
Title: Degeneration: A Chapter in Darwinism
Author: Sir E. Ray Lankester
Release date: March 31, 2019 [eBook #59171]
Language: English
Other information and formats: www.gutenberg.org/ebooks/59171
Credits: Produced by Turgut Dincer and the Online Distributed
Proofreading Team at http://www.pgdp.net (This file was
produced from images generously made available by The
Internet Archive)
DEGENERATION.

NATURE SERIES.
DEGENERATION.
A
CHAPTER IN DARWINISM.
BY
PROFESSOR E. RAY LANKESTER, F.R.S.,
FELLOW OF EXETER COLLEGE, OXFORD.
London:
MACMILLAN AND CO.
1880.
The Right of Translation and Reproduction is Reserved
LONDON:
R. CLAY, SONS, AND TAYLOR,
BREAD STREET HILL, E. C.
TO MY FRIEND
ANTON DOHRN
THESE PAGES
Are Dedicated
IN MEMORY OF OUR COMPANIONSHIP
AT JENA AND NAPLES.
E. R. L.
January, 1880.
It is the misfortune of those who study that branch of science which our President has done so much to advance—I mean the science of living things—that they are not able, in the midst of a vast assembly,[1] to render visible to all eyes the actual phenomena to which their inquiries are directed. Whilst the physicist and the chemist are able to make evident to the senses of a great meeting the very things of which they have to tell, the zoologist cannot hope ever to share with those who form his audience the keen pleasure of observing a new or beautiful organism; he cannot demonstrate by means of actual specimens the delicate arrangements of structure which[2] it is his business to record, and upon which he bases his conclusions. It is for this reason that he who would bring to the notice of laymen some matter which at the moment is occupying the attention of biological students, must appear to be unduly devoted to speculation—hypothesis—to support which he cannot produce the facts themselves but merely the imperfect substitutes afforded by pictures. It is perhaps not altogether a matter for regret that there should be in one great branch of science, as there is in biology, so very marked a disproportion between the facilities for demonstrating facts and the general interest attaching to the theories connected with those facts. We may be thankful that at the present day we are not likely, in the domain of biology, to make the mistake (which has been made under other circumstances) of substituting the mere inspection and cataloguing of natural objects for that more truly scientific attitude which consists in assigning the facts which come under our observation to their causes, or, in other words, to their places in the order of nature. Though we may rightly object to the attempt which is sometimes made to decry the modern teachings of biology[3] as not being “exact science,” yet we may boldly admit the truth of the assertion that we biologists are largely occupied with speculations, hypotheses, and other products of the imagination. All true science deals with speculation and hypothesis, and acknowledges as its most valued servant—its indispensable ally and help-meet—that which our German friends[2] call “Phantasie” and we “the Imagination.” Our science—biology—is not less exact; our conclusions are no less accurate because they are only probably true. They are “probably true” with a degree of probability of which we are fully aware, and which is only somewhat less than the probability attaching to the conclusions of other sciences which are commonly held to be “exact.”
These remarks are addressed to an Association for the advancement of science—of science which flourishes and progresses by the aid of suppositions and the working of the imagination. The Association has been holding its annual sitting in various parts of the British Islands for more than thirty years, and yet it is still a very common and widely spread notion that science, that is to say, true science according to[4] those who hold the notion, does not countenance hypotheses, and sternly occupies itself with the exact record of fact. On the other hand, there are many persons who run to an opposite extreme, and call by the name of science any fanciful attempt to deal with or account for a certain class of phenomena. The words “science” and “scientific” are used so vaguely and variously that one might almost come to the conclusion that it would be well for our Association to plainly state what is that thing for the advancement of which its meetings are held. I cannot venture to speak in the name of my colleagues; and no doubt a review of the work done by the Association would most fitly explain what that body understands by the word “science.” At the same time it is permissible to take this opportunity of briefly stating what science is and what it is not, so far as I am able to judge of the fitting use of the word.
Science is certainly not any and every kind of knowledge. Knowledge of literature, of the beautiful things which have been written or otherwise produced by human ingenuity, is not science. Knowledge of the various manufacturing processes in use by civilised men is not science; nor knowledge of the names of[5] the stars, or of the joints of a beetle’s leg. Science cannot be identified with knowledge of any particular class of objects, however detailed that knowledge may be. It is a common mistake to consider all knowledge of raw products, of living objects or other natural objects, as necessarily “science.” The truth is, that a man may have great knowledge of these things as so many facts, and yet be devoid of “science.” And, on the other hand, that which is properly called science embraces not only such subject-matter as that just alluded to, but also may find its scope in the study of language, of human history, and of the workings of the human mind.
The most frequent and objectionable misuse of the word ‘science’ is that which consists in confounding science with invention—in applying the term which should be reserved for a particular kind of knowledge to the practical applications of that knowledge. Such things as electric lighting and telegraphs, the steam-engine, gas, and the smoky chimneys of factories, are by a certain school of public teachers, foremost among whom is the late Oxford Professor of Fine Art, persistently ascribed to science, and gravely pointed out as the pestilential products of[6] a scientific spirit. They are, in fact, nothing of the kind. American inventors and electric lamps, together with all the factories in Sheffield, might be obliterated without causing a moment’s concern to a single student of science. It is of the utmost importance for the progress and well-being of science that this should be understood; that the eager practical spirit of the inventor who gains large pecuniary rewards by the sale of his inventions should not be confounded with what is totally different and remote from it, namely, the devoted, searching spirit of science, which, heedless of pecuniary rewards, ever faces nature with a single purpose—to ascertain the causes of things. It seems to me impossible to emphasise too strongly in such a place and in such a meeting as this, that Invention is widely separate from, though dependent on, Science. Invention is worldly-wise, and despises the pursuit of knowledge for its own sake. She awaits the discoveries of Science, in order to sell them to civilisation, gathering the golden fruit which she has neither planted nor tended. Invention follows, it is true, the footsteps of Science, but at a distance: she is utterly devoid of that thriftless yearning after knowledge,[7] that passionate desire to know the truth, which causes the unceasing advance of her guide and benefactress.
We may, it seems to me, say that of all kinds and varieties of knowledge that only is entitled to the name “science” which can be described as Knowledge of Causes, or Knowledge of the Order of Nature. It is this knowledge to which the great founder of European science—Aristotle the Greek—pointed as true knowledge: τότε ἐπιστάμεθα ὅταν τὴν αἰτίαν εἴδωμεν. Science is that knowledge which enables us to demonstrate so far as our limited faculties permit, that the appearances which we recognise in the world around us are dependent in definite ways on certain properties of matter: science is that knowledge which enables, or tends to enable us, to assign its true place in the series of events constituting the universe, to any and every thing which we can perceive.
The method by which scientific knowledge is gained—knowledge of the causes of nature—is precisely the same as that by which knowledge of causes in every-day life is gained. Something—an appearance—has to be accounted for: the question in both cases is “Through what cause, in relation to what[8] antecedent is this appearance brought about?” In scientific inquiry, as in every-day life, a hypothesis—a provisional answer or guess—is the reply; and the truth of that guess or hypothesis is then tested. This testing is an essential part of the process. “If my guess be true, then so-and-so as to which I can decide by inspection or experiment, must be true also,” is the form which the argument takes, and the inquiry is thus brought to a point where observation can decide the truth of the hypothesis or first guess. In every-day life we have often to be content without fully testing the truth of our guesses, and hurry into action based on such unverified suppositions. Science, on the other hand, can always wait, and demands again and again the testing and verification of guesses before they are admitted as established truths fit to be used in the testing of new guesses and the building up of scientific doctrine.
The delicately-reared imaginations of great investigators of natural things have from time to time given birth to hypotheses—guesses at truth—which have suddenly transformed a whole department of knowledge, and made the causes of things quite clear which before seemed likely to remain always[9] concealed. So great is the value of hypothesis, so essential to scientific discovery, that the most skilled and highly-trained observer may spend his life in examining and scrutinising natural objects and yet fail, if he is not guided by hypothesis, to observe particular facts which are of the uttermost importance for the explanation of the causes of the things which he is studying. Nature, it has been said, gives no reply to a general inquiry—she must be interrogated by questions which already contain the answer she is to give; in other words, the observer can only observe that which he is led by hypothesis to look for: the experimenter can only obtain the result which his experiment is designed to obtain.
For a long time the knowledge of living things, of plants and of animals could hardly be said to form part of the general body of science, for the causes of these things were quite unknown. They were kept apart as a separate region of nature, and were supposed to have been pitched, as it were, into the midst of an orderly and cause-abiding world without cause or order: they were strangers to the universal harmony prevailing around them. Fact upon fact was observed and recorded by students of plants and[10] animals, but having no hypothesis as to the causes of what they were studying, the naturalists of twenty years ago, and before that day, though they collected facts, made slow progress and some strange blunders. Suddenly one of those great guesses which occasionally appear in the history of science, was given to the science of biology by the imaginative insight of that greatest of living naturalists—I would say that greatest of living men—Charles Darwin.
