of importance probably are: the radial ganglia (one experiment); the interradial ganglia (?); the suspensoria, as shown by their supplying stimuli to isolated pieces of the sides connected with them; the frenula and the velarium, the latter of which gave contractions when removed with the frenula or in pieces only. No evidence is given that the frenula or the velarium can impart their contractions to other tissue, though this seems probable for the former. The proboscis can also contract of itself.
Reflexes between the velarium, frenula, subumbrella, sensory clubs, nerve, and any one pedalium, on the one hand, and the pedalia on the other hand, are very common, and point to the pedalia with the tentacles as organs of defense and offense. The pedalia serve also as rudders in swimming.
Finally, as judged by the results in this paper, Charybdea seems to occupy, physiologically, a position intermediate between the Hydromedusæ and the Scyphomedusæ. In its great activity as a swimmer, in its response to light, and in its reflexes it is Hydromedusan, while in the paralysis and recovery following the removal of its marginal bodies, as also in its response with several pulsations instead of one, when a deganglionated bell is stimulated, it is Scyphomedusan.
The observations on the Discomedusæ, Aurelia, Polyclonia, Cassiopœa, demonstrate the existence of motor nerve centers in the marginal bodies; but that other centers are present is shown by the recovery of pulsation following the removal of the marginal bodies or the margin. These results are mainly confirmatory of those of Romanes and Eimer. They differ from these in the fact that margins of Polyclonia and Cassiopœa, with only one ganglion attached, originated contractions distant from the ganglion. Removing of a single lithocyst resulted in a slowing of pulsation, as did also the removal of the oral lobes, though the immediate effect in the latter case was an acceleration. Isolated pieces of the subumbrella could contract.
Below follow Dr. Conant’s notes. They are printed about as Conant left them. Their order of succession, however, has been[24] changed to bring similar experiments together, while useless and often repeated ones have been omitted, and short elliptical sentences completed. Where the present writer wished to add any explanation, the same has been placed in brackets.
Light and Darkness.—1. Eight medusæ, in a deep glass jar and covered by a black coat, except one inch around the top, were placed in the dark-room.
a. When light from a lamp was thrown on the surface (one inch) layer, the animals were active near the surface; when the light was withdrawn, one or two were on the bottom and not moving but were probably pulsating.
b. After four or five minutes in the dark, three or four besides a feeble one are on the bottom. It took about two minutes to get them all to swim [by the lamp]. Of the three on the bottom, one, at any rate, was not pulsating. [Three other attempts like a and b were made, with very similar results.]
2. Experiment No. 1 was repeated several weeks later. Four in a large round glass dish were placed in the dark-room. A lamp being held to the dish all but one were found to be on the bottom. That one quickly went to the bottom, while two of those on the bottom quickly came to the top. In two or three minutes the one that had gone to the bottom began to pulsate and at about the same time the other one that had remained on the bottom also began to pulsate, while the two that had gone to the top stayed there swimming very actively. [Repeated with like results.]
3. Fresh ones did not show the reaction to light after darkness so well as did those in the experiments previously recorded. They were experimented with about nine A. M., while usually they were tried later in the day. I had rather suspected from previous work that they would not react so well when fresh.
4. a. In walking with the jar (1) of jelly-fish of experiment 1 from the dark-room to the back porch of the laboratory (fifty steps), in the bright sun and a cool breeze, all were found upon entering the laboratory door to have settled to the bottom and most of them to have ceased active swimming. In five minutes two or three were swimming somewhat, and in five minutes more all but one or two (eight in all) were swimming.
Walking with the jar about the laboratory did not suffice to make any change in their swimming, nor did blowing on the surface make any appreciable change.
b. Upon taking the jar to the back porch and placing it on the stone or cement flags, in the shade and a cool breeze, in four minutes time all were on the bottom not even pulsating.
Upon replacing them on the laboratory table all began to swim about at once. [Repeated.]
c. The jar (1) was placed on the back porch again; in fifteen seconds three were on the bottom; in one-half minute all but one. In three or four minutes all were on the bottom, but two were swimming lively and the others pulsating. In another minute all were swimming.
d. The jar (1) was tried again, not resting it on the flags but holding it by my hands on the sides. The effect was just as quick; they stopped pulsating at once. By the time I had got back to my table in the laboratory, one was at the surface and another arrived just as the jar was set down.
[Several other experiments of an order similar to those just noted were tried, with very similar results.]
5. Two buckets stood side by side in the laboratory. One bucket (1) had more Charybdeas in it than the other bucket (2), and also had more since brought in (about an hour). The water of one (1) was also more discolored and with more organic matter (sea weed, etc.). In the laboratory the animals were active on the surface of both buckets. Placed in the sunlight on the porch, no breeze, the sun slanting so that one side of the water in the buckets was bright while the other side was shaded, the jelly-fish in (1) went mostly to the bottom, while those in (2) seemed unaffected though some showed a tendency to go to the bottom after a longer exposure. The experiment with (1) was repeated and it took some five minutes for them all to go to the bottom. In a few minutes after replacing them in the laboratory several were active again on the surface.
6. Jar (a) with five large ones stood on my table; they were quite active. Placed in the sun (no breeze), on the porch, one or two sank to the bottom at once and the others seemed to slow their activities somewhat but not very markedly. In a few minutes all were swimming, apparently more actively than before, in the bright sunlight.
[In other experiments Conant shows that it is not the stimulus of walking that causes them to swim when carried into the room, for they would not swim when he walked with them on the porch. Also, he shows how they may change, some swimming, others not, when left for some time in any one place.]
7. In a tumbler were two pulsating very vigorously. Placed in the bright sunlight, very little breeze now and then, they showed no change whatever.
8. Some in a jar were covered with a black coat. The coat was taken off, and almost immediately they stopped pulsating, or pulsated but feebly, and sank to the bottom. The coat was put on again with one part near the bottom of the jar exposed. Almost at once, the animals, which were quite motionless, pulsating but little, resumed pulsation, which became more and more vigorous, and quickly swam to the top again. It seems plainly to be a reaction to light. [Such experiments as this were repeated at different times with very like results.]
9. A bucket with several bobbing actively on the surface was set out in a smart shower, and the animals continued bobbing on the surface as before. I could not see that they made the slightest attempt to go below.
There can be no doubt but that there is an individual difference in sensitiveness to the reaction of light after darkness. E. g., I just removed the coat from a dish with four in it; one went to the bottom at once, another presently, a third remained active at the surface, the fourth when noticed was on the bottom.
There is also a difference in the length of time they stay on the bottom as well as in the quickness in the response to light. Some recover very quickly, should say in less than a minute, and at once become very active. Some stay for a long time and only resume activity upon the coat being placed over them. Perhaps this explains some of the observations in Experiment 1.
Sensory Clubs.—10. All four concretions were removed and the animal stood the operation well. It swam more restlessly, however, than others did in the same surroundings. It seemed at first to show a trace of loss of sense-perception. It swam up, and down again, more changeable than those intact, which stay rather more constantly either on the bottom or at the surface. This may, however, have been[27] due solely to the restlessness of the animal after the operation. Later it swam actively for by far the most part on the surface only, which points to the truth of the preceding statement.
It showed no reaction to light. A coat placed over the jar was removed, when it was found to be on the surface and it remained there. This was twice repeated. I noticed specially that on pushing the bell above the surface of the water it at once turned and went deeper as the normal animal does. Finally, given another a trial with removing the coat from the jar, it went to the bottom as the normal animal usually does. After this, when next seen, it was keeping to the bottom. [This experiment was repeated on another occasion with almost identical results, no loss of sense-perception being noticeable.]
Sometimes it seemed as if access of light at removing the coat acted as a stimulus to one or more of those that were quiescent on the bottom. This was noticed again on the following day.
11. Two more were operated upon. These did not stand the operation well and stayed on the bottom, one swimming, while eight hours later one was in better condition (pulsating) than two left in the same dish for comparison.
12. a. Three clubs were cut off leaving only the stalks. A temporary paralysis of the power to swim was the immediate effect. Later it partially recovered this power. The proboscis, which was previously quiet, now showed convulsive twitchings and movements. It continued for some time to move to one side and then the other (after short pauses of varied length) as if to grasp some object. The lips of the proboscis were also moving and at times expanding. Often the movements were towards the side on which the club was uninjured.
b. The fourth club was next removed. A temporary paralysis as before resulted, followed by a quick recovery of pulsation; but the animal was now much weakened. The movement of the proboscis continued—shortening, lips expanding, moving to this side or that. The pulsations of the bell were kept up even when too weak to swim.
c. The sensory niches of this same animal were treated with 2.5 per cent. acetic acid by means of a pipette. The stalks of all four clubs showed white. Pulsations ceased. The velarium showed feeble local contractions. The movements of the proboscis and suspensoria drawing down the stomach continued. Upon stirring the animal it[28] gave rather feeble, somewhat convulsive pulsations with local (fibrillar) contractions; the pulsations in some cases were pretty well coördinated, but were more on the twitching kind.
13. Three clubs were removed. The animal pulsated well, only a little less strongly, perhaps. After a minute or two the fourth club was removed. It pulsated almost immediately, perhaps thirty seconds after the operation. It swam very well and pulsated feebly five hours after the operation.
14. One from jar (a) (Experiment 6) was operated upon. When the first club was cut off there was a paralysis of pulsation followed by a quick recovery. Cutting off the second club seemed to stimulate pulsation, the third to diminish it; after cutting off the fourth club it still pulsated. When placed in a large jar it pulsated on the bottom, but not strong enough to swim. The pulsations were fairly regular and sometimes seemed to occur in groups of two, but these groups were not well marked.
15. Another one from jar (a) was taken. One club was cut out, upon which there was a very temporary paralysis followed by good pulsations afterwards. The proboscis, as in all cases noticed, gave active movements to this side and that side. These movements of the proboscis were often very quick and definitely directed as if a well defined stimulus were given. After the operation one pedalium contracted so as to be at a right angle to the main axis of the bell; shortly a second pedalium also contracted. Placed in a small round dish the animal swam actively.
A second club was removed, and it swam as well as before. After fifteen minutes it was not swimming but pulsating against the jar. Upon stirring it a little it swam vigorously ten to fifteen strokes and then stopped. It seemed weak and its movements appeared not so definite, though this might be due to weakness.
A third club was removed. The only change seemed to be rather greater weakness.
After about five minutes the fourth club was removed. Paralysis of pulsation followed. It had the power to contract its pedalia when these were rather vigorously stimulated with a needle. It also gave one feeble pulsation when so stimulated.
16. The sensory clubs were removed from another. After removal of the third one it still pulsated actively, but stopped completely and apparently for good after the removal of the fourth club. Another[29] one stopped pulsating apparently for good upon removing the third club.
17. All four sensory clubs were removed from one, cutting as high up as possible so as to remove the endodermal tract of nerve fibers of the peduncle. It pulsated afterwards apparently the same as if the stalks had been left intact.
18. A small piece surrounding a sensory club and including the margin can contract by itself. The piece observed pulsated with quick pulsations and rhythmically but intermittently. After a fresh cutting away of such a piece, the portion of the velarium attached was seen to contract rhythmically, while the rest of the subumbrella was not so seen. The part of the subumbrella above the radial ganglion that was cut off did not contract by itself. The same portion of the velarium cut off did give contractions.
19. A sensory club with the surrounding region cut out pulsated rhythmically; when the club was cut from the end of its stalk pulsation stopped. This observation was repeated on another, and contractions were seen after the removal of the club. A piece of the subumbrella wall from the same animal gave contractions now and then even after an hour.
20. The normal position of a sensory club seems to be with the concretion almost at the lowermost end; often with it certainly lowermost, but probably oftener with the perpendicular passing through the center of the attachment of the club to its peduncle and just by the inner edge of the concretion. The eyes point inwards.
When the animal is on its side the concretions are always quite lowermost. When the animal was inverted the tendency was for the concretions to be lowermost. In this position the eyes may point in several directions. In one instance those of one club pointed rather outwards, while of two other clubs they pointed more in the plane of the body wall. (See also Experiments 24, 29.)
Nerve.—21. Cutting the nerve eight times, once on each side of each sensory club, produced no loss of coördination in pulsating. The animal was weakened, however, by the operation, which was made drastic to insure cutting the nerve; but it was still able to swim. This experiment was repeated four times.
22. That coördination was continued after the nerve was cut was proved beyond doubt by cutting from the edge up (eight times)[30] so as to perfectly separate the sensory clubs and the pedalia. Pulsations continued synchronously in all four sides—not the slightest evidence that one side contracted out of time with the others.
23. The eight cuts were made as in the preceding experiment with no loss of coördination noted. When the cuts were carried up to the base of the stomach, however, coördination ceased. The four side pieces seemed to contract each in its own time. Only two sides could be observed at one time, and they at any rate did not contract synchronously. One side often gave two contractions while the other side rested or gave one.
Yet, a little later, three of the sides at any rate showed a pretty good coördination. The fourth was smaller and did not seem to get into the game much—it went more on its own schedule. The four pieces were then cut apart and placed together under a dissecting microscope. No coördination at all could be made out. No evidence, therefore, of any definite rate of pulsation inherent in the sensory clubs.
Cutting the nerve causes the pedalia to forcibly contract inwards.
Side, Subumbrella.—24. A whole side was cut out, the transverse cut being above the sensory organ so as to take off [leave off] the radial ganglion also. This pulsated, or rather contracted, nicely. The upper end had been cut just through the suspensorium. It especially gave twitchings like the twitchings of the stomach. The piece was then halved transversely, when the upper part containing the portion of the suspensorium twitched as before while the lower part was not seen to contract again. This was repeated with the same result, except that a portion of the lower part gave a slight contraction several times. The part that contracted was at the upper end of the piece, i. e., nearest the suspensorium. The contractions were also more longitudinal than transverse, as the regular contractions would be.
The piece connected with the sensory clubs of course pulsated nicely. Upon cutting off the sensory club from the stalk, pulsation ceased, but twitching of the velarium continued. This was repeated with the same effect.
In the same animal, in cutting off the sides, the stomach was left, the cut being through the gastric ostium. The floor of the stomach was now cut off by cutting out the four interradial points of[31] attachment. The stomach and the proboscis gave vigorous contractions and tied themselves all up so that I could not cut off the proboscis.
The four pieces of the floor of the stomach left on the interradii gave contractions nicely. The phacelli continued their squirming movements.
25. Cutting off the whole aboral end of the animal excites to very rapid pulsations of the remaining part. The stream, as shown by particles in the water, is apparently stronger out the aboral end than past the velarium.
It seems that I get no good evidence that the subumbrella is able to contract of itself without connection with special nerve centers. In the one case noted (Experiment 31) I could not be sure but that the part that contracted was intimately associated with the suspensorium or frenulum.
26. A piece of the subumbrella cut off and having, so far as I could determine, no connection with ganglia, frenula, or suspensoria, gave contractions. Another piece was not seen to contract.
A small piece of the subumbrella connected with a club can contract. The proboscis can give contractions of itself when cut off with the base of the stomach. Even a cut-off lip can twitch by itself. A portion of the subumbrella by itself also showed twitchings. (See also Experiments 18, 19, 25, 26, 29, 47, 49.)
Pedalia, Velarium, Radial and Interradial Ganglia.—27. The pedalia with their tentacles were cut off at their bases to insure cutting out the interradial ganglia. The animal could pulsate well enough, but intermittently and without much progress (the velarium, of course, being injured). Cutting one pedalium caused the others to contract.
28. When the pedalia were cut off from one, the power of direct motion was entirely gone. It swam in circles, turned summersaults, changed its course continually, the oral end getting ahead of the aboral end, or trying to do so. The whole power of balancing was gone. It seemed excited by the operation and swam continually. [Repeated.]
29. The pedalia can be made to contract inwards by stroking their outer edge with a needle. This was noted last year and has been seen several times this year. Their inner edge is not so sensitive.
Touching a sensory club caused the pedalia to contract inwards in two cases.
The pedalia could be made to contract by giving the subumbrella a prick,—generally a rather severe one was necessary. The upper part of the subumbrella seems not so sensitive as the lower part and the proboscis, and the base of the stomach did not give any reflex at all (two specimens). One of the two could be made to give the reflex only with much difficulty. This was a very lively one. It would even stand severe pricks on the nerve, or even through the region of the sensory clubs, without contracting the pedalia or stopping pulsations.
Cutting the frenula seemed not to affect the ability to swim well. Cutting in this region brings about the reflex of the pedalia.
In the preceding fish the velarium was cut away wholly in some places, in other places it was left only as ragged strips. The pedalia became very strongly contracted and the tentacles were brought inside the bell. Pulsations that seemed strong produced much less progress than with the velarium intact. [Repeated.]
30. One with the whole margin cut off still gave pulsations. Upon the removal of the region of the radial ganglia, however, pulsations were seen no more.
The velarium in the above continued to give twitchings. The four pedalia were cut off with plenty of the tissue at their bases to insure the removal of interradial ganglia, and twitchings of the velarium with irregular contractions continued. No full contraction all around the velarium was noticed. When all the tissue was trimmed off as nearly as possible down to the velarium, the latter still gave twitchings and irregular contractions as before,—even more so as if excited by the operation. The power of originating contractions evidently resides in the velarium or in the ganglion cells of the frenula just as it does in the proboscis and the floor of the stomach.
