Plate 1.

LOVE-MAKING.
Frontispiece.
Title: The Courtship of Animals
Author: W. P. Pycraft
Release date: October 18, 2019 [eBook #60517]
Most recently updated: October 17, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/60517
Credits: Produced by Turgut Dincer (This file was produced from
images made available by The Internet Archive)
Hutchinson’s
Nature
Library
THE COURTSHIP OF ANIMALS
BY
W. P. PYCRAFT
OF THE
ZOOLOGICAL DEPARTMENT OF THE BRITISH MUSEUM:
FELLOW OF THE ZOOLOGICAL SOCIETY OF LONDON;
ASSOCIATE OF THE LINNEAN SOCIETY: MEMBER
OF THE ROYAL ANTHROPOLOGICAL INSTITUTE;
MEMBER OF THE BRITISH
ORNITHOLOGISTS’ UNION; HON.
MEMBER OF THE AMERICAN
ORNITHOLOGISTS’ UNION;
ETC., ETC.
Author of “A History of Birds,” “The Natural History Museum,”
“Pads, Paws and Claws,” “The Infancy of Animals,”
etc., etc., etc.
With 40 Plates on art paper
Containing over 80 Illustrations
THIRD EDITION
LONDON
HUTCHINSON & CO.
PATERNOSTER ROW
I DEDICATE THIS VOLUME
TO
H. ELIOT HOWARD
WHOSE OBSERVATIONS OF
THE COURTSHIP OF BIRDS
RECORDED IN HIS “HISTORY
OF THE BRITISH WARBLERS”
CONSTITUTE A BEACON FOR
ALL ENGAGED IN THE STUDY
OF ANIMAL BEHAVIOUR
ix
That “one touch of Nature which makes the whole World kin” is surely nowhere more obvious than in the “Courtship” of Animals. For the “Beasts that Perish,” no less than Man himself, are stirred by the same emotions; the Fever of Love runs as high in them as in ourselves; and its modes of expression are not so different, though they may superficially appear to be so. The nature of these differences and their interpretation, it is the purpose of this book to set forth.
Charles Darwin laid the foundation for the study of this phase of Animal behaviour in his masterly work on the “Descent of Man,” a work which has been much criticized and much misunderstood since Carlyle’s crude abuse of it as the “Gospel of Dirt.” Darwin was the first to show us that the fierce battles, and strange antics, which characterize so many of the “Lower Orders of Creation” under the exaltation of the Sexual emotions are manifestations fraught with tremendous consequences to the race.
The facts which he brought to light, and the discussions to which they have given rise, have, however, unfortunately been too commonly regarded as merely interesting to those who have a liking for Natural History.
This is a most unfortunate mistake. For such facts have a vitally important bearing on the very problems of social well-being which now loom so largely among us. “Reform” is in the air. Its protagonists are busyx amongst us with schemes for our regeneration, among which “Sex-problems” are made to occupy a very conspicuous place. But no good can come of their cogitations so long as they fail to realize the springs of behaviour in this regard. The facts herein set down will, it is hoped, help much towards this end.
My labours in the preparation of these pages have been materially lightened by the help and counsel of many friends. To them I desire now to record my very grateful thanks. More especially am I indebted to my friends Mr. H. Eliot Howard, Professor Lloyd Morgan and Mr. John Cooke.
I must also thank those who have contributed towards the illustrations which enliven these pages. The delightful Frontispiece, and many of the plates scattered through this work, I owe to the generosity of Messrs. Rowland Ward, Limited. The excellent rendering of the Birds of Paradise adapted in part from the work of Mr. G. E. Lodge and the late J. G. Keulemans, and partly drawn from specimens in the British Museum, is the work of Mr. Roland Green. The very difficult, and less fascinating, technical figures I owe to the skill of Mr. Philip Whelpley. The wonderful photographs illustrating the “Display” of the Sun-bittern and the Kagu were taken by my friend Mr. D. Seth-Smith.
Finally I have to thank Mr. Roger Ingpen for the immense amount of trouble which he has taken in seeing these pages through the press.
W. P. Pycraft.
October, 1913.
xi
| CHAPTER | |
| INTRODUCTION | |
The nature of Life and its power of reproduction—The stuff of which Life is made—The Emotions—The simplest living things—Where is neither Birth nor Death yet the Population increases—The First Marriage—The beginning of sex—The two dominating instincts—The conditions of survival—The Oyster’s narrow world—“Fiddling work”—Amorousness—The superior Male—Where Death begins—“Germ-plasm” and what it means—Sex and “Secondary sexual Characters”—Some theories—“Hormones” what are they? |
1 |
| CHAPTER II | |
| “MANKIND IN THE MAKING” | |
The use of the term “Courtship”—Primitive Man and the Foundations of Society—“Amorousness” as a motive force—Polygamy—Our half human ancestors—Standards of Beauty—Disquieting signs |
21 |
| CHAPTER III | |
| MAN’S COUSINS THE APES | |
The Man-like Apes and their mode of Life—Their “Courtships”—Musical Chimpanzees—How the Orang-utan improves his voice—His likeness to Caliban—The truculent visage of the Gorilla—“Ornament” in the lower Apes—The Concerts of the Howler Monkeys |
40 |
| CHAPTER IVxii | |
| AT DAGGERS DRAWN | |
The Birth of Weapons—All Flesh is Grass—Utility and Ornament—The Fever of Love—The “Challenge” of the Deer—What it means-More about “Hormones”—“Hummel” Stags—The Age of Deer—The “Courtship” of the Moose—Types of Antlers—Antlered Females—Fighting Topi—The Lance of the Oryx in the Lion’s Flanks—Happiness and Hartebeestes—Odoriferous Suitors—The Bloody Sweat of the Hippopotamus—The Elephant in Love—Concerning Tusks—Polygamy |
49 |
| CHAPTER V | |
| THE LION AND HIS KIN | |
A Surprising Relationship—The Lion’s Mane—The Sabre-toothed Tiger—Some Theories about Origins—Sea-lions in Love—Some Strange Ornaments—Whales and Weapons |
77 |
| CHAPTER VI | |
| COURTSHIP AMONG BIRDS | |
Generalities—Darwin v. Wallace—The Peacock in his Pride—The “Display” of the Peacock Pheasant—The Splendour of the Argus Pheasant and the Marvel of its Eyes—The Frill of the Amherst Pheasant—Birds of Paradise in the Toils of Love—Inflated Suitors-Ruffs and Reeves—Fearsome Weapons and their Uses—Birds which dance-Musical Birds—The Bird’s Voice-box—The “Lek” of the Capercaillie—Instruments of Percussion—The Curious Performance of the Woodpecker |
92 |
| CHAPTER VII | |
| THE SEXUAL SELECTION THEORY AS APPLIED TO BIRDS | |
Where the Rôle of the Sexes is reversed—Polygamy and how it is brought about—Coloration and Courtship—Instinctive Actions—The Importance of Landed Possessions—The Meaning of “Display”—The Springs of “Behaviour”—A New Light on the Wild-duck—The “Display” of the Great-crested Grebe—Some xiiiNeglected Factors |
134 |
| CHAPTER VIII | |
| SOME “COLD-BLOODED” LOVERS | |
The Courtship of the Crocodile—Amorous Lizards—Horned Chameleons—A Flagellating Terrapin—The Frog that would a-wooing go—Some Musical Frogs—Some marvellous instincts in Newts |
161 |
| CHAPTER IX | |
| LOVE-MAKING AMONG THE FISHES | |
Germinal variations—Fishes and Mate-hunting—Some Remarkable Sexual Differences displayed by the Teeth of “Rays”—The Double-eyed Fish—The Coloration of the Dragonet—Some Curious Facts about Salmon—The Strange Use of the Kidney in the Stickle-back—The Stickle-back and Parental Duties—Siamese Fighting-fish |
175 |
| CHAPTER X | |
| SOME OF THE “LOWER ORDERS” | |
Butterflies and Moths, and the Coloration of their Wings—Female Choice and “Fine Feathers”—When Male Butterflies are Dominant—Sexual Selection among Butterflies—Abortive Experiments—Wallace and the Sexual Selection Theory—The Sense of Smell in Butterflies and Moths—Fragrant Butterflies—Wingless Moths and their Lures to Lovers—Methods of Pairing among Butterflies and Moths—More Experiments |
185 |
| CHAPTER XI | |
| BEETLES THAT “BLUFF” | |
The Coloration, and other Forms of Ornament in Beetles, and the Significance thereof in regard to the Sexual Selection Theory—The Courtship of Grasshoppers and their Kin—The Remarkable Ears of Locusts and Grasshoppers—The Field-cricket and the Katydid as Troubadours—The Wonderful Performances of the Cicadas—The Duels of Long-horned Locusts—Dragon flies—The May-flies’ “Dance of Death”—The Jaws of the Giant Alder-fly and their Strange Use—Some Curious Facts xivabout Stone-flies |
208 |
| CHAPTER XII | |
| SCORPIONS, SPIDERS AND CRABS | |
Musical Lovers among Spiders and Scorpions—Colour among Spiders, and its uses—The Spiders’ Dance of Death—Spiders and Conjugal Bliss—How Pairing is accomplished—Scorpions in Love—Musical Crabs—Quarrelsome Fiddler-crabs—Crabs and Courtship in the Deep Sea-Amazons among Prawns—Brine-shrimps and Water-fleas—“Natural” v. “Sexual” Selection |
236 |
| CHAPTER XIII | |
| SOME STRANGE MARRIAGE-CUSTOMS: AND VIRGIN BIRTHS | |
The Courtship of the Cuttle-fish—The Sumptuous Cradle of the Argonaut—The Love-darts of the Snail—Hermaphrodites and the Dangers of Self-fertilization—Oysters and Beauty—Sex reduced to its Lowest Terms—Parthenogenesis and Virgin Birth—The Story of the Hive-Bee—The Departure of the Queen—The New Queen and her Marriage-flight—The Celebration of the Nuptials and its Surprising Sequel—The Widowed Queen turns Executioner—The Queen as Mother—The Queen’s Daughters—Nursemaids’ Duties—Change of Work—The Drones and their Career—Food and Sex—The Bumble-bee and its Life-story |
265 |
| CHAPTER XIV | |
| PARTHENOGENESIS AND ITS SEQUEL | |
Courtship among the Ants—The Great Renunciation—Maternity carried to Extremes—Where Males are Superfluous—Degenerate Males—Keeping Death at Bay—Where Females are Unknown |
296 |
xv
| Love-making | Frontispiece |
| Facing page | |
| The Gorilla preparing for hostilities | 42 |
| The barometer of maleness—among the Apes | 44 |
| Weapons of offence | 52 |
| Manchurian Wapiti “calling” | 54 |
| Group of Beisa Oryx | 60 |
| Eland Cows | 64 |
| American Bison | 64 |
| Elephants | 70 |
| Head of male Wart-hog | 72 |
| Male and female Babirusa | 72 |
| Somali Zebras | 72 |
| Giraffe | 72 |
| Californian Sea-lions, or Eared Seals | 82 |
| Elephant Seal | 88 |
| Northern Elephant Seal | 88 |
| “The Peacock in his pride” | 96 |
| Peacock Pheasant | 96 |
| Patterns which puzzled Darwin | 98 |
| The “Strutting Turkey” | 100 |
| The display of the Great Bustard | 100 |
| Some of Fortune’s favourites | 104 |
| The love-making of the Prairie Hen | 110 |
Grades of evolution in the syrinx or organ of voice in the males of Surface-feeding and Diving-ducks | 126 |
| Fighting for territory | 140 |
| xviThe display of the Grasshopper Warbler | 142 |
| The display of the Sun-bittern | 142 |
| The Kagu in display | 142 |
| A male-Savi’s Warbler | 152 |
| Another aspect of the Kagu’s “display” | 154 |
| Some strange accompaniments of courtship: | |
| The White-headed Bell-bird | 156 |
| The Umbrella-bird | 156 |
| Skull of the American white-beaked Pelican | 156 |
| Head of a Puffin, showing the moulting of the beak sheath | 156 |
| The Satin Bower-bird and its bower | 158 |
| The “bower” of the Bower-bird | 158 |
| The Bearded Lizard | 166 |
| Bright colours which cannot be attributed to “sexual selection” | 200 |
| Stridulating organs, etc. | 218 |
| Crickets and May-flies | 220 |
| Male Astia displaying before the less brilliant female | 242 |
| Male Icius displaying | 242 |
| Scorpions | 252 |
| Death of the male Scorpion | 254 |
| The female Mantis devouring her mate | 254 |
| The “Fiddler-crab” among mangrove roots | 258 |
| The “Fiddler-crab” | 258 |
| Some remarkable devices | 262 |
| Some remarkable methods of “courtship” | 268 |
THE COURTSHIP OF ANIMALS
The nature of Life and its power of reproduction—The stuff of which Life is made—The Emotions—The simplest living things—Where is neither Birth nor Death yet the Population increases—The First Marriage—The beginning of sex—The two dominating instincts—The conditions of survival—The Oyster’s narrow world—“Fiddling work”—Amorousness—The superior Male—Where Death begins—“Germ-plasm” and what it means—Sex and “Secondary sexual Characters”—Some theories—“Hormones” what are they?
The nature of life is generally regarded as affording a theme which possesses no more than an academic interest: but there is one aspect of this great subject which must attract us all, and that is its power of reproducing itself. Life begets Life, as Love is said to beget Love. The nature of this mysterious power we can only dimly realize, and the forces which underlie its manifestations few even suspect, save perhaps in a vague way. Yet the tree of Knowledge bears no fruit more vitally important to our well-being, than that which will make us “as Gods, knowing good and evil” in all that concerns the processes of reproduction. But curiously enough, this is a forbidden fruit, and those who eat thereof are expected 2to maintain a discreet silence on the subject. These enlightened ones, however, cannot remain altogether dumb. But they speak, in the veiled language, of Art and Poetry, Literature and the Drama. They talk round the subject rather than of it. Love, Hate, Jealousy, and Envy, are but attributes thereof. We profess to believe that “Knowledge is Power” and to desire to increase its force among us by raising the standard of our system of education. But education which does not, of set purpose, reveal the sources of our being and of our emotions, good and evil, is no more than a travesty of education; and they who seek to foist upon the community Knowledge thus emasculated, are unworthy to wield the power which has been placed in their hands. If social well-being be the aim of the high-priests of Education, then something more than copybook maxims like “Be good and you will be happy” must henceforth be preached. Of what avail is it to exalt the name of Knowledge, while the straightest road thereto is barred across and marked “No thoroughfare!” These blind leaders of the blind seem to imagine that the social well-being they profess to desire can only be attained by side roads, leading anywhere, save in the direction of this Pool of Siloam.
The stuff of which living things are made is called “Protoplasm.” Text-books of Physiology give its chemical constituents with fearsome accuracy, and each of these constituents can be isolated in the laboratory, but “all the king’s horses and all the king’s men” cannot build these up again into living matter. Its consistent inconsistency defies us; every statement we make of it has to be qualified by reservations and saving clauses. 3Its permanency is attested by the fact that it has endured through millions of years, yet we are daily reminded of its evanescent nature. Its power of reproducing itself according to type, none can doubt, yet no two individuals are exactly alike.
The purely physical phenomena of life, to be rightly appreciated, must always be considered in relation to the psychical phenomena which are the soul of life. These subtle and intangible forces cannot be experimented with in the laboratory, or expressed in formulæ; we cannot denote their strength in horse-power. Just as the physical manifestations of life begin with lowly types, so the psychical begin, and they gather strength and complexity with the bodies they pervade. These manifestations we call behaviour, and in their more intense developments, “emotions.”
These emotions present an infinite range of variety in the higher animals, and they attain their maximum of intensity wherever the reproductive activities are concerned. The part which these activities play in controlling behaviour is by no means always apparent, and is commonly not even suspected. Even man himself is subject to this control. And it is this fact which lifts the “Courtship” of the lower animals out of the category of merely curious phenomena. For the springs of his conduct, his behaviour and “emotions” under varying circumstances, can only be understood, and even then but imperfectly, by comparison with other creatures lower in the scale, so far, of course, as comparison is possible.
This line of inquiry, then, takes one back to the simplest living things, among which there is neither marrying nor giving in marriage, neither birth nor death. Life is reduced to its simplest terms—a speck of animated jelly is all that confronts one, and this is only to be seen under a 4high power of the microscope. It has neither mouth nor organs of digestion, no visible means of locomotion are traceable, and the special senses of sight and hearing are wanting; but taste and smell, of a nebulous kind, are there. Shape it cannot be said to have, for its bodily outline is constantly changing, thereby it moves. A long tongue of its jelly-like substance, or “protoplasm” as it is called, is thrust forwards, and the rest of the body is, as it were, dragged after it. Whatever animal, or vegetable, matter it passes over, in the course of its wanderings, is drawn up into the semi-fluid substance of this diaphanous body, and its juices extracted, the undigestible residue is left behind in the course of the morning’s walk! In due time it becomes adult; further growth is impossible. When this stage is attained a strange thing happens. A certain minute, more solid portion of this body, which lies in the very centre of the mass and is known as the “nucleus,” begins to assume an hour-glass shape. Speedily the constriction becomes apparent across the whole body and rapidly increasing, cuts it in two, as if by the tightening of some invisible thread. Here Death is cheated, and records of births are unknown! And just as there are no parents so there are no children. But a foreshadowing of what is to be occurs even here. For every now and then two individuals, to all appearances identical, meet and promptly begin to merge the one into the other till they twain become one flesh in very truth. Here is the most primitive form of marriage in Nature. And here, in this union, or fusion, of separate entities of Germ-plasm, we have the beginning of sex. Such unions are common among these primeval forms of life. In many cases this “marriage” takes place 5between two particles of Protoplasm of which one is rather larger than the other. In such case the smaller is regarded as male, the larger as female. Here we have the first sign of “sexual differentiation” or the evolution of “male” and “female” individuals.
Some such union, some such process of “rejuvenation” by the importation of “fresh blood” seems to be imperative for the continuance of existence throughout the whole animal world, even though it may take place at rare intervals of time. Why should this be? Is this strange meeting and commingling a matter of chance, or is the one seeking the other possessed by a ravenous mate-hunger?
As we ascend higher in the scale it becomes apparent that life has gathered force. That primitive speck of jelly, the Amœba, with which we started, gave but two signs of animation—the power of movement, and hunger. Whether these responses to internal stimuli can be called instinctive is open to argument. But there can be no question about the instinctive nature of the behaviour of these higher animals. After the instinct to feed the two most powerful are the desire for self-preservation—the avoidance of danger—and the desire to mate. These two are the dominating instincts throughout the rest of the animal world, not even excepting man himself.
The tremendous power of “mate-hunger” has been overlooked by a strange confusion between cause and effect. Almost universally its sequel, the production of offspring, has been regarded as the dominant instinct in the higher animals. This view has no foundation in fact. “Desire” for the sake of the pleasure it affords, and not its consequences, is the only hold on life which any race possesses. And this is true both in the case of man 6himself and of the beasts that perish. Wherever this instinct becomes weak, or defective, extinction speedily and inevitably follows. This “Amorousness” is the motive power of “Courtship” wherever it is met with; manifesting itself in the eccentric, and often grotesque posturings, or in the loud and often musical cries which constitute the study of courtship. Intensity of desire is indispensable to survival.
Only the lowly and sedentary types, of which the Oyster may be taken as an example, lack this fire; and here because it is unnecessary. For the reproductive germs of this animal are discharged into the water, to take their chance of attaining their object. They are liberated unconsciously, discharged like the undigested residue of the food, without effort, and without cognizance of the act. This must be so, for the Oyster merely lives—vegetates. Sightless, and without power of movement, after its larval wanderings are over, it lives merely to eat. And even in this, choice is denied it. The currents of water mechanically brought to afford the necessary oxygen for the maintenance of life, bring with them the food which is to restore the slowly wasting tissues. To such a creature there can be no “outer-world,” no consciousness of the existence of individuality other than its own.
The desire for sexual intercourse is met with only where the co-operation of two individuals is necessary to ensure the production of offspring. Such individuals being free to roam, must have some incentive to seek one another at the time when their germ-cells have attained maturity. And this incentive is furnished by the glands in which these elements are produced: supplemented by the secretions of certain ancillary glands. These stimulating juices, 7known as the “Hormones,” will be presently described.
But if we owe our existence to the gratification of what may be called our lower instincts, it is no less certain that all that is best in us we owe to our offspring. We meet with the beginnings of altruism, which the begetting of offspring entails, far down in the animal kingdom, and it attains to its full perfection in the human race. Here only, in its best and truest sense, Love begins: though affection may be found, and in a high degree, in many of the lower animals.
Living things are as clay in the hands of the potter. But it is as if they made themselves, for the designer and the guiding hand are alike invisible. No vessel is exactly like its neighbour, either in the quality of its substance or in the details of its construction. And this because the clay of which it is made possesses that mysterious property we call life. A property which endows each new feature as it appears, with an individuality of its own, whose survival, or suppression, depends entirely on its relationship to surrounding parts; on its harmony with its environment, in short. Colour, size, shape, temperament, behaviour, may each be regarded as so many entities depending for survival on whether or not they can exist in harmony with their environment—the several parts which make up what we call the individual.
In like manner the individual—the complex bundle of parts and qualities—must attain, and maintain, a certain harmony with its environment—the outer world. The process of change, both in quality and quantity, which is for ever going on among the several parts of every separate individual, brings about the elimination of unfavourable variations; and “selects” those which 8vary in the right direction: that is to say, which serve to maintain a place in the sun for the individual in which these momentous changes are going on. But it is not enough that the individual should be in “working order”; it must be in harmony with all the conditions on which existence depends. And the standard of this harmony is set by that very exacting arbiter of life and death, “Natural Selection.” It is not enough that the instincts in regard to this or that habit should be keen, or that this or that particular organ of the body should be efficient—a certain minimum, all-round, standard of efficiency is demanded, or elimination follows. It is through this instability of “temperament,” this tendency to vary in infinite directions, that the balance between the individual and the environment is maintained. Evolution follows the line of least resistance.
The little boy who remarked that it must be “fiddling work, making flies,” was more sage than he knew. The complex web of factors which even a fly represents are beyond the grasp of human understanding. But it is clear that the reproductive instincts, and the emotions they beget, have played, and play, a tremendous part in the evolution of the higher animals.
Those whose business it is, for one reason or another, to study these emotions know well that “mate-hunger” may be as ravenous as food-hunger, and that, exceptions apart, it is immensely more insistent in the males than in the females. But for this, reproduction in many species could not take place: for the sexes often live far apart, and mates are only to be won after desperate conflict with powerful rivals no less inflamed. Thus it is idle to speak of an equality between the sexes in this matter, in regard to the human race. Dogmatism, and the frequent 9repetition of pretty platitudes, will not alter what Nature has ordained. The failure to realize this is painfully obvious in the utterances of many who speak in the name of the newly-founded “Eugenics” society, which seeks the means to ensure the well-being of the race by the spread of a more intimate knowledge of this all-important subject. The existence of what Mr. Heape has recently called a “sex-antagonism” is beyond dispute, for the instincts of the male and female are fundamentally different. The male is dominated by the desire to gratify the sexual appetite; in the female this is counteracted by the stimulation of other instincts concerned with the cares of offspring.
Amorousness, then, is the dominant feature of the males among all animals: and this sex presents yet another characteristic which is to be borne in mind. In all that concerns the evolution of ornamental characters the male leads. In him we can trace the trend which evolution is taking; the female and young afford us the measure of the advance along the new line which has been taken. Why this should be is inexplicable. But sooner or later the females assume, or will assume, all the features originally possessed by their lords; and finally the young also follow suit. That is to say, the females and young tend to retain the ancestral characters. In the course of time the ability to develop new features by the male loses its impetus, and not till then, apparently, do the females, and still later, the young, begin to share his glory. These remarkable features are strikingly illustrated among the birds, as these pages will show.
Nature is nothing, if not perverse. And hence it happens that there are many exceptions to every rule which one formulates. Among the birds, for example, 10there are species wherein the rule that the female follows her mate in the acquisition of new characters is, so to speak, set aside. She follows a line of her own. This is true, at any rate, of superficial characters, such as coloration. By some curious change in her “metabolism,” as the conversion into living tissue of the substances taken as food is called, this coloration may attain a brilliance in no way inferior to that of the male, but strikingly different. The beautiful Orange Fruit-pigeon (Chrysoenas victor) furnishes a case in point, the male being of a gorgeous orange-yellow, the female of a no less vivid green. But the differences are not so great as they appear at first sight. For the male was originally green, and the female has thus but intensified the ancestral livery. Green, it should be remarked, of a more or less olive shade, always precedes yellow in development; and yellow may yield to red, but this order is never reversed. A no less striking case is that of the Upland Goose (Cloephaga magellanica), the male of which is pure white, while the female wears a livery of chestnut and brown. But so sharply are the colours defined that it would be difficult to say that one was of a higher order of coloration than the other. To what causes or factors are these departures due?
Reproduction in the simplest living things takes place by a simple division of the body into two as soon as its maximum size or adult condition has been attained. In such simple types the body consists only of a single “blob,” or particle, of jelly. But a new era began when large numbers of such particles, or “cells,” began to form coherent masses, different parts of the mass performing different work for the mutual benefit of the community. Some have come to form what we call the 11body, which is born, and in due course dies. Others are alone concerned with the task of reproduction. They are nourished by the body, and on attaining maturity, give rise to new bodies. These reproductive cells are excessively small. The male, or “sperm” cell, can only be distinguished under the highest powers of the microscope. The female cell, or “Ovum,” is always larger than the male, because, in addition to the germinal matter which it contains, it is furnished with a store of food in the shape of yolk. This accounts for the relatively enormous size of the egg of the hen. Within the hardened shell the germ develops into the chick, deriving food for its growth from this generous store. Where this yolk is limited in quantity the growing body is hastily fashioned, and launched forth into the world in the form of a “larva,” when it must forage for itself till it has attained its adult form. Or it is retained within the body of the mother until development is complete.
The reproductive cells are the bearers of the Germ-plasm, the stuff of which man and the beasts of the field alike are fashioned. Only a portion of this germ-plasm gives rise to a new body; the rest is, as it were, held over and stored within the new body to give rise to another in due course. That which produces the body we call the “Somatoplasm,” because it is the “plasm” or stuff of which the “Soma,” or body, is made. As to the nature of this Germ-plasm and its mysterious properties, a wide divergency of opinion exists among savants. But the views which find most favour to-day are those of the veteran Professor August Weissmann, as set forth in his work on the “Germ-Plasm, a Theory of Heredity.”
The excessively minute quantity of this germ-plasm 12which suffices to form a new body is incredible. By what miracle of miracles is the essence of a man distilled? His body arises from the union or commingling of two particles of living matter so minute as to be invisible to the naked eye. One of these particles is the “sperm”—cell furnished by the male parent; the other, the “ovum,” furnished by the mother. True the ovum may measure as much as the one-hundred and fiftieth part of an inch, but the bulk of this is yolk-food necessary to furnish the tender germ with life and energy till it shall have attached itself to the walls of the womb, whence all its future nourishment is derived.
By no process of analysis known to us could the germ-plasm of man be distinguished from that of, say, a jelly-fish; and in the matter of quantity there is no more difference. Yet, identical to our senses, in potentiality how amazingly different are these two particles of jelly! In the lowliest animals, such as jelly-fish, one cannot distinguish male and female at sight. The appearance of separate male and female individuals begins somewhat high in the scale marking an epoch in the history of animal life. For the birth of sex inaugurated not merely individuals producing distinctive “male” and “female” germs, but individuals which, by virtue of their sex, developed differences of behaviour and mentality which were to be followed by tremendous consequences. Certain aspects of this behaviour are to furnish the theme of these pages; others, and no less important, those who will may discover in Professor Arthur Thomson’s “Evolution of Sex.”
We are far, indeed, from being able to explain the attributes of sex. At most, we can but endeavour to interpret the behaviour associated therewith. This was 13the task which Darwin set himself to achieve in his theory of sexual selection. He was influenced in the train of thought which he followed up with such brilliant success by what he had observed in the behaviour of highly-ornamented species, such as the Peacock and the Birds of Paradise. The strange antics of these birds when under the influence of sexual excitement persuaded him that they were at least dimly conscious of their splendour, and of its power to fascinate. The female, on the other hand, was supposed to be coy, and to bestow her person on the finest performer. In this way the dullest birds and the poorest performers were gradually eliminated. Here, indeed, was sexual selection. The frills thus begotten he called “Secondary Sexual Characters,” a term which is also used, and was used, by him, to include any feature whereby the sexes can be distinguished apart from the character of the genital organs.
Horns, tusks, and spurs are other forms of secondary sexual characters. And these stand for another form of sexual selection—that of selection by battle. Herein victory falls to the strongest and most pugnacious male who, as the spoils of victory, annexes the females which formed the subject of the duel. This theory, which must be discussed at greater length in the course of these pages, has had many critics, and among them men of mark. But whatever modifications may be deemed necessary, they will be such as are demanded by the results of later discoveries rather than to the force and subtlety of the arguments of his opponents.
One of the most formidable of the opponents of the Sexual Selection theory was Wallace. But his arguments were far from convincing, and often inconsistent. He attributed the more frequent occurrence in male animals 14of brilliant coloration and exaggerations of growth such as give rise to manes, beards, long plumes, and so on, to a “surplus of strength, vitality and growth-power which is able to expend itself in this way without injury,” or, as he sometimes expresses it, to superabundant vitality. He was evidently striving to find words for the faith that was in him, and he was nearer the truth than he knew or than his critics supposed. He was seeking facts which only the physiologist could furnish. And these made their appearance long years after with Professor Starling’s discovery of Hormones. We are far from understanding the origin of these mysterious juices which must be so frequently alluded to in these pages, but they are evidently intimately associated with the expenditure of energy. This may sometimes find an outlet in increased stature, sometimes in pure luxuries of growth. The force of Wallace’s arguments was crushed out by the weight of detail they were made to bear.
Mr. J. T. Cunningham a few years ago entered the lists and failed to achieve his purpose no less completely. His was a theory which assumed too much. In the first place it was based on the transmissibility of acquired characters, of the truth of which there is at present no evidence.
He contends, for example, that the vivid hues of scarlet, blue, yellow and violet which colour the naked skin of the neck of the cassowaries and of both sexes, and the curious horny casque which surmounts the head, are the outcome of the constant laceration of the skin inflicted by the males during their conflicts for the possession of the females. He assumes that such conflicts take place, and he assumes that such “acquired characters” are transmitted. Now, as a matter of fact, these birds do 15not fight with their beaks, but with their feet. And to this end the claw of the inner toe is enlarged to form a great spur. But there is no evidence that the skin of the neck is ever damaged in such conflicts as they may engage in. No scars are ever found, at any rate, to lend support to this theory. The casque, which is similarly supposed to be a mark of honourable conflict, is an “ornament” of great frailty, for it is composed of a delicate filigree-work of bone covered with a thin sheath of horn. In like manner, the long plumes which surmount the heads of birds like the Peacock, and many Birds of Paradise, and the wattle which surmounts the beak of the Turkey, are supposed to have had their origin in similar pugilistic encounters in the past. Mr. Cunningham is surely pushing the theory of the transmission of acquired characters a little far. For what has been transmitted in these cases is not a number of scarred surfaces, but a series of hypertrophied structures. An amazing array of ornamental characters, symmetrically disposed, and often vividly coloured, in short, has been produced from lacerated tissues which in kind and extent can have varied but little.
Evidence has been accumulating during the last few years which would have rejoiced the heart of Darwin. Had he known that birds of sober hues “display” with the same animation and with as much elaboration of posture as the Peacock and the Pheasant, his theory of “Sexual Selection “would probably have left little for those who came after him to criticize. Since his time it has been discovered that both permanent and recurrent secondary sexual characters, such as the antlers of deer and the temporary nuptial plumage of birds, such as the Ruff for example, are controlled as to their growth by 16the stimulating action of the “secretions or juices formed by certain of the ductless glands “; that is to say, of glands having no apparent connection with their surrounding tissues. We owe much of our knowledge of this subject to Professor Starling, who has called these secretions “Hormones.”
Darwin knew that the essential sexual glands, the testes and the ovaries, in some mysterious way controlled, in a large degree, the development of these “hall-marks” of sex, for it was known in his time that castrated stags failed to produce antlers, and that hen pheasants, for example, in extreme old age, or when the ovaries were damaged by disease or injury, at once assumed the plumage of the cock; but the part played by these ductless glands was quite unsuspected. They are the Thyroid, and the Thymus glands, which are attached to the outer walls of the trachea or windpipe. The Pituitary body, which forms part of the brain, and the Suprarenal bodies, attached to the kidneys. It would be foreign to the purpose of these pages to enter into the functions of these glands; suffice it to say, that the juices formed therein are taken up by the blood, and distributed over the system. Their action is only very imperfectly understood. We know that any derangement in their efficiency results in disease, and that they play a very important part in the reproductive system, as will become abundantly evident in the course of these pages. Much hitherto attributed to the action of “Sexual Selection” alone, it is now evident is largely due to their action.
The all-sufficiency of the “Sexual Selection” theory to account for the development of armature, such as horns, antlers, and the huge spine-like outgrowths 17which form so conspicuous a feature of many of the extinct Land-dragons, or Dinosaurs, has been by no means universally accepted. Some authorities like Dr. A. Smith Woodward and Professor Osborne interpret these after another fashion. They hold that these are the “expression points” of inherent growth forces, a process of concentration marking the final stages of evolution prior to extinction. From which it may be inferred that there is a term to the life of a species as there is to the life of the individual. In many cases it is suggested the very exuberance of growth has been the exterminating factor, as in the case of the huge antlers of the Irish “elk,” whose enormous weapons hampered his endeavours to escape his enemies. This is the theory of “Orthogenesis,” or direct development. According to this, new structures, arising in the germ-plasm as “variations,” will of their own inherent vitality go on increasing in each generation unless, and until, checked by “Natural Selection.” Changes in the character of the “Hormones” might very well bring about these excesses of growth. It is well known that the exuberance of growth which produces giants among the human race is due to a derangement of the secretions or hormones of the pituitary body which largely control growth.
Another factor of Sexual Selection which is commonly ignored, but which is of profound importance, is to be found in the part played by the emotions in regard to sexual relationships; the part which the “mind” has played, and plays, in the mating of animals, at any rate of the higher types.
Darwin touched but lightly on this theme. Later writers have almost entirely ignored it. Almost all that is worth knowing on the subject we owe to Professor 18Lloyd Morgan, who was one of the first to take up this difficult line of investigation, and to Professor Groos. Their researches have shown that there can be no doubt but that the emotions have played and are playing an important part in the phenomena we are striving to analyse. Sexual selection, in short, is concerned not merely with the evolution of the physical characters of the body, but also, and no less, with the psychological attributes thereof. Many new and extremely valuable facts in this regard have been brought to light by Mr. H. Eliot Howard in the course of his remarkable studies on our native warblers. Not until the psychology of sex in the lower orders of creation has been further investigated shall we have a properly balanced account of the part played by sexual selection in the scheme of evolution.
By now it will have become apparent that the study of the “Courtship” of animals is one of alluring interest and full of pitfalls for the unwary. And this because of the apparent difficulty in drawing any hard-and-fast line between the part played by “Natural” and the part played by “Sexual” Selection, at any rate in some cases.
To this aspect of the theme Professor Lloyd Morgan has drawn particular attention. “It is difficult,” he remarks, to accept the view that individual choice has played no part where the sexual instincts are concerned. But supposing that it has played its part ... the effects will be wrought into the congenital tissue of the race if, and only if, there are certain individuals which, through failure to elicit the pairing response, die unmated. Is preferential mating, supposing it to occur, carried to such a degree that some individuals 19fail to secure a mate? That is the question. If so, sexual selection is a factor in race progress; if not, though it may occur in nature, it is inoperative as a means of evolutionary development. The whole question, in itself a difficult one, is further complicated by the fact that the males which are possessed of the most exuberant vitality, and are therefore by hypothesis rendered the most acceptable through emotional suggestion, are likely to compete with other males of less exuberant vitality by direct combat. Such competition, by which the weakest are excluded from mating through no choice on the part of the female, falls under the head of natural selection, and not of sexual selection, if by that term we understand preferential mating.
“This serves to bring out the difference ... between natural selection through elimination and conscious selection through choice.... Sexual selection by preferential mating begins by selecting the most successful in stimulating the pairing instinct.... The process is determined by conscious choice. It is in and through such choice that consciousness has been a factor in evolution.”
Herein Lloyd Morgan, like Darwin, recognizes the existence of a dual machinery in determining survival, where this depends on the co-operation of two individuals leading separate existences—Natural, and Sexual, Selection—sometimes the one and sometimes the other prevailing. In the former, the females are seized by force; in the latter, won by displays.
But is this really so? In these pages it is contended that a sharp line must be drawn between all those attributes and characters which are necessary to achieve individual survival, the survival of the Ego, and all those which, 20on the other hand, are necessary to achieve reproduction and the survival of the race. The former are governed, or determined, by Natural, the latter by Sexual, Selection.
The sphere of influence of these two factors may be delimited, if we regard natural selection as the factor accountable only for the qualities necessary for the survival of the individual—necessary to ensure success in the struggle for existence. Then it will become apparent that the qualities and attributes necessary to achieve the survival of the race are of a different kind, and these are the factors which are embraced under the term “Sexual Selection.”
It is a mistake to regard animals in relation to the selection theory as if they were so many tailors’ “mannikins.” Yet a large number of the critics of the selection theory seem to fall into this error, ignoring all but the most superficial characters.
The peculiarities of colour, structure and behaviour, that is to say, the characters and qualities which distinguish the individuals of any given race, are due to inherent qualities of the germ-plasm. Each of such qualities, therefore, may be regarded as entities. Selection determines their survival. Intracellular selection is the first sieve through which they have to pass, natural and sexual selection are others, as circumstances may determine.
As a rule the sex of an individual is attested by more or less conspicuous external features. These are known as the “Secondary Sexual Characters.” But no hard-and-fast line can be established for these, at any rate, so far as colour and ornament are concerned, for such, as will become apparent in the course of these pages, tend to appear first in the male, and then, later, to be acquired by the female, until in many cases the two sexes become again indistinguishable.
The use of the term “Courtship”—Primitive Man and the Foundations of Society—“Amorousness” as a motive force—Polygamy—Our half human ancestors—Standards of Beauty—Disquieting signs.
Our ideas on the subject of the “Courtship” of animals are of necessity largely framed on what has been observed by each of us in regard to our own race; and without any very careful analysis of motives, or thought of what lies behind. But no real insight into this most tremendous subject can be gained which does not strive to penetrate beyond what is actually seen; which does not endeavour to get at the source of conduct in this regard.
“Courtship” is the word we commonly employ to describe the act of wooing; and in civilized human society at any rate, the intensity of the emotions which inspire the desire to woo are held in restraint by a variety of causes—and hence the “Courtship.” In the lower animals it is a moot point whether the term “Courtship” can be accurately applied. They are governed by no conventions, for them there is neither modesty nor immodesty. Desire with them is not made to walk 22delicately, veiled according to custom; nor is it artificially fostered as among civilized communities by stimulating food and the crowding together of large numbers of both sexes in artificial surroundings. Rather it is a natural, rhythmical, highly emotional state, which gathering force inhibits the ordinary emotions, or, rather, overrides them, begetting an intensity of passion which brooks no control. It demands, without parleying, or mincing matters, what is really the object of courtship among the civilized human communities—the consummation of the nuptial ceremony. The term “Courtship” is a Euphemism. Nevertheless, bearing this in mind, it may conveniently be used in these pages.
We cannot hope to understand the springs of courtship in the human race from what we observe in present-day society, or even from what we have gleaned thereon from the records of remote ages. We must get back, so far as is possible, to the very dawn of the human race: to that period of man’s evolution when his conduct was controlled by purely savage instincts. But even then the mark of the beast must have been fading out. His most valuable asset, his larger brain, even then gave him an advantage over the Apes, his near relations, and over the beasts of the field which he had begun to bring into subjection. We may assume that like his anthropoid relations, he was of a solitary, nomadic disposition, wandering in small parties from place to place as fancy or food determined. His advance to this stage started when, by the activity of his enlarging brain, he began to be oppressed by the gloom of the forest, and drawn by the fascination of more open country, and the ever-varying scenes which exploration brought him. But this life 23begot new needs and new desires. Hitherto, hunger, self-preservation and self-perpetuation were the only stimulants which roused his activities; and they were also the three forces, and powerful forces, which shaped his love of solitude. The proximity of his fellows threatened his three most vulnerable points—they competed for his food, they endangered his life, and threatened the possession of his family.
This more varied and adventurous existence roused new centres of activity in his brain; he began to perceive, though dimly, the possibilities of a larger life, though doubtless one which would minister to his own comfort rather than to that of his family—the natural and only road to better things. He began to devise more expeditious means of securing food, and circumventing his enemies, among whom the most formidable was his fellow-man, because in him he met his match. In the course of his wanderings he had learned the use of stones as weapons—which he could never have done in the forests—and he had also discovered the value of his family as ministers to his comfort, if only by setting them to collect such food as did not require strength and cunning in its capture. An inherent love of the chase for the sake of the excitement which this afforded probably made him nothing loth to regard hunting as his own peculiar duty. A little later the advantages of neighbourliness were borne in on him, largely for the sake of the greater ease wherewith the animals of the chase could be captured by their combined efforts; but this begat comradeship and some of the graces which follow therefrom.
Thus was laid the foundation of Society and “civilization” with all its attendant barbarities. Then, as now, 24whatever discordant notes were heard, were those struck by the twin Demons Envy and Jealousy. These disturbers of the peace are parasites on Society, their very existence depends on it. They have played a larger part in fashioning its rules and regulations than is generally realized. Their influence is as powerful to-day as ever in the past. It expresses itself in varying degrees in different individuals, and is roused by varying causes. But the most potent of all is jealousy in regard to sexual matters.
Amorousness, a word with a deep meaning, was, and is, the underlying factor which shaped, and is sustaining, human society; and is no less powerful among the beasts that perish. The motive force in this has not been the desire for offspring, but for the satisfaction of the elemental animal passion, the gratification of the purely sexual emotions which at their height are irresistible. There may be some who will see in this contention a degrading aspect of life. But this view will obtain only among those who prefer the man-made sophistries of life to its Divine mysteries. This dominance of what are popularly called the animal passions is the outcome of a perfectly natural process, whereby those in which these passions were defective died without offspring, while those who tended to excess were similarly eliminated. The desire for offspring for its own sake may exist among our own species to-day but, normally, offspring follows as an effect not as a cause. Many of our social problems would straighten themselves out if these facts were once faced and acknowledged; we are apt to concern ourselves with what should be—according to our ideals—rather than what is. Let it be granted that this rendering is true, and much else that mystifies becomes 25clear.
Whether primitive man was monogamous or polygamous, or whether he practised promiscuity, are themes which have exercised the minds of the most ingenious since the custom of making books began, and the most diverse conclusions have been arrived at. In coming to any conclusion on this subject probability based on what we know of the higher apes can be the only standard of argument. In these animals monogamy is the rule, the male and female with their young roaming at large in a family party. Occasionally, however, a male is seen accompanied by two females, and this is only what we should expect. The Apes are not very prolific animals nor are they numerous in individuals, hence, should any male be killed either in combat with a rival or by any other means, his mate probably wanders in search of another male, by whom, when found, she is probably readily adopted even if he should be already mated.
In like manner lived our half-human progenitors. But with them family parties no longer wandered aimlessly searching for food, but with a purpose. No longer forest dwellers, or vegetarians, food would require more zeal and discrimination in collecting, and shelter of some kind had probably to be devised, partly as a protection against predatory animals, and partly for personal comfort, since it would now have become apparent that this could be appreciably increased by the exercise of a little effort and ingenuity. This appreciation of creature comforts formed a cement holding the family together; a sense of safety in Society helped still further. Rude tools chipped from flints were among their earliest and most cherished possessions for the sake of the advantages they secured. Here was the earliest form of wealth and the birth of 26labour and a further step on the road to progress. Little would now occur to derange the harmonious routine of the daily life, save only the ever-present jealousy of the head of the family which was assailable both from within and without. His sons and daughters were probably now regarded as a portion of his wealth, for they ministered to his comfort, and aided in the daily work which had now become a necessity. As his sons attained to maturity, so they became rivals to be watched with a jealous eye, and finally driven off, while his daughters at the same time became potential mates. This danger of close inter-marriage was a real one, though it cannot be supposed that it was in any way realized. The risk was evaded by perfectly natural means. The jealousy of the head of the family which drove him to expel his sons as they attained maturity provided the means. These young bachelors sought their mates from neighbouring families, and it is probable that they would not be hard to lure from their parental control, but in such matters force was able to effect where persuasion failed.
These mate hunting excursions are to be regarded as extremely powerful factors in securing the betterment of the race. They were adventures in which all must fail who did not possess courage, cunning, and brawn, for, paradoxical as it may seem, evolution depends, not so much on the qualities of the individual as on the elimination of the unfit. As yet might was right. But the strife of combat, fierce and merciless, had its beneficial results not only in weeding out the physically and mentally deficient, but in stimulating affection between the victor and his prize.
As the advantages of neighbourliness dawned upon these children of nature, rules and regulations, for the control 27of the individual on behalf of the good order of the community, came into being; and among the earliest laws to be framed, we may be certain, were those for the regulation of marriage. These, as we may gather from the history of savage races to-day, did not concern themselves with chastity, at any rate before marriage, it was enough if they secured the right of possession, and excluded the dangers of close intermarriage. Promiscuity in the past was never the practice of any race, its existence to-day, among both savage and civilized people, is due in part to imperfections in the social scheme, and in part to the vagaries of individuals.
That the sexual instincts form the bed-rock on which depends the survival of all races of animals, which, for their propagation, require the co-operation of separate sexes, is beyond dispute. And it is no less certain that in so far as the evolution of man is concerned, jealousy has been a powerful integrating factor.
Among the higher animals apart from Man, both polygamy and polyandry are met with, and this with no apparent detriment to the race. It is significant, however, that polyandry is never met with among the mammals, and but rarely among the birds, when, as will be shown, this form of sexual relationship has been accompanied by a profound modification of the behaviour of the sexes in regard, not only to courtship, but to the offspring. The male has lost his masculinity, and the female her femininity. In human society both forms of marriage prevail, and there can be no doubt, from the history of such customs, that of the two types, polygamy is much to be preferred. It is certain that no race which practices polyandry can do more than hold its own, and that in a low grade of development. This cannot be 28said of polygamy, which might indeed be commended as a solution of some of our own social problems, were it not almost certain that the remedy would prove as bad as the disease.
The subject of “Courtship” in so far as it applies to the human race is one concerning which little can be said. Westermark, Letourneau, Sutherland, and last but by no means least, Darwin, have brought together a mass of facts bearing on the status of women among communities, savage and civilized, ancient and modern, and from these much may be inferred. To this harvest, however, Darwin himself still remains the most important contributor on all that directly concerns the “Sexual Selection” theory. Other writers seem to have paid more attention to the laws governing the possession of women than to the discussion of the motives which may have controlled the choice of mates. Instances of amatory dalliance, such as are met with among the inferior apes, and the birds, seem to be wanting. This negative evidence seems to show that, even among the most ancient, the most Ape-like, half-human races of man such dalliance was unknown. And this because primitive man, in his love-making as in everything else, was accustomed to take what he wanted, or die in the attempt. It is to this forcefulness of character that the human race owes its progress throughout the ages. But did he, when desire possessed him, exercise any sort of choice, when this was possible? What were his standards? These are unanswerable questions; at most we can but infer what his behaviour may have been from observations on existing races of mankind. These seem to demonstrate that while some races profess admiration for certain of their physical peculiarities, these 29cannot be attributed to the action of sexual selection.
It has been suggested that the low, beetling brows, protruding mouth, and flat, broad nose which characterized the earliest human peoples, were slowly eliminated by the æsthetic taste displayed by the females in their choice of mates. Now in the first place, it is highly improbable that they had any choice allowed them, and if they had, these are just the characters which were most marked in the males and might, or probably would, in consequence, have been deemed “manly” and desirable, for it is hardly to be supposed that such people would be capable of conceiving ideas of a possible refinement of their personal appearance if they could but add to the height of their foreheads and reduce the size of their faces. These graces settled down on them as the brain enlarged and habits changed. But the process of transformation must have been infinitely slow, and quite imperceptible from one generation to another.
The absence of secondary sexual characters in man, such as the brightly coloured areas which are so conspicuous a feature of many of the lower apes, is to be explained by his fundamentally different mode of life. Such vivid hues obtain only in species which live in troops, and they serve as aphrodisiacs, ensuring mating to every female forming a part thereof, which would be by no means certain were there no external signs of her condition. Primitive man, like the higher apes, was instinctively monogamous, and of necessity solitary, till he had acquired a tolerable measure of self-control and neighbourliness. When lust possessed him, he was obliged, in making his maiden venture to scour the country in the search for the object of his desire. This found, and won, probably only after desperate conflict with the 30head of the family, the nuptial ceremonies would be short.
The greater physical strength of the male and his higher brain capacity are probably the result of Natural, rather than of Sexual Selection. The former would weed out all the weakly and dull-witted in the ordinary course of the struggle for existence, the latter, during the early days of man’s development, would award the prizes of life to the most amorous and cunning, and to the most ambitious of the competitors.
The secondary sexual characters of the female are chiefly negative characters, the absence of those which are conspicuous in the male. She retains more of the primitive characters of the race. This is the rule in regard to the animal kingdom. Wherever we desire to find the onward tendency of evolution, the latest developments of the race, we turn to the male; when we desire to learn something of the past history of the species we turn to the female and young. This standard, of course, yields by no means uniform results, for we find every gradation of progress on the part of the latter, till male and female and young are externally indistinguishable. But the order is almost invariably the same—first the male, then the female, then the young. Thus progress is more or less automatic or “Orthogenic,” as the scientific text books have it, new characters, as they appear, tending to go on increasing in amplitude till checked by Natural selection. It is to be noted, however, that this transference is limited, for the female never inherits characters which are concerned with aggressiveness to the same degree as in the males, as witness, for example, the brow-ridges and huge canines in the case of the gorilla.
Darwin believed that the beards of men have developed by the selective choice of the women who preferred 31bearded men, while the secondary sexual characters of the women indicate the lines of male choice. There is, however, no evidence to show that in the past—for these characters are as old as man himself—woman had any choice whatever in the choice of her mate, save under exceptional circumstances. He was led to this conclusion by one or two striking instances apparently demonstrating this choice, and on these he seems to have based his version of the influence of sexual selection in man. The first of them is furnished by the Hottentots wherein, in both sexes, there is a marked “Steatopygy,” or accumulation of fat on the buttocks. In the female this is excessively developed, and it is said that such females are highly prized by the males. Darwin cites an instance of a woman in which this accumulation was so enormous, that she could only rise with the greatest difficulty from a sitting position. But there is no evidence to show that less favoured females remained unmarried.
In other tribes the breasts attain excessive proportions, so much so that they can be slung over the shoulder to feed the infant strapped to her back. These may have been increased by sexual selection, the preference of the males for such mates as possessed this feature in the most marked degree; but there is good reason to believe that such characters, which, it must be remembered, are the outward manifestation of germinal variations, once having appeared, would of themselves, of their own inherent vitality, have gone on developing. They won favour from long familiarity, which has imparted a semblance of increment from choice. These increments of growth in any given generation would be imperceptible, but variations in excess of the average would be conspicuous, 32and excite admiration from their very strangeness.
The part which sexual selection has played in determining the physical characters of the human race has without doubt been overestimated. Its influence may be said to have ceased with the development of the emotional side of his nature. This momentous process began with the male and had its roots in the ebullitions of his inherently amorous nature which has been the dominating factor in his career, and will be to the end, however much its influence may be disguised by the complex conditions of civilization.
These emotions, varying in kind and intensity, are such as are embraced in the term “Love” in the highest sense. They control the selection of mates, but this selection takes no account, save by accident, of qualities which have any value as factors of race-survival. In the lower animals these are determined by natural selection, and sexual selection adopts as it were the material furnished thereby. It “selects” only in so far as it eliminates the non-sexually inclined, and those which lack the qualities essential to ensure reproduction, such as weapons for example. In human communities natural selection is largely avoided, and “mate-hunger” seems now to be swayed by more than the mere desire for its satisfaction. With the development of human faculty new factors have been introduced, complex emotions have come into being, whose influences are as yet only vaguely understood. Whither are they tending? What will be their effect on race-progress? These are matters of grave importance to us all, and to the student of Eugenics in particular.
Of man’s higher emotions, which, it is contended, now govern his conduct, probably the earliest to assert itself 33was the æsthetic. His quickening mentality could not fail to be captivated by the bright hues of birds and butterflies, and flowers, the glorious colour-effects of dawn and sunset, the seasons in their changes and so forth. And as this sense of the beautiful slowly gathered force he would seek to decorate his naked body with such of the more brightly-coloured objects around him as were suitable or rather with such as could be affixed thereto.
As a signal mark of his favour and affection, he would occasionally transfer some one, or another, of his most lasting ornaments to his mate, and the additional charm this would give her ensured a continuance of such gifts, and paved the way for tribal fashions. But then, as now among savages, the males take the lead in this matter of ornamentation, but in proportion as affection grows, they are transferred from him to her, so that among civilized races to-day, the custom is entirely reversed, the women, not the men, wearing the finery. So soon as families began to be neighbourly and to combine for the sake of company and mutual help, the spirit of rivalry, so essential to progress everywhere, would tend to increase the number of such gifts, and to set “fashions.” With the foundation of society “selection”—by the elimination of the unsocial, would ensure, not only the survival of such fashions, but their multiplication and diversification, producing results which, to our eyes, have often been hideous. The immediate effect of this form of selection, however, was not a change in physical characteristics, but in the evolution of personal ornaments and development of the æsthetic sense. Progress in this direction must have been infinitely slow, and the lower races of to-day furnish us with instructive object-lessons in its course. In many cases uglification 34rather than refinement has attended their efforts.
It is indeed more than probable that the various types of ornamentation obtaining among savage races had their origin in outbursts of sexual exaltation. One of the earliest methods of personal decoration was probably to daub the body with paint, as is the custom during the performance of various religious and semi-religious rites among the Australian aborigines. A desire to find a permanent substitute for paint led to the practice of cicatrization, and the later and more refined custom of tattooing. But personal mutilation has taken many and strange forms, such as knocking out the front teeth, filing them to saw-like points, inserting gold or jewels, or staining them. No less extraordinary are the various types of lip and ear ornaments, and the suspension of ornaments from the nose. The various fashions of dressing the hair are also traceable to this origin.
That these modes of personal decoration designed for special occasions should in course of time become permanent, and should, in many cases, have lost their original associations is but natural. To-day among savage and barbaric races many of these modes of transfiguration have become associated with religious and semi-religious ceremonies, but many have been retained solely to enhance the personal appearance, even though in our eyes an exactly opposite effect has been attained. Among the natives of the Congo, for instance, the face is covered with raised patterns formed by cicatrization; that is to say, by cuts made with a knife, which are made to form scars on healing by means of pungent juices or heated iron. Further, the teeth are filed to form saw-like cutting edges, producing a revolting effect according to European ideals, but charming according to the standards of those thus 35patterns which adorn the tattooed face of the Maori present a result more nearly pleasing. Many of the natives of East Africa pierce the lobes of the ear and hang ornaments therein so heavy, that in due course a hole large enough to run the arm through results. These are mutilations of a purely ornamental character. Curiously enough, precisely similar forms of mutilation occur among people dwelling in different continents, as in the case of the lip and ear ornaments worn by natives of Africa and South America. There can have been no means of communication between these races, and hence we must conclude they were independently derived.
More striking still is the practice of deforming the head which prevailed among the Peruvians, the Caribs of the West Indies, and the natives of Vancouver, and the Chinook Indians, wherein it attained its maximum. Among some tribes, the head was depressed from above downwards, giving the skull a cone-shape, the apex pointing backwards; among others the pressure was applied to the back and front of the head, giving a more or less globular shape, and causing the sides of the head to bulge ominously. Now these distortions are to be attributed solely to the whim of Fashion. But how could this have arisen? No adult could have started it, for the form of the skull cannot be altered once its growth is completed. The conception of this diabolical custom apparently then arose in the brain of some fiendishly ingenious person, who realized that to effect its realization pressure must be applied to the head of the infant at its birth and for some considerable time after, by squeezing the head between boards, or tying it round with thongs of hide. That disastrous results would follow from this tampering with the brain would seem an 36unavoidable conclusion; yet such was not the case. During the moulding process, travellers who have witnessed it tell us, children display no sign of suffering, even though their eyes seemed to be starting from their sockets from the pressure. But they cried when the thongs were loosened. On attaining to man’s estate, such victims to parental folly seemed to be in every way as intelligent as the men of neighbouring tribes which had no such insane customs.
How deeply rooted was the prejudice in favour of this extraordinary fashion is shown by the fact that when, during infancy, from sickness, or other cause, the bandaging was neglected or omitted, and the child, in consequence, attained to man’s estate with a head of the shape designed by nature, he was seriously hampered in the struggle for existence, for no honours among his tribe were possible. Indeed, as often as not he was sold as a slave. But thus did Public Opinion bring disaster on its advocates, for those misguided people have been swept off the face of the earth by their own folly. Those who survived the ordeal, it is true, seemed in no way mentally deficient, but the infant mortality must have been great, and none of the adults could ever have attained to their full potentiality.
These people were, however, not the only lunatics at large. For this extraordinary practice found its devotees in many other widely sundered parts of the world. Deformed heads of various types have been found in rock-tombs near Tiflis, in the Crimea, Hungary, Silesia, in South Germany, Switzerland, and even in France, Belgium and England! How did it spread from one nation to another? Since means of communication were extremely limited 37centuries ago, one can only suppose that in most cases it arose independently. It is possible that the idea started with the unintentional deformations of the head which follow the practice of carrying the child during early infancy. It is well known that if a child be constantly carried on one arm, so that one side of the head continuously presses against the shoulder, a more or less marked asymmetry of the skull results. It would be enough for the head of one of the chief’s children to show a rather unusually marked asymmetry of this kind for every mother to endeavour to copy the defect, for imitation ever was the sincerest form of flattery!
To place these superficial, non-transmissible, artificially created features, such as deformed heads, mutilated teeth and ears, and so on, in the same category as the “secondary sexual characters” of the lower animals which are physical, inherent and transmissible features, is to ensure confusion of thought. The one represents a physical, the other an emotional development. The persistence of certain forms of mutilation esteemed beautiful in human society is not to be attributed to Sexual selection, or to “preferential mating,” for these things are not only non-transmissible features, but outside the sway of the amorous instincts, as is shown by the case of those individuals who, living in a community where deformed heads are de rigueur, have heads of normal shape. So soon as such perversions become a part and parcel of everyday life, they become essential to the general well-being and comfort of their possessors, enabling them to follow their normal avocations without exciting the dislike or wounding the prejudices of their neighbours. The absence of the “tribal sign” alienates the esteem and comradeship of his neighbours and brings an unenviable 38notoriety. In like manner albinos among birds, for example, are hunted down by their fellows and killed, and birds of exotic species conspicuous by reason of their unfamiliar appearance are treated in the same manner. The sexual instincts have no part in this.
It will have become obvious in the course of this chapter that Sexual selection as a factor in shaping the evolution of the human race has not played a very conspicuous part. Nevertheless, the balance of opinion to-day is probably in favour of the view that the physical peculiarities by which we distinguish one race from another are, for the most part, due to the influence of this form of selection. A more careful survey of the facts will show that this view is untenable. And there is no more striking demonstration thereof than that it has been inconsequently applied to account for features in one race, which in another are attributed to environment or to Natural Selection. It may safely be asserted that colour, the shape of the nose, the prominence of the jaws, and the character of the hair, are no more the result of “Sexual Selection” than stature, for example. These are the manifestations of inherent growth forces, or “tendencies,” which owe their survival, and development, to the influence of Natural selection.
Sexual selection has brought about the dominance of the male, by the struggle between males for mastery, originally for females. It “selected” for survival, in primitive races, those males with the thickest skulls and the strongest physique; it determined the survival of the keenest witted and most aggressive and most amorous males, and it eliminated those in which the latter features were too active. It assured victory, in short, to those only who possessed just those qualities on 39which life or death depend in moments of conflict. In the case of the females, it assured survival only to those who possessed strongly developed maternal instincts and submissiveness.
It is by no means realized that the incidence of moulding forces has changed and is changing with the environment of the race. So long as physical force, as between man and man, determined survival, as among savage races to-day, so long does it ensure to such races strong men and strong children, for in conflict with neighbouring tribes victory rests with the most powerful of physique and endurance and the most prolific. This last is an all-important concomitant if repeated conflicts are to be successfully waged. Among civilized peoples such contests began to lose their value in this regard when, by the introduction of arms, physical personality became a steadily diminishing factor. Victory now rests rather with those peoples who are most skilful in devising engines of destruction. The brain, not brawn, tells. But man cannot live by brains alone. With the inevitable decline in his physical nature man’s hold on existence is seriously imperilled. Civilization is making for extinction as much as over-specialization in the case of the lower animals. Hitherto, save in the case of decaying nations, women have played but a minor part in what we may call the “tribal” affairs of the race. Among the civilized nations of to-day, in proportion as the “maleness” of the community becomes more and more effete, the victims of sophistry, and the slaves of shibboleths, so the influence of the females asserts itself. And recent events among us show plainly enough that that influence is the reverse of good. Having its roots in personal vanity, and the love of notoriety, it is intolerant alike of reason and self-restraint, and that way madness lies.
The Man-like Apes and their mode of Life—Their “Courtships”—Musical Chimpanzees—How the Orang-utan improves his voice—His likeness to Caliban—The truculent visage of the Gorilla—“Ornament” in the lower Apes—The Concerts of the Howler Monkeys.
We are none of us given to boasting of our poor relations, and most of us indignantly repudiate our kinship with the Apes. But facts are stubborn things: the relationship is there, whether we admit it or not: and those who love truth for truth’s sake will not shirk the comparison between themselves and their remote cousins. Unhappily, from our present point of view, this cannot be carried very far, for the “Love idylls” of the Apes have yet to be written. Such facts, however, as have been gleaned are interesting. Of the higher, man-like, or “Anthropoid” species only the most meagre information is to be obtained; but this nevertheless is interesting. For the most part we have to be satisfied with inferences drawn from a study of the external differences between the sexes—from the “Secondary Sexual Characters,” in short, and from the records of travellers who have encountered these creatures in their native wilds.
The species which throw most light on this theme are 41the Gorilla, the Chimpanzee and the Orang-utan. Of these the Chimpanzee has most in common with the human race. But it may satisfy the qualms of many to know that between the Ape and the Man there is a great gulf fixed. The brain of the largest Ape is less than half the size of that even of the lowest of mankind. Man is a reasoning, and for the most part a reasonable, creature; he is a tool-making animal. This is more than can be said of any of the apes, even the most intelligent. Their teeth and immensely powerful arms must serve their every need. No ape ever fashioned for himself either a knife, a vessel to carry water, or any means of transport; and herein we have a measure of his brain capacity. The huge jaws and great canine teeth are no less conspicuous “marks of the beast.”
These, however, man himself has but recently lost, as was proved by the sensational discovery of the skull of an ape-like man at Piltdown, in Sussex, during 1912. Herein the jaw was essentially that of an ape, while the base of the skull was as markedly human. The cheek teeth, or molars, were of the human type; but the canine was ape-like, though much inferior in point of size. That the men of this remote age—which was possibly that of Pliocene times and certainly not later than early Pleistocene—had begun to use rudely-fashioned tools, is proved by the roughly-chipped flints found with the remains. With the invention of tools the decline in the size of his “eye” teeth began.
In all the large apes these “eye” teeth are of great size. Their purpose, it would seem, is primarily to serve as weapons in conflicts between rivals. Such conflicts are apparently unintentionally, and unavoidably, provoked 42by the loud cries uttered by the males in their endeavours to discover the whereabouts of females desiring mates. Of necessity roaming far in search of food, the unmated have no means of making their whereabouts known, save by thus giving tongue to desire. Evidently the normal methods of voice production do not suffice for their urgent needs, for the carrying power of the voice is immensely fortified by means of great air sacs, or chambers, formed in part by an enlargement of the body of the hyoid, or the bone which supports the tongue, and in part by dilatations of the inner walls of the larynx. The females, it is to be noted, are by no means so well equipped in this matter. It is not necessary that they should be. All that those desiring mates have to do is to follow up the cries of avid males, a by no means difficult task, especially when under the spell of the emotions which possess them. But the mechanism which serves the Chimpanzee and the Gorilla by no means fulfils the needs of the Orang-utan. In this uncouth creature the system of resonating chambers is immensely increased by great, thin-walled, membranous pouches extending round the neck and under the armpits, so that when inflated these areas have a most extraordinarily swollen appearance. When the Orang chooses to lift his voice even the deaf must hear.
Where fighting instead of fondling is the sequel to these impassioned cries the conflict is probably not of long duration, for it is certainly severe. This is attested by the fact that captured specimens, if adult, are commonly found to be minus one or more fingers, which have been bitten or torn off in these love affairs.
Plate 2.

