Anmerkungen zur Transkription
Der vorliegende Text wurde anhand der 1923 erschienenen
Buchausgabe so weit wie möglich originalgetreu wiedergegeben.
Typographische Fehler wurden stillschweigend korrigiert. Ungewöhnliche
und heute nicht mehr gebräuchliche Schreibweisen sowie Schreibvarianten
bleiben gegenüber dem Original unverändert, sofern der Sinn des Texts
dadurch nicht beeinträchtigt wird.
Alle relativen Größenangaben in den Bildunterschriften
beziehen sich ausschließlich auf die Abbildungen der gedruckten
Originalausgabe. Die Liste der Druckfehler
wurde bereits in den Text eingearbeitet.
Bei den systematischen Namen wurde die Schreibweise nach
dem heute üblichen Standard vereinheitlicht, bei dem das Art-Epitheton
zum Gattungsnamen stets klein geschrieben wird, es sei denn, dieses
wurde von einem Personennamen abgeleitet; also z.B. ‚Gnetum gnemon‘
statt ‚Gnetum Gnemon‘. Weiterhin wurden heute möglicherweise nicht mehr
übliche Schreibweisen der Pflanzennamen sowie inkonsistente Varianten
beibehalten, sofern diese im Text mehrmals auftreten.
Abhängig von der im jeweiligen Lesegerät installierten
Schriftart können die im Original gesperrt
gedruckten Passagen gesperrt, in serifenloser Schrift, oder aber sowohl
serifenlos als auch gesperrt erscheinen.
LEHRBUCH
DER
BOTANIK
FÜR HOCHSCHULEN
BEGRÜNDET 1894
VON
EDUARD STRASBURGER, FRITZ NOLL
HEINRICH SCHENCK, A. F. WILHELM SCHIMPER
SECHZEHNTE, UMGEARBEITETE AUFLAGE
BEARBEITET
VON
DR.
LUDWIG JOST
O. PROFESSOR AN DER UNIVERSITÄT BONN
DR.
HANS FITTING
O. PROFESSOR AN DER UNIVERSITÄT HEIDELBERG
DR.
HEINRICH SCHENCK
O. PROFESSOR AN DER TECHNISCHEN HOCHSCHULE DARMSTADT
DR.
GEORGE KARSTEN
O. PROFESSOR AN DER UNIVERSITÄT HALLE-WITTENBERG
MIT 844 ZUM TEIL FARBIGEN ABBILDUNGEN IM TEXT
JENA
VERLAG VON GUSTAV FISCHER
1923
Alle Rechte vorbehalten
Copyright 1911 by Gustav Fischer, Publisher, Jena
Druck von Ant. Kämpfe in Jena
D
Die Verfasser dieses Lehrbuches wirken seit Jahren als Dozenten
der Botanik an der Universität Bonn zusammen. Sie haben dauernd
in wissenschaftlichem Gedankenaustausch gestanden und sich in
ihrer Lehraufgabe vielfach unterstützt. Sie versuchen es jetzt
gemeinschaftlich, ihre im Lehren gesammelten Erfahrungen in diesem
Buche niederzulegen. Den Stoff haben sie so untereinander verteilt,
daß EDUARD STRASBURGER die Einleitung und die Morphologie,
FRITZ NOLL die Physiologie,
HEINRICH SCHENCK die
Kryptogamen, A. F. W. SCHIMPER die Phanerogamen übernahm.
Trägt auch jeder Verfasser die wissenschaftliche Verantwortung nur
für den von ihm bearbeiteten Teil, so war doch das einheitliche
Zusammenwirken aller durch anhaltende Verständigung gewahrt. Es darf
daher das Buch, ungeachtet es mehrere Verfasser zählt, Anspruch auf
eine einheitliche Leistung erheben.
Dieses Lehrbuch ist für die Studierenden der Hochschulen bestimmt
und soll vor allem wissenschaftliches Interesse bei ihnen erwecken,
wissenschaftliche Kenntnis und Erkenntnis fördern. Zugleich nimmt es
aber auch Rücksicht auf die praktischen Anforderungen des Studiums
und sucht den Bedürfnissen des Mediziners und Pharmazeuten gerecht zu
werden. So wird der Mediziner aus den farbigen Bildern die Kenntnis
derjenigen Giftpflanzen erlangen können, die für ihn in Betracht
kommen, der Pharmazeut die nötigen Hinweise auf offizinelle Pflanzen
und Drogen in dem Buche finden.
Nicht genug ist das Entgegenkommen des Herrn Verlegers zu rühmen,
der die Kosten der farbigen Darstellungen im Texte nicht scheute,
und der überhaupt alles aufgeboten hat, um dem Buche eine vollendete
Ausstattung zu geben.
Bonn, im Juli 1894.
Die Verfasser.
[S. iv]
Aus dem
Vorwort zur 5. Auflage.
E
Einen schweren Verlust hat unsere gesamte botanische Wissenschaft, und
im besonderen auch unser Lehrbuch, durch den inzwischen erfolgten Tod
von A. F. W. SCHIMPER erfahren. In der Blüte der Jahre wurde
er uns entrissen, ein Opfer seines rastlosen Forschungstriebes. Mit ihm
erlosch auch so mancher fruchtbare Gedanke, ohne in wissenschaftliche
Tat umgesetzt zu werden. In seinem Nachlaß fanden sich nur lose Blätter
als Vorbereitung für die neue Auflage unseres Lehrbuchs vor. Die drei
anderen Mitarbeiter hielten es für ihre Pflicht, diese Blätter zu
sichten, zu ergänzen, einzuordnen und die begonnene Arbeit, im Sinne
des Verstorbenen, zum Abschluß zu bringen, damit sein Name auch noch
auf dieser, nach seinem Tode erscheinenden Auflage unseres Lehrbuches
stehe.
Bonn, im Dezember 1901.
Die Verfasser.
Aus dem
Vorwort zur 6. Auflage.
A
Aus der gleichmäßig fortschreitenden Aufeinanderfolge neuer Auflagen
unseres Lehrbuchs schöpfen wir dauernd neue Anregung, es auf der Höhe
seiner Aufgabe zu halten. Die sechste Auflage hat ganz besonders viele
Verbesserungen und Änderungen aufzuweisen. Morphologie, Physiologie und
Kryptogamen wurden wiederum einer eingehenden Durchsicht unterworfen,
die Phanerogamen aber, durch G. KARSTEN, vollständig
umgearbeitet.
Der Herr Verleger scheute seinerseits kein Opfer, um zu erreichen, daß
das Buch in seiner technischen Ausstattung den ersten Rang behaupte. Er
selbst äußerte den Wunsch, wir möchten noch vorhandene, fremden Werken
entnommene Habitusbilder möglichst durch Originale ersetzen. Da die
Kunst des Farbendrucks seit dem Erscheinen der 1. Auflage dieses Werkes
wesentliche Fortschritte gemacht hatte, so veranlaßte der Herr Verleger
uns auch, neue Vorlagen für die farbigen Abbildungen zu schaffen. Diese
sind von Herrn Dr. ANHEISSER meist nach der Natur entworfen
worden.
Bonn, im Oktober 1903.
Die Verfasser.
[S. v]
Aus dem
Vorwort zur 10. Auflage.
M
Mit dieser 10. Auflage vollendet unser Lehrbuch sein fünfzehntes Jahr.
Da es sich alle anderthalb Jahre durchschnittlich in einer neuen
Auflage verjüngte, so hoffen wir, daß es keine Zeichen hohen Alters an
sich trägt. Von dieser 10. Auflage können wir sogar behaupten, daß sie
in besonders neuem Gewande vor die Öffentlichkeit tritt.
Einen schweren Verlust hat unser Lehrbuch durch den Tod eines seiner
Mitarbeiter, FRITZ NOLL, erfahren. Seine Physiologie trug
nicht wenig zu den ersten Erfolgen des Buches bei. Ein ehrenvolles
Andenken ist ihm in unserer Wissenschaft gesichert. An seine Stelle
trat LUDWIG JOST in den Verband unseres Lehrbuchs ein.
Möge auch diese neue Auflage des Lehrbuchs Nutzen stiften und geneigte
Aufnahme finden.
Im Oktober 1909.
Die Verfasser.
Aus dem
Vorwort zur 12. Auflage.
S
Seit die letzte Auflage unseres Lehrbuches hinausging, hat es durch den
Tod EDUARD STRASBURGER’S den schwersten Verlust erlitten, der
es bisher betroffen. Er hatte an dem zunehmenden Erfolg des Buches, an
dem er 17 Jahre mitarbeitete, seine stete Freude und war unausgesetzt
bemüht, seinen Teil, die Morphologie, an Form und Inhalt höchsten
Ansprüchen genügend zu gestalten.
Nach seinem Ausscheiden trat HANS FITTING als Mitarbeiter ein;
ihm fiel die Aufgabe zu, den ersten Teil neu zu bearbeiten. Da zugleich
einige Änderungen in der ganzen Stoffverteilung geboten erschienen,
ist diese Auflage auch in den anderen Teilen mehr oder minder stark
umgestaltet worden. Wir waren bestrebt, die Einheitlichkeit des Buches
nach Möglichkeit zu wahren und zu erhöhen.
Zum ersten Male sind alle Figuren — bis auf die von R.
ANHEISSER gezeichneten Habitusbilder und die von den Mitarbeitern
für ihre eigenen Teile angefertigten Bilder — mit den Namen ihrer
Autoren versehen worden, so daß der Anteil der bisherigen Verfasser an
der Illustrierung des Buches zum Ausdruck gelangt.
Im April 1913.
Die Verfasser.
[S. vi]
Aus dem
Vorwort zur 14. Auflage.
M
Mit dieser neuen Auflage vollendet unser Lehrbuch das fünfundzwanzigste
Jahr seines Bestehens! Es hat im verflossenen Vierteljahrhundert
zahlreiche Freunde an deutschen und ausländischen Hochschulen
gewonnen; wir hoffen, daß ihm diese Freundschaft auch in Zukunft
nach Wiedereintritt des Friedens und mit Wiederaufnahme gemeinsamer
Arbeit zur Förderung der Wissenschaft erhalten bleibt. Auf den
erzielten Erfolg kann der Herr Verleger, der in dankenswerter Weise
die vortreffliche Ausstattung des Buches stets im Auge behielt, mit
besonderer Genugtuung zurückblicken. Uns Verfassern aber soll dieser
Erfolg erneuten Ansporn geben, fortgesetzt den Ausbau des Buches nach
besten Kräften zu fördern und so stets auf dem neuesten Stand der
Wissenschaft zu halten.
Obwohl wir den hie und da geäußerten Wunsch, auch die
Pflanzengeographie zu behandeln, gerne erfüllen möchten, müssen wir
in Rücksicht auf den stattlichen Umfang des Buches davon absehen, ihm
einen neuen größeren Abschnitt anzufügen, da eine Kürzung des Inhaltes,
wie sie bei den „Samenpflanzen“ dieses Mal versucht ward, nicht den
nötigen Raum dafür gewinnen läßt. Für unzweckmäßig aber müssen wir
es halten, die wichtige Pflanzengeographie nur auf wenigen Seiten zu
erledigen. Es sei daher zunächst noch auf die bereits vorhandenen
pflanzengeographischen Werke verwiesen.
Im Juni 1919.
Die Verfasser.
Vorwort zur 16.
Auflage.
D
Der im Januar 1921 erschienenen 15. Auflage müssen wir — wiederum nach
2 Jahren — eine neue folgen lassen, in welcher die neueste wichtigere
Literatur, soweit sie uns erreichbar war, gebührend berücksichtigt
wurde.
Wesentliche Änderungen erfuhr die systematische Anordnung der
Samenpflanzen auf Grund der Ergebnisse der serodiagnostischen
Untersuchungsmethode, die nach Ansicht des Verfassers dieses
Abschnittes nicht außer acht gelassen werden durfte.
Dem Herrn Verleger sind wir für die gute Ausstattung des Buches auch in
seiner neuesten Gestalt besonders dankbar.
Im April 1923.
Die Verfasser.
|
|
Seite
|
|
Einleitung
|
|
|
|
|
|
|
Morphologie
|
|
|
Erster Abschnitt. Zellenlehre
(Zytologie)
|
|
|
I.
|
Gestalt und Größe der Zellen
|
|
|
II.
|
Der lebende Inhalt der Zellen (der Protoplast)
|
|
|
|
A.
|
Bestandteile
|
|
|
|
B.
|
Physikalische Eigenschaften
|
|
|
|
C.
|
Chemische Eigenschaften
|
|
|
|
D.
|
Bau der Teile
|
|
|
|
E.
|
Ursprung der Elemente
|
|
|
III.
|
Gröbere leblose Einschlüsse der Protoplasten
|
|
|
IV.
|
Die Zellmembranen
|
|
|
Zweiter Abschnitt. Gewebelehre
(Histologie)
|
|
|
I.
|
Gewebebildung
|
|
|
II.
|
Zellarten. Gewebearten und Gewebesysteme
|
|
|
|
A.
|
Die Bildungsgewebe
|
|
|
|
B.
|
Die Dauergewebe
|
|
|
Dritter Abschnitt. Organlehre
(Organographie)
|
|
|
I.
|
Vegetationsorgane
|
|
|
|
A.
|
Der Thallus
|
|
|
|
B.
|
Der Kormus
|
|
|
|
|
1.
|
Bau des typischen Kormus
|
|
|
|
|
|
a)
|
Der Sproß
|
|
|
|
|
|
|
α)
|
Der Vegetationspunkt
|
|
|
|
|
|
|
β)
|
Die Sproßachse
|
|
|
|
|
|
|
γ)
|
Die Blätter
|
|
|
|
|
|
|
δ)
|
Die Verzweigung der Sprosse
|
|
|
|
|
|
b)
|
Die Wurzel
|
|
|
|
|
|
c)
|
Sekundäres Dickenwachstum des Kormus
|
|
|
|
|
2.
|
Anpassungen des Kormus an Lebensweise und Umwelt
|
|
|
|
|
|
A.
|
Autotrophe Kormophyten
|
|
|
|
|
|
|
a)
|
Anpassungen an den Feuchtigkeitsgehalt der Umwelt
|
|
|
|
|
|
|
b)
|
Anpassungen an den Lichtgewinn
|
|
|
|
|
|
|
c)
|
Anpassungen der grünen Kormophyten an besondere Ernährungsweise
|
|
|
|
|
|
B.
|
Heterotrophe Kormophyten
|
|
|
II.
|
Fortpflanzungsorgane
|
|
|
Vierter Abschnitt. Die Deszendenzlehre
und die Entstehung der Anpassungen
|
|
|
|
|
Physiologie
|
|
|
Kennzeichen des Lebens
|
|
|
Erster Abschnitt. Stoffwechsel
|
|
|
I.
|
Die stoffliche Zusammensetzung der Pflanze
|
|
|
II.
|
Aufnahme und Bewegung der Nährstoffe
|
|
|
III.
|
Assimilation der Nährstoffe
|
|
|
IV.
|
Wanderung und Wandlung der Assimilate
|
|
|
V.
|
Atmung und Gärung
|
|
|
Zweiter Abschnitt. Entwicklung
|
|
|
I.
|
Vorbemerkungen
|
|
|
|
|
1.
|
Wachstumsmessung
|
|
|
|
|
2.
|
Phasen des Wachstums
|
|
|
II.
|
Die Faktoren der Entwicklung
|
|
|
|
|
A.
|
Äußere Faktoren
|
|
|
|
|
B.
|
Innere Faktoren
|
|
|
III.
|
Der Entwicklungsgang und seine Abhängigkeit von
äußeren und inneren Faktoren
|
|
|
|
|
A.
|
Ruhe und Wachstum
|
|
|
|
|
B.
|
Wachstum und Zellteilung
|
|
|
|
|
C.
|
Vegetative Gestaltung
|
|
|
|
|
D.
|
Lebensdauer
|
|
|
|
|
E.
|
Fortpflanzung
|
|
|
|
|
F.
|
Vererbung, Variabilität, Artbildung
|
|
|
Dritter Abschnitt. Bewegungen
|
|
|
I.
|
Lokomotorische Bewegungen
|
|
|
II.
|
Krümmungsbewegungen
|
|
|
|
|
A.
|
Hygroskopische Bewegungen
|
|
|
|
|
B.
|
Bewegungen an der lebenstätigen Pflanze
|
|
|
|
|
|
1.
|
Autonome Bewegungen
|
|
|
|
|
|
2.
|
Paratonische Bewegungen
|
|
|
|
|
|
|
a)
|
Tropismen
|
|
|
|
|
|
|
b)
|
Nastische Bewegungen
|
|
|
III.
|
Rückblick auf die Reizerscheinungen
|
|
|
[S. viii]
|
|
|
|
Thallophyta
|
|
|
|
Bacteria
|
|
|
|
Cyanophyceae
|
|
|
|
Flagellatae
|
|
|
|
Myxomycetes
|
|
|
|
Dinoflagellatae
|
|
|
|
Diatomeae
|
|
|
|
Conjugatae
|
|
|
|
Heterocontae
|
|
|
|
Chlorophyceae
|
|
|
|
Phaeophyceae
|
|
|
|
Characeae
|
|
|
|
Rhodophyceae
|
|
|
|
Phycomycetes
|
|
|
|
Eumycetes
|
|
|
|
Lichenes
|
|
|
Bryophyta
|
|
|
|
Hepaticae
|
|
|
|
Musci
|
|
|
Pteridophyta
|
|
|
|
Filicinae
|
|
|
|
Equisetinae
|
|
|
|
Sphenophyllinae
|
|
|
|
Lycopodinae
|
|
|
|
Pteridospermeae
|
|
|
|
|
Spermatophyta
|
|
|
Übergang von den Farnpflanzen zu den
Samenpflanzen
|
|
|
Übersicht des Generationswechsels
|
|
|
Morphologie und Ökologie
der Blüte
|
|
|
|
Morphologie
|
|
|
|
Blütenstände
|
|
|
|
Ökologie
|
|
|
Entwicklung der Geschlechtsgeneration
bei den Samenpflanzen
|
|
|
|
A.
|
Gymnospermen
|
|
|
|
|
a)
|
Cycadeen
|
|
|
|
|
b)
|
Koniferen
|
|
|
|
|
c)
|
Gnetineen
|
|
|
|
B.
|
Angiospermen
|
|
|
|
|
a)
|
Mikrosporen
|
|
|
|
|
b)
|
Makrosporen
|
|
|
|
|
|
Der Samen
|
|
|
|
|
|
Die Frucht
|
|
|
|
|
|
Verbreitung der Samen
|
|
|
|
|
|
Die Keimung
|
|
|
Anordnung der Klassen, Ordnungen
und Familien
|
|
|
|
I.
|
Klasse Gymnospermae
|
|
|
|
|
1.
|
Ordnung Cycadinae
|
|
|
|
|
2.
|
„ Ginkgoinae
|
|
|
|
|
3.
|
„ Coniferae
|
|
|
|
|
|
Familie Taxaceae
|
|
|
|
|
|
„ Pinaceae
|
|
|
|
|
4.
|
Ordnung Gnetinae
|
|
|
|
|
Die fossilen Gymnospermen
|
|
|
|
II.
|
Klasse Angiospermae
|
|
|
|
|
1.
|
Unterklasse Dicotylae
|
|
|
|
|
|
Choripetalae
|
|
|
|
|
|
1.
|
Ordnung Polycarpicae
|
|
|
|
|
|
2.
|
„ Hamamelidinae
|
|
|
|
|
|
3.
|
„ Rosiflorae
|
|
|
|
|
|
4.
|
„ Leguminosae
|
|
|
|
|
|
5.
|
„ Myrtiflorae
|
|
|
|
|
|
6.
|
„ Umbelliflorae
|
|
|
|
|
|
7.
|
„ Centrospermae
|
|
|
|
|
|
8.
|
„ Primulinae
|
|
|
|
|
|
9.
|
„ Polygoninae
|
|
|
|
|
|
10.
|
„ Loranthiflorae
|
|
|
|
|
|
11.
|
„ Juglandiflorae
|
|
|
|
|
|
12.
|
„ Piperinae
|
|
|
|
|
|
13.
|
„ Querciflorae
|
|
|
|
|
|
14.
|
„ Saliciflorae
|
|
|
|
|
|
15.
|
„ Urticinae
|
|
|
|
|
|
16.
|
„ Rhoeadinae
|
|
|
|
|
|
17.
|
„ Cistiflorae
|
|
|
|
|
|
18.
|
„ Columniferae
|
|
|
|
|
|
19.
|
„ Tricoccae
|
|
|
|
|
|
20.
|
„ Gruinales
|
|
|
|
|
|
21.
|
„ Frangulinae
|
|
|
|
|
|
22.
|
„ Ericinae
|
|
|
|
|
|
Sympetalae
|
|
|
|
|
|
A.
|
Pentacyclicae
|
|
|
|
|
|
|
|
1. Ordnung Diospyrinae
|
|
|
|
|
|
B.
|
Tetracyclicae
|
|
|
|
|
|
|
|
2. Ordnung Contortae
|
|
|
|
|
|
|
|
3. „ Tubiflorae
|
|
|
|
|
|
|
|
4. „ Personatae
|
|
|
|
|
|
|
|
5. „ Rubiinae
|
|
|
|
|
|
|
|
6. „ Synandrae
|
|
|
|
|
2.
|
Unterklasse Monokotylae
|
|
|
|
|
|
|
|
1. Ordnung Helobiae
|
|
|
|
|
|
|
|
2. „ Spadiciflorae
|
|
|
|
|
|
|
|
3. „ Liliiflorae
|
|
|
|
|
|
|
|
4. „ Enantioblastae
|
|
|
|
|
|
|
|
5. „ Glumiflorae
|
|
|
|
|
|
|
|
6. „ Scitamineae
|
|
|
|
|
|
|
|
7. „ Gynandrae
|
|
|
|
|
Die fossilen Angiospermen
|
|
|
Offizinelle und giftige Pflanzen
|
|
Die Organismen, die unsere Erde bewohnen, teilt man in Tiere und
Pflanzen ein. Dementsprechend zerfällt die Biologie oder Lehre
von den Lebewesen in Zoologie, die Wissenschaft von den Tieren,
und in Botanik, die Wissenschaft von den Pflanzen.
Unter Pflanzen pflegt man festgewachsene, grüne, blühende und
fruchtende Lebewesen sich vorzustellen, unter Tieren dagegen meist
frei bewegliche Organismen, die Nahrung aufsuchen oder einfangen
und fressen. So leicht es also bei oberflächlicher Kenntnis zu sein
scheint, das Reich der Pflanzen gegen das der Tiere abzugrenzen, so
schwer ist es doch in Wirklichkeit. Bei sehr einfach gebauten,
d. h. äußerlich und innerlich wenig gegliederten, Organismen, die
man als die niedrigsten bezeichnet, läßt sich oft nicht entscheiden, ob
man sie in das Pflanzen- oder Tierreich einreihen soll. Tiere und
Pflanzen haben nämlich die folgenden wichtigen Eigenschaften gemein:
1. Die Pflanze besteht aus einem oder vielen, mikroskopisch kleinen
Kämmerchen, den Zellen, die sich durch Teilung vermehren. Aus
Zellen ähnlichen Baues und entsprechender Herkunft ist das Tier
zusammengesetzt. So haben Pflanzen und Tiere im wesentlichen
gleichen inneren Bau.
2. Die Pflanze ist wie das Tier ein lebendes Wesen
und stimmt in ihren wichtigsten Lebensregungen völlig mit dem
Tiere überein: Die Vorgänge der Ernährung und des Wachstums, der
Entwicklung und der Fortpflanzung sind bei Pflanzen und Tieren im
großen und ganzen wesentlich gleich, z. B. atmet auch die Pflanze und
entwickelt dabei Wärme; ferner besitzt sie auch Bewegungsvermögen und
Reizbarkeit mannigfaltiger Art.
3. Diese weitgehende Übereinstimmung zwischen den Lebensäußerungen
der Pflanzen und der Tiere kann nicht wundernehmen, wenn man weiß,
daß bei Pflanzen und Tieren das Leben an eine sehr ähnliche
„Grundsubstanz“, an das Protoplasma, gebunden ist, das sich in den
Zellen befindet.
Solche und viele andere Tatsachen weisen darauf hin, daß die
Pflanzen untereinander und mit den Tieren blutsverwandt sind.
Diese in der Abstammungs- oder Deszendenzlehre zum
Ausdruck kommende Auffassung kann man als eine grundlegende Theorie
der Biologie bezeichnen. Die Vorstellung, daß die Lebewesen mit
zusammengesetzterem Bau, mit höherer Organisation, aus einfacher
gestalteten sich entwickelt haben, reicht bis auf die griechischen
Philosophen zurück; sie wurde zu Beginn des 19. Jahrhunderts vor
allem von dem französischen Zoologen LAMARCK vertreten.
Eine wissenschaftliche Begründung erhielt sie aber erst später.
Namentlich war es CHARLES DARWIN[1], der durch eine Fülle von
Beweismaterial das zuvor[S. 2] herrschende Dogma von der Unveränderlichkeit
der Arten endgültig erschütterte und dadurch die großen Probleme der
organischen Entwicklung in Fluß brachte. Die Paläontologie lehrt uns
aus Versteinerungen und Abdrücken von Tieren und Pflanzen, daß in
früheren Erdperioden andere Lebewesen als in der Gegenwart, aber zum
Teil den jetzigen ähnliche den Erdball bewohnten. Diese Beobachtung
legt schon den Schluß nahe, daß die jetzt lebenden Formen durch
Umbildungen ausgestorbener entstanden sind. Er führt zu der Folgerung,
daß solche sehr ähnliche Organismen, die wir als Arten in einer Gattung
vereinigen, miteinander blutsverwandt sind, und daß man durch
Vereinigung von Arten zu Gattungen, von Gattungen zu Familien und von
Familien zu noch höheren Einheiten in einem „natürlichen“ Systeme
Verwandtschaftsgrade zum Ausdruck bringt. Die Entwicklung,
d. h. die Umwandlungen, die ein Lebewesen im Laufe von Generationen
während der Jahrtausende durchgemacht hat, nennt man mit ERNST
HAECKEL[2] seine Stammesgeschichte oder Phylogenie, die
Entwicklung, die es während seines Einzeldaseins durchmacht, seine
Ontogenie. Die Deszendenzlehre nimmt nun an, daß die höher
organisierten Pflanzen und Tiere phylogenetisch in letzter Linie aus
gemeinsamen Wurzeln entstanden sind, nämlich aus sehr einfachen Formen,
die vielleicht den einfachsten, heute noch lebenden ähnlich waren, und
zwar so, daß von solchen die phylogenetische Entwicklung einerseits
in der Richtung auf die höheren Tiere, andererseits in der Richtung
auf die ausgeprägten Pflanzen fortschritt. Nach dieser Annahme, die
ihre Stütze außer in den vielen, allen Tieren und Pflanzen gemeinsamen
Eigenschaften, eben in der Tatsache findet, daß eine scharfe Grenze
zwischen Tier- und Pflanzenreich in den Gruppen der niedersten Formen
sich nicht ziehen läßt, bilden alle lebenden Wesen im Grunde genommen
ein einziges Naturreich, das Reich der Organismen.
Ausgeprägt pflanzliche Merkmale wurden im Laufe der
phylogenetischen Entwicklung: die Ausbildung der wichtigsten
Körperflächen, die der Nahrungsaufnahme dienen, nach außen
(während dafür beim Tiere eine von einem Munde ausgehende
innere Körperfläche durch Einstülpung entstand), ferner die
Zellulosezellmembranen, mit denen die Zellen sich umkleideten,
endlich die grünen Farbkörper, die sich im Inneren der Zellen
ausbildeten. Der grüne Farbstoff befähigte die Pflanze, aus der
Kohlensäure der Luft, aus Wasser und aus gewissen Bodensalzen, also aus
anorganischen Verbindungen, ihre organische Leibessubstanz aufzubauen
und dadurch selbständig und unabhängig von allen anderen Organismen
zu leben; das Tier dagegen blieb in seiner Ernährung, unmittelbar
oder mittelbar, auf die Pflanze angewiesen, also in seinem Bestehen
von ihr abhängig. Fast alle Unterschiede, die zwischen ausgeprägten
Pflanzen und Tieren bestehen, lassen sich aus diesen Besonderheiten
der Ernährung ableiten. Als bezeichnend für die Pflanzen kann ferner
ihre ontogenetische Entwicklung gelten, die niemals
abgeschlossen wird, vielmehr an den Vegetationspunkten unbegrenzt
fortdauert, so daß die Pflanze im Prinzip immer weiter wächst. Daß aber
keines dieser Merkmale für sich allein ausreicht, um eine Pflanze von
einem Tiere mit Sicherheit zu unterscheiden, lehrt uns beispielsweise
die ganze Pflanzengruppe der Pilze; sie enthalten den grünen Farbstoff
nicht und sind infolgedessen wie die Tiere in ihrer Ernährung auf
organische Stoffe angewiesen, die letzten Endes einmal von grünen
Pflanzen gebildet worden waren. Gleichwohl rechnen wir die Pilze
zum Pflanzenreich, wen sie sich von grünen Gewächsen phylogenetisch
ableiten lassen.
Eine strenge, allgemeingültige Definition der „Pflanze“ und des
„Tieres“ zu geben, ist aber ganz unmöglich. Wir müssen uns also
hier mit dem Hinweise[S. 3] begnügen, daß von bekannteren Lebewesen die
Bakterien, Algen, Pilze, Flechten, Moose, Farn- und Samenpflanzen
(Gymnospermen und Angiospermen) der Pflanzenwelt zugerechnet
werden und somit Gegenstände der botanischen Forschung sind.
Viel leichter als die Begrenzung der Tier- und Pflanzenwelt
gegeneinander scheint die Aufgabe zu sein, das Reich der Organismen
gegen das der leblosen Körper abzugrenzen. Wir kennen kein Lebewesen,
dem das Protoplasma fehlt, aber keinen leblosen Körper, worin sich
tätiges Protoplasma nachweisen ließe. Seit es in der organischen
Chemie EMIL FISCHER gelungen ist, Zuckerarten synthetisch
darzustellen und die Synthese der Eiweißkörper anzubahnen, haben wir
aber mehr denn je Grund zu der Annahme, daß auch die Masse, die den
Ausgangspunkt der organischen Entwicklung bildete: das Protoplasma,
einen anorganischen Ursprung gehabt habe; denn diese Masse enthält nur
Elemente, die auch in der anorganischen Natur vorkommen. Eine solche
Urzeugung oder „Generatio spontanea“ hielt man im Altertume sogar bei
hochorganisierten Pflanzen und Tieren für möglich; weit verbreitet
war die Meinung, die selbst von ARISTOTELES geteilt wurde,
daß solche Lebewesen aus Schlamm und Sand hervorgehen könnten. Heute
wissen wir freilich durch tausendfältige Erfahrung, daß auch die
allerkleinsten und am einfachsten gebauten Organismen nicht in solcher
Weise entstehen, sondern nur aus ihresgleichen hervorgehen. So mag
die lebende Substanz aus lebloser vielleicht nur in einem bestimmten
Entwicklungszustande unserer Erde oder anderer Weltkörper entstanden
sein, als besondere Bedingungen zu ihrer Bildung sich eingestellt
hatten. Diese Annahme beseitigt freilich nicht alle Schwierigkeiten,
die der Vorstellung einer Urzeugung erwachsen. Damit aus solcher
lebenden Substanz die Welt der Organismen hervorgehen konnte, müßte
sie nämlich von vornherein die Fähigkeit gehabt haben, sich zu
erhalten, zu wachsen, fremde in ihren Körper aufgenommene Stoffe in
Körpermasse zu verwandeln, sich fortzupflanzen, d. h. sich durch
Teilung zu vervielfältigen, endlich neue Eigenschaften den vorhandenen
hinzuzufügen und sie erblich festzuhalten; kurz gesagt, es müßten in
dieser durch Urzeugung entstandenen Substanz alle wesentlichen Merkmale
des Lebens bereits ausgeprägt vorhanden gewesen sein[3].
Die Botanik zerfällt in eine Anzahl von Teilen. Die Morphologie
lehrt uns die äußere Gestalt und den inneren Bau der Pflanzen im
fertigen Zustande und während der ontogenetischen Entwicklung kennen
und verstehen. Die Physiologie erforscht die Lebenserscheinungen
der Gewächse. Beide Forschungszweige untersuchen auch die Beziehungen
der Bau- und der Lebenseigentümlichkeiten jeder Pflanze zu ihrer
Umgebung, zu ihren Außenbedingungen; sie sind bestrebt, festzustellen,
ob und wie weit diese Besonderheiten für ihren Träger nützlich
sind, also zu seiner Selbstbehauptung dienen, d. h. ob sie als
Anpassungen gedeutet werden können. Diese Teile der Morphologie
und Physiologie, die oft von den übrigen gesondert behandelt werden,
faßt man wohl auch als Ökologie zusammen. Die Systematik
beschäftigt sich mit der Beschreibung der Einzelformen und mit der
Klassifikation der Pflanzenwelt. Die Pflanzengeographie
hat zur Aufgabe, die Verteilung der Gewächse auf unserer Erde
festzustellen und die Ursachen dieser Verteilung zu ermitteln. Die
Paläophytologie erforscht die ausgestorbenen Pflanzen und die
zeitliche Aufeinanderfolge der Gewächse, mit anderen Worten, die
historischen Veränderungen der Pflanzenwelt auf[S. 4] der Erde. Alle diese
Gebiete rechnet man der reinen oder theoretischen Botanik
zu.
Aber nicht nur theoretische Ziele verfolgt die Botanik. Sie will
auch die gewonnenen Erkenntnisse für die Menschheit nutzbar machen:
die für den Haushalt des Menschen wertvollen Nutzpflanzen genau
kennen und immer zweckmäßiger ausnutzen lernen, die Schädlinge aus
dem Pflanzenreich erforschen, Verfälschungen der Handelsstoffe, die
aus dem Pflanzenreich stammen, nachweisen und dergleichen mehr. So
kommen zu den Teilgebieten der reinen Pflanzenkunde zahlreiche Zweige
der angewandten Botanik: z. B. die Pharmakognosie oder Lehre
von den Arzneipflanzen und ihren Produkten, den Pflanzendrogen; die
Lehre von den Giftpflanzen; die Lehre von den pflanzlichen Nahrungs-,
Genußmitteln und Gewürzen; die Lehre von den technisch wertvollen
Gewächsen und ihren Produkten; die landwirtschaftliche Botanik;
die Forstbotanik; die gärtnerische Botanik; ferner ein Teil der
Pflanzenpathologie, soweit sich nämlich dieser Wissenschaftszweig mit
der Bekämpfung der Pflanzenkrankheiten beschäftigt, und andere. —
Man kann die Botanik ferner auch, wie in diesem Buche, das in
allererster Linie die reine Botanik behandelt, in einen
allgemeinen und in einen speziellen Teil zerlegen.
Aufgabe und Ziel der allgemeinen Botanik ist es alsdann, aus
planmäßigen Untersuchungen an den einzelnen Pflanzen durch Vergleichung
die Eigenschaften zu ermitteln, die für die ganze Pflanzenwelt
oder ihre Hauptgruppen besonders bezeichnend sind. Die allgemeine
Botanik haben wir wieder in zwei Abschnitte, in Morphologie und
Physiologie, geteilt.
Aufgabe der speziellen Botanik ist es, die
Baueigentümlichkeiten, Fortpflanzungsverhältnisse und Lebensweise der
einzelnen Gruppen und Formen zu schildern, ferner die näheren
und ferneren Verwandtschaftsbeziehungen, die zwischen ihnen bestehen,
durch die Anordnung in einem möglichst „natürlichen“ Systeme zum
Ausdruck zu bringen. In diesen speziellen Teil des Lehrbuches haben
wir einige besonders wissenswerte Tatsachen aus manchen Zweigen der
angewandten Botanik, namentlich der Pharmakognosie, eingeflochten. Die
Ergebnisse der paläophytologischen Forschung sind an die Schilderung
der Einzelgruppen angeschlossen. Auch die Pflanzengeographie ist nicht
ganz unberücksichtigt geblieben, wenn davon auch keine zusammenhängende
Darstellung gegeben wurde.
[S. 5]
ERSTER
TEIL
Allgemeine Botanik.
Erste Abteilung.
Morphologie.
Die Morphologie der Pflanzen lehrt uns die äußere Gestalt und den
inneren Bau der Gewächse sowie die ontogenetische Entwicklung
ihres Körpers und seiner Glieder kennen. Sie strebt darüber hinaus
ein wissenschaftliches Verständnis der Pflanzenformen an,
indem sie sich bemüht, die Bedeutung und die phylogenetische
Herkunft der Pflanzenteile festzustellen und die Ursachen für die
Gestaltungsvorgänge zu ermitteln.
1. Man lernt nämlich den äußeren und inneren Bau eines Lebewesens nur
dann verstehen, wenn man sich erstens klar bewußt wird, daß das Tier
oder die Pflanze ein lebender Organismus ist, d. h. ein Gebilde,
dessen wichtigste Glieder nicht bedeutungslose Anhängsel, sondern für
das Leben notwendige Organe sind, durch deren harmonisches
Zusammenarbeiten erst das Leben des Ganzen zustande kommt[4]. Fast
alle äußeren Teile der Pflanze sind, wie beim Tiere, solche Werkzeuge
für bestimmte Lebensverrichtungen. Aber nur dann können die Teile des
Körpers ihre Leistungen im Dienste des ganzen Organismus erfüllen, wenn
sie äußerlich und innerlich im großen und ganzen leistungsfähig
gebaut sind, oder, wie man auch sagt, wenn ihr Bau einigermaßen
ihren Funktionen entspricht, ihnen angepaßt ist. Da nun die
einzelnen Teile bei höheren Pflanzen verschiedene Funktionen haben,
so wird es verständlich, daß sie auch ganz verschiedenen äußeren und
inneren Bau besitzen.
Um ein volles Verständnis vom Bau eines Organismus zu gewinnen, muß
man sich weiter über die Lebensverhältnisse klar werden, unter denen
er zu leben gewohnt ist: man muß seine Umwelt kennen. Jede Pflanze
besitzt nämlich, wie das Tier, Baueigentümlichkeiten, die es ihr
nur ermöglichen, unter bestimmten Außenbedingungen zu leben, wie
sie nicht überall da vorhanden sind, wo wir auf der Erde Gewächse
antreffen. Im Wasser z. B. sind die Lebensbedingungen ganz anders als
in der Wüste beschaffen. Dementsprechend sind die Wüstenpflanzen und
die Wassergewächse völlig verschieden gebaut und nur imstande, in
ihren gewohnten Verhältnissen oder solchen zu gedeihen, die diesen
einigermaßen ähnlich sind; jedenfalls können aber die Wüstengewächse
nicht im Wasser und die Wasserpflanzen nicht in der Wüste leben. Ein
Organismus ist also nur dann lebensfähig, wenn sein[S. 6] äußerer und
innerer Bau auch genügend auf die Umwelt abgestimmt, wenn er an
seine Lebensverhältnisse angepaßt ist.
Freilich lehrt eindringende morphologische Forschung alsbald, daß
zwar fast ein jedes Körperglied der Pflanze seine Funktionen hat, daß
aber längst nicht alle Eigentümlichkeiten seines äußeren und inneren
Baues als Anpassungen an diese Funktionen oder als Anpassungen an
die Umwelt gedeutet werden können: nur ein Teil der Merkmale eines
Pflanzengebildes steht in solchen Beziehungen zu seinen Verrichtungen
oder zur Umgebung, so z. B. bei den Laubblättern der Reichtum
an grünem Farbstoff und die flächenförmige Ausbildung zu ihrer
Hauptfunktion, der Kohlensäureassimilation. Solche Eigenschaften
bezeichnet man wohl auch als nützlich für den Organismus
oder als Anpassungsmerkmale. Daneben gibt es aber genug
gleichgültige, so an vielen Blättern wohl die Beschaffenheit ihrer
Ränder (Ganzrandigkeit, Sägung, Kerbung der Ränder u. dgl.), ja selbst
ungünstige (wie das Fehlen des grünen Farbstoffes in größeren Teilen
der Blätter, z. B. bei manchen wegen solcher „Weißbuntheit“ gern
kultivierten Ahornrassen), sofern sie den Organismus nicht unfähig
zum Leben machen. Eine Eigenschaft kann ferner bei einer Art
mehr oder weniger nützlich, bei einer anderen gleichgültig oder gar
schädlich sein. Solche Tatsachen zeigen nachdrücklich, wie vorsichtig
man bei der Beurteilung der Bedeutung aller organischen Formen und
Strukturen sein muß, zumal viele Annahmen über ihren Nutzen sich nur
sehr schwer durch Versuche auf ihre Richtigkeit prüfen lassen[5].
2. Aber noch in einer zweiten Richtung strebt die Morphologie
wissenschaftliches Verständnis der Pflanzenformen an. Alle lebenden
Pflanzen betrachten wir als mehr oder weniger blutsverwandt. Aus
einfachen, ungegliederten Formen, aus Einzelzellen, sind phylogenetisch
allmählich die am höchsten organisierten Wesen mit zahlreichen
verschiedenen Organen hervorgegangen. Dabei haben die Organismen und
ihre Teile mannigfaltige Weiter- und Umbildungen erfahren, indem z. B.
einzelne Organe durch Veränderungen ihres Baues neue Funktionen
übernahmen oder neuen Lebensverhältnissen angepaßt wurden. Eine sehr
wichtige Aufgabe der Morphologie ist es nun, diese phylogenetischen
Umbildungen zu erforschen. Da die stammesgeschichtliche Entwicklung
aber meist nicht direkt verfolgt, sondern nur erschlossen werden
kann, so ist die Morphologie zur Lösung dieser Aufgabe auf indirekte
Methoden angewiesen. Die wichtigsten Aufschlüsse in dieser Hinsicht
gewinnt sie 1. durch das Studium der Ontogenie der Organismen, ferner
2. durch die Vergleichung der jetzt bestehenden Lebewesen untereinander
und mit solchen, die in früheren Erdperioden gelebt haben. Die
Ontogenie eines Organismus durchläuft nämlich häufig innerhalb gewisser
Grenzen Entwicklungsstadien, die man als phylogenetische betrachten
darf; sie kann daher zur Ermittelung der stammesgeschichtlichen
Entwicklung beitragen. Und die vergleichende Forschung bemüht sich,
die verschiedenen Gestaltungen durch Zwischenglieder zu verbinden. Da
aber die Ontogenie die Phylogenie niemals vollständig oder unverändert
wiederholt und die Zwischenglieder zwischen den verschiedenen Formen
vielfach fehlen, so bleiben freilich die Ergebnisse auch dieser
Richtung der morphologischen Forschung entsprechend unvollkommen.
Haben wir durch eingehende Untersuchungen die Überzeugung gewonnen,
daß verschieden gestaltete Glieder des Pflanzenkörpers einen
gemeinsamen phylogenetischen Ursprung haben, so bezeichnen wir
die hypothetische Ursprungsform, von der wir sie ableiten, als ihre
Grundform, die verschiedenen Umbildungen aber, die die
Organe im Laufe der Stammesgeschichte erfahren haben, auch wohl als
ihre Metamorphosen. Eines der allerwichtigsten Ergebnisse der
Morphologie besteht in dem Nachweise, daß die mannig[S. 7]faltig
gestalteten äußeren und inneren Teile selbst der am reichsten
gegliederten Pflanzen sich auf ganz wenige Grundformen zurückführen
lassen, nämlich bei den höher organisierten Pflanzen die äußeren
Teile auf Wurzel, Stengel und Laubblatt, ferner bei allen Pflanzen die
inneren Teile auf die Zellen und ihre Bestandteile. Die Organe, die
sich aus einer gemeinsamen Grundform phylogenetisch weiter entwickelt
haben, nennen wir homolog, mögen sie auch noch so verschieden
aussehen. Ihnen schreiben wir gleichen morphologischen Wert zu.
Homolog sind z. B. die Laubblätter und die Blätter der Blüten (die
Kelch-, Kron-, Staub- und Fruchtblätter), ferner auch die Laubblätter,
die Blattranken (Fig. 207) und die Blattdornen (Fig. 195). Organe
völlig verschiedenen Baues und ganz verschiedener Funktion können
also doch homolog sein, also den gleichen morphologischen Wert
besitzen. Andererseits haben sich oft Organe mit gleichem Bau und
mit gleicher Funktion (z. B. Knollen Fig. 201,
203, 204, Dornen Fig.
195, 197, 198,
Ranken Fig. 206–208) phylogenetisch aus verschiedenen
Grundformen entwickelt. Solche Organe nennt man analog
(zahlreiche Beispiele vgl. S. 141 ff.). Wenig differenzierte Gebilde
ohne deutlich ausgeprägte Funktionen, die sich aus vollkommeneren
rückgebildet haben, bezeichnen wir als reduziert.
3. Schließlich setzt sich die Morphologie die Aufgabe, die Ursachen
oder Bedingungen zu ermitteln, die wie jedem Naturgeschehen, so auch
den Vorgängen der äußeren und inneren Ausgestaltung der Pflanze und
ihrer Teile, ferner ihren vererbbaren (phylogenetischen) Umwandlungen
zugrunde liegen, und darüber Klarheit zu gewinnen, wie sich im Laufe
der stammesgeschichtlichen Entwicklung die Eigenschaften ausbilden
konnten, die wir als Anpassungsmerkmale bezeichnet haben. Den Teil
der Morphologie, der sich mit solchen Fragen beschäftigt, kann man
experimentelle Morphologie nennen. Die Mehrzahl ihrer Probleme
findet man aber zweckmäßiger meist, so auch in unserem Lehrbuche, als
besonderen Abschnitt der Physiologie, d. h. des Zweiges der Botanik
behandelt, der sich überhaupt mit den Lebensvorgängen der Pflanzen
beschäftigt (Entwicklungsphysiologie).
Die Morphologie kann man zerlegen in die Lehre vom äußeren Bau (äußere
Morphologie) und in die Lehre vom inneren Bau (innere Morphologie
oder Anatomie). Für unser Buch aber wäre eine solche Gliederung nicht
zweckmäßig. Wollen wir doch die Glieder als Organe mit bestimmten
Lebensverrichtungen begreifen lernen. Dafür aber ist es notwendig,
zusammenhängend zu zeigen, in wie inniger Beziehung vielfach nicht
nur der äußere, sondern auch der innere Bau eines Organes zu seinen
Funktionen steht. Nicht die Pflanze als totes Gebilde, sondern als
lebender Organismus soll uns ja in erster Linie beschäftigen.
Die erste Frage, die es da zu beantworten gilt, ist die nach dem
Träger der Lebenserscheinungen. Woran ist das Leben gebunden? Nur
an einen Teil der ganzen Substanz einer Pflanze, nämlich an das
Protoplasma. Das Protoplasma aber ist in der Regel eingeschlossen in
die Zellen, die man als Elementarteile des Organismus ansehen
kann. Infolgedessen müssen wir den Bau der Zellen vor allem kennen
lernen. Den Teil der Morphologie, der dieser Aufgabe dient, nennt man
Zellenlehre oder Zytologie. Die Zellverbände (Gewebe) des
Pflanzenkörpers bilden alsdann den Gegenstand eines zweiten Teiles der
Morphologie, der Gewebelehre oder Histologie. Endlich
mit den äußeren Gliedern als Organen der Pflanze, und zwar mit ihrem
äußeren und inneren Bau, beschäftigt sich die Organlehre oder
Organographie.
[S. 8]
Erster Abschnitt. Zellenlehre (Zytologie).
I. Gestalt und Größe der Zellen.
Die Pflanzen werden, gleich den Tieren, aus Elementarteilen aufgebaut,
die wir als Zellen bezeichnen. Das sind bei den Pflanzen
meist mikroskopisch kleine Kämmerchen, deren Wände im Gegensatze zu
den Zellen der Tiere von besonderen Häuten gebildet werden.
Die Zellformen entsprechen im einfachsten Falle Kugeln, meist aber
kleinen Würfeln, Polyëdern oder Prismen, die bei vielzelligen Organen
in großer Menge aufeinander geschichtet sind; auch langgestreckte,
ja faser- oder schlauchförmige Zellen kommen häufig vor. Diese
Kämmerchen, von denen jedes aus den Kammerwänden, der Zellhaut
oder Zellmembran, und aus seinem Innenraum, dem Zellraum
oder Zelllumen, besteht, sind im allgemeinen so klein, daß
man sie erst bei stärkerer Vergrößerung erkennen kann. Ihr mittlerer
Querdurchmesser pflegt nämlich nur ein Hundertstel bis ein Zehntel
Millimeter zu betragen. Infolgedessen wurden die Zellen erst spät, in
der Neuzeit, entdeckt. Hier und da freilich werden Zellen auch viel
größer; manche an besondere Funktionen angepaßte faserförmige Zellen
(Sklerenchymfasern) werden bis zu 20 cm, Milchröhren sogar meterlang.
Das wichtigste an diesen Zellen ist ein Teil ihres Inhaltes, der
Zellenleib oder Protoplast. Er ist nämlich der
eigentlich lebende Teil der Zelle. Deshalb denkt man bei dem
Begriff Zelle heutzutage mehr an ihn als an das Gehäuse, das zudem
vielen „nackten Zellen“ ganz fehlt. In toten Zellen findet man freilich
nur noch Reste der Zellleiber, die aber auch vollständig geschwunden
sein können; alsdann enthalten die Zellräume nur Wasser oder Luft.
Ihre Bedeutung für den Pflanzenkörper brauchen die Zellen mit dem Tode
der Protoplasten aber nicht einzubüßen, ja ohne tote Zellen könnte ein
höher organisiertes Gewächs nicht auskommen; denn solche Zellen bilden
z. B. seine Wasserbahnen und tragen zu seiner mechanischen Festigung
bei.

Fig. 1. H
OOKES Bild des Flaschenkorkes, von ihm
als Schematism or Texture of Cork bezeichnet; vgl. dazu die
Fig. 58.
Die Entdeckung der Zellräume glückte bei den Pflanzen früher als
bei den Tieren; sie wurde dort durch die Zellhäute erleichtert.
Ihr Entdecker, der englische Mikrograph ROBERT HOOKE,
nannte sie Zellen wegen ihrer Ähnlichkeit mit den Zellen der
Bienenwaben und bildete sie in seiner Mikrographie 1667 zum ersten
Male ab (Fig. 1). Die eigentlichen Begründer der pflanzlichen
Histologie sind aber der Italiener MARCELLO MALPIGHI und
der Engländer NEHEMIAH GREW, deren Werke kurz nacheinander
von 1671 an, also wenige Jahre nach HOOKES Mikrographie,
erschienen. Der lebende Inhalt der Zellen, der Zellenleib, wurde
in seiner Bedeutung nicht vor der Mitte des vorigen Jahrhunderts
erkannt. Alsdann erst wandte man sich auch eingehend seiner
näheren Untersuchung zu, die u. a. SCHLEIDEN, HUGO V.
MOHL, NÄGELI, FERDINAND COHN und MAX
SCHULTZE anbahnten und besonders STRASBURGER förderte.
II. Der lebende Inhalt der Zellen (der Protoplast)[6].
A. Bestandteile des Protoplasten. Untersuchen wir bei starker
Vergrößerung zarte Längsschnitte durch die äußerste Stengelspitze
einer Samenpflanze, so finden wir, daß sie sich aus annähernd
rechteckig begrenzten Zellen zusammensetzen (Fig. 2), die mit Inhalt
dicht angefüllt und durch zarte strukturlose Wände, die Zellhäute,
voneinander getrennt sind. Die Zellen[S. 9] sind hier annähernd würfelförmig
oder prismatisch, weshalb sie eben im Schnitte als Quadrate oder
Rechtecke erscheinen.
Im Inhalte jeder Zelle fällt ein runder Körper (k) von Kugel- oder
Eiform besonders auf, der einen großen Teil des Zellraums ausfüllt: der
Zellkern (Kern, Nucleus). Die feinkörnige Masse,
die den Raum zwischen Zellkern (k) und Zellwand (m) einnimmt,
bezeichnet man als Zytoplasma (pl) oder Plasma. Um
den Zellkern herum findet man, in dem Zytoplasma verteilt, stark
lichtbrechende, farblose Körperchen, die Plastiden oder
Chromatophoren (ch). Zellkern, Plasma und Chromatophoren
sind die lebenden Inhaltsbestandteile der Zelle; sie zusammen
bilden das Protoplasma, den lebenden Zellenleib oder
Protoplasten. Zellkern und Chromatophoren, die stets im Plasma
eingebettet sind, kann man als Organe des Protoplasten ansehen,
denen besondere Lebensverrichtungen zukommen. Freilich kennen wir
die besonderen Funktionen des Kernes noch nicht; wir wissen nur, daß
zur Erhaltung des Lebens der Zelle eine Wechselwirkung zwischen Kern
und Plasma notwendig ist. Doch ist es für die niedersten Gewächse,
Spaltalgen (Cyanophyceen) und Bakterien, noch immer eine ungelöste
Frage, ob bei ihnen eine solche Arbeitsteilung im Protoplasten, d. h.
ob bei ihnen ein Zellkern vorkommt[7]. Im Protoplasma der Bakterien
fehlen auch die Chromatophoren, ebenso in den Zellen der Pilze und in
denen der Tiere.
Dagegen hat man in tierischen Zellen in unmittelbarer Nähe des
Kerns noch kleine lebende Gebilde, die Zentriolen, als
Bestandteile des Protoplasten nachgewiesen. Ähnliche Gebilde kommen
im Pflanzenreiche fast nur in Zellen von Kryptogamen vor, ohne aber
allgemeine Verbreitung zu besitzen (Fig. 21 A).
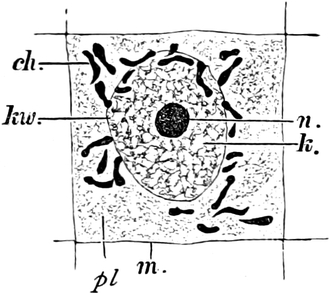
Fig. 2. Embryonale Zelle aus der Wurzelspitze des
Hafers. k Zellkern, kw Kernwandung, n Kernkörperchen, pl
Plasma, ch Chromatophoren, m Zellwandung. Etwas schematisiert.
Vergr. ca. 1500. Nach LEWITSKY.
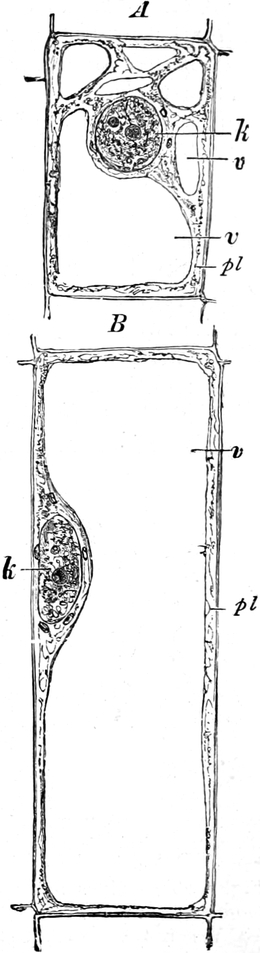
Fig. 3. Zwei Zellen der Stengelspitze einer
Samenpflanze, in verschiedener Entfernung von ihrem obersten Ende
entnommen. k Kern, pl Plasma, v Vakuolen (Safträume). Etwas
schematisiert. Vergr. ca. 500. Nach STRASBURGER.
Bei den Pflanzen sind nur die embryonalen Zellen, wie man
sie z. B. an den äußersten Stengel- und Wurzelspitzen findet, in
der eben beschriebenen Weise mit Protoplasma dicht gefüllt. Das ist
dagegen nicht der Fall in den ausgebildeten Körperzellen
(Dauerzellen), die durch Größenwachstum und mannigfaltige
Formveränderungen aus jenen hervorgehen. Während dieser Umwandlung zu
Dauerzellen sieht man nämlich die embryonalen Zellen bei den Pflanzen,
aber nicht bei den Tieren, immer plasmaärmer werden, weil das Plasma
während der Vergrößerung der Zellräume nicht wesentlich vermehrt
wird. Diese Umwandlung läßt sich an jedem Längsschnitt durch eine
Stengelspitze verfolgen. In einiger Entfernung von[S. 10] seinem oberen Ende
enthalten die heranwachsenden Zellen in ihrem Plasma bereits eine
größere Anzahl Hohlräume, Vakuolen (v in A Fig. 3), die mit
wäßrigem Saft, Zellsaft, gefüllt sind. Die Zellen fahren alsdann
noch fort, an Größe zuzunehmen, wobei die Vakuolen verschmelzen.
Schließlich wird meist ein Zustand erreicht, wo nur noch ein einziger,
großer, mit Zellsaft gefüllter Hohlraum, der Saftraum (v in
B Fig. 3), im Plasma der Zelle besteht, das Plasma aber nur einen
dünnen Belag an der Zellwandung bildet, in dem auch der alsdann
wandständige Kern eingebettet ist (Fig. 3 B k). Der Saftraum kann
aber auch in einer ausgewachsenen Zelle von Lamellen und Strängen oder
Fäden aus Plasma durchsetzt bleiben, worin oft der Kern, aber stets vom
Plasma allseitig umhüllt, aufgehängt ist (Fig. 5, 10). In jeder
noch lebenden Zelle ist die Zellwandung auf ihrer Innenseite von einem
ununterbrochenen Plasmabelag ausgekleidet, der der Zellwand überall
dicht anliegt, in älteren Zellen aber so dünn werden kann, daß man ihn
nicht unmittelbar sieht (Fig. 10). Erst wasserentziehende Mittel, die
ihn veranlassen, sich von der Zellwandung zurückzuziehen und abzulösen
(Plasmolyse, vgl. S. 192), wie etwa stärker konzentrierte Salz- oder
Zuckerlösungen, machen ihn alsdann sichtbar.
B. Physikalische Eigenschaften des Protoplasten. Um die
physikalischen Eigenschaften, insbesondere den Aggregatzustand, des
Protoplasmas kennen zu lernen, wenden wir uns zunächst zu einer
Gruppe von Organismen, die an der Grenze zwischen dem Pflanzen- und
dem Tierreiche stehen, zu den Schleimpilzen oder Myxomyceten. Sie
sind durch einen Entwicklungszustand ausgezeichnet, während dessen
ihr Protoplasma größere nackte Massen, die Plasmodien, bildet.
Ihr Plasma (Fig. 4) besteht aus netzartig verbundenen, dickeren und
dünneren Strängen einer glashellen Grundmasse, die Körnchen enthält
und zäh- oder dünnflüssig ist. In diesen Strängen sieht man nämlich
das Plasma innerhalb von festeren und dichteren ruhenden Hüllschichten
lebhaft nach Art einer Flüssigkeit strömen. Diese inneren Ströme
bewegen sich nach den Rändern des Plasmodiums hin oder von ihnen hinweg
und wechseln häufig ihre Richtung. An den Rändern des Plasmodiums
selbst werden Plasmafortsätze vorgestreckt oder schon vorhandene
eingezogen. Dadurch kann das Plasmodium sich kriechend fortbewegen. Wo
solche Protoplasmamassen fremden Körpern begegnen, sind sie befähigt,
sie in ihr Inneres aufzunehmen, in Vakuolen einzuschließen und, soweit
das möglich ist, auch zu verdauen.
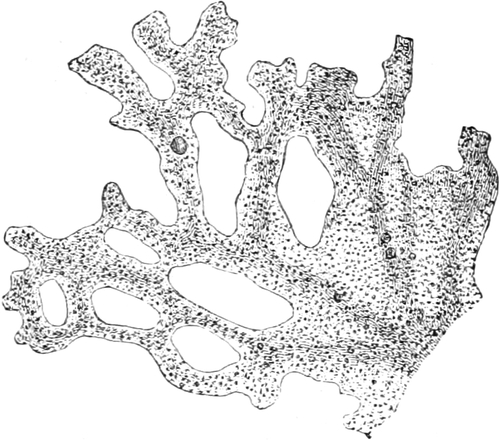
Fig. 4. Teil eines ausgewachsenen Plasmodiums von
Chondrioderma difforme. Vergr. 90. Nach STRASBURGER.
Wie in den nackten Plasmodien der Schleimpilze, so läßt sich auch
bei behäuteten pflanzlichen Zellen oft strömende Bewegung
im Plasma erkennen, solange es lebt. Meist stellt sie sich erst
in annähernd fertigen Dauerzellen, und zwar vielfach nur dann in
auffälliger Weise ein, wenn durch eine Verwundung, etwa das Schneiden
bei Herstellung des Präparats, ein Reiz auf die Protoplasten ausgeübt
worden ist[8]. Sie scheint den Transport von Nährstoffen nach der
Wundstelle zu beschleunigen. Schon an diesen Bewegungen kann man sehen,
daß auch hier das Protoplasma meist eine dünn-[S. 11] oder zähflüssige
Masse ist; aus seiner Hülle befreit, nimmt es dementsprechend die
Form eines runden Tropfens an. In behäuteten Zellen, in denen solche
Plasmabewegung vorkommt, sieht man das Protoplasma, abgesehen
von seiner stets ruhenden äußersten Schicht, die an die Zellwand
angrenzt, entweder in einem einzigen Strome von konstanter Richtung
oder in verschiedenen Strömen mit wechselnder Richtung sich bewegen.
Man hat danach zwischen Rotation und Zirkulation des
Plasmas unterschieden. In den Zellen, in denen Rotationsbewegung
vorkommt, z. B. bei vielen Wasserpflanzen, ist das Protoplasma auf
einen Wandbelag beschränkt. Der Rotationsstrom folgt der Zellwandung
und beschreibt eine kreisende, in sich zurücklaufende Bahn. Die bei
Landpflanzen häufige Zirkulation findet sich besonders in Zellen,
deren Innenräume von Plasmasträngen oder -lamellen durchsetzt sind;
das Plasma strömt in ihnen außerdem in bandförmigen, meist verzweigten
Streifen des Wandbelages, und zwar hier wie dort nach verschiedenen
Richtungen hin.
Von dem in Rotation befindlichen Plasma werden Zellkern
und Chromatophoren meist mitgeführt. Doch können letztere an der
ruhenden peripheren Schicht haften und infolgedessen unbeweglich
sein. So ist es beispielsweise bei den Characeen, Süßwasseralgen,
deren lange Gliederzellen in der Gattung Nitella besonders günstige
Beispiele für die Beobachtung starker Rotationsströmung sind. Ein
sehr gutes Objekt für das Studium der Zirkulation sind
die Staubblatthaare von Tradescantia virginica (Fig. 5). Die den
Saftraum durchsetzenden Plasmastränge verändern dabei langsam ihre
Gestalt und Lage und veranlassen dadurch auch Lageänderungen des
Kerns.
Bewegungen an eng umgrenzten Stellen des Plasmas beobachtet man
auch in den Protoplasten vieler niederer Algen, besonders ihrer
Schwärmzellen: In der Nähe des vorderen Körperendes umschließt
das Plasma eine oder mehrere kleine pulsierende Vakuolen,
die in kürzeren Zeitabschnitten rhythmisch verschwinden, d. h.
plötzlich sich entleeren, darauf aber wieder erscheinen und langsam
zur alten Größe heranwachsen (Fig. 335 1 v). Ferner besitzt
ihr Plasmakörper einen oder mehrere fadenförmige, kontraktile,
plasmatische Fortsätze, Geißeln, Zilien, die sehr
lebhaft schwingen und die Bewegungsorgane der Schwärmzellen sind.
Der Protoplast ist nur innerhalb ziemlich enger Temperaturgrenzen aktiv
lebenstätig (also auch strömungsfähig) und innerhalb etwas weiterer
lebensfähig. Er stirbt, d. h. gerinnt, erstarrt in der Regel rasch bei
Temperaturen, die nicht weit über +50° liegen. Auch durch Alkohol,
durch Säuren von bestimmter Konzentration, durch Lösungen von Sublimat
und vielen anderen Schwermetallsalzen wird das Protoplasma zum raschen
Erstarren gebracht, fixiert. Solche Gerinnungs- und Fixierungsmittel
spielen jetzt eine große Rolle in der mikroskopischen Technik[9].
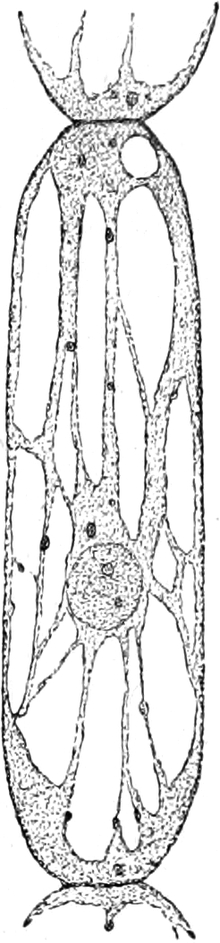
Fig. 5. Eine Zelle aus einem Staubblatthaare
von Tradescantia virginica. Innerhalb der Stränge als dunkle
Körner Leukoplasten und der runde Zellkern. Vergr. 240. Nach STRASBURGER.
C. Chemische Eigenschaften des Protoplasten[10]. Das in
Tätigkeit befindliche Protoplasma reagiert gewöhnlich alkalisch,
unter Umständen auch neutral, niemals aber sauer. Es ist nicht
ein einheitlicher chemischer Körper, sondern besteht aus einem
Gemische einer großen Zahl chemischer Verbindungen, die zum
Teil in Wasser gelöst, zum Teil fest sind. Ein Teil davon erfährt
im aktiv lebenstätigen Protoplasma fortdauernd Veränderungen,
auf denen ohne Zweifel viele wichtige Lebensäußerungen des
Protoplasten beruhen. Die wichtigsten Bestandteile in diesem
Gemische sind wohl die Eiweißkörper (Eiweißstoffe,
Proteïne, und Eiweißverbindungen,[S. 12] Proteïde). So gibt
das Protoplasma Eiweißreaktion und läßt beim Verbrennen infolge
seines Stickstoffgehaltes Ammoniakdämpfe entweichen. Und zwar ist
in dem Protoplasma eine ganze Reihe von Eiweißstoffen aufgefunden
worden. Im Zellkerne herrschen die Nukleoproteïde, phosphorhaltige
Eiweißverbindungen, vor, die von Pepsinlösung nicht aufgelöst werden.
Weiter enthält das Protoplasma wohl stets Spaltungsprodukte der
Eiweiße, vor allem Amide; außerdem Enzyme, Kohlehydrate und ölartige
Körper (Lipoide), wie Fette und Lezithine (vgl. S. 222); ferner
Phytosterine (aromatische Alkohole von der Formel C27H45OH)
und unter Umständen Alkaloide (heterozyklische, stickstoffhaltige
Pflanzenbasen) oder Glykoside (esterartige Verbindungen der Zucker
meist mit aromatischen Verbindungen). Daß auch Mineralstoffe im
Protoplasma nicht völlig fehlen, geht daraus hervor, daß es Asche
hinterläßt.
Durch verdünnte Kalilauge werden alle Teile des Protoplasten
gelöst, ebenso durch Chloralhydrat oder JAVELLEsche Lauge.
Durch Jod werden sie bräunlichgelb gefärbt, durch eine Lösung von
salpetersaurem Quecksilberoxydul (dem sog. MILLONschen
Reagens) ziegelrot. Die Reagenzien töten das Protoplasma, worauf
sich erst die charakteristische Reaktion einstellt. Diese
Reaktionen weisen auf Eiweißkörper hin, sind ihnen aber nicht
ausschließlich eigen.
D. Bau der Teile des Protoplasten. Sehr wichtige Hilfsmittel
für die Erforschung der Teile des Protoplasten bilden die
Fixierungs- und Färbeverfahren. Gewisse Gerinnungsmittel
fixieren und härten nämlich das Protoplasma anscheinend wenig
verändert. Man hat aber stets darauf zu achten, daß bei der Fixierung
auch Strukturen auftreten, die erst durch die Gerinnung entstehen[11].
Der Wert der Färbungen beruht darauf, daß die verschiedenen
Bestandteile des Protoplasmas mit ungleicher Begierde Farbstoffe
aufnehmen und mit größerer oder geringerer Kraft festhalten, wenn
man ihnen die Stoffe durch Lösungsmittel wieder zu entziehen
sucht. Viele Farbstoffe werden erst vom toten Protoplasma merklich
gespeichert. Zur Färbung der fixierten pflanzlichen Protoplasten
bedient man sich vornehmlich der Karminlösungen, des Hämatoxylins,
Safranins, Säurefuchsins, Gentianavioletts, Orange, Methylenblaus
u. a.
1. Das Plasma (Zytoplasma). In einer anscheinend glasklaren,
also optisch homogenen, dünn- oder zähflüssigen Grundmasse des Plasmas,
dem Hyaloplasma, sieht man bei stärkeren Vergrößerungen
gewöhnlich winzig kleine Körnchen und Tröpfchen in kleinerer oder
meist größerer Zahl, die Mikrosomen, eingebettet, die offenbar
aus verschiedenartigen Stoffwechselprodukten des Plasmas bestehen.
Körnerreiches Plasma bezeichnet man wohl als Körner- oder
Polioplasma. Das Hyaloplasma, das selbst mit dem Ultramikroskop
optisch nahezu leer erscheint, ist eine wäßrige Lösung der Art, die
die physikalische Chemie kolloidale Lösungen oder Sole
nennt[12]. Der Nachweis, daß das Protoplasma meist eine kolloidale
Lösung, und zwar ein Emulsoid, ist, hat, wie es scheint,
grundlegende Bedeutung. Dadurch dürften viele Lebensäußerungen des
Plasmas einer physikalisch-chemischen Erklärung zugänglich werden.
An seiner Peripherie ist das Plasma von einer äußerst dünnen,
körnchenfreien und oft zäheren Hyaloplasmaschicht umgeben, deren
äußerster Saum, die Hautschicht oder Plasmahaut, seine
eigentliche äußere Begrenzung bildet. Gegen den Saftraum und andere
Vakuolen grenzt es sich ebenfalls durch solche Hyaloplasmaschichten
und Plasmasäume, die Vakuolenwände, ab. Diese peripheren
Hautschichten und die Vakuolenwände können sich jederzeit neu bilden,
sind aber sehr wichtige Bestandteile des Protoplasten; denn sie
entscheiden über die Aufnahme von Stoffen in das Protoplasma. Sie sind
semipermeabel; d. h. lassen zwar Wasser durch, sind aber für
viele andere Stoffe undurchlässig oder schwer durchlässig.
[S. 13]
Ob das lebende Plasma außer der Emulsionsstruktur stets noch eine
andere bestimmte und bezeichnende, aber mit dem Mikroskope und
Ultramikroskope unsichtbare Struktur besitzt, wissen wir nicht. In sich
teilenden Protoplasten treten fadenförmige Sonderungen auf, die in
ruhenden Protoplasten wieder unkenntlich werden. Plasma, das fixiert
und gefärbt wurde, kann homogen sein oder bildet (bei schlechter
Fixierung), wie andere geronnene kolloidale Lösungen, ein Netz-,
Gerüst- oder Wabenwerk, in das Körnchen eingelagert sind.
Außer solchen Strukturen sind aber neuerdings im Plasma von
embryonalen und Dauerzellen, vor allem nach besonderem Fixierungs-
und Färbungsverfahren, noch korn-, stäbchen-, faden-, spindel-
oder hantelförmige Gebilde nachgewiesen worden, die in ihrem
Aussehen und in ihrem Verhalten gegenüber den Fixierungs-
und Färbungsmitteln so sehr mit den Chondriosomen
(Mitochondrien) embryonaler tierischer Zellen übereinstimmen, daß
man diese Bezeichnung auf sie ausgedehnt hat[13]. Wahrscheinlich
sind es verschiedenwertige Dinge, zum Teil kleine, an besonderen,
vielleicht nukleïnsäurehaltigen Eiweißkörpern reiche Vakuolen,
zum Teil fadenförmige Plasmastränge, zum Teil auch jugendliche
Chromatophoren; man hat sie auch bei Pilzen beobachtet und bei
gewissen Moosen in den embryonalen Zellen neben den
Chromatophoren.
2. Der Zellkern (Nukleus)[14] ist im allgemeinen kugel-, ei-
oder linsenförmig gestaltet, kann aber manchmal auch absonderliche,
z. B. gelappte Gestalt annehmen oder in gestreckten Zellen selbst
fadenförmig werden. In embryonalen Zellen beträgt sein Durchmesser etwa
zwei Drittel des Gesamtdurchmessers des Protoplasten. In ausgewachsenen
Dauerzellen dagegen macht er, da er nicht mitwächst, in dem größeren
Zellraume einen viel kleineren Eindruck. Große Kerne findet man bei
den meisten Koniferen und manchen Monokotylen, sowie bei Ranunculaceen
und Loranthaceen unter den Dikotylen. Mit besonders großen Kernen sind
meist Drüsenzellen ausgestattet. Dagegen sind die Kerne der meisten
Pilze (Fig. 6) und vieler Schlauchalgen sehr klein.

Fig. 6. Mehr-(5) kernige Zelle des Pilzes Hypholoma
fasciculare. Vergr. 500. Nach KNIEP.
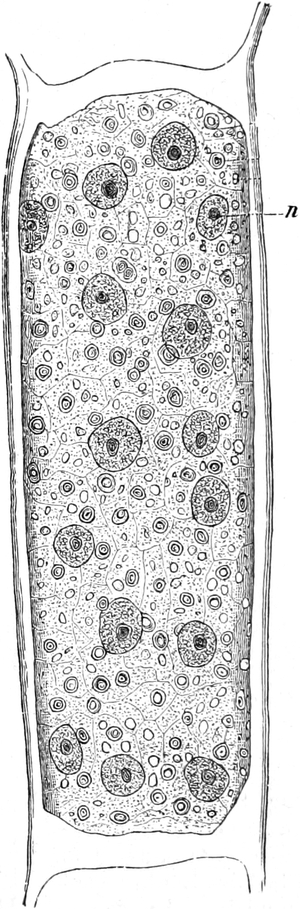
Fig. 7. Eine Zelle von Cladophora glomerata, nach einem
mit 1%iger Chromsäure fixierten und mit Karmin gefärbten Präparate. n
Kerne. Vergr. 540. Nach STRASBURGER.
Bei höheren Pflanzen kommen fast ausschließlich einkernige
Zellen vor. Bei den niederen Pflanzen sind aber vielkernige
Zellen sehr verbreitet; ja bei vielen Pilzen (Fig. 6) und bei den
Schlauchalgen herrschen sie vor. Der ganze Organismus wird alsdann
entweder von einer einzigen solchen vielkernigen Zelle gebildet, die
bei einigen Schlauchalgen, wie Caulerpa, äußerlich ungewöhnlich reich
gegliedert ist (Fig. 348); oder er besteht aus einer größeren Zahl
vielkerniger Zellen, so bei vielen Pilzen (Fig. 6) und z. B. bei der
Süßwasseralge Cladophora (Fig. 7).
[S. 14]
Der Zellkern sieht, solange er lebt, fein punktiert aus. Außerdem
fallen in ihm meist ein bis mehrere größere, runde, glänzende
Körner oder Tropfen auf: die aus Eiweißkörpern bestehenden
Kernkörperchen oder Nukleolen (Fig. 2 n), deren
Bedeutung wir noch nicht genau kennen. Der Kern, dessen Inhalt
zähflüssig zu sein scheint, ist von einer Kernwandung
umgeben (Fig. 2 kw), einer Hautschicht, mit der der Kernraum, die
Kernhöhle, gegen das umgebende Plasma abgegrenzt ist.
Einen Einblick in die feinere Kernstruktur erhält man nur an
entsprechend fixierten und gefärbten Präparaten. Man erkennt alsdann
im Kern meist ein stark gefärbtes wabig-netzartiges Gerüstwerk oder
Körner aus Chromatin (Fig. 13, 1 n), das vornehmlich aus
phosphorhaltigen Eiweißverbindungen (und zwar Nukleoproteïden) zu
bestehen scheint. In den Maschen des Gerüstes befinden sich die
Nukleolen, die sich ebenfalls intensiv, jedoch meist anders als das
Chromatin färben, weil sie meist nicht aus Chromatin bestehen. Das
Gerüstwerk und die Nukleolen des Kerns liegen innerhalb der Kernhöhle,
die mit Kernsaft, wohl einer Eiweißlösung, gefüllt ist.
In vielen Kernen scheint das Kerngerüst aus einer wenig färbbaren
Grundmasse, dem Linin, gebildet zu werden, dem das Chromatin
als kleine Körnchen eingelagert ist.
Bei Spirogyra unter den Algen, gewissen Flagellaten und Pilzen
enthalten vielleicht auch die Kernkörperchen einen Teil
des Chromatins, sind also denen der höheren Pflanzen nicht
gleichwertig, wie auch ihr Anteil an den Kernteilungsvorgängen
zeigt[15]. Solche Kerne werden wohl auch als Karyosomkerne
bezeichnet.
Welchen Anteil der Zellkern an den Lebenserscheinungen des Protoplasten
hat, ist noch ganz unbekannt; jedenfalls aber ist er zum Bestande des
Lebens in kernhaltigen Zellen nötig. Sehr große Bedeutung hat er als
hauptsächlicher Träger der erblichen Anlagen.
3. Die Chromatophoren[16]. In den embryonalen Zellen sind die
Chromatophoren kleine, farblose, stark lichtbrechende Gebilde von
Tropfen-, Körner-, Spindel- oder Fadenform, die sich vornehmlich
in der Nähe des Zellkerns (Fig. 2 ch) aufhalten. In Dauerzellen
sind sie meist zu Chloroplasten, Leukoplasten oder
Chromoplasten umgebildet, die man wegen dieses gleichen
Ursprunges ebenfalls Chromatophoren nennen kann.
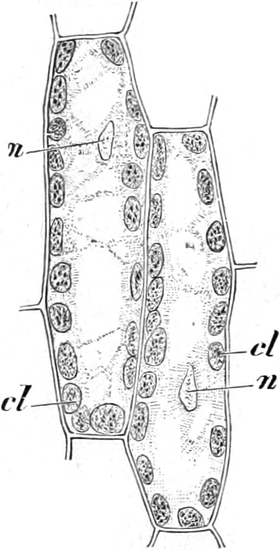
Fig. 8. Zwei Zellen mit Chlorophyllkörnern (cl) aus
dem Blatt des Laubmooses Funaria hygrometrica. n Zellkerne. Vergr.
300. Nach SCHENCK.
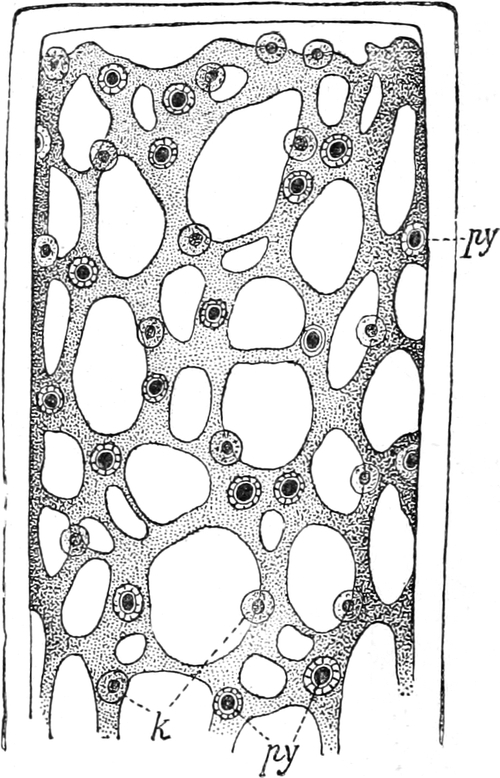
Fig. 9. Netzförmiger Chlorophyllkörper der Süßwasseralge
Cladophora arcta mit py Pyrenoïden, k Kerne. Nach SCHMITZ.
a) Chloroplasten. In peripherischen, dem Lichte
ausgesetzten Teilen der Pflanze gehen aus den Chromatophoren
der embryonalen Zellen in der Regel grüne Chloroplasten oder
Chlorophyllkörper hervor. Ihnen verdanken die grünen Pflanzen
ihre Farbe und auch ihre Befähigung zur Kohlensäureassimilation.
Die Chloroplasten liegen immer im Plasma, meist im[S. 15] plasmatischen
Wandbelage der Zellen, und besitzen bei allen höher organisierten
Pflanzen die Gestalt ellipsoidischer, etwas abgeflachter Körner
(Fig. 8 cl) Chlorophyllkörner. Bei den Algen sind die
Chlorophyllkörper aber oft anders gestaltet, nämlich bandförmig
(Fig. 329 C), sternförmig oder plattenförmig, häufig auch
netzartig durchbrochen (z. B. Cladophora Fig. 9). Alsdann sind
ihnen meist Pyrenoïde (Fig. 9 py) ein- oder angelagert:
runde Proteïnkörper, die in bestimmten Fällen einen Eiweißkristall
enthalten und mit kleinen schalenförmigen Stärkekörnern sich
umhüllen, weshalb die Pyrenoïde auch Stärkeherde genannt werden.
Nach längerer Belichtung findet man in den Chloroplasten der meisten
Pflanzen eine geringere oder größere Zahl sehr kleiner Stärkekörnchen
(Assimilationsstärke Fig. 15) und außerdem oft ölartige Tröpfchen, die
vielleicht aus Aldehyden bestehen. Die Grundmasse der Chloroplasten
läßt aber eine feinere Struktur in lebensfrischem Zustande selbst bei
den stärksten Vergrößerungen nicht erkennen; sie ist gleichmäßig grün
gefärbt.
Der grüne Farbstoff, das Chlorophyll, ist für die
Kohlensäurezerlegung in den Chloroplasten unentbehrlich.
Die Untersuchungen aus letzter Zeit[17], namentlich von
WILLSTÄTTER und seinen Schülern, haben ergeben, daß in den
Chloroplasten im ganzen vier Farbstoffe vorhanden sind: Zwei
sehr nahe verwandte grüne Pigmente, das Chlorophyll a und b,
im Mengenverhältnis von etwa 3 : 1, und, in wesentlich geringerer
Menge, zwei gelbe Farbstoffe. Die Chlorophylle sind Ester des
Phytols, eines Alkohols von der Formel C20H39OH, und einer
Trikarbonsäure, also hochmolekulare Verbindungen von Kohlenstoff,
Sauerstoff und Wasserstoff, in deren Aufbau noch Stickstoff und
Magnesium, entgegen früheren Annahmen aber kein Phosphor und kein
Eisen eingeht. Das blaugrüne Chlorophyll a entspricht der Formel
C55H72O5N4Mg + 1⁄2 H2O; das gelbgrüne Chlorophyll
b hat die Zusammensetzung C55H70O6N4Mg. Die gelben
Pigmente sind orangerote, kristallisierende Karotine
(Kohlenwasserstoffe von der Zusammensetzung C40H56), von
denen eines z. B. auch in den Möhrenwurzeln vorkommt, und gelbe,
ebenfalls kristallisierbare Xanthophylle (Oxyde der
Karotine: C40H56O2). An der Assimilation der Kohlensäure
sind aber nur die Chlorophylle beteiligt.
Alle vier Farbstoffe lassen sich aus den frischen oder getrockneten
Chloroplasten mit verschiedenen Lösungsmitteln ausziehen, z. B. mit
Azeton oder 80–90% Alkohol; am schnellsten kann man eine intensive
Lösung aller Pigmente aus frischen Laubblättern erhalten, wenn man
sie mit siedendem Alkohol übergießt. Solche Lösungen sind infolge
des Gehaltes an Chlorophyllen im durchfallenden Lichte smaragdgrün,
bei auffallendem Lichte durch Fluoreszenz blutrot[18].
Ihr Spektrum (Fig. 245) ist durch vier Absorptionsbänder im
weniger brechbaren (roten) Teile und drei im stärker gebrochenen
(blauen) ausgezeichnet. Die einzelnen Pigmente lassen sich durch
Ausschütteln dieser Lösungen voneinander trennen. Wird z. B. die
alkoholische Lösung mit Benzol geschüttelt, so nimmt dieses die
Chlorophylle auf und sammelt sich als grüne Lösung über dem nun
gelb gefärbten Alkohol an. Die Menge des in grünen Pflanzenteilen
vorhandenen Chlorophyllgrüns ist nur gering; sie macht nach
WILLSTÄTTER 0,5–1% der Trockensubstanz aus.
Bei manchen viel kultivierten Gewächsformen, den weißbunten
(panaschierten) Pflanzen, haben kleinere oder größere Teile
der Blätter nicht grüne, sondern weiße oder gelbliche Färbung; ihre
Zellen enthalten an Stelle der grünen Chloroplasten farblose oder
gelbliche Chromatophoren.
Viele Algen sind nicht grün, sondern anders gefärbt. In den
blaugrünen, spangrünen, blauen, seltener violetten Spaltalgen
und in den roten, violetten oder rotbraunen Chloroplasten
der Rotalgen sind nämlich neben den vier Farbstoffen der
grünen Chlorophyllkörper auch noch ein blauer Farbstoff, das
Phykozyan, und ein roter, das Phykoerythrin, allein
oder seltener nebeneinander vorhanden. Beide sind nach Abtötung
der Zellen schon in Wasser löslich, dem ein wenig Alkali oder
Neutralsalz zugesetzt ist, und fluoreszieren sehr schön. Um
Spaltalgen, die man auf Papier trocknet, bildet das Phykozyan oft
einen blauen Saum. Beide Farbstoffe sollen Proteïde sein. Über ihre
Bedeutung ist wenig Sicheres bekannt[19]. Bei den Braunalgen kommt
die Farbe der braunen oder gelben Chloroplasten dadurch zustande,
daß in ihnen außer[S. 16] Chlorophyll a und sehr wenig Chlorophyll b
etwa gleich viel gelbe Farbstoffe, nämlich Karotin, Xanthophyll und
außerdem in überwiegender Menge auch noch das dem letzten verwandte
rotbraune Phykoxanthin (C40H54O6), vorhanden
sind[20].
Die Verfärbungen[21], die die Blätter unserer Holzgewächse im
Herbste vor dem Blattfall erfahren, sind mit einer Zersetzung der
Chloroplasten und des Chlorophyllfarbstoffes verbunden. In den
Protoplasten findet man alsdann außer wäßriger, oft rotgefärbter
Flüssigkeit nur noch einige Öltröpfchen, Kristalle und gelbe, stark
lichtbrechende Kugeln. Anders steht es bei solchen Nadelhölzern,
deren Blätter im Winter sich bräunen, um im nächsten Frühjahr
wieder zu ergrünen; hier gehen in den Farbstoffen der Chloroplasten
Umwandlungen vor, die im Frühling wieder rückgängig gemacht
werden. Die Bräunung absterbender Laubblätter ist eine postmortale
Erscheinung, bei der braune wasserlösliche Farbstoffe auftreten.
In den nicht grünen phanerogamen Schmarotzern werden die Chloroplasten
nicht ausgebildet, sondern durch farblose, auch wohl bräunliche oder
rötliche Chromatophoren ersetzt, die übrigens bei manchen dieser
Gewächse noch Spuren von Chlorophyll enthalten können. Bei den Pilzen
fehlen die Chromatophoren ganz, wie schon hervorgehoben wurde.
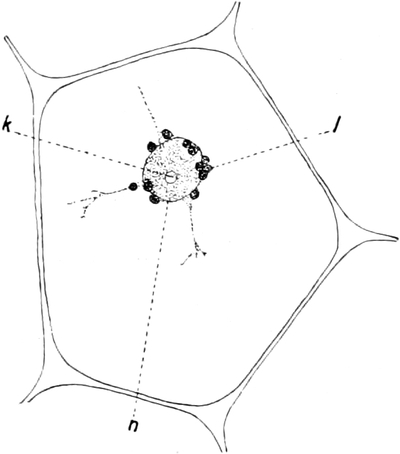
Fig. 10. Zelle aus der Epidermis der Commelinacee Rhoeo
discolor. n Kern mit Kernkörperchen k, umgeben von Leukoplasten
l. Vom Kern gehen Plasmastränge nach dem unsichtbaren wandständigen
Plasmabelag aus. Vergr. 240.
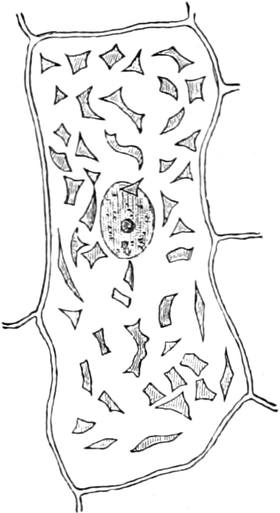
Fig. 11. Zelle mit Chromoplasten von der Oberseite des
gelb gefärbten Kelches der Kapuzinerkresse (Tropaeolum majus). Vergr.
540. Nach STRASBURGER.
Fig. 12. Chromoplasten aus der Möhrenwurzel, zum Teil
mit Stärkeeinschlüssen. Vergr. 540. Nach STRASBURGER.
b) Leukoplasten. In vielen Teilen der Pflanzen, namentlich
solchen, zu denen das Licht nicht gelangt, werden die Chromatophoren
zu farblosen Leukoplasten. Sie sind in vielen Zellen winzig
klein (Fig. 5, 10 l), kugelig, eiförmig oder nicht selten durch
einen eingeschlossenen Eiweißkristall gestreckt (Fig. 28 B kr). Dem
Lichte ausgesetzt wandeln sie sich häufig in Chloroplasten um, so
in den äußeren Partien belichteter Kartoffelknollen und in manchen
Erdwurzeln. Auch die Leukoplasten haben, wenigstens in vielen Zellen,
besondere Funktionen, nämlich die, Zucker in Stärke umzuwandeln, die
in ihnen als Körner auftritt, weshalb man die Leukoplasten auch als
Stärkebildner bezeichnet.
c) Chromoplasten gehen entweder direkt aus den farblosen
Chromatophoren der Embryonalzellen oder aus zuvor ausgebildeten
Chloroplasten hervor und bedingen die gelbe und rote Färbung vieler
Pflanzenteile, besonders von Blüten und Früchten. Sie können wie die
Chloroplasten rundliche Körner sein; doch sind sie oft kleiner und
stets gelb oder orangerot gefärbt. Diese Färbung rührt entweder von
gelben Xanthophyllen oder von leicht auskristallisierenden,
orangeroten Karotinen her. Die Farbstoffe[S. 17] sind nicht
gleichmäßig in ihrem Körper gelöst; vielmehr findet man in einer
farblosen, plasmatischen Grundsubstanz (dem Stroma) des
Chromoplasten viele winzige Tröpfchen (Grana) davon[22]. Die
Farbstoffe, namentlich die Karotine, kristallisieren aber auch oft aus;
alsdann sind die Chromoplasten nadelförmig oder zu dreieckigen oder
rhombischen Plättchen gestreckt (Fig. 11, 12).
Nach Herkunft und Bedeutung noch nicht genügend bekannt ist der
rote Augenfleck, den man in den Zellen vieler Algen,
besonders in ihren Schwärmzellen, neben dem Chloroplasten und
meist mit ihm verbunden findet (Fig. 335 1 a). Manche Forscher
glauben, daß er als Chromoplast aufzufassen ist und zur Wahrnehmung
des Lichtes, gewissermaßen als Auge, dient. Der rote Farbstoff,
Hämatochrom genannt, ist nichts anderes als Karotin.
E. Ursprung der Elemente des Protoplasten[6]. Alle lebenden
Elemente des Protoplasten, das Plasma, die Zellkerne und die
Chromatophoren, stammen ab von gleichnamigen Elementen; eine freie
Neubildung findet nirgends statt. Sie nehmen an Masse zu
durch Wachstum; sie vermehren sich aber an Zahl, ebenso wie
die Protoplasten selbst, nur durch Teilung oder Abspaltung aus
ihresgleichen. Dadurch werden die Eigenschaften der lebenden
Bestandteile einer Keimzelle auf alle Zellen des Organismus, so auch
wieder auf seine Keimzellen übertragen, und die ununterbrochene
Fortdauer des Lebens bleibt erhalten. Die Teilung der Protoplasten wird
gewöhnlich eingeleitet durch die Kernteilung. Das Ineinandergreifen der
Kern- und Zellteilung in einkernigen Zellen ist notwendig, um jeder
Tochterzelle einen Kern zu sichern. In vielkernigen Zellen (z. B. von
Algen und Pilzen) ist es nicht notwendig, wenn die Querwand so angelegt
wird, daß jedem Tochterprotoplasten die nötigen Kerne ohnedies bei der
Teilung zufallen; tatsächlich ist hier oft die Zellteilung nicht von
der Kernteilung abhängig.
Es kommt übrigens vor, daß der Protoplast einer Zelle als Ganzes
ohne Teilung seine alte Zellhülle aufgibt. Dieser Vorgang,
Zellverjüngung genannt, hat mit Zellteilung nichts zu tun.
Solche Zellverjüngung liegt z. B. vor, wenn in der grünen Alge
Oedogonium der Protoplast sich abrundet und aus einer Öffnung der
alten Zellhaut als nackte Schwärmspore heraustritt, oder wenn die
Protoplasten der Sporen von Moosen oder Farnen und der Pollenkörner
von Samenpflanzen innerhalb ihrer Zellhäute sich mit neuen
Membranen umgeben und mit diesen neuen Hüllen selbständig werden,
während die ursprünglichen Zellwände der Zerstörung anheimfallen.
1. Typische Teilung des Protoplasten. a) Kernteilung.
Von wenigen Fällen abgesehen vermehren sich die pflanzlichen Zellkerne
durch mitotische oder indirekte Teilung, einen Vorgang,
der auch als Karyokinese bezeichnet wird. Er spielt sich in
ziemlich verwickelter Weise ab und ist vor allem an fixierten und
gefärbten Schnitten näher studiert worden.
Indirekte Kernteilung[23]. Sie stimmt in ihren Hauptzügen bei
höher organisierten Pflanzen und Tieren überein. Fig. 13 stellt ihre
Stadien in etwas schematisierten Bildern so dar, wie sie in embryonalen
vegetativen Zellen solcher Pflanzen aufeinander folgen.
Das feine Gerüstwerk des ruhenden Zellkerns aus (Linin und)
Chromatin (Fig. 13, 1 n) sehen wir auf einzelne Punkte
des Chromatinnetzes sich zusammenziehen (Spiremstadium) und in
eine bestimmte Anzahl von fadenartigen Gebilden sich sondern (2
ch), die zunächst unregelmäßig begrenzt sind, aber allmählich
dichter werden und nun bestimmte Farbstoffe noch stärker speichern
(3, 4). Wir bezeichnen diese fadenförmigen Gebilde des Kerns als
Chromosomen. Sie spalten sich längs (5); etwas später werden
sie dicker,[S. 18] kürzer und glattrandig (6), worauf sie nach der Mitte
der Zelle befördert werden und sich hier zur Kernplatte oder
Äquatorialplatte (7 kp), einer sternförmigen Figur (dem Aster),
anordnen, die meist in der künftigen Teilungsebene der Zelle liegt
(Flächenansicht Fig. 14).
Während das Kerngerüst in die einzelnen Chromosomen sich sondert, legen
sich der Kernwandung von außen Plasmafäden an und umgeben sie mit einer
faserigen Schicht, die sich mehr und mehr an zwei gegenüberliegenden
Seiten der Kernwand sammelt und hier die Polkappen (6 k)
bildet. In ihnen neigen die Fasern nach den Endpolen der ganzen
Teilungsfigur zusammen, indem sie sich zu zugespitzten Büscheln
strecken. Alsdann wird das Kernkörperchen (1 nl) und die Kernwandung
(1 w) aufgelöst, worauf die Fasern der Kappen in die Kernhöhle
hineinwachsen (7). Sie endigen dort entweder, wie es scheint, an den
Chromosomen oder treffen mit den Enden aufeinander und verlaufen dann
als ununterbrochene Fäden von einem Pole zum anderen. Hiermit ist die
Kernspindel (7 s) aus den Spindelfasern fertiggestellt.
Fig. 13. Aufeinanderfolgende Stadien der Kern- und
Zellteilung in einer embryonalen Zelle einer höheren Pflanze.
Etwas schematisiert. Als Vorlage dienten Längsschnitte mit
Chromosmiumessigsäure fixierter Wurzelspitzen der monokotylen
Wasserpflanze Najas marina, nach Färbung mit Eisenhämatoxylin, n
Kern, nl Nucleolus, w Kernwandung, pl Plasma, ch Chromosomen,
k Polkappen, s Spindel, kp Kernplatte, t Tochterkern,
v Verbindungsfäden, z Zellplatte, m neue Scheidewand. Die
Chromatophoren sind bei solcher Fixierung und Färbung nicht sichtbar.
Vergr. ca. 1000. Nach CLEMENS MÜLLER.
Die beiden Längshälften jedes Chromosoms, die
Tochterchromosomen, rücken hierauf in entgegengesetzter Richtung
auseinander (8, 9), um die beiden Tochterkerne (10–12 t) zu bilden.
Während dieser Wanderung (Diasterfigur) sind die Chromosomen meist
U-förmig nach den Polen hin gekrümmt. An den Spindelpolen angelangt,
drängen sie sich aneinander. Darauf grenzt[S. 19] sich das umgebende Plasma
mit Hautschichten gegen die neuen Kernanlagen ab und bildet ihre
Kernwandungen. Innerhalb der Tochterkerne werden die Chromosomen wieder
wabig (Dispiremstadium 11) und vereinigen sich miteinander zu einem
Gerüstwerk (12), worin ihre Grenzen meist nicht mehr zu erkennen
sind, wenn sie auch, wie man annehmen muß, ihre Selbständigkeit nicht
einbüßen. Beide Tochterkerne werden dabei größer; es treten auch wieder
Nukleolen in Ein- und Mehrzahl in ihnen auf (12).
Durch diesen Teilungsmechanismus ist erreicht, daß die Substanz
des Kerns, besonders die der Chromosomen, bei jeder Kernteilung
ganz gleichmäßig auf die beiden Tochterkerne, und zwar infolge
der Längsspaltung der Chromosomen, so verteilt wird, daß
von den einzelnen, in der Längsrichtung aufeinanderfolgenden
Chromatinabschnitten eines jeden Chromosoms die eine Hälfte dem
einen, die andere Hälfte dem anderen Tochterchromosom mit Sicherheit
zufällt. Man schließt daraus, daß das Chromatin für das Leben der
Zelle und des ganzen Organismus besonders wichtig ist, nämlich daß die
Chromosomen die hauptsächlichsten Träger der vererbbaren Anlagen sind,
und daß in jedem Chromosom eine Anzahl verschiedener solcher
Anlagen perlschnurartig aneinander gereiht sind.
Die Zahl der Chromosomen in den Kernen einer Pflanzenart ist
übrigens eine bestimmte. Abweichende Zahlen kommen aber vor[24];
kleinere rühren zum Teil daher, daß einzelne Chromosomen mit ihren
Enden vereinigt bleiben; größere Zahlen kommen zum Teil dadurch
zustande, daß sich einzelne Chromosomen quer teilen. Bei den
verschiedenen Pflanzenarten ist dagegen die Chromosomenzahl verschieden
groß; die kleinste Zahl, die man bisher in den Zellen der höher
organisierten Gewächse angetroffen hat, ist sechs; meist aber ist sie
viel größer (bis über 130). Sehr häufig sind die Chromosomen eines
Kerns untereinander verschieden in Größe und Form (Fig. 14). Solche
Unterschiede werden bei allen Kernteilungen festgehalten. Diese
Tatsache deutet mit großer Sicherheit darauf hin, daß die Chromosomen
ihre Individualität auch im Ruhekern bewahren. Man nimmt jetzt
an, daß die untereinander verschieden gestalteten Chromosomen Träger
verschiedener Gruppen erblicher Anlagen der betreffenden
Pflanzenart sind.
In den untersten Abteilungen des Pflanzenreichs, bei manchen Algen und
Pilzen, verläuft die indirekte Kernteilung nicht selten einfacher,
indem die Chromatinmasse weniger sorgfältig auf beide Tochterkerne
verteilt zu werden scheint und die Spindel ganz oder wenigstens
teilweise dem Kern entstammt[14].
Fig. 14. Embryonale Zelle, dem Querschnitt einer
Wurzelspitze der Liliacee Galtonia candicans entnommen, mit einer
Kernplatte in Polansicht. Die Chromosomen zu Paaren angeordnet. Vergr.
1600. Nach STRASBURGER.
Die Vorgänge, die sich in einem Mutterkerne während der
Vorbereitung zur Teilung abspielen, werden als Prophase
der Teilung bezeichnet. Sie reichen bis zur Bildung der
Kernplatte, vor deren Fertigstellung sich auch die Längsspaltung
der Chromosomen vollzieht. Das Stadium der Kernplatte heißt die
Metaphase. Das Auseinanderweichen der Tochterchromosomen
erfolgt in der Anaphase, die Bildung der Tochterkerne in der
Telophase der Teilung. Der Höhepunkt der ganzen Kernteilung,
der Vorgang, der zur Bildung quantitativ und qualitativ gleicher
Teilungsprodukte führt, liegt offenbar in der Längsspaltung
der Chromosomen. Die fortschreitenden Vorgänge der Kernteilung
gehen mit dem Auseinanderweichen der Tochterchromosomen in die
rückläufigen über. Daher dauert wohl das Stadium der Kernplatte
meist längere Zeit an.
Wodurch die Chromosomen während der Karyokinese in der
beschriebenen Weise so regelmäßig bewegt werden, wissen wir
noch nicht. STRASBURGER nimmt an, daß die[S. 20]
Spindelfasern, die an den Chromosomen zu endigen scheinen, durch
Verkürzung die Tochterchromosomen aus der Kernplatte nach den Polen
ziehen (Zugfasern), während die von Pol zu Pol laufenden Fasern
(Stützfasern) gewissermaßen als Stützen der Kernteilungsfigur
dienen. Diese Annahmen erklären aber nicht die Bewegungen der
Chromosomen nach der Kernplatte hin.
In bestimmten Zellen der Pflanzen und Tiere, die der Fortpflanzung
dienen, vollzieht sich als notwendige Folge einer Befruchtung die
Kernteilung in besonderer, von der typischen Teilung abweichender
Art, die man als Reduktions- oder meiotische Teilung
bezeichnet (vgl. S. 172).
Direkte Kernteilung[25]. Außer der mitotischen oder indirekten
gibt es wenn auch selten, eine direkte oder amitotische
Kernteilung, auch Fragmentation genannt. Sie stellt sich meist
als Alterserscheinung an Kernen ein, die aus indirekter Teilung
hervorgegangen sind, und ist im wesentlichen eine Durchschnürung
des Kernes, wobei die Teilstücke durchaus nicht in ihrer Größe
übereinzustimmen brauchen. Lehrreiche Beispiele für direkte Kernteilung
sind die Kerne in den langen Gliederzellen der Characeen.
Bei den Characeen folgen die direkten Teilungen der Kerne in
den wachsenden Gliederzellen so rasch aufeinander, daß oft
perlschnurförmige Reihen zusammenhängender Teilstücke entstehen.
Auf die direkte Kernteilung folgt keine Zellteilung. Die direkte
Kernteilung kommt übrigens auch bei Samenpflanzen, z. B. bei
Tradescantia, der Liliacee Funkia, Impatiens balsamina (der
Balsamine) vor.
b) Vermehrung der Chromatophoren. Auch sie erfolgt durch
Teilung, und zwar auf direktem Wege durch Einschnürung. Man kann sie
am besten an den Chlorophyllkörnern verfolgen. Jedes Chlorophyllkorn
liefert dabei zwei gleich große Körner (Fig. 15).
c) Teilung des Plasmas. In den einkernigen Zellen der
höher organisierten Gewächse pflegen Kern- und Zellteilungen
ineinander zu greifen. Während die Tochterchromosomen sich trennen,
bleiben die von Pol zu Pol reichenden Fasern der Kernspindel als
Verbindungsfäden zurück (Fig. 13, 9 v), ja sie werden
durch Einschaltung neuer sogar noch vermehrt (Fig. 13, 10, 11)
und bilden schließlich zusammen einen tonnenförmigen Körper, den
Verbindungsfadenkomplex (Fig. 13, 11). Jeder Verbindungsfaden
schwillt alsdann in der Äquatorialebene zu einem Körnchen an (Fig. 13, 11);
dadurch entsteht die Zellplatte, die also in
Seitenansicht wie eine Körnchenreihe aussieht. Ist die Zelle sehr
plasmareich oder schmal, so erreicht der Komplex der Verbindungsfäden
an der Peripherie allseitig ihre Seitenwände. Aus den verschmelzenden
Körnchen der Zellplatte geht alsdann eine plasmatische Schicht hervor,
die sich spaltet und in der Spaltungsfläche eine Scheidewand aus
Zellhautstoff ausscheidet. Letztere teilt annähernd gleichzeitig,
simultan, den Mutterprotoplasten in zwei Tochterzellen (Fig. 13,
12 m).
Fig. 15. Chlorophyllkörner aus dem Blatte des Laubmooses
Funaria hygrometrica; ruhend und in Teilung. Im Innern der Körner
kleine Stärkekörnchen. Vergr. 540. Nach STRASBURGER.
Fig. 16. Drei Teilungszustände in derselben Zelle der
Orchidee Epipactis palustris. Nach dem Leben entworfen. Vergr. 365.
Nach TREUB.
Ist dagegen in der Zelle ein größerer Saftraum vorhanden oder ist
die Zelle sehr groß, so vermag der Komplex der Verbindungsfäden
sie nicht mit einem Male zu durchsetzen; vielmehr bildet er die
Scheidewand dann allmählich, succedan, aus (Fig. 16):
zunächst etwa einen Teil, der an eine Seitenwand der Mutterzelle
anschließt (Fig. 16 A), sodann einen folgenden, wobei er an seinem
freien Rande die Zellplatte ergänzt, aber[S. 21] sich von den schon
gebildeten Teilen der Scheidewand zurückzieht (B), und so fort
und fort, bis der ganze Protoplast durchschnitten und seine Teilung
vollendet ist (C). In langen Zellen, die sich längs teilen, z. B.
denen des Kambiums, schreitet die Zellwandbildung dagegen von der
Zellmitte aus, wo der Kern liegt, succedan allseits nach der
Peripherie fort[26].
Es gibt übrigens Fälle, wo die Verbindungsfäden klein an Zahl sind;
alsdann werden die Knötchen durch Plasmaplatten zur Zellplatte
verbunden.
Bei den Thallophyten werden die Scheidewände der vielkernigen und der
einkernigen Zellen dagegen fast niemals in Verbindungsfadenkomplexen
gebildet. Sie entstehen vielmehr entweder simultan und zwar
in Plasmaplatten, die auf einmal in der ganzen Teilungsebene
ausgebildet werden, oder succedan, indem eine ringförmige Leiste aus
Membransubstanz allmählich von der Mutterzellwand aus, einer Irisblende
ähnlich, in das Zellinnere immer tiefer vordringt (Fig. 17, 18) und es
schließlich durchschnürt. In einkernigen Zellen geht auch bei diesem
Teilungsvorgang die Teilung des Kerns der Zellteilung voraus; die neue
Scheidewand entsteht hierauf in gleichen Entfernungen von den beiden
Tochterkernen, und zwar in der Zone, wo ursprünglich der Kern gelegen
hatte.
Fig. 17. Eine Spirogyrazelle in Teilung. n Einer der
beiden Tochterkerne, w die wachsende Scheidewand, ch ein durch
letztere nach innen gedrängtes Chlorophyllband. Vergr. 230. Nach STRASBURGER.
Fig. 18. Stück einer sich teilenden Zelle von Cladophora
fracta. w Die wachsende Scheidewand. ch Chromatophoren, k Kerne.
Vergr. 600. Nach STRASBURGER.
Bei den nackten Zellen der Myxomyzeten und Flagellaten ist die
Teilung eine aktive Durchschnürung des Plasmas.
In vielkernigen Zellen folgt nicht auf jede Kernteilung eine
Zellteilung; ja unter den Algen und Pilzen gibt es sogar große,
äußerlich nicht selten reich gegliederte Formen, deren Inneres nur von
einem einzigen, sehr vielkernigen Plasmaleib gebildet, also
überhaupt nicht durch Zellwände gekammert wird.
2. Abarten der typischen Zellteilung. Hier und da im
Pflanzenreiche kommen Abweichungen von der typischen Zellteilung
vor, so die Vielzellbildung, die Zellsprossung und die
freie Zellbildung.
a) Freie Kernteilung und Vielzellbildung. Die Kernteilungen
in den vielkernigen Zellen der Thallophyten können bereits
als Beispiele für freie, d. h. von Zellteilungen nicht
begleitete, Kernteilungen angeführt werden. Aber auch
in Pflanzen mit typisch einkernigen Zellen kommen solche freie
Kernteilungen vor; besonders lehrreich in bestimmten, sehr großen
Zellen der Phanerogamen, den Embryosäcken, in denen der Embryo
ausgebildet wird. In den meisten Embryosäcken sieht man den
sekundären Embryosackkern sich in zwei Kerne teilen, die samt ihren
Nachkommen den Vorgang wiederholen. So entstehen schließlich nicht
selten Tausende von Kernen, die sich mit gleichen Abständen in dem
plasmatischen Wandbelag des Embryosackes verteilen. Zellteilungen
begleiten diese Teilungen nicht. Hört die Größenzunahme des
Embryosackes auf, so zerfällt sein protoplasmatischer Wandbelag
simultan oder fortschreitend in meist so viele Zellen, wie er Kerne
enthält. Dieser Vorgang, der als Vielzellbildung bezeichnet
wird, vollzieht sich folgendermaßen: Die Kerne umgeben sich in
ihrem ganzen Umkreis[S. 22] mit Verbindungsfäden, so daß sie strahlenden
Sonnen gleichen (Fig. 19); in diesen Fadenkomplexen treten gleich
weit von den Kernen Zellplatten und in diesen Zellwände auf. Die
Vielzellbildung läßt sich von der Zweiteilung ableiten und als ein
verkürzter Vorgang auffassen, der durch besondere Verhältnisse
(manchmal etwa durch ungewöhnlich rasche Größenzunahme einer
Zelle) bedingt sein kann. Durch Vielzellbildung entstehen auch die
Fortpflanzungszellen bei vielen Algen und Pilzen.
b) Zellsprossung. Eine Abart der typischen Teilung der
Protoplasten, aber mit ihr durch Zwischenstufen verbunden, ist auch
die Sprossung. Die Mutterzelle wird dabei nicht halbiert;
sie treibt vielmehr einen Auswuchs, der an seiner Ursprungsstelle
später durch eine Zellwand abgetrennt wird. So vermehren sich die
Zellen der Hefe (Fig. 20), und so entstehen auch die als Konidien
und als Basidiosporen bezeichneten Fortpflanzungszellen zahlreicher
Pilze (Fig. 398).
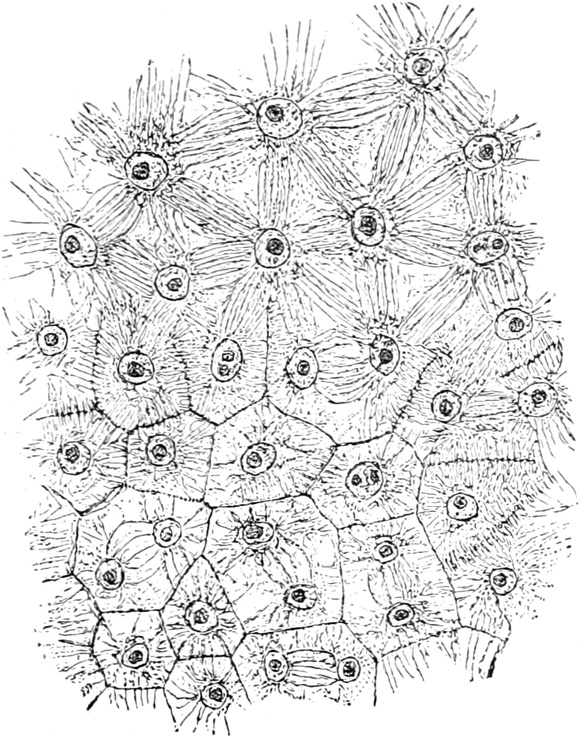
Fig. 19. Stück des protoplasmatischen Wandbelags aus dem
Embryosack von Reseda odorata, bei beginnender Vielzellbildung. Der
Vorgang schreitet von unten nach oben fort. Nach einem fixierten und
gefärbten Präparate. Vergr. 240. Nach STRASBURGER.
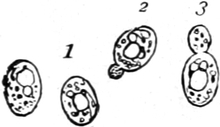
Fig. 20. Saccharomyces cerevisiae, 1 nicht sprossende,
2 und 3 sprossende Zellen. Vergr. 540. Nach STRASBURGER.
Fig. 21. Aufeinanderfolgende Stadien der Abgrenzung
einer Spore im Askus von Erysiphe communis. s Kerngerüst, n
Nucleolus. Vergr. 1500. Nach HARPER.
c) Freie Zellbildung. Dieser Vorgang entfernt sich von der
gewöhnlichen Zweiteilung der Zellen schon weiter; denn dabei folgt
auf die freie Kernteilung eine Bildung von Zellen, die einander
nicht berühren und nicht das gesamte Plasma ihrer Mutterzelle in
sich aufnehmen. Freie Zellbildung ist z. B. bei der Sporenbildung
der Ascomyceten, auch in der Keimanlage einiger nacktsamiger
Samenpflanzen (Gymnospermen), wie von Ephedra, und bei der
Bildung des Eiapparates und der Antipoden bei den Angiospermen zu
beobachten. Bei den Ascomyceten verläuft sie in folgender Weise.
Durch freie Teilung des in dem jungen Askusschlauch vorhandenen
Kernes und seiner Nachkommen werden in dem Plasma acht Kerne
gebildet. Um jeden Kern wird hierauf eine bestimmte Plasmamenge
der Zelle gegen die periphere Plasmamasse (Periplasma) durch eine
Plasmahautschicht abgegrenzt, die sich mit einer Zellhaut umgibt,
so daß acht voneinander getrennte Sporen entstehen (vgl. Fig. 382).
Wie die Untersuchungen von HARPER[27] gezeigt haben, geht
die Bildung der Hautschichten hierbei von einer zentriolenartigen[S. 23]
Plasmaansammlung aus (Fig. 21 A), die dem Spindelpol der
vorausgegangenen Teilungsfigur entspricht. Nach dieser Plasmamasse
hin ist der Kern schnabelartig lang vorgestreckt. Von ihr aus
werden springbrunnenartige Plasmastrahlen entsandt (kp), von
denen ein Teil schließlich zu der Hautschicht verschmilzt (B, C,
D).
III. Gröbere
leblose Einschlüsse der Protoplasten[28].
Abgesehen von den winzigen Mikrosomen, die im Plasma stets vorhanden
sind, treten bei der Umwandlung der embryonalen Zellen zu Dauerzellen
in allen Protoplasten, vor allem im Plasma und in den Chromatophoren,
gröbere leblose Einschlüsse auf. Erwähnt wurde ja schon der
Zellsaft, der in kleineren oder größeren Tröpfchen kaum einer
pflanzlichen Dauerzelle fehlt. Neben diesen Tröpfchen, die aus wäßrigen
Lösungen bestehen, kommen nicht selten auch Fett- oder Öltröpfchen und
feste Körper in amorpher Form oder als Kristalle vor. Viele dieser
Einschlüsse sind als Reservestoffe für das Leben der Pflanze
von großer Bedeutung; namentlich in den Zellen der Speicherorgane
(Knollen, Zwiebeln, Samen) häuft die Pflanze solche in großer Menge
auf, um sie im Falle des Bedarfs wieder zu verbrauchen. Andere sind
Endprodukte des Stoffwechsels, die aber ökologisch noch von
großer Wichtigkeit sein können. Von manchen Einschlüssen kennen wir die
chemische Zusammensetzung noch nicht.
A. Einschlüsse des Plasmas. 1. Flüssige Einschlüsse des Plasmas.
a) Der wäßrige Zellsaft. Wie wir schon wissen, wird als Zellsaft
die wäßrige Flüssigkeit in den größeren Vakuolen oder im
Saftraume ausgewachsener Pflanzenzellen bezeichnet (Fig. 3 v). Sie
ist reicher oder ärmer an sehr verschiedenen gelösten Substanzen, teils
Reservestoffen, teils Zwischen- oder Endprodukten des Stoffwechsels;
auch feste Einschlüsse, besonders in Form von Kristallen, kommen darin
vor. Der Zellsaft kann die gleichen, aber auch andere Stoffe gelöst
enthalten wie das Protoplasma und selbst in den Vakuolen einer
Zelle verschieden zusammengesetzt sein.
Jeder Zellsaft enthält zunächst anorganische Salze in Lösung,
besonders Nitrate, Sulfate und Phosphate. Er reagiert gewöhnlich
sauer, und zwar durch die organischen Säuren (Äpfelsäure,
C4H6O5, z. B. überall in den Blättern der Fettpflanzen;
Weinsäure, C4H6O6; Oxalsäure, C2O4H2 u. a.) oder
organischsauren Salze, die in ihm vorkommen.
Zu besonders häufigen Bestandteilen des Zellsaftes zählen ferner die
löslichen Kohlehydrate, die vielfach als Reservestoffe angehäuft
werden. Unter ihnen herrschen vor die Zuckerarten, vor allem die
Disaccharide (C12H22O11) Rohrzucker (Saccharose), Malzzucker
(Maltose) und von Monosacchariden (C6H12O6) der Traubenzucker
(Glykose). Oft wird Rohrzucker als Reservestoff gespeichert, z. B. in
der Mohrrübe, vor allem aber in der Zuckerrübe und dem Stengel des
Zuckerrohrs, woraus man ihn infolgedessen gewinnt. Eine ähnliche Rolle
spielen andere im Zellsaft gelöste Kohlehydrate, so bei den Kompositen
das Inulin, bei Pilzen das Glykogen. Als Zucker
wandern auch die Kohlehydrate innerhalb des Pflanzenkörpers.
Glykose oder Maltose haltige Schnitte, die in
Kupfersulfatlösung gelegt, dann abgespült und in Kalilauge
und Seignettesalzlösung erwärmt worden sind, reduzieren das
Kupferoxyd, so daß ein ziegelroter Niederschlag von Kupferoxydul
entsteht. Bei Vorhandensein von Rohrzucker wird der
Zellsaft nur blau gefärbt. Das Inulin, ein Polysaccharid
(C6H10O5)n, kann man mit Alkohol in Form kleiner
Kügelchen niederschlagen und in Wasser durch Erwärmen wieder
auflösen. Wenn inulinreiche Pflanzenteile, z. B. die Wurzelknollen
der Georgine (Dahlia variabilis), in Alkohol oder Glyzerin gelegt
werden, so fällt das Inulin in kugeligen Gebilden, vielleicht
Sphäriten (Sphärokristallen), aus, die von radialen Spalten
durchsetzt sind, leicht in keilförmige Stücke zerfallen und
manchmal auch deutlich konzentrisch geschichtet sind.
[S. 24]
Das bei Tieren als Reservestoff sehr verbreitete Kohlehydrat
Glykogen, ein Polysaccharid von der Zusammensetzung
(C6H10O5)n, kommt als Einschluß des Plasmas im
Pflanzenreich nur bei den Pilzen, Myxomyceten und Cyanophyceen
in Form von Tröpfchen vor. Bei den Pilzen tritt es an die Stelle
anderer Kohlehydrate, z. B. der Stärke und des Zuckers. Jodlösungen
färben das Glykogen rotbraun. Die Färbung schwindet größtenteils
beim Erwärmen, um bei der Abkühlung wieder aufzutreten.
Schleim, der aus Kohlehydraten besteht, als Reservestoff
enthält der Zellsaft häufig in den Zellen von Zwiebeln, z. B. von
Allium Cepa und Urginea (Scilla) maritima, ferner in denen der
Orchisknollen, doch auch in Zellen oberirdischer Pflanzenteile
(Fig. 22), besonders der Fettpflanzen-(Sukkulenten-)Blätter.
Schleim kommt aber auch außerhalb der Protoplasten in Zellmembranen
vor (vgl. S. 32).
Weiter enthält der Zellsaft als Reservestoffe oder als Zwischenprodukte
des Stoffwechsels ganz allgemein auch Amide, vor allem das
Asparagin, vielfach auch Eiweißstoffe (für deren Reaktionen vgl.
S. 12).
Mit konzentrierten Lösungen von Gerbstoffen[29] gefüllte,
stark lichtbrechende und unter Umständen sehr große Vakuolen sind
im Plasma vieler Zellen, besonders Rindenzellen, vorhanden; auch
Alkaloide, Glykoside (vgl. S. 12) und den Glykosiden
verwandte Bitterstoffe sind nicht selten im Zellsafte gelöst.
Das alles sind meist Endprodukte des Stoffwechsels.
Als Gerbstoffe werden Gemische sehr verschiedenartig
zusammengesetzter aromatischer Verbindungen bezeichnet, die oft
Glykoside sind. Besonders verbreitet bei den Pflanzen kommen
in glykosidischer Bindung die Gallussäure, die Gallusgerbsäure
(Digallussäure oder Tannin) und die Ellagsäure vor. Die dunkelblaue
oder grüne Färbung mit Ferrichlorid- oder Ferrisulfatlösung, der
rotbraune Niederschlag mit wäßriger Kaliumbichromatlösung gelten im
allgemeinen als Gerbstoffreaktionen. Freilich reagieren auch einige
andere Stoffe so. Die Gerbstoffe werden in den Pflanzen meist nicht
weiter verarbeitet. Infolge ihrer fäulniswidrigen Eigenschaften
dienen sie öfters zur Imprägnierung von Zellhäuten, die länger
ausdauern sollen.
Vielfach ist der Zellsaft gefärbt, besonders durch Anthozyane,
eine Gruppe stickstoffreier Glykoside. Sie sind rot in sauren,
blau in schwach alkalischen Zellsäften; unter Umständen sind sie
auch dunkelrot, violett (so in neutralem Zellsaft), dunkelblau,
selbst schwarzblau gefärbt. Alkalien wandeln die Farbe oft in grün
um. Bei einer sehr großen Anzahl intensiv gefärbter Pflanzen sind
die Anthozyane auch kristallinisch oder amorph ausgeschieden.
Seltener findet man, im Zellsaft gelöst, auch gelbe Farbstoffe, die
Anthochlore[30], z. B. in den Zellen der gelben Blütenblätter
der Primeln, des gelben Fingerhutes, der Löwenmäulchen, der
Königskerze; oder auch ein braunes Pigment, das Anthophaein, z. B.
in den Zellen der schwarzbraunen Flecken in den Saubohnenblüten.
Einsicht in die chemische Konstitution der Anthozyane verdankt
man vor allem den Untersuchungen von WILLSTÄTTER und
seinen Schülern[31]. Danach sind es meist Glykoside, in denen an
Zucker aromatische Farbstoffkomponenten, die Zyanidine,
gebunden sind, z. B. bei der Kornblumenblüte das Zyanidin
(C15H10O6), bei der Blüte des Rittersporns das
Delphinidin (C15H10O7). Die Zyanidine, die auch frei
in Zellsäften vorkommen können, sind Hydroxylverbindungen eines
Phenylbenzopyryliums; sie sind den Flavonen verwandt, die in
Pflanzen sehr weit verbreitet sind. In roten Blüten sind die
Zyanidine an Säuren gebunden, in blauen an Alkalien; in violetten
sind es neutrale Farbstoffe. Auch die Anthochlore sind Glykoside
mit aromatischen Farbstoffkomponenten, die zu den Flavonen gehören,
oder solche freien Flavone[30].
„Blutfarbige“, d. h. braune Laubblätter, z. B. die der
Blutbuchen, Bluthaselnüsse u. a., verdanken ihre eigenartige Färbung
dem Zusammenwirken von rotem Anthozyan und grünen Chlorophyllkörnern.
Auch die Rötung der Laubblätter im Herbste beruht auf Anthozyanbildung.
Bei den Blüten und Früchten kommen die verschiedenen
Farben, die im allgemeinen der Anlockung von Tieren dienen
und deshalb als Lockfarben bezeichnet werden, durch die Farben der
Zellsäfte, die Verteilung[S. 25] der farbstoffhaltigen Zellen, durch
Chromoplasten, endlich auch oft durch die Kombination der gelösten
Farbstoffe mit gelben, gelbroten oder roten Chromoplasten und grünen
Chloroplasten zustande.
b) Fettvakuolen. Als Reservestoffe sind die Fette (fetten Öle)
im Pflanzenreiche so verbreitet, daß ungefähr neun Zehntel aller
Phanerogamen sie im Plasma ihrer Samen und zwar als feinste, optisch
nicht nachweisbare Emulsion speichern. In besonders fettreichen Samen
macht das Öl bis zu 70% der Trockensubstanz aus. Fette können aber
auch als stark lichtbrechende Tröpfchen (Fettvakuolen) im Plasma
auftreten, so z. B. in den keimenden Samen. Die Fette sind Gemische
vieler Glyzerinester von Fettsäuren, besonders der Palmitinsäure
(C16H32O2), der Stearinsäure (C18H36O2), der Ölsäure
(C18H34O2) u. a. Mit diesen Reservestoffen wird der Raum der
Speicherorgane am besten ausgenutzt, da das Fett einen besonders großen
Energievorrat gegenüber anderen Speicherstoffen hat.
c) Vakuolen mit ätherischen Ölen und Harzen[32]. Auch sie
bilden stark lichtbrechende Tröpfchen; z. B. im Zellinhalt
zahlreicher Blumenblätter, in Rhizomen verschiedener Pflanzen
(Acorus calamus, Zingiber officinale), in Rinden (Cinnamomum),
in Blättern (Laurus nobilis), endlich in Fruchtschalen und Samen
(Piper nigrum, Illicium anisatum). Die Wände solcher Zellen sind
nicht selten verkorkt. Die ätherischen Öle sind vor allem Gemische
von Terpenen (C10H16)1 bis n und Terpenderivaten nebst
gewissen Estern, Phenolen, Phenolderivaten und höheren Alkoholen;
die Harze sind Gemische von Terpenen und Harzsäuren, die durch
Oxydation aus den Terpenen entstehen. Ätherische Öle und Harze
haben fäulniswidrige Eigenschaften. Die ätherischen Öle der
Blüten locken durch ihren Duft die bestäubenden Insekten an.
Unter Umständen nimmt das Öl auch Kristallform an, z. B. in den
Blumenblättern der Rose.
2. Feste Einschlüsse des Plasmas. a) Kristalle von
Kalziumoxalat, Ca(CO2)2 mit zwei oder sechs Mol.
Kristallwasser, kommen in sehr vielen Pflanzen vor. Sie werden, als
Endprodukte des Stoffwechsels, wohl meist im Zytoplasma (oder seltener
im Zellsafte kleinerer oder größerer Vakuolen) angelegt, liegen später
aber sehr oft im Zellsaftraum und nehmen unter Umständen schließlich
fast die ganze Zelle ein. In letzterem Falle sind die übrigen
Bestandteile der Zelle sehr reduziert, die Zellwände nicht selten
verkorkt. Es bilden sich entweder große Einzelkristalle (Fig.
132 k, 173 Bk, 182 k), deren Formen leicht zu erkennen sind, oder
viele winzige Kriställchen, die so zahlreich sein können, daß sie als
Kristallsand die Zelle anfüllen, oder viele, Rhaphiden
genannte Kristallnadeln, die parallel nebeneinander liegen und in
der Zelle Rhaphidenbündel bilden (Fig. 22), oder schließlich
morgensternförmige Kristalldrusen (Fig. 132 k′, 184 k). Bei
jeder Pflanzenart herrschen bestimmte Kristallformen vor.
Die großen Einzelkristalle gehören dem tetragonalen oder dem
monosymmetrischen Kristallsystem an. Im ersteren Fall enthalten sie
6 Mol., im letzteren 2 Mol. Kristallwasser. Der Konzentrationsgrad
der Lauge, aus der die Kristalle entstehen, soll es oft bedingen,
ob sie sich nach dem einen oder nach dem anderen System
bilden. Besonders häufig begegnet man den morgensternförmigen
Kristalldrusen, aus vielen Kristallen zusammengesetzt, die
von einem organischen Kern ausstrahlen. Bei monokotylen Gewächsen,
doch auch bei zahlreichen Dikotylen, sind die nadelförmigen,
monoklinen Rhaphiden verbreitet (Fig. 22). Ein solches
Bündel ist stets in eine große, mit Schleim gefüllte Vakuole
eingeschlossen. Die Oxalatkristalle sind ohne Aufbrausen löslich in
Salzsäure, aber unlöslich in Essigsäure.
Fig. 22. Eine mit Schleim und einem Rhaphidenbündel
gefüllte Zelle aus der Rinde von Dracaena rubra. r das
Rhaphidenbündel. Vergr. 160. Nach SCHENCK.
[S. 26]
Auch Kieselkörper, die sich nur in Fluorwasserstoffsäure
lösen lassen, werden in manchen Zellen, besonders bei Gräsern,
Palmen und Orchideen, gebildet. Sie füllen oft fast die ganze Zelle
aus.
b) Kleber und Eiweißkristalle. In saftigen Reservestoffbehältern
werden vor allem gelöste Eiweißkörper als Reservestoffe
im Zellsaft gespeichert. Man kann solche z. B. in den Zellen der
Kartoffelknolle mit Alkohol als feinkörnigen Niederschlag fällen. In
trockenen Reservestoffbehältern aber, so namentlich in zahlreichen
fetthaltigen Samen, werden die Eiweißkörper zu festen Körnern, den
Kleber-, Proteïn- oder Aleuronkörnern (Fig.
23), die in fettreichen Samen besonders groß sind. Sie gehen aus
Vakuolen hervor, deren Eiweißgehalt allmählich steigt, schließlich bei
Wasserverlust in Form eines rundlichen Korns oder in einzelnen Fällen
eines unregelmäßigen, sogar gelappten Gebildes erstarrt, und bestehen
vornehmlich aus Globulinen[33]. Diese Eiweißstoffe kristallisieren
in vielen Fällen teilweise aus und bilden einen, selten mehrere, im
Aleuronkorn eingeschlossene Kristalle (Fig. 23 k). Besonders groß
werden diese Kristalle in den Aleuronkörnern der Para„nüsse“ (der Samen
von Bertholletia excelsa). In Aleuronkörnern mit Eiweißkristallen
kommen meist noch rundliche Körner, die Globoide (Fig. 23 g),
vor, die wohl ebenfalls aus Eiweißkörpern bestehen, doch verbunden
mit dem Kalzium- und Magnesiumsalz (dem Phytin) der organischen
Inosithexaphosphorsäure C6H6[O2P(OH)2]6. Globoide liegen
übrigens bei manchen Samen auch frei im Plasma. Ferner können Kristalle
von Kalziumoxalat in Aleuronkörnern eingeschlossen sein. In den Körnern
unserer Getreidearten enthält die äußerste Zellschicht relativ kleine,
einschlußfreie Aleuronkörner (Fig. 24 al), das innere Gewebe dagegen
fast nur Stärke. Die Aleuronschicht bleibt, bei der Verarbeitung der
Körner zu Mehl, an den Körnerschalen haftend in der Kleie
zurück, geht also für das Mehl verloren.
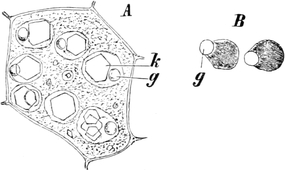
Fig. 23. A Zelle aus dem Endosperm des Rizinussamens
unter Wasser beobachtet. B Einzelne Aleuronkörner unter Olivenöl, k
Eiweißkristall, g Globoid. Vergr. 540. Nach STRASBURGER.
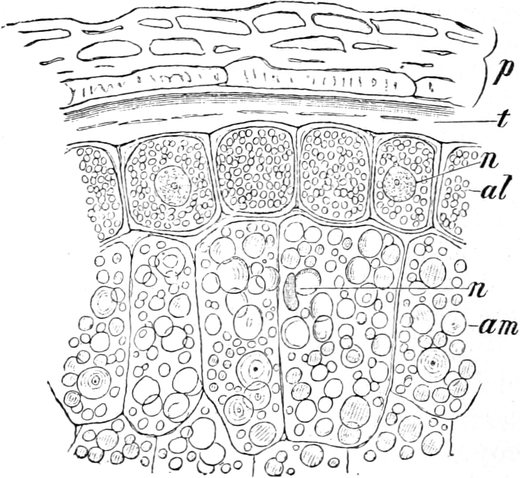
Fig. 24. Äußerer Teil eines Querschnittes durch ein
Weizenkorn (Triticum vulgare). p Fruchthülle, t Samenhaut. An die
Samenhaut grenzt das Endosperm. In diesem al Aleuronkörner, n
Zellkern, am Stärkekörner. Vergr. 240. Nach STRASBURGER.
Die Reaktionen des Klebermehls sind im wesentlichen die nämlichen,
die wir früher schon für Eiweißkörper kennen gelernt haben.
Mit Jodlösung färbt sich zum Beispiel die Aleuronschicht des
Weizenkorns gelbbraun.
Die Eiweißkristalle, die quellbar sind und sich ebenfalls mit
Jod gelbbraun färben, gehören dem regulären oder dem hexagonalen
Kristallsystem an. Solche Eiweißkristalle können aber auch
unmittelbar im Plasma vorkommen, so in peripherischen, stärkearmen
Zellen der Kartoffelknollen, ferner in Chromatophoren (Fig. 28) und
in Zellkernen, so[S. 27] nicht selten bei der Schuppenwurz (Lathraea) und
vielen anderen Scrophulariaceen, sowie den Oleaceen.
B. Einschlüsse der Chromatophoren. Eiweiß- und
Farbstoffkristalle haben wir schon als Einschlüsse der Chromatophoren
kennen gelernt (Fig. 28 kr). Sehr viel wichtiger aber ist
die Stärke[34]. Fast alle höher organisierten Pflanzen
bilden nämlich am Licht in ihren Chloroplasten Stärke, und zwar
in Körnerform aus. Die Körner treten hier in Mehrzahl auf (Fig.
15), werden aber nur ausnahmsweise groß, weil sie bald nach ihrer
Entstehung wieder aufgelöst werden, und sind meist aus noch kleineren
Körnchen zusammengesetzt. Große Stärkekörner findet man nur in
den Reservestoffbehältern, also dort, wo Stärke aus zugeführter,
assimilierter Substanz gebildet wird. Man bezeichnet solche Stärke als
Reservestärke im Gegensatz zu der Assimilationsstärke
der Chloroplasten. Auch sie entsteht, und zwar aus Zucker, nur in
Chromatophoren, den uns schon bekannten Leukoplasten (S. 16), die man
daher auch als Stärkebildner bezeichnet.
Alle Stärke des Handels ist Reservestärke. Ihre Menge in einem
Reservestoffbehälter ist oft sehr groß: sie macht etwa bis 20% des
Gesamtgewichts bei der Kartoffelknolle und sogar bis 70% beim Weizen
aus. Reines Stärkemehl, das nur aus Stärkekörnern besteht,
gewinnt man durch Auswaschen aus zerkleinerten Reservestoffbehältern.
Im gewöhnlichen Mehl aber sind auch die zermahlenen Zellhäute und
Protoplasten dieser Behälter enthalten.
Fig. 25. Stärkekörner aus der Kartoffelknolle. A Ein
einfaches, B ein halb zusammengesetztes Stärkekorn, C und D ganz
zusammengesetzte Stärkekörner. c Der Bildungskern des Stärkekornes.
Vergr. 540. Nach STRASBURGER.
Fig. 26. Stärkekörner aus den Kotyledonen von Phaseolus
vulgaris. Vergr. 540. Nach STRASBURGER.
Fig. 27. Stärkekörner des Hafers (Avena sativa). Ein
zusammengesetztes Korn und Teilkörner aus einem solchen. Vergr. 540.
Nach STRASBURGER.
Die Reservestärke besteht aus flachen oder rundlichen (eiförmigen oder
kugelrunden) Körnern, die in den Speicherorganen verschiedener Pflanzen
sehr ungleich groß sind, wie schon ein Vergleich der gleich stark
vergrößerten Figuren 25–27 zeigt; ihre Größe schwankt zwischen 0,002
und 0,17 mm. Die größten sind bereits mit dem bloßen Auge als helle
Körperchen zu erkennen. Verhältnismäßig große Stärkekörner, im Mittel
mit einem Durchmesser von 0,09 mm, enthalten die Kartoffelknollen.
Sie sind (Fig. 25) hier deutlich geschichtet. Die Schichtung
wird durch die verschiedene Dichte der Kornsubstanz verursacht und
ist exzentrisch: es wechseln dickere, dichtere Lagen, die im
durchfallenden Lichte heller sind, mit dünneren, weniger dichten
und dunkleren ab, und zwar ist der organische Initialpunkt oder
Bildungskern, um den die Schichten sich gelagert haben, dem einen
Rande[S. 28] des Kornes bedeutend genähert. Dagegen sind die Stärkekörner
der Hülsenfrüchte und der Getreidearten zentrisch geschichtet:
ihr Bildungskern liegt in der Mitte. Die deutlich geschichteten
Stärkekörner der Bohne (Phaseolus vulgaris, Fig. 26) werden außerdem
meist von radialen Spalten durchsetzt. Beim Weizen sind sie in
einer und derselben Zelle von zweierlei, sehr verschiedener Größe
als undeutlich geschichtete linsenförmige Großkörner und winzige
kugelförmige Kleinkörner ausgebildet. Die bisher betrachteten
Reservestärkekörner sind einfach. Es gibt aber auch halb
zusammengesetzte und ganz zusammengesetzte. Die ersteren
enthalten zwei oder mehr Teilkörner, die von gemeinsamen Schichten
umgeben sind: die letzteren bestehen nur aus Teilkörnern ohne
gemeinsame Schichten. Halb zusammengesetzte (Fig. 25 B) und ganz
zusammengesetzte (Fig. 25 C, D) Stärkekörner kommen in der
Kartoffelknolle vereinzelt zwischen den einfachen vor. In anderen
Fällen sind ganz zusammengesetzte Stärkekörner fast allein vorhanden,
so z. B. im Haferkorn (Fig. 27) oder im Reiskorn. 4–100 Teilkörner
setzen die Stärkekörner beim Reis, bis 300 beim Hafer, gelegentlich bis
30000 bei Spinacia glabra zusammen. Die Stärkekörner haben also bei
jeder Pflanzenart eine für sie bezeichnende Form.
Der Bau der Stärkekörner erklärt sich aus ihrer Bildungsgeschichte.
Bleibt das Stärkekorn während seines Wachstums von der Substanz der
Leukoplasten gleichmäßig umhüllt, so wächst es gleich stark nach allen
Seiten und erhält zentrischen Bau. Gelangt es während seines Wachstums
an die Peripherie des Stärkebildners, so wächst es dort stärker, wo
die Substanz des Leukoplasten es in größerer Dicke umgibt, und wird
exzentrisch (Fig. 28). Zusammengesetzte Körner bilden sich dann, wenn
in einem Leukoplasten gleichzeitig mehrere Stärkekörner nebeneinander
entstehen, die bei weiterem Wachstum zusammenstoßen. Werden um die
Teilkörner noch gemeinsame Schichten abgelagert, so kommt ein halb
zusammengesetztes Korn zustande.
Die Stärkekörner sind aus Kohlehydraten von der Zusammensetzung
(C6H10O5)n aufgebaut. Soll die Stärke im Stoffwechsel
weiter verwertet werden, so löst die Pflanze sie durch ein Enzym, die
Diastase, wieder auf; die Stärke wird dabei in Zucker (Maltose)
umgewandelt.
Fig. 28. Leukoplasten aus der oberirdischen Knolle der
Orchidee Phajus grandifolius. A, C und D von der Seite, B
von oben gesehen. st Stärke, kr Eiweißkristall. Vergr. 540. Nach
STRASBURGER.
Die Stärkekörner hält man für kristallinische Gebilde,
Sphärokristalle oder Sphärite, die aus miteinander verwachsenen,
feinen, radial angeordneten und büschelig verzweigten
Kristallnadeln der α- und β-Amylose aufgebaut sein sollen. Die
Schichtung ist der Ausdruck von Form- und Mengenverschiedenheiten
der Kristallnadeln in den aufeinander folgenden Schichten.
Im polarisierten Lichte zeigen die Stärkekörner, ähnlich wie
anorganische Sphärite, ein dunkles Kreuz. Auch Röntgenogramme der
Stärkekörner sprechen vielleicht für deren kristallinischen Bau.
Die Stärkekörner werden meist durch wasserhaltige Jodlösungen
zunächst blau, schließlich fast schwarz gefärbt; weinrot
färben sich aber z. B. die des Klebreises. Sie verquellen bei
gewöhnlicher Temperatur leicht in Kali- oder Natronlauge und in
Chloralhydratlösung, außerdem unter Kleisterbildung in Wasser von
60–80° C. Lösung, d. h. Umwandlung in Zucker ohne vorausgehende
Quellung, erfolgt in konzentrierter Schwefelsäure. Ohne Zusatz
von Wasser erhitzt, d. h. geröstet, geht Stärke in wasserlösliche
Stoffe („Röstgummi“, technisches Dextrin) über.
[S. 29]
Mit Jod rötlich färbt sich auch die (Florideen-)„Stärke“ der
Rotalgen. Diese rundlichen Körner haben ähnlichen Bau wie die
Stärkekörner der höheren Pflanzen, scheinen aber außerhalb der
Chromatophoren, jedoch in inniger Berührung mit ihnen zu entstehen
und sollen chemisch dem Glykogen näher stehen als echter Stärke[35].
IV. Die Zellmembranen[36].
Wie schon erwähnt, ist jeder Protoplast bei den Pflanzen in der Regel
von einem festen Gehäuse, der Zellhaut oder Zellmembran,
umgeben. Sie ist ein Außenprodukt des Protoplasten, das wir nicht als
lebend betrachten. Viele Gewächse beginnen freilich ihre Entwicklung
mit nackten Protoplasten, entweder als unbehäutete Schwärmsporen oder
Eizellen. Diese Zellen scheiden aber, ehe sie sich zu entwickeln, zu
teilen beginnen, an ihrer Oberfläche eine dünne Zellhaut aus. Bei
der Vermehrung der Zellen werden, wie wir gesehen haben, gewöhnlich
nach der Teilung des Plasmas Scheidewände zwischen die neu gebildeten
Zellen eingeschaltet, so daß auch dann alle Protoplasten von Zellhäuten
umhüllt bleiben.
Da nacktes Protoplasma meist Kugelform annimmt, so ist es die Zellhaut,
die die Gestalt der umhäuteten Zellen bedingt. Die Zellen, die
embryonal verhältnismäßig klein und ziemlich einförmig gestaltet sind,
wachsen nämlich zu ihren endgültigen Größen und zu ihren besonderen
Formen nur durch das Flächenwachstum ihrer Zellmembranen
heran. Bald ist dieses Wachstum ringsum überall gleich, bald auf die
Spitze oder eine Kante der Zelle oder einen die Zelle rings umlaufenden
Gürtel oder anders gestaltete, eng umschriebene Stellen beschränkt. Es
kommt entweder zustande durch Dehnung der vorhandenen Membran,
oder es erfolgt durch Einlagerung (Intussuszeption) neuer
Substanz zwischen die Teilchen der schon vorhandenen Haut.
Fig. 29. A Runde, gestielte Zelle von Saprolegnia
mit runden Tüpfeln in der Zellmembran. B Ein Tüpfel derselben, bei
stärkerer Vergrößerung im optischen Querschnitt.
Fig. 30. Steinzelle aus der Walnußschale mit
Membranschichtung und verzweigten Tüpfelkanälchen. Die unvollständig
gezeichneten Tüpfelkanäle verlaufen schräg zur Ebene der Zeichnung.
ROTHERT, frei nach REINKE.
Die Zellwand dient auch dem Schutze und ferner vor allem der
Festigung des Protoplasten. Diese wird durch Spannung der
Membran (Turgor, vgl. S. 191) und durch Dickenwachstum
der Zellhaut erreicht. Wie die Zelle durch das Flächenwachstum der
Membran ihre endgültige Form erhält, so bekommt die Membran
durch das Dickenwachstum ihre endgültige, bezeichnende Struktur.
Die Zellmembranen, die zuerst sehr zarte, dünne und strukturlose Häute
sind, werden nämlich weiterhin gewöhnlich ringsum überall gleich oder
nicht überall gleichmäßig verdickt, und zwar in der Weise, daß sie
an einzelnen Stellen verhältnismäßig dünn bleiben, während sie an
anderen viel stärker in die Dicke wachsen. In vielen Zellen wird die
ganze Zellhaut mit Ausnahme kleiner rundlicher (kreisförmiger,
elliptischer) oder spindelförmiger Stellen,[S. 30] Tüpfel, verdickt;
so entstehen in verdickten Zellmembranen Grübchen (Fig. 29) oder
röhrenförmige Kanäle (Fig. 30), die Tüpfelkanäle, die die
Verdickungsschichten durchsetzen, an einem Ende aber, zumeist dem
äußeren, durch unverdickte Zellhautteile, die Schließhaut
des Tüpfels, abgeschlossen sind (Fig. 29 B). Nicht selten werden
in gewissen Zellen mehrere Tüpfelkanäle bei weiter fortschreitender
Verdickung der Membranen zu einem einzigen Kanale vereint. Solche
verzweigte Tüpfel pflegen sehr eng zu sein und kommen
vornehmlich stark verdickten und harten Zellwänden zu, so denen der
Steinzellen oder Sklereïden (Fig. 30). In anderen
Zellen nimmt dagegen die Zellhaut im allgemeinen nur wenig
an Dicke zu, indem die Verdickung nur auf eng umgrenzte Teile
beschränkt bleibt, die dadurch die Form von Höckern, Warzen,
einfachen oder verzweigten Zäpfchen (Fig. 31), Stacheln (Fig. 32),
Leisten, Netzen oder Bändern (Fig. 67, 68) von charakteristischem
Bau erhalten. Solche Verdickungen sitzen der Zellhaut bald außen,
bald innen auf (zentrifugale, zentripetale Verdickungen). Kleine nach
außen vorspringende Höcker kommen z. B. an den meisten Haaren vor;
besonders mannigfaltig werden solche Verdickungen ausgebildet auf den
Außenflächen von Sporen und Pollenkörnern (Fig. 32) und in vielen
wasserleitenden Zellen der höheren Pflanzen (Fig. 67, 68).
Fig. 31. Stück einer schlauchförmigen Zelle (Rhizoid)
des Lebermooses Marchantia mit okalen, zapfenförmigen Wandverdickungen.
Vergr. 240.
Wandverdickungen können sehr seltsame Form annehmen, wenn sie auf
kleine Stellen in einer Zelle beschränkt sind, so ganz besonders
bei den Zystolithen, z. B. in den Blättern von Ficus
elastica (Fig. 33): zentripetalen Wandverdickungen von der Form
traubenförmiger gestielter Körper, in die sehr viel Kalziumkarbonat
eingelagert ist.
Das Dickenwachstum, das schon während des Flächenwachstums der Zellhaut
zu beginnen pflegt, aber auch nach dessen Beendigung noch fortdauern
kann, erfolgt meist durch Substanzanlagerung (Apposition)
von dem Protoplasma aus an die bereits vorhandenen dünnen Häute,
und zwar in Form neuer Membranlamellen. So entsteht in Zellen, in
denen der größte Teil der Zellhaut verdickt wird, gewöhnlich eine
schalenförmige Schichtung der Zellmembranen (Fig. 30): in den
Verdickungsschichten wechseln meist dickere, dichtere Lamellen mit
dünneren, weniger dichten, wasserreicheren und oft auch chemisch von
den dichteren verschiedenen Lamellen ab. Die dichteren brechen das
Licht stärker als die dünneren, erscheinen infolgedessen heller und
leuchtender. Auch viele scheinbar homogene Zellhäute lassen nach
Quellung mit starken Säuren oder Alkalien solche Schichtung deutlich
erkennen.
Fig. 32. A Pollenkorn des Kürbis in Flächenansicht
und zum Teil auch im optischen Durchschnitt. Das Präparat war mit
Zitronenöl durchsichtig gemacht worden. Vergr. 240. B Teil eines
Querschnittes durch die Pollenhaut von Cucurbita verrucosa. Vergr. 540.
Nach STRASBURGER.
Fig. 33. Zystolithenzelle von Ficus elastica. c
Zystolith. Vergr. 240.
[S. 31]
Nicht selten beruht das Dickenwachstum aber auch auf
Substanzeinlagerung (also Intussuszeption).
Besonders zentrifugale Wandverdickungen kommen oft durch
Intussuszeptionswachstum zustande. Solches kann auch fern vom
Protoplasma stattfinden und mit chemischen und strukturellen
Differenzierungen der Zellhäute verbunden sein, so daß solche
Membranen fast wie lebende Gebilde erscheinen. An Zellen aber,
die durch freie Zellbildung entstanden sind, wie z. B. bei den
Askosporen, werden die zentrifugalen Wandverdickungen von dem
Periplasma ausgebildet, aus dem die Zellen herausgeschnitten worden
sind (vgl. S. 22). Ebenso werden die zentrifugalen Verdickungen bei
Pollenkörnern und vielen Sporen von außen her durch die Tätigkeit
von Tapetenzellplasma aufgelagert, das die Behälter der Sporen
oder Pollenkörner innen auskleidet. Nach Auflösung der Tapete
verschmelzen nämlich ihre Protoplasten zu einem Periplasmodium, das
die Sporen- oder Pollenanlagen allseitig umgibt[37].
In manchen Fällen sieht man in den Verdickungsschichten einer
Membran bei Betrachtung von der Fläche feine Streifen (Fig.
34), die schräg zur Längsachse der Zelle verlaufen. Diese
Streifung beruht entweder auf einer Sonderung jeder
Verdickungslamelle in abwechselnd verschieden dichte Streifen,
wovon die dichteren oft in das Zellinnere vorspringen, oder,
bei vielen Algen (z. B. Cladophora), auf einer wellblechartigen
Fältelung der einzelnen Lamellen. Ist die Wandung deutlich
geschichtet, so sind die Streifen in den aufeinander folgenden
Verdickungslamellen meist entgegengesetzt geneigt (Fig. 34).
Chemie der Zellmembranen[38]. Trotz diesen Wachstumsvorgängen
ist die Zellmembran von Anfang an kein lebender Teil des Protoplasten,
sondern ein Ausscheidungsprodukt von ihm, das sich im Laufe der Zeit
noch in verschiedener Weise, auch chemisch, verändern kann. In lebenden
Zellen ist sie stets von Wasser durchtränkt und gequollen,
schrumpft infolgedessen bei Wasserentziehung mehr oder weniger
zusammen. Ihre Lamellen bestehen aus Kohlehydraten, vor allem
aus Zellulosen, doch teilweise auch aus Hemizellulosen
und Pentosanen, meist aus mehreren dieser Verbindungen zugleich.
In keinem Falle also sind die pflanzlichen Membranen nur aus reinen
Zellulosen aufgebaut, auch nicht, wenn man kurz von Zellulosemembranen
spricht. Die Zellulosen kommen in den Membranen aller Pflanzen vor,
mit Ausnahme der meisten Pilze; es sind Polysaccharide von der
Zusammensetzung (C6H10O5)n, die sich in Jodlösung nicht,
mit Chlorzinkjodlösung aber intensiv blau färben. Die gleiche Reaktion
gilt übrigens für viele Hemizellulosen, die ebenfalls Polysaccharide
sind. Die Zellhäute enthalten fast stets in größeren Mengen auch noch
andere Substanzen, z. B. auch solche, die sich mit Chlorzinkjod nicht
bläuen, sondern anders färben. Unter ihnen sind die Pektinstoffe
besonders wichtig, die mit diesem Reagens gelbbraune Färbung annehmen.
Darauf beruht es, daß viele „Zellulosemembranen“ sich mit Chlorzinkjod
nicht rein blau, sondern violett, braunviolett oder braun färben.
In den Membranen der meisten Pilze und Bakterien ist Chitin
vorhanden, das früher als spezifisch tierischer Membranstoff galt; es
soll bei den Pilzen die Zellulose vertreten[39].

Fig. 34. Teil einer Sklerenchymfaser von Vinca major
bei oberer Einstellung. Auch die inneren Grenzen der Wand wurden
bei tieferer Einstellung in das Bild eingetragen. Vergr. 500. Nach
STRASBURGER.
Die Zellulosen sind unlöslich in verdünnten Säuren, in
Alkalien, selbst konzentrierter Kalilauge. Dagegen sind sie unter
schwacher Hydrolyse löslich in Kupferoxydammoniak und, unter
Umwandlung in Dextrose, in konzentrierter Schwefelsäure oder[S. 32]
sehr stark konzentrierter Salzsäure. Ferner werden sie durch ein
besonderes Enzym, die Zellulase, das die Pflanze bildet, und
zwar in diesem Falle über das Disaccharid Zellobiose in Dextrose
übergeführt. Nach vorausgegangener Behandlung mit Schwefelsäure
oder Phosphorsäure werden sie durch wäßrige Jodlösung blau gefärbt,
ebenso bei gleichzeitiger Einwirkung der konzentrierten Lösungen
bestimmter Salze, wie Chlorzink oder Chloraluminium, mit Jod. Daher
ist das gebräuchlichste Reagens, um Blau- oder Violettfärbung der
Zellulosen zu erzielen, eben Chlorzinkjodlösung. Eine Reihe von
Stoffen, die den Zellulosen nahe stehen, aber schon durch verdünnte
Säuren in lösliche, von der Dextrose verschiedene Zuckerarten
(z. B. Mannose, Galaktose) umgewandelt werden, faßt man als
Hemizellulosen zusammen. Besonders reich daran sind die
Pflanzenschleime und die Reservezellulosen (vgl. S. 36). Einige von
ihnen sind in Kupferoxydammoniak unlöslich. So wie die Zellulosen
hochmolekulare Polysaccharide von Hexosen (C6H12O6)
sind, so sind die Pentosane (C5H8O4)n
entsprechende hochmolekulare Kondensationsprodukte von Pentosen
(C5H10O5), z. B. von Arabinose, Xylose. Die Pektine
sind durch die Leichtigkeit ausgezeichnet, womit sie sich, nach
vorhergegangener Behandlung mit verdünnten Säuren, in Alkalien
lösen. Sie färben sich im Gegensatz zur Zellulose mit Safranin
und Methylenblau intensiv. Die Pektine sind verwickelt gebaute
Verbindungen, worin an Tetragalakturonsäure (C24H34O25,
einem Kondensationsprodukt der Galakturonsäure C6H10O7)
Monohexosen, Pentosane, ferner esterartig Methylalkohol und
salzartig Kalzium und Magnesium gebunden sind[40]. Anwesenheit von
Pektinen bedingt die Gelatinierung von Fruchtdekokten (also die
Bildung von Fruchtgelees).
Das Chitin ist ein stickstoffhaltiges Polysaccharid
(C30H50O19N4), das Azetylessigsäure in säureamidartiger
Bindung enthält.
Die Zellhäute erfahren im Laufe des Lebens einer Zelle oft
mannigfache chemische Umwandlungen dadurch, daß die bereits
ausgebildeten Schichten ihre Beschaffenheit ändern oder die neuen
Verdickungsschichten in ihrer Zusammensetzung von den vorhandenen
abweichen. Diese Umwandlungen stehen oft in engster Beziehung zu
den Anforderungen, die an die Zellen gestellt werden. Was zunächst
die „Zellulose“membranen betrifft, so sind sie, ganz jung, wenig
elastisch, dagegen, wie es mit Rücksicht auf das bevorstehende starke
Längenwachstum günstig erscheint, verhältnismäßig stark dehnbar; später
pflegt sich das umzukehren. Sie setzen der Diffusion von Wasser und
gelösten Substanzen kaum Widerstand entgegen.
Nicht selten verschleimen Zellulosemembranen durch Umwandlung
ihrer Substanz in gallertige oder schleimartige, in Wasser stark
quellende Massen. Besonders oft tritt aber Verholzung,
Verkorkung und Kutinisierung der Zellmembranen ein.
Verholzung der Membranen verringert die Dehnbarkeit der Zellen ganz
bedeutend, erhöht also die Starrheit, ohne die Durchlässigkeit für
Wasser und darin gelöste Stoffe aufzuheben. Verkorkte und kutinisierte
Membranen aber sind verhältnismäßig undurchlässig für Wasser und
Gase und setzen die Verdunstung stark herab. Häufig werden auch die
Zellhäute nachträglich durch Derivate von Gerbstoffen sehr
dunkel gefärbt und gegen Fäulnis geschützt, so in Samenschalen und
in älterem Holz. In jede ältere Membran sind ferner anorganische
Stoffe unter Umständen in bedeutender Menge eingelagert, sehr
häufig Kieselsäure, seltener Kalziumkarbonat, ferner
organische Salze, z. B. besonders häufig Kalziumoxalat.
Die Verholzung beruht auf der Einlagerung von Ligninen in
die Kohlehydratlamellen. Die chemische Zusammensetzung der Lignine
ist aber noch wenig geklärt. Wahrscheinlich sind Benzolderivate an
ihrer Zusammensetzung beteiligt. In den Zellen, deren Membranen
verholzt sind, bestehen aber die innersten Membranschichten in
vielen Fällen aus Zellulose. Als besonders charakteristische
Reaktionen verholzter Zellwände gelten: Gelbfärbung mit
schwefelsaurem Anilin, Rotfärbung mit Phloroglucin und Salzsäure.
Diese Reaktionen werden wohl durch aromatische Stoffe bewirkt,
die in den verholzten Membranen vorkommen. Mit Chlorzinkjodlösung
färben sich verholzte Membranen gelb, nicht blau. Der Holzstoff
läßt sich technisch aus den verholzten Membranen durch längeres[S. 33]
Kochen mit Kalziumbisulfit- oder Natronlauge unter Druck
herauslösen (in mikroskopischen Schnitten auch durch Eau de
Javelle), so daß nur die Kohlehydratlamellen zurückbleiben. In
dieser Weise stellt man aus Holz „Zellulosezellstoff“ her.
Die Verkorkung beschränkt sich in der Regel auf die
mittleren Verdickungsschichten einer Membran. Die verkorkten
Lamellen bestehen nur aus Suberinen, enthalten
also keine Kohlehydrate; sie werden den unverkorkten
Membranlamellen angelagert. Mit der Verkorkung nicht völlig
übereinstimmend, wenn ihr auch nahe verwandt, ist die
Kutinisierung. Sie besteht in einer nachträglichen
Auflagerung von Kutinen auf Zellulosemembranen oder einer
Einlagerung in solche. Zwischen Kutinen und Suberinen bestehen
keine scharfen Unterschiede. Beide nehmen mit Chlorzinkjodlösung
gelbbraune, mit Kalilauge annähernd gleiche gelbe Färbung an,
färben sich mit Sudanglyzerin rot, und beide werden durch
konzentrierte Schwefelsäure oder Kupferoxydammoniak nicht gelöst.
Doch widerstehen die Kutine besser der Kalilauge. Die Kutine und
die Suberine verhalten sich übrigens je nach ihrer Abstammung gegen
Reagenzien etwas verschieden. Die Suberine sollen nach VAN
WISSELINGH[41] fettartige Körper sein aus Glyzerinestern
und anderen zusammengesetzten Estern der Phellon-, Suberinsäure
und anderen höheren Fettsäuren; den Kutinen soll dagegen die
Phellonsäure, die in den Suberinen stets vorhanden ist, immer
fehlen.
Kalziumkarbonat kommt bei manchen Pflanzen, wie den meisten
Characeen unserer Seen und Teiche, so massenhaft in den Membranen
vor, daß diese starr und brüchig werden. Kieselsäure ist
in den peripherischen, dadurch sehr harten Zellwänden der Gräser,
Schachtelhalme und vieler anderer Pflanzen, z. B. der einzelligen
Diatomeen, vorhanden. Das Kalziumoxalat ist meist in
Kristallen ausgeschieden.
Auch die zur Flavongruppe gehörenden Farbstoffe der
technisch benutzten Farbhölzer haben ihren Sitz in den Membranen.
Feste Zellmembranen können nachträglich in Gummi umgewandelt
werden, so bei der Gummosis in einem Holzkörper. Bei Prunus- oder
Citrus-Arten spielt sich dieser Vorgang so ab, daß nacheinander die
einzelnen Verdickungsschichten der Zellwände zu Gummi verquellen.
Schließlich wird auch der Zellinhalt zu einem Bestandteil der
Gummimasse[42]. Der Gummi ist chemisch den Pflanzenschleimen sehr
ähnlich (vgl. S. 32).
Röntgenspektroskopische Untersuchungen der letzten Zeit machen
es wahrscheinlich, daß die Zellulosemembranen wie die Stärke
kristallinische Struktur haben. Sie bestehen aus Kristalliten,
die mit einer ihrer Hauptachsen parallel zur Längsachse der Zelle
angeordnet sind[43].
Zweiter Abschnitt. Gewebelehre
(Histologie)[44].
I. Die Gewebebildung.
A. Begriffsbestimmung und Bedeutung des Zellgewebes. Jeder
innigere Verband umhäuteter Protoplasten wird als Zellgewebe
bezeichnet.
Nur die niedersten Organismen bestehen aus einem ein- oder
mehrkernigen Protoplasten; sie sind zeitlebens einzellig.
Meist ist aber der Körper einer Pflanze vielzellig, d. h.
er setzt sich aus vielen, voneinander durch Zellwände
getrennten Protoplasten, also aus Zellgewebe, zusammen. Höhere
äußere Organisation und größeres Körpervolumen sind nämlich bei den
weitaus meisten Pflanzen an die Ausbildung vieler umhäuteter
Protoplasten gebunden. Freilich gibt es, wie wir schon sahen, auch
äußerlich hochorganisierte Algen (die Schlauchalgen), bei denen eine
solche Kammerung nicht vorkommt, sondern das Innere von einem
vielkernigen Protoplasten eingenommen wird. Man kann sie als nicht
zellige Organismen den zellulären gegenüberstellen. Solche Organismen
gibt es aber nur wenige. Ausbildung von Zellgewebe war eben für
die Entwicklung höher organisierter Pflanzenformen offenbar von
größter Bedeutung. Sie ermöglichte es, eine Arbeitsteilung
im Protoplasma[S. 34] des Körpers durchzuführen. Durch die Zerteilung des
Protoplasmas in viele Protoplasten entstanden ebensoviele einzelne
Elementarteile, die verschiedene Aufgaben übernehmen konnten,
und vergrößerte sich außerdem die Plasmaoberfläche ganz
erheblich. Durch die Ausbildung der trennenden Zellwände wurde es
ferner möglich, diese Elementarteile mehr oder weniger gegeneinander
zu isolieren, zugleich den Zusammenhalt und die innere Aussteifung des
ganzen, aus vielen weichen Protoplasten bestehenden Körpers zu erhöhen.
Ein, freilich sehr unvollkommenes, lockeres Gewebe bildet sich
schon bei solchen Organismen, deren Zellen nach jeder Teilung
sich voneinander trennen, aber durch eine Gallerte aus ihren
verquellenden Zellhäuten zusammengehalten werden. Man nennt
derartige Verbände mehr oder weniger selbständiger Zellen
gemeinsamen Ursprungs Zellfamilien oder Zellkolonien.
Mannigfaltige Beispiele dafür bieten uns die Spaltalgen (Fig. 35)
und die Ordnungen der Volvocales und der Protococcales unter den
Grünalgen (vgl. diese). In den Zellfäden und Zellflächen niederer
Algen, deren Zellen bereits fest verbunden sind, beginnen
allmählich die Merkmale geschlossener Gewebe hervorzutreten. Diese
Merkmale werden mit wachsender Zellenzahl im Organismus und mit
der fortschreitenden Arbeitsteilung zwischen den Zellen immer
deutlicher.
Fig. 35. Gloeocapsa polydermatica. A Beginn einer
Teilung. B Links: kurz nach der Teilung. C Im Ruhezustande. Vergr.
540. Nach STRASBURGER.
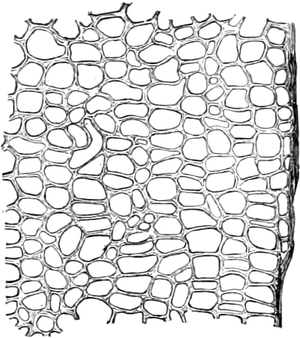
Fig. 36. Längsschnitt durch das Mutterkorn Vergr. 300.
Nach SCHENCK.
Fig. 37. Längsdurchschnitt durch den Stiel des
Fruchtkörpers des Steinpilzes. Vergr. 300. Nach SCHENCK.
B. Ursprung der Gewebe. Die Entstehung pflanzlicher Gewebe ist
im allgemeinen auf Zellteilungen zurückzuführen. Bei manchen
niederen Algen, z. B. bei Hydrodictyon, werden aber Gewebe durch
Anlagerung freier Zellen aneinander und durch Verwachsung ihrer
Zellmembranen, bei den Pilzen und Schlauchalgen (Siphoneen) durch
Verflechtung schlauchförmiger Zellen oder Zellfäden gebildet
(Flechtgewebe oder Plectenchym, Fig. 37). Kommt es dabei zu einer
innigen Verwachsung der verflochtenen Zellen und ist dieses Geflecht
besonders dicht, so macht es in dünnen Schnitten einen ganz ähnlichen
Eindruck wie ein Gewebe der höher organisierten Pflanzen (Fig. 36);
bei nachträglicher Verdickung der Wände können sogar die Tüpfel in den
nachträglich miteinander verwachsenen Zellhäuten aufeinander treffen.
C. Die Zellmembranen im Zellgewebe. Betrachtet man Schnitte
durch pflanzliches Zellgewebe bei schwächeren Vergrößerungen, so sind
es meist ausschließlich oder vor allem die Zellmembranen, die ins
Auge fallen; sie scheinen bei weniger genauem Zusehen ein Netzwerk
von Fäden, ähnlich einem Stoffgewebe, zu bilden, woraus sich der Name
Zellgewebe erklärt.
1. Schichtung. Alle Scheidewände, die bei den
Zellteilungen im Zellgewebe auftreten, sind, wie auch sonst bei
Zellteilungen, zunächst einfache und sehr dünne, meist
plattenförmige Membranlamellen, die den benachbarten Zellen
gemeinsam zukommen. In diesem Zustande bleibt die Zellmembran[S. 35]
aber nie. Sie wird schon in den embryonalen Zellen verdickt,
während die Zellen durch Flächenwachstum der Membranen sich
vergrößern. Die Verdickung erreicht aber ihr Ende erst, nachdem die
Zellen längst zu ihren endgültigen Größen herangewachsen sind. Sie
fällt verschieden aus, je nach den Funktionen, die die ausgebildeten
Dauerzellen übernehmen: besonders dicke Zellmembranen findet man
bei Zellen, die der mechanischen Festigung dienen (Fig. 38). In der
Regel wird die Verdickung einer Scheidewand von beiden angrenzenden
Protoplasten aus, und zwar meist durch beiderseitige Anlagerung
einer gleichen oder ungleichen Zahl neuer schalenförmiger
Membranlamellen an die dünne Scheidewand besorgt (Fig. 38,
40, 62). So bekommt eine jede Zelle nachträglich ihre eigenen
Membranschichten. Die den benachbarten Zellen gemeinsame
mittlere Membranlamelle einer Zellhaut bezeichnet man alsdann als
Mittellamelle (Fig. 38 m). Sie ist meist sehr dünn, nur an
den Zellecken und Kanten etwas dicker (Zwickel, Fig. 71 C m*) und
besteht hauptsächlich aus kalziumhaltigen Pektinstoffen, die
verhältnismäßig leicht löslich sind; in verholzten und verkorkten
Geweben ist sie außerdem oft verholzt.
In weichen Geweben lassen sich die Zellen schon durch Kochen in
Wasser voneinander trennen, das die Mittellamellen zum Verquellen
bringt, z. B. bei vielen Kartoffelknollen. In vielen reifen
Früchten tritt eine solche Isolierung von selbst ein. Durch das
SCHULZEsche Mazerationsgemisch (chlorsaures Kali
und Salpetersäure) oder durch heiße konzentrierte Ammoniaklösung
gelingt es, auch andere Zellen durch Auflösung der Mittellamellen
voneinander zu lösen, durch das Mazerationsgemisch z. B. die des
Holzes. Auch gibt es pektinvergärende Bakterien, die mittels des
Enzyms Pektinase die Mittellamellen zerstören und gewisse Zellen,
z. B. bei der Flachsrotte die mechanischen Zellen der Flachsstengel
von den anderen Zellen, trennen.
Die Verdickungsschichten unterscheiden sich meist optisch und chemisch
von der Mittellamelle; da sie sich in der Regel zu beiden Seiten der
Mittellamelle gleichmäßig anlagern, so erhält die Scheidewand zwischen
zwei Zellen mehr oder weniger symmetrischen Bau (Fig. 38–40,
41, 62), der sich selbst auf die Tüpfel erstreckt. Nicht selten ist
in stärker verdickten Zellhäuten, besonders den Zellen des Holzes,
beiderseits der Mittellamelle eine Sonderung in drei, ihrem
optischen und chemischen Verhalten nach verschiedene Schichten zu
erkennen, die sich als primäre, sekundäre und tertiäre
Verdickungsschichten unterscheiden lassen und selbst wieder aus
vielen Lamellen bestehen können. Am stärksten pflegt alsdann die
sekundäre Verdickungsschicht entwickelt zu sein; sie bildet die
Hauptmasse der Wandung. Die innerste Verdickungsschicht einer Zellhaut
ist meist stärker lichtbrechend; sie wird als Grenzhäutchen oder
Innenschicht bezeichnet (Fig. 71 Ci) und besteht meist aus Zellulose.
Zellwände, die nicht an andere Zellen grenzen (Fig. 40, 44), vor
allem also die Außenwände an der Peripherie des Pflanzenkörpers,
sind dagegen asymmetrisch gebaut. Bei solchen Wänden können
ja nur einseitig nach dem Zellinnern hin Verdickungslamellen an die
zunächst allein vorhandene dünne Zellhaut angelagert werden.
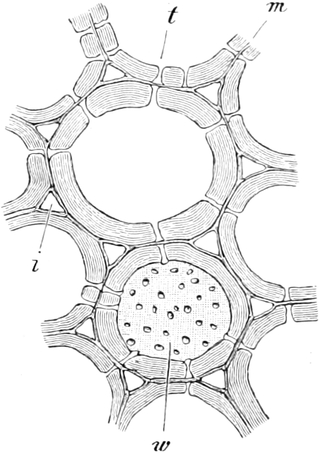
Fig. 38. Stark verdickte Zellen aus dem Marke eines
älteren Stammstückes der Waldrebe Clematis vitalba. m Mittellamelle,
i Interzellularraum, t Tüpfelkanäle in Seitenansicht. In der einen
Zelle ist die untere Wand w mit den Tüpfeln in Aufsicht zu sehen.
Vergr. 300. Nach SCHENCK.
[S. 36]
2. Tüpfelung. Die Zellmembranen, die die Protoplasten
voneinander trennen, erschweren begreiflicherweise den Stoffaustausch
zwischen den Zellen, und zwar um so mehr, je dicker sie sind. Ohne
regen Stofftransport von Zelle zu Zelle kann aber das Leben des ganzen
Organismus nicht bestehen. So muß also dafür gesorgt werden, daß die
Verdickung der Membranen, die der Festigung dient, die Stoffbewegung
nicht allzusehr hemmt. Diesem Bedürfnis wird meist durch Ausbildung von
Tüpfeln in den Scheidewänden zwischen den Protoplasten Rechnung
getragen. In den freien Wänden findet man dagegen in der Regel keine
Tüpfel.
Diese Tüpfel, die in stärker verdickten Zellmembranen Kanäle mit
kreisrundem (Fig. 38 bei w und 39 bei m), elliptischem oder
spaltenförmigem Querschnitte sind, treffen in den benachbarten
Zellen aufeinander; sie sind also beiden Zellen gemeinsam, werden
aber in der Mitte von der unverdickten Zellhaut, ihrer Schließhaut,
durchsetzt (Fig. 38 t, 39 p, 40 t). Spaltenförmige Tüpfel pflegen
schräg gerichtet zu sein und in den Verdickungsschichten benachbarter
Zellen einander zu kreuzen.
Der Bau der Tüpfel läßt sich besonders leicht untersuchen an den
stark verdickten und reichlich getüpfelten Zellwänden der Samen
verschiedener Palmen, zahlreicher Liliaceen, z. B. von Ornithogalum
(Fig. 39), und anderer Monokotylen. Die Verdickungsschichten
bestehen hier aus einer Hemizellulose, die ein Reservestoff des
Samens ist und bei der Keimung durch ein Enzym (Zytase) aufgelöst
wird. Sie sind glänzend weiß und können so hart werden, daß solche
Samen, z. B. die der Palme Phytelephas macrocarpa, technisch als
„vegetabilisches Elfenbein“ zur Anfertigung von Knöpfen u. dergl.
verwertet werden.
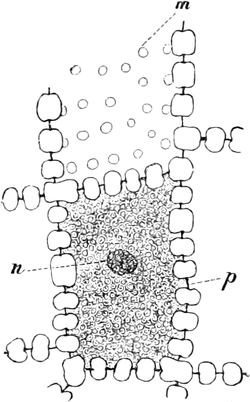
Fig. 39. Zellen aus dem Samen (Endosperm) der Liliacee
Ornithogalum umbellatum. m Tüpfel von oben, p Tüpfelkanal in
Seitenansicht mit der Schließhaut. n Zellkern. Vergr. 240. Nach
STRASBURGER.
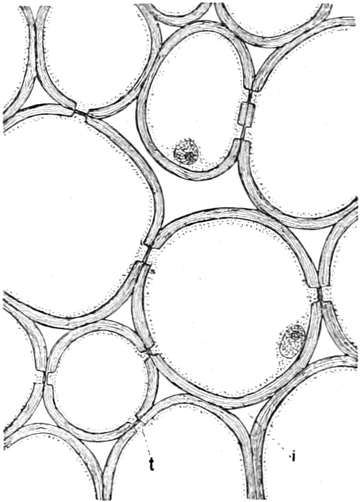
Fig. 40. Zellen aus der Wurzelrinde von Iris florentina.
t Tüpfel in den geschichteten Zellmembranen, i Interzellularen.
Vergr. gegen 400.
3. Verbindung der Protoplasten im Gewebe. Ein harmonisches
Zusammenarbeiten aller lebenden Teile des Körpers, wie es in den
Lebensäußerungen des gesamten Organismus so auffällig zutage tritt,
wäre freilich wohl kaum möglich, wenn die lebenden Protoplasten
im Zellgewebe des Körpers durch die Zellwände völlig getrennt
nebeneinander liegen würden. Tatsächlich sind sie nicht ganz
gegeneinander abgeschlossen, sondern durch zahlreiche, äußerst feine
Protoplasmafäden verbunden, die von ihren Hautschichten ausgehen
und die Zellhäute durchsetzen. Meist sind diese Plasmaverbindungen
oder Plasmodesmen[45], die sämtliche lebende Zellen
des Körpers verbinden, auf die Schließhäute der Tüpfel beschränkt
(Fig. 41 s); doch können sie auch die Zellhäute außerhalb der
Tüpfel durchsetzen (Fig. 42 pl). Sie erheben die Protoplasten des
ganzen Pflanzenkörpers trotz der Ausbildung der Zellwände zu einer
organischen, lebenden Einheit und dienen wohl ebenfalls zur
Stoff- und außerdem zur Reizleitung von Protoplast zu Protoplast.
[S. 37]
4. Zellfusionen. Die Lebenstätigkeit des vielzelligen Organismus
macht es aber auch nötig, Stoffe noch schneller, als es selbst
durch weite Tüpfelkanäle möglich ist, innerhalb seines Körpers von
einem Organ in ein anderes, etwa aus den Wurzeln in die Blätter, zu
schaffen. Die Diffusion von Stoffen durch die Zellmembranen oder
die Stoffbewegung in den äußerst feinen Plasmodesmen genügt dazu
vielfach nicht, wenn sie auch durch die Ausbildung der Tüpfelkanäle
sehr erleichtert wird, die ja in den Scheidewänden zwischen
benachbarten Zellen stets korrespondieren. Dementsprechend verschmelzen
viele, besonders dem Stofftransporte dienende, außerdem aber auch
manche andere Zellen, nachträglich durch breite offene Löcher zu
zusammenhängenden Röhren (vgl. S. 54, 55 u. 59), zu Zellfusionen
miteinander. Solche Löcher in Ein- oder Mehrzahl entstehen alsdann
durch entsprechende Auflösung der Wandsubstanz in den Zwischenwänden,
namentlich in den Endwänden benachbarter Zellen.
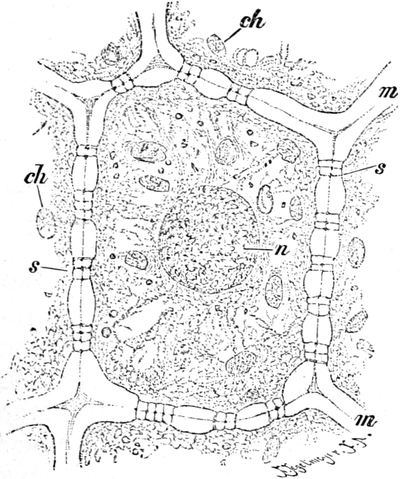
Fig. 41. Eine Zelle aus der Rinde der Mistel (Viscum
album) nach entsprechender Härtung und Färbung der Protoplasten und
Quellung der Wände (m). Die Schließhäute (s) der Tüpfel von
Plasmodesmen durchsetzt, ch Chloroplasten, n Zellkern. Vergr. 1000.
Nach STRASBURGER.
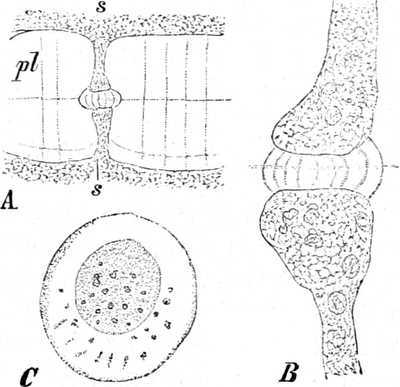
Fig. 42. A Ein etwas gequollenes Wandstück aus dem
Endosperm der Elfenbeinpalme (Phytelephas macrocarpa). Bei s und
s die aufeinander treffenden, mit Plasma gefüllten Tüpfelkanäle der
beiden angrenzenden Zellen, in der Schließhaut zarte Plasmodesmen,
außerdem die ganze Dicke der Zellwand durchsetzende Plasmodesmen pl.
Vergr. 375. B Die Tüpfelkanäle und die Plasmodesmen der Schließhaut
1500mal vergrößert. C Tüpfelkanal und Schließhaut von der Fläche
gesehen, bei 1500facher Vergrößerung. Der kleinere Kreis ist der
Tüpfelkanal, der größere die Schließhaut, die dunkleren Punkte darin
sind die Plasmodesmen. Nach STRASBURGER.
5. Bildung von Interzellularen und Durchlüftung der Gewebe.
Fast immer, wenn sich embryonale Zellen in Dauerzellen umwandeln,
werden die Mittellamellen nach Ausbildung von Verdickungsschichten
lokal, namentlich an den Ecken und Kanten der Zellen, gespalten,
so daß hier die Wände benachbarter Zellen auseinander weichen
können. So entstehen im Zellgewebe schon sehr frühzeitig, bis in
die äußersten Stengel- und Wurzelspitzen hinein, lufterfüllte
Zwischenzellräume (Interzellularen, Fig. 38 i, 40 i).
Meist haben sie im Querschnitt die Form kleiner Drei- oder Vierecke,
die der erwähnten Spaltung der Zellwand ihre Entstehung verdanken und
daher schizogen genannt werden. Die Interzellularen bilden ein
zusammenhängendes System (Interzellularsystem) reich verästelter
feiner Kanäle, die den Zellkanten entlang streichen und das
Gewebe allseitig durchziehen. Bei bevorzugtem Wachstum bestimmter
Zellwandstellen können derartige schizogene Interzellularen benachbarte
Zellen nachträglich völlig voneinander trennen und zu größeren
Kammern oder Gängen von mehr oder weniger regelmäßiger[S. 38] Gestalt
erweitert werden. Auch durch Zerstörung von Zellen kann die Bildung
von Zwischenräumen veranlaßt werden; solche auf Zerreißung von
Zellen zurückzuführende Interzellularräume heißen rhexigen,
durch Auflösung von Zellwänden entstehende lysigen. In
manchen Fällen werden schizogene Interzellularen weiterhin rhexigen
oder lysigen vergrößert. Ungleich verteiltes Wachstum führt oft zu
einer Dehnung und Zerreißung ganzer Gewebegruppen. Auf diese Weise
entstehen z. B. die hohlen Stengel. In Geweben, die sich durch
Verflechtung von Zellfäden gebildet haben, sind die Zwischenzellräume
von vornherein vorhanden (Fig. 37).
Die Interzellularen enthalten gewöhnlich Luft und sind für die lebenden
Gewebezellen von größter Bedeutung. Eine Zelle, die allseits von Wasser
oder Luft umspült ist, findet jederzeit in ihrer Umgebung leicht die
Gase, die sie zum Leben braucht. Die zahllosen Protoplasten in den
Geweben der vielzelligen Pflanzen würden aber nicht lebensfähig sein,
wenn nicht dafür gesorgt wäre, daß auch zu ihnen solche Gase gelangen
können. Diese Aufgabe, die Gase im Innern der Gewebe zirkulieren zu
lassen, erfüllen die Interzellularräume.
II. Zellarten,
Gewebearten und Gewebesysteme.
Nur bei den niederen mehrzelligen Pflanzen besteht das Zellgewebe des
Körpers aus lauter ziemlich gleichartigen kugelförmigen, polyëdrischen
oder zylindrischen Zellen (vgl. z. B. Fig. 84), die sämtlich in
fast gleicher Weise allen Lebensfunktionen dienen. Man kann diese
Gewebe Parenchym nennen. In dem Maße, wie mit fortschreitender
äußerer Organisation und mit Zunahme der Größe des Organismus die
Arbeitsteilung zwischen den Protoplasten zunimmt, erhalten Zellen
einzeln oder gruppenweise verschiedene Form, verschiedenen Bau und
besondere Aufgaben. So entsteht namentlich bei den höheren Pflanzen
eine Sonderung der gleichartigen Zellen in eine Anzahl verschieden
gebauter Zellarten, zwischen denen es aber immer Übergänge gibt.
Untersucht man vergleichend die verschiedensten Organe einer Pflanze
und aller höher organisierten Pflanzen miteinander, so findet man, daß
die Zahl dieser verschiedenen Zellarten klein ist und daß bestimmte
Zellformen überall wiederkehren.
Meist sind gleichartige Zellen zu Gruppen verbunden. Einen
solchen Verband aus lauter gleichartigen Zellen nennt man eine
Gewebeart. Die Gewebearten unterscheiden sich durch die
Formen, den Inhalt und den Membranbau der Zellelemente, aus denen sie
bestehen; eine jede Gewebeart hat ihre besonderen Aufgaben, die in
einer Hauptfunktion oder in mehreren Funktionen bestehen können. Je
höher die Pflanze organisiert ist, um so mehr Gewebearten setzen ihren
Körper zusammen. Doch ist entsprechend den Zellarten auch die Zahl der
Gewebearten klein, da sie in gleicher Weise bei den verschiedensten
Gewächsen immer wieder auftreten. Nicht selten kommt es vor, daß in
ein Gewebe aus sonst gleichartigen Zellen einzelne Zellen (wohl auch
Idioblasten genannt) oder Zellgruppen mit ganz abweichendem Bau und
Inhalt eingeschaltet sind, die also einer anderen Zellart angehören.
Bei den höheren Pflanzen bilden ferner einzelne Gewebearten größere
Gewebemassen, die auf weite Strecken oder durch den ganzen
Pflanzenkörper in ununterbrochenem Zusammenhange stehen. Man
nennt solche Verbände, die oft auch aus verschiedenartigen
Gewebearten zusammengesetzt sind, morphologische Gewebesysteme.
Auch derartige zusammengesetzte Gewebeverbände können durch ihre
Baueigentümlichkeiten sehr auffallen und bestimmte Hauptfunktionen
haben; und[S. 39] zwar pflegen die verschiedenen Gewebearten einander in
ihren Funktionen zu ergänzen oder zu unterstützen.
Zu einem physiologischen Gewebesysteme endlich faßt man
alle Zellen zusammen, die in ihren Hauptfunktionen übereinstimmen,
gleichgültig ob und wie sie morphologisch verbunden und
woraus sie ontogenetisch entstanden sind. Solche Systeme sind also
etwas ganz anderes als die morphologischen Gewebesysteme.
Wir können sämtliche Gewebearten der höher organisierten Pflanzen
in zwei Hauptgruppen teilen, nämlich 1. in die embryonalen oder
Bildungsgewebe und 2. in die fertigen oder Dauergewebe.
A. Die Bildungsgewebe.
Sie werden auch Meristeme genannt. Sie bestehen aus
verhältnismäßig kleinen, nach allen Richtungen ungefähr gleich großen,
also isodiametrischen, würfelförmigen oder aus prismatischen,
plattenförmigen oder langgestreckten, embryonalen Zellen mit
dünnen Zellmembranen, reichlichem Plasma, großen Zellkernen und wenigen
kleinen Vakuolen (vgl. Fig. 2). Bezeichnend für sie sind meist die
zahlreichen Zellteilungen, die man in ihnen, wenigstens so lange sie
tätig sind, findet. Diese Bildungsgewebe, aus denen die Dauergewebe
hervorgehen, zerfallen nach den Orten ihres Vorkommens und der Art
ihrer Entstehung in Urmeristeme und sekundäre Meristeme.
1. Urmeristeme. Sie entstehen durch die Teilung der Keimzelle
und setzen zunächst den Embryo allein zusammen. Später sind sie
hauptsächlich an den Spitzen der Zweige und Wurzeln, an den
Vegetationspunkten dieser Organe (Fig. 102, 154) vorhanden. Hier
findet alsdann die Vermehrung der embryonalen Zellen und die Anlage
vieler Seitenorgane statt (apikales oder Spitzenwachstum).
Eine oder einige dieser Meristemzellen an der äußersten Spitze des
Vegetationspunktes bleiben dauernd embryonal und vermehren durch
Wachstum und darauffolgende Zellteilungen fortgesetzt die Zellen des
Meristems, während die embryonalen Zellen, die durch diese Teilungen
entstanden sind, sich meist nach weiteren Teilungen allmählich in
Dauerzellen umwandeln. Ist eine solche Spitzenzelle vorhanden
(Fig. 100, 101, 153), die alsdann meist durch Form und besondere Größe
von den übrigen Meristemzellen abweicht, so spricht man von einer
Scheitelzelle, sind mehrere in einer oder mehreren
Schichten (Fig. 102, 154) vorhanden, von Initialzellen. Diese
sind meist von den übrigen Meristemzellen der Form nach nicht zu
unterscheiden; bei manchen Pflanzen ähneln sie jedoch mehr oder weniger
den Scheitelzellen.
Dicht hinter dem Vegetationspunkte beginnen die annähernd
gleichartigen, lückenlos verbundenen Urmeristemzellen verschieden zu
wachsen und sich in Stränge und Schichten verschiedenartig gestalteter
Bildungszellen zu sondern, die aber sonst zunächst die Eigenschaften
der embryonalen Zellen noch behalten (Fig. 100, 102, 154). Schon
hier treten Interzellularen auf. Erst in größerer Entfernung vom
Vegetationspunkte bilden sich allmählich die Merkmale der verschiedenen
Dauergewebe aus, basalwärts fortschreitend in immer stärkerem
Maße, bis der fertige Zustand schließlich erreicht ist. Bei dieser
Gewebedifferenzierung kommt es sehr häufig vor, daß Gruppen,
Stränge oder Schichten von Zellen ihre meristematische Beschaffenheit
beibehalten und zu Ausgangspunkten für weitere Neubildungen von
embryonalen und fertigen Geweben werden. In vielen Fällen stellen sie
vorübergehend eine Zeitlang ihre Teilungstätigkeit ein.
Bei vielen Monokotylen bleiben die basalen Abschnitte der
Stengelglieder lange Zeit meristematisch und dienen außer dem Meristem
der Vegetationspunkte als Bildungsherde für Dauergewebe, aber nicht
wie dieses[S. 40] Meristem auch zur Bildung von Seitenorganen. Dadurch kommt
das interkalare Wachstum dieser Sprosse und vieler anderer
Pflanzenteile zustande.
2. Sekundäre Meristeme sind Meristeme, die aus untätig
gewordenen Resten von Urmeristemen durch neue Teilungen ihrer
Zellen oder als Neubildungen aus Dauerzellen entstehen,
letzteres dadurch, daß diese Dauerzellen einen Funktionswechsel
durchmachen und durch neue Zellteilungen sich in embryonale Zellen
zurückverwandeln (Folgemeristeme). Ihre Elemente gleichen denen
der Urmeristeme, haben aber in der Regel die Gestalt langgestreckter
oder plattenförmiger Prismen (Fig. 167). Solche sekundären Meristeme
geben den Anlaß zur Korkbildung und zum sekundären Dickenwachstum der
Baumstämme und heißen Kambien. Diese Kambien sind mehr oder
weniger dünne Meristemschichten (Fig. 165, 166), die als mantelförmige
Hohlzylinder parallel zur Organoberfläche im übrigen Gewebe verlaufen.
In den Kambien pflegt eine mittlere Schicht von Meristemzellen, die
Initialzellen, durch fortgesetzte tangentiale, also
gleichgerichtete Teilungen nach außen oder innen oder nach beiden
Seiten in radialer Richtung Tochterzellen (Gewebemutterzellen)
abzugeben, die sich ihrerseits, manchmal erst nach weiteren Teilungen,
zu Dauerzellen umbilden.
Die in Meristemen bei den Zellteilungen entstehenden neuen
Zellwände sind sehr häufig ebenflächig und werden in der Regel,
doch nicht immer, senkrecht zu den schon vorhandenen älteren
Zellwänden gestellt (Regel der rechtwinkligen Schneidung),
wobei die Richtungen: mehr oder weniger parallel zur
Organoberfläche (perikline Zellwände) und senkrecht dazu
(antikline Wände) bevorzugt werden.
B. Die Dauergewebe.
Die Dauergewebszellen unterscheiden sich von den embryonalen Zellen
dadurch, daß in ihnen im allgemeinen keine Zellteilungen mehr
stattfinden, daß sie meist viel größer, verhältnismäßig plasmaarm und
reich an großen Vakuolen oder gar tot sind, und daß ihre Zellhäute
verschiedenartig verdickt und oft chemisch verändert sind. Meist wird
das Dauergewebe von Interzellularen durchzogen. Es besteht gewöhnlich
aus verschiedenen Zell- und Gewebearten, die auch ganz verschiedenen
Funktionen dienen.
Das Dauergewebe geht dadurch aus den embryonalen Zellen hervor,
daß letztere sich strecken, stellenweise auseinander weichen,
ihre Zellwände verdicken und chemisch verändern, ihren Zellinhalt
eigenartig weiter ausgestalten, oft auch einbüßen, und unter Umständen
miteinander durch Auflösung der trennenden Wände verschmelzen. Das
Streckungswachstum vollziehen die embryonalen Zellen oft ganz
unabhängig voneinander (Fig. 172), so daß einzelne, die sich
besonders stark strecken, sich mit ihren Enden oder ihren Kanten
zwischen andere und aneinander vorbeischieben (gleitendes
Wachstum[46].
Man kann die Dauergewebe in verschiedener Weise einteilen, einmal nach
ihrer Herkunft. Primäre Dauergewebe sind solche, die aus
den Urmeristemen hervorgehen; die sekundären sind solche, die
den sekundären Meristemen ihre Entstehung verdanken.
Eine morphologisch brauchbarere Einteilung der Dauergewebe gewinnen
wir, wenn wir sämtliche Verschiedenheiten der Dauerzellen, der
Zellarten, in Betracht ziehen, die sie zusammensetzen.
Früher berücksichtigte man dabei hauptsächlich die
Dimensionen der Zellen und unterschied Parenchyme
und Prosenchyme. Parenchym nannte man ein Zellgewebe,
dessen Zellen isodiametrisch sind oder, falls sie in einer
Richtung gestreckt sind, mit queren Wänden aneinander grenzen;
als Prosenchyme dagegen bezeichnete man solche Dauergewebe, die
aus gestreckten, beiderseits zugespitzten, also spindelförmigen
oder faserförmigen und mit ihren zugespitzten Enden fest
zwischeneinander greifenden Elementen[S. 41] bestehen. Diese beiden
Gruppen genügen aber nicht, um eine Übersicht über die
Mannigfaltigkeit der Gewebearten zu ermöglichen. Ja selbst ihre
Begriffsbestimmungen sind veraltet.
Untersucht man vergleichend die höheren Gewächse, so findet man
zunächst Gewebe, die, wie die Gewebe der niedersten vielzelligen
Pflanzen, aus lebenden, nährstoffhaltigen Zellen mit meist ziemlich
dünnen Zellulosewänden bestehen und vielseitige Funktionen haben;
wir wollen sie auch als Parenchyme bezeichnen. Daneben
finden wir aber Gewebe, die sich durch ihre Baueigentümlichkeiten
scharf von den Parenchymen unterscheiden und besonderen
Funktionen dienen. Die auffälligsten Gewebe sind nach ihren
Hauptfunktionen die Abschlußgewebe, die mechanischen
oder Festigungsgewebe und die Leitgewebe. Als
besondere Gewebe kann man weiter noch die Sekretgewebe und
die Drüsengewebe betrachten. Die Parenchyme bilden das
parenchymatische System, das primäre Abschlußgewebe das
Hautgewebesystem, die Festigungsgewebe das mechanische
und die Leitgewebe das Leitbündelgewebesystem.
Nicht selten begnügt man sich auch mit der Einteilung aller
Dauergewebe in Haut-, Leitbündel- und Grundgewebesystem.
1. Parenchym, Parenchymsystem. Bezeichnend für diese Gewebeart,
die zu den phylogenetisch ältesten Geweben (vgl. S. 38) gehört, ist
die Parenchymzelle. Sie kann isodiametrisch oder gestreckt
sein, hat verschiedenen Umriß und wird (vgl. Fig. 3 B, Fig. 40,
41) durch die folgenden Merkmale gekennzeichnet: Die wenig verdickte
Zellmembran bereitet dem Stoffaustausche keine Schwierigkeiten; sie
besteht in der Regel aus Kohlehydratlamellen. Sie besitzt außerdem
einfache runde oder elliptische Tüpfel, die den Stoffverkehr
zwischen den Zellen noch mehr erleichtern. Lebendes Protoplasma
ist meist vorhanden; es umschließt große Vakuolen, die in Menge
Nährstoffe enthalten können. Die Chromatophoren, die meist als Leuko-
oder Chloroplasten ausgebildet sind, enthalten oft mehr oder weniger
Stärke. Das Parenchym ist zur Durchlüftung meist von Interzellularen
durchzogen. Es kann ebensogut sekundäres wie primäres Dauergewebe
sein. Die Parenchymzelle dient, wie schon erwähnt, noch vielerlei
Funktionen. In ihr spielen sich die wichtigsten Lebensvorgänge der
ausgewachsenen Pflanze ab: die Nährstoffbereitung, -leitung und
-speicherung, die Atmung und die Wasserspeicherung; außerdem dient
sie infolge ihrer prallen Füllung mit Zellsaft der allgemeinen
Festigung des Pflanzenkörpers. Entsprechend der Vielseitigkeit der
Leistungen ergeben sich kleine Bauverschiedenheiten zwischen den
Parenchymzellen, je nach der Funktion, der sie hauptsächlich
dienen. Sind die Zellen reich an Chloroplasten, so nennt man das
Parenchym Assimilationsparenchym (Fig. 8), entsprechend seiner
wichtigsten Aufgabe: aus Kohlensäure organische Substanz zu bilden. In
dieser Weise ist das Parenchym vielfach in oberirdischen Pflanzenteilen
ausgebildet, soweit das Licht in sie einzudringen vermag. Weiter
im Innern liegen farblose Parenchyme. Sind sie besonders reich an
organischen Inhaltsstoffen, wie Zucker, Stärke, fetten Ölen und
Eiweißstoffen, oder in den alsdann stark verdickten Zellmembranen an
Hemizellulosen (Fig. 39), lauter Stoffen, die aufgespeichert
sind, um später wieder in den Stoffkreislauf gezogen zu werden, so
spricht man von Speichergewebe (Fig. 23 A, 24). Parenchym, das
der Wasserspeicherung dient, bezeichnet man als parenchymatisches
Wassergewebe; es besteht gewöhnlich aus besonders großen,
plasmaarmen, aber sehr zellsaftreichen, häufig mit dünnem Schleime
gefüllten, dünnwandigen Zellen, die sich bei Wasserabgabe stark
verkleinern. Die Leitung organischer Nährstoffe, namentlich der
Kohlehydrate, findet in Parenchymzellen statt, die zur Erleichterung
des Stofftransportes meist in der Hauptleitungsrichtung langgestreckt
sind, dem Leit[S. 42]parenchym, das oft in mantelförmigen lückenlosen
Scheiden um andere Gewebemassen auftritt. Parenchym, das von besonders
großen Interzellularen zur starken Durchlüftung oder Luftspeicherung
durchzogen ist, heißt Aërenchym (Durchlüftungsgewebe).
2. Abschlußgewebe. In vielzelligen Gewebekörpern besteht,
namentlich bei Landpflanzen, die Notwendigkeit, den ganzen Körper
oder einzelne Gewebe gegen schädliche Wasserverluste, gegen
mechanische Verletzungen, gegen zu hohe Erwärmung[47] und oft auch
gegen Verluste von diffusiblen Nährstoffen zu schützen. Diese Aufgabe
haben Zellen, die meist in Schichten (Scheiden) angeordnet
sind, durch besondere Baueigentümlichkeiten erhalten. Dadurch ist
eine Reihe weiterer Gewebearten entstanden, deren Hauptelemente die
Epidermiszellen und die verkorkten Zellen sind. Die
Epidermiszellen bilden die Epidermis und zusammen mit anderen
Zellarten das Hautgewebesystem.
a) Hautgewebesystem. 1. Epidermis. Die Epidermis
oder Oberhaut geht stets aus oberflächlich gelegenen
Urmeristemschichten (dem Dermatogen, vgl. S. 74) hervor, ist also
ein primäres Dauergewebe. Sie schließt als eine schützende Hülle
den Pflanzenkörper nach außen ab, vermittelt aber zugleich den
Stoffaustausch mit der Außenwelt. In typischer Ausbildung ist sie
fast immer einschichtig (Fig. 45 B) und besteht aus lückenlos zu
einer abziehbaren Haut miteinander verbundenen, in Flächenansicht
tafelförmigen oder auch langgestreckten lebenden Zellen, deren
seitliche Umrisse meist wellig (Fig. 43) oder zackig sind, was die
Festigkeit ihres seitlichen Verbandes erhöht. Im Querschnitt sind die
Zellen tafel-, linsen- oder pflastersteinförmig. Die Protoplasten der
Oberhautzellen sind gewöhnlich auf dünne Wandbeläge beschränkt, ihre
großen Safträume meist mit farblosem, manchmal aber auch gefärbtem
Zellsafte gefüllt. Die Epidermis am Lichte wachsender Teile der meisten
Farne, doch auch einer großen Zahl von Schatten liebenden Phanerogamen,
ist mit Chlorophyllkörnern ausgestattet und alsdann auch an der
Assimilationsarbeit der Pflanze beteiligt. Bei fortgeschrittenerer
Arbeitsteilung unterbleibt aber auch an oberirdischen Organen die
Ausbildung der Chlorophyllkörner in der Epidermis.
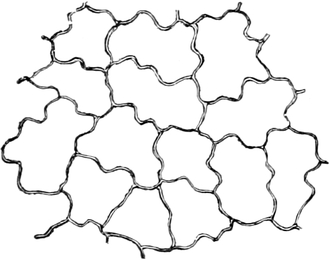
Fig. 43. Flächenansicht der Epidermis auf der
Blattoberseite von Mercurialis perennis. Vergr. 300. Nach H.
SCHENCK.
Alle für längere Lebensdauer eingerichteten, oberirdischen
Pflanzenteile haben, im Gegensatz zu den vergänglichen Blumenblättern,
verdickte Epidermisaußenwände. Diese Verdickung kommt
durch Anlagerung von Zelluloseschichten zustande, von denen namentlich
die äußeren hierauf meist, doch nicht immer, mehr oder weniger stark
kutinisieren (Fig. 188). Die Kutinisierung kann sich auch auf die
mittleren Lamellen der Seitenwände erstrecken. An den unter Wasser
lebenden und unterirdischen Pflanzenteilen, vor allem also den Wurzeln,
bei denen die Oberhaut oft besondere Funktionen, z. B. die Absorption
von Wasser und Salzen hat, sind die Außenwände dagegen dünn und nicht
kutinisiert.
Die Außenwände der Epidermen, mögen sie verdickt oder unverdickt
sein, sind außerdem, abgesehen von den Wurzeln, auf ihrer Außenseite
von einem zarten Kutinhäutchen, der Kutikula, bedeckt, das
ununterbrochen über sie fortläuft und auf den primären Außenwänden
der Epidermiszellen entsteht. Häufig ist die Kutikula ohne
Rücksicht auf die Zellgrenzen etwas[S. 43] gefältelt; sie sieht alsdann
in Flächenansicht gestreift aus. Die Kutikula und die kutinisierten
Schichten sind infolge ihres Gehaltes an Kutinen für Wasser und Gase
schwer durchlässig, und zwar um so schwerer, je dicker sie sind;
sie verhindern in wirksamster Weise schädliche Wasserverluste des
Gewebekörpers durch Verdunstung. Die Verdickung erhöht zugleich die
mechanische Festigkeit der Oberhautzellen. Den Wurzeln wird durch den
Mangel der Kutikula die Aufnahme des Bodenwassers und der Bodensalze
erleichtert.
In die Kutikula und in die kutinisierten Membranschichten ist oft
noch Wachs eingelagert, das sie noch weniger durchlässig für
Wasser macht. Von solchen Epidermen fließt das Regenwasser ab, ohne
sie zu benetzen. Tritt das Wachs aus der Kutikula nach außen hervor,
so entstehen Wachsüberzüge, die an Früchten, so besonders
auffällig an Pflaumen, Weinbeeren und anderen Organen einen hellgrauen,
abwischbaren Reif bilden. Sie können aus Körnchen (so z. B. bei den
Pflaumen und Weinbeeren), kürzeren oder längeren Stäbchen (Fig. 44)
oder Krusten bestehen und lösen sich in Äther oder in heißem Alkohol.
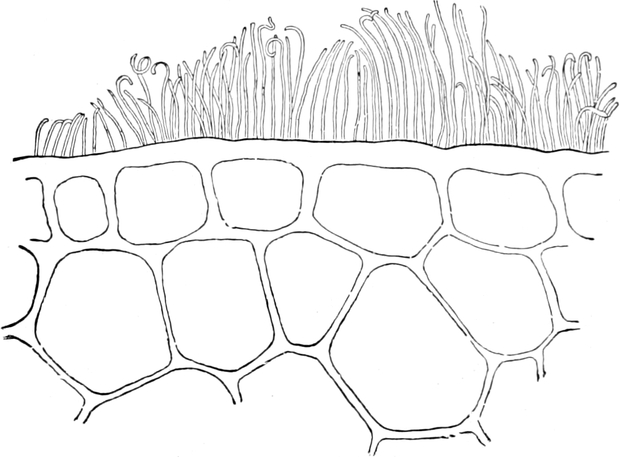
Fig. 44. Querschnitt durch einen Stengelknoten des
Zuckerrohrs mit stäbchenförmigen Wachsausscheidungen. Vergr. 540. Nach
STRASBURGER.
Das Pflanzenwachs besteht gleich den Suberinen und Kutinen aus
Fettsäureestern des Glyzerins, denen aber freie höhere Fettsäuren
und Fettsäureester anderer Alkohole beigemischt sind.
Vielfach schützt die Epidermis das umschlossene Gewebe
nicht nur gegen schädliche Wasserverluste durch Erschwerung
der Wasserdampfabgabe, sondern auch dadurch, daß sie ein
Wasserbehälter ist: Die meist unverdickten Seitenwände
legen sich bei Abgabe des Wassers, das im Zellraume gespeichert
ist, in Falten, um sich wie in einem Blasebalge bei neuer Füllung
der Zellen wieder zu strecken. Solche Epidermen sind manchmal auch
mehrschichtig.
Die Widerstandsfähigkeit der Epidermisaußenwände wird in bestimmten
Fällen durch Einlagerung von Kalk oder Kieselsäure erhöht. Bei
den Schachtelhalmen (Equiseten) ist die Verkieselung so stark,
daß man sie zum Polieren von Zinngefäßen nutzbar machen kann. Die
Fruchtschale der Graminee Coix Lacryma ritzt sogar Opal.
Eine besondere Mannigfaltigkeit in der Art der Verdickung und
in dem Verhalten der Verdickungsschichten findet man an den
Epidermiszellen der Früchte und häufiger noch der Samen. Die
Epidermen besorgen hier nicht allein den Abschluß der inneren
Teile, sondern fördern vielfach auch die Verbreitung und
Befestigung der Früchte und Samen, oder sie bilden druckfeste
Samenschalen.
2. Spaltöffnungsapparate[48]. Bei den höher organisierten
Gewächsen sind sehr bezeichnend für die Epidermen der meisten
oberirdischen, namentlich grünen Teile, die in der Luft und nicht
in Wasser wachsen, Paare gekrümmter, halbmondförmiger Zellen, die
eine Lücke, einen Spalt (Porus), zwischen sich lassen. Sie heißen
Schließzellen und samt den Spalten Spaltöffnungen
(Stomata) oder Spaltöffnungsapparate (Fig. 45 A, 46). Die
größten Spaltöffnungsapparate hat man bei den Gräsern beobachtet, so
beim Weizen von 0,079 mm Länge und 0,039 mm Breite mit einer 0,038 mm[S. 44]
langen und 0,007 mm breiten Spalte. In der Epidermis der Wurzeln kommen
dagegen solche Spaltöffnungen niemals vor.
Der Spalt unterbricht die lückenlose Schicht der
Epidermiszellen; er ist ein mit Luft gefüllter Interzellulargang,
der unter der Oberhaut in einen großen Interzellularraum (Fig. 45
B) mündet, den man Atemhöhle genannt hat, obwohl er mit
Atmung nichts zu tun hat. Dieser Raum steht mit den Interzellularen
des Parenchyms in Verbindung. Die Spalten sind für die Pflanzen von
größter Bedeutung; denn sie setzen das Interzellularsystem, das der
Durchlüftung der Gewebe dient, in Verbindung mit der Außenluft.
Solche Verbindungen sind aber wegen des lückenlosen Verbandes
der Epidermiszellen, wenn diese infolge der Ausbildung einer
Kutikula für Gase schwer durchlässig sind, unbedingt nötig, damit
die Interzellularenluft genügend schnell erneuert, in den grünen
Pflanzenteilen namentlich die verbrauchte Kohlensäure ersetzt werden
kann. Dagegen vermag der in der Luft reichlich vorhandene Sauerstoff
auch durch die Epidermiszellen, selbst wenn sie von einer Kutikula
überzogen sind, meist in hinreichender Menge in die Pflanzenteile
einzudringen.
Die Schließzellen enthalten stets Chlorophyll und zeichnen
sich, abgesehen von ihrer Form, durch ihre Wandverdickungen aus,
die an Querschnitten (Fig. 45 B, 47 B) kenntlich werden.
Diese Verdickungen bestehen meist aus je einer oberen und
unteren Verdickungsleiste an der Spaltseite (der Bauchseite) der
Schließzellen, während die Wand dort in halber Höhe ebenso wie die
ganze Rückenwand dünn bleibt (Fig. 45 B). Die Verdickungen haben
zu den Gestaltsänderungen, zu denen die Schließzellen befähigt
sind, Beziehung. Die Schließzellen haben nämlich die Aufgabe, durch
solche Gestaltsveränderungen die Spaltenweite zu verändern: durch
Verringerung ihrer Krümmung den Spalt z. B. etwa zu Zeiten allzu
großen Wasserverlustes zu schließen, durch Zunahme ihres Volumens und
Verstärkung ihrer Krümmung ihn zu anderen Zeiten weit zu öffnen. Sie
sind Regulatoren des Gasaustausches und der Transpiration.
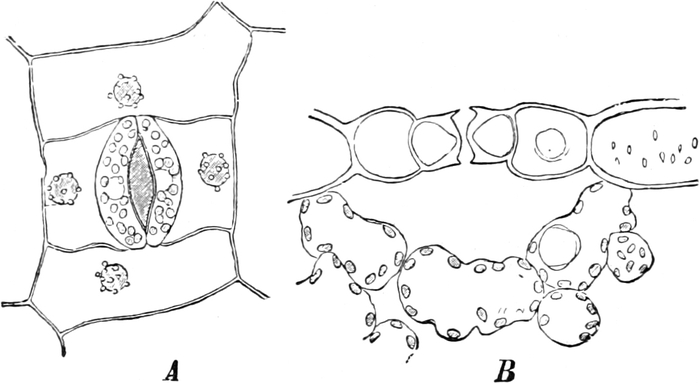
Fig. 45. Epidermis der Blattunterseite von Tradescantia
virginica. A In Flächenansicht von außen, B im Querschnitt. Vergr.
240. Nach STRASBURGER.
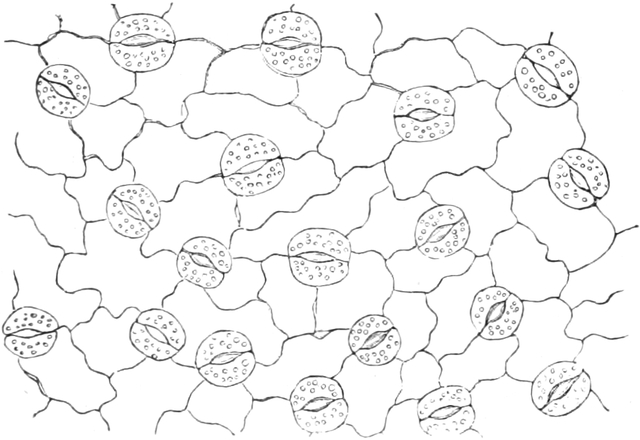
Fig. 46. Epidermis mit Spaltöffnungen auf
der Blattunterseite von Helleborus niger. Vergr. 120. Nach STRASBURGER.
Wie unser Querschnitt in Fig. 45 B lehrt, springen die
Verdickungsleisten der Schließzellen über und unter der Spalte
vor. So erweitert sich die Spaltöffnung meist oberhalb der
Zentralspalte zum sog. Vorhofe, unter ihr zum
Hinterhofe. An den Einfügungsstellen der Schließzellen
verdünnt sich die verdickte Außenhaut der angrenzenden
Epidermiszellen oft plötzlich, wodurch gewissermaßen Scharniere,
Haut[S. 45]gelenke, entstehen, die den Schließzellen die
Gestaltsveränderungen erleichtern (vgl. Fig. 47 B). Häufig werden
die Schließzellen, wie in Fig. 45 A zu sehen ist, von besonderen,
schwächer verdickten oder weniger hohen Epidermiszellen umgeben,
den Nebenzellen der Spaltöffnungsapparate.
Der Bau der Schließzellen und in Abhängigkeit davon auch die
Mechanik ihrer Öffnungs- und Schließbewegungen ist übrigens
verschieden. Zwei Hauptarten der Spaltöffnungsapparate lassen sich
dementsprechend unterscheiden, die aber durch Übergänge miteinander
verbunden sind: beim ersten Typus ändern die Schließzellen ihre
Form hauptsächlich parallel zur Epidermisoberfläche,
beim zweiten dagegen senkrecht zur Epidermis. Typus
I. Je nach dem Bau der Schließzellen kann die Spalte wieder
in verschiedener Weise geöffnet werden. a) Amaryllideentypus
(Fig. 47, bei den meisten Mono- und Dikotylen ausgebildet). Die
Rückenwand jeder Schließzelle (Fig. 47 B) ist unverdickt, die
Bauchwand dagegen verdickt, und zwar meist durch eine obere und
untere Verdickungsleiste. Nimmt der Turgor der Zelle zu, so wird
die dünne Rückenwand stärker gedehnt als die verdickte Bauchwand,
und die Zelle, die im wenig gedehnten Zustand kaum gekrümmt
war, wird in tangentialer Richtung halbmondförmig gekrümmt. b)
Gramineentypus (Fig. 48, bei den Gramineen und Cyperaceen). Die
Schließzelle hat hantelförmige Gestalt. Die erweiterten Enden sind
dünnwandig, das schmalere mittlere Verbindungsstück dagegen hat
sehr stark verdickte Außen- und Innenwände (Fig. 48 B). Bei der
Turgorzunahme der Zelle werden die eiförmigen dünnwandigen Enden
gedehnt und dadurch die starren Mittelstücke der Schließzellen
in tangentialer Richtung voneinander entfernt. Typus II.
Mniumtypus (Fig. 49, bei manchen Moosen und Farnen). Die Bauchwände
der Schließzellen sind dünn, dagegen die Rückenwände verdickt,
Außen- und Innenwände entweder verdickt oder unverdickt. Nimmt der
Turgor in der Zelle zu, so entfernen sich die Außen- und Innenwände
voneinander, wobei die Krümmung der dünnen Bauchwand abnimmt und
die Spalte sich erweitert, während die Rückenwand ihre Lage nicht
ändert. Einen Übergang zwischen Typus I und II stellen z. B. die
Spaltöffnungsapparate von Helleborus dar (vgl. Fig. 238); hier
werden die Schließzellen durch Dehnung der dünnen Rückenwände
stärker halbmondförmig gekrümmt und zugleich die verdickten Außen-
und Innenwände voneinander entfernt.
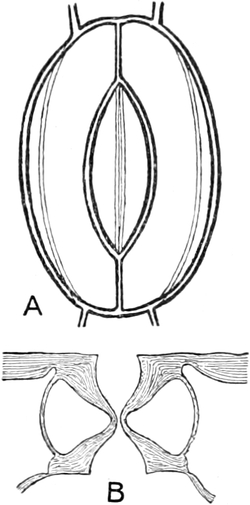
Fig. 47.
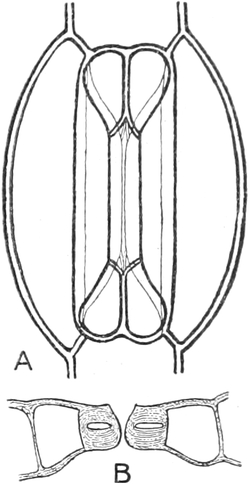
Fig. 48.
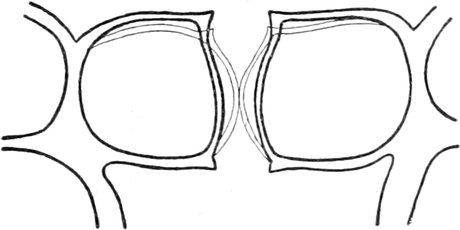
Fig. 49.
Fig. 47 bis 49. Typen von Spaltöffnungsapparaten. Die starken Linien
geben die Formen der Schließzellen bei geöffneten Spalten, die dünnen
Linien bei geschlossenen Spalten wieder.
Fig. 47. Amaryllideentypus, A in Flächenansicht, B im Querschnitt.
Fig. 48. Gramineentypus mit den beiden Nebenzellen, A in
Flächenansicht, B im Querschnitt.
Fig. 49. Mniumtypus im Querschnitt, nach HABERLANDT.
Im übrigen vgl. den Text.
Die Schließzellen entstehen durch Teilung junger Epidermiszellen.
Im einfachsten Falle zerfällt eine solche Zelle dabei in eine
kleinere inhaltsreichere Zelle, die zur Schließzellenmutterzelle
wird, und in eine größere inhaltsärmere, die sich zu einer
gewöhnlichen Epidermiszelle oder zu einer Nebenzelle ausbildet.
Die Mutterzelle rundet sich ellipsoidisch ab und teilt sich durch
eine Längswand in die beiden Schließzellen. In der Längswand bildet
sich hierauf der Spalt als schizogener Interzellulargang aus. Bei
Spaltöffnungsapparaten mit Nebenzellen folgen mehrere Zellteilungen
innerhalb einer jungen Epidermiszelle[S. 46] in bestimmter Weise
aufeinander, bevor die Schließzellenmutterzelle entsteht, oder die
Nebenzellen entstehen durch Teilungen von jungen Epidermiszellen,
die an die Spaltöffnungen angrenzen.
3. Haare. Der Epidermis fast keiner Pflanze fehlen Haare
(Trichome). Die Haare sind entweder einzellige Gebilde,
und zwar papillen- (Fig. 50), schlauch- (Fig. 51) oder pfriemförmige
(Fig. 52, 53, 56 links) Ausstülpungen der Epidermiszellen. Oder sie
sind mehrzellig, nämlich Zellreihen, gestielte und
ungestielte Zellflächen (Schuppenhaare, Fig. 55), die die
Gestalt von Blättchen haben können, wie z. B. die Spreuschuppen
der Farne, oder Zellkörper, die in der Epidermis befestigt
sind. Auch die mehrzelligen Trichome gehen ausschließlich aus
jungen Epidermiszellen, und zwar meist je aus einer solchen, der
Initialzelle des Haares, durch Wachstum und Teilung hervor.
Die einzelligen und mehrzelligen Haare sind ferner unverzweigt
oder verzweigt (Fig. 54, Sternhaare); sie haben dünne
und weiche Membranen oder stark verdickte, häufig verkalkte oder
verkieselte Seitenwände und stechende Spitzen (Borstenhaare,
Fig. 52 rechts); ihre Protoplasten sind entweder lebend, denen
der Epidermiszellen ähnlich, oder abgestorben. Abgestorbene
Haarzellen, die (infolge totaler Lichtreflexion) meist weiß aussehen,
sind mit Luft gefüllt oder seitlich zusammengedrückt (so an den langen
einzelligen Haaren der Baumwollsamen, Fig. 53, aus denen man das
Baumwollgewebe und die Watte macht). Stets wird an den Haaren der in
der Epidermis steckende Teil als Fußstück von dem herausragenden
Haarkörper unterschieden. Die Epidermiszellen, die das Fußstück
umgeben, sind oft ring- oder strahlenförmig angeordnet; man nennt sie
Nebenzellen des Haares. Sehr eigenartig gebaut sind die borstenförmigen
Brennhaare, die sich bei den Brennesseln (Fig. 52) und den
Loasaceen finden.
Fig. 50. Oberhaut vom Blumenblatte des Stiefmütterchens.
Die Zellen mit faltenartigen Leisten an den Seitenwänden und mit
vorspringenden Papillen. Vergr. 250. Nach H. SCHENCK.
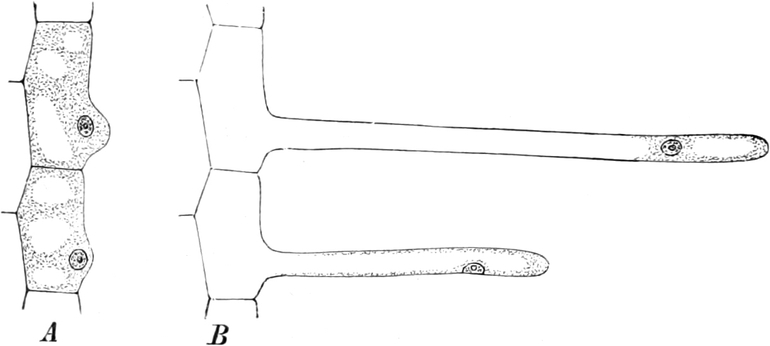
Fig. 51. Wurzelhaare (B) und ihre Entstehung
(A), halb schematisch. Wurzelepidermis im Längsschnitt. Nach
ROTHERT.
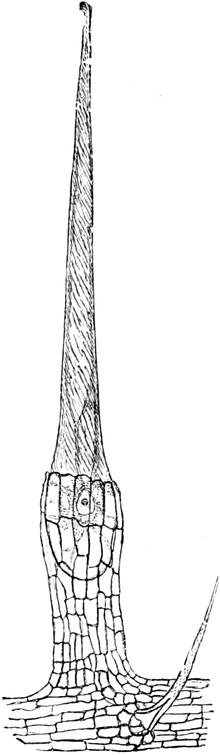
Fig. 52. Brennhaar von Urtica dioica, nebst einem Stück
Epidermis, auf dieser rechts ein kleines einzelliges Borstenhaar.
Vergr. 60. Nach STRASBURGER.
Sie gehen aus einer Epidermiszelle hervor, die während der
Haarbildung stark anschwillt und von benachbarten Epidermiszellen
becherförmig umwachsen wird. Durch Zellvermehrung[S. 47] in dem Gewebe,
das an den Haarfuß angrenzt, erhält das Haar gleichzeitig einen
säulenförmigen Stiel. Die Haarzelle spitzt sich oben zu und endet
mit einem kleinen, schräg aufgesetzten Köpfchen. Unter diesem
Köpfchen bleibt die Haarwandung unverdickt. Das glasartig spröde
Ende des Haares ist verkieselt, die übrigen Wandteile bis auf die
untere Anschwellung sind verkalkt. Wird das Köpfchen des starren
Haares leise berührt, so bricht es ab; die Haarspitze erhält nun
die Form einer Einstechkanüle und dringt in die Haut ein, in
die der Inhalt des Haares sich ergießt. Dieser enthält ein sehr
giftiges Toxin, das nach G. HABERLANDT eine eiweißähnliche
Substanz ist und in der Wunde eine mit brennendem Schmerz
verbundene Entzündung hervorruft. Sie kann, durch gewisse tropische
Brennesseln verursacht, gefährlich werden und starrkrampfähnliche
Zustände zur Folge haben.
Fig. 53. Samenhaare der Baumwollpflanze. A Ein
Stückchen der Samenschale mit Haaren, dreimal vergrößert. B1
Ansatzstelle und unterer Teil, B2, mittlerer Teil, B3 oberer
Teil eines Haares. Vergr. 300. Nach STRASBURGER.
Fig. 54. Sternhaar in Flächenansicht, von der
Blattunterseite der Levkoje (Matthiola annua). Vergr. 90. Nach
STRASBURGER.
Fig. 55. Schuppenhaar, A in Flächenansicht, B
im Längsschnitt von der Blattunterseite der Elaeagnacee Shepherdia
canadensis. Vergr. 240. Nach STRASBURGER.
Die Haare haben also sehr verschiedenen Bau und sie haben auch recht
verschiedene Funktionen. Vielfach unterstützen sie die Epidermis
in ihren schützenden Aufgaben. Alsdann bilden sie Überzüge an
ausgebildeten oder wachsenden Pflanzenteilen, besonders häufig aber
in der Knospe an den jungen Pflanzenteilen, die sie umhüllen. Solche
Überzüge, die seidig oder filzig sind, wenn sie aus abgestorbenen
Wollhaaren bestehen, können die Transpiration herabsetzen und
als Schirm gegen direkte Besonnung schützen. Die Wurzelhaare,
schlauchförmige Ausstülpungen der lebenden Wurzelepidermiszellen (Fig.
51 B,[S. 48] 155 r), dienen der Wasseraufnahme. Drüsenhaare (Fig.
75, 76, 77) scheiden Stoffe sehr verschiedener Art aus.
In den Dienst der Aufnahme mechanischer Reize treten nach G.
HABERLANDT[49] bestimmte plasmareiche Haare. Sie kommen an
Staubgefäßen, Blumenblättern und Blattgelenken vor und haben
eigenartigen Bau; sie werden als Fühlpapillen, Fühlhaare oder
Fühlborsten bezeichnet.
4. Emergenzen. Ähnliche Funktionen wie viele Haare haben
Auswüchse auf der Epidermis, an deren Bildung sich im Gegensatz zu
den Haaren außer der Oberhaut noch mehr oder weniger tief reichende
Teile des darunter liegenden Gewebes beteiligen. Man nennt sie
Emergenzen. Bald sind es Haftorgane, bald Drüsen.
Nur wenige subepidermale Zellreihen setzen sich beispielsweise
in die, sonst bloß aus Epidermis aufgebauten Auswüchse
(Drüsenzotten) fort, die sich an den Enden und an den Zähnen
der Nebenblätter des Stiefmütterchens (Viola tricolor) befinden
(Fig. 56). Tiefer reichende Zellschichten sind hingegen am Aufbau
jener Auswüchse beteiligt, die als ankerförmige Haftorgane den
Früchten der Hundszunge (Cynoglossum) aufsitzen, über 1 mm Länge
erreichen und der Verbreitung der Früchte durch Tiere dienen.
Noch größere Emergenzen sind die Stacheln, die z. B. dem
Rosenstrauch und den Brombeersträuchern beim Klettern behilflich
sind.
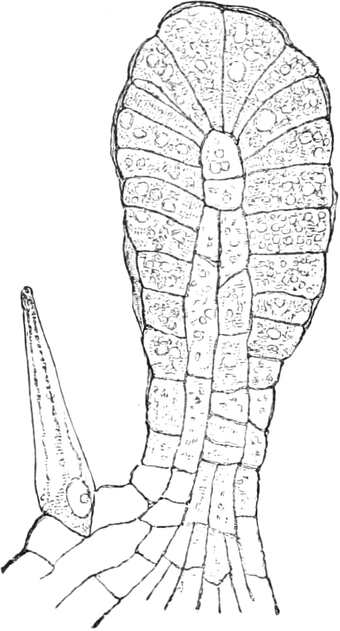
Fig. 56. Eine Drüsenzotte vom Nebenblatt der
Viola tricolor, neben ihr ein einzelliges Haar. Vergr. 240. Nach
STRASBURGER.
b) Abschlußgewebe aus verkorkten Zellen. In sehr vielen Fällen,
namentlich wo die Epidermis nicht während der ganzen Lebensdauer
des umschlossenen Organes lebt und tätig ist, wird der Abschluß des
Körpergewebes nach außen, z. B. auch der Schutz gegen zu starke
Erwärmung, noch viel wirksamer von Schichten verkorkter Zellen
übernommen. Solche Zellen werden in Form von Schichten oder Scheiden
sehr oft auch zur Abgrenzung und zum Abschlusse lebender Gewebemassen
gegen andere im Innern des Körpers verwendet. Sie können primären oder
sekundären Ursprungs sein. Die Verkorkung kommt dadurch zustande, daß
in jeder Zelle an die zunächst unverkorkten Kohlehydratmembranen eine
Suberinlamelle als sekundäre Verdickungsschicht angelagert wird, auf
die noch tertiäre, nicht verkorkte Verdickungsschichten folgen können.
Einige Schichten der Membranen sind oft verholzt. Wir können drei Arten
verkorkten Abschlußgewebes unterscheiden: 1. das Kutisgewebe, 2.
die Endodermis, 3. den Kork.
1. Das Kutisgewebe ist ein primäres Dauergewebe. Es entsteht
durch manchmal schon sehr frühzeitige Verkorkung von Zellen, z. B.
der Epidermis, oder dünnerer oder dickerer oft lückenlos verbundener
Parenchymschichten. Kutisgewebe letzterer Art schließt in Form einer
ein- oder mehrschichtigen Scheide nicht selten ältere Pflanzenteile
(z. B. Wurzeln, deren Epidermis frühzeitig zugrunde geht, Fig. 156
ex) nach außen oder Gewebestränge im Innern des Pflanzenkörpers gegen
das umliegende Gewebe ab. Die Kutiszellen behalten in der Regel ihren
lebenden Inhalt.
An Stelle der Verkorkung kann manchmal auch die Einlagerung von
Kutin oder anderen chemisch noch wenig erforschten Substanzen die
Membranen schwer durchlässig für Wasser machen.
2. Endodermis. Diese Gewebeart wird aus den
Endodermiszellen gebildet[50]. Die Endodermis tritt in
Scheidenform auf; sie dient ebenfalls, und zwar besonders häufig, als
einschichtige Schutzscheide der Abgrenzung[S. 49] und dem Abschlusse
lebender Gewebemassen gegen andere im Innern des Körpers, doch auch als
Abschluß nach außen. Sie ist bald primären, bald sekundären Ursprungs.
Die prismatischen, langgestreckten Endodermiszellen sind lebende
Zellen, die lückenlos zusammenschließen. In jugendlichem Zustande sind
ihre Membranen noch nicht verkorkt; in den radialen Seitenwänden
ist aber ein schmaler Membranstreifen in Form eines Bandes, das die
Zelle rings umläuft und für gewisse, in Wasser gelöste Stoffe schwer
durchlässig ist, durch Einlagerung eines noch nicht näher erkannten
(holzähnlichen?) Stoffes eigenartig verändert (Fig. 57 A). Dieser
Streifen erscheint im Querschnitte als dunkler Punkt oder als dunkles,
linsenförmiges Gebilde (CASPARYscher Punkt, Fig. 57 B und
Fig. 158 bei S), im radialen Längsschnitte als meist etwas welliges
Band. In älteren Endodermiszellen wird wie bei den Kutiszellen
an die Zellhaut ringsum eine sekundäre Verdickungsschicht aus
Korksubstanz angelagert, der noch dicke tertiäre, oft stark verholzende
Kohlehydratschichten folgen können.
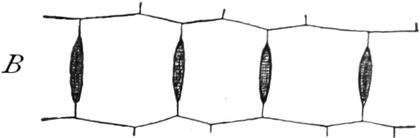
Fig. 57. A Schematische räumliche Darstellung einer
Endodermiszelle mit dem CASPARYschen Streifen in den radialen
Zellwänden. B Endodermis im Querschnitt. Der CASPARYsche
Streifen erscheint in Form einer dunklen linsenförmigen Figur.
Fig. 58. Querschnitt durch Flaschenkork. Vergr. 120.

Fig. 59. A Tangentialer Schnitt, B Querschnitt durch
die Korkhaut eines Lindenzweiges. Vergr. 120. Die Zellmembranen in B
weiß, der abgestorbene Zellinhalt punktiert gezeichnet.
In einschichtiges Kutisgewebe und in die Endodermis sind
nicht selten einzelne Zellen von abweichender Form mit
unverkorkten Zellwänden zerstreut eingeschaltet, die man als
Durchlaßzellen bezeichnet.
3. Kork. Wie die Epidermis und das Kutisgewebe stets primäre
Dauergewebe sind, so ist der Kork immer ein sekundäres Gewebe, das
der Tätigkeit eines sekundären Meristems, des Korkkambiums,
seine Entstehung verdankt. Das Korkgewebe bildet wie die Epidermis in
der Regel peripherische, aber mehrschichtige Scheiden in Form
dünner grauer und glatter Korkhäute oder dicker, außen rissiger
Korkkrusten aus regelmäßigen radialen Zellreihen (Fig. 58,
59) da, wo an ober- oder unterirdischen Pflanzenteilen die Epidermis
abgestoßen oder lebendes Parenchym durch Verwundungen freigelegt wird.
Die Korkzellen sind meist lufthaltige und durch den abgestorbenen
Inhalt gebräunte Zellen. Sie haben meist prismatisch-tafelförmige
Gestalt mit tangential gestellter Grundfläche (Fig. 59 A), schließen
lückenlos zusammen und sind mit verkorkten sekundären Membranschichten[S. 50]
versehen, während die Mittellamellen zwischen ihnen oft verholzt
sind. Tertiäre Verdickungsschichten fehlen in ihnen oder bestehen aus
Zellulose und bilden die sog. Zelluloseschicht, die übrigens auch
verholzt sein kann. Schon dünne Korkhäute, die aus wenigen
Zellschichten bestehen (Fig. 59), vermindern die Transpiration an der
Oberfläche der Pflanzenteile sehr, und zwar infolge der allseitigen
Verkorkung der Zellhäute begreiflicherweise viel stärker als die
Epidermis; dickere Korklagen (Krusten) verhindern außerdem das
Eindringen von Schmarotzern. Zudem besitzt Korkgewebe ein geringes
Wärmeleitungsvermögen, schützt also wirksam gegen zu hohe Erwärmung.
Korkhäute überziehen viele ältere Stengel, Stämme, Knollen,
Knospenschuppen und Früchte; aus einer Korkhaut besteht z. B. die
Kartoffelschale. Die Korkeiche besitzt eine Korkkruste, aus der die
Flaschenkorke hergestellt werden.
Die sehr selten getüpfelten Wände der fertigen Korkzellen bleiben
entweder verhältnismäßig dünn (Fig. 58) oder werden mehr oder
weniger stark verdickt (Fig. 59, 183 p). Stark verdickte
Korkzellen bilden den Steinkork. Die Korkzellen können mit
dem abgestorbenen, meist braunen Inhalt ganz angefüllt sein (Fig.
59 B).
Häufig wechseln im Korkgewebe Schichten verkorkter und unverkorkter
Zellen miteinander ab. Letztere Schichten, deren Elemente sich nach
Bau und Inhalt sonst nur wenig von den Korkzellen unterscheiden,
dünn- oder dickwandig sind und ebenso wie diese entstehen,
bezeichnet man als Phelloidgewebe.
Gewebemassen von noch verwickelterem Bau als der Kork bilden die
Borke, die an älteren Stämmen und Wurzeln als Abschlußgewebe an
Stelle des Korkes tritt (vgl. S. 139).
Fig. 60. Querschnitt durch eine Lentizelle von Sambucus
nigra. e Epidermis, pc Korkkambium, pd aus dem Korkkambium nach
innen abgeschnittene Parenchymzellen, pl Korkkambium der Lentizelle,
l Füllzellen. Vergr. 90. Nach STRASBURGER.
Lentizellen. Durch die Bildung eines von Interzellularen freien
Korkmantels an Stelle einer Epidermis würde der Gasaustausch zwischen
der Atmosphäre und dem Innern des Stammes aufgehoben werden, wenn
nicht für einen Ersatz der Spaltöffnungen gesorgt würde. Das geschieht
bei manchen Pflanzen, z. B. Clematis-, Vitis-, Lonicera-Arten, durch
Porenkork, d. h. dadurch, daß in die Korkhaut ovale oder
rundliche, eng umschriebene Flecke aus etwas kleineren verkorkten
Zellen eingeschaltet sind, die Interzellularen zwischen sich lassen,
meist aber durch die Lentizellen: längliche oder spindelförmige,
rauhe und poröse vorspringende Warzen, die man schon mit bloßem Auge
auf den Korkhäuten der Zweige unserer Holzgewächse sehen kann. Sie
bestehen aus abgestorbenem, meist unverkorktem und an Interzellularen
reichem[S. 51] Gewebe (Füllzellen), das pfropfenartig in das
Korkgewebe eingesetzt ist (Fig. 60). Die Interzellularen münden in
die Außenluft und setzen sich in das Interzellularsystem des lebenden
Gewebes fort.
Die Lentizellen entstehen oft unter den Spaltöffnungen, und zwar
sogleich zu Beginn der Korkbildung. Das Korkkambium, das auch
unter den Spaltöffnungsapparaten auftritt, hier aber radial
verlaufende Interzellularen zwischen seinen Zellen enthält, bildet
an diesen Stellen (Fig. 60 pl) nach außen die Füllzellen mit
Interzellularen (Fig. 60 l). Die Lentizellen durchbrechen alsbald
die Epidermis und heben sie lippenförmig empor. Abwechselnd mit den
Füllzellen erzeugt das Korkkambium in den Lentizellen namentlich
zur Herabsetzung ihrer Durchlässigkeit während des Winters
Schichten fester verbundener, verkorkter und verholzter Zellen,
Zwischenstreifen oder Verschlußschichten, die später (im Frühling)
gesprengt werden.
3. Mechanische oder Festigungsgewebe[51]. Ohne eine gewisse
Festigkeit könnte die Pflanze ihre Gestalt nicht beibehalten, die
meist für ihre Lebenstätigkeit unentbehrlich ist. Bei einzelnen
Zellen und bei wachsenden Geweben wird die nötige Festigkeit durch
die Zellmembranen, den Turgor (vgl. S. 191) und die Gewebespannung
(vgl. S. 248) erzielt. Da indes die Zellhäute dünn sind, Turgor
und Gewebespannung aber schon durch jeden stärkeren Wasserverlust
aufgehoben werden (Welken der Pflanze), so reicht beides nicht
aus, um der Pflanze, namentlich der Landpflanze, auf die Dauer die
nötige Festigkeit zu verleihen. So sehen wir denn bei sehr vielen
Pflanzen besondere Gewebe aus Zellen mit stark verdickten
Membranen, das Sklerenchym und das Kollenchym, mit
mechanischen Aufgaben betraut. Diese Gewebe werden auch Stereome
genannt.
Welche Ansprüche an die Festigkeit und den Zusammenhang der Teile
bei Pflanzen gestellt werden, das leuchtet sofort ein, wenn man sich
z. B. einen Roggenhalm vergegenwärtigt, der, aus Hunderttausenden
einzelner Zellen zusammengesetzt, bei einer Höhe von 1500 mm kaum 3
mm Durchmesser an seiner Basis mißt. Bis zu 3000 mm erheben sich die
schlanken Schäfte des Pfeilrohrs bei einer Grundfläche von nur 15
mm Durchmesser. Die Höhe des Pfeilrohres beträgt das 200fache, die
des Roggenhalmes gar das 500fache des Grunddurchmessers. Dabei trägt
aber der Roggenhalm an seiner Spitze noch die schwere Last der Ähre,
der schlanke Palmstamm die schweren und im Winde wie Segel wirkenden
Blätter, die bei Raphia-Arten 15 m Länge und entsprechende Breite
erreichen, und zeitweise noch die große Last der Früchte.
Neben seiner Festigkeit verfügt der Pflanzenkörper aber auch über
eine Eigenschaft, die wir unseren Bauten nicht entfernt im gleichen
Maße geben können; das ist seine außerordentliche Elastizität.
Der Roggenhalm weicht der Gewalt des starken Windes aus, indem er
seine Spitze bis zum Boden hinabbeugt, schnellt aber in die frühere
Lage zurück, wenn die Wirkung des Windes aufhört. Die technischen
Leistungen des Pflanzenkörpers sind also einzig in ihrer Art und höchst
vollkommen. Von dem festen und zugleich elastischen Baumaterial, das
die Pflanze sich herstellt, macht ja auch die Technik aller Völker
den ausgedehntesten Gebrauch, indem sie Holz zu Stützen und Trägern,
„Bastfasern“ zu Fäden, Tauen und Geweben (z. B. Leinwand) verwendet.
a) Als Sklerenchym bezeichnet man die Festigungsgewebe
der ausgewachsenen Pflanzenteile. Sie sind
aus Sklerenchymzellen (Steinzellen) oder aus
Sklerenchymfasern („Bastfasern“) zusammengesetzt, beides Zellen
mit sehr stark verdickten Zellmembranen aus Kohlehydratlamellen,
die oft zugleich verholzt sind. Die Sklerenchymzellen oder
Steinzellen (Fig. 30) sind mehr oder weniger isodiametrisch,
polyëdrisch und haben runde, unverzweigte oder verzweigte Tüpfel in
ihren fast stets stark verholzten Wänden.[S. 52] Die Sklerenchymfasern
(Fig. 61) dagegen sind schmal spindelförmige, sehr langgestreckte
Zellen mit zugespitzten Enden und mit spärlichen schräg aufsteigenden,
spaltenförmigen Tüpfeln und haben polygonalen Querschnitt (Fig. 62);
ihre Zellwände sind nahezu unverholzt (z. B. beim Lein) oder mehr oder
weniger verholzt (z. B. beim Hanf). Die Sklerenchymfasern haben immer
eine für Pflanzenzellen sehr bedeutende Länge, durchschnittlich von
1–2 mm. Sie können aber bei manchen Gewächsen noch sehr viel länger
werden: beim Lein 20–40 mm, bei der Brennessel bis 77 mm, ja bei der
Urticacee Boehmeria bis 220 mm. Solche langen Fasern sind für Gespinste
besonders brauchbar. Sie werden erst nach vollendeter Streckung der
Pflanzenorgane, vielfach unter Beteiligung von gleitendem Wachstum,
fertiggestellt.
Die Sklerenchymzellen und -fasern können einzeln für sich vorkommen,
so letztere z. B. in manchen Blättern, wo sie auch nicht selten
verzweigt sind. Meist aber sind sie, namentlich die Fasern, ohne
Interzellularen zwischen sich zu lassen, zu Sklerenchymsträngen,
-bändern und -scheiden gruppenweise recht verschieden, aber so
angeordnet, wie es die Ansprüche an die Biegungs-, Zug- oder
Druckfestigkeit des ganzen Organs oder seiner Gewebegruppen unter
Aufwand von verhältnismäßig wenig Festigungsmaterial erfordern.
Druckfestigkeit, z. B. in den Schalen von Nüssen und von Steinen
der Steinfrüchte, kommt meist durch Steinzellengewebe, Biegungs-
und Zugfestigkeit, z. B. von Stengeln und Wurzeln, dagegen durch
Sklerenchymfasergewebe zustande; beide Sorten mechanischer Zellen
bedingen außerdem den Widerstand, den viele Organe dem Schneiden und
anderen mechanischen Eingriffen entgegensetzen.
Die Festigkeit der einzelnen Zellen beruht auf der Verdickung
ihrer Zellmembranen, die manchmal noch durch mineralische
Einlagerungen verhärtet sind, die Zerreißungsfestigkeit der
Sklerenchymfasergewebe außerdem auf der Verzahnung der Fasern
miteinander. Infolge ihrer Faserform und der spindelförmigen Zuspitzung
ihrer Enden ist nämlich die Verwachsung benachbarter Fasern eine sehr
viel innigere als die anders gestalteter Zellen.
SCHWENDENERs Untersuchungen haben bestimmte
Vergleichszahlen für die mechanischen Eigenschaften dieser Zellen
ergeben. Danach kommt die Tragfähigkeit der Sklerenchymfasern
innerhalb ihrer Elastizitätsgrenze (ihr Tragmodul) im allgemeinen
der des besten Schmiedeeisens gleich; sie erreicht bei einzelnen
Pflanzen sogar die des Stahls. Dabei ist die Dehnbarkeit gegen
10–15mal größer als die des Schmiedeeisens. Nach Überschreiten der
Elastizitätsgrenze tritt alsbald Zerreißen ein, während bei dem
Eisen die Festigkeitsgrenze erst bei etwa dreifacher Belastung
erreicht wird. Für die Bedürfnisse der Pflanze hat es aber große
Bedeutung, daß bei ihr die Elastizitätsgrenze bis nahe an die
Festigkeitsgrenze reicht.

Fig. 61. Eine Sklerenchymfaser etwa 100mal vergr. Nach
STRASBURGER.
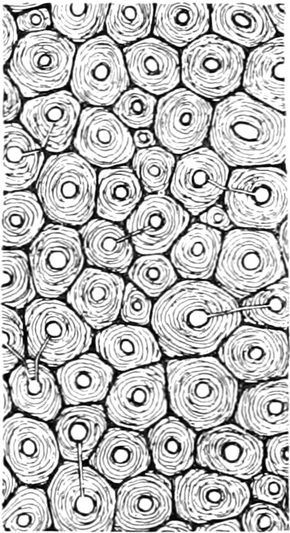
Fig. 62. Querschnitt durch das Sklerenchym im Blatte der
Liliacee Phormium tenax. Vergr. 240.
b) Das Kollenchym. Die Elemente des Sklerenchyms sind nicht
mehr imstande, zu wachsen; sie können deshalb in Pflanzenteilen, die
noch in lebhafter Streckung begriffen sind, keine Verwendung finden.
Bedürfen solche Pflanzenteile außer der Festigkeit, welche Zell- und
Gewebespannung ihnen[S. 53] verleihen, noch einer besonderen Verstärkung, so
wird sie durch Kollenchym erreicht.
Die Kollenchymzelle, die isodiametrisch oder meist sehr langgestreckt
(zugespitzt oder mit rechteckigem Umriß Fig. 64) ist, gleicht der
Parenchymzelle und enthält wie diese oft Chlorophyll, unterscheidet
sich aber von der Parenchymzelle wesentlich dadurch, daß ihre
Zellulosemembran ungleich, besonders an den Zellkanten
(Kantenkollenchym, Fig. 63) oder an den tangentialen Wänden
(Plattenkollenchym) stark verdickt ist. Ferner fehlen in ihrem
Plasma, abgesehen von großen Zellsaftvakuolen, leblose Einschlüsse.
Auch die Interzellularen fehlen oder sind sehr klein. Das Kollenchym
besitzt trotz hohem Wassergehalte infolge der Wandverdickungen seiner
Elemente eine ansehnliche Festigkeit gegen Zerreißen. Dabei gibt es
dem Wachstum seiner Umgebung dadurch nach, daß es selbst daran noch
teilnimmt. Die großen unverdickten Wandflächen, in denen noch rundliche
oder spaltenförmige Tüpfel vorhanden sind, ermöglichen zugleich einen
schnellen Transport von Baustoffen innerhalb dieser Gewebeart. Auch das
Kollenchym ist entsprechend seiner mechanischen Funktion angeordnet.
4. Die Leitgewebe. Je größer der Körper einer Pflanze wird und
aus je mehr Zellen er sich zusammensetzt, vor allem aber je mehr Teile
er aus dem Wasser oder aus dem Boden in den Luftraum streckt,
umso mehr entsteht die Notwendigkeit, Stoffe schnell von einem Organe
in ein anderes, etwa von den Wurzeln in die Blätter und umgekehrt,
zu schaffen. Die Diffusionsbewegung durch die Querwände selbst
langgestreckter Parenchymzellen genügt dazu vielfach nicht, auch wenn
die Stoffbewegung durch Ausbildung von Tüpfelkanälen sehr erleichtert
wird. So sind besondere, Leitungszwecken dienende Gewebearten,
die Leitgewebe, entstanden mit sehr charakteristischen
Zellelementen, die in der Hauptleitungsrichtung meist langgestreckt
sind, oft vergrößerte Diffusionsflächen besitzen, ja meist zu
eigenartigen Leitungskanälen (Fusionen) verschmelzen und stets zu
einem zusammenhängenden Systeme verbunden sind, das die ganze Pflanze
durchzieht. Bezeichnend für die Leitgewebe ist ferner der Mangel an
Interzellularen.
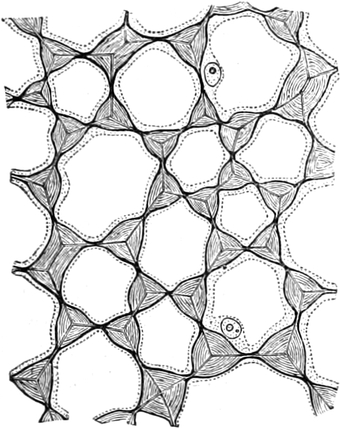
Fig. 63. Querschnitt durch das Kollenchym von Cucurbita
Pepo. Vergr. 240.

Fig. 64. Eine Kollenchymzelle in Seitenansicht. Vergr.
240.
a) Siebröhren. Durch offene Poren, die dem Transporte von
Eiweißstoffen und Kohlehydraten zu dienen scheinen, werden die in
Längsreihen angeordneten Glieder der Siebröhren[52] verbunden.
Die quergerichteten oder steilen Endwände (manchmal auch die
Seitenwände) jedes dieser übrigens langgestreckten, im Querschnitt
polygonalen Glieder besitzen nämlich siebartig durchlöcherte Stellen,
die von Plasmasträngen ausgefüllt werden. Man nennt diese Stellen
Siebplatten (Fig. 65 A, B). Bei vielen Pflanzen, z. B. dem
Kürbis (Fig. 65 A), ist die ganze Querwand eines Siebröhrengliedes
eine einzige Siebplatte mit verhältnismäßig groben Poren; auf den
Längswänden sind[S. 54] dagegen nur eng umgrenzte runde Stellen mit viel
feineren Poren als Siebplatten ausgebildet (Fig. 65 C c*), da wo
zwei Siebröhren seitlich aneinander grenzen. In anderen Fällen, z. B.
auf den alsdann meist steilen Endwänden der Siebröhren, findet man
dagegen mehrere solche durch nicht perforierte Membranteile
getrennten tüpfelartigen Siebplatten (Fig. 66) mit meist sehr feinen
punktförmigen Poren. Die Siebröhrenglieder, deren jedes einer Zelle
entspricht, enthalten lebende, dünne plasmatische Wandbeläge mit je
einem Zellkern, mit Leukoplasten und oft mit Stärkekörnern und als
Zellsaft eine wäßrige alkalische, mehr oder weniger konzentrierte
gerinnbare Flüssigkeit, die reich an Eiweißstoffen, oft auch an
Kohlehydraten und anorganischen Salzen (Phosphaten) ist (Fig. 65
D). Die Siebröhrenwandungen sind fast stets unverholzt, bestehen
aus Zellulose und sind durch den Inhalt elastisch gespannt. Im
allgemeinen funktionieren die Siebröhren nur während einer
Vegetationsperiode. Bevor sie untätig werden, überziehen sich ihre
Siebplatten mit stark lichtbrechenden Kallusplatten (Fig.
65 C), die den Stoffaustausch zwischen den Siebröhrengliedern
herabsetzen oder wohl ganz unterbrechen. Soll die Siebröhre in der
nächsten Vegetationsperiode nochmals tätig sein, so werden diese
Kallusplatten wieder gelöst.
Fig. 65. Teile von Siebröhren des Kürbis (Curcurbita
Pepo) in Alkohol gehärtet. A Eine Siebplatte von oben gesehen. B
und C Je zwei aufeinanderfolgende Siebröhrenglieder im Längsschnitt;
s Geleitzellen, u Schleimstrang, pr plasmatischer Wandbelag, c
Kallusplatte, c* kleine seitenständige Siebplatte mit Kallusbelag.
D Die Inhaltsmassen von zwei Siebröhrengliedern nach Auflösung der
Zellhäute mit Schwefelsäure. Vergr. 540. Nach STRASBURGER.
Fig. 66. A Grenze zweier Siebröhrenglieder vom
Weinstock (Vitis) mit schräger, längsdurchschnittener Querwand. Vergr.
600. Nach DE BARY. B Eine solche Querwand in Flächenansicht
mit den Siebplatten. Von ROTHERT schematisiert nach
DE BARY.
[S. 55]
Die Kallusplatten bestehen aus Kallose, einem seiner
chemischen Zusammensetzung nach noch unbekannten Körper, der
sich durch seine Unlöslichkeit in Kupferoxydammoniak, aber seine
Löslichkeit in 1%iger kalter Kalilauge auszeichnet. Sie färbt
sich in Chlorzinkjodlösung rotbraun, glänzend blau mit Anilinblau
und glänzend rot mit Korallin (Rosolsäure). — Kallose überzieht
übrigens in dünnen Schichten auch schon die Leisten der Siebplatten
zwischen den Poren, solange diese noch offen sind. Diese Schichten
werden allmählich dicker, die Poren mehr und mehr verengt und
schließlich ganz verstopft.
b) Gefäße. Besondere und zwar tote Zellarten,
die meist langgestreckt röhrenförmig sind, rundlichen oder
polygonalen Querschnitt haben und in Längsreihen, entsprechend der
Hauptleitungsrichtung, angeordnet sind, mit sehr charakteristischen
und auffälligen Verdickungen in den meist verholzten Wänden dienen
der Leitung, manchmal auch der Speicherung des Wassers in der Pflanze
(Gefäße). Die Gefäße enthalten dementsprechend, solange sie
tätig sind, Wasser, außerdem aber oft auch begrenzte Mengen Luft.
Man teilt sie ein in Tracheïden und Tracheen. Die
Tracheïden sind Einzelzellen, die mit eigenartig getüpfelten Zellhäuten
aneinander grenzen, meist zugespitzte Enden und in der Regel kleinen
Querdurchmesser (Fig. 70 B) besitzen; sie dienen oft zugleich als
Festigungszellen, z. B. im Stamme der Nadelhölzer. Die Tracheen
dagegen sind verhältnismäßig weite oder enge Membranröhren, die aus
Längsreihen vieler Zellen (ihren Gliedern) durch Auflösung
der Endwände entstanden sind. Die Endwände werden dabei, wenn sie
quer stehen, meist bis auf einen schmalen Rand aufgelöst, der als
ringförmige Membranleiste stehen bleibt und verdickt wird (Fig. 67
C bei s, Fig. 69 I bei q und q′); schräge Endwände sind
dagegen meist nicht von einem einzigen runden, sondern von mehreren,
übereinanderliegenden, spaltenförmigen oder elliptischen Löchern
(leiterförmige Perforation, Fig. 69 II; 171 tg) durchbrochen.
Einige der Endwände freilich sind nicht durchbrochen, sondern bloß mit
Tüpfeln versehen. Infolgedessen haben die Tracheen eine begrenzte Länge.
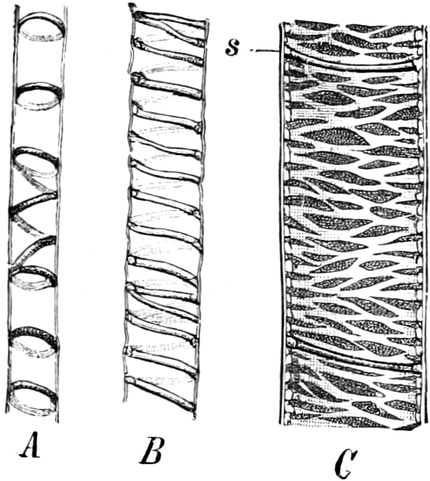
Fig. 67. Teile von Tracheïden und von einer Trachee. A
Ring- und Schraubentracheïde. B Schraubentracheïde. C Netztrachee
halb aufgeschnitten, bei s eine der beiden durchlöcherten Querwände.
Vergr. 240. Nach H. SCHENCK.
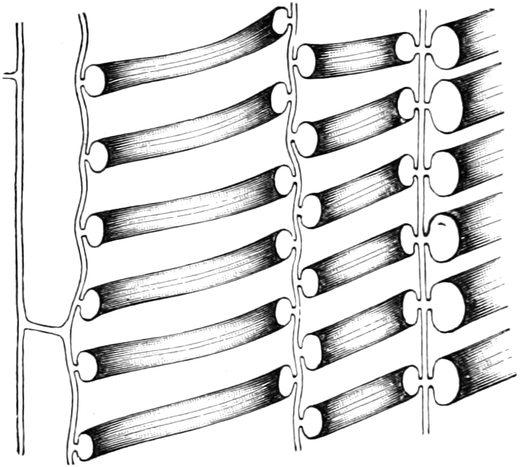
Fig. 68. Teil eines Längsschnittes durch drei
Schraubengefäße und eine Reihe von Parenchymzellen des Kürbis
(Curcurbita Pepo). Vergr. 560. Nach ROTHERT.
Einzelne Tracheen können zwar, im besonderen bei den kletternden
Holzgewächsen, den Lianen, einige Meter lang sein. Auch
bei unseren Eichen sind 2 m lange Tracheen noch sehr zahlreich.
Im allgemeinen beträgt aber ihre Länge weniger als 1 m, nämlich
meist nur gegen 10 cm. Wie die längsten, so sind auch die weitesten
Tracheen bei den Kletterpflanzen zu finden; ihr Durchmesser kann
hier 0,7 mm erreichen, während er bei unseren Eichen im Mittel
0,25, unserer Linde 0,06 mm beträgt.
Die Bezeichnungsweise der wasserleitenden Elemente ist in der
Literatur leider recht verschiedenartig. Meist wird unterschieden
zwischen Tracheïden und Tracheen oder[S. 56] Gefäßen. DE BARY
dagegen nannte alle Elemente Tracheen und unterschied zwischen
Tracheïden und Gefäßen. Am zweckmäßigsten scheint der Vorschlag
ROTHERTs, dem wir gefolgt sind, als Sammelbegriff von
Gefäßen zu reden und die Gefäße in Tracheïden und Tracheen
einzuteilen.
Die Wandverdickungen in vielen Gefäßen sind auf schmale Leisten
(Fig. 67 u. 68) in den sonst wenig verdickten Zellwänden beschränkt.
Diese Leisten können isolierte Ringe, zusammenhängende Schraubenbänder
oder ein Netzwerk mit großen queren Maschen bilden (Fig. 67, 68);
man unterscheidet danach zwischen Ring-, Schrauben-
und Netztracheïden oder -tracheen. In anderen Gefäßen
umfassen die Verdickungen den größeren Teil der Zellmembranen; alsdann
bleiben aber zahlreiche kreisförmige, polygonale oder in querer
Richtung mehr oder weniger gestreckte elliptische oder spaltenförmige
Tüpfel zwischen den verdickten Teilen ausgespart (Fig. 69, 70):
Tüpfelgefäße. Stehen an den Seitenwänden quergestreckte Tüpfel
regelmäßig in geraden Reihen übereinander, so wird das Gefäß als
Treppen- oder Leitergefäß bezeichnet (Fig. 69 II, 70
A). Zwischen sämtlichen Gefäßformen gibt es Übergänge.
Die Tüpfel sind in allen Tüpfelgefäßen zweiseitig oder
einseitig behöft (Hoftüpfel). Hoftüpfel heißt ein Tüpfel,
dessen Kanal sich nach der Schließhaut hin trichterartig erweitert
(Fig. 71 C). Sehr häufig sind die Hoftüpfel in der Flächenansicht
kreisförmig; alsdann sieht man in ihrer Mitte einen kleinen
konzentrischen Kreis (Fig. 71 A). Der kleinere innere Kreis ist die
enge Mündungsstelle des Tüpfelkanals in den Zellraum, der große äußere
Kreis (der „Hof“) seine weiteste Stelle, womit er an die Schließhaut
grenzt. Zwischen beiden Kreisen überwölben die Verdickungsschichten
der Membran die Schließhaut als Tüpfelwandung. Die Schließhäute sind
in der Mitte oft zu dem Torus verdickt (Fig. 71 C), vermögen
sich nach der einen oder anderen Seite vorzuwölben und mit den Tori die
engen Ausgänge der Tüpfel auf einer Seite nach Art von Klappenventilen
zu verschließen (Fig. 71 B, t). Die Hoftüpfel lassen infolge ihrer
großen Schließhäute sehr rege Flüssigkeits-(Wasser)bewegungen von
einem Zellraum in den anderen zu, ohne daß die toten Zellen auf die
notwendige Festigkeit ihrer Membranen zu verzichten brauchten, die eben
durch die einem Gewölbe ähnliche Tüpfelwandung gewährleistet wird.
Fig. 69. A Schemata von Tracheen in medianem
Längsschnitt. I. Weite Trachee mit kleinen elliptischen Hoftüpfeln
und mit einfacher Perforation der Querwände q q. Die Hinterwand
der Trachee ist im oberen Teil der Figur weggeschnitten. II. Enge
Treppentrachee mit leiterförmiger Perforation der Querwände q.
B Die Querwände beider Tracheen, von der Fläche gesehen. Nach
ROTHERT.
Die Hoftüpfel sind, wie es die Fig. 71 C zeigt, zweiseitig
behöft, wenn sie zwischen zwei wasserleitenden Elementen
ausgebildet sind. Sie sind einseitig behöft, wenn sie die
Zellwand eines wasserleitenden Elementes durchsetzen, das an eine
lebende Zelle grenzt. In diesem Falle erweitert sich[S. 57] der Tüpfelkanal
nämlich bloß von dem wasserleitenden Elemente aus nach der Schließhaut
hin und hat keinen Torus in der Schließhaut; auf der anderen Seite der
Schließhaut dagegen, also in den Wandschichten der lebenden Zelle, ist
ein gleich weiter (einfacher) Tüpfelkanal ausgebildet.
Auch die Tüpfel in den Endwänden der Tracheen und Tracheïden sind
stets Hoftüpfel. In den Tracheen sind es ihre Schließhäute, die
aufgelöst werden (vgl. Fig. 69 A I und II).
Auch in den Ring- und Schraubengefäßen können übrigens die
Verdickungsleisten gelegentlich den Wänden von Hoftüpfeln gleichen[53],
und zwar dadurch, daß sie T-förmigen Querschnitt haben und mit den
verschmälerten Kanten der Gefäßwandung angefügt sind (Fig. 68).
Die dünnen Membranstellen zwischen den Leisten entsprechen alsdann
den Schließhäuten von Hoftüpfeln; manchmal sind sie sogar, wenn
sie zwei wasserleitende Elemente trennen, wie diese Schließhäute,
in ihrer Mitte etwas stärker, als Tori, verdickt. Bei der
Anfertigung von Schnitten lösen sich die fast stets verholzten
schraubenförmigen Verdickungsbänder oft leicht von den unverdickten
(und nicht immer verholzten) Gefäßwandungen ab und werden weithin
aus den Gefäßen herausgerissen. Diese Erscheinung findet aber
weder in der Befestigungsweise der Bänder noch in der chemischen
Beschaffenheit der Schraubengefäßwände eine hinreichende Erklärung.
Fig. 70. A Unteres Drittel einer Treppentracheïde
aus dem Rhizom des Adlerfarns (Pteridium aquilinum). t Die
quergestreckten Tüpfel an den Seitenwänden, q die leiterförmig
getüpfelte Endfläche. Vergr. 95. Nach DE BARY. B Eine
Tracheïde mit runden Hoftüpfeln. Vergr. 100. Nach STRASBURGER.
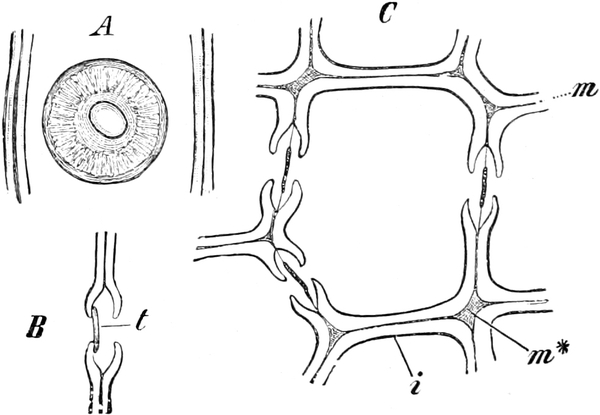
Fig. 71. Tracheïden aus dem Holze der Kiefer (Pinus
silvestris). A Radialer Längsschnitt mit Hoftüpfel in Flächenansicht.
B Tangentialer Längsschnitt mit Hoftüpfel im Querschnitt, t der
Torus. C Querschnitt durch eine Tracheïde, m Mittellamelle,
m* ein Zwickel in dieser, i das Grenzhäutchen. Vergr. 540. Nach
STRASBURGER.
Nur die mit ring- oder schraubenförmigen Verdickungsleisten
versehenen Gefäße sind noch streckungsfähig und dehnbar; daher
werden nur solche in wachsenden Pflanzenteilen ausgebildet.
Die Wandverdickungen erhöhen die mechanische Festigkeit der
wasserleitenden Elemente, indem sie es verhindern, daß die benachbarten
lebenden Zellen sie zusammendrücken. Der lebende Inhalt der Gefäße
wird, während die Zellmembranen sich verdicken, immer ärmer;
schließlich schwindet er, in den Tracheen nach Durchbrechung der
Querwände, ganz.
Leitbündelgewebesystem. Die Siebröhren treten fast nie für
sich allein auf, sondern sind meist mit Leitparenchym zu Strängen
oder Bündeln (Siebsträngen) verbunden, die die ganze Pflanze
durchziehen. Gleiches gilt für[S. 58] die Tracheïden und Tracheen
(Gefäßstränge); Tracheïden finden sich freilich auch einzeln
oder in kleineren Gruppen nicht selten als Wasserspeicher im Parenchym
zerstreut (Speichertracheïden).
Im primären Gewebe verbinden sich Sieb- und Gefäßstränge meist
zu gemeinsamen Strängen oder Bündeln, vollständigen
Leitbündeln, die meist parallel zur Längsachse eines Organes
verlaufen, durch Querzweige zu einem Netzwerk verbunden und
so auffällig sind, daß man für dieses Strangsystem den Namen
Leitbündelgewebesystem geprägt hat. Darin sind also die Elemente
der Wasserleitung mit denen der Leitung organischer Stoffe verbunden,
so daß das Wasser und diese Stoffe auf nahe benachbarten Wegen,
wenn auch oft in entgegengesetzter Richtung geleitet werden. Dieses
Gewebesystem kann primären oder sekundären Ursprungs sein. In jedem
vollständigen Leitbündel, das sich durch seine engen Elemente und den
Mangel an Interzellularen schon bei schwächster Vergrößerung von dem
übrigen weniger dichten Gewebe abhebt, ja manchmal sogar mit bloßem
Auge sichtbar ist, z. B. in den durchscheinenden Stengeln von Impatiens
parviflora, lassen sich also Gewebestränge zweierlei Art unterscheiden:
Gefäßstränge bilden den Gefäßteil oder das Xylem,
und Siebstränge bilden den Siebteil oder das Phloëm.
Xylem und Phloëm können in den Bündeln verschieden angeordnet sein;
infolgedessen sind auch die Querschnittsbilder der Bündel recht
verschieden (vgl. S. 85 ff.).
Für das vollständige Leitbündel und seine Teile werden noch andere
Namen in der Literatur gebraucht. Statt Leitbündel sagt man
auch Gefäßbündel, Fibrovasalbündel oder Mestom, statt Gefäßteil
Holzteil, Vasalteil oder Hadrom, statt Siebteil Bastteil,
Kribralteil oder Leptom.
Fig. 72. Stück einer Milchröhre der Asclepiadacee
Ceropegia. Vergr. 150. Nach STRASBURGER.
Sieb- oder Gefäßstränge kommen aber nicht selten auch für sich allein
vor. Solche Sieb- oder Gefäßstränge kann man als unvollständige
Leitbündel bezeichnen. Sie sind besonders im sekundären Dauergewebe
weit verbreitet, und zwar Gefäßstränge im Holz, Siebstränge im Bast
(vgl. S. 133, 136).
5. Sekretzellen und Sekretgewebe. 1. Einzelzellen.
In den verschiedensten Geweben findet man besonders häufig
Sekretzellen, einzeln für sich oder oft in längs verlaufenden
Reihen (so z. B. bei Liliaceen, Amaryllidaceen, Commelinaceen). Sie
sind isodiametrisch oder schlauchförmig (Schläuche) und unterscheiden
sich von den übrigen Zellen vornehmlich durch ihren Inhalt.
In dem oft stark geschwundenen, nicht selten schließlich
abgestorbenen Protoplasten liegen als Endprodukte des
Stoffwechsels sehr große Mengen Sekrete verschiedenster Art, die
als Schutzstoffe ökologische Bedeutung haben können. Als solche Sekrete
sind besonders verbreitet: Schleime, Gummi, ätherische Öle, Harze,
Gummiharze, Gerbstoffe, Alkaloide oder Oxalatkristalle (Fig. 22). Die
Wandungen dieser Zellen sind oft verkorkt.
Zu den Sekretzellen gehören auch die ungegliederten Milchröhren,
die als Sekrete Milchsäfte enthalten. Es sind reich verzweigte
Schläuche ohne alle Querwände, Röhren, die eine meist
unverdickte, glatte, elastische Zellulosewand (Fig. 72), einen lebenden
Wandbelag aus Plasma mit zahlreichen Zellkernen, manchmal auch mit
Stärkekörnern (bei vielen Euphorbien von knochenförmiger Gestalt)
besitzen[54] und als Zellsaft eine milchige, meist weiße, wäßrige
Flüssigkeit enthalten, die an der Luft rasch gerinnt. Der Milchsaft hat
ökologische Bedeutung; er dient zum Wundverschluß und als[S. 59] Schutzmittel
gegen Tierfraß. Leitfunktion haben die Milchröhren dagegen nicht.
In dem Milchsaft kommen gelöst vor: Gerbstoffe, Glykoside, manchmal
giftige Alkaloide und besonders Kalkmalat, ferner bei Ficus Carica
und Carica Papaya auch peptonisierende Enzyme; weiter als Tröpfchen
in Emulsion: Gummiharze, d. h. Gemenge von Gummi und Harz,
Kautschuk (C25H40), Guttapercha, Fett und Wachs; als feste
Bestandteile: vielfach Proteïnkörner.
Solche Milchröhren findet man bei vielen Euphorbiaceen (z. B.
Euphorbia), Moraceen, Apocynaceen und Asclepiadaceen. Sie gehen aus
Zellen hervor, die schon in der Keimpflanze kenntlich sind und mit der
ganzen Pflanze weiterwachsen, sich fort und fort verzweigen, in alle
ihre Glieder eindringen und so viele Meter lang werden können.
2. Zellfusionen. Mehrere Sekretzellen können auch durch
Auflösung der trennenden Querwände zu einem geräumigeren
Sekretbehälter verschmelzen. Am auffälligsten ist das der Fall
bei den gegliederten Milchröhren oder Milchgefäßen. Sie
sehen ganz ähnlich aus und besitzen auch ganz entsprechenden Inhalt
wie die ungegliederten Milchröhren, unterscheiden sich von ihnen nur
dadurch, daß sie aus Zellverschmelzungen hervorgehen und meist zu einem
Netzwerk verbundene Schläuche sind (Fig. 73). Infolgedessen findet man
in ihnen manchmal Reste von Querwänden.
Fig. 73. Tangentialer Längsschnitt aus der Peripherie
der Wurzel des Löwenzahns (Taraxacum), die netzförmig verbundenen
Milchgefäße zeigend. Vergr. 240.
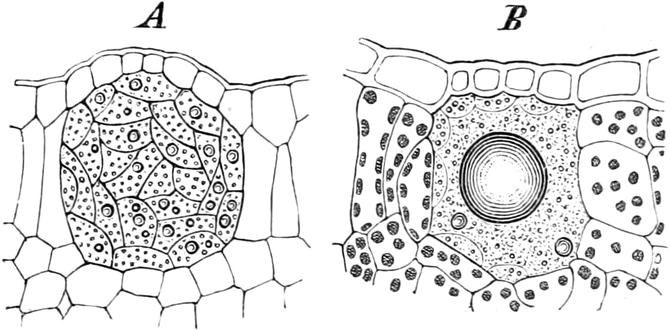
Fig. 74. Lysigener Ölbehälter im Querschnitt des Blattes
von Dictamnus Fraxinella. A Jung. B Nach Auflösung der Zellwände
fertig ausgebildet. ROTHERT frei nach RAUTER.
Wie die Milchröhren, so sind auch die Milchgefäße auf bestimmte
Pflanzenfamilien beschränkt, so auf gewisse Euphorbiaceen (z. B. bei
dem wichtigsten Kautschukbaum Hevea), die Papaveraceen mit Papaver
und dem durch orangerote Färbung seines Milchsaftes ausgezeichneten
Chelidonium, ferner auf die Campanulaceen und die Cichorieen unter
den Kompositen, etwa mit den Gattungen Cichorium, Taraxacum, Lactuca,
Scorzonera, Hieracium, Tragopogon. Ihre Funktionen entsprechen denen
der ungegliederten Milchröhren.
Den Milchgefäßen in vieler Beziehung ähnlich sind die
Schleimröhren, die bei vielen Monokotylen vorkommen. Ihr
Schleimsaft besteht aus Eiweiß, Stärke, Glykose, Gerbstoffen und
anorganischen Stoffen.
3. Lysigene Interzellularräume. Häufig entstehen ferner
Sekretbehälter auch lysigen, d. h. durch Auflösung der ganzen
Sekretzellen (Fig. 74)[S. 60] (lysigene Sekretbehälter). Sie
stellen rundliche Gebilde, unregelmäßige Hohlräume und schlauchförmige
Gänge dar. Sie gehen aus Zellgruppen hervor, in denen die Sekrete
entstanden sind und deren Wände allmählich aufgelöst wurden. Solchen
Ursprung haben unter anderem die mit ätherischem Öl gefüllten
Sekretbehälter der Orangen, Zitronen und anderer Rutaceen sowie vieler
Myrtaceen.
6. Drüsenzellen und Drüsengewebe. Außer Sekretzellen findet
man, ebenfalls einzeln oder auch zu Gruppen vereint, in der Epidermis,
im Parenchym oder in anderen Gewebearten oft Drüsenzellen, d. h.
Zellen, die Sekrete, also meist Endprodukte des Stoffwechsels,
aus ihren Protoplasten durch die Zellwände nach außen, aus
dem Pflanzenkörper oder in Interzellularen, ausscheiden. Auch die
Drüsenzellen, die immer lebend sind, gleichen den Parenchymzellen,
sind aber wie die Meristemzellen meist mit viel Plasma und mit großen
Zellkernen ausgestattet. Die ausgeschiedenen Stoffe haben oft eine
ökologische Bedeutung. Gruppen von lückenlos verbundenen Drüsenzellen,
die eine Zellschicht bilden, nennt man Drüsenepithelien.
Besonders häufig findet man in der Epidermis Drüsenepithelien
oder einzelne Drüsenzellen. Sie sind oft von einer porösen Kutikula
überzogen, oder eine Kutikula fehlt ganz. Hier kommen auch Drüsenhaare
vor, so auch Köpfchenhaare, deren als Köpfchen ausgebildete
Endzelle (Fig. 75) die Drüsenzelle ist. Andere solche Haare sind
schuppenförmig gestaltet (Fig. 76); auch Drüsenzotten (Fig. 56) kommen
vor. Das Sekret besteht sehr oft aus harzigen Stoffen; in diesem Falle
tritt es zunächst zwischen der Außenwand der Drüsenzelle und der
Kutikula auf, die Kutikula emporhebend und schließlich zersprengend.
Ähnliches gilt für andere klebrige Stoffe und Schleim.
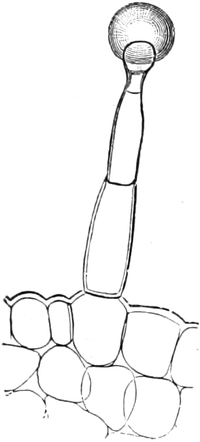
Fig. 75. Drüsenhaar vom Blattstiel der Primula sinensis,
oben das Sekret. Vergr. 142. Nach DE BARY.
Fig. 76. Gelbe Drüsenschuppen von den weiblichen
Blütenständen des Hopfens im senkrechten Durchschnitt. A Vor Beginn
der Sekretbildung. B Die Kutikula durch das Sekret emporgehoben, das
Sekret durch Alkohol entfernt. Vergr. 142. Nach DE BARY.
Fig. 77. Sitzende Digestionsdrüse der Blattoberseite
von Pinguicula vulgaris. A Im Längsschnitt. B In Aufsicht.
ROTHERT frei nach FENNER.
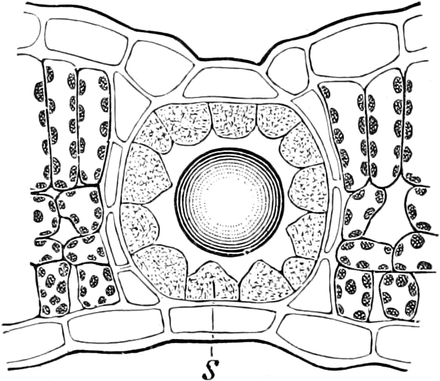
Fig. 78. Schizogener Ölbehälter im Blattquerschnitt von
Hypericum perforatum. s Das Drüsenepithel. Nach HABERLANDT.
Nach den Ausscheidungsprodukten, die recht verschiedene
ökologische Bedeutung haben können, unterscheidet man unter den
epidermalen Drüsen: Schleim-, Öl-, Harz-, Digestions- (Fig. 77),
Salzdrüsen, Wasserdrüsen (Hydathoden) und Nektarien[55].
Die letztgenannten scheiden zuckerreiche Sekrete aus, die Insekten
anlocken; sie finden sich als Drüsenflächen oder Drüsenhaare vor
allem innerhalb der Blüten (nuptiale Nektarien), oder außerhalb
(extranuptiale Nektarien, vgl. Fig. 141 n) und haben recht
verschiedenen Bau (vgl. auch S. 99).
[S. 61]
Die im Parenchym oder in anderem Gewebe eingeschlossenen
Drüsenzellen oder Drüsenepithelien grenzen stets an rundliche
oder unregelmäßig begrenzte Interzellularräume oder an gang- und
röhrenförmige, unverzweigte oder verzweigte Interzellularkanäle, die
manchmal die ganze Pflanze als kommunizierende Röhren durchziehen
können. Diese Interzellularen, die durch Auseinanderweichen der
Drüsenzellen, also schizogen, entstehen, sind es, in die die Sekrete
ausgeschieden werden; sie bilden die schizogenen Sekretbehälter
(Fig. 78). Ihr Inhalt besteht aus ätherischen Ölen, Harzen, Gummi,
oder Schleim; dementsprechend unterscheidet man zwischen Öl-, Harz-
(Fig. 133 A, h), Gummi- und Schleimgängen oder -kanälen. Solche
Harzkanäle finden sich bei vielen Coniferen, Ölgänge z. B. bei den
Umbelliferen, Schleim- und Gummigänge bei den Cycadeen und Araliaceen
(wie dem Epheu). Runde oder längliche schizogene Höhlungen (Lücken) mit
ätherischen Ölen kommen bei Hypericum-Arten vor (Fig. 78).
Übrigens gibt es auch schizolysigene Sekretbehälter.
Dritter Abschnitt. Organlehre (Organographie)[56].
Die Organismen, die wir zum Pflanzenreiche rechnen, sind sehr
verschieden gestaltet und gegliedert. Teils sind sie zeitlebens
einzellig, teils sind sie vielzellig. Einzellige wie Vielzellige können
sehr einfache und regelmäßige oder unregelmäßige Umrißformen haben
und äußerlich ganz ungegliedert sein oder einen durch Auszweigungen
mannigfaltigster Art reich und mehr oder weniger symmetrisch
gegliederten Körper besitzen.
I. Symmetrieverhältnisse. Die ganze Gestalt eines ungegliederten
oder irgendwie gegliederten Organismus und ebenso die Form und die
innere Ausbildung seiner Teile wird beherrscht durch die Eigenart
der Symmetrieverhältnisse, d. h. durch die mehr oder weniger
gesetzmäßige Verteilung der organischen Massen, die das Lebewesen oder
seine Organe zusammensetzen. Die Symmetrieverhältnisse stehen wie fast
alle Eigenschaften organischer Formen in engster Beziehung zu der
Lebensweise des Organismus und zu den Funktionen seiner Organe, vor
allem der Wuchsrichtung der Pflanze und ihrer Glieder. Meist entspricht
deshalb den äußeren Symmetrieverhältnissen eines Pflanzenteiles auch
die Symmetrie seines inneren Baues.
Von wenigen, sehr einfachen Pflanzen abgesehen, deren
Symmetrieverhältnisse hier unerörtert bleiben sollen, finden wir am
Körper pflanzlicher Organismen und jedem seiner Teile fast stets einen
polaren Gegensatz seiner durch die Längsachse verbundenen
Körperenden, seiner Spitze und Basis ausgebildet. Ein solcher
Unterschied kommt vor sowohl bei frei beweglichen Formen, bei denen die
Fortbewegungsrichtung meist durch die polare Ausbildung des Körpers
bestimmt wird, als auch bei festgewachsenen Arten, bei denen der Körper
mit dem unteren Pole, der Basis, am Substrate festgeheftet ist.
Jeder Schnitt parallel zur Längsachse, gleichgültig ob er diese in
sich aufnimmt oder nicht, ist ein Längsschnitt durch den
Pflanzenteil; die rechtwinklig zur Längsachse geführten Schnitte sind
Querschnitte. Ein polar gebauter Organismus oder Pflanzenteil,
der rings um seine Längsachse annähernd gleich gebaut ist, wird als
radiär, polysymmetrisch oder aktinomorph bezeichnet (Fig.
529 A). Er läßt sich durch mehrere, in der Längsachse sich
schneidende Längsschnitte in jeweils zwei spiegelbildlich ungefähr[S. 62]
gleiche Teile zerlegen; er hat also mehrere Symmetrieebenen.
An solchen Körpern heißen Längsschnitte radial, wenn sie durch die
Längsachse gehen (wie es z. B. bei seinen Symmetrieebenen der Fall
ist), tangential, wenn sie senkrecht auf einem Radius stehen und nicht
durch die Längsachse hindurch gehen. Sind bloß zwei aufeinander
senkrecht stehende Symmetrieebenen vorhanden, die sich ebenfalls in
der Längsachse schneiden, so spricht man von bilateralen oder
bisymmetrischen Gebilden (Fig. 107). Gibt es schließlich bloß eine
einzige Symmetrieebene, so liegt ein dorsiventraler,
monosymmetrischer oder zygomorpher Körper vor, bei dem nur
die beiden Flanken einander entsprechen, Rücken- und Bauchseite aber
verschieden sind (Fig. 529 B); die Symmetrieebene solcher Körper
nennt man ihre Medianebene oder Mediane. Pflanzen oder
Pflanzenteile, die in der Lotrichtung nach aufwärts oder abwärts
(orthotrop) wachsen, sind meist radiär oder wohl auch bilateral
symmetrisch; wenn sie dagegen senkrecht oder schräg zur Lotlinie
(plagiotrop) wachsen, so sind sie oft dorsiventral. Schließlich
gibt es auch ganz asymmetrische organische Gebilde, bei denen
sich der Körper überhaupt nicht in spiegelbildliche Hälften teilen
läßt. Manche sonst dorsiventrale Gebilde, wie z. B. manche Blätter,
werden dadurch asymmetrisch, daß die eine Hälfte sich anders ausbildet
als die andere. Das ist z. B. bei den Blättern von Begonia der Fall,
weshalb man diese Pflanzen auch „Schiefblätter“ nennt, ferner in
geringerem Grade u. a. bei den Blättern der Ulme.
Die Symmetrieverhältnisse sind für das Verständnis der pflanzlichen
Gestaltungsverhältnisse von sehr großer Bedeutung. Oft werden
die besonderen Symmetrieverhältnisse der Seitenglieder sofort
verständlich, wenn man den Aufbau der ganzen Pflanze in Betracht
zieht. So sind die asymmetrischen Blätter, z. B. bei den
Begonien[57], die Folge der dorsiventralen Symmetrie der ganzen,
meist mehr oder weniger plagiotropen Pflanze oder, wie bei der Ulme
und vielen anderen Gewächsen, ihrer Zweige.
II. Bedeutung der äußeren Gliederung für den Organismus. Bau und
Gliederung zeigen meist enge Beziehungen zu den Lebensbedürfnissen und
zur Lebensweise der Einzelformen. Äußere Gliederung ist meist gerade so
wie die innere der Ausdruck einer Arbeitsteilung, die zwischen
den Teilen einer Zelle oder eines vielzelligen Körpers eingetreten
ist. Die äußeren Glieder sind nämlich meist zu Organen mit
bestimmten Lebensfunktionen geworden. Der phylogenetische Fortschritt
von einfacheren zu reicher gegliederten organischen Formen besteht zum
guten Teile in der Zunahme dieser Arbeitsteilung.
III. Hauptgruppen von Organen. Jeder Organismus betätigt sich
in doppelter Weise: Er muß sich ernähren, um sich selbst
zu behaupten; und er muß sich fortpflanzen, um die Art zu
erhalten, da sein Leben begrenzt ist. Diesen beiden fundamentalen
Lebensregungen dient der Körper. Nur bei primitiven Pflanzen ist er
in gleicher Weise mit seiner ganzen Masse beiden Aufgaben dienstbar;
sonst besorgen bestimmte Teile die Ernährungsvorgänge, andere die
Fortpflanzung. So finden wir meist eine scharfe Arbeitsteilung zwischen
den Vegetationsorganen und den Fortpflanzungsorganen,
die wie in ihren Funktionen, so auch in ihrem äußeren und inneren Bau
fundamental verschieden sind. Beide Gruppen von Organen müssen wir
getrennt betrachten.
I. Vegetationsorgane.
Die höchste Gliederung, die die Pflanze in ihren
Vegetationsorganen erfahren hat, ist die Gliederung in
Wurzeln, Stengel und Laubblätter. Stengel und
Laubblätter faßt man auch als Sproß zusammen. Einen aus Sproß
und Wurzeln bestehenden Körper nennen wir Kormus. Die Gewächse[S. 63]
solchen Baues bezeichnet man wohl als Kormophyten; dazu gehören
die farnähnlichen Gewächse oder Pteridophyten und die aus ihnen
hervorgegangenen, noch reicher gegliederten Samenpflanzen.
Die Kormophyten sind phylogenetisch entstanden aus einfacher
organisierten Gewächsen, bei denen der Körper noch nicht eine so
weitgehende Gliederung erfahren hat: bei denen die Wurzeln und echten
Blätter noch fehlen, wenn bei manchen auch blattähnliche Zweige
vorkommen können. Solche Gebilde bis herab zu ganz einfachen, völlig
ungegliederten Pflanzenkörpern hat man Thalli genannt. Gewächse,
die einen Thallus besitzen, kann man als thallöse
Pflanzen den Kormophyten gegenüberstellen. Einen Thallus haben die
Algen, Pilze, Flechten und alle Moose.
Mit den thallösen Pflanzen darf man die Thallophyten nicht
verwechseln. Alle thallösen Pflanzen haben zwar einen Thallus,
aber nicht alle sind Thallophyten. Unter diesem Namen faßt die
Systematik nur die Algen, Pilze und Flechten zusammen.
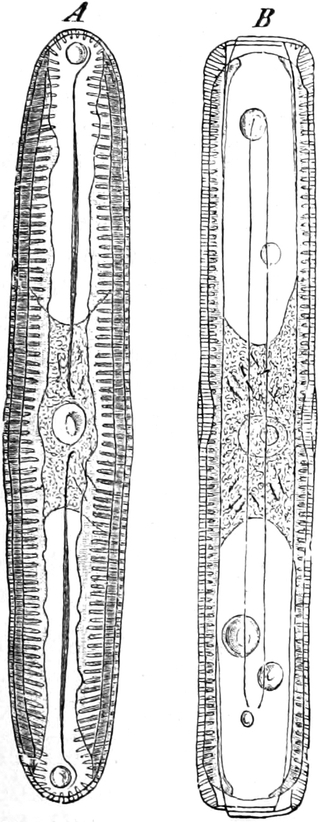
Fig. 79. Die Kieselalge Pinnularia viridis in zwei
Ansichten. A Die Schalenansicht, B die Gürtelbandansicht. Vergr.
540. Nach STRASBURGER.