In the form in which Mr. Darwin presented his view to the world it was no longer a mere guess. He had already tried it and proved it in an immense series of observations; it had already been converted by twenty years’ labour on his part into an established doctrine, and the twenty years which have passed since he published the Origin of Species have only served to confirm, by thousands of additional tests, the truth of his original guess.
Space will not allow me to go fully into the history of the Darwinian theory, but it is necessary for my present purpose that I should state precisely what that theory is. It involves a number of subordinate hypotheses which, together with the main hypothesis, furnish us with a complete “explanation,” as it is[11] called, of the facts which have been ascertained as to living things; in other words, it assigns living things to their causes, gives them their place in the Order of Nature.
It is a very general popular belief at the present day that the Darwinian theory is simply no more than a capricious and anti-theological assertion that mankind are the modified descendants of ape-like ancestors.
Though most of my readers, I do not doubt, know how imperfect and erroneous a conception this is, yet I shall not, I think, be wasting time in stating what the Darwinian theory really is. In fact, it is so continuously misrepresented and misunderstood, that no opportunity should be lost of calling attention to its real character. Bit by bit, naturalists had succeeded in discovering the order of nature—so far that all the great facts of the universe, the constitution and movements of the heavenly bodies, the form of our earth, and all the peculiarities of its crust, had been successfully assigned to one set of causes—the properties of matter, which are set forth in what we know by the name of the “laws of physics and chemistry.” Whilst geologists, led by Lyell, had[12] shown that the strata of the earth’s crust and its mountains, rivers, and seas were due to the long-continued operation of the very same general causes—the physico-chemical causes—which at this moment are in operation and are continuing their work of change, yet the living matter on the crust of the earth had to be excluded from the grand uniformity which was elsewhere complete.
The first hypothesis, then, which was present to Mr. Darwin’s mind, as it had been to that of other earlier naturalists, was this: “Have not all the varieties or species of living things (man, of course, included) been produced by the continuous operation of the same set of physico-chemical causes which alone we can discover, and which alone have been proved sufficient to produce everything else?” “If this be so,” Mr. Darwin must have argued (and here it was that he boldly stepped beyond the speculations of Lamarck and adopted the method by which Lyell had triumphantly established Geology as a science), “these causes must still be able to produce new forms, and are doing so wherever they have opportunity.” He had accordingly to bring the matter to the test of observation by seeking for[13] some case of the production of new forms of plants, or of animals, by natural causes at the present day. Such cases he found in the production of new forms or varieties of plants and animals, by breeders. Breeders (the persons who make it their business to produce new varieties of flowers, of pigeons, of sheep, or what not) make use of two fundamental properties of living things in order to accomplish their purpose. These two properties are, firstly, that no two animals or plants, even when born of the same parents, are exactly alike; this is known as Variation: secondly, that an organism, as a rule, inherits, that is to say, is born with the peculiarities of its parents; this is known as Transmission, and is simply dependent on the fact that the offspring of any plant or animal is only a detached portion of the parent—a chip of the old block, as the saying is. The breeder selects from a number of specimens of a plant or animal a variety which comes nearest to the form he wishes to produce. Supposing he wished to produce a race of oxen with short horns, he would select from his herd bulls and cows with the shortest horns, and allow these only to breed; they would transmit their relatively short[14] horns to their offspring, and from these again the cattle with the shortest horns would be selected by the breeder for propagation, and so on through several generations. In the end a very short-horned generation would be obtained, differing greatly in appearance from the cattle with which the breeder started.
Now we know of no facts which forbid us to suppose that could a breeder continue his operations indefinitely for any length of time—say for a few million years—he could convert the short-horned breed into a hornless breed; that he could go on and thicken the tail, could shorten the legs, get rid of the hind limbs altogether by a series of insensible gradations, and convert the race into forms like the Sirenia, or sea-cows. But if he could do this, you have only to give him a longer time still and there is no obstacle remaining to the conversion, by the same kind of process, of a polyp into a worm, or of a worm into a fish, or even of a monkey into a man.[3]
So far we have supposed the interference of a breeder who selects and determines the varieties which shall propagate themselves; so far we have[15] not got a complete explanation, for we must find a substitute in nature for the human selection exercised by the breeder. The question arises, then, “Is there any necessary selective process in nature which could have operated through untold ages, and so have represented the selective action of the breeder, during an immense period of time?” Strangely enough, Mr. Darwin was led to the discovery of such a cause existing necessarily in the mechanical arrangements of nature, by reading the celebrated book of an English clergyman, the Rev. Mr. Malthus, On Population. On happening to read this book, Mr. Darwin himself tells us that the idea of “natural selection” flashed upon him. That idea is as follows. Not only among mankind, but far more largely among other kinds of animals and of plants, the number of offspring produced by every pair is immensely in excess of the available amount of the food appropriate to the particular species in question. Accordingly, there is necessarily a struggle for existence—a struggle among all those born for the possession of the small quantum of food. The result of this struggle is to pick out, or select, a few who survive and propagate the species, whilst the majority perish[16] before reaching maturity. The fact that no two members of a species are alike has already been shown to be the starting-point which enables the breeder to make his selection. So, too, with natural selection in the struggle for existence; the fact that all the young born of one species are not exactly alike—but some larger, some smaller, some lighter, some darker, some short-legged, some big-eyed, some long-tongued, some sharp-toothed, and so on—furnishes the opportunity for a selection. Those varieties which are best fitted to obtain food and to baffle their competitors, gain the food and survive, the rest perish.
We have, then, to note that the hypothesis that there must be a selection—which was framed or deduced as a “test hypothesis” from the earlier hypothesis that species have arisen by the action of causes still competent to produce new forms—led Mr. Darwin to the discovery of this great cause—the “natural selection,” or “survival of the fittest,” in the struggle for existence. Just as the breeder can slowly change the proportions of the animals or plants on which he operates, so in inconceivably long periods of time has this struggling of varieties, and the consequent[17] natural selection of the fittest, led to the production, from shapeless primitive living matter, of all the endless varieties of complicated plants and animals which now people the world. Countless varieties have died out, leaving only their modified descendants to puzzle the ingenuity of the biologist.
Of the tens and hundreds of thousands of intermediate forms we know nothing by direct observation. They have perished as better fitted forms ousted them in the never-ending conflict. But we feel sure that they once were in existence, and can infer what was their structure, and what were their peculiarities, by the study of the structure and attributes of their now living descendants.
If all the forms of life at present living are the modified offspring of a smaller number of ancestral forms which have died out, and if these again were the modified descendants produced by ordinary parentage of a single original living thing—then the whole series of forms that have ever lived could, if we had them before us, be arranged in the form of a great family-tree—the various branches presenting a perfect gradation of forms arranged one after another, leading down from the terminal twigs (which would represent the latest[18] forms produced) to larger and larger branches, until the common trunk representing the original ancestor would be reached. Our actual means of observing the genealogical affinities of different kinds of animals and plants may be understood by a further use of the metaphor of a genealogical tree in shape like an elm or an oak. Suppose the genealogical tree completely written out—a perfect record—to be sunk in muddy water so that only its topmost branches and twigs are here and there visible—then you have a fair notion of the present condition of the great family of organisms. Only the topmost twigs remain visible, the rest of the great family-tree of living beings is hidden from view, submerged beneath the muddy waters of time. Naturalists have, however, undertaken to reconstruct this great genealogical tree. It is a main object now in the study both of zoology and of botany to find out what are the cousinships, what the exact genetic relationships of all the various species of plants and animals, and so to show, even to the minutest detail, in what particular ways physico-chemical causes have brought about and modified the forms of living things.
The task is not quite so difficult as the comparison[19] to a submerged forest-tree would lead one to expect; at the same time it is more difficult than those who have boldly attempted it appear to believe. We have one great help in the carefully worked out systematic classification of animals and plants according to their structure. We are justified in assuming as a general law that animals or plants of like structure have descended from common ancestors—that is to say, that the same kind of organisation (especially where a number of elaborate details of structure are involved) has not been twice produced by natural selection. Thus we are entitled to conclude that all the animals which have a backbone and pharyngeal gill-slits combined—the Vertebrates, as we call them—have descended from a common parent; that all the animals with a muscular foot-like belly and lateral gill filaments, the Molluscs, have also had a common parent, and so on.