From a drawing by I. Thornton.
THE GORILLA PREPARING FOR HOSTILITIES.
Note the “beetling” brows, the large size of the canine teeth, and the great development of the arms in these arboreal creatures, which play an even more important part in locomotion than the legs. The latter in this illustration are, however, relatively too small.
[Face page 42.
An added ferocity of expression is given to the male Gorilla by the development of enormous brow ridges and the huge canines. The former are regarded by some 43authorities as adaptations to afford increased powers of mastication. But if this were so, then such ridges should be equally developed in both sexes, and this is far from being true. Hideousness, rather than ferocity, has been given to the Orang-utan by the out-growth of enormous ridges on each side of the face, and these, when the great wind-bags encircling the neck are inflated, impart a repulsiveness of expression attained by no other animal living.
Of the normal every-day life of the great Apes but little is known. It would seem, however, that they live in family parties—an adult male accompanied by a female and one or more young of different ages, of which one is commonly an infant in arms. It is difficult to procure positive evidence on the point, but it is commonly believed that the young remain with their parents till they are several years old, when they are gradually driven off to fend for themselves. This is a common procedure with all animals. The dominant impulse in this is something akin to greediness, an indefinable perception that too large a family party will entail too great a strain on the food supply, hence the now no longer helpless young are regarded as a danger to the safety of the family, and are turned adrift. Incidentally this procedure is of immense benefit to the race, for it ensures its distribution, enlarges its chances of survival, and lessens the danger of in-breeding.
Attention must now be turned to the lower Apes. In these it is to be remarked the secondary sexual characters differ conspicuously from those of the man-like species. Manes and beards and brightly-coloured areas of bare skin are now the dominant feature. But canine teeth, 44in proportion rivalling those of the Gorilla, are found in the Baboons, while in some of the New-world monkeys voice production of quite remarkable power takes the place of ornament.
The precise part played by ornament among these animals can only be inferred from Darwin’s observations on captive animals, and then only in so far as they refer to colour. Manes, beards and moustaches, such as are shown in the adjoining illustrations, are borne only by the males, and sometimes take extravagant forms.
Darwin suggested that the mane of the Baboons, for example, served as a shield when fighting with rivals, protecting the great blood-vessels from injury. Incidentally this end may be attained, but from what we know of similar developments in other animals, this cannot be regarded as the primary function of the mane. One is tempted to look upon it as a protective device because of its position, but it is probably no more so than is the long flowing hair which adorns the flanks of the Guereza. This is of a purely ornamental character, although, according to some, it is to be reckoned as an instance of protective coloration, the long white hair matching the long pendant masses of lichen which hang from the boughs of the trees in the damp forests where these creatures live, and so concealing them from their enemies. Of beards and moustaches many examples might be cited, but the most striking must suffice. These are furnished by the Satan Monkey or Black Saki (Pithecia satanas), and the little Tamarin Monkey (Midas imperator)—one of the Marmosets. In the first-named the beard is thick and full, but in the latter scanty. This, however, is atoned for by the enormous upwardly curled moustache giving the face a most comically human appearance.
Plate 3.

From drawings by I. Thornton.
THE BAROMETER OF MALENESS—AMONG THE APES.
All the Man-like Apes possess great canine teeth and powerful voices. In the Orangutan the Compass of the voice is enormously heightened by means of a huge wind-bag which encircles the neck. The wind-bag is seen in fig. 1, which also shows the great folds of skin developed by adult males on each side of the face. In other species, as in the Tamarin Marmoset (Midas imperator) (fig. 2), and the Satan monkey (Pithecia satanus) (fig. 3), “ornaments” in the shape of beards and moustaches are developed, while in the Mandrill (fig. 4) the face is vividly coloured.
[Face page 44.
In the development of brilliantly-coloured areas of 45bare skin the monkeys stand alone among the Mammalia. The hues displayed are remarkable for their brilliancy, and this varies in intensity, waxing and waning with the varying moods of their possessors, and attaining their maximum during periods of sexual excitement. Blue, green, red, and violet are the dominant colours, and these are confined to the face, buttocks, and genital organs. The same hues are commonly present in both sexes, though in the female they are less brilliant. Normally the male appears to be unconscious of the conspicuous patches of colour, but when under the irrepressible stimulus of sexual excitement he seems to endeavour to make the utmost possible capital out of such adornments, more especially presenting his buttocks to his mate in an apparent endeavour to stimulate her desire. In some species, as with the Baboons for example, the naked area of this hinder part of the body is a much more conspicuous feature in the female than in the male, becoming enormously swollen and carunculated, and from its vivid red colour presents a positively revolting appearance, according to our standard of what is beautiful. The most vividly coloured species of all is the Mandrill, which, in this matter exceeds all other living Mammals. The face, in the male, is produced forward to give the head a dog-like shape, while the whole of the upper surface of the muzzle has been transformed into a swollen, deeply fluted mass by the excessive inflation of the underlying bone. The bare skin covering this is of a brilliant cobalt blue, with lines of violet in the furrows, while the nose is of a bright scarlet. The naked skin of the buttocks, and the genital organs, are suffused with brilliant tints of scarlet and blue. In spite of the purity and brilliance of the 46coloration the effect is to make the creature really hideous.
Of the display Cuvier writes: “La partie postérieure du corps n’est ni moins extraordinaire ni moins révoltante. Sous une courte queue sans cesse relevée est un anus entouré d’un gros bourrelet d’écarlate; de larges fesses nues, que l’animal semble montrer sans cesse avec autant de lascivité que d’impudence, sont colorées d’un rose vif nuancé sur les côtés de lilas et de bleu. Les parties genitales enfin sont d’un rouge de feu d’autant plus tranché qu’elles sont absolument nues, et qu’elles viennent a la suite d’un abdomen revêtu de poils blancs.”
While we cannot suppose these animals to possess any standard of beauty or ugliness, it must not be forgotten that they are more or less conscious, not only of the existence of these brightly-coloured areas, but of the effect they produce, as Darwin showed long since in the cases of a captive Mandrill, and some other smaller species of Monkeys, among them a Rhesus Monkey. These, when shown a looking-glass, at once presented their hinder ends to what they supposed to be the new arrival. A similar mark of friendliness was shown towards their keeper, and visitors introduced by him. Periodically, under the sexual stimulus, this desire becomes intensified and becomes an invitation to mating.
In this connection it is interesting to note that in some of the Macaque Monkeys we have signs of a reversal of the usual sequence of coloration. For in the Pigtailed Macaque the young of both sexes are more brilliantly coloured than the adults, in regard to the bare skin areas, while in the Hairy-eared Macaque (M. lasiotis) and the Rhesus Monkey (M. rhesus) the face of the female is brighter than that of the male. This surely means that this coloration is in process of suppression, for according 47to the rule the male is the first to develop new characters, then the female, and finally they are transmitted to the young. The extra brightness in the young, then, is to be regarded not as an incipient, but as an ancestral character in process of elimination.
As a rule, among the Mammals at any rate, brilliant coloration and weapons of offence are not associated in the same animal. The Baboons, and the Mandrill in particular, are exceptions, for these animals are provided with most formidable “tusks,” the canines of both upper and lower jaws being of great size, and opposed one to another in such a way that they wear away to form sharp, angular cutting-edges, more murderous than the fangs of the Tiger.
Reference has been made already to the existence of large sound resonators for the purpose of increasing the volume of the voice in the Orang, Gorilla and Chimpanzee. Some of the Gibbons are also well provided in this direction. But the most striking instances of the kind are furnished by the Orang, and the monkeys known as Howlers. In these last the base of the hyoid, as the skeleton for the support of the tongue is called, is fashioned into a deep bony cup, which has the effect of intensifying the volume of the voice to a most surprising extent. But more than this, apparently for the protection of this bony voice-bowl the upright branches of the lower jaw have become remarkably deepened, and widened, a correlation of growth between unrelated parts which is fraught with deep significance. “Terrific,” “terrible” and “harrowing” are terms which have been used by travellers like Bates, Belt and Wallace in describing the cavernous roar of these animals, a roar which will easily carry two miles. 48It would seem that these vocal efforts are not merely confined to what we may call the “Courting” season, as is the roar of the stag, but that they are heard nightly at dusk. They may be resumed again at dawn, and re-awakened when thunder-clouds gather. They have become the normal method of giving vent to excitement, and probably are intensified when isolated males are desirous of discovering the whereabouts of females equally anxious to find a mate.
Among the Apes we meet, as with the human species, with both monogamy and polygamy. But it would be dangerous to assume that the reasons for polygamy are the same in both. Polygamy, indeed, has by no means always the same significance. In the most primitive, half-human races of the past, as with the man-like Apes to-day, polygamy is determined by accident rather than choice. These extinct peoples, like the great anthropoids, were normally monogamous, but on the death of a male in conflict with his neighbour, or from other causes, his mate would probably of her own free will seek out the nearest male and even if he were already mated would be at once adopted into the family circle. This certainly happens in the case of the Gorilla and Chimpanzee to-day. But among living races of mankind, both savage and civilized, multiplicity of wives is a matter of choice on the part of the male, and in many cases to achieve this females from other tribes have to be secured—either by purchase or conquest. With the lower apes, or “monkeys,” polygamy only obtains among gregarious species; and either because the birth-rate of the females exceeds that of the males, or because a considerable number of young males are killed annually by exciting the jealousy of the older males, who are exceedingly pugnacious.
The Birth of Weapons—All Flesh is Grass—Utility and Ornament—The Fever of Love—The “Challenge” of the Deer—What it means—More about “Hormones”—“Hummel” Stags—The Age of Deer—The “Courtship” of the Moose—Types of Antlers—Antlered Females—Fighting Topi—The Lance of the Oryx in the Lion’s Flanks—Happiness and Hartebeestes—Odoriferous Suitors—The Bloody Sweat of the Hippopotamus—The Elephant in Love—Concerning Tusks—Polygamy.
From Apes to Antelopes is a far cry, but contrasts are always helpful. Antelopes and Deer, Zebras and Elephants, Rhinoceroses and Swine, are types, taken at random, of that great and important group of animals known as the “Ungulates,” or “Hoofed” animals. These illustrate in a very striking manner what is meant by the term “Secondary Sexual Characters.” They demonstrate no less forcibly what is meant by the term “Sexual Selection.” They are valuable in this connection, because of the often formidable weapons, in the shape of horns and tusks, which so many species have developed during the struggle for mates.
But “Sexual Selection” will not explain their origin, and it is difficult, in the present state of our knowledge, to discover any clues which will reveal this. In seeking these there are certain broad aspects of the problem which 50are not to be lost sight of. In the first place, horns, at any rate, are confined to the hoofed animals. That the various types of hoofed animals, living and extinct, have had a common ancestry, no one at the present day will probably call in question. The relationship, however, of the various living types, one to another, is by no means always apparent: the missing links are to be sought in the records of the rocks.
When the whole of the evidence comes to be surveyed, and not till then, it becomes apparent that this wonderful diversity is the result of complex factors. That the conditions of existence have controlled the results is beyond question; but it is equally certain that these conditions have been merely controlling and not causative. In other words, we must regard each of these different groups or types—Deer, Antelopes, Horses, Elephants, Swine, and so on—as witnesses of what we call “Heredity.” They are so many “Diathetic types.” That is to say, the forms, or individuals, belonging to each type have inherited certain peculiarities in common; they display a “Diathesis” as the doctors call it: an inherent, inborn tendency, or habit of growth, in a definite direction: a tendency which, ever and anon, develops new qualities, takes new directions. And thus it is that we get Oxen—using this term in its widest sense and not in its special sense—Antelopes, Goats and Sheep, for example. These have, among other things, a “diathesis” in the direction of horn production, and each, too, of a different type. What is meant by this apparently mystifying term “diathesis” will perhaps be made clear by taking the case of the Ox and the Sheep. While very different in appearance, these live on precisely similar food; yet no one has any difficulty in 51discriminating between the taste of beef and mutton. In the marvellous chemical laboratory of the body the grass gathered in the same field is converted into flesh which even in its uncooked state is easily distinguishable. Though for the purposes of this illustration domesticated animals have been used, the same is true of their wild relations. Sportsmen tell us that the various types of Antelopes and the Zebra, which may be seen feeding together, have yet flesh of very different qualities. These qualities are to be attributed neither to “Natural” nor to “Sexual” selection; they are “accidents.” Similarly, their horns are the witness of a horn-producing “diathesis”: the various divergencies in curvature, and in the form of their spirals, or the number of their encircling rings—as in the horns of Antelopes—are to be interpreted in like fashion. These twists and turns vary in the same way that the taste of the flesh varies, and for the same reason; that is to say, they are not the outcome of “Sexual Selection,” nor have they been brought about by “Natural Selection” to serve the purpose of “Recognition marks,” as Wallace would have us believe.
But horns, as horns, apart from their “accidents” of curvature and ornament, must certainly be regarded as the product of Sexual selection, for having once started into being those individuals had the best chance of leaving descendants which were best armed. The possession of horns was not necessary to the maintenance of the species; but such armature was essential among the males in securing possession of the females. Other things being equal, the male with the biggest horns wins the prize. Since these are also used as weapons of offence, or rather of defence, in warding off the attacks of beasts of prey, it might be contended that they are as much the product of Natural selection as of Sexual selection.
52It soon becomes apparent that this interpretation must fail. In the first place, if it were true, the females should be similarly armed. In the second, in the presence of many of their enemies they are useless. The Cape hunting-dog, for example, is more than a match for any antelope. This ferocious animal kills his victim by running it down, persistently tearing at its flanks, until at last the entrails protrude and the horrid chase is ended. Furthermore, the horns are a comparatively late acquirement of the species, as is shown in the case of the Deer; for the earliest known fossil species were hornless. That the females among the Oxen and many of the Antelopes possess horns is an interesting fact, but it can only be regarded as another instance of a character first acquired by the male and later, in successive generations, transferred to the female. And it is to be noticed that this transference is never found save in the cases where the character in question has attained its maximum in the male. The transference of weapons to the female is the more remarkable because there is no evidence that they play any part in the struggle for existence, either in securing mates or in warding off the attacks of enemies. Moreover, these weapons in the female may exceed those of the male, in length, though they are never so massive. They are to be regarded solely in the light of ornaments. There are few more striking instances indeed where the purely ornamental and the strictly utilitarian are so closely associated.
Plate 4.

By the courtesy of Rowland, Ward, Ltd.
WEAPONS OF OFFENCE.
Horns of various types furnish the most conspicuous of the “Secondary Sexual Characters” of the ruminants. In the Deer only are these branched. In the “hollow-horned” ruminants they are either lance-like or more or less spirally curved, or they may form more or less open loops.
1. Black-tailed Deer. 2. Hangul or Kashmir Barasingha Deer. 3. Greater Kudu. 4. Black-buck. 5. Saiga Antelope, remarkable also for its curiously swollen nose. 6. Marco-Polo’s Sheep.
[Face page 52.
Attention may now profitably be turned to the behaviour of these interesting tribes when under the alluring influences of love.
Tradition and the poets have contrived to persuade us 53that the fever of Love becomes epidemic in the spring. This, however, is by no means true, at any rate in so far as what we are pleased to call the “lower animals” are concerned. For with many, as for example the Deer and the Bats, this fever is not aroused till the time of autumn plenty. With regard to the deer, we can find a reason for this. It is determined in part by the period of gestation, and in part by the peculiar character of the most conspicuous of the male secondary sexual characters—the antlers. The deer, at any rate of the northern hemisphere, carry their young about eight months. Now it is important that they should make their entry into the world just as the food supply is increasing and the temperature is rising. With the summer before them the young have time to gather strength for the encounter with their first winter. We have a striking witness to the truth of this contention in the fact that when the Indian Spotted Deer, or Chital, was first introduced into Europe, nearly all the fawns perished owing to having been born in winter; later, the females took to calving in spring, and from thence onwards the species has held its own among us.
As touching the stags. The antlers, as everybody knows, are shed annually, and their renewal entails a very considerable strain on the system. As a consequence, it is necessary that this period of stress should fall after the trial of winter is overpast, and with the genial summer before them. From the end of March, when the old weapons are shed, till July, the masterful males of the community wander at large, seeking seclusion and avoiding all occasion of quarrel; for they are not only defenceless, but threatened with disaster should any accident 54befall the growing horns, which, during their formation, are exceedingly sensitive. Even a slight blow would not only spoil their shapely proportions, but, further, might render them useless in the warfare that is before them.
With some species this desire to go into retreat is more marked than in others. The Red-deer, and the Wapiti, on the one hand, and the Moose on the other, well illustrate this. The two first-named pass the winter in herds, in the case of the Wapiti numbering many thousand individuals; no other species, indeed, is so markedly gregarious. With the advance of the spring, however, all is changed, for the males withdraw from their companions to suffer humiliation in seclusion. As chill October arrives, a striking alteration in their demeanour becomes apparent, at any rate in the case of the older males. The new antlers are now hardened, and the blood supply, which has hitherto been building up the new weapons, is cut off. As a consequence, the “velvet,” which till now has been directly concerned with the growth of the antlers, dies, and peels off the underlying bone. To facilitate this work of cleaning, the animal rubs them, first against the stems of saplings, and, later, against larger trees, and even rocks, till at last they are ready for “battle, murder and sudden death.” The “rutting” season, in short, has commenced. And with the final completion of the antlers other signs of that approaching frenzy, which is soon to establish itself, become apparent. The most striking of these are the swelling of the neck, and a marked increase in the mane thereof; while the voice enlarges its compass enormously, whereby the females, so long neglected, are now feverishly sought for.
Plate 5.

Photo by G. IF. Wilson Co. Ltd., from “The Living Animals of the World.”
MANCHURIAN WAPITI “CALLING.”
The “stags” do not begin to call for mate’s until the horns have more or less completely shed their velvet.
[Face page 54.
The Red-deer, maddened with desire, scours the country, calling as he travels with a loud musical roar, ever and anon impatiently listening for the tremulous response of 55females hardly less anxious to mate than himself. One after another is speedily added to his harem, but not without conflict. For sooner or later he catches the call of another stag in like case. A jealous fury at once takes possession of him, and the call, intended as a message to mateless hinds, becomes translated into a challenge to fight for the mates possessed. Each of the now infuriated challengers makes all haste to come to blows, and speedily they are rushing headlong on one another to meet in a crash of antlers. Then follows a test of strength, a sort of tug-of-war reversed, for each strives to push the other to his knees, and succeeding, to deal a deadly sideways thrust at the kneeling adversary’s heart with the spike-shaped brow-tines. This attempt, however, is rarely achieved. Yet not seldom such encounters become a duel to the death, and one in which both die, for in the remorseless tilt at one another the antlers of one may spring apart, and then close in on those of the other. Once this happens, it seems to be rare indeed that they can be extricated from this close embrace. With heads thus locked, they sway, and twist, and tug, not now for the mastery, but for life itself. But as the hours run they become more and more exhausted by their efforts, weaker and weaker from loss of food and rest, till finally death releases both.
A male having once succeeded in forming a harem, will commonly contrive to repeat his success year after year, withstanding all comers. But sooner or later his vigour wanes and he is ousted by another and younger male. Not else would the stamina of the race be preserved. It is considered a moot point, however, whether physical strength and sexual potency run at the same pace; 56for it is believed by some that a stag will often contrive to hold a harem against all rivals after his fertility has declined. This, however, is extremely improbable. A lowering of fertility means a decline in the potency of the hormones, and in the development of the secondary sexual characters, among which are the antlers, which are by no means negligible factors. That they are not all-important, however, seems to be shown by the fact that, occasionally, stags appear in a herd which are congenitally unable to produce antlers—a reversion to the ancestral condition—and such are said, occasionally at any rate, to be able to oust their formidably armed rivals. This may be so, but the fact that “hummel” stags, as they are called, are so rare is surely to be regarded as eloquent testimony of the disadvantages of their unarmoured state. They become speedily eliminated, in short, by “Sexual Selection.”
After this outburst of sexual activity has spent itself, the various harems, with their lords, amalgamate; all living in peace through the winter. The stags retain their antlers at this season, partly as a protection against predatory enemies, such as wolves, and other large carnivores, which would otherwise play havoc in their ranks, and partly because the cold of winter and scanty fodder would inhibit the growth of new antlers or reduce their size. With the return of spring the dangers of attack are lessened, temperature rises, and food becomes once more plentiful. Then the inevitable disarmament takes place.
The Red-deer, though mature at six, does not reach his prime till his eleventh year, and from thence till his fifteenth or sixteenth year is at his best. The hinds mature earlier, and appear to be fertile for a much longer 57period. At any rate, a wild hind in Jura, known by certain peculiarities of its ears, during twenty-one years produced twenty calves. She was killed at last with a calf at her side, but was thin and haggard-looking. She was, therefore, not less than six-and-twenty at her death. The calves, it may be mentioned, are born in May and June.
Old stags shed their antlers, it is remarked, earlier than young ones. And this is an advantage to the species, since it prevents premature breeding on the part of sexually precocious but immature males, and limits competition to the adults.
What obtains in the case of the Red-deer obtains also with minor variations due to environment, climate, and so on, in the case of all other deer. The life-history of the Wapiti, as might be supposed, differs only in detail from that of the Red-deer. But during the winter they form vast herds, numbering thousands. It may be that in primitive times the Red-deer was no less numerous. But in this country, at any rate, conditions favourable to the maximum development, either in bodily size, or in the massiveness of the antlers, have long since passed away. Even in the Highlands of Scotland the conditions of existence have entirely changed owing to disafforestation. Deer are essentially forest dwellers. But the “deer forests” are such only in name, and for the most part the wild stags of to-day must get what shelter they can from rocks and inequalities of the ground. From this cause, and from the very natural desire of the owners of such “forests” to secure the finest heads in each year, the whole race has deteriorated. How great a change has come over it may be seen by comparing the heads of British Stags with those from German forests, where the 58conditions of existence are more favourable. If we turn to the records of the past we find that the antlers found in the fens, turbaries, and caverns of our islands are vastly larger, heavier, and carry a greater number of points on the sur-royals, than do those of the existing Scotch stags.
Having regard to the fact that hundreds, and in the distant past thousands, of antlers were shed annually, the comparative rarity of these weapons in the haunts of deer excites comment. This is accounted for by the fact that they are greedily eaten by their late owners, apparently, though unconsciously, for the sake of their bone-producing qualities.
By way of contrast with the Red-deer and Wapiti, we may take the Moose (Alces machlis), which at no time, and nowhere, attains to large herds. This is explained by the relatively restricted food supply which obtains in the haunts of these creatures. For they frequent the margins of streams, feeding largely on willows and birch. From the shortness of their necks, and the great length of their legs, they cannot crop grass and other short herbage, for unless they kneel they cannot reach the ground. Hence it is obvious that though their geographical range may be wide, their numbers are kept rigidly in check. They would be fewer still but for the fact that, unlike other deer, they glean no small amount of food from the water, wading out to feed upon aquatic vegetation. The roots of water-lilies are especially sought for, and to obtain these the animal will often disappear entirely under water.
As a consequence of the limited food supply the Moose lead solitary lives. On the Eastern side of America, where the winter is severe, a few individuals, generally a family party, will “yard up,” or make a fortress for their mutual 59protection by trampling down the snow over a restricted area. But in the Yukon district, my friend Mr. F. C. Selous tells me this is never done.
The rutting season of the bulls begins as soon as the antlers begin to “peel.” What follows is practically a repetition of what has already been related in regard to the Red-deer and Wapiti. And in this connection it is interesting to note that the natives take advantage of the period of desire in the bull to entice him to his death. Generally this is done by imitating the call of the cow in response to the bull’s anxious bellowing. But in Southern Alaska the opposite side of his nature is played upon. This is done by scraping or beating the bushes with the shoulder-blade of a Moose in such a way as to reproduce the sound of a bull cleaning his horns. The very suspicion of a rival enrages him, and, rushing in a blind fury in the direction of the tell-tale sounds, he speedily falls a victim to the trick which has been played him.
That the mating period is the most critical, and most searching in the whole life-history there can be no doubt. Every faculty during this time is put to the test, and from the time of sexual maturity until old age is at last attained it is an annual test. Alertness is all important. Other things being equal, success falls most certainly to those individuals with the keenest perception, and quickest interpretation of sight, sound and smell.
One is puzzled at what seems a concession of Darwin’s to the Lamarckian theory of the inherited effects of use in this connection. For in discussing the bellowing of the stag in “The Descent of Man,” he remarks that it “does not seem to be of any special service to him, either during courtship or battles, or in any other way. But may we not believe that the frequent use of the voice, under the 60strong excitement of love, jealousy and rage, continued during many generations, may at last have produced an inherited effect on the vocal organs of the stag, as well as of other male animals?” All the evidence goes to show that the production of sound, and the instant interpretation of its significance, is a matter of the highest importance. In the case of the Moose, for example, the noise occasioned by the cleaning of antlers provokes the same frenzy as at another time is aroused by the voice. Dullness of perception not only in these matters, but at all times, is fatal.
As touching the less conspicuous secondary sexual characters of Deer more must be said presently. For the moment the antlers must retain our attention. Time was when the Deer lacked these appendages. When they first appeared, in the now extinct species of the Middle Miocene period, they were no more than short prongs. Later, one of the prongs became elongated, and developed short branches or “tines,” which, in succeeding species, became more numerous, while at the same time, with the gradual evolution of more and more species, these antlers assumed new features both in the matter of size and in the character and number of the “tines,” a development which has reached its maximum to-day. But apart from these specific variations, which have given us such types as those of the Roe-deer, Red-deer, Wapiti, Caribou, Moose, Fallow-deer, Sambar, Schombergk’s deer, the strange Milou-deer, Elds-deer and Mule-deer, each species displays a quite remarkable range of variation in regard to its particular type of antler. Nowhere, perhaps, is this more strikingly marked than in the case of the Caribou and Moose. No doubt this feature is due largely to the fact that the horns are shed annually, and that the variations are due, in part at any rate, to temporary environmental conditions, such as food and weather. But these apart, individual peculiarities are constant, reappearing with more or less exactness each year.
Plate 6.

Photo by Lord Delamere, from “The Living Animals of the World.”
GROUP OF BEISA ORYX.
The lance-like horns of these animals can be used with deadly effect, even against lions.
[Face page 60.
In contemplating these facts one asks: What are the underlying factors of this variability? What is the significance of the branching? What end is attained by the annual shedding? That the antlers constitute very effective weapons of offence there can be no doubt, and one is inclined to regard the branching as the outcome of natural selection, on the assumption that branched antlers would be less deadly than lance-like weapons. It would perhaps be tempting to accept this interpretation as all sufficient were it not for the evidence afforded by the hollow-horned ruminants. The Oryx and the Kudu, for example, are lance-bearers, and therefore show conclusively that stags similarly armed might well have continued to survive in spite of the foils which the “tines” provide. Darwin, long since, guardedly suggested that while these weapons primarily served for offensive purposes, their elaborate systems of branching might have been brought about by sexual selection. That is to say, the extreme beauty of the weapons may excite the admiration of the females as well as our own. Granting this, he inferred they might have played an important part in elaborating the branching by constantly displaying a preference to mate with those males possessed of the largest and most branched antlers. But there are many and serious objections to this suggestion, and the most important of all is the fact that the female is allowed no choice in the selection of her lord and master. We can, then, only regard the antlers of deer as another instance of the survival of a “fortuitous” but inherent variation, which survived because, whatever the defects thereof, they 62proved advantageous in the struggle for existence.
Having regard to the fact that so many of the females among the hollow-horned ruminants have acquired horns, it is somewhat remarkable that in the Reindeer alone among the deer are these weapons normally possessed by the female. The gradual transference to the female of features which were originally secondary sexual characters in the male is an occurrence which is met with in every group of animals. In writing “The Infancy of Animals” I gave a number of instances of this kind. But the case of the Reindeer affords a more than usually striking illustration of this curious sequence; and this because rudiments of antlers are to be met with among the females in several different species of Deer to-day. They have been found in the females of both Roe- and Red-deer, though such cases are rarely met with. As a rule this assumption of the male secondary sexual characters by the female occurs only in very aged animals, or as one of the sequelæ of diseased ovaries and consequent sterility. But at least one instance is on record of a doe Roe-deer which possessed small antlers while pregnant. Thus, then, we gain a further insight into the process by which the female slowly assumes the outward attributes of the male; that is to say, the secondary sexual characters appear first in the male, and as seasonal characters. Sooner or later they become permanently established. By the time they have become firmly fixed in the male, and apparently not till then, they appear in a dilute form during senility, or in consequence of ovarian disease, in the female. Having once started, however, they appear earlier and earlier in the life-history of succeeding generations of females, and at last in the juvenile stages of both sexes.
63The hollow-horned ruminants, which must now be considered, afford some very striking facts in regard to these “secondary sexual characters,” more especially in so far as horns are concerned. In the first place these weapons are permanent structures, taking the form of a bony core ensheathed in horn, with which we may compare the temporary covering of velvet in the deer: in the second, they are unbranched. The only exception to this rule is furnished by the Prong-horned Antelope, wherein the sheath is both annually shed, and branched. The branching, however, is very slight, taking the form of a short forwardly directed prong about half-way up the sheath, which is borne on a long bony pedicle recalling that of the Muntjac. The shedding is due to the formation of new horn material at the base of the old sheath, which is gradually forced off by the growth of the new tissue. Structurally the horn of this remarkable Antelope differs somewhat from that of its relatives.
As may be seen in Plate 4, in the form of the horns the typical hollow-horned ruminants present an exceedingly varied range, and one often of great beauty in the matter of curvature. That they serve as formidable weapons of offence was demonstrated during 1912, when, according to the Annual Report of the Government Game Reserves, published by the Pretoria Government, the game warden, Major Stevenson Hamilton, reported of the Antelopes that “many carcases of males of almost all species, killed in single combat with rivals, were found during the mating season, untouched by anything except vultures.” As a rule, however, these animals, like the Sheep and Goats, and their larger relatives the Cattle, seem to avoid a duel to the death. One or two instances as to the general 64character of these combats for the possession of mates must suffice. Thus the late A. H. Neumann, a hunter of experience, remarks that he once or twice saw conflicts between the Topi (Damaliscus jimela), an ally of the Hartebeestes. The two rivals would stand a little apart, affecting, apparently, to be unaware of one another’s presence. Suddenly they would rush headlong at one another, bringing their heads together with a clash, each, at the same moment, falling on his knees.
Major Powell Cotton, again, once witnessed an affray between two Beisa Oryx. Here the master bull of the herd was infuriated by the advent of an intruder in his harem. Time after time they dashed at each other, their foreheads meeting with a thud; then, with horns interlocked, they wrestled fiercely; then, separating, they charged again. Yet neither, he remarks, tried to use his lance-points, as they do when cornered by man or beasts of prey. Nevertheless, encounters of a more sanguinary character appear to be by no means rare, for it is no uncommon experience of hunters to kill bulls of this species in which one eye has been burst by a horn-thrust. Another peculiarity of these animals is the extreme thickness of the hide of the neck and withers, which seems to afford a shield against such spear-thrusts during these battles. How powerful is the thrust of these weapons, and how efficiently they can be used, is shown by the fact that lions in making an attack on an old bull are often severely wounded, or even killed. And there are many instances on record of cases where both the lion and his intended victim have died together, the Antelope having been unable to withdraw his horns from his adversary’s body. The beautiful Pala Antelope fights furiously with rival rams, and the vanquished, as with so many of the Antelopes, form herds by themselves, till one by one they gather strength and skill enough to establish their right to mate.
Plate 7.

Photo by courtesy of the Duchess of Bedford, Woburn Abbey.
ELAND COWS.
Among antelopes the females commonly bear horns, which may be even longer than in the males, though less massive.
[Face page 64.
Plate 8.

Photo by courtesy of the Duchess of Bedford, Woburn Abbey.
AMERICAN BISON
The “Secondary Sexual Characters” of the male are here conspicuously developed, and are seen in the massive fore-quarters and enormous head.
The Elands present some puzzling features, for both sexes bear large horns, and they are very massive in the bulls. Yet these animals are generally described as the most inoffensive of all the horned ruminants. That the horns are used to any extent in conflicts between rival males seems doubtful, inasmuch as this species is remarkable for the development of an enormous “dewlap,” a thin pendulous fold of skin which runs from the throat to the chest. Such a form of “ornament”—for in this light we must regard it—would be dangerous, indeed, when much fighting was to be done. Nevertheless, it would be contrary to all our experience to conclude that weapons so well developed as are the horns of the bull Eland were entirely useless. This is a matter which decidedly calls for further investigation.
That our knowledge of that most important period of life of the larger mammals, the period of sexual exaltation, is lamentably incomplete will be realized by anyone who seeks enlightenment on this subject. Most of the meagre information we possess has been collected by travellers and sportsmen, neither of whom have the time to devote to the long and laborious watches that a fuller history demands. Every now and then a glimpse is afforded of this period of the life-history which brings home in a very convincing fashion, how little is really known. It seems certain that the fighting hitherto described is to be regarded as but a phase of a cycle of events which takes place at this time. Thus, for example, the old naturalist and traveller Schweinfurth tells how he once encountered a herd of Hartebeest which were apparently effervescing with animal spirits, for they kept running around in 66couples, like horses in a circus, using a clump of trees as a pivot. Others, in groups of three or four, stood by, interested spectators. After a time these, in turn, took their places and ran round, two at a time, in their own circuit, and in the same fashion. Their evolutions, he says, were so regular as to suggest the guidance of some invisible ring-master. These gyrations may be regarded as an erotic dance. The Sambar, under like excitement, will stalk about with erected tail, outstretched muzzle and everted face glands, and the Black-buck, among the antelopes, behaves in like fashion.
It cannot be supposed that these quaint performances are peculiar to the species in which they have been observed, but rather it may be inferred that similar antics, besides others yet to be discovered, are performed by all. Their purpose seems plain enough, for they must be regarded surely as aphrodisiacs, excitants to pairing. They recall the erotic dances of savages, or the ceremonial orgies of ancient civilizations. Such performances, on an even more elaborate scale, are to be met with among the birds.
So far, in describing the horned ruminants, the horns only have been considered; but these animals display yet other secondary sexual characters, which, while less conspicuous, are yet no less important during this critical period of life. Some, as for instance the canine teeth possessed by some of the deer, are decidedly puzzling. While absent, or vestigial, in most, in a few they are greatly developed, and this, too, in species which possess relatively large horns, as in the Muntjac. It seems difficult to believe that the co-existence of these very different kinds of weapons can be of 67vital importance to their possessors; yet unless this be so, one or other would surely have degenerated. It is significant that in the hornless Musk-deer these teeth attain to a very considerable length, at their maximum as much as three inches. That they are used by rival males, and with effect, is shown by the fact that the hides of these animals are often found scored by deep lines cut by these tusks. In those aberrant ruminants, the Camels, quite formidable tusks are present both in the upper and lower jaws, and these are used with effect whenever occasion demands, and often when it does not.
The armoury necessary for successful love-making contains yet other weapons, evolved to supplement physical force, and more subtle in their effect. Such are certain skin glands which, at the rutting season, secrete a copious flow of a creamy, or semi-fluid matter, and pungent odour. In the deer the more important of these are found in the deep pit, or “larmier,” which opens in front of the eye. In the Musk-deer, however, this secretion has a most powerful odour of musk, and is formed in a pouch, or “pod,” of about the size of a small orange, under the skin of the abdomen. The secretion, which is formed by the male only, is of a chocolate colour, and of about the consistence of moist gingerbread. It has a most pungent scent, and when diluted forms the basis of many of our most powerful and most highly-prized perfumes, on which account, it may be mentioned, this animal has for generations been submitted to a most unrelenting persecution. But that is another story.
In most of the antelopes the principal scent gland is seated in a pit in front of the eye, as in the deer. In some, as in the Gnu, it forms instead a swollen, tumid area, oblong in shape, instead of lying in a pit. In the Reedbuck it is placed around the bases of the horns; and in 68the Rocky-Mountain Goat it forms a great bare cushion behind the horns. All have more or less well-developed glands seated in the skin between the toes. But, wherever placed, the secretions thereof are more or less completely suspended save during the breeding season, when they are poured forth abundantly. The precise rôle they play is by no means certainly known. It seems reasonable to suppose that, in the first place, the odour they disperse enables the males to announce their whereabouts to the females seeking mates, should they fail to hear their bellowing. But the antelopes, for the most part, unlike deer, do not, the year round, lose touch with one another; so that it must be concluded that these odours serve as excitants to the act of pairing, and we know that the sense of smell plays a very important part at this time, which, so far as these animals are concerned, is the only period which comes more or less exactly within the meaning of the term “courtship.”
That scent among the antelopes holds a really important place is shown by the fact that the bull of the common Eland intensifies his natural odours by micturating upon the mass of long hair which grows upon the forehead. To do this the head is bent down and turned tailwards, in order that the tuft should receive its due urinary spray! And goats in captivity exhibit the same curious habit. In them, indeed, it is often pushed to such an excess that blindness results, so that the animal has to be slaughtered.
While in many cases these odours are imperceptible to human nostrils, in others this is far from being the case. Among the ruminants the goat is particularly odorous. So also are the giraffe and the water-buck, 69both of which may be detected by their smell at considerable distances. And these emanations are most noticeable in the males and at the breeding season. The bull elephant, both in the Indian and African species, during the breeding season produces a copious flow of aromatic matter from a gland which opens above the eye in the form of a tubular aperture large enough to admit a pencil. This aperture in the African elephant is remarkable for the fact that it is invariably found to be “plugged” with numerous spines of the acacia, which have from time to time found their way in as the animal was forcing its way through the dense undergrowth. This extraordinary fact was first noticed by Mr. F. C. Selous, and has since been confirmed by Dr. Einar Lonnberg.
It is probable that the “bloody sweat,” which at times covers the hide of the Hippopotamus just after leaving the water, is associated with the period of rut. This mysterious exudation is accompanied by small crystals; but though red in colour, it contains no blood. So far no reasonable explanation for this remarkable phenomenon has ever been given, but probably it will be found to be associated with the sexual activities and is possibly odoriferous. A precisely similar exudation occurs in the neck of the male of the Red Kangaroo.
That these secretions play an important and perhaps variable part in the selection of mates seems demonstrated in the case of an incident related to me by my friend Mr. John Cooke, who some time ago was watching a flock of some three hundred sheep while it was being driven by the shepherd and his dogs into a field. As soon as they were securely shut in, and the shepherd had gone, three rams who were included in the flock at once began a three-cornered 70fight. One, presumably the youngest, was soon vanquished. The other two soon settled their differences, and the clashing of horns was at once followed by a very different performance. The master ram began to run in and out among the ewes, sniffing at each, and driving out those whose odour most pleased him. Having at last satisfied himself with a harem of about one hundred, the second ram was allowed to make a like choice, and behaved in a like manner, leaving the remainder to the ram which was first vanquished. May we take it that the strongest and oldest rams selected the youngest ewes, and the oldest were left to the youngest, and first conquered ram? By some such rough and ready method of selection Nature may contrive that the immature male shall do as little harm to the race as possible by mating with the oldest, and in many cases barren females.
Our survey of the “hoofed” animals has so far been confined to the ruminants. Space must now be found for a brief review of what obtains under like circumstances in the case of the great pachyderms—the Elephant, Rhinoceros and Hippopotamus; the Pig and the Camel.
Plate 9.

Photo by Lord Delamere, from “The Living Animals of the World”
ELEPHANTS.
The sexes differ but little in general appearance: and chiefly in the superior size of the male and his more massive tusks.
[Face page 70.
As to actual “courtship” among these animals practically nothing is known; but the varied and formidable weapons which they possess are enough to show that the secondary sexual characters play a very important part in the preliminary capture of mates. That they may also be used for the more prosaic purpose of securing food is nothing to the point. In the Elephant, for example, the tusks are sometimes of enormous size and weight, specimens of eleven feet in length and weighing as much as two hundred and fifty pounds are on record. They are used for cutting through the bark of machabel trees, which is then seized by the trunk and torn off, for elephants are extremely fond of this bark; and they are 71also turned to account in breaking up roots which have been exposed by digging with the fore-feet. But this is certainly not the main purpose of such weapons. On the contrary, their use is primarily as weapons of offence between rival bulls. As one would expect, they never attain to a very large size in the female, but that they are large enough to serve her at need is shown by the fact that a portion of a tusk, evidently of a cow-elephant, was once found embedded in the jaw of a bull. There can be little doubt but that this was broken off in an endeavour to repel the advances of a too amorous male, for, as with all animals, pairing is impossible without the consent of the female, and this is never accorded until she is desirous that it should take place. As a preliminary to this, an amorous dalliance is perhaps the invariable rule among animals, and this takes many and often strange forms. The Elephant affords a case in point. For the late A. H. Neumann once came upon a pair which were evidently, as he says, “love-making.” Creeping upon them noiselessly, he found the male fondling his mate with his trunk, and then, standing side by side, they crossed their trunks, and put the tips thereof into each other’s mouths, the elephantine form of kissing. Deer, cattle and horses, cats and dogs, constantly lick one another under like circumstances.
Superficial secondary sexual characters are wanting both in the Hippopotamus and the Camel. Both, however, possess a formidable armature of teeth which are capable of inflicting very severe wounds. In the Hippopotamus the canines are of enormous size, and their punishing power is further strengthened by the fact that they work in opposition to a pair of similar teeth in the lower 72jaw; they cut like a pair of shears, the upper closing upon the lower pair with the precision of scissors-blades. In addition, the lower jaw develops two long, blunt-pointed, ivory spikes, which are scarcely less to be dreaded. With these weapons the bulls fight furiously, and it is no uncommon thing to find vanquished males frightfully mauled, the hide being lacerated from head to tail. Protection, in a measure, is afforded by its enormous thickness, but the great folds and pleats of skin seen in the Rhinoceros are never developed. The females, however, are similarly armed, and the teeth are nearly as large as in the males, which is a rather unusual occurrence.
The Swine, which are near relations of the Hippopotamus, in like manner develop huge pointed canines, and these reach their maximum in the great Wart-hogs of Africa. But in the swine the mechanism differs, for although the canines are closely opposed, the shaft of the upper teeth curves upwards, and the lower teeth are much smaller than the upper. In fighting, these animals do not bite, like the Hippopotamus, but use the upper canines to rip up their antagonist with a sudden, swift upward and sideways movement of the head. How dangerous is the wound thus inflicted those who have hunted the wild-boar know well. A curious exaggeration of this arrangement of the teeth is seen in the Babiroussa. Herein the upper canines grow directly upwards, actually piercing the upper lip as in the case of the downwardly growing tusks of the elephant. That these teeth, however, are of any service in fighting is doubtful, for the upper tooth curves upwards and backwards in a semicircle so that the points are harmless. The tusks of the lower jaw, however, are extremely long and pointed, though their wounding power is limited by reason of the upper teeth. This may account for the fact that 73the head, the part mostly attacked by enraged boars, presents no sort of armature designed for defence; while in the Wart-hog, on the other hand, great solid bucklers of hide stand out on either side of the head below the eyes, giving the animal a most repulsive appearance, but affording him a very present help in time of trouble. In the wild-boar, where the tusks are shorter, no such protective armature is needed.
Plate 10.

HEAD OF MALE WART-HOG.
In the “Swine” family the canine teeth are always greatly developed, but they attain to their maximum, relatively, in the Wart-hog.