Small pieces cut from between the pedalium corners and the frenula, so as to have tissue on them from neither, could contract by themselves. (See also for Pedalia, Experiments 15, 23, 41b; Velarium 18, 41c.)
Tentacles.—31. A cut-off tentacle can contract by itself, sometimes with squirming contractions. A prick at either end can produce a forcible contraction. A slight prick at the distal end may produce a local contraction. The proximal end is more sensitive, but this difference is not very marked. One with only the tentacles removed seemed to be a little less able to guide itself well.
Proboscis, Stomach, Phacelli.—32. The lips of the proboscis are highly contractile by themselves. The movement of the stomach and the phacelli goes on, after the lips are cut off, with increased vigor, due to the stimulus of shock. The vigor and frequency of their contractions, however, diminish quicker than that of the cut-off lips. (See for Proboscis, 12, 15, 18, 26, 29; Stomach, 18, 24, 29, 31; Phacelli, 18, 24, 31.)
Temperature.—33. Temperature does not seem to have much effect. Some placed in a tumbler half full of water, in the bright sunlight, swam vigorously over three-fourths of an hour. The water was quite warm to the hand.
34. The above experiment was repeated with the same results. A thermometer placed in the water with them showed 92° F.; hung in the sun near by, it showed 94° F.
Ice in the water did not stop their pulsating temporarily or permanently, except that it did for a short time after being held against one. Even then it took some time (fifteen to twenty pulsations) before it produced any effect.
35. Ice placed in the water again showed no marked effect. They swam as lively as ever. Some, after pulsating against the ice for a little while, seemed to be less vigorous, but quickly recovered in another part of the jar. Others did not seem to be the least bit affected by contact with the ice.
Food and Feeding.—36. I tried to feed one. A red and a white copepod were put into the subumbrella cavity. No attempt to eat it was observed in either case, though the copepods remained in the subumbrella cavity for some time.
Animals found in the stomach of Charybdea: small fish were most frequently seen; at another time a small stomatopod; again, a small polychæte; small shrimps; amphipod.
Those taken on August 16th (3 to 4 P. M.) seemed to have, for the most part, food in the stomach, and this more so than those taken in the morning.
Occurrence of Charybdea.—37. In the first tow on the bottom (with a net made of mosquito-netting and weighted with rocks in order to sink it) the haul was forty. I do not think that we could have been towing more than four or five minutes. The time was[34] about seven A. M. A light breeze was blowing and there had been a heavy shower a half-hour previous.
38. The usual time of towing was about 6.30 to 7.30 A. M. The water was four to five feet (1.2 to 1.5 m.) nearest shore but deeper farther out. At this time of day one could count on getting plenty of the larger sized (15 to 20 mm.), many small ones, but very few of the smallest. This was the experience of several mornings.
On August 12th I towed about nine A. M., and got but few of the larger sized, many small ones, and very many of the smallest.
The next day (7.00 to 7.45 A. M.) those obtained were mostly of the larger size. On the same day (3 P. M.) others of the party towed at the same place and obtained but few.
On another day I towed in the afternoon (3 to 4 P. M.) and obtained great numbers as I usually did in the morning.
39. We towed about 7.30 to 8.30 at night. Very few Charybdeæ were taken. On this evening we towed five times in the same locality, and obtained but seven or eight specimens. Towing with the same net on our way home, it was filled with Aureliæ and five or six Charybdeæ. It seems as if Charybdea came to the surface at night. Those towed in the evening were dead the next morning.
The next morning Richard, our colored attendant, towed from 5.30 to 6.30. There were heavy showers. The usual find of large and medium ones was obtained. There were only two with planulae.
40. The material of September 2nd was obtained about six A. M. They were mostly of large size. In all only fifteen or twenty were taken. Richard explained the small number by saying that the bottom had changed in the locality where we usually towed and that he got no weeds in his net, but mud.
The next day more were brought in by Richard (6.30 A. M.) There were rather more than yesterday but the quality was the same. There were three with planulae.
On another morning Richard brought in a great many, about a hundred. Among these there were three with planulae (light-colored and budding); on a previous day there was one with the reddish-brown kind and with a mouth.
Activity of Charybdea.—41. a. About five o’clock in the morning a Charybdea was taken in the tow. It was in good condition swimming incessantly round and round without change of direction,[35] in a jar of about twenty centimeters in diameter. It came to the surface every now and then, after eight to fifteen pulsations. The tentacles and the phacelli were of a lilac shade. If a pencil was placed in its way it would pulsate against it repeatedly without any effort to dodge around it.
| 6.58 | A. M., | 124 | pulsations | were | counted | to the | minute. |
| 8.00 | “ | 124 | “ | “ | “ | “ | “ |
| 9.25 | “ | 136 | “ | “ | “ | “ | “ |
| 10.15 | “ | 131 | “ | “ | “ | “ | “ |
| 11.00 | “ | 146 | “ | “ | “ | “ | “ |
At 10.15 it went around the dish in eight seconds, taking eighteen or nineteen pulsations. If a bright platinum spatula or a black pencil was placed in its circuit it would repeatedly butt against it each time it came around. After the second or third pulsation against it, however, it seemed to have some sense to change its direction.
b. The pedalia have no perceptible action of their own. They move inwards slightly toward the axis at each pulsation, but scarcely as much as one would suppose from their attachment to the pulsating margin. It seems as if they were for “winging” the moving animal more than for anything else.
c. The velarium is loose and it flaps. It seems to take part in swimming something more than the passive diaphragm function,—i. e., it straightens out during the recovery after each contraction of the bell.
[The following experiments were performed at Port Henderson, Jamaica, in 1896.]
42. May 12th. An Aurelia was pulsating normally at the rate of twenty-five or twenty-six pulsations to the half-minute. One lithocyst was cut out, when a few contractions, evidently caused by the stimulus of cutting, followed; then, rest. In the first minute there were only about five pulsations. In two or three minutes rhythmic pulsations were resumed. Four minutes after the cutting there were nineteen pulsations to the half-minute. About twenty minutes after there were nine to the half-minute, in groups of six and three.
A Polyclonia, about four and one-half inches (115 mm.) in diameter, gave twenty-six or twenty-seven regular pulsations to the half-minute. After one otocyst was removed, pulsations continued, but in groups with intervals of pause: e. g., thirteen, pause; ten, pause; six. Three minutes after the removal of the lithocyst there were 5, 3, 1, 3, 5, or seventeen pulsations to the half-minute. Eleven minutes after the operation there were fifteen to the half-minute. The removed lithocyst and surrounding tissue gave contractions.
43. May 13th. The Aurelia was in rather poor condition but would pulsate upon being stirred. The other seven lithocysts were removed when only a few contractions originated thereafter.
The Polyclonia was in good condition, but was pulsating only intermittently when first seen in the morning. When the remaining seven lithocysts were cut out and no more pulsations were observed, the oral arms could still move.
May 14th. Both were found dead upon returning in the evening.
44. May 15th. An Aurelia and a Polyclonia were taken in the morning. The Aurelia was two and one-half to three inches (62.5-75 mm.) in diameter, with three tufts of phacelli, three oral arms and seven lithocysts. The Polyclonia was normal and seven or eight inches (175-200 mm.) in diameter.
In the Aurelia all the lithocysts were removed. Spontaneous and coördinated contractions could still occur after time had been allowed for the shock from the operation to pass away. The next day the animal was still alive and pulsating, but ragged, and the next day following was quite dead.
In the Polyclonia the normal rhythm was fourteen pulsations to the minute. Some pulsations were apparently quicker than others and the intervals were not the same. Thirteen, ten, and twelve pulsations were also counted. After putting the animal into fresh sea-water, it pulsated thirty-three to the minute. Six minutes later it was still pulsating at the same rate, while in four minutes more eleven pulsations, many of which were in groups of two, were noted. In five minutes more it pulsated eleven times to the minute with only one double pulsation. One oral arm was then cut off and the rhythm counted about one minute afterward—fourteen pulsations, then a pause of fifteen seconds, then two pulsations, in all sixteen to the minute were counted. About ten minutes later there were eight pulsations, two or three minutes later only three, while in two or three[37] minutes more only three. There was a long latent period—two or three seconds—before the stimulation of cutting off the arm made itself evident in the rhythm.
A second oral lobe was removed. Then there followed twenty-four pulsations, a pause of two seconds, and two pulsations, in all twenty-six pulsations to a minute. The rate of pulsation soon fell to the previously abnormal low rate.
Third lobe removed: 21 pulsations in first half minute and then 16, or 37 per minute.
Fourth lobe removed: 17 pulsations in first half-minute plus 13 gives 30 for the minute.
No difference in the coördination of the animal was shown as a result of the removal of one-half the number of oral arms.
Fifth lobe removed: 17 pulsations plus 15 equals 32 to the minute.
Sixth lobe removed: 17 in first half-minute plus 4 in the second half-minute gives 21 pulsations for the minute.
Seventh lobe removed: 17 plus 9, or 26 per minute.
In all these instances the rhythm in the second half of the first minute was irregular and intermittent.
Seventeen and then seven pulsations were provoked after the animal had become quiescent, or nearly so, by merely handling it.
45. Eighth oral lobe was removed and pulsations stopped. The next day the animal was in good condition. The pulsations counted in the evening were 12, 14, 14, 11, per minute. The rhythm was not regular; there was a tendency to groups of twos, threes, or more, but no prolonged intervals of rest were observed. When placed into fresh sea-water, the pulsations were fourteen to the half-minute or twenty-six to the minute; seventeen to the half-minute, and thirty-three to the minute were also counted. This specimen gave spontaneous contractions during two weeks, after which it was thrown out, the aboral end being eaten through and little or no regeneration having taken place.
46. Two more were operated upon: A. Its rhythm was 18, 14, 17. Its entire margin was cut off. The separate pieces of the margin pulsated, 6, 7, 4, 6, 7, 9. The animal seemed paralyzed by the operation; it responded by a contraction now and then to stimulation but gave no spontaneous pulsations. B. Its rhythm was 17, 15, 12, 12. All its oral arms were removed. Its rhythm was only raised to seventeen and not perfect. In twenty-five minutes it had fallen to eleven, in four hours to ten pulsations [per minute].
May 22nd. A and B are living as also the pieces of the margin of A; all are giving spontaneous pulsations now and then at comparatively long intervals—even A, with its margin removed.
May 26th. Everything is still living. The one with the margin cut (A) counted sixteen and nineteen pulsations per minute, though this was not kept up all the time.
June 2nd. A and B and pieces are still living and contracting spontaneously. It is now two weeks, and they were thrown out eaten through at the aboral end with little or no regeneration.
47. The margin was cut off another one (C) and it was then paralyzed. The margin contracted vigorously by itself. The margin was next split, but a connection of about one-half an inch wide was left between the two rings. Over this bridge the contractions passed from the outer and inner ring. The inner ring did not originate any contractions. Both rings were then cut near their connecting bridge of tissue and the larger ring with the marginal bodies was split longitudinally so as to separate the exumbral from the subumbral portion. It was found that the contractions started only from the subumbral portion while the exumbral portion did not contract at all.
June 5th. Five of the eight small pieces of C were not seen to contract either to-day or yesterday. A slow rotary motion was observed in some of the pieces suggesting ciliation, but no cilia or currents pointing to ciliation were seen with a low power. C was seen to pulsate spontaneously. Possibly it did yesterday but it was not watched closely. A piece of the subumbral surface of C broken off (not from the margin) was found to contract spontaneously.
48. June 6th. In a fresh one (D) from Port Royal, the eight lithocysts of one side were removed in order to compare its movements with an intact one. Coördination was apparently unaffected.
June 9th. The margin of C is still pulsating vigorously. Parts of the subumbrella broken loose from the strip pulsated by themselves now and then. Fifteen lithocysts were removed, leaving only one at the end of the strip. It was found that with this single ganglion (lithocyst) left, and originating most of the contractions, now and then a contraction would originate at another part of the strip where there was no ganglion. Three days later contractions originated as often from other parts as from the ganglion.
[The remaining experiments were all performed in 1897, at Port Antonio.]
49. Removal of the sixteen marginal bodies caused paralysis for a time; then recovery followed.
Contraction was limited to the subumbrella.
A portion of the subumbrella not from the margin can contract by itself as well as a portion of the margin with the marginal bodies (lithocysts).
In the margin cut off as a strip with only one marginal body attached at one end, contractions sometimes started from the opposite end.
50. Size, seventeen or eighteen millimeters. Pulsations, thirty-two. Lithocysts, nine. The operation consisted in the removal of the concretions with as little injury to the pigmented parts of the marginal bodies as possible. One whole marginal body, however, was removed in the operation. Soon after the operation the pulsations were 28, 26, 20, 20, per minute.
Another one; size fifteen millimeters. Pulsations were forty per minute. The operation consisted in the removal of the concretions and pigmented parts of the marginal bodies with as little injury to the adjoining parts as possible. After the operation it seemed as if the intervals between the pulsations were irregular,—not a series at regular intervals. An hour or so after the operation the pulsations were very intermittent. During the afternoon it was not seen to pulsate except when it was stirred up, when six or seven vigorous pulsations followed. These, however, were rather aimless.
51. One sensory club (marginal body) was cut out, including its basal part also. In one or two other cases more or less injury was done to adjoining parts also. Pulsations ceased upon the removal of the last club, but upon placing it in an aquarium and allowing it to come to rest for two or three minutes, pulsations were now and then seen. In the evening, this one and another did not pulsate except when stirred, when they pulsated with good progress.
52. A circular cut, about two inches in diameter, was made through the epithelium of the subumbrella around the base of the[40] oral lobes. The animal pulsated well enough, but the contractions seemed not so simultaneous in all parts of the margin as normally. After a few days it had partly regenerated but died. One of the oral lobes cut off had some power of contraction, and this some time after the operation. A similar cut, but semicircular, made no difference between the contractions of the two halves.
53. The whole region of the sensory clubs was cut out when the animal was not seen to pulsate again, except in the evening, when pulsations were observed. The oral lobes also moved.
Method.—The following results on the histology of the sensory clubs, their eyes, and the tentacles, as already noted, were obtained from some of Dr. Conant’s preserved material. These results relate almost wholly to Charybdea, with only a few references to Tripedalia, noted in their proper place.
A portion of this material was killed after keeping the animals in the dark for some time, for the purpose of discovering any changes in the pigment of the eyes. I believe that a retraction of the pigment of the long pigment cells that project between the prisms and pyramids of the vitreous body in the retina of the distal complex eye is very evident in eyes killed in the dark. (But more on this below.)
I obtained my best results from the material preserved in saturated corrosive sublimate, to which had been added (5 to 10 per cent.) acetic acid. This also was Conant’s experience in his previous work on Charybdea and Tripedalia.
My best sections were obtained by embedding the sensory clubs in celoidin, passing the little blocks of celoidin with the sensory clubs into chloroform until perfectly transparent, and then into paraffine. I then cut sections as we ordinarily cut paraffine sections, mounted and stained them on the slide. My purpose in using this method was to avoid the displacement of the vitreous bodies of the eyes during embedding and cutting. This object was fully realized and more besides. Since the sections cut by the celoidin-paraffine method gave me so decidedly the best differentiation of the axial fibers of the retinal cells, as also of the cilia, basal bodies, etc., I am inclined to believe that the celoidin was in part responsible for this differentiation.
Most of my series were cut 4 µ in thickness. All in all I cut sixty-five clubs besides making some maceration preparations from material preserved for that purpose. These sixty-five series represent material from fourteen bottles. As a whole, my material was good, but the material from one bottle was decidedly superior for showing the axial fibers of the prisms and pyramids of the retinal cells. This shows the advantage of plenty of material. It will be evident that I had plenty of material.
I found iron-hæmatoxylin the most satisfactory stain. I stained for a shorter or a longer time—one-half to several hours and longer—and then washed out the sections until under a low power of magnification they appeared quite unstained, the nuclei and a few other parts only appearing darkly stained.
Depigmentation I practiced but little. I obtained many of my series almost wholly unpigmented, especially those I cut last. Others, of course, were very heavily pigmented. I am not certain but that alcohol slowly dissolves out the pigment after a long period of preservation. Slight variations in the technique of killing and preserving may also, perhaps, determine the stability or solubility of the pigment, as, of course, also the condition of the pigment at the time of killing.
Anatomy.—For a short epitome of the anatomy of a Cubomedusa and of a Cubomedusan sensory club see p. 2 of the Introduction.
The Distal Complex Eye—General.—The distal (larger) complex eye (Fig. 7) and the proximal (smaller) complex eye (Fig. 13) are so named to distinguish them from the lateral simple eyes of the clubs. The distal complex eye consists of the following parts: a cellular cornea, continuous with the epithelium of the sensory club; a cellular lens (externally cellular and internally often quite homogeneous) immediately beneath the cornea; a homogeneous capsule just internal from the lens, and evidently a secretion from the lens cells; a vitreous body composed primarily of prisms and pyramids just beneath the capsule; and a retina of pigmented cells, with subretinal nerve tissue, ganglion cells and fibers. To my knowledge all observers (except Carrière, who missed the capsule) are quite agreed on the anatomical structure of the distal complex eye as also[42] on the proximal complex eye and the lateral simple eyes.[d] It is on the histological structure of some of the various parts that differences exist.