A classification according to structure goes then a long way towards mapping out the main lines of the family-tree of organisms. We are further assisted in the task by the fossil remains of extinct organisms which sometimes give to us the actual ancestors of forms now living. But the most remarkable aid to[20] the correct building-up of the pedigree of animals at least (and the remarks which follow are confined to that division of the organic world), is afforded by the changes—the phases of development—which every animal exhibits in passing from the small shapeless egg to the adult condition. The aid which we here obtain depends on the following facts. Just as we suppose any one animal—say a dog—to have developed by slow change through an immense series of ancestors which become simpler and simpler as we recede into the past until we reach a small shapeless lump of living matter devoid of structure, so do we find actually as a matter of fact, which any one can see for himself, that every individual animal begins its individual life as a structureless particle which is thrown off from its parent, and is known as the egg-cell (Fig. 1). Gradually passing through a series of more and more elaborated conditions of structure, that egg grows into the adult dog. The changes which have taken countless ages in the one case, are accomplished in a few weeks in the other.
And now we have to note the important fact which makes this process of development so intensely interesting in relation to the pedigree of the animal[21] kingdom. There is very strong reason to believe that it is a general law of transmission or inheritance, that structural characteristics appear in the growth of a young organism in the order in which those characteristics have been acquired by its ancestors. At first the egg of a dog represents (imperfectly, it is true,) in form and structure the earliest ancestors of the dog; a few days later it has the form and structure of somewhat later ancestors; later still the embryo dog resembles less remote ancestors; until at last it reaches the degree of elaboration proper to its immediate forefathers.
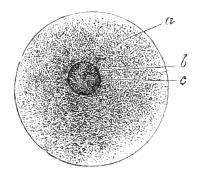
Fig. 1.—An egg: a single corpuscle of protoplasm with nucleus b c, and body a.
Accordingly the phases of development or growth of the young are a brief recapitulation of the phases of form through which the ancestors of the young creature have passed. In some animals this recapitulation is more, in others it is less complete. Sometimes the changes are hurried through and disguised, but we find[22] here and there in these histories of growth from the egg most valuable assistance in the attempt to reconstruct the genealogical tree. The history of the development of the common frog is a good illustration of the kind of evidence in question.

Fig. 2.—Tadpoles and young of the Common Frog. 1, Recently hatched (twice natural size); 2 and 2a, same enlarged to show the external gills; 3 and 4, later stages with gill slits covered by a membrane leaving only the spiracle (see Fig. 16) as an exit for the respired water; 5, with hind legs appearing; 6, with both fore and hind legs; 7, atrophy of the tail; 8, young frog.
The frog’s egg first gives rise to a little aquatic creature with external gills and a tail—the tadpole—which gradually loses its gills and its tail and acquires in their place lungs and four legs (Fig. 2), so as now to be fitted for life on dry land. From what we otherwise know of the structure of the frog and the animals to which it is allied, we are justified in concluding that the tadpole is a recapitulative phase of development, and represents to us more or less closely an ancestor of the frog which was provided with gills and tail in the adult state, and possessed neither legs nor lungs.
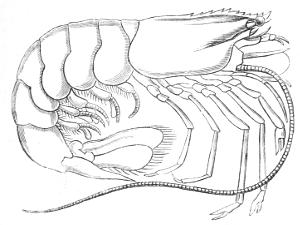
Fig. 3.—Adult shrimp of the genus Peneus.
A less familiar case is that of a certain kind of[24] shrimp, which is illustrated in the wood-cuts (Fig. 3 and right lower corner of Fig. 4). The little creature which issues from the egg of this shrimp is known as the “Nauplius form.” Many animals very different in appearance from this shrimp make their first appearance in the world as Nauplii; and it appears probable that the Nauplius-phase is the recapitulative re-presentation of an ancestor common to all this set of animals, an ancestor which was not exactly like the Nauplius, but not very different from it.
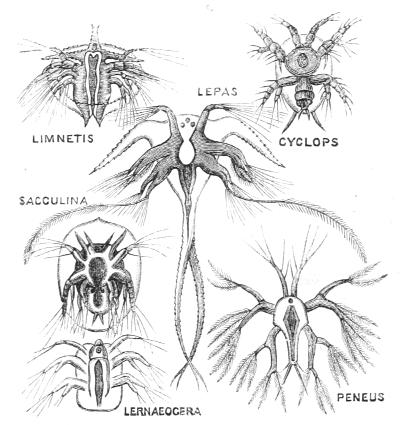
Fig. 4.—Nauplius larval-form of various Crustacea (Shrimps, Water-fleas, Barnacles, &c.).
The Nauplius of our shrimp gradually elongates.[25] At first it has but three pair of limbs, but it soon acquires additional pairs, and a jointed body, and thus by gradually adding to its complexity of structure as seen in Figs. 5 and 6, it approximates more and more to the adult form from the egg of which it originated.
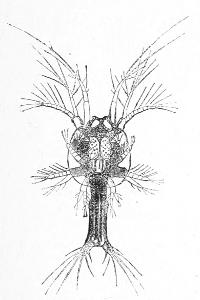
Fig. 5.—Larva of the Shrimp Peneus.
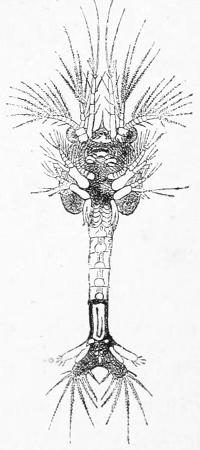
Fig. 6.—More advanced larva of the Shrimp Peneus.
And now we are approaching the main point to which I wish to draw the reader’s attention. In attempting to reconstruct the pedigree of the animal kingdom and so to exhibit correctly the genetic[26] relationships of all existing forms of animals, naturalists have hitherto assumed that the process of natural selection and survival of the fittest has invariably acted so as either to improve and elaborate the structure of all the organisms subject to it, or else has left them unchanged, exactly fitted to their conditions, maintained as it were in a state of balance. It has been held that there have been some six or seven great lines of descent—main branches of the pedigree—such as that of the Vertebrates, that of the Molluscs, that of the Insects, that of the Starfish, and so on; and that along each of these lines there has been always and continuously a progress—a change in the direction of greater elaboration.
Each of these great branches of the family-tree is held to be independent—they all branch off nearly simultaneously from the main trunk like the leading branches of an oak. The animal forms constituting the series in each of these branches are supposed to gradually increase in elaboration of structure as we pass upwards from the main trunk of origin and climb further and further towards the youngest, most recent twigs. New organs have, it is supposed, been gradually developed in each series, giving their[27] possessors greater powers, enabling them to cope more successfully with others in that struggle for existence in virtue of which these new organs have been little by little called into being. At the same time here and there along the line of march, certain forms have been supposed to have “fallen out,” to have ceased to improve, and being happily fitted to the conditions of life in which they were long ago existing, have continued down to the present day to exist in the same low, imperfect condition. It is in this way that the lowest forms of animal life at present existing are usually explained, such as the microscopic animalcules, Amœbæ and Infusoria. It is in this way that the lower or more simply-made families of higher groups have been generally regarded. The simpler living Mollusca or shellfish have been supposed necessarily to represent the original forms of the great race of Mollusca. The simpler Vertebrates have been supposed necessarily to represent the original Vertebrates. The simpler Worms have been supposed necessarily to be the stereotyped representatives of very ancient Worms.
That this is, to a certain extent, a true explanation of[28] the existence at the present day of low forms of animals is proved by the fact that we find in very ancient strata fossil remains of animals which differ, ever so little, from particular animals existing at the present day; for instance, the Brachiopods (lamp-shells), Lingula and Terebratula, the King-crabs, and the Pearly Nautilus are found living at the present day, and are also found with no appreciable difference in very ancient strata of the earth’s crust; strata deposited so long ago that most of the forms of life at present inhabiting the earth’s surface had not then been brought into existence, whilst other most strange and varied forms occupied their place, and have now for long ages been extinct.
Whilst we are thus justified by the direct testimony of fossil remains in accounting for some living forms on the hypothesis that their peculiar conditions of life have been such as to maintain them for an immense period of time in statu quo unchanged, we have no reason for applying this hypothesis, and this only, to the explanation of all the more imperfectly organised forms of animal or plant-life.
It is clearly enough possible for a set of forces such as we sum up under the head “natural selection”[29] to so act on the structure of an organism as to produce one of three results, namely these; to keep it in statu quo; to increase the complexity of its structure; or lastly, to diminish the complexity of its structure. We have as possibilities either Balance, or Elaboration, or Degeneration.
Owing, as it seems, to the predisposing influence of the systems of classification in ascending series proceeding steadily upwards from the “lower” or simplest forms to the “higher” or more complex forms,—systems which were prevalent before the doctrine of transformism had taken firm root in the minds of naturalists, there has been up to the present day an endeavour to explain every existing form of life on the hypothesis that it has been maintained for long ages in a state of Balance; or else on the hypothesis that it has been Elaborated, and is an advance, an improvement, upon its ancestors. Only one naturalist—Dr. Dohrn, of Naples—has put forward the hypothesis of Degeneration as capable of wide application to the explanation of existing forms of life;[4] and his arguments in favour of a general[30] application of this hypothesis have not, I think, met with the consideration which they merit.