Fig. 80. Bakterien des Zahnschleims. a Leptothrix
buccalis, bei a* nach Jodbehandlung, b Mikrokokken, c Spirochaete
dentium nach Jodbehandlung, d Spirillum sputigenum. Vergr. 800. Nach
STRASBURGER.
A. Der
Thallus[58].
a) Algen, Pilze, Flechten. 1. Einfachste (Kugel-)Formen.
Äußerlich ganz ungegliedert ist bloß eine Reihe mikroskopisch kleiner
einzelliger oder vielzelliger Gewächse. Die einfachste Form, die ein
Organismus annehmen kann, ist die Kugel. Aus solchen Kugelzellen
bestehen z. B. manche Algen, die an feuchten Mauern grüne Überzüge
bilden (Fig. 35), und viele Bakterien (Fig. 80 b), die bei weitem
kleinsten Organismen, die wir kennen.
2. Relative Oberflächenvergrößerung. Ausbildung einer
Längsachse. Die Kugel hat von allen geometrischen Figuren gleichen
Rauminhaltes die kleinste Oberfläche; und zwar ist die Oberfläche der
Kugel um so kleiner im Verhältnis zu ihrem Volumen, je größer dieses
wird, und umgekehrt (gleiches gilt übrigens auch für alle anders
geformten Gebilde). Bei den winzig kleinen Bakterienzellen ist also
die Oberfläche im Verhältnis zu ihrem Rauminhalt ganz außerordentlich
groß; ja man darf die auffallend geringe Größe dieser Organismen
in dieser Hinsicht wohl geradezu als Anpassung an ihre Lebensweise
bezeichnen. Alle Abweichungen von der Kugelgestalt sind mit einer
mehr oder weniger ausgiebigen relativen Vergrößerung der Oberfläche
verbunden. Namentlich wenn das Körpervolumen zunimmt, im Verhältnis
dazu seine Oberfläche also sich verringert, wird meist auf diese Weise
die Oberfläche des Körpers vergrößert. Alsdann finden wir Zylinder-,
Stäbchen-, Faden-,[S. 64] Band- und Scheibenformen, sowie schließlich
äußerlich gegliederte, mit Fortsätzen ausgestattete Körper, also
zumeist Gebilde, die bereits eine deutliche Längsachse erkennen lassen.
Die freie Oberfläche des Körpers nämlich ist bei jeder Pflanze
von allergrößter Bedeutung für die Aufnahme der zur Ernährung unbedingt
notwendigen flüssigen und gasförmigen Stoffe aus der Außenwelt. So ist
eben die Oberflächenvergrößerung das wichtigste Prinzip der
Oberflächendifferenzierung.
Von ellipsoidischer Form sind die einzelligen Individuen der
Bierhefe (vgl. Fig. 20); scheibenförmig oder zylindrisch sind
die Zellen vieler Algen, z. B. vieler Diatomeen-Arten. In dieser
Algengruppe gibt es auch spindel-, schiff- (Fig. 79), helm-,
fächer-, faden-, band- und kettenförmige Gebilde. Stäbchen- und
schraubenförmige Gestalten finden wir auch bei den Bakterien
(Fig. 80 a, c, d). Die Bakterienzellen besitzen natürlich
auch dann, wenn sie nicht Kugelgestalt haben, eben infolge ihrer
außerordentlichen Kleinheit gegenüber ähnlich gestalteten Zellen
anderer Organismen eine ungewöhnlich große freie Oberfläche.
Solche Lebewesen können mit Gallerte oder Schleim auf einer
Unterlage festsitzen oder auch frei in Flüssigkeiten, vor allem
in Wasser, flottieren. Die flottierenden Organismen des Wassers,
der Binnengewässer sowohl wie der Meere, bezeichnet man als
Plankton im Gegensatze zu den Wasserorganismen, die
festgeheftet sind, dem Benthos. Die Planktonflora, die
sehr reich an eigenartigen Formen ist, enthält fast lauter solche
Gestalten, wie wir eben genannt haben. Sie können mit aktivem
Bewegungsvermögen begabt sein (Schwimmer). Der Fortbewegung
dienen alsdann in der Regel besondere Organe: sehr häufig
fadenförmige kontraktile Geißeln oder Zilien, die
Fortsätze des Plasmakörpers sind. Ihr Besitz erlaubt es solchen
Planktonten, durch Reizbewegungen die Stellen mit den günstigsten
Ernährungsbedingungen aufzusuchen, ungünstige Stellen aber zu
fliehen. Andere Planktonorganismen schweben dagegen ohne eigenes
Bewegungsvermögen im Wasser (Schweber); viele von ihnen und
andere Planktonten besitzen besondere Schwebeeinrichtungen.
Die Oberflächen ihrer Körper sind durch lange Stacheln, Leisten,
fallschirmartige Platten außerordentlich vergrößert (Fig. 319, 321,
322); dadurch erhöht sich der Reibungswiderstand des Körpers am
Wasser bedeutend, und das Sinken wird erschwert[59].
Haben alle diese einfach organisierten Pflanzen starre
Körperformen, so gibt es auch niedere Pflanzen, die ständig ihre
Gestalt wechseln, also ohne feste Umrisse sind, z. B. die Myxamöben
und die Plasmodien der Schleimpilze.
3. Ausbildung des polaren Gegensatzes. Als nächste Stufe der
fortschreitenden Gestaltung kann bei Formen mit Längsachse jene gelten,
bei der sich ein Unterschied zwischen Basis und Spitze
oder Scheitel einstellt. Bei freibeweglichen Formen ist
alsdann der eine Pol oft Träger der Fortbewegungsorgane
(Geißeln). Bei festgewachsenen dient er meist als Haft- oder
Befestigungsorgan (als kreisrunde Haftscheibe oder als krallen-
oder fingerförmig verzweigte Haftlappen usw.) der Anheftung, während
das Wachstum sich auf eine eng umgrenzte Stelle des Körpers, einen
Vegetationspunkt, beschränken kann, nämlich bald auf eine
Zone zwischen Basis und Spitze (interkalares Wachstum, interkalarer
Vegetationspunkt), bald mehr und mehr auf den Scheitel (apikales oder
Spitzenwachstum, apikaler Vegetationspunkt). Ein Keimling der grünen
Meeresalge Ulva Lactuca mag für den letzteren Fall als Beispiel dienen
(Fig. 81).
Fig. 81. Keimling von Ulva Lactuca. Oben der Scheitel,
unten die Basis. Vergr. 220. Nach STRASBURGER.
4. Abplattung. Bei vielen Algen und Flechten ist der Thallus
bandförmig oder scheibenförmig abgeplattet (Fig. 83). Dadurch wird
die freie Oberfläche, worauf es der Pflanze ankommt, weiter
wesentlich vergrößert. Deshalb darf man diese Ausbildung wohl
als eine Anpassung an die Ernährungsverhältnisse[S. 65] dieser Organismen
ansehen. Sie bauen sich nämlich ihre organische Substanz aus dem
Kohlenstoffe der Kohlensäure auf, die sie zerlegen. Diese Zerlegung
und Assimilation aber erfolgt wie bei allen Pflanzen, die Chlorophyll
enthalten, nur am Lichte. Soll sie also in größerem Maße stattfinden,
so müssen möglichst viele Chlorophyllkörner dem Lichte ausgesetzt
werden. Das aber wird eben bei voluminösen Körpern vielfach durch
Abflachung erreicht.
5. Ausbildung von Dorsiventralität. Die Mehrzahl der bisher
besprochenen Formen ist radiär oder bilateral symmetrisch. Bei manchen,
namentlich solchen, die mit ihrem Thallus auf einer Unterlage sich
ausbreiten, also plagiotrop wachsen (z. B. bei vielen Flechten), ist
der Körper aber auch dorsiventral ausgebildet. Dorsiventrale Symmetrie
ist namentlich solchen Formen eigentümlich, deren Oberseite mehr Licht
erhält als die Unterseite. Alsdann ist vor allem die obere Seite für
die Assimilation eingerichtet.
6. Ausbildung von Verzweigungen. Noch höher organisiert sind
Fäden, Bänder und Scheiben, die durch Fortsätze verzweigt
sind. So ist die Mehrzahl der Thalli bei Algen, Pilzen und Moosen
gestaltet. Durch die Verzweigung wird die freie Oberfläche meist noch
weiter bedeutend vergrößert und zugleich eine bessere Raumausnutzung
ermöglicht. Es können dadurch schließlich busch-, strauch- und
baumförmige Thalli entstehen, bei den Algen vielfach mit Zweigen sehr
großer Biegsamkeit und Geschmeidigkeit, die dem bewegten Wasser keinen
Widerstand entgegensetzen, sondern in ihm fluten.
Fig. 82. a Schema der dichotomischen, b der
seitlichen, razemösen Verzweigung. K Keimlingsachse, H Hauptachse,
1, 2, 3, 4 Tochterachsen 1., 2., 3., 4. Ordnung.
Fig. 83. Dictyota dichotoma (braune Meeresalge).
2⁄3
nat. Gr. Nach SCHENCK.
Bei der Verzweigung kann sich der Scheitel der Keimlingsachse selbst
in zwei neue, gleichmäßig weiterwachsende Glieder teilen, gabeln
(dichotome Verzweigung), so bei dem sich fortdauernd gabelnden,
da durch fächerförmigen Thallus der braunen Meeresalge Dictyota
dichotoma (Fig. 83 und das Schema Fig. 82 a). Bei anderen verzweigten
Formen wachsen dagegen durch Neubildung von Vegetationspunkten
Seitenzweige hervor, oft mit gesetzmäßiger Anordnung (seitliche
Verzweigung). An höher organisierten Formen schränkt sich auch bei
dieser Art der Verzweigung die Bildung solcher Vegetationspunkte immer
mehr und mehr auf den Scheitel des Thallus ein; die dem Scheitel
nächsten, jüngsten[S. 66] Seitenzweige sind alsdann die kürzesten. Eine
solche scheitelwärts fortschreitende, akropetale Anlage neuer
Seitenglieder ist bereits bei der grünen Fadenalge Cladophora deutlich
(Fig. 84, vgl. auch Fig. 89). Bei der einfachsten Ausbildung der
seitlichen Verzweigung geht eine einheitliche Hauptachse, die
an der Spitze immer weiter wächst, ein Monopodium, durch das
ganze Verzweigungssystem. Sie ist die Mutterachse für eine
größere Zahl nacheinander meist ringsum entstandener und schwächer
wachsender Seitenachsen, die sich in gleicher Weise verzweigen können.
Man nennt diese Verzweigung die razemöse Verzweigung (vgl. das
Schema Fig. 82 b).
Alle Seiten- (oder Tochter-)achsen, die unmittelbar an der
Keimlingsachse entstehen, nennt man 1. Ordnung; solche, die
an Tochterachsen erster Ordnung durch Verzweigung entstehen, 2.
Ordnung und so fort (vgl. Fig. 82). Jede Achse, an der eine
Tochterachse irgendeiner Ordnung entsteht, wird mit Bezug auf diese
Auszweigung Mutterachse des Tochtergliedes genannt. Die
Seitenachsen können unbegrenztes Wachstum haben, Langtriebe
sein, oder es sind Kurztriebe mit begrenztem Wachstum.
An Achsen mit interkalaren Vegetationspunkten entstehen die
Seitenzweige entweder akropetal oder basipetal.
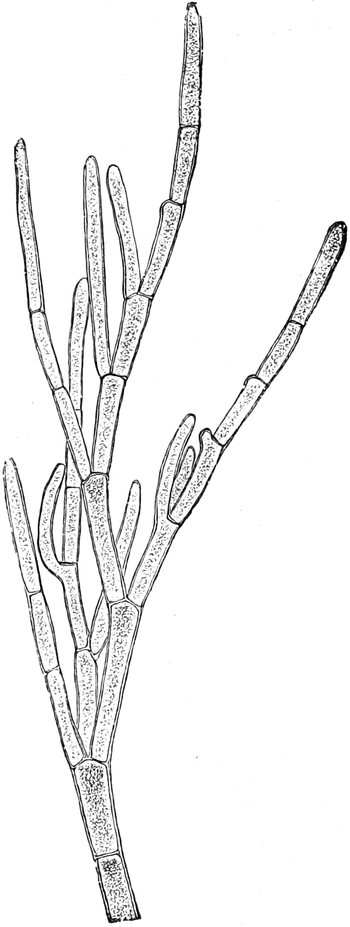
Fig. 84. Stück einer Cladophora glomerata
(Süßwasseralge). Vergr. 48. Nach SCHENCK.
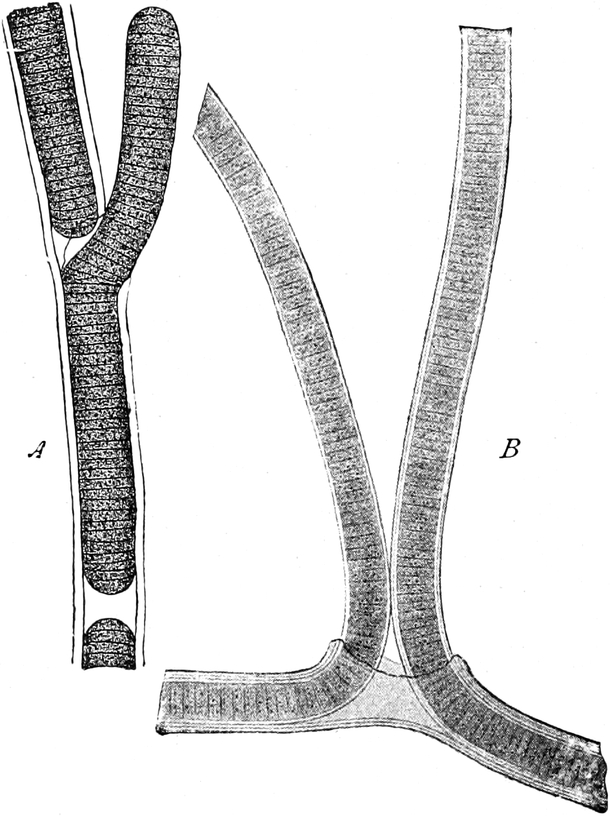
Fig. 85. Unechte Verzweigung bei Spaltalgen. A
Plectonema Wollei: nur das obere Ende des zerbrochenen Fadenstückes
wächst als Ast aus. B Pl. mirabile: beide Enden wachsen weiter.
OLTMANNS nach KIRCHNER
u. BORNET.
Übrigens kommt bei Thallophyten auch die zymöse Verzweigung vor,
die wir später beim Kormus näher kennen lernen werden.
Diese Verzweigungsarten nennt man echte. Im Gegensatz dazu
findet sich bei einigen niederen, fadenförmigen Algen und Bakterien
unechte Verzweigung. Sie kommt dadurch zustande, daß der
Faden in zwei Stücke zerbricht, die aber durch eine Gallertscheide
auch ferner zusammengehalten werden, und daß jedes durch den
Bruch entstandene neue Fadenende zu einer fadenförmigen Zellreihe
auswachsen kann (Fig. 85). Nicht von Ver[S. 67]zweigung, sondern von
Zerteilung spricht man dagegen, wenn ein unverzweigter Thallus
nachträglich in eine Anzahl Lappen zerteilt wird, wie es
z. B. bei dem bandartigen Thallus von Laminaria (Fig. 352) der Fall ist.
Bei den Pilzen, die keine Kohlensäure assimilieren, sondern
sich von organischen Stoffen ernähren, hat der Thallus dementsprechend
ein besonderes Aussehen. Man nennt ihn Myzelium. Er besteht
meist nur aus sehr dünnen in reichverzweigten, farblosen,
zylindrischen Fäden (Fig. 86 und Fig. 6), den Hyphen, die das
Substrat, z. B. den Waldhumus, allseitig durchziehen, so daß sie mit
außerordentlich großer Oberfläche die nötigen Nährstoffe aufnehmen
können. Schmarotzerpilze treiben meist Ausstülpungen von Hyphen als
Saugfortsätze (Haustorien) in die lebenden Zellen der
Wirtspflanzen, sofern sie nicht in den Zellen leben, sondern etwa mit
ihren Hyphen das Interzellularsystem durchziehen (Fig. 87).
Fig. 86. Teil eines Myzeliums von dem Schimmelpilz
Penicillium. Vergr. ca. 35.
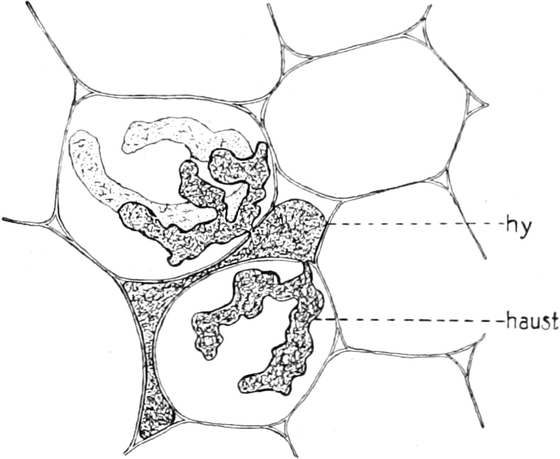
Fig. 87. Haustorien (haust) von Peronospora parasitica
in Parenchymzellen von Capsella. hy Interzellulare Hyphe. Vergr. 240.
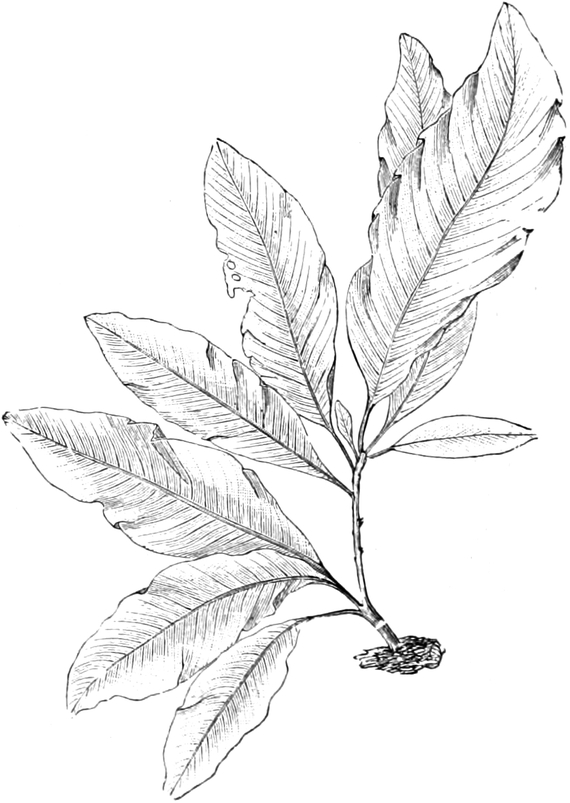
Fig. 88. Thallus der Rotalge Delesseria sanguinea.
1⁄2
nat. Größe. Nach SCHENCK.
7. Arbeitsteilung zwischen den Thalluszweigen. Am reichsten
gegliedert ist der Thallus in einigen Abteilungen der Schlauchalgen
(Siphoneen), der braunen und der roten Meeresalgen (Phaeophyceen und
Rhodophyceen). Die äußere Gliederung mancher solcher mit Haftscheiben,
Haftlappen oder verzweigten Strängen befestigten Formen, die zum
Teil sehr groß werden können (der Thallus der Braunalge Macrocystis
wird über 45 m lang), erinnert auffallend an die des Sprosses der
Kormophyten, so z. B. bei der roten Meeresalge Delesseria sanguinea
(Fig. 88): an zylindrischen, verzweigten Thallusästen sitzen
Seitenzweige, die blattähnlich gestaltet sind. Der Thallus hat bei
vielen solchen Formen außer der Ausbildung eines Haftorgans (einer
Haptere) und der Zweige eine weitere Arbeitsteilung zwischen
seinen Gliedern eintreten lassen: einige Zweige sind zylindrisch
und dienen[S. 68] dazu, das Wachstum und die Verzweigung des Thallus
als Langtriebe fortzusetzen und die übrigen Triebe zu tragen. Die
letzteren dagegen sind zu blattartigen Assimilationsorganen
(Assimilatoren) mit begrenztem Wachstum, zu Kurztrieben, geworden.
Ja, diese Kurztriebe zeigen manchmal unter sich nochmals eine
Arbeitsteilung. Solche Formen sind morphologisch von höchstem
Interesse, weil sie uns zeigen, wie die Blätter der Kormophyten aus
Kurztrieben entstanden sein könnten.
Die Ausbildung blattähnlicher Kurztriebe an den Körpern von
thallösen Pflanzen ist offenbar selbständig in jeder der
genannten Reihen entstanden, nämlich da, wo Thallusstücke zu
besonderen Assimilationsorganen wurden. Alle diese Gebilde nahmen
annähernd gleiche Form, eben die Blattform, an. Die blattartigen
Triebe der Siphoneen und Braunalgen sind, mit anderen Worten, denen
der Rotalgen nicht homolog, sondern nur analog.
8. Innerer Bau der Thalli. Alle diese Thalli, mögen sie
gegliedert oder ungegliedert sein, können aus einem einzigen
Protoplasten bestehen (z. B. Schlauchalgen: Caulerpa, Fig. 348) oder,
wie es meist der Fall ist, aus vielen Zellen sich zusammensetzen.
Bestehen sie aus vielen Zellen, so sind diese entweder in einer Reihe
zu einem Zellfaden (Fig. 84), in einer Fläche oder zu einem Zellkörper
angeordnet. Die einfachsten mehrzelligen Thalli setzen sich aus
lauter gleichförmigen und in gleicher Weise teilungsfähigen Zellen
zusammen. Sobald ein Vegetationspunkt sich ausbildet, tritt aber eine
Sonderung ein zwischen embryonalen, teilungsfähigen,
und Dauerzellen. Die äußersten Spitzen der apikalen
Vegetationspunkte werden bei vielzelligen Thalli fast stets von einer
einzigen Zelle, der Scheitelzelle, eingenommen, die bei manchen
Formen nur wenig von den anderen Zellen abweicht, so bei der Fadenalge
Cladophora glomerata (Fig. 84). An den vielzelligen Langtrieben der
büschelig verzweigten braunen Meeresalge Cladostephus verticillatus
fallen die großen kuppenförmig gestalteten Scheitelzellen aber sofort
auf (Fig. 89).
Jede solche an der Spitze fortwachsende Scheitelzelle teilt sich
durch quere, einander parallele Wände, die von ihrem unteren Ende
scheibenförmige Segmente abschneiden. Diese teilen sich
in gesetzmäßiger Weise weiter zunächst durch Längswände,
hierauf durch Querwände in eine größere Anzahl zunächst noch
embryonaler Zellen. Aus bestimmten Randzellen der Segmente wachsen,
spitzenwärts fortschreitend, die Seitenzweige (meist
als Kurztriebe) hervor, die das Aussehen der Pflanze bestimmen
(Fig. 89). Auch flache bandartige Körper können eine
ähnlich gestaltete, nur entsprechend abgeflachte Scheitelzelle
besitzen, so die in Fig. 90 dargestellte braune Meeresalge
Dictyota dichotoma[60]. Von ihr (Fig. 90 A) werden durch
grundwärts vorgewölbte Querwände flache Segmente abgeschnitten,
die sich weiterhin durch Längswände teilen. Gelegentlich wird die
Scheitelzelle aber auch durch eine Längswand in zwei nebeneinander
liegende gleichgroße Scheitelzellen geteilt (B, a, a), deren
jede einen Seitenzweig bildet. Dadurch kommen die Gabelungen des
Körpers zustande.
Die Dauerzellen des Thallus sind fast immer, selbst bei den am
reichsten gegliederten Thalli, nur Parenchymzellen. Ist der Thallus
ein vielzelliger Körper, so kann wohl eine Sonderung eintreten in
peripher gelegenes chlorophyllreiches Assimilationsparenchym, in
Speicherparenchym, das an Reservestoffen reich und farblos ist, und in
Leitparenchym aus langgestreckten Zellen.
Eine Veranlassung zur Ausbildung einer typischen Epidermis fehlt
bei den vielzelligen Algen, da sie im Wasser eines Schutzes
gegen Austrocknung nicht bedürfen und durch Schleimüberzüge vor
zu starkem Wasserverlust bewahrt bleiben, wenn sie bei der Ebbe
etwa an die Luft gelangen. Doch besitzen die Algen an ihren
Oberflächenzellen schon eine äußere Zellmembranlamelle, die
sich mit Chlorzinkjod braun färbt. Für genügende Festigung des
Thallus, besonders bei den in der Brandung wachsenden Arten,
wird durch starke Verdickung der Wände in den äußeren Zellagen,
unter Umständen auch durch Inkrustationen mit kohlensaurem Kalk
gesorgt. Beim Blasentang (Fucus vesiculosus) sind zudem besondere
mechanische, durch ihre Dickwandigkeit, große Dehnbarkeit und[S. 69]
Elastizität ausgezeichnete Zellen vorhanden. Den relativ höchsten
Grad innerer Differenzierung zeigen die ebenfalls zu den braunen
Algen gehörenden Laminarien. In den stammartigen Achsen, die bei
diesen Pflanzen sehr dick werden, läßt sich Rinde, Zentralkörper
und ein lockeres Mark unterscheiden. Die Rinde enthält vielfach
Schleimgänge, das Mark sogar Züge siebröhrenartiger Zellen, die
vielleicht der Stoffleitung dienen; solche Zellen kommen übrigens
auch bei manchen Rhodophyceen vor. Die Laminarienachsen wachsen
durch fortgesetzte Teilung der Rindenzellschicht in die Dicke. Die
Produkte dieser Teilungstätigkeit bilden eine Art sekundäres Gewebe
mit konzentrischen Zonen, die an Jahresringe der Samenpflanzen
erinnern.
Die Thalli der Flechten kommen durch Verflechtung von
Pilzhyphen zustande und können parenchymatische Struktur annehmen.
Bei vielen Arten werden die peripherischen Schichten durch sehr
dichte Verfilzung der Hyphen und sehr starke Verdickung der
Hyphenwände zu schützenden Rinden über den assimilierenden Algen.
Fig. 89. Endtrieb von Cladostephus verticillatus. Vergr.
30. Nach N. PRINGSHEIM.
Fig. 90. Der Vegetationspunkt von Dictyota dichotoma und
seine Gabelung. a Die Scheitelzellen. Nach
E. DE WILDEMAN.
Vergr. ca. 500.
Fig. 91. Riccia fluitans. Nat. Gr. Nach SCHENCK.
Fig. 92. Blasia pusilla. r Rhizoide. Vergr. 2. Nach SCHENCK.
b) Moose[61]. Im äußeren und inneren Bau der Moospflanzen
(Bryophyten) kommt wie bei den Algen wieder zum Ausdruck, daß sie
Kohlensäure assimilieren. Es gibt zunächst Lebermoosarten, deren
Körper bandartig ist, sich gabelig verzweigt und auffällig dem Körper
von Algen, wie Dictyota (Fig. 83), gleicht, so das Lebermoos Riccia
fluitans (Fig. 91). Bei dem Lebermoos Blasia pusilla (Fig. 92) ist
der bandartige Körper, der, wie viele andere thallöse Moose, eine
Mittelrippe besitzt, seitlich gelappt, als ob blattartige
Gebilde sich zu sondern begännen. Die am reichsten gegliederten
Lebermoose, wie Plagiochila asplenioides (Fig. 93), und alle Laubmoose
tragen solche an einem zylindrischen, verzweigten Stengel als besondere
Assimilationsorgane.[S. 70] Seitenzweige sitzen an den Mutterachsen
unter den Blättern. Auch diese dorsiventralen, bilateralen oder
radiären sproßähnlichen Körper, die bei den Laubmoosen oft Polster
bilden, sind den Sprossen der höheren Pflanzen nur analog. Man faßt
sie wohl am besten als hoch differenzierte Thalli auf. Den Moosen, die
im Gegensatze zu den meisten Algen in der Regel Luftorganismen sind,
fehlen nämlich noch die Wurzeln; sie befestigen sich am Boden nur
durch Rhizoiden: einzellige, an ihrer Basis mit einer Querwand
abgegrenzte Haare oder verzweigte Zellfäden, die den Körper auch mit
Wasser versorgen. Viele Formen können aber noch mit der ganzen
Oberfläche ihrer Vegetationsorgane Wasser aufnehmen.
Liegt der Thallus der Unterlage auf, so ist er wie bei entsprechend
lebenden Flechten meist dorsiventral ausgebildet und zeigt bei vielen
Lebermoosen oft nur an seiner dem Lichte ausgesetzten Oberseite
reicheren Chlorophyllgehalt (Fig. 95). Die Rhizoiden entspringen
alsdann ausschließlich der Unterseite.
Auch bei den Moosen, die immer vielzellig sind, wird die Spitze
des stets apikalen Vegetationspunktes oft von einer einzigen
Scheitelzelle eingenommen.
Fig. 93. Plagiochila asplenioides mit jalousieähnlich
übereinander greifenden Blättern. Nat. Gr. Nach SCHENCK.
Fig. 94. Schema des Vegetationspunktes von Metzgeria
furcata im Augenblick der Verzweigung. Von der Rückenfläche gesehen.
a Die Scheitelzelle der Mutterachse, b des Tochterzweiges. Vergr.
ca. 370. Nach KNY.
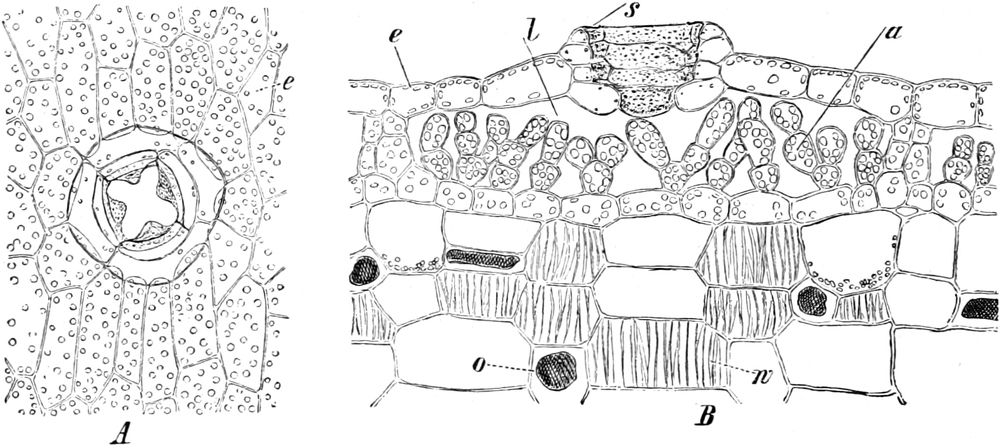
Fig. 95. Oberflächenansicht und Querschnitt des Thallus
der Marchantia polymorpha. In A eine Atemöffnung von oben, in B
im Querschnitt, e Epidermis, s Randzellen der Atemöffnung, l
Luftkammer, a Assimilationszellen, o Ölkörper, w Wassergewebe.
Vergr. 240. Nach STRASBURGER und KOERNICKE.
Diese Zelle hat bei bandartigen Lebermoosen, wie Metzgeria
und Aneura, ebenso schon bei ähnlich gestalteten Algen,
keilförmige Gestalt (Fig. 94) und ist meist
zweischneidig, seltener vierschneidig. Die zweischneidige
gibt durch aufeinanderfolgende, abwechselnd nach rechts und
links geneigte und schräg aufeinander stehende Wände nach
zwei Seiten hin Segmente ab, die durch weitere Teilungen
den Pflanzenkörper aufbauen; die vierschneidige gibt dagegen auch
noch nach oben und unten Segmente ab. Die scheinbar rein gabelige
Verzweigung der Lebermoose mit solchen Vegetationspunkten ist auf
die frühzeitige Anlage neuer Scheitelzellen aus der randständigen
Hälfte junger Segmente[S. 71] (Fig. 94 bei b) zurückzuführen. Bei
den aufrecht wachsenden, radiär gebauten Thalli der Laubmoose
hat die Scheitelzelle die Gestalt einer dreiflächig
zugespitzten Pyramide. Man kann sie dreischneidig nennen.
Auch die Blattanlagen der Laubmoose wachsen zuerst mit einer
Scheitelzelle, und zwar mit einer zweischneidigen, zeigen also
Spitzenwachstum; später wachsen sie interkalar.
Die Dauergewebe sind wesentlich vollkommener als bei den
Algen gesondert. Das ist durch das Landleben bedingt, das andere
Lebensbedingungen für die Moose als z. B. für die Algen brachte. Zur
Abgrenzung einer Epidermis kommt es gleichwohl auch bei den Moosen
nur ausnahmsweise, wenn auch die oberirdischen Teile von einer Art
Kutikula überzogen sind. Doch setzt sich am Thallus der Marchantien
eine äußerste Zellschicht von dem nächst inneren Gewebe deutlich ab.
Sie ist von Öffnungen (Fig. 95) durchbrochen, die als Atemöffnungen
bezeichnet werden und gleich den Spaltöffnungen der höheren Gewächse
Luftspalten sind. Auch haarähnliche, Schleim absondernde Bildungen in
Form von Papillen oder blattähnlichen Schuppen sind bei den Moosen weit
verbreitet.
Typische Spaltöffnungsapparate mit zwei Schließzellen, die eine
Spalte umschließen, findet man aber, wie GOEBEL[61]
gezeigt hat, beachtenswerterweise im Thallus der Lebermoosgattung
Anthoceros; freilich sind die Spaltöffnungen hier keine Luft-,
sondern Schleimspalten.
Ein eigenartiger kapillarer Apparat im Dienste der Wasserversorgung
ist bei den Torfmoosen (Sphagnaceen) ausgebildet. Die Rinde der
Stämmchen besteht aus drei bis vier Schichten inhaltsleerer
Zellen, die begierig Wasser aufsaugen, weil ihre ring- und
schraubenförmig verdickten Längs- und Querwände mit runden Löchern
versehen sind. In den Blättern liegen solche Zellen einzeln in den
Maschen eines einschichtigen Netzes aus langgestreckten, lebenden,
chlorophyllhaltigen Zellen.
Manche Lebermoose verfügen auch schon über besondere, der Stoffleitung
dienende Stränge aus langgestreckten Zellen, die ihren Körper, bei
bandartigen Formen in der Mittelrippe, durchziehen. Gegen das umgebende
Gewebe deutlich abgegrenzt treten uns die Leitstränge aber erst bei den
Laubmoosen entgegen.
Einen relativ einfach gebauten Leitstrang (l) dieser Art
im Stämmchen von Mnium undulatum führt im Querschnitt die
Fig. 96 vor. Am vollkommensten ist er in den Stämmchen der
Polytrichaceen ausgebildet. Dort verläuft ein zentraler Strang aus
langgestreckten, dünnwandigen und plasmaleeren, der Wasserleitung
dienenden, aus dickwandigen, der Festigung dienenden Zellen und aus
gestreckten Zellen, die Eiweiß und Kohlehydrate enthalten. Auch
die einschichtige Blattspreite besitzt oft einen mehrschichtigen
Mittelnerv, der einen Leitstrang der geschilderten Art enthalten
kann. Dieser setzt sich dann in das Gewebe des Stengels hinein bis
zu einem Leitstrang fort. Ferner kommen bei einigen Laubmoosen auch
mechanische Zellen vor, die langgestreckt und zugespitzt sind und
völlig Sklerenchymfasern gleichen.
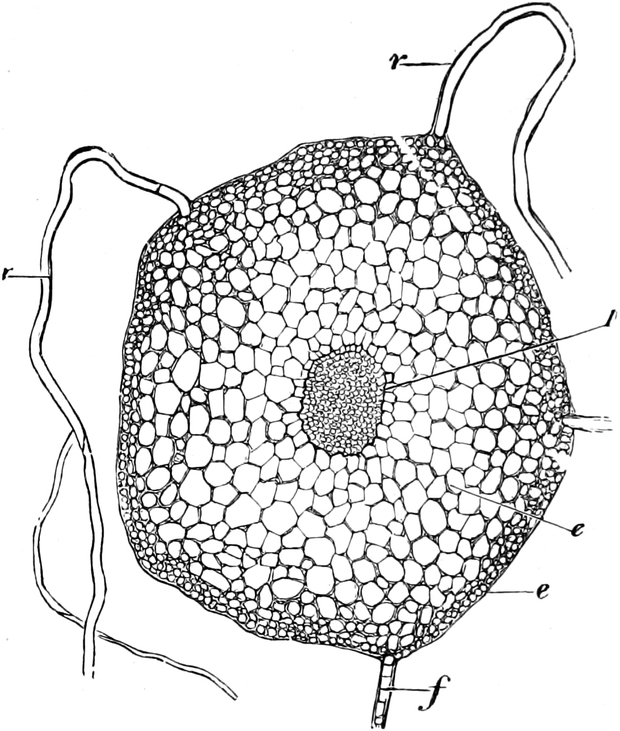
Fig. 96. Querschnitt durch das Stämmchen, von Mnium
undulatum. l Leitstrang, c Rinde, e die äußerste Zellschicht
der letzteren, f Blattflügel, r Rhizoiden. Vergr. 90. Nach STRASBURGER.
c) Gametophyt der Kormophyten[61]. Auch in den Entwicklungsgang
der Kormophyten, für die die Ausbildung des Kormus bezeichnend[S. 72]
ist, ist ein thallöser Vegetationskörper eingeschaltet: Bei ihnen
nämlich wechseln regelmäßig zwei Generationen von Vegetationskörpern
miteinander ab, von denen nur die eine, die Sporenpflanze
(Sporophyt), als Kormus, die andere aber, die Geschlechtspflanze
(Gametophyt), als meist sehr einfach gegliederter und gebauter
Thallus ausgebildet ist (Prothallium). Diese Generation lebt bei
den Farnpflanzen meist selbständig als ein grünes, mit in der Regel
einzelligen Rhizoiden am Boden befestigtes, flaches Gebilde (Fig. 97),
das nur einige Zentimeter lang wird und einem kleinen Lebermoosthallus
gleicht, aber auch aus verzweigten Zellfäden bestehen kann.
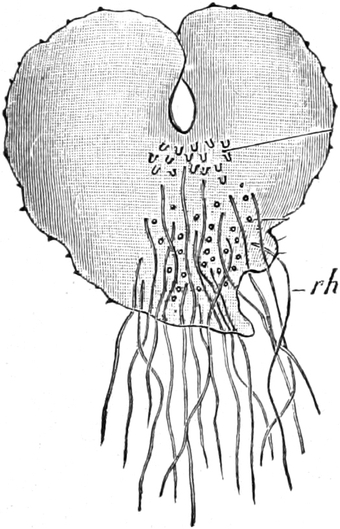
Fig. 97. Aspidium filix mas. Prothallium von der
Unterseite. rh Rhizoiden. Vergr. ca. 8. Nach SCHENCK.
B. Der
Kormus[62].
Die Vegetationsorgane des Sporophyten der Farnpflanzen (Pteridophyten)
und der Samenpflanzen, die wir Kormus nennen wollen, gliedern
sich, wie schon gesagt, noch viel weiter als die Thalli, nämlich in
Sprosse und Wurzeln, die Sprosse in Sproßachsen und Blätter. Stengel,
Blätter und Wurzeln sind die Grundformen des Kormus. Der Kormus
zeigt in seinem äußeren und inneren Bau augenscheinliche Anpassungen an
das Landleben.
Ebenso wie bei sehr vielen Thalli wird beim Kormus die Oberfläche
durch Verzweigungen fast stets bedeutend vergrößert. Die
Sproßachse bildet Seitensprosse (Seiten-, Tochterzweige), die Wurzel
Seitenwurzeln (Neben-, Tochterwurzeln). Durch die Verzweigung, die bei
vielen Gewächsen schon früh an der Keimpflanze beginnt; entsteht ein
Sproß- und ein Wurzelsystem.
Den Ausdruck Kormus gebraucht man meist als gleichbedeutend mit
Sproß und versteht darunter einen beblätterten Stengel ohne
die Wurzeln. Auch den beblätterten Moosen schreibt man dann
vielfach einen Sproß oder Kormus zu. Diese Auffassung stammt aus
einer Zeit, wo man den Entwicklungsgang der Moose noch nicht genau
kannte. Wir haben jetzt Grund zu der Annahme, daß der „Sproß“ der
Moose mit den Sprossen der Farn- und Samenpflanzen nicht
homolog ist. Also ist es zweckmäßiger, bei den Moosen, wie bei den
„beblätterten“ Algen, noch nicht von Sproß oder Kormus zu sprechen.
Es steht wohl nichts im Wege, den Begriff Kormus weiter zu fassen
als den Begriff Sproß und mit diesem Ausdruck die in Sproß und
Wurzeln gegliederten Vegetationsorgane der Kormophyten
zu bezeichnen. Übrigens gibt es Übergänge zwischen Wurzeln und
Sprossen (z. B. die Wurzelträger von Selaginella), wie auch
zwischen Blättern und Sprossen (z. B. bei Utricularia).
1. Bau des typischen Kormus.
Wir wollen zunächst solche Kormi betrachten, denen wir typischen Bau
zusprechen können. Die Besonderheiten der Grundformen treten nur in
typischer Ausbildung, wie wir sie etwa bei unseren Bäumen oder vor
allem bei vielen einheimischen Kräutern finden, deutlich zutage. Die
Grundorgane können nämlich mancherlei Umbildungen erfahren, die so weit
gehen können, daß ihre Unterschiede sich in extremen Fällen mehr oder
weniger verwischen.
[S. 73]
a) Der Sproß[63].
Der Sproß, der bei Landpflanzen ganz in der Luft oder teilweise
in der Luft (als Luftsproß), teilweise in der Erde (als
Erdsproß, Fig. 143) lebt, letzteres bei sehr vielen
ausdauernden krautigen Gewächsen (vgl. Fig. 125, 143), besteht aus
dem Stengel, den man auch Sproßachse nennt, und aus
den Blättern, die am meist grünen Luftsprosse hauptsächlich
als grüne Laubblätter (Laubsproß), an den farblosen (weißen)
Erdsprossen (Wurzelstöcken oder Rhizomen) aber als
blasse Schuppen ausgebildet sind. Die Sproßachse ist der Träger
der Blätter, sorgt für die Vergrößerung des Sproßsystems: für die
Verlängerung des Stengels, für die Neubildung von Blättern und von
Seitenzweigen, stellt die Verbindung der Blätter mit den Wurzeln her
und dient der Stoffleitung zwischen diesen Organen. Die Sproßachse
der meisten Erdsprosse dient ferner noch der Speicherung von
Reservestoffen. Die Laubblätter sind wie die blattähnlichen Kurztriebe
der thallösen Pflanzen die Assimilationsorgane und zugleich
die Transpirationsorgane der Kormophyten. Diesen Funktionen
entspricht der äußere und innere Bau der Laubblätter und des Stengels.
α) Der Vegetationspunkt. Der Sproß
zeigt Scheitelwachstum mittels eines apikalen
Vegetationspunktes, der sich an der äußersten Spitze, dem
Scheitel des Stengels, befindet. Da der Vegetationspunkt
gewöhnlich klein, dem bloßen Auge kaum sichtbar ist, so bekommt man
ihn erst zu Gesicht, wenn man Längsschnitte durch den Sproßscheitel
bei Lupenvergrößerung betrachtet (Fig. 98). Man sieht alsdann, daß er
flach (Fig. 99) oder vorgewölbt (Fig. 98 v), manchmal auch steil
kegelförmig ist (Vegetationskegel Fig. 100, 102), und daß an
seiner Oberfläche, exogen, seitlich oder unterhalb seiner Spitze
Höcker oder Wülste (f) dicht gedrängt in großer Zahl vorspringen: die
Blattanlagen und zwischen ihnen die Anlagen der Seitenzweige
(g). Die Blattanlagen entstehen in akropetaler Reihenfolge, sind
daher um so größer, je weiter sie vom Scheitel entfernt sind. Ihre
Gestalt wird auf Querschnitten durch den Vegetationspunkt besonders
deutlich (Fig. 99).
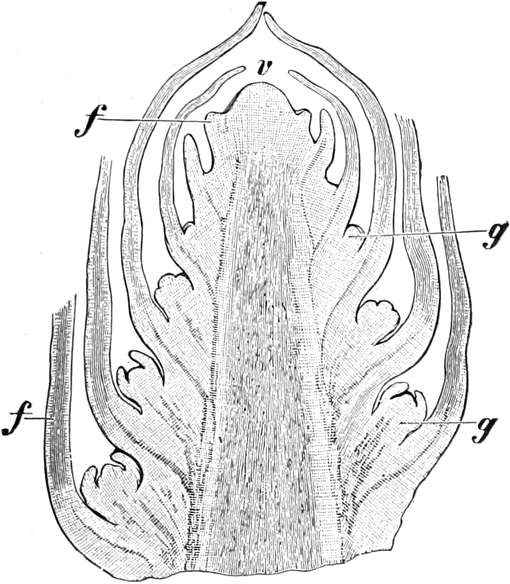
Fig. 98. Sproßscheitel einer Samenpflanze. Bei v
Vegetationspunkt, f Blattanlagen, g Seitenzweiganlagen. Vergr. 40.
Nach STRASBURGER.
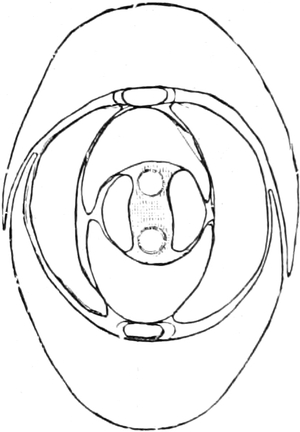
Fig. 99. Scheitelansicht eines Sproßvegetationspunktes
von Evonymus japonica. Vergr. 12. Nach STRASBURGER.
Der Vegetationspunkt und die ganz jugendlichen Blattanlagen, die
sich immer nur aus den embryonalen Teilen des Scheitels
bilden, bestehen aus embryonalem Gewebe. Bei den meisten Farnen und
den Schachtelhalmen liegt an der Spitze des Vegetationspunktes eine
Scheitelzelle (Fig. 100 t). Sie ist dreischneidig, hat also die Gestalt
einer dreiseitigen Pyramide (eines Tetraëders) mit vorgewölbter
Grundfläche als Außenseite.
Die Scheitelzelle (Fig. 100 t und 101 A) an den Hauptsprossen
des Ackerschachtelhalmes (Equisetum arvense) kann als Beispiel
dienen. Sie erscheint, vom Scheitel aus gesehen (Fig. 101 A),
als gleichseitiges Dreieck, in dem neue Scheidewände nacheinander[S. 74]
nach drei Seiten, parallel zu jeder der Seitenwände (p), angelegt
werden. Jedes Segment (S′, S″) wird durch Scheidewände (m)
weiter zerlegt. Bei den Farnpflanzen mit Scheitelzellen beginnen
auch die Blattanlagen (f, f′, f″) meist noch ihre Entwicklung
mit einer solchen, und zwar mit einer zweischneidigen (f).
Weiterhin büßen sie aber die Scheitelzelle meist ein und vollenden
ihre Ausbildung durch „Randwachstum“ mittels vieler gleichwertiger
zweischneidiger Randzellen. Ein solches Randwachstum findet sich
z. B. bei den Blattanlagen von Equisetum. Auch die Anlagen der
Seitenknospen (g) bilden sich aus einer Zelle, die zur
Scheitelzelle der Anlage wird.
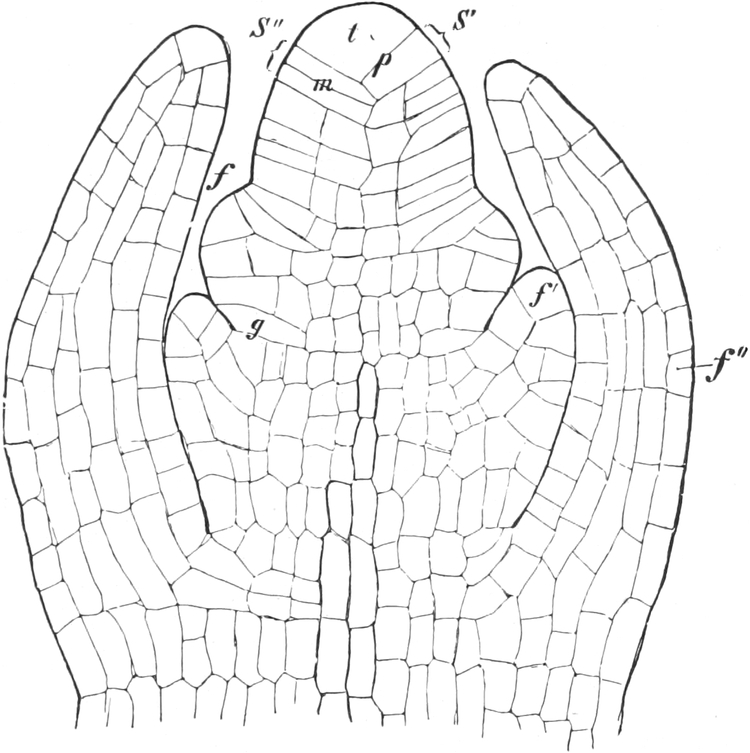
Fig. 100. Medianer Längsschnitt durch den
Sproßvegetationspunkt des Schachtelhalms Equisetum arvense.
Die Erklärung der Buchstaben im Text. Vergr. 240. Nach STRASBURGER.
Fig. 101. A Scheitelansicht des Vegetationskegels
von Equisetum arvense. B Optischer Durchschnitt desselben
Vegetationskegels unterhalb der Scheitelzelle. l Seitenwände der
Segmente. Die Erklärung der übrigen Buchstaben im Text. Vergr. 240.
Nach STRASBURGER.
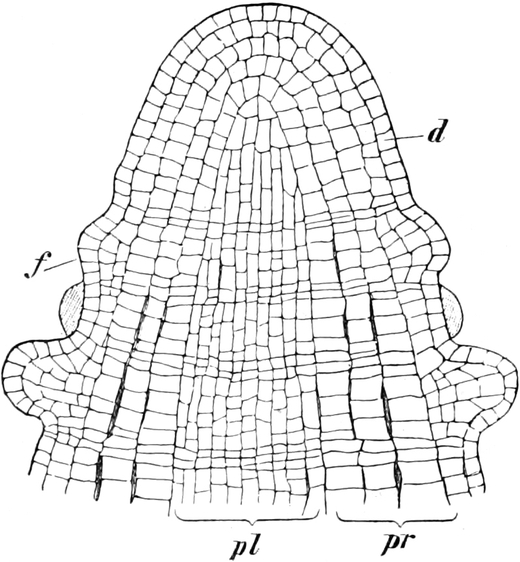
Fig. 102. Medianer Längsschnitt durch den
Vegetationskegel von Hippuris vulgaris (Tannenwedel). d Dermatogen,
pr Periblem, pl Plerom, f Blattanlagen. Vergr. 240.
Nach STRASBURGER.
Bei den Bärlappgewächsen (Lycopodiaceen) unter den Pteridophyten
und bei den Phanerogamen gibt es keine solche Scheitelzelle am
Vegetationspunkte. Hier treten an die Stelle der Scheitelzelle mehrere
gleichwertige embryonale Zellen, die oft regelmäßig in schalenförmigen
Schichten angeordnet sind (Fig. 102).
Die äußerste Zellschicht, die den Vegetationspunkt deckt, als
einfache Zellschicht auch die jungen Blattanlagen überzieht und
sich im Gegensatze zu den anderen Schichten meist nur durch
antikline Wände teilt, heißt Dermatogen (d), weil sie
meist ausschließlich die Epidermis der Pflanze liefert; die
Zellen, mit denen der zentrale Gewebestrang des Stengels, der
Zentralzylinder, im Vegetationspunkt endet, heißen Plerom
(pl), die zwischen beiden gelegenen Zellschichten Periblem
(pr). Plerom und Periblem lassen sich aber oft nicht
unterscheiden. An solchen Vegetationspunkten ohne Scheitelzellen
entstehen die Blätter und die Seitenzweige als vielzellige
Höcker (Fig. 102).[S. 75] Ihre Anlage pflegt durch örtliche Vermehrung
der äußersten Periblemschichten eingeleitet zu werden, während das
Dermatogen sich auch hier nur rechtwinklig zur Oberfläche teilt. An
den Anlagen der Blätter beteiligt sich außer dem Dermatogen nur das
Periblem, an den Anlagen der Seitenzweige auch noch das Plerom[64].
Da auch für diese Vegetationspunkte die Regel der rechtwinkligen
Schneidung der jungen Zellwände gilt, so bilden die Zellhäute in
ihrer Gesamtheit auf Längsschnitten durch die Spitzen mancher
kegelförmiger Vegetationspunkte auffallend symmetrische Figuren:
die Periklinen sowohl wie die Antiklinen je eine Schar von
Parabeln mit gemeinsamem Brennpunkte (Fig. 268). Die Elemente der
einen Schar sind entgegengesetzt gerichtet wie die der anderen
und schneiden diese annähernd rechtwinklig (SACHS). Auf
Querschnitten durch solche Scheitel bilden die Periklinen aber
konzentrische Kreise.
Knospe. Auf die Entwicklungsvorgänge, wodurch am Scheitel des
Sprosses aus embryonalem Gewebe neue Glieder angelegt werden, folgt
deren Größenzunahme, äußere und innere Ausbildung. Dieses Wachstum
pflegt meist mit einer ausgiebigen Streckung der Blattanlagen zu
beginnen. Dabei eilen die Blattanlagen also in ihrem Wachstum
dem Wachstume der Stengelspitze voraus, und zwar wachsen ihre
Unterseiten besonders stark. Infolgedessen schließen die älteren
über dem Vegetationspunkt domartig zusammen (Fig. 98) und decken
die jüngeren. Auf diese Weise bilden die größeren und älteren
Blattanlagen einen sehr wirksamen Schutz des zarten Vegetationspunktes
und der jüngsten Blattanlagen gegen Austrocknung, indem sie mit dem
Vegetationspunkte eine Knospe bilden. Die Knospe ist also nichts
anderes als das jugendliche, noch nicht fertig entwickelte Ende eines
Sprosses.
Knospenlage und Knospendeckung. Wie Querschnitte durch
Knospen lehren, fügen sich die Laubblattanlagen in verschiedener
Weise den engen Raumverhältnissen in der Knospe: Knospenlage
(Vernation). Sie können flach ausgebreitet oder auch der Länge nach
zusammengelegt, gefaltet, gerollt (Fig. 103 l) oder zerknittert
sein. Andererseits sieht man die aufeinanderfolgenden Blattanlagen
entweder mit ihren Rändern sich nicht erreichen oder nur berühren
oder, was gewöhnlicher ist, mit ihnen übereinander greifen (Fig.
103 k): Knospendeckung (Ästivation). Sie heißt im ersten
Falle offen (aperte Ä.), im zweiten klappig (valvate
Ä.), im dritten deckend oder dachziegelig (imbrikate Ä) (Fig.
103 k). Wenn alle Blätter einer Knospe mit dem einen Rande das
nächste Blatt decken, an dem anderen Rande vom vorhergehenden
Blatte gedeckt werden oder umgekehrt, so heißt die Knospendeckung
gedreht (kontorte Ä.).
Fig. 103. Querschnitt durch eine Laubknospe von Populus
nigra. Die Knospenschuppen k zeigen dachziegelige Deckung, die
Laubblätter l haben eingerollte Knospenlage; zu jedem Laubblatt
gehören zwei Nebenblätter ss. Vergr. 15. Nach STRASBURGER.
β) Die Sproßachse. A. Äußerer Bau. Der Stengel wächst erst in
einiger Entfernung vom Vegetationspunkte durch Streckung ausgiebig
in die Länge. Zugleich lösen sich hier die jugendlichen Blätter von
der Knospe. Bezeichnend für den Stengel, namentlich der Luftsprosse,
ist, daß dieses Streckungswachstum nicht auf ein kurzes Stengelstück
dicht hinter der Knospe beschränkt bleibt, sondern auch noch in
Stengelstücken stattfindet, die viele Zentimeter (bis über 50 cm) von
der Knospe entfernt sind. Freilich ist es in den aufeinanderfolgenden
Stengelzonen nicht gleich stark. Es kann überhaupt so gering sein, daß
die Blätter des Sprosses auch im fertigen Zustande aneinanderstoßen,
ohne freie Stammteile zwischen sich zu lassen. Meist aber ist es so
stark und zugleich so verschieden verteilt, daß die Ansatzstellen der
Blätter[S. 76] von nackten Stengelstücken getrennt werden (Fig. 115). Die
zwischen den Befestigungsstellen der Blätter dabei sich ausbildenden,
zylindrischen Stengelstücke nennt man Stammglieder,
Stengelglieder oder Internodien, die Stengelzonen
dagegen, an denen die Blätter befestigt sind, Knoten,
Nodi. Das Streckungswachstum des Stengels ist in den Knoten viel
geringer als in den Internodien und in diesen oft auf schmale Zonen,
z. B. auf die Basis der Internodien, beschränkt, so bei den Gräsern
(interkalares Wachstum); infolgedessen gibt es alsdann nicht mehr eine
einheitliche Streckungszone im Stengel, sondern deren mehrere, die
von ausgewachsenen Stengelstücken getrennt werden. Die Knoten können
angeschwollen sein (siehe Labiaten).
Bei den Luftsprossen sind die Internodien meist dünn, bei den
Erdsprossen dagegen oft sehr dick.
Die Länge der aufeinanderfolgenden Internodien an einer Achse (z. B.
einem Jahrestrieb) zeigt oft eine bestimmte Gesetzmäßigkeit. Am
häufigsten nehmen an der Hauptachse die Längen der Internodien in
aufsteigender Richtung zunächst zu und dann wieder ab.
Blattstellung[65]. Besonders bezeichnend für die Sprosse ist
die Blattstellung, d. h. die Verteilung ihrer Blätter. Sie kann
recht verschieden sein. An einem Knoten können ein bis mehrere Blätter
entspringen. Sind mehrere an einem Knoten vorhanden, so bilden sie
einen Wirtel oder Quirl; sie sind die Glieder
des Wirtels. In diesem Falle spricht man von wirteliger oder
quirlständiger Blattstellung. Ist an jedem Knoten bloß ein Blatt
ausgebildet, so liegt eine wechselständige Blattstellung vor.
Fig. 104. Querschnitt durch eine Laubknospe der Konifere
Tsuga canadensis, dicht über dem Sproßscheitel geführt,
5⁄13 Divergenz.
Vergr. etwa 20. Nach HOFMEISTER.
Fig. 105. Schema der 2⁄5-Stellung. Die Blätter
ihrer genetischen Aufeinanderfolge nach mit Zahlen versehen.
Nach STRASBURGER.
Untersucht man an aufrechten Sprossen mit allseitig ausgebreiteten
Blättern die Verteilung der Blätter, so findet man auffällige,
sehr beachtenswerte und eigenartige Gesetzmäßigkeiten. Unmittelbar
fällt die Regelmäßigkeit der Blattstellungen an Scheitelansichten
von Vegetationspunkten auf (Fig. 99, 104). Man sieht daran, daß
die jüngsten Anlagen in gesetzmäßiger Weise unter Ausnutzung des
vorhandenen Raumes sich den älteren anschließen. Am deutlichsten aber
treten die Stellungsverhältnisse der Blätter hervor, wenn man einen
schematischen Grundriß davon entwirft. Zu dem Zwecke zeichnet
man, wie bei einem Gebäudegrundriß die Teile des Gebäudes, so die
Lage der Blätter am Stengel auf eine zur Stengelachse rechtwinklige
Ebene ein, indem man die Blätter durch die schematisierten
Querschnittsfiguren ihrer Spreiten andeutet. Die Stengelachse
denkt man sich kegelförmig; so wird es möglich, Organe, die senkrecht
über tieferen stehen, innerhalb[S. 77] der unteren aufzuzeichnen.
Solche Grundrisse von Blattstellungen nennt man Diagramme
(Fig. 105). In ihnen ist das Zentrum der Stengelvegetationspunkt; die
dem Zentrum nächsten Blätter sind die jüngsten und zugleich obersten
Blattanlagen, die nach außen folgenden die jeweils im Alter nach unten
folgenden Blätter. Zweckmäßig deutet man jeden Knoten durch einen
Kreis an; auf die größeren dieser konzentrischen Kreise trägt man die
älteren, auf die kleineren die jüngeren Blätter ein, mehrere Blätter
an jedem Knoten natürlich auf die Peripherie eines Kreises.
Übrigens bilden solche Diagramme oft ähnliche Figuren wie Querschnitte
durch die Stengelknospe in der Nähe des Vegetationspunktes, die man bei
Vergrößerung betrachtet (Fig. 99, 104).
An radiären aufrechten Sprossen werden die Blätter möglichst
gleichmäßig rings um den Stengel verteilt. Durch diese Gesetzmäßigkeit
wird erreicht, daß die ausgewachsenen Blätter sich nur wenig
beschatten, also das Licht möglichst ausnutzen können. Diese
Verteilung ist so gleichmäßig, daß der Winkel, den die Medianen am
Stengel aufeinanderfolgender und in diesem Sinne benachbarter
Blätter miteinander einschließen (z. B. in Fig. 105, Blatt 1 und 2,
2 und 3 usw.), überall oben und unten am Stengel in der Regel der
gleiche ist. Man nennt ihn Divergenzwinkel oder, wenn man ihn in
Bruchteilen des Stengelumfanges ausdrückt, Divergenz. Er ist bei
verschiedenen Arten verschieden.
Fig. 106. Diagramm der dekussierten Blattstellung.
Die punkt. Linien sind die Orthostichen. Nach STRASBURGER
verändert.
Fig. 107. Diagramm der zweizeiligen Blattstellung.
Die punkt. Linien sind die Orthostichen. Nach STRASBURGER
verändert.
Bei wirteliger Blattstellung entspricht der Divergenzwinkel
der Blätter eines Wirtels (Fig. 106), dem Kreisumfange
dividiert durch die Anzahl der Wirtelblätter, die in der Regel bei
allen Wirteln konstant ist. Die Blätter der aufeinanderfolgenden
Wirtel stehen nicht übereinander, wechseln vielmehr von Wirtel
zu Wirtel miteinander so ab, daß die Glieder des nächst höheren
Wirtels in die Mitten der Lücken zwischen den Gliedern des
nächst tieferen Wirtels fallen (Fig. 99, 106); man sagt,
die Blätter aufeinanderfolgender Wirtel wechseln ab,
alternieren. Folge dieses regelmäßigen Wechsels und
der Gleichheit der Divergenzwinkel in allen Wirteln ist, daß
sämtliche Blätter an einem Stengel mit Quirlstellung in
Längsreihen angeordnet sind, deren Zahl doppelt so groß
ist wie die Zahl der Blätter eines Wirtels (Fig. 106).
Diese Längs- oder Geradzeilen heißen Orthostichen.
Verhältnismäßig häufig ist bei Wirtelstellungen die Ausbildung
zweigliedriger Quirle (Fig. 99, 106). Bei dieser Blattstellung,
die man dekussiert nennt, ist der Divergenzwinkel 180°
(die Divergenz also 1⁄2), und gibt es vier Orthostichen. Bei
dreigliedrigen Wirteln ist der Divergenzwinkel 120° (die Divergenz
1⁄3), bestehen sechs Orthostichen usw.
Fig. 108. Halbschematische Ansicht des
Fichtenzapfens von unten. Schuppen in 8⁄21-Stellung.
I–VIII System gleichartig im Sinne des Uhrzeigers den Zapfen umlaufender
Parastichen, 1–5 System entgegengerichtet den Zapfen umlaufender
Parastichen. Im übrigen vgl. den Text.
Bei wechselständigen Blattstellungen kann die Divergenz,
auf dem kürzesten Wege gemessen, 1⁄2, 1⁄3, aber auch z. B. 2⁄5,
3⁄8, 5⁄13 sein. Das Diagramm Fig. 107 führt uns die 1⁄2-Stellung,
Fig. 148 die 1⁄3-, Fig. 105 die 2⁄5-, Fig. 104 die 5⁄13-Stellung
vor. Auch bei wechselständigen Blattstellungen müssen die Blätter
infolge der Gleichheit der Divergenzwinkel in Längszeilen,
Orthostichen, am Stengel angeordnet sein: bei 1⁄3-Stellung[S. 78] fällt
augenscheinlich Blatt 4 senkrecht über Blatt 1 (Blatt 5 über
2, 6 über 3, 7 über 1 usw.); bei 2⁄5-Stellung (Fig. 105) fällt
Blatt 6 über Blatt 1, 7 über 2, 8 über 3 usw. Denkt man sich die
Ansatzstellen der am Stengel aufeinanderfolgenden Blätter auf
dem kürzesten Wege des Stengelumfanges durch eine Linie
verbunden (also in Fig. 105 von Blatt 1 über 2, 3, 4, 5 usw.),
so erhält man eine den Stengel umlaufende Schraubenlinie, die
als Grundspirale bezeichnet wird. Deshalb nennt man die
wechselständigen Blattstellungen wohl auch Schrauben-
oder Spiralstellungen. Jeder Abschnitt der Grundspirale,
den man von Blatt zu Blatt fortschreitend durchlaufen muß, um
von einem Blatte zu dem ersten senkrecht darüberstehenden zu
gelangen (in Fig. 105 z. B. von 1 bis 6, oder 3 bis 8), heißt
Zyklus der Grundspirale. Bei 1⁄3-Stellung besteht der
Zyklus aus drei Blättern; man muß einmal den Stengelumfang
durchlaufen, um den Zyklus zurückzulegen. Bei 2⁄5-Stellung (wie
in Fig. 105) besteht der Zyklus immer aus fünf Blättern; man
muß zweimal den Stengelumfang umkreisen. Der Zähler des Bruches
einer Divergenz gibt also stets an, wie oft ein Zyklus die
Sproßachse umkreist; der Nenner dagegen, wie viele Blätter der
Zyklus enthält, infolgedessen auch, wie viele Orthostichen es
gibt und welches Blatt als nächst höheres in der Orthostiche über
einem irgendwie bezeichneten steht. Bei 5⁄13-Stellung z. B. muß
man fünfmal die Sproßachse umkreisen, um das nächst höhere Blatt
zu erreichen, gibt es 13 Orthostichen, steht über Blatt 3 Blatt
16 (3 + 13), über Blatt 8 Blatt 21 (8 + 13). Da der Nenner des
Bruches stets die Anzahl der Orthostichen angibt, so nennt man
die 1⁄2-Stellung auch die zweizeilige, die 1⁄3-Stellung
die dreizeilige usw. Stehen die Blätter am Stengel so
gedrängt, daß sie sich berühren, so fallen nicht die Orthostichen,
sondern mehr oder weniger steil aufsteigende Schraubenlinien auf,
die als Schrägzeilen oder Parastichen bezeichnet
werden. Sie entstehen durch die Berührung derjenigen Blätter,
deren seitlicher Abstand voneinander an der Sproßachse am
kleinsten ist. Sehr deutlich sieht man die Schrägzeilen z. B.
am Fichtenzapfen, wovon in Fig. 108 eine etwas schematisierte
Ansicht von unten gegeben ist. Die Parastichen sind in
dieser Grundansicht Schraubenlinien. Mehrere Systeme untereinander
gleichsinnig verlaufender Parastichen treten deutlich hervor: eines
(mit ungebrochenen Linien I-VIII bezeichnet) umläuft den Zapfen im
Sinne des Uhrzeigers; zwei entgegengerichtete kreuzen dieses; davon
ist das eine (mit gestrichelten Linien 1–5 bezeichnete) flach, das
andere (mit fein punktierten Linien bezeichnete) steil gewunden.
Man kann zwei beliebige sich kreuzende Systeme gleichartiger
Parastichen dazu benutzen, die Divergenzen solcher Blattstellungen
zu bestimmen. Bezeichnet man irgendein Blatt mit 1 (vgl. dazu
die Fig. 108), so erhält man die Nummer des in der Parastiche
nächst folgenden Blattes dadurch, daß man zu 1 die Gesamtzahl der
gleichartigen Schrägzeilen des Systems addiert, die es rings um
den ganzen Stengel gibt. Parastichen mit ungebrochenen Linien gibt
es, wie man ohne weiteres abzählen kann, 8; also ist das nächste
Blatt in dieser Parastiche 1 + 8 = 9, das nächste 9 + 8 = 17 usw.
Gleichartig verlaufende Schrägzeilen von entgegengesetzter Neigung
gibt es z. B. gebrochen gestrichelte 5 (fein punktierte aber 13);
also sind die auf 1 in der gestrichelten Parastiche folgenden
Blätter 1 + 5 = 6, 6 + 5 = 11 usw. (in der punktierten Parastiche
dagegen 1 + 13 = 14, 14 + 13 = 27 usw.). Diese Gesetzmäßigkeit
rührt daher, daß in jedem System gleichartig verlaufender
Parastichen zwischen den benachbarten Blättern einer
Parastiche noch so viele Blätter am Stengel befestigt sein müssen,
als es außer dieser Parastiche noch weitere Schrägzeilen
in dem System gibt[S. 79] (z. B. in dem System mit ungebrochenen Linien
7; 7 Blätter liegen also zwischen 1 und dem nächsten Blatt der
Parastiche, demnach muß dieses auf 1 + 7 folgen, also das 9.
sein); das gleiche gilt natürlich auch für die Orthostichen.
Nummeriert man in dieser Weise alle Blätter, so ergeben die
aufeinanderfolgenden Zahlen 1, 2, 3, 4 usw. die Grundspirale und
die Divergenz. Der Fichtenzapfen in Fig. 108 hat die Blattstellung
8⁄21: dementsprechend liegen die Blätter 1, 22, 43 usw. in
einer Orthostiche übereinander. — Bestimmt man nun bei den
verschiedensten Pflanzen mit wechselständigen Blattstellungen die
Divergenzen, so fällt auf, daß gewisse Divergenzen ganz besonders
häufig sind; sie bilden die Reihe 1⁄2, 1⁄3, 2⁄5, 3⁄8, 5⁄13, 8⁄21,
13⁄34 usw. Diese Brüche haben merkwürdige Beziehungen zueinander:
Zähler und Nenner eines jeden sind die Summen der Zähler und Nenner
der beiden vorausgehenden Brüche. Die Divergenzen dieser Reihe
bewegen sich sämtlich zwischen 1⁄2 und 1⁄3 des Stengelumfanges. Sie
weichen um so weniger voneinander ab, je mehr sie sich vom Anfang
der Reihe entfernen, und nähern sich immer mehr einem Winkel von
137° 30′ 28″. Man hat diese Reihe als die Hauptreihe der
Blattstellungen bezeichnet. Daneben gibt es auch noch andere Reihen
ähnlicher Art. Die Hauptreihe ist aber vielleicht allen anderen
Reihen dadurch überlegen, daß bei ihren Brüchen mit der kleinsten
Zahl von Blättern die möglichst gleichmäßige Verteilung aller
an der Sproßachse erreicht wird. Die Entdecker der Reihen waren
CARL SCHIMPER und ALEXANDER BRAUN.
Aufrechte radiäre Stengel mit langen Internodien oder mit breiten
Blättern haben oft wenige Orthostichen, solche mit kurzen
Internodien oder mit schmalen Blättern meist viele. Man findet also
in diesem Falle bei Schraubenstellung stets Divergenzen, die den
höheren Gliedern der Reihen entsprechen.
An geneigten dorsiventralen Stengeln sind die Stellungsverhältnisse
der Blätter relativ einfach. Am häufigsten ist hier 1⁄2-Stellung
oder eine ähnliche Anordnung, wobei sich die Blattflächen parallel
zum Horizont stellen; dadurch werden die günstigsten Verhältnisse
für die Beleuchtung geschaffen. Die 1⁄2-Stellung wird überaus
häufig durch Drehung der Internodien erreicht, so bei der
verbreiteten dekussierten Blattstellung, die bei geneigten Achsen
durch solche Drehung zu einer zweireihigen Anordnung mit einer
Blattreihe rechts, der anderen links von der Achse wird. Auch bei
wechselständigen Stellungen kommt ähnliches vor und ermöglicht es
den Blattspreiten, das volle Oberlicht auszunutzen. So ist die
Stellung der Laubblätter eine Anpassung an die Lichtbedürfnisse
der Pflanzen. Bei manchen horizontal wachsenden Erdsprossen (z. B.
von Farnen) stehen die Blätter in einer oder zwei Reihen auf der
Oberseite.
Über die Ursachen der Blattstellungsgesetzmäßigkeiten wissen
wir noch gar nichts. SCHWENDENERs Annahme, daß rein
mechanische Ursachen die Anordnung der Blätter bestimmen, hat sich
als unbegründet erwiesen[66]. Jedenfalls brauchen die Blätter
durchaus nicht etwa in der Reihenfolge ihrer Grundspirale oder
als Glieder eines Wirtels gleichzeitig am Scheitel zu entstehen;
manchmal kann sogar eine Seite des Vegetationspunktes in der
Erzeugung von Blattanlagen wesentlich gefördert sein. Ebensowenig
nehmen sie bei Spiralstellungen als Anlagen am Scheitel stets
die gleichen Stellungen ein wie am ausgewachsenen Stengel; ihre
Divergenzen an letzterem werden vielmehr oft erst durch sekundäre
Verschiebungen hergestellt.
B. Primärer innerer Bau des Stengels[67]. Der Stengel zeigt
eine viel weitergehende Gewebedifferenzierung als die Langtriebe
selbst der am reichsten gegliederten Thalli. Zu äußerst finden wir
als Abschluß eine typische Oberhaut oder Epidermis. Darunter
liegt in den Internodien (die verwickelter gebauten Knoten lassen wir
außeracht) meist ein mehrschichtiger leitbündelfreier Gewebemantel, die
Rinde, die das übrige leitbündelhaltige Gewebe des Stengels, den
Zentralzylinder (Fig. 109), umschließt.
Wenn sich auch bei manchen Monokotylen eine Rinde von einem
Zentralzylinder nicht unterscheiden läßt, weil die Leitbündel bis
dicht unter die Epidermis gerückt sind, und auch sonst oft eine
scharfe Grenze zwischen beiden fehlt, so scheint es doch praktisch,
an dem viel gebrauchten Begriff Zentralzylinder festzuhalten.
Die Rinde besteht hauptsächlich aus Parenchym, und zwar
bei den grünen Luftsprossen an der Peripherie vorwiegend aus
chlorophyllhaltigem Parenchym, das in dicken Rinden weiter innen in
farbloses (Speicher-)Parenchym übergehen kann; bei den farblosen
Erdsprossen, die oft viel dicker als jene[S. 80] sind, besteht es nur
aus farblosem Parenchym, das, ebenso wie das übrige Parenchym der
Rhizome, reich an Reservestoffen ist. Häufig ist ein Teil der Rinde
als Festigungsgewebe ausgebildet. Die Stengel der Luftsprosse als
die Träger der Blattlast sind namentlich unter dem Einfluß des
Windes der Gefahr der Knickung ausgesetzt; sie müssen allseitig
biegungsfest gebaut sein. Dafür sorgt meist Festigungsgewebe,
in Form von Lagen oder Strängen aus Kollenchym oder Sklerenchym, die
möglichst peripher, nicht selten in vorspringenden Kanten des Stengels
direkt unter der Epidermis ausgebildet zu sein pflegen (Fig. 111, 1
u. 2).
Fig. 109. Querschnitt durch ein Stengelglied des Mais
(Zea mays). pr Rinde, pc Perizykel, cv Leitbündel, gc Parenchym
des Zentralzylinders. Vergr. 2. Nach SCHENCK.
Fig. 110. 1 Längsschnitt durch einen elastischen
Zylinder, vor der Biegung (punktiert) und danach (ausgezogen). F Die
Füllung. Vor der Biegung alle Längskanten gleich lang; nach der Biegung
die Außenkante a′ verlängert, die Außenkante a verkürzt. 2 Bei
ungenügender und unwirksamer Füllung f krümmen sich die Gurtungen a
und a′ leicht jede für sich, da sie dabei gleichlang bleiben können.
Nach NOLL.
Fig. 111. Biegungsfeste Konstruktionen des Stengels. 1
Querschnitt durch einen jungen Sambucussproß. c Kollenchymbündel.
Die innere punktierte Figur ist der Ring von Leitbündeln. 2 Teil
eines Halmquerschnittes vom Pfeifengras (Molinia coerulea). Sc
Sklerenchymrippen, ScR Sklerenchymring als tangentialer Verband
dazwischen. A Grünes Assimilationsgewebe. MH Markhöhle. Nach
NOLL. 3 Zusammengesetzter Träger, stärker vergr. u.
schematisiert. g′ g″ Gurtungen. f Füllung (in Form eines
Leitbündels).
Die Biegungsfestigkeit wird nämlich bei sparsamer
Verwendung von Festigungsmaterial am besten durch seine
periphere Anordnung erreicht. Wenn man einen geraden
Stab biegt, so wird die konvexe Seite notwendig verlängert,
die konkave verkürzt. Wie die Fig. 110 zeigt, müssen dabei die
äußersten Kanten a, a und a′, a′ des gebogenen Stabes am
meisten beeinflußt: a′, a′ am stärksten gedehnt, a, a
am stärksten zusammengedrückt werden, während die Längsstreifen
i, i und i′, i′ im Innern sich nur wenig verlängern oder
verkürzen. Wenn man also nicht den ganzen Stab aus fester Masse[S. 81]
aufbauen, sondern mit dem Festigungsmaterial sparsam umgehen will,
so wird man es möglichst an der Peripherie anbringen müssen;
denn hier wird es einerseits Biegungen den größten Widerstand
entgegensetzen, andererseits bei stärkeren Biegungen infolge
seiner Festigkeit weniger leicht zerrissen oder zerquetscht
werden als widerstandsloseres Material. Allbekannt ist ja, wie
groß die Biegungsfestigkeit von Eisenröhren selbst mit ganz
dünnen Wänden, sog. Mannesmannröhren, ist. Einen hohen Grad von
Festigkeit erreicht der Techniker schon dadurch, daß er an der
Peripherie biegungsfester Konstruktionen parallel zueinander und
zur Längsachse des Gebildes einzelne Stäbe aus Festigungsmaterial,
sog. Gurtungen, spannt. Von wesentlicher Bedeutung ist
dabei, daß diese Gurtungen durch widerstandsfähige,
gleichfalls elastische Füllungen (Fig. 111, 1) in ihrem
wirksamen Abstande und in ihrem Verbande erhalten werden. Jeder
Stab (Gurtung) bildet alsdann mit dem ihm auf der Gegenseite
gegenüberliegenden einen Träger; das Material, das quer
durch die Konstruktion hindurch zwischen den Stäben liegt, ist die
Füllung dieses Trägers (Fig. 110). Wenn solche Füllungen
fehlen, würde jeder Stab sich einzeln leicht biegen lassen. Sie
können aber in hohlen Gebilden durch tangentialen Verband
der Gurtungen, sei es durch Festigungsmaterial, sei es durch
anderes ersetzt werden. Bei größeren biegungsfesten Konstruktionen
ersetzt der Techniker die peripher gespannten Stäbe ihrerseits
meist durch Träger, die wiederum aus zwei Gurtungen und einer
Füllung bestehen (die Eisen-I-Träger der Technik).
Wie SCHWENDENER[51] zuerst gezeigt hat, sind die
mechanischen Gewebe, die dem Pflanzenstengel Biegungsfestigkeit
verleihen, etwa so angeordnet, wie es der Techniker tun würde, um
sie mit wenig Aufwand von Festigungsmaterial widerstandsfähig zu
machen. Bei vielen Gewächsen bildet das Festigungsgewebe einen
peripheren Hohlzylinder, der direkt auf die Epidermis
folgen oder tiefer ins Gewebe eingesenkt sein kann (Fig. 112
pc); bei anderen ein System entsprechend angeordneter, isoliert
nebeneinander verlaufender Stränge (System der einfachen
Träger Fig. 111, 1), die manchmal einem Hohlzylinder außen
noch aufgesetzt sind (Fig. 111, 2); bei noch anderen ist jeder
dieser peripheren Stränge selbst wieder in Form eines Trägers
gestaltet (Fig. 111, 3), dessen Gurtungen alsdann allein aus
mechanischem Gewebe bestehen, dessen Füllung aber meist aus
einem Leitbündel gebildet wird (System der zusammengesetzten
Träger). Monokotylenstengel sind im allgemeinen viel
vollkommener biegungsfest gebaut als die Dikotylen- und
Gymnospermenstengel in ihren primären Geweben; bei diesen wird die
Festigkeit durch das sekundäre Dickenwachstum nachträglich noch
erhöht. In grüngefärbten Stengeln, die an der Assimilationsarbeit
beteiligt sind, liegen die mechanischen Gewebe entweder
nicht direkt unter der Epidermis, so daß die Peripherie den
lichtbedürftigen grünen Geweben überlassen bleibt, oder sie teilen
sich an der Peripherie mit ihnen in den Raum (Fig. 111, 2).
Die innerste Zellschicht der Rinde pflegt in oberirdischen
Stengelteilen von Landpflanzen, namentlich wenn sie völlig ausgewachsen
sind, nicht besonders ausgebildet zu sein. In diesem Falle gibt es
also keine scharfe Grenze zwischen Rinde und Zentralzylinder. Sie kann
in ihnen aber auch als Stärkescheide, ferner namentlich in den
Erdsprossen von Landpflanzen und in den Stengeln von Wasserpflanzen
als typische Endodermis oder als eine Kutis ausgestaltet sein.
Ist sie eine Stärkescheide (st Fig. 112 A und B), so zeichnen
sich ihre Zellen durch den Gehalt an großen, leicht beweglichen
Stärkekörnern aus.
Vielfach ist eine Stärkescheide nur in jungen Pflanzentrieben
vorhanden, schwindet aber in älteren oder bleibt dort nur auf
bestimmte Stellen beschränkt. Statt gemeinsamer Stärkescheiden
oder Endodermen kann es auch solche um die einzelnen Leitbündel
geben (Fig. 119 pp) oder an ihrer Stelle einzelne Zellreihen, die
leicht bewegliche Stärkekörner enthalten.
Der Zentralzylinder besteht ebenfalls vor allem aus Parenchym,
das seiner Lage entsprechend farblos oder nur schwach grün ist und
hauptsächlich der Speicherung und Leitung von Stoffen dient; daneben
kommt oft auch Sklerenchym in ihm vor. Seine wichtigsten Bestandteile
aber sind die Leitbündel. Sie sind es, die die Blätter von den
Wurzeln her mit Wasser und mit den notwendigen Nährsalzen aus dem
Boden versorgen und umgekehrt organische Substanzen aus den Blättern
zu dem Wurzelsystem schaffen. Die Bündel[S. 82] sind in das übrige Gewebe
des Zentralzylinders eingebettet, wovon sie sich durch ihre engen
Elemente und den Mangel an Interzellularen schon bei ganz schwacher
Vergrößerung abheben. Sondert sich der Zentralzylinder gegen die
Rinde durch eine Scheide scharf ab, so pflegen die Leitbündel
nicht direkt an die Scheide anzugrenzen; den peripheren, ein- bis
mehrschichtigen, leitbündelfreien Gewebemantel des Zylinders, der nicht
selten aus Parenchym besteht (Fig. 112 A, B, pc), kann man als
Perizykel bezeichnen.
Entweder ist nur ein zentrales Leitbündel im Stengel vorhanden,
wie bei manchen Farnen und bei Lycopodium, oder es verlaufen im Stengel
mehrere Leitbündel. In diesem Fall, der die Regel bildet,
haben die Leitbündel einen bestimmten Verlauf und auf den
Stengelquerschnitten infolgedessen eine eigenartige Anordnung. Auf den
Querschnitten durch die Internodien sind sie nämlich im Zentralzylinder
bei den Schachtelhalmen (Equiseten), den meisten Farnen, Gymnospermen
und Dikotylen (Fig. 165) in einem Kreise angeordnet, dagegen bei den
Monokotylen (Fig. 109) ohne Ordnung zerstreut. Bilden die Leitbündel
einen Kreis, so gelten die von ihnen umschlossenen Gewebe des
Zentralzylinders, die meist aus lebenden oder auch aus frühzeitig
absterbenden Parenchymzellen bestehen, als Mark (Fig. 112 A,
m), die die Bündel seitlich trennenden Gewebe als Markstrahlen
(ms). Bei zerstreuter Verteilung der Bündel (Fig. 109) fehlt diese
Sonderung.
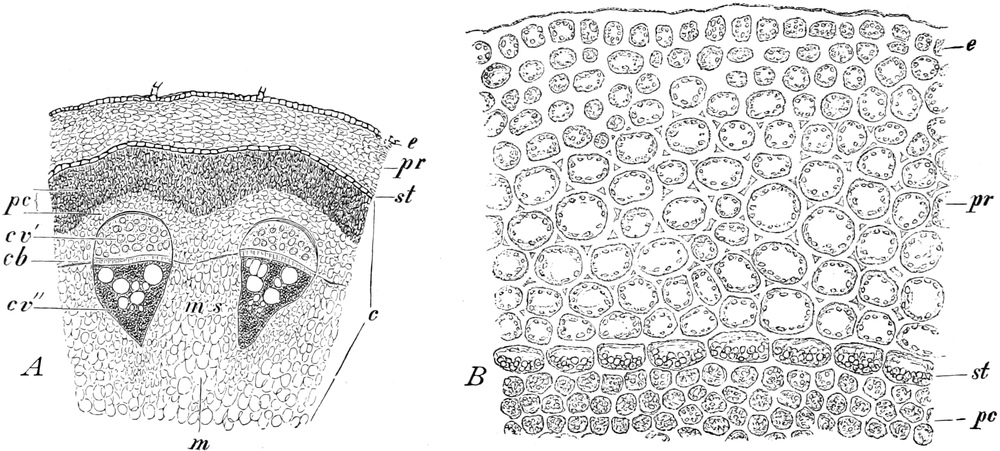
Fig. 112. A Teil eines Querschnittes durch einen
jungen Stamm von Aristolochia Sipho. e Epidermis, pr Rinde, st
Stärkescheide, c Zentralzylinder, pc Perizykel, in diesem Falle
mit einem Ring von Sklerenchymfasern, cv Leitbündel, und zwar
cv″ Gefäßteile, cv′ Siebteile, cb Kambiumring, m Mark, ms
Markstrahl. Vergr. 48. B Kleiner Teil eines Querschnittes aus dem
Umkreis eines noch jüngeren Stammteils. e Epidermis, pr Rinde,
st Stärkescheide mit leicht beweglichen Stärkekörnern, pc äußere
Zellschichten des Perizykels. Vergr. 350. Nach STRASBURGER.
Doch gibt es auch Farne (z. B. Pteris) und Dikotylen, bei
denen die Leitbündel zwei (Cucurbita, Phytolacca, Piper)
oder mehr Kreise (Amarantus, Papaver, Thalictrum) bilden.
Die inneren Kreise pflegen wenig regelmäßig zu sein. Ferner gibt es
in beiden Gruppen Gewächse, bei denen außer dem Bündelring und den
Markbündeln noch kleine akzessorische Rindenbündel vorkommen.
Die Markstrahlen können aus Parenchym bestehen; nicht selten, z. B.
bei vielen Kräutern, werden aber ihre inneren Teile, zwischen
den Gefäßsträngen der Leitbündel, aus Sklerenchym gebildet,
wogegen sich die äußeren parenchymatischen Teile zwischen den
Siebsträngen der Bündel alsdann scharf absetzen.
Unterirdische Stengelteile (Erdsprosse) und submerse
Wasserpflanzen, die zugfest gebaut sein müssen, haben die
mechanischen Gewebe oft in ihrer Mitte, also im Marke.
Leitbündelverlauf. Ihren Funktionen entsprechend bilden die
Leitbündel in den Pflanzen ununterbrochene Stränge, die sich,
namentlich an Ma[S. 83]zerationspräparaten, von den Wurzelspitzen bis in
die Blattspitzen verfolgen lassen. Solche Präparate kann man aus
krautartigen Pflanzenteilen gewinnen, die man in Wasser so lange liegen
läßt, bis die Gewebe mit Ausnahme der resistenteren Leitbündel verfault
sind.
Im Stengel ist der Verlauf besonders verwickelt. In die Basis der
Sproßachse tritt das Leitbündel der Wurzel ein, das sich mit den
Stengelbündeln vereinigt (vgl. S. 118). Die Stengelbündel können bis
zur Stengelspitze verlaufen, ohne an ihrem Ende in Blätter überzugehen.
Man nennt solche Leitbündel stammeigene Bündel. Umgekehrt sind
blatteigene solche, die gleich nach ihrem Eintritte aus den
Blättern in den Stengel mit stammeigenen Bündeln verschmelzen.
So bilden bei den Pteridophyten stammeigene Leitbündelstränge im
Stengel ein netzartiges Bündelrohr oder auch ein einziges zentrales
Leitbündel (Lycopodium u. a.), während die aus den Blättern
kommenden blatteigenen Bündelstränge sich mit diesen stammeigenen
Bündeln vereinigen.
Meist aber biegen die Bündel der Sproßachse an ihren Spitzen in die
Blätter aus: gemeinsame Bündel, die also mit ihren unteren
Teilen im Stengel, mit ihren oberen im Blatte verlaufen. In jedes Blatt
kann ein oder können mehrere solche Bündel eintreten, die man in ihrer
Gesamtheit als Blattspur bezeichnet. Es gibt also ein-
und mehrsträngige Blattspuren. Bei den Samenpflanzen besteht das
Leitbündelsystem des Stengels vor allem aus solchen Blattspuren.
In den Stengeln mancher Dikotylen (Begonien, Aralien) sind aber
die innerhalb des Kreises der Blattspurstränge verlaufenden Kreise
von Markbündeln stammeigene Bündel, die in den Knoten mit den
Blattspursträngen durch Querzweige verbunden sind.
Die Blattspurbündel können im Stengel dauernd voneinander getrennt
bleiben. Meist aber vereinigt sich jedes Bündel einer Blattspur bei
seinem Abwärtsverlaufe schließlich mit einem anderen Bündel, das aus
einem tiefer an der Achse befestigten Blatte stammt. Dieser Vereinigung
kann eine Spaltung (Gabelung) des Bündels vorausgehen. Durch einen
solchen netzartigen Verlauf der Bündel wird eine gleichmäßige
Versorgung der Pflanze mit Wasser erreicht, da ein jedes Bündel des
Stengels infolge seiner Verzweigungen größeren Sproßabschnitten
Wasser liefert. Je nach der Länge des Weges, den die einzelnen Bündel
im Stengel frei zurücklegen, der Richtung, die sie verfolgen, und
der Spaltung, die sie unter Umständen erfahren, ist das Bild des
Bündelverlaufes bei den verschiedenen Arten ein anderes. Natürlich ist
die Blattstellung für die Eintrittsstellen der Blattspuren in
den Stengel bestimmend; der Verlauf im Stengel ist aber
von der Blattstellung ganz unabhängig, so daß er bei ein und
derselben Blattstellung ganz verschieden sein kann.
Bei den Schachtelhalmen, den Koniferen und den
Dikotylen dringen alle Blattspurstränge gleich tief in
den Stengel ein, um im Stengel auch gleich weit von der
Stengelmitte, also auf dem Querschnitt zu dem charakteristischen
Kreise geordnet, nach abwärts zu laufen. Infolgedessen kann man
den Bündelverlauf in den Internodien auf einer Zylinderfläche
darstellen, die sich in eine Ebene ausbreiten läßt. In den Knoten
freilich ist der Bündelverlauf meist viel verwickelter, weil hier
die Blattspurstränge noch durch stammeigene Querverbindungen
miteinander verbunden sind. Nachträgliche seitliche
Verbindungen findet man übrigens oft auch in den Internodien.
Ein relativ einfaches Beispiel eines Bündelverlaufs liegt in den
jungen Zweigen von Juniperus nana vor (Fig. 113). Ihre Blätter
stehen in dreigliedrigen Quirlen. Aus jedem Blatt tritt eine
einsträngige Blattspur, somit ein einziges Bündel in den
Stengel ein. Etwa in der Mitte des nächst unteren Internodiums
gabelt es sich in zwei Schenkel, die je rechts und links mit
benachbarten Blattspuren verschmelzen. Weniger einfach erscheint
das Bild des in Fig. 114 dargestellten Bündelverlaufs in einem
jungen Zweige[S. 84] von Taxus baccata, obwohl auch hier die Blattspuren
einsträngig sind. Jede Blattspur läßt sich frei durch 12
Stengelglieder abwärts verfolgen, worauf sie mit einer anderen
verschmilzt. Zunächst läuft sie durch vier Internodien gerade
abwärts, dann biegt sie seitlich aus, um einer eintretenden
Spur Platz zu machen und sich mit ihr zu vereinigen. Bei Taxus
stehen die Blätter nach 5⁄13; dementsprechend zeigen auch die
Eintrittsstellen der Blattspuren in den Stengel 5⁄13 Divergenz.
Ein Beispiel dreisträngiger Blattspuren sei aus einem
jungen Zweige der italienischen Waldrebe (Clematis Viticella)
vorgeführt. Die Blattpaare an diesen Zweigen stehen dekussiert.
Die Medianstränge der Blattspuren (a u. d, g u. k, n u.
q, t u. x Fig. 115) laufen durch ein Internodium abwärts,
teilen sich im nächsten Knoten in zwei Schenkel und fügen diese
den ihnen zugekehrten Lateralsträngen der Blattspuren des dortigen
Blattpaares an. Die zwei Lateralstränge jeder Blattspur (b u.
c, e u. f, h u. i, l u. m, o u. p, r u. s)
lassen sich ebenfalls durch ein Internodium frei abwärts verfolgen,
biegen im nächsten Knoten zusammenneigend nach außen und legen sich
den nämlichen Lateralsträngen wie die Schenkel des Medianstranges
an.
Fig. 113. Schematische Darstellung des
Leitbündelverlaufs in einem jungen Zweige von Juniperus nana, auf
der eben gelegten Zylinderfläche entworfen. Bei k, k die in die
Achselsprosse tretenden Bündel. Nach GEYLER.
Fig. 114. Schematische Darstellung des Bündelverlaufs in
einem jungen Zweige von Taxus baccata. Das Bündelrohr ist einseitig bei
1 längs aufgeschlitzt und in einer Ebene ausgebreitet.
Einem ganz anderen Typus folgt der Bündelverlauf bei den
Monokotylen (Fig. 116). Hier befinden sich die
einzelnen Blattspurstränge im Zentralzylinder ungleich weit
von der Oberfläche des Stengels entfernt, sind also auf dem
Stengelquerschnitte zerstreut. Diese Anordnung kommt dadurch
zustande, daß das Dickenwachstum des Stengelvegetationspunktes
nach Anlage der ersten medianen Bündel des Blattes noch längere
Zeit anhält. Infolgedessen gelangen die später, und zwar
nacheinander erzeugten Bündel der Blattflächen nicht so weit,
und zwar verschieden tief nach innen. Besonders ausgeprägt
kommt diese Anordnung bei den Palmen (Palmentypus) vor. Jede
Blattspur besteht hier aus zahlreichen Strängen, die aus einem
stengelumfassenden Blattgrund im ganzen Umkreise in den Stengel
eintreten. Die in dem Blattgrund medianen Leitbündel (vgl. das
Bild des medianen Längsschnittes durch den Stengel Fig. 116, in
das für ein jedes Blatt A, B, C nur das mediane und ein
seitliches Leitbündel eingezeichnet ist) dringen fast bis zur
Mitte, die seitlich angrenzenden (a, b, c) immer weniger
tief in den Zentralzylinder ein. In ihrem Abwärtsverlaufe nähern
sich die Bündel langsam der Peripherie des Zentralzylinders, wo
sie mit anderen verschmelzen. Die Zahl der Internodien, die jedes
durchläuft, ist verschieden, für die medianen besonders groß.
Leitbündelbau[68]. Die Stengelbündel sind Gewebestränge von
kreisrundem, breit- oder schmalelliptischem Querschnitte, und zwar
sind es fast[S. 85] stets vollständige Leitbündel, d. h. solche
Bündel, worin Sieb- und Gefäßstränge zu gemeinsamen Strängen verbunden
sind (vgl. S. 58). Die Siebstränge, deren wichtigster Bestandteil
die Siebröhren sind, bilden das Phloëm (den Siebteil),
die Gefäßstränge mit den wasserleitenden Gefäßen das Xylem
(den Gefäßteil) des Bündels. Die Stengelbündel können bei den
einzelnen Kormophyten recht verschieden gebaut sein. In den Sproßachsen
findet man alle die verschiedenen Bündeltypen, die sich in den Organen
der Kormophyten überhaupt unterscheiden lassen, nämlich radiale,
konzentrische und kollaterale Bündel. Diese
Leitbündelformen unterscheiden sich voneinander durch die Anordnung und
die Ausbildung ihrer Sieb- und Gefäßstränge.
Fig. 115. Clematis Viticella. Zweigende durch Entfernung
der Oberfläche und Einwirkung von Kalilauge durchsichtig gemacht, den
Verlauf der Blattspuren zeigend. Die austretenden Enden der Stränge
infolge leichten Druckes etwas verschoben. Die jungen Anlagen der
beiden obersten Blattpaare bl1 und bl2 noch ohne Blattspuren,
v Vegetationskegel. Nach NÄGELI.
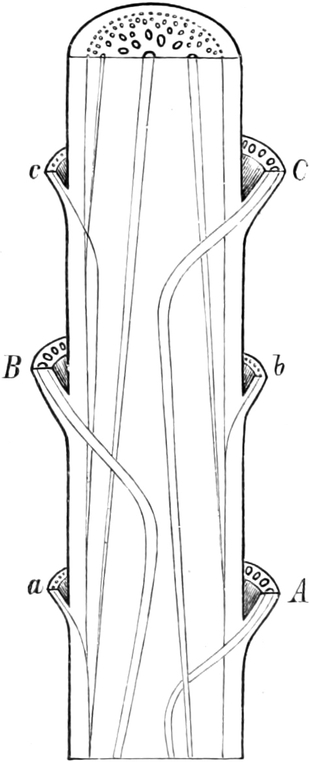
Fig. 116. Schematische Darstellung des Bündelverlaufs
nach dem Palmentypus, innerhalb eines medianen Längsschnittes durch
den Stengel in der Ebene der Blattmedianen. Zweizeilig alternierende,
stengelumfassende Blätter sind vorausgesetzt. Die Blätter Aa, Bb,
Cc sind nahe ihrer Basis abgeschnitten; die großen Buchstaben
bezeichnen ihre Medianen. Oben der Stengel im Querschnitt.
ROTHERT frei nach ROSTAFINSKI.
Im radialen Leitbündel (Fig. 117, vgl. auch Fig. 158, 160)
gibt es mehrere Gefäß- und Siebstränge, die auf dem meist
kreisrunden Bündelquerschnitt wie die Radien eines Kreises neben- und
miteinander abwechselnd angeordnet sind und in Seitenansicht parallel
zur Längsachse des Pflanzenteils verlaufen. Stoßen die Gefäßstränge
im Zentrum des Leitbündels zusammen, so bilden sie eine sternförmige
Querschnittsfigur; die Enden der Zacken werden von den engsten Gefäßen
(den Gefäß- oder Xylemprimanen, vgl. dazu S. 90) eingenommen, während
die Gefäße nach dem Zentrum des Bündels hin immer weiter werden (Fig.
117). In den Buchten zwischen den Zacken liegen die Siebstränge[S. 86] und
in ihnen außen die engsten Siebröhren (die Sieb- oder Phloëmprimanen).
Radiale Bündel, die für die Wurzeln bezeichnend sind, kommen in
Sprossen freilich nur selten, und zwar stets in Einzahl, vor, z. B. in
manchen Lycopodienstengeln.
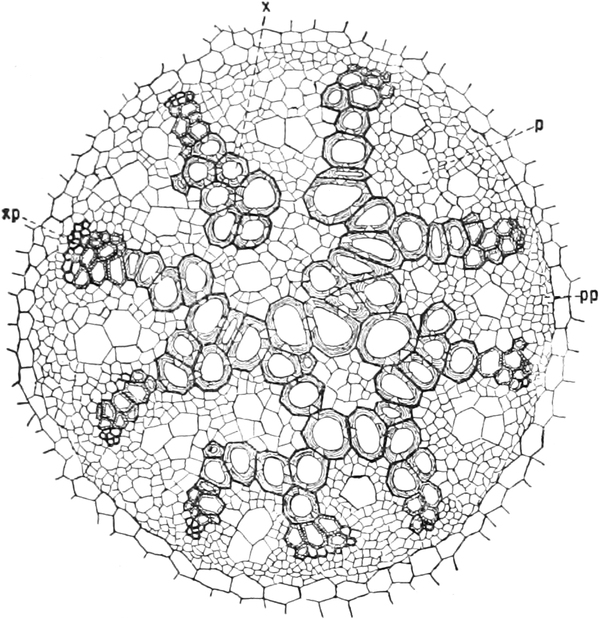
Fig. 117. Radiales Leitbündel aus dem Stengel von
Lycopodium Hippuris, p Phloëm, pp Phloëmprimanen, x Xylem, xp
Xylemprimanen. Vergr. 30.
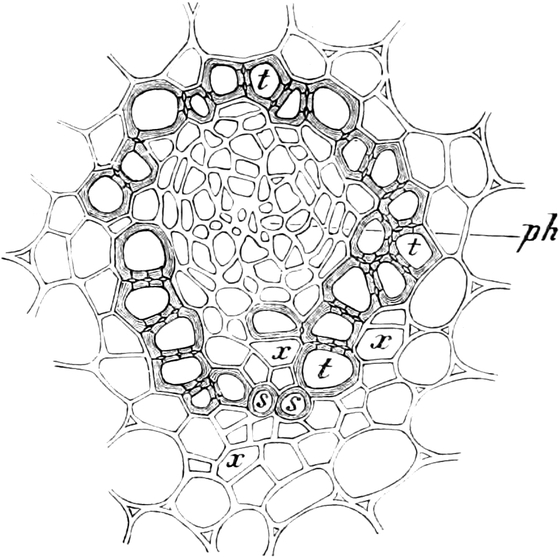
Fig. 118. Konzentrisches Leitbündel mit Außenxylem aus
dem Wurzelstock von Convallaria majalis. ph Phloëm, x, t Xylem,
s Xylemprimanen. Nach ROTHERT.
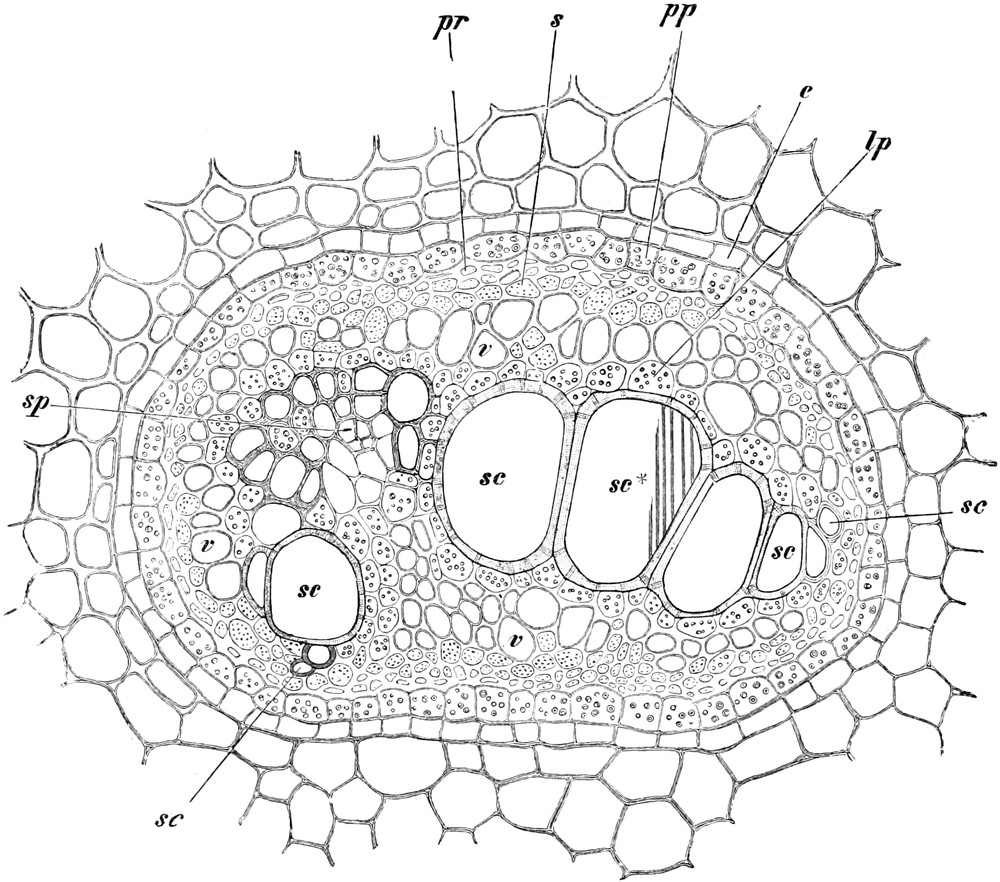
Fig. 119 Querschnitt durch ein konzentrisches Leitbündel
aus dem Blattstiel des Adlerfarns (Pteridium aquilinum). sc
Treppentracheïden, sp Xylemprimanen (Schraubentracheïden); in der
Treppentracheïde sc* Stück einer leiterförmig verdickten schrägen
Endwand, lp Xylemparenchym, v Siebröhren, s Phloëmparenchym, pr
Phloëmprimanen, pp Stärkeschicht, e Endodermis. Vergr. 240. Nach
STRASBURGER.
[S. 87]
Im konzentrischen Bündel wird ein zentraler Gefäß- oder
Siebstrang allseits von einem hohlmantelförmigen Sieb- oder
Gefäßstrang konzentrisch umgeben. Liegt das Xylem zentral, so kann man
das Bündel als konzentrisch mit Innenxylem, liegt es dagegen außen,
als ein solches mit Außenxylem bezeichnen. Konzentrisch mit Innenxylem
sind die Bündel bei den meisten Farnen (Fig. 119) und bei bestimmten
Dikotylen das Stengelbündel (viele Holzpflanzen). Konzentrisch mit
Außenxylem sind sie z. B. in gewissen Erdsprossen und Stämmen von
Monokotylen (Fig. 118); solchen Bau haben ferner die markständigen
Bündel z. B. bei Piperaceen, Begonia, Campanula u. a.
Bei den Pteridophyten liegen die engsten Gefäße (die Gefäßprimanen)
(sp) in dem Xylemstrang entweder gruppenweise peripher oder
zentral oder zwischen den älteren Gefäßen. Die Gefäßstränge
werden von einer Parenchymschicht (lp) umhüllt. Daran schließt
im Umkreis der aus Siebröhren (v) und aus Parenchym (s)
bestehende Mantel, an dessen Außenrand die engsten Siebröhren (die
Siebprimanen) gelegen sind.
Im kollateralen Leitbündel endlich (Fig. 120 A), das auch nur
einen Gefäßstrang und meist nur einen Siebstrang enthält,
liegt der Gefäßteil neben oder besser hinter dem Siebteil, so
daß Xylem und Phloëm sich nur einseitig berühren. Die Medianebenen
solcher Bündel sind in den Stengeln immer radiär gerichtet,
so daß diese Bündel im allgemeinen ihre Gefäßteile nach innen, ihre
Siebteile nach außen kehren. Die engsten Gefäße (die Xylemprimanen)
liegen im kollateralen Bündel gewöhnlich am Innenrande des Gefäßteiles
(bezogen auf den Stengelquerschnitt), die Phloëmprimanen am Außenrande
des Siebteiles. Solche kollaterale Leitbündel sind den Sprossen der
Samenpflanzen und der Schachtelhalme eigentümlich. Doch kommen auch
bikollaterale Leitbündel vor, die nicht nur außen, sondern auch
innen einen Siebstrang besitzen, so in den Stengeln der kürbisartigen
Gewächse (Cucurbitaceen). Die kollateralen Bündel sind bei den
Monokotylen, wie die radialen und die konzentrischen Leitbündel, meist
geschlossen, d. h. das ganze Bündel besteht aus Dauergewebe, und
der Gefäßteil grenzt unmittelbar an den Siebteil (Fig. 120 A). Bei den
Gymnospermen und Dikotylen sind sie dagegen meist offen, d. h.
die Sieb- und die Gefäßteile bleiben dauernd durch eine Schicht
meristematisches Gewebe, das Kambium der Bündel, getrennt (Fig.
121).
Bei sämtlichen Leitbündelformen bestehen die Gefäßstränge
vor allem aus engen oder weiten, verholzten Elementen, die der
Wasserleitung dienen: Tracheïden und Tracheen (Fig.
120 a, sp, m; Fig. 122 rp, sp, s, n, t) oder
Tracheïden allein, die sämtlich einzeln für sich oder zu Gruppen
ohne Interzellularen zwischen lebende, enge, langgestreckte und oft
unverholzte Leitparenchymzellen, Xylemparenchym, eingebettet
oder von ihnen in Form einer lückenlosen Scheide umgeben werden (Fig.
119 lp). Auch Sklerenchymfasern sind manchmal in den Gefäßsträngen
vorhanden. Bei den Farnpflanzen sind sämtliche Gefäße ausschließlich
als Tracheïden ausgebildet; in den Bündeln der Samenpflanzen
kommen dagegen meist Tracheïden und Tracheen nebeneinander vor. In
allen Bündeln (vgl. Fig. 122) sind die engsten Gefäße Ring- und
Schraubengefäße, die übrigen aber meist Netz- und Tüpfelgefäße, bei den
Pteridophyten, abgesehen von den Primanen, nur Treppengefäße (Fig. 70
A).
In den Siebsträngen der Leitbündel (Fig. 119 und 120)
verlaufen die der Eiweißleitung dienenden Siebröhren (v).
Sie sind stets von anderen lebenden Zellen begleitet, entweder nur
von Geleitzellen (Fig. 120 s), die meist kürzer als die
Siebröhrenglieder und mit diesen durch Siebplatten verbunden sind,
oder von Geleitzellen und von anderen gestreckten Parenchymzellen
(Phloëmparenchym) oder von letzteren allein (Fig. 119 s).
Ist Phloëmparenchym vorhanden, so sind die Siebröhren einzeln oder
gruppenweise lückenlos darin eingebettet.
[S. 88]
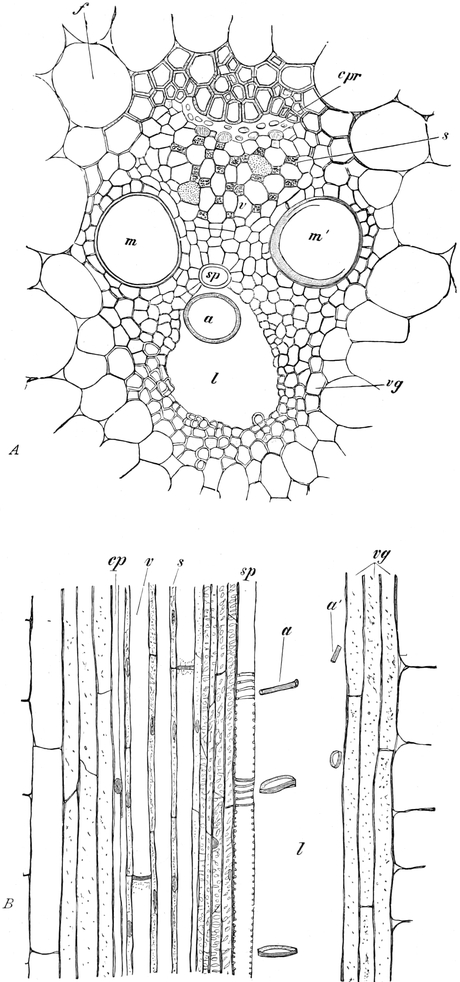
Fig. 120. A Querschnitt, B Längsschnitt durch
ein geschlossenes, kollaterales Leitbündel aus dem Stengel von Zea
mays. a Ring einer Ringtracheïde, sp Schraubentracheïde, m und
m′ Tüpfeltracheen, v Siebröhre, s Geleitzelle. cpr und cp
zerdrückte Phloëmprimanen, l Gefäßgang, vg Scheide. Vergr. 180.
Nach STRASBURGER.
[S. 89]
Geleitzellen kommen nur den Siebröhren der Angiospermen zu. Sie
sind Schwesterzellen der Siebröhrenglieder, gehen mit ihnen durch
Längsteilung aus derselben Mutterzelle hervor, erfahren aber
meist noch Querteilungen. Ihre Weite ist geringer als die der
Siebröhrenglieder, sie zeichnen sich vor letzteren auch durch ihren
reichlichen plasmatischen Inhalt aus. In einzelnen Fällen findet
man im Phloëm auch Milchsaft- oder Schleimröhren.
Das vollständige Bündel ist seinerseits gewöhnlich noch von einer
Bündelscheide mehr oder weniger umschlossen, die aus
interzellularenfreiem Parenchym (oft reich an großen Stärkekörnern:
Stärkescheide), aus Sklerenchym oder aus einer Schicht von
Endodermiszellen (manchmal auch aus Kutisgewebe) bestehen kann. Sie
wird nicht zum Leitbündel gerechnet. Die Scheiden dienen wohl
vielfach dazu, die Stoffleitung auf die Bündel zu begrenzen. Scheiden
aus Sklerenchym sind besonders häufig den Außenseiten der Siebteile als
halbmondförmige Sklerenchymschicht (Fig. 120 A, 121 vg) vorgelagert
und bei zerstreuter Bündelanordnung namentlich an den äußeren
Leitbündeln ausgebildet.
Wo eine sklerenchymatische Scheide ein kollaterales Bündel
umgibt, ist sie oft an jeder Seite des Bündels, an der Grenze
von Gefäß- und Siebteil, durch parenchymatische oder schwächer
verdickte und schwächer verholzte Elemente unterbrochen. Diese
Stellen erleichtern den Austausch von Wasser und Nahrungsstoffen
zwischen dem Bündel und dem Parenchym; sie werden als
Durchlaßstreifen bezeichnet.
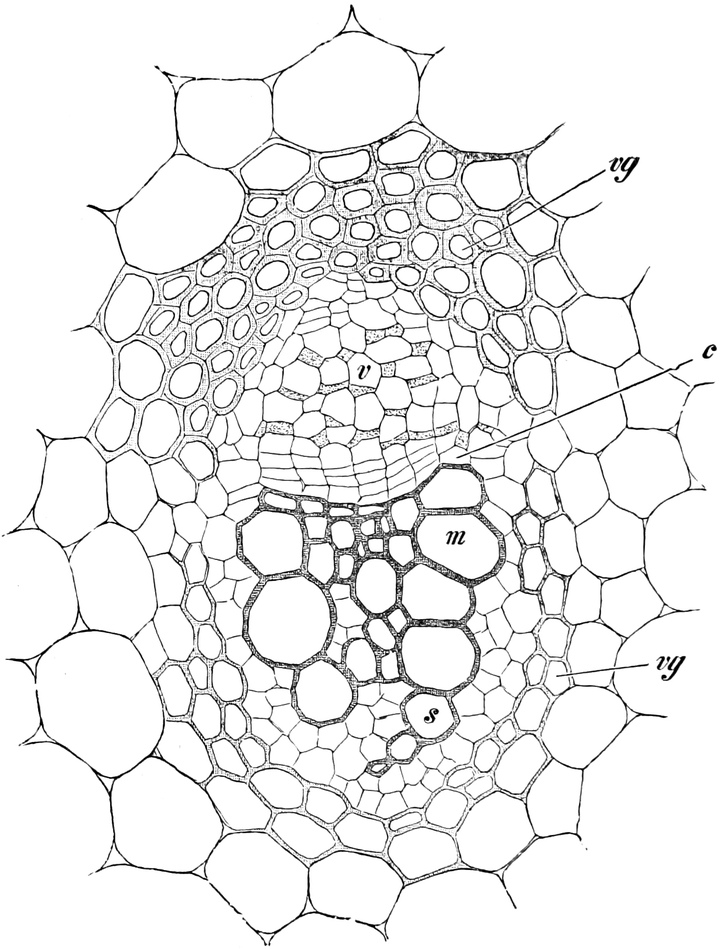
Fig. 121. Querschnitt durch das offene, kollaterale
Leitbündel eines Ausläufers von Ranunculus repens. s
Schraubentracheïden, m Tüpfeltracheen, c Kambium, v Siebröhren,
vg Scheide. Vergr. 180. Nach STRASBURGER.
Um den Bau der Bündel und die Unterschiede zwischen den Leitbündeltypen
vollständig zu verstehen, ist auch noch ein Einblick in ihre
ontogenetische Entwicklung erforderlich. Die primären Leitbündel
gehen aus Strängen langgestreckter Urmeristemzellen hervor. In
ihnen vollzieht sich die Gewebesonderung nur allmählich, und zwar
ungleichzeitig in den Zellen eines Stranges. Solange nämlich
ein Pflanzenteil noch stark in die Länge wächst, bleiben die Stränge
der Hauptmasse nach undifferenziert. Nur an engbegrenzten Stellen,
die meist an den Rändern jedes Stranges gelegen sind, wandeln sich
einzelne Zellenzüge in Dauergewebe um, und zwar auf Streckung oder
Dehnung eingerichtete Elemente: ring- und schraubenförmig verdickte
Tracheïden einerseits, Siebröhren oder Siebröhren nebst Geleitzellen
andererseits.[S. 90] Diese Tracheïden werden deshalb als Erstlinge des
Gefäßteils, Xylemprimanen (Protoxylem), die Siebröhren als
Erstlinge des Siebteils, Phloëmprimanen (Protophloëm),
bezeichnet. Erst nach vollendetem Längenwachstum des Pflanzenteils
werden die Leitbündel fertiggestellt, wobei die Differenzierung
der meristematischen Gewebe von den Primanen aus fortschreitet und
im Gefäßteil nacheinander zuerst Ring-, dann Schrauben-, Netz-,
schließlich Tüpfelgefäße ausgebildet werden (Fig. 120 B, 122). Die
Xylemprimanen sind im fertigen Leitbündel vielfach zerdrückt, auch
wohl durch Dehnung zerrissen (a und a′ Fig. 120 B, l; rp
Fig. 122); in manchen Fällen ist ein lysigener, von Wasser erfüllter
Interzellulargang („Gefäßgang“) an ihrer Stelle ausgebildet (Fig.
120 l), der aber noch der Wasserleitung dient[69]. Die Wände der
Phloëmprimanen (Fig. 120 B, cp) sind im fertigen Bündel verquollen,
ihre Siebplatten durch Kallusbeläge verschlossen.
Beim radialen Bündel schreitet die Ausbildung der Elemente entsprechend
der Lage der Primanen in den Gefäß- und Siebsträngen von der Peripherie
des Bündels nach dem Zentrum fort, beim kollateralen Bündel dagegen im
Siebteil vom Außenrande, im Gefäßteile aber vom Innenrande des Bündels
gegen die Mitte des Bündels; wird dabei alles Meristem aufgebraucht,
so entsteht ein geschlossenes kollaterales Bündel, bleibt etwas davon
erhalten, ein offenes. In den konzentrischen Bündeln vollzieht sich die
Ausbildung des Phloëms und Xylems nicht nach einem einheitlichen Typus;
dementsprechend ist die Lage der Primanen verschieden.
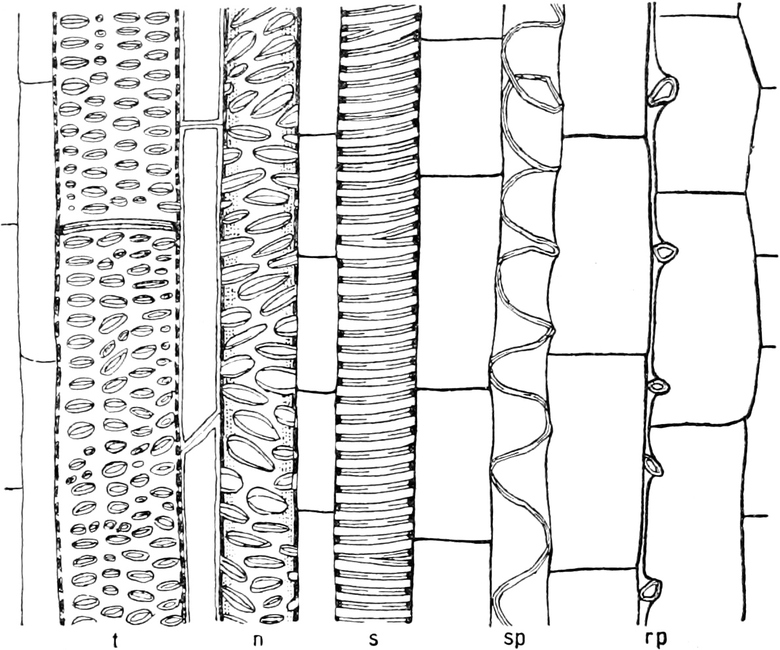
Fig. 122. Längsschnitt durch den Gefäßteil des
kollateralen Leitbündels von Impatiens parviflora. rp Durch
Längenwachstum des Stengels weit auseinander gerückte Ringverdickungen
einer stark gedehnten Ringprimane; sp gedehnte Schraubenprimane;
s Schrauben-, n Netz-, t Tüpfelgefäß. s, n, t erst nach
Beendigung des Streckungswachstums ausgebildet. Vergr. 120.
Bündel, in denen man das Protoxylem am Innenrande des Xylems
(wie bei den kollateralen und den konzentrischen Bündeln
der Monokotylen) oder im Zentrum des Xylems (wie oft bei
konzentrischen) findet, nennt man auch wohl endarch. Die
Xylemprimanen können aber auch an anderer Stelle ausgebildet sein,
z. B. am Außenrande des Xylems, exarch, wie beim radialen Bündel;
oder in einer oder in mehreren Gruppen mesarch, d. h. zwischen
Außen- und Innenrand des Gefäßteiles, z. B. in den Blattstielen
der Cycadeen und im konzentrischen Bündel vieler Farne zwischen
Peripherie und Zentrum des Xylems, also zwischen weitere Gefäße
eingebettet.
Ob irgendwelche Beziehungen zwischen der Anordnung der Sieb- und
Gefäßteile und den Leitungsbedürfnissen der Pflanzenteile bestehen,
darüber wissen wir noch gar nichts[67],[68].
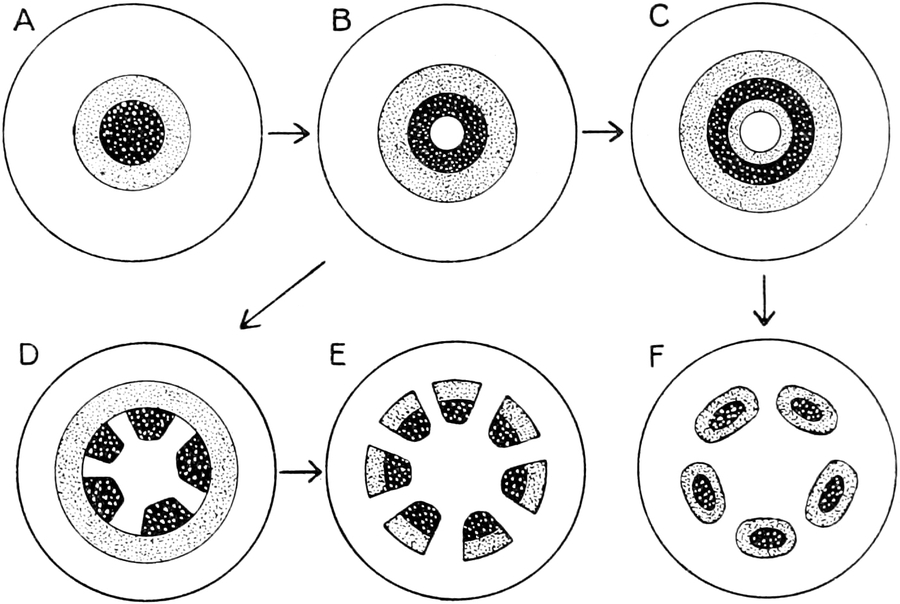
Fig. 123. Phylogenie der Leitbündeltypen,
schematisch. Weiß: Rinde und Mark, schwarz punktiert: Phloëm, weiß
punktiert auf schwarz: Xylem. Erklärung im Text.
Ebensowenig sehen wir über die Phylogenie der Bündeltypen
klar. Allen Grund haben wir freilich zu der Annahme, daß ein
Stengel mit einem einzigen zentralen Leitbündel als primitiv
anzusehen ist. Ein solches finden wir in den Stengelteilen mancher
lebender und ausgestorbener Farnpflanzen und in sämtlichen Wurzeln.
Zu den einfachsten und phylogenetisch ältesten Leitbündeln dürfte
das konzentrische Bündel mit einem soliden zentralen Xylemstrange
gehören; wenigstens ist ein solches, wie es scheint,[S. 91] in den
Keimsprossen fast aller lebenden Farne vorhanden (vgl. Fig. 123
A). Aber auch das radiale Bündel dürfte phylogenetisch sehr
alt sein. Dafür spricht, daß es allen Wurzeln lebender
und fossiler Kormophyten, soweit wir wissen, zukommt und daß es
außerdem auch in den Stengeln einiger Kormophyten ausgebildet
ist, während kein anderer Bündeltypus in Stengeln und
Wurzeln sich findet. Die Vielförmigkeit, die man im Bau und in der
Anordnung der Sproßbündel bei den Farnpflanzen im Gegensatz zu
den Samenpflanzen findet, erlaubt auch Vermutungen darüber, wie
aus einem Stengel mit einem konzentrischen Bündel Stengel
mit anderem Bau entstanden sein können. Einmal nämlich finden
wir Formen (Fig. 123 B), bei denen der Gefäßteil des einzigen
zentralen Bündels zu einem Hohlzylinder mit zentralem Strang von
Parenchym („Mark“) geworden ist (Gleicheniaceen, Schizaeaceen),
weiter solche (Fig. 123 C), wo auf den Xylemhohlzylinder noch ein
innerer Phloëmmantel und dann erst ein Mark folgt (z. B. Marsilia);
endlich solche (Fig. 123 F), bei denen dieses Bündelrohr von
rhombischen Lücken, den Blattlücken (d. h. den Ausgangsstellen der
Blattbündel), zu einem netzartigen Maschenwerk durchbrochen ist
(z. B. Aspidium Filix mas). In diesem Falle findet man auf dem
Stengelquerschnitte einen Kranz von zahlreichen, übrigens
typisch gebauten konzentrischen Bündeln, jedes mit einem soliden,
zentralen Xylemstrange. Ferner kommen aber auch Formen vor (Fig.
123 D), bei denen in dem zentralen, konzentrischen, markhaltigen
Bündel der an das Mark anschließende Xylemhohlzylinder in eine
Anzahl nebeneinander liegender, längs verlaufender Xylemstränge
aufgelöst ist, die durch radiale Parenchymplatten voneinander
getrennt und von einem geschlossenen Phloëmaußenmantel umhüllt
werden (z. B. Osmunda). Und schließlich gibt es Formen (Fig. 123
E), bei denen auch dieser Phloëmmantel noch in entsprechende
Stränge zerlegt ist, so daß die radialen Parenchymplatten als
„Markstrahlen“ die nunmehr kollateralen Xylem-Phloëmstränge völlig
voneinander trennen (Rhizom von Ophioglossum). Diese Reihen zeigen
uns, wie aus einem zentralen konzentrischen Bündel durch
Zerteilung erstens ein netzartiges Bündelrohr, das im Querschnitt
aus vielen konzentrischen, kreisförmig angeordneten Bündeln
besteht, zweitens ein Hohlrohr aus kollateralen Bündeln entstehen
kann. Nimmt man an, daß die phylogenetische Entwicklung sich
in dieser Weise vollzogen hat, so würde also nicht ein
kollaterales Bündel der Samenpflanzen oder eines der vielen
konzentrischen Bündel, die man auf den Stengelquerschnitten
vieler Farne findet, dem einen zentralen konzentrischen
Bündel „primitiv gebauter“ Farnpflanzen homolog sein, sondern die
Gesamtheit aller kollateralen oder konzentrischen Bündel im
Stengel, d. h. also der Zentralzylinder wäre homolog zu setzen dem
einen zentralen konzentrischen oder radialen Leitbündel.
Bezeichnet man nach dieser Auffassung, der Stelärtheorie[70],
das radiale Bündel oder das einzige zentral gelegene
konzentrische Bündel als Stele, so hätte die Gesamtheit der
konzentrischen oder kollateralen Bündel, die zu einem Bündelrohr
verbunden sind, samt dem umschlossenen Mark ebenfalls als Stele
zu gelten, da sie ja aus[S. 92] der Urstele entstanden gedacht ist.
Danach kann also ein „Einzelbündel“ eine ganze Stele oder nur
ein Teil einer solchen sein. Die Stele (der Zentralzylinder) der
Samenpflanzen liegt gewöhnlich als zentraler Gewebestrang im
Stengel (Monostelie). Doch kommen auch Fälle vor, wo er in mehrere
Teilzylinder zerlegt ist (Polystelie), so in den Stengeln der
Aurikeln und von Gunnera.
γ) Die Blätter[71]. 1. Anlage der Blätter. Wir sahen
die Blattanlagen am Vegetationspunkte des Sprosses exogen als
seitliche Höcker oder Wülste auftreten (Fig. 98, 102 f), die
zunächst ungegliedert sind. Man kann sie als Blattprimordien
bezeichnen (Fig. 127 A, b). Gewöhnlich nimmt eine Blattanlage nur
einen Teil des Vegetationspunktumfanges ein; doch kommt es auch vor,
daß sie als ringförmiger Wall den ganzen Vegetationspunkt umfaßt.
Auch die Anlagen mehrerer quirlständiger Blätter können zunächst als
einziger ringförmiger Wall auftreten und sich erst weiterhin sondern;
entstehen sie sogleich als selbständige Anlagen, so bilden sich diese
entweder gleichzeitig oder in der Regel[65], [114] nacheinander aus.
In freilich seltenen Fällen kann ein Blatt auch terminal am
Vegetationspunkte entstehen.
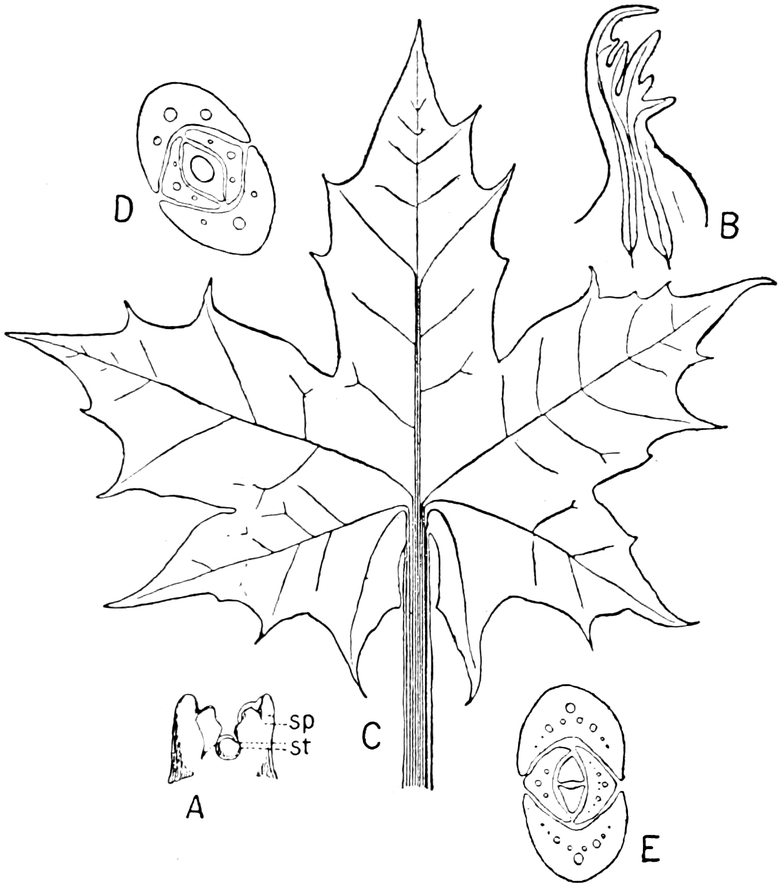
Fig. 124. Ahorn (Acer platanoides). A Außenansicht
einer Knospe mit zwei Blattanlagen, zwischen denen der Vegetationskegel
des Sprosses zu sehen ist. In sp die Blattspreite, an der fünf
Abschnitte angelegt sind, von denen der oberste Abschnitt zuerst
ausgebildet wurde. In st die Zone, durch deren Wachstum der
Blattstiel später entstehen soll. B Älteres Blatt von der Seite, die
Stränge in seinem Innern stellen die Leitbündelanlagen vor, die später
äußerlich die Nervatur bestimmen sollen. C Ausgewachsenes Blatt mit
schematischer Darstellung des Leitbündelverlaufs. D Querschnitt durch
den basalen Teil einer Knospe, mit je drei Bündelanlagen in jedem
Blatt. E höher geführter Querschnitt durch eine Knospe, die Zahl
der Bündelanlagen durch Verzweigung innerhalb der Blätter vermehrt.
A, B und E schwach vergrößert.
Nach DEINEGA aus Goebels Organographie.
Während der Sproß im allgemeinen mittels seines Vegetationspunktes
unbegrenzt, und zwar an der Spitze weiterwächst, ist das
Wachstum der Blattanlagen, die meist nur kurze Zeit an ihrer
Spitze wachsen, in der Regel begrenzt. Ausgesprochenes und
langdauerndes Scheitelwachstum zeichnet dagegen z. B. die Blätter
der Farne aus. Ist das Wachstum begrenzt, so wird die Spitze, die
in ihrer Entwicklung dem übrigen Blatte voranzueilen pflegt, zuerst
in Dauergewebe umgewandelt. Das geschieht zum Schutze der jüngsten
Knospenteile, den die Blätter zunächst zu übernehmen haben, wie wir
schon sahen. Das weitere Wachstum des Blattes vollzieht sich in der
Regel durch Streckung interkalar, und zwar meist so, daß
von der Spitze nach der Basis fortschreitend das Blattmeristem in
Dauergewebe übergeht, das Wachstum also am längsten und stärksten
in der Blattbasis bis zur fertigen Ausgestaltung des Blattes
fortdauert.
[S. 93]
Besonders auffällig ist das Vorauseilen in der Ausbildung der
Blattspitzen bei verschiedenen tropischen Gewächsen, besonders
Kletterpflanzen. Dort fällt diesen Blattspitzen nach M.
RACIBORSKI, der sie Vorläuferspitzen nennt, die Aufgabe
zu, vor der Fertigstellung des Blattes schon die Funktionen der
Blattspreite zu übernehmen.
Sehr eigenartig und abweichend von allen anderen Kormophyten
verhält sich Welwitschia mirabilis. Sie bildet über den
Keimblättern nur ein einziges Paar von Laubblättern, deren basale
Zonen in jährlicher Periode einen Zuwachs erfahren, während die
Blattenden allmählich verwittern.
2. Verschiedene Ausgestaltung der Blätter. Die Blätter des
Sprosses haben sehr verschiedene Funktionen und werden dementsprechend
an einem und demselben Stengel ganz verschieden ausgebildet, obwohl
sie alle der Anlage nach gleich angelegt sind. Den Wechsel in der
Ausbildung aufeinanderfolgender Blätter am Stengel bezeichnet man als
seine Blattfolge.
Fig. 125. Maiblume (Convallaria majalis). nd
Niederblätter, lb Laubblätter, hb Hochblätter, b Blüte,
ws Wurzelstock, aw Adventivwurzeln. Etwas verkleinert. Nach
SCHENCK..
Fig. 126. Vogelkirsche (Prunus avium). Knospenschuppen
1–3 und die Übergangsformen 4–6 zum Laubblatt 7, sp
Blattspreite, s Stiel, nb Nebenblätter. Etwas verkleinert. Nach
SCHENCK.
Der Hauptsproß der Keimpflanze trägt zu unterst die Keimblätter
oder Kotyledonen, die der Keimstengel, das Hypokotyl (Fig. 155), eines
Keimes schon im Samen besitzt. Nur ein einziges solches Blatt kommt
bei den Monokotylen vor; zwei Keimblätter sind bei den Dikotylen(Fig.
155) und bei manchen Gymnospermen, mehr als zwei bei anderen
Gymnospermen vorhanden. Auf die Keimblätter folgen an den Erdsprossen
und oft auch an den Luftsprossen mehr oder weniger schuppenförmige
Niederblätter (Fig. 125 nd), an den Luftsprossen hierauf die
Laubblätter (lb) und weiter oben wieder einfacher gestaltete
Hochblätter (hb). Wir wollen zunächst die Laubblätter
betrachten, da die übrigen Blätter nur Umbildungen von ihnen sind.
A. Die Laubblätter. Die Laubblätter sind äußerlich recht
verschieden gebaut. Ihre Gestalt und ihre Gliederung werden deshalb bei
der[S. 94] Diagnose der Pflanzen häufig verwertet. Meist ist das Laubblatt
gegliedert, und zwar in die lebhaft grüne, flächenförmige und
sehr dünne Blattspreite (Lamina, Fig. 126 sp), meist ungenau
Blatt genannt, in den stengelartigen Blattstiel (Petiolus, Fig.
126 s) und an seiner Basis in den Blattgrund. Dieser kann
als Blattpolster (Blattkissen) oder als Blattscheide
(Vagina, Fig. 138 v) ausgebildet sein, die den Stengel oberhalb
des Blattknotens mehr oder weniger umhüllt oder Nebenblätter
(Stipulae, Fig. 126 nb) ausgliedert. Bei vielen Blättern fällt der
Blattgrund aber nicht durch besondere Form und Größe auf, sondern geht
allmählich in den Blattstiel über, oft auch in die Nebenblätter
(Stipulae, nb). Fehlt der Stiel, so heißt das Blatt sitzend; ist
er vorhanden, gestielt. Die Anfänge der Blattgliederung pflegen sich
an den Laubblattanlagen schon sehr frühzeitig bemerkbar zu machen:
die Primordialblätter sondern sich sehr bald, nachdem sie als Wülste
am Vegetationspunkte sichtbar geworden sind, in das Oberblatt
(Fig. 127 A und B, o) und in den Blattgrund (A und B,
g). Aus dem Oberblatte geht die Blattspreite (Fig. 124 A, sp)
und, wo vorhanden, auch der Blattstiel hervor (A, st). Dieser
wird erst nachträglich, verhältnismäßig spät, durch interkalares
Wachstum zwischen die schon vorhandene Blattspreite und den Blattgrund
eingeschaltet, ist also niemals direkt am Stengel befestigt.
a) Die Blattspreite. Äußerer Bau (Fig. 129). Die in
der Regel ausgesprochen dorsiventrale, oberseits meist dunkler grün
gefärbte Blattspreite kann ungeteilt oder geteilt (Fig.
124 C), ja sogar aus Teilblättchen zusammengesetzt sein.
Solche zusammengesetzten Blätter entstehen entweder durch Verzweigung
aus den Rändern der Anlagen (Fig. 124 A) oder in seltenen Fällen (z. B.
Palmen) durch Zertrennung der jugendlichen unverzweigten Spreiten
bei ihrer Entfaltung. Den Monokotylen kommen vorwiegend einfache
Blätter zu, während die zusammengesetzten bei Dikotylen häufig sind.
Fig. 127. Blattentwicklung bei der Feldulme (Ulmus
campestris). A Der Vegetationskegel v, mit zwei Blattanlagen. Die
jüngste Blattanlage b noch ungegliedert, die nächstältere gegliedert
in Oberblatt o und Blattgrund g. B Das ältere Blatt von A,
schräg von außen gesehen. Vergr. 58. Nach STRASBURGER.
Die Blattspreite heißt schildförmig, wenn der Blattstiel
in ihrer Mitte zu entspringen scheint (Fig. 241). An der
ungeteilten Spreite (Fig. 125 lb, 126 sp) ist der Rand
nicht oder nur seicht eingeschnitten; er kann gesägt,
gezähnt, gekerbt oder ausgeschweift sein.
Sind tiefere Einschnitte vorhanden, so heißt das Blatt, wenn sie
nicht bis zur Mitte der Spreitenhälften reichen, gelappt;
wenn sie bis zu ihrer Mitte gehen, gespalten (Fig. 140
sb); wenn sie darüber hinaus sich fortsetzen, geteilt
(Fig. 142 l). Nur wenn die einzelnen Abschnitte der Spreite
so selbständig sind, daß sie als gesonderte Teilblättchen, kurz
Blättchen genannt, an dem gemeinsamen Blattstiel oder an der
ursprünglichen Mittelrippe der Spindel eingefügt sind, heißt das
Blatt zusammengesetzt (Fig. 141, 1–5); in allen anderen
Fällen gilt es als einfach. Die Teilung der Spreite ist
fingerartig (Fig. 142) oder fiederartig (Fig. 141,
1–5), je nachdem die Einschnitte gegen den Grund der Spreite
zusammenlaufen oder gegen deren Mittelrippe gerichtet sind.
Die Blättchen eines zusammengesetzten Blattes können während
ihrer Entwicklung ähnlich wie die Anlage gegliedert werden, aus
der sie hervorgingen; und an ihren Teilen kann sich die nämliche
Erscheinung wiederholen. Alsdann spricht man von doppelt, dreifach
und mehrfach zusammengesetzten Blättern. Häufig sind einfach und
doppelt gefiederte (Fig. 141) Blätter, deren Blättchen beiderseits
an Spindeln erster oder zweiter Ordnung eingefügt sind. Die
Blättchen eines zusammengesetzten Blattes können ganzrandig oder
mehr oder weniger tief eingeschnitten sein. Sie sitzen unmittelbar
der Spindel an, oder[S. 95] sie sind gestielt und unter Umständen sogar,
wie z. B. bei Phaseolus (Fig. 135 fg), Robinia, Mimosa, mit
angeschwollenen „Gelenkpolstern“ an den Einfügungsstellen versehen.
An Spreitenanlagen, die nicht einfach bleiben, vielmehr während
ihrer weiteren Entwicklung sich mehr oder weniger verzweigen,
werden die seitlichen Abschnitte meist in basipetaler Richtung,
also von der Spitze gegen die Basis fortschreitend, angelegt
(Fig. 124 A); doch ist auch eine entgegengesetzte, akropetale
Entwicklungsrichtung oder eine Kombination von beiden nicht
ausgeschlossen.
Die gelappten und oft auch durchlöcherten Blattspreiten der
Aracee Monstera kommen dadurch zustande, daß an den jugendlichen
Blättern inselartige Gewebemassen zwischen den Rippen absterben
und sich herauslösen. Auch die Abschnitte der fächerförmigen
und fiederförmigen Palmenblätter entstehen durch nachträgliche
Zertrennung der Blattspreiten, und zwar in der Weise, daß innerhalb
der ungeteilt angelegten und gefalteten jugendlichen Blattspreiten
vor deren Entfaltung entweder Gewebestreifen an den Oberkanten der
Falten absterben oder die Zellwände hier verschleimen und sich
voneinander trennen (so z. B. bei Cocos und Chamaerops)[72].
Die Blattspreiten ungestielter Blätter sitzen meist mit
breiter Basis an dem Stengel. Greift diese Basis jederseits noch
um den Stengel, so ist das Blatt stengelumfassend: z. B.
bei Papaver somniferum; umgibt sie geschlossen den Stengel, dann
heißt es durchwachsen (Bupleurum-Arten). Sind die Spreiten
zweier gegenüberstehender Blätter am Grunde verbunden, dann sind
sie verwachsen (Lonicera caprifolium). Setzt sich die
Spreite abwärts am Stengel flügelartig fort, so wird das Blatt als
herablaufend bezeichnet (Verbascum thapsiforme).

Fig. 128. Blatt des Rotdorns (Crataegus) mit
Netznervatur. 3⁄4
nat. Gr. Nach NOLL.
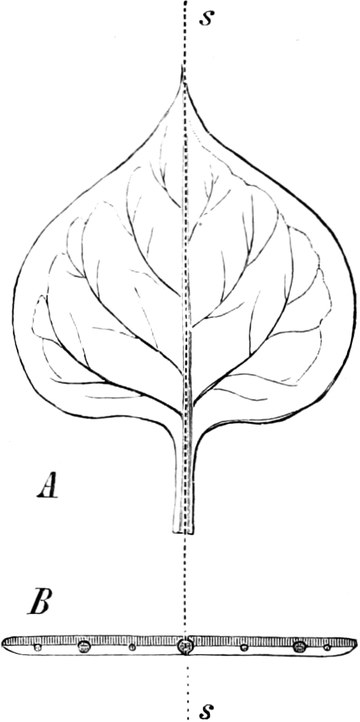
Fig. 129. Schematisches Bild eines dorsiventralen
Laubblattes. Bei A in Flächenansicht. Bei B im Querschnitt. ss
Die Symmetrieebene (Mediane). Nach STRASBURGER.
Die Blattspreiten sind von meist heller grünen Nerven (Adern)
durchzogen, die ein reichverzweigtes Netzwerk bilden. Diese Blattnerven
springen mit ihren dickeren Ästen (den Rippen) gewöhnlich
auf der Blattunterseite mehr oder weniger hervor, während ihnen auf
der Oberseite oft Furchen entsprechen. Die feineren Verästelungen
werden erst sichtbar, wenn man die Spreite im durchscheinenden Lichte
betrachtet. Vielfach ist ein in der Mediane der Spreite verlaufender
Nerv besonders kräftig entwickelt; er heißt Mittelnerv oder
Hauptnerv. Es können aber auch mehrere gleich starke Nerven
als Hauptnerven ausgebildet sein (Fig. 124). Von allen solchen Nerven
entspringen Seitennerven (Fig. 128).
Die Benennung der Nervatur richtet sich nach der Art des
Nervenverlaufs. Die Blätter der meisten Nadelhölzer sind
einnervig. Bei mehrnervigen Blättern stellt die
gabelige Nervatur einen besonderen Typus dar, wobei ein
Mittelnerv nicht zustande kommt. Sie ist für verschiedene Farne,
außerdem für Ginkgo biloba bezeichnend. Im übrigen unterscheidet
man bei den mehrnervigen Blättern die streifige Nervatur,
bei der mehrere Hauptnerven annähernd parallel oder im Bogen in
der Längsrichtung der[S. 96] Spreite verlaufen (Fig. 138 s) und gegen
deren Ende zusammenneigen, und die netzartige Nervatur
(Fig. 128), bei der die Nerven, an Stärke abnehmend, auseinander
entspringen und schließlich in einem feinen Maschenwerk ihren
Abschluß finden. Aber auch bei streifiger Nervatur pflegen die
Hauptnerven durch schwache Quernerven (Anastomosen) verbunden
zu sein. Bei netzartiger Nervatur unterscheidet man weiter
zwischen fiedernervigen Blättern (Fig. 128), wenn einem
medianen Hauptnerven schwächere Seitennerven entspringen, und
handnervigen Blättern (Fig. 124, 140 sb), wenn mehrere
annähernd gleich starke Hauptnerven an der Basis der Spreite
auseinandergehen. Die streifige Nervatur kennzeichnet im
allgemeinen die Monokotylen, die netzartige einige Farne und die
meisten Dikotylen.
Innerer Bau. Der innere Bau der Laubblätter ist sehr
mannigfaltig, aber meist ausgesprochen dorsiventral (bifazial):
die Gewebe auf der Oberseite sind anders ausgebildet als auf der
Unterseite (Fig. 129, 132).
Viele Blätter, besonders solcher Arten, die stark besonnte,
verhältnismäßig trockene Standorte bewohnen, doch z. B. auch vieler
submerser Wasserpflanzen, sind aber oben und unten gleich, also
isolateral (äquifazial, zentrisch) gebaut (Fig. 185, 191).
a) Nerven. Sie enthalten in Ein- oder Mehrzahl die Leitbündel
der Blätter. Die reiche Verzweigung, die Ausbildung der
Leitbündel als ein sehr feines Netzwerk, das sich über die ganze
Blattspreite ausdehnt, ist für die Blattspreiten besonders bezeichnend.
Es läßt sich durch Mazeration der Blätter als sehr zierliches
Blattskelett isolieren.
Der Bau der Leitbündel in der Blattspreite entspricht meist dem
im Stengel. Bei den Phanerogamen sind die Blattbündel gewöhnlich
kollateral. Da sie die Fortsetzungen der Blattspurbündel des Stengels
sind, so wenden sie ihren Gefäßteil nach oben, ihren Siebteil nach
unten.
Das Xylemparenchym der Blattbündel ist meist zu Platten angeordnet,
die auf den Bündelquerschnitten als radiale Zellreihen im
Gefäßteile erscheinen.
Im dem Maße, wie die Bündel sich in der Blattspreite mehr und mehr
verzweigen und schwächer werden, vereinfacht sich ihr Bau. Zunächst
schwinden die Tracheen; nur netz- und schraubenförmig verdickte
Tracheïden bleiben als wasserleitende Bahnen im Gefäßteile zurück.
Zugleich wird der Siebteil reduziert. Bei den Angiospermen,
deren Siebröhren von Geleitzellen begleitet werden, nehmen die
Siebröhren an Weite ab, während die Geleitzellen ihren früheren
Durchmesser behalten. Schließlich unterbleibt in den Zellen,
die Siebröhren fortsetzen, die Teilung in Siebröhrenglieder und
Geleitzellen: es werden Übergangszellen gebildet. Mit diesen
hört der Siebteil auf, während der Gefäßteil noch durch kurze
Schraubentracheïden vertreten ist und schließlich blind endigt
(Fig. 130).
Fig. 130. Leitbündelendigung im Blatt von Impatiens
parviflora. Vergr. 240. Nach SCHENCK.
Den Nadeln der Koniferen, die meist nur von 1–2 medianen längs
verlaufenden Leitbündeln durchzogen werden, fehlen alle feineren
Bündelverzweigungen. Den Außenrändern des Xylems folgt ein Saum
eigenartiger, toter, tracheïdaler Zellen mit Hoftüpfeln, den
Außenrändern des Phloëms ein entsprechender Saum eiweißreicher
Zellen. Dieses Transfusionsgewebe, das mehr oder weniger
tief in das lebende Blattgewebe beider Blatthälften eindringt,
vermittelt bei den Koniferen offenbar den Stoffverkehr zwischen
Nerv und diesem Blattgewebe.
Die Leitbündel werden von Parenchymscheiden umgeben, die in
den dickeren Nerven vielschichtig sind, einschichtig aber selbst ihre
feinsten Verzweigungen umschließen. Die Zellen dieser Scheiden sind
meist gestreckt[S. 97] und stets lückenlos verbunden. Den Leitbündeln folgen
auf einer oder beiden Seiten (Fig. 131, 1) häufig auch Stränge
von Sklerenchymfasern. Sie bilden namentlich an den Siebteilen der
stärkeren Bündel im Querschnitte sichelförmige Beläge, veranlassen
vorwiegend das Vorspringen der Blattrippen an der Spreitenunterseite
und machen die Spreite biegungsfest. Stränge aus Sklerenchym kommen
bei manchen Blättern auch zwischen den Nerven vor (Fig. 131,
1), ferner auch am Blattrande; solche sklerenchymatischen oder
kollenchymatischen Verstärkungen des Randes dienen zum Schutze gegen
scherende Kräfte, die die Blattflächen zu zerreißen suchen (Fig. 131,
2). Große Blattspreiten, denen ein solcher Schutz am Rande fehlt,
werden im Freien vom Winde zerfetzt (Banane).
b) Epidermis und Mesophyll. Das Laubblatt wird allseits
von einer typischen Epidermis umschlossen. Sie ist auf der
Blattunterseite besonders reich an Spaltöffnungen, die der
Oberseite nicht selten ganz fehlen (z. B. bei fast allen Laubbäumen).
Fig. 131. Blatt der Liliacee Phormium tenax (des
neuseeländischen Flachs). 1 Blattquerschnitt. Sc Sklerenchymplatten
und -stränge. A Grünes Assimilationsparenchym. H Epidermis
(Wasserspeicher). W Farbloses Mesophyll (innere Wasserspeicher).
2 Randpartie desselben Blattes. E Stark verdickte und gebräunte
Epidermis. R Randbündel aus Sklerenchymfasern. Nach NOLL.
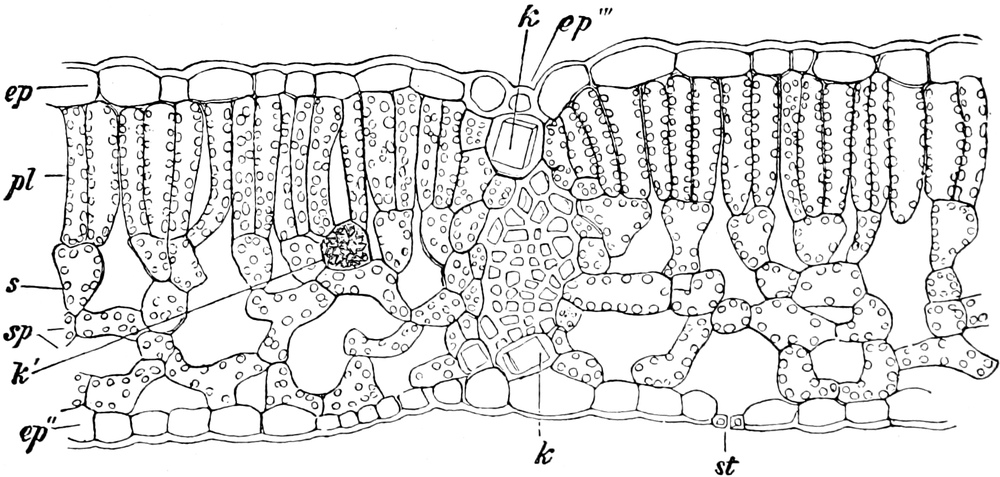
Fig. 132. Querschnitt durch das Blatt der Buche
(Fagus silvatica). ep Epidermis der Oberseite, ep″ Epidermis der
Unterseite, ep‴ längsgestreckte Epidermiszellen über einem der
Leitbündel, die der Querschnitt durch das Blatt der Quere nach trifft,
pl Palisadenparenchym, s Trichterzellen, sp Schwammparenchym,
k kristallführende Sekretzellen, k′ eine Kristalldruse, st
Spaltöffnung. Vergr. 360. Nach STRASBURGER.
Man findet an der Unterseite durchschnittlich 100–300
Spaltöffnungen auf dem Quadratmillimeter; doch kann diese Zahl in
einzelnen Fällen bis über 700 steigen. Isolaterale Blätter pflegen
auf beiden Seiten, Schwimmblätter nur oberseits Spaltöffnungen zu
besitzen.
Die Blattepidermis kann auch ein Wasserspeicher sein.
Besonders in diesem Falle ist sie nicht selten mehrschichtig.
[S. 98]
Das von der Epidermis umschlossene Gewebe der Blattspreite zwischen
den Rippen besteht hauptsächlich aus Parenchym, das man als
Mesophyll bezeichnet. Darin sind die feineren Nervenäste
eingebettet. Auf die Epidermis der Oberseite (Fig. 132 ep)
pflegen eine bis drei senkrecht zur Oberfläche gestreckte Lagen
zylindrischer (schlauchförmiger) Parenchymzellen zu folgen, die
Palisadenzellen (pl). Sie sind besonders chlorophyllreich,
bilden also ein Assimilationsparenchym und sind meist seitlich
voneinander durch enge Interzellularen getrennt. Oft neigen die
Palisadenzellen mit ihren unteren Enden deutlich zu Büscheln zusammen
(Fig. 132) und schließen an trichterförmig erweiterte Zellen (die
Trichterzellen s) an.
Die Dicke der Palisadenschicht ist bei den Blättern mancher
Bäume, z. B. der Rotbuche, verschieden: in den „Schattenblättern“
viel geringer als in den „Sonnenblättern“. Ein direkter
Einfluß der Belichtung liegt da aber nach den Untersuchungen
NORDHAUSENs[73] nicht vor. Es gibt auch Pflanzen (z. B.
Lactuca scariola), die nur in stark beleuchteten Laubblättern
Palisaden ausbilden.
In manchen Blättern kommen an Stelle der Palisadenzellen Schichten
von Zellen vor, deren Elemente nicht senkrecht, sondern parallel
zur Oberfläche, längs oder quer gestreckt sind, so z. B. bei vielen
Monokotylen. In den Nadeln der Kiefer und bei anderen Pflanzen
findet man ferner an Stelle der Palisaden große plattenförmige
Zellen, deren innere Oberfläche durch Einfaltungen der Zellmembran
bedeutend vergrößert ist (Faltenparenchym, Fig. 133 A, B fp,
C).
An das Palisadenparenchym schließt ein viel chlorophyllärmeres
Parenchym aus vorwiegend unregelmäßig gestalteten Zellen mit weiten
Interzellularen an, das Schwammparenchym (Fig. 132 sp),
das bis an die Epidermis der Unterseite (ep″) reicht. Die weiten
Interzellularen stehen mit den Spaltöffnungen in der Epidermis der
Blattunterseite in unmittelbarer Verbindung und dienen dem Gastransport
zu den Palisadenzellen.
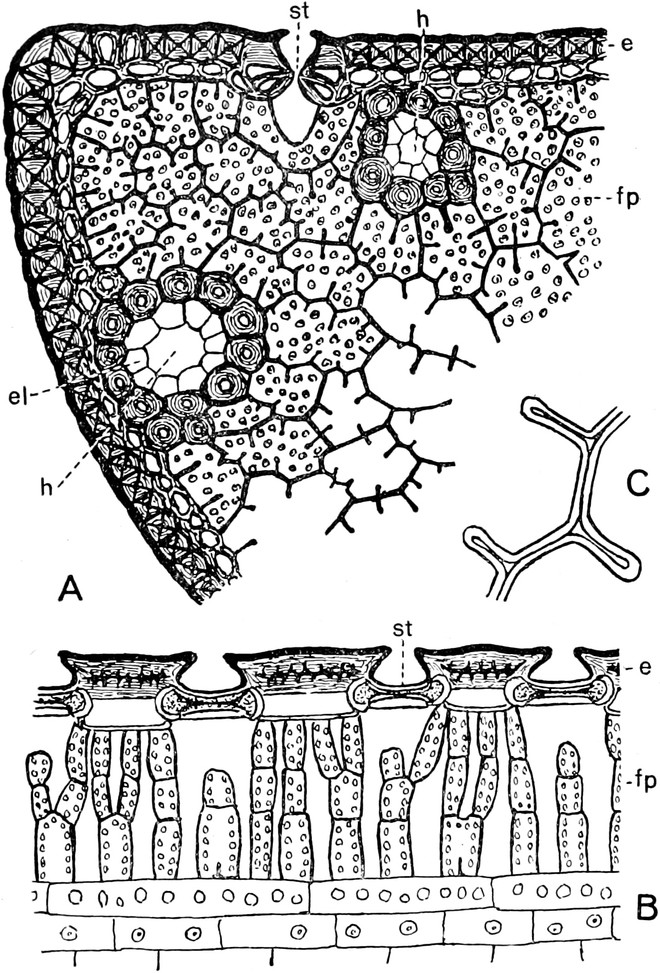
Fig. 133. Nadel von Pinus silvestris quer und längs.
A Querschnitt, B medianer Längsschnitt. Vergr. 160. e Epidermis,
st Spaltöffnung darunter, fp Assimilationsfaltenparenchym,
h Harzkanäle, das dünnwandige Drüsenepithel (el) von einer
Sklerenchymscheide umgeben. C Stück der Zellmembranen aus dem
Faltenparenchym. Vergr. 380.
HABERLANDT berechnet für einen Quadratmillimeter
Blattfläche bei Ricinus communis durchschnittlich im
Palisadenparenchym der Oberseite 403200, im Schwammparenchym
der Unterseite 92000 Chlorophyllkörner. Somit würden in diesem
Falle 82% der Chlorophyllkörner der Blattoberseite, 18% der
Blattunterseite angehören.
Nicht selten liegt auch im Mesophyll farbloses Wassergewebe
(Fig. 131 W).
Epitheme und Wasserspalten[74]. Bei gewissen Familien
der Mono- und Dikotylen bildet das Mesophyll der Blattlamina
lokal zwischen besonders angeschwollenen[S. 99] Leitbündelendigungen
und der Epidermis eigenartige Gewebepfropfen aus. Sie bestehen
aus kleinen lebenden Zellen mit farblosem Zellsaft, die mit
Wasser gefüllte Interzellularen zwischen sich lassen. Diese
Gewebepfropfen nennt man Epitheme. Sie vermitteln die
Ausscheidung von Wasser in tropfbar flüssiger Form. Dabei verhalten
sie sich der Hauptsache nach passiv; sie sind nur Stellen
geringsten Filtrationswiderstandes. Über solchen Epithemen, in die
Tracheïden münden, liegen in der Epidermis eigenartig ausgebildete
Spaltöffnungsapparate: die Wasserspalten (Fig. 134), die
größer als die Luftspalten sind. Ihre beiden Schließzellen sind
entweder lebend und können den Spalt, gleich den Luftspalten,
öffnen und schließen, oder sie verlieren (in anderen Fällen)
frühzeitig den lebenden Inhalt; der Spalt zwischen ihnen steht dann
unverändert weit offen. Die für die Schließzellen der Luftspalten
so bezeichnenden Verdickungsleisten werden bei den Wasserspalten
gewöhnlich nicht ausgebildet. Die ausgeschiedene Flüssigkeit
ist oft reich an kohlensaurem Kalke, der alsdann, wie an den
Blatträndern vieler Steinbrech- (Saxifraga-) Arten, in weißen
Schüppchen die Wasserspalten überdeckt.
Vielfach haben junge Blätter an ihren Spitzen und den Spitzen
ihrer Zähne über Epithemen Wasserspalten, die am fertigen Blatte
vertrocknet sind. Selbst bei submersen Pflanzen, bei denen keine
Luftspalten ausgebildet werden, kommen an den Blattspitzen nicht
selten Wasserspalten vor. Sie pflegen frühzeitig abzusterben,
werden auch wohl mit dem angrenzenden Gewebe zerstört, so daß
offene Grübchen, die Apikalöffnungen, entstehen, durch die Wasser
und darin gelöste Stoffe hervorgepreßt werden.
Auch in vielen Nektarien innerhalb und außerhalb der Blüten wird
die zuckerhaltige Flüssigkeit aus Wasserspalten („Nektarspalten“)
ausgeschieden.
Funktionen der Blattspreiten. Die Blattspreiten sind, wie schon
gesagt, die wichtigsten Ernährungs-, d. h. Assimilations- und zugleich
die Transpirationsorgane der Kormophyten. Ihr äußerer und innerer Bau,
ihre Anordnung und ihre Richtung zum Lichteinfall entsprechen diesen
Funktionen. Die Zerlegung der Kohlensäure ist an das Licht, außerdem an
das grüne Chlorophyll gebunden. So versteht man die grüne Farbe, die in
der Regel sehr große Oberflächenentwicklung der Blattspreiten, ihre bei
geringer Dicke flächenförmige Ausbildung und ihren meist dorsiventralen
Bau. Je größer die Oberfläche ist, um so mehr chlorophyllhaltige
Zellen werden ohne gegenseitige Beschattung dem Lichte ausgesetzt,
um so leichter kann die Kohlensäure aus dem spärlichen Vorrate der
Luft aufgenommen werden, und um so lebhafter wird zugleich die
Verdunstung der Spreite, ihre Transpiration, sein müssen. Da schon
wenige chlorophyllreiche Zellschichten das von außen in das Blatt
einströmende Licht so verändern, daß es in tieferen Schichten der
Kohlensäurezerlegung nicht mehr ausreichend zu dienen vermag, so ist
das Assimilationsgewebe vorzugsweise oberseits gelegen. Die Kohlensäure
der Luft wird aber hauptsächlich von der Unterseite des Blattes
durch die Spaltöffnungen aufgenommen; infolgedessen kann sie um so
schneller durch die weiten Interzellularen des Schwammparenchyms, das
im wesentlichen ein Durchlüftungsgewebe ist, zu den assimilatorisch
besonders tätigen oberen Gewebeschichten vordringen, je dünner das
Blatt ist.
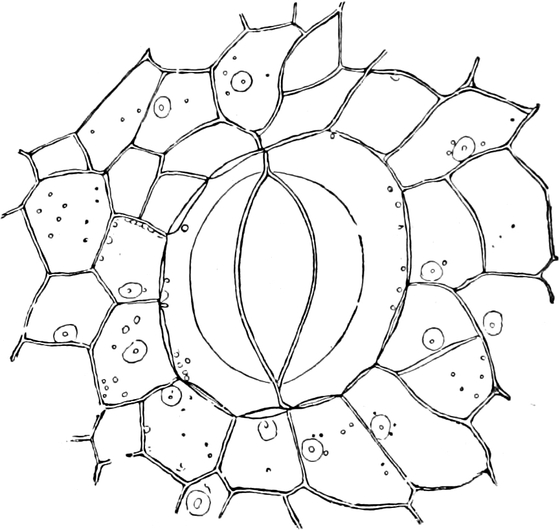
Fig. 134. Wasserspalte am Blattrande der Kapuzinerkresse
(Tropaeolum majus) nebst angrenzenden Epidermiszellen. Vergr. 240. Nach
STRASBURGER.
Das reich verzweigte Leitbündelnetz ermöglicht es, die Assimilate
von allen Teilen des Blattes schnell nach den Stengeln abzuleiten,
nachdem sie[S. 100] aus den Mesophyllzellen zu den feineren Verästelungen
des Nervennetzes hingeschafft worden sind; zugleich versorgt es durch
seine feine Zerteilung, die im Gegensatze zu dem wasserleitenden
Stengel für die wasserabgebende Blattspreite bezeichnend ist,
auf kürzestem Wege alle Teile der transpirierenden Blattspreite mit
Wasser. Schließlich erhöht die Aderung auch noch die Festigkeit der
Spreite.
Wie wir sahen, sind die Blätter am Stengel so angeordnet, daß die
Blattspreiten, die an aufrechten Sprossen nahezu horizontal stehen,
ohne allzu starke gegenseitige Beschattung dem Lichte möglichst
ausgesetzt werden. Viele Blätter besitzen außerdem Bewegungsvermögen
und können ihre Spreiten gegen das einfallende Licht einstellen.
Häufig, so namentlich an dorsiventralen plagiotropen Zweigen, fügen
sich die Blattspreiten, die sämtlich ihre Oberseiten gegen das Licht
wenden, bei Betrachtung von oben mehr oder weniger dicht zu einem
Blattmosaïk aneinander.
b) Der Blattstiel ist meist stengelartig ausgebildet. Sein
innerer Bau gleicht alsdann dem der Hauptrippe der Spreite oder
wohl auch der Sproßachse. Die Leitbündel sind jedoch bei den
Angiospermen oft in einem nach oben offenen Bogen angeordnet; typisches
Assimilationsgewebe fehlt dem Stiele ganz. Der Blattstiel dient dazu,
die Blattspreite von der Stengelachse weg in den Raum hinaus, also dem
Licht entgegen zu strecken. Sind Blattstiele vorhanden, so führen Teile
von ihnen auch meist die Einstellung der Spreiten gegen das Licht aus.
Fig. 135. Unpaarig gefiedertes Blatt von Phaseolus
mit Blattstielgelenken (Polstern). hg Polster des Blattstiels
(Hauptgelenk), fg Gelenk eines der Fiederblätter.
1⁄4 nat. Gr.
Manchmal werden die Einstellungsbewegungen durch besondere
örtliche Anschwellungen an der Basis oder an der Spitze oder an
beiden Stellen des Blattstieles ausgeführt, die wie Gelenke
arbeiten: Blattkissen oder Blattpolster, so besonders
bei vielen Leguminosen (Fig. 135).
An den Jahrestrieben der Holzgewächse kommt das Blattmosaïk außer
durch die Blattbewegungen nicht selten auch durch die verschiedene
Länge der Blattstiele (und durch die verschiedene Größe der
Blattspreiten) zustande: die unteren Blätter haben viel längere
Stiele (und größere Spreiten) als die oberen; so sehr auffallend
z. B. beim Ahorn oder der Roßkastanie. Auch bei solchen Gewächsen,
bei denen die Laubblätter eine Rosette bilden, kann man dies
beobachten, so besonders schön an den schwimmenden Rosetten der
Wassernuß (Trapa natans).
Gestielte Blattspreiten, die bei den Dikotylen weit häufiger als
bei den Monokotylen sind, setzen sich von den Blattstielen entweder
scharf ab, oder sie laufen an ihnen herab, so daß die Stiele
geflügelt erscheinen.
c) Der Blattgrund[75]. Ist der Blattgrund des Laubblattes in
besonderer Weise ausgebildet, so trägt er meist zum Schutze der Knospe
und des nächst jüngeren Blattes dadurch bei, daß er auch dann noch die
Knospe umhüllt, wenn die Blattspreite sich von der Knospe losgelöst und
entfaltet hat.
Vielfach werden aus dem Blattgrunde Nebenblätter oder
Stipulae gebildet, in typischen Fällen in Zweizahl, also je
eines zu beiden Seiten des zugehörigen Blattes. Sie können ganz
unscheinbar (Fig. 126 nb) oder ansehnlich (Fig. 136), gelblich oder
grün gefärbt sein. Haben sie nur die Knospen zu schützen, so sind sie
meist gelblich oder bräunlich gefärbt, im Innern viel einfacher als die
Blattspreiten gebaut und fallen frühzeitig ab.
[S. 101]
Wenn sich aber die Nebenblätter an der Kohlensäureassimilation der
Pflanze beteiligen (Fig. 207), so sind sie grün gefärbt und wie die
Blattspreiten gebaut.
Die Nebenblätter sind sehr verschieden ausgebildet. Bei vielen
Gewächsen sind es zwei freie Blättchen (Fig. 126 nb). Bei anderen
sind sie je mit ihrem einen Rande dem Blattstiel angewachsen
(Vaginalstipeln, Fig. 136 A), bei wieder anderen in
verschiedener Weise miteinander verwachsen, nämlich entweder
zu einem zungenförmigen Gebilde in oder oberhalb der Blattachsel
(zu der Axillarstipel, Fig. 136 B) oder zu einem dem
Blatte opponierten Gebilde (zu der opponierten Stipel).
Bei gegenständiger Blattstellung können die Nebenblätter der
Blattpaare paarweise mit ihren einander zugekehrten Rändern zu
Interpetiolarstipeln (Fig. 137) verwachsen. Die Nebenblätter
können aber auch an ihren beiden Blatträndern miteinander
verwachsen und den Stengel als vollständig geschlossene Tüte
umfassen, die den Stengel und die nächst jüngere Blattanlage in der
Knospe umhüllt; die Tüte ist aus einer opponierten Stipel bei dem
in Zimmern oft kultivierten Ficus elastica hervorgegangen, wo sie
durch das neu sich entfaltende Blatt aufgeschlitzt und an ihrem
Grunde abgesprengt wird; bei den Polygonaceen ist sie dagegen eine
Axillarstipel, die, von den Blättern an ihrer Spitze durchbrochen,
als trockene Scheide (Ochrea, Fig. 676) am Stengel zurückbleibt.
Fig. 136. Nebenblätter an den Blattstielbasen von
Jugendblättern der Seerose (Nymphaea alba). A Vaginalstipeln, B
Axillarstipel. Vergr. 12. Nach GLÜCK.
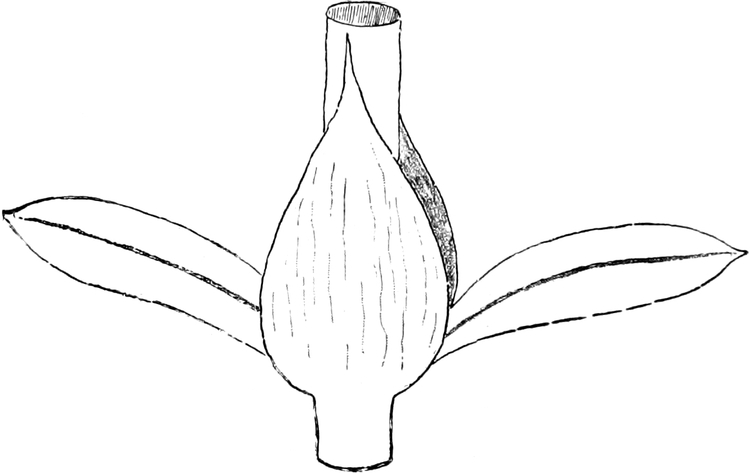
Fig. 137. Stengelknoten von Paronychia argentea
(Caryophyllacee). Rechts und links: ein Blattpaar. Vorn und hinten: je
eine interpetiolare Stipel. Vergr. 5. Nach GLÜCK.
Bei manchen Arten von Galium, wo die Nebenblätter vollständig dem
Oberblatt gleichen, glaubt man vier-, bei anderen sechs- oder
achtblättrige Blattquirle vor sich zu haben, während tatsächlich
nur zwei Blätter in dekussierter Stellung mit einer je nach der Art
verschiedenen Zahl von Nebenblättern den Wirtel bilden: nur zwei
dieser Blattgebilde nämlich tragen Achselknospen.
Sehr häufig bei Monokotylen, seltener bei den Dikotylen (z. B.
Umbelliferen) ist aus dem Blattgrund eine Scheide geworden. Bei
den Gräsern ist sie (Fig. 138 v) auf der einen Seite gespalten, bei
den Riedgräsern dagegen völlig geschlossen. Die Scheide der Gräser, die
den unteren Teil des noch wachsenden und weichen Internodiums schützt
und stützt, setzt sich am Grunde der ungestielten Blattspreite in einen
häutigen Auswuchs, die Ligula (l), fort; an ihrer Basis aber
ist sie unmittelbar oberhalb des Stengelknotens zu einem „Gelenk“ (dem
Gras„knoten“) angeschwollen (Fig. 138 k).
Die Ligula entspricht nach GLÜCK den miteinander
verwachsenen Spitzen der Vaginalstipeln, aus denen die Blattscheide
hervorgegangen ist.
Heterophyllie und Anisophyllie. Manche Pflanzen bilden
verschieden gestaltete Laubblätter aus, entweder in verschiedenen Zonen
des Stengels[S. 102] (Heterophyllie, Fig. 139, 140) oder in einer
und derselben Zone auf den beiden Seiten des dorsiventralen Sprosses
(Anisophyllie, Fig. 141). Mit Anisophyllie ist oft Asymmetrie
der Blattspreiten verbunden. Heterophyllie zeigen viele Wasserpflanzen
mit bandförmigen oder zerteilten untergetauchten Wasserblättern, die
an das Leben im Wasser angepaßt sind, und mit viel weniger zerteilten,
gestielten Luftblättern (Fig. 139). Die Blätter, die der Efeu zur Zeit
der Blütenreife entwickelt, sind wesentlich anders gestaltet als die,
die er vorher ausgebildet hat. Noch auffälliger ist dieser Unterschied
bei Eucalyptus globulus, der zunächst ovale und sitzende, später
sichelförmige Blätter ausbildet. Häufig sind die untersten Blätter
von Keimpflanzen (Jugend- oder Primärblätter) einfacher
geformt als die übrigen (Folgeblätter).
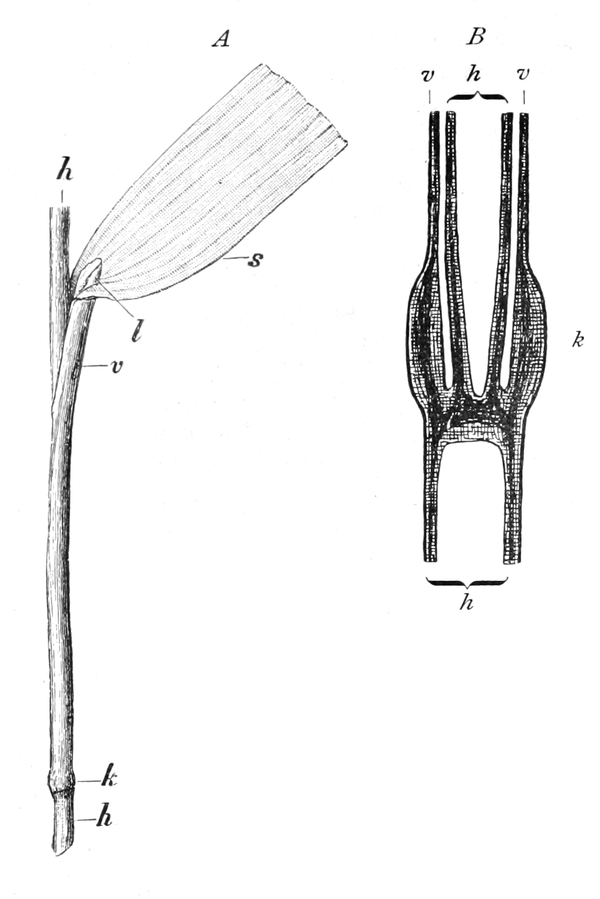
Fig. 138. A Stengel und Blattstück einer Graminee.
Nach SCHENCK.
B Gras„knoten“ im Längsschnitt, etwas schematisiert. h Halm, v
Blattscheide, k Anschwellung der Blattscheide über dem Stengelknoten,
s Stück der Blattspreite, l Ligula. Nat. Gr.
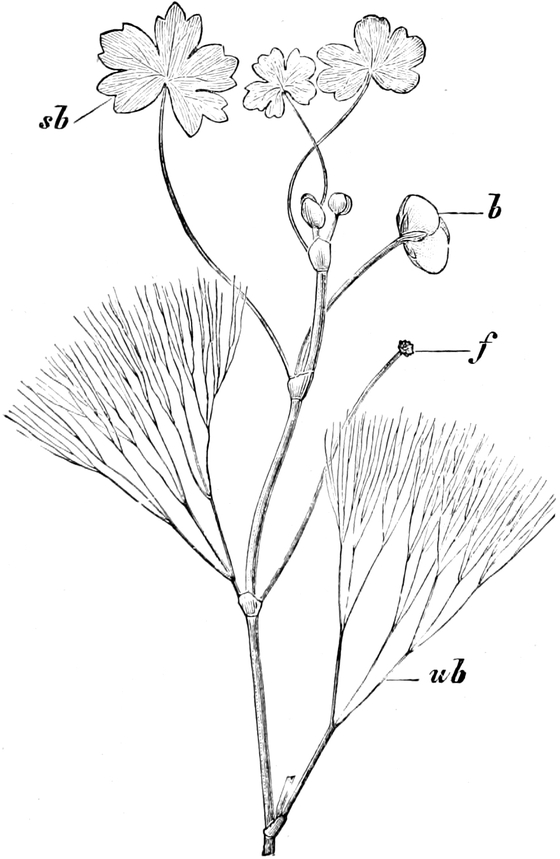
Fig. 139. Batrachium aquatile. Wasserhahnenfuß. ub
Untergetauchte Blätter, sb schwimmende Blätter, b Blüte, f
Fruchtanlage. Verkleinert. Nach SCHENCK.
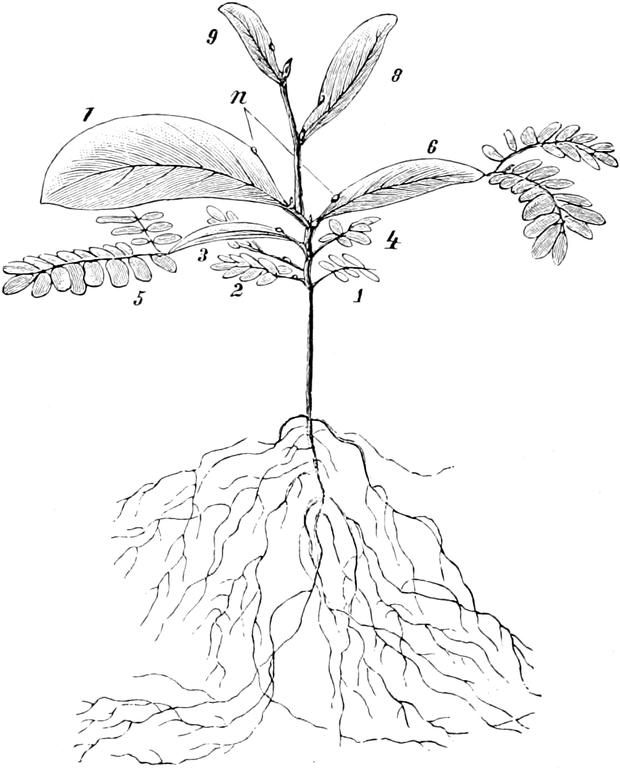
Fig. 140. Keimpflanze von Acacia pycnantha. Die
Keimblätter schon abgeworfen. 1–6 Jugendblätter, 1–4 einfach-,
die folgenden doppelt gefiedert. An den Blättern 5 und 6 sind die
Blattstiele bereits senkrecht abgeflacht. Bei den folgenden Blättern
(7, 8, 9) sind sie als Phyllodien ausgebildet. n Nektarien an
den Phyllodien. Vergr. ca. 1⁄2.
Nach SCHENCK.
B. Die Keimblätter. Die Keimblätter oder Kotyledonen, die
gestielt oder ungestielt sein können, sind[S. 103] fast immer viel einfacher
gestaltet als die Laubblätter, wenn sie auch oft im wesentlichen
dieselbe Gliederung wie diese erkennen lassen.
Sie können dauernd von der Samenschale umschlossen und unter der
Erde verborgen bleiben (hypogäische). In diesem Falle sind
sie gewöhnlich fleischige Reservestoffbehälter und bauen sich
hauptsächlich aus Speicherparenchym auf. Die epigäischen,
die die Samenschale sprengen und über der Erde erscheinen, pflegen
zu ergrünen und alsdann einige Zeit wie die Laubblätter Kohlensäure
zu assimilieren. Bei den Monokotylen, wo nur ein Keimblatt
ausgebildet wird, verläßt gewöhnlich nur der Scheidenteil des
Kotyledo den Samen; er kann unterirdisch und farblos bleiben oder
aus der Erde hervorwachsen und ergrünen.
C. Die Nieder- und Hochblätter sind in ihren Anlagen von
Laubblattanlagen nicht zu unterscheiden, stehen aber fertig ausgebildet
in ihrer Gliederung den Laubblättern bedeutend nach, haben gewöhnlich
Schuppenform und keinen Stiel. Sie bilden sich durch Vergrößerung von
Primordialblättern, und zwar vornehmlich aus deren Blattgrund aus,
während die Spreite mehr oder weniger unentwickelt bleibt (Fig. 126,
1–6, 142). Die Niederblätter, farblose oder grüne Schuppen, gehen am
Luftsprosse oft der Bildung der Laubblätter voraus (Fig. 125 nd). Sie
sind ferner als farblose, größere oder kleinere, oft kaum sichtbare
und meist kurzlebige Schuppen vielfach die einzigen Blattgebilde der
Rhizome, denen, entsprechend ihrem Leben im Dunkeln, die Laubblätter
meist fehlen (Fig. 125 ws, 143). Die Hochblätter dagegen, von
gleichem Bau und gleicher Beschaffenheit wie die Niederblätter des
Luftsprosses, manchmal aber andersfarbig, pflegen oben am Stengel auf
die Laubblätter als Deckblätter oder Brakteen für die Blüten
und Blütensprosse zu folgen. Der innere Bau beider Blattarten ist
wesentlich einfacher als der der Laubblätter. Nieder- und Hochblätter
sind an der Ernährung der Pflanze nicht oder kaum beteiligt,
sondern meist Schutzorgane für die jungen Blattspreiten oder die
Stengelknospen. Sie sind aber meist durch Zwischenformen mit den
Laubblättern verbunden (Fig. 126, 142).
Fig. 141. Anisophyllie der dorsiventralen Sprosse von
Selaginella Martensii; auf der Oberseite des Stengels zwei Reihen sehr
kleiner asymmetrischer grüner Blätter, auf jeder Flanke eine Reihe viel
größerer asymmetrischer Laubblätter. Etwas vergr.
Fig. 142. Helleborus foetidus. Laubblatt (l) und
Übergänge zum Hochblatt (h). Verkl. Nach SCHENCK.
Fig. 143. Rhizom von Polygonatum multiflorum. a Knospe
für den nächstjährigen oberirdischen Trieb, b Narbe des diesjährigen
Triebes, c, d und e Narben der drei vorausgegangenen Jahre. w
Wurzeln. Auf 3⁄4 verkleinert.
Nach SCHENCK.
[S. 104]
Daß die Niederblätter und Hochblätter der Hauptsache nach als
Hemmungsbildungen von Laubblättern aufzufassen sind, lehrt nicht
nur ihre Entwicklungsgeschichte, sondern auch die Möglichkeit,
ihre Anlagen zu Laubblättern werden zu lassen. So gelang es
GOEBEL, Blattanlagen, welche Niederblätter erzeugt hätten,
zur Laubblattbildung dadurch zu bewegen, daß er die Sprosse
entgipfelte und entblätterte. Unterirdische Stengel, die man
zwingt, sich im Tageslichte zu entwickeln, bilden Laubblätter aus
denselben Anlagen, die unter der Erde zu Niederblättern geworden
wären. Im inneren Bau sind Nieder- und Hochblätter aber nicht
ausschließlich Hemmungsbildungen von Laubblättern, sondern zeigen
oft diesen gegenüber besondere Differenzierungen, die mit ihren
Aufgaben zusammenhängen können[76].
3. Lebensdauer der Blätter. Die Blätter haben bei vielen
Gewächsen eine kürzere Lebensdauer als die Sproßachsen, an denen sie
entstanden sind. In diesem Falle werden sie bei den meisten Bäumen
und Sträuchern von den Sproßachsen abgestoßen (Blattfall) oder
verfaulen am Stengel (bei Erdsprossen); an den Luftsprossen der Kräuter
sterben sie meist mit den Stengeln ab. Blattnarben am Stengel
geben die Stellen an, wo früher Blätter gesessen haben. Pflanzen,
deren Laubblätter mehrere Vegetationsperioden tätig bleiben, nennt man
immergrün im Gegensatze zu den sommergrünen, bei denen
sie nur eine Vegetationsperiode dauern.
Der Blattfall der phanerogamen Holzgewächse wird durch eine
parenchymatische Trennungsschicht vermittelt, die am Grunde
des Blattstiels meist erst kurz vor dem Blattfall, mit oder ohne
vorausgehende Zellteilungen, ausgebildet wird. Alle mechanischen
Gewebe des Blattstiels sind an dieser Stelle sehr reduziert;
verholzt sind dort nur die Gefäße. Die Blattablösung erfolgt in der
Trennungsschicht meist durch Abrundung der Zellen gegeneinander
und durch Verschleimung ihrer Mittellamellen, während die Gefäße
und die Siebröhren zerrissen werden. Die Blattnarbe wird dadurch
abgeschlossen, daß die äußersten Zellschichten der Wundfläche sich
in (verholzendes) Kutisgewebe umwandeln, worunter meist noch durch
ein Korkkambium eine Korkschicht gebildet wird, die sich an die
Korkschicht des Stengels anschließt.
δ) Die Verzweigung der Sprosse[65] u. [77]. Je mehr Laubblätter
der Sproß im Sonnenlichte ausbreiten kann, um so mehr organische
Substanz vermag er im Assimilationsvorgange zu bilden. In dieser
Hinsicht ist, wie leicht ersichtlich, ein verzweigtes Sproßsystem einem
aufrechten Einzelsprosse weit überlegen. Jenes kann Blattflächen dem
ungeschwächten Lichte allseits über einen größeren Raum darbieten.
Wie bei den thallösen Pflanzen kommen die Verzweigungen der
Sprosse in zweierlei Weise zustande: entweder, doch nur
selten, durch Gabelung, Dichotomie, einer Mutterachse in
zwei Tochterachsen, oder meist durch seitliche Neubildungen
von Tochterachsen an einer weiter wachsenden Mutterachse, also durch
seitliche Verzweigung.
A. Die dichotome Verzweigung. Sie ist auf die Sprosse einiger
Lycopodiaceen beschränkt.
Bei solchen Bärlappgewächsen gabelt sich ein Sproß folgendermaßen
in zwei gleichwertige Teile: Der kreisförmige Querschnitt des
Vegetationspunktes, der gewöhnlich keine Scheitelzelle mehr
erkennen läßt, wird elliptisch. Den beiden Brennpunkten der Ellipse
entsprechend wölben sich die zwei neuen Vegetationskegel vor (Fig.
144). Die aufeinanderfolgenden Gabelungen können in rechtwinkligen
Ebenen zueinander stattfinden; in diesem Falle breitet sich das
Verzweigungssystem nicht in einer Ebene, wie in dem Schema (Fig. 82
a), sondern allseits im Raume aus.
Nicht selten weicht bei diesen Gewächsen, z. B. bei Selaginella,
das Verzweigungssystem in seinem Aussehen stark von dem Typus
dadurch ab, daß immer nur der eine Gabelast jedes
Zweigpaares einer Ordnung weiter wächst und sich wieder
gabelt oder, wie man auch sagt, die Verzweigung fortsetzt
(Fig. 145). Stellen sich alsdann alle die Zweigstücke, die jedesmal
die Verzweigung fortsetzen, annähernd in eine Richtung[S. 105] ein,
die anderen aber schräg dazu, so entsteht ein Verzweigungssystem,
das einem razemösen (Fig. 82 b) zum Verwechseln ähnlich werden
kann. Doch wird es nicht von einer einheitlichen Hauptachse,
sondern von einer nur scheinbaren Hauptachse durchzogen,
an der jedes Stück eine Tochterachse des vorausgehenden ist. Eine
solche Scheinachse bezeichnet man zum Unterschied von der echten
Hauptachse (Monopodium) als Sympodium, die Verzweigung als
sympodiale Verzweigung auf dichotomer Grundlage.
Übrigens kann man bei den Bärlappgewächsen alle Übergänge von
dichotomer zu seitlicher Verzweigung beobachten. Viele Arten
bilden bei der Gabelung eines Vegetationspunktes sofort zwei
Vegetationspunkte von verschiedener Größe aus, von denen der
kleinere sehr schnell gegen den größeren seitlich verschoben wird
(Fig. 146).
Fig. 144. Ein in zwei gleichstarke Gabeläste (p′
und p″) sich fortsetzender Sproß (p) von Lycopodium alpinum, im
Längsschnitt. b Blattanlage, c Rinde, f Leitbündel. Vergr. 60.
Nach HEGELMAIER.
Fig. 145. Sympodium auf dichotomer Grundlage.
Fig. 146. Ein in zwei ungleiche Gabeläste (p′ und
p″) sich teilender Sproß von Lycopodium inundatum. b Blattanlagen.
Vergr. 40. Nach HEGELMAIER.
B. Die seitliche Verzweigung. a) Ort der Entstehung der
Seitenknospen. An dem aus Sproßachse und Blättern bestehenden
Sprosse bilden sich Seitenzweige auch bei seitlicher Verzweigung in
der Regel nur an der Sproßachse oder an der untersten Basis
der Blattanlagen aus, und zwar meist schon am Vegetationspunkte des
Muttersprosses in akropetaler Reihenfolge als Auswüchse an seiner
Peripherie, also exogen wie die Blattanlagen (Fig. 98 g). Die
Orte der Seitensproßentstehung sind in der Regel fest bestimmt.
Bei Pteridophyten entspringen sie oft neben den Blatthöckern, bei den
Samenpflanzen aber in der Regel da, wo die Oberseite der höckerförmigen
Blattanlage in das Gewebe des Vegetationspunktes übergeht, mit
anderen Worten in der Blattachsel, bald mehr auf der Basis der
Blattanlage, bald mehr am Stengel.
Fig. 147. Schema für die entwicklungsgeschichtlichen
Beziehungen zwischen Achselsproß und Blattanlage; im Längsschnitt. Nach
GOEBEL.
Die Anlage eines Seitenzweiges kann 1. aus dem Gewebe der
Sproßachse dicht oberhalb der Blattanlage und nach ihr (Fig. 147
I) oder vor der Blattanlage entstehen; im letzteren Falle wölbt
sich die Blattanlage aus dem basalen Gewebe an der Unterseite der
Zweiganlage hervor (Fig. 147 III); 2. kann die Zweiganlage aus
dem Gewebe der ganz jugendlichen Blattanlage sich bilden (Fig. 147
II). Bei dorsiventralen Sprossen von Blütenpflanzen gibt es auch
extraaxilläre Seitenknospen seitlich von den Blattanlagen.
[S. 106]
An dem Längsschnitte durch einen Vegetationspunkt in Fig. 98 sieht
man die jüngste Anlage eines Seitensprosses (g) bereits in der
Achsel einer der allerobersten Blattanlagen sich vorwölben. In den
Achseln nächstälterer Blatthöcker sind die Sproßanlagen, da sie in
akropetaler Folge entstehen, schon größer und beginnen ihrerseits
Blatthöcker hervorzubringen. Solche in den Blattachseln erzeugte
Knospen werden als Achsel- (oder Seiten-) Knospen, die
aus ihnen hervorgehenden Sprosse als Achselsprosse bezeichnet;
die Knospe, die das fortwachsende Ende eines Sprosses abschließt, heißt
im Gegensatz dazu End- oder Terminalknospe. Das Blatt, in dessen Achsel
eine Knospe steht, ist ihr Tragblatt, Stützblatt oder Deckblatt
(Fig. 149 db). Die durch die Mittelrippe dieses Blattes und die
zugehörige Mutterachse gelegte Ebene heißt die Mediane des
Blattes. Im allgemeinen steht die Achselknospe in der Mediane
ihres Deckblattes: nur selten ist sie seitlich dagegen verschoben.
Regel ist bei den Angiospermen, daß jedes Laubblatt eine
Achselknospe trägt und daß nur eine Achselknospe in der Achsel ihres
Deckblattes entsteht; bei manchen Gymnospermen dagegen bilden nicht
alle Blätter Achselknospen aus.
Doch gibt es auch Fälle, wo auf die erste Achselknospe die
Bildung anderer, der Beiknospen, folgt. Entweder stehen
diese übereinander (seriale Beiknospen), so z. B. bei
Lonicera, Robinia, Gleditschia, Gymnocladus, oder nebeneinander
(kollaterale Beiknospen), z. B. bei manchen Liliaceen, wie
Allium- und Muscari-Arten.
Fig. 148. A Cuphea lanceolata (Lythracee). Der
(vegetative) Achselsproß in der Achsel des linken unteren Blattes nicht
verschoben; der des rechten unteren Blattes (eine Blüte bildend) dem
Sproß bis zu dem nächst oberen Blattpaar angewachsen. 1⁄2 nat. Größe.
B Samolus Valerandi (Primulacee). Die Tragblätter t an den
Achselsprossen a emporgerückt. Fruktifizierende Pflanze. Jeder
Achselsproß schließt mit einer Frucht ab. Nat. Größe. Nach
SCHENCK.
C Blatt der ostasiatischen Cornacee Helwingia: Der kleine männliche
Blütenstand mit dem Laubblatt bis zur Mitte der Spreite verwachsen.
Nach SIEBOLD und ZUCCARINI.
[S. 107]
Interkalare Wachstumsvorgänge in dem Gewebe an der Basis der
Achselknospe können Verschiebungen bewirken, wodurch die
ursprünglichen Beziehungen zwischen Deckblatt und Achselknospe
geändert werden. So gibt es Fälle, wo die Knospen den Achseln ihrer
Deckblätter durch Streckung des Gewebes der Mutterachse
unterhalb der Achselknospen entrückt werden, die einzelne Knospe
also viel höher am Stengel als ihr Deckblatt befestigt ist (Fig.
148 A). Das Deckblatt kann auch durch eigenes basales
Wachstum unterhalb der auf ihm sitzenden Knospe diese mitnehmen,
so daß der Achselsproß auf ihm sitzt (Fig. 148 C); oder es wird
selbst von der sich streckenden Basis des Achselsprosses,
wie in Fig. 148 B, mitgenommen und scheint ihm anzugehören.
Daß schon am Vegetationspunkte die Anlagen der Seitenzweige
sichtbar werden, ist bei den Phanerogamen Regel. Treten
Seitensproßanlagen erst in größerer Entfernung vom Scheitel auf,
so läßt sich meist nachweisen, daß embryonale Substanz für ihre
Bildung an den entsprechenden Orten aufgespart blieb.
Sproßanlagen, die in solcher Weise an vorbestimmten
Stellen meist noch jugendlicher Pflanzenteile entstehen, werden
als normale bezeichnet und solchen gegenübergestellt, die
beliebigen anderen Stellen jüngerer oder älterer Pflanzenteile,
nämlich Stämmen, Wurzeln und Blättern, entspringen und meist aus wieder
teilungsfähig gewordenem Dauergewebe hervorgehen. Solche Anlagen pflegt
man als adventive Bildungen zusammenzufassen. Adventivsprosse
können auch inneren oder endogenen Ursprung haben; sie müssen
in solchem Falle die äußeren Gewebe der Mutterpflanze durchbrechen,
um nach außen zu gelangen. An Stamm- und an Wurzelteilen auftretende
Adventivsprosse sind vornehmlich endogenen, die an Blättern erzeugten
exogenen Ursprungs.
Adventivsprosse brechen oft als Wurzelbrut aus den Wurzeln von
Kräutern (z. B. bei Convolvulus arvensis, Rumex Acetosella) oder
von Sträuchern (Rubus, Rosa, Corylus) oder von Bäumen (Populus,
Ulmus, Robinia) hervor, werden selbst an den Blättern mancher
Gewächse, so des Schaumkrautes (Cardamine partensis), der
Brunnenkresse (Nasturtium officinale), verschiedener Farnkräuter
hervorgebracht. Bei anderen Pflanzen regt erst eine Verwundung
des Pflanzenkörpers ihre Bildung an. So treten sie häufig als
Stockausschlag an den Stümpfen gefällter Bäume auf. Gärtner
verwerten vielfach Adventivknospen, die an abgeschnittenen
Stammstücken, Wurzelstücken oder abgeschnittenen Blättern
(Stecklingen) entstehen, um Pflanzen zu vermehren[78]. Gehen
die Knospen nicht aus vorhandenen Vegetationspunkten, sondern
aus Dauergewebe durch Neubildung hervor, so spricht man von
Restitution (vgl. Physiologie).
Fig. 149. A Grundriß (Diagramm) und B Seitenriß
einer Seitenknospe von einer Monokotyle mit 1⁄3-Blattstellung. M
Mutter- (Abstammungs-) Achse, db Deckblatt daran. t Tochterachse,
vb adossiertes Vorblatt daran. Verbindungslinie db–t–m Richtung
der Mediane der Seitenknospe, punktierte Linie: Richtung der
Transversalebene. h: was an der Tochterachse als hinten, v: was
daran als vorn bezeichnet wird.
b) Blattstellungsanschluß der Seitenknospen. Will man die
Stellungsverhältnisse an einem Seitenzweige beliebiger Ordnung
untersuchen, so orientiert man ihn stets so, daß sein
Deckblatt nach vorn (Fig. 149 db), d. h. nach dem Beobachter
hin gerichtet, seine Mutterachse (m) aber nach hinten, d. h.
von ihm weggewendet ist, und zugleich in der Weise, daß die Mediane des
Deckblattes mit der Mediane des Beschauers zusammenfällt. Die Mediane
des Deckblattes ist alsdann zugleich die Mediane des axillären
Seitensprosses (vgl. Fig. 149 t).[S. 108] Die Ebene, die man durch die
Längsachse des Seitensprosses senkrecht zu seiner Mediane legen kann,
heißt die Transversalebene des Seitensprosses (vgl.
Fig. 149 A). An dem Seitenzweige nennt man alles vorn (v),
was zwischen seiner Transversalebene und seinem Deckblatt gelegen
ist, hinten (h), was zwischen seiner Transversalebene und
seinem Muttersproß gelegen ist, seitlich rechts, was
sich an ihm rechts, links, was sich an ihm links von seiner
Mediane befindet. Median heißt ferner alles, was am Seitenzweig
in die Mediane, transversal, was in Richtung der Transversalen
fällt, diagonal endlich, was schräg nach vorn oder hinten (also
zwischen der Medianen und der Transversalen) an ihm liegt.
An den Seitenknospen pflegen die untersten Blätter, die
direkt auf das Deckblatt folgen, unabhängig von der Anordnung der
höheren Blätter eine ganz bestimmte Stellung zu dem Deckblatt und
zu der Mutterachse einzunehmen. Sie vermitteln den Anschluß
der Blattstellung des Seitenzweiges zu der am Muttersprosse. Bei den
Monokotylen gibt es ein solches Blatt (Fig. 149 vb), bei
den Dikotylen meist deren zwei von bezeichnender Stellung,
die Vorblätter. Es sind häufig Nieder- oder Hochblätter. Bei
den Monokotylen steht das Vorblatt median an der der Mutterachse
zugekehrten oder hinteren Seite des Zweiges. Man nennt es deshalb
adossiert. Häufig kommen ihm zwei als Kiele bezeichnete
Seitennerven zu, dagegen fehlt der Mittelnerv (Fig. 149 A). Es
dürfte durch Verwachsung zweier seitlicher Vorblätter entstanden
sein[79]. Bei den Dikotylen stehen die beiden Vorblätter (α und β) an
den Achselknospen gewöhnlich rechts und links transversal, worauf die
anderen Blätter oft in abweichenden Stellungen folgen.
Die Seitenknospen können im übrigen die gleiche oder eine andere
Blattstellung wie die Mutterachse zeigen.
Liegt schraubige Blattstellung vor, so ist die Grundspirale bei
manchen Gewächsen an den Seitenknospen gleichläufig (homodrom), bei
anderen gegenläufig (antidrom) zu der an den Muttersprossen.
c) Ausbildung des Sproßverzweigungssystems. Jedes Sproßsystem
erhält sein Aussehen, seinen Habitus, abgesehen von der
Wuchsrichtung seiner Hauptachse, durch die Zahl der Ordnungen von
Seitenachsen, die zur Ausbildung gelangen, durch die Stellung der zu
Seitenzweigen austreibenden Knospen an ihren Mutterachsen sowie
durch die Wachstumsintensität und die Orientierung der Seitenzweige
verschiedener Ordnungen im Verhältnis einerseits zu ihresgleichen
und andererseits zu ihren Mutterachsen. Auch die Verschiedenheiten
im Aussehen der Sproßsysteme lassen oft deutliche Beziehungen zur
Lebensweise der Pflanzen erkennen.
1. Wuchsrichtung der Hauptachse des Sproßsystems. Das
Sproßsystem erhält sein Gepräge zunächst durch die Wuchsrichtung der
Hauptachse.
Erhebt sich die Hauptachse senkrecht vom Boden, so nennt man die
Pflanze aufrecht, den Sproß orthotrop. In diesem
Falle pflegt die Hauptachse bei freiem Wuchse des Systems ihre
mehr oder weniger plagiotropen und dorsiventralen Seitenzweige
in radiärer Verteilung auszubilden. Wächst die Hauptachse
schräg oder horizontal, also plagiotrop, so sind die
Seitenzweige daran meist dorsiventralsymmetrisch angeordnet; bleibt
die Hauptachse samt den Seitenzweigen auf der Oberfläche des Bodens
oder horizontal unter dem Boden, ohne sich zu erheben, so entstehen
kriechende Pflanzen, deren Sprosse meist, und zwar auf ihren
Unterseiten, bewurzelt sind. Bei kriechenden Pflanzen pflegen die
Seitenzweige den Flanken der Sprosse zu entspringen; erheben sich
solche Seitenzweige senkrecht vom Boden, so verhalten sie sich
hinsichtlich ihrer Verzweigung oft wie aufrechte Pflanzen.
2. Sproßfolge. Ist schon der Vegetationspunkt der Hauptachse
(d. h. also der Keimlingsachse) nach entsprechender Erstarkung der
Pflanze zur Bildung der Fortpflanzungsorgane befähigt, so wird die
Pflanze einachsig (haplokaulisch) genannt. Einachsig[S. 109] ist
der Mohn, der schon seinen ersten, aus dem Keim hervorgegangenen
Sproß mit einer Blüte abschließt. Meist kommt aber erst Achsen
zweiter, dritter, vierter oder n-ter Ordnung die Fähigkeit zu,
eine Blüte auszubilden. Alsdann ist die Pflanze zweiachsig
(diplokaulisch), dreiachsig (triplokaulisch) oder n-achsig. Eine
dreiachsige Pflanze ist der große Wegerich, Plantago major, der
an seiner ersten Achse nach den Niederblättern nur Laubblätter,
an den Achsen zweiter Ordnung nur Hochblätter trägt und aus den
Achseln der letzteren die mit Blüten abschließenden Achsen dritter
Ordnung erzeugt. An unseren Bäumen sind erst Sprosse n-ter Ordnung
befähigt, Blüten zu bilden. In den meisten Verzweigungssystemen
gibt es viele Seitensprosse, die sich nicht bis zu den
blütenbildenden Ordnungen weiter verzweigen. Entweder treten sie
als Bereicherungssprosse auf, so bei vielen einjährigen
Pflanzen, oder sie entfalten sich als Erneuerungs- oder
Innovationssprosse alljährlich am Pflanzenstock, wie bei den
mehrjährigen Gewächsen. So pflegt in der Sproßordnung verzweigter
Pflanzen eine Arbeitsteilung zwischen den Seitensprossen
vorzukommen, die sich oft auch in der Stellung und in der
Ausgestaltung der Seitensprosse zu erkennen gibt; die Seitensprosse
sehen verschieden aus, je nachdem sie vornehmlich im Dienste der
Ernährung, der Speicherung oder der Fortpflanzung stehen.
3. Stellungen der austreibenden Knospen. Nur selten treiben
sämtliche Seitenknospen, die an einer Mutterachse entstanden sind,
sofort aus und werden zu Seitensprossen; das ist z. B. bei Kräutern der
Fall. Regel ist indes, daß viel mehr Seitenknospen angelegt werden, als
zur Entfaltung kommen, daß also nur ein ganz kleiner Teil zu Sprossen
auswächst. Die übrigen bleiben ruhende, schlafende Augen oder
verkümmern frühzeitig. Entfaltung aller Knospen wäre für die Pflanze
eine ganz unnötige, ja sogar schädliche Materialverschwendung. Die
Zweige würden sich gegenseitig so stark beschatten, daß ein Teil
absterben müßte.
Fast jeder Baum besitzt, namentlich im unteren Teile seiner
Jahrestriebe, solche „schlummernde Augen“, die kürzere oder längere
Zeit entwicklungsfähig bleiben und zur Entfaltung nur unter
besonderen Bedingungen gelangen. Bei der Eiche, Rotbuche u. a.
können schlummernde Knospen bis 100 Jahre alt werden. Vielfach sind
es daher Sprosse aus solchen Knospen und nicht Adventivsprosse, die
aus alten Stämmen hervorbrechen.
Das Austreiben der Seitenknospen kann regellos stattfinden oder
bestimmten Regeln unterworfen sein, akropetal oder basipetal erfolgen.
An reich verzweigten Sproßsystemen sind die peripheren bevorzugt; denn
hier besteht die größte Aussicht, die Blätter in günstiges Licht zu
bringen.
Fast alle einheimischen Bäume beschränken sich während einer
Vegetationsperiode darauf, die im Frühling aus den Winterknospen
hervorgegangenen Zweige an ihren Spitzen zu verlängern und ruhende
Knospen an diesen Ästen auszubilden. Meist erst bei Beginn einer
neuen Wuchsperiode lassen sie alsdann auf einmal Seitenzweige
aus den obersten Knospen, die im vergangenen Jahre an den Ästen
angelegt wurden, hervorgehen; etwa in einem echten oder Scheinquirl
(Araukarie, Tanne) oder meist so, daß die obersten Seitenknospen
zu Langtrieben, einige darunter befindliche zu Kurztrieben (Birne,
Apfel) werden. Bei anderen, namentlich aufrechten Sprossen treibt
von allen daran angelegten Knospen in streng gesetzmäßiger Weise
jede zweite oder dritte oder vierte usw. aus und zwar so, daß die
austreibenden Zweige seitlich oder longitudinal gleichen Abstand
voneinander erhalten.
Die Verteilung der austreibenden Knospen, ob wechselständig
oder quirlständig, bewirkt Unterschiede im Aussehen der
Verzweigungssysteme. Bei gegenständiger Stellung der Knospen kommt
eine Art unechte Gabelverzweigung zustande, wie bei der Roßkastanie
und dem Flieder.
4. Richtung und Wachstumsintensität der Seitenzweige im Verhältnis
zu ihresgleichen. Die seitlichen Winkel, die an orthotropen Ästen
die Längsachsen benachbarter Seitenzweige gleicher Ordnung
miteinander bilden, können bei einer Pflanzenart ziemlich beständig
sein (z. B. bei der Araukarie, Tanne).
[S. 110]
Dagegen ist die Wachstumsintensität der Seitenachsen an einer
Mutterachse oft recht verschieden. Oft bildet sich nämlich nur ein
Teil der Zweige als Langtriebe, der Rest zu gestauchten
Kurztrieben aus, vielfach als Ausdruck einer Arbeitsteilung
zwischen den Seitenzweigen. Die Kurztriebe haben meist kürzere
Lebensdauer, pflegen sich nicht zu verzweigen und nehmen bei Bäumen
am Aufbau des bleibenden Gerüstes keinen Anteil: so bei der Lärche,
die die Kurztriebe in Gestalt dichter Nadelbüschel an ihren älteren
Langtrieben trägt, und bei den Kiefern.
5. Richtung und Wachstumsintensität der Seitenzweige im Verhältnis
zu ihren Mutterachsen. Verschiedene Arten seitlicher Verzweigung.
Auch die Neigungswinkel der Seitenachsen zu ihrer Mutterachse pflegen
im allgemeinen bei einer Spezies ziemlich konstant, also für die Art
bezeichnend zu sein. Sie sind meist kleiner, selten größer als 90°.
Die Seitenachsen, die an einer Mutterachse entstanden sind, können
weniger intensiv als diese oder ebenso schnell, oft aber auch viel
schneller als die Mutterachse wachsen. Im letzten Fall sind sie also
gegenüber der Mutterachse gefördert. Die Mutterachse kann sogar
nach der Bildung von Seitenzweigen ihr Wachstum ganz einstellen und
einem oder mehreren Seitenzweigen die Ausbildung neuer Seitenzweige,
die Fortsetzung der Verzweigung überlassen. Es leuchtet ein, daß
die entstehenden Verzweigungssysteme durch solche Unterschiede im
Wachstum der Tochter- und Mutterachsen völlig verschiedenes Aussehen
erhalten müssen. Diese Differenzen haben Anlaß zur Unterscheidung
verschiedener Arten seitlicher Verzweigung gegeben, deren Kenntnis
für das Verständnis des morphologischen Aufbaues der höheren Pflanzen
unerläßlich ist. Besonders leicht kann man ihre Unterschiede an den
Blütenständen oder Infloreszenzen der Samenpflanzen
beobachten (vgl. speziellen Teil). Bezeichnend für viele Infloreszenzen
ist nämlich, daß die Achselknospen aller Hochblätter austreiben;
dadurch werden die Infloreszenzen im Gegensatze zu den vegetativen
Sproßsystemen zu außerordentlich dichten Zweigsystemen.
a) Wächst die Hauptachse stärker als die Seitenachsen I. Ordnung,
diese stärker als die an ihnen entstehenden Seitenzweige II.
Ordnung usw., oder wachsen die jeweiligen Mutterachsen ebenso
stark wie ihre Tochterachsen, so spricht man von razemöser
Verzweigung. Im ersteren Falle geht eine echte Hauptachse
(ein Monopodium) durch das ganze Verzweigungssystem hindurch
(vgl. das auch hierfür gültige Schema, Fig. 82 b). Diese typisch
monopodiale Verzweigung ist z. B. bei der Tanne und anderen
Koniferen mit pyramidenförmigen Gesamtumrissen ausgebildet: der radiäre
Hauptsproß wächst unter dem Einflusse der Schwerkraft (vgl. S. 299)
senkrecht nach oben, orthotrop; die meist dorsiventralen Seitenzweige
I. Ordnung strahlen in horizontaler oder schräger Richtung vom
Hauptsprosse allseitig aus. Wachsen auch die Seitenzweige I. Ordnung
steil aufrecht, wie z. B. bei der Zypresse und bei vielen Sträuchern,
so ist oft kein deutlicher Längenunterschied zwischen der Hauptachse
und den Seitenachsen I. Ordnung vorhanden. Das Verzweigungssystem hat
in diesem Falle ovalen oder runden Umriß.
b) Erlischt das Wachstum in den Mutterachsen rasch und geht es auf die
Tochtersprosse über, so liegen zymöse Verzweigungen vor. Sie
sehen verschieden aus, je nachdem mehrere gleichmäßig wachsende
Seitenachsen gleicher Ordnung oder nur eine Seitenachse
das Verzweigungssystem fortsetzen. Im letzteren Falle ist eine
scheinbare Hauptachse, ein Sympodium, ausgebildet.
Bei vielen zymösen Verzweigungen wachsen die jeweiligen
Mutterachsen nicht nur langsamer als die Tochterachsen, sondern
ihre Spitzen sterben sogar ab oder werden abgeworfen, wie bei
vielen unserer Laubbäume, z. B. den Weiden, der Linde.
[S. 111]
I. Setzen mehr als zwei Seitenzweige gleicher Ordnung die
Verzweigung fort, so spricht man von Pleiochasium. Diese
Seitenzweige pflegen dem oberen Ende ihres Muttersprosses genähert
zu sein und allseits, bei manchen Pflanzen quirlartig, schräg nach
außen in den Raum zu strahlen (z. B. Verzweigung von Euphorbia).
II. Setzen zwei Seitenzweige gleicher Ordnung, die in
spitzen oder rechten Winkeln einander gegenüber zu stehen pflegen,
die Verzweigung fort, so entsteht ein Dichasium. Schematisch
zeigt es Fig. 150 (vgl. dazu die dichasiale Infloreszenz Fig.
536). Die Seitenzweige breiten sich aber nicht, wie in dem Schema,
in einer Ebene aus, sondern allseits im Raume.
Dies wird dadurch erreicht, daß die Verzweigungsebenen in den
aufeinanderfolgenden Seitenzweigordnungen nicht zusammenfallen,
sondern rechte Winkel miteinander bilden. So kann nur der Grundriß
(Fig. 152 E) Aufschluß über die wahre Anordnung der Zweige des
Sproßsystems geben. Ein solches Verzweigungssystem, das z. B. auch
bei der auf unseren Bäumen wachsenden Mistel vorkommt, kann den
Anschein einer Dichotomie erwecken.
III. Setzt immer bloß ein Seitenzweig die Verzweigung
fort, so liegt ein Monochasium vor. Oft stellt sich dieser
Seitenzweig in die Verlängerung seines Muttersprosses, indem er
dessen Spitze zur Seite drängt (Fig. 151). So entsteht ähnlich, wie
es bei der dichotomen Verzweigung des Sprosses der Fall sein kann
(S. 104 ff.), ein Verzweigungssystem mit einer Scheinachse
(Sympodium), die sich aus Seitensprossen verschiedener
Ordnungen zusammensetzt. Ein solches Verzweigungssystem kann
einem monopodialen sehr ähnlich sehen, namentlich wenn die
Scheinachse, wie so oft, senkrecht emporwächst, die im
Wachstume zurückbleibenden Enden der Zweige, die die Scheinachse
zusammensetzen, dagegen Seitenzweigen ähnlich sich horizontal
oder schräg stellen. Von Seitenzweigen unterscheiden sich
solche Zweige aber stets dadurch, daß ein Deckblatt an ihrer
Basis fehlt, dafür aber ein Blatt ihnen gegenüber am Sympodium
befestigt ist, nämlich das Deckblatt des geförderten
Tochtersprosses (vgl. dazu Fig. 151). Nicht selten ist das
Sympodium weiter sympodial verzweigt. Stämme und Äste vieler
unserer Laubhölzer sind solche Sympodien, so bei der Linde oder
der Rotbuche. An ihrem Stamme und an ihren Ästen ist aber von dem
sympodialen Aufbaue nichts mehr zu erkennen. Dauernd erkennbar
bleibt dagegen der sympodiale Aufbau vielfach an unterirdischen
Stengelteilen, so an denen von Polygonatum multiflorum (Fig.
143). Jedes Jahr erhebt sich die jeweilige Endknospe dieses
unterirdischen Stammes als Sproß über den Boden, während eine
Achselknospe das Rhizom im Boden fortsetzt.
Fig. 150. Schema des Dichasiums. H Keimlingsachse,
1, 2, 3 Tochterachsen 1., 2., 3. Ordnung.
Fig. 151. Schema des Monochasiums vgl. Fig. 152.
Je nach der Stellung der Seitensprosse verschiedener Ordnung
zueinander entstehen monochasiale Verzweigungssysteme von sehr
verschiedenem und sehr bezeichnendem Aufbaue. Sehr oft setzt sich
die Verzweigung schon aus der Achsel eines Vorblattes fort.
A. Entweder fallen die Medianen aller Seitensprosse in
eine und dieselbe Ebene, nämlich in die Medianebene des
Seitensprosses I. Ordnung; sie stehen also median.
α) Alle aufeinanderfolgenden Seitenzweige fallen median nach
vorn von ihren Mutterachsen, d. h. zwischen die Mutterachse
und das Deckblatt der Mutterachse (vgl. S. 107 ff.), in der
Seitenansicht des Verzweigungssystems also auf ein und dieselbe
Seite: Sichel (Fig. 152 C, D).
β) Die aufeinanderfolgenden Seitenzweige fallen sämtlich median
nach hinten von ihren Mutterachsen (vgl. S. 107
ff.), in der Seitenansicht also abwechselnd nach links und rechts:
Fächel (Fig. 152 A, B).
B. Die Mediane jedes Seitensprosses (I., II., III. usw.
Ordnung) steht immer transversal, d. h. seitlich rechts oder
links zur Mediane des Deckblattes für seinen Muttersproß.
Solche Verzweigungssysteme lassen sich natürlich nur in Grundrissen
veranschaulichen.
α) Die aufeinanderfolgenden Seitenachsen stehen stets
nach der gleichen Seite transversal zu den Medianen ihrer
Muttersproßdeckblätter, entweder nach rechts oder
nach links: Schraubel (Fig. 152 F).
[S. 112]
β) Die aufeinanderfolgenden Seitenachsen stehen
abwechselnd nach rechts und links transversal zu den
Medianen ihrer jeweiligen Muttersproßdeckblätter: Wickel
(Fig. 152 G).
Schraubel und Wickel lassen sich aus dem Grundriß des Dichasiums
leicht ableiten (Fig. 152 E) und dadurch in ihrer Eigenart
verstehen.
Fig. 152. A Fächel im Seitenriß, B in Grundansicht.
C Sichel im Seitenriß, D in Grundansicht. E Dichasium in
Grundansicht und Ableitung der Wickel (blaue Linie) und Schraubel (rote
Linie) daraus. F Grundansicht der Schraubel. G Grundansicht der
Wickel. 1–9 Ordnungen aufeinanderfolgender Tochterachsen. (A–D
nach EICHLER, das übrige nach KARSTEN, verändert.)
Um das Verständnis zu erleichtern, sind in A–D und F, G die
aufeinanderfolgenden Tochterachsen je nächst niederer Ordnungen
mit verschiedenen Farben wiedergegeben. Das an jeder Tochterachse
befestigte Deckblatt hat die gleiche Farbe erhalten wie diese Achse, an
der es entstanden ist.
In einem Sproßsysteme sind nicht selten verschiedene
Verzweigungsarten miteinander verbunden. So können etwa auf dem
razemös verzweigten Keimsproß zymös verzweigte Seitensprosse
folgen. In besonders mannigfaltiger und wechselnder Verbindung sind
die verschiedenen Verzweigungsarten bei den Blütenständen vereinigt
(vgl. speziellen Teil).
b) Die Wurzel[80].
Den Wurzeln, die meist in der Erde (Erdwurzeln), seltener in
der Luft (Luftwurzeln) leben, fehlen immer die Blätter. Dadurch
haben sie ein ganz anderes Aussehen als die Sprosse, selbst als die
Erdsprosse. Ihre Aufgabe ist, die Pflanze im Boden zu befestigen,
aus dem Boden Wasser und Bodensalze aufzunehmen und zum Sproßsystem
hinzuleiten. Auch ihre Funktionen sind also völlig andere als bei den
meisten Sprossen, die hauptsächlich der Kohlensäureassimilation dienen.
1. Vegetationspunkt. Die Wurzel verlängert sich an der Spitze.
Sie zeigt Scheitelwachstum mittels eines kegelförmigen
Vegetationspunktes. Der Vegetationskegel bedarf für seine
dünnwandigen embryonalen Zellen eines besonderen Schutzes; denn
er wird bei dem Wachstum der Wurzel wie[S. 113] ein Nagel zwischen die
scharfkantigen Bodenpartikelchen vorwärts getrieben. Dieser Schutz wird
von einem besonderen Organe aus parenchymatischen Dauerzellen, der
Wurzelhaube oder Kalyptra, übernommen, die die Spitze
der Wurzel wie ein Däumling den Finger umhüllt, so daß der eigentliche
Vegetationspunkt im Innern des Gewebes der Wurzelspitze, also
interkalar, liegt. Die Verschleimung der äußeren Zellmembranen der
Haube erleichtert zugleich bei den Erdwurzeln das Vorwärtsdringen im
Boden. Die Wurzelhaube sieht man meist erst auf medianen Längsschnitten
durch die Wurzelspitzen (Fig. 153, 154); doch gibt es auch Fälle
(Pandanus), wo man sie schon an der unversehrten Wurzel als eine ihren
Scheitel deckende Kappe wahrnehmen kann.
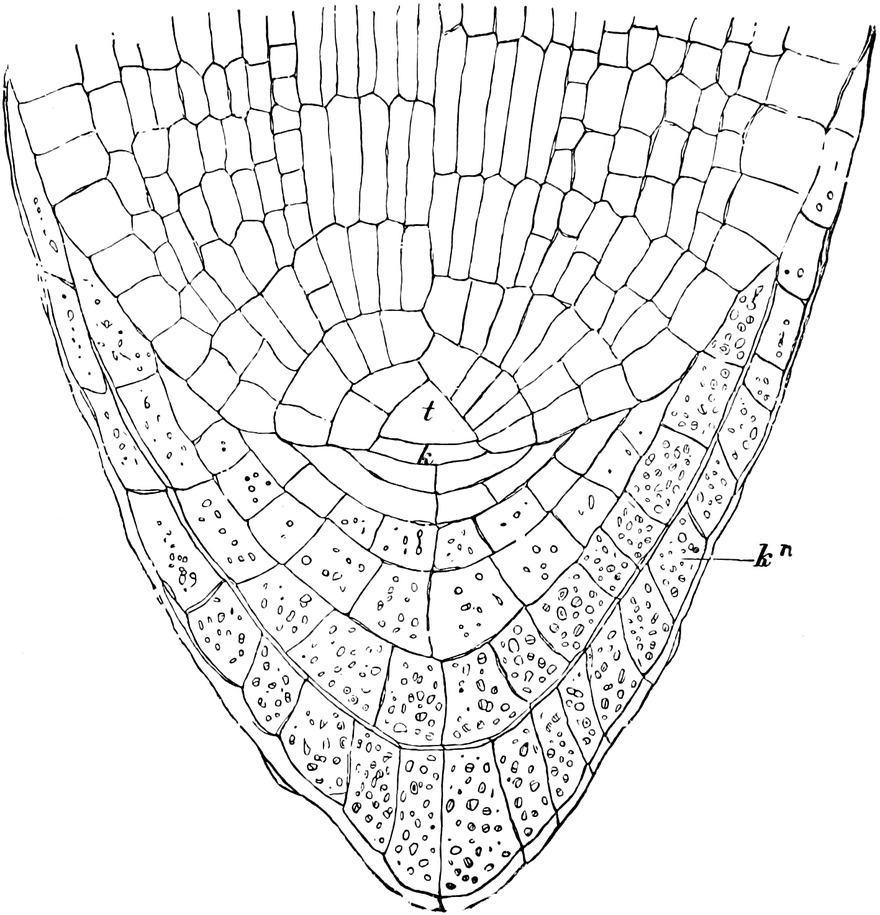
Fig. 153. Medianer Längsschnitt durch die Wurzel des
Farnes Pteridium creticum. t Scheitelzelle, k Haubeninitiale, kn
Wurzelhaube. Vergr. 240. Nach STRASBURGER.
Die besonders auffälligen Kappen an den Enden der Wasserwurzeln
unserer Wasserlinsen (Lemna-Arten) und mancher Hydrocharitaceen
gehören dagegen ihrem Ursprung nach nicht zur Wurzel; sie bilden
sich vielmehr aus einer die Wurzelanlage umgebenden Hülle, etwa
aus Sproßgewebe, und werden demgemäß als Wurzeltaschen
bezeichnet. Wurzeln ohne Wurzelhaube sind eine sehr seltene
Erscheinung; sie kommen z. B. vor bei den eben erwähnten
Wasserlinsen, bei denen die Wurzeltasche die Funktionen der
Wurzelhaube übernimmt. Ebenso fehlt eine Wurzelhaube der rasch
absterbenden Wurzel des parasitisch lebenden Teufelszwirns (vgl. S.
162).
In Zeiten der Ruhe, wo die Erdwurzeln nicht weiterwachsen, werden
die Wurzelhauben verkorkt; ihre parenchymatischen Zellen wandeln
sich in Kutisgewebe um, das die Wurzelspitzen wirksam nach außen
abschließt.[81]
Der Vegetationspunkt der Wurzel wird, wie schon gesagt, aus
Meristemzellen aufgebaut, von denen sich die basalwärts gelegenen in
Dauerzellen des Wurzelkörpers, die an der Spitze in die Dauerzellen der
Wurzelhaube umwandeln.
[S. 114]
Bei den meisten Pteridophyten haben die Wurzeln wie die Sprosse eine
dreischneidige Scheitelzelle (t Fig. 153) von der Gestalt einer
dreiseitigen Pyramide.
Außer den Segmenten, die sie parallel zu ihren drei inneren
Seitenwänden nach dem Wurzelkörper hin abgibt, bildet sie solche
auch nach außen (k). Letztere bauen die Wurzelhaube auf, indem
sie sich weiter teilen.
Die Vegetationspunkte der Phanerogamenwurzeln besitzen dagegen keine
Scheitelzellen. Sie bestehen aus gleichwertigen embryonalen Zellen, die
oft in regelmäßigen Schichten angeordnet sind.
Als Beispiel sei auf den Vegetationskegel einer Gramineen-Wurzel
(Fig. 154) hingewiesen: Die Meristemschichten, die das Dauergewebe
des Wurzelkörpers liefern, sondern sich hier in eine äußere
Zellschicht, das Dermatogen (d), in mehrere zentrale
Schichten, die einen Gewebestrang für sich bilden und sämtlich
oder teilweise in den Zentralzylinder der Wurzel übergehen,
das Plerom (pl), und in mehrere zwischen Dermatogen
und Plerom gelegene Schichten, das Periblem. Die
Schichten des Dermatogens (d Fig. 154) und Periblems (pr)
vereinigen sich am Scheitel zu einer einzigen Zellschicht.
Außerhalb davon liegt die Zellschicht, die die Wurzelhaube
bildet, das Kalyptrogen (k).
Bei vielen anderen Wurzeln (bei der Mehrzahl der Dikotylen)
wird dagegen die Wurzelhaube durch Vermehrung der Schichten des
Dermatogens gebildet, das auch an der Spitze des Scheitels vom
Periblem getrennt sein kann; bei manchen nimmt auch das Periblem,
ja unter Umständen selbst das Plerom an der Bildung der Haube teil,
so bei vielen Leguminosen und bei den Gymnospermen. Bei diesen sind
Periblem, Dermatogen und Kalyptrogen am Scheitel überhaupt nicht
gesondert; der Pleromzylinder schließt aber fast stets mit deutlich
abgegrenzten Initialen ab.
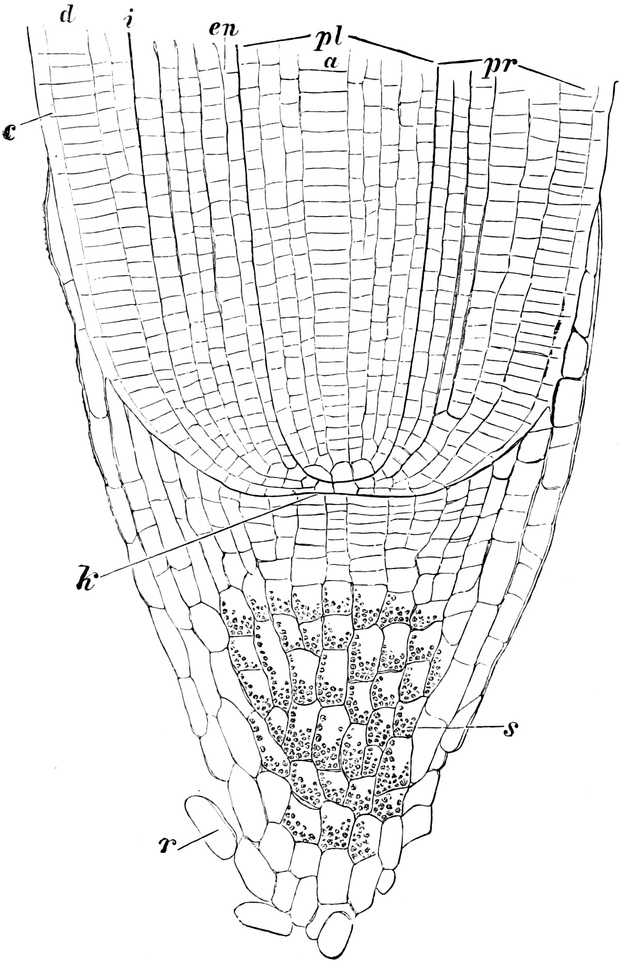
Fig. 154. Medianer Längsschnitt durch die Wurzelspitze
der Gerste. k Kalyptrogen, d Dermatogen, c dessen verdickte
Außenwand, pr Periblem, pl Plerom, en Endodermis, i mit Luft
sich füllende Interzellularen, a Zellreihe, aus der das zentrale
Gefäß hervorgehen wird, r abgestoßene Zellen der Wurzelhaube, s
große, leicht bewegliche Stärkekörner in den Haubenzellen. Vergr. 180.
Nach STRASBURGER u. KOERNICKE.
2. Äußerer Bau des Wurzelkörpers. Die embryonalen Zellen wandeln
sich an der Basis des Vegetationskegels unter starker Größenzunahme
allmählich in Dauerzellen um. Hiermit geht eine ausgiebige Verlängerung
des Wurzelkörpers Hand in Hand. Durch dieses Streckungswachstum, das
also erst hinter dem Vegetationskegel[S. 115] einsetzt und bei den Erdwurzeln
im Gegensatze zu den Luftsprossen auf eine sehr kurze, höchstens 5–10
mm lange Zone dicht hinter dem Vegetationspunkt beschränkt ist, wird
die Wurzel zu einem zylindrischen, fadenförmigen, farblosen Gebilde.
Bei Luftwurzeln kann die Streckungszone aber viele Zentimeter lang
werden. Ihre geringe Länge bei den Erdwurzeln hängt offenbar mit
der Lebensweise dieser Wurzeln im Boden zusammen.
In einiger Entfernung von der Wurzelspitze, etwa da, wo das
Streckungswachstum erlischt, entstehen an den Erdwurzeln wichtige
Anhangsgebilde der Wurzeln, die Wurzelhaare[82] (r in Fig.
155 und Fig. 51): lokale schlauchförmige, sehr dünnwandige und
schleimüberzogene Ausstülpungen der lebenden Epidermiszellen. Besonders
an Keimpflanzen, die in feuchtem Raume kultiviert werden, z. B. vom
Weizen, kann man sie in ungeheuer großer Menge (bei Zea mays etwa
420 pro qmm) mit bloßem Auge als zarten Flaum auf der Oberfläche der
Wurzeln gut erkennen. Ihre Länge schwankt je nach den Pflanzenarten
zwischen 0,15 und 8 mm. Sie vergrößern in sehr wirksamer Weise die
Oberfläche der Wurzeln (bei Pisum z. B. um das zwölffache). Diese Haare
dringen zwischen die Bodenpartikelchen ein, ja verwachsen sogar mit
ihnen. Im Boden bewahren sie infolgedessen nicht Zylinderform, wie in
feuchter Luft, sind vielmehr hin und her gekrümmt und an der Spitze
abgeplattet, keulig oder lappig (Fig. 237). Sie dienen der Wasser- und
Bodensalzaufnahme, haben aber nur wenige Tage Lebensdauer. In dem Maße,
wie spitzenwärts neue Wurzelhaare hinzukommen, sterben die älteren ab,
so daß immer nur ein begrenzter Teil der jungen Wurzel (einige Zenti-
oder Millimeter) von ihnen bedeckt ist. Der ältere kahle Teil dient
lediglich noch der Leitung, nicht mehr der Aufnahme von Wasser. An ihm
ist sehr häufig eine Querrunzelung der Oberfläche erkennbar, die durch
eine nachträgliche Kontraktion dieser Wurzelteile bedingt wird.
Durch diese Kontraktion verkürzt und spannt sich die Wurzel wie ein
gespannter Bindfaden straff und verankert den Sproß wesentlich fester
im Boden (vgl. Fig. 205, 6).
Manchen Pflanzen fehlen die Wurzelhaare, vor allem solchen, die
besonders leicht Wasser aufnehmen können, so vielen Wasser- und
Sumpfpflanzen. Die Wurzeln mancher Wasserpflanzen, z. B. von Nuphar
luteum, bilden aber dann Haare, wenn sie in den Boden eindringen;
die Wurzeln von Sumpfpflanzen, wie Carex paludosa, wenn es an
Wasser fehlt. Bei gewissen Wasserpflanzen, z. B. bei Hydrocharis,
tragen aber auch die Wasserwurzeln reichlich Wurzelhaare.
Fig. 155. Keimpflanze der Hainbuche (Carpinus betulus).
h Hypokotyl, c Kotyledonen, hw Hauptwurzel, sw Seitenwurzeln,
r Wurzelhaare, e Epikotyl, l und l′ Laubblätter.
Nat. Gr. Nach NOLL.
3. Primärer innerer Bau der Wurzel. Haben sich die embryonalen
Zellen des Vegetationspunktes in Dauerzellen umgewandelt, so sind in
der Wurzel die gleichen Gewebearten wie in der Sproßachse, und zwar
auch meist in radiär symmetrischer Anordnung gesondert.
Der äußere Abschluß wird an jüngeren Teilen der Wurzel durch die
dünnwandige Epidermis gebildet, die samt ihren Anhangsgebilden,
den Wurzelhaaren, der Stoffaufnahme dient. Bezeichnend für sie ist
der[S. 116] Mangel der Spaltöffnungen und der Kutikula. Die Epidermis
der Wurzel stirbt aber mit den Wurzelhaaren bald ab. Wo letztere
verschwunden sind, nimmt ein Kutisgewebe, nämlich die äußerste
Rindenschicht, deren Zellwände mehr oder weniger verkorken, als
Exodermis (Fig. 156 cx) die Oberfläche ein[83].
In einer solchen Exodermis werden oft unverkorkte Zellen
ausgespart, die als Durchlaßzellen gelten. Sie sind in bestimmten
Fällen regelmäßig zwischen den verkorkten verteilt und kleiner als
diese.
Das übrige Gewebe kann man wieder in Rinde und Zentralzylinder
einteilen.
Die Rinde der Erdwurzeln wird von farblosem Gewebe gebildet,
das fast immer parenchymatisch ist und nur in den inneren Teilen
Interzellularen enthält. In manchen Fällen erweitern sich die letzteren
zu Luftlücken oder Luftkanälen. In der Rinde vieler Luftwurzeln kommt
dagegen Chlorophyll vor. Manchmal werden Festigungsgewebe ausgebildet
(Fig. 159, 2), die die Epidermis oder Exodermis verstärken und die
Wurzeln biegungs- und druckfest machen. Die innerste Rindenschicht
pflegt eine Endodermis[84] zu sein (Fig. 156 e, 157 e,
158 S, 160 s), die eine scharfe Grenze zwischen Rinde und
Zentralzylinder zieht. Sie besteht aus rechteckig prismatischen,
gewöhnlich etwas gestreckten Zellen, die auf Querschnitten die dunklen
CASPARYschen Punkte in den radialen Wänden zeigen. Durch
diese Membranstreifen (vgl. S. 49) scheint ein gewisser Abschluß des
Zentralzylinders gegen die primäre Rinde erreicht zu werden; die
tangentialen Wände der jungen Endodermis aber erlauben dem Wasser den
Durchtritt aus der Rinde in den Zentralzylinder und umgekehrt.
Fig. 156. Querschnitt durch eine Wurzel der
Küchenzwiebel (Allium Cepa), ep Reste der Epidermis, ex Exodermis,
c Rinde, e Endodermis, cc Zentralzylinder. Vergr. 45. Nach M.
KOERNICKE.
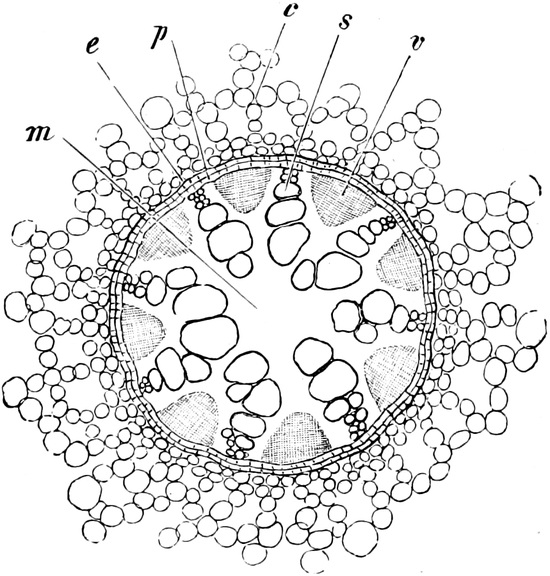
Fig. 157. Querschnitt durch die Wurzel von Acorus
Calamus. m Mark, s Gefäßstränge, v Siebstränge, p Perizykel,
e Endodermis, c Rinde. Vergr. 90. Nach STRASBURGER.
In etwas älteren Wurzelteilen werden die Endodermiszellen durch
Suberinlamellen verkorkt und, bei vielen Monokotylen, außerdem
durch tertiäre Verdickungsschichten meist einseitig nach dem
Zentralzylinder zu verdickt (Fig. 160). Treten solche Veränderungen
frühzeitig auf, so bleiben bestimmte, vor den Gefäßsträngen des
Leitbündels gelegene Endodermiszellen, die Durchlaßzellen, davon
ausgeschlossen (d Fig. 160).
Die äußerste parenchymatische Zellschicht des Zentralzylinders
der Wurzeln (Fig. 157 p, 158 pc, 160 p), also die Schicht
direkt unter der Endodermis, bildet den Perizykel, der
meist einschichtig ist, in seltenen Fällen auch fehlen kann. Im
Zentralzylinder verlaufen in gerader Längsrichtung die[S. 117] Leitungsbahnen,
die als Gefäß- und als Siebstränge ausgebildet sind und bei allen
Wurzeln ein radiales Leitbündel[61] bilden (vgl. S. 85). Die Gefäß-
und Siebstränge sind also in das übrige Gewebe des Zentralzylinders
radial nebeneinander und zwar so eingebettet, daß sie
voneinander durch eine bis mehrere Zellschichten (meist Leitparenchym)
getrennt bleiben. Die plattenförmigen Gefäßstränge sind in der Wurzel
umgekehrt wie in den kollateralen Leitbündeln des Stengels orientiert:
im Stengel haben sie ihre engsten Gefäße innen, die weitesten außen, in
der Wurzel dagegen die weitesten Gefäße innen, die engsten am Umkreis
des Leitbündels. Von außen nach innen folgen also Ring-, Schrauben-,
Netz- und Tüpfelgefäße aufeinander. Auch die Phloëmprimanen liegen
peripher, am Außenrande der Siebstränge, die rundlichen Querschnitt
haben. Nach der Zahl der vorhandenen Xylemstränge wird die Wurzel
als diarch, triarch usw., schließlich als polyarch bezeichnet. So
ist die in Fig. 157 dargestellte Wurzel oktarch, die der Fig. 160
pentarch. Die Gefäßstränge stoßen in der Mitte der Wurzel entweder
zusammen, wie in Fig. 158 u. 160; oder es ist dort, wie die Fig.
157 zeigt, ein zentraler Strang aus Parenchym oder Sklerenchym, oft
auch aus beidem vorhanden. Die meisten Wurzeln werden vor allem auf
Zugfestigkeit in Anspruch genommen. So ist das Festigungsgewebe
hauptsächlich in das Zentrum der Wurzel verlegt, wo es durch seine
geschlossene Masse die Wurzeln am besten vor Zerreißung schützt (Fig.
159).
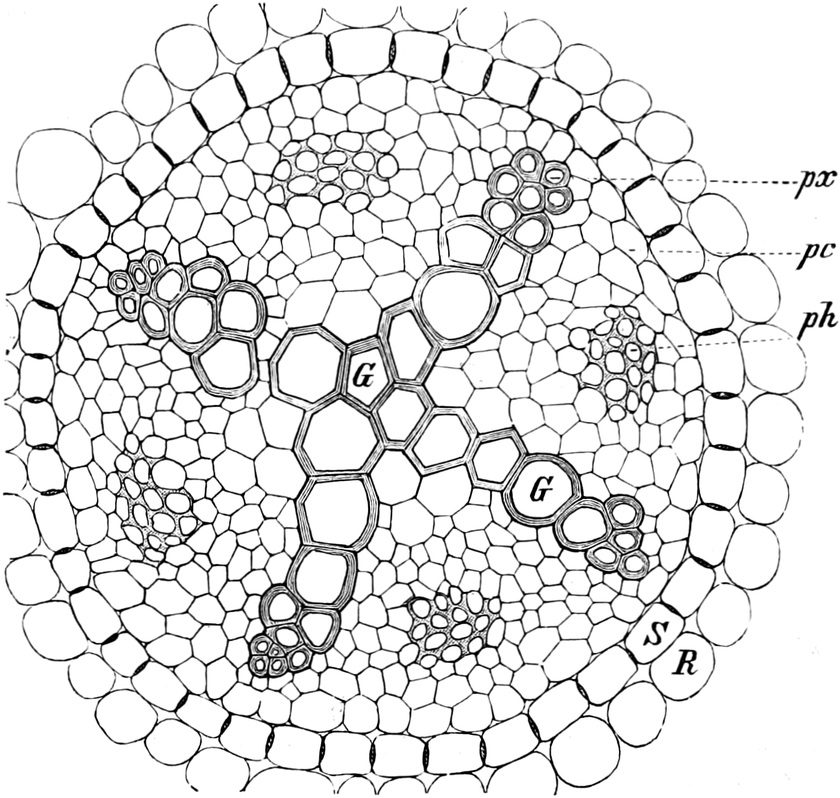
Fig. 158. Querschnitt durch das radiale Leitbündel der
Wurzel von Ranunculus acer. R Rindenparenchym, S Endodermis, pc
Perizykel, ph Phloëm, px Xylemprimanen, G Tüpfelgefäße. Vergr.
200. ROTHERT frei nach DIPPEL.
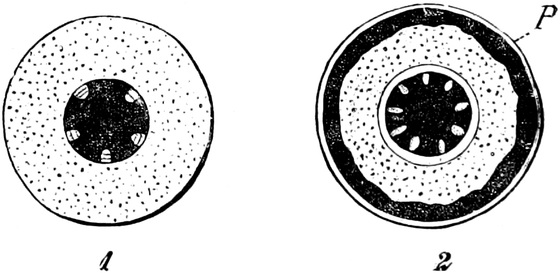
Fig. 159. Mechanisches Gewebe der Wurzel. 1 Zentral
angeordnet für Zugfestigung. 2 Neben dem zentralen Strang ein
peripherer Mantel P für Druck- und Biegungsfestigung (Stützwurzel).
Nach NOLL.
Für ein Organ, das zugfest sein soll, ist es an und für sich
ziemlich gleichgültig, wo auf dem Querschnitt die Festigungsmassen
liegen. Immerhin ist ihre Vereinigung im Zentrum zu einem einzigen
Strang jeder anderen Anordnung überlegen. Wären nämlich statt
dessen z. B. viele entsprechend dünnere Stränge an der Peripherie
verteilt, so würden einzelne bei einem einseitigen Zuge der Gefahr
der Zerreißung ausgesetzt sein.
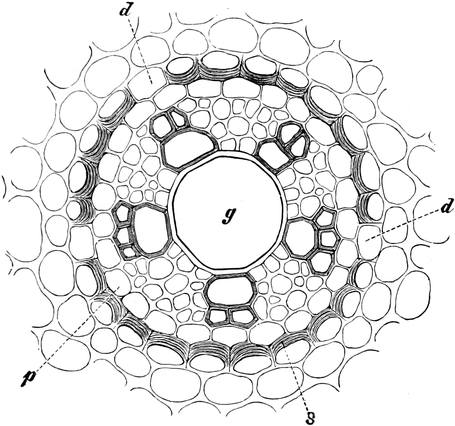
Fig. 160. Radiales Leitbündel der Wurzel von
Allium ascalonicum. s Endodermis mit verdickten Innenwänden,
d Durchlaßzellen, p Perizykel, g großes zentrales Gefäß.
ROTHERT nach HABERLANDT.
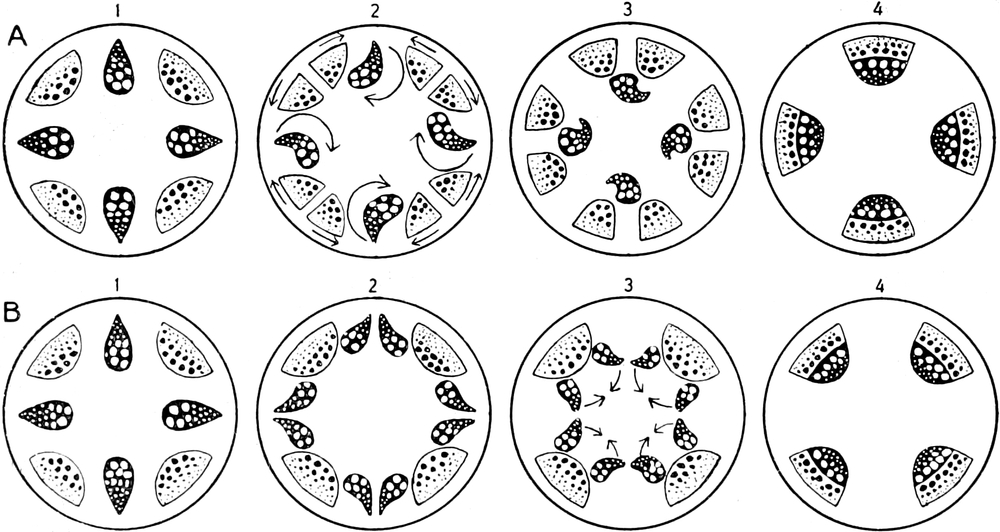
Fig. 161. Übergang des radialen Wurzelleitbündels in
die kollateralen Stengelbündel, schematisiert: nach Beschreibungen
VAN TIEGHEMs und CHAUVEAUDs. Weiß: Rinde und
Mark. Schwarz punktiert: Phloëm. Weiß punktiert auf Schwarz: Xylem.
Weitere Erklärung im Text.
Es bleibt noch zu erörtern, wie die Sieb- und Gefäßstränge des
radialen Wurzelbündels in die Sieb- und Gefäßstränge der anders
gebauten Stengelbündel übergehen[85].[S. 118] Das geschieht meist an der
Grenze der Keimwurzel und des Keimstengels; es sei nur für den
häufigsten Fall, für Pflanzen mit kollateralen Stengelbündeln,
kurz beschrieben. Das Wesentliche dieses Überganges besteht darin,
daß die Gefäßstränge des radialen Wurzelbündels sich in der
Übergangszone je um ihre eigene Längsachse, und zwar um annähernd
180° drehen (vgl. Fig. 161); dadurch werden aus den exarchen
Gefäßsträngen der Wurzel (vgl. S. 90) die endarchen Stränge
des Stengels. Die ältesten Xylemprimanen der Wurzel beteiligen
sich nach CHAUVEAUD an dieser Bewegung nicht, sondern
werden aufgelöst. Aus dem radialen Wurzelbündel werden nun
dadurch eine Anzahl kollateraler Bündel gebildet, daß die Sieb-
und Gefäßstränge, die in der Wurzel nebeneinander liegen, sich
gegeneinander verlagern. Diese Verlagerung vollzieht sich bei den
verschiedenen Arten verschieden. Zwei Haupttypen lassen sich nach
VAN TIEGHEM unterscheiden: 1. Die Gefäßstränge
verlaufen, abgesehen von ihrer Drehung, geradlinig aus der Wurzel
in den Stengel; die Siebstränge der Wurzel aber spalten sich durch
einen radialen Spalt je in zwei Hälften; die Hälften weichen
tangential auseinander und legen sich vor die benachbarten
gedrehten Gefäßteile, wo sie sich je mit der Hälfte des nächst
benachbarten Siebstranges vereinigen (Fig. 161 A). 2. Die
Siebstränge der Wurzel verlaufen geradlinig in den Stengel;
aber die Gefäßteile (Fig. 161 B) spalten sich durch einen
radialen Spalt in je zwei Hälften; diese Hälften drehen sich darauf
je um 180° mit den Primanen als Drehpunkt, weichen, ähnlich wie bei
den Siebsträngen unter 1, tangential auseinander und legen sich
alsdann hinter die benachbarten Siebstränge, wo sie sich je
mit der Hälfte des nächst benachbarten[S. 119] Gefäßstranges vereinigen.
Eine Abart dieses verbreitetsten Typus kommt durch gleichzeitige
Spaltung auch der Siebstränge (wie bei 1) zustande, so daß im
Stengel doppelt so viele kollaterale Bündel wie Gefäßstränge (oder
Siebstränge) in der Wurzel entstehen.
4. Verzweigung der Wurzel. Durch Verzweigungen, die immer wieder
Wurzeln (gleichnamige Organe) sind, wird es der Wurzel ermöglicht, den
Bodenraum nach allen Richtungen zu durchdringen und überall daraus
Wasser und Bodensalze aufzunehmen.
Dichotome Verzweigung durch Gabelung der Vegetationskegel in
je zwei Schenkel findet sich typisch bloß bei einigen Farnpflanzen
(Lycopodinae).
Im übrigen verzweigen sich die Wurzeln seitlich (Fig. 155).
Und zwar werden die Seitenwurzeln im Gegensatze zu den Seitenzweigen
am Sprosse meist erst in einiger Entfernung vom Vegetationspunkte der
Mutterwurzel, wo das Gewebe des Urmeristems bereits in Dauergewebe
übergeht, im Innern des Gewebekörpers der Mutterwurzel,
endogen (Fig. 162, 163), angelegt. Ihre Vegetationspunkte bilden
sich nämlich bei den Pteridophyten aus der innersten Rindenschicht,
bei den Phanerogamen dagegen aus der äußersten Zellschicht des
Zentralzylinders, d. h. aus dem Perizykel der Mutterwurzel, indem hier
Gruppen von Parenchymzellen sich zu teilen und in embryonalen Zustand
zurückzukehren beginnen. Die Seitenwurzelanlagen müssen also immer
die ganze Rinde ihrer Mutterwurzel durchbrechen. Demgemäß sind sie
nicht selten an der Austrittsstelle von dem vorgestülpten Rande der
durchbrochenen Rinde der Mutterwurzel wie von einem Kragen umgeben. Sie
entstehen in dieser Weise zunächst spitzenwärts fortschreitend, also
in akropetaler Reihenfolge. Sie pflegen auch in solcher, ihrem Alter
entsprechenden Reihenfolge zu Wurzeln auszuwachsen. Später werden sie
aber noch durch solche Seitenwurzeln vermehrt, die zwischen den bereits
vorhandenen selbst an älteren Wurzelteilen hervorkommen.
Fig. 162. Wurzelquerschnitt von Vicia Faba mit
Seitenwurzelanlage r, etwas schematisiert, e Endodermis, p
Perizykel, d Rinde, g Gefäßstrang, v Siebstrang des radialen
(tetrarchen) Leitbündels. Vergr. 40.
Fig. 163. Teil eines Längsschnittes durch eine Wurzel
von Amarantus mit Anlage einer Seitenwurzel. e Endodermis, vor der
Seitenwurzelanlage bereits resorbiert, d angrenzende Rinde, p
Perizykel, sp Schraubentracheïde, r Seitenwurzelanlage. Vergr. etwa
200. Nach PH. VAN
TIEGHEM.
Die Seitenwurzeln bilden immer gerade Reihen an der
Mutterwurzel[86]. Diese Anordnung wird dadurch bedingt, daß die
Nebenwurzeln entweder vor den längs verlaufenden Gefäßsträngen
der Mutterwurzel (Fig. 162) oder vor den Leitparenchymplatten
entstehen, die die Xylem- und Phloëmstränge trennen. Die Zahl der
Seitenwurzelreihen ist also entweder gleich der Zahl der Xylemstränge
oder doppelt so groß.
Die Seitenwurzeln haben den gleichen Bau wie die Hauptwurzel. Ihre
Gefäß- und Siebstränge setzen sich an entsprechende Stränge der
Mutterwurzel an.
5. Sproßbürtige Wurzeln. Außer an Wurzeln entstehen Wurzeln
sehr oft auch an ungleichnamigen Organen, d. h. an Teilen des
Sprosses, ebenfalls meist endogen aus Dauergewebe; bei den Farnen
schon aus dem embryonalen Gewebe der Sproßvegetationspunkte. Man nennt
solche Wurzeln je nach dem Orte ihrer Entstehung stengelbürtig oder
blattbürtig.
[S. 120]
Bei Sumpf- und Wasserpflanzen entspringen sie vielfach an den
unteren Stengelknoten zwischen den Blättern. Ein bevorzugter Ort
ihrer Entstehung, soweit die äußeren Bedingungen es zulassen,
sind überhaupt Stengelknoten; sie können das mit den unteren
Sproßpartien absterbende Hauptwurzelsystem ersetzen[87]. Besonders
zahlreich sind sie an den Unterseiten der im Boden wachsenden
Sproßteile (Rhizome, Fig. 143) oder kriechender Sprosse.
Abgeschnittene und in feuchten Boden gesteckte Sprosse oder
Sproßstücke bilden alsbald Wurzeln an ihrer Basis; solche brechen
auch aus der Basis mancher entsprechend behandelter Blätter, so der
Begonienblätter, hervor[88]. Die sproßbürtigen Wurzeln bezeichnet
man wohl auch als Adventivwurzeln.
Kommen vorhandene Wurzelanlagen nicht zur Entwicklung, so nennt man
sie schlafend. Solche ruhenden Anlagen von stengelbürtigen Wurzeln
sind z. B. an jedem Weidenzweige vorhanden; ihre Weiterentwicklung
läßt sich durch Feuchtigkeit und Verdunkelung leicht anregen.
6. Aussehen der Wurzelsysteme. Die Seitenwurzeln irgendeiner
Ordnung wachsen in der Regel schwächer und sind dünner als die
Mutterwurzeln, an denen sie entstanden sind. So wird das ganze
Wurzelsystem typisch razemös. Die Nebenwurzeln letzter Ordnung
bleiben gewöhnlich sehr kurz und haben nur eine begrenzte Lebensdauer;
man nennt sie Saugwürzelchen.
Das Wurzelsystem erhält sein Gepräge, wie das Sproßsystem, weiter
dadurch, daß Haupt- und Seitenzweige ganz verschiedene Lage zueinander
und im Raume annehmen, und zwar durch verschiedene Art von Geotropismus
(vgl. S. 299 ff.).
Viele Dikotylen (z. B. Lupine, Eiche) und Gymnospermen
(Edeltanne) haben eine radiäre Hauptwurzel oder Pfahlwurzel,
die schon als Keimwurzel den Hauptstamm nach unten fortsetzt und
senkrecht nach unten, orthotrop, in die Erde wächst (Fig. 155). An ihr
entstehen ebenfalls radiäre Seitenwurzeln I. Ordnung, die horizontal
oder schräg, also plagiotrop, in das Erdreich eindringen. Die an ihnen
entspringenden Seitenwurzeln II. Ordnung pflegen von denen I. Ordnung
allseits ausstrahlend das Erdreich rings um diese zu durchwachsen, so
daß also die Äste des Wurzelsystems den Boden nach allen Richtungen
möglichst gleichmäßig durchziehen und bei weitergehender Verzweigung
keinen Kubikzentimeter unausgenutzt lassen. Bei anderen Dikotylen und
Gymnospermen kann das Wurzelsystem auch mehr oberflächlich bleiben (z. B.
bei der Kartoffelpflanze und bei der Kiefer).
Den Monokotylen pflegt die Hauptwurzel zu fehlen, da sie
schon am Keimling zugrunde geht. An ihre Stelle treten zahlreiche
sproßbürtige Wurzeln aus der Stengelbasis, die senkrecht oder schräg
oder horizontal in den Boden eindringen. Sie verzweigen sich monopodial
und tragen Seitenwurzeln I. Ordnung, diese Seitenwurzeln II. Ordnung
usw., die das Erdreich in allen möglichen Richtungen durchwachsen
können. Hauptsächlich in horizontaler Richtung breitet sich das
Wurzelsystem z. B. beim Getreide über immer größer werdende Areale aus.
Addiert man die Längen sämtlicher Wurzeln einer Pflanze, so
bekommt man unerwartet hohe Werte; so kann die Gesamtwurzellänge bei
einer Getreidepflanze Hunderte von Metern sein.
Eigenartig wird ein Teil der Wurzeln bei vielen Bäumen der tropischen
Urwälder ausgebildet. Die außerordentlich hohen und dicken Stämme
vieler solcher Bäume sind an der Basis durch mächtige, strebenähnliche
Brettwurzeln oder durch stammartig verdickte, von den Ästen
zur Erde herabgewachsene Luftwurzeln (Säulen- oder
Stützwurzeln) vor dem Umfallen geschützt (z. B. bei vielen
Ficus-Arten, vgl. Fig. 694).
c) Sekundäres
Dickenwachstum des Kormus.
Wir sahen, daß die Sproß- und Wurzelteile, die in den
Vegetationspunkten durch Vermehrung der embryonalen Zellen neu angelegt
worden[S. 121] sind, durch Streckung fertig ausgebildet werden. Mit
diesem Längenwachstum ist meist auch ein gewisses Dickenwachstum
der Teile verbunden, das wie das Längenwachstum lediglich auf der
bedeutenden Vergrößerung der meristematischen Zellen während
ihrer Umbildung zu Dauerzellen, aber nicht auf Zellvermehrung
beruht (primäres Dickenwachstum oder Erstarkung, Fig. 98, 100,
102, 115). Es ist freilich meist verhältnismäßig gering. An diese
Dickenzunahme schließen sich in Sproßachsen und Wurzeln oft noch andere
Wachstumsvorgänge an, denen wir nun unsere Aufmerksamkeit zuwenden
müssen.
Je größer das Sproßsystem wird, um so besser vermag es der Beschattung
durch andere Gewächse zu entgehen und um so mehr organische Substanz zu
bilden. So sehen wir bei vielen Gewächsen aus einem zunächst kleinen,
blattarmen Keimpflänzchen durch Wachstum, oft auch durch Verzweigung
der Keimlingsachse, einen sehr blattreichen Kormus von riesigen,
baumartigen Größenverhältnissen sich entwickeln. Die Größenzunahme des
Sprosses im Luftraume stellt infolge Vermehrung der Blätter fortgesetzt
höhere Anforderungen an die Wasserversorgung durch die Wurzel, der
diese nur durch die Vergrößerung ihrer Oberfläche, meist verbunden
mit Verzweigung, genügen kann; oft werden sogar noch sproßbürtige
Wurzeln gebildet. Jede Vergrößerung des Wurzelsystems hat aber zur
Vorbedingung, daß dazu hinreichende Mengen organischer Nährstoffe
in den Blättern gebildet werden können. So stehen die Ausbildung
der Blattkrone und des Wurzelsystems in engsten Wechselbeziehungen
zueinander. Die Größenzunahme des Sproß- und Wurzelsystems hat ferner
zur Voraussetzung, daß in den Stengeln und Wurzeln die genügende
Anzahl von Leitungsbahnen einerseits für Wasser, andererseits für die
organischen Stoffe vorhanden ist oder ausgebildet werden kann, und daß
die Sproßachsen fest genug sind, um das größer und größer werdende
Gewicht der Blatt- und Zweigmassen, auch bei heftigem Winde, zu
tragen. Es bestehen also auch enge Beziehungen zwischen der Größe des
Kormus und der Ausbildung der Leitungsbahnen in seinen Achsen und der
Festigkeit der Sprosse.
Die Festigkeit muß um so größer sein, je größer die Pflanze wird und je
länger sie lebt. Pflanzen oder Sproßsysteme, die nur verhältnismäßig
kurze Zeit leben und nach Bildung der Fortpflanzungsorgane absterben,
bleiben meist „krautartig“: Kräuter. Große Kormi, die viele
Jahre leben und meist wiederholt fruktifizieren, erhöhen die Festigkeit
ihrer Sproßachsen und Wurzeln meist durch Holzbildung. Solche
Holzgewächse heißen Bäume[89], wenn sie einen Hauptstamm
ausbilden (der Säulenfestigkeit besitzen muß); sonst nennt man sie
Sträucher.
Dem Bedürfnisse nach der nötigen Anzahl von Leitungsbahnen und der
erforderlichen Festigung wird bei den Kormi der Sproßpflanzen in
verschiedener Weise Rechnung getragen: Erstens nämlich gibt
es Pflanzen, bei denen die Hauptachse des Keimlings und die etwa
entstehenden Seitenzweige schon, ehe sie in die Länge wachsen,
vor oder während ihrer Erstarkung, also primär, so verdickt und mit
so viel Leitungs- und Festigungsgewebe ausgestattet werden, daß sie
bei ihrem späteren Längenwachstum der ganzen künftigen[S. 122] Größenzunahme
des Kormus genügen; die Keimwurzel bleibt bei ihnen aber dünn, stirbt
meist frühzeitig ab und wird durch sproßbürtige Wurzeln aus der
Basis oder auch aus höheren Zonen der Keimlingssproßachse ersetzt.
Zweitens gibt es solche, bei denen zunächst lange, fadenförmig
dünne Stengel und Wurzeln mit wenigen Leitungsbahnen (und wenigen
mechanischen Elementen) entwickelt werden. Die Wasserversorgung der
Blätter und umgekehrt die Nahrungsversorgung des Wurzelsystems und
hiermit die Größenzunahme des Kormus würden bei ihnen meist sehr bald
eine Grenze in der geringen Zahl der Leitungsbahnen des Keimstengels
und der Keimwurzel finden, wenn nicht nach Umwandlung der Keimachse in
Dauergewebe, entsprechend den Bedürfnissen des heranwachsenden Kormus,
später für Vermehrung der Leitungsbahnen (und des Festigungsgewebes)
im Wurzelsysteme und im Keimstengel gesorgt würde. Das geschieht aber
durch nachträgliche Zellvermehrung, Bildung sekundärer
Gewebe (des Sekundärzuwachses), womit ein nachträgliches,
sekundäres Dickenwachstum der Stengel und Wurzeln verbunden
ist. Sekundäre Gewebe nennt man solche Gewebe, die durch Tätigkeit
sekundärer Meristeme, der Kambien (vgl. S. 40), den primären
Geweben hinzugefügt werden oder die primären Gewebe ersetzen.
Sekundäres Dickenwachstum findet sich sowohl bei krautigen als auch bei
Holzpflanzen.
Zum ersten Typus[91] gehören die meisten krautigen Pteridophyten
und Monokotylen, ja selbst fast alle ihre stammbildenden Formen
(Baumfarne, Palmen, Pandanaceen, bestimmte Liliifloren). Bei den
stammbildenden Monokotylen z. B. bleibt der Keimstengel nach der
Keimung zunächst lange Zeit sehr kurz. Das Urmeristem
seines flachen Vegetationspunktes wächst an der Peripherie des
Zentralzylinders durch Zellvermehrung stark in die Breite.
Infolgedessen erhält der Vegetationspunkt und die Sproßachse des
Keimlings, aus der der Stamm hervorgehen soll, von vornherein
einen sehr großen Durchmesser.
Bei solchen Formen, wie z. B. bei den Palmen und Pandanaceen,
kann der Stamm auch nach Ausbildung der Dauergewebe durch
Erweiterung der vorhandenen Dauergewebszellen noch längere
Zeit ein wenig weiter erstarken. Hauptsächlich die Zellen der
Sklerenchymfaserstränge, die die Siebteile der Leitbündel
begleiten, nehmen dabei an Weite zu, wodurch die ganzen Stränge
mächtiger werden. Hier und da freilich kann die Dickenzunahme von
Teilungen der Parenchymzellen begleitet sein (manche Palmen).
Zum zweiten Typus[92] gehören die meisten krautigen und
holzbildenden Gymnospermen, Dikotylen und einige baumförmige
Liliifloren. Bei ihnen folgt also auf das primäre Dickenwachstum (die
Erstarkung) der Stengel und Wurzeln durch Zellvergrößerung
meist ein sekundäres durch Zellvermehrung in einer besonderen
Meristemzone, dem Verdickungsring.
Einjährige, kletternde und windende Pflanzen beginnen oft erst in
ziemlich alten Internodien, lange nach Beendigung der Erstarkung,
sekundär in die Dicke zu wachsen. An den Zweigen der Bäume fängt
das sekundäre Dickenwachstum dagegen meist schon an, ehe die
primären Dauergewebe ausgebildet worden sind.
Sekundäres Dickenwachstum trat zuerst bei gewissen, jetzt nur aus
fossilen Überresten bekannten Pteridophyten auf; aber erst bei den
Gymnospermen und Dikotylen gelangte es zu allgemeiner Verbreitung.
Sekundäres Dickenwachstum monokotyler Stämme. Es gibt auch
einige baumartige Liliifloren (Dracaena, Cordyline, Yucca, Aloë),
deren Achsen befähigt sind, mit einem sekundären Meristem sekundär
in die Dicke zu wachsen. Dieses Meristem entsteht außerhalb
der primären Leitbündel, die im Zentralzylinder nach Monokotylenart
zerstreut sind, und zwar in der anschließenden Rinde, aus einer im
Querschnitte ringförmigen Zone von fertigen Rindenzellen, die sich
wieder zu teilen beginnen (bei den Dracaenen meist erst in größerer
Entfernung vom Stammscheitel, sonst schon in seiner unmittelbaren
Nähe).[S. 123] Es bildet einen Zylindermantel aus mehreren Schichten
prismenförmiger, lückenlos verbundener, embryonaler Zellen, die durch
tangentiale Wände längere Zeit nur Zellen nach innen, später auch
einige nach außen abgeben. Das weitere Schicksal dieser so durch
Teilung entstandenen embryonalen Zellen ist ein völlig anderes als
bei den Gymnospermen und Dikotylen: Die von dem Meristem nach außen
gebildeten Zellen werden nämlich zu sekundären Rindenzellen; die
nach innen abgegebenen aber teils zu vollständigen konzentrischen
Leitbündeln mit Außenxylem, teils zu Parenchym, dessen Zellwände
sich stark verdicken und verholzen (Fig. 164).
Die Meristemzellen haben im Querschnitt und im radialen
Längsschnitt rechteckige, im tangentialen Längsschnitt polygonale
Gestalt; es sind also Prismen mit tangential gerichteter
polygonaler Grundfläche (vgl. Fig. 167 A II). Solange das
Meristem einseitig tätig ist, werden seine Initialen aus dem
nach außen angrenzenden Dauergewebe der Rinde ergänzt. Sobald
es beiderseitig tätig wird, bleibt dauernd eine Zellschicht als
Initialschicht erhalten.
Ein echtes sekundäres Dickenwachstum monokotyler Wurzeln ist nur
für die Gattung Dracaena bekannt. Der Kambiumring, der es besorgt,
nimmt in der Wurzelrinde seinen Ursprung und zwar in den inneren
Teilen, die an die Endodermis grenzen.
Fig. 164. Cordyline (Dracaena) rubra. Querschnitt
durch den Stamm. f Leitbündel, und zwar f′ primäre, f″
sekundäre, f‴ ein aus einem Blatte kommendes, noch innerhalb der
Rinde befindliches Bündel, m parenchymatisches Grundgewebe, s
Leitbündelscheide, t Tracheïden, c Verdickungsring (Kambium), cr
Rinde, ph Korkkambium, l Kork, r Rhaphidenbündel. Vergr. 30. Nach
STRASBURGER.
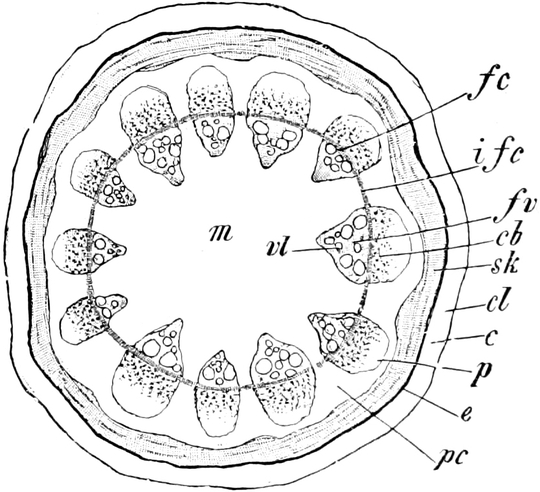
Fig. 165. Querschnitt durch einen 5 mm dicken Zweig von
Aristolochia Sipho. m Mark, fv Leitbündel, und zwar vl Gefäßteil,
cb Siebteil, fc Faszikularkambium, ifc Interfaszikularkambium,
p Phloëmparenchym an der Außenseite des Siebteils, pc Perizykel,
sk Sklerenchymring, e Stärkescheide, c Rinde, in dieser cl
Kollenchym. Vergr. 9. Nach STRASBURGER.
Sekundäres Dickenwachstum der Gymnospermen und Dikotylen. 1.
Bildung, Bau und Tätigkeit des Kambiums in den Stengeln. In den
offenen Leitbündeln der Gymnospermen und der Dikotylen kann die
sekundäre Gewebebildung an die Fertigstellung der primären Gewebe
anschließen oder schon früher beginnen. Nur der erstere Fall sei hier
besprochen, obwohl er keineswegs häufig ist. Die zwischen
den Gefäßteilen und Siebteilen der offenen[S. 124] Bündel vorhandenen
Reste von Urmeristem werden dabei zu Kambien, indem sie sich
von neuem lebhaft zu teilen beginnen. Die Leitbündel sind meist
im Kreise angeordnet. Nachdem die Kambiumtätigkeit innerhalb der
Bündel begonnen hat, bildet sich Kambium auch in den Markstrahlen.
Eine Zone von Dauerzellen nämlich, die die Kambien dar angrenzenden
Leitbündel verbinden, beginnt sich tangential zu teilen. Dieses
interfaszikulare Kambium ergänzt so die innerhalb der Bündel
gelegenen faszikularen Kambien zu einem vollen Zylindermantel
von Meristemgewebe. Seine Zellen, die in radialer Richtung
wachsen, teilen sich durch tangentiale und durch quere Scheidewände.
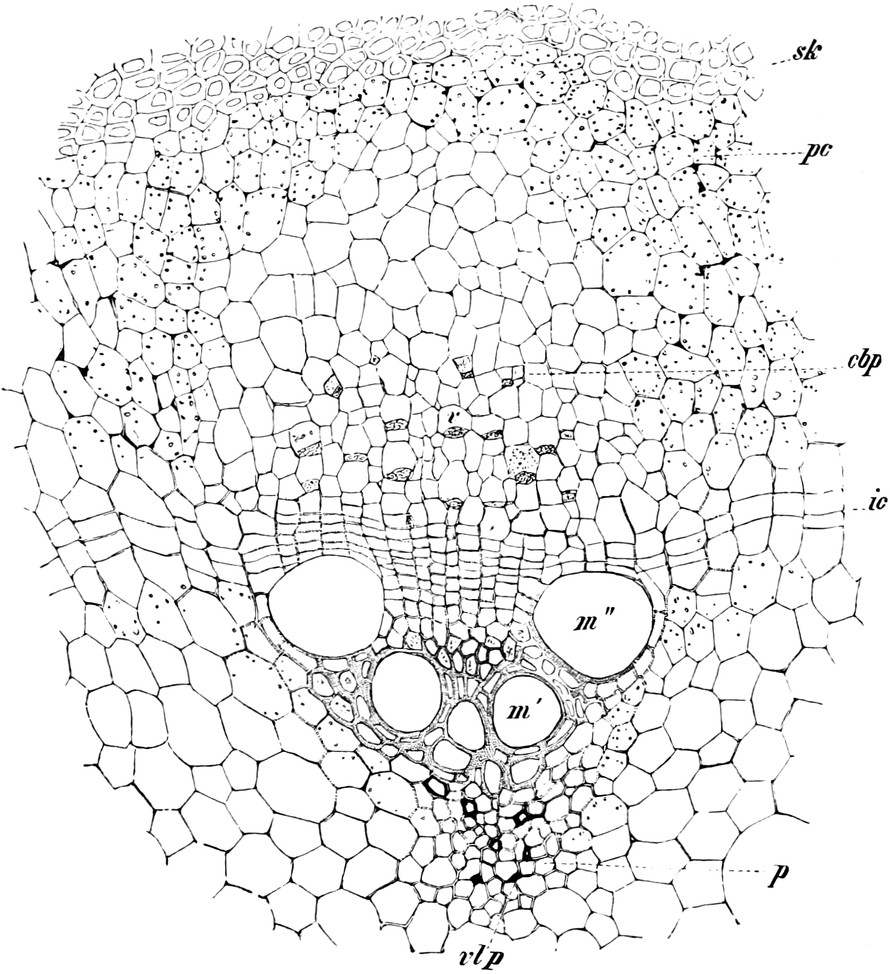
Fig. 166. Querschnitt durch ein Leitbündel eines Zweiges
von Aristolochia Sipho im ersten Jahre seiner Entwicklung, nach
begonnener Kambiumtätigkeit. p Xylemparenchym, an dem Innenrande des
Gefäßteils, vlp Xylemprimanen, m′ und m″ Tüpfeltracheen, ic
Interfaszikularkambium, sich in das Faszikularkambium fortsetzend,
v Siebröhren, cbp Siebprimanen, pc Gewebe des Perizykels, sk
innerer Teil des Ringes aus Sklerenchymfasern. Vergr. 130. Nach
STRASBURGER.
Die Fig. 165 und 166 stellen den Vorgang der Kambiumbildung für
den Stengelquerschnitt von Aristolochia Sipho dar, wo er sich
in möglichst einfacher und übersichtlicher Weise vollzieht. Die
Fig. 166 gibt ein einziges Leitbündel nebst dem angrenzenden
Interfaszikularkambium aus der Fig. 165 bei stärkerer Vergrößerung
wieder. In diesem Bündel ist die Kambiumtätigkeit bereits in
vollem Gange. Besonders fallen die in Ausbildung begriffenen
sekundären Gefäße (m″) auf. Innerhalb der Markstrahlen sind die
Parenchymzellen, die dem Interfaszikularkambium den Ursprung gaben,
noch deutlich zu erkennen.
[S. 125]
Die meristematischen Kambiumzellen des Verdickungsringes, die
lückenlos verbunden sind und radiale Reihen bilden, pflegen die
Gestalt langgestreckter, in tangentialer Richtung mehr oder weniger
abgeplatteter Prismen zu haben mit beiderseits meißelförmig
zugeschärften Enden, deren scharfe Kanten radial gestellt sind, so
daß die Zellform auf Tangential-, Radial- und Querschnitten ganz
verschieden erscheint (Fig. 167). Die tangentialen Wände, die die
polygonalen oder rhombischen Grundflächen der Prismen bilden, sind
dünn, die radialen, die senkrecht auf den Grundflächen stehen, dagegen
ziemlich dick und oft getüpfelt. In dem mehrschichtigen Kambiummantel
ist eine mittlere Zellschicht, die Initialschicht. Ihre
Zellelemente, die in radialer Richtung wachsen, bleiben dauernd
meristematisch und geben durch fortgesetzte Teilungen mittels
tangentialer Scheidewände Tochterzellen (Gewebemutterzellen) in
radialer Richtung nach außen, in viel größerer Zahl aber nach innen
hin ab. Diese Tochterzellen werden ihrerseits, meist nach weiteren
tangentialen Teilungen und nach oft starkem Längen- und Dickenwachstum
(Fig. 172), allmählich zu sekundären Dauerzellen, deren Formen vielfach
gar nicht mehr den embryonalen Kambiumzellen ähnlich sind.
Dadurch, daß das Kambium nach innen Zellen abgibt, wird es mit der
Dickenzunahme des Stammes selbst immer weiter nach außen geschoben;
dementsprechend muß sich der Umfang des Kambiummantels fortgesetzt
vergrößern. Das ist nur möglich durch Wachstum und Vermehrung der
Zellen in tangentialer Richtung. Auf Querschnitten durch
das Kambium sieht es so aus, als käme diese Vermehrung durch
gelegentliche radiale Teilungswände zustande. KLINKEN[93]
und NEEFF haben uns aber darüber belehrt, daß solche
Teilungen nicht vorkommen; soll die Zellenzahl tangential vermehrt
werden, so teilt sich vielmehr eine Kambiuminitialzelle quer,
worauf die Enden der Tochterzellen durch gleitendes Wachstum
tangential aneinander vorbei wachsen.
Alles durch die Kambiumtätigkeit nach innen erzeugte
Dauergewebe, das meist hart und fest ist und aus mehr oder weniger
verholzten Zellen besteht, wird als Holz, alles nach
außen gebildete Gewebe dagegen, das aus meist unverholzten
Zellen besteht, als Bast bezeichnet.
Die vom Kambium nach außen abgegebenen Gewebe heißen wohl auch
sekundäre Rinde.
Fig. 167. Schematische Darstellung der Form der
Kambiumzellen. A I und II die beiden vorkommenden Formen
körperlich, die tangentiale (Breit-)Seite zeigend; B Radialschnitt;
C Querschnitt. Nach ROTHERT.
Das sekundäre Gewebe, das vom faszikularen Kambium nach innen
gebildet wird, gleicht dem der Xylemteile, das nach außen gebildete
dem der Siebteile der primären Leitbündel. Durch die Tätigkeit des
Interfaszikularkambiums werden die Markstrahlen dauernd
auf der Holz- und der Bastseite verlängert, allerdings meist nicht in
ihrer ganzen Breite als Markstrahlen; denn der größere Teil auch des
interfaszikularen Kambiums bildet nach innen und nach außen in der
Regel ähnliche Gewebe wie das faszikulare. Da also in den Markstrahlen
nur an beschränkten, voneinander isolierten Stellen radiale Stränge von
Markstrahlgewebe gebildet werden, indem die Kambiumzellen eben nur hier
nach außen und innen Markstrahldauerzellen liefern, werden die zunächst
plattenförmigen Markstrahlen in zahlreiche neben- und übereinander
liegende kleinere, bandförmige, von spindelförmigem Querschnitt zerlegt
(Fig. 168), die das Mark mit der Rinde verbinden und als primäre
Markstrahlen des Holzes und des Bastes gelten. Bei zunehmender
Dicke des Holz- und Bastringes beginnen aber auch einzelne Streifen
des Faszikular[S. 126]kambiums, Markstrahlgewebe zu erzeugen. So werden die
sekundären Markstrahlen gebildet, die blind in dem Holze und
Baste endigen und um so weniger tief in beide hineinreichen, je später
sie angelegt wurden (Fig. 177).
Die Kambiumzellen, die Markstrahlzellen bilden, stehen den anderen
an Länge nach und haben auch weniger steile, ja zum Teil selbst
horizontale Endwände, weil die Kambiumzellen vor der Einschaltung neuer
Markstrahlen quer- oder schräggeteilt werden.
Man kann nach dem primären Bau der Stengel, der Entstehung des
Kambiums und der Art seiner Tätigkeit im wesentlichen drei Typen
unterscheiden[94]: 1. Im Stengel entsteht zunächst ein Ring von
kollateralen Leitbündeln, die durch breite Markstrahlplatten aus
Dauergewebe voneinander getrennt sind; die Markstrahlen
behalten auch während des sekundären Dickenwachstums im Stengel
ungefähr ihre Breite, indem das interfaszikulare Kambium fast nur
Markstrahlgewebe erzeugt, so bei vielen krautigen Pflanzen, unter
verholzten fast nur bei Lianen. Bei den Kräutern, bei denen der
innere Teil der Markstrahlen zwischen den primären Gefäßsträngen
aus Sklerenchym besteht (vgl. S. 82), bildet das interfaszikulare
Kambium in den Strahlen nach innen ebenfalls solches Gewebe. 2. Im
Stengel entsteht wie bei 1. ein Kranz kollateraler Blattspurbündel,
die durch sehr breite Markstrahlen voneinander getrennt sind. Noch
ehe das primäre Dickenwachstum beendigt ist, entstehen
aus dem zum Teil noch embryonalen Markstrahlgewebe, das
nun kambialen Charakter annimmt, in jedem Markstrahle ein bis
mehrere kleinere, tangential netzartig verbundene stammeigene
Zwischenbündelchen, die den Raum des Markstrahles fast völlig
ausfüllen und die in den Maschen zwischen sich sehr schmale
bandartige primäre Markstrahlen mit spindelförmigem Querschnitt
lassen (Fig. 168) (so bei vielen Kräutern und Holzgewächsen). 3. Im
Stengel entsteht bei der Umwandlung des primären Meristemgewebes
zu Dauergewebe von vornherein nicht ein Kranz kollateraler
Bündel, sondern ein ringförmiges Bündelrohr, das man
geradezu als konzentrisches Bündel mit Innenxylem und mit zentralem
Marke bezeichnen muß. Dieses Rohr hat eine Schicht von embryonalem
Gewebe zwischen Phloëm und Xylem, das spätere Kambium, und ist von
sehr schmalen, spindelförmigen primären Markstrahlen durchsetzt
oder kann auch alle primären Markstrahlen entbehren (so bei vielen
Bäumen).
Fig. 168. Auflösung eines primären Markstrahles in
viele kleine nach Beginn des sekundären Dickenwachstums. Tangentialer
Längsschnitt schematisch. l, l Benachbarte primäre Leitbündel; pm
primärer Markstrahl, durch die Tätigkeit des interfaszikularen Kambiums
zerlegt in viele kleine spindelförmige Markstrahlen und in netzartig
verbundene sekundäre Leitbündel.
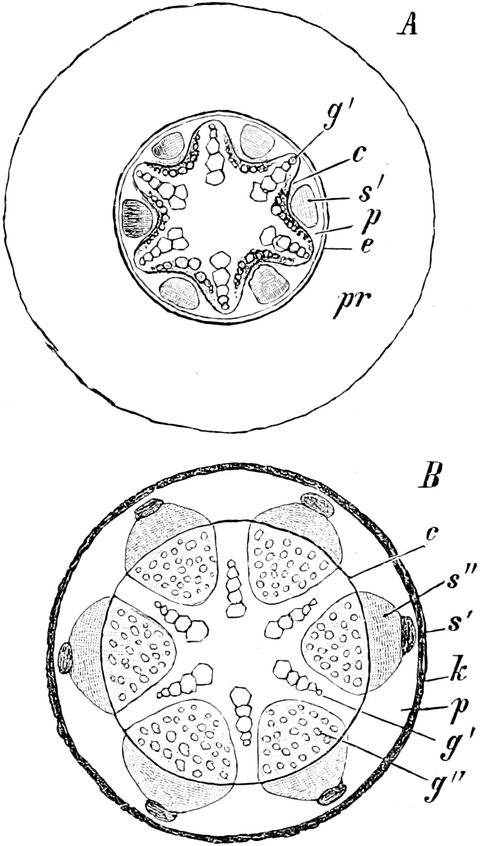
Fig. 169. Schematische Darstellung des Dickenwachstums
einer dikotylen Wurzel. In A bedeutet pr Rinde, e Endodermis. In
A und B sind c Kambiumring, g′ Gefäßstrang, s′ Siebstrang,
p Perizykel. In B bedeuten außerdem g″ Holz, s″ Bast, k
Periderm (vgl. S. 138). Nach STRASBURGER.
[S. 127]
Die primären Gefäßteile ragen im sekundär verdickten Stengel
als Vorsprünge in das Mark; man nennt sie in ihrer Gesamtheit
Markkrone.
2. Bildung und Tätigkeit des Kambiums in der Wurzel. Wie
wir gesehen haben (Fig. 157, 158), wechseln im Zentralzylinder der
Wurzeln die Gefäß- und Siebstränge miteinander ab; sie sind durch
Parenchymplatten voneinander getrennt. Beginnt nun eine solche
Gymnospermen- oder Dikotylenwurzel sekundär in die Dicke zu wachsen,
so bilden sich in diesen Parenchymplatten parallel zu ihren Flächen,
also zwischen den Gefäß- und Siebsträngen, durch Teilung der Zellen
Kambiumstreifen aus, die nach innen Holz, nach außen Bast erzeugen. Die
Ränder der Kambiumstreifen treffen im Perizykel vor den Gefäßsträngen
zusammen; hier werden die Perizykelzellen ebenfalls zu Kambiumzellen:
Nun ist ein vollständiger Kambiummantel mit welligem Querschnitt da,
der, wie beim Stengel, im ganzen Umkreis seine Tätigkeit ausüben
kann (vgl. die dunkle Linie der schematischen Fig. 169 A). Die
Einbuchtungen gleichen sich bald aus, so daß der Mantel kreisförmigen
Querschnitt erhält (Fig. 169 B). Dem Holzkörper und dem Baste fehlen
die eigentlichen primären Markstrahlen, doch werden wie im Stengel
sekundäre erzeugt; bei manchen Pflanzen besonders große, plattenförmige
vor den primären Gefäßsträngen (vgl. Fig. 169 B). Der Querschnitt
durch eine Wurzel, die jahrelang in die Dicke gewachsen ist, läßt
sich von einem Stammquerschnitt ohne eingehendere Untersuchung nicht
unterscheiden; erst durch solche kann man inmitten der Wurzel das
Vorhandensein ihrer eigenartigen primären Gefäßstränge feststellen.
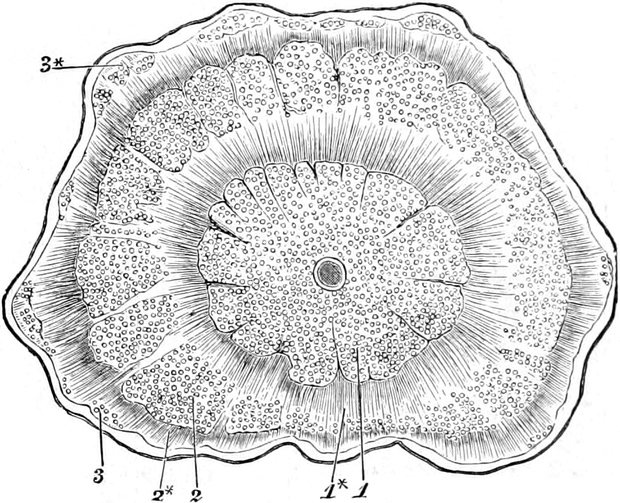
Fig. 170. Querschnitt durch einen Stamm von Mucuna
altissima. 1, 2, 3 aufeinanderfolgende Holzkörper. 1*, 2*,
3* aufeinanderfolgende Bastzonen. 3 und 3* innerhalb des
Perizykels in Bildung begriffen.
3⁄4 nat. Gr.
Nach SCHENCK.
Wiederholte Kambiumbildung in Stämmen und Wurzeln. Außer den
allgemein verbreiteten Vorgängen des sekundären Dickenwachstums
kommen in den Stämmen und in den Wurzeln bei Gymnospermen und
Dikotylen auch andere vor, die als Anomalien dem normalen
Typus gegenübergestellt werden. Sie kennzeichnen sich durch
abweichende Verteilung und Tätigkeit der Kambien.
Bei einigen Cycadeen und bestimmten Gnetum-Arten unter den
Gymnospermen, bei Chenopodiaceen, Amarantaceen, Nyctagineen,
Phytolaccaceen und noch anderen Familien unter den Dikotylen
hört der erste, in gewohnter Weise entstandene Kambiumring nach
längerer oder kürzerer Zeit zu funktionieren auf. Es bildet sich
ein neuer Kambiummantel meist im Perizykel, also außerhalb der
Bastzone, oder in einem vom ersten Kambiummantel abstammenden
Gewebe. Dieses Kambium erzeugt wiederum nach innen Holz, nach
außen Bast, beides mit Markstrahlen. Alsdann erlischt es; wieder
ein neues, außerhalb des neuen Bastmantels entstehendes Kambium
tritt an seine Stelle. Der Vorgang wiederholt sich und führt
zur Bildung konzentrischer Holz-Bastzonen. Solche treten uns z. B.
an dem Fig. 170 dargestellten Stammquerschnitt einer zu den
Papilionaceen gehörenden Liane, der Mucuna altissima, als Ringe
deutlich entgegen. Konzentrische Holz-Bastmäntel findet man auch an
manchen fleischigen Wurzeln, die zwei oder mehr Vegetationsperioden
ausdauern. So bei der Runkelrübe (Beta vulgaris), wo man sie schon
mit dem bloßen Auge auf Querschnitten sehen kann. Sie entstehen
wie eben beschrieben; nur herrschen in diesen Zuwachszonen, ebenso
wie im typischen sekundären Zuwachs anderer fleischiger Wurzeln,
die Parenchyme vor, die vermehrt werden, um der Speicherung von
Reservestoffen zu dienen.
[S. 128]
3. Das Holz. A. Gewebearten und deren Funktionen. Das Holz
ist ein Gewebekörper von verwickeltem Bau. Es setzt sich bei
den Dikotylen meist aus drei verschiedenen Gewebearten
mit mehr oder weniger verholzten Membranen zusammen: 1. aus
längsverlaufenden Strängen toter Gefäße (Fig. 171 g, tg),
2. aus längsverlaufenden Strängen meist toter Sklerenchymfasern:
Holzfasern (h) und 3. aus längs- und in den Markstrahlen auch
radialverlaufenden Strängen von Speicherparenchymzellen (hp):
Holzparenchym und Markstrahlparenchym. Dementsprechend dient
das Holz 1. zunächst wie die primären Gefäßteile der Leitbündel der
Wasserleitung, außerdem aber auch 2. der Festigung
der Stämme und der Wurzeln gegen Druck und Biegungen und 3. zur
Aufspeicherung organischer Substanzen. Die Eigenschaften, die
das Holz als Baumaterial für uns so wertvoll machen, werden aus seiner
Festigungsfunktion verständlich.
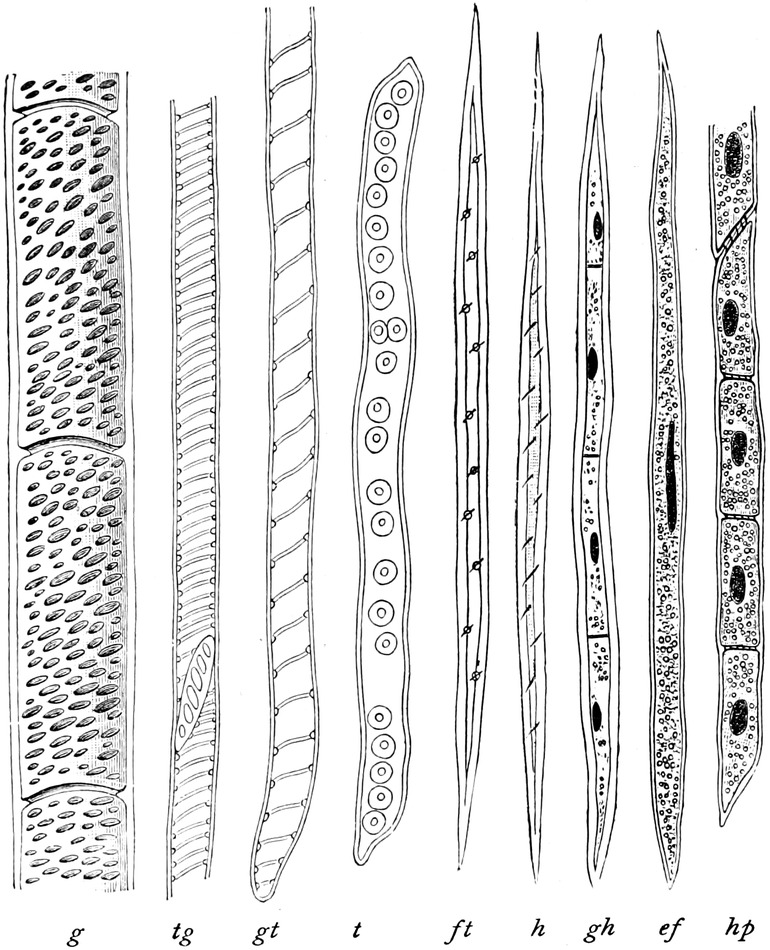
Fig. 171. Tracheen, Tracheïden, Holzfasern und
Holzparenchym der Dikotylen nebst ihren Übergangsformen, schematisch.
Erklärung im Text. Nach STRASBURGER. (Verändert.)
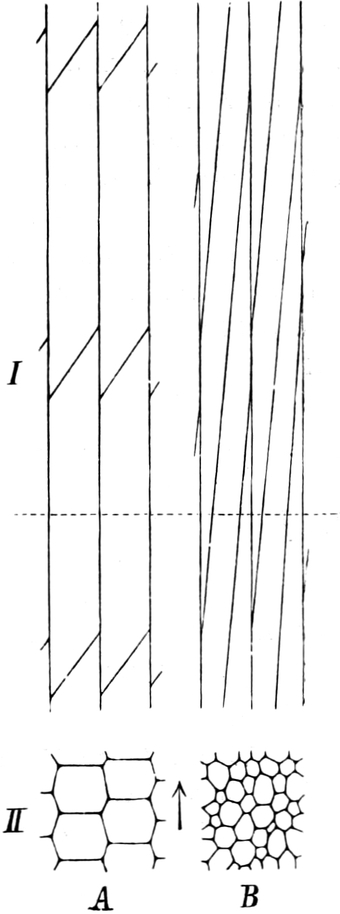
Fig. 172. Schematische Darstellung des gleitenden
Wachstums; für Holzfasern. A I und II jugendlicher Zustand der
Zellen, B I und II nach dem gleitenden Wachstum ausgewachsen. I
Im tangentialen Längsschnitte, II im Querschnitte in der punktierten
Linie von I. Nach ROTHERT.
Die Zellarten, aus denen das Holz sich aufbaut, lassen
sich am leichtesten untersuchen, wenn man das Holz mit
SCHULZEschem Gemisch mazeriert (vgl. S. 35).
Die Gefäße sind Tüpfel-, seltener Netzgefäße, und zwar in
der Regel teils sehr weite, kurzgliedrige und enge, mehr oder
weniger langgliedrige Tracheen (Fig. 171 g, tg), teils
enge, langgestreckte Tracheïden, die außer der Wasserleitung
zugleich zur Festigung des Stammes dienen. Die Holzfasern
(h) sind meist sehr eng und sehr langgestreckt, beiderseits
nadelförmig zugespitzt und dickwandig mit schrägen, schmalen,
spaltenförmigen[S. 129] Tüpfeln. Die Speicherparenchymzellen (hp)
endlich sind rechteckig prismatisch oder spindelförmig, in Richtung
des Strangverlaufs in der Regel gestreckt, dünn- oder dickwandig
mit meist kleinen, rundlichen, einfachen Tüpfeln und sind reich an
Reservestoffen (Stärke, Zucker oder Öl). Interzellularen kommen nur in
den Parenchymsträngen vor.
Bei zahlreichen Leguminosen, bei Weiden, Pappeln, Ficus-Arten
kommen im Holz als wasserleitende Elemente nur Tracheen vor.
Die Tracheïden und Holzfasern sind wesentlich länger als die
Kambiumzellen, aus denen sie hervorgehen, oft bis über 1 mm lang.
Diese größere Länge erreichen sie, ebenso wie die weiten Tracheen
ihren Durchmesser, durch gleitendes Wachstum (S. 40 und Fig. 172).
Bei der Bildung von Holzparenchym werden die Gewebemutterzellen des
Kambiums wiederholt quer geteilt. Dieses Parenchym besteht also
(vgl. Fig. 171 gh, hp) aus Zellreihen, denen man ihre Herkunft
aus Kambiumzellen ansieht, da sie oben und unten mit zugespitzten
Elementen endigen.
Die Scheidewände zwischen den Holz- oder Markstrahlparenchymzellen
und den Gefäßen sind einseitig behöft getüpfelt, soweit Tüpfel
vorhanden sind: die meist großen Tüpfel sind innerhalb der lebenden
Zellen ohne Hof, behöft dagegen in den Gefäßen, zudem im Gegensatze
zu den typischen beiderseits behöften Tüpfeln stets ohne Tori in
den Schließhäuten. Die Scheidewände zwischen den Gefäßen und den
Holzfasern und die zwischen den Holzfasern und den Parenchymzellen
sind dagegen meist gar nicht getüpfelt.
Bei den Hölzern, die aus Gefäßen, Holzfasern und Parenchymzellen
sich aufbauen, kann man nicht selten alle Übergänge zwischen
solchen typisch ausgebildeten Elementen beobachten; dementsprechend
gehen ihre Funktionen ineinander über. Neben mehr oder weniger
weiten Hoftüpfeltracheen (g) kommen enge Tracheen vor. Diese
(Fig. 171 tg) leiten über zu den Tracheïden (Fig. 171 gt, t).
Schmale, stark zugespitzte Tracheïden (Fasertracheïden ft), die
hauptsächlich der mechanischen Festigung dienen mögen, bilden einen
Übergang zu den Holzfasern (h). Wenig verdickte Holzfasern,
die ihren lebenden Inhalt behalten (die sog. Ersatzfasern ef),
ohne oder mit Querwänden (gh), vermitteln den Übergang zu den
Holzparenchymzellen (hp). Phylogenetisch sollen sich aber nach
STRASBURGER[68] die Fasertracheïden nur von den
Gefäßen, die Holzfasern dagegen nur durch Umbildung von
Holzparenchymzellen ableiten lassen.
Bei den Gymnospermen kommen im Holze außer wenig Holz- und viel
Markstrahlparenchym nur noch Tracheïden mit typischen Hoftüpfeln vor.
Hier ist also die Arbeitsteilung noch nicht so weit fortgeschritten;
die Festigung wird noch von den gleichen Elementen besorgt, die auch
der Wasserleitung dienen. Ebenso ist es bei der Magnoliaceen-Gattung
Drimys unter den Dikotylen.
B. Anordnung der Gewebearten im Holze. Bei den
Gymnospermen (Fig. 173–175) hat das Holz der Stämme und Wurzeln
also einen verhältnismäßig noch einfachen Bau. Die Tracheïden sind
entsprechend ihrer Entstehung (Fig. 173 B) in regelmäßigen radialen
Reihen angeordnet (Fig. 173 A), da sie nur in radialer Richtung, aber
so gut wie gar nicht in tangentialer und longitudinaler wachsen; sie
haben infolgedessen ähnliche Gestalt wie die Kambiumzellen (Fig. 167)
und besitzen oft nur in ihren radialen Wänden große runde Hoftüpfel, so
daß man diese vor allem in radialen Schnitten von der Fläche zu sehen
bekommt (Fig. 70 B, 71 A).
Holzparenchym ist in den meisten Gymnospermenhölzern nur
sehr spärlich vorhanden. Bei den Kiefern, Fichten und Lärchen
umgibt es ausschließlich die schizogenen Harzkanäle, die
das Holz zwischen den Tracheïden durchziehen (Fig. 173 Ah, 177
h) und mit radial in einzelnen breiten Markstrahlen verlaufenden
in Verbindung stehen. Daher können große Harzmengen aus einem
verwundeten Kiefern- oder Fichtenstamme ausfließen. Bei den
anderen Koniferen ist die Bildung des Holzparenchyms auf einfache
Zellreihen beschränkt, deren Zellräume sich weiterhin mit Harz
füllen.
Außer den Tracheïden findet man in den Gymnospermenhölzern, wie
gesagt, fast ausschließlich Markstrahlparenchym, das in Form
sehr zahlreicher, radial verlaufender, bandförmiger und meist nur eine
Zellschicht[S. 130] breiter Markstrahlen (Fig. 173 m,
175 sm, tm, 177
ms) das Holz durchsetzt. Eine jede Holztracheïde grenzt in ihrem
Längsverlaufe an einen oder auch an mehrere dieser Markstrahlen an.
Die Markstrahlzellen sind radial gestreckt, stärkereich, werden von
Interzellularen begleitet (Fig. 175 i) und dienen dazu, die in den
Blättern gebildeten und im Baste abwärts geleiteten Assimilate in
radialer Richtung dem Holz des Stammes und der Wurzeln zuzuführen und
hier zu speichern, umgekehrt Wasser aus dem Holzkörper nach außen zu
leiten. Diese Aufgaben können die Markstrahlen erfüllen, da sie, wie
wir sahen, mit ihren Enden gleicherweise in den Bast und in das Holz
eindringen (Fig. 173 B, 174, 177). Die Interzellularen münden in das
Interzellularsystem der Rinde und sichern den lebenden Zellen im Holze den
für ihre Lebensvorgänge notwendigen Gasaustausch mit der Atmosphäre.
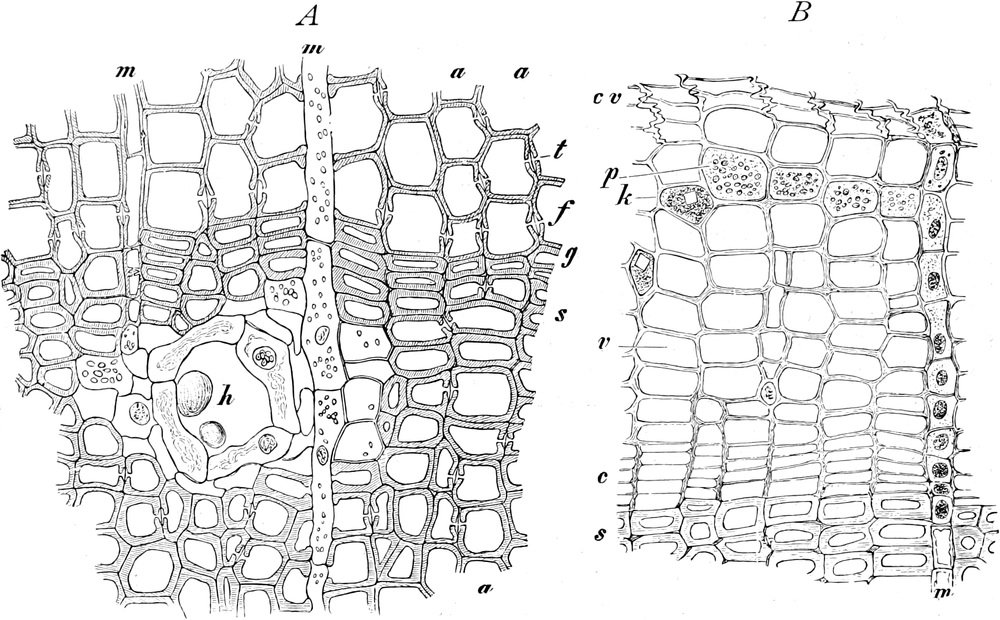
Fig. 173. A Partie eines Querschnittes durch das
Kiefernholz an einer Jahresgrenze. f Frühholz, s Spätholz, t
Hoftüpfel, a eine sich nach außen verdoppelnde Tracheïdenreihe,
h Harzgang, m Markstrahlen. Vergr. 240. B Querschnitt aus dem
Stamme der Kiefer, den äußeren Hand des Holzkörpers, das Kambium und
den angrenzenden Bast in sich fassend. s Spätholz, c Kambium,
v Siebröhren, p Bastparenchym, k Kristallzellen, cv außer
Funktion gesetzte Siebröhren, m Markstrahlen. Vergr. 240. Nach
SCHENCK.
Bei bestimmten Gymnospermen, vor allem den Abietineen (etwa der
Kiefer), sind einzelne Zellreihen der Markstrahlen des Holzkörpers,
gewöhnlich die oben und unten randständigen, ohne lebenden Inhalt,
tracheïdal ausgebildet, durch Hoftüpfel untereinander und mit
den Tracheïden verbunden (Fig. 174 tm). Vor der Zerdrückung
durch die turgeszenten lebenden Markstrahlzellen werden sie durch
besondere Verdickungsleisten der Wand geschützt. Sie erleichtern
den Wasseraustausch in radialer Richtung unter den Tracheïden,
die nur in den radialen Wänden getüpfelt sind. Bei den meisten
anderen Nadelhölzern dagegen, denen solche tracheïdalen Elemente
in den Markstrahlen fehlen, sind auch tangentiale Hoftüpfel in den
Tracheïden des Holzes vorhanden, wodurch eine radiale Bewegung des
Wassers gefördert wird. Auch die parenchymatischen Markstrahlzellen
des Holzes sind mit den Tracheïden, und zwar durch große einseitig
behöfte Tüpfel verbunden (Fig. 175 et).
Fig. 174. Radialer Längsschnitt durch den Kiefernstamm,
den Außenrand des Holzkörpers, das Kambium und den anschließenden
Bast sowie einen Markstrahl in sich fassend. s Spättracheïden,
t Hoftüpfel, c Kambium, v Siebröhren, vt Siebtüpfel, tm
tracheïdale Markstrahlzellen, sm stärkeführende Markstrahlzellen im
Holzkörper, sm′ im Bastkörper, em eiweißführende Markstrahlzellen.
Vergr. 240. Nach SCHENCK.
Fig. 175. Tangentialer Längsschnitt durch das Spätholz
der Kiefer. t Hoftüpfel, tm tracheïdale, sm stärkeführende
Markstrahlzellen, et einseitig behöfte Tüpfel, i Interzellularen am
Markstrahl. Vergr. 240. Nach SCHENCK.
Schon mit dem bloßen Auge nimmt man auf Stammholzquerschnitten, ebenso
wie bei den meisten Dikotylenhölzern, Jahresringe wahr (Fig.
176, 177). Betrachtung dünner Querschnitte durch Gymnospermenholz
bei stärkerer[S. 131] Vergrößerung (Fig. 173 A) belehrt darüber, daß
in jedem dieser Ringe die inneren Elemente (f) jeder radialen
Tracheïdenreihe weitlumig und dünnwandig, die äußeren aber (s)
englumig und dickwandig sind. Der Übergang der weiten zu den engen
ist im Jahresring ganz allmählich, der Übergang der englumigen zu den
weitlumigen des nächst äußeren Jahresringes aber unvermittelt. Die
Jahresringe[95] im Holze kommen durch die Periodizität der
Kambiumtätigkeit zustande, die mit dem Wechsel der Jahreszeiten
in Beziehung steht. Im Frühjahr, zur Zeit, wo die neuen Triebe sich
entwickeln, werden Tracheïden mit größerem Hohlraum ausgebildet als
in der Folgezeit. So entsteht ein weitlumigeres Frühholz
(Frühlings- oder Weitholz, Fig. 173 Af, 177 f), das vor allem
der Wasserzufuhr nach den Verbrauchsorten dient, und späterhin ein
englumiges Spätholz (Sommer- oder Engholz, Fig. 173 As, 177
s), das vor allem die Festigkeit des Stammes erhöht. In der zweiten
Augusthälfte hört die Holzbildung in unseren Breiten bei den Stämmen
meist auf. Sie beginnt von neuem im nächsten[S. 132] Frühjahr mit weitlumigen
Elementen. Demgemäß zeichnet sich zwischen dem letztjährigen Spätholze
und dem nächstjährigen Frühholze eine scharfe Grenze (Fig. 173 g,
177 i), die eben dem bloßen Auge schon kenntlich ist und die zur
Feststellung des Alters des betreffenden Pflanzenteils dienen kann.
In dem Holzkörper der Wurzeln ist die Grenze der Jahresringe meist
undeutlicher.
Je weiter spitzenwärts eine sekundär verdickte Sproßachse oder
Wurzel quer durchschnitten wird, um so weniger Jahresringe
findet man. Das folgt naturgemäß aus dem Scheitelwachstum dieser
Organe. Je älter die Jahresringe, um so früher müssen sie sich
ja scheitelwärts auskeilen. In ähnlicher Weise hören auch ältere
Bastschichten nach dem Scheitel zu auf.
Unter Umständen vermag die Zahl der Ringe im Holz die Zahl der
Altersjahre zu überschreiten, nämlich wenn Blattverlust durch
Frost, Raupenfraß oder andere schädliche Einflüsse das Austreiben
der für die nächstjährige Vegetationsperiode bestimmten Knospen
veranlaßt und Neubelaubung eine Wiederholung der Frühholzbildung
bedingt. Andererseits kann bei Holzgewächsen, die sonst die
Jahresringbildung streng einhalten, ausnahmsweise die Zahl der
nachweisbaren Jahresringe kleiner ausfallen, als das Alter des
betreffenden Individuums verlangt, weil sich die Jahresgrenzen
gelegentlich nicht deutlich markiert haben. So kann es auch
kommen, daß eine Zählung in einem Stammradius etwas weniger
Jahresringe als in einem anderen ergibt.
Fig. 176. Querschnitt durch einen im 4. Jahre
stehenden Zweig der Linde (Tilia ulmifolia). pr Rinde, c
Kambiumring, cr Bast, pm primäre Markstrahlen, pm′ äußeres,
durch Dilatation erweitertes Ende eines primären Markstrahls, sm
sekundärer Markstrahl, g Jahresgrenze, m Mark. Vergr. 6. Nach
SCHENCK.
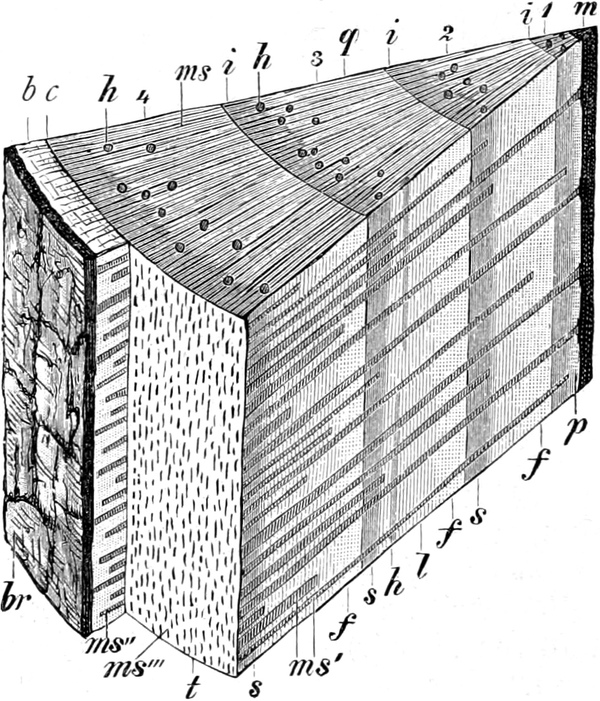
Fig. 177. Stück eines 4jährigen Stammteils der Kiefer
(Pinus silvestris) im Winter geschnitten. q Querschnitt-, l
radiale Längsschnitt-, t tangentiale Längsschnittansicht, f
Frühholz, s Spätholz, m Mark, p primäre Gefäßteile, 1, 2,
3 und 4 die vier aufeinanderfolgenden Jahresringe des Holzkörpers,
i Jahresgrenze, ms Markstrahlen in der Querschnittansicht
des Holzkörpers, ms′ in der radialen Längsschnittansicht des
Holzkörpers, ms″ innerhalb der Bastzone, ms‴ in der tangentialen
Längsschnittansicht, c Kambiumring, b Bastzone, h Harzgänge,
br die außerhalb der ersten Peridermlage befindliche, der Rinde
entsprechende Borke. Vergr. 6. Nach SCHENCK.
Das Holz der Stämme und Wurzeln von Dikotylen läßt sich
schon bei schwacher Vergrößerung leicht von einem Koniferenholze
unterscheiden (Fig. 178–180). Abgesehen davon, daß in den
Dikotylenhölzern außer Tracheïden und Parenchym stets noch Holzfasern
und fast immer auch sehr weite Tracheen vorhanden sind, wachsen die
verschiedenen Elemente, die Tracheen, Tracheïden, Holzfasern und
Holzparenchymzellen, nicht übereinstimmend; infolgedessen können sie
nicht ihre ursprüngliche radiale Anordnung und ihre ursprüngliche
Form beibehalten. Außerdem werden im Frühholze der Jahres[S. 133]ringe
hauptsächlich sehr weite Wasserbahnen, besonders Tracheen ausgebildet
(Fig. 178, 179 m), während im Spätholze vor allem die englumigen
Holzfasern (l) und die faserförmigen Tracheïden (t) vorherrschen.
Wir besitzen unter unseren dikotylen Holzgewächsen auch solche,
bei denen die Jahresringe sich deshalb nicht sehr deutlich
abheben, weil die verschiedenen Formelemente des Holzes annähernd
gleichförmig über den Jahreszuwachs verteilt sind, so bei der
Weide. Ja, es kann in diesem Falle die Abzählung der Jahresringe
ganz unmöglich werden, wie beim wilden Wein. Bei den Holzgewächsen
der feuchtwarmen Tropengebiete mit ununterbrochener Vegetationszeit
können die Jahresringe ebenfalls fehlen. Bei vielen sind aber auch
hier jahresringähnliche Zonen ausgebildet.
Nur die wasserleitenden Elemente des jüngsten Jahresringes
stehen in direkter Verbindung mit den Blättern der betreffenden
Vegetationsperiode. Da die Pflanze mit der Entfaltung der Blätter
im Frühjahr plötzlich viel Wasser durch Transpiration verbraucht,
so versteht man, daß im Frühholz eben zunächst für die nötigen
Wasserbahnen gesorgt wird. Bei vielen Holzgewächsen wird die
Laubmenge während des Sommers nicht weiter vermehrt, infolgedessen
kann das Kambium im Spätholze vorwiegend Festigungsgewebe bilden.
Fig. 178. Teil eines Querschnittes durch das Holz
von Tilia ulmifolia an einer Jahresgrenze. Vergr. 540. Nach
STRASBURGER.
Fig. 179. Tangentialer Längsschnitt aus dem Holz von
Tilia ulmifolia. Vergr. 160. Nach SCHENCK.
m Weites Tüpfelgefäß, t Tracheïden, l Holzfaser, p
Holzparenchym, r Markstrahl.
Trotz allen Verschiedenheiten, die in der Struktur dikotyler Hölzer
vorkommen, lassen sich für die Anordnung der verschiedenen Gewebearten
doch bestimmte Regeln angeben, die stets befolgt werden. Die aus
Tracheen oder Tracheïden bestehenden Gefäßstränge, reich
verzweigt in radialer und tangentialer Richtung, bilden in der
Längsrichtung des Holzes zusammenhängende Bahnen, die von den
Wurzeln ohne Unterbrechung bis zu den dünnsten Zweigenden verlaufen.
Nur so wird die ausreichende Wasserversorgung des Sproßsystems
gewährleistet.
Holzparenchym (Fig. 178, 179 p) ist bei den meisten
Dikotylenhölzern reichlich vorhanden, und zwar ebenfalls in Form längs
verlaufender[S. 134] Stränge oder Schichten, die aber oben und unten im Holze
früher oder später blind endigen. Das Holzparenchym bildet mit den
Markstrahlen stets ein zusammenhängendes System lebender Zellen.
Die Gefäße stehen immer mit diesen lebenden Holzparenchymzellen in
Verbindung; bald werden sie rings von ihnen umhüllt, bald einseitig von
ihnen begleitet (Fig. 178 p).
Das Holzparenchym umscheidet die Gefäße z. B. bei den Akazien u. a.;
es bildet tangentiale Binden im Holze, an die die Gefäße sich
einseitig anlehnen oder worin sie eingebettet sind, z. B. bei der
Walnuß, der Edelkastanie und bei Eichen; manchmal ist es auf die
Außenseiten der Jahresringe beschränkt.
Die Markstrahlen (Fig. 176 pm, sm: 178, 179 r) sind wie
bei den Gymnospermen radial verlaufende niedere oder hohe, eine oder
mehrere Zellschichten breite, unverzweigte oder meist verzweigte Bänder
(Fig. 180 tm, sm), die durch das Kambium hindurch in den Bast
eindringen (Fig. 176). Auch daran lehnen sich die Gefäßstränge hier
und da an. Das Markstrahlparenchym verbindet, wie schon hervorgehoben,
die Parenchymzellen des Bastes mit denen des Holzes, also alle
lebenden Gewebe des Stammes und der Wurzel zu einer Einheit. In den
Markstrahlen können auf diese Weise Assimilate, die im Baste abwärts
strömen, radial in den Holzkörper und dort im Holzparenchym eine
Strecke weit aufwärts oder abwärts geleitet werden. Diese Assimilate
werden in den Markstrahlen und im Holzparenchym meist als Stärke oder
auch als Fett aufgespeichert. Interzellularen, die den Markstrahl-
und Holzparenchymsträngen folgen, dienen dem Gaswechsel der lebenden
Elemente des Holzes.
Die Räume zwischen den Gefäß-, Holzparenchym-Strängen und Markstrahlen
werden von Holzfasersträngen (Sklerenchym) ausgefüllt.
Die Höhe und Breite der Markstrahlen läßt sich leichter an
tangentialen als an radialen Längsschnitten feststellen, weil man
alsdann ihre Querschnitte vor sich hat. An solchen tangentialen
Schnitten erscheinen die Markstrahlen spindelförmig (Fig. 179
r). Ihre Größe schwankt bei den meisten Hölzern nur innerhalb
relativ enger Grenzen, doch bei gewissen anderen, so der Eiche und
Rotbuche, sehr bedeutend. Die Eiche hat bis zu 1 mm breite und
fast 1 dcm hohe große Markstrahlen, dazwischen zahlreiche sehr
schmale niedrige. Bei der Pappel, der Weide, dem Buchsbaum sind
alle Markstrahlen so klein, daß man sie auch mit der Lupe kaum
unterscheiden kann. Besonders hoch und breit sind die primären,
sich über die Länge eines ganzen Internodiums erstreckenden
Markstrahlen bei vielen Lianen, so z. B. auch bei Aristolochia.
Fig. 180. Radialer Längsschnitt aus dem Holz von Tilia
ulmifolia mit einem kleinen Markstrahl. g Gefäß, l Holzfasern,
tm mit den Wasserbahnen durch Tüpfel verbundene, sm der Leitung
der Assimilate vornehmlich dienende Markstrahlzellen. Vergr. 240. Nach
SCHENCK.
Auch bei den Dikotylen, sehr ausgesprochen z. B. bei den Weiden,
sind im Holzkörper meist Zellreihen des oberen und unteren
Markstrahlrandes mit den Wasserbahnen durch einseitig behöfte
Tüpfel verbunden. Ihre lebenden Zellen sind höher als die der
mittleren Zellreihen; man hat sie daher auch als stehende
Markstrahlzellen bezeichnet (Fig. 180 tm). Die mittleren
Zellreihen sind in radialer Richtung stärker gestreckt, ohne
besondere Verbindung mit den Wasserbahnen und vornehmlich
auf Leitung und Speicherung der Assimilate eingerichtet:
liegende Markstrahlzellen (Fig. 180 sm).
Die Markstrahl- und Holzparenchymzellen, die an die Gefäße angrenzen,
nehmen Wasser aus den Wasserbahnen auf und geben es nach Bedarf an
andere lebende Zellen ab; andererseits pressen sie im Frühjahr,
zur Zeit der Knospenentfaltung, einen großen Teil der in ihnen
gespeicherten Assimilate, vor allem[S. 135] Glykose und geringe Mengen von
Eiweißstoffen, in die Gefäße ein, so daß diese Stoffe rasch nach den
Verbrauchsorten gelangen. Demgemäß kann man während des Winters und zu
Beginn des Frühjahrs Zucker und Eiweiß in den Gefäßen nachweisen. Diese
Stoffe sind auch in dem Blutungssaft enthalten, den man im Frühjahr
durch Bohrlöcher aus Birken, Ahornarten, Hainbuchen und anderen Bäumen
gewinnen kann.
Maserbildung. Bei vielen technisch verwendeten Hölzern
schätzt man vor allem außer der Farbe die Maserung. Sie
beruht zunächst auf der Ausbildung der Jahresringe und der
Markstrahlen, außerdem aber sehr oft, z. B. beim Nußbaumholz, auf
einem welligen Verlauf der Formelemente des Holzes, der durch
gedrängte Bildung von Seitenästen, Adventivknospen, Seitenwurzeln
oder auch durch Wundreize u. dgl. veranlaßt worden ist.
C. Nachträgliche Veränderungen des Holzes. Bei den meisten
Bäumen sterben die lebenden Elemente in den zentralen älteren
Teilen des Holzkörpers ab, und die Wasserbahnen werden verstopft
(Kernbildung), so daß nur die äußeren Holzschichten, die aus
wenigen, und zwar den zuletzt entstandenen Jahresringen bestehen, noch
lebende Zellen enthalten (Splint). Sie allein speichern noch
Reservestoffe. Auch die Wasserleitung ist auf den Splint, ja vielfach
sogar nur auf seine äußersten Schichten beschränkt; die peripheren
Gefäße sind es ja, wie wir sahen, allein, die direkt mit den Blättern
und mit den jüngsten Seitenwurzeln in Verbindung stehen. Der Kern dient
nur noch der Festigung. Viel seltener als solche Kernhölzer sind
Splinthölzer, deren gesamtes Holz Splint bleibt (Ahornarten,
Birke, Linde). Kernhölzer fallen meist unmittelbar durch die hellere
Farbe des Splintes auf; das dunklere Kernholz ist dichter, härter und
fester als das Splintholz und durch Einlagerung verschiedener Stoffe
gegen Zersetzung geschützt. Es gibt aber auch Kernhölzer, bei denen
sich der Kern vom Splint durch die Farbe nicht unterscheidet und leicht
zersetzt wird. Solche Stämme, wie die der Weiden, werden im Alter
leicht hohl.
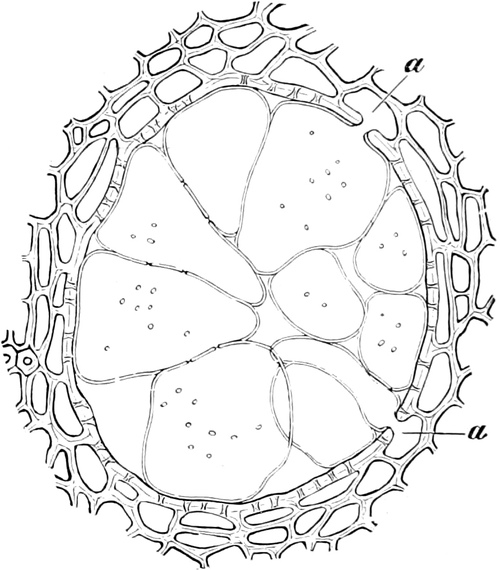
Fig. 181. Ein mit Thyllen erfülltes Gefäß, nebst
den angrenzenden Elementen aus dem Kernholz der Robinie (Robinia
Pseudacacia) im Querschnitt. Bei a und a ist der Zusammenhang
der Thyllen mit ihren Ursprungszellen zu sehen. Vergr. 300. Nach
SCHENCK.
Am schärfsten setzt sich der weißlichgelbe Splint vom Kernholz
dort ab, wo letzteres eine dunklere Färbung zeigt, so bei
unserer Eiche mit braunem Kern oder noch ausgesprochener beim
Ebenholz (Diospyros), dessen Kern schwarz ist. Je dunkler das
Kernholz, um so dauerhafter pflegt es zu sein. Vor dem Absterben
bilden die lebenden Zellen des Holzes, die ihre Reservestoffe
auflösen, meist verschiedene organische Stoffe, darunter besonders
Gerbstoffe, die in die Membranen der umgebenden Elemente
eindringen, ferner harz- und gummiartige Körper, die als Schutz-
und Kerngummi ihre Hohlräume zum Teil verstopfen. Meist verleihen
die Oxydationsprodukte der Gerbstoffe den toten Holzteilen die
dunkle Färbung, und die Gerbstoffe schützen den toten Holzkörper
vor Zersetzung. Die Gefäße in dem toten Holze werden teils durch
Ansammlungen von Kerngummi, teils durch „Zellen“ verstopft, die die
Gefäßlumina mehr oder weniger ausfüllen und als Thyllen[96]
bezeichnet werden (Fig. 181); sie entstehen dadurch, daß lebende
Holzparenchymzellen in die angrenzenden Gefäße durch die Tüpfel
unter Dehnung der Schließhäute hineinwachsen. Thyllen können ferner
in verwundeten Gefäßen entstehen; sie verschließen auch hier den
Gefäßhohlraum. Auch anorganische Stoffe sind nicht selten in den
Kernhölzern abgelagert, so bei Ulmus campestris und Fagus silvatica
kohlensaurer[S. 136] Kalk, und zwar hauptsächlich in den Gefäßen; in den
Gefäßen des Teakholzes (Tectona grandis) amorphe Kieselsäure. Die
Kerne mancher Hölzer dienen Färbezwecken, z. B. liefert das Blau-
oder Campecheholz (Haematoxylon campechianum L.) mit rotem Kern das
Hämatoxylin.
4. Der Bast. A. Gewebearten und deren Funktionen.
Auch den Bast (Fig. 173 B, 182) setzen meist drei Gewebearten
zusammen: 1. längs verlaufende Stränge von Siebröhren (v),
bei den Dikotylen mit Geleitzellen (Fig. 182 c), 2. bei vielen
Gewächsen längs verlaufende Stränge meist toter Sklerenchymfasern,
Bastfasern (Fig. 182 l) und 3. längs (p) und in den
Markstrahlen (Fig. 173 Bm, 182 r) radial verlaufende Stränge
von Parenchym mit Interzellularen: Bast- und
Markstrahlparenchym. Dazu kommen oft noch Sekretzellen
verschiedener Art (Kristallschläuche k, Milchröhren). Der Bast dient
wie das Phloëm der Leitbündel vor allem dazu, die Assimilate zu leiten
und vorübergehend zu speichern. Daneben trägt er zur Speicherung
organischer Substanzen und oft zur Festigung bei. Die Siebröhren
des Bastes haben bei vielen Pflanzen schräge Endwände (Fig. 182 v*);
sie sind eiweißreich, dünnwandig, unverholzt und meist nur kurze
Zeit tätig. Die Bastfasern sind sehr lang und englumig, stark
verdickt, verholzt oder unverholzt; die Parenchymzellen sind
in der Strangrichtung gestreckt, lebend, reich an Reservestoffen,
dünnwandig und meist nicht verholzt.
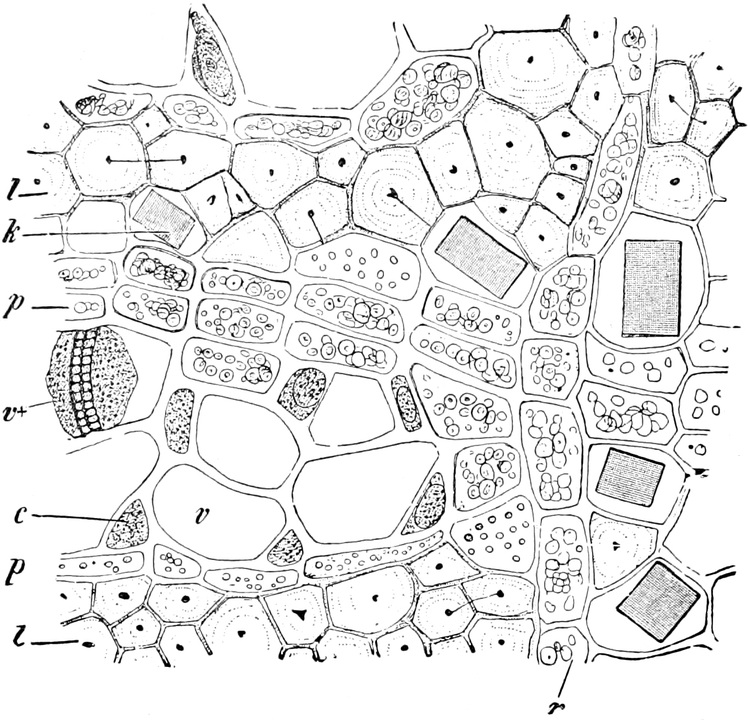
Fig. 182. Durchschnitt durch den Bast der Linde (Tilia
ulmifolia). v Siebröhren, bei v* eine Siebplatte getroffen, c
Geleitzelle, p Bastparenchym, k Kristallzellen des Bastparenchyms,
l Bastfasern, r Markstrahl. Vergr. 540. Nach
STRASBURGER.
In einiger Entfernung vom Kambium erhalten die Siebplatten der
Siebröhren Kallusbeläge. Noch weiter außen werden die Siebröhren
und ihre Geleitzellen zerdrückt (Fig. 173 B cv). In seltenen
Fällen, so beim Weinstock, sind sie mehr als 1 Jahr tätig; in
diesem Falle wird ihr Kallusbelag für die Zeit ihrer Tätigkeit
wieder entfernt. Bei solchen Koniferen, die eiweißhaltige
Bastparenchymzellen im sekundären Baste besitzen, werden diese
wie die angrenzenden Siebröhren zerdrückt. Die stärkehaltigen
Bastparenchymzellen hingegen bleiben jahrelang am Leben; sie werden
sogar zwischen den zerdrückten Siebröhren vergrößert.
B. Anordnung der Gewebearten im Baste. Die einzelnen Gewebearten
des Bastes verlaufen ähnlich wie im Holze. Die Siebstränge bilden
verzweigte Bahnen, in denen die Siebröhren ohne Unterbrechung von
der Wurzel bis in die Laubblattkrone ausgebildet sind. Außerdem
grenzen die Siebstränge hier und da ebenso wie die längs verlaufenden
Bastparenchymstränge an die auch im Baste bandförmigen (Fig. 177
ms″) Markstrahlen an, die, wie wir schon sahen, die radialen
Fortsetzungen der Holzmarkstrahlen sind. So können die Assimilate aus
der Laubblattkrone einesteils innerhalb des Bastes nach den Wurzeln
abströmen, andernteils in der schon geschilderten Weise aus dem[S. 137] Baste
durch die Markstrahlen in die lebenden Zellen des Holzes gelangen, wo
sie gespeichert werden.
Die Gewebearten des Bastes sind oft sehr regelmäßig zu tangentialen,
nur von den Bastmarkstrahlen unterbrochenen Binden angeordnet (Fig.
182). Die Kambiumperiodizität spricht sich im Baste aber nicht aus;
Jahresringe kommen also nicht vor. Das Kambium fährt auch nach
Abschluß der Spätholzbildung im Sommer und Herbst, solange es die
Witterungsverhältnisse gestatten, noch fort, nach außen Bast zu
erzeugen.
So wechseln bei der Linde Bänder von (Fig. 182) Siebröhren (v)
nebst Geleitzellen (c), stärkeführendem Bastparenchym (p),
Kristallzellen (k), Bastfasern (l), flachen Bastparenchymzellen
(p) und endlich wiederum Siebröhren miteinander ab. Der
Unterschied im Aussehen der Baststränge bei verschiedenen
Holzgewächsen wird vornehmlich durch die größere oder geringere
Weite der Siebröhren, das Vorhandensein oder das Fehlen von
Bastfasern, endlich durch die Art der Verteilung aller dieser
Elemente bedingt.
Bei den Kiefern und verschiedenen anderen Abietineen unter den
Gymnospermen sind eiweißreiche Zellreihen an den oberen und
unteren Markstrahlrändern ausgebildet (em Fig. 174). Sie sind
den Siebröhren dicht angeschmiegt, mit ihnen durch Siebtüpfel
verbunden, werden zugleich mit den Siebröhren entleert und
hierauf zusammengedrückt. Bei den Dikotylen sind die Markstrahlen
im Baste einfacher gebaut als im Holzkörper. Die Aufgabe der
Markstrahlzellen, die innerhalb des Bastes abwärts geleiteten
Stoffe aufzunehmen, zeigt sich durch ihre Tüpfel an, die bei
den Dikotylen nicht nur die Zellreihen des Markstrahls mit dem
Bastparenchym, sondern auch mit den Geleitzellen der Siebröhren
verbinden.
Folgen des sekundären Dickenwachstums für die Gewebe außerhalb des
Kambiummantels. 1. Dilatation. Indem das Kambium nach innen immer
mehr Holz, nach außen immer neuen Bast bildet, wächst der Stengel
oder die Wurzel sekundär in die Dicke. Die Dauergewebe, die außerhalb
des Kambiummantels gelegen sind: die Epidermis, die Rinde, die
primären Phloëmstränge und der Bast, bleiben von diesem Dickenwachstum
natürlich nicht unbeeinflußt; sie werden tangential gedehnt, zerrissen,
zerdrückt, verschoben oder auch durch tangentiales Wachstum verbreitert
(Dilatation). Zu solchem Dilatationswachstum sind natürlich vor
allem die lebenden Parenchymzellen der Rinde, des Phloëms, des Bastes
(namentlich des Markstrahlparenchyms), bei einigen Holzgewächsen sogar
die Epidermiszellen befähigt[97]. Alle diese Zellen wachsen dabei sehr
stark in tangentialer Richtung und werden meist durch radiale Wände
geteilt. Im Baste ist dieses Wachstum häufig bei den Markstrahlen
sehr auffällig: es kann in ihnen, z. B, bei der Linde, geradezu
zur Ausbildung sekundärer Meristeme kommen, die durch Teilungen
Parenchymzellreihen in tangentialer Richtung nach beiden Seiten
abgeben, wodurch die Bastmarkstrahlen von Jahr zu Jahr nach außen mehr
erweitert werden (Fig. 176 pm′). Die Siebröhren und ihre Geleitzellen
aber, die nur kurze Zeit tätig sind und dann absterben, werden samt den
Sekretzellen zerdrückt; auch die Sklerenchymfasern der Rinde und des
Bastes beteiligen sich an der Dilatation meist nicht. War in der Rinde
ein Hohlzylinder von Sklerenchymfasern vorhanden (Fig. 184 sc), so
wird er in tangentialer Richtung zerrissen; in die Risse wachsen die in
Dilatation begriffenen Parenchymzellen ein, füllen die Lücken aus und
werden darin bei vielen Gewächsen zu dickwandigen Steinzellen (Fig. 184
s). Auch sonst wandeln sich während der Dilatation Parenchymzellen
der Rinde und des Bastes einzeln, gruppen- oder schichtenweise
nachträglich in Sklerenchymzellen um.
Ein jahrelanges Dilatationswachstum der Epidermis findet
sich bei verschiedenen Rosen, Akazien, Ilex-, Ahornarten, den
Misteln u. a. m. Solche Epidermen haben meist mächtig verdickte
Außenwände. Sie vermögen diese Wände in dem Maße, wie sie an der
Oberfläche Risse bekommen und zerstört werden, von innen durch neue
Verdickungsschichten zu verstärken.
[S. 138]
2. Periderm. Gewöhnlich nimmt aber die Epidermis an der
Dilatation nicht teil; sie wird passiv gedehnt und schließlich
zersprengt. Schon lange vor dieser Zersprengung bildet sich ein neues
Abschlußgewebe, der Kork aus, der die Epidermis später ersetzt und
die Teile, die im Dickenwachstum begriffen sind, gegen Austrocknung
schützt. Der Kork entsteht durch die Tätigkeit eines besonderen
sekundären Meristems, das sich an der Peripherie der Organe bildet
(Fig. 183).
Meist beginnt die Bildung dieses Korkkambiums
(Phellogens) schon in der ersten Vegetationsperiode, bald nach
oder gar schon vor Beginn des sekundären Zuwachses. Es kann aus der
Epidermis selbst, und zwar durch tangentiale Teilungen ihrer Zellen
hervorgehen. Meist aber bildet es sich aus der Rindenschicht, die auf
die Oberhaut folgt, seltener aus tieferen Rindenschichten oder aus dem
Perizykel. Das letzte ist meist bei den Wurzeln der Fall (Fig. 169 B
k). Das Korkkambium erzeugt hauptsächlich nach außen Zellen, und zwar
in radialen Reihen. Dieses Meristem mit sämtlichen Produkten seiner
Tätigkeit nennt man Periderm. Die nach außen abgegebenen
Zellen werden zu Korkzellen, die nach innen gebildeten zu
chlorophyllreichen, unverkorkten Rindenzellen, die sich abrunden und
die Rinde ergänzen. Hat die Bildung des Periderms begonnen, so bräunt
sich die Stengeloberfläche.
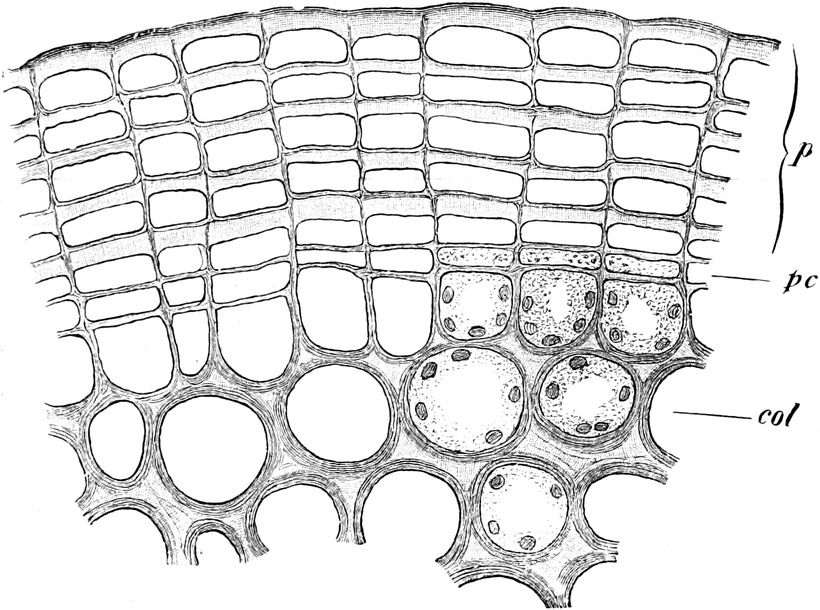
Fig. 183. Querschnitt durch die äußeren Teile eines
einjährigen Zweiges von Pirus communis im Herbst. Beginn der
Peridermbildung. p Kork, pc Korkkambium, col Kollenchym.
Die Korkzellen haben verdickte Außenwände. Vergr. 500. Nach
SCHENCK.
Die Gesamtheit der Zellen, die aus dem Phellogen nach innen
entstehen, bezeichnet man als Phelloderm.
Das Korkkambium ist in der Regel ein typisches Initialenkambium
(vgl. S. 39), zumal wenn es Kork und Phelloderm liefert. Eine
Initialschicht kann aber auch, z. B. bei fast allen Monokotylen,
fehlen; alsdann teilen sich die Dauerzellen, aus denen das
Korkkambium hervorgeht, in eine Anzahl Tochterzellen, die zu
Korkzellen werden, worauf der gleiche Vorgang auf angrenzende
Dauerzellen übergreift (Etagenkork).
Peridermbildung pflegt auch in solchen Pflanzen späterhin
einzutreten, deren Epidermen zunächst jahrelang dilatiert werden.
Sie geht nur den Mistelarten ab.
Echter Kork fehlt aber noch den Kryptogamen, selbst den
Pteridophyten. Einen Ersatz hierfür bietet, wo ein entsprechender
Schutz nötig ist, die Imprägnierung der Zellwände mit einem
sehr widerstandsfähigen braunen Stoff oder die Anlagerung von
Suberinlamellen in unverkorkte Zellen, also die Umwandlung von
Zellschichten in Kutisgewebe[98].
Durch die Tätigkeit des Korkkambiums können dicke, außen rissige
Korkkrusten gebildet werden, wie z. B. bei der Korkeiche, aus
deren Kork die Flaschenkorke geschnitten werden. Die Schichtung,
die sie zeigen, entspricht Jahresproduktionen. In anderen Fällen
werden Korkhäute mit glatter Oberfläche von nur ganz wenigen
Zellagen Dicke gebildet (Fig. 59, 183), deren äußerste Schichten dem
Dickenwachstum des Stengels durch passive Dehnung lange Zeit folgen,
schließlich zerreißen und abschilfern.
[S. 139]
Der Flaschenkork (Fig. 58) wird von breiten Lagen weicher, weiter
Korkzellen gebildet, mit denen schmale Lagen, die Grenzen der
Jahresproduktion bezeichnender, flacher Korkzellen abwechseln,
wie man bei Korkstöpseln erkennen kann. Die mit lockerem Pulver
angefüllten Poren, die den Flaschenkork in ganzer Dicke radial
durchsetzen, sind seine Lentizellen (vgl. S. 50). Die erste
Korkschicht der Korkeiche wird nach dem 15. Jahre bis auf
das Korkkambium künstlich vom Stamm entfernt, worauf einige
Zellagen tiefer sich ein neues Kambium bildet, das den technisch
verwertbaren Flaschenkork liefert. Dieser wird alle 6–8 Jahre
geschält. Da solche toten Korkkrusten der Dilatation nicht folgen
können, so müssen sie außen allmählich rissig werden.
3. Borkebildung. Was vom Korkkambium nach außen zu liegen
kommt, wird von weiterer Nahrungs- und Wasserzufuhr abgeschnitten
und muß absterben. Alle auf solche Weise abgetrennten toten Gewebe
werden mitsamt den Peridermschichten als Borke bezeichnet. So
können je nach dem Entstehungsorte des Periderms die Epidermis oder
kleinere oder größere Teile der Rinde zu Borke werden. Gewöhnlich
stellt das erste Korkkambium in Stamm und Wurzel bald (aber z. B.
nie bei der Buche) seine Tätigkeit ein. Es wird ein neues tiefer im
Stamm angelegt, dessen Tätigkeit ebenfalls nach einiger Zeit erlischt.
Hierauf entsteht weiter innen wieder ein neues, wie es Fig. 184 für die
Steineiche zeigt. Schließlich sind es nicht mehr primäre, sondern
sekundäre Gewebe, nämlich die lebenden Parenchyme der Bastzone, in
denen das neue Korkkambium sich bildet, so daß an älteren Stämmen
das lebende Gewebe außerhalb des Kambiummantels nur noch sekundären
Ursprunges ist, und auch die Borke abgestorbene sekundäre Gewebe
enthält. Die Gewebe, die durch die Korkschichten vom übrigen Gewebe des
Körpers abgeschnitten worden sind, werden von Nährstoffen entleert und
enthalten nur Nebenprodukte des Stoffwechsels. Die Borke vermag der
weiteren Dickenzunahme des Stammes oder der Wurzel natürlich nicht mehr
zu folgen, blättert mit der Zeit entweder außen ab oder zerreißt durch
Längsrisse. Sie stellt einen noch viel vollkommeneren Schutzmantel als
der Kork sowohl gegen Wasserverlust wie gegen Erwärmung dar.
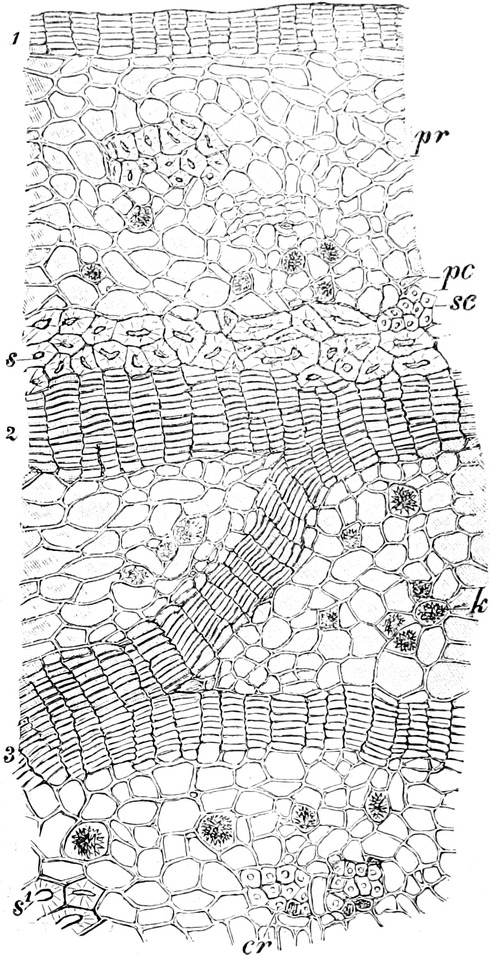
Fig. 184. Querschnitt aus der Stammoberfläche der
Steineiche (Quercus sessiliflora). 1, 2 und 3 nacheinander
erzeugte Korkschichten. pr Durch nachträgliche Dilatation veränderte
Rinde, pc Grenze des Perizykels, sc Sklerenchymfasern aus einem
zersprengten Sklerenchymfaserringe des Perizykels, s Steinzellen,
nach der Zersprengung dieses Ringes aus Parenchym entstanden, s′
Steinzellen im sekundären Zuwachs, cr Bastfasern von Kristallzellen
begleitet, k Zellen mit Kristalldrusen. Alles Gewebe außerhalb der
innersten Korkschicht abgestorben und gebräunt, in Borke verwandelt.
Vergr. 255. Nach SCHENCK.
Durch die Borkebildung werden an älteren sekundär verdickten
Pflanzenteilen die äußeren, jeweils ältesten Teile des Bastes
abgestoßen. Die Folge davon ist, daß der Bastmantel immer nur schmal
bleibt. Festigungselemente[S. 140] können also nur dann zu dauernden
Bestandteilen der Stämme werden, wenn sie innerhalb des Kambiums, d. h.
im Holze entstehen.
Umfassen die Korkschichten und die Korkkambien, aus denen sie
entstanden sind, nur begrenzte Teile der Stammoberfläche, so
schneiden sie schuppenförmige Gewebestücke aus ihr heraus. Dabei
setzen jüngere Korklagen mit ihren Rändern an ältere an. In solcher
Weise erzeugte Borke heißt Schuppenborke, so bei der
Platane, der Eiche (Fig. 184), unseren Nadelhölzern. Bilden die
Korklagen dagegen geschlossene Mäntel, so werden stammumfassende
Rindenmassen als Ringelborke entfernt, so beim Weinstock,
Kirschbaum, Geißblatt und bei der Waldrebe.
Bei solchen Stämmen, bei denen die Borke abblättert, geschieht das
nicht in rein mechanischer Weise, sondern durch Vermittlung von
Trennungsschichten aus besonders dünnwandigen Kork- oder
Phelloidzellen (vgl. S. 50), die zwischen die übrigen, oft stark
verdickten Kork- oder Phelloidschichten des Periderms eingeschaltet
sind. Diese Trennungsschichten werden durch hygroskopische
Spannungen der Borke durchrissen. Schwer sich ablösende Borke wird
während der Dickenzunahme des Stammes nur rissig (rissiges
Aussehen der Rinde bei den meisten alten Bäumen).
Die braune oder rote Färbung, die die Borken meist zeigen, wird
durch ähnliche Derivate von Gerbstoffen wie die Färbung
vieler Kernhölzer veranlaßt. Diese antiseptisch wirksamen Stoffe
bedingen die große Widerstandsfähigkeit der Borke. Die weiße
Färbung der Birkenborke rührt von Betulin (Birkenharz) her, das als
feinkörniger Inhalt die Korkzellen füllt.
Wundenheilung[99]. Im einfachsten Falle sterben die
verletzten Zellen am Körper der Landpflanzen ab, bräunen sich und
vertrocknen, während die Wände der darunter liegenden, unverletzt
gebliebenen Zellen mit Schutzstoffen imprägniert, unter Umständen
auch mit Suberinlamellen versehen, also zu Kutisgewebe werden. Bei
größerer Ausdehnung der Wunde bilden aber die Phanerogamen unter
diesen Zellschichten ein Korkkambium aus, das Wundkork
erzeugt. So werden auch die Blattnarben, die beim Laubfall
entstehen (S. 104), zunächst durch Verholzung und Verkorkung der
freigelegten Zellen, hierauf meist noch durch Ausbildung einer
Korkschicht abgeschlossen, die an die des Stammteils ansetzt.
Die Enden der Gefäße an der Blattnarbe füllen sich entweder mit
sog. Schutzgummi oder mit Thyllen oder mit beidem; die Enden der
Siebröhren werden zerdrückt und verholzen.
Wenn eine Wunde ein noch junges Gewebe bloßlegt, kommt es
gewöhnlich zur Bildung eines Kallus. D. h. alle an die Wunde
grenzenden lebenden Zellen wuchern aus ihr hervor, teilen sich und
schließen zusammen. Diese Kalluswucherung kann an ihrer Oberfläche
unmittelbar verkorken und so den nötigen Schutz gewähren. In den
meisten Fällen bildet sich aber in der Peripherie des Kallus ein
Korkkambium aus, das Kork erzeugt. Ausgedehnte Wunden an älteren
Stammteilen der Gymnospermen und Dikotylen, die bis in den
Holzkörper reichen, werden überwallt: Das an die Wundränder
grenzende Stammkambium wächst wulstartig zu einem Kallus hervor;
der Wulst schließt sich durch den Kork nach außen ab, während in
seinem Innern eine Kambiumschicht differenziert wird, die mit
dem Stammkambium in Verbindung tritt. Dieses Kambium bildet, wie
das angrenzende, nach innen Holz, nach außen Bast. So vergrößern
sich die Überwallungswülste und decken allmählich die Wundfläche.
An der bloßgelegten Holzfläche sind, falls sie noch aus Splint
bestand, die toten Elemente durch Wundgummi verstopft, den die
lebenden Zellen des Holzes erzeugen; die ganze Stelle ist gebräunt.
Gelingt es den Überwallungswülsten, sich über der Wundfläche
mit den Rändern zu erreichen, so verschmelzen schließlich ihre
Kambien zu einer einheitlichen Meristemschicht, die weiter eine
zusammenhängende Holzschicht nach innen und eine Bastschicht nach
außen bildet. Das deckende Holz verwächst nicht mit dem bei der
Verwundung bloßgelegten. Letzteres ist ja gebräunt und abgestorben.
Daher lassen sich in Stämme eingeschnittene Zeichen, die bis auf
den Holzkörper reichen, nach ihrer Überwallung wiederfinden. Durch
Überwallung vom Kambiummantel aus werden auch Aststümpfe mehr oder
weniger vollständig abgeschlossen. Das über den Wunden erzeugte
Holz ist in seinem Bau von normalem Holz zunächst verschieden,
wird daher als Wundholz bezeichnet. Es besteht aus fast
isodiametrischen Zellen, auf die erst allmählich gestrecktere
Formen folgen. Beim Kirschbaum erzeugt das Kambium infolge von
Verwundungen statt normaler Holzelemente Nester dünnwandiger
Parenchymzellen, die in Gummibildung eintreten (S. 33).
[S. 141]
2.
Anpassungen des Kormus an die Lebensweise und an die Umwelt[100].
Der äußere und innere Bau des Kormus steht in engen Beziehungen zu
seiner Lebensweise und diese wieder zur Außenwelt. Wir finden, daß fast
jede Pflanze durch ihren äußeren und inneren Bau an ihre Lebensweise
und an ihre gewohnte Umwelt, an ihren Standort, angepaßt ist.
Darauf vor allem beruhen die gemeinsamen Züge, die einheitliche
Physiognomie, der Gewächse eines Standortes und darauf auch
die Unterschiede in der Physiognomie der Vegetation zwischen solchen
Pflanzenstandorten auf der Erde, wo die Außenfaktoren, z. B. das
Klima, sehr verschieden sind. Infolgedessen sind die Vegetationsorgane
nicht bei allen Kormophyten typisch ausgebildet. Oft sehen sie
ganz anders aus, da die Grundformen in verschiedenster Weise
abgeändert, metamorphosiert sein können. Erst eine sehr genaue
entwicklungsgeschichtliche oder anatomische Untersuchung ermöglicht
es, bei vielen Kormophyten nachzuweisen, daß auch ihre ganz abweichend
gestalteten Organe immer wieder nur Metamorphosen der drei Grundorgane
des Kormus: Wurzel, Stengel, Blatt sind, und festzustellen, welcher
Grundform sie homolog sind. Der äußere Bau und die Funktionen
der fertig ausgebildeten Organe können nämlich sehr leicht irreführen,
weil nicht selten ein Grundorgan, z. B. eine Sproßachse, Bau und
Funktionen eines anderen, z. B. eines Blattes, übernommen hat, oder
weil verschiedene Grundorgane zu Werkzeugen von gleichem Bau
und gleicher Funktion umgebildet worden sind, also bloß analog,
aber nicht homolog sind. Bei Berücksichtigung aller morphologischen
Eigenschaften eines abgeänderten Organes wird man aber in der Regel
keinen Zweifel über seine Abkunft hegen.
Die Form einer Pflanze und die Ausbildung ihrer Teile ist in
allererster Linie an die Ernährungsweise des
Gewächses angepaßt. Infolgedessen sind wichtige und sehr auffällige
morphologische Unterschiede zwischen solchen Kormophyten vorhanden,
die mit anorganischen Nährstoffen auskommen (autotrophe K.) und
solchen, die organische Nahrung nötig haben (heterotrophe K.).
A. Autotrophe Kormophyten.
An Autotrophie sind die grünen Pflanzen in ihrem Bau
angepaßt. Soweit sie Kormophyten sind, wurden ihre typischen
Baueigentümlichkeiten im letzten Abschnitte besprochen. Die grünen
Kormophyten können aber untereinander wieder sehr verschieden gebaut
sein; denn sie sind in ihrem Bau auch noch an ihre Umwelt angepaßt, in
der sie leben, und diese Umwelt kann recht mannigfaltig sein.
Von allen den vielen Faktoren, die in der Umwelt wechseln können,
hat den bei weitem größten Einfluß auf die Gestaltung der grünen
Pflanzen das Wasser, nächstdem das Licht. Das ist leicht
begreiflich. Nur wenn genügend Wasser verfügbar ist, vermag ja, wie
jedermann weiß, die Pflanze ihr Leben zu fristen. Und nur wenn sie
genügend Licht aufnehmen kann, kann sie autotroph sein, organische
Substanz aus anorganischer aufbauen.
a)
Anpassungen an den Feuchtigkeitsgehalt der Umwelt.
1. Wasserpflanzen, Hydrophyten[101]. Zu besonderen
Baueigentümlichkeiten führte das Leben im Wasser, wo dauernd mit der
ganzen Oberfläche der Pflanze, also auch mit Stamm und Blättern, Wasser
und Nährsalze und nur aus dem Wasser die nötigen Gase (Kohlensäure
und Sauerstoff) aufgenommen werden können. Zum Verständnis der
Lebensbedingungen im Wasser ist es unerläßlich, den Gehalt des Wassers
an diesen Gasen zu kennen[S. 142] und mit dem der Luft zu vergleichen. Das
Liter Luft enthält etwa 210 ccm Sauerstoff und 0,3 ccm Kohlensäure.
Im Liter Wasser sind dagegen z. B. bei 20° (im Falle der Sättigung)
nur etwa 6 ccm Sauerstoff, dagegen wiederum etwa 0,3 ccm Kohlensäure
vorhanden. Den Wasserorganismen steht also zwar ungefähr ebensoviel
Kohlensäure (oder meist sogar noch etwas mehr) wie in der Luft, aber
sehr viel weniger Sauerstoff zur Verfügung, wenigstens im unbewegten
Wasser, da der Sauerstoff im Wasser sehr langsam diffundiert.
Nicht selten sind die Wurzeln überhaupt nicht mehr (Utricularia,
Ceratophyllum, Wolffia) oder nur schwach ausgebildet. Der Sproß aber
ist hier anatomisch insofern der Wurzel ähnlich geworden, als er auf
seinen stets sehr dünnen Epidermisaußenwänden nur eine sehr zarte
Kutikula ausbildet, die dem Wasser- und Salzeintritt, wenigstens an den
Blättern[102], kaum Schwierigkeiten entgegenstellt. Zur Langsamkeit
der Gasdiffusion im Wasser und zur Armut des Wassers an Salzen steht
aber wohl in Beziehung eine Oberflächenvergrößerung der meist
sehr dünnen, submersen Blätter, häufig durch feine Zerteilung
der Blattflächen (Batrachium Fig. 139, Utricularia, Myriophyllum,
Ceratophyllum), während die Schwimm- und Luftblätter meist typisch
ausgebildet sind (Heterophyllie, vgl. S. 102). Anatomisch sind die
submersen Blätter von diesen Blättern dadurch verschieden, daß
ihrer chlorophyllhaltigen Epidermis meist die Spaltöffnungen und in
der Regel auch die Haare fehlen, ferner dadurch, daß ihr an großen
Interzellularen reiches Mesophyll aus gleichartigem Parenchym besteht,
also nicht in Palisaden- und Schwammparenchym differenziert ist;
infolgedessen zeigen die Blätter im inneren Bau bilaterale
Symmetrie (Fig. 185). Mit dem Mangel der Transpiration und eines
lebhaften Wassertransportes bei untergetauchten Wasserpflanzen
geht Hand in Hand die schwache Ausbildung der wasserleitenden
Elemente in den Stengeln und Blättern, ferner der Mangel eines
sekundären Dickenwachstums. Der Auftrieb im Wasser macht auch das
Festigungsgewebe unnötig; höchstens in rasch strömendem Wasser wird für
die nötige Zugfestigkeit durch zentrale Lagerung der Leitbündel gesorgt.
Eine schwache Wasserdurchströmung der Pflanze, verbunden mit
Wasserausscheidung durch die Apikalöffnungen der Blätter (vgl. S.
99), läßt sich aber selbst bei vielen submersen Wasserpflanzen
nachweisen.
Auffallend bei fast allen Wasser- und auch Sumpfpflanzen ist die
mächtige Entwicklung der Interzellularen. Sie sind ganz
besonders weit, bilden ein System von oft sehr regelmäßig geformten
Luftkammern und Luftkanälen und werden durch parenchymatische
Zellschichten getrennt, die meist nur eine Zellage stark sind; so in
den Stengeln von Papyrus, Potamogeton u. a., den Blattstielen der
Nymphaeaceen, den Wurzeln der Onagracee Jussieua. Man hat solche
Gewebe als Luftgewebe oder Aërenchym bezeichnet; denn solche weiten
Luftkanäle sind Luftspeicher, zugleich ermöglichen sie eine sehr rege
Gasdiffusion im Innern des Pflanzenkörpers: den schnellen Transport von
Sauerstoff aus den assimilierenden grünen Teilen in die nicht grünen;
sie begünstigen also die Atmung.
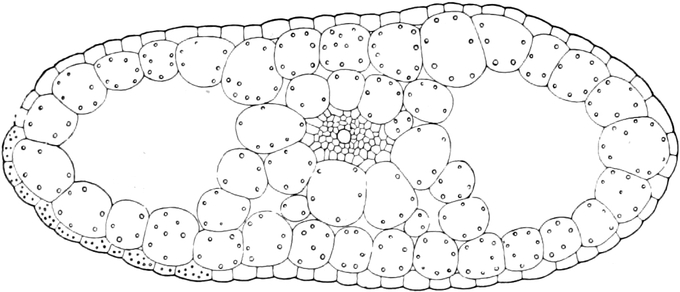
Fig. 185. Blattquerschnitt von Zanichellia palustris.
Vergr. 146. Nach SCHENCK.
[S. 143]
Bei manchen Sumpfpflanzen, die mit ihren unterirdischen Organen
in sehr sauerstoffarmem Sumpfboden leben, so z. B. bei gewissen
Palmen und bei Mangrovepflanzen der tropischen Küstensümpfe,
von denen einige durch ein System sproßbürtiger, stelzenförmiger
Luftwurzeln (Stelzwurzeln) sich im Schlamme verankern (Fig.
187)[100], werden besondere Organe zur Zuführung von Sauerstoff
ausgebildet: aus dem Sumpfboden aufsteigende Atemwurzeln
(Pneumatophoren Fig. 186), die aus der Luft durch
Lentizellen-ähnliche Öffnungen (Pneumathoden) Sauerstoff aufnehmen
und durch Luftgewebe zu den unterirdischen Teilen gelangen lassen.