Photos by Scholastic Photo Co., from “The Living Animals of the World.”
MALE AND FEMALE BABIRUSA.
A characteristic of this pig is the peculiar development of the tusks in the male, the upper pair of which grow through the lips and curve upwards.
[Face page 72.
Plate 11.

Photo by Lord Delamere, from “The Living Animals of the World.”
SOMALI ZEBRAS.
The Zebras, unlike their cloven-hoofed relations, have no weapons, save for inter-tribal conflicts. Yet they have been as successful in holding their own against lions and other predatory animals as species provided with horns.
[Face page 72
Plate 12.

Photo copyright by A. H. Bishop.
GIRAFFE.
The horns of this animal can prove formidable weapons of offence on occasion, though they are useless against predatory animals.
While the ungulates, or hoofed animals, are peculiar in the development of horns as weapons of offence, they are by no means singular in the use of teeth for this purpose. In some cases, as in the Muntjac, both forms of armature are present. The only other instances where teeth in this group of animals are used for offensive purposes are those furnished by the Camel and the Horse. But here they do not exhibit that excessive size which is met with in the Elephant, and some of the Swine. In both the Camel and the Horse it is the canine which is used, and both jaws are similarly armed. Since the camel has no upper incisors, the part played by the teeth is beyond dispute; but it has been contended that the horse uses his incisor or “front-teeth” alone when fighting. But this is not so; the canines can, and do, inflict ugly wounds, as is shown by the necks of zebras.
A further method of defence among the larger Ungulates, at any rate, is resorted to when hard pressed: and this is the use of the hoof in kicking. Giraffes kick both after the usual fashion and in striking downwards with the fore-foot. And an interesting demonstration of this has been furnished by Mr. F. C. Selous in his delightful “African Nature Notes.” He relates that on one occasion he came across a calf only a day or two old, with its back broken. 74 From scratches on the calf, and the footprints on the ground in its vicinity, he was at once enabled to gather the cause of its terrible plight. In a word, it had been attacked by two leopards, and the mother, in an endeavour to beat off the assailants with a blow of her fore-foot had accidentally struck her offspring. Horses, Cattle, Antelopes, Camels and Elephants can all kick with precision and effect. So far as the evidence goes, however, this is a method of defence used against beasts of prey, and is rarely, if ever, employed in conflicts between rival males. Females persecuted by the undesired attentions of amorous males, however, do, as we know from the case of domesticated animals, use this device to defend themselves.
It is not difficult to account for the origin of such secondary sexual characters as manes, beards, tusks, and brightly-coloured areas of skin, though whether our interpretations are really correct is another matter. But no attempt to explain the origin of horns has yet achieved a like degree of persuasiveness. These weapons appear only in the Ungulates, a group which has, in past times, given birth to some very extraordinary types of head armature of this kind. These must be excluded from the present discussion; suffice it to say that, as usual, they were the adjuncts of the males. According to current theories it is supposed that these weapons arose as the result of the action of sexual selection. It is assumed that the hornless ancestors of now horned ruminants fought for their mates by “butting” with the forehead. Naturally, other things being equal, the thickest skulled combatants obtained the mastery. Any tendency to develop frontal “bosses” of bone would further enhance the chances of success, and would, indeed, 75soon become necessary for survival. And from such “bosses” the passage to horns and antlers forms an easy transition. Just such incipient horns or “bosses” actually make their appearance in the domesticated horse: but these animals never butt at one another. If, however, we regard horn-production as an inherent diathesis of the ungulate somatoplasm, we have an intelligible basis for the explanation of horn development.
The formidable horns of the Rhinoceros are of a totally different character, being solid structures formed by hairlike agglomerations, firmly fixed upon a roughened area of the nasal region. These weapons play a very important part in settling disputes between rival males, but on other occasions demanding offensive tactics the Indian Rhinoceros at any rate seems to depend rather on his power of wounding by means of the chisel-shaped lower incisors. These, by means of a swift lateral movement of the head can be made to inflict most terrible gashes, as those who hunt with elephants well know. It is quite possible, however, that the teeth are also thus used during struggles for supremacy. And this may perhaps account for the enormous bucklers of skin developed by the Indian Rhinoceros, but only indicated in the case of the African species.
All the larger Ungulates, and many of the smaller species, are polygamous. The Rhinoceros, and all of the swine-group save the Hippopotamus, among the larger species are exceptions to the rule. The preponderance of females which this implies is generally supposed to be due to the losses sustained among the males by fighting during the struggle for mates. The case of horses, however, seems to militate against this view, for though they undoubtedly fight furiously, no evidence is forthcoming 76to show that such conflicts terminate fatally.
Were it possible to secure the necessary data it would probably be found that polygamy, and polyandry, are determined solely by the numerical proportions of the sexes: the excess of males or females being due neither to “Natural” nor “Sexual” Selection, but to inherent peculiarities of the germ-plasm tending to produce an excess of males, or females, as the case may be.
Finally, all the evidence goes to show that it is a mistake to suppose that polygamy is due to the excessive sexual avidness of the males, which impels them to first essay the overthrow of all possible rivals, and then to appropriate every female within their sphere of influence, holding them by force. On the contrary, this plurality of mates is thrust upon them. And this because the females, impelled by “mate-hunger,” attach themselves to the nearest male within call: the size of the harem depending on the number of available males. The battles which are fought between rival males are no more sanguinary than in the case of monogamous species. This contention is well illustrated by the African Wydah-birds (Vidua), which are markedly polygamous, though they have no special weapons of offence. In districts where males are numerous the harem will not exceed eight, or ten, females; where males are scarce this number may be increased to fifty. In like manner the varying number of hinds accompanying a stag are to be regarded, not as an index of his prowess, but of the scarcity or abundance of males in the neighbourhood.
A Surprising Relationship—The Lion’s Mane—The Sabre-toothed Tiger—Some Theories about Origins—Sea-lions in Love—Some Strange Ornaments—Whales and Weapons.
That the Lion and the Lamb could possibly have been derived from the same stock seems incredible: yet such is the case, though the pedigree is now well-nigh lost in the mists of a hoary antiquity. It is not surprising, then, that in their present-day garb they should show so little in common. Nor is it strange that among their many points of divergence the one should differ so conspicuously from the other in the matter of secondary sexual characters. For when these are conspicuous among the Ungulates they usually take the form of horns, of which the Carnivores have no need, for the teeth and claws whereby they win their daily portion of meat make equally serviceable weapons of offence when turned against their own kind.
Among the larger Carnivora, the Lion alone displays any obvious distinction between the sexes in the matter of ornament, and this in the form of the well-known mane. Darwin, and later authorities, have regarded this 78as a shield to protect the great blood-vessels from injury during battles between rivals. But it is not very clear that this alone is sufficient to explain its presence, inasmuch as the Tiger in this respect is defenceless. Mr. F. C. Selous long ago pointed out that the varying abundance of the mane is due to climatic causes. Lions which live in districts where the nights are very cold, as in high table-lands, have large manes; those which occupy lower ground, where the nights are relatively warm, have but a scanty mane. It is clear, however, that the abundance of the mane is not determined by the need for warmth, otherwise it would have been as well developed in the female. Rather we must regard a low temperature as conducive to the growth of long hair when a natural tendency to produce this is present.
There are few men who can claim to have so great a first-hand acquaintance with Lions as Mr. Selous, and he has pointed out to me one significant fact which seems to show not only that the mane has not been developed to serve as a shield when fighting, but that fights between rival males must be rare. And this because of the absence of any evidence in the shape of scars on the skin. With claws so formidable as those of the lion, ugly wounds would certainly be made in any prolonged conflicts, for the skin of this animal is very thin.
In the now extinct Sabre-toothed Tiger the upper canines were of enormous length, and it is not improbable that they, on this account, exceeded the bounds of usefulness; that, while as weapons of offence they may have proved exceedingly effective, yet they hampered the animal when feeding. In many ways one is reminded by these weapons of the huge tusks of the Walrus. These are 79blunt-pointed, and are said to be used very largely for digging up the large clams and other burrowing shell-fish on which this animal mainly feeds. They are also used as levers to drag the huge body out of the water on to the ice. As fighting weapons they are formidable, and the wounds they inflict are sometimes serious. The polygamous habits of this huge creature may account for the fact that they are so much larger in the males, wherein they may attain a length of thirty inches, and a weight of eight pounds a-piece.
In connection with the monstrous tusks of the Sabre-toothed Tiger there is a point which so far seems never to have attracted the attention it deserves. And this concerns two small flanges of bone which project from the lower border of the end of the lower jaw. In themselves they are unimportant: they lie, it is to be noticed, parallel with the points of the great upper teeth which descend on either side of them. Their full significance is not apparent till we turn to the skull of another extinct animal of quite another type—the huge Dinoceros, one of the Ungulates. This animal was also armed with an enormous pair of tusks, which also, when the mouth was closed, descended on either side of a flange. In this case, however, the flange was developed to such an extent that its free edge descended to the level of the point of the tusk, thus affording it protection against injury. The really striking feature of this curious down-growth is not apparent till an attempt is made to explain its presence. What determined its growth? It seems to furnish us with another of the many instances which are to be found of the correlation of growth between unrelated parts, for there is apparently no traceable connection between the growth of this pair of teeth in 80the upper jaw and the development of the flanges of the lower border of the jaw which are embraced by these teeth. In the Sabre-toothed Tiger the inciting cause to this flange growth, whatever it may have been, seems to have been much weaker than in the case of Dinoceros.
Naturally one asks, can the whole thing be explained by the theory of Kinetogenesis promulgated years ago by Cope? That is to say, are these curious down-growths the result of a response to a stimulus set up in the lower jaw by constant lateral blows dealt by the tusks against the side of the jaw during the lateral movements of the jaw when feeding or ruminating? Such movements in an Ungulate would be frequent and constant: hence perhaps the more striking result. On account of the scissor-like action of the jaws in the Sabre-tooth such lateral movements were far less extensive, and less powerful. But though this explanation sounds plausible, it presents many difficulties. In the first place it seems to commit one to the admission that the responses of the Somatoplasm during the life of the individual are transmitted to the germ-plasm: that, in short, the characters acquired by the individual during its lifetime are transmitted to its offspring. And there are insuperable difficulties in the way of such a theory. Yet, it must be admitted, it is no less difficult to believe that this correlation of growth is due solely to fortuitous variation, for one cannot really conceive of a variation of this kind taking place in two such different structures independently. Such a conception would have been less difficult if the case of Dinoceros alone were known to us. We could have supposed that, somehow, the lower jaw started to produce its flange just as the teeth began to develop an excess of growth which carried their points beyond the level of 81the jaw. But the Sabre-tooth shows that the tusks had assumed a growth relatively exaggerated as in Dinoceros, and yet the flange never attained to more than feeble development. We cannot rest content with the theory that the flange is due to the constant stimulus of blows struck against this region of the jaw during the lateral movements which take place when feeding. Were these animals alive to-day it could be tested by extracting the tusks during infancy, when, the stimulus being removed, the flanges should not appear.
There are yet other aspects of the skull of Dinoceros which may well be considered here. The first concerns the excessive armature of horns, there being no less than three pairs supported on massive bony cores; and the second the ridiculously small brain cavity which is proportionately smaller than that of any other known mammal, recent or fossil. This poverty of brain-power was probably one, if not the chief, factor among the causes which brought about the extinction of this strange beast. Even more formidable horns were borne by the extinct Arsinoetherium. But this animal did not display the double armature of horns and tusks.
Among the Carnivora monogamy is the rule, though the Lion is occasionally polygamous. But the Eared-seals (Otaria), or Sea-lions, and Sea-bears afford a striking example of polygamous species and of the ferocity they display when sexually excited. These animals, moreover, are capable of the most astonishing powers of endurance and vitality, exceeding indeed that of all other mammals. Since the habits of the Northern Fur-seal (Otaria ursina) have been more carefully studied than those of any others, it may serve as a sample of the rest.
Living for the greater part of the year in the open 82sea, the old bulls—animals of six or seven years old—are the first to seek the “rookeries,” or breeding grounds, taking up their territory a full month before the cows arrive. Later, the younger bulls appear, and the more daring endeavour to force their way through the ranks of those who have already taken up positions. This often leads to fighting, but more usually nothing further than “bluffing” is indulged in, though it is commonly supposed that very severe engagements take place. This seems, however, to be only occasionally true. In due course, generally about the second week in June, the cows begin to arrive, at first in straggling numbers, but soon the main body puts in an appearance, and before the end of the month many thousands of both sexes are crowded along the foreshore. But yet, contrary to the generally accepted belief, no serious fighting takes place. The bulls quietly seize the females as they arrive. It would seem that the first arrival serves as a focus of attraction for all later comers landing in the vicinity. The bull holding the most advantageous post—that is to say, that nearest the best landing-place—starts the collection and, unintentionally, the distribution of the cows. Having seized the first arrival, he places her by his side. As the later females arrive he gives each a most cordial welcome, and then proceeds to round up his harem. But soon he has more wives than he can continue to control. Do what he will, he cannot be in two places at once; and thus it is that in rushing off to chastise some covetous neighbour, one or more bulls on the opposite side of his harem proceed to make captures from his horde. And this system of abduction goes on over the whole rookery till all the cows have been appropriated, leaving a crowd of envious bachelors in the background who have not yet developed either courage or strength to secure mates for themselves.
Plate 13.

Photo by New York Zoological Society, from “The Living Animals of the World.”
CALIFORNIAN SEA-LIONS, OR EARED SEALS.
The “bulls” of the Eared Seal are much larger than the “cows”; they have otherwise no very conspicuous “Secondary Sexual Characters.”
[Face page 82.
But within forty-eight hours of their landing the cows give birth to their “pups.” And it is for this purpose, and not for mating, that they come to land. Within a few days of the birth, however, the females are “in use” again. This is the critical period in the life in the rookery. For the bulls now become frenzied with excitement and fight most viciously one with another, each hoping to possess himself of his opponent’s harem. Each tries to seize the other by the fore flipper, and, failing in this, the fangs are buried in the back. They hold tenaciously, each trying to force the other to relax his hold; but commonly this vice-like grip is maintained till the skin gives way, leaving great bleeding rents. Sometimes the contest rages till one or both is fatally wounded. Often during such duels an idle bull, hitherto unable to secure a harem, will rush in and capture that of one of the combatants!
In the management of the harem the bull is an adept. Whether he has five cows or fifty, he is, says Dr. Lucas, “master of the situation.” His will is law. Not that it is always tamely accepted as such, but the result is the same. If a cow becomes restless, and moves about, a warning growl usually quiets her. If the movement is persisted in and an attempt to escape evident, the bull is up at once with a show of fierceness and in chase. He may simply strike her down with his open mouth. Often in doing so his sharp canines tear a gash in her skin. He may even seize her in his mouth and deliberately throw her, or carry her back into the harem. If the cow thinks she has a chance to get away she may try to outrun him. If she miscalculates the distance he seizes her, after a few swift bounds, by the skin of the back, or by the hind flipper, and tosses her, often torn and bleeding, into the 84family circle. As a rule, however, she avoids this seizure by turning and facing her lord and master, and biting him in the breast and throat. But all to no purpose. In spite of her violent protests he pushes her backwards before him into the fold.
Sometimes in her efforts to improve her position she runs up to, and is seized by, a rival bull. Her lord speedily asserts his ownership by getting a grip wherever he can on the would-be truant. Then begins a tug-of-war between the two bulls, during which the wretched victim of their rage may be torn in pieces. By the elimination in each generation of the more querulous and discontented, the peculiarly gentle and passive nature so characteristic of the females has been developed.
After the first ten days’ sojourn ashore the female is allowed to go to sea to feed, returning presently to suckle her young. The bull, on the other hand, can enjoy no such privilege. For three long months he must keep watch and ward fasting—at first, in order that he may retain his territory; later, that he may retain his harem. This fast, having regard to the loss of energy and blood which this strenuous period entails, is wonderful; for in the case of all other animals fasts are always associated with absolute rest and sleep. Not so with the Sea-lion; he arrives at the breeding-ground fat and well-liking, he leaves a starved and battered wreck.
The foregoing summary of the habits of these most interesting and much persecuted animals is taken from the exhaustive report of Dr. F. A. Lucas and Mr. Charles Townsend. These two distinguished naturalists accompanied the United States contingent of the Fur-seal International Commission despatched in 1896–97 to 85inquire into the threatened extermination of these animals. Major Barrett Hamilton accompanied the British contingent, and also made a report. And it is curious to note that on some points he is diametrically opposed, not only to the American naturalists, but to all other writers on this theme. He contends, for example, that “nothing could better illustrate the fact that it is the cows, and not the bulls, which have the real control of the harem-system.” He traced the rapid growth of two harems from four or five to as many as eighty cows. And he tells these were completely out of control and free to move about as they wished. “The bulls, in spite of all their bluster, had the flimsiest of nominal dominion, and the cows were always able to, and frequently did, leave the harems daily to dally with the cowless bulls on the outside. Yet ... as long as they chose to sit massed together on the ground which had been appropriated by the two stronger bulls, no weaker rivals could approach to within ten yards of them. The master of the harem had no control over its occupants, but he was absolute lord of the ground on which they sat.” This is certainly curious, but more so is the fact that these females were allowed to return by the “cowless bulls” outside the charmed circle. Later in the season he tells us he witnessed an even better illustration of this singular behaviour. At this time “the division of the cows into harems was a very unequal one, the smaller bull being only able to keep a very few cows, while the larger one claimed the greater part of the rookery. But the cows could pass over to the smaller bull’s ground as often as they liked; and he probably was father to a great many more of the pups born in 1898 than those of the half-dozen cows over whom he claimed control.” In regard to two 86other bulls in another cart of the island, there came a time when the inequality of the harems reached such a pitch, that the newly-arriving cows “had to lie in scattered groups outside the main mass, and thus permitted the weaker bulls to form new harems out of the reach of the two strong old bulls.” But perhaps the most singular feature of all was the indifference which one old bull displayed towards a little bachelor, permitting him to enjoy the most intimate relations with one of his cows without displaying the least sign of annoyance, as if he could scarcely regard one so young as a rival.
There is much evidence to show that the erotic side of the male-seal develops early. “I saw,” he says, “the little black pups acting to each other in a way that made it certain that their sexual feelings had already made themselves felt.” This one can well understand, for only animals of strong sexual tendencies could survive the strenuous life which the period of sexual activity entails.
The very different interpretation of the behaviour of these animals at this very important stage of their life-history must be due to the fact that different colonies were studied which were living, too, under somewhat different conditions. It seems clear, for example, that the landing of the females so graphically described by Dr. Lucas was a landing under exceptional circumstances, the master bulls having taken up positions at the only spot where access to the desired breeding quarters was to be found; while Major Barrett Hamilton was probably fortunate in seeing phases which were wanting in the “rookeries” examined by Dr. Lucas. And both these observers again differ in the accounts they give of the life of such “rookeries” with those by Mr. Elliot, who explored these teeming colonies some years earlier when 87the number of animals forgathered there was far larger and the fighting, apparently in consequence, was far more severe.
In the matter of secondary sexual characters the most remarkable of the seal-tribe are those of the Elephant Seal and the Bladder-nosed Seal, and this because of the extraordinary development of inflatable tissue above the muzzle which these animals display. Of their life-history we know little enough, and this despite the fact that for generations the Elephant Seal was mercilessly hunted and slain for the sake of its oil. Millions were slaughtered during the last century, yet only scraps of information on the economy of the creatures has come down to us. All that is of any value, and especially in regard to the “Courting” period, we owe to Mr. Charles Townsend, of the New York Aquarium, and this in regard to the northern species, Macrorhinus angustirostris of Guadelupe, though it may safely be inferred that the Southern, Antarctic species, M. leoninus, differs in no essential respects.
According to Mr. Townsend, the adult bull, having taken possession of his territory and formed a harem, is constantly called upon to wage duels for both with less fortunate rivals. And the severity of such combats was attested by the deep wounds and festering sores of the necks of these old warriors—which, at their maximum, attained in the days of their prosperity a length of nearly thirty feet and a girth of sixteen feet; but the last survivors of the race to-day seem rarely to exceed twenty-two feet. The weapons used in fighting are the canines, and the only armour they possess is that formed by the thickening of the skin of the neck, which forms a great massive shield, so that really dangerous wounds are rare. The great fleshy proboscis, the most vulnerable part, is carefully 88guarded by the upturned position of the head. The use of this trunk-like organ, which may attain a length of about fifteen inches, is not clear; it seems to serve mainly as an “ornament,” at times, too, furnishing a very definite indication as to the temper of its owner. While the animal is slowly moving its great carcase from place to place, this remarkable organ is relaxed, and pendent; but when fighting it is closely contracted so as to be out of harm’s way. Whether it plays any useful part in the capture of food is not known; but it is probably much displayed during phases of sexual excitement. In young animals, it is significant to notice, as well as in the adult female this trunk is entirely wanting, which seems to suggest that this peculiar feature has only been recently acquired, the young and the adult female, as is the rule, standing nearer to the early forebears of this strange type. There is an enormous difference, it should be remarked, between the sexes in the matter of size, the female not attaining more than half the bulk of her lord. A further interesting point concerns the coloration of the young, which are black, while the adults are brown. Doubtless this is connected with the requirements of the young, the black coat attracting more heat than the lighter-coloured coat of the adult.
As touching that curious creature, the Crested, or Hooded Seal (Cystophora cristatus), a native of the colder regions of the North Atlantic. This animal is remarkable for the development, in the males alone, of a great crest or casque on the head, which is formed by a large inflatable air-sac over the ridge of the nose, and communicating with the nostrils. When fully inflated, it covers the head as far back as the eye. Its purpose is a matter of conjecture. It seems to be inflated either when he animal is greatly excited, as when challenging rival 89males, or when threatened with danger from other causes, as when attacked by man. The males are exceedingly pugnacious, and fight with one another for the possession of females with great ferocity, such contests being accompanied by cries which can be heard for miles. From the difficulty which Esquimaux and sealers find in killing the animal with clubs it certainly seems as if this strange wind-bag were more than merely ornamental.
Plate 14.

Photo copyright, W. P. Pycraft.
ELEPHANT SEAL.
This is a young animal. Note the great size of the eyes, and the general “seal-like” character of the head as compared with that of the adult.

Photo by courtesy of Charles Haskins Townsend, Director of New York Aquarium.
NORTHERN ELEPHANT SEAL.
Adult male and female, and yearling. The male shows the enormously inflated snout.
[Face page 88.
That those extraordinary creatures the Cetacea—the Whales and their kin—are derived from the same common stock as the typical carnivora there can nowadays be no doubt, widely as they have departed from their land-dwelling relatives in almost every possible feature of their organization. In the matter of their “Courtship” we know nothing, but we may infer certain incidents in this critical period of their life-history from the peculiar nature of the secondary sexual characters which some species display. Thus in the Pilot Whale (Globicephalus) and the Bottle-nose Whale (Hyperoödon) the forehead, in the bulls, is enormously swollen by a mass of fibrous tissue so dense as to turn the blade of the sharpest knife, as I know well from attempts to dissect this region. Now the only use, surely, for such a cushion is that of a battering-ram by rival males in charging one another, as rams and other horned animals will do. In the Bottle-nose Whale this cushion is backed up by an enormous mass of solid bone thrown up by the maxillæ. The origin of this bony growth is interesting, for it appears first as a slight swelling in the rare species Berardius; it is seen at a further stage of growth in the female “Bottle-nose” (Hyperoödon), and attains its maximum in the male, where it stands unique. There are two other species which demand 90notice here. The first is Layard’s Beaked Whale (Mesoplodon); the second the Narwhal. The former is the only vertebrate which in a wild state wears a muzzle! In this species the teeth have totally vanished save for a pair in the lower jaw, which are found towards the end of the jaw. These in the adult, or perhaps we should say senile, male grow upwards and inwards, finally meeting one another above the upper jaw, so as to make it impossible for the animal to open its mouth more than the fraction of an inch! Surely here we have a secondary sexual character carried to an excess, and so proving not only disadvantageous to the animal, but positively disastrous, for it seems clear that so hampered the creature can feed only on the most minute forms of animal life, which could only be captured and swallowed with difficulty. It is true that the Rorquals feed on excessively minute Crustacea, but they are able to take in enormous quantities at a time, the “whalebone” serving the office of a sieve to prevent their escape. The Mesoplodon has no such aids. One is tempted to believe that the skulls displaying this most curious feature are abnormal, comparable to those, say, of rabbits wherein the teeth have grown so excessively long as to close the mouth, on account of the displacement of the cutting surfaces by accident. But there is nothing to afford support to this view, and one must therefore fall back on the suggestion of senility.
The Narwhal has long been celebrated for the enormous size of the canine teeth, the only teeth present in the jaws. As a rule, only one leaves its bony socket, the other, commonly the right, remaining as a mere vestige, seven or eight inches long within the skull. The protruding tooth, which is spirally fluted, may attain a length of nine feet. Occasionally both teeth are developed, and in this 91case the spiral is the same, differing in a very striking manner from the spiral horns of ruminants, wherein one presents a right, the other left-handed spiral. But what purpose do these teeth serve? This question has never yet been definitely settled. Some hold that it is used to break open breathing holes in the ice, for the animal lives in the far north: others that it is used as a spear in hunting prey. Some aver that it serves as a weapon of offence, being used by rival males in their struggle for mates. Scoresby, the explorer, indeed, says he has seen young males in mock-battle, fencing with these remarkable weapons. But until we have more satisfactory data, we must regard this armature of the Narwhal as affording another instance of a secondary sexual character of doubtful value to its possessor.
Generalities—Darwin v. Wallace—The Peacock in his Pride—The “Display” of the Peacock Pheasant—The Splendour of the Argus Pheasant and the Marvel of its Eyes—The Frill of the Amherst Pheasant—Birds of Paradise in the Toils of Love—Inflated Suitors—Ruffs and Reeves—Fearsome Weapons and their Uses—Birds which dance—Musical Birds—The Bird’s Voice-box—The “Lek” of the Capercaillie—Instruments of Percussion—The Curious Performance of the Wood-pecker.
The fact that so little is known about the mammals during that period when the all-important work of securing mates is going on, and of the subsequent events, is largely due to the difficulties which close observation of this phase of their life-history entails. With the birds matters are far otherwise; their haunts are more accessible; they are far more numerous, and much more easily kept under observation. Consequently, we have a tolerably complete knowledge of the lives of some species, at any rate, during the reproductive period; that is to say, as to the sequence of events from the beginning of the reproductive activities onwards; but the interpretation of what is seen is another matter. No attempt which has yet been made to fathom the psychology of sex has yielded more than a slight insight into what is taking place. Nevertheless, this is 93an aspect of the subject which has a far more important bearing on the problems of evolution than is generally realized. But these pages are concerned rather with the relations between the sexes, than with the subtle forces which have fashioned and control conduct in this regard.
In all that concerns the problems of sex, which is to say of reproduction, birds, speaking generally, display a briefer and more condensed sequence of events than the mammals; and, moreover, many species compel the attention even of the most incurious, to their behaviour at this time, through the development, either of song, or of fantastic displays of their amorous feelings: while others force themselves no less conspicuously under notice by their habit of nesting in large, and often enormous colonies.
In the matter of the development of secondary sexual characters birds stand conspicuous among the Vertebrates, and easily eclipse the mammals; among which bright, strongly contrasted, colours are the exception. Among the birds they may almost be said to be the rule. Also, in this category we have to reckon song, and the production of more or less musical sounds by the agency of internal resonators or of specially modified feathers; as well as quaint forms of posturing which may be included under the head of dances. Further, some species have developed formidable weapons of offence. These things are interesting enough in themselves, but they become still more so when we reflect that they formed the corner-stone of Darwin’s theory of “Sexual Selection,” and that Wallace’s criticisms thereof were inspired by evidence from the same source.
The interests of this chapter will best be served if the evidence on which this theory was founded be first surveyed: 94when Darwin’s deductions and the criticism which they have aroused will be the more readily appreciated.
Definitions are always liable to exceptions; and concrete cases are better than abstract terms. Birds, then, perhaps better than any other group, illustrate what is meant by the term “secondary sexual characters,” if only because examples are so constantly at hand. Save among experts, sex among birds cannot be determined except by the differences in plumage, or sometimes in size, which the sexes display. But even here, it is only among species which occupy what we may call a mid-evolutionary phase in which this discrimination is possible. Among “generalized” species, wherein the plumage is of sombre hue, there is no external distinguishing mark between male and female; and the same is true with species which have attained to the maximum of resplendent plumage; as for example many Parrots and Kingfishers, where again both sexes, and at all ages, display the same vivid hues. Thus, in the case of either of the two extremes, the study of behaviour during the breeding season is one of great difficulty and no less uncertainty. Where the sexes are sharply distinguished by differences of coloration, however, as with the Peacock, the matter is otherwise. This bird, from time immemorial the symbol of vanity, illustrates in a singularly effective manner the broad features of what is commonly meant by “courtship” among birds, while it furnishes a no less striking example of the development of “secondary sexual characters.”
One might have supposed that birds, under the spell of that irresistible desire for sexual intercourse, would behave differently in regard to their “courtship” according 95to whether they were monogamous, polygamous, or polyandrous: but while their behaviour during this period of the life-history presents an extraordinary variety, it is only at any rate slightly determined by the plurality or otherwise of mates; and the same rule holds in regard to the brilliancy or otherwise of coloration.
The most common manifestation of sexual desire among birds takes the form of strange posturings which are, in some species, enormously exaggerated by the display of vividly coloured frills, tufts, or other conspicuous modifications of the normal plumage. The Peacock affords a most excellent example of this combination of the contortionist and the beau, though the nature of this display is by no means generally understood. This applies more particularly to artists, who from time immemorial to the present day, in essaying to paint the Peacock in his pride, have invariably fallen into the error of treating the great ocellated train as if it were the tail, placing it where, of course, the tail ought to be, at the end of the body! As a matter of fact it is nothing of the kind; these gorgeous plumes are really exaggerated tail-coverts which, when set on end, appear to arise from an oval shield of metallic green scales—the central back-feathers. When this trailing glory is erected, the bird throws the body forwards and downwards, so that the outermost train-feathers fall downwards on either side in front of the wings, which are more or less trailed: so that from the front only the head and neck are visible, the rest of the body being hidden behind the screen, as may be seen by a reference to the accompanying photographs. The manner of this display is extremely interesting, for the bird seems to be conscious of the effect produced: though it cannot be supposed that this is really the case.
When displaying, the bird gradually approaches the nearest female and slowly erects these extraordinary plumes. So soon as this is accomplished he begins to walk backwards towards the object of his attentions, presenting nothing but a great round shield of dull brown feathers, backed up by the tail-feathers, and the dull-coloured wings. So soon as he judges himself near enough, however, he suddenly swirls round, confronting her in all his splendour, and heightening the effect with a loud scream accompanied by a rapid, vibratory, motion of the train-feathers which produces sounds like the pattering of rain on leaves. Then he stands before her, with bowed head, as if to give her an opportunity of drinking in his splendour to the full. Commonly, however, she appears to be utterly indifferent, and either walks away or continues a real, or affected hunt for food, as if no such thing as a love-sick suitor were within a hundred miles of her! But sooner or later his suggestive attitudes beget an answering response, and pairing takes place.
The display of the beautiful Peacock Pheasant differs conspicuously from that of the Peacock, and recalls that of the Argus Pheasant. In the Peacock Pheasant, as will be seen from the adjoining photograph, the wings, and tail, are alike bedecked with ocelli. The display is made by the bird as it crouches close to the ground, with the wings and tail raised to form a continuous, patterned surface, the head being swiftly moved during the performance; hence its blurred outline in the photograph.
Plate 15.

Photo by D. Seth-Smith.
PEACOCK PHEASANT.
The display of this bird differs conspicuously from that of the Peacock and recalls that of the Pigeon in some respects. The “ocelli” on the wings afforded Darwin the interpretation he sought for as to the meaning of the notch in the “eye” of the Peacock’s tail-feather.
Plate 16.


Photos by the Author.
“THE PEACOCK IN HIS PRIDE.”
In the upper figure it will be noticed the “train,” when erected, encircles the base of the neck; the lower figure shows the train supported by the tail and dropping on each side in front of the wings.
[Face page 96.
The Argus Pheasant is an even more wonderful performer than the two preceding species. In this bird, it should be remarked, the tail and the secondary wing-feathers are enormously lengthened, the latter to an extent met with in no other bird, showing that the struggle for existence cannot be very severe with this species. For 97if long journeys had to be undertaken in search of food, or to avoid extremes of climate, or enemies had to be swiftly escaped, such cumbersome wings would lead to speedy extermination. But an even more remarkable feature of these wings is their wonderful coloration. The primaries have blue shafts, and a most delicately mottled pattern formed by spots of reddish chocolate on cream-coloured ground, while the secondaries have their broad webs ornamented with large ocelli, to be described in greater detail presently. When under the influence of sexual excitement Darwin tells us, the wings are so spread as to form a deep concavity, an effect which is gained by pressing the primaries close to the ground, and turning the elbows upwards. Within this concavity lie the ocelli, in radiating vertical rows. But to produce this effect the bird has to turn its head under its wing, so that it lies behind the screen. Hence it cannot see the female which is the object of these captivating antics. As a consequence, to discover whether he has an audience for she will often walk disdainfully away—he has constantly to thrust his head through the curtain, and hence many of the feathers in this region get much worn.
By nature it would seem the Argus Pheasant is a very solitary bird, though we must assume it is polygamous. As the breeding season advances, however, the male proceeds to choose some open space in the depths of the forest—which it never leaves—and therefrom to clear all the dead leaves, and twigs, for a space of some six or eight yards square, so that nothing but the bare earth remains, and thereafter this area is kept scrupulously clean. Here, in solitary state, for a short season he remains, calling at frequent intervals to advertise the fact 98that an eligible male is in the neighbourhood desiring mates. A dozen times in succession he will break the stillness of the forest gloom with a loud, “How-how, how, how, how!” Sooner or later comes a responsive, “How-owoo, how-owoo!” and in a short time, guided by the sound, one or more females discover the object of their quest. But the pairing desire has not yet reached its full intensity, and doubtless to kindle this the display just described is enacted, and not once, but a dozen times probably, before the desired state of frenzy has been aroused. Not seldom another male answers the cry, and this inevitably leads to a duel whereby the fittest and strongest male is speedily discovered.
A word as to these ocelli. This pattern is rare among birds, and Darwin brought to light some extremely interesting facts regarding it. He was led to investigate the matter by his curiosity as to the meaning of the notch in the ocelli of the Peacock’s train-feathers. At last he noticed that among the different species of Peacock Pheasants there was one (Polyplectron chinquis), in which the ocelli were paired, one lying on either side of the shaft, in another (P. malaccense) these approached and partly fused with one another. Now, to get the indented ocellus of the Peacock, we have only to imagine the fusion of two such ocelli, whose long axes inclined obliquely to one another, to get the “eye” of the Peacock with its indented lower edge; for such fusion would give a continuous upper and an indented lower border.
The “eyes” of the Argus Pheasant are more interesting still, for, as Darwin pointed out, these have the appearance, if the feathers are held more or less vertically, of a number of balls lying each within a socket, or cup: for each of these balls has a light area which exactly simulates the light glancing across the upper pole of a sphere, leaving 99the rest in shadow; and, singularly enough, this effect is produced in the living bird only when the feathers are erected for display. The probable steps in the evolution of these ocelli from simple spots, and through elliptical bars, Darwin traced with his usual skill and insight, and those who would follow this up should turn to that wonderful book, “The Descent of Man.”
Plate 17.
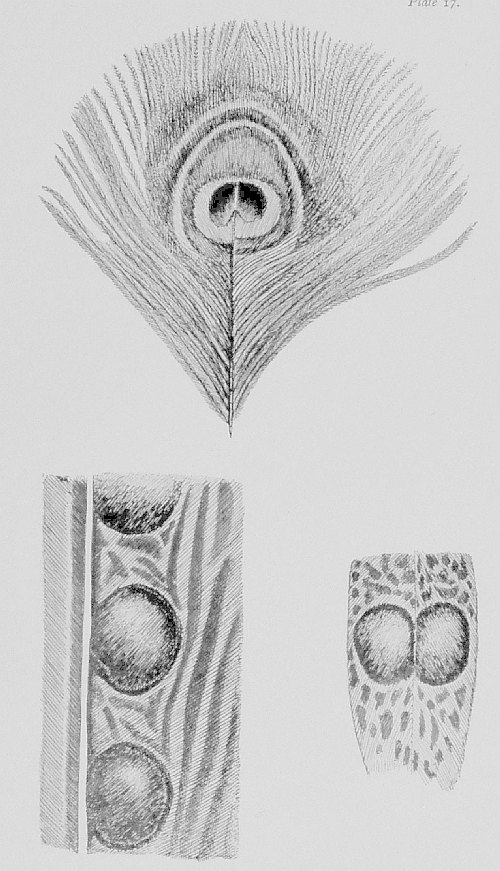
PATTERNS WHICH PUZZLED DARWIN.
The notch in the “eyes” of the Peacock’s train-feathers puzzled Darwin till he met with the ocelli of the Peacock-pheasant. The left-hand lower figure represents the ocellus of the Argus, the right-hand that of the Peacock-pheasant.
[Face page 98.
It is probable that the erroneous interpretation of the display of the Peacock is due to the more lasting and easily remembered impression of what obtains in the case of the Turkey under like emotions. This bird in his exultant moods, most people have seen. Herein the tail plays a very important part, being raised and spread to form a great half-circle, while at the same time the back-feathers, or at least those of the lower back, are set on end, and the wings are trailed on the ground. The effect is heightened by the suffusion of blood to the bare skin of the head and neck, and the sudden inflation of a long, pendent, fleshy wattle from the forehead, which hangs down over the beak. Great display is made with this, and an additional importance is added by the spasmodic vocal efforts which can best be described by the “gobble” rapidly repeated, as the bird struts about with mincing gait, turning the wheel-like tail now to one side now to the other. But the Turkey possesses yet another “ornament” which commonly escapes notice. This is the curious tuft of long, black, coarse, bristles which projects forward in front of the breast. It is difficult to discern what part this tuft may play, since it is quite inconspicuous. It seems as though this must be added to the number of structural characters which appear to survive without any apparent use.
The game-birds, it is significant to remark—and 100significant because they are commonly polygamous—afford a quite remarkable series of displays, only some of which can be summarized in these pages. In every case, too, they are accompanied by conspicuous coloration and a more or less excessive development of brightly-coloured plumes, or areas of bare skin. In some, as in those wonderful birds the Tragopans, the development of bare skin, vividly coloured, and produced into pendulous folds, has attained a degree met with nowhere else among this group. These flaps, or finger-like wattles, as the case may be, under the influence of sexual excitement become turgid, and their hues enormously intensified: though beyond this fact but little else is known of their performances. In Swinhoe’s Pheasant the face is bare, the skin being covered, as in the case of the common Pheasant, with tiny villi of a vivid red colour. But when excited by the presence of a female the upper part of this face area rises high above the head like a pair of horns. With these turgid, and erect, the bird makes a series of short, semicircular rushes around his prospective mate, accompanying each of these gyrations with an angry hissing sound. The Golden, and Amherst Pheasants are among the most gorgeously clad of birds. Not their least conspicuous ornament is a cape-like frill of long, highly coloured feathers of which the birds seem to be extremely conscious; for when endeavouring to excite the female nearest him to the necessary pitch of sexual desire, he places himself sideways before her, drawing the frill round to the side facing her, and dropping the wing, in order, as it would seem, that she may miss nothing of his resplendent livery. This side of his nature he reserves for her. Intruding rivals are treated after quite another fashion, for like most of the gallinaceous 101birds his legs are armed with formidable spurs which can, and do, inflict the most terrible wounds: as, indeed, has been shown from the evidence of the Cock-pit in the case of game-cocks.
Plate 18.

THE “STRUTTING TURKEY.”
This should be contrasted with the Peacock. Herein the tail itself is the principal ornament, the effect of which is heightened by the erection of the back-feathers, and the vivid play of colour of the “wattles” of the head.

Photo copyright by W. H. Quentin.
THE DISPLAY OF THE GREAT BUSTARD.
This is effected by the inflation of a great wind-bag in the neck, and the eversion of the wing and tail feathers as described in the text.
[Face page 100.
By way of contrast with the several displays just described, it would be hard to find a more striking illustration than that afforded by the Lesser Bird of Paradise (Paradisea minor), inasmuch as here the display is associated with rivalry between a number of individuals. For much of our knowledge on this subject we have to depend on the descriptions of natives; but happily this has now been supplemented by observations made by Mr. Ogilvie Grant on a captive in the Gardens of the Zoological Society of London.
Impelled by the surging wave of sexual desire, as yet only seeking consummation, these birds gather together at frequent intervals, on certain of the forest trees of the Aru Islands, selected apparently because they present an immense head of spreading branches, and large but scattered leaves. Here ample space is found for the revels, which take the form of “Sacaleli,” or dancing-parties, comparable to the erotic dances of many barbaric races.
By the time the ball opens, the birds, to the number of twenty or more, have worked themselves up into a state bordering on frenzy, and each commences his performance with quivering wings and loud, penetrating cries which may be syllabled as walk—walk—walk—walk—walk—walk, rapidly repeated. Then the wings are suddenly held out on either side, the tail is bent forward under the branch, and with a quick, barely perceptible rustle, the gorgeous, golden, diaphanous side-plumes are thrust upward and forward on each side of the body, forming 102an arched cascade above the back. With every muscle tense the performer will remain in this attitude from ten to twenty seconds, slightly quivering the wings, and from time to time imparting a tremor to the upraised plumes. Then follows a second phase. Each bird, seemingly possessed, commences to dance and hop wildly backwards and forwards along the bough, and with head bent forward, wings spread horizontally, and the side plumes raised to their utmost, he gives vent to a series of loud harsh cries—“ca! ca! ca! ca!” For some seconds he remains in a sort of ecstasy, rubbing his beak on the bough, and occasionally glancing backwards below his feet, and with the back fully arched. The climax passed, he reverts once more to the earlier, more erect stage of the display, when the paroxysm either gradually subsides or is renewed.
No less extraordinary is the behaviour of the King-bird of Paradise (Cicinnurus regius), which has been described by Sir William Ingram, who for a time had a captive in his aviaries. As the illustration shows, its posturing is quite remarkable. Before this is described, however, a brief description of its coloration should be given, which, it must be remarked, cannot possibly convey more than a very vague idea of its sumptuous character. Picture a bird no bigger than a thrush, but of a wonderful cinnabar red, with a gloss as of spun glass: the head clothed in short, velvety, orange-hued feathers; and with a white breast, having the softness and sheen of satin, and crossed by a band of deep metallic green, contrasting with the red of the throat. Add a yellow beak, and legs of cobalt blue, and you will have the features which catch the eye at the first glance. But a little closer examination will reveal yet other points for wonderment. 103Along each side of the body the upper flank-feathers become elongated and delicately tinted, and, furthermore, they are erectile: so that they can be raised up on each side of the body to form an almost circular shield of delicate ash grey, bordered with buff and emerald green. These play a most important part during the sexual frenzy, and the effect thereof is not a little heightened by the middle pair of tail-feathers, which have been modified to form a pair of slender stalks, some ten inches long, bearing at the ends a curious disc of emerald green formed by coiling upon itself—like a watch-spring—the only piece of the vane of the feather which remains.
So much for its fine feathers; now for the manner of their use. “He always commences his display,” writes Sir William Ingram, “by giving forth several short notes and squeaks, sometimes resembling the call of a quail, sometimes the whine of a pet dog. Next he spreads out his wings, occasionally quite hiding his head; at times, stretched upright, he flaps them, as if he intended to take flight, and then, with a sudden movement, gives himself a half turn, so that he faces the spectators, puffing out his silky-white lower feathers; now he bursts into his beautiful melodious warbling song, so enchanting to hear but so difficult to describe. Some weeks ago I was crossing a meadow and heard the song of a skylark high up in the heavens, and I exclaimed at once: ‘That is the love-chant of my King-bird.’ He sings a low bubbling note, displaying all the while his beautiful fan-like side-plumes, which he opens and closes in time with the variations of his song. These fan-plumes can only be expanded when his wings are closed, and during this part of the display he closes his wings and spreads out his short tail, pressing it close over his back, so as to throw the long 104tail-wires over his head, while he gently swings his body from side to side. The spiral tips of the wires look like small balls of burnished green metal, and the swaying movement gives them the effect of being slowly tossed from one side to the other, so that I have named this part of the display the ‘Juggling.’ The swaying of the body seems to keep time with the song, and at intervals, with a swallowing movement of his throat, the bird raises and lowers his head. Then comes the finale, which lasts only a few seconds. He suddenly turns right round and shows his back, the white fluffy feathers under the tail bristling in his excitement; he bends down on the perch in the attitude of a fighting cock, his widely-opened bill showing distinctly the extraordinary light apple-green colour of the inside of the mouth, and sings the same gurgling notes without once closing his bill, and with a slow dying-away movement of his tail and body. A single drawn-out note is then uttered, the tail and wires are lowered, and the dance and song are over.
“The King-bird has another form of display which he very rarely exhibits, and only on three or four occasions have I seen him go through this performance. Dropping under the perch, the bird walks backwards and forwards in an inverted position with his wings expanded. Suddenly he closes his wings and lets his body fall straight downwards, looking exactly like a crimson pear, his blue legs being stretched out to the full length and his feet clinging to the perch. The effect is very curious and weird, and the performance is so like that of an acrobat suddenly dropping on to his toes on the cross-bar of a trapeze that I have named this the ‘Acrobatic’ display. It has been witnessed on different days to his ‘Juggling’ display. While giving his ‘Acrobatic’ performance he sings the whole time, but never shows his side-plumes, 105and when he is in the pendulous position his body sways gently as if it were influenced by a fitful breeze. The whole of this performance takes but a very few seconds.”
Plate 19.

From a Drawing by Roland Green, Jun., adapted from G. E. Lodge and others.
SOME OF FORTUNE’S FAVOURITES.
The Birds-of-Paradise have few rivals in the matter of ornament. In the centre of this plate are seen the Lesser and the King Bird-of-Paradise displaying (after G. E. Lodge). The first-named is distinguished by the enormous development of the side plumes, which can be raised high above the back. In the second, the ornaments take the form of erectile frills on each side of the breast, and strangely modified tail-feathers which end in curious discs. At the top left-hand corner is the King of Saxony’s Bird-of-Paradise; on the right is the Long-tailed Bird-of-Paradise; at the bottom of the page, from left to right, are Hunstein’s, the Six-wired, and Superb Bird-of-Paradise.
[Face page 104.
Naturally one needs to witness such a display to appreciate its beauty and its weirdness; but the wonderful sketches which my friend, Mr. G. E. Lodge, made during one of these performances, should go far towards helping the reader to visualize what really takes place.
While it would be untrue to say that the Birds of Paradise are of a more amorous, or more excitable disposition than other less resplendent birds, one cannot but be impressed with the fact that they exhibit a range of variation in the matter of feather-ornament probably unequalled, and certainly unsurpassed, by any other group of birds. From what has been observed of the few species which have been kept in confinement, they seem to enjoy no less distinction in matters of display. On this latter subject no more of importance can be said, and exigencies of space forbid any attempt to describe the exquisite beauty of coloration which a survey of all the known species reveals. It would be hardly more profitable to attempt to describe the varied character of the shields, crests, frills, streamers, which are to be met with in different species: but a glance at the accompanying illustrations will show that it would be hard, indeed, to exaggerate the splendour of the ornamentation which these birds have developed. Even here, where no indication can be given of the glowing, vivid colours, often indescribably beautiful, it is obvious that these birds well deserve their name. St. John’s imaginary Paradise would probably have been described in far more enticing language had he known of the existence of these wonderful birds.
Among all the known species the dullest is Wallace’s Bird of Paradise, the general coloration being of a dull brown hue; but even here, a pair of wing-coverts are produced into long, broad streamers, unique among birds; while the feathers of the throat and flanks are of a marvellous metallic green, the flank-feathers being produced to form a long, pointed tuft.
To what factors must we attribute the growth of these wonderful colours, these strange outgrowths, frills, and tufts, and streamers, the like of which is almost unparalleled? In a group numbering some fifty or more species there is not one that does not display some strange feature. We cannot attribute it to the environment, for in such case the results should have produced uniformity; nor can we invoke the aid of sexual selection save in a very indirect manner, and in a sense other than generally understood by this term. It seems, then, not unreasonable to suggest that they are the expression points of the internal metabolism: the manifestations of that tendency to vary which is inherent in every fibre of the organism. But no attempt shall be made to elaborate this theory till more evidence has been taken. The humming-birds, and the game-birds, are perhaps the only other groups which exhibit quite such a prodigality of ornament; of the latter, instances have already been cited.
So far the displays which have been described have been such as are confined to the use of more or less resplendent plumage. There are, however, many species which contrive to secure most startling results, not so much by the parade of coats of many colours as by grotesque changes of shape produced by wind-bags of various kinds. The Pouter-pigeon affords a case in point. This bird possesses the power of inflating the gullet to an enormous size, so 107as to produce a strangely distorted form, at any rate, to our eyes. The “Pouter,” it is hardly necessary to mention, is an artificial product of the “fancier,” who has taken advantage of the natural tendency, seen in the Wild Pigeon, to inflate the neck during moments of excitement. By the selection from each generation of the finest performers in his stock, the Pouter of to-day has been developed. But there are many birds which, while not even remotely related, have developed the same strange device. The most striking illustration of this kind is furnished by the Great Bustard, a bird once common on the fen-lands of Great Britain, but now, unhappily, exterminated within these islands.
The means of inflation in this case is afforded by a large thin-walled sac of a very remarkable character. Opening by a small slit just under the tongue, it is continued down the front of the neck immediately under the skin, which in this region is thickened by an accumulation of fat and blood-vessels. Between the arms of the furcula, or merry-thought, its cavity is constricted, to expand again immediately to form a pear-shaped termination. How it is filled is something of a mystery. But once inflated, the bird draws its neck downwards and backwards, so that the head is brought to rest between the shoulders and is there almost buried, partly by pressure on this curious air-cushion and partly by the erection of a number of bristle-like feathers, which in calmer moments project backwards on each side of the head. At the same time the tail is drawn forwards to lie upon the back, thus exposing a billowy mass of white feathers forming the under tail-coverts. The tips of the wing-feathers are used to hold the tail in position. Meanwhile the scapulars are set on end, and the long inner secondary quills are 108similarly erected. The feathered contortionist, having completed his preparations, now approaches his partner with a mincing gait, then halts before her and solemnly utters a series of low grunts like “oak, oak, oak.” Having thereby declared his passion, and commonly without gaining any answering response, he returns to his normal shape again!
It is curious that a near relative of this bird, the Great Australian Bustard (Eupodotis australis), also captures the wind to declare his love; but it is disposed of after a quite different fashion, being drawn into the gullet, though the precise manner in which it is manipulated demands further investigation. The display Dr. Murie described many years ago. It begins, he tells us, with a swelling of the throat, while the head is thrown upwards. Immediately after, the neck swells, and the feathers of the lower part of the neck, set all on end, are carried downwards, apparently surrounding a huge bag which reaches nearly to the ground. During all this time the head and neck are held rigid and point skywards, the head surmounting a great feathery column. Meanwhile the tail, as with the Great Bustard, is drawn forwards over the back. In this peculiar attitude the bird struts about in a stiff, waddling manner, the elongated neck-bag swaying to and fro and the feathers of the throat standing out in the shape of a great rounded swelling. The acme of inspiratory effort completed, the bird begins to snap the jaws together, producing loud noises, which are accompanied by a soft dove-like cooing.
The Pectoral Sandpiper in like manner inflates its gullet. But, unlike the Pigeon and the bird just described, the neck is not markedly straightened, nor is the body raised. As the air is drawn in, the gullet expands, till 109it forms a great spherical drum. Then the excited performer runs along the ground uttering a resonant “too-u tooo-u” repeated seven or eight times in rapid succession, all the while he approaches nearer to the apparently very much-otherwise-engaged female. This effort failing, he will then often rise on quivering wings twenty or thirty feet into the air, and dive gracefully down again immediately afterwards, deflating this curious balloon to await a more favourable opportunity.
In some species where wind-bags are employed as aphrodisiacs the outer skin is brilliantly coloured and exposed during the display. The Prairie-hen affords a case in point. In this species the air-chamber is furnished, not by the gullet, but by the air-sacs of the neck. When these are inflated they appear as two large orange-coloured bodies standing boldly out among the feathers. Their effect is heightened by a tuft of long stiff feathers which are thrust forward like a pair of horns, on each side of the head, while at the same time the feathers of the back are set on end, the tail is spread like a great fan, and the wings are half opened and trailed like those of the Turkey.
The displays take place in the early hours of the morning, when parties of from a dozen to fifty, of both sexes, meet on some slight knolls where the grass is short. Having duly assembled, the more ardent cocks immediately begin to prepare for the morning revels, the first part of the performance apparently being of a comparatively passive nature—the parade of the air-sacs and the erection of the feathers.
Then some “proud cock, in order to complete his triumph, will rush forward at his best speed ... through the midst of the love-sick damsels, pouring out as he 110goes a booming noise ... which may be heard for at least two miles in the morning air. This sound is by no means harsh or unpleasant. When standing in the open prairie at early dawn listening to hundreds of different voices pitched in different keys, coming from every direction and from various distances, the listener is rather soothed than excited.
“Every few minutes this display is repeated. I have seen not only one, but more than twenty cocks going through this funny operation at once; but then they seem careful not to run against each other, for they have not yet got to the fighting point. After a little while the lady birds begin to show an interest in the proceedings by moving about quickly, a few yards at a time, and then standing still a short time.
“The party breaks up when the sun is half an hour high, to be repeated the next morning, and every morning for a week or two before all make satisfactory matches. It is towards the latter part of the love-season that the fighting takes place among the cocks, probably by two who have fallen in love with the same sweetheart....”
There is much that is extremely interesting in this account and a little that seems to have been misinterpreted. The fact that these antics are repeated during many days until at last the females are moved to display some interest is just what we should expect if this demonstrative behaviour on the part of the males acts, as we believe, as an aphrodisiac. And that actual fighting occurs is highly probable, but there can be no doubt that in such case the whole aspect of the bird must be changed, for anything in the nature of fighting with the delicate air-sacs inflated would greatly endanger the most important aid to success in achieving this object which these birds possess.
Plate 20.