The statement that the hypothesis of Degeneration has not been recognised by naturalists generally as an explanation of animal forms, requires to be corrected by the exception of certain kinds of animals, namely, those that are parasitic or quasi-parasitic. With regard to parasites, naturalists have long recognized what is called retrogressive metamorphosis; and parasitic animals are as a rule admitted to be instances of Degeneration. It is the more remarkable whilst the possibility of a degeneration—a loss of organisation making the descendant far simpler or lower in structure than its ancestor—has been admitted for a few exceptional animals, that the same hypothesis should not have been applied to the explanation of other simple forms of animals. The hypothesis of Degeneration will, I believe, be found to render most valuable service in pointing out the true relationships of animals which are a puzzle and a mystery when we use only and exclusively the hypothesis of Balance, or the hypothesis of Elaboration. It will, as a true scientific hypothesis, help us to discover causes.
We may now examine a few examples of undeniably degenerate animals, and first, I may call to mind the very remarkable series of lizard-like animals which exist in the south of Europe and in other countries, which exhibit in closely related genera a gradual loss of the limbs—a local or limited Degeneration. We have the common Lizard (Lacerta), with five toes on each of its well-grown fore and hind limbs; then we have side by side with this a lizard-like creature, Seps, in which both pairs of limbs have become ridiculously small, and are evidently ceasing to be useful in the way in which those of Lacerta are useful; and lastly, we have Bipes, in which the anterior pair of limbs has altogether vanished, and only a pair of stumps, representing the hinder limbs, remain.
No naturalist doubts that Seps and Bipes represent two stages of Degeneration, or atrophy of the limbs; that they have, in fact, been derived from the five-toed four-legged form, and have lost the locomotor organs once possessed by their ancestors. This very partial or local atrophy is not however that to which I refer when using the word Degeneration. Let us imagine this atrophy to extend to a variety of[32] important organs, so that not only the legs, but the organs of sense, the nervous system, and even the mouth and digestive organs are obliterated—then we shall have pictured a thorough-going instance of Degeneration.
Degeneration may be defined as a gradual change of the structure in which the organism becomes adapted to less varied and less complex conditions of life; whilst Elaboration is a gradual change of structure in which the organism becomes adapted to more and more varied and complex conditions of existence. In Elaboration there is a new expression of form, corresponding to new perfection of work in the animal machine. In Degeneration there is suppression of form, corresponding to the cessation of work. Elaboration of some one organ may be a necessary accompaniment of Degeneration in all the others; in fact, this is very generally the case; and it is only when the total result of the Elaboration of some organs, and the Degeneration of others, is such as to leave the whole animal in a lower condition, that is, fitted to less complex action and reaction in regard to its surroundings, than was the ancestral form with which we are comparing it (either actually[33] or in imagination) that we speak of that animal as an instance of Degeneration.
Any new set of conditions occurring to an animal which render its food and safety very easily attained, seem to lead as a rule to Degeneration; just as an active healthy man sometimes degenerates when he becomes suddenly possessed of a fortune; or as Rome degenerated when possessed of the riches of the ancient world. The habit of parasitism clearly acts upon animal organisation in this way. Let the parasitic life once be secured, and away go legs, jaws, eyes, and ears; the active, highly-gifted crab, insect, or annelid may become a mere sac, absorbing nourishment and laying eggs.
Reference was made above to the larval stage of a certain shrimp (Figs. 4, 5, 6). Let us now compare these with the young stages of a number of shrimp-like animals, viz., Sacculina, Lernæocera, Lepas, Cyclops, Limnetis, (all drawn in Fig. 4), some of which lead a parasitic life. All start equally with the recapitulative phase known as the Nauplius; but whilst the Nauplius of the free-living shrimp grows more and more elaborate, observe what happens to the parasites—they degenerate into[34] comparatively simple bodies; and this is true of their internal structure as well as of their external appearance. The most utterly reduced of these parasites is the curious Sacculina (Fig. 7) which infests Hermit-crabs, and is a mere sac filled with eggs, and absorbing nourishment from the juices of its host by root-like processes.
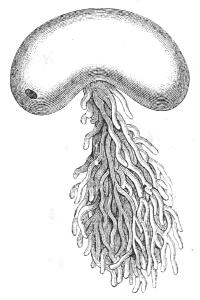
Fig. 7.—Adult Sacculina (female).
Lernæocera again, which in the adult condition is found attached to the gills of fishes, has lost the well-developed legs of its Nauplius childhood and become an elongated worm-like creature (Fig. 8), fitted only to suck in nourishment and carry eggs.
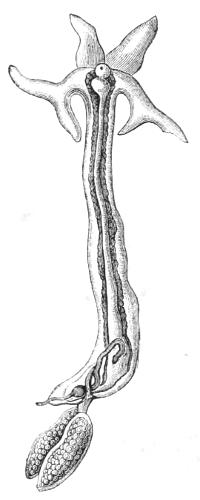
Fig. 8.—Adult female Lernæocera.
Amongst these Nauplii—all belonging to the great[35] group Crustacea, which includes crabs and shrimps—is one which gives rise to an animal decidedly degenerate, but not precisely parasitic in its habits. This Nauplius is the young of the ship’s Barnacle, a curious stalked body, inclosed in a shell of many pieces (Fig. 9). The egg of the Barnacle gives rise to an actively swimming Nauplius, the history of which is very astonishing. After swimming about for a time the Barnacle’s Nauplius fixes its head against a piece of wood, and takes to a perfectly fixed, immobile state of life (Fig. 10). The upper[36] figures represent the Nauplius stage of animals closely resembling the Barnacle: the lower figures show the transformation of the Nauplius into the young Barnacle. Its organs of touch and of sight atrophy, its legs lose their locomotor function, and are simply used for bringing floating particles to the orifice of the stomach; so that an eminent naturalist has compared one of these animals to a man standing on his head and kicking his food into his mouth.
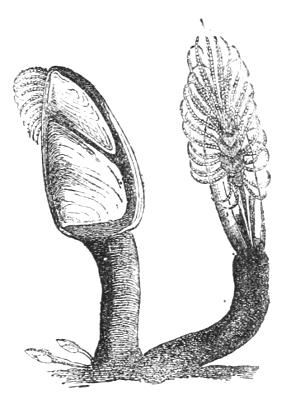
Fig. 9.—Adult Barnacle or Lepas (one of the Cirrhipedes). Natural size.
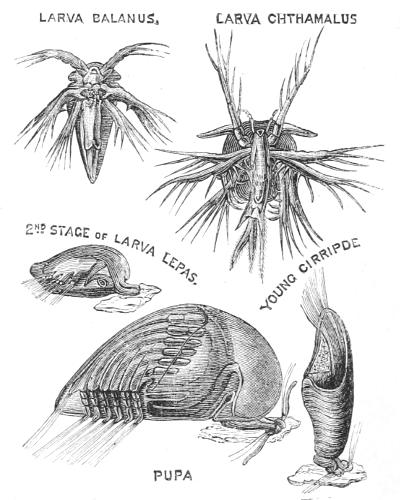
Fig. 10.—Development of Cirrhipedes (Barnacle and Sea-acorn). After Huxley.
Were it not for the recapitulative phases in the development of the Barnacle, we may doubt whether naturalists would ever have guessed that it was a degenerate Crustacean. It was in fact for a long time regarded as quite remote from them, and placed among the snails and oysters; its true nature was only admitted when the young form was discovered.
Other parasitic organisms, which exhibit extreme degeneration as compared with their free-living relatives, might be cited and figured in profusion, did our limits permit. Very noteworthy are the degenerate Spiders—the mites, leading to still more degenerate forms, the Linguatulæ.
We have two of these represented in Figs. 11 and 12. The one (Fig. 11), as compared with a spider is seen still to possess the eight walking legs,[38] small, it is true, whilst the palps and daggers of the spider have dwindled to a beak projecting from the front of the globular unjointed body. In the other the eight legs have become mere stumps, and the body is elongated like that of a worm.
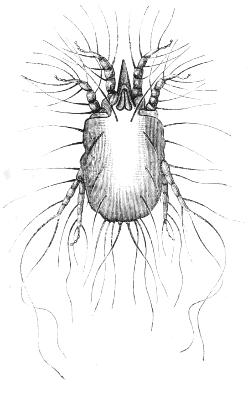
Fig. 11.—Acarus equi. A degenerate Spider or mite parasitic on the skin of the horse.

Fig. 12.—Degenerate Spider (Demodex foliculorum) found in the skin of the human face.