Fig. 186. Atemwurzel von Sonneratia alba. Verkleinerung
einer Abbildung in den „Vegetationsbildern“; nach JOH.
SCHMIDT.

Fig. 187. Stelzwurzeln bei Rhizophora mucronata.
Malayischer Archipel. Nach G. KARSTEN.
2. Die Landpflanzen. Sie nehmen meist nur aus dem Boden
Wasser und Nährsalze, aus der Luft Kohlensäure, dagegen
Sauerstoff sowohl aus dem Boden wie aus der Luft auf und geben mit
ihren Luftsprossen Wasser in Dampfform (durch Transpiration) an den
Luftraum ab.
Eine Ausnahme bilden nur einige Gewächse sehr feuchter
Örtlichkeiten, die mit ihrer ganzen Oberfläche Wasser
absorbieren; besonders die Hymenophyllaceen der tropischen
Regenwälder. Manche von ihnen bilden keine Wurzeln aus, besitzen
aber an den Wasser aufnehmenden Blättern oder Stengeln oder beiden
ein System von verschieden gestalteten Haaren, das eine gewaltige
Oberflächenvergrößerung bedingt.
Die Landpflanzen haben verschiedenen Bau, je nachdem es Pflanzen
immerfeuchter Standorte, Pflanzen trockener Standorte und Klimate oder
Pflanzen wechselfeuchter Klimate sind.
a) Anpassungen an immerfeuchte Standorte, Hygrophyten[103].
Landpflanzen, die dauernd in sehr feuchter Atmosphäre
leben (Hygrophyten, Hygrophile), z. B. viele tropische
Schattenpflanzen, bedürfen wie die Wasserpflanzen keine Einrichtungen,
die die Transpiration herabsetzen, ja haben sogar, scheint es, solche,
die die Wasserabgabe der Luftsprosse fördern (Hygromorphie). Nur
durch solche Einrichtungen dürfte die genügend starke Wasserbewegung
aus den Erdwurzeln in die oberirdischen Organe gesichert sein, die zur
Beschaffung und zur Fortbewegung der nötigen Nährsalzmenge erforderlich
ist. Im äußeren und inneren Bau gleichen viele Hygrophyten in mancher
Hinsicht den Wasserpflanzen, und zwar um so mehr, an je feuchteren
Standorten sie gedeihen.
[S. 144]
Wir finden bei den Hygrophyten mannigfaltige Einrichtungen, die
der Transpiration günstig sind: Ausbildung großer und dünner
Blattflächen, schwache Ausbildung der Kutikula, Verlegung der
Stomata auf exponierte Vorsprünge, wo sie über die Epidermis
emporgehoben sind, schließlich Baueigentümlichkeiten des Blattes,
die es, wie STAHL zeigte, nach Regengüssen möglichst
schnell wieder trocken werden lassen. Eine vorgezogene Spitze, mit
der die Spreiten oft abschließen, die Träufelspitze, oder
die Unbenetzbarkeit des Blattes durch Wachsüberzüge erleichtern
das Abfließen des Wassers; eine Samtoberfläche, durch Papillen
bedingt, breitet Wassertropfen kapillar zu einer äußerst dünnen,
schnell verdunstenden Schicht aus. Ferner hebt vielleicht, nach
STAHL, die Ausbildung Lichtstrahlen absorbierender
Farbstoffe, die Buntblättrigkeit, durch Wärmeabsorption die
Temperatur des Blattes über die Umgebung, so daß die Transpiration
am Tage auch noch in dampfgesättigter Luft unterhalten werden
kann. In der Abgabe tropfbar flüssigen Wassers, (der sog.
Guttation) aus wasserausscheidenden Organen der Blätter, den
Hydathoden, besitzen manche dieser Pflanzen vielleicht ein
Mittel, auch nach völliger Hemmung der Transpiration noch genügend
Wasser abzugeben. Diese Organe sind entweder Drüsenhaare oder
-flächen, die Wasser sezernieren, oder Wasserspalten, aus denen
Wasser von den Leitbündeln her ausgepreßt wird (vgl. Fig. 134).
b) Anpassungen an physiologisch trockene Standorte oder an
trockene Klimate, Xerophyten[100], [104]. Dagegen bedürfen die
Landpflanzen um so mehr Einrichtungen, die auf Hemmung der
Wasserabgabe berechnet sind, in je trockenerer Luft ihre Sprosse
wachsen und je schwerer zugleich ein hinreichend schneller und großer
Wassernachschub möglich ist. Denn die Einschränkung der Transpiration,
die unter ungünstigen Verhältnissen auch in allen anderen Klimaten
durch Verschluß der Stomata bewirkt wird, würde für Pflanzen besonders
trockener Klimate und Standorte nicht ausreichen. Austrocknungsfähig
aber (vgl. S. 189), wie Flechten und Moose, sind nur wenige
Kormophyten; die meisten werden schon durch starkes Welken getötet.
Pflanzen mit solchen Einrichtungen zur Hemmung der Wasserabgabe nennt
man Xerophyten (Xerophile). Man erkennt sie schon an ihrem
Habitus. Die Summe von morphologischen Eigenschaften, die solche
Einrichtungen bilden, fassen wir in dem Worte „Xerophytenbau“
(Xeromorphie) zusammen. Besonders extreme Xerophyten sind natürlich
viele Wüstenpflanzen, die Gewächse trockener Felsen und viele Epiphyten
(vgl. S. 156).
Sehr auffallend ist aber, daß mehr oder weniger ausgeprägter
Xerophytenbau auch bei Pflanzen von ganz anderer Lebensweise vorkommt,
wo er gar nicht oder wenigstens nicht auf den ersten Blick verständlich
erscheint, so bei den Pflanzen der Hochgebirge und des hohen Nordens,
bei vielen Sumpfpflanzen, bei den Pflanzen des Meeresstrandes,
namentlich den Salzpflanzen (Halophyten) (Fig. 192), selbst solchen,
die, wie die Mangrovegewächse tropischer Küsten, direkt im Wasser
wachsen. Wenn auch hier noch manche Frage ungelöst ist, wird man
doch annehmen dürfen, daß die Mehrzahl dieser Gewächse wenigstens
zeitweise in Gefahr ist, mehr Wasser durch Transpiration
abzugeben, als sie aus dem Boden aufzunehmen oder in die Blätter zu
schaffen vermögen. Wenn sie auch in verhältnismäßig feuchten Böden
leben, so scheinen diese für sie doch, zum mindesten vorübergehend,
mehr oder weniger physiologisch trocken zu sein, d. h. die
Wasseraufnahme zu erschweren.
Es gibt anatomische und morphologische Einrichtungen, die die
Transpiration herabsetzen. Ein Teil von ihnen mag zugleich als
Schutzmittel gegen zu starke Lichtbestrahlung oder gegen zu hohe
Erwärmung anzusehen sein.
Anatomische Einrichtungen, die die Transpiration
einschränken, sind: Verstärkung der Kutikula und der kutinisierten
Epidermisaußenwände; verschleimte Epidermiszellen; Ausbildung von
Wachs-, Harz- oder Kalküberzügen, bei Stämmen oder Wurzeln auch von
Korkschichten; Verringerung der Spaltöffnungszahl; Verengung der
Spaltöffnung oder ihre Verstopfung durch Harz- oder Wachsmassen;
Einsenkung der Spaltöffnungsapparate einzeln[S. 145] in Vertiefungen der
Epidermis (Fig. 188) oder in größerer Zahl in besondere krugförmige
Vertiefungen der Blattunterseite, die nur durch enge Kanäle nach außen
münden, wie beim Oleander, ferner ihre Überwölbung durch Nachbarzellen.
Endlich werden als Schirm gegen die Sonnenstrahlen Filzhaare, Stern-
und Schuppenhaare ausgebildet, die sich frühzeitig mit Luft füllen
und dadurch der Pflanze eine weiße oder graue Farbe verleihen
(Edelweiß, Proteaceen Australiens, Ölbaum). Oder die immergrünen
Blätter sind klein, dicklich, lederartig und verhältnismäßig saftarm
(Hartlaubgewächse, Sklerophyllen des Mediterrangebietes,
z. B. Ölbaum, Myrte). Bezeichnend für die Blätter ausgesprochener
Sklerophyllen (Fig. 191) ist die Kleinheit der Interzellularen
im Mesophyll, das meist sehr kleinzellig ist und vielfach kein
Schwammparenchym enthält, sondern oft nur aus Palisadenzellen besteht
oder doch zum mindesten ober- und unterseits mehrere typische Schichten
solcher Zellen aufweist, so daß die Blätter vieler Sklerophyllen
bilateral symmetrischen Bau besitzen. Einige Xerophyten
verzichten aber auf jeden Transpirationsschutz durch anatomische
Charaktere, weil sie durch ihren hochkonzentrierten Zellsaft zu einer
Wasseraufnahme auch aus verhältnismäßig trockenem Boden befähigt sind
(S. 195).
Meist verbinden sich diese Einrichtungen mit morphologischen
Besonderheiten des äußeren Baues, die ebenfalls die
Transpiration herabsetzen.
Viele kleinblättrige Xerophyten zeichnen sich durch Polsterwuchs
aus (Polsterpflanzen, z. B. viele Alpenpflanzen, Fig. 189),
wodurch schon die Transpiration eingeschränkt und zugleich ein Schutz
gegen zu starke Bestrahlung gewährt wird.
Ein sehr wirksamer Transpirations- und Lichtschutz wird durch
Einstellung der Blattflächen in die Vertikalebene erzielt,
vor allem bei neuholländischen Akazien und Myrtaceen; bei den
Akazien verbunden mit Reduktion der eigentlichen Blattflächen und
blattförmiger Ausbildung der Blattstiele (Phyllodien,
Fig. 140 und Fig. 190). Auch unter den einheimischen Pflanzen
findet sich ähnliche Einstellung der Blätter: so z. B. bei der als
Kompaßpflanze bekannten Lactuca scariola, die sämtliche Blätter
vertikal, und zwar in eine Nord-Süd-Ebene stellt. An solchen Blättern
gleiten bei höchstem Stand der Sonne die Lichtstrahlen mehr oder
weniger seitlich vorbei. Dadurch wird vielleicht eine übermäßige
Erwärmung und Transpiration verhindert.
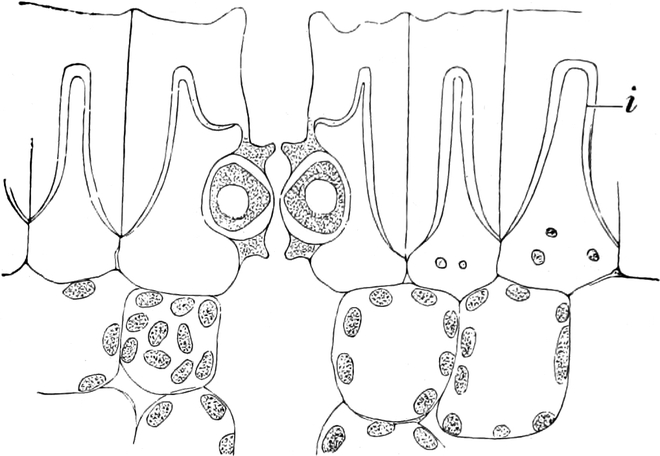
Fig. 188. Querschnitt durch die Epidermis von Aloë
nigricans. i Innere, nicht kutinisierte Verdickungsschicht. Vergr.
240. Nach STRASBURGER.
Fig. 189. Raoulia mamillaris (Composite) von
Neuseeland. Polsterförmiger Zusammenschluß der Einzelpflanzen. Aus
SCHIMPER, Pflanzengeographie.
[S. 146]
Sehr häufig werden die Blattspreiten reduziert. Bei den
Steppengräsern geschieht dies durch Einrollung ihrer freien Oberfläche
(Fig. 192); bei Ericaceen, Genisteen, Cupressaceen und gewissen
habituell ihnen ähnlichen Veronica-Arten Neuseelands (vgl. auch Fig.
193) durch Verkleinerung der Spreiten; bei Kakteen, baumartigen
Euphorbia-Arten, einigen Asclepiadaceen durch völliges Schwinden
der Spreiten. Mit der Verkleinerung der Blätter muß aber außer der
Transpiration auch die Assimilation des Kohlenstoffs abnehmen; so sehen
wir zur Kompensation des Verlustes Chlorophyllparenchym in den Stengeln
dieser Pflanzen auftreten. In diesem Falle sind die Sproßachsen grün
gefärbt; so z. B. bei Sarothamnus scoparius, der an seinen langen,
rutenförmigen Zweigen nur vereinzelte hinfällige, lanzettliche
Blättchen entwickelt (Rutengewächse, Sklerokaulen).
Fig. 190. Acacia marginata mit senkrecht gestellten
Phyllodien. Aus SCHIMPER, Pflanzengeographie.
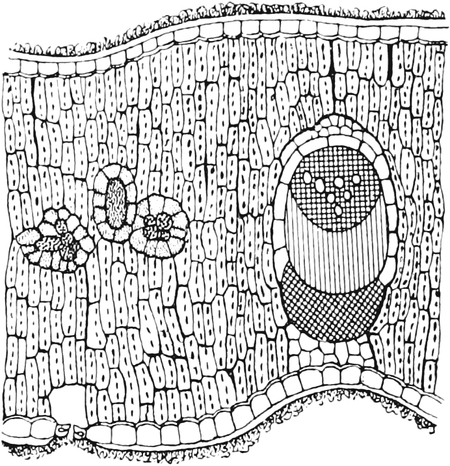
Fig. 191. Querschnitt des Blattes der Capparidacee
Capparis spinosa var. aegyptiaca. Vergr. 40. SCHIMPER nach
VOLKENS.
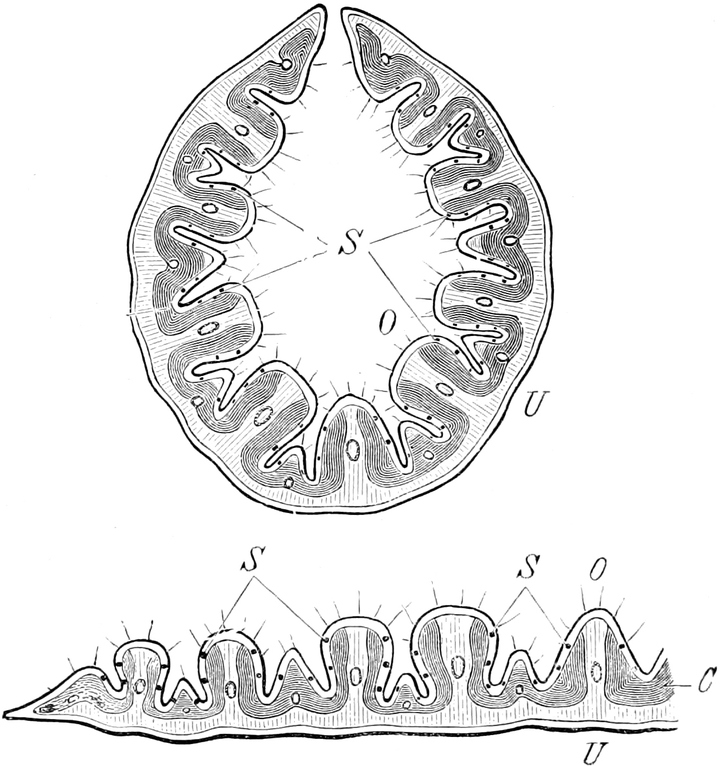
Fig. 192. Blattquerschnitte des Grases Stipa capillata.
Oben ein Blatt in geschlossenem Zustande, unten die Hälfte eines
offenen Blattes. U Spaltöffnungslose Unterseite. O Oberseite mit
Spaltöffnungen (S). C Chlorophyllhaltiges Assimilationsgewebe.
30fach vergr. Nach KERNER V. MARILAUN.
Oft geht Hand in Hand mit einer Reduktion der Blätter eine
Abflachung und blattähnliche Ausbildung der grünen Sproßachsen,[S. 147]
die alsdann weit vollkommener als die rutenförmigen die
Assimilationsfunktionen des Blattes übernehmen können (Fig. 194, 195),
aber natürlich zugleich auch wieder stärker transpirieren. Solche
blattartige Sprosse werden als Flachsprosse, Kladodien
oder Phyllokladien bezeichnet. GOEBEL schlägt vor, jene
Flachsprosse, die begrenzt wachsen und dadurch besonders blattähnlich
werden, Phyllokladien, andere verbreiterte Sproßachsen Kladodien
zu nennen. Ein lehrreiches Beispiel für Phyllokladien ist Ruscus
aculeatus, ein kleiner xerophytischer Strauch des Mittelmeergebietes,
der an seinen Zweigen (Fig. 194) in den Achseln schuppenförmiger
Blätter (f) breite, in eine scharfe Spitze auslaufende Phyllokladien
(cl) trägt, die durchaus den Eindruck von Blättern machen. Ihrer
Oberfläche entspringen manchmal in der Mittellinie, annähernd in ihrer
halben Länge, aus der Achsel eines schuppenförmigen Blattes eine bis
zwei Blüten. Diese Phyllokladien sind ein sehr schönes Beispiel für
Analogie von Organen: sie sehen aus und funktionieren wie
Blätter, sind aber Sproßachsen, wie ihre geschilderten morphologischen
Eigenschaften lehren. Eine blattartige Abflachung des gesamten massig
entwickelten Stammes, also Kladodienbildung, mit Verschmälerung an den
Verzweigungsstellen, zeigen uns die bekannten Opuntien (Fig. 195).
Fig. 193. Salicornia herbacea (Chenopodiacee), eine
ausgesprochene Salzpflanze. Aus SCHIMPER, Pflanzengeographie.
Fig. 194. Zweig der mediterranen Liliacee Ruscus
aculeatus. f Blatt, cl Phyllokladium, bl Blüte. Nat. Größe. Nach
SCHENCK.
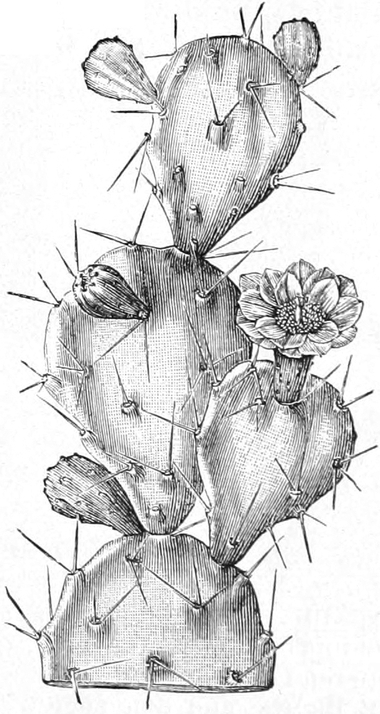
Fig. 195. Opuntia monacantha Haw. Kladodien mit
Blattdornen, Blüte und Früchten. Nach SCHUMANN. Etwa fünfmal
verkleinert.
Besonderes Interesse bieten schließlich einige Xerophyten, bei
denen nicht allein die Blätter, sondern auch die Sproßachsen ganz
schwinden. Dann übernehmen die abgeplatteten, grünen Wurzeln, aus
denen die Vegetationsorgane allein bestehen, die Funktionen der Blätter
(so bei der epiphytischen Orchidee Taeniophyllum, Fig. 196).
[S. 148]
Fig. 196. Taeniophyllum Zollingeri. Xerophytische
Orchidee ohne Blätter und ohne Sproßachsen, mit grünen, abgeflachten
Wurzeln. Nat. Gr. Aus SCHIMPER, Pflanzengeographie; nach
WIESNER.
Mit dem großen Reichtum an Sklerenchym, der für die Sprosse vieler
Xerophyten bezeichnend ist und ihre nötige Festigkeit auch im
Falle schwächeren oder stärkeren Welkens sicherstellt, steht oft
in Zusammenhang die Ausbildung von Dornen, die aber auch
bei einigen nicht xerophytisch gebauten Pflanzen unserer Klimate
vorkommen. Das sind sehr spitze, verholzte und infolgedessen starre,
verzweigte oder unverzweigte, pfriemförmige Gebilde, die durch
Umwandlung von Blättern oder Blatteilen (Blattdornen) oder
von Sprossen (Sproßdornen) oder in sehr seltenen Fällen von
Wurzeln (Wurzeldornen) entstanden sind. Beim Sauerdorn (Berberis
vulgaris) wird das ganze Blatt an den Hauptsprossen in einen meist
dreistrahligen Dorn umgestaltet; die Seitentriebe, die entwickelte
Laubblätter tragen, stehen in den Achseln dieser Dornen. Auch bei
den Kakteen (Fig. 195) gehen die Dornen aus Blattanlagen hervor.
Bei der Robinie (Fig. 197) und bei manchen sukkulenten Euphorbien
entwickeln sich die beiden Nebenblätter zu Dornen. Sproßdornen treten
bei Prunus spinosa, Crataegus oxyacantha und den in unseren Gärten
häufig kultivierten Gleditschien (Fig. 198) auf. Bei Colletia cruciata,
einer strauchartigen amerikanischen Rhamnacee, die an trockenen,
sonnigen Standorten wächst, sind alle Sproßachsen domartig ausgebildet,
zugleich etwas abgeflacht und grün; sie ersetzen die bald abfallenden,
kleinen Blätter. Wurzeldornen kommen bei einigen Palmen am Stamme vor
(z. B. Acanthorrhiza) und bei Myrmecodia unter den Dikotylen. Der
Dornsproß ist vor allem vielen Xerophyten der Wüsten und Steppen
eigentümlich.
Fig. 197. Stammstück von Robinia Pseudacacia mit dem
unteren Teile eines gefiederten Blattes und den zugehörigen, in zwei
Dornen verwandelten Nebenblättern n. g Gelenkpolster. 1⁄2 nat. Gr.
Nach SCHENCK.
Fig. 198. Sproßdorn von Gleditschia triacanthos. 1⁄2
nat. Gr. Nach SCHENCK.
Oft besitzen blattlose Xerophyten angeschwollene Blätter oder
Stämme: so die grünen säulenförmigen, prismatischen oder
zylindrischen, keulen- oder kopfförmigen Euphorbien und Kakteen. Viele
Xerophyten begnügen sich nämlich nicht damit, die Abgabe von Wasser
stark einzuschränken, sondern speichern Wasser in besonderen
Geweben während der Tage, wo es zur Verfügung steht, für die Zeiten der
Not. Solches Wassergewebe zeichnet sich bei typischer Ausbildung
durch große, chlorophyllfreie Zellen aus, die fast nur aus der riesigen
Vakuole bestehen. Im Grunde genommen[S. 149] ist jede Epidermiszelle ein
Wasserspeicher. Manchmal nehmen aber die Epidermiszellen ungewöhnliche
Dimensionen an, so daß sie die Hauptmasse eines Blattes ausmachen,
oder teilen sich auch parallel zur Oberfläche und werden zu einem
ein- bis mehrschichtigen (äußeren) Wassergewebe (verschiedene
Piperaceen, Begoniaceen, Ficus-Arten, Tradescantia). Sehr häufig ist
aber das Wassergewebe mehr zentral gelegen (inneres
Wassergewebe). Wo Wassergewebe sehr mächtig ausgebildet sind, bekommen
die betreffenden Organe dadurch einen fleischig-saftigen
Charakter. Daher nennt man solche Pflanzen „Sukkulenten“. Bei
gewissen Umbelliferen, Cucurbitaceen, Kompositen, Asclepiadaceen,
ferner Angehörigen der Gattungen Pelargonium und Oxalis der Steppen
und Wüsten sind Wurzeln zu Wasserspeichern ausgebildet.
Häufiger findet man Blattsukkulenten: Sedum, Sempervivum,
Agave, Aloë, Mesembryanthemum; oder Stammsukkulenten: Kakteen,
Euphorbia-Arten, Stapelia und andere Asclepiadaceen (Fig. 195, 199),
Kleinia unter den Kompositen. Ganz besonders bezeichnend für dürre
Gegenden in der neuen Welt sind die kugelförmigen oder säulenartigen
Kakteen, in der alten die säulenförmigen oder kandelaberartigen
Euphorbien und Asclepiadeen, die gewissen Kakteen zum Verwechseln
ähnlich sehen. Die Ähnlichkeit in der Lebensweise hat hier aus
ganz entfernt stehenden Pflanzen fast übereinstimmende Gestalten
geschaffen (vgl. Fig. 199 a–c), eine Erscheinung, die
außerordentlich häufig wiederkehrt (Konvergenz). Im Extrem
nähert sich der Stamm oder das Blatt der Sukkulenten der Kugel,
nimmt also bei gegebenem Volumen die kleinste Oberfläche an, wodurch
die Transpiration natürlich ganz besonders stark vermindert wird:
NOLL hat berechnet, daß die Verdunstung eines Kugelkaktus
600mal geringer ist als die einer gleichschweren Schlingpflanze
(Aristolochia sipho). Solche und ähnliche Sukkulentenstämme
sind ausgesprochene Sproßknollen, d. h. sehr geräumige
Speicherorgane für Wasser.

Fig. 199. Stammsukkulenten: a Stapelia grandiflora
(Asclepiadacee), b Cereus Pringlei (Cactacee), c Euphorbia erosa
(Euphorbiacee). 1⁄5 nat. Gr.
c) Anpassungen an wechselfeuchte Klimate. Tropophyten[100].
In einigen dauernd feuchtwarmen Tropengegenden ist das Klima jahraus
jahrein dem Pflanzenwachstume nahezu gleich günstig. Überall
sonst aber, wo eine ausgesprochene Klimaperiodizität, ein
regelmäßiger Wechsel zwischen einer dem Pflanzenwachstum günstigen
Vegetationsperiode und einer ihm mehr oder weniger schädlichen
Jahreszeit (Ruheperiode) besteht, die bald als trockne, bald als
kalte Zeit (Winter) ausgebildet sein kann, entspricht diesem Wechsel
eine auffallende Periodizität in den Lebensvorgängen der
Pflanzen. Darauf ist es auch zurückzuführen, daß viele Gewächse
solcher Klimate in mancher Hinsicht anders gebaut sind als diejenigen
gleichmäßig feuchtwarmer Tropengegenden. Nur solche Formen werden ja
dort gedeihen können, die in irgendeiner[S. 150] Weise den in der ungünstigen
Jahreszeit drohenden Gefahren gewachsen sind. Letztere bestehen aber in
allererster Linie auch da, wo ein sehr kalter Winter mit einem Sommer
abwechselt, in einem Tode durch Vertrocknung während der in den
Frostperioden sehr trockenen Winterszeit. Ein solcher Tod droht
freilich extremen Xerophyten nicht, weil ihre Standorte auch in der
günstigen Jahreszeit dauernd oder zeitweilig sehr trocken sind;
sie können oft ohne weiteres die Trockenperiode überdauern. Anders
steht es dagegen um die Gewächse, die in der günstigen Jahreszeit
nicht an trockenen Standorten gedeihen und nicht
xerophytischen Bau besitzen. Zunächst sind ihre Transpirationsorgane,
die Blätter, in der Trockenzeit gefährdet. So sehen wir denn als
sehr auffallende Eigentümlichkeit dieser Gewächse wechselfeuchter
Klimate, daß vor oder zu Beginn der schlechten Zeit die Blätter von
den Sproßachsen abgeworfen werden (laubabwerfende, sommergrüne
Holzgewächse) oder sogar mit einem Teil der Sproßachsen absterben
(sommergrüne Kräuter). Das genügt aber vielfach noch nicht, die Pflanze
vor dem Vertrocknungstode und die Art vor dem Aussterben zu bewahren;
denn auch die embryonalen Gewebe, die zu Beginn der guten
Jahreszeit die verloren gegangenen Teile ersetzen müssen, würden in der
Trockenzeit vertrocknen, wenn sie nicht irgendwie besonders geschützt
würden.
Solche Schutzeinrichtungen, bestimmt die Pflanze über eine
vegetationsfeindliche Zeit hinüber zu retten, kennzeichnen die Gewächse
wechselfeuchter Klimate, sofern sie nicht ausgesprochene Xerophyten
sind, so auch die meisten Kormophyten unserer Heimat. In der
günstigen Jahreszeit können sie einen wirksamen Transpirationsschutz
oft ganz entbehren und insofern Hygrophyten gleichen; in der
ungünstigen aber sind sie den extremsten Xerophyten ähnlich. Diesem
Wechsel im Aussehen verdanken die Tropophyten ihren Namen.
Entsprechend ist auch der anatomische Bau der ausdauernden Teile
xerophil, der nur in der feuchten Jahreszeit vorhandenen dagegen
hygrophil.
Die Gewächse wechselfeuchter Klimate können ausdauernde Holzgewächse:
Bäume und Sträucher, ausdauernde Krautgewächse (Perenne) und einjährige
Kräuter (Annuelle) sein. Jede dieser Gruppen hat ihre besonderen
Schutzeinrichtungen gegen Vertrocknung ihrer Transpirationsflächen und
ihrer embryonalen Gewebe.
1. Die Holzgewächse wechselfeuchter Klimate werfen mit
Ausnahme einiger weniger immergrüner, deren Blätter Xerophytenstruktur
besitzen (z. B. Koniferen, Ilex), am Ende der Vegetationsperiode
das Laub ab und schließen ebenso wie die erwähnten Immergrünen im
Gegensatze zu vielen, wenn auch nicht allen tropischen Formen, ihre
Sproßvegetationspunkte während der Trocken- oder Winterzeit in
schützende Gehäuse, in Winterknospen ein (Fig. 200).
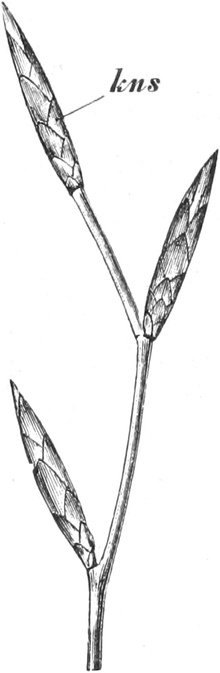
Fig. 200. Winterknospen der Rotbuche (Fagus silvatica). kns
Knospenschuppen. Nat. Gr. Nach SCHENCK.
Diese Gehäuse werden meist von fest zusammenschließenden
Knospenschuppen, Niederblättern, gebildet. Manchmal
werden dazu ganze Primordialblätter verwendet, die in
ungegliedertem Zustande verblieben sind. Am häufigsten aber
gehen die Schuppen aus dem Blattgrunde hervor, der sich
entsprechend vergrößert und ausgestaltet. Das Oberblatt kommt
alsdann entweder gar nicht zur Entwicklung oder sitzt in mehr oder
weniger reduziertem Zustande am Scheitel der Knospenschuppe. Eine
im Frühjahr austreibende Winterknospe der Roßkastanie (Aesculus
hippocastanum) zeigt dies unmittelbar; denn während ihre äußeren
Knospenschuppen an den Spitzen kaum etwas von einem[S. 151] Oberblatte
erkennen lassen, tragen die inneren Knospenschuppen oft schon
deutlich eine kleine Blattspreite. In anderen Fällen sind die
Knospenschuppen ihrem Ursprunge nach Nebenblätter, gehören also
auch dem Blattgrunde an (so bei der Eiche). Nicht selten ist auch
das Deckblatt an dem Schutz seiner Achselknospe beteiligt, indem
sein Blattgrund (oder die Basis seines Blattstieles) nach dem
Blattfall am Sproß sitzen bleibt und kappenförmig die Winterknospe
deckt. Bei Robinia ist dieser Blattgrund die einzige Schutzhülle
der Winterknospe.
Die Knospenschuppen werden lederartig dick und hart und färben
sich gewöhnlich braun. Kork- und Haarüberzüge, Harz-, Gummi-
und Schleimausscheidungen sowie eingeschlossene Luftschichten
machen sie zu sehr wirksamen Schutzorganen der Knospen gegen
Austrocknung. Der Abscheidung von Harz usw. dienen eigenartig
gestaltete Haargebilde, die Leim- und Drüsenzotten
oder Kolleteren (vgl. Fig. 56). So scheiden solche auf den
Deckschuppen sitzenden Drüsenzotten in Winterknospen vieler unserer
Bäume, z. B. der Roßkastanie, ein Gemenge von Gummi und Harz ab,
das durch Zersprengung der Kutikula frei wird und sich zwischen die
Deckschuppen ergießt und sie verklebt. Wenn die Knospen im Frühjahr
aufbrechen, so werden die Knospenschuppen gewöhnlich abgeworfen
und bedecken alsdann den Boden. An den Jahrestrieben der Bäume
sind die untersten Internodien, die zwischen den Knospenschuppen
lagen, besonders kurz. Sie lassen die dicht gedrängten
Schuppennarben und so die Grenzen der aufeinanderfolgenden
Jahrestriebe erkennen.
2. Die perennierenden Kräuter (Stauden) wechselfeuchter
Klimate opfern nicht allein die Blätter, sondern auch zum mindesten
die Teile der Laubsprosse, welche höher in die Luft ragen und
der Vertrocknung ausgesetzt sind, mit den daran sitzenden Knospen.
Sie überwintern mit oberirdischen Knospen, die dicht
über der Erde liegen, wo sie durch Schnee oder fallendes Laub gegen
Vertrocknung geschützt werden, oder „ziehen ganz ein“ und überwintern
mit unterirdischen Knospen (Geophyten), die in noch viel
wirksamerer Weise, von feuchter Erde umgeben, vor Vertrocknung und
zugleich vor dem Erfrieren bewahrt werden können.
Wo oberirdische Erneuerungsknospen vorhanden bleiben, sitzen
sie entweder an oberirdischen, niederliegenden Sproßstücken (z. B.
Saxifraga, Stellaria Holostea, Thymus u. a.) oder an unterirdischen
Sprossen (Rhizomen), so bei unzähligen Pflanzen Mitteleuropas,
z. B. den ausdauernden Rosettenpflanzen, wie Bellis, Taraxacum,
Primula; den Zweijährigen oder Biennen, die mit einer Blattrosette
überwintern, wie z. B. Verbascum, Digitalis und vielen anderen. Wie
bei den Geophyten können auch bei solchen Stauden unterirdische
Speicherorgane für organische Reservestoffe vorkommen (vgl. S. 153).
Bei den Kräutern mit unterirdischen Überwinterungsknospen,
den Geophyten[105], haben die Teile, die die Knospen tragen,
entsprechend ihrem Leben im Boden und ihren besonderen Aufgaben
einen eigenartigen Bau. Es sind metamorphosierte Sprosse:
Wurzelstöcke (Rhizome), Sproßknollen, Zwiebeln
oder metamorphosierte Wurzeln (Wurzelknollen). Die im
Frühjahr austreibenden unterirdischen Erneuerungsknospen brauchen
organische Nahrungsstoffe, und zwar um so mehr, je länger der Weg
ist, den sie bis zur Bodenoberfläche zurückzulegen haben. Diese
Nahrungsstoffe werden in der vorausgehenden guten Zeit gebildet, ehe
die Luftsprosse absterben. Da das Speicherungsvermögen mit dem Volumen
wächst, so wird es begreiflich, daß die unterirdischen, fast nur aus
Speicherparenchym bestehenden Überwinterungsorgane bei vielen solchen
Gewächsen angeschwollen, dick sind. Solche Speicherorgane können
Sproßachsen, Blätter oder Wurzeln sein. Sie entleeren sich bei Beginn
der guten Jahreszeit ihrer Bestimmung gemäß, gehen danach, mit Ausnahme
vieler Rhizome, meist zugrunde und werden oft in sehr eigenartiger
Weise durch neue ersetzt. Speicherorgane von Pflanzen sind es, die
wegen ihres Reichtums an ausnutzbaren organischen Reservestoffen
vielen Tieren und uns Menschen als besonders wertvolle vegetabilische
Nahrungsmittel dienen.
a) Die Wurzelstöcke und sehr viele Sproßknollen sind
unterirdische farblose Sprosse. Jene sind verhältnismäßig
dünn oder dick, mit[S. 152] langen oder kurzen Internodien (Fig. 125,
143); die Sproßknollen (z. B. die Kartoffelknollen, Fig. 201) aber
sind sehr dick. Ihre Blätter (Niederblätter) sind, wie meist an den
Erdsprossen, als Schuppen ausgebildet. Die Reservestoffe werden in den
Sproßachsen gespeichert, weshalb diese oft angeschwollen sind.
An solchen Schuppen, dem Vorhandensein regelmäßig verteilter Knospen,
dem Fehlen von Wurzelhauben, endlich an ihrem inneren Bau lassen sich
die Rhizome und unterirdischen Sproßknollen von Wurzeln unterscheiden.
Meist sind die Rhizome, die bei manchen Pflanzen senkrecht oder schräg,
bei anderen horizontal im Boden wachsen und verzweigt oder unverzweigt
sind, dauernd mit Wurzeln bedeckt, während die Sproßknollen nach ihrer
Bildung zunächst gewöhnlich keine Wurzeln ausbilden; doch kommen alle
Übergänge zwischen Rhizomen und Sproßknollen vor.
Fig. 143 stellt den Wurzelstock von Polygonatum
multiflorum dar, der auch als Beispiel eines Sympodiums (S. 111)
schon angeführt wurde. Die mit c, d und e bezeichneten
Stellen entsprechen den Narben der oberirdischen Triebe dreier
vorausgegangener Jahre. In b ist die Basis des Stengels zu sehen,
der in Blüte stand, als das Rhizom dem Boden entnommen wurde; a
ist die Knospe für den nächstjährigen Trieb.
Fig. 201. Unterer Teil einer Kartoffelpflanze (Solanum
tuberosum). Die mittlere dunklere Knolle ist die in die Erde gesteckte
Mutterknolle, aus der sich die Pflanze entwickelt hat. 1⁄3 nat Gr.
Nach der Natur mit Benutzung eines BAILLONschen Bildes. Nach
SCHENCK.
Fig. 202. Zwiebel der Tulpe (Tulipa Gesneriana) im
Längsschnitt. zk Achse, zs Zwiebelschuppen, v Terminalknospe,
k Anlage einer jungen Zwiebel, w Wurzeln. Nat. Gr. Nach
SCHENCK.
Die Knollen der Kartoffelpflanze, von Colchicum autumnale
oder Crocus sativus sind Beispiele für unterirdische
Sproßknollen. Die Knollen der Kartoffelpflanze (Fig. 201)
oder des Helianthus tuberosus sind unterirdische Sprosse mit
angeschwollenen Achsen und reduzierten Blättern. Sie entstehen in
Mehrzahl aus den angeschwollenen Enden verzweigter unterirdischer
Triebe, Ausläufer (Stolonen), und dienen gleichzeitig der
Vermehrung der Mutterpflanze. Die an jeder Kartoffelknolle
sichtbaren, regelmäßig verteilten Vertiefungen bergen Achselknospen
(die Augen), die bestimmt sind, im kommenden Jahr auszutreiben.
Die kleinen schuppenförmigen Blätter, in deren Achseln die Augen
entstehen, sind nur an ganz jungen Knollen kenntlich. Nach
Ausbildung der Knollen geht die Mutterpflanze zugrunde; die in
jenen angehäuften Nahrungsstoffe dienen zum Aufbau der aus den
Augen sich entwickelnden Triebe.
[S. 153]
Bei den Herbstzeitlosen (Fig. 812) entsteht die neue Knolle an der
alten, und zwar aus einer Achselknospe seitlich an ihrer Basis,
beim Safran (Fig. 821) aus einer Achselknospe nahe am Scheitel;
daher sitzt bei den Herbstzeitlosen die jüngere Knolle seitlich
neben der alten, während sie beim Safran ihr aufgesetzt erscheint.
Auch der Rettich und das Radieschen sind (unterirdische)
Sproßknollen, allerdings nur aus einem Teil eines einzigen
Internodiums, nämlich des hypokotylen Stengelgliedes der
Keimpflanzen. An der Bildung dieser Knollen nimmt aber auch der
oberste Teil der Keimwurzel teil.
Eine oberirdische Sproßknolle, die aus vielen Internodien
eines Laubsprosses hervorgeht und reich an organischen
Reservestoffen ist, ist z. B. der Kohlrabi.
b) Eine Zwiebel (z. B. die Küchen-, Tulpen-, Hyazinthenzwiebel)
ist ein unterirdischer Sproß, dessen Achse scheibenförmig (Fig.
202 zk) abgeflacht ist und dessen fleischig angeschwollene
Blätter, die Zwiebelschuppen (zs), mit organischen Reservestoffen
angefüllt sind. Aus dem Vegetationspunkt der Achse entwickelt sich der
oberirdische Trieb. Eine neue Zwiebel geht aus einer Knospe (k) in
der Achsel einer Zwiebelschuppe hervor.
c) Andere krautige Perennen wechselfeuchter Klimate wieder (z. B.
die Georginen und viele Orchideen) bilden Wurzelknollen aus
(Fig. 203, 204). Sie ähneln den Stammknollen, lassen ihre Wurzelnatur
aber an ihren Wurzelhauben, dem Fehlen von Blattanlagen und dem
inneren Bau erkennen. Eine knollige Hauptwurzel heißt Rübe
(so bei der Mohrrübe, Zuckerrübe; beides Bienne mit oberirdischen
Erneuerungsknospen).
Fig. 203. Wurzelknollen der Georgine (Dahlia
variabilis). Bei s die unteren Teile der abgeschnittenen Stengel. 1⁄5
nat. Gr. Nach SCHENCK.
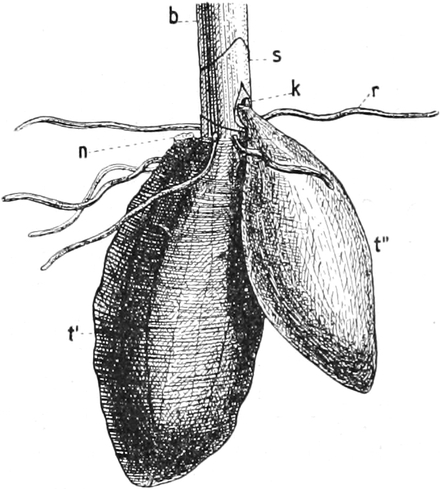
Fig. 204. Wurzelknollen von Orchis, etwas schematisiert.
t′ die alte, t″ die neue Wurzelknolle dieses Jahres. b
Blütensproß. k Die Achselknospe aus dem Deck- (Nieder-)Blatt s
daran, an der die neue Knolle t″ entstanden ist. r Adventivwurzeln,
unverdickt. n Die Narbe an der alten Knolle zeigt die Stelle, womit
die alte Knolle an ihrem Muttersproß befestigt war.
3⁄4 nat. Gr.
Eigenartig sind die Wurzelknollen der Orchideen aufgebaut: sie sind
eiförmig (Fig. 204) oder handförmig gestaltet (Orchis latifolia)
und entstehen stets nur aus einer Wurzel, die handförmigen
durch deren Gabelung. Man findet miteinander verbunden eine ältere
(t′) und eine jüngere Knolle (t″). Die ältere hat bereits
einen Blütensproß (b) getragen und ist im Schrumpfen begriffen.
Die jüngere ist an der Basis dieses Sprosses in der Achsel eines
Niederblattes (s) an dessen Achselknospe (k) entstanden, und
zwar durch Anschwellung einer sproßbürtigen Wurzel dieser Knospe.
Unverzweigte dünne Wurzeln entspringen über den Knollen aus der
Stengelbasis.
Viele Zwiebeln, Knollen und Rhizome haben eine spezifische
Tiefenlage, die freilich je nach der Beschaffenheit des Bodens
variieren kann. So[S. 154] liegen z. B. die Rhizome von Paris in 2–5 cm, die
von Arum bei 6–12, von Colchicum bei 10–16, von Asparagus officinalis
bei 20–40 cm Tiefe. Die Samen keimen aber auf oder dicht unter der
Erde. Die Erdsprosse der jungen Pflanzen müssen also tiefer und
tiefer in die Erde eindringen. Dies geschieht zum Teil durch die
Zuwachsbewegung des Stammes (vgl. S. 305), zum Teil aber auch durch
kontraktile Wurzeln (Zugwurzeln). Bei Lilium z. B. (Fig. 205)
sind alle Wurzeln stark kontraktil. Ihre Tätigkeit ist am besten aus
Fig. 205, 3 zu erkennen, wo die beiden untersten Wurzeln durch ihre
Kontraktion die Zwiebel so verlagert haben, daß die höheren an ihrer
Basis im Bogen gekrümmt worden sind. Hat die Zwiebel die richtige
Tiefe erlangt, so wird sie durch Wurzelkontraktion jährlich nur um so
viel tiefer gezogen, als sie durch Wachstum des Vegetationspunktes
aufsteigt. In anderen Fällen sind nicht alle Wurzeln kontraktil
(Arum), oder es sind überhaupt nur sehr wenige oder gar nur eine
einzige Kontraktionswurzel ausgebildet (Crocus, Gladiolus, Oxalis
elegans). Handelt es sich bei den bisher geschilderten Fällen durchweg
um Seitenwurzeln, die die richtige Tiefenlage sichern, so
kann z. B. bei manchen Rosettenpflanzen die Hauptwurzel durch
andauernde Kontraktion während ihres sekundären Dickenwachstums
dafür sorgen, daß der Vegetationspunkt jedes Jahr um so viel durch die
Wurzel in die Tiefe gezogen wird, wie er durch Wachstum in die Höhe
rückt, so daß die Rosette stets dem Erdboden anliegen bleibt (Gentiana
lutea).
Fig. 205. 1–4 Keimung von Lilium Martagon.
Verkleinert. Die horizontale Linie stellt die Erdoberfläche vor; die
vertikalen Marken haben Zentimeterabstand. 1 Keimling mit Samen; 2
Keimpflanze am Ende des 2. Jahres; 3 junge Pflanze noch im Absteigen
begriffen; 4 ausgewachsene Pflanze in der Normaltiefe; 5 Colchicum
autumnale (etwas verkleinert); punktiert ist die ursprüngliche Lage
der Knolle, ausgezeichnet die neue Lage, die durch Wurzelkontraktion
herbeigeführt wurde; 6 Kontrahierte Wurzel von Lilium. Vergr. 6fach.
Nach RIMBACH.
3. Die annuellen Kräuter schließlich geben für die
schlechte Jahreszeit ihre Vegetationsorgane überhaupt auf: sie
überdauern diese in der denkbar sichersten Weise mit ihren trockenen
Samen, in denen ebenfalls Reservestoffe reichlich gespeichert sind. —
Je gleichmäßiger günstig das Klima während des ganzen Jahres für
die Pflanzenwelt ist (wie in feuchten Gebieten der Tropen), um so
mehr überwiegen immergrüne Holzgewächse in der Vegetation, der
freilich immergrüne[S. 155] perennierende Kräuter, oft auch mit Erdsprossen,
nicht völlig fehlen. Umgekehrt, je extremer die Klimaperiodizität
ausgebildet ist, wie z. B. in den Steppen mit schroffer und sehr
langer Trockenzeit oder in Klimaten mit harten Wintern, um so
größer wird in der Vegetation der Prozentsatz an Tropophyten mit
hochgradigen Schutzeinrichtungen und bei den Kräutern an Geophyten und
Annuellen. Letztere treten aber in Gebieten mit sehr kurzen und kalten
Vegetationsperioden, z. B. im Hochgebirge und in der Arktis, wieder
stark zurück.
b)
Anpassungen an den Lichtgewinn.[100].
Je üppiger sich durch die Gunst des Klimas die Vegetation entfaltet,
um so riesenhaftere Pflanzenformen treten uns in ihr entgegen. In den
tropischen Regenwäldern sind, wie eben schon erwähnt, neben wenigen
niedrigen immergrünen Kräutern kleinere und größere immergrüne
Sträucher ausgebildet, die wieder von zum Teil riesigen ebensolchen
Bäumen (vgl. dazu auch S. 120) überschattet werden. Je höher die
Pflanzengestalten des Waldes sind, um so mehr direktes Sonnenlicht wird
von ihren Blättern aufgefangen, das kleineren Pflanzenformen verloren
geht.
Die Kutikula der Blätter solcher Tropenbäume ist oft besonders
glatt. Sie wirft einen Teil des Sonnenlichtes zurück, wodurch die
Glanzlichter entstehen, die für das Laub der Tropen bezeichnend
sind. Das ist vielleicht eine Schutzeinrichtung gegen zu starke
Bestrahlung. Andere Schutzmittel gegen zu starke Bestrahlung wurden
bei Besprechung der Xerophyten auf S. 145 ff. erwähnt.
Die kleinen und niedrigen Schattenpflanzen der Urwälder und auch
unserer Wälder sind angepaßt, die Kohlensäure mit oft großen
Blattspreiten in auffallend schwachem Lichte noch hinreichend zu
assimilieren.
Im Kampfe um das Licht sind außer Bäumen und Sträuchern zwei
Kormophytengruppen ganz eigenartigen Baues entstanden, die für die
tropischen Regenwälder besonders bezeichnend sind, ohne bei uns
ganz zu fehlen: die Kletterpflanzen (Lianen) und die
Epiphyten.
1. Lianen oder Kletterpflanzen[106]. Sie vermögen ohne großen
Materialaufwand, ohne säulenförmige Stämme, in kurzer Zeit ihr Laub
dem Schatten, etwa des Waldes, zu entziehen und es an die Peripherie
der Baumkronen oder der sonstigen Vegetationsdecke stärkerem Lichte
darzubieten, indem sie mit dünnen Stengeln an fremden Sprossen,
Baumstämmen und Ästen emporklettern. Die tauartigen Stämme
der Lianen sind es, die den Urwald der Tropen in ein vielerorts
undurchdringliches Dickicht verwandeln.
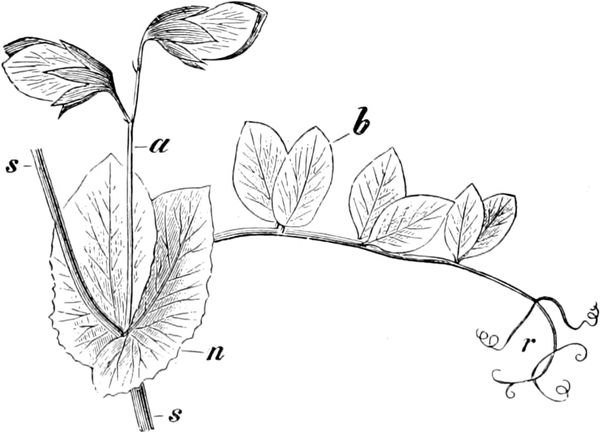
Fig. 206. Stengelstück und Blatt der Erbse (Pisum
sativum). s Stengel, n Nebenblätter, b Blättchen des einfach
gefiederten Blattes, r die zu Ranken ausgewachsenen Blättchen, a
der Blüten tragende Achselsproß. 1⁄2
nat. Gr. Nach SCHENCK.
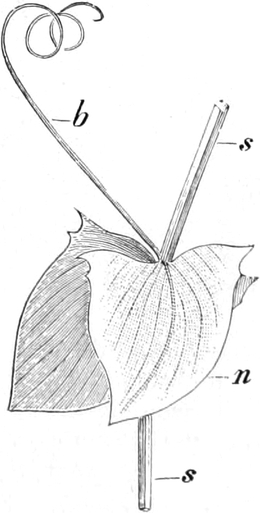
Fig. 207. Stengelstück der Rankenplatterbse (Lathyrus
Aphaca). s Stengel, n Nebenblätter, b Blattranke. 1⁄2 nat. Gr.
Nach SCHENCK.
[S. 156]
Das Klettern wird in sehr verschiedener Weise bewerkstelligt, z. B.
durch widerhakenförmige Seitensprosse oder Haare und Stacheln
oder durch beides, oder durch Dornen (Spreizklimmer:
Galium Aparine, Kletterrosen, Solanum Dulcamara), oder durch
Wurzeln (Wurzelkletterer: Efeu, viele Araceen), oder durch
Windebewegungen (Schlingpflanzen: Hopfen, Feuerbohne), oder
endlich durch besondere Befestigungsorgane, die Ranken
(Rankenpflanzen). Die Ranken sind fadenförmige, unverzweigte
oder verzweigte Organe, die, mit Kontaktreizbarkeit (vgl. S. 312)
ausgestattet, fremde Stützen umwickeln und den Sproß daran befestigen
können. Bei vielen Kletterpflanzen sind es metamorphosierte
Sproßachsen (Sproßranken), so beim Weinstock, wilden Wein (Fig.
208) und bei den Passionsblumen, bei anderen umgebildete Blätter
(Blattranken), z. B. beim Kürbis, bei der Gurke und bei Lathyrus Aphaca
(Fig. 207), wo die Funktionen der zur Ranke gewordenen Blattspreite
durch die blattartig ausgebildeten Nebenblätter übernommen worden sind,
oder es sind umgebildete Teile von Blattspreiten (z. B. bei der
Erbse, Fig. 206), wo sich die oberen Blättchen des Fiederblattes in
eine verzweigte Ranke umgewandelt haben).
Bei gewissen Arten des wilden Weins, Parthenocissus quinquefolia und P.
tricuspidata (Fig. 208), sind die Rankenzweige befähigt, an ihren Enden
Haftscheiben auszubilden und sich mit diesen auch an flachen Stützen zu
befestigen.
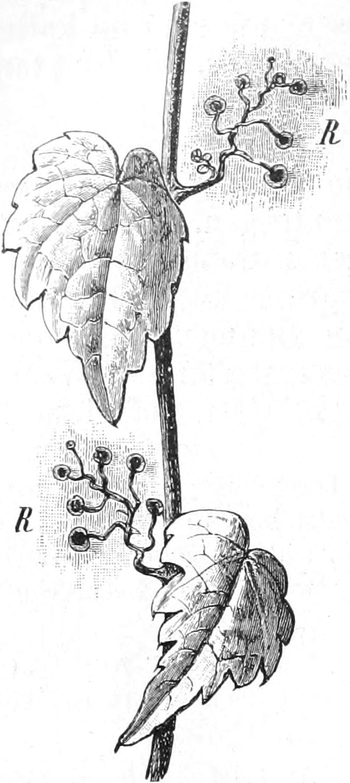
Fig. 208. Parthenocissus tricuspidata. R R
Sproßranken. 3⁄4
nat. Gr. Nach NOLL.
Fig. 209. Querschnitt durch den Stamm von Serjania
Laruotteana. sk Teile des zersprengten Sklerenchymringes des
Perizykels, l und l* Bastzonen, lg Holzkörper, m Mark. Vergr.
2. Nach STRASBURGER.
Fig. 210. Querschnitt durch den Stamm einer Bignoniacee
aus Blumenau. Nat. Gr. Nach SCHENCK.
Bezeichnend für fast alle Lianen sind ihre ungewöhnlich weiten
Tracheen und Siebröhren, ferner bei den tropischen Kletterpflanzen
Anomalien des sekundären Dickenwachstums, wodurch
gefurchte, zerklüftete oder geteilte Holzkörper entstehen, die
die tauförmigen, langen und schwanken Sprosse biegungs- und
torsionsfähig machen. Gefurchte Holzkörper sind bei vielen
Bignoniaceen-Lianen ausgebildet. Der Kambiummantel erzeugt zuerst
in gewohnter Weise nach innen Holz, nach außen Bast und bildet
einen normal gebauten Holzzylinder[S. 157] Hierauf beginnt er aber an
einzelnen Stellen nur noch wenig Holz nach innen, um so mehr Bast
nach außen zu bilden. Das hat zur Folge, daß tiefe Bastkeile, die
stufenförmig nach außen an Breite zunehmen, in das äußere Holz
hineinreichen (Fig. 210). Das Kambium ist dabei in Längsstreifen,
breitere an den Holzvorsprüngen, schmälere im Grunde der Bastkeile,
zerfallen. Das fortwachsende Holz muß an diesen Bastkeilen
vorbeigleiten, so daß seitlich zwischen diesen Holzteilen und den
Bastkeilen keine Verbindung besteht. Mehrere Holzzylinder
sind verschiedenen tropischen Lianen aus den Sapindaceen-Gattungen
Serjania und Paullinia eigen (Fig. 209). Sie kommen dadurch
zustande, daß die Leitbündel in den primären Stammteilen auf
dem Querschnitte nicht im Kreise angeordnet sind, sondern eine
ringförmige Figur mit mehr oder weniger tiefen Ausbuchtungen
bilden. Letztere werden quer durch den Stamm hindurch durch Kambium
verbunden, worauf mehrere Kambiumringe nebeneinander im Stengel
liegen.
2. Epiphyten[107]. Eine andere Gruppe von Kormophyten kann die
Blätter dadurch in stärkeres Licht bringen, daß ihre Vertreter nicht
im Boden wurzeln, sondern sich von vornherein in den Kronen der
höchsten Bäume auf ihren Ästen ansiedeln. Solche Pflanzen nennt man
Epiphyten. Diese Bäume dienen ihnen nur als Unterlagen;
sie können deshalb auch durch anorganische Substrate, z. B. Felsen,
ersetzt werden. Zu epiphytischer Lebensweise sind natürlich nur solche
Gewächse geeignet und befähigt, deren Samen durch Luftströmungen oder
durch Tiere immer wieder auf die Äste der Bäume gebracht werden.
Für die epiphytischen Pflanzen ist offenbar die Beschaffung des
nötigen Wassers und der Nährsalze recht schwierig. Infolgedessen
kommen sie nur in warmen Gebieten mit reichlichem Regenfall und
großer Luftfeuchtigkeit, deshalb also vor allem in den tropischen
Regenwäldern, vor. Außerdem findet man bei ihnen oft eigenartige
Einrichtungen, die diesen Schwierigkeiten begegnen: bei vielen
Epiphyten Sproßknollen als Wasserspeicher (z. B. Orchideen), die bei
Regenwetter gefüllt werden, oder Einrichtungen, um Wasser aufzufangen.
Fig. 211. Dischidia Rafflesiana mit Laub- (l) und
Kannenblättern (k). B Längsschnitt einer Kanne. ö Öffnung, st
Stiel der Kanne, w Wurzel. A ca. 1⁄3, B ca. 1⁄2 nat. Gr. Nach
TREUB.
In unseren Gegenden sind die Epiphyten nur durch rindenbewohnende
Algen, Flechten und Moose vertreten, die vollständige Austrocknung
vertragen. In den Tropen kommen aber auch sehr viele Kormophyten
als Epiphyten vor, z. B. in der Gruppe der Pteridophyten, in den
Familien der Orchideen, Bromeliaceen, Araceen u. a.
Die Schwierigkeiten der Wasserversorgung machen es
verständlich, daß die Epiphyten fast sämtlich ausgesprochene
Xerophyten sind (Fig. 196). Sie befestigen sich[S. 158] an ihren
Unterlagen mit Haftwurzeln; das sind verhältnismäßig
kurze, unverzweigte und das Licht fliehende Wurzeln, die fremde
Gegenstände umklammern und oft Drähten gleichen. Außer diesen
Haftwurzeln werden übrigens bei vielen Araceen auch viel längere
Nährwurzeln ausgebildet, die frei nach unten in die Luft
hängen und ohne jede Verzweigung nach abwärts wachsen, bis sie
den Boden erreichen, worin sie sich verzweigen. Die Mehrzahl
aber, die keine Wurzeln bis zum Erdboden treibt, ist auf das
Wasser der Regengüsse angewiesen, das sie meist mittels besonderer
Apparate aufnehmen und sammeln. Baumbewohnende tropische
Orchideen und auch einige Araceen besitzen zur
Aufnahme von Regen an ihren Luftwurzeln ein besonderes Gewebe:
die mehrschichtige Epidermis ist eine Hülle, Wurzelhülle
oder Velamen, die in manchen Fällen nicht unbeträchtlich
dick wird. Ihre Zellen, die den lebenden Inhalt einbüßen und in
den Zellwänden meist schrauben- oder netzförmige Verdickungen und
häufig auch Löcher besitzen, sind je nach dem Feuchtigkeitsgrade
der Umgebung mit Luft oder mit Wasser gefüllt. Sie saugen wie
Fließpapier das Wasser auf. Sind die Zellen mit Luft gefüllt, so
erscheint die Hülle weiß; enthalten sie Wasser, so schimmert das
innere Gewebe mit grüner Farbe durch. Bei anderen epiphytischen
Orchideen und Araceen bilden die nach aufwärts wachsenden
Luftwurzeln ein reichverzweigtes Geäst, in dem sich durch Verwesung
daraufgefallener Blätter Humus und darin auch Wasser ansammelt.
Namentlich unter den Farnen aber finden sich Epiphyten, deren
Blätter den Humus sammeln: Bei Asplenium nidus sitzen die
Blätter dem dichten Stamm in kurzer Rosette an; der trichterförmige
Raum über der Stammknospe, den sie einschließen, füllt sich mit
Humus. Bei Polypodium- und Platycerium-Arten aber
werden besondere Blätter als „Nischenblätter“, „Mantelblätter“ usw.
ausgebildet, die alle der Sammlung von Humus und von Wasser dienen.
Noch weiter geht in der Umbildung der Blätter die Asclepiadee
Dischidia Rafflesiana (Fig. 211): Einige Blätter sind als
tiefe Urnen mit engen Mündungen ausgebildet; in die Urnen,
in denen das Transpirationswasser kondensiert wird, wachsen Wurzeln
hinein, verzweigen sich darin und nehmen zusammen mit diesem Wasser
wertvolle stickstoffhaltige Substanzen auf; die Urnen enthalten
nämlich gewöhnlich Kolonien von Ameisen, deren Fäkalien und
abgestorbene Leiber.
Ein Extrem unter diesen Epiphyten sind die amerikanischen
Bromeliaceen, bei denen Wurzeln überhaupt fehlen können
(Tillandsia usneoides) oder nur Haftorgane sind. Die Wasseraufnahme
erfolgt hier ausschließlich durch eigenartige schildförmige
Schuppenhaare, die auf den Blättern sitzen. Nicht selten bilden
bei diesen Gewächsen die dicht aneinander schließenden Blattbasen
einen trichterförmigen Hohlraum, in dem sich Wasser sammelt
(Zisternenepiphyten).
c)
Anpassungen der grünen Kormophyten an besondere Ernährungsweise.
Solche kommen bei den insektenfressenden,
fleischfressenden oder karnivoren Pflanzen vor[108]. Das
sind Pflanzen, die zwar mit grünen Blättern organische Substanzen aus
Kohlensäure bilden und völlig autotroph leben können, außerdem aber
mit Einrichtungen zum Fangen und Festhalten kleiner Tiere, vor allem
Insekten, ausgestattet sind und durch ausgeschiedene Enzyme die Beute
so weit wie möglich auflösen, verdauen und als Nahrung resorbieren.
Für den Tierfang bestehen bei den karnivoren Pflanzen die
mannigfaltigsten Einrichtungen. Auf den Droserablättern stehen
wie Schneckenfühler gestaltete, von einem Leitbündel durchzogene
Auswüchse oder Emergenzen, die Tentakeln (Fig. 212, 213), deren
Drüsenköpfchen ein klebriges Sekret in Form glänzender Tröpfchen
absondern. Insekten bleiben an diesen Drüsen hängen, kommen bei ihren
Befreiungsversuchen mit noch mehr Drüsen in Berührung und werden
dadurch um so fester gehalten. Durch den Reiz veranlaßt, krümmen sich
sämtliche Tentakeln nach dem Opfer hin, wobei die Blattfläche selbst
hohl wird und das Insekt umfaßt.
Bei der ebenfalls in Deutschland heimischen Pinguicula legt
sich einfach der Blattrand um das kleine Tier, das an den winzigen
Hautdrüsen hängen bleibt.
[S. 159]
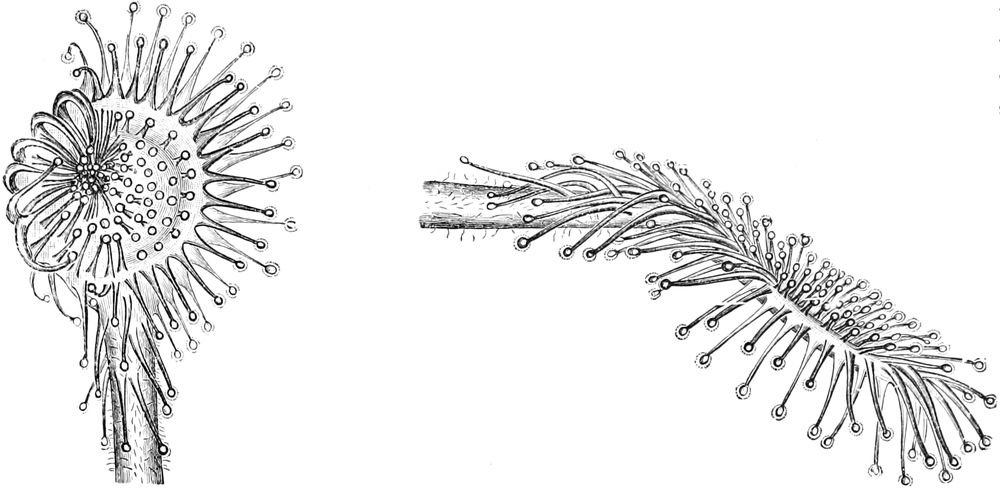
Fig. 212. Blätter von Drosera rotundifolia, links mit
teils eingekrümmten Tentakeln, von oben; rechts mit ausgebreiteten
Tentakeln, von der Seite gesehen. Vergr. 4fach. Nach DARWIN.
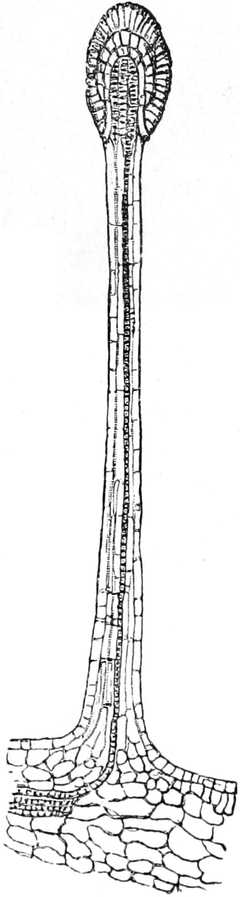
Fig. 213. Digestionsdrüse von Drosera rotundifolia.
Vergr. 60. Nach STRASBURGER.
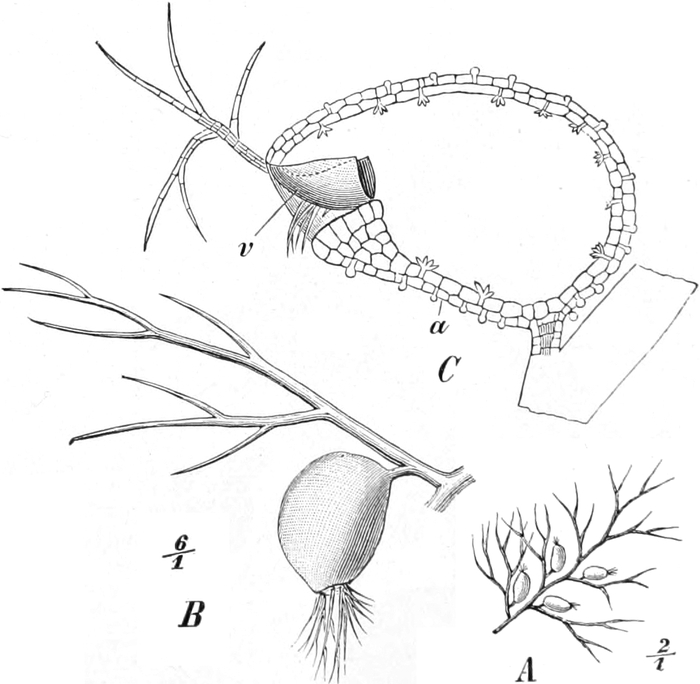
Fig. 214. Utricularia vulgaris. Bei A ein Blattstück
mit mehreren Blasen. Vergr. 2. Bei B ein Fiederteil des Blattes mit
Blase. Vergr. 6. Bei C eine Blase im Längsschnitt. Vergr. etwa 28.
In Cv Klappe, a Blasenwandung. A, B nach SCHENCK, C
nach GOEBEL.
Die auch bei uns, in stehenden Gewässern, vorkommenden
Utricularia-Arten tragen an den zerschlitzten Blättern (Fig.
214) in grüne Blasen umgewandelte Blattzipfel. Die Blasen sind mit
Wasser gefüllt und besitzen eine kleine viereckige Öffnung, die mit
einer elastischen, nur nach innen sich öffnenden Klappe verschlossen
ist. Berühren kleine Wassertiere, z. B. Krebse, eine der vier
hebelartig wirkenden Borsten, die am unteren Rande des Blasendeckels
(Fig. 214 C) nach außen hervorstarren, oder den Deckel selbst, so
werden sie durch eine eigenartige „Schluckbewegung“ der Blase, die
durch Entspannung der durch einen Kohäsionsmechanismus (vgl. S. 296)
elastisch gespannten Blasenwände zustandekommt, mit einem Wasserstrom
in die Blase gesogen; darauf springt die Klappe sogleich in ihre
Ausgangsstellung zurück[S. 160] und verwehrt ihnen den Austritt, so daß man
oft viele Tierchen in einer Blase gefangen findet. Haare, die von der
inneren Blasenwand entspringen, besorgen die Resorption der verdauten
tierischen Körper und eines Teiles des Wassers in den Blasen, wodurch
eben die Blasenwände elastisch gespannt und zu der Schluckbewegung
bereit gemacht werden[109].
Stattlicher und noch leistungsfähiger sind die Fangeinrichtungen
ausländischer Karnivoren. Geradezu überraschend ist die Schnelligkeit,
mit der die Venusfliegenfalle, Dionaea, auf den
Torfmooren Carolinas ihre gezähnten Blatthälften zusammenklappt und
das Insekt, das sich darauf wagt, gefangen nimmt. Fig. 215 stellt
ein Dionaeablatt in offenem Zustande, zum Fange bereit, dar. Ähnlich
gebaute Blätter besitzt die auch bei uns heimische, aber sehr seltene
Wasserpflanze Aldrovanda.
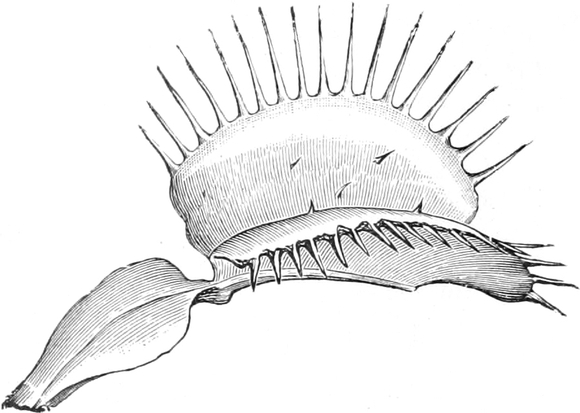
Fig. 215. Ein Blatt der Venusfliegenfalle (Dionaea
muscipula). Der schraffierte Teil der Innenfläche, dicht mit
Verdauungsdrüsen besetzt. Vergr. 4fach. Nach DARWIN.

Fig. 216. Nepenthes robusta. Gewächshauspflanze. 1⁄9
nat. Gr. Nach SCHENCK.
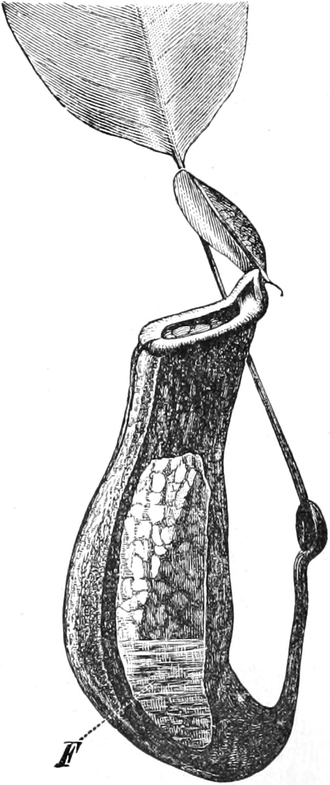
Fig. 217. Blattkanne einer Nepenthes. Am Grunde der
Kanne, aus der ein Stück herausgeschnitten gedacht ist, steht die von
den Blattdrüsen ausgeschiedene Flüssigkeit F, in der hineingefallene
Tiere verdaut werden. 1⁄2
nat. Gr. Nach NOLL.
Bei anderen ausländischen Karnivoren tritt vorherrschend die Kannenform
als Tierfalle auf, so bei Nepenthes, Cephalotus,
Sarracenia, Darlingtonia. Die Kannen entstehen, indem
ganze Blätter oder Teile dieser sich zu krugartigen Behältern ausbilden
(vgl. Fig. 216, 217), die meist[S. 161] von einem Deckel, wohl zur Abhaltung
des Regenwassers, überdacht sind. Bei Nepenthes z. B. geht jede Kanne,
wie Goebel nachgewiesen hat, aus einer entsprechend umgeformten
Blattspreite hervor; der Blattgrund verbreitert sich zugleich
spreitenförmig; der Blattstiel, der beide trennt, kann eine Ranke sein.
Im Grunde solcher Kannen steht eine von Drüsen ausgeschiedene wäßrige
Flüssigkeit. Tiere, die bei Nepenthes durch Honigabscheidungen am
Rande der Kanne angelockt werden und den Kannenrand betreten, gleiten
aus, fallen in die Flüssigkeit und werden im Innern der Kannen durch
kleine Haare, die alle nach abwärts gerichtet sind, am Zurückklettern
verhindert.
B.
Heterotrophe Kormophyten[110].
Neben den grünen Kormophyten, die das Licht brauchen, um
mittels des Chlorophylls aus Kohlensäure und Wasser die organischen
Substanzen aufzubauen, und wohl auch stark transpirieren
müssen, um die Bodennährsalze in genügender Menge in ihrem Körper
anzureichern, gibt es wie bei den Thalluspflanzen auch solche,
die alle oder einen Teil ihrer organischen Substanzen von außen
beziehen, und zwar meist aus lebenden Organismen (Schmarotzer oder
Parasiten). Diese Formen brauchen also kein Licht und keine
Transpiration. Ihrer besonderen Ernährungsweise entspricht ein ganz
eigenartiger, von den grünen Kormophyten völlig abweichender Bau. Er
führt vor Augen, wie sehr die Chlorophyllfunktion mit der Ausgestaltung
der grünen Pflanze in Beziehung steht. Mit dem Zurücktreten oder dem
Verschwinden des Chlorophylls in den Schmarotzern gehen die besonders
für die Assimilation und die Transpiration eingerichteten großen
Blattflächen verloren; die Blätter werden zu unscheinbaren gelblichen
Schuppen oder fehlen ganz. Auch ihre Träger, die Sproßachsen, sind
mehr oder weniger reduziert und gelblich, nicht grün gefärbt. Da
eine lebhafte Transpiration nicht mehr stattfindet, so schwinden
bei vielen Formen auch die Wurzeln. Infolgedessen bleiben auch die
Gefäßteile der Leitbündel schwach entwickelt, und eine Holzbildung
findet höchstens in ganz geringem Umfange statt. Diesem Fortfall
der Assimilationseinrichtungen steht aber die Ausbildung neuer
Einrichtungen gegenüber, die es den Parasiten gestatten, in den Körper
des befallenen Organismus bis zu den Leitungsbahnen[S. 162] einzudringen und
derart in dessen Stoffwechsel einzugreifen, daß er seinen Wirt
ausgiebig berauben kann.
Fig. 218. Ein Leguminosenzweig, dicht besetzt mit
hervorbrechenden Blüten des Schmarotzers Pilostyles Ulei Solms
(Rafflesiacee). Nach GOEBEL, Organographie.
Manche ausländische Schmarotzerpflanzen schließlich, so besonders
die Rafflesiaceen, sind derart in parasitischer Lebensweise
aufgegangen, daß ihre Vegetationsorgane äußerlich überhaupt nicht mehr
sichtbar und gar nicht mehr kormophytisch gegliedert sind, sondern als
hyphenähnliche Stränge ganz innerhalb der Wirtspflanze wachsen, aus der
die fremdartigen Blüten des Schmarotzers überraschend hervorbrechen, z. B.
Pilostyles-Arten (Fig. 218). Auch die größte der bekannten
Blüten, die bis zu 1 m Durchmesser erreichende Blüte der in Sumatra
lebenden Rafflesia Arnoldi, sitzt den Wurzeln ihrer rebenartigen
Nährpflanze, einer Cissus-Art, in dieser Weise unmittelbar auf.
Fig. 219. In der Mitte ein Weidenzweig, umwunden von der
schmarotzenden Cuscuta europaea. An den warzenförmigen Anschwellungen
des Cuscutastengels treten Haustorien in die Weide ein. b Reduzierte
Blättchen. Bl Blütenknäuel. Links: Verbindung des Schmarotzers
(Cus) mit einer Wirtspflanze W. Die Haustorien H dringen teils in
das Rindenparenchym ein, teils legen sie sich dicht an den Gefäßteil
v und den Siebteil c der Leitbündel an, deren Sklerenchymscheide
s sie zum Teil abheben. Rechts: Keimende Cuscuten: der längste
Keimling auf dem Boden kriechend, indem er vorn auf Kosten des
absterbenden Teils t weiterwächst. Nach NOLL.
Als Beispiel einer einheimischen parasitisch lebenden Phanerogame
sei hier die zur Familie der Convolvulaceen gehörige Cuscuta
europaea behandelt, obgleich der Chlorophyllgehalt ihrer
Sproßachsen, denen die Laubblätter fehlen, noch an die normal
assimilierenden Pflanzen erinnert, wenn er auch nur sehr gering
ist. Rechts unten in Fig. 219 sind Cuscutakeimlinge dargestellt,
wie sie sich im Frühjahr aus dem Samen entwickeln. Schon der
Keimling verzichtet auf die Ernährung mittels der Kotyledonen,
die verkümmert sind; auch sein Keimwürzelchen stirbt immer bald
ab. Das Keimstengelchen aber streckt sich sofort zu einem langen
dünnen Faden, dessen freies Ende sich in weitem Kreise herum
bewegt und dadurch eine in seinem Bereiche wachsende Nährpflanze
auffindet. Ist von dem Orte der Keimung aus keine Wirtepflanze zu
erreichen, so vermag der Keimling eine kurze Strecke weiter zu
kriechen, indem er am hinteren Ende (Fig. 219 t) abstirbt und
auf Kosten der diesem Teile entzogenen Nährstoffe am vorderen Ende
sich verlängert. Trifft das freie Fadenende bei seinem Kreisen
aber schließlich auf eine Nährpflanze, z. B. einen Weidenschoß
oder einen Brennesselstengel, so umschlingt er diese wie eine
Schlingpflanze. Nach kurzer Zeit entwickeln sich aus der dem
Wirte angeschmiegten Seite zunächst papillöse Wucherungen der
Epidermis, die in die Gewebe der Wirtspflanze eindringen. Finden
diese Prähaustorien dort zusagende Verhältnisse vor,[S. 163] so
folgt ihnen sehr rasch die Ausbildung der eigentlichen Saugorgane,
der Haustorien (H). Diese brechen aus dem Innern des
Parasiten hervor und besitzen in hohem Grade die Fähigkeit, in
die Wirtspflanze einzudringen. Sie breiten sich scheinbar ohne
Schwierigkeit im fremden Gewebe aus, legen sich eng an dessen
Leitbündel an, während einzelne, aus dem Körper des Haustoriums
hervorsprossende freie Zellreihen wie Pilzfäden in dem zarten
Parenchym vordringen und diesem weitere Nahrung entnehmen. Da, wo
sich das Haustorium an den Gefäß- und Siebteil eines Leitbündels
herangedrängt hat, bilden sich in ihm Elemente des Gefäßteils und
des Siebteils aus, die sich einerseits an die Gefäß- und Siebteile
des Wirtes organisch anschließen, andererseits aber mit den
Leitbündeln des Schmarotzerstengels in Verbindung treten (Fig. 219
links). Wie ein der Wirtspflanze angehöriges Seitenorgan entnimmt
alsdann der Schmarotzer dieser sein Transpirationswasser und seine
sämtlichen Nährstoffe.
Die Samen der ebenfalls bei uns einheimischen schmarotzenden
Orobanchen keimen erst bei Berührung mit den Wurzeln der
Wirtspflanze. Ihre Haustorien bleiben auf die Wurzeln ihrer
Nährpflanze beschränkt; es kommen nur die hellgelblichen,
rötlichbraunen oder amethystblauen, spargelartigen Blütensprosse
neben dem Wirte aus der Erde hervor. Auch die Orobanchen enthalten
noch geringe Mengen assimilierender Chromatophoren (Fig. 772).
Beide, Cuscuta und Orobanche, sind gefürchtete Feinde der
Landwirtschaft, die an Kulturgewächsen, erstere als „Teufelszwirn“,
„Flachs- und Kleeseide“, letztere als „Würger“, großen Schaden
anrichten und schwer auszurotten sind.
Ähnlichen Habitus wie die Orobanche-Arten haben einige Formen,
die man zunächst wohl nicht für Parasiten halten wird, weil sie
im Humus des Waldbodens leben: Orchideen (Neottia,
Corallorrhiza, Epipogon) und die Pirolacee
Monotropa. Der Mangel an Chlorophyll und die Reduktion der
Blätter zu Schuppen, bei Corallorrhiza auch die Reduktion der
Wurzeln (vgl. Fig. 220 das wurzellose Rhizom) beweisen aber schon,
daß auch diese Formen organische Substanz von außen beziehen. Sie
können aber den Humus nicht selbst ausnutzen, sondern sind darauf
angewiesen, daß humusbewohnende Pilze, die daraus ihre Nahrung
beziehen, in ihre unterirdischen Gewebe mit einem Teil ihres
Myzeliums eindringen; ein Teil der eingedrungenen Pilzhyphen wird
alsdann verdaut. Die Kormophyten sind also gewissermaßen Parasiten
auf Humuspilzen; ihre verpilzten Wurzeln nennt man Mykorrhizen.
Fig. 220. Rhizom von Corallorrhiza innata. Bei a
Blütenschaft, bei b Anlagen neuer Rhizomzweige. Nat. Gr. Nach
SCHACHT.
Gegenüber diesen Schmarotzern, die in die größte Abhängigkeit
von ihren Nährpflanzen geraten sind, gibt es aber auch solche,
die äußerlich noch sehr selbständig erscheinen, da sie mit
großen grünen Blättern assimilieren können. Trotzdem sind sie
zugleich Parasiten, da sie sich nur dann normal entwickeln,
wenn ihr Wurzelsystem mit fremden Wurzeln oder zur Not auch
mit denen der Artgenossen durch Haustorien verbunden ist
(„Halbschmarotzer“). So leben z. B. die Santalacee
Thesium und die Rhinanthaceen Rhinanthus,
Euphrasia, Pedicularis, Bartschia,
Melampyrum und Tozzia, von denen sich die letzte,
zumal in den ersten Entwicklungsstadien, am meisten ausgeprägt
parasitisch ernährt.
Auch unsere einheimische, auf Baumkronen schmarotzende
Mistel besitzt, wie viele ihrer ebenso lebenden
fremdländischen Verwandten unter den Loranthaceen, noch
stattliche Blätter, aber reduzierte Wurzeln und ist so reich an
Chlorophyll, daß sie wohl ihren Bedarf an Kohlehydraten vollständig
selbst zu decken vermag.
II.
Fortpflanzungsorgane[111].
A. Bedeutung der Fortpflanzung für den Organismus. Gewaltsamer
oder natürlicher Tod ist das Ende jedes Organismus. Für den Fortbestand
aller lebenden Wesen ist also die Fortpflanzung ebenso unbedingt
notwendig wie die Ernährung. Wie im Worte Fortpflanzung schon zum
Ausdruck[S. 164] kommt, ist ihr wichtigstes Merkmal, daß ein Individuum
Teilen von sich, die zu neuen Individuen werden, Fortdauer sichert.
Die einfachste Art der Fortpflanzung liegt bei vielen einzelligen
Pflanzen in der Zweiteilung ihrer Zellen und Trennung der Tochterzellen
voneinander vor. Solchen Formen schließen sich gewisse einfach
organisierte vielzellige Thallophyten (z. B. Spirogyra) aufs engste
an, insofern ihr Körper zu Zeiten in seine Einzelzellen zerfällt, die
alsdann durch wiederholte Teilung ebenso viele neue Tochterindividuen
liefern. Meist aber erfolgt die Fortpflanzung durch die Ausbildung
besonderer Keime, die sich von der Mutterpflanze ablösen und
später durch Keimung zu neuen Individuen werden. So entstehen
junge Tochterorganismen, Keimlinge, die die Entwicklung der
Mutterpflanzen wiederholen.
Während sich bei den Einzelligen und bei einfachen Vielzelligen
eine jede Zelle an der Fortpflanzung beteiligen kann, ist
bei allen höher organisierten Pflanzen eine Arbeitsteilung
zwischen Vegetationsorganen und Fortpflanzungsorganen
durchgeführt, d. h. solchen besonders gestalteten Organen, denen die
Aufgabe zufällt, Keime auszubilden. Diese Arbeitsteilung wird um so
auffälliger und zieht um so mehr Teile des Körpers in den Dienst der
Fortpflanzung, je höher organisiert der Organismus ist. In erster
Linie auf die Verschiedenartigkeit des Baues und der Stellung dieser
Fortpflanzungsorgane gründet sich die Einteilung des Pflanzenreiches in
Klassen, Ordnungen, Familien usw.
B. Allgemeine Eigenschaften der Keime. Auch der Bau der Keime
steht in engster Beziehung zu den Aufgaben, denen sie zu dienen haben.
Bezeichnend für die meisten Keime ist ihre geringe Größe im
Verhältnis zu dem Mutterorganismus, der sie hervorbringt; dadurch wird
den Mutterpflanzen ohne allzuviel Materialaufwand die Bildung sehr
zahlreicher Keime und den Keimen die Verbreitung erleichtert.
Bei der Fortpflanzung handelt es sich nicht nur um die Bildung
eines neuen Individuums an Stelle des Mutterindividuums,
sondern um eine Vermehrung der Individuen. Da viele, manchmal
die meisten Keime überhaupt nicht die zur Keimung und zum Wachstum
günstigen Verhältnisse finden, auch ein großer Teil der Keimlinge
abstirbt, ehe er sich fortpflanzen kann, so würde Ausbildung nur
eines Keimes gleichbedeutend mit dem baldigen Untergange der
Art sein. So ist die ungeheure, anscheinend verschwenderische Menge
von Keimen für die Fortpflanzung bezeichnend. Ein Hutpilz oder ein
Farnkraut kann Millionen von Keimen bilden; ein Pappelbaum soll nach
BESSEY jährlich 28 Millionen solcher reifen.
Weiter muß dafür gesorgt werden, daß die Keime von der Mutterpflanze
losgelöst und daß sie danach möglichst weit verbreitet
werden; denn an den Stellen, wo sie entstehen, sind meist nicht die
Bedingungen für ihre Keimung und ist kein Raum für die Entwicklung der
Tochterorganismen vorhanden.
Endlich ist es nötig, daß den Keimen vom Mutterorganismus
Reservestoffe mitgegeben werden, damit sie sich weiter entwickeln
können, bis sie sich selbst zu ernähren vermögen.
Sehr häufig haben die Keime die Aufgabe, den Organismus über Zeiten,
die ihr Leben gefährden, z. B. über eine Kälte- oder Trockenperiode,
hinüber zu retten. Sie befinden sich alsdann im Ruhezustand (S.
186), worin sie viel widerstandsfähiger gegen schädigende Einflüsse
(Austrocknung, Frost, Hitze) sind; dicke Hüllen gewähren solchen
Keimen oft außerdem Schutz. Die Keimung beginnt bei solchen
Keimen erst nach Eintritt günstiger Lebensbedingungen.
C. Einteilung der Keime. Die entwicklungsfähigen Keime selbst
sehr hochorganisierter Vegetationskörper sind einzellig
(Keimzellen, Sporen)[S. 165] oder mehrzellig
(Brutkörper und Samen). Unschwer lassen sich zwei
Typen der Fortpflanzung in fast allen Klassen des Gewächsreiches
erkennen.
Der eine zielt auf die Bildung von Keimzellen oder von Brutkörpern
hin, die meist nach ihrer Lostrennung von der Mutterpflanze, ohne
weiteres, entweder sofort oder nach einer Ruhezeit, keimen und zu
neuen selbständigen Einzelwesen heranwachsen. Diese Fortpflanzungsweise
hat man die vegetative, ungeschlechtliche oder
monogene genannt.
Der zweite bei der Fortpflanzung eingeschlagene Weg ist dagegen
viel umständlicher. Es werden dabei zweierlei Keime erzeugt.
Sie sind, jeder für sich, gewöhnlich unfähig, auszukeimen, gehen
vielmehr in kurzer Zeit zugrunde, wenn sie keine Gelegenheit
haben, sich paarweise zu vereinigen (Befruchtung,
Geschlechtszellen); erst wenn zwei Geschlechtszellen
miteinander zu einer Zelle verschmolzen sind, wird das
Produkt entwicklungsfähig. Diese Art der Fortpflanzung wird als die
sexuelle, geschlechtliche oder digene bezeichnet.
Die meisten Gewächse pflanzen sich auf beide Weisen fort; die
sexuelle Fortpflanzung fehlt nur bei niedrig organisierten Gruppen, so
den Bakterien, Cyanophyceen, manchen Algen und Pilzen.
Erst die Befruchtung befähigt, wie gesagt, die Geschlechtszellen zur
Weiterentwicklung. Doch gibt es Ausnahmen, wo eine Geschlechtszelle
auch ohne Befruchtung auskeimt. Erfolgt diese Entwicklung aus einer
solchen Zelle, also monogen, so spricht man von jungfräulicher Zeugung
oder Parthenogenesis[112]. Diese ist im Pflanzenreiche bisher
bei Algen, z. B. bei Chara crinita, festgestellt, sowie bei der
Keimbildung aus unbefruchteten Eizellen zahlreicher höher organisierter
Pflanzenfamilien, wie Kompositen, Ranunculaceen, Rosifloren,
Thymelaeaceen, Urticaceen, auch Marsiliaceen.
Bei allen diesen Pflanzen liegt habituelle Parthenogenesis
vor, d. h. die Eizellen entwickeln sich ohne Befruchtung ohne
weiteres. In anderen Fällen tritt Parthenogenese erst durch
experimentelle Eingriffe ein: experimentelle Parthenogenesis.
In bestimmten Fällen wird die Befruchtung von Geschlechtszellen
durch einen Verschmelzungsvorgang der Kerne benachbarter
vegetativer Gewebezellen ersetzt[113]. So ist es bei dem
Prothallium gewisser Kulturformen von Farnkräutern (z. B. von
Dryopteris [Lastrea] und Athyrium). Das Verschmelzungsprodukt
dient alsdann der Fortpflanzung, während die Geschlechtsorgane des
Prothalliums verkümmern.
Ein tieferes Verständnis der Fortpflanzungserscheinungen im einzelnen
kann man erst durch genaueres Studium des Entwicklungsganges der
Pflanzengruppen gewinnen. Diese Aufgabe fällt dem speziellen
Teile unseres Buches zu. Die im folgenden gegebene ganz knappe
Übersicht über die Bauvariationen der Keime und der
Fortpflanzungsorgane im Pflanzenreich soll nur die für die allgemeine
Morphologie wichtigen Tatsachen und Probleme hervortreten lassen.
1. Brutkörperbildung.
Mehrzellige monogene Fortpflanzungskörper, Brutkörper,
kommen bei vielen Moosen vor (z. B. bei Marchantia, wo sie in
besonderen Organen, den Brutbechern, auf dem Thallus, Fig. 446, 447,
erzeugt werden); ferner als Brutknospen in weiter Verbreitung
bei Farnpflanzen und Phanerogamen.
Nicht selten lösen sich bei ihnen Seitensprosse ab, die oft
durch besonderen Bau als Fortpflanzungskörper kenntlich sind, so
etwa die unter- oder oberirdischen Ausläufer (Stolonen).
Letztere wachsen z. B. aus der grundständigen Blattrosette der
Erdbeerpflanzen als fadenförmige Seitenzweige hervor und tragen
Endknospen, die sich bewurzeln und durch späteres Absterben
der Ausläufer selbständig werden. Solche der vegetativen
Vermehrung dienenden Gebilde bei höheren Pflanzen sind auch
viele Knollen und Zwiebeln, ferner die Brutknospen,
Brutzwiebeln (Bulbillen, Fig. 221) und die sich los[S. 166]lösenden
Überwinterungsknospen (Hibernakeln) zahlreicher
Wasserpflanzen (z. B. bei Hydrocharis, Stratiotes).
Fig. 221. Zwiebeltragende Zahnwurz (Dentaria bulbifera)
mit Brutzwiebel br. Nat. Gr. Nach SCHENCK.
Brutknospen können auch an Orten auftreten, wo gewöhnlich
keine Vegetationspunkte vorhanden sind; es sind alsdann
Adventivbildungen. Solche werden besonders häufig an Blättern,
zumal auf Blattspreiten, ausgebildet; z. B. in den Kerben der
Blattränder von Bryophyllum-Arten, auf den Blättern von Cardamine
pratensis. Die Blätter von Begonien, Drosera u. a. entwickeln erst
nach gewaltsamer Lostrennung von der Mutterpflanze Knospen.
Schließlich sei erwähnt, daß manche Pflanzen, z. B. viele krautige
Perennen, ohne besondere vegetative Fortpflanzungskörper auch
dadurch sich ungeschlechtlich vermehren, daß ihre verzweigten
Rhizome durch Verwesung der älteren, absterbenden Teile in die
einzelnen Zweige zerfallen. Auch bei gewissen Meeresalgen kann der
Thallus, z. B. durch die mechanischen Wirkungen der Brandung, in
Stücke zerlegt werden, die zu ebenso vielen Thalli auswachsen; ja
Caulerpa pflanzt sich, wie es scheint, nur auf diese Weise fort.
2. Keimzellenbildung.
a) Ungeschlechtliche Keimzellen (Sporen). Sie bilden sich in
zwei Formen aus: 1. Einzelne Zellen, die oft durch Sprossung entstehen,
werden als Keimzellen aus dem Zellverband des Körpers losgelöst oder
abgeschnürt: Exosporen oder Konidien, so bei vielen
Pilzen (Fig. 222). 2. Die Keimzellen entstehen als Endo- oder
Sporangiensporen in besonderen Behältnissen (Sporangien),
aus deren Wandungen sie durch ein Loch oder einen Riß ausschlüpfen
oder ausgestoßen werden (Fig. 223, 229 sp), so bei anderen Pilzen,
vielen Algen, den Moosen, Farn- und Samenpflanzen. Diese Sporangien
sind bei den Thallophyten Einzelzellen, deren Protoplast sich meist in
mehrere oder viele Endosporen teilt (Fig. 223). Bei den Moosen, Farn-
und Samenpflanzen sind es dagegen verwickelter gebaute, vielzellige
Gewebekörper, in denen eine bis mehrere äußere „sterile“ Zellschichten
die Wandung des Behältnisses bilden und nur das davon umschlossene
Gewebe, das sporogene Gewebe, Sporen liefert (Fig. 224 sg).
Fig. 222. Konidienbildende Hyphe des Schimmelpilzes
Aspergillus herbariorum. Vergr. 540. Nach KNY.
Fig. 223. Saprolegnia mixta. Sporangium, die zweiziligen
Zoosporen s2 entlassend. Nach G. KLEBS.
Die ungeschlechtlichen Sporen sind zum Teil angepaßt an die Verbreitung
im Wasser, so viele Sporangiensporen bei Algen und Pilzen.
Diese Sporen entbehren eine Zellmembran völlig, sind also nackt und
meist mit eigenem Bewegungsvermögen im Wasser durch Zilien begabt (Fig.
223, 227 A). Solche Sporangiensporen heißen Schwärmsporen
oder Zoosporen, die Behältnisse, in denen sie entstehen,
Zoosporangien.
[S. 167]
Die Sporen anderer Thallophyten, der Moose, Farn- und Samenpflanzen,
sind dagegen angepaßt an die Verbreitung in der Luft, durch den Wind
oder durch Tiere. Alsdann sind sie von einer dicken Sporenmembran
umgeben und sehr widerstandsfähig gegen Austrocknung.
Bei den Thallophyten entstehen die ungeschlechtlichen
Fortpflanzungsorgane an beliebigen oder an bestimmten Stellen des
Thallus.
Bei den Moosen sitzt der meist langgestielte, große
Sporenbehälter ebenfalls entweder auf dem Thallus oder bei den
beblätterten Formen an den Enden beblätterter Zweige (Fig. 456).
Bei den Farnpflanzen werden die in der Regel ziemlich kleinen
und unansehnlichen Sporangien in sehr großer Zahl meist an Blättern,
den Sporophyllen, ausgebildet (Fig. 473). Diese können den
Laubblättern völlig gleichen; häufig aber ist eine Arbeitsteilung
eingetreten zwischen Laubblättern und Sporophyllen. Letztere dienen
alsdann vorwiegend oder nur noch der Erzeugung von Sporangien und
haben dementsprechend einen ganz anderen Bau als die Laubblätter,
nämlich keine flächenförmig entwickelten und grünen Spreiten. Nicht
selten werden die Sporophylle in größerer Zahl an Enden von Zweigen,
meist begrenzten Wachstums, gebildet: so bei den Schachtelhalmen und
Bärlappgewächsen (vgl. Fig. 488, 493). Die Sproßenden, die mit den
Sporophyllen besetzt sind, haben ein ganz anderes Aussehen als die
vegetativen; sie dienen ebenfalls nur noch der Fortpflanzung und
sterben danach ab. Sie gleichen Zäpfchen oder Ähren, werden deshalb
auch Sporophyllzapfen oder Blüten genannt. Die
einfachste Blüte ist also das Ende eines Sprosses, das Sporophylle
trägt. Diese Zäpfchen können wohl an ihrer Basis von einer Hülle
aus einigen schuppenförmigen, aber sterilen Blättern umgeben werden.
Auch bei den Samenpflanzen werden die Sporangien in solchen
besonderen Sproßteilen, in Blüten gebildet, die mit allen ihren
Gliedern nicht mehr der Ernährung, sondern nur noch der Fortpflanzung
dienen. Diese Blüten sind denen der Farnpflanzen homolog, also wieder
stark metamorphosierte Teile von Laubsprossen, und zwar Enden von Lang-
oder meist Kurztrieben, deren Blattanlagen nicht zu Laubblättern
werden, sondern sich zu den dicht zusammengedrängt stehenden
andersartigen Blattgebilden der Blüte, zu Kelch-, Kron-,
Staub- oder Fruchtblättern umbilden.
Fig. 224. Schematisches Bild des Sporangiums einer
Farnpflanze mit steriler zelliger Wandung und mit sporogenem Gewebe
sg.
Die Staubblätter oder Staubfäden entwickeln in vielzelligen
Sporangien mit mehrschichtiger Wandung, den Pollensäcken,
in großer Zahl die Pollenkörner (den Blütenstaub): zunächst
einzellige Sporen (Fig. 32). An den Fruchtblättern dagegen, die
bei den Gymnospermen frei, bei den Angiospermen zum geschlossenen
Fruchtknoten verwachsen sind, entstehen kurzgestielte, ovale
Gewebekörper von verwickeltem Bau, die Samenanlagen, die in
ihrem Innern je eine Spore einschließen.
Bei den Gymnospermen[114] haben die zapfenähnlichen Blüten (vgl. Fig.
604–607), die nur aus zahlreichen, meist schraubig angeordneten,
schuppenförmigen Staub- oder Fruchtblättern bestehen, noch
die größte Ähnlichkeit mit den Sporophyllähren der Lycopodiaceen.
Bei den Angiospermen[114] erhält aber die Blüte eine meist völlig
abweichende Gestalt (Fig. 225): 1. durch die Beschränkung der
quirlständigen Blattgebilde auf eine oft kleine und meist
konstante Zahl; 2. durch die Blütenblätter, die in Form von
grünen und derben Kelchblättern und oft andersfarbigen und[S. 168]
zarten Kronblättern zur Ausbildung gelangt sind, 3. durch
die pfriemförmigen Staubblätter (Staubgefäße) und endlich
4. durch die Verwachsung der Fruchtblätter zu Behältern, den
Fruchtknoten. Alle diese Teile sind in regelmäßiger Weise in
der Blüte angeordnet. In der typischen Angiospermenblüte wechseln
nämlich fünf gleichgliedrige Wirtel von Blattgebilden alternierend ab
(Fig. 226): der äußerste Wirtel kommt den Kelchblättern zu, die die
jugendlichen Blütenteile in der Blütenknospe umhüllen und schützen,
der zweite wird durch die Kronblätter gebildet, der dritte und vierte
von den Staubblättern, der oberste fünfte von den Fruchtblättern[114],
[115]. Diese Blattgebilde, die der sehr kurzen, vielfach auch
abgeflachten oder ausgehöhlten Blütenachse entspringen, sind oft
untereinander und in manchen Fällen auch mit der Achse verwachsen;
eingehende vergleichende und entwicklungsgeschichtliche Untersuchungen
klären alsdann häufig erst den Sachverhalt auf.
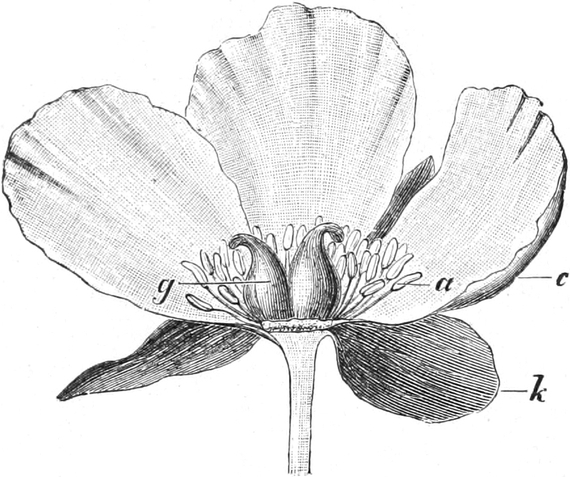
Fig. 225. Blüte von Paeonia peregrina. k Kelch, e
Krone, a Staubblätter, g Fruchtblätter. Die vorderen Kelchblätter,
Kronblätter und Staubblätter wurden entfernt, um die beiden, zwei
getrennte Fruchtknoten bildenden Fruchtblätter zu zeigen. 1⁄2
nat. Gr. Nach SCHENCK.
Fig. 226. Diagramm der Liliaceenblüte, darunter das
Deckblatt; darüber, durch einen schwarzen Punkt angedeutet, die
Mutterachse. Nach STRASBURGER.
b) Geschlechtliche Keimzellen (Sexualzellen oder Gameten). 1.
Verschiedene Formen der Sexualzellen und Sexualorgane. Auch
die sexuelle Fortpflanzung tritt uns im Pflanzenreiche in sehr
mannigfaltigen Formen entgegen, deren Extreme zwar sehr verschieden,
aber durch zahlreiche Übergänge verbunden sind.
Die Geschlechtszellen oder Gameten sind bei der einfachsten
Art der sexuellen Fortpflanzung, wie wir sie bei niederen Algen und
Pilzen finden, zwei völlig gleich große und gleich gestaltete, meist
nackte Protoplasten, die wie asexuelle Schwärmsporen aussehen,
aber miteinander kopulieren (Isogamie Fig. 227 B). Man nennt
die Zellen, in denen sie auch ganz wie Sporangienschwärmsporen in Ein-
oder Mehrzahl aus den Protoplasten entstehen, Gametangien,
das Kopulationsprodukt Zygote oder Zygospore (Fig. 227
B 4). Alles spricht dafür, daß solche Gameten den Schwärmsporen,
von denen sie sich oft nur durch geringere Größe unterscheiden, und
desgleichen die Gametangien den Sporangien homolog, d. h. durch
Umbildung aus ihnen phylogenetisch entstanden sind. Derartige Gameten
können mit Zilien aktiv beweglich sein. Sie suchen sich im Wasser
gegenseitig auf, um paarweise zu kopulieren (Fig. 227 B).
Nicht selten sind aber schon bei Algen und Pilzen und ganz allgemein
bei den Moosen, Farn- und Samenpflanzen die Gameten ungleich
groß (Heterogamie); alsdann bezeichnet man den größeren
Gameten, der gewöhnlich an Reservestoffen sehr reich ist, als
weiblichen (Ovium  ), den kleineren als männlichen
(Spermium
), den kleineren als männlichen
(Spermium  ). Der größere Gamet kann auch ganz unbeweglich[S. 169] sein
(Eizelle). In diesem Falle sucht das Spermium das Ovium auf und
befruchtet es (Eibefruchtung, Oogamie). Liegt Oogamie
vor, so werden die männlichen und weiblichen Keimzellen in verschieden
gestalteten Geschlechtsorganen (männlichen und weiblichen) ausgebildet.
). Der größere Gamet kann auch ganz unbeweglich[S. 169] sein
(Eizelle). In diesem Falle sucht das Spermium das Ovium auf und
befruchtet es (Eibefruchtung, Oogamie). Liegt Oogamie
vor, so werden die männlichen und weiblichen Keimzellen in verschieden
gestalteten Geschlechtsorganen (männlichen und weiblichen) ausgebildet.
Fig. 227. A Eine ungeschlechtliche Schwärmspore der
Grünalge Ulothrix zonata. B 1 ein Gamet, 2 und 3 kopulierende
Gameten, 4 eine durch Kopulation erzeugte Zygote. Vergr. 500. Nach
STRASBURGER.
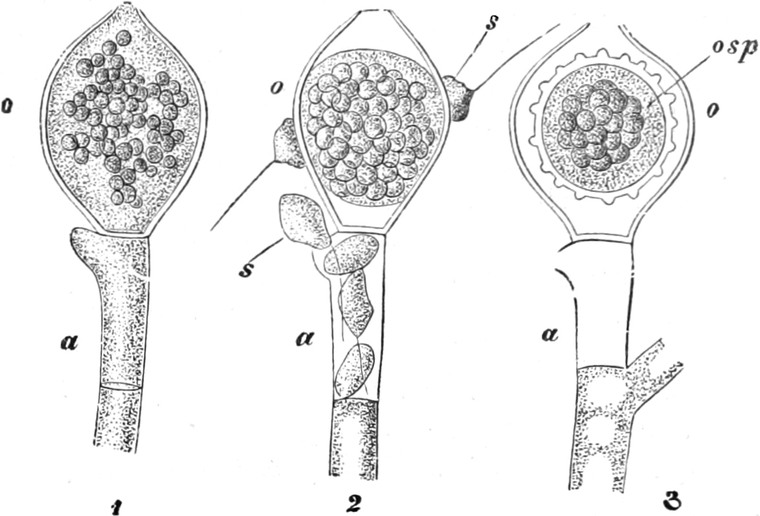
Fig. 228. Monoblepharis sphaerica. Ende eines Fadens mit
einem Oogonium o und dem darunter liegenden Antheridium a, in 1
vor der Bildung der Eizelle und der Spermien, in 2 die letzteren s
austretend und an dem Oogonium hinaufkriechend, in 3 reife Oospore
osp, das Antheridium entleert. Vergr. 800. Nach CORNU.
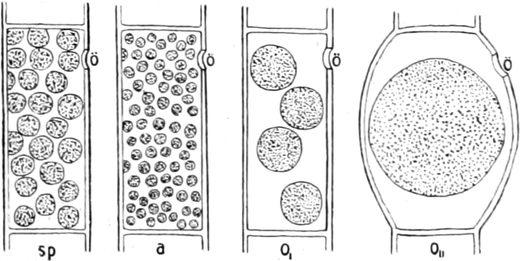
Fig. 229. Schematische Zeichnungen (nach Algen) für
Sporangium sp mit Sporen; Antheridium a mit Spermien; Oogonium
o′ mit mehreren, o″ mit einer Eizelle. ö Das Loch in der
Zellwand.
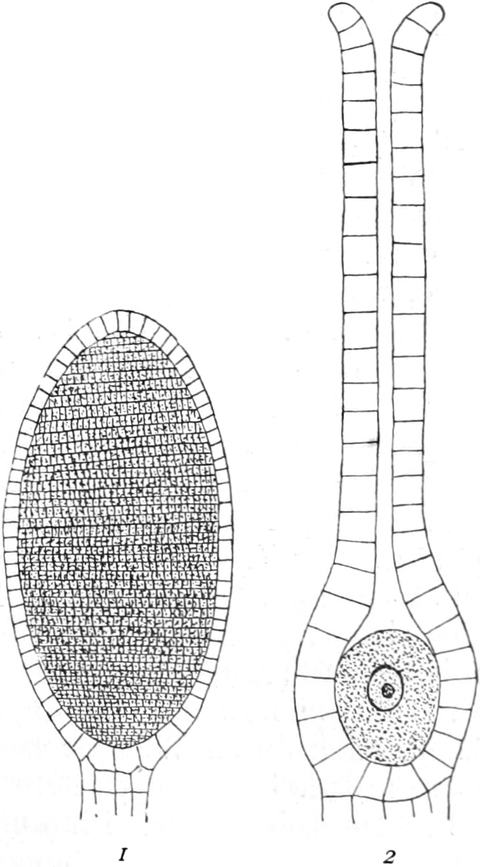
Fig. 230. 1 Antheridium mit steriler, zelliger Wandung
und spermatogenem Gewebe; 2 Archegonium mit entsprechender Wandung
und Eizelle. Beides von einem Lebermoos.
Die Behälter, in denen die fast stets sehr kleinen und nackten
Spermien meist in sehr großer Zahl entstehen, also die
männlichen Geschlechtsorgane, werden Antheridien genannt (Fig.
228 2a, 229 a, 230 I). Es sind bei den Thallophyten (Fig. 228
2a, 229 a) meist Einzelzellen, bei den Moosen und Farnpflanzen
dagegen Gewebekörper mit einer Wandung aus sterilen Zellen, die das
Spermien bildende, spermatogene, Gewebe umschließt (Fig. 230
I). Dagegen heißen die Behälter, in denen die Ovien in Ein-
oder Mehrzahl gebildet werden, bei den Thallophyten, wo sie
ebenso wie die Antheridien meist Einzelzellen sind, Oogonien
(Fig. 228 2 und 229 o′), o″), bei den Moosen und
Farnpflanzen, wo sie aus verwickelt gebauten Gewebekörpern
bestehen (Fig. 230 2), Archegonien. Die Eizelle, die ebenfalls
nackt zu sein pflegt, bleibt gewöhnlich in ihrem Behältnis liegen, in
[S. 170]dessen Wand eine Öffnung ausgebildet wird (Fig. 228 2, 229 o′
und o″ ö, 230 2). Die Befruchtung der empfängnisfähig
gewordenen Eizellen erfolgt in der Weise, daß die Spermien, die
durch ein Loch in der Antheridienwandung in das umgebende Wasser
ausschwärmen, also meist aktiv beweglich sind, von den Eizellen durch
ausgeschiedene Stoffe chemotaktisch (vgl. S. 292) angelockt werden.
Zahlreiche Übergänge zwischen Oogamie und Isogamie bei den Thallophyten
zeigen uns augenscheinlich, daß sich die Oogamie phylogenetisch durch
Weiterbildung aus der Isogamie entwickelt hat. Infolgedessen sind
bei ihnen die Antheridien und Oogonien einander und zugleich den
Gametangien und Sporangien der isogamen Pflanzen homolog (vgl. auch
Fig. 229).
Erst durch Aufnahme eines Spermiums hat die Eizelle die Fähigkeit
erlangt, sich, nachdem sie sich mit einer Membran umgeben hat, sofort
oder nach einer Ruhezeit als Oospore (Fig. 227 3 osp), weiter
zu entwickeln. Bei den Moosen und Farnpflanzen geht aus der Eizelle
alsbald nach der Befruchtung ein Gewebekörper, der Embryo,
hervor, der allmählich zu dem Keimling heranwächst.
Bei den Samenpflanzen entfernen sich die Sexualorgane am
weitesten vom einfachsten Typus. Die Gameten sind hier in den
Pollenkörnern und Samenanlagen der Blüten eingeschlossen,
und zwar enthalten die Pollenkörner die männlichen Sexualzellen,
die Samenanlagen in Mehr- oder Einzahl die Eizellen. Hiermit
hängt es zusammen, daß sich auch die Befruchtung der Eizellen
bei den Samenpflanzen in eigenartiger Weise vollzieht. Sollen
die Eizellen befruchtet werden, so müssen die Pollenkörner, die
ja die männlichen Keime enthalten, bei den Gymnospermen zunächst
auf die Samenanlagen, bei den Angiospermen auf ein besonders
ausgebildetes Organ des Fruchtknotens, die Narbe, übertragen
werden (Bestäubung). Die meisten und sehr mannigfaltigen
Weiterbildungen, die die einfachsten Blüten bis zu den am höchsten
organisierten der Angiospermen erfahren haben, sind Anpassungen an
die Art der Bestäubung[116]. Diese erfolgt stets durch besondere
Transportmittel der Pollenkörner. Wenn, wie sehr häufig,
männliche und weibliche Organe in einer Blüte vereinigt sind,
d. h. bei den Zwitterblüten, sollte man freilich meinen, daß
keinerlei besondere Einrichtungen nötig wären, um den Pollen auf die
Narbe zu bringen. Genauere Untersuchung hat aber gezeigt, daß auch
hier solche Einrichtungen in Hülle und Fülle bestehen, ja daß sie oft
sogar sehr verwickelt sind. Sie alle zielen nicht einfach darauf hin,
den Pollen der Blüte auf die zugehörige Narbe zu befördern, sondern
machen oft eine derartige Selbstbestäubung (Autogamie) ganz
unmöglich und bewirken eine Fremdbestäubung (Allogamie), d. h.
eine Übertragung des Pollens auf Narben benachbarter Blüten des
gleichen (Geitonogamie) oder womöglich eines anderen Individuums
(Xenogamie). Die Übertragung des Blütenstaubes wird, je nach dem
Bau der Blüten, durch Wind, durch Tiere oder, in seltenen
Fällen, auch durch Wasser besorgt; dementsprechend kann man die
Blüten in anemophile, zoidiophile und hydrophile
einteilen (vgl. den speziellen Teil). Die zoidiophilen Blüten
bilden bunte Blütenblätter, Duft oder Honig aus, wodurch die
Bestäubungsvermittler angelockt werden. Die meisten Samenpflanzen haben
sich also bei der Befruchtung von der Gegenwart von Wasser ganz
frei gemacht; sie konnten zu ausgeprägten Landpflanzen werden.
Neben Pflanzen mit Allogamie fehlt es aber auch nicht an solchen,
die Einrichtungen zur Beförderung der Autogamie haben,
mag diese erst, wenn die Allogamie nicht geglückt ist, oder, wie
bei den kleistogamen Blüten, von vornherein eintreten (vgl. den
speziellen Teil).
[S. 171]
Nach der Bestäubung treiben die Pollenkörner je eine schlauchförmige
Ausstülpung, den Pollenschlauch, der in die Samenanlage bis
zur Eizelle vordringt, nachdem er bei den Angiospermen zuvor durch
den Griffel in das Fruchtknotengehäuse hinabgewachsen ist (vgl.
Fig. 567, 569). Nachdem nun eine offene Verbindung zwischen dem
Pollenschlauch und der Eizelle hergestellt ist, wird die Eizelle
meist nur durch einen Kern des Pollenschlauches ohne
Plasmahülle befruchtet. Die befruchtete Eizelle entwickelt
sich in der anschwellenden Samenanlage zu einem vielzelligen
Gewebekörper, dem Embryo, der sich schon hier meist in die
Keimblätter, das Keimwürzelchen (Radicula) und oft in
die Keimknospe (Plumula) gliedert: die Samenanlage aber wird
durch Umbildung ihrer Gewebe zum Samen, wobei ihre äußeren
Zellschichten die Samenschale liefern. Der Same, der in
reifem Zustande abgeworfen wird und zur Vermehrung dient, ist also
eine weiterentwickelte Samenanlage, die einen Embryo umschließt.
Auch das Fruchtknotengehäuse entwickelt sich nach der Befruchtung der
Eizelle weiter, und zwar zur Frucht. Diese kann sich als Ganzes
mit den eingeschlossenen Samen von der Mutterpflanze ablösen (so bei
den Beeren, Nüssen und Steinfrüchten), oder sie bleibt an der Pflanze
sitzen, platzt auf und entläßt die Samen (so bei den Kapseln).
Wie bei allen Fortpflanzungskörpern wird auch bei den Samen und den
sich loslösenden Früchten für eine Verbreitung gesorgt[117].
Dies geschieht mit den gleichen Mitteln wie die Pollenübertragung:
durch Luft-, durch Wasserströmungen, durch Tiere, sowie endlich auch
durch eine Eigentätigkeit der Pflanze. Der Bau der Samen und Früchte
ist an ihre Verbreitungsmittel angepaßt (vgl. den speziellen Teil).
Früher oder später nach erfolgter Aussäung beginnt die
Samenkeimung[118]. In der Regel tritt am keimenden Samen
zunächst die Wurzel des Keimlings aus, indem sie die Samenschale
durchbricht. Da diese Schale oft außerordentlich fest gebaut ist,
so sind nicht selten besondere Austrittsstellen für die Keimwurzel
ausgebildet (z. B. bei der Kokosnuß). Der Keimling wächst zunächst
allein auf Kosten von Nährstoffen, die den Samen von der Mutterpflanze
in besonderen Speicherorganen mitgegeben worden sind.
Eine sehr eigenartige ungeschlechtliche Fortpflanzung, die sich
bei manchen Blütenpflanzen findet, täuscht eine geschlechtliche
Fortpflanzung vor. Hier entstehen nämlich innerhalb der Samenanlage
an Stelle der unterdrückten Eizelle aus anderen Zellen
ungeschlechtliche Embryonen, die also später im Samen
eingeschlossen sind[113] (Apogamie). Die Samen enthalten
alsdann kein Geschlechtsprodukt, sondern sind zu Organen der
vegetativen Vermehrung geworden. Diese „Adventivkeimbildung“ ist
in der Regel mit Polyembryonie, d. h. Ausbildung vieler
Keime in einem Samen verbunden (Funkia ovata, Citrus aurantium,
Caelebogyne ilicifolia u. v. a.).
2. Die Vorgänge der Zellverschmelzung bei der Befruchtung und ihre
Folgen. Die Befruchtungsvorgänge selbst, in ihrer einfachsten
Form, kann man am besten beobachten bei den Geschlechtszellen niederer
Organismen, wo diese Zellen gleich gestaltet sind, also an Isogameten
(Fig. 227). Bei solchen Formen läßt sich leicht feststellen, daß
nicht bloß die Plasmakörper der beiden Zellen, sondern früher oder
später auch ihre Kerne verschmelzen. Dagegen vereinigen sich
die Chromatophoren der beiden Geschlechtszellen nicht, wenn in
ihnen solche vorhanden sind, was bei manchen Algen (Florideen, Chara
u. a.) nicht der Fall ist; entweder leben sie nebeneinander in der
befruchteten Zelle fort, oder es geht ein entsprechender Teil zugrunde,
falls die Zahl der Chromatophoren in der Zelle konstant ist. Bei den
Angiospermen dringt aber, nach unseren jetzigen Kenntnissen, nur
ein männlicher Kern (Spermakern) ohne Plasma und
ohne Chromatophoren in die Eizelle ein,[S. 172] um hier mit dem Eikern
zu verschmelzen. Daraus hat man geschlossen, daß das Wesentliche
bei der Befruchtung der Übertritt des männlichen Kernes in die Eizelle
ist.
Wir haben bei der typischen Kernteilung gesehen, daß die Kerne
eines Individuums stets eine konstante, für die Art bezeichnende
Chromosomenzahl behalten. So bringt die männliche Zelle meist
ebensoviele Chromosomen wie die weibliche mit. Diese Chromosomen
verschmelzen bei der Kopulation der Geschlechtszellkerne nicht,
so daß der Kopulationskern die doppelte Chromosomenzahl besitzt wie die
Kerne der Geschlechtszellen[119]. Er ist diploid gegenüber den
haploiden Kernen dieser Zellen.
Auch die Kerne, die durch Teilung aus dem Kopulationskerne hervorgehen,
sind meist diploid. In jedem diploiden Kerne stammen meist ebensoviele
Chromosomen von dem männlichen wie vom weiblichen Kerne. Kommen in
den haploiden Kernen zwischen den einzelnen Chromosomen Größen- und
Formunterschiede vor, die bei jeder Kernteilung wieder sichtbar werden,
so sind die Chromosomen in den diploiden Kernen paarweise gleich groß
und gleich gestaltet. Diese paarweise gleichen Chromosomen, von denen
immer eines von dem Vater, das andere von der Mutter stammt, liegen in
den Kernplatten meist nebeneinander (Fig. 14).
Da die Kerne der Geschlechtszellen aller Individuen einer Rasse stets
haploid, die Kopulationskerne aber und meist auch ihre Teilungsprodukte
diploid sind, so müssen irgendwo im Entwicklungsgange des Individuums
aus den diploiden Kernen wieder haploide werden; sonst müßte ja von
Generation zu Generation die Zahl der Chromosomen sich verdoppeln!
Dieser Vorgang vollzieht sich in der Reduktionsteilung[120],
d. h. einer eigenartigen Kernteilung, wobei nicht die Längshälften
der Chromosomen, sondern die ganzen Chromosomen zur Hälfte auf
die beiden Tochterkerne gleichmäßig verteilt werden. Sie tritt im
Entwicklungsverlaufe an ganz bestimmter Stelle ein, die bei
verschiedenen Organismen freilich recht verschieden sein kann. So ist
für den Entwicklungsgang sexueller Organismen ein regelmäßiger Wechsel
von Haploidie und Diploidie der Kerne (ein Kernphasenwechsel)
bezeichnend.
Bei vielen Algen ist aber gleich die erste Teilung des befruchteten
Eikernes die Reduktionsteilung, so daß alle Zellen des
Organismus, also auch seine Geschlechtszellen, mit Ausnahme der
befruchteten Eizelle, haploid sind. Bei anderen (z. B. bei Fucus)
erfolgt die Reduktion erst bei Bildung der Geschlechtszellen,
so daß umgekehrt alle Zellen des Körpers mit Ausnahme der
Geschlechtszellen diploide Kerne haben. Oft ist aber der
Kernphasenwechsel mit einem Generationswechsel verbunden (vgl. S.
175).
Bei manchen Pflanzen wird indes der Reduktionsvorgang aus dem
Entwicklungsgange dadurch ausgeschaltet, daß sie diploide
Eizellen bilden[112], [113]. Ein solches Ei, das somit bereits
über die doppelte, sonst erst durch die Befruchtung geschaffene
Zahl von Chromosomen verfügt, entwickelt sich ohne Befruchtung,
also parthenogenetisch weiter. So ist es bei den S. 165 genannten
unbefruchteten Eizellen der Samenpflanzen, Marsiliaceen und von
Chara, während bei anderen Algen die haploide Eizelle
parthenogenetisch sich zu einer neuen Pflanze entwickeln kann.
Erwähnt sei hier, daß man vielfach dann nicht von Parthenogenesis,
sondern von Apogamie (vgl. S. 171) spricht, wenn diploide
und nicht mehr befruchtungsfähige Geschlechtszellen, also ohne
Befruchtung, sich weiter entwickeln.
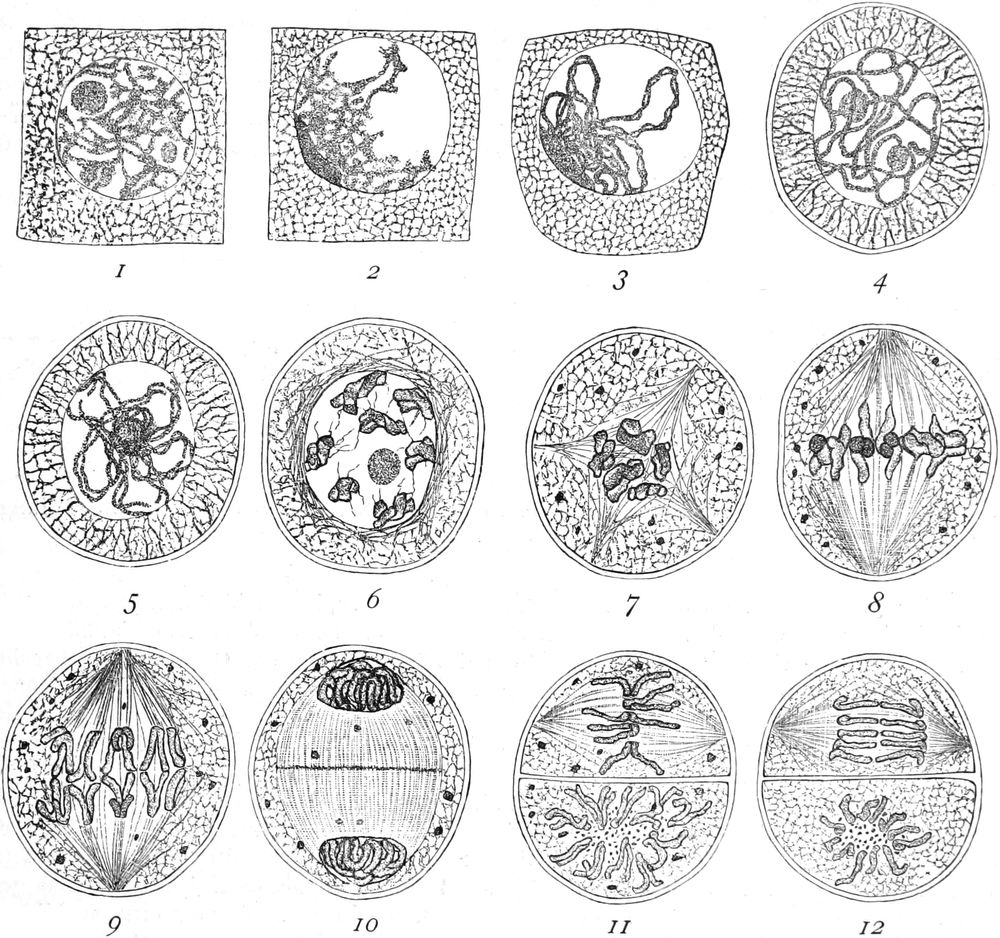
Fig. 231. Pollenmutterzellen einer Lilie in Teilung,
etwas schematisiert. Nach Fixierung mit Chromosmiumessigsäure und
Eisenhämatoxylinfärbung. Die Chromatophoren nach solcher Fixierung und
Färbung nicht sichtbar. Erklärung im Text. Nach STRASBURGER.
Eigentümlich ist es für die Reduktionsteilung, die man im Gegensatze
zu der gewöhnlichen Mitose oder typischen Teilung auch
heterotypische Teilung oder Meiosis nennt, daß der
Kerninhalt in der Prophase, und zwar im Spiremstadium vorübergehend
einseitig zusammengeballt ist (Synapsis, Fig. 231, 2, 3).
Besonders bezeichnend für den weiteren Verlauf der Teilung ist aber,
daß sich die väterlichen und mütterlichen Chromosomen paarweise[S. 173]
fest, zu den Gemini, aneinanderlegen oder gar vorübergehend
vereinigen. Die Zahl dieser Gemini, in denen die Chromosomen auffällig
kurz und dick werden, ist natürlich halb so groß wie die Zahl der
Chromosomen in den Gewebezellen derselben Pflanze. In dem Zustand der
sog. Diakinese (5, 6), der auf die Synapsis folgt, sind die
Gemini annähernd gleichmäßig an der Kernwand verteilt. Nun beginnen
Plasmafäden sich von außen der Kernwandung anzuschmiegen (6), worauf
die Kernwandung schwindet und aus den Plasmafäden die Anlage der
Kernspindel hervorgeht, die zunächst mehrpolig ist (7), allmählig
aber zweipolig (8) wird. Die Gemini werden nun zu einer äquatorialen
Kernplatte angeordnet (8). Bald erfolgt hier die Trennung der zu
Paaren vereinigten Chromosomen (9). Bei dieser Trennung werden
somit nicht Längshälften von Chromosomen, sondern ganze Chromosomen
voneinander geschieden, die zur Hälfte dem einen, zur Hälfte dem
anderen Tochterkern zugeteilt werden. Das hat zur Folge, daß die
Zahl der Chromosomen, die jedem Tochterkern zufällt, nur die Hälfte
jener Zahl ist, die die Gewebezellen derselben Pflanze in ihren
Kernen enthalten, und daß beide Tochterkerne teils männliche, teils
weibliche Chromosomen bekommen: Da sich bei der Bildung der Gemini
in der Reduktionsteilung[S. 174] immer gleichgestaltete Chromosomen
aneinanderlegen, von denen eines dem Vater, das andere der Mutter
entstammt, und da bei dieser Teilung die Chromosomen sich voneinander
trennen, um regellos in die beiden Tochterkerne einzugehen, so muß
jeder dieser haploiden Tochterkerne einige Chromosomen vom Vater, die
übrigen von der Mutter erben. Welche vom einen, welche vom anderen
Elter stammen, darüber scheint der Zufall zu entscheiden. Die Bildung
der Tochterkerne vollzieht sich wie bei einer gewöhnlichen Kernteilung
(10). Rasch folgt aber auf die Reduktionsteilung meist eine zweite
Teilung, die homöotypische Teilung, die im wesentlichen typisch
verläuft (11, 12). In der homöotypischen Teilung trennen sich also
Längshälften von Chromosomen wie in der typischen Teilung voneinander.
Ein Unterschied gegenüber der letzteren besteht aber darin, daß
die Chromosomen nicht in der Prophase dieser Teilung längs
gespalten werden, sondern, wie es scheint, bereits in der Prophase der
vorausgegangenen Reduktionsteilung, freilich ohne daß die gebildeten
Hälften getrennt werden. So bilden zwei schnell aufeinanderfolgende
Kernteilungen, die hetero- und die homöotypische, ein bezeichnendes
Merkmal der meisten Reduktionsvorgänge.
Fig. 232. In A schematische Darstellung
der Äquationsteilung, in B der Reduktionsteilung. Nach
STRASBURGER.
Der grundsätzliche Gegensatz, der zwischen der typischen Teilung
und der Reduktionsteilung besteht, soll auch schematisch deutlich
gemacht werden. Die Fig. 232 stellt in A eine typische
Kernteilung dar, die also auf Längsspaltung der Chromosomen
beruht. Es sind sechs längsgespaltene, zur Kernplatte angeordnete
Chromosomen eingezeichnet und durch verschiedene Schraffierung
kenntlich gemacht. Die zwei mittleren sind in Frontansicht, die
vier anderen in Seitenansicht gezeichnet. In Ab sieht man die
getrennten Längshälften dieser Chromosomen auf ihrem Wege nach den
Spindelpolen begriffen. Fig. 232 B ist dagegen ein Schema der
Reduktionsteilung. Die sechs in entsprechender Weise wie in Fig.
231 A gekennzeichneten Chromosomen sind in Ba zu drei Gemini
vereinigt. Die beiden seitlichen Gemini sind in Seitenansicht, der
mittlere Geminus ist in Frontansicht gezeichnet. In Bb haben sich
die beiden Chromosomen jedes Geminus getrennt. Sie werden nach den
Spindelpolen geschafft, um hier die Tochterkerne zu bilden. Diese
Teilung ist mit einer Reduktion der Chromosomenzahl von sechs auf
drei in den beiden Tochterkernen verbunden. Im Gegensatz zu der
Reduktionsteilung, die eine Verschiedenheit der Teilungsprodukte
bedingt, weil sie ganze, untereinander verschiedene
Chromosomen trennt, wird die typische Kernteilung wohl auch als
Äquationsteilung bezeichnet, da sie in den Längshälften der
Chromosomen völlig übereinstimmende Teilungsprodukte liefert.
Über die Fragen, wann und wie die Chromosomen bei der
Reduktionsteilung sich zu Paaren aneinanderlegen oder auch
vorübergehend zu einheitlichen Gebilden vereinigen, sind
die Ansichten noch geteilt. Möglicherweise erfolgt die
Reduktionsteilung überhaupt nicht nach einem einheitlichen Schema.
Meist scheinen sich die Chromosomen dabei paarweise nebeneinander
(Parasyndese), bei manchen Pflanzen dagegen mit den Enden
aneinander zu legen (Metasyndese).
Bei der Wiedertrennung der paarweise miteinander verkoppelten
Chromosomen scheint ein Austausch von Chromosomensubstanz zwischen
den Paarlingen möglich zu sein[121].
[S. 175]
D. Generationswechsel[122]. Mit der Fortpflanzung ist bei den
Pflanzen meist ein Generationswechsel, d. h. im typischen
Falle ein regelmäßiger Wechsel mindestens zweier, durch ihre
Fortpflanzungsweise voneinander verschiedener Generationen verbunden,
die häufig morphologisch ganz verschiedene und selbständige Individuen
sind. Der Entwicklungsgang eines solchen Gewächses setzt sich alsdann
meist aus zweierlei Individuen zusammen, die in regelmäßigem
Wechsel aufeinanderfolgen, häufig äußerlich und innerlich ganz
verschieden gebaut sind und verschiedene Fortpflanzungsorgane
erzeugen. Die eine Generation pflanzt sich nur vegetativ fort, man
nennt sie Sporophyt; die andere, der Gametophyt, auch
sexuell. Als typisches Beispiel für einen solchen Generationswechsel
sei der Entwicklungsgang der Farnpflanze erwähnt: Das beblätterte
Farnkraut erzeugt nur ungeschlechtliche Sporen in Sporangien, ist
also der Sporophyt. Die Sporen werden ausgestreut und keimen nicht zu
einem neuen beblätterten Farn, sondern zu einem kleinen, thallösen
Gebilde aus, dem Prothallium (Fig. 97), das die Antheridien
und Archegonien ausbildet und sich sexuell fortpflanzt, also der
Gametophyt ist. Erst die befruchtete Eizelle wird wieder zu einer
beblätterten Farnpflanze. Jede der genannten Fortpflanzungszellen der
einen Generation erzeugt also nur die andere Generation;
so folgen Sporophyt und Gametophyt (desgleichen ungeschlechtliche
und geschlechtliche Fortpflanzung) in regelmäßigem Wechsel
aufeinander. Sporophyt und Gametophyt können aber auch gleichen Bau
besitzen (z. B. bei der Braunalge Dictyota).
Sehr häufig kommt es vor, daß nicht beide Generationen selbständige
Individuen sind, sondern daß die eine dauernd mit der anderen,
wie ein Schmarotzer mit seinem Wirt, verbunden bleibt. In diesem
Falle läßt sich erst durch genaue Untersuchung feststellen, daß ein
Generationswechsel vorhanden ist. So ist es bei den Moosen und bei den
Samenpflanzen. Bei den Moosen ist nämlich das Moospflänzchen
die geschlechtliche Generation, der Gametophyt, der die Antheridien
und Archegonien erzeugt; die gestielte Mooskapsel, die mit der
Moospflanze dauernd verbunden bleibt, ist dagegen eine Generation für
sich, der Sporophyt, also nicht, wie es den Anschein hat, ein Glied
der Moospflanze. Bei den Samenpflanzen ist die beblätterte Pflanze wie
die Farnpflanze ein Sporen erzeugender Sporophyt; aus den Sporen gehen
stark reduzierte männliche und weibliche Prothallien hervor, von denen
die ersteren in die Pollenkörner und Pollenschläuche, die letzteren
dauernd in die Samenanlagen der Blüten eingeschlossen bleiben.
Wo ein Generationswechsel im Pflanzenreich vorkommt, pflegt mit ihm
der Kernphasenwechsel verbunden zu sein, so bei vielen Algen, Pilzen,
den Moosen, Farn- und Samenpflanzen. Der aus der befruchteten Eizelle
entstandene Sporophyt ist diploid; der Bildung seiner Sporen geht
die Reduktionsteilung unmittelbar voraus; infolgedessen sind die
Sporen selbst, der daraus sich entwickelnde Gametophyt und seine
Geschlechtszellen haploid. Dementsprechend ist z. B. die beblätterte
Generation der Farn- und Samenpflanzen ebenso wie die Mooskapsel
diploid; das Moospflänzchen, das Farnprothallium und die reduzierten
Prothallien der Samenpflanzen sind dagegen haploid.
Sehr merkwürdig sind einige Fälle, wo aus vegetativen Zellen
der einen Generation ohne weiteres sich die andere, dann
also mit gleicher Chromosomenzahl entwickeln kann. Bei
einer Form von Athyrium filix femina geht aus solchen vegetativen
Prothallienzellen, deren Kerne diploid sind, ohne Verschmelzung
die diploide Farnpflanze hervor, die unter Ausschaltung der
Sporen und der Reduktionsteilung aus ebenfalls diploiden Zellen
des Blattrandes wieder diploide Prothallien erzeugt
(Aposporie). Nach YAMANOUCHI kann aber auch (bei
Nephrodium molle) aus einer haploiden Prothallialzelle
ohne Verschmelzungsvorgänge eine haploide Farnpflanze
entstehen. Ferner kann man auf experimentellem Wege durch
Regeneration aus zerschnittenen Mooskapselstielen,[S. 176] die als
Sporophytengewebe diploid sind, eine diploide Moospflanze,
also einen diploiden Gametophyten erziehen, der auch diploide und
gleichwohl befruchtungsfähige Geschlechtszellen hervorbringt. Durch
Befruchtung entstehen tetraploide Mooskapseln, die wiederum auf
dem Wege der Regeneration selbst tetraploide Moospflanzen liefern
können. Eine direkte Beziehung zwischen der Chromosomenzahl und der
Ausbildung der Generation besteht also offenbar nicht[112], [113].
Vierter
Abschnitt. Die Deszendenzlehre und die Entstehung der Anpassungen.
A. Die Deszendenztheorie[123]. Eine der allerwichtigsten
theoretischen Fragen der Morphologie ist die, wie die organischen
Formen unserer Erde und ihre morphologischen Baueigentümlichkeiten
entstanden sind. Die frühere Annahme, die Einzelarten seien selbständig
und unabhängig voneinander erschaffen worden (Schöpfungstheorie), ist
allmählich, namentlich infolge der Vertiefung unserer morphologischen
Kenntnisse und unter dem Einflusse der Werke DARWINs, durch
die Deszendenztheorie völlig verdrängt worden, wie schon in der
Einleitung hervorgehoben wurde; d. h. durch die Vorstellung, daß die
Organismen, die jetzt die Erde bewohnen, aus anderen und oft einfacher
gebauten sich entwickelt haben, die in früheren Erdepochen gelebt haben
(vgl. S. 1 ff.). Die Deszendenz- oder Abstammungslehre durchdringt
heute als eine grundlegende Theorie der Biologie die morphologische
Forschung in allen ihren Teilen so vollständig, daß es für den
Morphologen unerläßlich ist, sich mit den zahlreichen Indizienbeweisen
vertraut zu machen, wodurch sie uns geradezu aufgezwungen wird.
Solche Beweise liefert vor allem die Systematik, die Morphologie, die
Pflanzen- und Tiergeographie und die Paläontologie.
1. Systematische Beweise. Nach der Schöpfungstheorie sind die
Arten unabhängig voneinander erschaffen worden und im wesentlichen
konstante, d. h. nur so wenig veränderliche Gebilde, daß aus einer
Art nicht andere Arten, sondern höchstens Varietäten als mehr oder
weniger erbliche Abänderungen hervorgehen können. Diese Lehre fordert
also scharfe Grenzen zwischen den Spezies und deutliche
Unterschiede im Wesen der Arten und in dem der Varietäten.
Dem Systematiker, der sich in das Studium irgendwelcher Lebewesen
vertieft, fällt es aber immer wieder auf, daß sich überhaupt keine
Merkmale auffinden lassen, wodurch sich Varietäten durchgreifend von
Arten unterscheiden. Das Maß der morphologischen Verschiedenheiten
zwischen den Arten einer Gattung, den Varietäten einer Art
oder zwischen Arten und Varietäten ist eben ganz und gar unbestimmt.
Auch hat sich im Laufe der Zeit gezeigt, daß die Arten gar keine
selbständigen morphologischen Einheiten sind, sondern in
vielen Fällen sehr umfangreiche Formenkreise von „Kleinarten“
(z. B. in den Gattungen Erophila, Rubus, Rosa, Hieracium, Quercus),
deren scharfe Abgrenzung gegen andere Arten, d. h. andere solche
Formenkreise, zudem oft sehr schwierig, ja manchmal kaum möglich
ist. Auch sind die erblich konstanten Kleinarten oft viel weniger
voneinander verschieden als viele sog. Varietäten. So ist es oft
lediglich Sache des Geschmackes, des „systematischen Taktes“, ob
man eine Form als Art oder[S. 177] Varietät auffassen und wie man eine Art
umgrenzen will. Denn auch die Regel, an die man früher wohl glaubte,
die Kreuzungen zwischen den unabhängig voneinander erschaffenen
Arten seien unfruchtbar, zwischen den Varietäten einer Art aber
fruchtbar, hat sich als nicht richtig erwiesen; es gibt fruchtbare und
unfruchtbare Bastarde zwischen zwei Varietäten und zwischen zwei Arten.
Aber nicht nur Übergänge zwischen Arten kommen vor, sondern oft selbst
zwischen Gattungen oder gar Familien, so daß es manchmal auch hier der
Willkür des Systematikers überlassen bleibt, die Grenzen zu ziehen.
Alle diese Tatsachen werden nur verständlich, wenn man annimmt, daß
die Arten nicht selbständig erschaffen, sondern erblich veränderlich
sind, d. h. daß aus einer Spezies durch erbliche Umwandlungen andere
hervorgehen können, durch stärkere Veränderungen aus den Arten einer
Gattung neue Gattungen, schließlich aus einer Familie andere Familien.
Ohne diese Annahme bleibt es auch völlig unbegreiflich, daß es möglich
ist, die Organismen überhaupt in Gruppen niederer und höherer Ordnung
(Arten, Gattungen, Familien, Klassen usw.) zu bringen, die sich teils
einander koordinieren (wie die Arten einer Gattung, die Gattungen einer
Familie), teils einander subordinieren lassen (wie die Gattungen der
Familie, die Arten der Gattung), und daß die Gruppen der ausgestorbenen
Organismen, die in früheren Erdepochen lebten, zwischen die der
lebenden meist zwanglos eingeordnet werden können. Alle Schwierigkeiten
fallen sofort weg, wenn man die Organismen als blutsverwandt betrachtet
und das natürliche System als den Ausdruck der näheren und ferneren
Verwandtschaft, also gewissermaßen als eine genealogische Anordnung
(Stammbaum) der Lebewesen ansieht.
Fig. 233. Blütendiagramme von Scrophulariaceen. A
Verbascum nigrum, B Digitalis purpurea, C Gratiola officinalis,
D Veronica Chamaedrys. A, C, D nach EICHLER. Die
unfruchtbaren Staubgefäße sind durch schwarze Punkte, vollständig
geschwundene durch Sterne angedeutet.
2. Morphologische Beweise. Unverständlich für die
Schöpfungstheorie, sofort einleuchtend bei Annahme der Deszendenzlehre
sind ferner einerseits die gemeinsamen, morphologischen
Baupläne, die den Angehörigen einer systematischen Gruppe,
einer Gattung, einer Familie, einer Klasse, in gewissem Sinne sogar
allen Organismen der Erde (im zelligen Bau, Protoplasma) zugrunde
liegen, andererseits das Vorkommen von einzelnen Eigenschaften bei
einer Gruppe, bei der man sie nach ihrem Bauplan eigentlich gar nicht
erwarten sollte (z. B. die Spermien im Pollenschlauch der Zykadeen).
Die großen Gruppen der Moose, Farne und Gymnospermen haben bei allen
morphologischen Verschiedenheiten doch im wesentlichen den gleichen
Entwicklungsgang, den gleichen Generationswechsel und die gleichen
Geschlechtsorgane. Nur mit der Annahme der Blutsverwandtschaft kann
man ferner verstehen, daß so oft die Organe verschiedener Arten
bei genauerer morphologischer Untersuchung trotz gleichem Bau und
gleichen Funktionen doch verschiedenen Grundformen entsprechen, also
nur analog sind, daß sie umgekehrt aber auch dann einander homolog
gefunden werden, wenn sie völlig verschieden gebaut sind und ganz
verschiedenen Funktionen dienen, oder daß auch die Organe eines und
desselben Organismus trotz ganz verschiedenem Bau und verschiedenen
Funktionen doch so oft homolog sind. Dornen[S. 178] und Ranken z. B. sind
„umgewandelte“ Blätter, Nebenblätter, Sproßachsen oder Wurzeln; die
Keimblätter, Nieder-, Hoch-, Kelch-, Kronblätter, die Staubgefäße
und die Fruchtblätter einer Pflanze sind sämtlich nichts anderes als
„umgewandelte“ Laubblätter. Alle diese Umwandlungen und Weiterbildungen
von Organen haben offenbar während der phylogenetischen Entwicklung
stattgefunden. Ebenso weisen die funktionslos gewordenen, reduzierten
Organe, die man bei manchen Formen findet, auf die Abstammung von
solchen Formen hin, bei denen die Organe noch wohl ausgebildet waren:
In der Familie der Scrophulariaceen (Fig. 233) finden wir Formen mit
fünf Staubgefäßen bei Verbascum bis zu solchen mit nur zwei halben
bei Calceolaria; in der Gattung Scrophularia ist eines der fünf zwar
noch vorhanden, aber reduziert; bei Digitalis fehlt das reduzierte;
bei Gratiola sind nur zwei fruchtbare vorhanden, außerdem noch zwei
reduzierte, bei Veronica nur zwei fruchtbare, bei Calceolaria nur
zwei halbe. Reduzierte, nutzlose Organe sind aber für die
Schöpfungstheorie schwer verständlich. Gelegentlich beobachtet man
wohl auch, daß bei einer Form plötzlich eine fremdartige Eigenschaft
auftaucht, die man nur als einen Rückschlag (Atavismus) zu
längst verloren gegangenen Eigenschaften von Vorfahren auffassen
kann, z. B. Fruchtbarwerden reduzierter Staubgefäße, Auftreten von
reduzierten oder fruchtbaren Staubgefäßen an Stellen, wo solche in
der phylogenetischen Entwicklung verloren gegangen waren. Auf eine
Blutsverwandtschaft der Organismen deutet ferner die weitgehende
Ähnlichkeit der Embryonen selbst sehr verschiedener Organismen
hin, wofür namentlich das Tierreich viele auffallende Beispiele
liefert; weiter die Tatsache, daß manchmal die Embryonen viel höher
gegliedert sind als die ausgebildeten Organismen (so bei manchen
reduzierten Organismen, z. B. vielen Parasiten), und daß die
Jugendblätter an den Keimlingen mancher Gewächse, die an extreme
Lebensbedingungen einseitig angepaßt sind, den Folgeblättern nicht so
einseitig angepaßter Arten der gleichen Gattung entsprechen (z. B.
bei Acacia, vgl. Fig. 140). Ja nicht selten wiederholt eine Art mehr
oder weniger in ihrer ontogenetischen Entwicklung die Entwicklung,
die man aus sonstigen Gründen als ihre phylogenetische ansehen muß
(biogenetisches Grundgesetz).
3. Geographische Beweise. Geographische Schranken,
Hindernisse für freie Wanderung (wie z. B. hohe Gebirge, Meere für
Landpflanzen oder Ländermassen für Meeresorganismen) stehen mit den
Verschiedenheiten der Flora und Fauna der einzelnen Wohngebiete,
Länder, Kontinente und Meere in auffälligem Zusammenhang. Die
Organismenwelten zweier Kontinente sind durchschnittlich in Familien,
Gattungen usw. um so verschiedener, je schwieriger die Verbindungen
sind und früher waren, weil die Formen in einem jeden solchen Land sich
selbständig phylogenetisch weiter entwickelten. Zwei Gebiete haben
dagegen um so mehr gemeinsame Formen, je leichter ein Formenaustausch
möglich war. Sehr allgemeine Regel ist, daß die Bewohner eines Gebietes
mit denen desjenigen nächsten Gebietes am engsten verwandt sind,
aus dem sich die Einwanderung aus geologischen und geographischen
Gründen mit Wahrscheinlichkeit annehmen läßt. Das gilt z. B. für
die Kap Verdischen Inseln und das afrikanische Festland, für die
Galapagosinseln oder Juan Fernandez und die Nachbargebiete von
Amerika. Je abgeschlossener aber ein Wohnbezirk, etwa eine Insel, von
der übrigen Welt ist, um so reicher pflegt sie an ihm eigentümlichen
Formen, Endemismen, zu sein. Das sind teils solche Formen, die
sich von anderen nicht endemischen oft nur wenig unterscheiden und
offenbar aus ihnen in solchen Gebieten erst nach deren Abschließung
entstanden sind, ohne sich darüber hinaus haben ausbreiten zu können
(progressive Endemismen), teils auch solche, die als[S. 179] Reste
früher weiter verbreiteter Formen in dem Gebiet ohne Verwandte sind,
vielfach als Zeugen untergegangener Floren angesehen werden können und
in diesem Falle hohes Alter aufweisen (Reliktendemismen).
4. Paläontologische Beweise. Die Paläontologie endlich zeigt
uns, daß Arten in der Geschichte unseres Planeten völlig erlöschen,
andere dafür auftauchen, ferner daß nicht selten in benachbarten
Erdschichten die Formen sich zu Formenreihen fortschreitender
Organisationshöhe anordnen lassen, und daß die Formengruppen,
deren Arten wir als die am reichsten gegliederten ansehen, erst
verhältnismäßig spät in der Erdgeschichte, die Angiospermen z. B.
erst in der Kreidezeit, erschienen sind. Vor allem aber hat diese
Wissenschaft uns heute fehlende, ausgestorbene Zwischenglieder zwischen
Gattungen, Familien und Klassen kennen gelehrt, d. h. Formen, die
in ihren Baueigentümlichkeiten zwischen diesen vermitteln. Daß sie
nicht häufiger sind, hängt offenbar mit der großen Unvollständigkeit
unserer geologischen Urkunden zusammen. Auf botanischem Gebiete ist
die wichtigste dieser Zwischengruppen die der Pteridospermeen oder
Cycadofilices. Das sind Gewächse der Karbonformation, die zwischen
Farnen und Zykadeen vermitteln, d. h. Blätter wie Farne, Samen aber
wie die Zykadeen und anatomischen Bau teils wie jene, teils wie diese
gehabt haben.
5. Direkte Beweise für die Veränderlichkeit der Arten. Alle
diese verschiedenartigen Indizienbeweise gewinnen aber noch dadurch
ganz wesentlich an Bedeutung, daß es gelungen ist, die Inkonstanz
mancher Arten direkt zu beobachten. Bei sorgfältiger Untersuchung
sehen wir nämlich in der freien Natur, viel häufiger aber in der
Kultur, bei Organismen gelegentlich erbliche Veränderungen auftreten,
die den systematischen Wert von Varietäten oder Arten haben. Auch ist
es im Experimente auf verschiedenen Wegen geglückt, solche erblich
konstanten Veränderungen, also die Erzeugung neuer Formen zu erzwingen.
Derartige Beobachtungen sind zugleich deshalb sehr wertvoll, weil sie
uns Einblicke in das Problem der Artbildung, der Entstehung neuer
morphologischer Eigenschaften erlauben.
B. Entstehung der Anpassungen. Alle bisherigen Beobachtungen
weisen darauf hin, daß die erblichen Änderungen der Organismen bald
diese, bald jene Eigenschaften betreffen und in kleineren oder wohl
auch größeren Sprüngen, und zwar ganz regellos erfolgen. Daraus
erklärt sich die große Mannigfaltigkeit der organischen Formen. Diese
sprunghaften Veränderungen können bald schädlich, bald gleichgültig,
bald nützlich für den Organismus sein. Sind sie so schädlich, daß
der Organismus damit nicht mehr lebensfähig ist, so verschwinden
derartige Varianten meist ebenso schnell, wie sie entstanden sind
(z. B. Keimlinge, die die Eigenschaft verloren haben, Chlorophyll
auszubilden). Wieweit solche erblichen Veränderungen unter dem
Einflusse äußerer erfaßbarer Bedingungen entstehen, bleibt im einzelnen
noch zu erforschen.
Seitdem man sich auf den Boden der Deszendenzlehre gestellt hat, hat
man erkannt, daß eine besondere Erklärung noch die Entstehung der für
die Organismen so bezeichnenden Anpassungsmerkmale bedarf.
Mit der Erkenntnis, daß die Lebewesen regellos erblich abändern, ist
ja noch keinerlei Einsicht in die auffällige Tatsache gewonnen, daß
der Organismus vielfach so ausgesprochen an seine Umwelt, ferner ein
Organ in seinem Bau mehr oder weniger an seine Funktionen angepaßt ist
und nützlich reagiert. Dieser Zustand der Anpassung, dieses
erbliche Angepaßtsein muß phylogenetisch irgendwie entstanden
sein; wie, darauf geben uns alle Beobachtungen und Versuche bis jetzt
keine unmittelbare Antwort. Erklärungen[S. 180] hat man auf ganz verschiedenem
Wege versucht. Die wichtigsten Hypothesen in dieser Hinsicht sind der
Lamarckismus und der Darwinismus.
1. Der Lamarckismus[124] geht von der Tatsache aus, daß manche
Organismen je nach der Umgebung, in der ihre Keimzellen auskeimen
und sich zum fertigen Organismus entwickeln, verschiedene Gestalt
annehmen, ohne daß diese Lebewesen dadurch die Fähigkeit verlören,
sich in anderer Umgebung dieser entsprechend zu entwickeln. So gibt
es Pflanzen, die sowohl auf dem Lande wie im Wasser leben können (man
nennt sie amphibische) und die je nach den Außenbedingungen, in
denen sie sich entwickeln, verschiedene Formen annehmen. Auf dem Lande
ausgesät, nehmen sie Gestalt und inneren Bau typischer Landpflanzen
an; bei Kultur im Wasser gleichen sie typischen Wasserpflanzen. Manche
Pflanzen bilden bei Trockenkultur xerophile Merkmale aus, bei Kultur in
feuchter Luft dagegen hygrophile. Diese Befähigung, in verschiedener
Umgebung mit Ausbildung verschiedener Merkmale zu reagieren, nennt man
Modifikationsfähigkeit. Solche Modifikationen (vgl. Physiologie
S. 285) sind nicht in dem Sinne erblich, daß die Samen z. B. einer
amphibischen Pflanze, die sich im Wasser zur Wasserpflanze entwickelt
hat, bei Aussaat auf dem Lande nun ebenfalls die Wasserform lieferte.
Vielmehr entsteht auf dem Lande immer die Land-, im Wasser
stets die Wasserform, von welcher Form auch immer die Samen
entnommen sein mögen.
Man hat nun in diesen Wirkungen der Außenwelt auf solche
modifizierbaren Pflanzen ein direktes „Sichanpassen“
erblicken wollen und dem Organismus die Fähigkeit zugeschrieben, auf
jede äußere Einwirkung, auch auf solche, die ihm in der Natur
nicht zu begegnen pflegen, mit einer nützlichen Reaktion zu
antworten. So wie an äußere Faktoren, so soll auch ein Sichanpassen
an neue Funktionen möglich sein; jedes „Bedürfnis“ nach einem
Organ soll auf die Bildung eines solchen hinwirken. Und weiter nimmt
der Lamarckismus an, daß jede einmal entstandene Veränderung,
insbesondere jede durch Außenfaktoren oder durch „Bedürfnisse“ bewirkte
Abänderung auch erblich sei oder zum mindesten mit der Zeit erblich
werden könne. Wenn also eine Pflanze Generationen hindurch sich immer
wieder an das Wasserleben oder an das Leben im Schatten oder Leben
auf Kosten eines anderen Organismus im obigen Sinne direkt anpaßt,
so sollen die so gewonnenen Baueigentümlichkeiten allmählich fixiert
werden; d. h. sie sollen auch dann auftreten, wenn die Anlässe dazu
nicht mehr vorhanden sind. Zu dieser Auffassung ist zunächst zu
bemerken, daß die Annahme, „ein Bedürfnis nach einem Organ könne
die Bildung eines solchen bewirken“, eine unklare Überlegung
ist. Sodann muß man betonen, daß von einem Erblichwerden solcher
Wirkungen äußerer Bedingungen, die wir Modifikationen genannt haben,
schlechterdings nichts zu bemerken ist. Schon aus diesem Grunde
ist der Lamarckismus abzuweisen. Zweitens aber hieße es an ein Wunder
glauben, wenn man annehmen wollte, daß der Organismus von vornherein
auf beliebige Außenfaktoren nützlich reagierte. In der Tat sehen wir
auch nicht ganz selten Reaktionen auf neue ungewohnte Reize
eintreten, die durchaus „gleichgültig“ oder gar schädlich erscheinen.
So krümmen sich Droseratentakeln auf hohe Temperatur hin so ein, als
wären sie mit einem Insekt in Berührung gekommen; abgeschnittene
Blätter können bei vielen Pflanzen durch Ausbildung von Wurzeln sich
manchmal jahrelang am Leben erhalten, auch wenn ihnen die Möglichkeit
der Sproßbildung abgeht. Wenn andererseits auf verschiedene Reize,
denen gewisse Organismen an ihren verschiedenen Standorten ausgesetzt
sind, wie Wasser, Luft, Licht, Schatten usw., ein direktes Sichanpassen
zu folgen scheint,[S. 181] so kann man diese Erfolge auch anders deuten. Man
kann annehmen, daß solche Organismen schon die Befähigung, d. h. die
Anlagen besitzen, die ihnen je nach den äußeren Bedingungen bald
diese, bald jene Entwicklung einzuschlagen erlauben. Die Außenfaktoren
erzeugen also nicht solche Befähigungen oder Anlagen, sondern
sie bewirken nur ihre Entfaltung oder Nichtentfaltung. Wie aber
jene Anlagen historisch entstanden sind, warum manche Lebewesen sie
besitzen, andere nicht, warum also z. B. nur manche angepaßt
sind, im Wasser in der Gestalt von Wasserpflanzen, auf dem Lande in der
Gestalt von Landpflanzen zu leben, das eben bleibt wie eine jede andere
Anpassung noch immer zu erklären. Und hier versagt der Lamarckismus
völlig.
2. Der Darwinismus[123], [125]. DARWIN geht von der
Tatsache aus, daß die beschränkten Lebensbedingungen auf unserer Erde
keine unbegrenzte Vermehrung der Organismenmenge erlauben. Fast jedes
Lebewesen liefert aber während seines Einzeldaseins so viele Keime,
daß, wenn alle aufkämen, schon nach ganz kurzer Zeit die Erde allein
von einer Art übervölkert sein würde. Nur wenige Nachkommen eines
Individuums bleiben aber am Leben, weil nämlich die Umwelt so viele
in jedem Entwicklungsstadium von der Keimzelle an vernichtet. Die
Nachkommen jedes Individuums unterliegen dem „Kampfe ums Dasein“
mit der Umwelt, wozu wir natürlich auch die anderen Organismen der
gleichen oder anderer Arten zu rechnen haben. Wären alle Nachkommen
ganz gleich, so könnte allein der Zufall darüber entscheiden, welche am
Leben bleiben. Solche Zufälle haben auch sicher eine große Bedeutung.
Da aber zuweilen unter den Nachkommen erbliche Verschiedenheiten
bestehen, so werden in der Regel diejenigen in diesem Kampfe bevorzugt
sein, die an dem Platze, wohin sie der Zufall verschlagen hat, durch
ihre Besonderheiten erhaltungsfähig oder erhaltungsfähiger als
die anderen sind. Es findet also eine Auslese (Selektion,
natürliche Zuchtwahl) statt. Und wenn nun die ausgelesenen
Varianten ihre Eigenschaften an ihre Nachkommen übertragen, bei
denen sich die erblichen Variationen und der Kampf wiederholen,
dann muß die Entwicklung zu immer besser angepaßten Formen führen.
Entstehen können Organismen mit ganz beliebigen Eigenschaften,
nützlichen, gleichgültigen oder schädlichen. Da alle mit schädlichen
Eigenschaften früher oder später wieder verschwinden müssen, bleiben
nur solche übrig, die besser angepaßt sind als die Besiegten, sich
aber außerdem auch noch in vielen gleichgültigen Eigenschaften von
diesen unterscheiden können. Nützlichkeit (d. h. Angepaßtsein) erklärt
sich also nach dem Lamarckismus überhaupt nicht, da dieser das
nützliche Reaktionsvermögen der Organismen auch gegenüber ihnen völlig
fremden, neuen Außenverhältnissen als gegeben hinnehmen muß, nach dem
Darwinismus dagegen aus den Vorzügen besser erhaltungsfähiger erblicher
Eigenschaften im Kampfe ums Dasein. Darin liegt der große Fortschritt
der DARWINschen Theorie gegenüber dem Lamarckismus. Sie wird,
wie wir sahen, durch alle Beobachtungen unterstützt, die wir über die
Entstehung neuer erblicher Eigenschaften bei Organismen bisher gemacht
haben, wenn auch bei Annahme des Darwinismus noch immer mancherlei
Schwierigkeiten zu überwinden bleiben.
Die Physiologie hat die Aufgabe, die Lebenserscheinungen zu
beschreiben, ihre Abhängigkeit von äußeren Faktoren zu studieren
und sie soweit wie möglich auf ihre Ursachen zurückzuführen. Wie
Chemie und Physik, so forscht also auch die Physiologie nach
den Ursachen des Geschehens, sie ist eine Wissenschaft mit
kausaler Fragestellung. Doch muß sie notwendigerweise auch die
Bedeutung des Geschehens für den Organismus mit in Betracht
ziehen. Wie in der Fragestellung, so folgt die Physiologie auch in
ihrer Arbeitsmethode der Physik und Chemie: sie bedient sich in erster
Linie des Experimentes.
Die Hauptergebnisse der physiologischen Forschung sind folgende:
1. Einen grundsätzlichen Unterschied zwischen den
Lebenserscheinungen der Tierwelt und der Pflanzenwelt gibt es
nicht. Das ist nicht überraschend, weil schon morphologisch
Tier und Pflanze nur in ihren extremen Ausbildungen scharf
unterscheidbar sind. Auf physiologischem Gebiet aber hat sich,
je weiter die Forschung vorschreitet, desto deutlicher gezeigt, wie
ähnlich das Leben in den beiden Hauptreichen verläuft. Dementsprechend
gibt es eigentlich nur eine Physiologie, die Physiologie der
Organismen. Ein Lehrbuch der Botanik hat aber selbstverständlich
nur die Physiologie der Pflanzen darzustellen; doch soll, wo es
nützlich erscheint, auf analoge Vorkommnisse im Tierreich kurz
hingewiesen werden.
2. In mancher Hinsicht reagiert die lebende Pflanze nicht anders als
beliebige tote Körper. Trotz ihres hohen Wassergehaltes ist sie im
allgemeinen ein fester Körper und hat die physikalischen Eigenschaften
eines solchen. Schwere, Festigkeit, Elastizität, Leitungsfähigkeit für
Wärme und Elektrizität kommen ihr in gleicher Weise zu wie leblosen
Körpern. So wichtig diese Eigenschaften nun auch für das Bestehen und
für das Leben der Pflanze sind, so bedingen sie doch noch nicht das
Leben selbst.
3. Die eigentlichen Lebenserscheinungen scheinen nämlich auf
den ersten Blick recht verschieden zu sein von den Vorgängen, die man
bei leblosen Körpern antrifft. So lange der Organismus in voller
Lebenstätigkeit ist, können wir eine ununterbrochene Kette von
Veränderungen an ihm wahrnehmen, die sich in dreifach verschiedener
Weise äußern:
I. Ein Organismus besteht selbst dann, wenn eine Vergrößerung durch
Wachstum nicht mehr erfolgt, keineswegs aus einer gleichbleibenden
Stoffmasse. Während die äußere Form konstant bleibt, finden im
Innern fortwährend Veränderungen statt; neue Stoffe werden von außen
aufgenommen, werden im Innern umgewandelt und auch wieder nach außen
abgegeben. Der Organismus besitzt einen Stoffwechsel.
II. Gewöhnlich aber verläuft dieser Stoffwechsel nicht in der Weise,
daß die Stoffaufnahme der Stoffabgabe gleichkommt, sondern es wird
mehr aufgenommen als abgegeben; die Masse nimmt zu, der
Organismus wächst. Wachstum kennen wir auch an chemischen
Ausfällungen (Niederschlägen) oder an Kristallen. Hier pflegt es indes
so zu verlaufen, daß eine wesentliche[S. 183] Veränderung der Form nicht
erzielt wird (bei Kristallen), oder daß die Gestaltsveränderung eine
zufällige, keine gesetzmäßige ist (Niederschläge). Der Organismus aber
nimmt unter gesetzmäßiger Gestaltsveränderung ganz bestimmte,
immer wiederkehrende Formen an; er macht eine Entwicklung durch,
und diese führt früher oder später zur Entstehung von neuen Organismen,
Tochterindividuen; es tritt Fortpflanzung ein. Wachstum, Entwicklung
und Fortpflanzung sind für die Lebewesen ungemein charakteristische
Vorgänge.
Manche Niederschläge haben unter Umständen eine gewisse
äußerliche Ähnlichkeit mit Pflanzen. Bringt man z. B. in
eine mit Gelatine versetzte Lösung von gelbem Blutlaugensalz und
Kochsalz etwas Kupfervitriol, dem Zucker zugegeben wurde, so
bildet sich ein Niederschlag von Ferrocyankupfer, der zusehends
wächst und in seiner Gestalt an Pflanzen erinnert. Es fehlt dieser
„künstlichen Pflanze“ aber nicht nur die innere Struktur der
wirklichen Pflanze, sondern vor allem auch die Fortpflanzung und
die gesetzmäßige Entwicklung.
III. Endlich zeigen die Organismen Bewegungsvermögen; sie
ändern entweder im ganzen den Ort, oder sie bringen kleinere
oder größere Teile in andere Lagen. Da auch Anorganismen und
tote Organismen Bewegungen ausführen können, so ist für die Lebewesen
nur die Art und Weise, wie die Bewegung zustande kommt und
unterhalten wird, charakteristisch.
In der Natur treten die drei genannten Prozesse, der Stoffwechsel,
die Entwicklung und die Bewegungen, fast stets gleichzeitig auf.
Der Stoffwechsel ist ohne Stoffbewegungen undenkbar, die
Entwicklung ist immer mit stofflichen Veränderungen und mit
Bewegungen verknüpft, die Bewegungen endlich können ohne
Stoffwechsel und ohne Gestaltswechsel nicht stattfinden. Dennoch können
wir in unserer Darstellung die drei Prozesse gesondert betrachten und
die Physiologie in drei Abschnitte einteilen, nämlich:
Die Lehre vom Stoffwechsel oder die chemische Physiologie, auch
wohl Physiologie der Ernährung genannt.
Die Lehre von der Entwicklung, Physiologie der Gestaltung,
Formwechsel.
Die Lehre von den Bewegungen.
4. Diese Lebenserscheinungen sind durchaus an das Protoplasma
gebunden und beruhen auf der eigentümlichen Art, wie diese Substanz
auf Einflüsse der Außenwelt reagiert, d. h. vor allem auf ihrer
Reizbarkeit und Regulationsfähigkeit.
a) Reizbarkeit. Bei den Reaktionen des Organismus ist in
der Regel der Zusammenhang zwischen Ursache und Wirkung nicht so
durchsichtig wie bei physikalischen und chemischen Vorgängen. Das rührt
daher, daß wir kaum jemals die nächsten, sondern meist nur ganz
indirekte Folgen einer äußeren Einwirkung zu sehen bekommen.
Auch kann je nach dem Zustand des Protoplasmas ein und derselbe Faktor
ganz verschiedenartige Wirkung haben. Hierfür ein Beispiel:
Befestigt man einen biegsamen Stab in horizontaler Lage an einem
Ende, so wird sich das freie Ende durch das Gewicht des Stabes
bis zu einem bestimmten Punkte abwärts biegen. Dasselbe tut jeder
Pflanzenteil, und bei toten Pflanzenteilen bleibt es dabei. Hat
man zu dem Versuche jedoch einen lebendigen, wachsenden Stengel
benutzt, dann zeigt sich nach einiger Zeit in diesem eine Wirkung
der Schwerkraft, die im Vergleich zu ihrer rein physikalischen
Wirkungsweise durchaus überrascht: Der wachsende Teil des Stengels
richtet sich durch eigene Tätigkeit wieder auf; er krümmt
sich dem Zug der Schwere entgegen. Macht man den Versuch mit einer
Pfahlwurzel, so wird diese, unter Entfaltung von Kraftäußerungen,
die ihr Eigengewicht erheblich übertreffen, sich senkrecht abwärts
krümmen; ein Rhizom von Scirpus dagegen würde in der Horizontallage
weiterwachsen und würde auch seine wachsende Spitze wieder
wagerecht stellen, wenn man es aus dieser Lage gebracht hätte.
Bei diesen drei Versuchen sind die physikalischen[S. 184] Bedingungen
jedesmal die gleichen: Die Erdschwere wirkt auf einen horizontalen
Pflanzenteil. Das Resultat aber fällt so verschieden wie nur
möglich aus.
Man muß in solchen Fällen annehmen, daß zwar die Einwirkung von außen
zunächst stets den gleichen Effekt hat wie in Anorganismen —
in unserem Beispiele würde also die Schwerkraft stets zunächst
einmal eine Druckwirkung bedingen —, daß aber diese primäre
Veränderung innere Tätigkeiten der Pflanze veranlaßt, wobei
vielfach der Außenfaktor als auslösendes Moment aufzutreten
scheint. Solche Vorgänge werden erst verständlich, wenn wir den
Organismus mit einem Mechanismus vergleichen. Der
Zusammenhang zwischen dem schwachen Fingerdruck, den wir auf den
Abzugsbügel eines Gewehres ausüben, und der Flugbewegung des Geschosses
ist nicht einfach. Der Druck bewirkt zunächst die Entspannung einer
Feder; die frei gewordene Energie treibt einen Schlagbolzen auf die
Zündmasse; diese explodiert und bringt das Pulver zur Explosion; die
dabei entstehenden Gase erst schleudern das Projektil aus dem Lauf.
Es leuchtet ein, daß die Kraft des Schlagbolzens keinerlei Beziehung
zur Kraft des Fingerdruckes hat, den der Schütze ausübt, und daß
ebensowenig eine Beziehung zwischen der Größe der Expansionskraft
des Pulvers und der Schlagbolzenkraft besteht. Es werden vorhandene
Energien, die der gespannten Feder und die des Pulvers, ausgelöst.
Solche Auslösungen, vor allem Auslösungen, die einander
folgen, die eine Kette von Prozessen bilden, treten uns bei allen
Lebenserscheinungen entgegen; sie sind freilich nicht so einfach
und übersichtlich wie bei den Mechanismen, weil sie sich durchweg im
Protoplasma abspielen. Auslösungen, die in der lebenden Masse
aufgespeicherte Spannkraft in kinetische Energie verwandeln, seien
als Reizerscheinungen, der auslösende Faktor als „Reiz“
bezeichnet (vgl. aber S. 322).
So wie die Leistung einer Maschine nur verständlich wird, wenn man
ihren Bau kennt, so ist auch die Kenntnis des äußeren und inneren Baues
der Pflanze Vorbedingung für physiologische Studien. Es hat sich aber
gezeigt, daß das Verständnis der Funktion aus dem Bau bei der Pflanze
nicht annähernd in dem Maße erzielt werden kann wie bei der Maschine;
es handelt sich im Organismus eben nicht um ein Ineinandergreifen von
Rädern oder anderen Mechanismen, sondern wesentlich um eine Folge
chemischer Reaktionen.
b) Regulationen. Maschinen können aber nicht nur das Wesen der
Auslösung versinnbildlichen, sondern sie können auch noch die
zweite weitverbreitete Eigenschaft des Organismus klarmachen, nämlich
seine Regulationsfähigkeit. So wie an einer Maschine z. B. die
Geschwindigkeit automatisch auf einer gewissen Höhe erhalten
wird, so sehen wir auch in der Pflanze bei zahllosen Prozessen eine
Selbststeuerung eintreten, die Qualität und Quantität eines
jeden Geschehens regelt. Wenn es auch in der anorganischen Welt nicht
an regulatorisch verlaufenden Vorgängen fehlt, so treten uns doch diese
nicht in der Häufigkeit entgegen wie im Organismus. Deshalb kann
man die Regulationsfähigkeit neben der Reizbarkeit als ein besonders
wichtiges Merkmal der lebenden Wesen betrachten.
Wenn nun auch die Lebenserscheinungen bisher noch nirgends gründlich
aufgehellt sind, so kann uns das doch nicht an der Überzeugung hindern,
daß sie sich lediglich durch ihre außerordentlich große Komplikation
von den Vorgängen in der leblosen Natur unterscheiden und im
Prinzip einer physikalisch-chemischen Erklärung zugänglich sind.
5. Die Lebenstätigkeit der Pflanze vollzieht sich nur, wenn zahlreiche
Bedingungen, die sog. allgemeinen Lebensbedingungen, erfüllt
sind, die[S. 185] man in zwei Gruppen, innere und äußere
Ursachen, einzuteilen pflegt[127]. Die inneren Ursachen des Lebens sind
an das Protoplasma geknüpft. Sein Bau, seine Organisation bedingen
nicht nur, daß das Geschehen im Organismus die Charakterzüge des
Lebendigen an sich trägt, sondern daß es auch je nach der Abstammung
der betreffenden Plasmamasse spezifisch verschieden ausfällt.
Demnach ist das Vorhandensein einer lebenden Plasmamasse die
fundamentalste Lebensbedingung. Alle anderen Lebensbedingungen
können wir nach Wunsch schaffen oder wegnehmen; das Protoplasma dagegen
können wir nicht synthetisch herstellen, es entsteht nur im Organismus
durch die Tätigkeit schon existierenden Plasmas.
Das Protoplasma kann aber nur bei ständiger Wechselwirkung mit der
Außenwelt etwas leisten. Dabei fällt der Außenwelt eine dreifache Rolle
zu: sie liefert erstens das Material zum Aufbau des Pflanzenkörpers;
sie wirkt zweitens als auslösendes Moment (Reiz, S. 184); sie
versorgt endlich die Pflanze mit der nötigen Energie, mag diese als
chemische Energie mit den Stoffen von außen aufgenommen werden oder in
Form von Ätherschwingungen eindringen.
Unter den äußeren Faktoren, die für das Leben von Wichtigkeit
sind, kann man die notwendigen oder unentbehrlichen von den
entbehrlichen unterscheiden. Als unentbehrliche Bedingungen
für die Lebenstätigkeit nennen wir vor allem eine gewisse Temperatur
und das Vorhandensein bestimmter Stoffe sowie das Fehlen anderer,
die schädigend oder tödlich wirken (Gifte); dagegen ist
das Licht keine solche allgemeine Lebensbedingung. Manche
Pflanzen verlangen freilich wenigstens für ihre oberirdischen Organe
direktes Sonnenlicht; andere meiden dies und suchen den Schatten auf
(Schattenpflanzen); wieder andere können ihren ganzen Lebenszyklus in
vollkommener Dunkelheit durchmachen.
Die notwendigen Faktoren sind ferner an ganz bestimmte Grenzen
gebunden. Ein Zuviel von ihnen (Überschreiten des „Maximums“) wirkt
ebenso schädlich auf die Pflanze wie ein Zuwenig (Überschreiten des
„Minimums“), und oberhalb des Minimums sieht man die Lebensäußerungen
an Intensität zunehmen bis zum „Optimum“ und oberhalb von diesem wieder
abnehmen. Die drei Punkte, Minimum, Maximum und Optimum, die uns bei
der Abhängigkeit jeder Lebenserscheinung von einem Außenfaktor
entgegentreten, nennt man die Kardinalpunkte der Einwirkung
dieses Faktors (Fig. 259). Sie sind indes keineswegs Konstanten;
sie sind vielmehr für die einzelnen Organismen und Lebenserscheinungen
verschieden: sie ändern sich ferner mit der Dauer der Einwirkung des
Faktors, und sie hängen auch vom Zustand der Pflanze sowie von anderen
äußeren Faktoren ab.
Jede Überschreitung des Minimums wie des Maximums eines äußeren
Faktors führt früher oder später zum Tod. Dieser kann also
z. B. durch zu hohe (Hitzetod) oder zu niedrige Temperatur
(Erfrieren) erfolgen; er kann durch ein Zuviel oder Zuwenig an
Licht, durch ein Zuviel oder Zuwenig eines Stoffes bedingt sein; ist z. B.
zu wenig Wasser gegeben, so spricht man zunächst von Welken
und dann von Vertrocknen; ist ein Stoff in zu großen,
schädigenden Mengen gegeben, von Vergiftung.
Die meisten Pflanzen werden durch bestimmte niedere
Temperaturen zum Erfrieren[128] gebracht, fast alle
werden durch hohe Temperaturen, die noch weit vom Siedepunkte des
Wassers entfernt sind, getötet; nur manche Cyanophyceen ertragen
die sehr hohe Temperatur gewisser heißer Quellen.
Empfindliche Pflanzen, insbesondere Pflanzen tropischer Herkunft,
sterben schon bei Temperaturen über Null. Andere werden durch die
Eisbildung im Innern getötet, und wieder andere können mehrfach
im Winter hart gefrieren, ohne Schaden zu nehmen. So erträgt in
Nordsibirien Cochlearia fenestrata eine Temperatur von -46° C
ohne Schädi[S. 186]gung, und gewisse Waldbäume halten sogar -60° aus.
Bemerkenswert ist die Widerstandskraft niederer Organismen
gegen extreme Kältegrade. So ertrugen in Versuchen PICTETs
Diatomeen längere Zeit eine Temperatur von -200° C.
Durch Steigerung der Lichtintensität kann man schließlich
wohl jede Zelle zum Absterben bringen, mag nun das Licht dabei im
einzelnen mehr thermisch oder mehr chemisch wirken. Viele Bakterien
sterben schon im hellen Tageslicht ab, weshalb diesem eine wichtige
hygienische Bedeutung in unseren Wohnräumen zukommt.
Das Lichtbedürfnis wechselt aber nicht nur bei verschiedenen
Pflanzenarten und je nach Gewöhnung auch wieder individuell,
sondern es kann sich das Optimum der Lichtwirkung bei demselben
Einzelwesen mit der Entwicklung verschieben. Viele tropische
Kulturpflanzen, z. B. Kakao und Kaffee, verlangen während ihrer
Jugend Schatten und müssen daher durch besonders angepflanzte
schattenspendende Pflanzen (Albizzia-Arten, Musa) zunächst
geschützt werden, während sie später die volle Tropensonne ertragen
oder gar nötig haben.
Unter den stofflichen Einflüssen kommt oft dem Wasser
eine besonders in die Augen fallende Bedeutung zu. Wenn Licht
und Temperatur in optimalem Maße geboten sind, wie das in den
Tropen zutrifft, hängt die Entwicklung der Pflanze vor allem
vom Wasser ab. Ist dieses reichlich geboten, und erfolgen die
Niederschläge auch gleichmäßig über das ganze Jahr verteilt,
dann sehen wir den üppigsten Pflanzenwuchs in der Formation
des tropischen Regenwaldes auftreten. Eine regelmäßig
wiederkehrende Trockenperiode bedingt den sommergrünen Wald,
geringere Niederschläge lassen die Formation der Savannen
entstehen und noch weniger ausgiebige Niederschläge endlich die
Wüsten[129].
Weitgehende, mit Austrocknung verbundene Wasserentziehung
ertragen nur wenige Pflanzen. — In vielen Fällen beruht der bei
niederer Temperatur eintretende Tod nicht auf einer direkten
Kältewirkung, sondern auf ungenügender Wasserversorgung; die
Wurzeln können dem kalten oder gar gefrorenen Boden nicht so viel
Wasser entziehen, wie die oberirdischen Organe verdunsten.
6. Die Überschreitung des Maximums oder des Minimums der äußeren
Faktoren führt aber keineswegs immer sofort zum Tod. Vielfach kann
der Organismus zunächst in einen Zustand übergehen, den man als
latentes Leben bezeichnet. Äußerlich kann man es dem Organismus
nicht immer ansehen, ob er sich im Zustand des aktiven
Lebens, des latenten Lebens oder des Todes befindet.
Das latente Leben hat mit dem toten Zustand das gemeinsam, daß jede
Lebenstätigkeit unterbleibt; aus dem latenten Leben kann aber
durch geeignete Einwirkungen das aktive Leben wieder erweckt werden,
was beim toten Zustand unmöglich ist.
Nicht immer freilich ist der Eintritt des latenten Lebens einfach die
Folge der Überschreitung der extremen Kardinalpunkte. Vielfach arbeitet
die Pflanze auch aus inneren Ursachen auf die Herstellung latenten
Lebens hin (Samen und Sporen).
Im latenten Leben befinden sich vor allem viele Dauerzustände der
Pflanzen, wie die Samen und Sporen. Sie sind in der Regel viel
widerstandsfähiger gegen Austrocknung, Hitze und Kälte als Organe,
die im aktiven Zustand sind. So ist bekannt, daß Bakteriensporen
im feuchten Zustand eine Zeitlang Temperaturen von 100° und
mehr ertragen können; genau so verhalten sich einige Samen
(Medicago-Arten). Andererseits widerstehen Sporen und Samen in
trockenem Zustand sogar einer Temperatur von -253°[130].
I. Die stoffliche
Zusammensetzung der Pflanze[132].
Jede Beschäftigung mit dem Stoffwechsel setzt die Kenntnis der
chemischen Zusammensetzung der Pflanze voraus. Diese studiert man mit
den Methoden der Chemie.
Wasser und Trockensubstanz. Ein gewisser Einblick in die
Zusammensetzung der Pflanze wird freilich schon ohne besondere
Hilfsmittel gewonnen. Daß die Pflanze aus Wasser und Trockensubstanz
besteht, weiß jeder, der Pflanzen getrocknet, für das Herbarium
hergerichtet hat. Er weiß auch, wie durch diesen Wasserverlust
fundamentale physikalische Eigenschaften der Pflanze, wie Festigkeit
und Elastizität, beeinflußt werden. Mehrjährige Zweige von
Schlingpflanzen z. B. (Cobaea, Passiflora), die in frischem Zustande
außerordentlich biegsam und elastisch sind, werden nach dem Trocknen an
der Luft starr und spröde; sie brechen wie Glas, wenn man sie zu biegen
versucht. Nimmt man die Wage zu Hilfe, so stellt man leicht fest, wie
groß der Anteil des Wassers an der Gesamtmasse der Pflanzensubstanz
ist. Dabei genügt es freilich nicht, die Pflanzen der Luft zu
exponieren: vielmehr muß ihnen im Exsikkator oder in einer Temperatur
von etwas über 100° auch der nicht unbeträchtliche Rest von Wasser
entzogen werden, den sie im sog. „lufttrockenen“ Zustand noch führen.
Dann zeigt sich, daß der Wassergehalt sehr groß ist; er beträgt bei
holzigen Teilen etwa 50%, bei saftigen Kräutern 70–80%, bei Sukkulenten
und Früchten 85–95% und endlich bei Wasserpflanzen, namentlich Algen,
95–98% des Frischgewichts. Daß Körper mit so hohem Wassergehalt noch
„fest“ sein können, erscheint wunderbar, findet aber seine Erklärung
durch nähere Studien über die Verteilung des Wassers (S. 188).
Asche. So wie uns die Austrocknung zur Unterscheidung von
Wasser und Trockensubstanz führt, so können wir mit Hilfe der
Verbrennung zwischen verbrennbarer oder organischer Substanz
und unverbrennbarer oder Aschensubstanz unterscheiden. Der
Aschengehalt der Pflanzen fällt beim Verbrennen von Holz, beim Rauchen
einer Zigarre ohne weiteres auf; daß auch winzige Zellhautfragmente
oder Stärkekörner Asche hinterlassen, weist das Mikroskop nach. Über
die quantitativen Verhältnisse gibt die Analyse Aufschluß, die vor
allen Dingen zeigt, daß die verschiedenen Organe einer Pflanze nicht
den gleichen Aschengehalt aufweisen; die Blätter z. B. pflegen mehr zu
enthalten als die Stengel. So hat man gefunden, daß in den Blättern von
Brassica rapa rund 20%, in den Stengeln 10% der Trockensubstanz aus
Asche besteht (vgl. S. 204).
Aber auch je nach dem Standort und anderen äußeren Einflüssen kann die
Aschenmenge variieren. Andererseits können wieder verschiedene Spezies,
selbst wenn sie den gleichen äußeren Bedingungen ausgesetzt sind, doch
ganz verschiedene Mengen von Asche führen.
Wenn auch die Mehrzahl der überhaupt auf der Erde vorkommenden
Grundstoffe in der Asche von Pflanzen gefunden worden ist, so
treten doch nur wenige Elemente in so großer Menge in ihr auf, daß man
sie quantitativ zu bestimmen pflegt. Es sind dies von Metalloiden Cl,
S, P, Si und von Metallen Na, K, Mg, Ca, Fe.
Organische Substanz. Die organische Substanz besteht in erster
Linie aus den Elementen H, O, N und C; sie werden bei völliger
Verbrennung in flüchtige Stoffe übergeführt, nämlich in Kohlensäure,
Wasser, Ammoniak oder freien Stickstoff. Auch in der Asche sich
findende Elemente können[S. 188] in der lebenden Pflanze organisch gebunden
sein. — Der Masse nach nimmt stets der Kohlenstoff die erste
Stelle in der organischen Substanz ein, denn die Trockensubstanz
besteht etwa zur Hälfte aus ihm. Auch ohne chemische Untersuchung
erkennt man den reichen Gehalt der Pflanze an diesem Element durch
das „Verkohlen“ jedes beliebigen Teiles, wenn er erhitzt wird.
Die Betrachtung eines Stückes Holzkohle, in dem sich die feinste
Holzstruktur erhalten hat, läßt weiterhin darauf schließen, wie
gleichmäßig die Verteilung des Kohlenstoffes in der Substanz und wie
vorherrschend seine Masse in dieser ist.
Herkunft der Stoffe. In größerer Menge finden sich somit nur
folgende 13 Elemente in der Pflanze:
H, Cl, O, S, N, P, C, Si und Na, K, Mg, Ca, Fe.
Während des Wachstums nimmt ihre Menge dauernd zu; sie werden demnach
fortwährend von außen aus dem Boden, dem Wasser, der Luft oder aus
anderen Organismen aufgenommen.
In der Regel werden nur Gase und Flüssigkeiten in die Pflanze
eindringen. Die starre Zellwand gestattet festen Körpern den Zutritt
ins Innere erst, nachdem sie gelöst sind. Wo die Zellwand aber fehlt,
wie bei Flagellaten und Schleimpilzen, da kann das nackte Protoplasma
auch feste Körper umfließen und so ins Innere aufnehmen.
Die Tiere stimmen in ihrer chemischen Zusammensetzung im wesentlichen
mit der Pflanze überein. Die Stoffaufnahme erfolgt aber bei ihnen
zunächst in das Darmsystem. Der Unterschied gegenüber den Pflanzen
ist indes nicht so groß, als er auf den ersten Blick zu sein scheint;
denn im allgemeinen treten die Stoffe auch erst, nachdem sie flüssig
geworden sind, in die Zellen ein.
II.
Die Nährstoffe, ihre Aufnahme und ihre Bewegung in der Pflanze.
Die aufgenommenen Stoffe können notwendige, überflüssige oder
schädliche sein. Das läßt sich in jedem einzelnen Fall nur durch
das Experiment feststellen, denn es wäre ein großer Irrtum, zu
glauben, daß alle konstant vorkommenden Stoffe auch notwendig sind.
Es hat sich vielmehr gezeigt, daß im allgemeinen nur 10 von
den eben genannten 13 Elementen wirklich unentbehrlich sind.
Die notwendigen Stoffe kann man als Nährstoffe bezeichnen.
Sie treten fast alle nicht als Elemente, sondern als
Verbindungen in die Pflanze ein. Nur eines von ihnen, der
Sauerstoff, findet sowohl in Verbindungen als auch in freiem Zustand
Verwendung in der Pflanze. Wir unterscheiden a) Wasser, b) in
Wasser gelöste Salze, c) Gase als die drei Hauptgruppen
von Nährstoffen.
Ohne dauernde Zufuhr von Nährstoffen kann die Pflanze
nicht existieren. Das ist für die wachsende Pflanze
selbstverständlich; denn hier tritt ja eine Körpervergrößerung
auf Kosten der von außen aufgenommenen Stoffe ein. Aber auch die
ausgewachsenen Teile bedürfen ständig neuer Stoffzufuhr, da ihr
„Stoffwechsel“ mit ständigen Stoffverlusten verbunden ist.
a) Das Wasser.
Die ganze Fülle von chemischen Prozessen, die den Stoffwechsel der
Pflanze ausmachen, vollzieht sich in wässeriger Lösung. Daher
ist das Wasser ein ganz unentbehrlicher Bestandteil der Pflanze;
alle lebenstätigen Pflanzenteile sind stark wasserdurchtränkt: die
Zellwände sind mit Wasser imbibiert, im Zellumen finden sich
die großen mit wässerigen Lösungen[S. 189] erfüllten Vakuolen vor,
und der Träger des Lebens, das Protoplasma, ist stets etwa zu 75%
oder mehr aus Wasser aufgebaut. Nur in diesem Zustand der annähernden
Wassersättigung vermag die Pflanze voll zu vegetieren; jeder stärkere
Wasserentzug vernichtet entweder das Leben völlig oder setzt wenigstens
alle Lebensäußerungen so sehr herab, daß man sie nicht mehr nachweisen
kann.
Bei den Vegetationsorganen, insbesondere der höheren Pflanzen,
führt meist schon ein weitgehendes Welken zum Tod; nur selten
können Pflanzen, wie gewisse Sukkulenten, etwa 9⁄10 ihres
Wassergehaltes einbüßen, ohne geschädigt zu werden. Um so
auffallender ist es, daß manche epiphytische und xerophile Farne,
gewisse Isoëtes-Arten Algiers und Selaginella lepidophylla, die
in den regenarmen Hochebenen Zentralamerikas zu Hause ist, durch
einen bis zur „Lufttrockenheit“ fortschreitenden Wasserverlust
nicht getötet werden, sondern in latentem Leben verharren,
um sofort bei Wiederbefeuchtung weiter zu wachsen. Bei niederen
Pflanzen, vor allem bei Flechten und Moosen, ist diese
Austrocknungsfähigkeit viel weiter verbreitet und gestattet
ihnen die Besiedelung von Felsen, Baumrinden und ähnlichen
Standorten, an denen sie nur unmittelbar während und nach einem
Regen wasserdurchtränkt sind, um bald darauf wieder, von der Sonne
ausgedörrt, zu pulverisierbaren Massen zu erstarren.
Bei Samen und Sporen ist in der Regel mit der Ablösung von der
Mutterpflanze eine Austrocknung verbunden, die fast überall gut
und lange ertragen wird. Auch hier hört während des wasserarmen
Zustandes jede Lebensäußerung auf.
Manche Samen verlieren ihre Keimkraft im trockenen Zustand erst
nach einem oder einigen Jahren, manche schon nach Tagen, und wieder
andere sollen ein Austrocknen überhaupt nicht vertragen. — Es muß
aber hervorgehoben werden, daß Pflanzenteile auch im Zustande
völliger Lufttrockenheit immer noch etwa 9–14% Wasser
enthalten. Selbst über der Schwefelsäure des Exsikkators
bewahren Samen noch wochenlang 6 und mehr Prozent Wasser. Aber auch
den weitgehenden Wasserverlust durch Trocknung bei 110° oder durch
Einwirkung von absolutem Alkohol vertragen manche Samen und Sporen
ganz gut.
Aufnahme des Wassers.
Wasseraufnahme der Zelle. Alle Teile der Zelle sind, wie
bemerkt, wasserdurchtränkt. Die Zellhaut führt das Wasser
zwischen ihren kleinsten Teilchen in so feiner Verteilung, daß
man auch mit Hilfe der stärksten Vergrößerungen Wasser und feste
Substanz nicht unterscheiden kann. Läßt man das Wasser verdunsten,
so treten nicht etwa an seiner Stelle lufterfüllte Hohlräume auf,
sondern es findet eine Zusammenziehung der Zellhaut statt.
Umgekehrt ist die Wasseraufnahme in trockene oder in nicht voll
wassergesättigte Zellhäute mit einem Aufquellen verbunden. Unter
„Quellung“[133] versteht man die Volumvergrößerung, die
ein Körper durch Einlagerung von Flüssigkeit erfährt. Bei gegebener
Temperatur ist die aufgenommene Menge Flüssigkeit eine begrenzte. —
Es gibt Substanzen, die in Alkohol oder Xylol usw. quellbar sind; die
pflanzliche Zellhaut aber quillt in Wasser. Dabei nehmen die Wände der
Holzzellen etwa 1⁄3 ihres Gewichts, die Membranen mancher Algen und
einzelner Samen- und Fruchtschalen das Mehrfache ihres Gewichts an
Wasser auf. Die Wasseraufnahme erfolgt mit beträchtlicher Energie und
dementsprechend auch gegen bedeutende Widerstände.
Das lufttrockene Protoplasma mancher Samen und Sporen nimmt gerade wie
die Zellwand Wasser durch Quellung auf. Ähnlich wie Gummi arabicum
verliert es aber dabei den Charakter des festen Körpers und geht in
eine kolloidale Lösung über. In der gewöhnlichen, lebenstätigen
Zelle hat dann das Protoplasma in der Regel die Eigenschaften einer
solchen, doch können auch einzelne Partien festere Konsistenz annehmen.
Kolloidale Lösungen haben ja stets die Neigung, aus dem flüssigen
(Sol-) Zustand in den Gelzustand überzugehen.
[S. 190]
Der Zellsaft endlich ist immer eine molekulare Lösung von Kristalloiden
in Wasser, denen auch Kolloide beigemengt sein können.
Nur eine Zelle, die nicht vollkommen wassergesättigt ist, kann ihrer
Umgebung Wasser entziehen. Es fragt sich also, wann ist die Zelle
wassergesättigt? Für die Zellwand ist diese Frage rasch beantwortet:
Sättigung ist eingetreten, wenn das Quellungsmaximum erreicht ist.
Viel schwieriger ist es, für das Protoplasma und den Zellsaft die
Grenzen der Wasserkapazität zu bestimmen. — Halten wir uns zunächst
an den Zellsaft! Wir nehmen der Einfachheit halber zunächst an, er
bestehe lediglich aus Kristalloiden, und er sei direkt von
einer Zellhaut umschlossen, ohne durch Protoplasma von
ihr getrennt zu sein. Wir füllen also einen Schlauch aus Zellulose
mit einer Lösung, z. B. von Kochsalz, und tauchen ihn in Wasser; dann
wird ein Diffusionsprozeß beginnen. Wasser bewegt sich ins
Innere des Schlauches, Salz dagegen tritt aus dem Innern aus. Und wenn
auch die Zellhaut dem Durchtritt der Salze einen größeren Widerstand
entgegensetzt als dem des Wassers, so wird der Diffusionsprozeß doch
so lange fortgesetzt werden, bis innerhalb und außerhalb überall
die gleiche Konzentration herrscht. Eine Wand, die für Wasser
und Salz durchlässig ist, wirkt also auf die Diffusionsbewegung,
die in jeder freien Flüssigkeitsmasse eintritt, nur insofern ein,
als sie die Geschwindigkeit dieses Vorganges verringert. Im
Endzustand aber herrscht gleiche Konzentration des Salzes auf
beiden Seiten der Membran.
Fig. 234. Osmometer. Schematisch. T Tonzelle, N
Niederschlagsmembran, R Manometer, Q Quecksilber, Z Zuckerlösung.
1 Osmometer nach Füllung mit der Lösung in Luft. 2 Dasselbe
nach Eintauchen in Wasser. Ein osmotischer Druck ist aufgetreten,
das Quecksilber dementsprechend im Manometer gestiegen. Nach
URSPRUNG.
Wesentlich anders verläuft der Diffusionsprozeß, wenn die Wand aus
einer Substanz besteht, die für Wasser wohl, für Salz dagegen gar nicht
permeabel ist. Solche Membranen nennt man semipermeabel, und man
erhält sie z. B., wenn man eine Kupfervitriollösung mit Ferrocyankalium
oder Leim mit Gerbsäure in Berührung bringt. Da die semipermeablen
Membranen nur eine geringe Festigkeit besitzen, kann man aus ihnen
nicht wie aus Zellulose Schläuche herstellen. Man verleiht ihnen
die nötige Festigkeit dadurch, daß man sie auf einer porösen Wand von
gebranntem Ton auflagert; man benutzt also die bekannten, bei gewissen
galvanischen Elementen zur Verwendung kommenden Tonzellen und
lagert ihnen auf der Innenseite eine Schicht von Ferrocyankupfer auf.
Füllt man dann die so vorbereitete Tonzelle mit einer Salzlösung,
verschließt sie und taucht sie in Wasser ein, so kann wohl das Wasser
nach innen, aber nicht das Salz nach außen wandern; die Diffusion
findet also einseitig statt und als Folge davon tritt ein
Druck im Zellinnern auf, den man als osmotischen Druck zu
bezeichnen pflegt. — Ähnlich wie Kochsalz können auch andere Salze,
doch auch andere Verbindungen, sofern sie nur wasserlöslich sind und
Ferrocyankupfer schlecht durchwandern können, einen osmotischen Druck
erzeugen.
[S. 191]
Zum Nachweis und zur Messung des osmotischen Druckes[134] bedient man
sich seit PFEFFER (1877) der eben geschilderten Tonzelle,
die eine Niederschlagsmembran von Ferrocyankupfer auf der Innenseite
erhalten hat. Sie wird mit Zuckerlösung gefüllt, verschlossen und mit
einem Quecksilbermanometer versehen. Das Quecksilber stellt sich dann
in beiden Schenkeln in gleiche Höhe ein (Fig. 234 1). Wird nun die
Tonzelle in Wasser eingetaucht, so dringt dieses ins Zellinnere
ein, und es entsteht ein Innendruck („osmotischer Druck“), der sich
am Steigen des Quecksilbers im rechten Schenkel bemerkbar macht (Fig.
234 2). Das Steigen erfolgt anfangs rasch, später langsam, und
schließlich — nach vielen Stunden — ist der Maximaldruck
erreicht. Er tritt dann auf, wenn durch den Druck der Quecksilbersäule
dem weiteren Eindringen von Wasser ein Ziel gesetzt wird. Die Höhe des
Quecksilberstandes gibt also direkt das Maß für den osmotischen Druck,
den man in Atmosphären anzugeben pflegt.
Messungen mit solchen „Osmometern“ haben ergeben, daß der
osmotische Druck innerhalb gewisser Grenzen der Konzentration
der angewandten Lösung proportional ist; für eine 1%ige
Rohrzuckerlösung fand PFEFFER 2⁄3 Atm. Vergleicht man
mit Rohrzucker andere nicht dissoziierte organische Substanzen,
so zeigt sich, daß allgemein die osmotische Wirkung der Zahl
der gelösten Moleküle proportional geht; äquimolekulare
Lösungen sind isosmotisch. Es ist deshalb zweckmäßig, statt
der prozentischen Lösungen molekular-normale Lösungen zu
verwenden, im allgemeinen volumnormale. Bei dissoziierenden
Verbindungen, z. B. den Salzen, wirken auch die einzelnen Ionen
neben den undissoziierten Molekülen. Eine Lösung von 0,1 Mol.
KNO3 hat deshalb eine viel größere osmotische Wirkung als 0,1
Mol. Rohrzucker.
Vergleichen wir diese Tonzelle mit einer Pflanzenzelle, so entspricht
die Zellhaut der Wand der Tonzelle, das Protoplasma dagegen dem
Ferrocyankupfer; denn das Protoplasma ist, wenigstens solange es
lebendig ist, annähernd semipermeabel, die Zellwand dagegen permeabel.
[Es wird freilich S. 207 gezeigt werden, daß die Impermeabilität
des Protoplasmas für viele gelöste Körper nicht besteht, und
es mag hier betont sein, daß es auch semipermeable Zellhäute
gibt[135].] Es wird also auch in die pflanzlichen Zellen Wasser
einströmen, ohne daß Salz austritt, und somit wird ein osmotischer
Druck entstehen. Das Protoplasma dehnt sich unter diesem Innendruck
ohne wesentlichen Widerstand aus; die Zellhaut dehnt sich zwar
ebenfalls aus, aber sie übt auch vermöge ihrer Elastizität einen
erheblichen Gegendruck aus. Dieser nimmt mit dem Anwachsen des
Innendruckes zu und führt schließlich dahin, daß weiteres Wasser nicht
mehr aufgenommen werden kann.
Auf den Wassergehalt des Protoplasmas wollen wir nicht näher eingehen;
es sei nur bemerkt, daß auch er begrenzt sein muß, weil
das Protoplasma sich einerseits unter dem Druck des Zellsaftes,
andererseits unter dem der Zellhaut befindet.
Die Dehnung der Zellwand unter dem Einfluß des Innendruckes ist oft
eine recht beträchtliche; sie hängt einerseits von der Größe des
Druckes, andererseits von den elastischen Eigenschaften der Zellhaut
selbst ab. Sehr häufig können Zellwände durch den Innendruck um
10–20%, manchmal sogar um 50% gedehnt werden. Sticht man die Zelle
an oder tötet man ihr Protoplasma, so hört der Druck auf, und die
Membran zieht sich elastisch auf ihre ursprüngliche Länge zusammen;
während der Dehnung war sie also gespannt, jetzt ist sie
entspannt (Fig. 235). Diese Spannung aber hat für die Pflanzenzelle
eine große Bedeutung; durch sie wird die Zellhaut fester, gerade wie
auch ein dünner Kautschukballon durch Einpressung von Luft gegen
Gestaltsänderung widerstandsfähiger wird. Eine Zelle, deren Haut durch
den Innendruck gespannt ist, heißt turgeszent. Dementsprechend
nennt man auch den Innendruck „Turgordruck“[136].
[S. 192]
Der Turgordruck kann bei embryonalen Zellen des Vegetationspunktes, die
noch keine Vakuolen führen, lediglich auf der Quellung des Protoplasmas
beruhen. Bei der typischen ausgewachsenen Zelle aber pflegt man von dem
dünnen Protoplasma ganz abzusehen und zu sagen, daß der osmotische
Druck der Vakuolenflüssigkeit direkt auf die Zellmembran einwirkt
und diese spannt.
Diese Festigung der Pflanze durch die elastische Dehnung der Zellhaut
ist sehr wichtig, sie ist die einfachste und vielfach alleinige Art
der Zellenfestigung. Sie ist aber natürlich von der Gegenwart einer
genügenden Wassermenge abhängig; nimmt man eine pralle Zelle aus dem
Wasser und läßt sie an der Luft Wasser abgeben, so verschwindet immer
mehr die Spannung ihrer Wand und damit die Festigkeit; die Zelle
wird schließlich welk, d. h. schlaff. Durch erneute
Wasserzufuhr kann der frühere turgeszente Zustand wieder hergestellt
werden. Solange eine Zelle das Maximum ihres Wassergehaltes
nicht besitzt, wirkt sie wie eine Saugpumpe, sie hat also ein
„Saugvermögen“. Um dieses Saugvermögen[137] oder diese
Saugkraft voll zu verstehen, kehren wir nochmals zu der
PFEFFERschen Zelle (Fig. 234) zurück. Diese entwickelt nach
der Füllung mit der Rohrzuckerlösung in dem Moment, wo wir sie in
Wasser tauchen, ein großes Saugvermögen; ihre Saugkraft nimmt aber
offensichtlich in dem Maße ab, als mit dem Eindringen von Wasser
sich der Innendruck steigert; schließlich wenn der volle Innendruck
eingetreten ist, der maximale „osmotische Druck“ sich ausgebildet hat,
ist die Saugkraft = Null geworden. Um die Größe der Saugkraft beim
Beginne des Versuches zu messen, wird man nach Eintauchen des
Osmometers (Fig. 234 1) in Wasser den Quecksilberdruck soweit erhöhen
müssen, daß überhaupt kein Wasser einströmen kann. Die Höhe dieser
Quecksilbersäule ist dann das Maß für die Saugkraft, und ein Versuch
zeigt, daß diese so groß ist wie der osmotische Druck, den die Lösung
entwickeln kann. Und von dieser Höhe sinkt dann, wenn Wasseraufnahme
ermöglicht wird, die Saugkraft bis auf den Wert Null. In den zwischen
den Extremen gelegenen Stadien findet man ihren Wert so, daß man
von der ursprünglichen Saugkraft des Inhaltes den jeweiligen
Manometerdruck abzieht. In ganz entsprechender Weise wird die
Saugkraft der pflanzlichen Zelle gefunden, indem man von der
Saugkraft des Zellsaftes den Druck der Zellhaut abzieht.
Fig. 235. Einzelliges Stammglied einer Nitella
(Characee) etwa 6mal vergrößert. F Frisch und durch Turgordruck
gespannt. P Dasselbe nach zerstörtem Turgordruck, schlaff, kürzer und
schmäler; ss Seitenglieder. Nach NOLL.
Da man an der Pflanzenzelle kein Manometer anbringen kann, ist auch
eine direkte Messung des osmotischen Druckes in ihr unmöglich. Doch
kann man auf Umwegen, nämlich durch Studium der plasmolytischen
Erscheinungen zu diesem Ziele gelangen. Bringt man eine turgeszente
Zelle in eine Salzlösung, die osmotisch wirksamer ist als der Zellsaft,
so tritt zunächst eine Verkürzung der Zelle ein, die bis zur völligen
Entspannung der Membran geht, sodann aber erfolgt eine Abhebung des
Protoplasmas von der Zellwand, weil ja die Zellwand sich nicht weiter
verkürzen kann, während das Protoplasma dauernd der sich weiter
verkleinernden Vakuole folgt. Diese Abhebung („Plasmolyse“)
beginnt an den Ecken und führt schließlich zu einer Ab[S. 193]rundung
des Plasmas im Innern der Zellhaut (Fig. 236). Es ist im Grunde
gleichgültig, welche Stoffe man zur Plasmolyse verwendet; sie dürfen
nur das Plasma nicht schädigen, und das Plasma muß für sie impermeabel
sein. Am besten eignet sich Rohrzucker, während der früher viel
benutzte Kalisalpeter doch sehr stark eindringt.
Hat man durch systematisches Ausprobieren diejenige Konzentration
des Plasmolytikums gefunden, die gerade die erste Abhebung des
Protoplasmas an den Ecken der Zelle bewirkt, so kann man sagen, daß
diese „plasmolytische Grenzkonzentration“ denselben osmotischen
Wert besitzt wie der Zellsaft dieser entspannten Zelle. Wenn z. B.
festgestellt ist, daß die Grenzkonzentration 0,2 Mol. Rohrzucker ist,
so ist der Zellsaft isosmotisch mit 0,2 Mol. Rohrzucker, man sagt
auch, der Zuckerwert des Zellsaftes beträgt 0,2 Mol. Da man
nun in physikalischen Versuchen (z. B. mit Osmometern, doch auch mit
anderen Methoden) den osmotischen Druck verschiedener Konzentrationen
von Rohrzucker bestimmt hat[138], so weiß man, wie groß der osmotische
Druck in einer solchen Zelle beim Eintauchen in Wasser werden
kann. Wenn die Zellwand stark verdickt ist und dementsprechend so
gut wie gar nicht dehnbar ist, dann kann auch der Zellsaft nicht durch
eindringendes Wasser verdünnt werden, und eine solche Zelle kann den
physikalisch ermittelten Wert als maximalen osmotischen Druck bekommen.
Bei dehnbarer Membran aber tritt mit dem Wachsen des Zellvolumens
bei der Wasseraufnahme stets eine Verdünnung des Zellsaftes
ein. Nehmen wir an, die plasmolysierte Zelle dehne ihr Volumen bei
Wasseraufnahme bis zur Sättigung auf das Doppelte aus, so wird
also die Konzentration ihres Zellsaftes und damit ihr osmotischer
Druck auf die Hälfte des Wertes sinken, den er bei fehlender
Volumzunahme erreichen könnte. Nur bei sorgfältiger Berücksichtigung
der Volumänderungen kann man also aus dem „Zuckerwert“ Schlüsse auf
den tatsächlichen osmotischen Druck in einer Zelle machen. In den
meisten Fällen ist der osmotische Wert nicht mit Zucker, sondern mit
KNO3 bestimmt worden. In gewöhnlichen Zellen ist der Salpeterwert
0,15–0,30 Mol.; er kann aber auch auf 3 und mehr Mol. ansteigen. Im
übrigen pflegt er selbst bei Nachbarzellen eines Gewebes um 0,1–0,2
Mol. zu differieren und je nach Außenbedingungen zum Teil periodisch zu
schwanken[139].
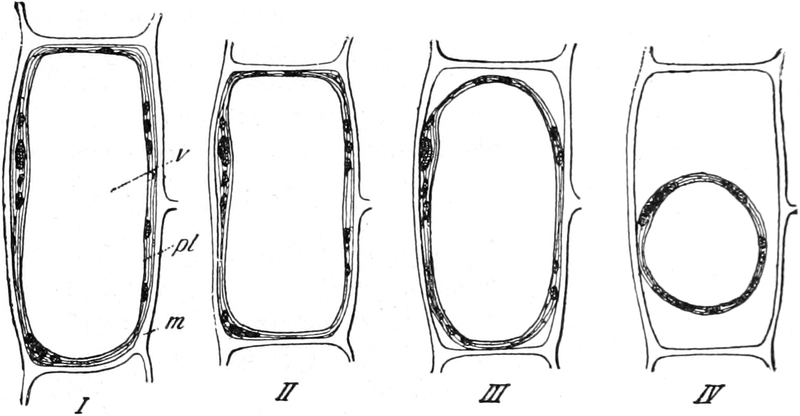
Fig. 236. Schema der Plasmolyse an einer jungen Zelle
aus dem Rindenparenchym des Blütenstiels von Cephalaria leucantha
(Compositae). m Zellhaut, pl Protoplasma, v Vakuole. I in
Wasser. II in 4%iger Salpeterlösung. III in 6%iger Salpeterlösung.
IV in 10%iger Salpeterlösung. Nach DE VRIES.
Die Ausdrücke osmotischer Druck, Turgordruck, Saugkraft werden
in der Literatur vielfach in verschiedenem Sinn gebraucht. Sie
sind hier durchweg im Anschluß an URSPRUNG[136], dessen
Ausführungen uns klar und folgerichtig scheinen, gefaßt. Besonders
betont sei noch, daß manche Chemiker von einem osmotischen Druck
auch bei Lösungen sprechen, die nicht durch eine semipermeable
Membran von Wasser getrennt sind, die also einen meßbaren
Druck nicht erzeugen. Diese Ausdrucksweise will kaum
etwas anderes bedeuten, als daß eben die Lösung den betreffenden
osmotischen Druck erzeugen kann, wenn sie durch eine
semipermeable Wand von Wasser getrennt ist.
[S. 194]
Die Abhebung der Protoplasten von der Zellwand erfolgt nicht
so glatt, wie die schematische Figur 236 das darstellt. In
Wirklichkeit pflegt das Plasma durch feine Fäden mit der Haut
verbunden zu bleiben; diese reißen später durch. Es ist wohl
möglich, daß, wie HANSTEEN ausführt, das Protoplasma
ursprünglich gar nicht scharf von der Zellhaut getrennt ist und daß
die bei der Plasmolyse entstehende Oberfläche eine künstliche ist,
die keineswegs dieselben Eigenschaften zu haben braucht wie die
ursprüngliche Hautschicht.
Die Plasmolyse kann durch Überführung der Zelle in reines Wasser
wieder rückgängig gemacht werden. Es tritt dann wieder die alte
Turgeszenz ein. Tötet man aber das Protoplasma, so verliert es
seine Semipermeabilität, und die Herstellung der Turgeszenz ist
unmöglich. Besonders anschaulich tritt eine derartige Veränderung
des Protoplasmas durch Abtöten bei Zellen mit gefärbtem Zellsaft
(z. B. rote Rüben) vor Augen, weil hier dann sofort der Farbstoff
in die Umgebung übertritt, während solche Zellen im lebenden
Zustand tagelang im Wasser liegen können, ohne es zu färben.
Der Salpeterwert der Zelle ist zunächst einmal spezifisch
verschieden. Besonders hohe Werte finden sich z. B. bei den
Grasknoten (0,5–1,0 Mol.) und bei Wüstenpflanzen (3,0 Mol.). Die
höchsten Werte aber treten in Pflanzen auf, die wie die Meeres-
und Strandpflanzen in Salzlösungen oder wie viele Pilze in
konzentrierten Zuckerlösungen gedeihen. In beiden Fällen muß, um
eine Turgeszenz zu ermöglichen, der osmotische Wert des Zellsaftes
den der Außenlösung übersteigen, und er paßt sich stets
dem letzteren an; er ist nicht ein für allemal gegeben, sondern
er ist regulationsfähig[140]. Man begreift, daß Zellen mit
so hochkonzentriertem Zellsaft sofort platzen, wenn sie aus ihrer
bisherigen Umgebung in reines Wasser oder in schwächere Lösungen
überführt werden.
Außerdem ist aber der Salpeterwert auch bei den einzelnen Organen
einer Pflanze verschieden. Das zeigt z. B. die folgende Tabelle
(nach BLUM):
|
|
Fagus
Mol.
|
Urtica
Mol.
|
|
Epidermis Blattunterseite
|
0,365
|
0,498
|
|
Schwammparenchym
|
0,571
|
0,635
|
|
Palisadenparenchym
|
1,017
|
1,015
|
|
Äußere Rinde (Stengel)
|
0,671
|
0,472
|
|
Kambium
|
0,634
|
0,548
|
|
Holzmarkstrahl
|
0,938
|
—
|
Wasseraufnahme vielzelliger Pflanzen. Bei vielen niederen
Pflanzen beteiligen sich alle lebenden Zellen an der
Wasseraufnahme; bei anderen komplizierteren dagegen grenzen nur
die „Außenzellen“ an die wasserhaltige Umgebung an, und sie
allein können demnach der Wasseraufnahme dienen. Beim Kormus ist die
Wasseraufnahme — wenigstens bei den typischen „Landpflanzen“ — auf
die Epidermiszellen der Wurzeln beschränkt. Die oberirdischen
Teile sind schon durch die mehr oder minder stark entwickelte
Kutikula ungeeignet für die Wasseraufnahme, so daß sie in der
Natur nie so viel Wasser aufnehmen, als die Pflanze nötig hat. Die
Wurzel dagegen ist durch ihre äußere Gestalt wie durch die feinere
Struktur ihrer Oberhaut hervorragend zur Wasseraufnahme geeignet.
Der Umstand, daß das Wasser im gewöhnlichen Boden in unsichtbar
feiner Verteilung auftritt und von den Oberflächenkräften der
Bodenpartikelchen festgehalten wird, macht eine große Oberfläche der
aufnehmenden Wurzel notwendig. Eine solche wird einerseits durch eine
reiche Verzweigung des Wurzelsystems, andererseits durch die Ausbildung
von Wurzelhaaren hergestellt, die mit den kleinsten Bodenteilchen
verwachsen (Fig. 237).
Fig. 237. Die Spitze eines Wurzelhaars mit Bodenteilchen
verwachsen. Vergr. ca. 240. Nach NOLL.
Durch die zahllosen feinen Seitenwürzelchen und die aus ihnen
entspringenden Wurzelhaare ist die Pflanze mit dem Boden verbunden
und kann[S. 195] das kapillar in diesem festgehaltene Wasser aufnehmen,
sobald sich in den Haaren durch Wasserverlust ein Saugvermögen
eingestellt hat. Auch einem Boden, der sich schon trocken anfühlt,
vermag die Pflanze noch Wasser zu entziehen. Bei dauernder
Wasseraufnahme aus so trockenen Böden erfolgt schließlich Welken der
Pflanze; aber auch in welkem Zustand geht die Wasseraufnahme noch
fort, freilich nie so weit, daß auch die letzten Spuren dem Boden
entzogen werden könnten. Weiter als unsere typischen Landpflanzen gehen
nach FITTING die Wüstenpflanzen, weil ihre Zellsäfte
ungewöhnlich hohe Konzentration besitzen und deshalb ein sehr starkes
osmotisches Saugvermögen entwickeln können[141].
Es leuchtet ein, daß Zellen mit hohem Salpeterwert eine höhere
Saugkraft entwickeln können als solche mit niedrigem
Salpeterwert. Allein die Bestimmung des Salpeterwertes gibt
kein Maß für die in der Zelle tatsächlich bestehende
Saugkraft ab; diese hängt ja von der Wassersättigung ab.
Andere Arten der Wasseraufnahme. Manche Pflanzen nehmen
das Wasser nicht aus dem Boden auf; sie gehören vor
allem zu zwei ganz verschiedenen ökologischen Gruppen: zu den
Epiphyten und zu den Wasserpflanzen. Über die
morphologischen und anatomischen Eigentümlichkeiten, die eine
Aufnahme von Regen oder Tau durch oberirdische Organe ermöglichen,
ist schon S. 157 berichtet.
Bewegung des Wassers in der Pflanze.
Eine Bewegung des Wassers von der Wurzel nach den oberirdischen
Teilen muß schon aus dem Grund erfolgen, weil Wasser zum Aufbau
neuer Zellen in den wachsenden Teilen unentbehrlich ist. Tatsächlich
braucht aber die Pflanze ungemein viel mehr Wasser, als das zu ihrem
Aufbau nötige, weil sie große Wassermassen in Dampfform, geringere
in tropfbar flüssiger Form an den oberirdischen Teilen abgibt. Der
erste Vorgang ist unter dem Namen Transpiration, der zweite als
Guttation bekannt.
Transpiration[142].
Wie jede freie Wasserfläche, wie jeder mit Wasser gequollene Körper,
z. B. Gelatine, Leim, muß auch die Pflanzenzelle Wasser an die
Luft abgeben, solange diese nicht völlig dunstgesättigt ist. Die
Wasserabgabe ist bei manchen Pflanzenteilen (z. B. bei Wurzeln,
bei submersen Teilen, bei Schattenpflanzen), wenn sie trockener
Luft ausgesetzt werden, so groß, daß sie kollabieren, schlaff, welk
werden und schließlich vertrocknen. Anders verhalten sich z. B. die
Blätter unserer gewöhnlichen Landpflanzen. An ihnen ist zunächst
nichts von Wasserabgabe zu bemerken. Doch auch sie welken, wenn
es lange nicht regnet, wenn also die Wasseraufnahme aus dem Boden
erschwert ist. Unterbindet man ihnen die Wasserzufuhr völlig, indem
man sie abschneidet, so tritt das Welken schneller ein. Stellt man
aber die abgeschnittenen Zweige in Wasser, so nehmen sie dieses mit
der Schnittfläche auf und welken nicht. Daß sie für gewöhnlich an
ihrem Standort nicht welken, beruht also offenbar darauf, daß
Wasser in demselben Maß von unten her nachgeschoben wird, wie es oben
verdunstet. Es läßt sich denn auch die Abgabe von Wasserdampf mit
einfachen Mitteln nachweisen.
In sehr anschaulicher Weise wird die Transpiration durch Verfärbung
von sog. Kobaltpapier demonstriert; Filtrierpapier, das
mit einer Lösung von Kobaltchlorür getränkt ist, zeigt im Zustand
völliger Trockenheit eine blaue Farbe; bei Wasserzutritt wird es
rot. Legt man nun ein Stückchen blaues Kobaltpapier z. B. auf ein
Blatt und bedeckt es zur Abhaltung der atmosphärischen Feuchtigkeit
mit einer Glasplatte, so weist die beginnende Rotfärbung auf
Transpiration hin, zugleich lassen sich aus der geringeren oder
größeren Geschwindigkeit des Eintretens und Fortschreitens der
Farbenänderung Schlüsse auf die Quantität des abgegebenen Wassers
ziehen. Exakte Nachweise in dieser Hinsicht[S. 196] kann freilich nur die
Wägung bringen. Sie muß an einer Pflanze ausgeführt werden, deren
Blumentopf durch passende Umhüllung völlig an jeder Wasserabgabe
verhindert ist. Es zeigt sich, daß die Wasserdampfabgabe der
Pflanze in der Regel so groß ist, daß schon im Verlaufe einer
Viertelstunde ein Gewichtsverlust eintritt, der beträchtlich genug
ist, um ihn ohne jede Schwierigkeit auf einer gewöhnlichen Wage
feststellen zu können. Über die Größe der Transpiration, die man
zweckmäßigerweise auf die Einheit der transpirierenden Fläche
beziehen wird, läßt sich nichts Allgemeines aussagen, da sie von
vielen äußeren Umständen, z. B. von Temperatur, Licht, Wasserzufuhr
usw., außerdem auch von der Struktur der Pflanze abhängt.
Der Vorgang der Transpiration vollzieht sich in folgender Weise. Eine
der Luft exponierte Zelle muß von dem Quellungswasser ihrer
Zellhaut dauernd Wasser an die Luft abgeben; dieser Prozeß würde bis
zur Lufttrockenheit der Zellhaut weitergehen, wenn nicht von innen
her Nachschub erfolgte. Ein solcher tritt nun aber in der Tat aus
dem Protoplasma ein. Die nicht völlig gesättigte Zellhaut übt einen
Zug auf das Wasser des Protoplasmas aus, und das Protoplasma sucht
Deckung für den Verlust aus dem Saftraum. So pflanzt sich also die
Wasserbewegung bis ins Zellinnere fort und bewirkt eine Verringerung
des Turgordruckes. Damit ist dann eine Saugkraft in dieser Zelle
hergestellt und die Bedingung zur Wasseraufnahme aus einer Nachbarzelle
gegeben, die selbst nicht transpiriert; und so setzt sich die
Wasserabgabe von der verdunstenden Oberflächenzelle in die Tiefe der
Gewebe fort. Die Größe der Transpiration hängt vor allem davon ab,
wie groß die Wasserdurchlässigkeit der Zellhaut ist. Handelt
es sich um eine gewöhnliche Zellulosewand, so ist die Transpiration
demnach groß; ist aber die Zellwand mit Wachs und Kutikula bedeckt
oder mit Kutikularsubstanz imprägniert, so gibt sie wenig Wasser ab.
Vergleichende Versuche mit Kobaltpapier lassen dementsprechend an
geeigneten Objekten leicht erkennen, wie die Transpiration mit der
Zunahme der Dicke der Kutikularschichten abnimmt, bis sie schließlich
so gut wie ganz verschwindet. Ähnlich wie Kutikularschichten wirken
Korkhäute. Jedermann weiß, wie lange Zeit Äpfel oder Kürbisse, d. h.
Früchte mit wohl ausgebildeter Kutikula, oder Kartoffelknollen, die mit
einer Korkhaut versehen sind, ohne Wasseraufnahme turgeszent bleiben.
Wenn aber die Laubblätter im allgemeinen derartige Schutzmittel
gegen Transpiration nicht in dem Maße besitzen, so ist das kein
Nachteil für die Pflanze; denn die Transpiration ist nicht
etwa ein Übel, sondern sie ist nötig, einerseits weil
sie die Temperatur der Pflanze herabsetzt, die in der prallen
Sonne leicht lebensgefährlich hoch werden könnte, andererseits
weil sie eine rasche Bewegung der sehr verdünnten Nährsalze
bewirkt und endlich weil sie diese verdünnten Lösungen eindickt. So
haben wir denn auch Einrichtungen kennen gelernt (S. 144), die man als
Förderungsmittel für die Transpiration bezeichnen kann. Dahin
gehört vor allem die große Flächenausdehnung der Blätter.
Nun ist aber die Transpiration nicht auf unmittelbar an die Luft
grenzende Zellen beschränkt; auch zahllose Binnenzellen können
Wasserdampf abgeben, wenn sie an Interzellularräume angrenzen. Die
lufterfüllten Interzellularen müßten freilich nach kurzer Zeit
völlig wasserdampfgesättigt sein, wenn sie ringsum abgeschlossen
wären. In Wirklichkeit aber finden sich, wie wir gesehen haben,
Kommunikationswege zwischen der Atmosphäre und den Interzellularen,
von denen die wichtigsten die Spaltöffnungen (S. 43) sind. Aus
ihnen kann der Wasserdampf austreten, und so wird erzielt, daß die
Wasserdampfsättigung der Interzellularenluft keine vollständige ist.
Den aus den Spaltöffnungen austretenden Wasserdampf kann man leicht mit
Kobaltpapier nachweisen. Legt man dieses gleichzeitig auf Ober- und
Unterseite eines Blattes, das nur auf der Unterseite Spaltöffnungen
führt, so zeigt[S. 197] sich, daß hier rasche Farbenänderung eintritt, während
an der Oberseite die blaue Farbe lange Zeit erhalten bleibt.
Diese Ausführungen zeigen, daß man zwischen kutikulärer und
stomatärer Transpiration unterscheiden kann. Bei der typischen
Landpflanze spielt zweifellos die stomatäre Transpiration die
Hauptrolle; bei Pflanzen, die feuchte Lokalitäten bewohnen, nimmt aber
auch die kutikuläre Transpiration beträchtliche Dimensionen an. Die
Spaltöffnungen sind zwar außerordentlich feine Poren in der Epidermis
— vgl. S. 43 —, aber sie sind in sehr großer Zahl vorhanden und zudem
außerordentlich zweckmäßig angeordnet. NOLL hat berechnet,
daß ein einziges mittelgroßes Kohlblatt mit etwa 11 Millionen, ein
Blatt der Sonnenblume mit etwa 13 Millionen Luftspalten versehen
ist. Die Transpiration erfolgt also durch zahllose, zwar sehr enge,
aber sehr dicht stehende Öffnungen in der Kutikula, die selbst so
gut wie kein Wasser durchläßt. BROWN und ESCOMBE
aber haben gezeigt, daß durch eine solche siebartig durchlöcherte
Membran, die über den transpirierenden Mesophyllzellen ausgespannt
ist, unter Umständen ebensoviel Wasser diffundieren kann, wie wenn
die Mesophyllzellen frei der Luft exponiert wären. Wenn das richtig
ist, muß man fragen, warum denn die Pflanze überhaupt einen derartig
komplizierten Apparat aufbaut und nicht die freie Transpiration
aus ungeschützten Zellen vorzieht. Der Grund liegt darin, daß die
Spaltöffnungen nicht nur die Transpiration fördern, sondern auch ganz
aufheben können, daß sie die Transpiration regulieren, was einer
Kutikula nicht möglich ist. Die Weite der eigentlichen Spalte kann
nämlich durch Vorgänge in den Schließzellen verändert werden; ist die
Spalte geschlossen, so ist die Transpiration fast Null, ist die Spalte
weit geöffnet, so erfolgt maximale Transpiration. Da nun Öffnen und
Schließen der Spalten nach Bedürfnis der Pflanzen erfolgt, so haben
wir es in den Spaltöffnungen mit Organen zu tun, die in wunderbar
zweckmäßiger Weise reagieren. Die Öffnung der Spalten wird vielfach
durch Beleuchtung und durch einen gewissen Feuchtigkeitsgrad der Luft
verursacht; umgekehrt wirken Verdunklung oder trockene Luft auf ein
Schließen der Spalten hin.
Fig. 238. Spaltöffnung von Helleborus spec. im
Querschnitt. Die dicken Linien zeigen die Form der Schließzellen bei
geöffnetem Spalt, die feineren Linien bei geschlossenem Spalt. Nach
SCHWENDENER. Der Saftraum der Zellen im geschlossenen Zustand
wurde hier schraffiert; er ist sichtlich kleiner als bei geöffneten
Schließzellen.
Die Bewegungen der Schließzellen sind Reizbewegungen und
erfolgen durch Turgordruck. Vermöge der eigenartigen
Verdickungen der elastischen Zellwände der Schließzellen (S. 45)
führen Druckschwankungen in ihnen nämlich zu Gestaltsveränderungen,
derart, daß eine Steigerung des Turgordruckes die Krümmung der Zellen
vermehrt, eine Abnahme sie vermindert. Der erste Vorgang hat demgemäß
das Öffnen, der zweite den Verschluß der Spalten zur Folge, wie das aus
der Abbildung Fig. 238 ohne weiteres hervorgeht (vgl. auch Fig. 47–49).
Eine Zunahme des Turgordruckes könnte schon dadurch zustande kommen,
daß der osmotisch wirksame Inhalt der Schließzellen mehr Wasser
aufzunehmen in die Lage käme; tatsächlich erfolgt er aber dadurch, daß
bei[S. 198] Zunahme der Beleuchtung und der Luftfeuchtigkeit der osmotische
Wert der Schließzellen ganz beträchtlich zunimmt. Dieser Wert nimmt
umgekehrt bei Verdunklung und Trockenheit ab.
Der maximale osmotische Wert der Schließzellen kann auf 1
Mol., ja selbst auf 2 Mol. NaCl steigen gegenüber 0,15 Mol.
der gewöhnlichen Epidermiszellen. Bei Wasserentziehung (durch
Austrocknung oder Zuckerlösung) oder durch Verdunklung sinkt der
osmotische Wert der Schließzellen unter Umständen noch unter den
der Epidermiszellen[143].
Die Spaltöffnungen sind vorzugsweise auf den Blattflächen zu finden,
die Blätter sind demnach als Organe der Transpiration (und
Assimilation S. 215) zu betrachten. Die Leistungen der
Blätter im Verdunsten von Wasser sind denn auch außerordentlich
überraschend[144]. Eine kräftige Pflanze, beispielsweise eine
Sonnenblume von Manneshöhe, verdunstet an einem hellen Tage über ein
Liter Wasser, und man hat festgestellt, daß ein Morgen mit Kohlpflanzen
in 4 Monaten zwei Millionen Liter, ein solcher mit Hopfen 3–4 Millionen
Liter Wasser verdunstet. Für eine einzeln stehende Birke mit etwa
200000 Blättern berechnete VON HÖHNEL das verdunstete
Wasser an einem heißen, trockenen Tage auf etwa 300–400 Liter, im
Tagesdurchschnitt auf 60–70 Liter. Ein Hektar Buchenhochwald verdunstet
im Durchschnitt 20000 Liter täglich. Auf je 100 g Blattsubstanz
berechnet, verbraucht in einer Vegetationsperiode die Rotbuche 75
Liter, die Tanne aber nur 7 Liter. Im Durchschnitt verdunstet eine
Pflanze für jedes Gramm Trockensubstanz, das sie in ihren oberirdischen
Teilen bildet, 250–900 g Wasser.
Die Flächenentwicklung und die feinere Organisation des Blattes
(Zahl und Weite der Spaltöffnungen, Ausbildung von Kutikula und
Behaarung) beeinflussen die Wasserabgabe außerordentlich stark;
deshalb versteht man es, daß verschiedene Pflanzen sehr ungleich
stark transpirieren. Aber auch ein und derselbe Sproß transpiriert
nicht immer gleichmäßig; das kommt daher, daß äußere und innere
Ursachen einerseits physiologische Folgen haben, indem sie die
Weite der Spaltöffnungen verändern, und daß sie andererseits
die Transpiration auch rein physikalisch ebenso beeinflussen
wie die Verdunstung eines freien Wasserspiegels. Wärme, Trockenheit
und Bewegung der Luft steigern die Verdunstung physikalisch,
während Beleuchtung und Feuchtigkeit sie physiologisch
fördern. Beide Momente wirken zusammen dahin, daß am Tage die
Verdunstung im allgemeinen stärker ist als des Nachts. Pflanzen,
die, wie Impatiens parviflora, an heißen trockenen Tagen leicht
welken, werden daher nachts wieder frisch.
Vergleicht man die Transpiration eines Blattes mit der Evaporation
einer gleichen Fläche reinen Wassers, die unter gleichen
Außenverhältnissen steht wie das Blatt, so gibt das Verhältnis
Transpiration : Evaporation, das immer kleiner als 1 ist und als
relative Transpiration[145] bezeichnet wird, ein Maß für die
physiologisch wirksamen Momente im Blatt, vor allem also für
die regulierende Tätigkeit der Stomata.
Dieser Begriff muß indes noch mit Vorsicht verwendet
werden, denn SIERP hat gezeigt, daß bei steigender
Windgeschwindigkeit die Evaporation der freien Wasserfläche
ganz anderen Gesetzmäßigkeiten folgt als die Wasserabgabe durch
multiperforate Membranen, die relative Transpiration
also bei verschiedenen Außenbedingungen auch ohne organische
Regulationen ganz ungleiche Werte haben kann. Der Einfluß des
Windes, der in den Studien von BROWN und ESCOMBE
ganz zurücktrat, ist erst neuerdings voll gewürdigt worden; er ist
um so wichtiger, als die Luft in der Natur niemals so unbewegt ist,
daß eine Diffusion aus einem Blatt in der Weise vor sich geht, wie
BROWN und ESCOMBE das annahmen. — Während bei
ganz ruhiger Luft auch die Gestalt des Blattes und die Gestalt
der ganzen Pflanze für die Größe der Transpiration von Bedeutung
sein könnten, ist bei bewegter Luft stets nur die transpirierende
Fläche wirksam (SIERP).
Über die Öffnungsweite[146] der Spaltöffnungen orientiert man
sich durch direkte Beobachtung mit dem Mikroskop, Anwendung von
Kobaltpapier (S. 195) oder durch die „Infiltrationsmethode“:
sind die Stomata geöffnet, so dringen Flüssigkeiten wie
Petroleum, Alkohol usw. leicht ein und injizieren das ganze
Interzellularensystem, das Blatt wird also[S. 199] durchsichtig. Legt man
quer über ein Blatt einen Streifen schwarzen Papiers, so schließen
sich unter diesem die Spaltöffnungen. Nach Behandlung mit Alkohol
ergibt sich das Bild der Fig. 239. Sehr anschaulich wird das
Offensein der Spaltöffnungen auch mit der „Gasdiffusionsmethode“
demonstriert. Werden z. B. rote, anthozyanhaltige Blätter mit
offenen Spalten in ammoniakhaltige Luft gebracht, so tritt in
wenigen Sekunden eine Blaufärbung ein, die bei geschlossenen
Spalten ausbleibt. In neuester Zeit ist ganz besonders das von
DARWIN zuerst konstruierte Porometer ausgiebig zu
Messungen über die Öffnungsweite der Stomata benutzt worden. Es
besteht aus einer kleinen Glasglocke (Fig. 240 G), die mit Leim
einem Blatt (B) aufgedichtet wird. Die Luft im Innern wird durch
Saugen bei Q etwas unter Atmosphärendruck gebracht; ihr Druck
kann an einem Wassermanometer abgelesen werden. Die Geschwindigkeit
des Ausgleiches zwischen dem Atmosphärendruck und dem Druck in der
Glocke gibt ein Maß für den Öffnungszustand der Spaltöffnungen.
Daß die Spaltöffnungen vortrefflich regulatorisch wirken, kann
man daraus entnehmen, daß in unserem Klima die Schwankungen im
Wassergehalt eines Blattes nur etwa 1% betragen[147]. Ein so
geringfügiger Wasserverlust führt freilich schon zum Welken.