Photo from The Museum of Natural History, New York.
THE LOVE-MAKING OF THE PRAIRIE HEN.
During the “display” large, yellow, air-sacs in the neck are inflated. The bird in the foreground shows one of these, and the ornamental feather frill, very clearly.
[Face page 110.
No less remarkable is the performance of the Frigate-bird (Fregata), a tropical species allied to the Pelicans and Boatswain-bird, and to our own more familiar Cormorants and Gannets. It might well be called a marine Swift, having excessively short legs and small feet, and a wonderful expanse of wing. As with the Swifts, of course most of its time is spent on the wing; the feet are only useful for supporting the body when ashore, they are never used for walking, at any rate, for more than a few steps. The wings afford the only means of locomotion. Our knowledge of these birds when under the stress of sexual excitement we owe to Dr. C. W. Andrews, who had the good fortune to study the species known as the Great Frigate-bird (Fregata aquila) during his task of surveying Christmas Island (Indian Ocean).
“About the beginning of January,” he remarks, “the adult males begin to acquire a remarkable pouch of scarlet skin beneath the throat; this they can inflate till it is nearly as large as the rest of the body, and a dozen or more of these birds sitting on a tree with outspread, drooping wings and this great scarlet bladder under their heads are a most remarkable sight. When a hen bird approaches the tree the males utter a peculiar cry, a sort of ‘wow-wow-wow-wow,’ and clatter their beaks like castanets, at the same time shaking their wings. When they take to flight the air is allowed to escape from the pouch, but occasionally they might be seen flying with it partly inflated.”
Here again there can be no doubt about the purpose, or perhaps one should say the stimulus, of this strange performance. This pouch, I have been enabled to ascertain 112from dissection, is not formed by inflating the gullet, but, as in the case of the Prairie-hen, by the enlargement of the air-sacs of the neck.
These air-sacs, which are present in all birds, are only enlarged to further the ends of sexual display in a few species, and, curiously enough, these are in no way related one to another. The Adjutant storks, it may be remarked in this connection, have used the air-sacs which are fed by the nasal system instead of those fed by the lungs, as in all the species so far described. When deflated this pouch forms a quite inconspicuous conical swelling in front of the neck; under the stimulus of excitement, it awakens as it were into activity, and is suddenly transformed into a great red or red-and-black bag, encircling the neck and projecting far downwards in front of it, only to be deflated an instant later with a speed which leaves one gasping.
The specialization of the air-sacs, that is to say their transformation to perform new functions subservient to the ends of sexual activities, is not exclusively confined to display. In at least one instance an air-sac has been specially developed to act as a voice resonator. This is furnished by the Emu, wherein the wind-pipe, near the middle of its length and on its anterior aspect, has a number of incomplete rings forming a long slit. The lining of the windpipe escapes from this slit in a hernia-like pouch, and takes up a position beneath the skin. Even when inflated this pouch gives no very obvious sign of its existence, but it serves to produce a curious hollow, drumming sound, like the boom of a big drum softly beaten. But why it should have been developed, when the Ostrich and the Cassowary produce similar but louder “music” without any special apparatus 113whatever, is a mystery. At least one species of Cassowary can emit a roar which would do credit to a lion.
In the males of all healthy animals the periodic stimulus to reproduction finds expression in more or less striking eccentricity of conduct. Sometimes, as the foregoing instances have shown, this has been exaggerated by the development of long, resplendent plumes: sometimes by brilliant coloration, displayed either by the plumage or by bare areas of skin, or by both, while in not a few cases attitudes, to our eyes grotesque and made still more so by the aid of inflatable pouches, are the outward and visible sign of the raging fires within. For the completion of this chapter yet other instances of this kind must be cited, instances which reveal a further elaboration of some of the more striking of these tricks of posturing; or which concern the growth of the aggressive instincts, which are proclaimed by the development of armature often of a very formidable character. As the sequel shows, however, there are no hard and fast dividing lines between these several modes of expression.
That remarkable bird, the Ruff (Machetes pugnax), now, alas! no longer to be met with in our fens, exhibits a curiously composite character in the phases of its love display.
Preparations for this are begun in the early spring by the assumption of what is called a “nuptial dress,” which is worn only by the male, and which contrasts in a very conspicuous manner with the plumage worn during the rest of the year. The most striking features of this dress are the great, erectile, Elizabethan ruff which encircles the neck immediately behind the head, and the long, tongue-shaped “ears” which surmount the head itself. These exhibit a most remarkable diversity 114in their coloration, and it is no exaggeration to say that no two are ever alike. Red, cream-colour, buff, black, white; spotted, streaked, freckled and barred are the only descriptions that can be applied to them, for the combinations of their hues and patterns seem infinite. Having grasped this fact, the eye next turns to the colouring of the rest of the body, and it will be found that here too is the same diversity, though less conspicuously so; and finally it will be noticed that at this time the feathers around the base of the beak have been replaced by yellow or orange-coloured papillæ. The females also now wear a dress differing from that of the so-called “winter plumage,” but it does not present any very striking features nor any form of ornamentation comparable to that of the males.
The Ruff is a polygamous bird, which, in its display, presents some curious and puzzling features, one of which consists of a sort of tournament between rival males. At the break of day the performers, selecting such eminences as the fen-lands afford, assemble apparently to display their finery, for a couple of males will often stand facing one another with frills erected and beaks touching the ground, silent and immovable, for perhaps half a minute. Sooner or later, however, they will commence to spar, and this presently leads to blows, during which one of the combatants will attempt to seize the other by the wings. However, no damage seems to be inflicted during such encounters, which are by no means aimless or profitless, for during such bouts the weaker, less vigorous birds are driven from the field, and the victor in consequence wins for himself a larger harem.
When the actual pairing time arrives the parade of the frills begins again. The amorous instincts, it is important to notice, are awakened earlier in the males, so that by the time the females have attained to a like condition the least mettlesome males have been driven off. What follows is not the selection by the females of the finest performers so much as a process of sorting out, whereby the females discover and cleave to those males which are readiest for mating. This display succeeds in revealing both the most mettlesome males and the most amorous females, who, however, would seem to require great persistence and much demonstration on the part of the males before they can be finally aroused to the pitch necessary for pairing. Again and again a male may be seen to approach an apparently very unconcerned female, and then to crouch down before her with his beak pressed to the ground and his frill and “ears” set off to their fullest. For some seconds he will remain lost in apparent contemplation, then with a dazed, far-off, expression he will look up, to find, as often as not, that she is still apparently feeding, quite unmoved by his protestations; or that she has even flown off and left him. Pursuit speedily follows, and the performance is repeated until at last she too catches the flame of passion and permits, or rather invites, the final act of sexual congress.
Though these birds on occasion will fight, and savagely, they cannot inflict serious damage on one another by reason of the relative feebleness of their beaks and legs, which are but ill-adapted for violent measures. Inasmuch as the Ruff is a polygamous species, these bloodless battles have a peculiar interest. They show that the preponderance of females, which polygamy implies, is not, as is commonly supposed to be the case, due to a 116high death-rate among the males by fighting. The same is true of the Wydah-birds, and their kin, the only polygamous species among the Passeres.
In this connection it is to be remarked that fighting, of a more or less sanguinary character, is apparently universal among birds, the conflicts being waged not so much in the way of squabbles for the possession of females as for the acquisition and retention of territory and all that this entails during the breeding season and, to a much less extent, in the defence of the eggs and young. But to this point we must return. For the moment it will be more profitable to focus attention on the character of this fighting. In the first place, it is by no means necessary that the combatants should be armed. The “dove of peace” at this time of the year appears in a new and not always pleasing light, for not only will he fight his neighbours, but he does not always show that gentleness towards his wife with which tradition has credited him. The little Humming-bird would seem to be as little capable of fighting as a bird could be, yet few are more pugnacious. The naturalist Gosse tells of a pair which had torn one another’s tongues out in their blind fury; and everybody knows that Robins and Tits fight savagely to preserve their chosen haunts from invasion by their neighbours. In some birds this pugnacity has become an overmastering passion. Some of the Quails, and a species of Rail (Gallicrex cristatus), a near relation of the Moorhen, are commonly kept by the natives of the East, as our forefathers kept Fighting-cocks, for the sake of seeing them fight one another. Yet, save in the case of the Fighting-cock, neither of these birds possesses any aggressive 117weapons.
Among the game-birds, however, powerful armature, in the shape of long, pointed, spurs on the legs are met with. In the Jungle-fowls and Pheasants only a single pair are found on each leg, but in other species, as in the Francolins, there are several pairs, and these birds, it is instructive to notice, are notorious for the ferocity of their encounters. It is said that in the Indian Swamp-Francolin (Francolinus gularis) nearly every individual is marked by scars and wounds received in duels with rivals.
Certain members of the Plover-tribe, and certain Anserine birds, have developed spurs of a very formidable character on the wings. Among the Plover-tribe the best example of such armoured species is the Egyptian Spur-winged Plover (Hoplopterus). This bird, after the fashion of its unarmoured relatives, such as the Common Lapwing, fights by turning suddenly in the air and striking with the wings. In the case of the formidably armed Egyptian bird the result is often fatal; but with our Lapwing a fatal result is rare, since but slightly swollen knobs take the place of spurs. In Hoplopterus and in the Jacana this spur arises from the base of the thumb, but in the Spur-winged Goose (Plectropterus) it is borne by one of the wrist bones (the radial) while in the aberrant Geese-like birds (Palamedea and Chauna) there are two spurs on each wring, one at each end of the metacarpus. That these weapons have come into being in response to need seems a very natural conclusion, but it is one which presents many difficulties when more closely examined. The wing spurs, differing widely in their nature as they do, in one case borne on a carpal bone, in others on the metacarpus, seem rather 118to owe their origin to fortuitous variations which have become, so to speak, adopted by selection, than to a response to the oft-repeated stimuli incidental to fighting. The latter explanation is Lamarckian and to-day finds favour with but few. The stimulus theory seems to be effectually discounted by the existence of the spurs on the legs of gallinaceous birds. That these owe their origin to impacts, or blows, seems more than doubtful: and one can hardly see how they could have served any useful purpose until they had attained a sufficient length to serve as weapons. Even if we suppose that the spurs of, say, the Jungle-fowl or the Francolin have been derived from tuberosities such as are found on the legs of the French Partridge (Caccabis rufa), we should still lack evidence that the use of the legs in fighting caused the origin of the tuberosities.
There is yet another puzzling feature in regard to the armature of the wings, and one which may yet help to a better understanding of the puzzles presented by spurs. A Jacana, one of the Plovers, has the radius broadened or flattened out from its middle onwards to form a flat plate or blade, but the use thereof is unknown. It may possibly serve as a weapon of offence, enabling the bird to beat its rivals with its wings, but from the nature of the structure, and of the effect such a use of the forearm would have upon the hand, it seems doubtful whether it serves any aggressive function. If used at all in fighting it is probably during fights in mid-air, when, after the fashion pursued by the Spur-winged Plover, and even in the case of our own Lapwing, a blow is struck by the uppermost bird at its rival, and often with fatal effect. It is significant to remark, by the way, that in the Lapwing a tubercle answers to the spur of Hoplopterus 119just as the tubercles of the French Partridge (Caccabis) answer to the spur of the Jungle-fowl or Pheasant: but the flattened radius of the wing of the Metopidius jacana has no parallel.
With birds, as with men, there must always remain the ability to appeal to force when some important end cannot otherwise be gained. The species which adopts the crazy tactics of the Quaker is doomed to extinction, sooner or later. The foregoing instances display force, as we may say, aggressively. But even the peacefully disposed birds can fight when aroused.
Reference has already been made to dancing in this chapter; but so far no very striking instances thereof as a form of sexual display have been cited. The subject has been deferred because this peculiar type of activity is not always directly associated with the furor amantium.
With some species, which, it should be remarked, also lack distinctive colouring, the erotic state is manifested apparently not so much by the display of expanded wings and tail as by frenzied dances. The Jacanas, aberrant members of the Plover tribe resident in South America, are expert performers, displaying moreover a curious spontaneity during such outbursts. A flock will be apparently sedulously feeding when suddenly and with quick, excited gestures all will cluster together in a group and go through a singular and pretty performance, holding their wings outstretched and agitated, some with a fluttering and others with more leisurely movement, like that of a butterfly sunning itself. The performance over, all scatter and feed again.. The Honourable Walter Rothschild, in his “Avi-fauna of Laysan” tells us of the stately Albatross, which breeds, or rather bred there—for 120the Japanese display a singular callousness in regard to animal life where commercial interests are concerned in thousands: “First they stand face to face, then they begin nodding and bowing vigorously, then rub their bills together with a whistling cry. After this they begin shaking their heads and snapping their bills with marvellous rapidity, occasionally lifting one wing, straightening themselves out and blowing out their breasts; then they put their bills under the wing or toss them in the air with a groaning scream, and walk round each other often for fifteen minutes at a time.”
Cranes are much given to dancing. Mr. Nelson, an American ornithologist, has described with much vigour the dancing of the Sandhill Crane in Alaska. As he lay in a “hunting-blind” he was suddenly aroused by the arrival of a crane, followed speedily by a second, uttering his loud note as he came, until he espied the first-comer on the ground, when he made a circuit and dropped close by. Both birds then joined in a series of loud rolling cries in quick succession. Suddenly, the last-comer, which seemed to be a male, wheeled his back towards the female and made a low bow, his head nearly touching the ground, and ending by a quick leap into the air. Another pirouette brought him facing his charmer, whom he greeted with a still deeper bow, his wings trailing loosely by his sides. She replied by an answering bow and hop, and then tried to outdo the other in a series of spasmodic hops and starts, mixed with a set of comically grave and ceremonious bows. The pair stood for some moments bowing right and left, when the legs appeared to become envious of the large share taken in the performance by the neck, and then would ensue a series of skilled hops and skips, like the steps of a minuet. Such 121antics are characteristic of the Cranes of all species, and sometimes a whole flock will join in such dances. But, it is to be noted, they are not necessarily signs of the furor amantium: they certainly always accompany this, but frequently they are indulged in, apparently, solely as an outlet for exuberance of feeling.
Before the theme of dancing can be dismissed the performance of a small species of perching bird, one of the South American Manakins, must be described. The natives call it the “Bailador,” or dancer. In an account of his travels in Nicaragua Mr. Nutting tells us: “I once witnessed one of the most remarkable performances it was ever my lot to see. Upon a bare twig ... at about four feet from the ground, two male ‘bailadors’ were engaged in a song and dance act that simply astonished me. The two birds were about a foot and half apart and were alternately jumping about two feet in the air and alighting exactly on the spot whence they jumped. The time was as regular as clockwork, one bird jumping up the instant the other alighted, each bird accompanying himself to the tune of to-le-do—to-le-do—to-le-do, sounding the syllable to as he crouched to spring, le while in the air, and do as he alighted. This performance was kept up without intermission for more than a minute, when the birds suddenly discovered they had an audience and made off.” Here again we have no evidence of the furor amantium; nor that any females were spectators of the scene.
It is important to notice that Mr. Howard, in the course of his study of the Warblers, witnessed a performance having some likeness to this on the part of three young Sedge Warblers but newly escaped from the nursery. And this not in some solitary instance, but on several 122occasions. Just after leaving the nest, he remarks, they are very playful, “their games sometimes taking the form of a tilting match. Three take part; two sit on convenient twigs facing one another, and the third, from the central position, might almost be called an umpire. Numbers One and Two lower their heads, each in anticipation of the other moving; one of them, call him Number One, then springs into the air and darts at Number Two: Number Two dodges and occupies the position vacated by Number One; each of them then faces round ready to continue the fray, the change of positions becoming quite rapid.” But no recurrence of these antics has been noted during the course of the adult sexual display, which is confined to posturing and displaying the outspread wings and tail. Nevertheless there can be no doubt but that such games in later life are incorporated, in the case of many species, with the love display.
That the reproductive glands have played, and still play, a by no means unimportant rôle in Evolution is shown by the history of the secondary sexual characters. Among the birds, at any rate, the early stages of physical changes belonging to this “figuration” are to be seen in various forms of posturing, which in their more elaborate developments we call “dances.” In many cases, as for example among the Warblers, the periods of sex-emotion are marked by posturing alone. But in a number of species, as has already been shown, the products of the sexual gland seem to have undergone some further elaboration which has resulted in the additional phenomena of gaudy coloration, in hypertrophied plumes, and in weapons of offence.
But not yet is the list of such sexual products exhausted, for no mention has so far been made of the 123development of the many wonderful devices for the production of peculiar and arresting sounds, musical and otherwise. These are of two kinds: one wherein certain feathers have been modified to produce rhythmical notes either by percussion or by vibration; the other wherein the internal organs have been modified to produce musical notes or loud, resonant cries.
Instances of the latter kind are innumerable, and as a consequence no more than one or two can be cited in these pages. The facts associated with the production of vocal, as distinct from instrumental, music are both curious and puzzling. To begin with, this music is produced by the lower end of the trachea or windpipe, which has become modified in various ways, though not so strictly in relation to the sounds produced as is commonly supposed. The anatomical details of these modifications cannot, or rather need not, be described now, save in the most general terms.
Briefly the syrinx, or organ of voice, of birds, is formed in part by the lowermost rings which form the tubular windpipe, and in part by the smaller pair of tubes which, running therefrom to the lungs, form the bronchi. These last are formed of semi-rings only, the inner wall of the tube being formed by very delicate translucent membranes. As air is forced from the lungs along the bronchi and up the windpipe, the modulation of the voice is effected by muscles which regulate the amount of air driven through the syrinx, and the height of the column in the tube; the latter being effected by muscles which alternately lengthen and shorten it.
So far so good. Next it is to be noted that this syrinx presents a great variety of modifications, or types, differing not only in plan, but also in the number and 124distribution of the muscles for its manipulation. The most accomplished performers are to be found among that great group of birds known as the Passeres, or perching birds, wherein the number of these muscles is never less than five pairs, and generally rises to seven. This association of musculature with performance is exactly what we should expect. In Nature, however, it is always the unexpected that happens. In the first place, the females are, so far as the dissecting-knife and the microscope can show, as well provided as the males, yet they do not sing. In the second, the Nightingale and the Crow are equally endowed, so far as we can discover, yet it is unnecessary to state that the talents which the Crow possesses are never used! More disconcerting still is the reflection that the Parrot, which is far less generously endowed by Nature in so far as singing muscles are concerned, is a much more skilful performer, inasmuch as it will reproduce with equal fidelity the human voice and the song of the Canary! The latter feat, at any rate, has been accomplished with amazing accuracy both by the little Budgerigar (Melopsittacus undulatus) and the Quaker Parrot (Myopsittacus monachus). In their wild state the Parrot family are notorious for their discordant cries. It is therefore the more remarkable that such feats should be capable of attainment. But wherefore the elaborate syrinx of the Nightingale, if the simple type seen in the Parrot is capable of the same result, and why the elaborate syrinx in the case of the Crow, which never attains to a greater perfection of vocal effort than the wild Parrot?
One speaks of the syrinx of the Parrot as of a simpler type because of its feebler musculature and the lesser complexity of its framework, but it is nevertheless a more 125efficient instrument, since it is capable of reproducing both the human voice and songs such as that of the Canary. This fact becomes still more remarkable when we reflect that the natural voice of the Parrot, as we have just remarked, attains to no more than a harsh screech. How is it that, capable of so much, it has achieved so little? The same question may be asked in the case of the Raven. This bird has a syrinx indistinguishable from that of the Nightingale, save in point of size; yet the Raven’s voice is never musical, nor can it be trained to such an achievement. Like the Parrot, however, it can be taught to speak, though its vocabulary is never so extensive. One would have imagined that when the syrinx of, say, the Raven, or any of the Crow tribe, was compared with that of the Nightingale or the Skylark, some structural differences, commensurate with the difference in performance, would be discovered; but such is not the case.
What interpretation are we to place on these paradoxical facts? One cannot help asking why seven pairs of muscles should have been produced by one group of birds to perform what can as easily be achieved in another by two? It is true that the more generously endowed species are musicians by birth, the others only by training. But one cannot make a silk purse out of a sow’s ear. In like manner one asks why male and female, possessing precisely similar voice-organs, should not sing equally well, but they do not. Evidently mere mechanism does not alone answer these questions.
Some, perhaps, may see in them instances of what is known as “Hypertely,” wherein the bounds of mere utility seem to be transcended. Hypertely, however, implies something more than this: it implies a shooting 126beyond the mark, the overdoing of a feature, where the momentum gained, from some obscure cause, keeps on being increased by cumulative inheritance: and not being checked by Natural Selection, causes the species in respect of such characters to pass beyond its congeners. Professor Lloyd Morgan’s theory of “over-production” would seem better to apply here, though in a somewhat different sense from that used by him. For in the instances just quoted there is a latent potentiality for response to new demands which the struggle for existence may make, but a potentiality varying in degree, and here selection finds its métier.
Yet further illustrations of secondary sexual characters, such as are concerned with vocal music, must now be considered. The discussion of these has been designedly deferred. They embrace instances of voice production more singular than any yet referred to, and if possible more difficult to interpret.
The facts first to be reviewed concern the syrinx of certain of the Anatidæ. It is noteworthy that each of the three divisions of this group—the Swans, Geese and Ducks—contains species in which either the syrinx or the windpipe has acquired some singular feature. In the surface-feeding Ducks, modifications of the syrinx are most frequently found. Commonly, as in the Mallard, this takes the form of a spherical bony case; in the diving Ducks this bony chamber has enormously increased in size. Furthermore it has conspicuously changed both in form and character: for it is now roughly trihedral in form, and its walls present large fenestræ closed only by delicate membrane, suggesting that the increased size of the chamber has not been accompanied by a corresponding increase of bony tissue for its construction. Hence all that is available is used for the construction 127of girders to form supports for the now membranous chamber walls. Some species seem to show that this fenestration has been pushed to excess, leaving only vestiges of this singular chamber, as is shown in PI. 21. In some species the bronchi are much swollen, and the syringeal chamber has entirely disappeared: in others, as in the Merganser and Goosander, a large syringeal chamber is supplemented by dilatations of the windpipe.
Plate 21.
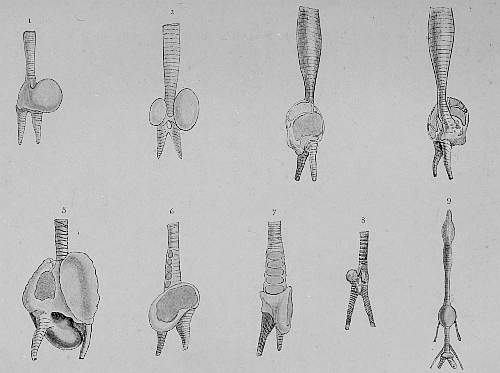
GRADES OF EVOLUTION IN THE SYRINX OR ORGAN OF VOICE IN THE MALES OF SURFACE FEEDING AND DIVING-DUCKS.
1. Wigeon. 2. Common Sheldrake. 3 and 4. Red-crested Pochard. 5. Red-crested Merganser. 6 and 7. Long-tailed Duck. 8. Steller’s Eider. 9. Common Scoter.
[Face Page 126.
Save in the case of the Goosander, these peculiar structures are found only in the male, but in the species first named the male, in addition to the syringeal chamber, has two fusiform swellings in the windpipe, one above the other: in the female one of these swellings is present, but there is no syringeal box.
This box is generally, and probably correctly, regarded as a sort of musical instrument. Nevertheless the males are far less vociferous than the females which have no such voice resonator. One has only to listen to, and compare the notes of the Mallard drake and duck to discover this fact. Here, then, we seem indeed to have a case of “Hypertely.” Before, however, we build too much on this we must discover whether the sibilant sounds uttered by the males do, or do not, play an important part in arousing the sexual passions of the females.
Certain of the Swans and Cranes afford illustrations of musical instruments of an even more remarkable kind. Herein the windpipe at the base of the neck enters a large chamber formed by the absorption of the diploe sandwiched between the outer walls of the keel of the breastbone and the enlargement of the space so created until it can accommodate the tubular windpipe. This, entering the cavity in the form of a loop, runs the whole 128length of the keel, the upper limb of the loop finally running to the lungs. That we have here an indubitable musical instrument there can be no question, for its possessor is enabled thereby to utter loud, trumpet-like, if harsh, sounds. Here again only the males are so provided.
The profound interest of this really extraordinary association of unrelated structures has never attracted the attention it deserves. Originally, no doubt, one would have met with nothing more than a loop of the windpipe impinging against the anterior border of a normal, blade-like keel: later there would have been formed a broad shallow surface on the keel at the point of contact with the loop, and gradually the depression must have deepened till the bony chamber came into being. By what nexus of sympathy were these reciprocal responses made?
Another very singular type of looped windpipe is that wherein the trachea forms a series of coils between the body and the skin. It is surely somewhat surprising to find that precisely similar coils are met with in widely different groups of birds. Among the Passeres they occur in the Manucode: among the Plovers in the Painted Snipe (Rhynchea rostratula): among the game-birds in some of the Curassows, and among the Anatidæ in the aberrant Australian Black-and-White Goose (Anseranas).
Very little is really known of the part played by these musical instruments of the Anatidæ, nor, for the matter of that, of most of the “musicians” among birds. Of some of the game-birds more has been gleaned, and among these surely the most interesting is the love-song or “lek” of the Capercaillie. With the advent of April the cock, just before dawn, repairs to some favourite tree—used 129year after year—and there performs a most astonishing if unmusical serenade; with outstretched neck, drooping wings and spreading tail he gives forth a weird, uncouth kind of song, more or less divisible into three parts. He begins with a series of notes which remind one of nothing so much as the sound made by two sticks knocked together at intervals of ten to fifteen seconds, getting quicker and quicker, and changing in key till at last they become bell-like. Then follows a series of sounds like the drawing of a cork out of a bottle, and these end with bird-like twitterings. By this time, however, the singer has worked himself up to an ecstasy of fervour and passion so intense as to deaden him to all that may be passing in the outer world. During these moments no sound disturbs him, partly, apparently, because the excitement of the “song” causes a turgid condition of the blood-vessels which for the time effectually deafens him. “Sportsmen,” in Swedish and other European forests, knowing this, select such performances as affording the most favourable time for Capercaillie shooting, only cocks being selected.
A survey must now be made of some of the more remarkable cases whereby more or less musical, or rhythmical, sounds are made by instruments of percussion; or by rapid vibrations. These are in almost every instance formed by varying grades of modification in the feathers of the wings or tail. Their presence, and their use, seem natural enough until we recall the fact that many other birds without any apparatus whatever, make sounds in no way less remarkable or less penetrating. Pigeons, Nightjars and Owls, for example, can produce at will curious snapping sounds by bringing the wings smartly together over the back. The White, and Shoebilled Storks make castanets of the beak, throwing the 130head backwards till the point of the beak touches the back, when the jaws are set rapidly clashing one against another, producing a sound comparable to the “bones” of negro minstrels, but without the varying rhythm. As this performance is proceeding, the head and neck are slowly moved through half a circle, till the tip of the beak touches the ground, when the music ceases. As with the wing-snapping just referred to, both sexes are equally skilful performers; but while they seem to indulge in such exercises much more frequently, and with more vim during the breeding season, they will break out after this demonstrative fashion at all times of the year. But why, then, the need for the yet more elaborate contrivances which are to be met with among the Snipe, the Game-birds, and certain of the Passeres?
However, be this as it may, in a large number of species a special mechanism has been evolved to produce sounds which, as has been remarked, in other species are no less effectually made without that mechanism.
One of the simplest of the cases is that furnished by the remarkable “bleating” or “drumming” performances of many species of Snipe, generally, if not only, when sexually excited, and especially of the Common Snipe (Gallinago cœlestis) during its love-flights. Mounting to a great height, this bird, at such times, suddenly turns, and descends with prodigious speed, meanwhile holding the tail fully expanded. The outermost pair of feathers are, however, specially modified so that, in the first place, during this descent they stand at right angles to the long axis of the body and well apart from the rest of the tail-feathers. This alone, however, would not produce these weird sounds, which owe their origin 131to the fact that these particular feathers have their shafts conspicuously thickened and peculiarly curved, while the vane or web of the inner side of the feather is of great width and structurally differs from the vanes of the other feathers, whereby the vane becomes more resistant to the rush of air caused by the wings during the descent.
But in the case of these Snipe it is to be noted this curious form of musical instrument is found in both sexes, and there is little difference in the quality of the sounds produced, but the bleating of the male is said to be the more resonant.
The Common Snipe is the best performer among several different species, and it is to be noted presents, to a casual examination, no remarkable or peculiar feature whatever—the structural differences just described are only to be discovered by very patient scrutiny. But in the Pin-tailed Snipe (Gallinago stenura) the number of the feathers has been greatly increased, while at the same time their webs have been so reduced that the outspread tail seems to consist of little more than spines. With such a transformation one expects to find a quite exceptional performance, far surpassing that of the Common Snipe. Yet so far as observation and experiment go they effect absolutely nothing! Here again we have a case where modification of structure has passed the bounds of need and passed so far as to make the whole tail useless as a sound-producing organ!
A contrast and a parallel are afforded by some of the gallinaceous birds of South America. The Black Penelope (Penelopina nigra) of Guatemala, while on the wing, will, during its “love-flights,” pitch suddenly earthwards with outstretched wings, and at such times a crashing, rushing sound is produced, which has been likened to the sound of a falling tree. Yet there is nothing in the 132shape of the wing which will account for this. On the other hand, a near relation of this bird, the Black-wattled Guan, Aburria (Penelope) aburri has the four outermost primaries deeply incised along their inner vanes, reducing the outermost portion of the feathers to mere spines. Yet, so far as is known, this wing makes no especial noise. However, the males of certain little South American Perching-birds known as Manakins have the shafts of the secondary quills thickened to an extraordinary degree so as to form solid, horny lumps, and these, when the wings are brought together smartly over the back, produce a noise not unlike the crack of a whip, so that here again structure and function are found together. In the contradictory cases just cited where specialized parts are found which are apparently functionless, we must suppose that the habit of using them has been supplanted by some new stimulant.
The part played by musical instruments of percussion would seem to be a variable one. In some cases, and possibly in all, it may serve as an excitant, or stimulant, to the rousing of a “sex-storm”; in many, at any rate, such sounds serve as calls to the sexes when separated. This much seems to be demonstrated in the case of certain of the Woodpeckers, which in this matter differ conspicuously from any other species yet referred to, in that they have developed no special sound-producing mechanism, but make use of hollow trees which serve them as drums, the beak being used as the drumstick. This is a very noteworthy fact, for one would have supposed that here at any rate, where the production of loud and far-reaching sounds is of vital importance, the means would have been provided by some such modification 133of the wing-feathers as we have already seen to obtain in the case, for example, of the Manakins. More closely examined, however, this apparent failure of the organism to produce its own mechanism becomes less remarkable, for Woodpeckers are forest-dwellers and but indifferent fliers; loud sounds produced by the rapid vibration of the wings or tail, as in the case of the Snipe, in mid-air, are thus impracticable, if not impossible, and sounds produced after the fashion of the Manakins would not have sufficient carrying power.
One of the most skilled performers among the Woodpeckers is the Great Spotted Woodpecker (Dendrocofus major), whose weird drumming once heard will never be forgotten. These sounds are produced by blows of the beak on a branch, delivered so rapidly that the bird’s head presents but a blurred appearance. The sounds thus made vary with the resonance of the wood and can be heard at a distance of half a mile. These strange vibrating notes are most frequently heard during the courting season, and they will commonly beget a speedy response from some more or less distant part of the wood, so that their purpose is clear. They attain the same end as the bellowing of the stag or the “lek” of the Capercaillie. They are, however, to be heard at other times, as when the birds are greatly alarmed or when the nest is being robbed.
Where the Rôle of the Sexes is reversed—Polygamy and how it is brought about—Coloration and Courtship—Instinctive Actions—The Importance of Landed Possessions—The Meaning of “Display”—The Springs of “Behaviour”—A New Light on the Wild-duck—The “Display” of the Great-crested Grebe—Some Neglected Factors.
The significance of the varied behaviour of birds—more especially of the males—during the period of reproductive activity must now be more minutely analysed. But before this analysis can be profitably begun, it will be necessary to recall the fact that there are several cases known wherein the rôle of the sexes is largely reversed. Herein the females do the “courting,” and fight one another as rivals for the males; while the males perform the duties of incubation and brooding, and feeding the young. This is really very remarkable, and demands more attention than it has yet received.
What factors have brought about this curious reversal? In any search for an explanation it must be borne in mind that in all such cases polyandry is the rule, and in all such cases the female is larger and more vividly coloured than the male. Here, then, we have exactly the opposite to what obtains in cases of polygamy. What 135is the reason for this preponderance of males? Why is it that when the males are in excess of the females the latter should be the more brilliantly coloured and the more amorous? These questions at present are unanswerable. When polygamy obtains it seems always to be assumed that it is explained by the excessive pugnacity of the males, which, after fierce contests for the mastery, take forcible possession of as many females as may be captured and held in durance; the same argument seems never to have been applied when polyandry obtains. There can be no doubt but that it applies in neither case.
When polygamy obtains, as we have already pointed out, the females are not seized and captured by the males, they are not victims of a lecherous lord. On the contrary, they seek the males, and the intensity of the desire to satisfy their natural cravings extinguishes any feeling of jealousy.
The same interpretation must obtain where the numerical values of the sexes is reversed. Failure to appreciate this accounts for one of the many futile suggestions made for the suppression of the rabbit plague in Australia, which was that large hauls of these pests should be made by netting, and that the females should be slain and the males released. This, it was held, would lead to the speedy reduction of the latter, which would kill one another in their fights for the remaining females. The plan was impracticable, but the suggestion demonstrated the prevalent belief as to the attitude of the male in this respect. Had it been well founded, surely polyandrous species, whether of birds or beasts, would never have existed; for, by the reduction of the males, monogamy would speedily have been restored. How, then, are 136we to explain polyandry? How are we to explain the fact, as it seems to be the fact, that the excess of males has brought about such a complete reversal in behaviour—the males, instead of the females, requiring the aphrodisiac? The solution of this problem probably lies with the physiologist. We now know that the problem of sex does not rest merely in the complete development of the primary sexual organs; we know that fertile unions do not depend merely on the act of pairing, but on the functional activity of those ancillary glands already referred to. And it may well be that some change in the character of the secretions has not only altered the numerical values of the sexes, but reversed the normal rôle of coloration and behaviour. That is to say, neither polygamy nor polyandry among the lower animals, at any rate, has been brought about or is maintained by the excessive death-rate due to combats for possession of mates, but must be explained as demonstrating inherent changes in the germ-plasm, disturbing the relative proportions of the sexes and correlated with a profound transformation, not only in the behaviour of the sexes during the period of reproductive activity, but also in their physical characteristics.
The action of the primary sexual glands and of the ancillary glands has, then, to be allowed for in all attempts to interpret behaviour in sexual matters. No less so must this be the case in regard to the development of coloration and other forms of ornament, and the genesis of weapons of offence. But at present we are, in this direction, dealing with an unknown quantity. The recognition of this, however, should not deter us from attempting to solve the riddle of sex from the phenomena 137which have so far been surveyed.
To-day the interpretation which holds the field is Darwin’s theory of “Sexual Selection.” But this was framed rather to account for the existence of conspicuous secondary sexual characters—the antlers of Deer, the train of the Peacock, and so on; it did not take cognizance of the unarmed, and the soberly-clad individuals. But whatever shortcomings we may discover, real or imaginary, in this theory, we must never forget that he had not only to analyse and present his facts, but he had first to collect them. This, in his case, was a more laborious task than most people seem to suppose. Our criticisms to-day are based, not so much on the revelations of new facts, as on the harvests of his gleaning. Yet when all is said and done, the theory of “Sexual Selection” remains, though perhaps in a new setting.
To attempt to epitomize this theory is to essay a very difficult task. But, in a condensed form, it may be said to be a theory which accounts for the development of secondary sexual characters, on the one hand through the agency of conquest by battle, whereby rival males strive for the possession of one or more females, who have no choice in the matter, or who may deliberately elect to follow the victor: and on the other by display of conspicuous ornamentation, or of more or less grotesque antics, or of some form of music, using this term in a very wide sense. Wherever display is the agent, however, its purpose seems to be to win the affections of the female to whom such attentions are addressed. She is supposed to elect to mate with the finest performers of a number of suitors. In this way, it is assumed, the intensity of the display, whatever its nature, has been gradually increased.
138Wallace strongly opposed this, contending that it assumed too much, that it assumed a common and uniform standard of perfection shared by all the females concerned in the selection, which is indeed assuming too much. But his own theory was no more satisfactory. Indeed it was very much less so, for he contended that these various exaggerations of colour and form are to be regarded simply as evidences of a superabundant vitality, though there is no evidence that “superabundant vitality,” if it exists, is a transmissible character.
The revised version of the Sexual Selection theory advanced in these pages is largely inspired by the work of Mr. H. Eliot Howard who, in his Monograph on the British Warblers, has not only added very materially to our knowledge of the life-histories of these birds, during the reproductive period, but has also done much—both in the direction of destructive, and constructive criticism, of generally accepted conceptions on this head—to set us on the right track for further research.
A study of his work leaves one with the conviction that, while these birds exhibit what we may call a nascent intelligence, their actions, on the whole, may be described as instinctive, or congenitally definite. That is to say, they follow one another in definite sequence. Hence we must regard each new phase in the chain of events appertaining to the reproductive cycle, as following one another in a definite sequence, so that any break therein throws the orderly performance of the necessary acts out of gear. There is no realization of what reproduction means, no deliberate striving to achieve that end. Each new phase brings its own set of associations and sets a new train of actions in motion, which are performed mechanically. For instance, these Warblers, like hosts of other species 139under similar circumstances, are scrupulously careful to remove the fæces of their young from the nest; thereby preserving it in a sanitary condition. It is certain that any neglect to do this would speedily end in the death of the young. This act is “instinctive”; it is not performed because the parents have evolved any views on sanitation, and any strain in whom this instinct was defective would speedily become eliminated. Mr. Howard has demonstrated the mechanical character of this sanitary measure by placing leaves in nests of young. The parents, having fed their offspring, at once seized upon the leaf and commenced to dispose of it after their usual fashion, first by trying to swallow it and then by carrying it away. They did not, evidently, realize the difference between the texture of the leaf and the milk-white, jelly-like envelope which always encloses the fæcal matter of the nestling. We shall probably never know how this most vitally important instinct came into being; nor can we hope to discover what chain of happenings begot the instinct, which each parent displays, to gently stimulate the cloacal lips of their offspring in order to induce the discharge of the fæces when this does not immediately follow the stimulus of swallowing food.
We cannot credit these birds with notions on the importance of the regular discharge of the evacuations. Equally mysterious is the development of the envelope enclosing the fæcal matter. This is jelly-like in substance, and of considerable thickness, and is enclosed within a very delicate skin or pellicle, enabling one to lift the whole in the fingers without soiling them. How and where it is formed should not long evade discovery. But how it has come to be is another matter. We can, at any rate, vaguely account for responses of the organism to 140internal stimuli reacting directly on the individual, but here is an elaborate mechanism evolved in response to extra-personal needs: and which cannot be regarded as of exactly the same configuration as the instinct to feed the young.
A return must be made to the nature of the early phases in the procession of the reproductive instincts. Mr. Howard’s study of the Warblers seems to show conclusively that these first manifest themselves in an overmastering desire to seize upon territory large enough to ensure an abundance of food for the offspring that are yet to be. To this end the males arrive from their far-distant winter quarters at least a week in advance of the females. Since each returns approximately to the scene of last year’s nursery, the arrivals are fairly distributed at the first; but nevertheless this distribution inevitably brings a conflict of interests between one or more males, perchance young birds about to start in life, and having therefore no definite objective. But whatever the reason, the competition is there. The strongest male remains in possession, and immediately commences to express the ecstasy of feeling which possesses him in continuous outbursts of song. Such, doubtless, answer to the bellowing of the male stag. They advertise the presence of a male to the female, who, as she arrives, would seem to be already stirred by the rising storm of sexual desire, for having once discovered a male in possession of the all-necessary site for the nest, and the equally necessary domain, each settles down to conjugal bliss: within twenty-four hours the task of building has begun. There is evidently here no sexual selection in Darwin’s sense: no choice from among a number of males of the individual which most excites desire within her; but the mating of the most mettlesome, most virile males has been determined before her arrival and by a double sieve. In 141the first place, the duller-witted birds fail to secure suitable territory, and in the second, the territory, having been taken, must be held by force, so that only the strongest males remain to mate when the females eventually arrive. So far as one can see, selection is less exacting in the case of the females, which apparently need do little more than respond to the advances of the males.
Plate 22.

From a drawing by H. Grönvold.
FIGHTING FOR TERRITORY.
Two Black-caps are here seen fighting for their annual breeding territory. A Chiff-chaff has been unable to resist the excitement of conflict.
Face page 140.
With the advent of the females the amorous instincts of the male speedily gather force; but for their satisfaction it is imperative that the female should be possessed by a like desire. To provoke this, for it is essential to the well-being of the race that offspring should be produced as early as possible, some form of aphrodisiac seems to be necessary. This fact has never been properly realized, though it is implied in Darwin’s theory of “Sexual Selection.” Here, however, it was used to account for the evolution of resplendent coloration, eccentric postures, and dances which, it was assumed, enabled or induced the female to choose the most mettlesome males. What obtained among sombre-clad species, appears to have excited no curiosity among the students of the evolution theory. Hence it comes somewhat as a surprise to find that the soberly-clad Warblers behave exactly as though they too wore coats of many colours. After what has been said in the last chapter on this head it will be unnecessary to describe these displays among the Warblers in detail, more especially as my friend Mr. Howard has kindly allowed me to use some of the illustrations from his book. These show convincingly enough that the wings and tail are made to play the same part as though they bore all the hues of the rainbow. To bring this fact home compare 142the figures of some of these small birds clad in sober russet and black with that of the Sun Bittern (Eurypyga helias) in like mood, whose wings and tail when spread, and only then, display bands of vivid chestnut-red, contrasting with bands of black, on a background of grey and buff, variegated with delicate mottlings and vermiculations of black and brown, and streaks of white. In the case of the Warblers, it is to be remarked, the male, in these ecstatic moods, will commonly hold a leaf, or a piece of stick, in his beak, as if suggesting the work of nest-building and its delightful sequence. This, or its equivalent, is a common phase, for the Great Crested Grebe, for example, in these paroxysms will dive and bring up weed, the nest material of the species, as an offering to his mate, or as a stimulant to her yet slumbering passion.
It seems clear, then, that the evolution of colour is not the stimulant to display, for this is present where conspicuous colours are wanting. Yet it can readily be understood how the association of ideas in regard to colour and display arose, for there are cases where this interpretation seems inevitable. Such are afforded by certain sea-birds like the Kittiwake, Guillemot, Fulmar and Cormorant, wherein the inside of the mouth is of a lurid orange-red in the case of the first-mentioned, and of flaming gamboge yellow in that of the others. During moments of sexual ecstasy the mouth is widely opened, as if to charm the beholder with its gaudy hue. Both sexes have the same colouring, and both behave alike. But it is doubtful whether either is conscious that its own mouth is like that exposed to its gaze: the action is sympathetic. No doubt it may play its part in stimulating desire, but we cannot contend from this that it has been evolved by sexual selection, that is to say, that 143the hues have undergone a process of gradual intensification owing to the deliberate rejection of the less gaily-coloured suitors. The tendency to develop colour in the mouth would appear to be latent in all birds.
Plate 23.

From a drawing by H. Grönvold.
THE DISPLAY OF THE GRASSHOPPER WARBLER.
The behaviour of this bird under the stimulus of sexual excitement is precisely similar to that of the Sun-bittern and the Kagu, yet it has no brilliant colours to exhibit by such actions.
Face page 142.
Plate 24.

THE DISPLAY OF THE SUN-BITTERN.
Quite inconspicuous in repose, this bird, in its moments of exaltation, becomes banded and blotched with vivid colours, revealed by spreading the wings and tail.

Photos copyright, D. Seth-Smith.
THE KAGU IN DISPLAY.
What is true of the Sun-Bittern is true also of the Kagu.
It is significant that whenever bright colours appear, they do so first in the males, the females and young retaining the dress common, up to this time, to the species at all ages. In the majority of instances, at any rate, it would seem that this accession of colour appears with the seasonal re-awakening of the reproductive activities: it forms a “nuptial” dress, and is discarded after the breeding season is over for a livery indistinguishable from that of the female, this forming the so-called “winter plumage.” But if all the available facts are taken into consideration there seems good reason to believe that the nuptial plumage tends to be assumed earlier and to be retained later, as this disposition to develop ornament gathers force, till finally only the head and neck go into “eclipse,” as in the case of the Black-cock, Jungle-fowl and Partridge.
In the Pheasant we have an instance—one of hundreds—where the resplendent dress is worn throughout the year. The next phase in the direction of the growth of colour occurs when the female, towards old age, develops a more or less well marked tendency to assume the hues of her lord, and this accession of colour makes its appearance earlier and earlier in succeeding generations, till finally the adults of both sexes are coloured alike, save that, as a rule, the female lacks the intensity of coloration which her mate displays. The original sombre dress is now only worn by the young. In due course the resplendent dress is assumed also by the young, as witness the numerous instances among the Kingfishers 144and among the Parrots, where adults and young are all habited in the same vivid hues. There are infinite variations of these changes which cannot be discussed here, for obvious reasons. All that matters now is the fact of such sequences, which inevitably raise the questions: Why, in so many cases, do the females show no disposition to assume resplendent colours? And to what factors can such coloration, when it occurs, be attributed? The second only of these questions is germane to the present discussion, and to this no very satisfactory answer can be returned.
To say that the development of brilliance in species hitherto sombrely clad is due to “changes in the metabolism” is only an affectation of wisdom. What we want to know is what induces the changes? Time was when no more than a guess could be hazarded as to this: a suggestion that ornament, of whatever kind, was one of the many modes of the expression of that instability of the organism which is characteristic of living things: that it was one of the outward and visible signs of that inward, intangible tendency to vary which is so familiar. Later research seemed to show, fairly conclusively, that ornament was one of those “secondary sexual characters” which was dependent on the stimulating juices, or “hormones,” emanating from the primary sexual glands. To-day it is manifest that this is only partly true, for it is certain that these glands are not alone concerned and they may only participate indirectly. It seems to have been clearly demonstrated that the thyroid and pituitary glands, or the “hormones” therefrom, play a large part in this matter of the “secondary sexual characters.”
Castration, it is true, profoundly affects these characters. In the case of Deer it inhibits the growth of antlers, 145in Cattle the horns are increased in length but reduced in thickness—they are longer than those of the female, but resemble them in appearance, and further, the whole stature is greatly increased, but it is at the same time conspicuously less massive, particularly at the neck and fore-quarters. In eunuchs it results in immense stature and the loss of the more characteristic male features, such as the beard and the bass voice. The removal of the testes in birds is always a difficult operation and is rarely successfully performed. Hence the accounts of changes in plumage consequent on this operation are inconclusive. It has generally been supposed that whenever, either by removal or by disease, the testes are rendered inoperative the plumage, when normally of a resplendent type, assumes the coloration of the female. This is probably an erroneous supposition, but what happens is a failure to secrete the more intense pigments and the more specialized forms of feathers, so that the resultant dress answers to the juvenile male dress. It is not a case of “reversion” to this livery, but a failure to assume the latest acquirements of the species. These, as has already been shown, are only very gradually developed. The intensity of pigmentation, or concentration of pigmentation, which results in sharply defined areas of colour, is a cumulative process. As it loses in intensity at any given moult, so the individual tends to reproduce the phases of the earlier and vanishing livery. Sooner or later, however, this earlier livery disappears more or less completely: is eliminated from the system, so to speak: and what is commonly called lack of “vigour” results, not in a return to the earlier, sombre dress, but in the later-acquired, resplendent plumage lacking intensity. The seasonal, temporary secondary sexual character has become, as some say, a “somatic” character. Highly probable as 146this view appears, it ought, it may be argued, to receive support from nestling plumages. Young gulls, for example, should occasionally revert from the mottled to the earlier striped livery. But we have no evidence of this; and it does not follow that this sequence of events should occur. The conditions of control are different.
What exactly are the factors which govern the evolution of resplendent plumage is not known. But they would seem to be more complex than was supposed. That the primary sexual glands play an important part, through the juices or “hormones” which they liberate, there can be no doubt but these are only partial factors. The “hormones” of the pituitary and thyroid glands are also necessary contributors, controlling as they do both fertility and the more superficial characters, such as colour and ornament. Evidence, indeed, is slowly accumulating to show that the problem of the behaviour of animals during the period of sexual activity, as well as the peculiarities of structure and coloration which they develop at this time, are all largely governed by the action of these secretions.
These, in their turn, are undoubtedly inhibited, or increased, by the control of the nervous system, though this control is of course involuntary. This much seems clear from the fact that birds will display when under the excitement of fear, though the character of that display is never the same as that in moments of sexual exaltation. If the nervous system, through the eye, by “suggestion,” played no part, there could be no use for display, but it is equally certain that for the realization of the sexual activities a number of other factors have 147to contribute.
The existence of this nexus of conditions is commonly overlooked, but it is extremely important. Normally, not only among birds, but other animals higher and lower in the scale of life, “suggestion” does not suggest until the “hormones” concerned with the sexual activities have, as it were, saturated the system and rendered it, so to speak, highly inflammable. Even then it commonly happens that, with the male at any rate, this inflammable state bursts into flame of its own accord. But for this, indeed, how could the consummation—of the period of sexual activity ever be realized? In many cases the sexes are sundered far apart. What, but the merest accident, could bring them together if it were not for this consuming fire of desire which impels each sex to seek out the other? This stage is manifested in the case of the Deer, where, we have seen, the stag wanders far and wide bellowing to advertise his errand and listening for a response to his call. He is possessed by a “male-hunger” which eventually attains to a state of frenzy. Here no “suggestion” is needed, but the necessity for this stimulus, for some form of aphrodisiac, occurs with him after the first relief of his pent-up state has been attained. This stimulus is applied, both through the eye and the sense of smell, by the females of his herd. The same conditions apply in the case of the birds. But it is to be noted that with the females, as in the case of mammals, sexual desire is commonly less intense than in the males, and hence, in their case the need for “suggestion” by display of some sort. But apart from this, a “display” of some kind is necessary. How else can desire be indicated? And here is “sexual selection.” For males, mate-hungry as they might be, which resorted to no 148means of expressing their condition would go mateless: and the same is true, though perhaps in less extent, with the females; hence, then, it is clear display is a product of sexual selection.
That sexual desire is less intense in the case of the females is to be regarded as another result of this form of selection. If they displayed the same intensity of passion the males would speedily become exhausted, for it is well known that the gratification of the sexual emotions is far more enervating in the case of the male. It may well be that polyandry has arisen from this transference to the females, or development by the females, of increased sexual hunger.
The fact that birds will repeat, albeit imperfectly, the phases of the sexual display under the stimulus of fear, or anger, and when no females are present, must be regarded as an indication, for we can scarcely call it a proof, that exaggerated movements have become the normal concomitants of great excitement, at any rate during the season of reproductive activity. They are purely nervous responses to external conditions. It must not be forgotten that, at this time, fear begets other movements, equally striking, such as feigning lameness, and death, which have no part in the sexual display.
Interpreted in this light one can understand that to the female not as yet sexually “ripe” or sexually “hungry,” these movements, when not interpreted as signs of fear or anger, fail to produce any response. So soon, however, as this period of “ripeness” arrives, the stimulus through the nervous system produces the desired response, begetting a complementary stimulus through the secretions of the sexual glands, by what we may call 149the flow of the hormones; just as the sight of food stimulates the flow of saliva, or “makes the mouth water” before we are conscious of feeling hungry. In due time hunger will assert itself without the stimulus of the nervous system through the senses. But there must in any case be some form of display, some form of communicating and stimulating desire between the sexes, to secure the consummation of the reproductive acts. How else could intimation of sex hunger be indicated and satisfied?
That the desire for sexual congress is inherently more avid, more intense, in the male than in the female is often called in question; and more especially so by those who imagine that they have a mission to carry on “social reforms” and to regulate the relations between the sexes of the human race. Such aims and ambitions are commonly those of the arrogantly ignorant. There are few people who possess a sufficiently wide knowledge of this theme, or of the factors which underlie it, to qualify them to become the mentors of their fellow-men in these matters. However much we may choose to seek refuge in sophistry, the fact remains that man is still an animal, and if the human race is to continue he must always remain so.
A lurid light has just been shed on the fierceness of the sexual passion in the male by Mr. Julian Huxley, who relates some facts pregnant with meaning to all who have understanding, in regard to what obtains among birds. These facts are primarily concerned with the Mallard (Anas boscas). This bird is ostensibly monogamous, and, on the whole, seems to be a fairly considerate mate. The normal period of pairing having passed, and the duties of incubation having begun, the female ceases to harbour any further desire for sexual intimacy. Her whole energies 150are devoted to nursing her embryonic young into life. Not so the male. He is yet far from satiated; in him the sexual fever still burns fiercely, but somehow he seems never to make any attempt to provoke in his mate a like condition, as in the days before brooding began. On the other hand, he does not scruple to savagely pursue every other female who ventures abroad in his neighbourhood. So soon as a duck takes wing for a brief relaxation from the arduous work of brooding she is pursued by ten or a dozen already mated males, till at last she is obliged to descend on the water, and with her descend her pursuers, now to mob her without mercy. Commonly at least half of these infuriated males will eventually succeed in treading her; leaving their victim only after she has become completely exhausted or killed outright. This is no unusual occurrence. On the reservoirs at Tring, where every spring from one thousand to one thousand two hundred pairs congregate to breed, from seven per cent, to ten per cent, of females are annually killed in this way.
It is just possible, however, that an error may have crept into these observations. One cannot help asking, may it not be possible that these pursuing males were actually unmated birds? The chief argument against this is the fact that there is no sort of attempt to “display” apparent with these birds, simply an overmastering, ravenous desire to satisfy the craving which possesses them.
Evidence is not wanting that the evolution of pigment intensification and the consequent development of vividly coloured liveries, or the equivalent development of ornament, has been accompanied by an intensification of the reproductive instincts. For there can be no doubt but 151that the display of species which are conspicuous for their ornamentation is more animated than those of duller hues. As an argument in favour of this view the case of the display of the Great Crested Grebe may be cited, wherein each sex has developed both colour and ornament to a high degree, and are distinguishable only to the expert.
The latest and the best exponent of the behaviour of this species under the spell of sexual exaltation is Mr. Julian Huxley, whose observations, in a condensed form, are now to be surveyed. The most conspicuous features in this bird are the great Elizabethan ruff of bright chestnut and dark Vandyke brown, and the long dark-brown tufts of feathers, or “ears,” which surmount the head. But the satin-like sheen of the white breast and the fore part of the neck and face add not a little to the general effect. These ornaments are worn only during the breeding season. So soon as the fires within begin to burn, the parade of this finery commences, and it would seem that a somewhat protracted dalliance takes place before any actual pairing. During the early phases of these performances much play is made with ruffs and “ears.” The courting pair will frequently face one another on the water, and go through a strange ceremony of head-shaking. To this is soon added a sort of ghost dance, wherein the male suddenly dives, leaving his mate swinging excitedly from side to side. In a moment or two, however, he appears, not suddenly, as usual, but arising gradually out of the water. He seems to “grow” out of the water. First his head appears, with ears and ruff extended, and beak pointed downwards; then his neck, and finally the body arises into view, till only the extreme tail end remains submerged, so that he looks 152more like a penguin than a grebe! All the while he is turning on his long axis, as it were, till he gradually displays before his mate the dazzling white sheen of his breast and neck, set off by the rich red chestnut and brown of his face and frills. A moment more and both subside into their normal attitude, shake their heads at one another, and then proceed to feed as if nothing had happened.
But these quaint antics are only the preliminaries to still stranger. A pair of birds, engaged, apparently, solely in fishing and feeding, will suddenly approach one another and begin head-shaking, each striving to outdo the other. Then the ears, till now erect, are thrust out laterally, and the ruff is still further erected till it forms, with the ears, a common disc. Then the hen dives: immediately after down goes the cock. After some fifteen seconds or so she appears at the surface again, speedily followed by the cock, who breaks out about five-and-twenty yards off. Each crouches low over the water, and each will be seen bearing a tuft of weed in the beak. As each sights the other a tremendous rush is made, as if they intend to charge. But when about a yard apart each springs up and assumes the penguin position, save that the beak, instead of pointing downwards, is now held horizontally and bears its burden of weed. Still approaching, they eventually touch one another, treading the water and swaying in a sort of ecstasy, all the while shaking their heads from side to side. Then they gradually settle down into the normal swimming pose, though still keeping up the head-shaking; then this, too, subsides, the weed is dropped, and the performers drift apart and begin feeding. But no actual pairing accompanies these strange performances. This final rite is associated with a quite different ceremonial, and was 153witnessed more than once by Mr. Huxley. On the particular occasion which he describes he was watching a male swimming along near the reeds, apparently on the look-out for something, and turning his eyes in the direction of the course, he saw, at some distance off, what he supposed was a dead grebe lying hunched up in the water, with outstretched neck, and ruff and ears depressed. Presently the male swam alongside the body and bent down his head as if to examine it. Then he swam to the tail end, and suddenly scrambled out of the water on to the body; and there, with bowed head and depressed ears and crest, he seemed to stand a moment. Then he waddled forward over its head and into the water. Instantly the supposed corpse raised its head and neck, gave a sort of jump, and was swimming by the side of its mate. They had been pairing on a half-made nest, whose surface lay level with the water.
Plate 25.