Cornea.—Little need be said on the cornea except that it consists of flattened cells applied to the outer surface of the lens. It is continuous with the epithelium of the club and evidently a modified portion of this epithelium (Fig. 7). All observers conform to this statement.
The Lens.—The lens is of cellular origin, but in its interior the cells are often so changed—absence of nuclei, cell walls, and protoplasmic structure—as to make a mass quite homogeneous and structureless. While this internal mass sometimes shows practically no structure, yet at other times it is found broken up into masses much the size and shape of cells but without nuclei, while again, cells with nuclei may be quite evident. This occasional breaking up of this mass is evidently predetermined by its original cell structure. Iron-hæmatoxylin stains this inner mass very dark and it is difficult to wash out the stain. Borax carmine and Lyons blue give the best results on the lenses. In figure 7 the lens of the distal complex eye is shown as quite homogeneous internally, while in figure 13 (proximal complex eye) it is drawn cellular. In this latter lens the inner cells are quite round and nucleated as they may also appear in the distal eye. What I have said applies equally to the lenses of both complex eyes, though the cellular nature of the inside of the lens is more readily demonstrated in the proximal eye.
It appears that it is in younger specimens that the central mass of the lens shows the cellular structure best, and that as the animal grows older this structure is more and more lost until no trace[43] of it remains. As concerns most of my series I could not well determine which were from younger and which from older individuals, yet, several series of quite small (5 mm.) and therefore young animals, in which the eyes were so small that the lenses were compassed into less than half a dozen sections, the cellular structure of the lens was very evident.
The external cells of the lens form a spherical shell (both complex eyes) which, in section, shows as a hollow ring (Figs. 7, 13). The thicker ends of these cells lie at the inner (toward the capsule) half of the sphere and the cells taper toward the corneal surface, dovetailing laterally with their immediate neighbors as also distally with those from the opposite side of the sphere. The thicker inner ends of the cells contain the large nuclei with nucleoli. At a point (* Figs. 7 and 13) on the inner (next the capsule) surface of the lens the cells only approximate each other and thus leave a place which is easily broken through, as is shown by portions (drops, probably representing cells or portions of cells) of the mass within the lens becoming squeezed out into the substance of the capsule and the vitreous body, and found occasionally also among the cells of the retina. A considerable portion of the inside of the lens may be found thus squeezed out, and its path can often be traced. This phenomenon is evidently brought about by a contraction of the shell of the lens during fixation and before the inside of the lens has become hardened.
In origin the lens is evidently ectodermal, originating from an ectodermal invagination which becomes pinched off as a hollow sphere, the outer (i. e. next the cornea) half of which becomes the lens, the inner half the retina (i. e. vitreous body plus the so called retina). (See Retina.) The transition from retinal to lens cells is quite readily made out at the lower side of Fig. 7, but the corresponding structure on the upper left side is not so manifest. It is further evident that the lens is again an invagination into this sphere, and the point at which the lens cells approximate (where the central mass of the lens may be squeezed out as above described) represents the place of pinching off of the original lens-retina sphere. It appears, then, that the lens is formed in the lens-retina sphere in the following manner: The cells of the secondary invagination going to form the lens begin to lengthen distally (i. e. toward the cornea) during their invagination to form a hollow sphere, at the[44] same time dovetailing with each other and budding off cells to form the inside of the lens (Figs. 7, 13).
At the lower side of the lens, near the margin of the retina, the cells of the lens are slightly indented or pushed inwards (Fig. 7, ind.). I believe this to be due to the weight of the lens in the normal position of the club, when the lens rests against the margin of the retina and the capsule and adjacent tissue.
Anticipating the description of the retina, it may here be added, that the retina is formed from the inner half of the lens-retina sphere. The cells of this portion of the sphere become differentiated into prism cells, pyramid cells, and long pigment cells, while laterally, beyond the margin of the vitreous body, they are differentiated into pigmented iris cells (Figs. 7, 6a).
Above are my results on the lens. Haake[2] speaks of the lens as consisting of a cellular “Kern” with a covering of lamellated cells. Carrière describes it as cellular and filled internally with a “Gerinsel,” or coagulation. Carrière and Haake are each in part right. Claus describes it as wholly cellular. Schewiakoff regards the lens as wholly cellular, and like Claus has not noted that internally this cell structure may be quite obliterated. Schewiakoff regards the lens and retina as formed from an invaginated sphere, and shows the transition from the lens cells into retinal cells as I have figured. Conant also gives the structure of the lens for the complex eyes as cellular but missed the change of structure that the interior of the lens may undergo.
The Capsule.—The capsule of the lens (Figs. 4, 7) lies immediately below (inward from) the lens. In structure it is homogeneous, except for certain fibers from the long pigment cells of the retina that traverse it, while sometimes also other fibers can be seen which, possibly, are branches from the fibers just mentioned or continuations from the fine fibers of the prism cells of the retina soon to be described. I have, however, no evidence that the fibers from the prism cells extend beyond the prisms in whose axis they lie. The capsule lies very closely applied to the lens, never becoming separated from it in sections, and is, hence, regarded as a secretion from the lens cells. Just what its function may be is difficult to surmise. The proximal complex eye possesses no capsule. I have thought, however, that if the lens should be adjustable, the capsule might[45] serve as a protection to the prisms of the vitreous portion of the retina during the adjusting movements of the lens. (But more on this below.) To my knowledge all previous observers are quite agreed on the structure of the capsule. Carrière and Haake, however, missed it altogether.
Retina.—While I have enumerated (following previous observers) the vitreous body and the so-called retina as distinct parts, yet, as the sequel will show, they are, histologically, different parts of the same thing—namely the sensorium proper of the eye—and I propose to use the term retina for both taken together, while I retain the expression vitreous body (as hitherto used) for the vitreous portion of the retina. This simplifies matters; and using a word that is already used for analogous structures of other eyes (vertebrates, anthropods, molluscs) is conducive to clearness. I have been tempted, furthermore, to use the words rods and cones for the prisms and pyramids that I find in the vitreous bodies of the retinas of the complex eyes. But since the prisms in reality approximate prisms and the pyramids pyramids, in their shape, I have decided to retain the words prism and pyramid for these structures. The former of these terms (prism) was first used by Conant in his description of the complex eyes.
What I shall call the retina, then, in the distal and proximal complex eyes of Charybdea, consists of three kinds of elements: the prism cells, the pyramid cells, and the long pigment cells. (Figs. 4, 7, 22, prc, pyrc, lp.) We may also describe the retina as composed of three zones: the vitreous zone (vitreous body of authors), the pigmented zone, and the nuclear zone. (Figs. 4, 7, 22, vb, pz, nz.)
The cells composing the retina form a single layer in the shape of a hollow cup, into which cup the lens with its capsule fits. (Fig. 7.) This single layer of cells takes in the thickness of the vitreous zone, the pigmented zone, and the nuclear zone. Indeed, the distinctions vitreous zone (vitreous body), pigmented zone, and nuclear zone characterize three topographical regions of the retinal cells.
That the retina is made up of three kinds of cells is most readily demonstrated in transverse sections through the vitreous body. Fig. 1 is such a section, taken quite near the pigmented zone (at about the level x, Fig. 4). Three different kinds of areas are readily made out in such a section. The more numerous areas[46] (pr) are transverse sections of the distal prisms of the prism cells, the less numerous and lighter areas (pyr) are transverse sections of the pyramids of the pyramid cells, and the large oval heavily pigmented areas (lp) are the transverse sections of the long pigment cells. The dots within the two first named areas represent fine fibers in the axes of the prism and pyramid cells, to be described below. The presence of three kinds of cells can again be readily seen in such Figs. as 4 and 7, in which the elements of the retina are cut parallel to their long axis. (Fig. 22.) Again, a transverse section through the most distal part of the pigmented zone of a slightly pigmented retina (Fig. 2) also shows us the presence of three kinds of elements. The larger and more heavily pigmented areas (lp) are the long pigment cells; the smaller, lighter areas (pyrc) with a central dot are the pyramid cells, and the more numerous dots, with no definite polygonal areas outlined about them (prc), belong to the prism cells. Thus, I believe, we have conclusive evidence of the existence of three kinds of cells in the retina of the distal complex eye.
(a) The prism cells are the more numerous, and, as the name implies, end distally in a vitreous polygonal prism (Figs. 4, 7, 22, pr). The prismatic structure of the vitreous body is also shown in Figs. 10 and 11, which are drawn from a macerated preparation of Conant’s. (See the descriptions of these figures.)
In Figs. 4 and 7 the prism cells correspond to the cells with the darker nuclei (npr); in Fig. 2 they are represented by the dots without defined polygonal areas about them (prc), and in Fig. 1 by the most numerous areas (pr). These cells, then, consist of a centrad portion with nucleus, a pigmented portion with granules of a dark-brown pigment, distal from the nucleus, and a distal vitreous prism which extends to the capsule of the lens.
In the axis of each prism is a fine darkly-staining fibril extending the entire length of the prism. I found no good evidence that this fiber extends into the capsule. Centrad this fiber is continued through the pigmented part of its cell and approaches to or near the nucleus (Fig. 2, dots without defined polygonal areas; Fig. 7, part of retina left unpigmented). In some instances I could trace this fiber quite to the nucleus, while in others it ended before reaching the nucleus or a little to one side of it. I am inclined to believe, however, that it extends past the nucleus and is continued as a nerve[47] fiber. I believe this to be so because the fiber is evidently sensory, and a priori we should expect it to be so continued. Further, I find decided evidence in sections of the simple eyes to show that the fibers there extend past the nucleus into the subretinal tissue where I could not trace them farther. (Fig. 16.) Again, that the flagella of the epithelial cells of the club are also continued into the cells, in some instances could be traced past the nuclei (Figs. 12 and 26), and the fact, too, that the retinal cups of the eyes represent invaginated epithelium (the axial fibers of the prisms are hence cilia?)—all this leads me to believe that the axial fibers of the prism-cells extend centrad past the nuclei through their cells and are continued as nerve-fibers. (See below under pyramid-cells and under epithelium). Immediately upon entering the pigmented part of its cell the axial fiber of a prism-cell has a dumbbell-shaped enlargement which lies quite at the distal edge of the pigmented part of the cell (Fig. 7, unpigmented part of figure). This, of course, can be seen only in unpigmented retinas. This dumbbell-shaped body, (Basalkörperchen of Apathy), which name I give it, since it evidently is homologous to the basal bodies described by others for the cilia of epithelia, can be most beautifully seen as two minute spheres lying close together and in line with the nucleus. These two little spheres of the basal bodies put to the test the highest powers of the microscope; but, when, after a prolonged and careful study, one satisfies himself of their existence and exact shape, the very difficulty with which they are resolved adds a zest to be appreciated. The length of a basal body is about one-fifth to one-fourth that of the nuclei of the prism-cells.
The structure of the nuclei of the prism-cells is that of a dense network (Figs. 4, 7, npr) which stains dark with hæmatoxylin. A nucleolus can often be seen in these nuclei. In some few series, again, these nuclei did not show a network-like structure, but the chromatin was arranged in masses (Fig. 5, npr). These nuclei can usually be distinguished from those of the other cells of the retina by their denser, darker-staining network (Figs. 4, 7, npr), or as shown in Fig. 5 (npr). Their denser structure and staining capacity are a distinguishing characteristic of the nuclei of the prism-cells. I must add, however, that not in every series is this apparent.
That portion of a prism-cell that contains the nucleus rarely contains any pigment; and when pigment is present, I believe that[48] it has been dissolved in from the pigmented zone. The nucleus, again, lies a little centrad from the pigmented part of its cell, so that an unpigmented zone is seen in the retina between the pigmented zone and the row of nuclei (Figs. 4, 7, 22).
Centrad the prism-cells are continued as a single process (Figs. 6, b, c, d, and 8a, b, c, d). In some sections I thought I could trace these processes to the basement membrane, but I could not satisfy myself that such appearances were not due to artificial splitting in the tissue. Schewiakoff makes a similar remark about his supporting cells, which cells I believe are the same as my long pigment cells, but these do not extend to the supporting lamella.
At the margin of the retina the cells do not develop prisms but remain pigmented and form an iris (Fig. 7), which was so named by Claus and also described by Schewiakoff. These cells also assume a somewhat different shape (Fig. 6a). This cell (Fig. 6a) is seen from its broader side with which it is applied to the capsule or the lens. Schewiakoff figures similar cells. That the cells of the iris are prism cells without the prisms does not necessarily follow. They simply represent cells of the retinal cup that have become differentiated to serve as an iris.
As to the exact origin of the prisms, and pyramids (to be described below), it is difficult to say anything definite. If the so-called basal bodies of the axial fibers are really homologous with the basal bodies of flagella, then it would seem that they (the prisms and pyramids) are secretions comparable to cuticular secretions.
(b) The pyramid-cells, like the prism-cells, are differentiated into three regions: a distal vitreous pyramid, a pigmented part, and a centrad part with nucleus. The pyramids are seen in transverse section in Fig. 1 (pyr) and in longitudinal section in Figs. 4 and 7 (pyr).[e]
Each pyramid extends between the bases of the prism-cells about one-third to one-half the depth of the vitreous body (Figs. 4, 7, 12 (pyr)). The pyramids are also a shade lighter than the prisms,[49] which fact is characteristic. In the axis of each pyramid is a darkly-staining fiber quite like the one described for the prism-cells (Figs. 1, 4, 7, 22). That this fiber extends distally beyond the limits of the pyramids I could not determine, but I do not think that it does. Centrad this fiber extends into the pigmented portion of its cell quite to or near the nucleus as was described for the fibers of the prism-cells (Figs. 7, 22). Whether or not these fibers extend past the nucleus and become continued as nerve fibers, the same course of reasoning holds as was given for the fibers of the prism-cells. Each of these fibers possesses a basal body just on its entrance into the pigmented part of the cell (Fig. 7), but I could not determine that it was dumbbell-shape. In form it represents an enlargement of the fiber itself, which gradually tapers again to its normal size. The continuations of these fibers within the pigmented parts of the pyramid-cells, as also the basal bodies, could only be demonstrated in unpigmented series.
Patten[5] describes axial fibers extending centrad through the rods (vitreous portions) of retinal cells (“retinophora”) into the region of the nucleus and past the nucleus (arthropods and molluscs). My retinal cells (prism and pyramid cells) evidently correspond to Patten’s retinophora, but I find no evidence that one of my retinal cells represents more than a single cell, while Patten gives evidence that his retinophora are made up of two cells closely applied to each other as twin cells. If this were also true for the retinal cells that I have described, I believe my macerated preparations would have shown it. Schreiner[12b] and Hesse[13] also figure and describe axial fibers for the rods of the visual cells in polychætous annelids, and Schreiner[12a] also for molluscs. Neither of these observers finds the fibers to extend distally beyond the rods nor centrad toward the nucleus as Patten and myself show. Neither Schreiner nor Hesse figures these cells as twin cells as Patten does, so that to my knowing Patten stands alone in this respect. Andrews[14] describes and figures rods for the visual cells of polychæte annelids but no axial fibers. He was the first to describe these rods in annelids.
The pigmented zone of the pyramid cells, in heavily pigmented series, is filled throughout with dark-brown pigment granules, and is quite like that of the prism cells (Figs. 4, 7). In transverse sections, however, through the most distal part of the pigmented zone, of unpigmented series (Fig. 2), lighter areas with central dots could[50] occasionally be demonstrated, which areas are the pyramid cells. In Fig. 2, the more definite polygonal outline as well as the lighter shade of these areas was a distinguishing feature. The difference in shade was not wholly due to a difference in pigmentation but to a structural difference.