Fig. 239. Ein Blatt der Syringe wird in der Mitte
verdunkelt, an den Enden dem Licht exponiert. Nur die beleuchteten
Spaltöffnungen bleiben offen und lassen absoluten Alkohol
eindringen. Nach MOLISCH.
Fig. 240. Porometer nach DARWIN. B Blatt,
G Glasglocke, T Manometer, Q Quetschhahn, W Wasser.
Erst nach Eintritt des Welkens schließen sich die Stomata; eine
ganz ideale Regulation vermögen sie also nicht auszuüben. Es
konnte mehrfach gezeigt werden[148], daß dem Spaltenverschluß bei
starker Transpiration zunächst eine ganz unzweckmäßige Erweiterung
der Spalten vorausgeht, die ihrerseits das Welken beschleunigt.
Wie alle Regulation hinkt also auch die der Stomata etwas nach.
— Nach den Angaben einiger Autoren[149] sieht es so aus, als ob
außer den Spaltöffnungen noch ein anderer Faktor die Transpiration
herabzusetzen vermöchte. Abgeschlossene Resultate liegen noch nicht
vor.
Pflanzen trockener Standorte, die mit dem aufgenommenen Wasser
haushälterischer verfahren müssen als die typische Landpflanze,
zeigen eine Fülle von Einrichtungen, die sie vor übermäßiger
Transpiration schützen (vgl. S. 144), können andererseits auch
einen viel größeren Wasserverlust ohne Schaden ertragen als unsere
einheimische Flora (LIVINGSTON). Umgekehrt finden sich bei
Pflanzen sehr feuchter Lokalitäten Einrichtungen zur Förderung
der Transpiration. Gelingt es dem Blatt durch Wärmeabsorption von
außen her oder durch Wärmeproduktion aus eigenen Mitteln (S. 241)
seine Temperatur über die der Umgebung zu heben, so ist selbst in
dunstgesättigter[S. 200] Luft noch eine Transpiration möglich. In der
Guttation liegt endlich ein Mittel der Pflanze vor, auch nach völligem
Aufhören der Transpiration noch Wasser abzugeben.
Guttation[150].
Die Ausscheidung tropfbar flüssigen Wassers aus unverletzten Pflanzen
ist ungleich seltener wahrzunehmen als die Transpiration, denn sie
findet nur unter engbegrenzten Bedingungen statt, nämlich dann,
wenn die Pflanze wassergesättigt, die Luft dunstgesättigt ist. In
der Natur bemerkt man sie dementsprechend bei uns nur am frühesten
Morgen nach einer regenlosen, aber feuchtwarmen Nacht. Man sieht
da an den Blatträndern und Spitzen mancher Pflanzen glitzernde
Wassertropfen ansitzen, die langsam an Größe zunehmen, abfallen und
durch neue Tröpfchen ersetzt werden. Es handelt sich aber nicht
etwa um Taubildung, sondern um Tropfen, die von den Blättern selbst
ausgeschieden werden. Mit der Zunahme der Temperatur verschwinden sie
dann, weil die Pflanze nicht mehr voll wassergesättigt ist. Erhöht man
aber den Wassergehalt der Pflanze, entweder indem man ihre Verdunstung
herabsetzt (z. B. durch Überstülpen einer Glasglocke), oder indem
man Wasser künstlich in abgeschnittene Zweige einpreßt, so tritt die
Tropfenausscheidung alsbald wieder auf.
Die Tropfen treten bei den Gräsern an der Blattspitze, bei
Alchimilla an den Blattzähnen, bei Tropaeolum (Fig. 241) an den
stumpfen Ausbuchtungen des Blattes auf; sie kommen entweder aus
sog. Wasserspalten (S. 99) oder aus gewöhnlichen Spaltöffnungen,
oder sie werden von Apikalöffnungen oder von Haaren (manchmal auch
von Brennhaaren) sezerniert. Alle solche wasserausscheidenden
Organe nennt man Hydathoden.
Ungleich verbreiteter als in unserem Klima findet sich die
Wasserausscheidung im feuchten, tropischen Urwald. Besonders
auffallend ist sie bei manchen großblätterigen Araceen, an deren
Blattspitzen das Wasser oft in kurzen Zwischenräumen abtropft.
Bei Colocasia nymphaeifolia kann es sogar auf kurze Strecken
fortgeschleudert werden; dabei scheidet ein einziges Blatt zuweilen
190 Tropfen in der Minute, in einer Nacht oft 1⁄10 Liter aus. —
Auch an einzelligen Pflanzen, so an Schimmelpilzen, ist oft eine
reichliche Ausscheidung von Wassertröpfchen durch die Zellwand
und die Kutikula hindurch wahrzunehmen, die hier, wie auch bei
Wasserpflanzen, durchlässig ist.
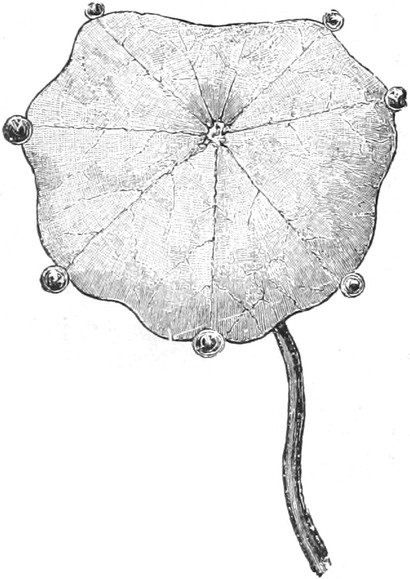
Fig. 241. Ausscheidung von Wassertropfen aus einem
Blatte der Kapuzinerkresse (Tropaeolum majus). Nach NOLL.
Die Ausscheidung flüssigen Wassers kann unter ungünstigen
Transpirationsbedingungen, vor allem bei submersen Wasserpflanzen, die
Verdunstung in gewissem Sinne vertreten oder ersetzen. Ihre Bedeutung
für die Pflanze deckt sich aber insofern nicht mit der Transpiration,
als in den Tropfen sehr häufig mineralische oder organische Substanzen
mit ausgeschieden werden. Diese Stoffe sind manchmal so reichlich
im ausgestoßenen Wasser enthalten, daß sie nach dessen Verdunstung
als kleine Krusten zurückbleiben. Auf diese Weise entstehen die
Kalkschüppchen der Steinbrecharten, die Salzmassen mancher Halophyten
(S. 206). STAHL hat wahrscheinlich gemacht, daß in der
Ausscheidung solcher gelöster Salze (Exkretion) die physiologische
Hauptrolle der Guttation liegen kann, da manche derartige Stoffe,
wenn sie sich in den Zellen anhäufen, schädlich wirken. Daß bei den
Aussonderungen der Nektarien, bei den Verdauungsdrüsen
der Insektivoren (S. 222) und bei den Ausscheidungen mancher
Narben nicht das Wasser, sondern der gelöste Körper das
wichtigste ist, versteht sich von selbst. Bei manchen Nektarien[S. 201] ist
die Ausscheidung weitgehend unabhängig vom Wassergehalt und kann auch
noch an der welkenden Pflanze weiter gehen.
Bluten. Einen Wasseraustritt aus der Pflanze kann man vielfach
nach Verwundungen beobachten; regelmäßig sieht man ihn an beschnittenen
Bäumen und Sträuchern im Frühjahr, z. B. besonders stark an der Rebe.
Auch an Stauden, die man dicht über der Erde abgeschnitten hat, läßt
sich leicht ein Wasseraustritt aus der Wunde zeigen. Man spricht vom
Tränen oder Bluten der Wunden und stellt leicht fest, daß das Wasser
aus den Gefäßen hervorquillt und daß es auch gegen hohen Widerstand
ausgepreßt wird (Blutungsdruck).
Die Figur 242 zeigt, in welcher Weise man den Blutungsdruck, den
ein Stammstumpf s produziert, mit Hilfe eines Manometers messen
kann. Die Quecksilbersäule wird bei gewissen Pflanzen bis zu 50 und
60, unter günstigen Bedingungen aber bis zu 140 Zentimeter (Birke)
oder noch höher emporgedrückt. Diese Druckkräfte könnten also eine
Wassersäule bis zu 6, 8 und 18 Metern halten.
Besonders reichlich ist der Wasseraustritt, wenn die Erde
feucht und warm ist; er dauert dann je nach der Pflanze und
ihrem Entwicklungszustand oft mehrere Tage bis Monate an,
und das ausgeschiedene Wasser beträgt unter Umständen bis zu
einigen Litern und mehr; bis zu 1 Liter am Tag bei der Rebe,
bis 5 Liter bei der Birke, 10 bis 15 Liter bei Palmen. Bei
längere Zeit hindurch blutenden Pflanzenteilen bemerkt man eine
gewisse Periodizität der Blutungsmenge; nachts wird mehr
ausgeschieden als am Tag.
Der „Blutungssaft“ führt außer mineralischen Salzen zuweilen
erhebliche Mengen von organischen Substanzen (gelöste Eiweißstoffe,
Asparagin, Säuren, besonders aber Kohlehydrate) mit sich; bei
einigen Pflanzen ist der Zuckergehalt dieses Saftes so groß, daß
Zucker technisch daraus gewonnen werden kann. So liefert der
Zuckerahorn Nordamerikas mit 1⁄2% Zucker im Saft in einem
Frühjahr etwa 2–3 Kilo von einem Baum. Solcher Saft kann auch
gleich Most oder Biermaische zu berauschenden Getränken vergoren
werden und liefert dann Birkenwein, Palmwein oder die „Pulque“ der
Mexikaner. Diese wird aus dem Safte blühreifer Agaven gewonnen;
eine einzelne Pflanze kann in 4–5 Monaten nahezu 1000 Liter Saft
ausscheiden.
Ursachen der Wasserausscheidung[151]. Bei der
Tropfenausscheidung an der intakten Pflanze handelt es sich nur zum
Teil um Wasserausscheidung aus Oberflächenzellen. In anderen
Fällen wird Wasser in die Gefäße eingepreßt und tritt dann
an Orten geringeren Widerstandes zutage (S. 99). Auch bei den
Erscheinungen des Blutens wird Wasser aus Parenchymzellen in die
Hohlräume der Gefäße gedrückt, und wenn das auch ganz besonders
häufig in der Wurzel stattfindet, so fehlt der Vorgang doch in den
Stengeln und Blättern durchaus nicht.
Allen geschilderten Vorkommnissen gemeinsam ist also in letzter
Linie eine einseitige Flüssigkeitsausscheidung aus lebenden Zellen.
Auf die Ursachen einer solchen Drüsentätigkeit der Zellen kann hier
nicht eingegangen werden.
Fig. 242. Demonstration des Wurzeldruckes an einer
Georgine. Auf den glatt abgeschnittenen Stumpf s ist mittels
Kautschukschlauchs c das gebogene Glasrohr g aufgesetzt. Das aus
der Erde durch die Wurzel aufgenommene Wasser W wird so kräftig
ausgepreßt, daß es den Druck der Quecksilbersäule Q überwindet. Nach
NOLL.
Leitung des
Wassers[152].
Das Wasser, das teils in Dampfform namentlich von den Blättern
abgegeben wird, teils durch Hydathoden oder Wunden in flüssiger
Form der Pflanze entströmt, ist im allgemeinen von der Wurzel
aufgenommen worden.[S. 202] Es hat also einen Weg zurückzulegen, der schon
bei einjährigen Pflanzen nach Metern messen kann, bei den Riesen des
Pflanzenreiches aber etwa 100 m beträgt (Eucalyptus amygdalina 110 m;
Sequoia gigantea 95 m Stammhöhe). Durch osmotische Saugung von Zelle
zu Zelle würde diese Wasserbewegung viel zu langsam stattfinden,
um die Verluste decken zu können. Tatsächlich erfolgt denn auch
die Strömung des Wassers zur Deckung der Transpiration, der sog.
Transpirationsstrom, im Gefäßteil der Leitbündel bzw. bei Bäumen
im Holzkörper. Das kann man schon aus einem uralten Versuch, dem sog.
Ringelungsversuch, entnehmen. Werden an einem Aste eines Baumes
eine Strecke weit die Gewebe bis auf den Holzkörper fortgenommen, so
bleiben zunächst, d. h. solange nicht eine Austrocknung oder Zersetzung
des Holzkörpers an der entrindeten Stelle eingetreten ist, die Blätter
dieses Astes ebenso frisch wie die eines anderen, nicht geringelten
Astes; das beweist, daß der Transpirationsstrom nicht durch die Rinde,
sondern durch den Holzkörper sich bewegt. Entfernt man dagegen aus
einem Zweige sorgfältig eine Strecke weit den Holzkörper und läßt die
Rinde größtenteils im Zusammenhang, so welken die Blätter über der
Operationsstelle so rasch wie an einem völlig abgeschnittenen Zweige.
Man kann diese Versuche ebensogut an intakten Pflanzen ausführen wie
an abgeschnittenen, in Wasser gestellten Zweigen; denn letztere nehmen
wenigstens eine Zeitlang (solange keine sekundären Änderungen an der
Schnittfläche eingetreten sind) das Wasser ebenso lebhaft direkt mit
dem Holzkörper auf wie intakte Pflanzen mit der Wurzel. Wird ein
abgeschnittener Zweig mit seiner Schnittfläche in eine Lösung von
Gelatine gestellt, und läßt man dann die Gelatine, die eine Strecke
weit in die Gefäße eingedrungen ist, erstarren, so hat der Holzkörper
seine Leitfähigkeit für Wasser verloren. Man sieht daraus, daß das
Lumen der Gefäße für die Leitung des Wassers unentbehrlich
ist. Aber freilich, in der lebenden Pflanze sind die Gefäße nicht nur
mit Wasser gefüllt, sondern sie führen wenigstens in Zeiten lebhafter
Transpiration immer auch Luft.
Entsprechend ihrer Aufgabe findet man die Gefäßteile in
Wasserpflanzen und Sukkulenten, in denen gar keine oder eine sehr
schwache Transpiration besteht, nur spärlich entwickelt. Dagegen
besitzen die transpirierenden Blattflächen ein außerordentlich
reiches Leitbündelgewebe, das zudem durch vielfache Anastomosen
dafür sorgt, daß jeder beliebige Punkt auch bei Nichtfunktionieren
des nächsten Verbindungsstranges doch genügend Wasser enthält. Die
Fig. 128 gibt von diesem Berieselungssystem eines Blattes noch
keine vollständige Vorstellung, denn die feinen, nur mikroskopisch
nachweisbaren Stränge sind in ihr gar nicht dargestellt. Auch
in den Zuleitungen zu den Blättern, in den Stämmen, findet
man, insbesondere bei den in die Dicke wachsenden Bäumen, ein
außerordentlich leistungsfähiges Wasserleitungssystem. Es ist
freilich nicht mehr das gesamte Holz eines dicken Stammes, welches
der Wasserleitung dient; sie ist vielmehr stets auf die jüngsten
Jahresringe beschränkt. Wo Kernholz (S. 135) gebildet wird, ist
dieses von der Leitung völlig ausgeschlossen.
Über die bewegenden Kräfte des Transpirationsstromes ist man
noch nicht im klaren. Man wird in erster Linie an eine Druckwirkung
von unten oder eine Saugwirkung von oben her denken und für erstere
den Blutungsdruck, für letztere den Vorgang der Transpiration
verantwortlich machen. Allein der Blutungsdruck kommt aus mehreren
Gründen nicht in Betracht, und ob die Saugkraft der Transpiration
ausreicht, um Wasser bis zum Gipfel hoher Bäume dauernd in
ausreichender Menge zu heben, erscheint zweifelhaft. Eine allseitig
anerkannte Lösung des vielumstrittenen Problems existiert demnach heute
nicht.
Bezüglich des Blutungsdruckes der Wurzel (Wurzeldruck) ist
folgendes zu bedenken: Bei vielen Pflanzen erreicht der Wurzeldruck
nur sehr geringe Höhe oder fehlt ganz. Aber auch bei Pflanzen
mit kräftigem Wurzeldruck ist die von der Wurzel ge[S. 203]lieferte
Wassermenge erheblich geringer als die bei der Transpiration
verbrauchte. Bei einigermaßen starker Transpiration kommt der
Wurzeldruck daher in der geschilderten Weise überhaupt nicht
zur Geltung. Wird eine kräftig verdunstende Pflanze am
Wurzelstumpf abgeschnitten, so tritt aus diesem zunächst gar
kein Wasser hervor, im Gegenteil, der Stumpf saugt dargebotenes
Wasser begierig in sich auf (es herrscht also ein Druck unter
Atmosphären-Größe). Erst nach seiner vollen Sättigung beginnt
eine Auspressung. Im Freien kommt der Wurzeldruck nur dann zur
Geltung, wenn bei feuchter kühler Luft, wie zumal des Nachts,
die Transpiration sehr vermindert ist. Am günstigsten liegen
die Umstände dazu im Frühjahr, wenn bei höchstem Wassergehalt
des Holzes das transpirierende Laub noch nicht entfaltet ist.
Bei Verletzungen des Holzkörpers quillt dann der „Saft“ aus
Tracheen und Tracheïden in Tropfen hervor. Ein Blutungsdruck über
Atmosphärengröße bei belaubten Bäumen dürfte nur im tropischen
Urwald vorkommen.
Daß die Transpiration eine Saugung von Zelle zu Zelle
bewirkt, ist schon hervorgehoben (S. 196), und es ist ohne weiteres
verständlich, daß diese sich dann von den Parenchymzellen in die
angrenzenden Gefäße fortsetzt. Auch ist diese Saugkraft leicht zu
demonstrieren.
Ein abgeschnittener, in Wasser gestellter Sproß zeigt durch
sein Frischbleiben an, daß er das Wasser bis in seine obersten
Zweigspitzen zu heben vermag. Er leistet aber noch mehr:
luftdicht mit einem langen wassergefüllten Rohre verbunden,
vermag er leicht eine Wassersäule von 2 Metern und mehr
emporzusaugen; wird das untere Ende des Rohres in Quecksilber
gebracht, dann wird selbst dieses bis zu ansehnlicher Höhe
gehoben. Kräftige, sonst unverletzte Koniferenzweige sind
imstande, das Quecksilber unter Umständen bis weit über
Barometerhöhe zu heben, ohne zu welken. Bedingung für eine
solche Saugung ist einerseits ein luftdichter Abschluß der
Wasserbahnen, andererseits eine nicht unbeträchtliche Kohäsion
der Flüssigkeiten, die gehoben werden. Tatsächlich erreicht denn
auch die Kohäsion des Wassers ganz gewaltige Werte; nach den
Messungen von URSPRUNG und RENNER etwa 300
Atmosphären. So konnte man zu der Vorstellung kommen, daß der
Zug der Transpiration durch die Kohäsion des Wassers bis in die
Wurzelspitzen sich fortpflanze. Die Versuche RENNERs, die
den Nachweis beträchtlicher negativer Drucke in den Leitbahnen
transpirierender Pflanzen erbracht zu haben schienen, sind
von anderer Seite stark angezweifelt worden[153]. Somit ist
die Kohäsionstheorie[154] noch nicht bewiesen. Wenn
wirklich die Saugung durch Kohäsion des Wassers nach unten
weitergegeben werden soll, dann müßten die Gefäße kontinuierlich
mit Wasser erfüllt sein, während man tatsächlich Luft- und
Wassersäulchen in ihnen gefunden hat. Mit Eintreten eines
Zuges müssen sich die Luftbläschen expandieren, und in der Tat
findet man in lebhaft transpirierenden Zweigen stark verdünnte
Luft in den Gefäßen. Werden solche Gefäße unter Quecksilber
durchschnitten, so stürzt dieses unter dem äußeren Luftdruck
auf weite Strecken in sie hinein. Jedenfalls erfolgt das Reißen
der Wasserfäden in Gefäßen durchaus nicht immer, weil die
Kohäsion überwunden, sondern auch, weil die Adhäsion an
die Wand dem Zug nicht mehr gewachsen ist, oder weil Luft durch
die Wand tritt. Dadurch wird es verständlich, daß das Reißen in
verschiedenen Elementen bei ganz verschiedener Spannung erfolgt,
während die Kohäsion ja stets gleich groß sein muß. Es ist auch
schon der Gedanke ausgesprochen worden, daß zweierlei Typen
unter den Gefäßen existieren: 1. leitende (Tracheïden) mit hoher
Inhaltsfestigkeit und 2. speichernde (Tracheen), die, weil in ihnen
das Wasser leicht reißt, dieses auch leicht abgeben können[155].
— Die Vertreter der Kohäsionstheorie nehmen an, daß neben
solchen lufthaltigen Bahnen immer auch völlig mit Wasser erfüllte
existieren, und daß nur diesen eine Leitfähigkeit zukomme, während
die anderen als Wassermagazine von Bedeutung sein sollen. —
Es ist aber keineswegs ausgeschlossen, daß auch den lebenden
Elementen, die in keinem Holzteil fehlen, eine Rolle bei der Hebung
des Wassers zukommt.
b) Die Nährsalze.
Die Nährsalze, die von einer Pflanze aufgenommen werden, finden sich
fast alle in der Asche wieder; nur die Stickstoffverbindungen fehlen.
Demnach kann uns die nachstehende Tabelle über den Gehalt einiger
Kulturpflanzen an Aschenbestandteilen schon einen gewissen
Einblick in die Menge und die Verteilung der Nährsalze geben.
[S. 204]
|
Pflanzenteile
|
Asche in
100 Tln.
der
Trocken-
substanz
|
In 100 Teilen der Asche sind enthalten:
|
|
K2O
|
Na2O
|
CaO
|
MgO
|
Fe2O3
|
Mn3O4
|
P2O5
|
SO3
|
SiO2
|
Cl
|
|
Roggen (Körner)
|
2,09
|
32,10
|
1,47
|
2,94
|
11,22
|
1,24
|
—
|
47,74
|
1,28
|
1,37
|
0,48
|
|
„ (Stroh)
|
4,46
|
22,56
|
1,74
|
8,20
|
3,10
|
1,91
|
—
|
6,53
|
4,25
|
49,27
|
2,18
|
|
Erbsen (Samen)
|
2,73
|
43,10
|
0,98
|
4,81
|
7,99
|
0,83
|
—
|
35,90
|
3,42
|
0,91
|
1,59
|
|
„ (Stroh)
|
5,13
|
22,90
|
4,07
|
36,82
|
8,04
|
1,72
|
—
|
8,05
|
6,26
|
6,83
|
5,64
|
|
Kartoffel (Knolle)
|
3,79
|
60,06
|
2,96
|
2,64
|
4,93
|
1,10
|
—
|
16,86
|
6,52
|
2,04
|
3,46
|
|
Weintraube (Beere)
|
5,19
|
56,20
|
1,42
|
10,77
|
4,21
|
0,37
|
—
|
15,58
|
5,62
|
2,75
|
1,52
|
|
Tabak (Blätter)
|
17,16
|
29,09
|
3,21
|
36,02
|
7,36
|
1,95
|
—
|
14,66
|
6,07
|
5,77
|
6,71
|
|
Baumwolle (Faser)
|
1,14
|
36,96
|
13,16
|
17,52
|
5,36
|
0,60
|
—
|
10,68
|
5,94
|
2,40
|
7,60
|
|
Fichte (Holz)
|
0,21
|
19,66
|
1,37
|
33,97
|
11,27
|
1,42
|
22,96
|
2,12
|
2,64
|
2,73
|
0,07
|
Zu diesen Zahlen ist zu bemerken, daß sie keine konstanten
Verhältnisse ausdrücken, daß vielmehr die Zusammensetzung der Asche
je nach dem Kulturboden wechseln kann. Auch ist es vielleicht nicht
ganz überflüssig, darauf aufmerksam zu machen, daß diese Oxyde
der Mehrzahl nach erst bei der Veraschung gebildet werden. In der
lebenden Zelle waren Salze gegeben und vor allem vielfach
die Metalle an organische Säuren gebunden.
Man sieht aus dieser Tabelle zunächst einmal, wie allgemein verbreitet
die Aschensubstanzen sind, erkennt aber andererseits auch sofort, daß
sie in verschiedenen Pflanzen, ja sogar in verschiedenen Teilen einer
Pflanze, in recht ungleicher Menge vorkommen. So findet sich z. B. in
den Samen viel Phosphorsäure, dagegen wenig Kieselsäure und Kalk; das
Stroh aber verhält sich gerade umgekehrt. Die Kartoffel enthält
viel K2O und wenig CaO, während Fichtenholz reichlich CaO und wenig
K2O besitzt.
Trotz ihrer allgemeinen Verbreitung wird man die Aschensubstanzen
zunächst für Ballast, für eine zufällige Verunreinigung der organischen
Pflanzensubstanz zu halten geneigt sein. Allein jeder Versuch, eine
Pflanze aschenfrei zu erziehen, zeigt die Unentbehrlichkeit dieser
Stoffe.
Daß die mineralischen Inhaltsstoffe der Pflanze wesentliche
Bestandteile der Pflanzennahrung ausmachen, sprach zuerst
BERTHOLLET 1803 aus; später (1832) betonte es KARL
SPRENGEL, dann auch LIEBIG. Exakt bewiesen wurde
diese wichtige Tatsache aber erst 1842 durch WIEGMANN und
POLSTORFF.
Fig. 243. Buchweizen in Wasserkultur. I In Nährlösung
mit Kali. II In Nährlösung ohne Kali. Beide gleich stark verkleinert.
Nach NOBBE.
Zwei Wege bieten sich, um diesen Nachweis zu führen und zugleich zu
zeigen, ob alle oder nur gewisse Aschensubstanzen notwendig sind.
Einmal kann man die Pflanze in künstlichem Boden kultivieren, der
aus unlöslichen Substanzen (wie Platin, reiner Kohle, reinem Quarz)
besteht und dem dann die in Frage stehenden Stoffe[S. 205] beigemengt werden.
Bequemer ist der andere Weg, die sog. Wasserkulturmethode. Es hat sich
gezeigt, daß viele Pflanzen ihr Wurzelsystem statt in Erde auch in
Wasser entwickeln können. Man hat es dann in der Hand, diesem Wasser
der Reihe nach alle Elemente der Asche in verschiedenen Verbindungen
zuzusetzen und so zu erproben, was nötig, was überflüssig ist. Ein
Blick auf Fig. 243 I zeigt, daß in einer solchen „Nährlösung“ von
passender Zusammensetzung die Pflanze (Buchweizen) gut gedeiht; sie
produziert Wurzeln, Sprosse, Blüten und Früchte und vermehrt dabei ihr
Trockengewicht auf das Hundert- oder Tausendfache, gerade wie wenn sie
in gutem Boden stände. In destilliertem Wasser dagegen wächst sie zwar
anfangs ganz normal, stellt aber bald ihr Wachstum gänzlich ein und
bleibt ein außerordentlich kümmerliches Gewächs.
Nährlösungen werden in recht verschiedener Zusammensetzung
verwendet[156]. Besonders häufig wird die KNOPsche
Lösung benutzt (Wasser 1000, Kalziumnitrat 1, Magnesiumsulfat
0,25, saures phosphors. Kalium 0,25, Kaliumnitrat 0,25,
Eisenchlorid Spur). Die V. D. CRONEsche Lösung (Wasser
1000, Kaliumnitrat 1, Kalziumsulfat 0,5, Magnesiumsulfat 0,5,
tert. Kalziumphosphat 0,25, Ferrophosphat 0,25), die statt des
Ferrisalzes ein Ferrosalz enthält und die Phosphorsäure als fast
ganz unlösliches Kalziumphosphat führt, wird manchmal mit Vorteil
verwendet. Neuerdings sind von amerikanischen Forschern zahlreiche
Salzkombinationen geprüft worden.
Der Erfolg einer solchen Wasserkultur lehrt, daß die typischen
Landpflanzen mit einigen Verbindungen auskommen, die sich
aus den Elementen K, Ca, Mg, Fe und H, O, S, P, N zusammensetzen
— vorausgesetzt, daß ihnen außerdem noch der Sauerstoff und die
Kohlensäure der Luft zur Verfügung stehen. Es sind also im ganzen 10
Elemente, aus denen sich die unentbehrlichen Nährstoffe aufbauen. Von
ihnen interessieren uns an dieser Stelle nur die nach Ausschluß von H,
O, C übrigbleibenden sieben, die als Nährsalze aus dem
Boden bzw. dem Wasser aufgenommen werden müssen. Sechs davon
finden sich in der Asche der Pflanze vor, eines, der Stickstoff,
geht bei der Verbrennung in flüchtige Substanzen über. Daß diese sieben
Elemente völlig unentbehrlich sind, ergibt sich daraus, daß das Fehlen
jedes einzelnen auch durch den größten Überschuß der anderen oder eines
ihm nahestehenden Elementes nicht kompensiert werden kann.
So kann z. B. Kalium im allgemeinen nicht durch Natrium, Lithium,
Rubidium ersetzt werden. Niedere Organismen (Algen, Bakterien,
Pilze) machen geringere Ansprüche als die höheren; sie können
das Ca entbehren. Der Mangel eines notwendigen Elementes macht
sich entweder nur in einer äußerst kümmerlichen Entwicklung der
Pflanze bemerkbar (Fig. 243 II, Kalimangel) oder er ruft höchst
charakteristische Veränderungen an ihr hervor. Am bekanntesten
in dieser Beziehung ist die Wirkung des Eisenmangels. Ohne Eisen
ergrünen die Pflanzen nicht (Chlorose). Bei Kalkmangel
machen sich schwere Schädigungen (Vergiftungen) bemerkbar.
Die genannten Elemente werden häufig auch schlechthin als
„Nährstoffe“ der Pflanze bezeichnet. Der Ausdruck ist indes nicht
ganz korrekt, weil die Pflanze mit wenigen Ausnahmen die Elemente
selbst gar nicht auszunützen vermag. H und O können das
Wasser durchaus nicht ersetzen, und metallisches Kalium ist
ebenso unverwendbar wie Schwefel. Die Pflanze bedarf ganz bestimmter
Salze oder — da diese im Wasser zum Teil ionisiert werden —
bestimmter Ionen. Von Kationen sind nötig: K+, Ca++, Mg++,
Fe++ (oder Fe+++), von Anionen SO4– –, H2PO4– und
NO3–. Während nun der Phosphor und der Schwefel in keiner anderen
Verbindung Verwertung finden kann, ist das beim Stickstoff anders; er
kann auch als Kation NH4+ in vielen Fällen wohl ebenso gute Dienste
leisten wie als NO3–.
So wie C, N, H und O beteiligen sich auch manche Nährsalzelemente
am chemischen Aufbau wichtiger Pflanzenstoffe: so S und P, die
im Eiweiß und[S. 206] in Eiweißderivaten vorkommen, so das Mg, das im
Chlorophyllfarbstoff auftritt. Möglich ist es, daß auch die
anderen Metalle in Verbindungen notwendiger Stoffe eingehen,
sicher aber spielen sie auch noch eine ganz andere, nämlich eine
physikalisch-chemische Rolle. Die Salze greifen in alle physiologischen
Prozesse regulierend ein und erhalten das kolloidale Protoplasma in
normalem Zustand.
Neben dem positiven Ergebnis der Notwendigkeit gewisser Salze hat die
Wasserkultur auch gezeigt, daß viele normalerweise von der
Pflanze aufgenommene Stoffe entbehrlich sind, so vor
allem Natrium, Chlor und Silizium.
Das Natrium ist selbst bei den Halophyten, bei denen
es in größter Menge gefunden wird, entbehrlich. Diese Pflanzen
leben auf kochsalzhaltigem Boden, nicht weil sie das NaCl zu ihrer
Entwicklung nötig haben, sondern weil sie es besser ertragen
als andere Pflanzen, deren Konkurrenz also an solchen Lokalitäten
ausgeschlossen ist. Ihre charakteristische sukkulente Ausbildung
(Fig. 193) verlieren die Halophyten freilich mehr oder weniger,
wenn ihnen das Kochsalz entzogen wird. Den Diatomeen und gewissen
Meeresalgen scheint Na unentbehrlich zu sein[157].
Silizium ist auch den Schachtelhalmen und den Gräsern,
die sehr reichlich SiO2 führen, entbehrlich; dagegen brauchen
es die Diatomeen, deren Schalen fast ganz aus Kieselsäure
bestehen. Diesem Umstande verdanken sie ihre Dauerhaftigkeit.
Diatomeenschalen bilden ja als Kieselgur mächtige geologische
Ablagerungen. Aluminium[158] ist zwar in kleinen Spuren
allgemein verbreitet, wird aber nur von wenigen Pflanzen in
beträchtlicher Menge aufgenommen (z. B. Lycopodium-Arten); nach
STOKLASA soll es den Wasserpflanzen unentbehrlich sein.
Jod, im Meerwasser kaum in Spuren analytisch nachweisbar,
wird in Meeresalgen trotzdem reichlich aufgespeichert, so daß diese
lange Zeit das Material für die Jodgewinnung bildeten. Ob es diesen
Pflanzen entbehrlich ist, wissen wir nicht.
Entbehrliche Aschenbestandteile können der Pflanze insofern
nützlich sein, als sie die unentbehrlichen Bestandteile in
allgemeineren Eigenschaften, so als Basen zur Neutralisation von
Säuren u. dgl., vertreten und damit die unentbehrlichen Stoffe
für ihre speziellen Wirkungen, in denen sie unersetzbar sind, in
vollem Umfange disponibel machen. So kann Na partiell das
K und Ca bis zu einem gewissen Grade das Mg ersetzen. Auch können
manche Stoffe und Stoffgemische der Pflanze nützlich und zu ihrem
Gedeihen förderlich werden, ohne gerade unentbehrlich zu sein. So
gedeihen viele Pflanzen, z. B. der Buchweizen besser, wenn ihnen
Chloride zur Verfügung stehen, und die Kieselerde wirkt u. a. in
hohem Grade nützlich durch die größere Widerstandsfähigkeit, die
sie den Geweben verleiht. Auch hat man gefunden, daß die Gegenwart
gewisser, an sich nicht wertvoller Stoffe die Giftwirkungen anderer
zum Teil auch nötiger Stoffe aufheben kann.
Daß auch die Tiere die Salze ebenso nötig haben wie die Pflanzen,
ist bekannt. Einzelerfahrungen scheinen hier aber nur sehr wenig
vorzuliegen. Man darf vielleicht sagen, daß im Tierreich dieselben
Elemente notwendig sind wie in der Pflanze, daß aber Na und Cl dazu
treten.
Aufnahme der Nährsalze. Die Nährsalze können nur in gelöstem
Zustande von Oberflächenzellen der Pflanze aufgenommen werden. Es
erhebt sich also die Frage, wie gelöste Stoffe durch die Zellwand
und das Protoplasma bis in die Vakuole gelangen können. Es wurde ja
bei Besprechung der Plasmolyse (S. 192) hervorgehoben, daß
das Protoplasma semipermeabel, d. h. wohl für Wasser, aber
nur schwer für gewisse, in Wasser gelöste Stoffe permeabel ist. Wäre
diese Impermeabilität eine vollständige, so könnten sich im Zellinnern
unmöglich Nährsalze vorfinden; tatsächlich ist sie aber vielleicht für
keinen Stoff eine absolute, und wir kennen stufenweise Übergänge von
Stoffen, die ebenso leicht durch das Plasma dringen wie das Wasser,
bis zu solchen, die fast gar nicht durchgehen. Nicht zu langsam
eindringende Stoffe bewirken eine vorübergehende Plasmolyse.
Außerordentlich rasch diffundierende Stoffe, z. B. Alkohol, Äther,
Chloralhydrat, verursachen keine Plasmolyse.
[S. 207]
Die Permeabilität des Protoplasmas ist keine konstante. Sie wird
wenigstens für manche Stoffe durch Außenumstände verändert[159];
Alkalisalze z. B. bewirken eine zunehmende Impermeabilität für
sie selbst. Aber auch Erdalkalisalze können die Permeabilität für
Alkalisalze vermindern. Nicht das Gesamtprotoplasma, sondern nur
die äußerste Hautschicht ist für Aufnahme oder Nichtaufnahme
eines Stoffes maßgebend. Auch der Übertritt der Stoffe in den
Zellsaft wird durch eine Plasmahaut (die Vakuolenwand) reguliert. Die
Eigenschaften dieser beiden Plasmahäute bedingen also zusammen mit
dem Speichervermögen, von dem alsbald die Rede sein soll, das sog.
Wahlvermögen der Zelle; dieses äußert sich darin, daß eine Pflanze
manche in ihrer Umgebung häufige Stoffe gar nicht oder in ganz geringer
Menge aufnimmt, und daß verschiedene Spezies in der Stoffaufnahme stark
differieren.
Aus demselben Boden nimmt die eine Pflanze vorzugsweise
Kieselsäure, die andere vornehmlich Kalk, eine dritte Kochsalz
auf. Besonders lehrreich in dieser Beziehung sind die Meeresalgen,
die von einer Lösung umspült werden, die etwa 3% Kochsalz,
dagegen wenig Kalisalze enthält. Trotzdem nehmen ihre Zellen
verhältnismäßig wenig Kochsalz, dagegen reichlich Kalisalze auf.
Jeder Stoff, für den die Grenzschichten des Protoplasmas permeabel
sind, kann so lange in die Vakuole diffundieren, bis die
Konzentrationsdifferenz zwischen dieser und der Umgebung ausgeglichen
ist. Mit der Herstellung dieses physikalischen Gleichgewichtes hört
aber die Diffusion keineswegs immer auf; in nicht wenigen Fällen
findet sich schließlich eine bestimmte Substanz in der Zelle in
relativ viel größerer Menge als in der Umgebung. Das trifft z. B.
für das Jod in den Meeresalgen zu; dieses ist ja im Meer selbst nur
spurenweise, in den Tangen aber in so ansehnlicher Menge vorhanden,
daß man es aus ihnen technisch gewinnen kann. Die Zelle hat also ein
Speichervermögen, und sie speichert die Stoffe dadurch, daß
sie sie in eine unlösliche oder in eine nicht diffusible Form
bringt.
Ganz vorzüglich eignen sich gewisse organische Farbstoffe[160],
wie z. B. Methylenblau, zum Nachweis des Eindringens und der
Speicherung. Viele Zellen enthalten in den Vakuolen Gerbstoffe und
andere Stoffe, die mit dem eintretenden Farbstoff eine Verbindung
eingehen, die nicht diffusibel oder gar unlöslich ist. Demnach
sieht man auch in äußerst verdünntem Methylenblau die Vakuole tief
blau werden oder in ihr blaue Niederschläge auftreten. — Besonders
zu betonen ist, daß das Protoplasma selbst ungefärbt bleibt und
nicht im mindesten geschädigt wird; totes Plasma würde ja den
Farbstoff stark speichern.
Ein genaues Studium des Durchtritts von Farbstoffen durch
das Protoplasma hat wertvolle Einblicke in die Ursachen der
Permeabilität gestattet, ohne z. Z. zu einer allseits anerkannten
Theorie zu führen. — Vorauszuschicken ist die Bemerkung, daß nicht
alle Zellen sich gleich verhalten. So nehmen z. B. die an Gefäße
angrenzenden die sauren Farbstoffe leicht auf, während sonst fast
nur basische Farben durchgelassen werden. Da nun die basischen
Farbstoffe in den Lipoiden, d. h. in Fetten und fettähnlichen
Körpern (Lezithin, Cholesterin) löslich sind, die Säuren aber
nicht, so hat man die Vorstellung entwickelt[161], die Hautschicht
des Protoplasmas bestehe aus einer Lipoidschicht, und diese
Ansicht hat mit einigen Modifikationen auch heute noch recht viel
für sich. Denn die anderen Hypothesen, die ausgesprochen wurden,
haben sich alle schwerwiegender Einwände zu erwehren, z. B. die
Ultrafiltertheorie[162], nach der das Eindringen der Farbstoffe
lediglich von ihrer Teilchengröße abhängen soll, das Protoplasma
also wie ein Sieb wirken würde; die Haftdrucktheorie[163], nach der
die Fähigkeit eines Stoffes, die Oberflächenspannung des Wassers zu
erniedrigen, maßgebend für seine Aufnahme in die Zelle sein soll.
Wie in unseren Wasserkulturen, so nehmen auch in der Natur manche
Pflanzen die Nährsalze aus dem Wasser auf. Dies ist der Fall bei
manchen Wasserpflanzen, bei denen dann die ganze Oberfläche in den
Dienst dieser Aufnahme gestellt sein kann. Da die Nährsalze nur
spärlich in den Gewässern vorkommen, so ist es verständlich, daß
die Wasserpflanzen eine recht große[S. 208] Oberfläche zu ihrer Aufnahme
benötigen; so erklärt sich zum Teil das häufige Vorkommen fein
zerteilter Blätter bei ihnen. Aber nicht allen Wasserpflanzen genügen
die im Wasser gelösten Nährsalze. Manche nehmen mit Hilfe ihrer Wurzeln
aus dem Boden am Grunde der Gewässer Stoffe auf und gedeihen nicht,
wenn sie der Wurzeln beraubt werden.
Ganz allgemein erfolgt bei den Landpflanzen die Nährsalzaufnahme
aus dem Boden. Im Wasser des Erdbodens sind die in unserer
Nährlösung gegebenen oder ähnliche Salze stets vorhanden, aber
manche von ihnen in so minimalen Mengen, daß sie nur für kurze Zeit
Pflanzenwachstum gestatten. Wo solches längere Zeit anhält, da müssen
andere Quellen für die Nährsalze bestehen. In der Tat gibt der
Nährsalzgehalt des Bodenwassers keinen Maßstab für die Fruchtbarkeit
des Bodens ab; denn die Pflanze ist nicht auf diesen allein angewiesen,
sie kann vielmehr auch im Boden „absorbierte“ Stoffe aufnehmen
und selbst Gesteine aufschließen. Die Erschließung erfolgt in erster
Linie durch Ausscheidung von Kohlensäure aus den Wurzelhaaren. In
kohlensäurehaltigem Wasser sind viele Stoffe ungemein viel leichter
löslich als in reinem Wasser.
Die Lösung fester Gesteine durch die Pflanze läßt sich besonders
bequem an glatt polierten Marmorplatten demonstrieren, auf denen
man Wurzeln hat wachsen lassen; der Verlauf der Wurzeln wird durch
Ätzfiguren wiedergegeben.
Es fehlt nicht an Angaben, wonach solche Gesteinslösungen auch
durch stärkere Säuren als Kohlensäure zustande kommen sollen; aber
es liegt keinerlei Beweis vor, daß höhere Pflanzen
aus lebenden Zellen andere Säuren ausscheiden als CO2. Wenn so
schwer lösliche Substanzen wie Feldspat und Glimmer von gewissen
Flechten gelöst werden, so ist hier an der Ausscheidung einer
stärkeren Säure[164] aus der Pflanze um so weniger zu zweifeln, als
ja Pilze und Bakterien sehr häufig in ihrem Stoffwechsel solche
Stoffe produzieren und ausscheiden. Da nun im Boden eine große
Menge von Bakterien lebt, so müssen auch diese eine Rolle bei der
Auflösung schwer löslicher Gesteine spielen.
Auch die aus toten Wurzelhaaren und Wurzelhaubenzellen austretenden
sauren Salze dürften bei der Lösung der Gesteine von Bedeutung sein.
Manche Böden, vor allem die tonigen und humösen, nicht aber die
Sandböden, haben die Fähigkeit, Kali- und Ammoniaksalze, in
geringerem Grade auch Kalk- und Magnesiumsalze, in der Weise
festzuhalten, daß diese Stoffe zwar nicht leicht ausgewaschen
werden, aber doch der Pflanze zugänglich sind. Man schreibt dem
Boden ein Absorptionsvermögen für die genannten Stoffe zu.
Dieses besteht aber nicht für alle Nährsalze; so werden z. B.
Sulfate und Nitrate nicht absorbiert.
Wenn das Substrat der Pflanze außer Wasser und Nährsalzen auch noch
andere Stoffe, z. B. gelöste organische Stoffe enthält,
so können diese in der gleichen Weise aufgenommen werden wie die
besprochenen Substanzen. Der Erfolg der Wasserkultur beweist aber, daß
wenigstens die typische grüne Pflanze auf solche Stoffe nicht
angewiesen ist. Anders ist das bei Pilzen und anderen Pflanzen, die den
Pilzen in ihrem Stoffwechsel gleichen (S. 219).
Neben Wasser und den Nährsalzen können auch gelöste Gase durch die
Wurzel aufgenommen werden; von diesen wird im allgemeinen nur dem
Sauerstoff eine Bedeutung zukommen. Hauptquelle für die Gasaufnahme
bildet die Atmosphäre.
Wanderung der Nährsalze. Die aufgenommenen Nährsalze bleiben
nicht in den Epidermiszellen der Wurzel bzw. des Sprosses, sondern sie
wandern von der Aufnahmestelle aus durch die ganze Pflanze. Für
diese Wanderungen gibt es zwei Wege, den der Diffusion
und den der Massenströmung. Bedingung für die Diffusion ist,
daß Zellwand und Protoplasma für die betreffende Substanz permeabel
sind, und daß ein Konzentrationsunterschied zwischen dem Ausgangspunkt
und dem Ziel der Wanderung besteht. Bei dem Übertritt von Stoffen
aus einer Vakuole in die Vakuole der Nachbarzelle müssen diese
zunächst ins Protoplasma, dann in die Zellwand, dann wieder[S. 209] ins
Protoplasma und endlich in die Vakuole übertreten. Eine besondere
Schwierigkeit bei dieser Wanderung scheint die Zellwand zu bieten,
wenigstens wenn sie dick ist. Dementsprechend sind alle dickeren
Zellhäute mit Tüpfeln versehen, und die Schließhäute der Tüpfel sind
noch von zarten Protoplasmafäden (Plasmodesmen, S. 36) durchsetzt;
ja bei gewissen Elementen der Pflanze, den Siebröhren, werden die
Tüpfelschließhäute resorbiert, und es gehen dementsprechend dickere
Stränge von Protoplasma in die Nachbarzelle hinüber. Die Untersuchungen
von BROWN und ESCOMBE haben ergeben, daß eine
feinperforierte Wand der Diffusion keinen wesentlichen Widerstand
leistet.
Auch innerhalb der Zelle können Diffusionsbewegungen eintreten, wenn
gelöste Stoffe nicht überall gleiche Konzentration aufweisen. Jede
Diffusionsbewegung verläuft nun recht langsam. Ein mg NaCl z. B.
braucht, um aus 10%iger Lösung einen Meter weit in Wasser zu
diffundieren, fast ein Jahr; andere Stoffe diffundieren noch erheblich
langsamer. Die Geschwindigkeit der Mischung kann aber erheblich
beschleunigt werden, wenn zu der molekularen Diffusionsbewegung
eine Massenströmung hinzutritt, wie wir sie im gewöhnlichen Leben
und im Laboratorium durch Umrühren herstellen. Eine solche Mischung
innerhalb der Zelle kann z. B. durch die Protoplasmabewegung erzielt
werden. — Je länger nun eine Zelle ist, desto geeigneter wird sie zum
Stofftransport durch die Pflanze, weil dann nur noch selten, nämlich
an den Enden der Zelle, die langsame Diffusionsbewegung eintreten muß,
während im übrigen Massenbewegungen Platz greifen können.
Wo es der Pflanze auf besonders lebhafte Stoffbewegung ankommt,
da verwendet sie Massenströmungen in Röhren, die sie sich
durch Auflösung von Querwänden in ganzen Zellzügen herstellt
(Tracheen), und so sehen wir, daß die Nährsalze innerhalb
der Pflanze mit dem Transpirationsstrom wandern. Bei der
„Wasserbewegung“ in den Leitbündeln wird demnach kein reines
Wasser, sondern eine sehr verdünnte Nährlösung transportiert, und
die Transpiration hat in erster Linie den Erfolg, diese Nährlösung
einzuengen, in zweiter Linie, sie rasch überallhin zu führen. Sonst
wäre die Transpiration schwer verständlich, und die Pflanze hätte sie
gewiß allgemein eingeschränkt. Da wo sie tatsächlich eingeschränkt ist
(vgl. S. 144), haben wir es mit Pflanzen zu tun, die wegen geringer
Versorgung mit Nährsalzen und auch wohl mit Kohlensäure nur langsam
wachsen. — Erwähnt muß freilich werden, daß offenbar auch gelegentlich
ein Überfluß an Nährsalzen im Innern der Pflanze auftritt, der
schädliche Wirkungen ausüben kann, und von dem die Pflanze vor allem
durch Guttation (S. 200) sich zu befreien sucht.
Nährsalze und Landwirtschaft. Wenn die Pflanze somit immer neue
Nährsalze aus dem Boden aufnimmt, so muß dieser allmählich an diesen
Stoffen verarmen, wenn nicht für Ersatz gesorgt wird. In der Natur
geschieht das dadurch, daß die absterbenden und abfallenden Teile
wieder auf und in die Erde gelangen und ihren Gehalt an Nährsalzen neu
aufkommendem Leben zur Verfügung stellen. Bei landwirtschaftlichem
Betrieb aber werden große Mengen Pflanzensubstanz mit der Ernte
entfernt, und ihr Nährsalzgehalt geht dem Acker verloren; höchstens
ein Bruchteil davon kehrt mit dem Miste der Tiere dahin zurück. Auf
dem Gehalte des Mistes an Nährsalzen beruht wenigstens zum Teil
seine wachstumsfördernde Wirkung, die dem Praktiker seit alten Zeiten
bekannt ist. Da indes diese Nährsalze der Quantität nach nicht genügen,
um den Verlust zu decken, muß die Landwirtschaft zur künstlichen
Düngung[165] greifen. Unter den Düngestoffen sind vor allen Dingen
solche wichtig, die Stickstoff, Kalium und Phosphorsäure enthalten.
Als stickstoffhaltige Substanzen finden neben dem Guano (der auch
Phosphorsäure enthält) Chilisalpeter, schwefelsaures Ammonium[S. 210]
und die neuerdings künstlich aus Luftstickstoff dargestellten
Verbindungen, Kalziumcyanamid, salpetersaurer Kalk und vor allem
Ammoniumbikarbonat und Harnstoff Verwendung. Kaliumsalze
verschiedener Art werden für landwirtschaftliche Zwecke in großen
Mengen bergmännisch gewonnen; das wichtigste unter ihnen ist wohl der
Karnallit. Als Phosphorsäurequellen dienen die Thomasschlacke und das
Superphosphat. Erstere ist tertiärer phosphorsaurer Kalk, der bei
Verhüttung phosphorhaltiger Erze entsteht und äußerst fein zermahlen
als „Thomasmehl“ verwendet wird. Superphosphat wird durch Aufschließen
von phosphorsaurem Kalk mit Schwefelsäure gewonnen.
Boden und Pflanzengeographie. Nach den mitgeteilten Erfahrungen
sollte man glauben, daß ein Boden, der eine Pflanzenspezies zu
ernähren vermag, auch für jede andere geeignet sein müßte. Tatsächlich
zeigt uns aber die Pflanzengeographie[166], daß die Beschaffenheit
des Bodens einen weitgehenden Einfluß auf die Verteilung der Pflanzen
ausübt. Das hängt damit zusammen, daß verschiedene Pflanzen an die
Menge und Löslichkeit der notwendigen Stoffe verschiedene Ansprüche
machen, ferner damit, daß neben den nötigen auch überflüssige
Verbindungen in einem Boden sein können, die auf die einzelnen Spezies
ganz verschieden wirken. CaCO3 z. B. und ebenso NaCl wirken auf
manche Pflanzen giftig, während andere große Dosen dieser Stoffe
ertragen können.
Es kommt indes bei der Pflanzenverteilung in ihrer Abhängigkeit
vom Boden keineswegs bloß auf chemische Verhältnisse an. Auch die
physikalischen Eigenschaften der Böden spielen eine große Rolle.
Endlich kann auch eine Pflanze an einem Ort, der ihr an sich
zusagen würde, deshalb fehlen, weil ihre Samen nie dahin gelangt
sind.
c) Gase.
Wasser und Nährsalze werden, wie wir gesehen haben, im allgemeinen
dem Boden entnommen, aber auch die Luft enthält Stoffe, die notwendig
zum Gedeihen vieler Pflanzen sind, die also Nährstoffe genannt werden
müssen. Es sind das die Kohlensäure und der Sauerstoff. Diese werden
denn auch im allgemeinen aus der Luft aufgenommen; nur die submersen
Wasserpflanzen beziehen sie aus dem Wasser. Im letzteren Falle erfolgt
ihre Aufnahme genau so wie die von anderen gelösten Stoffen.
Sauerstoff. Entzieht man einer Pflanze den Sauerstoff, so werden
meist alle Lebensäußerungen sistiert (vgl. S. 257). Diese Tatsache
erscheint nicht wunderbar, da auch dem menschlichen Organismus der
Sauerstoff unentbehrlich ist. (Weiteres s. S. 240.)
Kohlensäure. Viel weniger plausibel ist es dagegen für den
Laien, daß auch die Kohlensäure der Pflanze unentbehrlich sein
soll. Und doch ist das so. Wir setzen einer Wasserkultur keinerlei
Kohlenstoffquelle zu, die Pflanze vermehrt aber trotzdem ihren
Kohlenstoffgehalt; da bleibt keine andere Möglichkeit, als daß sie die
Kohlensäure der Luft benützt. Kohlensäure ist in der gewöhnlichen Luft
in einer Menge von etwa 0,3 pro Mille enthalten; ein Liter enthält
also 0,3 ccm. Läßt man diese Luft über grüne Pflanzen streichen, die
an hellem Licht stehen, so zeigt sich, daß ihr Kohlensäuregehalt
abnimmt oder verschwindet; farblose Teile, wie Wurzeln, oder
nichtgrüne Organismen, wie die Pilze, verhalten sich anders, sie
absorbieren keine Kohlensäure. Wird nun ein grüner Pflanzenteil in
einer Glasglocke gehalten, durch die man kohlensäurefreie
Luft hindurchleitet, so stellt er bald sein Wachstum ein, und jede
Vermehrung der Trockensubstanz hört völlig auf. Kohlensäure ist demnach
ein unentbehrlicher Nährstoff; sie ist offenbar die Quelle für den
Kohlenstoff der grünen Pflanze. Die in der Luft vorhandene prozentual
geringe Menge reicht zur Ernährung dieser völlig aus (S. 216). Es zeigt
sich[S. 211] ferner, daß die Zufuhr von organisch gebundenem Kohlenstoff zum
Boden oder zur Nährlösung die Kohlensäure der Luft nicht entbehrlich
macht; für die grüne Pflanze, die wir einstweilen allein im Auge
haben, ist jedenfalls CO2 die beste Kohlenstoffquelle. Auch genügt
es durchaus nicht, wenn wir einer solchen Pflanze die Kohlensäure im
Boden oder in der Nährlösung zur Verfügung stellen. Sie muß unbedingt
den Blättern geboten werden.
Andere Gase. Sauerstoff und Kohlensäure sind die einzigen Gase,
die den gewöhnlichen Pflanzen notwendig sind. Der atmosphärische
Stickstoff dagegen ist für die Mehrzahl der Pflanzen ohne jede
Bedeutung (vgl. S. 224).
Aufnahme der Gase. Kohlensäure und Sauerstoff dringen zum Teil
in die Epidermiszellen, zum größeren Teil aber durch die Spaltöffnungen
in die Interzellularen und von diesen aus in die inneren Gewebe.
Weder in der Zellwand noch im Protoplasma existieren lufterfüllte Räume
oder Kanäle, durch die Gase ins Zellinnere gelangen können. Deshalb
ist eine Aufnahme der Gase nur in dem Grade möglich, als sie in der
Zellhaut und im Protoplasma bzw. in dem diese Teile durchdringenden
Wasser löslich sind. Die gelösten Gase verhalten sich dann gerade
wie andere gelöste Stoffe, sie diffundieren in die Zelle. Je
wasserreicher eine Zellhaut ist, desto leichter diffundieren die Gase
durch sie. Die gewöhnliche Zellhaut ist in trockenem Zustande für Gase
auf dem Wege der Diffusion nur sehr wenig durchlässig[167]; allein in
der Natur findet sie sich ja immer mehr oder weniger wassergesättigt
vor. Dagegen ist die Kutikula sehr wenig imbibitionsfähig für Wasser
und stellt deshalb dem diosmotischen Gasdurchtritt erhebliche
Schwierigkeit entgegen; doch ist sie keineswegs völlig undurchlässig.
Der Hauptsache nach erfolgt also die Gasdiffusion gar nicht durch die
eigentliche Zellhautsubstanz, sondern durch die Stoffe, mit denen diese
imprägniert ist. Da nun Kohlensäure in Wasser viel leichter löslich
ist als Sauerstoff, so ist es begreiflich, daß sie auch schneller
die wassergetränkte Zellhaut durchsetzt als der Sauerstoff. Aller
Wahrscheinlichkeit nach gilt das gleiche auch bei der Kutikula. Da
aber die Partiärpressung des Sauerstoffes in der Luft eine relativ
ansehnliche, die der Kohlensäure dagegen eine recht geringe ist, so
kann tatsächlich zwar Sauerstoff, nicht aber Kohlensäure in genügender
Menge durch die Kutikula hindurch, und deshalb sehen wir, daß fast
alle Organe, die nur Sauerstoff aufzunehmen haben, ohne Spaltöffnungen
sind, daß dagegen an den Kohlensäure aufnehmenden Organen stets
Spaltöffnungen sich vorfinden.
Nicht nur in der Luft, sondern auch im Boden finden die Pflanzen in
der Regel so viel Sauerstoff, daß er in genügender Menge durch die
Epidermiszellen eindringen kann. Eine Ausnahme bilden nur solche
Organe, die in sauerstoffarmem Sumpfboden leben. Gehören diese
einer Pflanze an, deren Sprosse sich in der Luft ausbreiten, so
erfolgt die Sauerstoffaufnahme in der Regel in diesen Luftsprossen;
weite Interzellularen, die stets bei den Sumpfpflanzen ausgebildet
sind, sorgen dann dafür, daß der Sauerstoff auch den unterirdischen
Teilen zuströmen kann. Manchmal aber (besonders bei Palmen und
Mangrovepflanzen) werden besondere, aus dem Boden aufsteigende
Atemwurzeln (Pneumatophoren, Fig. 186) ausgebildet,
die an der Luft Sauerstoff aufnehmen.
Die Spaltöffnungen sind für den Gasaustausch je nach der
Öffnungsweite der Spalte sehr verschieden leistungsfähig. Ihr
Verschluß, der aus Gründen der Wasserversorgung unter Umständen
nötig ist, hat nicht nur die Aufhebung der Transpiration zur Folge,
sondern er unterbindet auch das Eindringen von CO2 in die
Pflanze.
Es ist schon bei Gelegenheit der Wasserdampfabgabe darauf
hingewiesen worden, daß die Spaltöffnungen trotz ihrer geringen
Größe durch die enorme Zahl und dichte Anordnung die Diffusion
fördern. Das gilt auch für die Aufnahme der Kohlensäure.
Dementsprechend konnte denn auch gezeigt werden, daß beispielsweise
ein Quadratmeter Blattfläche von Catalpa in der Zeiteinheit
etwa 2⁄3 der Kohlensäuremenge absorbiert,[S. 212] die ein Quadratmeter
3–10%iger Natronlauge, frei der Luft ausgesetzt, dieser
entreißt.
Die Bewegung der Gase von Zelle zu Zelle sowie ihr Austausch
zwischen den Zellen und den Interzellularen erfolgt durch
Diffusion; bei ihrer Bewegung in den Interzellularen können aber
auch Massenströmungen erfolgen, wenn Druckzustände bestehen. Solche
werden aber in der Interzellularenluft durch Erwärmung und Abkühlung
oder durch Deformation der Pflanze erzielt. Die Interzellularen
sind ein weitverzweigtes System untereinander und mit der
Atmosphäre kommunizierender Lufträume. Ihre Austrittsöffnungen
werden in erster Linie durch die Spaltöffnungen, daneben auch durch
Lentizellen und ähnlich funktionierende Organe gebildet (S. 43, 50);
durch sie erfolgt ebensowohl Diffusion wie Massenbewegung der Gase.
Der Zusammenhang der Interzellularen unter sich und mit der
Außenwelt läßt sich experimentell leicht feststellen. Es gelingt
nämlich, Luft aus den Spaltöffnungen oder den Lentizellen austreten
zu lassen, wenn man sie in die Interzellulargänge einpreßt;
auch umgekehrt treten bei mäßiger Saugung aus den freigelegten
Interzellularen dauernd große Mengen von Luft heraus, die nur durch
Spaltöffnungen und Lentizellen ihren Weg in die Pflanze gefunden
haben können. Das S. 199 geschilderte Porometer läßt die Wegsamkeit
der Spaltöffnungen und der Interzellularen für Luft besonders
deutlich hervortreten.
In hervorragendem Maße sind luftführende Interzellularräume bei
Wasserpflanzen und Sumpfgewächsen entwickelt (vgl. S. 142), wo
sie oft bis zu zwei Drittel des Gesamtvolumens einnehmen. Die
untergetauchten Wasserpflanzen verschaffen sich auf diese Weise
eine Binnenatmosphäre, mit der ihre Zellen einen lebhaften
Gaswechsel unterhalten können, da die Binnenatmosphäre durch
Diffusionsvorgänge wieder mit der Umgebung in langsamem Austausch
steht. Im übrigen sind diese Pflanzen bei ihrem Gaswechsel ganz
auf Diffusionsvorgänge angewiesen, da ihnen Spaltöffnungen usw. zu
fehlen pflegen. Aber auch Pflanzen, die Spaltöffnungen besitzen,
können daneben auf diosmotischem Wege Gase gewinnen, vorausgesetzt,
daß die Kutikula ihrer Epidermis gasdurchlässig ist.
III.
Die Assimilation der Nährstoffe.
Die Pflanze wächst, sie bildet beständig neue Organe, und zur
Herstellung dieser bedarf sie fortwährend neuer Nährstoffe. Die
Nährstoffe ihrerseits werden aber nach ihrer Aufnahme verändert, es
geht aus ihnen Pflanzensubstanz hervor; sie werden assimiliert,
wie man zu sagen pflegt. Unter Assimilation eines Stoffes versteht
man dementsprechend seine Überführung in Pflanzensubstanz. Besonders
interessant sind solche Assimilationsvorgänge, bei denen tiefgreifende
Veränderungen erfolgen, z. B. Überführung von anorganischen in
organische Verbindungen, namentlich dann, wenn es uns mit künstlichen
Mitteln noch nicht gelingt, die entsprechenden Reaktionen auch
außerhalb des Organismus herbeizuführen. Ein solcher Prozeß ist z. B.
die Assimilation des Kohlenstoffes bei der grünen Pflanze; hier geht
aus Kohlensäure organische kohlenstoffhaltige Substanz hervor.
A. Assimilation des Kohlenstoffes.
1. Assimilation der Kohlensäure bei grünen Pflanzen.
In Wasserkultur nimmt die Trockensubstanz einer grünen Pflanze zu;
diese aber besteht zur Hälfte aus Kohlenstoff, den wir der Nährlösung
nicht zugesetzt haben. Sie bezieht ihn aus der Kohlensäure der Luft;
sie führt die CO2 unter Mitwirkung des Sonnenlichtes in Zucker über.
Dabei wird auch Wasser mitverarbeitet und Sauerstoff ausgeschieden; der
Prozeß vollzieht sich also nach der Formel
6 CO2 + 6 H2O = C6H12O6 + 6 O2.
[S. 213]
Nimmt man an, daß die Kohlensäure der Luft (das Kohlendioxyd, CO2)
bei ihrer Lösung in der Zelle durch Wasseraufnahme zu H2CO3 wird,
so dürfte sich die Zuckerbildung in zwei Hauptschritten vollziehen;
beim ersten wird Sauerstoff abgespalten und Formaldehyd[168] gebildet:
H2CO3 = H2CO + O2,
beim zweiten wird der Aldehyd zu Zucker kondensiert:
6 H2CO = C6H12O6.
Es muß also für jedes verschwindende Volum Kohlensäure das gleiche
Volum Sauerstoff auftreten. In der Tat haben eudiometrische
Bestimmungen gezeigt, daß das zutrifft (WILLSTÄTTER). Der
auftretende Sauerstoff kann aber, auch wenn er nur qualitativ
nachgewiesen wird, als Indikator der Kohlensäurezerlegung dienen. So
kann man z. B. eine in sauerstoffreiem Raum befindliche Pflanze bei
Gegenwart von Phosphor dem Licht exponieren; die dann auftretenden
Phosphordämpfe beweisen die Bildung von Sauerstoff. Oder man kann
aus der Bewegung gewisser Bakterien, die zuvor unbeweglich im Wasser
lagen, auf die Sauerstoffproduktion einer grünen Pflanze schließen
(S. 290). Am anschaulichsten aber ist der Assimilationsnachweis mit
Hilfe gewisser Wasserpflanzen, wie Helodea oder Potamogeton. Werden
abgeschnittene, unter Wasser befindliche Sprosse oder Blätter dieser
Pflanzen dem Licht exponiert, so sieht man alsbald einen lebhaften,
kontinuierlichen Strom von Blasen der Schnittfläche enteilen. Fängt man
das Gas in einem Reagenzglas (Fig. 244) in größerer Menge auf, so zeigt
sich, daß es zwar keineswegs aus reinem Sauerstoff, aber doch immerhin
aus einem sauerstoffreichen Gemisch besteht; ein glimmender Holzspahn
beginnt in ihm zu brennen.
Das Auftreten von Sauerstoffblasen erklärt sich in folgender Weise.
Die im Wasser gelöste Kohlensäure gelangt durch Diffusion in die
grünen Zellen der Pflanze und wird da zerlegt. Der auftretende
Sauerstoff ist viel weniger löslich als die Kohlensäure, und
deshalb wird er in Gasform abgeschieden. Er tritt in die
Interzellularräume, bewirkt hier eine Vermehrung des Druckes,
und diese wird die Ursache des Austretens von Gasblasen an jeder
Schnittstelle.
Fig. 244. Ausscheidung von Sauerstoff durch
assimilierende Pflanzenteile. In dem Glaszylinder C befindet
sich in Wasser eine frisch abgeschnittene Wasserpflanze (Helodea
canadensis). Die Schnittflächen sind in ein ebenfalls mit Wasser
gefülltes Probierröhrchen R eingeführt. Die ausgeschiedenen
Sauerstoffbläschen B steigen darin auf und sammeln sich oben bei S.
Nach NOLL.
Die Grundlagen unserer Kenntnisse über die CO2-Assimilation der
grünen Pflanze wurden durch die Forschungen von PRIESTLEY,
INGENHOUSZ, SENEBIER und Th. de SAUSSURE
am Ende des 18. und am Anfang des 19. Jahrhunderts gewonnen. Diese
Entdeckung ist außerordentlich bedeutsam, weil die Bildung
organischer Substanz aus Kohlensäure durch die grüne Pflanze der Prozeß
ist, der allen anderen Organismen, vor allem also auch den Tieren, das
Leben auf der Erde ermöglicht (vgl. S. 220).
[S. 214]
Mit Hilfe der Gasblasenmethode läßt sich leicht der Beweis für die
oben aufgestellte Behauptung erbringen, daß nur chlorophyllführende
Pflanzenteile, und diese nur im Licht imstande sind, CO2 zu
assimilieren. So sieht man z. B. den Blasenstrom einer Helodea, der am
hellbeleuchteten Fenster lebhaft war, in dem Maße, als man die Pflanze
in die Tiefe eines Zimmers bringt, langsamer werden und schließlich
bei einer Lichtintensität erlöschen, die unserem Auge noch gut das
Lesen erlaubt. Innerhalb gewisser Grenzen läuft die Assimilation der
Lichtintensität proportional. Entsprechende Versuche kann man auch mit
künstlichen Lichtquellen ausführen; sie zeigen, daß alle gebräuchlichen
Beleuchtungsarten bei der CO2-Assimilation verwendbar sind. Die
Strahlen verschiedener Wellenlänge haben aber nicht alle die gleiche
Bedeutung für die CO2-Assimilation[169].
Um die Assimilation im Licht verschiedener Wellenlängen zu
untersuchen, bedient man sich entweder spektral zerlegten oder
durch Absorption monochromatisch gemachten Lichtes. Zur Absorption
können die von Schott u. Gen. hergestellten Rot- und Blaugläser
verwendet werden, oder man benutzt doppelwandige Küvetten, deren
Wandraum mit passenden Lösungen gefüllt ist.
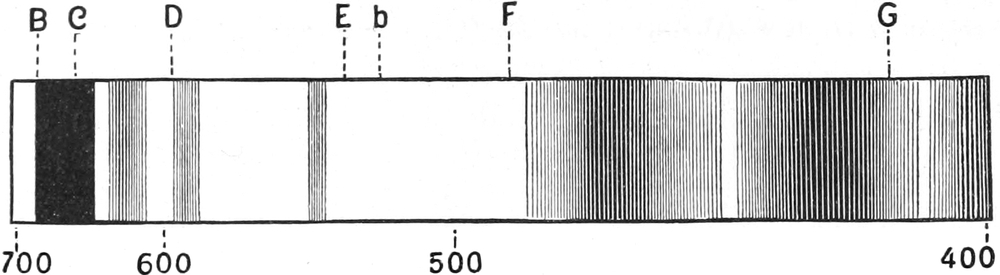
Fig. 245. Absorptionsspektrum des Chlorophylls nach
GR. KRAUS. Oben sind die FRAUNHOFERschen Linien B,
C usw. angegeben, unten die Wellenlänge 700–400 μμ. An den dunkeln
bzw. schraffierten Stellen ist das Licht ausgelöscht bzw. geschwächt.
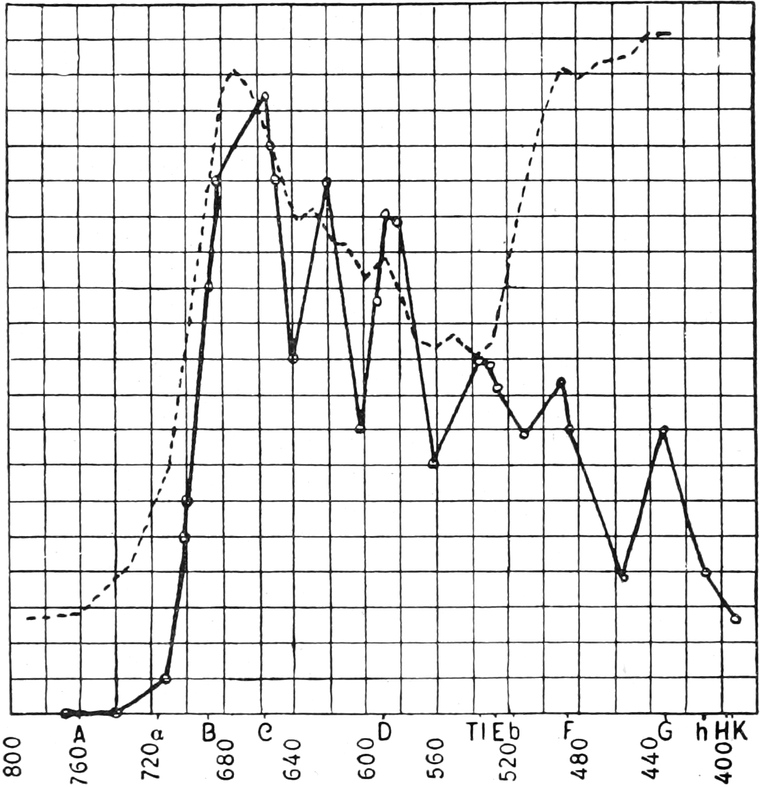
Fig. 246. Die ausgezogene Linie zeigt die Abhängigkeit
der Stärkebildung von der Wellenlänge des Lichtes, wenn die
auffallende Lichtenergie für alle Strahlen gleich ist. Die punktierte
Linie stellt die vom grünen Farbstoff lebender Blätter absorbierte
Strahlung dar, wobei die eintretende Strahlung = 100 gesetzt wird.
Auf der Abszisse sind die Wellenlängen des Lichtes in μμ und einige
FRAUNHOFERsche Linien angegeben (Gitterspektrum). Nach
URSPRUNG.
Im großen und ganzen ist die assimilatorische Wirksamkeit auf die
sichtbaren Strahlen von rund 0,4–0,8 μ beschränkt, doch ist
sicher nachgewiesen, daß auch ultrarote und ultraviolette verwertet
werden können. Wie sich die einzelnen Wellenlängen verhalten,
ist sehr viel untersucht, aber bis heute doch nicht ganz sicher
entschieden. Zweifellos hat das Licht von 0,68 μ, das maximal
absorbiert wird, auch eine besonders starke assimilatorische
Wirkung. Nach ENGELMANN soll allgemein eine weitgehende
Übereinstimmung zwischen Lichtabsorption im Chlorophyll und
Assimilation bestehen. URSPRUNG hat Bohnenblätter[S. 215] in
jeweils zwei Ausschnitten eines Spektrums assimilieren lassen,
die auf gleiche Lichtenergie abgestimmt waren, und hat so die
Stärkebildung in allen Wellenlängen untersucht. Seine Resultate
sind in der Fig. 246 dargestellt; es zeigt sich der erwartete
Parallelismus, der freilich in Blau und Violett aufhört, weil da
durch Verschluß der Spaltöffnungen die Zufuhr an CO2 zu gering
wurde. KNIEP und MINDER haben die Assimilation
in verschiedenfarbigem Licht mit der Gasblasenmethode untersucht:
sie fanden, daß blaues Licht ebenso wirksam ist wie rotes, wenn es
gleiche Energie liefert.
Auch gewisse als Purpurbakterien bezeichnete Organismen, die zwar
kein Chlorophyll, wohl aber andere eigenartige Farbstoffe führen,
zerlegen CO2; anscheinend nützen sie dabei gerade die Strahlen
am meisten aus, die das Chlorophyll am wenigsten absorbiert und
verwertet[170].
Wenn das Sonnenlicht in der Natur ein unentbehrlicher Faktor bei der
CO2-Assimilation ist, dann wird es erst voll verständlich, warum die
Laubblätter, deren wesentlichste Funktion in der CO2-Assimilation
besteht, flächenförmig ausgestaltet sind. Ihre große Oberfläche
gestattet ihnen, viel Licht zu absorbieren. Und nicht nur eine große
Fläche, sondern auch eine geringe Dicke müssen die Laubblätter haben,
wenn sie ihrer Funktion gut nachkommen sollen. In der Tat zeigte sich
in Versuchen WILLSTÄTTERs, daß Sonnenlicht, das durch zwei
Laubblätter hindurchgegangen ist, keine assimilatorische Wirkung mehr
auszuüben vermag. Die Blätter müssen aber außerdem viel Chlorophyll
enthalten. Daß das zutrifft, zeigt schon die dunkelgrüne Farbe der
Blätter. Ein Blick ins Mikroskop bestätigt es. Die Stengel haben
viel weniger Chloroplasten als die Blätter; die Wurzeln und andere
unterirdische Organe haben keine.
Jeder Versuch zeigt nun, daß chlorophyllfreien Organen das Vermögen
der CO2-Assimilation gänzlich abgeht. Das gilt nicht nur für die
äußerlich sichtbaren groben Organe der Pflanze, sondern auch für die
feinsten Teile der Zelle. Der Zellkern, das farblose Protoplasma
geben im Sonnenlicht keinen Sauerstoff ab — das kann man mit Hilfe
von Bakterien (S. 213) leicht nachweisen. Nur die Chloroplasten, und
diese nur wenn sie „Chlorophyll“ führen, sind die tätigen Organe
der CO2-Assimilation; etiolierte, panaschierte und chlorotische
Chloroplasten sind nicht funktionsfähig.
Bei rotblätterigen Abarten grüner Pflanzen (Rotbuche, Rotkohl)
ist das Chlorophyll ganz wie in den grünen Stammformen entwickelt
und nur durch einen roten Farbstoff in der Epidermis oder in
tieferliegenden Zellen verdeckt. Über die Farbstoffe der Rotalgen,
Braunalgen und Diatomeen vgl. S. 15.
Nur ein verhältnismäßig geringer Teil, wenige Prozente, des
einfallenden und absorbierten Lichtes wird übrigens bei der
CO2-Assimilation verwendet[171]. Daß aber Licht als solches bei
der CO2-Assimilation verschwinden muß, ist klar; denn woher
anders sollte der Energiegehalt der organischen Substanz, die im
Assimilationsprozeß erzeugt wird, stammen, als aus der Lichtenergie.
Tatsächlich dient diese in der organischen Substanz enthaltene Energie
der Pflanze zur Unterhaltung ihres Lebensgetriebes, und auch die durch
unsere Dampfmaschinen erzeugten Kräfte sind auf die Assimilationsarbeit
jener Pflanzen zurückzuführen, deren Holz oder deren verkohlte Reste
(Kohle, Braunkohle, Torf) unter dem Maschinenkessel verbrennen.
Denn beim Verbrennen dieser reduzierten Kohlenstoffverbindungen zu
Kohlensäure wird nur die Energie wieder frei, die zuvor bei der
Überführung der Kohlensäure in diese Brennstoffe gebunden wurde.
Demnach handelt es sich bei der Bildung organischer Substanz nicht
nur um den Gewinn von unentbehrlichen Baustoffen, sondern auch von
Energie in einer Form, die eine leichte spätere Verwendung auch
an anderen Orten ermöglicht (S. 237).
Da bei der CO2-Assimilation ein Energieaufwand nötig ist, so
wird die S. 212 aufgeführte Formel vollständiger so geschrieben:
6 CO2 + 6 H2O + 684000 cal = C6H12O6 + 6 O2.
[S. 216]
Nach den Untersuchungen von BROWN und PURIEWITSCH
kann man annehmen, daß ein Blatt in der Natur etwa 80% des
auffallenden Sonnenlichtes absorbiert; davon wird weitaus der
größte Teil zur Erwärmung verwendet, denn nur etwa 0,5–6%
dienen der Assimilation. Unter besonderen Versuchsbedingungen,
bei schwachem, gelbem Licht, das voll absorbiert wurde, konnte
WARBURG eine Ausnützung bis zu 70% für assimilatorische
Zwecke feststellen[172].
Wie jede Lebensfunktion ist auch die Assimilationstätigkeit eines
Chloroplasten von einer ganzen Menge von inneren und äußeren
Faktoren abhängig. Zu den inneren gehört das Vorhandensein des
Chlorophyllfarbstoffes und seine Einlagerung in einen lebendigen
Chloroplasten; der Farbstoff als solcher, aus der Pflanze herausgelöst,
vermag so wenig die Kohlensäure zu zerlegen wie ein Chloroplast, der
den grünen Farbstoff aus irgendwelchen Gründen nicht entwickelt hat (im
Dunkeln gebildeter, in eisenfreier Nährlösung entstandener Chloroplast,
Leukoplasten der unterirdischen Teile oder der Epidermis), oder ihn
verloren hat (Chromoplast). Da aber die Assimilation keineswegs
immer dem Chlorophyllgehalt proportional erfolgt, muß man mit
WILLSTÄTTER[173] annehmen, daß neben dem Chlorophyllfarbstoff
noch ein anderer Faktor wesentlich ist, mag das nun das Protoplasma der
Chloroplasten oder ein in ihm enthaltenes Enzym (S. 229) sein.
Von äußeren Faktoren ist vor allem das schon besprochene Sonnenlicht
zu nennen. Nächst ihm dann die Gegenwart von Kohlensäure. Da diese
nur in geringer Menge in der Luft vorhanden ist und durch andere
Verbindungen nicht ersetzt werden kann (auch nicht durch Kohlenoxyd),
so müßte das Leben der Pflanze und damit die Existenz aller Organismen
schließlich aufhören, wenn nicht fortwährend neue Kohlensäure auf der
Erde entstände. Man schätzt die Menge des Kohlensäurevorrates in der
Luft auf 2100 Billionen kg, die Menge der Kohlensäure, die jährlich von
den grünen Pflanzen des Festlandes konsumiert wird, auf 50–80 Billionen
kg; demnach würden die Pflanzen in einigen 30 Jahren diesen Vorrat
erschöpfen[174].
Die Luft erhält andauernd große Kohlensäuremengen durch die Atmung
von Organismen, durch Verbrennung von Holz und Kohle und durch
vulkanische Tätigkeit. Ein erwachsener Mensch atmet täglich
etwa 900 g Kohlensäure (245 g C) aus, die ganze Menschheit, zu
1400 Millionen gerechnet, also allein schon etwa 1200 Millionen
Kilo CO2 (340 Millionen Kilo C). Weitere große Massen von
CO2 liefern die übrigen Tiere, dann aber auch die Pflanzen,
insbesondere Pilze und Bakterien (vor allem die Bodenbakterien).
Die aus sämtlichen Schornsteinen der Erde entweichende Kohlensäure
liefert enorme Werte, da allein in Deutschland im Jahre 1911 neben
73 Millionen Tonnen Braunkohlen 161 Millionen Tonnen Steinkohlen
gefördert wurden; letztere würden schon etwa 400000 Millionen
Kilogramm Kohlensäure ergeben, also etwa 1⁄5000 der Gesamtmenge der
in der Atmosphäre enthaltenen CO2.
Die Festlegung der Kohlensäure durch die grüne Pflanze und die
Neuentstehung durch die angeführten Prozesse scheint annähernd zu
einem Gleichgewicht zu führen. Man findet fast stets etwa 3 Liter
CO2 in 10000 Liter Luft; im Winter ist bei uns die Menge etwas
größer (3,0–3,6 Liter) als im Sommer (2,7–2,9 Liter). (Die Luft des
Bodens ist wegen der Tätigkeit der Bakterien CO2-reicher.) Diese
3 Liter CO2 wiegen etwa 7 g; davon sind aber 8⁄11 Sauerstoff
und nur 3⁄11 Kohlenstoff. In den 10000 Liter Luft sind demnach
nur 2 g Kohlenstoff enthalten. In einem Baume von 100 Zentnern
Trockengewicht sind etwa 50 Zentner oder 2500 Kilo Kohlenstoff
angesammelt. Um diesen zu erlangen, muß der Baum also etwa 1250000
× 10000 Liter = etwa 12 Millionen Kubikmeter Luft von ihrer
Kohlensäure befreit haben. Bei der Berücksichtigung solcher Zahlen
findet man es begreiflich, daß die Entdeckung INGENHOUSS’
ungläubig aufgenommen, später ganz zurückgewiesen und vergessen
wurde. Erst LIEBIG brachte sie in Deutschland wieder zur
Geltung, und heute steht sie über allen Zweifel erhaben da. Die
angeführten Zahlen haben aber nichts Ungeheuerliches, wenn man
bedenkt, daß trotz des geringen Prozentgehaltes der Atmosphäre an
Kohlensäure sich der tatsächlich vorhandene Vorrat auf etwa 2100
Billionen Kilo berechnet, in denen 560 Billionen kg Kohlenstoff
enthalten sind. Den[S. 217] Pflanzen steht aber der ganze Vorrat des
Luftmeeres zur Verfügung, da sich die Kohlensäure durch Diffusion
und Luftströmungen immer wieder gleichmäßig ausbreitet.
Nach SCHRÖDER soll in den Landpflanzen der Erde der
Kohlenstoff von 1100 Billionen Kilogramm CO2 festgelegt sein,
also etwa die Hälfte des in der Luft enthaltenen; fast 90% davon
kommt auf das Holz der Bäume. Die Tierwelt scheint ganz
außerordentlich viel weniger Kohlenstoff zu enthalten, etwa 1% des
in den Pflanzen angesammelten.
Submerse Wasserpflanzen nehmen die im Wasser gelöste
Kohlensäure auf. Hier schwankt ihre Menge in sehr beträchtlicher
Weise je nach der Temperatur. Bei 15° C enthält der Liter
Wasser ungefähr ebensoviel CO2 als ein Liter atmosphärischer
Luft. Neben der Kohlensäure spielen aber auch die gelösten
Bikarbonate, die in Karbonat und CO2 dissoziieren, eine
wichtige Rolle für den Kohlenstoffgewinn der Wasserflora. Eine
künstliche Bereicherung des Wassers durch Einleiten von Kohlensäure
vermehrt bis zu einem gewissen Grade die Assimilation.
Die geringe Menge der in der Luft enthaltenen Kohlensäure macht eine
große aufnehmende Fläche notwendig. Somit sind auch aus
diesem Grunde, nicht nur wegen der Lichtabsorption, die Laubblätter
flächenförmig gestaltet. Eine Vermehrung des CO2-Gehaltes der
Luft hat, wenn sie nicht zu weit geht, eine vermehrte Assimilation zur
Folge. Demnach kann durch künstliche Anreicherung der Luft mit
CO2 eine ganz erhebliche Steigerung der landwirtschaftlichen
Produktion erzielt werden. Es unterliegt keinem Zweifel mehr,
daß die günstigere Wirkung der Stallmistdüngung gegenüber der Zufuhr
von mineralischem Dünger u. a. auch durch die fortgesetzte starke
CO2-Produktion von Bakterien[175] bedingt ist, die im Boden die
nötigen organischen Nährstoffe finden und die ihre Atmungskohlensäure
aus dem Boden in die Luft übertreten lassen.
Wie alle Lebensprozesse ist auch die CO2-Assimilation von der
Temperatur abhängig. Sie beginnt bei Temperaturen hart unter Null,
erreicht ihren größten Wert bei etwa 37° C und hört bei etwa 45° C
wieder auf.
Diese Kardinalpunkte haben nicht nur bei verschiedenen Pflanzen
eine differente Lage, sondern sie bleiben auch bei einer bestimmten
Pflanze nicht konstant. Insbesondere gilt das für das Optimum, da
dieses im Laufe einiger Stunden von 37° auf 30° sinken kann. —
Bei hellem, warmem Wetter erreicht die Assimilation ihren vollen
möglichen Wert nicht, weil dann der Zufluß von Kohlensäure nicht
genügend ist.
Andere weniger wichtige Faktoren sollen nicht im einzelnen
aufgeführt werden; erwähnt sei nur, daß zahlreiche Substanzen
eine temporäre oder schließlich auch dauernde Beschränkung oder
Vernichtung der Assimilationsfähigkeit bewirken können.
Bis vor kurzem nahm man mit BLACKMAN an, daß bei
gleichzeitiger Einwirkung der zahlreichen Faktoren, die für die
CO2-Assimilation wichtig sind, immer einer, nämlich
der im geringsten Ausmaß vorhandene, die Größe der Assimilation
bestimme (Gesetz des Minimums). Neuere Untersuchungen[176]
haben aber gezeigt, daß unter Umständen ebensowohl durch Steigerung
der Beleuchtungsstärke wie durch Zunahme der Kohlensäure eine
Vermehrung der Assimilation eintreten kann. — Entsprechende
Resultate sind übrigens auch bei den Nährsalzen gefunden worden, wo
früher nach LIEBIG das Minimumgesetz zu gelten schien.
Produkte der Assimilation der Kohlensäure. Wir nahmen oben an,
es werde Zucker aus der Kohlensäure gebildet. In der Tat zeigt die
Analyse eine Zunahme des Zuckergehaltes der Laubblätter bei Besonnung.
Nicht immer und nicht ausschließlich wird freilich Traubenzucker
nachweisbar sein, meist treten andere, kompliziertere Kohlehydrate auf.
Diese lassen sich alle auf Hexosen wie Traubenzucker zurückführen,
schließlich entsteht durch Zusammentreten mehrerer Hexosenmoleküle bei
gleichzeitigem Wasseraustritt Stärke [(C12H20O10)n]. Stärke
findet sich in Chloroplasten beleuchteter Laubblätter sehr häufig,
doch keineswegs allgemein. — Hält man die Blätter einige Zeit im
Dunkeln, so verschwindet die Stärke. Bringt man umgekehrt entstärkte
Pflanzenteile ans Sonnenlicht, so bilden sich in ihren Chloroplasten[S. 218]
oft in überraschend kurzer Zeit (5 Minuten) neue Stärkekörnchen,
die dann bald an Größe zunehmen und schließlich die Substanz des
Chloroplasten an Masse übertreffen. Da nun die Stärke mit Jodlösung
sich blau färbt, kann man das Eintreten der Assimilation leicht
makroskopisch demonstrieren (SACHSsche Methode).
Fig. 247. Assimilationsversuch mit dem Blatt der Aracee
Ariopsis peltata. Verkleinert.
Blätter, die sich am Licht befanden, werden zunächst durch Alkohol
ihrer grünen Farbe beraubt; sie bläuen sich dann nach Jodzusatz.
Bei größerem Gehalt an Stärke tritt tiefe Blaufärbung ein, bei
noch größerem endlich Schwarzfärbung. Die entstehende Farbennuance
erlaubt also zugleich eine gewisse Schätzung der Quantität
der Stärke. Um auch kleinere Mengen Stärke sichtbar zu machen,
empfiehlt es sich, die entfärbten Blätter vor der Anwendung der
Jodlösung in Kalilauge oder in Chloralhydratlösung zu legen, um
die Stärkekörnchen durch Aufquellen zu vergrößern. — Mit dieser
Methode des Assimilationsnachweises kann man auch sehr schön
zeigen, daß nur an beleuchteten Stellen Stärke auftritt.
Wird dem Blatt eine Schablone aus undurchsichtigem Material
aufgelegt, die etwa das Wort „Stärke“ als Ausschnitt besitzt, so
ergibt sich nach Beleuchtung und Behandlung mit Jod das Bild der
Fig. 247: das Wort Stärke erscheint blau auf hellem Grunde. Statt
einer Schablone kann man auch, wie MOLISCH gezeigt hat,
ein geeignetes photographisches Negativ dem Blatt auflegen; nach
Belichtung und Behandlung mit Jod erhält man dann eine positive
Photographie (Fig. 248). Mit der gleichen Methode läßt sich auch
unschwer der Nachweis erbringen, daß panaschierte Blattteile
nicht assimilieren.
Bei manchen Pflanzen, z. B. vielen Monokotylen, kommt es in den
Chloroplasten überhaupt nicht zur Bildung von Stärke, und das
Assimilationsprodukt tritt gelöst in den Zellsaft über. Sorgt man
aber für eine starke Anhäufung des Assimilationsproduktes, so
findet auch hier Stärkebildung statt; in anderen Zellen dieser
Monokotylen, in den Spaltöffnungsschließzellen und den Zellen der
Wurzelhaube wird stets Stärke gebildet. Übrigens wird bei den
anderen Pflanzen stets nur ein Bruchteil, bei Helianthus z. B. 1⁄6
der Assimilationsprodukte in Stärke umgewandelt, der Rest bleibt
als Zucker erhalten oder wird sonst irgendwie verwertet. Man sieht
daraus, daß man die Menge der gebildeten Stärke nicht ohne weiteres
als Maß der Assimilation betrachten darf.
Daß die Stärkebildung nicht mit der Kohlensäureassimilation direkt
zusammenhängt, sondern nur die Folge einer gewissen Anhäufung
von Zucker in der Zelle ist, ergibt sich auch daraus, daß man
Stärkebildung im Finstern erzielen kann, wenn man Blätter auf
Zuckerlösungen von geeigneter Konzentration schwimmen läßt. Statt
Zucker kann man mit gleichem Erfolg auch andere organische Stoffe
verwenden, auch solche, die den Kohlehydraten ferner stehen, wie z. B.
Glyzerin.

Fig. 248. Ein stärkefreies Blatt von Tropaeolum
wurde mit einem Negativ-Porträt bedeckt der Sonne exponiert.
Nach Assimilation ruft Jodlösung ein positives Bild hervor. Nach
MOLISCH.
Bei manchen Algen kommen nicht Zucker und Stärke, sondern andere
Assimilationsprodukte vor, z. B. Florideenstärke.
Was für Stoffe die sog. „Fettröpfchen“ sind, die vielfach in
assimilierenden Zellen auftreten, und was für eine Beziehung zum
Assimilationsprozeß sie haben, ist noch nicht aufgeklärt.
Die Quantität der Assimilate hängt einerseits von der
Pflanzenspezies ab, die wir untersuchen, andererseits von den äußeren
Umständen, unter[S. 219] denen die Pflanze weilt. Es hat sich gezeigt, daß
unter möglichst günstigen Verhältnissen der Quadratmeter
Blattfläche pro Stunde 0,5–1 g Trockensubstanz produzieren kann. —
Wenn man bedenkt, wie viele Quadratmeter Blattfläche auf der Erde Tag
für Tag assimilatorisch tätig sind, so bekommt man einen Begriff von
der ungeheuren Produktion von organischer Substanz in dieser größten
aller chemischen Fabriken. SCHRÖDER schätzt die organische
Substanz, die durch die Landpflanzen jährlich gebildet wird, im Mittel
zu 35 Billionen kg. Allein die deutsche Ernte enthielt im Jahre 1912
rund 9 Milliarden Kilo an Assimilaten im Getreide (Roggen, Weizen,
Spelt und Gerste).
Es gibt zwei Methoden[177] zur Bestimmung der
Assimilationsmenge. Die eine rührt von SACHS her. Nach
seinem Vorgang verfährt man in folgender Weise: Am Morgen werden
Teile von Blättern, gewöhnlich Hälften, sorgfältig ihrer
Fläche nach ausgemessen, dann getrocknet und gewogen. Am Abend
werden gleichgroße Flächen (die restierenden Hälften),
die den Tag über dem Licht exponiert waren, ebenfalls getrocknet
und gewogen. Die Gewichtszunahme bedeutet den Gewinn der Pflanze
an C-Assimilaten (SACHSsche Blatthälftenmethode).
— Eine ganz andere Methode zur quantitativen Bestimmung der
CO2-Assimilation rührt von KREUSLER her und ist auch
von GILTAY und BROWN benutzt worden. Man bringt
ein an der Pflanze befindliches Blatt in einen abgeschlossenen
Raum, der von einem konstanten Luftstrom durchflossen wird,
und bestimmt nun die Menge Kohlensäure, die von dem Blatt der
durchströmenden Luft entrissen wird. Die Menge von Zucker oder
Stärke, die aus einer gegebenen Menge von Kohlensäure gebildet
werden kann, läßt sich ja leicht berechnen.
2. Kohlenstoffgewinn einiger Bakterien[178].
Gewisse Bakterien, von denen auch später (S. 238) die Rede sein
wird, zeichnen sich dadurch aus, daß sie bei Gegenwart von
Karbonaten im Dunkeln und ohne Chlorophyll in rein
anorganischer Nährlösung ihre Substanz vermehren; so die Nitrit- und
Nitratbakterien, die Wasserstoff-, Eisen- und gewisse Schwefelbakterien
sowie die Methanbakterien. Manche von ihnen sind durchaus auf den
Kohlenstoffgewinn aus CO2 angewiesen, während andere, vor allem die
Wasserstoffbakterien, daneben unter Umständen auch organisch gebundenen
Kohlenstoff zu nutzen verstehen.
Über die Produkte der Kohlenstoffassimilation dieser Bakterien
wissen wir noch gar nichts. Auch ist der Gewinn an organisch
gebundenem Kohlenstoff gering. Es verdankt also jedenfalls nur ein
ganz minimaler Bruchteil organischer C-Verbindungen, die momentan
auf der Erde vorhanden sind, ihre Entstehung diesen Bakterien.
Die Kohlensäureassimilation bei ihnen bleibt darum nicht minder
interessant, zumal sie in wesentlich anderer Weise erfolgt als bei der
grünen Pflanze. Vor allem findet sie ohne Gegenwart von Licht statt.
Demnach muß hier eine andere Energiequelle beim Aufbau der organischen
Substanz tätig sein als bei der grünen Pflanze; es ist die Energie,
die durch Oxydation des Ammoniaks, des Nitrits, des Eisenoxyduls,
des Schwefelwasserstoffes, Methans oder Wasserstoffes gewonnen
wird. Man kann die Bildung der organischen Substanz bei der grünen
Pflanze eine Photosynthese, bei den angeführten Bakterien eine
Chemosynthese nennen.
3. Kohlenstoffgewinn der Heterotrophen.
Den Gewinn des Kohlenstoffes aus der Kohlensäure betrachtet man
als typische Kohlenstoffassimilation der Pflanze, doch ist
dieser Modus keineswegs der einzige im Pflanzenreich. Da er —
von den eben besprochenen Bakterien sehen wir jetzt ab — an die
Gegenwart von Chlorophyll und von Sonnenlicht gebunden ist, kann er
bei den unterirdischen Pflanzenteilen, bei[S. 220] allen nichtgrünen
Pflanzen und bei sämtlichen Tieren nicht in Betracht kommen.
In der Tat sind diese auf organische Kohlenstoffverbindungen
angewiesen, die direkt oder indirekt der Assimilationstätigkeit
grüner Pflanzenteile entstammen. Man nennt alle Organismen, die in
ihrer Ernährung von der Tätigkeit der grünen Pflanzen abhängig sind,
heterotroph; die grünen Pflanzen und die soeben besprochenen
Bakterien werden als autotroph bezeichnet. Doch auch die
Autotrophen hängen von anderen Organismen ab. Es wird noch zu zeigen
sein, daß das Leben auf der Erde nur dadurch sich kontinuierlich
erhält, daß jede einseitige Veränderung der Stoffe, die sich unter
dem Einfluß bestimmter Organismen ergibt, durch die Tätigkeit anderer
Organismen wieder kompensiert wird. — Schon durch ihre Lebensweise,
vor allem durch ihren Wohnort, weisen die heterotrophen Organismen
darauf hin, daß sie andere Ansprüche auf Nährstoffe machen als
autotrophe. Sie finden sich entweder als Parasiten auf oder in
lebenden Pflanzen und Tieren, oder sie leben als Saprophyten von
toten Organismen oder Derivaten derselben.
Die Ansprüche, die von heterotrophen Pflanzen an die Kohlenstoffquelle
gemacht werden, lassen sich am besten bei saprophytischen
Bakterien und Pilzen studieren. Man kann diese Organismen auf
verschieden zusammengesetzten Substraten kultivieren und aus ihrem
Gedeihen Rückschlüsse auf den Nährwert der dargebotenen Verbindungen
ziehen. Die zur Kultur dienenden Nährlösungen müssen neben den
unentbehrlichen Aschensubstanzen und einer Stickstoffquelle (etwa einem
Ammoniumsalz) im allgemeinen Zucker als C-Quelle enthalten; sie
sollen für Schimmelpilze schwach sauer, für Bakterien meistens schwach
alkalisch oder neutral reagieren und werden oft durch Zusatz von
Gelatine oder Agar-Agar in den festen Aggregatzustand übergeführt. In
der Regel kann der Zucker durch zahlreiche andere organische Substanzen
mehr oder minder gut ersetzt werden, z. B. durch andere Kohlehydrate,
durch Fette, Eiweiß und Eiweißabbauprodukte, durch organische Säuren
usw. usw. Während man nun für einen einzelnen Organismus diese
C-Quellen nach ihrem Nährwert in eine bestimmte Reihenfolge bringen
kann, ist das generell ganz unmöglich, denn es gibt nicht wenige
Saprophyten, die sich ganz besonderen Bedingungen angepaßt haben,
und die gerade solche C-Quellen mit Vorliebe verzehren, die bei der
Mehrzahl der anderen kaum einen Nährwert besitzen (z. B. Ameisensäure,
Kohlenwasserstoffe, Oxalsäure).
Aber selbst solche Saprophyten, die mit recht verschiedenen
Kohlenstoffverbindungen auskommen (Omnivore), haben doch ein
sehr feines Unterscheidungsvermögen für diese. So wird von dem
Schimmelpilz Penicillium aus gewöhnlicher Weinsäure zunächst
überwiegend die Rechtsweinsäure, von einer Bakterienart dagegen
zuerst die Linksweinsäure verarbeitet. In einem Gemisch von Glykose
und Glyzerin wird erstere von Aspergillus bevorzugt. Das Glyzerin
wird also durch den Zucker vor dem Verbrauch geschützt (Elektion
der Nährstoffe), während es, allein dargeboten, völlig
aufgebraucht wird.
Sehr bemerkenswert ist die Fähigkeit mancher Pilze, auch solche
organische Verbindungen auszunutzen, die wie Stärke, Zellulose usw.
in Wasser unlöslich sind und demnach erst nach zuvoriger Verwandlung
und Lösung aufnahmefähig werden. Diese Pilze und Bakterien scheiden
Stoffe besonderer Art (Enzyme; vgl. S. 229) aus, die imstande sind, die
betreffenden Stoffe zu spalten und damit löslich zu machen.
Die Saprophyten sind nach dem Gesagten durch die Art ihrer
Assimilationstätigkeit charakterisiert: sie können den ersten
Schritt der Assimilation des Kohlenstoffes, den die grüne Pflanze mit
Hilfe des Lichtes vollzieht, nicht ausführen. Dagegen besteht
aller Wahrscheinlichkeit nach im weiteren Verlauf der Assimilation, in
dem Aufbau komplizierter Verbindungen, die den Körper zusammensetzen,
aus der gebotenen[S. 221] einfachen organischen Verbindung kein Unterschied
gegenüber den Autotrophen.
Über heterotrophe Blütenpflanzen vgl. auch S. 226.
Den Saprophyten stehen dann Parasiten gegenüber, die wir
in reicher Auswahl bei Pilzen und Bakterien finden; doch fehlen sie
auch unter den Algen, Cyanophyceen und den höheren Pflanzen nicht ganz.
Daß nun diese Parasiten oder wenigstens viele von ihnen Nährstoffe
aus dem Wirte aufnehmen, sieht man dem letzteren oft direkt an; er
kann durch den Parasiten schwer geschädigt, ja sogar schließlich
getötet werden. Welche Stoffe aber im einzelnen es sind, die der
Parasit aufnimmt und zu seinem Gedeihen bedarf, ist schwer zu sagen.
Da vielfach nur Organismen einer bestimmten Verwandtschaft (Familie,
Gattung, Art, Kleinart) von einer Parasitenspezies befallen werden,
muß man annehmen, daß diese bezüglich Qualität und Quantität ihrer
Nahrung ganz spezifische Anforderungen macht. Eine solche Vermutung
wird noch dadurch unterstützt, daß man die Mehrzahl der Parasiten nicht
unabhängig von ihrem Wirt kultivieren kann.
A. Assimilation des Stickstoffes.
Wenn die grüne Pflanze den Kohlenstoff als Kohlensäure aus der Luft
bezieht, wo dieses Gas in relativ sehr geringer Menge vorhanden
ist, so könnte man glauben, daß der Riesenvorrat von Stickstoff in
der Atmosphäre die erste und die beste Quelle für den Bezug dieses
Baustoffes sei. Jede Wasserkultur lehrt indes auf das eindringlichste,
daß der Luftstickstoff von der typischen grünen Pflanze nicht
ausgenützt werden kann. Läßt man den gebundenen Stickstoff aus der
Nährlösung weg, so ist es mit dem Gedeihen der Pflanze vorbei.
In der KNOPschen Nährlösung war der Stickstoff als
salpetersaures Salz geboten, und diese Form gilt auch heute noch als
die beste für die höhere Pflanze. Immerhin muß zugegeben werden, daß
auch gewisse Ammoniumverbindungen, z. B. schwefelsaures oder salzsaures
Ammonium, den Nitraten gleichwertig sind, da sie nicht wie z. B.
kohlensaures Ammonium durch alkalische Reaktion die Pflanze schädigen.
Auch organische Stickstoffverbindungen können als Nährstoffe dienen,
so z. B. Aminosäuren, Säureamide, Amine usw., doch ist wohl mit keiner
von ihnen ein so guter Erfolg zu erzielen wie mit Salpetersäure. Auch
salpetrigsaure Salze können eine brauchbare Stickstoffnahrung abgeben,
wenn sie nicht durch zu hohe Konzentration schädigen.
Über die Assimilation der Salpetersäure und des Ammoniaks sind wir
nicht annähernd so gut orientiert wie über die der Kohlensäure. Wir
kennen den Ort der Assimilation nicht genau, wir wissen über die
mitwirkenden äußeren Umstände nicht so gut Bescheid, und wir sind
endlich über die auftretenden Produkte nicht ganz im klaren. In
letzter Linie handelt es sich vor allem um Bildung von Eiweiß,
also um eine sehr viel kompliziertere Substanz als die Kohlehydrate
sind, eine Substanz, die neben C, H und O stets ca. 15 bis 19% N und
außerdem auch noch S, eventuell auch P enthält. Einen Einblick in den
Bau des Eiweißmoleküls haben uns vor allem die methodischen Studien
über den Eiweißabbau gegeben. Sie zeigten, daß im Eiweiß
eine große Anzahl von Aminosäuren durch Wasserabgabe miteinander
verkettet sind. So wie nun EMIL FISCHER künstlich durch
Zusammenschweißen von Aminosäuren und darauffolgende Kondensation
eiweißähnliche Körper (Polypeptide) hergestellt hat, so wird es
aller Wahrscheinlichkeit nach auch in der Pflanze sich darum handeln,
daß zunächst solche Aminosäuren gebildet und dann gekoppelt werden.
Betrachtet man nun die einfachste Aminosäure, das Glykokoll NH2CH2 · CO2H, das freilich in der Pflanze nicht sehr verbreitet
[S. 222] ist, so
zeigt sich, daß diese sich von der Essigsäure ableiten läßt, wenn man
ein am Kohlenstoff hängendes H-Atom durch eine NH2-Gruppe ersetzt.
Es muß also die aufgenommene HNO3 reduziert werden, wenn ihr
Stickstoff zum Aufbau von Eiweiß verwendet werden soll. Diese Reduktion
ist unabhängig von Sonnenlicht und Chlorophyll; auch im
Dunkeln und von farblosen Teilen wird Salpetersäure assimiliert[179].
— Indirekt freilich wird das Chlorophyll und ebenso das Licht von
Bedeutung für die Eiweißsynthese sein können, insofern als auch
C-haltige organische Substanz für den Eiweißaufbau nötig ist, und
diese in der Sonne vom Chlorophyll gebildet wird. Wegen ihres
reichlichen Gehaltes an Kohlehydraten werden deshalb die Laubblätter
besonders zur Eiweißbildung geeignet sein; sie sind aber durchaus nicht
in dem Grade „Organe der Eiweißbildung“, wie sie Organe der
Kohlehydratbildung sind. Auch kann man bei vielen Pflanzen (vor allem
bei den Ruderalpflanzen, z. B. Chenopodium, Amarantus, Urtica) die
Salpetersäure noch in den Blättern nachweisen, bei der Mehrzahl scheint
sie schon sofort nach ihrer Aufnahme in der Wurzel verändert zu werden.
Ebensowenig wie über die Assimilation der Salpetersäure sind wir über
die Assimilation des Ammoniaks unterrichtet. Da hierbei nicht erst eine
Reduktion nötig ist, sollte man das Ammoniak für leichter assimilierbar
halten als die Salpetersäure; in den Fällen, wo bei der Wasserkultur
ein Zusatz von Ammoniak weniger günstig wirkt als Salpetersäure,
werden wahrscheinlich irgendwelche Nebenwirkungen des NH3 in
Betracht kommen.
Die hypothetischen Zwischenprodukte zwischen den aufgenommenen
Stickstoffverbindungen und dem fertigen Eiweiß, d. h. verschiedene
Aminosäuren und ihnen nahestehende Substanzen, fehlen wohl keinem
Pflanzenteil. Besonders verbreitet sind Leucin, Tyrosin und Asparagin.
Aber nur selten wird man mit Bestimmtheit entscheiden können, ob diese
Stoffe synthetisch aus Ammoniak oder Salpetersäure, oder ob sie
durch Eiweißabbau entstanden sind (vgl. S. 231).
Außer im Eiweiß finden wir den Stickstoff auch noch in den
Lezithinen und den organischen Pflanzenbasen. Erstere
sind komplizierte Ester, in denen sich Glyzerin mit zwei
Molekülen Fettsäure, einem Molekül Phosphorsäure und der N-haltigen
Base Cholin verbunden hat. Sie fehlen keinem lebenden Protoplasma. —
Die meisten organischen Basen (Alkaloide) dürften Nebenprodukte
der N-Assimilation sein und keine Verwendung mehr finden.
Wenn man im allgemeinen sagen kann, daß die typische autotrophe Pflanze
den Stickstoff als Salpetersäure ebensogut oder besser zu assimilieren
versteht wie als Ammoniak, so trifft das für die Mehrzahl der Pilze
nicht zu. Nur wenige von ihnen nehmen überhaupt mit Salpetersäure
vorlieb; in der Regel gelten Ammoniumsalze als beste anorganische
N-haltige Nahrung für sie. Es ist oft behauptet worden, daß manche
Pilze mit so einfachen Stickstoffverbindungen nicht auskommen, oder
daß sie wenigstens leichter und sicherer aus organischen
N-Verbindungen die komplizierteren Körpersubstanzen aufbauen.
Da aber offenbar viele Fehlerquellen bei diesen Untersuchungen
nicht berücksichtigt wurden, bedarf diese Frage einer erneuten
Untersuchung[180].
An dieser Stelle müssen auch die insektenfressenden,
fleischfressenden oder karnivoren Pflanzen genannt
werden[181] (vgl. S. 158). Es sind das Pflanzen, die mit Einrichtungen
zum Fangen und Festhalten kleiner Tiere, Insekten vor allen Dingen,
versehen sind, und die dann durch ausgeschiedene Enzyme die Beute
soweit als möglich auflösen, verdauen und resorbieren. Alle diese
Insektivoren besitzen Chlorophyll; es kann ihnen also gewiß nicht in
erster Linie auf den Gewinn organisch gebundenen Kohlen[S. 223]stoffes
ankommen. Es ist auch bekannt, daß sie ohne tierische Nahrung ganz
gut gedeihen, doch wird durch Zufuhr einer nicht zu großen Menge
animalischer Substanz eine entschiedene Förderung erzielt, die sich
in vermehrter Frucht- und Samenproduktion kenntlich macht. Es ist
sehr wahrscheinlich, aber keineswegs bewiesen, daß die Karnivorie auf
den Gewinn von brauchbarem Stickstoff hinausläuft. Ob dieser an den
Wohnorten der Insektivoren — sie wachsen vielfach in nährsalzarmem
Torfmoor oder Wasser — nur der Quantität nach unzureichend ist, oder
ob seine Qualität da nicht optimal ist, muß dahingestellt bleiben. Die
Möglichkeit besteht zweifellos, daß organisch gebundener Stickstoff
diesen Pflanzen besonders zuträglich ist. Daneben ist es freilich auch
nicht ausgeschlossen, daß die Insektivoren nicht allein Stickstoff,
sondern auch andere Nährsalze, vor allem Kali und Phosphorsäure, aus
ihrer Beute gewinnen. Ob diese Nährsalze dann in organischer Bindung
ausgenützt werden oder ob sie bei dem Verdauungsprozeß in anorganische
Form übergeführt werden, wissen wir nicht. Im letzteren Falle würde
der Nutzen der Insektivorie vor allem darin gesucht werden müssen, daß
mehr Nährstoffe gewonnen werden, als der Boden bietet.
Vollkommen geklärt ist die Karnivorie auch heute noch nicht.
Man hat darauf hingewiesen, daß die Insektivoren mehrfach
ein sehr schwaches Wurzelsystem besitzen; das trifft aber
für Nepenthes nicht zu. Eine Mykorrhiza (S. 226), die
wahrscheinlich eine vermehrte Aufnahme von Bodensalzen bewirkt,
fehlt den Insektivoren aber allgemein.
Die fleischfressenden Pflanzen scheinen dem Laien in ihrer
Ernährungsweise weit von den anderen Pflanzen abzuweichen und mehr
an die Tierwelt heranzukommen. Wie die Tiere, so nützen auch die
Insektivoren feste Nahrung aus, die sie durch ausgeschiedene
Enzyme verflüssigen, ehe sie sie in ihre Zellen aufnehmen. Daß bei
den Tieren die „Verdauung“ der Nahrung im Magen sich vollzieht, bei
den Insektivoren aber manchmal wenigstens in einem Gebilde, das
dem Magen vergleichbar ist (Kannen usw.), macht die Ähnlichkeit
der beiden Gruppen besonders groß. Es verdient aber hervorgehoben
zu werden, daß in physiologischer Hinsicht manche Pilze und
Bakterien den Tieren entschieden näher stehen. Denn sie können wie
Tiere ihre gesamte Nahrung durch Verdauung fester organischer
Körper gewinnen, während ja die Insektivoren wenigstens bezüglich des
Kohlenstoffes autotroph sind.
An die Insektivoren schließen sich gewisse phanerogame
Parasiten[182] an, die oben S. 221 nicht erwähnt worden sind,
weil sie durch den Besitz grüner Blätter schon anzeigen, daß
sie bezüglich des Kohlenstoffes autotroph sind. Trotzdem sind
sie ausgesprochene Parasiten, die sich nur dann normal entwickeln,
wenn ihr Wurzelsystem mit fremden Wurzeln oder zur Not auch mit
denen der Artgenossen durch Haustorien verbunden ist. So leben z. B.
die Santalacee Thesium und auch viele Rhinanthaceen; von letzteren
zeigt Tozzia, zumal in den ersten Entwicklungsstadien, die
parasitische Lebensweise am meisten ausgeprägt. Unsere einheimische, in
Baumkronen wurzelnde Mistel besitzt, wie viele ihrer fremdländischen
Verwandten (Loranthaceen), ebenfalls noch stattliche Blätter und
ist so reich an Chlorophyll, daß sie ihren Bedarf an Kohlehydraten
vollständig selbst zu decken vermag. Wasser und Nährsalze aber entnimmt
sie der Wirtspflanze. Dementsprechend ist ihr Wurzelsystem weitgehend
reduziert. Das gleiche trifft auch für die oben genannten Rhinanthaceen
zu. In welcher Form sie die Nährsalze aus dem Wirt entnehmen, ist indes
noch unbekannt.
In scharfem Gegensatz zu den Karnivoren, die vielleicht durch organisch
gebundenen Stickstoff gefördert werden, stehen gewisse Mikroorganismen,
die bezüglich ihres Stickstoffgewinnes autotroph, bezüglich ihrer
Kohlenstoffassimilation dagegen durchaus heterotroph sind. Wir sprechen
von den[S. 224] Organismen, die imstande sind, den atmosphärischen
Stickstoff zu binden. Der Nachweis ihrer Existenz wurde erst
am Ende des vorigen Jahrhunderts, vor allem durch die Arbeiten von
WINOGRADSKY, HELLRIEGEL und WILFARTH
erbracht[183].
Fig. 249. Eine aus dem Boden gehobene Wurzel der
Feldbohne (Vicia faba), dicht besetzt mit Bakterienknöllchen.
Verkleinert. Nach NOLL.
Fig. 250. 1 Junge Knöllchen K auf einer Wurzel
W von Vicia faba. B Großzelliges mit Bakterienmasse angefülltes
Gewebe, bei M dessen „Meristem“. T Tracheïdenzüge. Vergr. 60fach.
2 Eine mit Tausenden von Bakterien erfüllte Zelle des Knöllchens,
mit benachbarten nicht infizierten Zellen. Vergr. 320fach. 3 Ein
infiziertes Wurzelhaar mit dem „Infektionsfaden“ im Innern. Vergr.
320fach. 4 Formen von „Bakteroiden“. 5 Unveränderte Bakterien.
Vergr. 1200fach. Nach NOLL.
Es sind zunächst einmal gewisse heterotrophe Bakterien, wie Clostridium
Pasteurianum und Verwandte sowie Azotobacter chroococcum, die im
einzelnen unter recht verschiedenen äußeren Bedingungen im Boden
des Ackers freien Stickstoff binden und damit eine für ihr eigenes
Gedeihen wie für das vieler anderer Organismen außerordentlich wichtige
Eigenschaft besitzen, die auch für die Landwirtschaft von großer
Bedeutung ist. Im Wasser scheinen wieder andere Bakterien im gleichen
Sinne tätig zu sein. Eine nach neueren Untersuchungen immer größer
werdende Zahl niederer Pilze scheint die gleiche Befähigung, wenn auch
meist in geringerem Grade, zu besitzen. Neben diesen freilebenden gibt
es auch noch im Innern von höheren Pflanzen auftretende Mikroorganismen
von gleicher Wirksamkeit. Unter diesen sind am besten studiert die
verschiedenen Formen des Bacillus radicicola, die die Wurzeln von
Leguminosen befallen und an ihnen in oft großer Zahl gallenartige
Knöllchen (Fig. 249 u. 250) erzeugen. Soviel wir wissen, handelt es
sich hier um eine[S. 225] Art gegenseitigen Parasitismus, d. h. um eine
auf wechselseitiger Ergänzung beruhende Lebensgemeinschaft, wie sie
von DE BARY als Symbiose bezeichnet worden ist.
Die Leguminosen erscheinen dann, abweichend von den meisten grünen
Pflanzen, als Stickstoffsammler[184], was durch GILBERT und
LAWES in England, durch SCHULTZ-LUPITZ in Deutschland
zuerst festgestellt worden ist.
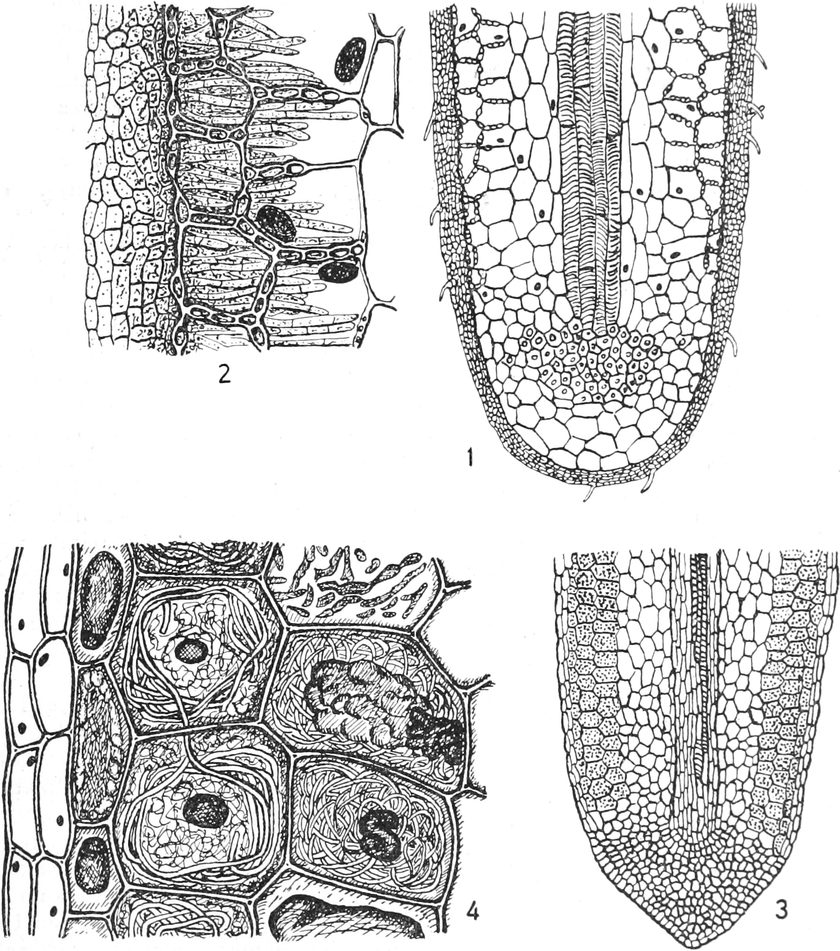
Fig. 251.
1 Längsschnitt durch die Wurzelspitze von
Pinus silvestris mit
ektotropher Mykorrhiza. Vergrößert.
2
Ein Teil der
Fig. 1 stärker vergrößert; vom äußeren Pilzmantel gehen
Hyphen aus, die zwischen die Zellen der Rinde dringen.
3 Längsschnitt
durch die Wurzelspitze von Neottia. Drei Zellreihen mit dunklem Inhalt
führen die Pilze (
endotrophe Mykorrhiza). Vergrößert.
4 Ein
Teil der
Fig. 3 stärker vergrößert. Die mittlere pilzbewohnte Schicht
besteht aus
Pilzwirtzellen, in denen die Pilze erhalten bleiben;
die äußere und die innere sind Verdauungszellen. Nach W
ERNER
M
AGNUS. (K
NY, Wandtafeln.)
Die stabförmigen Bakterien dringen durch die Wurzelhaare, deren
Wand sie lokal zur Aufquellung bringen, in die Rinde der Wurzel
ein, verursachen hier die genannten Wucherungen der Parenchymzellen
und füllen diese mit einer Bakterienmasse an, die schließlich in
abnorm gebildete vergrößerte und verzweigte, aber noch wachstums-
und teilungsfähige Formen, die sog. Bakteroiden, übergeht.
Während die Bakterien von dem Überschuß der Kohlehydrate ihres
Wirtes, in der ersten Zeit auch noch von dessen Eiweiß zehren,
kommt den Bakteroiden die Fähigkeit zu, den atmosphärischen
Stickstoff zu binden. Die Leguminose aber kann andauernd den
Bakteroiden den gebundenen Stickstoff entziehen. Man hat berechnet,
daß Lupinen pro Hektar auf diesem Wege bis[S. 226] zu 200 kg Stickstoff zu
gewinnen vermögen, also mehr als in 300 Doppelzentnern Stallmist
durchschnittlich enthalten ist. Demnach hat diese Stickstoffbindung
eine große Bedeutung für die Landwirtschaft, und man sucht sie
durch Impfung der Felder mit wirksamem Boden oder mit Reinkulturen
besonders virulenter Bakterien zu fördern. Bei der Kultur der
Seradella (Leguminose) ist denn auch durch Impfung eine erhebliche
Steigerung der Produktion erzielt worden; während der ungeimpfte
Boden 5 kg Frischgewicht pro Ar ergab, wurden nach Impfung 400 kg
pro Ar geerntet. — Stehen der Leguminose genügend Nitrate im Boden
zur Verfügung, so vermag sie mit diesen vorzüglich zu gedeihen; da
die Nitrate zugleich auf Bacillus radicicola schädlich wirken, so
kommt es unter diesen Umständen gar nicht zur Knöllchenbildung.
Außer bei den Leguminosen kommen auch bei Alnus, Elaeagnus
und Casuarina regelmäßig Wurzelknöllchen durch Infektion mit
niederen Pflanzen zustande. Elaeagnus und Alnus können den freien
atmosphärischen Stickstoff assimilieren, wenn ihre Wurzeln
Knöllchen besitzen. Auch Podocarpus hat die gleiche Befähigung.
Hier aber handelt es sich um die Verbindung der Wurzel mit einem
Pilz, um eine „Mykorrhiza“[185].
Eine Mykorrhiza findet sich bei zahlreichen Pflanzen, vor
allem solchen, die im Humus der Wälder und Heiden leben. Man
unterscheidet zwei Extreme, die durch Zwischenstufen verbunden
sind, als ekto- und endotrophe Mykorrhiza. Bei der
ersteren (Fig. 251, 1, 2) umgibt ein Mantel von Pilzhyphen
die Wurzel und umhüllt auch schon deren Vegetationspunkt (viele
unserer Bäume, Monotropa); bei der letzteren (Fig. 251, 3, 4)
lebt der Pilz in den Zellen gewisser Wurzelschichten und sendet
nur einzelne Fäden in den Boden (Orchideen, Ericaceen, viele
Liliaceen). — Die endotrophen Mykorrhizapilze werden von den
Zellen der Wurzel teilweise verdaut; es müssen also alle dabei
freiwerdenden Stoffe der Phanerogame zugute kommen. Wenn auch
noch nicht ganz sichergestellt ist, ob der Pilz die Fähigkeit
hat, atmosphärischen Stickstoff zu binden, so kann man doch wohl
sagen, die endotrophe Mykorrhiza sei einigermaßen
klargelegt. Sie ist auch in manchen Fällen als eine ganz
unentbehrliche Assoziierung erkannt, insofern als bei den Orchideen
vielfach schon die Keimung, bei den Ericaceen die Weiterentwicklung
des Keimlings unterbleibt, wenn der Pilz fehlt. Weniger klar liegen
die Verhältnisse bei der ektotrophen Mykorrhiza. Hier stehen
sich noch immer mehrere Anschauungen unvermittelt gegenüber: nach
der einen soll der Pilz ein reiner, also auch im Grunde schädlicher
Parasit sein; nach der zweiten ist die Blütenpflanze der Parasit;
die dritte nimmt eine wirkliche Symbiose zwischen beiden
Organismen an und stellt sich mit STAHL die Beziehungen
zwischen beiden etwa in folgender Art vor: der Pilz nimmt sehr
energisch Nährsalze aus dem Boden und versorgt mit diesen auch die
autotrophe Blütenpflanze; diese liefert dem Pilz dafür organische
Nahrung. Da aber eine ektotrophe Mykorrhiza sich auch bei farblosen
Phanerogamen (z. B. Monotropa) findet, so muß wenigstens hier an
einen Parasitismus der Blütenpflanze gedacht werden.
Das gleiche dürfte bei endotropher Mykorrhiza für die
nichtgrünen Orchideen (Neottia, Coralliorhiza, Epipogon)
gelten, die also nicht saprophytisch vom Humus leben, sondern
offenbar von den Mykorrhizapilzen ernährt werden.
Die Konsortien von Algen und Pilzen, die man Flechten nennt,
finden gewöhnlich eine Deutung, die der dritten bei der ektotrophen
Mykorrhiza angeführten entspricht[186].
Neuerdings sind in den Blättern von tropischen Pflanzen
(Rubiaceen und Myrsinaceen) Knöllchen gefunden worden, die einer
Infektion durch Bakterien ihren Ursprung verdanken. Während
aber bei den Leguminosen die Infektion immer von dem zufälligen
Zusammentreffen zwischen Bakterien und Blütenpflanze abhängt,
wird bei den genannten Pflanzenfamilien schon dem Embryo in
der Mutterpflanze eine gewisse Bakterienmasse mitgegeben; und
wenn diese künstlich ferngehalten wird, erfolgt die Entwicklung
bei Ardisia anomal. Daß auch hier eine Assimilation von freiem
Stickstoff stattfindet, ist wenigstens für die Rubiaceen
erwiesen[187].
C. Assimilation der übrigen Stoffe.
Die Schwefelsäure schließt sich im Stoffwechsel der Pflanze
am engsten an den Stickstoff an, da sie ja ebenfalls zum Aufbau von
Eiweißstoffen, die etwa
1⁄2–11⁄2%
S enthalten, Verwendung findet. Wo
und unter welchen Bedingungen ihre Assimilation stattfindet, ist noch
unklar; nur so viel ist bekannt, daß dabei eine Reduktion stattfinden
muß. Bei einigen Pflanzen[S. 227] findet Schwefel nicht nur im Eiweiß, sondern
auch in anderen Verbindungen Verwendung.
Die Phosphorsäure schließt sich insofern an die Schwefelsäure
an, als sie zum Aufbau wenigstens gewisser Proteïnsubstanzen
(Nukleoalbumine) und vor allem der Nukleoproteïde der Zellkerne in
einer Menge von 0,3–3% dient. Im Gegensatz zur Schwefelsäure wird die
Phosphorsäure bei ihrer Aufnahme in diese Moleküle nicht reduziert.
Auch die in keiner Pflanze fehlenden Lezithine (vgl. S. 222) enthalten
Phosphor; ebenso das besonders im Samen auftretende Phytin.
Die Metalle. Kalium, Kalzium, Magnesium und Eisen sind, wie z. B.
durch die Wasserkulturmethode gezeigt wurde, ebenso unentbehrlich
wie irgendeiner der bisher besprochenen Stoffe; es ist zum mindesten
für Kalium und Magnesium sehr wahrscheinlich, daß sie am Aufbau
gewisser Verbindungen teilnehmen, die für die Existenz der Pflanze
wesentlich sind. Vermutlich enthält z. B. das Protoplasma solche
Stoffe. Aber auch andere Substanzen können sie enthalten; so ist z. B.
für den Chlorophyllfarbstoff ein beträchtlicher Gehalt an Magnesium
nachgewiesen. Man glaubte früher irrtümlicherweise, das Chlorophyll
enthalte Eisen, weil in einer Nährlösung ohne Fe die Chloroplasten
gelb bleiben (Chlorose S. 205). Wir wissen jetzt, daß das
Chlorophyll eisenfrei ist und daß auch nichtgrüne Pflanzen Eisen nötig
haben; deshalb gewinnt die Vermutung an Wahrscheinlichkeit, daß das
Protoplasma selbst Eisen bedürfe, und daß die Chlorose bei
Fe-Mangel die Folge eines kranken Protoplasmas sei.
Kalium, Magnesium und Eisen müssen demnach, da sie in die
Pflanzensubstanz übergehen, irgendwo assimiliert werden. Wo und wie
das geschieht, ist noch unbekannt. — Etwas anders liegt die Sache
beim Kalzium. Es ist bei manchen Pflanzen (Algen) entbehrlich, bei
anderen hat es jedenfalls in erster Linie eine schützende
Wirkung, insofern als es Giftwirkungen, die von Eisen, Magnesium,
Kalium und Natrium, aber auch von Phosphorsäure, Schwefelsäure,
Salpetersäure und Salzsäure ausgehen, aufzuhalten vermag. Es ist
aber nicht wahrscheinlich, daß auch bei den höheren Pflanzen seine
Unentbehrlichkeit durch diese Schutzwirkung allein bedingt ist.
Bei Besprechung der Insektivoren und gewisser grüner Parasiten ist
darauf hingewiesen worden, daß diese vielleicht die Aschensubstanzen
bereits in organischer Bindung aufnehmen. Sicheres ist aber nicht
bekannt.
Wasser. Daß das Wasser unentbehrlich für die Pflanze ist,
wissen wir. Wenn es aber ohne chemische Umwandlung als Wasser in den
Pflanzenkörper eingelagert wird, werden wir nicht von „Assimilation“
sprechen. So z. B. bei dem Wasser, das die Vakuole füllt, oder das in
Plasma und Zellhaut eingelagert ist. Anders wenn das Wasser chemisch
gebunden wird. Das findet z. B. notwendigerweise statt, wenn aus
Kohlensäure Kohlehydrate entstehen und wahrscheinlich auch anderwärts.
In diesen Fällen kann man demnach von einer Assimilation des Wassers
mit dem gleichen Recht reden, wie man von Assimilation der Kohlensäure
spricht.
IV. Wanderung und Wandlung
der Assimilate.
Die Assimilate dienen in erster Linie als Baustoffe
zum Aufbau neuer Pflanzensubstanz, also zur Herstellung neuer
Zellen; daneben finden sie auch Verwendung als Reserven,
als Wanderstoffe, und ein Teil von ihnen wird als
Betriebsstoffe verbraucht, während andere in Exkrete oder
Sekrete übergeführt werden.
[S. 228]
Nur selten findet das Wachstum an den Stellen statt, die
assimilatorisch tätig sind; die Kohlensäureassimilation z. B. erfolgt
größtenteils in ausgewachsenen Laubblättern, während das Wachstum in
der Nähe der Vegetationspunkte, räumlich mehr oder minder weit von den
Blättern entfernt, sich vollzieht. Auch zeitlich fällt Organbildung
und Assimilationstätigkeit nur teilweise zusammen. Viele Pflanzen
haben Zeiten lebhafter Assimilation, die mit geringer Organbildung
verbunden ist; und abwechselnd damit dann umgekehrt Zeiten lebhaften
Wachstums bei geringer oder gänzlich fehlender Assimilationstätigkeit.
Unsere Bäume verlieren im Herbst die Blätter, unsere Stauden die
ganzen oberirdischen Organe; beide müssen im Frühjahr erst neue
Assimilationsorgane bilden, ehe sie mit erneuter Assimilation beginnen,
und zu dieser Organbildung brauchen sie gespeicherte Assimilate.
Auch jeder keimende Same lebt zunächst auf Kosten der Assimilate
einer älteren Generation. Solche gespeicherte Assimilate nennt man
Reservestoffe. Sie können am Ort ihrer Bildung deponiert
sein, oder sie können sich an sekundärer Lagerstätte befinden. Den
ersten Fall illustriert jedes Laubblatt, das am Abend eines hellen
Sommertages mit Stärke überfüllt ist. Den zweiten sehen wir bei Samen,
die in einem besonderen Nährgewebe (Endosperm) oder in den Kotyledonen
die Reserven beherbergen, doch auch an Vegetationsorganen, die als
Reservestoffbehälter schon an ihrer Gestalt kenntlich sind, so die
angeschwollenen Blätter der Zwiebeln, die angeschwollenen Stengel (z. B.
Kartoffel) oder Wurzeln (z. B. Rübe). Wenn Assimilate in diese
Reservestoffbehälter gelangen sollen, so müssen sie wandern, und
wenn sie aus diesen Behältern austreten und für Organbildung verwendet
werden sollen, so müssen sie abermals wandern. Viele Reservestoffe
oder Assimilate sind aber zunächst in einer Form gegeben, die ihnen
den Übertritt von Zelle zu Zelle ganz unmöglich macht, sie sind fest;
man denke an die Stärke. Andere sind zwar gelöst, aber sie besitzen
so große Moleküle, daß sie wenig diffusionsfähig sind (z. B. Inulin).
Aus diesen Gründen sieht man die Reservestoffe in der Regel einer
Umwandlung („Mobilisierung“) unterworfen, ehe ihre Wanderung
beginnt.
A. Mobilisierung der Reservestoffe.
Bei der Mobilisierung der Reservestoffe handelt es sich meist um eine
wenig tiefgreifende Veränderung, eine sog. Hydrolyse, d. h. um
eine unter Wasseraufnahme vor sich gehende Zerspaltung in kleinere
Moleküle. Diese ist bei den drei Grundtypen der Reservestoffe, den
Kohlehydraten, den Fetten, den Eiweißkörpern gesondert zu betrachten.
1. Hydrolyse der Kohlehydrate.
Die Stärke ist einer der wichtigsten Reservestoffe der
Pflanze. Sie bildet bei Samen sowie bei Knollen und Zwiebeln nicht
selten die Hauptmasse der Reserven. In der Kartoffelknolle besteht
20%, im Weizensamen 70% der frischen Substanz aus ihr. Auch im Mark,
dem Holzparenchym, den Markstrahlen und der Rinde der Bäume ist sie
in reicher Menge enthalten. Chemisch gilt heute die Stärke als ein
polymerisiertes Maltoseanhydrid, dem wahrscheinlich die Formel
(C12H20O10)n zukommt[188]. Sie ist völlig unlöslich;
um wanderungsfähig zu werden, muß sie abgebaut worden. In unseren
Fabriken geschieht das durch Behandlung mit Säuren. Der im Handel
befindliche Traubenzucker wird durch Behandlung von Kartoffelstärke mit
Schwefelsäure gewonnen. Entsprechend der Formel
(C12H20O10)n +
2n H2O = n C6H12O6
wird das Stärkemolekül unter Wasseraufnahme in n Glykosemoleküle
zerspalten. In der Pflanze geht aber die Hydrolyse unter dem Einfluß
eines[S. 229] besonderen Stoffes, der Diastase, vor sich. Es ist
das eine Substanz, die man mit Wasser oder Glyzerin aus der Pflanze
herauslösen kann. Durch Zusatz von Alkohol wird sie aus dem Extrakt
ausgefällt und kann nach abermaliger Lösung wieder die gleiche
Wirkung auf Stärke ausüben wie zuerst. Nach Erhitzung wird sie aber
unwirksam. WILLSTÄTTER hat durch Adsorption Enzyme, darunter
auch eine Diastase, weitgehend gereinigt und hat gezeigt, daß sie
weder zu den Kohlehydraten noch zu den Proteinen gehört. Von anderen
Enzymen unterschied sich dieses Präparat nur durch seine Wirkung auf
Stärke, chemisch aber gar nicht; nach dem Erhitzen war überhaupt jeder
Unterschied verschwunden. So birgt die Frage nach der chemischen Natur
der Diastase und anderer Enzyme noch viele Rätsel[189].
Diese Diastase hat nun eine ganz ähnliche Wirkung auf Stärke wie die
Schwefelsäure, aber sie baut freilich die Stärke nicht so vollständig
ab wie die Säure, sondern sie macht nach Bildung des Disaccharids
Maltose halt. Man bezeichnet die Schwefelsäure und ebenso
die Diastase als einen Katalysator. Katalysatoren aber nennt
man Stoffe, die die Geschwindigkeit einer freiwillig verlaufenden
chemischen Reaktion ändern. In erster Linie handelt es sich um die
Beschleunigung von Reaktionen. In unseren Laboratorien bedienen
wir uns zur Beschleunigung von Reaktionen vor allem einer erhöhten
Temperatur; dieses Mittel kann der Organismus nicht weitgehend
verwenden, weil sein Leben an enge Temperaturgrenzen gebunden ist.
In zweiter Linie beschleunigen wir Reaktionen durch anorganische
Katalysatoren. Da viele von diesen, z. B. die oben verwendete
Schwefelsäure, das Protoplasma schädigen, so begreifen wir, daß im
Organismus besondere, unschädliche Katalysatoren auftreten. Diese nennt
man Enzyme[190] und findet sie bei Pflanzen wie bei Tieren in
gleicher Weise vor. Während nun viele anorganische Katalysatoren auf
sehr verschiedenartige chemische Vorgänge Einfluß haben, sind die
organischen Katalysatoren von ganz spezifischer Wirkungsweise; Diastase
z. B. wirkt nur auf Stärke ein. Da der Katalysator nicht oder
nicht dauernd in die Reaktion eingeht, so kann eine kleine Menge von
ihm eine sehr große Masse hydrolysieren, wenn für dauernde Abfuhr der
Reaktionsprodukte gesorgt ist.
Die Diastase findet sich in zahlreichen Pflanzenorganen, insbesondere
solchen, die viel Stärke enthalten, z. B. in Laubblättern, in keimenden
Samen. Der Diastasegehalt eines Organs ist aber kein konstanter,
vielmehr wird er nach Bedürfnis der Pflanze reguliert, auch kann die
Diastase durch Bildung von anderen Enzymen in ihrer Wirkung gehemmt
werden (Antienzyme). Hier treffen wir also wieder einen der vielen
Regulationsvorgänge, die für den Organismus so charakteristisch sind.
Fig. 252. Korrodierte Stärkekörner aus keimender Gerste.
1, 2, 3, 4 aufeinanderfolgende Stadien der Auflösung, die sich
in einem Samen nebeneinander vorfinden. Nach NOLL.
In der Pflanze wirkt die Diastase auf die Stärkekörner; diese
werden unter ihrem Einfluß korrodiert, sie schmelzen von außen
her ab, doch geht dieses Abschmelzen meist etwas unregelmäßig vor
sich, so daß die Körner andere Gestalt annehmen. An einzelnen Punkten
frißt die Diastase rascher in die Tiefe und zerlegt so das Korn unter
Benutzung präexistierender Spalten und Kanäle in kleinere Stücke, die
dann weiter abschmelzen (Fig. 252). Außerhalb[S. 230] der Pflanzen kann man
die Diastasewirkung am besten an einem dünnen Stärkekleister
demonstrieren, der nach Zusatz von Diastase im Laufe von Minuten oder
Viertelstunden in Zucker übergeht. Dementsprechend bemerkt man an
Proben, die man mit Jod versetzt, daß die anfangs bläuliche Färbung
einer weinroten, endlich einer gelben Farbe weicht.
Auch Zellulose, die chemisch der Stärke sehr nahe steht[188],
tritt häufig als Reservestoff auf. In dem Endosperm vieler Samen,
besonders auffallend bei den Palmen (z. B. Elfenbeinpalme), sieht man
die Zellwände außerordentlich stark verdickt. Die Verdickungsschichten
werden bei der Keimung aufgelöst. Auch hier ist ein Enzym tätig (die
sog. Zytase), das nun aber freilich nicht jede beliebige
Zellulose anzugreifen vermag. Gerade typische Zellulose (S. 31) wird
nicht von ihm angegriffen, wohl aber von einem anderen Enzym, der
Zellulase.
Das Inulin, das namentlich bei den Kompositen und Campanulaceen
verbreitet ist, geht in ähnlicher Weise aus Fruktose hervor wie
die Stärke und die Zellulose aus Maltose[188]. Im Gegensatz zu
diesen beiden Kohlehydraten kommt es stets nur in gelöstem Zustand in
den Pflanzen vor. Trotzdem ist es nicht ohne weiteres wanderungsfähig
und wird bei der Keimung durch ein Enzym in Fruktose abgebaut.
Auch Rohrzucker findet sich sehr häufig als Reservestoff
und wird in großer Menge z. B. im Zuckerrohr und in der Zuckerrübe
gespeichert. Er wird durch das weit verbreitete Enzym „Invertin“ in
Dextrose und Lävulose gespalten.
2. Die Fette.
Wenn wir auch die unter den Reservekohlehydraten erwähnten Körper nicht
aus Dextrose bzw. Lävulose herstellen können, so begreifen
wir doch, daß es der Pflanze ebenso leicht wird, sie aufzubauen wie
abzubauen. Viel schwieriger ist es zu verstehen, wie die Pflanze Fette
(Glyzerinester verschiedener Fettsäuren vgl. S. 25) aus Kohlehydraten
zu bilden vermag. Fette fehlen wohl keinem lebenden Protoplasma; es
wurde ja schon mehrfach auf die allgemeine Verbreitung des Lezithins
hingewiesen, das ein Fettderivat ist. In besonders großen Mengen
aber treten die Fette als Reservestoffe auf. In den assimilierenden
Laubblättern freilich nicht, wohl aber in vielen reifen Samen, wo sie
an Stelle verschwindender Kohlehydrate treten. Bei der Keimung werden
sie durch das Enzym Lipase in Fettsäure und Glyzerin zerspalten.
Die Fettsäure als solche kann die wasserdurchtränkte Zellhaut schon
leichter durchwandern als das Fett, doch dürfte sie in der Regel keine
allzu großen Strecken in der Pflanze zurücklegen, vielmehr rasch wieder
in Kohlehydrate verwandelt werden. — Fettes Öl findet sich außerdem im
Fleisch (Perikarp) mancher Früchte, z. B. bei der Ölpalme und Olive,
und wird dann nicht mehr in den Stoffwechsel der Pflanze hereingezogen.
3. Die Eiweißkörper.
Das Eiweiß findet sich in den Reservestoffbehältern teils gelöst, teils
in kristallinischer oder amorpher Form. Die Kristalle kommen frei
im Plasma, Zellkern oder den Chromatophoren vor, in den Samen aber
besonders in den Aleuronkörnern, wo sie von den Globoiden (vgl. S. 26)
begleitet werden.
Die hydrolytischen Abbauprodukte der Eiweißkörper sind vor allem
Aminosäuren, deren weite Verbreitung in der Pflanze schon erwähnt
wurde. Namentlich wenn eiweißreiche Samen, wie z. B. Rizinus. Pinus
u. v. a. keimen, wird man die reichlich auftretenden Aminosäuren
als Abbauprodukte des Eiweißes betrachten dürfen. Aminosäuren,
die sich an anderen Stellen vorfinden,[S. 231] können ja immer auch beim
Eiweißaufbau entstanden sein. Das Eiweißmolekül zerfällt
nicht sofort und nicht ausschließlich in Aminosäuren. Der Abbau
erfolgt durch allmähliche Zertrümmerung des Riesenmoleküls, wobei
zunächst Albumosen auftreten, dann Peptone und dann erst Aminosäuren.
Neben letzteren treten Ammoniak, ferner auch schwefelhaltige und
eventuell phosphorhaltige Spaltungsprodukte und wohl auch allgemein
Kohlehydratgruppen auf.
Dieser hydrolytische Abbau des Eiweißes vollzieht sich unter dem
Einfluß von „proteolytischen“ Enzymen (Proteasen), die aller
Wahrscheinlichkeit nach große Ähnlichkeit mit den entsprechenden
Enzymen des Tierkörpers haben. Demnach hätten wir zu unterscheiden:
- Pepsine, die das Eiweißmolekül nur wenig tief,
bis zu Albumosen und Peptonen abbauen.
- Erepsine, die das Pepton in Aminosäuren überführen.
- Trypsine, die Eiweiß direkt bis zu den Aminosäuren abbauen.
- Desamidasen, die aus Aminosäuren Ammoniak abspalten.
Die Abbauprodukte des Eiweißes erfahren in der Pflanze rasch
Veränderungen; demnach ist das Gemisch stickstoffhaltiger
organischer Verbindungen, das man bei einer im Dunkeln gehaltenen
Pflanze erhält, nicht identisch mit den durch Hydrolyse
von Eiweiß außerhalb der Pflanze entstehenden Spaltungsprodukten.
Es folgen nämlich in der Pflanze auf die primäre Spaltung sofort
wieder Synthesen, die z. B. zur Bildung von Amiden führen.
Unter den Amiden steht an Verbreitung das Asparagin obenan.
Sehr reichlich findet es sich in Gramineen und Leguminosen;
im Liter Preßsaft von Bohnenkeimlingen z. B. ist es bis zu 15
g enthalten. Bei Cruciferen und Cucurbitaceen wird es durch
das Glutamin ersetzt, während in den Koniferen einer
Diaminosäure, dem Arginin, die entsprechende Rolle
zuzufallen scheint. Bei solchen Synthesen wird vor allem die
Anhäufung des giftigen Ammoniaks vermieden; die Amidbildung
in der Pflanze hat eine ähnliche Bedeutung wie im Tierkörper die
Harnstoffbildung: Asparagin und Harnstoff sind also entgiftetes
Ammoniak. Die Entstehung von Asparagin ist aber an die Gegenwart
von unoxydierten C-Atomen (also z. B. Kohlehydrat) gebunden,
während Harnstoff aus NH3 und oxydiertem Kohlenstoff, also auch
im Hungerzustande, gebildet werden kann[191]. Noch weitergehend
sind die Synthesen am Licht, wo aus den Eiweißspaltprodukten sofort
wieder Eiweiß gebildet wird.
B. Wanderung der mobilisierten Reservestoffe.
Wenn die Reservestoffe durch geeignete Enzyme in lösliche Form gebracht
oder in Substanzen von kleinerem Molekulargewicht verwandelt sind,
dann können sie wandern; sie sind „mobilisiert“. Bei diesen Bewegungen
muß wie bei anderen Stoffwanderungen vor allem ein Diffusionsgefälle
gegeben sein und dauernd unterhalten werden. Das geschieht dadurch,
daß in den Zellen, die sich an die Reservestoffbehälter in größerer
oder kleinerer Entfernung anschließen, ein lebhaftes Wachstum
einsetzt. Solange dieses andauert, wird jedes zutretende Molekül rasch
verwandelt, Zucker z. B. in Zellulose umgebildet, und dadurch Platz
für nachrückende Moleküle geschaffen. Doch auch in nicht wachsenden
Organen, z. B. Kotyledonen, Endospermen, stellt die Pflanze ein
Diffusionsgefälle her, z. B. dadurch, daß die Zellen, die das Ziel
der Wanderung sind, ein stärkeres Kondensationsvermögen für Zucker
(Stärkebildung) haben als andere. Man kann das Diffusionsgefälle auch
künstlich herstellen, wenn man Reservestoffbehälter unter geeigneten
Vorsichtsmaßregeln einseitig mit großen Wassermassen in Berührung
bringt. Auf diese Weise ist es z. B. gelungen, künstlich eine
Entleerung von Samen, Zwiebeln usw. zu erzielen.
[S. 232]
Da jede Diffusionsbewegung nur sehr langsam sich vollzieht, so muß
bei Stofftransporten auf größere Entfernung eine Unterstützung
durch Massenbewegung hinzutreten. So werden z. B. im Frühjahr die
Reservestoffe, die im Holzkörper unserer Bäume deponiert sind, mit
dem aufsteigenden Wasserstrom in den Gefäßen fortgerissen. Im Inhalt
der Gefäße läßt sich zu dieser Zeit reichlich Glykose nachweisen.
Umgekehrt wird sich der Strom von mobilisierten Reserven, der von den
Laubblättern abwärts wandert, mindestens zum Teil in den Siebröhren
bewegen. Im einzelnen bedarf hier aber noch vieles der Aufklärung[192].
Auch eine andere Stoffwanderung aus dem Laubblatt hat man
beobachtet. Kurz vor dem Laubfall „vergilben“ die Blätter: während
die grünen Farbstoffe des Chlorophylls gelöst und weggeführt
werden, bleiben die gelben in den Chloroplasten zurück. Es werden
dann wenigstens manche, keineswegs alle, der im Blatt enthaltenen
nützlichen Stoffe in den Stengel übergeführt und bleiben so der
Pflanze erhalten. Die Zellwände, ein protoplasmatischer Wandbelag
und osmotisch wirksame Vakuolenstoffe bleiben, das Blatt
fällt in turgeszentem Zustand ab; Kalium, Phosphorsäure und
Stickstoff aber werden in den Stamm übergeführt[193].
C. Weitere Stoffmetamorphosen.
Regeneration der Reservestoffe. Früher oder später werden
die durch Enzyme mobilisierten Reservestoffe wieder in feste oder
hochmolekulare Stoffe zurückverwandelt. Dies geschieht unter allen
Umständen am Ende ihrer Wanderung, einerlei, ob sie da wieder als
Reservestoffe deponiert oder als Baustoffe verwendet werden. So
kann z. B. im Blatt gebildete Glykose in einen Samen oder in eine
Knolle wandern und dort zu Stärke oder zu Zellhaut transformiert
werden. Bei Wanderung auf größere Strecken findet aber nicht nur am
Endziele, sondern auch unterwegs fortwährend Reservestoffbildung
statt. Das läßt sich besonders gut für Stärke zeigen. Auf den Bahnen
der Zuckerwanderung kann in jeder Zelle sog. transitorische Stärke
gebildet werden. Durch diese vorübergehende Stärkebildung wird das
Konzentrationsgefälle unterhalten, das für dauernde Bewegung notwendig
ist.
Andere Stoffwechselprodukte[194]. Mit den bisher genannten
organischen Stoffen ist nur ein ganz kleiner Teil der „Pflanzenstoffe“
erwähnt. Es mag hier genügen, an die organischen Säuren, die
Gerbstoffe, die Glykoside, die Alkaloide, Farbstoffe, ätherischen
Öle, Harze, Gummiharze und Federharze (Kautschuk und Guttapercha)
zu erinnern, um auf die Legion von Stoffen hinzuweisen, die aus den
Assimilaten gebildet werden. Von diesen werden nur die organischen
Säuren bei späterer Gelegenheit noch Besprechung finden (S. 235);
alle anderen sind weder nach ihrer Entstehung noch nach ihrer
physiologischen Bedeutung genügend erforscht. Bekannt ist nur, daß
sie nach ihrer Bildung in der Regel keine weitere Verwendung mehr
finden. Vermutlich handelt es sich also um Nebenprodukte des
pflanzlichen Stoffwechsels.
Sie brauchen nicht nutzlos zu sein. In der Tat nimmt man für manche
bittere oder giftige Stoffe an, daß sie Tiere vom Genuß der Pflanze
abhalten; auch manche Farbstoffe sollen nützlich werden dadurch,
daß sie Tiere anlocken, die Pollen, Samen oder Früchte verbreiten,
oder daß sie schädliche Tiere abschrecken (Schreckfarben). Harze
und Federharze endlich können, wenn sie austreten und erhärten,
einen Wundverschluß liefern.
Das Reifen fleischiger Früchte. Eine sehr auffallende
Umwandlung von Stoffen findet auch bei der Reife fleischiger
Früchte statt. Auf einen freilich seltenen Fall (Fettbildung) ist
schon oben hingewiesen worden. Viel häufiger ist die Umwandlung von
Stärke in Zucker sowie das Verschwinden der organischen Säuren und
der Gerbstoffe. Die Früchte hören also auf, sauer oder bitter zu
schmecken, sie werden süß. In diesem Zustand werden sie dann zur
Nahrung für Tiere, die die Samen verbreiten. Die ganze Bedeutung
der chemischen Vorgänge liegt hier also auf ökologischem Gebiet.
Bei der höheren Pflanze wird niemals die ganze Menge der im
Assimilationsprozeß erzeugten organischen Substanzen zu Bau- und
Reservezwecken verwendet; stets wird ein Teil davon abgebaut, in
anorganische Substanz rückverwandelt. Neben der Assimilation existiert
immer auch Dissimilation. Die Bedeutung dieses Prozesses,
der im allgemeinen unter Sauerstoffaufnahme vor sich geht und als
Atmung bezeichnet wird, liegt nicht in dem Auftreten gewisser
Stoffe, sondern in dem Freiwerden von Energie, die für die
Pflanze unentbehrlich ist. Bei gewissen niederen Pflanzen erfolgt
aber der Gewinn an freier Energie unter Umständen in anderer
Weise. Hier werden meist organische Substanzen aus dem Substrat
aufgenommen und (ohne erst weiter assimiliert zu werden) sofort
wieder abgebaut. Im einzelnen können bei diesem Abbau Oxydationen,
Reduktionen oder Spaltungen erfolgen; alle diese Prozesse werden als
Gärungen zusammengefaßt. Andere niedere Organismen vermögen die bei
der Oxydation gewisser anorganischer Stoffe freiwerdende Energie
auszunützen. Diese verschiedenen Arten des Energiegewinnes sind durch
Übergänge verknüpft.
Schienen früher die einzelnen Formen der Dissimilation zwar
biologisch gleichwertig aber chemisch außerordentlich
verschieden, so bricht sich nach den Ausführungen WIELANDs
mehr und mehr die Anschauung Bahn, daß sie auch chemisch im
Prinzip immer wieder auf die gleichen Vorgänge hinauslaufen. Denn
es hat sich gezeigt, daß nicht dem Sauerstoff die aktive Rolle
zukommt, die man ihm früher zuschrieb, sondern daß überall der
Wasserstoff primär eingreift, daß Hydrierungen und Dehydrierungen
zuerst einsetzen und daß die Rolle des Sauerstoffs auch von anderen
Stoffen übernommen werden kann[195].
A. Atmung.
Unter Atmung in ihrer typischen Form versteht man die Oxydation
organischer Substanz zu Kohlensäure und Wasser; dazu ist die Aufnahme
von Sauerstoff aus der Umgebung nötig (vgl. S. 210).
Die Atmung der Pflanzen bietet sich dem Beobachter nicht so augenfällig
dar wie die der höheren Tiere. Wie sich die Ernährung der grünen
Pflanzen nur durch das eigens angestellte Experiment feststellen
ließ, so bedurfte es gleichfalls besonderer Versuche, um zu
erkennen, daß auch die Pflanzen atmen müssen, um zu leben,
daß sie ganz wie Tiere Sauerstoff aufnehmen und Kohlensäure abgeben.
SAUSSURE und DUTROCHET haben dies in den Jahren 1822
bis 1837 durch eingehende Untersuchungen bewiesen. Später leugnete
man freilich die Existenz der Atmung bei den Pflanzen wieder unter
Hinweis auf die Kohlensäurezersetzung und Sauerstoffabscheidung
bei der Assimilation; man konnte sich nicht vorstellen, daß beide
Prozesse gleichzeitig stattfinden könnten. Erst SACHS hat
dann der richtigen Anschauung zum Sieg verholfen. Assimilation und
Atmung sind zwei Lebensvorgänge, die ganz unabhängig voneinander in der
Pflanze bestehen. Während nur die grünen Pflanzenteile, und zwar
nur im Licht, bei der Assimilation Kohlensäure zerlegen und Sauerstoff
ausscheiden, atmen alle Organe der höheren Pflanzen — genau
wie die der Tiere — Tag und Nacht Sauerstoff ein und Kohlensäure
aus. Wird bei der Assimilation organische Substanz gewonnen, so
geht umgekehrt bei der Atmung solche verloren. Wenn grüne Pflanzen im
Licht einen bedeutenden Überschuß organischer Substanz gewinnen, so
verdanken sie diesen ausschließlich dem Umstande, daß die zeitweilige
Produktion durch die Assimilationstätigkeit der Chlorophyllkörper
die Verluste durch die ständige Atmung aller Organe übertrifft. So
genügt nach BOUSSINGAULTs Schätzungen beim Lorbeer[S. 234] 1 Stunde
Assimilation, um das Material für 30 Stunden Atmung zu beschaffen.
Unterdrückt man die Assimilation, läßt man die Pflanze im Dunkeln
verweilen, so verliert sie ganz beträchtlich an Trockengewicht.
Die Pflanzen produzieren in 24 Stunden durchschnittlich das
5–10fache ihres Volumens an Kohlensäure. Bei Schattenpflanzen
ist die Produktion meist auf das 2fache Volumen beschränkt; die
bekannte Zimmerpflanze Aspidistra bringt es aber nur auf die
Hälfte ihres Eigenvolumens und kann sich eben deshalb auch mit der
geringen Assimilation in sehr gedämpften Lichte begnügen.
Zum Nachweis der Atmung kann man entweder die Aufnahme von Sauerstoff
oder die Abgabe von Kohlensäure durch die Pflanze benutzen. Läßt
man eine Hand voll gequollener Samen am Grunde eines zylindrischen
Glasgefäßes keimen und hält dessen Glasstöpsel einen Tag lang
verschlossen, so ist der Sauerstoff, der sich zuvor in dem Gefäß
befand, von den Keimlingen aufgezehrt; eine Kerze erlischt, wenn man
sie in den Raum einführt. Eine andere Versuchsanordnung wird durch Fig.
253 illustriert. In dem umgekehrten Kolben befinden sich Keimpflanzen,
Blüten oder Hutpilze; durch den Wattepfropfen W werden sie am
Herabfallen verhindert. Die Öffnung des Kolbens ist durch Quecksilber
gesperrt, im Hals befindet sich Kalilauge K. Die auftretende
Kohlensäure wird dann von der Kalilauge absorbiert und das Quecksilber
steigt. Bei quantitativer Ausführung dieses Versuches ergibt sich,
daß ein Fünftel des Luftvolumens verschwindet, daß also der ganze
Sauerstoff der Luft aufgenommen wurde. Da sich aber ein abgeschlossenes
Luftvolumen bei der Atmung von Pflanzen (wenn keine Kalilauge zugegen
ist) nicht ändert, muß für jedes Volumen absorbierten Sauerstoffes
ein gleichgroßes Volumen CO2 gebildet werden; das Verhältnis
der ausgeschiedenen Kohlensäure zum aufgenommenen Sauerstoff, der
Atmungsquotient, ist gleich eins
(CO2O2 = 1).
Da Glykose das gewöhnliche Material der Atmung ist, so erfolgt
diese nach der Formel:
C6H12O6 + 6 O2
= 6 CO2 + 6 H2O.
Bei diesem Prozeß, der genau entgegengesetzt der CO2-Assimilation
verläuft, muß also auch Wasser auftreten, das freilich nicht
so bequem wie der Verbrauch von Sauerstoff und die Produktion
von Kohlensäure nachzuweisen ist. Quantitative Bestimmungen des
Trockengewichtsverlustes und der Kohlensäure zeigen, daß letztere nicht
ausreicht, den ersteren zu erklären; ein Teil der Trockensubstanz muß
also zu Wasser geworden sein.
Fig. 253. Atmungsversuch. Der bauchige Teil des
Glasgefäßes B ist mit Wucherblumen gefüllt. Die durch ihre Atmung
erzeugte Kohlensäure wird von der Kalilauge K absorbiert und die
Absorption angezeigt durch das Steigen des Quecksilbers Q in der
Röhre. Nach NOLL.
Aber nicht unter allen Umständen bleibt ein abgeschlossenes Gasvolumen
unter der Atmungstätigkeit von Pflanzen unverändert, nicht immer ist
die[S. 235] auftretende Kohlensäure volumgleich mit dem verschwindenden
Sauerstoff. Kleine Abweichungen von diesem Verhältnis findet man wohl
bei allen Pflanzen, beträchtliche z. B. beim Keimen fetthaltiger
Samen und bei Blättern gewisser Sukkulenten (Crassulaceen). Das hängt
damit zusammen, daß bei diesen Samen Fette veratmet werden,
die viel sauerstoffärmer sind als die Kohlehydrate, und daß bei den
Crassulaceen aus Kohlehydraten nicht Kohlensäure und Wasser, sondern
bestimmte organische Säuren entstehen. Auch bei anderen Pflanzen treten
solche Säuren auf, wenn auch nicht in so großer Menge. Sie dürften
zum größten Teil beim Atmungsprozeß entstehen, doch können sie wohl
auch im aufbauenden Stoffwechsel erzeugt werden. Vor allem ist da die
Oxalsäure zu nennen, deren weite Verbreitung in Form von oxalsaurem
Kalk ja bekannt ist. Ihre Bildung geht in dem Maße weiter, als ihre
Neutralisation durch Kalk ermöglicht wird. Ohne solche Neutralisation
aber wird ihre Entstehung eingeschränkt. Selbst in oxalatfreien
Pflanzen kommt es zu einer vorübergehenden Bildung von Oxalsäure, die
aber dann rasch durch ein oxydierendes Enzym weiter oxydiert wird.
Bei der Keimung fettreicher Samen wird sehr viel mehr Sauerstoff
aufgenommen, als Kohlensäure abgegeben wird; oft so viel, daß
in den ersten Tagen im Dunkeln trotz bestehender Atmung eine
Zunahme des Trockengewichts erfolgt. Der Atmungsquotient ist also
kleiner als 1. Die Hauptmenge dieses Sauerstoffes wird zur
Umwandlung der sauerstoffarmen Fette in Kohlehydrate verbraucht;
nur ein kleiner Teil dient zur Atmung.
Bei den Crassulaceen ist die Oxydation der Kohlehydrate
eine unvollständige; nur ein Teil ihres Kohlenstoffs wird in
CO2 übergeführt, der übrige zur Bildung organischer Säuren
verwandt. Deshalb erscheint weniger Kohlensäure, als man nach
der Sauerstoffaufnahme erwarten sollte. Der Atmungsquotient ist
kleiner als 1. — Dieser eigenartige Atmungsprozeß, der mit
einer schon am Geschmack wahrnehmbaren Säurebildung im Zellsaft
verbunden ist, hat für sukkulente Pflanzen eine große ökologische
Bedeutung. Die auftretenden Säuren (Äpfelsäure und Oxalsäure vor
allem) geben nämlich am Licht Kohlensäure ab. Diese kann dann
sofort im Assimilationsprozeß wieder Verwertung finden, während
bei der typischen Atmung wenigstens alle in der Nacht entstandene
CO2 entweicht und für die betreffende Pflanze verloren geht. Die
Sukkulenten sparen also mit ihrem C-Vorrat, und das dürfte damit
zusammenhängen, daß sie sich nicht so leicht mit Kohlensäure aus
der Luft versehen können wie gewöhnliche Pflanzen. (Verringerung
des Gasaustausches wegen Einschränkung der Transpiration.)
Wie gesagt, ist die Atmung bei den höheren Pflanzen allgemein
verbreitet; sie findet sich nicht nur bei Pflanzenteilen, die
chlorophyllfrei sind (vgl. die Versuche S. 234), sondern sie läßt sich
auch in chlorophyllhaltigen Zellen nachweisen. Hier wird sie freilich
am Licht durch den quantitativ sehr überwiegenden Assimilationsprozeß
überdeckt; sie äußert sich also nur in einer Verminderung der
Assimilationsprodukte. Dämpft man das Licht mehr und mehr ab, so
verschwindet die Assimilation schließlich ganz, und die Atmung zeigt
sich in voller Klarheit.
Wenn demnach Atmung in jeder einzelnen Zelle stattfindet, so ist sie
in verschiedenen Organen und unter verschiedenen äußeren Bedingungen
doch ihrer Intensität nach außerordentlich verschieden. Lebhaft
wachsende Pflanzenteile, junge Pilze, keimende Samen, Blütenknospen
und vor allem die Infloreszenzen von Araceen und Palmen zeigen eine
sehr energische Atmungstätigkeit. Diese übertrifft bei manchen
Bakterien und Pilzen nicht unbeträchtlich die des Menschen, wenn man
sie an gleichen Gewichtsmengen Körpersubstanz mißt. In der Mehrzahl
der Fälle, insbesondere bei Pflanzenteilen, die ganz oder überwiegend
aus ausgewachsenen Geweben bestehen, ist aber der Sauerstoffkonsum
und entsprechend die Kohlensäureproduktion ganz bedeutend geringer
als bei den warmblütigen Tieren. — Unter den äußeren Einflüssen,
die von Wichtigkeit für die Intensität der Atmung sind, müssen vor
allem die Temperatur und der Sauerstoff genannt werden. Wie auf
alle[S. 236] Lebensvorgänge, so wirkt auch auf die Atmung eine Zunahme der
Temperatur zunächst beschleunigend. Die Produktion der Kohlensäure wird
genau in der gleichen Weise wie viele andere chemische Vorgänge durch
eine Temperatursteigerung um 10° ungefähr verdoppelt bis verdreifacht.
Bald folgt aber auch hier mit weiter steigender Temperatur eine Abnahme
der Atmung. Im Gegensatz zu anderen Erscheinungen ist der abfallende
Ast der Kurve bei der Atmung ganz außerordentlich steil, so daß Optimum
und Maximum fast zusammenfallen.
Man pflegt die Atmung als einen Verbrennungsprozeß zu bezeichnen.
Wenn das richtig ist, sollte man glauben, daß die Menge des zur
Verfügung stehenden Sauerstoffes von fundamentaler Wichtigkeit sei;
insbesondere sollte man erwarten, daß die Atmung in reinem Sauerstoff
enorm gesteigert, im sauerstoffreien Raum völlig sistiert werde.
Keines von beiden trifft zu. In reinem Sauerstoff ist die Atmung
nicht wesentlich vermehrt, und erst bei einem Sauerstoffdruck von 2–3
Atmosphären macht sich anfänglich eine Zunahme der Atmung bemerkbar,
der aber bald eine als Absterbeerscheinung zu deutende Abnahme
folgt. — Sehr viel auffallender ist die Tatsache, daß Pflanzen ohne
Gegenwart von Sauerstoff fortfahren, Kohlensäure zu produzieren. Hier
kann natürlich von einem „Verbrennungsprozeß“ nicht mehr die Rede
sein; man spricht von „intramolekularer Atmung“[196], weil die
auftretende Kohlensäure ihre Entstehung einer Umlagerung von Atomen im
Molekül des Atmungsmateriales (Zucker) verdankt. Dabei zerfällt das
Zuckermolekül und bildet neben Kohlensäure stets andere, nämlich stark
reduzierte Substanzen, manchmal z. B. Alkohol, nach der
Formel:
C6H12O6 =
2 C2H6O + 2 CO2.
Nimmt man statt dieser empirischen die Strukturformeln:
COH · CHOH · CHOH · CHOH · CHOH · CH2OH
= CO2 + CH3 · CH2OH + CH3CH2OH + CO2,
so sieht man, daß das Zuckermolekül in vier Stücke zerbricht, von denen
zwei sauerstoffärmer, die beiden anderen sauerstoffreicher sind als die
Molekülgruppen, aus denen sie hervorgehen. Es entziehen also bei dieser
Art von Atmung gewisse Molekülgruppen anderen den gebundenen
Sauerstoff.
Man nimmt an, daß Sauerstoffatmung und intramolekulare Atmung der
Ausdruck ein und derselben Befähigung der Pflanze sind, mit anderen
Worten, daß die Sauerstoffatmung bei Entziehung des Sauerstoffes in
eine intramolekulare Atmung übergeht. Wenn das zutrifft, dann wird
man sagen müssen, daß das Wesentliche bei der Atmung gar nicht
in einer Oxydation besteht, sondern in einer Veränderung des
Atemmaterials, bei der Produkte entstehen, die leicht Sauerstoff
aufnehmen können. Die Materialien, die in der Pflanze veratmet werden,
Kohlehydrate und Eiweiß, werden bei gewöhnlicher Temperatur nicht
leicht oxydiert. Fette freilich, die auch als Atmungsmaterial dienen
können, sind oxydabel; allein von ihnen wissen wir, daß sie in der
Pflanze zuerst in Kohlehydrate verwandelt werden, ehe sie der Atmung
verfallen. Die Pflanze muß also über Mittel besonderer Art verfügen, um
die Oxydation ihrer Reservestoffe zu bewerkstelligen.
Die Vorstellungen, die man sich über den Chemismus der Atmung
machen kann, haben sich in den letzten Jahren, vor allem durch
die chemischen Studien WIELANDs und die physiologischen
PALLADINs sehr geklärt[197]. Man darf jetzt annehmen, daß
bei der Verbrennung des Atmungsmaterials der primäre Vorgang nicht
etwa in einer Aktivierung des Sauerstoffs, im Auftreten von Ozon
besteht, denn sonst müßten zahllose Stoffe in der Zelle verbrannt
werden, und es könnte nicht eine „auswählende“ Verbrennung
bestehen derart, daß leicht oxydable Stoffe verschont bleiben,
schwer oxydable spielend oxydiert werden. Vielmehr besteht der
primäre Vorgang in einer Anlagerung von[S. 237] Wasser an das
Atmungsmaterial, z. B. an den Zucker. Hierauf erst findet eine
Oxydation statt, und zwar in Form von Wasserstoffentziehung
(Dehydrierung), wobei gleichzeitig CO2 frei wird.
WIELAND konnte nämlich zeigen, daß die Glykose in
Gegenwart von Palladiumschwarz als Katalysator auch unter
Sauerstoffabschluß bei niederen Temperaturen H und CO2
abgibt; der Wasserstoff wird an das Palladiumschwarz gebunden,
so daß sich der Prozeß mit zunehmender Wasserstoffsättigung des
Palladiums abschwächt, jedoch weitergeht bei Gegenwart eines
Wasserstoffakzeptors. Als solcher kann bei Luftabschluß
z. B. Methylenblau unter Bildung seiner Leukoverbindung dienen,
während bei Luftzutritt der Sauerstoff dessen Rolle übernimmt,
wobei dann Wasser entsteht.
In den höheren Pflanzen scheint nun das Palladiumschwarz durch
gewisse Enzyme ersetzt zu sein, die den Wasserstoff von der
Additionsverbindung des Zuckers mit Wasser auf leicht reduzierbare
Substanzen übertragen. Diese H-Akzeptoren hat PALLADIN
„Atmungspigmente“ genannt. Von den Atmungspigmenten aber geht
dann der Wasserstoff unter Einwirkung sog. Oxydasen an den
Luftsauerstoff über, und es entsteht Wasser. Die Oxydasen,
die auch Enzyme sind, übertragen also den Luftsauerstoff nicht
auf den Zucker, sondern auf den Wasserstoff, der auf dem
beschriebenen Wege aus dem Zucker frei gemacht wird. Sie sind in
den Pflanzen ganz allgemein nachgewiesen.
Im Schema würde sich also die Zuckerveratmung so vollziehen:
1.
C6H12O6 + 6 H2O
=
6 CO2 + 24 H
C6H12O6 + 6 O2
=
6 CO2 + 6 H2O
Fehlt der freie Sauerstoff, so muß natürlich die Gleichung einen
anderen Verlauf nehmen. Entweder wird, wie bei gewissen Gärungen
(S. 238), der Wasserstoff als solcher ausgeschieden, oder er wird
zur Bildung weniger hoch oxydierter Verbrennungsprodukte, z. B.
Alkohol, verwendet. Aller Wahrscheinlichkeit nach entstehen bei der
Atmung aus dem Zucker zunächst die Stoffe, von denen unten bei der
alkoholischen Gärung zu reden sein wird.
Auf den ersten Blick erscheint die Atmung als ein widersinniger Prozeß;
es wird ja doch organische Substanz, die im Assimilationsprozeß
aufgebaut wurde, wieder zerstört. Verständlich wird die Atmung
erst, wenn man von ihrer stofflichen Seite ganz absieht und die
energetische ins Auge faßt. In der Tat kommt es bei der Atmung
für die Pflanze gar nicht in Betracht, daß CO2 und Wasser entstehen;
wichtig ist einzig und allein, daß freie Energie auftritt.
Solche muß beim Abbau z. B. von Kohlehydraten entstehen, da ja zu deren
Aufbau, wie wir sahen, ein Energieaufwand nötig ist; und diese frei
werdende Energie ist für die Pflanze unentbehrlich, sie liefert die
Betriebskraft für zahlreiche Lebenserscheinungen. Dementsprechend
steht bald nach der Entziehung des Sauerstoffs die Protoplasmabewegung
still; das Wachstum und die Reizbewegungen werden eingestellt. Alle
diese Lebensäußerungen aber beginnen wieder, wenn nach nicht zu
langer Zeit Sauerstoff von neuem der Pflanze zur Verfügung gestellt
wird. Man kann sich wohl vorstellen, daß der Organismus Einrichtungen
besäße, mit deren Hilfe er die ihm direkt von außen zufließende
Energie, Licht und Wärme, als Betriebsenergie verwenden könnte.
Allein tatsächlich sehen wir, daß er es vorzieht, die Sonnenenergie
in Form von chemischer Energie zu speichern. Das hat für die Pflanze
vor allem den einen großen Vorzug, daß die gespeicherte Energie durch
Stoffwanderung außerordentlich leicht an andere Orte
geführt werden kann (auch an solche, die wie z. B. Wurzeln im Dunkeln
leben und nicht selbst die Speicherung ausführen können), und daß sie
auch zu Zeiten verwendet werden kann, in denen ein Energiegewinn
unmöglich ist (z. B. nachts).
Auch durch intramolekulare Atmung wird Energie frei; doch genügt diese
bei den meisten Organismen nicht zur Aufrechterhaltung des ganzen
normalen Lebensbetriebes, bei vielen nur zu kümmerlicher Fristung
des Lebens. Während manche Samen bei intramolekularer Atmung viele
Stunden oder[S. 238] Tage am Leben bleiben und einzelne sogar die gleiche
Kohlensäuremenge ausgeben wie bei Sauerstoffatmung — bei der Mehrzahl
vermindert sich diese Menge rasch —, tritt bei anderen Pflanzen
wahrscheinlich durch die als Gifte wirkenden reduzierten Stoffe bald
der Tod ein, und die Leistung der intramolekularen Atmung ist hier in
jeder Hinsicht ganz unbedeutend. Umgekehrt erlangt sie bei gewissen
Organismen, von denen alsbald die Rede sein soll, ganz außerordentlich
große Bedeutung.
B. Oxydation anorganischer Substanz[198].
Während die meisten Pflanzen in der geschilderten Weise organische
Substanz, vor allem Kohlehydrate, veratmen, haben sich gewisse
Bakterien ganz andere Energiequellen erschlossen. So oxydieren die im
Boden weitverbreiteten Nitritbakterien Ammoniak zu salpetriger Säure,
und die mit ihnen vergesellschafteten Nitratbakterien oxydieren die
salpetrige Säure weiter zu Salpetersäure. Mit Hilfe der so gewonnenen
freien Energie vermögen sie dann — wie S. 219 schon berichtet wurde —
auch Kohlensäure zu assimilieren; die frei gemachte chemische Energie
tritt an die Stelle der Sonnenenergie bei den typischen autotrophen
Pflanzen. Ein Abbau organischer Substanz fehlt hier völlig; es wird
also die Gesamtmenge assimilierter Nahrung dauernd festgehalten, so daß
diese Organismen außerordentlich ökonomisch arbeiten. Da aber das ihnen
zur Verfügung stehende Ammoniak nur in begrenzter Menge und nur von
anderen Organismen geliefert wird, so können die Nitrobakterien keine
so dominierende Stellung in der Natur einnehmen wie die grünen Pflanzen.
An die Nitrobakterien schließen sich die Schwefelbakterien an,
die Schwefelwasserstoff zu Schwefelsäure oxydieren, wobei sie
intermediär Schwefel bilden und als Reservestoff in ihrem Körper
speichern. In ähnlicher Weise gewinnen andere Bakterien bei
der Oxydation von Methan zu Kohlensäure und Wasser die nötige
Betriebsenergie. Daß diese überall in erster Linie zur Synthese
organischer Substanz aus CO2 dient, ist sehr wahrscheinlich.
Im Gegensatz zu diesen streng spezialisierten, autotrophen
Bakterien ist die Verbrennung von Wasserstoff nicht die Eigenschaft
ganz bestimmter „Wasserstoffbakterien“, sondern sie wird weit
verbreitet von gewöhnlichen, typisch heterotrophen Bakterien
ausgeübt, die daneben auch die Verbrennung organischer Substanz
vollziehen. Offenbar ist es bei den sog. Eisenbakterien
(z. B. Leptothrix ochracea) ähnlich, die vielleicht Eisen und
Mangan nur dann wirklich nötig haben, wenn ihnen wenig geeignete
organische Substanzen geboten werden.
C. Gärungen[199].
Mit der Entziehung des Sauerstoffes tritt intramolekulare Atmung ein;
diese vermag zwar nicht bei höheren Pflanzen, wohl aber bei niederen
die zur dauernden Erhaltung des Lebens nötige freie Energie zu liefern.
Viele Bakterien, Pilze, auch gewisse Algen (Characeen) sind in
auffallender Weise unabhängig vom Sauerstoff, sie nehmen mit geringen
Spuren von ihm vorlieb, oder sie fliehen ihn überhaupt gänzlich und
leben an sauerstoffreien Orten. Im Gegensatz zu dem verbreitetsten
Typus von Organismen, die man aërobe oder Aërobionten nennt, heißen sie
anaërobe oder Anaërobionten. Die beiden Extreme sind durch allerlei
Abstufungen verbunden. Die echten Anaërobionten zersetzen organische
Substanzen in sehr großen Mengen, und diese Zersetzung, die im Prinzip
mit den Vorgängen bei der intramolekularen Atmung identisch ist,
nennt man Gärung. Wie dort, so handelt es sich auch hier um Gewinnung
gebundenen Sauerstoffs.
Das Prototyp der Gärung ist die alkoholische Gärung, die ganz
besonders von Hefepilzen verursacht wird. Hier wird Zucker in Alkohol
und Kohlensäure[S. 239] zerspalten, und dieser Vorgang hat bekanntlich
eine große Bedeutung in der Technik (Bier-, Wein-,
Branntweinbereitung). Der chemische Prozeß selbst ist der
gleiche wie in der grünen Pflanze, die intramolekular atmet; im
Gegensatz zu dieser aber vermag die Hefe in der Gärung einen vollen
Ersatz für die Atmungstätigkeit zu finden. Sie ist aber nur so
lange unabhängig vom Sauerstoff, als ihr ein geeignetes Gärmaterial
(Zucker) zur Verfügung steht. Fehlt Zucker, so ist der Sauerstoff
unentbehrlich, und es findet dann normale Atmung statt; ist Zucker
und Sauerstoff gegeben, so tritt gleichzeitig Atmung und Gärung
ein, es wird also ein Teil des Zuckers zu Kohlensäure und Alkohol, ein
anderer zu Kohlensäure und Wasser verarbeitet. Es leuchtet ein, daß die
Verarbeitung von Zucker zu Alkohol und Kohlensäure viel weniger Energie
liefert als die vollständige Verbrennung zu Kohlensäure und Wasser.
Deshalb versteht man es, daß die Hefe ungeheure Massen von Zucker
verbraucht. Nur etwa 2% Zucker in der Nährlösung wird zum Aufbau von
Körpersubstanz verbraucht (assimiliert), der Rest wird vergoren. Zur
Ausführung einer so weitgehenden Spaltung des Zuckers besitzt die Hefe
spezifische Enzyme, vor allem die Zymase.
Wenn bei der normalen Atmung die gesamte Verbrennungswärme der
Glykose = 709 cal frei wird, so ist davon bei der Alkoholgärung
die Verbrennungswärme von 2 Molekülen Alkohol 2 × 326 = 652 cal
abzuziehen: somit werden nur 57 statt 709 cal frei.
Daß die chemische Tätigkeit der Hefe auf der Wirkung von Enzymen
beruht[200],
hat man schon lange vermutet. Erst E. BUCHNER
aber gelang es, diese vom lebenden Plasma zu trennen. Da die
„Zymase“ im Gegensatz zu anderen Enzymen nicht aus der Zelle
herauszudiffundieren vermag, mußte sie nach Aufreißen der Zellen
durch hohen Druck aus diesen herausgepreßt werden. Die Zymase ist
übrigens kein einheitliches Enzym, sondern ein Gemisch vieler
Enzyme, die zusammen schließlich zur Alkoholbildung führen.
Im einzelnen dürfte nach NEUBERG[201] dieser Prozeß
etwa so verlaufen, daß zunächst das Zuckermolekül in 2 Moleküle
Methylglyoxal übergeführt wird, die Brenztraubensäure liefern;
diese wird durch CO2-Abspaltung in Acetaldehyd übergeführt.
Acetaldehyd aber läßt sich als Zwischenprodukt der Gärung
nachweisen und wird seinerseits auch direkt von der Hefe zu Alkohol
reduziert.
Wenn auch die Hefe durch ihr Gärvermögen weitgehend unabhängig
vom Sauerstoff ist, so kann man sie doch nicht zu den streng
anaëroben Organismen rechnen, da sie in ihrem Wachstum durch
freien Sauerstoff stark gefördert wird. Dementsprechend wird, da der
chemische Umsatz von der Zahl der Hefezellen abhängt, mit der Zeit
mehr Alkohol bei Sauerstoffzutritt als ohne solchen gebildet werden
können. Andere Gärungsorganismen werden aber von freiem Sauerstoff
direkt geschädigt und leben dementsprechend in der Natur nur an Orten,
wo solcher fehlt. Zu diesen echten Anaërobionten gehören vor allem
die Buttersäurebakterien, die Kohlehydrate aller Art, höhere
Alkohole und Salze der Milchsäure in Wasserstoff und organische Säuren
überführen, unter denen die Buttersäure nie fehlt. Sie spielen, da sie
auch die sonst so schwer angreifbare Zellulose lösen, eine sehr große
Rolle in der Natur: sie führen den von den Pflanzen in ihren Membranen
festgelegten Kohlenstoff wieder in eine Form über, die eine weitere
Verwendung durch andere Organismen gestattet, sie verhindern also ein
Ausscheiden großer Mengen Kohlenstoffs aus dem Kreislauf der Stoffe (S.
240).
Auch bei der Buttersäuregärung soll das Kohlehydrat zunächst in
Brenztraubensäure übergeführt werden, aus der dann durch eine
Umlagerung Buttersäure neben freiem Wasserstoff auftritt. Indem für
letzteren der freie Stickstoff als „Akzeptor“ fungiert, soll es
zur Bildung von Ammoniak, also zur Stickstoffbindung kommen; vgl.
224[195].
Es können hier nicht alle Gärungen angeführt werden. Es sei nur noch
die Milchsäuregärung genannt, die in der Milchwirtschaft (Sauermilch,
Käsebereitung, Kumys, Kefir) und auch sonst in der Praxis (Säuerung
von Ge[S. 240]müsen, z. B. Sauerkraut) eine große Rolle spielt. Auch sie geht
anaërob vor sich, indem das Zuckermolekül entweder direkt in zwei
Moleküle Milchsäure gespalten wird oder neben Milchsäure auch CO2
und H2 entstehen läßt.
War bisher vorwiegend von der Vergärung der Kohlehydrate
die Rede, so muß jetzt noch hervorgehoben werden, daß alle,
auch die komplizierteren organischen Verbindungen des Pflanzen- und
Tierkörpers, vor allem also die Eiweißkörper, vergoren werden können.
Die Eiweißvergärung pflegt man als Fäulnis zu bezeichnen,
wenn sie ohne Sauerstoffzufuhr erfolgt, als Verwesung, wenn
Oxydationen möglich sind. In der Natur treten zunächst gewöhnlich
aërobe Bakterien bei der Eiweißvergärung auf, die den anaëroben den Weg
bahnen; eine scharfe Grenze zwischen Verwesung und Fäulnis existiert
also nicht. Unter allen Umständen wird das Eiweiß zunächst hydrolytisch
gespalten. Es treten also die früher erwähnten Spaltungsprodukte auf,
vor allem Aminosäuren. Diese werden weiter verändert, zumeist unter
Abspaltung ihrer NH2-Gruppe und auch durch noch tiefergreifenden
Abbau. Vielfach treten dann auch übelriechende Substanzen, wie Indol
und Skatol, auf, die aber nicht bei jeder Eiweißgärung sich bilden
müssen.
Es ist nicht möglich, eine scharfe Grenze zu ziehen zwischen den
Dissimilationsvorgängen, die mit Eingreifen des Luftsauerstoffes und
solchen, die ohne dieses sich vollziehen. Als Gärung müssen alle
Dissimilationsprozesse, die von der typischen Atmung abweichen,
bezeichnet werden. Demnach wäre auch die Entstehung von Äpfelsäure
und Oxalsäure bei den Crassulaceen so gut wie die Entstehung von
Oxalsäure bei Pilzen und Bakterien als Gärung zu bezeichnen. Und eine
ganz typische Oxydationserscheinung, die Überführung des Alkohols in
Essigsäure und Wasser, die durch die Essigbakterien bewirkt wird, muß
ebenfalls als Gärung betrachtet werden.
Wenn allen diesen Vorgängen im Grunde ein Gewinn von Sauerstoff
gemeinsam ist, bald von freiem, bald von gebundenem, so müssen
schließlich auch solche Prozesse, bei denen anorganische Stoffe
den Sauerstoff liefern, hier angeschlossen werden. So leben gewisse
Bakterien anaërob, wenn ihnen Nitrate als Sauerstoffquelle
dienen, wenn also diese Nitrate etwa bis zu freiem Stickstoff
reduziert werden (Denitrifikation), oder wenn Sulfate etwa in
Schwefelwasserstoff übergeführt werden (Sulfatreduktion). Es
gibt sogar Bakterien (Micrococcus selenicus), die zwar nicht anaërob
sind, aber doch den freien Sauerstoff nicht zu nützen vermögen,
vielmehr nur aus leicht reduzierbaren Stoffen, wie Natriumselenit,
Natriumthiosulfat, Indigkarmin oder Methylenblau Sauerstoff entnehmen
können[202].
Kreislauf der Stoffe. Werden organische Substanzen, wie das
in der Natur mit den Resten abgestorbener oder den Exkreten lebender
Organismen stets geschieht, verschiedenartigen Mikroorganismen
preisgegeben, so arbeiten sich diese wechselseitig in die Hand, und
Stoffwechselprodukte der einen werden von anderen weiter zersetzt, bis
nur anorganische Materie übrig ist, bis die organischen Verbindungen
„mineralisiert“ sind. Als Endprodukte treten Kohlensäure,
Wasser, Wasserstoff, Methan, Ammoniak, Stickstoff, Schwefelwasserstoff
auf.
Alle diese Endprodukte der Gärung können wieder von anderen Organismen
verwertet werden. Sehen wir von CO2 und H2O ganz ab, da
diese zur Genüge besprochen sind, so wäre hervorzuheben, daß der
Wasserstoff, das Methan, das Ammoniak und der Schwefelwasserstoff durch
spezifische Bakterien oxydiert werden, daß der Stickstoff durch
wieder andere Bakterien assimiliert wird. Durch dieses Zusammenarbeiten
aller Organismen wird die einseitige Bildung und Anhäufung eines
Stoffes verhindert; es entsteht ein fortwährender Kreislauf der Stoffe,
durch den sich das Leben dauernd auf der[S. 241] Erde erhält. Existierte nur
ein Typus von Organismen, so hätte dieser in kurzer Zeit durch
seinen einseitigen Stoffwechsel sich jede Lebensmöglichkeit unterbunden.
D. Entwicklung von Wärme und Licht bei
Atmung und Gärung.
Wärme[203]. Da die typische Atmung ein Oxydationsprozeß ist, so
wird es begreiflich, daß eine Wärmeentwicklung mit ihr verbunden ist.
Daß sich Pflanzen durch die Atmung aber meistens nicht fühlbar
erwärmen, rührt daher, daß diese nicht ausgiebig genug ist, und daß
die Transpiration der großen Flächen erhebliche Wärmeverluste
herbeiführt, wodurch transpirierende Pflanzen meist sogar kühler
als ihre Umgebung sind. — Auch bei einigen Gärungen werden nicht
unbeträchtliche Wärmemengen frei, z. B. bei der Alkoholgärung. Bekannt
ist ferner die Erhitzung, die in faulendem Mist eintritt und die in den
„Mistbeeten“ der Gärtner ausgenutzt wird.
Werden Transpiration und Wärmestrahlung verhindert und werden
zur Untersuchung lebhaft atmende Pflanzen ausgewählt, dann läßt
sich in der Tat eine Erwärmung nachweisen; sehr deutlich z. B.
mit angehäuften Keimpflanzen oder Blütenknospen. Eine starke
Erwärmung ist an blühenden Kolben von Araceen beobachtet worden,
die ihre Temperatur um 10–20° C durch intensive Atmung erhöhen.
Ein Gramm der Kolbensubstanz liefert dabei in einer Stunde bis
zu 30 Kubikzentimeter CO2, und in kurzer Zeit kann bei so
intensiver Atmung die Hälfte der Trockensubstanz, der ganze Vorrat
an Zucker und Stärke, veratmet werden. Auch in den großen Blüten
der Victoria regia sind 15° C Temperaturerhöhung gemessen worden.
Diese hohen Temperaturen in Blüten und Infloreszenzen locken
Bestäubung vermittelnde Insekten an. — Besonders hohe Temperaturen
erhält man, wenn Blüten, Blätter usw. in Dewargefäßen (z. B.
Thermosflaschen) vor Wärmeverlusten bewahrt werden. Die Temperatur
steigt dann unter Umständen auf 40–50° C und die Objekte sterben
ab. Nach ihrem Tode erhöht sich dann die Temperatur durch die
Tätigkeit von Mikroorganismen weiter.
Nach Verwundung wird die Atmung und auch die Erwärmung merklich
gesteigert; das Gegenteil wird bei Hungerzuständen beobachtet.
Auch bei der Tabakfermentation findet eine beträchtliche
Temperatursteigerung statt, und noch größer fällt diese aus, wenn
feuchtes Heu oder feuchte Baumwolle in größeren Massen aufgetürmt
sich selbst überlassen werden; es kommt dann durch die Entstehung
leichtentzündlicher Gase schließlich zur Selbstentzündung. Am
genauesten sind die Verhältnisse der Selbsterhitzung des Heus
untersucht. Hier tritt zunächst durch die Atemtätigkeit des
Bacillus coli ein Steigen der Temperatur bis auf 40° C ein; eine
ganze Reihe von thermophilen Schimmelpilzen und Bakterien, deren
Minimum etwa bei dieser Temperatur liegt, stellt sich dann ein;
unter ihnen ist es namentlich der Bacillus calfactor, der die
Temperatur bis zu 70° steigert. Schließlich gehen alle Organismen
an der durch sie selbst produzierten Temperatur zugrunde; das Heu
ist steril geworden.
Leuchten[204]. Unter denselben Bedingungen, die die Atmung
unterhalten, bemerkt man bei einer beschränkten Zahl von Pflanzen
ein Leuchten. Die bekanntesten leuchtenden Pflanzen sind
gewisse Bakterien und das früher als „Rhizomorpha“ beschriebene
Myzelium eines baumtötenden Hutpilzes, der Armillaria mellea.
Auf toten Fischen und Fleisch treten oft Überzüge unschädlicher
leuchtender Bakterien (Bacterium phosphoreum, Pseudomonas lucifera
u. a.) auf. Das Leuchten zahlreicher Tiere scheint durchweg von
Bakterien hervorgebracht zu werden, die in ganz bestimmten Organen
dieser Tiere regelmäßig zur Entwicklung kommen.
Das Leuchten verschwindet in sauerstoffreier Umgebung, stellt
sich aber bei Zutritt freien Sauerstoffs sofort wieder ein.
Leuchtbakterien lassen sich deshalb als empfindliches Reagens zum
Nachweis der Assimilation benutzen. Alle Umstände, die die Atmung
fördern, verstärken das Leuchten und umgekehrt. Ein Nutzen der
Lichtentwicklung für die Produzenten ist nicht bekannt.
Die Entwicklungsphysiologie soll hier in drei Abschnitten
behandelt werden. Der erste bringt Vorbemerkungen, die
rein deskriptiv manches näher ausführen, was schon
im morphologischen Teil besprochen ist. Dann erst folgt die
kausale Entwicklungsphysiologie, deren Ziel ist, die
aufeinanderfolgenden Vorgänge der Entwicklung nach ihren Ursachen zu
verstehen und willkürlich abzuändern. Die Ergebnisse sind freilich zur
Zeit noch weit vom Ziel entfernt, wir haben noch viel mehr Probleme als
Lösungen.
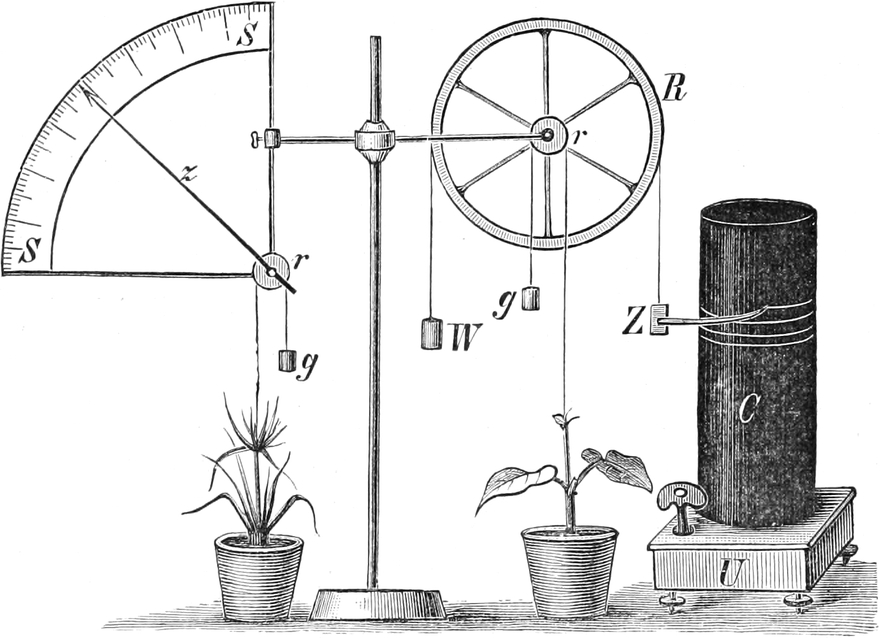
Fig. 254. Wachstumsmesser (Auxanometer). Links
ein einfaches Hebelauxanometer (Zeiger am Bogen), rechts ein
selbstregistrierendes Auxanometer. Nähere Erklärung im Text. Nach
NOLL.
Es empfiehlt sich, diese Probleme in doppelter Weise zur Anschauung
zu bringen: es sollen im 2. Abschnitt die Faktoren, die
von Einfluß auf die Entwicklung sind, in den Vordergrund der
Betrachtung gestellt werden, während umgekehrt im 3. Abschnitte
gerade die Entwicklungsvorgänge das Einteilungsprinzip
ergeben.
I. Vorbemerkungen.
Entwicklung, mit Gestaltsveränderung verbundenes Wachstum, ist eine
der auffallendsten Lebenserscheinungen. Unter Wachstum versteht man
nicht jede beliebige Volumvergrößerung. Wenn eine ausgetrocknete
verschrumpfte Rübe im Wasser schwillt, so ist das kein Wachstum. Nur
bleibende, nicht rückgängig zu machende Größenzunahme kann
Wachstum genannt werden, mag dabei die Pflanze im ganzen Substanzgewinn
oder -verlust erfahren. In der Regel ist freilich das Wachstum mit
Substanzgewinn verbunden; die im Keller treibende Kartoffel aber
erleidet durch Transpiration und durch Atmung Verluste, und doch
wachsen ihre Triebe.
1. Wachstumsmessung.
Gesamtverlängerung. — Handelt es sich darum, die
Zuwachsgröße einer Pflanze, d. h. die Gesamtverlängerung in
der Zeiteinheit zu bestimmen, so kann man bei raschwüchsigen Organen,
z. B. den Blütenschäften einer Agave, den Sprossen einer Bambusa,
in bestimmten Zeitabschnitten (Tagen, Stunden) einen gewöhnlichen
Maßstab anlegen und ablesen. — In der Regel aber ist es nötig, den
Zuwachs der Pflanzen zum Zweck der Messung zu vergrößern. Das
kann z. B. durch das Mikroskop geschehen, das den von der
Pflanze durchschrittenen Raum beliebig zu vergrößern gestattet. Die
bei gröberen Versuchsobjekten meist benutzte Methode der Vergrößerung
ist aber die mittels[S. 243] Hebelübertragung. Die darauf beruhenden Apparate
werden als Auxanometer bezeichnet.
Fig. 254 stellt links ein einfaches Auxanometer, den „Zeiger am
Bogen“ vor, mit dem der Zuwachs eines Blütenschaftes beobachtet
wird. Ein dicht unter der Gipfelknospe befestigter Faden läuft
über die kleine Rolle r und wird durch das Gewicht g straff
gehalten, ohne einen störenden Zug auf den Schaft auszuüben. z
ist ein mit der Rolle r fest verbundener Zeiger, der etwa 20mal
so lang ist als der Halbmesser der Rolle, den jeweiligen Zuwachs
des Schaftes also zwanzigfach vergrößert an der Skala S angibt.
Um die zu bestimmten Zeiten hier notwendigen Ablesungen zu
ersetzen, hat man selbstregistrierende Auxanometer verwandt,
deren Konstruktion in Fig. 254 rechts in einfacher Ausführung
dargestellt ist. Der große Hebelarm wird durch einen Radius der
größeren Rolle R gebildet, der kleine durch einen Radius der
kleinen Rolle r. Bei der durch den Sproßzuwachs erfolgenden
Drehung der Rollen hebt sich ein mit dem Zeiger Z versehenes
Metallstück, das durch das Gegengewicht W äquilibriert ist. Der
horizontale spitze Zeiger berührt rechts eine durch das Uhrwerk
U in gleichmäßige Drehung versetzte, mit einem berußten Papier
überzogene Trommel C, auf der der Zeiger einen weißen Strich
hinterläßt. Dreht sich die Trommel in je einer Stunde einmal,
dann gibt der senkrechte Abstand zwischen den Zeigerspuren
den jeweiligen stündlichen Zuwachs in bekannter Vergrößerung
selbsttätig an.
Im allgemeinen ist die Zuwachsgröße der Pflanzen so gering, daß man bei
kurzer Beobachtungszeit überhaupt kein Wachstum bemerkt. Nur gewisse
Pilze und die Staubfäden mancher Gräser wachsen so rasch, daß man die
Verlängerung mit bloßem Auge wahrnehmen kann. Der Fruchtkörper des
Gasteromyceten Dictyophora verlängert sich nach A. MÖLLER um
5 mm, die Staubfäden von Triticum (Weizen) nach ASKENASY um
1,8 mm in der Minute; das Ende der letzteren rückt also etwa
mit der gleichen Geschwindigkeit vor wie die Spitze des großen Zeigers
einer Taschenuhr. Die nach diesen Staubfäden am schnellsten wachsenden
Pflanzenteile, nämlich die Blattscheiden der Bananen, stehen mit 1,1
mm, die Bambusschößlinge mit 0,75 mm, kräftige Kürbissprosse mit 0,1
mm, die Hyphen von Botrytis mit 0,034 mm in der Minute schon erheblich
dagegen zurück; die allermeisten Pflanzen erreichen aber auch unter
günstigen Verhältnissen nur einen viel geringeren Zuwachs (0,005 mm und
darunter in der Minute).
Niemals bleibt die Zuwachsgröße eines Organs dauernd gleich: auch
bei konstanten äußeren Verhältnissen sieht man vielmehr die
Zuwachse zunächst von sehr kleinen Werten bis zu einem Maximum
ansteigen und dann wieder allmählich auf Null abklingen. Man nennt
diese Erscheinung „die große Periode des Wachstums“. Ein
Beispiel mag ihren Verlauf illustrieren:
Für das erste Stengelglied der Lupine wurden an
aufeinanderfolgenden Tagen bei konstanter Temperatur im Dunkeln
folgende Zuwachse (in Zehntelmillimeter) gefunden:
8, 9,
11, 12, 35, 43, 41, 50, 51, 52,
65, 54, 43, 37, 28,
18, 6, 2,
0.
Nicht immer freilich verläuft diese Periode so regelmäßig; vielfach
treten durch „stoßweise Änderungen“ des Wachstums erhebliche
Unregelmäßigkeiten in ihrem Verlauf ein.
Wachstumsverteilung. — In der Regel wächst ein Pflanzenteil
nicht in seiner ganzen Ausdehnung, es gibt vielmehr an ihm
ausgewachsene und wachsende Teile; und die wachsenden Abschnitte
verlängern sich auch nicht etwa gleichmäßig, sondern sie bestehen
aus verschieden rasch wachsenden Zonen, die allmählich ineinander
übergehen. — Die Lage und die Länge der Wachstumszonen
ist bei verschiedenen Organen nicht die gleiche. Die typische
Wurzel hat eine einzige Wachstumszone, und diese
liegt dicht hinter der Spitze und nimmt eine Länge von 5–10 mm ein;
Luftwurzeln freilich haben eine Wachstumszone, die erheblich länger
werden kann und im Extrem 1 m[S. 244] beträgt. Die Stengel verhalten
sich ungleich. Solche, die keine scharf differenzierten Knoten
besitzen, haben wie die Wurzeln bloß eine einzige Wachstumszone,
aber die Länge der Wachstumszone ist immer beträchtlich, oft bis zu
einem halben Meter. Wo aber typische Knoten ausgebildet sind, da
haben wir so viele Wachstumszonen, als Internodien sich strecken,
und diese sind entweder von weniger stark wachsenden Partien oder
von ganz ausgewachsenen getrennt. Man spricht im letzteren Falle von
interkalarem Wachstum; sehr schön ist dieses z. B, bei den Grashalmen
ausgebildet, wo an der Basis jedes Internodiums eine Wachstumszone
sich findet. Auch bei manchen Blättern, namentlich denen
der Monokotylen, haben wir an der Basis eine interkalare
Wachstumszone.
Die Verteilung des Wachstums wird dadurch festgestellt, daß man von
Zeit zu Zeit die Entfernung gewisser natürlicher oder künstlich
angebrachter Marken mißt.
So ist z. B. in Fig. 255 I eine Wurzelspitze dargestellt, die
vom Vegetationspunkt aus (Punkt 0; darunter die Wurzelhaube) mit
Tuschestrichen in Millimeterabstand versehen ist. 22 Stunden
später sind diese Striche in der Weise auseinandergerückt, wie das
Fig. 255 II zeigt. Das Wachstum ist also in der Wachstumszone
ungleich; am oberen und unteren Ende verläuft es langsam und
vermittelt so den Übergang zu der Partie, die das maximale
Wachstum aufweist. Untersucht man den Zuwachs einer
Querscheibe, z. B. des zwischen den Marken 0 und 1 gelegenen
Millimeters, an aufeinanderfolgenden Tagen, so zeigt sich, daß
sie erst langsam, dann schnell, dann wieder langsam wächst; mit
anderen Worten: ein jeder Abschnitt der Wachstumszone zeigt die
große Periode des Wachstums. Die verschiedenen von der Spitze
aus abgetragenen Millimeter befinden sich in verschiedenen Stadien
ihrer großen Periode; die 2 ersten sind im aufsteigenden Ast, 3
und 4 auf dem Gipfel, die übrigen im absteigenden Ast der Kurve.
Entsprechendes gilt für andere Organe.
Zeitlich getrennte Wachstumsperioden treten z. B. an
den Schäften von Taraxacum auf, die erste gelegentlich bei der
Blütenentwicklung, die zweite bei der Fruchtbildung. Ähnlich
verhalten sich andere Organe, deren Aufgabe zeitweise sich ändert
(Blütenstiele bzw. Fruchtstiele von Linaria Cymbalaria, Arachis
hypogaea, Tropaeolum).
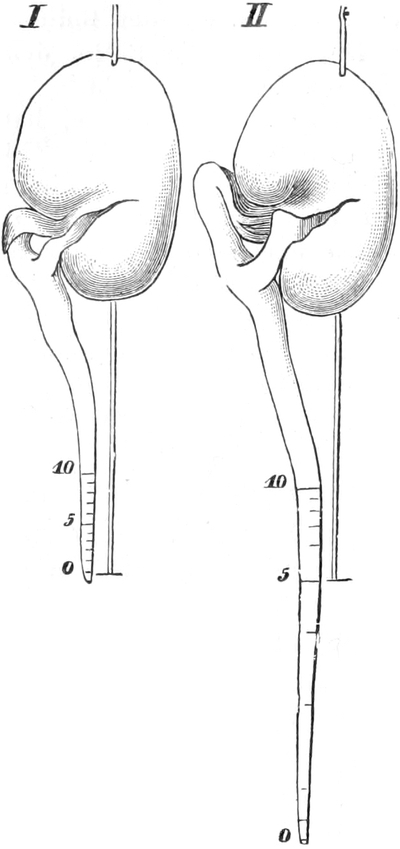
Fig. 255. Die Verteilung des Zuwachses an der
Wurzelspitze einer Feldbohne (Vicia Faba). Bei I die Wurzelspitze
durch Tuschemarken in 10 gleiche Querzonen von 1 mm geteilt. In II
dieselbe Wurzel nach 22 Stunden. Die Tuschestriche sind durch
ungleiches Wachstum der Zonen verschieden weit auseinandergerückt. Nach
SACHS.
Wachstumsgeschwindigkeit. — Aus der Tatsache, daß in
verschiedenen Organen ganz verschieden lange Zonen im Wachstum
begriffen sind, kann man entnehmen, daß die Angaben über den
Gesamtzuwachs eines solchen Organs, wie sie S. 243 gemacht wurden,
keinen Schluß auf die eigentliche Wachstumsgeschwindigkeit,
d. h. den Zuwachs der Längeneinheit in der Zeiteinheit
zulassen. Bei den Sprossen von Bambusa z. B. ist die wachsende Zone
viele Zentimeter, bei Botrytis nur 0,02 mm lang; wenn also Bambusa bei
gleichen Außenverhältnissen etwa einen 20mal so großen Zuwachs in der
Minute erfährt als Botrytis, so ist doch ihre Wachstumsgeschwindigkeit
eine viel geringere. Zur Charakterisierung der Wachstumsgeschwindigkeit
sind demnach Angaben der Verlängerung pro Minute in Prozenten der
Wachstumszone nötig. Da ergibt sich dann eine Geschwindigkeit von 83%[S. 245]
bei Botrytis, von nur 1,27% bei Bambusa; im Maximum hat man 220% bei
gewissen Pollenschläuchen beobachtet, während manche noch immerhin
ansehnlich wachsende Sprosse nur 0,5% ergeben.
Größe der Pflanze. Aus der Wachstumsgeschwindigkeit und der
Größe der wachsenden Zone kann man die definitive Verlängerung
eines Pflanzenteiles erst dann bestimmen, wenn man auch die
Wachstumsdauer kennt. Durch die Variation dieser Faktoren ist
die Größe der Pflanze wie auch ihrer Teile bestimmt, die,
wie jedermann weiß, zwar in mannigfacher Weise von äußeren Faktoren
abhängt, aber doch eine spezifisch verschiedene ist. Eine
bestimmte Größe gehört so gut zu den spezifischen Eigenschaften eines
Organismus wie seine Blattgestalt usw.; auch ist die ganze Organisation
einer Pflanze derart, daß sie nur mit einer gewissen Größe verträglich
ist.
2. Die Phasen des Wachstums.
Bei den einfachsten Pflanzen, niederen Algen, Pilzen, Bakterien,
besteht die Entwicklung lediglich im Wachstum der Zelle mit
darauffolgender Teilung. Diese Fälle sind in der Morphologie zur
Genüge besprochen. Bei komplizierten Pflanzen findet sich zwar auch
stets Zellenwachstum und häufig genug Zellteilung, aber beide Prozesse
erscheinen unter das Gesamtwachstum untergeordnet, und dieses setzt
sich vielfach aus drei verschiedenen, wenn auch zeitlich meist nicht
scharf getrennten Phasen zusammen, der embryonalen Anlage der
Organe, der Streckung und der inneren Ausgestaltung.
a) Embryonale Anlage. — Das embryonale Wachstum erfolgt
normalerweise an Vegetationspunkten, und neue Vegetationspunkte
entstehen im allgemeinen unmittelbar aus schon vorhandenen; nur bei den
Wurzeln erfolgt die Ausbildung der Vegetationspunkte der Seitenglieder
etwas verspätet aus Resten des Vegetationspunktes, die
embryonalen Charakter beibehalten haben. Die charakteristischen Züge
der Organbildung an Vegetationspunkten sind schon in der Morphologie
behandelt. Dort wurde auch auf S. 64 und 65 die Symmetrie und
die Polarität besprochen; auch diese werden häufig schon am
Vegetationspunkt ausgebildet; ja die Polarität, der Gegensatz zwischen
Basis und Spitze, wird bei den höheren Pflanzen schon in der Eizelle
angelegt und bleibt, einmal entstanden, gewöhnlich dauernd erhalten.
Hier aber muß noch betont werden, daß nicht alle Vegetationspunkte von
ihresgleichen abstammen. Nicht nur durch die normale Organogenese,
sondern auch durch Restitution kann die Entwicklung der Pflanze
vonstatten gehen.
Unter Restitution[206] versteht man die Neubildung von
Organen, die in der Regel nach Verstümmelung einer Pflanze
auftritt und die an Orten erfolgen kann, wo an der unverletzten Pflanze
keinerlei Wachstumstätigkeit eingetreten wäre. Man kann zwei Fälle
von Restitution unterscheiden, je nachdem die Ersatzbildung aus der
Wundfläche oder in einer gewissen Entfernung von ihr entsteht.
Die Wiederherstellung des verlorenen Organs von der Wundfläche aus
findet sich bei niederen Pflanzen, z. B. bei Algen und Pilzen, nicht
ganz selten, dagegen ist sie bei höheren Pflanzen von ganz beschränktem
Vorkommen. Nur Gewebe, die noch embryonal sind, aber bei weitem nicht
alle embryonalen Gewebe, sind dazu befähigt. Am häufigsten sind
derartige Restitutionen am Vegetationspunkt der Wurzel beobachtet;
hier wird nach Abtragung der Spitze durch einen Querschnitt diese
wieder gebildet, wenn der Schnitt nicht weiter als etwa 0,5 mm von
der Kuppe des Vegetationspunktes entfernt war. Längsgespaltene
Wurzelvegetationspunkte pflegen sich derart zu ergänzen, daß eine
solche Wurzel dann zwei Spitzen erhält. An Sproßvegetationspunkten
kommt diese Art von Restitution nicht vor, an Blattanlagen ist sie sehr
selten.
[S. 246]
Dagegen ist die andere Art von Restitution ganz außerordentlich
verbreitet im Pflanzenreich. Hier wird für das verlorene Organ
dadurch Ersatz geschaffen, daß in der Nähe der Wunde ein neues
gebildet wird oder ein in der Anlage schon vorhandenes auswächst.
Auch für diese Art der Restitution liefern Algen und Pilze, vor allem
aber die Laub- und Lebermoose, zahlreiche Beispiele, die hier nicht
angeführt werden können. Wir beschränken uns auf die Betrachtung der
Blütenpflanzen. Ganz besonders verbreitet ist bei diesen die Befähigung
zur Wurzelbildung. Bei den Pelargonien, bei der Weide und bei
vielen anderen Pflanzen hat man es durch Abtrennen der Sprosse in der
Hand, an jeder beliebigen Stelle Wurzeln entstehen zu lassen; bei
anderen Pflanzen sind es bevorzugte Orte, wie die älteren Knoten, an
denen sie sich entwickeln. Nach der Wurzelbildung ergänzt sich aber
der Stengel zu einer vollen Pflanze, wenn entweder vorhandene
Blattachselknospen austreiben oder wenn neue Sproßvegetationspunkte
auftreten. Auch abgeschnittene Blätter haben sehr oft die
Fähigkeit, sich zu bewurzeln, doch ist mit dieser nur selten auch
das Vermögen der Sproßbildung verbunden. Selbst aus abgeschnittenen
Wurzeln können — wenn sie imstande sind, Knospen zu erzeugen
— neue Pflanzen entstehen. Außer an Stengeln, Blättern, Wurzeln
hat man auch schon an Ranken, Blüten und Früchten, vielfach unter
Neubildung von Vegetationspunkten, Knospen, auftreten sehen. Wird der
Vegetationspunkt einer Blütenpflanze zerstört, so kann aus dem Meristem
oberhalb der jüngsten Blattanlagen ein Ersatzvegetationspunkt
geschaffen werden. Ist hier die Restitution auf ganz embryonale
Zellen beschränkt, so sehen wir in anderen Fällen ältere, zum Teil
schon ausgewachsene Zellen wieder anfangen zu wachsen und sich zu
teilen, also wieder embryonal werden. Vielfach bildet sich so zunächst
ein besonderes Gewebe an der Wunde: der Kallus; im Innern
dieser Zellwucherung treten dann Sproßanlagen auf. In wieder anderen
Fällen sehen wir ausgewachsene Zellen, mögen das nun Parenchymzellen
oder Epidermiszellen sein, direkt, d. h. ohne Kallusbildung, zu
Vegetationspunkten werden. So stellt z. B. die Fig. 256 die Entstehung
eines Begoniensprosses aus einer Epidermiszelle des Blattes dar.
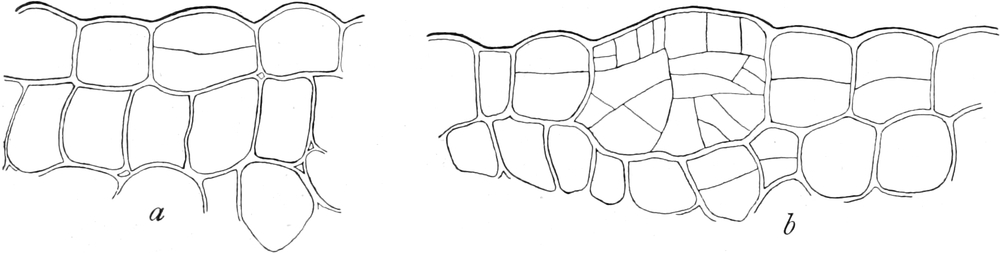
Fig. 256. Querschnitt durch das Blatt von
Begonia. Vergr. 200. Bildung eines Adventivsprosses aus einer
Epidermiszelle nach HANSEN. a Die Epidermiszelle hat sich
einmal durchgeteilt. b Aus der Epidermiszelle ist ein vielzelliges
Meristem geworden.
Nicht nur äußerlich sichtbare Organe, sondern auch Gewebe
können sich aus ausgewachsenen Parenchymzellen bilden.
So werden z. B., wenn die Leitbahnen unterbrochen worden
sind, aus dem Parenchym neue Gefäße gebildet, die wieder eine
Verbindung herstellen. Nicht immer freilich wird das entfernte
oder unterbrochene Gewebe wieder gebildet; häufig kommt es zu
Ersatzbildungen. So wird in der Regel die Epidermis durch
Kork ersetzt, und nur ausnahmsweise kommt es zu ihrer echten, mit
der Bildung von Spaltöffnungen verbundenen Wiederbildung.
Hier anzuschließen wäre auch die Neubildung der Epidermis, wie sie
im normalen Verlauf der Entwicklung bei bestimmten Araceen sich
einstellt, die Löcher in ihren Blattspreiten entstehen lassen.
Bei Monstera deliciosa sterben einzelne Stellen der Spreite[S. 247]
ganz junger Blätter ab. Um diese Stellen tritt das Mesophyll in
Teilung ein und bildet aus der äußersten Zellschicht eine sekundäre
Epidermis, die nunmehr die Löcher auskleidet und an die primäre
Epidermis der beiden Blattflächen anschließt. — Auch sonst finden
sich in der normalen Entwicklung vielfach Vorgänge, die man als
Restitutionen betrachten kann; z. B. die wiederholte Korkbildung
(S. 139).
Neben der Tatsache, daß Restitution eintritt, interessiert auch die
Frage, wo sie sich einstellt. Und da zeigt sich dann vielfach, daß
die Polarität, die an der intakten Pflanze auffällt, sich auch
an der restituierenden geltend macht. So pflegen an Stengeln die
Sprosse am Apikalende, die Wurzeln am Basalende aufzutreten, während
an der Wurzel genau die entgegengesetzte Verteilung besteht. Auch
bei niedrig organisierten Pflanzen zeigt sich oft eine Polarität in
den Restitutionsprozessen. Wird z. B. ein Zellfaden einer Cladophora
in Einzelzellen zerlegt, so bildet jede an der Basis ein farbloses
Rhizoid, an der Spitze einen grünen Faden.
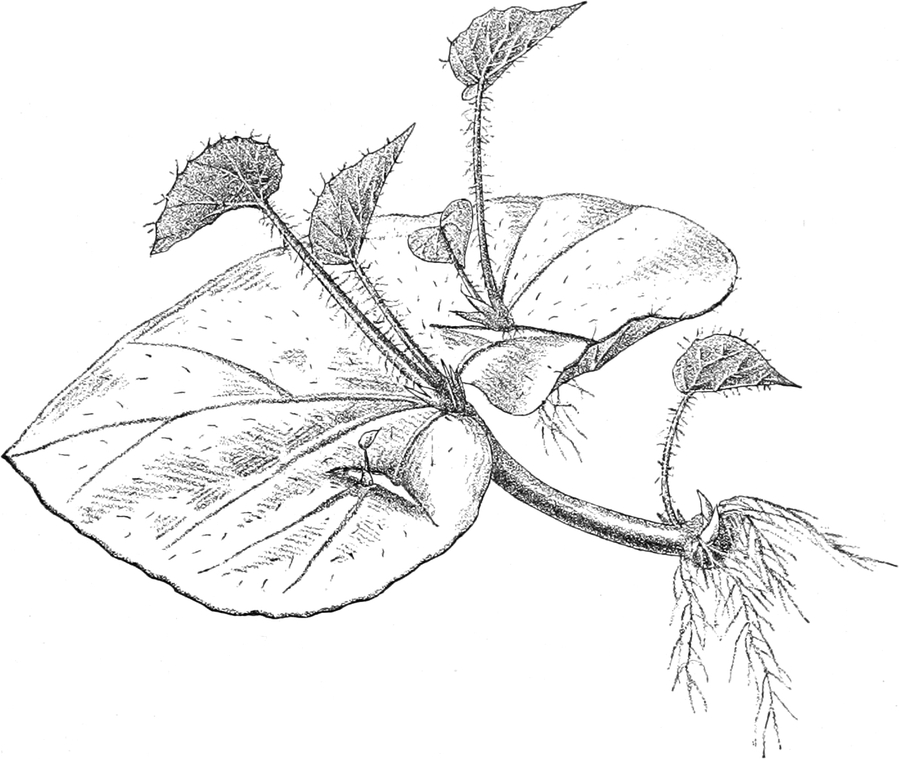
Fig. 257. Blatt von Begonia als Steckling
behandelt. Mit den Restitutionssprossen. Nach STOPPEL.
An Laubblättern tritt bei Restitutionen dieser Gegensatz von Basis
und Spitze nicht zutage. Das mag damit zusammenhängen, daß das
restituierende Blatt nicht in die Neubildung eingeschaltet wird;
es entsteht vielmehr an der Blattbasis eine völlig neue Pflanze,
worauf das Blatt selbst abstirbt. In einzelnen Fällen treten auch die
Neubildungen auf der ganzen Blattfläche ein (Torenia); manchmal läßt
sich aber der Ort ihrer Entstehung durch Einschnitte in die Lamina
beeinflussen: bei Begonia z. B. (Fig. 257) bilden sich die jungen
Pflänzchen oberhalb der Schnitte.
Die besprochenen Erscheinungen der Restitution haben eine
große Bedeutung für die Gärtnerei, da sie gestatten, Pflanzen
ohne Zuhilfenahme von Samen rasch zu vermehren. Bei dieser
künstlichen Vermehrung werden abgetrennte Teile von Pflanzen
benutzt, um aus ihnen wieder vollständige Pflanzen zu erzielen.
Das gelingt bei manchen Pflanzen leicht, bei anderen ist es
schwieriger, bei wieder anderen ist es überhaupt nicht möglich. Das
einfachste Verfahren ist die Fortpflanzung durch Stecklinge,
d. h. das Einsetzen abgeschnittener Zweige in Wasser, Sand oder
Erde, wo sie sich bewurzeln (Oleander, Pelargonien, Tradescantien,
Fuchsien, Weiden usw.). Durch abgetrennte Blätter[S. 248] werden
hauptsächlich die Schiefblätter (Begonien) vermehrt. Selbst im
Zusammenhang mit ihrer Mutterpflanze erzeugen die Blätter mancher
Gewächse jungen Nachwuchs (Bryophyllum). Auch aus Wurzeln und
Wurzelstücken können einzelne Pflanzen vermehrt werden, wie z. B.
der Löwenzahn (Taraxacum).
b) Streckung. Um in Funktion treten zu können, müssen sich die
embryonalen Anlagen vergrößern und entfalten; das geschieht durch den
Prozeß der Streckung in höchst eigenartiger, haushälterischer Weise.
Die Vergrößerung erfolgt nämlich vor allem durch Einlagerung von Wasser
(„Schwellwasser“), das von außen aufgenommen werden kann. Organische
Substanz muß nur zum Flächenwachstum der Zellhaut aufgewandt werden.
Dagegen bedarf es keiner Vermehrung des Protoplasmas bei der
Streckung; es wird also gerade die besonders kostbare N-haltige
Substanz gespart. In dieser Hinsicht besteht ein großer Unterschied
im Wachstum der Pflanzen gegenüber dem typischen Tier, dem eine der
„Streckung“ entsprechende Größenzunahme abgeht.
Schon die embryonale Zelle des Vegetationspunktes enthält in der
Zellhaut und dem Protoplasma reichliche Wassermengen; die gesamte
organische Substanz ist ja mit Wasser imbibiert. Bei fortgesetzter
Wasseraufnahme von außen tritt aber eine Sonderung zwischen dem
wasserdurchtränkten Protoplasma und den mit wäßriger Lösung gefüllten
Vakuolen auf, und schließlich kommt es durch Verschmelzen der
Vakuolen zur Ausbildung des einzigen zentralen „Saftraumes“ und des
peripheren Plasmaschlauches (vgl. S. 9, Fig. 3). Daß der Saftraum der
Sitz osmotischer Kräfte ist, wurde schon früher (S. 191) ausgeführt;
der Turgordruck aber ist eine unentbehrliche Bedingung für das
Flächenwachstum der Zellhaut.
Zellen, deren Turgeszenz (S. 191) durch Wasserentzug aufgehoben
ist, zeigen kein Wachstum mehr. Man nimmt deshalb an, daß die
mechanische Dehnung der Zellhaut ihr Wachstum erleichtert oder
erst ermöglicht. Man darf aber nicht glauben, daß irgendeine
Proportionalität zwischen Wachstum und Dehnung existiere. Auch
läßt sich die Dehnung durch den Turgordruck keineswegs durch eine
beliebige mechanische Dehnung ersetzen. Das Protoplasma spielt
eben beim Flächenwachstum der Zellhaut stets die Hauptrolle, und
mit seiner Betätigung muß es wohl zusammenhängen, daß manchmal
Zellhäute bei sehr schwacher Spannung ausgiebig wachsen.
Über die Vorgänge beim Zellhautwachstum, die man als Apposition
und Intussuszeption bezeichnet, ist S. 29 das Nötige gesagt
worden. Bei Flächenwachstum ohne Substanzaufnahme („plastische
Dehnung“), dem dann gewöhnlich Lamellenanlagerung folgt, ist die
Turgordehnung eine leichtverständliche Voraussetzung des Wachstums;
bei Intussuszeptionswachstum erscheint uns der Turgordruck weniger
notwendig.
Wenn durch Wachstum der Zellhaut erneute Wasseraufnahme ermöglicht
wird, muß der Zellsaft zunächst verdünnt werden. Tatsächlich
tritt eine solche Verdünnung nicht ein, weil das Protoplasma der
wachsenden Zelle fortwährend die Konzentration des Zellsaftes
zu regulieren vermag. Durch Überführung von Zucker in
organische Säuren kann der osmotische Wert erheblich vergrößert
werden; würde z. B. Oxalsäure aus Glykose gebildet, so könnte er
auf das Dreifache seines früheren Wertes steigen. Umgekehrt kann
er z. B. durch völlige Verbrennung von Zucker bei der Atmung ganz
erheblich verringert werden.
Neben der Streckung in der Längsrichtung existiert auch eine
Streckung in der Querrichtung (Dickenwachstum). Der
Durchmesser der fertigen Wurzel, des fertigen Stengels z. B.
ist größer, oft beträchtlich größer als unmittelbar am
Vegetationspunkt. Man unterscheidet, wie S. 121 auseinandergesetzt
ist, ein primäres vom sekundären Dickenwachstum. Nur das primäre
Dickenwachstum ist ein reines Streckungswachstum; beim sekundären
werden durch ein interkalares Meristem, das Kambium, zunächst neue
embryonale Zellen geschaffen, die erst allmählich in Streckung
übergehen.
Gewebespannung. Die Streckung der Zellen in die Länge und Weite
erfolgt in einem Querschnitt eines Organs nicht immer gleichmäßig und
gleichzeitig. Sehr verbreitet findet sich z. B. bei wachsenden Stengeln
die Erschei[S. 249]nung, daß das Mark ein stärkeres Streckungsbestreben
hat als die peripherischen Gewebe. Da eine Kontinuitätstrennung
zwischen beiden nicht eintreten kann, kommt es zu Spannungen
(„Gewebespannung“); das Mark dehnt die Rindengewebe, diese
komprimieren das Mark: die tatsächliche Länge des Organs ist eine
aus antagonistischen Bestrebungen resultierende. Trennt man die
Gewebe künstlich, so nimmt jedes seine spezifische Länge an, das Mark
verlängert sich, die Rinde kontrahiert sich: die Spannung hört auf.
Diese in wachsenden Organen weit verbreitete Gewebespannung
kann man in folgender Weise demonstrieren. Löst man in einem
Sonnenblumensproß mit Hilfe des Korkbohrers das Mark eine Strecke
weit aus seinem Zusammenhang mit den Nachbargeweben, so tritt es
nach Entfernung des Bohrers an der Schnittfläche weit hervor (Fig.
258, 1). Wird ein Sproß der Länge nach gespalten, so krümmen
sich die beiden Hälften nach außen, da das Mark sich verlängert,
die Epidermis sich verkürzt. Selbst an hohlen Sprossen, z. B.
den Blütenschäften des Löwenzahns (Taraxacum), besteht eine
Spannung zwischen inneren und äußeren Gewebeschichten. Wird ein
Taraxacum-Stengel der Länge nach gespalten, so krümmen sich seine
Teile nach außen (Fig. 258, 2a), und diese Krümmungen nehmen noch
erheblich zu (Fig. 258, 2b) wenn das Objekt in Wasser gelegt wird.
Auch in Blättern und Wurzeln finden sich Gewebespannungen. Die
Spannungen müssen nicht ausschließlich in der Längsrichtung
eintreten; es gibt auch Querspannungen. So wird z. B. die Rinde von
Bäumen, die sekundär in die Dicke wachsen, in tangentialer Richtung
ganz beträchtlich gedehnt; sie kontrahiert sich also nach Loslösung
vom Holzkörper.
Die Gewebespannungen bilden sich in geringer Entfernung vom
Vegetationspunkt mit dem Beginne der Streckung allmählich aus, und
sie verschwinden im allgemeinen wieder in der ausgewachsenen Zone. In
einigen Fällen bleiben sie aber dauernd erhalten (S. 297). Sie
sind von großer Bedeutung für die wachsenden Gewebe; sie vermehren die
schon durch die Turgeszenz der Einzelzellen bedingte Festigkeit. Die
Gewebespannung hat auch eine gewisse Ähnlichkeit mit der Turgeszenz der
Einzelzelle; das zeigt sich am deutlichsten beim typischen Stengel:
wie der Zellsaft durch den osmotischen Druck die Zellhaut dehnt, so
expandiert das schwellende Mark die Rindenpartien. Wie aus der Dehnung
der Zellhaut, so resultiert auch aus der der Rinde ein vermehrter
Widerstand gegen Deformationen, also eine vergrößerte Festigkeit.
Fig. 258. 1 Sproß von Helianthus annuus nach
Entfernung der Blätter; Mark mit dem Korkbohrer von der Peripherie
getrennt. 2 Blütenstand von Taraxacum. Stiel der Länge nach übers
Kreuz gespalten. a Sofort nach dem Spalten, b nach Einlegen in
Wasser.
Die Gewebespannung wird dadurch aufgehoben, daß sämtliche Zellen
schließlich die mittlere Länge dauernd annehmen, die ihnen durch
die antagonistischen Bestrebungen diktiert wird. Manchmal aber
zeigen gewisse Zellen, nachdem sie ihre größte Länge erreicht
haben, ein ganz beträchtliches, mit Änderung ihrer Form verbundenes
Kontraktionsbestreben. Sehr verbreitet kommt das z. B. bei
Wurzeln vor, wo die Zellen der Rinde und die zentralen Partien
durch zwischenliegende Gewebe, die sich kontrahieren, in Falten
gelegt werden. Die Bedeutung dieser Wurzelkontraktion (S. 154), die
eine Verkürzung der ausgewachsenen Teile um 10–70% herbeiführen
kann, ist sehr groß. Sie bewirkt es z. B., daß die Blätter vieler
„Rosettenpflanzen“ trotz des andauernden Längenwachstums des Stammes
doch immer dem Boden angedrückt bleiben;[S. 250] sie bedingt und reguliert
das Eindringen vieler Knollen und Zwiebeln in eine bestimmte Tiefe der
Erde; sie erhöht schließlich auch die Befestigung der Pflanze im Boden,
da durch straffe Wurzeln eine größere Stabilität erzielt wird als durch
schlaffe.
c) Innere Ausbildung. Die Zellen des typischen
Vegetationspunktes des Sprosses und der Wurzel behalten andauernd ihre
Wachstums- und Teilungsfähigkeit. Man nennt sie embryonale Zellen.
Alle Organe, die mit solchen Zellen versehen sind, haben im Prinzip
die Befähigung zu unbegrenztem Wachstum. Embryonale Gewebe finden
sich nicht nur am Vegetationspunkt, sondern auch in den sekundären
Meristemen (S. 40).
Ein Teil der embryonalen Zellen, in Organen mit begrenztem Wachstum
sogar alle, verwandelt sich in somatische Zellen,
Dauergewebe, die bald mit Wachstum und Teilung aufhören und
früher oder später dem Tode verfallen (S. 272).
Die innere Ausbildung der Organe beginnt direkt hinter dem
Vegetationspunkt und dauert sehr verschieden lang. Während Haare
vielfach außerordentlich rasch fertiggestellt werden, wird die
definitive Ausgestaltung von inneren Geweben oft erst nach Abschluß der
Streckung vollendet; falls sekundäres Dickenwachstum eintritt, ist sie
überhaupt nie abgeschlossen. Die Ausbildung von „Dauergeweben“
aus den Ur- und Folgemeristemen ist im morphologischen Teil schon
geschildert.
II. Die Faktoren der Entwicklung.
Wenn wir den Versuch machen, die Faktoren zu besprechen, die von
Einfluß auf die Entwicklung sind, so kann es sich da immer nur
um Beispiele handeln, die uns in möglichst charakteristischer
Weise den Einfluß eines einzelnen solchen Faktors vor Augen führen
sollen. An Vollständigkeit ist weder in der Aufzählung der Faktoren
noch ihrer Wirkung gedacht. Wie in anderen Fällen, so können wir auch
hier die Faktoren in zwei Gruppen bringen: äußere und innere.
A. Äußere Faktoren.
Als äußere Faktoren treten uns alle die Kräfte und Stoffe
entgegen, die wir schon beim Stoffwechsel als physiologisch wirksam
kennen gelernt haben, oder die bei den Bewegungen eine Rolle spielen.
Gewisse äußere Faktoren sind schon S. 185 genannt und als allgemeine
Lebensbedingungen bezeichnet worden; ohne diese gibt es überhaupt
keine Entwicklung. Neben der allgemeinen (formalen) Bedeutung können
aber dieselben Faktoren auch noch eine andere Bedeutung für das
Wachstum haben. Durch Wechsel in ihrer Intensität, Qualität, eventuell
auch Richtung, können quantitative, ja sogar auch qualitative
Änderungen in den Organen der Pflanzen hervorgerufen werden. Wir nennen
diese Wirkungen „formativ“ und stellen leicht fest, daß überall
der Zusammenhang zwischen Ursache und Wirkung ein sehr komplizierter
ist.
1. Temperatur[207]. Wie beim Stoffwechsel, so konstatieren wir
auch jetzt, daß eine gewisse Temperatur unerläßliche formale Bedingung
für das Wachstum ist.
Eine zu niedere Temperatur (etwa 0° oder weniger) und ebenso eine zu
hohe (40–50° oder mehr) heben das Wachstum völlig auf. Zwischen dem
Minimum und dem Maximum liegt dann, meist nicht in der Mitte, sondern
dem Maximum genähert bei etwa 22–37° das Optimum (Fig. 259). Im
einzelnen zeigen aber die Pflanzen, die verschiedene Klimate bewohnen,
recht große[S. 251] Differenzen in der Lage der Kardinalpunkte (vgl. S. 185)
der Temperatur. Daß auch die verschiedenen Individuen der gleichen
Art große Unterschiede in der Abhängigkeit ihres Streckungswachstums
von der Temperatur aufweisen, erkennt man z. B. an der ungleichen
Entwicklung der Knospen der Roßkastanien im Frühjahr. Aber auch
am einzelnen Individuum unterliegen die Wachstumsprozesse der
verschiedenen Organe einer ganz differenten Beeinflussung durch die
Temperatur.
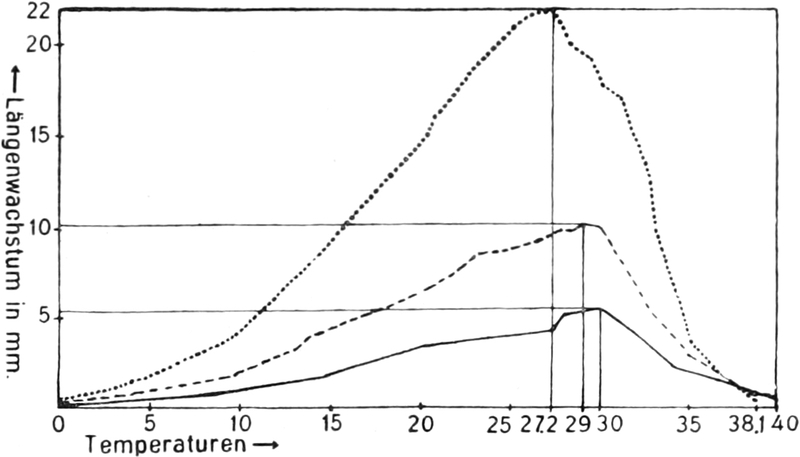
Fig. 259. Abhängigkeit des Wachstums von der Temperatur
nach TALMA. Die Abszisse gibt die Temperatur. Die Kurven geben
den Zuwachs der Wurzeln von Lepidium sativum an, und zwar die
ausgezogene Linie in 31⁄2 Stunden, die gestrichelte in 7 Stunden und
die punktierte in 14 Stunden. Man sieht, daß das Optimum mit der Dauer
des Versuchs auf niedrigere Temperaturen rückt.
Bei Pflanzen tropischer Klimate kann beispielsweise das Minimum bei
+10° C liegen, während unsere, oft die Schneedecke durchbrechenden,
ersten Frühjahrspflanzen ebenso wie die Bewohner der Hochalpen und
der polaren Regionen bei Temperaturen wenig über 0° noch kräftig
wachsen. — Zahlreiche unserer Frühlingspflanzen zeigen, daß die
Blütenentfaltung durch niedrigere Temperaturen gefördert werden
kann als die Laubblattentfaltung.
2. Licht[208]. Das Licht ist wenigstens nicht allgemein
eine so unentbehrliche Wachstumsbedingung wie die Temperatur. Es gibt
Pflanzen (Bakterien und Pilze), die ihre ganze Entwicklung normal auch
im Dunkeln vollziehen können. Wenn das bei der grünen Pflanze nicht der
Fall ist, so liegt das zum Teil schon daran, daß hier das Licht ja zur
Herstellung wichtiger Baustoffe unentbehrlich ist. Sind aber solche
Pflanzen reichlich mit Reservestoffen versehen (Samen, Rhizome), so
können sie meistens ohne Licht lange Zeit wachsen. Dabei nehmen sie
freilich abnorme Gestalten an, von denen S. 252 noch die Rede sein
soll. Es fehlt aber nicht an Organen, denen wenigstens vorübergehend
eine gewisse Lichtmenge geboten werden muß, damit sie überhaupt ihre
Entwicklung beginnen können (Samen und Sporen, S. 267). Andererseits
gibt es für jedes Organ eine gewisse Lichtstärke, die das Wachstum
sistiert. Diese ist bei Schattenpflanzen geringer als bei Lichtpflanzen.
Der Einfluß verschiedener Beleuchtungsstärke auf die
Wachstumsgeschwindigkeit wurde früher dahin gedeutet, daß „Licht
das Wachstum hemmt, Dunkelheit es fördert“. Es gibt aber nur
wenige einwandfreie Versuche, die über den Erfolg einer bestimmten
konstanten Beleuchtungsstärke bei konstanter Temperatur berichten. In
vielen Fällen muß jedoch diese alte Darstellung auch heute noch als
wahrscheinlich gelten, wenn auch in dem eingehend studierten
Fall von Avenakeimsprossen sich ergeben hat, daß das Licht am 1. Tag
eine fördernde, erst späterhin eine hemmende Wirkung ausübt. Wird ein
Avenakeimling eine Zeitlang bei konstanter Beleuchtung kultiviert und
dann in eine höhere, aber ebenfalls konstante Beleuchtung gebracht, so
zeigt sich auch hier die gleiche doppelte Wirkung des Lichtes, zuerst
Förderung und dann Hemmung des Wachstums. Umgekehrt wirkt eine Abnahme
der Beleuchtungsstärke: nach anfänglicher Abnahme folgt eine Zunahme
des Wachstums.
[S. 252]
Viel öfter ist in den letzten Jahren der Einfluß eines
Lichtwechsels auf das Längenwachstum studiert worden. Es hat
sich gezeigt, daß ebenso eine Zunahme wie eine Abnahme der Beleuchtung
das bisher gleichförmige Wachstum ungleichförmig macht. In manchen
Fällen tritt bei Beleuchtungszunahme zuerst Wachstumsbeschleunigung,
dann Hemmung ein, manchmal aber wird auch der umgekehrte Erfolg
beobachtet. Meist tritt bei dieser „Lichtwachstumsreaktion“
(BLAAUW) nicht nur ein Wellenberg und ein
Wellental auf, sondern es folgen einander mehrere Wellen von
abnehmender Amplitude, bis dann der stationäre Zustand erreicht wird,
der noch genauer zu erforschen ist.
Sehr groß sind die formativen Erfolge[209] des Lichtes, doch
verhalten sich die einzelnen Organe dabei sehr verschieden. Das tritt
nirgends deutlicher in Erscheinung als bei dauernder Verdunkelung. Wie
eingangs bemerkt, kann eine solche bei Autotrophen nur bei genügender
Versorgung mit Reservestoffen längere Zeit durchgeführt werden und
bedingt dann ein völlig verändertes Wachstum und deshalb ein ganz
fremdartiges Aussehen der Pflanzen. Diese als Etiolement oder
Vergeilung bezeichnete Erscheinung kommt dadurch zustande,
daß einzelne Organe im Wachstum gefördert, andere gehemmt werden.
Bei Dikotylen findet man z. B. die Stengel stark überverlängert
und schlaff. Überverlängert sind auch die Blattstiele, während die
Blattspreiten klein bleiben und lange in der Knospenlage verharren
(Fig. 260). Da im Dunkeln zwar die gelben Chloroplastenfarbstoffe,
nicht aber der Chlorophyllfarbstoff gebildet werden kann, so
erscheinen die Blätter etiolierter Pflanzen gelb, während die
chloroplastenarmen Stengel weiß aussehen. Etiolement kommt aber
auch bei nichtgrünen Pflanzen vor: gewisse Hutpilze z. B. verlängern
bei Verdunkelung ihren Stiel ganz beträchtlich unter gleichzeitiger
Verkleinerung der Hüte.
Auch im inneren Bau bestehen beträchtliche Differenzen zwischen der
etiolierten und normalen Pflanze. In etiolierten Pflanzen sind die
Gewebe wenig differenziert, es fehlen die verdickten Zellen.
Die Überverlängerung gewisser Organe bei gleichzeitiger Reduktion
anderer hat in der Natur bei allen im Dunkeln austreibenden Samen
und Rhizomen eine große ökologische Bedeutung. Die nur am Licht
funktionierenden Teile werden zunächst noch nicht ausgebildet, die
Baustoffe für sie, insbesondere auch für das Chlorophyll gespart;
die starke Streckung der anderen Organe, die vorzugsweise auf
Wassereinlagerung beruht, hat den Effekt, daß die lichtbedürftigen
Teile möglichst bald aus der Dunkelheit herausbefördert werden.
Fig. 260. Zwei gleichalte Keimpflänzchen des weißen
Senfs (Sinapis alba). E Im Dunkeln erwachsen, etioliert. N Bei
gewöhnlicher Tagesbeleuchtung gewachsen, normal. Nach NOLL.
Auch ohne zu dem Extrem völliger Verdunkelung zu greifen, kann man
weitgehende formative Erfolge durch verschiedene Beleuchtungsstärke
erzielen. Am genauesten sind solche bei den Farnprothallien durch
die Studien von KLEBS bekannt geworden. Er fand:
1. Bei schwachem Licht (Osramlampe von 27 MK) bilden die
Prothallien oft mehr als 2 mm lange, unverzweigte Fäden ohne
jede Zellteilung.
2. Bei etwas stärkerem Licht bleibt die Gestalt des Prothalliums
die gleiche, aber es treten Querwände auf.
[S. 253]
3. Bei etwa 250 MK hört der Zellfaden auf und wird durch die
Zellfläche ersetzt. Bei passender Beleuchtungsstärke kann schon die
zweite Zelle des Prothalliums zur Flächenbildung übergehen; die
Fadenbildung wird also völlig unterdrückt.
4. Bei 500–1000 MK endlich treten Zellkörper auf.
Weitere formative Erfolge der Beleuchtungsstärke treten uns in
der Gestalt und im Bau der Laubblätter entgegen. Schattenblätter
weisen einen ganz anderen Bau auf als die in voller Sonne
erwachsenen Blätter der gleichen Spezies. Sie sind dünner, ihre
„Palisadenzellen“ verjüngen sich nach unten trichterförmig,
lassen weite Interzellularen zwischen sich und treten auch nur in
einer Reihe auf, während am Licht die Palisaden höher werden
und auch in mehreren Schichten sich ausbilden können.
Fig. 261. Taraxacum officinale. 1 in der Ebene,
2 im Hochgebirge kultiviert. Beide gleichmäßig verkleinert. Nach
BONNIER.
Die Pflanzen der Alpen, die das Sonnenlicht länger, intensiver und
in anderer Zusammensetzung erhalten als die der Ebene, weichen
auch in ihrem ganzen Habitus (Fig. 261) weit von diesen ab. Ihre
Vegetationsorgane sind gedrungen, ihre Blüten groß und intensiv
gefärbt. Indes sind neben dem Licht auch andere Faktoren bei dieser
Abänderung beteiligt.
Wo im Laufe der Entwicklung zweierlei Blätter produziert werden,
wie z. B. bei Campanula rotundifolia, manchen Wasserpflanzen usw.,
da zeigen sich die Jugendblätter an die schwache, die Folgeblätter
an eine hohe Beleuchtungsstärke gebunden (S. 270). Auch die
definitive Größe einer Pflanze kann von der Beleuchtung
abhängen. Die[S. 254] Fig. 262 zeigt die große Periode von Avena bei
verschiedenen Beleuchtungen. Sie läßt ohne weiteres erkennen, daß
in starkem Licht die Pflanzen viel kleiner bleiben als in schwachem
Licht.
Die Wirkung der verschiedenen Strahlenarten[210], aus denen das weiße
Tageslicht besteht, ist nicht die gleiche. Wenn Licht die Streckung des
Stengels hemmt, so sind dabei die kurzwelligen blauen und violetten
Strahlen wirksam, während sich rote Strahlen wie Dunkelheit verhalten.
Bei den Farnprothallien wird durch rotes Licht das Wachstum
wie durch Dunkelheit gefördert, aber die Zellteilung gehemmt, während
umgekehrt die blauen und violetten Strahlen das Wachstum hemmen,
aber die Teilung fördern. Da das Licht nicht nur als Wachstumsreiz,
sondern auch als Energiequelle tätig ist, so ist die Komplikation
der Erscheinungen einigermaßen verständlich. — Ultraviolettes Licht
schädigt die Pflanze, Radium- und Röntgenstrahlen wirken hemmend auf
Wachstumsvorgänge ein, können aber wie Gifte (S. 257) in kleinen Mengen
das Wachstum fördern[211].
Neben der Intensität und der Qualität des Lichtes hat auch seine
Richtung eine große Bedeutung für die Ausgestaltung des
Pflanzenkörpers. Einseitig einfallendes Licht führt zu
Krümmungen (Phototropismus, vgl. S. 307). Aber auch auf Polarität
und Symmetrie hat das Licht Einfluß. So kann z. B. bei niedrig
organisierten Pflanzen an der Zelle, von der die Entwicklung ausgeht,
die stärker beleuchtete Seite zur Spitze, die andere zur Basis
werden. Oder es kann ein ursprünglich radiärer Vegetationspunkt durch
einseitige Beleuchtung zu einem bilateralen oder dorsiventralen werden.
Endlich kann auch ein Organ, nachdem es längst aus dem embryonalen
Stadium herausgetreten ist, noch durch einseitige Lichtwirkung
dorsiventral werden, z. B. indem es bloß auf der Schattenseite Wurzeln
bildet. — Und wenn es gelingt, experimentell die äußere Symmetrie
umzugestalten, so ist damit in der Regel auch der innere Bau verändert.
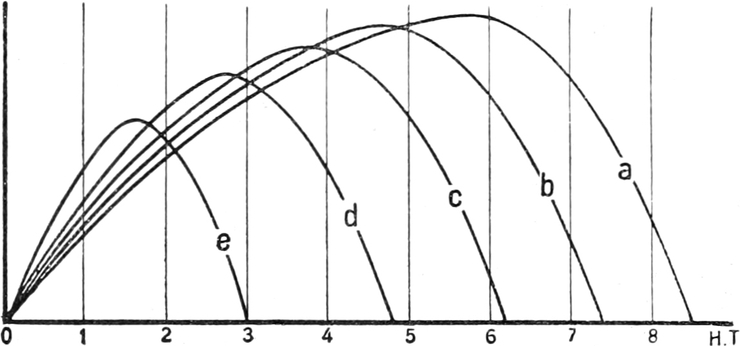
Fig. 262. Große Periode des Wachstums der
Avenakoleoptile nach SIERP. Die Abszisse gibt die Zeit in
halben Tagen an, die Ordinaten den Zuwachs. Kurve a bei Dunkelheit,
b, c .. bei zunehmender Beleuchtungsstärke.
Bei der Keimung der Sporen von Equisetum wird die Richtung der
ersten Scheidewand und damit die Lage von Basis und
Spitze durch die Lichtrichtung bestimmt. Ein ähnlicher
Einfluß des Lichtes auf die Polarität zeigt sich bei den Eizellen
von Fucus und Dictyota. — Antithamnion cruciatum (eine Floridee)
bildet in zerstreutem Licht aufeinanderfolgende Auszweigungen
ungefähr in gekreuzter Stellung aus; bei einseitigem Lichteinfall
aber stellen sie sich alle senkrecht zu den Strahlen in eine
Ebene. Weitere Beispiele für solche durch einseitige Beleuchtung
induzierte Dorsiventralität liefern die Zweige vieler
Laubmoose, die Thalli der meisten Lebermoose und endlich die
Prothallien der Farne, Gebilde, die ohne solche einseitige
Lichtwirkung teils radiär, teils bilateral werden. So wird z. B. an
den Farnprothallien und bei Marchantia die Oberseite durch
die stärkere Beleuchtung bestimmt. Während nun bei Farnprothallien
nach Umkehrung der Beleuchtungsrichtung der Neuzuwachs sich den
neuen Lichtverhältnissen anpaßt, also die bisherige Oberseite
zur Unterseite wird, ist der Marchantienthallus, wenn einmal die
Dorsiventralität induziert ist, nicht mehr veränderlich. — Als
Beispiel einer durch Beleuchtung induzierten Dorsiventralität
bei höheren Pflanzen seien die Sprosse des Efeus und anderer
Wurzelkletterer genannt, deren Kletterwurzeln auf der beschatteten
Seite entstehen.
[S. 255]
3. Schwerkraft. Dem Lichte können wir die Pflanze leicht
entziehen; die Schwerkraft aber wirkt überall auf sie ein. Was wir da
ändern können, ist nur die Richtung. Fällt die Schwerkraftrichtung
mit der Richtung der Hauptwurzel und des Hauptsprosses zusammen, so
bemerken wir keinerlei Einfluß: bildet sie aber einen Winkel mit
diesen Organen, so treten ähnlich wie bei einseitig einfallendem
Lichte Krümmungen auf (s. Geotropismus). Davon abgesehen, ist
vor allem ein Einfluß der Schwerkraft auf die Polarität der
Pflanzen konstatiert. Nirgends freilich in dem Maße, daß es gelänge,
durch Umkehrung (Inversstellung) einer Pflanze etwa den Sproß in eine
Wurzel zu verwandeln. Nicht einmal an der undifferenzierten Eizelle
läßt sich durch die Schwerkraft die Polarität verändern. Sie wird fast
überall durch innere Ursachen bedingt, und die Schwerkraft kann nur
modifizierend eingreifen.
Werden abgeschnittene Weidenzweige in einem feuchten Raum
aufgehängt, so bilden sich Wurzeln vorzugsweise nahe dem
unteren Ende, und von den angelegten Knospen treiben nur die
höchst stehenden aus (Fig. 263, 1). Werden die Zweige aber in
verkehrter Lage aufgehängt, so sind es die gleichen (jetzt nach
unten schauenden) Knospen, die austreiben, und am basalen (nach
oben schauenden) Ende bilden sich die größten Wurzeln (Fig. 263,
2). Somit zeigt also der Versuch, daß in erster Linie innere
Ursachen für den polaren Gegensatz bestehen. Da aber bei den
invers gestellten Zweigen die Wurzelbildung erheblich nach
abwärts, die Sproßbildung aufwärts verschoben ist, so muß auch die
Schwerkraft mitbeteiligt sein. Nur ausnahmsweise ist es gelungen,
durch Inversstellung eine vollkommene und eine dauernde Umkehrung
der Polarität einer Pflanze zu erzielen. In den meisten Fällen
sterben inverse Pflanzen bald ab; wo sie aber längere Zeit am
Leben bleiben, zeigen sie schwere Störungen in ihrem anatomischen
Bau[212].
Auch an schräg oder horizontal gelegten Zweigen macht sich ein
Einfluß der Schwerkraft auf die inneren Dispositionen geltend.
Letztere bringen es mit sich, daß die obersten Knospen am stärksten
austreiben und sich zu Langtrieben entwickeln. Aus der Vertikallage
gebrachte Zweige zeigen die basalen Knospen gefördert, die apikalen
gehemmt. Bogig gekrümmte lassen die stärkst wachsenden Triebe am
höchsten Punkt auftreten. In der Reben- und Obstzucht wird
durch Verbiegung der Äste auf die Bildung schwächer wachsender
Sprosse (Kurztriebe) hingewirkt, die erfahrungsgemäß leichter
Blüten produzieren.
Fig. 263. Ein Zweig einer Weide, 1 in normaler Lage,
2 in inverser Lage im feuchten Raum treibend. Nach VÖCHTING.
4. Mechanische Einflüsse. Druck und Zug wirken in zweierlei
Weise auf das Wachstum, einmal rein mechanisch, außerdem auch als
Reiz. Mechanische Widerstände verlangsamen zunächst das
Wachstum der Zellen und heben es schließlich ganz auf. Da aber das
Flächenwachstum der Zellwand noch weiter geht, schwindet ihre Spannung
schließlich ganz. Nach völliger Entspannung der Zellhaut aber wirkt
der ganze Innendruck der Zelle auf die Widerlage ein, in manchen
Fällen wird unter solchen Umständen der osmotische Wert des Zellsaftes
noch vergrößert. So kommt es, daß Wurzeln Felsen sprengen können.
Ist das Hindernis nicht zu überwinden, so veranlaßt die plastische
Nachgiebigkeit der Membranen ein enges Anschmiegen an dasselbe;
Wurzeln[S. 256] und Wurzelhaare, die in enge oder flache Hohlräume eindringen,
füllen diese daher oft so vollkommen aus, als ob sie als flüssige
Masse hineingegossen worden wären. — Von einem starken mechanischen
Zug sollte man erwarten, daß er durch Unterstützung und
Förderung der Dehnung das Längenwachstum beschleunige. Tatsächlich
macht sich aber eine Reizwirkung geltend, der Zug hat zunächst
geradezu eine Verzögerung des Wachstums zur Folge, und erst
später treten Beschleunigungen bis 20% auf.
Von anderen Reizwirkungen mechanischer Einflüsse nennen wir noch
folgende: An gekrümmten Wurzeln treten Seitenwurzeln ausschließlich
auf den Konvexseiten auf; die Ursache liegt wahrscheinlich in
den Spannungsverhältnissen, die an den antagonistischen
Seiten bestehen (Fig. 264). — Die Anlage der Haustorien von
Cuscuta und die Ausbildung der Saugnäpfe an den Ranken mancher
Parthenocissus-Arten (Fig. 207) wird durch Berührungsreiz
ausgelöst.
Steigern sich mechanische Einflüsse bis zur Verwundung, so treten
entweder einfache Heilungserscheinungen (S. 140) oder Restitutionen (S.
245) auf.
Fig. 264. Junge Lupine mit bogig gewachsener
Hauptwurzel. Die Seitenwurzeln s ausschließlich auf den Konvexflanken
entwickelt. Nach NOLL.
5. Stoffliche Einflüsse. Die Gegenwart der nötigen Nährstoffe
in genügender Menge ist ebenso wie die Abwesenheit von Giften formale
Bedingung für jedes Wachstum. Wenn auch, wie wir wissen, jeder einzelne
Nährstoff unentbehrlich ist, durch eine Überfülle der anderen nicht
ersetzt werden kann, so darf man doch für bestimmte Vorgänge
einem einzelnen eine besondere Wichtigkeit zuschreiben. — Da die
Streckung ganz überwiegend auf Wassereinlagerung beruht, so
ist die Bedeutung der Wasserzufuhr für die wachsende Pflanze
ohne weiteres einleuchtend. Nur die turgeszente Pflanze wächst, und
der Wassergehalt des Bodens beeinflußt weitgehend die Größe der
Pflanze und ihrer Zellen. Im trockenen Boden entstehen Zwerge[213].
Manche Pflanzen aber besitzen Wasserspeicher und sind dadurch etwas
unabhängiger von direkter Wasseraufnahme; sie wachsen auf Kosten des
gespeicherten Wassers. Vielfach kann auch das Wasser älteren Teilen
entnommen werden, die dann vertrocknen, während an der Spitze Wachstum
stattfindet; so bei der im Keller treibenden Kartoffel. Ganz anders
wie der Wassergehalt des Bodens wirkt der Feuchtigkeitsgehalt
der Luft. Durch ihn wird Form und anatomischer Bau weitgehend
geändert. In feuchter Atmosphäre entwickeln sich lange Internodien und
lange Blattstiele, die Blattfläche wird groß, aber sie bleibt dünn,
und ihr Rand ist wenig ausmodelliert, die Behaarung tritt zurück. Die
Gewebedifferenzierung ist sehr gering. Es besteht also in mehrfacher
Hinsicht Annäherung an die etiolierte Pflanze. Eine lokale Überfüllung
der Pflanze mit Wasser, wie sie nach Hemmung der Transpiration z. B.
durch Überzüge von Paraffinöl auftritt, führt zur Anschwellung der
Zellen und somit zur Intumeszenzbildung. Umgekehrt wird durch trockene
Luft die anatomische Differenzierung sehr gefördert; die Kutikula
wird verstärkt, Kollenchym, Sklerenchym und Gefäße werden vermehrt,
kurz es entstehen vielfach Abänderungen, die in feuchter Luft auf
Transpirationssteigerung, in trockener[S. 257] auf Transpirationshemmung
hinauslaufen, also zweckmäßige Abänderungen. Anpassungen.
Eine sehr auffallende Reizwirkung wird durch die dauernde Berührung
mit flüssigem Wasser bei denjenigen Pflanzen erzielt, die eine
solche überhaupt ertragen können; zweifellos handelt es sich da
freilich nicht um eine einfache stoffliche Wirkung des Wassers, sondern
um eine kombinierte Wirkung vieler Faktoren: neben dem Wasserüberfluß
dürfte z. B. die Aufhebung der Transpiration, doch auch die Änderung
der Beleuchtung und der Sauerstoffzufuhr wichtig sein.
Amphibische Pflanzen, d. h. solche, die befähigt sind,
sowohl auf dem Lande wie im Wasser zu leben, entwickeln im Wasser
oft ganz andere Formen als in der Luft. Dies tritt zumal in der
Gestalt der Blätter hervor, die im Wasser häufig lineal und
sitzend oder aber fein zerschlitzt, an der Luft dagegen gestielt
und mit breiter Spreite ausgebildet werden (vgl. Fig. 140). Aber
auch Blattstiele und Internodien zeigen im Wasser oft ein anderes
Wachstum als in der Luft, und es erinnert ihr Verhalten im Wasser
an die Überverlängerung im Finstern. Das ist besonders der Fall
bei untergetauchten Wasserpflanzen, von denen gewisse Teile die
Oberfläche erreichen müssen (Stengel von Hippuris, Blattstiele
von Nymphaea). Diese Pflanzenteile sind durch verschieden starke
Streckung unter Wasser imstande, ihre Länge der Höhe des über ihnen
stehenden Wasserspiegels anzupassen, in flachem Wasser kurz zu
bleiben, in tiefem sehr lang zu werden.
Auch die Gewebe der Wasserformen zeigen manche Abweichungen
von denen der Landformen: in den Stengeln fehlen die verdickten
Zellen, und auch die Leitbündel sind reduziert; die Blätter
erinnern an Schattenblätter. In schroffstem Gegensatz zu den
Wasserpflanzen stehen solche Landpflanzen, die bei geringer
Wasserzufuhr bzw. bei starker Transpiration entstanden sind; hier
sind die Leitbündel sehr kräftig entwickelt, und in der Epidermis
werden Einrichtungen ausgebildet, die wir unter den Mitteln zum
Schutz vor zu großer Transpiration kennen gelernt haben.
Neben den eigentlichen Nährstoffen, die zum Aufbau der Pflanzensubstanz
Verwendung finden, verdient auch der Sauerstoff genannt zu werden.
Obwohl sein Eintritt in die Pflanze mit einer Verminderung der
organischen Substanz verbunden ist, ist er — bzw. die durch ihn
bewirkte Atmung — doch ganz unentbehrlich für das Wachstum. Bei den
Aërobionten wenigstens steht das Wachstum nach Sauerstoffentziehung
völlig still; auch eine Verminderung oder Vermehrung des normalen
Sauerstoffgehaltes der Luft beeinflußt es schon.
Aber auch Reize mannigfacher Art gehen von Stoffen aus, die auf die
Pflanze einwirken.
Da sind zunächst die Gifte zu nennen, Stoffe, die schon in sehr
geringer Konzentration das Wachstum und schließlich überhaupt das
Leben sistieren. So tötet Kupfervitriol Algen, zumal Spirogyren,
aber auch Erbsen in Wasserkultur, schon in der Verdünnung
1 : 25000000. Auffallenderweise wirken aber viele Gifte in starker
Verdünnung geradezu stimulierend auf das Wachstum und bewirken
eine bessere Ausnützung der Nährstoffe. Chemische Reizung anderer
Art spielt vor allen Dingen bei der Keimung vieler Samen, Sporen
und Pollenkörner sowie bei der Entwicklung der Früchte eine große
Rolle. Viele parasitische Pilze und auch parasitische Phanerogamen
(Orobanche, Lathraea) werden durch noch unbekannte, von ihren
Wirten ausgehende Stoffe zum Wachstum gereizt. Bei Algen und Pilzen
kann namentlich durch hohe Konzentration gewisser Nährstoffe eine
beträchtliche Gestaltsveränderung hervorgerufen werden.
6. Einflüsse fremder Organismen. Pilze und Bakterien, die
parasitisch in einer Pflanze leben, veranlassen nicht selten
weitgehende Deformationen in ihr, die man Gallen[214] nennt. In
einfacheren Fällen entsteht dabei nur eine Vergrößerung der Zellen,
in komplizierteren treten qualitative Veränderungen der Organe ein.
Noch viel auffallender sind die Gallenbildungen, die durch Tiere, vor
allem Insekten an den Pflanzen hervorgebracht werden. Es entstehen
Wucherungen, die dem Parasiten zur Wohnung und Nahrung[S. 258] dienen und
die in ihrem Bau oft als durchaus nützlich für den Parasiten
erscheinen: wir treffen Schutzschichten, Nährschichten und
Deckelbildungen, die für die Pflanze selbst ganz ohne Bedeutung sind.
Die von dem Rostpilz Aecidium Euphorbiae infizierte
Zypressenwolfsmilch (Euphorbia Cyparissias) wird unfruchtbar,
bleibt unverzweigt, erhält kürzere aber breitere Blätter und
erscheint in ihrem ganzen Aussehen so verändert, daß man
sie kaum wieder erkennt. Blattläuse können Vergrünungen der
befallenen Blütensprosse veranlassen, wobei statt Blumenblättern
laubblattähnliche Gebilde entstehen. Besonders eigenartig sind
die Gallen (Cecidien), deren Bildung durch Pilze und
vornehmlich durch Tiere (Würmer und Arthropoden) bedingt wird.
Der Eingriff dieser Organismen in die normale Entwicklung der
befallenen Pflanze ist mehr oder weniger bedeutend, je nachdem
er die embryonale Substanz der Vegetationspunkte oder ein schon
angelegtes, jedoch noch in Ausbildung begriffenes, oder endlich
ein schon fertiges Glied des Pflanzenkörpers trifft. Es können
histoide Gallen entstehen, bei welchen es sich um Produkte
abnormer Gewebebildung handelt, oder organoide, die auf
Umbildung oder Neubildung der Glieder des Pflanzenkörpers beruhen.
Die letzteren sind besonders lehrreich. Die Larve der Gallmückenart
Cecidomyia rosaria, die über dem Vegetationspunkt der befallenen
Weidensprosse sich entwickelt, veranlaßt die Verbildung der
ganzen Sprosse, die Entstehung jener Gallen, die als Weidenrosen
bekannt sind und Blattschöpfe aus stark veränderten Blättern
und gestauchten Achsenteilen darstellen. Verschiedene Fliegen
(Diptera) legen ihre Eier in noch unfertige Blätter und regen
um so stärkere Anschwellungen und Rollungen an, je jünger die
Blätter im Augenblick der Infektion waren. Unverändert in seiner
Gesamtausbildung bleibt hingegen das Blatt der Eiche, das erst
im schon weit entwickelten Zustand von Gallwespen der Gattung
Cynips angestochen wird; hier wird nur eine lokale Wucherung des
Blattgewebes gebildet, die zur Bildung der kugeligen gelben oder
roten, an den Seitenrippen der Blattunterseite sitzenden Gallen
führt.
Auch „Symbionten“, d. h. friedlich zusammenlebende Organismen, von
denen nicht der eine die Rolle des Wirtes, der andere die des Parasiten
spielt, können sich formativ beeinflussen. Das sieht man z. B. bei den
Flechten.
Es ist wahrscheinlich, daß bei der Beeinflussung durch fremde
Organismen neben der Verwundung stoffliche Wirkungen die
Hauptrolle spielen. Freilich konnte bisher nur in seltenen Fällen
eine der Gallbildung ähnliche Deformation schon durch Einwirkung
toter, aus dem Gallenbewohner extrahierter Substanzen erzielt werden.
Auch Parasiten, die keine Gallbildung zur Folge haben, wirken durch
ihre Gifte stofflich auf die Wirtpflanze ein. Umgekehrt kann die
Wirtpflanze durch Bildung von Antikörpern den Parasiten schädigen
oder ihn ganz am Eindringen hindern. So hat HEINRICHER
gezeigt, daß manche Birnsorten leicht, andere schwer von der Mistel
infiziert werden, und er hat wahrscheinlich gemacht, daß eine einmalige
Infektion mit dem Schmarotzer den Wirt bei künftigen Infektionen
widerstandsfähiger macht. Es kommen also auch im Pflanzenreich
Immunitätserscheinungen vor, sie sind aber hier bei weitem nicht
so eingehend untersucht wie bei Tieren[215].
7. Rückblick auf die äußeren Faktoren. Es hat sich gezeigt,
daß eine Menge von äußeren Faktoren in einfacheren Fällen nur auf die
Wachstumsgeschwindigkeit, in komplizierteren dagegen auch auf die
schließliche Gestalt der Pflanze von Einfluß ist. Wir pflegen die
Gestalt der Pflanze, die uns gewöhnlich in der Natur entgegentritt,
als ihre normale Gestalt zu bezeichnen. Jede Abweichung von
dieser, die unter dem Einfluß äußerer Faktoren entsteht, nennen
wir eine Modifikation dieser normalen Gestalt. Es ist aber
klar, daß auch die Normalform lediglich der Ausdruck ganz bestimmter
Außenfaktoren ist, nämlich derjenigen, die gewöhnlich die Pflanzen
während ihrer Entwicklung treffen. Mit anderen Worten, Normalform
und Modifikation sind beide gleich gesetzmäßige Bildungen. Nicht die
Modifikation, sondern nur die Konstellation von Außenbedingungen,
unter denen sie entstanden ist, ist abnorm. Wenn auch manche dieser
Veränderungen für die betroffene Pflanze[S. 259] ganz bedeutungslos
sind oder gar, wie das bei den Gallen betont wurde, nur für den
Veranlasser der Veränderung von Nutzen sind, so ist doch eine große
Zahl von Reaktionen auf äußere Reize derart, daß die Pflanze einen
Nutzen von ihnen hat. So haben wir z. B. die Überverlängerung beim
Etiolement, die charakteristische Ausbildung der amphibischen Pflanzen
im Wasser bzw. auf dem Lande, die Steigerung des Transpirationsschutzes
bei Zunahme der Lufttrockenheit usw. usw. als nützliche Reaktionen, als
Anpassungen zu bezeichnen, Woher es kommt, daß die Pflanze oft
mit Anpassungen reagiert, ist an dieser Stelle nicht zu besprechen (S.
179). Im Gegensatz zu den Anpassungen nennen wir solche Modifikationen,
die für die Pflanze nutzlos oder gar schädlich sind Mißbildungen.
B. Innere Faktoren.
Auf innere Faktoren müssen wir stets dann schließen, wenn eine
Veränderung am Organismus bei Konstanz aller Außenfaktoren
eintritt. Wir können die Innenfaktoren bei weitem nicht so gut
analysieren wie die Außenfaktoren, und deshalb ist die Zurückführung
mancher Erscheinungen auf „innere Ursachen“ oft nicht viel mehr als ein
Geständnis unserer Unkenntnis.
1. Anlagen. Innere Ursachen sind zunächst einmal die
„Anlagen“, die eine Pflanze von ihren Eltern mitbekommen hat;
diese bewirken, daß aus der Spore eines Pilzes immer wieder dieser
Pilz, aus dem Samen der Bohne wieder eine Bohne hervorgeht. Sie
bewirken auch im einzelnen, daß eine bestimmte „Art“ bei gleichen
Außenbedingungen in allen Individuen die gleichen Eigenschaften
hat, wie z. B. Blütenfarbe, Blattgestalt, Größe usw. Den Bestand
einer Spezies an Anlagen kann man im allgemeinen nicht experimentell
verändern, ja man kann ihn nicht einmal ohne weiteres durch Beobachtung
feststellen; erst das Vererbungsexperiment gibt über ihn Aufschluß.
Deshalb kann auch erst bei Behandlung der Vererbung mehr von den
„Anlagen“ gesagt werden (S. 280).
2. Korrelationserscheinungen[216]. Wenn auch äußere Faktoren
einen weitgehenden Einfluß auf den inneren Bau der Pflanzen haben, so
findet die Gewebedifferenzierung doch auch bei ganz konstanten
äußeren Bedingungen statt — sie ist also durch innere Ursachen
bewirkt. Was für Ursachen das im einzelnen sind, die eine embryonale
Zelle in eine bestimmte Entwicklungsrichtung hineindrängen, das wissen
wir nicht. Nur eins ist uns ganz sicher bekannt: aus jeder Zelle
des Vegetationspunktes kann noch alles werden, alle Zellen sind ihren
Anlagen nach gleich. Daß die eine sich so, die andere sich anders
entwickelt, ist einzig und allein durch die Wechselbeziehungen oder
Korrelationen bedingt, die zwischen den Zellen bestehen. Heben
wir diese Beziehungen auf, heben wir den Zusammenhang zwischen den
Zellen auf, so sehen wir bei den Erscheinungen der Restitution (S. 245)
in der Tat ganz andere Leistungen von den Zellen vollführt, als sie
im Zusammenhang zuvor zeigten. Und nicht nur embryonale Zellen, auch
ausgewachsene können noch nach Aufhebung des Zusammenhangs mit den
Nachbarzellen neue Leistungen übernehmen und dann auch neue Gestalten
ausbilden. So können bei Restitutionsprozessen Zellen, die ausgewachsen
waren und die bald abgestorben wären, wieder jung werden, und wir sehen
z. B. aus einer Epidermiszelle (Fig. 256) die ganze Fülle von
Zellarten sich bilden, die für die betreffende Pflanze charakteristisch
sind. Es ist klar, daß ein Organismus, bei dem solche gegenseitige
Beeinflussung der Zellen fehlt, nicht die Arbeitsteilung aufweisen
kann, wie wir sie bei den höheren Pflanzen zu sehen gewohnt sind. Es
sind also, mit anderen Worten, die[S. 260] Korrelationen mit zu den
„Regulationen“ zu rechnen, ohne die ein Organismus undenkbar ist.
Nicht nur zwischen den Zellen, auch zwischen den äußeren Organen
der Pflanze bestehen solche Korrelationen. Nimmt man diese auch für
gewöhnlich nicht wahr, so treten sie doch sofort zutage, wenn man
entweder ein Organ entfernt und nun die Reaktion des isolierten Organs
sowie der Pflanze, von der es genommen ist, studiert, oder wenn man ein
Organ künstlich an einen Platz bringt, an dem es vorher nicht war.
Im ersten Fall können zunächst einmal sog. Kompensationen
auftreten: Durch Entfernung eines Organs werden andere
größer. Die Blätter, die am Vegetationspunkt entstehen,
verhindern ältere Blätter, ihre maximale Größe zu erreichen; wird
der Vegetationspunkt entfernt, so nimmt demnach (z. B. beim Tabak)
die Blattgröße zu. Das Austreiben einzelner Achselknospen
verhindert zahlreiche andere am Streckungswachstum; werden aber
die dominierenden Sprosse entfernt, so treiben die bisher ruhenden
Knospen sofort aus. Man wird daraus schließen, daß auch in der
normalen Entwicklung die Größe der Organe durch korrelative
Einflüsse der Nachbarorgane mit bestimmt wird. In anderen Fällen
sehen wir einen mehr qualitativen Erfolg der Entfernung eines
Organs. Wird der Gipfel einer Tanne entfernt, so tritt einer der
nächststehenden Seitenzweige an seine Stelle: er nimmt den aufrechten
Wuchs des Hauptsprosses an und ordnet auch seine Nadeln so an, wie
der Hauptsproß das tut. Wir dürfen also vermuten, daß für gewöhnlich
die schräge Lage und die dorsiventrale Anordnung der Benadelung am
Seitenzweig sich unter dem Einfluß des Hauptsprosses vollzieht. In
diesen und in zahllosen anderen Fällen korrelativer Beeinflussung ist
es übrigens keineswegs notwendig, ein Organ zu entfernen; meist
genügt es, wenn es z. B. durch Eingipsung oder durch andere Mittel in
seiner normalen Funktion gestört wird.
Daß an isolierten Organen je nach Umständen neue Wurzeln, neue
Sprosse entwickelt werden, ist bei der Besprechung der Restitution
S. 246 ausgeführt worden. So wie jede Zelle ursprünglich zu
vielfacher Gestaltung befähigt ist, so sind es auch die Glieder.
Nur die gegenseitige Beeinflussung hindert sie an so vielseitiger
Betätigung.
Korrelationen können endlich noch dadurch nachgewiesen werden, daß
man ein Organ an eine neue Stelle verpflanzt. Mit den Methoden der
Transplantation, die aus der Praxis der Gärtnerei hervorgegangen
sind, gelingt es bei manchen Pflanzen leicht, einen abgetrennten
Teil an eine Wunde anwachsen zu lassen. Man nennt den zugefügten das
„Pfropfreis“, den anderen Komponenten die „Unterlage“. Das Reis kann
der gleichen Spezies entstammen wie die Unterlage oder von einer
verwandten Pflanze entnommen sein. Bei solchen Transplantationen können
auch korrelative Beeinflussungen auftreten. Eine solche ist z. B. schon
darin gegeben, daß durch die Verwachsung die Restitution sowohl an der
Unterlage wie am Reis unterbleibt. Es adoptiert gewissermaßen der eine
Teil die Wurzeln des anderen, der andere die Sprosse des ersten, und
damit entfällt die Notwendigkeit, solche Organe neu zu bilden.
Die künstliche Verwachsung spielt ähnlich wie die künstliche
Vermehrung eine große Rolle in der Gärtnerei, bei der
Veredelung. Es werden abgetrennte knospentragende Teile
einer Pflanze, das sog. Edelreis, auf einer bewurzelten
Pflanze, der sog. Unterlage, zur Verwachsung gebracht, weil es
so rascher als durch künstliche Vermehrung oder gar durch Samen
gelingt, hochwüchsige Exemplare einer Rasse oder Art herzustellen.
In der gärtnerischen Praxis haben sich verschiedene Methoden für
die Verbindung der Pflanzenteile bei der Veredelung herausgebildet,
von denen hier nur die wichtigsten[S. 261] erwähnt sein mögen. Unter
Pfropfen versteht man solche Transplantationen, bei denen
mit Knospen besetzte Zweige mit dem abgestutzten
Stamm der Unterlage zur Verwachsung gebracht werden. Es wird dabei
beachtet, daß wenigstens stellenweise die Kambialregionen der zu
verbindenden Teile aufeinander stoßen. Man pfropft im übrigen
entweder in den Spalt, in die Rinde oder in die
Seite. Im ersten Falle wird das Ende der Unterlage längs
gespalten, und in den Spalt wird entweder ein einziges Pfropfreis
eingesetzt, das dann so stark sein kann wie die Unterlage, oder
es wird auf jeder Seite peripherisch ein Reis zur Verwachsung
gebracht; unter allen Umständen wird die Basis des Pfropfreises
keilförmig zugespitzt. Bei dem Pfropfen in die Rinde wird das
flach zugeschnittene Edelreis zwischen die durch einen Längsspalt
klaffend gemachte Rinde und den Splint eingeschoben (Fig. 265 I).
Bei dem Pfropfen in die Seite werden die Edelreiser, keilartig
zugeschnitten, in seitliche Einkerbungen der Unterlage eingesetzt.
— Wird die Unterlage schräg abgeschnitten und über ihr ein
ebenfalls schräg zugeschnittenes Pfropfreis von gleicher Dicke
so angesetzt, wie das Fig. 265 II zeigt, so spricht man wohl
von Kopulieren. — Eine besondere Art der Transplantation
ist das Okulieren (Fig. 165 III), wobei nicht ein
Zweigstück, sondern eine Knospe, ein „Auge“, unter
die Rinde der Unterlage eingeschoben wird. Das Edelauge bleibt
im Zusammenhang mit einem schildförmigen Rindenstück, das sich
leicht vom Splint ablöst, wenn die Pflanzen „im Saft“ sind. Durch
einen T-förmigen Schnitt wird dann die Rinde der Unterlage
gelockert, das Rindenschild des Edelauges eingeschoben und das
Ganze dicht abgeschlossen. Auf das „treibende Auge“ okuliert man im
Frühjahr, auf das „schlafende“, erst im nächsten Jahr austreibende
Auge, im Sommer.
Fig. 265. Verschiedene Veredelungsweisen. I Pfropfen.
II Kopulieren. III Okulieren. In allen Figuren bedeutet W
Unterlage (Wildling) E Edelreis bzw. Edelauge. Nach NOLL.
Die Verwachsung erfolgt mit Hilfe eines an beiden Pflanzenteilen
entstehenden Wundgewebes, des sog. Kallus (vgl. S. 140). In
dem Kallus bilden sich nachträglich Gefäße und Siebröhren aus und
verbinden die gleichartig funktionierenden Elemente beider Stücke.
— Derartige Verwachsungen sind aber nur bei nahe verwandten
Pflanzen möglich. So lassen sich die zu den Amygdalaceen gehörigen
Obstbäume leicht verbinden, also Pfirsich, Aprikose, Mirabelle,
Pflaume und Schlehe untereinander, ebenso die Pomaceen Birne und
Quitte, Quitte und Weißdorn, nicht aber Apfel mit Pflaume oder gar,
wie auch behauptet worden ist, mit Eiche u. dgl.
Die Polarität, die sich bei der Restitution so bemerkbar macht,
tritt auch bei künstlich herbeigeführter Verwachsung
sehr auffallend zutage. Ungleichnamige Pole einer Pflanze sind
leicht zum Verwachsen zu bringen, gleichnamige verwachsen nicht
oder schwieriger, und ihre Verbindung trägt dann dauernd einen
schadhaften Charakter.
Unterlage und Pfropfreis beeinflussen sich in mannigfacher Weise. So
gewinnen z. B. Teile einjähriger Pflanzen, wenn sie auf ausdauernde
gepfropft werden, eine längere Lebensdauer. Ebenso läßt sich auch der
entgegengesetzte Erfolg, Verkürzung der Lebensdauer durch Pfropfung,
erzielen. Aber auch qualitative Erfolge können eintreten, die im
Extrem so[S. 262] weit gehen, daß eine Laubknospe durch Aufpfropfung auf eine
passende Unterlage zu einem Blütensproß umgewandelt werden kann. Die
spezifischen Eigenschaften der beiden Komponenten bleiben indes
bei der Transplantation erhalten; gewisse als „Chimären“ bezeichnete
Fälle scheinen dem auf den ersten Blick zu widersprechen, doch zeigt
sich bei näherer Betrachtung, daß auch bei ihnen, obwohl sie äußerlich
als Mittelbildungen zwischen den beiden Pfropfsymbionten
erscheinen, dennoch keine „Vermischung“ zweier Spezies vorliegt.
Chimären[217]. In unseren Gärten finden sich unter dem Namen
Laburnum Adami und Crataegomespilus einige Pflanzen, die in mehrfacher
Hinsicht an Bastarde (S. 278) erinnern, die aber zweifellos
nicht durch sexuelle Vereinigung der Mutterpflanzen entstanden sind.
Laburnum Adami (Fig. 266) ist eine Mittelbildung zwischen
Laburnum vulgare und Cytisus purpureus und bildet sehr häufig Zweige
aus, die man als „Rückschläge“ zu Laburnum vulgare betrachten
muß, seltener solche, die ganz wie Cytisus purpureus aussehen.
Als Crataegomespilus oder Bronveauxhybriden werden einige
Mittelformen zwischen Crataegus monogyna und Mespilus germanica
bezeichnet, die ebenfalls sehr häufig Rückschläge zu den Stammformen
bilden. Von den Bronveauxhybriden ist nun der Ursprung bekannt. Man
weiß, daß die Mittelformen — es existieren deren mehrere differente
— in der Nähe einer Pfropfstelle von Mespilus auf Crataegus zuerst
in einem Garten in Bronveaux bei Metz entstanden sind. Daß Laburnum
Adami ähnlichen Ursprungs ist, muß jetzt als sicher gelten. Man hat
demnach die beiden Pflanzen als „Pfropfhybriden“ betrachtet, d. h.
als Bastarde die nicht durch Verschmelzung von Sexualzellen, sondern
durch eine irgendwie beschaffene Beeinflussung vegetativer Zellen
hervorgebracht worden sind.
HANS WINKLER hat solche „Pfropfbastarde“ experimentell
hergestellt. Er pfropfte Solanum nigrum (Nachtschatten) auf Solanum
Lycopersicum (Tomate) und schnitt nach erfolgter Verwachsung den
Stengel des Pfropfreises quer durch die Pfropfstelle ab. Unter den
Adventivsprossen, die sich unmittelbar aus der Verwachsungsstelle
der beiden Komponenten bildeten, befanden sich auch ausgesprochene
Mittelbildungen zwischen Tomate und Nachtschatten. Und zwar traten
zunächst solche Formen auf, die aus verwachsenen Längshälften
der beiden aufeinander gepfropften Pflanzen bestanden; WINKLER
nannte sie Chimären. Später aber stellten sich auch andere,
äußerlich einheitlich erscheinende Mittelbildungen ein (Fig.
267), die zunächst als die gesuchten Pfropfbastarde gedeutet wurden.
Nähere Untersuchung zeigte aber, daß auch sie als Chimären zu
betrachten sind, da sie aus verwachsenen, sonst aber unveränderten
Teilen der Tomate und des Nachtschattens bestehen. Es sind aber
nicht Längshälften, die miteinander verwachsen sind, sondern Innen- und
Außenschichten derart, daß die Schichten des Vegetationspunktes
(vgl. S. 75 u. 269) aus Geweben verschiedener Spezies aufgebaut
sind. Man nennt sie „Periklinalchimären“ (Mantelchimären);
im Gegensatz dazu heißen die der Länge nach verwachsenen Chimären
„Sektorialchimären“.
Auch Cytisus Adami und die Crataegomespili sind Periklinalchimären;
echte Pfropfbastarde, bei denen durch Pfropfung eine
Vermischung der Arteigenschaften in der einzelnen Zelle erfolgt wäre,
sind bisher noch nicht bekannt.
[S. 263]

Fig. 266. Laburnum Adami Poit. (Cytisus Adami Hort.)
mit Rückschlagszweigen in seine beiden Stammformen: Laburnum vulgare
(links) und Cytisus purpureus (rechts). Nach NOLL.
[S. 264]
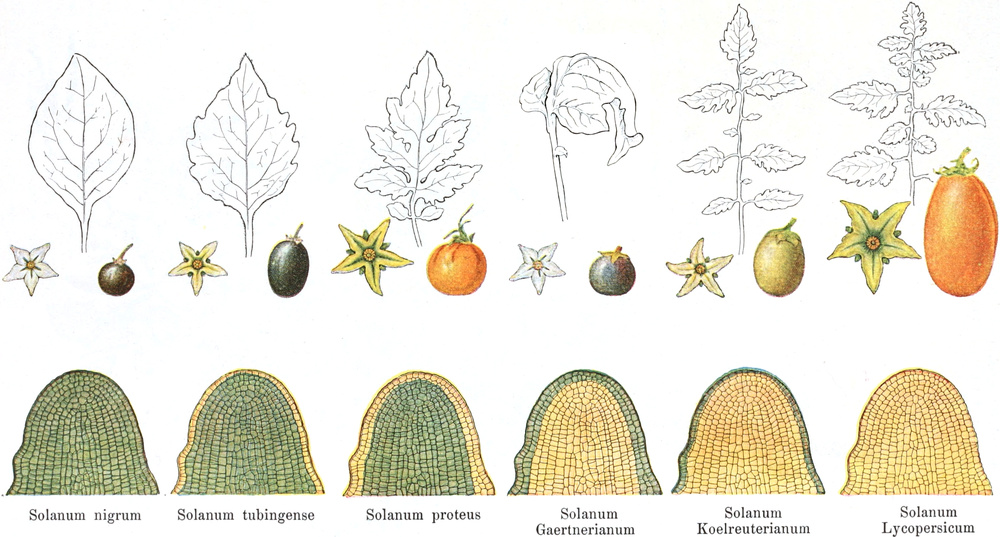
Fig. 267. Pfropfchimären zwischen Solanum nigrum und Solanum
Lycopersicum. Nebst den Eltern. Original von H. WINKLER.
Dargestellt ist jeweils ein Blatt, eine Blüte und eine Frucht sowie
ein Schema des Vegetationspunktes. In letzterem sind die von Solanum
nigrum stammenden Schichten grün, die von der Tomate herrührenden gelb
gehalten.
Solanum tubingense hat das Dermatogen der Tomate, das Innere des
Nachtschattens; bei Solanum Kölreuterianum ist es gerade umgekehrt;
bei S. proteus sind die zwei äußeren Lagen von der Tomate,
die übrigen vom Nachtschatten, und bei S. Gaertnerianum ist es
wahrscheinlich umgekehrt (Fig. 267). Entsprechend stammt bei
Cytisus Adami das[S. 265] Dermatogen von Cytisus purpureus, das Innere
von Laburnum vulgare. Bei einer der Bronveauxhybriden, der
forma „Asnieresii“, ist ein Crataeguskern von einer Mispelepidermis
bedeckt, die andere Form „Dardari“ hat zwei oder mehr
Mantelschichten von Mespilus. Diese Angaben beziehen sich auf den
Bau des Vegetationspunktes und der aus diesem entstehenden Achsen.
Die Blätter dagegen können nur bei Formen mit einschichtigem
Mantel Chimären sein; wo aber der Mantelsymbiont zweischichtig
auftritt, da muß er nach NOACK[218] das ganze
Blatt aufbauen, da dieses nur aus Epidermis und Hypodermis des
Vegetationspunktes sich herleitet. Demnach können diese Blätter
keine Periklinalchimären sein; auffallenderweise zeigen sie aber
doch Zwischenbildungen zwischen den Komponenten.
Wenn es gelingt, aus der einzelnen Schicht Adventivsprosse zu
erzielen, so haben diese den reinen Speziescharakter dieser Schicht
ohne jede Spur eines Einflusses des anderen Symbionten. — Über die
Entstehung der Periklinalchimären wissen wir noch nichts sicheres,
doch ist kaum daran zu zweifeln, daß die Vegetationspunkte der
Adventivsprosse aus Zellen der beiden Komponenten sich aufbauen,
und zwar derart, daß die eine Spezies die Bildung des Kernes, die
andere die des Mantels übernimmt.
III.
Der Entwicklungsgang und seine Abhängigkeit von äußeren und inneren Faktoren.
Der Entwicklungsgang besteht in einer Folge von Vorgängen, die sich
bei einer bestimmten Spezies immer wieder in der gleichen Weise zu
wiederholen pflegt. Doch lehren schon gelegentliche Beobachtungen in
der Natur, erst recht aber Versuche, daß diese Folge veränderungsfähig
sein muß. Die Aufgabe der Entwicklungsphysiologie ist es, den
Entwicklungsgang experimentell zu beherrschen und so Einblick in
seine Ursachen zu gewinnen. Von großer Wichtigkeit ist es auch, die
Teilprozesse der Entwicklung voneinander zu unterscheiden und durch
ihre ungleiche Abhängigkeit von Außenfaktoren voneinander zu trennen.
Wenn wir auch sehr viele Einzelheiten eines Entwicklungsganges
heute noch nicht experimentell beherrschen, so drängen uns doch
die bisherigen Erfolge die Überzeugung auf, daß der typische
Entwicklungsgang nur eine von vielen Möglichkeiten ist, die stets dann
eintritt, wenn ein bestimmter Komplex von Ursachen gegeben ist, und
daß jede Abweichung von diesem Ursachenkomplex auch in der Entwicklung
der Pflanze ihren Ausdruck findet.
Alle Umgestaltungen aber fallen um so extremer aus, je jünger die
Zellen sind, die beeinflußt werden. Wird bereits die embryonale
Substanz eines Vegetationspunktes von ihrem normalen Entwicklungsgang
abgelenkt, so kann ein ganz anderes Gebilde an Stelle des erwarteten
treten. Oder es bilden sich Mittelformen von mehr oder weniger
monströser Gestalt aus. Die embryonale Substanz der Vegetationspunkte
ist noch zur Bildung aller Anlagen befähigt, die den Formenkreis
der Spezies umfassen, und so kann z. B. an Stelle einer Blüte ein
vegetativer Sproß entstehen, ja in seltenen Fällen kann sogar der
Vegetationspunkt einer Wurzel sich unmittelbar als Sproß weiter
entwickeln. Hingegen werden schon angelegte Blätter in ihrer
Veränderung sich vornehmlich innerhalb des Formenkreises der
Blattmetamorphose halten, Blumenblätter beispielsweise an Stelle von
Staubblättern oder Fruchtblättern sich bilden. Je später der ändernde
Einfluß auf die Anlage sich geltend macht, um so unvollkommener wird
ihre Umwandlung sein.
Die Entwicklung der Organismen vollzieht sich nicht in ewig gleicher
Tätigkeit, in andauernd gleichem Wachstum. Vielmehr sehen wir überall,
meist periodisch, Veränderungen auftreten. So wechseln Perioden der
Ruhe mit solchen der Wachstumstätigkeit; und während der letzteren
sehen wir periodisch Zellteilungen auftreten, periodisch andere
Blattformen und Sproßformen[S. 266] entstehen und Fortpflanzungsorgane sich
bilden, periodisch auch kleinere und größere Teile absterben, bis
schließlich der Tod des ganzen Organismus erfolgt.
A. Ruhe und
Wachstumsbeginn[219].
Es ist schon früher ausgeführt worden, daß wir drei verschiedene
Zustände in der Pflanze unterscheiden: den des aktiven Lebens,
den des latenten Lebens und den des Todes. Es wurde auch erwähnt,
daß im latenten Leben alle Lebensäußerungen stillstehen: alle
Stoffwechseltätigkeit, selbst die Atmung, ist sistiert, und von
Wachstum und Bewegung ist nichts zu bemerken. Im Gegensatz zum toten
Zustand aber bleibt bei latentem Leben die Entwicklungsfähigkeit
gewahrt.
Ruhezustände. Solches latentes Leben treffen wir bei
den Samen, bei manchen Sporen der niederen Pflanzen, aber auch
bei vielen ausgewachsenen Teilen und Knospen in ungünstiger
Jahreszeit (Kälteperioden, Trockenperioden). Es wird aber nicht
unbegrenzt von der Pflanze ertragen, selbst Samen und Sporen,
die es am vollkommensten zeigen, büßen früher oder später die
Entwicklungsfähigkeit ein, sie sterben. — In anderen Fällen sehen
wir nicht alle Lebensfähigkeit unterdrückt, sondern einzig und
allein das Wachstum sistiert. So z. B. bei unbefruchteten Eizellen.
Auf den ersten Blick möchte man glauben, daß die Ruhezustände
während der ungünstigen Jahreszeit unmittelbar von dieser veranlaßt
werden. Tatsächlich tritt aber auch bei vielen tropischen Bäumen
periodisch ein Wachstumsstillstand ein; bei andauernd günstiger
Temperatur und Wasserzufuhr sehen wir die Blattbildung nicht
kontinuierlich vor sich gehen, sondern von Ruhepausen unterbrochen,
so daß im Laufe eines Jahres mehrfaches Austreiben erfolgt. Und bei
den einheimischen Pflanzen ist der Eintritt der Ruheperiode keineswegs
durch die zu niedere Temperatur bedingt. Viele Bäume hören schon im Mai
oder Juni mit der Blattentfaltung völlig auf. Auch zeigen unsere Bäume,
in ein tropisches Klima versetzt, vielfach eine ähnliche Periodizität
wie dort einheimische Pflanzen. Diese Erscheinungen werden noch nicht
von allen Forschern gleichartig gedeutet. Auf der einen Seite wird
angenommen, daß jede Periodizität im Wachsen der Pflanze durch eine
Periodizität in der Außenwelt bedingt sei, die freilich nicht nur in
der Zufuhr von Wärme und Wasser, sondern auch z. B. in der Aufnahme
von Nährsalzen gegeben sein kann. Auf der anderen Seite nimmt man eine
Periodizität aus inneren Ursachen an, die sich in Ländern mit
wechselnden Jahreszeiten diesen anpaßt: die Bäume verlegen bei uns die
Ruhezeit in den Winter, in anderen Ländern in die trockene Jahreszeit.
Das ist durchaus nicht bei allen Pflanzen so. Es gibt bei uns einige
Kräuter, wie z. B. Senecio vulgaris, die das ganze Jahr hindurch
wachsen, wenn nur die äußeren Bedingungen es zulassen, und in den
Tropen fehlt es erst recht nicht an dauernd wachsenden Pflanzen.
Eichen-, Buchen-, Apfel- und Birnbäume behalten in dem
subtropischen Klima von Madeira ihre Ruheperiode bei; unter den
noch gleichmäßiger günstigen Verhältnissen gewisser Tropengegenden,
wie in den Bergen Javas, besteht eine Ruheperiode fort, aber sie
verschiebt sich bei verschiedenen Individuen, ja sogar an den Ästen
eines und desselben Baumes zeitlich derart, daß Eichen, Magnolien,
Obst- und Mandelbäume wie gewisse dort einheimische Bäume zu
gleicher Zeit belaubte und blattlose Zweige tragen.
Entwicklungsbeginn. Der Schluß des Ruhezustandes, der
Wiederbeginn des Wachstums, ist vielfach lediglich mit der Herstellung
der allgemeinen Wachstumsbedingungen gegeben. In anderen Fällen aber
ist der Ruhezustand ein sehr fester. Die Herstellung der üblichen
Wachstumsbedingungen[S. 267] genügt dann nicht, um ihn zu überwinden, doch
läßt er sich manchmal durch besondere Reizmittel abkürzen.
Die Keimung der Samen erfolgt in der Regel nach Herstellung der
allgemeinen Wachstumsbedingungen, vor allem also der nötigen
Temperatur, Zufuhr von Sauerstoff und Wasser; doch fehlt es nicht
an Beispielen, die spezielle Reize erfordern. Solche spezielle Reize
heben auch die Entwicklungshemmung der Eizellen auf; für gewöhnlich
werden sie mit der Befruchtung gegeben; doch können manche Eizellen
auch ohne Befruchtung, z. B. durch Einwirkung von Chemikalien, zur
Entwicklung angeregt werden (vgl. auch S. 277 Parthenogenesis).
Manche Samen machen eine längere Ruheperiode durch, ehe sie zu
keimen beginnen. Zum Teil liegt das daran, daß sie erst nach der
Loslösung von der Mutterpflanze voll ausreifen [„Nachreife“[220]],
zum Teil aber auch daran, daß sie nur langsam Wasser aufnehmen. Vor
allem fallen bei der Keimung große individuelle Differenzen auf.
Es können einzelne Samen einer Pflanze oft jahrelang unverändert
im Boden liegen, während gleichalte längst gekeimt haben; das
hängt gewöhnlich mit der sog. Hartschaligkeit zusammen, d. h.
mit einer außerordentlich erschwerten Quellungsfähigkeit der
Schale. Diese scheint auch einen Grund dafür abzugeben, daß die
Samen vieler Wasserpflanzen[221] in reinem Wasser nicht keimen
können, während nach Zusatz von Säuren und Alkalien die Keimung
beginnt. In einzelnen Fällen aber können auch völlig in Wasser
gequollene Samen nicht keimen, solange ihnen nicht gewisse
Stoffe oder Licht[222] geboten werden. Bei den
sog. „Lichtkeimern“ sind meistens die roten oder gelben
Strahlen wirksamer als die stark brechbaren, und es genügt oft
eine erstaunlich kurze Belichtungszeit (Lythrum Salicaria 1⁄10
Sekunde bei 370 Hefner-Kerzen). Nicht selten vermögen eine gewisse
hohe Temperatur oder chemische Einflüsse das Licht zu ersetzen.
Chemische Reize spielen die Hauptrolle bei gewissen Parasiten
(Orobanche, Tozzia), die nur in der Nähe ihrer Nährpflanzen
keimen. — Als Gegenstück zu den Lichtkeimern mögen noch
die Dunkelkeimer genannt sein (z. B. Amarantus), bei denen
gerade das Licht die Keimung hindert oder verzögert.
Auch bei Sporen kann die Keimung entweder durch besondere Reize
oder schon durch Herstellung der formalen Wachstumsbedingungen
eingeleitet werden.
Ein auffallender und recht fester Ruhezustand findet sich bei
unseren Laubbäumen. In einer bestimmten Jahreszeit, im Herbst oder
früher, können ihre Knospen in keiner Weise zum Treiben gebracht
werden (Stadium der Vollruhe). Späterhin aber (Nachruhe) bewirken
nicht nur höhere Temperatur, sondern auch zahllose Reize (Frost,
Wärme, Trockenheit, Dunkelheit, Beleuchtung, Ätherdämpfe, Azetylen,
Tabakrauch, Verwundung, Wasserinjektion usw.) eine beträchtliche
Verkürzung der Ruheperiode[223]; fast ebensogut kann aber auch
kurz nach eingetretener Wachstumshemmung, in der Vorruhe,
getrieben werden. Mit diesen Verhältnissen muß die gärtnerische
Frühtreiberei rechnen.
Restitutionsreiz[224]. Wie bei den besprochenen Fällen von
Wachstumsbeginn, so kann man auch bei den Restitutionen nach
den Ursachen ihres Beginnens fragen. Und hier scheint die Antwort
sehr einfach zu sein; sehen wir doch diese Erscheinungen in der Regel
nach Verwundungen eintreten. Schon der Umstand, daß manchmal im Laufe
der normalen Entwicklung Prozesse auftreten, die auffallend an
Restitutionen erinnern, muß uns aber vorsichtig machen. So können z. B.
in den Blattkerben von Bryophyllum junge Pflänzchen sich entwickeln,
und bei gewissen Begonien entstehen nicht nur am abgeschnittenen Blatt,
sondern auch auf der intakten Lamina Sproßbildungen. Durch Experimente
ist denn auch für viele echte Restitutionen festgestellt worden, daß
nicht die Entfernung eines Organs, sondern vielmehr die Störung seiner
normalen Funktion ihre Bildung auslöst (S. 260).
Polarität. Die befruchtete Eizelle der Blütenpflanzen
bildet, sowie ihre Wachstumshemmung aufgehoben ist, sofort zwei
verschiedene Vege[S. 268]tationspunkte, einen, der den Sproß,
einen anderen, der die Wurzel erzeugt. Entsprechende polare
Differenzierungen, Ausgestaltung von Basis und Spitze finden sich
auch bei viel einfacher gebauten Pflanzen. Wenn auch schon gezeigt
worden ist, daß in manchen Fällen ein äußerer Faktor bestimmt,
welcher Teil der jungen Pflanze Basis und welcher Spitze wird, so ist
doch bei allen höheren Pflanzen das polare Wachstum eine durchaus
spezifische, auf inneren Ursachen beruhende Erscheinung: so
wenig wir eine apolare kuglige Alge zu polarem Wachstum veranlassen
können, so wenig gelingt es, eine polar gebaute höhere Pflanze apolar
wachsen zu lassen.
Die in der Eizelle entstandene Polarität bleibt auch im großen
und ganzen dauernd erhalten. Bei einzelnen Pflanzen aber sehen
wir aus inneren Gründen eine Veränderung eintreten. So bilden
sich bei Platycerium- und Adiantum-Arten unter den Farnen,
Neottia nidus avis unter den Orchideen Sprosse direkt aus dem
Wurzelvegetationspunkt. Bei den Natterzungen (Ophioglossum), deren
vegetative Vermehrung ganz auf der Bildung von Wurzelsprossen
beruht, rücken die Sproßanlagen fast bis auf die Wurzelspitze. Auch
der Scheitel mancher Farnblätter (Adiantum Edgeworthii)
vermag unmittelbar in der Bildung einer Sproßanlage aufzugeben.
Symmetrie. Jeder Vegetationspunkt baut dann in der für ihn
charakteristischen Weise das betreffende Organ weiter und sorgt
zugleich für Anlage von Seitenorganen, die in einer ganz bestimmten
Verteilung (radiär, bilateral, dorsiventral) auftreten. So sehen wir
also eine gewisse Symmetrie schon am Vegetationspunkt gegeben,
und diese kann wenigstens in vielen Fällen durch rein innere Ursachen
bestimmt werden; in anderen Fällen haben äußere Faktoren einen
maßgebenden Einfluß.
B. Wachstum
und Zellteilung.
Auch das in Gang gesetzte Wachstum verläuft nicht immer gleichmäßig.
Wohl sehen wir manche Algenzellen, wie Vaucheria, oder Pilze,
wie Saprolegnia, eine gegebene Zelle durch Spitzenwachstum immer
weiter verlängern. In der großen Mehrzahl der Fälle aber ist dieser
Verlängerung ein Ziel gesetzt, indem nach Überschreitung einer gewissen
Größe durch Teilung das Normalmaß der Zelle wieder hergestellt
wird. Dieses ist freilich kein konstantes. Es hängt nicht nur von
äußeren, sondern maßgebend auch von inneren Umständen ab. So sehen
wir z. B. meistens unmittelbar am Vegetationspunkt die Teilungen
rasch eintreten, während sie später bei andauerndem Wachstum seltener
werden. Dementsprechend nimmt die Zellgröße vom Vegetationspunkt bis
in die Zone der Dauergewebe im allgemeinen recht beträchtlich zu. Von
großer Bedeutung für die Zellgröße ist auch die Kernmasse, die sie
enthält. Es ist in einigen Fällen durch gewisse Eingriffe gelungen,
das Doppelte oder gar Vierfache der normalen Kernmasse in einer Zelle
anzusammeln, und es zeigte sich, daß vielfach alle Derivate solcher
Zellen beträchtlich größer waren als die Norm[154].
Immerhin kann man aber ein mittleres Maß der Zelle als eine
erbliche Eigentümlichkeit jeder Pflanzenspezies betrachten. Und wenn
man verschieden große Arten vergleicht, so fällt es auf, daß die
Zellgröße nicht so variiert wie die Außendimensionen; mit anderen
Worten: große Pflanzen sind in erster Linie (nicht ausschließlich)
durch eine große Zahl von Zellen bedingt[225].
Was im einzelnen die Ursachen der Teilung sind, ist noch wenig
bekannt[226]. Zweifellos handelt es sich um eine recht komplizierte
Folge von Erscheinungen; sehen wir doch, daß häufig nicht nur das
Protoplasma, sondern auch der Zellkern bei diesem Prozeß eine Rolle
spielt, insofern als er die Teilung einleitet.
[S. 269]
Zweifellos spielen zunächst äußere Faktoren eine große Rolle.
So haben wir schon gesehen, daß die Intensität und Qualität des
Lichtes z. B. bei Farnprothallien einen maßgebenden Einfluß auf die
Zellteilung ausübt (S. 252). Daneben wirken auch innere Faktoren,
vor allem das Alter der Zelle mit: während die junge Zelle
teilungsfähig ist, nimmt diese Fähigkeit mit dem Alter mehr und
mehr ab — doch können Außeneinflüsse von neuem die Teilung
ermöglichen; das sieht man vor allem bei den Restitutionen. Wie
HABERLANDT[227] gezeigt hat, sind es die aus verwundeten
oder auch nur mechanisch etwas geschädigten Zellen stammenden
Stoffe, die von neuem zur Teilung anregen. Auch bei der Plasmolyse
entstehen solche Stoffe, vielleicht durch Zerreißung von
Plasmafäden, und auch körperfremde Gifte können im gleichen Sinne
wirken. — In den Sproßvegetationspunkten der höheren Pflanzen
und auch bei manchen Algen ist eine gewisse Periodizität der
Zellteilung[228] nachgewiesen, sie tritt z. B. bei Spirogyra des
Nachts ein, offenbar weil sie vom Licht gehemmt wird. Bei Wurzeln
aber finden wir zu allen Tageszeiten Kernteilungen, doch nicht
gleich viele. Ein Maximum liegt in den Vormittagsstunden. Hier
müssen noch unbekannte Außenfaktoren oder innere Ursachen bei
dieser Periodizität mitwirken.
Nicht nur die Tatsache, daß eine neue Zellwand auftritt,
auch die Richtung, in der sie entsteht, ist ein Problem der
Entwicklungsphysiologie. Man hat schon lange bemerkt, daß die Stellung
der neuen Scheidewände auffallende Ähnlichkeit mit dem Verhalten
dünner gewichtsloser Flüssigkeitslamellen (z. B. Seifenlamellen)
zeigt. Diese haben die Tendenz, sich auf die kleinste mögliche Fläche
zusammenzuziehen, und setzen sich deshalb im allgemeinen möglichst
unter rechtem Winkel an etwa vorhandene Wände an. Trotz der großen
Ähnlichkeit in der Anordnung von Zellwänden einerseits und Flächen
„minimae areae“ andererseits darf man doch nicht behaupten, die junge
Zellwand werde aus den gleichen Ursachen wie jene in die beobachteten
Lagen gebracht; denn die Zellwand ist nie flüssig.
Sehr auffällig ist das Prinzip der rechtwinkligen Schneidung im
Vegetationspunkt der Phanerogamen gewahrt. Hier treten uns, wenn
wir das von SACHS herrührende Schema eines Längsschnittes
betrachten (Fig. 268), die Zellwände in zwei Systemen von
Parabeln entgegen, die sich rechtwinklig durchschneiden
und einen gemeinsamen Brennpunkt besitzen. Das erste System
(I–VI in Fig. 268) verläuft mehr oder weniger parallel mit der
Oberfläche des Vegetationspunktes; wir nennen diese Zellwände
Periklinen. Die senkrecht zu ihnen stehenden Wände (I-II)
heißen Antiklinen.
Die Zellen des Vegetationspunktes sind noch alle gleich. Erst in einer
gewissen Entfernung von ihm beginnt der Prozeß der Differenzierung.
Die Zellen nehmen ungleiche Funktion und Hand in Hand damit ungleiche
Formen an. Während die embryonalen Zellen noch befähigt sind,
jedes beliebige Gewebe zu erzeugen, nimmt diese Befähigung mit dem
Fortschreiten der Differenzierung ab, die Zellen werden mehr und
mehr in eine bestimmte Entwicklung gedrängt (Determination). Im
allgemeinen ist aber bei der Pflanze die Determination keine so feste,
wie beim Tier. Das sieht man vor allem aus der weiten Verbreitung der
Restitution.
Fig. 268. Schema eines Sproßvegetationspunktes. Nach
SACHS.
Was im einzelnen die Ursachen der Differenzierung und der
Determination sind, ist unbekannt. Daß aber die Korrelationen bei
diesen Prozessen eine große Rolle spielen, ist S. 259 ausgeführt
worden.
[S. 270]
C. Weitere
periodische Änderungen in der vegetativen Gestaltung.
Während der Wachstumstätigkeit treten uns vielfach auch noch
ganz andere periodische Erscheinungen entgegen. So sieht man z. B.
periodische Änderungen in der Ausgestaltung der Blätter und
Stengel, die nicht nur quantitativer, sondern auch qualitativer
Art sind; es wechseln Laubblätter mit Nieder- und Hochblättern,
Laubsprosse mit Rhizomen ab, wobei der Übergang ein allmählicher oder
ein sprungweiser sein kann. Bei diesen Erscheinungen tritt uns wieder
die Korrelation des Wachstums, von der S. 259 die Rede war, entgegen.
Die Existenz oder besser: die Tätigkeit einer gewissen Menge
von Laubblättern übt auf die weiter am Vegetationspunkt entstehenden
Anlagen einen Einfluß dahin aus, daß diese zu Niederblättern,
Knospenschuppen werden. Entfernt man die Laubblätter im Frühsommer,
so bilden sich aus diesen Anlagen keine Niederblätter, sondern
Laubblätter. In entsprechender Weise wirkt die Entfernung von
Laubsprossen oft auf die unterirdischen Rhizome ein; sie veranlaßt
diese aus dem Boden herauszuwachsen und an Stelle von Niederblättern
Laubblätter zu bilden.
Außer Laub- und Niederblattbildung treffen wir bei manchen Pflanzen
auch noch eine andere Art von Heterophyllie. Vielfach treten in der
Jugend Blätter anderer Gestalt auf als später, und manchmal gelingt
es, eine Pflanze zur erneuten Produktion der Jugendform zu zwingen,
wenn die äußeren Bedingungen, unter denen sie sich bildete, wieder
hergestellt werden. So folgen z. B. bei Campanula rotundifolia auf
die schmalen Folgeblätter wieder runde Jugendblätter, wenn die
Beleuchtungsintensität herabgesetzt wird. In ähnlicher Weise sind
auch bei manchen Wasserpflanzen die Wasserblätter Jugendformen,
die Schwimmblätter und die Luftblätter Folgeformen. Auch hier kann
künstlich die Jugendform wieder hervorgerufen werden. Das ist aber
nicht immer so. Manchmal wird der Vegetationspunkt selbst so stark
alteriert, daß er nur noch Folgeblätter zu produzieren vermag.
Die „Schattenblätter“ (S. 253) unserer Laubbäume gelten heute in
erster Linie als Jugendblätter, denn sie hängen nicht so sehr direkt
von der Beleuchtungsstärke ab, als von der Stellung am Sproß. Jeder
Sproß beginnt mit Schattenblättern und bildet erst später Lichtblätter.
Offenbar sind also Ernährungseinflüsse bei Ausbildung dieser
Blattformen maßgebend.
Wie das Blatt, so kann auch der Stengel weitgehende Umbildung erfahren.
So sehen wir ihn im Laubsproß aufrecht wachsen, im Rhizom horizontal
im Boden oder auf dem Boden kriechen: bei den Windepflanzen streckt er
seine Internodien über das sonst übliche Maß, bei Rosettenpflanzen ist
er ungewöhnlich gestaucht; auch im Dickenwachstum haben wir weitgehende
Differenzen: im Extrem erscheint der Stengel als Knolle. Alle diese
verschiedenen Gestalten oder Wachstumsweisen sind die Folgen ganz
bestimmter Einwirkungen, sie lassen sich demnach wenigstens zum Teil
willkürlich erzielen, auch zu einer Zeit und an einem Ort, wo sie in
der „normalen“ Entwicklung nicht erwartet werden können.
Als ein Beispiel für die Beeinflußbarkeit des Stengels wollen wir die
Knollenbildung bei der Kartoffel betrachten. Wie Fig. 201 darstellt,
entwickeln sich gewöhnlich die Knollen am Ende von horizontalen
Ausläufern, die ihrerseits aus der Basis des Laubtriebes im Boden
entspringen. Die Knollenbildung erfolgt durch starkes Dickenwachstum
am Ende der Ausläufer unter Sistierung des Längenwachstums. Wird aber
der Laubtrieb rechtzeitig entfernt, so gehen statt der Knollen aus
den Enden der Ausläufer gestreckte aufrechte Zweige hervor, die über
den Boden gelangen und grüne Laubblätter[S. 271] erzeugen. Man kann also
den typischen Entwicklungsgang der Kartoffel offenbar so abändern,
daß es gar nicht zur Knollenbildung kommt. Andererseits lassen sich
auch die Knollen willkürlich an ganz anderen Orten erzeugen;
so kann z. B. bei niederer Temperatur die Hauptachse einer
bestimmten Kartoffelsorte ganz kurz bleiben und sich sofort in eine
Knolle verwandeln; bei anderen Sorten können Knollen nahe der Spitze
der Laubsprosse, also in der Luft erzwungen werden, wenn man die
Sproßspitze verdunkelt. Noch plastischer als die Kartoffel hat sich
die Basellacee Boussingaultia baselloides erwiesen: hier kann man jede
beliebige Knospe in eine Knolle umgestalten, und wenn Knospen fehlen,
können Internodien oder Wurzeln knollig anschwellen. Es sieht so aus,
als ob die Produktion einer gewissen Menge von Reservestoffen als Reiz
wirke, dem die Bildung eines Reservestoffbehälters folge.
D.
Lebensdauer[229].
Eine weitere hier zu besprechende Erscheinung äußert sich im Werden und
Vergehen, in der Lebensdauer der Pflanze. Es gibt Pflanzen, wie
Stellaria media, Senecio vulgaris, die in wenigen Wochen ihre ganze
Entwicklung von der Samenkeimung bis zur Samenreife durchlaufen, bei
denen auch jeder Same sofort keimen kann, so daß mehrere Generationen
ohne Ruhe und unbekümmert um die Jahreszeit in einem Kalenderjahre
auftreten. Mit der Ausbildung einer gewissen Anzahl von Samen stirbt
die Pflanze ab; die Samen sorgen aber für die Forterhaltung ihres
Typus. Ähnlich, aber doch mehr an die Jahreszeiten gebunden, sind
zahlreiche „einjährige Pflanzen“; an sie schließen sich andere einmal
fruchtende (monokarpische) Pflanzen an, bei denen der Ausbildung der
Samen ein zwei- bis vieljähriges Stadium rein vegetativen Wachstums
mit oder ohne eingeschobene Ruhezeiten vorausgeht. Vermutlich liegt
bei ihnen allen in der Fruchtbildung die Ursache des Absterbens der
Vegetationsorgane; denn man kann durch Verhinderung des Samenansatzes
ihr Leben bedeutend verlängern. In einem Gegensatz zu ihnen stehen die
Pflanzen, deren Prototyp unsere Bäume sind, die mehrfach fruchten,
bei denen der Fortbestand des einzelnen Exemplars nicht mit der
Samenbildung erlischt. Bei allen mehrjährigen Typen tritt nun neben der
Jahresperiode noch eine andere Periodizität zutage: Ein Baum hat im
ersten Jahre, als Keimpflanze, eine viel geringere Wachstumsintensität
als manche Annuelle; er „erstarkt“ aber allmählich, und sein
Längenwachstum, sein Dickenwachstum, selbst die Elementarorgane seines
Holzkörpers nehmen immer mehr an Größe zu. Manche Bäume erreichen ein
hohes Alter und sind an sich zu unbegrenztem Leben befähigt. Von einem
gewissen Höhepunkt ihrer Entwicklung an, werden aber ihre Jahrestriebe
wieder kleiner, offenbar weil der Stoffaustausch zwischen Wurzeln und
Blättern sich immer schwieriger gestaltet. Schließlich stirbt der
Baum aus diesem Grunde oder weil Parasiten oder sonstige von außen
kommende Störungen ihm ein Ende bereiten. Sorgt man dafür, daß in
der Nähe der Vegetationspunkte der Sprosse immer neue Wurzeln sich
bilden, so werden die Sprosse mit stets gleicher Intensität wachsen,
und unter diesen Umständen ist ein Ende des Wachstums nicht abzusehen.
Man kann dieses Experiment nicht bei jedem Baum ausführen, weil nicht
alle leicht Wurzeln bilden; mit der Weide aber gelingt es leicht,
wenn man Zweige als Stecklinge behandelt. Ist somit der Tod für den
Vegetationspunkt abwendbar, so treten uns bei ausgewachsenen Zellen
ganz andere Verhältnisse entgegen. Lange bevor der ganze Baum zugrunde
geht, sind ihm schon einzelne Teile im Tode vorausgegangen. So werden
die Blätter abgestoßen, nachdem sie ein oder mehrere Jahre funktioniert
haben. Manchmal[S. 272] werden auch ganze Äste abgeworfen. In anderen Fällen
freilich gehen sie ohne Abstoßung zugrunde und zerfallen allmählich
am Ort ihrer Entstehung. Aber auch alle älteren Gewebe des Stammes
gehen dem Tod entgegen; die peripherischen werden in Borke verwandelt,
fallen ab oder bilden eine schützende Hülle für die übrigen Teile; im
Zentrum geht das Holz in Kernholz über, wobei die letzten lebenden
Elemente absterben. Nur die Vegetationspunkte und die Kambien sowie
ihre jüngsten Derivate sind an einem älteren Baume noch am Leben. —
So sehen wir also, daß jede Zelle, die ihren embryonalen Charakter
verloren hat, aus Gründen, die wir nicht kennen, nach kürzerer oder
längerer Frist dem Tode verfällt. Dieses Absterben aber läßt sich
im allgemeinen nicht aufhalten. Trotzdem können wir nicht sagen,
daß der Tod notwendig sei. Nur dadurch, daß gewisse
Zellen sich entwickeln, sterben andere ab; der Tod ist also eine
Korrelationserscheinung. Entfernen wir bei restitutionsfähigen
Pflanzen, bevor die Dauergewebszellen zu alt geworden sind, die
Vegetationspunkte, so können ausgewachsene und somit normalerweise
dem Tode verfallene Zellen wieder embryonal werden und weiter leben.
Bei niederen Pflanzen, denen der Gegensatz von embryonalen und
ausgewachsenen Zellen fehlt, die nur aus embryonalen Zellen bestehen,
kann demnach das Wachstum ein unbegrenztes sein, der Tod kann hier nur
durch ungünstige Außenfaktoren herbeigeführt werden. Es fehlt indes
nicht an Beispielen dafür, daß innere Faktoren tödlich wirken;
so ist z. B, eine gelegentlich beobachtete Pflanzenrasse, die kein
Chlorophyll bilden kann, dem Tod verfallen. In der Vererbungslehre sind
vielfach letale Faktoren bekannt geworden, d. h. innere Anlagen, die —
wenn sie zur Entfaltung kommen — den Tod des Organismus herbeiführen
(vgl. S. 288, Oenothera).
Am besten bekannt oder abgeschätzt ist das Alter von Bäumen[230],
über deren Anpflanzung zum Teil geschichtliche Daten vorliegen,
während das Alter anderer Bäume oft Jahrhunderte weit über die
geschichtliche Zeit zurückreicht. Die berühmte Linde bei Neuenstadt
am Kocher in Württemberg ist annähernd 700 Jahre alt, eine Littauer
Linde von 25,7 m Umfang hatte 815 Jahresringe, und ein Taxus in
Braburn (Kent) wurde bei 18 m Umfang auf 2880 Jahre geschätzt.
Sequoia gigantea, der Mammutbaum Kaliforniens, wird nach H.
MAYR 4000 Jahre alt. Berühmt durch ihr Alter ist auch eine
Wasserzypresse (Taxodium mexicanum) auf dem Friedhofe von St. Maria
Tule bei Oaxaca in Mexiko (Fig. 603). Einer der ältesten Bäume
Deutschlands dürfte eine ungefähr 1200jährige Eiche sein, die auf
dem Gut Ivenak in Mecklenburg steht und in Brusthöhe einen Umfang
von fast 13 m hat. — Auch Bryophyten erreichen oft ein hohes
Alter; die an der Spitze fortwachsenden verkalkten Gymnostomumrasen
und die Stengel der metertief ins Torfmoor hinabreichenden
Sphagnaceen zählen ihr Dasein sicher nach vielen Jahrhunderten.
E. Fortpflanzung.
Über die Organe, die der Fortpflanzung dienen, ist das Nötige schon in
der Morphologie gesagt; hier sind nur die Bedingungen und die Bedeutung
der Erscheinung zu berücksichtigen und dann die Eigenschaften der
Nachkommen zu besprechen.
1. Die Bedingungen der
Fortpflanzung[231].
In der Natur scheint die Fortpflanzung mit einer gewissen Notwendigkeit
auf das vegetative Wachstum zu folgen. Sie tritt gewöhnlich ein,
wenn das vegetative Wachstum nachläßt, wenn die Pflanze ein
gewisses Alter erreicht hat. Tatsächlich läßt sich aber zeigen,
daß diese Aufeinanderfolge nicht notwendig ist, daß der natürliche
Entwicklungsgang nur dann eintritt, wenn ganz bestimmte Bedingungen
gegeben sind, und daß er durch andere Einflüsse sehr stark abgeändert
wird.
[S. 273]
Die Hauptfrage, die wir hier zu studieren haben, ist diese: unter
welchen Bedingungen findet das vegetative Wachstum, unter welchen
die Bildung von Fortpflanzungsorganen statt? Da diese Frage noch
verhältnismäßig wenig studiert ist, so läßt sich nicht gut eine
allgemeine Antwort auf sie geben. Wir müssen uns also darauf
beschränken, an einigen Beispielen das Wesentlichste klar zu machen.
Niedere Pflanzen. Als Beispiel betrachten wir einen Pilz aus der
Gattung Saprolegnia. Diese Pilze besitzen einen einzelligen,
verzweigten, chlorophyllfreien Vegetationskörper. Sie finden sich in
der Natur gewöhnlich auf toten Insekten, die ins Wasser gefallen sind,
und ihr Thallus durchwuchert zunächst den Insektenleib. Nach einiger
Zeit wächst er aber auch allseits aus diesem heraus in das Wasser. Das
Ende dieser ausstrahlenden Fäden wird in der Regel durch eine Querwand
abgegliedert, und sein Inhalt zerfällt in viele Schwärmsporen, die dann
austreten, sich bewegen und schließlich an einem anderen Orte zu einer
neuen Saprolegnia auskeimen. Später bilden sich an der alten Pflanze
Eier und Spermazellen, und die ersteren entwickeln sich wenigstens bei
manchen Spezies erst nach der Befruchtung weiter. Mit der Ausbildung
der befruchteten Eier pflegt die Tätigkeit des Saprolegniapflänzchens
beendet zu sein, es geht allmählich zugrunde.
G. KLEBS hat nun gezeigt, daß man diesen Entwicklungsgang von
Saprolegnia völlig verändern kann; unter anderem ist es gelungen, die
Entwicklung in folgender Weise zu lenken:
1. Der Thallus kann jahrelang rein vegetativ weiter wachsen, wenn stets
für frische, gute Nährstoffe gesorgt wird.
2. Ein derartig gut ernährter Thallus geht bei Übertragung in reines
Wasser rasch und vollständig in der Bildung von Sporangien auf.
3. In Lösungen von Leucin (0,1%) und Hämoglobin (0,1%) erfolgt zuerst
kräftiges Wachstum, dann Bildung von Geschlechtsorganen; Schwärmsporen
werden nicht gebildet. Diese treten aber auf, und zwar nach den
Geschlechtsorganen, wenn man verdünntes Hämoglobin (0,01%) verwendet.
Es zeigt sich also, daß ganz bestimmte, wohl zu definierende
Bedingungen für vegetatives Wachstum, andere für die Bildung
von Geschlechtsorganen, wieder andere für das Auftreten von
ungeschlechtlicher Fortpflanzung existieren.
Bedingungen der Blütenbildung[232]. Bei den Phanerogamen
tritt die ungeschlechtliche Vermehrung durch besondere Brutknospen
usw. stark zurück gegenüber der geschlechtlichen Fortpflanzung. Diese
aber ist an die Ausbildung von Blüten geknüpft. Das Auftreten von
Blüten scheint in der Regel an ein bestimmtes Alter gebunden. Es
liegen aber Beobachtungen genug vor, daß auch schon an ganz jungen
Exemplaren Blüten auftreten können. Demnach ist zu erwarten,
daß die Bildung der Blüten geradeso wie andere Organbildung durch ganz
bestimmte Bedingungen gekennzeichnet ist. In einzelnen Fällen ist es
denn auch gelungen, diese näher zu erforschen. Für Sempervivum Funkii
z. B. hat KLEBS folgendes festgestellt:
1. Bei lebhafter Kohlenstoffassimilation in hellem Licht und bei
starker Aufnahme von Wasser und Nährsalzen wächst die Pflanze
unbegrenzt rein vegetativ.
2. Bei lebhafter Kohlenstoffassimilation in hellem Licht, aber bei
Einschränkung der Wasser- und der Nährsalzaufnahme tritt
Blütenbildung auf.
3. Bei einer mittleren Wasser- und Nährsalzaufnahme hängt es von der
Intensität der Beleuchtung ab, ob Blühen oder vegetatives Wachsen
eintritt. Bei schwächerer Lichtintensität (bzw. bei Verwendung von
blauem Licht)[S. 274] erfolgt nur Wachsen, bei stärkerer Beleuchtung
(bzw. bei Verwendung roten Lichts) Blühen.
KLEBS unterscheidet bei der Blütenbildung von Sempervivum
drei Phasen: 1. die Herstellung des Zustandes der Blühreife, 2.
die Bildung der mikroskopisch nachweisbaren Blütenanlagen, 3. die
Streckung der Infloreszenz. Diese drei Phasen sind an ganz verschiedene
Bedingungen geknüpft und hängen deshalb in ganz verschiedener Weise
von den Außenfaktoren ab. Die Blühreife ist bedingt durch ein starkes
Überwiegen der C-Assimilation über die Prozesse, bei denen
Kohlehydrate konsumiert werden, wie Atmung und vegetatives Wachstum. Da
hohe Temperatur die Atmung steigert, Nährsalze das vegetative Wachstum
fördern, so ergibt sich neben starker Beleuchtung niedrige Temperatur
und Einschränkung der Nährsalze als notwendig für die Erzielung der
Blühreife. Schon bestehende Blühreife kann durch hohe Temperatur
vernichtet werden, sie kann durch niedere Temperatur selbst im Dunkeln
lange konserviert und noch gesteigert werden. Wirkt somit in diesem
Fall das Licht anscheinend nur, insofern es die Assimilation der CO2
bedingt, so hat es bei der zweiten Phase eine ganz andere Bedeutung:
eine gewisse Dauer der Beleuchtung ist hier ganz unerläßlich,
und zwar sind ausschließlich die langwelligen Strahlen wirksam,
während kurzwellige sogar den Zustand der Blühreife wieder vernichten.
— In der Natur ist der blühreife Zustand schon im Herbst erreicht,
es fehlt aber an einer genügend langen und intensiven Beleuchtung.
Wird eine Dauerbelichtung mit Osramlampen, die reich an roten
Strahlen sind, durchgeführt, so tritt die Blütenbildung um Monate
verfrüht ein; je intensiver die Lichtquelle ist, desto kürzer kann die
Belichtung sein. Je früher im Winter man diese Bestrahlung durchführt,
desto länger muß sie sein; gegen Ende des Winters ist durch die
dauernde Abkühlung der Zustand der Blühreife so weit gediehen, daß eine
kurze Bestrahlung die Blüte bewirkt. Pausen, die in die Dauerbelichtung
eingeschaltet werden können, dürfen nicht zu lang sein, sonst heben sie
die Wirkung der Lichtperiode auf. — Die letzte Phase der Streckung
endlich hängt wieder wie die erste nur von der ernährenden Wirkung des
Lichtes ab; dementsprechend kann sie bei genügender Vorernährung auch
im Dunkeln wenigstens zum Teil eintreten.
Eine ähnlich tiefdringende Analyse der Bedingungen des Blühens
liegt bei anderen Objekten bisher noch nicht vor, doch lassen
zahlreiche Beobachtungen und Versuche erkennen, daß allgemein
das Licht, die Temperatur, die Nährsalze
in erster Linie von Bedeutung für die Blütenbildung sind. Da
dieselben Faktoren auch im vegetativen Leben unentbehrlich sind,
so ist es nur das Maß, in dem sie gegeben sind, und vor allem ihre
relative Menge, die darüber entscheidet, ob eine bestimmte
Knospe zur Blüte wird oder vegetativ wächst.
Die Bedeutung des Lichtes für die Blütenbildung ergibt
sich z. B. aus der lange bekannten Tatsache, daß der Efeu nur
an hellem Standort, nicht aber im Schatten des Waldes blüht,
obwohl er an letzterem Orte gut gedeiht. Das gleiche beweisen
VOECHTINGs Versuche mit Mimulus Tilingii. Diese
Pflanze stellt bei einer gewissen niederen Lichtintensität,
die ihr vegetatives Wachsen noch gut erlaubt,
die Blütenbildung gänzlich ein. Entsprechende Versuche hat
KLEBS z. B. mit Veronica chamaedrys ausgeführt, und
er gibt an, daß bei allen Pflanzen, die keine nennenswerten
Massen von Reservestoffen führen, eine Lichtverminderung die
Blütenbildung unterdrückt. Er betrachtet die vom Licht bewirkte
Kohlenstoffassimilation als in erster Linie maßgebend für die
Blütenbildung. — Bei einer gewissen Lichtintensität, die zur
Ausbildung normaler Blüten nicht mehr ausreicht, entstehen bei
manchen Pflanzen kleistogame Blüten.
Auch die Temperatur spielt offenbar eine wichtige Rolle.
Eine andauernd hohe Temperatur verhindert das Blühen. So wachsen
Pflanzen unserer Klimate in den Tropen eventuell dauernd vegetativ
(Kirsche), und zweijährige einheimische Pflanzen, wie Rübe,
Digitalis, kann man auch im zweiten Jahr am Blühen hindern, wenn
man sie im Winter[S. 275] warm hält und weiter wachsen läßt. So ist es
KLEBS gelungen, die Rübe mehrere Jahre lang rein vegetativ
zu erhalten. Auch Glechoma, Sempervivum wuchsen, wenn sie an der
Winterruhe verhindert waren, jahrelang vegetativ.
Endlich sind noch die Nährsalze zu erwähnen. Keimpflanzen
können durch Nährsalzentziehung zu kümmerlichen Hungerpflänzchen
gemacht werden, bei denen oft nach wenigen winzigen Laubblättern
schon die Blütenbildung beginnt. Auch Versuche von MOEBIUS
haben gezeigt, daß Gräser und Borrago bei geringer Salzzufuhr
besser blühen als bei starker Düngung. Die Erhöhung der
Fruchtbarkeit der Obstbäume, die man durch Beschneiden der
Wurzeln erzielt, dürfte ebenfalls auf einer Einschränkung der
Nährsalzaufnahme beruhen. Daß aber nicht alle Nährsalze in gleicher
Weise wirken, hat besonders BENECKE betont, der nachweisen
konnte, daß Verminderung der Stickstoffnahrung, Vermehrung
des Phosphors zur Blütenbildung führt.
Werden, nachdem die Blütenbildung begonnen hat, wieder die
Bedingungen für vegetatives Wachstum hergestellt, so kann ein
schon zur Blüte prädisponierter Sproß zu vegetativem Wachstum
zurückkehren. So bleiben, wenn Mimulus Tilingii in schwaches
Licht gebracht wird, die bereits angelegten Blütenknospen
unentwickelt, und ruhende Achselknospen der Brakteen entwickeln
sich zu Laubsprossen. Es tritt eine Gestaltung der Pflanze ein, die
außerordentlich stark vom gewohnten Aussehen abweicht.
Geschlechtsbestimmung. Die große Mehrzahl der Blüten
sind Zwitterblüten, produzieren also sowohl männliche
als auch weibliche Sexualzellen. — In anderen Fällen sehen wir
eingeschlechtige Blüten allein oder neben den zweigeschlechtigen
auftreten. Schon der Umstand, daß dann die weiblichen Blüten meist
an anderen Stellen der Pflanzen sich finden als die männlichen,
weist darauf hin, daß jede dieser Formen ihre spezifischen
Bildungsbedingungen hat; welche diese sind, ist freilich unbekannt.
Ein Problem der Entwicklungsphysiologie wird die
Geschlechtsbestimmung aber ganz besondere dann, wenn wir es mit
sog. zweihäusigen Pflanzen zu tun haben, d. h. mit solchen,
die in nur männlichen oder nur weiblichen Exemplaren vorkommen.
Vgl. S. 284.
Die Befruchtung. Wenn Eizelle und Spermazelle verschmolzen sind,
umgibt sich das Produkt allgemein mit einer Membran. Bei niederen
Pflanzen entsteht so eine Zygospore oder Oospore, die gewöhnlich
erst nach einer Ruheperiode keimt. Bei den höheren Pflanzen tritt
sofort Wachstum und Zellteilung ein; es entsteht ein Embryo, der bei
Moosen und Farnen alsbald in bestimmter Weise sich weiter entwickelt,
während er bei den Phanerogamen nach kurzer Zeit in einen Ruhezustand
verfällt. Zuvor geht von ihm aber noch eine Fülle von Reizwirkungen
aus, die namentlich bei den Angiospermen hoch entwickelt sind. Die
Samenanlage, in der der Embryo eingeschlossen ist, fängt an zu wachsen,
sie vergrößert sich beträchtlich und erhält eine ganz charakteristische
Struktur; sie wird zum Samen, der in der Regel erst nach Ablauf
einer Ruheperiode keimt. Aber auch der Fruchtknoten fängt nach der
Befruchtung lebhaft zu wachsen an und entwickelt sich zur Frucht.
Diese durch die Embryobildung ausgelösten Wachstums- und
Gestaltungsvorgänge in Samenanlage, Fruchtknoten und
eventuell auch noch in anderen Teilen der Blüte sind als
Korrelationserscheinungen zu deuten; denn falls die
Befruchtung verhindert wird, unterbleiben meist alle die
Veränderungen, die aus der Blüte eine reife Frucht hervorgehen
lassen: es tritt vielmehr eine andere korrelative Wirkung, die
Abstoßung des ganzen, nun nutzlosen Organs, ein. Einzelne Pflanzen,
zumal solche, die schon lange in Kultur genommen sind, können
freilich eine Ausnahme machen; bei fast allen Varietäten von
Bananen, bei der echten kernlosen Mandarine und den als Sultaninen
bezeichneten Rosinen z. B. unterbleibt die Bildung der Embryonen;
trotzdem aber entwickeln sich die Früchte. Der Anstoß zu dieser
Entwicklung kann auch hier entweder von der bloßen Bestäubung der
Narbe oder doch von der Befruchtung der Samenanlagen ausgehen,
welch letztere dann aber früher oder später verkümmern, ohne der
sonstigen Entwicklung der Frucht Eintrag zu tun. In manchen Fällen
entwickeln sich sog. „taube“ Früchte aber auch ganz ohne den Anstoß
der Bestäubung, wie die parthenokarpen[233] Früchte der
Feige, der Gurke und gewisser Apfel- und Birnsorten.
[S. 276]
Auch von Pollenkörnern und Pollenschläuchen, die sich auf der Narbe
befinden, können Einflüsse ausgehen, die weithin geleitet werden.
So tritt nach Bestäubung der Narbe der Orchideen vielfach ein
Verschwellen der Narbe und des Gynostemiums ein, und es wird auch
das Perianth plötzlich in seinem Wachstum gehemmt und zum Welken
gebracht. Wie FITTING[234] zeigen konnte, geht diese
Wirkung von löslichen, organischen, hitzebeständigen Substanzen
aus, die leicht von der ungekeimten Pollenmasse getrennt werden
können.
Mag nun eine einfache Spore oder ein komplizierter Embryo die Folge der
Befruchtung sein, so unterscheidet sich dieses Produkt von den Zellen,
die es bildeten, stets dadurch, daß es doppelte Kernmassen, also die
doppelte Anzahl von Chromosomen aufweist (S. 172). Deshalb ist eine
bald früher, bald später eintretende Reduktionsteilung, die wieder die
einfache Zahl der Chromosomen herstellt, stets die notwendige Folge der
Befruchtung.
2. Die Bedeutung der sexuellen Fortpflanzung.
Die Bedeutung der sexuellen Fortpflanzung ist nicht ohne weiteres zu
erkennen. Viele Pflanzen kommen in der Natur oder in der Kultur ganz
ohne sie aus und begnügen sich mit vegetativer Fortpflanzung.
Auf niedere Pflanzen, die noch gar keine sexuelle Fortpflanzung
besitzen, ist schon hingewiesen (S. 165). Von höheren Pflanzen
bringen z. B. die kultivierten Bananen, manche Dioscoreaceen, der
Kalmus, Spielarten der Weinrebe, der Apfelsinen, Gartenerdbeeren
überhaupt keine sexuellen Nachkommen mehr hervor. Der Knoblauch,
der an Stelle der Blüten kleine Zwiebelchen bildet, die weiße
Lilie und der durch Wurzelknöllchen sich vermehrende Ranunculus
Ficaria u. a. bilden, falls sie ungestört ihre vegetativen
Vermehrungsorgane zu entwickeln vermögen, kaum noch keimfähige
Samen aus. Solche können aber unter Umständen, so z. B. manchmal
an abgeschnittenen Blütenständen jener Pflanzen, noch gewonnen
werden. Für gewöhnlich vermehren sie sich ausschließlich durch
ungeschlechtliche Sprossung. Irgend eine Degeneration,
die man früher bei ausschließlich vegetativer Vermehrung für
unvermeidlich hielt, ist dabei nicht zu bemerken[235].
Wenn demnach die monogene Fortpflanzung für die Erhaltung der Art
genügt, so muß doch die digene irgend etwas bieten, was die monogene
nicht leistet; es wäre sonst unbegreiflich, daß die digene überhaupt
entstanden ist, da sie ja viel komplizierter eingerichtet ist und viel
weniger sicher zum Ziel führt als die vegetative Fortpflanzung.
Bei einseitiger Betrachtung der Algen und Pilze könnte man
glauben, die sexuelle Fortpflanzung führe zur Ausbildung besonders
widerstandsfähiger Keime, die demnach eine längere Ruhezeit unter
ungünstigen Vegetationsverhältnissen überdauern können; denn
tatsächlich sind die Zygosporen und Oosporen häufig sehr viel
resistenter als die Schwärmsporen und Konidien. Aber schon bei den
Farnen kehrt sich das Verhältnis um, die befruchtete Eizelle muß
sofort die Bedingungen zur Weiterentwicklung finden, widrigenfalls sie
zugrunde geht; dagegen ertragen die ungeschlechtlichen Sporen eine
lange Ruhe.
Sehr häufig finden wir bei der digenen Fortpflanzung, daß die
einzelne Sexualzelle entwicklungsunfähig ist. Erst
durch die Verschmelzung wird früher oder später die Entwicklung
ausgelöst. Demnach liegt eine Bedeutung der Befruchtung in der
Aufhebung einer bestehenden Wachstumshemmung. Man kann sich aber nicht
vorstellen, daß das ihre ursprüngliche und wesentliche Bedeutung ist.
Viel wahrscheinlicher ist es, daß die Sexualzellen erst allmählich
die Entwicklungsunfähigkeit erworben haben, weil durch diese die
Verschmelzungsmöglichkeit gesichert wurde. Wenn jede Sexualzelle sofort
nach ihrer Bildung wüchse, so würde sie in der Mehrzahl der Fälle zu
wachsen beginnen, ehe sie mit einer anderen Sexualzelle verschmelzen
konnte.
[S. 277]
Diese Auffassung wird gestützt durch das Verhalten mancher
Algen, bei denen nicht selten die Sexualzellen auch ohne
Verschmelzung zu keimen vermögen; insbesondere können sich die
Eizellen bei ihnen ohne Befruchtung weiter entwickeln.
Im Anschluß an ähnliche Vorgänge im Tierreich nennt man diese
Erscheinung Parthenogenesis. Bei den primitiven Algen ist
Parthenogenesis möglich, weil hier die Entwicklungsunfähigkeit
der Eizelle noch nicht erworben ist oder weil sie
durch verschiedene Einwirkungen leicht behoben werden kann.
So tritt z. B. bei der Alge Protosiphon durch hohe Temperatur
parthenogenetische Entwicklung ein, und bei den Eiern niederer
Tiere (Echinodermen) findet sich das gleiche z. B. nach Behandlung
mit Lösungen von einer gewissen Konzentration. Man wird vielleicht
vermuten dürfen, daß in den Fällen, wo die Entwicklung nur
nach Befruchtung erfolgt, eine in der Spermazelle enthaltene
Substanz die Entwicklung anregt.
Es fehlt freilich auch bei höheren Pflanzen nicht an Vorkommnissen,
die man als Parthenogenesis[236] bezeichnen kann. So entwickelt
sich die Eizelle mancher Kompositen, ferner bei Alchimillen,
Thalictrum purpurascens, Wickstroemia indica, Ficus hirta, Marsilia
Drummondii und Chara crinita ohne vorhergehende Befruchtung. Von
den eben besprochenen Vorkommnissen unterscheiden sich aber diese
Fälle dadurch, daß die betreffenden Eizellen in der Chromosomenzahl
ihrer Kerne wie vegetative Zellen sich verhalten, diploid (S. 172)
sind und überhaupt nicht für die Befruchtung eingerichtet zu sein
scheinen (vgl. den speziellen Teil).
Wir kommen also zu der Überzeugung, daß in der Aufhebung einer
bestehenden Entwicklungshemmung das Wesentliche der sexuellen
Fortpflanzung nicht bestehen kann. Somit werden wir es in der
Verschmelzung der Substanz zweier Zellen, in der dadurch bedingten
Mischung väterlicher und mütterlicher Eigenschaften zu suchen
haben. In diesem Punkt liegt jedenfalls der Hauptunterschied
zwischen den beiden Fortpflanzungsarten: die vegetativ erzeugte
Nachkommenschaft besteht aus unvermischten Abkömmlingen, die sexuell
erzeugte aus Mischlingen. Der in den Nachkommen entfaltete Komplex
von Eigenschaften weicht deshalb bei vegetativer Vermehrung von
dem der Stammform in der Regel nicht ab. Die Varietäten, Sorten
und Rassen vieler Kulturpflanzen können deshalb in der Regel nur durch
Pfropfung oder Stecklingsbildung vermehrt werden. Im Gegensatz zu den
vegetativen Abkömmlingen wird der sexuelle Nachkomme gewöhnlich
der Mutterpflanze nicht völlig ähnlich sein können, sondern
die Eigenschaften beider Eltern in sich vereinigen müssen. Je
abweichender diese voneinander waren, desto größer wird der sichtbare
Erfolg der Vermischung ausfallen.
F.
Vererbung, Variabilität, Artbildung.
Vererbung[237]. Unter Vererbung versteht man die allbekannte
Erscheinung, daß die Eigenschaften der Eltern sich bei den Nachkommen
wieder finden. Eine solche Vererbung tritt uns ebenso bei der
Zweiteilung einer Zelle, der einfachsten Form der Fortpflanzung,
entgegen wie beim komplizierten Sexualakt. Daß die Tochterzellen
der Mutterzelle gleichen, bedarf keiner Erklärung: ein Problem wird
die Vererbung erst dann, wenn aus kleinen Teilen eines kompliziert
gebauten Mutterorganismus, aus Keimen, die Nachkommen durch
Entwicklung hervorgehen. Man schreibt solchen Keimen
bestimmte „Anlagen“ oder „Gene“ zu, die es bewirken,
daß ein Organismus in ganz bestimmter, spezifischer Weise auf äußere
Faktoren reagiert. Daß diese Anlagen an die Chromosomen der Zellkerne
gebunden sind, ist wahrscheinlich; über die Art und Weise aber, wie sie
den Entwicklungsgang beeinflussen, wissen wir nichts.
Auch in den Sexualzellen der höheren Pflanzen müssen solche Anlagen
vorhanden sein, und zwar sowohl in den männlichen wie in den
weiblichen. Demnach muß also die befruchtete Eizelle die doppelte
Anzahl von Anlagen[S. 278] besitzen — und dennoch geht nur ein einziger
Organismus aus ihr hervor. Doch nicht nur in den Keimzellen,
sondern in allen Zellen der Pflanze finden sich wenigstens
ursprünglich die gleichen Anlagen; das zeigen uns die
Erscheinungen der Restitution.
Zweifellos sind die Probleme der Vererbung am interessantesten bei der
sexuellen Fortpflanzung, wo vor allem die Frage nach dem Anteil der
beiden Eltern an der Gestaltung der Nachkommen von Bedeutung
ist. Diese Frage aber kann nur an den Bastarden gelöst werden,
weil die einzelnen Individuen einer reinen Art die gleichen
Anlagen haben.
Bastarde[238]. In der Regel ist eine erfolgreiche Vereinigung
der Sexualzellen nur dann möglich, wenn sie beide der gleichen
„Art“ angehören. Unter Umständen können aber auch Sexualzellen
differenter Rassen, Arten, ja selbst Gattungen verbunden werden.
Die Produkte solcher Befruchtung werden Bastarde (auch
Hybriden oder Blendlinge) genannt. Man bezeichnet sie
auch als Heterozygoten, aus zwei ungleichen Sexualzellen
entstandene Individuen, im Gegensatz zu den Homozygoten, die aus
Sexualzellen mit ganz identischen Anlagen entstehen. Je näher sich die
Formen stehen, desto leichter bilden sie im allgemeinen auch Hybriden,
doch ist das keine durchgängige Regel.
Seitdem man weiß, daß nicht nur die Eizelle, sondern auch der
Embryosackkern mit einer Spermazelle verschmilzt, kann man auch
die sog. Xenienbildung verstehen. Xenien sind aus verschmolzenen
heterozygotischen Kernen entstandene Bastardendosperme.
Manche Familien neigen sehr zur Bastardbildung (Solanaceen,
Caryophyllaceen, Iridaceen usw.), andere bilden nur schwierig oder
überhaupt keine Bastarde (Papilionaceen, Koniferen, Convolvulaceen
usw.). Auch verwandte Gattungen und Arten verhalten sich oft recht
verschieden; Arten von Dianthus, Nicotiana, Verbascum, Geum sind
leicht, die Arten von Silene, Solanum, Linaria, Potentilla dagegen
schwer untereinander zu bastardieren. Eine Hybridisierung von nahe
verwandten Arten will oft nicht gelingen, während fernerstehende
gekreuzt werden können.
Auch in der freien Natur findet man Bastarde; insbesondere in den
Gattungen Salix, Rubus, Hieracium und Cirsium ist das der Fall.
Daß hybride Formen hier nicht häufiger sind, liegt einmal am
Mangel zeitlicher oder räumlicher Gelegenheit zur Bastardierung,
andererseits aber auch daran, daß der Pollen der eigenen Art bei
gemischter Bestäubung meist allein zur Wirkung kommt.
Bastarde kann man oft daran erkennen, daß sie Zwischenformen der
beiden verschiedenen Stammeltern sind; sie halten entweder zwischen
beiden genau die Mitte, z. B. Nicotiana rustica × Nic. paniculata
und Sorbus Aria × aucuparia (Fig. 269), oder gleichen in manchen
Eigenschaften mehr dem Vater, in anderen mehr der Mutter. Selten nur
kommt es vor, daß der Bastard bis auf verschwindende Merkmale wieder
ganz dem Vater (gewisse Erdbeerbastarde) oder ganz der Mutter ähnlich
ist. In weitaus den meisten Fällen fällt der Bastard ganz gleich
aus, einerlei ob die Pflanze A Vater oder Mutter ist; in einzelnen
Fällen aber ist der Bastard A × B von A × B deutlich
verschieden.
Häufig ist die Durchdringung der Eigenschaften bei den Bastarden
eine vollständige. Hat die eine Art ganzrandige Blätter,
die andere Art gefiederte, so kann der Bastard eine Mittelform,
nämlich eichenähnlich (Fig. 269) gebuchtete Blätter besitzen.
Hat die väterliche Art rote Blüten, die mütterliche aber gelbe,
so kann der Bastard orangefarbige Blüten hervorbringen. Wird
eine frühblühende Form mit einer spätblühenden gekreuzt, so kann
die Blütezeit des Bastards in der Mitte liegen usw. — Es gibt
aber auch, freilich seltener, eine andere Art von Bastarden,
die man Mosaikbastarde genannt hat, bei denen Teile mit
den Eigenschaften der Mutter abwechseln mit solchen, die die
Eigenschaften des Vaters aufweisen.
[S. 279]