From a drawing by H. Grönvold.
A MALE-SAVI’S WARBLER
—in one of his “courtship” attitudes. Note the leaf held in the beak.
Face page 152.
Mr. Edmund Selous seems to have witnessed some almost incredible behaviour on the part of the owners of a nest he had under observation, inasmuch as, on more than one occasion, he declares the male lay prone upon the nest and the female assumed the position of the male. After this pantomime both would leave the nest, but commonly the female would speedily return and pairing would be duly performed.
This brief summary of Mr. Huxley’s observations, which he was generous enough to give me the privilege of seeing in manuscript, taken in conjunction with many other facts of a like kind given in these pages, seems to lend support to the view that an excessive amorousness is commonly associated with conspicuous ornamentation, as if these stood in the relation of cause and effect.
Finally, it is contended, the facts garnered during recent years show that the theory of Sexual Selection, as Darwin propounded it, especially in so far as birds are concerned, is no longer tenable: but it is not an exploded theory, it has only undergone modification. So far as the evidence goes, it would seem that the first of the series of events in the sexual cycle is performed by the already avid male, when he proceeds to secure a “territory” large enough for his needs. In insectivorous and carnivorous species this area is fairly extensive. No other male will be allowed within its confines. The perfection of this instinct is vitally important, if sufficient food for the offspring that are to be is to be assured. Where the food is inexhaustible, as with the Auk-tribe, only a ledge large enough to hold the egg is required. Only avid males will develop and respond to this stimulus. The second stage occurs with the arrival of a female in the area. She does not at once proceed to “select” her mate, passing on if he fails to provoke her admiration. Her sexual condition is apparently as yet but half awakened: to rouse this, the male supplies an aphrodisiac in some form of display to which, in the normal course of things, she responds, often also with some form of display, or indication of the desire which has been aroused. The intensity of the performance seems to vary with the intensity of the sexual passion, which appears to be greater in some species than others, and especially so with such as have conspicuously ornamental plumage. There is, indeed, a variation in the sexual appetite as there is in the ornamentation. The two are reciprocal, and are determined in degree by the stimulatory qualities of the hormones of the sexual glands. Where these have been developed in like intensity by the females, they also display. Diminution in the quality and quantity of the 155stimulating secretions of the ancillary sexual glands, the hormones of the pituitary and thyroid, or the primary glands—testis and ovary—decreases fertility, or induces sterility. Where these stimulants are lacking there will be no desire, no display, and no pairing, and consequently an end to this defective strain. Here then is Sexual Selection.
Plate 26.

Photo copyright by D. Seth-Smith.
ANOTHER ASPECT OF THE KAGU’S “DISPLAY.”
Herein two birds are seen facing one another with the great head-crest fully erected. While in this mood these birds will strut up and down with mincing gait and drooping wings. This is a posture commonly assumed during momentary excitement, whereas the posture shown in plate 24 is apparently only assumed during moments of sexual excitement.
[Face page 154.
Instances of such impotency on the part of either sex are wanting, and we can only speculate as to how such cases would be met. Would a female who had chanced to settle in the territory of a male whose sexual impulses carried him no further than seizing territory remain with him throughout the mating season, held by an imperfectly roused, ill-defined, sexual instinct? Or, eventually becoming mate-hungry, and failing to stimulate him to perform his part, would she desert him and seek another mate? On the other hand, would a male, failing to arouse response in the female he had secured, drive her away and supplant her?
In other words, are we then justified in postulating differential effects in regard to display: a minimum of intensity to ensure mating? A display of some sort is essential. It may be feeble as compared with that of another species—that of the Sparrow, for instance, compared with that of the Peacock—but it must be sufficiently good of its kind to effect its purpose, which is to “hustle” up the production of offspring. A phlegmatic but virile male, or a too feeble performer, is almost as certainly doomed to extinction as an impotent male; for his offspring will probably be eliminated by the adverse conditions of existence to which their late appearance exposed them. Where a female settles down with a male which does not attain to the standard of display 156characteristic of his race, it is conceivable she may sooner or later seek a mate elsewhere, deserting the phlegmatic bird as if under the impression that she had made the mistake of settling down with one of her own sex. There is no need that the female should have to “select” the best performer of a number of males displaying at the same time and place as a number of rivals.
Finally, the ornamental crests and frills, and the vivid hues which so many birds display have not arisen, as is generally supposed, as a direct result of the selection, by the females, of the most vividly coloured, or ornamented, from among a number of suitors presenting varying degrees of intensity in ornamentation. Such “frills and furbelows” are to be regarded as “expression points” of internal variations in the germ-plasm, which have been free to develop along their own lines because they have not proved in disharmony with the conditions of the birds’ environment. Their development is to be traced to the stimulating action of the “hormones” which control both pigmentation and structure, as is shown by the fact that both are modified by any interference with the glands in question. Such ornamental features then are the concomitants not the results of Sexual selection.
The development of ornament, whether of colour or structure, may be taken then as an index of specialization, and as one of the many manifestations of that variation which is going on in every part of every living organism.
So long as the continued increments in the development of these characters do not hamper their possessors in the struggle for existence, they are free to go on developing. Sexual selection, other things being equal, operates by according the greatest number of descendants to the most amorous, and not necessarily to those of the brightest hues.
Plate 27.

Photos copyright, G. Herring.
SOME STRANGE ACCOMPANIMENTS OF COURTSHIP.
THE WHITE-HEADED BELL-BIRD.
This species is remarkable for the enormous, erectile wattles which arise from the base of the beak of the male at the courting season.

THE UMBRELLA-BIRD.
The crest which adorns the crown of the head has many counterparts, but the long-feather clad wattle which depends from the fore-part of the breast is unique.
[Face page 156.
Plate 28.
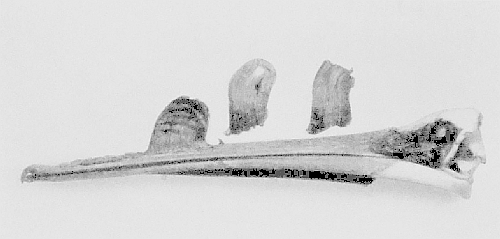
SKULL OF THE AMERICAN WHITE BEAKED PELICAN.
The beak of this bird develops at each breeding season an irregular horny plate which falls off at the end of this period, It is difficult to regard this as a sexual “ornament,” yet it comes under this category.
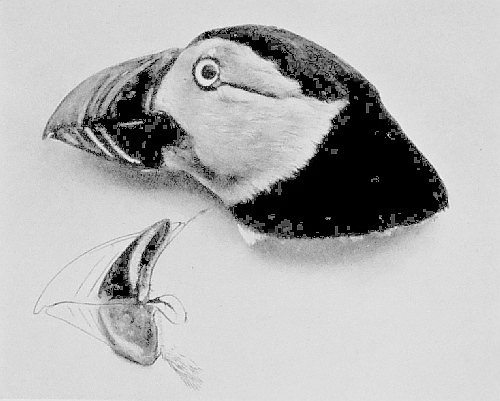
Photos copyright, G. Herring.
HEAD OF A PUFFIN, SHOWING THE MOULTING OF THE BEAK SHEATH.
At the breeding season, in both sexes, a triangular horny plate is developed over the eye, an oblong plate below it, while the sides of the beak become deepened by means of larger triangular horny plates. All these embellishments are highly coloured, and they are shed at the end of the breeding season. A further ornament is developed at the gape, in the shape of a fleshy rosette of a bright orange colour.
Face page 156.]
But Sexual selection does not begin, and end, with the evolution of frills and furbelows. “Behaviour” counts for more than is generally supposed. This is as specific as “structure,” that is to say, it is as constant for each species as is its coloration, and it is also as variable. That Evolution may be determined by variation in behaviour, no less than through structural variations, is a possibility which has received but little consideration at the hands of students of Evolution.
The singular history of the Australian Bower-birds lends additional support to this view, and at the same time provides an additional argument against the generally accepted opinion that bright colours have been evolved by reason of the preference shown by the females for the most vividly coloured of their suitors. For while the males affect all the tricks and turns which are the common accompaniment of courtship, they, in addition, introduce very extraordinary features in the shape of “bowers” cunningly constructed and often gaily decorated, as will be seen presently. Eight of the total number of species of this group exhibit this behaviour, and while they differ very conspicuously in coloration among themselves, they agree very closely in the type of the bower they build. If the coloration is determined by the female, then in this they display very different standards, and if they do select, each according to the standard of the species, then we must suppose that they also must exercise a choice in regard to the character of the bower, the favoured male being the best builder. But why, in this case, is there not as much diversity in the form of the bowers as in the coloration of the feathers? A survey of the facts will perhaps make this 158point clear.
One of the best known of these bowers is that of the Satin Bower-bird (Ptilonorhynchus violaceus). On either side of a platform of small twigs a fence of similar twigs is reared, sloping inwards to form a more or less complete tunnel. At the entrance to this is placed a platform of sticks, which is strewn with a miscellaneous assortment of brightly coloured feathers, bleached bones, and occasionally flowers. The work of construction is almost entirely performed by the male: it is indeed a little curious, having regard to the circumstances, that the female should bear any share in its construction at all.
Really this is a more wonderful piece of architecture than would appear from the mere description of its main features: for it represents psychical activities which are difficult to fathom. It does not take the place of display, but is an extension of this. During his amorous moments the cock becomes greatly excited, chasing his mate in and out of the bower, carrying the while, in his beak, a brightly coloured feather or a leaf.
At the same time he sets all his feathers on end and every now and then drops first one wing, then the other, accompanying these actions with curious whistling notes and pretences of picking up food.
The Regent-bird (Sericulus melinus) differs conspicuously from the Satin Bower-bird, for while this is of a uniform, deep, metallic steel-blue, the Regent-bird is jet black, with a golden yellow crown and hind-neck and a great blaze of golden yellow on the wing. Yet the bowers of the two species—which belong to different genera—are practically identical, save that brightly coloured berries are used more frequently by the Regent-bird.
Plate 29.

Photo by W. P. Dando.
THE SATIN BOWER-BIRD AND ITS BOWER.
The “Bowers” of the “Bower-builders” are the most remarkable variants on “Secondary Sexual Characters” yet brought to light.

Photo by L. Medland.
THE “BOWER” OF THE BOWER-BIRD.
The “Bower” must not be confused with the nest, which is placed in a tree and bears no sort of likeness to the bower.
[Face page 156.
The Spotted Bower-birds (Chlamydodera maculata and 159C. nuchalis) are quite dull-coloured species save for a vivid semicircular crest of pink and mauve feathers which arise from the nape of the neck. Their bowers differ from those just described in having a longer run and for the immense quantities of shells which are deposited at each end of the run. Some of them are brought from long distances, as is shown by the large number of sea shells which are to be found in the collections made by birds living far from the sea.
By far the most remarkable of all are the bowers of Newton’s Bower-bird (Prionodura newtoni) and the Gardener Bower-bird (Amblyornis inornata). The first of these, a native of the Mountains of Queensland, is somewhat strikingly coloured, at any rate so far as the male is concerned, for he is of an oil green above and has a small yellow crest, while his breast is of a bright yellow; the female, on the other hand, is brown above and grey below.
The Gardener Bower-bird, on the other hand, is of a sombre olive-brown, but the male boasts an enormous crest of a flaming orange yellow. Yet, widely dissimilar as are these two species, in the matter of their bowers they display much in common.
That of the Gardener Bower-bird takes the form of a hut-like structure of twigs, arranged around a central support, commonly a very young sapling. As a rule the thin stems of an orchid (Dendrobium) are used in the construction of this curious hut, whose diameter is about three feet. Before the entrance is a carpet of moss, which is kept clear of leaves or debris of any sort, and on this the most vividly coloured fruit, seed-pods, fungi, and flowers are laid, being constantly replaced as they 160wither. Newton’s Bower-bird, in like manner, forms a hut around a central column: a hut which may attain to a height of as much as six or even eight feet, and the walls of the pyramid thus raised are generally gaily decorated with flowers and fruit. Around the central a number of subsidiary huts are not infrequently found, and in and out of these the birds pursue one another in ecstasies of excitement.
We have in these facts some extremely puzzling features, which at present, at any rate, permit of no more than a very rough analysis. Probably the whole of these bower-building instincts have their origin in the habit, which the males of so many birds exhibit, of carrying a leaf in the beak when under the excitement of love-making. This is suggestive of nest-building, and in many species this is actually begun before the arrival of a female in the breeding territory, while others build what are known as “cock-nests” which are never used. Among the Bower-birds these “cock-nests” have taken a new and more elaborate form, and are placed on the ground instead of in the trees, the normal site for the nest in all these birds. Furthermore, stages in the evolution of such strange fabrications can be found. These are furnished by the Tooth-billed Bower-bird (Scenopaeetes dentirostris), the Cat-bird (Aeluredus viridus) and the gorgeous Lawe’s Bird of Paradise (Parotid lawesi)—which is not perhaps a Bird of Paradise. These build no bowers, but are content with clearing a patch of ground, of about ten feet in diameter, on which to disport themselves. But while the “displays” of these birds closely resemble one another, in the matter of coloration and ornament they present the most striking contrasts.
The Courtship of the Crocodile—Amorous Lizards—Horned Chamæleons—A flagellating Terrapin—The Frog that would a-wooing go—Semo musical Frogs—Some marvellous instincts in Newts.
The measure of the vitality of animals may be estimated by their response to stimuli; and their behaviour increases in variety and complexity as the nervous system develops. Our interpretation of that behaviour commonly leaves out of account the character of this responsiveness: we are apt to see proof of intelligence in acts which should be read as instinctive. And instinct is to be regarded as a co-ordinated response to stimulus, independent of prior experience.
The complexity of this response stands in very close relation to the structural complexity of the organism in which it occurs, and this because an ever-increasing number of mechanisms and actions must be set in motion to carry out the fulfilment of any given stimulus, as this is traced from the lower to the higher groups of animals: till at last we have to distinguish between movements that are merely reflexes, and those which are “instinctive.” The latter must be fulfilled by the former—the reflex 162actions are the agents of the instinctive. Indifferent performance in either, endangers the existence of the individual, and in some directions of the race itself.
The sexual instincts, with which alone these pages are concerned, are primarily stimulated and sustained by internal forces, generated, as we have already seen, by the juices of certain glands whose relation to the reproductive system has only recently been discovered. Though not commonly realized, and though denied by some, the sexual instincts are the dominant factors in the animal world. Even Man himself, the lord of Creation, knowing good and evil, cannot escape their overmastering rule. Commonly he is by no means inclined to rebel against this control But there be some who, in their arrogance, imagine that its overthrow is an end to be desired. Having scaled some slight intellectual eminence they fondly imagine this feat was accomplished by virtue of some spiritual grace of their own cultivation, and call to their fellow-men to emulate their example. But such preceptors are labouring under a strange delusion: they are suffering from a disease they wot not of, a “Disharmony,” as Metschnikoff calls it, a disease which blinds their perception of the motive power which has given them all that they believe themselves to have created. For these same despised instincts are the sacred fires of our being, and when they are quenched all that makes us human, love, ambition, and life itself will be extinguished. If the continuance of the race be a thing to be desired it is well that the choice should not be left to us.
Truisms are sometimes trite, and while it is a truism to say that no race can continue which does not reproduce its kind, it is more exact to say that, other things being 163equal, the race depends for its existence, primarily, on the efficient working of the sexual instincts. In the higher animals, the phenomena which these present are so complex that they often assume something more than a semblance of intelligent, purposeful behaviour. It is therefore necessary, for their right understanding, that they should be analysed in animals lower and lower in the scale of life till at last we come to the very simplest types of organisms wherein instinct can be said to play a part.
The lower we descend in the scale of animal life in our survey of behaviour during the reproductive period, the more the evidence seems to grow in favour of the interpretation of the Sexual selection theory adopted in these pages—the view that neither the formal displays nor the exaggerations of colour and ornament which so commonly accompany them, are due to female choice; a choice not necessarily conscious, but rather to be interpreted as the final abandonment to the finest performer of a number of suitors. On the contrary, this ornamentation, of whatever kind, is the expression of an intensification of the gland secretions which is manifested by the process of pigment concentration and a consequent intensification of coloration. Hand in hand with these developments it would appear goes an exaggeration of the normal movements which characterize the species when under the influence of great excitement, whether of fear or pleasure. At any rate, the displays of gaudily coloured and highly ornamental species are commonly more striking than those of sober hue.
On this rendering, the behaviour of Reptiles, Amphibia and Fishes, is much more readily interpreted, and this is even truer of the more lowly groups of animals such as Spiders, Butterflies and Beetles.
164Among the Reptiles, as among the birds and beasts, the desire to obtain territory seems to be strong. But the information to be gathered as to their behaviour in the search for mates, and after, is exceedingly small.
Sluggish by nature, all become animated under the stimulus of mate-hunger, and this is especially true of the males. As one would have expected, from what has just been said, desire is most demonstrative in brightly coloured and highly ornamented species. But even the dullest hued and most phlegmatic display quite surprising agility and animation under the fever of Love. Thus among the Crocodiles fierce battles are fought by rival males for the possession of some coveted female: and later the victor strives to dispel the apathy of his mate by caperings most undignified in a Crocodile. He will twist and turn, or rather twirl, round on the surface of his chosen pool, with head and tail raised high in air, and his capacious barrel of a body swollen out to bursting point. These antics are performed to the accompaniment of loud bellowings and roars heard at no other season of the year. But more than this, an appeal is made to the nose as well as to the eyes of his apathetic mate, for during all this parade of love he exudes from glands in the lower jaw, and tail, an almost overpowering smell of musk. At last, however, these antics have their reward, for sooner or later apathy awakens into interest, and interest ends in desire.
The Crocodile is colourless, or at least is monochromatic; not so many of the Lizards, which rival the birds in the vividness of their hues. With the birds the colours undergo no changes save such as are due to the incidence of light; with the Lizards, however, the bare 165skin is exposed and this can, as it were, be made to blush with all the colours of the rainbow. Having regard to what has been said already as to the sources of this coloration it is not surprising to note that here also the males are the more vividly coloured whenever the sexes differ in this particular. And further, it is among the most vividly coloured males that most animated displays take place when the endeavour is being made to excite the amorous instincts of the females.
The males of the genus Sitana are very brightly garnished. They possess a large throat pouch, coloured blue, black, and red when expanded, and this occurs only during moments of excitement, whether this is due to fear or pleasure. And at the same time the vividness of the coloration is greatly increased. No such secondary sexual characters are present in the female.
A variant on the throat pouch, of a much more striking character, is displayed by the Frilled Lizard (Chlamydosaurus kingi), wherein the tongue bones have become enormously elongated so as to project backwards on each side of the body almost as far as the base of the tail. With them they have carried a thin fold of skin, so that whenever the mouth is opened these bones stand out at right angles to the head and display a circular fold of skin stretched as it were on rods; or they may be compared to the ribs of an umbrella. The great Elizabethan frill thus formed, is displayed only during moments of great excitement, and the open mouth, at such times, is flushed with a vivid red, which, contrasting with the teeth, gives a very terrifying aspect to prospective enemies, and doubtless also proves a valuable asset as a “secondary sexual character.”
The display of a vividly coloured mouth during moments 166of sexual excitement, it may be remembered, occurs in some birds. Among reptiles it is a common feature. A good illustration of this is furnished by the Moustached Lizard (Phrynocephalus mystaceus), a native of Southern Russia. When violently excited it raises itself on its hind legs, curls and uncurls its tail, and opens its mouth to its widest extent, presenting, to our eyes, a quite fearsome aspect. This effect is immensely increased by the fact that the corners of the mouth are provided with flanges of skin, which at this time swell up into crescentic plates, the inner borders of which pass gradually into the rosy lining of the mouth, thereby causing it to appear much wider than it really is. So far this display has been witnessed only when the animal is under the influence of fear. But since we find that birds will make similar displays, both when under the stimulus of fear and that of sex, we may assume, with no little degree of certitude, that the same applies in the case of the reptiles, for the origin of the ornaments is almost certainly to be attributed to the same gland secretions which produce the secondary sexual characters of birds and beasts.
This, however, is no mere assumption, for we have some positive evidence as to the association of bright coloration with “courtship,” which has been furnished by Mr. Annandale, a naturalist of long experience and having a first-hand acquaintance with tropical life. He has given us a lively description of the courtship of the Malayan Lizard (Calotes emma). “The males,” he says, “are very pugnacious, and change colour as they fight. At the time of courtship a curious performance is gone through by the male, the female remaining concealed in the foliage hard by. He chooses some convenient station, such as a banana-leaf, or the top of a fence, and advances slowly towards the female. His colour is then pale 167yellowish flesh colour, with a conspicuous dark spot on each of the gular pouches, which are extended to their utmost. He stands upright, raising the fore-part of the body as high as possible, and nodding his head up and down. As he does so the mouth is rapidly opened and shut, but no sound is emitted. When he is driven away, caught, or killed, the dark spot disappears entirely from the neck.”
Plate 30.