The nuclei of these cells are usually a little larger than those of the prism cells and are filled with a finer and less dense network (Figs. 4 and 7, npyr), in consequence of which they present a lighter appearance in sections when examined with a high power. It will be seen in the figures (4, 7) with what regularity these lighter nuclei lie opposite the pyramids. Some few exceptions occur. These are probably due to the fact that a nucleus or pyramid was not differentiated by the technique. If this opposition between the pyramids and the lighter nuclei were all, I believe it would be sufficient evidence for associating these lighter nuclei with the pyramid cells.[f]
(c) The long pigment cells are about as numerous as the pyramid cells. In these cells, as in the prism and pyramid cells, three regions can be distinguished: the region of the nucleus, a pigmented region (the distal half of which extends between elements of the vitreous body), and a distal rod-like portion, or fiber, which is continued between the prisms into the capsule of the lens (Figs. 4, 7, 9). The pigmented portion is about twice the length of that described for the other cells, and also often of greater diameter, so that in transverse sections (Figs. 1, 2, 3) these cell-areas are larger than those of the other cells. As nearly as I could determine, these cells are pigmented just like the other retinal cells described. In quite unpigmented series, however, they often contain more pigment than the other cells do[51] (Fig. 2). Distally, the pigmented part becomes narrowed to a strong pigmentless fiber (Figs. 3, 4, 7). This fiber stains quite dark with iron-hæmatoxylin and appears homogeneous. It passes between the prisms into the capsule, where it usually bends in a direction toward the margin of the capsule (Fig. 7) and passes diagonally across this to the lens. In sections, a space is often seen about these fibers in the vitreous body, which I regard as a shrinkage space (Figs. 3, 4), since it is not evident in all series (Fig. 1). In Fig. 7, I have assumed that these spaces are due to shrinkage and have not indicated them. Also, in this same figure I have assumed that the spiral appearance of the fibers (Fig. 4) is due to a shortening of the prisms during fixation, and have drawn them straight. At the lens these fibers seem to end. In a few instances they were seen to branch upon reaching the capsule (Fig. 4). In Fig. 9, also, which shows some of these cells from a macerated preparation by Conant, the rods show evidence of branching at their distal terminations. In the same preparation I thought I could see that a fiber became expanded into a membrane spreading over one of the lens-cells. I could not satisfy myself, however, that this was the actual condition of things. Judging from Fig. 9, one might conclude that all the fibers are branched distally; yet, if such were the case I should have seen more of it in sections, but branching as seen in Fig. 4 is the exception. Hence, if all these fibers do branch, I am inclined to believe that it must be among the bases of the lens-cells. Or, if the fibers do expand into membranes to cover the lens-cells (I could not explain purpose), the evidence in Fig. 9 may be nothing more than fragments of this membrane left attached to the ends of the fibers. As is seen in Fig. 7, most of these rods end opposite the cells of the lens, and not usually between two adjacent cells as Schewiakoff has described for Charybdea marsupialis. The nuclei of these cells are like the nuclei of the pyramid cells (Figs. 4, 5, 7, 9) and often have a nucleolus.[g] Centrad these cells are continued into a number of processes as is seen in Figs. 5, 7 and 9. How far the several centrad processes extend and where they end I cannot say; but, as seen in Fig. 5, they soon taper to a thin end which I suppose may be continuous with a nerve fiber. I believe Schewiakoff was mistaken when he stated that these cells extend to the basement membrane.
I have found no evidence in these cells of the existence of an axial fiber such as I have described for the prism and pyramid cells. I find no definite arrangement of the nuclei of the retina into definite layers, but the nuclei of the three kinds of cells lie quite mixed, sometimes one kind lying deeper than the other as can be seen in the figures. Again, they may lie quite at the same level. (This point will be referred to later.)
It is these long pigment cells that I believe retract their pigmented part from between the prisms and pyramids when the medusæ are placed in the dark, protruding with their pigment when placed in the light. Fig. 5 is a section from a slightly pigmented retina killed in the dark. The parts of the cells projecting beyond the pigmented zone, and which would lie between the prisms and pyramids (here not shown) of the vitreous body are seen to be narrower than in sections from retinas killed in the light (Figs. 1, 3, 4, 7) and the cells themselves appear in a condition of retraction as is shown by their large centrad portions with the nuclei, which latter, also, here lie at quite a lower level than the other nuclei. (The pyramid cells were not shown in this series.) I occasionally found appearances like Fig. 5 in retinas killed in the dark (indeed, in some the pigmented portions in the vitreous body were much thinner and more retracted than in Fig. 5). Yet this appearance was not of sufficiently general occurrence to leave no doubt as to its significance. As positive evidence, however, I cannot give it any other interpretation than the one given—that the cells retract themselves with their pigment when in the dark. Again, it must be added that the nuclei of these cells may occasionally lie quite deep even in retinas killed in the light. Indeed, like structures in different retinas may vary considerably in size and shape. None of my darkness retinas, however, showed such a large proportion of the pigmented parts of the long pigment cells projected between the prisms and pyramids as did the light retinas. I examined and tabulated all my series with respect to the extent the long pigment cells were projected into the vitreous body, and I found that those which showed these cells with their pigment least projected between the prisms and pyramids to be those that had been killed in the dark. I thus feel satisfied that the pigmented parts of these cells become in part or quite completely retracted from between the prisms and pyramids of the vitreous body when in the[53] dark, but just how this is accomplished—whether the whole cell with its nucleus takes up a deeper position, the cell substance at the same time collecting in the region about the nucleus, as shown in Fig. 5 and the diagram (Fig. 22), I cannot with certainty state. It would seem, too, as though the pigment became less in the cells exposed to darkness, for I rarely, even in the most retracted heavily pigmented series, saw the pigment to extend farther towards the nucleus than commonly. The time of keeping in the dark, prior to fixing, varied from three-fourths of an hour to one and one-half hours. I could not bring the amount of retraction into relation with the time of exposure, except that in general the retinas longest exposed showed the greater amount of retraction.
(d) The tissue underlying the retina is described by former observers (Claus, Schewiakoff, Conant) as composed of nerve-fibers and ganglion cells. I cannot give it any other interpretation, but I must add that the supposed ganglion cells are seen only as nuclei, no cell bodies ever being demonstrable in any of my sections. Conant also recognized no cell bodies. Occasionally, as in Fig. 7, long fibers could be traced for some distance in this subretinal tissue, in some instances quite to or from a visual cell. Pigment was not regularly observed in this tissue, as Schewiakoff describes, and when present I believe it has been dissolved in from the pigmented zone.
(e) Schewiakoff describes the retina (my pigmented and nuclear regions) as composed of spindle-shaped visual cells (my pyramid cells?) alternating with pigmented supporting cells (long pigment cells), with the nuclei of the former lying more centrad than those of the latter. The visual cells are pigmented only at their periphery, or surface, leaving an unpigmented axis, while the supporting cells have pigment throughout their whole substance within the pigmented zone. Distally, the visual cells have hyaline rods, or fibers, which extend into spaces in the vitreous body, and pass through this and the capsule to the lens. The vitreous body is described as homogeneous, except the spaces for the visual rods, and a secretion from the retinal cells.
It will thus be seen that my results are quite different from those just described. I find the vitreous body to be composed of prisms and pyramids with axial fibers, while the long pigment cells (supporting cells of Schewiakoff) are continued into the vitreous body, and becoming narrowed into a non-pigmented fiber,[54] extend to the lens as described. The prisms and pyramids are, further, the distal continuations of cells whose pigmented and nuclear parts lie in the so-called retina, but which, together with the vitreous body, I have named the retina proper. Conant has so summarily disposed of Schewiakoff’s distinction between retinal cells based on pigmentation and location of nuclei, that I need not say more. Schewiakoff’s Fig. 18 corresponds to my Fig. 1. In this figure he shows the vitreous body as homogeneous with pigmented areas (my long pigment cells) and with spaces with his visual rods. It is quite evident that his spaces with the visual rods correspond to my lighter areas with central dots; i. e. my pyramids of the vitreous body are the same as the spaces shown in his Fig. 18. It is quite evident that Schewiakoff mistook the lighter areas for spaces. That they are not spaces can readily be seen by comparing them with real spaces. It is, of course, possible, too, that the reagents had dissolved the pyramids, leaving only the axial fibers with a little pyramid substance about them, and that this is what Schewiakoff saw. I often found small circular spaces in the centers of the pyramid areas, as also in the prism areas (Fig. 3), which might be taken for hyaline visual rods, fibers, in transverse section, but in such spaces I could usually see a small dot to one side of the space that I take to be the rod (fiber) proper. Fig. 14 also shows such small circular spaces that have very much the semblance of hyaline rods. This figure is a transverse section of the vitreous body of the proximal complex eye, in which no long pigment cells or pyramid cells are present, but it serves well to illustrate the point. The above explanation also accounts for the large size of the visual rods (fibers) in Schewiakoff’s figures. That the fibers of the pyramid cells (visual rods of Schewiakoff) do not extend to the lens is quite evident in my Figs. 4 and 7.
Again, since the long pigment cells are often not seen to terminate in a fiber, but a part of the fiber can often be seen in the distal part of the vitreous body and in the capsule, it will be quite readily seen how Schewiakoff should associate his visual rods, or fibers, with these distal parts of the fibers of the long pigment cells and suppose his visual rods to extend to the lens.
Again, since the long pigment cells sometimes cannot be seen to terminate distally in a fiber, while the vitreous body at the same time may be broken away from the pigmented zone (Fig. 4), it is[55] quite evident how Schewiakoff should have interpreted the parts of the long pigment cells in the vitreous body as conical pigmented caps placed opposite his supporting cells (long pigment cells).
Finally, since Schewiakoff had only twelve marginal bodies to study, and since this tissue is difficult to preserve properly, I do not believe that I am doing Schewiakoff any injustice by explaining away his results as I have done. This fact remains, that Conant and myself agree in all points in which we differ from Schewiakoff.
To Conant belongs the credit of having first demonstrated the prismatic structure of the vitreous body, and he also regarded the prisms as a part of the retinal cells. H. V. Wilson[15, 8b] suggested, however, some years prior to Conant, that the vitreous body might be of a prismatic structure. Conant had evidence also of both the prism and pyramid fibers, as is well shown in his figures of transverse sections but he found his evidence too meager to make any very definite statements. Indeed, Conant concludes that there are three kinds of fibers in the vitreous body and complains of finding but two kinds of cells in the so-called retina (pigmented and nuclear zones) to which to refer them. He saw the pyramids with their axial fibers as lighter areas in transverse sections of the vitreous body (his Figs. 64 and 68, and my Figs. 1, 4 and 7), but suggests that they may be the same as the long pigment cells, the cells having only to project themselves or their pigment in order to become long pigment cells. This suggested to him to preserve material both in the light and in the dark. I do not think Conant’s supposition to be a fact, for I find the pyramids in specimens preserved in the light as well as in the dark. It is, of course, possible that the pyramid cells are in a stage of structural transition to the long pigment cells, for, besides their pigmentation, they also have like nuclei. Furthermore, I held for a long time with Conant that there may be only two kinds of cells in the retina, but I soon found the pyramids so definitely shown as to leave no doubt but that they represented a third kind of cell. For me it remained to first definitely see all the fibers in the vitreous body as also the pyramids in sagittal sections.
Conant describes the long pigment cells with their fibers extending between the prisms of the vitreous body quite as I have described, and in this my work is only confirmatory of his. Conant does not, however, describe the several centrad processes of these cells, nor is[56] he clear that their distad processes extend to the lens, though he speaks of fibers within the capsule.
(f) What, now, is the function of these three varieties of cells of the retina? Schewiakoff regards his visual cells (pyramid cells), as the name implies, as having a visual function. That they have such it seems reasonable to suppose, since they have an axial fiber in their pyramids. If the pyramid cells are visual cells, it appears that the prism cells also are such. Indeed, since these are the only ones present in the proximal eye and the more numerous ones in the distal eye, and like the pyramid cells have an axial fiber in their prisms, it seems that they are the visual cells par excellence of the Cubomedusan eye. Also, the analogy between the prisms and pyramids on the one hand, and the rods and cones of the vertebrate eye on the other hand, does not seem to be so far fetched. It may be of interest, here, to briefly consider Patten’s theory of color vision.[5b]
The gist of Patten’s theory is this: In the eyes of certain molluscs and arthropods, in the parts of the retinal cells corresponding to my prisms and pyramids, he not only finds an axial fiber (or fibers) but finer fibrils that extend at right angles from these axial fibers to the surface of the rods (I shall here, for convenience, call the prisms, pyramids, etc., rods) where they probably become continuous with other fibrils in the surface of the rods. These fibrils from the axial fibers are arranged in superimposed planes, and if I understand rightly, an axial fiber with its radiating fibrils may be compared to the axial wire with its radiating bristles of a brush used for cleaning bottles, provided the bristles of such a brush be arranged in superimposed planes. The lateral arrangement of the fibrils will, of course, be modified according whether a rod is circular, hexagonal, square, etc., in transverse section. It will also be remembered (p. 49) that Patten describes the retinal cells studied by him as composed of twin cells, and he gives the name retinophora to a pair. The system of fibers and fibrils in the rods he names a retinidium. Centrad the axial fibers are continued past the nucleus as a nerve fiber. The fibrils extending laterally in superimposed planes from the axial fiber of a rod, Patten supposes to be the ones stimulated by the incoming rays of light, the retinophora being so arranged that the light rays entering them are parallel to the axial fibers or perpendicular to the lateral fibrils of the[57] retinidium. Again, since the rods are usually the shape of truncated pyramids or cones the lateral fibrils, which are perpendicular to the axial fibers, are of different lengths accordingly as they are situated at the larger or smaller end of a rod. Patten assumes similar fibrils to exist in the rods and cones (particularly the cones) of the vertebrate eye, and he thus makes a general application of his theory. He supports himself in this rather sweeping generalization by the claim to have demonstrated the twin-cell nature of the cones in amphibia and fishes.
For illustration, Patten supposes that if red light only were admitted to the retinophora this would stimulate the fibrils near the broader end of the cone (but that all the fibrils of the retinidium would be stimulated a little) and that we would thus have the sensation of red light. Likewise, if violet light only were admitted, the fibrils at the narrower end of the cone would be stimulated, and we should have violet light. Similarly, if light including all the different wave lengths of the spectrum were admitted, all the lateral fibrils would be stimulated and the sensation of white light produced. The method of stimulation need not be that of a vibration of the fibrils.
Certain grave objections may be raised against such a theory, the most serious, perhaps, being the fact that no such fibrils as Patten has described have as yet been demonstrated for the eyes of those animals that we know have color vision. Yet, as a whole, the objections are perhaps no more serious than any that can be brought against other theories of color vision. What Patten’s theory does do,—it gives us a definite mechanical basis to work from, and if these fibrils should be demonstrated for the rods and cones of vertebrates, physiologists would then have a mechanical basis for color vision quite as they now have for hearing. As Patten says, the problem is primarily a mechanical one. However, the theory cannot well pass for more than a suggestion, a stimulus for future work, and in this lies its present value.
It is quite evident that my results for the retinal cells of Charybdea are, if any thing, a support to Patten’s theory. While I have not been able to demonstrate the fibrils that are the essential to Patten’s theory, yet I have demonstrated the axial fibers of the rods, and if these fibers should be continued as a nerve fiber to some central ganglion (as I believe is reasonable to suppose, see p. 47), I[58] do not see how we can avoid the conclusion that these axial fibers of the prism and pyramid cells are somehow concerned in vision. In Patten’s theory these fibers would represent a conducting element, the real sensory element (fibrils perpendicular to these axial fibers) not having been demonstrated by me.
I have recently read in a short review of Patten’s theory[9] that the evidence we at present have points to the tips of the cones (vertebrate eye) as being the seat of the sensation of red. This would be exactly the converse of what Patten’s theory supposes. Whether or not this objection is a real one, future investigation only can determine.
Hesse[13] regards the axial fibers that he describes for the rods in worms as the primitive fibers of Apathy. In this I agree with him, regarding the axial fibers I have described as “Primitivfibrillen.” Further, I believe, if I understand Apathy rightly, that the fibrils described by Patten as extending laterally from the axial fibers correspond to Apathy’s “Elementarfibrillen.”
It is the long pigment cells that are the puzzling element. Since there can be little doubt but that these cells can project and retract their pigmented parts (as already described), it would seem that a part of their function is to check the diffusion of light in the vitreous body when exposed to strong light. This function would be quite analogous to that of the pigmented cells of the vertebrate retina, which in light become projected between the rods and cones. Similar observations have also been made on the compound eyes of arthropods by Herrick[10] and by Parker[7], who find that the distal retinula cells of Palæmonites project themselves distad in the dark, thus surrounding the vitreous cones with a cylinder of pigment, while (Parker) the pigment of the proximal retinula cells migrates centrad and the accessory cells move distad; in light the reverse takes place. Other observations of this kind are not wanting for crustacea, insects and arachnids. To my knowledge, the pigment changes that I have described are the first of their kind for medusæ.
I suggested while describing the capsule, that the lens might be adjustable. That the fibers of the long pigment cells extend to the lens is my principal reason for this. May these cells not represent ganglion cells and their distad fibers nerve fibers? That they are not sensory (i. e. are stimulated by light waves) seems to be suggested by their not having any axial fiber and in having several centrad processes.[59] These facts suggest that they are not sensory but the center of a reflex mechanism.[h] When the sensory cells proper are stimulated, the impulses are conducted centrad into some nerve center (it may be the nerve tissue underlying the retina, or other nerve centers such as the two groups of ganglion cells in the upper part of the club, or the radial ganglia) from which center, again, impulses return over fibers leading to the long pigment cells causing them to project their pigment, and conducting the impulse to the lens, to produce a change in its adjustment. Since these cells are not so numerous as the prism and pyramid cells taken together, but in turn have a number of processes continued centrad (the sum of which processes approximates the number of sensory cells, prism and pyramid cells) it appears that these cells are admirably adapted to function in just such a mechanism as I have described,—each long pigment cell serving a number of its immediate neighbors.
Further, we may conceive each of the centrad processes of the long pigment cells as receiving a fiber from one of the sensory cells directly as well as indirectly, as just described. While I have been able to demonstrate only a single centrad process for the sensory cells (prism and pyramid cells), yet this does not exclude the possibility of a nerve fibril passing out from such a centrad process to one of the processes of the long pigment cells, and it seems possible that this constitutes the reflex mechanism. That nerve fibrils ramify in ganglion and sensory cells, and may even leave these cells to join those of other cells, has been well demonstrated by Apathy,[6] so that my finding only a single process of the visual cells leading centrad without giving off lateral fibers cannot be a serious objection. Again, fine nerve fibers coming off from the main centrad process of sensory cells in medusæ have been figured by other observers, among whom I mention the Hertwigs. Careful macerations at the seashore would probably demonstrate them for Charybdea.