The instances of degeneration which we have so far examined are due to parasitism, except in the example of the Barnacle, where we have an instance of degeneration due to sessile and immobile[39] habit of life. We may now proceed to look at some sessile or immobile animals which are not usually regarded as degenerate, but which, I think, there is every reason to believe are the degenerate descendants of very much higher and more elaborate ancestors. These are certain marine animals, the Ascidians, or sea-squirts. These animals are found encrusting rocks, stones, and weeds on the sea bottom. Sometimes they are solitary (Fig. 13), but many of them produce buds, like plants, and so form compound masses or sheets of individuals all connected and continuous with one another, like the buds on a creeping plant (Fig. 14).
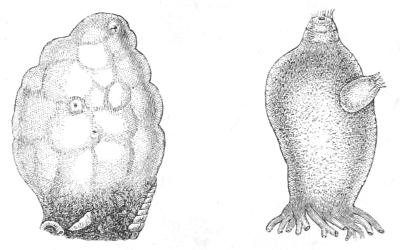
Fig. 13.—Two adult Ascidians: to the left Phallusia—to the right Cynthia: the incurrent and excurrent orifices are seen as two prominences. Half the natural size.
We will examine one of the simple forms—a tough mass like a leather bottle with two openings; water is continually passing in at the one and out at[40] the other of these apertures. If we remove the leathery outer-case (Fig. 15), we find that there is a soft creature within which has the following parts:—Leading from the mouth a great throat, followed by an intestine. The throat is perforated by innumerable slits, through which the water passes into a chamber—the cloaca: in passing, the water aërates the blood which circulates in the framework of the slits. The intestine takes a sharp bend, which causes it to open also into the cloaca. Between the orifice of the mouth and of the cloaca there is a nerve-ganglion.
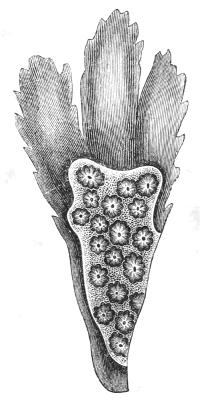
Fig. 14.—A colony of compound Ascidians (Botryllus) growing on a piece of sea-weed (Fucus). Each star corresponds to eight or more conjoined Ascidians. Natural size.
My object in the next place is to show that the structure and life-history of these Ascidians may be best explained on the hypothesis that they are instances of degeneration; that they are the modified descendants of animals of higher, that is more elaborate structure, and in fact are degenerate Vertebrata, standing in the same relation to fishes, frogs, and men, as do the barnacles to shrimps, crabs, and lobsters.
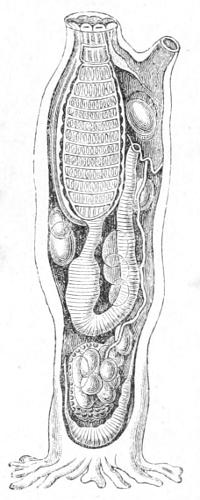
Fig. 15.—Anatomy of an Ascidian (Phallusia). At the top is the mouth, to the right the orifice of the cloaca. In the cloaca lies an egg, and above it the oblong nerve ganglion. The perforated pharynx follows the mouth and leads to the bent intestine which is seen to open into the cloaca. The space around the curved intestine is the body-cavity; in it are seen oval bodies, the eggs, and quite at the lower end the curved heart. The root-like processes at the base serve to fix the Ascidian to stones, shells, or weed.
The young of some, but by no means of all these[42] Ascidians, have a form totally different from that of their parents. The egg of Phallusia gives rise to a tadpole, a drawing of which placed side by side with the somewhat larger tadpole of the common frog is seen in the adjoining figure (Fig. 16). The young Ascidian has the same general shape as the young frog, but not only this; the resemblance extends into details, the internal organs agreeing closely in the two cases. Further still as shown by the beautiful researches of the Russian naturalist, Kowalewsky, the resemblance reaches absolute identity when we examine the way in which the various organs arise from the primitive egg-cell. Tail, body, spiracle, eye, and mouth, agree in the two tadpoles, the only important difference being in the position of the two mouths and in the fact that the Ascidian has one eye while the frog has two.
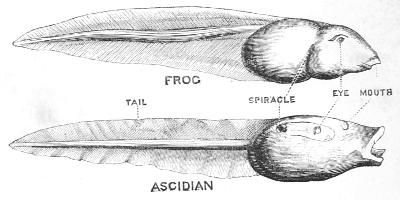
Fig. 16.—Tadpole of Frog and of Ascidian. Surface view.
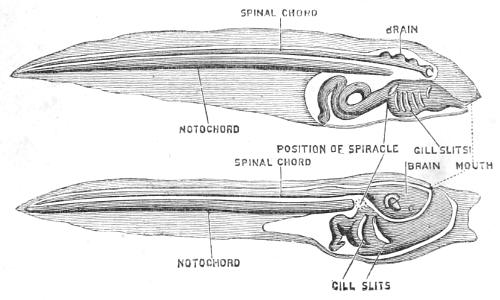
Fig. 17.—Tadpole of Frog and of Ascidian. Diagrams representing the chief internal organs.
Now let us look at the internal organs (Fig. 17). There are four structures, which are all four possessed at some time of their lives by all those animals which we call the Vertebrata, the great branch of the pedigree to which fishes, reptiles, birds, beasts, and men belong. And the combination of these marks or structural peculiarities is an overwhelming piece of evidence in favour of the supposition that the creatures which possess this combination are derived from one common ancestor. Just as one would conclude that a man whom one might meet, say on Salisbury Plain, must belong to the New Zealand race, if it were found not only that he had the colour, and the hair, and the shape of head of a New Zealander, but also that he[44] was tattooed like a New Zealander, carried the weapons of a New Zealander, and, over and above in addition to these proofs, that he talked the Maori language and none other; so here, in the case of the vertebrate race, there are certain qualities and possessions, the accumulation of which cannot be conceived of as occurring in any animal but one belonging to that race. These four great structural features are—first, the primitive backbone or notochord; second, the throat perforated by gill-slits; third, the tubular nerve-centre or spinal cord and brain placed along the back; and lastly, and perhaps most distinctive and clinching as an evidence of affinity, the myelonic or cerebral eye.
Now let us convince ourselves that these four features exist not only in the frog’s tadpole, as they do in all fishes, reptiles, birds, and beasts, but that they also exist in the Ascidian tadpole, and, it may be added, co-exist in no other animals at all.
The corresponding parts are named in Figs. 16 and 17, in such a way as to render their agreement tolerably clear, whilst in Fig. 18 a more detailed representation of the head of an Ascidian tadpole is given.
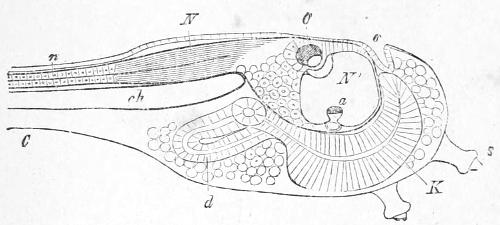
Fig. 18.—Ascidian Tadpole with a part only of the tail C. N, nervous system with the enlarged brain in front and the narrow spinal cord behind n; N´, is placed in the cavity of the brain: O, the single cerebral eye lying in the brain; a, similarly placed auditory organ; K, pharynx; d, intestine; o, rudiment of the mouth; ch, notochord or primitive backbone. (From Gegenbaur’s “Elements of Comparative Anatomy.”)
It is clear then that the Ascidians must be admitted to be Vertebrates, and must be classified in that great sub-kingdom or branch of the animal pedigree. The Ascidian tadpole is very unlike its parent the Ascidian, and has to go through a process of degeneration in order to arrive at the adult structure. The diagrams which are reproduced in Figs. 19 and 20, show how this degeneration proceeds. It will be observed, that in somewhat the same manner as the young barnacle, the young Ascidian fixes itself to a stone by its head: then the tail with its notochord and nerve-chord atrophies. The body grows and gradually changes its shape, whilst the cloacal chamber forms. The brain remains quite small and undeveloped, and the remarkable myelonic eye (the eye in[46] the brain) disappears. The number of gill-slits increases as the animal grows in size and its outer skin becomes tough and leather-like.
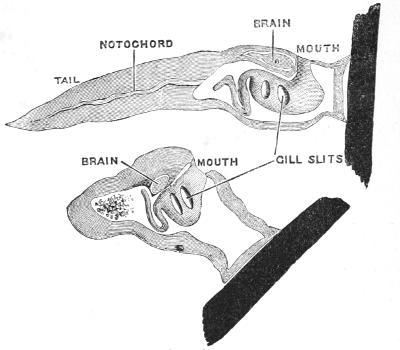
Fig. 19.—Degeneration of Ascidian Tadpole to form the adult. The black pieces represent the rock or stone to which the Tadpole has fixed its head.