Photo by W. Saville-Kent.
THE BEARDED LIZARD.
Paring moments of excitement the Bearded Lizard opens the mouth widely displaying a vividly coloured interior.
[Face page 166
Normally sluggish, the Lizards display, it will have been remarked, a quite surprising degree of animation when maddened by mate-hunger. Some exhibit a considerable degree of pugnacity. In Anolis carolinensis, for example, when two males meet they face one another, bob the head up and down two or three times, expand the throat pouch, lash their tails from side to side, and then, worked up to the requisite degree of fury, rush at one another, rolling over and over and holding firmly with the teeth. The conflict generally ends in one of the combatants losing his tail, which is eaten by the victor.
The Chamæleons include among their number species which have developed quite formidable horns, recalling those of the Rhinoceros or, better, of the extinct Arsinoetherium, since they are placed side by side instead of one behind the other. In Owen’s Chamæleon there are three such horns, two on the forehead and a median horn on the snout, and these are borne only by the males.
The marvellous play of colour which many Lizards display is commonly attributed indifferently to “protective coloration” and to “sexual selection.” It is unlikely that both have played equally important parts in their development. If the case of certain of the 168Geckoes alone is taken, then there would seem to be no doubt but that “Natural Selection” was the agent which had determined their elaborations for protective purposes, and in such and similar cases this may be largely true. But the material which “Natural Selection” has worked upon has been furnished by the secretions of the sexual glands to which reference has so frequently been made already. These seem to possess a very marked tendency to contain an excitant which promotes the formation of intense pigmentation, or an excess of tissue which may assume the form of weapons of offence, or of excrescences in the form of spines, or other ornamental features. Animals in whom this tendency to pigmentation and ornament has developed must, so to speak, obtain a licence from “Natural Selection” if they are to retain it. That is to say, if such ornament whenever it appears makes the wearer conspicuous to its enemies, or hampers it in escaping therefrom, or in fulfilling the ordinary avocations of life, then its further progress will be inhibited, or the wearer will be exterminated. But the tendency to produce colour, a by-product of the sexual gland secretions, may incidentally serve to afford it a protective garb, and in this event its further elaboration in the required direction is assured.
In certain abnormal, sexually poisoned individuals among the human race it is well known pleasure is derived from flagellation. There is but one instance known to me where this obtains as a normal accompaniment of desire among the lower animals, and this occurs in one of the Painted Terrapins (Chrysemys picta), whose finger-nails are produced into long, whip-like ends. I had the good fortune to witness their use one day when in the 169Reptile House at the Zoological Gardens in London.
The unusual activity of a male of this species was the first thing to attract attention to his movements. Watched more closely, he was found to be dodging a female and making frantic efforts to swim round so as to oppose her path. This done, he closed up and immediately commenced to apply the bastinado to her head. The movements were so rapid that nothing more than a blurred image of these strange whips was visible. As soon as she escaped his attentions, he set about circumventing her again, and again succeeded: and this most extraordinary performance was repeated many times during my watch.
Turning to the Amphibia, the descendants of that stock which must be regarded as the ancestors of the Reptiles, the version of the sexual rôle which is adopted in these pages, that “Sexual Selection” in the older, Darwinian sense, does not exist, finds further support.
Among the tailless Batrachians—the Frogs and their kind—there is no “display” immediately preceding the act of pairing. The males seize upon the females and hold them in a close embrace which lasts for a very prolonged period, covering many days or even weeks, until the extrusion of the eggs, which he impregnates by successive emissions of the fertilizing element. What controls the orgasm no one has yet succeeded in discovering, but this is an important point, for it is essential that the seminal fluid should not be emitted until the moment the eggs are set free. The pairing act is here purely instinctive, as is shown by the fact that if a Frog in embrace be removed and replaced on some inanimate body, this will be treated as though it were a female.
170With the tailed Batrachians—the Newts and Salamanders—the male commonly executes a very animated display which is followed by behaviour of a quite remarkable character. The display, which is always associated with vivid coloration, or the development of fin-like frills along the back, takes the form of amorous writhings and other gesticulations. At times he will hit his mate with his snout, and at others he will simply rub sides with her, as if to entice her to respond to his advances. These evolutions may be followed by an amplexus, an embrace. In some species, however, these performances are followed by behaviour which leaves one gasping with astonishment.
To begin with, there is no act of pairing, no coitus, but the male discharges a number of conical or bell-shaped “spermatophores,” each of which is crowned by a bunch of spermatozoa, the male germs necessary to ensure fertilization of the ova. These spermatophores adhere to the bottom of the stream, and are gathered up by the female, either directly, by placing herself in such a position that they can be seized by the lips of the genital opening, or by seizing the spermatophore, with its fertilizing germs, between her hind legs and pressing it home! The more one contemplates this extraordinary proceeding the more one marvels at the evolution of a departure from the normal sexual relations so inconceivably strange. Here one sees the purpose of the aphrodisiac in its true light. But for these facts it would have seemed certain that its primary object was to enable the male to relieve desire and at the same time to accomplish its end—the fertilization of ova—without undue waste. And this, in all the cases so far discussed, is possible only when the female has become inflamed with a like 171desire for coitus. But here the male finds relief, without more ado, by depositing the precious germs upon the ground. The display then is indeed to serve as an aphrodisiac. For the continuation of the race now rests entirely on the female. Any defect in the orderly working of her sexual appetite means the waste of the spermatozoa and the failure to effect fertilization. We cannot suppose that there is any realization of these facts, or any deliberate action on her part, but rather that she derives pleasurable feelings from the necessary passage of the spermatophore, which, probably, she recognizes by its smell.
The statement that the Frogs and their kind dispense with a display requires some qualification. For in the first place they, like their tailed relatives the Newts, develop secondary sexual characters, but these are of a quite peculiar kind. Among the Newt tribe, as has been mentioned, these characters take the form of frills and crests and vivid colours. They are intended to stimulate through the sense of sight, and arouse emotion, as a city is beflagged to welcome those it may delight to honour. The Frog tribe appeal to the musical sense, even though that music be of a barbaric kind. But, it would seem, when once the errant females have been drawn to the spot chosen by the males, no further aphrodisiac is used, the male simply seizing upon the female nearest at hand and, having once embraced her, she is not released again until the eggs have been extruded and fertilized. To maintain his hold, the forearm is often excessively muscular, while one or more of the fingers may be armed with pads. In some cases, as with the Himalayan Rana liebigi, the inner side of the arm and each side of the breast are studded with small conical 172spines. But the absence of ornament in these cases, as with such of the Newts where there is no amorous display before embrace, is significant.
And now, as touching the musical performances of these troubadours. These commence in the early spring. With many species, as with our Common Frog (Rana temporaria) nothing more than loud croakings are attained. But with others this “music” is enormously increased in volume by resonators in the form of air-sacs or wind-bags. We may surely, with some show of certainty, liken this “music” to the song of birds, and assign its primary purpose to the same cause—a device to advertise their presence to wandering females seeking mates. That birds sing after mates have been found, and later, is no doubt due to a general feeling of “fitness,” which finds expression in what has become the usual mode for such emotional states. Most people must have heard the spring concerts of our Common Frog; but these are incomparably surpassed in volume by the Edible Frog and the Bull Frog, which are provided with large, globular, inflatable, air-sacs in the throat, serving as voice-resonators. Such performances, however, are mere bawlings compared with some other species, which mew like cats, or bark like dogs. The most famous of all is the Brazilian “Ferreiro” or “Smith” (Hylodes faber), whose voice is one of the most characteristic sounds to be heard in Tropical South America. “Fancy,” says Dr. Gadow, “the noise of a mallet, slowly and regularly beaten upon a copper plate, and you will have a pretty good idea of the concert given generally by several individuals at the same time and with slight variations of tone and intensity.” When seized, the performer utters a “loud and shrill, most startling cry, somewhat similar to that 173of a wounded cat.” Another, a Paraguayan species, Phryniaxus nigricans, at the breeding season, utters a call note which consists of two clear, musical “rings,” followed by a long descending “trill” like that of our British Greenfinch. But, it is to be noted, both sexes in this case perform.
The period of sexual activity with perhaps the majority of animals is intermittent and extends over but a short period annually; with others potency is continuous, at least with the males, though desire becomes clamant only when aroused by external stimuli. But whenever this condition be aroused it invariably finds expression in exaggerated movements or vocal demonstrativeness. It uses the normal channels of expression, in short, but intensifies them. Now this period of sexual activity represents the maximum of “fitness” in animals, and it is not surprising, therefore, to find that when the barometer of vitality stands high some approach to the maximum of activity is indicated. In many birds this is revealed in song, though the earlier stimulating cause is absent. Among the cold-blooded frogs the same obtains. In the Edible Frog (Rana esculenta), for example, the males, which “are great musicians,” remarks Dr. Gadow, “go on singing for sheer enjoyment not only during the pairing time, but throughout the months of June and July. Warm, moonlight nights are the favourite times for the concert, which takes place in the water, beginning at sunset and continuing till early dawn. A few individuals utter a single note, ‘gwarr-oo-arr’ or ‘coarx’ but these are only preliminaries. The precentor ... begins with a sharp-sounding ‘brekeke’ and this is the signal for all the others to chime in with the same note, varied with all sorts of other sounds, bass, 174tenor and alto, each performer filling its resounding vocal sacs to bursting size, and these bags then look as if they acted as floats. When there are several hundred of these sociable creatures the din is continuous, and may be heard more than a mile off.”
From what has been said of the Amphibia, and especially of the Newts, it would seem that, among the land vertebrates at any rate, the sexual instincts in this lowest or simplest form are satisfied with the discharge of the germinal products. Many, however, have advanced a stage further and reveal the rudiments of that instinctive care for offspring which develops to higher and higher grades as we ascend in the animal kingdom, till at last, in the human race, where the offspring is desired for its own sake, we ascend to the highest plane of all. The varied means of expression which these rudimentary instincts take in the Amphibia have already been discussed in “The Infancy of Animals,” which preceded this present volume, and hence no more need be said on this head in these pages.
Germinal Variations—Fishes and mate—hunting—Some remarkable Sexual differences displayed by the Teeth of Rays—The Double-eyed Fish—The Coloration of the Dragonet—Some curious facts about Salmon—The strange use of the kidneys in the Stickle-back—The Stickle-back and parental duties—Siamese Fighting-fish.
Mammals, Birds, Reptiles and Amphibia, as has already been shown, all exhibit practically the same line of conduct in regard to their mate-hunting instincts; all use like modes of expression. And this is a very significant fact. It becomes more so when we turn to the fishes, for here again we meet with the same behaviour, and here again we meet with the same rules in “secondary sexual characters.”
An instance or two of the latter distinction between the sexes should suffice. As a rule, among fishes, the males are smaller than the females: commonly there is no other external distinguishing feature between them. In many cases, however, the males are more or less strikingly different, thereby showing a departure in the nature of a higher degree of complexity, or “specialization,” just as obtains among the birds. And the same 176sequence to this also obtains. That is to say, as has already been remarked in the case of the Mammals and the Birds, the new features first appear in the males, leaving the females and young of both sexes unmodified. A singular illustration of this is afforded by some of the Rays, or Skates, as they are often called. In the Thorn-backed Ray (Raia clavata) the teeth of the adult male are sharp-pointed and directed backwards, while those of the female are broad and flat, forming a sort of mosaic or pavement. The young male agrees with the female in this respect. In the Common Blue Skate (Raia batis) the teeth are pointed in both sexes, though more so in the adult male. In the Spotted Skate (Raia maculata) the teeth are fully pointed in both sexes. Here, then, the normal course in the evolution of new characters is followed, but it is remarkable that the teeth, which are so intimately related to the capture of food, should be thus affected. Whether the change of teeth is associated with a change of food, or whether neither pointed nor pavement teeth affect the feeding, is unknown.
Still more remarkable is the case of the Double-eyed Fish (Anableps). In this fish there is an intromittent organ in the shape of a tube which is formed by a continuation of the urinogenital ducts down the front of the anal fin. In the hinder half of this organ a bend is made either to the right or left. Out of seventeen males, this bend was to the right in eleven, to the left in six. Further, there is a small fleshy tubercle at the side of the anal fin-ray, at the middle of its length. When this prominence is on the left side, the organ bends to the right; when it is on the 177right, the bend is to the left. In the females the genital opening is covered by a special scale, which is free on one side, left or right, and not on the other. Thus copulation is possible only from the side, and a left-sided male can only conjugate with a right-sided female, and vice versa. Here is one of the most extraordinary cases of specialized secondary sexual characters known. How do the sexes distinguish their complemental mates? It is important that they should, for unions are otherwise impossible.
In the Dragonet (Callionymus lyra) the male differs conspicuously from the female in being much the larger—an exception to the rule—and in having the fin-rays enormously elongated. Further he wears a conspicuously resplendent livery, but this is strictly a “nuptial” livery, the colours waning as soon as the period of sexual activity is past. That these colours play the same part as with the birds is clear from the observations of the late Saville Kent. “The male,” he says, “resplendent in his bridal livery, swims leisurely round the female, who is reclining quietly on the sand, his opercula distended, his glittering dorsal fins erect and his every effort being concentrated upon the endeavour to attract the attention of his mate.... The female, at first indifferent, becomes at length evidently dazzled by his resplendent attire and the persistency of his wooing. She rises to meet him, the pair so—far as is practicable with fishes—rush into each other’s arms, and with their ventral areas closely applied, ascend perpendicularly towards the surface of the water.” In the course of this ascent the ova and sperms are shed, and fertilization takes place.
The difficulties in the way of the study of the behaviour of fishes during the critical period of mate-hunting are many and obvious. Something may be inferred from the 178nature of the secondary sexual characters which they exhibit, and more definite information can be obtained from such species as can be kept in aquariums. From these two sources enough has been gleaned to show that these cold-blooded creatures, in many cases, exhibit the same emotions and the same means for their fulfilment as the higher vertebrates. And it is significant that wherever anything approaching what may be called “Courtship” obtains, the males commonly exhibit secondary sexual characters, whether in the form of ornament or of armature; while among species which consort in shoals during the breeding season no such distinctions are present. The ova and milt are shed and fertilization takes place as they escape.
Comment is frequently made in works on Natural History on the fact that among fishes the males are commonly smaller, often conspicuously so, than the females. Among mammals the males are the larger; but among birds this is by no means always the case. It is somewhat surprising to find this discrepancy among the birds of prey, where, as in the case of the Sparrow-hawk, the male is little more than half the size of his mate; commonly, however, there is little or no difference. Among the fishes the differences are often much more marked, as for example in the Conger-eel, wherein the male never exceeds a length of two feet six inches or a weight of one pound; females, on the other hand, may exceed eight feet in length and attain a weight of one hundred and twenty-eight pounds, though such giantesses are rare, but specimens of fifty pounds and upwards are frequently met with. The explanation of this may lie in the fact that among fishes it is no uncommon thing to 179and males becoming sexually mature long before they have attained their full stature. With the Salmon, for instance, ripe spermatozoa have been found in individuals of not more than a few inches in length, and in this species also the male is the smaller. Ova take longer to attain maturity, for in addition to the germ-plasm they must be provided with a more or less extensive amount of food-material in the shape of yolk. The formation of this is inhibited until the demands on the system for the building up of the body have begun at least to lessen.
Mate-hunger among the fishes seems generally to find peaceable modes of expression, either in “display” or in consorting in vast shoals, though, so far, the factors which govern their conduct in this matter are as yet unknown. But here, as with the higher vertebrates, there are some species which adopt more violent methods. A good illustration of such conduct is furnished by the Salmon, which, during the period of sexual activity, develops a curious modification of the lower jaw, which is produced forwards and upwards to form a hook-shaped projection of fibrous tissue. When the mouth is closed this hook is received into a cavity formed within the fore-part of the roof of the mouth. It has been described as a weapon of offence. But this it can hardly be. On the other hand it has been suggested that it serves to protect the jaws when charging a rival, for the shock on such occasions is considerable. It answers, in short, like to the fibrous mass of tissue which protects the fore-part of the head in Whales like the Black Whale (Globicephalus) and the Bottle-nose Whale (Hyperoodon), serving as a battering-ram. In the Pacific Salmon (Onchorhynchus) both jaws are hooked, so that when the mouth is closed 180the hooks cross one another as in the beak of the Crossbill. In this Salmon, too, the front teeth attain a considerable length, while the body becomes laterally compressed and a hump forms at the shoulders. Little, however, seems to be known as to the nature of their battles.
The combats of the Salmon of our own islands, however, are evidently severe, and this has long been known, for Darwin speaks of as many as three hundred, all with one exception males, being found dead in the Tyne during the month of June, killed by fighting. Such battles are fought, it is to be noticed, not so much for the possession of females—for it is a polygamous fish—as for the privilege of fertilizing the eggs as they are shed. The absence of a “display” here is a noticeable feature, and it is on this account, probably, that the reproductive period is not associated with the appearance of any form of resplendent livery. On the contrary, the marvellous silvery sheen which adorned both sexes on their arrival at the spawning ground from the sea has entirely vanished by the time that the consummation of the journey has been attained, and in its place is naught but a slimy, dingy copper-coloured hue. But no sooner has the reproductive period passed than the silver lustre makes its appearance once more.
These facts are the more interesting when contrasted with what obtains among other fighting species which must woo the females. Take the case of the common freshwater Stickle-back. In this species the body is invested with an armature of bony plates and spines in place of scales, while the males are arrayed in vivid hues of red and blue. Any survey, however, of the reproductive activities of this little fish must take into account 181certain quite remarkable prenuptial actions and instincts. Briefly, before the male commences his search for a mate he constructs a nest of fine fragments of aquatic weeds, which are held together, not by interweaving as with birds’ nests, but by a sticky and copious secretion from the kidneys. According to some authorities, this secretion is to be regarded as a pathological product caused by the undue pressure of the ripening testes. It is difficult to accept this interpretation, for it might with as much reason be argued that the copious secretions of the salivary glands of the edible Swift—which builds a nest constructed entirely of hardened saliva—are also pathological in character. But be this as it may, the nest completed, the male seeks a mate, or mates, for polygamy is the rule of his tribe. In his search for these he has constantly to do battle with other males, whom he endeavours to disembowl by swift rushes contrived to t rip open his rival as he passes, by means of one or other of the erectile spines which project from his back and belly. With the females whom he desires he uses the arts of peaceful persuasion, swimming backwards and forwards before her in his endeavour to excite her amorous instincts. At last he persuades her to enter his bower and deposit a few eggs, fertilizing them immediately they are laid. The first to enter leaves by forcing a passage through the opposite wall of the nest, a happy contrivance, for thereby a current of water can be constantly driven through, leaving fresh oxygen to the developing eggs. One female after another is inveigled into the bower, until the complement of eggs is complete. These, singularly enough, are now taken charge of by the male. He it is who creates life-sustaining currents which bathe the eggs, by the rapid vibrations of his breast-fins, 182and he it is who protects them from their most persistent enemies—the females who laid them. As soon as the fry appear the duties of the male are still further increased. He must guard them from their mothers, and other foes, and he must prevent their too extensive wanderings. Such as stray too far afield are sucked into the mouth and brought back again to the nursery, where they are set at liberty by a reversal of the sucking action. That the male of a polygamous species, and with all the attributes of a polygamous species—pugnacity and vivid coloration—should take upon himself the duties which under like circumstances among the higher vertebrates are undertaken by the female is a very remarkable and puzzling feature. In this species, in short the male plays successively a polygamous and a polyandrous rôle.
Strange as these facts are, they are not apparently without parallel among fishes, for certain of the labyrinth-gilled fish present many features in common, though as yet proof seems to be wanting. Thus the small Siamese “Fighting Fish” (Betta pugnax) is endowed with so ferocious a nature that it is kept, as the Malays keep fighting cocks, for the amusement of native sportsmen, two fish being pitted against one another and large bets being made on the result. In a state of quiescence it presents no very remarkable coloration, but if two be brought together, or if one sees its image in a looking-glass, it becomes thrown into a paroxysm of rage, the fins are raised and the whole body becomes irradiated with metallic colours of dazzling beauty. There can be no doubt but that a like play of colour occurs during moments of sexual excitement; it is highly probable that it is polygamous. Of its breeding habits, however, little or 183nothing seems to be known. Not so, however, in the case of a closely-related species, less pugnacious in disposition, but almost as vividly coloured, in so far as the male is concerned. Now in this species a nest of froth is made and the eggs, after deposition therein, are jealously guarded by the male. Hence, on these facts, we may assume with a fair amount of certainty that the closely-related “Fighting Fish” displays like habits.
That the Reptiles, Amphibia and Fishes have much in common with one another, and with the higher vertebrates, in the manner of their love-making is indisputable. We find no evidence anywhere that the first faint throbbings of the sexual pulse in the female are quickened to fever beats after the efforts of several successive wooers, each more demonstrative than the last, to arouse this state—the conditions required by the Sexual Selection theory. But successful mating depends, in each year, on the sexual fitness of the male himself, and the mate, or mates, which for that year he has taken “for better or worse.” It is possible, of course, that a male, ambitious but impotent, will be forsaken by his mate; it is possible that a female of low sexual vitality may fail to respond to the most impassioned displays; in either case no offspring result, and thus the failures are eliminated. It is possible that here, as with the higher vertebrates, coition may by no means always be immediately preceded by display. But the “display” has done its work. It has stimulated the sexual appetite, as the sight of tempting food stimulated the bodily appetite.
But both the Amphibia and the Fishes reveal a lower plane of the sexual instincts, when the sexes, dominated by some imperious instinct, gather in hordes, commingling to shed their precious germs into the surrounding water, 184there to effect the work of fertilization and the achievement of new birth. The all-important union of these germs is no mere work of chance, as it might seem, but the sperms seek the ova with unerring surety, guided, in this case, by that very efficient substitute for instinct, chemotaxis, or the attraction which certain chemical substances have for lowly organized living bodies. In this case the allurement is furnished by the ova. It is surely no unreasonable surmise that here we have the beginnings of the complex phenomena which the earlier chapters have revealed. On this lower plane we are probably confronted by instinct alone, but from this level upwards intelligence plays an increasingly important part.
Butterflies and Moths, and the Coloration of their Wings—Female Choice and “Fine Feathers”—When Male Butterflies are Dominant—Sexual Selection among Butterflies—Abortive Experiments—Wallace and the Sexual Selection Theory—The Sense of Smell in Butterflies and Moths—Fragrant Butterflies—Wingless Moths and their Lures to Lovers—Methods of Pairing among Butterflies and Moths—More Experiments.
Not the least impressive feature met with in the study of animal behaviour under the spell of the Sexual Instincts is its uniformity. This fact becomes the more apparent as one turns to the lower grades of life. Whether one starts with the vertebrates and works downwards, or vice versa, the same problems arise and the same interpretation is demanded. That is to say, the theory of “Sexual Selection” leads one to the same conclusions whether it be tested by the evidence afforded by the Butterflies and Moths, or that furnished by Birds or Mammals.
The accessory phenomena, the vehicles which give expression to these internal fires, are in like manner curiously similar. These “vehicles” are the “secondary sexual characters”—colour, and armature, and scent. These very tangible signs are the phenomena in the 186Mystery Play of Sex which first catch the attention of the investigator. To account for these the theory of “Sexual Selection” was first devised.
After the birds, probably the group most conspicuous for its splendour is that which contains the Scale-winged Insects or Lepidoptera, and it has always been allowed that any explanation of the one must apply also to the other. It seems impossible to avoid this conclusion. But before going further it would be well to take note of one or two interesting features in regard to coloration that have so far not been touched upon in these pages.
The Coloration of Animals is generally regarded as a by no means fortuitous feature, but one, on the contrary, controlled and determined by various factors. Hence are recognized various kinds of coloration: Obliterative or Protective-resemblance Coloration; Warning Coloration; Mimetic Coloration; and Epigamic Coloration, or the colours associated with courtship. These various types have been subdivided and accorded technical labels by Professor E. B. Poulton, in his “Colours of Animals,” but these need not be enlarged upon here. Suffice it to say that it is generally held that all forms of coloration can be explained, and all can be labelled, as to their origin, with more or less certainty. There are those who doubt the warranty for this classification. Commonly, it must be admitted, the arguments of these sceptics are not impressive; they are sometimes even stupid. That such coloration, however it be labelled, is subjected to some control seems to be shown in the case of the Lepidoptera, for, generally, in the Butterflies, the upper surface of the wings is much more vividly coloured than the under surface, and this, apparently, because when the 187creature is at rest the wings are brought up over the back like the leaves of a book, so that the brightly-tinted, and therefore conspicuous, area is concealed, as, for example, in the “Red Admiral.” With the Moths the wings, while the creature is at rest, are held horizontally, and it is the upper instead of the under surface which is exposed, but the hind-wing is covered by the fore-wing. The coloration is here very different; for while the exposed surfaces of the fore-wings are commonly soberly tinted, the hind-wings may be quite glaringly coloured. These bright colours are exposed only during flight, or during moments of unusual excitement, as in the case of the Eyed Hawk-moth. According to Weismann, this insect when alarmed raises the fore-wings so as to expose the “eye-spots “on the hind-wings, which, with the increased area of the wings, impart a terrifying appearance to the body to would-be assailants. This is as it may be, but for the moment the feature to be insisted upon is that the bright colours are almost invariably hidden when the insect is at rest, and by quite different means, determined, apparently, by the different carriage of the wings. Now, according to some, bright colours are begotten by strong light, but in the Moth and Butterfly the surface area of the wing which is most exposed is the surface turned to the light during rest, and this is the least coloured. The curious relation between this coloration and the resting position is strikingly illustrated by the case of one of the “small Blues” (Lyccenæ), cited by Weismann. Herein the male, which has the upper surface of the wings of a bright blue, rests in the position common to Butterflies—with the wings raised and concealing the bright colour—while the female, which has the upper surface of a dull brown, rests with the wings expanded. 188As, however, the concealed under surface is not brightly coloured, it is difficult to believe that these different postures and conspicuously different colours can have been brought into existence solely by the action of Natural Selection, which, it is generally contended, has brought about the extinction of those individuals which neglected, when resting, and therefore liable to be “caught napping,” to conceal their arresting colours. There is, indeed, no apparent reason why the female, which has nothing to conceal, should depart from the custom common to Butterflies, of resting with the wings closed and raised, this position effectively protecting the male. The facts seem to show that the coloration of the exposed surfaces of the wings is determined primarily by some physiological factor rather than by the incidence of Natural Selection directly through external agencies. Thus, for example, the action of light on the surface of the wings when in the resting posture may well inhibit the production of vivid pigment owing to some inherent physiological idiosyncrasy. But any individuals which lack this inhibiting factor—as some species which, though resting, are brightly coloured, appear to do—will be eliminated, if they live in an environment harbouring eliminating factors, which the exceptions to the rule we must suppose do not. But on this interpretation the fundamental factor in the determination of the coloration is the action of light. Selection imposes a bar only to certain types of coloration.
Some Butterflies and Moths, it has just been hinted, when resting exhibit bright colours. Our “Swallow-tail” the under surface of the wings is as brightly tinted as the upper. Among the Moths may be cited many of the 189gorgeous Atlas Moths, the Hawk Moths, the beautiful Indian Dysphania militaris—wherein the whole of the exposed surface is of a beautiful and vivid violet and yellow—and the tropical members of the Burnet Moths, belonging to the family Syntomidæ. In all these cases it is not the under but the upper surface of the fore-wings which has thus departed from the usual rule of the tribe. Not the least remarkable feature of these insects is the fact that while the Atlas and Hawk Moths are crepuscular in habits, the Dysphanias and Syntomids and Burnet Moths are diurnal, and revel in the sunlight.
To revert for a moment to the factors to which these and other bright and often conspicuous hues are due. That all highly-coloured animals are descendants of dull-coloured ancestors there can be no room for doubt. The vivid tints they now display are to be regarded as due to some change in the metabolism, some clarifying process of the organism whereby the various pigments became segregated, concentrated and intensified. But many of the most vivid hues are not due to pigment at all, but to changes in the surface structure of the coloured areas. Such are the wonderful metallic colours which all kinds of animals display. The iridescence is due to the breaking up of the light by reflection from finely-grooved surfaces.
Whatever their nature, one still asks what is their origin, what brought them into being. They cannot be regarded simply as adaptations which have arisen to meet the demands of the environment, as are the structural peculiarities of the skeleton for example; for in this case both sexes, and all stages of growth, should display the same hues, and this is rarely the case. Furthermore, we should not in this case be left with a vast assemblage 190of forms which certainly cannot be “pigeon-holed” as to the nature of their coloration. Such, for example, as the marine types of birds.
The metallic and iridescent tints to which reference has just been made, occur among animals to which they can be of but doubtful value, as in the Golden Mole, for example, or the inside of the Oyster shell. Their existence in such places well illustrates what we may call the fortuitous, or apparently fortuitous, beginning of colour of whatever kind, regarded from an analytical point of view. That is to say, we are not concerned with the fact that animals are coloured—that is inseparable from their existence; but with why this coloration should, in some cases, assume so conspicuous a brilliancy and vividness—a coloration varying in its character with every species, but apparently unchanging among the individuals of that species.
No answer to this, likely to find general acceptance, seems to be forthcoming at present. But it is significant to remark that all coloration of the kind now under consideration has its origin, as have most other structural characters, in the male. It is as true of coloration as of, say, skeletal characters. One turns to the male for what is new in the history of a species, to the female and young for indications of past history. It is equally true that in their coloration one finds the same sequence of development—the male first, then the female, then the young, till both sexes, and all stages, are once more alike in hue. And this rule seems to apply to coloration of all kinds—Protective—Warning—Epigamic.
The tendency to develop brilliant colours is associated with some physiological diathesis with which we are not yet acquainted. But once having started, this tendency gathers force with each succeeding generation and continues 191to exhibit an almost kaleidoscopic capacity for change, unless, and until, checked by Natural Selection, whereby its further progress in any given direction may be barred, or some other element or aspect of the coloration may be introduced.
Given this controlling factor, all the various types of coloration would seem to be interpretable. By almost common consent, however, the resplendent coloration of the males among many species of birds, a coloration often apparent only during the reproductive period, and the more conspicuous ornamentation of the males of many other groups, higher and lower in the scale of organization, are supposed to be governed by an entirely different factor—female choice, or preference. The exercise of this, it is contended, has gone on for countless generations, and the tendency has ever been to heighten the intensity of the ornament by the rejection of the less favoured suitors in favour of their more resplendent rivals. Birds and Butterflies alike are supposed to be swayed by the same irresistible desire to mate, and mate only with what we may call the smartest and best—groomed of their many suitors; and these, of course, being the most vigorous, most virile, sustain the stamina of the race and so attain Nature’s end.
So long as attention was focused alone, or mainly, on birds conspicuous for the highly ornamental character of their plumage, this theory seemed reasonable and probable enough, for one may admit in their courtships an element, at least, of intelligence and keenness of perception. But it has now been abundantly demonstrated that the animated displays so characteristic of these gaily-bedecked gallants, are enacted with no less persistence and vim by species which exhibit a Quaker-like soberness 192of dress. Thus, then, the champions of the Sexual Selection theory have been dazzled by the tinsel, and have missed the essential elements—the physical and psychological side of the display—the contortions, prancings, and so on, and they have missed the even more important element, the preliminary struggle for territory.
In this new light, the gaily-bedizened individuals of the Insect world may be surveyed afresh. The explanation of such of their features as are commonly attributed to Sexual Selection in terms of female choice, whereby only the most favoured from among a crowd of suitors could hope to succeed, may now be replaced by that which obtains also in the case of the higher animals. It seems to fit the facts better. One cannot understand, for example, how, on the interpretation of Sexual Selection, the extraordinary disparity in numbers between the sexes of some species of Butterflies came about. Thus in that marvellously beautiful genus Ornithoptera there is one species (O. brookiana) in which the females are excessively rare; so much so that the collector Kunstler could only obtain fifteen females to one thousand males. Though the males, among the Butterflies, are commonly much more numerous than the females, the disparity is rarely so great as with this species; but there are many in which the proportion of males to females is as fifty to one. As with the higher vertebrates selection affords no explanation of this curious disproportion. Though according to Weismann it fulfills “the first postulate in ‘Sexual Selection’ namely, that there be an unequal number of individuals in the two sexes.” But Sexual Selection here has a little over-reached itself, for surely one hundred suitors seems an embarrassing number for 193an inexperienced female to have to choose from! To say nothing of the ninety-nine males doomed to perish without leaving offspring.
That the beauty of colour and form which the Lepidoptera, and especially the diurnal Lepidoptera, or Butterflies, exhibit is due to the choice by the females—albeit an unconscious choice—of the most resplendent of her suitors, that is, in other words, that she yields at last to the most ravishing member of the crowd—there is no evidence to show. There would seem to be no possibility of a differential selection from among a number of males, for there is no “display” comparable to that, say, of birds. And what is more, it is unlikely that, if there were, she would find anything to choose between them, for the range of variation in, say, one hundred males of any given species is very slight. Finally we have no trustworthy evidence to show that the eyes of Butterflies and Moths are sufficiently good to enable them to make nice distinctions between slightly different males. We have no evidence that the eyes of Insects are capable of discriminating the details of the often intricate patterns which their own wings, and those of their suitors, exhibit.
In the matter of “Secondary Sexual Characters,” indeed, the Lepidoptera exhibit very little difference between the sexes. As a rule the females are larger, often strikingly so, but in the matter of coloration they show far less disparity. But there are exceptions to every rule. A striking illustration of this is afforded by the genus Ornithoptera. The butterflies of this superb group are of huge size, and the females are larger than their consorts, and commonly are extremely different therefrom both in coloration and habits. In Ornithoptera paradisea this disparity attains its maximum. The female, remarks 194Mr. David Sharp, “is a large, sombre creature of black, white and grey colours, but the male is brilliant with gold and green, and is made additionally remarkable by a long tail of unusual form on each wing.” But a glance at the two sexes will show that the female, though less gorgeously arrayed, still disports a livery which is of a highly specialized or elaborated character. How are we to account for her differences in shape, size and coloration on the older interpretation of Sexual Selection? The perceptual powers, the mentality, of a Butterfly are surely of a far lower grade than those of a bird, or even a fish. Here, therefore, we cannot attribute the same possibilities of response to form and colour which we can ascribe with tolerable safety to the vertebrates. Yet the Sexual Selection theory as generally understood demands this.
So far so good. And now as to the part played by Sexual Selection among the Lepidoptera. Darwin, in formulating this, found its application to the Lepidoptera a very disconcerting problem, being naturally disposed to regard the extraordinary wealth of colour which these insects exhibit as the outcome of a process of female selection, in every way comparable to that which he held to obtain among the birds. He did not postulate a conscious, deliberate, selection; but a final abandonment on the part of the female to the male which, by his beauty and demonstrativeness, pleased her most. He assumed that at this critical time she would always be surrounded by rival suitors, offering varying if slight degrees of difference: and, indeed, in many cases she is thus surrounded. He remarks, in discussing the case of Butterflies: “The males sometimes fight together in rivalry, and 195many may be seen pursuing or crowding round the same female.” But in the case of the Silk-moths—and here is another illustration of the merciless criticism to which he submitted his own theories—he remarks: “The females appear not to evince the least choice in regard to their partners.” This fact, which is certainly true in the case of both Butterflies and Moths, and these gorgeous hues, disconcerted him, as is shown in the passage: “Unless the female prefer one male to another, the pairing must be left to mere chance, and this does not appear probable.” The facts which have come to light in regard to the “Courtship” of Butterflies since Darwin wrote are meagre enough, but such as have been recorded give no support to the supposition that the females are really influenced by, or even perceive the colours of, their mates. Just on five-and-twenty years ago the naturalist Skertchly published some observations on the Courtship of that magnificent Bornean Butterfly Ornithoptera brookiana. He one day came on a male sipping honey from the flowers of a tree, vibrating its wings with the rapidity of a Hawk-moth, and the vivid green of the wings flashing in the sunlight, though the crimson areas thereof were invisible. The female came “and did all the wooing.” They circled about in flight with the female above and somewhat behind, so that she could see, we are told, the emerald markings; but there was no real evidence here that she was really influenced by his coloration, and if this really were the case then the coloration of the female equally demands an explanation, for this, though less gorgeous than that of the male, is far from a primitive type; on the contrary, it is of a highly differentiated character. Furthermore, in this genus, as has already been remarked, the males outnumber the females by, roughly, one hundred to one. Again, Moseley, the naturalist on the Memorable Voyage of the Challenger 196in 1872, when in the Aru Islands, was once “lucky enough to find a flock of about a dozen males fluttering round and mobbing a single female. They were then hovering slowly, quite close to the ground, and were easily caught.” But he was by no means convinced that any choice was exerted. And he suggests “a series of experiments, in which, in the case of highly-coloured and decorated Butterflies, the colours should be rubbed off the wings of a few among a number of males, or painted over of a black or brown colour. It might be tested whether the females would always prefer the highly-coloured ones.” Such experiments are foredoomed to failure, for the removal of the scales would remove the only source of communication between the sexes.
Wallace, always a strenuous opponent of the Sexual Selection theory, found in the behaviour of Butterflies and Moths when mate-hunting a particularly powerful countervailing weapon. He assumes that Darwin postulated a conscious selection on the part of the female, and with some show of reason, though it is probable that Wallace was mistaken in this. “The weakness of the evidence for conscious selection among these insects,” he remarks, “is so palpable, that Mr. Darwin is obliged to supplement it by the singularly inconclusive argument, ‘Unless the female prefer one male to another the pairing must be left to mere chance, and this does not appear probable’ But he has just said, ‘The males sometimes fight together in rivalry, and many may be seen pursuing or crowding round the same female’ While in the case of the Silk-moths—‘the females appear not to evince the least choice in regard to their partners.’ Surely the plain inference from all this is, that the males fight 197and struggle for the almost passive female, and that the most vigorous and energetic, the strongest-winged or the most persevering wins her. How can there be chance in this? Natural Selection would here act, as in birds, in perpetuating the strongest and most vigorous males; and as these would usually be the more highly coloured of their race, the same results would be produced as regards the intensification and variation of colour in the one case as the other.”
Commenting on Darwin’s interpretation of those cases wherein the females are more brilliantly coloured than the males, he insists that on his (Darwin’s) theory “throughout the whole animal kingdom the males are usually so ardent that they will accept any female, while the females are coy, and choose the handsomest males, whence it is believed the general brilliancy of males as compared with females has arisen.”
“Mr. Darwin admits,” he continues, “that these bright colours have been acquired for protection [because they resemble those of species which from their disagreeable taste are avoided by birds and other insect-eating enemies]; but as there is no apparent cause for the strict limitation of the colour to the female, he believes that it has been kept down in the male by its being unattractive to her. This appears to me to be a supposition opposed to the whole theory of Sexual Selection itself. For this theory is, that minute variations of colour in the male are attractive to the female, have always been selected, and that thus the brilliant male colours have been produced. But in this case he thinks that the female Butterfly had a constant aversion to every trace of colour, even when we must suppose it was constantly recurring during the successive variations which resulted in such a marvellous 198change in herself. But the case admits of a much more simple interpretation. For if we consider the fact that the females frequent the forests where the Heliconidæ abound [the distasteful species already referred to] while the males fly much in the open and assemble in great numbers with other white and yellow Butterflies on the banks of rivers, may it not be possible that the appearance of orange-stripes or patches would be as injurious to the male as it was useful to the female, by making him a more easy mark for insectivorous birds among his white companions? This seems a more probable supposition than the altogether hypothetical choice of the female, sometimes exercised in favour of, and sometimes against, every new variety of colour in her partner.”
Wallace’s arguments are not so crushing as he supposed them to be, and they contribute nothing towards the solution of the problem to be faced. But if colour played the part which Darwin believed, and colour alone be concerned, it is curious that the males should recognize their mates in a guise so unlike their own. How is it that they do not pass them by as members of the totally different distasteful species? Whenever, indeed, the female is more or less brightly liveried than the male, how do the sexes recognize one another, and how, when they live in environments so different as those referred to by Wallace, do they find one another when possessed by the insistent demands of the “sex-hunger” which is the all-essential stimulant to secure the continuation of the race?
The factors which assure the satisfaction of this hunger differ in some important features from those which obtain among the higher animals—birds, for example. In the first place there is no necessity to find and hold territory, 199which is an imperative necessity where there are eggs to be brooded and young to be fed. In the second, the males, as has just been remarked, must search for the females, often indeed, in the case of many Moths, because they are wingless.
This search is conducted by the sense of smell. This fact, familiar enough to-day to the entomologist and the student of Evolution, was unknown to the earlier naturalists. Neither Darwin nor Wallace suspected it. It would have been wonderful if they had, for there is nothing in the general appearance of these insects which suggests an organ of smell, nor is there anything in the structure of the nervous system which would indicate this subtle sense. During recent years, however, the number of workers engaged on the investigation of the senses of animals has increased immensely, and great strides have been made in perfecting instruments of research. To the efforts of these workers we owe the discovery of the seat of the scent-detecting organs and the source of the scent. The former are furnished by the antennæ, which lodge also the senses of taste and touch.
Among the Lepidoptera these constitute important secondary sexual characters, the antennæ, among the Moths at any rate, presenting striking differences in male and female. The scent-producing organs are very elusive structures, and so far have been definitely traced, among Butterflies, only in the males, where they are formed by certain peculiarly modified scales known as “androconia.” They may be either irregularly scattered over the wing, or may form complex structures. Sometimes they are arranged in the form of brightly-coloured, bristle-like tufts on the hind-wings, sometimes in a fringe 200along the edge of the hind-wing. In some of the Moths they are arranged to form a thick, glistening white felt, which fills a folded-over portion of the edge of the hind-wing, and in many cases “the perfume can be retained,” Weismann remarks, “and then, by a sudden turning out of the wing-fold, be allowed to stream forth.” In the Ghost-moth (Hepialus humuli), the hind-legs of the male have become pressed into service and have become transformed into scent-bottles, since they are swollen and filled with glands for the manufacture of odorous matter.
The naturalist Fritz Müller discovered the fact that some of the Butterflies which haunted his Brazilian garden exhaled a flower-like fragrance. Anyone can test this curious trait for himself who will take the trouble to brush his finger over the wing of a newly-caught male Garden-White Butterfly (Pieris napi). The white powder which will adhere to the finger will be found to be made up of the wing-scales, which will exhale a delicate perfume of lemon or balsam! Among the Moths the strong odour of musk is exhaled by the Convolvulus Hawk-moth (Sphinx convolvuli).
It is, however, only in the males that these odours can be detected, and, though palpable enough to human nostrils, their power of diffusion is apparently extremely limited. They would seem to serve as aphrodisiacs for the stimulation of the female, and, as a consequence, there is no need that they should start into activity until the male has arrived at the immediate neighbourhood of his prospective mate.
Plate 31.
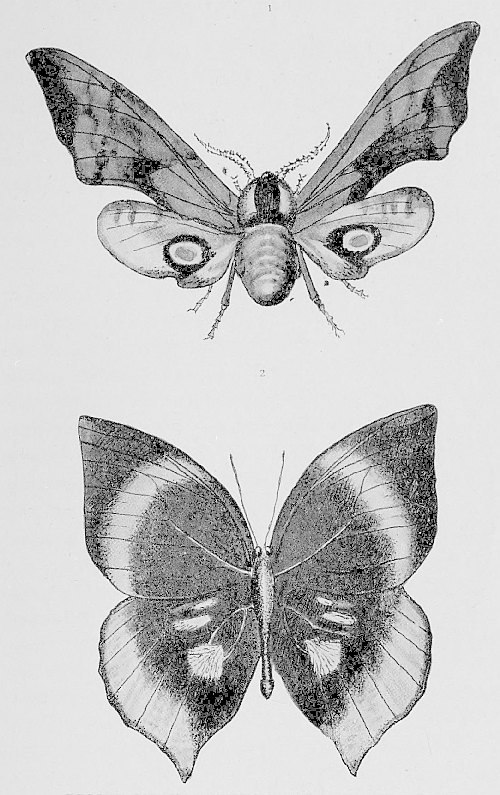
BRIGHT COLOURS WHICH CANNOT UK ATTRIBUTED TO “SEXUAL SELECTION.”
1. Eyed Hawk-moth, under the influence of excitement.
2, A Butterfly, Zeuxidia horsfieldi, Feld, showing tufts of scent-diffusing scales on the hind-wings.
[Face page 200.
With the females of the Moths, however, matters are otherwise. For the most part Moths are nocturnal, and hence could not distinguish one another when on the search for mates, and in many species the females are wingless, and consequently are unable to move 201from the immediate neighbourhood in which they emerged from the pupal stage. In either case some means of informing the males of the presence of females is an imperative necessity for the continuation of the race. This is provided by means of a subtle odour exhaled by the females which, though imperceptible to human nostrils, must possess an extraordinarily penetrating power. Weismann gives an instance of this in the case of the nocturnal Eyed Hawk-moth (Smerinthus ocellatus). He placed some females, without any special intention, in a covered vessel near an open window. “The very next morning several males had gathered, and were sitting on the window-sill, or on the wall of the room close to the vessel, and by continuing the experiment I caught, in the course of nine nights, no fewer than forty-two males of this species, which I had never believed to be so numerous in the gardens of the town....” To this power of exhaling odours we may attribute the wingless condition of many Moths, for otherwise the loss of flight would have brought about extinction long before any perceptible reduction in the wings had taken place. The odour which such prisoners emit seems to possess an irresistible attractiveness, and this fact is commonly taken advantage of by entomologists. The Common Vapourer Moth (Orgyia antiqua) affords a good illustration of this. The female is wingless, and little more than a pouch for eggs, but in certain seasons it is very abundant, even in the midst of London. That experienced entomologist Prof. Selwyn Image, in a letter to my friend Mr. John Cooke, remarks, on this theme, that the Caterpillars may be seen crawling by hundreds in and around the squares, while the males may be seen flying up and 202down New Oxford Street or Tottenham Court Road. If a virgin female be put in a box placed outside the window, within a very short space of time, often not more than a few minutes, several males will be fluttering round her. This device for attracting males is commonly known as “assembling.”
More striking is the case of the Oak-eggar Moth (Lasiocampa quercus). Mr. Richard South, in his most useful “Moths of the British Isles,” relates that on one occasion he had a number of pupæ in a cage in a cottage on the edge of a moor near Lynton, North Devon, and these attracted quite a number of males into the room containing the precious casket, and he was enabled to capture several. The next day he placed a female which had meanwhile emerged, in a “roomy chip-box, and carried it, in a satchel, to the moor, where it was placed on the ground; the males began to arrive soon afterwards, and some fine examples were secured.” But the sequel is even more remarkable; for, he remarks: “Although the female was taken on the moor only on one occasion, that satchel continued to be an object of interest to the male Eggars for several days afterwards.” That this scent is capable of being transferred to foreign objects, and of retaining its power for several days, is a striking proof of its pungency, yet it is quite impalpable to human nostrils! The Kentish Glory Moth (Endromis versicolor) affords yet another instance of this curious attraction by scent, the effectiveness of which is not even lessened by exhalations of the human body, for if a virgin female be placed in a box, and this be placed in one’s pocket, the males will often swarm round one and even endeavour to gain access to the box. In all such cases the females, even when capable of flight—the 203female Vapourer is wingless—never fly until after impregnation has taken place. Hence males with defective scent—detecting powers inevitably fail to leave offspring.
Selection, then, here lies between males of the most active scent-detecting powers, and not between those of the most brilliant colours. Nevertheless, both males and females—where the females are winged—exhibit a remarkably beautiful coloration, and this is especially true of the Kentish Glory, wherein both sexes wear a resplendent dress. That of the male—which is much smaller than the female—differs in that the fore-wings are darker, but bear the same pattern as in the female, while the hind-wings are chestnut-red instead of cream colour as in the female. If this scent-factor has replaced colour as an inciting agent to pairing, then these Moths should be of sombre hues. That such is not the case seems sufficient to show that the colour is not due to Sexual Selection, for it is highly improbable that scent and colour are both of equal importance, and this being so, one would expect to find the negligible factor eliminated.
The existence, then, of bright colours in this and other species in like case, seems to show that it has nothing to do with Sexual Selection, directly at any rate. The males having assembled, their presence is probably communicated to the female by the characteristic male odour, which is never of the same penetrating quality as that of the female. There is no need that it should possess this, for the females never seek their mates. The successful male, where several rivals are competing, is probably not simply the strongest, but he who also disperses the right odour necessary to provoke the 204pairing response. These illustrations furnished by the scent-hunting, scent-dispersing males and females are of the highest importance to students of the Sexual Selection theory, for they seem to show conclusively that coloration plays at any rate but a minor part therein. The importance of the scent-detecting organs is shown in the very different types of antennæ which obtain between male and female Moths, those of the male taking the form of huge feather-like structures, as in some Saturniidæ, and far exceeding those of the female in size.
The methods of pairing which obtain among Butterflies and Moths, it is not surprising to find, are very different; for whereas in the former it takes place on the wing, in the latter the female is always in a resting position. Where the females are winged, long flights are often taken for the purpose of depositing and distributing the eggs: the flightless forms make no such excursions. A few, as in the case of some of the Psychidæ are not only wingless, but limbless and maggot-like. They never leave the chrysalis case, but deposit their eggs inside it. Though there is undoubtedly much that is wonderful about the mating of these scent-distributing species, the history of the Moths of the genus Acentrophus is more wonderful and more mysterious still. For the females are aquatic. The males may sometimes be found in crowds fluttering over the surface of large but shallow sheets of water. The females, which are wingless, come to the surface and, like sirens, draw the males under water, where coupling takes place; after which they probably immediately die. But how do they discover their submerged mates? The escape from the water of any odour which the females may possess seems 205well nigh impossible.
Whether display, such as birds appear to delight in ever takes place among the Lepidoptera seems doubtful Nevertheless, something closely akin thereto seems to have been found in the case of certain species of Butterflies (Heliconius melpomene and H. rhea), which have been seen dancing in the air like gnats, and when some of them withdrew others took their places. Again, having regard to the fact that birds, when alarmed or excited, will perform the display which is more or less characteristic of periods of sexual excitement, it is possible that the position of alarm assumed by some of the Hawk Moths may also be used in Courtship (Fig. 1, Plate 31). But we have no evidence on this point, and from the part played by scent in the mating of Butterflies it seems improbable that such displays take place.
A serious attempt to test the Sexual Selection theory by experiment—to test the extent, if any, of female choice in mating—was made some years ago by Mayer, an American naturalist, on the large Bombycid Moth (Callosamia promethea). This species exhibits striking dissimilarity between the sexes in regard to colour and pattern. “The females,” remarks Professor Kellog, “are reddish brown in ground colour, while the males are blackish, and in the two sexes the pattern is distinctly different....” Mayer took four hundred and forty-nine pupæ, in cocoons, of this moth and endeavoured to discover, first of all, whether the males found the females by sight or smell. Enclosing females in jars, some of which were covered and some of which were uncovered, he found that males paid no attention to females enclosed in transparent jars so closed as to prevent the escape of odours, while such as were enclosed in boxes or wrapped 206in cotton-wool, so as to be invisible, but yet capable of exhaling odour, were besieged by males. To locate the organs of scent in the female he cut off the abdomen of several and placed the abdomens and their late owners at some distance apart. Males came to the abdomens and not to the thorax and wings. Males whose antennæ were covered with shellac, photographic paste, glue, paraffin, etc., showed no response to the female exhalations, until the covering medium was removed.
Mayer next tested the selective action of the females. He began by removing their wings and affixing to the stumps the wings of males. The males mated with the females quite as readily as under normal conditions, though the most conspicuous female characters had been exchanged for those of the male. After this he affixed female wings upon the males, but mating took place as usual. The females did not seem to detect anything unusual in their suitors, nor did normal males attempt to pair with males bearing female wings. Later he tried the experiment of dyeing the wings of three hundred males scarlet or green, and matched these against three hundred which were left untouched. The disguised, dyed males succeeded in pairing as easily as their normally-coloured brethren. The females exhibited no choice whatever. Hence, then, we have further reason to believe that with the Lepidoptera scent, not sight, is the channel by which mates are found. So far as the evidence goes, it seems to show conclusively that in all that concerns sexual relationships, scent is the guiding and determining factor. By scent the females attract the males, and by scent of another kind the males sharpen the procreative appetites of the females.
If the interpretation adopted in these pages is correct, 207these manifestations and emanations of colour and scent are readily accounted for; for they are manifestations of inherent growth changes which, having started, are free to go on increasing in amplitude unless, and until, checked by natural selection. There is nothing unreasonable or improbable in this interpretation; on the contrary, it embraces also many other features hitherto ignored, but no less demanding an explanation. Such, for example, as the infinite variety of form and sculpture which the scales of the wings and the eggs display. These are details visible only by the aid of the microscope, but they demand explanation as much as the more obvious characters. Moreover they have the advantage of belonging to a set of characters which cannot in any way influence the choice, if choice there be, in the selection of mates, nor are they of a nature likely to affect the results of the struggle for existence. Of these characters, then—the sculpturing of the egg-shell and of the scales, the “nervation” of the wings, and coloration—we can say no more than that they are idiosyncrasies of growth, free to develop in any direction unless, and until, checked by natural selection, which will speedily eliminate disharmonies with the environment.
The Coloration, and other Forms of Ornament in Beetles, and the Significance thereof in regard to the Sexual Selection Theory—The Courtship of Grasshoppers and their Kin—The Remarkable Ears of Locusts and Grasshoppers—The Field-cricket and the Katydid as Troubadours—The Wonderful Performances of the Cicadas—The Duels of Long-horned Locusts—Dragon-flies—The May-flies’ “Dance of Death”—The Jaws of the Giant Alder-fly and their Strange Use—Some Curious Facts about Stone-flies.
In these pages it is contended that neither brilliant coloration nor any other form of ornamentation is to be ascribed to the direct action of “Sexual Selection.” That is to say, such conspicuous features have not been dependent on the action of female choice for their survival and development, but are rather the “expression points” of the internal, inherent growth variations, which, not being inimical to the welfare of the species, have been free to pursue their development in any direction which apparent chance may dictate.
The Butterflies and Moths well illustrate this in regard to coloration, for scent, not colour, would seem to be their principal source of information as to the outer world. The Beetles are no less instructive; for these 209creatures, though they contain numerous highly-coloured and some exquisitely beautiful species, are more remarkable for their bizarre shapes, and it seems impossible to regard these as the products of sexual selection. Yet this is the interpretation of their origin which, in the judgment of Darwin, we must adopt. He evidently had misgivings as to the correctness of this view; but it must be remembered that in reviewing the facts relating to these lower orders of Creation he was biased by the evidence which he had brought together in regard to the behaviour of the higher groups under the stimulus of sexual emotion. Convinced that female choice obtained here, he was but following the logical result of such conclusions in postulating the same factor wherever it could conceivably be applied. The most formidable critic of the Darwinian theory of Sexual Selection was Darwin himself. The dominant ambition in all his work was to explain his facts, not to establish his theory; and he was convinced that his theory of Sexual Selection did achieve that end; though there were cases where the evidence he was analysing seemed less clear than in others. That the Beetles presented difficulties is evident from his comments thereon. He was puzzled by the vivid coloration which some species present. “They may serve,” he remarks, “as a warning or means of recognition ... as with Beetles the colours of the two sexes are generally alike, we have no evidence that they have been gained through sexual selection; but this is at least possible, for they may have been developed in one sex and then transferred to the other; and this view is even in some degree probable in those groups which possess other well-marked secondary sexual characters....
“Some Longicorns, especially certain Prionidæ, offer 210an exception to the rule that the sexes of Beetles do not differ in colour. Most of these insects are large and splendidly coloured. The males of the genus Pyrodes ... are generally redder but rather duller than the females, the latter being coloured of a more or less splendid golden-green. On the other hand, in one species the male is golden-green, the female being tinted with red and purple. In the genus Esmeralda the sexes differ so greatly in colour that they have been ranked as distinct species: in one species both are of a beautiful shining green, but the male has a red thorax. On the whole, as far as I could judge, the females of those Prionidæ in which the sexes differ are coloured more richly than the males, and this does not accord with the common rule in regard to colour when acquired through sexual selection.”
While there is nothing very remarkable in the two sexes being coloured alike, it is certainly strange to find the female more brilliantly coloured than the male. And this because among the higher vertebrates, as among the birds, the female exceeds in brilliance only where she also plays the rôle of wooer instead of wooed; leaving to the male the whole responsibility of rearing the family. With the Beetles the family has to rear itself, parental care being limited to the right disposal of the eggs. By some change in the character of the germ-plasm the females may have, in these cases, acquired more “maleness,” more of the qualities which are answerable for the secondary sexual characters of the male, or, what seems rather to be the case here, a result like that which has been reached in certain of the Pigeons and Parrots has been arrived at. That is to say, the tendency to intensification of pigment in the female struck out a new line, instead of following 211that of the male. This rather rare form of sexual dimorphism is also met with, it will be remembered, among the Butterflies and Moths.
While brilliant colour is the more usual form of ornament among the Beetles, there are many species wherein the males have developed enormous horns, or have greatly exaggerated the length of the jaws; and these outgrowths give the impression of a formidable armature, but so far as the evidence goes this is by no means the case. They must therefore be relegated to the category of “ornaments,” though the term “excrescences” would more fittingly apply to them, for they are “ornaments” only from a human standpoint. At any rate, there is no evidence whatever that they serve to enhance their possessors in the eyes of the females.
In relation to the Sexual Selection theory these excrescences are of quite exceptional interest, for they throw a strong light on the meaning of ornament, such as obtains among birds, which seem to show a consciousness of its existence and effectiveness. Darwin argued from the birds to the Beetles. Convinced that the gorgeous crests and trains and vivid colours were appreciated by the females of the former, he was impelled to believe that the ornaments of the latter had developed in like case by the fostering influences of the females. Similarly, from the evidence as to the use of horns in the case of mammals, and spurs in the case of birds, he was induced to believe that the horn-like outgrowths of Beetles had been attained by like influences. But in both kinds of cases, he could only infer their action, for he could discover no really decisive instances of conquest either by display or by battle, such as he was able to produce in the case of the higher animals. Had chance directed his attention in the beginning either to the Warblers 212among the birds, or the beetles among the insects, his interpretation of the action of sexual selection, it is more than probable, would have been materially different from that developed in the “Descent of Man.” No additions of any importance have been added to the facts he so laboriously collected.
As touching the “horns” it should be remarked that these may arise either from the head or from the thorax, or from both, and sometimes even from the under surface of the body.
One of the most remarkable instances of these singular outgrowths is that of the Hercules Beetle (Dynastes hercules) of the West Indies and tropical America. Herein the roof of the head is prolonged into a great upturned beam bearing tooth-like prominences, and the top of this is opposed to a still more massive beam, whose base covers the whole roof of the thorax, and whose tip extends far beyond that projecting from the head. A pair of “teeth” point downwards from the middle of this beam, whose under surface is thickly covered with short chestnut-coloured hairs forming a brush-like surface. In another, Copris isidis, the head bears two short, rhinoceros-like horns, and the thorax a short, triangular overhanging ledge: in Phanœus jaunus there is a single horn on the head, and the thorax bears two short, forwardly-projecting blades, one on each side; while in Onthophagus rangifer—the Reindeer Beetle—the head bears a pair of horns curiously like the antlers of a deer. One might cite many such instances, all varying in detail, but these will suffice.
Darwin, in commenting on these structures, remarked: “The extraordinary size of the horns and their widely 213different structure in closely-allied forms indicate that they have been formed for some purpose; but their excessive variability in the males of the same species leads to the inference that this purpose cannot be of a definite nature. The horns do not show marks of friction, as if used for any ordinary work. Some authors suppose that as the males wander about much more than the females, they require horns as a defence against their enemies; but as the horns are often blunt, they do not seem well adapted for defence. The most obvious conjecture is that they are used by the males for fighting together, but the males have never been observed to fight, nor could Mr. Bates, after a careful examination of numerous species, find any sufficient evidence, in their mutilated or broken condition, of their having been thus used. If the males had been habitual fighters, the size of their bodies would probably have been increased through sexual selection, so as to have exceeded that of the females; but Mr. Bates, after comparing the two sexes in above a hundred species of the Copridæ, did not find any marked difference in this respect amongst well-developed individuals. In Lethrus, moreover, a Beetle belonging to the same great division of the Lamellicorns, the males are known to fight, but are not provided with horns, though their mandibles are much larger than those of the female.
“The conclusion that horns have been acquired as ornaments is that which best agrees with the fact of their having been so immensely, yet not fixedly, developed—as shown by their extreme variability in the same species This view will at first appear extremely improbable, but we shall ... find with many animals standing much higher in the scale, namely, fishes, amphibians, 214reptiles and birds, that various kinds of crests, knobs, horns and combs have been developed apparently for this sole purpose.”
The assumption that these “animals standing much higher in the scale” owe their weapons to the selective action of the females forms the crux of the whole Sexual Selection theory in regard to the significance of ornament. The evidence that the intensification of pigment and the eccentricities of growth in the shape of crests and frills have a fascinating effect on the female is more than under suspicion; it is discredited by the facts which have come to light in regard to behaviour during the periods of sexual exaltation. And there is a growing conviction that this is so. No better proof could be found that “ornaments” can, and do, exist in spite of, rather than because of, the action of “sexual selection.” They are the accidents of this selection, not a part of its machinery.
Incipient horns are found in not a few cases among the females of these insects, while in others, as in the case of the Reindeer Beetle, they are almost as well developed as in the males. This is what one would expect to find if these outgrowths were the result of inherent variations restrained as to their size by natural selection, which eliminates only when this growth penalizes, by increasing the struggle for existence.
As to the actual behaviour of Beetles when sexually excited but very little information is obtainable; but there are records of species the males of which fight with rivals for the possession of females. Wallace saw two males of Leptorhynchus augustatus, a Beetle with no name in common speech and a long beak, “fighting for a female, who stood close by busy at her boring. They pushed at each other with their rostra, and clawed 215and thumped in the greatest rage.” The smaller male, however, “soon ran away, acknowledging himself vanquished.” In this case, it is to be noted, the combatants lacked weapons. With the Stag Beetle it is otherwise, and this species is said to engage in fierce conflicts. Darwin cites an instance where two males were enclosed with one female in a box, when the larger severely pinched the smaller one, until he resigned his pretensions. This being so, it is curious to find that the female, which makes no display of pugnacity, has the stronger jaws. The fact that there are so few records of fighting among male Beetles, and the absence of injury to the highly-polished surfaces of the horns or jaws where these are conspicuously large, seem to indicate that at most no more than a semblance of fighting ever takes place. In a North American Stag Beetle (Lucanus elaphus) the jaws, which are greatly developed, are used, Darwin tells us, for seizing the female, but they do not appear to be employed for this purpose in any other species. It might be held that they play the part of terrifying agents, as the eye-spots of Caterpillars and adult Lepidoptera are believed to do. At any rate, they seem to be so used in the case of a Beetle of South Chile (Chiasmognathus grantii) wherein the jaws are of great size and have their inner edges toothed. When threatened “he faces round, opens his great jaws, and at the same time stridulates freely.” But this parade of force is evidently no more than “bluffing,” for Darwin, who describes this behaviour, remarks, “the mandibles were not strong enough to pinch my finger so as to cause actual pain.” In the female, it may be remarked, the jaws are quite small.
That too much stress has been laid upon the significance 216of the enlarged jaws and other hypertrophied developments in the Beetles seems to be shown by the case of certain carnivorous Beetles, of which one species (Taphroderes distortus) may serve as an example. Herein the left jaw takes the form of a long, crooked strap-shaped outgrowth, whose purpose cannot even be conjectured. And in this connection one may cite the case of certain species of Homoptera—Bugs—which occur in tropical South America. Here, in both sexes, as may be seen in Plate 40, Fig. 4, the neck-shield is produced backwards far beyond the body, to form a most elaborate superstructure which appears to confound the most ingenious attempts at interpretation.
It is to be noted that wherever special structures are necessary for the performance of specific acts such as are of vital importance to the well-being of the race, they are developed to perfection: there is little or no variation in their size, and no doubt as to their purpose. Thus in many species means are necessary to enable the male to seize and hold the female during the sexual embrace. In the Water Beetle of our ponds and ditches (Dytiscus marginalis) the male bears a very remarkable sucker on each fore-leg, the adhesive surface of which, under the microscope, reveals an extraordinary complexity and wondrous beauty. This sucker forms a very conspicuous “secondary sexual character,” and is used in embracing his mate, whose back is deeply grooved to enhance the hold of the suckers. In some species punctures take the place of grooves. Suckers, like those of Dytiscus are met with again in a Wasp (Crabro cribrarius). In another genus of Beetles (Penthe) cited by Darwin, the antennæ of the male have a few of the middle joints dilated and their under surfaces furnished 217with a cushion of hairs to aid in the sexual embrace.
Beetles are creatures of solitary habits; how, then, do they find their mates when, by the insistence of the reproductive desires, they are driven forth to begin the search? Though we have no direct evidence, it seems more than probable that, as with the Butterflies and Moths, scent furnishes their most reliable guide. At any rate, in a large number of species, as among the Lamellicornia, the antennæ bear leaf-like plates, which are much more developed in the males, in which they probably serve as scent-detecting organs.
In some species stridulating organs occur such as are met with in even greater perfection among the Crickets and Grasshoppers, and among the Spiders and Scorpions. That these “musical-boxes” provide a means of communication between the sexes there can be no doubt, even if, as some contend, they are commonly used only to frighten enemies. This purpose may well be the explanation of their presence in the larval Stag Beetle, for it cannot be claimed that they have any relation to the acts of courtship at this stage of development.
Stridulating organs, wherever they are met with, are fashioned on the same principle. The mechanism for sound-production differs conspicuously from that which produces the voice in the vertebrates. For where there are no lungs or breathing apparatus, comparable to that of birds and beasts, there can be no internal voice-mechanism. Instead, the skeleton which in these creatures forms the external surface of the body—that is to say, it encloses the muscles, whereas in the vertebrates it is internal and overlain by the muscles—produces the necessary sounds. And this by means of rubbing two opposed surfaces against 218one another, one of which is ridged, the other toothed. In the details both of position and structure a wonderful variety will be discovered when all the known types are surveyed, and it is possible that they perform different functions in different groups.
The Locusts and Grasshoppers are among the finest performers of these “harpists,” and it would seem that in this group, at any rate, the music they make is of an erotic character. In one of our native Grasshoppers (Stenobothrus melanopterus) these high-pitched and somewhat strident notes are produced by rubbing the roughened inner surface of the hindmost thigh, which forms the base of the great leaping leg, against one of the libs of the wing-case which is specially enlarged and has a sharp edge. Thereby the wing is thrown into a state of vibration and the musical sound produced. The roughening of the inner surface of the thigh just referred to is produced by a row of bead-like projections whose appearance under the microscope is depicted in the adjoining illustration. This apparatus is well developed in the males, and but feebly, or not at all, in the females. That the music it produces is appreciated by the performers and their mates there can be no doubt, for they are provided with a special apparatus which fulfils the purpose of an ear. In the short-horned Grasshoppers (Acridiidæ) this is placed in the middle of the body just above the base of the great thigh. It differs in the details of its construction. In some cases it is formed by a delicate sheet of membrane surrounded by a rim, in others the membrane may be slightly depressed, and in some very much so, the rim closing up to form a broad slit. Such ears, it is to be noted, exist in both sexes, while the stridulating organs do not. That such sound-producing organs serve as stimulants to the sexual passions of the 219females is but a natural inference. Some authorities, however, regard this as doubtful, since there are species which appear to lack these stridulating instruments, though possessing ears. But closer observation will probably show that these apparently dumb species are not really so, as Dr. David Sharp, commenting on this fact remarks: “It is well known that sounds inaudible to some human ears are perfectly distinct to others. Tyndall, in his work on Sound, has illustrated this by a fact that is of special interest from our present point of view. ‘Crossing the Wengern Alp with a friend’ he says, ‘the grass on each side of the path swarmed with insects which, to me, rent the air with their shrill chirruping. My friend heard nothing of this, the Insect world lying beyond his limit of audition!’ If human ears are so different in their capacity for perceiving vibrations, it of course becomes more than probable that auditory organs so differently constituted as are those of insects from our own may hear sounds when the best human ear can detect nothing audible. On the whole, therefore, it would appear most probable that the Orthoptera provided with acoustic organs, and which we consider dumb, are not really so, but produce sounds which we cannot hear, and do so in some manner unknown to us. If this be the case, it is probable that these ears are special organs for hearing particular sounds.”
Plate 32.
STRIDULATING ORGANS. ETC.
1. The stridulating mechanism of the Red Ocypode Crab.
2. The stridulating apparatus of a Grasshopper—highly magnified.
3. The head of a Gnat with the compound eyes split up.
4. The “ear” of a Grasshopper.
[Face page 218.
Certain of the Grasshoppers of Africa, known to entomologists as Pneumorides, have undergone a most extraordinary transformation of their bodily shape, as if in response to the demands of these musical performances. They have entirely lost the power of leaping, and the abdomen, in the male, has become transformed into a 220huge, pellucid, inflated bag or bladder, apparently to serve the purpose of a resonator, increasing the volume of the sound produced by the stridulating organ, which consists of a series of ridges placed on each side. The noise which this mechanism produces is, as might be supposed, considerable. It is curious to remark that in this group the females are more vividly coloured than the males. In the case of one South African species—Pneumora scutellaris—this coloration is so extravagant that she has been said to look as if “got up” for a fancy-dress ball (Plate 33, Fig. 1). Her ground colour is of a light green, with pearly-white markings, surrounded by an edging of magenta; the white areas are very numerous. The face has magenta patches, and numerous, tiny, pearly-white tubercles, each of which, when placed on a green part, is surrounded by a ring of mauve. This scheme of coloration distinguishes her as one of the most remarkably coloured of insects. But to what are we to attribute these hues? Sexual Selection will not explain them, and it seems unreasonable to regard them either as affording a protective or a warning coloration. They may then, perhaps, be allowed to rank as another instance of unchecked variation in the direction of vivid colouring, such as has been already described as occurring both among birds and other animals lower in the scale.
Plate 33.
CRICKETS AND MAY FLIES.
1 and 2 afford illustrations of the excessive development of “ornament “; fig. 3 of devices for seizing the female; figs. 4 and 5 of unaccountable differences in the development of wings.
1. The Pneumatic Cricket (Pneumora scutellaris), showing the strange markings on the female.
2. The Cleft-footed Burrowing Cricket (Schizodactylus monstrosus).
3. The Giant Alder-fly (Corydalis crassicornis), with its huge jaws for grasping the female.
4. The Stone-fly (Perla maxima), the large-winged Continental form.
5. Loch Tanna Stone-fly (Isogenus nubecula), male, with vistigial wings.
[Face page 220.
In the Locustidæ the ear is placed on the side of the front leg and the rim surrounding it may either take an oval shape or close up to form a slit. The air necessary for the efficient action of the acoustic apparatus is admitted through a gaping hole in the side of the body, above the base of the leg, an arrangement not met with among any other insects. The musical apparatus of these insects differs from that of the Acridiidæ, for it is formed only by the wing-cases, and not by the wing 221case and the leg. One of the wings bears a file on its inner surface, the other, the right wing, is furnished with a sharp edge placed on a prominent part of its inner margin. By slightly tilting the fore-wings, or wing-cases, and vibrating them rapidly, the edge passes under the file and a musical sound is produced. By this means one of our native long-horned Grasshoppers (Locusta viridissima) produces a shrill, but not unpleasant, sound, capable of being sustained continuously for a quarter of an hour. But a species encountered by Bates during his travels in the Amazons is a much more efficient performer. Known by the name of Tanana by the natives, it is so much admired by them for its singing powers that it is kept in little cages as we keep Canaries. That these organs are of importance to the species may be gathered from the case of a Bulgarian long-horned or Green Grasshopper (Poecilimon affinis), wherein the wings have so degenerated as to be useless in flight, but in the male they have been retained solely as musical instruments. In some species both sexes have a music-producing apparatus, but as a rule this is present only in the male.
That these curious and complex stridulating organs do indeed primarily act as aphrodisiacs seems to have been clearly demonstrated by the naturalist Bates, who, in speaking of the European Field Cricket remarks: “The male has been observed to place himself at the entrance to his burrow, and stridulate until a female approaches, when the louder notes are succeeded by a more subdued tone, whilst the successful musician caresses with his antennæ the mate he has won.”
Among the most efficient and most celebrated performers of all on these instruments of percussion are 222the “Katydids” of North America. The sounds they produce are said to form the words “Katy-did, O-she-did, Katy-did-she-did.” The first of these extraordinary concerts is heard about mid-July; by mid-August they are in full song. By others the sounds have been likened to those produced by the slow turning of a child’s rattle, ending in a sudden jerk; and this prolonged rattling, which is peculiar to the male, is always answered by a single, sharp “chirp” or “tschick” from one or more females, who produce the sound by a sudden upward jerk of the wings.
Pride of place, however, among insect performances of this kind must surely be awarded to the Cicadas, which are notoriously the noisiest members of the Insect world, far eclipsing the shrill calls of the Grasshoppers and even of the Crickets. Darwin remarks that the noise they made could “be plainly heard on board the Beagle when anchored off Brazil at a quarter of a mile from the shore; and Captain Hancock says it can be heard at a distance of a mile. The Greeks formerly kept, and the Chinese now keep, these insects in cages for the sake of their song, so that it must be pleasing to the ears of some men.” Only the males sing, the females being completely dumb, and this prompted the Greek poet Xenarchus to make the now famous remark, “Happy the Cicadas’ lives, for they have voiceless wives.” Another naturalist, Riley, says: “The general noise, on approaching the infested woods, is a combination of that of a distant threshing-machine and a distant frog-pond.” Another species, Tympanoterpes gigas, also Brazilian, is said to make a noise equal to the whistle of a locomotive: recalling that of a nest of young snakes, or young birds, when disturbed—a sort of scream. They can also produce a 223chirp like that of a Cricket and a very loud, shrill screech prolonged for fifteen or twenty seconds, gradually increasing and decreasing in force.
Curiously enough, no special auditory organs have yet been discovered, and it has been suggested that these insects do not hear in our sense of the word, but feel rhythmical vibrations. But whether the males “sing in rivalry,” as Dr. David Sharp suggests, is another matter. The purpose of the “song” in the first place is no doubt intended as a guide to the females seeking mates. But it is quite conceivable that the call of one male may stimulate every other male in the neighbourhood. Darwin, commenting on this aspect of the music, gives a quotation from Dr. Hertman, who says: “The drums are now ... heard in all directions. This I believe to be the marital summons from the males. Standing in thick chestnut sprouts about as high as my head, where hundreds were around me, I observed the females coming around the drumming males.... This season a dwarf pear-tree in my garden produced about fifty larvæ of Cicada pruinosa; and I several times noticed the females alight near a male while he was uttering his clanging notes.”
The structures, he remarks, from which these sounds proceed, “must be ranked amongst the most remarkable voice-organs in the animal kingdom. They are totally different from the stridulating organs that are found in many other insects.... Some difference of opinion has existed as to the manner in which the structures act, but the account given by Carlet ... will, we believe, be found to be essentially correct.” They are partly thoracic and partly abdominal. On examining a male Cicada there will be seen, on the under surface, two plates, meeting in the middle line of the body and 224overlapping the base of the abdomen. They can be slightly moved away from the abdomen, and thereby a wide fissure will be produced, displaying the mechanism beneath. If the whole operculum be removed, three membranes will be seen, an external, called the “timbal,” an anterior folded and soft membrane, and a posterior “mirror.” This last is a very beautiful object, tensely stretched and pellucid, yet reflecting all the hues of the rainbow. The sound is primarily produced by the vibrations of the timbal, to which a muscle is attached; the other membranes are probably also set vibrating, and the whole skeleton helps to increase or modify the sound, which is probably also influenced by the position of the operculum, which varies in different species. A further control of the tension of the air is exerted by “stigmata” or pores, and certain air-chambers connected therewith.
Throughout these pages comment has been made on the apparently “fortuitous” character of complex patterns and structures. The “musical-box” of the Cicada affords yet another instance. Nevertheless there is an impressive harmony between the several parts; an interdependence which is not fortuitous. There is obviously a nexus of growth-controlling factors preserving harmony between each separate part which as yet has escaped all endeavour to discover.
While it is difficult to picture the initial stages of growth of so complex an organ as that of the Cicada, the beginnings of simpler structures such as the stridulating organs of Beetles and Grasshoppers are more easily discernible. “It is probable,” remarks Darwin, “that the two sexes of many kinds of Beetles were at first enabled to find each other by the slight shuffling noise produced by the rubbing together of the adjoining hard parts of their 225bodies; and that as those males or females which made the greatest noise succeeded best in finding partners, rugosities on various parts of their bodies were gradually developed by means of sexual selection into true stridulating organs.”
Structures to which we can ascribe a use are commonly supposed to have been evolved for the purpose which we assign to them. The “horns” of Beetles afford a case in point; but there are many other equally remarkable and extravagant developments among the insects which seem to defy explanation. And they will continue to do so until it is realized that they are but exaggerations of the normal processes of growth, which is not limited to definite areas but may produce extensions and excrescences of an almost infinitely varied character. The only controlling factor is that imposed by Natural Selection when these growth-changes tend to impair the well-being of the organism as a whole. Often such changes confer benefits, giving rise to new organs, and in this case Natural Selection encourages the new departure. Nothing, indeed, “succeeds like success.” New departures in one direction may be promptly suppressed, in another they spell fortune: there is no “Socialism” in Nature. Often these “new departures” neither help nor hinder, and instances of this kind are commonly afforded by “ornaments.” One of the most singular illustrations of this kind is furnished by that extraordinary Long-horned Grasshopper of India (Schizodactylus monstrosus), wherein the wings, when at rest, have their tips coiled up like a watch-spring, while the appendages to the legs are scarcely less remarkable. It is a burrower, driving long tunnels in the banks of rivers. But little is known of its habits, save that it does not emerge from its burrow till night, when it takes long flights. This being 226so, the bizarre character of its wings and legs is the more difficult to explain on any Sexual Selection theory. But regarded as spontaneous variations which have not fallen under the ban of Natural Selection, they are somewhat less puzzling, though, having regard to the extraordinary transformation which the burrowing Mole-cricket and the allied Cylindrodes have undergone, in adaptation to fossorial habits, the legs of this insect are remarkable indeed.
While there can be no doubt that the musical performances of the Crickets and Locusts play an important part in courtship, in some of the Long-horned Locusts, at any rate, the males fight furiously when mate-hunting, and to this end the head and jaws are greatly enlarged. During the progress of the duel the wings are extended and held erect, which is hardly what one would have expected, since in this position they would seem to be more exposed to danger.
All the insects so far surveyed have been more or less conspicuous for their vivid hues, yet in none of these have elaborate “displays” been recorded. To find demonstrativeness of this kind one must turn apparently to a group of minute, lowly organized, dull-coloured, wingless insects with ugly misshapen heads and bodies. The sexes do not differ in appearance, but they are interesting on account of the sedulous court which the males pay to the females. The late Lord Avebury, in a Communication to the Linnean Society, remarked of them: “It is very amusing to see these little creatures (Smynthurus luteus) coquetting together. The male, which is much smaller than the female, runs round her, and 227they butt one another, standing face to face and moving backwards and forwards like two playful lambs. Then the female pretends to run away and the male runs after her. With a queer appearance of anger, he gets in front and stands facing her again; then she turns coyly round, but he, quicker and more active, scuttles round too, and seems to whip her with his antennæ; then for a bit they stand face to face, play with their antennas and seem to be all in all to one another.”
The Dragon-flies are among the most beautiful of insects; they are also relatively long-lived, and they are conspicuous. Yet this beauty must be attributed to some inherent inward grace rather than to the æsthetic instincts of the female. Moreover, in the matter of size and beauty there is little to choose between the sexes; where any difference occurs the males have the advantage. Though the mode of copulation is well known, nothing has been discovered as to the means whereby male and female discover one another. It is doubtful whether this can be done by sight, for with all the beauty of their shimmering suits of mail and gauzy wings, their vision is limited to a field of a few inches. Possibly scent is their guide; at any rate, dead Dragon-flies have a vile odour.
It is worth noting that there are no wingless Dragon-flies, and that none have developed unnecessary ornament in the form of spines, horns, or frills of any kind, such as are so commonly met with among groups of more sedentary habits like the Phasmidæ and the Beetles, for example. In other words, there is clearly a direct relation between ornament and the mode of life. It is also clear that some modes of subsistence are very inelastic, allowing of no more than very slight structural variations, for the Dragon-flies are an extremely ancient group. Fossil species of large size are known from the Lower Lias, and the remains 228of a giant measuring two feet in expanse of wings has been found in the Carboniferous. This species, however, seems to have stood near the parting of the ways between the May-flies and the Dragon-flies. But be this as it may, undoubtedly Dragon-flies hovered over the backs of sleeping Ichthyosaurs and furnished food for Pterodactyles millions of years ago, as they now hover over lazy kine for the sake of the flies forgathered there, or dodge to avoid the stoop of the Hobby, and in all this vast space of time they have not appreciably changed.
And what is true of the Dragon-fly is true also of the May-fly, for it is clear that they are of the same stock. It is true at any rate in so far as the conformation of the body is concerned. The possibility that it may be equally true in regard to the details of their life-history almost staggers one, because these are, in many respects, of a quite remarkable character. As with the Dragon-fly, there is a prolonged period of larval life, lasting from one to two years, which time is passed in streams and pools where a luxuriant vegetation ensures a plentiful supply of food. Some are carnivorous, but in the majority of species minute plants only are eaten. More than forty species are to be reckoned as natives of the British Islands, the commonest being the “Green Drake” and “Grey Drake,” beloved by the fisherman. These names are applied, it may be mentioned, to the phase known as the sub-imago which precedes the fully-adult stage, of Ephemera vulgata and E. danica.
Save that it is curious that while the larvæ of some species are carnivorous those of others are vegetarians, there is nothing very remarkable about what may be called the infantile period. But when this is ended the 229span of life remaining to them as adults is brief indeed. Instinctively realizing that the time of transfiguration is at hand, the erstwhile crawling grub rises to the surface of the stream, and almost in the twinkling of an eye it mounts into the air on gauzy wings, there for a brief space to execute an aerial dance which in its every phase is amazing. Some species never see the sun. They emerge as the sky begins to redden, and as its glory fades they, too, expire. This brief space is all that Nature has allowed them in which to fulfil her behest to all living things—to increase and multiply. And myriads die without even a chance to effect this consummation of existence.
The dance is a Dance of Death, and it is performed by a host so vast as to surpass the bounds of belief save to those who have had the good fortune to witness a scene so amazing.
D’Albertis tells of a gathering which he witnessed on the Fly River, New Guinea—for these insects have a world-wide distribution—wherein countless myriads were assembled. “For miles the surface of the river, from side to side, was white with them as they hung over it on gauzy wings; at certain moments, obeying some mysterious signal, they would rise in the air and then sink down anew like a fall of snow.” And in this assemblage he estimated that there was but one female to every five or six thousand males. It is during this flight that the act of mating is performed. The fortunate male from the host of rivals, in this mid-air embrace is aided by the foremost pair of legs, which are especially curved to effect this purpose. The embrace is momentary. Thereafter he dies; to the female a somewhat longer span of life remains, for she has yet to deposit her eggs, and 230this being done en masse, she, too, expires.
It is curious that these creatures, which in their winged state have never seen the sun, should be attracted by light. But such is the case. I well remember witnessing an instance of this years ago, while staying, one August, at Bingen on the Rhine. Dinner was served in the open air, and just as the soup was served May-flies in myriads swarmed round the lamps and fell on the tables as thick as snow-flakes. Some of these were in copula, and I succeeded in bottling a few specimens for the British Museum, where they still remain to remind me of this amazing scene.
About three hundred species of May-flies are known, and some enjoy a somewhat longer span of life than others. In no case, however, do they emerge till just before sunset; but in some species it is believed life may be prolonged for as much as three or four days, or even longer, if the weather be cold and wet, so as to keep them in a state of enforced rest, which amounts to a state of coma.
That their hold on life during this final stage of existence is brief there can be no gainsaying, for it is passed fasting. Jaws are wanting, and the whole alimentary canal has been transformed into one long air-chamber. Its walls are now of extreme tenuity, and by changes in the interior of the tube, valves are formed which convert the stomach into a capacious air-sac.” When movements,” remarks Dr. David Sharp, “tend to increase the capacity of the body cavity then air enters into the stomachic sac by the mouth orifice, but when muscular contractions result in pressure on the sac they close the orifices of its extremities by the valve-like structures just referred to; the result is, that as the complex movements of the body are made the stomach becomes more 231and more distended by air.” It was known even to the old naturalists that the dancing May-fly is a sort of balloon, but they were not acquainted with the exact mode of inflation. Palmen says that in addition to the valve-like arrangements we have described, the entry into the canal is controlled by a circular muscle with which are connected radiating muscles attached to the walls of the head. The canal thus strangely transformed performs the functions of a balloon, and at the same time aids the functions of the reproductive organs.
Where vast numbers of individuals set out simultaneously to achieve their nuptials there would seem to be no need for special devices on the part of either sex to call attention to their whereabouts. Nevertheless, it is highly probable that the female exhales some distinctive odour; otherwise, having regard to the fact that she is overwhelmingly outnumbered by suitors, her discovery in such a crowd would be impossible, and it is of vital importance that no time should be lost in effecting conjugation, for the time for its accomplishment is perilously short. But there is another possible means of discrimination—the males may distinguish the females by the very different appearance of the head in the latter. At any rate, this may be true of some species wherein the males have no less than seven eyes, and these of three different kinds! The compound eyes, characteristic of insects, are, in these, divided, one half being set upon the summit of a pillar raised high above the level of the head, the other part remaining in its normal place at the side of the head; and in front of these, on what may be called the forehead, are three separate simple eyes, or “ocelli.” A reference to Plate 32, Fig. 3, will make this clear.
That the history of the later life of the May-fly is 232remarkable no one will deny: in many respects it is unique. Yet for all its strangeness it enables us to set our compasses, so to speak, in regard to the phenomena of sex in other groups. The extraordinary disparity in the proportions between male and female, for example, is full of significance, for it shows, as has been suggested more than once in these pages, that, in the case of polygamous species, we are probably in error in supposing that the excess of females is due to the reduction in the number of males by reason of the elimination of males by fighting. The excess of males, or females, as the case may be, is due to an inherent quality in the germ-plasm. The May-fly might be regarded as an excessively polyandrous species if the number of males in relation to females alone be regarded: but actually it is monogamous. After a prosaic infancy they are suddenly transformed into gay lovers, dancing a marriage-dance. But for them is no marriage feast, nor any later sequence of domesticity. One in ten thousand may find a mate, and only in this is he more fortunate than his neighbours, for, like them, he too must die before the dawn. Theirs is not even a sleep and a forgetting, but “one splendid hour of Life, and then—oblivion.” It may be urged that even these which might seem to have been fooled, have not really lived in vain, for hosts of animals feast upon their bodies. Myriads, indeed, are snapped up by fishes even before they have opened their wings, while birds rudely invade the swarms as they dance in mid-air, feasting on these fasting ones. But this is, after all, an inglorious end, and leaves us still asking Cui bono?
Is this amazing life-history a thing of yesterday, a new phase, or an order of things as old as the origin of the species, dating back some millions of years?
233So far as one can profitably speculate on such a theme it would seem more likely to be a relatively recent innovation. The nearly-related Alder-flies (Sialidæ), so well known to anglers, seem like to meet with a similar fate, for the female lives but for a few days only and the male has an even briefer existence as a winged insect. The family to which the Alder-flies belong contains a few species which attain gigantic proportions, as, for example, in the case of the North American members of the genus Corydalis, which are giants. The males thereof are remarkable for the fact that they are armed with enormous jaws, which may be likened to a pair of callipers whose limbs have been crossed. These weapons serve as claspers, enabling the males to seize and hold the females during the act of mating. But even here the same brief span of life has to suffice them, for death follows swiftly on the fulfilment of the nuptial rites.
The Perlidæ, or Stone-flies, which, like the Sialidæ, are aquatic Neuroptera, the larval stages being passed in streams, present very puzzling features in regard to the adult males which, so far, have baffled all attempts at solution; yet they seem to have a very important bearing on the all-important work of reproduction. They are among the earliest insects to appear in spring, and possess an extraordinary power of resisting cold. One species, Capina vernalis, common in the Albany River, in Canada, frequently comes up through cracks in the ice and casts its skin there! Another, Nemoura glacialis, which appears at about the same time, actually performs the nuptial rites in crevices in the dissolving ice! Happily reason is denied them, or they would find life a mockery indeed; for having attained their final development, when the joyous and exhilarating exercise of 234flight should be theirs, they are compelled forthwith to fulfil their reproductive functions and die—in an ice-chamber! The males have wings which are rarely or never unfurled; as a rule they present nothing more than a crumpled mass of gauzy tissue, as if glued together. Such species as attain to flight are most indifferent performers, travelling but slowly, with laboured movements and settling after a few yards have been traversed.
As a rule, among insects, where there is a difference in the power of flight, it is the male which is superior. The case of Nemoura, just referred to, affords an instance where the contrary is the case, and Mr. J. J. Lister records the case of one of these flies—Isogenus nubecula—taken at Loch Tanna in Arran, wherein the wings of the female were greatly reduced, while those of the male were so much so as to be mere useless vestiges. Similar facts have been recorded of more than one species in Scotland, but in all such cases the phenomenon seems to be associated with the appearance of the insect in very early spring. In another species—Nemoura trifasciata—only the front wings are reduced, the hind pair being large enough to cover the body. In male specimens of Perla maxima taken in Scotland, the wings are so short as to be useless for the purposes of flight, yet, in the same species taken in Central Europe, they are of ample proportions.
These facts are puzzling indeed, but they seem to show that flight is not essential to attain the ends of reproduction. As to whether these flies secure their mates by any kind of “courtship,” or how they find one another, seems not to be known. But the female is remarkable for the fact that she carries her eggs about with her, to the number of five or six thousand, attached to the 235end of the abdomen.
Having regard to the fact that three thousand species of Perlidæ are known, and that they have a wide distribution over the earth’s surface, one might have expected that more would be known of their singular life-history. They are, however, flies of very unattractive appearance and great frailty, hence, save to anglers, by whom they are esteemed as bait for trout, they attract but little attention.
Musical Lovers among Spiders and Scorpions—Colour among Spiders, and its uses—The Spiders’ Dance of Death—Spiders and Conjugal Bliss—How Pairing is accomplished—Scorpions in Love—Musical Crabs—Quarrelsome Fiddler-crabs—Crabs and Courtship in the Deep Sea—Amazons among Prawns—Brine-shrimps and Water-fleas—“Natural” v. “Sexual” Selection.
It is a curious and significant fact that in the most brilliantly coloured of the Invertebrates—the Butterflies and Moths—” courtship” in the sense of “wooing” is extremely rarely met with; and this is quite contrary to what the Sexual Selection theory of Darwin demands, for, according to this, the colours are the result of that selection. On the other hand, Spiders, which are for the most part dull-coloured creatures, and the Scorpions, which are also dull-coloured, are commonly extraordinarily demonstrative during the early stages of “mate-hunger.” Some practise the form of instrumental music known as “stridulation,” others dance and indulge in other forms of posturing.
In the Spiders the stridulating apparatus is formed either between the limb-bearing portions of the body, or “cephalothorax,” and the abdomen; between the 237palps or leg-like feelers, and the jaws; or between these feelers and the front legs. But the construction is similar in all. In some Spiders the abdomen bears a horny collar, which is toothed, and these teeth, as the abdomen is raised and depressed, scrape against a number of delicate ridges on the thorax, or “chest,” which form a surface recalling that of a file. The grating of these opposing surfaces against one another produces shrill rasping or chirping sounds, which, in some cases at any rate, seem to be designed to inform the female of the presence of a suitor. Those who will, may examine this strange instrument for themselves if they will take the trouble to seek for it in one of our commonest English Spiders (Steatoda bipunctata). That it serves as a sexual excitant, or as an aid to mate-hunting, is indicated by the fact that it is found in males only, or in a very rudimentary condition in the female. There is a large Spider in Assam (Chilobrachys stridulatus) which produces a sound like the drawing of the back of a knife along the edge of a strong comb; and there are others which, by the friction of the feelers against the jaws, produce sounds like the buzzing of bees. One of the Wolf-spiders (Lycosa kochy) is known as the “purring” or “drumming” Spider from its custom, at mating-time, of rapidly drumming on dead leaves with its feelers. It is a wood-haunting species, and runs hither and thither over the ground as if searching for something, and pausing frequently to “purr.” This singular method of producing sound recalls that of the drumming of Woodpeckers on the hollow branches of trees, and similarly is produced without any special mechanism.
That the Scorpions should possess similar stridulating organs is only what we should expect, having regard to 238their kinship with the Spiders. In the great Rock scorpions of India and Africa the stridulating apparatus lies between the basal segment of the pincers and that of the first pair of legs, and consists of a set of tubercles and a cluster of curved, hair-tipped spines. During moments of excitement the pincers are waved up and down so that the spinules scrape against the tubercles, emitting a rustling sound, which has been compared to that produced by rubbing a stiff tooth-brush with one’s finger-nails. In the South African Opisthophthalmus the mechanism differs, consisting of leaf-like hairs placed on the inner surface of the jaws. But since both sexes possess these strange sound-producing mechanisms it has been suggested that their main, if not their only purpose, is to serve as a warning to enemies to keep their distance. Some of the great bird-eating Spiders (Aviculariidæ) produce a kind of whistle; others, sounds like the dropping of shot upon a plate.
These stridulating contrivances present some curious and puzzling features. In the first place the sounds they produce are never loud to human ears; therein they differ from the shrill piercing sounds produced by like mechanism by the Crickets and Grasshoppers, though even with some of these the notes are, to us, inaudible. In the second, it has been suggested that where both sexes possess a stridulating apparatus its purpose is solely to warn off enemies, and this because the performers have no sense of hearing, and are thus, we presume, unaware of the sounds they produce. There is something unsatisfactory about this line of argument. There seems to be no evidence either that the sounds produced are loud enough to terrify an enemy, or that the performers are really deaf.
239In cases where the males alone stridulate it is always supposed that this “music” serves the purpose of a lure, or acts as an excitant, to the female, even though inaudible to human ears. But there are many people who are unable to hear the shrill squeal of our native bats. Yet no one doubts but that all bats hear it. The argument as to the absence of any sense of hearing in certain Spiders is based on their failure to respond to the vibrations of a tuning-fork, but this evidence is not conclusive. Neither is it safe to infer that the presence of stridulating organs in the adult and immature stages of both sexes, in some species, precludes their recognition as secondary sexual characters. They may serve the double purpose of sexual excitants and terrifying enemies, their motive being expressed by the quality of the sound as certainly as the timbre of the human voice may express rage or pleasure.
Neither Spiders nor Scorpions exhibit any very striking secondary sexual characters. As a rule the female is the larger, often strikingly so. Bright colours are rare, and are met with only among the Spiders, wherein sometimes the male, sometimes the female, is the more resplendent; where bright colours—apple-green, red and yellow—do occur, they seem rather to be of the type known as Anti-cryptic, or aggressive resemblance colours. That is to say, they are hues developed to deceive prey by reason of the likeness they afford the wearer to its surroundings. Thus, for example, one of our native Spiders (Tibellus oblongus) is straw-coloured, and has an elongated body, which is therefore seen with difficulty amid dry grass and rushes which are the haunts of the species. Misumena vatia, one of the Crab-spiders, resembles the flowers on which it is accustomed to lurk for its prey. It is of a 240variable hue, commonly yellow or pink, and a favoured lurking-place is near the blooms of the great mullein (Verbascum thapsus), where it seizes upon bees coming for honey. Exotic relatives of this species afford far more striking illustrations of this kind. One has a pink, three-lobed body which bears a striking likeness to a withered flower, and it exhales a sweet odour of jasmine. Insects attracted by the smell are thus readily pounced upon. Dr. Trimen, of Cape Town, describes a rose-red species which exactly matches an oleander flower, and to complete the deception the abdomen is marked with white. The same observer, approaching a bush of the yellow-flowered Senecio pubigera, noticed that two of the numerous butterflies settled upon it did not fly away with their companions. Each of these he found to be in the clutches of a spider whose remarkable resemblance to the flower lay not only in its colour, but in the attitude it assumed. “Holding on to the flower-stalk by the two hinder pairs of legs, it extended the two long front pairs upwards and laterally. In this position it was scarcely possible to believe that it was not a flower seen in profile, the rounded abdomen representing the central mass of florets, and the extended legs the ray florets; while to complete the illusion the femora of the front pair of legs, addressed to the thorax, have each a longitudinal red stripe which represents the ferruginous stripe on the sepals of the flower.” But more remarkable still is the case cited by my friend Dr. H. O. Forbes. This came under his notice while butterfly hunting in Java. The butterflies of the family Hesperidæ have a habit of settling on the excreta of birds. Forbes noticed one on a leaf apparently enjoying a feast. Creeping up, he seized hold of this victim 241of a depraved taste and found it mysteriously held down. On further examination of this “excreta” he found that it was really a spider! Later, when in Sumatra, the same species once more in like manner deceived him. The deception is more than usually remarkable, for it is not due to the coloration of the body, but to what may almost be described as a diabolically ingenious display of intelligence. For the creature weaves upon a leaf a small white patch of web exactly resembling the fluid excrement of a bird sliding down the smooth surface of the leaf. Having completed this, the weaver lies on its back in the middle of the web holding on by the spines with which the legs are furnished. It then awaits its victim with the disengaged portions of the legs ready to close in a deadly embrace the moment the lure has done its work. Though somewhat in the nature of a digression, these facts show that colour often plays a vital part in well-being; though in the matter of courtship its rôle has probably been overestimated. Colour as an aid to “mate hunting “probably nowhere plays so important a part as was at one time believed. The Warblers among the birds, and the Spiders among more lowly animals, seem to demonstrate this fact.
The actual mating of Spiders, the act of coition, is peculiar, and demands notice, for the orgasm is not accomplished at the moment of the ejection of the sexual products. The male discharges the seminal fluid upon a small web woven for the purpose, and the liberated spermatozoa are then sucked up into a tube—the receptaculum seminis—which lies coiled up within a hollow bulb attached to the base of the last joint of the leg-like feeler, or “pedipalp” at the base of the head. The precious fluid is there stored and retained until the moment arrives when these palps can be thrust into die 242genital aperture of the female, and their contents discharged for the second and last time. This is the critical moment of the Spider’s life, and it is noteworthy that it should occur now, instead of at the moment of the discharge from the body. The ejection from the palpal organ is effected by means of a fibro-elastic bag, in its normal, collapsed, state spirally disposed round the base of the bulb which contains the sperm tube. Immediately preceding copulation this elastic bag or “hæmatodocha” becomes turgid with blood, and it is probably the pressure thus exerted on its base which affords the final fury of desire without which, indeed, one might well imagine the necessary courage for copulation would never be raised, at any rate, in the case of some species.
Strange as these facts are, the nice adjustment of the instincts for their effectual performance is, by comparison, stranger still. By what subtle sense is the male Spider informed of the importance of the fertilizing fluid which escapes his body? What prompts him before its escape to prepare a web for its reception? What prompts him after its deposition to collect it within the palp till it shall be needed? The least defect in the instincts appertaining to these vitally important acts would mean the extinction of the race. We cannot suppose that the nature of their performance is in any way realized by the performer, and this makes their orderly execution the more wonderful.
Plate 34.