Hesse thinks that the eyes of the Alciopidæ are adjustable. He[60] describes what he supposes to be muscle fibers just exterior (distal) to the lens, and believes that a contraction of these fibers would have the effect of forcing the lens nearer the retina, or vice versa. His supposition, like mine, needs experimental verification. Hitherto the only instance known of accommodation in the eyes of invertebrates was that described by Beer[17] for Cephalopods.
The Proximal Complex Eye.—With four exceptions, the description and discussion given for the distal complex eye also holds good for the proximal complex eye (Fig. 13). The four exceptions are: the absence of a capsule to the lens; the absence of the long pigment cells; the absence of the pyramid cells; and the different relative position of the lens and retina. This eye, then, has a cornea continuous with the epithelium of the sensory club, a lens, in structure and probable origin quite like that described for the distal complex eye, and a retina of prism cells with axial fibers for the prisms. Since Conant[8b] has described this eye quite fully, and discussed Schewiakoff’s conclusions at length, I shall be brief. Suffice it to say, that Schewiakoff describes two kinds of cells (supporting cells and spindle-shaped visual cells) for the retina of this eye just as he described for the distal complex eye. The vitreous body he likewise describes as being homogeneous and with spaces for the visual rods (fibers) of the visual cells. It is evident that Schewiakoff has interpreted the structure of this eye from analogy with his results on the distal complex eye. Claus likewise has described two kinds of cells for the retina, and the vitreous body as homogeneous. Conant and myself find only one kind of cells in the retina of this eye. The pigmentation that Schewiakoff describes for the vitreous body I believe to have been dissolved in from the pigmented zone of the retina, for I find no regular pigmentation in the vitreous body. Haake’s observation, previously noted (p. 42), applies also to the proximal complex eye.
Conant’s evidence for the axial fibers of the prisms was clearly insufficient, so that he did not in this respect complete his Fig. 69. I republish this figure with the prism fibers drawn (Fig. 13).
Since the long pigment cells are absent my reasons for supposing the lens of this eye to be adjustable vanish.
Finally, a word on the origin of the lens and the relative position of the lens and retina. The lens and retina in this eye[61] are evidently not developed from an outer and an inner half, respectively, of the invaginated and pinched-off lens-retina sphere (as is true for the distal complex eye) but from proximal and distal halves respectively. It is also quite easy to understand the connection of the lens in this eye with the supporting membrane. Since the cells of the ectoderm of the club can in many instances be seen to extend to the basement membrane, or supporting lamella, the cells of the lens, which arise from the ectoderm, simply remain in connection with the basement membrane, this becoming thickened to form a support for the lens. That the lens of the distal complex eye has lost its connection with the basement membrane is evidently due to the fact that the lens is formed from the outer half of the lens-retina sphere. The cells of the lens are by this so far separated from the basement membrane as to lose their connection with it. Schewiakoff also notes the fact that the lens and retina of the proximal complex eye are developed from proximal and distal halves of the lens-retina sphere. He further supposes that the portion of the basement membrane that acts as a support to the lens takes the place of the capsule in the distal complex eye. This latter supposition I do not think probable, since the supporting lamella does not form a distinct covering to the lens on its retinal side.
The Simple Eyes.—Since the shape and position of these eyes have already been described (Claus, Schewiakoff, Conant), I shall not tarry long in this respect. Speaking generally, these eyes are flask-shaped (Fig. 12), the proximal pair quite so, while the distal pair are drawn out in the transverse diameter of the club. These eyes are invaginations of the surface epithelium and the shape of the cells lining these invaginations is quite like that of the epithelial cells, except that their distal portions (bordering the lumen of the invagination) are heavily pigmented. The proximal walls (Fig. 12, left side) of the distal pair are heavier pigmented than the distal walls and the proximal pair of eyes. Schewiakoff calls attention to this point. The pigmentation is, furthermore, not only heavier, but the pigmented portion of each cell is much longer in the proximal walls of the distal eyes (indeed, the cells are longer) than in the distal walls. The significance of this I do not understand. Indeed, I am inclined to believe that in life all these eyes are pigmented quite alike and that it is the reagents used that alter or dissolve the pigment in[62] certain places. Yet, the fact that the cells of the proximal walls of the distal eyes have their pigmented portions nearly double the usual length, shows some deeper significance.
I also note here the small secondary, non-pigmented invagination into the tissue of the clubs from each of the distal simple eyes. Schewiakoff describes this invagination, and it extends in a proximal and dorsal direction (dorsal-side of club opposite complex eye) from the dorsal sides of the distal simple eyes. The cells of these invaginations are not pigmented, but quite like the other pigmented cells in shape, and like these with distal flagellate fibers. I do not see the necessity of assuming, however, that these secondary invaginations are the real sensitive parts of these eyes, while the pigmented parts serve as an iris, as Schewiakoff does in his general discussion.
The histological structure of both pairs of simple eyes is the same. Sections and macerations give me evidence of only one kind of cells, all pigmented alike (except, of course, the non-pigmented secondary invaginations just noted). The cells in these eyes are very closely crowded so that their nuclei lie at several different levels. That they all extend to the lumen of the eyes and are all pigmented could be demonstrated with certainty in many sections, when some of these cells whose nuclei lay most centrad could be followed with the greatest nicety to the lumen (Fig. 12). Macerations (Figs. 8, unlettered cells 21) also show cells with very long cell bodies pigmented at their distal ends and occasionally with a distal process or fiber. While there are, therefore, spindle-shaped cells found, yet they are in every other respect alike, and their differences of shape and position of nuclei are simply the result of crowding. There is, therefore, no evidence of supporting (pigmented) cells and spindle-shaped visual cells (pigmented only externally) as Claus and Schewiakoff have described and which Conant and myself cannot corroborate.
Distally, the retinal cells of the simple eyes have each a fiber (flagellum) that extends into the lumen (Figs. 12, 15, 16, 21). Each flagellum has a dumbbell-shaped basal body just on its entrance into its cell quite like the basal bodies described for the visual cells of the complex eyes (Fig. 12, part left unpigmented). Each flagellum, or fiber, can usually be seen to extend into the cell. In one series I found appearances like Fig. 16, which is a drawing of a part of a section through one of the proximal simple eyes. This section is[63] quite in the angle between the proximal complex eye and the group of network cells in the upper part of the club. In this series I could very definitely trace the distal fibers of the retinal cells centrad, past the nucleus and into the subretinal nerve-tissue. These fibers could be so easily followed that no doubt can exist as to the fact noted. It thus appears that the axial fibers just described pass centrad through the cells and are continued as nerve fibers. On the evidence of such sections as Fig. 16 I have indicated these fibers as extending centrad through their cells. The lumen of the simple eyes is filled with a homogeneous vitreous secretion. This is often incomplete in some parts; occasionally the secretion shows a formation of globules, but all this I believe to be due to the action of reagents. Indeed, I have found simple eyes in which hardly any secretion was present, while others showed an almost completely filled cavity. In that portion of the vitreous secretion just outside the mouth of the distal eyes I occasionally found numbers of very darkly staining granules. I suspect that these are either bacterial or algal organisms.
As already noted, Claus and Schewiakoff describe two kinds of cells for the retinas of these eyes which neither Conant nor myself can demonstrate. Further, I believe I have shown that only one kind exists. If any doubt should still exist, a section like Fig. 25 (which is from the epithelium of the club, but similar smaller areas with central dots could often be demonstrated in transverse sections of the retinal cells of the simple eyes) I believe should be convincing. Schewiakoff further describes flagella for the retinal cells (his visual cells) of the simple eyes quite as I have described them for all the cells. The pigmentation that Schewiakoff mentions as occurring in the secretions within the lumina of these eyes I believe to have been dissolved in from the pigmented zones. I find no definite pigmentation in these vitreous secretions. These secretions are evidently products of the retinal cells and have been so regarded by former observers.
Lithocyst and Concretion.—The cavity filled by the concretion is lined in places by a single layer of cells, two of which are shown in Fig. 7. This fact has been noted by both H. V. Wilson and Conant. Such cells are evidently remnants of the cells that formed the concretion. The supporting lamella completely surrounds the cavity of the concretion.
The concretion filling the lithocyst has the shape of a hemiprolate spheroid cut in the plane of the axis of revolution. Whether it is of endo- or of ectodermal origin, I believe developmental studies only can determine. Tests made in the Chemical Laboratory show the presence of calcium sulphate with perhaps a very small trace of phosphate.[i] Nitric acid slowly dissolves these concretions, but I believe Claus was mistaken when he said that they dissolve with an evolution of gas. I watched them dissolve under the microscope, and never could see the least bit of gas formed. If Claus’s observation is correct, then the composition of the concretions of C. marsupialis is different from that of the concretions of C. Xaymacana. The concretions, further, were dissolved out of the material preserved in formaline and in osmic acid solutions. For dissolving them in situ I used either nitric or hydrochloric acid, or both. A slight husk remains after all the lime is dissolved.
The Epithelium of the Clubs.—The epithelium is thickest on the dorsal side of a club. The thickening here, as in several other places, seems to be due to a crowding of the cells, in consequence of which the nuclei come to lie at different levels, but I believe that all the cells quite reach the surface. The cells with their nuclei nearest the surface are pyramidal in shape, with the bases of the pyramids toward the surface, while those cells whose nuclei lie deeper (where several layers of nuclei occur) may be spindle-shaped (Figs. 12, 23, 24, 26). Centrad these cells are continued into a single process, which often seems to extend to the basement membrane (Figs. 7, 12, 13, 23, 24). Where the epithelium covers the region of the concretion, the cells become flattened and with the long axis of their nuclei parallel with the surface of the club (Fig. 7). The same holds true for the corneal epithelium (Figs. 7, 13).
It is a significant fact that in many places the nuclei form only a single layer, and in such places one cannot speak of spindle-shaped cells. I cannot find any evidence of sensory and supporting cells as Schewiakoff describes. The fact that spindle-shaped cells may exist is simply a physical consequence of their being closely crowded. Conant arrived at the same conclusion.
But I have another and better reason for supposing the existence[65] of only one kind of cells in the epithelium. In a tangential section taken just through the tips of the epithelial cells (Fig. 25) I find polygonal areas with a central dot. This section does not at all agree with Schewiakoff’s Fig. 8, in which he figures two kinds of cells. In Fig. 25 there can be no evidence of two kinds of cells, unless both kinds have like flagella, for these dots are the transverse sections of flagella continued within the cells (Fig. 26).
The epithelium, then, is flagellate, a flagellum to a cell. Whether there are flagella on the epithelium covering the region of the concretion, I could not determine. But I believe that in all other parts, excepting, of course, the corneas, it is flagellated. The fibers (flagella) of the simple eyes are evidently the flagella of the invaginated epithelium. Each flagellum has a basal body, and I could in many instances determine that it was dumbbell-shaped (Fig. 12). This fact was not always evident, however, and it was only occasionally that I felt sure of it. Often the flagella showed only a general thickening within the cells (Fig. 26) while, again, the thickening (basal body) might be quite localized near the surface of the cell. Each flagellum extends into its cell, and occasionally I could trace one clear past the nucleus into the subepithelial nerve-tissue (Fig. 26), just as I did for the axial fibers of the retinal cells of the simple eyes. In those instances in which I could do this, the fibers could so clearly be traced that little if any doubt can exist. I have thus made bold and have drawn the flagella as continued through their cells into the subepithelial nerve-tissue for all the cells of the epithelium of Fig. 12.
A word on the epithelium covering the network cells of Fig. 13. Conant and Schewiakoff here describe fibers from the supporting lamellæ that pass in bundles in among the network cells. These fibers are supposed to be a part of the supporting lamella which reaches out to be a support for the epithelial cells. (Schewiakoff also describes similar fibers for other parts of the epithelium.) Now, as Conant himself shows in Fig. 13, these coarse fibers are not of the same consistency and staining capacity as the supporting lamella. I found them to stain just like the intracellular parts of the flagella or like the central continuations of the axial fibers of the cells of the simple eyes. I could, also, occasionally trace them to the surface of the epithelium, and beyond, when they became continued as short blunt processes or flagella (Fig. 13). I, therefore, conclude that they are sensory fibers like those I have described for the other epithelial[66] cells. Yet, that they pass to the supporting lamella, just as Conant shows in Fig. 13, would seem to indicate that they are fibers from the supporting lamella or processes of the epithelial cells. While this stands as an objection to their being sensory fibers, yet I cannot explain away their being continued distally as a flagellum, except I assume this continuation to be an artefact. This does not seem probable. Perhaps they serve both purposes; namely, that the cell body with its axial fiber is continued to the supporting lamella, the cell proper ending there, while the axial fiber is continued as a nerve fiber. I believe this to be the proper explanation.
The epithelium of the peduncle is quite like the epithelium of the club just described. Sections through the tips of the epithelial cells of the peduncle and also sections sagittal to the axis of these cells give sections like Figs. 25 and 26. I, therefore, conclude that this epithelium is a sensory flagellate epithelium like that of the clubs. Nerve tissue and unstriped muscle fibers underly the epithelium of the peduncles. Claus and Conant also describe a small ventral endodermal tract of nerve tissue, which according to Conant is connected with the endodermal nerve tissue found in the region of the radial ganglia.
To sum up, the epithelium of the club and the peduncle is a flagellate sensory epithelium whose flagella are continued through the cells as nerve fibers into the nerve tissue below. A priori, judging from the mass of nerve tissue underlying the epithelium, we should expect the epithelium to be one strictly sensory. What sense it serves is difficult to surmise. In the physiological part of this paper I suggested that it might be tactile, serving in connection with the lithocysts in giving the animal sensations of space relations.
Claus mentions having seen patches of flagella on the epithelium of the clubs. Schewiakoff supposes that his spindle-shaped sensory cells have only a single flagellum, while his supporting cells have many cilia. In the latter supposition he was evidently mistaken. Conant (from an unpublished note) saw the flagella of the epithelium on the living object and does not think that there could be more than a single one to each cell. He also concludes from living specimens squeezed out under a cover-glass, that there is only one kind of cells in the ectoderm.
Cilia and flagella extending into the cells to which they are attached are described by a number of observers.
I shall not endeavor to discuss the subject further, but shall append the literature on the subject that has come to my notice. (See Literature). Some of these observers ascribe a nervous function to these centrad continuations. I am inclined to believe that they represent the primitive fibrils of Apathy, whether the cilia or flagella are motile or sensory. I should mention, however, that Apathy has traced the “Primitivfibrillen” to be continuous with cilia, and also traces them into the sensory rods of the sensory cells in the sense organs of leeches. Eimer also describes cilia as continued centrad.
The Network Cells and the Multipolar Ganglion Cells.—Conant is the first to accurately describe the true structure of the network cells (Fig. 13) that fill the upper part of the club between the proximal complex eye and the attachment of the peduncle. I cannot add anything to Conant’s description. As their name implies, they are filled with a coarse network-like structure with a central nucleus and nucleolus. Schewiakoff erroneously described them as ganglion cells and Claus as supporting cells. I have sometimes thought that they are not made up of a network, but of a vesicular structure; i. e. the network we see is really produced by the sections of planes that intersect to form little polyhedral cavities. I could not, however, satisfy myself on this point. I further saw similar but smaller cells, with a finer network, disposed in small groups laterally and distally from the attachment of the peduncle to the club.
What the function of these network cells is can only be guessed. In size and shape they somewhat resemble some of the cells found in luminous organs. Conant, however, nowhere mentions that Charybdea is luminous.
Lateral to the larger group of network cells lie two groups of large multipolar ganglion cells (a group on each side). Claus describes these cells, but Schewiakoff does not specially note them, and evidently considered them a part of the network cells, which he erroneously described as ganglion cells.
The Nerve Tissue.—I cannot add anything new on this. It consists of fine fibers and ganglion cells, quite as described by Claus, Schewiakoff, and Conant, and fills the club between the ampulla and the epithelium, except the spaces occupied by the eyes, lithocyst, and network cells. It is likewise present under the ectoderm of the[68] peduncle, where also a small tract is found under the endoderm. (See preceding head, or Claus[3], and Conant[8b]). As already noted, under the distal complex eye, I find only large nuclei to represent the ganglion cells. By saying this, however, I do not wish to dispute their ganglionic nature. The large multipolar ganglion cells I have noted under the preceding topic.
The Supporting Lamella.—The supporting lamella is a continuation, through the peduncle, of the jelly of the bell. It completely surrounds the ampulla and the lithocyst, and also forms a partition between them, so that, as already noted, the lithocyst becomes completely surrounded by it. It also sends a partition ventrally between the complex eyes (Figs. 7, 13). Its thickening to form a support for the lens of the proximal complex eye has already been noticed. I shall limit myself in the discussion of the supporting lamella to the above short resumé, since Schewiakoff gives further detail.