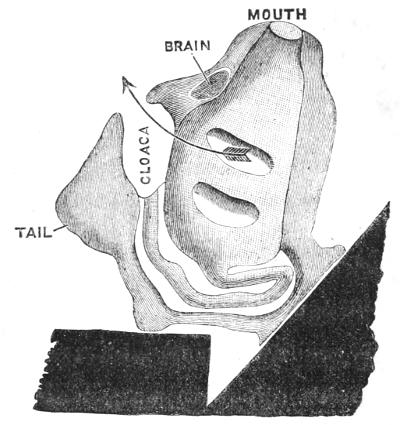
Fig. 20.—Very young Ascidian with only two gill-slits. Compare with Fig 15; which is, however, seen from the other side, so that left there corresponds to right here.
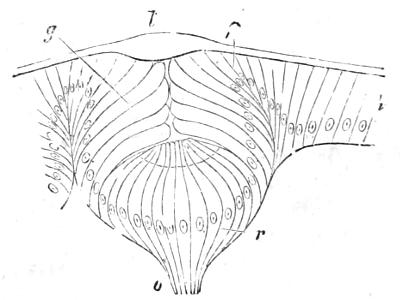
Fig. 21.—Section through the eye (“surface-eye”) of a Water-beetle’s larva. All the cells are seen to be in a row continuous with h, the cells of the outermost skin or ectoderm. p, pigmented cells; r, retinal cells connected at o with the optic nerve; g, transparent cells (forming a kind of “vitreous body”); l, cuticular lens. (From Gegenbaur’s “Elements of Comparative Anatomy,” after Grenacher.)
Before saying anything further on the subject of degeneration, it seems desirable once more to direct attention to the myelonic or cerebral eye which the Ascidian tadpole possesses in common with all Vertebrates. All other animals which have eyes develop the retina or sensitive part of the eye from their outer skin (See Figs. 21 and 22, and explanation.) It is easy to understand that an organ which is to be affected by the light should form on the surface of the body where the light falls. It has long been known as a very puzzling and unaccountable peculiarity of Vertebrates, that the retina or sensitive part of the eye grows out in the embryo as a bud or vesicle of the brain, and thus forms deeply below the surface and away from[48] the light (see Fig. 23, and explanation). The Ascidian tadpole helps us to understand this, for it is perfectly transparent and has its eye actually inside its brain. The light passes through the transparent tissues and acts on the pigmented eye, lying deep in the brain. We are thus led to the conclusion—and I believe this inference to be now for the first time put into so many words—that the original Vertebrate must have been a transparent animal, and had an eye or pair of eyes inside its brain, like that of the Ascidian tadpole. As the tissues of this ancestral Vertebrate grew denser and more opaque, the eye-bearing part of the brain was forced by natural selection to grow outwards towards the surface, in order that it might still be in a position to receive the influence of the sun’s rays. Thus the very peculiar mode of development of the Vertebrate eye from two parts, a brain-vesicle (Fig. 23, A a, and B p r), and a skin-vesicle (Fig. 23, B e, l), is accounted for.
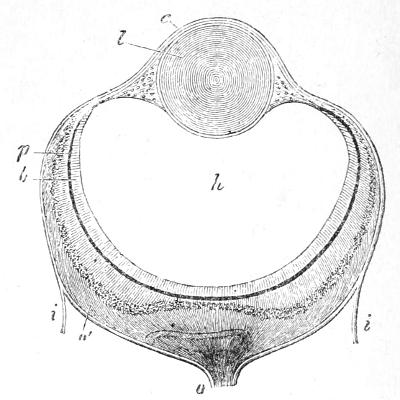
Fig. 22.—Section through the eye (“surface-eye”) of a Marine Worm (Neophanta). i, integument spreading over the front of the eye c; l, cuticular lens; h, cavity occupied by vitreous body; p, retinal cells; b, pigment; o, optic nerve: o´, expansion of optic nerve.
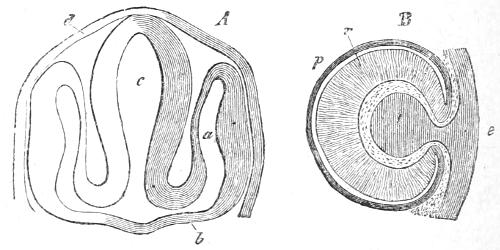
Fig. 23.—A. Vertical section through the head of a very young fish, showing in the centre the cavity of the brain c. On each side is a hollow outgrowth (a) which will form the retina of the fish’s eye (“cerebral eye”); b, will become the optic nerve connecting the brain and the retina; d, integument.—B. Later condition of the hollow outgrowth (a) of A. Its outer wall r is pressed against its deeper wall p by an ingrowth (l) from the outer skin (ectoderm) e; r, gives rise to the retinal cells, whilst only l, the cellular lens, is derived from the surface of the skin.
The cases of degeneration which I have up to this point brought forward, are cases which admit of very little dispute or doubt. They are attested by either the history of the individual development of the organisms in question, as in Sacculina, in the Barnacle, and in the Ascidian, or they are cases[50] where the comparison of the degenerate animal, with others like it in structure, but not degenerate, renders the hypothesis of degeneration an unassailable one. Such cases are the Acarus or mite, and the skin-worm (Demodex).
We have seen that degeneration, or the simplification of the general structure of an animal, may be due to the ancestors of that animal having taken to one of two new habits of life, either the parasitic or the immobile. Other new habits of life appear also to be such as to lead to degeneration. Let us suppose a race of animals fitted and accustomed to catch their food, and having a variety of organs to help them in this chase—suppose such animals suddenly to acquire the power of feeding on the carbonic acid dissolved in the water around them just as green plants do. This would lead to a degeneration; they would cease to hunt their food, and would bask in the sunlight, taking food in by the whole surface, as plants do by their leaves. Certain small flat worms, by name Convoluta, of a bright green colour, appear to be in this condition. Their green colour is known to be the same substance as leaf-green; and Mr. Patrick Geddes has recently shown[51] that by the aid of this green substance they feed on carbonic acid, making starch from it as plants do. As a consequence we find that their stomachs and intestines as well as their locomotive organs become simplified, since they are but little wanted. These vegetating animals, as Mr. Geddes calls them, are the exact complement of the carnivorous plants, and show how a degeneration of animal forms may be caused by vegetative nutrition.
Another possible cause of degeneration appears to be the indirect one of minute size. It cannot be doubted that natural selection has frequently acted on a race of animals so as to reduce the size of the individuals. The smallness of size has been favourable to their survival in the struggle for existence, and in some cases they have been reduced to even microscopic proportions. But this reduction of size has, when carried to an extreme, resulted in the loss or suppression of some of the most important organs of the body. The needs of a very minute creature are limited as compared with those of a large one, and thus we may find heart and blood-vessels, gills and kidneys, besides legs and muscles, lost by the diminutive degenerate descendants of a larger race.[52] That this is a possible course of change all will, I think, admit. It is actually exemplified in Appendicularia—the only adult representative of the Ascidian tadpole—still tadpole-like in form and structure, but curiously degenerate and simplified in its internal organs. This kind of degeneration is also exemplified in the Rotifers, or wheel animalcules, in the minute Crustacean water-fleas (Ostracoda), and in the Moss-polyps, or Polyzoa. Roughly then we may sum up the immediate antecedents of degenerative evolution as, 1, Parasitism; 2, Fixity or immobility; 3, Vegetative nutrition; 4, Excessive reduction of size. This is not a logical enumeration, for each of these causes involves, or may be inseparably connected with, one or more of the others. It will serve for the present as well as a more exhaustive analysis. (See Note C.)
And now we have to note an important fact with regard to the evidence which we can obtain of the occurrence of this process of degeneration. We have seen that the most conclusive evidence is that of the recapitulative development of the individual. The Ascidian Phallusia shows itself to be a degenerate Vertebrate by beginning life as a[53] tadpole. But such recapitulative development is by no means the rule. Quite arbitrarily, we find, it is exhibited in one animal and not in a nearly allied kind. Thus very many animals belonging to the Ascidian group have no tadpole young—just as some tree-frogs have no tadpoles. It is quite possible, and often, more often than not, occurs, that the most important part of the recapitulative phases are absent from the developmental history of an animal. The egg proceeds very rapidly to produce the adult form, and all the wonderful series of changes showing the animal’s ancestry are absolutely and completely omitted; that is to say, all those stages which are of importance for our present purpose. Just as certain bodies pass from the solid to the liquid state at a bound, omitting all intermediate phases of consistence, but giving evidence of “internal work” by the suggestive phenomenon of latent heat—so do these embryos skip long tracts in the historically continuous phases of form, and present to us only the intangible correlative “internal work” in place of the tangible series of embryonic changes of shape.
Now I want to put this case—a supposition—before[54] the reader who has so far followed me in these pages. Suppose, as might well have happened, that the Barnacles, one and all, instead of recapitulating in their early life, were to develop directly from the egg to the adult form, as so many animals do; should we have ever made out that they were degenerate Crustaceans? Possibly we should: their adult structure still bears important marks of affinities with crabs and shrimps; but as a matter of fact before their recapitulative development had been discovered they were classed by the great Cuvier and other naturalists with the Molluscs, the mussels and snails.