MALE ASTIA DISPLAYING BEFORE THE LESS BRILLIANT FEMALE.
From drawings, T. Carreras, in “Marvels of the Universe.”
MALE ICIUS DISPLAYING.
The “courtship” of the male spider takes the form of a “display” recalling that of birds. He commonly ends in being eaten by his mate.
[Face page 242.
Our knowledge of Spiders under the afflatus of sexual desire has been immensely increased by the long and patient observations of Mr. and Mrs. Peckham. The fact that their investigations were carried on with captive specimens, and therefore under artificial conditions both as to environment and the number of individuals placed 243together at one time, must not be lost sight of; nor must we forget that they worked under the firm conviction that the Sexual Selection theory of choice by the females was an indisputable fact. Wherever colour was present they looked for, and saw, evidence that the female appreciated such hues, though from their observations it would seem that dull-coloured species behaved as though they were suffused with resplendent hues. In the course of their studies the courtship of several species was investigated, but a summary of their results is all that can be given here. Saitis pulex formed the subject of one of their experiments. A male was placed in a box containing a mature female. “He saw her as she stood perfectly still, twelve inches away; the glance seemed to excite him and he moved towards her; when some four inches from her he stood still, and then began the most remarkable performance that an amorous male could offer to an admiring female. She eyed him eagerly, changing her position from time to time so that he might be always in view. He, raising his whole body on one side by straightening out the legs, and lowering it on the other by folding the first two pairs of legs up and under, leans so far over as to be in danger of losing his balance, which he only maintains by sidling rapidly towards the lowered side. The palpus, too, on this side was turned back to correspond to the direction of the legs nearest to it. He moved in a semicircle for about two inches, and then instantly reversed the position of the legs and circled in the opposite direction, gradually approaching nearer and nearer to the female. Now she dashes towards him, while he, raising his first pair of legs, extends them upwards and forwards as if to hold her off, but withal slowly retreats. Again and again he 244circles from side to side, she gazing towards him in a softer mood, evidently admiring the grace of his antics. This is repeated until we have counted one hundred and eleven circles made by the ardent little male. Now he approaches nearer and nearer, and when almost within reach whirls madly around and around her, she joining and whirling with him in a giddy maze. Again he falls back and resumes his semicircular motions with his body tilted over; she, all excitement, lowers her head and raises her body so that it is almost vertical; both draw nearer; she moves slowly under him, he crawling over her head, and the mating is accomplished. After they have paired once the preliminary courtship is not so long. On one occasion a female was the more eager of the two, but this is evidently very exceptional. The female always watches the antics of the male intently, but often refuses him in the end, even after dancing before her for a long time.”
Of another species—Epiblemum scenicum—they write: “The females seemed to have some difficulty in choosing from among the males, but after a decision has been reached and a male accepted, there appeared to be complete agreement.” A species of the genus Iritis, which seems to have baffled identification, was watched for hours under natural conditions as well as in confinement.” A dozen or more males, and about half as many females, were assembled together within the length of one of the rails. The males were rushing hither and thither, dancing opposite now one female, now another; often two males met each other, when a short passage of arms followed. They waved their first legs, sidled back and forth, and then rushed together and clinched, but quickly separated, 245neither being hurt, only to run off in search of fairer foes.”
These most patient observers seem to have been convinced that whenever Spiders possess vividly coloured areas on their bodies they are not only conscious of this fact, but desire to make the most of such splendour during the period of love-making. Thus they interpret the behaviour of a curiously ant-like Spider—Synageles picata—which has the first pair of legs especially thickened, flattened on the anterior surface, and of a highly iridescent steel-blue colour. As he approaches the female he pauses “every few moments to rock from side to side, and to bend his brilliant legs so that she may look full at them; ... he could not have chosen a better position than the one he took to make a display.” And similarly they interpret the movements of another species—Dendryphantes capitatus—which has a bronze-brown face, rendered conspicuous by snow-white bands. The attitude he assumes when sexually excited is one which seems, to them at any rate, to serve admirably to expose this feature to the watchful female. But he has other charms, and his “antics are repeated for a very long time, often for hours; when at last, the female, either won by his beauty or worn out by his persistence, accepts his addresses.” Habrocestum splendens—unhappily these creatures have no names in common speech—possesses an abdomen of a magnificent purplish red, and the attitude which he assumes at courtship they regard as one designed to display this to the full. Another case of quite remarkable interest is that of Astia vittata, because the males appear to be dimorphic. That is to say, they appear under two quite distinct forms, the one red, like the female; and the other black, with three tufts of hair just behind the head. The attitudes and 246the movements of courtship, it is significant to remark, are entirely different in the two varieties: the black form, assumed to be the later development, “is much the more lively of the two, and whenever the varieties were seen to compete for a female, the black one was successful.”
Professor Poulton, commenting on this particular case, contends that “it must be admitted that these facts afford the strongest support to the theory of Sexual Selection.” But do they? A further examination of the facts will probably show that the red “form” is but an immature example, and this being so, the difference in performance and the invariable success of Othello is at once accounted for. The fact that the “two forms pass into each other” and that the “tufts only occur in the fully developed niger form” is an additional reason for regarding the red form as immature.
Professor Poulton remarks: “When the males possess any special adornment they make a point of displaying them as fully as possible.” If this be so it seems to be a very foolhardy proceeding, akin to holding the proverbial “red rag” to a bull: for it is well known that the male Spider seeking a mate carries his life in his hands, at any rate in the case of many species. Mr. and Mrs. Peckham observed several instances of this remarkable sequel to Love’s embraces. In describing the female of Phidippus morsitans they remark that she was “a savage monster. The two males we provided for her had offered her only the merest civilities, when she leaped upon them and killed them.” The first pair of legs in the males of this species possess “special adornments” in the shape of long white hairs, and it was “while one of the males was waving these handsome 247legs over his head that he was seized by his mate and devoured.” Again, in the case of a male of Phidippus rufus, the display of his “ornaments” was his undoing, for he was “caught and eaten when he insisted upon showing off his fine points too persistently.” Thus the females seem to “select” the more resplendent males as much for eating as for mating! The ogre-like habits of the females in this regard, indeed, are almost without parallel in the animal kingdom.
Anyone who cares to take the trouble to watch the web of the large Garden Spider (Epeira diademata) may witness one of these connubial tragedies. In this species, the males are conspicuously smaller than the females, and it is possible that this disparity has been brought about by Sexual Selection, the largest and least active males having been exterminated. In some species the discrepancy in size is most striking, as for example in Nephila chrysogaster, the female of which measures two inches in length, the male not more than one-tenth of an inch, and less than one one-thousand-three-hundredth part of her weight.
The males, apparently, fully realize the perils which their amours may lead them into. They haunt the borders of the webs of unmated females, but exhibit a hesitating, irresolute manner. For hours they will linger near her, feeling the silken carpet cautiously with their legs, and apparently trying to ascertain the nature of the welcome likely to be extended to them. The odds are against them: for even if allowed to mate, unless they are extraordinarily agile in slipping away the moment they have attained their object, the chances are they will be slain and eaten!
Among some species, however, matters are otherwise: 248for the males of the genus Linyphia, for example, are generally to be found living peacefully with their consorts.
More rarely the male weaves a small nuptial tent, into which he partly leads and partly drives the female: though the “driving” would appear to be merely for form’s sake! The habits of the Cellar Spider (Tegenaria parietina), a long-legged species fairly common in the South of England, affords a yet further interesting and instructive contrast with the foregoing accounts. The pairing habits of this species have been studied by many observers, but perhaps the best account is that of Mr. F. M. Campbell. He found, to begin with, that in this species the tender ties of mating are at any rate rarely violated by the horrible aftermath of cannibalism so common a feature with so many other Spiders.
One or two illustrations from Mr. Campbell’s work must suffice. On one occasion he placed together a male and a female. For four days they took no notice of one another; then the female cast what proved to be her last skin, and within three hours after, the male began to show signs of interest in her presence—which is a fact of some significance, for not till then had she attained maturity. “After a few convulsive twitchings of the legs, the male pressed forwards, moving his palpi”—the leg like “feelers” on each side of the head which form the genital organs (page 241)—“up and down, when, as they touched the palpi of the female, the pair played with these organs like two friendly bees with their antennæ. After a few minutes the female raised herself, leaning a little on her left side, and the male crept forward until his head was under the breast of his mate, while his first pair of legs were resting upon hers. He now 249advanced his right palpus, leaning a little to the left and using the left palpus as part of his support. The right palpus was slightly twisted so as to bring the surface (containing the fertilizing germs) opposite the sexual organs of the female.... He now rapidly raised his palpus up and down for four or more seconds, and with such energy as to compel her to assume a vertical position. He then retired and again approached her, repeating the movements ... occasionally pausing before he withdrew his palpus.... At times he would leave the female for five minutes, and strut with straightened legs round the vase, wagging his abdomen. Now and then he would remain perfectly still with the palpus withdrawn, or play with the palpi of the female, while she seemed in a comatose state. He would then renew the union with undiminished vigour, appearing on each occasion less desirous of changing his position. I left them at 12–30 a.m. and returned at 7 a.m. The male was still using his right palpus. I saw no application of the left palpus, but have no doubt that it was employed during the night, as in other cases. I have not observed the pairing ever interrupted by a fresh collection of semen, although there is no reason to think this may not occur. The duration of pairing is long; but I am inclined to think it is more dependent on the difficulty in inserting the palpus than on sexual endurance. The impregnation appeared to take place when the male retained his palpi in front of the bursa copulatrices for about thirty seconds, which was frequently the case.” There are occasions, however, when a very different sequel attends this dalliance. In one instance, for example, Mr. Campbell placed a pair together, and at once the male began to pay his addresses. “Shortly 250afterwards he rapidly applied one of his palpi to the female ... apparently with her consent.” Five hours later “he charged her, tore away two legs ... and began to suck one, using the mandibles to hold the limb as a human being would a stick of asparagus.” It is not surprising to find she died an hour afterwards. An examination of her remains brought to light the fact that she was not mature. But this does not apparently explain the ferocity of her partner, for this investigator on two other occasions saw males similarly dismember their spouses an hour or so after impregnation. This horrid feast cannot have been prompted by hunger, for one of these males had, but a few hours previously, eaten a daddy-long-legs and two blow-flies. Only twice did this investigator see a female of this species drive away a male, and in each case immediately after union. “On the other hand,” he says, “I have kept an adult pair together from the 22nd of August to the 28th of October, and they lived in perfect amity. The male never ceased paying unrequited attentions except to feed.” It will have been remarked that the behaviour of this species in regard to mating differs conspicuously from the accounts of observations on other species, wherein the aggressive instincts are displayed by the female. Mr. Campbell, commenting on these facts, remarks that such conduct is just what one would expect from creatures which lead solitary lives, and must have “come to regard weaker forms of animal life as food, or as an inconvenience, if we except its young or its mate when in the act of pairing.” Instincts which are habitually practised throughout the greater portion of the life of the species, and on which existence depends, would scarcely be suspended for a longer period than necessary for sexual 251union. Spiders frequently eat one another, and such an occurrence after pairing is only curious if considered apart from their habits. When the sexual desire is satisfied, their actions would again be directed by the dominant instinct of destruction.
It is to be noticed that the attack, when made by a female, often immediately follows the sexual union, while in the case where males assume the aggressive it takes place some time afterwards. Mr. Campbell explains this by the supposition that the action of the female, when satiated, would be precipitated by the threatened and unacceptable continued application of the hard, spiny palpus, while the more lasting desire of the male would have to subside before he became directed by another instinct. By that time, other attractions, if not his wandering disposition, would take him away from the web.
The fact that male Spiders are comparatively rare is perhaps explained by the fact that they are very short-lived; they probably die soon after pairing—even if they are not eaten! The snares they spin, it is to be noted, are very imperfect, though curiously enough, when young they make perfect snares on a small scale.
It will have been noticed, in the course of the foregoing descriptions, that Spiders display a more or less conspicuous wariness, a cool, deliberate “counting on the cost” in their matrimonial ventures that is often wanting in such matters in the human race. But, then, the risks involved are more patent, more imminent. Mr. Campbell comments on this intelligent behaviour in the case of the Cellar Spider, remarking that they measure “each other’s strength when on the same web by the tension and motion of the threads.”
252A word as to the Scorpions. These creatures are near relations of the Spiders, and in many things resemble them, notably in regard to their ferocity. One does not meet here, however, with the same disparity in size between the sexes, nor are vivid colours ever developed. This, according to some, would be accounted for by the fact that though these creatures possess numerous eyes they are practically blind, and depend for their information as to what is going on around them by their sense of touch, which is excessively delicate. They are morose in disposition and always solitary. It has been said that if two are found under the same stone—a favourite lurking-place—one is engaged in eating the other! Nevertheless, they are of abstemious habits, for the naturalist Fabre found that from October to March they last, though throughout this time they remain alert, and always ready to resent disturbance. In April they exhibit more activity, though even then they eat but little. But now they begin to wander in search of mates.
Fabre’s observations on their mating habits are exceedingly interesting, and they have brought to light some very extraordinary phenomena. His notes were made on the species common in the South of France—Buthus occitans. Mr. Cecil Warburton, referring to the distinguished Frenchman’s work, quotes the following noteworthy passage in the Cambridge Natural History: “After some very curious antics, in which the animals stood face to face with raised tails, which they intertwined ... they always indulged in what Fabre calls a ‘promenade à deux’ hand in hand, so to speak, the male seizing the chelæ of the female with his own and walking backwards, while the female followed, usually without any reluctance. This promenade occupied an 253hour or more, during which the animals turned several times. At length, if in the neighbourhood of a suitable stone, the male would dig a hole, without for a moment entirely quitting his hold of the female, and presently both would disappear into the newly-formed retreat.”
Plate 35.

Photo by P. H. Fabre.
SCORPIONS.
The early stages in the courtship of the scorpions are full of romance. The two prospective partners for life engaging in a kind of waltz, holding each other’s “hands.”
[Face page 252.
After the mating, as with the Spiders, the male is often devoured by the female. After any combat with an enemy, such as a Lycosa or a Scolopendra, it seems to be de rigueur to eat the vanquished.
If the mating period in the case of the higher animals rouses the males to the pitch of frenzy, that frenzy is dangerous only to possible rivals. With the more lowly Spiders and Scorpions ferocity of disposition is a normal feature, and one which can with difficulty be held in check long enough to permit the all-important act of mating to take place. In how far this is accounted for by the extremely deficient senses of sight and hearing, which are such marked features in these animals, it would be difficult to estimate. But that the manner of their display is governed by these deficiencies there can be no doubt. The Spider, having a more or less efficient vision at short range, executes more or less elaborate antics in front of the female, designed, as in the case of the birds, to serve as excitants to fan sexual desire, already smouldering, to a flame. With the purblind Scorpion the Spider-antics are useless; he must proclaim his desire by a pressure of the hand, and by intertwining his tail with that of his prospective mate as they “walk out” together. But Scorpions at one time were credited with a very acute sense of hearing; later investigations, however, fail to yield any evidence whatever that they possess this sense, though experiment has proved that 254their sense of touch is excessively delicate and seems to reside in the hairs which are thickly distributed over the legs and body. Now, hearing and touch are senses near akin, and the vibrations produced by stridulation may be, and probably are, received by, and interpreted through, the medium of these hairs. For though the Scorpion may not respond to sounds made by curious investigators, it may be that they can perceive notes of a low pitch imperceptible to our ears, such as are made by stridulating organs, as in the case of the Spiders.
Perchance certain comb-like structures known as the “pectines” may play a part in mate-hunting. These are placed on either side of the under-surface of the body between the last two pairs of legs. The fact that they are larger in the male, and sometimes strangely modified in the female, seems to show that they have some function in relation to sex. They also appear to serve as sources of information as to the nature of the ground traversed by the animal, since they are long in species which walk with the body raised high off the ground and short in such as adopt a more grovelling posture. That the Scorpions possess but a very limited means of gleaning information of the outer world there can be no doubt. How, then, do they find one another when that insistent desire to mate begins to make itself felt? Are the “pectines” their informants through the sense of smell? Do the hairs scattered over the body act as sound-collectors responding to the notes emitted by the stridulating organs? These are points on which information is much to be desired.
Plate 36.

Photo by P. H. Fabre.
DEATH OF THE MALE SCORPION.
But by the time the nuptial rites have been performed the female has thrown off her “sweetness,” and ends by eating her lover!
[Face page 254.
Plate 37.

Photo by Paul H. Fabre.
THE FEMALE MANTIS DEVOURING HER MATE.
With these insects, as with the spiders and scorpions, the male is often eaten by the female.
Our survey of the “Arthropoda,” as those limb-bearing jointed animals invested in a horny, or, more exactly, a “chitinous” external skeleton are called has so far 255been confined to such as, during adult life, at least, are land-dwellers. But the aquatic types known as the “Crustacea” furnish some extremely interesting facts in regard to the problems of sex. In the first place, they too possess a stridulating apparatus. This is curious, but not surprising, because, although the skeleton of such creatures is of a harder and almost stone-like character, the development of roughened surfaces working in opposition to one another might well have been foretold to occur, at least in some individuals. Colonel Alcock—a naturalist who has contributed largely to our knowledge of marine animals by his researches in the Indian Ocean—in his most delightful book “A Naturalist in Indian Seas,” describes what he calls a “musical crab.” This is the great-horned Coromandel Strand Crab (Ocypoda macrocera). In both sexes of this remarkable genus he says, “the nippers, or chelipeds, are singularly unequal in size, and in all the species but one there is present on the inner surface of the ‘hand’ of the larger cheliped a transverse row of five teeth, which, when the cheliped is flexed, can be made to play against a ridge or another row of teeth on its ‘arm’ ... much as a man might rub one side of his chest with the palm of the corresponding hand. The whole mechanism, except that it is on a larger scale and has a more finished appearance, is very much like that by means of which crickets and grasshoppers produce their shrill music, and no one has ever doubted that it is used for the same purpose, though very few people have actually heard it in action. I myself ... was beginning to think that the structure must, after all, have some quite other function, when one morning ... on the sandy wastes 256of the Godavari delta, I at last, like Ancient Pistol, heard with ears that which I had been so long waiting for. That is to say, I heard a noise very much like that which an angry squirrel makes, and discovered that it came from a red ocypode crab into whose burrow another individual had trespassed.
“In order to understand the matter it should be known that these crabs ... are gregarious, and that each one has a burrow of its own. Though they may be seen marching in battalions across the sand, yet as a rule they stay close to their burrows, methodically searching and sifting the surrounding sand for any food that may have been thrown up by the tide, and flying to their burrows with headlong speed when alarmed. At first sight one does not understand the necessity for so much wariness, and for such a deep system of entrenchment, for the creatures seem to hold undisputed possession of the whole shore; but as a matter of fact they are preyed upon all day long by Brahminy kites, and when the jackals come out in the evening, by them. Now, although each crab may on ordinary peaceful occasions know its own home, yet when a crowd of them are running for their lives they may sometimes, one would think, act on the devil take the hindmost principle and try to squeeze into the nearest burrow. But as ancient philosophers do report, things may be done upon occasion which it is inexpedient to make a habit of doing, and this seems to be one of those things; for if many Crabs made a practice of crowding into one small burrow they would certainly run the risk of being suffocated, if not crushed to death outright. It seems probable, therefore, that it would be advantageous to the species as a whole if the rights of property in burrows were rigidly respected, and if each individual 257member possessed some means of giving notice that its burrow was occupied ... and I think that this consideration gives us a clue to the use of the stridulating mechanism. At any rate, I was often able, after my first accidental discovery, to elicit the sound, by catching one of these crabs and forcing it into a burrow which I knew was already occupied: the intruder would never go far in, but would crouch just inside the mouth of the burrow, and if it were made to travel deeper, then the voice of the rightful owner would be heard in indignant remonstrance from the depths.” Another species, the Grey Ocypode Crab (Ocypoda ceratophthalmus), possesses a similar instrument, and makes therewith a loud, croaking noise. But it does not often burrow deeply. Colonel Alcock therefore suggests that in this case it may be used for scaring enemies.
That these curious musical instruments may also be used in mate-hunting seems highly probable. If the stridulation is produced on one occasion to announce the fact that callers are not desired, it may on another signify an equally emphatic invitation to enter, the mood of the occupant being expressed by the character of the sounds emitted. It is significant, at any rate, that there are no external sexual differences in these species; hence the probability that it is by stridulation that the sexes distinguish one another.
This view seems to obtain confirmation from the fact that the Crabs of the genus Gelasimus, or “Fiddler-crabs,” which are near relations of the ocypode Crabs, and, like them, live in burrows in large companies, and are exposed to the same enemies, which they avoid in the same way by burrowing, have no stridulating mechanism, but the sexes are strikingly different. This is especially so in the case of the nippers, or chelipeds. These, in the 258female, are slender and much shorter than the legs, being used mainly for feeding. In the adult male one of these “hands” is often twice as big as the body itself! “Many uses,” remarks Colonel Alcock, “have been assigned to this enormous, lop-sided organ: some say that it is used as a stopper to barricade the mouth of the burrow, others that it is a sort of cradle or bridal-couch upon which the female reclines—the male, in this case, literally bestowing his hand upon the female; but from observations of Gelasimus annulipes, the species which most frequents the Godavari mud-flats, I believe that it primarily serves as a war-club, for the males indulge in interminable tournaments for the hand of the female; and secondarily, for it is of a most beautiful cherry-red colour, as an ornament to attract and delight the latter capricious sex.
“Landing one afternoon in March upon a cheerful mud-flat of the Godavari sea-face, I was bewildered by the sight of a multitude of small pink objects twinkling in the sun, and always, like will-o’-the-wisps, disappearing as I came near to them, but flashing brightly on ahead as far as the eye could reach. It was not until I stayed perfectly quiet that I discovered that these twinkling gems were the brandished nippers of a host of the males of Gelasimus annulipes. By long watching, I found out that the little creatures were waving their nippers with a purpose—the purpose apparently being to attract the attention of an occasional infrequent female, who, uncertain, coy, and hard to please, might be seen unconcernedly sifting the sand at the mouth of her burrow. If this demure little flirt happened to creep near the burrow of one of the males, then that favoured individual became frantic with excitement, dancing round his 259domain on tip-toe and waving his great cherry hand as if demented. Then, if another male, burning with jealousy, showed a desire to interfere, the two puny little suitors would make savage back-handed swipes at one another, wielding their cumbrous hands as if they had no weight at all. Unfortunately, though I spent many a precious hour on the watch from time to time, I could never see that these combats came to anything; the males seemed always to be in a state of passionate excitement, and the females to be always indifferent and unconcerned; and though the dismembered chelipeds of vanquished males could often be seen lying on the battle-field, I never had the satisfaction of beholding a good stand-up fight, fought out to the sweet end, or a female rewarding a successful champion with her heartless person.”
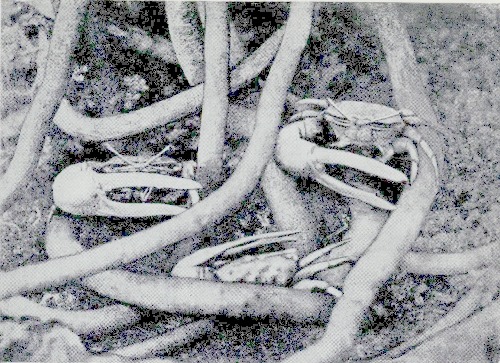
Photos by W. Saville-Kent.
THE “FIDDLER-CRAB.”
This “strong right arm” is used in conflicts with rivals for the possession of the females.
Plate 38.