Fig. 269. 1 Sorbus aucuparia, 2 Sorbus Aria und
3 ihr Bastard. Nach SCHLECHTENDAL, LANGETHAL und
SCHENCK. (Flora v. Deutschl. 5. Aufl. von HALLIER.
[S. 280]
Bei Bastarden treten auch bemerkenswerte neue Eigenschaften auf,
verringerte Fruchtbarkeit, größere Neigung zu Abänderungen
(Varietätenbildung) und oft eine üppigere Gestaltung
(„luxurierendes Wachstum“).
Die Fruchtbarkeit ist oft so weit herabgesetzt, daß Bastarde zum
Teil überhaupt nicht blühen (Rhododendron, Epilobium) oder doch
unfruchtbar sind (daher „Blendlinge“) und sich sexuell nicht
fortpflanzen; die Unfruchtbarkeit ist im allgemeinen um so größer,
je weiter die Stammeltern in der Verwandtschaft auseinander
standen; andere, wie Salix- und gewisse Hieraciumbastarde, bleiben
dagegen fruchtbar.
Bastarde auch von nahe verwandten Arten bilden häufig kräftigere
Vegetationsorgane, blühen früher, länger und reichlicher als die
Stammeltern; dabei sind die Blüten oft größer, prächtiger und zur
Füllung geneigt. Dieses luxurierende Wachstum und die gesteigerte
Neigung zu Abänderungen machen die Bastarde ganz besonders
wertvoll für die Gärtnerei und die Landwirtschaft.
Vererbung von Bastarden[239]. Dem ersten Forscher, der Bastarde
experimentell zu wissenschaftlichen Zwecken erzog, JOSEPH GOTTLIEB
KÖLREUTER (1761), waren diese aus ganz anderen Gründen von
Interesse als uns heutzutage. Sie waren ihm ein unwiderleglicher Beweis
für die Sexualität der Pflanzen, an der man so oft gezweifelt
hatte. Heute sind die Vererbungsfragen, die sich an die Bastarde
knüpfen, in erster Linie wichtig. Für das Studium der Vererbung aber
sind Artbastarde viel zu kompliziert. Dagegen ist es bei Benutzung
nächst verwandter Formen zuerst 1866 GREGOR MENDEL in Brünn
gelungen, gewisse Gesetzmäßigkeiten zu entdecken, die unbeachtet
blieben und erst, nachdem sie dann 1900 gleichzeitig von DE
VRIES, CORRENS und TSCHERMAK wiederentdeckt
worden waren, die wissenschaftliche Welt bewegten. Um zu diesen
Gesetzen oder Regeln zu gelangen, mußte MENDEL die Bastarde in
mehreren aufeinander folgenden Generationen verfolgen und mußte
alle auftretenden Individuen statistisch betrachten.
1. Uniformität der Bastarde. In der ersten Generation
sind alle Bastardpflanzen untereinander völlig gleich. Im Einzelfall
können sie dabei entweder eine genaue Mittelbildung zwischen
den Eltern vorstellen (intermediäre Vererbung), oder sie können
ganz oder vorwiegend dem einen Elter gleichen (dominierende
Vererbung).
Fig. 270. Mirabilis Jalapa alba und rosea nebst ihrem
Bastard in erster und zweiter Generation. Schema nach CORRENS.
Ein Beispiel für intermediäre Vererbung liefern die Bastarde von
Mirabilis Jalapa rosea und alba, zwei Rassen der Wunderblume, die sich
nur darin unterscheiden, daß die eine rote, die andere weiße Blüten
besitzt. Der Bastard hat hellrote Blüten (Fig. 270). — Der andere
Fall wird z. B. vom Bastard zwischen zwei Brennesseln geliefert,
Urtica pilulifera und[S. 281] Dodartii, von denen die erste scharf gesägte,
die zweite nur schwach gezackte Blätter besitzt. Hier ist die erste
Bastardgeneration nicht von Urtica pilulifera zu unterscheiden,
da alle Pflanzen mit scharf gesägten Blättern vorsehen sind (Fig.
271). Es bedurfte einer besonderen, gründlichen Untersuchung von
CORRENS, um nachzuweisen, daß wenigstens in der Jugend
ein kleiner Unterschied zwischen diesen Bastardpflanzen und reiner
pilulifera besteht. In diesem Falle sagt man, Urtica pilulifera
„prävaliert“ oder „dominiert“, während Dodartii sich
„rezessiv“ verhält. — Welche Merkmale bei der Kreuzung
prävalieren werden, läßt sich nicht voraussagen, sondern nur durch
Erfahrung bestimmen; meist sind es phylogenetisch ältere
Merkmale, die prävalieren.
Fig. 271. Der Bastard von Urtica pilulifera und
Urtica Dodartii in drei Generationen nebst den Eltern. Schema nach
CORRENS.
2. Spaltungsregel. In der zweiten Generation der
Bastardpflanzen, die durch Bestäubung der Blüten erster Generation mit
ihrem eigenen Pollen oder dem von gleichen Pflanzen entsteht, tritt
nun aber eine große Überraschung auf. Die zweite Generation
ist nicht mehr gleichförmig, sondern besteht aus verschiedenen
Pflanzen. In den einfachen Fällen von Mirabilis oder Urtica, wo die
Eltern nur in einem einzigen Merkmal sich unterschieden, ist
die Gesetzmäßigkeit der zweiten Bastardgeneration leicht
zu erkennen: es treten bei Mirabilis dreierlei Pflanzen in ganz
bestimmten Zahlenverhältnissen auf, nämlich 50%, die so aussehen wie
die I. Bastardgeneration, 25% die der einen und 25% die der
anderen Elternpflanze gleichen. Es blühen demnach 50% rosa,
25 rot und 25 weiß. Der Bastard ist also wenigstens zum Teil, hier
zur Hälfte, wieder auseinander gefallen, indem ein Viertel reine
Vaterpflanzen, ein anderes Viertel reine Mutterpflanzen und nur noch
die Hälfte Bastarde sind. Daß diese Deutung richtig ist, ergibt die
III. und alle folgenden Generationen, in denen bei weiter fortgesetzter
Inzucht in der Tat alle Nachkommen der in Generation II weiß oder rot
blühenden Pflanzen unverändert bleiben, während die Nachkommen
der rosablütigen weiter im gleichen Verhältnis zerfallen. Diesen
Zerfall nennt man das Spalten der[S. 282] Bastarde. In den folgenden
Generationen treten die Mischlinge mit rosa Blüten der Zahl nach
natürlich immer mehr gegen die roten und weißen Pflanzen zurück, so daß
sie in der 8. Generation nur noch 0,75% ausmachen; dieser kleine Rest
aber „spaltet“ weiter.
Bei Dominanz des einen Merkmals gestaltet sich das Spalten, wie
das Schema der Fig. 271 für Urtica zeigt, etwas anders. In der II.
Generation gleichen 25% Urtica Dodartii; da sie auch in den folgenden
Generationen unverändert bleiben, so sind sie reine U. Dodartii.
75% aber gleichen Urtica pilulifera. Sie sind indes nicht einheitlich;
ein Drittel von ihnen ist reine Urtica pilulifera, zwei Drittel sind
Mischlinge, die in den folgenden Generationen in der gleichen Weise
spalten wie in der II., also im Verhältnis 3 : 1.
Theoretisch sucht man seit MENDEL die Spaltungsregel durch
die Annahme zu verstehen, daß in den Mischlingen beiderlei
Anlagen unverändert weiter bestehen und sich bei der Ausbildung
der Geschlechtszellen zu gleichen Teilen auf diese verteilen. Die
Geschlechtszellen der rosa blühenden Mischlinge besäßen demnach
keinen Mischlingscharakter mehr, sondern hätten sich in rein „rote“ und
rein „weiße“ Geschlechtselemente geschieden. Bei der Befruchtung wird
dann die zu neuen Mischlingen führende Verbindung rot × weiß (weiß ×
rot , rot × weiß ) doppelt so oft möglich sein als die Verbindung
rot × rot oder weiß × weiß.
In sehr anschaulicher Weise werden die Tatsachen der Vererbung durch
Symbole dargestellt. Man bezeichnet die einzelnen Anlagen oder
Gene mit Buchstaben. Hat also eine bestimmte Pflanze die Gene A,
B, C, D,... X, so wird ein durch Reinzucht erzeugter Nachkomme
die „Erbformel“ AA, BB, CC, DD... XX besitzen, denn er hat
ja jeweils von Vater und Mutter die gleichen Anlagen mitbekommen.
Unterscheiden sich aber die zwei geschlechtlich verschiedenen
Keimzellen in einem Gen, so wird in diesem dann der Nachkomme
heterozygot sein. Nennen wir die Anlage für Rotfärbung bei Mirabilis R
(rot), so wird durch r das entsprechende nichtrote, also weiße
Gen bezeichnet.
In diesem Fall könnte man ebensogut der Anlage für weiß
den großen Buchstaben geben und rot als nicht weiß mit
kleinen Buchstaben bezeichnen. Wenn aber Dominanz besteht,
bezeichnet man stets das dominierende Merkmal mit großem, das
rezessive Merkmal mit kleinem Buchstaben.
Indem man dann alle Merkmalspaare, in denen Gleichheit besteht,
wegläßt, nimmt das Symbol für die Vererbung folgende Gestalt an:
|
Eltern
|
|
RR (rot)
|
rr (weiß)
|
|
Deren Geschlechtszellen
|
|
100% R
100% R
|
100% r
100% r
|
|
Bastard I. Generation
|
|
Rr (rosa)
|
|
Dessen Geschlechtszellen
|
|
50% R
50% R
|
50% r
50% r
|
|
Deren Kombinationen
|
|
RR, Rr, rR, rr.
|
Aus der letzten Zeile kann man also die Spaltung, die in der II.
Generation eingetreten ist, direkt ablesen: 25% RR müssen rot, 25% rr
weiß blühen; beide sind Homozygoten; 50% aber sind Heterozygoten rR,
blühen rosa und spalten weiter. Wenn aber rot über weiß dominierte, so
würde man ebenfalls aus dieser Zeile ablesen können, daß 25% weiße und
75% rote Blüten in der II. Generation auftreten müssen, und daß von
letzteren 1⁄3 homozygotisch, 2⁄3 heterozygotisch sind.
3. Autonomie der Merkmale. Waren die Eltern in zwei
Merkmalen statt in einem voneinander verschieden, liegen also
statt Monohybriden[S. 283] sog. Dihybriden vor, so zeigt es sich, daß die
einzelnen Merkmale beim Spalten unabhängig voneinander sich verteilen
können (Autonomie der Merkmale). Dementsprechend können neue
Kombinationen der Merkmale in den Nachkommen auftreten, was
ebenso für die Pflanzenzüchtung wie für das Verhalten der Pflanze in
der Natur von großer Bedeutung ist. Aus der Kreuzung einer Maissorte
mit glatten, weißen und einer zweiten mit runzligen,
blauen Körnern erhält man als neue Kombinationen glatte
blaue und runzlige weiße Körner.
Dieses Resultat läßt sich leicht aus der nachstehenden symbolischen
Darstellung entnehmen. Da glatt über runzlig dominiert, bezeichnen
wir glatt mit G, runzlig mit g; da blau über weiß dominiert,
schreiben wir B blau und b weiß.
|
Eltern:
|
Gb
|
|
gB
|
|
I. Generation:
|
Gg
glatt,
|
Bb
blau
|
|
Keimzellen:
|
GB
|
Gb
|
gB
|
gb
|
|
Kombinationen dieser Keimzellen:
|
|
GB
GB
glatt
blau
|
GB
Gb
glatt
blau
|
GB
gB
glatt
blau
|
GB
gb
glatt
blau
|
|
Gb
GB
glatt
blau
|
Gb
Gb
glatt
weiß
|
Gb
gB
glatt
blau
|
Gb
gb
glatt
weiß
|
|
gB
GB
glatt
blau
|
gB
Gb
glatt
blau
|
gB
gB
runzlig
blau
|
gB
gb
runzlig
blau
|
|
gb
GB
glatt
blau
|
gb
Gb
glatt
weiß
|
gb
gB
runzlig
blau
|
gb
gb
runzlig
weiß
|
Man entnimmt diesem Schema, daß eine Spaltung in folgenden
Verhältnissen eingetreten ist:
9 glatt blau: 3 glatt weiß: 3 runzlig blau: 1 runzlig weiß,
auch zeigt das Symbol, daß nur 4 von den 16 Kombinationen
homozygotisch sind, nämlich
Die 12 anderen sind Heterozygoten und spalten in der Folge.
Das wichtigste Resultat, das man diesen Vererbungsforschungen entnehmen
kann, ist: daß zwei Organismen bei ganz verschiedenem Gehalt an
Anlagen doch ganz gleich aussehen können; nicht das Aussehen, sondern
nur die Vererbungsanalyse kann also den Gehalt an Anlagen aufdecken.
Rückkreuzung. Das Ergebnis einer Rückkreuzung eines Bastardes
mit einer der Stammarten läßt sich am einfachsten unter Verwendung
unserer Symbole verstehen. Wenn wir eine Pflanze AA mit einem Bastard
Aa kreuzen, so bildet AA nur einerlei Geschlechtszellen A, während
der Bastard Aa Geschlechtszellen A und a bildet. Demnach werden in
50% der Fälle A mit A,[S. 284] in anderen 50% A mit a zusammenkommen. Die
Hälfte der Pflanzen sind wieder Bastarde, die andere Hälfte artreine
Mutterpflanzen.
Geschlechtsbestimmung[240]. Es hat sich nun gezeigt, daß
die Vererbung des Geschlechtes bei diözischen Pflanzen, von der
S. 275 schon die Rede war, nach der MENDELschen Regel,
genauer nach dem eben erörterten Schema der Rückkreuzung erfolgt.
In Versuchen von CORRENS wurde die monözische
Bryonia alba mit der diözischen Bryonia dioeca gekreuzt.
Bryonia dioeca-Weibchen mit dem Pollen von Bryonia alba belegt,
ergaben ausschließlich Weibchen; dagegen ergab die
Kreuzung alba × dioeca 50% Männchen und 50% Weibchen.
CORRENS deutet dieses Resultat so: es bestehen bei
diözischen Pflanzen zwei Sippen, die nur durch Kreuzung sich
erhalten können. Bei der Kreuzung dominiert das Merkmal
männlich; weiblich ist rezessiv. Die Weibchen
müssen im Geschlechtsfaktor homozygotisch sein (aa), die Männchen
heterozygotisch (Aa). Es müssen sich in der Hälfte der Fälle die
Faktoren aa kombinieren und das gibt Weibchen; in der anderen
Hälfte aber A und a, und das gibt, weil A dominiert, Männchen. Bei
den Versuchen mit Bryonia kommt zu dem geschlechtsbestimmenden
Faktor A noch ein zweiter hinzu, den wir Z nennen und der aus einer
monözischen eine diözische Pflanze macht. Die Erbformeln sind also:
|
Bryonia
|
alba zzaa
|
Geschlechtszellen
|
za
|
|
„
|
dioeca Männchen ZZAa
|
„
|
ZA und Za
|
|
„
|
„ Weibchen ZZaa
|
„
|
Za.
|
Die beiden reziproken Kreuzungen lauten dann so:
dioeca + alba
|
=
|
|
Za + za
|
=
|
Zzaa; alle weiblich
|
alba + dioeca
|
=
|
|
za + ZA
|
=
|
ZzAa; 50% männlich
|
|
za + Za
|
=
|
Zzaa; 50% weiblich.
|
Die Annahme, daß die weibliche Pflanze homozygot mit der Erbformel
aa, die männliche heterozygot mit der Erbformel Aa ist, macht
auch verständlich, warum in der Natur die Männchen annähernd
in gleicher Menge vorzukommen pflegen wie die Weibchen. Da man
ferner im allgemeinen nicht in der Lage ist, einen Einfluß auf
die Paarung der Gameten auszuüben, so kann man das Verhältnis
der beiden Geschlechter nicht ändern. In einigen Fällen ist das
CORRENS aber doch geglückt, z. B. bei Melandrium. Wird
hier die Narbe mit wenig Pollen bestäubt, so erhält man 43%
Pflanzen, wird aber viel Pollen aufgetragen, so treten nur 30%
auf. CORRENS erklärt dieses Resultat so: Die Pollenkörner
mit der Anlage a, die also Weibchen geben, sind raschwüchsiger. Bei
Aufbringen von viel Pollen findet eine Konkurrenz zwischen Eizellen
und Pollenschläuchen statt, und die raschwüchsigen Schläuche haben
mehr Wahrscheinlichkeit, zur Paarung zu kommen, als die langsam
wachsenden. Auch in anderen Eigenschaften, wie z. B. Resistenz
gegen Alkohol und gegen Austrocknung, unterscheiden sich die
zweierlei Pollenkörner.
Gültigkeit der Mendelschen Regeln. — Diese Regeln sind
nun nicht nur auf die Bastarde im gewöhnlichen Sinne des Wortes
beschränkt, sondern sie beherrschen die Vererbung im Tierreich und
Pflanzenreich weitgehend. Daß es keine Vererbung gäbe, die anderen
Gesetzen unterliegt, kann man zur Zeit gewiß nicht sagen, denn es
sind tatsächlich auch schon gut untersuchte Fälle bekannt, die sich
nicht nach den Mendelschen Regeln richten[241]. Wohl aber kann
man betonen, daß schon viele Erscheinungen, die anfangs den Mendelschen
Regeln zu widersprechen schienen, bei näherer Betrachtung sich ihnen
völlig unterordneten.
Auch manche auf den ersten Blick schwer verständliche Tatsache wird
durch die Mendelschen Regeln erklärt. So hat man z. B. beobachtet, daß
der Bastard zwischen einem weißblühenden und einem hellgelbblühenden
Löwenmäulchen nicht etwa eine sehr hellgelbe Blüte, sondern eine
rote Blüte besitzt. In der zweiten Generation treten Spaltungen
ein; es entstehen auf 3 rote 6 blaßrote, 3 hellgelbe und 4 weiße
Pflanzen. Wir können die Erklärung dieses Falles hier nicht bringen und
benützen ihn nur, um darauf hinzuweisen, daß die rote Farbe zweifellos
die Farbe der Stammform ist, von der sowohl die weißen wie
die hellgelben Formen abstammen. Bei der Bastardierung können also
Ahnenmerkmale (Atavismen) auftreten. Und das[S. 285] trifft
nicht nur in diesem Beispiel zu, sondern ist eine häufige Erfahrung der
Züchter.
Die Chromosomen als Träger der Gene[242]. Die Hypothese, daß
die Chromosomen die Träger der Gene sind, nimmt mehr und mehr an
Wahrscheinlichkeit zu. Bei der Befruchtung bringt die Eizelle genau
ebenso viele Chromosomen mit, wie die männliche Zelle, und der
diploide Organismus baut also seine Zellkerne aus väterlichen und
mütterlichen Chromosomen in gleicher Zahl auf. Bei der Ausbildung
neuer Sexualzellen aber erfolgt die Reduktionsteilung,
bei der nach den Gesetzen des Zufalls die väterlichen und die
mütterlichen Chromosomen auf die Tochterzellen verteilt werden.
So wird die wichtige Tatsache, daß die Geschlechtszellen immer
rein sind, nie Bastardnatur haben, in der einfachsten
Weise erklärt. Im Bastard ist keine Vermischung der
Gene eingetreten, sie blieben nebeneinander liegen; in den
Geschlechtszellen werden sie wieder getrennt. Wenn wirklich die
Chromosomen die Gene tragen, dann kann es in einem Organismus
nur so viele unabhängig spaltende Gene geben, als Chromosomen
vorhanden sind. Indes ist die Zahl der Chromosomen viel zu gering,
als daß man annehmen könnte, jedes trage nur ein einziges Gen.
Alle Gene aber, die in einem Chromosom vereinigt sind, spalten für
gewöhnlich nicht, sie sind aneinander gekoppelt. In
der Tat kennt man Faktoren genug, die in der Regel miteinander
gekoppelt bleiben, und für ein tierisches Objekt, die Fliege
Drosophila, ist durch MORGAN und seine Schule bis ins
einzelne hinein festgestellt, daß wirklich nur so viele unabhängig
spaltende Gruppen von Genen existieren, als Chromosomen vorhanden
sind.
Variabilität[243]. Unter Variabilität versteht man die
Tatsache, daß die Individuen, die zu einer Spezies gehören, nicht
alle gleich sind. Vielfach ist die Variabilität überhaupt nur eine
scheinbare, weil man die Spezies nicht eng genug begrenzt
hat. So existieren bei Rosen, Brombeeren, Draba verna usw. viele
einander nahestehende Arten nebeneinander. Der Eindruck, daß hier eine
„variierende“ Art vorliege, ist völlig falsch; jede der „elementaren
Arten“, aus denen sich die „Sammelart“ zusammensetzt, erweist sich als
konstant, macht keine Übergänge zu den anderen elementaren Arten.
Von solchen Fällen ist hier natürlich ganz abzusehen. Wir halten uns
ausschließlich an möglichst eng begrenzte Arten, wenn möglich an
die Nachkommenschaft einer sich selbst bestäubenden Pflanze, eine
sog. reine Linie (JOHANNSEN). Und da zeigt sich,
daß auch sie variiert. Wir können den Vorgang der Veränderung, die
Variationen, auf zwei Ursachen zurückführen und demnach auch mit
zweierlei Namen benennen: Modifikationen und Mutationen. Dazu kommen
bei Kreuzung noch die Kombinationen.
Modifikationen. Darunter versteht man Variationen, die durch
äußere Faktoren erzeugt sind. Es ist (S. 250 ff.) darauf hingewiesen
worden, daß zahllose Außenfaktoren die Gestalt der Pflanze weitgehend
beeinflussen. Groß sind z. B. die Unterschiede, die zwischen der
Landform und der Wasserform einer amphibischen Pflanze oder der
Ebenenform und der alpinen Form einer Art bestehen. Die in Fig.
261 (S. 253) abgebildeten Pflanzen sind Teile, eines und desselben
Individuums, 1 wurde in der Ebene, 2 im Hochgebirge erzogen. Um die
ganze Modifikationsfähigkeit einer Pflanze kennen zu lernen, muß man
sie unter allen Kulturmethoden erziehen, unter denen sie überhaupt
existieren kann; solche Versuche hat namentlich KLEBS in
größerem Maßstabe mit Erfolg ausgeführt. Wenn es möglich wäre, zwei
Pflanzen gleicher Abstammung unter ganz identischen äußeren Umständen
zu erziehen, so müßten diese ununterscheidbar sein. Tatsächlich
gelingt das nie, und dementsprechend weisen die homozygotischen
Individuen einer reinen Linie auch bei möglichst gleichartiger Kultur
doch zahlreiche quantitative Unterschiede auf. Betrachten wir z. B.
die Samen einer reinen Linie von Bohnen, so können wir diese nach
dem Gewicht in verschiedene Kategorien bringen und feststellen, wie
viele Exemplare zu jeder Kategorie gehören. Das Resultat einer[S. 286]
solchen Untersuchung gibt die Kurve Fig. 272. Es zeigt sich, daß
diejenigen Gewichtskategorien am häufigsten vertreten sind, die dem
Durchschnittsgewicht am nächsten stehen, und daß, je weiter
eine Kategorie vom Durchschnitt entfernt ist, desto weniger Individuen
zu ihr gehören. Das gleiche Resultat haben fast alle statistischen
Aufnahmen von Variationen ergeben. Die Variationskurven, die
man erhalten hat, stimmen immer mehr oder minder genau mit der sog.
Zufallskurve überein. Das erscheint begreiflich; denn es
sind stets mehrere äußere Faktoren tätig, die entweder eine
Vergrößerung oder Verkleinerung der Größe, der Zahl oder des Gewichts
bewirken können. Nur der Zufall entscheidet, welche Einwirkung
stattfindet. Dann werden sehr selten alle Faktoren auf Verkleinerung,
ebenfalls sehr selten alle Faktoren auf Vergrößerung hinwirken, und am
häufigsten werden Kombinationen eintreten müssen, die ein mittleres Maß
bewirken.
Sät man einen Samen einer reinen Linie aus, so ist es gleichgültig,
ob man von einem kleinen, mittleren oder großen ausgeht. Die
Variationskurve der nächsten Generation sieht nicht anders aus als
die, von der man ausging. Auch die durch Kultur im Hochgebirge
erzielten Veränderungen (Fig. 262) sind nicht erblich. Somit sind diese
Modifikationen durchaus keine erblichen Veränderungen; sie
dauern nur so lange oder wenig länger, als ihre Ursachen wirken.
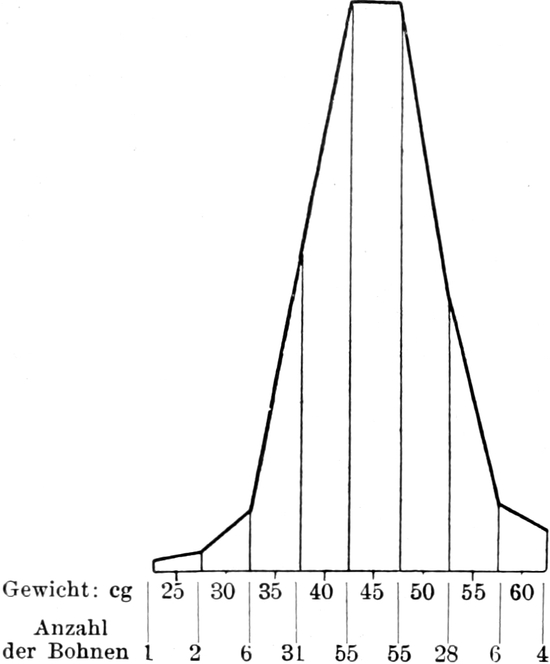
Fig. 272. Variationskurve der Gewichte der Samen
einer reinen Linie von Bohnen. (Johannsens Linie K.). Nach
BAUR.
Die Erfahrungen der Praxis scheinen diesem Resultat zu widersprechen.
Bei dem sog. Selektionsverfahren greift man eine Pflanze mit
besonderen Eigenschaften aus einer großen Menge heraus und sieht
vielfach in ihren Nachkommen die gleichen Eigenschaften wieder
auftreten. Das liegt daran, daß man in diesem Fall aus einem Gemisch
von verschiedenen Rassen oder Linien eine einzelne isoliert
hat, die ihre charakteristischen Eigenschaften auf ihre Nachkommen
überträgt. Bei wirklich reinem Ausgangsmaterial hat die Selektion
keinen Erfolg.
Kombinationen. Reine Linien können sich nur bei Selbstbestäubung
oder vegetativer Vermehrung erhalten. Tritt dagegen Fremdbestäubung
zwischen den einzelnen Linien einer Art ein, so werden homozygotische
Individuen selten, Heterozygoten die Regel sein. Diese werden die
Eigenschaften teils nur des einen, teils des anderen, teils auch
beider Eltern besitzen; sie werden also verschieden aussehen. Diese
Form von Variation ist äußerlich oft von der Modifikation
nicht zu unterscheiden, denn auch sie kann in Form der Zufallskurve
auftreten; innerlich aber unterscheidet sie sich sehr wesentlich
von ihr, denn sie ist erblich. Die Nachkommen variieren nach
den MENDELschen Regeln. Diese Form der Variation wird
Kombination genannt.
Unter Mutationen[244] endlich versteht man Variationen, die
sich von den Kombinationen dadurch unterscheiden, daß sie keinesfalls
durch Bastardierung entstehen, die ihnen aber darin gleichen, daß
sie erblich sind. Mit Sicherheit können Mutationen nur im Experiment
erkannt werden, wenn in[S. 287] den Nachkommen einer reinen Linie
Individuen auftreten, die eine neue Eigenschaft besitzen oder eine
Eigenschaft des Mutterorganismus nicht besitzen, und wenn ihre
Nachkommen die Abweichung beibehalten. In der Tat hat man im Experiment
das Auftreten solcher Mutationen sehr häufig beobachtet. BAUR
fand z. B. bei seinen Antirrhinumkulturen im Durchschnitt 2‰ mutierte
Sämlinge. Aber auch für viele in der Natur gefundene Variationen
ist die Wahrscheinlichkeit groß, daß sie als Mutationen betrachtet
werden dürfen. So z. B. das Chelidonium laciniatum, eine Mutation von
Chelidonium majus mit gezackten Blättern, die zuerst 1590 in Heidelberg
gefunden wurde (Fig. 273); Fragaria monophylla, eine zuerst im Jahre
1761 beobachtete Erdbeere, die sich durch einfache Blätter von der
Stammform mit dreizähligen Blättern unterscheidet. Auch die merkwürdige
kronblattlose Nicotiana tabacum virginica apetala, die während
der Versuche von KLEBS auftrat, gehört hierher. Viele solcher
Mutationen sind nicht einmal, sondern mehrfach aufgetreten.
Das gleiche hat man bei den in Kulturen entstehenden Mutanten
beobachtet. Alle diese Formen unterscheiden sich von den Mutterformen
nur in einem einzigen Merkmal. Und in weitaus den meisten
Fällen verhält sich dieses Merkmal bei der Kreuzung mit der Stammform
rezessiv; doch fehlt es nicht an Mutationen, die sich als
dominant erwiesen haben. Bei Selbstbestäubung bleiben die
Mutanten unbegrenzt konstant.
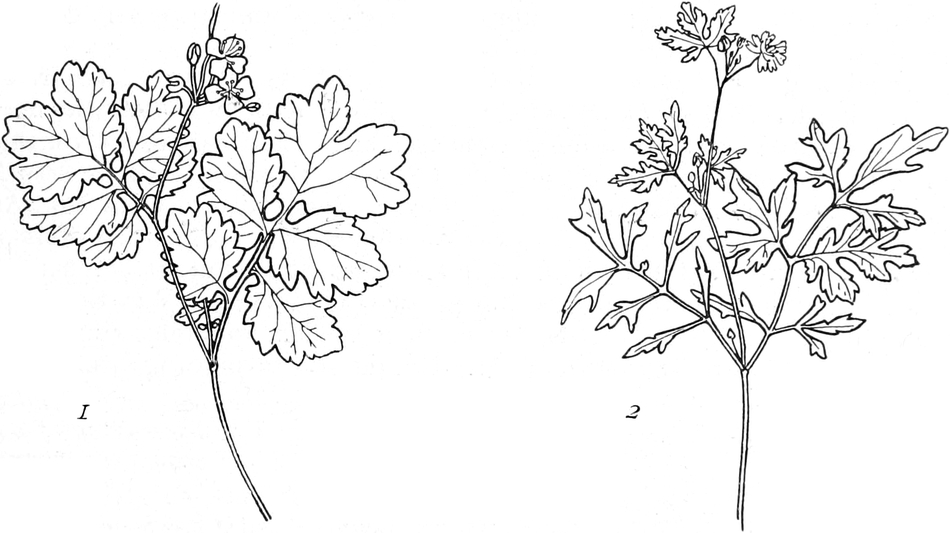
Fig. 273. Habitus von Chelidonium majus (1) und von
Chelidonium majus laciniatum (2). Nach LEHMANN.
Zahlreiche „Spielarten“ unserer Gärten beruhen auf derartigen
Mutationen in einem Merkmal. Dahin gehören nicht nur die schon
genannten geschlitztblättrigen Formen, sondern auch die Rassen mit
Trauerwuchs, Pyramidenwuchs, die schmalblättrigen,
krausblättrigen und rotblättrigen Abarten. Sie treten vor allem
an Sämlingen auf, in manchen Fällen aber auch rein vegetativ an
Seitenzweigen (vegetative Mutation). Verständlich ist, daß
sie gewöhnlich heterozygotisch sind; homozygotisch können sie
ja bei geschlechtlicher Entstehung nur dann sein, wenn beide zur
Verschmelzung kommende Sexualzellen gleichartig mutiert hatten, und das
ist offenbar ein seltener Zufall. In den genannten Antirrhinumkulturen
BAURs kamen auf 1000 Sämlinge nur 0,05 homozygotische
Mutanten.
[S. 288]
Außer den besprochenen hat man auch andere Mutationen gefunden, die
sich zum Teil weitgehend von diesen unterscheiden. Hier sei nur
erwähnt, daß manchmal ein Mutant sich durch eine veränderte Anzahl von
Chromosomen, z. B. durch Verdoppelung derselben, von der Mutterart
unterscheidet, was sich dann äußerlich in der Größe der Pflanze geltend
macht. Eine besondere Kategorie von Mutation findet sich bei Oenothera
Lamarckiana[245] und einigen anderen Arten dieser Gattung. Historisch
sind sie von besonderem Interesse, weil hier durch DE VRIES
zum erstenmal exakt die Entstehung der neuen Formen beobachtet wurde.
Aber echte Mutationen können wir heute diese Varianten schwerlich
mehr nennen, da Oenothera Lamarckiana offenbar ein Bastard mit
der besonderen Eigentümlichkeit ist, daß seine Eltern nicht mehr
existenzfähig sind. Endlich wären noch die Mutationen der Bakterien zu
nennen. Diese vermehren sich ja nur durch Zweiteilung; die Mutationen
sind also vegetative, und sie treten hier nach Behandlung mit
Giften auf, während wir bei den Mutationen höherer Pflanzen die
Ursachen gar nicht kennen. Sollten sie in einzelnen Fällen auch hier
durch äußere Faktoren bedingt sein, so wären sie deshalb doch scharf
von den Modifikationen zu unterscheiden; denn bei Mutationen tritt
eine Veränderung in den Anlagen auf, bei den Modifikationen
nicht.
Artbildung. Eine Reihe von Gründen, auf die schon S. 176
ff. hingewiesen wurde, hat zu der Vorstellung geführt, daß die
Organismen, die heute unsere Erde bewohnen, sich aus anderen entwickelt
haben, die vor ihnen lebten. Diese Vorstellung, die unter dem Namen
Deszendenztheorie[246] bekannt ist und eine große Wichtigkeit
besitzt, nimmt also an, daß die „Art“ nichts Konstantes, sondern
etwas Veränderliches sei. Unter Hinweis auf das früher Gesagte (S.
176 ff.) haben wir an dieser Stelle nur zu betonen, daß von den
bisher beobachteten Variationen ausschließlich die Mutationen und
Kombinationen eine Rolle bei der Artbildung spielen konnten. In
neuester Zeit mehren sich die Anzeichen dafür, daß die Bastardierung
bei der Artbildung wichtig ist.
Dritter Abschnitt. Bewegungen.
Nicht minder allgemein wie Stoffwechsel und Entwicklung treten uns
Bewegungserscheinungen an der lebenden Pflanze entgegen. Der
Stoffwechsel ist mit einer fortgesetzten Bewegung der aufgenommenen
rohen Nahrung sowie der Stoffwechselprodukte verknüpft. Diese
Bewegungen sind zwar nicht direkt wahrzunehmen, aber darum nicht
weniger sicher festgestellt; von ihnen war im ersten Abschnitt der
Physiologie genügend die Rede. Daneben existiert eine Fülle von auch
äußerlich sichtbaren, oft freilich nur langsam verlaufenden, manchmal
aber auch ganz plötzlich eintretenden Ortsveränderungen, die entweder
von der ganzen Pflanze oder von ihren einzelnen Organen ausgeführt
werden.
Das Protoplasma selbst ist zu verschiedenen Bewegungsformen befähigt,
sowohl im nackten Zustand (ohne Zellmembran) als auch eingeschlossen in
einer Membran. Nackte Plasmakörper zeigen fast stets Bewegungen, aber
auch behäutete Zellen besitzen das Vermögen der freien Ortsveränderung
oft in hohem Grade. Vielzellige höhere und niedere Pflanzen befestigen
sich jedoch meist am Orte ihrer Keimung durch Wurzeln und andere
Haftorgane und verzichten damit ein für allemal auf einen Ortswechsel.
Statt dessen[S. 289] haben sie aber sehr allgemein das Vermögen, die Lage
und Richtung ihrer Organe durch Krümmung zu verändern.
Dabei bedienen sie sich nicht nur ungleichen Wachstums, sondern sie
lassen auch andere Prozesse eingreifen, die zu einer Gestaltsänderung
führen. Durch solche Krümmungen werden in der Regel ihre Teile in
Stellungen gebracht, die für ihre Funktion notwendig oder vorteilhaft
sind; so werden die Stengel aufwärts, die Wurzeln abwärts, die Blätter
mit der Oberseite nach dem Lichte hin gerichtet: Schlingpflanzen und
Ranken umfassen die tragenden Stützen, und die Stengel vieler junger
Keimpflanzen werden rückwärts so umgebogen, daß sie ohne Verletzung der
Endknospe das feste Erdreich durchbrechen können.
Wir haben also zu unterscheiden zwischen lokomotorischen
Bewegungen einerseits, Krümmungsbewegungen andererseits.
I.
Lokomotorische Bewegungen[247].
A. Mechanik der lokomotorischen Bewegungen.
Ortsveränderungen kommen vor allem durch amöboide Bewegung,
Zilienbewegung oder durch Protoplasmabewegung in der
behäuteten Zelle zustande.
Die amöboide Bewegung freier Protoplasten ist eine
kriechende Fortbewegung auf festem Substrat, bei der der nackte
Plasmakörper der Amöben und Plasmodien einzelne Fortsätze nach einer
oder mehreren Seiten austreibt; diese werden entweder alsbald wieder
eingezogen, oder es folgt ihnen der ganze Plasmaleib in fließender
Bewegung nach. Die Bewegung erinnert äußerlich an das Fließen eines
zähen Flüssigkeitstropfens auf nicht benetzter Unterlage, und man nimmt
an, daß die Arbeit der Bewegung wie dort von Oberflächenspannungen
geleistet werde, die das reizbare Plasma lokal verändern kann.
Durch lokale Änderung der Oberflächenspannung können auch
an Tropfen lebloser Substanzen (Öltropfen in Seifenlösung;
Ölseifenschaum in Wasser; Quecksilbertropfen in 20% Salpetersäure
und in Wechselwirkung mit Kaliumbichromatkristallen) ähnliche
amöboide Bewegungen auftreten.
Die Zilienbewegung[248] ist eine Schwimmbewegung, bei
der eigenartige Bewegungsorgane in Gestalt äußerst feiner, oft bei
starker Vergrößerung kaum wahrnehmbarer Fädchen in Tätigkeit treten.
Diese „Geißeln“ oder „Zilien“ sind kontraktile Fortsätze
des Protoplasmas und finden sich zu 1, 2, 4 oder sehr zahlreich und
in verschiedener Anordnung an den beweglichen Zellen (Fig. 222 und
226). Sie durchsetzen, wenn die Zelle nicht nackt ist, die Zellhaut
und treiben durch ihre komplizierten Bewegungen den Plasmaleib oft
mit ansehnlicher Geschwindigkeit durch das Wasser fort. Die winzigen
Schwärmer von Fuligo varians legen dabei in 1 Sekunde 1 mm, das 60fache
der eigenen Länge, zurück, die von Ulva noch 0,15 mm; andere sind
träger. Einer der schnellsten Bazillen, der Choleravibrio, gebraucht 22
Sekunden für die Millimeterstrecke.
Die Geißeln bewirken nicht nur eine Vorwärtsbewegung, die
geradlinig oder in einer Schraubenlinie erfolgt, sondern sie
versetzen gleichzeitig auch den Organismus in eine Rotation
um seine Längsachse. Bei Spirillum fand METZNER 40
Umdrehungen der Geißeln und 13 Umdrehungen des Bakterienkörpers in
der Sekunde. Im einzelnen ist die Tätigkeit der Geißeln eine recht
verschiedene. Schon ein einfaches Schwingen einer bogig gekrümmten
Geißel in einem Kegelmantel kann die Schwimmbewegung herbeiführen.
In anderen Fällen aber hat die Geißel die Gestalt einer Schraube,
die ähnlich wie die Schiffsschraube oder ein Propeller je nach der
Drehungsrichtung eine Vorwärts- oder Rückwärtsbewegung bewirkt.
Während aber bei Mechanismen die Schraube starr ist, fest an
einer Achse sitzt und durch Rotation der letzteren sich betätigt,
muß die fest[S. 290] mit dem Zelleib verbundene Geißel eines Organismus
ihre Schraube während ihrer Bewegung immer neu bilden. Nach
BÜTSCHLIs Ausführungen nimmt man an, daß das in der Weise
geschieht, daß eine schraubig verlaufende Linie größter Kontraktion
fortgesetzt den Körper der Geißel umwandert. Auf andere und
kompliziertere Bewegungen der Geißeln kann hier nicht eingegangen
werden.
Die Diatomeen zeigen andere Formen der Bewegung. Die Arten,
die einen Spalt (Raphe) in ihrer Kieselschale führen, gleiten
oder schwimmen gewöhnlich in der Richtung ihrer Längsachse hin
und her und ändern ihre Bewegungsrichtung durch Schwenkungen
und Oszillationen. Aus der Art, wie sie kleine Körnchen ihrer
Umgebung in Bewegung setzen, hat man schon früher auf einen aus
der Raphe nach außen tretenden Plasmastrom geschlossen, der nach
O. MÜLLER die Ursache der Bewegung sein soll[249]. —
Die Zellen der Desmidiaceen führen mit Hilfe lokaler
Gallertabscheidungen ihre eigenartigen Bewegungen aus; die
Oszillarien scheinen sich ähnlich zu verhalten[250].
Neben solchen Ortsveränderungen der ganzen Zelle kennt man auch
Bewegungen des Protoplasmas innerhalb der Zellhaut, bei denen
vor allem die Rotations- und Zirkulationsbewegung zu
unterscheiden sind (vgl. S. 11).
Bei diesen Bewegungen ist die äußerste Schicht des Protoplasmas,
die an die Zellwand grenzt, stets in Ruhe. Das Protoplasma
stützt sich also bei seiner Strömung nicht etwa auf die
Zellwand, und deshalb kann die Bewegung nicht mit der einer
Amöbe verglichen werden, die in die Zelle eingeschlossen ist.
Auch dauert nach Abhebung des Protoplasmas von der Wand die
Strömung noch eine Zeit lang fort; demnach müssen die Änderungen
in den Oberflächenspannungen zwischen Protoplasma und
Zellsaft die Ursache für diese Bewegungen sein.
Die Protoplasmaströmungen sind 1772 von CORTI entdeckt
worden. Gute Beispiele für ihre Demonstration sind die Haare
mancher Pflanzen, die Blattzellen mancher Wasserpflanzen, die
langen Zellen der Characeen und Siphoneen.
B. Die Bedingungen der Lokomotion.
Da diese Bewegungen vom Protoplasma und seinen Organen ausgehen, so
ist es begreiflich, daß sie an ein gewisses Ausmaß derjenigen Faktoren
gebunden sind, die wir als allgemeine Lebensbedingungen kennen gelernt
haben.
Das Auftreten und die Lebhaftigkeit aller dieser Bewegungen hängt
demnach vor allem von günstiger Temperatur und bei den Aërobionten
auch von der Gegenwart freien Sauerstoffes ab. Fakultative Anaëroben
(Nitella) können auch bei Ausschluß des Sauerstoffs die Plasmabewegung
wochenlang unterhalten. Obligat anaërobe Bakterien verlieren ihre
Beweglichkeit bei Sauerstoffzutritt; umgekehrt gewinnen aërobe
Bakterien, die bei Sauerstoffmangel unbeweglich waren, bei erneutem
Sauerstoffzutritt ihre Beweglichkeit wieder (vgl. S. 213).
Überschreitung des „Minimums“ oder „Maximums“ dieser Faktoren bedingt
zunächst Bewegungslosigkeit, Starrezustände (so Kälte-,
Wärmestarre usw.), die anfangs noch durch Wiederkehr günstiger
Bedingungen aufgehoben werden können, bei längerer Dauer aber
schließlich zum Tode führen. In manchen Fällen genügt es, daß diese
allgemeinen Lebensbedingungen erfüllt sind, in anderen aber
muß die Bewegung durch einen besonderen Reiz ausgelöst werden.
So ist z. B. bekannt, daß die Protoplasmabewegung vielfach erst nach
Verwundung auftritt oder wenigstens durch einen solchen Eingriff stark
beschleunigt wird. Bei gewissen Bakterien wird erst durch das Licht
oder durch eine bestimmte Konzentration des Substrats die Bewegung
erweckt. Ebenso kann aber auch durch äußere Einflüsse die Beweglichkeit
aufgehoben werden, während bei den nur temporär bewegungsfähigen
Objekten (Schwärmsporen, Spermien) auch innere Ursachen die
Bewegung sistieren.
Eine ganz besondere Rolle spielen äußere Reize aber bei den
lokomotorischen Bewegungen insofern, als sie diesen eine bestimmte
Richtung[S. 291] geben. Ohne solche richtenden Reize bewegen sich die
Plasmodien ziellos; die Richtung der Schwimmbewegungen und der
Zirkulationsbewegung wechselt häufig und nur die Rotationsströme sind
durch eine konstante Richtung ausgezeichnet.
C. Taxien.
Als richtende Reize kommen vor allem einseitig einwirkende Beleuchtung
und ungleich im Wasser verteilte, gelöste Stoffe in Betracht. Die durch
solche Faktoren erzielten Richtungsbewegungen werden als Taxien
bezeichnet; durch Licht bewirkte heißen Phototaxis, durch gelöste
Stoffe hervorgerufene Chemotaxis. (Weitere Taxien s. S. 293.)
Richtungsbewegungen führen die freibewegliche Pflanze oder das
bewegliche Organ einer Zelle entweder zum Reizmittel hin oder von ihm
weg; im ersten Falle spricht man von positiver, im zweiten von
negativer Taxis. Welche von diesen verschiedenen Reaktionsweisen
eintritt, hängt vielfach nicht nur vom Objekt, sondern auch von äußeren
Umständen ab. Man unterscheidet ferner eine topische Reaktion,
bei der eine Einstellung und Bewegung in der Richtung des wirksamen
Reizes stattfindet, und eine phobische Reaktion, bei der ein
Übergang zu einer anderen Intensität des Reizmittels die
Reizbewegung auslöst.
1. Phototaxis.
Phototaktische Bewegungen[251] kann man am besten wahrnehmen, wenn man
Wasser mit Volvocineen und Chlamydomonaden oder Schwärmsporen von Algen
in einem Glasgefäß der einseitigen Beleuchtung etwa in der Nähe eines
Fensters aussetzt. Nach kurzer Zeit ist die gleichmäßige Grünfärbung
des Wassers verschwunden, da sich die beweglichen Organismen alle an
der Lichtseite des Gefäßes angesammelt haben. Dreht man das Gefäß um
180°, so eilen die Algen sofort an die nunmehr belichtete Seite. Läßt
man aber stärkeres Licht, etwa direktes Sonnenlicht einfallen, so
sieht man dieselben Organismen, die bisher positiv reagierten, negativ
phototaktisch werden und von der Lichtquelle wegschwimmen. Auch andere
äußere Faktoren können eine solche „Umstimmung“ bewirken.
Bei manchen chlorophyllfreien Organismen, so den Plasmodien der
Schleimpilze, kommt es auch bei niedrigen Lichtintensitäten
gewöhnlich nur zu negativer Reaktion. Es gibt aber auch farblose
Organismen, die positiv phototaktisch reagieren.
Man kennt phobische und topische Reaktionen bei der Phototaxis.
Manche Organismen können sowohl phobisch als topisch reagieren,
andere zeigen nur eine dieser Reaktionsweisen. —
Phobotaktisch reagieren vor allem gewisse Bakterien, die auf den
Übergang von Licht zu Dunkelheit mit einer Rückwärtsbewegung
antworten. An einer stark beleuchteten Stelle werden sie dadurch
gefangen, daß sie jedesmal, wenn sie durch ihre Bewegungen ins
Dunkle geführt werden, zurückprallen (Lichtfalle). — Die
topotaktischen Organismen stellen vor allem ihre Längsachse in
die Lichtrichtung ein, um sich dann nach der Lichtquelle hin zu
bewegen, wenn sie positiv reagieren, oder von ihr wegzuschwimmen,
wenn sie negativ phototaktisch sind. Fallen Lichtstrahlen
verschiedener Richtung gleichzeitig auf solche Organismen ein, so
bewegen sie sich in der Resultierenden. Dieses „Resultantengesetz“
gilt nicht nur in Beziehung auf die Richtung, sondern auch in
Beziehung auf die Intensität.
Im Experiment kann man Bedingungen herstellen (konvergentes Licht),
die z. B. negativ topotaktische Schwärmer zwingen, indem sie sich
von der Lichtquelle entfernen, in immer hellere Zonen zu eilen. In
der Natur aber führen zweifellos die phototaktischen Bewegungen die
Organismen an Orte optimaler Helligkeit.
Sehr auffallende Phototaxis besitzen die Chlorophyllkörner[252], deren
Bewegungsmechanismus freilich noch ganz unbekannt ist. Diese Bewegungen
bringen das Chlorophyllkorn in eine derartige Lage, daß es eine
optimale[S. 292] Lichtmenge aufnehmen kann. Diese Lage wird bald durch
Drehungen an Ort und Stelle, bald durch Wanderung an andere Stellen
erreicht.
In den zylindrischen Zellen der Alge Mesocarpus befindet sich ein
einziger Chloroplast, der die Gestalt einer rechteckigen Platte
hat. Licht niedriger Intensität sucht er möglichst auszunutzen,
indem er sich senkrecht zu der Richtung der Strahlen stellt
(Flächenstellung); bei höherer Lichtintensität dreht sich die
Platte um ihre Längsachse und bildet einen spitzen Winkel mit den
Strahlen oder wendet ihnen schließlich gar die schmale Kante zu
(Profilstellung), nimmt dann also sehr wenig Licht auf.
In den Blättern der Moose und der höheren Pflanzen sowie in
den Prothallien der Farne wird eine Stellungsänderung der
zahlreichen Chlorophyllkörner durch Verschiebung auf
den Wänden der Zelle erreicht. In gemäßigtem Lichte werden
die Chlorophyllkörner an denjenigen Wänden verteilt, die quer
die Richtung der Lichtstrahlen schneiden (Fig. 274 T); sie
gleiten aber alsbald an die den Lichtstrahlen parallel laufenden
Seitenwände und werden der Lichtwirkung damit möglichst entzogen,
wenn das Licht anfängt, zu stark zu werden (Fig. 274 S). Im
Finstern oder bei sehr schwachem Licht kann eine dritte, aus
der Fig. 274 N ersichtliche Art der Gruppierung eintreten, deren
Ursache und Bedeutung hier nicht erörtert werden kann.
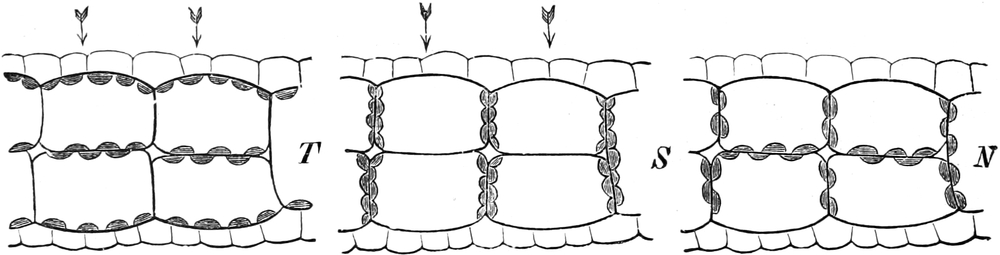
Fig. 274. Wechselnde Stellung der Chlorophyllkörner
in den Zellen der untergetauchten Wasserlinse (Lemna trisulca) bei
verschiedener Beleuchtung. T In diffusem Tageslicht. S In direktem
Sonnenlicht. N Des Nachts. Die Pfeile geben die Richtung des
einfallenden Lichtes an. Nach STAHL.
Die Chlorophyllkörner erfahren zudem bei Beleuchtungswechsel
Formveränderungen; in gemäßigtem Lichte sind sie abgeflacht,
in zu starkem und zu schwachem Lichte werden sie dicker und
entsprechend kleiner. — Durch die Änderungen in der Anordnung der
Chlorophyllkörner erscheint die Farbe grüner Organe in wechselnder
Abtönung. In starker Besonnung sehen sie meist heller, in
zerstreutem Licht dunkler grün aus.
2. Chemotaxis[253].
Eine Chemotaxis kommt, wie bemerkt, durch ungleiche Verteilung von im
Wasser gelösten Stoffen zustande. Positive Chemotaxis führt zu einer
Ansammlung der reizbaren Pflanze in der höheren Konzentration des
Chemotaktikums.
Als Chemotaktikum funktionieren nicht beliebige, sondern ganz
bestimmte Substanzen. So werden z. B. viele Bakterien von gewissen
Nährstoffen, anorganischen wie organischen, z. B. Pepton, Zucker,
Fleischextrakt, Phosphaten usf., „angelockt“; andere Stoffe,
vor allem Säuren, Alkalien, „stoßen sie ab“. Steht hier die Chemotaxis
im Dienste des Nahrungserwerbes, so sehen wir sie bei den Spermien
eine ganz andere Bedeutung gewinnen; diese männlichen Sexualzellen
finden durch chemotaktische Anlockung die Eizellen. Fig. 356 zeigt die
chemotaktische Anlockung der Spermien durch die weibliche Sexualzelle
bei Ectocarpus. Auch Zellkerne und Chloroplasten können chemotaktische
Bewegungen ausführen.
Chemotaxis freibeweglicher Organismen wird seit PFEFFER
gewöhnlich in der Weise nachgewiesen, daß man den chemotaktisch
empfindlichen Organismus in Wasser auf dem Objektträger hält und
mit Deckglas bedeckt,[S. 293] während eine Glaskapillare, die mit dem
Chemotaktikum gefüllt ist, unter das Deckglas geschoben wird. Nach
kurzer Zeit erfolgt eine Ansammlung der chemotaktischen Organismen
vor oder in der Kapillare. Manche stellen sich, wenn sie in das
Diffusionsfeld eingetreten sind, in die Richtung des Diffusionsgefälles
ein und steuern direkt auf das Diffusionszentrum los (topische
Reaktion). Andere kommen nur zufällig an die Mündung der Kapillare
und werden dann durch phobische Reaktion am Verlassen dieser Stelle
gehindert.
Die Samenfäden der Farne werden durch Äpfelsäure bzw. äpfelsaure
Salze in den Hals der Archegonien gelockt; bei den Lycopodien spielt
Zitronensäure, bei den Laubmoosen Rohrzucker, bei den Marchantien
Eiweiß die gleiche Rolle. Es sind oft äußerst geringe Substanzmengen,
die eine kräftige Reizbewegung auslösen; so genügt schon eine
0,001prozentige Lösung von Äpfelsäure zur Anlockung der in reinem
Wasser ziellos umherschwärmenden Farnspermien.
Die chemotaktischen Bewegungen kommen nur dann zustande, wenn das
Chemotaktikum ungleich verteilt ist, wenn also Diffusion
stattfinden kann. Aber auch homogene Lösungen der Chemotaktika sind
nicht ohne Einfluß auf die chemotaktisch empfindlichen Organismen; sie
setzen kurz gesagt deren Empfindlichkeit herab. Man bezeichnet
diejenige minimale Konzentration des Chemotaktikums, die bei Benutzung
der Kapillarenmethode gerade noch zu einer sichtbaren Ansammlung führt,
als Reizschwelle. Durch Verwendung einer homogenen Lösung des
Chemotaktikums wird nun die Reizschwelle erhöht. Es hat sich
gezeigt, daß diese Erhöhung in streng gesetzmäßiger Weise verläuft. Für
Farnspermien z. B. fand PFEFFER folgende Werte:
|
|
Reizschwelle
|
|
In
|
Wasser
|
|
0,001%
|
Äpfelsäure
|
|
„
|
Äpfelsäure
|
0,0005
|
0,015%
|
„
|
|
„
|
„
|
0,001
|
0,03 %
|
„
|
|
„
|
„
|
0,01
|
0,3 %
|
„
|
Man sieht also, daß immer das gleiche Verhältnis zwischen der
einseitig und der allseitig wirksamen Lösung bestehen muß; erstere
muß 30mal so konzentriert sein wie letztere, wenn es zu einer
Ansammlung kommen soll. Diese Gesetzmäßigkeit ist unter dem Namen
WEBERsches Gesetz bekannt. Das WEBERsche Gesetz
der Psychophysik hat wenigstens eine große Ähnlichkeit mit dem hier
konstatierten. Es darf aber nicht vergessen werden, daß es sich dort
um das Verhältnis zwischen Reiz und Empfindung, hier um das Verhältnis
zwischen Reiz und Reaktion handelt.
Ist die Chemotaxis durch Sauerstoff bedingt, so spricht man von
Aërotaxis. Sie findet sich z. B. bei Bakterien, die aus diesem
Grunde zum Nachweis der Kohlensäureassimilation Verwendung finden
können (S. 213).
Findet sich hier eine positive Aërotaxis typischer Aërobionten,
so zeigen umgekehrt echte Anaërobionten negative Aërotaxis, und gewisse
Bakterien, die an bestimmte niedere Sauerstoffspannungen angepaßt sind,
können bald positive, bald negative Bewegungen ausführen und so die
optimale Sauerstoffspannung aufsuchen.
An die Chemotaxis läßt sich die Erscheinung der Hydrotaxis
anschließen, eine Richtungsbewegung, die durch ungleiche Verteilung
des Wasserdampfes in der Luft veranlaßt wird. Eine positive
Hydrotaxis kommt den Plasmodien der Myxomyceten zu, und sie geht
zur Zeit der Sporenbildung in negative Hydrotaxis über.
Außer den genannten sind noch viele andere Taxien mehr oder weniger
genau bekannt. Von Osmotaxis spricht man, wenn nicht die chemische
Natur, sondern lediglich die Konzentration einer Lösung zu einer
Ansammlung beweglicher Organismen führt. Bei[S. 294] der Thermotaxis
wird durch ungleiche Wärmeverteilung, bei der Galvanotaxis durch
galvanische Ströme, bei der Rheotaxis durch Wasserströme die
Bewegung ausgelöst.
II. Krümmungsbewegungen.
Die Arten der Krümmung, die an den Organen festsitzender
Pflanzen auftreten können, werden durch Fig. 275 illustriert. Ein
vierkantiges Prisma hat gleich lange Kanten; wird es aber in einer
Ebene gekrümmt, so müssen die Kanten der Konkavseite notwendig kürzer
werden als die der Konvexseite. Eine Verlängerung der einen Seite,
oder eine Verkürzung der anderen Seite, oder endlich gleichzeitige
Verlängerung und Verkürzung antagonistischer Seiten müssen zur Krümmung
führen. Wenn bei dieser Einbiegung der Stab in einer Ebene bleibt,
spricht man von Krümmung (II) schlechthin, wenn aber der Stab
aus der Ebene in den Raum hinaustritt (dadurch, daß die Biegung um
schief zu der Längsachse gelegte Linien erfolgt), dann nennt man ihn
gewunden (IV). Wenn endlich der Stab im ganzen zwar gerade
bleibt, seine Kanten aber Schraubenlinien beschreiben, dann nennen wir
ihn gedreht oder tordiert (III); die Torsion kommt
durch eine Längendifferenz zwischen der Mittellinie und sämtlichen
(untereinander gleichbleibenden) Kanten zustande.
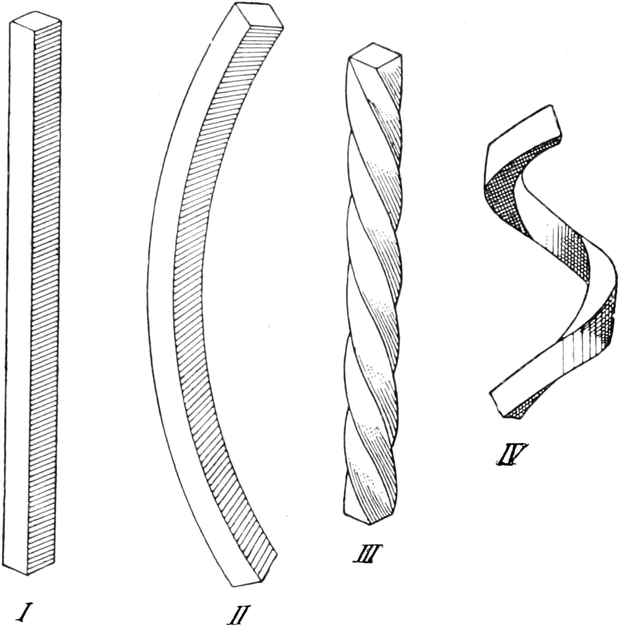
Fig. 275. Vierkantiges Prisma. I gerade, II
gekrümmt, III gedreht, IV gewunden.
Mittel zur Ausführung der Krümmungen. Bei der Ausführung
der Krümmungen handelt es sich, wie gezeigt wurde, stets um
ungleiche Verlängerung oder Verkürzung eines Organs, also um
Dimensionsänderungen. Zur Ausführung von Dimensionsänderungen aber
stehen der Pflanze folgende Mittel zur Verfügung:
1. Wachstum. Es dient fast stets nur zur Verlängerung.
2. Turgordruck. Er kann, je nachdem er zu- oder abnimmt, sowohl
eine Verlängerung als auch eine Verkürzung herbeiführen.
3. Wasserschwankungen in der Membran oder in toten Zellen.
Sie können wieder ebensogut im Sinne einer Verlängerung wie einer
Verkürzung wirken.
Nach den Mitteln, die zur Realisierung einer Dimensionsänderung
Verwendung finden, kann man die pflanzlichen Krümmungsbewegungen
einteilen in 1. Wachstumsbewegungen (Nutationen),
2. Turgeszenz- oder Variationsbewegungen und 3.
hygroskopische Bewegungen. Da Wachstum und Turgordruck
Lebenserscheinungen sind bzw. vom lebenden Protoplasma wesentlich
beeinflußt werden, so sollen sie von den hygroskopischen Bewegungen
getrennt behandelt werden. Denn diese letzteren sind
keine Lebenserscheinungen; sie können zwar vereinzelt auch
an lebenden Organen beobachtet werden, aber sie treten ebensogut an
absterbenden oder an toten Organen auf, und sie werden ausschließlich
von äußeren Faktoren bewirkt. Das Protoplasma ist nur insofern an
diesen Bewegungen beteiligt,[S. 295] als es die Organe so aufbaut, daß sie
bei Schwankungen des Wassergehaltes nicht einfache Längenänderungen,
sondern Krümmungen erfahren.
A. Hygroskopische Bewegungen.
Bei den hygroskopischen Bewegungen kann man zwei recht verschiedene
Fälle unterscheiden. Im ersten handelt es sich darum, daß die Zellwände
durch Quellen sich verlängern oder durch Schrumpfen sich
verkürzen. Bewegungsapparate, die auf diesem Prinzip beruhen, nennt man
Quellungsmechanismen[254].
Die Quellung der Membranen hängt damit zusammen, daß das
Imbibitionswasser nicht in vorgebildete Hohlräume aufgenommen wird, wie
das bei dem Kapillarwasser eines porösen Körpers (Schwamm, Gips) der
Fall ist, sondern daß es bei seiner Aufnahme die kleinsten Teilchen der
Zellhaut auseinander drängt. Umgekehrt nähern sich diese wieder, wenn
das Quellungswasser bei der Schrumpfung verdunstet. Wenn nun in einem
Organ auf verschiedenen Seiten verschieden stark quellbare Schichten
abgelagert sind, so müssen notwendig mit jeder Anfeuchtung und mit
jedem Wasserverlust Krümmungen zustande kommen. Obwohl es sich dabei
um rein physikalische Erscheinungen handelt, so können diese doch eine
große Bedeutung für die Pflanze haben.
Das Aufspringen und Aufreißen reifer Samenbehälter ist die
Folge ungleicher Kontraktionen beim Austrocknen. Hierbei werden
oft Spannungen erzeugt, die bei plötzlicher Überwindung des
Hindernisses die Samen weit fortschleudern (Euphorbiaceen,
Geranium u. a.). Man nennt dieses Aufspringen beim Austrocknen
Xerochasie. Im Gegensatz dazu findet umgekehrt bei manchen
Wüstenpflanzen ein Öffnen der Früchte und die Ausstreuung der
Samen bei Befeuchtung statt (Hygrochasie). Das beste
Beispiel hierfür sind die Früchte von Mesembryanthemum linguiforme
(„Auferstehungssterne“). Ebenso verhält sich die „Jerichorose“
(Anastatica hierochuntica). Diese zeichnet sich auch noch
dadurch aus, daß die in frischem Zustand wie ein gewöhnliches
niederliegendes Kraut erscheinende Pflanze im Zustand der
Fruktifikation beim Austrocknen durch ungleiche Längenveränderung
der Ober- und Unterseite der Äste zu einer Kugel sich einkrümmt.
Bei Wasseraufnahme nimmt die Pflanze ihre ursprüngliche Gestalt
wieder an und öffnet ihre Früchte; die Samen werden also nur
ausgestreut, wenn sie Keimungsbedingungen finden. An Anastatica
schließen sich einige andere Pflanzen an, die zum Teil den gleichen
Namen „Jerichorose“ führen (Odontospermum). Einzelne Früchte
führen beim Wechsel ihres Wassergehaltes nicht nur Krümmungen,
sondern auch Torsionen und Windungen aus, wie besonders die
Teilfrüchtchen von Erodium gruinum (Fig. 276), die Früchte von
Stipa pennata und von Avena sterilis. Diese Bewegungen führen dazu,
den Samen in die Erde zu vergraben. Liegt eine Erodiumfrucht
von der Gestalt der Fig. 276 A mit beiden Enden dem Boden auf,
so macht ihre Spitze bei wechselndem Wassergehalt der Luft eine
bohrende Bewegung. Die an ihr befindlichen schräg gerichteten Haare
wirken dann so, daß nur ein Eindringen nach unten möglich ist.
Eine wichtige Rolle fällt den Quellungskrümmungen auch bei der
Entleerung der Mooskapseln zu: an diesen sind es die
Zähne des Peristoms, die durch ihre Bewegungen die Kapselöffnung
hygroskopisch verschließen oder öffnen. Bei den Schachtelhalmen
führt die in Form zweier paralleler Bänder sich ablösende Außenwand
der Sporen sehr lebhafte hygroskopische Bewegungen aus.
Fig. 276. Teilfrüchtchen von Erodium gruinum. A in
trockenem Zustande, aufgerollt. B in feuchtem Zustande, gerade
gestreckt. Nach NOLL.
[S. 296]
Um die Quellungsbewegungen hervorzurufen, ist eine direkte
Benetzung nicht notwendig; die Membranen kondensieren bei
wechselndem Feuchtigkeitsgehalt der Luft wechselnde Mengen Wasser;
sie sind hygroskopisch. Deshalb benutzt man z. B. die
Grannen von Erodium zur Messung der Luftfeuchtigkeit in Hygrometern
und „Wetterhäuschen“.
Den Quellungsmechanismen werden die Kohäsionsmechanismen[255]
gegenübergestellt. Sie unterscheiden sich von jenen dadurch, daß die
Zellwände während der Ausführung der Bewegung mit Wasser imbibiert
bleiben. Bei eintretendem Wasserverlust verkleinert sich hier der
Innenraum der Zelle. Als Beispiel für eine solche Krümmung
betrachten wir die Bewegung des Polypodiaceensporangiums beim
Austrocknen. Diese Sporangien sind gestielte, bikonvexe Körper, die
innerhalb einer einschichtigen Wand die Sporen enthalten. Während im
allgemeinen die Zellwände dünn sind, umgibt den Rand des Sporangiums
etwa in der Ausdehnung eines Halbkreises ein sog. Ring
(Annulus) von eigenartig verdickten Zellen (Fig. 277 R). Sie
haben (Fig. 277, 2) dünne Außenwände, nach innen sich verdickende
Seitenwände und dicke Innenwände. Trockener Luft ausgesetzt, verlieren
die Annuluszellen allmählich ihr Füllwasser. Es kommt aber nicht zu
einer Loslösung des wäßrigen Inhaltes von der Zellwand und auch nicht
zu einem Zerreißen der Flüssigkeit in sich, weil die Adhäsion an die
Wand und die Kohäsion der Wassermoleküle untereinander sehr groß ist,
mehr als 300 Atmosphären beträgt. Dagegen folgt die Zellwand unter
Deformation dem abnehmenden Füllwasser: unter Einstülpung der zarten
Außenmembranen (Fig. 277, 3) nähern sich die derben seitlichen
Zellwände einander und so entstehen sehr energische einseitige
Verkürzungen des Kohäsionsgewebes, die zur Öffnung des Sporangiums
führen. Dabei bleibt die Mehrzahl der Sporen an der Sporangiumwand
haften. Nun aber tritt bei fortgesetzter Wasserverdunstung der
Augenblick ein, da der Wasserrest dem wachsenden Zug der gespannten
Membranen nicht mehr Widerstand zu leisten vermag. Die Flüssigkeit
in den Annuluszellen reißt plötzlich in sich selbst, oder sie löst
sich von der Wand los, und der Annulus springt unter Ausschleudern
der Sporen in seine Anfangsstellung zurück. Er stellt also eine
Wurfmaschine dar, welche die Sporen ziemlich weit ausstreut und
ihnen so günstigere Entwicklungsbedingungen schafft, als wenn alle an
einer Stelle niederfielen.
Fig. 277. 1 Sporangium von Polypodium falcatum nach
CAMPBELL. R Ringzellen. St Stomium. 2 Ringzellen in
ursprünglichem Zustand, gerade. 3 Nach teilweiser Verdunstung des
Füllwassers W; die oberen Zellwände o eingestülpt, die unteren u
in ihrer ursprünglichen Länge. 2, 3 nach NOLL.
Kohäsionsmechanismen sind auch bei anderen Sporangien höherer
Kryptogamen sowie in der Wandung der Staubbeutel ausgebildet und
bewirken deren Öffnen. — Manche hygroskopische Krümmungen kommen durch
gemeinsame Wirkung von Quellung und Kohäsion zustande.
B. Krümmungsbewegungen an der lebenstätigen
Pflanze.[256]
Wie bei der lokomotorisch tätigen, so treten auch an der
festgewachsenen Pflanze die Bewegungserscheinungen einesteils schon
ein, wenn alle allgemeinen[S. 297] Bedingungen für die Lebenserscheinungen
gegeben sind, andernteils aber erst dann, wenn ein bestimmter Faktor
(Reiz) sich geltend macht, der entweder nur das Maß der Krümmung
oder auch ihre Richtung bedingt. Man nennt die Bewegungen, die ohne
solche spezifische äußere Reize erfolgen, autonome, die anderen
induzierte oder paratonische.
1. Autonome Krümmungsbewegungen.
Wie bemerkt, genügt ein gewisses Ausmaß der äußeren Faktoren,
an die das Leben gebunden ist (S. 185), um diese Krümmungen zu
ermöglichen. Überschreitung ihres Minimums oder ihres Maximums führt
zu Starrezuständen, macht die Pflanze bewegungslos. So kennen wir
Kälte-, Wärme-, Dunkel-, Trockenstarre usw. Auch chemische Schädigungen
(Giftwirkungen) führen zu Starrezuständen.
Eine autonome Bewegung ist auch das geradlinige Wachstum des
Sprosses und der Wurzel mit seiner charakteristischen, rein aus
inneren Ursachen entspringenden großen Periode. Eine ganze Reihe
von Wachstumskrümmungen (Nutationen) schließt sich
ihm an; ja, man kann wohl sagen, es gibt überhaupt kaum irgendwo
wirklich geradliniges Wachstum. Die Spitzen der Organe beschreiben
vielmehr eine außerordentlich unregelmäßige Kurve im Raume, sie führen
die von DARWIN entdeckten „Zirkumnutationen“ aus. Sind
diese Krümmungen auch gewöhnlich so unbedeutend, daß man sie ohne
besondere Hilfsmittel nicht wahrnehmen kann, so fehlt es doch nicht
an Organen, die sehr auffallende und regelmäßig verlaufende autonome
Wachstumskrümmungen zeigen.
So ist die Entfaltung der meisten Laub- und Blütenknospen eine
Nutationsbewegung, die durch stärkeres Wachstum der Oberseite
(Epinastie) der jugendlichen Blätter erfolgt. Besonders auffällig
tritt das bei den in der Jugend durch verstärktes Wachstum der
Unterseite (Hyponastie) eingerollten Blättern der Farne und mancher
Cycadeen hervor. Der Keimstengel zahlreicher Pflanzen nimmt bei
seinem Austritt aus dem Samen häufig eine scharfe Krümmung an,
die ihm beim Durchbrechen des Bodens zustatten kommt, und eine
ähnliche, mit dem Zuwachs der Triebe nach vorn weiterrückende
Nutationskrümmung zeigen u. a. die Sprosse des wilden Weins
(Parthenocissus quinquefolia).
Besonders auffällig werden die Nutationsbewegungen, wenn das
Wachstum nicht eine Seite bevorzugt, sondern abwechselnd
verschiedene Seiten fördert. Sehr schön läßt sich das an
Blütenschäften der Küchenzwiebel beobachten. Diese zuletzt
senkrecht gestellten Sprosse krümmen sich im halb erwachsenen
Zustande oft derart, daß der Gipfel den Boden berührt. Solche
Krümmungen sind aber nicht von langer Dauer, der Schaft streckt
sich vielmehr wieder gerade, um bald darauf nach einer anderen
Seite sich zu beugen.
Rückt die im Wachstum geförderte Seite in bestimmter Richtung
rings um den Stengel herum, so wird dieser eine gleichsinnige
kreisende Bewegung mit seinem Gipfel ausführen (kreisende oder
rotierende Nutation). Diese Bewegungsform tritt vornehmlich
ausgeprägt bei Ranken und Sprossen von Kletterpflanzen auf und
ermöglicht es ihnen, Stützen in ihrem Bereiche sicher aufzufinden.
In beiden Fällen wird neuerdings eine wesentliche Beteiligung von
Geotropismus und Autotropismus angenommen und die reine Autonomie
der Bewegung bezweifelt.
Neben diesen durch Wachstum bedingten Nutationen finden sich auch,
aber freilich ungleich seltener, autonome Variationsbewegungen
(S. 294); sie sind fast ganz auf Laubblätter beschränkt, und zwar auf
solche, die an der Basis des Stieles, oder auch an der Basis ihrer
weiteren Auszweigungen, Gelenkpolster besitzen. Sie finden sich vor
allem bei Leguminosen und Oxalideen, doch auch bei Marsilia, und
zeichnen sich durch einen Bau aus, der ihrer Funktion sehr zustatten
kommt.
In den gewöhnlichen Parenchymzellen wird die Zellhaut allmählich fast
ganz entspannt; deshalb kontrahieren sich ausgewachsene Zellen bei[S. 298]
Plasmolyse nicht in dem Maße wie wachsende Zellen (vgl. Fig. 237), und
sie werden umgekehrt bei einer Steigerung des Binnendruckes nur wenig
gedehnt, weil sie derbwandig sind. In einzelnen Fällen aber, und zu
diesen gehören gerade die Parenchymzellen der Gelenkpolster, bleibt
die Zellhaut auch im ausgewachsenen Zustande durch den Turgordruck
ganz beträchtlich gedehnt. Das sieht man nicht nur bei Ausführung der
Plasmolyse, sondern man merkt es schon an der bestehenden lebhaften
Gewebespannung.
Ein solches Gelenk einer Leguminose, z. B. der Bohne, zeigt
die Leitbündel und das Sklerenchym, die im Blattstiel (Fig.
278, 1) peripher angeordnet sind, zu einem zentralen, leicht
biegsamen Strange vereinigt, der von einer mächtigen Hülle von
Parenchym umgeben ist (Fig. 278, 2 u. 3). Wird aus dem durch
zwei Querschnitte isolierten Gelenk eine mittlere Gewebeschicht
herausgespalten (Fig. 278, 4), so zeigt sich schon an der
Vorwölbung des Rindenparenchyms, oben und unten, die starke
Spannung. Längsspaltungen, wie sie in Fig. 278, 5 angedeutet
sind, lassen das Expansionsbestreben des Parenchyms gegenüber dem
Leitbündel auf das deutlichste erkennen.
Es ist nun leicht einzusehen, daß eine allseitige Zunahme
des Turgordruckes die Spannung zwischen Leitbündel und Parenchym
steigert, somit die Festigkeit des Gelenkes erhöht. Dagegen wird
eine einseitige Zunahme des Druckes, oder eine Abnahme des
Turgordruckes auf der Gegenseite, oder endlich das Eintreten beider
Prozesse zugleich eine Verlängerung der einen, eine Verkürzung der
anderen Seite herbeiführen, wobei sich natürlich das Gelenk krümmt.
Das Leitbündel wird dabei ebenfalls gebogen, es erfährt indes keine
Veränderung in der Länge. Mit der Krümmung des Gelenkes ist aber eine
passive Bewegung des ansitzenden Blattteiles verbunden.
Fig. 278. 1 Querschnitt durch den Blattstiel der
Buschbohne. 2 Desgl. durch das Gelenkpolster. 3 Längsschnitt
durch das Gelenkpolster mit Übergang zum Blattstiel. 4 Mittlere
Lamelle, in Wasser liegend. 5 Dieselbe nach Abtrennung der Rinde vom
Leitbündel. 4 und 5 nach SACHS. Alles schwach vergrößert.
Autonome Variationsbewegungen fehlen wahrscheinlich keinem mit Gelenk
versehenen Blatte; auffällige Dimensionen nehmen sie aber nur bei
wenigen Pflanzen an.
So schwingen z. B. die kleinen Seitenblättchen von Desmodium gyrans
in lang gestreckten Ellipsen bald gleichmäßig, bald mehr ruckweise.
Bei relativ hoher Temperatur (30–35°) wird ihre Bewegung äußerst
lebhaft, ein Umgang kann in einer halben Minute vollendet werden.
Noch lebhafter schwingen die Blättchen von Oxalis hedysaroides,
ihre Spitzen können einen Weg von 0,5 bis 1,5 cm in einer oder in
wenigen Sekunden zurücklegen. Während die autonomen Bewegungen
beider Pflanzen vom Licht gar nicht beeinflußt erscheinen, werden
diejenigen von Trifolium pratense am Licht fast völlig unterdrückt;
im Dunkeln aber macht das Endblatt Schwingungen von oft über 120
Bogengraden, die sich in 2–4stündigem Rhythmus wiederholen.
2. Paratonische Bewegungen
(Reizbewegungen)[257].
Bei den induzierten oder paratonischen Bewegungen wirkt
stets ein äußerer Faktor als Reiz, der die Bewegung auslöst. Nur
durch solche[S. 299] Reizbewegungen bringen festgewachsene Organismen ihre
einzelnen Organe in diejenige Stellung, in der sie ihre Funktion am
besten ausüben können. Wenn die Organe einer Keimpflanze einfach in
der Richtung weiter wüchsen, die sie bei der Ausstreuung des Samens
zufällig einnehmen, müßten nicht selten die Wurzeln in die Luft, die
Stengel in die Erde gelangen.
Licht, Wärme, Schwerkraft, stoffliche und mechanische Einflüsse der
verschiedensten Art geben der Pflanze Mittel, sich in der Welt zu
orientieren. Dabei verhalten sich verschiedene Organe einer Pflanze oft
ganz verschieden gegen ein und dieselbe äußere Einwirkung. Die Stengel
z. B. wachsen zum Licht hin, die Wurzeln vom Licht weg; beide wachsen
in der Richtung der Strahlen weiter; die Blätter dagegen stellen
sich mit ihren Flächen ungefähr senkrecht zu den einfallenden
Strahlen. — Aber diese Reaktionsweise ist keine ein für allemal
gegebene, sondern sie kann sich weitgehend ändern. Man sagt dann wohl,
die „Stimmung“ der Pflanze habe sich geändert, und konstatiert,
daß solcher Stimmungswechsel teils durch innere Zustände, teils auch
durch Außenfaktoren bestimmt wird.
Als Richtungsbewegungen oder Tropismen werden wir
diejenigen Bewegungen zusammenfassen, die eine bestimmte Lage zu
der Richtung des wirkenden Reizes herbeiführen. — Die übrigen
Krümmungsbewegungen nennen wir Nastien; es sind durchweg
Bewegungen, die zu einer bestimmten Lage in Beziehung auf die
Pflanze, nicht in Beziehung auf die Richtung des Reizmittels führen.
a) Tropismen.
Bei den Richtungsbewegungen hat man orthotrope (parallelotrope)
und plagiotrope Organe zu unterscheiden. Erstere stellen sich
in die Richtung des Reizes, indem sie sich der Reizquelle nähern
(positive Reaktion) oder sich von ihr entfernen (negative
Reaktion). Die plagiotropen Organe stellen sich senkrecht oder schief
zur Richtung des Reizes. Die Reaktionsweise eines bestimmten Organs
kann durch äußere und innere Einflüsse eine Änderung erfahren.
Je nach dem wirksamen Reiz werden diese Richtungsbewegungen als
geotropische, phototropische usw. bezeichnet.
Die Tropismen der festgewachsenen Pflanzen entsprechen den Taxien
der freibeweglichen. Ihre Bedeutung liegt wie dort in dem Aufsuchen
günstiger Lebensbedingungen. Die wirksamen Reize, die positive bzw.
negative Reaktionsweise sowie der Wechsel zwischen beiden sind
vollkommen analog den bei den Taxien geschilderten Erscheinungen.
1. Geotropismus[258].
Es ist eine Erfahrungstatsache, daß die Stämme eines Tannenwaldes
exakt lotrecht und demnach untereinander parallel stehen; die Äste
und Blätter solcher Bäume aber nehmen andere Lagen ein. Betrachten
wir statt eines Baumes die Keimpflanze von Zea, so finden wir,
zunächst wenigstens, ausschließlich Organe, die sich in die Lotlinie
einstellen. Zugleich aber bemerken wir hier leichter als an einem
Baume das total verschiedene Verhalten der Wurzel und des Sprosses.
Beide stehen in der Lotlinie, aber der Sproß verlängert sich in ihr
aufwärts, die Wurzel abwärts. Bringen wir die Keimpflanze aus dieser
ihrer natürlichen Lage heraus, legen wir sie z. B. horizontal, so
sehen wir in beiden Organen eine Krümmung eintreten; die Wurzel krümmt
sich abwärts, der Keimsproß aber aufwärts. Da diese Krümmungen nicht
an der Stelle ausgeführt werden, wo Sproß und Wurzel zusammenstoßen,
vielmehr in der Nähe der Spitze beider Organe, so bleibt ein je nach
Umständen verschieden großes Stück der Achse horizontal gerichtet, und
nur die beiden Enden werden durch die Krümmung in die natürliche[S. 300] Lage
zurückgebracht, in der dann auch der weitere Zuwachs erfolgt. Daß die
Schwerkraft es ist, die diesen aufrechten Wuchs der Hauptachse
und der Hauptwurzel bedingt, das ergibt eigentlich schon die direkte
Beobachtung, die zeigt, daß diese Organe auf unserer ganzen Erdkugel in
der gleichen Weise orientiert sind, eben in der Richtung der Erdradien;
denn außer der Schwerkraft kennen wir keine überall gegebene,
in der Radiusrichtung wirkende Kraft. Doch nicht auf Grund dieses
Gedankenganges, sondern durch die Versuche von KNIGHT (1806)
hat sich in unserer Wissenschaft diese Erkenntnis Bahn gebrochen.
KNIGHTs Versuche beruhen auf folgender Überlegung: Offenbar
kann die Schwerkraft nur dann die Wurzel zum Abwärtswachsen, den
Stamm zum Aufwärtswachsen veranlassen, wenn der Same in Ruhe und in
derselben relativen Lage zur Richtung der Erdanziehung verbleibt;
deshalb vermutet KNIGHT, „daß eine solche Beeinflussung durch
stetigen und schnellen Wechsel der Lage des keimenden Samens aufgehoben
werden könne, und daß man ferner in der Lage wäre, durch das Mittel der
Zentrifugalkraft eine Gegenwirkung auszuüben“.
Er befestigte also am Rande eines Rades eine Anzahl von keimenden
Samen in möglichst verschiedenen Lagen, so daß die austretenden
Wurzeln nach außen, nach innen und zur Seite hervorwachsen mußten,
und ließ dieses Rad um eine horizontale Achse rotieren. Da er
aber dem Rade eine sehr ansehnliche Geschwindigkeit erteilte, so wurde
nicht nur die einseitige Wirkung der Schwerkraft aufgehoben, sondern
gleichzeitig eine recht beträchtliche Zentrifugalkraft erzeugt, die nun
ihrerseits die Keimlinge beeinflußte. Als Resultat ergab der Versuch,
daß sämtliche Wurzeln radial nach außen, sämtliche Sprosse radial
nach dem Zentrum des Rades wuchsen. Die Pflanze reagiert also auf die
Zentrifugalkraft gerade so wie auf die Schwerkraft.
Ein anderes Experiment, das ebenfalls KNIGHT schon ausgeführt
hat, läßt Schwerkraft und Zentrifugalkraft gleichzeitig, aber in
verschiedener Richtung auf die Keimlinge einwirken. Die Pflanzen
werden in Vertikalstellung auf einer horizontalen Scheibe
befestigt, die um eine vertikale Achse rotiert, und wenn nun
der Abstand der Pflanzen vom Zentrum und die Rotationsgeschwindigkeit
so gewählt wird, daß der mechanische Effekt von Schwerkraft und
Zentrifugalkraft gleich ist, dann wachsen die Wurzeln nach außen und
unter 45° nach unten, die Stengel nach innen und unter dem gleichen
Winkel nach oben; wird aber die Rotation gesteigert, so nehmen die
Keimachsen immer mehr eine der Horizontalen sich nähernde Lage an.
Daraus muß man schließen, daß die senkrecht nach unten wirkende Kraft
nicht nur in ihrer Richtung, sondern auch in ihrer Größe mit der
Schwerkraft übereinstimmt, d. h. daß sie eben wirklich die Schwerkraft
ist. Die Pflanze vermag also zwischen Schwerkraft und Zentrifugalkraft
keinen Unterschied zu machen. Beide Kräfte aber haben das miteinander
gemein, daß sie den Körpern eine Massenbeschleunigung erteilen. Für
die experimentelle Erforschung des Geotropismus ist diese Tatsache von
größter Bedeutung, da man nur die Zentrifugalkraft, nicht aber die
Schwerkraft in ihrer Intensität variieren kann.
Eine sehr wesentliche Ergänzung zu den KNIGHTschen
Fundamentalversuchen brachten dann erst sehr viel später (1874) die
Experimente von SACHS. Wie im ersten KNIGHTschen
Versuche wurden auch in den Versuchen von SACHS die Pflanzen
an der horizontalen Achse gedreht, aber die Geschwindigkeit der
Umdrehung wurde sehr gering gewählt, so daß eine Umdrehung in
10–20 Minuten erfolgte. Dabei konnten nennenswerte Zentrifugalkräfte
nicht entwickelt werden; da aber durch die fortwährende Drehung jede
einseitige Schwerewirkung eliminiert ist, so wachsen Wurzeln
und[S. 301] Sprosse in jeder beliebigen Richtung, die man ihnen zu Anfang
gegeben hat. SACHS benutzte zu solchen Versuchen ein Drehwerk,
das er Klinostat genannt hat.
Die Eigenschaft der Pflanze, unter dem Einfluß der Erdschwere eine
bestimmte Lage einzunehmen, bezeichnet man als Geotropismus. Es
hat sich gezeigt, daß es nicht nur orthotrope Organe gibt, die sich
in die Richtung der Schwerkraft einstellen und dabei positiv
geotropisch (nach unten) oder negativ geotropisch (nach
oben) wachsen, sondern auch plagiotrope, die eine horizontale oder
schräge Ruhelage besitzen. Die Stellungen der Seitenorgane sind also
gleichfalls — wenn auch gewöhnlich nicht ausschließlich — von der
Schwerkraft bedingt.
Negativ geotropisch sind alle gerade lotrecht nach oben
wachsenden Pflanzenteile, seien es Stengel, Stämme, Blätter (von
Liliifloren), Blütenschäfte, Blütenteile oder Wurzeln (wie die
senkrecht aus dem Schlamm oder der Erde aufsteigenden Atemwurzeln von
Sonneratien [Fig. 186, S. 143], Palmen u. a.). Werden derlei Organe aus
ihrer aufrechten Lage herausgebracht, dann richten sie sich, soweit
sie noch wachstumsfähig sind, wieder auf. Die Krümmung erfolgt
durch das gesteigerte Wachstum der erdwärts gerichteten Flanke
und das verminderte Wachstum der Gegenseite; die Folge ist eine
Aufrichtung des fortwachsenden Endes.
Fig. 279. Verlauf einer geotropischen Bewegung.
Die Figuren 1–16 bezeichnen aufeinanderfolgende Stadien der
geotropischen Krümmung einer im Halbdunkel erwachsenen Keimpflanze.
Diese bei 1 horizontal gelegt, bei 16 wieder völlig aufgerichtet.
Für die Zwischenstadien vgl. den Text. Schematisiert. Nach
NOLL.
Der tatsächliche Verlauf der geotropischen Krümmung eines Stengels wird
durch Fig. 279 dargestellt; sie zeigt, daß dieser Vorgang ein recht
komplizierter ist. Eine im Halbdunkel erwachsene Pflanze wird in Nr. 1
horizontal gelegt. Ihr Wachstum ist dicht hinter den Keimblättern am
lebhaftesten; deshalb tritt an dieser Stelle die erste geotropische
Krümmung auf (Nr. 2, 3). Allmählich greift dann die Krümmung immer
weiter basal um sich, geht also immer mehr auf die langsamer wachsenden
Teile über. An der Grenze der Wachstumszone macht sie dann halt.
Durch die Krümmung der Basalstücke werden (Nr. 7, 8) die Apikalstücke
über die Vertikallinie hinausgeführt, es tritt eine „Überkrümmung“
ein. Eine solche muß sich aber auch schon deshalb ergeben, weil jede
geotropische Reizung nicht mit dem Moment des Einrückens in die
Ruhelage aufhört, sondern noch lange nachwirkt. — Die Überkrümmung muß
aber aus einem doppelten Grunde wieder verschwinden (Nr. 13–16).
Einmal muß in dem übergekrümmten Teil eine neue, der bisherigen
entgegengesetzte geotropische Krümmung ausgelöst werden; außerdem aber
kombiniert sich mit dieser ein Bestreben, das man Autotropismus
nennt (S. 315).
[S. 302]
In einzelnen Fällen sind negativ geotropische Krümmungen auch an
„ausgewachsenen“[259] Sprossen möglich, d. h. an solchen, die
ohne einen geotropischen Reiz kein Längenwachstum mehr zeigen.
So wird an verholzten Zweigen und Stämmen, die aus der Ruhelage
gekommen sind, durch einen geotropischen Reiz das Kambium der
Unterseite zu einem Längenwachstum veranlaßt, das dann zu einer
Aufrichtung des Organs führt. Diese erfolgt um so langsamer und
unvollkommener, je größer der Widerstand des passiv zu krümmenden
Teils ist. Auch die sog. Knoten der Grashalme, die in Wirklichkeit
Blattpolster (Fig. 138) sind, werden durch geotropischen Reiz zu
neuem Wachstum angeregt. Erfolgt dieser Reiz allseitig, dreht
man also den Grasknoten um seine horizontal gelegte Längsachse
auf dem Klinostaten, so fangen alle Parenchymzellen gleichmäßig
an, sich zu verlängern; wird aber der Knoten einfach horizontal
gelegt, so beschränkt sich das Wachstum auf die Unterseite, während
die Oberseite passiv komprimiert wird (Fig. 280). Durch solche
Krümmungen in einem oder in mehreren Knoten werden die durch Wind
oder Regen gelagerten Grashalme wieder aufgerichtet.
Positiver Geotropismus wird vornehmlich bei Pfahlwurzeln,
vielen Luftwurzeln und den Keimsprossen mancher Liliaceen sowie den
Rhizomen von Yucca beobachtet. Diese Organe erreichen die senkrechte
Richtung nach abwärts aus jeder anderen Stellung und behalten sie
dauernd bei. Die positiv-geotropischen Bewegungen werden ebenso wie
die negativ-geotropischen durch aktives Wachstum ausgeführt.
Die Wurzel sinkt also nicht etwa dem Gewichte ihrer Spitze folgend
passiv in den Boden, sondern sie vermag einen das eigene Gewicht
weit übertreffenden Gegendruck zu überwinden, kann also z. B. in das
spezifisch viel schwerere Quecksilber eindringen. Bei der Krümmung wird
das Wachstum auf der Oberseite gefördert, auf der Unterseite gemindert,
während die Mittellinie mit unveränderter Geschwindigkeit weiter
wächst[260]. Fig. 281 stellt den Verlauf der geotropischen Krümmung an
der Wurzel dar.
Plagiogeotropisch sind viele Seitenzweige und Seitenwurzeln
erster Ordnung. Diese Organe sind in der Ruhelage, wenn
ihre Längsachse einen bestimmten Winkel mit der Lotrichtung
bildet. Sehr häufig wird übrigens die natürliche schiefe Stellung
von Pflanzenteilen nicht durch Geotropismus allein bewirkt.
— Ein besonderer Fall von Plagiogeotropismus liegt in der
horizontalen Ruhelage von Organen vor. Es sind besonders
Rhizome und Stolonen, die solchen Transversalgeotropismus
(Diageotropismus) zeigen und die aus jeder anderen Stellung
mit der fortwachsenden Spitze immer wieder in die wagerechte
Richtung zurückkehren, vorausgesetzt, daß sie sich in der
richtigen Tiefenlage befinden. Ist das nicht der Fall, so wird
zunächst durch aufwärts oder abwärts gerichtete Bewegungen diese
erstrebt, und dann erst folgt horizontales Wachstum. Seitenzweige
und Seitenwurzeln höherer Ordnung sind oft gar nicht geotropisch
und stehen nach allen Seiten vom Mutterorgan ab.
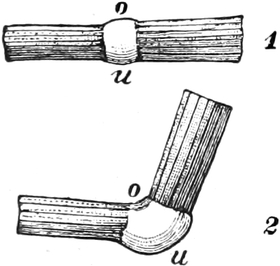
Fig. 280. Geotropische Aufrichtung eines
Grasblattpolsters. 1 Der vorher aufrechte Halm horizontal gelegt. 2
Die Unterseite u des Polsters stark verlängert, die Oberseite o
unverlängert (sogar etwas verkürzt). Die dadurch bedingte Krümmung hat
das jüngere Halmstück um etwa 75° emporgerichtet. Nach NOLL.
Fig. 281. Geotropische Krümmung einer Wurzel (Keimwurzel
von Vicia Faba). I Die vorher senkrecht abwärts gewachsene Wurzel
wagerecht gelegt und mit Tuschemarken versehen. II Dieselbe Wurzel
nach 7 Stunden. III Dieselbe Wurzel nach 23 Stunden, wieder senkrecht
abwärts gerichtet. Z Ein fester Index. Nach SACHS.
Eine besondere Art der geotropischen Orientierung tritt bei
dorsiventralen Organen (Laubblättern, zygomorphen Blüten,
S. 62) auf. Alle diese Organe bilden, ebenso[S. 303] wie die radiären
plagiotropen, einen bestimmten Winkel mit der Lotlinie, sind aber
nur dann in der Ruhelage, wenn auch die Dorsalseite nach oben,
die Ventralseite nach unten schaut, während es bei radiären
Organen nicht darauf ankommt, welche Flanke gerade oben liegt,
wenn nur die Organ-Achse die richtige Neigung hat. Bei der
Orientierung dorsiventraler Organe reichen dementsprechend einfache
Krümmungen häufig nicht aus, sondern es kommt zu Torsionen.
Die Drehung der Fruchtknoten vieler Orchideen, der Blüten von
Lobeliaceen, der Blattstiele an allen hängenden oder schräg
gestellten Zweigen, wie auch die Umdrehung der Blätter der
Alstroemerien und des Allium ursinum sind bekannte Beispiele für
regelmäßig auftretende Orientierungstorsionen.
Unter den dorsiventralen Organen verdienen die mit Gelenkpolstern
versehenen Laubblätter wieder besonders hervorgehoben zu werden,
weil sie auch im ausgewachsenen Zustande durch geotropische
Variationsbewegungen ihre Lage verändern können.
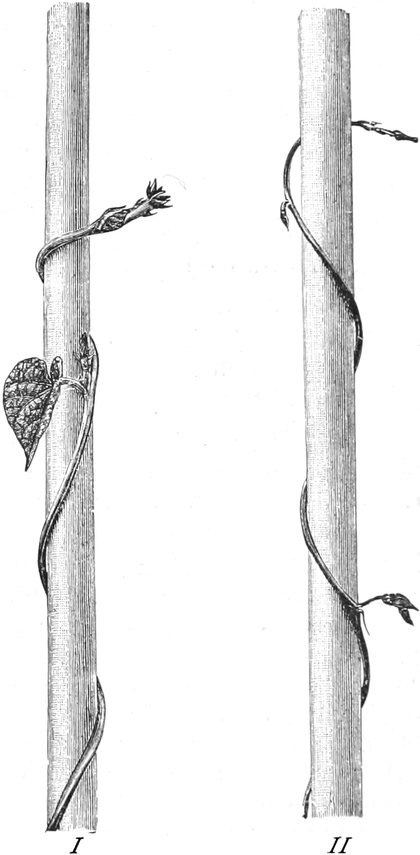
Fig. 282. I Linkswindender Sproß von Pharbitis.
II Rechtswindender Sproß von Myrsiphyllum asparagoides. Nach
NOLL.
Die Schlingpflanzen[261]. Die Schlingpflanzen, die in den
verschiedensten Pflanzenfamilien auftreten, besitzen Sprosse, die
sich nicht aus eigener Kraft zu halten vermögen, aber dennoch
aufwärts wachsen. Die Stengel und Stämme anderer Pflanzen,
die sich mit Aufwand großer Mengen von assimilierter Substanz (Holz,
Sklerenchym) zu aufrechtem Wuchs gefestigt haben, werden von den
Schlingpflanzen benutzt, um an ihnen die eigenen Assimilationsorgane
in freier Luft und in vollem Licht auszubreiten. Die Ausnutzung
fremder Assimilationsgerüste haben die Schlingpflanzen mit anderen
Kletterpflanzen, wie den Rankenpflanzen und Wurzelkletterern,
gemein. Sie erreichen ihr Ziel aber nicht durch die Ausbildung
seitlicher Haftorgane, sondern durch schlangenartiges Winden ihrer
Hauptachsen an den Stützen hinauf. Die ersten aus dem Samen oder aus
unterirdischen Reservestoffbehältern sich entwickelnden Stengelglieder
der Schlingpflanzen stehen in der Regel noch aufrecht. Bei weiterem
Wachstum krümmt sich das freie Ende aber aktiv seitwärts über und
nimmt eine mehr oder weniger schräge oder wagrechte Stellung an.
Zugleich beginnt der so geneigte Gipfel wie ein Uhrzeiger sich
im Kreise zu drehen, vgl. S. 297. Diese Bewegung dauert von dem
Moment ihres Entstehens an so lange, als der betreffende Sproß im
Wachstum verbleibt, und sie behält in der Regel eine bestimmte Richtung
dauernd bei; bei der Mehrzahl der Windepflanzen erfolgt die kreisende
Bewegung, von oben her gesehen, in der Richtung entgegengesetzt
der Uhrzeigerbewegung (nach links, wie man gewöhnlich zu sagen
pflegt); in der Richtung des Uhrzeigers, also nach rechts,
kreist z. B. der Hopfen und das Geißblatt; verschiedene Winderichtung
bei verschiedenen Individuen und selbst Wechsel der Richtung beim
Einzelindividuum hat man z. B. bei Polygonum Convolvulus und Loasa
lateritia beobachtet. Die linkskreisenden Pflanzen winden
auch links (Fig. 282 I),[S. 304] d. h. die „Wendeltreppe“, die sie bilden,
steigt (von außen gesehen) von links unten nach rechts oben, von oben
gesehen entgegen dem Uhrzeiger; die rechtskreisenden Pflanzen
winden auch rechts (Fig. 282 II). Es besteht also eine enge
Beziehung zwischen kreisender Bewegung und Winden.
Mit dem Beginn der kreisenden Bewegung ist noch nicht ohne weiteres
eine Windebewegung gegeben; diese beginnt erst dann, wenn wir dem Sproß
eine mehr oder minder lotrechte, nicht zu dicke Stütze bieten. Eine
solche wird dann in lockeren und anfangs sehr flachen Schraubenlinien
umwunden, die sich erst allmählich steiler aufrichten. Die Aufrichtung
erfolgt durch negativen Geotropismus und geht bei nachträglicher
Entfernung der Stütze — unter sonst geeigneten Umständen — in eine
völlige Geradestreckung der Schraubenwindung über, wobei der Stengel
dann gedreht erscheint; wird die Stütze nicht entfernt, so tritt nur
ein Engerwerden der Windungen und demnach ein Druck auf die Stütze
ein. Durch kreisende Bewegung und negativen Geotropismus kommt also
die Windebewegung zustande. Die Stütze spielt insofern eine Rolle, als
sie die sonst unvermeidliche Geradestreckung unmöglich macht. Sie muß
mehr oder minder lotrecht stehen, weil sie sonst von dem überhängenden
Gipfel gar nicht dauernd erfaßt werden kann.
Erleichtert wird das Winden noch dadurch, daß die Sprosse der
Windepflanzen zuerst die Internodien stark strecken und ihre
Blätter unentwickelt lassen. Sie ähneln hierin den etiolierten
Pflanzen, und sie erreichen durch die späte Entfaltung der Blätter
das regelmäßige Kreisen des Gipfels, das andernfalls durch
Anstoßen von Blättern an die Stütze unmöglich gemacht würde.
Der feste Halt an der Stütze wird vielfach durch Rauheit der
Stengeloberfläche, durch Haare, Borsten, Riefen noch erhöht. Auch
Torsionen, auf deren Ursache hier nicht eingegangen werden kann,
wirken oft im gleichen Sinne.
Änderung der geotropischen Ruhelage. Die Ruhelage, die
ein Organ nach einer bestimmten geotropischen Reizung einnimmt, ist
nicht ein für allemal gegeben; vielmehr ändert sie sich durch innere
und äußere Einflüsse. Man spricht von einer „Umstimmung“
der geotropischen Reizbarkeit. Eine gewisse „Stimmung“ der Pflanze
betrachtet man demnach als die normale, und die bei ihr erfolgenden
Reaktionen haben zur Einteilung in orthotrope und plagiotrope, in
positiv und negativ geotropische Organe geführt.
Von den äußeren Faktoren, die Einfluß auf die geotropische Stimmung
haben, nennen wir hier Licht und Temperatur, die Zentrifugalkraft, den
Sauerstoff; von inneren die Entwicklungsphase, in der sich ein Organ
befindet.
Die Veränderung der geotropischen Reaktion durch die Beleuchtung
hat eine große Bedeutung für die Tiefenlage der Rhizome. Wenn ein
Rhizom von Adoxa, etwa an einem Abhang wachsend, mit der Spitze ins
Licht gerät, geht sein bisheriger Transversalgeotropismus sofort in
positiven Geotropismus über, der das Rhizom wieder in den Erdboden
führt. Indessen genügt offenbar auch der Lichteinfluß auf den
oberirdischen Sproßteil oft schon, um ein unterirdisches
Rhizom zu dirigieren. Wird das Rhizom von Polygonatum zu hoch
im Boden eingepflanzt, doch immer noch so, daß es ganz von Erde
bedeckt und verdunkelt ist, so wendet sich der Neuzuwachs schräg
nach unten; wird es aber zu tief gesetzt, so wendet er sich
nach oben (Fig. 283); bei richtiger Tiefenlage verhält er sich
dagegen transversal-geotropisch. — Auch auf den Geotropismus der
Seitenwurzeln wirkt das Licht sehr stark ein: bei Beleuchtung
nähern sich die Seitenwurzeln erster Ordnung viel mehr der
orthotropen Ruhelage als im Dunkeln.
Eine Wirkung der Temperatur läßt sich an den Stengeln mancher
Frühjahrspflanzen beobachten. Bei Temperaturen in der Nähe von 0°
legen sich diese vielfach dem Boden an, um sich erst bei höherer
Temperatur orthotrop aufzurichten. — Durch Sauerstoffmangel werden
manche Wurzeln und Rhizome negativ geotropisch und gelangen so in
Regionen, wo ihnen mehr Sauerstoff zur Verfügung steht.
Umstimmungen durch innere Ursachen sehen wir z. B an Rhizomen,
die in einem gewissen Entwicklungsstadium ihre diageotrope Lage
aufgeben und orthotrop werden, oder an Blütenstielen, die nach
der Befruchtung positiv geotropisch werden[262]. So werden die
Früchte von Trifolium subterraneum und von Arachis hypogaea in die
Erde ein[S. 305]gegraben. — Auch am windenden Stengel haben wir eine
Umstimmung kennen gelernt; er windet in der Jugend noch nicht.
Geotropismus als Reizerscheinung. Der Entdecker des
Geotropismus, KNIGHT, suchte die geotropischen Bewegungen in
rein mechanischer Weise zu erklären, was insbesondere für die positiv
geotropischen Organe nicht schwierig erschien. Er stellte sich vor, daß
diese einfach dem Zug der Schwere folgend in ihre Ruhelage gelangen.
Später hat noch HOFMEISTER ähnliche Ansichten vertreten. Die
richtige Auffassung, daß wir es mit komplizierten Reizbewegungen zu
tun haben, bei denen die Erdschwere nur die Rolle des auslösenden
Faktors spielt, verdankt man vor allem DUTROCHET,
FRANK und SACHS. Schon die einzige Tatsache, daß die
Wurzel auch gegen den Widerstand von Quecksilber eine geotropische
Krümmung auszuführen vermag, hätte genügen müssen, um jede rein
mechanische Auffassung zu widerlegen.
Fig. 283. Rhizome von Polygonatum. Nach
RAUNKIAER. Die gestrichelte Linie stellt die Erdoberfläche
vor. Die Blütenstengel sind abgeschnitten. Rhizom 1 war zu hoch
eingepflanzt; seine Verlängerung geht abwärts (nur die
Endknospe, aus der sich wieder ein Blütenstengel bildet, ist aufwärts
gerichtet). Rhizom 2 war aufrecht und zu tief eingepflanzt; sein
Zuwachs geht schräg nach oben.
Erst in neuerer Zeit hat man den Versuch gemacht, zu ergründen,
welches die primäre Wirkung der Schwerkraft in der Pflanze
ist[263]. Es kann keinem Zweifel unterliegen, daß es sich da um eine
Druckwirkung handeln muß. Die Ersetzbarkeit der Schwerkraft
durch Zentrifugalkraft spricht schon dafür. — Weiter aber zeigt
sich, daß dieser Druck bei orthotropen Organen nur so weit zur
Geltung kommt, als er senkrecht zur Längsachse steht; schief
angreifende Schwerkraft wirkt also nur im Verhältnis ihrer senkrechten
Komponente. („Sinusgesetz“, weil die Größe dieser Komponente
durch den Sinus des Einfallwinkels bestimmt wird.) Zwei unter einem
Winkel angreifende Kräfte (Schwerkraft und Zentrifugalkraft) treten
nach dem Parallelogramm der Kräfte zu einer Resultante zusammen
(Resultantengesetz). — Endlich wissen wir, daß der Druck
durchaus im Innern der Zellen wirken muß und in keiner Weise
ersetzbar ist durch von außen kommende Wirkungen.
F. NOLL hat zuerst die Vorstellung entwickelt, daß
irgendwelche Zellteile, die spezifisch schwerer sein müssen als das
sie umgehende sensible Plasma, unter dem Einfluß der Schwerkraft
einen einseitigen Druck auf dieses ausüben, worauf dann das
Plasma die Wachstumsvorgänge im Sinne der Schwererichtung lenkt.
NĚMEC und HABERLANDT haben dann die Vermutung
ausgesprochen, daß solche spezifisch schwereren Körperchen
(„Statolithen“) in gewissen Stärkekörnern zu suchen seien,
die relativ rasche Fallbewegungen in der Zelle auszuführen vermögen
und sich deshalb immer der nach unten[S. 306] schauenden Zellwand anlegen.
Sie finden diese Stärkekörner bei den Stengeln in der Stärkescheide
(S. 81), bei den Wurzeln in den Zellen der Haube. Sie nehmen an,
daß nur in den mit solchen Stärkekörnern versehenen Teilen ein
Schwerereiz direkt von Wirkung sei, daß er aber von diesen Punkten
aus weiter geleitet werde. In der Tat hat man nachgewiesen, daß in
der Wurzel vorzugsweise die Spitze den Schwerereiz aufnimmt. Nach
Versuchen von STAHL und ZOLLIKOFER gelingt es
in manchen Fällen, die Statolithenstärke zu lösen und damit auch
den Geotropismus zum Verschwinden zu bringen, während Wachstum
und phototropische Reaktionsbefähigung fortdauern. Damit ist
gezeigt, daß die Stärkekörner hier für die Reizaufnahme nötig
sind; aber freilich die leichte Beweglichkeit dieser
Körner, die die Hypothese so eindrucksvoll gemacht hat, ist nach
gewissen physiologischen Erfahrungen für die Reizperzeption ganz
gleichgültig. Auch ist in anderen Fällen (Moosrhizoiden) nach
Verschwinden der Stärke noch Geoperzeption möglich. Da müssen also,
wie ja auch bei Pilzen, andere Statolithen tätig sein.
Im allgemeinen können wir auf geotropische Reizbarkeit eines Organs
nur aus der eintretenden Krümmung schließen. In einzelnen Fällen aber
läßt sich auch ohne solche Reaktion eine geotropische Reizbarkeit
erkennen. So ist z. B. bei manchen Graskeimlingen, die ein wohl
ausgebildetes Internodium unterhalb des Scheidenblattes besitzen
(Paniceen), das Scheidenblatt in einem gewissen Moment
ausgewachsen und deshalb nicht mehr geotropisch krümmungsfähig;
daß es aber noch geotropisch reizbar ist, erkennt man daraus,
daß nach einseitiger Einwirkung der Schwerkraft auf diese Scheide das
Internodium sich krümmt, obwohl es selbst nicht geotropisch
reizbar ist. Der geotropische Reiz muß ihm also von der Scheide her
zugeleitet worden sein. Bei anderen Graskeimlingen (Poaeoideen)
hat man bemerkt, daß die Spitze des Scheidenblattes viel stärker
geotropisch reizbar ist als die Zone maximalen Wachstums, und eine
ähnliche Abnahme der geotropischen Sensibilität von der Spitze nach
der Basis zu liegt auch bei Wurzeln vor. Man kann nun mit Hilfe eines
besonderen Apparats bei solchen Objekten Spitze und Wachstumszone durch
Fliehkräfte entgegengesetzt geotropisch reizen und bemerkt dann, daß
die Wachstumszone sich im Sinne der gereizten Spitze krümmt. Es
findet also eine Reizleitung von der Spitze basalwärts statt,
und der geleitete Reiz überwindet den in der Wachstumszone direkt
induzierten vollkommen. — Es lassen sich also in solchen Fällen
deutlich drei Prozesse trennen: Reizaufnahme (Perzeption), Reizleitung
und Reizreaktion. Ein Organ kann perzipieren, ohne selbst zu reagieren,
und umgekehrt kann auch ein Organ, das selbst nicht perzipiert,
geotropisch reagieren. Wir haben allen Grund anzunehmen, daß diese drei
Teile des Reizprozesses auch da unterschieden werden müssen, wo sie
nicht so scharf sich trennen lassen.
Wir entnehmen schon diesen Ausführungen, daß der Grad der geotropischen
Krümmung und die Geschwindigkeit, mit der sie eintritt, keineswegs
ein Maßstab für die Größe der Reizung ist, da sie weitgehend von der
Wachstumsbefähigung abhängen. Die Größe der geotropischen Reizung hängt
zunächst einmal von der spezifischen Empfindlichkeit der gereizten
Organe, außerdem aber auch von der „Reizmenge“ ab, die es getroffen
hat. Die Größe der Reizung eines gegebenen Organs ist direkt der
„Reizmenge“ proportional. Unter Reizmenge aber versteht man das Produkt
aus der Intensität des Reizes und der Dauer seiner Einwirkung. Es ist
also für den Erfolg gleichgültig, ob wir eine hohe Fliehkraft für kurze
Zeit oder eine geringe entsprechend längere Zeit einwirken lassen.
Diese Gesetzmäßigkeit, das „Reizmengengesetz“[264], das
enge Beziehungen zu den früher erwähnten Gesetzmäßigkeiten, dem
Sinusgesetz und dem Resultantengesetz aufweist, gilt
freilich nur innerhalb gewisser Grenzen. Es hat sich gezeigt,
daß ein orthotropes Organ, horizontal gelegt,[S. 307] unter konstanten
Außenbedingungen nach einer ganz bestimmten Zeit anfängt, sich zu
krümmen. Die Zeit, die vom Beginn der Reizung bis zum Beginn der
Reaktion verstreicht, nennt man die Reaktionszeit. Es ist aber
zur Erzielung einer geotropischen Reaktion nicht nötig, ein Organ
während der ganzen Reaktionszeit zu reizen. Es genügt vielmehr eine
sehr viel kürzere Zeit, um an dem weiterhin senkrecht gestellten Organ
durch Nachwirkung eine geotropische Krümmung zu erhalten. Die minimale
Reizzeit, nach der noch eine gerade sichtbare Krümmung erfolgt, nennt
man „Präsentationszeit“. Nur für Reize, die solange oder etwas
länger währen als die Präsentationszeit, gilt zunächst einmal das
Reizmengengesetz; die Präsentationszeit ist also umgekehrt proportional
der Reizintensität. Größere Reizmengen haben keine entsprechende
Steigerung der geotropischen Krümmung zur Folge.
Wie andere Eigenschaften der Pflanze ist auch die Reaktionszeit und
die Präsentationszeit weitgehenden Schwankungen unterworfen, so
daß man bei statistischer Untersuchung typische Variationskurven
erhält. TRÖNDLE fand als mittlere Reaktionszeit für
Hafersprosse 32 Min., für Kressewurzeln 21 Min. Einzelne Haferkeime
reagierten schon nach weniger als 14 Min., andere erst nach mehr
als 49 Min. Bei den meisten Pflanzen sind die Reaktionszeiten aber
größer. Die Präsentationszeiten sind häufig zu 2, 3 bis 10 und mehr
Min. gefunden worden.
Reize unter Präsentationsdauer bleiben indes nicht wirkungslos. Bei
Wiederholung summieren sie sich und führen schließlich zu einer
Krümmung, wenn die Summe der Einzelreize die Präsentationszeit
erreicht, und wenn die Pausen zwischen den Einzelreizen nicht zu groß
waren. Eine untere Grenze für die Dauer des Einzelreizes konnte bis
jetzt nicht gefunden werden.
2. Phototropismus [Heliotropismus][265].
Phototropische Krümmungen kommen bei einseitigem Lichteinfall
zustande. Man beobachtet sie leicht bei Pflanzen, die am Waldrande
wachsen oder die im Zimmer gehalten werden. Die Stengel der im Zimmer
aufgestellten Pflanzen wachsen nicht wie im Freien bei allseitiger
Beleuchtung aufrecht, sondern sind dem nächsten Fenster zugeneigt;
sie sind orthotrop und positiv phototropisch. Im
Gegensatz zu diesen Teilen findet man ihre Blattflächen senkrecht
zu den einfallenden Lichtstrahlen gestellt, um möglichst viel Licht
aufzufangen; die Blattflächen sind transversal phototropisch
(lichtfangend). Weniger häufig hat man Gelegenheit,
negativen Phototropismus, also ein Wegwachsen von der
Lichtquelle, zu sehen. — In Fig. 284 sind die verschiedenartigen
phototropischen Krümmungen, die an einer Wasserkultur des weißen Senfes
bei einseitiger Beleuchtung eintreten, dargestellt.
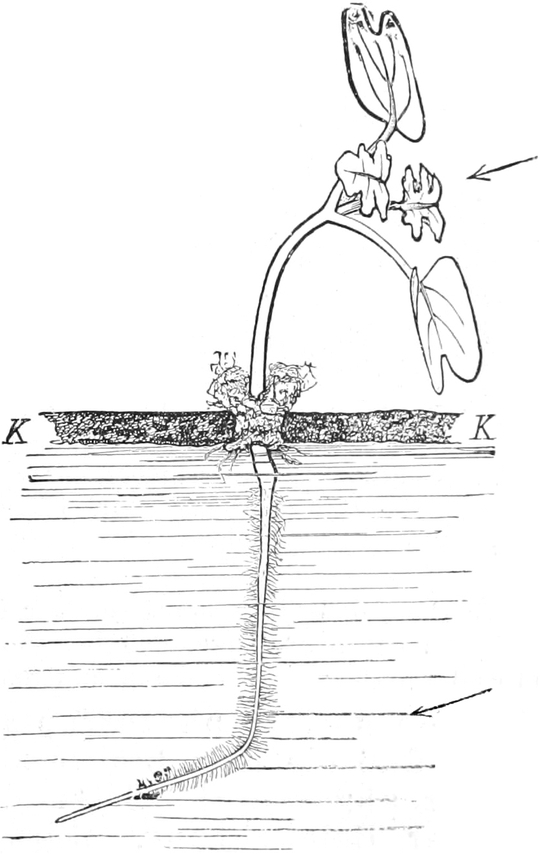
Fig. 284. Keimling des weißen Senfs in Wasserkultur,
ursprünglich allseitig, dann einseitig beleuchtet. Stengel dem Lichte
zugekehrt, die Wurzel abgewendet, die Blattflächen senkrecht zum Lichte
ausgebreitet. K K Korkplatte (Schwimmer). Nach NOLL.
[S. 308]
Phototropismus ist im Pflanzenreich weit verbreitet. Am häufigsten
kommt der positive Phototropismus zur Beobachtung, er
bildet die Regel bei den oberirdischen Vegetationsachsen. Viel
seltener zeigt sich der negative Phototropismus, z. B.
bei Luftwurzeln, zumal Kletterwurzeln (Ficus stipulata, Begonia
scandens u. a.), beim hypokotylen Stammglied der keimenden
Mistel, bei manchen, aber nicht allen Erdwurzeln (Sinapis,
Helianthus), Ranken (zumal den mit Haftscheiben versehenen), am
Stengel einzelner Kletterpflanzen. Haftwurzeln und Haftranken von
Kletterpflanzen haben, wie auch die Keimwurzel der Mistel, von
ihrem negativen Phototropismus den Vorteil, der dunklen Unterlage
zugeführt zu werden. Wie später (S. 310) zu zeigen sein wird, kann
die phototropische Reaktionsweise abgeändert werden, also z. B. ein
sonst positiv reagierendes Organ zu negativen Krümmungen veranlaßt
werden.
Zur genaueren Beobachtung der phototropischen Erscheinungen ist es
notwendig, das einseitig einfallende zerstreute Tageslicht durch eine
enger begrenzte Lichtquelle zu ersetzen. Dann zeigt sich vor allem,
daß, wie bei den topophototaktischen Bewegungen, die Richtung
des einfallenden Lichtes maßgebend ist für die phototropische Ruhelage.
Jede Änderung der Strahlenrichtung hat auch eine Stellungsänderung
der phototropischen Organe zur Folge. Das Gipfelende mancher positiv
phototropischen Organe findet man völlig in der Richtung der Strahlen
eingestellt.
Mit welcher Genauigkeit dies bei einzelnen Pflanzen geschieht,
zeigt ein Versuch mit dem kleinen Pilze Pilobolus crystallinus.
Seine Sporangienträger kommen aus feucht gehaltenem Pferde- und
Kuhmist nach kurzer Zeit zahlreich hervor; sie sind positiv
phototropisch und richten alle das schwarze Sporangium der
Lichtquelle zu. Zur Zeit der Reife wird das Sporenköpfchen mit
großer Gewalt geradeaus fortgeschleudert. Hat man nun das Licht
nur durch ein kleines verglastes Rundfenster seitlich in die
Versuchskammer einfallen lassen, so findet man die klebrigen
Sporangien alle dicht um das Zentrum der kleinen Lichtscheibe
angeschossen, ein Zeichen, daß die Sporangienträger genau dorthin
gerichtet waren (Fig. 285).
Es ist anzunehmen, daß bei gleichzeitigem Einfall zweier oder
mehrerer Strahlenbüschel verschiedener Richtung und Stärke das
Resultantengesetz gilt (vgl. S. 305).
Fig. 285. Pilobolus crystallinus (P), seine Sporangien
nach der Lichtscheibe abschießend. G Glasscheibe. B Blechschieber
mit rundem Fenster F. M Kulturkasten mit Pferdemist gefüllt. Vgl.
den Text. Nach NOLL.
Die positiv phototropischen Krümmungen kommen dadurch
zustande, daß die dem Lichte zugewandte Seite langsamer, die vom
Lichte abgewandte Seite dagegen stärker wächst als bei allseitiger
Beleuchtung. Beim negativen Phototropismus besteht die umgekehrte
Wachstumsverteilung. Im allgemeinen treten Krümmungen nur in der
Strecke auf, die noch im Wachsen begriffen ist, und der Ort des
lebhaftesten Wachstums pflegt zugleich derjenige der schärfsten
Krümmungen zu sein.
Der Verlauf der phototropischen Krümmung entspricht vollkommen
dem der geotropischen (S. 301). — Man hat früher versucht, das
geförderte Wachstum der Schattenseite bei positiver phototropischer
Reaktion durch beginnendes Etiolement, das gehemmte der Lichtseite
durch die verzögernde Wirkung zu erklären, die das Licht auch auf
das geradlinige Wachstum der Stengel ausüben sollte (S. 251). Diese
Vorstellung, die lange Zeit verlassen war, ist neuerdings vor allem
durch BLAAUW[266] in modifizierter Form wieder aufgenommen
worden. Nach ihm sollen die Veränderungen des Längenwachstums,[S. 309] die
man nach einer Änderung der Beleuchtungsstärke beobachtet, wenn
sie an verschiedenen Flanken eines orthotropen Organs ungleich
sind, direkt zu einer phototropischen Krümmung führen. Diese
Veränderungen sind nicht bei allen Pflanzen gleich (vgl. S. 252)
und bestehen bald in einer primären Wachstumsförderung,
bald in einer Hemmung. So sucht BLAAUW, indem er
ferner auch die verschiedene Durchsichtigkeit der Organe und die
Lichtbrechung im Innern berücksichtigt, die bald positive, bald
negative phototropische Reaktion oder auch das Ausbleiben jeglichen
Phototropismus verständlich zu machen. Es muß anerkannt werden, daß
in neuerer Zeit immer mehr ein weitgehender Parallelismus zwischen
der Lichtwachstumsreaktion und dem Phototropismus festgestellt
werden konnte. Daneben muß aber auch heute noch betont werden,
daß manche Lücken in der Beweisführung der BLAAUWschen
Theorie bestehen und daß diese bei der Übertragung auf andere
Reizerscheinungen (Geotropismus und noch mehr Haptotropismus)
großen Schwierigkeiten begegnet.
In einzelnen Fällen hat man[267] auch an Organen, deren
Längenwachstum abgeschlossen ist, noch phototropische Krümmungen
festgestellt. Dies trifft einmal für mehrjährige Bäume zu, bei
denen wahrscheinlich das Kambium eine große Bedeutung für die
beobachteten Krümmungen hat, andererseits finden sie sich auch an
Knoten, wie z. B. denen der Gramineen und Commelinaceen, wo durch
Licht allein oder nur bei gleichzeitiger Schwereeinwirkung das
Längenwachstum wieder aufgenommen wird.
Hat sich ein Organ durch eine phototropische Krümmung in die Richtung
des Lichtes eingestellt, so ist es auch allseitig gleich stark
beleuchtet und befindet sich in seiner phototropischen Ruhelage.
Sorgt man, ohne den Lichteinfall oder die Lichtintensität zu ändern,
dafür, daß die Pflanze durch ein Uhrwerk (Klinostat) in dauernde
Rotation um ihre vertikale Längsachse versetzt wird, so heben sich die
auf verschiedene Seiten orthotroper Organe nacheinander einwirkenden
phototropischen Reize auf; es bleiben also die phototropischen
Krümmungen aus.
SACHS hat seiner Zeit die Vorstellung entwickelt, daß beim
Phototropismus — ähnlich wie beim Geotropismus die Richtung der
Schwerkraft — die Lichtrichtung maßgebend sei. Eine große
Anzahl von Tatsachen spricht heute dafür, daß die phototropische
Reizung vielmehr durch ungleiche Helligkeit an verschiedenen
Stellen des phototropisch-empfindlichen Organs zustande kommt[268].
Wenn auch in einzelligen Schläuchen von einer bestimmten
Lichtrichtung gesprochen werden kann, so dürfte das doch für
vielzellige Gewebe, mit ungleich brechendem Zellinhalt und
reichlichen lichtführenden Interzellularen bestimmt nicht möglich
sein. Es lassen sich aber auch experimentell Bedingungen schaffen,
wo zweifellos die phototropische Krümmung nicht in der Richtung der
Strahlen verläuft. So z. B. wenn man Avena-Koleoptilen halbseitig
von oben beleuchtet: da tritt die Krümmung senkrecht zu der
Strahlenrichtung nach der beleuchteten Seite zu auf: oder wenn man
Avena-Koleoptilen von innen her beleuchtet: die Krümmung erfolgt
genau in entgegengesetzter Richtung wie die Strahlen; es ist also
gleichgültig, ob die helle Hälfte der Koleoptile das Licht von
außen oder von innen erhält.
Phototropische Krümmungen können im Licht aller
Wellenlängen, die das sichtbare Spektrum bilden, ausgeführt
werden. Bei gleicher Energie wirken aber die blauen und violetten
Strahlen ungleich stärker phototropisch, wie sie ja auch
phototaktisch sich wirksamer erweisen.
Der Transversalphototropismus findet sich bei Blättern
und blattartigen Assimilationsorganen (wie Farnprothallien,
dem Thallus von Lebermoosen, Algen) also bei dorsiventralen Gebilden.
Bei diesen Organen überwiegt der transversale Phototropismus alle
anderen Bewegungsreaktionen. Solche Organe stellen sich also im
allgemeinen senkrecht zu dem hellsten diffusen Licht, das sie
während ihrer Entwicklung trifft; bei dieser Einstellung führen
einfache Krümmungen meist nicht mehr zum Ziel, es kommt zu
Torsionen der Blätter oder der Internodien.
[S. 310]
In sehr hellem, direktem Licht kann die transversale Stellung
der Blattflächen durch andere, den intensiven Lichtstrahlen
ausweichende Lagen ersetzt werden. Es hat also das Laubblatt
ähnlich wie die Chlorophyllplatte von Mesocarpus die Fähigkeit,
eine Profil- oder eine Flächenstellung einzunehmen und dadurch
die Menge des einfallenden Lichtes zu regulieren. Die
Blätter unserer Lactuca Scariola und des nordamerikanischen
Silphium laciniatum, die blattartigen Sprosse mancher Kakteen,
stellen sich auf sehr sonnigem Standort in die Meridianebene ein
(„Kompaßpflanzen“), so daß nur die Morgen- und Abendsonne ihre
Fläche trifft, während die heißen Strahlen der Mittagssonne sie nur
streifen.
Eine Reihe von Laubblättern besitzt an der Basis des Stieles bzw.
auch am Grunde sekundärer und tertiärer Ausgliederungen Blattpolster
(Fig. 134, 278), die Variationsbewegungen auszuführen vermögen.
Dadurch sind diese Blätter in den Stand gesetzt, zeitlebens ihre
Lage zu ändern und in jedem Moment die Stellung einzunehmen, die
ihnen den optimalen Lichtgenuß verschafft. Sie erreichen also nicht
eine „fixe“ Lichtlage, die durch das stärkste während ihrer
Entwicklung einfallende diffuse Licht bedingt ist, sondern sie nehmen
bald Flächen-, bald Profilstellung zu dem gerade herrschenden Licht ein.
Stimmungswechsel[269]. Ein bestimmter Pflanzenteil reagiert
nicht immer in gleicher Weise auf ein und denselben Reiz; vielmehr kann
die Reaktionsweise durch das Alter oder durch äußere Einflüsse geändert
werden. Man spricht in diesem Sinn auch hier von einer „Stimmung“ und
einer „Umstimmung“ der Pflanze.
So sind die Blütenstiele der Linaria Cymbalaria zunächst
positiv phototropisch; nach der Bestäubung werden sie aber bei
gleichzeitiger starker Verlängerung (S. 244) negativ phototropisch
und führen die jungen Samenkapseln zur Aussaat in Mauer- und
Gesteinsspalten ein.
Von umstimmenden Außenfaktoren ist vor allen Dingen
die Lichtmenge selbst zu nennen. Kleine einseitig auf
Avena einfallende Lichtmengen führen ausschließlich zu positiv
phototropischer Krümmung; größere Mengen bewirken eine schwächere
positive Krümmung, der bald eine negative folgt; noch größere
Mengen bedingen eine rein negative Reaktion. Bei weiterem
Anwachsen der Reizmenge bemerkt man zunächst wieder positive und
späterhin mindestens abgeschwächt positive, wenn nicht negative
Reaktion. Inwieweit auch die Intensität der Beleuchtung die
Ergebnisse beeinflußt, kann hier nicht besprochen werden.
Der Phototropismus ist gerade wie der Geotropismus eine
Reizerscheinung[270]. Auch bei ihm lassen sich Reizaufnahme,
Reizleitung und Reizreaktion unterscheiden. Auch bei ihm gibt es
eine Präsentationszeit und eine Reaktionszeit. Ferner ist die
Gültigkeit des Reizmengengesetzes sowohl bei der positiven wie bei
der negativen Reaktion von Avena nachgewiesen, während es bei der
nach noch stärkerer Beleuchtung auftretenden abermaligen positiven
Reaktion nicht zutrifft. Weiter gilt ein Gesetz, das freilich nur
einen Spezialfall des Reizmengengesetzes bildet, das aber deshalb
von großer Bedeutung ist, weil es zuerst für die Lichtempfindung im
menschlichen Auge nachgewiesen wurde. Dieses sog. TALBOTsche
Gesetz[271] sagt aus, daß unterschwellige einseitige Lichtreize
sich summieren können und daß die Summe der in den Einzelreizen der
Pflanze zugeführten Lichtmengen genau den gleichen Effekt
hat, wie wenn sie auf einmal, ohne Unterbrechungen gewirkt
hätte. Vorausgesetzt ist dabei nur, daß die Einzelreize von nicht zu
langen Verdunkelungen getrennt sind, da sonst die Wirkung des ersten
abgeklungen ist, wenn der zweite einsetzt.
Lokalisation der phototropischen Reizung. Sehr häufig erfolgt
die Aufnahme des Lichtreizes an derselben Stelle, an der auch die
Bewegung ausgeführt wird. Bei gewissen Laubblättern aber ist die
Lamina imstande, einen phototropischen Reiz zu perzipieren, ohne
daß sie selbst auch die entsprechende Bewegung auszuführen vermag;
diese erfolgt vielmehr erst, wenn der Reiz[S. 311] weiter geleitet
ist, im Blattstiel. Der Blattstiel kann freilich auch auf direkte
Reizung reagieren. Noch auffallendere Verhältnisse treffen wir bei
den Keimlingen gewisser Gramineen an; bei manchen Paniceen ist
nur die Spitze des sog. Kotyledons phototropisch reizbar, und nur
das abwärts auf ihn folgende hypokotyle Glied ist phototropisch
krümmungsfähig. Hier haben wir also eine ausgesprochene Trennung in
Perzeptionsorgan und Bewegungsorgan; die Ähnlichkeit mit entsprechenden
Vorkommnissen beim Geotropismus ist somit sehr auffallend. Aber auch
an die Verhältnisse im Tierreich erinnert diese Differenzierung in
Perzeptions- und Bewegungsorgan. Ein wesentlicher Unterschied liegt
aber in der Art der Reiztransmission[272]. Denn es fehlen in
der Pflanze spezifisch reizleitende Zellen („Nerven“) völlig, und
der Reiz wird von Zelle zu Zelle weitergegeben, ja er kann sogar
nach Unterbrechung des organischen Zusammenhanges durch seitliche
Einschnitte auch dann weitergeleitet werden, wenn in den Einschnitt ein
Blättchen von Gelatine gelegt wird; dagegen hört die Leitung auf, wenn
in gleicher Weise ein Stanniolblättchen angebracht wurde. Schneidet
man bei Gramineenkeimlingen die Spitze ab und setzt sie wieder dem
Stumpf auf, so kann ein Reiz von der Spitze aus über die Wundstelle weg
zur Basis geleitet werden; der Versuch gelingt selbst dann, wenn man
die Spitze einer anderen Grasart dem Stumpf aufgelegt hat. Somit muß
wohl ein diffundierender Stoff bei der Reizleitung eine Rolle
spielen, und es liegt nahe anzunehmen, daß derselbe Stoff die Leitung
besorgt, der bei der phototropischen Reizung entsteht. Mit anderen
Worten, man muß sich vorstellen, daß die Pflanze über Stoffe verfügt,
die unter dem Einfluß des Lichtes entstehen oder vergehen können und
deren ungleiche Verteilung Phototropismus bewirkt. Warum deren Leitung
aber nur in der Längsrichtung der Organe und nur basipetal erfolgt, ist
noch ganz ungeklärt.
3. Chemotropismus[273].
Chemotropisch nennt man Richtungsbewegungen, die durch ungleichmäßige
Verteilung von gelösten oder gasförmigen Stoffen in der Umgebung
der Pflanze veranlaßt werden. Bei Pilzen und bei Pollenschläuchen
sind solche Bewegungen konstatiert, die den Organismus in eine
bestimmte Konzentration des betreffenden Stoffes, wohl die optimale,
hineinführen. Dementsprechend fallen diese Bewegungen bei dem gleichen
Organismus und beim gleichen Reizstoff bald positiv, bald negativ
aus; positiv, wenn es gilt, eine höhere Konzentration zu gewinnen;
negativ im entgegengesetzten Falle. — Als Reizstoffe kommen für die
Pollenschläuche Zucker und Proteïne in Betracht; bei Pilzen außer
diesen noch Pepton, Asparagin, Ammoniumverbindungen und Phosphate. Es
fehlt auch nicht an Stoffen, die schon in sehr schwacher Konzentration
stets nur abstoßende Wirkung ausüben, so z. B. freie Säuren und bei
Pilzen Stoffwechselprodukte unbekannter Natur, die aus den Zellen
heraus diffundieren. Auch bei Wurzeln ist eine chemotropische
Reizbarkeit nachgewiesen, ohne daß man sagen könnte, diese spiele eine
große Rolle in ihrem Leben.
Bei den bisher angeführten Beispielen von Chemotropismus waren die
Reizstoffe feste Körper, die in Wasser gelöst wurden. Wenn dagegen
die Pflanze durch ungleich im Raume verteilten Wasserdampf
oder Gase zu einer Richtungsbewegung veranlaßt wird, so hat
man dieser einen besonderen Namen zu geben für nötig befunden,
obwohl im Prinzip kein Unterschied gegenüber den gelösten
Substanzen besteht. Reizbewegungen durch Feuchtigkeitsdifferenzen
nennt man hydrotropische, durch Gasdifferenzen bedingte
aërotropische. Aërotropismus ist bei Pollenschläuchen,
Wurzeln und Sprossen nachgewiesen, Hydrotropismus bei Wurzeln und
Schimmelpilzen;[S. 312] positiv hydrotropisch sind z. B. die Wurzeln, die
mit Hilfe dieser Reaktionsweise die feuchten Stellen in der Erde
auffinden, negativ hydrotropisch verhalten sich z. B. Sporangienträger
der Mucorineen, die aus dem feuchten Substrat herauswachsen. Diese
Reaktionen können so energisch sein, daß sie eventuell andersartige (z. B.
geotropische) Reize überwinden.
4. Traumatotropismus[274].
Eine einseitige Verwundung eines Pflanzenteils wird leicht zu
einer Wachstumshemmung an der Wundstelle führen, so daß eine
Krümmung entsteht, die auf ihrer Konkavseite die Wunde trägt. Eine
solche Krümmung wäre keine tropistische Reizkrümmung. Unter
Traumatotropismus versteht man demnach eine ganz andere
Erscheinung, die vor allem bei Wurzeln leicht zu beobachten
ist. Macht man an ihrem Vegetationspunkt einseitig einen
Einschnitt, oder ätzt oder sengt man ihn einseitig an, so tritt in
einiger Entfernung in der darüber liegenden Wachstumszone
eine Krümmung ein, die man negativ traumatotropisch nennt,
weil durch sie die Spitze der Wurzel von dem schädigenden Agens
entfernt wird. Die senkrecht oberhalb der Wunde gelegene Stelle
der Wachstumszone wächst also hier stärker als die Gegenseite. Das
besondere Interesse dieses Tropismus liegt demnach darin, daß wir
eine scharfe Trennung zwischen dem Ort der Reizaufnahme und dem
Ort der Reizreaktion und dementsprechend auch eine ausgesprochene
Reizleitung feststellen können.
Auch bei oberirdischen Organen ist gelegentlich ein negativer
Traumatotropismus nachgewiesen worden, meistens aber zeigen diese
einen positiven Traumatotropismus. Daß dieser nicht mit
der eingangs erwähnten rein mechanisch erfolgenden einseitigen
Wachstumshemmung verwechselt werden darf, daß er vielmehr eine
echte Reizbewegung ist, ergibt sich schon daraus, daß er
vielfach unter beträchtlicher Beschleunigung des mittleren
Wachstums erfolgt, und daß Reizleitung auf weite Strecken bei ihm
eine gewöhnliche Erscheinung ist. STARK hat gezeigt,
daß ähnlich wie bei der phototropischen Reizleitung auch hier
diffundierende Stoffe eine Rolle spielen, Stoffe also, die bei der
Verwundung entstehen und die bei verschiedenen Spezies gleich oder
ähnlich sind, so daß man den traumatischen Reiz aus einer
Art in eine andere weiterleiten kann.
Es ist jetzt eine Anzahl von Tropismen besprochen, aber die
ganze Reihe der bisher bekannt gewordenen tropistischen Erfolge
ist damit lange nicht erschöpft. Vielmehr kennt man außerdem
noch einen Thermotropismus, bei dem die Wärme, einen
Rheotropismus, bei dem das fließende Wasser und einen
Galvanotropismus, bei dem galvanische Ströme den Reiz
abgeben. Auf diese Tropismen kann hier nicht eingegangen werden.
5. Haptotropismus [Thigmotropismus][275].
Eine Einkrümmung nach einseitiger Berührung findet namentlich
bei Kletterpflanzen statt, die mit Hilfe solcher Greifbewegungen
eine Stütze umfassen und sich an ihr befestigen. Die Einrichtung
läuft also auf das gleiche hinaus wie bei den Windepflanzen, doch
sind die Bewegungen hier durchaus keine geotropischen. Es sind
Seitenorgane verschiedenen morphologischen Charakters (vgl. S.
155), die die Befestigung besorgen, und die entweder ihrer normalen
Leistung und Ausbildung (als Laubblätter, Laubsprosse, Blütensprosse)
treu geblieben, oder aber, wie dies meist der Fall ist, zu typischen
„Ranken“ geworden sind, um ausschließlich und in vollkommenster
Weise der Umklammerung zu dienen. Die einseitige Berührung mit einem
festen Körper bewirkt eine starke Wachstumsbeschleunigung, die maximal
der Berührungsstelle gegenüber erfolgt und von da aus bis zur berührten
Stelle allmählich ausklingt, so daß sie auch noch in der Mittellinie
einen nennenswerten Betrag erreicht. So wird die berührte Stelle
konkav, und der berührende Körper, die Stütze, wird umfaßt. Dies
geschieht natürlich um so rascher und leichter, je dünner die Ranke
und je kräftiger ihre Reaktion ist. Einige Ranken fassen sehr rasch
(Passiflora, Sicyos, Bryonia), andere sind sehr träge (Smilax, Vitis).
Da die Krümmung nach dem Anlegen der Ranke an die Stütze sich noch zu
verengern strebt, so wird die letztere oft derart[S. 313] fest umwickelt, daß
an weichen Stengeln und dgl. tiefe Eindrücke entstehen können.
Von großer Wichtigkeit für die Funktion der Ranken ist der Umstand, daß
sie, wie PFEFFER gezeigt hat, nicht durch jeglichen Kontakt,
sondern nur durch die Berührung mit den Unebenheiten eines festen
Körpers zum Einkrümmen gereizt werden. Auch der heftigste Regenfall
wirkt daher nicht als Berührungsreiz, und selbst der bis zur Quetschung
gesteigerte Anprall reinen Quecksilbers geht ohne eine spezifische
Reizwirkung vorüber, wogegen ein vom Luftzug bewegtes Baumwollfäserchen
von 0,00025 mg Gewicht sie bereits auslösen kann.
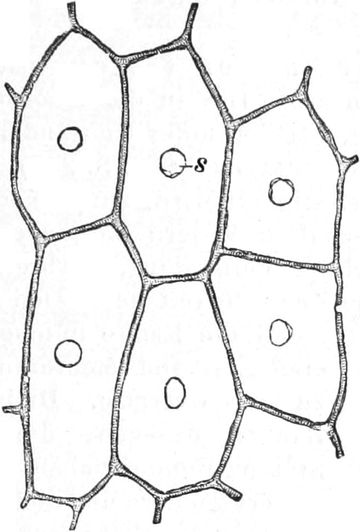
Fig. 286. Oberflächenansicht einiger Epidermiszellen der
Rankenunterseite von Cucurbita Pepo mit Fühltüpfeln s. Vergr. 540.
Nach STRASBURGER.
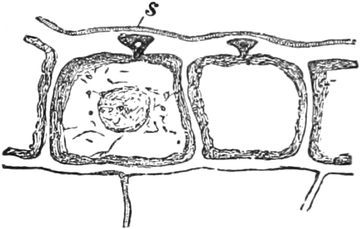
Fig. 287. Querschnitt durch ebensolche
Epidermiszellen wie in Fig. 286; im Fühltüpfel s ein sehr kleiner
Kalziumoxalatkristall. Vergr. 540. Nach STRASBURGER.
Aller Wahrscheinlichkeit nach wird die Aufnahme so schwacher Reize
durch die sog. „Fühltüpfel“ (Fig. 286, 287) begünstigt. Es sind
das protoplasmaerfüllte und nach außen stark erweiterte Tüpfel in
der Außenwand der Epidermiszellen. Sie finden sich z. B. bei den
Cucurbitaceen, können aber selbst bei sehr reizbaren Ranken (z. B.
bei Passiflora) auch fehlen.
Die Ranken mancher Pflanzen sind allseitig reizbar und krümmungsfähig
(Cobaea, Eccremocarpus, Cissus), andere, so die der Cucurbitaceen
u. a. mit eingekrümmtem Gipfel, sind nach FITTING zwar
allseitig gegen Berührung empfindlich, krümmen sich aber nur
auf eine einseitige Berührung der Unterseite ein, während
eine gleichzeitige Reizung der Oberseite in der Hemmung der
Einkrümmung zum Ausdruck kommt.
Manche einseitig empfindliche Ranken haben die Fühltüpfel nur auf
der reizbaren Unterseite. Überhaupt sind die Ranken mehr oder
weniger dorsiventral, und deshalb sind ihre Reaktionen oft schon
mehr nastisch als tropistisch.
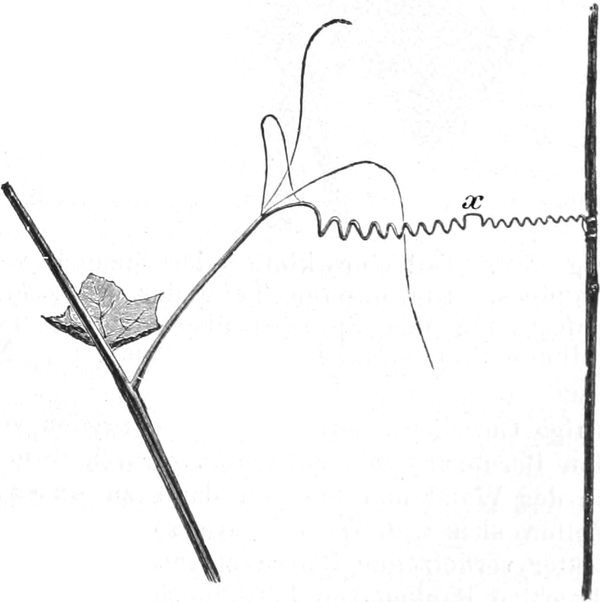
Fig. 288. Stengelstück mit Ranke von Sicyos angulatus,
einer Cucurbitacee. Ein Rankenast hat mit seiner Spitze die aufrechte
Stütze rechts erfaßt und seine freie Strecke bereits spiralig
aufgerollt. Bei x Wendepunkt der Aufrollung. Nach NOLL.
Bei den vollkommensten Ranken bleibt die Krümmung nicht auf den
unmittelbar gereizten Teil beschränkt. Abgesehen davon, daß durch
das Umwickeln der Stütze immer weitere Stellen der Ranke gereizt
werden, pflanzt sich die Krümmung auch auf Regionen fort, die
unberührt blieben. Dadurch wird zunächst einmal die Rankenspitze um
die Stütze gerollt. Später krümmt sich auch der übrige, zwischen
Stütze und Muttersproß[S. 314] ausgespannte Teil der Ranke kräftig
ein, so wie ja auch alternde Ranken, die keine Stütze gefunden
haben, sich schraubig einrollen. Während aber in diesem Falle eine
einzige Schraube entsteht, müssen bei Ranken, die sich
oben an einer Stütze befestigt haben, im basalen Teil mindestens
zwei einander gegenläufige Schrauben auftreten, die
durch einen Wendepunkt (Fig. 288 x) getrennt werden. Durch diese
schraubige Rollung wird der Stengel der Kletterpflanze nicht nur
näher an die Stütze herangezogen, sondern auch elastisch daran
aufgehängt und vor dem Abreißen durch plötzliche Erschütterung
geschützt.
Auch in der anatomischen Ausbildung der Ranken treten,
nachdem eine Stütze erfaßt ist, vorteilhafte Veränderungen
auf. Die in der Knospenlage meist gerollte junge Ranke macht
während ihrer Streckung, bei der unter Umständen eine tägliche
Verlängerung um 90% erreicht wird, sehr lebhafte Nutationen (vgl.
S. 297). Dabei ist sie dünn, biegsam und leicht zerreißbar;
ihre Turgorfestigkeit wird, zumal nach der Spitze zu, nur von
Kollenchym unterstützt. Diese Eigenschaften ändern sich aber bald,
nachdem die Ranke gefaßt hat. Dann tritt oft eine erhebliche
Verdickung, Verbreiterung und Erhärtung der umklammernden Teile
ein, während die Zugfestigkeit des freien Teiles durch Verholzung
und Sklerenchymbildung so erhöht wird, daß die Ranke nunmehr oft
eine Last von mehreren Kilo zu tragen vermag. Diejenigen Ranken
dagegen, die keine Stütze gefunden haben, pflegen zu verkümmern und
abzufallen, nachdem sie sich zuvor oft von selbst noch eingerollt
haben.
Ein Vorteil, den die Rankenkletterer vor den Schlingpflanzen voraus
haben, ist der, daß sie nicht auf vertikale Stützen angewiesen
sind. Die Art der Einrollung ihrer Ranken weist sie aber auch
auf dünne Stützen an, da sie an dickeren abgleiten. Nur
vereinzelte Rankenpflanzen haben sich von dünnen Stützen unabhängig
gemacht und vermögen mittels besonderer Einrichtungen selbst an
glatten Wänden emporzuklettern. Ihre Ranken sind nämlich negativ
phototropisch und besitzen an oder dicht unter der Spitze kleine
knopfartige Gewebewucherungen, die entweder von vornherein
vorhanden sind oder erst durch den Berührungsreiz entstehen. Durch
ihre Klebrigkeit haften diese Knöpfchen zunächst an der Wand und
wachsen dann zu saugnapfähnlichen Scheibchen aus, deren
Zellen sich wie Wurzelhaare so eng mit dem Substrat verbinden,
daß man eher die später verholzende Ranke zerreißen, als die
Haftscheiben ablösen kann. Fig. 208 zeigt derartige Ranken von
Parthenocissus tricuspidata. Die Haftscheibchen sind hier an
jungen Ranken schon als Knöpfchen vorgebildet. Bei anderen Arten
des wilden Weins bilden sich die Haftscheiben aber erst nach der
Berührung aus; ihre Ranken vermögen auch dünne Stützen zu umfassen.
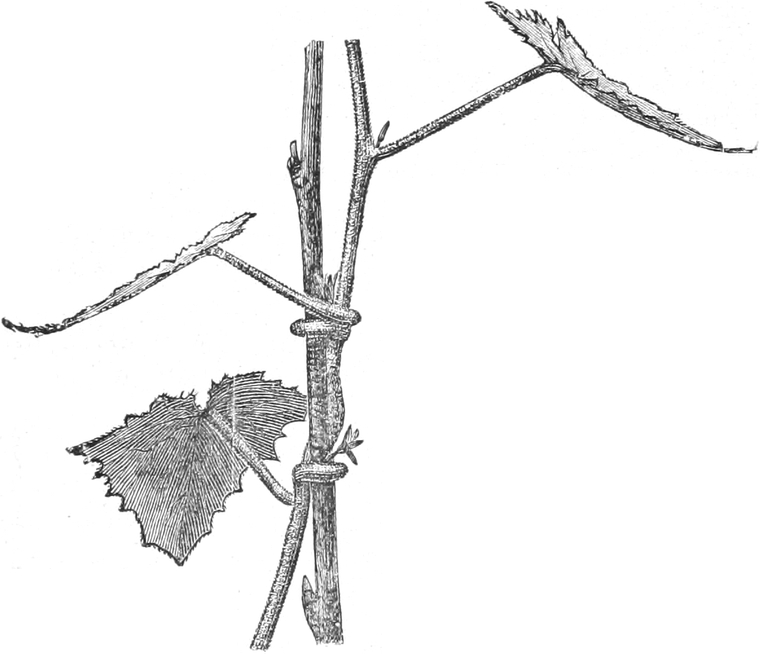
Fig. 289. Teil eines kletternden Stengels von Maurandia
scandens. Die unteren Teile der Blattstiele haben die Befestigung des
Sprosses übernommen, indem sie die dünne Stütze rankenartig umklammern.
Nach NOLL.
Fig. 289 zeigt ein Stück einer Kletterpflanze (Maurandia scandens),
deren Blattstiele gleich Ranken reizbar sind.
Hier wird die Stütze aber nur etwa einmal oder nur teilweise
umfaßt. Ähnlich klettern mittels ihrer Blattstiele die bekannte
Kapuzinerkresse und andere Tropaeolum-Arten, Solanum jasminoides,
Nepenthes u. a. Bei manchen Tropenpflanzen sind Achselsprosse
oder Nebenblätter zu rankenartigen „Kletterhaken“ geworden. Bei
Gloriosa, Littonia und Flagellaria ranken fadenförmige Blattenden,
und bei manchen Fumaria- und Corydalis-Arten umwickelt außer dem
Fiederstielchen auch die Fiederspreite dünne Stützen.
Selbst bei niederen Kryptogamen (Florideen)[S. 315] kommen rankende
Thallusteile vor. — Die schmarotzenden Sprosse der Cuscuta (Fig.
218) sind zum Winden wie zum Ranken befähigt und machen von beiden
Eigenschaften beim Erklimmen und Umklammern ihrer Nährpflanze
abwechselnd Gebrauch.
Neuere Untersuchungen[276] haben gezeigt, daß der Haptotropismus
sehr viel weiter verbreitet ist, als man bisher angenommen
hatte. Etiolierte Keimlinge sind stets haptotropisch, aber auch
ältere Sprosse grüner Pflanzen, insbesondere von Schling- und
Kletterpflanzen verhalten sich häufig ebenso. Die Empfindlichkeit
dieser Pflanzen weicht übrigens von der der Ranken insofern ab,
als sie auch durch beliebige Erschütterungen (z. B. auch durch
Wasserstrahlen) gereizt werden. Einen Nutzen scheinen sie alle aus
diesen Befähigungen nicht zu ziehen.
STARK zeigte, daß beim Haptotropismus der Keimlinge das
Resultantengesetz gilt; die Krümmung erfolgt also im Sinne der
Resultierenden z. B. aus zwei an verschiedenen Flanken angreifenden
Reizen. Er wies ferner nach, daß auch das WEBERsche
Gesetz gilt. Werden opponierte Flanken verschieden stark gereizt,
so erhält man nur bei gleichem Verhältnis der Reizgrößen
die gleiche Einkrümmung. Auch wenn man zwei opponierte Flanken
gleichstark reizt und eine einseitige Reizung senkrecht dazu
einwirken läßt, gilt das WEBERsche Gesetz[277].
6. Autotropismus[278].
Es ist nötig, hier noch einer Erscheinung von allgemeiner
Verbreitung zu gedenken, bei der nicht äußere, sondern
innere, in der Pflanze selbst liegende Veränderungen als
Reiz wirken. Ganz allgemein kann man nämlich beobachten, daß jede
tropistische Krümmung, mag sie nun durch die Schwerkraft, das
Licht oder einen anderen Faktor ausgelöst sein, nach Aufhören der
Reizwirkung von einer Geradstreckung gefolgt wird. In den
meisten Fällen wird sie durch Wachstumsprozesse bewerkstelligt,
bei gekrümmten Wurzeln aber kann diese auch durch Kontraktion
(S. 249) erfolgen. Dieses Bestreben eines ursprünglich geraden
Organs, sich wieder gerade zu strecken, ebenso aber auch das
Bestreben eines ursprünglich gekrümmten Organs, seine normale
Gestalt wieder anzunehmen, nennt man Autotropismus. Auf
manche solche autotropische Erscheinungen mußte früher schon
hingewiesen werden. So wurde beim Geotropismus erwähnt, daß die
über das Lot hinausgehende Krümmung autotropisch zurückreguliert
wird. Es muß aber jetzt gesagt werden, daß jede geotropische
Krümmung wieder mehr oder weniger rückgängig gemacht wird, wenn
man die einseitige Schwerewirkung aufhebt, das Objekt also auf dem
Klinostaten rotieren läßt, und daß ebenso phototropische Krümmungen
zurückgehen, wenn die ungleiche Lichtverteilung beseitigt wird
usf. Auch Ranken, die sich haptotropisch gekrümmt haben, können
sich nach Aufhebung des Kontaktreizes wieder gerade strecken.
Ja selbst bei Fortdauer der einseitigen Einwirkung läßt sich
vielfach ein Rückgang in den tropistischen Krümmungen erkennen.
Die physiologische Wirksamkeit der Reize nimmt also offenbar
mit der Dauer ihrer Wirkung ab. Der Krümmungsausgleich kann zur
völligen Geradstreckung führen, ja er kann sogar gelegentlich
über das Ziel hinausschießen, zu einer neuen Krümmung in
entgegengesetzter Richtung führen, die dann ihrerseits autotropisch
ausgeglichen wird; so kann einer tropistischen Reizung ein Hin-
und Herpendeln eines Sprosses folgen. Voraussetzung für das
Eintreten des autotropischen Ausgleiches ist nur, daß die Pflanze
noch wachstumsfähig ist. Und nicht nur nach tropistischer Reizung,
sondern auch nach Krümmungen, die man durch mechanische Biegung
einer Pflanze aufgezwungen hat, macht sich der Autotropismus
geltend. Was im einzelnen der wirksame Reiz ist, der zur
Wiederherstellung der ursprünglichen Körperform führt, wissen
wir nicht, doch darf man vermuten, daß die Pflanze da auf innere
Spannungen reagiert. So sieht man auch, daß wenn eine
tropistische Krümmung mechanisch verhindert wird, nur
Spannungen entstehen, auch ein Ausgleich dieser Spannungen
autotropisch erfolgt.
b) Nastische Bewegungen[279].
Im Gegensatz zu den tropistischen und taktischen Reizbewegungen, bei
denen die Richtung des Reizes in einer ganz bestimmten Beziehung
zur Richtung der Bewegung steht, handelt es sich hier um Reaktionen,
die entweder überhaupt durch nicht bestimmt gerichtete, also durch
diffuse Reize veranlaßt werden, oder bei denen doch eine eventuelle
Reizrichtung ohne Einfluß ist. Nastien sind dementsprechend auf
dorsiventrale Organe be[S. 316]schränkt, und die Richtung der Bewegung wird
hier stets allein von dem reagierenden Organ bestimmt und nicht von
der Außenwelt; demnach dienen auch die Nastien nicht der Orientierung
im Raum, wie das für die Tropismen der Fall ist. Ihre ökologische
Bedeutung ist vielfach noch gar nicht klar; nach GOEBEL sollen
sie z. T. völlig nutzlose Bewegungen sein.
Typische nastische Variationsbewegungen liegen bei den
Spaltöffnungen[279] vor, deren Bau es mit sich bringt, daß auf
Turgorschwankungen Veränderungen der bestehenden Krümmung der
Schließzellen, Öffnen oder Schließen, eintritt. Man nimmt freilich
wohl vielfach an, daß das bei Wasserverlust eintretende Schließen
und das nach Beleuchtung eintretende Öffnen rein mechanisch
erfolge; Wasserverlust wird ja eine Minderung der Zellhautspannung
zur Folge haben, Lichtzufuhr wird, wenigstens wenn die Möglichkeit
der Wasseraufnahme gegeben ist, diese Spannung durch Produktion
von Assimilaten steigern. Es kann aber kein Zweifel bestehen, daß
neben rein physikalischen Beeinflussungen vielfach auch echte
Reizbewegungen eine Rolle spielen. So kann das Licht, so
können auch manche andere Faktoren als Reize wirken, auf die hin
das Protoplasma die Produktion osmotisch wirksamer Substanzen in
bestimmtem Sinne lenkt.
Wie bei den Spaltöffnungen, so können auch bei anderen nastischen
Bewegungen Licht und Wärme, chemische Substanzen, außerdem aber auch
Erschütterungen die Rolle von Reizen spielen. Meist erfolgt die
Bewegung eines bestimmten Organes auf mehrere dieser Reize hin in
gleicher oder ähnlicher Weise.
1. Nyktinastische Bewegungen[280].
Viele Laubblätter und Blütenblätter nehmen bei Tag eine andere Ruhelage
ein als bei Nacht. Je nachdem der Übergang von der einen zur anderen
durch Schwankungen in der Lichtintensität, in der Temperatur oder in
beiden Faktoren gleichzeitig bewirkt wird, unterscheidet man zwischen
Photo-, Thermo- und Nyktinastie. Nach den
mechanischen Mitteln bei ihrer Ausführung sind die Bewegungen teils
Wachstums-, teils Variationsbewegungen.
1. Thermonastie. Durch Temperaturschwankungen bedingte
Wachstumsbewegungen finden sich namentlich bei Blüten, so z. B. bei
Tulipa, Crocus, Ornithogalum, Colchicum und Adonis. Diese Blüten bzw.
ihre Perigonblätter oder Kronblätter zeigen nach Temperatursteigerung
eine plötzlich einsetzende und nur eine gewisse Zeit währende
Wachstumssteigerung, die auf der Innenseite dieser Blätter maximal ist.
Die Blüten öffnen sich also. Umgekehrt schließen sie sich bei Abkühlung.
Äußerst empfindlich gegen Temperaturschwankungen erweisen sich
die Blüten der Tulpe und des Crocus. Geschlossene Tulpen- und
Crocusblüten, die man aus dem Freien ins warme Zimmer bringt,
öffnen sich hier in kurzer Zeit, bei einer Temperaturdifferenz
von 15–20° C schon in 2–4 Minuten. Empfindliche Blüten von Crocus
reagieren schon auf eine Schwankung von 1⁄2° C, die der Tulpe auf
2–3° C.
Fig. 290. Blütenköpfchen einer Composite (Leontodon
hastilis), links durch Verdunkelung geschlossen, rechts nach Belichtung
geöffnet. Aus DETMER, Physiol. Prakt.
2. Photonastie. In ähnlicher Weise öffnen sich andere Blüten
(Nymphaea, Kakteen) und auch die Blütenköpfchen der Kompositen (Fig.
290) auf Erhellung und schließen sich nach Verdunkelung.
Umgekehrt verhalten sich die „Nachtblüher“, z. B. Silene noctiflora,
Victoria regia, Nicotiana-Arten.
Die Bedeutung dieser Bewegungen dürfte darin liegen, daß die
Geschlechtsorgane nur dann exponiert werden, wenn Insektenbesuch
zu erwarten ist, zu anderen Zeiten aber[S. 317] vor den Unbilden der
Witterung, vor allem vor Regen geschützt werden. Die Nachtblüher
sind bei ihrer Bestäubung auf Nachtfalter angewiesen.
3. Nyktinastie. Viele Laubblätter führen nyktinastische
Bewegungen aus, die in der Regel mehr vom Licht als von der Temperatur
beeinflußt werden. In manchen Fällen, so z. B. bei Chenopodiaceen,
Caryophylleen, Balsaminen, manchen Kompositen, sind es, wie bei den
Blütenblättern, ausschließlich Wachstumsbewegungen; bei den
Leguminosen, Oxalideen und anderen mit Gelenkpolstern versehenen
Pflanzen sehen wir dagegen Variationsbewegungen, eintreten.
Die ersteren sind natürlich von kurzer Dauer; sie hören auf, wenn
die Blätter ausgewachsen sind; die anderen aber werden lange Zeit
fortgesetzt. Bei den Variationsbewegungen tritt eine Erhöhung
des Turgors mit der Verdunkelung wahrscheinlich in beiden
Gelenkhälften ein, jedoch in der Konkavseite schwächer oder langsamer.
Überall ist die Nachtstellung — die man auch „Schlafstellung“ nennt,
obwohl diese Erscheinungen durchaus nichts mit dem „Schlaf“ der Tiere
zu tun haben — durch die vertikale Lage der Blattspreiten
charakterisiert, wobei der Stiel oder das Polster entweder nach oben
oder nach unten sich krümmt, die Lamina selbst also entweder ihre
Unterseite oder ihre Oberseite nach außen kehrt. In der Tagstellung
stehen die Flächen ungefähr horizontal oder senkrecht zum Lichteinfall
(S. 309) (Fig. 291).
Fig. 291. Sproß von Amicia zygomeris (Leguminose). Links
in zerstreutem hellem Tageslicht mit ausgebreiteten Blättchen, rechts
nach Verdunkelung in Schlafstellung.
Daß es sich bei den geschilderten Erscheinungen nicht um
Phototropismus handelt, ergibt sich daraus, daß die Tagstellung
eintritt, einerlei, ob die Unterseite oder die Oberseite stärker
beleuchtet ist, oder ob beide gleich hell gehalten werden.
Entsprechendes gilt bei der Verdunkelung.
Die Bedeutung der nächtlichen Senkrechtstellung der Laubblätter
liegt nach STAHL darin, daß durch sie die Taubildung
vermindert und damit die Transpiration gefördert wird. Als
Förderung der Transpiration kann auch die Tatsache gedeutet werden,
daß vielfach die Spaltöffnungen auf der in der Schlafstellung
geschützten Seite der Blätter liegen.
Übermäßig hohe Beleuchtung oder Temperatur veranlaßt die
Blätter, die übliche Tagstellung zu verlassen und eine neue
Lage aufzusuchen, die entweder mit der Nachtstellung äußerlich
übereinstimmt oder ihr diametral entgegengesetzt ist. So sind
die Fiederblättchen der Robinie nachts abwärts geschlagen,
in zerstreutem Tageslicht flach ausgebreitet; in der heißen
Mittagssonne richten sie sich steil aufwärts. — Dieser sog.
„Tagesschlaf“ findet sich nur bei den Gelenkblättern und vollzieht
sich hier in mechanischer Hinsicht anders als die abendliche
Lageveränderung: es tritt keine Turgeszenzsteigerung, sondern eine
Erschlaffung ein, die auf zwei Seiten eines Gelenkes ungleich ist.
Periodische Bewegungen[281].
Wenn Blätter längere Zeit hindurch regelmäßige nyktinastische
Bewegungen infolge des täglichen Lichtwechsels ausgeführt haben, so
setzen sie diese Periodizität einige Tage[S. 318] lang auch in konstantem
Licht oder konstanter Dunkelheit fort. Bei manchen Blättern läßt
sich durch geeignete experimentelle Maßnahmen auch eine kürzer
oder länger als 24 Stunden dauernde Schwingungsperiode herstellen,
die dann ebenfalls eine Zeitlang nachwirkt. — Andererseits
ist bei gewissen Blüten (Calendula) und Blättern (Phaseolus)
festgestellt, daß es auch Bewegungen mit 24 stündiger Periode gibt,
die nicht durch den Rhythmus der Beleuchtung und Verdunkelung
sowie deren Nachwirkung bedingt sind. Daß diese Bewegungen autonom
sind, ist ausgeschlossen. Man muß erwarten, daß noch irgendein
Faktor gefunden wird, der für die Periodizität dieser Bewegungen
verantwortlich gemacht werden kann. Die von STOPPEL
ausgesprochene Ansicht, daß die Schwankungen in der elektrischen
Leitfähigkeit der Atmosphäre maßgebend seien, konnte nicht
bestätigt werden.
2. Chemonastie[282].
Im gleichen Verhältnis wie die Photonastie zum Phototropismus steht die
Chemonastie zum Chemotropismus. Einerlei, von welcher
Seite her ein chemischer Reiz (z. B. Dampf von Äther, Chloroform,
Ammoniak) empfindliche Ranken trifft, immer wird diejenige Seite
konkav, die wir als vorzugsweise haptotropisch empfindlich erkannt
haben.
Diese chemonastischen Einkrümmungen der Ranken sind offenbar
ohne jeden Nutzen für die Pflanze. Das gleiche gilt von den
nastischen Bewegungen der Ranken, die nach Verwundung eintreten
(Traumatonastie) oder die sich in ganz gleicher Weise ebenso bei
Abkühlung wie bei Erwärmung zeigen (Thermonastie). Dagegen spielen
die chemonastischen Bewegungen bei gewissen Insektivoren eine große
Rolle.
Sehr ausgeprägte chemonastische Bewegungen sieht man an den Tentakeln
von Drosera, die sich nach chemischer Reizung so einkrümmen, daß
ihre Oberseite konkav wird, das Drüsenköpfchen also nach der Mitte der
Blattscheibe gelangt (Fig. 212). Als Reizmittel dienen einmal solche
Stoffe, wie Eiweiß, Phosphate usw., die Drosera zu ihrer Ernährung
gebrauchen kann (S. 223), andererseits aber auch gleichgültige und
sogar giftige Substanzen. Oft genügen minimale Spuren von ihnen (z. B.
0,0004 mg Ammoniumphosphat), um die Reizbewegung einzuleiten, und zwar
müssen diese auf das Köpfchen des Tentakels gelangen, damit die
Basis des Tentakels die Einkrümmung ausführt; es besteht also
hier, ähnlich wie bei manchen anderen Reizbewegungen, ja noch schärfer
als dort, eine Trennung zwischen dem Perzeptionsorgan, das den Reiz
aufnimmt, und dem Bewegungsorgan, das die äußerlich sichtbare Reaktion
ausführt. Der im Köpfchen aufgenommene Reiz muß zur Tentakelbasis
geleitet werden.
Ein Insekt, das sich auf einem randständigen Tentakel niedergelassen
hat, wird also durch dessen Krümmung in die Mitte der Blattscheibe
gebracht. Die hier stehenden kurzgestielten Tentakel aber senden einen
Reiz nach allen randständigen, die sich daraufhin sämtlich nach innen
krümmen. So wird also das Insekt immer von vielen Drüsenköpfchen
berührt und mit deren verdauendem Sekret übergossen.
Die durch Wachstum erfolgende Krümmung wird ganz in der gleichen
Weise ausgeführt wie bei den Ranken; nach der Einkrümmung sind die
Tentakel erheblich länger geworden. Mit dem Abschluß des Wachstums
hört also die Bewegungsfähigkeit der Tentakel auf, diese können nur
einige wenige Male die Krümmung ausführen. Mit den Ranken haben die
Droseratentakel ferner noch gemeinsam, daß sie ebenfalls thigmo-,
traumato- und thermonastisch reagieren. Zweifellos ist aber ihre
chemonastische Reizbarkeit die größte und wichtigste. Chemische
Reize lösen auch bei anderen Insektivoren Bewegungen aus, z. B. bei
Dionaea und Pinguicula.
3. Seismonastie und
Traumatonastie[283].
Bei der soeben erwähnten Insektivore Dionaea muscipula klappen die
beiden Blatthälften (Fig. 215) nicht nur nach chemischen, sondern
auch nach mechanischen Reizen zusammen. Im Gegensatz zu den nur nach
Berührung[S. 319] mit festen Körpern erfolgenden haptotropischen Bewegungen
der Ranken oder von Drosera wirkt hier jede durch einen Stoß erzielte
Erschütterung als Reiz; die Bewegung kann also auch durch Regentropfen
ausgelöst werden. Solche Bewegungen nennt man seismonastische.
Das bekannteste Beispiel für seismonastische Bewegungen liefert die
Leguminose Mimosa pudica, ein tropischer Halbstrauch, der wegen seiner
hohen Empfindlichkeit den Namen „Sinnpflanze“ erhalten hat. Die
Blätter tragen am Ende eines langen Stieles 4 (eventuell 6) fingerartig
auseinanderspreizende Sekundärstiele, die mit zahlreichen Paaren
feiner Blättchen in fiederförmiger Anordnung besetzt sind (Fig. 292).
An der Basis des primären und der sekundären Blattstiele sowie der
Fiederblättchen finden sich Gelenkpolster, so daß alle Teile beweglich
sind. Die Lichtlage der ungereizten Blätter zeigt Fig. 292 links. Wird
das Blatt erschüttert, dann führen alle seine Teile bei günstiger
Temperatur (25–30° C) und Feuchtigkeit sehr rasche Bewegungen aus, die
dem Blatt ein völlig verändertes Aussehen geben. Die Fiederblättchen
klappen nach oben und vorn zusammen, die Sekundärstiele legen sich nach
vorn zu seitlich aneinander, und der Blattstiel senkt sich scharf nach
unten (Fig. 292 rechts). Die gereizte Pflanze erholt sich aber bald
wieder und nimmt ihre frühere Blattstellung wieder an, wenn ihr Ruhe
gelassen wird.
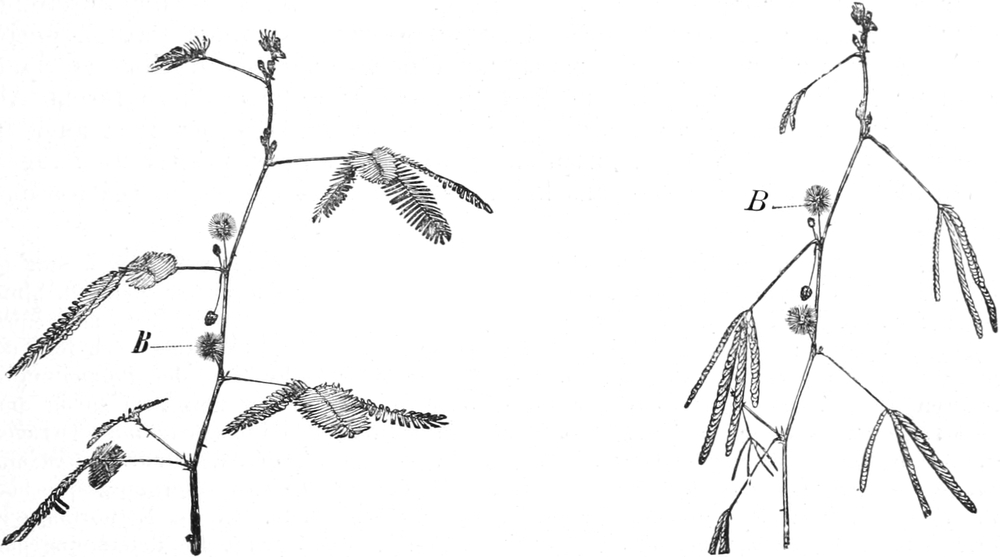
Fig. 292. Mimosa pudica. Links ungereizt in der
Tagstellung, rechts durch Erschütterung gereizt, wodurch sich
die Blätter zusammengefaltet und abwärts geschlagen haben. B
Blütenköpfchen.
Der Stellung eines erschütterten Blattes äußerlich ähnlich ist
seine Schlaf- oder Nachtstellung, doch sind die
Spannungszustände, die zu den beiderlei Stellungen führen, in
dem Gelenkpolster verschieden; auch in der Nachtstellung ist das
Blatt noch mechanisch reizbar. — Wie die Schlafstellung, so wird
auch die seismonastische Reizstellung durch Turgorschwankungen
bewirkt, allein es handelt sich hier um eine Abnahme des
osmotischen Druckes und eine Erschlaffung in den konkav werdenden
Polsterhälften. Besonders am Hauptstielpolster kann man diese
Erschlaffung der übrigens allein reizbaren Unterseite sehr deutlich
erkennen; sie ist hier mit einem Austritt von Flüssigkeit aus
den Zellen und einer Injektion der Interzellularen mit dieser
Flüssigkeit verbunden.
Die gleiche Reizstellung wie nach Erschütterung nimmt
das Blatt der Mimosa auch nach Verwundung (Einschneiden,
Anbrennen, Anätzen) ein, und die Vorgänge im Gelenk sind bei einer
solchen traumatonastischen[S. 320] Reizung die gleichen wie nach
seismonastischer Reizung. Auch auf andere Reize: elektrische Schläge,
plötzliche Temperaturänderungen, chemische Reize können ganz die
gleichen Bewegungen an den Blättern beobachtet werden wie nach Stoßreiz.
Alle diese Reize sind in ihrer Wirkung aber nicht auf das zunächst
betroffene Gelenk beschränkt, sondern werden weit fortgeleitet, im
Extrem über die ganze Pflanze, überall zu einer Krümmung in den
Gelenken führend. Am schnellsten und weitgehendsten erfolgt diese
Reizleitung[284] bei den traumatischen Reizen; hier wird
sie deshalb am besten näher zu schildern sein. Reizt man etwa ein
einzelnes Fiederblättchen am Ende eines Sekundärstieles durch Anbrennen
mittels eines Zündholzes, so klappen augenblicklich beide
Endfiederchen nach oben, und es folgen ihnen rasch der Reihe nach
sämtliche Fiederpaare bis zum Ansatz des Sekundärstieles. Von hier
überträgt sich die Erscheinung auf die anderen Sekundärstrahlen,
an denen jetzt die Blättchen in umgekehrter Reihenfolge, also
spitzenwärts fortschreitend, zusammenschlagen. Haben sich auch die
Sekundärstrahlen seitlich aneinander gelegt, und wähnt der Beobachter
den Bewegungsvorgang abgeschlossen, dann überrascht ihn plötzlich noch
einmal der Hauptstiel durch rasche Abwärtsbeugung. Von diesem einen
Blatte aus kann sich der Reiz auch im Stengel, oft bis 50 cm weit,
fortpflanzen und auf andere Blätter übertragen. Doch auch von der
Wurzel aus kann sich der Reiz bis in die Blätter fortpflanzen.
Die Geschwindigkeit der Reizleitung kann nach Verwundung 10
cm, nach Stoß bis zu 3 cm in der Sekunde betragen, also recht
ansehnlich sein. Immerhin steht sie hinter der Reizleitung in den
Nerven des Menschen weit zurück. Wenn auch noch nicht sicher
nachgewiesen ist, wie die Reizleitung bei Mimosa erfolgt,
so steht doch fest, daß sie von der Leitung einer Erregung in den
Nerven oder selbst von der Reizleitung in anderen pflanzlichen
Fällen wesentlich verschieden ist. Denn der Reiz geht sicher
auch über tote Strecken weg und bewegt sich auch in der
lebenden Pflanze in toten Organen, nämlich in den Gefäßen des
Holzes. Nach RICCA gelingt es, den Reiz auch durch eine
mit Wasser gefüllte Glasröhre weiterzuleiten, und die von diesem
Autor ausgesprochene Vermutung, daß es sich um eine Fortleitung von
Stoffen handle, die bei Berührung oder Verwundung entstehen, hat
manches für sich. Im übrigen scheint die Reizleitung nach Berührung
etwas anders zu erfolgen als die nach Verwundung.
Ähnlich, doch weniger reizbar wie die Mimose sind manche andere
Leguminosen und gewisse Oxalideen. So führen z. B. Robinia
Pseudacacia und Oxalis acetosella auf kräftige mechanische Reize
hin Bewegungen aus, die freilich viel unbedeutender sind als die
der Mimose. In der Regel bedarf es bei diesen Pflanzen mehrerer
Stöße, um den Reiz auszulösen, während bei Mimosa ein einziger
genügt; auch vergrößert sich bei ihnen der Reizerfolg mit Zunahme
der Stöße, während bei Mimosa jeder überhaupt wirksame Reiz unter
normalen Bedingungen gleich die maximale Bewegung auslöst. Auch die
Blattbewegungen nach Verwundung sind nicht auf Mimosa beschränkt.
Die Reaktionsfähigkeit der Mimose auf Stoßreiz hängt selbstverständlich
weitgehend von Außenfaktoren ab, und jeder dieser Faktoren kann,
wenn er im Übermaß oder in zu geringem Maß geboten wird, zu einem
Starrezustand führen. Die Reizbewegungen der Mimose treten nur
bei genügend hoher Temperatur lebhaft ein; unter einem gewissen
Wärmegrad (15°) findet überhaupt keine Bewegung statt, es ist
dann Kältestarre eingetreten. Bei etwa 40° Lufttemperatur
tritt dagegen Wärmestarre, bei längerem Aufenthalt im
Finstern Dunkelstarre ein. Im Vakuum, in Wasserstoff, in
Chloroformdämpfen, Leuchtgas usw. tritt ebenfalls Bewegungslosigkeit
ein, die teils durch Sauerstoffmangel, teils durch unmittelbare
Giftwirkungen bedingt ist. Wirken die zu Starrezuständen führenden
nachteiligen Verhältnisse nicht zu lange ein, dann kann der frühere
reizbare Zustand wiedergewonnen[S. 321] werden. — Auch bei anderen
Reizerscheinungen treten uns ähnliche Starrezustände entgegen.
Die Variationsbewegungen, die bei den Staubfäden einiger
Berberideen (Berberis, Mahonia) und Kompositen (besonders schön bei
Centaurea americana) auftreten, schließen sich in ihrer Mechanik
an die der Blätter an. Die Staubfäden solcher Kompositen verkürzen
sich nach mechanischer Reizung unter Geradestreckung auf ihrer
ganzen Länge oft um 10–20% und lassen so den mit Pollen
bedeckten Griffel aus der Staubbeutelröhre heraustreten (Fig.
293). Die Verkürzung erfolgt bei mäßiger Dickenzunahme, unter
Wasseraustritt in die Interzellularen, durch die elastische
Kontraktion der Membranen. Die Staubfäden von Berberis und Mahonia
sind nur an der Innenseite ihrer Basis gegen Berührung empfindlich.
Hier tritt dann auch die Verkürzung ein und verursacht, daß sich
die Antheren auf die Narbe hinbiegen.
Die zweilippigen Narbenlappen der Griffel von Mimulus,
Goldfussia, Martynia, Torenia und anderen Pflanzen schließen sich,
wenn sie berührt werden, zusammen. Kurze Zeit darauf öffnen sie
sich wieder und sind dann von neuem seismonastisch empfindlich. Ein
Öffnen erfolgt auch dann, wenn Pollen auf die Narben gebracht wird
und auf ihnen keimt. Unter seiner zerstörenden Einwirkung auf das
Narbengewebe kommt es dann aber zu einer Schließbewegung, die keine
Reizerscheinung ist.
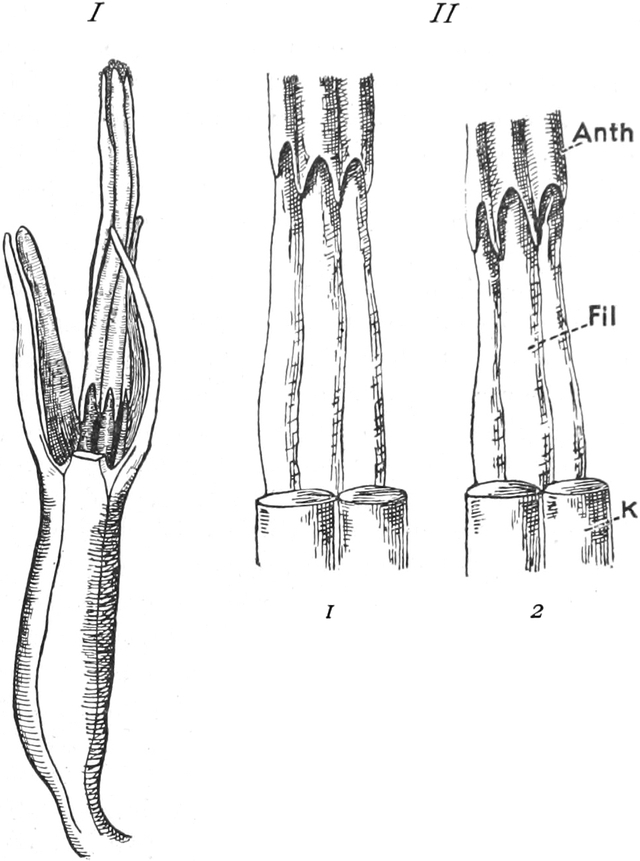
Fig. 293. Centaurea dealbata. I Blüte nach Abtragung
eines Kronblattes. Vergr. 4. II Unteres Ende der Antherenröhre und
die Filamente (nach Abtragung des Kronblattes). 1 ungereizt, 2 nach
Reizung verkürzt. Vergr. 21.
Wenn auch die Extreme von Haptotropismus einerseits, Seismonastie
andererseits eine ganz verschiedenartige Reizbarkeit aufweisen, so
fehlt es doch nicht an Pflanzen, die in ihrer Empfindlichkeit zwischen
Stoßreizbarkeit und Kontaktreizbarkeit vermitteln; namentlich gewisse
etiolierte Keimlinge, deren Haptotropismus oben (S. 315) erwähnt wurde,
sind dadurch ausgezeichnet, daß sie auch durch Wasserstrahl und durch
Stoß — freilich schwächer als durch Streichen mit festen Körpern
— gereizt werden können[285]. In bezug auf die Mechanik der
Bewegung folgen nicht alle seismonastisch empfindlichen Pflanzen dem
Beispiel der Mimose. Bei Dionaea soll die Krümmung in ähnlicher Weise
wie bei den Ranken durch Wachstum erfolgen.
III. Rückblick auf die Reizerscheinungen[286].
Reizerscheinungen sind bei den Organismen ganz
außerordentlich häufig. Bei den Bewegungen, die in diesem
dritten Abschnitt der Physiologie behandelt wurden, spielen sie
eine so große Rolle, daß hier fast ausschließlich von Reizen
die Rede war. Aber auch im zweiten Abschnitt (Formwechsel) konnten
die Reize nicht übergangen werden, und es zeigte sich, daß die
gleichen Faktoren, die Bewegungen auszulösen vermögen, auch in der
Formbildung der Pflanze die Rolle von Reizen spielen. Am wenigsten
traten die Reize bei Behandlung des Stoffwechsels hervor, weil hier
schon manche Erscheinung ihre exakte chemische Erklärung gefunden
hat, die man früher als Reizwirkung zu betrachten geneigt war. Wenn
auch zu hoffen ist, daß der Reizbegriff mehr und mehr eingeschränkt
werden kann, so sind wir doch heute gewiß noch nicht so weit, ihn
entbehren zu können, wie manche etwas stürmische Autoren glauben.
Bei der Wichtigkeit[S. 322] des Gegenstandes empfiehlt es sich aber, das
zerstreut Vorgebrachte hier am Schlusse kurz zusammenzufassen und
dabei auch einen Blick auf die Reizerscheinungen im Tierreich zu
werfen.
Zunächst ist hervorzuheben, daß im großen und ganzen die gleichen
äußeren Faktoren in der Pflanze und im Tier eine Reizwirkung
entfalten, nämlich das Licht, die Wärme, mechanische Einflüsse
(Schwerkraft, Druck, Stoß), chemische Einflüsse, galvanische
Ströme. Ein Unterschied zwischen den extremen Vertretern der
zwei Reiche besteht nur insofern, als die Pflanzen nicht auf
Schallwellen ansprechen.
Wenn man diesen Faktoren nun eine auslösende Wirkung
zuschreibt (S. 184), so soll damit nur gesagt sein, daß sie
nicht die Energie für das weitere Geschehen liefern, sondern
daß sie die Verwendung anderer, in der Pflanze disponibler Energie
veranlassen. Durchaus nicht soll etwa behauptet werden, daß
notwendigerweise jede Proportionalität zwischen Reizgröße und
Reizeffekt fehlen müsse. Es gibt ja Auslösungen, wie z. B. bei
Mimosa, wo jeder überhaupt wirksame Stoßreiz die volle Senkung des
Blattes herbeiführt, und solche Fälle zeigen die bloß auslösende
Bedeutung des Reizes besonders klar. Allein wenn, wie vielfach bei
geotropischen und phototropischen Reizen, eine Zeitlang wenigstens
eine Proportionalität zwischen Reizgröße und Krümmungsgröße
besteht, so spricht das in keiner Weise gegen Auslösung.
Sehr viel größere Schwierigkeiten stehen dem Auslösungsbegriff
auf anderen Gebieten gegenüber. So hat KLEBS betont,
daß bei vielen Entwicklungsvorgängen nicht ein einzelner
Außenfaktor maßgebend sei, sondern daß eine quantitative
Veränderung aller Lebensbedingungen eintreten müsse. Solche
quantitative Änderung der Lebensbedingungen führt z. B. Blühen
nach vorhergehendem vegetativem Wachstum herbei; rein quantitative
Änderungen in der Lichtintensität können bei Farnprothallien
weitgehend die Gestalt bedingen. Da ist es, ähnlich wie bei den
unten noch zu nennenden stationären Reizen, schwer, an
dem Begriff der Auslösung festzuhalten. Nach dem heutigen Stand
unserer Kenntnisse kann man also nur sagen, daß eine einheitliche
Definition für „Reize“ nicht mehr möglich sei, es sei denn, daß
man mit SACHS etwas unbestimmt sagt: „unter Reizbarkeit
versteht man die eigentümliche Art, wie lebende Organismen auf
Außenfaktoren antworten“. Allenfalls kann man noch hinzufügen, daß
die Eigentümlichkeit darin besteht, daß zwischen erstem Anlaß und
letzter Reaktion sich zahllose Vorgänge einschieben. Gewiß wird der
Reiz sehr häufig nur auslösend wirken, aber es ist möglich, daß
er gelegentlich auch energetisch oder auch katalytisch, d. h. nur
beschleunigend, wirkt.
Damit nun der Reiz vom Protoplasma aufgenommen werden kann, muß
dieses im reizempfänglichen Zustand sich befinden. Es ist
sehr wichtig zu konstatieren, daß dieser Zustand unter Umständen
fehlen kann, unter denen andere Funktionen des Protoplasmas völlig
normal ausgeübt werden. Die Reizempfänglichkeit („Empfindlichkeit“)
des Protoplasmas ist nämlich sehr weitgehend von Außeneinflüssen
abhängig, und es können zum Teil dieselben Faktoren, die
reizauslösend wirken, zum Teil auch andere die Empfindlichkeit
hemmen oder auch steigern. Unter den hemmenden Stoffen
sind besonders erwähnenswert die sog. Narkotika (Äther,
Chloroform, Chloralhydrat), weil sie ja auch in der Tierphysiologie
eine vergleichbare Rolle spielen. Sie können im Extrem eine
bestimmte Reizbarkeit, z. B. die phototaktische, ganz
aufheben, während Bewegungsfähigkeit erhalten bleibt. —
In anderen Fällen fehlt dem Protoplasma die Reizbarkeit aus
inneren Gründen. Endlich kann auch die Reizempfänglichkeit
gegeben sein, aber sie kann doch nicht zu einem sichtbaren
Erfolg führen, weil die Reaktionsbefähigung fehlt. So haben wir
z. B. vielfach Grund zur Annahme, daß auch ausgewachsene Organe
geotropisch reizbar sind; aber zu einer Krümmung kann es bei ihnen
nicht kommen.
Jeder Reiz muß nun eine unsichtbare, für ihn charakteristische
Veränderung im Protoplasma herbeiführen, die von einer ganzen
Kette von weiteren Veränderungen gefolgt wird, deren letzte als
„Reaktion“ bezeichnet werden; man versteht darunter die äußerlich
sichtbaren Erfolge der Reizung. Die primäre Veränderung muß aber
vor allem eine gewisse Größe haben, damit schließlich ein äußerlich
sichtbarer Effekt zutage treten kann. Es hat sich indes gezeigt,
daß auch Reize, die diese „Schwelle“ nicht erreicht haben,
sog. unterschwellige Reize, wenn sie in passender Folge
wiederholt werden, sich addieren können und schließlich doch zu
einem sichtbaren Effekt führen. Somit muß man betonen, daß auch
jeder einzelne unterschwellige Reiz einen Eindruck im Protoplasma
hinterläßt.
In vielen Fällen genügt das einfache Vorhandensein des Reizfaktors,
um an der reaktionsfähigen Pflanze eine Reizerscheinung auszulösen.
Alle für das Leben unentbehrlichen Faktoren müssen in diesem Sinn
als Reize betrachtet werden.
[S. 323]
Für gewöhnlich sieht man aber von den durch die normalen
Lebensbedingungen erzielten „stationären“ Reizen ab und
betrachtet nur diejenigen Einwirkungen als Reize, die
durch Veränderungen im bisherigen Zustand erzielt werden.
Die Veränderungen aber können entweder im Raum oder in
der Zeit erfolgen. Wenn eine Veränderung nur in der Zeit
stattfinden soll, muß ein bisher schon wirkender Faktor fernerhin
seine Qualität oder Intensität ändern, oder ein neuer Faktor muß
zu den bisherigen hinzutreten; dabei kann die Veränderung an
dem reizbaren Organ allseits gleichmäßig erfolgen. Wir
nennen solche Reize diffuse oder homogene. Besteht
dagegen die Veränderung darin, daß ein Faktor einseitig
ein Organ trifft (räumliche Änderung) oder wenigstens die eine
Seite mehr als die anderen, so spricht man von einseitigen
oder anhomogenen Reizen. Als Beispiele für diffuse Reizung
führen wir an: das Fallen oder Steigen der Temperatur, die
Veränderung der Lichtintensität und Lichtqualität, oder überhaupt
das Auftreten von Licht nach zuvoriger Dunkelheit. Als Beispiele
für anhomogene Reizung seien eine einseitige Erwärmung, eine
einseitige Beleuchtung, eine einseitige Einwirkung von Chemikalien
genannt. Es kann also ein und dasselbe Reizmittel zu diffuser wie
zu anhomogener Reizung führen, und deshalb hat man nicht nur vom
Reizmittel, sondern auch vom Reizanlaß, d. h. der Art
der Einwirkung des Mittels zu reden. — Unter Umständen kann
übrigens auch ein an sich homogener Reiz zu anhomogener Reizung
führen, nämlich dann, wenn die Pflanze durch ihren Bau eine nur
einseitige Wirkung des Reizes gestattet oder wenn sie einseitig
empfindlicher ist. Dies trifft vor allem bei dorsiventralen Organen
zu.
Von Reizen, die unseren eigenen Körper treffen, erlangen wir
wenigstens vielfach dadurch Kenntnis, daß sie uns „zum Bewußtsein
kommen“. Bei allen anderen Organismen aber können wir die Existenz
der Reizbarkeit nur daran erkennen, daß einem Reiz früher oder
später eine Reaktion folgt. Diese Reaktion kann darin
bestehen, daß eine schon im Gange befindliche Aktion beschleunigt,
gehemmt oder in andere Bahnen gelenkt wird, oder daß neue Aktionen
ausgelöst werden. Die Aktionen selbst aber können sich entweder
im Stoffwechsel oder im Formwechsel oder im Ortswechsel äußern,
genauer gesagt, sie können ihren Hauptausdruck, den uns am meisten
auffallenden Effekt, auf einem der genannten Gebiete haben.
Sicher liegt zwischen der Perzeption eines Reizes und der
Reaktion immer eine Kette zahlloser Einzelvorgänge. Von
diesen Teilvorgängen des Reizprozesses wissen wir im allgemeinen
nicht viel. Manchmal aber, nämlich dann, wenn die Reaktion
an anderer Stelle erfolgt als die Perzeption, ist ein
Glied dieser Teilprozesse, nämlich die Reizleitung ohne
weiteres bemerkbar. Die Unterscheidung dreier Einzelvorgänge:
Reizperzeption, Reizleitung und Reizreaktion erinnert uns um so
mehr an die Verhältnisse bei den höheren Tieren, je mehr besondere
Organe für sie ausgebildet sind. So ist das Tentakelköpfchen
von Drosera das Perzeptionsorgan, der obere Teil des Stieles das
Leitungs-, der untere Teil das Reaktionsorgan.
Wenn der Physiologe von Empfindung, von Stimmung, von Sensibilität
usw. redet, wenn er also Ausdrücke verwendet, die dem Seelenleben
des Menschen entnommen sind, so will er damit zweifellos auf
Ähnlichkeiten[287] hinweisen, die auf dem Gebiete der
Reizerscheinungen in den beiden Reichen bestehen, aber es liegt
ihm doch im allgemeinen ganz fern, der Pflanze eine „Seele“
zuzuschreiben. Wenn es erlaubt ist, von der Empfindlichkeit einer
Wage und von der Empfindlichkeit eines Menschen zu sprechen, so
ist auch nichts dagegen einzuwenden, wenn man von „Empfindung“ bei
Tier und Pflanze redet, auch wenn weitgehende Differenzen bestehen.
Es ist ja ganz allgemein üblich, alte Namen auf neue Begriffe zu
übertragen, dem alten Wort also einen neuen Sinn unterzuschieben.
Nötig ist dabei freilich zur Vermeidung von Mißverständnissen eine
scharfe Definition. Daran hat es aber in der Regel gefehlt. Und so
kommt es, daß der Ausdruck die Pflanze „empfindet“ die Reize dahin
geführt hat, ihr eine Seele zuzuschreiben.
Es fragt sich nun, ob Gründe für eine solche Annahme sich finden
lassen. In der Tat wird diese einerseits von ernsthaften Forschern,
andererseits aber auch von populären Schriftstellern vertreten,
welch letzteren es oft mehr auf die erzielte Sensation als auf
die Exaktheit ankommt. — So reden dann solche Schriftsteller von
„subjektiver Empfindung“, von „Wollen“ und „Denken“ bei der
Pflanze; sie schreiben ihr also die ganze komplizierte Psyche des
Menschen zu. Die Beweise für eine solche sind aber äußerst dürftig
und beschränken sich schließlich auf einige Fälle von nützlicher
Reaktion, denen man mit Leichtigkeit gegenteilige Beispiele an
die Seite stellen kann. Namentlich bei den raschen Bewegungen
der niederen Organismen entsteht leicht der Eindruck[S. 324] von
„vernünftigem“ Handeln. Daß die Pflanze denkt, wird niemand
ernstlich begründen können, und wenn man fragt, ob sie so empfindet
wie wir, ob sie Bewußtsein hat, so kann man nur antworten:
Darüber fehlt uns jede Kenntnis.
Wir können vom phylogenetischen Standpunkt aus nur sagen, daß
Pflanzen und Tiere von denselben einfachen Organismen aus sich
entwickelt haben. In beiden Reichen finden wir eine „Beantwortung“
von Reizen. Im Tier hat sich ein besonderes Organsystem
ausgebildet, dessen einzige Funktion es ist, die Reize aufzunehmen
und weiterzugeben. Ein solches Nervensystem mit seinen Zentren
(Gehirn und Rückenmark) fehlt der Pflanze zunächst einmal in
morphologischem Sinn; aber auch der Funktion nach kann man bei ihr
ein Zentralorgan nicht finden. Daraus kann man aber nicht
mit Sicherheit schließen, daß die Pflanze keine Psyche
habe. Geht man im Tierreich abwärts zu immer einfacheren Formen,
so schwindet ja auch mehr und mehr die zuverlässige Kenntnis der
psychischen Befähigung. Es sind stets nur Analogieschlüsse, die uns
die Annahme einer Psyche bei anderen Organismen als bei uns selbst
nahelegen. Sie werden um so unsicherer, je weiter wir uns vom
Menschen entfernen. Jedenfalls kann man gegen die Annahme
einer primitiven Psyche bei der Pflanze keine ernstlichen
Einwände machen.
Für die prinzipielle Gleichheit der Grundlage aller
Reizerscheinungen kann man nun die Tatsache anführen, daß gewisse
Gesetzmäßigkeiten ebenso bei der Reizbarkeit der einfachen
Pflanze wie des komplizierten Tieres festgestellt worden sind:
das Reizmengengesetz einerseits, das WEBERsche
Gesetz andererseits. Das Reizmengengesetz sagt aus, daß weder die
Intensität des Reizes noch seine Dauer allein maßgebend
ist, daß es vielmehr auf das Produkt beider ankommt. Dieses
Gesetz ist beim Geotropismus und Phototropismus nachgewiesen
und ist uns dort auch in der Form des Sinusgesetzes,
des Resultantengesetzes und des Talbotgesetzes
entgegengetreten. Auch bei Tieren ist gelegentlich das
Reizmengengesetz gültig befunden, und es ist ferner bei manchen
anderen Vorgängen an der Pflanze beobachtet worden, z. B. bei der
wachstumshemmenden Lichtwirkung an der Koleoptile von Avena und bei
der Bildung des roten Farbstoffes in den Zellen des Buchweizens.
Gerade weil solche verhältnismäßig einfachen Vorgänge diesem
Gesetz gehorchen, und weil es ferner auch bei der Lichtwirkung auf
Silbersalze gilt, kann man sagen, hier haben wir es mit rein
physikalischen und chemischen Dingen zu tun, die vor den
eigentlichen physiologischen Erscheinungen liegen.
Wichtiger ist deshalb das WEBERsche Gesetz, das bei der
Unterschiedsempfindung für Drucke beim Menschen aufgefunden wurde,
nun auch bei den chemotaktischen und haptotropischen Bewegungen
sicher nachgewiesen ist. — Aber freilich, es muß betont werden,
daß das WEBERsche Gesetz durchaus nicht bei allen
Reizerscheinungen weder bei der Pflanze noch beim Tiere gilt, und
daß ihm selbst da, wo es zutrifft, gewisse Grenzen gezogen sind.
Als weitere Ähnlichkeit der Reizerscheinungen im Tier- und
Pflanzenreich kann man noch die Tatsache anführen, daß man auch
bei der Pflanze von verschiedenen Sinnen reden kann. Denn
es hat sich gezeigt, daß, wenn auch die Reaktionen oft nach
verschiedener Reizung, z. B. nach geotropischer und phototropischer
ganz gleich ausfallen, doch die ersten physiologischen Wirkungen
von Schwerkraft und Licht, ja selbst von verschiedenen Stoffen bei
der Chemotaxis, ganz verschieden sind. Das muß man erstens
daraus schließen, daß eine Summierung solcher Reizwirkungen
nicht immer möglich ist, was doch der Fall sein müßte, wenn sie
die gleichen primären physiologischen Wirkungen besäßen, und
zweitens, daß durch homogene Reizung mit dem einen Agens keinerlei
Abstumpfung der Empfindlichkeit für einseitige Wirkung der anderen
erzielt werden kann.
Endlich mag noch darauf hingewiesen sein, daß auch die
Erscheinungen der „Stimmung“ in beiden Reichen vorkommen. Äußere
Faktoren aller Art wirken so auf den Reizprozeß ein, daß die
Reaktion weitgehend geändert werden kann. Das kann schließlich
dahin führen, daß die Richtung einer Reaktion sich ändert,
eine sonst positive Taxis z. B. in eine negative übergeht.
Es mag sein, daß diese Analogien zwischen pflanzlicher und
tierischer Reizbarkeit vielleicht nicht so tiefgehend sind, wie
manche Forscher glauben. Jedenfalls muß man aber zugeben, daß die
Annahme einer prinzipiellen Gleichheit beider einen sehr großen
heuristischen Wert gehabt hat, da sie eben zur Auffindung vieler
Gesetzmäßigkeiten geführt hat.
[S. 325]
ZWEITER
TEIL
Spezielle Botanik.
Die spezielle Botanik ist die spezielle Morphologie, Physiologie
und Ökologie der Gewächse. Während der allgemeinen Botanik die
Aufgabe zufiel, uns mit den Gesetzen bekannt zu machen, welche
die Gestaltung, die Lebensvorgänge und die Anpassungen im ganzen
Pflanzenreiche beherrschen, soll uns die spezielle Botanik in die
Kenntnis der einzelnen Gruppen des Pflanzenreichs einführen. Auf Grund
morphologischen Vergleichs der mannigfaltigen einzelnen Pflanzentypen
miteinander versucht die spezielle Morphologie einen Einblick in die
Phylogenie des Pflanzenreichs zu gewinnen. Die Lösung dieser
Aufgabe würde uns den Schlüssel zur Aufstellung eines natürlichen
Systems der Organismen liefern; denn als ein natürliches kann
nur jenes gelten, das auf ihrer wirklichen Verwandtschaft fußt.
Freilich wird jedes von uns aufgestellte „natürliche“ System nur ein
sehr unvollkommenes sein, da wir den phylogenetischen Zusammenhang
nicht direkt feststellen können, sondern ihn nur indirekt aus dem
morphologischen Vergleich erschließen müssen.
Einem solchen natürlichen System der Organismen, das nach dem
wirklichen Zusammenhang zwischen den Lebewesen sucht, stehen die
künstlichen Systeme gegenüber, die von vornherein nur ein
praktisches Ziel ins Auge fassen und die Wesen so gruppieren wollen,
daß man den Namen eines jeden derselben möglichst leicht auffinden
oder bestimmen könne. Von allen künstlichen Systemen früherer Zeiten
hatte für solche Zwecke das von CARL LINNÉ im Jahre 1735
aufgestellte Sexualsystem die größte Bedeutung gewonnen.
LINNÉ verwertete ausschließlich Merkmale, die sich auf
die Verhältnisse der Geschlechtsorgane beziehen, und unterschied
danach in seinem Sexualsystem im ganzen 24 Klassen von Pflanzen. In
der letzten, 24. Klasse vereinigte er alle Gewächse ohne deutlich
sichtbare Geschlechtsorgane und nannte sie Kryptogamen; von
diesen waren damals nur verhältnismäßig wenige Formen bekannt,
und die mannigfaltigen Fortpflanzungsverhältnisse dieser großen
Gewächsgruppe lagen noch in tiefem Dunkel. Den Kryptogamen stehen
die übrigen 23 Klassen als Phanerogamen oder Pflanzen mit
deutlich sichtbaren Geschlechtsorganen, „in Blüten“, gegenüber.
Die Phanerogamenklassen unterschied LINNÉ zunächst nach
der Verteilung der Geschlechter in den Blüten in solche mit
Zwitterblüten (Klasse I-XX) und solche mit eingeschlechtigen
oder mit teils eingeschlechtigen, teils zwitterigen Blüten
(XXI-XXIII). Die zwitterblütigen teilte er weiter in drei Gruppen
ein: Pflanzen mit freien Staubblättern (I-XV), solche mit
verwachsenen Staubblättern (XVI-XIX) und solche, deren Staubblätter
mit dem Fruchtknoten verwachsen sind (XX); die erste dieser
drei Gruppen weiterhin nach der Zahl, der Insertion[S. 326] und den
Längenverhältnissen der Staubblätter. Jede der 24 Klassen gliederte
er in Ordnungen nach ähnlichen Gesichtspunkten. Manche der so
von ihm erhaltenen Klassen und Ordnungen entsprechen natürlichen
Verwandtschaftsgruppen, die indessen in seinem System bunt
durcheinander gewürfelt sind; die meisten aber enthalten Pflanzen,
die phylogenetisch einander sehr fern stehen.
Bereits LINNÉ hatte indessen auch versucht, natürliche
Pflanzenfamilien aufzustellen (im Jahre 1738) und sie nach ihrer
„Verwandtschaft“ anzuordnen. Solange aber an die Unwandelbarkeit der
Spezies geglaubt wurde, hatte die Bezeichnung Verwandtschaft und
Familie in einem System der Organismen nur eine mystische Bedeutung.
Sie bedeutete tatsächlich weiter nichts als eine Zusammenstellung
ähnlich aussehender Wesen. Erst durch die Deszendenzlehre hat das
natürliche System der Organismen seine reale Grundlage gewonnen.
Das System, das für die nachfolgende Darstellung zugrunde gelegt
werden soll, ist das von ALEXANDER BRAUN aufgestellte, von
EICHLER, ENGLER, WETTSTEIN, MEZ und
anderen weiter ausgebildete natürliche System.
Wir gliedern das Pflanzenreich in folgende vier Hauptgruppen:
- Thallophyta, Thalluspflanzen.
- Bryophyta, Moospflanzen.
- Pteridophyta, Farnpflanzen.
- Spermatophyta, Samenpflanzen.
Erste Abteilung.
Thallophyten, Bryophyten, Pteridophyten.
Thallophyten, Bryophyten und Pteridophyten werden seit LINNÉ
allgemein als Kryptogamen, die Samenpflanzen als
Phanerogamen bezeichnet. Diesen beiden Hauptgruppen kommt
indessen vom streng systematischen Standpunkt aus ungleicher Wert
zu, denn die tiefer stehenden Vertreter der Phanerogamen nähern
sich den höchstentwickelten Kryptogamen, nämlich den Farnpflanzen,
aus denen sie hervorgegangen sind, entschieden mehr als diese den
Moosen. Die Moose zeigen wie die Thallophyten noch einen Aufbau
aus mehr oder weniger gleichartigen Zellen; beide Gruppen werden
daher auch als Zellenpflanzen zusammengefaßt und den
Gefäßpflanzen, die durch die Farnpflanzen und Samenpflanzen
vertreten werden, gegenübergestellt. Da aber andererseits die Moose
in mancherlei Beziehungen den Farnpflanzen näher stehen und mit
diesen aus gemeinsamer Wurzel als gesondert weiter entwickelte Stämme
hervorgegangen zu sein scheinen, so kann auch die Unterscheidung von
Zellenpflanzen und Gefäßpflanzen nicht zur obersten Einteilung des
Pflanzenreichs Verwendung finden.
Die Samenpflanzen oder Spermatophyten unterscheiden sich wesentlich
durch ihre Verbreitung mittels Samen von den Kryptogamen, bei
denen allgemein Sporen, das sind einzellige Keime,
von der Mutterpflanze sich loslösen und den Ausgangspunkt für die
Entwicklung eines neuen Individuums abgeben. Die Kryptogamen verdienen
daher richtiger den Namen Sporenpflanzen. Bei den Samenpflanzen
ist allerdings auch Sporenbildung vorhanden, allein die Sporenbehälter
und Sporen, aus denen die Samen als vielzellige eigenartige Gebilde
hervorgehen, entwickeln sich in Verbindung mit der Mutterpflanze
weiter, und erst die Samen lösen sich von dieser ab.
[S. 327]
Die Unterschiede zwischen Thallophyten, Bryophyten und Pteridophyten
sind in Kürze folgende:
Die Thallophyten enthalten mannigfaltige Pflanzengestalten,
die einen einzelligen oder einen mehrzelligen, mehr oder weniger
reich verzweigten Thallus vorstellen. Die Fortpflanzung geschieht
ungeschlechtlich oder geschlechtlich, meist ohne bestimmte
Aufeinanderfolge beider Reproduktionsarten. Ein Generationswechsel ist
nur bei höher stehenden Vertretern vorhanden.
Die Bryophyten und die Pteridophyten legen ihre Entwicklung in
zwei regelmäßig miteinander abwechselnden Generationen zurück. Die
ungeschlechtliche Generation bildet Sporen und wird daher Sporophyt
genannt. Aus den Sporen geht die geschlechtliche Generation, der
Gametophyt, hervor, der Geschlechtsorgane von charakteristischem Bau
erzeugt; die männlichen heißen Antheridien, die weiblichen Archegonien.
Aus der befruchteten Eizelle der Archegonien geht wiederum der
Sporophyt hervor.
Bei den Bryophyten oder Moospflanzen stellt der Gametophyt stets
einen Thallus vor, auch bei den höher stehenden Moosen, wo er zwar eine
Gliederung in Stämmchen und Blättchen, aber noch keine echten Wurzeln
und keine Leitbündel mit Gefäßen aufweist; der Sporophyt dagegen ist
eine gestielte oder ungestielte, auf der geschlechtlichen Moospflanze
halbparasitisch heranreifende Sporenkapsel.
Bei den Pteridophyten oder Farnpflanzen ist der Gametophyt ein
kleiner Thallus, während der Sporophyt Gliederung in Stengel, Blätter
und Wurzeln, mit echten Leitbündeln, also schon den Aufbau der
Spermatophyten besitzt.
Die Moose und Farnpflanzen werden auf Grund des übereinstimmenden
Baues ihrer weiblichen Sexualorgane, der Archegonien, auch als
Archegoniaten vereinigt. Indessen treten diese Organe
in etwas vereinfachter Form auch noch bei tiefer stehenden
Spermatophyten (bei den meisten Gymnospermen) auf, so daß sich
eine scharfe Abgrenzung der Archegoniaten von den höher stehenden
Pflanzengruppen nicht durchführen läßt.
Die Thallophyten unterschied man früher in Algen oder Algae,
Pilze oder Fungi und Flechten oder Lichenes. Algen sind
Thallophyten, die Chromatophoren mit Farbstoffen, vor allem
stets Chlorophyll, besitzen und dementsprechend zu selbständiger
Ernährungsweise oder Autotrophie befähigt erscheinen, während die
Pilze als heterotrophische Gewächse keine solchen Farbstoffe enthalten
und saprophytische oder parasitische Lebensweise führen. Jedoch gibt
es auch mixotrophe Algen, die nicht ausschließlich auf autotrophe
Lebensweise angewiesen sind, sondern auch in mehr oder weniger hohem
Maße organische Substanzen in ihrem Stoffwechsel verwerten können,
also in fauligem Wasser gut gedeihen. Die Einteilung in Algen und
Pilze hat nur physiologischen, keinen phylogenetischen Wert, da sie
die Verwandtschaftsbeziehungen der einzelnen Klassen, in die beide
Gruppen zerlegt werden müssen, nicht zum Ausdruck bringt. Der Thallus
der früher als einheitliche Organismen betrachteten Flechten stellt
eine Symbiose von Fadenpilzen mit Algen vor. Vom streng systematischen
Standpunkt aus müssen die Flechtenpilze und die Flechtenalgen in die
Klassen der ihnen nächstverwandten Organismen eingereiht werden;
andererseits zeigen die Flechten so viel Übereinstimmendes in Bau und
Lebensweise, daß eine zusammenfassende Behandlung im Anschluß an die
Fadenpilze für unsere Zwecke vorzuziehen ist.
Die verwandtschaftlichen Beziehungen der 14 Klassen, in die wir die
Thallophyten gliedern, bedürfen noch in vielen Punkten der Klärung und[S. 328]
sollen nur mit Vorbehalt in folgender Zusammenstellung veranschaulicht
werden:
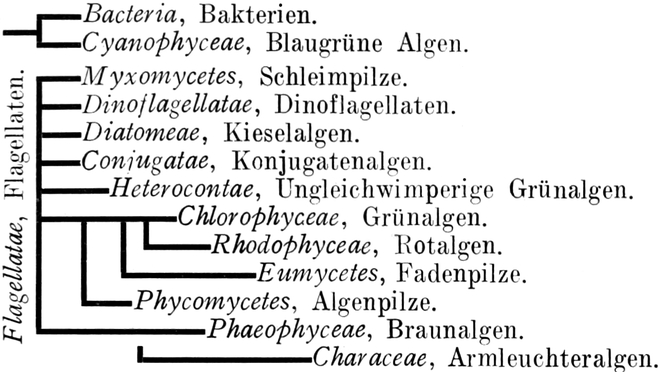
Zu den einfachsten Thallophyten gehören die Bakterien und die
Cyanophyceen, die untereinander nähere Beziehungen aufweisen
und die man daher auch als Schizophyta oder Spaltpflanzen
zusammengefaßt hat. Obwohl einige Gründe für die Angliederung der
Schizophyten an Flagellaten sprechen, erscheint es einstweilen noch
angemessen, sie in besonderer Stellung den übrigen einfacheren
Thallophyten gegenüber zu belassen, die mit mehr oder weniger
großer Wahrscheinlichkeit auf Flagellaten als Ausgangsformen
hinweisen. Die Flagellaten wurden und werden vielfach auch zu den
niedersten Tieren gerechnet; in der Tat vereinigen sie pflanzliche
und tierische Charaktere und sind auch als Ausgangsformen für
niedere Tiere zu betrachten. Die Schleimpilze dürften als farblose
Saprophyten ebenfalls von ihnen abstammen. Die Dinoflagellaten
stellen nur einen weiter entwickelten Zweig der Flagellaten dar.
Die einfachsten Formen der Heteroconten, der Grünalgen und der
Braunalgen schließen an Flagellaten an, während dagegen eine
direkte Verbindung der letzteren mit den Diatomeen und mit den
Konjugaten auf größere Schwierigkeiten stößt.
Von der Hauptreihe der Grünalgen haben sich die Algenpilze
abgezweigt; zweifelhaft ist die Anfügung der Rotalgen und der
Fadenpilze, welche beide auf gemeinsame Wurzel zurückzugehen
scheinen. Eine vorgerückte Stellung nehmen die Armleuchteralgen
ein, die meist als höchst entwickelte Grünalgen aufgefaßt werden,
in wichtigen Charakteren aber auch zu den Braunalgen einige
Beziehungen aufzuweisen scheinen; ihr Ursprung ist uns rätselhaft.
Allgemein verbreiten und vermehren sich die Thallophyten durch
ungeschlechtlich erzeugte Sporen, deren Bildungsweise große
Mannigfaltigkeit aufweist. In vielen Fällen entstehen die Sporen durch
Zellteilung im Innern von Zellen, die dann als Sporangien
bezeichnet werden, in anderen Fällen entstehen sie durch Umbildung
und Loslösung von Thalluszellen oder durch Zellsprossung. Die Sporen
heißen Schwärmsporen, Zoosporen, wenn sie kontraktile Zilien
besitzen, deren Schwingungen ihre Fortbewegung im Wasser bewirken; sie
heißen Aplanosporen, wenn sie keine Zilien tragen. Im letzteren
Falle können sie nackt sein bei Verbreitung im Wasser, oder mit Membran
umgeben bei Verbreitung in der Luft.
Außer der ungeschlechtlichen ist auch geschlechtliche Fortpflanzung
sehr verbreitet. Im einfachsten Falle besteht sie in Isogamie,
d. h. in der Verschmelzung von zwei zwar äußerlich gleichgestalteten,
physiologisch aber verschieden differenzierten Sexualzellen oder
Gameten zu einer einzigen Zelle, der Zygote oder
Zygospore. Die Organe, in denen die Gameten gebildet werden,
heißen Gametangien. Planogameten sind mit Zilien begabt,
Aplanogameten dagegen zilienlose Gametenzellen. Auf höheren
Stufen ist die Isogamie in Heterogamie übergegangen; die Gameten
sind in kleinere männliche und größere weibliche differenziert. Ist
diese Differenzierung[S. 329] so weit vorgeschritten, daß wir männliche,
meist mit Zilien versehene Zellen, Spermien (Spermatozoiden,
Spermatozoen, Samenzellen), die in Antheridien erzeugt werden,
und weibliche größere, reservestoffreichere, zilienlose, unbewegliche
Zellen, Ovien (Oosphären, Eizellen), die in Oogonien
entstehen, unterscheiden können, so sprechen wir von Eibefruchtung
oder Oogamie. Die befruchtete Eizelle, die Zygote, wird
als Oospore bezeichnet, wenn sie einen Dauerzustand eingeht;
sie kann aber auch bei gewissen Gruppen ihre Entwicklung sofort
beginnen. Man nimmt an, daß die Gameten aus ungeschlechtlichen Sporen
phylogenetisch hervorgegangen sind. Gametangien, Oogonien, Antheridien
sind bei den Thallophyten den Sporangien homologe Gebilde.
Die sexuelle Fortpflanzung hat sich in verschiedenen Gruppen, also
unabhängig mehrmals, herausgebildet. Parthenogenesis ist unter den
Thallophyten verbreitet.
Während bei gewissen Thallophyten ausschließlich ungeschlechtliche,
bei anderen nur geschlechtliche Fortpflanzung stattfindet,
kommen bei vielen beide Formen der Fortpflanzung vor, sei es an
derselben Pflanze neben- oder nacheinander, oder in einander
ablösenden getrennten Generationen. Im allgemeinen ist aber
bei den Thallophyten keine regelmäßige Aufeinanderfolge von
ungeschlechtlichen und geschlechtlichen Generationen vorhanden, da
äußere Faktoren auf die Art der Fortpflanzung von großem Einfluß
sind[289]. Nur bei wenigen Gruppen (bei manchen Braunalgen, bei
den Rotalgen und bei gewissen Fadenpilzen) folgt regelmäßig
auf eine geschlechtliche Generation (Gametophyt) eine
ungeschlechtliche (Sporophyt); ein solcher regelmäßiger
Generationswechsel ist auch bei allen Archegoniaten vorhanden.
Bei der Kopulation zweier Sexualzellen erhält der Keimkern
die doppelte Anzahl von Chromosomen; er wird diploid,
während die Sexualzellen haploide Kerne besitzen (vgl.
S. 172). Im Laufe der ontogenetischen Entwicklung muß also
eine Reduktionsteilung der diploiden Kerne zu haploiden
erfolgen. Wir unterscheiden somit eine haploide und eine
diploide Phase im Entwicklungsgange der Gewächse. Die
Reduktionsteilung erfolgt bei vielen Thallophytengruppen in der
keimenden Zygote, kann aber auch selbst innerhalb eines
Verwandtschaftskreises verschieden, an anderen Stellen einsetzen
und ist somit ein Vorgang, der mit dem Beginn einer neuen
Generation nicht notwendig verbunden zu sein braucht. Bei gewissen
Braunalgen, Rotalgen und allen Archegoniaten erfolgt die Reduktion
in den Sporangien, so daß hier regelmäßig der Gametophyt haploid,
der Sporophyt diploid ausfällt. Die Wertigkeit der Kerne kann aber
nicht als maßgebend für die spezifische Gestaltung der Generationen
erachtet werden.
Die Keimzellen der von den Flagellaten abzuleitenden
Thallophytenklassen, sowohl Schwärmzellen, als auch Gameten,
sind in vielen Fällen zilientragende nackte Protoplasten von
Flagellatencharakter. Selbst bei den Moosen und Farnen, sogar
auch bei Cycadeen und Ginkgoaceen zeigt sich noch in der Gestalt
der männlichen Gameten, wenn sie auch sekundär verändert sind,
dieses Zurückgehen in der Ontogenese auf die phylogenetischen
Ausgangsformen.
Die Bakterien stellen sehr einfach gebaute, einzellige oder
fadenförmige Organismen dar, die des Chlorophyllfarbstoffs ermangeln
und meist saprophytische oder parasitische Lebensweise führen. Sie
sind in enormer Arten- und Individuenzahl über die ganze Erde, in
der Atmosphäre, im Wasser, im Boden, ferner auf und in toten oder
lebenden Pflanzen und Tieren verbreitet. Man bezeichnet sie auch als
Spaltpilze oder Schizomycetes, weil die Vermehrung ihrer
einzelligen Formen nur durch Zweiteilung oder Spaltung der Zellen sich
vollzieht, eine Vermehrungsweise, die übrigens auch bei den anderen
einzelligen Pflanzen vorkommt.
Die Zellen der Bakterien sind von einer dünnen Membran umgeben und
enthalten ein meist farbloses Protoplasma, das bei Plasmolyse sich
von der[S. 330] Wand ganz oder teilweise zurückzieht und das im Innern
des Wandbelag seinen einzigen Saftraum oder auch mehrere Vakuolen
umschließen kann. In den Protoplasten sind zwar körnige Gebilde in
Ein- oder Mehrzahl, sog. Chromatinkörner, beobachtet, die sich durch
Farbstoffe intensiv färben lassen und von verschiedenen Autoren als
Zellkerne gedeutet werden; indessen ist es bis jetzt nicht gelungen,
unzweifelhafte Kernteilung (Karyokinese) an ihnen nachzuweisen, so daß
das Vorhandensein von Kernen noch nicht sichergestellt ist.
Die Bakterien sind zum größten Teil außerordentlich winzige Organismen,
und es gehören zu ihnen überhaupt die kleinsten bekannten Lebewesen.
So messen die kugeligen Zellen der kleinsten Arten im Durchmesser nur
0,0008 mm, die stäbchenförmigen Zellen des Tuberkelbazillus haben nur
0,0015 bis 0,004 mm Länge, die meisten Arten etwa 0,001 mm Breite und
0,005 mm Länge.
Die einfachsten Formen der Spaltpilze sind winzige kugelrunde
Zellen, Kokken. Formen mit stäbchenförmigen Zellen werden als
Bakterium oder als Bazillus bezeichnet, Stäbchen mit
kurz schraubiger Krümmung heißen Vibrio, längere Schrauben
Spirillum. Die einzelligen Kokken, Stäbchen, Vibrionen können
nach der Teilung in Zellketten vereinigt bleiben. Häufig kommt es vor,
daß die äußeren Zellmembranschichten gallertartig aufquellen und so
die Zellen oder Zellketten in Gallerte eingebettet werden. Ein solcher
Zustand heißt Zoogloea.
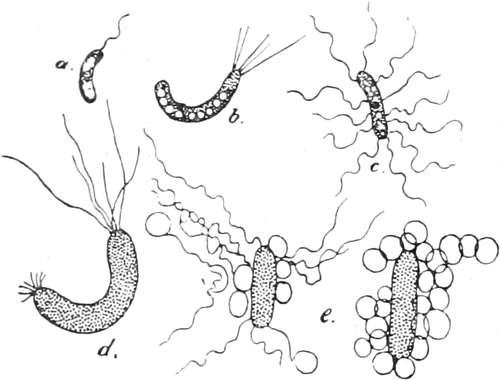
Fig. 294. Geißeltypen. a Vibrio cholerae. b, d
Spirillum undula. d Entwicklung eines neuen Geißelbüschels bei der
Teilung, c Bacillus Typhi, e Bacillus subtilis. Vergr. 2250. Nach
A. FISCHER.
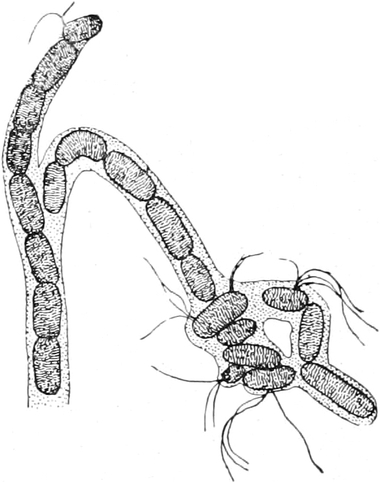
Fig. 295. Cladothrix dichotoma. Bildung der
Schwärmstäbchen aus den Fadenzellen. Vergr. 1000. Nach A.
FISCHER.
Diesen einzelligen, als Haplobakterien bezeichneten Formen stehen
die Trichobakterien gegenüber, die meist einfache Zellfäden
aufweisen (Leptothrix, Beggiatoa, Crenothrix),
bei Cladothrix aber eine unechte Verzweigung der Fäden
erreichen, die dadurch zustande kommt, daß eine Fadenzelle sich in der
Längsrichtung geradlinig weiter teilt und dabei den über ihr gelegenen
Teil des Fadens beiseite drängt.
Viele Bakterien sind durch Eigenbewegung ausgezeichnet, welche durch
Schwingungen von feinen Plasmazilien vermittelt wird[291]. Diese
Geißeln sind entweder über die Oberfläche verteilt (Heubazillus Fig.
296a, d; Typhusbazillus 294c; Tetanusbazillus 299e) oder sie
entspringen von einem Punkte aus, entweder als Einzelgeißel oder als
Geißelbüschel. Danach unterscheidet man peritriche, monotriche und
lophotriche Bakterien. Eine polare Einzelgeißel hat der Cholerabazillus
(Fig. 294a), ein polares Geißelbüschel Spirillum undula (Fig. 294b,
d); ein seitenständiges Geißelbüschel die Schwärmzelle von Cladothrix
(Fig. 295). Die Geißelbüschel können sich zu zöpfchenartigen Gebilden
zusammendrehen; sie werden niemals eingezogen, sondern gehen[S. 331] vor der
Sporenbildung oder durch ungünstige Einflüsse, oft unter vorheriger
Einrollung (Fig. 294e), zugrunde.
Die Vermehrung und Verbreitung geschieht auf vegetativem Wege durch
eine sehr ausgiebige Zweiteilung der Zellen, die bei gestreckten
Formen stets quer zur Längsachse erfolgt, die Erhaltung der Art
durch ungeschlechtliche Bildung von Dauersporen, die als
Endosporen (Fig. 296c, 297e, f) entstehen, wohl überall
in der Weise, daß die Spore im Zellplasma entweder in der Mitte oder
näher einem Ende der Zelle sich abgrenzt und mit neuer Membran umgibt.
Die Mutterzellmembran geht nach der Reife der Sporen durch Verquellen
zugrunde. Sporen sind aber nicht bei allen Arten nachgewiesen.
1. Ordnung Haplobacteria. Einzellige Bakterien. Sie umfassen die
Hauptmasse der Arten.
Obwohl ihr Formenkreis ein sehr einfacher ist, weisen ihre Arten
eine ungemeine Mannigfaltigkeit in ihrem Stoffwechsel auf. Die
meisten Bakterien haben Sauerstoff zu ihrer Atmung nötig wie die
übrigen Pflanzen, sind also aërob; manche können aber auch
ohne Sauerstoff sich weiterentwickeln, während gewisse Arten, wie
z. B. die Buttersäurebakterien, der Starrkrampfbazillus, streng
anaërob nur bei Abschluß von Sauerstoff gedeihen. Manche
Bakterien erzeugen durch ihre Atmung beträchtliche Wärmemengen;
darauf beruht die Selbsterhitzung von feuchtem Heu, Mist, Tabak,
Baumwolle (Bacillus coli und calfactor. (Vgl. S. 241.)
Wir unterscheiden autotrophe, saprophytische und
parasitische Arten, obwohl eine scharfe Trennung oft nicht
möglich ist und die letztgenannten in Kulturen auf geeigneten
Substraten auch saprophytische Lebensweise führen können.
Eine sehr bekannte saprophytische Art ist der Heubazillus,
Bacillus subtilis (Fig. 296), der sich in dem Extrakt, den man
durch Kochen von Heu gewinnt, in der Regel einstellt. Die Sporen
bleiben trotz der Hitze dabei lebensfähig und keimen zunächst zu
peritrich begeißelten, schwärmenden Stäbchen, die sich teilen
und auch in kurzen Ketten zusammenhaften. An der Oberfläche der
Flüssigkeit gehen die schwärmenden Stäbchen über in ruhende
geißellose, die sich zu langen geschlängelten Ketten weiter teilen.
Die Zellketten legen sich zu einer sog. Kahmhaut, einer besonderen
Form von Zoogloeabildung zusammen. Nach Erschöpfung der Nährstoffe
tritt dann Sporenbildung ein.
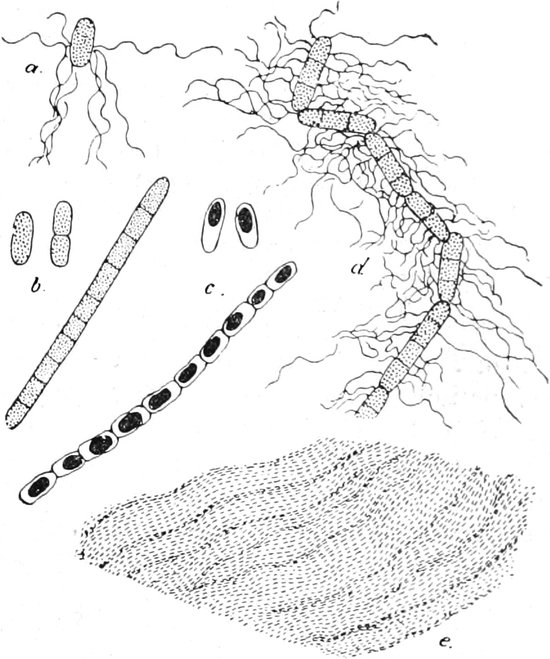
Fig. 296. Bacillus subtilis. a, d Bewegliches
Stäbchen und Kette, b unbewegliche Stäbchen und Kette, c Sporen aus
der Kahmhaut e. Vergr. a-d 1500, e 250. Aus A. FISCHER,
Vorlesungen über Bakterien.
Zu den saprophytischen Bakterien gehören als wichtige
Vertreter die zymogenen oder Gärungsbakterien und die
saprogenen oder Fäulnisbakterien. Erstere oxydieren oder
vergären hauptsächlich Kohlehydrate; letztere dagegen spalten
stickstoffhaltige tierische und pflanzliche Substanzen, Eiweiß,
Fleisch usw. unter Abscheidung übelriechender Gase. (Vgl. S. 238.)
Die Essigbakterien (Fig. 297a, b, e) oxydieren den Alkohol
zu Essigsäure. Die Vergärung von Milchzucker zu Milchsäure
wird durch den Formenkreis des Bacillus acidi lactici (Fig.
297d) bewirkt; die Bildung von Buttersäure aus verschiedenen
Kohlehydraten bei Abschluß von Sauerstoff erfolgt durch
Clostridium butyricum (Fig. 297 e) u. a., während gewisse
Sumpfbakterien (Fig. 297 f) bei Sauerstoffabschluß die Vergärung
der Zellulose zu Methan oder auch zu Kohlensäure und Wasserstoff
besorgen. Der häufigste Fäulniserreger auf Fleisch, Eiweiß usw. ist
Bacillus proteus.
[S. 332]
Streptococcus (Leuconostoc) mesenterioides (Fig. 298), der
Froschlaichpilz der Zuckerfabriken, wandelt den Zucker in Schleim
um; seine kugeligen Zellen teilen sich zu rosenkranzartigen Ketten
und umgeben sich mit dicken Gallerthüllen, deren Bildung in
zuckerfreien Substraten unterbleibt.
Die Purpurbakterien, die sich auf in Wasser faulenden
Substanzen bei Sauerstoffmangel und bei Zutritt von Licht
entwickeln, enthalten nach MOLISCH[292] einen grünen und
einen roten Farbstoff (Bakteriochlorin und Bakteriopurpurin).
Letzterer spielt nach BUDER eine Rolle für
photosynthetische Vorgänge; die Purpurbakterien scheinen die
absorbierten Lichtstrahlen zu einer Assimilation der Kohlensäure
unter Einbeziehung des freiwerdenden Sauerstoffs in den
Stoffwechsel auszunutzen. Auch andere Pigmentbakterien
scheiden Farbstoffe in ihren Zellen oder nach außen ab. Letzteres
ist der Fall bei Bacillus prodigiosus, dessen ellipsoide,
peritriche Stäbchen auf Milch oder Gebäck fuchsinrote Kolonien
bilden und so die Veranlassung zu dem Wunderglauben an blutende
Hostien gegeben haben.
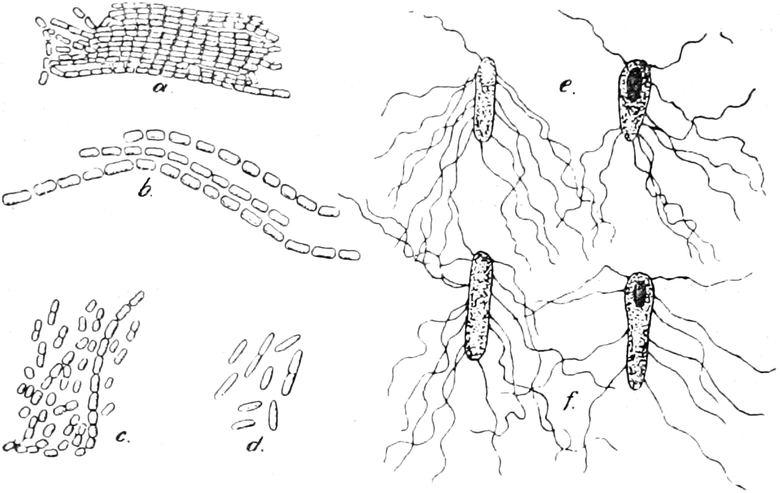
Fig. 297. Gärungsbakterien a-c Essigbakterien, a
Bacillus aceti, b Bac. Pasteurianus, c Bac. Kützingianus, d
Bac. acidi lactici, Milchsäurebazillus, e Clostridium butyricum,
Buttersäurebakterie, f Plectridium paludosum, Gärungsbakterie aus
Sumpfwasser. Vergr. 1000. Aus A. FISCHER, Vorles. über Bakt.
Die photogenen Bakterien erzeugen in ihren Zellen eine
Substanz, die bei Oxydation leuchtet. Bacterium phosphoreum
ist die verbreitetste, auf Fleisch vorkommende und sein Leuchten
verursachende Leuchtbakterie[293].
Gewisse Bodenbakterien (Clostridium Pasteurianum, Azotobacter
chroococcum) und Meeresbakterien sind imstande, den freien
Stickstoff zu assimilieren.[294] Zu diesen stickstoffbindenden
Formen gehören auch die symbiotisch in den Wurzelknöllchen der
Leguminosen lebenden, unter der Bezeichnung Bacillus radicicola
zusammengefaßten Arten von Bazillen, die im freien Zustand
bewegliche, lophotrich begeißelte Stäbchen vorstellen (Fig.
249 u. 250), ferner Mycobacterium Rubiacearum, das analoge
Bakteriengallen an den Blättern tropischer Rubiaceen erzeugt und
mit diesen Pflanzen in dauernder Symbiose lebt[295]. Andererseits
gibt es auch im Boden und sogar im Meere denitrifizierende
Bakterien, die Nitrate und Nitrite unter Abscheidung von freiem
Stickstoff zersetzen (vgl. S. 240).
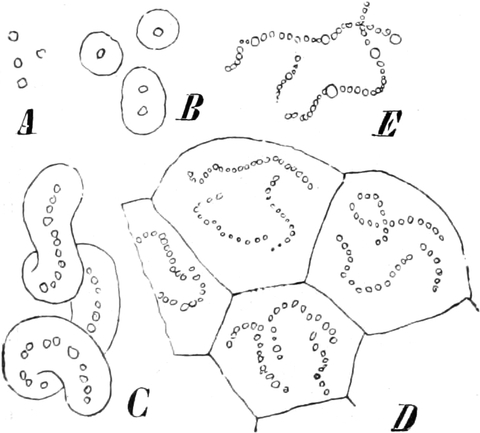
Fig. 298. Streptococcus mesenterioides. A Zellen
ohne Gallerthülle. B, C Bildung der Gallertkörper. D Teil einer
erwachsenen Zoogloea. E Rosenkranzartige Fäden der Zoogloea. Vergr.
620. Nach VAN TIEGHEM.
Die parasitischen Bakterien leben entweder in Pflanzen oder
in Tieren. Als Bakteriosen erkannte Pflanzenkrankheiten sind
z. B. die krebsartigen Geschwülste, die das von SMITH
entdeckte, auch für Menschen pathogene Bacterium tumefaciens an
höheren Pflanzen erzeugt, ferner die durch Bacillus phytophthorus
hervorgerufene Schwarzbeinigkeit der Kartoffel[296].
Von den zahlreichen pathogenen Bakterien, deren schädliche
Einwirkung auf die Gewebe und das Blut des tierischen und
menschlichen Körpers durch Abscheidung von giftigen Substanzen,
Toxinen, bedingt ist, sind als wichtigste Erreger von
Infektionskrankheiten folgende zu nennen:
[S. 333]
Staphylococcus pyogenes (Fig. 299 a), regellose oder
traubenförmige Haufen von Kokken bildend, ist der häufigste
Eitererreger, ebenso der regelmäßig bei Wundrose oder Erysipel und
anderen Eiterungen auftretende, in Ketten wachsende Streptococcus
pyogenes (Fig. 299 b), während Micrococcus (Diplococcus)
Gonorrhoeae (Fig. 299 c u. 300 a), dessen semmelförmige Kokken
paarweise nebeneinander liegen, den Tripper verursacht. Im Blut
und in den Organen milzbrandiger Tiere findet sich der durch
R. KOCH bekannt gewordene Bacillus Anthracis (Fig.
299 d, 300 c), dessen relativ große Stäbchen auch in kurzen
Ketten vorkommen und in Kulturen reichlich Endosporen, ähnlich
wie der Heubazillus bilden. Der im Erdboden verbreitete Bacillus
Tetani (Fig. 299 e) ist der Erreger des Wundstarrkrampfes. Seine
geraden, peritrich begeißelten Stäbchen wachsen nur in den Wunden
selbst; sie bilden die Sporen in ihren keulig angeschwollenen
Enden. Bacillus influenzae zeigt sehr zarte Kurzstäbchen,
Bacillus pestis kleine dickere unbewegliche Stäbchen. Der
LÖFFLERsche Bacillus Diphtheriae (Fig. 299 f) besteht
aus kleinen, zuweilen an den Enden kolbig verdickten Stäbchen. Der
KOCHsche Tuberkelbazillus Mycobacterium tuberculosis
(Fig. 299 g, 300 b), der sich in allen tuberkulösen Geweben
und Sekreten und auch im Sputum findet, ist für gewöhnlich ein
schlankes Stäbchen, kann aber auch verzweigte Formen bilden; er
ist unbeweglich, bildet keine Sporen und wird daher mit gewissen
anderen Arten zu einer besonderen Familie der Mycobacteriaceen
vereinigt[297]. Der Unterleibstyphus wird durch die peritrich
begeißelten Stäbchen des Bacillus typhi (Fig. 299 h) verursacht.
Die größte Ähnlichkeit mit letzterem hat der meist unschädliche,
stets im Darm des Menschen anwesende Kolonbazillus, Bacillus coli
(Fig. 299 i). Ebenfalls durch R. KOCH entdeckt wurde
der Kommabazillus der asiatischen Cholera Vibrio cholerae (Fig.
299 k). Dieser findet sich nur im Darm in Form kurzer, schraubig
gekrümmter Stäbchen mit polarer Einzelgeißel, nicht selten auch von
längeren Schraubenketten.
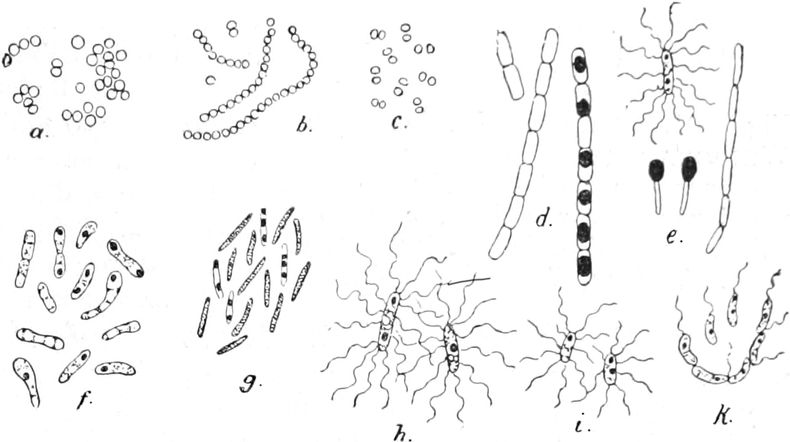
Fig. 299. Pathogene Bakterien. a Eiterkokken,
b Erysipelkokken, c Tripperkokken, d Milzbrandbazillen, e
Starrkrampfbazillen, f Diphtheriebazillen, g Tuberkelbazillen, h
Typhusbazillen, i Kolonbazillen, k Choleravibrionen. Vergr. etwa
1500. Aus A. FISCHER, Vorles. über Bakt.
Fig. 300. Färbungspräparate (aus ZIEGLERs
Lehrbuch der allgemeinen Pathologie). a Tripperkokken
im Trippersekret, Schleim- und Eiterkörperchen mit Kokken
(Methylenblau-Eosin), Vergr. 700; b Tuberkelbazillen im Sputum eines
Lungenkranken (Fuchsin-Methylenblau), Vergr. 400; c Milzbrandbazillen
in Milzbrandpusteln (Methylenblau-Vesuvin), Vergr. 350. Aus A.
FISCHER, Vorles. über Bakterien.
[S. 334]
Außer diesen bösartigen Parasiten gibt es aber auch zahlreiche mehr
oder weniger harmlose, auf den Schleimhäuten, in der Mundhöhle
(Fig. 80), im Darm lebende Arten, so z. B. die im Magen und
Darm des Menschen auftretende Sarcina ventriculi, welche aus
würfelförmigen Klumpen von Kokken besteht.
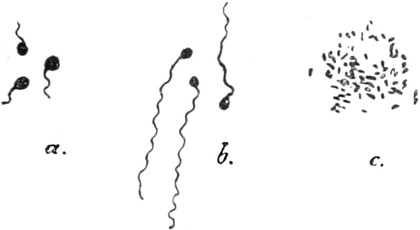
Fig. 301. Salpeterbakterien nach WINOGRADSKY.
a Nitrosomonas europaea von Zürich, b Nitrosomonas javanensis von
Java, c Nitrobakter aus Quito. Vergr. 100. Aus A. FISCHER,
Vorl. über Bakt.
Den saprophytischen und parasitischen Formen stehen
die autotrophen gegenüber, die sich trotz ihres
Mangels an Chlorophyll ganz selbständig aus anorganischen
Verbindungen ernähren. So verhalten sich die im Boden lebenden
Nitritbakterien (Nitrosomonas) und Nitratbakterien
(Nitrobacter), von denen erstere Ammoniak zu salpetriger Säure
und letztere die salpetrige Säure zu Salpetersäure oxydieren.
Beide benutzen als Kohlenstoffquelle die Kohlensäure, kommen also
gänzlich ohne organische Substanzen aus (Fig. 301, vgl. auch S.
219).
2. Ordnung Trichobacteria. Fadenbakterien. Die Fadenbakterien
umfassen nur einige Gattungen. Sie stehen in ihrer Organisation den
fadenförmigen Cyanophyceen nahe und können, wenigstens zum Teil,
von diesen als farblose Formen abgeleitet werden. Die meisten Arten
leben saprophytisch im Wasser, einige auch autotroph.
Überall verbreitet in unreinen Gewässern ist die morphologisch
am höchsten stehende Cladothrix dichotoma. Ihre feinen, aus
stäbchenförmigen Zellen bestehenden, unecht verzweigten (vgl. S.
67), festsitzenden Fäden bilden schleimige Überzüge an Algen,
Steinen oder Holzwerk. Sie vermehren sich durch zilientragende
Schwärmzellen, die durch Teilung aus den Fadenzellen entstehen und
durch Verquellen der Fadenscheide frei werden (Fig. 295). Nach dem
Schwärmen setzen sich die Zellen fest und wachsen zu neuen Fäden
heran.
Sehr häufig ist ferner der Brunnenfaden, Crenothrix polyspora,
mit unverzweigten, festsitzenden, aber leicht zerbrechlichen
Fäden, die Eisenoxydhydrat in ihren Scheiden speichern können. Er
entwickelt sich oft in solchen Massen in Wasserleitungen, daß die
Röhren sich verstopfen und das Trinkwasser ungenießbar wird. Bei
Crenothrix zerfallen die Fadenzellen in der Scheide durch Teilung
in zahlreiche geißellose, rundliche Zellen, welche die Vermehrung
besorgen.
In Schwefelquellen und am Boden von Gewässern, wo durch Fäulnis
organischer Stoffe Schwefelwasserstoff auftritt, siedeln sich
zahlreiche Schwefelbakterien[298] an, unter denen die
fadenförmige Beggiatoa alba (Fig. 302) am verbreitetsten ist.
Die Schwefelbakterien ernähren sich auch autotroph, also ohne
organische Substanzen, indem sie Ammoniumsalze und Kohlensäure
in ihrem Stoffwechsel verwerten. Den für ihre Lebenstätigkeit
nötigen Schwefelwasserstoff oxydieren sie zu Schwefel, den sie
in rundlichen Tröpfchen gleichsam als Reservestoff aufspeichern
und zu Schwefelsäure oxydieren, die weiterhin durch Karbonate
neutralisiert wird. Bei fortschreitendem Schwefelwasserstoffmangel
werden immer mehr Schwefeltröpfchen gelöst, die Querwände in
den Fäden treten deutlicher hervor, die Fadenzellen runden sich
schließlich ab und lösen sich voneinander los. So können die
Beggiatoen sich leicht nach neuen Standorten verbreiten. Zu den
Schwefelbakterien gehören auch einige Haplobakterien. (Vgl. S. 238.)
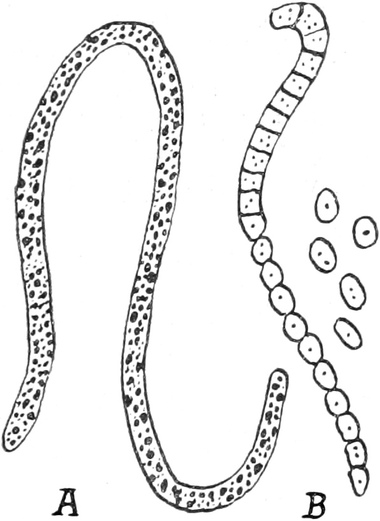
Fig. 302. Beggiatoa alba. A Faden mit
Schwefeltröpfchen, Querwände nicht sichtbar. B Faden bei Mangel an
Schwefelwasserstoff, in die Gliederzellen zerfallend. Vergr. 600. Nach
DÜGGELI.
Zu den Eisenbakterien[299] gehört die in Wiesensümpfen
und Bächen häufige, fadenförmige Chlamydothrix (Leptothrix)
ochracea. Sie oxydiert kohlensaures Eisenoxydul zu
Eisenoxydhydrat, das in den Fadenscheiden aufgespeichert wird,
und kommt bei dieser Ernährungsweise mit Spuren organischer
Nahrung aus. Andererseits gedeiht sie aber auch ohne Eisen recht
gut in organischen Nährlösungen. Andere Eisenbakterien, so nach
LIESKE Spirophyllum ferrugineum, sind rein autotroph.
Sie gedeihen nur in[S. 335] Wasser, in dem außer geringen Mengen
anorganischer Salze Eisenoxydulkarbonat gelöst ist. Sie oxydieren
das Eisenoxydul zu Eisenoxydhydrat und speichern dieses auf.
Diese Oxydationen scheinen die Energie für die Assimilation der
Kohlensäure zu liefern.
Klasse II.
Cyanophyceae, Blaugrüne Algen oder
Spaltalgen[288],
[300].
Die Cyanophyceen sind einfach organisierte, teils einzellige, teils
fadenförmige, blaugrün gefärbte Thallophyten, deren Zellen oder
Fäden häufig durch Gallerte, die durch fortgesetzte Verquellung
der Außenschichten ihrer Membranen entsteht, zu Kolonien vereinigt
erscheinen. In zahlreichen Arten über die ganze Erde verbreitet,
bewohnen sie die Gewässer oder vegetieren auf feuchtem Schlammboden,
an feuchten Felsen oder Baumrinden in gallertartigen Massen oder
feinfädigen Überzügen. Sie sind wie die rein grünen Algen autotroph.
Der Protoplast besitzt ein hohlzylindrisches oder hohlkugeliges
peripherisches Chromatophor, welches außer Chlorophyll einen blauen
Farbstoff, das Phykocyan, nach dem die Gruppe ihren Namen
trägt, bei gewissen Arten außerdem auch rotes Phykoerythrin
in verschiedener Menge enthält und als Assimilationsprodukte
Glykoproteïde, die aus Glykose gebildet werden, liefert. Das
innerhalb des Chromatophors befindliche, als Zentralkörper
bezeichnete, farblose Plasma enthält chromosomenähnliche Gebilde;
es ist als Zellkern gedeutet worden. Besondere Einschlüsse
der Zellen sind ferner die aus Proteïnsubstanzen bestehenden
Cyanophycinkörner, die vorwiegend in den Chromatophoren liegen. Die
Membranen bestehen aus Zellulose und Pektinstoffen.
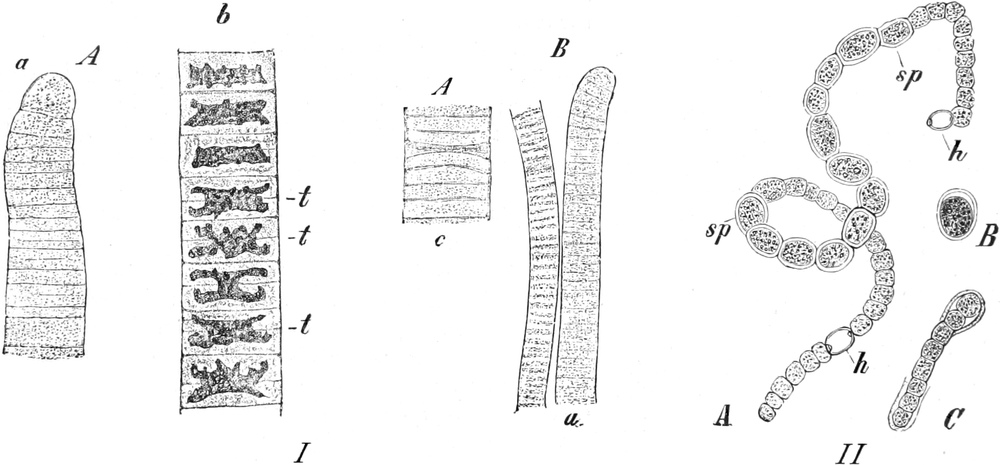
Fig. 303. I A Oscillaria princeps. B Oscillaria
Froelichii. a Fadenenden, b und c Stücke aus dem inneren Teil des
Fadens, t Zellen in Teilung. Nach Fixierung und Färbung. Vergr. von
A 1080, von B 540. Nach STRASBURGER. II Nostoc Linckii,
in Wasser schwimmende Art. A Fadenstück mit zwei Heterozysten h und
einer Anzahl von Sporen sp. B Spore in Keimung. C Junger Faden
aus der Spore hervorgegangen. Vergr. 650. Nach BORNET.
Die Vermehrung geschieht ausschließlich durch Zellteilung. Bei
vielen Arten werden Sporen als Dauerzustände gebildet durch
Vergrößerung und starke Wandverdickung einzelner Zellen (Fig. 303
II), also in anderer Weise als bei den Bakterien.
Wie die Bakterien als Spaltpilze, Schizomyceten, so werden die
blaugrünen Algen als Spaltalgen, Schizophyceen, infolge
ihrer Vermehrung durch Teilung oder Spaltung, bezeichnet. Beide
Gruppen werden vielfach zu einer Klasse der Spaltpflanzen,
Schizophyta, vereinigt. In der Tat zeigen sie vieles
Gemeinsame, indessen ist zu bemerken, daß die für Bakterien
charakteristischen Geißeln und Endosporen den Spaltalgen fehlen.
Die einfachsten Cyanophyceen bestehen aus blaugrünen rundlichen
Zellen, so die Arten der Gattung Chroococcus. Bei Gloeocapsa
(Fig. 35), deren Arten meist in gallertigen[S. 336] Überzügen an feuchten
Felsen und Mauern auftreten, bleiben die Zellen nach der Teilung
durch geschichtete Gallerthüllen zu mehrzelligen Kolonien verbunden.
Unter den fadenförmigen Arten, die teils unverzweigt sind,
teils unechte Verzweigung aufweisen (Fig. 85) und die kein
Spitzenwachstum zeigen, sind die überall im Wasser oder auf
Schlammboden häufigen Oscillaria-Arten die einfachsten, da
sich hier die meist von einer dicken Scheide eingeschlossenen
Fäden aus gleichartigen scheibenförmigen Zellen zusammensetzen
(Fig. 303 I). Die Fäden zergliedern sich in kurze Fadenstücke,
Hormogonien, die aus der Scheide nach außen gelangen, der
Vermehrung dienen und zu neuen Fäden heranwachsen. Die Gattung
Nostoc besitzt rosenkranzförmige Fäden (Fig. 303 II); ihre
Arten leben auf feuchtem Boden oder im Wasser in Form von
unregelmäßig gestalteten oder kugeligen Gallertkolonien. Bei
manchen fadenförmigen Cyanophyceen kommt es zur Ausbildung
besonderer, wohl als Reservestoffbehälter dienender Zellen mit
gelblichem Inhalt, Grenzzellen oder Heterozysten (Fig. 303
II, h), die nach einer Ruhezeit unter günstigen Umständen
wieder ergrünen und keimen können.
Die fadenförmigen Blaualgen, besonders die Oscillarien, aber
auch die Hormogonien von Nostoc und verwandten Gattungen, führen
auf festen Unterlagen photo- und chemotaktische Kriechbewegungen
aus. Das gewöhnlich unter Drehung um die Längsachse erfolgende
Vorwärtsgleiten der Fäden kommt dadurch zustande, daß die Zellen
aus Poren ihrer Längswände Schleim, ein nicht doppelbrechendes
Kohlehydrat, nach rückwärts abscheiden[301].
Gewisse Cyanophyceen leben an der Oberfläche von Teichen und
bedecken sie mit einer „Wasserblüte“; sie verdanken ihr
Schwimmvermögen nach KLEBAHN winzigen Gasvakuolen,
die im Zellplasma gelagert sind[302]. Eine in wärmeren Meeren
schwimmende Art ist Trichodesmium erythraeum, das als „Seeblüte“
dem Wasser rote Färbung verleiht (Rotes Meer).
Manche Blaualgen beteiligen sich an der Zusammensetzung der
aus Pilzen und Algen bestehenden Flechten. Einige Arten leben
endophytisch in Gewebehöhlungen anderer Pflanzen, so Anabaena in
Azolla; Nostoc-Arten in gewissen Lebermoosen, in Wasserlinsen
(Lemna), in den Wurzeln von Cycas; Nostoc punctiforme als
fakultativer Parasit im Rhizom von Gunnera[303].
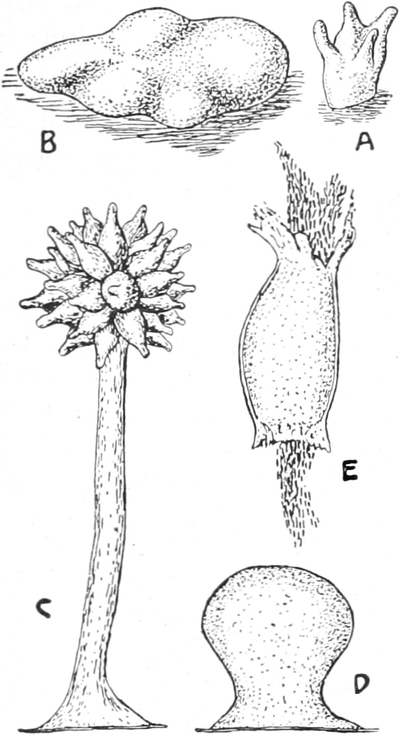
Fig. 304. A Myxococcus digitatus, hellroter
Fruchtkörper auf Mist. Vergr. 120. B Polyangium primigenium, roter
Fruchtkörper auf Kaninchenmist. Vergr. 40. C Chondromyces apiculatus,
rotgelbe Fruchtkörper auf Antilopenmist. D Fruchtkörperanlage.
Vergr. 45. E Einzelne Zyste in Keimung. Vergr. 200. A, B nach
QUEHL, C-E nach THAXTER.
Unter Vorbehalt reihen wir hier die Polyangideae
(Myxobacteriaceae)[312] an, die zuerst durch THAXTER
genauer bekannt geworden sind und von ihm zu den Bakterien
gerechnet wurden, nach VAHLE den Schleimpilzen näher
stehen sollen, von JAHN aber wieder zu den Schizophyten
gestellt werden und vielleicht als farblos gewordene Organismen
aus dem Formenkreis der blaugrünen Algen hervorgegangen sein
mögen. Sie sind sehr verbreitet und leben saprophytisch auf Mist
von Tieren. Im vegetativen Stadium sind sie plasmodienähnliche
Schwärme von stäbchenförmigen kleinen Zellen, die, in Schleim
eingebettet, zusammenhalten, kriechende Bewegungen ausführen
und schließlich meist lebhaft gefärbte Fruchtkörper bilden, in
denen die Stäbchenzellen zu kugeligen Sporen sich umwandeln. Die
Fruchtkörper, mit Wandung versehene Zysten, sind ungestielt oder
erheben sich einzeln oder in Gruppen auf Stielen, deren Substanz
ebenso wie die Zystenwände aus dicht nebeneinander gelagerten, von
der Sporenbildung ausgeschalteten Stäbchenzellen hervorgeht. Bei
der Keimung treten die aus den Sporen entstehenden Stäbchenschwärme
aus der aufbrechenden Zyste hervor (Fig. 304).
Die Flagellaten bilden eine ungemein formenreiche Gruppe einzelliger,
wasserbewohnender Organismen, die pflanzliche und tierische
Eigenschaften in sich vereinigen und als Ausgangsformen einerseits für
niedere Thallophyten, andererseits für Protozoën betrachtet werden
können.
Der kontraktile oder amöboid sich bewegende Protoplast dieser
Organismen ist nach außen durch eine dichtere Plasmahautschicht
abgegrenzt. Er besitzt eine oder mehrere Zilien
(Geißeln, Flagellen) als Bewegungsorgane, einen Zellkern, pulsierende
Vakuolen und bei vielen Vertretern wohlausgebildete grüne
oder gelbbraune Chromatophoren, vielfach auch einen roten
Augenfleck. Als Assimilationsprodukt tritt meist Öl auf, aber
auch Stärke oder andere Kohlehydrate. Andere Formen sind farblos
und ernähren sich ausschließlich saprophytisch oder auch zugleich
animalisch. Der Protoplast mancher Flagellaten, besonders der
farblosen, aber auch gefärbter Arten, kann nämlich Formänderungen und
kriechende Bewegungen ausführen (Amöbenzustand) und auch fadenförmige
dünne Fortsätze, Pseudopodien (Rhizopodien), aussenden und wieder
einziehen, durch welche die Aufnahme von Nahrungskörpern vermittelt
wird (Fig. 305, 311).
Die meisten Vertreter leben als nackte freie Zellen, andere in
mehr oder weniger komplizierten, durch Gallerte zusammengehaltenen
Zellkolonien; gewisse Gattungen zeichnen sich aus durch die Bildung von
eigenartigen, abstehenden, gestielten oder ungestielten, hornartigen
Gehäusen, durch Kieselskelett oder durch Kalkhüllen.
Die Vermehrung geschieht auf rein vegetativem Wege durch
Teilung der Zellen allgemein der Länge nach. Bei vielen Arten werden
als Ruhestadien geißellose, mit Membran umgebene Dauerzellen oder
Zysten erzeugt, die vielfach bei ihrer Keimung nach Teilung
ihres Inhalts mehrere Tochterzellen ausschwärmen lassen (Fig. 310).
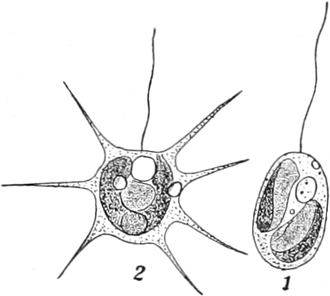
Fig. 305. Chrysamoeba radians. Im Süßwasser; mit 1 Zilie
und 2 gelbbraunen Chromatophoren. 1 Gewöhnliche Form. 2 Amöboider
Zustand mit Pseudopodien. Nach KLEBS.
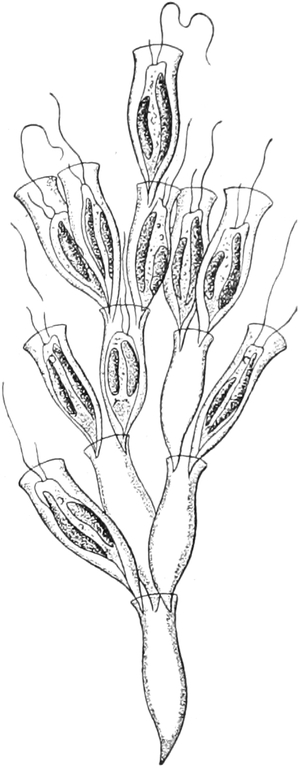
Fig. 306. Dinobryon Sertularia. Gehäuse und Kolonie
bildend. Im Süßwasserplankton. Vergr. 600. Nach SENN.
Eine wichtige Flagellatengruppe sind die Chrysomonadinen, zu
denen u. a. Chrysamoeba (Fig. 305) und das Kolonien bildende
Dinobryon (Fig. 306) gehören. Sie zeichnen sich durch radiären
Bau und durch meist gelbbraune, seltener durch rote oder
blaugrüne Chromatophoren aus, die ein im Plasma abgelagertes
besonderes Kohlehydrat (Leucosin) bilden. Als eine höher stehende
Chrysomonadine gilt Hydrurus foetidus (Fig. 307), der in Form
von Zellkolonien, bis 30 cm langer verzweigter, gallertartiger
Fäden, an Steinen, in fließendem Wasser lebt. In der Fadengallerte
liegen zahlreiche geißellose, durch Längsteilung sich vermehrende
Zellen eingebettet. Diese liefern schließlich ausschlüpfende,
tetraëdrisch gestaltete, einzilige Flagellatenschwärmer, die
sich wieder[S. 338] festsetzen und zu neuen Gallertfäden heranwachsen.
Auch einige andere verwandte Flagellaten sind zu solchen
algenartigen Lebensformen vorgeschritten, bei denen das ruhende
Stadium vorherrscht, während das bewegliche der Vermehrung und
Ausbreitung dient. An die Chrysomonadinen schließen sich zwei
eigenartige Familien mariner sehr kleiner Planktonorganismen
an, die Silicoflagellaten[306], die sich durch ringförmige
oder hutförmige, durchbrochene Kieselskelette auszeichnen (Fig.
308 A), und die Coccolithophoriden[307], die eine Hülle aus
Kalkplättchen und Kalkstachelchen besitzen und sich durch meist
zu 4 gebildete Schwärmsporen vermehren (Fig. 308 B, C).
Aus dem Verwandtschaftskreise der Chrysomonadinen scheinen die
Diatomeen und auch die Heteroconten hervorgegangen zu sein, so daß
diese drei Gruppen auch als Chrysophyta zusammengefaßt werden
können. Ihre gemeinsamen Merkmale sind u. a. die sehr häufige
oder stete Zusammensetzung der Gehäuse- und Zellmembranen aus
zwei Schalenstücken, die Verkieselung der Membran und die häufige
Ungleichheit der Geißeln.
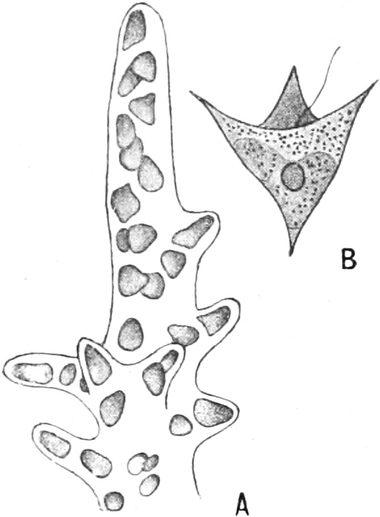
Fig. 307. Hydrurus foetidus. A Spitze eines
Gallertlagerzweiges nach BERTHOLD. B Schwärmzelle nach
KLEBS. Aus PASCHER, Flagellaten.
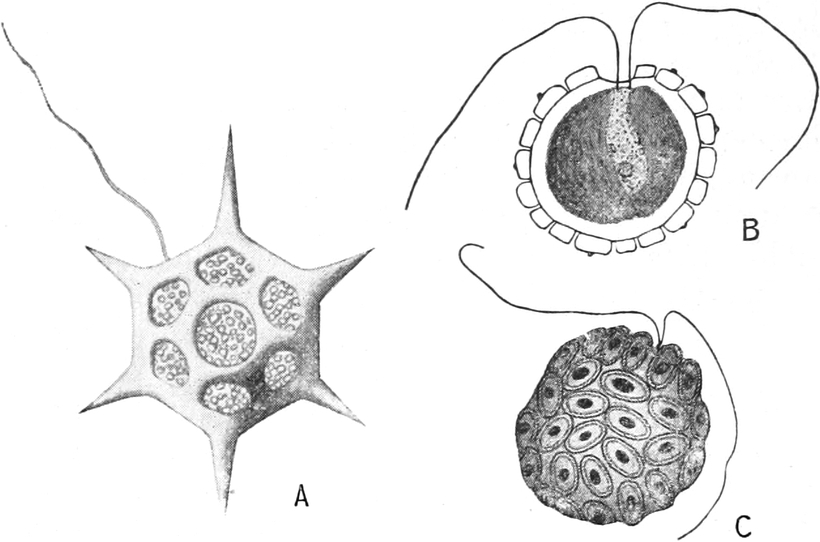
Fig. 308. A Distephanus speculum nach
BORGERT. B, C Calyptrosphaera insignis aus der Adria,
B im optischen Durchschnitt mit 2 braungelben Chromatophoren, C
Außenansicht. Vergr. 1600. Nach SCHILLER.
Die etwas höher als die Chrysomonadinen differenzierten, meist
gelbbraun, doch auch grün oder blaugrün gefärbten Cryptomonadinen
unterscheiden sich von ihnen durch dorsiventrale, vorn schräg
abgestutzte Protoplasten mit zwei ungleich langen, aus einer
furchenartigen Vertiefung entspringenden Geißeln (Fig. 309). Zu
ihnen gehört auch Chrysidella (Zooxanthella). Sie lebt in
Symbiose mit zahlreichen Meerestieren (Radiolarien, Actinien usw.),
in deren Plasma ihre gelben Ruhezellen liegen. Aus der Ruhezelle
schlüpft später der Protoplast als begeißelter Flagellat aus. In
die Verwandtschaft der Cryptomonadinen gehören die Dinoflagellaten.
Auch schließen sich an sie einige Gattungen (Phaeocystis,
Phaeothamnion u. a.) an, die den größten Teil ihres Lebens in
unbeweglichen Gallertkolonien oder Zellverbänden verbringen und
bereits an Braunalgen erinnern, aber nur mit Zweifel als Vorstufe
zu solchen gerechnet werden können.
Fig. 309. Cryptomonas erosa. Vergr. 650. Nach
STEIN.
Unter den mit grünen Chromatophoren versehenen Flagellaten
ist der Ausgang für die Chlorophyceen zu suchen.
Die Eugleninen[308] sind eine hochstehende grüne
Flagellatengruppe. Euglena-Arten (Fig. 310 A) treten oft in
Dorfteichen in Form einer grünen „Wasserblüte“ auf. Nahe verwandt
mit ihnen sind ähnlich gestaltete, aber farblose saprophytische
Formen. Euglena gracilis kann sogar durch Kultur in organischen
Lösungen unter Lichtabschluß in eine hyaline Dunkelform mit
Leukoplasten übergeführt werden. In dieser Gruppe[S. 339] scheint zum
ersten Male bei Flagellaten sexuelle Fortpflanzung, Kopulation
zweier gleichgestalteter Gameten vorzukommen; jedoch bedürfen die
Angaben noch weiterer Bestätigung.
An die Flagellaten mit gefärbten Chromatophoren schließen wir
die zahlreichen farblosen, saprophytisch oder animalisch
lebenden Formen an, die aus ersteren hervorgegangen sein dürften.
Für gewisse Vertreter läßt sich sogar noch nahe Verwandtschaft mit
gefärbten auf Grund übereinstimmender Zellstrukturen nachweisen.
Bei den Pantostomatinen geschieht die Aufnahme von
Nahrungskörperchen durch die ganze Oberfläche des Protoplasten
meist mittels Pseudopodien (Fig. 311), bei den Protomastiginen
meist nur an einer Mundstelle, bei den Distomatinen an zwei
Mundstellen. Zur zweiten Gruppe gehören auch gewisse im Blut und im
Darm von Tieren lebende Arten, die zum Teil Erreger gefährlicher
tropischer Krankheiten sind, so Trypanosoma brucei, das die
Tsetsekrankheit des Rindviehs, und T. gambiense (Fig. 312), das
die Schlafkrankheit beim Menschen verursacht; beide werden durch
Fliegen der Gattung Glossina übertragen.
Wir dürfen annehmen, daß aus farblosen Flagellaten die Myxomyceten
ihren Ausgang genommen haben, ferner daß auch die niederen
Protozoën (Rhizopoden) hier ihren Anschluß nach unten finden.
Fig. 310. Euglena gracilis. A Form mit grünen
Chromatophoren ch, Zellkern n, Vakuole nebst rotem Augenfleck
v, Geißel g. B Halbsaprophytische Form mit kleinen grünen
Chromatophoren. C Farblose saprophytische Form, bei Lichtabschluß in
Nährlösung gezogen. D Dauerzyste der Form C, r roter Augenfleck.
E Keimung der Dauerzyste der Form A in vier später austretende
Tochterzellen. A, C Vergr. 360, B 650, D, E 1000. (Nach
ZUMSTEIN.)
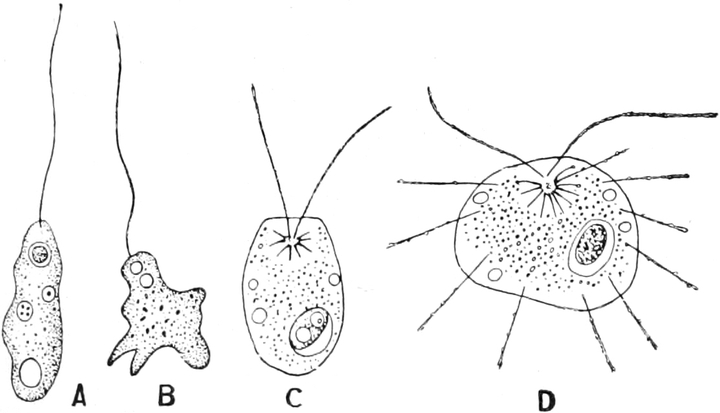
Fig. 311. Mastigamoeba invertens. A freischwimmend,
B Amöbenzustand. Vergr. 1033. Dimorpha mutans, C mit eingezogenen,
D mit ausgestreckten Pseudopodien. In einer Vakuole ein aufgenommenes
Nahrungskörperchen. Vergr. 666. Nach LEMMERMANN.
Fig. 312. Trypanosoma gambiense. A aus dem Blut
infizierter Affen. Geißel eine undulierende Membran bildend. B aus
der Fliege Glossina. Geißel im Innern. Nach MINCHIN.
Die Schleimpilze bilden eine eigenartige, von Flagellaten abgeleitete
Gruppe niederer Thallophyten; sie nehmen ebenfalls eine Mittelstellung
zwischen Pflanzen und Tieren ein und werden daher auch als Mycetozoa
oder[S. 340] Pilztiere bezeichnet. Zunächst sei das Verhalten der
umfangreichsten Ordnung, der Myxogasteres, dargestellt, die in
zahlreichen Arten über die ganze Erde verbreitet sind. Im vegetativen
Zustande bestehen diese Schleimpilze aus nackten, saprophytisch
sich ernährenden Protoplasmamassen, den Plasmodien, welche
zahlreiche kleine Zellkerne enthalten, des Chlorophylls vollständig
ermangeln und als Reservestoff keine Stärke, sondern Glykogen bilden.
Die Plasmodien (Fig. 4) finden sich mit Vorliebe auf dem Boden der
Wälder, auf abgefallenen Blättern, auf und in faulendem Holz. Sie
nehmen auch feste Nahrung auf, kriechen unter Formänderung und
Verzweigung im Substrat umher und gelangen dort zu den für ihre
Ernährung günstigsten Stellen hauptsächlich vermöge ihrer Befähigung
zu chemotaktischen, hydrotaktischen und negativ phototaktischen
Bewegungen. Vor der Sporenbildung aber ändern sich diese Reizbarkeiten;
das Plasmodium kriecht dann aus dem feuchten Substrat dem Licht
entgegen und wandelt sich je nach den Gattungen in einen einzigen oder
in zahlreiche, dicht nebeneinander stehende Fruchtkörper um. Jeder
Fruchtkörper (Sporangium) bildet an seiner Peripherie eine
Hülle (Peridium) und in seinem Innern zahlreiche kleine, mit
Membran umkleidete Sporen. Bei vielen Gattungen kommt es auch
zur Ausbildung eines Capillitiums (Fig. 313 A, B, 314 B),
das entweder aus freien oder aus netzförmig verbundenen feinen Röhrchen
oder Fasern besteht und aus dem zwischen den Sporen befindlichen Plasma
hervorgeht. Bei der Fruchtreife bricht das Peridium des Sporangiums
auf; das Capillitium lockert sich, streckt sich hervor und die Sporen
werden durch seine hygroskopischen Bewegungen und durch den Wind
ausgestäubt. Die Gattung Ceratiomyxa verhält sich insofern einfacher,
als ihre Fruchtkörper keine Hüllen besitzen, sondern die Sporen an der
Oberfläche auf kleinen Stielchen tragen.
Fig. 313. Reife geöffnete Fruchtkörper nach Entleerung
der Sporen. A Von Stemonitis fusca. Vergr. 10. B Von Arcyria
punicea. Vergr. 12. C Von Cribraria rufa. Vergr. 32.
Fig. 314. A-C Trichia varia. A Sporangien
geschlossen und geöffnet. Vergr. 6. B und C Capillitiumfaser und
Sporen. Vergr. 240. D Leocarpus fragilis. Gesellige Einzelsporangien
auf Moos. Nat. Gr.
Die Sporen (Fig. 315, Chondrioderma) keimen im Wasser oder
auf nassem Substrat. Der aus der Sporenhaut austretende
Protoplast erzeugt an seinem vorderen Ende nur eine einzige lange
Zilie oder Geißel als Bewegungsorgan und wird so zu einer
Schwärmspore (Fig. 315 e–g); sie besitzt einen Zellkern
am vorderen Ende und eine pulsierende Vakuole am hinteren Ende.
Schon innerhalb der Sporenhaut kann eine Zellteilung erfolgen,
so daß dann zwei Schwärmsporen aus ihr entlassen werden. Die
Schwärmsporen können sich bei gewissen Arten noch durch Zweiteilung
vermehren. Nach einiger Zeit verlieren sie ihre Zilien und gehen
in den Zustand der Myxamöben (Fig. 315 i, k) über. Die
Amöben vermehren sich ebenfalls durch Teilung (Fig. 316 A, B).
Unter ungünstigen Lebensbedingungen umgeben sie sich mit Membran
und bilden Ruhe[S. 341]zustände, sogenannte Mikrozysten, die unter
günstigen Bedingungen wieder Schwärmsporen austreten lassen.
Die Myxamöben verschmelzen nach JAHN[310] paarweise
miteinander, wobei auch ihre haploiden Kerne kopulieren (Fig. 316
C).
Die so durch einen Sexualakt entstandenen einkernigen
Amöbozygoten vereinigen sich zu größeren mehrkernigen Plasmodien.
Diese nehmen auch noch weitere haploide Amöben auf, verdauen sie
aber in Vakuolen(Fig. 316 D). Schließlich schreiten sie zur
Fruchtkörperbildung. Die Kerne der Plasmodien sind diploid und
erfahren wiederholte mitotische Teilungen (Fig. 316 E). Ihre
letzte Teilung vor der Sporenabgrenzung ist eine Reduktionsteilung,
wodurch die Zahl der Chromosomen wieder auf die Hälfte verringert
wird. Jeder so entstandene haploide Tochterkern wird zum Kern
einer Spore. Die nicht zur Sporenbildung verwendeten Kerne gehen
zugrunde. Bei Ceratiomyxa können die Sporen außer ihrem normalen
Kern auch noch einen degenerierenden enthalten. Aus dem ersteren
entstehen hier durch zweimalige Teilung in der reifen Spore vier
Kerne, die sich bei der Keimung nochmals teilen, so daß schließlich
acht Schwärmsporen aus einer Spore hervorgehen.
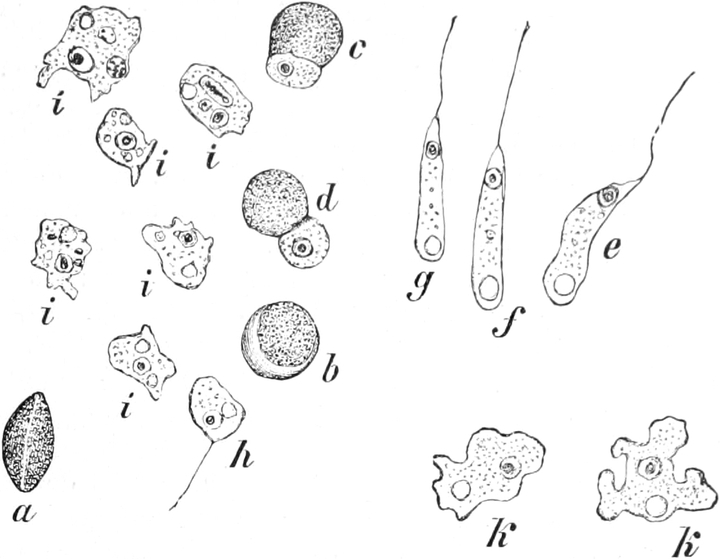
Fig. 315. Chondrioderma difforme. a eine trockene
Spore, b eine geschwellte Spore, c und d Austritt des Inhaltes
aus der Spore, e, f und g Schwärmsporen, h Übergang der
Schwärmspore zur Myxamöbe, i jüngere, k ältere Myxamöben. Vergr.
540. Nach STRASBURGER. (Vgl. Fig. 4, S. 10.)
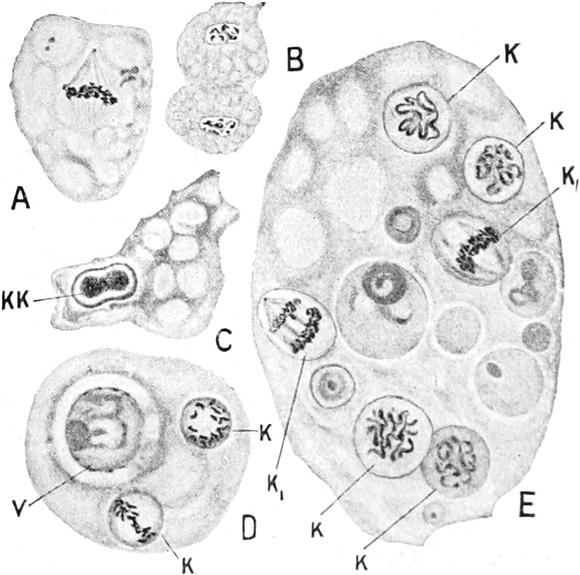
Fig. 316. Physarum didermoides. A, B Amöben in
Teilung. C Kopulation zweier haploider Amöben, kk die beiden Kerne
in Verschmelzung. D Zweikerniges Plasmodium, in der Verdauungsvakuole
eine haploide Amöbe. E Sechskerniges Plasmodium in Kernteilung
(k1). Mit Verdauungsvakuolen. Nach JAHN.
In ihren Schwärmsporen und Myxamöben weisen die Myxomyceten auf
flagellatenartige Organismen als ihre Ausgangsformen hin; auch sind
plasmodienartige Zellfusionen bereits bei gewissen Flagellaten
nachgewiesen.
Die stattlichsten Plasmodien, oft von über 30 cm Durchmesser, von
lebhaft gelber Farbe und rahmartiger Beschaffenheit, bildet Fuligo
varians (Aethalium septicum), die als sog. Lohblüte im Sommer
auf feuchter Gerberlohe sehr verbreitet ist. Auf trockenem Substrat
können diese Plasmodien unter Zerfall in zahlreiche, behäutete
Zellen zu kugeligen oder strangartigen Dauerzuständen, sog.
Sklerotien, sich umwandeln, die bei Zutritt von Feuchtigkeit
wieder in die bewegliche Form übergehen. Die Plasmodien der
meisten Schleimpilze besitzen solches Eintrocknungsvermögen und
können somit ungünstige Perioden überdauern. Schließlich wird
das Plasmodium zu einem gelblichen oder braunen, kuchenförmigen
Fruchtkörper, der eine stark kalkhaltige Hülle besitzt, im Innern
durch zahlreiche Wandungen gefächert ist, von einem fädigen
Capillitium mit unregelmäßigen, Kalkkörnchen enthaltenden Blasen
durchzogen wird und zahlreiche violettschwarze Sporen umschließt.
Dieser Fruchtkörper ist somit aus zahlreichen verschmolzenen
Einzelsporangien zusammengesetzt, während bei den meisten übrigen
Schleimpilzen die Sporangien getrennt ausgebildet werden.
[S. 342]
Bau und Beschaffenheit der Sporangien geben die Merkmale zur
Unterscheidung der einzelnen Formen ab. Die meist braunen oder
ockergelben Sporangien sind kugelig, oval oder auch zylindrisch,
gestielt (Fig. 313, 314 D) oder ungestielt (Fig. 314 A).
Gewöhnlich öffnen sie sich durch Absprengung oder Zerfall
des oberen Teiles der Wandung, während der untere als Becher
zurückbleibt (Fig. 313 B, 314 A); bei Cribraria (Fig. 313
C), deren Fruchtkörper kein Capillitium enthält, wird der
obere Teil gitterartig durchbrochen, bei Stemonitis (Fig. 313
A) hingegen zerfällt das ganze Peridium, und das Capillitium
entspringt einer Columella, der Fortsetzung des Stieles.
Die Ordnung der Plasmodiophoraceae[311] enthält einige wenige
parasitäre Pilze, als Typus die Plasmodiophora Brassicae, die
die sog. Kohlhernie an Brassica-Arten: knollenförmige Verdickungen
am Strunk und an den Nebenwurzeln der befallenen Kohlpflanzen
verursacht. Ihre Myxamöben leben in den Zellen dieser Wucherungen,
und zwar in den Vakuolen des lebendigen Plasmas; sie zehren den
Inhalt der Wirtszelle auf und verschmelzen zu Plasmodien, die
schließlich die zahlreichen von Chitinmembranen umhüllten Sporen
liefern. Im Plasmodium vollzieht sich vor der Sporenbildung
eine der Reduktion der Chromosomenzahl dienende Kernteilung,
die die Kerne für die Sporen liefert. Die Sporen werden bei der
Verwesung der Pflanze frei und keimen wie bei Chondrioderma; die
Myxamöben dringen wieder in die Wurzeln junger Pflanzen ein. Eine
Peridiumbildung findet also nicht statt, so daß der Pilz einen
einfacher organisierten oder infolge der parasitären Lebensweise in
der Sporangienbildung reduzierten Schleimpilz vorstellt.
Die systematische Stellung der Ordnung ist noch zweifelhaft, da
sie in einigen zytologischen Merkmalen Ähnlichkeit mit den zu den
Phycomyceten gerechneten Chytridiaceen aufweist.
Die Dinoflagellatae oder Peridineen schließen sich an gewisse
Flagellaten als weiter entwickelte Gruppe enge an, so daß sie auch
zu diesen selbst gestellt werden könnten. Sie leben als einzellige,
freischwimmende Organismen, teils im Süßwasser, überwiegend aber im
Meere, wo sie zusammen mit Diatomeen einen wichtigen Bestandteil des
Phytoplanktons abgeben. Ihre Zellen sind ausgezeichnet durch den Besitz
von zwei langen Zilien, die in der Mitte der Bauchseite in einer
Längsfurche entspringen; die eine Zilie ist nach hinten gestreckt, die
andere dagegen legt sich wellig gebogen in eine den Körper umziehende
Querfurche (Fig. 317). Der Protoplast besitzt einen Zellkern, Vakuolen
verschiedener Art, zahlreiche gelbbraune Chromato[S. 343]phoren, die
mehrere Farbstoffe enthalten. Als Assimilationsprodukt tritt Stärke
oder Öl auf. Während bei den Gymnodiniaceen (Fig. 318 d) die Zellen
nackt sind, ist dagegen bei den Peridiniaceen eine Zellulosewand
vorhanden, die sich aus polygonalen, meist zierlich gezeichneten, von
Poren durchbrochenen Platten zusammensetzt; die Querfurche wird
von einer gürtelförmigen Plattenreihe eingenommen (Fig. 317).
Fig. 317. Peridinium tabulatum.
Nach SCHILLING.
Fig. 318. Cystodinium Steinii. a Zyste, b Teilung
in zwei Schwärmzellen, c Zyste in Quellung, d befreiter Schwärmer.
Vergr. 480. Nach KLEBS.
Bei vielen Planktonperidineen (Fig. 319) zeichnen sich
die Platten durch besondere Flügelbildungen aus, oder die Zellen
besitzen lange hornförmige Fortsätze, Einrichtungen, die das
Schweben und Steuern im Wasser ermöglichen[314].
Unter den Dinoflagellaten gibt es auch farblos gewordene, somit
saprophytisch lebende Formen, deren Chromatophoren noch
als Leukoplasten nachweisbar sind. Bei einigen Formen ist sogar
tierische Lebensweise nachgewiesen, so bei dem im Süßwasser
lebenden Spirodinium hyalinum, dessen Protoplast zum Zwecke
der Nahrungsaufnahme die Geißeln verliert und zu einer, kleine
Algenzellen aufnehmenden und verdauenden Amöbe wird.
Fig. 319. Planktonperidineen. A Ceratocorys horrida
var. africana, Ind. Ozean. Vergr. 250. B Ceratium tripos intermedium
var. aequatoriale, Ind. Ozean. Vergr. 62. C Ceratium tripos gibberum
und D Ceratium palmatum, Atl. Ozean. Vergr. 62. E Ceratium furca,
Atl. Ozean. Vergr. 125. Nach G. KARSTEN.
Gewisse marine Peridineen (z. B. Ceratium tripos, Peridinium
divergens) besitzen Leuchtvermögen; sie haben einen
Hauptanteil am Meeresleuchten[293].
Die Vermehrung geschieht durch Zweiteilung meist im
beweglichen Zustande der Zellen. Bei gewissen Gattungen
(Peridinium, Cystodinium, [Fig. 318]) gehen die beweglichen
Zellen kürzere oder längere Ruhezustände ein, bilden geißellose
Zysten, in denen die Teilung erfolgt; die Tochterzellen
treten dann aus der verquellenden Zyste als Schwärmzellen aus.
Endlich kann auch das bewegliche Stadium ganz unterdrückt werden,
und die beiden anfangs nackten Tochterzellen werden als behäutete
geißellose und unbeweglich bleibende Zellen aus der Mutterhülle
entlassen (Hypnodinium).
Einige Gattungen (Ceratium) bilden dickwandige Dauerzysten
innerhalb des alten Membranpanzers.
Sexuelle Fortpflanzung ist bei Dinoflagellaten nicht mit
Sicherheit nachgewiesen[315].
Fossile Dinoflagellaten sind zuerst aus der Kreideformation
bekannt geworden.
Die Diatomeen (Bacillariaceae) stellen eine ungemein reichhaltige
Klasse von einzelligen Algen vor, die teils im Süßwasser, teils
im Meere, teils auf nassem Boden, meist in großer Menge gesellig
vegetieren.
[S. 344]
Die Zellen leben einzeln oder in Kolonien, entweder freischwimmend oder
auf dünnen, aus Poren ausgeschiedenen Gallertstielchen festsitzend
(Fig. 320). Bei gewissen Arten bleiben die Zellen in Bändern oder
Zickzackketten durch kurze Gallertpolster vereinigt, oder sie sind in
festsitzende schlauchförmige Gallertröhren eingeschlossen, die bei
der im Meere lebenden Gattung Schizonema sich büschelig verzweigen.
Der Umriß der Zellen ist höchst mannigfaltig, kreisrund, elliptisch,
stabförmig, keilförmig, gerade oder gebogen, oft regelmäßig bilateral
symmetrisch. Sehr charakteristisch ist die Verkieselung der aus
Pektinstoffen bestehenden Zellmembran und ihre Zusammensetzung aus
zwei Schalen, von denen die eine wie der Deckel einer Schachtel
über die andere übergreift. Die Zelle bietet daher zwei verschiedene
Ansichten dar, je nachdem man sie von der Schalenseite oder von
der Gürtelseite betrachtet (Fig. 79).
Die Seitenwände beider Schalen sind durch die unter ihren Rändern
sich ansetzenden Gürtelbänder gebildet; bei gewissen Gattungen
wird die Gürtelseite noch durch Einfügung von ring- oder
schuppenförmigen Zwischenbändern verlängert.
Häufig ist die Membran der Schalenseiten mit Querrippen, Warzen oder
Gruben besetzt und vielfach auch von offenen Porenkanälen durchbohrt,
welche der Gallertausscheidung dienen. Beim Glühen der Zelle auf
einem Glimmerplättchen bleibt das Kieselskelett der Membran mit allen
Strukturen zurück, beim Herauslösen der Kieselteile mit Flußsäure aber
ebenso das Pektingerüst.
Fig. 320. Licmophora flabellata. Diatomeen-Kolonie mit
verzweigten Gallertstielen. Nach SMITH, aus GOEBEL,
Organographie.
Fig. 321. Planktoniella Sol. Atl. Ozean. Scheibenförmige
Planktondiatomee mit hohlem, von der Gürtelseite entspringendem
Schwebeflügel. Plasma mit Kern und zahlreichen Chromatophoren. Vergr.
322. Nach G. KARSTEN.
Die Diatomeenzelle enthält einen Zellkern (Fig. 79) und entweder
ein oder zwei bis vier (Fig. 324) große, flache, oft gelappte oder
auch zahlreiche (Fig. 321) kleinere, durch Chlorophyll und gelbes
Phykoxanthin braungelb gefärbte Chromatophoren, die häufig
Pyrenoide führen. Im Zellplasma finden sich gewöhnlich einige Tropfen
von fettem Öl, das an Stelle der Stärke als Assimilationsprodukt
auftritt.
[S. 345]
Die Diatomeen vermehren sich durch Zweiteilung, die sich
immer nur nach einer Richtung hin vollzieht. Die beiden Schalen werden
dabei durch den sich vergrößernden Plasmakörper an den Gürtelbändern
auseinander geschoben: jede der beiden Tochterzellen erzeugt eine
neue Schale, welche unter die von der Mutterzelle übernommene Schale
mit ihren Rändern eingreift; dann trennen sich die Tochterzellen
voneinander. Die beiden Schalen einer Zelle sind somit ungleichaltrig.
Diese Art der Membranbildung hat, da die verkieselten Wände nicht
wachstumsfähig sind, zur Folge, daß stets eine der Tochterzellen
fortschreitend kleiner wird, und dies geht so fort bis zur Erreichung
eines gewissen Minimums der Zellgröße. Alsdann tritt in der Regel
die Bildung von Auxosporen ein, die mehrmals größer sind als
die Zellen, aus denen sie hervorgegangen sind, und die bei ihrer
Weiterentwicklung somit die Anfangsgröße der Zellen wieder herstellen.
Die sexuelle Fortpflanzung besteht in Kopulation
gleichgestalteter Gameten.
Die Diatomeen umfassen zwei Ordnungen, Centricae und Pennatae.
Die Auxosporen wachsen bei den Centricae, die sich durch
zentrischen Bau der Schalen auszeichnen, ohne vorhergehenden
Kopulationsvorgang aus vegetativen Zellen heran, dagegen bei den
mit fiedriger Schalenskulptur versehenen Pennatae aus den durch
Gametenkopulation hervorgegangenen Zygoten. Die Pennaten haben
diploide vegetative Zellen, und ihre Reduktionsteilung erfolgt bei
der Bildung der Gameten, während die Centricae haploid sein dürften
und schon bei der Teilung der Zygoten, die hier, soweit bekannt,
durch Verschmelzung begeißelter Gameten gebildet werden, ihre
einfache Chromosomenzahl erhalten. Die beiden Kieselalgengruppen
zeigen somit scharfe Unterschiede.
1. Ordnung. Diatomeae centricae. Die Schalenseite ist
zentrisch gebaut, mit radialer oder konzentrischer Anordnung der
Wandskulpturen. Die überwiegende Mehrzahl der hierher gehörigen
Arten lebt im Meere und beteiligt sich in hervorragendem
Maße an der Zusammensetzung des Planktons[314].
Die Planktondiatomeen sind mit besonderen Schwimm- und
Schwebeeinrichtungen, oft mit hornförmigen Fortsätzen oder
Membranflügeln ausgestattet (Fig. 321 u. 322).
Fig. 322. Corethron Valdiviae. Antarktisches Plankton.
a Zelle mit Schwebeborsten und Fangarmen, b Auxosporenbildung,
Protoplast nach Abwerfen einer Schale aus der anderen hervortretend und
zu etwa vierfachem Durchmesser herangewachsen, vom Perizonium umgeben,
c Protoplast innerhalb des Perizoniums kontrahiert und die neue obere
Schale ausbildend, d Perizonium oben aufgelöst. Die Auxospore bildet
die untere Schale und tritt aus dem Perizonium hervor. Nach G.
KARSTEN.
Die Auxosporenbildung der Centricae vollzieht sich als
reiner Wachstumsvorgang in der Weise, daß der sich von den
Schalen befreiende Plasmakörper einer Zelle zu einer vergrößerten
Zelle heranwächst, die zunächst von einer schwach verkieselten[S. 346]
Hülle (Perizonium) umgeben, in dieser die beiden neuen Schalen
nacheinander ausscheidet (Fig. 322, 323 B).
Den Pennaten gegenüber zeichnen sich die Centricae durch
bewegliche, mit Zilien versehenen Gameten aus. Die Bildung
dieser bisher als Mikrosporen bezeichneten Sexualzellen wurde
zuerst von BERGON bei Biddulphia mobiliensis (Fig. 323
A–D) untersucht. Hier teilt sich die Zelle zunächst in zwei
sich gegeneinander abrundende Gametangien, deren Inhalt durch
wiederholte Zweiteilungen in viele (32) Gameten zerlegt wird.
Diese sind mit zwei gleich langen Geißeln versehen und schlüpfen
als nackte Schwärmzellen aus den Behältern aus (Fig. 323 C–E).
KARSTEN beobachtete an konserviertem Material von
Corethron Valdiviae, daß die Mikrosporen paarweise zu Zygoten
kopulieren, die heranwachsen und sich in je zwei Tochterzellen
teilen. Jede Tochterzelle hat erst zwei Kerne, von denen einer
später verschwindet; sie wächst allmählich zu einer fertigen
Corethronzelle heran. Diese Vorgänge erinnern an das für Closterium
zu schildernde Verhalten der Desmidiaceen. Auch bei einigen
anderen Gattungen sind solche zweiwimperigen Gameten, und zwar von
zweierlei Form, größere chromatophorenführende und etwas kleinere
farblose, beobachtet worden (Fig. 323 E). Ihre Kopulation wurde
in neuester Zeit von P. SCHMIDT an lebendem Material von
Melosira festgestellt[317]. Die Geißeln der Gameten deuten auf
phylogenetische Beziehungen der Diatomeen zu den Flagellaten, unter
denen in erster Linie die Chrysomonadinen als Ausgangsgruppe in
Betracht zu ziehen sind.
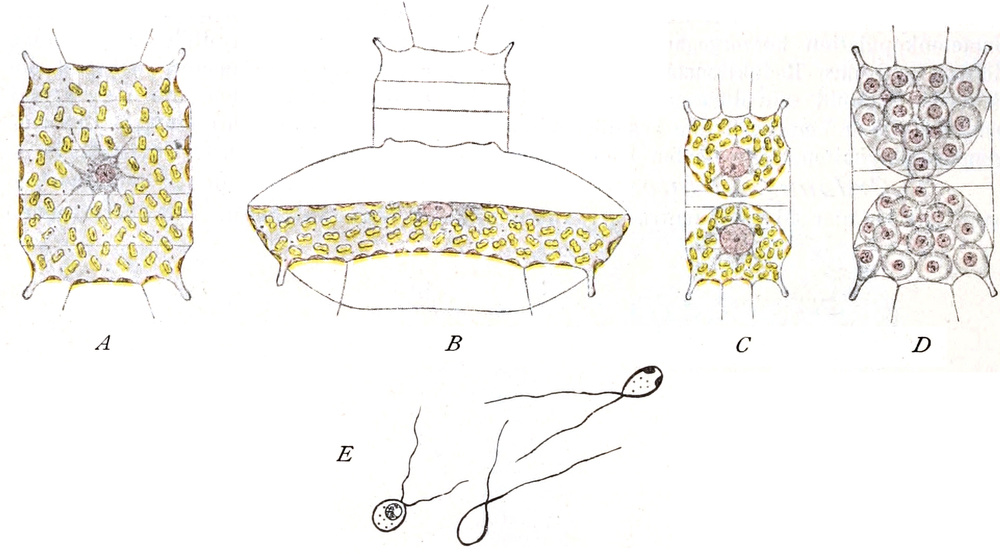
Fig. 323. A-D Biddulphia mobiliensis. A
Gürtelbandansicht. B Auxosporenbildung. C Zelle in zwei Gametangien
geteilt. D Gametenbildung in diesen. Vergr. 228. Nach P.
BERGON. E Coscinodiscus, Gameten. Nach PAVILLARD.
2. Ordnung. Diatomeae pennatae. Sie sind meist grundbewohnende
Kieselalgen. Die meist langelliptische oder stabförmige oder
schiffchenförmige oder auch keilförmige Schalenseite zeigt fiedrige
Anordnung ihrer Wandskulpturen (Fig. 320, 324, 325). Bei manchen
Pennaten (Fig. 79) verläuft über die Schalenseite eine von zwei
Endknoten ausgehende und in der Mitte zu einem Mittelknoten
anschwellende Längsleiste oder Raphe, in welcher schraubig
verlaufende Längsspalten die Zellwand durchsetzen. Die Formen mit
Raphe zeichnen sich durch eine eigentümliche, ruckweise erfolgende
Fortbewegung aus, deren Zustandekommen auf das in den
Spalten befindliche und aus ihnen hervortretende strömende Plasma
zurückgeführt wird. Die Bewegungen dieser Diatomeen werden durch
äußere Reize veranlaßt.
Die Bildung der Auxosporen vollzieht sich in mannigfaltiger
Weise. Wir gehen aus von dem Verhalten von Navicula,
Pleurosigma u. a., auf das sich die anderen Typen zurückführen
lassen; zwei Zellen legen sich hier nebeneinander und scheiden eine
Gallerthülle aus. Ihre Kerne erfahren eine mit Chromosomenreduktion
verbundene Tetradenteilung in je vier Kerne, nämlich zwei Großkerne
und zwei Kleinkerne. Dann teilt sich jede Zelle in zwei Gameten,
von denen ein jeder einen Großkern und einen Kleinkern mitbekommt.
Die aus den Schalen hervortretenden Gameten kopulieren paarweise
zu anfangs vierkernigen Zygoten; in diesen verschmelzen die
Großkerne, während die Kleinkerne[S. 347] schwinden. Jede Zygote wächst
innerhalb einer dünnen Hülle (Perizonium) zu einer mehrmals
größeren Auxospore heran, die schließlich ihre beiden neuen Schalen
ausbildet und die Reihe der vegetativen Zweiteilungen beginnt (Fig.
324).
Bei Surirella und Cocconeïs (Fig. 325) teilen sich die
kopulierenden Zellen nicht in zwei Tochterzellen, sondern
verschmelzen direkt miteinander; aber die Zellkerne erfahren noch
Teilungen, und zwar teilt sich bei ersterer Gattung der Kern
einer jeden Zelle zweimal, so daß ein großer Sexualkern und drei
Kleinkerne entstehen, bei letzterer Gattung aber nur einmal in
einen Großkern und einen Kleinkern. Die Großkerne verschmelzen, die
Kleinkerne gehen zugrunde.
Bei Achnanthes subsessilis teilt sich der Inhalt einer Zelle in
zwei austretende einkernige Tochterzellen, die sich alsdann zu
einer Auxospore vereinigen.
Bei manchen Pennaten tritt die Sexualität zurück, und die
Auxosporen entstehen parthenogenetisch. Bei Synedra teilt
sich eine Mutterzelle in zwei zu Auxosporen heranwachsende
Tochterzellen, deren Kerne zwar noch eine zweite Teilung ausführen,
aber wieder verschmelzen; ebenso verhält sich Rhabdonema
arcuatum, nur ist die zweite Kernteilung nicht mehr nachzuweisen.
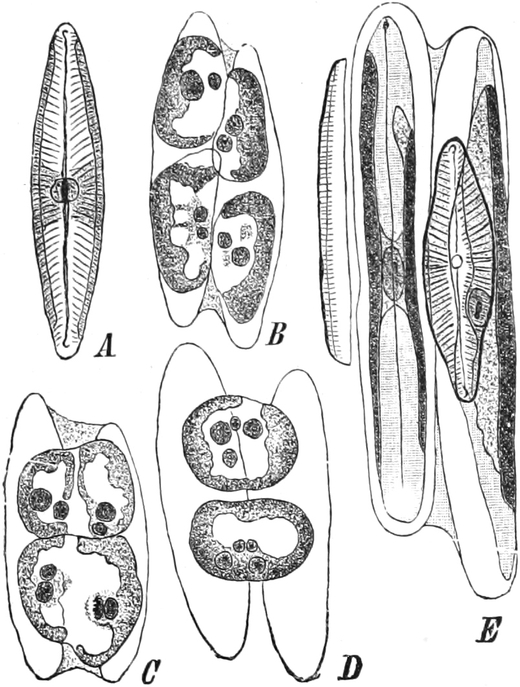
Fig. 324. Auxosporenbildung von Navicula viridula. A
Zelle von der Schalenseite. B Zwei Zellen in je zwei Tochterzellen
mit zwei Kernen geteilt. C, D Kopulation der Tochterzellen zu
zwei anfangs vierkernigen Auxosporen. E Die beiden herangewachsenen
Auxosporen. Vergr. 500. Nach G. KARSTEN.
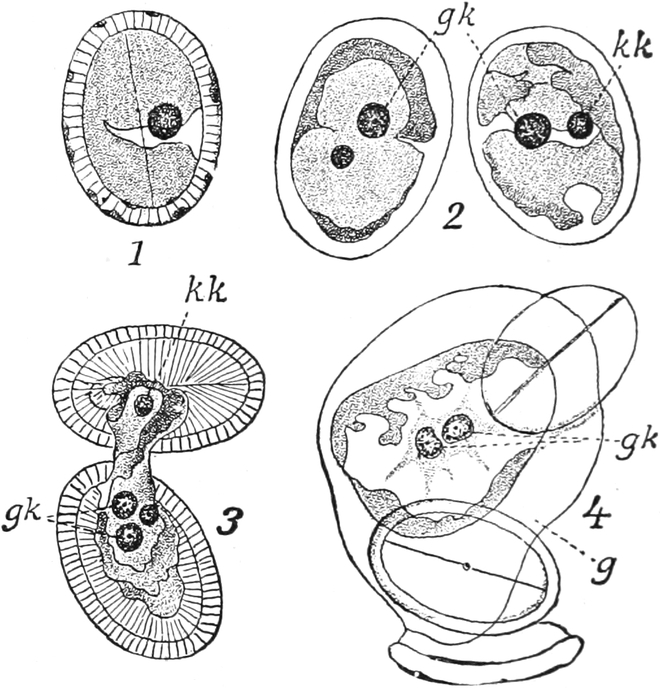
Fig. 325. Cocconeïs Placentula. 1 Vegetative
Einzelzelle, 2 Zellenpaar vor der Kopulation, 3 und 4 in
Kopulation. gk Großkern, kk Kleinkern, g Gallerte. Nach G.
KARSTEN, aus OLTMANNS, Algen.
Rhabdonema adriaticum geht noch einen Schritt weiter; es stößt
einen der beiden Tochterkerne aus dem Plasmakörper aus und
entwickelt aus der ganzen Mutterzelle nur eine Auxospore.
Viele Pennaten siedeln sich mit Vorliebe an Stellen an, wo
verwesende Substanzen reichlich vorhanden sind. Solche Arten
können zu saprophytischer Lebensweise übergehen.
Ihre Chromatophoren erleiden dabei unter Umständen eine
bedeutende Verkleinerung und eine Entfärbung. Für einige
marine Nitzschia-Arten ist sogar ausschließliche Ernährung
aus organischen Substanzen und vollständiges Schwinden der
Chromatophoren und Farbstoffe nachgewiesen[318].
Navicula ostrearia, eine im Mittelmeer vereinzelt auftretende,
in den Austernparks der französischen Küsten aber sehr verbreitete
und den Austern als Nahrung dienende Diatomee, zeichnet sich durch
einen im Plasma enthaltenen himmelblauen Farbstoff, Marennin, aus.
Das Marennin bewirkt Grünfärbung der Austern, in denen es
sich unter Änderung seiner Farbe ablagert. Bei Neapel kommt sie auf
Padina (S. 365) vor und verleiht ihr grüne Färbung[319].
[S. 348]
Wegen ihrer außerordentlich feinen Membranskulptur dienen
gewisse pennate Diatomeen als Testobjekte zur Prüfung von
Mikroskopobjektiven, so namentlich Pleurosigma angulatum.
Fossile Diatomeen sind bis in den oberen Lias hinab nachgewiesen
worden, in besonders reicher Fülle im Tertiär, wo ihre Schalen
vielfach in größeren Massen abgelagert wurden, als Hauptbestandteil
der Kieselgur (Bergmehl, Diatomeenerde), die zur Dynamitfabrikation
Verwendung findet. Die tertiären Arten sind zum Teil mit jetzt noch
lebenden identisch oder gehören meist denselben Gattungen an.
Die Konjugaten bilden eine formenreiche Gruppe von einzelligen oder von
unverzweigt fadenförmigen, im Süßwasser lebenden grünen Algen, die sich
von den Chlorophyceen scharf unterscheiden. Ihre Zellen vermehren sich
nur durch Zweiteilung, sind einkernig und haben im Gegensatz zu
den Diatomeen eine kieselfreie Membran sowie große, verwickelt
gebaute grüne Chloroplasten. Ungeschlechtliche Vermehrung durch
Schwärmsporen fehlt ihnen ebenso wie den Diatomeen, mit denen sie
auch in ihrer sexuellen Fortpflanzung zum Teil übereinstimmen. Diese
besteht in der Kopulation oder Konjugation zweier gleichgestalteter,
geißelloser Gameten zu einer Zygospore oder Zygote.
Die Konjugaten und Diatomeen sind daher auch zu einer nunmehr
als unnatürlich zu betrachtenden Gruppe der Zygophyceen
zusammengefaßt und in Rücksicht auf den Mangel von Zilien an den
Gameten auch als Acontae, Geißellose, bezeichnet worden. Letztere
Bezeichnung trifft indessen für gewisse Diatomeen, wie wir sahen,
nicht mehr zu. Die Reduktionsteilung erfolgt bei den Konjugaten
nach der Kopulation in den keimenden Zygoten, bei den Pennaten
dagegen bei Bildung der Gameten. Beide Gruppen haben offenbar
getrennt voneinander ihren Ausgang aus Flagellaten genommen.
1. Die Mesotaeniaceae umfassen als einfachste einzellige
Konjugaten nur wenige Gattungen; sie besitzen eine einfache, nicht
wie bei den Desmidiaceen aus zwei Hälften bestehende Membran ihrer
kurzzylindrischen, in Gallerte an feuchten Orten lebenden Zellen.
Die Kopulation zeigt Verschiedenheiten. Bei Cylindrocystis
(Fig. 326) verschmelzen die Protoplasten von zwei Zellen als
Gameten zu einer Zygote, beide Kerne vereinigen sich, während
die vier Chloroplasten erhalten bleiben. Vor der Keimung teilt
sich die Zygote schrittweise in vier Keimzellen, die dann aus
ihr heraustreten. Im ersten Teilungsschritt vollzieht sich die
Reduktion der Chromosomen. Bei Spirotaenia dagegen teilen sich
erst die Protoplasten der beiden kopulierenden Zellen in zwei
Tochterzellen, die dann paarweise zu Zygoten verschmelzen. Auch
sollen hier aus diesen nur zwei Keimlinge hervorgehen, indem die
beiden anderen unterdrückt werden.
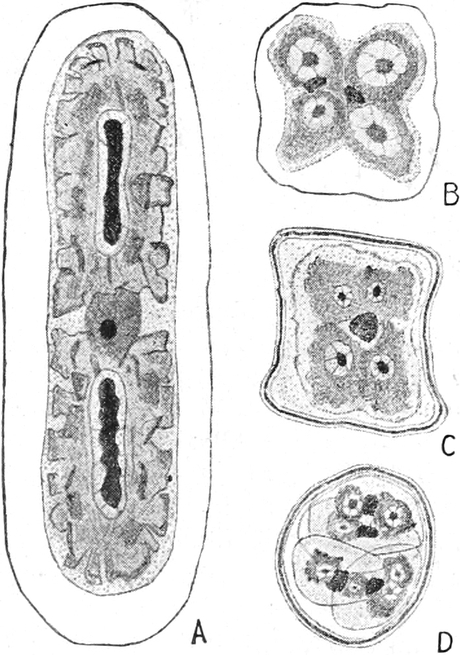
Fig. 326. A Cylindrocystis Brebissonii. Der Zellkern
in der Mitte zwischen zwei großen viellappigen Chloroplasten mit langen
Pyrenoiden. B Zygote vor, C nach der Verschmelzung der Kerne, D
Zygote vor der Keimung mit vier Tochterzellen. Nach KAUFFMANN.
2. Die Desmidiaceen sind ebenfalls einzellige oder in Zellketten
erscheinende Konjugaten; sie gehören zu den zierlichsten Algen
der Torfmoore und Sümpfe und weisen ebenso wie die Diatomeen eine
ungemeine Mannigfaltigkeit der Gestalt auf. Ihre Zellen bestehen
aus zwei symmetrischen Hälften, die meist durch eine Einschnürung
begrenzt sind. Jede[S. 349] Hälfte enthält einen großen Chloroplasten
mit einigen Pyrenoiden oder Stärkeherden; in der Mitte der Zelle
ist der Kern gelegen. Die Gesamtform ist sehr verschieden, bald
abgerundet eckig (z. B. Cosmarium, Fig. 327 A, B), bald
sternförmig (Micrasterias, Fig. 327 D). Häufig ist die Membran,
die wie bei den Diatomeen aus zwei Hälften besteht, mit stachel-
oder warzenartigen Verdickungen versehen und meist von Poren,
die der Gallertausscheidung dienen, durchsetzt. Einige Gattungen
zeigen keine Einschnürungen zwischen den beiden Hälften der Zelle,
so z. B. Closterium moniliferum (Fig. 328 F)[321], dessen
zwei Chromatophoren die Gestalt von kegelförmigen, mit Rippen
besetzten Körpern haben, und das an den Zellenden je eine Vakuole
mit winzigen, in Bewegung befindlichen Gipskristallen aufweist.
Manche Desmidieen vollziehen phototaktische Bewegungen; sie stoßen
an ihren Enden durch Poren der Membran Schleimfäden aus, mittels
deren sie sich fortschieben und in die Richtung der einfallenden
Lichtstrahlen stellen können.
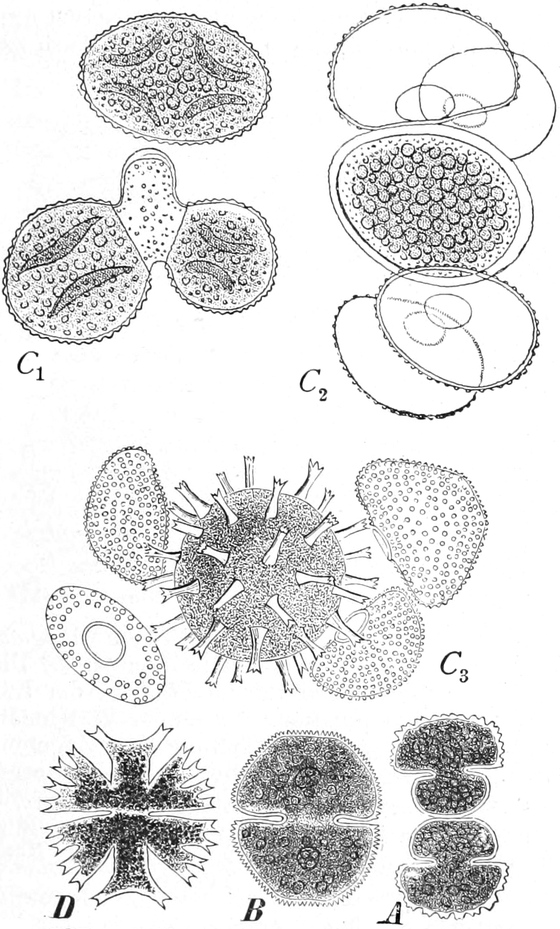
Fig. 327. A Cosmarium coelatum in Teilung. B, C
Cosmarium Botrytis. C1 Zwei gekreuzt stehende Zellen im Beginn
der Kopulation, die untere Zelle zeigt den Kopulationskanal. C2
Gameten zur jungen Zygote verschmolzen. C3 Fertige Zygote. D
Micrasterias Crux melitensis. Nach RALFS, C2 und C3
nach DE BARY.
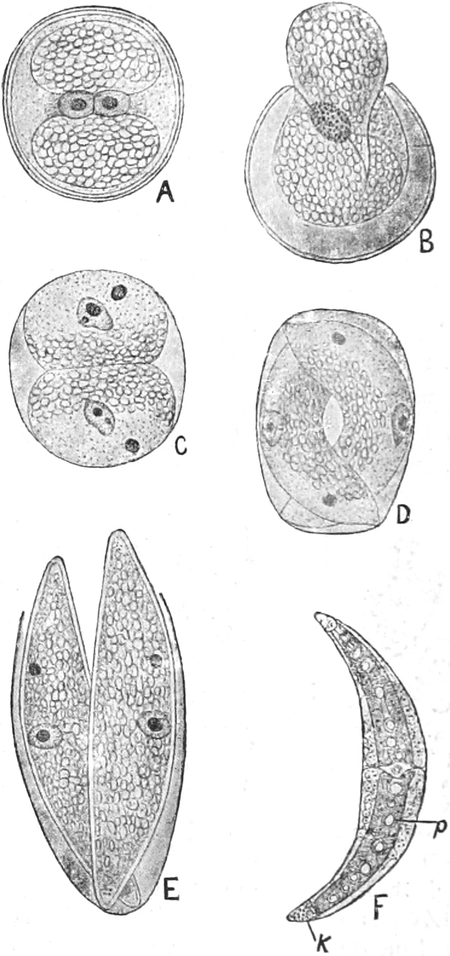
Fig. 328. Closterium. A Zygote vor der Keimung mit den
beiden noch nicht verschmolzenen Gametenkernen. B Zygote in Keimung,
Kerne zu einem vereinigt. C Teilung in zwei Zellen, jede mit einem
Großkern und einem Kleinkern. D Weiterentwicklung der Keimlinge.
E Heraustreten derselben aus der Membran. F Fertiges Stadium von
Closterium moniliferum. A–E nach KLEBAHN.
Die Vermehrung geschieht durch Zweiteilung, die nach der
Kernteilung durch eine in der Einschnürung der Zelle auftretende
und dann sich spaltende Querwand vollzogen wird. Die Tochterzellen
bilden sodann neue Zellhälften aus, indem sie sich nach der
Teilungsfläche zu ausstülpen (Fig. 327 A).
Zur Kopulation legen sich zwei Zellen nebeneinander
und umgeben sich mit Gallerte; die Zellwand bricht in der
Einschnürung auf, die Protoplasten treten in die sich vorwölbenden,
bald verschleimenden Kopulationskanäle und vereinigen sich
zur Zygospore,[S. 350] deren Wandung häufig durch Stachelbildungen
ausgezeichnet ist (Fig. 327 C). Neben oder an den reifen Sporen
liegen die vier Membranhälften. Bei einigen Desmidieen werden die
kopulierenden Zellen erst in je zwei Tochterzellen geteilt, die
dann paarweise sich vereinigen.
Die in die Zygoten eingetretenen beiden Gametenkerne verschmelzen
erst bei beginnender Keimung zu einem einzigen, der sich
darauf nacheinander in vier Kerne, zwei große und zwei kleine,
voraussichtlich unter Reduktion der Chromosomenzahl, teilt. Es
werden aber nur zwei Keimzellen gebildet, von denen jede zwei
ungleich große Kerne mitbekommt, deren kleinerer später schwindet
(Fig. 328). Die zweizellige Keimung erscheint so als Reduktion der
vierzelligen von Cylindrocystis. Die vier Chromatophoren der Zygote
gehen zur Hälfte zugrunde, die beiden übrigbleibenden teilen sich
vor der Keimung in je zwei.
3. Unter den fadenförmigen Konjugaten, den Zygnemaceen[322] ist
am bekanntesten die Gattung Spirogyra, deren zahlreiche Arten
als frei schwimmende, fädige grüne Watten im Frühjahr in ruhigen
Gewässern häufig auftreten. Die aus längeren oder kürzeren Zellen
bestehenden Fäden wachsen in die Länge unter Teilung und Streckung
aller Zellen. Auch ist der Fadenverband kein inniger, denn die
Zellen können sich unter Umständen leicht aus ihm lösen. Jede
Zelle besitzt einen Kern und einen oder mehrere wandständige,
bandförmige, schraubige Chloroplasten (Fig. 329 C und 17).
Die Zellwand ist glatt und porenlos. Bei der Gattung Zygnema
sind zwei sternförmige Chloroplasten vorhanden, bei Mesocarpus
ein axiler bandförmiger. Die Fäden können sich in einer noch
unbekannten Weise fortbewegen.
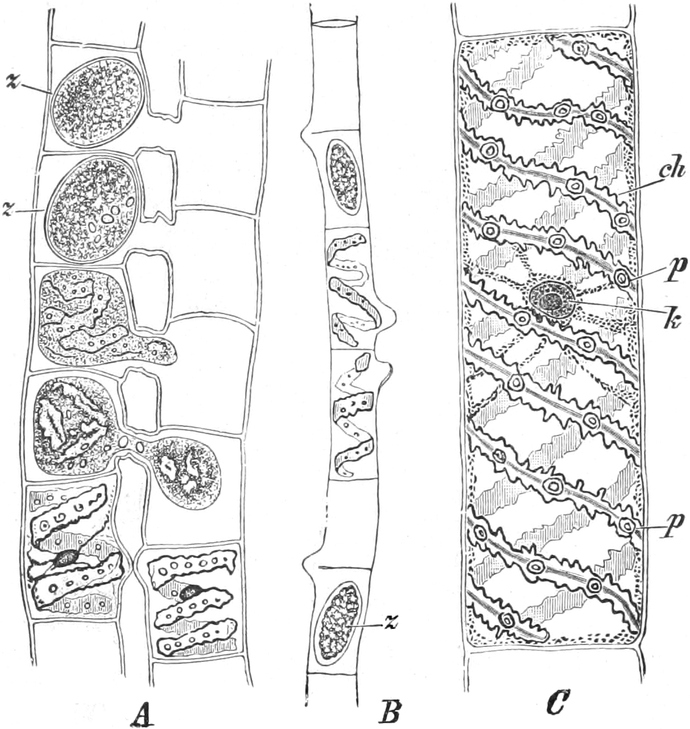
Fig. 329. A Kopulation von Spirogyra quinina. z
Zygosporen. Vergr. 240. B Desgleichen von Sp. longata. Vergr. 150.
C Zelle von Sp. jugalis, k Kern, ch Chromatophor, p Pyrenoide
oder Amylumherde. Vergr. 256.
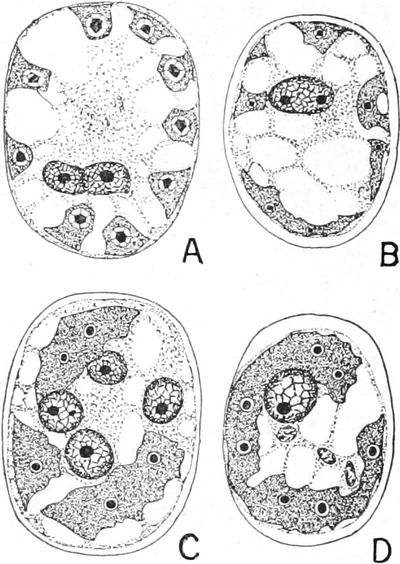
Fig. 330. Spirogyra longata. Junge und alte Zygoten. A
Die beiden Sexualkerne vor der Kopulation, B nach der Verschmelzung,
C Teilung des Zygotenkerns in vier haploide Kerne. D Die drei
Kleinkerne in Zerfall. Chlorophyllbänder wandständig, durchschnitten
gezeichnet. Nach TRÖNDLE.
Die Kopulation der Gameten vollzieht sich innerhalb der
Membranen der Zellen, die durch Kopulationskanäle miteinander in
offene Verbindung treten (Fig. 329). Der Protoplast der einen Zelle
wandert dann durch den Kanal zu dem anderen hinüber. Ersterer kann
als männlich, letzterer als weiblich bezeichnet werden. Plasma und
Kerne verschmelzen miteinander, nicht aber die Chlorophyllkörper,
von denen die in der männlichen Zelle befindlichen zugrunde gehen.
So entsteht eine sich abrundende mit dicker dreischichtiger Membran
umkleidete, dicht mit Fett und rotbraunen Schleimkugeln erfüllte
Zygospore. Bei den meisten Spirogyren erfolgt die Kopulation
von zwei, zuweilen auch von mehreren Fäden leiterförmig
(Fig. 329 A). Zuerst legen sich diese dicht[S. 351] nebeneinander;
dann wölben sich die gegenüberstehenden Zellen aufeinander zu und
werden an den Berührungsstellen fest verbunden. Hier strecken sich
die Wände zu kurzen Kanälen, wodurch die Fäden etwas voneinander
geschoben werden. Schließlich wird die trennende Wand im Kanal
aufgelöst.
Sind, wie es meist der Fall ist, die Fäden getrennt geschlechtig,
so liegen die Zygosporen sämtlich im weiblichen Faden. Es gibt aber
auch gemischt geschlechtige Arten, bei denen sie dann teils im
einen, teils im anderen Faden liegen. Bei diesen letzteren Arten
kann auch seitliche Kopulation von aufeinanderfolgenden
geschlechtlich verschieden differenzierten Gameten eintreten (Fig.
329 B). Nach HEMLEBEN findet diese geschlechtliche
Differenzierung in der Regel vor der letzten Zweiteilung der
Fadenzellen statt und dem entspricht auch die Lagerung der
Zygosporen.
Es gibt übrigens auch Gattungen, bei denen die Zygote mitten in den
Kopulationskanal gelagert wird.
Der Kopulationskern der jungen Zygospore erfährt eine mit
Chromosomenreduktion verbundene Tetradenteilung. Von den vier
Kernen wird einer zum Kern des Keimlings, die drei übrigen werden
zu Kleinkernen und gehen zugrunde (Fig. 330). Von den Chloroplasten
der beiden Gameten werden die des übertretenden aufgelöst. So
entsteht nur ein Keimling, der schlauchförmig auswächst und durch
Zellteilung den Faden bildet.
Die Heterocontae umfassen eine Anzahl Gattungen grüner Algen,
die zu den Chlorophyceen gerechnet wurden, besser aber als
selbständiger, aus Chrysomonadinen hervorgegangener Formenkreis zu
betrachten sind.
Sie zeichnen sich aus durch gelbgrüne Färbung ihrer
plattenförmigen Chromatophoren, die außer Chlorophyllgrün einen
gelben, mit Säuren sich blau färbenden Farbstoff enthalten und
als Assimilationsprodukt nie Stärke, sondern fettes Öl bilden,
ferner fast stets durch zwei ungleich lange, etwas seitlich
eingefügte Zilien ihrer Schwärmzellen, endlich in vielen
Fällen durch Zusammensetzung ihrer pektinhaltigen und meist
verkieselten Zellmembran aus zwei ineinander geschachtelten Teilen.
Teils sind die Heteroconten einzellig, teils leben ihre Zellen in
gallertigen Zellkolonien oder auf Gallertstielen, teils stellen sie
Zellfäden vor.
Die Vermehrung erfolgt durch Schwärmsporen, bei gewissen Gattungen
aber auch an deren Stelle durch endogene Aplanosporen; auch werden
endogene Zysten gebildet, die wie die zuletzt genannten Sporen
meist mit zweischaligen Kieselmembranen versehen sind. Bei einigen
Gattungen sind außer Schwärmsporen auch diesen ähnliche, paarweise
kopulierende Gameten beobachtet worden.
Die Heteroconten beteiligen sich mit mehreren einzelligen Gattungen
(Meringosphaera, Halosphaera) an der Zusammensetzung des
marinen Planktons.
An den Beginn der Klasse stellen wir flagellatenartige Formen, wie
die im Süßwasser lebende Chloramoeba heteromorpha (Fig. 331),
deren nackte, amöboid veränderliche Zellen einen Zellkern, 2–6
gelbgrüne Chloroplasten und am Vorderende über einer Vakuole zwei
sehr ungleich lange Geißeln besitzen. Chloramoeba gehört zu den
niederen grünen Organismen, die bei Kultur im Dunkeln in Nährlösung
farblos werden und zu unselbständiger Ernährung übergehen.
Ruhestadien werden von derbwandigen Dauerzellen gebildet.
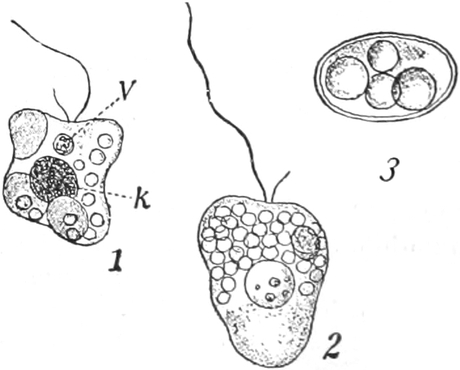
Fig. 331. Chloramoeba heteromorpha. 1 Grüne Form.
2 Farblose Form. v Vakuole, k Kern. 3 Dauerzelle. Nach
BOHLIN, aus OLTMANNS, Algen.
Unter den höher stehenden Heteroconten ist besonders die im
Süßwasser sehr verbreitete Gattung Conferva (Tribonema) (Fig.
332) zu nennen, deren Arten unverzweigte Zellfäden vorstellen
mit eigenartig gebauter Membran, die aus zwei in der Mitte der
Zelle durch einen schräg-ringförmigen Spalt getrennten Stücken
besteht und bei der Teilung[S. 352] durch Einschiebung eines neuen, im
Längsschnitt H-förmigen Membranstückes verlängert wird.
Die Zoosporen entstehen zu 1 oder 2 aus den Fadenzellen. Außer
Zoosporen werden auch, unter Zerfall der Fäden, geißellose
derbwandige Aplanosporen erzeugt.
Die Mehrzahl der Heteroconten besitzt einkernige, nur gelegentlich
zweikernige Zellen. Doch gehören auch Gattungen mit mehrkernigen
Zellen wie Ophiocytium und Sciadium in diese Klasse.
Zu den Heteroconten stellen wir mit Vorbehalt auch Botrydium
granulatum (Fig. 333), eine auf feuchtem Lehmboden sehr
verbreitete Alge, deren einzelliger, aber vielkerniger Thallus die
Form von einer etwa 2 mm großen, birnförmigen, grünen, unterwärts
mit farblosem verzweigtem Rhizoïd festgeankerten Blase aufweist.
Die Zoosporen werden in großer Zahl erzeugt, treten aus einem Loch
am Scheitel aus, besitzen zwei Chloroplasten, eine endständige
Geißel, umgeben sich nach dem Schwärmen mit Membran und keimen
zu neuen Blasen heran. Geschlechtliche Fortpflanzung ist nicht
bekannt[289].
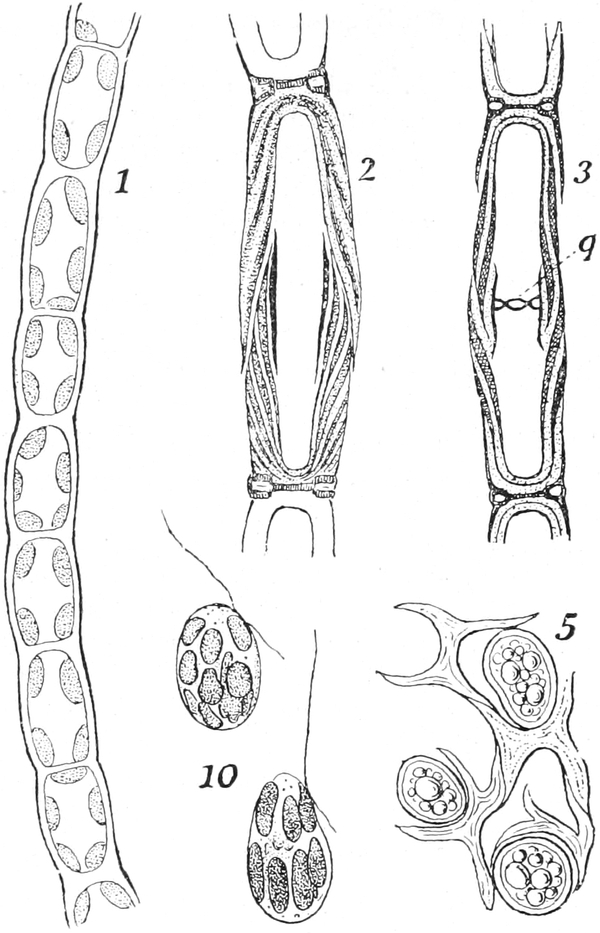
Fig. 332. Conferva bombycina. 1 Faden, 2, 3
Zellwandbildung, q Querwand, 5 Aplanosporenbildung unter
Zerfall der Fäden, 10 Zoosporen mit ungleich langen Geißeln. Nach
GAY (1, 5), BOHLIN
(2, 3), LUTHER
(10). Aus OLTMANNS, Algen.
Fig. 333. Botrydium granulatum. A Vergr. 28. B Eine
Schwärmspore. Vergr. 540. Nach STRASBURGER.
Nach Ausscheidung der ebenfalls grünen Konjugaten und Heteroconten
verbleiben die Chlorophyceen als eine natürliche, in mehreren
Reihen aufsteigende Gruppe von Algen, von denen der größere Teil in
Süßwasser oder an feuchten Orten, manche größere Arten aber auch an
den Meeresküsten leben, während sie am marinen Plankton keinen Anteil
haben. Ihre mit Membran umhüllten Zellen zeichnen sich aus durch
rein grün gefärbte, fast stets Stärke bildende und häufig
Pyrenoide führende Chloroplasten. Ihre ungeschlechtlichen
Schwärmsporen haben birnförmige Gestalt und besitzen bei den
typischen Vertretern zwei oder vier gleich lange Geißeln
(daher[S. 353] auch die Bezeichnung Isocontae für die Gruppe), und im unteren
Teil einen gebogenen oder auch becherförmigen Chloroplasten. Bei
einigen Gattungen treten an Stelle der Schwärmsporen unbewegliche
Aplanosporen auf; bei gewissen höher stehenden Gattungen
(Oedogonium, Vaucheria) sind die Schwärmsporen weniger einfach gebaut,
lassen sich aber auf die ursprünglicheren Formen zurückführen.
Die Schwärmsporen gelangen durch phototaktische Bewegungen an Orte,
die ihnen für ihre Keimung die günstigsten Lichtbedingungen bieten
(vgl. S. 291).
Die sexuelle Fortpflanzung geschieht meist durch Kopulation von
Gameten, die den Zoosporen ähnlich sind. Bei den Endgliedern aller
Gruppen mit Ausnahme der Protococcales hat sich aber an Stelle dieser
Isogamie bereits Oogamie herausgebildet. Die Reduktionsteilung erfolgt,
soweit untersucht, bei der Keimung der Zygoten.
Von den fünf Ordnungen der Chlorophyceen schließen sich die
Volvocales am nächsten an die Flagellaten an, von denen sie sich aber
durch den Besitz von Zellmembranen unterscheiden; sie umfassen ebenso
wie auch die Protococcales einzellige oder Zellkolonien bildende
Formen. Die Ulotrichales und Siphonocladiales stellen Zellfäden
vor, die bei ersteren aus einkernigen, bei letzteren aus großen
vielkernigen Zellen bestehen und in den einfachsten Fällen unverzweigt,
bei höher stehenden Formen oft vielfach verzweigt sind. Die
Siphonales endlich haben einen aus einer einzigen schlauchförmigen,
vielkernigen Zelle aufgebauten Thallus.
Die Grünalgen stellen sicher eine sehr alte Gruppe
niederer Pflanzen vor. Mit Sicherheit sind aber nur die
durch Kalkabscheidungen widerstandsfähigen Thalli mariner
Siphonocladiales bis in das Silur abwärts nachgewiesen worden.
Diese Kalkalgen spielten namentlich in der Trias eine bedeutende
Rolle und erreichten dort eine große Formenfülle.
1. Ordnung. Volvocales. Sie zeichnen sich dadurch aus, daß
die Zellen, die einen Kern und einen Chloroplasten
enthalten, auch während ihres vegetativen Zustandes die Geißeln
behalten, also ständig beweglich bleiben. Darin verhalten
sie sich wie Flagellaten.
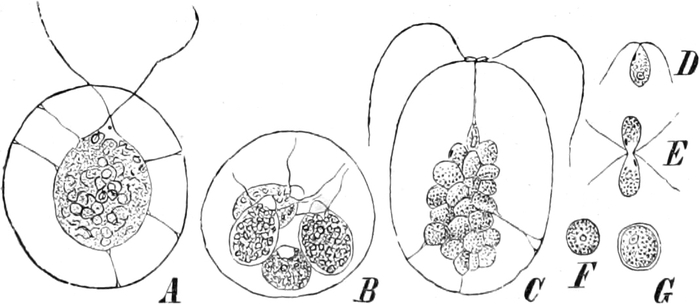
Fig. 334. A, B Haematococcus pluvialis. A
Schwärmende Zelle. B Bildung der Schwärmsporen. Vergr. 360. C-G
Haematococcus Bütschlii. C Gametenbildung. Vergr. 400. D Gamet. E
Kopulation zweier Gameten. F, G Zygoten. Vergr. 800. C-G nach
BLOCHMANN.
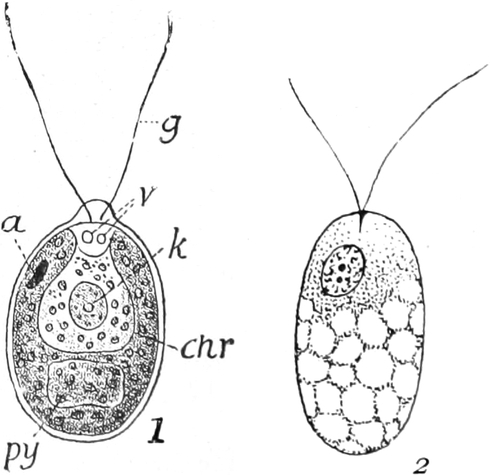
Fig. 335. Chlamydomonas angulosa, nach DILL.
g Geißeln, v Vakuole, k Kern, chr Chromatophoren, py
Pyrenoid, a Augenfleck. 2 Polytoma uvella, nach DANGEARD.
Aus OLTMANNS, Algen.
Chlamydomonas (Fig. 335) und Haematococcus (Fig. 334) gehören
zu den freischwimmenden einzelligen Formen[325]. Bei
ersterer Gattung liegt die Membran dicht dem Protoplasten an, der
vorn zwei Geißeln und einen roten Augenfleck besitzt; bei letzterer
dagegen ist der Protoplast bis auf einige Fortsätze von der Membran
durch eine Gallertschicht getrennt. Haematococcus pluvialis
tritt häufig in Regenpfützen auf und zeichnet sich ebenso wie die
auf Firnfeldern den „roten Schnee“ verursachende Chlamydomonas
nivalis durch einen roten Farbstoff, Hämatochrom (= Karotin),
in seinen Zellen[S. 354] aus. Die Vermehrung geschieht ungeschlechtlich
durch Schwärmsporen, die zu 2–8–16 in einer Mutterzelle gebildet
und durch Zerreißen der Membran frei werden, und außerdem
geschlechtlich durch Kopulation kleiner, zweiwimperiger,
gleichartiger Gameten, die in größerer Menge (64 oder noch mehr)
in der Mutterzelle entstehen und paarweise mit den Vorderenden zu
ruhenden Zygoten sich vereinigen. Bei Chlamydomonas coccifera ist
nach GOROSCHANKIN[326] im Gegensatz zu den übrigen Arten
eine weitgehende Differenzierung der Gameten eingetreten. Einzelne
Zellen werden zu großen, zilienlosen weiblichen Gameten oder
Eizellen, andere teilen sich und liefern je 16 kleine, zweizilige
männliche Gameten. Der Übergang zur Oogamie vollzieht sich hier
also schon bei einzelligen Algen.
Polytoma uvella, die wie Chlamydomonas gebaut ist, verdient
Erwähnung, weil sie eine farblose, saprophytisch sich ernährende
Form darstellt (Fig. 335, 2).
Mehrere Chlamydomonaden verlieren unter gewissen Bedingungen ihre
Geißeln. Die Zellen teilen sich weiter, werden aber durch die
gallertig aufquellenden Zellwände noch in Kolonien zusammengehalten
(sog. Palmella-Stadium). Unter günstigen Bedingungen werden die
Zellen wieder beweglich.
Die Gattungen Pandorina, Eudorina, Volvox u. a. vereinigen
ihre zweiwimperigen Zellen zu freischwimmenden Kolonien
oder Cönobien. Volvox (Fig. 336) als höchststehende Form
zeigt hohlkugelige Kolonien, deren zahlreiche Protoplasten durch
feine Fortsätze noch in Verbindung bleiben, so daß eine solche
Kolonie bereite als ein Individuum oder auch als eine einfache Form
eines Plasmodiums aufgefaßt werden kann. Bei Volvox sind die
Geschlechtszellen in Eier und Spermatozoiden differenziert.
Die Eizellen entstehen durch Vergrößerung einzelner Koloniezellen,
sind groß, grün, unbeweglich und von Gallerte umgeben, während
die viel kleineren, langgestreckten, nackten hellgrünen Spermien
unter ihrem schmalen farblosen Vorderende seitlich zwei lange
Zilien tragen und durch Teilung von Koloniezellen in zahlreiche
Tochterzellen entstehen. Nach Kopulation der Eizelle mit einem
Spermium wird die Eizelle zu einer derbwandigen, ruhenden
Oospore, in der sich bei der Keimung die Reduktionsteilung des
Zygotenkerns abspielt[327]. Volvox vermehrt sich vegetativ durch
Teilung einzelner Koloniezellen zu neuen Tochterkolonien, ein
Vorgang, welcher der Schwärmsporenbildung der einfacheren Gattungen
entspricht. Außer Volvox zeichnet sich auch noch Eudorina durch
Oogamie aus.
Fig. 336. Volvox globator. A Kolonie mit Eizellen
und Spermiengruppen in verschiedenen Entwicklungsstadien. Vergr. 165.
B Spermienbündel, aus einer Zelle durch Teilung entstanden. Vergr.
530. C Spermien. Vergr. 530. D Eizelle, in der Gallertmembran von
Spermien umschwärmt. Vergr. 265. Nach F. COHN.
2. Ordnung. Protococcales. Einzellige oder in Zellkolonien
verschiedener Form lebende grüne Algen, deren vegetative
Zellen aber keine Zilien tragen, also unbeweglich
sind. Meist ist nur ein Kern und ein Chloroplast in
der Zelle vorhanden.[S. 355] Die Vermehrung geschieht durch Zoosporen,
an deren Stelle aber bei manchen Gattungen auch zilienlose
Aplanosporen treten. Die geschlechtliche Fortpflanzung besteht in
Kopulation gleichgestalteter Gameten; sie ist aber nur bei einigen
Gattungen nachgewiesen und scheint bei den einfacheren Formen
überhaupt noch nicht zur Ausprägung gekommen zu sein.
Zu den einfachsten Formen gehören die einzelligen Gattungen
Chlorococcum und Chlorella[328], [329]. Erstere bildet kugelige
Zellen, die in Süßwasser, aber auch auf feuchten Substraten
vorkommen. Die Vermehrung geschieht ungeschlechtlich durch Teilung
der Zellen in eine Anzahl von ausschlüpfenden Zoosporen (Fig.
337), an deren Stelle unter gewissen Umständen auch zilienlose
Aplanosporen gebildet werden können. Chlorella vulgaris (Fig.
338) ist eine ungemein verbreitete Alge, deren kleine grüne Zellen
häufig symbiotisch im Plasma von niederen Tieren, Infusorien,
Hydra, Spongilla, Planarien leben und sich ausschließlich durch
Teilung der Zellen in 2, 4 oder 8 austretende Aplanosporen
vermehren, die sich mit Membran umgeben und heranwachsen.
Fig. 337. Chlorococcum (Chlorosphaera) limicola.
1 Vegetative Zelle und Zelle in acht Zoosporen geteilt. 2
Freie Zoosporen. 3 Dieselben nach Umhüllung mit Membran. Nach
BEIJERINCK, aus OLTMANNS, Algen.
Fig. 338. Chlorella vulgaris. 1 Zelle. 2, 3
Teilung in vier Aplanosporen. 4, 5 Desgleichen in acht. Nach
GRINTZESCO.
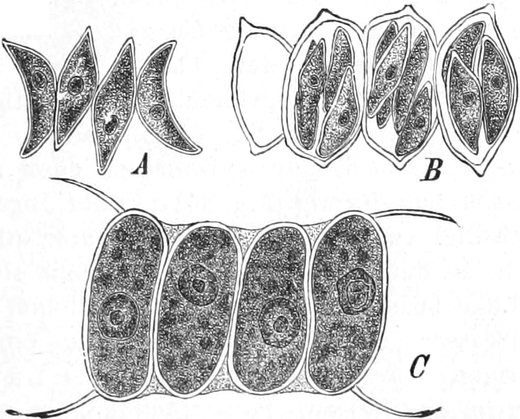
Fig. 339. A Scenedesmus acutus. B Desgleichen in
Teilung. C Scenedesmus caudatus. Vergr. 1000. Nach SENN.
Fig. 340. Pediastrum granulatum. A Alte Zellfamilie,
entleert bis auf die drei Zellen a; die Zelle b entläßt 16
Schwärmzellen. B Zellfamilie nach dem Austritt. C Zellfamilien
41⁄2
Stunden später. Vergr. 300. Nach AL. BRAUN.
An Chlorella schließen wir die im Süßwasser sehr verbreitete
Gattung Scenedesmus (Fig. 339) an, welche Zellkolonien
einfachster Art, meist aus je vier Zellen in einer Querreihe
vorstellt. Die häufigste Art, S. acutus, hat spindelförmige
Zellen, während S. caudatus an jeder Endzelle zwei lange,
hornförmige Membranfortsätze besitzt. Jede Zelle teilt sich der
Länge nach in vier zilienlose und sich mit Membran umgebende
Tochterzellen, die nach Verlassen der alten Membran eine neue
Kolonie bilden. Reicher zusammengesetzte Zellkolonien treffen
wir bei Pediastrum (Fig. 340) an in Form von zierlichen,
freischwimmenden Täfelchen. Die Bildung ungeschlechtlicher,
mit zwei Zilien versehener Schwärmsporen findet in der Weise
statt, daß der Inhalt einer Zelle sich in[S. 356] eine Anzahl (bei dem
abgebildeten P. granulatum in 16) Schwärmsporen teilt, welche,
von einer gemeinsamen Blase umgeben, durch einen Riß in der Wandung
austreten, sich sodann in der Blase bewegen und schließlich zu
einer neuen heranwachsenden Zellfamilie zusammenlegen. Neben der
ungeschlechtlichen tritt bei Pediastrum auch geschlechtliche
Fortpflanzung auf. Die Gameten sind den Schwärmsporen ähnlich,
nur kleiner, und entstehen in den Zellen in größerer Zahl; sie
schwimmen frei im Wasser und kopulieren paarweise zu Zygoten. Beide
Gameten sind gleich gestaltet. Die Weiterentwicklung der Zygoten zu
den Zellfamilien ist noch nicht ganz lückenlos bekannt.
Ähnlich ist auch der Entwicklungsgang des Wassernetzes,
Hydrodictyon utriculatum[289], [330], einer der zierlichsten
freischwimmenden Süßwasseralgen, deren zylindrische vielkernige
Zellen zu einer Kolonie in Form eines sackförmigen,
langgestreckten, vielmaschigen Netzes verbunden sind.
Hierher gehört auch Protosiphon botryoides, eine auf Schlammboden
meist gemeinsam mit dem habituell ähnlichen Botrydium (S.
352) lebende Alge, deren blasenförmige bis 5 mm breite, nach
unten in einen farblosen Wurzelschlauch sich verlängernde Zelle
einen netzförmig durchbrochenen Chloroplasten und zahlreiche
Kerne enthält und sich durch Sprossung und Teilung vermehrt.
Fortpflanzung erfolgt durch Isogamie. Protosiphon zeigt, daß der
Thallus schon bei den Protococcales siphonalen Charakter annehmen
kann.
Die Protococcales leiten wir ebenso wie die Volvocales von
Flagellaten ab. Im Gegensatz zu den Volvocales hat aber bei
ihnen, wie auch weiterhin bei allen höherstehenden Algen, der
unbewegliche zilienlose Zustand der Zellen die Oberhand gewonnen,
so daß bei einigen Gattungen sogar auch die Sporen keine Geißeln
mehr erzeugen, obwohl im allgemeinen gerade in den Keimzellen der
Algen der Flagellatencharakter sich recht zähe erhält. Hand in Hand
mit der Aufgabe der Beweglichkeit erfolgt dann bei den Arten mit
ruhenden Einzelzellen eine fortschreitend reichere Ausgestaltung
der äußeren Zellform.
3. Ordnung. Ulotrichales. Sie bezeichnen den einzelligen
Grünalgen gegenüber einen Fortschritt in der äußeren Gliederung
des Thallus, der stets mehrzellig erscheint und meist aus
einfachen oder verzweigten Zellfäden besteht. Die Zellfäden
sitzen entweder mit einer farblosen Fußzelle am Substrat unter
Wasser fest (Fig. 342 A) oder schwimmen frei. Bei der im Meere
lebenden Gattung Ulva (Ulva lactuca, Meersalat) besteht der
Thallus aber aus großen blattartigen, grünen, zweischichtigen
Zellflächen (Fig. 81, Keimpflanze), bei Enteromorpha ist
er zylindrisch oder abgeflacht bandförmig (Fig. 341); in der Jugend
auch hier zweischichtig, wird er später innen hohl, so daß die
Wandschicht nur aus einer Zellage besteht. Die Ulotrichales leben
im Süßwasser oder im Meere. Nur einige Formen (Chroolepideen)
wachsen als Luftalgen an Felsen, Baumstämmen, in den Tropen
auch auf Blättern. Hierzu gehört die auf Steinen in Gebirgen
wachsende Trentepohlia (oder Chroolepus) Jolithus, deren
Zellfäden infolge Hämatochromgehaltes rot erscheinen und einen
veilchenartigen Geruch besitzen (Veilchenstein).
Die Zellen besitzen stets nur einen Zellkern und meist auch
nur einen Chloroplasten. Die ungeschlechtliche Fortpflanzung
vollzieht sich durch zilientragende Schwärmsporen, die
geschlechtliche besteht entweder in Kopulation von Planogameten,
oder die Geschlechtszellen sind in Eizellen und Spermien
differenziert.