Suppose again that all the existing Ascidians, as many of them actually have, had long ago lost their recapitulative history in growth from the egg: suppose that no such a thing as an Ascidian tadpole existed, but that the Ascidian’s egg grew as directly as possible into an Ascidian, in every living species of the group. This might easily be the case. Then most assuredly we should not have the least notion that the Ascidians were degenerate Vertebrates. We should still class them where they used to be classed before the Russian naturalist Kowalewsky discovered the true history and structure of the Ascidian tadpole.[55] I believe that I shall have the assent of every naturalist when I say that the vertebrate character of the Ascidians and the history of their degeneration would never have been suspected, or even dreamed of, had the Ascidian tadpoles ceased to appear in the course of the Ascidian development at a geological period anterior to the present epoch.
This being the case, it must be admitted that it is quite possible—I do not say more than possible—that other groups of animals besides parasites, Barnacles, and Ascidians, are degenerate. It is quite possible that animals with considerable complexity of structure, at least as complex as the Ascidians, may have been produced by degeneration from still more highly-organized ancestors. Any group of animals to which we can turn may possibly be the result of degeneration, and yet offer no evidence of that degeneration in its growth from the egg.
Accordingly, wherever we can note that a group of organisms is characterized by habits likely to lead to degeneration, such as I have enumerated, viz., parasitism or immobility, or certain special modes of nutrition, or again, by minute size of its representatives—there we are justified in applying the[56] hypothesis of degeneration, even in the absence of any confirmatory evidence from embryology. When we so apply this hypothesis we find in not a few cases, in working over the details of the organization of many different animals by the light which it affords—that much becomes clear and assignable to cause which, on the hypothesis either of “balance” or of “elaboration,” is quite hopelessly obscure. As examples of groups of animals which can thus be satisfactorily explained I may cite first of all the Sponges: as only somewhat less degenerate, we have all the Polyps and Coral-animals, also the Starfishes. Amongst the Mollusca—the group of headless bivalves, the oysters, mussels and clams, known as the Lamellibranchs, are, when one once looks at their structure in this light, clearly enough explained as degenerated from a higher type of head-bearing active creatures like the Cuttle-fish; whilst the Polyzoa or Moss-polyps stand in precisely the same kind of relation to the higher Mollusca as do the Ascidians to the higher Vertebrates: they have greatly degenerated, and become minute encrusting organisms which, like some of the Ascidians, build up colonies by plant-like budding growth. The Rotifers, or wheel animalcules,[57] I have already mentioned as best explained by the supposition that they are the descendants of far larger and more fully-organized animals provided with locomotive appendages or limbs: they have dwindled and degenerated to their present minute size and curiously suggestive structure.
Besides these there are other very numerous cases of animal structure which can best be explained by the hypothesis of degeneration. A discussion of these, and a due exposition of the application of the hypothesis of degeneration to the various groups just cited, would involve a complete treatise on comparative anatomy and embryology, and lead far beyond the limitations of this little volume.
All that has been, thus far, here said on the subject of Degeneration is so much zoological specialism, and may appear but a narrow restriction of the discussion to those who are not zoologists. Though we may establish the hypothesis most satisfactorily by the study of animal organization and development, it is abundantly clear that degenerative evolution is by no means limited in its application to the field of zoology. It clearly offers an explanation of many vegetable phenomena, and is already admitted by[58] botanists as the explanation of the curious facts connected with the reproductive process in the higher plants. As a further example of its application in this field, the yeast-plant may be adduced, which is in all probability a degenerate floating form derived from a species of mould (Mucor). In other fields, wherever in fact the great principle of evolution has been recognised, degeneration plays an important part. In tracing the development of languages, philologists have long made use of the hypothesis of degeneration. Under certain conditions, in the mouths and minds of this or that branch of a race, a highly elaborate language has sometimes degenerated and become no longer fit to express complex or subtle conceptions, but only such as are simpler and more obvious. (See Note D.)
The traditional history of mankind furnishes us with notable examples of degeneration. High states of civilisation have decayed and given place to low and degenerate states. At one time it was a favourite doctrine that the savage races of mankind were degenerate descendants of the higher and civilised races. This general and sweeping application of the doctrine of degeneration has been proved to be erroneous by[59] careful study of the habits, arts, and beliefs of savages; at the same time there is no doubt that many savage races as we at present see them are actually degenerate and are descended from ancestors possessed of a relatively elaborate civilisation. As such we may cite some of the Indians of Central America, the modern Egyptians, and even the heirs of the great oriental monarchies of præ-Christian times. Whilst the hypothesis of universal degeneration as an explanation of savage races has been justly discarded, it yet appears that degeneration has a very large share in the explanation of the condition of the most barbarous races, such as the Fuegians, the Bushmen, and even the Australians. They exhibit evidence of being descended from ancestors more cultivated than themselves.
With regard to ourselves, the white races of Europe, the possibility of degeneration seems to be worth some consideration. In accordance with a tacit assumption of universal progress—an unreasoning optimism—we are accustomed to regard ourselves as necessarily progressing, as necessarily having arrived at a higher and more elaborated condition than that which our ancestors reached, and as destined[60] to progress still further. On the other hand, it is well to remember that we are subject to the general laws of evolution, and are as likely to degenerate as to progress. As compared with the immediate forefathers of our civilisation—the ancient Greeks—we do not appear to have improved so far as our bodily structure is concerned, nor assuredly so far as some of our mental capacities are concerned. Our powers of perceiving and expressing beauty of form have certainly not increased since the days of the Parthenon and Aphrodite of Melos. In matters of the reason, in the development of intellect, we may seriously inquire how the case stands. Does the reason of the average man of civilised Europe stand out clearly as an evidence of progress when compared with that of the men of bygone ages? Are all the inventions and figments of human superstition and folly, the self-inflicted torturing of mind, the reiterated substitution of wrong for right, and of falsehood for truth, which disfigure our modern civilisation—are these evidences of progress? In such respects we have at least reason to fear that we may be degenerate. Possibly we are all drifting, tending to the condition of intellectual Barnacles or Ascidians. It is possible[61] for us—just as the Ascidian throws away its tail and its eye and sinks into a quiescent state of inferiority—to reject the good gift of reason with which every child is born, and to degenerate into a contented life of material enjoyment accompanied by ignorance and superstition. The unprejudiced, all-questioning spirit of childhood may not inaptly be compared to the tadpole tail and eye of the young Ascidian: we have to fear lest the prejudices, pre-occupations, and dogmatism of modern civilisation should in any way lead to the atrophy and loss of the valuable mental qualities inherited by our young forms from primæval man.
There is only one means of estimating our position, only one means of so shaping our conduct that we may with certainty avoid degeneration and keep an onward course. We are as a race more fortunate than our ruined cousins—the degenerate Ascidians. For us it is possible to ascertain what will conduce to our higher development, what will favour our degeneration. To us has been given the power to know the causes of things, and by the use of this power it is possible for us to control our destinies. It is for us by ceaseless and ever hopeful labour[62] to try to gain a knowledge of man’s place in the order of nature. When we have gained this fully and minutely, we shall be able by the light of the past to guide ourselves in the future. In proportion as the whole of the past evolution of civilised man, of which we at present perceive the outlines, is assigned to its causes, we and our successors on the globe may expect to be able duly to estimate that which makes for, and that which makes against, the progress of the race. The full and earnest cultivation of Science—the Knowledge of Causes—is that to which we have to look for the protection of our race—even of this English branch of it—from relapse and degeneration.
[1] These pages formed a discourse delivered before the British Association at Sheffield on the evening of August 22nd, 1879, under the presidency of Professor Allman, LL.D., F.R.S.
[2] See Note A.
[3] See Note B.
[4] Der Ursprung der Wirbelthiere und das Princip des Functionswechsels. Leipzig, 1875.
“Die Phantasie ist ein unentbehrliches Gut; denn sie ist es, durch welche neue Combinationen zur Veranlassung wichtiger Entdeckungen gemacht werden. Die Kraft der Unterscheidung des isolirenden Verstandes sowohl, als der erweiternden und zum allgemeinen strebenden Phantasie sind dem Naturforscher in einem harmonischen Wechselwirken nothwendig. Durch Störung dieses Gleichgewichts wird der Naturforscher von der Phantasie zu Träumereien hingerissen, während diese Gabe den talentvollen Naturforscher von hinreichender Verstandesstärke zu den wichtigsten Entdeckungen führt.”—Johannes Müller, Archiv für Anatomie, 1834.
To many persons the conclusion that man is the naturally modified descendant of ape-like ancestors appears to be destructive of the belief in an immortal soul, and in the teachings of Christianity; and accordingly they either reject Darwinism altogether, or claim for man a special exemption from the mode of origin admitted for other animals.