The Endothelium of the Ampulla and the “Floating Cells.”—The ampulla is lined by a secreting epithelium. This is shown by the large masses of a secretion within the bases of the cells, and by smaller masses scattered in the central and more distal parts (Figs. 7, and 27, lower half). The section of the cells is such in Fig. 7, that the bases of some (those nearest the supporting lamella) are taken, the central nuclear region of others, and the tips of those farthest from the supporting lamella. The section may be said to be taken diagonally through the bases and central parts of some of the cells, but owing to the curvature of the ampulla wall, through the tips of others. The secretion is a colloid substance, staining yellowish gray with iron-hæmatoxylin, blue with Lyons blue, and reddish with borax-carmine. Sometimes darkly staining rods and fibers of unknown origin could be seen within the larger masses of the secretion (Fig. 7). These rods and fibers could also be seen in spaces within the cells, from which the secretion had evidently been dissolved. I think there can be no question but that the masses described are a secretion. Many series, however, do not show it; indeed, an examination of Conant’s slides gave me little evidence of a secreting function, though I could demonstrate it in his sections both within the endothelium and also the floating bodies. The[69] presence or absence of this secretion is evidently correlated with the feeding habits of the animals, or else it would be more generally present.
The endothelium is thickest (the cells are longest) in the upper part of the ampulla where the supporting lamella approaches the lens of the proximal complex eye, and in the lower portion of the ampulla (Fig. 7), in the angle between the concretion cavity and the region of the distal complex eye. In general, the cells are longest in the upper part of the ampulla, while in the lower part, especially where they cover the concretion cavity and the dorsal wall, they may be quite cubical instead of columnar. Often they present a vacuolated appearance at their bases (Fig. 27). Claus and Schewiakoff describe and figure this endothelium, but not in detail. No one, to my knowledge, has described this secretory function.
The nuclei of these cells are peculiar. They may contain a network with a nucleus (Fig. 27). Again, they may show evidence of amitotic division (Fig. 20, h, i, j). Indeed, Remak’s scheme (Wilson[18] “The Cell,” p. 46) can be quite readily demonstrated. It is, however, such dumbbell-shaped, elliptical, or ringed nuclei as seen in Figs. 7 and 20 that are of special interest.
I have spoken of some of these nuclei as dumbbell-shaped, elliptical, or ringed. This is so, however, only in sections. They are really flattened spheres with a rod of tissue, of the same structure as the nuclear wall, stretching between the poles. One may conveniently compare the shape of these nuclei with that of an apple, the core of the apple representing the rod connecting the two opposite flattened or slightly hollowed poles of the nucleus. For convenience I shall call the rod connecting the two poles the axis of the nucleus. The dumbbell or elliptical shape would be obtained by a meridional section through the axis (Figs. 20, a, b, c, e, g, k, l, m, n, o, 7). Likewise a ringed appearance with a central dot would be obtained by a section parallel with the flattened surfaces or perpendicular to the axis (Figs. 20, d, 7). In a section not strictly meridional the axis would be cut as in Fig. 29, a, or not show at all. As nearly as I could determine, the inside of these nuclei is a vacuole, which the axis penetrates.
The walls and axis of these nuclei have the structure of a very fine and dense network that stains very dark with iron-hæmatoxylin. It stains quite like the reticulum of any nucleus, but is very dense,[70] as though all the reticulum of the nucleus had been crowded together at the surface. Judging from appearances like p (Fig. 20), the hollowing out, so to speak, of these nuclei, would seem to be a process of vacuolation, the reticulum becoming crowded aside to the surface. But how, on this view, to amount for the formation of the axis, I do not know. Perhaps the axis is formed by a pushing in of two opposite poles of a nucleus, the two invaginations meeting and fusing. On this supposition one might expect the axis to be hollow (cylindrical), but I could not determine that it was. Perhaps the centrosphere (or spheres) (see the next paragraph) has something to do with the formation of the axis (Fig. 20, b, g, e, etc.).
In the nuclei of Fig. 20 with the dark outlines, and of Fig. 7 a small reticular body is seen just opposite one end of the axis, or opposite both ends in g. In d (Fig. 20) this body is seen next the axis just below (outside) the hollow cup represented by the hollow ring. In this instance a central granule is seen in the reticular body, as also in c. I take this reticular body to be the centrosphere, and the central granule in c and d the centrosome. In k, l, m, n, and o (Fig. 20), which are from another series, in which the walls of the nuclei did not stain so dark as in the other nuclei of the same figure, a nucleolus could be definitely seen, indeed, sometimes quite perched upon the wall of the nucleus (k, l). In several instances I could see two nuclei, as in o. But besides these nucleoli, I could in several instances see quite definitely a reticular body (centrosphere) opposite the axis (m, n, o) quite as I described for the nuclei with the dark outlines. In a, b, c, d, e and g the nuclei could not be so readily demonstrated, but I could occasionally see a darker stained body as in a, c and g, that I have no doubt is the nucleolus, which here, again, is perched quite upon the surface of the nucleus. This position of the nucleolus is perhaps due to its having been crowded to one side by the nucleus becoming hollow. It is no uncommon thing, either, to find several nuclei in a single cell, sometimes in process of division or just divided as o and e (Fig. 20), also h, i and j. The whole nuclear phenomenon that I have described seems to be one of division. Perhaps it is somehow associated with the giving off of the secretion of the cells, for these nuclei seem to be found in greatest abundance in those cells in which the secretion is most abundant. In Conant’s sections I found but little evidence of these nuclear phenomena as also little secretion, which all goes to[71] show the association of the nuclear phenomenon with the secretion. I have failed to find any descriptions in the literature of nuclei to which I could refer my observations.
The endothelium of the ampulla is flagellated (Figs. 7, 17, 27). It will be seen that there are two slender flagella to a cell. Each pair of flagella has a pair of basal bodies that are longer than thick, and which are continued as a thin fiber towards the nucleus of the cell. That these centrad continuations of the basal bodies extend to or past the nucleus I could not determine. Sometimes the basal bodies with the centrad continuations are pushed quite to one side of the cell (Fig. 27), while in other cells they are applied quite to the distal surface (Figs. 7, 17, 27). Fig. 17, and the part of Fig. 7 that shows these points, are taken just through the tips of the cells. The darker lines within the polygonal areas are the intracellular basal bodies with their centrad continuations, while the thinner lines are the flagella, and are supposed to lie in the plane just below the plane of the figure. In those instances in which the centrad continuations are applied to the distal surface of the cells they could occasionally be seen to bend centrad (Fig. 27b). While these cilia with their basal bodies and centrad continuations are usually separate, as shown in the figures, yet they are at times applied quite closely to each other so that the double nature of the basal bodies and their centrad continuations is not evident. When the intracellular continuations of the cilia become pushed to one side or applied to the distal surface of the cells, I believe this to be due to the turgor of the cells consequent upon the deposition of large masses of secretion within them. But I must add that this explanation is not altogether satisfactory, since in the endoderm cells of the pedalia of both Charybdea and Tripedalia I found like conditions with no evidence of a secreting function. (See below, under tentacles.) No one, to my knowledge, has described the flagellation in detail, although both Claus and Schewiakoff state that the endoderm is ciliated.
The “floating cells” in the stomach pockets and in the ampulla, described by Conant, I believe are in part derived from the endothelial cells of the ampulla. That a portion of them may arise from the ovary, as Conant explains, I do not doubt; I have, further, found a mass of floating cells in a small Charybdea quite as Conant describes for Tripedalia (his Fig. 71). In this Charybdea, however, I could find no traces of any ovary. Conant speaks of larger and smaller floating[72] cells, and that the smaller ones are also found in the males. This latter fact agrees with what I have suggested, that some of the floating cells arise in the ampulla. My chief reasons for my supposition, however, are the following: I find globules of the secretion of the ampulla cells in some of the floating cells and also scattered loosely among them (Fig. 19). These globules in and among the floating cells have the same general appearance and a similar staining capacity as the secretion in the ampulla cells. Again, in spaces within some of the ampulla cells I find bodies resembling the floating cells with lumps of the secretion within them (Fig. 18). The conclusion, therefore, lies near that some of the floating cells originate within the cells of the ampulla, engulf within them some of the secretion, and are then expelled into the lumen of the ampulla. Better said, perhaps, they represent portions of the ampulla cells with some of the secretion. I also found several instances in which a floating cell had the appearance of being expelled from an ampulla cell. Conant suggests for a similar observation that the cells were about to be swallowed by the ampulla cells. I believe, however, that my finding a secretion similar to that within the cells of the ampulla, in some of the floating cells, as also bodies very much like them and filled with secretion within the ampulla cells, together with Conant’s finding floating cells in males, and finally the observation that the floating cells are usually quite dilapidated, never showing a healthy cell structure—all this leads me to conclude that some of the floating cells originate from the ampulla cells, and that they have a nutrient function in distributing the secretion. This is quite the reverse of what Conant supposed,—that they were taken in as nourishment by the ampulla cells. I also find what appears to be a secretion in the endoderm of the tentacles of both Charybdea and Tripedalia, and believe this is another source of the floating cells. (See below, under tentacles.)
I also found other very darkly staining bodies (Fig. 19) both within the floating cells and free in the ampulla cavity, and more numerous in the ampulla cells themselves. This again goes to show that floating cells take their origin from the ampulla cells. What these darkly staining bodies are, I cannot say. Perhaps they are something akin to the “Chromatoider Nebenkörper” described by Lenhossek (L), or they represent another kind of secretion. If these floating cells are derived from the cells of the ampulla, the active[73] nuclear division within these also receives an explanation. Some nuclear matter can usually be observed in the floating cells.
The Endothelium of the Peduncle.—The endothelium of the peduncle consists of flagellate columnar cells (Fig. 27, upper half). The cells are vacuolated at their bases like some of the cells of the ampulla, and contain a comparatively large nucleus with nucleolus. The flagella are long and slender, quite like those described for the cells of the ampulla, except that there is only one to each cell. The basal bodies of the flagella are of a peculiar shape. They may be described as a bent spindle, continuous at their distad ends with the cilia and at their centrad ends with a fiber that can be traced quite to the neighborhood of the nucleus. I could not trace these fibers into the basal parts of the cells, except in one instance, and I could not be sure of that (Fig. 27a).
Another interesting observation in connection with the basal bodies is that they are bent in one direction on one side of the canal and in an opposite direction on the other side. In Fig. 27, which represents a longitudinal section of the endoderm and the supporting lamella of the dorsal (i. e. farthest from the eyes) side of the peduncle, the distal ends of the basal bodies are bent towards the ampulla, while on the ventral side they would be bent away from the ampulla. This seems to suggest that the flagella move the contents of the canal in one direction on the dorsal side of the canal and in an opposite direction on the ventral side. Conant observed in living material that bodies in the ampulla and the canal were moving about, and that bodies within the tentacles were moving in opposite directions at the same time. This last observation and the histological facts just described, I believe, are mutually corroborative. Again, a priori, we should expect some such mechanism as the one described to bring about an exchange between the contents of the ampulla and that of the stomach pockets. I have not as yet been able to demonstrate a similar flagellate mechanism in the tentacles. Flagella and basal bodies are present in the tentacles, but I could not determine that the basal bodies had any definite arrangement like that shown in Fig. 27. (See under tentacles.) I may add, yet, that the cells in the canal of the manubrium have cilia, similar to the ones just described, with large basal bodies, and with centrad continuations. Finally, I am not certain but that these cells form buds at their ends quite[74] like those I describe for the endothelial cells of the tentacles (see below), and that they aid in the formation of the floating cells. I thought I saw such buds just at the entrance of the lumen of the peduncle into the ampulla, but could not find conclusive evidence.
The Tentacles and the Pedalia.—My observations on the tentacles were begun with the object of demonstrating a flagellate mechanism similar to the one described above for the endothelium of the peduncle. While I have failed to demonstrate such a mechanism for the tentacles, yet several interesting points came to my notice. It will be remembered that the tentacles of the Cubomedusæ are not directly attached to the bell, but that a blade-like portion, the pedalium, intervenes between the tentacles and the bell. For figures of the pedalia and the tentacles the works of Haake, Claus, Conant and Maas[22] may be consulted.
The Ectoderm.—The ectoderm of the tentacles is the seat of a number of differentiations. It is quite thick, as the figures (28 and 29) show, and in this respect is very different from the pedalia, on which the ectoderm cells are quite cubical. I found evidence of cilia here and there, but I can add nothing definite about them. Neither can I add any definite statements regarding the ectoderm cells proper, but what I have to say relates to their differentiations.
(a) The thread cells are of two kinds, larger ones and smaller ones. This is well shown in Fig. 29, which is part of a transverse section of a tentacle of Tripedalia. Two kinds of nettle-cells are also present in the tentacles of Charybdea, but they were specially well shown in Tripedalia. The structure of these thread-cells seems to be typical, and I have little more to say about them. I wish, however, to call attention to the five or six unstriped muscle-fibers that are attached to their basal lateral parts, and which connect them with the basement membrane (Figs. 28, 29). Claus describes these muscle-fibers and mentions that Fr. Müller has described them before him, but I have not found them mentioned elsewhere in the literature of nettle-cells. Professor Brooks tells me, however, that he has often found them. It would appear from Fig. 29 that they serve to retract the thread-cells from the surface. Claus suggests that the muscles are developed from the cnidoblasts.
(b) The plain subectodermal muscle-fibers are of interest. In[75] Charybdea they lie wholly enclosed within canals of the supporting lamella (Fig. 32, upper part). They run longitudinally, and near the base of each tentacle pass out of their canals and become strictly subectodermal (Figs. 31, 32). This is for Charybdea. In Tripedalia they rarely come to lie in closed canals as in Charybdea. These facts show beyond doubt that these muscles are developed from the ectoderm. Claus has suggested their ectodermal origin, but did not demonstrate it. He also suggested that they become inclosed in canals by the supporting lamella pushing up around them and finally fusing above them. This, I believe, is demonstrated by the conditions in Tripedalia (Fig. 29). Here the canals usually remain open, but occasionally, as in the left-hand canal, one may become completely inclosed. This condition of things suggests the intra-lamellar muscles found in actiniarians. The nuclei found in the canals with the muscle-fibers probably belong to the cells from which the muscles become differentiated. Claus figures these muscle-fibers and nuclei, and it may be added that the supporting lamella he figures, for C. marsupialis, is much thicker than I have figured it for C. Xaymacana and Tripedalia cystophora. The number of muscle-canals also is greater and occupies a much greater depth of the thickness of the lamella. Since Claus gives a figure of a transverse section showing the muscles in their enclosed canals, I have not deemed it necessary to duplicate his figure. In the transition from a tentacle to a pedalium, the muscles are most strongly developed toward and at the edges of the pedalium. This is true for the pedalia in general, and accounts for the readiness with which they can be bent inwards, as noted in the physiological part of this paper.
(c) I have found a single ganglion-cell among the cells of the ectoderm of the tentacles. This showed so plainly that I have figured it (Fig. 28). Other ganglion-cells no doubt exist, but could probably not be distinguished from other cells. In its position in Fig. 28 it appears to be associated with the nettle-cell shown just above it. Its position is very much the same as that figured by Lendenfeld (25a).
The Endoderm.—The cells of the endoderm of a tentacle are long and quite slender (Fig. 31). At their bases they are vacuolated quite like the cells of the ampulla and the canal of the sensory clubs. They contain a well-formed nucleus with a nucleolus. In their distal half small light bodies with a dark center are very evident. These bodies are evidently a secretion.
Another peculiar phenomenon presents itself in these cells. The distal part of each cell becomes separated off from its body by what appears to be the formation of a transverse cell-wall (Fig. 31, c-d). I have found the ends of these cells quite separated off in some series. The formation of the walls seems to begin as a thickening at the sides of the cells, and a section through this region, transverse to the cells, would appear like Fig. 30. The dots in the centers of the polygonal areas of this figure are the centrad continuations of the cilia to be described below. As already remarked in describing the endoderm of the ampulla, I believe we here have another place of origin of the “floating cells.” The secretion just described moves into the distal parts of the cells prior to their separation (Fig. 31). In some series I could see these secretion bodies much more numerous within the distal ends of the cells than in Fig. 31.
As will be seen in Fig. 31, each of the endoderm cells of the tentacles has a flagellum that extends into the lumen of the tentacle. Each flagellum has a thickening just within its cell, which may be regarded as a basal body. From this basal body, again, a small fiber extends centrad into each cell. It does not appear that the flagella are thrown off with the distal parts of the cells; at all events, I never found them connected with any of the floating cells except in a few doubtful instances.
What I have said for the endoderm of the tentacle of Charybdea applies equally to Tripedalia.
Claus, in his figure of a transverse section of a tentacle of C. marsupialis shows the endoderm as cubical. I cannot explain why there should be such a difference between the endoderm of the tentacles of C. marsupialis and that of the tentacles of C. Xaymacana and Tripedalia cystophora. Claus does not describe the endoderm in detail.
The endoderm cells of the pedalia of both Charybdea and Tripedalia are cubical and possess flagella, basal bodies, and centrad continuations, quite like those I have described for the endoderm cells of the ampulla. The double nature of the basal bodies and the centrad continuations is, however, not so evident. A secretion I did not find. Histologically, therefore, the endothelium of the pedalia corresponds rather with that of the ampulla, and that of the tentacles with that of the peduncle of the clubs.
The most important results in the histological part of this paper relate to the structure of the retinas of the eyes of the sensory clubs.