Fig. 341. Enteromorpha compressa. Häufige Grünalge der
Nordseeküsten. 1⁄2 nat. Gr.
Ulothrix zonata[297] (Fig. 342 A), der typische Vertreter
der Gruppe, ist eine der häufigsten Fadenalgen. Sie besteht aus
unverzweigten, mit einer Rhizoïdzelle festsitzenden[S. 357] Fäden ohne
Spitzenwachstum; ihre kurzen Zellen enthalten einen bandförmigen
Chloroplasten. Die ungeschlechtliche Fortpflanzung geschieht
durch vierzilige Schwärmsporen (Zoosporen) (C), die
einzeln oder durch fortgesetzte Zweiteilung zu mehreren in einer
Fadenzelle gebildet werden und durch ein seitlich entstehendes Loch
aus der Zellmembran ausschlüpfen (B), umherschwärmen und dann
zu neuen Fäden auswachsen. Die geschlechtlichen Schwärmzellen,
Planogameten, bilden sich in gleicher Weise aus Fadenzellen
oder Gametangien, aber in viel größerer Zahl (D, E); sie
sind kleiner und besitzen nur zwei Zilien, außerdem einen roten
Augenfleck und einen Chloroplasten, wie die Schwärmsporen;
Ulothrix ist morphologisch isogam, physiologisch aber heterogam,
denn Gameten, die aus ein und demselben Faden stammen, kopulieren
nicht miteinander, sondern nur solche von getrennter Herkunft; sie
verschmelzen dann paarweise zu Zygoten (F-H), welche die Zilien
einziehen, sich abrunden und mit Membran umkleiden. Die Zygote
stellt einen Ruhezustand dar, sie wird zu einem kleinen einzelligen
Keimpflänzchen (J); ihr Inhalt teilt sich in mehrere Zellen, aus
denen die neuen Ulothrixfäden wieder heranwachsen. Übrigens können
sich die Planogameten unter Umständen auch parthenogenetisch ohne
Kopulation direkt weiter entwickeln. Damit ist die Mannigfaltigkeit
der Schwärmerbildung noch nicht erschöpft, denn die Fäden können
außer den oben genannten Schwärmsporen mit vier Wimpern auch
kleinere ungeschlechtliche, aber gametenähnliche Mikrozoosporen
mit vier oder zwei Wimpern erzeugen, welche bei Temperaturen über
10° meist zugrunde gehen, bei solchen unter 10° nach einigen Tagen
zur Ruhe kommen und dann langsam keimen. Die Alge ist insofern von
Interesse, als bei ihr die sexuelle Differenzierung der Gameten
noch in einem Anfangsstadium steht.
Fig. 342. Ulothrix zonata. A Junger Faden mit
Rhizoïdzelle r. Vergr. 300. B Fadenstück mit ausschlüpfenden
Schwärmsporen, zu zwei in jeder Zelle. C Einzelne Schwärmspore. D
Gametenbildung und Entleerung eines Fadenstückes. E Gameten. F, G
Kopulation der Gameten. H Zygote. J Zygote nach der Ruheperiode.
K Zygote, deren Inhalt in mehrere Zellen sich geteilt hat. B-K
Vergr. 482, nach DODEL.
Fig. 343. A, B Oedogonium. A Schwärmsporen
beim Ausschlüpfen. B Schwärmspore. C, D Oed. ciliatum. C
Vor der Befruchtung. D Während der Befruchtung. o Oogonien,
a Zwergmännchen, S Spermatozoid. Vergr. 350. Nach N.
PRINGSHEIM.
Als Beispiel oogamer Ulotrichales sei die Gattung
Oedogonium[331] genannt, an die sich Bulbochaete anschließt.
Während letztere verzweigte Zellfäden aufweist, hat erstere
Gattung unverzweigte Fäden, deren Zellen einen aus zahlreichen
zusammenhängenden Bändern bestehenden Chloroplasten besitzen.
Die ungeschlechtlichen Schwärmsporen (Fig. 343 B) sind
bei Oedogonium besonders groß, haben ein farbloses Vorderende,[S. 358]
an dessen unterem Rande zahlreiche Zilien in Form eines Kranzes
entspringen. Sie entstehen in Einzahl aus dem ganzen Inhalt einer
Fadenzelle (Fig. 343 A) und schlüpfen unter Aufbrechen dieser
Zelle aus, um alsbald zu neuen Fäden auszukeimen, nachdem sie sich
mit ihrem farblosen Ende festgesetzt haben. Die Oogonien
gehen aus einzelnen Fadenzellen hervor, indem diese tonnenförmig
anschwellen und ihren Inhalt zu einer großen Eizelle ausbilden.
Am oberen Ende des Oogoniums entsteht in der Membran ein Loch und
unter diesem ein farbloser Empfängnisfleck an der Eizelle. An
anderen Stellen desselben oder eines anderen Fadens werden die
Spermien erzeugt, und zwar meist zu je zwei in relativ niedrig
bleibenden Fadenzellen, den Antheridien. Die Spermien sind
kleiner als die ungeschlechtlichen Schwärmsporen, aber wie diese
mit einem Zilienkranz versehen. Sie schlüpfen durch die Öffnung in
das Oogonium und verschmelzen mit der Eizelle, die dann zu einer
großen derbwandigen Oospore wird. Bei der Keimung teilt sich
ihr Inhalt in vier große Schwärmsporen, welche ausschlüpfen und
neue Fäden bilden. Fig. 344 stellt die Bildung dieser Sporen für
Bulbochaete dar.
Bei gewissen Oedogonien liegen die Verhältnisse weniger einfach.
Die Spermien werden nämlich bei diesen in kleinen, nur aus wenigen
Zellen bestehenden Pflänzchen, sog. „Zwergmännchen“
erzeugt. Diese Pflänzchen entwickeln sich aus spermienähnlichen
Schwärmsporen (Androsporen), die sich nach dem Ausschwärmen auf
den weiblichen Fäden, ja sogar auch direkt auf den Oogonien
festsetzen und zu den wenigzelligen Zwergmännchen heranwachsen,
deren obere Zellen die Spermien erzeugen. Fig. 343 C zeigt ein
reifes Zwergmännchen auf einem noch geschlossenen Oogonium, D den
Eintritt der Befruchtung, das Spermium auf dem Empfängnisfleck, die
obere Wandung des Zwergmännchens deckelartig abgesprengt.
Fig. 344. Bulbochaete intermedia. A Oospore. B
Bildung von vier Schwärmsporen aus der keimenden Oospore. Vergr. 250.
Nach N. PRINGSHEIM.
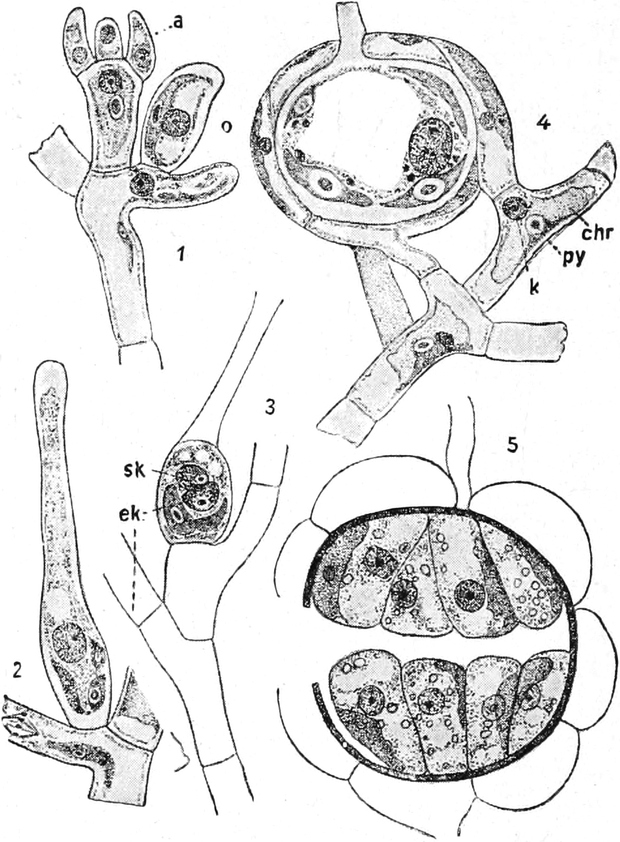
Fig. 345. Coleochaete pulvinata. 1 Antheridium a
und junges Oogon o, 2 Oogonium kurz vor der Öffnung, 3 dasselbe
befruchtet, ek Eikern, sk Spermakern, 4 Oospore durch Umwachsung
zur „Frucht“ entwickelt, 5 keimende Oospore. Nach OLTMANNS.
Die Gattung Coleochaete[332] zeichnet sich ebenfalls durch
Oogamie aus (Fig. 345). Ihr flaschenförmiges Oogonium
hat einen farblosen Hals, der sich an der Spitze zur Aufnahme
des Spermiums öffnet. Die Antheridien sind endständig und
erzeugen nur je ein mit zwei Geißeln versehenes
Spermium. Die heranwachsende, kugelige Oospore wird in ein
einschichtiges, pseudo-parenchymatisches Gewebe eingeschlossen,
indem von der Tragzelle des Oogoniums und benachbarten Zellen
Zellfäden aussprossen und sie dicht umhüllen. So entsteht eine
Oosporenfrucht. Bei der Keimung führt der Kern der Oospore
eine Reduktionsteilung aus. Hierauf entsteht ein 16–32zelliger
Körper, der die Oosporenhülle durchbricht und aus jeder Zelle eine
Schwärmspore entläßt, die sich zu einem neuen Thallus entwickelt.
Coleochaete vermehrt sich ungeschlechtlich durch zweiwimperige
Schwärmsporen, die in Einzahl in Thalluszellen gebildet werden.
[S. 359]
4. Ordnung. Siphonocladiales. Die hierher gehörigen, meist
reich verzweigten Algen unterscheiden sich von den Ulotrichales
durch große, vielkernige Zellen, deren Chloroplasten
entweder in Einzahl groß und netzförmig gestaltet erscheinen oder
als zahlreiche kleine Plättchen auftreten.
Die Gattung Cladophora mit zahlreichen Arten im Süßwasser und
im Meere kann als wichtigster Vertreter genannt werden. Cl.
glomerata (Fig. 84) ist eine der häufigsten Flußalgen, oft
fußlange verzweigte Fadenbüschel bildend, die an der Basis mit
rhizoïdartiger Zelle festsitzen und Spitzenwachstum aufweisen,
das bei anderen Vertretern der Ordnung noch nicht zur Ausbildung
gelangt ist. Der Bau der Zellen ist aus Fig. 7, 9 und 18 zu
ersehen. Die Verzweigung erfolgt an den oberen Enden der Zellen
durch Ausstülpung und Abgrenzung von Astzellen. Die Vermehrung
geschieht durch zweiwimperige (Fig. 346), bei den marinen
Arten der Gattung durch vierwimperige Zoosporen, die in
großer Zahl meist aus den oberen Zellen der Fäden entstehen und
durch ein seitliches Loch aus diesen Sporangien austreten. Die
geschlechtliche Fortpflanzungsweise ist Isogamie wie bei
Ulothrix.
Nur bei einer Gattung, der im Süßwasser in Form einfacher
Fäden vorkommenden Sphaeroplea annulina, ist die sexuelle
Fortpflanzung zur Oogamie vorgeschritten.
Manche Vertreter der Ordnung leben im Meere, z. B. Siphonocladus,
und haben zum Teil einen reich gegliederten, aber stets aus
verzweigten Zellfäden aufgebauten Thallus, der bei gewissen
Gattungen durch Kalkinkrustationen eine korallenartige
Beschaffenheit annimmt. Als Beispiel solcher Kalkalgen sei die
zierliche, im Mittelmeer heimische Acetabularia mediterranea
(Fig. 347) genannt. Der dünne Stiel des Thallus sitzt mittels
einiger Rhizoiden fest, der Schirm besteht aus zahlreichen, dicht
zu einer Fläche zusammenschließenden radialen Schläuchen, deren
jeder als ein Gametangium aufzufassen ist. Ihr Inhalt aber bildet
die zweiwimperigen Gameten nicht direkt, sondern zerfällt erst in
eine große Zahl derbwandiger Zysten, welche ruhend überwintern und
dann zahlreiche, paarweise kopulierende Gameten entlassen. Die
Zygoten keimen sehr bald und wachsen schrittweise zu neuen Pflanzen
heran.
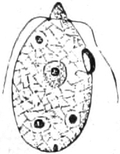
Fig. 346. Cladophora glomerata. Schwärmspore. Vergr.
500. Nach STRASBURGER. Hierzu Fig. 84.
Fig. 347. Acetabularia mediterranea. Kalkalge. Nat. Gr.
Nach OLTMANNS.
5. Ordnung. Siphonales. Die Siphoneen oder
Schlauchalgen unterscheiden sich von den vorhergehenden
Gruppen dadurch, daß in ihrem äußerlich mehr oder weniger
reich gegliederten Thallus zunächst keine Querwände
vorhanden sind. Die Zellhaut umschließt somit eine einzige
Plasmamasse, in welcher zahlreiche Zellkerne und
zahlreiche kleine grüne Chloroplasten enthalten sind. Ähnliche
Thallusformen kehren bei den Phycomyceten oder Algenpilzen wieder.
Die meisten Siphoneen leben im Meere und gehören infolge
ihres eigenartig gegliederten Thallus zu den interessantesten
Algentypen. So besitzt die in vielen Arten in wärmeren Meeren
vertretene Gattung Caulerpa[333] eine kriechende, an der Spitze
fortwachsende Hauptachse, die nach unten farblose Rhizoiden in
den Boden entsendet, nach oben dagegen grüne, bei den einzelnen
Arten sehr verschieden gestaltete Thallusglieder trägt. Bei
der mediterranen C. prolifera (Fig. 348) sind diese Glieder
blattartig, entweder einfach oder durch wiederholte Aussprossungen
verzweigt. Dabei umschließt die ganze Pflanze nur einen einzigen
Zellraum, der von netzförmig verbundenen Zellstoffbalken
durchsetzt wird. In den farblosen Thallusteilen enthält das Plasma
stärkebildende Leukoplasten.
Die Gattung Bryopsis hat einen zierlich federförmig verzweigten
Thallus[334]. Bei den in wärmeren Meeren lebenden Halimeda-Arten
setzt er sich aus scheibenförmigen[S. 360] Gliedern, ähnlich wie eine
Opuntia im kleinen, zusammen und erhält durch Kalkablagerungen eine
korallenartige Beschaffenheit. Die Glieder sind aus verzweigten und
miteinander verflochtenen Schläuchen gebildet.
Bei Bryopsis hat sich eine Differenzierung der kopulierenden
zweiziligen Gameten in größere, mit einem grünen Chromatophor
versehene weibliche und dreimal kleinere, nur ein kleines
gelbliches Chromatophor führende männliche vollzogen; bei
Vaucheria und Dichotomosiphon ist bereits Oogamie scharf
ausgeprägt[335]. Diese letzteren Algen kommen im Süßwasser vor
oder bilden auf feuchtem Boden kleine Rasen. Ihr Thallus besteht
aus einem fadenförmigen, verästelten und im Substrat mit farblosen
Rhizoïden befestigten Schlauch (Fig. 349 D).
Die Bildung der ungeschlechtlichen Schwärmsporen geschieht
bei Vaucheria in anderer Weise als bei den übrigen Siphonales.
Einzelne Zweigenden schwellen zur Bildung des Sporangiums etwas
an und grenzen sich mit einer Querwand ab (Fig. 349). Der Inhalt
der Endzelle verwandelt sich nun in eine einzige, sehr große,
grüne, mit bloßem Auge schon sichtbare Schwärmspore, die einen
farblosen, zahlreiche Kerne enthaltenden Saum besitzt und vor jedem
Kern je zwei Zilien hervorstreckt. Bei der Entleerung reißt der
Sporangiumscheitel auf, und die Spore zwängt sich unter Drehung um
ihre Längsachse aus der Öffnung hervor. Morphologisch entspricht
die Vaucheriaspore der Gesamtheit der Zoosporen eines gewöhnlichen
Sporangiums.
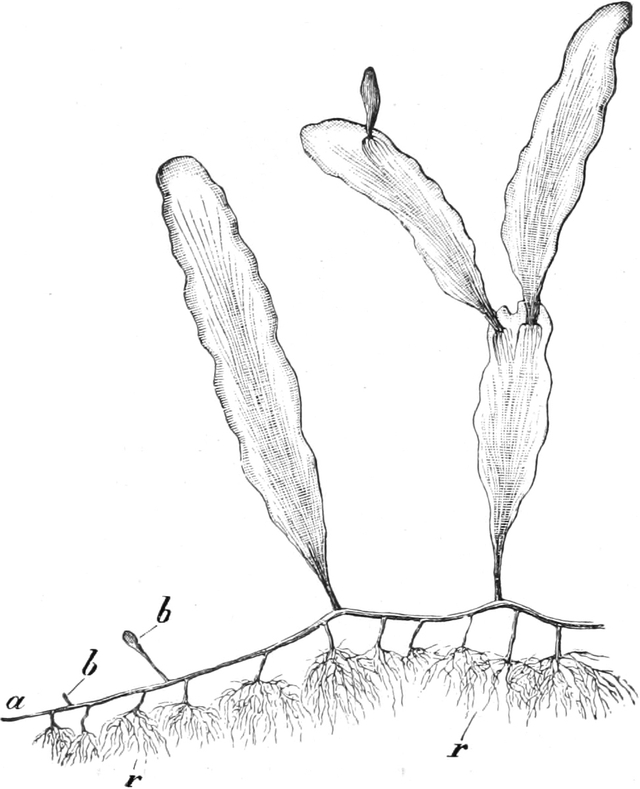
Fig. 348. Caulerpa prolifera. Die feinen Linien auf den
Thallusblättern bezeichnen die Plasmaströme. a Fortwachsende Spitze
der Thallusachse, bb junge Thalluslappen, r Rhizoïden.
1⁄2 nat. Gr.
Fig. 349. Vaucheria sessilis. A Anlage des
Sporangiums. B Aus dem Sporangium ausgeschlüpfte Zoospore. C Ein
Stück der Peripherie einer Zoospore. D Keimpflanze aus einer Zoospore
mit Rhizoïd. A, B nach GÖTZ, D nach SACHS, aus
OLTMANNS Algen, C nach STRASBURGER.
Die sexuelle Fortpflanzung von Vaucheria weicht bedeutend
von der Gametenkopulation der übrigen Siphoneen ab, ist aber
von dieser als der ursprünglicheren Befruchtungsart abzuleiten.
Oogonien und Antheridien entstehen an den Thallusfäden als
Ausstülpungen, die durch eine Scheidewand abgegrenzt werden (Fig.
350 o und a).[S. 361] Die Oogoniumanlage enthält anfangs zahlreiche
Kerne, die aber nach OLTMANNS und HEIDINGER
alle bis auf den zurückbleibenden einzigen Eikern vor der
Scheidewandbildung wieder in den Tragfaden zurückwandern. Im reifen
Zustande besitzt das Oogon eine schnabelartige, mit farblosem
Plasma angefüllte Vorstülpung, an welcher es sich öffnet. Ein
Ballen Plasma tritt zunächst aus, und die Eizelle rundet sich ab.
Das in seiner Anlage ebenfalls vielkernige Antheridium ist mit
seinem Tragast ein hornförmig gekrümmtes Gebilde (a); es bildet
einkernige Spermien, öffnet sich bei der Reife an seiner Spitze und
entleert seinen schleimigen Inhalt, aus dem die winzigen, völlig
farblosen, mit zwei seitlich inserierten Zilien versehenen Spermien
herausschwärmen, um an dem farblosen Empfängnisfleck des Oogoniums
sich anzusammeln. Ein Spermium dringt ein und vollzieht die
Befruchtung durch Verschmelzung seines Kerns mit dem Eikern. Die
befruchtete Eizelle umgibt sich mit einer Membran, geht als Oospore
in einen Ruhezustand über und keimt dann zu einem neuen Faden aus.
Fig. 350. Vaucheria sessilis forma repens. Fadenstück
mit Oogonium o, Antheridium a, ch Chromatophoren, n Zellkerne,
ol Öltropfen. Vergr. 240. Nach STRASBURGER.
Die Phaeophyceen scheinen wie die Chlorophyceen ihren phylogenetischen
Ausgang von Flagellaten genommen zu haben; in ihrer vegetativen
Gestaltung erreichen sie aber höhere Stufen als die Grünalgen.
Sie sind mit Ausnahme einiger Süßwasserarten Meeresalgen, die ihre
größte Entwicklung in den kälteren Ozeanen finden. In der Gestalt
ihres Thallus herrscht eine ungemeine Mannigfaltigkeit. Die
einfachsten Vertreter (z. B. Ectocarpus) zeigen unverzweigte
oder verzweigte, festsitzende Fäden, die aus einfachen Zellreihen
bestehen. Sodann gibt es Formen mit zylindrischem, reich verzweigtem,
vielzelligem Thallus, z. B. Cladostephus, dessen Hauptzweige mit
dichtem Filz von kurzen vielzelligen Seitenzweigen bedeckt sind (Fig.
89), oder mit bandförmigem, dichotomisch verzweigtem, vielzelligem
Thallus (z. B. Dictyota, Fig. 83). Diese Vertreter wachsen an ihren
Thallusenden vielfach mittels großer Scheitelzellen weiter (Fig. 89 und
90). Andere Arten haben scheiben- oder blasenförmigen Thallus.
Die höchste Entwicklung erfahren die Braunalgen in den Ordnungen
der Laminariaceen und Fucaceen. Zu den ersteren gehört die in
den nördlichen Meeren verbreitete Gattung Laminaria, deren Thalli
einem großen einfachen oder gefingerten, gestielten Blatt gleichen,
das an seiner Stielbasis mittels eines verzweigten, wurzelähnlichen
Haftorganes befestigt ist.
Bei Laminaria digitata und L. Cloustoni (Fig. 352) zeigt das
handförmig geteilte Thallusblatt ein sehr eigenartiges Wachstum,
indem es an seiner Basis eine interkalare wachstumsfähige Zone
besitzt, die jährlich gegen Ende des Winters ein neues Blatt auf
dem perennierenden Stiel erzeugt. Das alte wird dabei emporgehoben
und stirbt allmählich ab; das neue spaltet sich in mehrere
zugespitzte Lappen. Die Laminarien erreichen riesige Dimensionen;
so wird der Zuckertang L. saccharina (Nordsee) mit ungeteiltem,
ebenfalls sich jährlich erneuerndem Thallusblatt bis 3 m lang und
der Stiel über 1 cm dick.
Die größten Thalli unter den Phaeophyceen besitzen gewisse
Laminariaceen der kälteren Ozeane, vor allem die Macrocystis
pyrifera (Fig. 351). Ihr in einer Tiefe von 2–25 m festsitzender
Thallus verzweigt sich nach SKOTTSBERG[336] anfangs
dichotomisch. Einzelne Thallussprosse wachsen zur Wasseroberfläche
und an dieser flottierend zu großer[S. 362] Länge heran; ihre Achsen
tragen einseitig lange herabhängende Thalluslappen, die an der
Basis je eine große Schwimmblase besitzen und durch Spaltung der
endständigen weiterwachsenden Thallusfläche gebildet werden.
SKOTTSBERG maß in der Antarktis Exemplare von 70 m Länge,
FRYE, RIGG und CRANDALL bestimmten an
der kalifornischen Küste die Thalluslänge bis zu 45,7 m. Sehr
bemerkenswert sind ferner die antarktischen Lessonia-Arten, die
eine schenkeldicke verzweigte Hauptachse mit überhängenden langen
Thallusblättern an den Zweigen entwickeln und mehrere Meter Höhe
erreichen, also Algen mit baumartigen Habitus vorstellen.
Fig. 351. Macrocystis pyrifera Ag. a Junger, b
älterer Thallussproß. 1⁄80
nat. Gr. Nach Skottsberg.
Fig. 352. Laminaria Cloustoni, Nordsee. Sporophyt im
April-Stadium. Auf 1⁄3
verkleinert. Offizinell.

Fig. 353. Fucus serratus, Helgoland. 1⁄3 nat. Gr. Links
Endstück eines älteren Thallus mit Konzeptakeln.
[S. 363]
Die Fucaceen[337] bleiben hinter den Laminarien an Größe
zurück. Am bekanntesten sind von nordeuropäischen Formen Fucus
vesiculosus, der Blasentang, mit runden luftführenden Blasen in
dem bandförmigen, gabelig verzweigten Thallus, F. platycarpus
ohne Blasen, F. serratus mit gezähntem Thallus (Fig. 353). Sie
sitzen mit Haftscheiben dem Substrat an und wachsen gesellig
in der Brandungszone; ihr Thallus erreicht über 1 m Länge. Die
höchste Gliederung erlangt der Thallus bei Sargassum durch
eine scharfe Sonderung in zylindrische Achsen und in Seitenäste,
die je nach ihrer Funktion laubblattartig, hochblattartig oder
als fruktifizierende Seitenäste oder endlich als Schwimmblasen
ausgebildet erscheinen.
Bemerkenswert sind die Sargassum-Arten, die in wärmeren
Zonen der Ozeane oft in großen, lebhaft bräunlichgelben Massen
freischwimmend auftreten. In dem nach ihnen benannten Sargasso-Meer
des Atlantischen Ozeans treten nach BÖRGESEN zwei Arten,
S. natans (= S. bacciferum) und S. fluitans mit solcher
ausschließlich pelagischer Lebensweise auf; sie vermehren sich seit
altersher nur durch vegetative Sprossung und stammen ursprünglich
wohl von festsitzenden Arten der westindischen und tropisch
amerikanischen Küsten ab[338]. Auch im Pazifischen Ozean kommt S.
natans vor.
Die Zellen der Phaeophyceen[339] enthalten meist nur einen Zellkern
und mehrere scheibenförmige, ovale oder gelappte, braune
Chromatophoren, die den Algen eine gelbbraune oder dunkelbraune
Gesamtfärbung verleihen; außer den allgemein verbreiteten
Chlorophyllfarbstoffen enthalten sie noch einen besonderen
gelben Farbstoff, das Phykoxanthin (Fucoxanthin). Als
Stoffwechselprodukt entsteht hieraus Glykose das Polysaccharid
Laminarin, das als Reservestoff dient, ferner auch
Mannit. In den Zellen findet man allgemein verbreitet
blasenartige Vakuolen, die einen gerbstoffartigen Stoff,
Fucosan, ein Nebenprodukt des Assimilationsprozesses enthalten.
Bei den höheren Formen zeigt sich bereits eine ziemlich weitgehende
anatomische Differenzierung des Thallus. Die äußeren Zellschichten
sind in der Regel als Assimilationsgewebe ausgebildet, die
inneren als Speicherzellen. Bei den Laminariaceen und Fucaceen
finden sich im Thallus Schläuche, die wie die Siebröhren der
Kormophyten gebaut sind und wohl auch der Leitung eiweißartiger Stoffe
dienen (vgl. S. 69).
Selbst bei den stattlichsten Meeresalgen, auch bei den
Rotalgen, fehlen in den Geweben allgemein die lufterfüllten
Interzellulargänge. Nach KNIEP ist sowohl der
assimilatorische Gaswechsel als auch besonders der
Atmungsstoffwechsel dickfleischiger Algen dementsprechend sehr
träge; andererseits sollen Gase sehr leicht durch Algenmembranen
diffundieren können.
Wir unterscheiden vier Ordnungen der Braunalgen. Die Phaeosporeen
umfassen die Formen mit einfacherem Bau ihres meist fädigverzweigten
Thallus; sie vermehren sich ähnlich wie die einfacheren fadenförmigen
Grünalgen ungeschlechtlich durch Zoosporen, geschlechtlich durch
zilientragende Gameten. Auf höherer Stufe stehen zunächst die
Tilopteridaceen und Dictyotaceen, bei denen die Sexualzellen
in große unbewegliche Eizellen und kleine zilientragende Spermien
geschieden sind. Die Sexualorgane werden bei ihnen an besonderen
Individuen erzeugt, den Geschlechtspflanzen oder Gametophyten;
aus der befruchteten Eizelle geht eine der geschlechtlichen
gleichgestaltete ungeschlechtliche Generation, der Sporophyt, hervor,
der ungeschlechtliche Sporen erzeugt. So ist also hier ein regelmäßiger
Generationswechsel ausgeprägt, der übrigens bei gewissen Phaeosporeen
auch schon hervortritt. Auch bei den Laminariaceen herrscht Oogamie
und regelmäßiger Generationswechsel; Sporophyt und Gametophyt sind aber
wesentlich voneinander verschieden, der letztere nur ein kleines fädig
verzweigtes Zwergpflänzchen. Die Fucaceen zeichnen sich durch Oogamie
aus, erzeugen keine Sporen und entbehren also des Generationswechsels.
[S. 364]
Die Zoosporen, Gameten und Spermien besitzen spindelförmige
Gestalt, stets einen roten Augenfleck und zwei seitlich eingefügte
Geißeln, eine nach vorn, die andere nach hinten gerichtet. Sie
haben große Ähnlichkeit mit gewissen gelbbraunen Flagellaten.
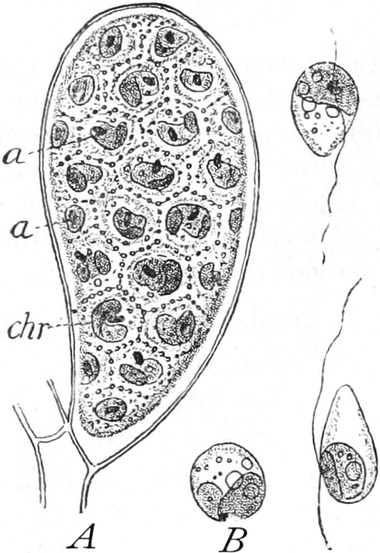
Fig. 354. A Pleurocladia lacustris. Unilokuläres
Sporangium mit noch eingeschlossenen Zoosporen, a Augenfleck, chr
Chromatophor nach KLEBAHN. B Chorda filum, Zoosporen nach
REINKE. Aus OLTMANNS, Algen.
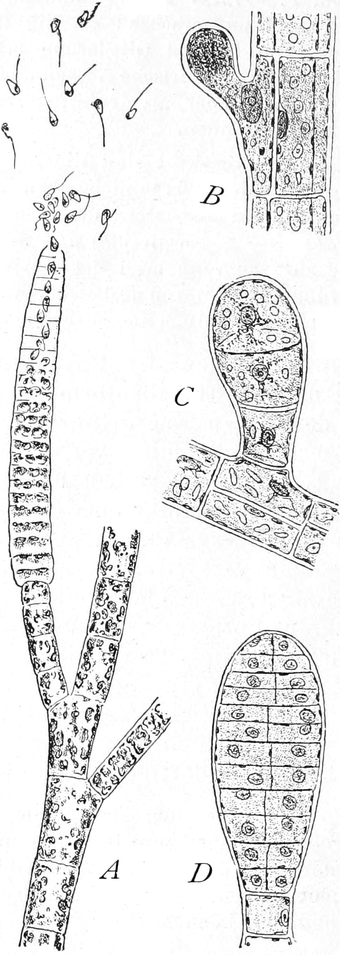
Fig. 355. A Ectocarpus siliculosus. Plurilokuläres
Sporangium in Entleerung (nach THURET). B, C, D
Sphacelaria cirrhosa, desgl. in Entwicklung nach REINKE. Aus
OLTMANNS, Algen.
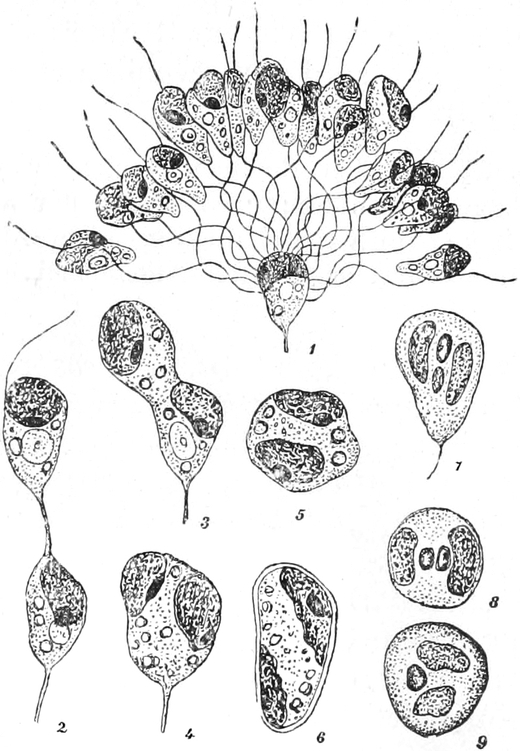
Fig. 356. Ectocarpus siliculosus. 1 Weiblicher
Gamet von vielen männlichen Gameten umgeben, von der Seite gesehen.
2–5 Verschmelzung der Gameten. 6 Keimling nach 24 Stunden.
7–9 Vereinigung der Zellkerne bei der Kopulation, nach fixiertem
und gefärbtem Material, 1–5 nach BERTHOLD, 6–9 nach
OLTMANNS.
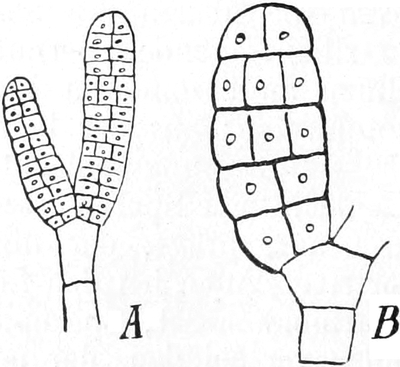
Fig. 357. A Antheridium. B Oogonium von Cutleria
multifida. Vergr. 400. Nach REINKE.
1. Ordnung. Phaeosporeae. Hierher gehört die Mehrzahl der Formen. Sie vermehren sich
durch ungeschlechtliche Schwärmsporen, die in großer Anzahl in[S. 365]
einfächerigen (unilokulären) Sporangien erzeugt
werden und nach dem Ausschwärmen bald keimen (Fig. 354).
Außer einfächerigen werden von den Phaeosporeen auch
mehrfächerige (plurilokuläre) Sporangien erzeugt
(Fig. 355), die in jedem Zellenfach nur eine, selten mehrere
Schwärmsporen bilden. Bei einigen Gattungen ist Kopulation dieser
Schwärmsporen beobachtet worden. Wir haben sie demnach als
Planogameten und ihre Sporangien als Gametangien
zu bezeichnen. Allerdings ist die Sexualität verschieden stark
ausgeprägt, und die aus plurilokulären Sporangien entlassenen
Schwärmer keimen vielfach auch ohne Kopulation zu neuen Pflanzen,
wie dies unter den Chlorophyceen auch für Ulothrix bemerkt wurde.
Als Beispiel für Kopulation sei Ectocarpus siliculosus (Fig.
355) genannt, bei dem bereits ein Unterschied in dem Verhalten der
im übrigen gleichgestalteten Gameten festzustellen ist, derart,
daß männliche und weibliche, beide in besonderen, diözisch oder
monözisch verteilten Gametangien erzeugt, zu unterscheiden sind.
Die weiblichen Gameten setzen sich fest und zahlreiche männliche
Gameten berühren sie mit ihren Zilien. Schließlich verschmilzt
ein männlicher Gamet mit dem weiblichen zu einer Zygote (Fig.
356 2–9), die zuletzt nur einen Kern, aber zwei Chromatophoren
enthält, sich mit einer Membran umgibt und dann ohne Ruhezustand zu
einer neuen Pflanze heranwächst.
Bei anderen Phaeophyceen ist der Unterschied zwischen den Gameten
auch in Form und Größe ausgeprägt, und besonders in der Familie
der Cutleriaceen ist ein entschiedener Übergang von Isogamie zur
Oogamie, eine Differenzierung der Gametangien in Antheridien
und Oogonien (Fig. 357), festzustellen.
Bei gewissen Phaeosporeen hat sich bereits ein regelmäßiger
Generationswechsel herausgebildet, so u. a. bei den
Cutleriaceen[340]. Die haploiden geschlechtlichen Pflanzen
alternieren mit diploiden ungeschlechtlichen, an denen die
Reduktionsteilung in den Zoosporangien sich vollzieht. Bei
Zanardinia sind beide Generationen gleich gestaltet, besitzen
einen scheibenförmigen Thallus, bei Cutleria dagegen sind
sie verschieden, indem die Sexualpflanzen aufrechte gabelig
zerschlitzte Sprosse, die Sporangienpflanzen (Aglaozonia)
dagegen flache, gelappte, niederliegende Scheiben vorstellen. Der
Generationswechsel wird indessen bei Cutleria nicht überall strenge
eingehalten; aus Sporangienpflanzen oder aus Geschlechtspflanzen
können auch die gleichnamigen Generationen wieder hervorgehen. Die
Eizellen entwickeln sich öfters parthenogenetisch zu Sporophyten.
2. Ordnung. Tilopteridaceae[341]. Nur wenige Vertreter, die sich
in vegetativer Hinsicht an einfachere Phaeosporeen (Ectocarpus)
anschließen, so die nordatlantische Tilopteris Mertensii. Die
Oogonien erzeugen nur je eine große Eizelle, die Antheridien kleine
zweizilige Spermien. In den ungeschlechtlichen Sporangien tritt
keine Sonderung in einzelne Sporen ein, sondern ihr Inhalt wird zu
einer einzigen großen, mit Membran versehenen Monospore mit meist 4
Kernen, während bei den Dictyotaceen sich an ihrer Stelle 4 freie
Sporen bilden.
3. Ordnung. Dictyotaceae[342]. Hierher gehören nur wenige in
vegetativer Hinsicht weiter als vorige Ordnung vorgeschrittene
Gattungen, z. B. die mediterrane, fächerförmig gestaltete Padina
Pavonia, die in europäischen Meeren verbreitete bandförmige,
typisch gabelig verzweigte Dictyota dichotoma (Fig. 83). Die
Sporen entstehen ähnlich wie bei den Rotalgen, meist zu vier als
sog. Tetrasporen, seltener zu acht, und sind membranlos,
unbeweglich, ohne Zilien, also als Aplanosporen zu bezeichnen
(Fig. 358 1). Man kann die Tetrasporangien von den unilokulären
der Phaeosporen ableiten. Oogonien und Antheridien
stehen bei Dictyota je gruppenweise zusammen; sie bilden sog.
Sori (Fig. 358 3, 2), die aus nebeneinander stehenden
Rindenzellen nach Abgliederung je einer Stielzelle hervorgehen. An
den Antheridiengruppen bleiben bei Dictyota die peripheren Zellen
steril und bilden eine Art Hülle. Die Oogonien erzeugen nur je
eine große einkernige, ausschlüpfende Eizelle, die vielzelligen
Antheridien, die mit den plurilokulären Gametangien übereinstimmen,
aus jeder Zelle je ein Spermium, das im Unterschied zu den übrigen
Braunalgen nur eine einzige lange, seitlich inserierte Zilie
besitzt. Die Zygoten keimen, ohne einen Ruhezustand zu erfahren.
Dictyota ist zweihäusig. Die haploiden männlichen und weiblichen
Sexualpflanzen gehen hervor aus den ungeschlechtlich erzeugten
Tetrasporen; aus den befruchteten Eizellen entstehen diploide
Pflanzen, welche ungeschlechtliche Tetrasporen erzeugen. Die
Reduktion der Chromosomenzahl von 32 auf 16 findet bei der
Tetradenteilung in den[S. 366] Sporangien statt. So liegt hier ein
regelmäßiger Generationswechsel vor; Gametophyt und
Sporophyt zeigen aber im Bau ihres Thallus keinen morphologischen
Unterschied.
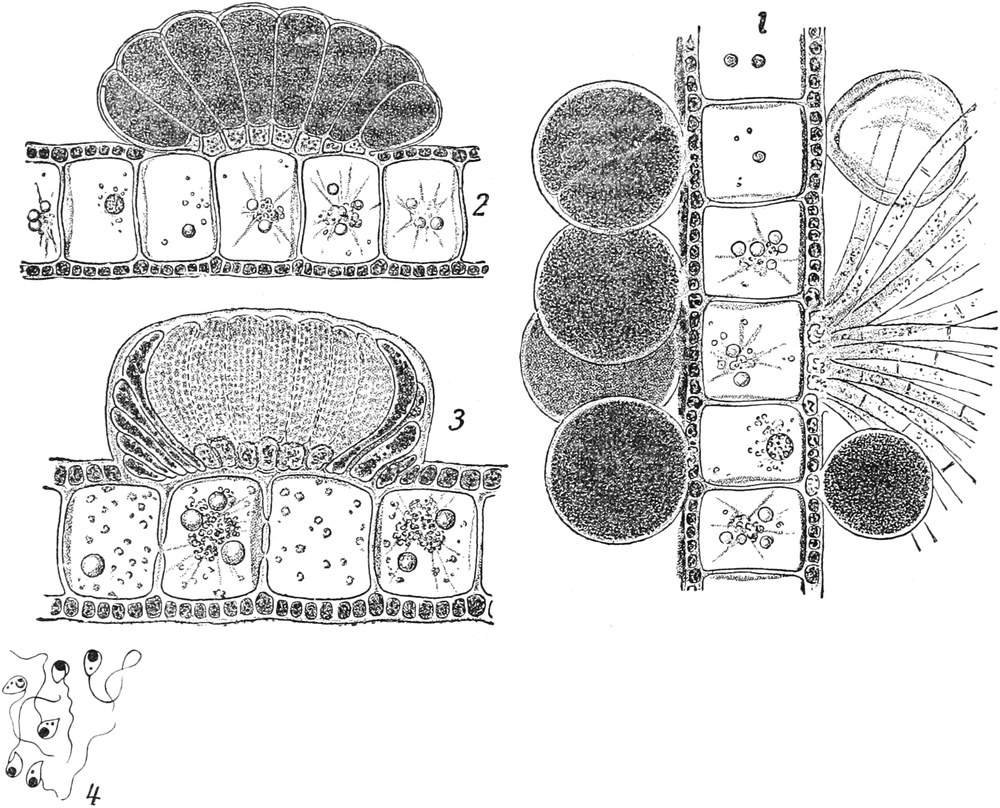
Fig. 358. Dictyota dichotoma. Thallusquerschnitte.
1 mit mehreren Tetrasporangien, davon eins entleert, 2 mit
Oogoniumgruppe, 3 mit Antheridiengruppe, nach THURET, 4
Spermien, nach WILLIAMS. Aus OLTMANNS, Algen.
Fig. 359. Laminaria digitata. A Männlicher Gametophyt,
a entleerte Spermatangien. B Größerer, C kleiner, D auf das
Oogonium reduzierter weiblicher Gametophyt, og Oogonium, o Eizelle.
E Junger Sporophyt, noch auf dem entleerten Oogonium sitzend. F
Weiteres Stadium mit zwei Rhizoïden. A Vergr. 600, B 292, C 322,
D 625, E 322, F 390. Nach H. KYLIN.
4. Ordnung. Laminariaceen[343]. Der erst in den letzten Jahren
entdeckte regelmäßige Generationswechsel dieser stattlichsten
aller Braunalgen entspricht dem[S. 367] der Farne insofern, als auch
hier die Gametophyten, durchaus verschieden von den Sporophyten,
nur winzige Zwergpflänzchen vorstellen. Der diploide Sporophyt
erzeugt keulenförmige oder zylindrische Sporangien, die
in ausgebreiteter oberflächlicher Schicht angeordnet sind. Jede
Oberflächenzelle des Thallus setzt sich fort in eine mittlere
keulenförmige sterile Zelle oder Paraphyse, neben denen die
Sporangien als kürzere Zellen entspringen. In den Sporangien
vollzieht sich die Reduktionsteilung. Aus den zweiziligen
Schwärmsporen entwickeln sich winzige männliche und weibliche,
haploide Geschlechtspflänzchen (Fig. 359), die aus Zellfäden sich
aufbauen, die ersteren reicher verzweigt, die letzteren dagegen
nur wenigzellig und im Extrem sogar nur aus einer schlauchförmigen
Zelle gebildet. Die männlichen Gametophyten erzeugen die
Antheridien an den Zweigspitzen neben- oder auch hintereinander
als einzellige Gebilde, die nur je ein Spermium liefern; an den
weiblichen kann jede Zelle zu einem eineiigen Oogonium werden. Die
nackte Eizelle tritt durch ein Loch an der Spitze des Oogoniums
hervor, bleibt aber vor der Mündung liegen und wächst dann nach der
Befruchtung zum Keimling des Sporophyten (Fig. 359 E, F) heran.
Oogonien und Antheridien sind den Gametangien der Phaeosporeen
homolog.
Für Laminaria saccharina hat PASCHER an kultivierten
jungen Keimpflänzchen des Sporophyten eine ausnahmsweise frühzeitig
eintretende Sporangienbildung beobachtet. Es kann also auch der
für gewöhnlich stattlich ausgebildete Sporophyt unter bestimmten
äußeren Bedingungen auf einer einfacheren Entwicklungsstufe stehen
bleiben. Wir gewinnen so einen Gesichtspunkt für die Erklärung des
Zustandekommens des auffallenden Dimorphismus der Generationen.
5. Ordnung. Fucaceae[344]. Bei den Fucaceen fehlt die
ungeschlechtliche Sporenbildung, dagegen ist sexuelle
Fortpflanzung vorhanden und als Oogamie ausgeprägt.
Bei Fucus sitzen die Oogonien und Antheridien in besonderen
krugförmigen Vertiefungen, sog. Conceptacula, die zu vielen in
die letzten angeschwollenen Auszweigungen des Thallus eingesenkt
und für die ganze Familie sehr charakteristisch sind (vgl.
Fucus serratus, Fig. 353). Bei F. platycarpus enthalten die
Konzeptakeln (Fig. 360) sowohl Oogonien als Antheridien, bei
F. vesiculosus dagegen herrscht Diözie. Der Innenwand der
Conceptacula entspringen zahlreiche, unverzweigte, sterile Haare,
sog. Saftfäden oder Paraphysen, die zum Teil als Büschel nach außen
hervortreten, und zwischen ihnen die Oogonien und Antheridien.
Die Antheridien sitzen als ovale Zellen dicht gedrängt an reich
verzweigten, kurzen Fäden (Fig. 380 a, 361 1). Der Inhalt des
Antheridiums teilt sich in 64 Spermien; er wird als Ganzes, von
der dünnen inneren Wandschicht umgeben, entleert (Fig. 361 2)
und entläßt dann die birnförmigen, hauptsächlich aus Kernsubstanz
bestehenden, mit zwei verschieden langen seitlichen Zilien, einem
orangegelben Chromatophor und einem roten Augenfleck versehenen
Spermien. Die Oogonien (Fig. 360 o, 361 3) sind große
rundliche, auf einzelligem Stiel sitzende, gelbbraune Gebilde,
deren Zellhaut acht große, aus der Oogoniummutterzelle durch
Teilung entstandene Eizellen umschließt. Der Oogoniuminhalt tritt,
von einer dünnen Hülle umgeben, aus der aufplatzenden Oogoniumwand
heraus; die Hülle verquillt am oberen Teil, stülpt sich teilweise
zurück, und die nackten braunen Eier werden nun frei ins Wasser
entleert (Fig. 361 4, 5). Dort haften die Spermien in großer
Menge ihnen an, versetzen sie durch ihre Zilien in rotierende
Bewegung, wobei die Befruchtung durch ein Spermium erfolgt (Fig.
361 6). Nach der Befruchtung umgibt sich das Ei mit Membran,
setzt sich fest und wächst unter Teilung zu einer neuen Pflanze
heran.
Fig. 360. Fucus platycarpus. Monözisches Conceptaculum
mit Oogonien verschiedenen Alters o und Antheridienbüscheln a,
Paraphysen p. Vergr. ca. 25. Nach THURET.
Bei anderen Fucaceen werden in dem Oogonium nur vier, zwei oder
schließlich auch nur ein befruchtungsfähiges Ei erzeugt; immer aber
teilt sich der eine Kern der[S. 368] Oogoniumanlage nach OLTMANNS
in acht Kerne, wovon dann in diesen Fällen vier, zwei oder einer zu
Eikernen ausgebildet werden, die übrigen als befruchtungsunfähige
Eikerne zugrunde gehen.
Da die Fucaceen der ungeschlechtlichen Sporenbildung ermangeln,
so fehlt ihnen der für Laminaria charakteristische
Generationswechsel. Der aus der befruchteten Eizelle
hervorgehende Fucus-Thallus, der durchaus dem Laminarien-Thallus
gleichwertig ist, hat diploide Zellkerne, deren Reduktion erst in
den Anlagen der Oogonien und Antheridien, und zwar in den ersten
zwei Teilungsschritten erfolgt, so daß vier haploide Kerne erzielt
werden, die im Oogonium dann noch eine, im Antheridium noch vier
Teilungen bis zur Bildung der Sexualzellen erfahren. Bei Fucus
ist also nur eine ganz kurze haploide Phase zu erkennen. Die
Entwicklung und Beschaffenheit der Sexualorgane, namentlich der
Oogonien, läßt die Deutung zu, daß sie morphologisch den Sporangien
der Laminarien homolog sind, daß aber in ihnen an Stelle von Sporen
Gameten differenziert werden. Die Fucaceen bezeichnen somit das
Endglied in der Reduktion des Gametophyten, die bei den Laminarien
noch nicht ganz so weit vorgeschritten erscheint.
Nutzpflanzen der Phaeophyceen sind: die früher
offizinellen Laminaria digitata und L. Cloustoni (Pharm.
germ.), deren Thallusstiele getrocknet als Quell-Stifte (stipites
Laminariae) in der Chirurgie Verwendung fanden. — Verschiedene
Laminariaceen und Fucaceen lieferten früher aus ihrer Asche
(Varec, Kelp) Jod; auch wird Soda aus ihnen gewonnen. Viele
Laminarien sind reich an Mannit (z. B. Lam. saccharina),
dienen zur Gewinnung seiner und werden auch, besonders von Chinesen
und Japanern, als Nahrung genossen.
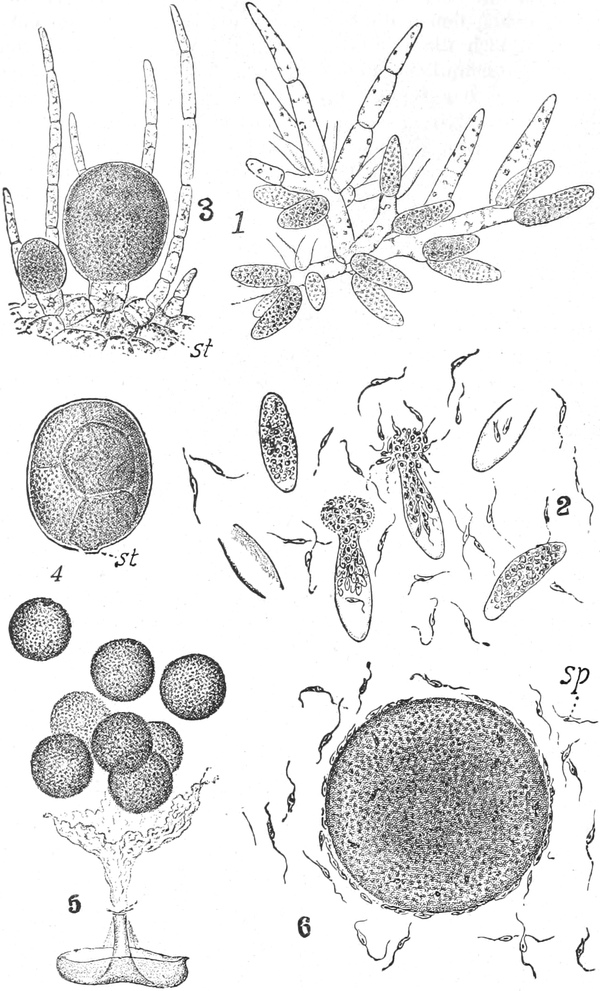
Fig. 361. Fucus. 1 Antheridiumstand. 2 Antheridien
in Entleerung. 3 Oogonium in acht Eizellen geteilt. 4 Nach der
Ablösung vom Stiel st. 5 Befreiung der Eier. 6 Ei von Spermien
umschwärmt. Nach THURET, aus OLTMANNS, Algen, Hdw. d.
Nat.
Die Characeen oder Charophyten bilden eine nach oben und unten hin
isoliert stehende Gruppe von hochentwickelten grünen Thallophyten.
Ihr Ursprung ist zwar bei den Chlorophyceen gesucht worden, jedoch
gestatten sie infolge des eigenartigen Baues ihrer Sexualorgane
keinen unmittelbaren Anschluß an oogame Grünalgen, verraten dagegen
in einigen morphologischen Charakteren Analogien mit Braunalgen,
von denen sie aber durch rein grüne Färbung der Chromatophoren
abweichen. Man kann sie nicht etwa als Vorläufer der Moose ansehen,
wenn auch ihre Karyokinese mit derjenigen der Archegoniaten
weitgehende Übereinstimmung zeigt.
Die Characeen vegetieren in Form von oft über fußhohen Wiesen in
Teichen und Bächen. Sie sind ausgezeichnet durch ihren regelmäßigen
Aufbau; ihre Hauptachsen bestehen aus langen Internodien und kurzen
Knoten, an denen aus wenigen Gliedern bestehende Seitenäste und Quirle
entspringen (Fig. 362). Diese Seitenachsen sind entweder einfach
oder tragen an ihren Knoten kurze Ausstrahlungen zweiter Ordnung. In
der Achsel eines Seitenastes entspringt in jedem Quirl eine
der Hauptachse ähnliche Seitenachse. So kommt ein armleuchterartiger
Habitus zustande. Am Grunde sind die Achsen mittels farbloser,
verzweigter, aus den Knoten entspringender fädiger Rhizoïdwurzeln
im Substrat befestigt. Diese zeigen ebenfalls eine Gliederung in
lange Fadenzellen und zwischen ihnen stehende kurze, hier einseitig
gerichtete Knotenzellen, aus denen die Verzweigung erfolgt.
Haupt- und Seitenachsen wachsen an ihren Spitzen mittels
Scheitelzellen, die durch Querwände Segmente abteilen;
jedes Segment teilt sich nochmals durch eine Querwand, und nun
entwickelt sich aus der unteren Zelle die langgestreckte, ungeteilt
bleibende Internodienzelle; aus der oberen Zelle dagegen unter
weiterer Teilung die Knotenscheibe, ferner die Seitenachsen und an
dem unteren Teil der Hauptachsen auch die Rhizoïden. Während bei
Nitella die Internodiumzellen nach außen hin frei bleiben, werden
sie bei den meisten Arten von Chara mit einer Schicht längs
verlaufender Zellreihen, die an den Knoten aus den Basilarzellen
der Seitenachsen hervorwachsen, dicht umschlossen; diese
morphologische Eigentümlichkeit wird in ähnlicher Art bei gewissen
Braunalgen (z. B. Spermatochnus) angetroffen.
Die Zellen enthalten je einen normalen, auf karyokinetischem
Wege entstandenen Zellkern. In den langen Internodialzellen aber teilt
sich dieser amitotisch in zahlreiche Kerne, die in dem einen
großen Saftraum umschließenden und lebhafte Strömung aufweisenden
Plasmabelag verteilt liegen. Die grünen Chloroplasten sind
rundlich und in großer Zahl vorhanden.
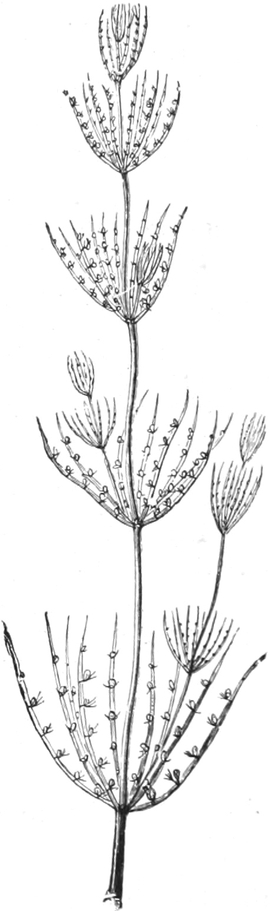
Fig. 362. Chara fragilis. Ende eines Hauptsprosses.
Nat. Gr.
Ungeschlechtliche Fortpflanzung mittels Sporen fehlt
vollständig; die sexuelle Fortpflanzung besteht in
Eibefruchtung. Die weiblichen Organe, hier als Eiknospen
bezeichnet, sind eiförmige, mit bloßem Auge sichtbare Gebilde und
stehen ebenso wie die kugeligen, rotgefärbten männlichen Organe an
den Knoten der Seitenachsen. Meist sind die Pflanzen monözisch,
einige Arten auch diözisch. Die befruchtete Eizelle wird zu einer[S. 370]
Oospore. Die Characeen zeigen also keinen Generationswechsel,
sondern stets Aufeinanderfolge von Gametophyten.
Das männliche Organ (Fig. 363 A) geht aus einer sich
zunächst in 8 Zellen teilenden Mutterzelle hervor. Jeder Oktant wird
durch 2 tangentiale Wände in 3 Zellen zerlegt. So ergeben sich 8
äußere flache Wandzellen (Schilder), die durch einspringende Wände
unvollständig gefächert werden: 8 mittlere Zellen (Griffzellen), die
sich radial strecken, und 8 innere Zellen (primäre Köpfchenzellen),
die rundliche Form annehmen. Infolge stärkeren Flächenwachstums
der 8 Schilder entsteht im Innern des Organs ein Hohlraum, in den
die Griffzellen mit ihren aufsitzenden Köpfchenzellen hineinragen.
Letztere bilden 3–6 sekundäre Köpfchenzellen, und aus diesen sprossen
nun in den Hohlraum hinein je 3–5 lange, aus scheibenförmigen Zellen
bestehende, einfache, spermatogene Zellfäden, die aus jeder
Zelle ein spiralig gewundenes, mit zwei Zilien versehenes Spermium
(Fig. 363 C) entlassen. Morphologisch erinnern diese Zellfäden oder
Antheridien an die plurilokulären Braunalgengametangien, die z. B. bei
Stilophora ebenfalls aus einfachen Zellreihen bestehen und in Gruppen
(Sori) beieinanderstehen. Das männliche Organ der Characeen, das als
Ganzes zwar gewöhnlich Antheridium genannt wird, enthält also 8
Antheridiengruppen von endogener Entwicklung und sollte demnach als
Antheridienstand bezeichnet werden.
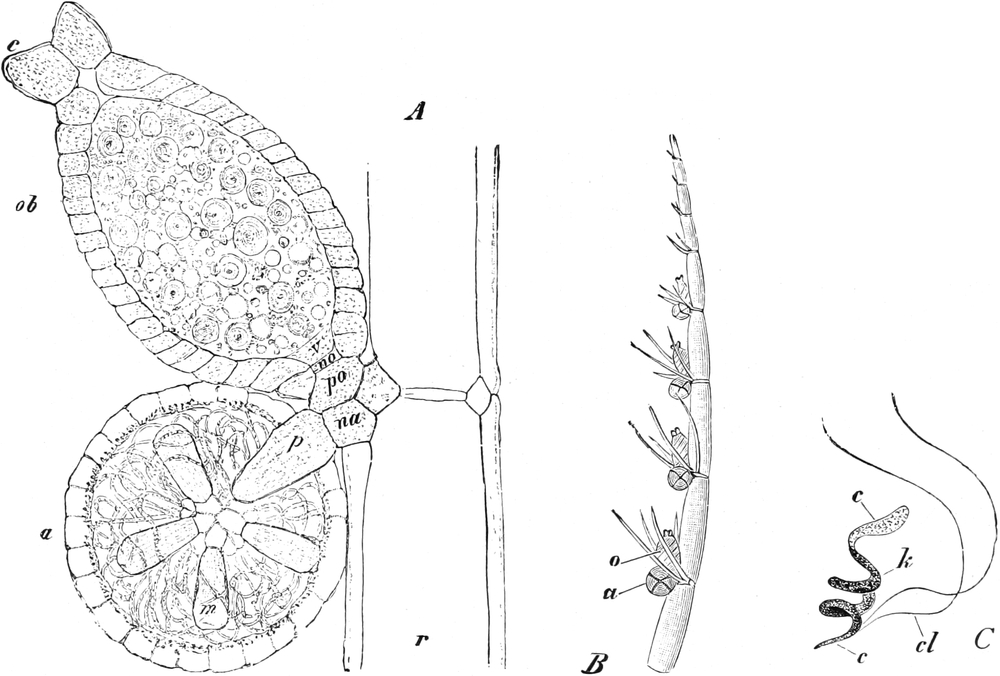
Fig. 363. Chara fragilis. A Medianer Längsschnitt
durch eine Seitenachse r, a Antheridiumstand, und zwar na
Basilarknotenzelle, p Stiel, m die Griffzellen, aus deren
aufsitzenden Zellen die spermatogenen Zellfäden entspringen, ob
Eiknospe, und zwar po Stielzelle, no die Knotenzelle, v die
Wendungszelle, c das Krönchen. Vergr. 60. B Ganze Seitenachse.
Vergr. 6. C Spermium, k Kern, cl Zilien, c Plasma. Vergr. 540.
Nach STRASBURGER.
Die Eiknospe (Fig. 363 ob) besteht aus einem Oogonium,
das nur eine einzige mit Öltropfen und Stärkekörnern dicht gefüllte
Eizelle enthält, anfangs frei hervorragt und dann von 5
schraubig gewundenen Hüllschläuchen dicht umschlossen wird. Letztere
endigen in den Krönchen c, zwischen dessen Spalten die Spermien
eindringen. Am Grunde des Oogoniums finden sich bei Chara eine,
bei Nitella drei kleine sog. Wendungszellen, die von der anfangs
einzelligen Oogoniumanlage abgeteilt wurden. Diese Teilungen
entsprechen den ersten Teilungen in den Mutterzellen des männlichen
Organs.[S. 371] Die Eiknospe kann daher als ein auf ein einziges Oogonium
reduzierter Oogoniumstand betrachtet werden.
Nach der Befruchtung umgibt sich die Eizelle mit einer dicken
farblosen Haut, und auch die Innenwände der Hüllschläuche verdicken
sich, werden braun und mit einer Schicht von amorphem kohlensaurem
Kalk bedeckt, während die äußeren weichen Zellwände der Schläuche
bald nach dem Abfallen der Frucht vergehen.
Bei der Keimung der Oospore teilt sich nach OEHLKERS und
ERNST der Kern schrittweise in vier Kerne: die erste
Teilung ist eine heterotypische. Dann wird die Membran an der
Spitze durch die sich hier vorwölbende Zygotenzelle geöffnet. Drei
Kerne verbleiben im Bauchteil der Zygote, wo sie degenerieren, der
vierte, sich vergrößernde Kern aber wandert in die hervortretende
Ausstülpung, die dann durch eine Querwand abgegrenzt wird. Aus
dieser Zelle geht unter weiterer Teilung ein einfach gestalteter,
fadenförmiger, mehrgliedriger Keimling hervor, an dessen erstem
Knoten Rhizoïden entspringen, während am zweiten einige einfache
Seitenachsen stehen, sowie eine oder mehrere Hauptachsen, aus
deren weiterer Verzweigung die fertige Pflanze heranwächst. Das
diploide Stadium beschränkt sich also bei den Characeen nur auf die
Oosporen, während die Armleuchterpflanzen selbst haploid sind.
Bemerkenswertes Verhalten zeigt Chara crinita. Nach
ERNST kommen von dieser diözischen Art die haploiden, 12
Chromosomen führenden männlichen und weiblichen Geschlechtspflanzen
nur sehr zerstreut vor, weit verbreitet dagegen diploide 24
Chromosomen führende weibliche Pflanzen, die sich vermittelst
diploider Eizellen parthenogenetisch vermehren und die aus der
Kreuzung der Ch. crinita mit anderen Arten hervorgegangen
sein sollen. Nach WINKLER aber könnte diese Diploidie
durch Verdoppelung der Chromosomenzahl in der Scheitelzelle
einer weiblichen haploiden Pflanze zustande gekommen sein oder
eher dadurch, daß von den bei der Reduktionsteilung in der
Zygote entstehenden 4 Kernen die beiden weiblich differenzierten
wieder miteinander verschmelzen. Es liegt also hier somatische
Parthenogenese und nicht, wie man früher annahm, generative
vor[346].
Einige Characeen zeichnen sich durch die Bildung besonderer, mit
Stärke dicht gefüllter Knöllchen in der unteren Partie
der Achsen aus. Diese dienen als Überwinterungsorgane und
gehen entweder aus Knoten mit verkürzten Astquirlen hervor (so bei
Tolypellopsis stelligera, wo sie sternförmige Gestalt haben) oder
entsprechen modifizierten Rhizoïden (z. B. bei Chara aspera, wo
sie kugelige, weiße Gebilde vorstellen).
In fossilem Zustand sind Characeen (in Form von Zygoten) schon
aus dem Jura sicher nachgewiesen. Vielleicht gehören auch gewisse
devonische Fossilien hierher.
Die Rotalgen, Rhodophyceen oder Florideen, bilden eine scharf umgrenzte
Gruppe höherer Algen, deren phylogenetischer Ursprung vielleicht bei
höher stehenden Grünalgen zu suchen ist, die jedoch auch Beziehungen zu
den Braunalgen zeigt. Sie sind wie die letzteren fast ausschließlich
Meeresalgen und bewohnen vorzugsweise die tieferen Algenregionen an
den Küsten. Nur wenige Gattungen (Batrachospermum z. B.) wachsen im
Süßwasser, am Boden fließender Gewässer.
Der Thallus der Rotalgen weist große Mannigfaltigkeit auf.
Die einfachsten Formen stellen verzweigte Zellfäden dar (z. B.
Callithamnion). Bei anderen baut sich der verzweigte, zylindrische
Thallus aus mehreren Zellen im Querschnitt auf. Zahlreiche Formen
besitzen einen breiter oder schmäler bandförmigen und oft reich
fiederig oder gabelig verzweigten Thallus (z. B. Chondrus crispus
Fig. 364, Gigartina mamillosa Fig. 365). Sodann gibt es Arten,
die in Form von Zellflächen der Unterlage aufliegen. Alle Florideen
sitzen an der Basis mittels Haftfäden oder Haftscheiben fest. Eine
der reicher gegliederten Formen ist z. B. die Delesseria sanguinea
(Hydrolapathum Fig. 88) des Atlantischen Ozeans. Der blattartige,
einer Basalscheibe entspringende[S. 372] Thallus ist hier mit Mittel- und
Seitenrippen versehen. Im Herbst gehen die Spreiten zugrunde; die
Hauptrippen aber bleiben als Achsen stehen, um im nächsten Frühjahr
neue Thallusblätter zu treiben. Die Familie der Corallinaceen, deren
Gattungen teils einen zierlich verzweigten, teils einen krustenförmigen
Thallus aufweisen, zeichnet sich dadurch aus, daß in und um die
Membranen kohlensaurer Kalk massenhaft abgelagert wird, so daß diese
Algen etwa den Eindruck von Korallen machen. Die Kalkflorideen
vegetieren hauptsächlich an Küsten mit starker Brandung, besonders in
den Tropen.
Die Rotalgen sind meist rot oder violett, auch purpurschwarz oder
braunrot gefärbt. Ihre scheibenförmigen, ovalen oder gelappten
Chromatophoren treten in größerer Zahl und dichter Lagerung in den
Zellen auf und enthalten außer Chlorophyll einen roten Farbstoff,
das Phykoërythrin, bei gewissen Arten außerdem auch blaues
Phykozyan; sie gehen aus farblosen spindelförmigen Leukoplasten
der Scheitelzellen und Keimzellen hervor. Als Assimilationsprodukt
wird aus Glykose nicht echte Stärke erzeugt, sondern ein anderes
Kohlehydrat, die Florideenstärke, in Form von rundlichen,
oft geschichteten, mit Jod sich rötlich färbenden Körnchen. Auch
Öltröpfchen kommen vor. Die Zellen sind einkernig oder auch mehrkernig.

Fig. 364. Chondrus crispus. Nach einem Exemplar von
Helgoland. 1⁄2
nat. Gr. Offizinell.
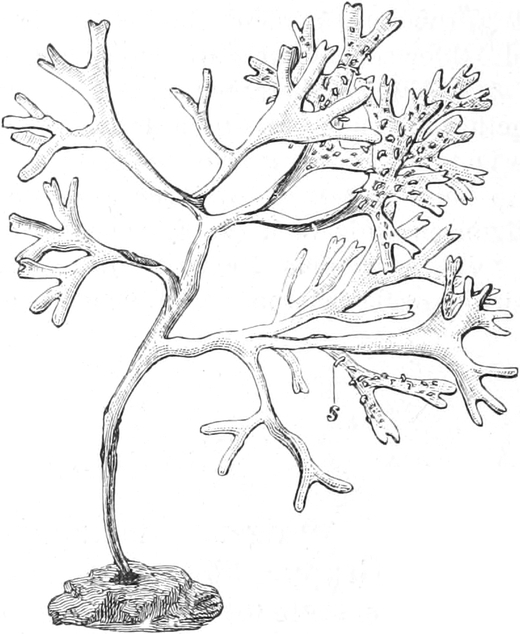
Fig. 365. Gigartina mamillosa. s Warzenförmige
Fruchtkörper. 3⁄4
nat. Gr. Offizinell.
Die Fortpflanzung geschieht bei den Florideen einerseits
ungeschlechtlich mittels Sporen, andererseits geschlechtlich mittels
Befruchtung weiblicher Organe durch männliche Zellen.
Die ungeschlechtlichen Sporen werden auf zweierlei Weise
gebildet. Im ersten Falle sind es nackte, runde, unbewegliche,
einkernige Zellen ohne Zilien, die zu vier durch Teilung in
einem Sporangium entstehen. Die Sporangien sitzen als rundliche
Körper an den Thallusfäden oder sind dem Thallus eingesenkt;
sie entlassen die vier Sporen aus einem Querriß ihrer Wandung.
Infolge ihrer Entstehung zu vieren nennt man die Florideensporen
Tetrasporen (Fig. 366). Sie vertreten die Rolle der
Schwärmsporen der übrigen Algen und finden sich in ähnlicher
Weise nur bei den Dictyotaceen unter den Braunalgen wieder. Die
Tetrasporangien führen in der Regel anfangs[S. 373] einen
Kern, der sich dann in die vier Sporenkerne teilt. In einigen
Fällen (Martensia, Nitophyllum) aber sind sie zwar anfangs
vielkernig, dann aber degenerieren alle Kerne bis auf einen, der
die vier Sporenkerne liefert. Den Tetrasporangien gleichwertig sind
die in der Gruppe der Nemalionaceen auftretenden, nur je eine
Spore entlassenden Monosporangien, sowie auch die zahlreiche
Sporen erzeugenden Polysporangien der Ceramiaceen.
Die zweite Form ungeschlechtlicher Sporen der Rotalgen sind die
Karposporen (vgl. S. 374), die in Einzahl aus endständigen
Karposporangien als kugelige, anfangs nackte, zilienlose
Protoplasten entleert werden, also äußerlich sich den Monosporen
nähern.
Die Ausbildung der Sexualorgane, besonders der weiblichen,
ist sehr eigenartig und von dem Verhalten der übrigen Algen
sehr abweichend. Sie sei an dem Beispiel von Batrachospermum
moniliforme, einer einheimischen Süßwasserfloridee, erläutert.
Diese Alge besitzt einen in Gallerte gehüllten, bräunlichen, aus
wirtelig verzweigten Fäden bestehenden Thallus, der im Herbst
fruktifiziert. Die Sexualorgane sitzen in den Zweigquirlen an
kurzen, dicht zusammengedrängten Seitenästchen.
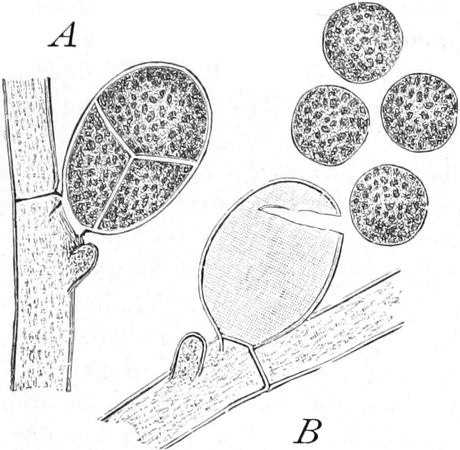
Fig. 366. Callithamnion corymbosum. Tetrasporenbildung.
A Geschlossenes, B entleertes Sporangium mit den vier ausgetretenen
Tetrasporen. Nach THURET.
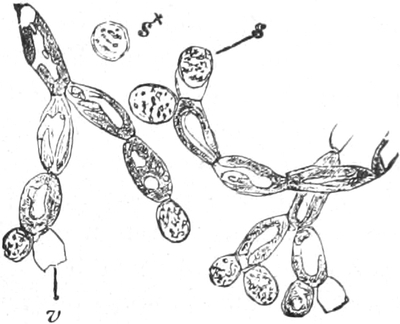
Fig. 367. Batrachospermum moniliforme. Wirtelzweige mit
Antheridien. Bei s* ein Spermatium, bei s ein solches im Augenblick
der Entleerung, bei v ein leeres Antheridium. Vergr. 540. Nach
STRASBURGER.
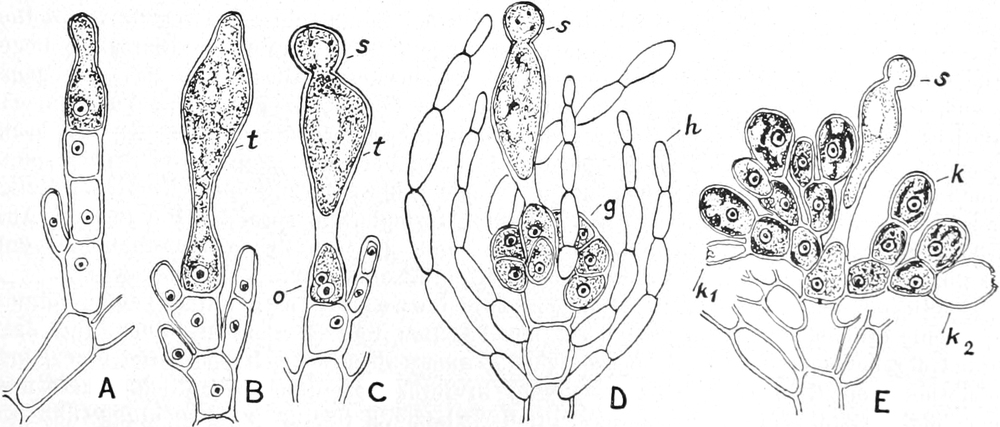
Fig. 368. Batrachospermum moniliforme. A Anlage eines
Karpogons am Ende eines Zweiges. B Reifes Karpogon, t Trichogyn.
C Nach Befruchtung durch ein Spermatium s, Eizelle o mit den
beiden kopulierenden Sexualkernen. D Gonimoblasten g und Hüllfäden
h. E Teil eines reifen Gonimoblasten mit den Karposporen k,
k1 und k2 entleert. A–D Vergr. 960, E Vergr. 720. Nach
H. KYLIN.
Die Antheridien, auch Spermatangien genannt (Fig. 367),
sprossen meist in Zweizahl aus den Endzellen der Wirtelzweige
hervor. Jedes Antheridium besteht aus nur einer Zelle, deren
gesamtes Plasma in die Bildung eines einzigen Spermatiums
aufgeht. Die Spermatien werden aus der zurückbleibenden Zellhaut
entleert, sind rundlich, farblos, anfangs einkernig, nur bei
Batrachospermum und Nemalion später zweikernig. Sie sind
nackte Zellen, können sich nicht selbständig bewegen wie die mit
Zilien versehenen Spermien der übrigen Algen und verdanken diesem
Unterschied ihre besondere Bezeichnung. Die weiblichen Organe,
hier Karpo[S. 374]gonien genannt, sitzen ebenfalls an Zweigenden,
zwischen den Antheridien tragenden Ästen. Das Karpogon (Fig.
368) besteht aus einer langen, im unteren Teil flaschenförmig
angeschwollenen, im oberen Teil keulenförmig gestalteten Zelle.
Der Basalteil enthält Eikern und Chromatophoren, der obere wird
als Trichogyn bezeichnet und fungiert als Empfängnisorgan
für die Spermatien, die zu einem oder mehreren an seiner Spitze
festhaften, sich mit einer Membran umgeben und dann ihren Inhalt
durch eine entstehende Öffnung in das Karpogon unter Zurücklassung
ihrer Membran entlassen. In der Regel enthält das junge Karpogon
der Florideen zwei Zellkerne, von denen der in dem Trichogyn
vorhandene später degeneriert. Bei Batrachospermum aber ist nur
ein Kern im Bauchteil sicher nachgewiesen, und bei Nemalion
soll die Bildung des Trichogynkernes nach CLELAND meist
unterbleiben. Trotz ihres auffallend verschiedenen Baues sind beide
Sexualorgane der Rotalgen als homolog zu erachten[348].
Nachdem Spermakern und Eikern miteinander verschmolzen sind,
grenzt sich die Eizelle vom Trichogyn durch eine Querwand ab,
wird aber nicht zu einer Oospore, sondern aus den Seitenflächen
des Bauchteils des Karpogons wachsen vielmehr sich weiter
verzweigende Zellen oder Zellfäden hervor, die sporogenen Fäden
oder Gonimoblasten. Zugleich sprossen aus den Tragzellen des
befruchteten Karpogons Hüllzweige, die sich um diese Gonimoblasten
lagern und mit ihnen eine sog. Hüllfrucht, Zystokarp,
bilden. Die sporogenen Fäden erzeugen in ihren anschwellenden
Endzellen die kugelrunden, einen Kern und ein Chromatophor
führenden Sporen, die man hier als Karposporen bezeichnet.
Sie werden aus den zurückbleibenden Hüllen der Endzellen entleert.
Aus den Karposporen entwickelt sich bei Batrachospermum zunächst
ein aus Zellfäden bestehender Vorkeim, der aus seinen Endzellen
ungeschlechtliche, einzellige, nackte, erst nach ihrer Festsetzung
sich mit Membran umgebende Sporen (Monosporen) erzeugt. Diese
dienen der Vermehrung des Vorkeims. Schließlich wachsen einzelne
Zweige des Vorkeims zu den geschlechtlich differenzierten
Thallusfäden heran. Die Sporenbildung am Vorkeim entspricht der
Tetrasporenbildung der übrigen Florideen.
Die Bildung der Antheridien[349] sowohl, wie auch der oft
sehr verwickelt gebauten Zystokarpien folgt bei den Florideen
verschiedenen Typen. Überall aber lassen sich die Karposporen
nach OLTMANNS in ihrer Entstehung als Abkömmlinge der
befruchteten Eizelle nachweisen.
Als Beispiel für reichere Ausbildung der karposporenbildenden
Generation sei die an den wärmeren europäischen Küsten verbreitete
Dudresnaya coccinea, mit büschelig reich verzweigtem Thallus,
gewählt (Fig. 369). Die Karpogonäste bestehen aus etwa sieben
Zellen; die endständige Karpogonzelle trägt ein sehr langes
Trichogyn. Nach der Befruchtung treibt die Karpogonzelle einen
Zellfaden nach unten, der sich weiterhin verlängert und verzweigt
und schrittweise mit bestimmten, dichten Inhalt führenden
vegetativen Zellen, den Auxiliarzellen, durch Fusion
in Verbindung tritt. Die ersten Auxiliarzellen liegen in dem
Karpogonast, die folgenden in anderen Seitenästen. Alle Kerne des
sporogenen Fadens sind durch Teilung des befruchteten Eikerns
hervorgegangen. Die Fusionen mit Auxiliarzellen führen nicht
zu Kernverschmelzungen, sondern dienen nur der Ernährung der
sporogenen Fäden. Aus der Karpogonzelle kann noch ein zweiter und
dritter sporogener Faden in gleicher Weise entspringen. Aus den
blasenförmig angeschwollenen Zellen der sporogenen Fäden, die mit
den Auxiliarzellen fusionierten, sprossen nun je zwei Ausstülpungen
hervor, welche sich weiter teilen und zu rundlichen Sporenhaufen
entwickeln, aus denen die Karposporen schließlich entlassen werden.
Wir können somit bei allen Rotalgen zwei Generationen
unterscheiden, einmal die geschlechtliche, den Gametophyt,
der Eizellen und Spermatien bildet, und dann die aus der
befruchteten Eizelle hervorgehende ungeschlechtliche, Karposporen
erzeugende Generation, den Sporophyt. Beide sind in ihrer
Gestaltung wesentlich voneinander verschieden, erstere eine
selbständige Pflanze, letztere in ihrer Entwicklung und Ernährung
von dieser abhängig, und vielleicht als eine eingeschobene, aus der
Zygote neu hervorgegangene Bildung aufzufassen.
Die in Tetrasporangien oder in ihnen gleichwertigen Monosporangien
oder Polysporangien erzeugten Sporen stellen eine zweite Form
ungeschlechtlicher Sporen vor, durch die eine Vermehrung der
Geschlechtspflanzen bewirkt wird. Sie können in einzelnen Fällen
(Nemalion) übrigens ganz fehlen.
Bei gewissen Gattungen treten diese Sporangien ausschließlich auf
den Geschlechtspflanzen selbst auf; bei der Mehrzahl der Gattungen
aber gehen aus den Karposporen zunächst Pflanzen hervor, die nur
Tetrasporen erzeugen, und aus den Tetrasporen erst die[S. 375] meist
diözischen Geschlechtspflanzen, so daß dann die Entwicklung in
drei Generationen, Gametophyt, Karposporophyt und Tetrasporophyt,
verläuft. Von der geschlechtlichen Hauptgeneration hat sich hier
also eine rein ungeschlechtliche, ihr im übrigen morphologisch
gleiche und ihr vorangehende Generation abgespalten. Bei manchen
Rotalgen mit solchem Entwicklungsgang tritt noch eine weitere
Komplikation hinzu, indem an dem Gametophyten auch noch Tetrasporen
oder diesen gleichwertige Monosporen erzeugt werden, aus denen dann
wiederum Gametophyten hervorgehen.
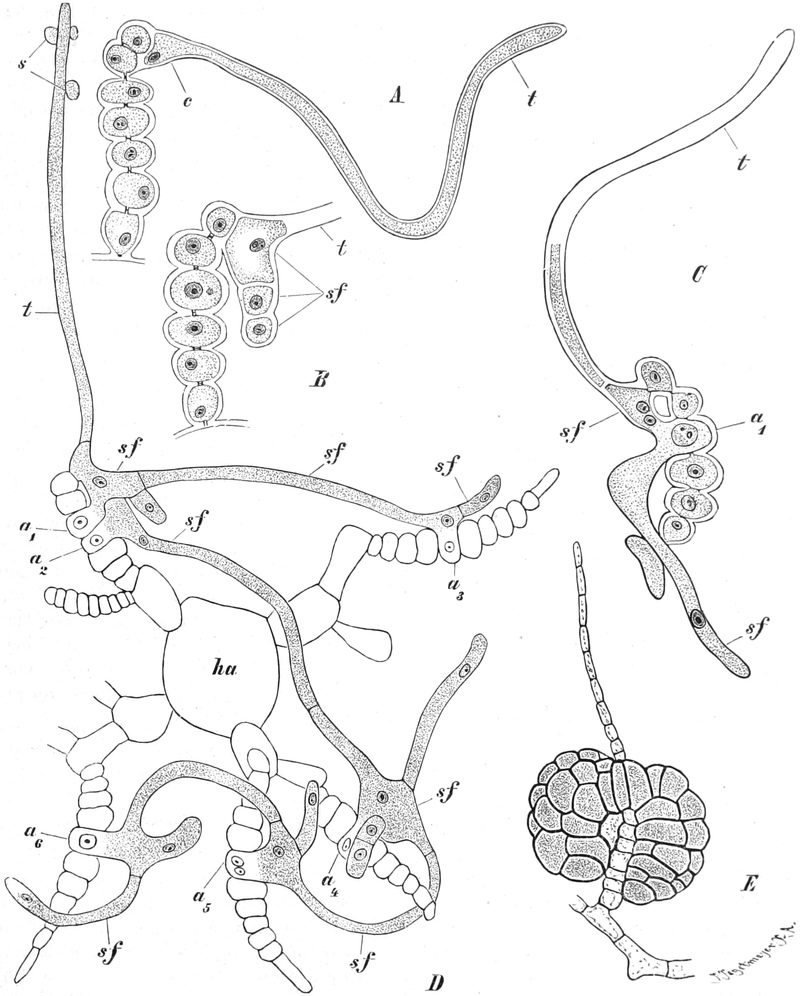
Fig. 369. Dudresnaya coccinea. A Karpogon-Ast,
Karpogon c mit Trichogyn t. B Nach der Befruchtung, Karpogon zum
sporogenen Faden sf ausgewachsen. C Verbindung dieses Fadens mit
der ersten Auxiliarzelle a1. D Die Verzweigung des sporogenen
Fadens und Verbindung mit sechs Auxiliarzellen a1–a6.
Die Zellen a3–a6 sind Ästen eingefügt, die von der Achse
ha entspringen. Schema. E Reifer Karposporenknäuel aus einem Ast
hervorgegangen. Vergr. A–C ca. 500, D 250, E 300. A–D nach
OLTMANNS, E nach BORNET.
Das Verhalten der Zellkerne und ihre Reduktionsteilung ist erst
für eine kleine Zahl von Arten untersucht worden, wobei sich
prinzipiell bemerkenswerte Verschiedenheiten in der Verteilung der
haploiden und diploiden Phasen auf die Generationen herausgestellt
haben[350].
[S. 376]
Bei Scinaia tritt nach SVEDELIUS die Reduktion der
Chromosomenzahl bei der ersten Teilung des befruchteten Eikernes
auf, so daß nur dieser diploid ist, der Karposporophyt, die
Karposporen und die aus letzteren hervorgehenden Gametophyten
aber haploid werden. Der Gametophyt vermehrt sich hier außerdem
noch ungeschlechtlich durch haploide Monosporen an Stelle der
fehlenden Tetrasporen. Ebenso verhält sich nach KYLIN
Batrachospermum, ferner Nemalion, bei welchem aber weder
Monosporen noch Tetrasporen am Gametophyten vorkommen.
Es steht zu erwarten, daß bei allen Rotalgen mit einfachem
Generationswechsel die Reduktionsteilung nach dem Scinaia-Typus
verläuft, der als der primitive angesehen werden darf.
Bei denjenigen Rotalgen, die ihre Entwicklung in drei
Generationen zurücklegen, ist dagegen die Reduktionsteilung in
das Tetrasporangium verlegt, so daß die aus den Tetrasporen
hervorgehenden Gametophyten haploid sind, während die
Karposporophyten, die Karposporen und die aus letzteren
hervorgehenden Tetrasporophyten hier die diploide Phase darstellen.
So verhalten sich Polysiphonia, Griffithia, Delesseria,
Nitophyllum, Rhodomela. Tritt nun in solchen Fällen an dem
haploiden Gametophyten außerdem noch eine ungeschlechtliche
Sporenbildung auf, so muß diese naturgemäß ohne Reduktionsteilung
erfolgen. So verhält sich nach SVEDELIUS der Gametophyt
von Nitophyllum punctatum, der außer Sexualorganen auch noch
Monosporangien bildet, durch deren Sporen er eine weitere
Vermehrung erfährt. Diese Monosporangien entsprechen den an den
Tetrasporophyten auftretenden Tetrasporangien, sind anfangs
vielkernig, erzeugen aber unter Degeneration aller Kerne bis
auf einen und unter Wegfall einer Reduktionsteilung nur je eine
haploide Monospore.
Besonderes Interesse verdient eine kleine Nordseefloridee
Harveyella mirabilis[351], welche auf einer anderen Rotalge,
der Rhodomela subfusca, parasitisch in Form von kleinen
weißlichen Polstern auftritt. Chromatophoren sind in ihren Zellen
nicht mehr nachweisbar, so daß diese Floridee sich wie ein Pilz
ernährt. Ebenso verhalten sich noch einige andere Arten.
Offizinell sind Gigartina mamillosa (Fig. 365) mit
zäpfchenförmigen, 2–5 mm langen, dem Thallus aufsitzenden
Zystokarpien und Chondrus crispus (Fig. 364) mit ovalen, der
Thallusfläche eingesenkten, etwa 2 mm langen Zystokarpien und
mit Tetrasporangienlagern an den Thallus-Endsegmenten. Beide
leben an den Nordseeküsten als purpurrote Algen; getrocknet sind
sie von hellgelblicher Farbe und liefern, besonders Chondrus,
das offizinelle Carrageen oder irländische Moos (Pharm.
germ., austr., helv.), das zur Gallertbereitung verwendet wird. —
Verschiedene Florideen liefern das ebenfalls zur Gallertbereitung
benutzte Agar-Agar; so Gelidium corneum und
cartilagineum das Agar von Japan; Sphaerococcus (Gracilaria)
lichenoides das Agar von Ceylon (auch Fucus amylaceus genannt);
Eucheuma spinosum das Agar von Java und Makassar.
Im Bau des Thallus und auch der Sexualorgane zeigen die meisten
Algenpilze nähere Beziehungen zu den Schlauchalgen oder Siphoneen,
unter denen vielleicht ihr phylogenetischer Ursprung zu suchen
ist; indessen deuten gewisse Formen auch auf andere niedere grüne
Algen hin, so z. B. die Basidiobolaceen auf Conjugaten und die
Chytridiaceen als einfache Algenpilze auf Protococcales und
Flagellaten. Wenn zurzeit die Phykomyceten noch als geschlossene
Klasse aufgeführt werden, so geschieht dies mit dem Vorbehalt ihrer
späteren Auflösung in mehrere an die entsprechenden Algenklassen
anzugliedernde Reihen.
Im einfachsten Falle, bei Chytridiaceen, besteht der Thallus
aus einer einfachen Zelle, die in den ersten Stadien sogar einen
membranlosen Protoplasten vorstellt. Bei den höher stehenden Formen
weist der schlauch- oder fadenförmige, meist reich verzweigte Thallus,
wie bei Vaucheria, bis zur Bildung der Fortpflanzungsorgane keine
Querwände auf und umschließt ein einheitliches Plasma, in welchem
zahlreiche sehr kleine Zellkerne, aber keine Chromatophoren enthalten
sind. Die Pilzfäden bezeichnet man als Hyphen,
[S. 377]
den ganzen Thallus als Myzelium. Meist sind die Hyphen
ungegliedert, nur in wenigen Fällen tritt eine Gliederung der Hyphen
durch Querwände ein. Die Algenpilze leben teils saprophytisch im Wasser
auf faulenden Pflanzen oder Tieren oder an der Luft auf verwesenden
organischen Substanzen, teils parasitisch in den Geweben von höheren
Pflanzen oder auch von Insekten.
Die ungeschlechtliche Vermehrung geschieht durch Sporen,
die meist in Sporangien erzeugt werden. Das gesamte
Protoplasma der Sporangien zerklüftet sich in zahlreiche Sporenzellen,
die bei den wasserbewohnenden Gattungen als zilientragende
Schwärmsporen entlassen werden (Fig. 372), bei den terrestrischen
dagegen vor der Entleerung bereits mit Membran umkleidet und so
der Verbreitung in der Luft angepaßt sind (Fig. 378). Bei gewissen
Gattungen werden neben den Sporangien oder auch ausschließlich
Konidien gebildet durch Hervorsprossung und Abschnürung von
Sporenzellen aus Hyphen, die dann meist als Konidienträger sich vom
Substrat erheben. Die Verbreitung der Konidien findet ebenfalls in der
Luft statt.
Die Sexualorgane bieten viel Eigenartiges. Nach ihrer
Beschaffenheit unterscheiden wir bei den höher stehenden Phykomyceten
die beiden Gruppen der Oomyceten und Zygomyceten.
Bei den Oomyceten, die den Siphonales noch am nächsten stehen,
differenzieren sich Oogonien und Antheridien, und letztere entleeren
durch schlauchartige Ausstülpungen ihren Inhalt in erstere, die dann
nach der Befruchtung Oosporen liefern. Nur bei Monoblepharis werden
noch freie Spermien gebildet. Bei den Zygomyceten sind die beiden
Sexualorgane von gleicher Beschaffenheit und liefern eine Zygospore.
Sie sind aber meist vielkernig und entsprechen dann morphologisch je
einem ganzen Gametangium der isogamen Grünalgen. Bei den einfacheren
Phykomyceten, den Archimyceten, sind nur in wenigen Fällen
Sexualorgane, Antheridien und Oogonien oder auch einander gleiche
Gameten beobachtet worden.
Vielkernige Gametangien, Oogonien und Antheridien, die ihre Gameten
nicht trennen und einzeln ausschlüpfen lassen, sondern direkt
miteinander kopulieren, werden insgemein als Cönogameten
bezeichnet.
1. Ordnung. Archimycetes[355].
Die hierher gehörigen Chytridiaceen sind mikroskopisch kleine,
in Algen, Wasserpilzen, Wassertieren oder in den Zellen von
Landpflanzen schmarotzende Pilze, deren Thallus nur bei einigen
höher stehenden Gattungen die Form eines kleinen verzweigten
Myzeliums aufweist, bei den meisten Vertretern aber eine
einfache, in der Wirtszelle lebende Zelle vorstellt. Die
ungeschlechtliche Vermehrung erfolgt durch ein- oder
zweizilige Schwärmsporen, die sich auf der Nährpflanze
festsetzen und ihren Protoplasten in die Wirtszelle eintreten
lassen. Der Protoplast wächst heran, bleibt anfangs membranlos,
umgibt sich dann mit Membran und wird zu einem vielkernigen
Sporangium, das zahlreiche einkernige Schwärmsporen, meist aus
einem halsförmigen Fortsatz, nach außen entläßt. Auch werden
dickwandige Dauersporangien gebildet, die erst nach längerer
Ruhe Schwärmsporen erzeugen. So verhält sich u. a. Olpidium
Brassicae (Fig. 370), das in der Stengelbasis junger Kohlpflanzen
schmarotzt und sie zum Absterben bringt.
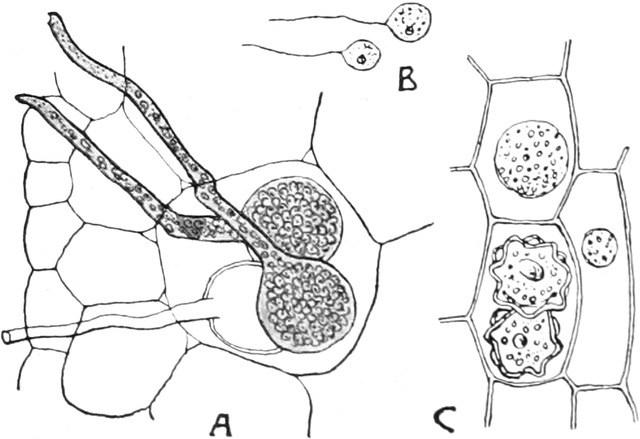
Fig. 370. Olpidium Brassicae. A Drei Zoosporangien,
eines entleert. Vergr. 160. B Zoosporen. Vergr. 520. C
Dauersporangien. Vergr. 520. Nach WORONIN.
[S. 378]
Geschlechtliche Fortpflanzung, der Bildung der
Dauersporangien vorausgehend, ist nur bei wenigen Vertretern sicher
nachgewiesen. Bei dem auf Vicia unijuga schmarotzenden Olpidium
Viciae sind die eingeißeligen Schwärmsporen teils asexuell, und
aus ihnen gehen nach ihrem Eindringen in eine Wirtszelle nach
wenigen Tagen wieder Zoosporangien hervor, teils aber kopulieren
sie als Planogameten paarweise miteinander zu einer dann mit zwei
Geißeln versehenen nackten Zygote, die sich auf der Nährpflanze
festsetzt, mit Membran umgibt und ihren Protoplasten in eine
Epidermiszelle entleert. Hier entwickelt sich die Zygote zu einem
Dauersporangium, in welchem erst die beiden Gametenkerne kopulieren
und dessen Inhalt zahlreiche Zoosporen liefert.
Ähnlicher Entwicklungsgang ist von CURTIS auch für
Synchytrium (Chrysophlyctis) endobioticum nachgewiesen.
Dieser sehr schädliche Pilz, der als Erreger des Kartoffelkrebses
warzenförmige, später zerfallende und verfaulende Wucherungen an
Knollen und Sprossen seiner Nährpflanze hervorruft, breitet sich
in neuerer Zeit in Europa aus. Seine mehrere Jahre lebensfähig
bleibenden derbwandigen Dauersporangien entlassen im Frühjahr
einzilige Zoosporen, die in junge Knollenzellen eindringen und
hier zu größeren, mit orangegelber äußerer und hyaliner innerer
Membran versehenen Zellen (Prosorus) heranwachsen. Ihr Protoplast
tritt dann in das Plasma der Wirtszelle aus, zehrt es auf und
teilt sich in eine Gruppe (Sorus) von rundlichen, dünnwandigen
Einzelsporangien, die je 200–300 einzilige kleine Schwärmsporen
bilden und nach Verquellung des umliegenden Knollengewebes nach
außen entlassen. Diese Zoosporen entwickeln sich in gleicher
Weise wieder zu neuen Sori. Außer dieser ungeschlechtlichen
Vermehrung kommt aber auch geschlechtliche vor. Die den Zoosporen
gleichgestalteten Gameten kopulieren paarweise zu Zygoten, die
als nackte Zellen wieder in die Kartoffeln eindringen und in
ihren Wirtszellen zu Dauersporangien heranwachsen. Ob Sporangien
oder Gametangien in den Sori gebildet werden, scheint von äußeren
Lebensbedingungen abzuhängen.
Bei der auf Saprolegniafäden schmarotzenden Olpidiopsis vollzieht
sich die Kopulation in anderer, mehr an das Verhalten der höheren
Phykomyceten erinnernder Weise. Größere weibliche und kleinere
männliche Protoplasten, beide anfangs noch membranlos, liegen in
den Nährzellen nebeneinander, wachsen heran, werden vielkernig,
umgeben sich mit Zellulosewänden, und nun tritt der Inhalt des
männlichen in den weiblichen über, der zu einer dickwandigen
Oospore wird; die Kerne scheinen paarweise zu verschmelzen. Das
Schicksal der Oospore ist unbekannt. Außerdem vermehrt sich
Olpidiopsis durch zweigeißelige, in Sporangien gebildete Zoosporen.
2. Ordnung. Oomycetes.
1. Nur bei der kleinen primitiven Familie der
Monoblepharidaceen[356] entlassen die Antheridien freie
Spermien, während bei den übrigen Oomyceten der vielkernige
Antheridiuminhalt sich nicht mehr in freie Spermien sondert.
Die Monoblepharis-Arten finden sich an Pflanzenresten im
Wasser, vermehren sich ungeschlechtlich durch einzilige
Schwärmsporen, die in größerer Zahl in Sporangien erzeugt
werden. Die meist terminal stehenden Oogonien enthalten
nur eine Eizelle (Fig. 371); die den Sporangien ähnlichen
Antheridien entlassen eine Anzahl von einziligen Spermien.
Diese gelangen zu den Oogonien, deren Wandung sie an der Spitze
auflösen, und dringen so zur Eizelle vor, die dann nach der
Befruchtung zu einer stachelig behäuteten Oospore wird.
Fig. 371. Monoblepharis sphaerica. Ende eines Fadens mit
einem Oogonium o und dem darunter liegenden Antheridium a, in 1
vor der Bildung der Eizelle und der Spermien, in 2 die letzteren s
austretend und an dem Oogonium hinaufkriechend, in 3 reife Oospore
osp, das Antheridium entleert. Vergr. 800. Nach CORNU. Aus
V. TAVEL, Pilze.
Fig. 372. Saprolegnia mixta. Sporangium die zweiziligen
Zoosporen s2 entlassend. Nach G. KLEBS.
Fig. 373. Saprolegnia mixta. Faden mit
Geschlechtsorganen. a Antheridium, das einen Befruchtungsschlauch
in das Oogonium hineingetrieben hat, o1 Eizelle, o2 Oospore mit
Membran umgeben, op parthenogenetische Oosporen, g junges Oogonium.
Nach G. KLEBS.
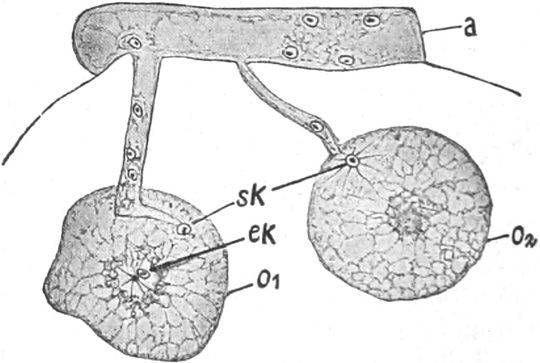
Fig. 374. Achlya polyandra. Befruchtung zweier Eizellen
o eines Oogoniums durch zwei Schläuche des Antheridiums a. ek
Eikern, in o2 der Eikern nicht im Schnitt getroffen, sk
Spermakerne. Nach TROW.
[S. 379]
2. An die erste Familie schließen sich die Saprolegniaceen[357]
an, die mit ihren Myzelien ebenfalls im Wasser meist saprophytisch
an der Oberfläche faulender Pflanzen, Insekten und sogar auf
lebenden Fischen vegetieren. Sie vermehren sich ungeschlechtlich
durch keulenförmige Sporangien (Fig. 372), aus denen zahlreiche,
mit zwei Zilien begabte Schwärmsporen nach außen entleert
werden. Bei Saprolegnia umgeben sich diese mit endständigen
Geißeln versehenen Sporen nach dem Schwärmen unter Einziehung ihrer
Geißeln mit kugeliger Hülle, schlüpfen aber bald aus ihr von neuem
aus, in anderer Gestalt, nämlich als bohnenförmige Schwärmer mit
seitlicher Geißeleinfügung. Diese kommen schließlich zur Ruhe,
werfen die Zilien nun ab und entwickeln sich zum Myzel. Bei anderen
Gattungen tritt nur eine Form von Schwärmsporen auf. An
älteren Myzelfäden entstehen die Geschlechtsorgane (Fig. 373, 374).
Die Oogonien bilden mehrere oder viele Eizellen (bis 50),
selten nur eine einzige aus. Sie enthalten anfangs viele Kerne,
die aber zum größten Teile zugrunde gehen; die übrig bleibenden
teilen sich nochmals mitotisch in Tochterkerne, von denen ein
Teil wiederum degeneriert, während um die übrigen sich die stets
einkernigen Eizellen abgrenzen. Die Antheridien enthalten
ebenfalls mehrere, sich nochmals mitotisch teilende Kerne. Sie
treiben einfache oder sich verästelnde Befruchtungsschläuche in die
Oogonien bis zu den Eizellen, in die nun je ein männlicher Kern
eintritt, um mit dem Eikern zu verschmelzen. Hierauf bilden sich
die Eizellen zu derbwandigen Oosporen um, bei deren Keimung[S. 380]
die Reduktionsteilung vollzogen wird. Bei einigen Formen dieser
wie auch der vierten Familie kann die Bildung der Antheridien
gelegentlich oder auch stets ausbleiben; die Eisporen reifen dann
ohne Befruchtung (Fig. 373 op). Eine in Abwässern massenhaft
vegetierende wasserreinigende Saprolegniacee ist Leptomitus
lacteus, von dem nur Vermehrung durch Schwärmsporen bekannt ist.
3. Die Peronosporaceen[358] sind parasitische Pilze, deren
Myzel in den Geweben höherer Landpflanzen lebt (Fig. 87). Gewisse
Arten bewirken in nassen Jahren epidemische Erkrankungen von
Kulturgewächsen, so vor allem die Phytophthora infestans, der
um 1830 aus Amerika bei uns eingeschleppte Pilz der bekannten
Kartoffelkrankheit (Kraut- und Knollenfäule). Seine
Myzelfäden leben interzellular, senden kurze Saugschläuche oder
Haustorien in die Zellen hinein und verursachen die Braunfärbung
und das Absterben der von ihnen befallenen Organe. Bis jetzt
sind bei dieser Art Geschlechtsorgane, die man bei Kultur des
Pilzes auf gewissen Nährböden zur Entwicklung bringen kann,
auf der Kartoffelpflanze selbst noch nicht beobachtet, sondern
nur ungeschlechtliche Sporangien, die als ovale Gebilde
auf verzweigten, vorzugsweise auf der Blattunterseite aus den
Spaltöffnungen herauswachsenden Sporangienträgern gebildet werden
(Fig. 375). Diese erscheinen dem bloßen Auge als weißer Schimmel.
Die Sporangien werden endständig angelegt und durch eine Querwand
abgegliedert; dann wächst der Träger neben dem Sporangium vorbei,
so daß es eine seitliche Stellung erhält. Noch vor der Teilung des
Inhaltes lösen sich die Sporangien (B) ab, werden durch den Wind
verbreitet und tragen so zur raschen Ausbreitung der Epidemie bei.
Die Entwicklung der Schwärmsporen aus den Sporangien erfolgt
nur in Wasser, somit nur bei nassem Wetter. Der Sporeninhalt
teilt sich in mehrere, mit zwei Zilien versehene, ausschlüpfende
Schwärmsporen (C, D), die zu einem neuen, in das Blatt
eindringenden Myzelfaden auskeimen. Die Sporangien können auch
direkt ohne Teilung des Inhaltes und ohne Bildung von Schwärmsporen
zu Keimschläuchen auswachsen. Es läßt sich somit bei dem
Kartoffelpilz, wie auch bei anderen Peronosporeen eine Umbildung
von Sporangien zu Konidien verfolgen, die hier mit dem Übergang
von der aquatischen zur terrestrischen Lebensweise zusammenhängen
dürfte.
Plasmopara viticola, mit reich verästelten Sporangienträgern, ist
ein sehr schädlicher, aus Amerika stammender Parasit, der Pilz des
falschen Mehltaues der Blätter und Beeren des Weinstockes. Eine
sehr häufige Art ist Albugo candida (= Cystopus candidus) auf
Cruciferen, besonders Capsella, weiße Auftreibungen der Stengel
verursachend. Die Sporangien werden bei dieser Art in Ketten am
Myzelium unter der Epidermis der Nährpflanze erzeugt und entleeren
bei Regenwetter zahlreiche Schwärmsporen.
Fig. 375. A Blattepidermis von Solanum tuberosum
mit Sporangienträgern der Phytophthora infestans. Vergr. 90.
B Sporangium. C Ein solches mit geteiltem Inhalt. D Eine
Schwärmspore. B–D Vergr. 540. Nach STRASBURGER.
Die Sexualorgane der Peronosporeen, die an Vaucheria
(Fig. 350) erinnern, entstehen meist im Innern der Nährpflanze,
die Oogonien als kugelige Anschwellungen von Hyphenenden, die
Antheridien als schlauchförmige Ausstülpungen meist dicht unter
den Oogonien (Fig. 376). Beide Organe werden durch Querwände
abgegrenzt und enthalten viele Kerne, die dann eine einmalige
Teilung erfahren. Im Verhalten der Kerne zeigen sich bei den
einzelnen Arten interessante Verschiedenheiten. Bei Peronospora
parasitica, Albugo candida und lepigoni, Pythium,
Plasmopara, Sclerospora differenziert sich im Plasma des
Oogoniums eine einzige große zentrale Eizelle oder Oosphäre, die
im reifen Zustand[S. 381] nur einen Eikern in der Mitte enthält, während
die übrigen Kerne sämtlich in dem peripherischen sog. Periplasma
liegen. Die Antheridien treiben in das Oogonium einen Fortsatz, der
sich an der Spitze in die Eizelle öffnet und nur einen männlichen
Zellkern eintreten läßt. Die Oosphäre grenzt sich sodann durch eine
Membran ab, die Kerne verschmelzen aber erst nach einiger Zeit,
und das Periplasma wird zur Bildung der äußeren Sporenmembran,
des Episporiums, verbraucht. Bei Peronospora parasitica ist die
reife Oospore einkernig, bei Albugo durch Teilung des Keimkerns
vielkernig. Albugo Bliti und A. Portulacae legen zwar ebenfalls
eine zentrale Oosphäre, von Periplasma umgeben, an; sie enthält
aber zahlreiche Kerne, und auch der Antheridiumschlauch führt
zahlreiche Kerne in sie ein, die paarweise mit den weiblichen
Kernen kopulieren. Aus dieser zusammengesetzten Eizelle geht dann
eine vielkernige Oospore hervor. Das Verhalten beider Arten kann
als ein ursprüngliches betrachtet werden, während die einkernigen
Eizellen der zuerst genannten Vertreter sich von den vielkernigen
ableiten lassen. Eine vermittelnde Stellung nimmt nun Albugo
Tragopogonis ein, deren Eizelle zwar vielkernig angelegt ist,
aber schließlich doch nur einen weiblichen Kern infolge
Schwindens der übrigen enthält. Die überzähligen Kerne in den
Oogonien und Antheridien können als funktionslos gewordene
Gametenkerne betrachtet werden, ähnlich wie die überzähligen
Eikerne bei gewissen Fucaceen. Die Oosporen keimen entweder direkt
zu einem Myzelium aus oder erzeugen zunächst Schwärmsporen. Die
Keimkerne in den Oosporen sind diploid; ihre erste Teilung, die
schon vor der Keimung erfolgen kann, ist eine Reduktionsteilung.
Fig. 376. Befruchtung der Peronosporeen. 1 Peronospora
parasitica. Junges vielkerniges Oogonium og und Antheridium an. 2
Albugo candida. Oogonium mit der zentralen einkernigen Oosphäre und
dem Befruchtungsschlauch a des Antheridiums, der den männlichen Kern
einführt. 3 Desgl. Befruchtete Eizelle o umgeben von dem Periplasma
p. Vergr. 666. Nach WAGER.
Fig. 377. Rhizopus nigricans. Teil eines Myzeliums
mit Sporangien, das rechts befindliche die Sporen entleerend mit
stehenbleibender, halbkugeliger Columella. Vergr. 38.
3. Ordnung. Zygomycetes.
Zu der ersten Familie, den Mucoraceen[359] gehören terrestrische
Schimmelpilze, die auf pflanzlichen und tierischen Stoffen
saprophytisch vegetieren. Die ungeschlechtliche Vermehrung
geschieht durch unbewegliche behäutete Sporen aus Sporangien
oder durch Konidien. Die sexuelle Fortpflanzung besteht in der
Kopulation zweier gleichwertiger Cönogameten zu einer
Zygospore.
Eine der verbreitetsten Arten ist der Kopfschimmel, Mucor
mucedo, dessen Myzel weiße Schimmelrasen auf feuchtem Brot, Mist,
Fruchtsäften bei Abschluß von frischer Luft bildet, ferner auf
gleichen Substraten der Ausläufer treibende Mucor stolonifer (=
Rhizopus nigricans) mit bräunlichem Myzelium. Bei den Mucor-Arten
entstehen die kugeligen Sporangien (Fig. 377) an den Enden
von senkrecht sich erhebenden Myzelschläuchen[S. 382] durch Abgrenzung
mittels Querwand, die sich kegelförmig als sog. Columella (Fig.
378) in das Sporangium vorwölbt. Das Plasma des Sporangiums
zerfällt durch fortgesetzte Zerklüftung in zahlreiche Sporen, die
durch Zerfließen der Sporangienwandung unter Aufquellen einer
zwischen ihnen liegenden Zwischensubstanz entleert werden. Bei
den auf Mist häufig sich entwickelnden Pilobolus-Arten wird
das Sporangium durch den stark turgeszierenden und unterhalb der
Columella schließlich aufplatzenden Träger weit abgeschleudert.
(Vgl. S. 308.) Nach HARPER sind bei Pilobolus die Sporen
zweikernig, bei Sporodinia vielkernig (Fig. 379).
Unter gewissen Bedingungen wird die ungeschlechtliche Fortpflanzung
abgelöst durch die sexuelle, die darin sich zeigt, daß an den
Myzelschläuchen keulenförmige Äste hervorkommen, paarweise mit
den Enden aufeinander stoßen (Fig. 380) und dann erst dort
die vielkernigen Cönogameten durch je eine Querwand
abgrenzen. Die Gameten verschmelzen nun zu einer Zygospore
mit warzenbesetzter Membran. Über das Verhalten der Kerne bei
der Kopulation liegen bis jetzt nur wenige Angaben vor. Bei
Sporodinia, Phycomyces und anderen Gattungen kopulieren die in
die Zygospore eintretenden Gametenkerne paarweise miteinander. Die
Zygosporen keimen nach längerer Ruhe; in der Regel schließen die
Keimschläuche ihre Entwicklung mit der Bildung von Sporangien ab
(Fig. 380, 5). Die Reduktionsteilung soll nach BURGEFF
bei Phycomyces in der Anlage des Sporangiums, das den aus der
Zygospore hervorgehenden Keimschlauch abschließt, stattfinden.
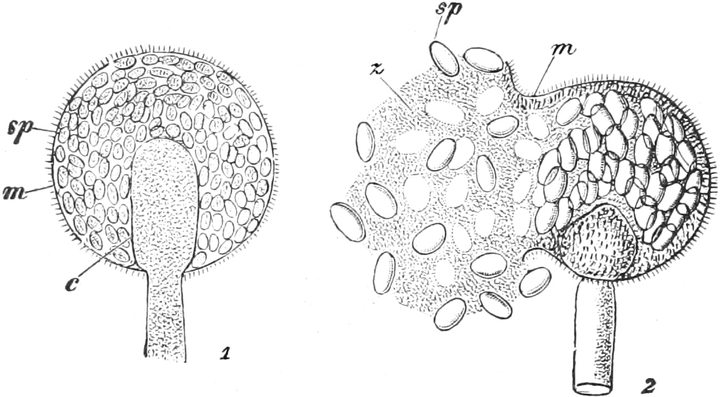
Fig. 378. 1 Mucor mucedo. Sporangium im optischen
Längsschnitt, c Columella, m Membran, sp Sporen. 2 Mucor
mucilagineus. Sporangium in der Sporenentleerung begriffen, die Membran
m zerfließend, die Zwischensubstanz z stark aufquellend. 1 Vergr.
225. 2 Vergr. 300. Nach BREFELD. Aus V. TAVEL,
Pilze.
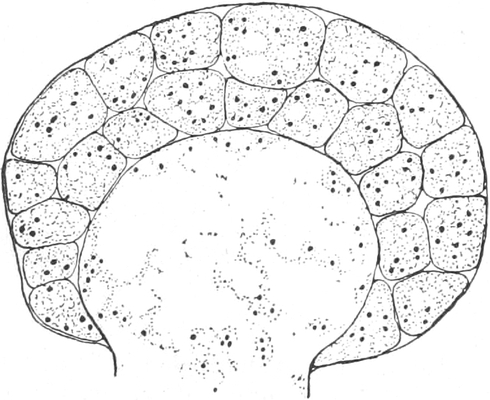
Fig. 379. Sporodinia grandis. Schnitt durch ein reifes
Sporangium. Sporen mehrkernig. Vergr. 425. Nach HARPER.
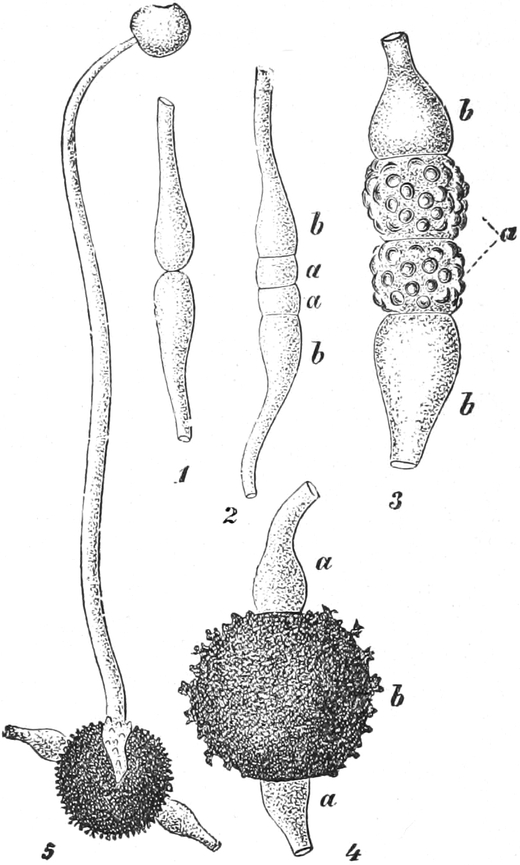
Fig. 380. Zygosporenbildung von Mucor mucedo. 1 Die
Konjugationsäste. 2 Abgrenzung der Gameten a von den Suspensoren
b. 3 Weiteres Stadium, die Warzen der Membran beginnen ihre
Bildung. 4 Reife Zygospore b zwischen den Suspensoren a. 5
Keimung der Zygospore mit einem Sporangium. 1–4 Vergr. 225, 5
Vergr. ca. 60. Nach BREFELD. Aus
V. TAVEL, Pilze.
Interessant ist der von BLAKESLEE für die meisten
Mucorineen erbrachte Nachweis von diözischen
(heterothallischen) Myzelien, so bei Mucor Mucedo und
Rhizopus nigricans; Zygosporen werden nur dann gebildet, wenn
solche mit + und - bezeichneten Myzelien, die zwar äußerlich nicht
unterscheidbar, physiologisch aber verschieden differenziert sein
müssen, aufeinander stoßen. Bei einigen Mucorineen (Sporodinia
grandis)[S. 383] dagegen entstehen die kopulierenden Gameten auf
ein- und demselben, also homothallischen Myzelium. Bei sonst
heterothallischen Arten, wie Phycomyces nitens, können unter
gewissen Umständen homothallische Myzelien auftreten oder auch
neutrale, die nur Sporangien bilden[360].
Innerhalb der Gruppe der Zygomyceten ist eine Rückbildung der
Sexualität zu verfolgen: Bei gewissen Mucorineen werden zwar
die Konjugationsschläuche paarweise angelegt; es findet aber keine
Verschmelzung mehr statt, sondern die Endzellen bilden sich direkt
zu Sporen aus, die man dann als Azygosporen bezeichnet;
endlich bei anderen Formen werden die an ihren Enden Azygosporen
bildenden Schläuche einzeln am Myzelium angelegt. Bei vielen Arten
stellt sich die Zygosporenbildung nur selten ein.
Bei Mucor mucedo unterliegt die Größe und die Sporenzahl der
Sporangien auffallenden Schwankungen. Bei der Gattung Thamnidium
hat sich ein Dimorphismus der Sporangien ausgebildet,
ein größeres vielsporiges steht am Ende des Trägers und eine
größere Anzahl keiner, wenigsporiger, sog. Sporangiolen, an
wirtelig verzweigten Seitenästen des Trägers. Letztere können
sogar unter bestimmten Ernährungsbedingungen einzellig bleiben
und auf diese Weise zu Konidien werden. Bei der tropischen
Gattung Choanephora ist der Dimorphismus am weitesten
gegangen, indem hier neben den großen Sporangien auf besonderen
Trägern Konidien erzeugt werden. Endlich gibt es Zygomyceten
(z. B. Chaetocladium), bei denen ausschließlich Konidien als
ungeschlechtliche Fruktifikation auftreten. So lassen sich also in
derselben Pilzgruppe alle Übergänge vom vielsporigen Sporangium bis
zur einzelligen Konidie feststellen.
Rhizopus nigricans enthält in seinem Zellsaft ein auf Tiere
tödlich wirkendes Gift[361].
2. Die Familie der Entomophthoraceen[362] stellt eine kleine
Gruppe meist parasitisch in den Leibern von Insekten und
Raupen lebender Pilze vor, deren vielkernige, meist verzweigte
Myzelschläuche einzellig bleiben oder später in Zellen sich
zergliedern, die dann im Blut durch den ganzen Tierkörper
verbreitet werden. Sie vermehren sich ungeschlechtlich durch
vielkernige oder einkernige Konidien, die einzeln an den
Enden der Myzeläste entstehen und bei der Reife abgeschleudert
werden, geschlechtlich durch Zygosporen, an deren Stelle
aber auch häufig Azygosporen gebildet werden.
Am bekanntesten ist Empusa Muscae, der Schimmelpilz der
Stubenfliege (Fig. 381). Die abgeschleuderten, vielkernigen
Konidien umgeben die Fliegen, die durch den Pilz infiziert und
getötet wurden, mit einem weißen Hofe, der besonders auffällt, wenn
die Fliegen an Fensterscheiben haften.
3. Basidiobolaceen[363]. Basidiobolus ranarum, ein auf
Froschmist saprophytischer Pilz, muß von der vorhergehenden Gruppe
abgetrennt werden. Sein verzweigtes Myzel ist vielzellig, die
einzelnen Zellen enthalten je einen großen Kern. Die im reifen
Zustand abgeschleuderten, einkernigen Konidien entstehen
einzeln an ihren Tragzellen. Eigenartig verläuft die Bildung der
Zygosporen. Zwei benachbarte Zellen eines Fadens schwellen
an und kopulieren, nachdem sie vorher zwei schnabelartige Fortsätze
getrieben und diese durch Querwände als vergängliche und bei
der Kopulation nicht beteiligte Zellen abgegrenzt haben. In der
Zygospore teilen sich die beiden Gametenkerne in vier Kerne, von
denen ein Paar zugrunde geht, das andere Paar aber verschmilzt. In
diesen Vorgängen und auch in der Struktur der Zellkerne zeigen sich
Ähnlichkeiten mit Konjugaten.
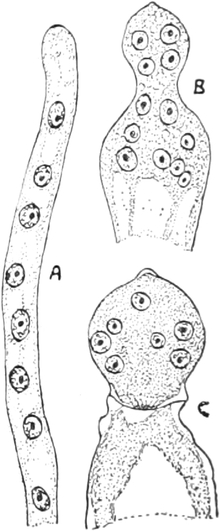
Fig. 381. Empusa Muscae. A Myzelschlauch aus einer
Fliege. B Junger, aus solchem Schlauch entstandener und nach außen
hervorgebrochener Konidienträger. C Bildung der Konidie, in welche
die zahlreichen Kerne des Trägers hineingewandert sind. Vergr. 450.
Nach OLIVE.
Zu dieser ungemein formenreichen Klasse gehören zwei große Gruppen
terrestrischer Pilze, die Askomyceten und die Basidiomyceten,
über deren[S. 384] phylogenetische Ableitung noch Unsicherheit herrscht. Es
ist versucht worden, sie von den Phykomyceten abzuleiten; indessen
spricht nicht nur die Beschaffenheit des Thallus dagegen, sondern auch
der Bau der Sexualorgane und die Entwicklung der Fruchtkörper, die
im besonderen bei den Askomyceten einige Beziehungen zu den Rotalgen
zeigen, während zu den Grünalgen keinerlei Anknüpfungspunkte erkennbar
sind. Die Basidiomyceten lassen sich durch eine ihrer einfacheren
Ordnungen, die der Uredineen oder Rostpilze, auf die Askomyceten
zurückführen. Andererseits scheinen die Basidiomyceten durch eine
ihrer einfacheren Ordnungen, die der Uredineen oder Rostpilze, mit den
Askomyceten verknüpft zu sein.
Wie bei den Phykomyceten besteht der vegetative, an saprophytische
oder parasitische Lebensweise vortrefflich angepaßte Thallus der
Eumyceten aus reichverzweigten, farblosen Zellfäden oder Hyphen,
deren Gesamtheit als Myzelium (Fig. 86) bezeichnet wird. Die
Hyphen sind aber gegliedert, sie bestehen aus Zellreihen. In diesen
weisen die einzelnen Zellen eine meist dünne, chitinhaltige Membran
auf und in ihrem farblosen Plasma winzige Zellkerne, die oft zu
mehreren (Fig. 6), doch in bestimmten Fällen zu zweien oder auch nur
in Einzahl vertreten sind. Die Zellen enthalten keine Chromatophoren
und nie echte Stärke, an deren Stelle vielmehr Glykogen, oft
in sehr beträchtlicher Menge, und Fett treten. Die Hyphen durchziehen
das Substrat und entnehmen ihm die für ihren Aufbau erforderlichen
Substanzen. Bei manchen Pilzen bilden sie durch reiche Verzweigung
Gewebekörper (Flechtgewebe oder Plektenchym). Wenn die Fäden dabei
dicht zusammenlagern und sich in kurze Zellen teilen, so entsteht
ein dem echten Parenchym überaus ähnliches Gewebe (Pseudoparenchym)
(Fig. 36), das bei gewissen Arten erzeugt wird, wenn sie aus ihren
Myzelien vegetative Ruhezustände, sog. Sklerotien bilden; diese
sind knollige oder strangartige feste Körper, die unter bestimmten
Bedingungen wieder auskeimen. Ferner bestehen die Fruchtkörper aus
lockerem oder dichterem Flechtgewebe (Fig. 37).
Gewisse Fadenpilze bilden an den Wurzeln humusbewohnender grüner
Pflanzen die weit verbreiteten Mykorrhizen (S. 226). Die
Bestimmung der hier in Betracht kommenden Pilzarten, die wohl
beiden Unterklassen entstammen, ist nur auf experimenteller
Grundlage sicher zu stellen. Von MELIN wurde auf diese
Weise nachgewiesen, daß Boletus-Arten (S. 406) Mykorrhizen an
Kiefern und Lärchen liefern[354].
Die beiden Unterklassen unterscheiden sich durch die Art der
ungeschlechtlichen Sporenbildung. Für sämtliche Askomyceten ist der
Askus charakteristisch, ein keulenförmiger Sporenschlauch, der
eine bestimmte Zahl von Sporen (meist 8) in eigenartiger Weise durch
freie Zellbildung erzeugt (Fig. 382), während die Basidiomyceten an
Stelle des Askus die Basidie aufweisen, die von verschiedener
Gestalt, entweder schlauchförmig einzellig oder vierzellig die Sporen
in bestimmter Zahl (meist 4) durch Zellsprossung abgliedert (Fig. 398,
399, 401, 402, 404, 411).
1. Unterklasse. Ascomycetes,
Schlauchpilze[288],
[296],
[352],
[353],
[364]–[378].
Die Schlauchpilze besitzen in ihren typischen Vertretern
Sexualorgane, Oogonien, die hier als Askogone oder wie bei den
Rotalgen als Karpogone bezeichnet werden, und Antheridien. Nur für eine
geringe Zahl von Gattungen sind die Sexualorgane näher untersucht; sie
zeigen eine auffallende Mannigfaltigkeit in ihrer Gestaltung. Unter
Hinweis auf die spezielle Darstellung seien hier nur einige Typen kurz
erwähnt.
1. Bei den Laboulbenieen (Fig. 397) zeigen das Karpogon mit seinem
Trichogyn und die Antheridien, von denen Spermatien gebildet werden,
große Ähnlichkeit mit den gleichnamigen Gebilden der Rotalgen.
[S. 385]
2. Die Flechten bildenden Askomyceten (Fig. 431, 432) schließen sich
am nächsten an die erstgenannte Gruppe an; ihr Karpogon ist aber
ein schraubig gewundener, vielzelliger Faden mit Trichogyn; die
Spermatien werden in besonderen krugförmigen Einsenkungen des Thallus,
den Spermogonien, erzeugt. Auch bei einigen nicht flechtenbildenden
Schlauchpilzen kommen solche Organe vor.
3. Einem anderen Typus folgen Pyronema[372] (Fig. 391) und verwandte
Gattungen, Ascodesmis (= Boudiera CLAUSSEN)[372],
Monascus[366], Aspergillus[369], bei denen die Befruchtung eines
vielkernigen, mit Trichogyn versehenen Karpogons durch ein vielkerniges
Antheridium stattfindet, die beide als Cönogameten (S. 377) zu
bezeichnen wären. Hier schließt sich auch Lachnea[367] an.
4. Bei den Erysibeen (Fig. 383) kopuliert ein einkerniges Antheridium
direkt mit einem einkernigen Oogonium.
Andere Gattungen weisen in der Gestaltung der Karpogone und Antheridien
Übergänge von dem Verhalten der Flechtenpilze zu dem von Pyronema und
dem der Erysibeen auf, so daß wir vielleicht erstere als primitive,
letztere als reduzierte Schlauchpilze betrachten können[365].
Bei gewissen Schlauchpilzen sind die Sexualorgane zwar noch vorhanden,
aber eine Befruchtung der Karpogone findet nicht mehr statt, oder
sie sind mehr oder weniger rückgebildet. Öfters kommt es vor, daß
Antheridien nicht mehr ausgebildet werden.
Das Karpogon liefert nicht eine ruhende Oospore, sondern
verbleibt im Zusammenhange mit der Mutterpflanze und läßt askogene
Zellfäden aussprossen, die sich verzweigen und schließlich an ihren
Enden die Sporenschläuche oder Asci bilden. Die aus
einem Karpogon, in gewissen Fällen auch aus einer Gruppe benachbarter
Karpogone hervorgegangenen Fäden und Asci vereinigen sich zu einem
Fruchtkörper, an dessen Zusammensetzung und Umhüllung auch
vegetative, dem Myzel der Mutterpflanze entsprossene und stets von
den askogenen scharf geschiedene Zellfäden sich beteiligen. Das die
Geschlechtsorgane erzeugende, aus den Askosporen hervorgegangene Myzel
stellt die geschlechtliche Generation (Gametophyt) vor, das aus dem
Karpogon hervorgegangene, in den Sporenschläuchen endigende Fadensystem
dagegen die ungeschlechtliche (Sporophyt).
In oder an den Fruchtkörpern gewisser Gruppen der Schlauchpilze stehen
die Asci parallel nebeneinander in einer Schicht (Hymenium), und
zwischen ihnen finden sich in der Regel Saftfäden (Paraphysen),
die dem sterilen Fadensystem der Fruchtkörper entstammen.
Bei einigen Ordnungen der Schlauchpilze fehlen die Sexualorgane und
Fruchtkörper, vielleicht infolge von Rückbildung, vollständig; die Asci
entstehen dann direkt aus den Myzelien.
Der Askus geht aus der endständigen Zelle eines askogenen
Fadens hervor, die bei ihrer Anlage zwei Kerne mitbekommt. Beide Kerne
verschmelzen miteinander, und aus dem Verschmelzungsprodukt gehen dann
durch dreimalige Teilung acht Kerne hervor, um die sich auf dem Wege
freier Zellbildung nach der in Fig. 21 dargestellten Weise die acht
Sporen mittels Membranen abgrenzen (Fig. 382 und 392).
Im Gegensatz zu den Sporangien der Phykomyceten wird nicht das
gesamte Plasma des Sporenschlauches zur Bildung der Askosporen
verbraucht; diese liegen vielmehr meist in einer Längsreihe angeordnet
in dem übriggebliebenen glykogenhaltigen Periplasma.
Das Ausstäuben der Sporen geschieht bei den Ordnungen der
Diskomyceten und Pyrenomyceten aktiv infolge Verquellens des
Epiplasmas, wodurch die Sporen aus den[S. 386] Schläuchen einige
Millimeter weit herausgeschleudert werden. Bei ersterer Ordnung
erfolgt sie als Wirkung des Reizes, den Licht- und Wärmestrahlen
auf das Hymenium ausüben. Die Sporen werden durch aufsteigende
Luftströmungen verbreitet. Bei den übrigen Askomyceten werden die
Sporen durch Zerfall der Fruchtkörper frei[367].
In seltenen Fällen teilen sich die acht Kerne vor der Abgrenzung
ihrer Zellen weiter; so entstehen zahlreiche freie Askussporen,
bei Thecotheus z. B. 32; häufiger aber vollziehen sich Teilungen
der acht Zellen nach dieser Abgrenzung und führen zur Bildung von
acht zwei- oder mehrzelligen Sporenkörpern an Stelle von acht
Einzelsporen.
Das Verhalten der Sexualkerne bei und nach der Befruchtung
der Karpogone ist nur für wenige Fälle sicher erkannt. Für
einige Askomyceten (Pyronema, Monascus) ist in neuerer Zeit
festgestellt, daß im Karpogon die Sexualkerne sich nur dicht
nebeneinander lagern, daß sie sich in den askogenen Fäden
konjugiert weiterteilen und daß erst in der Askusanlage die
Abkömmlinge je eines männlichen und je eines weiblichen Kerns
miteinander verschmelzen. So erscheint die Kopulation der
Sexualkerne vom Karpogon in den Askus hinausgeschoben und der
Befruchtungsvorgang in zwei Stadien zerlegt.
Soweit sich aus den bisherigen Angaben schließen läßt, scheint bei
den Schlauchpilzen allgemein die Reduktion der Chromosomen
nach der Vereinigung der beiden Kerne in der Askusanlage bei
der ersten Teilung des Fusionskerns sich abzuspielen.
In dem Entwicklungsgange der Schlauchpilze tritt vor der
Fruchtkörperbildung vielfach noch eine ungeschlechtliche Vermehrung
mittels Konidien auf; diese sind behäutete Sporen, die durch
Zellsprossung an den Enden einfacher oder verzweigter Hyphenäste,
der Konidienträger (z. B. Fig. 385), entstehen und sich
abgliedern.
Von zahlreichen Fadenpilzen ist bis jetzt nur solche
Konidienvermehrung bekannt, nicht aber die Askusfrüchte. Man
bezeichnet sie daher als Fungi imperfecti, wobei die Frage
offen steht, ob nicht in gewissen Fällen die Askusbildung ganz in
Wegfall gekommen ist.
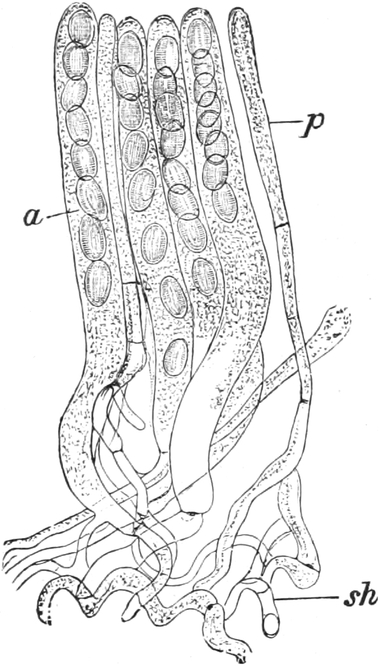
Fig. 382. Teil des Hymeniums von Morchella esculenta.
a Asci, p Paraphysen, sh subhymeniales Gewebe. Vgr. 240. Nach
STRASBURGER.
Nach der Beschaffenheit der Fruchtkörper unterscheiden wir zunächst
die Ordnungen der Erysibeen, Plectascineen und Pyrenomyceten mit
geschlossenen oder mit krugförmigen Fruchtkörpern (Perithecium), der
Discomyceten mit offenen Fruchtkörpern (Apothecium), der Tuberineen
mit anfangs offenen, später fast geschlossenen Fruchtkörpern.
Diesen Ordnungen reihen wir die Exoasceen an, bei denen die Asci ohne
Fruchtkörperbildung aus den Hyphenzellen hervorgehen, ferner die sehr
einfachen Saccharomyceten oder Hefepilze. Beide Gruppen können als
reduzierte Askomyceten aufgefaßt werden.
Die Laboulbenieen endlich, deren Asci in kleinen Perithecien
eingeschlossen sind, nehmen eine isolierte Stellung ein.
Die verwandtschaftlichen Beziehungen dieser Ordnungen zueinander
sind noch nicht geklärt.
1. Ordnung. Erysipheae, Mehltaupilze[296],[352],[368]. Die
kleinen kugeligen Perithecien besitzen eine geschlossene
Hülle (Peridie), die schließlich unregelmäßig aufplatzt und die
Askosporen entläßt. Die Asci stehen im Zentrum der Frucht in
Einzahl oder zu mehreren büschelig angeordnet.
Die Mehltaupilze überziehen als Parasiten die Blätter höherer
Pflanzen spinnwebartig und entsenden aus ihren Hyphen Haustorien
oder Saugfortsätze in die Epidermiszellen. In einzelnen Fällen
lebt das Myzel auch interzellulär im Blattgewebe. Die Peri[S. 387]thecien
sind in den weißen Überzügen als kleine, mit eigentümlichen
Anhängseln versehene, kugelige, schwarze Körperchen zu erkennen. Im
einfachsten Falle (z. B. bei der Gattung Sphaerotheca) umschließt
das Perithecium nur einen einzigen Askus mit acht Sporen, der
von sterilen Hyphen oder Hüllfäden in mehreren Schichten dicht
umwachsen ist. Bei den Gattungen Erysibe und Uncinula dagegen
finden sich in jedem Perithecium mehrere, bei Phyllactinia 12
bis 25 Asci vor. Die Zahl der Sporen beträgt meist vier oder auch
nur zwei, indem nicht alle acht Kerne des Askus zur Sporenbildung
verwendet werden. Wie HARPER nachgewiesen hat, besteht
die erste Anlage des Peritheciums aus einem Oogonium
und einem Antheridium. Beide werden an Hyphenästen als
einkernige Sexualzellen abgegrenzt, stehen dicht nebeneinander,
und der männliche Kern tritt durch ein Loch in der Zellwand in
das Oogon über (Fig. 383, 1–4). Nach der Befruchtung wird das
Oogonium von Hüllfäden, die aus seiner Stielzelle oder auch aus
derjenigen des Antheridiums entspringen, umgeben (5) und zu einem
mehrzelligen Gebilde weiterentwickelt (6), aus dessen vorletzter,
zweikerniger Zelle bei Sphaerotheca nach vorausgegangener
Kernvereinigung und Reduktionsteilung der achtsporige Askus
entsteht, während bei Erysibe und bei Phyllactinia diese
Zelle ausschließlich oder wenigstens vorwiegend askogene
Schläuche treibt, die ihrerseits die hier in Mehrzahl vorhandenen
Asci bilden. Die Mehltaupilze vermehren sich, bevor sie zur
Peritheciumbildung übergehen, zunächst durch Konidien, die an
kleinen aufrechten Myzelzweigen in Form von Ketten von der Spitze
nach abwärts fortschreitend, abgegliedert und durch den Wind
verbreitet werden.
Fig. 383. Sphaerotheca Castagnei. 1 Oogonium og
und Antheridiumzweig az. 2 Abgrenzung des Antheridiums an. 3
Übertritt des Antheridiumkernes zum Oogoniumkern. 4 Befruchtetes
Oogonium, in 5 mit zwei Lagen Hüllfäden aus der Stielzelle st.
6 Mehrzelliges Askogon durch Teilung des Oogoniums hervorgegangen,
die vorletzte zweikernige Zelle, as, liefert den Ascus. Nach
HARPER.
Fig. 384. Uncinula necator. A Konidienbildung,
c Konidie, b Konidienträger. B Myzelfaden m, der auf
Epidermiszellen die Anheftungsscheiben a und ein Haustorium h
gebildet hat. C Perithecium mit Anhängseln. Aus SORAUER,
LINDAU, REH. Handb. der Pflanzenkrankheiten, II1,
S. 246, 1921.
Der auf Blättern und Beeren von Vitis in Amerika, seit 1845 auch in
Europa erschienene, sehr schädliche Mehltau des Weinstocks
ist die als Oidium Tuckeri bezeichnete Konidienfruktifikation
von Uncinula necator (= U. spiralis), deren Perithecien mit
an der Spitze spiralig eingerollten Anhängseln versehen sind.
Sie erscheinen nur selten bei uns, während sie in Nordamerika
regelmäßig im Spätherbst auftreten (Fig. 384). Auch der bei uns
seit 1907 sehr verbreitete Eichenmehltau, Microsphaera quercina
vermehrt sich hauptsächlich durch Konidien und erzeugt nur sehr
selten die Perithecien, deren lange Anhängsel an der Spitze
mehrfach gegabelt sind.
[S. 388]
2. Ordnung. Plectascineae. Die Perithecien sind rundlich,
haben eine geschlossene Peridie und enthalten die Asci im Innern in
unregelmäßiger Anordnung.
1. Aspergillaceae[369]. Fruchtkörper klein, nicht unterirdisch.
Hierher gehören zwei der gemeinsten Schimmelpilze, Aspergillus
(Eurotium) herbariorum und Penicillium crustaceum, die
saprophytisch auf organischen Stoffen leben. Beide vermehren sich
anfangs in reichlichem Maße nur durch Konidien, bevor sie zur
Bildung der Perithecien übergehen.
Die Konidien von Aspergillus herbariorum sind unter dem
Namen Gießkannenschimmel bekannt; sein Konidienträger zeigt am
oberen Ende eine kugelige Anschwellung, von der radial angeordnete
Konidienketten entspringen (Fig. 385). Die Träger stehen
reihenweise nebeneinander und erscheinen als anfangs weißer, später
blaugrüner Schimmel auf feuchten Vegetabilien, Früchten, Brot
usw. Einige Arten von Aspergillus können auch als pathogene Pilze
Erkrankungen bei Wirbeltieren und Menschen hervorrufen, so bewirkt
A. fumigatus, der in gärenden Heuhaufen vorkommt und bei 40° C
sein Temperaturoptimum erreicht[370], die Mykosen des Ohrgangs, des
Rachens und der Lunge.
Die ebenfalls blaugrünen Schimmelrasen von Penicillium
crustaceum, dem überall verbreiteten Pinsel- oder Brotschimmel,
bestehen dagegen aus verzweigten Konidienträgern (Fig. 385).
Penicillium brevicaule, auf modrigem Papier vorkommend, wird zum
Nachweis von Arsen benutzt, da es auf arsenhaltigen Substraten
(Tapeten) in seinem Stoffwechsel das stark riechende Diäthylarsin
abscheidet.
Die Perithecien von Aspergillus und Penicillium
erscheinen später am Myzel, bei letzterer Gattung treten sie nur
selten auf. Sie sind verwickelter gebaut als bei den Erysibeen.
In ihrer ersten Anlage sind Sexualorgane, ein Antheridium und ein
mit Trichogyn versehenes Karpogon, nachgewiesen. In den reifen
Früchten erscheinen die Schlauchwandungen und das sterile, die
Asci umgebende Hyphengewebe bis auf die unregelmäßig aufplatzende
Fruchtwand aufgelöst.
Fig. 385. Konidienträger von Aspergillus herbariorum
(links), von Penicillium crustaceum (rechts).
2. Die Elaphomycetaceae besitzen unterirdische, knollenförmige,
trüffelähnliche Fruchtkörper, deren Peridie gegen den zu einer
pulverigen Sporenmasse reifenden Askuskomplex scharf abgesetzt ist.
Offizinell war früher und wird jetzt noch zu Tierarzneien
gebraucht die in Nadelwäldern Europas häufige Hirschtrüffel,
Elaphomyces cervinus (Boletus cervinus) mit walnußgroßen,
gelbbraunen, bitter schmeckenden Fruchtkörpern.
3. Die Terfeziaceae unterscheiden sich von den vorigen durch eine
nicht scharf abgesetzte Peridie ihrer Fruchtkörper. Hierher gehören
die besonders im Mittelmeergebiet verbreiteten Terfezia-Arten mit
eßbaren trüffelähnlichen Fruchtkörpern.
3. Ordnung. Pyrenomycetes, Kernpilze. Außerordentlich
formenreiche Gruppe von Pilzen, die teils parasitisch auf
Pflanzenteilen, teils saprophytisch auf faulem Holz, Mist usw.
leben. Einige wenige Gattungen befallen parasitisch verschiedene
Insektenlarven. Die Pyrenomyceten charakterisieren sich
durch die krugförmige Gestalt ihrer Askusfrüchte oder
Perithecien, die an ihrer Spitze eine offene Mündung und in
ihrem Grunde ein Hymenium aus Sporenschläuchen und haarförmigen,
oft verzweigten Saftfäden oder Paraphysen (Fig. 386)
besitzen. Die Seitenwände des Peritheciums sind bis zur Mündung
ausgekleidet mit ähnlichen Hyphenhaaren, den Periphysen. Die
Askosporen werden aus der Mündung nach außen entleert.
Die einfachsten Pyrenomyceten besitzen freie, dem Myzel
aufsitzende, meist schwarz gefärbte, kleine Perithecien (Fig. 386),
so die Gattungen Sphaeria und Podospora. Bei anderen Kernpilzen
aber erscheinen die Perithecien zu mehreren oder vielen dicht
nebeneinander eingebettet in einen rundlichen, polsterförmigen
oder keulenförmigen, zuweilen[S. 389] verzweigten Myzelkörper von
pseudoparenchymatischer Struktur. Man bezeichnet diesen als
Stroma.
Der Peritheciumbildung gehen in dem Entwicklungsgang der meisten
Kernpilze mannigfaltige Nebenfruktifikationen, hauptsächlich
Konidien voraus, die in verschiedener Weise von den
Myzelfäden teils direkt, teils auf besonderen Trägern abgegliedert
werden und zur Ausbreitung des Pilzes beitragen. Häufig erscheinen
die Konidienträger zu Fruchtkörpern vereinigt. Eine besondere
Form solcher Früchte sind die bei manchen Gattungen auftretenden
Pykniden, kleine kugelige oder flaschenförmige Gebilde, die
als Auskleidung verzweigte Hyphenfäden besitzen, an deren Spitzen
die Konidien, hier Pyknosporen (oder Pyknokonidien) genannt,
abgegliedert werden (Fig. 387). Die Pykniden und ihre Sporen
stimmen in ihrer Bildung mit den Spermogonien und Spermatien der
Flechtenpilze überein und mögen wohl auch als die ursprünglichen
männlichen Organe der Askomyceten zu betrachten sein.
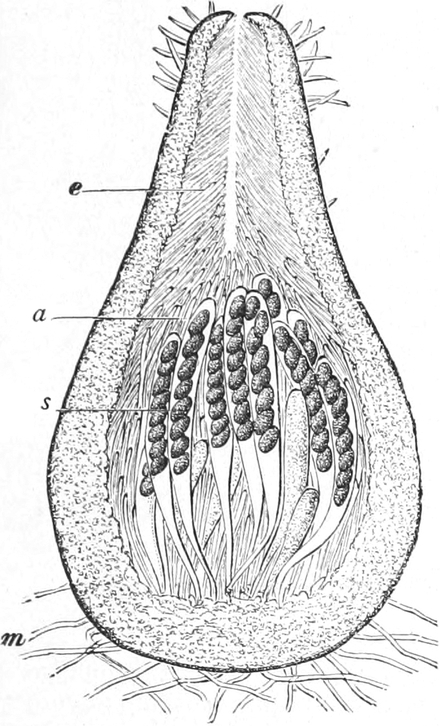
Fig. 386. Perithecium von Podospora fimiseda im
Längsschnitt. s Die Asci, a die Paraphysen, e die Periphysen, m
Myzelfäden. Vergr. 90. Nach V. TAVEL.
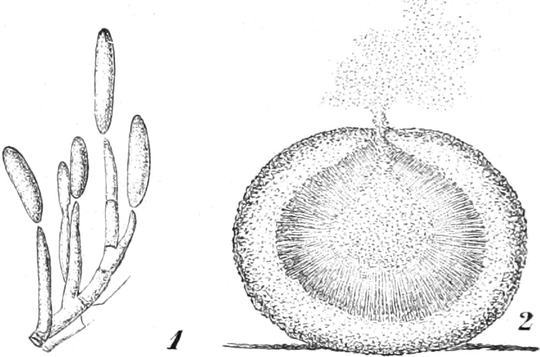
Fig. 387. 1 Konidienabschnürung an den Konidienträgern
aus der Pyknide von Cryptospora hypodermia. Vergr. 300. Nach
BREFELD. 2 Pyknide von Strickeria obducens, im Durchschnitt.
Vergr. 70. Nach TULASNE. Aus V. TAVEL, Pilze.
Wichtig als offizinelles Gewächs und als Schädling
der Roggenfelder ist Claviceps purpurea, der Pilz des
Mutterkorns. Er lebt parasitisch in den jungen Fruchtknoten
von Gramineen, hauptsächlich des Roggens. Diese werden im
Frühsommer durch die Askosporen infiziert. Das Myzel überwuchert
die Fruchtknoten und geht bald zur Bildung von Konidien über,
die auf kurzen seitlichen Trägern in kleinen Köpfchen vereinigt
abgegliedert werden (Fig. 388 A). Zugleich findet Ausscheidung
eines süßen Saftes statt, mit dem die massenhaft erzeugten
Konidien zu Tropfen zusammenfließen. Dieser sog. Honigtau
des Getreides wird von Insekten aufgesucht, auf andere Blüten
übertragen und so der Pilz verbreitet. Das Myzel geht nach
Aufzehrung des Fruchtknotengewebes schließlich in ein Sklerotium
über, dadurch, daß die Hyphenfäden dicht zusammenwachsen und
namentlich an der Peripherie unter Querteilung ein geschlossenes
Pseudoparenchym bilden (Fig. 36). Diese langgestreckten,
schwarzviolett gefärbten, aus der Kornähre mit schwach
hornförmiger Krümmung hervorragenden Sklerotien werden als
Mutterkorn, Secale cornutum, bezeichnet (Fig. 388
B). Die mit Reservestoffen (Fett) dicht angefüllten Sklerotien
fallen zu Boden und keimen erst im nächsten Frühsommer zur Zeit
der Roggenblüte. Es kommen Hyphenbündel aus ihnen hervor, die zu
langgestielten, blaßrot gefärbten Köpfchen heranwachsen (C).
In letzteren werden zahlreiche eingesenkte Perithecien,
gleichmäßig über die Oberfläche verteilt, erzeugt (D, E).
Jedes Perithecium enthält eine Anzahl Asci mit acht langen,
fadenförmigen Askosporen, die aus der Mündung hervorgepreßt werden
und, durch den Wind verbreitet, auf die Grasähren gelangen.
Nectria galligena, ein sehr schädlicher Parasit in der Rinde
einiger Laubbäume, besonders Apfel- und Birnbaum, verursacht den
sog. Krebs der Obstbäume; sie bildet im Winter und Frühjahr
zahlreiche kleine, rot gefärbte Perithecien[371].
Offizinell ist Secale cornutum (Pharm. germ., austr.,
helv.), Mutterkorn, das Sklerotium von Claviceps purpurea.
[S. 390]
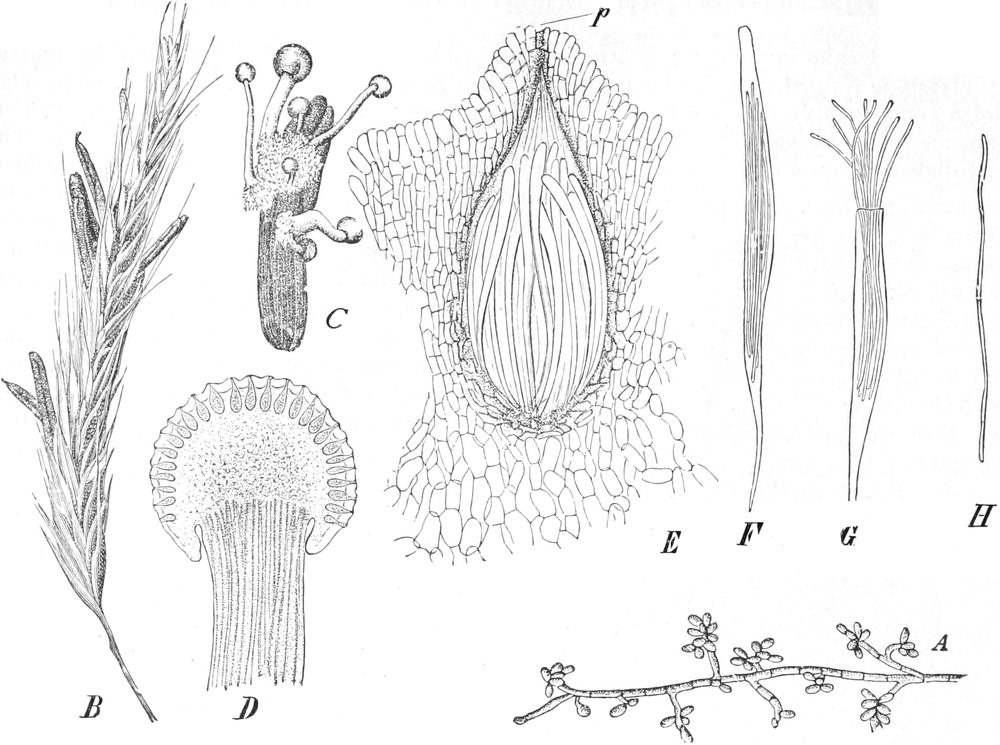
Fig. 388. Claviceps purpurea. A Konidienbildender
Myzelfaden. B Roggenähre mit mehreren reifen Sklerotien. C
Gekeimtes Sklerotium mit gestielten, zusammengesetzten Fruchtkörpern.
D Längsschnitt durch einen Fruchtkörper mit zahlreichen Perithecien.
E Einzelnes Perithecium stärker vergrößert. F Geschlossener Askus
mit acht fadenförmigen Sporen. G Austreten der Sporen. H Einzelne
Spore. A nach BREFELD, C–H nach TULASNE, B
phot. nach der Natur. — Offizinell und giftig.
4. Ordnung. Discomycetes, Scheibenpilze[372]. Die formenreichen
Diskomyceten unterscheiden sich von den übrigen Ordnungen dadurch,
daß ihre reifen Schlauchfrüchte das aus Sporenschläuchen und aus
Saftfäden oder Paraphysen bestehende Hymenium offen an ihrer
Oberseite tragen (Fig. 382, 390).
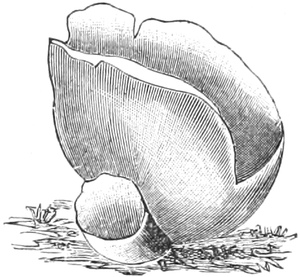
Fig. 389. Peziza aurantiaca. Nat. Größe. Nach
KROMBHOLZ.
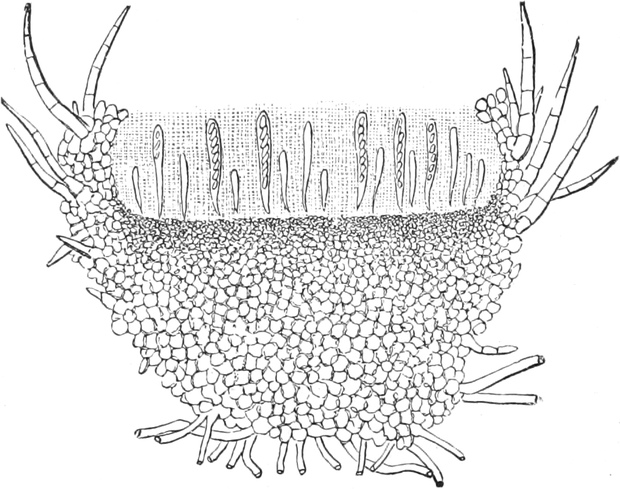
Fig. 390. Lachnea pulcherrima. Sporenreifes geöffnetes
Apothecium. Zwischen den Paraphysen sind alte und junge Schläuche
verteilt. Nach WORONIN. Aus V. TAVEL, Pilze.
Die überwiegende Mehrzahl der Diskomyceten, als deren Typus die
Gattung Peziza gelten kann, vegetiert auf lebenden oder toten
Pflanzenteilen, besonders auf altem Holz, zum Teil aber auch als
Erdpilze in Humusboden. Sie besitzen napf- oder becherförmige,
fleischige oder lederartige Askusfrüchte, meist von geringem
Durchmesser. Eine der größten Formen ist die erdbewohnende Peziza
aurantiaca (Fig. 389)[S. 391] mit bis 7 cm breiten, unregelmäßig
becherförmigen Früchten, die lebhaft orangerot gefärbt sind,
während die Mehrzahl der Arten graue oder braune Färbung aufweist.
Solche Becherfrüchte bezeichnet man als Apothecien.
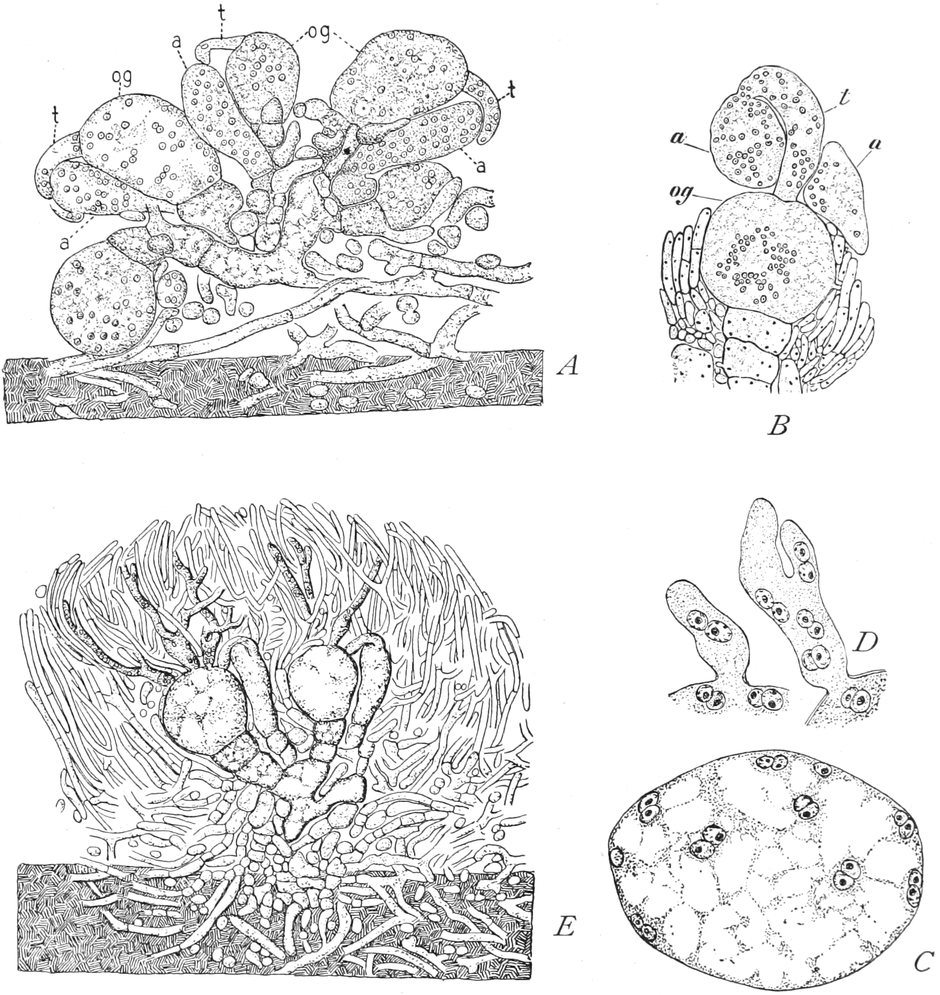
Fig. 391. Pyronema confluens. A Anlage eines
Apotheciums, Oogonien og mit Trichogynen t, Antheridien a. Vergr.
450. B Fusion des Antheridiums mit der Trichogynspitze. Vergr.
300. C Querschnitt, Paarung der männlichen und weiblichen Kerne im
Oogonium. Vergr. 1000. D Einwanderung der Paarkerne in die askogenen
Schläuche des Oogoniums. Vergr. 1000. E Junges Apothecium. Die den
Oogonien entspringenden askogenen Schläuche verzweigen sich und werden
von sterilen Hyphen eingehüllt. Vergr. 450. B nach HARPER,
A, C, D, E nach CLAUSSEN.
Die Apotheciumentwicklung sei an dem Beispiel des zuerst
von R. HARPER eingehend untersuchten Pyronema
confluens dargestellt, dessen etwa 1 mm breite, fleischige,
gelbliche oder rötliche, gesellig beisammenstehende Fruchtkörper
häufig auf Brandstellen in Wäldern gefunden werden. Das Myzelium
erzeugt eine Rosette von mehreren größeren Karpogonen und kleineren
Antheridien als Anlage eines Apotheciums (Fig. 391 A). Das
Karpogon oder Askogon ist ein kugeliges, vielkerniges
Oogonium, dessen Scheitel eine vielkernige, schnabelförmig
gebogene Zelle, das Trichogyn, aufsitzt. Aus einem
benachbarten Myzelfaden entspringt das keulenförmige, vielkernige
Antheridium, dessen Spitze mit dem Trichogynscheitel
mittels Durchbrechung der Wandung in offene Verbindung tritt.
Die männlichen Kerne wandern zunächst in die[S. 392] Trichogynzelle
ein (B), dann nach Durchbrechung der Basalwand des Trichogyns
in das Oogonium, während die Trichogynkerne zugrunde gehen. Nun
grenzt sich die Eizelle wieder ab und treibt zahlreiche askogene
Schläuche, die die Kerne aus ihr aufnehmen, sich verzweigen und
schließlich die Asci bilden (E), während die sterilen Hyphen
und die Paraphysen zwischen den Schläuchen aus den Hyphenzellen
unterhalb der Sexualorgane entspringen. Nach HARPER
sollen die männlichen und weiblichen Kerne im Karpogon
paarweise miteinander kopulieren; nach neuerer Untersuchung von
CLAUSSEN aber legen sie sich nur dicht nebeneinander
(C) und bleiben auch in den askogenen Fäden, in denen
sie sich konjugiert weiter teilen, deutlich voneinander getrennt
(D). Die Zellen der askogenen Fäden führen in der Nähe des
Askogons bis zu acht Kernpaare, weiter oben aber nur ein Kernpaar.
Erst in der zweikernigen Anlage des Askus findet die Kopulation
der beiden Kerne, also eines männlichen mit einem weiblichen
Sexualkernabkömmling, zum Askuskern statt (Fig. 392).
Bei manchen Diskomyceten ist eine mit Geschlechtsverlust
verbundene Rückbildung der Sexualorgane eingetreten.
Entweder funktionieren die Antheridien nicht mehr oder sind ganz
unterdrückt, und in extremen Fällen fehlen auch die Askogone,
an deren Stelle nur Hyphenknäuel sich erkennen lassen. Stets
aber lassen sich die askogenen Hyphen in den Fruchtkörperanlagen
nachweisen.
Die Asci entstehen an den Enden der askogenen Hyphen in
verschiedener Weise, entweder direkt aus den zweikernigen
Endzellen, meist aber indem diese je einen seitlichen, rückwärts
gerichteten, hakenförmigen Auswuchs bilden, worauf sich das
Kernpaar konjugiert teilt (Fig. 392 A). Seine beiden unteren
Tochterkerne liefern die Kerne für die Hakenzelle h und die
Stielzelle s, die sich durch Querwände von der endständigen, die
beiden oberen Tochterkerne aufnehmenden Askusanlage a abgrenzen
(B). Hierauf verschmelzen die beiden Kerne des jungen Askus
(C). Dieser wächst heran und bildet nach dreimaliger Teilung
seines Kernes die acht Askosporen (D). Die Hakenzelle tritt in
offene Verbindung mit der Stielzelle s, so daß eine zweikernige
Fusionszelle entsteht, die nun ebenfalls zu einer neuen Askusanlage
schreiten kann. So entstehen komplizierte askogene Hyphensysteme.
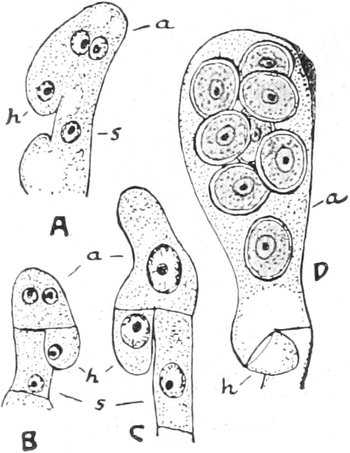
Fig. 392. Askus-Entwicklung. A–C Pyronema confluens.
Nach HARPER. D Junger Askus mit acht Sporen von Boudiera.
Nach CLAUSSEN. Erklärung im Text.
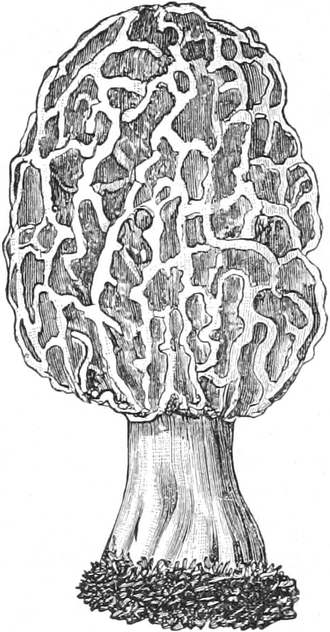
Fig. 393. Morchella esculenta. 4⁄5 nat. Gr.
Die eigenartigste Ausbildung erfährt der Fruchtkörper
der Diskomyceten in der Gruppe der Helvellaceen oder
Morchelpilze, die mit ihrem Myzel im Humusboden vegetieren.
Bei der Gattung Morchella, Morchel (Fig. 393), besteht der
große Fruchtkörper aus einem aufrechten dicken Stiel, auf dem
ein kegelförmiger oder abgerundeter Hut mit grubiger Oberfläche
sich erhebt. Sexualität ist bei den Morcheln noch nicht
nachgewiesen. Das Hymenium (Fig. 382) mit seinen achtsporigen
Ascis überzieht die Oberfläche des Hutes. Die Morcheln sind
vorzügliche Speisepilze[373], besonders M. esculenta,
die Speisemorchel, mit blaßgelbbraunem, eiförmigem Hut, bis
12 cm hoch, M. conica, die Spitzmorchel, mit dunkelbraunem,
kegelförmigem Hut, bis 20 cm hoch, u. a. Verwandt sind die
ähnlich gestalteten Lorcheln, deren Hut aber mützenförmig
herabgeschlagen, unregelmäßig gelappt und blasig aufgetrieben ist,
so Gyromitra esculenta, mit schwarzbraunem Hut und weißlichem
Stiel. Die Lorcheln sind eßbar, enthalten aber in frischem Zustand
die Helvellasäure, die unter Umständen giftig wirken kann; starke
Erhitzung beim Braten der Pilze oder längeres Abbrühen ist
anzuraten[374].
[S. 393]
5. Ordnung. Tuberineae, Trüffelpilze[375]. Die Trüffelpilze sind
saprophytische, im Humus der Wälder unterirdisch (hypogäisch)
lebende Askomyceten, die zu den Mykorrhizen bildenden Pilzen
gehören. Die Askusfrüchte, unter der Bezeichnung Trüffeln
bekannt, stellen knollenförmige Körper vor. Sie sind von einer
dicken Hülle umgeben und von nach außen mündenden Gängen
durchzogen, deren Wandungen von Hymenien aus keulenförmigen Ascis
bedeckt sind (Fig. 394). Die Sporen werden zu wenigen in den Ascis
erzeugt, bei den echten Trüffeln (Tuber) meist zu vier und meist
mit stacheligem oder netzförmig verdicktem Epispor versehen. Sie
gelangen durch Zerfall der Asci und der Fruchtkörperwandung in den
Boden.
Manche Tuberaceen liefern eßbare Fruchtkörper[373] von
aromatischem Geschmack. Sie werden mit Hilfe dressierter Hunde
oder Schweine besonders in Frankreich und Italien gesammelt.
Die wichtigsten sind die schwarzen Trüffeln, Tuber brumale,
melanosporum (Perigord-Trüffel), aestivum und mesentericum,
die außen schwarzbraun gefärbt und mit Warzen versehen sind,
ferner die weiße Trüffel, Choiromyces maeandriformis, die außen
blaßbraun, innen weiß gefärbt ist.
Die Fruchtkörper werden in frühester Jugend offen angelegt, wie bei
den Diskomyceten, mit denen die Trüffeln verwandt sind.
6. Ordnung. Exoasceae[376]. Die wichtigste Gattung ist Taphrina
(einschl. Exoascus), deren Arten als parasitische Pilze auf
verschiedenen Bäumen leben und teils als einjährige Pilze sich
nur in den Blättern entwickeln und fleckiges Erkranken derselben
bewirken, teils mit ihrem Myzel im Gewebe der Nährpflanzen
überwintern, somit jährlich wiederkehrende Krankheiten an diesen
verursachen. Das Myzel veranlaßt dann häufig die befallenen
Sprosse zu reichlichen anomalen Verzweigungen, die man als
Hexenbesen bezeichnet. So erzeugt Taphrina Carpini
Hexenbesen auf der Weißbuche, Taphrina Cerasi solche auf
Kirschbäumen. Taphrina deformans bewirkt die Kräuselkrankheit der
Pfirsichblätter. Taphrina Pruni dagegen schmarotzt in den jungen
Fruchtknoten der Pflaumen, die sie zu hohlen Pilzgallen, sog.
Taschen, umbildet; ihr Myzel überwintert in den Zweigen.
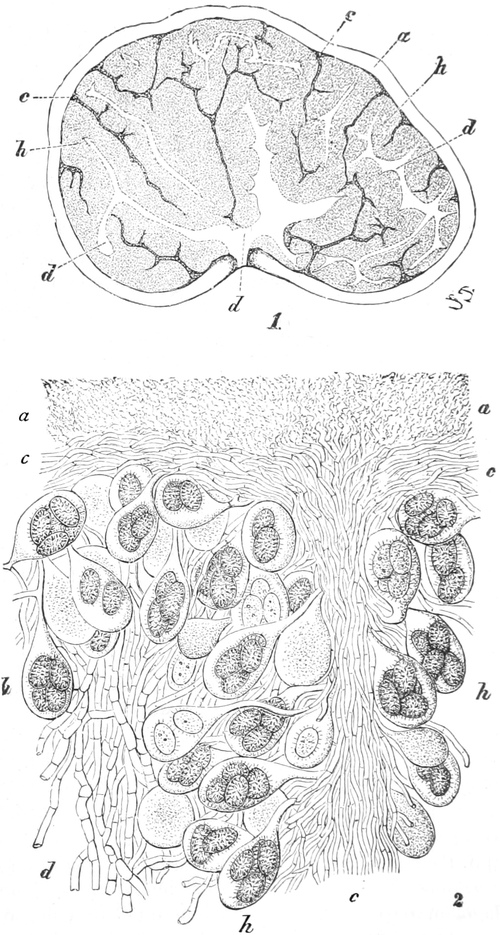
Fig. 394, Tuber rufum. 1 Ein Fruchtkörper im
Vertikalschnitt. Vergr. 5. a Die Rinde, d lufthaltiges Gewebe, c
dunkle Adern lückenlosen Gewebes, h das askusbildende Gewebe. 2 Ein
Stückchen des Hymeniums. Vergr. 460. Nach TULASNE. Aus V.
TAVEL, Pilze.
Die Askusbildung vollzieht sich, soweit bisher bekannt, in
der Weise, daß das Myzelium zwischen die Epidermis und die Kutikula
der Blätter oder der Fruchtknoten eindringt und sich hier reichlich
verzweigt. Die einzelnen Hyphenzellen schwellen an und bilden
meist unter Abgliederung einer basalen Stielzelle je einen die
Kutikula nach außen durchbrechenden Askus, der wie bei den übrigen
Schlauchpilzen anfangs zwei[S. 394] Kerne aufweist und nach Verschmelzung
beider in drei Teilungsschritten acht Sporen erzeugt (Fig. 395).
Die zahlreichen Asci stehen dicht nebeneinander.
Die Sporen sprossen in Wasser oder zuckerhaltiger Nährlösung,
häufig sogar schon in den noch geschlossenen Schläuchen direkt
zu Konidien aus, eine Form der Konidienvermehrung, die als
Hefesprossung bezeichnet wird; so bei Taphrina Pruni.
Die Exoascaceae sind vielleicht als reduzierte Askomyceten
aufzufassen, bei denen die Sexualorgane vollständig rückgebildet
wurden.
7. Ordnung. Saccharomycetes, Hefepilze, Sproßpilze[377]. Die
zur Gattung Saccharomyces vereinigten Bier-, Branntwein- und
Weinhefen stellen sehr einfache, einzellige Pilze vor, die nur in
Form von kugeligen, ovalen oder zylindrischen Zellen auftreten.
Sie enthalten einen Kern und vermehren sich fortgesetzt durch
Zellsprossung (Fig. 396). Myzelbildung fehlt, höchstens
bleiben die Zellen in Ketten eine Zeitlang vereinigt. Nach
Erschöpfung des Substrats, bei freiem Zutritt von Sauerstoff
und bei günstiger Temperatur bilden die Hefen Asci,
die äußerlich den Hefezellen gleichen, im Innern aber einige
wenige Sporen erzeugen. Bei manchen Hefepilzen ist eine mit
Kernverschmelzung verbundene Kopulation zweier Zellen
beobachtet, so bei Saccharomyces Ludwigii, dessen vier Sporen
im Askus keimen und dann paarweise durch je einen schnabelartigen
Kopulationskanal fusionieren, worauf dieser zum Keimschlauch
sich verlängert und mit der Abgliederung von Hefezellen beginnt,
ferner bei der Ingwerbierhefe Zygosaccharomyces und bei
Schizosaccharomyces, deren Hefezellen mittels langer Schläuche
vor der Sporenbildung kopulieren. Diese Kernfusionen entsprechen
vielleicht derjenigen im jungen Askus der übrigen Askomyceten.
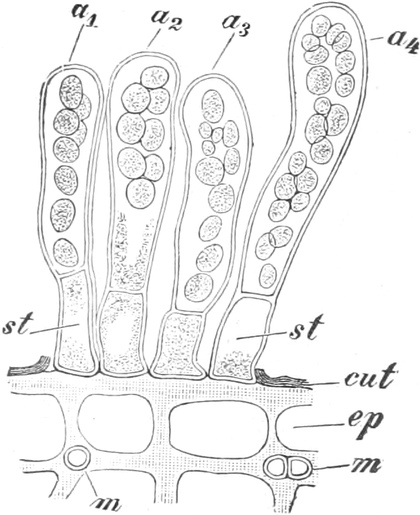
Fig. 395. Taphrina Pruni. Querschnitt durch die
Epidermis einer infizierten Pflaume. Vier reife Asci. a1 a2,
mit acht Sporen, a3, a4 mit Konidiensprossung aus den Sporen,
st Stielzelle des Askus, m Myzel quer durchschnitten, cut
Kutikula, ep Epidermis. Vergr. 600. Nach SADEBECK.
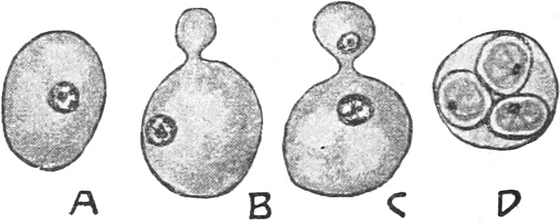
Fig. 396. Saccharomyces cerevisiae. A Hefezelle. B,
C In Sprossung. D Askus mit Sporen (in Vierzahl gebildet). Vergr.
1125. Nach GUILLIERMOND.
Die Hefepilze sind in physiologischer Beziehung als
Gärungserreger bemerkenswert; sie bewirken vermittels
der von ihnen erzeugten Zymase die Spaltung von Trauben- und
Fruchtzucker in Alkohol und Kohlensäure (vgl. S. 238). Die
Bierhefe, Saccharomyces cerevisiae, ist nur in der kultivierten
Form bekannt; der Weinhefenpilz, S. ellipsoideus, dagegen tritt
in der Natur schon im Boden der Weinberge in Sporenform auf und
gelangt von dort auf die Trauben und so in den Most. Beide Arten
kommen in verschiedenen Rassen vor, die in Reinkulturen gezüchtet
werden, um so in den Gärungsbetrieben Verwendung zu finden.
Außer Saccharomyces gehören zu den Hefepilzen noch einige andere
Gattungen, die zum Teil auch Myzelien bilden.
Die Hefen sind selbständige Pilze, wenigstens ist bis jetzt der
Nachweis nicht geführt, daß sie in den Entwicklungsgang anderer
Fadenpilze gehören, wenn auch bei verschiedenen Gattungen der
Exoasceen und Ustilagineen ebensolche Hefesprossung zu beobachten
ist. Ob sie reduzierte Askomyceten vorstellen oder zusammen mit
einigen Gattungen sehr einfacher Schlauchpilze an den Beginn der
Klasse gehören, bleibt noch unentschieden.
Infolge ihres Gehaltes an leichtverdaulichen Nährstoffen,
besonders Eiweiß und Glykogen, auch etwas Fett, hat Hefe einen
bedeutenden Nährwert und findet daher gereinigt und bei 125° C
getrocknet als Nährhefe Verwendung. Aus Hefe läßt sich eine
plastische hartgummiartige Masse, Ernolith, gewinnen, die
zu technischen Zwecken (Druckstöcken) unter Heißpressung gebraucht
wird.
[S. 395]
8. Ordnung. Laboulbenieae[378]. Diese durch THAXTER
genauer bekannt gewordenen eigenartigen Askomyceten sind winzige,
auf Insekten, besonders Käfern, parasitisch lebende Pilze mit zwei-
bis vielzelligem Thallus, der mittels eines spitzen Fortsatzes
der untersten Zelle in der Chitinhaut befestigt ist oder auch
mittels Rhizoiden in den Körper des befallenen Insektes eindringt.
Als Beispiel sei der auf Stubenfliegen in Europa verbreitete
Stigmatomyces Baerii genannt. Die zweizellige, mit Schleimhülle
versehene Spore (Fig. 397 A) setzt sich mit der unteren Spitze
fest (B); beide Zellen teilen sich sodann (C), und aus
der oberen Zelle geht ein Anhängsel mit mehreren einzelligen,
flaschenförmigen Antheridien (D, an) hervor, aus welchen
die nackten, zilienlosen, kugeligen Spermatien entleert
werden, während die untere Zelle zunächst in vier Zellen (D a,
b, c, d) sich teilt, von denen a sich vorwölbt und einen
vielzelligen weiblichen Apparat liefert. Die eigentliche Eizelle
(Eac), Karpogon genannt, ist von einer Hülle umgeben und
über ihr stehen zwei Zellen (Etp, t), von denen die oberste als
Trichogyn frei hervorragt und als Empfängnisorgan für die
Spermatien fungiert. Das Karpogon teilt sich nach der Befruchtung
in drei Zellen, von denen die obere verschwindet, die untere (F,
st) steril bleibt, die mittlere aber zu den Sporenschläuchen
auswächst, die je vier zweizellige spindelförmige Sporen erzeugen
(G). Die Sexualkerne treten im Karpogon nur paarweise dicht
zusammen und teilen sich dann konjugiert weiter; erst in den
Askusanlagen vollzieht sich die Kopulation der Kerne. Bei gewissen
Arten fehlen die Antheridien; nach FAULL erhält das
Karpogon in diesen Fällen einen zweiten Kern aus der Tragzelle des
Trichogyns.
Actinomycetes, Strahlenpilze[379].
Die Stellung der Strahlenpilze im System ist zur Zeit noch durchaus
unsicher. Im vegetativen Zustand bestehen sie aus sehr feinen
monopodial verzweigten Fäden (Hyphen) von Bakteriendicke
(0,5–0,8 μ). Der Fadeninhalt erscheint homogen, doch zerfallen
die Fäden leicht in Bruchstücke und könnten daher doch wohl in
Zellen gegliedert sein. Stärker färbbare Körnchen im Plasma
sind vielleicht Zellkerne. Chlorophyll fehlt. Rundliche oder
zylindrische Sporen entstehen reihenweise durch Teilung und Zerfall
der Hyphenenden in kurze Zellen. Da weder Sexualorgane noch die
für Algenpilze oder für Fadenpilze besonders charakteristischen
Sporenbildungen bei den Strahlenpilzen nachgewiesen sind,
fragt es sich, ob sie primitive Organismen oder ob
sie als reduzierte Fadenpilze etwa den Ascomyceten
anzuschließen sind. Von den Bakterien unterscheiden sie sich
durch die Verzweigung der Hyphen und andere Sporenbildung,
andererseits stimmen sie vielfach mit ihnen in Vorkommen und
Lebensweise überein. In zahlreichen Rassen sind sie überall im
Erdboden, an Pflanzen und Tieren, auf allen möglichen Substraten
verbreitet, meist als Saprophyten, einige auch als Parasiten. So
ist Actinomyces scabies der Erreger der Schorfkrankheit der
Kartoffeln und Rüben. Eiterige Geschwüre beim Rind erzeugt A.
bovis, ähnliche Krankheiten, besonders Eiterungen im Kiefer, beim
Menschen A. hominis. Übrigens ist die Unterscheidung bestimmter
Arten bei dem jetzigen Stand unserer Kenntnisse und bei der großen
Variabilität der Rassen in Kulturen noch nicht durchführbar.
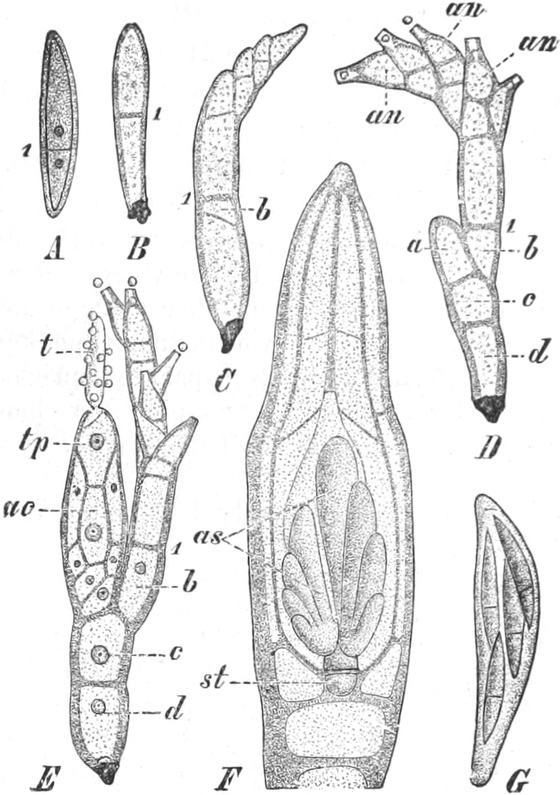
Fig. 397. Stigmatomyces Baerii. Erklärung im Text.
A Spore, B, F Aufeinanderfolgende Stadien. D Mit Antheridien
an, aus denen Spermatien austreten. E Oben mit Antheridien, links
mit ausgebildetem weiblichem Apparat. F Perithecium mit sich
entwickelnden Sporenschläuchen. G Reifer Sporenschlauch. Nach
THAXTER.
[S. 396]
2. Unterklasse. Basidiomycetes[288],
[352],
[353],
[380]–[392].
Die Basidienpilze scheinen keine Sexualorgane mehr zu besitzen;
nur bei den Uredineen oder Rostpilzen treten noch Gebilde auf, die
man als überkommene, aber funktionslos gewordene männliche Organe
deuten kann, und sogar auch noch Zellen, die den Karpogonen der
Askomyceten zu entsprechen scheinen. An Stelle der Asci treten die
ihnen gleichwertigen Basidien, die aber durch Zellsprossung
die Basidiosporen, bei den meisten Vertretern in der Zahl
vier, erzeugen. Ihre Anlagen zeigen insofern Übereinstimmung
mit denen der Asci, als in sie ebenfalls zwei Zellkerne eintreten, die
miteinander verschmelzen. Auf diese Kernverschmelzung (Karyogamie)
scheint allgemein eine Reduktionsteilung zu folgen. In zwei
Teilungsschritten entstehen so die vier Sporenkerne, die in die
Aussprossungen der Basidie hineinwandern (Fig. 398).
Der bei den Askomyceten vorhandene Generationswechsel läßt
sich bei den Basidiomyceten innerhalb der Gruppe der Uredineen
noch in ähnlicher Form erkennen, bei den übrigen Gruppen aber
mangels der Sexualorgane nicht mehr feststellen. Als Ersatz einer
Kopulation von Sexualzellen kommen Zellfusionen in Betracht, die
zweikernige Zellen liefern. Jedes Kernpaar entspricht zwar einem
diploiden Kern, aber erst in der Anlage der Basidien kommt eine
Verschmelzung der beiden haploiden Kerne eines Paares zu einem
wirklich diploiden zustande, der dann bei seiner Teilung gleich
wieder haploide Kerne liefert.
Die Basidien treten in drei verschiedenen Formen auf. Bei den
Ordnungen der Uredineen und Auricularieen ist ihr oberer Teil durch
Querwände in vier Zellen geteilt, von denen eine jede an ihrem oberen
Ende eine auf einem dünnen Stielchen (Sterigma) sitzende Spore erzeugt
(Fig. 404, 409). Bei den Tremellineen dagegen teilt sich die Basidie
durch zwei Längswände in vier mit langen schlauchförmigen Sterigmen
versehene Zellen (Fig. 399). Bei den Exobasidiinen, Hymenomyceten
und Gasteromyceten ist der Basidienträger einzellig, ungeteilt; er
bildet an seinem Gipfel vier Sporen, die meist auf Sterigmen sitzen,
seltener ungestielt sind (Fig. 398, 421 2). Von Interesse ist das
Verhalten der Ustilagineen, indem bei der einen Familie dieser Pilze
quergeteilte Basidien, bei der anderen dagegen ungeteilte Basidien
auftreten. Die Zahl der gebildeten Sporen ist hier nicht scharf
begrenzt, sondern oft sehr groß.
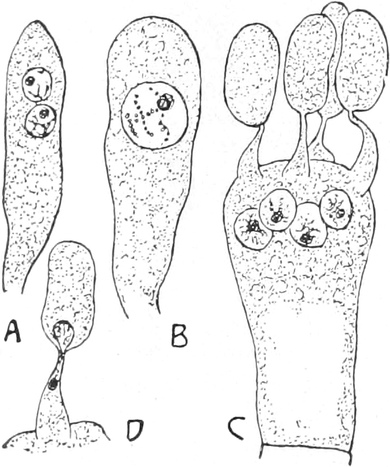
Fig. 398. Armillaria mellea. A Junge Basidie mit
den beiden primären Kernen. B Nach Verschmelzung der beiden Kerne.
— Hypholoma appendiculatum. C Basidie vor dem Übertritt der aus
dem sekundären Basidienkern entstandenen vier Kerne in die jungen
scheitelständigen Basidiosporen. D Übertritt des Zellkernes durch das
Sterigma in die Basidiospore. Nach RUHLAND.
Fig. 399. Basidie einer Tremellinee (Tremella
lutescens). Vergr. 450. Nach BREFELD. Aus V. TAVEL,
Pilze.
Außer den Basidien treten wie bei den Askomyceten auch hier
Konidien als Nebenfruktifikationen in dem Entwicklungsgang
mancher Arten auf. Von der Konidienbildung verschieden ist
die Entstehung ungeschlechtlicher Sporen durch Abrundung,
Membranverdickung und schließliche Abtrennung der Hyphenzellen
(Chlamydosporen nach BREFELD). So entstehen
reihenweise die Brandsporen der Ustilagineen, endständig[S. 397] die
Rostsporen der Uredineen. Bei ersteren gehen die Basidien
unmittelbar aus den keimenden Brandsporen hervor (Fig.
401), bei letzteren aus bestimmten Rostsporen (Fig. 404 2).
Beide Gruppen unterscheiden sich dadurch scharf von den übrigen
Basidiomyceten, bei denen, abgesehen von einigen einfacheren
Formen, von den Myzelien durch Hyphenverflechtung Fruchtkörper
angelegt werden, an deren Oberfläche oder in deren Innerem
dann die Basidien, meist in besonderen Schichten oder Hymenien
zur Bildung gelangen. Diese Fruchtkörper entsprechen denen der
Askomyceten, nur fehlen in ihren Anlagen die Sexualorgane.
Die den Rost- und Brandsporen entsprechenden Basidienanlagen
gehen hier aus Hyphenzellen der Fruchtkörper hervor, ohne daß
Chlamydosporenbildung eintritt.
1. Ordnung. Ustilagineae, Brandpilze[381]. Die Brandpilze leben
parasitisch in höheren Pflanzen. Besonders dienen die Gramineen
als Nährpflanzen. Gewisse Arten sind dem Getreide in hohem Maße
schädlich: sie erzeugen in den Fruchtständen von Hafer, Gerste,
Weizen, Hirse, Mais die als Getreidebrand bekannten
Krankheiten.
Das Myzelium bildet die Brandsporen, indem seine Hyphen
sich durch Querwände in kurze Zellen teilen, die anschwellen,
sich abrunden, ihre Membran aufquellen lassen und sich als Sporen
innerhalb der später verschwindenden Gallerthüllen mit einer neuen
Membran umgeben. So zerfällt das Myzelium in eine dunkelbraune oder
schwarze Masse von Brandsporen. Diese sind Dauersporen, werden
von den Wirtspflanzen aus durch den Wind zerstreut und keimen
nach der Winterruhe zu den Basidien aus, deren Bildung bei
den beiden Familien der Brandpilze, den Ustilaginaceen und den
Tilletiaceen, nach verschiedenen Typen erfolgt.
Fig. 400. Ustilago. A In Nährlösung keimende
Brandspore cl mit der Basidie (Promyzel) t, den Basidiosporen
(Sporidien) c. Vergr. 450. B In Nährlösung sprossende Konidien.
Vergr. 200. C Sproßverband von Konidien. Vergr. 350. Nach
BREFELD. Aus V. TAVEL, Pilze.
Fig. 401. Ustilago scabiosae. A Aus der Brandspore
gekeimte junge Basidie mit vier Kernen. B Sporenbildung an der
vierzelligen Basidie. Nach HARPER.
Als wichtigster Vertreter der Ustilaginaceen ist die Gattung
Ustilago zu erwähnen. Ust. Avenae, Hordei, Tritici, die
früher als U. Carbo zusammengefaßt wurden, verursachen den
Staubbrand in den Fruchtknoten von Hafer, Gerste, Weizen, Ust.
Maydis an den Halmen, Blättern und Infloreszenzen des Mais die
Bildung von großen, mit schwarzem Brandsporenpulver erfüllten,
geschwürartigen Beulen und Blasen. Ust. violacea lebt in
Caryophyllaceen (Lychnis, Saponaria), deren Staubbeutel von ihr an
Stelle des Pollens mit Brandsporen erfüllt werden. An weiblichen
Lichtnelkenpflanzen bewirkt der Pilz in den Blüten die Entwicklung
von Staubgefäßen mit brandigen Antheren.
Die Brandsporen von Ustilago keimen nach der Ruhezeit auf dem Boden
zu einem kurzen Schlauch (Promyzel), der sich durch drei bis vier
Querwände teilt (Fig. 401) und die Basidie vorstellt; diese
bringt seitlich am oberen Ende ihrer einzelnen Zellen sowie an
ihrer Spitze die eiförmigen, hier einkernigen Basidiosporen
(Sporidien) hervor. Wenn reichlich Nährstoffe dem Pilz zur
Verfügung stehen, wie bei Kultur in Nähr[S. 398]lösungen, werden
beständig neue Sporen in großer Zahl abgegliedert (Fig. 400),
und diese vermehren sich dann, indem sie fortgesetzt in Konidien
weitersprossen. Sind keine reichlichen Nährstoffe im Substrat
vorhanden, so erfolgen bei manchen Brandpilzen paarweise Fusionen
zwischen den Konidien oder zwischen den Zellen des Promyzels (Fig.
403). Auf den Getreideäckern findet die Konidienbildung im feuchten
gedüngten Boden statt, also bei saprophytischer Ernährungsweise,
und die aus Konidien oder Fusionszellen schließlich hervorgehenden
Fäden gehen zur parasitischen Lebensweise über, indem sie in
Getreidekeimlinge eindringen und diese bis zur Vegetationsspitze
durchwachsen, wo später die Infloreszenzen angelegt werden. In
letzteren entwickelt sich das Myzel weiter und schließt mit der
Erzeugung der Brandsporen ab.
Außer der Infektion junger Pflanzen kommt auch eine
Infektion der Blüten vor, indem Brandsporen oder am Boden gebildete
Konidien auf die Narben gelangen und dort zu Myzelien auskeimen,
die in die Samenanlagen eindringen und schließlich im Keimling
überwintern. Entweder findet ausschließlich Blüteninfektion statt,
so bei Ustilago Tritici, Hordei, oder vorwiegend Infektion der
Keimpflanzen, so bei U. Avenae, Sorghi, Panici miliacei,
Crameri, während der Maisbrand mit seinen Konidien alle Teile
noch junger Maispflanzen infizieren kann und sich auf die
infizierte Stelle beschränkt.
Die Tilletiaceen führen ähnliche Lebensweise wie die
Ustilaginaceen. Am bekanntesten sind Tilletia Tritici (auch T.
Caries genannt) und Till. laevis, die Pilze des Stein- oder
Stinkbrandes des Weizens. Die Weizenkörner werden vom Pilz mit
schwärzlichen, nach Heringslake riechenden Brandsporen
erfüllt, die bei ersterer Art netzförmig verdickte, bei letzterer
glatte Wandung besitzen. Im Gegensatz zu den Ustilaginaceen erzeugt
der Keimschlauch nach vorausgegangener mehrmaliger Teilung seines
Kernes die hier fadenförmigen Basidiosporen nur an seinem
Scheitel, in wirteliger Anordnung zu 8 bis 16 (Fig. 402 A). Sie
zeigen die Eigentümlichkeit, daß sie paarweise miteinander in
Verbindung treten (B) und somit auch paarweise abfallen. Die
Sporenpaare keimen leicht aus und erzeugen an ihren Keimschläuchen
wiederum je eine Konidie, aber von sichelförmiger Gestalt
(Fig. 402 C). Bei reichlicher saprophytischer Ernährung wachsen
die Keimschläuche zu größeren Myzelien heran, an denen in reichem
Maße solche sichelförmigen Konidien an der Luft abgegliedert werden
(D). Tilletia weist somit im Gegensatz zu Ustilago zweierlei
Formen von Konidien auf. Im übrigen stimmt die Entwicklung bei
beiden Gruppen überein.
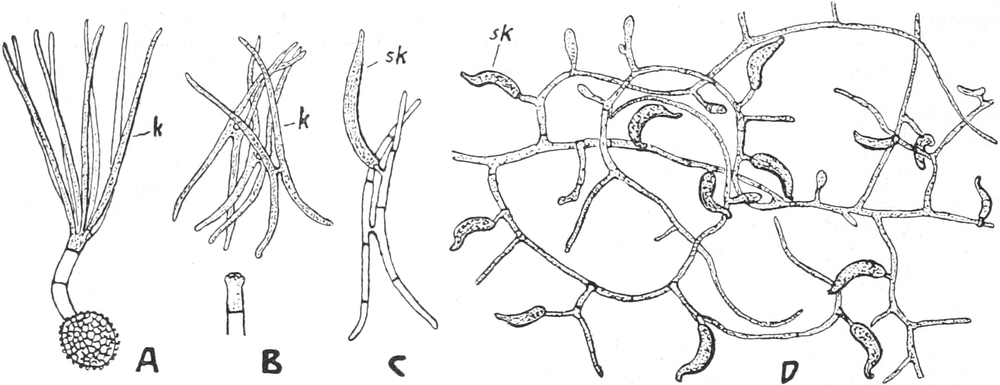
Fig. 402. Tilletia Tritici. A Die aus der Brandspore
hervorgegangene Basidie mit vier Paaren endständiger Sporen k.
Vergr. 300. B Ablösung der fusionierten Sporenpaare. Vergr. 250.
C Ein solches in Keimung mit sichelförmiger Sekundärkonidie sk.
Vergr. 400. D Myzelium mit sichelförmigen Konidien. Vergr. 350. Nach
BREFELD.
Bezüglich des Verhaltens der Kerne der Brandpilze ist zu
bemerken, daß allgemein in die jungen Brandsporen zwei Kerne
eintreten, die dann verschmelzen. Bei der Keimung der Brandsporen
ist eine Reduktionsteilung zu erwarten. Die Zellen des Promyzels
und die Sporidien sind einkernig und bezeichnen den Beginn
der haploiden Phase. Die Zweikernigkeit der Zellen wird nun
auf verschiedene Weise erreicht. Bei U. Maydis, Doassansia
Sagittariae besteht das parasitische Myzel aus einkernigen Zellen,
und erst kurz vor der Sporenbildung fusionieren benachbarte
Hyphenzellen durch Lösung der Querwand paarweise miteinander,
wodurch zweikernige, die Sporen liefernde Zellen[S. 399] entstehen.
Dagegen erhält bei U. carbo und wohl den meisten Brandpilzen das
parasitische Myzel zweikernige Zellen dadurch, daß die Sporidien
oder die aus ihnen hervorgehenden Konidien oder sogar schon die
Promyzelzellen oder auch die Zellen der aus ihnen oft entstehenden
verzweigten Myzelien paarweise fusionieren (Fig. 403). Ähnlich
verhält sich auch Tilletia, bei der die Sporidien bereits vor
ihrer Loslösung paarweise in Verbindung treten; aus der einen
Sporidie tritt der Kern in die andere über. Die bei der Keimung aus
ihr hervorgehenden Hyphenzellen und sekundären Sporidien, ebenso
die Zellen des parasitischen Myzeliums sind demgemäß sämtlich
zweikernig.
Für Ustilago violacea hat KNIEP nachgewiesen, daß
die aus Sporidien hervorgehenden Konidien nicht miteinander
kopulieren, wenn sie Abkömmlinge nur eines Sporidiums sind.
Physiologische Geschlechtsdifferenzierung führt hier zur Bildung
zweier zwar äußerlich gleicher, innerlich aber heterogener Sorten
von Sporidien. Solche Geschlechtsdifferenzierung bei Ustilagineen
dürfte sich nach KNIEP und RAWITSCHER bereits
bei der Reduktionsteilung den Brandsporenkerns in die zwei ersten
Promyzelkerne vollziehen, von denen einer und seine Abkömmlinge das
eine Geschlecht (+), der andere das andere Geschlecht (–) erhält.
Bei den Brandpilzen sind somit haploide und diploide Phasen nicht
übereinstimmend festgelegt.
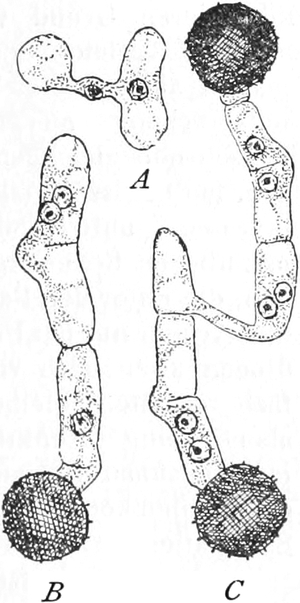
Fig. 403. Ustilago Carbo. A Kopulierende Sporidien.
B Die zwei obersten Zellen eines Promyzels zu einer zweikernigen
Zelle kopuliert. C Kopulation zwischen zwei Promyzelien. Vergr. 1000.
Nach RAWITSCHER.
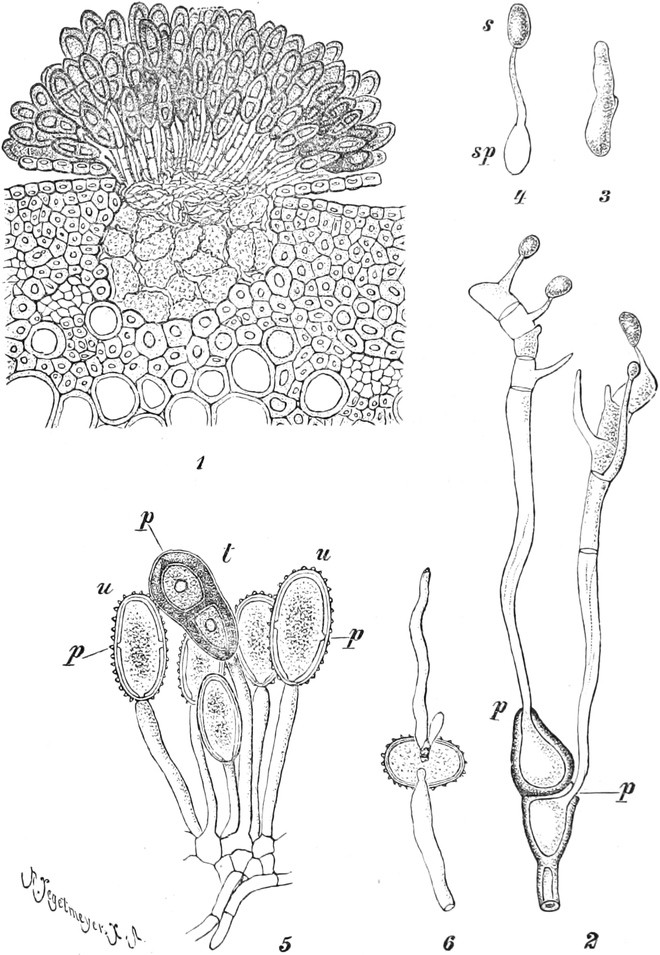
Fig. 404. Puccinia graminis. 1 Querschnitt durch einen
Getreidehalm mit einem Teleutosporenlager. 2 Keimende Teleutospore
mit zwei Basidien. 3 Vegetativ, 4 fruktifikativ keimende
Basidiospore. Letztere mit Sekundärspore, welche gebildet wird, wenn
zur Infektion einer Pflanze keine Gelegenheit geboten ist. 5 Gruppe
von Uredosporen u, untermischt mit einer Teleutospore t, p die
Keimsporen. 6 Keimende Uredospore. 1 Vergr. 150. 2 Vergr. ca.
230. 3, 4 Vergr. 370. 5 Vergr. 300. 6 Vergr. 390. 2, 3,
4 nach TULASNE. 5, 6 nach DE BARY. Aus V.
TAVEL, Pilze.
2. Ordnung. Uredineae, Rostpilze[382]–[386].
Die Rostpilze leben als Parasiten mit ihren Myzelien in den
Interzellularräumen hauptsächlich der Blätter höherer Pflanzen und sind
die Erreger der sehr verbreiteten Rostkrankheiten. Von den Brandpilzen
unterscheiden sie sich durch viel mannigfaltigere Sporenbildungen.
[S. 400]
Auch hier entstehen Basidien nicht direkt am Myzel, sondern
bei der Keimung besonderer Sporen, der Teleuto-
oder Wintersporen, die für fast sämtliche Rostpilze
charakteristisch sind. Diese Sporen werden in kleinen Gruppen unter
der Epidermis der Nährpflanze aus den Endzellen dicht nebeneinander
stehender Hyphenäste gebildet, entweder einzeln oder häufig zu
zwei oder mehr in kurzer Kette; sie sind dickwandige, den Winter
überdauernde Ruhesporen (Fig. 404, 1 und 5 t). Die Sporenlager
durchbrechen gewöhnlich die Epidermis. Die Sporen besitzen anfangs,
wie auch alle Zellen des sie bildenden Myzels, zwei Kerne; in
reifem Zustande aber sind letztere zu einem Kern verschmolzen.
Bei der Keimung wächst aus der Teleutosporenzelle eine
Basidie (Promyzel) hervor, die sich, wohl allgemein unter
Reduktionsteilung ihres Kernes, in vier Zellen quer teilt
und aus jeder Zelle auf einem Sterigma eine Basidiospore
(Sporidie) erzeugt, die nur einen Zellkern enthält (Fig. 404,
2). Die Sporen werden durch den Wind verbreitet und keimen im
Frühjahr auf den Blättern von Nährpflanzen derselben Art oder
einer anderen Art zu einem interzellulären Myzelium, dessen Zellen
sämtlich einkernig bleiben. Aus diesem Myzelium entwickeln sich nun
zweierlei Gebilde, und zwar Spermogonien an der Blattoberseite,
Äcidien an der Blattunterseite.
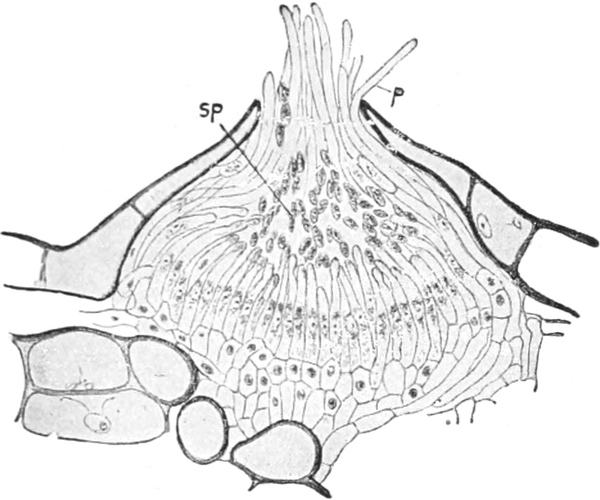
Fig. 405. Gymnosporangium clavariaeforme. Spermogonium
auf Crataegusblättern, die Epidermis durchbrechend. sp Spermatien,
p sterile Saftfäden oder Paraphysen. Nach BLACKMAN.
Die Spermogonien (Fig. 405) sind krugförmige Gebilde, deren
Grund von Hyphenenden ausgekleidet wird, welche die mit je einem
Zellkern ausgestatteten Spermatien abgliedern. Sie entsprechen
morphologisch den gleichnamigen, bei gewissen Askomyceten
auftretenden männlichen Sexualorganen, die unter den Basidiomyceten
nur bei Uredineen zwar noch vielfach erhalten bleiben, aber keine
Funktion mehr ausüben und auch ganz fehlen können. Die Spermatien
vermögen in Nährlösungen kurze Keimschläuche zu treiben, sind aber
nicht imstande, auf Blättern neue Infektionen hervorzurufen.
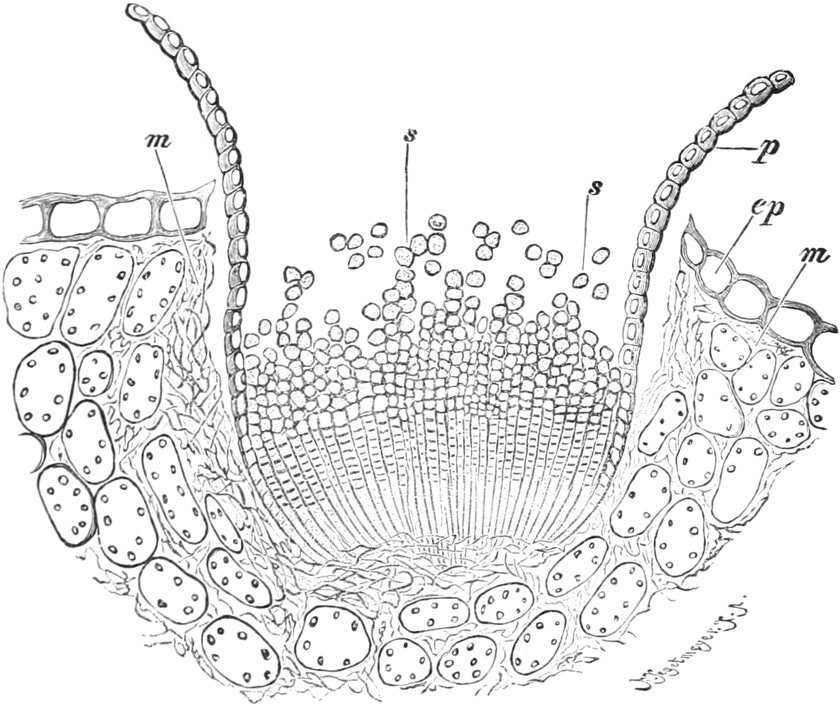
Fig. 406. Puccinia graminis. Äcidium auf Berberis
vulgaris. ep Epidermis der Blattunterseite, m interzellulares
Myzel, p Peridie, s Sporenketten. Vergr. 142.
Die Äcidien (Fig. 406) sind anfangs geschlossene, später
sich öffnende, becherförmige Fruchtkörper, die zahlreiche,
dicht nebeneinander stehende, von basalen Trägern abgegliederte
Sporenketten enthalten. Meist umgibt eine einschichtige, aus
dickwandigen Zellen bestehende Hülle (Peridie) das Äcidium.
Die von BLACKMAN[382] an dem auf Brombeerblättern
vorkommenden Rost, Phragmidium viola[S. 401]ceum, untersuchte Entstehung
eines solchen Organs erfolgt in der Weise, daß die unter der
Blattoberhaut gelegenen Hyphenenden zunächst eine sterile, bald
vergängliche Zelle an ihrer Spitze abgrenzen, und daß die darunter
gelegene fertile Zelle anschwillt (Fig. 407 A). Sie enthält
nur einen Kern; aber sie wird zweikernig dadurch, daß der
Zellkern einer anstoßenden Myzelzelle in sie hinüberwandert,
ohne daß Kernverschmelzung eintritt (konjugierte Kerne). Diese
zweikernige „basale“ Zelle teilt sich nun in eine Kette von
zweikernigen Sporenmutterzellen, die nochmals je in eine obere
zweikernige Äcidiospore und eine untere, ebenfalls zweikernige,
aber steril bleibende, bald zusammenschrumpfende Zwischenzelle
zerlegt werden (B, C).
Nach CHRISTMAN[383] vollziehen sich diese Vorgänge in
etwas anderer Weise bei dem auf Rosen schmarotzenden Phragmidium
speciosum (Fig. 408) und ebenso nach neueren Untersuchungen auch
bei Puccinia und anderen Gattungen. Die Äcidiumentwicklung
dieser Formen darf als die typische gelten. Zwar teilen sich auch
hier die Hyphenenden (A) in eine obere, sterile, vergängliche
Zelle und in eine untere fertile Zelle (B); die fertilen Zellen
verschmelzen aber paarweise miteinander (C), indem ihre Wandung
im oberen Teile aufgelöst wird. Die beiden Kerne teilen sich darauf
gleichzeitig und nebeneinanderliegend (konjugierte Teilung) in je
zwei Tochterkerne, von denen zwei in den unteren Teil, zwei in den
oberen Teil der Zelle rücken (D). Dieser grenzt sich nunmehr als
erste Sporenmutterzelle durch eine Querwand ab. Die Bildung der
Äcidiosporenketten erfolgt im übrigen wie zuvor dargestellt. Eine
Peridie wird bei Phragmidium nicht angelegt, wohl aber bei Puccinia
u. a.; sie geht hervor aus den steril bleibenden peripherischen
Sporenketten und den die Decke des Peridiums bildenden Endzellen
der mittleren Sporenreihen.
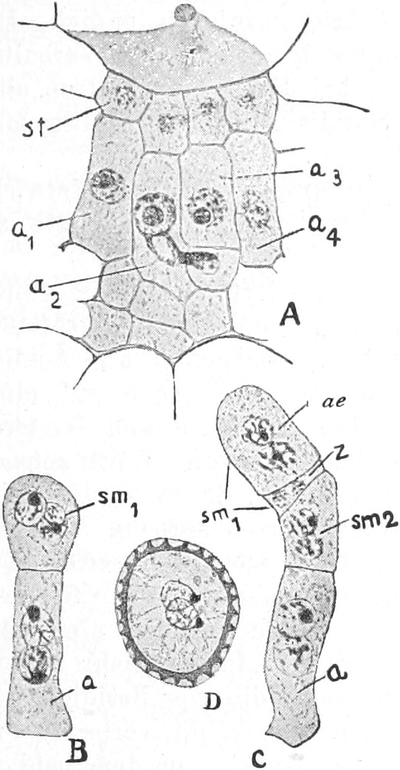
Fig. 407. Phragmidium violaceum. A Teil einer
Äcidiumanlage, st sterile Zelle, a fertile Zellen, bei a2
Übertreten des Kernes einer benachbarten Zelle. B Bildung der ersten
Sporenmutterzelle sm1 aus der Anlage a einer Sporenkette. C
Weitere Teilung, aus sm1 die Äcidiospore ae und die Zwischenzelle
z gebildet, sm2 die zweite Sporenmutterzelle. D Reife
Äcidiospore. Nach BLACKMAN.
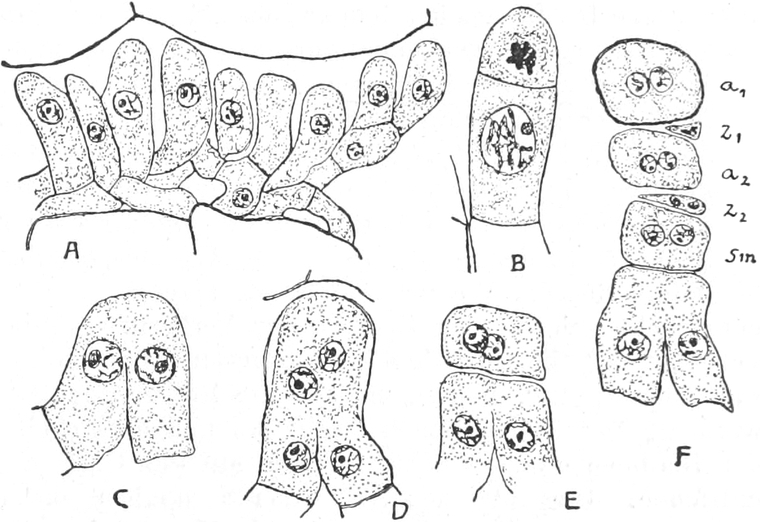
Fig. 408. Phragmidium speciosum. A Erste Anlage
eines Äcidiums unter der Blattepidermis von Rosa. B Teilung einer
Hyphenendzelle in die obere sterile vergängliche und die untere fertile
Zelle. C Kopulation zweier benachbarter fertiler Zellen. D Weiteres
Stadium, erste Kernteilung vollendet. E Abgliederung der ersten
Äcidiosporenmutterzelle. F Kette von Äcidiosporen a1 a2,
Zwischenzellen z1 z2, die zuletzt gebildete Mutterzelle sm
noch nicht geteilt. Nach CHRISTMAN.
Die reifen zweikernigen Äcidiosporen (Fig. 407 D) stäuben aus,
infizieren eine neue Wirtspflanze, und aus jeder Spore geht wieder
ein interzellulares Myzelium hervor, das dann bald im Sommer zur
Bildung von Uredo- oder Sommersporen übergeht.
Diese entstehen in kleinen rundlichen oder strichförmigen Lagern,
einzeln aus den anschwellenden Endzellen ihres Trägers (Fig. 404,
5 und 6) und enthalten, wie auch die Zellen des ganzen, aus
der Äcidiospore hervorgehenden Myzels, zwei Kerne. Sie besorgen
im[S. 402] allgemeinen die Ausbreitung des Pilzes im Sommer. Später
werden dann in denselben oder in anderen Lagern die Teleutosporen
gebildet, in denen erst die Kopulation der beiden Kerne zu einem
einzigen stattfindet, wie dies überhaupt in den Basidienanlagen zu
geschehen pflegt.
Die beiden Formen der Zellfusionen in den Anlagen der Äcidien
sind auch noch bei anderen Uredineen nachgewiesen und können nur
als Ersatz ehemaliger Befruchtungsvorgänge angesehen werden. Wenn
wir die Uredineen von den Askomyceten ableiten wollen, müssen wir
die Spermatien als funktionslos gewordene männliche Sexualzellen
ansehen und die sog. fertilen Zellen der Äcidienanlagen mit den
Karpogonen vergleichen. Zieht man den Vergleich noch weiter,
so würden in dem Entwicklungsgang der Uredineen diese fertilen
Zellen den Anfang von diploiden ungeschlechtlichen Generationen
vorstellen, die nach Bildung von Äcidio-, Uredo- und Teleutosporen
in den den Ascis entsprechenden Basidien ihren Abschluß finden,
während die haploide geschlechtliche Generation aus den
Basidiosporen hervorgeht und in den Äcidiumanlagen endigt. Die
Übereinstimmung zwischen Askomyceten und Uredineen zeigt sich auch
in dem Verhalten der Sexualkerne, die in den Sexualorganen nur sich
paaren, bei den Schlauchpilzen aber erst in den Askusanlagen, bei
den Rostpilzen erst in den Basidienanlagen endgültig miteinander
kopulieren.
Die drei Sporenformen der Uredineen zeigen nach CHRISTMAN
in ihrer Entwicklung aus den sie bildenden „basalen“ Zellen
gewisse Übereinstimmung, so daß sie als morphologisch gleichwertig
angesehen werden können.
Der Entwicklungsgang der Rostpilze ist also höchst kompliziert.
Entweder treten die verschiedenen Sporenformen im Laufe des Jahres
an ein und derselben Nährpflanze auf; solche Uredineen bezeichnet
man als autözische. Oder Spermogonien und Äcidien finden
sich auf der einen Nährspezies, Uredo- und Teleutosporen dagegen
auf einer anderen, der ersteren im System oft sehr ferne stehenden
Pflanze. Bei diesen letzteren heterözischen Arten liegt
also ein Wirtswechsel des Parasiten vor. Neben solchen sehr
wählerischen gibt es aber auch pleophage heterözische Rostpilze,
deren Äcidien oder deren Uredo-Teleutosporen auf zahlreichen
verschiedenen Nährpflanzen auftreten[384].
Als Beispiel für Heterözie sei Puccinia graminis, der
häufigste Getreiderostpilz, erwähnt, der seine Uredo-
und Teleutosporen an Blättern und Halmen von Gräsern, besonders
Roggen, Weizen und Gerste erzeugt. Die Äcidien und Spermogonien
dieser Art entwickeln sich auf den Blättern der Berberitze
(Berberis vulgaris). Im Frühjahr treiben zunächst die überwinterten
Teleutosporen ihre Basidien, von denen die vier Basidiosporen
sich ablösen (Fig. 404, 2), um auf die Berberitzenblätter durch
den Wind verbreitet zu werden. Der Keimschlauch dringt ein und
entwickelt sich zum Myzel, aus dem bald an der Blattoberseite die
Spermogonien, auf der Unterseite die Äcidien (Becherrost, Fig. 406)
entstehen. Die rotgelben Äcidiosporen stäuben aus der Peridie aus
und gelangen auf Gräser. Das aus ihnen hervorgehende Myzel bringt
im Sommer zunächst die Uredosporen (Fig. 404, 5) hervor. Die
Uredosporen sind einzellig, mit vier äquatorialen Keimporen in der
außen mit kleinen Warzen bedeckten Wandung versehen und enthalten
rotgelbe Fettröpfchen in ihrem Plasma. Sie keimen auf Getreide
und verbreiten rasch die Rostkrankheit. Gegen Ende des Sommers
werden in denselben Lagern (Fig. 404, 1) die dunkelbraunen,
stets zu zwei vereinigten, dickwandigen Teleutosporen mit je einem
Keimporus erzeugt, von denen im nächsten Jahr die Entwicklung
von neuem anhebt. Auch kann in dem durch Uredosporen infizierten
Wintergetreide der Pilz überwintern und mit Übergehung von
Basidiosporen und Berberis-Äcidium im nächsten Sommer seine Uredo-
und Teleutosporen bilden[385].
Nicht alle Uredineen weisen einen so reich gegliederten
Entwicklungsgang auf. Rostpilze, die sämtliche Sporenarten
erzeugen, heißen Eu-Formen, solche ohne Uredo: Opsis-Formen,
solche ohne Äcidien: Brachy-Formen, solche ohne Äcidien und
Uredo: Mikro-Formen. Bei denjenigen Rostpilzen, die keine Äcidien
besitzen, finden wir in den aus den Basidiosporen hervorgehenden
Myzelien einkernige Zellen, später aber, vor der Bildung der
Teleutosporen, zweikernige Zellen. Die Zweikernigkeit der Zellen
wird bei ihnen, wie bereits für einzelne Fälle festgestellt
wurde, in den Anlagen der ersten Uredosporen oder, wenn solche
fehlen, in den Anlagen der Teleutosporen (z. B. bei Puccinia
Malvacearum) durch Kopulation zweier Zellen ähnlich wie in den
oben geschilderten Äcidiumanlagen erreicht, ein Verhalten, das die
Homologie der drei genannten Sporenformen bestätigt.
[S. 403]
Abweichend von allen übrigen Rostpilzen und einfacher verhält sich
Endophyllum[386], dessen Arten auf Sempervivum und auf Euphorbia
schmarotzen und weder Uredo- noch Teleutosporen bilden. Das aus
den Basidiosporen hervorgehende, aus einkernigen Zellen bestehende
Myzel bildet Spermogonien und Äcidien; in den Anlagen der
Äcidiosporenketten erfolgen Zellfusionen nach Art von Phragmidium,
wodurch sie zweikernig werden. Die reifen Äcidiosporen verhalten
sich nunmehr wie die Teleutosporen der übrigen Rostpilze; ihre
beiden Kerne verschmelzen zu einem einzigen; dann keimen die Sporen
aus zu Basidien mit vier einkernigen Basidiosporen und erfahren
vorher eine Reduktionsteilung ihrer Kerne (Fig. 409). Vielleicht
kann Endophyllum als eine primitive Form betrachtet werden.
Ebenso verhält sich auch Caeoma nitens[387].
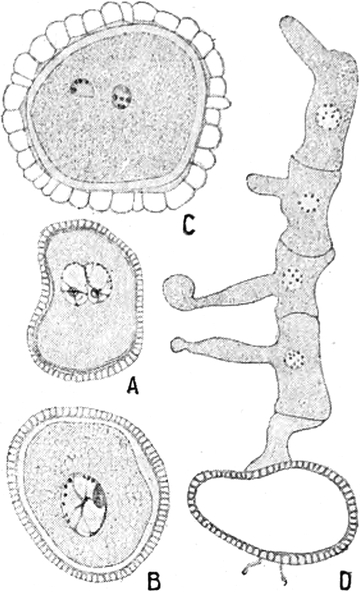
Fig. 409. Endophyllum Sempervivi. A Junge noch
zweikernige Äcidiospore. B Reife einkernige Spore. C Keimende
Spore, deren Kern sich bereits in zwei Kerne geteilt hat. D
Äcidiospore zu einer jungen vierzelligen Basidie ausgekeimt. Nach
HOFFMANN.
3. Ordnung. Auricularieae. Basidien wie bei den Uredineen
quergeteilt, mit vier Sporen. Hierher nur wenige Formen,
unter denen am bekanntesten der als Volksheilmittel noch vielfach
verwendete Holunderschwamm oder das Judasohr, Auricularia
sambucina ist, mit gallertartigen, dunkelbraunen, muschelförmigen
Fruchtkörpern, die aus alten Holunderstämmen hervorbrechen und auf
ihrer Innenseite das Basidienhymenium tragen.
4. Ordnung. Tremellineae, Zitterpilze. Basidien der Länge
nach geteilt (Fig. 399). Die Fruchtkörper der Zitterpilze sind
von gallertartiger Beschaffenheit, lappig oder runzlig gefaltet
und auf ihrer Oberseite mit dem Basidienhymenium überkleidet. Nur
wenige Gattungen, saprophytisch in faulenden Baumstämmen, aus deren
Oberflächen die Fruchtkörper hervorkommen.
5. Ordnung. Exobasidiineae. Die Fruchtkörperbildung fehlt hier,
vielmehr entstehen die keulenförmigen, ungeteilten, an ihrem Gipfel
vier Sporen auf dünnen Sterigmen tragenden Basidien frei aus den
Myzelfäden, so bei Exobasidium Vaccinii, einem auf Ericaceen,
besonders Preißel- und Heidelbeeren auftretenden Pilz, dessen
Myzel Auftreibungen der befallenen Pflanzenteile verursacht. Die
Basidien werden in Lagern unter der Epidermis gebildet und brechen
durch diese nach außen hervor (Fig. 410). Als Nebenfruktifikationen
treten bei dieser Gattung, wie bei manchen anderen, Konidien
auf, die als schmal spindelförmige Zellen vom Myzel abgegliedert
werden und der Basidienbildung vorausgehen.
6. Ordnung. Hymenomycetes[373].
Die Basidien sind
ungeteilt und tragen an der Spitze auf dünnen Sterigmen
vier Sporen (Fig. 411 sp). Sie werden an Fruchtkörpern erzeugt,
die durch Verflechtung von Hyphen entstehen, und sind in Schichten
oder Hymenien angeordnet. An der Zusammensetzung dieser
beteiligen sich die Saftfäden oder Paraphysen (Fig. 411 p)
und vielfach auch die ebenfalls sterilen Zystiden (c),
einzellige Schläuche, die sich durch größeren Umfang auszeichnen.
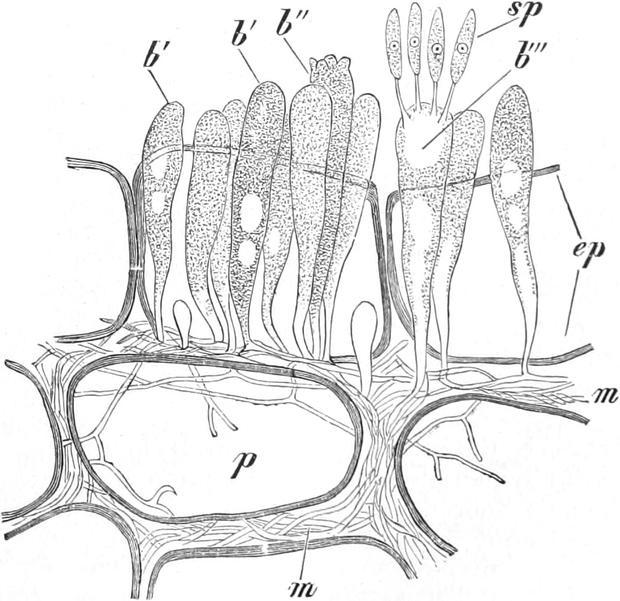
Fig. 410. Exobasidium Vaccinii. Querschnitt durch die
Stengelperipherie von Vaccinium. ep Epidermis, p Rindenparenchym,
m Myzelfäden in den Interzellularräumen, b die nach außen
hervorbrechenden Basidien, b′ noch ohne Sterigmen, b″ Anlage der
Sterigmen, b‴ mit vier Sporen. Vergr. 620. Nach WORONIN.
[S. 404]
Die vier Sporen werden vermittels des osmotischen Druckes der
Basidien von den Sterigmen nacheinander eine kurze Strecke
weit abgeschleudert; sie haften überall leicht an. Ihr freies
Herabfallen von den Hymenien wird aber durch die Paraphysen
begünstigt, die die Basidien voneinander trennen. Die Zystiden sind
nach KNOLL Wasser und Schleim abscheidende Organe. In
einzelnen Fällen mögen sie auch noch andere Funktionen erfüllen, so
bei Coprinus-Arten das Auseinanderhalten der Lamellen und somit
die Sicherung des Sporenfalles[388].
Da bei den Hymenomyceten wie auch bei den ihnen nächst verwandten
Ordnungen besondere Sexualorgane fehlen, die Basidien aber den
Schläuchen der Askomyceten entsprechen und anfangs wie diese
zwei miteinander kopulierende Kerne aufweisen, so entsteht die
Frage, ob in dem Entwicklungsgang beider Pilzgruppen noch weitere
Homologien bestehen, und wie bei ersteren die Zweikernigkeit der
Basidienanlage zustande kommt[389].
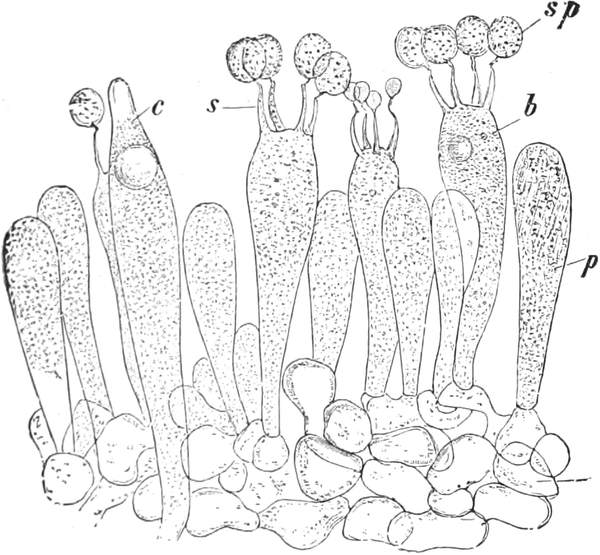
Fig. 411. Russula rubra. Teil des Hymeniums. sh
Subhymeniale Schicht, b Basidien, s Sterigmen, sp Sporen, p
Paraphysen, c ein Zystide. Vergr. 540. Nach STRASBURGER.
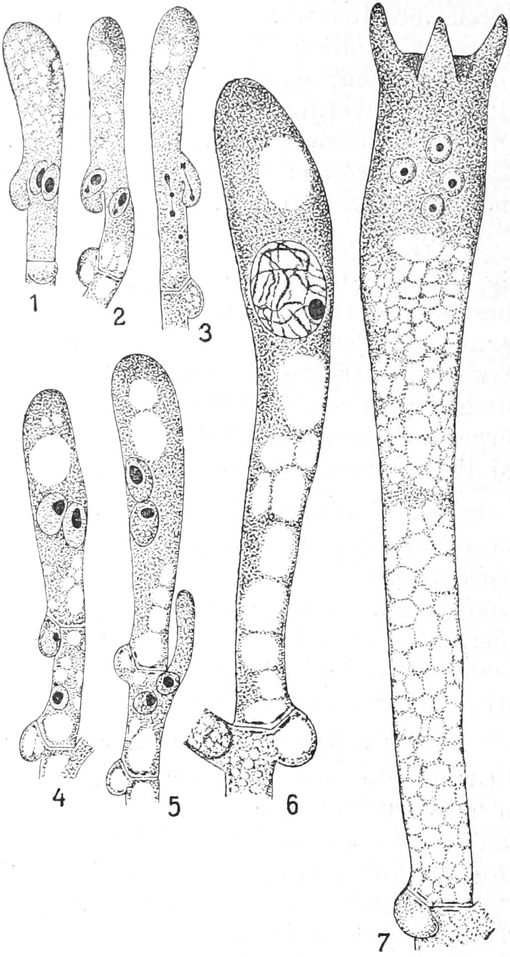
Fig. 412. Armillaria mucida, Schnallenbildung
und Basidienentwicklung. 1 Beginn der Schnallenbildung in der
zweikernigen Endzelle. 2 Ein Zellkern nach der Schnallenausbuchtung
gerückt. 3 Konjugierte Kernteilung. 4 Schnallenzelle und Stielzelle
abgegrenzt von der Basidienanlage. 5 Fusion dieser beiden Zellen.
6 Die beiden Kerne der Basidienanlage zu einem vereinigt. 7 Junge
Basidie mit den vier Basidiosporen, oben mit vier Sterigmenanlagen
(eine davon verdeckt). Nach H. KNIEP.
Neuere Untersuchungen, besonders von KNIEP, ergaben,
daß wohl bei den meisten Hymenomyceten die aus den Basidiosporen
hervorgehenden Myzelien aus einkernigen Zellen bestehen, daß
aber dann früher oder später schon vor und unabhängig von der
Fruchtkörperbildung weiterhin Zweikernigkeit sich einstellt,
daß die Kernpaare in den Zellen sich konjugiert weiterteilen
und daß diese Paarkernigkeit mit der eigenartigen Bildung von
Myzelschnallen Hand in Hand geht bis zu den Anlagen der
Basidien. Nur wenige Hymenomyceten haben vielkernige Hyphenzellen
und erst kurz vor der Basidienbildung paarkernige. Die
Myzelschnallen entstehen sowohl an den langzelligen vegetativen
Hyphen als auch an den kürzeren und dickeren Hyphen, aus denen die
Hymenien hervorgehen, in gleicher Weise als seitliche, hakenförmig
nach unten gerichtete, kurze Ausstülpungen, je eine etwa in der
Mitte einer endständigen Hyphenzelle (Fig. 412, 1). Hierauf
wandert der eine der beiden Kerne zum Teil in diese Ausstülpung
ein und teilt sich hier (2, 3); gleichzeitig teilt sich
auch der andere Kern, zwischen dessen Tochterkernen darauf eine
Querwand dicht unterhalb der Schnalle entsteht. In die Endzelle
des Fadens wandert auch der obere Tochterkern des Schnallenkerns
ein, während der untere in der Schnalle[S. 405] selbst verbleibt. Diese
grenzt sich durch eine Querwand von der Endzelle ab, fusioniert mit
der tieferstehenden Zelle und läßt ihren Kern in sie übertreten.
Durch diese Schnallenbildung erhalten also beide Fadenzellen ihre
neuen Kernpaare als Abkömmlinge des ursprünglichen Kernpaares.
Vielleicht liegt die Bedeutung dieses umständlichen Vorganges in
der Sicherung einer solchen Verteilung der Schwesterkerne auf zwei
getrennte Fadenzellen. Die zweikernige Endzelle wird zur Anlage
der Basidie. Beide Kerne kopulieren miteinander und teilen sich
dann in vier Sporenkerne (Fig. 412, 5, 6, 7). Wie bei den
Askomyceten vollzieht sich also auch bei den Basidiomyceten der
Befruchtungsvorgang in zwei Stufen, Zellkopulation bei der ersten
Schnallenbildung und Kernkopulation in den Basidien.
Die Schnallenbildung entspricht nach KNIEP genau der
Hakenbildung vieler Askomyceten, bei denen sie aber nur auf die
askogenen Hyphen beschränkt ist.
KNIEP hat den Nachweis erbracht, daß die Myzelien wohl
der meisten Hymenomyceten heterothallisch sind, wie
bei Mucorineen (S. 382). Schnallenbildung und als Folge davon
Paarkernigkeit, ferner Fruchtkörperbildung tritt nur ein, wenn
sexuell verschieden differenzierte Myzelien miteinander, jedenfalls
durch Anastomosen, in Verbindung treten. Die beiden Kerne einer
jeden Myzelzelle entstammen dann verschiedenen Myzelien. Diese
sexuelle Differenzierung wird nach KNIEPs Feststellung bei
Aleurodiscus polygonius bei der ersten Reduktionsteilung in der
Basidie vollzogen, so daß von ihren vier Basidiosporen zwei das
eine Geschlecht (+), zwei das andere (–) erhalten.
Nicht alle Basidiomyceten sind getrenntgeschlechtlich. Es gibt
auch solche mit zweikernigen homothallischen Myzelien, die
Fruchtkörper mit normalen Basidien bilden. Die Paarkernigkeit der
Hyphenzellen stellt sich dann an ein und demselben Einspor-Myzelium
ein, bei Hypochnus nach KNIEP bereits durch Teilung des
Basidiosporenkerns.
Das Paarkernmyzel bezeichnet die diploide Phase, die haploide
beginnt in der Basidie, endigt unbestimmt mit dem Beginn der
Schnallen, bei Hypochnus aber bereits mit dem einkernigen
Anfangsstadium der Basidiospore. Der Generationswechsel ist also
infolge Unterdrückung der Sexualorgane nicht mehr bei allen
Vertretern in gleicher Weise ausgeprägt.
Übrigens gibt es auch einzelne sexuell reduzierte Basidiomyceten
(z. B. Coprinus fimetarius), bei denen die Schnallenbildung
unterbleiben kann und die Zellen der aus den Sporen hervorgehenden
Myzelien stets einkernig sind. So entstehen haploide Fruchtkörper,
in denen die Kernverschmelzung in der einkernigen Basidienanlage
ausfällt, trotzdem aber Basidiosporen erzeugt werden.
Die meisten Hymenomyceten leben mit ihrem Myzelium im humushaltigen
Boden der Wälder oder im faulenden Holz, in absterbenden
Baumstämmen und erheben ihre Fruchtkörper, die gemeiniglich
als Schwämme bezeichnet werden, über die Oberfläche des
Substrats. Das Myzel der im Boden vegetierenden Formen breitet
sich an der Peripherie immer weiter aus und nimmt eine von Jahr zu
Jahr immer größer werdende, ringförmige Zone ein. Infolgedessen
erscheinen dann auch die jährlich hervorkommenden Schwämme bei
ungestörter Entwicklung in Ringen angeordnet, die vom Volk
Hexenringe genannt werden. Weniger zahlreiche Hymenomyceten
vegetieren parasitisch in der Rinde und dem Holze von Holzgewächsen.
Die fortschreitend reichere Gestaltung der mannigfachen
Basidienfruchtkörper dient zur Einteilung der Hymenomyceten.
1. Die Thelephoraceen erzeugen einfach gestaltete
Fruchtkörper meist von korkig lederartiger Beschaffenheit; sie
bilden auf Baumstümpfen teils flache Krusten von rundlichem oder
getapptem Umriß, und das Basidienhymenium überzieht die glatte
Oberseite dieser Krusten; oder die flachen Fruchtkörper heben sich
in horizontaler Richtung vom Substrat ab, bilden halbkreisförmige,
oft dachziegelartig gruppierte Hüte, und das Hymenium ist auf
ihrer Unterseite entwickelt, so bei dem an Laubholzstämmen
häufigen Stereum hirsutum. Eigenartige trichterförmige schwarze
Fruchtkörper hat Craterellus cornucopioides, die Totentrompete,
ein wohlschmeckender Speisepilz.
2. In der Gruppe der Clavariaceen haben die von dem
Hymenium an ihrer Oberfläche bedeckten Fruchtkörper die Form von
fleischigen Keulen oder sind korallenartig verzweigt.
Die größeren, reich verästelten Formen liefern minderwertige
Speiseschwämme, so Clavaria flava mit gelbem Fruchtkörper, und
Clavaria Botrytis (Fig. 413), von blaßrötlicher Farbe, beide
als Hahnenkamm oder Korallenschwamm bezeichnet, ferner der krause
Ziegenbart, Sparassis crispa, auf Sandboden in Nadelwäldern
auftretend, mit blattförmigen, reich verzweigten Ästen, bis 1⁄2 m
im Durchmesser erreichend.
[S. 406]
3. Die Hydnaceen oder Stachelschwämme besitzen
Fruchtkörper mit stachelartigen Auswüchsen, auf denen die
Hymenien als Überzug entwickelt werden. Die einfachsten Hydneen
haben krustenförmige Fruchtkörper, auf deren Oberseite diese
Stacheln stehen, andere dagegen gestielte, hutförmige, fleischige
Fruchtkörper, die auf der Hutunterseite die abwärts gerichteten
Stacheln tragen. Zu letzteren gehören verschiedene eßbare Schwämme,
so Hydnum imbricatum, der Habichtsschwamm, in Kiefernwäldern,
mit braunem, oben schwärzlich beschupptem, bis 15 cm breitem
Hut, ferner Hydnum repandum, der Stoppelschwamm (Fig. 414) mit
gelblichem Hut.
Fig. 413. Clavaria Botrytis. Verkl.