It seems worth while, in order to secure a calm and unprejudiced consideration for the teachings of Darwinism, to point out to such persons that, as a matter of fact, whatever views we may hold with regard to a soul and the Christian doctrines, they cannot be in the smallest degree affected by the admission that man has been derived from ape-like ancestors by a process of natural selection, so long as the demonstrable fact, not denied by any sane person, is admitted, namely, that every individual man grows by a process of natural modification from a homogeneous egg-cell or corpuscle. Assuredly it cannot lower our conception of man’s dignity if we[67] have to regard him as “the flower of all the ages,” bursting from the great stream of life which has flowed on through countless epochs with one increasing purpose, rather than as an isolated, miraculous being, put together abnormally from elemental clay, and cut off by such portentous origin from his fellow animals, and from that gracious Nature to whom he yearns with filial instinct, knowing her, in spite of fables, to be his dear mother.
A certain number of thoughtful persons admit the development of man’s body by natural processes from ape-like ancestry, but believe in the non-natural intervention of a Creator at a certain definite stage in that development, in order to introduce into the animal which was at that moment a man-like ape, something termed “a conscious soul,” in virtue of which he became an ape-like man. It appears to me perfectly legitimate and harmless for individuals to make such an assumption if their particular form of philosophy or of religion requires it. Such an assumption does not in any way traverse the inferences from facts to which Darwinism leads us; at the same time zoological science does not, and cannot be expected to, give any support to such an assumption. The gratuitous and harmless nature of the assumption so far as zoological science is concerned, and accordingly the baselessness of the hostility to Darwinism of those who choose to make it, may be seen by the consideration of a parallel series of facts[68] and assumptions, which puts the matter clearly enough in its true light.
No one ventures to deny, at the present day, that every human being grows from the egg in utero, just as a dog or a monkey does; the facts are before us and can be scrutinised in detail. We may ask of those who refuse to admit the gradual and natural development of man’s consciousness in the ancestral series, passing from ape-like forms into indubitable man, “How do you propose to divide the series presented by every individual man in his growth from the egg? At what particular phase in the embryonic series is the soul with its potential consciousness implanted? Is it in the egg? in the fœtus of this month or of that? in the new-born infant? or at five years of age?” This, it is notorious, is a point upon which Churches have never been able to agree; and it is equally notorious that the unbroken series exists—that the egg becomes the fœtus, the fœtus the child, and the child the man. On the other hand we have the historical series—the series, the existence of which is inferred by Darwin and his adherents. This is a series leading from simple egg-like organisms to ape-like creatures, and from these to man. Will those who cannot answer our previous inquiries undertake to assert dogmatically in the present case at what point in the historical series there is a break or division? At what step are we to be asked to suppose that the order of nature was stopped, and a non-natural[69] soul introduced? The philosopher or theologian of this or that school may arbitrarily draw an imaginary line here or there in either series, and the evolutionist will not raise a finger to stop him. As long as truth in the statement of fact, and logic in the inference from observed fact are respected, there need be no hostility between evolutionist and theologian. The theologian is content in the case of individual development from the egg to admit the facts of individual evolution, and to make assumptions which lie altogether outside the region of scientific inquiry. So, too, it would seem only reasonable that he should deal with the historical series, and frankly accept the natural evolution of man from lower animals, declaring dogmatically, if he so please, but not as an inference of the same order as are the inferences of science, that something called the soul arrived at any point in the series which he may think suitable. At the same time, it would appear to be sufficient, even for the purposes of the theologian, to hold that whatever the two above-mentioned series of living things contain or imply, they do so as the result of a natural and uniform process of development, that there has been one “miracle” once and for all time. It should not be a ground of offence to any school of thinkers, that Darwinism, whilst leaving them free scope, cannot be made actually contributory to the support of their particular tenets.
The difficulties which the theologian has to meet[70] when he is called upon to give some account of the origin and nature of the soul, certainly cannot be said to have been increased by the establishment of the Darwinian theory. For from the earliest days of the Church, ingenious speculation has been lavished on the subject. As to the origin of the individual soul, Tertullian tells us as follows:—De Anima, ch. xix.—“Anima velut surculus quidam ex matrice Adami in propaginem deducta, et genitalibus semine foveis commodata. Pullulabit tam intellectu quam et sensu.”
Whilst St. Augustine says:—“Harum autem sententiarum quatuor de anima, utrum de propagine veniant, an in singulis quibusque nascentibus mox fiant, an in corpora nascentium jam alicubi existentes vel mittantur divinitus, vel sua sponte labantur, nullam temere affirmari oporteret: aut enim nondum ista quæstio a divinorum librorum catholicis tractatoribus, pro merito suæ obscuritatis et perplexitatis, evoluta atque illustrata est; aut si jam factum est, nondum in manus nostras hujuscemodi litteræ provenerunt.”
A very important form of degeneration, not touched on in the text, is that exhibited in the Mexican axolotl, where the larval form of a Salamander develops generative organs, and is arrested in its further progress to the adult parental form. It is not possible to class this with the other phenomena which I have enumerated as Degeneration, since there is no modification of an adult structure, but simple arrest, and retention of the larval structure in all its completeness. I should call the phenomenon exhibited by axolotl “arrest” or “super-larvation” rather than degeneration.
The result of super-larvation is in so far similar to that of those changes to which it is desirable to restrict the term “degeneration,” that it may be classed under “simplificative evolution” as opposed to “elaborative evolution.” That there is a very real difference between super-larvation and degeneration may best be seen by taking a case of each process and instituting a comparison. Axolotl proceeds[72] regularly on its course of development from the egg, but instead of passing from the aquatic gilled condition to the terrestrial gill-less adult form of the Salamander, it remains arrested in the earlier condition, develops its reproductive organs, and propagates itself. There is no loss or atrophy in this case, but simply a dead stop in a progressive course. On the other hand, as we have seen, the Ascidian loses, by a process of atrophy and destruction, a powerful locomotive organ, a highly-developed eye, a relatively large nervous system. The former may be compared to a permanent childishness, the latter to the second childhood, which is really atrophy and decay. It is highly probable that super-larvation has taken place at various epochs and in various groups of the animal kingdom, just as it does in axolotl, and yet we cannot hope for evidence fitted to establish its occurrence in any one case, where it is no longer possible by exceptional conditions to recover (as in the case of axolotl, which can experimentally be made to advance to the Salamander phase by proper treatment), the discarded, more developed adult form. By super-larvation it would be possible for an embryonic form developed in relation to special embryonic conditions and not recapitulative of an ancestry, to become the adult form of the race, and thus to give to the subsequent evolution of that race a totally and otherwise improbable direction.
It seems also exceedingly probable that “super-larvation” does not occur only as in axolotl through premature maturation of the reproductive organs, but the phenomenon may develop itself more slowly by a gradual creeping forward, as it were, of larval features. Just as the adaptations acquired in, and having relation to, later life tend to show themselves in an early period of the development of the individual and out of due season; so do characters acquired by the early embryo, and having relation only to this early period of life tend to remain as permanent structures, and by their invasion to perturb the adult organization. Such perturbation may tend either to simplification or elaboration.
The term (degeneration of language) includes two very distinct things; the one is degeneration of grammatical form, the other degeneration of the language as an instrument of thought. The former is a far commoner phenomenon than the latter, and, in fact, whilst actually degenerating so far as grammatical complexity is concerned, a language may be at the same time becoming more and more serviceable, or more and more perfect as an organ having a particular function. The decay of useless inflexions and the consequent simplification of language may be compared to the specialization of the one toe of the primitively five-toed foot of the horse, whilst the four others which existed in archaic horses are, one by one, atrophied. Taken by itself, this phenomenon may possibly be described as degeneration, but inasmuch as the whole horse is not degenerate but, on the contrary, specialized and elaborated, it is advisable to widely distinguish such local atrophy from general degeneration. In the same way language[75] cannot, in relation to this question, be treated as a thing by itself—it must be regarded as a possession of the human organism, and the simplification of its structure merely means in most cases its more complete adaptation to the requirements of the organism.
True degeneration of language is therefore only found as part and parcel of a more general degeneration of mental activity. To some extent the conclusion that this or that language, as compared with its earlier condition, exhibits evidence of such degeneration, must be matter of taste and open to discussion. For instance, the English of Johnson may be regarded as degenerate when compared with that of Shakspeare. There is less probability of a difference of opinion as to the degeneracy of modern Greek as compared with “classical” Greek; or of some of the modern languages of Hindustan as compared with Sanskrit, and I am informed that the same kind of degeneration is exhibited by modern Irish as compared with old Irish. Degeneration, in the proper sense of the word, so far as it applies to language, would seem to mean simply a decay or diversion of literary taste and of literary production in the race to which such language may be appropriate.
LONDON:
R. CLAY, SONS, AND TAYLOR,
BREAD STREET HILL, E.C.