The retina of the distal complex eye is composed of three kinds of cells: two kinds of sensory cells (the prism and pyramid cells), and the long pigment cells (Figs. 1-9). The prism and pyramid cells have each an axial nerve fiber in their prisms and pyramids respectively. These fibers I could, however, trace only to the neighborhood of the nuclei. But since I could trace similar fibers in the retinal cells of the simple eyes (Fig. 16) past the nucleus into the subretinal nerve tissue, I believe that the axial fibers in question also extend centrad as nerve fibers into the subretinal nerve tissue. Other observers also figure such fibers as extending centrad as nerve fibers. The axial fibers of the prism cells have each a dumbbell-shaped basal body at their entrance into the pigmented part of a cell. The evidence for a body of such shape in the pyramid cells was not conclusive, though a basal body for the axial fiber exists. The long pigment cells project or retract their pigment in light or darkness respectively and thus seem to serve to check the diffusion of light in the retina. I have also supposed that these cells may serve for conducting impulses to the lens, and that the latter is adjustable.
The proximal complex eye (Fig. 13) has only the prism cells present in its retina, and not two kinds of cells as Schewiakoff has described (see text, pp. 53, 60, 63) for all the eyes.
The simple eyes (Fig. 12), two on each side of a club, four in all, also have only one kind of cells in their retinas, and each cell has a flagellum extending into the vitreous secretion of the lumen. These flagella could be traced centrad as a nerve fiber (Figs. 12, 16). Similarly, a nerve fiber could be traced centrad from the flagella of the epithelial cells of the clubs. Dumbbell-shaped basal bodies for the flagella of the simple eyes could also be demonstrated, but the evidence for this in the epithelial cells of the clubs was not so satisfactory.
Other points of interest are: A secretory epithelium lining the ampulla of the clubs, and a somewhat similar epithelium lining the canals of the tentacles (Figs. 7, 27, 31); the partial origin of the “floating bodies” in the canals of the clubs and tentacles and the stomach pockets from these epithelia (Figs. 18, 19); two flagella to[78] each cell of the endothelium of the ampulla and of the pedalia (Figs. 7, 17); the peculiar nuclei in the endothelial cells of the ampulla (Fig. 20); the longitudinal muscles of the tentacles being completely inclosed within canals of the supporting lamella, but near the base of a tentacle becoming subectodermal. This demonstrates their ectodermal origin. In Tripedalia it is seldom that any of these muscles become enclosed as in Charybdea (Fig. 29).
If to the reader my results seem to embody a somewhat heterogeneous detail, he must remember that the work consists partly in corroborating and partly in supplementing the work of previous observers, and that, in general, histological detail does not usually make the most readable paper.
Biological Laboratory, Johns Hopkins Univ., May 1899.
[a] It was at one time supposed that the concretions in the marginal bodies of medusæ represented lenses and the surrounding nerve tissue the optic nerve, a supposition so highly improbable that it never gained any acceptance. (Ib., p. 41, note.)
[c] By no means do I wish to attribute intelligence to these animals.
[d] Haake[2] says that in the adult Charybdea Rostonii the vitreous bodies of the complex eyes are absent but present in the young. It is difficult to explain this observation except on grounds of imperfect preservation of the adult material, for in all observations on other forms a vitreous body is described. Haake evidently did not use sections, and for this reason his results must be regarded as of doubtful accuracy. Haake also says that the simple lateral eyes of the clubs are absent in the adult, but present in the young.
[e] In the series from which Fig. 3 is taken the pyramid-cells are not so readily demonstrated. Indeed, I missed them altogether at first in this and some other series and supposed that there were only two kinds of cells (19), but upon a careful re-examination I could demonstrate them to my satisfaction. They did not show, however, in the particular section of Fig. 3, so that they are not indicated in this figure.
[f] I go into this at some length because the cell-walls in the series that showed the nuclei best differentiated as lighter and darker ones did not show well, and there might be some doubt that these lighter nuclei belonged to the pyramid cells. I could, however, in many instances, trace the axial fibers of the pyramids through the pigmented zone to these lighter nuclei (as already noted) which fact can leave no doubt but that some of these nuclei belong to the pyramid cells. (Similar nuclei, however, are found to belong to the long pigment cells, to be described below.) Centrad these pyramid cells are continued into a single process just as the prism cells were shown to be (Fig. 7). Figures 6, 8, 9, and 21 show samples of all the pigmented cells found in macerated preparations, and none of these (except Fig. 9, long pigment cells) show more than a single centrad process. Hence, I conclude that centrad both the pyramid cells and prism cells are continued as a single prolongation.
[g] I have been able to demonstrate nucleoli in all the different nuclei of the cells of the sensory clubs.
[h] It may be objected that my criterion, the presence of axial fibers, is not necessarily characteristic of visual cells. However, the great general occurrence of such axial fibers (Patten,[5] Grenacher,[16] Schreiner,[12] Hesse,[13] myself, in simple complex eye, see below, and perhaps others) in eyes in which the retina has only one kind of cells, would seem to indicate that they are quite characteristic of visual cells. Note again that in the proximal eye of Charybdea there is only one kind of cells and with axial fibers.
[i] Mr. J. C. Olsen, of the Chemical Laboratory, kindly made these tests for me.
I. Romanes, G. J. a. ’75, ’77. The Locomotor System of Medusæ. Philosophical Transactions. London. Vol. CLXVI, pt. 1. Vol. CLXVII, pt. 2.
b. ’85. Jelly-fish, Star-fish and Sea-urchins. London.
II. Murbach, Louis. ’95. Preliminary Notes on the Life-history of Gonionemus. Journal of Morphology. Vol. XI.
III. Hesse, R. ’95. Über das Nervensystem und die Sinnesorgane v. Rhizostoma Cuvieri. Zeit. Wis. Zool., B. LX.
IV. Eimer, Th. Zoologische Untersuchungen. ’74. Würzburg Verhandlungen. VI. Bd.
V. Haeckel, E. ’79. Monographie der Medusen. Jena.
VI. Berger, E. W. ’98. Abstract of Dr. F. S. Conant’s Notes on the Physiology of the Medusæ. Johns Hopkins University Circulars. Vol. XVIII, No. 137.
VII. (See also 8, below.)
1. Carrière, J. ’85. Die Schorgane der Thiere. München u. Leipzig.
2. Haake, W. ’87. Scyphomedusen des St. Vincent Golfes. Jen. Zeit. f. Naturwis., Bd. XX., pp. 596-597, 602-604.
3. Claus, C. ’78. Über Charybdea marsupialis. Arb. aus dem Zool., Inst. Univers. Wien., Bd. I.
4. Schewiakoff, W. ’89. Beiträge zur Kenntniss des Acalephenauges. Morph. Jahrb., Bd. XV, H. 1.
5. Patten, William. a. ’89. Studies on the eyes of Arthropods. II. Eyes of Acilius. Journal of Morphology. Vol. II.
b. ’98. A Basis for a Theory of Color Vision. American Naturalist. Vol. XXXII, No. 383.
6. Apathy, St. ’97. Das Leitende Element des Nervensystems u. seine topographischen Beziehungen zu den Zellen. Mitt. Zool. Stat. Neapel., Bd. XII, H. 4.
7. Parker, G. H. ’97. Photomechanical Changes in the Retinal Pigment Cells of Palæmonites, and their Relation to the Central Nervous System. Bull. Mus. Comp. Zool. Harvard Coll. Vol. XXX, No. 6.
8. Conant, F. S. a. ’97. Notes on the Cubomedusæ. Johns Hopkins University Circulars. Vol. XVII, No. 132.
b. ’98. The Cubomedusæ. Memoirs Biological Laboratory Johns Hopkins Univ. Vol. IV, No. 1.
9. A Review of 5b. ’99. A Theory of Color Vision. Natural Science. Vol. XIV, No. 85.
10. Herrick, F. H. ’91. The Embryology and Metamorphosis of the Macroura (Brooks and Herrick). Natl. Acad. Sciences. Vol. V, p. 454.
11. Hertwig, O. & R. ’78. Das Nervensystem und die Sinnesorgane der Medusen. Leipzig.
12. Schreiner, K. E. a. ’96. Die Augen bei Pecten und Lima. Bergens Museums Aarbog.
b. ’97. Histologische Studien über die Augen der freilebenden marinen Borstenwürmer. Bergens Museums Aarbog.
13. Hesse, R. ’99. Untersuchungen über die Organe der Lichtempfindung bei niederen Thieren. V. Die Augen der Polychäten Anneliden. Zeit. Wis. Zool., B. LXV, H. 3.
14. Andrews, E. A. ’92. On the Eyes of Polychætous Annelids. Journal of Morphology. Vol. VII.
15. Wilson, H. V. ’78. Unpublished Notes.
16. Grenacher, H. ’84. Abhandlungen zur vergleichenden Anatomie des Auges. I. Die Retine der Cephalopoden. Abhandl. der Naturf. Gesellsch. zu Halle. Bd. XVI.
17. Beer, Theodore. ’98. Die Accomodation des Auges in der Thierreihe. Wiener klinische Wochenschrift. Nr. 42.
18. Wilson, E. B. ’96. The Cell.
19. Berger, E. W. ’98. The Histological Structure of the Eyes of Cubomedusæ. The Journal of Comp. Neurology. Vol. VIII, No. 3.
20. Lendenfeld, R. Die Nesselzellen der Chidarier. (Review and bibliography.) Biol. Centralbl. Bd. XVII, Nr. 13.
21. Schneider, K. ’90. Histologie von Hydra fusca mit besonderer Berücksichtigung des Nervensystems der Hydropolypen. Arch. Mik. Anat. Vol. XXXV.
22. Maas, O. ’98. Die Medusen. (Charybdea arborifera, Systematic.) Mem. Mus. Comp. Zool., Harvard Coll. Vol. XXIII, No. 1.
A. Haeckel, E. ’72. Die Kalkschwämme. Vol. I, p. 141; Vol. III, Pl. 25, Figs. 3-5.
B. Schultze, F. E. ’75. Rhizopodien Studien. V. Arch. Mik. Anat. Bd. II, p. 583.
C. Eimer, Th. ’77. Weitere Nachrichten über d. Bau des Zellkerns, nebst Bemerkungen über Wimperepithelien. Arch. f. Mik. Anat. Bd. XIV, Taf. VII, p. 114.
D. Bütschli, O. ’78. Beiträge zur Kenntniss der Flagellaten, u. s. w. Zeit. f. Wis. Zool. Bd. XXX, p. 269.
E. Engelmann, Th. W. ’80. Zur Anatomie u. Physiologie d. Flimmerzellen Pflüger’s Arch. Bd. XXIII.
F. Hatschek, B. ’85. Entwickelung der Trochophora von Eupomatus uncinatus Arb. Zool. Inst. Wien., Bd. VI, p. 139.
G. Heider, K. ’86. Zur Metamorphose der Oscarella lobularis. Arb. Zool. Inst. Wien., Bd. VI, pp. 189-194.
H. Schneider, K. C. ’92. Einige histologische Befunde an Coelenterata. Jen. Zeit. f. Nat. 27, N. F. 20.
I. Hecht, Emile. ’95. Contribution a l’Étude des Nudibranchs. Memoirs de la Société Zool. de France. T. 8, Pl. IV, Fig. 45.
J. Minchin, E. A. ’96. Notes on the Larva and Postlarval Development of Leucolosolemia variabilis, etc. Proc. R. Soc., London. Vol. LX.
K. Henneguy, L. F. ’98. Sur le rapports des ciles vibrales avec les centrosomes. Arch. d’anat. micros., T. 1.
L. Lenhossek, H. ’98. Über Flimmerzellen. Anat. Anz. (Supplement.) Bd. XIV.
M. Petre, Carl. ’99. Das Centrum für die Flimmer u. Geisel-bewegung. Anat. Anz. Bd. XV, Nos. 14 and 15.
N. See also 6.
| a | = | flagellum in Fig. 27, that is supposed to extend centrad beyond the nucleus. |
| b | = | twin flagella in Fig. 27, of which the centrad continuation is seen applied against the distal surface of the cells and to be continued centrad. |
| c | = | capsule of lens. |
| cf | = | axial fibers of cells extending centrad. |
| co | = | cornea. |
| concr | = | concretion cavity. |
| ec | = | ectoderm. |
| en | = | endoderm. |
| f | = | flagella. |
| flp | = | distal fiber of a long pigment cell. |
| fpr | = | axial nerve fiber of a prism cell. |
| fpyr | = | axial nerve fiber of a pyramid cell. |
| frc | = | axial nerve fiber of the retinal cells of the simple eyes. |
| gc | = | ganglion cells. |
| ind | = | impression of the lens probably due to the pressure of weight against the surrounding tissue. |
| l | = | lens. |
| lp | = | long pigment cells. |
| m | = | muscle fibers. |
| namp | = | nuclei of ampulla cells. |
| nc | = | network cells (Figs. 13 and 16), and nettle cells (Figs. 28, 29). |
| nf | = | nerve fibers and tissue. |
| nlp | = | nucleus of long pigment cell. |
| nm | = | nucleus of muscle cells. |
| nprc | = | nucleus of prism cell. |
| [81]npyrc | = | nucleus of pyramid cell. |
| nz | = | nuclear zone. |
| pr | = | prism of prism cell. |
| prc | = | prism cell. |
| pyr | = | pyramid of pyramid cell. |
| pyrc | = | pyramid cell. |
| pz | = | pigmented zone. |
| r | = | retina. |
| s | = | secretion in endo. of tent. and ampulla. |
| sh | = | shrinkage space. |
| sec | = | vitreous secretion in the lumen of the simple eyes. |
| sla | = | supporting lamella. |
| vb | = | vitreous body or zone. |
| x | = | (1) the approximate level at which Fig. 4 should be cut transversely to give Figs. 1 and 3. (2) the thickening of the supporting lamella in Fig. 13 to support the lens. |
| * | = | Point of approximation of cells of lenses in Figs. 7 and 13. |
ALL FIGURES, UNLESS OTHERWISE STATED, ARE FROM CHARYBDEA.
Fig. 1. This figure represents a transverse section through a portion of the vitreous body of the distal complex eye at about the level x of Fig. 4. Three kinds of areas are seen, namely, the prisms and pyramids with their axial fibers and the distal continuations of the long pigment cells. Towards the lower left of the figure the section is a little more distal than at the right and the transverse areas of the long pigment cells are no more so large as at the right of the figure. The dark granules in the areas of the long pigment cells represent pigment. Camera lucida sketch. ×920. pp. 45, 46, 48, 49, 50, 51, 52, 54.
Fig. 2. This figure is a camera lucida sketch from a section taken transverse through the most distal part of the pigmented zone of a slightly pigmented retina of a distal complex eye. The presence of three kinds of elements is again evident. The dots without the polygonal areas represent the centrad continuations of the axial fibers of the prism cells. The lettering explains the other areas. ×920. pp. 46, 48, 50.
Fig. 3. This is from a section similar to that of Fig. 1, but a little more distal. At the right, the section is more distal than at the left of the figure, in consequence of which the long pigment cells are there taken through their distal fibers. Note the small shrinkage spaces about the axial fibers of the prisms. The white lines bounding the prism areas appear as in nature. The pyramid cells are not shown in this figure. ×950. Camera sketch. pp. 50, 51, 52, 54.
Fig. 4. This figure is from a section taken parallel to the long axis of the cells of the retina of a distal complex eye. It is from a camera sketch, and nothing has been put into the figure except what could be clearly seen. The lateral boundary lines of the prisms are not shown. Note the evidence for the existence of three kinds of cells. ×920. pp. 44-52, 54.
Fig. 5. This figure represents a sagittal section through the nuclear and pigmented zones and the subretinal nerve tissue of a slightly pigmented retina of a distal complex eye, that had been killed in the dark. Camera sketch. The pyramid cells are not shown. ×900. pp. 47, 51, 52, 53.
Fig. 6. These cells are from a preparation by Conant of a sensory club, macerated[82] in acetic acid. Cell a is evidently an iris cell. The others are probably prism cells from the proximal complex eye. ×900. pp. 44, 48.
Fig. 7. In this figure I represent a sagittal section through the distal complex eye. In the middle half of the section, the nuclei, the prism and pyramid cells with their axial fibers, and the long pigment cells with their large distal fibers are all strictly camera lucida sketched. A portion of the pigmented zone has been left unpigmented to better show its structure. At the right and above the concretion cavity is shown a portion of the endoderm of the ampulla. The section is not strictly in a dorsoventral plane of the club, in consequence of which the cells of the ampulla are cut diagonally and through their tips. Note the dumbbell-shaped nuclei of the ampulla cells, as also the masses of secretion. A part of the retina of the proximal complex eye is shown in the upper part of the figure. ×920. pp. 41-54, 63, 64, 68-71.
Fig. 8. These cells are from a macerated preparation. Cells a, b, c, d may be either prism or pyramid cells from the distal complex eye or prism cells from the proximal complex eye. Cells e and f are probably from the right fourth (Fig. 13) of the retina of the proximal complex eye or from the simple eyes. The unlettered cells are probably from the simple eyes. Some of these show a distal process. ×900. pp. 48, 62, 65.
Fig. 9. The cells here figured are long pigment cells from the same preparation as Fig. 6. ×900. pp. 50, 51.
Fig. 10. This drawing shows an end view of a group of prisms from the same preparation as Fig. 6. ×900. pp. 46.