THE “FIDDLER-CRAB” AMONG MANGOE ROOTS.
This species is remarkable for the enormous size of the right “arm,” which exceeds that of the body.
[Face page 258.
The fascinating tale of Colonel Alcock’s observations does not end here, however, for he has brought to light some extremely interesting facts in regard to the sexual aspect of Crustacean life in the deep sea; information gathered during his exploration work on board the Investigatory much of which was done to enlarge our knowledge of the abysses of the ocean where the light of day never penetrates. Here, he remarks, the conditions of life might seem to be reduced to a minimum of simplicity, yet evidences are not wanting that, among the higher Crustacea, they are complicated, much as they are everywhere else, by the play of the sexual instincts.
In these awful depths, where reigns eternal night, most of the inhabitants, of whatever kind, from fishes downwards, are blind and eyeless, or they possess 260enormous eyes and a purblind vision responding to the only light these regions display, that of phosphorescence, which is generated by so large a number of those creatures which are condemned by Fate to live this sunless life.
“It is written,” he remarks, “that the male must exert himself to find a mate, and where sight cannot help him in his search, a kind of blind-man’s buff is the only alternative. In this serious game many deep-sea Crustacea, especially those of the Shrimp-tribes, trust to the sense of smell, as the greatly developed outer, or olfactory, branch of the first pair of antennæ bears witness. These antennæ, again, seem to be used by the males of some species for catching their partners, and in Parapeneus rectacutus ... they are turned into a sort of crook for this purpose. This has long been thought to be their function in the Prawns of the oceanic genus Sergestes.” In the male of certain other deep-sea Prawns, the hind pair of foot-jaws are modified in a way which can only mean that they are used for hooking on to a partner of the opposite sex. In the deep-sea Hermit-lobsters of the genus Munida the nippers are greatly enlarged, as in many Shore-crabs, for the purpose of subjugating rivals and embracing the females; and in all such cases these are much smaller in the female and immature male.
Mention of numerous cases has already been made where the female is larger than the male, and is the more pugnacious, and in such cases the females are generally more numerous than the males. Some of the deep-sea Prawns exhibit the same peculiarity. And in these the sword-like forward prolongation of the head-shield is far larger than in the male. Now this rostrum is the most formidable weapon which the Prawn possesses, 261so that we may, with tolerable certainty, conclude that the females fight their rivals for the possession of the males, which are, in these species, far less numerous than the females.
Among the lower Crustacea, such as the “Fairy-shrimps,” “Brine-shrimps,” the “Water-fleas,” and the “Copepoda,” which play so important a part in furnishing food for many of the fishes which in turn feed us, secondary sexual characters of an extremely interesting kind are met with. These, however, are never such as appeal to the eye, for the vision in these creatures is but feebly developed. Scent, as is usual where sight is defective, plays an important part in enabling the sexes to discover one another. Selection here secures success only to such as have the proper odour and the most sensitive organs of smell. In these creatures, as with the butterflies and moths, the odour emanating from the female is most powerful, while the sense of smell is most developed in the male. One of the most striking illustrations of these facts is furnished by that very beautiful species Leptodora hyalina—a veritable giant among these small Crustacea—wherein the antennæ of the male are produced into enormously elongated comb-like structures, the teeth of the comb being formed by delicate olfactory filaments. In the female these antennæ are extremely short and their olfactory filaments are limited to a small terminal tuft to the antennæ, answering to the larger tuft at the base of the comb of the male.
To the majority of species, however, delicate odours seem to make little or no appeal, since excessive development of the olfactory apparatus, such as is seen in the aberrant Water-flea (Leptodora), is rare. This is perhaps explained by the fact that Leptodora is a species which 262does not herd together in vast numbers, hence, probably, the need of some exceptional means whereby the males may discover the whereabouts of the females, while in the case of the swarming hosts formed by Water-fleas and Brine-shrimps, for example, no such highly specialized aid is necessary. Instead, the males have developed powerful arms for the capture and retention of the females. In the case of the Brine-shrimp these arms are of quite formidable proportions. The males of the Copepoda, remarks Weismann, “possess on their anterior antennæ an arrangement which enables them to throw a long, whip-like structure like a lasso round the head of the female as she rapidly swims away. The antennæ of the male Daphnids, too, are in one genus (Moina) developed into a grasping apparatus; ... the first antennæ ... are not only much longer and stronger than those of the female, but they are also armed with claws at the end, so that the males can catch their mates as with a fork, and hold them fast. And even that was not enough, for, in addition, the males of most Daphnids possess a sickle-shaped but blunt claw on the first pair of legs, which enables them to cling to the smooth shell of the female, and to clamber up on to it to get into the proper position for copulation.
Plate 40.
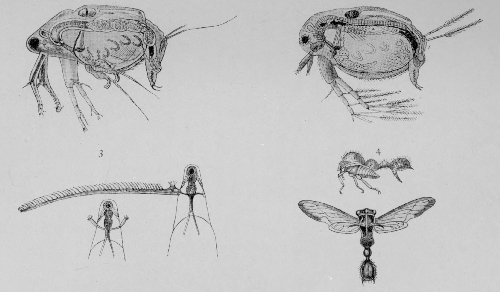
SOME REMARKABLE DEVICES.
1. A Water-flea (Moina rectirostris): male showing the claspers-the front pair of “legs,” for grasping the female.
2. The female of the same, in which the “claspers” appear as mere stumps.
3. The aberrant Water-flea (Leptodora kindlii): the male showing the long comb-like antenna for the discovery of the female (the left only is drawn), and the female, just beneath, lacking this olfactory organ.
4. An extraordinary species of Bug in which the upper surface of the thorax has been produced backwards to form an overhanging pent-house, of unknown function, and illustrating the theory of “Hypertely.”
[Face page 262.
“If we inquire into the manner of the origin of secondary sexual characters of this kind, we shall find that both may have been increased by sexual selection, for a male with a better sickle will succeed more quickly in getting into the proper position for copulation than one with a less perfect mechanism. This assumption does not rest on mere theory, for I was once able ... to observe for a considerable time, under the microscope, a female to whose shell two males were clinging, each trying to push 263the other off. Nevertheless, it seems to me very questionable whether the origin of this sickle-claw can be referred to sexual selection, for without this clamping-organ copulation in most Daphnids would not be possible. It was thus not as an advantage which one male had over another that the clamping-sickle evolved, but rather as a necessary acquisition of the whole family, which must have developed in all the species at the same time as the other peculiarities, and notably those of the shell. The competition of the males among themselves is thus in this case simply an expression of the struggle for existence on the part of the species as such, and it is not a question merely of a character which makes it easier for the males to gain possession of the females, but of one which had necessarily to arise lest the species should become extinct. In other words, in this case Natural Selection and Sexual Selection coincide.
“The case of the antennæ of Moina, which have been modified into grasping organs is quite different; these owe their origin, not to natural selection, but to sexual selection, for antennæ of that kind are not indispensable to the existence of the species, as we can see from the closely related genera, Daphnia and Simocephalus, where the males have quite short, stump-like antennæ, furnished with olfactory filaments not much more numerous than the females possess. Just as these supernumerary olfactory filaments were produced by sexual selection and not by the ordinary natural selection, because those males with the more acute sense of smell had an advantage over those in which it was blunted, so the males of the genus Moina which could grasp most securely had an advantage over those that gripped less firmly, and thus arose these two different kinds of male characteristics. 264Neither of them is of advantage to the species as such, but only to the males in their competition for the possession of the females.”
Much uncertainty would seem to exist in regard to two very extraordinary marine species of Copepoda. In one, Calocalanus pavo the male possesses enormous antennæ, and a remarkable development of iridescent feather-like structures at the end of the body, arranged in a sort of open fan-work; the female has what may be called “normal” antennæ, and a brush-like tuft at the end of the body. In the other species—Calocalanus plumulosus—of which the female only is known, there is a similar arrangement of plume-like structures at the end of the body, but all but one are extremely small; the single plume differs from the rest in being of enormous length. Commonly these structures are regarded as mechanisms to reduce the expenditure of energy necessary to keep at the surface of the water, for these creatures inhabit the surface-waters of the open ocean. Many larval Crustacea inhabiting similar areas are in like manner kept afloat, or at any rate aided in keeping afloat, by the excessive development of spines. But if this be the purpose of these strange excrescences of Calocalanus it seems curious that the female of C. pavo should not be similarly provided. If they are to be regarded as secondary sexual characters it is curious that the females of C. pavo and C. plumulosus should be so utterly dissimilar. The male of C. plumulosus is unknown. On the whole, it seems more reasonable to regard these strange structures as mechanical aids to swimming rather than as secondary sexual characters.
The Courtship of the Cuttle-fish—The Sumptuous Cradle of the Argonaut—The Love-darts of the Snail—Hermaphrodites and the Dangers of Self-fertilization—Oysters and Beauty—Sex reduced to its Lowest Terms—Parthenogenesis and Virgin Birth—The Story of the Hive-bee—The Departure of the Queen—The New Queen and her Marriage-flight—The Celebration of the Nuptials and its Surprising Sequel—The Widowed Queen turns Executioner—The Queen as Mother—The Queen’s Daughters—Nursemaids’ Duties—Change of Work—The Drones and their Career—Food and Sex—The Bumble-bee and its Life-story.
That the psychical emotions sway the goad of sexual instincts in the higher animals there can be no doubt; and there can be as little uncertainty that this stimulating and controlling factor gradually loses force as we descend in the scale of animal life. Just where it ceases it is impossible to say. A vague, nebulous intelligence doubtless persists after these more subtle emotions have ceased, and this, probably, in turn, gives place to purely instinctive behaviour. These various phases of the sexual problem grade one into the other. But they are all parts of a continuous sequence, beginning, apparently, in relatively simple responses to chemical interactions of the kind known as chemotaxis and ending with the passion 266which, in the human race, may become a consuming fire, purifying and ennobling, or exactly the reverse—according to the nature of the inflammable material. That is to say, in the phenomena of sex one sees emotions in the making. The begetting of children becomes the underlying goal of life, the hidden heart and soul of animated nature.
This being so, one cannot but feel surprised at the discovery that, in certain groups of the animal kingdom one meets with a strange exception to this great rule. And this is furnished by the phenomenon of parthenogenesis, wherein sexual desire has been dethroned. Offspring result from Virgin births: parental care is non-existent. This anomalous condition must be regarded as an offshoot of the normal course of events traced in these pages, and not as a primitive condition. This interpretation seems to be shown clearly enough in that almost every case where parthenogenesis obtains, males, sooner or later, make their appearance—periodically or sporadically. Every stage between the normal, seasonal appearance of males and their entire suppression can be traced, and an analysis of these cases demonstrates unequivocally the uplifting character of the bi-sexual state, if only by the fact that the uni-sexual condition makes no demands on the parent, and does nothing to foster the growth of the higher emotions.
No attempt need now be made to discover the origin of parthenogenesis. Let it be assumed, for the moment, that it is a condition derived from hermaphroditism, wherein each individual is monœcious or bi-sexual. In all diœcious or uni-sexual animals, that is to say, where the individuals composing the species are either male or 267female, each contains a leaven of the opposite sex, even when adult. It is still a moot point whether, in the earlier stages of development, chance decides whether the sex shall be male or female, or, at any rate, whether the growing body is potentially male or female, till the die is cast by some as yet undiscovered factor; or whether this is determined from the very beginning of germinal life. In many of the lower animals, as among the Mollusca and some of the insects, each individual is as much male as female, and it is from a condition such as this that parthenogenesis probably had its rise.
These two groups are selected here because they, more than any others in like case, afford some extremely interesting gradations in this strange phenomena of what is to be regarded as the degeneration of sexual individuality, for each contains some members wherein the sexes are separate, and in these cases sexual desire is present in varying degrees. In some it is associated with very remarkable phenomena.
Among the Mollusca the Octopuses afford one of the most striking illustrations of such phenomena. In these creatures one of the sucker-bearing arms is more or less completely transformed to subserve the ends of sexual congress. Without entering into the technical details of the changes, it will suffice to remark that it is modified in such a way as to allow the transference of the spermatozoa from the body cavity wherein they are formed, to the arm near, or at, the tip of which they are stored in a special sac or “spermatophore,” and such modified arms are said to be “hectocotylized.” This extraordinary modification attains its maximum development in the celebrated Argonaut, and one or two of the more typical Octopuses. In the Argonaut this arm does not make its 268appearance until sexual maturity has been attained, when a large more or less globular swelling appears, enclosing the third arm of the left side, coiled upon itself. Having attained its full development the sac bursts and releases the arm. The folds which formed the sac now bend back to form a new receptacle into which the spermatophore is passed. But this is not all. The tip of the newly released arm bears another sac, which sooner or later bursts, forming a long, slender penis, and along the central tube of this the spermatozoa pass from the spermatophore to their destination. Their conveyance thereto forms the last and most amazing feature of this strange history. The male, eager with pent-up desire, and glowing with all the colours of the rainbow, gradually approaches the female of his choice, who apparently awaits him with no little palpitation, and then, with a sudden rush flings himself upon her, and apparently thrusts the penis into her mantle cavity, when at once the whole arm breaks off from his body and remains attached to her person, retaining its vitality, strange as it may seem, for some considerable time, during which, no doubt, the spermatozoa are slowly making their way out of the spermatophore and along the channel prepared for their reception. That the Cuttle-fish are polyandrous there seems to be little room for doubt, inasmuch as no less than four such detached arms have been found beneath the mantle of one female. With the majority of the Cuttle-fish and Octopus tribe the arm is not detached, but when it is so, and this occurs in all the species belonging to three different genera, a new arm is grown.
Plate 41.
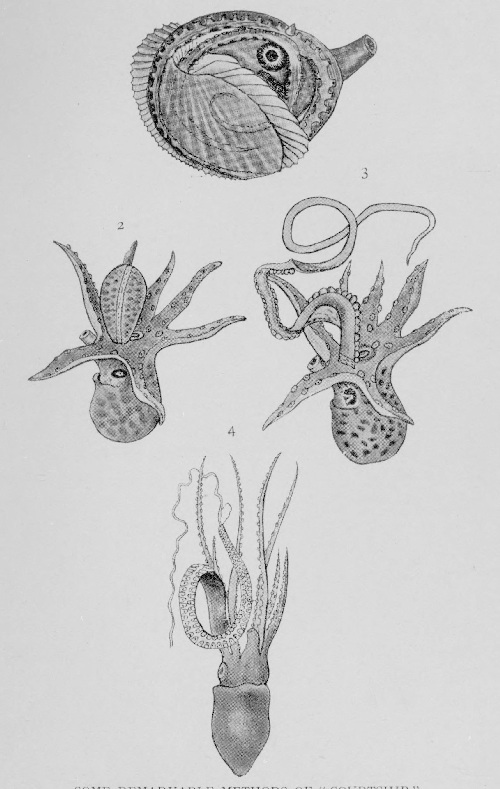
SOME REMARKABLE METHODS OF “COURTSHIP.”
1. The female Argonaut and her egg-casket.
2 and 3. The male Argonaut and his “hectocotylized” arm.
4. A Cuttle-fish (Ocyhöe catenulata ♂), showing the “hectocotylized” arm described in the text, and the “spermatophore” at the base of the long filament.
[Face page 268.
As a rule, among these animals the males are smaller than the females. In the case of the Argonaut there is a yet more striking difference, for the female possesses a very beautiful shell in which she carries her eggs. This 269remarkable cradle, translucent and beautifully sculptured, she attaches to her person by means of a pair of arms which are expanded to form great lobes, almost but not quite completely covering the shell. The earlier naturalists believed that this shell served as a boat, and that the lobated arms were spread as sails! This supposed fact naturally caught the fancy of the poets, who seized upon it to point a moral and adorn a tale. Byron celebrated these imaginary feats of seamanship in the familiar lines:
and Pope bids us:
Sir Richard Owen years ago, however, dispelled these pretty fancies, though the facts are surely as wonderful as the fables they have replaced. They afford, too, one of the most striking secondary sexual characters to be met with among the Mollusca; nowhere else, indeed, among the members of this group is so strange a cradle to be met with.
But little, unfortunately, is known of the behaviour of these animals, which are by far the most active of the Mollusca, and which also display no small degree of intelligence. Their eyes, which are of great size and complex structure, are undoubtedly far more effective organs of vision than are possessed by any other Molluscs. It is possible, therefore, that the sexes discover one another by sight; and it is certain that something in the nature270 of a “Courtship” takes place. The majority of the species, also, possess the most extraordinary powers of changing their coloration, especially during moments of great excitement. The magnificence of the hues which succeed one another, like a series of variegated blushes suffusing the whole body, may be one of the weapons in the armoury of Cuttle-fish love-making. In how far the “courtship” of the Cuttle-fish resembles that of terrestrial animals, however, is a matter on which at present nothing is really known. That even the comparatively sedentary species, like the Octopus, seize upon and hold territory is very improbable, for there is no need of such landed estates, inasmuch as the offspring are not tended and fed by the parents—this would indeed be a laborious task in the case of some of the “Squids” which lay between thirty thousand and forty thousand eggs! Having regard to the fact that the records of the reproductive habits of the Octopus tribe date back to the time of Aristotle, more than two thousand two hundred years ago—for he first drew attention to the hectocotylized arm—it is curious that so little has been gleaned during this vast space of time.
There are facts in regard to the sexual relationships of some of the Snails that are in nowise less remarkable than those just related of the Octopus tribe. Unlike the Octopuses, the Snails are hermaphrodite, nevertheless sexual congress takes place as with unisexual species: the eggs of the one being fertilized by the spermatozoa of the other. During this process the orgasm of the sexual act appears to be brought about by stabbing one another by means of a little dart formed of carbonate of lime, the dart burying itself in the flesh and apparently promoting a pleasurable, tingling sensation in the course271 of its journey. Speedily, no doubt, it becomes absorbed, the material being then available for the formation of a new dart.
This remarkable instrument, which is known as a “Love-dart,” or Spiculum amoris, assumes a different form in each species in which it occurs. In some the shaft is ridged like a bayonet, as in the case of the Garden Snail, in others the form assumed is that of an awl. These darts are formed within a special receptacle, or “dart-sac,” but so far no explanation as to the origin of these remarkable structures has even been hinted at. They do not seem to have been derived by the modification of some pre-existing organ serving a different function, as wings, for example, are derived from walking limbs, or as lungs are derived from air-sacs. Their origin is as mysterious as their use: for they are not found in all Snails, though they occur in one or two Slugs—which are degenerate Snails. But no other Molluscs save the Snails and one or two of their immediate allies are so armed.
The hermaphrodite conditions of these animals, as with other Mollusca in like case, present some knotty points for consideration, and especially in regard to the problem of sex-attraction. Where each individual is as much male as female, which is the dominating factor in desire, the maleness or the femaleness? Though each individual contains both ova and sperm cells, probably these ripen at different times, to avoid danger of self-fertilization. In this case the sex impulses are on the same footing as in the case of animals wherein the sexes are not thus combined. That is to say, the individual which is for the moment only potentially male mates with another for the moment only potentially272 female. But this being so, how does each discover the condition of the other?
Many of the Snails, like Helix nemoralis, are gaily coloured. Are these hues, these bands of black and yellow, the product of “sexual selection”—the outcome of a process of selection from among the most conspicuously coloured individuals as postulated by the Darwinian theory of Sexual Selection? If so, then this choice must be regarded as a periodic recurrence coinciding with the period during which the individual is dominated by its female attributes. In due course it becomes, for the time, a male, and may find itself rejected, owing to a lack of intensity in its coloration, or, on the other hand, it may vanquish a rival by its very splendour. Each, in short, would help materially in this process of beautification. If the choice of mating for it is this rather than a choice of mates—proceeds on these lines, the bright coloration of the members of this species becomes easy to understand. But does it? It is more than doubtful whether the eyes of Snails are sufficiently good to distinguish the coloration of their neighbours’ shells, or for the matter of that of their own, for their eyes being carried on long mobile stalks, they should have no difficulty in contemplating their own charms. And what of Snails of more sober hues? It seems highly probable that here, as in so many cases, scent is the selecting factor, and the coloration is an “accidental” feature. That the colour of the shell plays no such part as that just postulated may be gathered from the evidence afforded by many marine species, whose shells, though conspicuously marked, are, during life, completely enveloped and concealed by the all-investing, fleshy mantle. In like manner the273 exquisite beauty in the form and sculpturing of the shell which so many species exhibit, are characters which cannot be regarded as due to sexual selection.
As touching the danger of self-fertilization to which reference has been made. That this is real is shown by the fact that the ova and spermatozoa are rarely ripe in one individual at the same time. However, among the pulmonata, or air-breathing gastropods, it seems to have been established that self-fertilization can, and does, occur. That in some species, at any rate, where cross-fertilization, for some reason, is impossible, the individual thus isolated can store up its own spermatozoa to be used in fertilizing its own eggs. But the fact that this rarely happens is testimony enough that such occurrences are inimical to well-being.
The Lamellibranch, or bivalve Mollusca, e.g., Oyster, Mussel, and Cockle, afford valuable evidence as to excrescences and extravagances of growth which appeal to our eyes as ornamental, and therefore likely to be due to the influence of sexual selection. And this because such ornamentation is a very conspicuous feature among these animals. Yet, save in a few cases, locomotion is impossible, and sight is wanting. Light-distinguishing organs, and therefore eyes, are possessed by some, but in no case probably are they strong enough to appreciate form. Even if they did, such revelations of beauty would play no part in mate selection from among the most ornamental; for these creatures are commonly fixed throughout life in one position, often, indeed, buried in mud or sand. Some move laboriously: a few, like the Cockles and Pectens, swim by rapidly opening and closing the shell. The Pectens are brilliantly coloured, not only as regards the shell, which is also beautifully sculptured, 274but the foot also is of a vivid scarlet, and the Pecten have numerous minute eyes. But the Cockles and Mussels possess like attributes as to colour and sculpture, yet they are blind. More to the point is the fact that these animals do not mate after the fashion of higher animals, but the males, where the sexes are distinct, discharge immense quantities of spermatozoa into the water, and these find their way to the ova of the female through the action of the inhalent currents set up by the animal for the purpose of drawing in fresh supplies of water containing food and oxygen. There are no “secondary sexual characters,” that is to say, that even where the sexes are separate, and many, like the Oysters, are hermaphrodite, they are externally indistinguishable. Nevertheless, many, as has been already remarked, have shells of great beauty. As, for example, the giant Tridacna and the strangely spinous valves of the “Thorny Oysters” (Spondylidæ).
The fact that the Lamellibranch, or bivalve molluscs, are far less numerous in point of species than the univalve tribes is accounted for by the fact that in the first place they are of necessity aquatic, and in the second their means of locomotion is extremely limited. Some few species swim spasmodically: some crawl: many are incapable of movement when once the motile larva settles down and the shell-bearing adult stage is attained. Such species can extend their range only by means of larval wanderings. Enormous numbers, millions, of young have to be produced and set adrift each year by every adult in the community, and yet but a few of each brood can ever attain to maturity. Life, for such species, must be a dull, monotonous business: the only opportunity for excitement is that which is preliminary to being eaten275, and the only purpose in life is to be eaten. But happily Oysters don’t think. They and their kind are simply semi-conscious living things, responding mechanically to stimuli. Any approach, then, to beauty, either of form or coloration, or both, must be regarded as due to innate, inherent changes in the germ-plasm affecting the parts so made conspicuous: the only form of selection to which such “ornaments” can be subjected is Natural Selection. If, and when, such ornaments penalize their possessor either by their cumbrousness or their conspicuous characters, or by increasing the difficulty of feeding or distributing offspring, then the further development of such excrescences is checked by the death of all individuals which have passed the bounds of endurance in this respect.
Sex, and all that appertains thereto, in short, is in these creatures reduced to its lowest terms. There are not wanting, to-day, both men and women, who affect to believe that all would be well for the human race could a similar slowing-down, or strangulation, of the sexual instincts be brought about. Such blind leaders might profitably contemplate the Oyster: but such contemplation, to be profitable, requires intelligence of a higher order than these protagonists of folly appear to possess.
In justice to Darwin it should be remarked that he himself fully realized, and carefully points out, the inconceivability of the application of the Sexual Selection theory to the Mollusca. In commenting on the beauty of colour and shape which many species display, he remarks: “The colours do not appear in most cases, to be of any use as a protection; they are probably the direct result, as in the lowest classes, of the nature of the 276tissues1: the patterns and the sculpture of the shell depending on the manner of growth.” Just so: and this is surely the fundamental explanation of ornament, using this term in its widest sense, everywhere in the Animal Kingdom. The peculiarities and eccentricities of behaviour, which occur among the higher groups, act as “aphrodisiacs” to hasten reproduction because this confers an advantage, the earliest to produce offspring—so soon as the conditions for their nurture are favourable—having the best chance of survival. Premature sexual activity is checked by the death of the offspring.
1 Italics mine.
It has been contended that the hermaphrodite condition represents the primitive mode of reproduction among the multicellular animals—that is to say, all animals above the level of those whose bodies are composed of but a single cell, or particle, of protoplasm—but this view is probably erroneous, and the hermaphrodite state must be regarded as a secondary condition, a later innovation.
More remarkable are the facts concerned with that singular form of reproduction known as parthenogenesis, or the production of offspring by virgin females. This is undoubtedly a degenerate sexual condition occurring as a normal mode of reproduction, among the microscopic “Rotifers,” e.g. the “Wheel-animalcule,” Crustacea, and Insects, and in varying degrees of intensity.
The most familiar instances of Parthenogenesis are furnished by the Hymenoptera, and notably by the Bees and the Aphides.
There are certain cases among the Rotifers where no males have ever been found, and it is possible that they 277have become entirely suppressed, but in every other case the periodical advent of males is an absolute essential for the continuation of the race. Perhaps the least degenerate of these types are the Bees, wherein we meet with well-developed, highly-organized males and females, which, in their sexual relationships, are perfectly normal. But in the fulfilment of the mating instincts in these insects, a most amazing sequence of events is revealed such as are without parallel in the rest of the Animal Kingdom. The story has been charmingly told already by Maeterlinck, in his delightful “Life of the Bee,” and it has been told again by Tickner Edwardes, with less of poetry, perhaps, but still fascinatingly: and it must be told again now, but in a condensed fashion.
Briefly, a community of hive-bees harbours both male and female individuals only for a very short space. During the greater part of the year it consists only of a vast concourse of infertile females, the daughters of one mother; the “queen” of the hive. The males of that hive are the brothers, not the fathers, of the workers, as some have supposed, and their sojourn there is brief. To gain a clear idea of the facts in regard to the life-history of these insects it is necessary to trace some of the incidents which lead up to the manner in which the population of the hive is regulated, and its continuance ensured. These may well begin with the time when the number of the inhabitants consonant with the well-being of the hive has reached its limit. This occurs during the early part of June, when the queen leaves the hive, accompanied by several thousands of her daughters; they settle at some distance from their late abode in a “swarm” for the purpose of founding a new colony. Here we may leave them. The house just vacated is, however, not entirely deserted.278 A few of the inhabitants, the infertile sexless workers, degenerate females—degenerate so far as the power of reproduction is concerned at any rate—are left behind, and there remain also in their cradles a variable number of unhatched queens, and drones or males. One of these potential queens and the males now speedily emerge, and for a day or two remain within the seclusion of the hive, feeding upon the honey stored in the combs.
The males are the first to leave, making daily excursions abroad in the search for mates. They display in this a very leisurely behaviour, rising late and not venturing out till the day is well aired. Returning early in the afternoon with sharpened appetites, they feed to repletion and soon fall asleep.
In about three days, however, the young queen ventures abroad, timidly at first, to stretch her wings in the sunshine. She is preparing for the great moment of her life, the nuptial flight. So far, though drones may swarm on every side of her, no sign of recognition is given, nor do the males evince any consciousness of her presence. She behaves warily and demurely throughout. Her first excursions abroad are very brief; they are not so much trial flights, apparently, as efforts to locate the exact position of the hive in relation to the outer world. To this end the flights are rapidly extended in ever-widening circles, till at last, with lightning speed, she makes for the blue sky, to return to the gloom of the hive almost immediately after. During all this time the stimulus of sexual desire has been gathering force, and now, being no longer controllable, she darts off, and up into the sky; almost at once she is recognized by the swarms of males from neighbouring hives, some thousands in number, which for days have been279 seeking this event. Instantly they give eager chase, mounting after her higher and ever higher. But as they ascend so their numbers decrease. Some, the feeble, the ill-fed from impoverished hives, are speedily left behind; many endure to the end, but only one secures the prize, and this great moment of his life is also his last, for the fact of impregnation is no sooner completed than Death claims him. He falls earthwards, as if struck by lightning, and in his fall the intromittent organ is dragged from his body, to be removed by the survivor of this mad flight, on her descent.
She leaves a bride and returns a widow, filled with murderous intentions. There are captive queens in the hive, and she can tolerate no rivals. So soon as she has removed from her person the embarrassing souvenir of her nuptial flight she makes for the Royal cells. Accompanied by attendant workers she proceeds to tear off their waxen coverings and put their occupants to death with a thrust of her stiletto. No sooner is the work of execution over than the dead bodies are seized by the workers and borne out of the hive. This awful task is soon over, however, and henceforth for four or five long years she remains a prisoner within the walls of her own palace. Craving neither the air nor the light of the sun, she will die without once having sipped the nectar from a flower. And during all this time, save during the winter sleep, her sole duty is to produce sons and daughters. In the prime of her maternity she may lay as many as three thousand eggs a day. But strangely enough the number of eggs produced is determined for her by the workers, who are the real rulers in this constitutional state. By varying the amount and quality of the food they give her they can increase or check the280 number of eggs produced; while even the sex of the resultant larva is apparently also under their control.
During that brief, weird honeymoon in the clouds she received a store of spermatozoa, the fertilizing male germs, sufficient for all the eggs she can ever lay, and they may amount to nigh on a million. Incredible as this may seem, their purpose is yet more so; for they are destined to be expended solely in the production of female offspring doomed for the most part to perpetual spinsterhood. One youngster in ten thousand may attain to a higher state, may, if Fate wills, become a queen and mother. And because of this need for mothers to carry on the race, this extraordinary state of affairs has been brought about. All is under the control of her daughters—the spinster-workers. As she proceeds on her rounds of egg-laying an attendant crowd waits upon her, controlling her actions by gentle caresses. As she passes from cell to cell, the cradles of the young that are to be, she thrusts down her abdomen and lays an egg in each. The cells destined to produce the workers are the smallest, those for drones are larger, and those for queens are largest of all, and the walls are formed of pure pollen, not of wax as are those of the workers and drones. But it would seem that she never lays an egg in any of the last named. The sight of a queen-cell rouses her to fury. These cells, then, are filled by the workers, who remove the requisite number of worker—eggs from the cells in which they were laid and deposit them in the queen-cradles. The larvæ at hatching, and for the first three days of life, differ in no wise from their sisters around them. Their Royal state is determined solely by the food which is administered to them. This consists of “bee-jelly,” which is furnished in abundance: a white, shining liquid, regurgitated by the ever-zealous281 nurse-bees. These superfed babies cease feeding at about the fifth day, and each spins for herself a silken vestment in which to undergo the pupal state. This done, the door of each cell is sealed up with pollen. During the following sixteen days strange transformations take place: the queen that is to be is taking shape. But the cradle now becomes a prison, for at the end of the sixteenth day each of the four or five young queens begins to clamour for release. But this cannot be, for such as succeeded in emerging would immediately be slain by the reigning queen. A small hole is bored through the roof of the cell, and through this each is fed, and a close guard is kept night and day to ensure that they shall not emerge till the moment is ripe. Soon each captive begins to gnaw away the roof of her prison chamber, and as rapidly more material is placed by her guards on the outer surface. Not until the old queen leaves the hive with thousands of her daughters to “swarm” and found a new colony will freedom be allowed; and then only to one. The rest must remain till the new queen either also “swarms,” or returns from her nuptial flight, and in this case all will be slaughtered in their cramped quarters, unable to resist.
But what of the drone? He, as has already been mentioned, is reared in a larger cradle than that of his sisters—save such as are destined to be queens—and for the first three days of his life is fed on “bee-milk” of a special kind and more generous quality than that of his worker—sisters, the Cinderellas of the hive; but this generous diet is diminished at the end of three days, when a mixture of honey and pollen is given him. In about three weeks or rather more he emerges, a great, lazy drone, and for a fortnight more he wanders about the hive alternately soliciting bee-milk from his sisters282 and helping himself to honey from the comb, and when full to repletion he seeks some snug corner in which to sleep off his surfeit. In due time, however, he ventures abroad, his hour is at hand. He takes his daily flights abroad in search of a mate, returning home early in the afternoon for his rations, being too indolent or too stupid to draw nectar from the flowers for himself. Thus for many days he and his brothers disport themselves in riotous living, till one or other of them attains the end for which he was born; and after a few delirious moments drops earthwards a mutilated corpse.
But so far only a part of the story of the drone’s life-history has been told. Though the son of a queen, he has never had a father; and should he ever attain to the dignity of fatherhood his posthumous children are all daughters, most of whom die spinsters within six or seven weeks of their birth, worn out by a life of ceaseless toil and drudgery!
The queen, it will be remembered, cohabits with the male but once in her life. The sperm-cells then received are stored in a special receptacle and are released during the passage of the egg down the oviduct. In this act of releasing the fertilizing germs a singular economy is practised. In the case of most other creatures myriads of sperm-cells are released for the fertilization of a single egg, and of these but one can possibly attain its goal, the minute aperture or “micropyle” which is the doorway to the germ liberated, in the form of an egg, by the female. The rest die. In the case of the queen bee but one of these precious sperm-cells is liberated at a time. Hence her prolonged ability to produce fertilized eggs. But eggs destined to produce males, or drones, are never thus fertilized: they are born without the283 intervention of a father. A queen which has never mated will lay only male-producing eggs. This is an astounding thing, but it is true. No less remarkable is the fact that the sperm-cells should survive in their encapsuled state for periods extending over several years: it seems almost incredible, but it is nevertheless true.
One cannot suppose that the queen in coming to a drone cell deliberately withholds the male germ as the egg passes down her oviduct; some inhibitory factor preventing the release of the sperm-cell must be brought into play which as yet we have not discovered. This production of males from unfertilized eggs, or “parthenogenesis” as it is called, is a common feature among the hymenoptera, and some other groups of insects, and it occurs also among other lowly creatures to be described later.
Having regard to the importance of the workers, a brief summary of their life-history must be given. These, it has already been indicated, are all, at any rate till three days old, potential queens. Their development into, or degradation to, the lower grade is determined, apparently, solely by the quality of the food, for the fact that queens are reared only in specially constructed cells of large size with walls of pollen instead of wax is explained by the larger size of the queen and the need for a more porous, air-permeated cell-wall on account of the longer time which must be spent in confinement. The worker is certainly the most “intellectual” member of the hive, but this superiority has been gained at a great price. Emerging from the chrysalis skin at about three weeks from the time that the egg from which she emerged was laid, she begins forthwith to gnaw her way through the mass of wax and pollen which forms the door of her284 prison. Rather, she eats her way through, for the material removed is swallowed as it is detached, thus the young bee, as Mr. Tickner Edwardes remarks, is caused to effect her own release by the promptings of her appetite. Hunger-strikes in the bee community are unknown. Speedily the youngster steps out, distinguishable from her elder sisters only by her weak, grey-hued, flaccid appearance. Her first act on gaining freedom is to groom herself down, after which she proceeds to explore the gloomy, busy, crowded thoroughfares of the hive. A day or two is thus passed in gathering strength. On the second appetite returns, and she proceeds to help herself from the vats of honey and pollen bins scattered here and there among the cradles of her sisters yet prisoners. But speedily she is caught and thrust, so to speak, on to the treadmill of work which is to know no cessation during her short span of life-some six or seven weeks. Her first duties are those of nursemaid. Without instruction, or previous experience, she begins to feed her younger sisters and brothers yet in the larval stage. But besides, during her first fortnight, before she is allowed to leave the hive she and her sisters of the same age have to fulfil a variety of tasks. All the indoor work of the house falls on these Cinderellas. Not only do they, and they alone, feed the young, but they have to produce the wax and build the combs and attend to the sanitary arrangements: “they are the brewers of the honey and the keepers of the stores; they feed the queen bee on her ceaseless rounds and give the drones, their brothers, their daily rations of bee-milk”—what else these lazy creatures need they take for themselves from the honey-vats. But this is not all. They have to meet their older sisters returning from the fields and gardens laden with nectar. This is regurgitated285 and transferred to the pouches of the youngsters, by whom it is transformed into honey and stored in the combs in the upper region of the hive. At the end of about a fortnight these little drudges are allowed a brief respite, during the heat of the day, to emerge into the outer air and gather ideas on the world which is yet to be explored. Soon a measure of freedom is allowed, the indoor work ceases, and each takes up the new and more agreeable task of gathering pollen, and after a few days of this the more responsible task of gathering nectar is undertaken, which is continued till death ends one of the most crowded, surely, of existences. Such as are born near “swarming-time” may have the good fortune to take part in the exodus and the settling down in the new home, and some may taste yet other moments of excitement, but they are moments only. The worker bee knows no leisure for the improvement of her mind and morals. She needs none, for she has neither: she is a creature of routine, a living automaton apparently. Yet there are incidents in this wonderful community which seem too complex to be merely the result of instinct unaided, uninspired, by intelligence albeit of a nebulous kind.
The worker-bees, it has been remarked, are barren: their reproductive organs are atrophied, and by the decree, not of the queen-mother of the hive, nor of the males, but of their own caste. In spite of the fact that they are incapable of producing offspring, they, and they alone, determine who shall undertake this task; and they decree the fate that awaits those thus appointed when they can no longer fulfil this purpose.
When the queen, waxing old, and waning in fecundity, lays fewer and fewer eggs, and these only producing males, they take silent note of the fact, and at the286 appointed time decree the death of their Sovereign-mother. Yet they hesitate to lay violent hands on her. She, as queen, claimed the right in her early youth to slay her sister-queens, and sped them with a dagger-thrust; now her turn comes to die. But it must be a bloodless death, carried out with due ceremonial. So her daughters cluster about her, and in a mock embrace, that tightens every moment, her breath is squeezed out of her body. There are no State pensions for those who are past work, but a State execution instead. This is vastly more economical, and it may yet commend itself to some would-be social “reformers,” who will doubtless contrive to make exceptions to the rule!
The execution of a queen is not an event of common occurrence; but that of male members of the hive forms part of the ordinary routine, though coming only within the larger cycle of the year. As the summer wanes and the harvest of nectar grows perceptibly less, visions of a possible famine, and its attendant horrors, seem to arise. So heads are counted and occupations are scrutinized, when it is discovered that the only members of the community who are contributing nothing to the general well-being are the males, who are now but useless drains on the hive. None of the neighbouring hives are now likely to send forth a virgin queen to her nuptials, to which end each hive is obliged to contribute—for no hive utilizes the services of its own drones; these idle fellows, then, are “eating their heads off”—and males, too; perish the thought! While they had anything to gain from him their motto was “Feed the brute”; but now, on each, doom is pronounced. It must be admitted that a live drone at the end of summer is one of life’s failures. Notoriously unable to feed himself save upon287 the honey made by his sisters, and having no function in life to perform save that of mating, his very existence now is a damning witness against himself.
When the mother of the hive ceases to maintain the standard of fertility set by her exacting daughters, she is put to death stealthily, as if in an excess of devotion: she is smothered under their embraces. Towards the drones now under sentence no such consideration is to be shown. When the word goes forth, the slaughter begins, and it gathers in ferocity. It begins in a massacre of the innocents—every helpless larval drone is ruthlessly dragged from its cot and thrown out of the hive to die: there is now no crime in infanticide, nor in the most gruesome massacre that is presently to follow. The drones, all unsuspecting, are to be tolerated a brief spell longer. The cool, calculating spirit of these unsexed ones seems to realize that there is even yet a remote possibility that the services of these doomed ones may be wanted. No sooner, however, does it become clear that this chance is past, than the decree of death is made absolute, and the poor drones are suddenly and viciously attacked by half a dozen frenzied spinsters at once. Each tries to bite through the base of the victim’s wings, and succeeding in this, he is speedily pushed towards the door of the hive and out into the open, whence return is impossible, so that nothing is left but death by starvation. Some of the victims will escape in the mêlée, but only for a brief season. Such as find their way, unmaimed, to the open air, are still faced by inevitable death. To remain out is to die of starvation or cold, to return is to fall a prey to the now infuriated guards, who, strongly reinforced, stand at the doorway of the hive to intercept and dispatch these unlucky288 fugitives. It will be remarked that these executioners make no use of their stings; these they might be unable to withdraw from their victim’s body, in which case they, too, would die. But there is no need to run this risk, for the males, their brothers, whom they so cheerfully slay, are unarmed; they may be attacked without risk. The dreadful work, however, is soon over, and the survivors, the queen and her daughters, have the house to themselves to make the final preparations for the winter sleep, which is apparently undisturbed by qualms of conscience.
There are certain structural differences distinguishing the three types in such a hive—the queen, the drone and the worker—which must now be referred to. The queen is larger than the worker; she has a larger and longer abdomen, a longer and much-curved sting, and her eyes have fewer facets. Only vestiges remain of the wax-secreting organs, and no trace is to be found of the wonderful pollen-baskets which perform so important a function in the worker; and finally, her instincts are of a very different kind.
The “pollen-basket” of the worker is a strange contrivance. The pollen is mainly collected by the hairs which clothe the under surface of the body, from which it is scraped by special brushes of hairs which clothe the inner surface of the “metatarsus “—the big, flat joint to which are attached a series of small triangular joints, the last of which bears the claws. When the brushes are “clogged up,” the legs are crossed and the pollen is combed out by specially stiff hairs on the “tibia”—the joint immediately above the metatarsus—and the bolus thus formed is then transferred to the outer surface of289 the tibia, which is trough-shaped, forming the “corbiculum,” or pollen-basket. The next, or middle, pair of legs are then employed to ram the pollen well into the basket, for safe conveyance to the hive. On arrival at the combs, the bee pushes its hind-legs into a cell, or “pollen-tub,” and with a special spur dislodges the pellet of pollen and lets it fall into the tub. These are complex movements, performed without instruction and, we must suppose, without any intelligent conception of their purpose.
The drone is larger than either queen or worker, and has enormous eyes, which meet one another over the top of the head; he has no wax-secreting organs, no pollen-basket, no sting. His antennæ are longer, his hum is deeper, his sole function is to fertilize a queen, and this done, he promptly dies. Failing in his first flight, he may make yet other ventures, but the chances are that he will die without attaining the only purpose for which he exists.
The fact that he lives for some days in the hive with the queen, before her nuptial flight, apparently unaware of her presence, would seem to indicate some special “trigger” for the release of the sexual instincts. But it must be remembered that he does not attain to maturity until after his first flight, and this it is, probably, which arouses the mate-hunger. More than this, however, it is probable that coitus is possible only when on the wing, when the air-sacs become inflated, and exert pressure on the genital organs. How he recognizes the queen when on her wild flight heavenwards is unknown: possibly by scent, but more probably by the very different vibrative note of her wings, that of the male being much stronger and deeper. His continued return to the hive 290is a proof of his failure to justify his existence, for no drone ever experienced Love’s embrace and lived to tell the tale: hence, when the time comes, he is slain without compunction.
These differences between the fully-developed male and female present nothing very striking; but how are the singular peculiarities of structure and instinct in the “workers” to be accounted for? They are present in neither queen nor drone, yet by them they are transmitted to their offspring from one generation to another! It is true that every worker, for a time, is a potential queen, and every queen, but for the grace of Chance, might have been a worker. All depends on the food. It is remarkable, but apparently the fact, that a more generous diet, or, rather, a more stimulating diet, should so profoundly modify the organism, but, it is to be noted, this sleight-of-hand is only successfully practised on a larva during its first three days of existence. Thus the royal bee jelly stimulates the growth of the sexual organs and inhibits the development of the structures peculiar to the worker—the basket, and pollen-hairs, and so on. These structures are not made by the food; they are simply nourished or inhibited, as the case may be. Nevertheless, one cannot help being mystified by the fact that the mere difference in the quality of the food, or, rather, in the chemical constituents thereof, should cause the inhibition, or, rather, the suppression, of relatively complex structures like the corbiculum and the reduction of the number of the facets of the eye. To say that the structures inhibited, in the case of the queen, are just those which will be of no service when in her royal state, is by no means to explain the mystery. And what is true of the physical side is291 no less true of the psychical, for with this change of diet the behaviour of the insect, throughout its whole life, is most profoundly changed. If the pollen-basket is wanting, no less so are the instinctive actions associated with its use; if the genital organs are atrophied, so also are the instinctive acts associated therewith. This nexus between instinct and structure is not to be lost sight of.
How—and the question has often been asked—are the experiences of the infertile females, the workers, transmitted to the germ-plasm? For the workers, it has been contended, being sterile, are incapable of handing on such acquirements: this is so. These workers hold the same position in regard to the species that structures essential to well-being hold in regard to the individual. These last are not under the control of the individual, but are determined by a plus or minus quality in its germ-plasm. The worker-bees are products of the germ-plasm, committed to the care of the queens. Any strain, so to speak, of that germ-plasm which gives rise to defective workers brings about its own extinction, or elimination, sooner or later. Any strain of germ-plasm which contains, so to speak, a spark of that quality which in the individual is expressed by intelligent behaviour, will gain advantages in the struggle for existence.
The complex, the extraordinarily complex, behaviour of the worker-bees on any interpretation is still mysterious. This interpretation can be tested only by a reference to the life-history of other social-bees which have attained to a less complexity. This shows us that the sterile worker is not to be regarded as a newly-evolved type so much as an arrested stage of a more complete ancestral condition, and the fact that the worker is potentially a queen is further evidence of this.
A clue to many of the more puzzling features presented by the domestic economy of the Hive-bee may be obtained by a study of the life-history of other species of social-bees which have not attained to so high a degree of specialization. The Bumble-bees afford illustrations of the stages through which Apis mellifica, the Hive-bee, must have passed.
In the stone Bumble-bee (Bombus lapidarius), a queen, who has passed the winter in blissful sleep, will lay the foundation for a new colony on some bright May morning by collecting a small quantity of moss. This done, she starts forth to gather pollen, with which, under cover of the moss, she forms a waxen cell, mixing the newly-gathered pollen with the wax so mysteriously formed within her body, as in the case of Hive-bees of the worker type. Slowly and laboriously this waxen cradle grows. Fashioned like a globe, its inner surface is lined with pollen soaked in honey, and with the last pellet of this a number of eggs are laid arid the nursery is sealed up. By the time these labours are completed the queen is worn out; she therefore rests awhile, clinging to the outer wall of this cunningly-wrought cradle. After a few days’ rest she adds another and commonly yet a third cell to the first, joining each to the other with wax. But before the third cradle is finished the eggs in the first have hatched. The youngsters will have consumed the layer of honey-soaked pollen placed there for this purpose. They therefore require feeding, and thus the labours of this very industrious queen are still further increased. Divining the needs of her imprisoned first-born, she bites a small hole through the nursery wall and pours in a quantity of honey for their sustenance. In due time they are “full-fed,” and each spins for itself a silken293 vestment wherein to undergo its transformation into a worker-bee. The careful mother, during this period of transition, now scrapes away an opening through which the young bees may creep when they awake. This event takes place in the course of a few days, when her work is materially lightened, for these newly-hatched workers at once take over the duties of building nurseries and feeding the further batches of young which, for a time, follow one another in quick succession. The queen, indeed, has now nothing else to do but to lay eggs in the nurseries as they are ready. So far all the children born to her are daughters. The earliest-born, it is to be noted, were “workers”; those which follow and are tended by the workers are also females, and supplement their mother’s labours by producing fertile eggs, though they have never even seen the male of their own species. Thus, if the queen-mother die her virgin daughters carry on the colony. But it sometimes happens that she may have left no descendants capable, for the time, of laying fertile eggs. In this case, if there be larvæ still in the nursery, the workers feed them assiduously as if in the hope that some may prove fertile. But if there be no infants to be fed they apparently abandon work, become despondent, and spend the greater part of their time sitting at home by the empty cradles, till at last death comes to their rescue and the colony is extinct.
Much that baffles one in the history of the Hive-bee becomes clear in the light of the facts revealed by the life-story of the Bumble-bee. In the first place it will be remembered her first eggs produced only workers, which appeared at a time when her energies were severely strained, and their food allowance was no more than barely sufficient to sustain life. The females which294 appeared later produced fertile eggs, having been more abundantly fed by their infertile elder sisters. The number of fertile females which appear at this stage of the colony seems again to be regulated by the abundance of food, which varies in amount with fine, or cold, weather. Even among the worker broods fertile females may appear. They owe their fertility apparently to good luck, which afforded them the opportunity of securing more food than their sisters. The birth of young from females about whose virginity there can be no question is certainly remarkable, but it would seem that this parthenogenetic state is one of limited endurance, for towards the end of summer males appear, and these mating with some of the later-born females, lead again to the appearance of a queen, who, being fertilized, alone survives the winter to carry on the race with the succeeding summer.
Thus, then, the mysterious existence of the workers among the Hive-bees, displaying structural peculiarities and instincts so different from those of the queen-mother, is explained. For the queen, in this case, is evidently the product of a more intensified, more perfected, social system, relieved, from the first, of the labours of building and the care of her offspring, duties which the queen Bumble-bee has at first to perform for herself, because all her children die at the end of the summer. Among Hive-bees fertile workers also occasionally occur; they are probably bees which in their larval state received a more than usually abundant supply of food, or food approximating to the “bee jelly” which produces young queens. The difference, then, between the individuals of a colony of Hive-bees and one of Bumble-bees lies in the greater abundance of fertile workers and in the295 fact that the queen of the Hive-bees is relieved of all work from the first, and so is enabled to devote her whole energies to the duties of reproduction. She is the descendant of a race of queens which in earlier times, like the Bumble-bee queen, had to perform the duties now relegated to her daughters, who inherit not only her house-building and child-nurturing instincts, but also her potentiality for child-bearing, though this potentiality is commonly inhibited by the starvation of the reproductive activities. Selection secures survival of this state of affairs by the elimination of any tendency to lose any of these qualities on the part of the queen. The workers of the Hive-bee, in short, have not evolved their peculiarities of structure and instinct by some mysterious process of natural selection confined to the workers individually, for these, being infertile, could not transmit any of their inherent qualities or tendencies to variation in the direction of more efficient workers. On the contrary, all that they possess they inherit from the queen-mother, who transmits to her offspring the qualities and characteristics her forebears in the female line possessed in their own person.
Courtship among the Ants—The Great Renunciation—Maternity carried to Extremes—Where Males are Superfluous—Degenerate Males—Keeping Death at Bay—Where Females are Unknown.
The phenomenon of virgin birth is one of profound mystery. The existence of so astonishing a mode of reproduction was an established belief among the ancients, though they could have had no means of demonstrating the faith that was in them. But these men saw no difficulty in ascribing to the females of their own race this faculty of producing offspring without the intervention of a male. One suspects, indeed, that there was no solid foundation whatever for this belief in these miraculous powers: they lived in credulous times, and the recorded occurrences of these, even to them, irregular births are to be regarded as devised to afford a convenient means of escape from the consequences of lapses from the path of virtue. Yet, incredible as it may appear, there are not wanting to-day both men and women who affect to believe that this mode of reproduction obtains still among the human race, in certain exceptional cases; and further, they profess a conviction 297that in the future it may become the normal mode, males, in consequence, becoming unnecessary! Such professions of faith are made only by the ignorant, or by those who trade on human credulity. Parthenogenesis not only does not occur in the human race, but it does not even occur in any member of the great group of vertebrates of which man himself stands at the head, and it never will occur.
Those near relations of the Bees, the Ants, afford a further insight into this strange method of reproduction. Each community in the case of these insects harbours not one, but many queens. The nuptial flight, like that of the Bees, takes place in mid-air; but myriads of both sexes participate therein, forming a filmy, ever-shifting cloud, now rising, now falling, in the shimmering sunlight. At no time do they seek to attain the altitude, or the privacy, so strenuously striven for by the Bees. But in the case of the latter there is but one female, and her life is precious. She must seek sanctuary for the consummation of her marriage in the highest heavens, beyond the risk of instant destruction by insect-eating birds; for though thousands of suitors accompany her, she rises above them all, save one or two, and hence would form an easy mark. With the Ants there are thousands of queens, and the destruction of a few hundreds more or less is rather an advantage to the species than otherwise. On their return to earth the males die: their life’s work is accomplished. The females, or as we must call them, the queens, on the other hand, have a long life before them; far longer than that of the queen bee. But for them the joys of flight are restricted to this one brief revel, for, no sooner have they reached298 terra firma, than they renounce, as it were, the pleasures of life to devote themselves entirely to the work of reproduction. And as if to make all regrets vain, to stamp out all possible temptation to desert their vows, they tear off their gauzy wings, and with them goes all hope of fertile repentance: for the rest of this life their home is underground.
Each queen, on her descent, departs a separate way, and hard is the road before her. She left the parental nest well-fed, and in good liking, her body well, stored with food in the shape of fat and the now useless, bulky, wing-muscles, and with this, her only dowry, she starts the formation of a new colony out of her own substance. Her first task is to form a burrow, and at the end of this she fashions a small chamber. This done, she closes the mouth of the burrow and cuts herself off from the world. The labour of this burrowing is so severe that it often wears away her teeth, her only tools, and the hairs from her body. In this retreat she now waits patiently for the eggs within her to ripen, which may take months to accomplish: she is still fasting, or, rather, feeding upon herself. When at last the eggs are laid and hatched, she feeds her children on saliva, the very juice of her body, for she is still fasting. Nor is the strain relaxed till the larvæ undergo their transformation into pupæ, and, after a brief sleep, emerge as “worker” Ants, puny in stature owing to the poorness of their food during larval life. In some species this fast may last for ten long months. So soon, however, as these little workers emerge, like dutiful daughters they make their way to the outer world, and go forth in search of food, which they share with their now exhausted mother. But, besides, they enlarge the original chamber, and drive galleries in all299 directions to provide accommodation for the vast population that is soon to crowd the thoroughfares. Meanwhile the queen resumes her task of producing more and yet more daughters, in whom she now displays not the slightest interest. Her elder children now bear away the eggs, and feed the young as they hatch. In course of time, as with the Bees, the task of wet-nurse falls on the youngest of the Ants, those who have just attained to anthood. For ten or fifteen years this queen-mother may continue her work of reproduction, a slave, indeed, to domesticity, with monotonous regularity, checked only by the chill of autumn and the sleep of winter.
Those among our own race who profess to hail the prospect of a time when parthenogenesis shall be the normal mode of reproduction may well take the Ant as an awful warning. Their ambitions may overreach the mark. The poor queen becomes a slave to reproduction; children in myriads are born to her; even if she would she could not sustain her interest in them, she could not even recognize them as the fruit of her body. Her daughters are born to a lifelong drudgery, her sons are mere fertilizing agents: for their only purpose in life is to perpetuate this awful thraldom, this appalling prolificness; and having accomplished this, they die forthwith. If there be any joy in this life it is drunk by the males alone. Thus does the female rule overreach itself. It is well, indeed, that the participants of the joyous nuptial flights dancing deliriously on gauzy wings in the glare of a summer day, have no foreknowledge of the long night that is to follow.
Unlike the Bees, the Ants may produce as many as five grades of workers, each of which have different duties towards the community. But the nature of those300 duties and the manner of the evolution of these types, are themes foreign to these pages: enough has been said already to indicate the nature of the problems they present when discussing the life-history of the Bees.
The subject of parthenogenesis need be pursued no further in this volume than is sufficient to bring out its retrograde character. It is a form of reproduction which may be limited to a small number of generations, as with the Aphides, or to a single generation alternating with normal sexual generations, as in many Cynipidæ or Gall-flies, or it may be the only mode of reproduction, as in some other Gall-flies, some Saw-flies and some Crustacea, wherein no males have ever been seen. In some species this form of reproduction gives rise to females only—the Thelyotokous parthenogenesis of scientific text-books—as in the Saw-flies and Gall-flies, and the parasitic Tomognatbous. In some other Saw-flies, unfertilized queens and workers of Ants, Bees, and Wasps, which occasionally produce offspring, the progeny is always male, and this is known as Arrhenotokous parthenogenesis. In one or two species of Saw-fly, e.g. Nematus curtispina, both males and females may be produced, when the species is said to be Deuterotokous.
In the case of the Aphides, winged males normally appear in large numbers at the end of the summer, and these fertilize the females; but if kept in a warm green-house, parthenogenetic reproduction may be sustained for as long as four years. Under quite normal circumstances these tiny insects show a singular range of variability, for egg-laying and viviparous individuals are met with; while winged and wingless generations appear301 sporadically, apparently according to the abundance of food. The winged form is sometimes so abundant as to float about in swarms that darken the air. There are at least three kinds of males-winged males, wingless males with a functional mouth, and small wingless males which have no mouth, and, one need hardly say, are very short-lived. The Aphides are a feeble folk, individually, but collectively a power in the land, causing at times incalculable loss to the farmer and gardener; but on this head and on the subject of their strange habits, and sometimes adventurous lives as slaves in the service of Ants, no more than a hint may be dropped in these pages. But some such aids to faith seem to be necessary when those who are not tolerably familiar with these insects are told of their amazing fertility. Linnæus long since estimated, in regard to one species, that in the course of one year a single Aphis will give rise to a quintillion of descendants—all produced without the aid of a male. Every one of these females begins to reproduce within from ten to twenty days of her birth, but even this statement does not bring home the result of such an astounding fecundity like Huxley’s calculation which was carefully worked out. He estimated that the produce of a single female would, in the course of ten generations, supposing all the individuals to survive—and possess the normal fertility of their race—“contain more ponderable substance than five hundred millions of stout men: that is, more than the whole population of China.”
To explain such a riot of reproduction one might almost suppose these insects to be imbued with a dread of the impending dissolution of their race, and endowed with the power to avert such a calamity by these302 stupendous efforts; for it is evident that parthenogenesis confers quite extraordinary powers of raising the birth-rate. But then the normal mode of procreation is capable of achieving results quite as remarkable. The queen Termite or White Ant, for instance—which, by the way, is no Ant, but a near relation of the Stone-flies—when in her prime will lay eggs at the rate of sixty a minute, or eighty thousand and upwards in the course of a day of twenty-four hours. But this unenviable mode of breaking the record is attended, surely, with some little inconvenience; for to attain to such fertility her abdomen increases until it attains something like two thousand times that of the workers of the community in which she lives. That the history of the queen Termite is unique of its kind is not surprising: indeed, such an amazing story could only be told of creatures which enjoyed the seclusion of a subterranean existence. Here, on a bare couch, with her Royal spouse beside her, she lies, a bloated, heaving mass, incapable of movement, depositing eggs with the rhythm of a machine, the mother of offspring which she will never see. A more unsightly picture of maternity it would be impossible to conceive: it is well, indeed, that it is hidden from the light of day. No such state of affairs could ever arise among creatures living an outdoor life, with enemies to avoid, and food to find.
The instances just surveyed, these extremes of the potentiality of procreation, are instructive in more ways than one. They are to be regarded as “excrescences” of reproduction, comparable to those “excrescences” of individual growth which we call “ornament,” for example. Individuals on whom this fertility has settled, so to speak, are the victims of the machinery of sex and reproduction. Their amazing powers of multiplication303 are not of their own seeking, they are inherent manifestations of variations of growth, uncontrollable save by the machinery of Natural Selection. Incidentally such victims serve a useful purpose, for their myriad hosts afford food for hordes of other animals, which in turn are eaten. Little though we realize it, the well-being of the human race would suffer if these prolific creatures—the uncomplaining victims of that inexorable law which bids all living things “increase and multiply” or die—should cease to be; for with them would disappear a host of animals on whose existence man’s comfort more or less depends.
During the millions of years that have rolled by since the first appearance of life on the earth, who shall count the number of types which have been exterminated without leaving the faintest trace of their having ever existed? The survivors which have contrived to maintain a place in the sun present an infinite range of variation in colour, size, habit, and structure, as well as in emotions. These varied aspects are all so many facets of the mysterious phenomenon we call Life: and they are so many witnesses of the versatility of Life. Not the least mysterious feature of this Life is its faculty of reproduction, which expresses itself in an infinite variety of ways, defying all but the crudest forms of analysis. The evolution of sex has exercised the speculative ingenuity of some of the acutest students of Nature from the earliest times, and we are still far from a satisfactory solution of the problems it presents. Hermaphroditism and Parthenogenesis are commonly regarded as degenerate forms of reproduction, but it would probably be more correct to see in them exceptional modes of adaptation enabling such individuals to occupy304 niches in the world untenable to creatures of more conservative habit. That the peculiar “strains” of animal life have turned into backwaters which offer no opportunity or possibility of further advancement seems clear enough, but they are nevertheless interesting and instructive.
The parthenogenetic Crustacea and the Rotifers afford some good evidence of this adaptability—of the way in which creatures manage to cling to the skirts of life by reason of their power to survive the extremest tests of endurance. And this success has largely been due to some mysterious property of the germ-plasm enabling reproduction to take place through the female line alone, or in some cases with an occasional fillip from the intervention of males. Of the many marvellous things that could be related of these creatures but few instances can be cited here.
The case of the Brine Shrimp (Artemia salina) will afford an exceptionally good illustration because the facts can be tested by anyone who will take the trouble to make a simple experiment for himself. Those anxious to do this should dissolve eight ounces of Tidman’s sea-salt in a glass jar containing five pints of water, keeping the mixture well stirred till the salt is dissolved. It should be allowed to stand and be carefully watched. In about three days, with a pocket-lens, or even without, minute white specks will be seen moving with a jerky motion up and down the water. These are larval Brine Shrimps. Now they must be fed. Take a piece of lettuce-leaf or any green stuff, and pound it up, or grind it up with a knife-blade on a plate with a little water, till the whole is reduced to the consistency of green paint; then empty this into the water. This must be305 done daily, or at any rate frequently. Quickly these tiny specks will grow into Brine Shrimps, translucent creatures nearly half an inch long, swimming about back downwards with a marvellously rhythmical movement of delicate feet. In all probability no males will be found, but, on the other hand, both sexes in almost equal numbers may be present. The males may readily be distinguished by their massive arms immediately behind the head, for the purpose of embracing the females.
Whence came these wonderful animals? The mystery is easily explained. The salt is genuine sea-salt, formed in brine-pans, chiefly in the Mediterranean. As the water evaporated the Shrimps it contained gradually died; but the eggs in the females became encapsuled in the salt-crystals to hatch out long months after. In one of my own experiments I succeeded with salt that I had kept for more than a year. Of course, every sample of salt experimented with will not yield successful results, but failures are not expensive. Now in this brine-pan there were myriads of other animals which were killed outright: the Brine Shrimp is at least able to pass on descendants by reason of the vitality of its eggs. Some near relations of the Brine Shrimps live in fresh water and possess similar powers of resistance to adverse conditions. The Fairy Shrimp (Chirocephalus) is one of these. Not unlike its cousin the Brine Shrimp in appearance, it lives in shallow pools, such as have muddy bottoms and are constantly liable to dry up. Birds hunting by the margins of the pool where the retreating water has left a fringe of mud bear away more or less of this on their feet and transport it to similar pools, or even puddles. Such transplanted samples may easily contain numbers of eggs of this tiny creature. Only a year or 306two ago Fairy Shrimps were found in abundance in rain pools at Eton, and some, indeed, were discovered swimming gaily about in a rain-filled cart-rut!
Another very singular Crustacean, known as Apus, bears a curious superficial likeness to the King Crab (Limulus), having a large back-shield and a long tail. This little creature, a giant compared with his nearest relations, is an inhabitant of wayside ponds and ditches. Thousands of females may be taken for years in succession without the advent of a single male. Then, for some strange reason which we cannot even guess at, males appear. Like its freshwater cousin, the Fairy Shrimp, Apus can withstand drought: its favourite haunts may be transformed into sun-baked hollows, but with a heavy fall of rain and a few hours’ soaking the eggs left by dead females develop, and once more the pool and its inhabitants are established again. Having regard to the extraordinary vitality of these small creatures, it is curious that they should ever disappear from their favoured haunts. But they do. Not many years ago Apus could be found in abundance in many parts of the South of England. It is now extinct; its last resorts were the ponds at Hampstead: now one may search in vain for them. “No British specimens,” remarks Dr. Caiman, a great authority on the Crustacea, “had been recorded for over forty years, and the species was believed to be extinct in this country, when it was found in 1907 by Mr. F. Balfour Browne in a brackish marsh near Southwick, in Kirkcudbrightshire.” These had probably developed from eggs accidentally transported by some bird from the Continent. The extinction of the race throughout the British Islands can only be attributed to the too long absence of males, and the consequent307 inability to restore vigour by the more normal method of reproduction by sexual congress.
Among the Rotifers the little Wheel-animalcules exhibit an even greater vitality, for not only can their eggs withstand prolonged desiccation, but in some the body of the animal survives even harsher treatment. If specimens be enclosed within a chamber containing a little sand or moss the contents may be dried over sulphuric acid, or heated up to 200° F., or left to the neglected dust of years, and will yet revive if a little fresh water be added to the sand. Males are rare, and when they do occur are little more than animated receptacles for semen, for they are incapable of feeding, the gullet and digestive tract being reduced to a solid cord. A certain amount of nourishment, however, may be absorbed through the delicate body wall.
The degeneration of the males in these parthenogenetic species irresistibly reminds one of the smile of the Cheshire cat; they grow smaller and smaller, and their functions less and less, till finally nothing is left. The “complemental males” discovered years ago by Darwin in the Barnacles well illustrate this process. In dissecting adult specimens of the stalked Barnacle (Scalpellum) he found, just inside the valves, in a pocket of the mantle, a varying number of “complemental males,” tiny organisms which Mr. Geoffrey Smith describes as “little more than bags of spermatozoa,” and they apparently serve to fertilize the ripe ova of the larger animal—one cannot say of the female, for Scalpellum, like most of the Barnacles, is hermaphrodite. But it is believed that these complemental males are really arrested hermaphrodites. At any rate, if it so be noted that with some of the Barnacles, as with some other Crustacea, the larvæ are males, but 308when adult life is attained female glands appear and hermaphroditism is established. Such hermaphrodites have the singular distinction of being males which have acquired female attributes, true females being unknown among them!
In one of the parasitic Crustacea (Chondracanthus) infesting the gills of Gurnard, Plaice, Skate and other fish, the adult female is about half an inch long, and very unlike a Crustacean in appearance; the male is an extremely minute maggot-like object—a few millimetres in length—and lives permanently attached to the belly of his mate just at the base of the egg masses. More remarkable still is the case of another nearly related parasitic species—Lernea—which becomes sexually mature in its childhood. The males perform their part and die; their mates arrive at maturity and settle down to a comfortable life as parasites on fish, reproducing without further mating.
That Parthenogenesis and Hermaphroditism are but specialized forms of reproduction, leading sooner or later to degeneration and extinction, there can be no doubt. They are, so to speak, failures in the evolution of sex, demonstrating in a very forcible fashion the impossibility of progress—as we understand it—where the sexual functions are thus combined.
To the differentiation of sex, resulting in separate male and female individuals, we must attribute the marvellous complexity of the pageant of life which confronts us to-day. The story of the Courtship of Animals is only one of an infinite number of incidents in this pageant, and one which is by no means easy of interpretation.
In these pages an attempt has been made to show that this differentiation of sex has, throughout, been309 accompanied by, and largely moulded by, common instincts and behaviour, and this interpretation is only to be reached by a study of the phenomena in their simplest form among the lower grades of animal life. Colour and the various sexual differences in form have been allowed to dominate this investigation of the problem of sex, and have diverted attention from more profitable and fruitful channels.
The lower we descend in the scale of animal life the less convincing becomes the argument that the colour, ornament or armature of the males is the result of sexual selection in the older, Darwinian sense. The argument of Geddes and Thomson and others that the males are more “katabolic,” the females more “anabolic,” seems no less unsatisfactory, for in many cases the female is just as highly ornamented as the male, and in others she is considerably large. Further, in their less specialized species the sexes are almost or quite indistinguishable externally, and are sombrely clad, just as at the opposite extreme we find them equally ornamented and equally active.
We shall be nearer the truth if we regard these secondary sexual characters as expression points of germinal variations. Though we seem hopelessly ignorant as to the inciting cause of the variations, at least we seem to be able to lay a finger on the agents by which they are effected. And these are the hormones of the primary and secondary sexual glands, whose functions affect more than the merely sexual side of the organism. They profoundly affect the coloration of animals, giving rise on the one hand to purely ornamental “secondary sexual characters,” and on the other to changes of coloration which achieve the ends of protective resemblance310 colours, or of “warning coloration,” as circumstances may demand. There is nothing more remarkable in this than the control which the pituitary body exercises over stature, either when in a pathological condition, or when the controlling action of the other gland secretions is removed, as by castration.
Hitherto much has been made of trophic nerves, which control growth; but it is probable we have overlooked the still more important action of “trophic” glands, such as the thyroid. This apparently controls growth in many directions. Adaptations to environment which are effected by changes in bodily shape-as in the transformation of land-dwelling mammals into Seals and Whales—are probably largely controlled by these glands. Their activity is as great as their manifestation is varied.
Why their action should be more stimulating in the case of the male, why he should lead the way in all the new acquirements of the species, both in non-sexual as well as in sexual characters, is by no means plain. But the fact remains that this is so. Remove any one of these glands and the machinery of growth is thrown out of gear; it is not merely the secondary sexual characters which are affected.
But these glands are concerned no less intimately with the behaviour of animals. This is most obvious in all that concerns sexual appetite as the preceding chapters have already shown. Having regard to the immense variety of animals concerned, this behaviour presents an underlying uniformity of expression which must not be lost sight of: and the same is no less true of what we may call the physical manifestations of these glandular activities.
THE END
Printed at The Chapel River Press, Kingston, Surrey.
For the first line of page 16, instead of “by certain glands of the ductless glands,” read: “by certain of the ductless glands.”