Fig. 414. Hydnum repandum. Verkl.
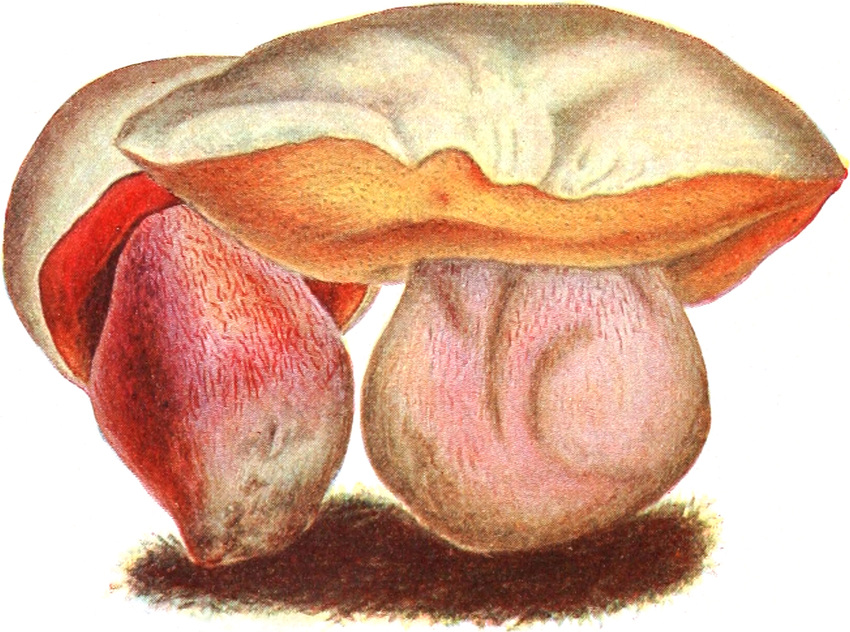
Fig. 415. Boletus Satanas, Satanspilz. 1⁄2 nat. Gr. Nach
KROMBHOLZ. Giftig. Verkl.
4. Bei den artenreichen Polyporaceen oder Löcherschwämmen
besitzen die gestielten oder sitzenden Hüte in der Regel
auf ihrer Unterseite röhrenförmige Vertiefungen oder
tiefgewundene Gänge oder dicht zusammenstehende Röhrchen, und
das Basidienhymenium ist in diesen auf der Innenseite
entwickelt. Hierher gehört die Gattung Boletus, Röhrling, mit
großen, fleischigen, auf Waldboden auftretenden, gestielten Hüten,
deren Unterseite mit einer dicken Schicht von feinen Röhrchen
bekleidet ist. Die Arten sind teils vorzügliche Speisepilze,
so u. a. B. edulis, der Steinpilz, B. badius, der Maronenpilz,
B. elegans, der schöne Röhrling, und B. luteus, der Butterpilz,
teils aber sehr giftig, wie der Satanspilz, B. Satanas
(Fig. 415), mit fahlweißlichem, bis 20 cm breitem Hut, gelb bis
purpurrot gefärbtem und mit roter Netzzeichnung versehenem Stiel
und erst blutroter, dann orangeroter Hutunterseite. Wegen seines
bitteren Geschmacks ungenießbar ist der[S. 407] dem Steinpilz ähnliche
Gallenröhrling, B. felleus, der sich durch hellrosa Röhren
statt weißer von ihm unterscheidet. Von den zahlreichen Arten der
Gattung Polyporus ist offizinell der südeuropäische, an
Lärchen vorkommende Polyporus officinalis, dessen unregelmäßig
knollige, weiße Fruchtkörper einen bitteren harzartigen Bestandteil
enthalten. Verwandt mit Polyporus ist der ebenfalls offizinelle
Feuer- oder Zunderschwamm, Fomes fomentarius. Sein Myzelium
lebt parasitisch in Laubbäumen, besonders Buchen, und erzeugt
große konsolförmige, bis 50 cm breite und 35 cm dicke, mehrjährige
Fruchtkörper mit harter, grauer Rinde und wergartiger, den
Zunderschwamm liefernder Innenmasse. Auf der Unterseite stehen die
engen Hymeniumröhren in übereinander lagernden Jahresschichten.
Der ähnliche Fomes igniarius, unechter Zunderschwamm (Fig. 416),
besonders an Eichen auftretend, ist rotbraun gefärbt, viel härter
und liefert nur einen schlechten Zunder.
Fig. 416. Fomes igniarius. Durchschnitt durch einen
mehrjährigen Fruchtkörper mit Zuwachszonen. a Befestigungsstelle des
halbkreisförmigen Hutes. 1⁄2 nat. Gr.

Fig. 417. Psalliota campestris. Champignon, rechts
junger Fruchtkörper. Verkleinert.
Manche Polyporeen sind sehr schädliche Parasiten der
Waldbäume, so Fomes annosus, an Kiefern und Fichten. Eine
sehr schädliche saprophytische Art ist Merulius lacrymans,
der Hausschwamm[390], dessen Myzelium in feuchtem Bauholz,
in erster Linie in Nadelholz, vegetiert und dieses zerstört; an
der Oberfläche des Holzes und an Mauerwerk bildet sein Myzel große
grauweiße Watten mit derben, sich verzweigenden Strängen, die
neben gewöhnlichen Hyphen siebröhrenartige Hyphen zur Leitung von
Wasser und Nährstoffen und verdickte Faserhyphen enthalten. Die
Hyphen zeichnen sich durch schnallenförmige Verbindungen ihrer
aufeinanderfolgenden Zellen aus. Die aus Ritzen hervorkommenden,
im jungen Zustand weißen, unregelmäßig lappigen Fruchtkörper
tragen das Hymenium auf ihrer grubigen Oberfläche und sind bei
der Reife mit rostbraunen Sporenmassen bedeckt. Trockenlegung und
gute Durchlüftung der infizierten Räume ist das sicherste Mittel
zur Bekämpfung des Hausschwammes. Nahe verwandt mit ihm ist der in
Wäldern vorkommende Merulius silvester.
5. Als artenreichste Gruppe sind schließlich die
Agaricaceen oder Blätterschwämme zu nennen, deren Hüte auf
der Unterseite radiale Lamellen tragen, die mit dem Hymenium
überzogen sind. Die Fruchtkörperanlagen sind rundliche Körper, in
denen sich bald der Stiel und der Hut differenzieren. Bei vielen
Blätterpilzen spannt sich eine dünne Hyphenhaut (Velum)
in dem jungen Fruchtkörper vom Hutrand quer zum Stiel; sie
reißt später ein und kann als ringförmiger, festsitzender oder
verschiebbarer Hautlappen (Annulus) am Stiele verbleiben
(Fig. 417). Manche Arten besitzen auch eine oben am Stiel hängende
Haut (Manschette, Armilla), die sich unter dem Hut
von der Stieloberfläche ablöst. Bei Amanita (Fig. 418–420) und
Verwandten ist eine gemeinsame Hülle vorhanden, die am Grunde
des Stiels als Volva und auf dem Hut in weißen Fetzen
zurückbleibt.
Manche Blätterschwämme Mitteleuropas werden als vorzügliche
Speiseschwämme geschätzt, so vor allem der auch in Kultur
genommene Champignon oder Egerling, Psalliota campestris (Fig.
417), mit weißlichem Hut und erst weißen, dann rosenroten, zuletzt
braunschwarzen Lamellen; ferner der Pfifferling oder Eierschwamm,
Cantharellus cibarius, mit dottergelbem, kreiselförmigem Hut;
der Reizker, Lactaria deliciosa, mit rotgelbem Hut und rotgelbem
Milchsaft in besonderen Hyphenschläuchen; Lactaria volema, der
Brätling, mit rotbraunem Hut, dickem Stiel und weißem Milchsaft;
Tricholoma equestre, der Grünling, mit braungelber Hutoberseite,
sonst schwefelgelb gefärbt; der Parasolschwamm, Lepiota procera,
mit weißem braunbeschupptem Hut; der Kaiserling, Amanita
caesarea, mit orange[S. 408]farbigem, oben anfangs einige dicke lose
weiße Hautfetzen tragendem Hut und gelben Lamellen. Eßbar sind
auch die bräunlichen Fruchtkörper des Hallimasch, Armillaria
mellea, der als sehr verderblicher Baumparasit an Laub- und
Nadelhölzern auftritt; sein Myzel zeichnet sich aus durch Bildung
photogener Substanzen, die das Leuchten des infizierten
Holzes im Dunkeln bedingen[293]. Bemerkenswert sind ferner die
als Rhizomorphen bezeichneten Dauerzustände seines Myzels,
schwarze verzweigte Stränge unter der Rinde oder zwischen den
Wurzeln der Nährbäume.
Verhältnismäßig gering ist die Zahl der giftigen Blätterschwämme.
In erster Linie sind zu nennen der Fliegenschwamm, Amanita
muscaria (Fig. 418), mit weißen Lamellen; die oft mit dem
Champignon verwechselten, tödlich wirkende Gifte enthaltenden
Knollenblätterschwämme, nämlich die besonders giftige, auf der
Hutoberfläche heller oder dunkler grünlich gefärbte A. phalloides
(Fig. 419), die rein weiße A. verna und die gelblichweiße
A. mappa (Fig. 420), alle drei mit weißen Lamellen und mit
dickknolligem Stielfuß, der bei den zwei erstgenannten Arten eine
große gelappte Scheide trägt, bei der dritten Art aber nur kantig
gesäumt ist; der Speiteufel, Russula emetica, mit rötlichem Hut
und weißen Lamellen; der Giftreizker, Lactaria torminosa, mit
rotbraunem, zottigem Hut und weißem Milchsaft.

Fig. 418. Amanita muscaria, Fliegenpilz. 1⁄2 nat. Gr. —
Giftig.

Fig. 419. Amanita phalloides. Grünlicher
Knollenblätterschwamm. 1⁄2 nat. Gr. — Sehr giftig.

Fig. 420. Amanita mappa. Gelber Knollenblätterschwamm.
1⁄2 nat. Gr. — Giftig.
Ökologisch sehr interessant ist die südbrasilianische Agaricinee
Rozites gongylophora,[S. 409] deren Myzel nach A. MÖLLER
von den Blattschneiderameisen in ihren Nestern auf
herbeigeschleppten und zerkauten Blattstückchen regelrecht
kultiviert wird. Das Myzel erzeugt hier kugelige, dicht
mit Plasma erfüllte Anschwellungen seiner Hyphenenden, die
sog. Kohlrabiköpfchen, welche den Ameisen als Nahrung dienen.
Die Ameisen verhindern die Entwicklung der Konidien, die als
Nebenfruktifikation dem Pilze eigen sind und nur bei Kultur des
Myzels ohne Ameisen gebildet werden, erhalten also den Pilz
in ihren Nestern stets in seinem vegetativen Zustande. Die
Fruchtkörper finden sich nur selten auf den Nestern; sie haben in
ihrer Form Ähnlichkeit mit denen des Fliegenschwamms, zu dessen
Verwandtschaft Rozites gehört. Im tropischen Asien wird nach
HOLTERMANN das Myzel von Agaricus Rajab von Termiten in
ihren Nestern kultiviert[391].
Offizinell: Fomes fomentarius, liefert Fungus
Chirurgorum (Pharm. germ. austr.). — Polyporus officinalis
(= Boletus laricis) Lärchenschwamm liefert Agaricus albus
(Pharm. helv.) oder Fungus Laricis (Pharm. austr.),
Agaricinum (Pharm. germ.) und Acidum agaricinum
(Pharm. helv.).
7. Ordnung. Gasteromycetes, Bauchpilze[373].
Die Gasteromyceten haben geschlossene
Fruchtkörper, die sich erst nach der Sporenreife öffnen,
wobei die als Peridie bezeichnete äußere
Hyphenrinde in charakteristischer Weise aufplatzt. Die von der
Peridie umschlossene sporenbildende Innenmasse wird insgesamt als Gleba bezeichnet. Die Gleba ist von zahlreichen
Kammern durchsetzt, die von dem Basidienhymenium ausgekleidet werden,
oder sie ist erfüllt von locker verflochtenen Hyphen, deren Zweige in
Basidien endigen.
Das Myzel lebt saprophytisch im Humusboden der Wälder und Wiesen.
Die Fruchtkörper aber erheben sich über die Oberfläche. Nur die
Gruppe der Hymenogastreen besitzt unterirdische, trüffelähnliche
Fruchtkörper.
Verhältnismäßig einfach gebaut ist der Fruchtkörper von
Scleroderma vulgare, dem Hartbovist, dessen breitkugelige, meist
5 cm dicke Basidienfrucht eine weißlich braune, lederartige, am
Scheitel rissig gefelderte Peridie besitzt (Fig.421, 1). Die im
reifen Zustande schwarze Gleba ist von zahlreichen sterilen Adern
durchzogen und erfüllt mit birnförmigen Basidien, die vier sitzende
kugelige Sporen tragen (Fig. 421, 2). Der Hartbovist gilt als
giftig und wird zuweilen mit Trüffeln verwechselt.
Fig. 421. 1 Scleroderma vulgare, Fruchtkörper. 2
Basidien aus demselben. Nach TULASNE. 3 Lycoperdon gemmatum.
4 Geaster granulosus. 1, 3, 4 in nat. Gr. 2 vergrößert.
Die Gattungen Bovista und Lycoperdon (Fig. 421, 3), Boviste
und Stäublinge, haben ebenfalls kugelige, bei letzterer Gattung
auch gestielte, anfangs weißliche, später bräunliche Fruchtkörper.
Sie erreichen bei dem Riesenbovist Lycoperdon Bovista sogar bis
1⁄2 m Durchmesser. Ihre Peridie ist in Form von zwei Schichten
entwickelt, von denen sich die äußere bei der Reife gewöhnlich
ablöst und die innere sich am Scheitel öffnet. Die Kammern der
Gleba werden hier von einem regelmäßigen Hymenium aus Basidien
ausgekleidet. Eine Eigentümlichkeit der Boviste besteht ferner
in dem Auftreten von Capillitiumfasern in den Kammern der
Gleba; das sind braune dickwandige, verästelte Hyphen,[S. 410] welche die
Auflockerung der Sporenmasse besorgen. Die jungen, noch weißen
Boviste sind eßbar, die reifen dagegen enthalten Harnstoff.

Fig. 422. Ithyphallus impudicus.
1⁄2 nat. Gr.
Bei Geaster (Fig. 421, 4), Erdstern, ist die Peridie der
Fruchtkörper ebenfalls als doppelte Hülle ausgebildet. Die äußere
Hülle breitet sich in sternförmigen Lappen aus, die innere öffnet
sich am Scheitel mit einem Loch.
Die höchste Ausbildung erreichen die Phallaceen[392], als deren
bekanntester Vertreter Ithyphallus impudicus, die Stinkmorchel,
in Wäldern Deutschlands einheimisch, zu nennen ist. Dieser Pilz
galt vielfach als giftig. Früher wurde er zu Gichtsalben verwendet.
Sein Fruchtkörper ähnelt den echten, zu den Diskomyceten gehörenden
Morcheln, hat aber eine ganz andere Entwicklungsgeschichte. Er
ist etwa 15 cm hoch, hat einen langen, innen hohlen, netzförmig
gekammerten, weißen Stiel und einen glockenförmigen, mit
der braungrünen, im reifen Zustand zu Schleim verflüssigten,
sporenhaltigen Glebamasse überzogenen Hut, unter dem
am Stiel noch die Reste eines zarthäutigen Schleiers
(Indusium) haften, der den Hut auf seiner Innenseite anfangs
bedeckte (Fig. 422). Der junge Fruchtkörper bildet einen eiförmigen
weißen Körper (Hexenei oder Teufelsei genannt) und wird von
einer doppelwandigen Hülle mit gallertartiger Mittelschicht ganz
umschlossen. Im Innern der Hülle oder Peridie (auch Volva
genannt) differenziert sich das Hyphengewebe in den axilen Stiel
und in den glockenförmigen Hut. Im Umkreise des Hutes wird die
Gleba ausgebildet. Bei der Reife streckt sich der Stiel rasch
in die Länge, sprengt dabei die an seiner Basis als Scheide
zurückbleibende Hülle und hebt den glockenförmigen Hut mit der
Gleba empor. Letztere erzeugt aasartig riechende Stoffe und lockt
dadurch Aasinsekten an, die die Sporen verbreiten.
Die Flechten sind symbiotische Organismen; sie bestehen aus
Fadenpilzen, und zwar aus Askomyceten, nur in ganz vereinzelten
Fällen aus Basidiomyceten, die mit einzelligen oder fädigen Algen,
Cyanophyceen oder Chlorophyceen, gemeinsam vegetieren und so einen
eigenartigen zusammengesetzten Thallus, ein Konsortium, bilden.
Die Flechtenalgen und die Flechtenpilze sind im natürlichen System in
die Gruppen der nächstverwandten Algen und Pilze einzureihen. Doch
besitzen die Flechten untereinander so viel Übereinstimmendes in Bau
und Lebensweise und haben sich als Konsortien phylogenetisch weiter
entwickelt, daß sie zweckmäßiger als besondere Klasse behandelt werden.
Der Pilz ernährt sich von den durch die assimilierenden Algenzellen
erzeugten organischen Stoffen; er umspinnt mit seinen Hyphen die
Algen (Fig. 423), kann aber auch Haustorien in sie hinein entsenden
und als Parasit sogar ihren Inhalt aufzehren, auch als Saprophyt
abgestorbene Algenzellen ausnutzen. Andererseits gewährt der Pilz
den Algen bestimmte Vorteile, liefert ihnen die anorganischen
Stoffe und Wasser, wahrscheinlich auch organische Verbindungen.
Der größere Vorteil in dieser mutualistischen Symbiose mag
auf seiten des Pilzes liegen, vor allem bei solchen Flechten,
die auf sterilem, jeglicher organischen Stoffe barem Gestein
wachsen, während der Pilz bei Humusboden- oder Baumrindenbewohnern
wenigstens einen Teil seiner Nahrung saprophytisch aus dem Substrat
beziehen kann. Wenn[S. 411] auch die Algen in manchen Fällen vom Pilz
stärker ausgenützt werden, so erfahren sie doch eine ausgiebige
Vermehrung, und beide können infolge ihres Zusammenlebens auch
an Orten gedeihen, wo weder Pilz noch Alge für sich allein leben
könnte.
Den Flechten eigene Stoffwechselsekrete sind die zahlreichen, nur
den Gallertflechten fehlenden Flechtensäuren, deren Bildung
durch das chemische Zusammenwirken von Pilz und Alge bedingt ist
und deren Ablagerung an der Oberfläche der Hyphenmembranen in Form
von Kristallen oder Körnchen erfolgt. Ihre Rolle als Schutzmittel
gegen Tierfraß (Schnecken) scheint nach ZOPF nicht von
allgemeiner Bedeutung zu sein[394].
Fig. 423. Cetraria islandica. Querschnitt durch den
Thallus. or Rindenschicht der Oberseite, ur der Unterseite, m
Markschicht mit den grünen Algenzellen von Cystococcus humicola. Vergr. 272.
Fig. 424. Parmelia physodes. Soral (a) und Soredium
(b). Nach BITTER und NIENBURG.
Die Flechten sind in großer Artenzahl über die ganze Erde
verbreitet; selbst in den Polarländern und auf den höchsten
Berggipfeln dringen sie viel weiter vor als die Moose. Ihre
Hauptentwicklung erfahren sie in feuchten Gebirgsregionen, wo
sie oft in geselliger Vegetation den Boden, die Felsblöcke und
Baumstämme überziehen oder in Form langer Bärte massenhaft von den
Baumästen herabhängen. In der Arktis bilden sie auf trockenem Boden
ausgedehnte Flechtentundren.
Die einfachsten Flechten sind die Fadenflechten; sie bestehen
aus Algenfäden, die der Länge nach von Pilzhyphen umsponnen sind. Als
Beispiel sei Ephebe pubescens genannt, deren vielästige verzweigte
Fäden an feuchten Felsen in Form schwärzlicher Filzlager auftreten.
Sodann unterscheidet man Gallertflechten, mit gallertigem,
laubartigem Lager. Ihre Algen sind Chroococcaceen und Nostocaceen mit
aufquellenden Membranen. In der Algengallerte verlaufen die Pilzhyphen.
Von einheimischen Gattungen gehört z. B. Collema hierher.
Sowohl bei den Faden- als Gallertflechten sind Algen und Pilzhyphen
gleichmäßig im Thallus verteilt, und dieser wird daher als
ungeschichtet oder homöomer bezeichnet. Die Algen bestimmen hier
in erster Linie seine Form.
Die übrigen Flechten weisen dagegen einen geschichteten oder
heteromeren Thallus auf. Die Flechtenalgen, unter denen die
einzellige Protococcacee Cystococcus humicola die häufigste, von
fadenförmigen die zu den Ulotrichales gehörige Trentepohlia umbrina
zu nennen ist, treten hier in Schichten auf, die von einer
aus dicht verflochtenen Pilzhyphen bestehenden Rindenschicht
bedeckt werden (Fig. 423). Man unterscheidet unter den heteromeren
Flechten Krustenflechten, deren Thallus in Form von Krusten
an Baumstämmen, an Felsen oder auf dem Erdboden festgewachsen ist
oder mittels Pilzhyphen etwas in das Substrat eindringt; ferner
Laubflechten (Fig. 428), deren Thallus laubartig, klein- oder
großlappig ist und auf der Unterseite entweder nur in der Mitte
oder bis auf die freien Ränder mittels rhizoïdartiger Pilzhyphen
(Rhizinen) angewachsen ist; endlich Strauch[S. 412]flechten (Fig. 425,
427), mit verzweigtem, fadenförmigem oder bandförmigem, an der Basis
angeheftetem, zuweilen auch frei auf dem Substrat liegendem Thallus.
Das Wachstum der Flechtenthalli ist im allgemeinen ein recht langsames.
So beträgt der jährliche Zuwachs an den Thalluslappen gewisser
Laubflechten nur wenige Millimeter; Parmelia furfuracea erreicht bei
günstiger Belichtung in 10 Jahren nur einen Durchmesser von 31 × 60
mm[395].
An den natürlichen Standorten entwickeln sich die Flechtenpilze nur
dann aus ihren Sporen weiter, wenn sie die ihnen zusagenden Algenzellen
zur Verfügung haben. Nur für wenige Flechten ist festgestellt, daß
ihr Pilz auch ohne Algen in der Natur existenzfähig ist, so für die
tropische Cora pavonia (Fig. 433), deren Pilz zu den Basidiomyceten
gehört und auch algenfreie Fruchtkörper erzeugen kann, die denen der
Pilzgattung Thelephora gleichen. Wohl aber ist es gelungen, aus den
Sporen gewisser flechtenbildender Askomyceten unter Zufuhr geeigneter
Nährlösung Myzelien zur Entwicklung zu bringen.
Fig. 425. Usnea florida. ap Apothecium. Nat. Gr.
Fig. 426. Cetraria islandica. ap Apothecium. Nat. Gr.
— Offizinell.
Viele Flechten vermehren sich vegetativ dadurch, daß Teile des Thallus
sich lostrennen und wieder mit Rhizinen festsetzen. Die meisten
heteromeren Flechten besitzen ferner in der Bildung von Soredien
ein ausgezeichnetes Mittel vegetativer Vermehrung: Kleine Gruppen von
sich teilenden Algenzellen werden dicht von Myzelfäden umsponnen, lösen
sich los und bilden Körperchen, die unter Aufreißen von Thallusrinde
als staubartige Masse frei werden, um durch den Wind verbreitet,
anderswo wieder zu einer Flechte heranzuwachsen. Häufig entstehen
die Soredien am Thallus in scharf umschriebenen Brutstätten, den
Soralen (Fig. 424, 427).
Im Flechtenthallus fruktifizieren nur die Flechtenpilze, nicht
aber die stets vegetativ bleibenden Flechtenalgen.
Nur bei einigen Gattungen (Endocarpon) finden sich besondere
klein ausgebildete Algenzellen auch in den Früchten vor, werden mit
den Sporen gemeinsam ausgeworfen und von den Keimschläuchen des
Flechtenpilzes alsbald umsponnen.
1. Ascolichenes. Nur wenige Flechtengattungen haben
krugförmige Perithecien; ihre Pilze gehören daher zu den
Pyrenomyceten, so die Laubflechte Endocarpon, die Krustenflechte
Verrucaria. Die meisten Gattungen aber besitzen schüssel-
oder scheibenförmige Apothecien, die wie die Fruchtkörper der
Diskomyceten gebaut[S. 413] sind. Von Strauchflechten gehört hierher
als eine der häufigsten Arten die an Baumstämmen festsitzende
Usnea florida, die Bartflechte, mit großen, am Rande bewimperten
Apothecien (Fig. 425); die breit bandförmige, verzweigte, an
Bäumen wachsende Ramalina fraxinea; ferner die an Küstenfelsen
der warmen Zone verbreiteten Roccella-Arten mit gabelig
verzweigtem, drehrundem oder bandförmigem Thallus (Fig. 427).
Eine Mittelstellung zwischen Strauch- und Blattflechten nimmt
die auf den Gebirgen und im Norden der nördlichen Hemisphäre
weitverbreitete offizinelle Cetraria islandica, das
isländische Moos (Fig. 426), ein, mit gegabelten, blattartigen
Thalluslappen, welche braun, auf der Unterseite weißlich
gefärbt sind und die Apothecien an ihren Rändern tragen. Zu
den Laubflechten gehören die zahlreichen, an Bäumen und Felsen
wachsenden Arten von Parmelia (Fig. 428). Eine eigenartige
Krustenflechte ist die Schriftflechte, Graphis scripta, deren
grauweißer Thallus auf Rinde, besonders von Buchen, lebt und deren
Apothecien die Form von schwarzen schmalen, strichförmigen oder
gegabelten, an Schriftzüge erinnernden Rinnen haben.
Fig. 427. Roccella tinctoria DC. Kanarische Inseln. Mit
randständigen Soralen. Nach WIESNER, Rohstoffe.
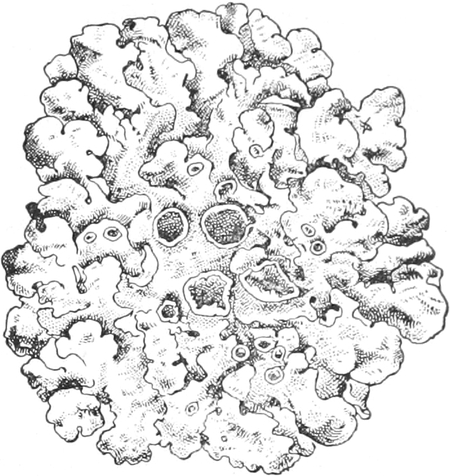
Fig. 428. Parmelia acetabulum, an Bäumen. Nach
REINKE.
Eine besondere Entwicklung erfährt der Flechtenthallus bei der
vielgestaltigen erdbewohnenden Gattung Cladonia. Auf einem aus
horizontalen, dem Substrat aufsitzenden, gekerbten Schüppchen
bestehenden primären Thallus erheben sich vertikale Gebilde
(Podetien) von sehr verschiedener Gestalt und Größe. Sie sind bei
manchen Arten, so bei Cladonia pyxidata, der Becherflechte, und
bei Cladonia coccifera (Fig. 429) gestielt kreiselförmig und
tragen am Becherrand oder seinen Aussprossungen, die bei ersterer
Art braunen, bei letzterer roten rundlichen Apothecien. Bei anderen
Arten sind die Podetien zylindrisch, einfach oder gegabelt; bei
Cladonia rangiferina, der Renntierflechte, die über die ganze
Erde verbreitet ist und rasenbildend in den nordischen Tundren
auftritt, sind die Podetien (Fig. 430) zierlich verästelt; ihr
Primärthallus geht frühzeitig zugrunde.
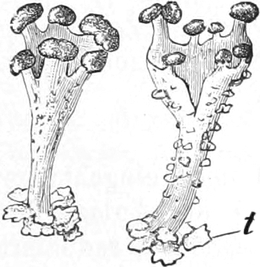
Fig. 429. Cladonia coccifera. t Thallusschüppchen.
Nat. Gr.
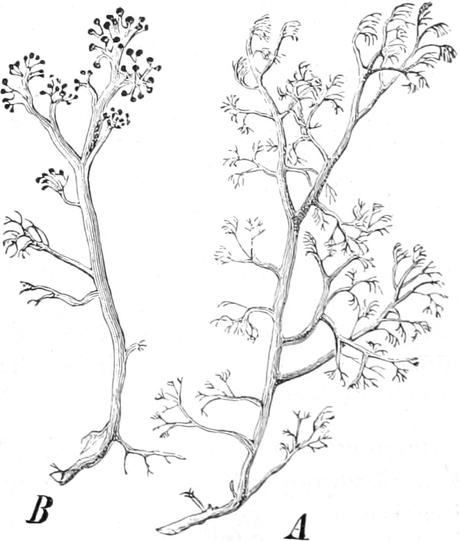
Fig. 430. Cladonia rangiferina. A Steril. B Mit
Askusfrüchtchen an den Astenden. Nat. Gr.
Die Askusfrüchte, Apothecien und Perithecien, nehmen,
wie zuerst Stahl und in neuerer Zeit besonders
BAUR[396] nachgewiesen hat, ihren Ursprung aus
befruchteten[S. 414] Karpogonen, weiblichen Sexualorganen, die
im jungen Thallus oft in sehr großer Anzahl angelegt werden. Das
Karpogon (Fig. 431) ist hier ein vielzelliger, im unteren
Teile mehrfach schraubig gewundener Faden, der sich in ein
langzelliges, mit seiner Spitze aus dem Thallus hervorragendes
Trichogyn fortsetzt. Die Zellen enthalten je einen Kern,
führen im unteren Teile des Karpogons dichteres Plasma und sind
durch Tüpfel verbunden. Abgesehen von der Vielzelligkeit erinnern
diese Gebilde an die Karpogone der Florideen. Als männliche
Sexualzellen erscheinen die in krugförmigen Behältern, den
Spermogonien (Fig. 432), erzeugten Spermatien, deren
Entwicklung auf verschiedene Art erfolgt[397]. Entweder ist die
Innenwand der Spermogoniumhöhlung mit einfachen oder verzweigten
Hyphenästen ausgekleidet, die an ihren Enden die Spermatienzellen
abgliedern (Peltigera, Parmelia), oder das Spermogonium ist
anfangs von Hyphengewebe dicht erfüllt, später aber durch dessen
Auseinanderweichen von Hohlräumen durchsetzt, aus deren Wandzellen
die Spermatien auf sehr kleinen und dünnen Stielchen hervorsprossen
(Anaptychia, Physcia, Sticta). Die Spermatien werden, in
Schleimmassen eingebettet, aus ihren Behältern entleert, kopulieren
mit den klebrigen Spitzen der Trichogyne (Fig. 431 B) und
erscheinen dann leer, ohne Kern; darauf kollabieren die Zellen des
Trichogyns, gehen später zugrunde, während die mittleren Zellen des
schraubigen Karpogons anschwellen, sich auch noch weiter teilen
und nun zu den askogenen Hyphen aussprossen, die an ihren Enden
die Asci bilden. Die vegetativen Hyphen und die Paraphysen der
Früchte entspringen aus den unter dem Karpogon befindlichen Hyphen.
Entweder nur ein oder auch mehrere Karpogone zusammen liefern eine
Frucht. Das Verhalten der Sexualkerne bedarf noch eingehender
Untersuchung. Solche Karpogone sind bereits bei manchen Gattungen
als Anlage der Früchte nachgewiesen, indessen erscheinen sie bei
einigen (Peltigera, Solorina) reduziert und besitzen kein
Trichogyn mehr; diese Flechten sind anscheinend apogam geworden.
Spermogonien werden bei ihnen überhaupt nicht mehr gebildet, oder
sind, wie bei Nephromium, deutlich in Rückbildung begriffen. Von
A. MÖLLER wurde festgestellt, daß die Spermatien auch
vegetativ auskeimen können; indessen kann dies nicht als Beweis
gegen die ursprünglich sexuelle Natur dieser Gebilde gelten, da es
sich möglicherweise um Funktionswechsel handelt.
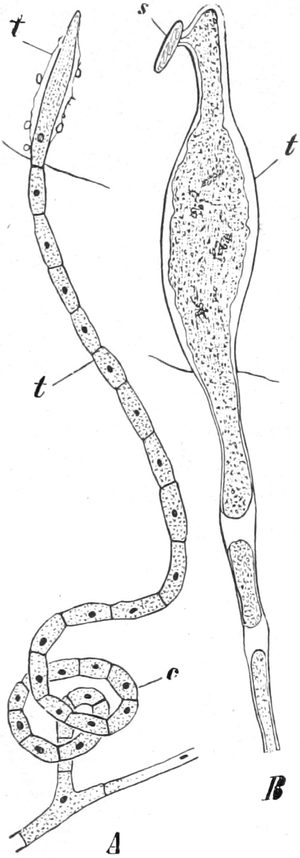
Fig. 431. Collema crispum. A Karpogon c mit
Trichogyn t. Vergr. 405. B Spitze des Trichogyns mit Spermatium
s. Vergr. 1125. Nach E. BAUR.
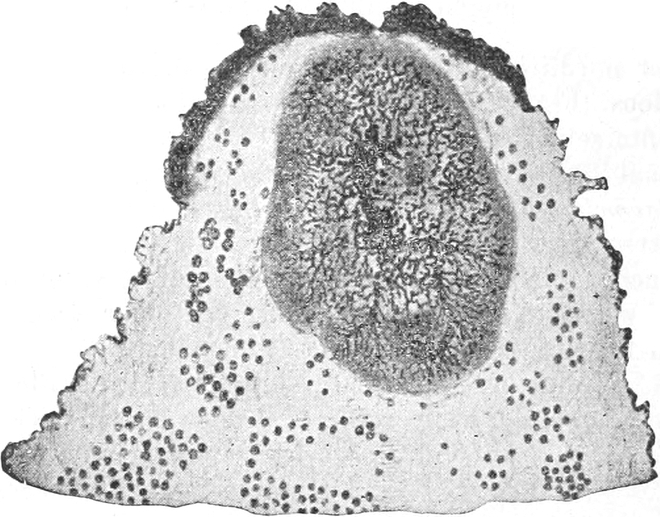
Fig. 432. Anaptychia ciliaris. Reifes Spermogonium.
Vergr. 192. Die runden Zellen im Thallusgewebe stellen die grünen
Algenzellen vor. Nach GLÜCK.
Sehr merkwürdig verhält sich nach F. BACHMANN Collema
pulposum. Die Spermatien entstehen hier im Innern des Thallus zu
wenigen gruppenweise an den Hyphen und lösen sich nicht ab; die
langen Endzellen der Trichogyne bleiben im Thallus, wachsen auf sie
zu und fusionieren mit ihnen[398].
[S. 415]
2. Basidiolichenes (Hymenolichenes)[399]. Als Typus der
Basidiolichenen ist die in den Tropen weitverbreitete, auf
dem Erdboden oder an Bäumen lebende Cora pavonia zu nennen.
Der Pilz der Cora ist eine Thelephoree (vgl. S. 405),
deren halbkreisförmige, gelappte, dachziegelartig gruppierte
Fruchtkörper auch ganz ohne Algen gefunden werden. Tritt der
Pilz in Symbiose mit einzelligen Chroococcusalgen, so resultiert
als Fruchtkörper die typische Cora pavonia (Fig. 433), die wie
ein Thelephorafruchtkörper auf ihrer Unterseite ein durch Risse
gefeldertes Basidienhymenium entwickelt. Tritt dagegen derselbe
Pilz mit den Fäden der blaugrünen Alge Scytonema in Symbiose, so
bildet sich, wenn der Pilz überwiegt, die Flechte zu strahlig
fädigen, an Baumästen abstehenden Scheiben mit dem Hymenium
auf der Unterseite aus (Dictyonema-Form), und wenn die Alge
formbestimmend ist, in Form von feinfädigen filzigen Überzügen auf
Baumrinde mit unregelmäßigen, an den vom Lichte abgewandten Stellen
des Thallus erscheinenden Hymenien (Laudatea-Form).
Fig. 433. Cora pavonia. A Von oben. B Von unten.
hym Hymenium. Nat. Gr.
Offizinell ist Cetraria islandica, Lichen
islandicus (Pharm. germ., austr., helv.), die beim Kochen
die bittere Licheningallerte liefert. Als Volksmittel wird auch
noch die Lungenflechte Lobaria pulmonaria (Muscus pulmonarius),
die stattlichste einheimische, an Bäumen wachsende Laubflechte,
gebraucht.
Die Mannaflechte, Lecanora esculenta, ist eine ursprünglich auf
Gestein lebende alpine Krustenflechte des Orients. Sie zerfällt
in Stücke, die zu kugeligen Knollen von 7–12 mm Durchmesser
heranwachsen und vom Winde weithin in die Steppen und Wüsten
Südrußlands, Kleinasiens und Nordafrikas verbreitet werden. Hier
dienen sie den Eingeborenen als Nahrungsmittel, den Tartaren
zur Bereitung von Erdbrot. Auch Cetraria islandica (Fig. 426)
kann nach Auswässerung ihrer Bitterstoffe wegen ihres reichen
Gehalts an Kohlehydraten (Flechtenstärke) als Nahrungsmittel zu
Brot und zu Gallerte verwendet werden. Cladonia rangiferina (Fig.
430) bildet die wichtigste Nahrung der Renntiere, kann auch nach
Entbitterung als Futter für Vieh und Geflügel benutzt werden und
dient in Norwegen zur Gewinnung von Alkohol. In Japan wird eine an
Stärke und Gallerte reiche felsbewohnende Laubflechte, Gyrophora
esculenta, mit kreisrundem, 3–13 cm großem, in der Mitte dem
Substrat angewachsenem bräunlichen Thallus, als wohlschmeckendes
Nahrungsmittel viel benutzt.
Einige an Flechtensäuren besonders reiche Arten werden zur
Darstellung der Farbstoffe Orseille, Persio,
französischer Purpur und Lackmus verwertet, in erster
Linie Roccella-Arten (besonders R. Montagnei, R. tinctoria
(Fig. 427), R. fuciformis und R. phycopsis, die an Küstenfelsen
wärmerer Länder wachsen, ferner die Krustenflechte Ochrolechia
tartarea im nördlichen Europa und Amerika.
Die Moospflanzen umfassen die Lebermoose (Hepaticae) und die
Laubmoose (Musci). Sie sind in ihrer äußeren Gliederung zwar
noch Thallophyten, unterscheiden sich aber von diesen durch den
eigenartigen Bau ihrer Geschlechtsorgane, der Antheridien
und Archegonien, die in ähnlicher Ausbildung auch bei den
Pteridophyten wiederkehren. Bryophyten und Pteridophyten werden daher
den Thallophyten gegenüber auch als Archegoniaten bezeichnet.
[S. 416]
Außer der sexuellen Fortpflanzung findet allgemein bei den Moosen wie
auch bei den Pteridophyten eine ungeschlechtliche durch einzellige,
mit Membran umkleidete, an die Verbreitung in der Luft angepaßte
Sporen statt. Beide Fortpflanzungsweisen wechseln stets
miteinander ab und sind auf zwei scharf geschiedene Generationen
verteilt, eine geschlechtliche (Gametophyt), welche die
Sexualorgane erzeugt, und eine ungeschlechtliche (Sporophyt),
welche die Sporen hervorbringt. Die geschlechtliche Generation geht
aus der Spore hervor, die ungeschlechtliche aus der befruchteten
Eizelle. Die Zahl der Chromosomen der Zellkerne ist im Sporophyt
doppelt so groß als im Gametophyt. Bei der Vereinigung der Sexualkerne
wird die doppelte Chromosomenzahl gewonnen, bei der Teilung der
Sporenmutterzellen dagegen die Reduktion auf die Hälfte vollzogen.
Dieser regelmäßige Generationswechsel ist charakteristisch
für alle Archegoniaten. Bei den Bryophyten stellt die Moospflanze
den haploiden Gametophyten, die gestielte Sporenkapsel den diploiden
Sporophyten dar; bei den Pteridophyten ist der Gametophyt eine kleine
Thalluspflanze, der Sporophyt hingegen ein stattlicher Kormophyt.
Fig. 434. Funaria hygrometrica. A Keimende Spore, ex
Exine. B Protonema mit Knospen kn und Rhizoïden r, s Spore.
Vergrößert. Nach MÜLLER-THURGAU.
Die die geschlechtliche Generation liefernde Spore, deren Wand
aus zwei Häuten, einer äußeren kutinisierten Exine und einer inneren
zarten Intine besteht, keimt unter Sprengung ihrer Exine zu einem
Schlauche aus, der bei den Lebermoosen alsbald an seinem vorderen
Ende zur Ausbildung der Moospflanze schreitet, während er bei den
meisten Laubmoosen zunächst ein aus verzweigten Zellfäden bestehendes
Protonema erzeugt. Dieses gliedert sich in chlorophyllführende
aufwärts wachsende Fäden und in farblose, in den Boden eindringende
Rhizoïden (Fig. 434). An dem Protonema entstehen aus seitlichen
Knospen die beblätterten Moospflanzen. Protonema und Moospflanze
stellen aber, auch wo sie in solcher Weise voneinander abgesetzt
sind, nur die eine, geschlechtliche Generation der Pflanze vor. Alle
Moose wachsen mittels Scheitelzellen heran (S. 70). Viele Lebermoose
weisen noch einen aus dichotomisch verzweigten Lappen bestehenden
Thallus (Fig. 448 u. 449) auf, der an seiner Basis oder an seiner
Unterseite mittels Rhizoïden festgeheftet ist; sie wiederholen somit
den vegetativen Aufbau mancher Algen. Bei höheren Lebermoosen und
bei allen Laubmoosen ist dagegen eine scharfe Gliederung der Pflanze
in Stämmchen und kleine Blättchen durchgeführt (Fig. 451 u. 458). An
den unteren Teilen der Stämmchen entspringen Rhizoïden; echte Wurzeln
fehlen den Moosen, die sich somit in ihrer äußeren Gestaltung nicht
über die Stufe eines reichgegliederten Thallus erheben, wie wir
ihn beispielsweise bei Sargassum unter den Braunalgen antreffen. Die
Moosstämmchen und Blätter sind von einfacher anatomischer Struktur;
sie[S. 417] werden, wenn überhaupt, nur von sehr einfachen, aus gestreckten
Zellen gebildeten Leitbündeln durchzogen. Die Sexualorgane, Antheridien
und Archegonien entspringen, in der Regel zu mehreren, bei thallösen
Formen dem Rücken des Thallus, bei den in Stengel und Blätter
gegliederten Formen auf dem Scheitel des Stämmchens oder seiner Äste.
Die Antheridien[401] (Fig. 435) oder männlichen Organe sind
auf einem mehrzelligen Stiele sitzende, kugelige oder keulenförmige
Gebilde, deren dünne, meist einschichtige Wandung zahlreiche kleine
kubische Zellen umschließt, von denen eine jede sich schließlich
in zwei Spermienzellen diagonal oder quer teilt[402]. Das Öffnen
des Antheridiums vollzieht sich bei den Laubmoosen an seiner Spitze
in einer aus einer oder mehreren Zellen bestehenden besonderen
Öffnungskappe, deren schleimhaltiger Inhalt aufquillt und so die
Kutikula aufsprengt (Fig. 440 A); bei den Lebermoosen ist die Kappe
nicht scharf abgegrenzt, die schleimhaltigen aufquellenden Wandzellen
reißen im oberen Teile des Antheridiums unregelmäßig auseinander. Nun
werden die Spermienzellen entleert, aus denen durch Verquellung ihrer
Wandung die Spermien als kurze, etwas gewundene Fäden frei werden, die
nahe am Vorderende zwei lange feine Zilien tragen (Fig. 435).
Fig. 435. Marchantia polymorpha. A Ein fast reifes
Antheridium im Durchschnitt, p Paraphysen. Vergr. 90. B Spermien.
Vergr. 600. Nach STRASBURGER.
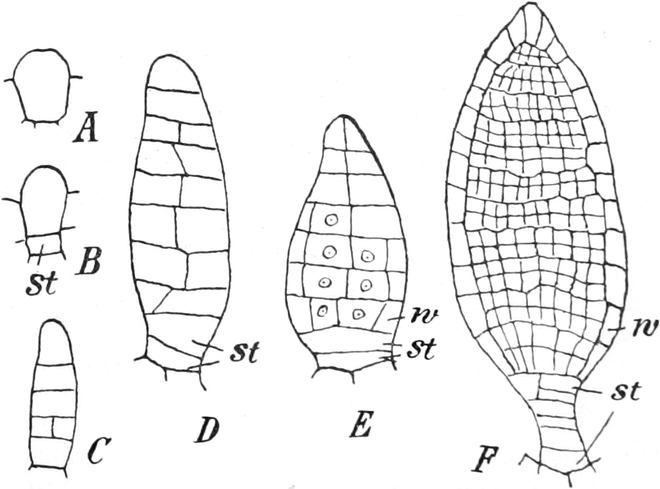
Fig. 436. Antheridiumentwicklung von Fegatella conica,
einem Marchantiaceen-Lebermoos. A Einzellige Anlage. B Die
Stielzelle st abgetrennt. C, D Querscheibenzellen abgeteilt,
die sich durch senkrechte Wände fächern. E Anlage der Wandschicht
w. F Halbreifes Stadium. A–E Vergr. 400. F Vergr. 220. Nach
BOLLETER.
Die Entwicklung des Antheridiums erfolgt durch Teilung
einer Oberflächenzelle; nur bei Anthoceros (Fig. 445) wird es
endogen angelegt. Bei den übrigen tiefer stehenden Lebermoosen
(Marchantiales) teilt sich diese Zelle in scheibenförmige
Quersegmente, die durch senkrechte Wände in je vier Zellen zerlegt
werden, worauf in diesen Quadranten durch tangentiale Wände die
peripherischen Wandzellen sich von den inneren, das spermatogene
Gewebe liefernden Zellen abteilen (Fig. 436 A–F). Bei den
höheren Lebermoosen (Jungermanniales) wird die Ausgangszelle
in drei Quersegmente zerlegt, von denen nur das oberste den
Antheridienkörper liefert, indem es sich zunächst senkrecht in
zwei Zellen teilt: dann wird in diesen durch je zwei schiefe
aufeinanderfolgende Längswände die Anlage der Wandung und die des
spermatogenen Gewebes geschieden. Bei den Laubmoosen (Musci)
hingegen baut sich das Antheridium auf aus Segmenten einer
zweischneidigen Scheitelzelle, die aus der obersten Querscheibe
durch zwei schräge Wände herausgeschnitten wird. Die einzelnen
Segmente werden dann in peripherische Wandzellen und je eine,
spermatogenes Gewebe liefernde Innenzelle zerlegt (Fig. 437
A–F).
[S. 418]
Die Archegonien (Fig. 438)[401] stellen kurzgestielte,
flaschenförmige Organe vor, deren Wandung einen Bauchteil und
einen Hals unterscheiden läßt. Der Bauchteil umschließt eine große
Zentralzelle, deren Inhalt kurz vor der Reife in die Eizelle und in
eine am Grunde des Halses gelegene Bauchkanalzelle zerfällt. An diese
schließt im Halse eine zentrale Reihe von Halskanalzellen an, deren
Zahl bei den Lebermoosen geringer (4 oder 8), bei den Laubmoosen größer
(etwa 10–30 oder noch höher) ist. Die Öffnung des Halses geschieht
dadurch, daß seine obersten schleimhaltigen Zellen aufquellen, die
Kutikula zerreißen und sich, oft in Form von vier Lappen, zurückrollen
(Fig. 440 B)[403]. Die Kanalzellen verquellen alsdann zu Schleim. Da
der Befruchtungsvorgang sich nur im Wasser vollziehen kann, so erfolgt
er bei den Landformen nur nach Benetzung durch Regen oder Tau. Die
Bewegungsrichtung der Spermien, die auf die Archegonien zusteuern und
den Hals hinab zum Ei gelangen, wird bestimmt durch besondere Stoffe,
die aus dem weiblichen Organ herausdiffundieren.
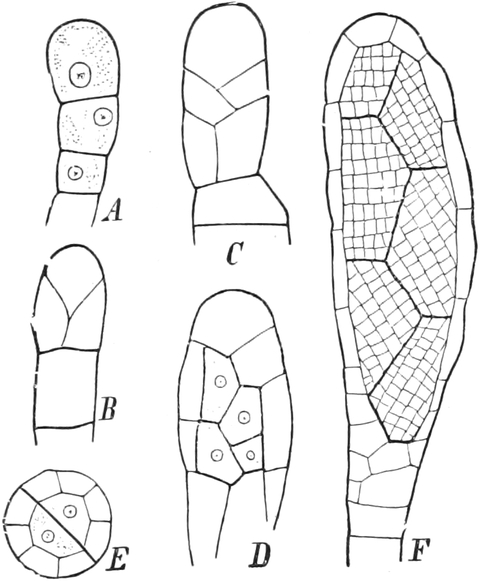
Fig. 437. Antheridiumentwicklung von Funaria
hygrometrica, einem Laubmoos. A Querteilung der Anlage. B
Bildung der Scheitelzelle aus der obersten Zelle. C Teilungen der
Scheitelzelle. D Scheidung in Wandung und Anlage des spermatogenen
Gewebes. E Desgleichen im Querschnitt. F Älteres Stadium. Nach
D. CAMPBELL.
Fig. 438. Marchantia polymorpha. A Junges,
B geöffnetes, C befruchtetes Archegonium mit achtzelligem
Keimling, k′ Halskanalzellen, k″ Bauchkanalzellen, o Ei, pr
Pseudoperianth. Vergr. 540. Nach STRASBURGER.
Fig. 439. Entwicklung des Lebermoosarchegoniums.
A (Längsschnitt) und B (Querschnitt) Teilung der oberen Zelle
durch drei Wände. C Die mittlere Zelle in Deckelzelle d und
Innenzelle i geschieden. D Die Innenzelle geteilt in Anlage
der Halskanalzellen hk und Zentralzelle c, die die Eizelle und
Bauchkanalzelle liefert. st Stielanlage. Nach GOEBEL.
Die Samenfäden der Laubmoose werden von Rohrzuckerlösung
angelockt, diejenigen des Lebermooses Marchantia von
Proteïnstoffen, außerdem bei Versuchen auch von Kalium-,
Rubidium- und Cäsiumsalzen[404].
Die Entwicklung des Archegoniums vollzieht sich aus einer
Oberflächenzelle. Bei den Lebermoosen teilt sie sich in eine den
Stiel liefernde untere Zelle und in[S. 419] eine obere Zelle, die durch
drei Längswände in drei äußere und eine mittlere zerlegt wird,
diese wiederum durch eine Querwand in eine Deckelzelle und eine
Innenzelle. Aus den äußeren Zellen geht die Wandung des Hals-
und Bauchteiles hervor, aus der Innenzelle die Halskanalzellen,
Bauchkanalzelle und Eizelle (Fig. 439). Bei den Laubmoosen
dagegen wird in der Ausgangszelle durch zwei schiefe Wände eine
zweischneidige Scheitelzelle abgeteilt, deren Segmente den Stiel
aufbauen. Dann wird die endständige zweischneidige Zelle durch
drei schräge Wände und eine Querwand in eine dreischneidige, unten
abgestutzte Scheitelzelle, eine unter dieser liegende Zentralzelle
und drei peripherische Wandzellen zerlegt. Die Zentralzelle liefert
die Eizelle, Bauchkanalzelle und Halskanalzellen, die Scheitelzelle
dagegen aus ihren Segmenten die Wandung des Halsteils und durch
Querteilung auch noch die obersten Halskanalzellen. Die Sphagnaceen
nehmen nach MELIN eine Mittelstellung ein, insofern der
Stiel wie bei den Laubmoosen, der Archegoniumkörper aber ähnlich
wie bei den Lebermoosen ohne dreiseitige Scheitelzelle gebildet
wird.
Antheridien und Archegonien sind, wie aus gelegentlich auftretenden
intermediären Gebilden hervorgeht, homologe Organe; Bauchkanal- und
Halskanalzellen würden demnach funktionslos gewordene Gametenzellen
vorstellen. Die Bauchkanalzelle ist in der Regel kleiner als
die Eizelle, kann aber zuweilen gleichwertig mit der Eizelle
ausgebildet werden. Auch können sich gelegentlich mehrere Eizellen
in einem Archegoniumbauch entwickeln, so bei Sphagneen vier oder
noch mehr.
Nach der Befruchtung teilt sich die Zygote und entwickelt sich
im Archegonium weiter zum Embryo, ohne erst einen Dauerzustand
durchzumachen (Fig. 438 C). Der Embryo wächst heran zum
Sporogon, das die ungeschlechtliche Generation vorstellt,
die aber zeitlebens mit der anderen Generation verbunden bleibt und
wie eine halbparasitische Pflanze von dieser zum Teil die zu ihrer
Entwicklung nötigen Substanzen bezieht. Das Sporogon ist ein
kürzer oder länger gestielter, rundlicher oder ovaler Sporenbehälter,
in welchem zahlreiche Sporen erzeugt werden. Allgemein entstehen die
Sporen der Moose, wie auch die der Farnpflanzen und Samenpflanzen zu
vier, in Tetraden, durch zweimalige, mit Reduktion der
Chromosomenzahl verbundene Teilung der Sporenmutterzellen, die sich
vorher voneinander loslösen und abrunden.
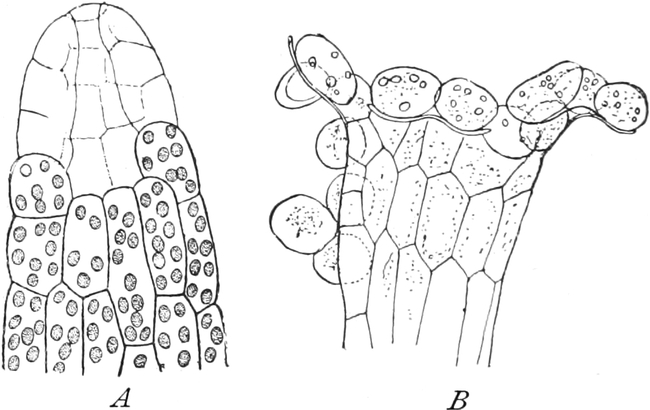
Fig. 440. A Spitze des entleerten Antheridiums von
Polytrichum mit Resten der Öffnungskappe, halbiert gezeichnet. Nach
GOEBEL. B Geöffneter Hals des Archegoniums von Mnium
undulatum. Nach ZIELINSKI.
Bei den Laubmoosen dringt der untere, als Saugorgan dienende
Teil des Embryos in das sich oft stark vergrößernde Gewebe des
Archegoniumstieles, in manchen Fällen sogar bis in das Gewebe des
Stämmchens ein. Dieses Gewebe und der mitwachsende Archegoniumbauch
bilden dann zusammen die Hülle, die schließlich von dem
sich streckenden Embryo durchbrochen wird. Der oberste, aus dem
Archegoniumbauch hervorgegangene Teil der Hülle wird dabei als
Calyptra oder Haube emporgehoben, während der untere als
Scheide den Sporogonstiel an seiner Basis umgibt. In ähnlicher
Weise wird auch die Hülle bei einem Teil der Lebermoose (so in
der Regel bei den Marchantiales) gebildet; bei den übrigen aber
wächst der Embryo mit seiner Basis in das unter dem Archegonium
befindliche Thallus- oder Stämmchengewebe mehr oder weniger
tief hinein; auch kann in bestimmten Fällen das dem Archegonium
benachbarte Gewebe zu einem beutelartigen, meist in den Boden
eindringenden Gebilde, dem Marsupium, heranwachsen, in
welches das Archegonium mit seinem Embryo hineinversenkt erscheint.
So kommt es schließlich für diesen zur Bildung eines eigenartigen
Schutz- und Ernährungsorganes.
[S. 420]
Die Entwicklung der Sporogone[405] zeigt bemerkenswerte
Verschiedenheiten. Bei den niederen Lebermoosen
(Marchantiales) teilt sich die Zygote quer und längs in 8,
dann radial weiter in 16 Zellen und diese durch perikline Wände
in äußere und innere Zellen (Fig. 441). Aus der unteren Hälfte
des Embryos geht der Fuß und der kurze Stiel hervor, aus der
oberen die Kapsel, deren zentrale Zellen, das Archespor,
das vielzellige sporogene Gewebe liefern. Diese Zellen werden
zum Teil zu Sporenmutterzellen; zum Teil bleiben sie steril und
dienen zunächst als Nährzellen für die ersteren (Fig. 441 C).
Dann wachsen die Nährzellen meist zu faserförmigen Gebilden
mit schraubenbandförmigen Wandverdickungsleisten heran, zu
Schleudern (Elateren), die nach der Öffnung ihrer
Kapsel durch ihre hygroskopischen Bewegungen die Sporen auflockern
und ausstreuen. Nur bei den Ricciaceen werden sämtliche Zellen
des inneren Gewebes zu Sporenmutterzellen, und das ganze Sporogon
ist hier zu einer rundlichen ungestielten Kapsel mit einschichtiger
Wand vereinfacht.
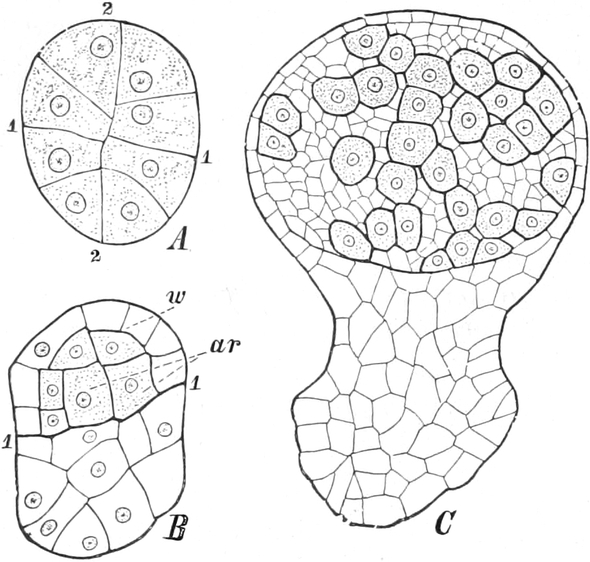
Fig. 441. Sporogonentwicklung von Corsinia
marchantioides, einem Marchantiaceen-Lebermoos. A Die Zygote in
16 Zellen geteilt. B Die untere Hälfte des Embryos entwickelt
sich zum Fuß, die obere zur Kapsel, w Wandzellen, ar Archespor.
Vergr. 170. C Älteres Sporogon. In der Kapsel die aus dem Archespor
hervorgegangenen Sporenmutterzellen und kleineren sterilen Zellen.
Letztere erfahren bei Corsinia keine Weiterentwicklung zu Elateren.
Vergr. 90. Nach K. MEYER.
Bei den höheren Lebermoosen (Jungermanniales) teilt sich
die Zygote erst einigemale quer, die unterste Zelle wird meist
zu einem ein-, selten mehrzelligen Saugorgan, die oberen Zellen
liefern Fuß, Stiel und Kapsel. Aus dem sporogenen Gewebekomplex
gehen auch hier neben den Sporenmutterzellen sterile Zellen
hervor, die bei den meisten Gattungen wiederum zu Elateren sich
weiterentwickeln.
Die Lebermoosgruppe der Anthocerotales weicht im Bau des
Sporogons bedeutend von den vorher genannten ab und nähert sich in
einigen Eigentümlichkeiten den Laubmoosen (vgl. S. 423).
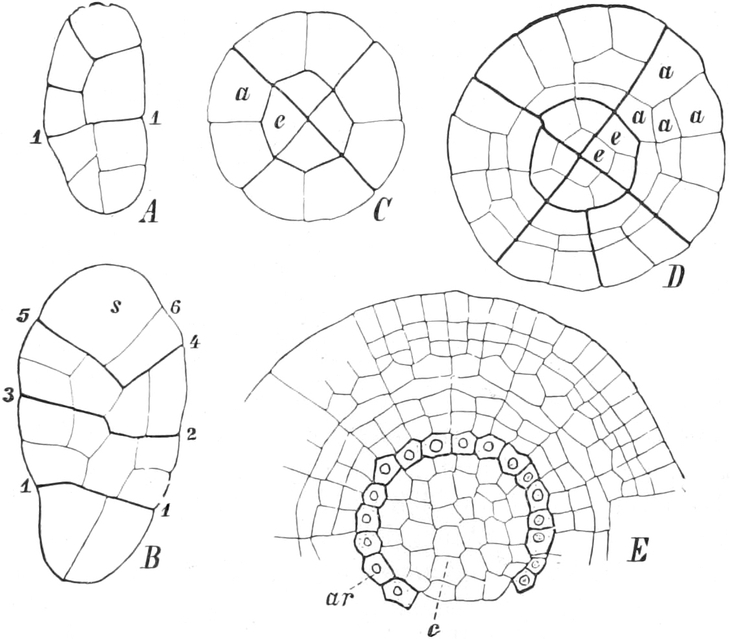
Fig. 442. Sporogonentwicklung von Funaria hygrometrica,
einem Laubmoos. A, B Längsschnitt. Erste Teilungen der Zygote, s
Scheitelzelle. C–E Querschnitt. C Teilung in Endothecium e
und Amphithecium a. D Weitere Teilungen. E Älteres Sporogon, im
Endothecium die äußerste Zellschicht, das Archespor ar, abgeteilt von
der Columella c. Nach CAMPBELL.
Bei den Laubmoosen besitzt das Sporogon einen axilen,
der Stoffleitung dienenden Strang sterilen Gewebes, eine
Columella, in deren Umkreis das Archespor in Form einer
meist einfachen Zellschicht angelegt wird. Bei den Sphagnales
(Fig. 454 C) und den Andreaeales überwölbt die Archesporschicht
kuppelförmig die Columella, bei den Bryales (Fig. 460) dagegen
hat sie die Form eines Zylindermantels. Der langgestreckte Embryo
baut sich aus Quersegmenten auf, die bei den Sphagnales durch
Querteilung der Zygote entstehen; bei den übrigen Laubmoosen aber,
deren Zygote zuerst auch quergeteilt wird, treten in der oberen
Zelle schiefe Wände auf und die von ihnen gebildete zweischneidige
Scheitelzelle scheidet[S. 421] nach beiden Seiten hin Quersegmente
ab, die sich dann weiter teilen. In den Quersegmenten, die die
Mooskapsel liefern, findet zunächst eine Längsteilung statt,
dann in den so entstandenen Quadranten durch perikline Wände die
Zerlegung in äußere Zellen (Amphithecium) und innere Zellen
(Endothecium) (Fig. 442). Das Archespor entsteht nur
bei den Sphagnales als innerste Zellschicht des Amphitheciums, bei
allen übrigen Laubmoosen dagegen als äußerste Zellschicht aus dem
Endothecium. Es liefert ausschließlich Sporen, aber keine sterilen
Zellen (Fig. 443).
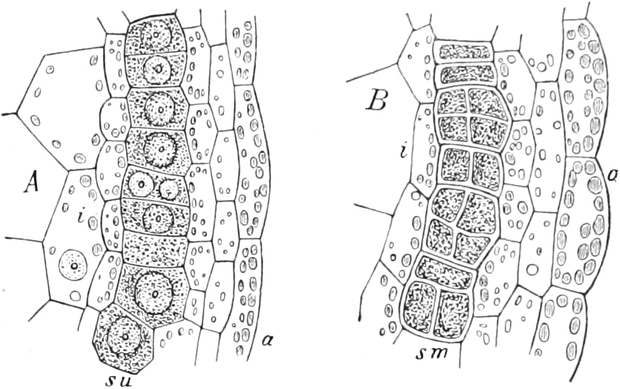
Fig. 443. Funaria hygrometrica. Querschnitt durch
das Archespor (A su) und die aus ihm hervorgegangenen, noch nicht
isolierten Sporenmutterzellen (B sm). Nach GOEBEL.
Die Moose sind bis auf wenige, sekundär zum Leben im Wasser
übergegangene Formen Landpflanzen im Gegensatz zu den Algen
und zeigen dementsprechende Anpassungen in ihrer anatomischen
Struktur. So sind alle ihre oberirdischen Teile von einer Kutikula
überzogen. Die bemerkenswerte Kleinheit der Moose im Vergleich zu
den Farnpflanzen steht in Zusammenhang mit ihrem einfachen zelligen
Aufbau, mit dem Mangel echter Gefäße und echter Wurzeln. Manche
Arten sind winzige Pflänzchen, die größten Laubmoose sind die
neuseeländischen Dawsonien, deren einfache beblätterte Stämmchen
bis 50 cm Höhe erreichen.
Viele Moose besitzen rötliche, bräunliche, einige sogar fast
schwarze Färbung, die in den meisten Fällen durch besondere
Membranfarbstoffe (Anthocyane und Phlobaphene), seltener durch
gefärbten Zellsaft bedingt ist[406].
Die Moose weisen eine ungemeine Regenerationsfähigkeit aus
abgeschnittenen Stücken aller ihrer Organe auf; ferner ist bei
ihnen vegetative Vermehrung durch Brutkörper und Brutknospen sehr
verbreitet, die am Thallus, an den Stämmchen, an den Blättern, am
Protonema in verschiedener Art entstehen und sich loslösen[407].
Die phylogenetische Ableitung der Bryophyten von einer
bestimmten Algengruppe begegnet Schwierigkeiten. Zwischen
den Moosen einerseits, den höherstehenden Grünalgen und den
Characeen andererseits sind keine einen Übergang vermittelnde
Formen bekannt. Der morphologische Vergleich ergibt vielmehr eher
eine Anknüpfung der Moose an Braunalgen, deren vielfächerige,
bei einigen Gattungen bereits in Antheridien und Oogonien
differenzierte Gametangien als homologe Vorläufer der
Antheridien und Archegonien der Archegoniaten gelten dürfen.
So zeigt das Antheridium der niederen Lebermoose noch ganz
übereinstimmenden Zellenaufbau mit Braunalgengametangien (vgl. Fig.
436, 355, 357); es unterscheidet sich von ihnen durch den Besitz
einer sterilen, schützenden Wandschicht, deren Differenzierung als
eine Anpassung an terrestrische Lebensweise angesehen werden kann.
Ferner zeigt sich unter den Braunalgen, z. B. bei Dictyota, ein
mit dem der Moose übereinstimmender Generationswechsel. Gametophyt
und Sporophyt haben allerdings dort in ihrem vegetativen Bau
gleiche Form. Den Tetrasporangien des Dictyotasporophyten
entsprechen die Sporenmutterzellen des Moossporophyten,
deren endogene Anlage in zusammenhängenden Schichten auf den
Einfluß terrestrischer Lebensweise sich zurückführen ließe. Der
Sporophyt der Moose geht frühzeitig zur Entwicklung seiner Sporen
über, beschließt damit sein Wachstum, ohne erst eine Gliederung in
vegetative Organe zu erfahren, und wird so wesentlich verschieden
von dem Gametophyten, dessen Gestaltung bei den thallösen
Lebermoosen mit dem Thallus gewisser Braunalgen manche Analogien
aufweist[408].
Die beiden scharf geschiedenen Klassen der Bryophyten charakterisieren
sich kurz folgendermaßen:
1. Hepaticae, Lebermoose. Geschlechtliche Generation mit schwach
entwickeltem und meist nicht scharf abgesetztem Protonema, ihr
Thallus entweder flächenförmig und gabelteilig oder in Stämmchen mit
dorsiventral angeordneten Blättchen gegliedert. Der Sporenbehälter
erzeugt bei den meisten außer Sporen auch Elateren. Nur bei einer
Ordnung, den Anthocerotales, wird in der Kapsel eine Columella
ausgebildet.
[S. 422]
2. Musci, Laubmoose. Vorkeim der geschlechtlichen Generation meist
kräftig entwickelt, scharf abgesetzt, Thallus stets in Stämmchen
und Blättchen gegliedert. Die Blätter in spiraliger, mehrzeiliger,
seltener in zweizeiliger Anordnung, Stämmchen also poly- oder seltener
bisymmetrisch beblättert. Sporenbehälter stets ohne Elateren, aber mit
Columella.
Fossile Moose. Die Lebermoose sind primitiver organisiert
als die Laubmoose, scheinen auch älter zu sein, da ihre fossilen
Reste vereinzelt bis zum Karbon hinab, mit einiger Sicherheit aber
erst im Mesozoikum gefunden werden, während die ersten Laubmoose
aus der oberen Kreide bekannt wurden. Die meisten fossilen
Moosreste entstammen dem Tertiär und zeigen größte Ähnlichkeit mit
heutigen Gattungen.
Die Mehrzahl der Lebermoose bewohnt feuchte Standorte und besitzt
dementsprechend hygrophile Struktur. Echte Wasserpflanzen sind
unter ihnen nur spärlich vertreten. Manche zarte Jungermanniaceen
leben versteckt in Laubmoosfilzen. Weniger zahlreich sind Formen,
die extrem trockene Standorte auf Baumrinden und Felsen oder auf
dem Boden bewohnen und xerophilen Bau sowie Einrichtungen zum
Wasserspeichern besitzen. Unter den epiphytischen Arten sind
die kleinen, epiphyllen, d. h. auf Blättern lebenden Lebermoose
feuchter Tropenwälder bemerkenswert. Im allgemeinen spielen
die Lebermoose keine bedeutende Rolle in der Zusammensetzung
kryptogamer Pflanzenformationen.
Die Rhizoïden vieler Lebermoose, namentlich der Jungermanniaceen,
sowie auch die chlorophyllfreien Thallusgewebe bei Marchantiaceen
werden häufig von endophytischen Pilzfäden (u. a. des Mucor
rhizophilus) besiedelt, die den Moosen keinen besonderen Nutzen,
aber auch keinen bedeutenden Schaden bringen[410].
Die Lebermoose zerfallen nach dem Bau der Sporogone und der Gliederung
der geschlechtlichen Generation in drei Ordnungen, von denen die
Anthocerotales und Marchantiales ausschließlich Thalluspflanzen,
die Jungermanniales teils ebensolche, teils aber Formen mit
dorsiventral beblättertem, seltener, und zwar nur in der Gruppe der
Haplomitrieen, mit radiär gebautem, beblättertem Stämmchen umfassen.
1. Ordnung. Die Anthocerotales[411], eine isoliert stehende
Gruppe, die nur wenige Formen umfaßt, können als eine primitive
Moosordnung gelten. Ihr Sporogon zeichnet sich durch einen
reicheren inneren Bau aus, als das der übrigen Lebermoose, bei
denen es eine fortschreitende Vereinfachung erfahren hat.
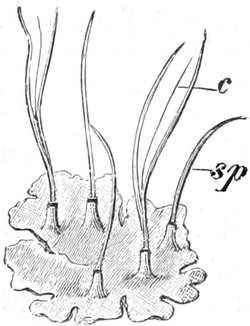
Fig. 444. Anthoceros laevis. sp Sporogon, c
Columella. Nat. Gr.
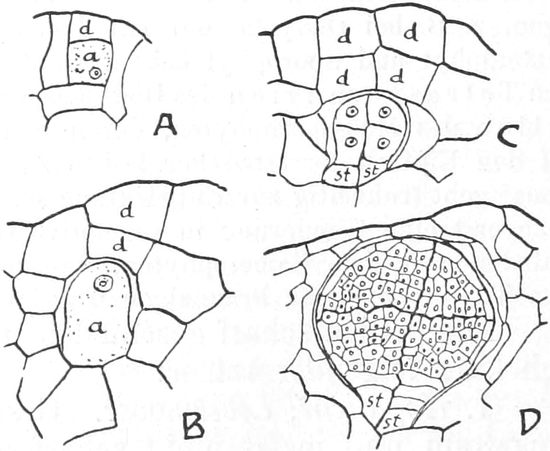
Fig. 445. Anthoceros Pearsoni. Entwicklung des endogenen
Antheridiums. d Deckzellen, st Stielzellen, a Anlage des
Antheridiums. Nach D. CAMPBELL.
Der Gametophyt hat die Gestalt eines gelappten, am Boden mittels
Rhizoïden festgewachsenen Thallus. Seine Zellen enthalten
zum Unterschied von allen anderen Moosen nur einen einzigen
großen pyrenoidführenden Chlorophyllkörper. Auf der Unterseite,
seltener auf der Oberseite finden sich Spaltöffnungen. Die
Antheridien stehen einzeln oder zu vier im Innern
geschlossener Höhlungen unter der Oberseite des Thallus (Fig.
445). Die Höhlung wird erst bei der Reife der Antheridien
vermittels Schleimbildung in den Deckzellen[S. 423] geöffnet. Die Anlage
der männlichen Organe ist also hier abweichend von allen übrigen
Archegoniaten sekundär zu einer endogenen geworden, indem eine
Oberflächenzelle sich zunächst in eine die Decke liefernde äußere
(d) und eine innere (a), die Mutterzelle der Antheridien,
teilt. Die Archegonien sind in die Oberseite des Thallus
eingesenkt und werden nach der Befruchtung von einer durch
Wucherung des Thallusgewebes entstehenden, mehrschichtigen Hülle
(Marsupium) überwölbt, die später von der Kapselfrucht durchbrochen
wird und eine Scheide an deren Basis bildet. Die befruchtete
Eizelle teilt sich im Gegensatz zu den übrigen Moosen zunächst
durch eine Längswand in zwei Zellen, diese dann weiter durch
Querwände. Das aus dem Embryo hervorgehende Sporogon besitzt
einen angeschwollenen, mit rhizoïdähnlichen Schläuchen im Thallus
befestigten Fuß und eine ungestielte, lang schotenförmige, mit
zwei Längsklappen aufspringende Kapsel, in deren Längsachse eine
aus wenigen Zellreihen bestehende Columella sich befindet (Fig.
444). Diese wird kappenförmig von der schmalen sporenbildenden
Zellschicht bedeckt. Außer Sporen erzeugt letztere auch Schleudern;
sie sind hier mehrzellig, vielgestaltig, oft gegabelt. Im Gegensatz
zu allen übrigen Lebermoosen reift der Kapselteil dieses Sporogons
nicht gleichzeitig heran, sondern von der Spitze ausgehend unter
andauernder Fortentwicklung an seiner Basis. Auch enthält die
Sporogonwand Chlorophyll und besitzt Spaltöffnungen, die sonst bei
Lebermoosen nicht auftreten.
An der Unterseite des Thallus der Anthocerotaceen werden durch
Auseinanderweichen benachbarter Zellen Spalten erzeugt, die in
Höhlungen führen, welche Schleim enthalten. In diese dringen
häufig Nostocfäden ein, um sich dort zu endophytischen Kolonien zu
entwickeln[412].
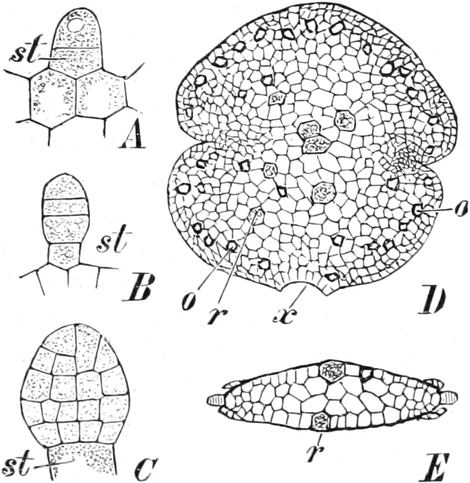
Fig. 446. Marchantia polymorpha. A–C
Brutkörperentwicklung, st Stielzelle, D Brutkörper von der
Fläche, E im Querschnitt, x Ablösungsstelle, o Ölzellen,
r Rhizoïdanlagen. A–C Vergr. 275. D–E Vergr. 65. Nach
KNY.
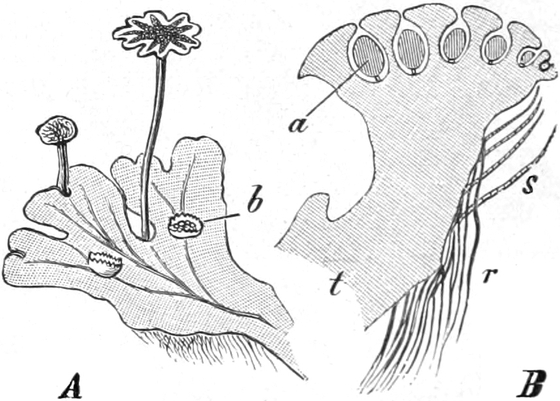
Fig. 447. A Männliche Pflanze von Marchantia
polymorpha, b Brutkörbchen. Nat. Gr. B Antheridiumstand mit den
eingesenkten Antheridien a, t Thallus, s Ventralschuppen. r
Rhizoïden. Etwas vergr.
2. Ordnung. Die Marchantiales[413] besitzen zum Teil recht
komplizierten Aufbau. Als Typus sei die an Quellen häufige
Marchantia polymorpha geschildert. Sie bildet bis 2 cm breite,
sich gabelig verzweigende Thalluslappen (Fig. 447 A, Fig. 448
A) mit schwachen Mittelrippen. An der Unterseite entspringen
einschichtige Zellamellen oder Ventralschuppen und die
Rhizoïden, die den Thallus befestigen und ihm Wasser
zuführen; sie sind zum Teil glattwandig, zum Teil aber mit
zapfenförmigen, nach innen vorragenden Wandverdickungen versehen
(Fig. 31). Diese Zäpfchenrhizoïden verflechten sich längs der
Mittelrippe zu einem Strange. Die Dorsiventralität des Thallus
macht sich auch im anatomischen Aufbau geltend. Auf der Oberfläche
des Thallus bemerkt man schon mit bloßem Auge eine rhombische
Felderung. Jedes Feld entspricht einer unter der obersten
Zellschicht befindlichen, von geschlossenen seitlichen Wänden
abgegrenzten Luftkammer, die durch eine Atemöffnung in
der Mitte des Feldes nach außen führt (Fig. 95 A, B). Die
Öffnung besteht aus einem kurzen Kanal mit einer aus mehreren
ringförmigen Stockwerken von je vier Zellen gebildeten Wandung.
Vom Boden der Kammer erheben sich zahlreiche kurze, aus rundlichen
Zellen bestehende Fäden, die Chlorophyllkörner enthalten und das
Assimilationsgewebe vorstellen. Auch in den[S. 424] Kammerwänden
und in der Epidermis befindet sich Chlorophyll, aber in geringerer
Menge. Im übrigen besteht der Thallus aus großen chlorophyllarmen,
als Speicherzellen dienenden Parenchymzellen, die an der
Unterseite von einer einschichtigen geschlossenen Zellschicht
bedeckt werden. Auf die Ausbildung der Luftkammern ist die
Belichtung von großem Einfluß. Bei sehr schwacher Belichtung kann
ihre Bildung ganz unterbleiben.
Auf den Mittelrippen der Oberseite des Thallus treten in der Regel
becherförmige Auswüchse mit gezähntem Rand, die Brutbecher
oder Brutkörbchen (Fig. 447 b) auf, in denen eine Anzahl
von flachen Brutkörperchen sich befinden. Sie entstehen, wie Fig.
446 zeigt, durch Hervorwölbung und weitere Teilung einzelner
Oberflächenzellen und sitzen mit einer Stielzelle (st) fest,
von der sie sich (D bei x) ablösen. Sie haben an den beiden
Einschnürungsstellen zwei Vegetationspunkte und bestehen aus
mehreren Schichten von Zellen, von denen eine Anzahl mit Ölkörpern
erfüllt ist (D, o), andere, farblose, als Anlagen der späteren
Rhizoïden dienen. Ölhaltige Zellen treten auch im fertigen Thallus
zerstreut auf und sind überhaupt bei Lebermoosen sehr verbreitet.
Mit Hilfe der Brutkörperchen kann sich Marchantia in reichlichem
Maße vegetativ vermehren. Die Dorsiventralität des aus dem
isolateralen Brutkörper hervorgehenden Thallus wird durch den
Einfluß des Lichtes bedingt.
Die Sexualorgane, Antheridien und Archegonien, werden von
besonderen aufstrebenden Zweigen des Thallus getragen. Im unteren
Teile sind diese Zweige stielartig zusammengerollt, im oberen Teile
verzweigen sie sich sternförmig. Antheridien und Archegonien sind
diözisch verteilt. Die männlichen Zweige schließen mit einer
lappig gerandeten Scheibe ab, an deren Oberseite die Antheridien
eingesenkt sind, und zwar ein jedes in einen flaschenförmigen
Hohlraum, der mit einer engen Öffnung nach außen mündet (Fig. 447
B u. 435). Diese Höhlungen werden von Luftkammern führendem
Gewebe getrennt. Die Spermien sammeln sich oben auf dem Hut in
einem Wassertropfen, der durch den welligen Hutrand gehalten wird.
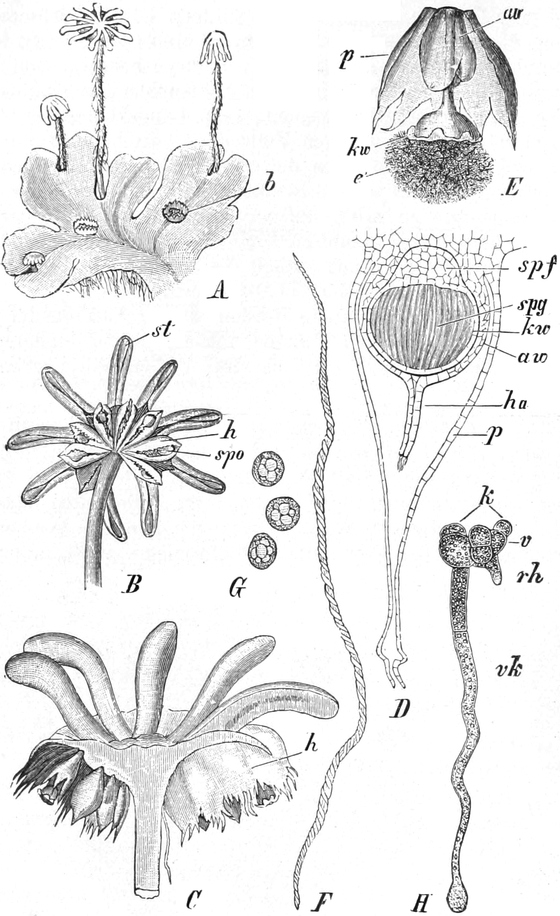
Fig. 448. Marchantia polymorpha. A Weibliche
Pflanze mit vier verschiedenalterigen Archegoniumständen, b
Brutkörbchen. Nat. Gr. B Archegoniumstand von unten, st Strahlen,
h Hülle, spo vortretende Sporogone. Vergr. 3. C Derselbe halb
durchschnitten. Vergr. 5. D Junges Sporogon im Längsschnitt, mit dem
Fuß spf, dem sporenbildenden Gewebe spg, der Kapselwandung kw,
der Archegoniumwandung aw, dem Archegoniumhals ha, dem Perianth
p. Vergr. 70. E Aufgesprungenes Sporogon, Sporen mit Elateren e.
Vergr. 10. F Ein einzelner Elater. G Reife Sporen. Vergr. 315. H
Gekeimte Spore mit Keimschlauch vk und Keimscheibe k, letztere mit
der Scheitelzelle v und dem Rhizoïd rh. Vergr. 100. C, E nach
BISCHOFF. B, D, F–H nach KNY.
Die weiblichen Zweige (Fig. 448 A) schließen mit einem
meist neunstrahligen Schirm ab. Die Oberseite des Schirmes ist
zwischen den Strahlen umgeschlagen und trägt[S. 425] hier die Archegonien,
die somit der Unterseite des Schirmes zu entspringen scheinen. Sie
bilden radiale Reihen zwischen den Strahlen. Jede dieser Reihen
wird von einer zierlich gezähnten Hülle (Perichaetium) (B,
C, h) umgeben. Die Gestalt der Archegonien ist aus Fig. 438
ersichtlich.
Die Befruchtung erfolgt bei Regenwetter, indem Regentropfen
die Samenfäden enthaltende Flüssigkeit von den männlichen Hüten
auf die weiblichen Schirme spritzen, deren Epidermiszellen
papillenförmig vorspringen und ein oberflächliches Kapillarsystem
darstellen, in welchem die Samenfäden zu den Archegonien
hinabgeleitet werden.
Nach der Befruchtung entwickelt sich die Eizelle zu einem
vielzelligen Embryo (Fig. 438 C), dieser zu einem gestielten
ovalen Sporogon. Seine Kapsel hat eine einschichtige
Wandung, deren Zellen Ringfaserverdickungen aufweisen. Nur am
Scheitel ist die Wandung zweischichtig, hier beginnt auch das
Einreißen der Kapsel, indem das Deckelstück zerfällt und die
Wandung in Form mehrerer Zähne sich zurückkrümmt. Die reife
Kapselfrucht ist anfangs noch bedeckt von der eine Zeitlang
mitwachsenden Archegoniumwandung (Fig. 448 DE, aw), die aber
bei der Streckung des Stieles durchbrochen wird und an der Basis
als Scheide zurückbleibt. Außerdem wird die Kapsel von einer
vier- bis fünfspaltigen, dünnhäutigen Hülle, dem Perianth,
umgeben, das schon vor der Befruchtung aus dem kurzen Stiel des
Archegoniums ringsum als sackartige Hülle hervorzusprossen beginnt
(Fig. 438 C, pr, 448 D, E, p). Die Kapsel entläßt
Sporen und Elateren (Fig. 448 F, G).
Marchantia war früher als Mittel gegen Leberkrankheiten offizinell,
daher auch die Bezeichnung Lebermoose.
Die Ricciaceen[414] weisen zwar eine weitgehende
Vereinfachung ihrer Sporogone auf, schließen sich aber an
einfacher gebaute Marchantiaceen als reduzierte Formen an. Ihr
dichotomisch gelappter Thallus bildet auf Schlammboden am Ufer der
Gewässer oder auf feuchten Äckern kleine Rosetten. Riccia natans
schwimmt mit ihren breiten Thalluslappen auf der Oberfläche des
Wassers nach Art der Lemnaceen (Fig. 449 C). Riccia fluitans
(Fig. 449 A), vielleicht eine Sammelart von submersen Formen
verschiedener terrestrischer Arten, hat dagegen schmale, reicher
verästelte Thalluslappen. Diese beiden wasserbewohnenden Arten
können aber auch auf Schlammboden niederliegende Rosetten bilden
(Fig. 449 B). Der Thallus besteht an der Oberseite ähnlich wie
bei Marchantia aus einem von schizogen entstandenen Luftkammern
durchzogenen, assimilierenden Gewebe; an seiner Unterseite trägt
er feine Rhizoïden und außerdem eine Reihe von quergestellten
Ventralschuppen, die wie erstere sich an der Nährstoffaufnahme
beteiligen. Beide Organe fehlen vollständig der submersen Form von
Riccia fluitans.
Antheridien und Archegonien sind auf der Oberseite eingesenkt. Aus
der Eizelle entwickelt sich nach der Befruchtung ein ungestieltes
kugeliges Sporogon mit einschichtiger Wandung. Die Wandung wird vor
der Sporenreife aufgelöst, und die Sporen werden durch Verwitterung
der sie umgebenden Zellen des Thallus frei. Elateren fehlen.
Fig. 449. A Riccia fluitans, untergetaucht
schwimmende Form. B Riccia natans, Landform. Nat. Gr. C Riccia
natans. Schwimmform mit langen Ventralschuppen. Vergr. 2. B Nach
GOEBEL, C nach BISCHOFF.
3. Ordnung. Die Jungermanniales, meist kleine, auf Erde oder
an Baumstämmen, in den Tropen auch auf Blättern von Waldpflanzen
lebende Lebermoose, weisen in ihren einfacheren Formen einen
breitlappigen Thallus wie Marchantia auf, z. B. die auf
feuchtem Erdboden häufige Pellia epiphylla, oder einen schmal
bandförmigen, dichotom verzweigten, ähnlich wie Riccia fluitans,
so die an Baumstämmen oder Felsen lebende Metzgeria furcata
(vgl. Fig. 94). Sodann gibt es Formen, deren breitlappiger, mit
Mittelrippe versehener Thallus bereits eine schwache Ausbildung
von blattähnlichen Gliedern an seinem Rande aufweist, so die
erdbewohnende Blasia pusilla (Fig. 450). Die Mehrzahl aber
besitzt eine deutliche Gliederung in ein niederliegendes oder
aufstrebendes,[S. 426] reichverzweigtes, dorsiventrales Stämmchen und in
einschichtige Blättchen ohne Mittelnerv, die in zwei Zeilen an
den Flanken des Stämmchens mit schiefer Stellung ihrer Spreite
angeordnet sind (Fig. 451). Bei gewissen Gattungen tritt zu diesen
zwei Zeilen von Flankenblättern auch noch eine bauchständige Reihe
von kleineren und anders beschaffenen Blättchen, Amphigastrien oder
Bauchblättern, hinzu, so bei Frullania Tamarisci (Fig. 452),
einem zierlich verzweigten, an Felsen und Baumstämmen häufigen
Lebermoos von bräunlicher Farbe. Die Flankenblätter gliedern sich
häufig in einen Oberlappen und einen Unterlappen. Der letztere
erscheint bei gewissen Arten, die an ihren Standorten zeitweise der
Gefahr des Vertrocknens ausgesetzt sind, sackartig ausgebildet und
dient als kapillarer Wasserbehälter, so bei Frullania Tamarisci.
Die Flankenblätter sind entweder oberschlächtig, wenn der
Hinterrand eines Blattes von dem Vorderrand des nächstunteren
überdeckt wird (Fig. 452 Frullania), oder unterschlächtig,
wenn der Hinterrand eines Blattes über dem Vorderrand des
nächstunteren liegt (Fig. 451).
Fig. 450. Blasia pusilla mit Sporogon s, r
Rhizoïden. Vergr. 2.
Fig. 451. Plagiochila asplenioides mit Sporogon s.
Nat. Gr.
Das Sporogon besitzt einen langen zarten und weichen
Stiel; es ist schon fertig ausgebildet, ehe es bei der rasch
erfolgenden Streckung des Stiels die Archegoniumwand durchbricht
und als häutige Scheide an seinem Grunde zurückläßt; es weist eine
kugelige, meist in vier Klappen sich öffnende Kapsel auf, bildet
keine Columella aus und erzeugt Sporen und Elateren. Bei einigen
Gattungen (Pellia, Aneura) sind in der Kapsel Elaterenträger
vorhanden, die aus Gruppen steriler elaterenähnlicher Zellen
bestehen. Die Kapselwandzellen sind mit ringförmigen oder
leistenartigen Verdickungen versehen oder gleichmäßig verdickt
bis auf die dünnen Außenwände. Das Aufspringen erfolgt durch die
Kohäsion des schwindenden Füllwassers unter Einbiegung der dünnen
Außenwände.
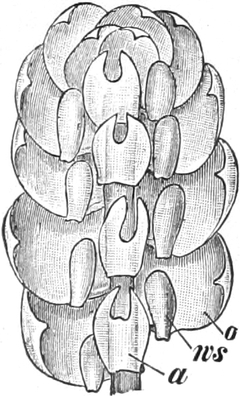
Fig. 452. Frullania Tamarisci, von unten. o
Flankenblatt, ws als Wassersack ausgebildeter Unterlappen des
Flankenblattes, a Amphigastrien. Vergr. 35.
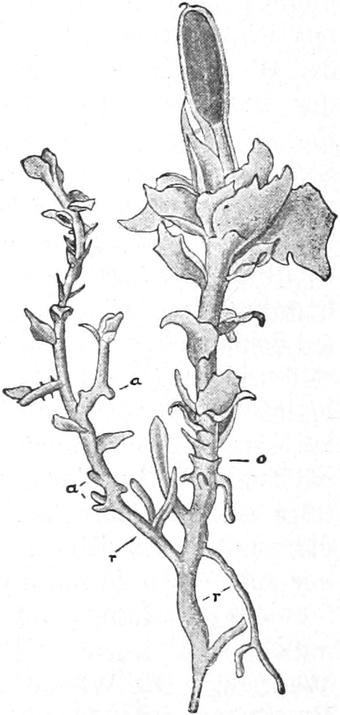
Fig. 453. Haplomitrium Hookeri. a Anlagen neuer
Sprosse, r Rhizome, o untere Grenze des oberirdischen Sprosses.
Nach GOTTSCHE.
Nach der Stellung der Sporogone gliedert sich die Mehrzahl der
Jungermanniales in zwei Gruppen. 1. Bei den Anacrogynae
wird der Scheitel zur Archegoniumbildung nicht mit verwendet, die
Sporogone stehen rückenständig und sind an ihrer Basis von einem
scheidenartigen Auswuchs des Thallus, einem Perichaetium, umgeben.
Hierher gehören die thallösen Formen (Pellia, Metzgeria) und
die Übergangsformen zu den beblätterten Formen (Blasia). 2. Bei
den Acrogynae dagegen stehen die Archegonien und somit
auch die Sporogone am Ende des Stengels oder seiner Äste und sind
von einem aus besonders gestalteten Blättchen gebildeten, nach
der Befruchtung heranwachsenden Perianth umhüllt. Hierher die
dorsiventral beblätterten Formen (z. B. Plagiochila, Frullania
und die artenreiche Gattung Jungermannia). 3. Eine vorgerückte[S. 427]
Stellung nehmen die Haplomitrieae ein, die noch gewisse
Beziehungen zu den Anacrogynen aufweisen. Sie werden nur durch
zwei Gattungen vertreten, von denen Calobryum in den Tropen,
Haplomitrium Hookeri (Fig. 453) als einzige Art in Europa,
vielleicht als Überbleibsel präglazialer Lebermoose, vorkommt. Sie
weichen von allen übrigen Lebermoosen ab durch radiären Bau ihrer
dreizeilig oder ringsum beblätterten Sprosse. Die Sexualorgane sind
bei ersterer Gattung in terminalen Ständen vereinigt, bei letzterer
zwischen den oberen Blättern verteilt.
Die Laubmoose sind in ungemeiner Formenfülle in allen Zonen
verbreitet, sie wachsen auf trockenem Erdboden, in Sümpfen, an
Felsen, an Baumstämmen, in tropischen Wäldern auch als Epiphyten
auf den Baumästen, seltener im Wasser, und zeigen dementsprechend
sehr verschiedene Strukturen. Für sehr trockene Standorte sind
besonders dichte Polster oder Rasen charakteristisch, während die
typischen Bodenbewohner unserer Wälder in ausgebreiteten lockeren
oder dichteren Filzen vegetieren. In feuchten Bergwäldern der
Tropen und Subtropen besiedeln Laubmoose oft in unglaublichen
Mengen die Äste in Form schwellender Polster oder in Form lang
herabhängender Schleier[416]. Ausgedehnte Bestände bilden die
Torfmoose in Mooren, ferner rasenbildende Arten (besonders
Polytrichum) auf feuchtem Boden in den arktischen Moostundren.
Das reich verzweigte Protonema der Laubmoose erscheint dem
bloßen Auge als ein feiner grüner Filz (Fig. 434). Es entsendet in
den Boden Rhizoïden, chlorophyllfreie verzweigte Fäden, die sich
durch schräge Stellung ihrer Querwände auszeichnen. Am Protonema
entstehen die Knospen der Moospflänzchen als seitliche Ausstülpungen
einzelner Zellen des Hauptfadens, meistens aber der Anfangszellen
der Protonemazweige. Diese Ausstülpungen werden durch eine Querwand
abgetrennt, teilen sich weiter in eine oder auch in zwei Stielzellen
und eine anschwellende Endzelle, die bei ihrer weiteren Teilung
die dreiseitig pyramidale Scheitelzelle des Moospflänzchens
liefert[417]. Letzteres ist stets in Stengel und Blättchen gegliedert.
Die Laubmoose unterscheiden sich leicht von den beblätterten
Jungermanniaceen durch die spiralige Anordnung ihrer Blättchen, die
nur selten zweizeilig gestellt sind. Bei solchen Laubmoosen, die
niederliegende Stengel haben, sind die Blättchen bei spiraliger
Anordnung häufig einseitswendig oder gescheitelt, so daß zwar ein
Gegensatz von Ober- und Unterseite, aber in anderer Weise als bei den
Lebermoosen, zustande kommt.
Der Moosstengel wird von Zellen aufgebaut, die nach der
Oberfläche zu enger und dickwandiger werden. Bei verschiedenen
Gattungen, z. B. bei Polytrichum, bei Mnium (Fig. 96), findet
sich in der Achse des Stengels ein zentrales Leitbündel aus
langgestreckten Zellen vor. Diese Wasser und organische Substanzen
leitenden Bündel stehen auf niederer Stufe der Differenzierung:
sie führen weder echte Siebröhren noch echte Gefäße, enthalten
aber neben lebendigen Elementen auch plasmaleere wasserhaltige
Zellen. Sie fehlen ganz den Sphagnaceen oder Torfmoosen, die an
sumpfigen Standorten leben. Der Stengel dieser Formen zeigt eine
eigentümliche Ausbildung der peripherischen Zellschichten, die
plasmaleer sind, mit großen offenen Poren untereinander und mit der
Atmosphäre in Verbindung stehen und spiralige Verdickungsleisten
als Aussteifungen an ihren Wandungen besitzen, somit einen Bau
aufweisen, der sie befähigt, Wasser mit Leichtigkeit aufzusaugen
und als kapillare Wasserbehälter und Leitungsbahnen zu dienen.
Die Blätter mancher Moose bestehen nur aus einer Schicht von
polygonalen chlorophyllführenden Zellen; meist aber sind sie in
der Mittellinie mehrschichtig und von einem aus dem Zentralstrang
des Stengels entspringenden Bündel langgestreckter Zellen
durchzogen. Den Torfmoosblättern geht letzteres ab, dagegen sind
sie eigenartig differenziert, indem ihre einschichtige Blattfläche
ähnliche plasmaleere wasserspeichernde Zellen führt wie die
Stengelperipherie. Diese Zellen sind hier groß, langgestreckt und
ebenfalls mit queren Verdickungsleisten und offenen Poren versehen.
Zwischen ihnen bilden die chlorophyllhaltigen, schmalen Zellen
ein zusammenhängendes Netz. Außer den Torfmoosen zeigt auch noch
die Familie der Leucobryaceen eine ähnliche Differenzierung der
Blattzellen (z. B. Leucobryum glaucum).
[S. 428]
Eigenartigen Blattbau, der sich als Anpassung an die Wasseraufnahme
und Schutz gegen Trockenheit darstellt, besitzt unter den
Laubmoosen u. a. Polytrichum commune, der gemeine Widerton,
dessen mehrschichtige Blätter auf der Oberseite zahlreiche
einschichtige, dichtstehende Längslamellen aus chlorophyllhaltigen
Zellen tragen, die das assimilierende Gewebe vorstellen und in den
Zwischenräumen Wasser leiten und festhalten. Bei Trockenheit faltet
sich das Blatt mittels Kohäsionsmechanismus zusammen und
legt sich dicht dem Stamm an, wodurch die zarten Lamellen in eine
vor übermäßiger Transpiration geschützte Lage gebracht werden[418].
Überhaupt können viele Laubmoose unbeschadet große Trockenheit
vertragen.
Am Grunde des Stengels entspringen die fadenförmigen, verzweigten
Rhizoïden (Fig. 456, 458), die chlorophyllfrei sind,
sonst aber den gleichen Bau aufweisen wie das Protonema und auch
gelegentlich zu solchem auswachsen und neue Moospflänzchen in
derselben Weise wie dieses erzeugen können.
Die Sexualorgane stehen bei den Laubmoosen in Gruppen an
den Enden der Hauptachsen oder kleiner Seitenzweiglein, umgeben
von den obersten Blättchen, die oft als besondere Hüllblättchen,
Perichaetium, ausgestaltet sind (Fig. 458). Zwischen den
Sexualorganen steht gewöhnlich eine Anzahl von mehrzelligen,
oft mit kugeligen Endzellen versehenen Safthaaren oder
Paraphysen. Die Stände sind entweder zwitterig oder einhäusig
oder zweihäusig. Bei gewissen getrenntgeschlechtlichen Laubmoosen
erscheinen die aus besonderen Sporen hervorgehenden männlichen Pflanzen
im Vergleich zu den weiblichen als winzige Zwergpflänzchen, die nach
Bildung einiger weniger Blättchen bereits zur Erzeugung der Antheridien
übergehen[419]. Die Antheridien und Archegonien der Laubmoose
unterscheiden sich entwicklungsgeschichtlich von denen aller übrigen
Archegoniaten durch den Aufbau ihres Körpers aus Segmenten einer bei
ersteren Organen zweischneidigen, bei letzteren aber dreischneidigen
Scheitelzelle.
Das Sporogon der Laubmoose[420] weist in seiner Kapsel ein
zentrales Säulchen aus sterilem Gewebe, die Columella, auf,
in deren Umkreis der Sporensack mit den Sporen liegt (Fig. 460).
Die Columella fungiert als Nährstoffzuleiter und Wasserspeicher
für die sich bildenden Sporen, denen die plasmareichen Zellen der
Sporensackwandung die Nährstoffe zuführen. Elateren werden nie
gebildet. Im jungen Sporogon liegt außerhalb des Sporensackes ein
wohlentwickeltes Assimilationsgewebe, das von einer Epidermis
bedeckt wird. Bei den meisten Laubmoosen sind im unteren Teile der
Kapselwandung Spaltöffnungen ausgebildet. Die reife Kapsel zeigt eine
Fülle eigenartiger Strukturen, die zu ihrer Öffnung dienen und das
Ausstreuen der Sporen vermitteln. Der Kapselstiel, die Seta,
hebt die Kapsel empor, so daß der Wind die Sporen leicht weithin
verbreiten kann. Im einzelnen weist die Gestaltung des Sporogons
bei den drei Ordnungen der Laubmoose, nämlich den Sphagnales, den
Andreaeales und den Bryales, mancherlei Verschiedenheiten auf.
1. Ordnung. Sphagnales[421]. Sie umfassen nur die Familie der
Sphagnaceen oder Torfmoose mit der einzigen, allerdings sehr
formenreichen Gattung Sphagnum. Sie leben an sumpfigen Orten
und bilden große Polster, die an ihrer Oberfläche von Jahr zu
Jahr weiterwachsen, während die tieferen Schichten absterben und
schließlich in Torf übergehen. Die Stämmchen verzweigen sich
reichlich; ein Teil ihrer Zweige wächst aufwärts und bildet das
gipfelständige Köpfchen, ein anderer abwärts und umhüllt den
unteren Teil des Stämmchens (Fig. 454 A). Diese abwärtswachsenden
Zweige sind peitschenförmig gestreckt. Ein Zweig unter dem Gipfel
entwickelt sich alljährlich ebenso stark wie der Muttersproß, der
damit eine falsche Gabelung erhält. Indem nun die Stämmchen von
untenher allmählich absterben, werden die nacheinander erzeugten
Tochtersprosse zu selbständigen Pflanzen. Einzelne Zweige des
Köpfchens fallen durch ihre besondere Gestalt und Färbung auf;
sie erzeugen die Geschlechtsorgane. Die männlichen Zweige tragen
neben den Blättern die runden gestielten Antheridien, die
weiblichen Zweige an[S. 429] ihrer Spitze die Archegonien. Die Sporogone
entwickeln nur einen kurzen Stiel mit angeschwollenem Fuß, sind
längere Zeit von der Archegoniumwand oder Kalyptra eingeschlossen
und sprengen diese an der Spitze, lassen sie also an ihrer
Basis als Scheide zurück (B, C). In der kugeligen Kapsel
wird die hier halbkugelige Columella von dem sporenbildenden
Gewebe (spo) kuppelförmig überlagert. Die Kapsel öffnet sich
mittels eines abspringenden Deckels. Das reife Sporogon ist mit
seinem erweiterten Fuß in das angeschwollene obere Ende einer
nach der Befruchtung des Archegoniums sich emporstreckenden
stielförmigen Verlängerung der Stengelspitze, des Pseudopodiums
(ps), eingesenkt. Auf den eigentümlichen Bau der Blätter und
der Stengelrinde ist bereits oben hingewiesen. Eigenartig sind
die Vorkeime der Torfmoose gestaltet. Die Spore keimt zu einem
kurzen Faden aus, der dann in eine Zellfläche übergeht, auf der die
Stammknospen entstehen.
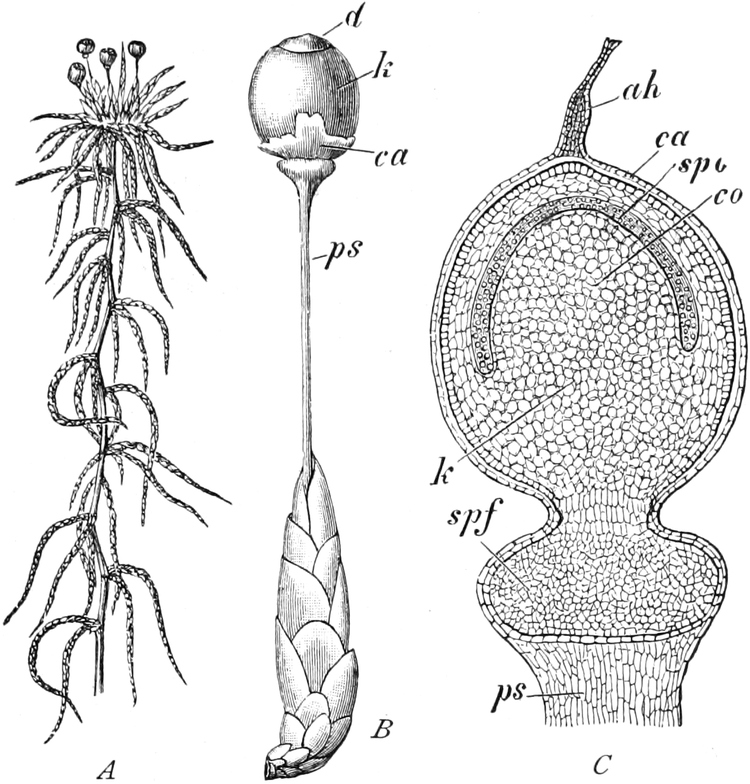
Fig. 454. A Sphagnum fimbriatum. Mit vier reifen
Sporogonen. Nat. Gr. — B Sphagnum squarrosum. Reifes Sporogon am
Ende eines kleines Zweiges, ca durchrissene Kalyptra, d Deckel.
Vergr. — C Sphagnum acutifolium. Junges Sporogon im Längsschnitt,
ps Pseudopodium, ca Archegoniumwand oder Kalyptra, ah
Archegoniumhals, spf Sporogonfuß, k Kapsel, co Columella, spo
Sporensack mit Sporen. B, C nach W. P. SCHIMPER.
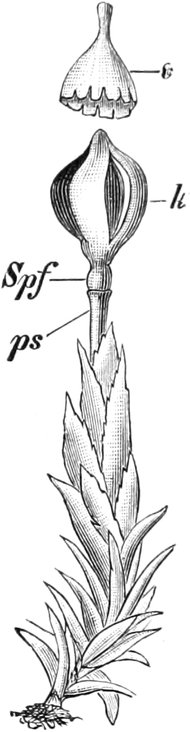
Fig. 455. Andreaea petrophila. ps Pseudopodium, Spf
Sporogonfuß, k Kapsel, c Kalyptra. Vergr. 12.
2. Ordnung. Andreaeales. Sie umfassen nur die Gattung Andreaea,
deren Arten kleine bräunliche Moospolster an Felsen vorstellen.
Die Sporogone stehen an der Spitze des Stengels. Die anfangs
von einer mützenförmigen Kalyptra bedeckte Kapsel öffnet sich
in eigentümlicher Weise mittels vier an der Spitze und Basis
verbundener Klappen (Fig. 455); der Stiel bleibt kurz und besitzt
an seiner Basis einen erweiterten Fuß (Spf), welcher, wie bei
Sphagnum, in ein Pseudopodium (ps) eingesenkt ist. Die Columella
ist wie bei Sphagnum von dem Sporensack kuppelförmig überlagert.
Das Protonema ist anfangs ein kleiner Zellkörper, der zu einem
verzweigten bandförmigen Gebilde auswächst.
3. Ordnung. Bryales[422]. Hierzu gehört die Mehrzahl
der Familien. Die Moosfrucht erreicht bei ihnen höchste
Differenzierung. Das Sporogon besteht aus einem elastischen Stiel,
der Seta (Fig. 456 s), die am Grunde mit ihrem Fuß
in das Gewebe der Mutterpflanze eingesenkt ist, und aus der
Kapsel, die anfangs von der später abfallenden Haube
oder Kalyptra bedeckt wird. Die Haube geht aus dem den
oberen Teil des Sporogonembryos umschließenden Archegoniumbauch
hervor, während der Hals vertrocknet und als Spitze noch auf ihr
sitzen bleibt. Die Bauchwand löst sich an ihrer[S. 430] Basis an einer
vorgebildeten Trennungslinie mit dem Beginn der Sporogonstreckung
los und dient als Schutz für die heranreifende Kapsel. Sie besteht
aus mehreren Schichten von Zellen, erzeugt bei manchen Moosen,
namentlich trockenen Standortes, Haare, die ihrem Bau nach
Protonemafäden begrenzten Wachstums entsprechen. Bei gewissen
Moosen (z. B. Funaria) erweitert sich die junge Haube bauchig und
dient als Wasserspeicher für die junge Kapsel[423]. Der oberste
Teil der Seta unter der Kapsel wird als Apophyse bezeichnet.
Sie ist bei Mnium klein und nur durch eine ganz schwache
Einschnürung von der Kapsel abgesetzt (Fig. 462 A, ap), dagegen
bei Polytrichum commune in Form eines Ringwulstes (Fig. 456 ap)
und am auffälligsten, als rot oder gelb gefärbter Kragen, bei den
nordischen Splachnum-Arten entwickelt, bei denen sie durch ihre
Färbung und zugleich auch durch Abscheidung aasartig riechender
Duftstoffe im Dienste der Anlockung von sporenverbreitenden Fliegen
steht[424]. Die Kapsel wird der Länge nach von der Columella
durchzogen, in deren Umkreis der Sporensack liegt. Der obere Teil
der Kapselwandung ist in Form eines Deckels mit oder ohne
schnabelartige Spitze ausgebildet. Unterhalb des Deckelrandes
ist eine schmale Zone der Kapselwandungszellen als sog. Ring
differenziert. Der Ring, dessen Zellen aufquellenden
Schleim enthalten, vermittelt das Absprengen des Deckels bei der
Reife. Am Rande der Kapselöffnung, zunächst von dem Deckel bedeckt,
befindet sich bei den meisten Laubmoosen ein in der Regel von
Zähnen gebildeter Mundbesatz, das Peristom, das den übrigen
Moosen fehlt.
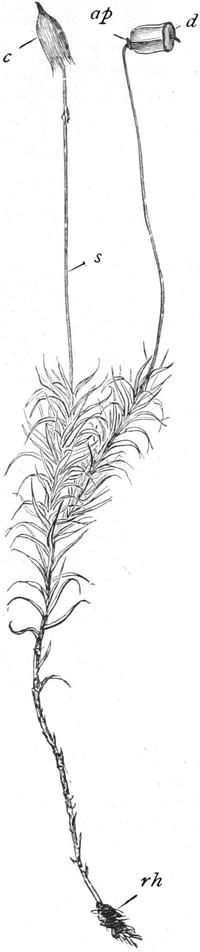
Fig. 456. Polytrichum commune, verzweigtes Exemplar
(Stengel in der Regel einfach), rh Rhizoïden, s Seta, c Kalyptra,
ap Apophyse, d Deckel. Nat. Gr.
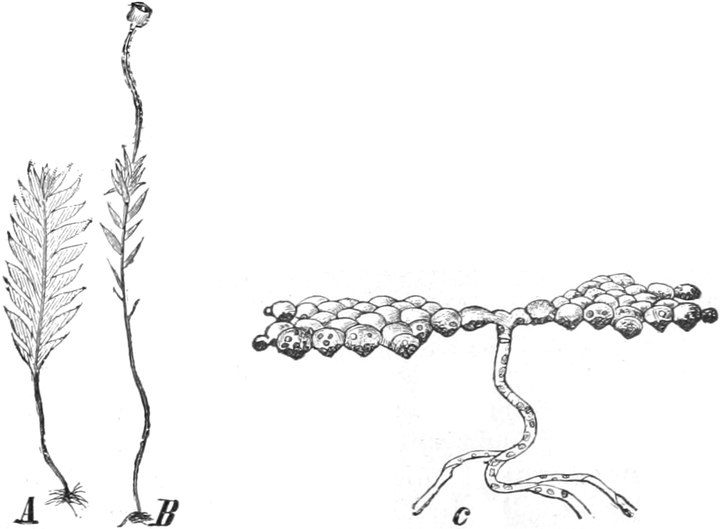
Fig. 457. Schistostega osmundacea. A Sterile,
B fertile Pflanze. Vergr. 5. C Protonema. Vergr. 90. C Nach
NOLL.

Fig. 458. Mnium undulatum. Orthotroper Sproß mit
endständigem, von Hüllblättchen umgebenem Antheridiumstand.
Seitensprosse plagiotrop. Nach GOEBEL.
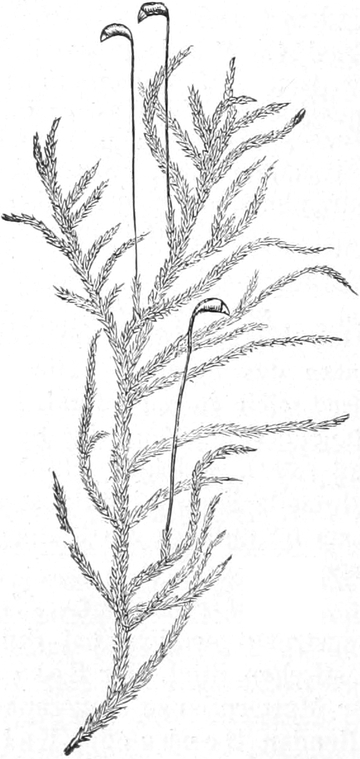
Fig. 459. Scleropodium purum. Nat. Gr.
[S. 431]
Das Peristom von Mnium hornum (Fig. 462) möge als Beispiel
dienen; es ist doppelt. Das äußere besteht aus 16 am Innenrande
der Kapselwandung inserierten, quergestreiften Zähnen. Das innere
Peristom liegt dem äußeren dicht an und setzt sich zusammen aus
schmalen Lamellen und Fäden, die mit Querleisten an der Innenfläche
besetzt und in ihrem unteren Teile zu einer gemeinsamen Membran
verschmolzen sind. Zwischen zwei äußeren Peristomzähnen stehen
jedesmal zwei Wimpern des inneren Peristoms.
Die Entwicklungsgeschichte ergibt, daß die Zähne und Wimpern
aus einer der an die Innenseite des Deckels anschließenden
Zellschichten durch stellenweise Verdickung der gegenüberstehenden
Wände angelegt werden (Fig. 461), und zwar die Zähne aus den
Außenwänden, die Wimpern aus den inneren Wänden dieser Zellschicht.
Die Querleisten entsprechen den Ansatzstellen der Querwände. Bei
dem Öffnen der Kapsel trennen sich die Zähne und Wimpern in den
dünnbleibenden Wandungsstellen.
Bei den Polytrichaceen entstehen die Peristomzähne nach einem
besonderen Typus; sie bauen sich hier aus langgestreckten
verdickten ganzen Zellen auf.
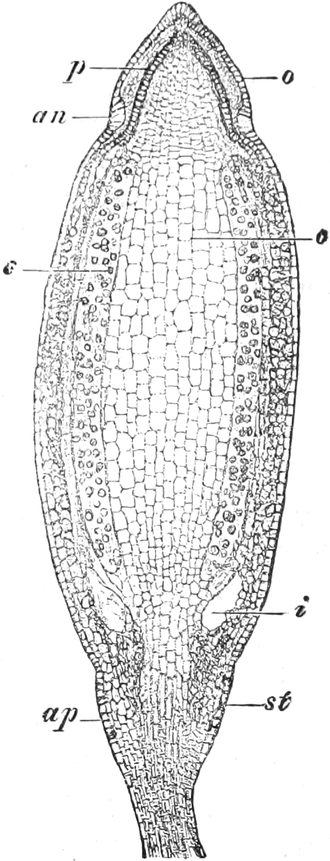
Fig. 460. Mnium hornum. Medianer Längsschnitt durch
ein halbreifes Sporogon. o Deckel, p Peristom, an Ring, c
Columella, s Sporensack mit Sporen, i ringförmiger Hohlraum, ap
Apophyse, st Spaltöffnung. Vergr. 18. Nach STRASBURGER.
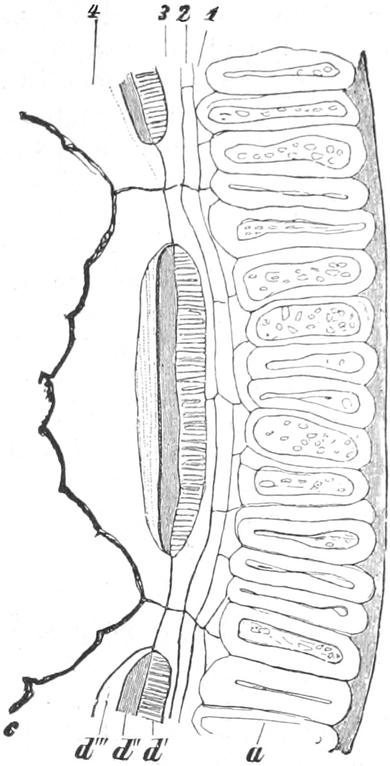
Fig. 461. Mnium hornum. Querschnitt durch den
Kapselrand. a Zellen des Ringes, 1–4 aufeinanderfolgende
Zellschichten, d′ die in der dritten, d″ die in der vierten
Zellschicht entstandene Verdickungsmasse der Zähne, d‴
vorspringende Querleisten, c verschmolzene Wimpern. Vgr. 240. Nach
STRASBURGER.
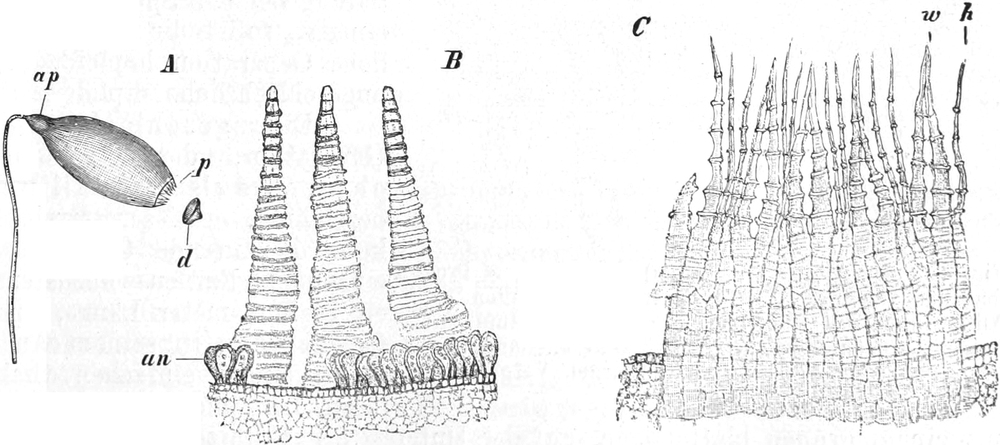
Fig. 462. Mnium hornum. A Kapsel mit einem Stück der
Seta, ap Apophyse, p Peristom, d abgesprengter Deckel. B Drei
Zähne des äußeren Peristoms von außen gesehen, an Ring. C Inneres
Peristom, w die breiteren, h die schmäleren Wimpern, von innen
gesehen. A Vergr. ca. 4. B, C Vergr. 60.
[S. 432]
Im Bau des Peristoms herrscht große Mannigfaltigkeit. Seine Zähne
führen hygroskopische Bewegungen einwärts und auswärts aus und
bewirken so ein allmähliches Ausstreuen der Sporen aus der
Kapsel.
Gestalt der Kapsel, des Peristoms, des Deckels und der Haube geben
die wichtigsten Gattungsunterschiede ab. Die Bryales teilt man
bislang in zwei übrigens kaum natürliche, große Unterordnungen nach
der Stellung der Archegonien oder der Kapseln ein.
a) Bei den Acrocarpi stehen die Archegonien und somit
auch die Sporogone am Ende des Hauptstengels. Von häufigeren
Arten gehören hierher Mnium undulatum (Fig. 458) und hornum,
Polytrichum commune (Fig. 456), Funaria hygrometrica. Eine
sehr eigentümliche Ausbildung des Protonemas treffen wir bei dem
in Erdlöchern oder in Höhlen lebenden Leuchtmoos Schistostega
osmundacea (Fig. 457). Die fertilen Sprosse dieses Mooses sind
einfach oder verzweigt, spiralig beblättert und tragen auf langer
Seta eine peristomlose Kapsel; die sterilen Sprosse dagegen sind
zweizeilig beblättert. Der Vorkeim allein leuchtet smaragdgrün,
indem seine nach unten linsenförmig ausgebauchten Zellen die durch
die Chlorophyllkörner hindurch gehenden Lichtstrahlen reflektieren.
Bei einigen winzigen Moosen (Archidium, Phascum, Ephemerum)
erfährt das Sporogon bedeutende Vereinfachung seiner Struktur;
Deckel-, Ring- und Peristombildungen unterbleiben, und die
Kapselwand öffnet sich unregelmäßig durch Verwesen.
b) Bei den Pleurocarpi wachsen die Hauptachsen unbegrenzt
weiter, und die Archegonien, somit auch die Sporogone, stehen
auf besonderen, kurzen Seitenzweigen (Fig. 459). Hierher gehören
zahlreiche, meist reich verzweigte, Rasen oder Pilze bildende
Arten, darunter unsere größten Waldmoose, die den Familien der
Neckeraceen und Hypnaceen entstammen, ferner auch die in Bächen
und Flüssen flutende Fontinalis antipyretica.
Die Pteridophyten umfassen die Farne nebst den Wasserfarnen, die
Schachtelhalme und die Bärlappgewächse und stellen die am höchsten
entwickelten Kryptogamen vor. Wie bei den Bryophyten vollzieht sich
auch hier der Entwicklungsgang in zwei miteinander abwechselnden
Generationen. Die geschlechtliche Generation trägt Antheridien und
Archegonien, die ungeschlechtliche geht aus der befruchteten Eizelle
hervor und erzeugt ungeschlechtliche einzellige Sporen. Aus der
Keimung der letzteren entsteht wieder die geschlechtliche Generation.
Da die Reduktionsteilung bei der Sporenbildung eintritt, so ist die
geschlechtliche Generation haploid, die ungeschlechtliche diploid.
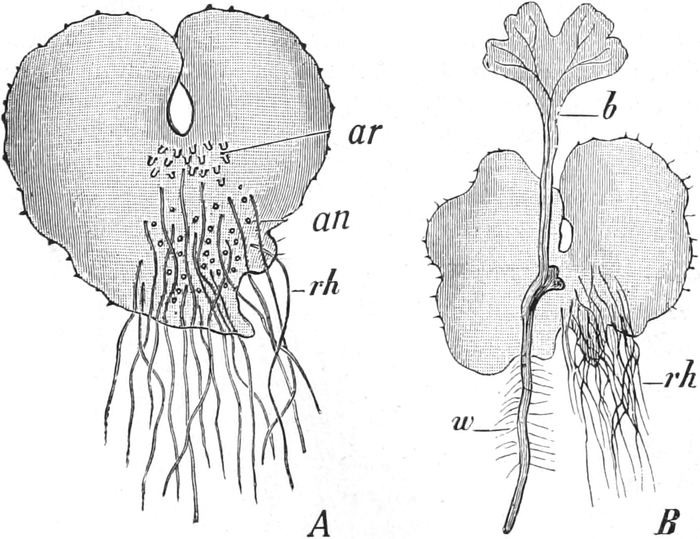
Fig. 463. Dryopteris (Aspidium) filix mas. A
Prothallium von der Unterseite mit Archegonien ar, Antheridien an,
Rhizoïden rh. B Prothallium mit jungem, aus einer befruchteten
Eizelle entstandenem Farnpflänzchen, b erstes Blatt, w Wurzel.
Vergr. ca. 8.
Die geschlechtliche Generation, der Gametophyt, wird
als Prothallium bezeichnet. Dieses erreicht keine bedeutende
Größe, bei einzelnen Farnen höchstens einige Zentimeter Länge, und
gleicht dann in seinem Aussehen einem einfachen thallösen Lebermoos; es
besteht aus einem grünen blattartigen, auf der Unterseite mit Rhizoïden
am Boden befestigten Thallus (Fig. 463 A). In einigen Fällen ist
das Prothallium verzweigt fadenförmig ausgebildet, in anderen Fällen
halb oder ganz unterirdisch in Form von knollenförmigen, farblosen
Gewebekörpern mit saprophytischer[S. 433] Lebensweise und mit endophytischen
Fadenpilzen nach Art der Mykorrhizen; in gewissen Abteilungen der
Pteridophyten endlich erleidet es eine Rückbildung und bleibt in der
Spore mehr oder weniger eingeschlossen. An dem Prothallium entstehen
die Geschlechtsorgane, Antheridien (Fig. 470, 477), in denen
zahlreiche, meist schraubig gewundene, vielzilige oder nur zweizilige
Spermien erzeugt werden, und Archegonien (Fig. 471, 478), die
je eine Eizelle ausbilden. Die Befruchtung ist wie bei den Moosen nur
in Wasser, also bei Benetzung der Prothallien möglich. Eine aus dem
Archegonium in das umgebende Wasser ausgeschiedene Substanz induziert
den Spermien die Bewegungsrichtung nach der Eizelle. Das spezifische
Reizmittel der Samenfäden ist für Farne, Salvinia, Equisetum,
Selaginella und Isoëtes Äpfelsäure oder deren Salze, für Lycopodium
Zitronensäure. Aber auch andere Dikarbonsäuren, gewisse Metallsalze,
sogar auch einige Alkaloide vermögen als Lockmittel zu dienen, und
die einzelnen Gattungen zeigen dabei verschiedenes Verhalten. Die
chemotaktischen Sensibilitäten der Samenfäden können sich auf mehrere
Stoffe erstrecken[427].
Nach der Befruchtung der Eizelle durch ein Spermium entwickelt sich aus
ihr, wie bei den Bryophyten, die ungeschlechtliche Generation, hier das
kormophytische Farnkraut.
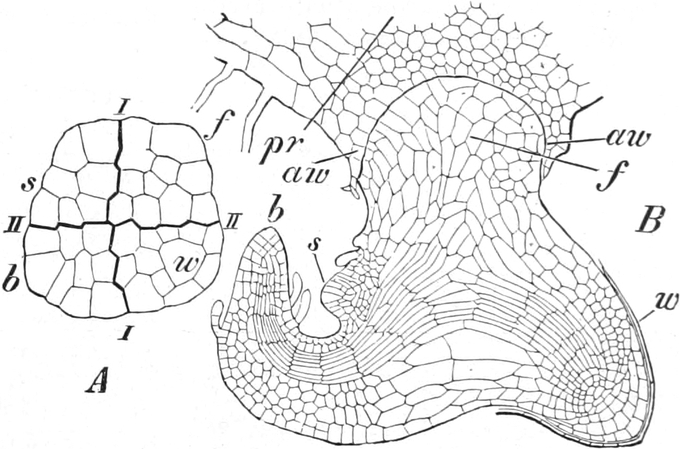
Fig. 464. A Pteris serrulata. Aus dem Archegonium
befreiter Embryo im Längsschnitt. I Basalwand, II senkrecht
zu dieser stehende Quadrantenwand, f Anlage des Fußes, s des
Stammscheitels, b des ersten Blattes, w der Wurzel. Nach
KIENITZ-GERLOFF. B Pteridium aquilinum. Weiter entwickelter
Embryo, mit dem Fuß f im erweiterten Archegoniumbauch, aw,
steckend, pr Prothallium. Vergr. Nach HOFMEISTER.
Die ungeschlechtliche Generation, der Sporophyt, ist
bei den Pteridophyten eine in äußerer Gliederung und innerer Struktur
hochdifferenzierte Pflanze mit Stengel, Blättern und Wurzeln. Bei der
Mehrzahl der Pteridophyten, so bei den Farnen und Schachtelhalmen,
teilt sich die befruchtete Eizelle, nachdem sie sich mit einer
Zellulosemembran umgeben hat, zunächst durch eine Basalwand in zwei
Zellen und jede dieser dann durch zwei zur Basalwand senkrecht
stehende, sich kreuzende Wände in Oktanten. Unter weiterer Teilung
dieser acht Zellen entsteht ein noch im Archegonium eingeschlossener
Gewebekörper, an welchem der Stammscheitel, das erste Blatt, die
erste Wurzel und neben dieser ein der Keimpflanze der Pteridophyten
eigentümliches Organ, der sog. Fuß, angelegt werden (Fig. 464
f). Der Fuß ist ein höckerartig vorspringender Gewebekörper, mit
welchem die junge Keimpflanze in dem durch Wachstum sich erweiternden
Archegoniumbauch eingefügt bleibt; er sorgt als Saugorgan für ihre
Ernährung, bis die Wurzel in den Boden gedrungen ist, die ersten
Blätter sich entfaltet haben und die Keimpflanze somit selbständig
sich ernähren kann. Bei manchen Bärlappgewächsen (Lycopodium,
Selaginella) wird in verschiedener Weise ein ein- oder wenigzelliger
Embryoträger oder Suspensor gebildet. Das Prothallium
geht nach Entwicklung der Keimpflanze in der Regel bald zugrunde. Aus
dem Stammscheitel des Embryo entwickelt sich ein einfacher oder sich
gabelig, ohne Beziehung zu den Blättern verzweigender, aufrechter,
radiärer oder niederliegender, dorsiventraler Stamm, der in schraubiger
oder quirliger Anordnung die Blätter trägt. Die Farnpflanzen bilden
echte, aus verschiedenartigen Geweben aufgebaute[S. 434] Wurzeln, wie wir
sie auch bei den Samenpflanzen vorfinden. Auch die Blätter stimmen im
wesentlichen in ihrer Struktur mit denen der Phanerogamen überein.
Die drei Grundorgane wachsen bei den meisten Pteridophyten mittels
Scheitelzellen heran (Fig. 100, 101, 153); bei Lycopodium und Isoëtes
läßt sich im Vegetationskegel keine solche mehr erkennen, während
Selaginella Scheitelzellwachstum und Übergänge zu Wachstum mittels
zahlreicher Initialzellen zeigt. Stämme, Wurzeln und Blätter werden von
wohl differenzierten Leitbündeln durchzogen, die hier zum ersten Male
im Pflanzenreich erscheinen und als wasserleitende Elemente vorwiegend
Treppentracheïden führen. Die Leitbündel im Stamm und im Blatt der
Pteridophyten sind nach verschiedenen Typen, überwiegend aber nach dem
konzentrischen und nach dem radialen, gebaut (vgl. S. 85 ff., Fig. 465,
466). Sekundäres Dickenwachstum durch Kambiumtätigkeit kommt bei den
jetzt lebenden Familien nur ganz vereinzelt vor, zeichnete aber die
Stämme von gewissen fossilen Pteridophytengruppen aus.
[S. 435]
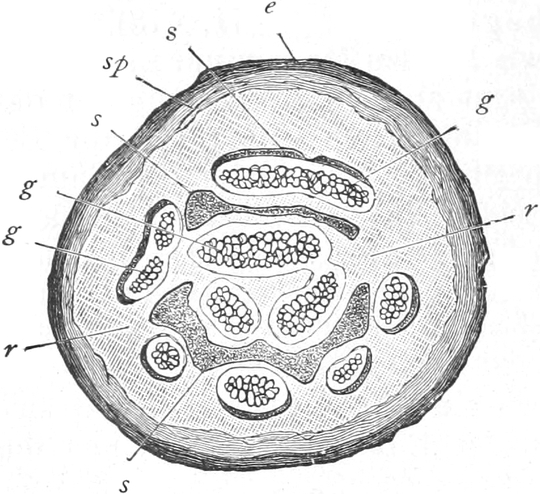
Fig. 465. Querschnitt durch das Rhizom von Pteridium
(Pteris) aquilinum. g Leitbündel, s Sklerenchymplatten, sp
peripherischer Sklerenchymfaserring, r Rinde, e Epidermis. Vergr. 7.
Fig. 466. Querschnitt durch den Stengel von Lycopodium
complanatum. ep Epidermis, ve, vi und pp äußere, innere und
innerste Partie der primären Rinde. Diese umgibt das aus mehreren
Holzteilen und Siebteilen zusammengesetzte zentrale Leitbündel, in
welchem sc Treppentracheïden, sp Ring- und Schraubentracheïden, v
Siebteile sind. Vergr. 26. Nach STRASBURGER.
Fig. 467. Blattaderung bei Farnen. A Adiantum capillus
veneris (Venatio Cyclopteridis). B Asplenium adiantum nigrum (V.
Sphenopteridis). C Asplenium esculentum (V. Goniopteridis). D
Polypodium serpens (V. Marginariae). E Polypodium nereifolium (V.
Goniophlebii). F Onoclea sensibilis (V. Sageniae).
Der Leitbündelverlauf in den Blättern, die Venation, liefert
wichtige Merkmale für die systematische Gruppierung namentlich
bei den Farnen (Fig. 467). Während in den einfachen Blättern der
Schachtelhalme und Bärlappe nur ein Mittelnerv vorhanden ist,
verzweigen sich in den Farnblättern die Nerven in mannigfaltigster
Weise, entweder gabelig oder fiederig und mit freien Ästen, oder
sie anastomosieren zum Teil zu einem System von Maschen. In diesen
polygonalen Maschen können die letzten Auszweigungen blind endigen.
An den Blättern, in einzelnen Fällen in den Blattachseln, werden auf
ungeschlechtlichem Wege die Sporen in besonderen Behältern oder
Sporangien erzeugt (Fig. 468). Die sporangientragenden Blätter
heißen Sporophylle. Die Sporangien umschließen das sporogene
Gewebe, dessen Zellen sich abrunden, voneinander loslösen und die
Sporenmutterzellen darstellen, die eine Reduktionsteilung
ihrer Kerne ausführen und je vier oft tetraëdrisch angeordnete Sporen
(Sporentetraden) liefern. Im Umkreis des sporogenen Gewebes befinden
sich plasmareiche, die Ernährung der Sporen vermittelnde Zellen,
sog. Tapetenzellen, die bei den Lycopodinen erhalten bleiben,
bei Farnen und Schachtelhalmen aber ihre Membranen auflösen und sich
zu einem die Sporenmutterzellen umgebenden Periplasmodium
vereinigen, dessen Zellkerne eine Vermehrung durch amitotische
Teilungen erfahren. Es wandert dann zwischen die sich aus dem
Tetradenverband lösenden jungen Sporen ein, ernährt sie, beteiligt
sich an der Bildung der Sporenhäute und wird dabei aufgebraucht[428].
Die Wand der reifen Sporangien ist entweder einschichtig oder
mehrschichtig. Die jungen Sporenzellen umgeben sich bei ihrer Lösung
aus dem Tetradenverband zunächst mit einer kutinisierten Membran,
dem Exospor, innerhalb dessen eine dünne Zellulosehaut, das
Endospor, abgeschieden wird. In vielen Fällen wird dem Exospor
von dem Periplasmodium noch ein Perispor aufgelagert, so
bei den Schachtelhalmen, Wasserfarnen und gewissen Farnkräutern.
Fig. 468. Entwicklung des Sporangiums von Asplenium.
A Erste Teilungen der aus einer Oberflächenzelle hervorgehenden
Anlage. B Teilung in peripherische Wandanlage w und zentrale Zelle
ar (Archespor), die bereits eine Tapetenzelle t abgeteilt hat. C
Älteres Stadium. Das Archespor hat sich in Tapetenzellen und sporogenes
Gewebe sp geteilt. Vergr. 300. Nach SADEBECK.
Bei der Mehrzahl der Pteridophyten sind die Sporen von gleicher
Beschaffenheit, und bei der Keimung geht aus ihnen ein Prothallium
hervor, an dem sowohl Antheridien als auch Archegonien entstehen.
In gewissen Fällen können aber auch die Prothallien diözisch
sein. Diese Trennung der Geschlechter erstreckt sich bei einigen
Pteridophytengruppen auch schon auf die Sporen und führt zur
Ausbildung von zweierlei Formen von Sporen, Makrosporen, die
in Makrosporangien entstehen und bei der Keimung nur
weibliche Prothallien liefern, und Mikrosporen, die in
Mikrosporangien erzeugt werden und männlichen Prothallien den
Ursprung geben. Danach hat man also zwischen gleichsporigen oder
isosporen (oder homosporen) und verschiedensporigen oder
heterosporen Ordnungen zu unterscheiden, ein Unterschied, der
aber nicht zur Gesamteinteilung verwertet werden kann, da er sich in
gleicher Weise in systematisch getrennten Klassen, also mehrmals,
herausgebildet hat.
[S. 436]
Die Übereinstimmungen in der Struktur der Antheridien,
der Archegonien und der Sporenmutterzellen sprechen für
Verwandtschaft der Farnpflanzen mit den Moosen.
Obwohl beide Gruppen ihren phylogenetischen Ausgang aus einer
gemeinsamen Algengruppe (vgl. S. 421) genommen haben mögen, müssen
wir getrennte Weiterentwicklung annehmen. Vor allem kann der
Farnsporophyt nicht von dem ihm entsprechenden Moossporophyten,
dem Sporogon, abgeleitet werden. Während dieses, ohne vegetative
Ausgliederungen, frühzeitig mit der Sporenbildung seinen Abschluß
erreicht, gliedert sich der Farnembryo in Stamm, Blatt und
Wurzel. Als eine ganz neue Struktur erscheinen in den Geweben
der Farnpflanze die Tracheïden, deren Besitz sie befähigte, zu
größeren, reich gegliederten und sogar zu baumartigen Landpflanzen
sich weiter zu entwickeln, im Gegensatz zu den Moosen, die
infolge ihres einfachen zelligen Aufbaus und des Mangels an
ausgiebigen Wasserbahnen keine großen Dimensionen erreichen
können. Die Farnpflanze geht erst in einem späteren Stadium zur
Erzeugung der Sporen über. Die Sporenmutterzellen werden im Innern
besonderer Blattausgliederungen angelegt; diese heißen zwar
allgemein noch „Sporangien“, sind aber den Thallophyten-Sporangien
nicht homolog. Daher würde es sich empfehlen, für die
sog. Farnsporangien eine neue Bezeichnung (Sporotheken) zu
wählen. Den Thallophytensporangien entsprechen vielmehr bei den
Farnen und Moosen die Sporenmutterzellen, die am ehesten mit den
Tetrasporangien der Braunalgen und Rotalgen verglichen werden
können.
Der Gametophyt der Farnpflanzen schließt seine Entwicklung
frühzeitig mit der Bildung von Geschlechtsorganen ab. Die typischen
Farnprothallien erheben sich nicht über das Jugendstadium
eines Thallus, während umgekehrt bei den Moosen gerade die
geschlechtliche Generation eine fortschreitende Entwicklung
aufweist[408].
Die Pteridophyten gliedern sich in folgende Klassen:
1. Filicinae, Farne. Stengel einfach oder verzweigt, mit
wohlentwickelten, abwechselnden, meist reichgegliederten Blättern,
die hier als Wedel bezeichnet werden. Sporophylle mit zahlreichen,
der Unterseite entspringenden Sporangien, die entweder frei oder
zu mehreren in Soris vereinigt oder in besonderen Blattabschnitten
eingeschlossen sind. Spermien mit vielen Zilien.
- Unterklasse Filicinae eusporangiatae. Reife Sporangien mit
derber mehrschichtiger Wandung ohne Ring. Isospor.
- Unterklasse Filicinae leptosporangiatae. Reife Sporangien mit
einschichtiger Wandung.
- Ordnung. Filices, Farne im engeren Sinne. Isospor. Sporangien
mit Ring.
- Ordnung. Hydropterides, Wasserfarne. Heterospor. Sporangien
ohne Ring.
2. Equisetinae, Schachtelhalme. Stengel einfach oder quirlig
verzweigt, mit quirlig gestellten, einfachen, selten gabelig geteilten,
oder schuppenartigen, zu geschlossenen Scheiden verwachsenen Blättern.
Sporophylle am Ende der Zweige zu einem ährenförmigen Sporangienstand
vereinigt, schildförmig, auf der Unterseite mit mehreren Sporangien.
Spermien mit vielen Zilien.
- Ordnung. Equisetaceae. Schachtelhalme. Isospor. Krautige
Pflanzen.
- Ordnung. Calamariaceae. Schachtelhalmbäume. Isospor oder
Heterospor. Baumartige Pflanzen. Ausgestorben.
3. Sphenophyllinae. Keilblattgewächse.
- Ordnung. Sphenophyllaceae. Stengel schlank, quirlig
beblättert. Sporophylle mit 1–4 Sporangien, in ährenförmigen
Ständen. Isospor, vereinzelt heterospor. Ausgestorben.
4. Lycopodinae. Bärlappartige Gewächse. Stengel einfach oder
gabelig verzweigt. Wurzeln gabelig verzweigt. Blätter einfach, meist
abwechselnd[S. 437] gestellt. Sporangien derbwandige Kapseln, stets einzeln in
den Achseln oder am Grunde der Oberseite der Sporophylle.
- Ordnung. Lycopodiaceae. Bärlappe. Isospor. Spermien zweizilig.
Kräuter mit gegabelten Stengeln.
- Ordnung. Selaginellaceae. Selaginellen. Heterospor. Spermien
zweizilig. Kräuter mit gegabelten Stengeln und kleinen Blättchen.
- Ordnung. Psilotaceae. Homospor. Spermien vielzilig.
Stengel krautig, gegabelt, mit abwechselnden einfachen oder
schuppenförmigen Blättern. Statt Wurzeln Rhizome mit Wurzelhaaren.
Sporophylle gegabelt, auf der Oberseite nahe der Basis je
ein 2- oder 3-fächeriges Sporangium tragend. Isospor.
- Ordnung. Isoëtaceae. Brachsenkräuter. Heterospor. Spermien
vielzilig. Stengel knollig, mit Dickenzuwachs, einfach, Blätter
pfriemlich.
- Ordnung. Sigillariaceae. Siegelbäume, Heterospor. Baumartig.
Stamm einfach oder wenig gegabelt. Ausgestorben.
- Ordnung. Lepidodendraceae. Schuppenbäume. Heterospor. Reich
gabelig verzweigte Bäume. Ausgestorben.
5. Pteridospermeae. Samenfarne. Pflanzen vom Habitus
großer Farne, heterospor, mit Mikrosporangien und mit samenartigen
Makrosporangien. Aus eusporangiaten Farnen hervorgegangen. Ausgestorben.
Zu den Farnen im weiteren Sinne gehört die Hauptmasse der heute
lebenden Pteridophyten. Nach dem Bau ihrer Sporangien werden sie in
zwei Unterklassen unterschieden. Bei den Eusporangiaten besteht
die derbe Sporangienwand aus mehreren Zellschichten und öffnet sich
mittels eines Längsrisses, bei den Leptosporangiaten dagegen ist
sie im reifen Zustand nur einschichtig und reißt quer oder längs auf.
Erstere haben am Grunde der Wedel Nebenblätter oder Scheiden, die den
letzteren fehlen. Auch in der Beschaffenheit der Prothallien und im Bau
der Sexualorgane zeigen sich Unterschiede. Die Sporen besitzen nur bei
einigen Gruppen der Leptosporangiaten eine dem Exospor aufgelagerte
äußere Hülle, das Perispor.
In früheren Erdperioden waren die Eusporangiaten reich vertreten;
heute umfassen sie nur zwei Familien mit wenigen Gattungen.
Allem Anschein nach stellen sie ältere Farntypen dar, die
den Ausgangsformen der Filicinen noch am nächsten zu stehen
scheinen. Neben ihnen erscheinen bereits in paläozoischer Zeit
die Leptosporangiaten, von denen später, in der Kreideperiode
und im Tertiär, die Hydropterides als kleine Gruppe sumpf- und
wasserbewohnender Farne sich abzweigten. Bei diesen haben sich die
bei allen übrigen Farnen noch gleichartigen Sporen in Mikro- und
Makrosporen differenziert.
1. Unterklasse. Eusporangiatae.
Die 1. Ordnung, die Marattiaceen[430], vielleicht die
primitivsten aller heutigen Farne, umfaßt etwa 20 stattliche
tropische Farne mit dicken Stammknollen und meist sehr großen,
an der Basis mit je zwei Nebenblättern versehenen Wedeln. Die
Sporangien stehen an der Wedelunterseite in Gruppen, Sori, entweder
frei (Angiopteris) oder zu einem in Fächern aufspringenden,
kapselartigen, ovalen Gebilde verwachsen. Das Prothallium hat die
Form eines lebermoosähnlichen, grünen, herzförmigen, zuweilen
gegabelten, mehrschichtigen und langlebigen Thallus, dessen
unterseits entspringende, fast ganz eingesenkte Sexualorgane wie
bei folgender Ordnung beschaffen sind; seine Zellen beherbergen
endophytische Fadenpilze.
[S. 438]
Die 2. Ordnung, die Ophioglossaceen[431], enthält ebenfalls nur
wenige Arten. Bei uns heimisch sind die Natterzunge, Ophioglossum
vulgatum (Fig. 469 E) und verschiedene Arten der Mondraute,
Botrychium (Fig. 469 A). Beide haben einen kurzen Stamm, an
dem jährlich meist nur ein einziges, mit Blattscheide versehenes
Blatt sich entfaltet, das bei ersterer Gattung zungenförmig, bei
letzterer gefiedert ist. Diese Blätter tragen an ihrer Oberseite
einen im oberen Teile des Stieles entspringenden fertilen
Blattabschnitt, der bei Ophioglossum einfach zylindrisch ist und
die Sporangien in zwei Reihen in das Gewebe eingesenkt trägt, bei
Botrychium dagegen fiederartig verzweigt und mit großen rundlichen
Sporangien auf seiner Innenseite dicht besetzt ist. Aus dem Verlauf
der Leitbündel und gelegentlichen Rückschlagsbildungen läßt sich
schließen, daß der fertile Blattabschnitt aus der Vereinigung
zweier basaler Blattfiedern hervorgegangen ist.
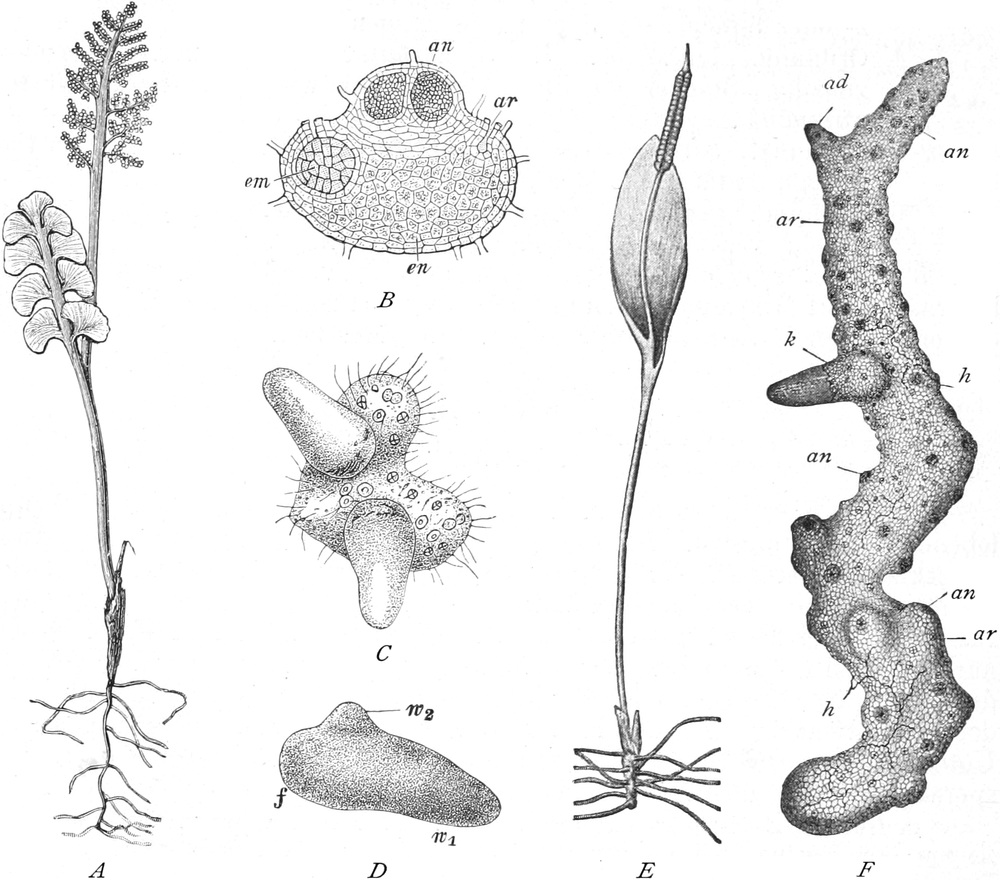
Fig. 469. Botrychium Lunaria. A Sporophyt. 1⁄2 nat.
Gr. B Prothalliumquerschnitt mit Antheridium an, Archegonium ar,
Embryo em, Pilzfäden en. Vergr. 45. C Prothallium mit zwei
Embryonen, deren Wurzeln hervortreten. Vergr. 16. D Embryo mit Wurzel
1 und 2 und Fuß. Vergr. 16. — Ophioglossum vulgatum. E Sporophyt
mit Knospe für das nächste Jahr. 1⁄2 nat. Gr. F Prothallium. an
Antheridien, ar Archegonien, k junge Keimpflanze mit erster Wurzel,
ad Adventivsproß, h Pilzhyphen. Vergr. 15. B–D, F nach
BRUCHMANN.
Sehr eigenartige und von den Marattiaceen abweichende
Beschaffenheit zeigen die besonders durch BRUCHMANN
bekannt gewordenen monözischen Prothallien, unterirdische,
chlorophyllfreie, saprophytische, wie die Mykorrhizen gewisser
Samenpflanzen von Pilzfäden durchzogene, langlebige Knöllchen, bei
Ophioglossum (Fig. 469 F) zylindrisch einfach oder verzweigt
und radiär gebaut, bei Botrychium oval oder herzförmig und
dorsiventral (Fig. 469 B, C). Antheridien (Fig. 470) und
Archegonien (Fig. 471) sind in das Gewebe eingesenkt; erstere
umschließen einen großen Komplex von Samenzellen und öffnen sich
bei der Reife dadurch, daß eine mittlere Deckzelle wohl infolge
Verschleimung[S. 439] ihrer Wände abgeworfen wird. Das Spermium besteht
aus einem schraubig gewundenen Körper mit anhaftendem Bläschen
und zahlreichen Zilien (Fig. 470 E). Die Antheridien gehen aus
oberflächlich gelegenen Zellen (Fig. 470 A–C) hervor, ebenso
auch die Archegonien (Fig. 471 A–C), deren kurzer Halsteil
sich etwas hervorstreckt und nach Verquellung der Halskanalzelle
(hk) sich öffnet, während der Bauchteil eingesenkt bleibt und die
Eizelle (o) umschließt. Der Embryo führt bei manchen Arten eine
Reihe von Jahren hindurch ein unterirdisches Dasein; seine erste
Wurzel wird zunächst angelegt und tritt bald aus dem Archegonium
hervor (Fig. 469 C, F, k), während erst viel später das
erste Blatt und die Scheitelzelle des Stammes zur Differenzierung
kommen. Bei einigen Botrychium-Arten liefert die sich teilende
Eizelle einen langen mehrzelligen Embryoträger oder Suspensor, an
dessen Spitze erst der eigentliche Embryokörper gebildet wird. In
dieser Eigentümlichkeit zeigt sich eine Übereinstimmung mit den
Lycopodinen (vgl. Fig. 495 u. 500), die mit den Eusporangiaten im
übrigen in keiner engeren Verwandtschaft stehen.
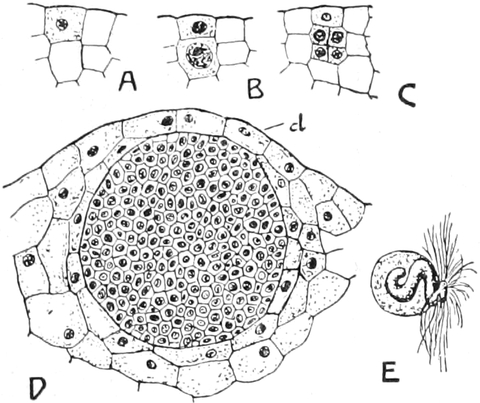
Fig. 470. Ophioglossum vulgatum. A–C Entwicklung
des Antheridiums aus einer oberflächlichen Zelle, die obere Zelle
in C liefert die Deckzellen, die untere die Spermienzellen. D
Antheridium noch geschlossen, d Deckzellen. E Spermium. Nach
BRUCHMANN.
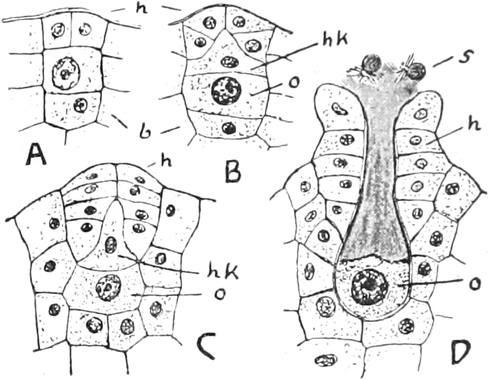
Fig. 471. Ophioglossum vulgatum. A–C Entwicklung des
Archegoniums. D Reifes Archegonium geöffnet mit zwei Spermien s vor
der Mündung. h Halszellen, hk Halskanalzellen, o Eizelle, b
Basalzelle. Nach BRUCHMANN.
2. Unterklasse. Leptosporangiatae.
1. Ordnung. Filices.
Die Filices sind in außerordentlicher Fülle von Arten in allen
Erdteilen verbreitet; ihre Hauptentwicklung erreichen sie in den
Tropen. Hier treffen wir auch die stattlichsten Vertreter an, die
Baumfarne (Cyathea, Alsophila, Dicksonia), welche die
besondere Familie der Cyatheaceen bilden. Der holzige, meist etwa
armdicke Stamm der Baumfarne (Fig. 472) ist unverzweigt und trägt
an seinem Ende eine Rosette von sehr großen, mehrfach gefiederten
Blättern oder Wedeln, die nach dem Absterben große Blattstielnarben
hinterlassen. Der Stamm ist mittels zahlreicher Adventivwurzeln im
Boden befestigt und ist auch von solchen dicht umhüllt. Die meisten
Farne leben als krautartige bodenständige Pflanzen, besitzen ein
wagerechtes oder aufsteigendes, wenig verzweigtes Rhizom
und meist an seinem Ende eine Rosette reichgefiederter Blätter. So
verhält sich u. a. der in Wäldern sehr häufige Wurmfarn Dryopteris
(Aspidium) filix mas, dessen Rhizom als wurmtreibendes Mittel
offizinell ist (Fig. 473). Bei dem gewöhnlichsten einheimischen
Farnkraut, dem Engelsüß, Polypodium vulgare, sind die Blätter einfach
gefiedert und entspringen einzeln auf der Oberseite des kriechenden,
verzweigten Rhizoms. Auch gibt es manche Farne, welche ungeteilte
Blätter aufweisen, so die Hirschzunge, Scolopendrium vulgare
(Fig. 474). In den Tropen wachsen zahlreiche krautartige Farne als
Epiphyten auf den Bäumen.
[S. 440]
Wie Fig. 472 zeigt, sind die Blätter in der Knospe eingerollt,
eine Eigentümlichkeit, die sämtlichen Farnen und auch den Wasserfarnen
zukommt. Im Gegensatz zu der Mehrzahl der Phanerogamenblätter bleibt
bei den Farnblättern der an ihrer Spitze befindliche Vegetationspunkt
länger tätig.
Die meisten Farne sind an ihren Stämmen, Blattstielen und zum Teil
auch den Blättern mit bräunlichen, einschichtigen, oft gefransten
Spreuschuppen (Schuppenhaaren oder Paleae) bekleidet.
Die Sporangien werden in großer Zahl auf der Unterseite der Blätter
erzeugt. Die Sporophylle sind in der Regel nicht von den sterilen
Laubblättern in ihrer äußeren Form verschieden. Nur bei einigen
Gattungen sind sie wesentlich anders gestaltet. Als einheimische
Vertreter sind hier der Straußfarn, Struthiopteris germanica, ferner
Blechnum spicant zu nennen, bei denen gedrungene, dunkelbraune
Sporophylle zu mehreren im Innern der Rosette grüner Wedel stehen.
Im Bau der Sporangien zeigen die einzelnen Familien Unterschiede.
Es sei zunächst das Verhalten der Mehrzahl unserer einheimischen
Farne dargestellt, die zu der umfangreichen Familie der
Polypodiaceen gehören. Die Sporangien erscheinen hier in
verschieden gestalteten Häufchen, sog. Sori, vereinigt.
Sie entspringen auf einem hervortretenden Blattgewebepolster, dem
Receptaculum (Fig. 473 A), und werden bei vielen Arten vor der
Reife von einem häutigen Auswuchs der Blattfläche, dem sog. Schleier,
Indusium, bedeckt und geschützt (Fig. 473 B, C). Das
einzelne Sporangium (Fig. 475) geht aus einer einzigen Epidermiszelle
durch Teilung hervor (Fig. 468), besteht im reifen Zustande aus einer
kleinen Kapsel mit mehrzelligem, dünnem Stiel und mit einschichtiger
Wandung und enthält eine größere Anzahl von Sporen, die nur bei einigen
Gattungen (Asplenium, Aspidium, Acrostichum und Verwandte)
ein sackartiges Perispor aufweisen. Sehr charakteristisch für die
Polypodiaceen ist der Ring, Annulus, der über den Rücken und
Scheitel des Sporangiums bis zur Mitte der Bauchseite als vortretende
Zellenreihe mit stark verdickten Radial- und Innenwänden verläuft.

Fig. 472. Alsophila crinita, Baumfarn von Ceylon. Verkl.
Beim Austrocknen der Kapselwand werden durch den Kohäsionszug des
schwindenden Wassers in den Annuluszellen die dünnen Außenwände
nach innen eingestülpt, der Ring also an seiner Außenseite verkürzt
und dadurch das Aufreißen der Sporangien in einer[S. 441] Querspalte
zwischen den breiten Endzellen des Ringes verursacht. Ist der
Kohäsionszug des Wasserrestes schließlich überwunden, so erfolgt
ein elastisches, die Sporenausstreuung beförderndes Zurückschnellen
des Ringes, worauf das Sporangium infolge Austrocknens und
Kontraktion der dünnen Membranteile dauernd geöffnet bleibt (vgl.
Fig. 277)[432].
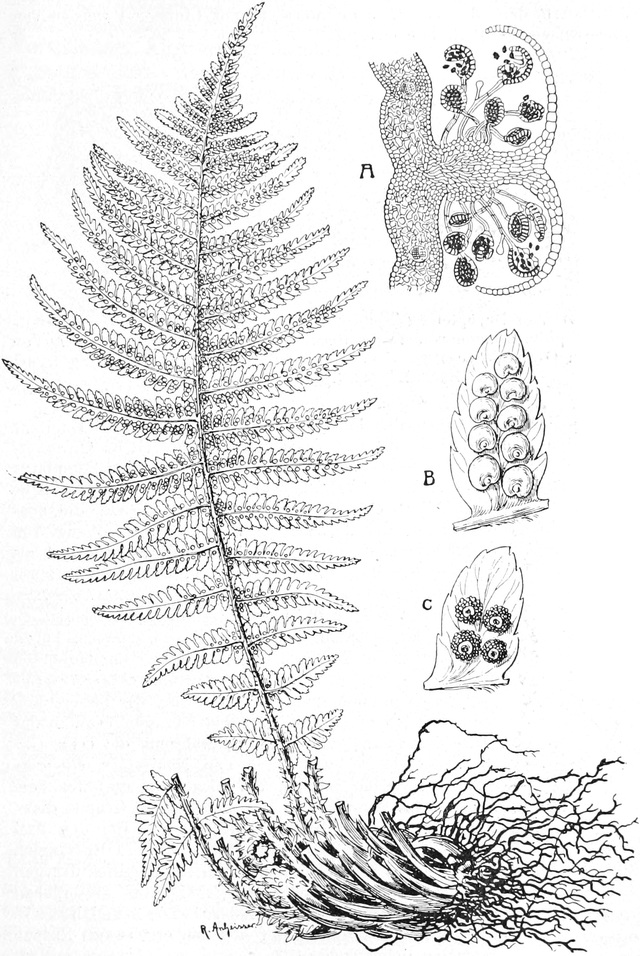
Fig. 473. Dryopteris (Aspidium) filix mas. 2⁄5 nat.
Gr. A Sorus, quer durchschnitten. Vergr. 20. Nach KNY.
B Fiederchen mit jungen, noch vom Schleier bedeckten Soris. C
Desgleichen in älterem Stadium mit geschrumpftem Schleier. Schwach
vergrößert. — Offizinell.
Fig. 474. Scolopendrium vulgare. 1⁄4 nat. Gr.
Die Form und Insertion der Sori, das Vorhandensein und die
Gestalt oder das Fehlen der Indusien geben die wichtigsten
Gattungsunterschiede ab. Bei Scolopendrium (Fig. 474) sind
die Sori strichförmig, bestehen aus zwei parallel über je einen
Blattnerven laufenden Streifen und werden an beiden Seiten von
einem lippenförmigen, einschichtigen[S. 442] Indusium bedeckt, das bei
der Reife zurückklappt. Bei Dryopteris (Aspidium) (Fig. 473)
dagegen treffen wir zahlreiche rundliche Sori, bedeckt mit einem
weißlichen, nierenförmigen, dem Receptaculumscheitel eingefügten
Indusium, und die Sporangien tragen öfters an ihrem Stiel ein
gestieltes, köpfchenförmiges Drüsenhaar. Bei Polypodium vulgare
sind die rundlichen Sori ganz ohne Schleier. Bei dem Adlerfarn,
Pteridium aquilinum, stehen die Sporangien an den Rändern der
Blattfiedern in ununterbrochener Linie und werden von dem nach
unten eingeschlagenen Blattrande bedeckt.
Fig. 475. Sporangien. A Von Dryopteris (Aspidium)
Filix mas. Am Stiel ein Drüsenhaar. B und C von Alsophila armata,
von zwei entgegengesetzten Seiten gesehen. D Von Aneimia caudata. E
Von Osmunda regalis. A–D Vergr. 70, nach der Natur. E Vergr. 40.
Nach LÜRSSEN.
Außer den Polypodiaceen umfassen die Farne noch andere, vorwiegend
tropische Familien, deren Sporangien in der Ringbildung und
dementsprechend auch im Öffnungsmechanismus Verschiedenheiten
zeigen. So besitzen die Cyatheaceen oder Baumfarne
Sporangien mit vollständigem, in schiefem Verlauf über den Scheitel
ziehendem Ring (Fig. 475 B, C); die Hymenophyllaceen,
die ausgezeichnet sind durch zierliche dünnhäutige Wedel von
einfachster anatomischer Struktur und überwiegend als Epiphyten in
tropischen Wäldern, in wenigen Vertretern auch noch im atlantischen
Europa, Hymenophyllum tunbridgense sogar noch in Luxemburg
und in der sächsischen Schweiz an feuchten Sandsteinfelsen,
vorkommen, haben einen vollständigen, schief über das Sporangium
laufenden Ring; die tropischen Schizaeaceen und
Gleicheniaceen dagegen einen quer gestellten Ring, der bei
ersteren nahe der Spitze (Fig. 475 D), bei letzteren etwa in der
Mitte des Sporangiums verläuft, während die Osmundaceen,
die bei uns durch den Königsfarn, Osmunda regalis, vertreten
werden, auf dem Rücken unter dem Scheitel des Sporangiums nur eine
kleine Gruppe dickwandiger Zellen aufweisen (Fig. 475 E). Bei
den drei zuletzt genannten Familien öffnen sich die Sporangien
mittels Längsspalte, bei den drei ersten Familien dagegen
mittels quergestellter oder schiefer Spalte. So ergeben sich
zwei Hauptgruppen, die longiciden und breviciden
Leptosporangiaten, von denen erstere den Eusporangiaten näher
stehen[433].
Alle Filices sind homospor. Ihr Prothallium hat meist die
Gestalt eines flachen, herzförmigen, kleinen Thallus von der für
Dryopteris in Fig. 463 dargestellten Form. Antheridien und
Archegonien entstehen an der dem einfallenden Licht abgewandten Seite,
normal also an der Unterseite.
Fig. 476. Trichomanes rigidum. Teil eines Prothalliums
mit Archegonienträgern A, davon einer mit Keimpflanze. Nach
GOEBEL.
Bei gewissen Hymenophyllaceen (Trichomanes) aber ist das
Prothallium fädig verzweigt und trägt an seinen Ästen die
Antheridien und auf besonderen mehrzelligen Seitenästen[S. 443] die
Archegonien (Fig. 476). Im Aufbau erinnern diese Prothallien an das
Protonema der Laubmoose.
Die Antheridien und Archegonien[434] zeigen denjenigen
der Eusporangiaten gegenüber einige Unterschiede. Die Antheridien (Fig.
477) werden an jungen Prothallien angelegt und sind kugelig vorgewölbte
Gebilde, die ohne Stiel mitten auf einer Prothalliumzelle sitzen und
aus dieser durch papillenartige Vorwölbung, Abgrenzung durch eine
Querwand und weitere Teilung hervorgegangen sind. Ihre Wand besteht
aus zwei ringförmigen Zellen und einer Deckelzelle, die bei einigen
Familien in zwei oder mehr Zellen sich teilt, bei den Polypodiaceen
aber meist einfach bleibt. Die Spermienzellen gehen aus der zentralen
Zelle durch Teilung hervor. Die Entleerung der Antheridien geschieht
durch den Druck der schleimerfüllten und aufquellenden Ringzellen,
welche die ebenfalls aufquellende Deckelzelle absprengen. So gelangen
die rundlichen Spermienzellen ins Wasser und entlassen nach einiger
Zeit je ein pfropfenzieherartig gewundenes, mit zahlreichen Zilien an
den vorderen Windungen besetztes Spermium, an dessen Hinterende ein
Bläschen befestigt ist, das einige kleine Körnchen enthält und einen
unverbrauchten Rest des Inhaltes der Mutterzelle darstellt.
Fig. 477. A Reifes Antheridium von Woodsia ilvensis.
Die Kutikula c ist geplatzt. B Geöffnetes Antheridium, d
Deckelzelle, r aufgequollene Ringzelle. Nach SCHLUMBERGER.
— C Spermium von Struthiopteris germanica. k Zellkern, cl
Geißeln, b Blase aus einer Vakuole hervorgegangen, c Plasma. Vergr.
850. Nach SHAW.
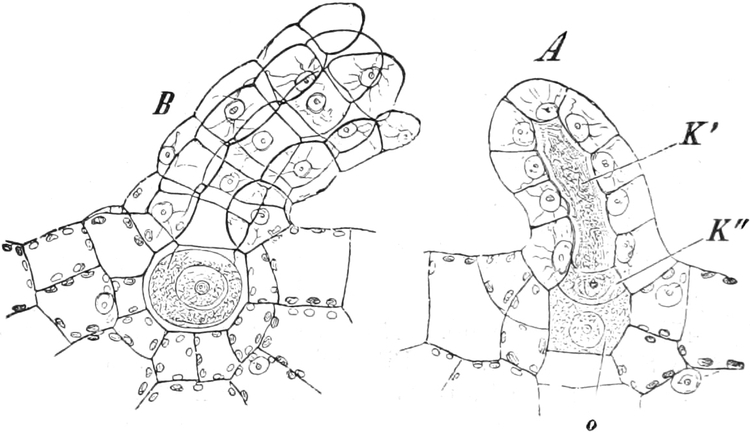
Fig. 478. Polypodium vulgare. A Junges Archegonium.
K′ Halskanalzelle, K″ Bauchkanalzelle, o Ei. B Geöffnetes
Archegonium. Vergr. 240. Nach STRASBURGER.
Die Archegonien (Fig. 478) entstehen in dem mehrschichtigen mittleren
Teile älterer Prothallien durch Teilung einzelner Zellen. Ihr Halsteil
ist länger als bei den Eusporangiaten, besteht aus vier Zellreihen und
schließt eine zentrale langgestreckte Halskanalzelle ein. Im Bauchteile
befindet sich die große Eizelle, über ihr die Bauchkanalzelle. Die
Kanalzellen werden aufgelöst und erfüllen den Kanal mit einer stark
lichtbrechenden Substanz, die bei Wasserzutritt stark aufquillt; das
Archegonium öffnet sich an seiner Spitze. Die Embryoentwicklung ist aus
Fig. 464 zu ersehen.
Ausnahmsweise kann bei gewissen Farnkräutern der Sporophyt auf dem
Prothallium durch vegetative Knospung sich entwickeln, ohne daß
Sexualorgane mitwirken oder ausgebildet werden (Apogamie), und
umgekehrt kommt es auch vor, daß an den Farnwedeln keine Sporen,
sondern direkt Prothallien erzeugt werden (Aposporie).
Giftig: Pteridium aquilinum, der Adlerfarn, enthält
einen Giftstoff, der bei Pferden Erkrankungen und selbst den Tod
herbeiführt.
[S. 444]
Offizinell ist Dryopteris (Aspidium) filix mas,
Rhizoma Filicis (Pharm. germ., austr., helv.), ferner
das südeuropäische Adiantum Capillus Veneris, Frauenhaar,
dessen Blätter benutzt werden: Folium Adianti seu Herba
Capilli Veneris (Pharm. helv.). Auch das nordamerikanische
Adiantum pedatum liefert Folium Adianti (Pharm. helv.).
Die seidenähnlichen, glänzendbraunen Gliederhaare am Grunde der
Blattstiele verschiedener Baumfarne, besonders von Cibotium
Baranetz, im tropischen Asien und auf den pazifischen Inseln,
liefern die als Wundwatte und auch als Polstermaterial benutzten
Paleae haemostaticae (Pennawar Djambi auf Sumatra,
Pakoe-Kidang auf Java, Pulu in Amerika) (Pharm. austr.).
2. Ordnung. Hydropterides, Wasserfarne[435]–[438].
Zu den Wasserfarnen gehören nur wenige Gattungen wasser- oder
sumpfbewohnender Kräuter. Sie sind sämtlich heterospor. Die
Makro- und Mikrosporangien werden in besonderen, an der Basis
der Blätter sitzenden Behältern, sog. Sporangienfrüchten
oder Sporokarpien, eingeschlossen. Auch besitzt ihre
einschichtige Wandung keinen Ring. Die Sporen sind von eigenartigen
Perisporien umgeben.
Die Wasserfarne umfassen die beiden Familien der Marsiliaceen
(mit drei Gattungen) und der Salviniaceen (mit zwei Gattungen).
Zu ersterer gehört die Gattung Marsilia, die bei uns durch M.
quadrifolia vertreten ist (Fig. 479 A). Sie hat eine kriechende,
verzweigte Achse mit einzeln stehenden, langgestielten Blättern,
deren Spreite aus zwei nahe beieinander stehenden Fiederblattpaaren
sich zusammensetzt. Über der Basis des Blattstiels entspringen
paarweise, bei anderen Arten in noch größerer Anzahl, die
gestielten ovalen Sporokarpien, von denen ein jedes seiner
Anlage nach dem assimilierenden sterilen, hier aber ungegliedert
bleibenden Blatteil entspricht. Die jungen Blätter sind, wie bei
den Farnen, an der Spitze schneckenförmig eingerollt.
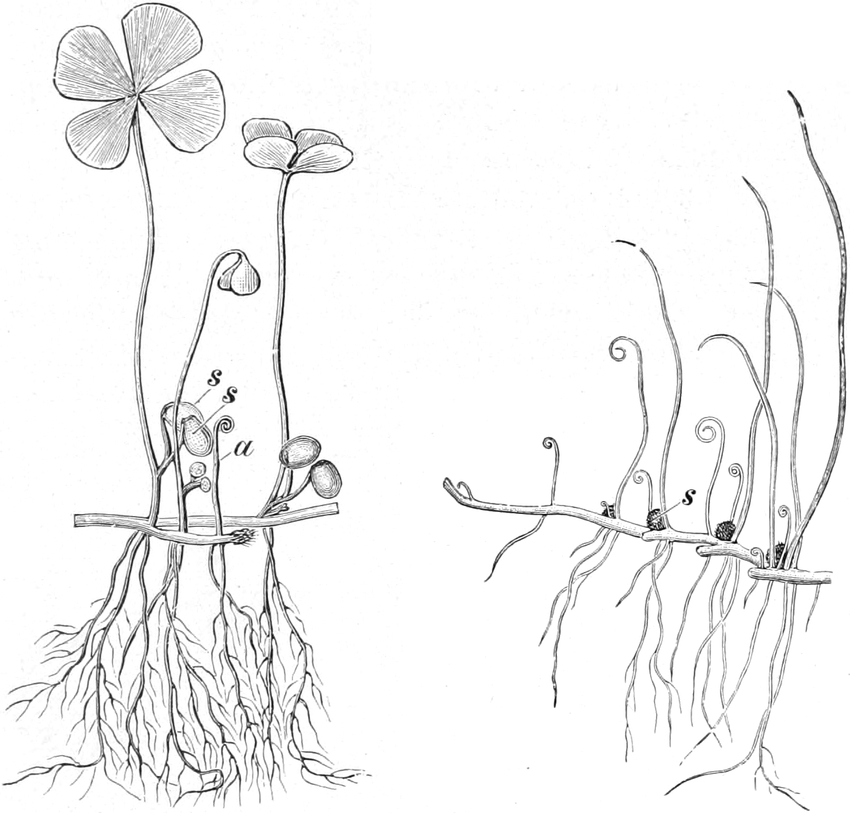
Fig. 479. Wasserfarne. A Marsilia quadrifolia.
a Junges Blatt, s Sporokarpien. B Pilularia globulifera. s
Sporokarpien. Verkleinert. Nach BISCHOFF.
Die Gattung Pilularia, zu der als einheimische Art P.
globulifera, ebenfalls auf sumpfigen Wiesen wachsend, gehört,
unterscheidet sich von Marsilia durch einfache lineale Blätter, an
deren Grunde die kugeligen, in der Anlage dem sterilen Blattstiel
entsprechenden Sporokarpien einzeln entspringen (Fig. 479).
Die zweite Familie, Salviniaceen, enthält frei schwimmende
Wasserpflanzen. Die erste Gattung Salvinia ist in unserer
Flora durch S. natans vertreten, deren wenig verzweigter
Stengel an jedem Knoten drei Blätter trägt; die beiden oberen
sind als ovale Schwimmblätter ausgebildet, das untere dagegen
ist in zahlreiche, in das Wasser herabhängende, fadenförmige,
behaarte Zipfel geteilt und übernimmt die Funktion der fehlenden
Wurzeln. An diesen Wasserblättern sitzen am Grunde der basalen
Zipfel zu mehreren die kugeligen Sporokarpien (Fig. 480 A),
die bei den Salviniaceen eine andere Entwicklungsgeschichte
zeigen als bei den Marsiliaceen. Die Sporangien entspringen auf
einem säulenförmigen Receptaculum, das seiner Anlage nach einem
modifizierten Wasserblattzipfel entspricht. Die Hülle dagegen ist
als Indusium aufzufassen; sie entsteht als Neubildung in Form
eines Ringwalles, der krugförmig und schließlich hohlkugelförmig
über das Receptaculum und seinen Sporangiensorus emporwächst, am
Scheitel aber dicht zusammen[S. 445]schließt. Die zweite Gattung Azolla
ist vorwiegend tropisch und stellt zierliche, reichverzweigte
Schwimmpflänzchen vor mit dicht aufeinanderfolgenden Blättchen in
zweizeiliger Anordnung. Jedes Blatt hat zwei Lappen, von denen der
obere schwimmt und assimiliert, der untere ins Wasser taucht und
an der Wasseraufnahme sich beteiligt. Der obere Lappen enthält
eine Höhlung, die mit enger Öffnung nach außen mündet und stets
Fäden der blaugrünen Alge Anabaena Azollae beherbergt. Zwischen
diesen wachsen aus der Wand der Höhlung Haare hinein, eine
Erscheinung, die auf das Bestehen eines symbiotischen Verhältnisses
zwischen Azolla und Anabaena hindeutet. Azolla besitzt zarte lange
Würzelchen an der Unterseite des Stengels und Sporenfrüchte, die
meist zu zweien am Unterlappen des Blattes einzelner Seitenzweige
entspringen.
Der Bau der Sporangien und Sporen und die Entwicklung der
Prothallien zeigen manche Unterschiede den Filices gegenüber.
Sie mögen zunächst für die Salviniaceen an dem Beispiel von
Salvinia natans[435] erläutert werden. Die Sporokarpien enthalten
entweder Mikrosporangien in größerer Zahl oder Makrosporangien in
geringerer Zahl (Fig. 481 A, ma, mi). Beiderlei Sporangien
erinnern in ihrem Aufbau und ihrer Entwicklung an die Sporangien
der leptosporangiaten Farnkräuter; sie sind gestielt, besitzen
im reifen Zustande eine einschichtige dünne Wandung, aber keinen
Ring (B, D). Die Mikrosporangien umschließen 64
Mikrosporen, die in einer schaumigen, erhärteten Zwischensubstanz
eingebettet liegen, und zwar, ihrer Entstehung in Tetraden
aus den Sporenmutterzellen entsprechend, zu je vier genähert
(C). Die schaumige Zwischensubstanz geht aus dem Plasma der in
einschichtiger Lage gebildeten Tapetenzellen hervor.
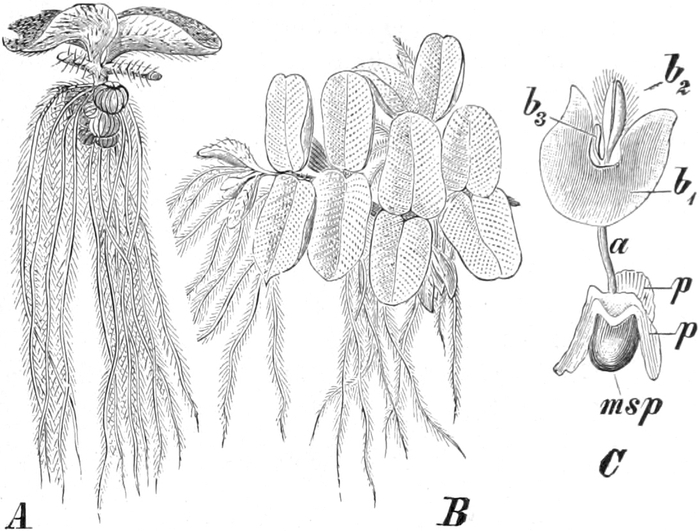
Fig. 480. Salvinia natans. A von der Seite. B Von
oben. Verkleinert. Nach BISCHOFF. C Keimpflanze, msp
Makrospore, p Prothallium, a Stengel, b1 b2 b3 die drei
ersten Blätter. b1 das sog. Schildchen. Vergr. 15. Nach N.
PRINGSHEIM.
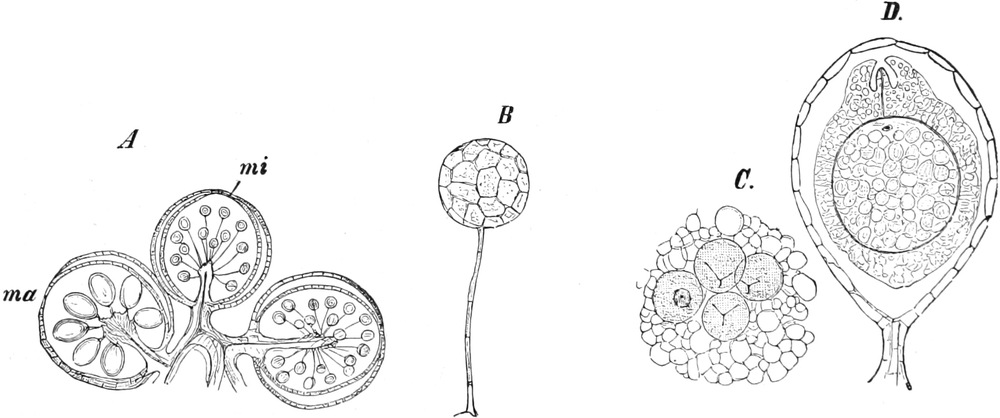
Fig. 481. Salvinia natans. A, ma Makrosporokarp,
mi Mikrosporokarp im Längsschnitt. Vergr. 8. B Ein Mikrosporangium
von außen. Vergr. 55. C In schaumige Zwischensubstanz eingebettete
Mikrosporen. Vergr. 250. D Makrosporangium und Makrospore, im
Längsschnitt. Vergr. 55. Nach STRASBURGER.
Die Mikrosporangien platzen nicht auf; die Mikrosporen entwickeln
nur je ein kurzes schlauchförmiges männliches Prothallium,
das nach außen durch die Sporangienwand hervortritt, nur aus
wenigen Zellen sich aufbaut und nur zwei Antheridien enthält (Fig.
482). Jedes Antheridium erzeugt im ganzen vier Spermien, die durch
Aufbrechen der Zellwände nach außen gelangen. Obwohl somit dieses
Prothallium sehr vereinfacht erscheint, läßt es sich unschwer auf
die Prothallien der Filices zurückführen.
Die Makrosporangien sind größer als die Mikrosporangien
und besitzen ebenfalls eine einschichtige Wandung (Fig. 481 D),
enthalten aber nur eine einzige große[S. 446] Makrospore, da nur eine der
32 angelegten Sporen auf Kosten der übrigen sich weiter entwickelt.
Die Makrospore ist mit eckigen Proteïnkörnern, mit Öltröpfchen und
Stärkekörnern dicht erfüllt; an ihrem Scheitel liegt dichteres
Plasma und der Kern. Ihre braune Sporenwand (Exospor) ist von
einer dicken schaumigen Hülle, dem Perispor, überlagert, die der
Zwischensubstanz des Mikrosporangiums entspricht und wie diese aus
dem Tapetenzellplasma hervorgeht. Die Makrospore bleibt von der
Sporangiumwand umschlossen, wird mit dieser von der Mutterpflanze
frei und schwimmt an der Wasseroberfläche. Bei ihrer Keimung
teilt sie sich in eine scheitelständige kleinere Zelle, aus der
ein kleinzelliges weibliches Prothallium hervorgeht, und in
eine darunter liegende große Zelle, die mit ihrem Reichtum an
Reservestoffen zu dessen Ernährung dient und sich nicht weiter
teilt, obwohl ihr Kern durch freie Kernteilung zahlreiche,
wandständige Tochterkerne liefert. Die Sporenhaut platzt in drei
Klappen auf, ebenso springt die Sporangienwand auf, und das grüne
Prothallium ragt nun als kleines sattelförmiges Gebilde etwas
hervor. Es entwickelt drei bis fünf Archegonien; aber nur die
befruchtete Eizelle eines von ihnen kommt zur Weiterentwicklung
und zur Bildung eines Embryo, der mit seinem Fuß im erweiterten
und schließlich gesprengten Archegoniumbauch steckt (Fig. 483).
Das erste Blatt der Keimpflanze (Fig. 480 C) hat schildförmige
Gestalt, es schwimmt auf der Oberfläche des Wassers.
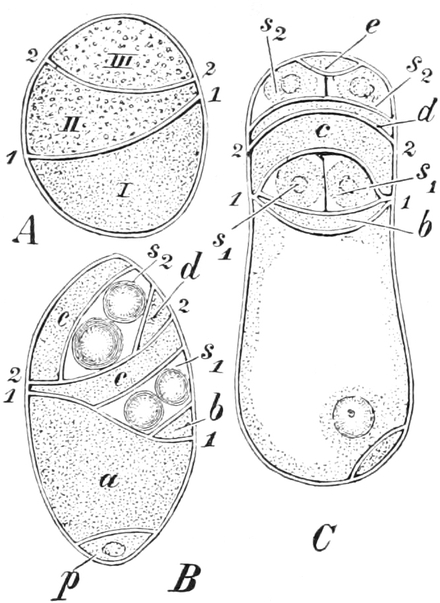
Fig. 482. Salvinia natans. Männliche Prothallien. A
Teilung der Mikrosporen in die drei Zellen I–III. Vergr. 860. B
Fertiges Prothallium von der Flanke. C Von der Bauchseite. Vergr.
640. Zelle I hat sich in die Prothalliumzellen a und p geteilt,
p ist als Rhizoïdzelle zu deuten; Zelle II in die sterilen Zellen
c, b und die beiden spermatogenen Zellen s1, von denen jede
zwei Spermien bildet; Zelle III in die sterilen e, d und die
beiden spermatogenen Zellen s2. Die Zellen s1 s1 und
s2 s2 stellen zwei Antheridien vor, die Zellen b, c, d,
e deren Wandungszellen. Nach BELAJEFF.
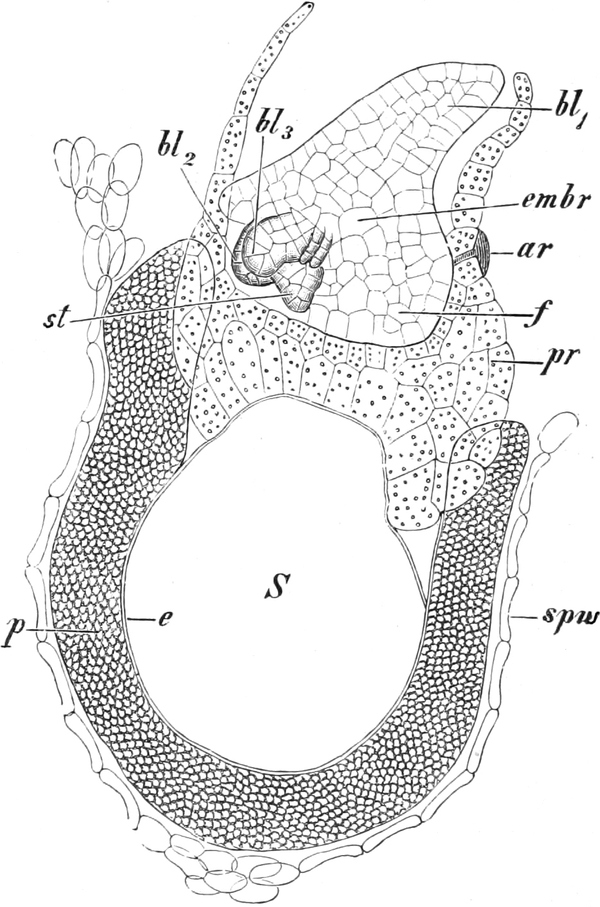
Fig. 483. Salvinia natans. Embryo im Längsschnitt,
Prothallium pr, s Sporenzelle, e Exospor, p Perispor, spw
Sporangiumwand, embr Embryo, f Fuß, bl, bl2 bl3
die drei ersten Blätter, st Stammscheitel. Vergr. 100. Nach
PRINGSHEIM.
Bei Azolla[436] verläuft der Entwicklungsgang in ähnlicher Weise,
aber die Sporangien und Sporen zeigen einige Besonderheiten. Die
Mikro- und Makrosporocarpien entwickeln sich anfangs in gleicher
Weise; ein jedes legt ein einziges Makrosporangium an, das von
der Hülle umwallt wird, und aus dessen Stiel dann innerhalb der
Hülle die Mikrosporangien hervorsprossen. Im Mikrosporokarp
schrumpft die Anlage des Makrosporangiums, und nur die
Mikrosporangien entwickeln sich weiter; im Makrosporokarp
gelangt dagegen nur das Makrosporangium zur Ausbildung. In den
Mikrosporangien werden die[S. 447] 64 Sporen durch eine schaumige
Zwischensubstanz, die von dem Periplasmodium geliefert wird, zu 5–8
rundlichen Ballen, den Massulae, vereinigt. Jede Massula ist
an der Oberfläche mit gestielten Widerhäkchen, Glochiden,
Auswüchsen der Zwischensubstanz besetzt. Die Sporangiumwand platzt
auf und entläßt die Massulae, die im Wasser zu den Makrosporen
gelangen. In den Makrosporangien werden 32 Makrosporen angelegt,
aber nur eine Spore wächst weiter, verdrängt alle anderen und preßt
schließlich auch die Wandung des Makrosporangiums zusammen, so daß
diese dicht an die eiförmige Sporenfruchtwandung zu liegen kommt.
Das Perispor umgibt die Makrospore als schaumige, mit Vertiefungen
und fadenförmigen Verlängerungen versehene Haut und bildet an ihrem
Scheitel einen Aufsatz von drei birnförmig gestalteten Gebilden,
den Schwimmkörper. Die Massulae haken sich in das Perispor
fest. Die Sporenfrucht reißt am unteren Teile auf, ihr Scheitel
verbleibt an der freigewordenen Makrospore in Form eines Schirmes.
Die Prothalliumbildung stimmt im wesentlichen mit Salvinia überein,
an den wenigzelligen männlichen Prothallien, die aus den Massulae
hervorwachsen, entsteht aber nur je ein einziges Antheridium mit
acht Spermien.
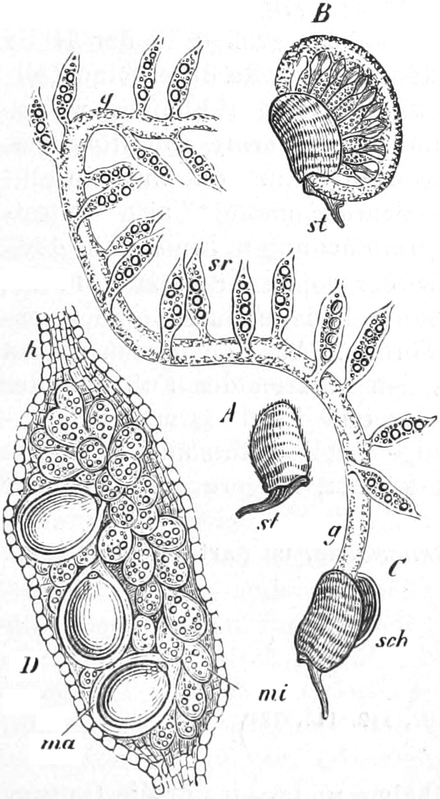
Fig. 484. Marsilia salvatrix. A Sporokarp in nat.
Gr., st Stiel. B In Wasser aufgesprungenes Sporokarp mit dem
hervortretenden Gallertring. C Dieser (g) zerrissen und gestreckt
mit den Sorusfächern sr, Sporokarpschale sch. D Unreifer Sorus.
m Makrosporangien, mi Mikrosporangien. Nach J. SACHS und
J. HANSTEIN.
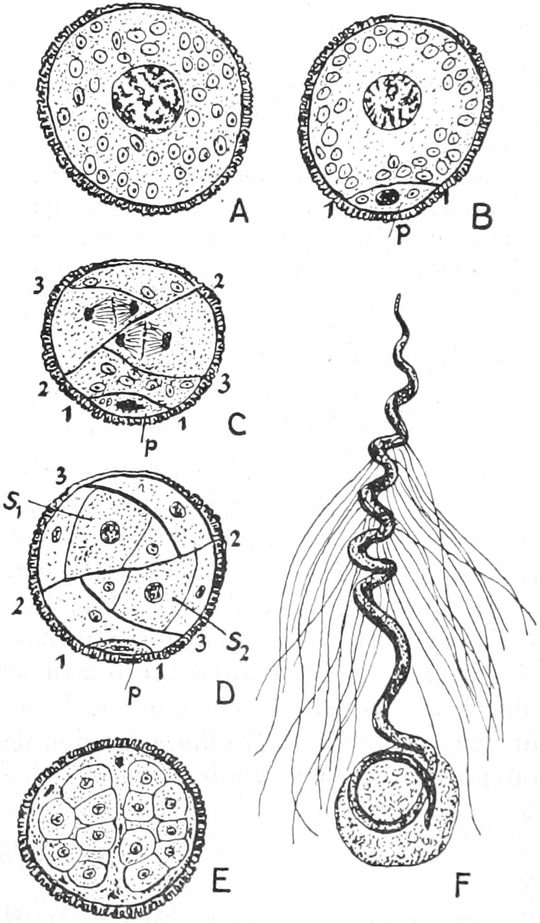
Fig. 485. Marsilia quadrifolia. Entwicklung des
männlichen Prothalliums aus der Spore A. In B durch Wand 1 eine
kleine Prothalliumzelle (rudimentäre Rhizoïdzelle) p abgeteilt,
C und D weitere Teilungen. s1 und s2 die Mutterzellen
der spermatogenen Zellen in beiden Antheridien. E Reifes Stadium,
die beiden Gruppen von je 16 Spermienzellen, aus s1 und s2
hervorgegangen, liegen in dem durch Auflösung der peripheren sterilen
Zellen gebildeten Plasma. F Ein Spermium stark vergrößert, die
Zilien entspringen einem bandförmigen, dem schraubig gewundenen Kerne
anliegenden Blepharoplasten. Nach LESTER W. SHARP.
Die Sporokarpien der Marsiliaceen[437] sind komplizierter
gebaut, enthalten bei Pilularia globulifera vier Fächer, jedes
mit einem Sorus, bei Marsilia zahlreiche Fächer (14–18) mit
je einem Sorus, in zwei Reihen nebeneinander gelagert. Die Sori
beider Gattungen führen zugleich Makro- und Mikrosporangien. Sie
entstehen wie bei manchen Farnen am Blattrande der Sporophyllanlage
aus Oberflächenzellen, die dann nachträglich vom umgebenden
Gewebe überwallt werden. So liegen sie schließlich eingebettet
im Sporophyllgewebe, dessen äußere Wandschichten zu einer harten
Schale sich differenzieren. Nach[S. 448] einer Ruhezeit keimen die
Sporokarpien in Wasser. Bei Pilularia quillt das die
Sori umgebende Gewebe stark auf, sprengt die Schale am Scheitel
vierklappig auf und tritt als Schleimmasse mit den Sporangien
hervor, aus denen ebenfalls durch Verquellung der Wandungen
die Sporen frei werden. Prothalliumentwicklung und Befruchtung
erfolgen in dieser mehrere Tage lang sich haltenden Schleimmasse.
Die Sporokarpschale von Marsilia dagegen wird zweiklappig
aufgesprengt. Eine den Sorusfächern an Rücken- und Bauchnaht des
Sporokarps ringförmig anliegende knorpelige Gewebemasse quillt
stark gallertig auf, sprengt die Bauchnaht, tritt hier hervor,
zieht die Sori, die von häutigen Hüllen umgeben sind, mit heraus,
verlängert sich wurmförmig und reißt schließlich durch (Fig. 484).
Aus der Mikrospore geht innerhalb der Sporenhülle ein
reduziertes männliches Prothallium hervor, das im fertigen Zustand
nur zwei Antheridien mit je 16 Spermienzellen umschließt und nach
dem Aufplatzen der Hülle die korkzieherartig gewundenen, mit
zahlreichen Zilien versehenen Samenfäden entläßt (Fig. 485).
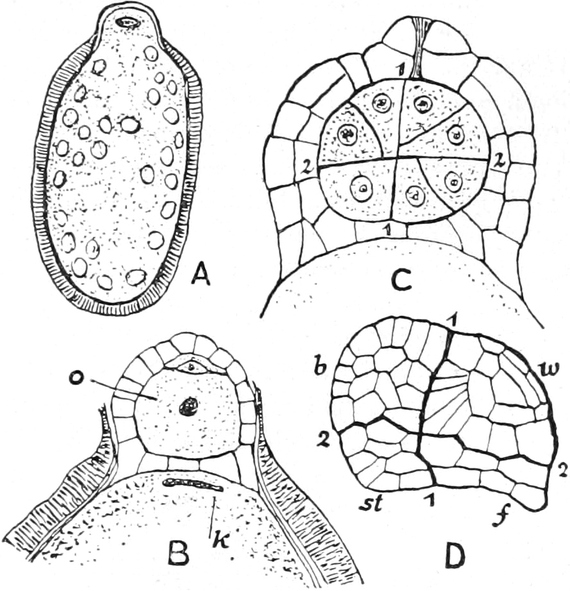
Fig. 486. Marsilia vestita. A Makrospore, am Scheitel
der Kern und das Plasma, aus dem das in B dargestellte weibliche
Prothallium hervorgeht; o Eizelle des Archegoniums, darüber
Bauchkanal- und Halskanalzelle; k Kern der in der Sporenhülle
verbleibenden großen Sporenzelle. C Junger Embryo im Archegonium nach
den ersten Teilungen, 1 Basalwand, 2 Quadrantenwand. D Späteres
Stadium, w Wurzelanlage, b erstes Blatt, st Stamm, f Fuß. A
Vergr. 60, B 360, C 525, D 260. Nach D. CAMPBELL.
Die dickwandige Makrospore entwickelt in ähnlicher Weise
wie bei Salvinia an ihrem aufspringenden Scheitel aus dem hier
befindlichen dichteren Plasma, das von der großen, in der Hülle
verbleibenden Sporenzelle durch eine Zellwand abgeteilt wird, ein
kleines, wenigzelliges, im Längsschnitt sattelförmiges, ergrünendes
Prothallium, das nur ein einziges Archegonium erzeugt, also
weitestgehende Vereinfachung aufweist (Fig. 486).
Die Entwicklung des Embryos folgt dem Typus der
leptosporangiaten Farne, indem die Eizelle sich erst durch eine
längsgerichtete Basalwand, dann durch eine Querwand in Quadranten
teilt, die darauf in Oktanten zerlegt werden. Aus den beiden
oberen Oktantenpaaren geht die Wurzel und das erste Blatt, aus den
unteren der Fuß und der Stammscheitel hervor (Fig. 486 C D). Das
Prothallium wächst eine Zeitlang mit und umhüllt den Embryo; es
treibt aus seinen unteren Zellen einige farblose Rhizoïden. Findet
keine Befruchtung statt, so entwickelt sich aus ihm ein kleiner,
längere Zeit lebender Thallus, der keine neuen Archegonien mehr
erzeugt.
Für gewisse neuholländische Marsilien der Gruppe Drummondi ist
parthenogenetische Keimbildung nachgewiesen worden[438].
1. Ordnung. Equisetaceae. Die Schachtelhalme umfassen nur
die Gattung Equisetum, die in vergangenen Erdperioden bis
in die Trias zurückreicht und heute nur noch in 20 weit
verbreiteten Arten vertreten ist. Diese sind teils Land-, teils
Sumpfpflanzen. Sie zeigen einen sehr charakteristischen Aufbau
ihrer ungeschlechtlichen Generation. Aus einem im Boden
kriechenden Rhizom entspringen aufrechte Halme von meist nur
einjähriger Lebensdauer. Bei Equisetum arvense (Fig. 488),
dem Ackerschachtelhalm, sowie auch bei anderen Arten werden
seitliche kurze Rhizomäste in Form von rundlichen Knollen als
Reservestoffbehälter und Überwinterungsorgane ausgebildet. Die
oberirdischen Halme bleiben entweder einfach, oder sie verzweigen
sich in quirlig gestellte Äste zweiter, dritter usw. Ordnung. Alle
Achsen sind aus gestreckten Internodien zu[S. 449]sammengesetzt, innen von
einem zentralen und von peripherischen Luftgängen sowie von einem
Kreis von kollateralen Leitbündeln durchzogen (Fig. 487).
An den Knoten sitzen abwechselnde Quirle von zugespitzten,
unterwärts in eine den Stengel umschließende Scheide verwachsenen
Schuppenblättern. Die Internodien sind mit ihrer Basis in
diese Scheiden eingeschachtelt. Die Seitenzweige werden in den
Blattachseln angelegt und brechen quer durch die Scheiden nach
außen hervor. Entsprechend der geringen Größe und ungeeigneten
Beschaffenheit der Blattspreiten übernehmen die Halme die Funktion
der Assimilation und bilden das chlorophyllführende Gewebe aus.
Die Sporangien werden von besonders gestalteten Blättern,
Sporophyllen, erzeugt. Diese sind in mehreren Quirlen an den
Enden der Sprosse angeordnet und bilden somit zapfenförmige
Sporophyllstände (Fig. 488), die in ihrem Aufbau den männlichen
Blüten der Koniferen gleichen und auch als Blüten zu bezeichnen
sind. Der unterste Quirl ist steril, bildet einen kurzen Kragen.
Die Sporophylle selbst haben die Form eines gestielten Schildes,
an dessen Unterseite 5–10 sackförmige, mit Längsriß aufspringende
Sporangien sitzen (Fig. 488 B, C). Das sporenbildende Gewebe
ist im jüngeren Sporangium von einer mehrschichtigen Wandung
umgeben. Während die inneren Lagen als Tapetenzellen ihre Wandung
auflösen und ihr Plasma zum Periplasmodium wird, das zwischen
die sich abrundenden Sporen eindringt und bei der Bildung der
Sporenwand aufgebraucht wird, bleibt bei der Reife nur die
äußerste Zellschicht als definitive Wandung erhalten; ihre Zellen
erhalten Spiral- und Ringfaserverdickungen; die Sporangien
gleichen darin den ihnen homologen Pollensäcken der Phanerogamen.
Das Aufspringen geschieht durch Kohäsionszug des schwindenden
Füllwassers und durch die Kontraktion der dünnen Membranteile
beim Austrocknen. Das geöffnete Sporangium entleert zahlreiche
grüne Sporen mit eigenartig gebauter Wand. Der aus Endospor und
Exospor zusammengesetzten eigentlichen Sporenwand wird von dem
Periplasmodium ein Perispor aufgelagert. Diese äußere Schicht
besteht aus zwei schraubig gewundenen, parallel laufenden, an ihren
Enden löffelförmigen Bändern (Elateren), die sich beim Austrocknen
der Sporen ablösen, aber an einer Stelle in ihrer Mitte miteinander
und mit dem Exospor verbunden bleiben; sie breiten sich aus, legen
sich bei Zutritt von Feuchtigkeit aber wieder zusammen und mögen
durch ihre hygroskopischen Bewegungen dazu dienen, die Sporen, die
meist diözische Prothallien bilden, gruppenweise zu verketten und
zu verbreiten (Fig. 488 D, E).
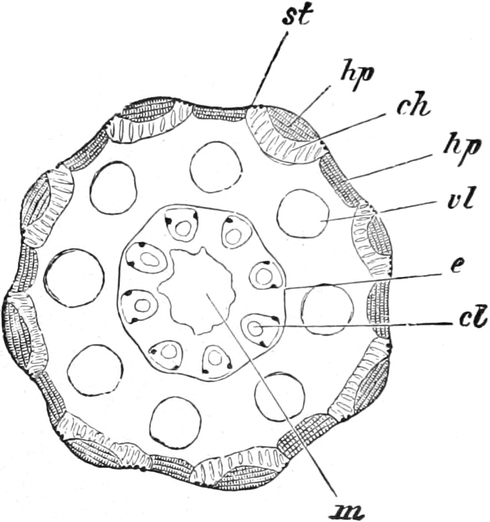
Fig. 487. Equisetum arvense. Stengel quer, m lysigene
Markhöhle, e Endodermis, ci Karinalhöhlen in den kollateralen
Leitbündeln, vl Vallekularhöhlen, hp Sklerenchymstränge in den
Riefen und Rippen, ch chlorophyllführendes Gewebe der Rinde, st
Spaltöffnungsreihen. Vergr. 11. Nach STRASBURGER.
Bei gewissen Schachtelhalmen hat sich ein Unterschied in der
Ausgestaltung der oberirdischen Halme herausgebildet. Teils bleiben
sie steril und verzweigen sich reichlich, teils tragen sie an ihrem
Ende die Blüten und verzweigen sich dann später sparsamer oder
überhaupt nicht in unfruchtbare Seitenzweige. Am ausgeprägtesten
ist dieser Unterschied bei Equisetum arvense und E. Telmateja,
bei denen die fertilen Halme ganz einfach sind, an ihrem Ende mit
einer einzigen Blüte abschließen (Fig. 488) und sich auch durch
den Mangel des Chlorophylls und ihre blaßrötliche Färbung von den
vegetativen Halmen unterscheiden. Sie verhalten sich also gleichsam
wie parasitische Sprosse, die aus dem Rhizom ihre Nahrung beziehen.
Das im tropischen Amerika einheimische Equisetum giganteum ist
die größte Art der Gattung, sie erhebt sich kletternd im Gesträuch
mit ihren 2 cm dicken, quirlig verzweigten Halmen bis über 12 m
Höhe.
Die Sporen sind sämtlich von gleicher Beschaffenheit und keimen zu
thallösen Prothallien aus. Diese sind meist diözisch (Fig.
489); weibliche bedürfen zu ihrer Entwicklung guter Ernährung, bei
schlechter bilden sie Antheridien. Die weiblichen Prothallien sind
bedeutend größer als die männlichen und verzweigen sich reichlicher
in dorsiventrale krause Lappen, an deren Grunde die Archegonien
sitzen. Diese sind ganz[S. 450] ähnlich wie bei den Farnen beschaffen, nur
sind die obersten Zellen des aus vier Zellreihen bestehenden Halses
stark verlängert und biegen sich bei der Öffnung des Archegoniums
stark nach außen um. Die Spermien besitzen wie bei den Farnen
zahlreiche Zilien (Fig. 489 III). Am Embryo treten die
ersten Blätter gleich in einem Quirl angeordnet auf und umwallen
ringförmig den Stammscheitel, der mit dreiseitiger Scheitelzelle
weiterwächst (Fig. 489 IV, 100, 101).
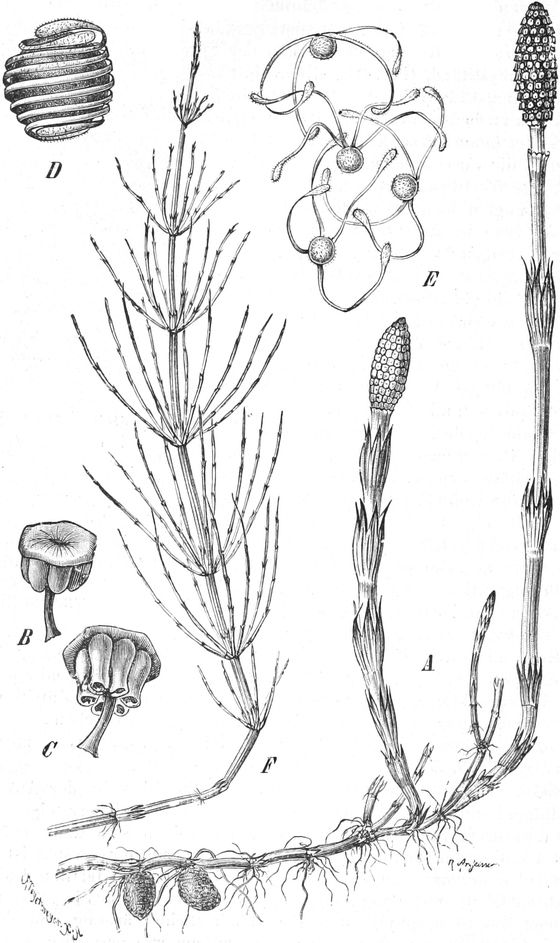
Fig. 488. Equisetum arvense. A Fertile Halme, dem
knollentragenden Rhizom entspringend, mit vegetativem Halm noch in der
Knospe. F unfruchtbarer vegetativer Halm. B und C Sporophylle
mit Sporangien, in C aufgesprungen. D Spore mit den beiden
Spiralbändern (Elateren) des Perispors. E Sporen mit den in trockenem
Zustand ausgebreiteten Spiralbändern. A, F 1⁄2 nat. Gr. B, C,
D, E Vergr.
Die äußeren Membranen der Stengelepidermis sind bei den
Schachtelhalmen mehr oder weniger stark mit Kieselsäure
imprägniert, in besonderem Maße bei Equisetum hiemale, das ebenso
wie auch E. arvense infolgedessen zum Scheuern von metallenen
Gefäßen, zum Polieren von Holz und zu ähnlichen Zwecken Verwendung
findet.
[S. 451]
Giftig: In einigen Equisetum-Arten sind giftig wirkende
Stoffe nachgewiesen. Heu mit viel Schachtelhalmen untermischt ist
für das Vieh schädlich.
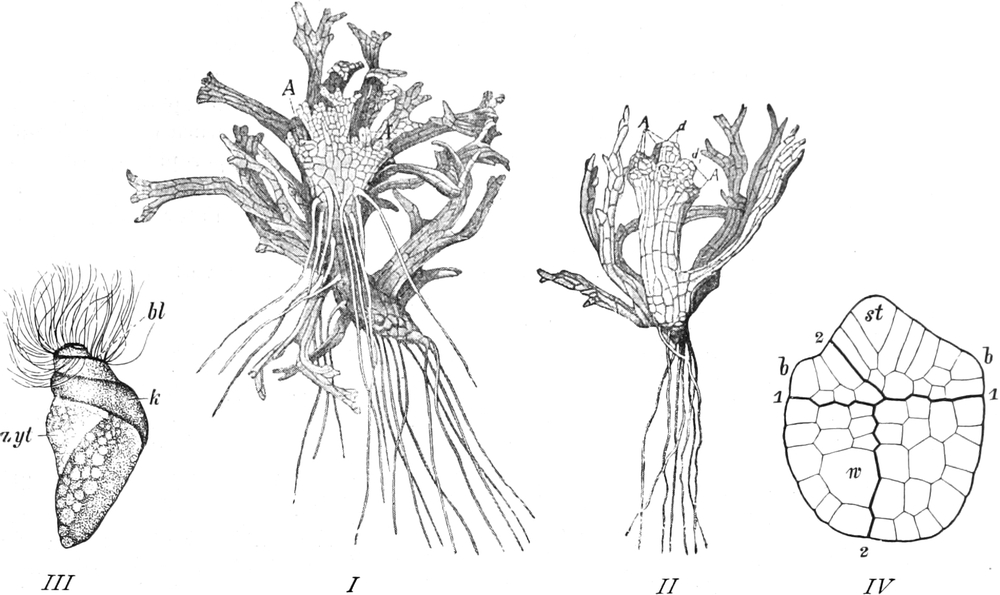
Fig. 489. I, II Equisetum pratense. I Weibliches
Prothallium von der Unterseite, mit Archegonien A. II Männliches
Prothallium mit Antheridien A, d deren Deckzellen. I Vergr.
17. II Vergr. 12. Nach GOEBEL. III Equisetum arvense.
Spermium, k Kern, bl Blepharoplast (Zilienbildner) und Zilien,
zyt Zytoplasma. Vergr. ca. 1250. Nach SHARP. IV Equisetum
arvense. Embryo. 1, 2 Oktantenwände. Aus der über der Basalwand 1
liegenden Hälfte entsteht der Stamm st und der erste Blattquirl b,
aus der unteren Hälfte die Wurzel w und der Fuß. Vergr. 165. Nach
SADEBECK.
Fig. 490. 1 Asterocalamites radiatus, nach
STUR. 2 Annularia stellata, nach SEWARD. Aus
LOTSY, Botan. Stammesgeschichte.
Offizinell: Die sterilen Halme von Equisetum arvense
liefern Herba Equiseti (Pharm. austr.).
2. Ordnung. Calamariaceae[450]. Die ausgestorbenen, im
Paläozoikum sehr reich entwickelten, besonders im Karbon sehr
häufigen Calamariaceen waren habituell den Schachtelhalmen
ähnliche, in einzelnen Arten wohl bis 30 m hohe, baumartige
Gewächse, deren mit Periderm bedeckte, hohle, monopodiale,
quirlig verzweigte Stämme (Calamites) sekundäres Dickenwachstum
aufwiesen. Ihre Blätter (Annularia, Fig. 490) standen in
abwechselnden Quirlen, waren schmal-lanzettlich, anfangs zu einer
Scheide verbunden, später sich trennend und in dem ältesten
Typus Asterocalamites (Fig. 490) noch dichotom geteilt, also
farnblattähnlich. Die Sporangienstände hatten bei dieser Gattung
ähnlichen Bau wie bei Equisetum, bei den übrigen Vertretern
aber waren sie zusammengesetzt aus abwechselnden Quirlen von
Schuppenblättern, zwischen denen superponierte Quirle von
besonderen Sporangienträgern standen. Jeder solcher Träger
hatte die Form eines gestielten Schildes und trug unter diesem[S. 452]
vier Sporangien (Fig. 491). Bei Calamostachys waren die
Sporangienträger ein Stück über die zugehörigen Sporophylle in
die Höhe gerückt, bei Palaeostachya standen sie in den Achseln
dieser; sie können morphologisch als besondere Auswüchse der
schuppenförmigen Sporophylle angesehen werden. Interessant ist
die Tatsache, daß unter den Calamarien neben isosporen bereits
heterospore Arten vertreten waren.
Klasse III.
Sphenophyllinae,
Keilblattgewächse[450].
Diese kleine, ausschließlich paläozoische Klasse nimmt eine
vermittelnde Stellung zwischen Equisetinen und Lycopodinen ein,
schließt sich aber im vegetativen Aufbau an erstere näher an.
Fig. 491. 1 Calamostachys Binneyana. Sporangienstand
im Längsschnitt, 2 im Querschnitt. 3 Calamostachys Casheana.
Querschnitt durch den Stiel eines Sporangienträgers, das drei
Makrosporangien und ein Mikrosporangium trägt. 4 Palaeostachya.
Sporangienstand mit axillären Sporangienträgern, Längsschnitt. Nach
SCOTT und HICKLING. Aus LOTSY, Botanische
Stammesgeschichte.
Fig. 492. 1 Sphenophyllum, verzweigte Stengel mit
linearen und keilförmigen wirteligen Blättern, der rechte Zweig mit
längerem Sporangiumstand. Nach SCOTT. 2 S. emarginatum. Nach
SEWARD. Aus LOTSY, Bot. Stammesgeschichte.
Die Sphenophyllinen waren durch zwei Gattungen vertreten.
Cheirostrobus aus dem untersten Karbon besaß reich gegliederte
Blüten von ähnlichem Bau wie bei den Calamarien. Die vom Devon
bis Perm verbreiteten Sphenophyllum-Arten waren krautige
Landpflanzen, anscheinend Kletterpflanzen, deren langgliedrige
verzweigte Stengel ein axiles dreistrahliges markloses Leitbündel
mit sekundärem Zuwachs aufwiesen. Sie waren mit übereinander
stehenden, meist sechszähligen Quirlen keilförmiger oder gabelig
geteilter Blätter, und mit ährenförmigen Equisetum-ähnlichen
Blüten, deren Sporophylle ein bis vier Sporangien trugen,
ausgestattet (Fig. 492).
Klasse IV.
Lycopodinae,
Bärlappgewächse[288],
[400],
[426],
[429].
Die Lycopodinen unterscheiden sich durch den Habitus ihrer Sporophyten
und ihre Sporangienentwicklung scharf von den übrigen Pteridophyten.
Sie waren bereits in der paläozoischen Periode reich vertreten und
umfaßten auch baumartige Vertreter[S. 453] hauptsächlich in den ausgestorbenen
Ordnungen der Sigillariaceae und Lepidodendraceae.
Die heute noch lebenden zahlreichen Arten sind sämtlich krautige
Gewächse und gehören den Gattungen Lycopodium, Selaginella,
Psilotum und Tmesipteris, Isoëtes an, die sich auf vier Ordnungen
verteilen.
Charakteristisch für die Sporophyten ist die gabelige Verzweigung ihrer
Wurzeln und Stengel (Fig. 144 u. 146), ferner die einfache Form ihrer
Blätter. Die beiden erstgenannten Gattungen besitzen gestreckte Stengel
und kleine Blättchen, Isoëtes dagegen gestauchte knollige Stengel und
lange pfriemliche Blätter. Während bei Filicinen und Equisetinen die
Sporophylle stets zahlreiche Sporangien erzeugen, tragen sie hier
diese Organe stets in Einzahl am Grunde der Blattoberseite oder in
der Blattachsel. Bei manchen Lycopodinen sind die Sporophylle von
den sterilen Blättern kaum verschieden, bei den meisten aber anders
gestaltet und an den Sproßenden zu ährenförmigen Sporophyllständen
oder Blüten, ähnlich wie bei Equisetum, vereinigt. Die Sporangien sind
im Verhältnis zu den Blättern relativ groß und besitzen eine derbe
mehrschichtige Wand ohne Ring. Die innerste Schicht der Wandung, die
Tapetenschicht, wird nicht aufgelöst. Daher weist auch die Membran der
Sporen kein ihr aufgelagertes Perispor auf. Der Raum zwischen den sich
entwickelnden Sporenzellen ist nur mit schleimiger, ihre Ernährung
vermittelnder Flüssigkeit erfüllt. Die Sporangien öffnen sich meist
mit zwei Klappen, die in einer über den Scheitel laufenden Spalte
sich voneinander trennen. Die Spalten sind durch zwei Reihen dünn
bleibender Zellen vorgebildet. Nur bei Isoëtes werden die Sporen durch
Verwesung der Sporangiumwand frei. Während Lycopodium und Psilotum
isospor sind, treffen wir bei Selaginella und Isoëtes Heterosporie
an und zugleich eine weitgehende Reduktion und sehr eigenartige
Ausbildung der Prothallien; bei Lycopodium und Psilotum dagegen sind
die Prothallien wohl entwickelt und zeigen gewisse Ähnlichkeiten mit
denen der Ophioglossaceen. Man kann die vereinfachten Prothallien der
Selaginellen und Isoëten mit Jugendzuständen der Lycopodium-Prothallien
vergleichen, die dann keine vegetative Weiterentwicklung erfahren
hätten, sondern frühzeitig zur Gametenbildung gelangt wären.
Lycopodiaceen und Selaginellaceen stehen einander näher in
der Gliederung des Embryos, der sich bei beiden durch den Besitz
eines Embryoträgers auszeichnet, und in der primitiveren Struktur
ihrer nur zwei Zilien tragenden Spermien. Die Isoëtaceen
dagegen zeichnen sich durch trägerlosen Embryo und ebenso wie die
Psilotaceen durch vielzilige Spermien aus. Danach unterscheiden
wir Lycopodinae biciliatae und pluriciliatae. Schon im
Karbon waren krautige Lycopodinen vorhanden, die Vorläufer von
Lycopodium und Selaginella, während Isoëtes erst aus der
unteren Kreide sicher bekannt ist, die Psilotaceen reichen bis in
das Devon zurück.
1. Ordnung. Lycopodiaceae[440]. Die zahlreichen Arten der Gattung
Lycopodium, Bärlapp, sind krautige, meist erdbewohnende Gewächse;
in den Tropen gibt es auch viele epiphytische Formen mit schlaff
herabhängenden Sproßbüscheln. Eine der häufigsten Arten unserer
Flora ist Lycopodium clavatum (Fig. 493). Der Stengel dieser
wie auch anderer Arten kriecht weit über den Boden hin, gabelt
sich in aufsteigende Äste und ist dicht mit linealpfriemlichen
kleinen Blättchen besetzt. Auf der Unterseite der Stengel
entspringen dichotom verzweigte Wurzeln. Die ährenförmigen
Blüten stehen zu zwei oder mehreren an den Enden von aufrechten,
dichotom verzweigten Stengeln und tragen breit schuppenförmige,
lang zugespitzte Sporophylle, die am Grunde ihrer Oberseite je
ein großes nierenförmiges, zweiklappig aufspringendes Sporangium
mit zahlreichen winzigen Sporen erzeugen (Fig. 493 H). Das
einheimische Lycopodium Selago weicht in seinem Habitus von den
übrigen Arten ab: seine gegabelten Stengel stehen alle aufrecht,
und die Sporophyllstände sind von der vegetativen Region der Zweige
nicht abgesetzt.
[S. 454]
Die Sporangien sind isospor. Da die Sporen bis zu
ihrer Reife in Tetraden verbunden bleiben, behalten sie ihre
kugeltetraëdrische Gestalt bei. Ihr Exospor ist mit netzförmigen
Verdickungsleisten versehen oder mit rundlichen Grübchen getüpfelt
(Fig. 493 J, K).
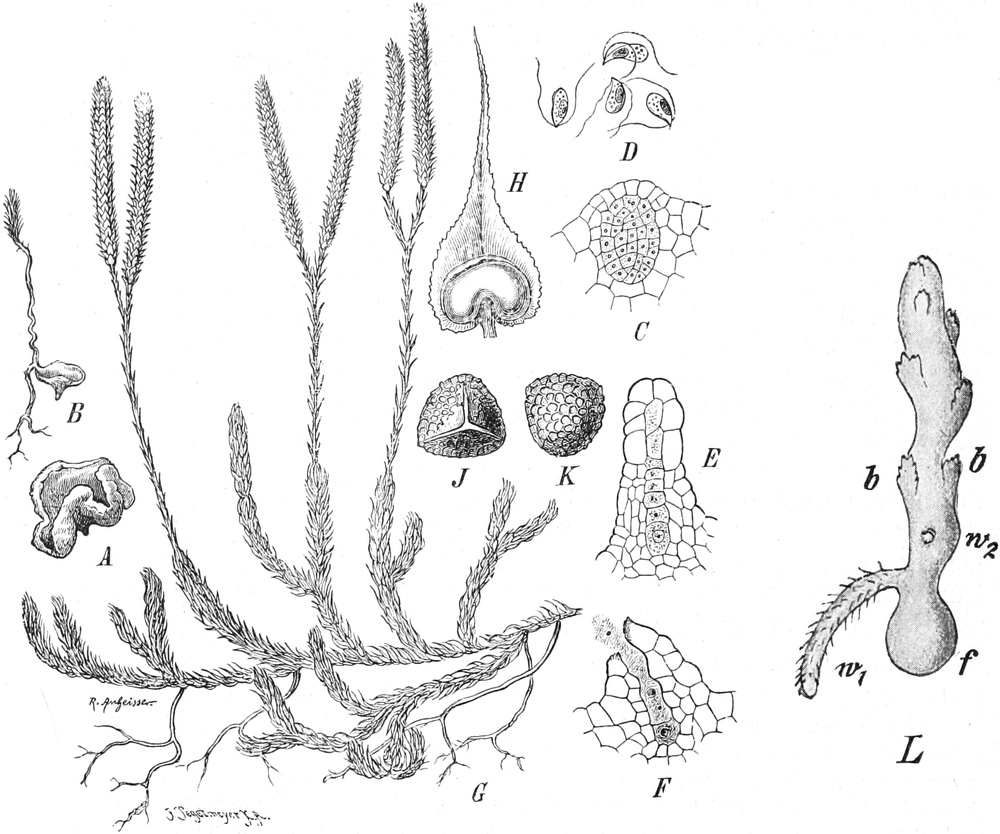
Fig. 493. Lycopodium clavatum. A Älteres Prothallium.
B Prothallium mit junger Pflanze. C Antheridium, noch geschlossen,
Längsschnitt. D Spermien. E Jüngeres, noch geschlossenes, F
befruchtungsreifes geöffnetes Archegonium. G Sporangientragende
Pflanze. 1⁄2 nat. Gr. H Ein Sporophyll mit aufgesprungenem
Sporangium. J und K Sporen in zwei Ansichten. L Junge
unterirdische chlorophyllfreie Keimpflanze, mit Fuß f, Wurzel w und
Schuppenblättern b. Vergr. 10. A–F, L nach BRUCHMANN.
Während die Sporophyten der Lycopodien im wesentlichen
übereinstimmenden Bau aufweisen, zeigen dagegen die aus den Sporen
hervorgehenden Prothallien bemerkenswerte Verschiedenheiten.
Bei L. clavatum (Fig. 493 A, B) und dem nahe verwandten
L. annotinum stellen sie unterirdische, saprophytisch lebende,
weißliche Knöllchen dar, die anfangs kreiselförmig gestaltet,
später durch Auswachsen der Randpartien zu vielgestaltigen,
becherförmigen, wulstig gelappten, bis etwa 2 cm großen
Gewebekörpern werden, die mit langen, der Wasseraufnahme dienenden
Haaren oder Rhizoïden besetzt sind und auf ihrer oberen Fläche
zahlreiche Antheridien und Archegonien tragen. Die Sporen keimen
erst nach 6–7 Jahren und liefern auf Kosten ihrer Reservestoffe
zunächst einen fünfzelligen Keimling, der nach einer Ruhezeit
erst dann sich weiterentwickelt, wenn Pilzfäden in seine unteren
Zellen eintreten (Fig. 494 A, B). Die endophytischen Pilzfäden
bewohnen auch in älteren Prothallien nur die peripheren Gewebe, zum
Teil treten sie aber wieder ins Freie aus den als Durchlaßzellen
dienenden Fußzellen der Wurzelhaare und umspinnen diese [441].
Nach 12–15 Jahren tritt erst die Geschlechtsreife ein, und die
gesamte Lebensdauer der Prothallien mag etwa 20 Jahre betragen.
Bei L. complanatum (Fig. 494 C) sind diese Gewebekörper
rübenförmige, bei L. Selago an der Basis kegelförmige, dann
zylindrische dorsiventrale Knöllchen, die bei letzterer Art auch
an der[S. 455] Oberfläche des Erdbodens sich entwickeln können und dann
ergrünen. Anders dagegen verhält sich das auf feuchtem Torfboden
lebende kleine L. inundatum unserer Flora und das tropische mit
aufrechten, reichverzweigten Sprossen versehene L. cernuum, deren
Prothallien kleine, im Boden steckende und mit Rhizoïden befestigte
chlorophyllarme Gewebekörper vorstellen, die am oberen Ende grüne,
oberirdische Thalluslappen entsenden. Die Archegonien entspringen
am Grunde dieser Thalluslappen, die Antheridien auch auf den Lappen
selbst. Alle Lycopodiumprothallien enthalten in ihren peripheren
Geweben endophytische Pilzfäden nach Art der Mykorrhizen.
Die Prothallien sind sämtlich monözisch. Die Antheridien
(Fig. 493 C) sind in das Gewebe etwas eingesenkt und umschließen
zahlreiche Spermienzellen; jede Zelle entläßt ein ovales, unter
seiner Spitze zwei Zilien tragendes Spermium (Fig. 493 D). Die
Archegonien (Fig. 493 E, F) sind ähnlich wie bei den
Farnen beschaffen, an ihrem Halsteil gehen die obersten Zellen beim
Öffnen zugrunde. Die Zahl der Halskanalzellen ist bei den einzelnen
Arten verschieden (1, 3 bis 5, oder 6 bis 10, sogar bis 20).
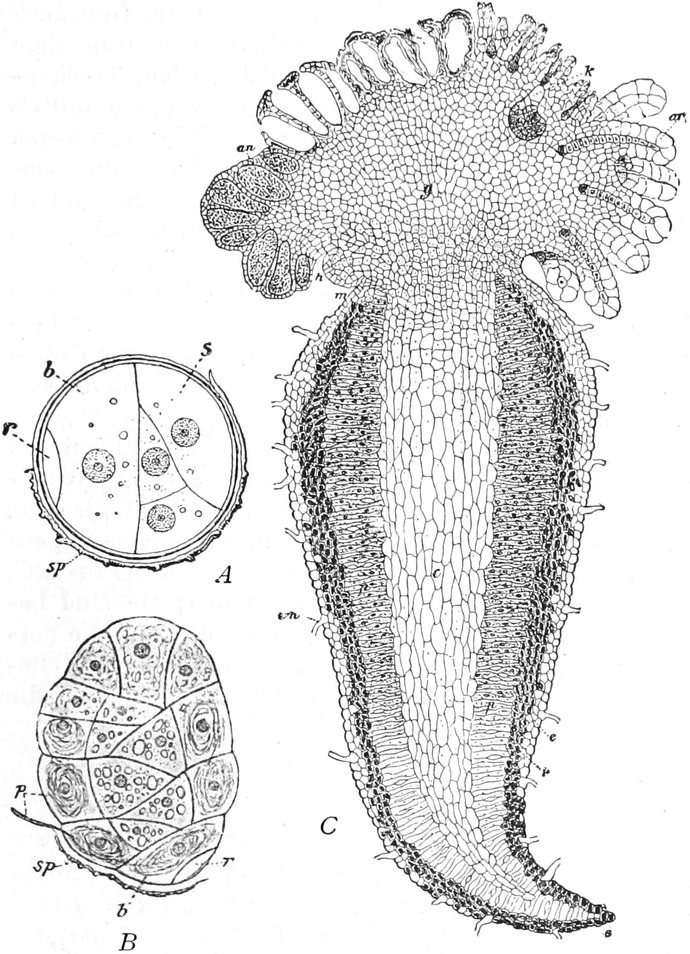
Fig. 494. Lycopodium annotinum. A Fünfzelliger
farbloser Sporenkeimling, mit Rhizoïdzelle r, Basalzelle b,
Scheitelzelle s, Sporenhaut sp. Vergr. 580. B Junger Keimling,
in dessen unteren Zellen der endophytische Pilz p wohnt. Die
Scheitelzelle in drei Scheitelmeristemzellen geteilt. Vergr.
470. — Lyc. complanatum. C Reifes Prothallium mit Antheridien
an, Archegonien ar, k junger Embryo. Vergr. 26. Nach
BRUCHMANN.
Fig. 495. Lycopodium complanatum. Embryoentwicklung.
A Embryo mit den ersten Teilungen; die Basalwand I teilt die
Anlage des Embryoträgers et von der Anlage des Embryokörpers ab,
die Transversalwände II und III (letztere in der Ebene des
Schnittes) sowie die Querwand IV liefern zwei vierzellige Stockwerke,
von denen das zwischen I und IV gelegene den Fuß liefert, das
unterste den Sproßteil. Vergr. 112. B Mittleres Stadium, s
Stammscheitel, b Blattanlage, f Fuß. Vergr. 112. C Embryo kurz
vor dem Herauswachsen aus dem Prothallium. bb die beiden ersten, den
Stammscheitel bedeckenden Blätter, w die erste Wurzel. Vergr. 40.
Nach BRUCHMANN.
Der Embryo bleibt während seiner Entwicklung (Fig. 495) im
Prothallium eingeschlossen. Er besitzt einen kugeligen, bei L.
complanatum keulenförmigen und warzigen Fuß als Saugorgan für
die Keimpflanze; unter dem Fuße differenziert sich die Anlage des
Sprosses, dessen erste Blätter schuppenartig sind und aus dessen
basalem Teile die erste Wurzel hervorkommt. Zwischen Sproß und Fuß
befindet sich der Embryoträger[S. 456] oder Suspensor, ein Gebilde, das
als anfängliches Saug- und Ernährungsorgan des Embryos dient.
Offizinell sind die Sporen von Lycopodium clavatum und anderen
Arten (Lycopodium, Pharm. germ., austr., helv.). Sie werden als
Hexenmehl bezeichnet.
Fig. 496. A Selaginella helvetica. Nat. Gr. Nach der
Natur. B S. Kraussiana, Keimpflänzchen mit der Makrospore. Vgr. Nach
BISCHOFF.
Fig. 497. Selaginella helvetica. A Makrosporangium
von oben mit Dehiszenzlinie d. B Geöffnet von der Seite, die vier
Makrosporen C ausgeschleudert. D Mikrosporangium in der Achsel des
Schuppenblattes von innen. E Geöffnet. F Mikrosporen. Vergr. ca. 15.
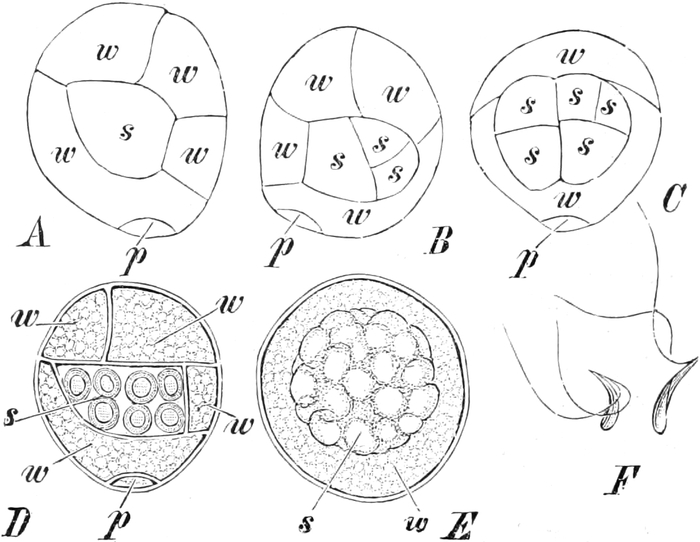
Fig. 498. A–E Selaginella stolonifera.
Vergr. 640. Keimung der Mikrosporen, aufeinander folgende
Stadien, p Prothalliumzelle, als Rhizoïdzelle aufzufassen, w
Antheridiumwandzellen, s spermatogene Zellen, A, B, D von der
Seite, C vom Rücken. In E die Prothalliumzelle nicht sichtbar, die
Wandzellen, aufgelöst, umgeben die Spermienzellen. F Sel. cuspidata.
Spermien. Vergr. 780. Nach BELAJEFF.
2. Ordnung. Selaginellaceae[442]. Die Gattung Selaginella ist
bei uns nur durch wenige, in den Tropen dagegen durch zahlreiche
Formen vertreten. Diese besitzen teils niederliegende, teils
aufrechte, reich gabelig, mit sympodialer Ausbildung verzweigte
Stengel; einige sind rasenbildend, andere klettern sogar mit
mehrere Meter langem Stengel im Gesträuch empor. Gewisse xerophile
Arten (S. lepidophylla im tropischen Amerika u. a.) können
monatelang, ja mehrere Jahre lang Trockenheit ertragen, wobei sie
ihre rosettig angeordneten Sprosse mittels Kohäsionsmechanismus
zusammenschließen; bei Eintritt von Regen breiten sie sich wieder
aus[443]. Im allgemeinen haben die Selaginellen ähnlichen Habitus
wie die Lycopodien. Der Stengel ist mit kleinen, schraubig oder
dekussiert in vier Zeilen stehenden, schuppenartigen Blättchen,
und zwar meist in dorsiventraler Ausbildung besetzt, so bei der
in den Alpen heimischen Selaginella helvetica (Fig. 496), deren
Stengel zwei Reihen kleiner, sog. Oberblätter und zwei Reihen
diesen gegenüberstehender größerer Unterblätter trägt (vgl. auch
Fig. 141). Eigentümlich ist den Selaginellen, daß ihre Wurzeln
nicht unmittelbar dem beblätterten Stengel, sondern zu je einer
oder mehreren an den Enden von kürzeren oder längeren, verzweigten
oder unverzweigten Wurzelträgern[444] endogen entspringen.
Diese sind zylindrische, blattlose nach unten wachsende,
wurzelähnliche Sprosse, eigenartige Organe, die paarweise stets
an Gabelungsstellen der Stengel, gekreuzt mit dessen Gabelästen
exogen entstehen; sie sind befähigt, zu beblätterten Sprossen
weiterzuwachsen, wenn die normalen Sprosse zurückgeschnitten
werden. Schon an dem Keimling entstehen ganz kurze Wurzelträger,
aus deren Spitzen die[S. 457] ersten Wurzeln endogen sich bilden. Die
Blätter der Selaginellen sind ausgezeichnet durch eine am Grunde
der Blattoberseite entspringende kleine häutige Ligula,
die als Organ der Wasseraufnahme ein sehr rasches Aufsaugen von
Regentropfen durch die beblätterten Sprosse vermittelt[445]. Die
epidermalen Assimilationszellen der Blätter führen bei manchen
Arten, ähnlich wie bei Anthoceros, nur je einen großen
muldenförmigen Chloroplasten[446].
Fig. 499. Selaginella Martensii. A Aufgesprungene
Makrospore von oben. Prothallium mit drei Rhizoïdhöckern und mehreren
Archegonien. Vergr. 112. B Längsschnitt, zwei Archegonien mit sich
entwickelnden Embryonen. Vergr. 112. Nach BRUCHMANN.
Die endständigen Sporophyllstände oder Blüten sind einfach
oder verzweigt, vierkantig radiär, seltener dorsiventral. Jedes
Sporophyll trägt nur ein über der Blattachsel aus dem
Stengel entspringendes Sporangium. In ein und derselben Blüte
treten sowohl Makro- als auch Mikrosporangien auf. In den ersteren
(Fig. 497 A–C) gehen die angelegten Sporenmutterzellen alle
zugrunde bis auf eine, welche die vier großen, paarweise gekreuzten
und die Sporangienwand buckelig vorwölbenden Sporen liefert. Das
durch einen Kohäsionsmechanismus erfolgende Aufspringen vollzieht
sich auf vorbezeichneter Dehiszenzlinie mit zwei auf einem basalen,
kahnförmigen Teile stehenden, sich zurückkrümmenden Klappen; durch
den Druck des sich verengenden Kahnteils werden die Sporen dann
herausgeschleudert. In den flachen Mikrosporangien sind zahlreiche
kleine Sporen vorhanden. Die Öffnung geschieht hier in ähnlicher
Weise, nur ist der kahnförmige Teil viel kürzer, die Klappen
reichen fast bis zur Basis.
Fig. 500. Selaginella Martensii. Embryo vor dem
Hervorbrechen aus dem Prothallium im Längsschnitt. f Fuß. wt
Wurzelträger, et Embryoträger, k Keimblätter mit Ligula. Vergr.
150. Nach BRUCHMANN.
Die Mikrosporen beginnen ihre Weiterentwicklung schon
innerhalb des Sporangiums. Die Sporenzelle teilt sich zunächst
in eine kleine linsenförmige, der Rhizoïdzelle von Salvinia
entsprechende Zelle, und in eine große Zelle, die nacheinander
in acht sterile Prothallien- oder Wandzellen und zwei oder vier
zentrale spermatogene Zellen zerlegt wird (Fig. 498 A). Durch
weitere Teilung der letzteren Zellen, die ein einziges Antheridium
vorstellen, entstehen die sich abrundenden Spermienmutterzellen
in größerer Anzahl (B–D). Die Wandzellen lösen alsdann ihre
Wände auf und werden zu einer Schleimschicht, in welcher die
zentrale[S. 458] Masse der Spermienzellen eingebettet liegt (E). Die
kleine Prothalliumzelle (p) bleibt hingegen erhalten. Das
ganze männliche Prothallium ist bis zu diesem Stadium von der
Mikrosporenhaut noch umschlossen; schließlich bricht diese auf
und die Spermienzellen werden frei, um die keulenförmigen, an der
Spitze mit zwei langen Zilien versehenen Samenfäden zu entlassen.
Auch die Makrosporen beginnen, allerdings nicht bei allen
Arten, ihre Weiterentwicklung schon, wenn sie noch im Sporangium
eingeschlossen liegen. Der Zellkern teilt sich in Tochterkerne, die
in dem Wandplasma am Scheitel sich verteilen, und nun erfolgt hier
die Ausbildung von Zellwänden. So wird vom Scheitel bis zur Basis
fortschreitend die Spore mit großen Prothalliumzellen angefüllt;
zugleich beginnt aber auch in derselben Richtung die weitere
Teilung dieser Zellen in kleinzelliges Gewebe. Bei einigen Arten
wird zunächst nur am Scheitel eine kleinzellige Scheibe von Zellen
angelegt, die durch verdickte getüpfelte Innenwände (Diaphragma)
sich von der großen unteren Zelle abgrenzen, und diese teilt sich
erst später in großzelliges Gewebe. In dem kleinzelligen Gewebe
am Scheitel werden einige wenige Archegonien angelegt, und zwar
manchmal bereits, wenn die Spore noch nicht vom Prothalliumgewebe
ganz ausgefüllt ist. Meist werden die Archegonien erst gebildet,
wenn die Sporen aus dem Sporangium entleert sind; öfters findet
aber die Befruchtung auch noch auf der Mutterpflanze statt.
Die Sporenwand springt am Scheitel in den drei Sporenkanten auf;
das kleinzellige farblose Prothallium tritt etwas hervor und bildet
auch einige Rhizoïden auf drei Gewebehöckern, die zum Sprengen
und Aufsperren der Wand wie zum Festhalten von Wasser dienen. Es
erfolgt dann die Befruchtung von einem oder wenigen Archegonien und
die direkte Weiterentwicklung der befruchteten Eizellen zum Embryo
(Fig. 499).
Die Entwicklung des Embryos, an dem wir den ein- oder
mehrzelligen Embryoträger, den Sproßvegetationskegel mit den ersten
Blattanlagen, die ersten Wurzelträger und den Fuß unterscheiden,
verläuft in auffallend verschiedener Weise innerhalb der Gattung.
Die befruchtete Eizelle teilt sich zunächst quer; aus der
oberen, hypobasalen Zelle geht bei S. Martensii und Verwandten
der Embryoträger allein hervor, aus der unteren, epibasalen
Zelle dagegen der Hauptkörper des Embryos (Fig. 500); bei S.
denticulata, helvetica und den meisten anderen Arten aber
liefert die obere Zelle außer dem Embryoträger auch noch den Fuß
und Wurzelträger. Der Sproßscheitel mit dem ersten Blattpaar
wächst nach oben, die Wurzel abwärts; die junge Keimpflanze bleibt
mit ihrem Fuß in dem Prothalliumgewebe der Makrospore stecken
(Fig. 496 B). Bei einigen Arten (S. rubricaulis, spinulosa,
helvetica) bleiben die Archegonien geschlossen. und die Eizelle
entwickelt sich parthenogenetisch zum Embryo. Bei S. Kraussiana
und Verwandten ist der Embryoträger nach BRUCHMANN
rückgebildet, aber in seiner Funktion ersetzt durch einen
eigenartigen Embryoschlauch, der aus der Membran der Eimutterzelle
hervorgehen soll; in diesem soll sich die Anlage des Embryos
abgrenzen und dann in das Nährgewebe hinabwandern.
3. Ordnung. Psilotaceae[447]. Von jetzt lebenden Formen gehören
zu dieser anscheinend sehr primitiven Ordnung nur Psilotum mit
zwei tropischen Arten und Tmesipteris mit einer australischen
Art. Bemerkenswert ist das gänzliche Fehlen von Wurzeln. Die
gabelig verzweigten Sprosse tragen in schraubiger Anordnung kleine
einfache Blätter und in ihren oberen Teilen die Sporophylle, die
stets tief gegabelt sind und auf ihrer Innenseite am Grunde je ein
derbwandiges zwei- oder dreifächeriges Sporangium besitzen.
Die in neuerer Zeit erst bekannt gewordenen Prothallien beider
Gattungen sind im Substrat verborgene, zylindrische, verzweigte,
farblose, von einem Phykomycetenmyzel nach Art der Mykorrhizen
durchzogene, nur einige Millimeter große Thalli, die auf ihrer
Oberfläche zahlreiche Antheridien und Archegonien entwickeln. Die
Spermien haben zahlreiche Zilien.
Die im Devon vertretenen Rhyniaceen scheinen zu den Anfangsformen
der Psilotaceen zu gehören und sind insofern besonders
bemerkenswert, als ihr wurzelloser, gabelig verzweigter Stengel
noch keine blattartigen Ausgliederungen zeigte, äußerlich also noch
einem Algenthallus glich, und an seinen Enden Sporangien trug, die
bei Hornea eine Columella, ähnlich wie die Laubmoossporogone,
aufwiesen[448].
4. Ordnung. Isoëtaceae[449]. Hierher gehört nur die Gattung
Isoëtes, Brachsenkraut, (Fig. 501), die als selbständiger Zweig
der in früheren Erdperioden viel formenreicheren Klasse anzusehen
ist. Die Isoëtes-Arten sind teils untergetauchte, teils[S. 459] auf
feuchtem Boden lebende, perennierende Kräuter mit knolliger,
gestauchter, selten dichotom gegabelter Achse, die nach unten
ein Büschel von dichotom verzweigten Wurzeln, nach oben eine
dichte Rosette von langen pfriemförmigen, von vier Luftkanälen
durchzogenen Blättern trägt. Die Stammknolle zeichnet sich aus
durch sekundäres Dickenwachstum mittels eines Kambiums, das
nach außen Rinde, nach innen Tracheiden und aus Prismenzellen
bestehendes Parenchym, das das hier fehlende Phloëm vertritt,
abscheidet. Die Blätter verbreitern sich am Grunde zu einer breiten
Scheide und sind an der Innenseite über ihrer Basis mit einer
länglichen grubenartigen Vertiefung, der Fovea, versehen.
Die Sporophylle tragen im Grunde der Fovea je ein Sporangium; im
übrigen gleichen sie den Laubblättern, die den innersten Teil
der Rosette einnehmen. Über der Fovea ist die Ligula, die hier
als schleimbildendes Organ dient, als dreieckiges Häutchen mit
eingesenkter Basis eingefügt. Im Habitus weicht also Isoëtes von
den übrigen Ordnungen bedeutend ab, mit Selaginella ist ihr die
Ligula gemeinsam; daher werden auch Isoëtes und Selaginella
als Ligulatae bezeichnet, zu denen auch die fossilen Sigillariaceen
und Lepidodendraceen gehören.
Fig. 501. Isoëtes lacustris. 1⁄2 nat. Gr.
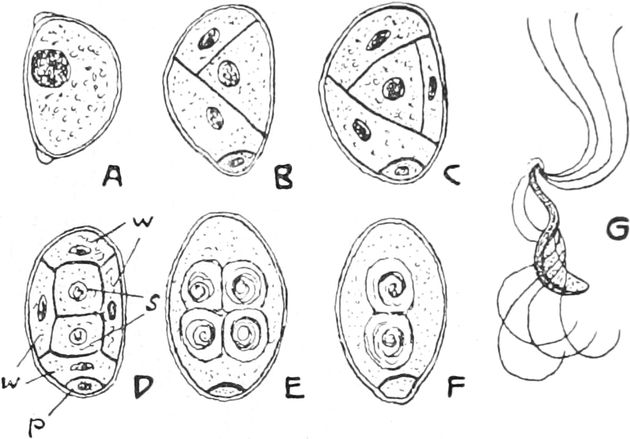
Fig. 502. A–F Isoëtes setacea. Vergr. 640.
A Mikrospore von der Seite. B–D Teilung der Spore, p
Prothalliumzelle, w die vier Wandzellen, s spermatogene Zellen.
E Die Wandzellen aufgelöst, umgeben die vier Spermienzellen,
Flächenansicht. F Desgleichen, Seitenansicht. G Isoëtes
Malinverniana, Spermium. Vergr. 780. Nach BELAJEFF.
Fig. 503. Isoëtes echinospora. A Weibliches
Prothallium mit Archegonium ar, Eizelle o. B, C Entwicklung
des Archegoniums aus einer oberflächlichen Zelle, h Halszellen, hk
Halskanalzelle, b Bauchkanalzelle, o Eizelle. Vergr. 250. Nach
CAMPBELL.
Fig. 504. Isoëtes echinospora. Embryo vor dem
Herauswachsen aus dem Prothallium; Längsschnitt. cot Keimblatt, l
Ligula, v Scheide an der Basis des Keimblattes, in dessen Achsel
der Stammscheitel sich bildet, w Wurzel, f Fuß. Vergr. 200. Nach
CAMPBELL.
Die Makrosporangien, die hier im Unterschied zu Selaginella
noch zahlreiche Makrosporen ausbilden, sitzen an den äußeren
Blättern der Rosette, die ihnen ähnlichen Mikrosporangien
an den auf diesen folgenden Blättern. Beide sind hier von
querverlaufenden[S. 460] sterilen Gewebesträngen, Trabeculae
genannt, durchsetzt und unvollständig gefächert. Die Sporen werden
erst durch Verwesung der Behälter frei.
Die Entwicklung der geschlechtlichen Generation geschieht in
ähnlicher Weise wie bei Selaginella. Das reduzierte männliche
Prothallium entwickelt sich bereits in der Spore (Fig. 502).
Auch hier wird die Sporenzelle in eine kleine linsenförmige
Prothalliumzelle (p) und eine größere, die Anlage eines einzigen
Antheridiums, zerlegt. Die große Zelle teilt sich weiter in
vier sterile Wandzellen (w), welche allseitig zwei zentrale
spermatogene Zellen umschließen. Aus beiden entstehen je zwei
Spermienzellen (E), im ganzen also vier, die nach dem Auflösen
der Wandzellen und Aufplatzen der Sporenhülle nach außen gelangen;
eine jede entläßt nun ihren schraubig gewundenen und am vorderen
Ende mit einem Zilienbüschel besetzten Samenfaden (G).
Fig. 505. 1 Lepidodendron, Rekonstruktion, entworfen
von POTONIÉ. 2 L. aculeatum. Schaftstück, Hohldruck
nach STERNBERG. 3 und 4 Lepidodendron, Blattpolster,
nach POTONIÉ. 5 Rindenstück, nach SEWARD. Aus
LOTSY, Botanische Stammesgeschichte.
Wie bei Selaginella bleibt auch hier das einfach gebaute
weibliche Prothallium (Fig. 503) in der Makrospore
eingeschlossen und ist nicht zu selbständigem Wachstum befähigt.
In seiner Bildung zeigt es wie bei Selaginella Annäherung an die
Koniferen, indem zunächst der Kern der Makrospore in zahlreiche,
freie, wandständige Tochterkerne sich teilt, bevor die Zellwände,
vom Scheitel der Spore zur Basis längs der Wandung fortschreitend,
angelegt werden. Die ganze Spore wird so mit einem Prothallium
gefüllt, an dessen Scheitel einige wenige Archegonien zur
Entwicklung kommen. Der Embryo (Fig. 504) besitzt im
Gegensatz zu den übrigen Lycopodinen keinen Embryoträger.
5. Ordnung. Sigillariaceae[450]. Die Siegelbäume sind vom Kulm
ab nachgewiesen, waren im Karbon am artenreichsten und scheinen im
Rotliegenden ausgestorben[S. 461] zu sein. Sie waren stattliche Gewächse
mit mächtigen, in die Dicke wachsenden, säulenförmigen, einfachen
oder nur wenig gegabelten Stämmen, oben mit langen pfriemlichen,
einfachen, oberseits am Grunde mit Ligula versehenen Blättern
besetzt und am Schafte bedeckt mit den Längszeilen sechseckiger
Blattnarben, mit stammbürtigen, langgestielten, zapfenförmigen
Blüten, deren Sporangien in Einzahl auf jedem Sporophyll gebildet
wurden und heterospor waren.
6. Ordnung. Lepidodendraceae[450], [451]. Die Schuppenbäume,
vom Oberdevon bis in das Rotliegende, besonders aber im Karbon
verbreitet, waren ebenfalls baumartige bis etwa 30 m hohe Pflanzen,
aber mit dichotom verzweigten, rhombisch gefelderten, in die Dicke
wachsenden Stämmen, an denen oben die meist schraubig angeordneten,
schmalen, bis 15 cm langen, mit Ligula versehenen Blätter auf
rhombischen Blattkissen saßen (Fig. 505). Die zapfenförmigen Blüten
(Lepidostrobus, Fig. 506) entsprangen endständig oder am Stamme
selbst und enthielten Makro- und Mikrosporangien, je eins auf
jedem Sporophyll. Die Zahl der Sporen im Makrosporangium war eine
größere als bei Selaginella. In der Makrospore entwickelte sich ein
Prothallium von ähnlichem Bau wie bei dieser Gattung.
Fig. 506. Lepidostrobus Veltheimianus. 1
Mikrosporangien, Querschliff, rechts Tetraden von Mikrosporen.
2 Sporangienstand, Längsschliff, oben mit Mikro-, unten mit
Makrosporangien. 3 Makrosporangien, Querschliff. 4 Makrospore
im Längsschliff. 5 Makrospore, wahrscheinlich infolge der Keimung
aufgesprungen. 1–5 nach SCOTT, KIDSTON,
BINNEY. Aus LOTSY, Bot. Stammesgeschichte.
Von hohem Interesse ist die Auffindung samenähnlicher
Gebilde bei einigen paläozoischen Lycopodinen (Lepidocarpon,
Miadesmia), die daher auch als Lepidospermeae von den übrigen
Ordnungen abgesondert werden können. Das Makrosporangium war
bei ihnen bis auf eine enge Furche von einem Integument umhüllt
und zum Teil auch von dem Sporophyll bedeckt. Nur eine einzige
Makrospore kam in ihm zur Entwicklung; das Prothallium blieb wie
bei Isoëtes in der Spore eingeschlossen. Die Mikrosporen wurden in
Lepidostrobus-ähnlichen Blüten erzeugt. Wahrscheinlich gelangten
die ausstäubenden Mikrosporen zu den Makrosporangien bereits auf
der Mutterpflanze, von der sich später die Makrosporophylle samt
ihren Sporangien loslösten.
Klasse V.
Pteridospermeae,
Samenfarne[450],
[452].
Soweit unsere Kenntnisse reichen, stellen die Equisetinae und
Sphenophyllinae Seitenzweige des Pteridophytenstammes dar, die keine
Weiterentwicklung[S. 462] zu höheren Pflanzen erfuhren. Aus den Lycopodinen
aber scheinen die Koniferen sich entwickelt zu haben, während aus den
Filicinae bereits im Paläozoicum die ersten Samenpflanzen, die
Pteridospermeae (Cycadofilices) hervorgegangen sind, die, auf einer
höheren Stufe der Organisation als sämtliche übrigen Pteridophyten
stehend, ein verbindendes Glied zwischen den Farnkräutern einerseits
und den Cycadeen andererseits vorstellen. Sie sind bereits im Perm
wieder ausgestorben.
Fig. 507. Lyginodendron. Wedel (Sphenopteris
Hoeninghausii). Verkl. 1⁄3.
Nach POTONIÉ.
Fig. 508. Lyginodendron oldhamium. Stammquerschliff.
Vergr. 21⁄2.
Nach SCOTT.
[S. 463]
In ihren stattlichen vegetativen Organen trugen sie den Charakter
von eusporangiaten Farnen, im besonderen von Marattiaceen, zur
Schau. Ihre Wedel (Sphenopteris Fig. 507, Neuropteris) waren
reich gefiedert, der Blattstiel über der Basis gegabelt. Die
axillär verzweigten Stämme (Lyginodendron) zeichneten sich
durch sekundäres Dickenwachstum mittels eines Kambiums aus,
das nach innen im Umkreis des großen zentralen Marks einen aus
radial gereihten Elementen bestehenden Holzkörper, nach außen
eine Phloëmzone abschied (Fig. 508). In der Rinde verliefen
Blattspurbündel, die den Holzkörper durchsetzten und an die
primären Gefäßstränge in der Markperipherie anschlossen. Auch die
Wurzeln wurden sekundär verdickt.
Die Samenfarne waren heterospor. Ihre Sporangien entsprangen an
Wedeln, die kaum von gewöhnlichen Farnwedeln verschieden waren.
Mikrosporangien sind noch nicht bekannt; die als Crossotheca
bezeichneten und hierher gerechneten Sporangien scheinen nicht
zu Lyginodendron zu gehören. Die Makrosporangien (Lagenostoma,
Fig. 509), bis zur Anheftungsstelle an der Basis frei, mit
einem Integument versehen, waren von einer Cupula umhüllt
und hatten ähnlichen Bau wie die Samenanlagen der Cycadeen;
die Makrosporophylle waren aber noch nicht wie bei diesen zu
zapfenartigen Blüten angeordnet.
Fig. 509. Lyginodendron oldhamium. Makrosporangium
(Lagenostoma). Die offene Cupula mit gestielten Drüsen besetzt,
Rekonstruktion. Nach SCOTT.
[S. 464]
Zweite Abteilung.
Spermatophyta oder Samenpflanzen.
Übergang von den Farnpflanzen zu den Samenpflanzen[453]. Der
scharf ausgeprägte Generationswechsel ist eines der wesentlichen
Merkmale der Pteridophyten: Aus der Spore entsteht der selbständig
lebende haploide Gametophyt, das meist nur kurzlebige Prothallium,
aus dessen befruchteter Eizelle der ebenso selbständige diploide
Sporophyt, das Farnkraut, der Schachtelhalm oder Bärlapp hervorgeht.
Das Auftreten der Heterosporie führte zu weiterer Reduktion der
Prothalliumpflanze, welche damit der Aufgabe, beiderlei Sexualorgane
hervorzubringen, überhoben ward und nur verminderter Ausbildung
bedurfte. So hatte das männliche Prothallium, in der Mikrospore
entwickelt, nur noch Antheridien zu bilden, welche die Spermien
lieferten; daneben blieb lediglich eine kleine funktionslose vegetative
Zelle des Prothalliums erhalten. Auch das weibliche, in der Makrospore
sich ausbildende Prothallium, bei Salvinia noch ergrünend und aus
der Makrospore hervortretend, hat bei Selaginella und Isoëtes die
Fähigkeit selbsttätiger Ernährung bereits verloren. Es beginnt seine
Entwicklung schon innerhalb des mütterlichen Makrosporangiums, und
die frei gewordenen Makrosporen öffnen sich schließlich nur, um den
Spermien Zugang zu den scheitelständigen Archegonien zu gewähren. Aus
der befruchteten Eizelle wächst ohne Ruhepause der Embryo zu dem jungen
Sporophyten heran.
Von diesen am höchsten differenzierten Archegoniaten unterscheiden sich
die einfachsten Samenpflanzen (Spermatophyten) nur unwesentlich.
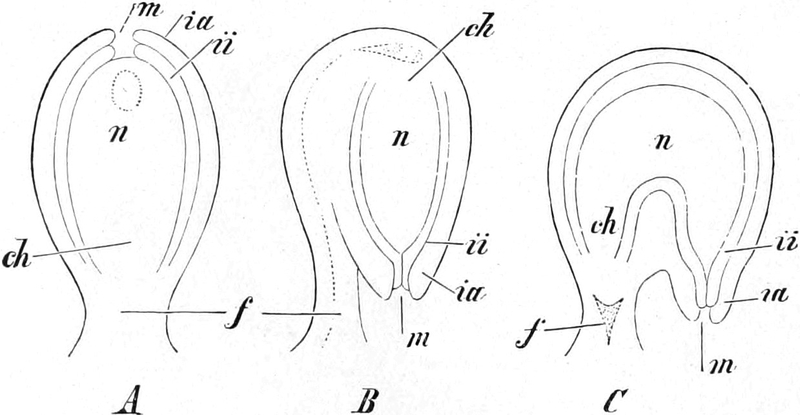
Fig. 510. A atrope, B anatrope, C kampylotrope
Samenanlage. Schematisch und vergrößert. Modifiziert nach A. F. W.
SCHIMPER. Erklärung im Text.
Die Makrospore, hier von altersher als Embryosack
bezeichnet, bleibt stets im Makrosporangium, der
Samenanlage (Fig. 510), eingeschlossen. Diese besteht aus
dem Nucellus (n) und einer oder zwei von seinem Grunde,
der Chalaza (ch), aus rings emporwachsenden Hüllen, den
Integumenten (ii, ia), welche nur über dem Nucellusscheitel
einen schmalen Zugang, die Mikropyle (m), freilassen. Mit
einem oft sehr kurzen Stiele, dem Funiculus (f), ist ein
solches Makrosporangium dem Sporophyll, hier Makrosporophyll
oder Fruchtblatt genannt, angefügt. Die Ansatzstelle einer oder
mehrerer Samenanlagen trägt den Namen Placenta. Ist der Nucellus
die geradlinige Fortsetzung des Funiculus, so nennt man die Samenanlage
gerade, atrop. Sehr viel häufiger erleidet der Funiculus
unterhalb der Chalaza eine scharfe Krümmung, so daß die Samenanlage
mit dem äußeren Integumente an ihm entlang zurückläuft; sie ist dann
umgewendet, anatrop. Die in solchem Falle noch am reifen Samen
kenntliche Verwachsungslinie von Funiculus und äußerem Inte[S. 465]gument
heißt die Samennaht oder Raphe. Endlich kann der Nucellus
selbst gekrümmt sein, man spricht dann von einer kampylotropen
Samenanlage (Fig. 510 C).
Innerhalb einer solchen Samenanlage findet sich in der Regel nur ein
einziger Embryosack vor. Wie im Makrosporangium (von Selaginella
z. B.) durch Tetradenteilung vier Makrosporen entstehen, so
werden auch in den Samenanlagen von der Embryosackmutterzelle vier
Tochterzellen gebildet, von denen aber drei zugrunde gehen, während
die vierte sich zur Makrospore, dem Embryosacke, entwickelt. Der
Embryosack der einfachsten Samenpflanzen gleicht der Makrospore auch
darin, daß er mit Prothalliumgewebe, hier Endosperm genannt, ausgefüllt
ist, an dessen Scheitel ein oder mehrere Archegonien stehen, deren
Eizelle besondere Größe besitzt. Die befruchtete Eizelle wächst zum
Keim oder Embryo noch innerhalb der Makrospore und auf
Kosten der Mutterpflanze heran. Erst nachdem der Embryo eine bei den
verschiedenen Pflanzen verschieden hohe, aber für die betreffende
Art charakteristische Stufe der Entwicklung erreicht hat, stellt
er das Wachstum ein und geht nach Trennung von der Mutterpflanze
einer Ruheperiode entgegen. Er ist von den entsprechend weiter
entwickelten und in ihrer Ausbildung ihm angepaßten übrigen Teilen des
Makrosporangiums umgeben, nämlich dem Prothallium oder Endosperm, dem
Nucellus und der aus den Integumenten hervorgegangenen Schale. Das
ganze aus der Samenanlage entstandene Gebilde wird Samen genannt. Diese
Weiterentwicklung des geschlossenen Makrosporangiums zum Samen ist das
charakteristische Merkmal der Samenpflanzen oder Spermatophyten.
Die Mikrosporen der Samenpflanzen heißen seit altersher
Pollenkörner. Sie entstehen in großer Anzahl in den
Mikrosporangien oder Pollensäcken, die einzeln oder in
Mehrzahl von den Mikrosporophyllen, den Staubblättern
(Stamina), erzeugt werden. Der die Pollensäcke führende Teil der
Staubblätter heißt Anthere.
Die Bildung der Pollensäcke wird durch eine Teilung der auf die
Epidermis folgenden Zellschicht der Anthere eingeleitet, die parallel
zu der Oberfläche erfolgt und das primäre Archespor von den
Schichtzellen trennt. Diese zerfallen in drei verschiedene
Zellagen (Fig. 511), deren direkt unter der Epidermis liegende
äußerste Lage bei den Angiospermen die Faserschicht gibt,
die sich bei den Gymnospermen aus der Epidermis selbst bildet. Die
mittlere Zellschicht geht später zugrunde, während die innerste die
Tapetenschicht liefert, und das Archespor in weiteren Teilungen
die Pollenmutterzellen gibt, deren jede, wie bei den Bryo- und
Pteridophyten durch eine Tetradenteilung in vier Tochterzellen, die
Pollenkörner, zerfällt. Diese sind kugelige bis ellipsoidische
Körper; sie werden von einer Membran umhüllt, die eine äußere
kutinisierte Exine und eine aus pektinreicher Zellulose
bestehende Intine erkennen läßt.
Für die Beförderung der Pollenkörner zu den Samenanlagen wird bei den
Samenpflanzen der Wind oder Beihilfe von Tieren in Anspruch genommen,
während alle Farnpflanzen sich des Wassers als Transportmittel ihrer
männlichen Sexualzellen bedienen. Die fortschreitende Rückbildung des
männlichen Prothalliums, das schon bei den heterosporen Pteridophyten
auf einzelne Zellen beschränkt war, führt bei den Samenpflanzen
dahin, daß bei der Keimung des Pollenkornes, wenn von den bei vielen
Gymnospermen noch gebildeten, sehr vergänglichen Prothalliumzellen
(Fig. 512) abgesehen wird, nur noch zwei Zellen zu beobachten sind:
eine vegetative und eine Antheridiumzelle. Die erstere
wächst zu einem langen Schlauch,[S. 466] dem Pollenschlauche, aus,
der durch chemotropische Reizbarkeit (vgl. S. 311) zu den Eizellen
geleitet wird; letztere, die Antheridium-Mutterzelle, wandert
in den Pollenschlauch ein und bildet dort früher oder später zwei
generative oder Spermazellen, welche im Schlauche (daher die
Bezeichnung Siphonogamen) vorwärtsgleitend in den Embryosack eindringen
und zur Eizelle gelangen.
Das Ergebnis dieser Übersicht läßt sich also dahin zusammenfassen,
daß die Samenpflanzen die Reihe der Pteridophyten fortsetzen und
ihnen im Besitze eines Generationswechsels gleichen. (Vgl. Schema S.
467.[454]) Die immer reicher und mannigfaltiger werdende Gestaltung
und Ausgliederung des Sporophyten ist aber von einer fortschreitenden
Rückbildung des Gametophyten begleitet. Der Sporophyt hält die
weibliche Geschlechtsgeneration während ihrer ganzen Entwicklung
eingeschlossen; erst in und mit dem fertigen Samen, der den Beginn der
nächsten ungeschlechtlichen Generation als Embryo in sich birgt, wird
sie von dem Sporophyten getrennt.
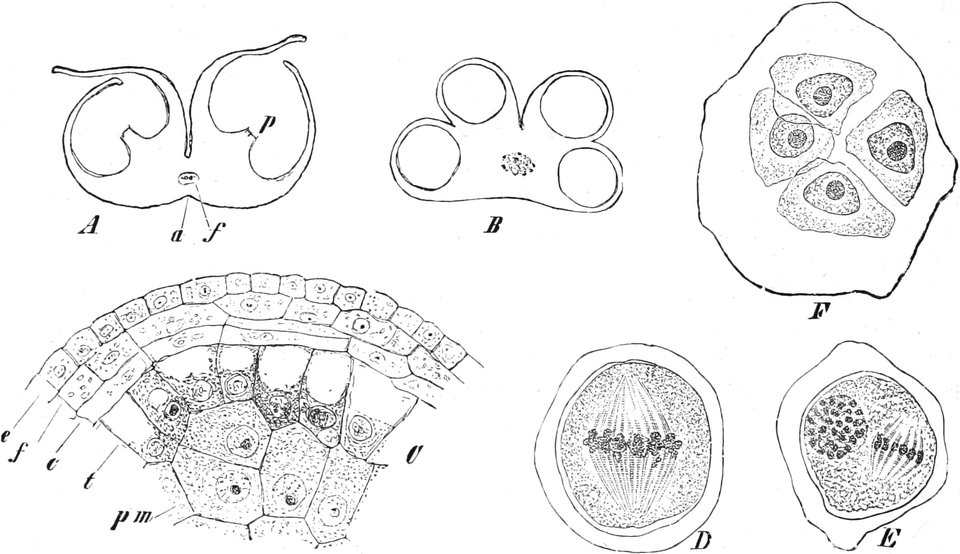
Fig. 511. Hemerocallis fulva. A Querschnitt durch eine
fast reife Anthere, mit ihren durch den Schnitt geöffneten Fächern,
p die Scheidewand zwischen den Fächern, a Furche am Konnektiv, f
Leitbündel. Vergr. 14. B Querschnitt durch eine junge Anthere. Vergr.
28. C Teil des Querschnittes aus einem Fache, e Epidermis, f
die spätere Faserschicht, c später verdrängte Wandschicht, t die
sich später auflösende Tapetenschicht, pm Pollenmutterzellen. Vergr.
240. D, E Pollenmutterzelle von Alchimilla speciosa in Teilung
begriffen. Vergr. 1125. F fertige Pollentetrade von Bryonia dioica.
Vergr. 800. Nach E. STRASBURGER.
In voller Übereinstimmung mit dieser Darlegung des innerhalb der
Samenpflanzen weiter zu verfolgenden Generationswechsels stehen die
Ergebnisse der Untersuchungen über die Reduktionsteilung[455] der
Sporenmutterzellen (vgl. S. 173). Die einer Pflanze zukommende Zahl von
Chromosomen verringert sich bei den zur Bildung der Sexualgeneration
führenden Teilungen auf die Hälfte, und erst der Befruchtungsakt stellt
für den Embryo die volle Chromosomenzahl wieder her. So verfügt der
Sporophyt stets über die diploide, der Gametophyt über die haploide
Zahl der Kernfadensegmente.
[S. 467]
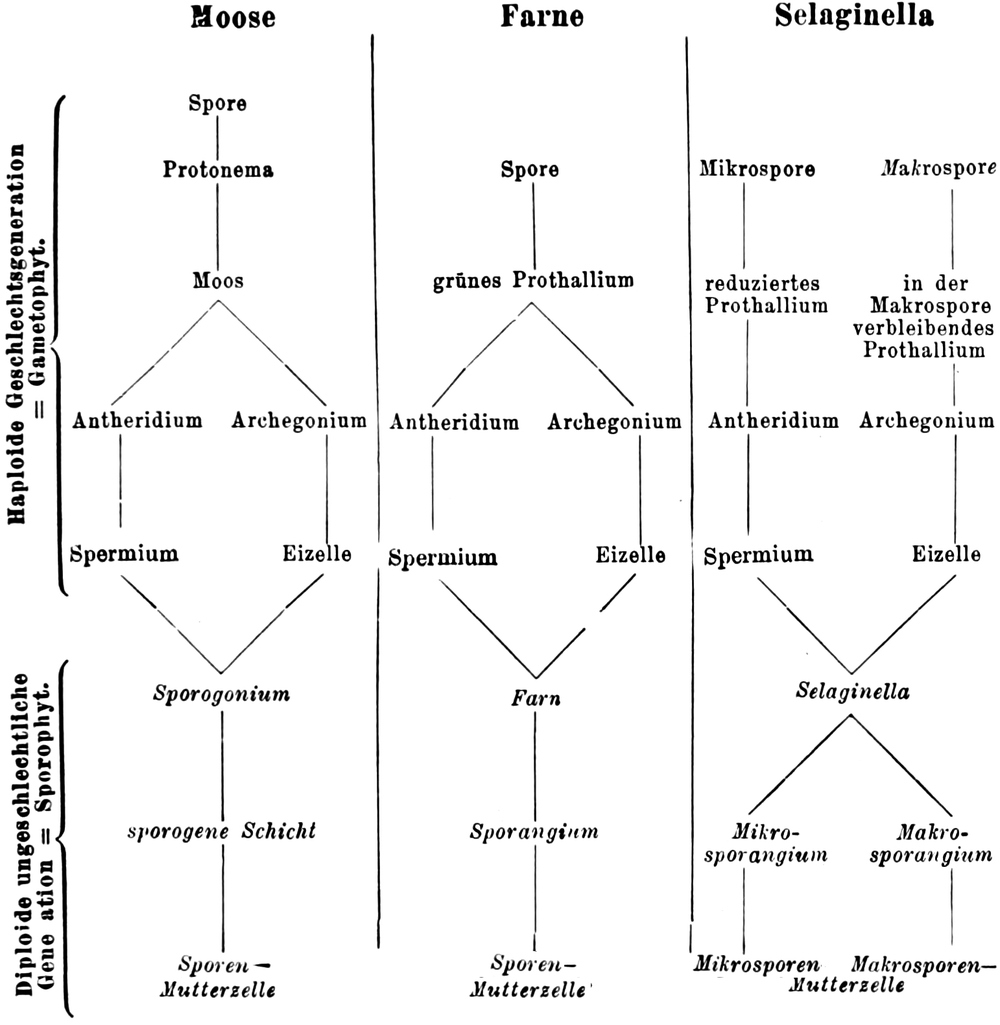
Generationswechsel; linker Teil des Schemas.
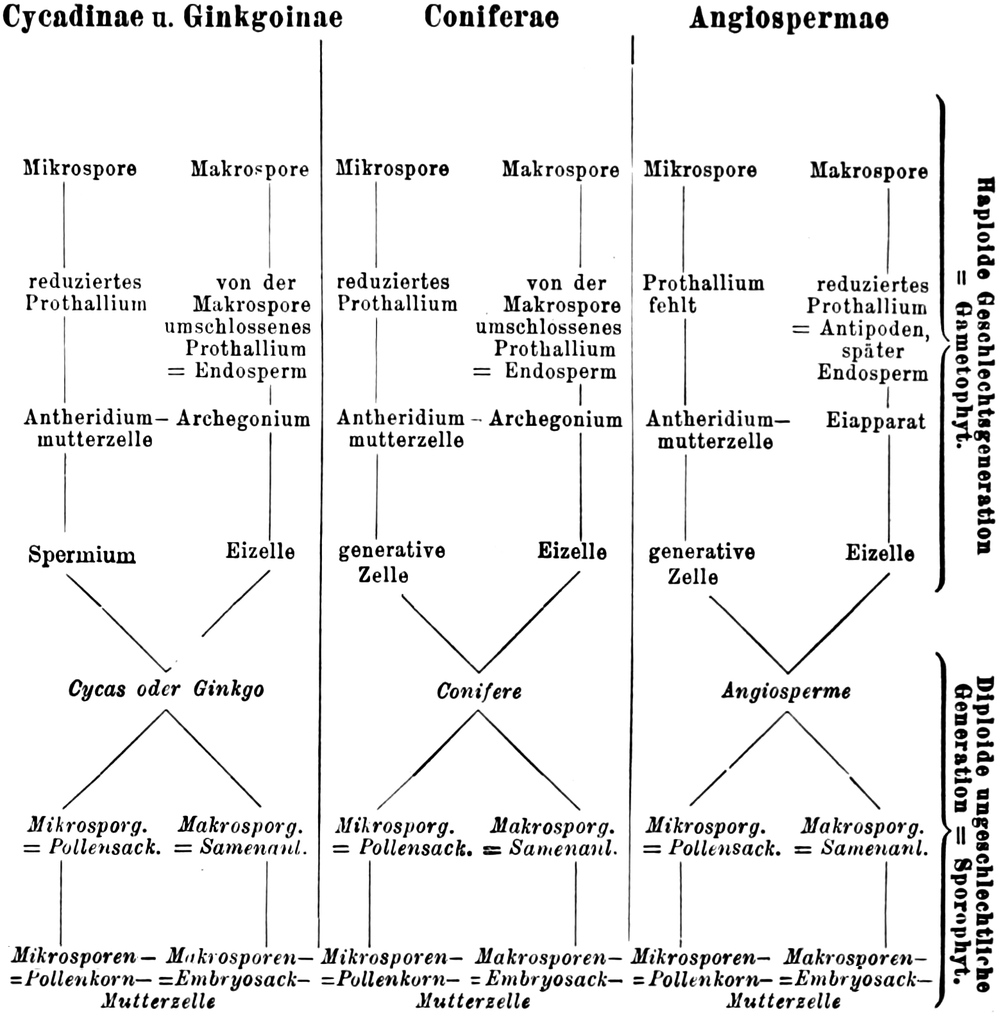
Generationswechsel; rechter Teil des Schemas.
[S. 468]
Die Samenpflanzen zerfallen in zwei Klassen, die ihrem ganzen
Aufbau nach erheblich verschieden sind: 1. die Gymnospermen oder
nacktsamige Pflanzen, und 2. die Angiospermen oder bedecktsamige
Pflanzen. Die Fruchtblätter der Angiospermen treten zur Bildung
geschlossener Hohlräume, der Fruchtknoten, zusammen, in denen die
Samenanlagen sich entwickeln; die Fruchtblätter der Gymnospermen
bleiben offen, sie tragen die Samenanlagen frei auf ihrer Oberseite
oder an ihren Rändern.
Die Gymnospermen sind die phylogenetisch älteren Pflanzenformen;
sie sind einfacher gebaut und schließen im Verhalten ihrer
Geschlechtsgeneration unmittelbar an heterospore Farnpflanzen an.
Die Angiospermen zeigen eine viel größere Mannigfaltigkeit im
morphologischen wie anatomischen Aufbau. Ihre Entwicklung weicht von
jener der Gymnospermen erheblich ab und läßt die Beziehung zu den
Archegoniaten nicht mehr so deutlich erkennen.
Paläontologische Befunde zeigen uns dementsprechend
Gymnospermenähnliche oder ihnen gleichende Formen neben Übergängen
von den Farnpflanzen in den Gesteinsschichten des Devon, in der
Steinkohlenformation und im Perm, während die Angiospermen erst viel
später, in der Kreideformation, sicher nachweisbar werden.
Morphologie und Ökologie der Blüte[456].
1. Morphologie. Die Gymnospermenblüten sind
stets eingeschlechtig, diklin, Mikro- und Makrosporophylle
bleiben getrennt und bilden die männlichen oder die weiblichen Blüten.
Beide Geschlechter finden sich entweder auf einem Individuum:
die Pflanze ist einhäusig, monözisch, oder sie sind
auf verschiedene verteilt: dann ist die Art zweihäusig,
diözisch. Gymnospermenblüten haben außer den Sporophyllen, also
den Staubblättern und den Fruchtblättern, nur bei den Gnetaceen andere,
als Hülle dienende Blattgebilde aufzuweisen.
Die männlichen Blüten der Gymnospermen stellen Sprosse
begrenzten Längenwachstums dar, deren Achse mit Sporophyllen in meist
schraubiger Anordnung dicht besetzt ist. Ihre Knospenschuppen bleiben
oft noch an der Basis der entfalteten Blüte erhalten (Fig. 512).
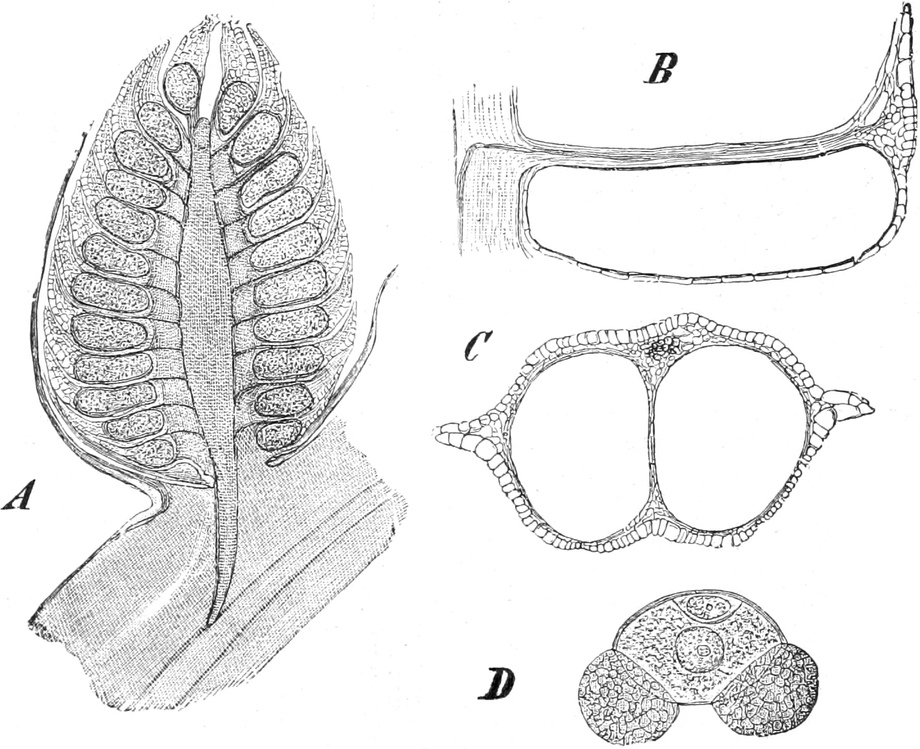
Fig. 512. Pinus montana. A Längsschnitt durch eine
fast reife männliche Blüte. Vergr. 10. B Längsschnitt durch ein
einzelnes Staubblatt. Vergr. 20. C Querschnitt durch ein Staubblatt.
Vergr. 27. D Ein reifes Pollenkorn von Pinus silvestris. Vergr. 400.
Nach E. STRASBURGER.
[S. 469]
Die Mikrosporangien stehen in Zwei- oder Mehrzahl auf der Unterseite
der Sporophylle. Ihre Öffnung wird, wie bei den Sporangien der
Pteridophyten, durch eigenartige Ausbildung meist der äußersten,
als „Exothecium“ bezeichneten Zellage der Wandung herbeigeführt.
Die Pollenkörner sind rundlich, häufig mit zwei Luftsäcken versehen,
welche die Verbreitung durch den Wind erleichtern (Fig. 512 A–D),
Bei der Keimung entledigen sie sich ihrer äußeren derben Wandschicht,
der Exine, vollständig; diese wird durch Dehnung des Plasmakörpers zum
Platzen gebracht.
Die weiblichen Blüten sind bei zahlreichen Gymnospermen den
männlichen ähnlich, insofern auch sie aus einer Achse und zahlreichen,
in schraubiger Anordnung daran sitzenden Sporophyllen bestehen, also
Zapfen darstellen. In anderen Fällen weichen sie aber erheblich von
ihnen ab und sind überhaupt von einer weit größeren Mannigfaltigkeit;
so kann über die Einzelheiten erst im speziellen Teil berichtet werden
(S. 511 ff.).
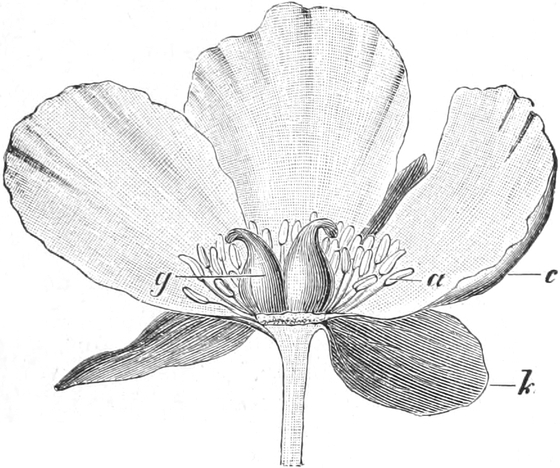
Fig. 513. Blüte von Paeonia peregrina. k Kelch, c
Krone, zusammen das Perianth, a Andröceum, g Gynäceum, vorderer
Teil der Blüte entfernt. 1⁄2
nat. Gr. Nach H. SCHENCK.
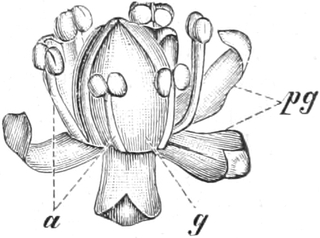
Fig. 514. Blüte von Acorus Calamus nach A.
ENGLER. pg Perigon. a Andröceum, g Gynäceum. Vgr.
Die Angiospermen zeigen dagegen in der Regel die Vereinigung
von Mikro- und Makrosporophyllen in einer Blüte, die danach zwittrig
oder hermaphroditisch heißt. (Ausnahme z. B. Querciflorae.)
Daneben sind in den Angiospermenblüten meist farbige, d. h. vom
grünen Laubblatt verschiedene Hüllblätter, insgesamt Perianth
genannt, vorhanden (Fig. 513, 514). (Ausnahme z. B. Querciflorae.)
Der schraubigen Anordnung gymnospermer Sporophylle an den
langgestreckten Achsen ihrer Blüten gegenüber fällt die meist
wirtelige Stellung der Hüllblätter wie der Sporophylle um
sehr verkürzte Achsen in den meisten angiospermen Blüten
auf. (Ausnahme z. B. Polycarpicae.) Wirtelige Stellung der
verschiedenen Blütenblätter, farbiges Perianth und Zwittrigkeit
sind also charakteristisch für Angiospermenblüten, ohne aber
allen Angiospermenblüten eigen zu sein. Der wichtige Faktor der
Pollenübertragung ist für diese Verschiedenheiten verantwortlich
zu machen. Wo der Wind diese Funktion übernommen hat, bietet die
Streckung der Achse, die freie Stellung der weiblichen Empfängnisorgane
nur Vorteile. Sind aber Tiere, Insekten oder Vögel, für die Bestäubung
in Anspruch genommen, so ist Ausbildung des Perianths als eines
weithin sichtbaren Schauapparates oder anderer Lockmittel notwendig,
die etwa auf Formgefühl oder Geruchsinn wirken, und die Form der
Blüte, Anordnung ihrer Sporophylle wie der Orte für Absonderung süß
schmeckenden Nektars müssen sich den Körperformen oder den Gewohnheiten
der betreffenden Tiere anbequemen. Diesem Umstande ist der unendliche
Farben- und Formenreichtum angiospermer Blüten zum großen Teil
zuzuschreiben.
Bei gewissen Angiospermen findet man hermaphrodite und
eingeschlechtige Blüten an demselben Individuum, man nennt
diese Erscheinung[S. 470] Polygamie; und zwar liegt Andro-
oder Gynomonözie vor, je nachdem männliche oder weibliche
Blüten neben Zwitterblüten vorkommen. Sind verschiedene
Individuen einer Art teils mit Zwitterblüten, teils mit
eingeschlechtigen Blüten versehen, so ist Andro- (bzw.
Gyno-)diözie vorhanden.
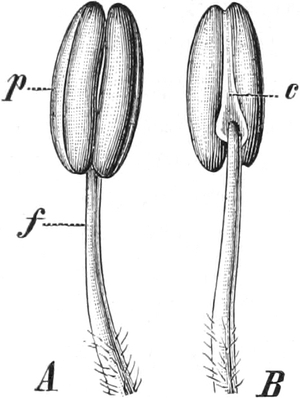
Fig. 515. Staubblätter von Hyoscyamus niger. A Von
vorn, B von hinten. f Filament, c Konnektiv, p eine Theca.
Vergr. Nach A. F. W. SCHIMPER.
Das Perianth angiospermer Blüten besteht aus meist zwei Wirteln,
welche gleich geformt und gefärbt sein können (z. B. Lilium) und
dann als Perigon bezeichnet werden, oder sie gliedern sich
in einen äußeren grünen Kelchblattkreis und einen inneren
gefärbten Kreis, die Krone oder Korolle (z. B.
Rosa). Der äußere Wirtel umhüllt und schützt die Blütenknospe.
In jeder vollzähligen Blüte folgen zwei Kreise von
Mikrosporophyllen oder Staubblättern auf das Perianth, und ein
Wirtel von Fruchtblättern, Makrosporophyllen, schließt die Blüte. Die
Wirtel alternieren der Regel nach. Die gesamten Staubblätter pflegt
man als Andröceum, die Fruchtblätter als Gynäceum
zusammenzufassen.
Jedes Staubblatt besteht aus einem fadenförmigen Träger,
dem Filament, und der Anthere, welche aus den
beiden Thecae zusammengesetzt ist; sie werden durch das
Konnektiv miteinander und dem Filament verbunden (Fig.
515). Jede Theca umfaßt ein Paar von Pollensäcken. Die
Anthere kann ihre Thecae der Innenseite der Blüte, also dem
Fruchtblattkreise, zuwenden oder sie nach außen kehren; sie heißt
danach intrors oder extrors. Dem Öffnen einer reifen
Theca dient meist (Ausnahme Ericaceae) die eigenartige Ausbildung
der hypodermalen Faserschicht (Fig. 511 C f), „fibröse
Schicht“ oder Endothecium genannt, während die Gymnospermen,
bis auf Ginkgo (vgl. S. 511), gleich den Farnen ein „Exothecium“
besitzen. In der Regel wird die Scheidewand zwischen den beiden
Pollensäcken beim Öffnen zerrissen, so daß sie dann nur ein Fach
bilden (Fig. 511 A). Die Mikrosporen selbst sind in ihrer Ausrüstung
verschieden; bei anemophilen Pflanzen auf die Verbreitung durch Wind
angewiesen, sind sie glatt, trocken und leicht. Bei den von Tieren
besuchten Blüten dagegen pflegen sie klebrig oder mit Stacheln und
Vorragungen der Exine versehen zu sein, die ein besseres Haften am
Haarkleid der bestäubenden Tiere ermöglichen. Sie unterscheiden
sich von denen der Gymnospermen auch dadurch, daß bei ihrer Keimung
die Exine nicht völlig abgeworfen zu werden braucht, weil sie von
vornherein mit mehr oder weniger zahlreichen Durchtrittstellen
für die Pollenschläuche ausgerüstet wird (Fig. 516). Sterile
Staubblätter, welche keinen fruchtbaren Pollen hervorbringen, werden
Staminodien (vgl. z. B. Scitamineen) genannt.
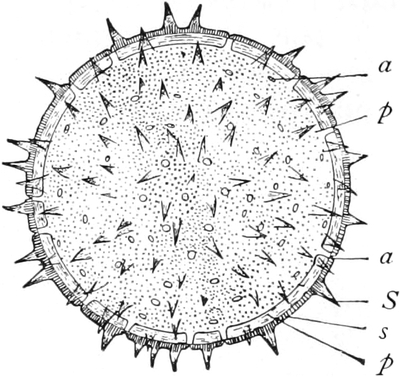
Fig. 516. Pollenkorn von Malva silvestris. S Stacheln
der Exine, s Stäbchenschicht der Exine, a Austrittstellen der
Pollenschläuche, p Stäbchen von oben. Nach A. MEYER.
Den Abschluß der Blüte bildet stets das Gynäceum. Seine
Fruchtblätter, oder Karpelle, können frei bleiben und
jedes für sich eine Frucht bilden (Fig. 517, 520 A), man spricht
dann von einem apokarpen Fruchtknoten, oder sie verwachsen
miteinander und stellen ein synkarpes Gynäceum[S. 471] (Fig. 518)
dar. Die Fruchtblätter tragen ihre Samenanlagen meist am Rande auf
mehr oder weniger deutlichen Wucherungen, den Placenten
(Fig. 517 p). Apokarpe Fruchtknoten werden demnach an ihren
beiden zusammenschließenden Rändern, an der sog. Bauchnaht,
je eine Reihe von Samenanlagen führen; die ihr gegenüberstehende
Rückennaht wird von der Mittelrippe des Blattes gebildet. In
synkarpen Fruchtknoten treten die Samenanlagen ebenfalls meist an den
Rändern der miteinander verwachsenen Fruchtblätter auf (Fig. 519 pl).
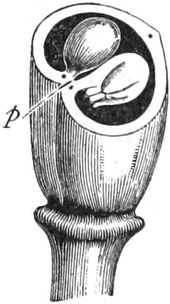
Fig. 517. Delphinium consolida. Querschnitt durch
den Fruchtknoten mit Samenanlagen an der Plazenta p. Nach
ENGLER-PRANTL.
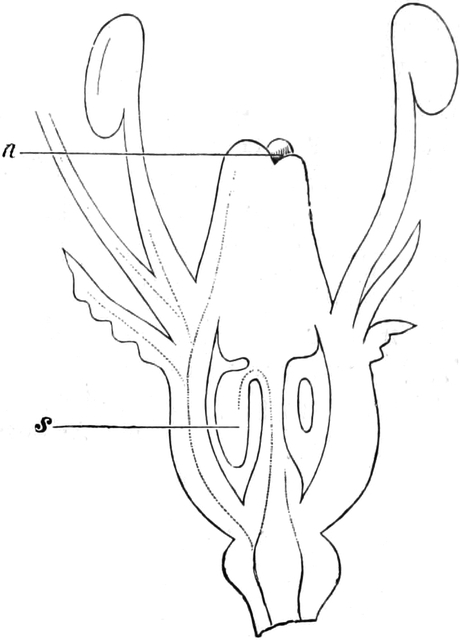
Fig. 518. Sambucus nigra. Blütenlängsschnitt. s
Samenanlage, n Narbe. Nach TSCHIRCH-ÖSTERLE.
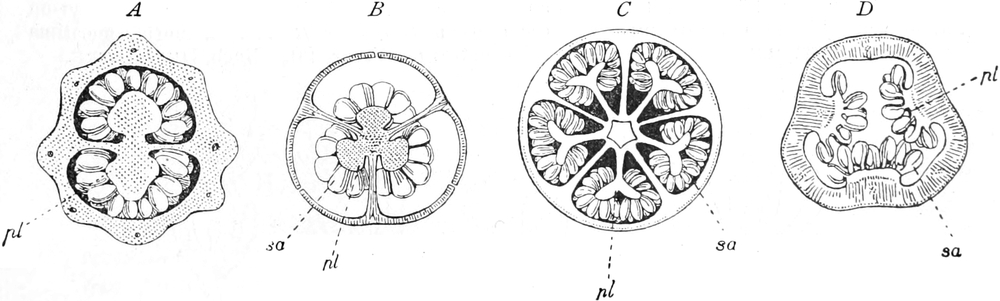
Fig. 519. Querschnitt von Fruchtknoten. A Lobelia.
B Diapensia. C Rhododendron. D Passiflora. pl Plazenta, sa
Samenanlagen. Nach LE MAOUT
et DECAISNE.
Die Samen werden parietal genannt, wenn die Placenten der
Wandinnenseite als flache Wülste entspringen (Fig. 519 D). Tritt
durch weiteres Eindringen der Karpellränder eine Fächerung des
Fruchtknotens ein, so rücken die Placenten und ihre Samenanlagen
gleichfalls nach innen vor; sie werden zentralwinkelständig,
wie Fig. 519 B zeigt. Nur die aus den Karpellrändern selbst
gebildeten Scheidewände pflegt man als echte zu
bezeichnen gegenüber den z. B. bei den Cruciferen vorhandenen
falschen (Fig. 701), die aus Wucherungen der Fruchtblätter
oder ihrer Nähte hervorgehen. Durch Beteiligung der Blütenachse,
welche im Zentrum zwischen den Fruchtblättern emporwächst und
sich von dem Karpellgewebe nicht scharf trennen läßt, kommt eine
sog. Zentralplacenta zustande (z. B. Primulaceen). Die
ursprünglich vorhandenen Scheidewände bleiben schon in früherem
Alter im Wachstum zurück oder werden aufgelöst, so daß die mit
dem Karpellgewebe emporgehobenen Samenanlagen an einer zentralen,
von den Fruchtblättern überkleideten Achse sitzen, während die
Außenteile der Fruchtblätter sie als einheitliche Wand umgeben.
[S. 472]
Jedes einzelne Fruchtblatt im apokarpen Gynäceum setzt sich nach oben
in einen stielartigen Griffel (Stylus) fort und endet
in einer sehr verschieden gestalteten Narbe (Stigma),
die als Empfängnisapparat dient und dementsprechend papillös, feucht
und klebrig zu sein pflegt (Fig. 520 D). In einem völlig synkarpen
Fruchtknoten ist nur ein einheitlicher Griffel und eine Narbe
vorhanden. Fig. 520 stellt ein apokarpes Gynäceum (A), ein synkarpes
(C) und einen nur im unteren Teil verwachsenen Fruchtknoten mit
freien Griffeln (B) dar.
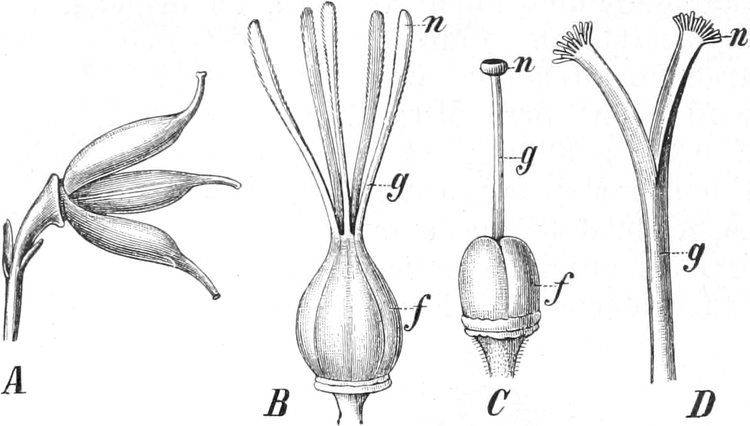
Fig. 520. Verschiedene Formen des Gynäceums. A Von
Aconitum Napellus. B Von Linum usitatissimum. C Von Nicotiana
rustica. D Griffel und Narbe von Achillea millefolium. f
Fruchtknoten, g Griffel, n Narbe. Vergr. Nach BERG und
SCHMIDT.
Fig. 521. Fruchtknoten von Conium maculatum mit
hängenden Samenanlagen im Längsschnitte. Raphe ventral. Nach
TSCHIRCH-ÖSTERLE.
Fig. 522. Fruchtknoten mit aufrechter Samenanlage, im
Längsschnitte. A Von Fagopyrum esculentum (atrop). B Von Armeria
maritima (anatrop). Vergr. 20. Nach DUCHARTRE.
Fig. 523. A Oberständiger Fruchtknoten (hypogyne
Blüte). B, B′ Mittelständiger Fruchtknoten (perigyne Blüten).
C Unterständiger Fruchtknoten (epigyne Blüte). Nach A. F. W.
SCHIMPER.
Die Lage der Samenanlagen im Fruchtknoten kann aufrecht,
hängend, wagerecht oder schräg zur Achse sein (Fig. 521, 522). Bei
anatropen Samenanlagen ist die Raphe ventral gelegen, wenn sie
der Bauchseite des Fruchtblattes zugekehrt ist, dorsal, wenn sie
gegen die Rückenseite sieht.
Eine große Mannigfaltigkeit der Blütenformen wird ferner durch
verschiedene Gestaltung der Blütenachse und entsprechend
geänderte Lage[S. 473] des Gynäceums bedingt. Die schematischen Figuren
(Fig. 523 A–C) geben einige der häufigeren Fälle wieder. Eine
Verdickung des Achsenendes gegenüber dem Tragstiele ist meist
vorhanden, häufig eine Verbreiterung und Vorwölbung oder Aushöhlung und
Vertiefung. Stehen an einfach kegelförmiger Achse die alternierenden
Quirle der Blüte übereinander, so ist das Gynäceum als Abschluß
der oberste Kreis, es ist oberständig, die übrige Blüte selbst wird
also unterständig oder hypogyn (Fig. 524 1). Wird aber durch
starke Verbreiterung des Achsenendes zu einem flachen Blütenboden,
Receptaculum (Hypanthium), oder zu einem ausgehöhlten
Blütenbecher (Fig. 523 B, B1), ein breiter Rand zwischen
Andröceum und Gynäceum eingeschoben, so spricht man von einer
perigynen Blüte, einem mittelständigen Fruchtknoten (Fig.
524 2). Schließt sich endlich die ausgehöhlte Blütenachse mit ihren
Kelch, Krone und Andröceum tragenden Rändern oben wieder zusammen,
so daß die Fruchtblätter mit ihr verwachsen, so ist das Gynäceum
unterständig, die Blüte oberständig oder epigyn (Fig. 524 3).
Der Deutlichkeit halber soll im folgenden stets die Bezeichnung
von der Stellung des Gynäceums hergeleitet werden, also nur vom
oberständigen, mittelständigen oder unterständigen Fruchtknoten die
Rede sein.
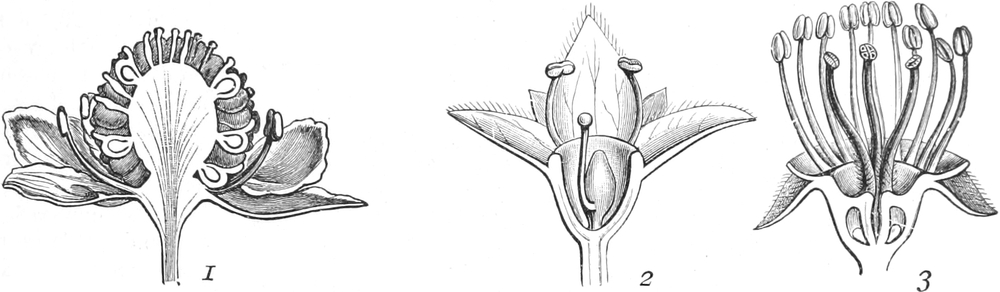
Fig. 524. Blütenlängsschnitte. 1 Ranunculus
sceleratus mit zahlreichen oberständigen apokarpen Fruchtblättern
auf kegelförmiger Blütenachse (hypogyne Blüte). Vergr. (Nach H.
BAILLON). 2 Alchimilla alpina, mittelständiger Fruchtknoten
(perigyne Blüte). 3 Pirus malus, unterständiger Fruchtknoten (epigyne
Blüte). Vergr. Nach W. O. FOCKE.
Ökologisch wichtige Gebilde der Blüte sind endlich die
Nektarien, bestimmte Stellen der Achse oder anderer Blütenteile,
die zuckerreiche Flüssigkeit (Nektar, Honig) zur Anlockung der
bestäubenden Tiere ausscheiden.
Eine typische Angiospermenblüte führt ihre Organe also in fünf
miteinander alternierenden Wirteln, von denen zwei auf das Perianth,
zwei auf das Andröceum, einer auf das Gynäceum entfallen. Sie ist
fünfwirtelig, pentazyklisch. Die Zahl der Glieder ist
entweder für alle Wirtel dieselbe, so z. B. für eine regelmäßige
Monokotylenblüte drei, eine Dikotylenblüte fünf, oder es tritt
besonders innerhalb der zum Andröceum und Gynäceum gehörenden
Wirtel eine Vermehrung oder Verminderung der Zahl ein. Auch kann
z. B. im Andröceum ein Wirtel ganz ausfallen oder die Wirtelzahl
vermehrt werden. Blüten mit nur einem Andröceumwirtel heißen
haplostemon, solche mit zwei Wirteln diplostemon.
Steht der äußere Andröceumwirtel (und dementsprechend derjenige der
Fruchtblätter) nicht alternierend mit dem Kronblattwirtel, sondern
direkt vor ihm, so ist das Andröceum obdiplostemon.
Das Diagramm (S. 77) einer fünfwirteligen Monokotylenblüte,
derart orientiert, daß der Querschnitt der Infloreszenzachse oben,
derjenige des[S. 474] Deckblattes (vgl. S. 106) sich unten befindet, ergibt
Fig. 525, dasjenige einer ebensolchen Blüte dikotyler Art Fig. 526.
Fig. 525. Diagramm einer pentazyklischen
Monokotylenblüte (Lilium).

Fig. 526. Diagramm einer pentazyklischen Dikotylenblüte
(Viscaria).
Fig. 527. Theoretisches Diagramm einer Irisblüte. Der
fehlende Staubblattquirl ist durch Kreuze bezeichnet.
Nach A. W. EICHLER.
Beide Diagramme sind sog. empirische Diagramme. Von einem
theoretischen Diagramm spricht man dagegen, wenn nicht
nur die tatsächlich vorhandenen Organe eingetragen werden,
sondern auch solche, deren früheres Vorhandensein man aus
phylogenetischen Gründen annehmen muß; so ist z. B. bei den mit
den Liliaceen nahe verwandten Iridaceen nur einer, und zwar der
äußere Staubblattwirtel vorhanden, der innere, dessen Gegenwart
der Verwandtschaft nach zu erwarten wäre, ist ausgefallen. Somit
erhalten wir, wenn die Stellung der fehlenden Glieder mit Kreuzen
in das empirische Diagramm eingetragen wird, das beistehende
theoretische Diagramm der Iridaceen (Fig. 527). Gelegentlich kommen
derartige dem theoretischen Diagramm entsprechende „vollständige“
Irisblüten vor, und es ist HEINRICHER[454] gelungen, eine
solche Blütenform durch Generationen hindurch aus Samen weiter
zu ziehen. Ein derartiges auf Merkmale der Vorfahren weisendes
Rückschlagen wird als „Atavismus“ bezeichnet (vgl. S. 178).
Zu einem kurzen Ausdruck eines solchen Diagramms bedient man sich
der sog. Blütenformeln, d. h. man setzt für jeden Wirtel
ein Buchstabenzeichen, etwa Kelch = K, Corolle = C, oder aber
Perigon = P, Andröceum = A, Gynäceum = G und die Zahl der Glieder
im Wirtel dahinter. Bei starker Vermehrung der Glieder kann das
Zeichen ∞ gesetzt werden. Verwachsung der Glieder eines Wirtels
wird durch eine Klammer um die betreffende Zahl, die Stellung
des Fruchtknotens durch einen unter (oberständig) oder
über (unterständig) der Zahl angebrachten Strich angedeutet.
Die Blütendiagramme (Fig. 525, 526) wären also zu schreiben: P3
+ 3, A3 + 3, G (3) für Monokotylen und K5, C5, A5 + 5, G(5) für
Dikotylenblüten, für eine Ranunkel K5, C5, A ∞, G ∞, für den
Schierling: K5, C5, A5, G (2), für Artemisia endlich: KO, C (5), A
(5), G (2).