Fig. 11. This group of prisms are from the same preparation as Fig. 6. Two of them are broken off. The fibers seen at the lower end are probably some of the axial fibers. The fiber at the upper end I believe is interprismatic and the distal fiber of a long pigment cell. ×900. pp. 46.
Fig. 12. This figure is a summary of my results on the simple eyes. It is from a camera sketch of one of the distal eyes, but somewhat diagrammatic. The left side of the figure is proximal, the right side distal. ×920. pp. 61, 62, 64, 65.
Fig. 13. Sagittal dorsoventral section of a proximal complex eye. Conant drew and published this as his Fig. 69. Conant’s evidence regarding the axial fibers of the prism cells was incomplete; so that, in this respect, he left his figure unfinished. I have drawn in these fibers and republish the figure. At the right of the retina and next the lens (the white space) the vitreous body is incomplete and the fibers from the retinal cells project freely into the space. This part of the retina also remains unpigmented. Like my Fig. 7, this figure evidently represents a section somewhat to one side of a sagittal dorsoventral plane of the club, so that the endoderm cells of the ampulla are cut diagonally or transversely. pp. 41-44, 60, 64-68.
Fig. 14. This is drawn to show how regularly small shrinkage spaces may occur in transverse sections of the vitreous bodies. This figure is from a transverse section of the vitreous body of a proximal complex eye. I believe that these spaces are determined by the axial fibers of the prisms. Prism outlines are not shown. ×950. pp. 54.
Fig. 15. This figure is a drawing of a portion of a transverse section of one of the simple eyes. Note the flagella from the retinal cells. pp. 62.
Fig. 16. The section of the lower left hand corner of this figure is through a portion of one of the proximal complex eyes, and shows the centrad continuation of the axial nerve fibers of the retinal cells. The section is such, that, besides the simple eye, the nuclei of the proximal complex eye (upper part of figure) and two network cells are cut. ×920. pp. 47, 62, 63.
Fig. 17. A transverse section through the tips of the ampulla cells is here shown. To the left is towards the upper end of the ampulla. The basal bodies with the centrad fibers are in the plane of the section, while the flagella are supposed to extend below the plane of the section. ×1350. pp. 71.
Fig. 18. These bodies, from within the ampulla cells, contain some of the secretion of the ampulla cells, and resemble the “floating bodies.” ×1350. pp. 72.
Fig. 19. The “floating bodies” here represented are from the ampulla. Globules of a secretion similar to that found in the ampulla cells are seen both within and without the bodies. Note also the two black bodies without the cells and two or three similar ones within the cells. These latter bodies are of doubtful nature. ×1320. pp. 72.
Fig. 20. This figure represents sections of the various nuclei found within the ampulla cells. ×1350. pp. 69, 70.
Fig. 21. These cells are from the same preparation as Fig. 6. They are evidently retinal cells from the simple eyes. The tendency of their pigmented ends to become globular, I believe, is due to their having become isolated before they hardened during maceration. ×920. pp. 62.
Fig. 22. This diagram illustrates the retraction of the long pigment cells. The dotted lines in the vitreous body mark the outlines of the prisms, while the continuous lines represent the axial fibers of the prism and pyramid cells. pp. 45, 46, 48, 49, 53.
Fig. 23. These cells are from the epithelium of a sensory club. They are from the same preparation as Fig. 6. Flagella are not shown. ×900. p. 64.
Fig. 24. This group of epithelial cells of a club are from the same preparation as Fig. 6. ×850. p. 64.
Fig. 25. This sketch is a transverse section through the tips of the epithelial cells of a club. The polygonal areas are the cells, while the central dots are the centrad continuations (nerve fibers) the flagella of the cells. ×920. pp. 63, 65, 66.
Fig. 26. The flagella of the epithelium of a club are in this figure seen to extend centrad, some beyond the nuclei. Cell outlines are not shown. ×920. pp. 64, 65, 66.
Fig. 27. The cells of the lower half of this figure belong to the ampulla, those of the upper half to the canal of the peduncle. The right side of the figure is towards the eyes (the ventral side) of the club. Globules of secretion are seen within the ampulla cells, as also a globule without. The ring above the latter globule is probably an empty shell of a floating cell. ×1320. pp. 68, 69, 71, 73.
Fig. 28. This figure is from a transverse section of a tentacle of Charybdea.[84] The mass with darkly stained granules is the remains of a thread cell. The ectoderm and a small part of the supporting lamella only are figured. Note the large ganglion cell. ×920. pp. 74, 75.
Fig. 29. Part of a transverse section of a tentacle of Tripedalia. The endoderm is not figured. The supporting lamella is seen to be considerably thinner than in Charybdea. Note the subectodermal muscles, as also the muscle fibers to the thread cells. ×920. pp. 69, 74, 75.
Fig. 30. This is a transverse section through the endothelium of a tentacle of Charybdea in the line c d of Fig. 32. The dark lines bounding the polygonal areas are the thickenings of the sides of the walls of the cells in the line indicated. The central dots are the centrad continuations of the flagella. ×920. p. 76.
Fig. 31. This figure is a transverse section through a tentacle of Charybdea at about the middle of Fig. 32, i. e. so near to where the tentacle joins the pedalium, that the muscles within the lamella have all come to lie under the ectoderm. The ectoderm is not shown. ×920. pp. 75, 76.
Fig. 32. A longitudinal section through the supporting lamella only, of a tentacle of Charybdea, is here shown. In the upper part of the figure the muscle fibers are seen wholly enclosed by the supporting lamella. In the middle of the figure they are seen to pass out of their canal. In the lower part of the figure, the supporting lamella is seen to bend to the right where it becomes continuous with the lamella of the pedalium. ×920. p. 75.
CUBOMEDUSÆ. PLATE I.
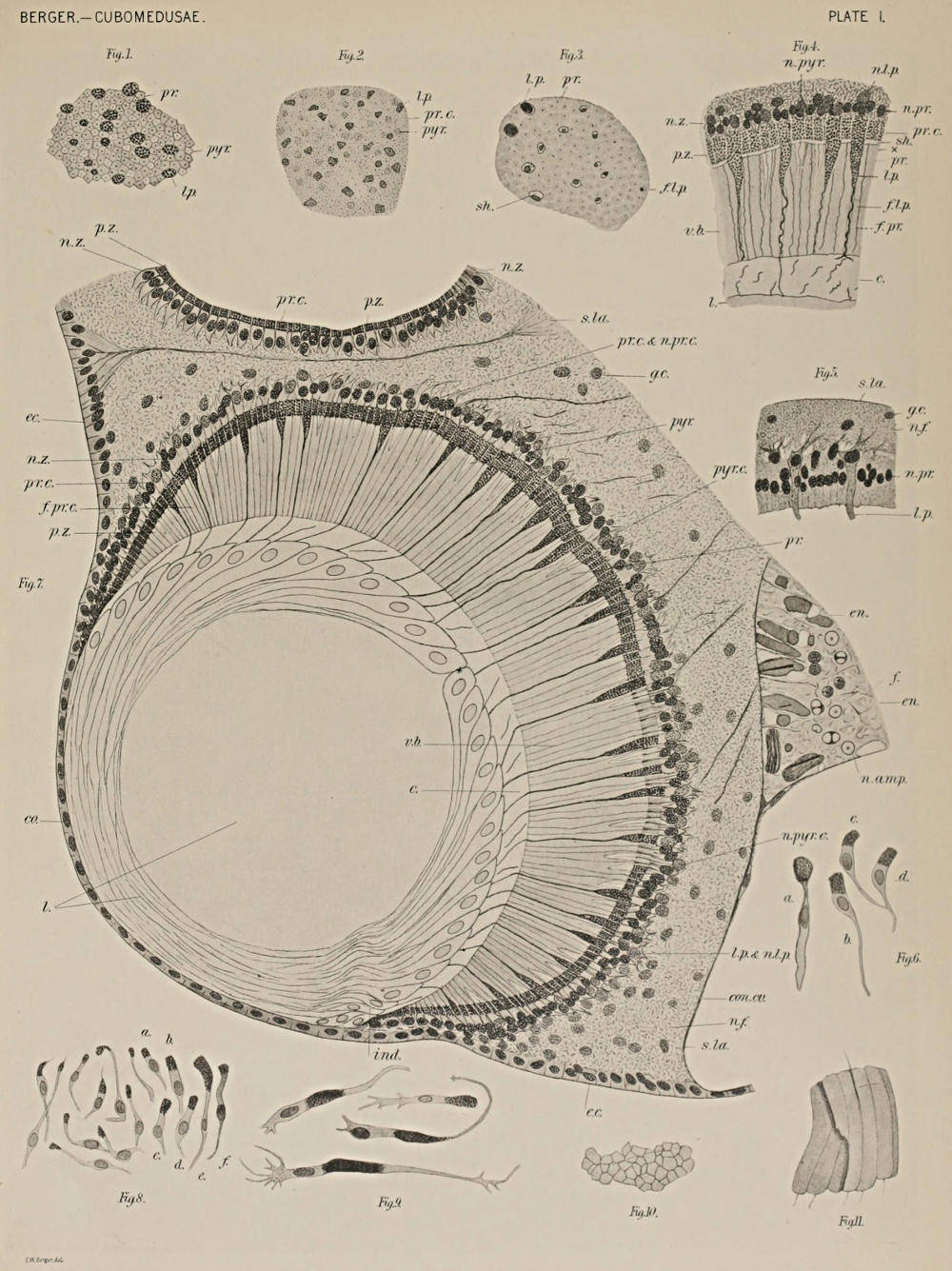
E. W. Berger, del.
CUBOMEDUSÆ. PLATE II.
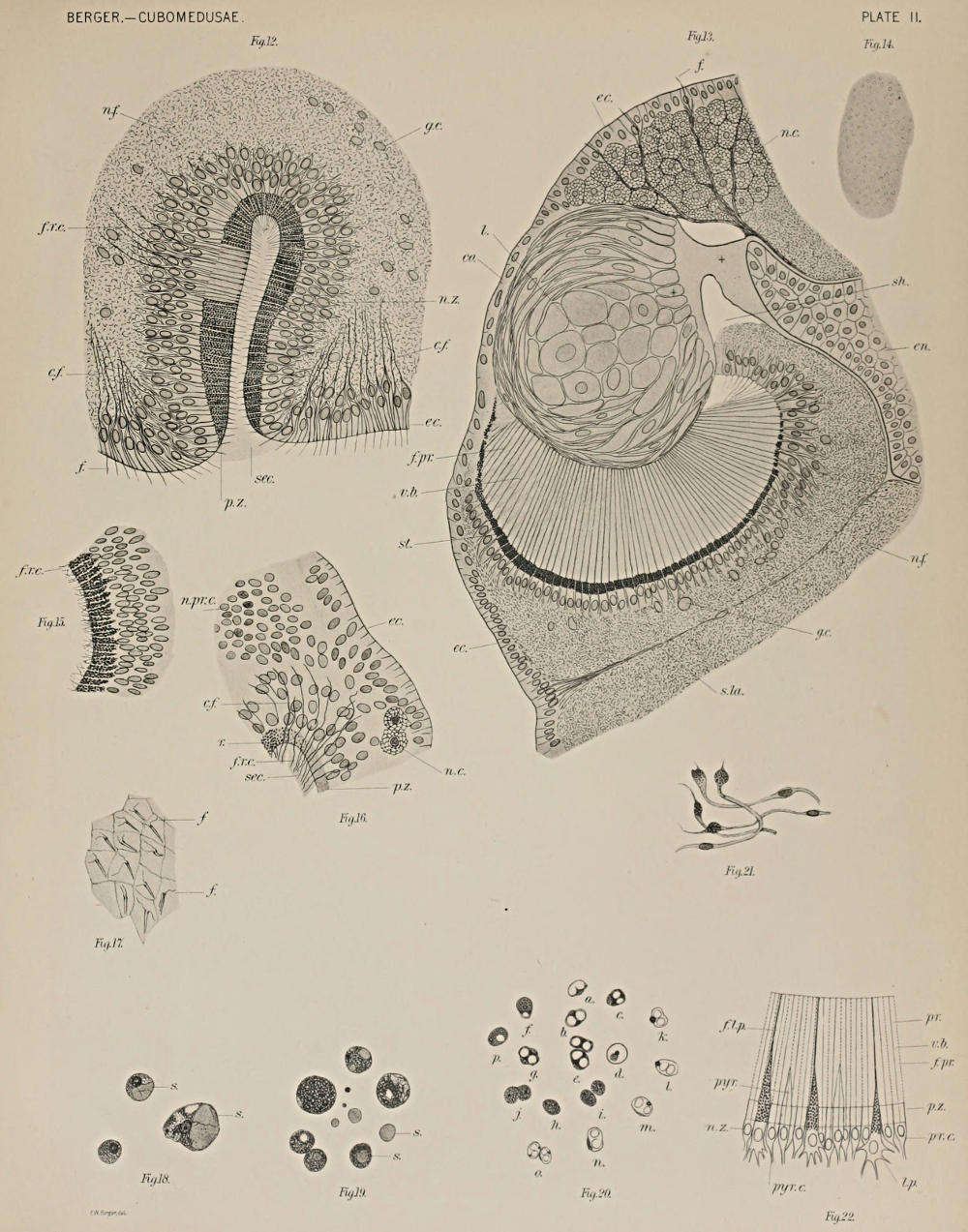
E. W. Berger, del.
CUBOMEDUSÆ. PLATE III.
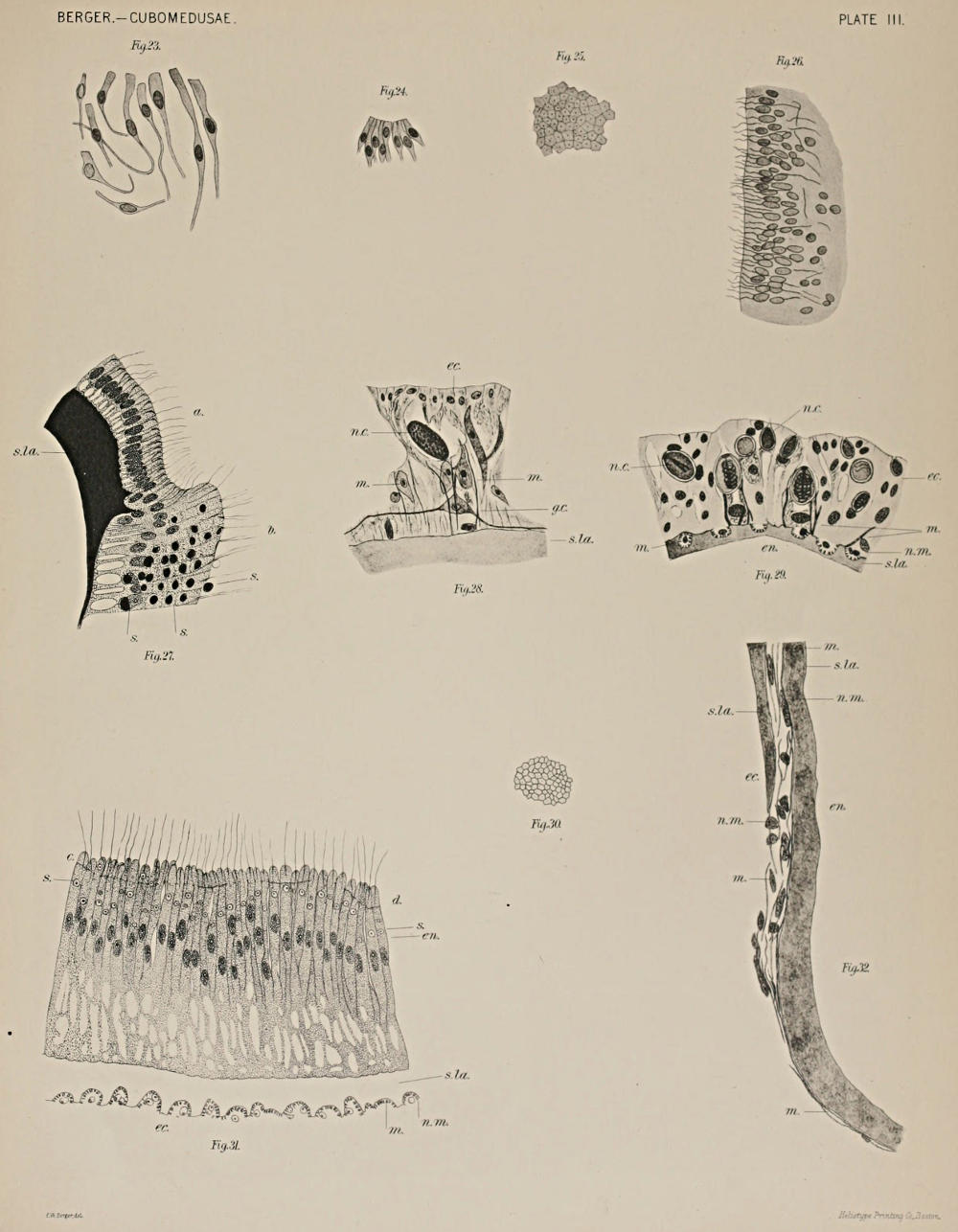
E. W. Berger, del. Heliotype Printing Co., Boston.