Fig. 528. Atavistische Form von Iris pallida Lam.
abavia. Nach E. HEINRICHER.
[S. 475]

Fig. 529. A Radiäre (aktinomorphe) Blüte von Geranium
sanguineum. B Dorsiventrale (zygomorphe) Blüte von Viola tricolor.
Nach A. F. W. SCHIMPER. C Asymmetrische Blüte von Canna
iridiflora. f Fruchtknoten, k Kelch, c Krone, l Labellum, st
1–3 die übrigen Staminodien, a fertile Anthere, g Griffel. 1⁄4,
nat. Gr. Nach H. SCHENCK.
Fig. 530. Schemata razemöser Blütenstände. A Traube.
B Ähre. C Dolde. D Köpfchen. E Rispe. F Zusammengesetzte
Dolde oder Doppeldolde.
Fig. 531. Blütenähre von Plantago lanceolata. Nach
DUCHARTRE.
Fig. 532. Blütenkätzchen von Corylus americana. Nach
DUCHARTRE.
[S. 476]
Durch Verschiebung der Blütenglieder, ungleiche Größe oder
Unterdrückung einzelner sind aus dem ursprünglich strahligen, radiären
(aktinomorphen) Bau (Fig. 529 A) abweichende Gestalten hervorgegangen
(vgl. S. 65 f.), die entweder dorsiventral (zygomorph)
(Fig. 529 B), oder aber völlig asymmetrisch geworden sind
(Fig. 529 C). In den Blütenformeln werden radiäre Blüten mit  ,
dorsiventrale mit ↓ gekennzeichnet, z. B.: für den Goldregen ↓ K(5), C5
A(5 + 5), G1. Dorsiventrale Blüten pflegen immer eine ganz bestimmte
Lage zur Richtung der Schwerkraft anzunehmen. Unter Pelorien
versteht man radiäre Bildungsabweichungen sonst dorsiventraler Bluten.
,
dorsiventrale mit ↓ gekennzeichnet, z. B.: für den Goldregen ↓ K(5), C5
A(5 + 5), G1. Dorsiventrale Blüten pflegen immer eine ganz bestimmte
Lage zur Richtung der Schwerkraft anzunehmen. Unter Pelorien
versteht man radiäre Bildungsabweichungen sonst dorsiventraler Bluten.
Blütenstände (Infloreszenzen).
Die im vorstehenden beschriebenen Blüten der Angiospermen stehen in
verhältnismäßig seltenen Fällen einzeln, viel häufiger sind sie zu
mehreren oder vielen auf Verzweigungssysteme verteilt, die man als
Blütenstände oder Infloreszenzen bezeichnet. Von den
vegetativen Verzweigungen unterscheiden sich die Blütenstände durch
die gedrängte, dichte Stellung ihrer Seitenzweige, durch die meist
schuppenförmige Ausbildung der Deckblätter oder Brakteen und der
Vorblätter, in deren Achseln die Seitenzweige oder Einzelblüten stehen,
und durch das Austreiben aller Achselknospen, wenigstens in vielen
Fällen. In den Infloreszenzen der Cruciferen fehlen die Hochblätter
vollkommen.
Fig. 533. Traube von Linaria striata. Blüten mit
Deckblättern d. Nach A. F. W. SCHIMPER.
Fig. 534. Doldiger Blütenstand der Kirsche. Nach
DUCHARTRE.
Fig. 535. Blütenrispe von Yucca filamentosa. Verkl. Nach
A. F. W. SCHIMPER.
A. Die Hauptachse wächst stärker als
die Seitensprosse.
I. Die Blütenstände können, wie die vegetativen Verzweigungen,
Monopodien (vgl. S. 110) sein, so daß die Hauptachse dem Wachstum
ihrer Seitenzweige stets überlegen[S. 477] bleibt (oder die Seitensprosse
wachsen ebenso stark); solche razemösen Blütenstände treten
in verschiedenen Formen auf (Schemata Fig. 530):
α) Seitenachsen unverzweigt.
1. Traube, gestielte Einzelblüten an der verlängerten
Hauptachse, Schema 530 A. Fig. 533.
2. Ähre, sitzende Einzelblüten an der verlängerten
Hauptachse, Schema 530 B. Fig. 531.
Ist die Achse fleischig verdickt, so heißt die Ähre Kolben
wie bei den Araceen. Fällt die Ähre nach dem Verblühen (der
Fruchtreife) als Ganzes ab, so heißt sie Kätzchen Fig. 532.
β) Seitenachsen verzweigt.
3. Rispe, eine verlängerte Hauptachse, deren Seitenachsen Trauben
sind. Schema 530 E. Fig. 535.
B. Die Haupt- und Seitenachsen wachsen gleich stark.
4. Dolde, eine Anzahl von Seitenachsen, die alle gleichmäßig
mit der in eine Einzelblüte ausgehenden Hauptachse wachsen.
Schema 530 C. Fig. 534.
5. Zusammengesetzte Dolde, eine Dolde, die an Stelle der
Einzelblüten wiederum Dolden trägt. Schema 530 F. Fig. 666.
6. Köpfchen, sitzende Einzelblüten an einer verkürzten und
oben verbreiterten Hauptachse. Schema 530 D (Compositae). Fig.
799.
II. Oder die jeweilige Hauptachse wird jedesmal von ihren
Seitenachsen überholt; dann liegen zymöse Blütenstände vor,
die sich nach Zahl und Stellung der Seitensprosse in Pleiochasium,
Dichasium und Monochasium unterscheiden. Diese Verzweigungen sind
S. 111 bereits ausführlich beschrieben und im Grundriß dargestellt
(Fig. 152). Auch sind dort die monochasialen Verzweigungssysteme,
Sichel und Fächel, wie die aus dem Dichasium ableitbaren Schraubel
und Wickel behandelt, so daß hier nur auf die Abbildung eines
typischen Dichasiums (Fig. 536) und diejenige des Wickels (Fig.
537) hingewiesen zu werden braucht.
Fig. 536. Zymöser Blütenstand (Dichasium) von
Cerastium collinum. t–t⁗ die aufeinanderfolgenden Achsen. Nach
DUCHARTRE.

Fig. 537. Blütenwickel von Heliotropium curassavicum.
Nach ENGLER-PRANTL.
2. Ökologie. Die Blütenbestäubung[457] (vgl. S. 170).
Viele sonst rätselhaft bleibende Unterschiede im Bau der Blüten und
in der Anordnung ihrer[S. 478] Organe werden verständlich, wenn man sie
ökologisch betrachtet. Gemeinsam ist es allen Blüten, Nachkommenschaft
auf sexuelle Weise hervorzubringen; die Wege, die zu diesem Ergebnisse
führen, sind aber sehr verschieden. Gegenüber den Bryophyten und
Pteridophyten, bei welchen die Vereinigung der Geschlechtszellen unter
Zuhilfenahme des Wassers sich vollzieht, sind die Samenpflanzen, die
keine freibeweglichen männlichen Geschlechtszellen entlassen, und deren
Eizellen dauernd im Gewebe der Mutterpflanze eingeschlossen bleiben,
gezwungen, andere Wege einzuschlagen. Eine besondere Art der Zuführung
der Pollenkörner zu den von ihren Makrosporangien umschlossenen
Makrosporen und der darin befindlichen Eizelle wird notwendig. Es
müssen die zum Empfange des Pollens bereiten Narben (oder Mikropylen)
bestäubt werden.
Eine sehr große Zahl von Samenpflanzen bedient sich der bewegten Luft,
des Windes, als Übermittler des Pollens, wie z. B. unsere
gesamten Koniferen und auch die Mehrzahl der einheimischen Laubbäume:
die Ulme, Eiche, Buche, Hainbuche, ferner unsere Gräser, besonders die
Getreidearten. So einfach die Verhältnisse in diesem Falle zu liegen
scheinen, so bedarf es doch mancherlei notwendiger Voraussetzungen für
einen sicheren Erfolg dieser Übertragungsart.
Vor allem müssen solche windblütigen oder anemophilen Pflanzen
eine ungeheuere Masse von Pollen erzeugen, da naturgemäß nur ein
geringer Bruchteil den Ort seiner Bestimmung erreichen kann. So sieht
man wohl zur Zeit, da unsere Nadelwälder in Blüte stehen, ungezählte
Mengen des in die Luft entführten Pollenstaubes bei eintretendem
Regenwetter als dichten gelben „Schwefelregen“ niederfallen.
Fig. 538. Alnus glutinosa. 1 Blühender Zweig mit
kleinen aufrechten weiblichen und hängenden männlichen Kätzchen und
Blatt. 2 Eine Deckschuppe mit männlichen Blüten. 3 Weibliches
Kätzchen. 4 2 weibliche Blüten mit Deckschuppe. 5 Fruchtstand. 6
Frucht. 3⁄4
nat. Gr., 2–6 vergr.
Die anemophilen Pflanzen und Blüten haben nun einige Charakterzüge
gemein, die in Beziehung zu der Windbestäubung stehen und keineswegs
als bloße Zufälligkeiten gelten können. Das gilt in erster Linie für
den Bau der männlichen Blütenstände. Sie haben oft die Form
mehr oder minder lang herabhängender Kätzchen (Fig. 538) angenommen,
die eine große Zahl von Mikrosporophyllen vereinigen und so orientiert
sind, daß der Wind nach Öffnung der Sporangien alle Sporen ausfegen
oder ausschütteln kann; z. B. Eichen (Fig. 690), Birken (Fig. 683),
Erlen (Fig. 538), Haselnuß, Hainbuchen (Fig. 684), die auffallend
langen Walnußkätzchen (Fig. 679), dasselbe gilt für die aufrechten
Koniferenblüten(Fig. 604). Gleiche Bedeutung kommt der Befestigung[S. 479]
der Gramineen-Antheren auf langen schwanken Filamenten zu (Fig. 539).
Einige Urticaceen (Pilea-Arten, Brennesseln usw.) schleudern ihren
Pollen bei Öffnung der Fächer mit Hilfe der elastisch gespannten
Filamente als leichte Staubwölkchen in die Luft. Wie die Infloreszenzen
und Blüten, so haben auch die Pollenkörner der Anemophilen
charakteristische Merkmale. Sie sind leicht und glatt, bei einigen
Nadelhölzern sogar mit zwei lufthaltigen Flugblasen (Fig. 512 D)
versehen, die ein längeres Schweben in der Luft ermöglichen.
Die weiblichen Blüten pflegen der lebhaften Farben zu
entbehren und besitzen keine Nektarien. Ihre Narben, die den Pollen
auffangen, sind jedoch besonders mächtig entwickelt und mit langen
Federhaaren versehen (Fig. 539) oder pinselförmig, federförmig oder
lang fadenförmig ausgezogen. Bei vielen Gymnospermen, besonders
deutlich bei Taxus, scheidet die Mikropyle des Makrosporangium zur
Blütezeit einen Flüssigkeitstropfen aus, an dem die Pollenkörner
hängen bleiben und beim Eintrocknen des Tropfens auf den Scheitel
des Nucellus niedergesogen werden; in anderen Fällen gleiten die
Pollenkörner zwischen den auseinander gespreizten Sporophyllschuppen
der zapfenförmigen, aufgerichteten Blüten hinab, bis sie am Grunde
zwischen die Integumentfortsätze gelangen und an den feuchten
Mikropylenöffnungen haften.
Endlich ist aber auch der Zeitpunkt des Blühens nicht bedeutungslos.
Die Ulme blüht lange vor ihrer Belaubung bereits im Februar oder März,
die Haselnuß, Pappel und Erle (Fig. 538) machen es ebenso, und bei
Walnuß, Eiche, Hainbuche und Birke, ja auch bei der Rotbuche sieht man
die Blüten mit den ersten Blättern hervorbrechen und ihre Blütezeit
vollenden, bevor die Belaubung voll entwickelt ist. Es würde im anderen
Falle eine ungeheuere Menge von Pollenkörnern an der Behaarung und
in den Falten der großen Laubblätter hängen bleiben und damit noch
mehr Blütenstaub, als so schon der Fall ist, seiner eigentlichen
Bestimmung entzogen werden. Bei den Nadelhölzern ist die Blattform viel
weniger hinderlich, und außerdem sehen wir bei ihnen die weiblichen
Blütenzapfen in den Spitzen ihrer Kronen (Abies) oder am Gipfel der
Baumpyramiden (Picea) hervorsprossen, während die männlichen Blüten
sich mehr an den unteren Zweigen entwickeln. Der bei trockenem, warmem
Wetter entleerte Pollen wird von der bei Sonnenschein sich erwärmenden
und an den Bäumen entlang aufsteigenden Luft mit emporgeführt und
erreicht so die hoch über seiner Bildungsstätte stehenden weiblichen
Zapfen mit ihren zahlreichen Samenanlagen.
Sehr gering ist die Zahl derjenigen Samenpflanzen, bei denen das
Wasser eine Rolle bei der Pollenübertragung zu spielen hat;
es handelt sich ausschließlich um submerse Wasserpflanzen, die sich
ihrem Medium nicht entziehen können und als wasserblütige oder
hydrophile Gewächse neben die anemophilen treten (z. B. Zostera,
das Seegras Vallisneria spiralis und Helodea, die Wasserpest).
Fig. 539. Anemophile Blüte von Festuca elatior. Nach
H. SCHENCK.
[S. 480]
Für die große Mehrzahl der Samenpflanzen jedoch kommen weder Wind
noch Wasser, sondern Tiere, und zwar in erster Linie Insekten als
Pollenüberträger in Betracht. Auf Insekten zur Pollenübertragung
angewiesene Pflanzen heißen entomophil.
Seit KONRAD SPRENGEL in seiner berühmten Schrift „Das
entdeckte Geheimnis der Natur im Bau und in der Befruchtung der Blumen
1793“ die Wechselbeziehungen zwischen den die Blumen besuchenden
Insekten und den Formen und Farben der Blumen aufgedeckt hatte,
ist kein anderes Gebiet der Biologie so eifrig durchforscht worden
wie die Blütenökologie, die man schon begann mit dem Begriff der
Pflanzenbiologie überhaupt zu verwechseln. Um so merkwürdiger ist
es, daß sich niemand die Frage vorgelegt hatte, ob denn die unserem
Auge sich darbietenden Farben auch von den Insekten in gleicher
Art wahrgenommen werden könnten? Freilich erschien es schwer, sich
das Auftreten der ganzen Farbenpracht unserer Wiesen und Obstbäume
anders zu denken, denn als „Schauapparat“ für die Nahrung suchende
Insektenwelt, deren Hunger zu befriedigen die Blumen eigene
Vorkehrungen in ihren Nektarien besitzen, obwohl ja auch nicht
entomophile Blüten, wie diejenigen unserer Koniferen, die Narben von
Corylus u. a., intensive Farben entwickeln.
Es ist das Verdienst von C. HESS[458], dieser Frage
nachgegangen zu sein. Auf Grund seines Nachweises, daß die Bienen,
die wichtigsten Blütenbestäuber, in einem Merkmal mit farbenblinden
Menschen übereinstimmen, darf man eine Rot-Grün-Farbenblindheit bei
Bienen im allgemeinen annehmen, während HESS für völlige
Farbenblindheit eintrat.
Durch eine Reihe sorgfältiger, experimenteller Arbeiten hat V.
FRISCH[459] diese Meinung zu entkräften gewußt und höchst
interessante innigere Beziehungen zwischen Bienen und Blütenbesuch
festgestellt. Er unterscheidet bei den Blütenbesuchen der Bienen
zwischen „Suchern“ und „Sammlern“. Die Sucher finden neue ergiebige
Nektarquellen mittels der Farben (es wurde mit gelb und blau
experimentiert) aus beträchtlicher Entfernung auf und erkennen
in der Nähe an dem Blütendufte mit großer Genauigkeit die für
sie geeigneten Blüten heraus. Sie übermitteln ihre Befunde den
Stockgenossen mittels einer charakteristischen Zeichensprache und
kehren mit einem oder mehr Sammlern zu der Fundstelle zurück, die
nun stetig weiter ausgebeutet wird.
Unzweifelhaft bleibt also die Wechselwirkung zwischen Blumen und
Insekten bezüglich des Farben-, Geruch- und Formensinns bestehen.
Wenn eine Fernwirkung des Duftes für Bienen im allgemeinen nicht
nachzuweisen war, so scheinen Falter, Sphingiden und Eulen darin
abzuweichen. Denn welche Vorstellung sollte man sich von dem starken,
gegen Abend sich steigernden Duft von Lonicera, Philadelphus usw.
machen, wenn nicht die eines Anlockungsmittels für Nachtfalter,
die dem Dufte entgegenfliegen und damit ihre Nahrungsquelle zu finden
vermögen? Wie wollte man sich die Nektarien und die Ausgabe
von großen Mengen eines so wichtigen Pflanzenreservestoffes wie
Zucker erklären, wenn die Gäste, die ihn gierig aufsuchen, den Blüten
nicht unentbehrlich wären? Wie endlich kann man den Blütenbau
einer Salvia, einer Orchis, ja irgendeiner dorsiventral
gebauten Blüte verstehen wollen, wenn man ihn nicht in Beziehung
bringt zu den sie aufsuchenden, ihren Nektar saugenden und sie dabei
bestäubenden Insekten? Die wechselseitigen Anpassungen der Blütenformen
und der Insektenkörper sind so zahlreich, und die Erfahrung, daß sonst
wohlgedeihende Pflanzen fern ihrer Heimat aus Mangel an den ihnen
angepaßten Blütenbestäubern unfruchtbar bleiben — wie es z. B. bei der
Vanille der Fall war, bis man die Blüten durch Menschenhand bestäubte
—, nachgerade so häufig gemacht worden, daß man an dem Angepaßtsein
von Blüten an bestimmte Insekten und umgekehrt nicht zweifeln kann.
[S. 481]
Meist ist die Lage der vom Insekt aufgesuchten Nektarien derart,
daß das Haarkleid der Tiere an ganz bestimmten Stellen Pollenkörner
mitnehmen muß, die beim Besuche anderer Blüten auf die Narbe
übertragen werden. Da ist es denn wichtig, daß solche entomophilen
Pflanzen ihre Pollenkörner ganz anders ausrüsten als die vorher
betrachteten anemophilen. Viele stachlichte Hervorragungen, rauhe
oder klebrige Oberflächen oder das Zusammenbleiben in Tetraden, ja
in ganzen Pollenfächern wie bei Orchis (Fig. 842) und Asclepias
sind für den Pollen entomophiler Pflanzen charakteristisch. Daß
solcher Pollen neben dem Nektar für manche Insekten ein besonders
wertvolles, weil stickstoffhaltiges Nahrungsmittel bildet, wie für
die Bienen, die ihn zum „Bienenbrot“ verkneten, ist ebenfalls von
Bedeutung.
Fig. 540. Blütenbestäubung bei Salvia pratensis.
(Erklärung im Text.) Nach F. NOLL.
Ein sehr inniges Zusammenwirken von Blütenbau und Insektenkörper
bietet die Bestäubung von Salvia pratensis durch Hummeln. Fig.
540 1 zeigt eine Salviablüte mit einer Nektar suchenden Hummel
auf der Unterlippe. Salvia hat nur zwei Staubblätter, deren
Antherenhälften ganz verschieden entwickelt sind: die eine ist
steril und bildet eine Verschlußklappe in der Blumenkronröhre
(Fig. 540 4), die andere sitzt am Ende eines langgestreckten
Konnektivs, das diese fertile Hälfte unter die Wölbung der
Oberlippe hinaufhebt. So ist ein ungleichseitiger Hebel gebildet,
der drehbar an der Filamentspitze befestigt ist. Bei Einführung
des Rüssels übt die Hummel einen Druck auf den kürzeren Arm, die
fertile Antherenhälfte wird damit durch die Hebelwirkung des
Konnektivs (c) um den Ansatzpunkt am Filament (f) gedreht und
fest auf dem Haarkleid des Hummelhinterleibes abgestreift (Fig. 540
1, 3). Beim Besuche einer älteren Blüte findet sie den Griffel
weiter unter der Oberlippe hervorgewachsen (vgl. S. 483 Dichogamie)
und seine Narbe genau an der Stelle jener Antherenhälfte, so daß
von dort mitgenommene Pollenkörner auf dem sichersten Wege der
Narbe übermittelt werden müssen.
Jedoch nicht die Nahrung allein zieht die Insekten zu den Blüten
hin, auch der Fortpflanzungsinstinkt führt sie in einigen
Fällen zum Blütenbesuch. Das mißfarbige Aussehen und der Aasgeruch
mancher Araceen, Asclepiadaceen- und Aristolochiaceenblüten
veranlassen Aasfliegen zum Besuch der Blüten, in denen sie ihre
Eier ablegen und gleichzeitig Pollen anderer vorher besuchter
Blüten übertragen. Die Bestäubung der Feigenblüten (Fig. 541)
wird durch eine Gallwespe besorgt, welche in die krugförmigen
Blütenstände einkriecht, ihre Eier in die kurzgriffeligen
Gallblüten ablegt und dabei den vom „Caprificus“, den männlichen
Blütenpflanzen, mitgebrachten Pollen den langgriffeligen weiblichen
Samenblüten zuführt (Fig. 541). Yucca filamentosa endlich ist für
ihre Fortpflanzung gänzlich auf eine Motte (Pronuba) angewiesen,
welche ihre Eier in die Yucca-Fruchtknoten ablegt und die Narbe
gleichzeitig bestäubt. Wenn auch die Raupen einen großen Teil der
Samenanlagen fressen, so bleiben doch stets Samen übrig, während
ohne Bestäubungsvermittler, wie die bei uns kultivierten Exemplare
zeigen, überhaupt kein Samenansatz erfolgen kann.
Fig. 541. Ficus carica. A Längsschnitt durch einen
Blütenstand. B Samenblüte. C Gallenblüte. D Männliche Blüte.
B–D vergr. D nach KERNER. B, C nach H. Graf
SOLMS-LAUBACH.
[S. 482]
Neben der Entomophilie spielt die auf amerikanische Kolibris und
die Honigvögel der alten Welt sich beziehend Ornithophilie
eine weit bescheidenere Rolle. Einen besonders merkwürdigen Fall
von Anpassung zwischen einer Blüte und ihrem Bestäuber stellt die
in unseren Gewächshäusern häufig kultivierte Strelitzia reginae
dar (Fig. 542). Ihre drei äußeren Perigonblätter (t) sind
lebhaft orangerot gefärbt; das große azurblaue Labellum (p)
entspricht einem der inneren Perigonblätter, während die beiden
übrigen (p) unscheinbar bleiben und den Zugang zum Nektarium
überdachen, das reichlich Nektar austräufeln läßt. Staubblätter
(st) und Griffel (g) liegen in einer das Labellum der Länge
nach durchziehenden Rinne, deren Ränder leicht auseinanderklappen;
die Narbe (g) ragt frei über das Labellum hinaus. Der in
den gleichen Farben prangende Vogel, Nectarinia afra, fliegt
zuerst die Narbe an und streift nachher, auf dem Labellum weiter
vordringend, den Pollen der Staubblätter ab, den er auf eine
nächste Narbe übertragen kann. Ebenso eigenartig ist der Bau der
hängenden Marcgravia-Infloreszenzen mit Deckblättern, die zu Nektar
haltenden Kannen umgewandelt sind; doch steht dieser nach neueren
Beobachtungen nicht mit der Entomophilie in Beziehung[460]. Durch
die angeführten Untersuchungen von HESS (S. 480) wird
es erklärlich, daß die meisten ornithophilen Blüten intensiv rot
gefärbt sind (Aloë, Clianthus, epiphytische Loranthaceen usw.),
da die Empfindlichkeit für Rot beim Tagvogel jener für unser Auge
ähnlich ist.
Fig. 542. Ornithophile Blüte von Strelitzia reginae
und Querschnitt durch das große Labellum p. t Äußere, p innere
Perigonblätter, g Griffel bzw. Narbe, st Staubblätter. Aus A.
F. W. SCHIMPER, Pflanzengeographie.
Der Vollständigkeit halber seien auch noch die
chiropterophilen Blüten erwähnt, z. B. diejenigen der
Pandanacee Freycinetia, deren innere Deckblätter den großen
Fledermäusen des Malayischen Archipels, den bekannten „fliegenden
Hunden“, ein beliebter Leckerbissen sind, wofür sie sich durch die
Pollenübermittelung dieser diözischen Pflanze nützlich erweisen.
Endlich sollen Calla palustris, Chrysosplenium und Aspidistra durch
Schnecken bestäubt werden, sie wären demnach als malakophile
Pflanzen zu bezeichnen.
Da die Mehrzahl angiospermer Pflanzen hermaphrodite Blüten besitzt,
könnte es merkwürdig erscheinen, daß so mannigfache und verwickelte
Anpassungen der Übertragung des Pollens auf andere Blüten, also
der Kreuzung[S. 483] dienen müssen. Zunächst aber liefert oft
die Bestäubung mit eigenem Pollen, die Selbstbestäubung oder
Autogamie, minder guten Samenansatz, z. B. bei dem Roggen, oder
bleibt ganz ohne Erfolg. Derartige „selbststerile“ Pflanzen sind z. B.
Lobelia fulgens, Corydalis cava, Cardamine pratensis. Ebenso wird
Fremdbestäubung, Allogamie, eintreten müssen, wenn der Pollen
nur nach Verwundung der Narbe zu keimen vermag, wie bei Laburnum
vulgare; hier erfüllt nur Insektenbesuch, der in der Regel fremden
Pollen mitbringen dürfte, die Keimungsbedingung und schließt damit
blüteneigenen Pollen aus. Bei Orchideen aber wirkt der eigene Pollen
vielfach direkt schädigend und läßt die damit belegte Blüte alsbald
absterben.
Fig. 543. Blüte von Anthriscus silvestris. Schwach
vergr. 1 im männlichen, 2 im weiblichen Stadium. Nach H.
MÜLLER.
Aber auch wo keine Selbststerilität besteht, gibt es zahlreiche
und sehr mannigfaltige Einrichtungen, die eine Selbstbestäubung
hermaphroditer Blüten völlig unmöglich machen und Fremdbestäubung
begünstigen. Daß Diözie die Selbstbestäubung ausschließt, daß
monözische Blütenverteilung wenigstens die Bestäubung mit blüteneigenem
Pollen verhindern muß, bedarf ja keiner Erörterung. Ein gleiches
Verhältnis wird dort vorliegen, wo die beiden Geschlechter einer
hermaphroditen Blüte ungleichzeitig reifen. Dieser sehr häufige
Fall wird als Dichogamie bezeichnet. Naturgemäß sind bei
dichogamen Pflanzen zwei verschiedene Möglichkeiten vorhanden; entweder
reifen die Staubblätter zuerst, und der Pollen wird entleert, bevor
die Narben derselben Blüte belegungsfähig sind, die Pflanze ist
protandrisch, oder umgekehrt der Griffel mit seinen Narben
ist reif, bevor der Pollen verbreitet werden kann, die Pflanze ist
protogyn.

Fig. 544. Blütenstand von Plantago media mit protogynen
Blüten. Aus den mittleren noch geschlossenen Blüten ragt der
bestäubungsfähige Griffel hervor (
). Die unteren Blüten haben den
Griffel bereits verloren, dafür aber die langen Staubblätter entfaltet
(
). Nach F. N
OLL.
Die Protandrie ist der weitaus häufigere Fall der Dichogamie.
So sind bei den Blüten der Kompositen, Campanulaceen, Lobeliaceen,
Umbelliferen (Fig. 543), Geraniaceen, Malvaceen (Fig. 709) u. a. die
Narben noch unentwickelt, wenn die Staubblätter ihre Pollenmassen
entlassen. Auch bei Salvia (Fig. 540) ist Protandrie notwendige
Voraussetzung der Fremdbestäubung. Bei der Protogynie dagegen
wird die Bestäubung nur von seiten älterer Blüten möglich sein, deren
Pollen nach Abblühen des Griffels und seiner Narben freigeworden ist.
Hierher gehören die Plantaginaceen[S. 484] (Fig. 544), Scrophularia nodosa,
Aristolochia Clematitis, Arum maculatum, Helleborus, Magnolia.
In gleichem Sinne wirkt die von DARWIN zuerst aufgedeckte
Heterostylie, die freilich TISCHLER zufolge durch
Ernährungseinflüsse verändert werden kann. Halten wir uns an das
abgebildete Beispiel (Fig. 545) von Primula sinensis, so zeigt sich
beim Vergleich von Blüten verschiedener Individuen, daß sie sich in
der Länge ihrer Staubblätter und Narben unterscheiden. Man findet
„langgriffelige“ Blüten, deren Narben den Eingang der Kronröhre
verengern, deren Antheren dagegen tief unten in der Röhre sitzen; ein
anderes „kurzgriffeliges“ Individuum zeigt die Antheren in Höhe der
Narbe jener erstbetrachteten Blüte, die Narbe in Höhe ihrer Antheren.
Ein und dasselbe Insekt kann natürlich nur gleich hochstehende
Blütenorgane mit derselben Körperstelle berühren, also nur die
sich ihrer Lage nach entsprechenden Blütenteile bestäuben, so daß
Fremdbestäubung gesichert ist. Die Betrachtung der Pollenkörner und
der Narbenpapillen läßt leicht erkennen, daß ihre Größenverhältnisse
wechselseitige Bestäubung bedingen.
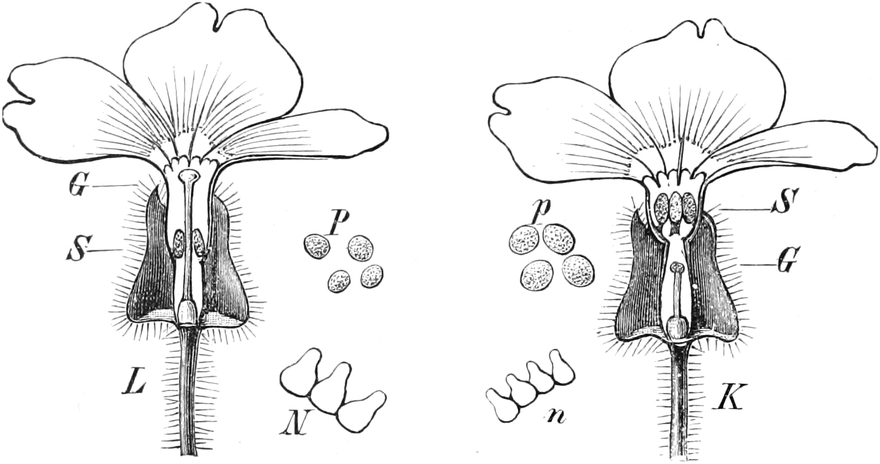
Fig. 545. Primula sinensis. Zwei heterostyle Blüten
von verschiedenen Stöcken. Schwach vergrößert. L Langgriffelige,
K kurzgriffelige Blütenform, G Griffel, S Staubbeutel. P
Pollenkörner und N Narbenpapillen der langgriffeligen, p und n
Pollenkörner und Narbenpapillen der kurzgriffeligen Form. P, N,
p, n bei 110facher Vergrößerung. Nach F. NOLL.
Derartige „dimorphe“ Heterostylie finden wir noch bei Hottonia,
Pulmonaria, Linum, Menyanthes; dagegen besitzen Lythrum salicaria und
Oxalis-Arten dreierlei verschiedene Stellungen für Narben und Antheren,
sie sind „trimorph heterostyle“ Pflanzen.
Fig. 546. Blüten von Aristolochia Clematitis, längs
durchschnitten. I Junge Blüte. N Narben, S Staubbeutel. II
Ältere Blüte, vgl. den Text. 2/1. Nach F. NOLL.
Bei zahlreichen Blüten ist endlich die Anordnung derartig, daß der
Pollen durch seine Lage vollkommen verhindert wird, überhaupt mit
der eigenen Narbe in Verbindung zu kommen. Dies Verhalten heißt
Herkogamie. So trägt Iris ihre drei Antheren unter den
Griffelwölbungen, Orchis heftet die beiden Pollinien[S. 485] über der Narbe
fest, Asclepias schließt die fünf Pollinien an Griffelschwellungen mit
Klemmkörperchen paarweise zusammen (vgl. Fig. 755).
Bisweilen wirken Herkogamie und Dichogamie zusammen: Die protogyne
Blüte von Aristolochia Clematitis (Fig. 546) steht im ersten
Blütenstadium mit geöffnetem Schlunde aufgerichtet. Kleine Insekten
vermitteln die Bestäubung. Beim Einkriechen in den aufrecht
stehenden Trichter können sie zwischen abwärts gerichteten
Reusenhaaren hindurch in den Kessel vordringen. Ihrer Flucht aus
diesem Gefängnis stehen aber eben jene Haare so lange entgegen, bis
im zweiten Blütenstadium die Bestäubung der Narbe durch aus älteren
Blüten mitgebrachten Pollen vollzogen ist. Dann öffnen sich die
unter dem säulenförmigen Griffel befindlichen Antheren, die Blüte
sinkt schlaff herab und die Insekten können mit frischem Pollen
versehen über die gleichzeitig vertrocknenden Reusenhaare hinweg
ins Freie gelangen und neue, jüngere Blüten aufsuchen. Alle diese
mannigfaltigen und zum Teil direkt raffinierten Einrichtungen zur
Erzielung der Kreuzung weisen darauf hin, daß es bei der
Befruchtung darauf ankommen dürfte, derartige Geschlechtszellen
zu vereinigen, die in ihren vererbbaren Eigenschaften weiter
voneinander differieren, als es bei Abkömmlingen derselben Blüte
der Fall sein könnte. Auch pflegen allogam erzeugte Nachkommen
kräftiger zu sein als autogam entstandene.
Wenn nun trotzdem bei gewissen Pflanzen neben den für Wind- oder
Insektenbestäubung eingerichteten großen chasmogamen Blüten
kleine unscheinbare Blüten vorkommen, die sich niemals öffnen und
nur der Selbstbestäubung dienen können, so lassen sich solche
kleistogamen Blüten[461] wohl damit verständlich machen,
daß diesen Pflanzen ein weiteres Propagationsmittel gegeben ist,
das ihren Nachkommen wieder zu gelegentlicher Kreuzung mit Hilfe
der großen chasmogamen Blüten verhelfen kann. Kleistogamie ist
häufig oder regelmäßig vorhanden bei Impatiens-, Viola-, Lamium-,
Stellaria-Arten, bei Specularia perfoliata, den unterirdischen
Infloreszenzen von Lathraea squamaria, Juncus hufonius u. a.;
Polycarpon tetraphyllum besitzt nur kleistogame Blüten.
Entwicklung
der Geschlechtsgeneration bei den Samenpflanzen.
A. Bei den Gymnospermen[462] enthalten die
Mikrosporen ein wenigzelliges Prothallium, das sich innerhalb
der großen, später zum Pollenschlauch auswachsenden Zelle, deren
Kern in Fig. 547 mit k bezeichnet ist, der Außenwand anlegt. Die
ältesten (p) stellen den Rest vegetativer Prothalliumzellen
dar. Auf sie folgt, als letzte abgegebene Zelle, die spermatogene
Zelle (sp). Diese zerfällt früher oder später in die
Mutterzelle des Antheridiums (Fig. 548 B[m]) und eine
sterile Schwesterzelle, die jene an die übrigen Zellen des
Prothalliums anheftet (s). Nur durch Ab- oder Auflösung der sterilen
Schwesterzelle, die GOEBEL daher Dislokatorzelle nennt, kann
also die Antheridium-Mutterzelle frei werden und in den Pollenschlauch
einwandern. Sie bildet dabei, oder schon solange sie noch festsitzt,
zwei Tochterzellen, die generativen Zellen, Spermazellen oder
männlichen Geschlechtszellen.
Fig. 547. Pollenkorn von Ginkgo biloba noch innerhalb
des Mikrosporangiums. Vergr. 300. Nach E. STRASBURGER.
a) Cycadeen.
Bei den Cycadeen und bei Ginkgo erhalten diese Zellen noch die Form
von Spermien, so daß sie sich hierin direkt an die spermiumbildenden
heterosporen Archegoniaten anschließen. Die Entwicklung ist in Fig.
548 an Zamia dargestellt, und die Figurenerklärung gibt über die
Einzelheiten Auskunft. Wie die Fig. 549 (a) weiter zeigt, bleiben die
beiden Rücken an Rücken ausgebildeten Spermien eine Zeitlang an der
sterilen Schwesterzelle des Antheridiums haften, nach ihrer Ablösung
(b) runden sie sich ab und zeigen ihr verjüngtes Vorderende mit
einem schraubig den Körper umlaufenden Zilienkranze versehen, der ihre
Schwimmbewegung ermöglicht (Fig. 552).
[S. 486]
Fig. 548. Zamia floridana. Spermienbildung nach H.
J. WEBBER. A Reifes Pollenkorn, 800. B, C, D Verschiedene
Stadien der Antheridiumentwicklung, B, C 400, D 200. k
Kern des Pollenschlauches, sp spermatogene Zelle, p erhaltene
vegetative Prothalliumzelle, die (B) in die sterile Schwesterzelle
s des Antheridiums hineinwächst, m Antheridium-Mutterzelle, d. h.
Mutterzelle der Spermien. e Exine. In der Mutterzelle sind die
großen sternförmigen Blepharoplasten bl sichtbar, welche die Zilien
bilden werden, die in D als kleine Körnchen die Querschnitte des
Zilienbandes zeigen. Stärkekörner sind im Pollenschlauch vorhanden,
in C treten sie auch in der vegetativen Zelle und in der sterilen
Schwesterzelle auf, in D erscheinen beide mit Stärke vollgepfropft,
D zeigt die beiden aus der Spermienmutterzelle hervorgehenden
Spermien sp durch eine Wand voneinander getrennt.
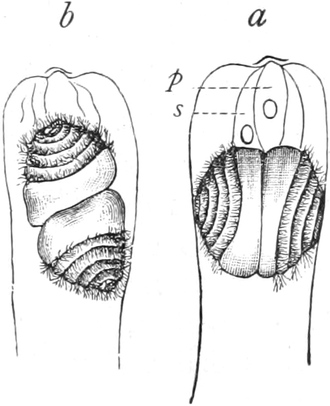
Fig. 549. Oberes Ende des Pollenschlauches von Zamia
floridana mit vegetativer Prothalliumzelle p. steriler Schwesterzelle
s und den beiden Spermien, a Vor Beginn der Bewegung, b nach
Eintritt der Zilienbewegung. Die Prothalliumzellen sind zerrissen,
die Trennung der beiden Spermien ist fortgeschritten. Nach H. J.
WEBBER. Vergr. ca. 75.
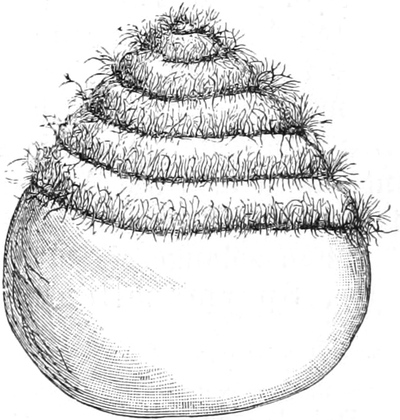
Fig. 550. Zamia floridana. Frei schwimmendes reifes
Spermium. Nach H. J. WEBBER. Vergr. 150.
Weibliche Zapfen von Zamia tragen eine Anzahl von Sporophyllen,
deren sechseckige Oberflächenbilder genau aneinander passen. Jedes
Sporophyll führt zwei Makrosporangien. Sie bestehen aus einem
Nucellus und einem Integument. Zwischen den Integumenträndern bleibt
über dem Nucellusscheitel die Mikropyle als offener Kanal erhalten.
Zur Zeit des Stäubens der männlichen Zapfen weichen die einzelnen
sechseckigen Makrosporophyllschilder auseinander, so daß der vom
Winde herbeigeführte Pollen freien Zutritt findet. Auf dem Scheitel
des Nucellus bildet sich zu dieser Zeit eine mehr[S. 487] oder minder tiefe
Höhlung — die sog. Pollenkammer (Fig. 551) —, während die
dabei aufgelösten Zellen, vielleicht in Gemeinschaft mit flüssiger
Ausscheidung der angrenzenden Nucelluszellen, eine schleimige Masse
darstellen, welche den Mikropylenkanal füllt und als Tropfen aus ihm
hervorquillt. In diesen Tropfen gelangen die zwischen die Sporophylle
eingedrungenen Pollenkörner und werden mit der eintrocknenden
Flüssigkeit durch den Mikropylenkanal auf den Nucellus und in die
Pollenkammer niedergesogen.
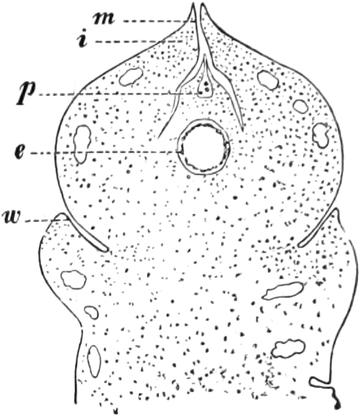
Fig. 551. Längsschnitt durch ein junges Makrosporangium
von Ginkgo biloba nach COULTER und CHAMBERLAIN.
Vergr. 35. m Mikropyle, i Integument, p Pollenkammer. e
Embryosack, w Wucherung des Sporophylls.
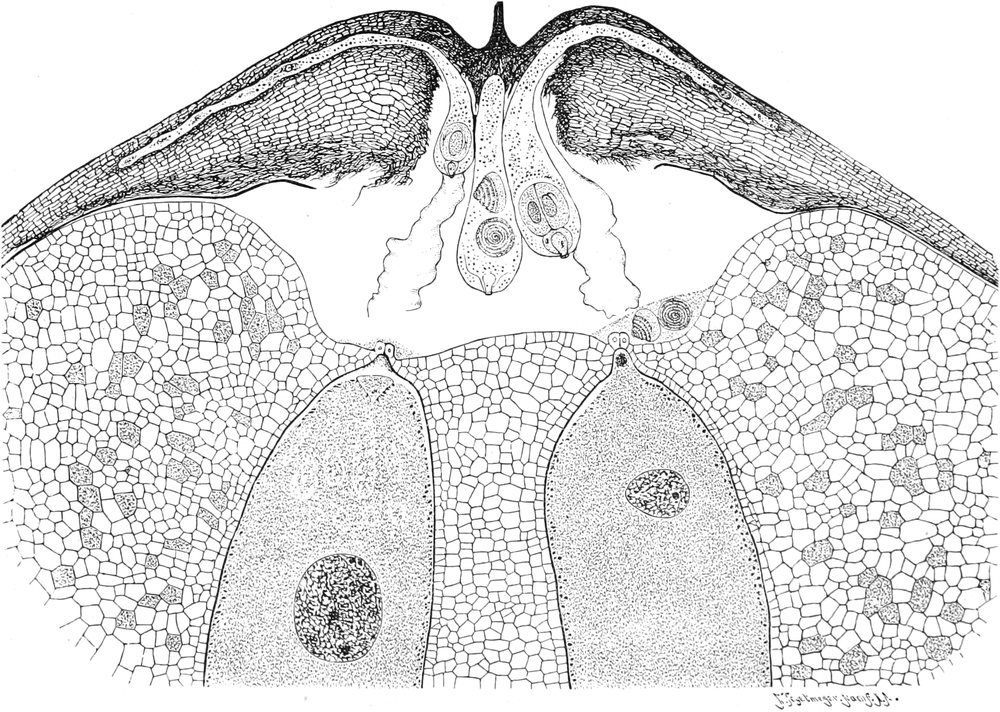
Fig. 552. Dioon edule. Oberer Teil eines Nucellus zur
Zeit der Befruchtung. Die Pollenschläuche haben sich zunächst durch
seitliche Auszweigungen im Nucellusgewebe fest verankert und sind dann
aus der bereits zugewachsenen Pollenkammer heraus in den Nucellus
tiefer eingedrungen. Sie haben die Archegonienkammer erreicht, und aus
zweien ist der Inhalt bereits entlassen. Zwei große Archegonien ragen
mit ihren Halszellen in die Archegonienkammer vor. Nach CH. J.
CHAMBERLAIN.
Während der geschilderten Entwicklung der Mikrosporen zu
Pollenschläuchen und der Bildung ihrer Spermien (Fig. 550)
wächst der im Grunde des Nucellus liegende, mit Prothalliumgewebe
bereits gefüllte Embryosack mächtig heran. Wie bei den Koniferen (Fig.
558) geht er aus der Tetradenteilung einer Embryosackmutterzelle
hervor, die meist wie im Makrosporangium von Selaginella alle übrigen
gleichen Anlagen verdrängt hat, und von deren vier Tochterzellen nur
eine Makrospore, der Embryosack übrig bleibt. Der Nucellus
schwindet fast bis zum Gipfel, und der Embryosack gelangt so[S. 488] in die
Nähe der Pollenkammer. Am Scheitel des Embryosackes sind die großen
Archegonien meist in Vierzahl vorhanden, je durch einige Gewebelagen
voneinander getrennt. Jedes Archegonium besitzt einen Halsteil und gibt
schließlich auch eine Kanalzelle ab. Gerade über den Archegonien findet
sich eine Einsenkung im Prothallium, die Archegonienkammer (vgl.
Fig. 552), bei Dioon z. B. von 2 mm Durchmesser und 1 mm Tiefe. In
diese Höhlung wachsen die Pollenschläuche hinein und entlassen hier,
sich vielleicht unter Mitwirkung der mit Reservestoffen gefüllten
Prothallium- und Dislokatorzelle apikal öffnend, ihre Spermien zugleich
mit einem Tropfen wäßriger Flüssigkeit, der ihnen einige Bewegung
gestattet. Sie müssen beim Eindringen in das Archegonium ihre breite
Form erheblich zusammenpressen, um die schmale Pforte zwischen den
aufgebrochenen Halszellen hindurch zu passieren. Das Spermium streift
im Eiplasma vordringend das Zilienband ab und vereinigt sich mit dem
Eikern, womit die Befruchtung vollzogen ist. (Vgl. jedoch S. 491.) Aus
der Vereinigung der Kerne entsteht der Keimkern (Fig. 553), der
alsbald in Teilung eintritt. In seinen Tochterkernen werden in rascher
Folge stets gleichzeitig verlaufende Weiterteilungen durchgeführt,
bis nach der achten Teilung etwa 256 freie Kerne den Zellraum füllen.
Sie drängen ins untere Ende des befruchteten Eies, wo Zellwandbildung
zwischen ihnen eintritt.
Damit ist ein sog. Proëmbryo entstanden (Fig. 554), dessen
fortwachsender Scheitel von dem zunächst aus nur wenig Zellen
bestehenden Embryo gebildet wird. Die weiter zurückliegenden
Zellen strecken sich stark und schieben als Embryoträger
oder Suspensor den Embryo in das Prothallium hinein,
das bei den Spermatophyten Endosperm genannt wird und als
Nährgewebe für den heranwachsenden Embryo dient. Dieser besitzt
schließlich an seinem in das Prothallium vorgeschobenen Ende zwei
mächtige Keimblätter oder Kotyledonen, zwischen denen
sich die Anlage der Stammknospe, die Plumula, birgt.
Der unterhalb der Kotyledonen befindliche Teil des Stammes heißt
Hypokotyl; er geht allmählich in die Hauptwurzel oder
Radicula über, die stets gegen die Mikropyle gekehrt ist.
Fig. 553. Zamia floridana. Eizelle unmittelbar nach
der Verschmelzung des eingedrungenen Spermakernes mit dem Eikern. Das
abgeworfene Zilienband in der Spitze des Eies. Ein zweites Spermium
versucht ins Ei einzutreten. Nach H. WEBBER. Vergr. 18.
Fig. 554. Zwei junge Proembryonen von Dioon edule in
ihrer Lage an der Archegonkammer. s Suspensor. e Embryo. Nach
CH. CHAMBERLAIN.
b) Koniferen[463].
Die Koniferen zeigen eine vom geschilderten Entwicklungsgange
abweichende Ausbildung ihrer keimenden Mikrosporen. Der Pollenschlauch
entwickelt sich an der morphologischen Basis der Mikrospore. Die
Prothalliumzellen, deren Zahl bei der sehr alten Gattung Araucaria
(Fig. 555) größer als bei den übrigen Koniferen und Cycadeen ist,
vergehen sehr bald (Fig. 556 A), und die generativen Zellen sind
niemals mehr in Form von Spermien ausgebildet.
[S. 489]
Fig. 555. Pollenkorn von Araucaria brasiliensis
mit mehrzelligem Prothallium pc, pc′ und sich teilender
Antheridium-Mutterzelle sp. k Pollenschlauchkern. 616. Nach L.
BURLINGAME.
Fig. 556. Entwicklung des Pollenschlauches.
A, B Pinus Laricio. 300. Nach COULTER und
CHAMBERLAIN. C Picea excelsa. 250. Nach K. MIYAKE.
p Reste der Prothalliumzellen. sp Spermatogene Zelle. m
Antheridium-Mutterzelle, s deren sterile Schwesterzelle. g
Generative Kerne von ungleicher Größe in gemeinsamer Plasmahülle. k
Pollenschlauchkern.
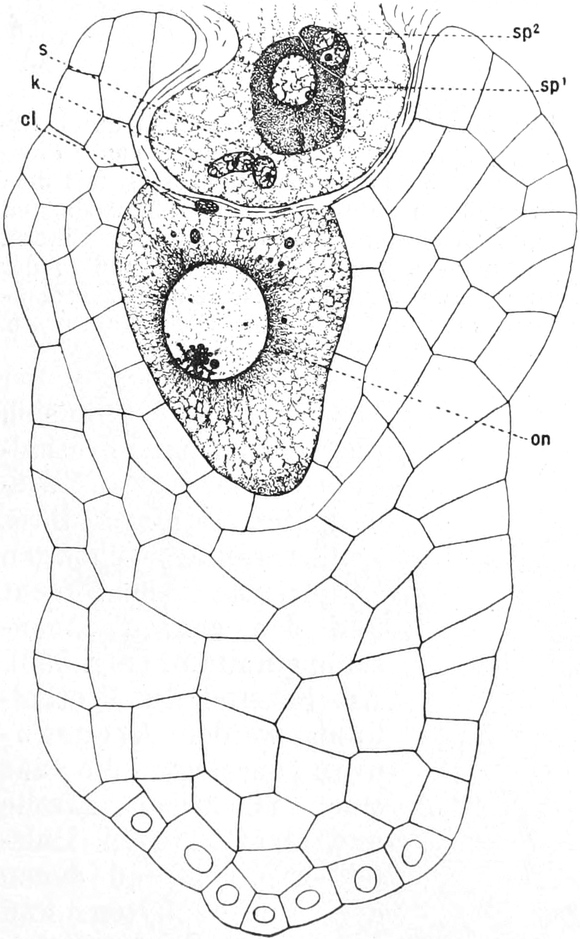
Fig. 557. Torreya taxifolia. Längsschnitt durch ein
weibliches Prothallium mit großer Eizelle und Eikern (on); abgegebene
Bauchkanalzelle (cl) und ein aufliegender Pollenschlauch mit
Pollenschlauchkern (k), Kern der sterilen Schwesterzelle (s) und
zwei Spermazellen (sp1), (sp2), von denen die größere (sp1)
allein funktionsfähig ist. Nach COULTER und LAND.
Die Teilung der spermatogenen Zelle ergibt bei Araucaria neben
der sterilen Zelle, dem Dislokator GOEBELS, die
Antheridium-Mutterzelle, die durch eventuelles Platzen des Dislokators
losgelöst wird. Sie liefert zwei zunächst gleichgroße Spermakerne in
gemeinsamer Plasmahülle, doch scheint einer von ihnen häufig nach
und nach zu schwinden. Das ist bei den Taxaceen zur Regel geworden.
Fig. 557 von Torreya taxifolia, einer nordamerikanischen Taxacee,
zeigt im Pollenschlauchende neben den Kernen des Schlauches (k) und
der sterilen Schwesterzelle (s) einen sehr großen funktionsfähigen
(sp1) und einen um mehr als die Hälfte kleineren, nicht fertilen
Spermakern (sp2), jeder von eigener Plasmamasse umhüllt. Die
Größendifferenz ist bei Taxus selbst noch erheblicher. Während nun
die Cupressineen durchweg zwei gleiche Spermazellen besitzen, haben
die Abietineen, ähnlich den Araucarien und Taxaceen zwei ungleich
große generative Kerne in[S. 490] gemeinsamer Plasmamasse (Fig. 556). Der
vorangehende größere allein ist fruchtbar.

Fig. 558. Taxus baccata. Längsschnitt durch das
sporogene Gewebe mit einer Embryosack-Mutterzelle, nach stattgehabter
Tetradenteilung. Vergr. 250. Nach E. STRASBURGER.
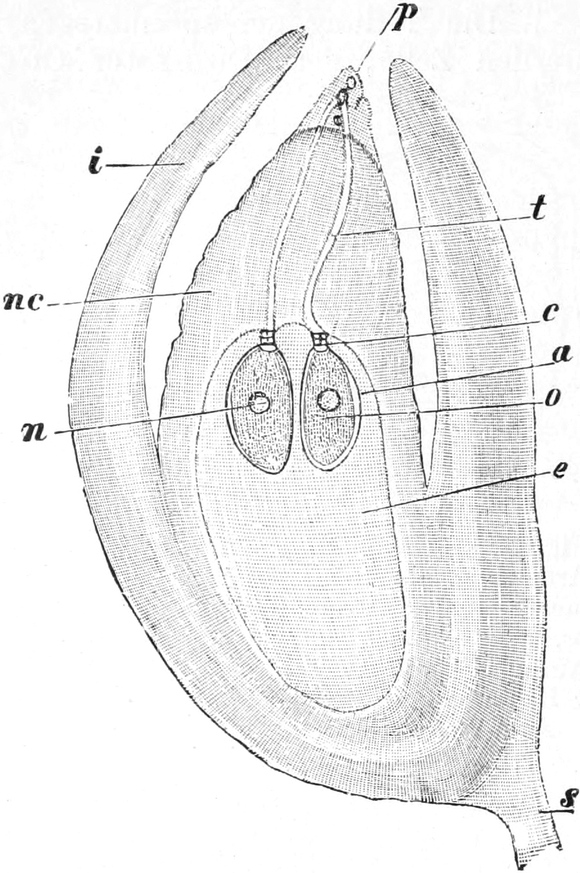
Fig. 559. Medianer Längsschnitt durch die
empfängnisreife Samenanlage von Picea excelsa. Vergr. 9. e Embryosack
mit dem Prothallium gefüllt, a Bauchteil, c Halsteil eines
Archegoniums, o Eizelle, n Eikern, nc Nucellus, p Pollenkörner
auf und in der Knospenwarze, t Pollenschläuche, i Integument, s
Samenflügel. Nach E. STRASBURGER.
Die Makrosporophylle tragen in der Regel zwei
Makrosporangien. Die meist nur in Einzahl vorhandene
Makrosporenmutterzelle geht eine Tetradenteilung ein (Fig. 558);
doch entwickelt sich von ihren vier Tochterzellen nur eine
zum Embryosacke, der Makrospore. Sie verdrängt die
Schwesterzellen und nach und nach den gesamten sporogenen Zellkomplex.
Die Makrospore füllt sich unterdessen mit Prothalliumgewebe, das aus
wiederholten Teilungen ihres Zellkernes und zugehörigen Zellplasmas
hervorgeht und den ganzen Innenraum einnimmt (Fig. 559). Am Scheitel
des Prothalliums werden Archegonien angelegt, die aus einer
mächtigen Eizelle und einem kurzen Halsteil bestehen und denen der
Pteridophyten und Cycadeen auch darin gleichen, daß kurz vor der
Befruchtung eine kleine Bauchkanalzelle von der Eizelle abgegeben wird
(Fig. 560), die bald zugrunde geht.
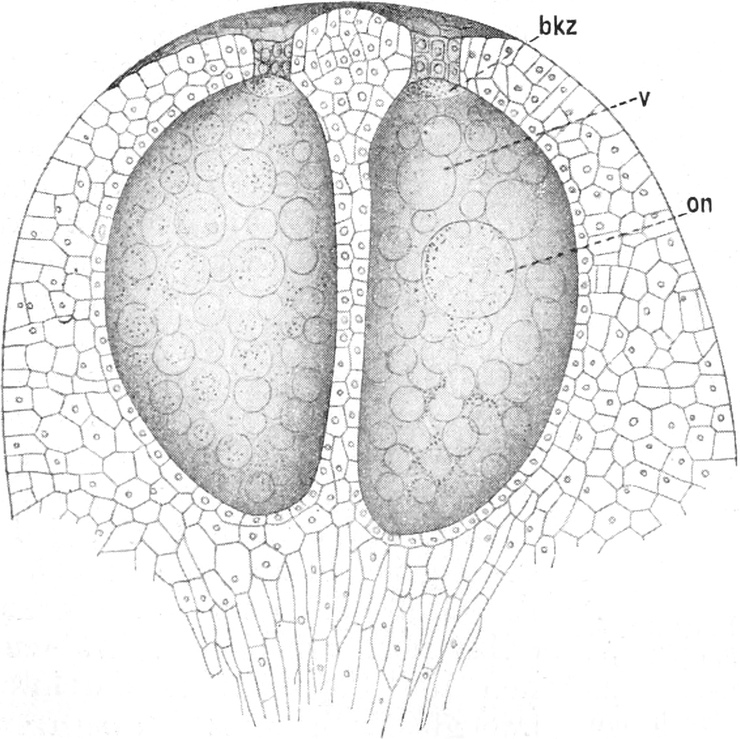
Fig. 560. Längsschnitt durch den Scheitel eines
Embryosackes von Picea excelsa mit zwei Archegonien, deren
Bauchkanalzelle bkz bereits abgeteilt ist, on Eikern, v Zahlreiche
Eiweiß-Vakuolen. Nach E. STRASBURGER. 80.
[S. 491]
Die Befruchtung selbst sei bei Torreya taxifolia dargestellt. Der
Pollenschlauch ist nach Durchbrechung der oberen Abschlußwandung in
die Eizelle eingebrochen, und der fertile Spermakern hat sich auf den
Eikern gelegt, während das Plasma der Spermazelle beide Kerne umhüllt.
Der übrige Inhalt des Pollenschlauches, wie er in Fig. 557 vorhanden
war, findet sich in einer oberen Ecke der Eizelle zusammengedrängt.
Die weitere Verschmelzung der Kerne zum Keimkern zeigt Fig. 562 A für
Picea excelsa.
In einigen Fällen (vielleicht aber bei allen Cycadeen und
Koniferen) ist die Kernvereinigung weit komplizierter als bisher
angenommen. So schildert HUTCHINSON für Abies balsamea
den Vorgang der Verschmelzung derart, daß die beiden miteinander
vereinigten Kerne sich jeder für sich teilen und dabei die haploide
Chromosomenzahl erkennen lassen. Die Chromosomen treten dabei aber
paarweise zusammen, was der eigentlichen Verschmelzung entsprechen
dürfte, gerade wie es im Diakinesestadium der heterotypischen
Teilung zu sehen ist. Dann wird durch Querteilung der Paare und
Auseinanderweichen der beiden Längshälften die zu fordernde
diploide Zahl hergestellt. CHAMBERLAIN schließt sich
dieser Darstellung für die Cycadee Stangeria an.
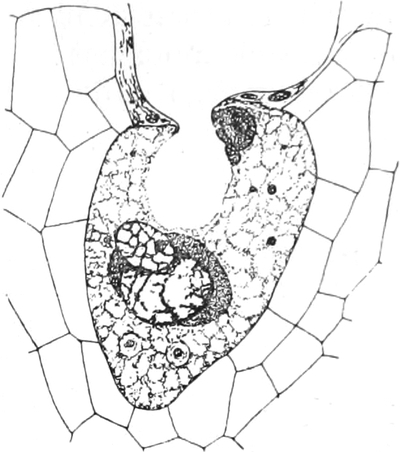
Fig. 561. Befruchtung bei Torreya taxifolia. Nach
COULTER und LAND. Beschreibung im Text.
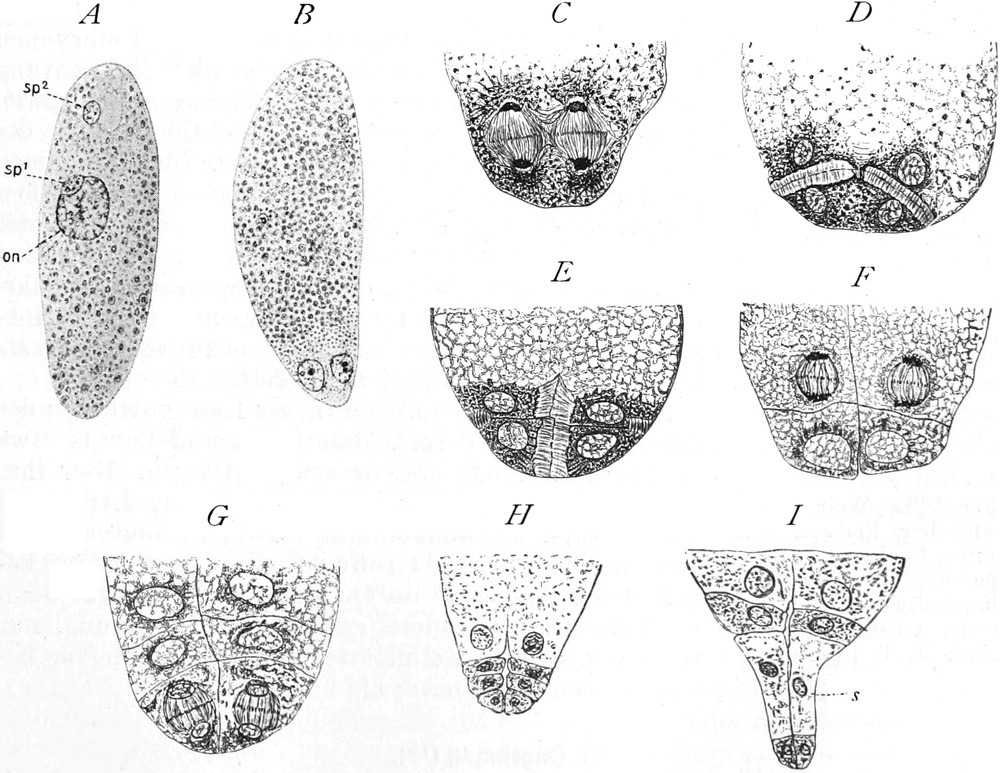
Fig. 562. A, B Picea excelsa. Vergr. 73. Nach K.
MIYAKE. C–I Pinus Laricio. C–G Vergr. ca. 200. Nach
N. I. KILDAHL. H, I Vergr. 104. Nach COULTER und
CHAMBERLAIN. on Eikern, sp1, sp2 Spermakerne, s
Suspensor. Beschreibung im Text.
Nach doppelter Teilung des Keimkernes wandern die vier Kerne ins
untere Ende des Eies und ordnen sich in einer Ebene nebeneinander an
(Fig.[S. 492] 562 B); da die weitere Entwicklung des Embryos nicht für
alle Gattungen die gleiche ist, so sei sie hier zunächst für Pinus
dargestellt.
Bei Pinus Laricio (Fig. 562) wandern die aus der zweifachen
Teilung des Keimkernes entstandenen vier Kerne ebenfalls in die Basis
der Keimzelle, ordnen sich in einer Fläche nebeneinander an und
teilen sich wieder (C). Zwischen diesen acht Kernen, die in zwei
Stockwerken übereinander liegen, bilden sich zunächst Querwände,
dann Längswände aus; so entsteht ein achtzelliger „Proëmbryo“
(DE). Die vier oberen Zellen bleiben jedoch gegen die Keimzelle hin
offen, so daß ihr Plasma mit dem Keimzellplasma in Verbindung steht.
Diese vier oberen Zellen treten zunächst in Teilung ein (F). Darauf
folgt Teilung der unteren vier Zellen (G). Der Proëmbryo besteht
demnach jetzt aus vier Etagen von je vier Zellen. Das oberste Stockwerk
bildet den Abschluß der Keimzelle. Die drei übrigen beteiligen sich an
der weiteren Entwicklung in der Art, daß das obere, wohl als Rosette
bezeichnete, durch eine stärkere Wand, die Basalplatte, völlig gegen
das Ei abgeschlossen wird (Fig. 563 p). Das mittlere wächst zum
Embryoträger oder Suspensor (Fig. 662 I s) aus und
schiebt die letzte, zur Embryobildung bestimmte Etage vor sich
her in das mit Nährstoffen gefüllte Prothalliumgewebe hinein.
Nach neueren Untersuchungen von BUCHHOLZ spalten sich die
Suspensorzellen mit den jungen Embryoanlagen bei Pinus ausnahmslos
auseinander (Fig. 563), so daß jedes befruchtete Archegonium vier
Embryonen liefert und bei der durchweg erfolgenden Befruchtung von
mehreren Archegonien eine außergewöhnlich starke Polyembryonie die
Regel ist. Eine Querteilung der Suspensorzellen findet nicht statt,
doch nehmen die an den Suspensor stoßenden oberen 1–3 Reihen von
Embryozellen als „Embryonalschläuche“ an der weiteren Verlängerung
lebhaften Anteil, was insofern von Bedeutung ist, als nur der am
weitesten ins nährstoffreiche Prothallium eingedrungene Embryo Aussicht
hat, im Konkurrenzkampfe obzusiegen und als alleiniger Embryo erhalten
zu bleiben.
Die Verschiedenheiten in der Embryobildung der Abietineen sind
derart, daß Pinus und Cedrus etwa gleichaltrig, Tsuga erst in etwas
späterem Alter ihre Embryoanlage spalten; bei Abies, Picea, Larix und
Pseudotsuga unterbleibt eine Spaltung, und es wird stets nur ein
einheitlicher Embryo aus jedem befruchteten Archegonium entwickelt.
Er besitzt dann die vorher für Cycadeen geschilderte Gliederung,
nur ist die Zahl der Keimblätter bei den Koniferen, besonders den
Abietineen, oft höher als zwei.
Fig. 563. Weiter entwickelter Embryo von Pinus
Banksiana. s Suspensor, e1 erster Embryonalschlauch, r
Rosette, p Basalwand. 80. Nach J. T. BUCHHOLZ.
c) Gnetineen[464].
Die letzte Ordnung der Gymnospermen, die Gnetineen, zeigt
eine abweichende und eigenartige Entwicklung. Zwar ist die
Mikrosporenbildung und -keimung nicht erheblich von derjenigen
der anderen Gymnospermen verschieden, wenn auch die Ab[S. 493]grenzung
der generativen Zellen minder deutlich zu werden, ja teilweise
zu fehlen scheint, so daß zwei gleich große Kerne in
gemeinsamer Plasmahülle vorliegen. Aber bei den Makrosporen zeigen
sich größere Abweichungen. Die Makrosporen von Ephedra und
Welwitschia besitzen ein wohlausgebildetes Prothallium.
Ephedra entwickelt am Scheitel Archegonien, die etwa denen der
Koniferen gleichen. Welwitschia läßt 2–5 kernige Schläuche
vom Prothalliumscheitel aus in das Nucellusgewebe hinein den
eindringenden Pollenschläuchen entgegenwachsen; ihre Deutung
als Archegonien wird durch die bauchige Erweiterung der
Basis wahrscheinlicher. Gnetum endlich bildet kein
Prothalliumgewebe, sondern zeigt den Embryosack (Fig. 564)
lediglich mit zahlreichen im Plasma verteilten Eikernen gefüllt.
Die beiden generativen Kerne des Pollenschlauches verschmelzen mit
je einem weiblichen Kern, dann erst beginnt die Endospermbildung.
Auf die Verwendung beider Spermakerne im Embryosack sei mit
Rücksicht auf die „doppelte Befruchtung“ der Angiospermen
hingewiesen. Von allen beim Eindringen mehrerer Pollenschläuche in
größerer Zahl entstandenen Keimzellen kommt nur ein Embryo
zu voller Entwicklung.
B. Angiospermen[465].
a) Die Mikrosporen der Angiospermen
bilden noch vor ihrem Verstäuben eine Antheridiummutterzelle
(Fig. 565 A m), die sich uhrglasförmig von der großen vegetativen
Pollenzelle abtrennt, ohne eine Zellulosehaut auszuscheiden. Sie
löst sich allmählich von der Außenwandung und liegt zur Zeit
der Pollenverbreitung als spindelförmiges Gebilde in der Mitte
der Mikrospore neben dem vegetativen Pollenkern (k). Bei der
Keimung auf der Narbe wandert der vegetative Kern und hinter ihm
die Antheridiummutterzelle in den Pollenschlauch ein. Diese tritt
in Teilung, und die beiden generativen Tochterkerne (g) bleiben
ohne besondere Abgrenzung einer zugehörigen Plasmamasse frei im
Pollenschlauch. Sie sind von länglich ovaler oder ellipsoidischer
Form und wandern hintereinander im Schlauch abwärts. Den Gymnospermen
gegenüber ist also das Fehlen sowohl der Prothalliumzellen
und der sterilen Schwesterzelle des Antheridiums, wie einer
Zellulosemembran für dieses, endlich das Auftreten nackter
generativer Kerne statt generativer Zellen im Pollenschlauch
hervorzuheben. Die Rückbildung des männlichen Prothalliums ist demnach
so weit gegangen, daß nur die unumgänglich notwendigen Teile erhalten
geblieben sind.
Ob das von HERRIG beobachtete Auftreten zweier
Spermazellen bei künstlicher Keimung von monokotylen
Pollenkörnern sich bei normaler Keimung ebenfalls zeigt, wäre zu
untersuchen.
Fig. 564. Embryosackscheitel von Gnetum rumphianum. Kurz
vor der Keimzellenbildung. wk Weibliche Kerne, mk männliche Kerne,
pk Pollenschlauchkern, ps Pollenschlauch. Vergr. 500.
[S. 494]
b) Makrosporen. Die für die Angiospermen charakteristische
Abweichung von dem vorher geschilderten Entwicklungsgange des
gymnospermen Makrosporangiums beginnt erst, wenn nach der
Tetradenteilung der Makrosporenmutterzelle der allein zur Ausbildung
gelangende Embryosack seinen Kern wiederum teilt (Fig. 566 1–5).
Der „primäre Embryosackkern“ teilt sich, die Tochterkerne
weichen auseinander und teilen sich abermals und zum dritten Male.
Hierauf erst tritt teilweise Zellbildung um diese Kerne ein (Fig. 566
6–8). Im oberen, der Mikropyle zugekehrten, wie im unteren Ende
des Embryosackes entstehen drei nackte nur von Plasmahaut umgebene
Zellen; die beiden übrigbleibenden „Polkerne“ wandern gegeneinander
in die Mitte der Zelle und vereinigen sich zum „sekundären
Embryosackkern“. Die Zellen des unteren Zellendes heißen
Antipoden oder Gegenfüßlerinnen; sie entsprechen den
vegetativen Prothalliumzellen, wie solche bei den Gymnospermen bis auf
Gnetum die ganze Makrospore füllen.
Bei Peperomia hispidula und Pandanus u. a. ist die Zahl der
Antipoden eine erheblich größere; ob dieses Verhalten vielleicht
als primitives Merkmal zu deuten ist, muß bis auf weiteres
dahingestellt bleiben.
Die drei oberen Zellen dagegen stellen den „Eiapparat“ dar (Fig.
568). Zwei einander gleichende seitliche werden als Synergiden
oder Gehilfinnen bezeichnet, die dritte, tiefer in den
Embryosack hineinragende ist die Eizelle selbst. Die Gehilfinnen
vermitteln den Übertritt des Pollenschlauchinhaltes in den Embryosack.
Auch hier ist also die Reduktion fast bis an die Grenze des Möglichen
gegangen; an Stelle der mehr oder minder zahlreichen Archegonien in
der gymnospermen Makrospore ist nur eine Eizelle vorhanden. Die
Synergiden kann man entweder als steril gewordene Archegonien oder mit
TREUB und PORSCH als Halszellen des zum Eiapparat
gewordenen Archegoniums ansprechen (Fig. 568).
Fig. 565. Pollenkorn von Liliummartagon und seine
Keimung. Vergr. 400. k Pollenkornkern, m Mutterzelle des
Antheridiums, g generative Kerne. Nach E. STRASBURGER.
In manchen Fällen erfährt die Embryosackmutterzelle keine
Tetradenteilung mehr, sondern liefert nur drei oder zwei
Tochterzellen, oder sie wird ohne Teilung direkt zum Embryosack,
der bei Cypripedium und Plumbagella seinen Inhalt unter Ausfall
der letzten Teilung auf Eizelle, zwei Polkerne und einen
Antipodenkern, oder bei Cypripedium in anderer Verteilung auf
Eizelle, zwei Synergiden und einen Polkern, also jedesmal
auf[S. 495] vier Kerne beschränkt, während bei den sich sonst ebenso
verhaltenden Lilium-Arten die normale Achtzahl erreicht wird.
Die Reduktionsteilung findet dann im Embryosack statt, ist
also vom Ende der Sporophytengeneration in den Anfang der
Gametophytengeneration verschoben. Für weiter sich findende
Abänderungen der Embryosackbildung und -ausstattung vgl.[466].
Da ein direkter Zutritt der Mikrosporen zu den im Fruchtknoten
eingeschlossenen Makrosporen hier ausgeschlossen ist, müssen sie
auf der Narbe keimen (Fig. 567). Ihre Pollenschläuche
durchwachsen die ganze Länge des Griffels, und in der Regel dringt
dann ein Schlauchende durch die Mikropyle zum Nucellusscheitel
vor. Dieser häufigste Fall des Pollenschlauchzutrittes wird als
Porogamie bezeichnet. Doch sind neuerdings zahlreiche
Abweichungen davon bekannt geworden.
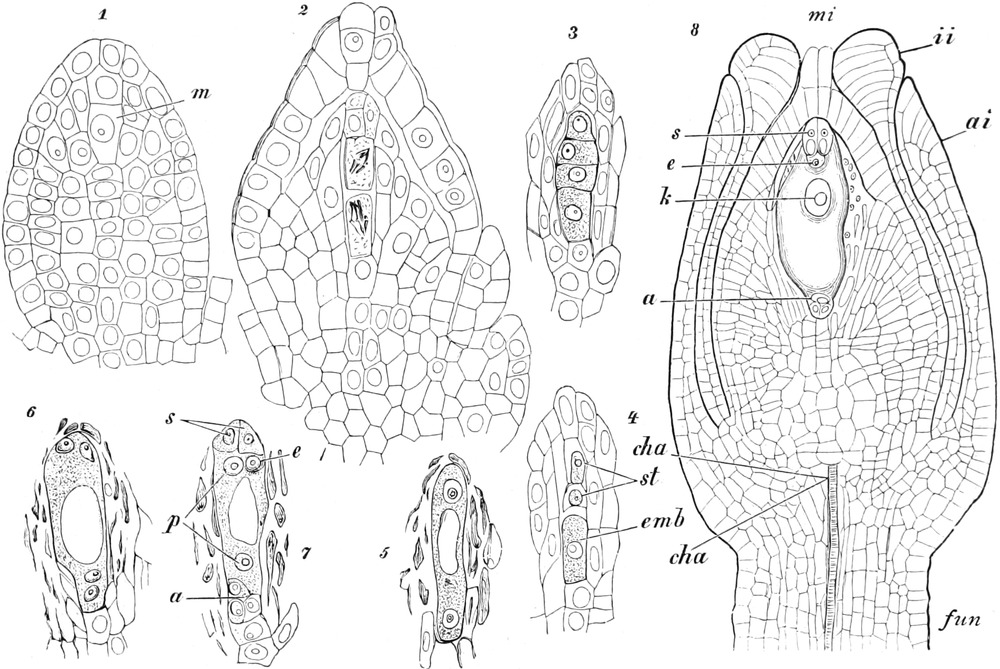
Fig. 566. Embryosackentwicklung von Polygonum
divaricatum. 1–7 Vergr. 320, 8 Vergr. 135. m
Embryosackmutterzelle, emb Embryosack, st sterile Schwesterzellen,
e Eizelle, s Synergiden, p Polkerne, a Antipoden, k
sekundärer Embryosackkern, cha Chalaza, mi Mikropyle, ai,
ii äußeres und inneres Integument, fun Funiculus. Nach E.
STRASBURGER.
Nachdem M. TREUB zuerst für Casuarina nachgewiesen
hatte, daß die Pollenschläuche hier von der Chalaza her zu
dem höchst eigenartig entwickelten, zahlreiche Makrosporen
bergenden sporogenen Gewebe gelangen, ist eine größere Zahl
derartiger, als Chalazogamen den normalen Porogamen
gegenübergestellter Formen bekannt geworden. Es gehören
hierher vor allem die Casuarinaceen, dann die Juglandaceen,
Betulaceen, Ulmaceen, Celtoideen, Urticaceen, Cannabinaceen,
Euphorbiaceen, die alle das gemeinsame Merkmal haben, daß sie
ihre Pollenschläuche innerhalb der Gewebe der Samenanlage
verlaufen lassen. Sie verschmähen also den Durchtritt durch die
Mikropyle, die in einigen Fällen auch geschlossen wird (bei den
Urticaceen) oder die bei den Euphorbiaceen durch den Obturator
gedeckt ist, — und bahnen sich den Weg zum Embryosack teils
von seinem Chalazaende (Fig. 569), teils von der Seite her zum
Eiapparate, indem sie das zwischenliegende Gewebe durchbrechen.
Da die genannten Familien nach der Auffassung vieler Autoren
am unteren Ende der Dikotylenreihe stehen sollen, hat man in
dieser Art des Pollenschlauchwachstums innerhalb der Gewebe
eine Annäherung an die ursprünglicheren Verhältnisse bei den
Gymnospermen erblicken wollen, wo ebenfalls zur Erreichung des
Embryosackes das ganze darüberliegende Gewebe des Nucellus vom
Pollenschlauche durchwachsen[S. 496] werden muß (Fig. 559). Weiter
findet nun NAWASCHIN, daß auch bei der Entwicklung
des Pollenschlauchinhaltes dieser Pflanzen deutliche
Anzeichen einer gegenüber der Mehrzahl der Angiospermen
niedrigeren Entwicklungsstufe vorhanden sind, indem die beiden
generativen Kerne (bei Juglans), von einer gemeinsamen Plasmahülle
umgeben, in den Embryosack eintreten, um hier erst allmählich
nackt hervorzutreten und ihre Funktionen zu erfüllen. Aller
Wahrscheinlichkeit nach handelt es sich in den genannten Familien
aber um stark reduzierte Formenkreise, nicht, wie der genannte
Autor meint, um aufsteigende; dem Aussehen nach können ja beide
übereinstimmen.
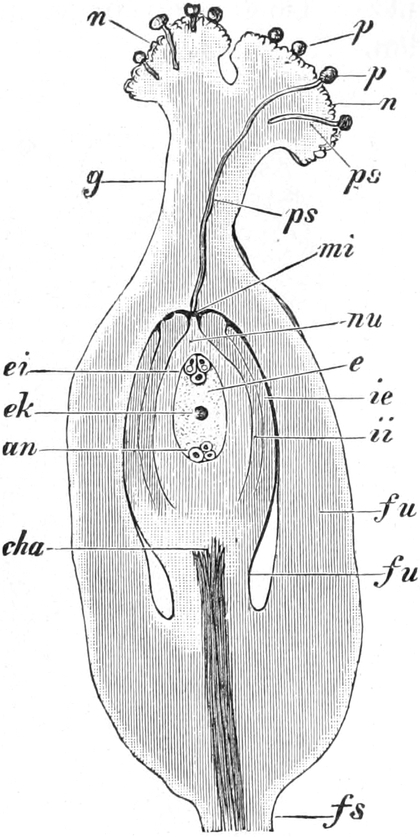
Fig. 567. Fruchtknoten von Polygonum convolvulus mit
atroper Samenanlage (schematisiert). fs Stielartige Basis, fu
Funiculus, cha Chalaza, nu Nucellus, mi Mikropyle, ii inneres,
ie äußeres Integument, e Embryosack, ek Embryosackkern, ei
Eiapparat, an Antipoden, g Griffel, n Narbe, p Pollenkörner,
ps Pollenschläuche. Vergr. 48. Nach H. SCHENCK.
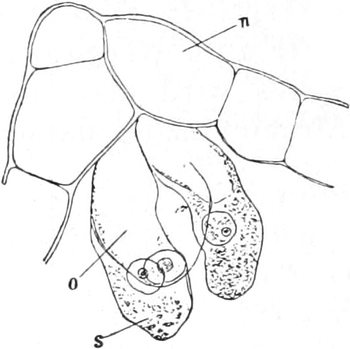
Fig. 568. Funkia ovata. Nucellusscheitel mit Eiapparat
vor der Befruchtung, o Eizelle, s Synergide. Vergr. 390. Nach
E. STRASBURGER.
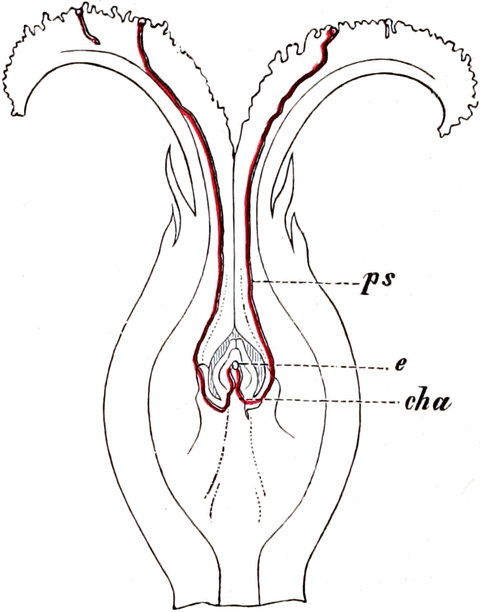
Fig. 569. Längsschnitt durch einen Fruchtknoten von
Juglans regia zur Darstellung der Chalazogamie. Vergr. 6. ps
Pollenschlauch, e Embryosack, cha Chalaza (schematisiert).
So gelangt der Pollenschlauch mit den beiden generativen Kernen an den
Embryosack. Er entläßt seinen Inhalt, welcher durch eine der Synergiden
zur Eizelle vordringt. Die betreffende Synergide stirbt ab. Einer
der beiden generativen Kerne dringt in die Eizelle ein und verschmilzt
mit dem Eikerne: die befruchtete Eizelle umgibt sich mit einer
Zellulosemembran. Der zweite generative Kern ist am Ei vorbeigewandert
und vereinigt sich mit dem großen „sekundären Embryosackkern“ zum
„Endospermkern“ (Fig. 570, 571). Beide männlichen Kerne haben oft
pfropfenzieherartig gewundene Form, so daß NAWASCHIN, der
das Eindringen und Verbleiben des zweiten generativen Kernes zuerst
beobachtet hatte, sie direkt mit den Spermien der Sporenpflanzen
vergleicht. Die Weiterentwicklung pflegt sodann mit der Teilung[S. 497]
des Endospermkernes einzusetzen, der zunächst eine große Zahl im
plasmatischen Wandbelag verteilter Kerne liefert. Durch Ausbildung
der Querwände zwischen den einzelnen von je einem Kern beherrschten
Plasmabezirken und weitere Vermehrung dieser Zellen zu einem massiven
Gewebekörper entsteht das Endosperm (Fig. 579 A).
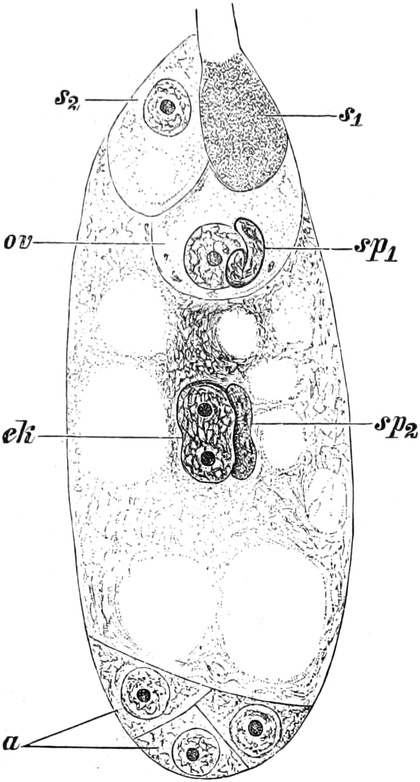
Fig. 570. Befruchtung von Lilium martagon. Einer der
männlichen Kerne ist neben dem Eikern, der andere neben den gerade
verschmelzenden Polkernen sichtbar. Schematisiert. (Bezeichnung wie
Fig. 571.)
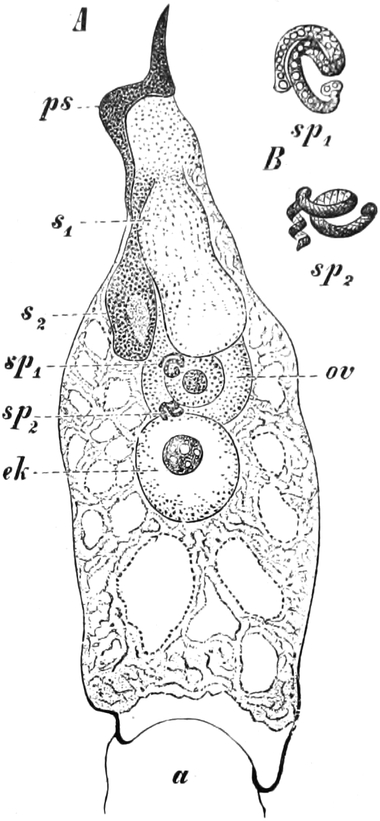
Fig. 571. A Embryosack von Helianthus annuus nach
S. NAWASCHIN. B Die männlichen Kerne daraus stärker
vergrößert. ps Pollenschlauch, s1 s2 Synergiden, sp1
sp2 männliche Kerne, ov Eizelle, ek Embryosackkern, a
Antipoden.
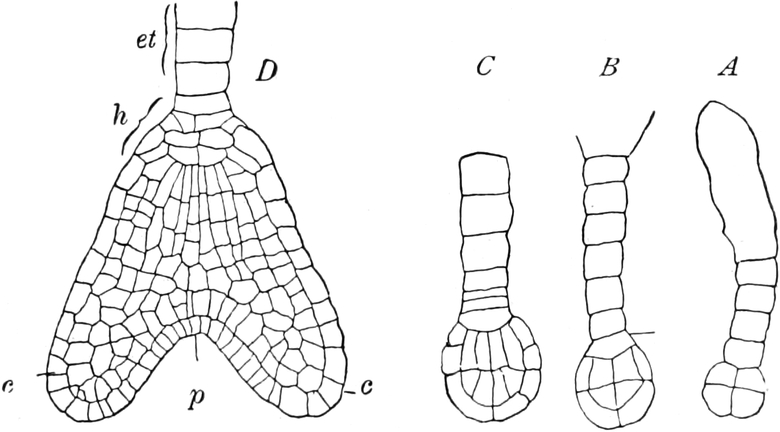
Fig. 572. Entwicklung des Keims von Capsella bursa
pastoris. h Hypophyse, et Embryoträger, c Kotyledonen, p
Plumula. Vergr. Nach J. HANSTEIN.
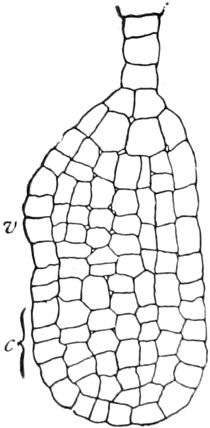
Fig. 573. Junger Keim von Alisma plantago. c
Kotyledon, v Vegetationspunkt. Vergr. Nach J. HANSTEIN.
Übrigens ist bei zahlreichen Kompositen, ebenso bei Aponogeton,
Potamogeton usw., das Endosperm von der ersten Teilung des
sekundären Embryosackkernes an zellulär ausgebildet. Das Stadium
der frei im Wandbelag verteilten Kerne fällt hier aus, wie übrigens
auch schon W. HOFMEISTER es für verschiedene Familien
angegeben hatte.
[S. 498]
Das unterscheidende Merkmal im Entwicklungsgange des
Angiospermen-Endosperms gegenüber dem Gymnospermen-Prothallium liegt
demnach in der Unterbrechung seiner Bildung, in der „fraktionierten
Endospermbildung“. Zunächst wird in dem der Befruchtung harrenden
Embryosack nur eine Andeutung des Prothalliums, die Antipoden,
gegeben. Die eigentliche Endospermbildung dagegen ist von der
Weiterentwicklung des betreffenden Embryosackes abhängig gemacht
und damit jeder Materialvergeudung vorgebeugt. Den Ausgangspunkt
dazu bildet der sekundäre Embryosackkern, welcher einer Anregung zur
Weiterentwicklung durch Vereinigung mit dem zweiten generativen Kern
des Pollenschlauches zum Endospermkern bedarf. (Vgl. dazu das bei den
Gnetaceen Gesagte.)
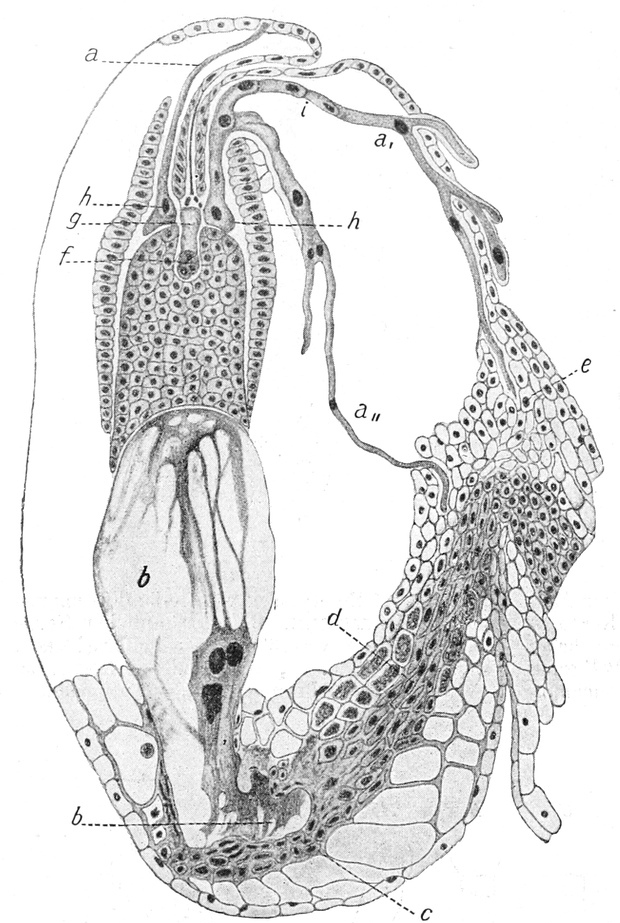
Fig. 574. Haustorien des befruchteten Embryosackes von
Melampyrum nemorosum nach BALICKA-IWANOWSKA. b Haustorien
des Chalazaendes, c Nährgewebe, d Leitbündelzweig, e Funiculus,
f Embryo, g sein Suspensor, a, a′, a″ am Mikropylenende
schon früh entstandene Haustorienschläuche, deren innere sich im
Funiculus weit ausbreiten, zum Teil die Epidermis durchbrechen, h
ihre Ansatzstellen, i Querwände im Schlauche.
Aus der befruchteten, alsbald mit fester Zellulosehaut bekleideten
Eizelle, der Keimzelle, geht eine Zellreihe, der Vorkeim,
hervor, welcher aus seiner Gipfelzelle die Hauptmasse des
Embryos[467] entwickelt. Der Rest des Vorkeimes bildet den
Embryoträger oder Suspensor. An der Grenze von Embryo und
Suspensor liegt eine als Hypophyse (Fig. 572 D h) bezeichnete
Zelle, die an dem Aufbau des unteren Embryoendes, der Radicula,
in geringem Maße beteiligt zu sein pflegt. Je nach der Zugehörigkeit
der betreffenden Pflanze zu der Unterklasse der Monokotyledonen oder
Dikotyledonen ist die Gliederung des Embryokörpers verschieden.
Die Dikotyledonen lassen zwei Keimblätter, Kotyledonen, am Scheitel
des heranwachsenden Embryos hervorsprossen (Fig. 572) und bilden
am Grunde des Spaltes zwischen beiden den Sproßvegetationspunkt, die
Plumula; die Monokotyledonen dagegen besitzen der Regel nach einen
scheitelständigen großen Kotyledon und einen seitlich gelegenen
Vegetationspunkt (Fig. 573).[S. 499] Die Wurzel, Radicula, geht in beiden
Fällen aus dem der Mikropyle zugekehrten Teile des Embryokörpers
hervor; ihre Abgrenzung wird an älteren Embryonen deutlich.
Da der Embryosack nach eingetretener Befruchtung für den Aufbau
des Keimlings, wie zur Füllung der Reservestoffbehälter im
jungen Samen sehr erheblicher Zufuhr von Nährstoffen bedarf,
so ist es verständlich, daß besondere Wege dafür eingeschlagen
werden. Während im einfachsten Falle das Endosperm sich mächtig
vergrößert und den ganzen Nucellus nach und nach verdrängt, sind
häufig die Antipodenzellen, als der Chalaza nächst gelegene Teile,
mit der Funktion der Embryosackernährung betraut. Sie erfahren
dann starke Vermehrung und bisweilen eine mächtige Entwicklung.
Aber auch andere Teile des Embryosackes selbst können zu langen
Saugorganen, Haustorien, auswachsen, die bald aus der Mikropyle
hervordringen, bald in der Chalazaregion das umliegende Gewebe weit
durchziehen und das in vielen Fällen, besonders bei insektivoren
und halbparasitisch lebenden Pflanzen, an diesem Orte vorher
angehäufte, reiche Reservestoffmaterial der Makrospore zuführen
(Fig. 574).
Völlig abweichende Entwicklung des Embryosackes und Embryos
ist endlich bei Pflanzen zu finden, die unter ganz besonderen
Lebensbedingungen vorkommen, wie die nur in reißenden
Gebirgswässern tropischer und subtropischer Gebiete lebenden
Podostemaceen. Hier kommt alles darauf an, daß während der
kurzen Trockenzeit die Blütenanlage und -entwicklung auf Kosten
vorher angesammelten Ernährungsmaterials vor sich gehe, und daß
Bestäubung, Befruchtung und Samenentwicklung in abgekürztem
Verfahren rasch durchlaufen werden, damit bei Wiederkehr des
Wassers die reifen Samen ihre Keimungsbedingungen finden und der
Vermehrung und Ausbreitung der Pflanzen dienen können.
Fig. 575. Vegetative Sprossung aus dem Nucellus von
Funkia ovata. (Apogamie.) n Nucellus, dessen Zellen am Scheitel
Adventivkeime (ae) bilden, s Synergide, o aus der Eizelle
entstandener Embryo mit Eikern und Spermakern, ii inneres Integument.
Nach E. STRASBURGER.
In einzelnen Fällen ist Pflanzen die Fähigkeit sexueller Fortpflanzung
mehr oder minder verloren gegangen[468] und durch eine oft nur
bei genauerer Untersuchung als davon verschieden festzustellende
anderweitige Vermehrung ersetzt worden. So unterlassen einige
Pflanzen, wie Alchimilla, Thalictrum, Taraxacum, einige Urticaceen u. a.
eine Reduktionsteilung bei der Anlage ihres Embryosackes. Daraus
ergibt sich, daß die „Eizellen“ nicht befruchtungsfähig sind, sie
sind keine normalen Geschlechtszellen, sondern besitzen vegetativen
Zellcharakter. Derartige diploide „Eizellen“ sind nun ohne weitere
Kernverschmelzung zur Fortentwicklung befähigt, und in Verbindung damit
— oder als Ursache davon? — ist meist auch der Pollen unfruchtbar.
Um die Übereinstimmung mit der zoologischen Terminologie zu bewahren,
soll dies Verhalten, d. h. die Weiterentwicklung einer Eizelle ohne
Befruchtung als „Parthenogenesis“ oder „jungfräuliche Zeugung“
bezeichnet werden.
Da nun neben der Weiterentwicklung solcher an Stelle von Eizellen
stehender diploider Zellen auch Verdrängung normaler, haploider
Eizellen durch vegetative Sprossungen, sog. „Adventivembryonen“ aus
Nucelluszellen bekannt geworden ist, so mag diese Vortäuschung einer
sexuellen Fortpflanzung[S. 500] die Bezeichnung „Apogamie“ erhalten.
In einzelnen Fällen solcher Apogamie ist die Bestäubung der Narbe als
Vorbedingung nachgewiesen worden, so bei Funkia (Fig. 575) und Citrus
aurantium, während bei Caelebogyne ilicifolia und den Calycanthaceen
ohne derartige in ihrer Beziehung nicht aufgeklärte Mitwirkung des
Pollens die Adventivembryonen zustande kommen.
Unter „Aposporie“ endlich versteht man den völligen Ausfall
der Sporenbildung oder aber Ausschaltung der Makrospore aus dem
Entwicklungsgang; wenn z. B. bei Farnen aus diploiden Zellen des
Blattrandes diploide Prothallien direkt hervorsprossen, oder bei
Samenpflanzen Makrosporangien nicht entwickelt werden, sondern
beliebige vegetative Zellen an ihre Stelle treten (Fig. 576).
Besonderes Interesse verdient nach den Untersuchungen von
OSTENFELD und ROSENBERG die Gattung Hieracium,
deren Samenanlagen die verschiedenartigsten Wege zur Embryobildung
einschlagen können. Die Mehrzahl führt Tetradenteilung mit
Chromosomenreduktion aus, aber nur einige solcher Samenanlagen gelangen
in den Besitz eines normalen befruchtungsfähigen Embryosackes,
weil dieser meist durch eine vegetative Zelle verdrängt wird. Eine
derartige Verdrängung der Sporen — hier der Makrospore — wäre also
Aposporie (Fig. 576). Außerdem werden auch Embryosäcke mit
parthenogenetischer Eizelle ausgebildet.
Fig. 576. Apospore Entstehung des Embryosackes von
Hieracium flagellare. a Normale Makrosporentetrade, b und c ihre
Desorganisation. Der diploide Embryosack geht aus einer bereits in a
kenntlichen Integumentzelle hervor. Nach O. ROSENBERG aus
A. ERNST 1918.
Der Samen.
Das ganze aus der Samenanlage nach stattgehabter Befruchtung
entstehende Gebilde heißt Samen. Jeder Samen besteht aus
dem mehr oder minder entwickelten Embryo, dem umgebenden
Nährgewebe und der schützenden Schale. Diese geht
in der Regel aus dem oder den Integumenten hervor, deren Zellen
sich durch Verdickung, Verkorkung, Verholzung ihrer Wände zu einem
wirksamen Schutzorgan gegen Austrocknen und Verletzungen für die darin
schlummernde junge Pflanzenanlage umgestaltet haben. Ausbildung der
Samenepidermis als Schleimzellen findet sich recht häufig, z. B. bei
Quitten, Linum, zahlreichen Cruciferen u. a. Der Schleim dient den
Samen als erstes Befestigungsmittel und gleichzeitig zum Festhalten
einer bei der Keimung nötigen Wassermenge. Sonstige Strukturen der
Oberfläche wie Haare, Stacheln usw. haben ebenfalls die Bedeutung, das
Festhaften zu sichern, wenn sie nicht zur Samenverbreitung in Beziehung
stehen.
Besondere morphologisch bemerkenswerte Stellen der Schale sind:
1. Mikropyle, 2. Nabel oder Hilum, d. h.
Funiculusabbruchstelle, endlich 3. die Raphe.[S. 501] S.
464 ist gezeigt worden, daß Mikropyle und Nabel an den beiden
Polen einander gegenüber liegen, falls die Samenanlage atrop war,
daß sie nebeneinander sich befinden bei Samen, die aus anatropen
Samenanlagen hervorgegangen sind, und daß nur diese letzteren eine
Raphe, also eine die Chalaza mit dem Hilum verbindende, bei der
Trennung des Samens vom Funiculus entstehende Narbstelle besitzen
können. Kampylotrope Samenanlagen ergeben den anatropen ähnliche
Samen, doch ist der Embryo hier gekrümmt.
In gewissen Fällen ist die Funktion der Samenschale dadurch
verändert, daß die Fruchtschale selbst den Schutz des oder der
Samen übernimmt, indem sie ihre Wand oder deren innerste Schicht zu
Steinzellen umbildet, also zu einer Nuß oder Steinfrucht wird. In
solchen Fällen, wie Mandel, Lorbeer, Kirsche, Pfeffer u. a., pflegt
die Samenschale reduziert zu sein; ihre verschiedenen Zellschichten
werden zusammengedrückt, und eine Verstärkung oder Veränderung der
Wandungen unterbleibt, da die schützende Fruchtwand eine besondere
Ausrüstung der Samenschale unnötig macht.
Das Nährgewebe ist bei den Gymnospermen, abgesehen von Gnetum,
schon zur Zeit der Befruchtung als Prothallium ausgebildet
(vgl. S. 487). Es füllt den Embryosack aus und dient der Ernährung
des hineinwachsenden Embryo; die umgebenden Reste des Nucellus werden
verdrängt, so daß das Prothallium bis an die Samenschale reicht.
Seine Zellen sind mit Stärke, Fett und Eiweiß als Reservestoffen
vollgepfropft, die zur Entwicklung des Embryo bei der nach Ablauf einer
Ruheperiode eintretenden Keimung Verwendung finden.
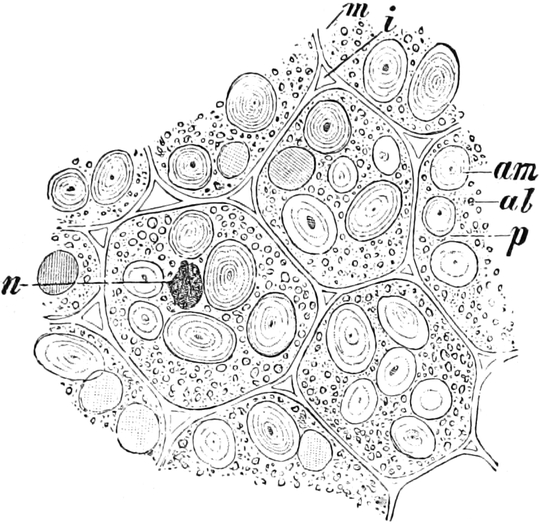
Fig. 577. Zellen der Erbsenkeimblätter mit
Reservestoffen. am Stärkekörner, al Aleuronkörner, p Protoplasma,
n Zellkern. Vergr. 160. Nach E. STRASBURGER.
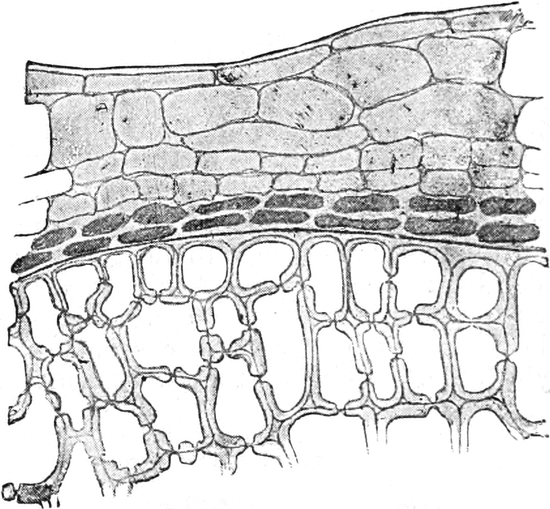
Fig. 578. Querschnitt des Colchicum-Samens. Innerhalb
der Samenschale Reservezellulose des Endosperms.
Das Nährgewebe der Angiospermen (und von Gnetum) entsteht
dagegen erst nach der Befruchtung der Eizelle aus der Vereinigung
der beiden Polkerne zum sekundären Embryosackkern, der durch
Aufnahme des zweiten Spermakernes den Anreiz zu weiterer Teilung
erhält (vgl. S. 498). Seine Tochterkerne verteilen sich in der Regel
rings im wandständigen Plasmaschlauch, und wenn die Zahl der Kerne
eine hinreichend große geworden ist, zerfällt dieser Plasmakörper in
zahlreiche Zellen, die für weitere Ausfüllung des ganzen Embryosackes
mit Endospermgewebe sorgen.
Auch bei den Angiospermen verdrängt das Endosperm meist den
ganzen Rest des Nucellus und häuft Reservestoffe, wie Stärke, fettes
Öl, Aleuronkörner, in den Zellen an (Fig. 577) oder speichert in den
sich stark verdickenden Wänden Zellulose als Reservezellulose (Fig.
578). In selteneren Fällen, so bei Piperaceen, Scitamineen usw., bleibt
ein Rest des Nucellus vorhanden, der dann ebenfalls als Nährgewebe
fungiert und den Namen Perisperm führt (Fig. 579 B). Wenn
Lamellen des Perisperms oder auch der[S. 502] Samenschale in das Endosperm
einwachsen, von dem sie in Farbe und Inhalt abweichen, so spricht man
von ruminiertem Endosperm (Myristica Fig. 617, Areca).
In sehr zahlreichen Fällen aber, so bei den Leguminosen, Cruciferen
u. a., wird nicht nur der Nucellus vom Endosperm, sondern auch
dieses bereits vom Embryo völlig verdrängt, die Reservestoffe werden
dann in den Keimblättern allein oder im ganzen Körper des Embryos
aufgespeichert (Fig. 580).
Endlich ist noch eine meist wohl zur Samenverbreitung in
Beziehung stehende Bildung, der Arillus, zu erwähnen, auch
Samenmantel genannt. Er entsteht als fleischiger (Taxus) oder
auch trockener (Pahudia javanica, Strelitzia reginae), meist lebhaft
gefärbter Wulst am Funiculus und wächst an der Samenanlage bereits in
ziemlich frühem Alter empor, indem er sie endlich mehr oder minder
umhüllt (Fig. 581 D, 582). — Einen der Mikropyle benachbarten
Auswuchs, der unter anderem besonders den Euphorbiaceen eigen ist,
nennt man Caruncula (Fig. 581 C, B.).
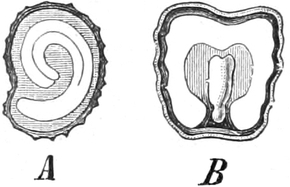
Fig. 579. A Samen von Hyoscyamus niger. Der dikotyle
Keim in Endosperm eingebettet. B Samen von Elettaria cardamomum.
Innerhalb der dunklen, von einem dünnen Arillus umhüllten Samenschale
liegt zunächst weißes mehliges Perisperm, dann (schraffiert) ein öliges
Endosperm und in der Mitte der monokotyle Keim. Nach BERG und
SCHMIDT.
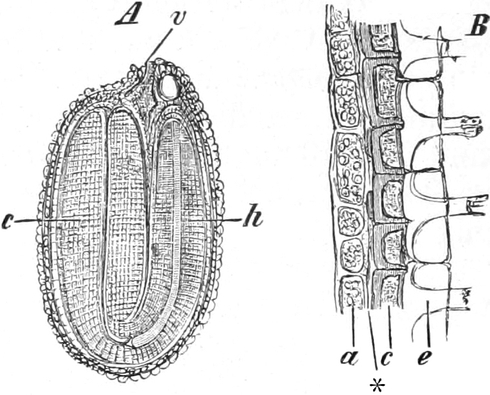
Fig. 580. Capsella bursa pastoris. A Längsschnitt
durch den reifen Samen, h hypokotyles Glied, c Kotyledonen, v
Leitbündel des Funiculus. Vergr. 26. B Partie aus dem Längsschnitt
durch die Samenschale, e die gequollene Epidermis, c braune,
stark verdickte Schicht, * zerdrückte Zellagen, a Aleuronschicht,
einzige erhaltene Zellage des Endosperms. Vergr. 250. Nach E.
STRASBURGER.
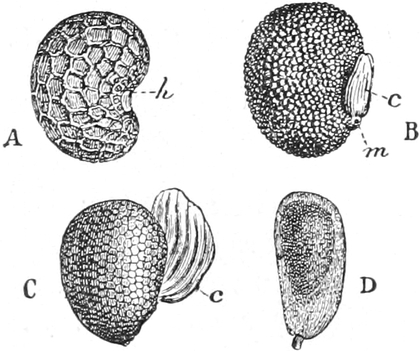
Fig. 581. A Samen von Papaver Rhoeas, h Hilum. B
Samen von Corydalis ochroleuca, m Mikropyle, c Caruncula. C Samen
von Chelidonium majus. D Samen von Nymphaea alba mit Samenmantel
(Arillus). Nach DUCHARTRE.
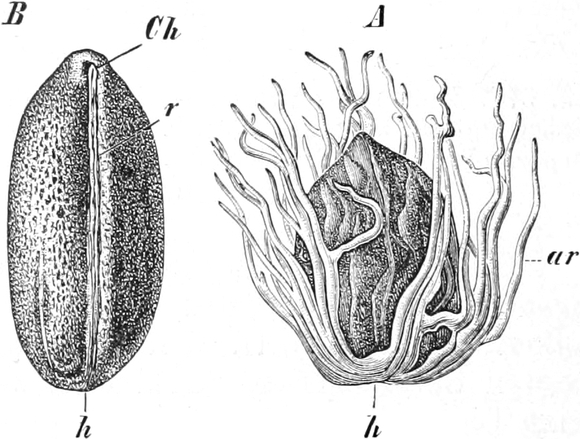
Fig. 582. A Myristica fragrans. Samen mit abgelöstem
Arillus ar. B Myristica argentea. Samen nach Entfernung des
Arillus. Ch Chalaza, r Raphe, h Hilum (Nabel). 3⁄4 nat. Gr. Nach
O. WARBURG.
Die Frucht[469].
Doch nicht auf die Makrosporangien allein erstrecken sich die Folgen
der Befruchtung, auch die Makrosporophylle, die Fruchtblätter, werden
in Mitleidenschaft gezogen. Das aus ihnen, oft unter Mitwirkung
des noch erhaltenen[S. 503] Kelches und der Blütenachse hervorgehende,
außerordentlich verschieden gestaltete Gebilde, welches die Bedeutung
hat, den in Entwicklung begriffenen Samen Schutz zu gewähren,
nennt man die Frucht. Bei den Gymnospermen freilich, wo die
Samenanlagen nackt auf den Fruchtblättern sitzen, hat man Früchte im
eigentlichen Sinne nicht, da ja ein Fruchtknoten fehlt. So kann man
bei Cycas, Ginkgo, Taxus, Podocarpus, Gnetum, Ephedra nur von Samen,
nicht von Früchten sprechen. Wenn aber die Fruchtblätter nach der
Befruchtung zusammenschließen, wie bei den verholzende Zapfen tragenden
Gymnospermen und den Beerenzapfen von Juniperus, so ist ein der
Angiospermenfrucht entsprechendes Gebilde gegeben, auf das man auch die
Bezeichnung Frucht wird anwenden dürfen.
Eine große Mannigfaltigkeit in der Entwicklung der Angiospermenfrüchte
läßt ja schon die verschiedenartige Ausbildung des Gynäceums erwarten.
Denn die einfachste Definition der Frucht ist: der reife Fruchtknoten.
Schwierigkeiten bereiten dabei die apokarpen Gynäceen.
Die zu vielen beisammenstehenden, aus apokarpen Gynäceen
hervorgegangenen Einzelgebilde, z. B. der Rosaceen, sollen hier
als Früchtchen, das ganze Gynäceum als Frucht
(eventuell als Sammelfrucht) bezeichnet werden. So ist
z. B. die Erdbeere eine durch Fleischigwerden des Blütenbodens
entstandene Sammelfrucht, deren einzelne Körnchen je einem
Früchtchen, und zwar einer Nuß, entsprechen; ebenso wäre beim
Apfel nur das Kerngehäuse die Frucht, das fleischige Gewebe um das
Gehäuse entstammt der um die Fruchtblätter ausgehöhlten und mit
ihnen verwachsenen Blütenachse. Bei den Hagebutten hat man ebenso
Sammelfrüchte vor sich; die Früchtchen sind die von dem fleischig
gewordenen Blütenboden umhüllten harten Nüßchen (Fig. 583). Bei
den aus synkarpen Gynäceen entstandenen Früchten kommt weiter die
Ausbildung der Fruchtwand, des Perikarps, besonders
in Betracht. Ihre äußere, mittlere und innerste Schicht werden als
Exo-, Meso- und Endokarp unterschieden.
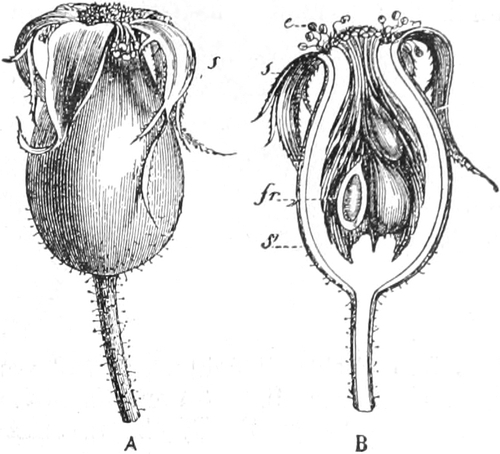
Fig. 583. Sammelfrucht von Rosa alba. Auf dem
fleischigen Blütenboden s′ sitzen die erhaltenen Kelchblätter und
die als Nüßchen ausgebildeten Einzelfrüchte fr. e Vertrocknete
Staubblätter. Nach DUCHARTRE.
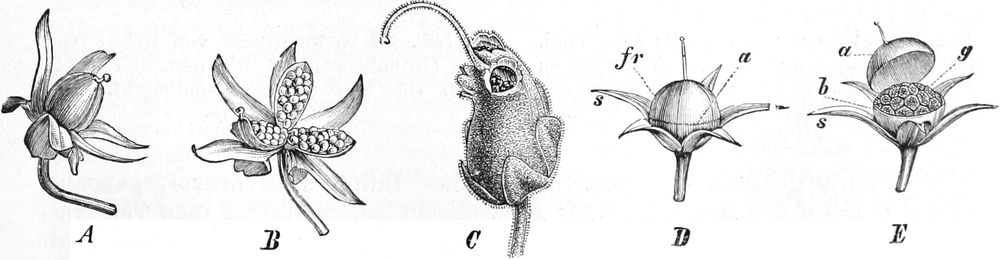
Fig. 584. Aufspringen der Kapselfrüchte. A Kapsel von
Viola tricolor, vor dem Aufspringen. B Dieselbe nach dem Aufspringen.
C Poricide Kapsel von Antirrhinum majus. Vergr. D Deckelkapsel von
Anagallis arvensis, geschlossen. E Dieselbe geöffnet. Nach A. F.
W. SCHIMPER.
Nach der Beschaffenheit dieses Perikarps ist folgende Einteilung der
Fruchtformen aufgestellt:
1. Eine Frucht mit meist trockenem, bei der Reife sich öffnendem
Perikarp heißt Kapsel (capsula) (Fig. 584). Geschieht
die Öffnung durch Trennung der Karpelle in den Nähten,
so ist die Kapsel septicid wie bei Colchicum (Fig. 812), tritt
ein Längsspalt im einzelnen Fache ein, so heißt sie
loculicid, z. B.[S. 504] Ornithogalum (Fig. 814), und werden bestimmt
umschriebene Löcher dabei gebildet, so haben wir die poricide
Kapsel, wie bei Papaver. Spezialfälle der Kapsel sind die
Balgfrucht (springt an der Naht des einblätterigen Fruchtknotens auf
[z. B. Aconitum, Fig. 520]), die Hülse (springt an der Naht und in der
Mittelrippe auf), z. B. die Erbse, endlich die Schote der Cruciferen
(öffnet sich durch Abspringen der Fruchtblätter von der falschen
Scheidewand, an der die Samen hängen bleiben), wie bei Cheiranthus
(Fig. 701 A).
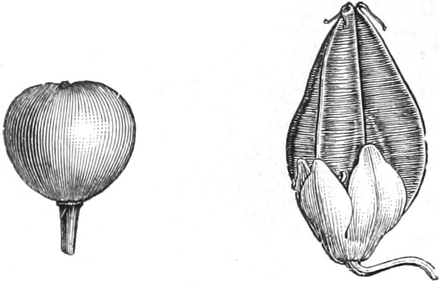
Fig. 585. Schließfrüchte. A Nuß von Fumaria
officinalis. Vergr. 6. B Von Fagopyrum esculentum. Vergr. 2. Nach
DUCHARTRE.
Fig. 586. Spaltfrucht von Galium Mollugo. Vergr. 6. Nach
DUCHARTRE.
2. Alle andersgearteten Früchte, die sich bei der Reife nicht öffnen,
kann man als Schließfrüchte der Kapsel gegenüberstellen.
Nach der Ausbildung des Perikarps unterscheidet man dann:
a) Die Nuß (nux), eine Schließfrucht mit trockenem,
hartem Perikarp, wie z. B. Haselnuß, Linde, Helianthus, Fumaria,
Fagopyrum (Fig. 585).
b) Zerfällt eine trockene, aus mehreren Fruchtblättern bestehende
Frucht bei der Reife in ihre Teilfrüchtchen, ohne daß diese sich
öffnen, so wird sie Spaltfrucht (schizocarpium) genannt.
Solche besitzen die Umbelliferen, auch Malva, Galium (Fig. 586).
c) Wird das Perikarp völlig und in allen Schichten
fleischig, so ist die Frucht eine Beere (bacca),
wie bei Vaccinium, Vitis, Physalis (Fig. 587).
d) Ist unter einem fleischigen Exokarp ein hartes, geschlossen
bleibendes Endokarp vorhanden, so haben wir eine Steinfrucht
(drupa). Prunus (Fig. 641) und Juglans (Fig. 671) sind
bekannte Beispiele.
Fig. 587. Frucht von Physalis Alkekengi. Sie besteht
aus der aus dem Fruchtknoten hervorgegangenen Beere fr, die vom
herangewachsenen Kelch s umhüllt wird. Nach DUCHARTRE.
Fig. 588. A Sammelfrucht von Rubus fruticosus
(Brombeere). B Scheinfrucht (Fruchtstand) von Morus nigra
(Maulbeere). Nach DUCHARTRE.
Nimmt dagegen ein ganzer, aus einer Infloreszenz hervorgegangener
Fruchtstand das Aussehen einer Einzelfrucht an, so spricht man
von einer Scheinfrucht. Die Feige ist das bekannteste Beispiel
einer solchen, wie sie überhaupt bei den Urticaceen und Moraceen
besonders häufig sich findet. Der Vergleich einer Brombeere,
die als Sammelfrucht aus einer Blüte hervorgeht, und einer
Maulbeeren-Scheinfrucht wird die sehr große äußere Ähnlichkeit
beider Gebilde erkennen lassen (Fig. 588).
[S. 505]
Verbreitung
der Samen[470].
Das wichtigste Mittel der Spermatophyten, ihre Art den unter gleichen
Bedingungen mit ihnen zusammenlebenden Gewächsen gegenüber zu erhalten,
ist möglichst reichliche Samenerzeugung; denn mit der Masse
der Nachkommen steigt die Wahrscheinlichkeit, daß wenigstens einige
davon ans Ziel gelangen. Aber die Zahl allein würde nur geringen
Einfluß haben können, wenn alle Samen an den Ort ihrer Entstehung
gebunden blieben; so ist neben der Zahl auch eine die Verbreitung
begünstigende Ausrüstung von der größten Bedeutung, und Form
wie Beschaffenheit von Frucht und Samen zeugen zur Genüge von dem
tiefgreifenden Einfluß dieses Faktors auf ihre Ausgestaltung.
Als Verbreitungsmittel stehen den Samen der Pflanzen dieselben
Agenzien zur Verfügung wie für die Pollenübertragung, also neben
Luft- und Wasserströmungen die Tiere, und endlich menschliche
Verkehrseinrichtungen; doch ist insofern ein Unterschied zu beachten,
als die Pollenkörner fast durchweg von überaus geringem Gewicht
und von minimaler Größe sind, während im Samen doch meist eine
verhältnismäßig größere Menge von Reservestoffen abgelagert sein muß,
die ihm ein höheres Gewicht und größere Masse verleihen. Trotzdem
ist Samentransport durch Wind die alle anderen Möglichkeiten weitaus
überwiegende Verbreitungsart.
Fig. 589. Geflügelter Samen von Pithecoctenium echinatum
in natürlicher Größe. Nach F. NOLL.
Die Ausrüstung der Samen für die Aussäung durch den Wind
besteht vielfach nur in ihrem außerordentlich geringen Ausmaß
an Größe und Gewicht; so zählen sie z. B. bei Stanhopea
gewiß nach Millionen in jeder Kapsel, und ihr Gewicht ist für
Dendrobium attenuatum auf etwa 1⁄200 mg bestimmt worden. So
können diese Orchideen vermöge der Leichtigkeit und Menge ihrer
Samen eine so hervorragende, nur von den mit ebenso leichten
Sporen versehenen Farnen erreichte Rolle unter den epiphytischen
Bewohnern feuchter Tropenwaldungen spielen. Weit häufiger ist eine
Oberflächenvergrößerung und Darbietung einer Angriffsfläche für den
Wind bei nicht ganz so leichten Samen zu beobachten, indem entweder
die ganze Samenoberfläche kürzere oder längere Haare trägt, wie
bei den Weiden (Fig. 691), Pappeln (Fig. 692) und der Baumwolle
(Fig. 712), oder ein längerer Haarschopf dem Samen an einem
Ende aufgesetzt ist, wie bei den Asclepiadaceen und Apocynaceen
(Strophanthus, Fig. 754), vielen Gesneriaceen und Bromeliaceen.
Ebenso häufig findet sich bei anderen Familien die Ausbildung
einer richtigen Segelfläche aus einer äußerst dünnen und leichten
Membran, wie sie bei unseren Fichten (Fig. 605) und Kiefern (Fig.
606) von der Fruchtschuppe abgespalten wird, bei Rhododendron,
Bignoniaceen und bei Rubiaceen (Cinchona, Fig. 774) sich an jedem
Samen ausbildet; nirgends wohl schöner als bei der Cucurbitacee
Zanonia und der ihr fast gleichkommenden Bignoniacee Pithecoctenium
echinatum, deren leichte seidig glänzende Flügel den fallenden
Samen eine fast wagerechte Stellung einzunehmen erlauben und damit
ein Hinschweben über weite Strecken bei dem leichtesten Luftzuge
ermöglichen (Fig. 589).
[S. 506]
Mannigfaltige andere Blüten- bzw. Fruchtorgane sind besonders
in solchen Fällen, wo es sich um einsamige Früchte handelt, zu
Flügeln oder Windsegeln umgestaltet worden, so die Kelchblätter
bei Dipterocarpaceen, das große Hochblatt der Lindeninfloreszenz
(Fig. 714), das Deckblatt mit den Vorblättern bei Carpinus (Fig.
684), meist aber die Fruchtknotenwandung, wie bei Betula (Fig.
683), Alnus, Ulmus (Fig. 693), Polygonaceen (Fig. 677 D). Acer
(Fig. 736), Fraxinus (Fig. 749) oder den Früchtchen der Typhaceen,
Eriophorum (Fig. 824), Dryas und Anemone (Fig. 630). Gleiche
Bedeutung hat endlich ein Haarkelch (Pappus), der am oberen Rande
der einsamigen Frucht entwickelt ist, wie bei Valerianaceen (Fig.
777) und Kompositen (Fig. 793 B, 791), besonders wenn er durch
nachträgliche Verlängerung des Fruchtendes fallschirmähnlich
wirken kann, wie bei Taraxacum (Fig. 794), Tragopogon u. a. Nach
DINGLER beträgt die Fallverzögerung z. B. für die mit
Haarschuppen versehenen Früchte von Cynara Scolymus bereits in
der ersten Sekunde das Sechsfache im Vergleich zum freien Fall
im luftleeren Raume, für Pinus silvestris das Siebenfache, für
Pithecoctenium das Dreißigfache.
Weit beschränkter, aber für viele Gewächse von ganz hervorragender
Bedeutung ist die Samen- oder Fruchtbeförderung durch
Meeresströmungen. Die ganze Strand- und Küstenflora z. B. im
Malayischen Archipel besteht nach SCHIMPERs Untersuchungen
ausschließlich aus Pflanzen mit schwimmfähigen Früchten oder Samen,
deren Ausrüstung dann stets mehr oder minder derjenigen der überall
an Tropenküsten verbreiteten Kokosnuß (Fig. 806) entspricht. Ein
dickes, aus grobem Fasergewebe bestehendes, von pergamentartigem
Exokarp bedecktes Mesokarp erhält die Frucht schwimmfähig und
schützt das spröde steinharte Endokarp vor dem Zertrümmertwerden
beim Anprall an Felswände und Ufersteine. Derselbe oder doch
ein sehr ähnlicher Bau findet sich bei Barringtonia speciosa
und anderen Arten, bei Cerbera Odollam (Fig. 590), Terminalia
Catappa, Nipa fruticans, den Pandanusarten und zahlreichen
weiteren Vertretern auch der strauchigen und krautigen Dünen-
und Strandflora; bei allen aber ist lange Schwimmfähigkeit ohne
Beeinträchtigung ihrer Keimfähigkeit Bedingung für Verbreitung und
Erhaltung der Art. Die Notwendigkeit beider Bedingungen
zeigt z. B. die an allen Küsten des indischen Ozeans gelegentlich
angespülte große Doppelfrucht von Lodoicea Seychellarum, die zwar
schwimmfähig ist, aber den Transport auf dem salzigen Meere nicht
zu ertragen vermag und so auf das geringe Verbreitungsgebiet einer
kleinen Inselgruppe beschränkt bleiben mußte.
Fig. 590. Frucht von Cerbera odollam aus der Drift.
Nach A. F. W. SCHIMPER. Hautartiges Exokarp und das Fleisch
des Mesokarps fehlen, so daß das von groben Fasersträngen durchzogene,
etwas abgeriebene Schwimmgewebe frei liegt.
Die Verbreitung von Früchten oder Samen durch Tiere beruht
in der Regel darauf, daß fleischige Früchte Vögeln als Nahrung
dienen, von denen die Samen unverdaut wieder abgegeben werden.
In Mitteleuropa ist besonders der Holunder, Sambucus nigra, ein
bekanntes Beispiel; für manche Samen scheint die Wanderung durch
den Darm von Tieren die Keimfähigkeit zu bedingen. Besondere
Anpassungen an die Samenverbreitung durch Tiere dürften in vielen
Fällen die Arillusbildungen (vgl. S. 502) darstellen. Der
Arillus von Taxus mit seiner intensiv roten Farbe, der einzige
nicht giftige Teil des Baumes, wird von Amseln mit Vorliebe
verzehrt, die roten Früchte von Evonymus europaea lassen geöffnet
ihre mit hochrotem Arillus versehenen vier Samen hervorschauen, die
von Rotkehlchen aufgesucht werden. Die Verbreitung der Muskatnüsse
wird von einer großen Taubenart, die dem Arillus eifrig nachstellt,
über die ganzen Molukken vermittelt. Endlich werden unsere
Misteln im Winter, wenn wenig andere Nahrung zu finden ist, von
Amseln und anderen Vögeln gern aufgesucht; die Samen der Misteln[S. 507]
bleiben, vermöge ihres Viscinklebstoffes, beim Abwetzen des
Schnabels an Zweigen haften und können dort zur Keimung gelangen.
Die Verbreitung der Klettpflanzen Galium aparine, Lappa-Arten
(vgl. Fig. 790 a), Bidens, Xanthium usw. durch das Wollkleid
unserer Vierfüßler, die allgemeine Verbreitung der Wasserpflanzen
von einem See oder Tümpel zu weit entfernten anderen durch die
Wasservögel, die Verstreuung der Haselnüsse usw. durch Eichhörnchen
bedürfen keiner weiteren Erklärung. Schließlich sei noch auf die
Samenverbreitung durch Ameisen hingewiesen; diese Tiere werden
durch ölreiche Anhängsel gewisser Samen und Früchte, wie die
Caruncula, veranlaßt, sie in ihre Behausungen zu verschleppen.
Daß der Mensch durch Handel und Erwerbstätigkeit gewaltigen
Einfluß auf die Verbreitung der Nahrungs- und Nutzpflanzen gewonnen
hat, und daß mit diesen zugleich eine Menge der als Unkraut
dazwischen lebenden Gewächse in ihren Samen über die bewohnten
Teile der Erde verbreitet sind, ist durch zahlreiche Beispiele
belegt und allgemein bekannt.
Die
Keimung[471].
Samen, die alle Fährlichkeiten der Reise gut überstanden haben,
bedürfen zur Keimung eines geeigneten Standortes. Kleinen Samen
ist es nicht schwer, in Ritzen oder Rillen des Bodens Unterkunft
zu finden, wo sie häufig, durch besondere Eigenschaften ihrer
Oberfläche unterstützt, leicht festhaften bleiben. Größere Samen
werden durch das fallende Laub die genügende Bedeckung erhalten.
Die Früchtchen von Erodium und anderen Geraniaceen, von Avena
sterilis, Stipa-Arten und sonstigen Gramineen gelangen mit Hilfe
ihrer hygroskopischen Krümmungen in den Boden (vgl. S. 295, Fig.
276); rückwärts gerichtete Haare ihrer Oberfläche verhindern, daß
sie wieder hinausgedrängt werden. Geotropisch abwärts wachsende
Fruchtstiele (vgl. S. 304) sorgen für hinreichend tiefes Eingraben
der Früchte bei Arachis hypogaea, Trifolium subterraneum, Okenia
hypogaea u. a., negativer Phototropismus führt diejenigen von
Linaria cymbalaria in die Ritzen der von der Pflanze bewohnten
Mauern ein (vgl. S. 310).
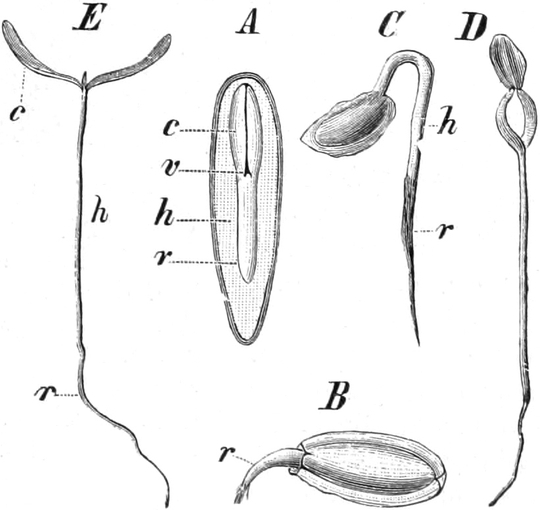
Fig. 591. Thuja occidentalis. Bei A
medianer Längsschnitt durch den reifen Samen. In B bis E
Keimungsstadien. h Hypokotyl, c Kotyledonen, r Radicula, v
Stammvegetationskegel, A 5 mal, B und C 2 mal vergr. D und E
nat. Größe. Nach H. SCHENCK.
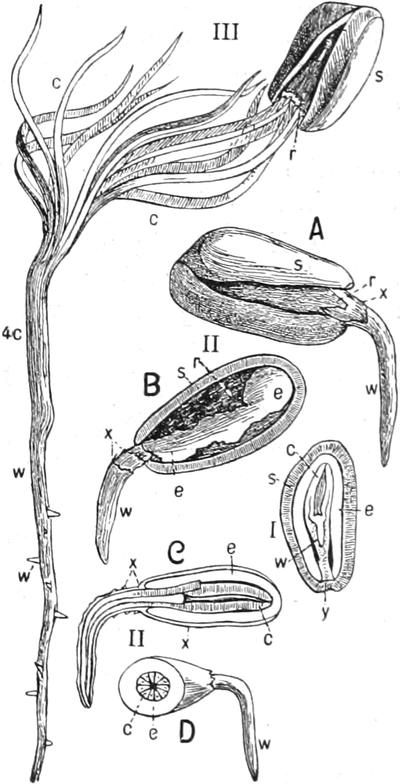
Fig. 592. Pinus pinea. Keimung nach J. SACHS.
I Samenlängsschnitt. y Mikropylenende. II Keimungsbeginn.
s Samenschale, e Endosperm, w Hauptwurzel, x der von ihr
ausgestülpte Embryosack (zerrissen), r rote Samenhaut in der Schale.
III Kotyledonen c verlassen den ausgesogenen Samen, hc Hypokotyl,
w′ Nebenwurzeln.
Finden die so geborgenen Samen dann hinreichend Feuchtigkeit,
so quellen sie zunächst stark auf. Damit geht ihnen freilich
bereits ein Teil ihrer großen Unempfindlichkeit gegen alle
Fährlichkeiten, speziell Temperaturextreme und Trockenheit,
wieder[S. 508] verloren, die sie nur ihrem sehr geringen Wassergehalt
verdankten. Die nächste Aufgabe für den Samen ist alsdann die
Sprengung der Samenschale, die meist von der Wurzel besorgt wird.
Da die Orientierung der Wurzelspitze stets gegen die Mikropyle
gerichtet ist, wird diese Stelle geringeren Widerstandes von der
Wurzelspitze durchbohrt und durch Herauswachsen der stärkeren Teile
des Hypokotyls auseinandergesprengt (Fig. 591, 592). Die aus dem
Samen austretende Wurzel wendet sich alsbald geotropisch abwärts
und sorgt durch Wurzelhaare, die am Wurzelhals, d. h. der
Grenze von Wurzelanlage und Hypokotyl, besonders lang und zahlreich
hervorbrechen, für schleunige Befestigung des Keimlings im Boden.
Inzwischen wächst das Hypokotyl nach und nach aus der Samenschale
heraus, während die Kotyledonen zum großen Teil noch darin stecken
und die Reste der Reservestoffe aufsaugen (Fig. 591, 593 a). So ist
eine mehr und mehr bis an die Insertionsstelle der Keimblätter sich
hinaufziehende scharfe Krümmung des Hypokotyls unausbleiblich,
und die bei weiterem Aufwärtswachsen sich steigernde Spannung muß
schließlich die Kotyledonen aus der Samenschale herausziehen,
worauf der Keimling eine aufrechte Lage einnimmt und die Blätter
zur Assimilation ausbreitet, um ein selbständiges Leben zu
beginnen. Die Zahl der Keimblätter ist meist 2, bei einigen
Gattungen der Koniferen aber wechselt sie von 3–∞ (Fig. 592).
Die häufigste Form der Keimung ist charakterisiert durch
die oberirdische Ausbreitung der Kotyledonen, sie heißt
epigäische Keimung. Man findet sie bei den kleineren Samen
fast ausschließlich.
Die hypogäische Keimung findet sich hauptsächlich
bei großsamigen Dikotyledonen, deren Keimblätter als
Reservestoffbehälter dienen, wie Vicia faba, Pisum, Aesculus,
Juglans, Quercus u. a. Ihr Charakteristikum besteht darin, daß
die Kotyledonen, nachdem die Wurzel sich im Boden befestigt hat,
in der Samenschale verbleiben, während das epikotyle Stammende
sich zwischen den Keimblättern emporrichtet und die Folgeblätter
entwickelt. So scharf morphologisch die Trennung beider
Keimungsformen ist, so wenig ist dieser Unterschied systematisch
verwertbar; schon innerhalb der Papilionaceen z. B. finden sich
mannigfache Übergangsstufen; Phaseolus vulgaris keimt epigäisch,
Phaseolus multiflorus hypogäisch.
Die Keimung monokotyler Samen weicht von den beschriebenen
Fällen dadurch ab, daß nach der Hauptwurzel der Kotyledon mit
seinem Scheidenteil aus dem Samen austritt (Fig. 593 b), während
das andere Ende des Keimblattes sehr lange im Samen verbleibt und
als Saugorgan die im Endosperm aufgespeicherten Reservestoffe
dem Keimling zuführt, dessen erstes Blatt alsbald aus dem
Scheidenteil des Kotyledons hervortritt. Sehr harte Samenschalen
weisen besondere Vorkehrungen auf, welche das Austreten der Wurzel
erleichtern. So finden sich an der Kokosnuß drei Keimlöcher vor,
den drei Fruchtblättern entsprechend. Dasjenige, vor welchem der
einzige zur Entwicklung gelangende Keimling mit seinem Wurzelende
liegt, ist von nur sehr dünner Schale überdeckt, die beiden
anderen dagegen sind fest verschlossen. Die harte Steinschale der
Palme Acrocomia sclerocarpa (Fig. 594) besitzt an der der Wurzel
vorgelagerten Stelle einen leicht herausschiebbaren Pfropf; in der
ganzen Familie der Scitamineen sind ähnliche Deckel vorhanden.
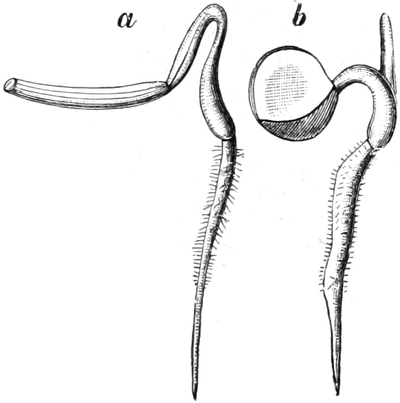
Fig. 593. Keimlinge von a Scorzonera humilis. b Iris
pseudacorus. Nach G. KLEBS.
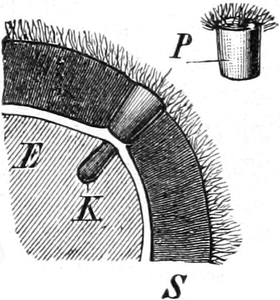
Fig. 594. Schnitt durch den oberen Teil der Frucht von
Acrocomia sclerocarpa. S Steinschale, P Pfropf, K Keimling, E
Endosperm. Nach E. PFITZER.
Ganz abweichende Verhältnisse, die hier nur kurz erwähnt werden
können, zeigen die sog. „viviparen“ Pflanzen (Fig. 659). Die
Viviparie stellt eine zum Standort in Beziehung stehende
ökologische Anpassung der tropischen Mangrovepflanzen dar. Sie
lassen ihre einsamigen Früchte bereits an der Mutterpflanze
keimen, d. h. die Fruchtwandung wird vom mächtig heranwachsenden
Keimling, der zunächst mit dem Radikularende aus der Mikropyle der
Samenschale hervordringt, ebenfalls durchbrochen, und das ins Freie
tretende Hypokotyl erreicht z. B. bei Rhizophora mucronata und Rh.
mangle bis[S. 509] über 1 m Länge (vgl. Fig. 187 u. 659), so daß die vom
Baum fallenden Keimlinge mit dem zugeschärften Wurzelende bei ihrem
ansehnlichen Gewicht tief in den Schlamm eindringen, dort festen
Fuß fassen und sich sogleich weiterentwickeln können.
Anordnung der
Klassen, Ordnungen und Familien.
I. Klasse. Gymnospermae[472].
Die Gymnospermen sind nach morphologischen Gesichtspunkten
(wie nach den Ergebnissen der Serodiagnostik (S. 524) nicht
einheitlicher Abstammung. Vielmehr gehen die Cycadeen zwar auf
die Cycadofilices (vgl. S. 461 Pteridospermeae) zurück, wohl
vermittelst der Benettitaceae (vgl. S. 522, Cycadeoidea), aber die
Coniferae würden auf die Lycopodinae ligulatae zurückzuführen sein,
wobei die Deckschuppe dem Blatte, die Fruchtschuppe vielleicht der
Ligula? entsprechen könnte. Die Abietineenzapfen bestehen demnach
aus zahlreichen Makrosporophyllen, und jeder Zapfen entspricht
einer Blüte, nicht einem Blütenstand, so daß sich ein direkter
Anschluß an die spiralig aufgebauten Magnoliaceenblüten ergibt.

Fig. 595. Cycas revoluta-Gruppe aus einem Tempelgarten
in Japan. Die mächtigsten Stämme in Japan von 8 m Höhe und 2 m Umfang.
Nach E. R. WIELAND.
Die 1. Ordnung, Cycadinae, enthält als einzige
Familie die Cycadaceae. Die Cycadeen sind den Tropen und
Subtropen angehörige Holzgewächse. Und zwar ist Cycas in Asien
heimisch, Macrozamia und Bowenia in Australien, Encephalartos
und Stangeria gehören Afrika an; dagegen besitzt Amerika die
Gattungen Dioon, Zamia, Ceratozamia und Microcycas. Den
Habitus eines Cycadeenhaines gibt Fig. 595 wieder, wo zahlreiche
ansehnliche Exemplare von Cycas revoluta, darunter auch
verzweigte, beisammenstehen.
Der Stamm ist mit Dickenwachstum begabt, er bleibt entweder
unverzweigt oder stellt ein Sympodium dar und bringt große,
gefiederte Laubblätter von mehrjähriger Dauer und lederiger
Beschaffenheit in gipfelständiger Rosette abwechselnd mit
schuppenförmigen Niederblättern hervor. Diese bekleiden gemeinsam
mit den Basen der abgeworfenen Laubblätter die Oberfläche des
säulenartigen oder knollenförmigen Stammes. In allen Teilen der
Pflanzen finden sich Schleimgänge. Die Leitbündel sind kollateral,
führen jedoch im Gefäßteil nur Tracheïden.
[S. 510]
Die Blüten der Cycadaceae sind diözisch verteilt. Fig. 596 stellt
eine blühende weibliche Pflanze von Cycas revoluta dar. Der
Vegetationskegel des Stammes bildet hier abwechselnd Laubblätter
und Makrosporophylle. Die Laubblätter sind in der Jugend gleich
denen der Farne eingerollt. Ein Sporophyll ist in Fig. 596 a
genauer wiedergegeben. Es zeigt noch die gefiederte Form der
Laubblätter, entbehrt aber des Chlorophylls und ist dicht mit
braunen Haaren bedeckt. Der Basis genähert trägt es zwei bis acht
randständige Makrosporangien an Stelle von Fiedern. Jede blühbare
weibliche Cycaspflanze durchläuft also in regelmäßigem Wechsel eine
vegetative und eine Blütenperiode; die Blüte, von der Gesamtheit
der Sporophylle dargestellt, wird stets durchwachsen, da dem Stamme
die Fähigkeit sich zu verzweigen im allgemeinen fehlt. Dagegen
vereinigen die männliche Cycaspflanze und die übrigen Cycadaceae
ihre Sporophylle in terminalen Zapfenblüten von oft riesigen
Dimensionen; eine Seitenknospe des Stammes schiebt sodann diese
terminalen Blüten beiseite und setzt in sympodialem Aufbau das
Wachstum in ursprünglicher Richtung fort.

Fig. 596. Cycas revoluta, weiblich, blühend. Nach einer
Photographie. a Makrosporophyll (Fruchtblatt) von Cycas revoluta
(nach SACHS). b Mikrosporophyll (Staubblatt) von Cycas
circinalis (nach RICHARD).
Derartige Zapfen bestehen aus zahlreichen Sporophyllen, die
in spiraliger Stellung an der Achse aufeinander folgen.
Mikrosporophylle sind auf ihrer Unterseite mit zahl[S. 511]losen
Mikrosporangien übersät, wie Fig. 596 b für Cycas darstellt.
Makrosporophylle der übrigen zapfenblütigen Cycadaceae
beschränken die Zahl der Makrosporangien auf je zwei, welche dem
Cycas gegenüber erheblich abgeänderten Sporophyll am Rande
eingefügt werden, wie Fig. 597 für Ceratozamia robusta zeigt;
Entwicklungsgeschichte vgl. S. 485.
Der einzige Vertreter der 2. Ordnung, Ginkgoinae,
die auf die Familie der Ginkgoaceae beschränkt ist, ist
der aus Japan stammende, bei uns jetzt vielfach angepflanzte
Baum Ginkgo biloba. Ginkgo biloba verliert jährlich seine
langgestielten, ein- bis mehrfach zweilappigen, Adiantum ähnlichen
Blätter. Bei diözischer Blütenverteilung sind die hüllblattlosen
Staubblätter zahlreich an einer gestreckten Achse vereinigt, ihre
Mikrosporangien besitzen ein „Endothecium“ (S. 470). Die
Makrosporangien stehen zu zweien auf dem Gipfel kurzer Sprosse
in einer kragenartig die Sporangien umgebenden Wucherung, dem
Sporophyll (Fig. 598). Entwicklungsgeschichte vgl. S. 485.
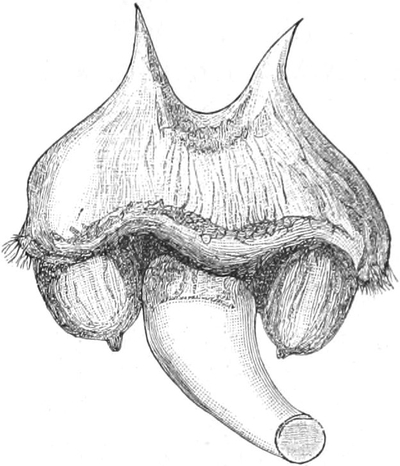
Fig. 597. Ceratozamia robusta. Ein Makrosporophyll mit
zwei Makrosporangien. Nach K. GOEBEL.
In der 3. Ordnung Coniferae, den Koniferen, sind die unter
dem Namen der „Nadelhölzer“ bekannten Pflanzen vereinigt.
Stattliche Gewächse von baumförmigem oder strauchartigem Habitus, mit
holzigen Stämmen versehen, zeichnen sie sich durch den gemeinsamen
Charakter kleiner, ungeteilter, fester, flacher oder kantiger, meist
xerophil gebauter Nadelblätter von meist mehrjähriger Dauer aus, so
daß Nadelhölzer, bis auf wenige Vertreter, wie die Lärche, zu der
„immergrünen“ Vegetation zählen. Eine reiche Verzweigung in
Lang- und Kurztriebe typischer Form findet sich bei den Gattungen
Pinus, Larix und Cedrus. In allen Fällen aber sind Hauptstamm und
Seitenzweige durch Wachstumsrichtung und -stärke scharf unterschieden,
wenigstens an jüngeren Individuen. Im Alter nehmen die Bäume oft
Schirmform an.
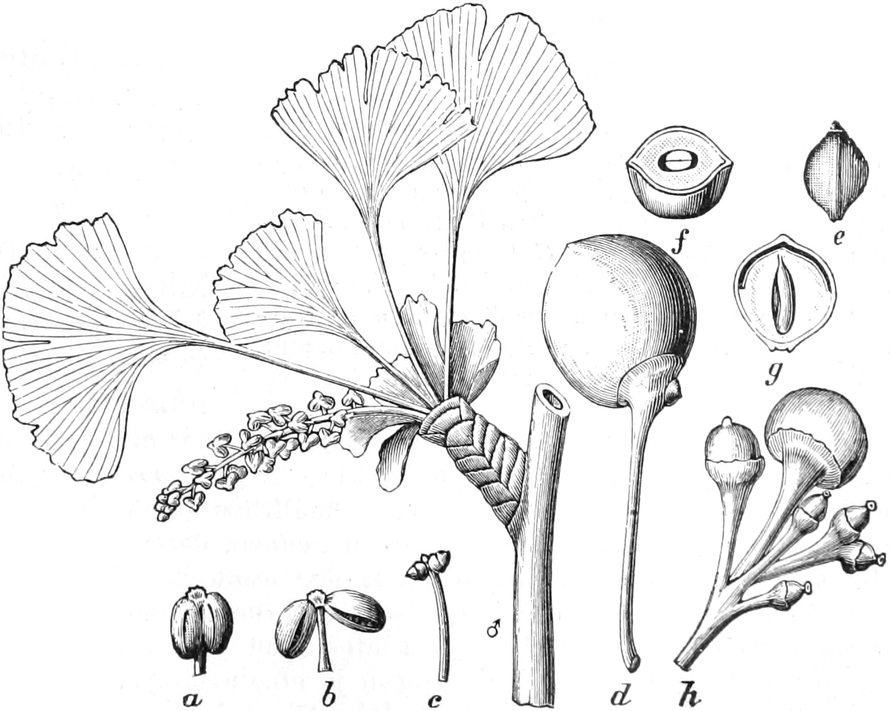
Fig. 598. Ginkgo biloba. Männlicher Kurztrieb mit
Blüte und jungen Blättern. a, b Staubblätter, c weibliche
Blüte, d Frucht, e Steinkern daraus, f derselbe im Querschnitt,
g im Längsschnitt nach Ausbildung des Embryo, h weibliche Blüte
mit ausnahmsweise zahlreichen gestielten Samenanlagen. — Männliche
Blüte und c in nat. Gr., d ein wenig verkleinert, die übrige
Figur vergrößert. Nach L. C. RICHARD, a–d
nach A. W. EICHLER.
[S. 512]
Der Mangel an Tracheen im Holz, wie in den Gefäßteilen der jungen
Pflänzchen, ist in anatomischer Hinsicht charakteristisch. Große
Tracheïden mit eigenartigen Hoftüpfeln besonders auf den radialen
Längswänden vertreten ihre Stelle und bilden Holzkörper von
außerordentlicher Gleichmäßigkeit. Der Mehrzahl der Koniferen ist
reicher Harzgehalt in allen Teilen eigen.
Die Nadelhölzer sind im Gegensatz zu den Cycadinae meist Bewohner
gemäßigter Zonen, auch gehören sie mit zu den am weitesten gegen
den Pol vordringenden Baumformen. Wo sie innerhalb der Wendekreise
auftreten, handelt es sich meist um Arten, die hoch ins Gebirge
hinaufsteigen.
Nach den Verschiedenheiten ihrer Blütenbildung werden die Koniferen auf
zwei Familien verteilt.
Die Taxaceae sind charakterisiert durch weibliche Blüten mit einem
oder wenigen Makrosporangien, die in der Regel mit einem Arillus
versehen sind. Diese Blüten pflegen nicht zapfenförmig zu sein. Die
Mehrzahl der Vertreter hat diözische Geschlechtsverteilung.
Die Pinaceae dagegen führen mehrere Samenanlagen in jeder
weiblichen Blüte, welche aus zahlreichen, an einer Spindel zu Zapfen
vereinigten Sporophyllen gebildet wird. Ein Arillus fehlt, und die
Geschlechtsverteilung ist meist monözisch.
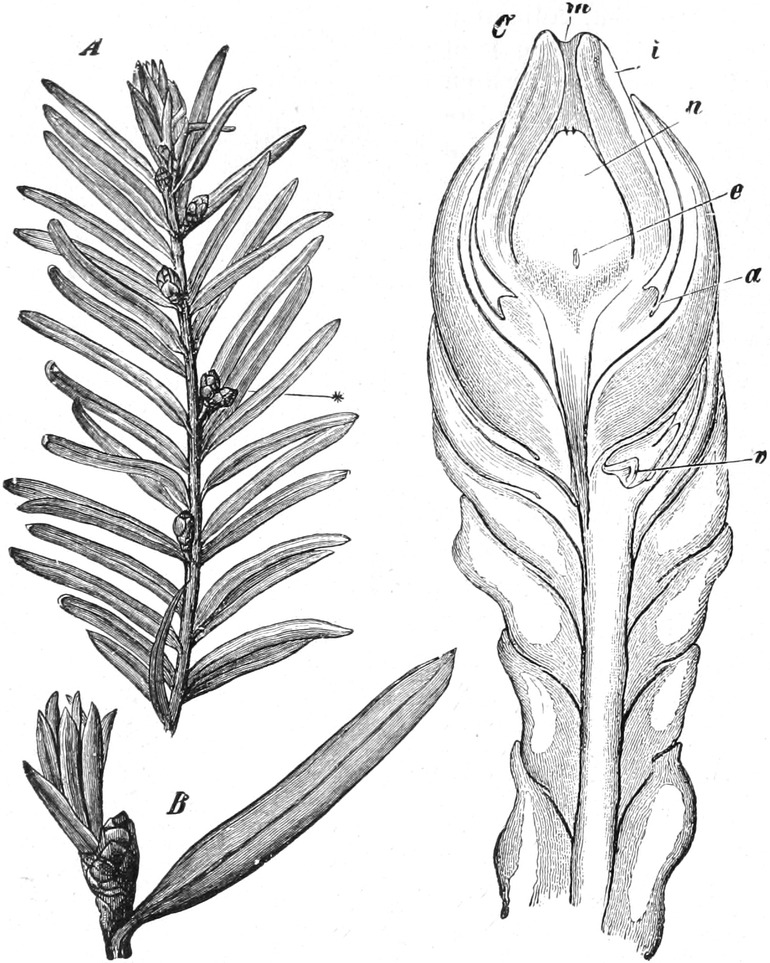
Fig. 599. Taxus baccata. A Habitusbild eines
Zweiges mit weiblichen Blüten, bei * zwei Samenanlagen an demselben
Primansprößchen. Nat. Gr. B Ein Blatt mit achselständiger
Samenanlage. Vergr. 2. C Längsschnitt durch die gemeinsame
Mediane des Priman- und Sekundansprößchens, v Vegetationskegel
des Primansprößchens, a Arillusanlage, e Embryosackanlage, n
Nucellus, i Integument, m Mikropyle. Vergr. 48. — Nach E.
STRASBURGER. Giftig.
Familie Taxaceae. Die Angehörigen der Taxaceen sind in
verschiedenen kleinen Gattungen teils auf der nördlichen
Hemisphäre, wie Taxus, Torreya, Cephalotaxus, vorwiegend
aber in zahlreichen Gattungen auf der südlichen Hemisphäre
verbreitet. Als wichtigste von diesen ist die Gattung Podocarpus
zu nennen, deren zahlreiche Arten sich als stattliche Bäume in
den Bergwaldungen meist der asiatischen Tropen finden und in den
gemäßigten Zonen Ostasiens und der australisch-neuseeländischen
Inselwelt eine weite Verbreitung besitzen. Ihre weiblichen Blüten
sind kleine Sprosse, deren Sporophylle fleischig anschwellen; 1
oder 2 davon tragen je eine anatrope, von fleischig werdendem
Arillus umhüllte Samenanlage auf dem Gipfel. Die auf demselben
oder auf einem anderen Individuum vorhandenen männlichen Blüten
stellen kleine aufgerichtete Zäpfchen dar, die mit zahlreichen
Mikrosporophyllen besetzt sind. Jedes Sporophyll trägt unterseits
zwei Sporangien, deren Mikrosporen mit Flugblasen versehen sind.
Die Eibe, Taxus baccata, ist die einzige in Europa
vorhandene Art der Familie (Fig. 599, 600). Jetzt vielfach
in Anlagen gepflanzt, hat sie früher eine große Ver[S. 513]breitung
als immergrünes Unterholz unserer Wälder besessen. Einzelne
mächtige Exemplare finden sich in Nord- und Mitteldeutschland
noch vor, größere Bestände sind wohl nur spärlich erhalten
geblieben. Der Baum erreicht eine Höhe von etwa 10 m. Seine
Zweige sind sämtlich als Langtriebe ausgebildet und tragen
an den aufstrebenden Hauptästen allseitig, an den wagerecht
ausgebreiteten Seitenzweigen nach rechts und links gescheitelt,
flache Nadelblätter von mehrjähriger Dauer. Der Baum ist diözisch.
Die Blüten sitzen auf der Unterseite der Zweige und entstehen als
Achselsprosse vorjähriger Nadeln. Männliche Blüten werden von
einer Anzahl Schuppenblätter am Grunde umhüllt und enthalten etwa
10 schildförmige Staubblätter, mit je 5–9 Pollensäcken (Fig. 600
A). Bei ihrer Öffnung wird die Außenwand nach Lösung an der
Basis und den Seiten zurückgeschlagen; so ähnelt das Staubblatt
einem nach unten aufgespannten Schirm, in dessen Höhlung die
ausgefallenen Pollenmassen liegen bleiben, bis der Wind sie
entführt. Die Pollenkörner von Taxus entbehren der Flugblasen.
Weibliche Blüten (Fig. 599) entstehen meist einzeln als sekundärer
Achselsproß der obersten Schuppe eines primären Triebes,
dessen zur Seite gedrängter Vegetationskegel sich nicht weiter
entwickelt. Sie bestehen aus einer einzigen atropen Samenanlage,
die von einem Integument umhüllt wird. Die den Gymnospermen
eigene Tropfenausscheidung aus der Mikropyle ist hier besonders
gut zu beobachten. Während der Samenentwicklung bildet sich ein
fleischiger Arillus aus, der als hochroter Becher den reifen Samen
umgibt. Das Laub und der Samen, nicht aber der ihrer Verbreitung
durch Vögel dienende süße Arillus, sind giftig.

Fig. 600. Taxus baccata mit Früchten. 1⁄2 nat. Gr. A
Männliche Blüte nach RICHARD. — Giftig.
Familie Pinaceae. In der Familie der Pinaceen sind
die wichtigsten Nadelholzbäume enthalten, die sich nach
Verschiedenheiten der Blattstellung und Lage der Samenanlagen auf
zwei Unterfamilien verteilen lassen. Alle Formen mit gegenständigen
oder quirlständigen Blättern fassen wir als Cupressineae
zusammen. Diesen kommt auch eine aufrechte Stellung ihrer
Samenanlagen zu.
Alle Formen mit wechselständigen Blättern bilden die Unterfamilie
der Abietineae, welche fast ausnahmslos umgewendete Samenanlagen
besitzen.
Unterfamilie Cupressineae. Die Cupressineen haben
teils quirlständige Nadeln, wie unser Wacholder (Fig. 601),
teils schuppenförmige und dann dekussiert stehende Blätter wie
Thuja-Arten und Juniperus sabina (Fig. 602). Doch zeigen die
Keimpflanzen von Thuja noch nadelförmige Blätter, und einzelne
Zweige der schuppig beblätterten Juniperus-Arten fallen ebenfalls
häufig auf Nadelblätter und dreizählige Blattquirle zurück, so daß
man diese als ältere Blattform aller Cupressineen zu betrachten
berechtigt ist. Taxodium distichum trägt zweizeilig beblätterte
hinfällige Kurztriebe.
[S. 514]
Die Blüten der Cupressineen sind monözisch, nur bei Juniperus
meist diözisch verteilt. Die männlichen sind bei Juniperus
communis blattachselständig. Der kleine Sproß beginnt mit einer
Anzahl anliegender Schuppenblätter (Fig. 601 Aa) und endet mit
einigen Wirteln von schildförmigen Sporophyllen c, die auf der
Unterseite je 2 bis 4 Mikrosporangien (d) tragen, welch letztere
sich mit einem vertikalen, in Längsrichtung des Sporophylls
verlaufenden Riß öffnen. Die Stellung der weiblichen Blüten ist
die gleiche. Auf die Hülle der Schuppenblätter (Fig. 601 B)
folgt ein Quirl Fruchtblätter (Fig. 601 Cb); jedes trägt schräg
vor der Mittellinie eine aufrechte Samenanlage c. Nach der
Befruchtung geht hauptsächlich aus dem Grunde der Sporophylle
fleischiges Parenchym hervor, welches die drei Makrosporangien
emporhebt und sich zwischen sie eindrängt, ohne jedoch den freien
Raum ganz auszufüllen. Die drei Fruchtblätter wachsen dann über
den Samenanlagen völlig zusammen. Eine Verwachsungsnarbe auf dem
Scheitel kennzeichnet noch an der reifen Beerenfrucht die Grenzen
der Fruchtblätter. Übrigens ist Juniperus die einzige Gattung der
Cupressineen mit Beerenfrüchten; die anderen wie Cupressus,
Thuja, Taxodium tragen Zäpfchen und führen ihre Samenanlage auf
einer kaum abgegliederten Wucherung der Zapfenschuppen.

Fig. 601. Juniperus communis. 2⁄3 nat. Gr. — Zweig mit
Früchten und Zweig mit männlichen Blüten. — Offizinell. A
Männliche Blüte, B fertiler Zweig mit weiblicher Blüte, C weibliche
Blüte, ein Fruchtblatt zurückgeschlagen, D Frucht nach BERG
und SCHMIDT.
Juniperus communis, der Wacholder, ist ein auf der ganzen
nördlichen Hemisphäre verbreiteter, stellenweise sehr häufiger
Strauch. Juniperus sabina, der Sadebaum (Fig. 602), findet sich
in den Alpen und sonstigen mittel- und südeuropäischen Gebirgen als
niederliegender Strauch. Die Zypresse, Cupressus sempervirens,
ist im Mittelmeergebiet zu Hause. Thuja-Arten, Lebensbäume
genannt, werden als Ziersträucher und Bäume bei uns angepflanzt.
Das sommergrüne Taxodium distichum bildet ausgedehnte Sumpfwälder
an den Nordküsten des mexikanischen Golfes von Florida bis
Galveston, während das immergrüne Taxodium mexicanum auf dem
mexikanischen Hochplateau weit verbreitet ist, in zum Teil
mächtigen Exemplaren, wie „dem großen Baum von Tule“, der von
HUMBOLDT auf 4000 Jahre geschätzt, bei 50 m Höhe 44 m
Stammumfang besitzt (Fig. 603).
[S. 515]
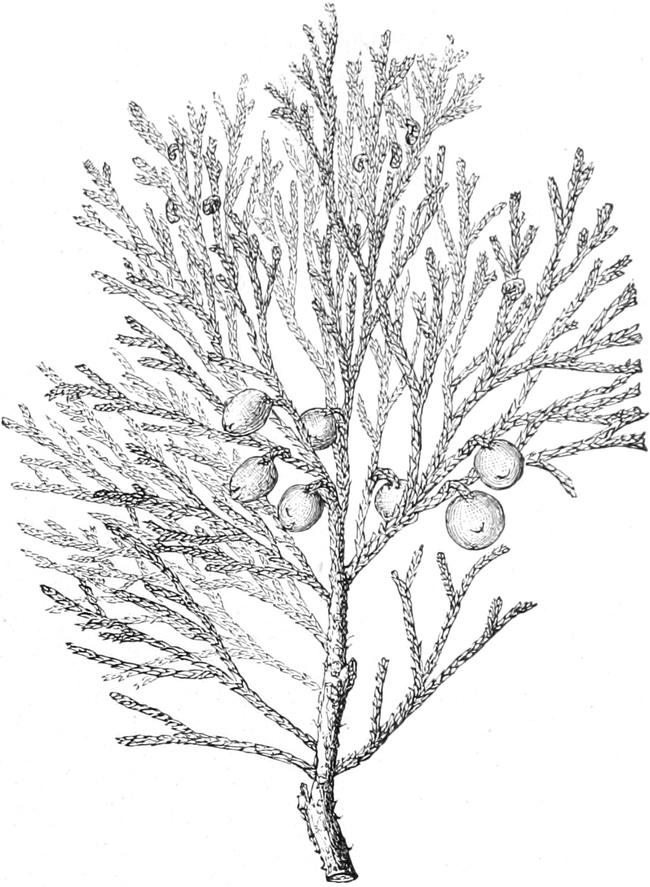
Fig. 602. Zweig von Juniperus Sabina mit Früchten. —
Offizinell und giftig. Gez. H. SCHENCK.

Fig. 603. Taxodium mexicanum auf dem Kirchhofe Sa. Maria
de Tule bei Oaxaka. Der große Baum von Tule, einer der ältesten Bäume
der Erde. Nach einer Photographie.
Unterfamilie Abietineae. Die männlichen Blüten (vgl. S.
479) bestehen lediglich aus einer am Grunde mit Schuppenblättern
besetzten Achse, an der zahlreiche Staubblätter sich finden, die
auf ihrer Unterseite Pollensäcke tragen. Ihre Zahl ist bei den
Abietineen im engeren Sinne auf zwei beschränkt, sie steigt
bei Agathis und Araucaria auf 5–15. Die Mikrosporen haben
meist Flugblasen. Weibliche Blüten stellen durchweg Zapfen
dar, sie bestehen also aus einer Achse und Zapfenschuppen, den
Makrosporophyllen. Bei Agathis und Araucaria trägt jede Schuppe
eine anatrope Samenanlage an der Basis. — Ähnlich liegen
die Verhältnisse bei Sequoia und Sciadopitys, deren Schuppen
aber stets mehrere (4–9) anatrope Samenanlagen auf einem
sich schärfer abhebenden Auswuchs tragen. Bei den Abietineen im
engeren Sinne ist die Trennung dieses Auswuchses von der Schuppe
weiter durchgeführt; man findet eine zweite, mit der Zapfenschuppe
am Grunde verbundene innere Schuppe, welche die beiden, stets
anatropen Samenanlagen trägt. Die äußere Schuppe heißt
Deckschuppe, die innere Fruchtschuppe (Fig. 604 C, 605 5).
Die Fruchtschuppe wird erheblich stärker ausgebildet. Sie ist der
verholzende, die Samenanlage schützende Teil. Schon zur Zeit der
Blüte wird meist die Deckschuppe von[S. 516] der Fruchtschuppe überdeckt
und erst bei genauer Untersuchung kenntlich. Doch gibt es einige
Arten, z. B. Abies-Arten (Fig. 604), Pseudotsuga Douglasi
usw., welche die Deckschuppe stets und in jedem Alter des Zapfens
zwischen den Fruchtschuppen deutlich hervortreten lassen.
Wichtige Gattungen und Arten: Agathis (Dammara) ist im
Malayischen Archipel und bis nach Neuseeland hinüber verbreitet.
Agathis australis und A. Dammara liefern Kauri-Kopal, jedoch
nicht das Dammaraharz. Araucaria brasiliana und A. imbricata
sind stattliche Waldbäume Südamerikas. Zur Gattung Sequoia
gehören die gewaltigen Baumgestalten der Mammutbäume aus der
kalifornischen Sierra Nevada, Sequoia gigantea, deren Stämme
über 100 m Höhe und 12 m Durchmesser erreichen, und die schöne,
jener an Höhe und Dicke kaum nachstehende S. sempervirens der
Küstengebirge.
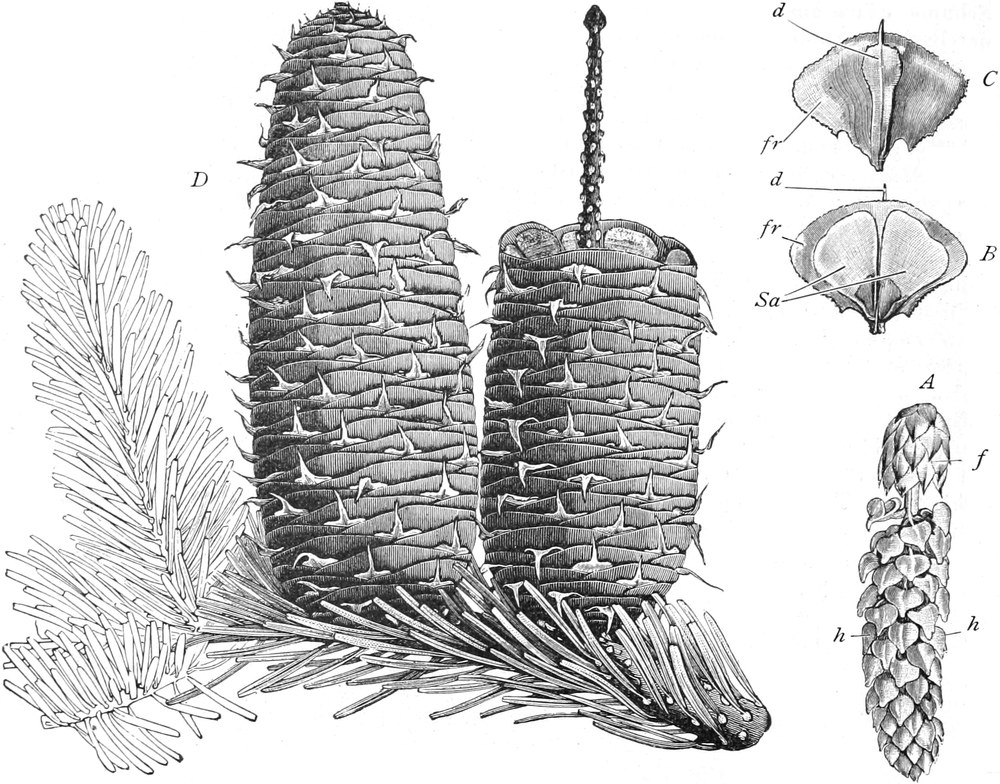
Fig. 604. Abies pectinata. A Männliche Blüte, f
Schuppen, h Sporophylle. B Deck- d und Fruchtschuppe fr
von oben, C dieselben von unten. Sa Geflügelte Samen. Nach
BERG und SCHMIDT. Offizinell. D Abies
Nordmanniana mit reifen, zum Teil zerfallenen Zapfen, verkl. aus
ENGLER-PRANTL.
Heimische Waldbäume: Die Edeltanne (Fig. 604 A–C),
Abies pectinata, ihrer hellen Rinde wegen auch Weißtanne
genannt, ist in den mittel- und südeuropäischen Gebirgen zu Hause.
Der Baum führt in der vegetativen Region nur Langtriebe. Flache,
unterseits mit zwei weißen Streifen versehene Nadeln, deren
Spitze ausgerandet ist, stehen mehr oder weniger allseitig an
dem Zweige, werden aber an den wagerechten, von oben belichteten
Seitenästen durch Drehung ihres Grundes gescheitelt. Sie bleiben
6–8 Jahre erhalten, sollen bisweilen sogar 15 Jahre alt werden.
Männliche Blüten entstehen achselständig auf der Unterseite oder
den Flanken des Sprosses. Sie wachsen abwärts, so daß ihre der
Sporophyll-Unterseite eingesenkten Pollensäcke aufwärts schauen.
Durch eine schräge Längsspalte geöffnet, klafft die Wandung weit
auseinander, und die Mikrosporen können leicht herausfallen.
Weibliche Blüten entspringen oberseits und sind steil aufgerichtet.
Die Deckschuppe ist hier etwas länger als die viel breitere
Fruchtschuppe. Befruchtete Zapfen behalten ihre aufrechte
Stellung; die[S. 517] Schuppen fallen bei der Reife von der Spindel
ab, wobei die Samen frei werden. Ihre Ausbildung erfordert ein
Jahr. Abies Nordmanniana aus dem Kaukasus (Fig. 604 D), A.
concolor, A. balsamea, A. nobilis, alle drei aus Nordamerika,
sind in Parkanlagen anzutreffen.
Picea excelsa, die Fichte (oder Rottanne) (Fig.
605), eine stattliche pyramidale Baumgestalt unserer Wälder.
Kurztriebe fehlen. Langtriebe allseitig mit vierkantigen spitzen
Nadeln besetzt, die 5–7 Jahre, an Haupttrieben bis 12 Jahre alt
werden. Männliche Blüten meist an den Flanken vorjähriger Triebe.
Sie krümmen sich zur Blütezeit aufwärts. Beide Pollensäcke öffnen
sich durch einen Längsriß. Weibliche Zapfen stehen endständig
an vorjährigen Trieben desselben Individuums, meist dem Gipfel
genähert; sie sind zur Blütezeit gerade aufgerichtet.
Die reifen Zapfen hängen und entlassen die Samen zwischen
den geöffneten Schuppen hindurch, fallen dann als Ganzes ab;
ebenso verhält sich Tsuga canadensis, die „Hemlokstanne“. Die
Samenentwicklung ist einjährig. Picea orientalis aus Kleinasien,
die serbische Picea omorica und Picea alba aus Nordamerika,
ferner Picea engelmannii und Picea pungens; beide als
„Blautannen“ bekannt, werden häufig angepflanzt.
Larix europaea, die Lärche (Fig. 607), gehört zu den
wenigen laubabwerfenden Koniferen. Sie erneuert ihre zarten,
kaum xerophilen Nadeln jährlich. Ihre Triebe sind in Langtriebe
und Kurztriebe gegliedert. Erstere, allseitig mit schmalen,
linealen Laubblättern bestanden, setzen die Verzweigung des
pyramidenförmigen Baumes, dessen Seitenzweige oft tief herabhängen,
fort. Kurztriebe entstehen in den Blattachseln an vorjährigen
Langtrieben; sie bestehen aus einer Rosette von 30 bis 40
Blättern, denen der Langtriebe ähnlich, nur etwas kürzer. Die
Blüten entsprechen in ihrer Stellung einem Kurztriebe. Männliche
Blüten sind nach völliger Entwicklung abwärts gekrümmt, sie
öffnen ihre aufwärts schauenden Pollensäcke wie bei Abies. Die
weiblichen Zapfen stehen aufgerichtet, sie reifen in einem Jahr.
Cedrus-Arten, immergrüne Waldbäume des Atlas, Libanon und
Himalaya, sind in West- und Süddeutschland in Anlagen anzutreffen.
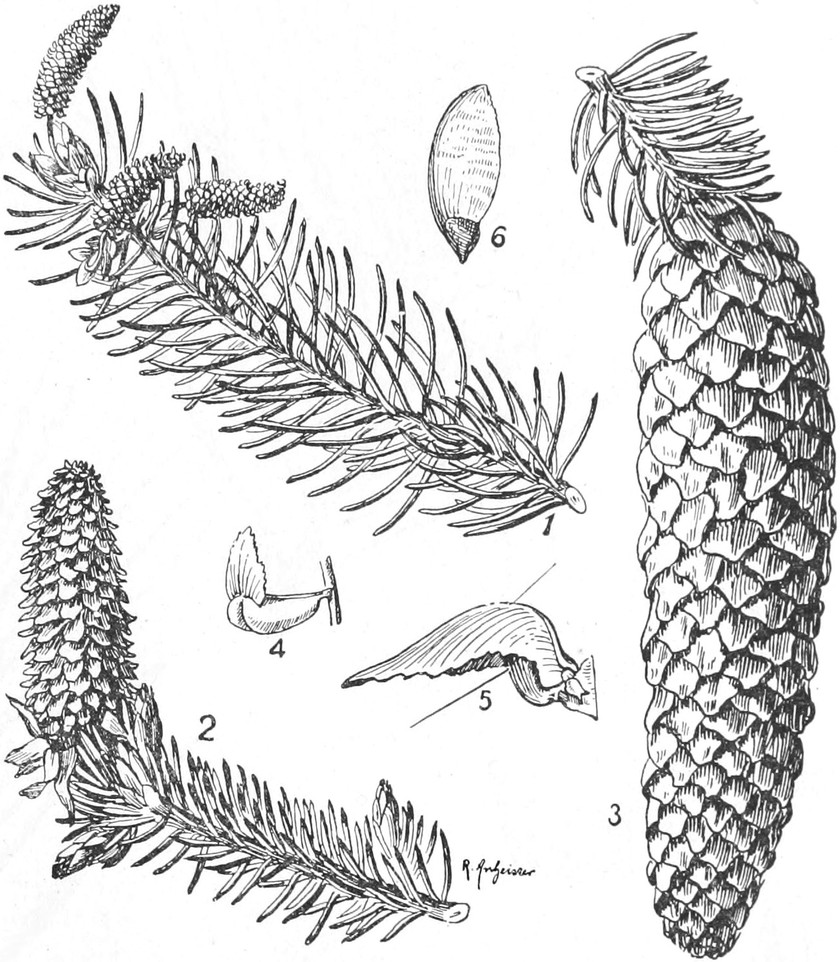
Fig. 605. Picea excelsa. 1⁄2 nat. Gr. 1 Zweig mit
männlichen Blüten. 2 Endständige weibliche Blüte. 3 Hängender
Fruchtzapfen. 4 Mikrosporophyll. 5 Makrosporophyll, Deckschuppe
d von der nach außen umgeschlagenen größeren Fruchtschuppe fr
überdeckt, eine Samenanlage am Grunde der letzteren. 6 Reifer Samen
mit Flughaut aus einer abgelösten Lamelle der Fruchtschuppe gebildet.
4–6 vergr. — Offizinell.
[S. 518]
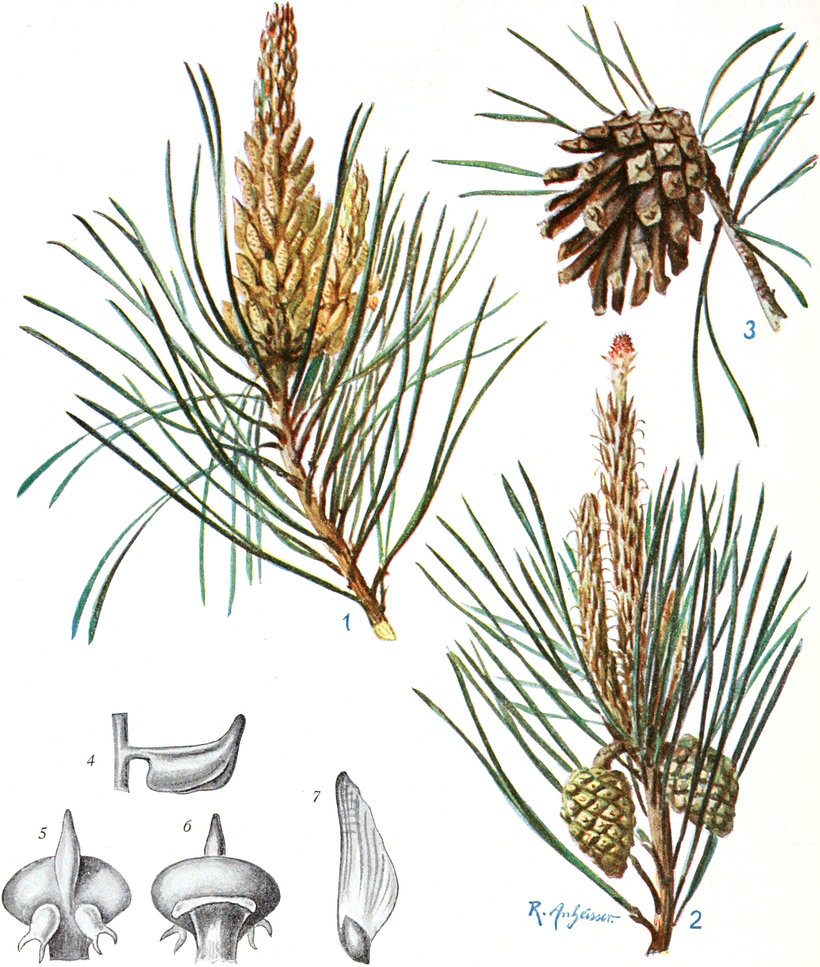
Fig. 606. Pinus silvestris. 2⁄3 nat. Gr. 1 Neuer
Jahrestrieb am Gipfel eines mit zahlreichen Kurztrieben besetzten
Langtriebes. Am Grunde des jungen Langtriebes viele männliche
Blüten je an Stelle eines Kurztriebes; weiter oben bräunliche
Schuppenblätter mit je einer Kurztriebanlage in der Achsel. 2
Ebenso, jedoch eine weibliche aufgerichtete Blüte am Gipfel an
Stelle eines Langtriebes. Zwei vorjährige, unreife, grüne Zapfen,
umgebogen, am Gipfel des vorjährigen Langtriebes. 3 Zapfen des
vorvorigen Jahres aufgesprungen, um die reifen Samen zu entlassen.
4 Ein Mikrosporophyll. 5 Makrosporophyll von der Oberseite, zeigt
Fruchtschuppe mit zwei Samenanlagen am Grunde. 6 Dasselbe von der
Unterseite, die kleine Deckschuppe unter der Fruchtschuppe freigelegt.
7 Reifer Samen mit dem von den Oberflächenschichten der Fruchtschuppe
abgespaltenen Flügel. 4–7 vergr. Offizinell.
Die höchste Differenzierung der vegetativen Organe ist in der
Gattung Pinus zu finden. Pinus silvestris, unsere gewöhnliche
Kiefer (Fig. 606), mag als Beispiel dienen. Junge, 1
bis 2jährige Keimpflanzen führen nach Verlust der zahlreichen
Keimblätter (Fig. 592) regelmäßig benadelte Langtriebe. An den
älteren Exemplaren geht diese Benadelung völlig verloren und wird
durch bräunliche Schuppenblätter ersetzt, in deren Achseln die
von häutigen Niederblättern umscheideten zweinadeligen Kurztriebe
stehen[S. 519] (vgl. Figurenerkl. 606). Die Nadeln fallen im 3. Jahre
ab. Die Samen reifen im 2. Jahre. Sie werden durch Öffnung
ihrer bis dahin fest aufeinander gepreßten Fruchtschuppen frei.
Der Zapfen fällt dann als Ganzes vom Baume. Pinus montana,
Zwergkiefer unserer Gebirge. P. Laricio, die Schwarzkiefer, aus
Niederösterreich. P. Pinea, Pinie, und P. Cembra, Arve, beide,
wie auch P. Lambertiana, Zuckerkiefer aus Nordamerika mit eßbaren
Samen. Arve, Zuckerkiefer und die amerikanische P. Strobus,
Weymouthskiefer mit fünfblätterigen Kurztrieben.
Giftig: Juniperus sabina, der Sadebaum und Taxus
baccata, die Eibe.
Offizinell: Juniperus communis: Fructus Juniperi
(Pharm. germ., austr., helv.). Oleum Juniperi (Pharm. germ.,
austr., helv.) und Lignum Juniperi (Pharm. austr., helv.),
Juniperus oxycedrus: Oleum cadinum (Pharm. helv.). —
Juniperus Sabina: Herba Sabinae (Pharm. austr., helv.).
— Larix europaea: Terebinthina veneta (Pharm. helv.).
— Larix sibirica (Nordrußland, Sibirien): Pix liquida
(Pharm. germ.). Verschiedene Pinus-Arten, wie P. silvestris,
Laricio, Pinaster, Taeda, australis, cubensis
usw., wie auch Abies pectinata und Picea excelsa liefern
Terebinthina, Kolophonium, Ol. Terebinthina,
Pix liquida (Pharm. germ., austr., helv.); Pinus montana
liefert Ol. Pini Pumilionis (Pharm. austr., helv.). —
Pinus silvestris: Turio Pini (Pharm. helv.). — Callitris
quadrivalvis: Sandaraca (Pharm. austr.).
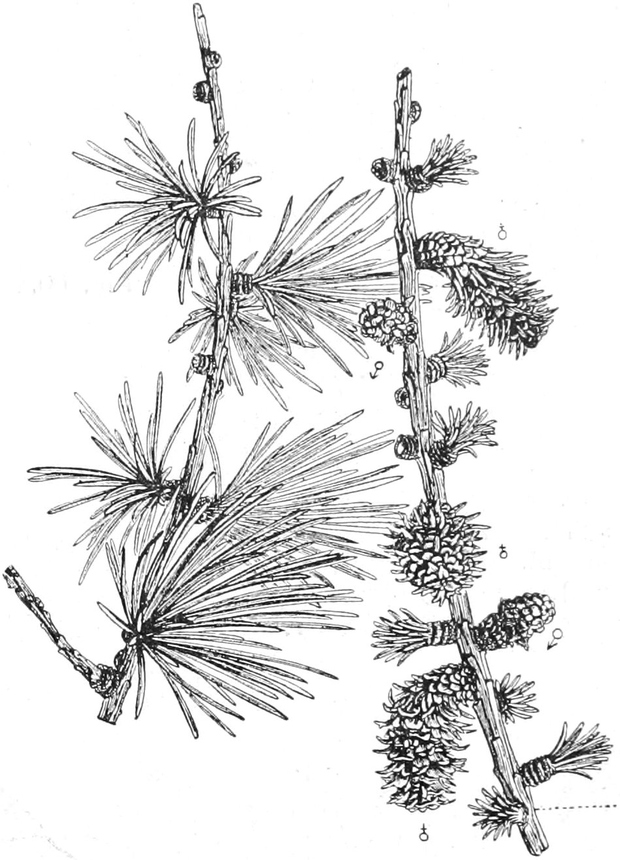
Fig. 607. Larix europaea. Vorjährige Langtriebe, rechts
vegetative Kurztriebe tragend, links männliche und weibliche Blüten an
ihrer Stelle. Offizinell. Aus ENGLER-PRANTL.
Die einzige Familie der 4. Ordnung, Gnetinae, ist die der
Gnetaceae. Nur drei Gattungen gehören ihr an: Ephedra (Fig.
608), blattlose Sträucher wärmerer, trockener Gegenden, vorwiegend
der nördlichen Hemisphäre, Welwitschia (Fig. 609) mit der
einzigen Art W. mirabilis aus den Wüsten Südwestafrikas, welche
außer den hinfälligen Kotyledonen zeitlebens nur ein einziges Paar
meterlanger, am Grunde fortwährend nachwachsender Blätter an dem
nur gerade über die Erdoberfläche ragenden, keulig angeschwollenen
Stammscheitel hervorbringt, und Gnetum (Fig. 611) mit paarig
gestellten, breiten, netzadrigen Laubblättern versehene Bäume und
Lianen der Tropen. Diese so verschieden aussehenden Gattungen
stimmen überein in dem Besitze gegenständiger Blätter (bei Ephedra
auf Schuppen beschränkt), in der Entwicklung echter Tracheen im
sekundären Holz und Fehlen der Harzgänge, endlich in dem Auftreten
einer Hülle in den meist diözisch verteilten Blüten (Fig. 610).
Der Besitz dieser Blütenhülle verbietet es, die Gnetaceenzapfen
als Blüten anzusprechen; es liegen hier also Infloreszenzen vor,
und die Gnetaceenzapfen wären den Koniferenzapfen nicht homolog zu
setzen. Wegen naher Beziehungen ihrer Entwicklung sowohl zu den
Gymnospermen wie zu den Angiospermen ist die Familie geeignet,
den Übergang zu vermitteln. Auch ist bei allen drei Gattungen
Insektenbesuch der Blüten beobachtet worden, der zur Zeit freilich
nur bei Ephedra campylopoda zur Bestäubung zu führen scheint.
Über die Entwicklung der Geschlechtsgeneration vgl. S. 492.
Die fossilen Gymnospermen[473].
Reste von Gymnospermen sind, im Gegensatz zu Pteridophyten, in
den ältesten paläozoischen Schichten, dem Kambrium und Silur,
bisher nicht gefunden; sie treten zuerst im Devon auf, jedoch nur
spurenweise, und erreichen erst in der Steinkohlenflora größere
Bedeutung. Von den Cycadofilices, Stämmen mit sekundärem
Dickenwachstum und farnähnlicher[S. 520] Belaubung, welche bisher immer
den Pteridophyten zugerechnet wurden, trennen OLIVER und
SCOTT neuerdings die Pteridospermeae ab, die sie kurz
als farnähnliche Samenpflanzen charakterisieren. Diese sind
im Anschluß an die Pteridophyten S. 461 behandelt.
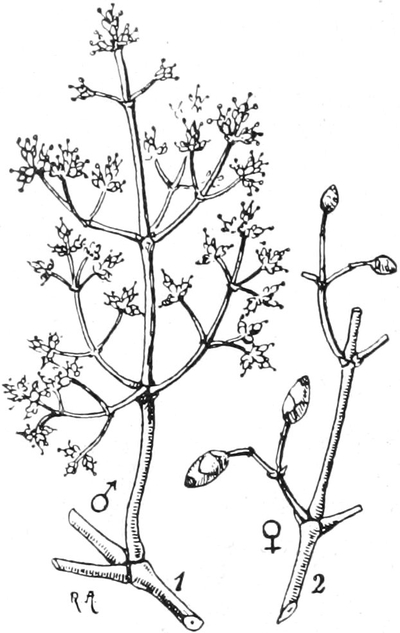
Fig. 608. Ephedra altissima. 1 Habitus eines
männlichen Blütenstandes und 2 eines unreifen Fruchtstandes.
2⁄3 nat. Gr.
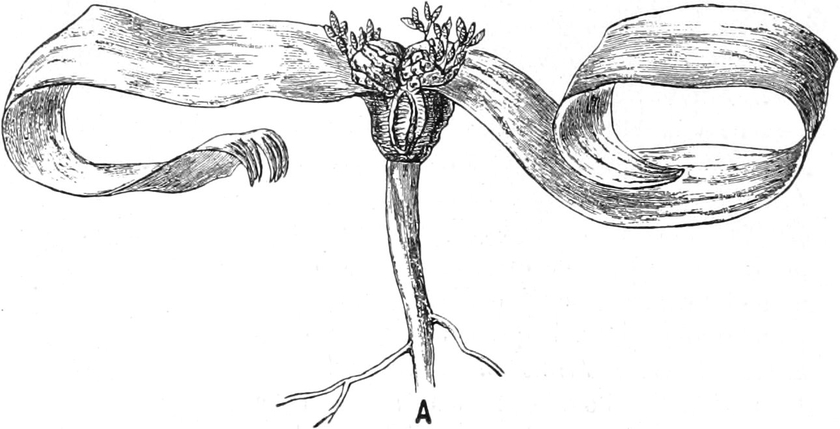
Fig. 609. Welwitschia mirabilis. Jüngere Pflanze nach
ENGLER-PRANTL, stark verkleinert.
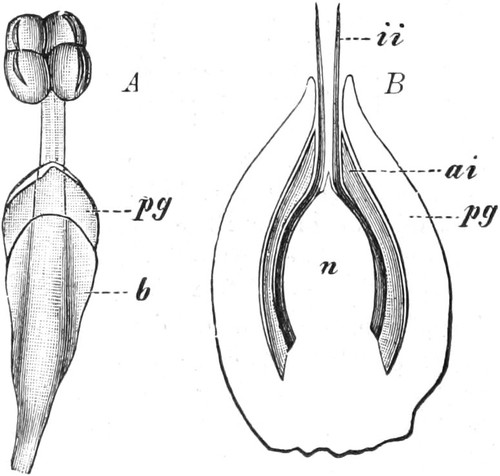
Fig. 610. A Ephedra altissima. Männliche Blüte. Vergr.
16. pg Perigon, b Blatt. Nach E. STRASBURGER. — B
Längsschnitt durch eine weibliche Blüte von Gnetum gnemon. Nach J.
P. LOTSY. Vergr. 32. n Nucellus, ii inneres, ai äußeres
Integument, pg eine weitere integumentartige Hülle, die als Perigon
gedeutet worden ist.
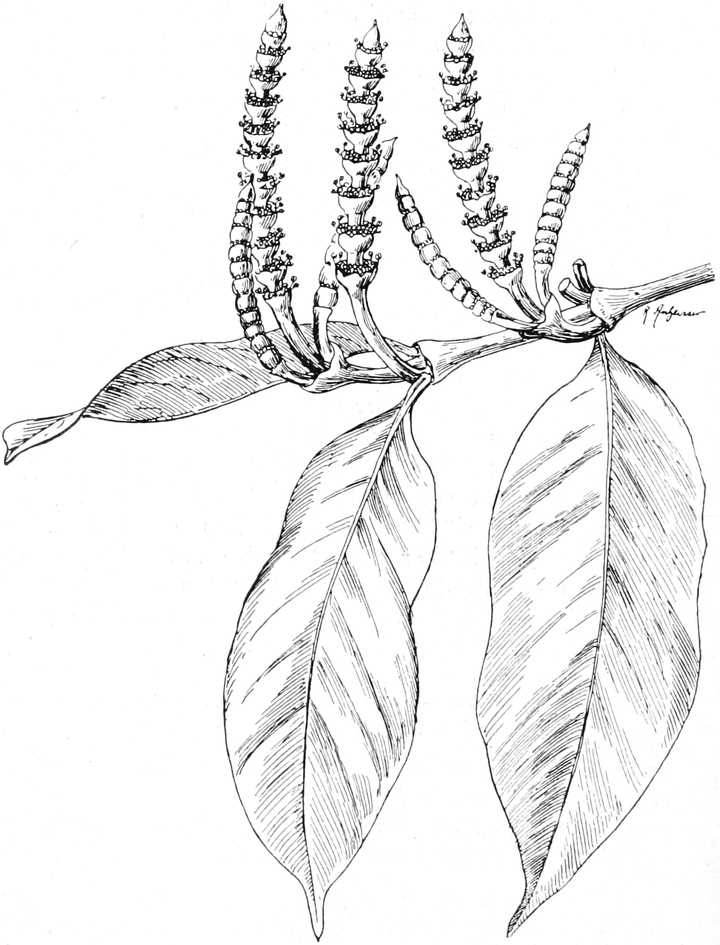
Fig. 611. Gnetum gnemon. Zweig mit männlichen
Blütenständen aus zahlreichen akropetal aufsteigenden Wirteln
bestehend, deren jeder über den in Spiralen angeordneten männlichen
Blüten eine Reihe steriler weiblicher Blüten trägt.
1⁄2 nat. Gr.
Cordaitaceae. Ein auf die paläozoischen Epochen beschränkter,
höchst eigenartiger Typus ist Cordaites. Dank der vorzüglichen
Erhaltung ist Cordaites morphologisch fast ebenso genau bekannt,
wie die jetzt lebenden Gymnospermen. Es waren hohe verzweigte Bäume
mit handförmig schmalen oder breiteren, ganzrandigen oder wenig
gelappten parallelnervigen Blättern, die am Zweigende schopfig
gehäuft stehen, und mit Blüten, die von[S. 521] denen der jetzigen
Gymnospermen sehr abweichen. Männliche wie weibliche Blüten sind
je in ährenförmigen, achselständigen Blütenständen zu mehreren
vereinigt. Die weiblichen Blüten bestehen lediglich aus einer
atropen Samenanlage, die das in der Achsel eines Hochblattes
befindliche Fruchtblatt aufbraucht. Die Hochblätter gleichen
den vegetativen Laubblättern (Fig. 612, 3, 4). Am Scheitel des
Nucellus ist eine tiefe Pollenkammer eingesenkt, in der vielfach
Pollenkörner angetroffen werden. Die männlichen Blüten schließen
kleine Sprosse ab, die von zahlreichen sterilen Hochblättern
umhüllt sind und am Vegetationspunkt nacheinander zahlreiche mit
2–4 Antheren gekrönte Staubblätter hervorbringen (Fig. 692, 1,
2). Phylogenetisch wichtig ist der Umstand, daß das männliche
Prothallium einen mehrzelligen Gewebekörper darstellt. Die
Struktur der Samenanlagen und der Samen zeigt ebenfalls große
Ähnlichkeit mit Cycas. Neben wenig zahlreichen Resten (Cycadites,
Dicranophyllum), welche man in dieselbe Verwandtschaft
rechnen mag, stellt Cordaites im ganzen Karbon den am reichsten
entwickelten Gymnospermentypus dar. Erst im unteren Rotliegenden
zeigen sich zweifellose Cycadophyten.
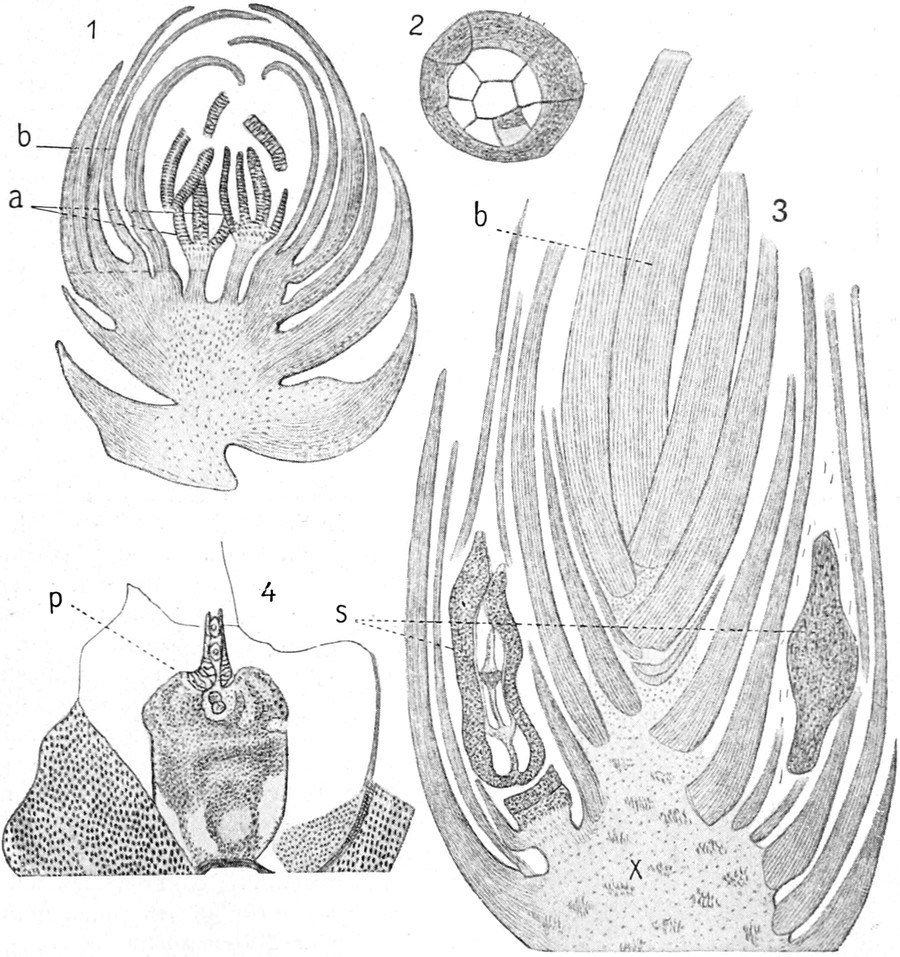
Fig. 612. 1. Cordaites subglomeratus. Längsschliff
einer männlichen Blütenknospe. b Hüllblätter. a Staubblätter mit
mehreren Antheren. 2. Ein Pollenkorn. Die Prothalliumzelle durch
gebogene Wand abgeteilt; der Rest des Kornes in zahlreiche Zellen
zerlegt. 3. C. Williamsoni. Längsschliff eines Fruchtsprosses. b
Blätter, s Samenlängsschliff. 4. G. Grand’Euryi. Längsschliff durch
eine Samenanlage mit tiefer Pollenkammer im Nucellus, die mehrere
Pollenkörner enthält. Nach B. RENAULT.
Mit dem Beginn der mesozoischen Schichten schwinden die Cordaiten.
Die Gymnospermenflora geht mit Cycadophyten, Ginkgoinen und
Koniferen erloschener Typen durch die Trias hindurch und findet im
Jura eine mächtige Entwicklung. Die Ginkgoinen und die Cycadophyten
erreichen hier ihren Höhepunkt.
Bennettitaceae. Über das Aussehen und die hohe Entwicklungsstufe
mesozoischer Cycadophyten berichtet auf Grund reichen, in
Nordamerika gefundenen und von WIELAND[S. 522] bearbeiteten
Materiales SCOTT. Es handelt sich um Bennettites-Arten,
deren aus Europa früher bekannt gewordene Früchte hermaphrodite
Blüten bereits hatten vermuten lassen; die von dem amerikanischen
Autor angewandte Benennung Cycadeoidea ist also mit Bennettites
synonym. Niedrige, zum Teil verzweigte Stämme, im Aussehen
und der Beblätterung den lebenden Cycadaceen ähnlich, tragen
12 cm lange hermaphrodite Blüten. Hundert oder mehr spiralig
angeordnete Perianthblätter umschließen einen Wirtel von 18–20
Mikrosporophyllen, die am Grunde zu einer tief ausgehöhlten
Schüssel verwachsen sind, in deren Mitte sich das Gynäceum erhebt
(Fig. 613). Die gefiederten, 10 cm langen Mikrosporophylle erinnern
an Farnblätter, ebenso ihre Mikrosporangien an die Sporangien der
Marattiaceen. Das Gynäceum besteht aus zahlreichen, langgestielten,
atropen Samenanlagen, die von Schuppenblättern umhüllt und durch
nach oben stark verdickte Wände voneinander getrennt werden, jedoch
die Mikropyle frei nach außen münden lassen. Die reifen Samen
enthalten einen hochentwickelten dikotylen Embryo und entbehren
des Endosperms; sie werden von den an ihren äußeren Enden sich
zusammenfügenden Schuppen wie von einem Fruchtknoten eingeschlossen
(Fig. 614). Wie die paläozoischen Pteridospermeen Charaktere der
Farne und Gymnospermen in sich vereinigen, so finden sich in den
mesozoischen Bennettites- bzw. Cycadeoidea-Blüten solche
der Angiospermen mit denen von Gymnospermen und Farnen zugleich
verbunden.
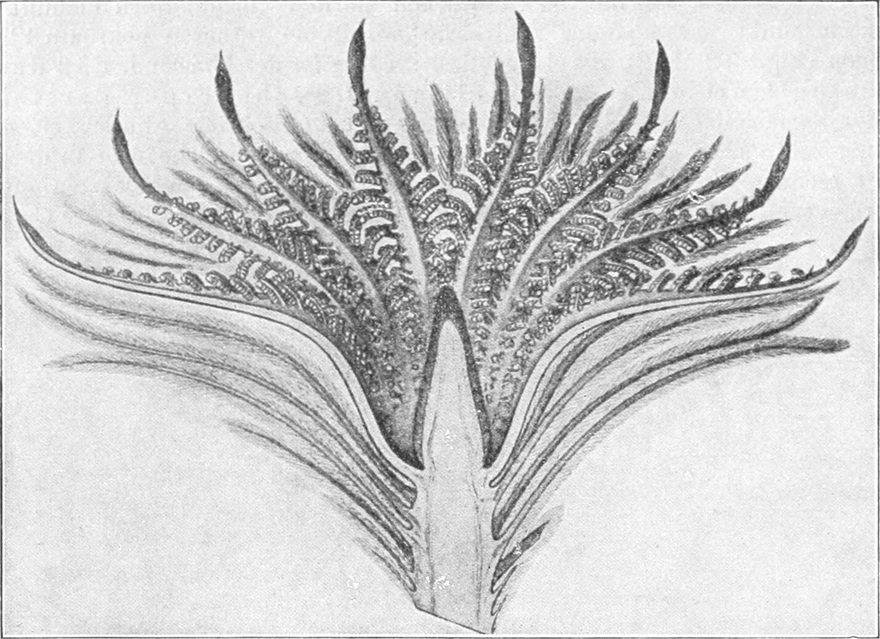
Fig. 613. Rekonstruierter Blütenlängsschnitt von
Cycadeoidea (Bennettites) ingens nach G. R. WIELAND aus D.
H. SCOTT.
Im Jura zeigen sich echte Araucarien, welche zu den älteren
Koniferen gehören. Im Wealden herrschen unter den Gymnospermen
noch die Cycadophyten und Ginkgoinen mit einigen Koniferen; in der
eigentlichen Kreide treten die altertümlichen Typen immer mehr
zurück, während die Koniferen immer zahlreicher werden. Unter ihnen
zeigen sich bereits jetzt noch lebende Gattungen, wie Dammara,
Sequoia, Pinus, Cedrus, Abies, Callitris usw. Auch die
Taxaceen scheinen vertreten zu sein, doch ist die Zugehörigkeit
der Reste zweifelhaft.
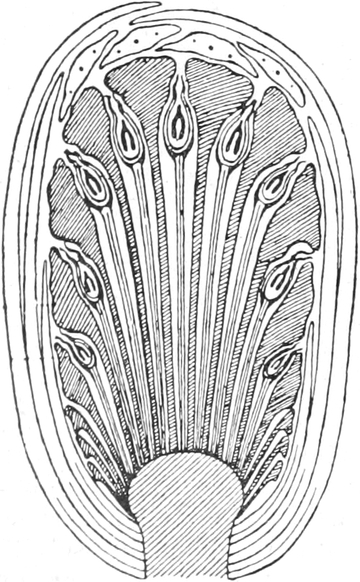
Fig. 614. Längsschliff durch eine Frucht von Bennettites
Gibsonianus nach D. H. SCOTT.
Die Gymnospermen des Tertiärs gehören durchaus noch lebenden
Typen, zum großen Teil noch lebenden Arten an. Koniferen sind
vorherrschend; von Ginkgoinen ist nur Ginkgo biloba vorhanden,
und zwar auch in Europa, zusammen mit anderen jetzt auf[S. 523] Ostasien
oder Nordamerika beschränkten Arten, wie Cryptomeria japonica,
Taxodium distichum, Sequoia gigantea und sempervirens, Pinus
strobus usw. Auch eine Angehörige der Cycadaceae (Encephalartos)
ist gefunden.
II. Klasse. Angiospermae[474].
Bei den Angiospermen ist die lange strittige Frage, ob die
Monokotyledonen oder die Dikotyledonen voranzustellen seien,
zur Zeit wohl dahin entschieden, daß man die Monokotyledonen
von den Polycarpicae unter den Dikotyledonen ableitet, die
in Blütenorganisation, anatomischem Aufbau und verschiedenen
morphologischen Merkmalen mit monokotylen Gewächsen übereinstimmen.
So wird man jetzt die Dikotylen in der systematischen Reihenfolge
voranstellen und die Monokotylen ihnen folgen lassen.
Dafür ist außerdem entscheidend, daß ein Übergang von den Gymnospermen
direkt zu den Monokotylen völlig ausgeschlossen erscheint, daß dagegen
eine Anknüpfung dikotyler Pflanzen an die Gymnospermen nicht ganz
aussichtslos sein dürfte. Wie sich in der Entwicklung der männlichen
und weiblichen Organe Parallelen und Weiterbildungen zwischen
Gymnospermen und Angiospermen erkennen lassen, ist vorher S. 486 f.
dargestellt; doch auch in der Ausgestaltung der ganzen Blüten ergeben
sich Ableitungsmöglichkeiten.
Der Versuch WETTSTEINs, die einfachsten Angiospermenblüten,
etwa diejenigen der Gattung Casuarina, von den ebenso einfachen
Infloreszenzen von Ephedra abzuleiten, leidet daran, daß, wie
wohl immer mehr die Überzeugung durchdringt, in den „einfachen“
Angiospermenblüten durchweg reduzierte, nicht aufsteigende Reihen
vorliegen. So kann daran nicht mehr angeknüpft werden, trotz mancher
verlockend erscheinenden Einzelheiten, wie Insektenbestäubung bei
Ephedra.
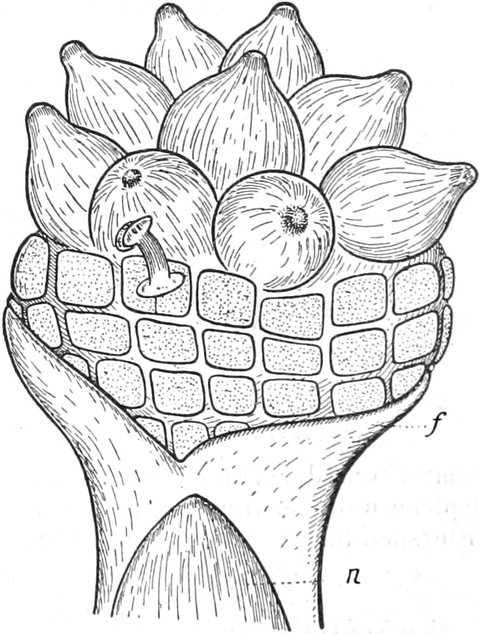
Fig. 615. Androgyne Infloreszenz von Gnetum hypothetisch
auf einen Wirtel beschränkt (vgl.
Fig. 611,
S. 520). Seitenansicht
halb von oben. Sie beginnt mit einem sterilen Brakteenpaar
n, zu
dem das einzige fertile Paar
f dekussiert steht. Gez. von N.
P
ATSCHOVSKY.
Nach Ablehnung dieses Ableitungsversuches wird man sich an die
Polycarpicae erinnern müssen, von denen ja zweifelsohne die Reihe der
Monokotyledonen abgeleitet werden konnte. Zunächst ist der spiralige
Blütenaufbau der typischen Polycarpicae ein starker Hinweis darauf,
daß hier die Verbindung zu den spiraligen Zapfenblüten der Koniferen
liegen müsse. Die stets monözische Koniferenblüte könnte man sich mit
Hilfe der androgynen Blütenstände von Gnetum etwa in die hermaphrodite
Anordnung der angiospermen Polycarpicaeblüte übergeführt denken,
wie Fig. 615 dies zu veranschaulichen sucht. Sie bietet direkt das
Vorbild einer Polycarpicaeblüte mit zahlreichen spiralig stehenden
Staubblättern und einigen apokarpen Fruchtblättern, ohne daß daraus
auf die phylogenetischen Beziehungen geschlossen werden soll. Eine
wesentliche Stütze dieser Anschauung wurde durch blütenbiologische
Befunde von DIELS[475] gegeben, der nachweisen konnte, daß
ebenso wie gewisse südafrikanische Encephalartos-Arten auch einige zu
den Polycarpicae gehörige Pflanzen durch Käfer bestäubt werden. Da nun
die Coleopteren die phylogenetisch ältesten blütenbesuchenden Insekten,
an der ältesten lebenden Gymnospermenfamilie[S. 524] als Bestäuber auftreten,
läßt sich auf ähnliches Alter einiger ebenfalls von Käfern bestäubter
Polycarpicae schließen.
1. Unterklasse. Dicotylae.
Die Dikotylen sind bis auf vereinzelte Ausnahmen mit zwei Keimblättern
versehen. Die Unterschiede epigäischer und hypogäischer Keimung sind S.
507 f. besprochen.
Der Stamm besitzt in der Regel kreisförmig angeordnete offene
Leitbündel (vgl. S. 123 u. Fig. 165), die Wurzel abwechselnd
gelagerte Gefäß- und Siebteile auf dem Querschnitte. Das in den
Leitbündeln des Stammes und auf der Innenseite der Siebteile der Wurzel
enthaltene Meristem wird bald zu einem geschlossenen Ringe ergänzt,
der als Kambium ein regelrechtes Dickenwachstum der Stämme und Wurzeln
vermittelt.
Das typische Dikotylenblatt ist mit mehr oder minder langem
Stiel versehen, es besitzt häufig Nebenblätter als Auszweigungen
des Blattgrundes, entbehrt aber meist einer Scheide (Ausnahme
Umbelliferen). Seine Spreite ist einfach oder zusammengesetzt; ihre
Gliederung kommt nur durch seitliche Verzweigung der Blattanlage
zustande. Der Blattrand ist von sehr verschiedenartiger Form, die
Nervatur in der Regel netzartig (Fig. 128, S. 95).
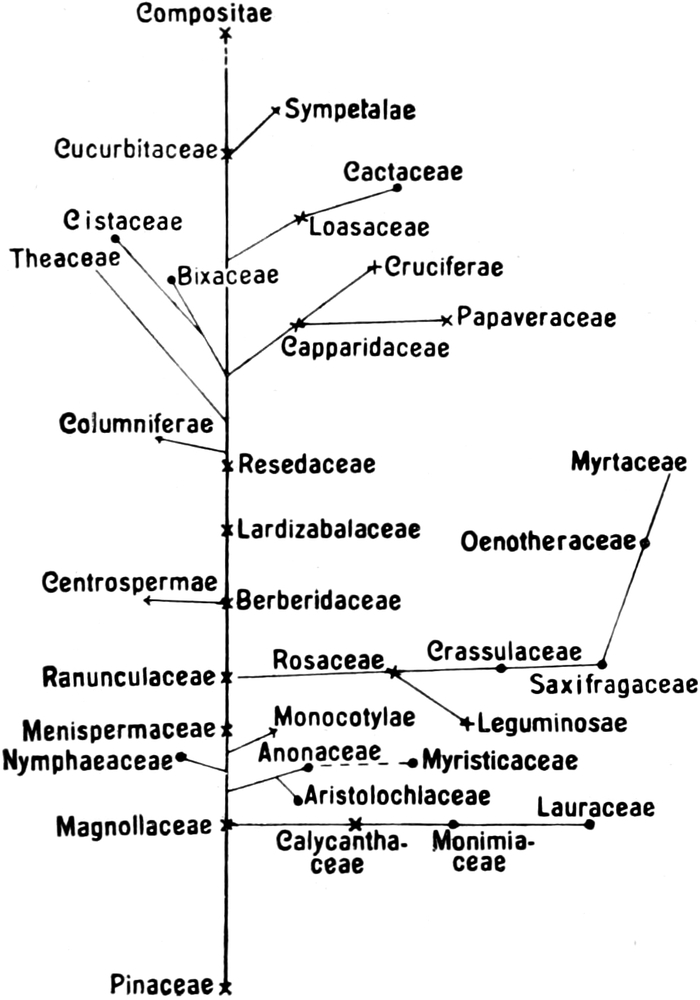
Stammbaum der Dikotylen, besonders der Polycarpicae und Verwandten,
nach serodiagnostischen Untersuchungen von CARL MEZ
(modifiziert).
Die Dikotylenblüten sind in den typischen Fällen fünfzählig und
fünfwirtelig, doch finden sich abweichende Formen in großer Menge. Sie
entsprechen in regelmäßig gebauten Vertretern der Formel K 5 C 5 A5 + 5
G 5.
Neben diesen morphologischen und biologischen Gesichtspunkten
erfordert aber auch die von der Königsberger Schule durchgeführte
serodiagnostische Methode[476] eine Berücksichtigung, die von sich
behauptet, daß sie sich „nicht auf unsichere und den Einflüssen
der Außenwelt direkt ausgesetzte, deshalb eventuell konvergent
ausgebildete Eigenschaften der Lebewesen bezieht, sondern auf
wirkliche Verwandtschaftsverhältnisse, nämlich auf die chemischen
Verwandtschaften der Eiweißstoffe“. Diese Methode beruht auf
den Erfahrungen der Immunitätslehre. Wenn im tierischen
Blut, oder besser Serum, durch gewisse eingeführte giftige
Eiweißstoffe automatisch antitoxisch wirkende Eiweißverbindungen oder
Antigene gebildet werden und diese Antigene innerhalb größerer
Verwandtschaftsreihen bei gleichem Eingriff die gleichen sind, so wird
man auch umgekehrt aus einer Antigengleichheit auf eine Verwandtschaft
der betreffenden eingeführten Eiweißkörper zurückschließen dürfen. Und
wenn es auf diese Weise gelungen ist, die Eiweißstoffe verschiedener
Vogeleier voneinander zu unterscheiden und die Blutsverwandtschaft
der Menschen mit den Menschenaffen serodiagnostisch festgestellt
werden konnte, so müßte[S. 525] es auch möglich sein, auf gleichem Wege
(durch Tierversuche) die Beziehungen verschiedener Pflanzeneiweiße
zu bestimmen. Wenn dann die Vorsicht beobachtet wird, daß stets
erst beim Gelingen auch des reziproken Versuches das Resultat als
gültig angenommen wird, so ist damit ein gewisser Grad von Sicherheit
erreicht; d. h. also, wenn etwa die Antigene von Pinus und den
Magnoliaceen eine Eiweißgleichheit ergeben, so muß verlangt werden,
daß ebenso das Magnoliaceen-Serum die Abietineen oder Pinusarten
als verwandte Gruppen anzeigt. Wegen aller Einzelheiten muß auf die
einschlägige Literatur verwiesen werden. Wenn wir also den Versuch
machen, die Ergebnisse dieser serodiagnostischen Untersuchungen in
der Form von Stammbäumen dem System unterzulegen, so wird das um so
unbedenklicher dort geschehen können, wo die Morphologie schon vorher
dieselbe Richtung eingeschlagen hatte, während dort, wo das nicht der
Fall ist, die nötige Reserve beobachtet werden muß. Da der Plan dieses
Buches eine Berücksichtigung nur der offizinell und wissenschaftlich
wichtigen Familien vorsieht, so sollen die vollständig mitgeteilten
serodiagnostischen Stammbäume nur dazu dienen, das vorliegende Material
gleichsam als Gerippe zu geben, das der fraglichen Familie ihren
Standplatz anweist.
A. Choripetalae
(getrennt blättrige Blumenkrone).
1. Ordnung.
Polycarpicae.
Hermaphrodite, meist lebhaft gefärbte Blüten. Eine starke Verlängerung
der Blütenachse mit spiralig daran aufgereihten freien einzelnen
Blütenteilen des Perianths, der Staubblätter und der apokarpen
Fruchtblätter, deren Zahl unbestimmt und sehr erheblich sein kann,
zeichnet die typischen Vertreter der Polycarpicae aus. Die Sonderung
von Kelch und Krone ist vielfach nicht durchgeführt, und in einzelnen
Fällen, wie bei Calycanthus, schließen sogar die Laubblätter mit
spiraliger Stellung direkt an die Blütenhochblätter an. Die Form der
Staubblätter ist häufig blattartig mit einem die Antheren überragenden
Konnektiv oder blattartiger Verbreiterung des Filaments oberhalb der
Anthere. Die Narben bilden das Ende der Fruchtblätter ohne stielartigen
Griffel, Insektenbestäubung (und zwar in einigen primitiveren Formen
durch Käfer) ist allgemein verbreitet. Auch der Aufbau des Holzkörpers
ist primitiv und kommt in einzelnen Fällen dem Koniferenholze nahe.
Zu dieser Ordnung zeigen die einfachsten Formen der Monokotyledonen,
die Helobiae, unverkennbare Verwandtschaft, da sie mit ihnen in
der Vermehrung der Staubblätter und der apokarpen Fruchtblätter gut
übereinstimmen. Außerdem lassen sich zahlreiche Familien unter die
Polycarpicae einreihen, welche in vielen der oben als typisch
angeführten Eigenschaften abweichen können, in anderen aber sich
trotzdem als Angehörige der Reihe erkennen lassen. Hier muß es genügen,
die wichtigsten anzuführen:
1. Familie Magnoliaceae. Die Magnoliaceae sind durchweg
Holzgewächse mit großen endständigen Blüten, deren Perianth,
ohne Scheidung in Kelch- und Kronblätter, ebenso wie die
zahlreichen Staubblätter und apokarpen Fruchtblätter an einer
langen Blütenachse spiralig aufsteigen. Die Narbe ist direkt
den Fruchtblättern ohne Griffel aufgesetzt. Ölzellen in Stamm
und Blättern, Pollenkörner mit einer Austrittstelle
charakterisieren die Familie innerhalb der Reihe. Drimys
und Zygogynum besitzen koniferenartiges tracheenloses Holz.
Magnolia und der Tulpenbaum, Liriodendron sind vielfach
angepflanzt. Offizinell: Fructus Anisi stellati (Pharm. austr.,
helv.) von Illicium anisatum, Früchte von Illicium religiosum
sind giftig. 2. Anonaceae. Durch spiralige Anordnung der
Staubblätter und apokarpe Fruchtblätter hierher gehörige tropische
Holzgewächse, deren Samen durchweg ruminiertes Nährgewebe besitzen.
Darin gleicht ihnen die 3. Familie der Myristicaceae,
deren diözisch verteilte Blüten wesentlich einfacher gebaut sind.
Offizinell: Semen Myristicae, Oleum Nucistae, Oleum
Macidis (Pharm. germ., austr., helv.) und Macis (Pharm. austr.),
alles abstammend von dem Samenkerne von Myristica fragrans
(Fig. 616, 617). 4. Calycanthaceae zeigen direkten
Anschluß der Laubblätter an die durchaus spiralig[S. 526] gebaute Blüte
mit zahlreichen freien Perianth-, Staub- und Fruchtblättern im
vertieften Blütenboden. Hier und bei Familie 10 ist die Abzweigung
der Rosaceae zu vermuten.
Weitere Familien der Polycarpicae zeigen eine Beschränkung
auf dreizählige einfach oder doppelt vorhandene Perianth- und
Staubblattkreise, wobei die 5. Familie der Berberidaceae
nur ein Fruchtblatt besitzt, während die 6. der
Menispermaceae deren drei führt. An die
Berberidaceen soll nach den Resultaten der serodiagnostischen
Untersuchungen der Centrospermenast Anschluß finden.
Die dornblättrige Berberis vulgaris ist der einheimische
Vertreter der weit verbreiteten Gattung. Offizinell ist
Podophyllinum (Pharm. germ., helv.) von der nordamerikanischen
Berberideenstaude Podophyllum peltatum (Fig. 618) und Radix
Colombo (Pharm. germ., austr., helv.) von der schlingenden
Menispermacee Jatrorrhiza palmata.
Ebenfalls aus dreizähligen Doppelkreisen der Perianth-
und Staubblätter besteht die Blüte der 7. Familie der
Lauraceae, deren einzelnes einsamiges Fruchtblatt der
dreizähligen Narbe nach als ein aus drei Blättern verwachsener
Fruchtknoten gedeutet wird. Die Frucht wird beerenförmig oder
zur Steinfrucht. Die Lauraceen sind aromatische immergrüne Bäume
oder Sträucher mit lederigen, ganzrandigen Blättern, die nur bei
dem nordamerikanischen blattwechselnden Sassafras bisweilen
dreizählig sind (Fig. 619). Charakteristisch sind klappig sich
öffnende Antheren.
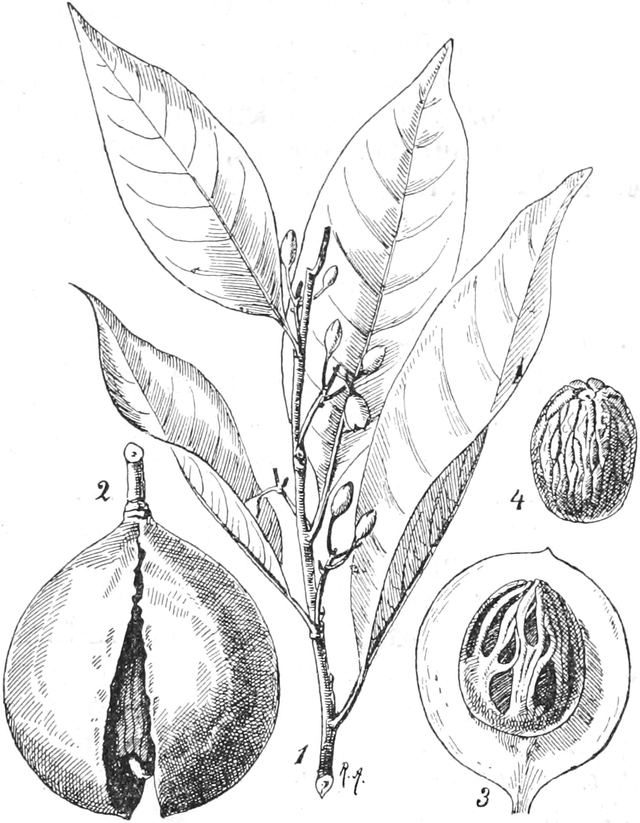
Fig. 616. Myristica fragrans. 1 Blühender männlicher
Zweig. 1⁄2 nat. Gr. 2 Reife Frucht hängend, aufgesprungen. 3
Dieselbe Frucht nach Entfernung der halben Fruchtschale, zeigt den
dunkelbraunen Samen vom durchbrochenen Arillus (Macis) umhüllt. 4
Samenkern (Droge) von der Samenschale befreit. — Offizinell.
Fig. 617. Myristica. Samenquerschnitt. sa sch
Samenschale. end Endosperm. pe Perisperm. — Offizinell.
Laurus nobilis, der Lorbeer, diözischer immergrüner
Baum der Mittelmeerländer (Fig. 621, 622), wird bei uns
vielfach als Kalthauspflanze kultiviert. Größere Pflanzungen
zur Ölgewinnung finden sich schon am Gardasee, wo der Baum vom
Oktober ab seine länglich-ovalen, schwarzblauen Steinfrüchte
reift. Cinnamomum umfaßt eine Anzahl wichtiger Nutzbäume,
so den japanisch-chinesischen Kampferbaum, den chinesischen
und den Ceylon-Zimtbaum, stattliche immergrüne Bäume, mit
lederig-glänzenden Blättern, aber unansehnlichen grünlichen Blüten
in blattachselständigen Infloreszenzen. Persea gratissima
(Fig. 620), Baum des tropischen Mexiko, liefert als Frucht die
wohlschmeckende Aguacate oder Avocadobirne. Arten von Cassytha,
einzige Gattung mit krautigen Vertretern sind überall in den Tropen
häufige, Cuscuta-ähnliche Parasiten.
Offizinell: Fructus Lauri (Pharm. germ., austr.)
und Oleum Lauri (Pharm. germ., austr., helv.) von L.
nobilis. Camphora. Kampfer (Pharm. germ., austr., helv.),
von Cinnamomum Camphora; Cortex Cinnamomi und Oleum
Cinnamomi Ceylon-Zimt (Pharm. germ., austr., helv.) von C.
Cassia und C. Zeylanicum. Lignum Sassafras (Pharm. germ.,
austr.), Cort. Sassafras (Pharm. helv.) von Sassafras
officinale.
[S. 527]
Fig. 618. Podophyllum peltatum. 1⁄2 nat. Gr. Aus Natürl.
Pflanzenfamilien. — Offizinell.
Fig. 619. Sassafras officinale. Nach BERG und
SCHMIDT. 1⁄2 nat. Gr. 1 Männliche Infloreszenzen am noch
unbelaubten Zweig. 2 Früchte am beblätterten Zweig. 3 Männliche
Blüte. 4 Weibliche Blüte. 5, 6 Geschlossene Staubblätter der
beiden äußeren Kreise. 7 Geöffnetes Staubblatt des inneren Wirtels.
8 Fruchtknoten mit Griffel und Samenanlage. — Offizinell.
Fig. 620. Blütendiagramm von Persea. Nach A. W.
EICHLER.
[S. 528]
8. Familie Aristolochiaceae. Die zygomorphen Blüten
(Fig. 546) haben ein einfaches verwachsenes Perianth und ein zum
Gynostemium vereinigtes Andröceum und Gynäceum.
Die parasitischen Rafflesiaceae und die insektivoren
Familien der Cephalotaceae, Sarraceniaceae,
Nepenthaceae und auch der Droseraceae schließen sich
am besten an die Polycarpicae an.

Fig. 621. Laurus nobilis mit männlichen Blüten. 1⁄2 nat.
Gr. — Offizinell.

Fig. 622. Laurus nobilis mit Früchten. 1⁄2 nat. Gr. —
Offizinell.
Die 9. Familie der Nymphaeaceae enthält durchweg
Wasserpflanzen mit untergetauchten oder schwimmenden Blättern
von oft riesiger Größe. Die vegetativen Organe führen Milchsaft
(Fig. 623, 625). In der Nähe der Nymphaeaceen ist der Anschluß der
Monokotyledonen zu suchen.
Nymphaea alba, unsere heimische Seerose (Fig. 623, 624),
breitet zwischen ihren großen schwimmenden Blättern die weißen, von
vier starken grünen Kelchblättern geschützten Blüten aus, deren
vielzähliges weißes Perianth und leuchtend gelbe Staubblätter
spiralig an dem unterständigen, aus zahlreichen Fruchtblättern
verwachsenen Fruchtknoten angeordnet sind. Bei Nuphar mit
oberständigem Fruchtknoten und unscheinbaren, Nektarien tragenden
Kronblättern dient der Kelch als Schauapparat.
Freie apokarpe Fruchtblätter besitzen dagegen die amerikanische
Gattung Cabomba (Fig. 625), durch untergetauchte,[S. 529] vielfach
zerteilte Blätter neben den ganzrandigen Schwimmblättern
ausgezeichnet, und Nelumbium, dessen schildförmiges Laub sich
gleich den Blüten über das Wasser erhebt. Die käferblütige
Victoria regia und die autogame Euryale ferox sind ihrer
Riesenschwimmblätter wegen bekannt und werden in unseren
Warmhäusern viel kultiviert; diese ist in den asiatischen Tropen,
jene im Amazonas heimisch.

Fig. 623. Nymphaea alba. 1⁄2 nat. Gr. Der entblätterte
Fruchtknoten zeigt die spiralige Stellung der Staubblätter und
Kronblätter.
Fig. 624. Nymphaea. Diagramm. Nach F. NOLL.
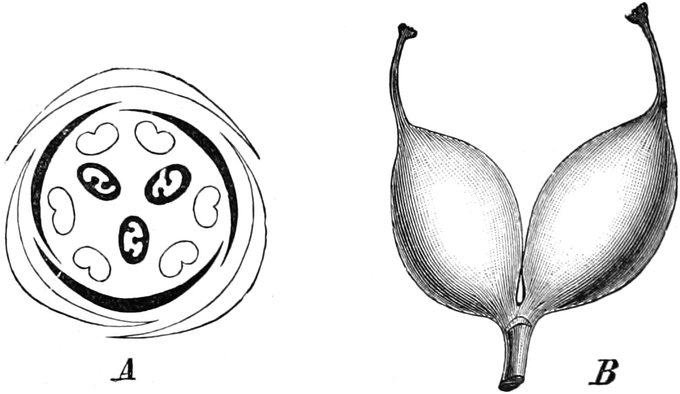
Fig. 625. A Diagramm. B Frucht von Cabomba aquatica.
Nach H. BAILLON. Vergr. 4.
Fig. 626. Blütendiagramme von Ranunculaceen. A
Adonis autumnalis. B Aconitum Napellus. C Aquilegia vulgaris. D
Cimicifuga racemosa. Nach A. W. EICHLER.
10. Familie Ranunculaceae. Die Angehörigen der Ranunculaceen
sind einjährige Kräuter (Myosurus), häufiger Stauden (Caltha) oder
ganz selten Holzgewächse (Paeonia-Arten), mit wechselständigen,
nebenblattlosen Blättern. Die spiralige Anordnung der Teile ihrer
Zwitterblüten tritt an der langen Blütenachse von Myosurus,
Mäuseschwänzchen, am deutlichsten in Erscheinung, ist aber fast
überall da zu beobachten, wo Staub- oder Fruchtblätter in großer Zahl
auftreten (Fig. 626, 627). Das[S. 530] Perianth ist einfach oder doppelt,
perigonartig (Aconitum) oder in Kelch und Krone gesondert (Ranunculus);
das vielgliedrige Andröceum birgt Pollenkörner mit mindestens
drei Austrittstellen. Die Fruchtblätter sind oberständig,
frei; sie stehen auf der gewölbten Blütenachse (Fig. 627) zu drei
bis vielen beisammen, enden in eine griffellose Narbe und tragen die
Samenanlagen an der Bauchnaht (Fig. 626 D) einzeln oder in Mehrzahl.
Früchtchen der Sammelfrucht sind Balgfrüchte (Paeonia), Nüßchen
(Anemone) oder Beeren (Hydrastis), deren Samen den
kleinen Embryo in großem, ölhaltigem Endosperm führen (Fig. 628).
An die Ranunculaceen schließen sich nach den serodiagnostischen
Untersuchungen die ihnen auch morphologisch nahestehenden Rosifloren an.
Fig. 627. a Blüte von Ranunculus sceleratus. b im
Längsschnitt, vergr. Nach H. BAILLON.
Fig. 628. Ranunculus arvensis. Nach H. BAILLON.
Fruchtblatt im Längsschnitt, vergr.

Fig. 629. Ranunculus sceleratus. 1⁄2 nat. Gr. —
Giftig.
[S. 531]
Die Ranunculaceen liefern zahlreiche unserer häufigsten Wald- und
Wiesenpflanzen. Sie sind alle in mehr oder minder hohem Grade
giftig. Die Gattung Ranunculus, an ihren meist gelben glänzenden
Blüten und hochgewölbten Blütenachsen mit zahlreichen Nüßchen
kenntlich, ist in vielen Arten bei uns verbreitet. Ihre Kronblätter
sind mit einem Honiggrübchen am Grunde versehen. R. sceleratus
ist eine gefährliche Giftpflanze (Fig. 629, 627), R. arvensis
durch die Größe der stacheligen Nüßchen auffallend (Fig. 628). Im
Wasser lebende R.-Arten (Batrachium) sind vielfach heterophyll
(Fig. 139); die Schwimmblätter dienen, wie bei Cabomba, den sich
über den Wasserspiegel erhebenden Blüten als Stütze.

Fig. 630. Anemone pulsatilla. 1⁄2 nat. Gr. —
Giftig.
[S. 532]
In allen Laubwäldern bildet Anemone nemorosa mit den ersten
Schmuck des Waldbodens im Frühjahr. Die Pflanze besitzt ein flach
im Boden kriechendes Rhizom, das mit einer terminalen Blüte
abschließt und durch einen Seitensproß fortgesetzt wird. Das
Perianth der Blüte ist einfach, kronartig gefärbt; doch kommt
allen Anemonen ein mehr oder minder tief darunter sitzender, meist
dreiblättriger Wirtel von grünen Hüllblättern zu, der bei A.
Hepatica, der Leberblume, unmittelbar an das Perianth heranrückt
und dadurch einem Kelche ähnlich sieht. Alle Arten sind etwas
giftig, besonders A. Pulsatilla (Fig. 630), die Küchenschelle.
Die Gattung Clematis enthält meist Sträucher und stellt in
Cl. Vitalba eine der wenigen einheimischen Lianen; sie liefert
zahlreiche Zierpflanzen und weicht durch gegenständige Blätter
von allen übrigen R. ab. Clematis-Arten und viele Anemonen
versehen ihre kleinen Nüßchen mit Haar- oder Federanhängen, welche
der Verbreitung durch den Wind dienen. Caltha palustris (Fig.
631), ein als Butter- oder Dotterblume bekannter
Frühjahrsblüher, hat einfaches, intensiv gelbes Perianth und
glänzende herz- oder nierenförmige, kurzgestielte Blätter. Die
Früchte entwickeln sich, ebenso wie bei den im Winter blühenden
giftigen Helleborus-Arten, zu Balgfrüchtchen. Der Eisenhut,
Aconitum Napellus (Fig. 632, 633), eine stattliche, mit jährlich
sich erneuernden Knollen ausdauernde, stark giftige Staude, ist
besonders auf Gebirgswiesen häufig. Seine Blätter sind handförmig
geteilt, die einzelnen Zipfel tief fiederspaltig eingeschnitten,
der Blütenstand dicht traubig mit dorsiventralen Blüten. Eines
der fünf dunkelblauen Kelchblätter ist helmförmig emporgewölbt,
es birgt zwei langgestielte, röhrig-zweilappige Nektarien, die,
wie bei Helleborus und Eranthis, Kronblättern entsprechen.
Die übrigen Kronblätter fehlen oder sind zu unscheinbaren Fädchen
verkümmert. Aconitum Lycoctonum besitzt kleinere gelbe Blüten
desselben Baues. Aquilegia, Delphinium und Paeonia (Fig.
513) liefern beliebte Zierpflanzen mit lebhaft gefärbten, zum
Teil eigenartig geformten Blüten. Actaea und Hydrastis haben
Beerenfrüchte.
Offizinell: Tubera Aconiti von Aconitum Napellus
(Pharm. germ., helv.). Folia Aconiti von derselben
Pflanze (Pharm. helv.). Rhizoma Hydrastis (Pharm. germ.,
austr., helv.) von der nordamerikanischen Staude Hydrastis
canadensis (Fig. 634), die jedes Jahr ihre mit scharf gekielten,
zweizeilig stehenden Niederblättern beginnenden Zweige über die
Erde emporsendet und einzelne gipfelständige Blüten auf den
zweiblättrigen Sprossen bringt. Das einfache weiße Perianth fällt
nach Öffnung ab, wie das auch bei Thalictrum-Arten geschieht.
Sammelfrucht aus zahlreichen kleinen Beeren mit je 1–2 Samen. Aus
dem Rhizom wird Hydrastin dargestellt. Herba Adonidis
von Adonis vernalis (Pharm. austr.).

Fig. 631. Caltha palustris. 2⁄3 nat. Gr. —
Giftig.
Die 2. Ordnung umfaßt die beiden Familien der
Hamamelidinae, Fam. Hamamelidaceae und Platanaceae. Beide
Familien enthalten Holzpflanzen mit unansehnlichen anemophilen
Blüten und einfachem Perianth, seltener entomophile Blüten mit
gefärbtem, eventuell doppeltem Perianth. Zwei Fruchtblätter.
Offizinell: Styrax liquidus (Pharm.[S. 533] germ., austr., helv.)
von Liquidambar orientalis. Folia Hamamelidis von Hamamelis
virginiana (Pharm. austr.). Platanen sind beliebte Alleebäume.
Beide Familien sollen nach den Ergebnissen der serodiagnostischen
Untersuchungen nähere Beziehungen zu den verschiedenen Familien der
Rosifloren besitzen.

Fig. 632. Aconitum Napellus. 1⁄2 nat. Gr. —
Offizinell und giftig.
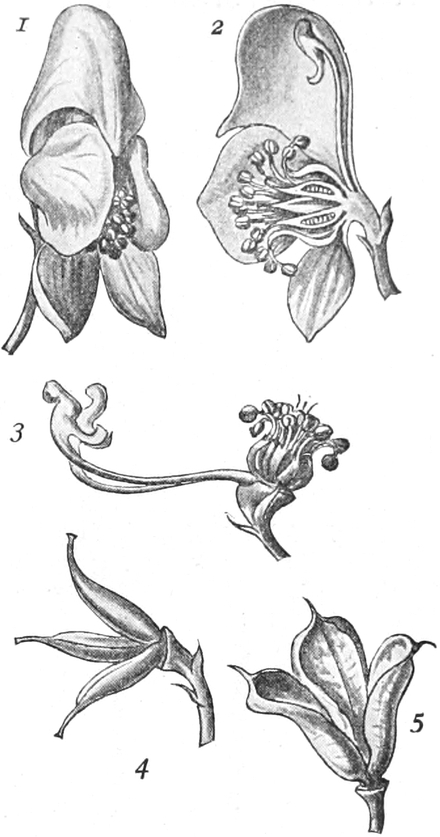
Fig. 633. Aconitum Napellus, nat. Gr. 1 Einzelblüte
schräg von vorn. 2 Einzelblüte längsdurchschnitten. 3 Die zu
Nektarien umgebildeten Kronblätter und das Andröceum nach Entfernung
des Perigons. 4 Frucht aus drei apokarpen Fruchtblättern. 5 Frucht,
aufgesprungen.
3. Ordnung.
Rosiflorae.
Die wirtelig gebauten Blüten sind im übrigen denen der Polycarpicae
ähnlich: besonders eng ist der Anschluß der Rosaceen an die
Calycanthaceen und[S. 534] Ranunculaceen. Einzahl der Fruchtblätter bei den
Pruneen und dorsiventrale Blüten der Chrysobalaneen leiten zu den
Leguminosen über.
Die Ordnung umfaßt Pflanzen mit wechselständigen Blättern von sehr
verschiedener Form und Ausbildung. Ihre fast immer strahligen Blüten
sind wirtelig angeordnet, haben 5, 10 oder viele Staubblätter und
Fruchtblätter, die Neigung zu apokarper Fruchtbildung zeigen.
Charakteristisch ist die starke Beteiligung der Blütenachse an der
Blüten- und Fruchtbildung. K5, C5, A5-∞ G1-∞.
Sukkulente Kräuter (vgl. S. 149) oder Halbsträucher mit
vielgliedrigen cymösen Blütenständen enthält die 1. Familie
Crassulaceae. Sedum, die Fetthenne (Fig. 635), mit
fünfzähligen und Sempervivum mit 6-∞zähligen Blüten sind häufige
und artenreiche einheimische Gattungen. Bryophyllum-Arten mit
vierzähligen Blüten sind ausgezeichnet durch die regelmäßige
Bildung von Adventivknospen in allen Kerbstellen des Blattrandes.
Crassula, Südafrika, bildet Mimikryformen[477], die in ihrer
klobigen Form Steine nachahmen.
Die 2. Familie Saxifragaceae enthält Kräuter neben
Holzgewächsen mit zwitterigen obdiplostemonen Blüten. Die Kapsel-
oder Beerenfrüchte der Saxifragaceen werden in der Regel aus zwei
Fruchtblättern gebildet und enthalten viele endospermhaltige Samen.
Die Saxifraga- (Steinbrech-)Arten sind kleine Rosettenpflanzen,
die im Gebirge ihre Hauptverbreitung finden. Ihre oft ansehnlichen
Infloreszenzen tragen lebhaft gefärbte Einzelblüten mit stets
zwei, im oberen Teile freien Fruchtblättern. Parnassia palustris
tritt im Hochsommer auf unsern Wiesen auf, ihre fünfzählige
Blüte hat vier Fruchtblätter und zu handförmig zerschlitzten
Staminodien umgebildete Kronstaubblätter, die als Nektarien
fungieren. Ribes-Arten, mit unterständigem Fruchtknoten, liefern
Beerenfrüchte. R. rubrum (Fig. 636), Johannisbeere, R. nigrum,
schwarze Johannisbeere, R. grossularia, Stachelbeere. Andere
Saxifragaceen sind als Zierpflanzen beliebt, wie Ribes aureum und
R. sanguineum, Hydrangea, Hortensie, Philadelphus, Deutzia.
Offizinell: Syrupus ribium (Pharm. austr.) von Ribes
rubrum.

Fig. 634. Hydrastis canadensis. 1⁄2 nat. Gr. Apokarpe
Sammelfrucht daneben. — Offizinell.
Fig. 635. Sedum Telephium. Vergr. 4. a Blüte, b
Blüte im Längsschnitt. Gez. H. SCHENCK.
3. Familie Rosaceae[478]. Die charakteristischen Merkmale[S. 535] der
Rosaceen sind der stete Besitz von Nebenblättern; ferner die
Endospermlosigkeit der meisten Rosaceen-Samen, endlich die apokarpen
Früchte und meist auch die Vielzähligkeit des Andröceums (Fig. 637).
Letztere beiden Eigenschaften kommen auch den Polycarpicae zu und
sprechen für engere verwandtschaftliche Beziehungen, doch sind die
Blütenglieder bei den Rosaceen streng wirtelig und perigyn gestellt.
In vielen Fällen geht die Vermehrung der Andröceum- und
Gynäceumglieder von einer an der schüsseligen Vertiefung der Achse
interkalar gelegenen, gürtelförmigen Vegetationszone aus längere
Zeit nebeneinander her. Über die Einschiebung neuer Glieder
entscheiden räumliche Verhältnisse, so daß bei verschiedenen
Individuen derselben Art Differenzen in den Zahlen vorkommen.
Zahlreiche Arten der Gattung Spiraea mit typisch fünfzähligen
Blüten und oberständigen Fruchtknoten finden als Ziersträucher
Verwendung (Fig. 637 E). Quillaja Saponaria (Fig. 638),
der Seifenrindenbaum Chiles, ist ein immergrüner Baum mit
kurzgestielten, wechselständigen, lederartigen Blättern und
gipfelständigen, meist dreiblütigen Dichasien. Die Blüten sind
denen von Spiraea ähnlich, aber durch ihre fünfzackige, auf
die großen Kelchblätter hinaustretende, honigabsondernde Scheibe
charakterisiert, an deren eingekerbten Vorsprüngen die fünf
Kelchstaubblätter stehen, während sich die Kronstaubblätter am
inneren Rande befinden. Die schmalen Kronblätter sind weiß. Nur
die Mittelblüte des Dichasiums ist zwittrig und fruchtbar, die
seitlichen mit verkümmertem Fruchtknoten sind männlich. Der
oberständige Fruchtknoten liefert eine sternförmige Sammelfrucht.
Jedes Fruchtblatt springt in zwei Klappen auf und entläßt seine
geflügelten Samen.
Fig. 636. Ribes rubrum. 2⁄3 nat. Gr. —
Offizinell.
Fig. 637. Blütendiagramme von Rosaceen. A Sorbus
domestica. B Prunus Padus. C Rosa tomentosa. D Sanguisorba
officinalis. E Spiraea hypericifolia. Nach A. W. EICHLER.
Durch den unterständigen, meist aus fünf Fruchtblättern
bestehenden, rings mit der ausgehöhlten fleischig anschwellenden
Blütenachse verwachsenen, gefächerten, pergamentenen Fruchtknoten
und freie Griffel von den sonstigen Rosaceen unterschieden
sind die Gattungen Pirus, Cydonia u. a. Pirus malus,
Apfelbaum (Fig. 524 3), Pirus communis, Birnbaum,
unsere wichtigsten einheimischen Obstbäume, alte Kulturpflanzen in
zahlreichen Varietäten. Cydonia vulgarus,[S. 536] die Quitte,
mit einzeln stehenden großen hellrosa Blüten. Ihre apfel- oder
birnförmigen, wollig-filzig behaarten Früchte sind roh ungenießbar,
doch von höchst angenehmem Aroma. Mespilus germanica, der
Mispelbaum; seine Früchte haben eine tiefe Grube am
Scheitel, die rings von den Überresten der Kelchblätter umgeben
ist. Die immergrüne japanische Mispel, Eriobotrya
japonica, wird im Mittelmeergebiete häufig angepflanzt. Sorbus
aucuparia, der Vogelbeerbaum, und Crataegus-Arten,
Rotdorn, Weißdorn, sind teils als Alleebäume, teils
in Hecken und als Zierbäume verbreitet (vgl. S. 262).
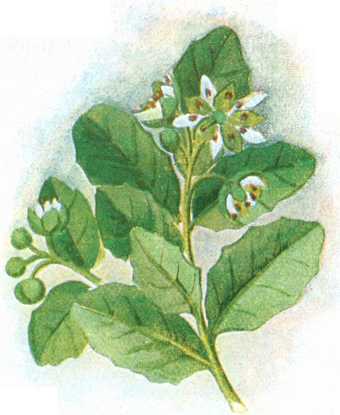
Fig. 638. Quillaja Saponaria. 4⁄5 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Fig. 639. Hagenia abyssinica. 1 Weibliche Blüte, e
Außenkelch, f Kelch, g Korolle. Vergr. 4.
2 Frucht mit vergrößertem Außenkelch. Nat. Gr. Nach BERG und
SCHMIDT. — Offizinell.

Fig. 640. Hagenia abyssinica. Zweigstück mit
Blütenstand. 1⁄2
nat. Gr. Nach BERG und SCHMIDT. —
Offizinell.
Eine krugförmig vertiefte Blütenachse, auf der ein
bis viele freie Fruchtblätter stehen, die zu nußartigen, vom
Blütenbecher umschlossenen Früchtchen werden, eignet der Gattung
Rosa (Fig. 637 C). Ihr Blütenbecher wird fleischig (Fig.
583), und die kronblattartige Ausbildung zahlreicher Staubblätter
gibt staminodial gefüllte Formen.[S. 537] Dagegen ist bei Agrimonia
und Hagenia abyssinica der Blütenbecher hart. Hagenia, ein
diözischer Baum Abessyniens, mit unpaarig gefiederten Blättern,
deren Blattstiel durch Nebenblätter rinnig geflügelt wird, trägt
reichverzweigte Blütenrispen. Die Einzelblüten mit zwei Vorblättern
und Außenkelch werden durch Fehlschlagen eingeschlechtig; ihre
Krone ist nach der Blüte hinfällig, der Kelch eingerollt, der
Außenkelch vergrößert. Aus den zwei freien Fruchtblättern mit je
einer Samenanlage wird eine einsamige Frucht (Fig. 639, 640).
Die kronblattlose Alchimilla (Fig. 524, 2) und Sanguisorba
officinalis mit köpfchenförmig gehäuften, polygamen, vierzähligen
Einzelblüten ohne Außenkelch und Krone mit ein bis zwei
Fruchtblättern (Fig. 637 D) sind stark reduzierte Formen.
Potentilla mit flachem Blütenboden, apokarpen Fruchtblättern und
Außenkelch hat zahlreiche einheimische Arten. Geum und Dryas
verbreiten ihre weichhaarigen, langauswachsenden Früchtchen durch
den Wind. Fragaria, die Erdbeere, trägt kleine Nüßchen auf der
fleischig geschwollenen Achse. Rubus, die Brombeere, ist in
zahlreichen meist kletternden Arten verbreitet. Ihre Blätter sind
stets dreizählig, ihre Früchte kleine Steinfrüchtchen, die in
Sammelfrüchten beisammenbleiben. Eine der wenigen nicht kletternden
Arten ist die Himbeere R. Idaeus.
Ein einziges Fruchtblatt, mittelständig auf flachschüsselförmig
vertieftem Blütenbecher (Fig. 637 B) haben die Steinobstbäume:
Prunus Cerasus, Sauerkirsche (Fig. 641). Prunus avium,
Süßkirsche, Pr. domestica, Pflaume (Zwetsche); Pr.
armeniaca, Aprikose, chinesischen Ursprungs, ebenso wie
Pr. persica, Pfirsich; Pr. Amygdalus, Mandel, aus
dem östlichen Mittelmeergebiete. Das fleischige Exokarp der Mandel
trocknet beim Reifen und platzt auf, so daß das steinige Endokarp
freigelegt wird.
Giftig: Die Samen vieler Rosaceen sind amygdalinhaltig;
durch die entstehende Blausäure wirken Preßrückstände, z. B. von
bitteren Mandeln, häufig giftig. Die Blätter des Kirschlorbeers,
Prunus Laurocerasus, können aus gleichem Grunde ebenfalls
Vergiftungen hervorrufen.
Fig. 641. Prunus Cerasus 2⁄3 nat. Gr. 1 Blühender
Zweig. 2 Einzelblüte halbiert (etwas vergr.). 3 Fruchtstand. 4
Einzelfrucht halbiert. — Offizinell.
Offizinell: Pirus Malus: Extractum ferri pomati
(Pharm. germ., helv.). — Cydonia vulgaris liefert Semen
Cydoniae (Pharm. helv.). — Hagenia abyssinica: Flores
Koso (Pharm. germ., austr., helv.). — Rosa centifolia und
R. gallica: Flores Rosae (Pharm. germ., austr., helv.);
dieselben und andere Arten: Ol. Rosae (ibid.). — Rubus
Idaeus: Syrupus R. Idaei (Pharm. germ., austr., helv.). —
Prunus Amygdalus: Amygdalae dulces und A. amarae,
Oleum Amygdalarum (Pharm. germ., austr., helv.). — Pr.
domestica: Pulpa prunorum (Pharm. austr.). — Pr.
Laurocerasus: Aqua Laurocerasi (Pharm. austr., helv.). —
Pr. Cerasus: Syrupus Cerasorum (Pharm germ.). — Quillaja
Saponaria: Cortex Quillajae (Pharm. germ., austr.). —
Spiraea Ulmaria: Flos Spiraeae Pharm. helv.). — Rubus
fruticosus: Folium Rubi fruticosi (Pharm. helv.). —
Potentilla Tormentilla: Rhizoma Tormentillae (Pharm.
helv.).
[S. 538]
4. Ordnung.
Leguminosae.
Das gemeinsame Kennzeichen aller Leguminosen ist in der Beschaffenheit
ihres Fruchtknotens gegeben, der die Ordnung von den sonst nahe
verwandten Rosifloren scharf unterscheidet. Er geht stets aus einem
einzigen Fruchtblatt hervor, ist einfächerig und trägt die Samenanlagen
in einer oder zwei Reihen an der nach hinten gekehrten Bauchnaht (Fig.
642, 645, 650). Meist wird die Frucht eine Hülse (Legumen), die an der
Bauchnaht und in der Rückenlinie in zwei Klappen aufspringt (wie die
Erbsen). Wechselständige, zusammengesetzte Blätter mit Nebenblättern
sind fast allen Leguminosen eigen. Sehr viele besitzen Blattgelenke
(Fig. 643), welche Variationsbewegungen ermöglichen.
Die 1. Familie Mimosaceae enthält Bäume und aufrechte oder
kletternde Sträucher mit paarig doppeltgefiederten Blättern und
strahligen. fünf- oder vierzähligen Blüten (Fig. 642). Die Knospenlage
von Kelch und Krone ist klappig. Die freien, halb-, voll- oder meist
überzählig vorhandenen Staubblätter bedingen durch ihre bedeutende
Länge oder große Zahl die Blütenfarbe. Ihre Pollenkörner bleiben häufig
in Tetraden oder zu mehreren vereinigt. Die Blüten stehen in Ähren oder
Köpfchen zu vielen beisammen. ihre Krone ist unscheinbar, der Embryo
gerade.
Fig. 642. Blütendiagramme von Mimosaceen. A Von Mimosa
pudica. B Von Acacia lophantha. Nach A. W. EICHLER.
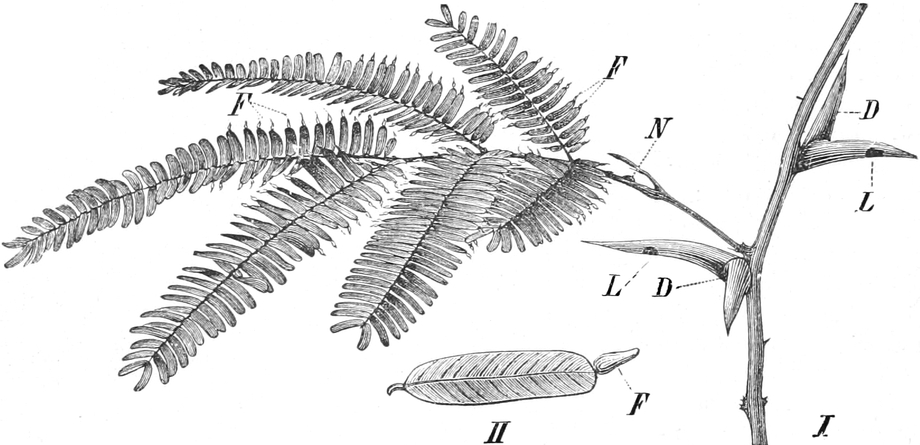
Fig. 643. Acacia nicoyensis aus Costa Rica. I
Stammstück mit Dornen (D) und einem Blatte. Die hohlen Dornen werden
von Ameisen angebohrt und bewohnt. L Eingangsöffnungen. An den
basalen Blattfiederchen die Futterkörper F. Auf dem Blattstiel bei
N ein Nektarium. Verkleinert. II Einzelnes Blattfiederchen mit dem
Futterkörper F, etwas vergrößert. Nach F. NOLL.
Die in den Tropenwaldungen reich entwickelte Familie besitzt
in Europa keinen einheimischen Vertreter. Mimosa pudica, die
Sinnpflanze (Fig. 292), mit hochgradiger Empfindlichkeit
gegen Berührung, ist ein pantropisches Unkraut. Die Gattung
Acacia ist in zahllosen Arten in den Tropen und Subtropen der
alten und neuen Welt verbreitet. Ihre australischen Formen sind
durch den Besitz von Phyllodien ausgezeichnet deren vertikale
Stellung wesentlich zum eigenartigen Habitus der australischen
Wälder mit beiträgt (vgl. Fig. 190). Einige amerikanische
Akazien-Arten werden von Ameisen bewohnt (Fig. 643), denen sie
in großen Nebenblattdornen Wohnung, in den BELTschen
Körperchen[479] an den Enden der Fiederblättchen Nahrung bieten,
ohne daß ein symbiotisches Gegenseitigkeitsverhältnis nachweisbar
ist. Durch Lieferung von Gummi und reichen Gehalt an Gerbstoffen,
teils in der Rinde, teils im Extrakt des Kernholzes, teils in den
Hülsen, gewähren zahlreiche Acacia-Arten einen erheblichen Nutzen.
[S. 539]
Offizinell: Durch Desorganisation des Stammparenchyms
liefern Acacia Senegal (Nilländer und Senegambien) und andere
Arten: Gummi arabicum (Pharm. germ., austr. helv.), das
aus Wunden als dicke Flüssigkeit herausfließt und erhärtet. —
Catechu (Pharm. germ., austr., helv.) ist ein Dekokt aus dem
Kernholze der ostindischen Acacia Catechu (Fig. 644) und Ac.
Suma.
Die 2. Familie Caesalpiniaceae umfaßt Bäume oder Sträucher
mit einfach oder doppelt paarig gefiederten Blättern. Ihre Blüten sind
meist etwas dorsiventral mit dachig aufsteigender Knospendeckung
der Krone, deren Blattzahl ebenso wie die der freien Staubblätter
häufig unvollzählig ist (Fig. 645). Typisch: K5, C5, A5 + 5, G̱1. Auch
hier ist der Embryo gerade. In den Tropen und Subtropen ist die
Familie reich ausgebildet.

Fig. 644. Acacia Catechu. 2⁄3 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Fig. 645. Blütendiagramme von Caesalpiniaceen.
A Cercis siliquastrum. B Tamarindus indica. Nach A. W.
EICHLER.
Bei Cassia angustifolia sind Kelch und Kronblätter frei und in
Fünfzahl vorhanden (Fig. 646), die unteren deckenden Kronblätter
etwas größer als die oberen. Von zehn[S. 540] Staubblättern sind die drei
oberen kurz und unfruchtbar, sieben nach unten bogig gestreckte
nehmen von oben nach unten an Länge zu. Ihre Antheren öffnen
sich mit endständigen Poren. Der Fruchtknoten ist eine breite
und flach zusammengedrückte Hülse. Derartige Blüten stehen in
blattachselständigen Trauben beisammen an dem etwa meterhohen, mit
lebhaft grünen, paarig gefiederten Blättern besetzten Strauche.
Am Grunde des Blattstieles stehen zwei kleine Nebenblättchen.
Tamarindus indica (Fig. 647), ein schöner Baum des tropischen
Afrika, wird in den gesamten Tropen jetzt vielfach angepflanzt.
Mit einfach paarig gefiederten Blättchen besetzte Seitenzweige
bringen die Blütentrauben endständig. Aus den stark dorsiventralen
Einzelblüten geht eine vom Typus abweichende Frucht hervor. Ihre
Wandung differenziert sich in ein äußeres brüchiges Exokarp, ein
fleischiges musartiges Mesokarp M und ein festes, die einzelnen
mehr oder minder zahlreichen Samen umhüllendes, aus Steinzellen
bestehendes Endokarp (Fig. 648). Kaum merklich dorsiventral
sind die kronblattlosen Blüten von Copaifera, in denen auf
4 Kelchblätter sogleich 8–10 freie Staubblätter folgen. Die
einsamige Frucht öffnet sich bei der Reife. Ein fleischiger,
unregelmäßig begrenzter Arillus umhüllt den Samen einseitig.
Einheimische Caesalpiniaceae fehlen; der im Mittelmeer beheimatete
Johannisbrotbaum Ceratonia Siliqua, wird im Kalthause, die
kauliflore (vgl. S. 573) Cercis siliquastrum (Fig. 645 A)
und Gleditschia triacanthos (N.-Am., Fig. 198) werden als
Zierpflanzen bei uns kultiviert.

Fig. 646. Cassia angustifolia. 2⁄3 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Offizinell: Folia Sennae (Pharm. germ., austr.,
helv.), Fiederblättchen von Cassia angustifolia; Pharm. helv.
läßt auch Fol. S. Alexandrinae von C. acutifolia zu.
— Cassia obovata und C. acutifolia: Fructus Sennae
(Pharm. austr., helv.). — Cassia Fistula (tropisches Amerika):
Fructus Cassiae Fistulae (Pharm. austr., helv.). —
Copaïfera-Arten (Bäume des tropischen Amerika) enthalten in
Balsamgängen des Holzes: Balsamum Copaïvae (Pharm. germ.,
austr., helv.). — Rad. Ratanhiae (Pharm. germ., austr.,
helv.) von Krameria triandra, einem Strauch der Kordilleren mit
silberweißen, einfachen Blättern, dessen Blüten völlig abweichen;
sie haben innen lebhaft gefärbte Kelchblätter, eine kleine Krone
und drei Staubblätter mit Porenöffnung am Scheitel. Ihre Früchte
sind kugelig und mit Stacheln besetzt (Fig. 649). — Lignum
Haematoxyli (Pharm. austr.), Kernholz von Haematoxylon
campechianum (tropisches Amerika). — Pulpa[S. 541] Tamarindorum
(Pharm. germ., austr., helv.), das musartige Mesokarp der Frucht
von Tamarindus indica (Fig. 648).

Fig. 647. Tamarindus indica. 4⁄5 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Fig. 648. Tamarindus indica. Frucht im Längsschnitt. M
Das fleischige Mesokarp. Nach BERG und SCHMIDT. —
Offizinell.

Fig. 649. Krameria triandra. 4⁄5 nat. Gr. Nach A.
MEYER u. SCHUMANN. — Offizinell.

Fig. 650. Blütendiagramme von Papilionaceen. A Von
Vicia Faba. B Von Laburnum vulgare. Nach A. W. EICHLER.

Fig. 651, Lotus corniculatus. 1⁄2 nat. Gr. Blühender
Sproß. Blüte. Schiffchen, Staubblätter. Fruchtblatt (nat. Gr.) und
Frucht, 1⁄2 nat. Gr.

Fig. 652. Myroxylon Pereirae. 2⁄3 nat. Gr. Nach
BERG und SCHMIDT. — Offizinell.
Fig. 653. Myroxylon Pereirae (vgl. Text). Vergr. Nach
BERG und SCHMIDT. — Offizinell.
Fig. 654. Frucht von Myroxylon Pereirae. 2⁄3 nat. Gr.
Nach BERG und SCHMIDT. — Offizinell.
Die Angehörigen der 3. Familie Papilionaceae haben in der
Regel unpaarig gefiederte Blätter und stets stark dorsiventrale
Blüten mit absteigender Knospendeckung (Fig. 650). Auf[S. 542] den
fünfblättrigen Kelch folgt die dorsiventrale fünfblättrige Krone
und 10 Staubblätter, deren Filamente alle miteinander zu einer,
den Fruchtknoten umhüllenden Röhre verwachsen sind (Lupinus),
oder die neun vorderen sind verwachsen und das hintere Staubblatt
bleibt frei (Lotus) oder alle sind frei (Myroxylon, Fig. 653). Der
Keimling ist im Samen stets gekrümmt[S. 543] eingelagert. Die Familie ist
weniger in den Tropen als in den gemäßigten Zonen reich vertreten.
Die Bestandteile einer Schmetterlingsblüte sind in Fig. 651 einzeln
auseinandergelegt. Das hintere, in der Knospenlage (Fig. 650)
beiderseits übergreifende Kronblatt heißt Fahne (vexillum), zwei
darauffolgende seitliche die Flügel (alae), und die mit ihren
abwärts gekehrten Rändern in der Regel verwachsenen beiden unteren
bilden das Schiffchen oder den Kiel (carina). Die Staubblätter
sind an ihren oberen Enden frei und aufwärts gekrümmt, ebenso der
Griffel mit seiner Narbe.
Fig. 655. Astragalus gummifer. 4⁄5 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Myroxylon balsamum var. Pereirae, ein Baum von mäßiger Höhe,
besitzt unpaarig gefiederte Blätter (Fig. 652) in wechselständiger
Anordnung. Die Blüten, in endständigen Trauben stehend, sind nur
mit einem großen Vexillum versehen, alle übrigen Blätter der
Krone bleiben unansehnlich und schmal. Die Staubblätter, nur am
Grunde miteinander verwachsen (Fig. 653), tragen ansehnliche
rot-gelbe Antheren. Der langgestielte Fruchtknoten enthält zwei
Samenanlagen, eine davon wird zum Samen der geschlossen bleibenden,
zusammengedrückten Hülse, deren Form durch eine an der oberen Naht
breite, an der unteren schmale Flügelung, wie durch zwei seitliche
Balsamblasen sehr charakteristisch ist (Fig. 654). Oben am Stiel
bleibt der glockige Kelch erhalten. Amicia, Gattung der Anden
für nyctinastische Untersuchungen bevorzugt, hat paarig gefiederte
Blätter. — Genista, Sarothamnus, Lupinus, Cytisus zeigen
alle 10 Staubblätter verwachsen (Fig. 650 B). Ihre Blätter sind
ganzrandig, gefiedert oder einfach. Der Goldregen (Fig.
266) Laburnum vulgare, einer der häufigsten und beliebtesten
Zierbäume unserer Gärten mit dreiteilig gefiederten Blättern und
lang herabhängenden gelben Blütentrauben, wächst in den Alpen
wild. Ulex ist eine Charakterpflanze Englands, Spartium
im Mittelmeergebiet verbreitet. — Trifolium, der Klee,
hat bleibenden Kelch und Krone, dreiteilige Blätter, kopfig
gehäufte Blüten mit (9) + 1 Staubblättern und Schließfrüchten.
Medicago, Schneckenklee, hat hinfällige Krone und sichelförmige
oder schraubig gekrümmte Früchte. Melilotus, Steinklee, mit
traubigen Blütenständen. Trigonella, der Bockshornklee,
bringt lang auswachsende Hülsenfrüchte. Ononis, Hauhechel,
mit 10 verwachsenen Staubblättern. Bei den im großen angebauten
Trifolium-, Medicago- und Lupinus-Arten (wie der unten
genannten Seradella) kommt die Stickstoffanreicherung des Bodens
durch die „Leguminosenknöllchen“ (vgl. S. 224, Fig. 249, 250) für
europäische Verhältnisse am meisten zur praktischen Verwertung.
— Lotus, der Hornklee (Fig. 651), trägt die üblichen unpaarig
gefiederten Blätter, doch wird das unterste Fiederpaar durch Fehlen
des Blattstieles nebenblattähnlich. Anthyllis, Wundklee. — Bei
Astragalus-Arten, den Traganthsträuchern, niedrigen Sträuchern
des östlichen Mittelmeergebietes und westlichen Asiens, fallen die
Fiederchen der Blätter ab, und die Blattspindeln bleiben lange
Jahre als scharf stechende Dornen erhalten und dienen zum Schutz
der jungen Triebe, Blätter und Blüten (Fig. 655). Einheimische
Arten sind krautig. Robinia (Fig. 181), rasch wachsender Baum
Nordamerikas mit sehr sprödem, windbrüchigem Holz, wird vielfach
angepflanzt als „Akazie“. Glycyrrhiza,[S. 544] Süßholz, Stauden des
südlichen Europa. Wistaria sinensis (Glycine), eine schön
blau blühende Schlingpflanze, findet sich häufig an Häusern u. dgl.
— An Gliederhülsen kenntlich sind Coronilla,
Ornithopus sativus, Seradella, Arachis hypogaea,
Erdnuß, eine wichtige ölfrucht der Tropen und Subtropen.
Ihre Blütenstiele dringen nach der Befruchtung geotropisch in
den Boden ein; dort reifen die Früchte. — Endranken, an Stelle
der unpaaren Fiederblättchen und hypogäische Keimung besitzen
Pisum, Erbse (Fig. 206), Lens, Linse, Lathyrus,
Platterbse (Fig. 207), Vicia, Wicke, und als
aufrechte, rankenlose Pflanze Vicia Faba, Saubohne,
deren Endblättchen zu einem borstenförmigen Stummel verkümmert.
— Windende Pflanzen mit dreiteilig gefiederten Blättern sind
Phaseolus, Bohne; Physostigma, die Kalabarbohne.
Giftig: Durch hochgradige Giftigkeit ausgezeichnet ist
unter den einheimischen Papilionaceen nur Laburnum vulgare und
die verwandten Arten der Gattung Cytisus. — Als giftig gelten
auch Coronilla varia, ein wildwachsendes Kraut mit rosaroten
Blütendolden, und die windende Wistaria sinensis unserer Gärten.
Offizinell: Astragatus-Arten liefern Tragacantha
(Pharm. germ., helv.). — Glycyrrhiza glabra in ihren Wurzeln
und Ausläufern: Rad. Liquiritiae (Pharm. germ., austr.,
helv.) — Melilotus officinalis: Herba Meliloti, Steinklee
(Pharm. germ., austr.). — Trigonella Foenum graecum: Semen
Foenugraeci, Bockshornsamen (Pharm. germ., austr., helv.).
— Ononis spinosa: Rad. Ononidis (Pharm. germ., austr.,
helv.). — Spartium scoparium: Sparteïnum (Pharm.
helv.). — Physostigma venenosum (bohnenähnliche Schlingpflanze
Westafrikas): das aus den Samen (Semen Calabar) dargestellte
Alkaloid Physostigminum (Pharm. germ., austr., helv.).
— Andira Araroba, ein brasilianischer Baum, enthält in
seinem Stamm eine pulverige Exkretmasse: Chrysarobinum
oder Araroba genannt (Pharm. germ., austr., helv.). —
Pterocarpus santalinus, ein ostindischer Baum, liefert in
seinem Kernholze Lignum Santali rubrum (Pharm. aust.).
— Pterocarpus marsupium, ein ostindischer Baum, in seinem
eingetrockneten Safte: Kino (Pharm. helv.). — Myroxylon
balsamum var. genuinum, ein südamerikanischer Baum, Balsamum
tolutanum (Pharm. germ., austr., helv.); M. balsamum var.
Pereirae (San Salvador): Balsamum peruvianum (ibid.).
5. Ordnung.
Myrtiflorae.
Die Ordnung weicht durch unterständigen Fruchtknoten und Fehlen der
Nebenblätter von den Rosifloren ab.
1. Familie Thymelaeaceae. Daphne Mezereum (Fig. 656).
Giftiger Strauch unserer Wälder, blüht im Februar bis
April vor Entfaltung der Blätter aus den vorjährigen Knospen.
Die rosenroten stark duftenden Blüten sind kronenlos, im übrigen
vierzählig, haben aber nur ein Fruchtblatt und eine hängende
Samenanlage im Fruchtknoten, der sich zu einer hochroten Beere
entwickelt. Die Blätter stehen zunächst schopfförmig, bis die
Achse sich streckt. In den Alpen und im Mittelmeergebiet mehrere
Daphne-Arten, die alle giftig sind.
[S. 545]
Offizinell: Cortex Mezereï (Pharm. helv.) von D.
Mezerum.

Fig. 656. Daphne Mezereum. 1⁄2 nat. Gr.
Offizinell und giftig.
Fig. 657. Diagramm von Oenothera (Onagraceae). Nach
F. NOLL.
Zu der 2. Familie Elaeagnaceae, die durch aufrechten
Stand ihrer Samenanlage unterschieden ist, gehören der Sanddorn,
Hippophaë, und die Ölweide, Elaeagnus. Blätter und junge Zweige
sind bei beiden Pflanzen mit glänzenden Schildhaaren bedeckt,
ebenso bei Shepherdia (S. 47).
3. Familie Lythraceae. Der einheimische Weiderich,
Lythrum Salicaria, hat eine typisch sechszählige, durch trimorphe
Heterostylie (S. 484) ausgezeichnete Blüte mit zwei bis sechs
Fruchtblättern.
Die Angehörigen der 4. Familie Onagraceae haben stets
vierzählige Blüten und obdiplostemones Andröceum. Epilobium, das
Weidenröschen, ist in mehreren Arten bei uns verbreitet, seine
Kapselfrüchte enthalten Samen mit Flughaaren. Oenothera-Arten
(Fig. 657) sind bekannt als Versuchspflanzen zur experimentellen
Begründung der Mutationstheorie durch DE VRIES.
Einheimisch sind ferner Circaea, Hexenkraut, und Trapa,
Wassernuß. Die in Amerika beheimateten Fuchsia-Arten werden ihrer
auch durch petaloiden Kelch lebhaft gefärbten Blüten wegen viel
kultiviert. Sie tragen Beerenfrüchte.
Fig. 658. Blütendiagramme von Myrtaceen. A Von Myrtus
communis. B Von Eugenia aromatica. Nach A. W. EICHLER.
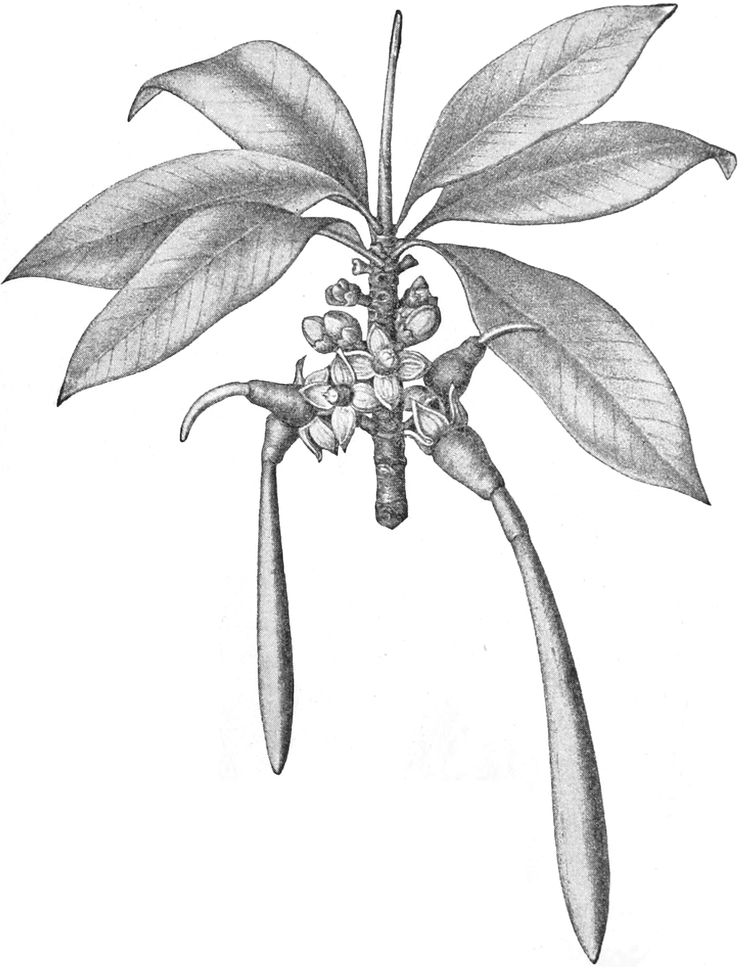
Fig. 659. Rhizophora conjugata. 1⁄4 nat. Gr.
Die 5. Familie Rhizophoraceae umfaßt tropische
Küstenpflanzen der Mangroveformation, die durch ihre, den
Standortseigentümlichkeiten angepaßte Viviparie, wie durch
Atemwurzeln und Stelzwurzeln eigenartigen Habitus aufweisen (Fig.
187). Rhizophora (Fig. 659), Bruguiera, Ceriops, Kandelia
sind alle an den Küsten des Indischen Ozeans, Rhizophora-Arten
auch sonst an tropischen Meeresküsten verbreitet.
Die 6. Familie der Myrtaceae enthält durchweg immergrüne
Pflanzen mit gegenständigen, lederartigen, oft aromatischen Blättern
und strahligen, vier- oder fünfzähligen Blüten, die im Andröceum
vielzählig sind und deren Staubblätter häufig in Bündeln stehen, die
durch Spaltung entstanden sind. Fruchtblätter sind zwei bis viele
vorhanden (Fig. 658), die mit der Blütenachse zum unterständigen
Fruchtknoten verwachsen, der sich zur Beere oder Kapsel entwickelt.
Ihre Hauptverbreitung finden die Myrtaceen im tropischen Amerika
und in Australien.
[S. 546]
Myrtus communis, die Myrte, ist im Mittelmeergebiet zu Hause
und bildet die einzige europäische Art. Häufig angepflanzt finden
sich in wärmeren Ländern die Eucalyptus-Arten[480] Australiens,
besonders E. Globulus, welche ihres schnellen Wachstums und
brauchbaren Nutzholzes halber von Wert ist. Junge Exemplare haben
sitzende gegenständige Blätter, ältere Bäume dagegen gestielte
wechselständige, sichelartig gebogene, die vertikal herabhängen. In
ihrer australischen Heimat rührt die Schattenlosigkeit der Wälder
teils von dieser eigenartigen Stellung der Blätter, teils von dem
weiten Abstande der einzelnen Baumindividuen her. E. amygdalina
ist bei 150 m gemessener Höhe und 30 m Umfang am Grunde der
gewaltigste Baumriese, den man kennt; Psidium guajava und einige
Jambosa-Arten liefern eßbare Früchte. Jambosa caryophyllus,
der Gewürznelkenbaum der Molukken (Fig. 660), gibt in den
ungeöffneten Blütenknospen die „Nägelchen“ oder „Nelken“. Der
Baum wird in den Tropen vielfach kultiviert. Fig. 660 zeigt auch
den unterständigen zweifächerigen Fruchtknoten im Längsschnitt.[S. 547]
Sonneratia-Arten sind vielfach die am weitesten ins Meer
vordringenden Angehörigen der Mangrovevegetation, so daß ihre
Pneumatophoren besondere Höhe erreichen müssen (Fig. 186, S. 143).
Offizinell: Caryophylli, Oleum Caryophyllorum
(Pharm. germ., austr., helv.) von Jambosa caryophyllus, Folia
Eucalypti (Pharm. helv.) von Eucalyptus Globulus, Oleum
Cajeputi (Pharm. austr., helv.) von Melaleuca Leucadendron.
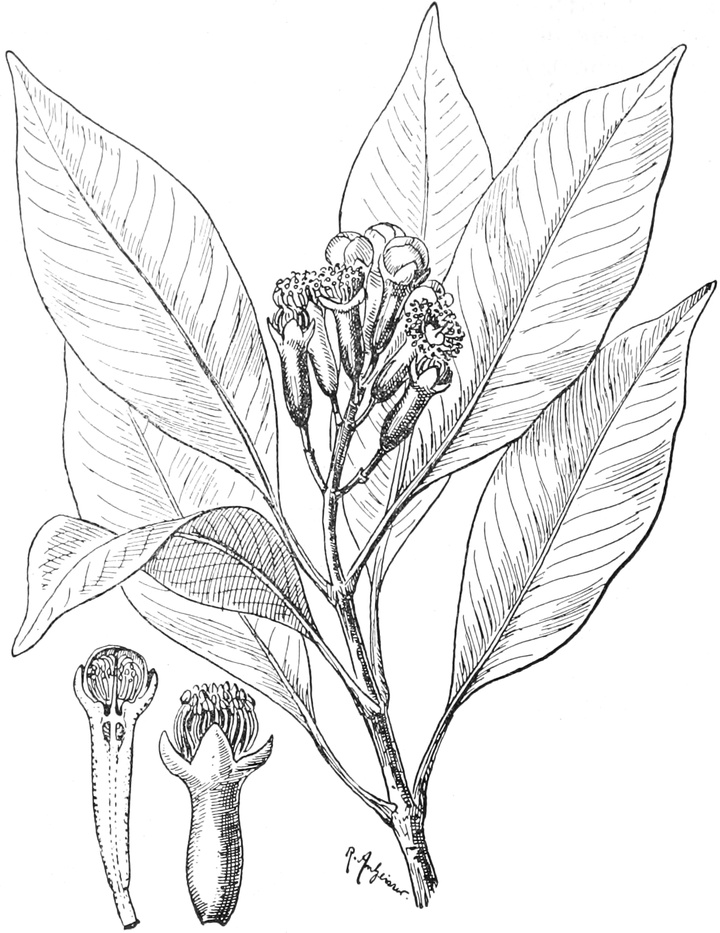
Fig. 660. Jambosa caryophyllus. 2⁄3 nat. Gr. Blühender
Zweig. Eine Knospe halbiert und eine geöffnete Blüte daneben, etwa nat.
Gr. — Offizinell.

Fig. 661. Punica granatum. 1⁄2 nat. Gr. 1 Zweig mit
Blüte und Knospe. 2 Blütenlängsschnitt. 3 Frucht (vgl. Text).
Offizinell.

Fig. 662. Blütendiagramm von Punica granatum. Nach
A. W. EICHLER.
Die einzige Gattung der 7. Familie der Punicaceae
ist Punica. Punica granatum, ein aus dem Orient stammendes
Bäumchen, das seiner säuerlichen, äußerst erfrischenden Früchte
wegen vielfach im Mittelmeergebiet gezogen wird (Fig. 661), hat
kleine Blätter und ansehnliche Blüten mit steifem, rotem Kelch, der
unbestimmt viele Kron- und zahlreiche Staubblätter umschließt. 7–14
Fruchtblätter sind in zwei Stockwerken angeordnet, deren oberes
der vollen Kelchblattzahl, das untere ihrer Hälfte entspricht
(Fig. 661 2, 662). Die Frucht wird von lederiger Wand umhüllt,
mit zahllosen Samen in den Fächern beider Etagen. Die äußeren,
fleischigen Schichten der Samenschale stellen den genießbaren Teil
der Frucht dar.
Offizinell: Cortex Granati (Pharm. germ., austr.,
helv.), Stamm- oder Wurzelrinde des Bäumchens.
6. Ordnung.
Umbelliflorae.
Das gemeinsame Merkmal der in dieser Ordnung vereinigten Pflanzen
sind die doldenförmigen Blütenstände, aus zwittrigen, strahligen
Einzelblüten, mit nur einem Staubblattwirtel und unterständigem,
zweifächerigem, aus zwei Fruchtblättern gebildeten Fruchtknoten,
dessen obere Diskusfläche als Nektarium dient; jedes Fach enthält eine
hängende Samenanlage.
[Nach den neuesten sero-diagnostischen Untersuchungen sollen
die Umbellifloren den Abschluß des Rosiflorenastes bilden.
Morphologisch wäre ihre nächste Verwandtschaft unter den
Rubiaceen-Caprifoliaceen zu suchen.]
1. Familie Cornaceae. Cornus mas. Die
Kornelkirsche (Fig. 662) entwickelt ihre kleinen Dolden
vierzähliger gelber Blüten bereits vor Erscheinen der einfachen
Blätter; jede Dolde von vier Hochblättern gestützt. Ihre
Steinfrüchte von säuerlich-herbem Geschmack werden besonders in
den Balkanländern vielfach verwendet. Nächstjährige Blütenstände,
schon zur Zeit der Fruchtreife in den Blattachseln ausgebildet,
überdauern den Winter unter dem Schutze ihrer als Knospenschuppen
ausgebildeten Hochblätter. C. sanguinea, häufiger Strauch. C.
suecica, nordische Staude, die in Norddeutschland ihre Südgrenze
erreicht.
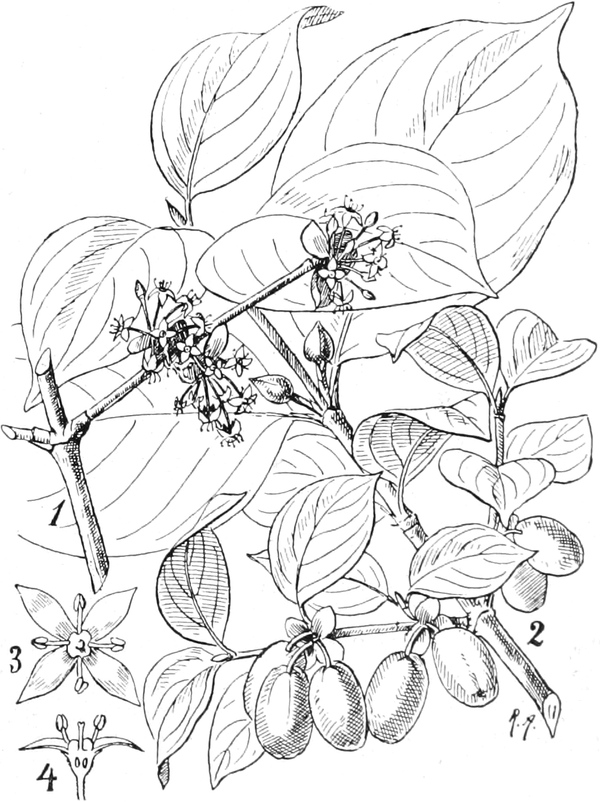
Fig. 662. Cornus mas. 1⁄2 nat. Gr. 1 Blühender, 2
fruchttragender Zweig. 3 Blüte von oben. 4 Blütenlängsschnitt. 3,
4 vergr.
Von der 2. Familie, Araliaceae, ist nur der bekannte
wurzelkletternde Efeu, Hedera Helix[481], in Deutschland
heimisch. Seine elliptisch zugespitzte Blattform tritt erst an
den orthotropen Trieben älterer Pflanzen auf, welche alsdann auch
(im Spätsommer oder Herbst) zur Blüte gelangen. Die jugendlichen
Blätter der kriechenden oder kletternden plagiotropen Triebe sind
gelappt und wenigstens bei wildwachsenden Individuen sehr kurz
gestielt. Ein fünfzipfeliger Kelch entspricht den fünf Rippen des
Fruchtknotens. Die Krone ist von grünlicher Färbung; ihr großer
Diskus auf der Fruchtknotenoberfläche lockt reichlichen Besuch von
Fliegen und Bienen herbei. Die Früchte reifen während des Winters
oder Frühjahrs zu blauschwarzen Beeren; die Samen werden durch
Amseln und andere Vögel, die den Früchten eifrig nachstellen,
verbreitet.
[S. 548]
Die 3. Familie, Umbelliferae, ist bei weitem die
umfangreichste und wichtigste der Ordnung. Sie enthält ausschließlich
Kräuter oder Stauden von bisweilen mächtiger Größe. Der Stengel
trägt wechselständige Blätter, welche ihn mit ihren oft stark
entwickelten Scheiden völlig umfassen; er ist in massive Knoten und
hohle Internodien gegliedert und trägt meist reich zusammengesetzte,
nur in wenigen Fällen einfache Blätter. Seine Infloreszenzen sind
gipfelständige Dolden oder häufiger Doppeldolden mit Stützblättern,
die eine „Hülle“ bzw. „Hüllchen“ bilden; die Enddolden werden vielfach
durch nächstjüngere Achselsprosse übergipfelt. Ihre Einzelblüten sind
weiß, grünlich oder gelb, andere Farben sind sehr selten. (Fig. 663.)
K5 (meist nur in Form kurzer Spitzen), G5, A5, G(̅2). Die Samenanlage
hängt an der medianen Scheidewand (Fugenfläche) herab und kehrt ihre
Mikropyle aufwärts und auswärts. Die Scheitelfläche der Fruchtblätter
wird von einem geschwollenen Diskus, dem Nektariumpolster, eingenommen,
das in den Griffeln mit kugeligen Narben endet. Durch Trennung in
der Fugenfläche zerfällt die Spaltfrucht in zwei Teilfrüchtchen,
die in vielen Fällen nach ihrer Trennung noch eine Zeitlang von dem
in der Mittellinie der Scheidewand befindlichen, aus mechanisch
wirksamem Gewebe bestehenden Karpophor, dem Fruchtträger,
festgehalten werden. Hauptverbreitungsgebiete der Umbelliferen sind
das westasiatische Steppengebiet, das mittlere Nordamerika, Chile und
Australien.

Fig. 663. Umbelliferae, Diagramm (Siler). Nach F.
NOLL.
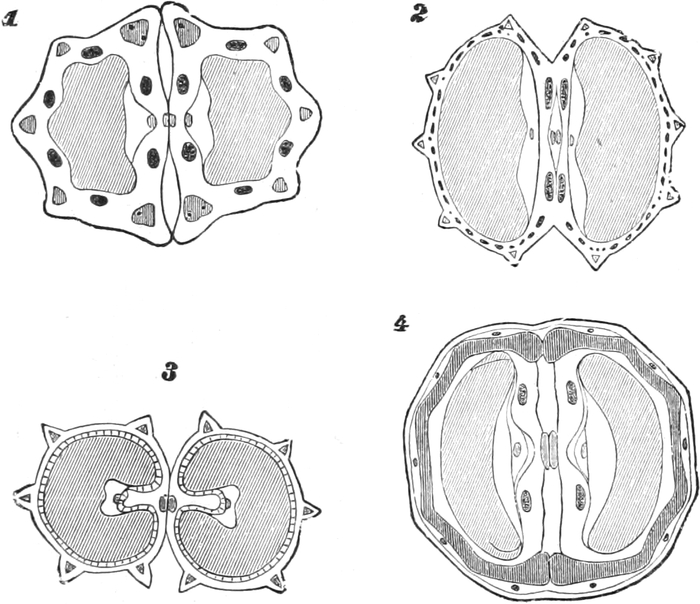
Fig. 664. Umbelliferenfrüchte im Querschnitt. 1
Foeniculum capillaceum. 2 Pimpinella Anisum. 3 Conium maculatum.
4 Coriandrum sativum. (4 mit Benutzung einer Figur von O.
DRUDE) Vergr. — Offizinell.
Fig. 665. Carum Carvi. 1⁄2 nat. Gr. Fruchtstand,
Einzelblüte und Teilfrüchte am Karpophor. Vergr. — Offizinell.
Für die systematische Unterscheidung sind vor allem die Früchte von
Bedeutung. Jede Teilfrucht ist mit fünf Rippen ausgestattet,
welche die Leitbündel führen. An[S. 549] der Fugenfläche liegen die
Randrippen der beiden Teilfrüchte, die häufig von den
übrigen drei Rückenrippen verschieden sind. Zwischen den
fünf Hauptrippen finden sich bisweilen noch vier Nebenrippen
(Coriandrum). In der Regel folgt jedoch ein Tälchen auf
eine Rippe, und in jedem Tälchen verläuft ein großer, die Frucht
der Länge nach durchziehender, als Ölstrieme bezeichneter
Sekretgang. Die Fugenfläche führt beiderseits des Karpophors je
eine Ölstrieme, so daß sechs davon jeder Teilfrucht zukommen
(Fig. 664, 1). Daneben finden sich bei einzelnen Formen noch
anderweitige Sekretbehälter (Fig. 664, 2, 3). Verschiedene
Querschnittsform der Früchte, je nachdem der quer zur Fugenfläche
genommene oder der in ihr liegende Durchmesser größer ist, Form von
Rand- und Rückenrippen, Fehlen oder Vorhandensein von Ölstriemen
oder Nebenrippen ermöglichen eine gute und sichere Erkennung der
Früchte; diese sind für die Bestimmung der Arten unentbehrlich.
Da zahlreiche Umbelliferenfrüchte als Arzneimittel und Gifte,
andere als Gewürze usw. benutzt werden, ist ihre Unterscheidung
von erheblicher Bedeutung. Das Endosperm der Umbelliferensamen hat
fettes Öl als Reservestoff gespeichert.
Bei Pimpinella, Bibernell, und den folgenden Gattungen ist
das Endosperm an der Fugenseite flach oder schwach konvex
(Fig. 664, 1, 2). Pimpinella Anisum, der einjährige
Anis. Carum Carvi, Kümmel, eine alte Kulturpflanze (Fig.
665), deren Blätter doppelt gefiedert-fiederspaltig sind, mit
nebenblattartigen untersten Fiederchen; die folgenden größeren
werden an der vertikal aufsteigenden Hauptspindel horizontal
gestellt. Die letzten Fiederchen endlich sind einfach lineal.
Jede gipfelständige, erstblühende Dolde wird von den aus oberen
Blattachseln entwickelten seitlichen übergipfelt. Die Pflanze
ist zweijährig. Foeniculum, Fenchel, Pastinaca, Pastinak und
Levisticum, Liebstöckel, blühen gelb; Petroselinum, Petersilie,
Daucus, Möhre oder Karotte, Apium, Sellerie, Anethum,
Dill, sind Gemüsepflanzen. Die Wasser- oder Sumpfpflanzen:
Cicuta, Wasserschierling (Fig. 666), Sium, Merk, Oenanthe,
Pferdekümmel, Berula, Berle, ebenso die sog. Hundspetersilie
Aethusa Cynapium (Fig. 667), ein Gartenunkraut mit gekielten
Fruchtrippen und drei langlinealen außenwendigen Hüllblättchen
am Döldchen, sind sämtlich giftig. Archangelica officinalis,
Engelswurz, eine bis etwa 2 m hohe stattliche Pflanze, hat doppelt
gefiederte große Blätter mit sackförmigen Scheiden; ihre grünlichen
Blüten sind auffallend stark protandrisch; die Pflanze dient im
hohen Norden als Nahrungsmittel.
Fig. 666. Cicuta virosa. 1⁄2 nat. Gr.
Rhizom-Längsschnitt. Frucht vergr. — Giftig.
Bei Scandix und Anthriscus, dem Kerbel, sind die Früchte
geschnäbelt, und die Fugenseite des Endosperms wird, wie bei
den folgenden Gattungen, von einer Längsrinne[S. 550] durchzogen.
Chaerophyllum, Kälberkropf. Conium maculatum (Fig. 668),
der Schierling, ist eine häufige zweijährige Pflanze, oft von
stattlicher Höhe. Diese bekannte Giftpflanze ist völlig unbehaart,
die roten Flecken am Stengel wie den Blattstielen rechtfertigen
den Artnamen. Die mattgrünen Blätter sind doppelt bis dreifach
gefiedert-fiederteilig, und ihre äußersten Spitzen laufen stets in
einen farblosen stachelartigen Fortsatz aus. Diese Blattform und
die wellig gekerbten Längsrippen des der Ölstriemen in den Tälchen
entbehrenden Fruchtknotens sind neben dem eigenartigen unangenehmen
Geruch die Hauptkennzeichen der Pflanze (Fig. 664, 3).
Bei Coriandrum sativum ist die Fugenseite des Endosperms
ausgehöhlt (Fig. 664, 4). Der einjährige Koriander hat durch
starke Verlängerung der nach außen gekehrten Kronblätter
dorsiventrale Blüten. Durch feste Verwachsung der an der Fugenseite
tief ausgehöhlten zwei Teilfrüchte wird die Gesamtfrucht kugelig
mit geschlängelten, kaum vortretenden Hauptrippen und dazwischen
etwas schärfer markierten geraden Nebenrippen.
Fig. 667. Aethusa Cynapium. 2⁄3 nat. Gr. B
Einzeldolde. C Frucht. Vergr. — Giftig.
Offizinell: Archangelica officinalis liefert Rad.
Angelicae (Pharm. germ., austr., helv.). — Levisticum
officinale: Rad. Levistici (Pharm. germ., helv.),
Pimpinella magna und P. Saxifraga: Rad. Pimpinellae
(ibid.). — Imperatoria Ostruthium: Rhizoma Imperatoriae
(Pharm. helv.). — Pimpinella Anisum: Fructus Anisi,
Oleum Anisi (Pharm. germ., austr., helv.). — Foeniculum
capillaceum: Fruct. Foeniculi (ibid.). Carum Carvi:
Fruct. Carvi (ibid.). — Coriandrum sativum: Fruct.
Coriandri (Pharm. austr.). — Petroselinum sativum: Fruct.
Petroselini (Pharm. helv.) und Rad. Petroselini (Pharm.
austr.). — Conium maculatum: Herba Conii (Pharm.
germ., austr.), Fructus Conii (Pharm. helv.). — Dorema
Ammoniacum (Persien): Ammoniacum (Pharm. germ., austr.,
helv.). — Ferula galbaniflua und andere Arten (Persien):
Galbanum (Pharm. germ., austr., helv.). — Ferula Narthex
(Tibet) und F. Asa foetida (Persien): Asa foetida (ibid.).
[S. 551]

Fig. 668. Conium maculatum. 1⁄2 nat. Gr. —
Offizinell und giftig.
7. Ordnung.
Centrospermae.
Pflanzen meist mit Zwitterblüten von dem fünfzähligen Dikotylentypus.
An die Polycarpicae, und zwar die Berberidaceae, sollen nach den
sero-diagnostischen Untersuchungen die Centrospermae anschließen, denen
nach ihrer zentralen Placenta auch die Primulinae angehören würden.
Während ich die hierher zugewiesenen Lentibulariaceae am alten Platze
bei den Scrophulariaceae belasse, weise ich den Cactaceae die ihnen von
jeher angewiesene Stelle neben den Aïzoaceae zu, da ihr Hinaufrücken
fast bis an den Sympetalenast kaum morphologisch zu rechtfertigen ist.
1. Familie Aïzoaceae. Gattung Mesembryanthemum.
Perenierende, meist krautige, stark xerophile und blattsukkulente
Pflanzen Südafrikas. Die hermaphroditen Blüten haben eine
vielblättrige, aus Staubblättern umgebildete Krone, zahlreiche
Staubblätter und 2-∞ Fruchtblätter, die hygroskopische Kapselfrüchte
liefern. Die Gattung enthält steinähnliche „Mimicry“-Arten wie M.
truncatellum, pseudotruncatellum, Bolusii und calcareum[477].
2. Familie Cactaceae. Diese auf Amerika beschränkte
Familie umfaßt blattlose Stammsukkulenten von sehr geringen bis
zu gewaltigen Dimensionen, deren Zwitterblüten strahlig, seltener
dorsiventral sind mit vielzählig spiraligem, langsam von Kelch in
Krone[S. 552] übergehendem Perianth und vermehrtem Andröceum und Gynäceum.
Der unterständige Fruchtknoten ist einfächerig und enthält
zahlreiche wandständige Plazenten mit langgestielten Samenanlagen.
Die Früchte werden zu Beeren, deren Fleisch wesentlich aus diesen
Samenstielen hervorgeht.
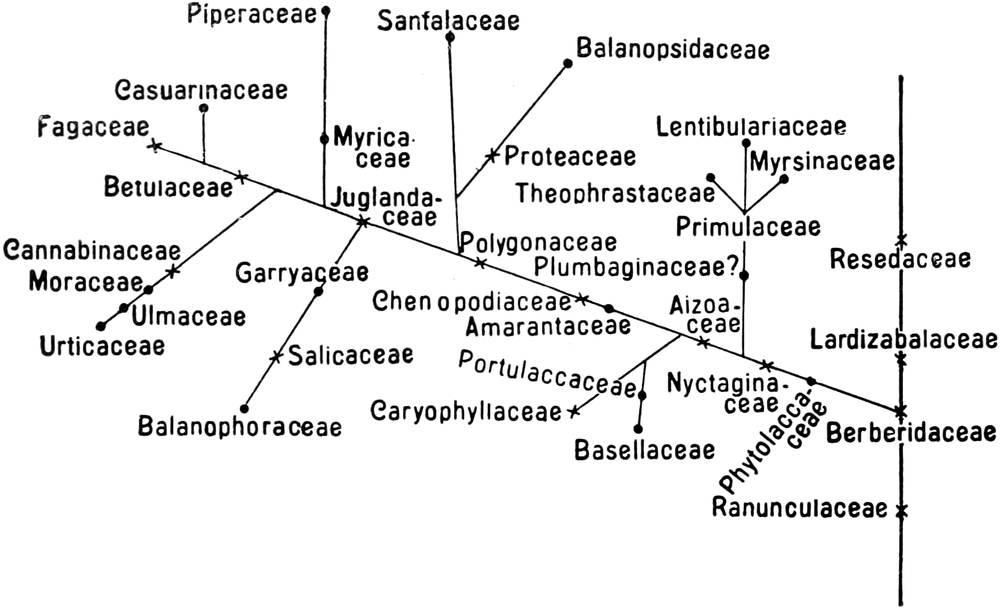
Stammbaum des Astes der Centrospermen und der anschließenden Familien
nach den sero-diagnostischen Untersuchungen von F. MALLIGSON.
Nur Peireskia und einzelne Opuntia-Arten haben noch Blätter,
andere Opuntien nur flache Sproßglieder (Fig. 195). Cereus
(Fig. 199), Echinocactus u. a. haben Längsrippen, Mamillaria
freie Höcker (Mamillen); die zahlreichen Dornbüschel auf den
Gliedersprossen, den Rippen oder den einzelnen Mamillen entsprechen
je einem Achselsproß, dessen Tragblatt verkümmert ist, während die
Blattanlagen der verbreiterten Sproßachse in Dornen umgebildet
werden (Fig. 669).
Cactaceae bilden einen Hauptbestandteil der Vegetation im
regenarmen Südwesten der Vereinigten Staaten von Nordamerika,
Mexiko und den Anden Südamerikas. Ihre eigenartige Gestaltung kehrt
bei Euphorbiaceen und Asclepiadaceen unter ähnlichen klimatischen
Bedingungen wieder (vgl. S. 149). (Konvergenzerscheinung.) Die
Gattungen Rhipsalis, Epiphyllum und Phyllocactus enthalten
zahlreiche epiphytische Formen. Im Mittelmeergebiet verwildert:
Opuntia Ficus indica, deren Früchte genießbar sind, wie auch
zahlreiche andere Arten der Familie in ihrer Heimat als Obstbäume
geschätzt werden. Einige Cactaceae, wie Anhalonium u. a.,
führen stark giftige Alkaloide und Saponine. Opuntia und
Nopalea-Arten (Nopalea coccinellifera) dienen zur Kultur der
Cochenille-Schildlaus, die den Karminfarbstoff liefert.
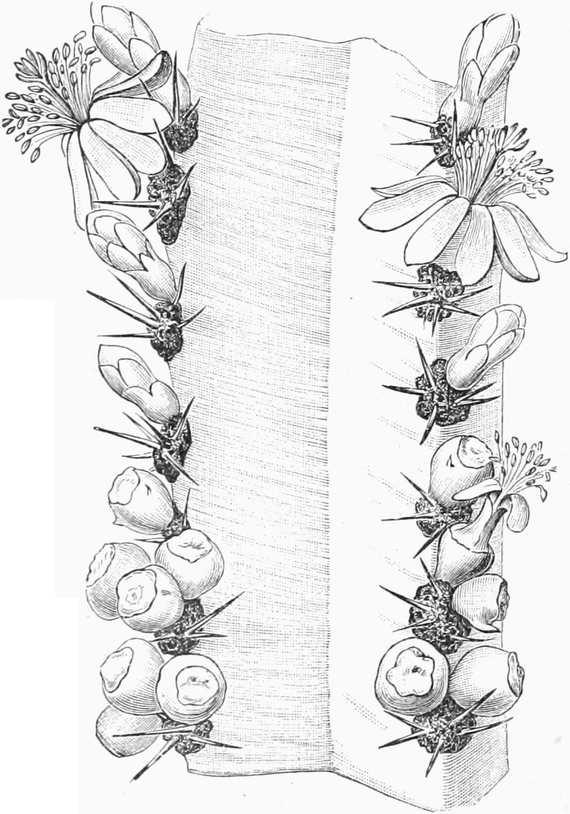
Fig. 669. Cereus geometrizans. Zwei Rippen eines
fünfrippigen Stammes mit Blüten und Früchten. 3⁄4 nat. Gr.
3. Familie Caryophyllaceae. Hierher gehören Kräuter mit
ein[S. 553]fachen, lineal-länglichen, in der Regel gegenständigen Blättern
und pentazyklischen fünfzähligen Blüten mit Kelch und Krone; die
Andröceumkreise sind obdiplostemon, und der einfächerige Fruchtknoten
ist oft unvollständig gefächert. K5, C5, A5 + 5, G(5) (Fig. 671).
Die Kapselfrüchte bergen zahlreiche Samen, die einen um das mehlige
Perisperm herumliegenden gekrümmten Embryo enthalten.

Fig. 670. Saponaria officinalis. 1⁄2 nat. Gr. —
Giftig.
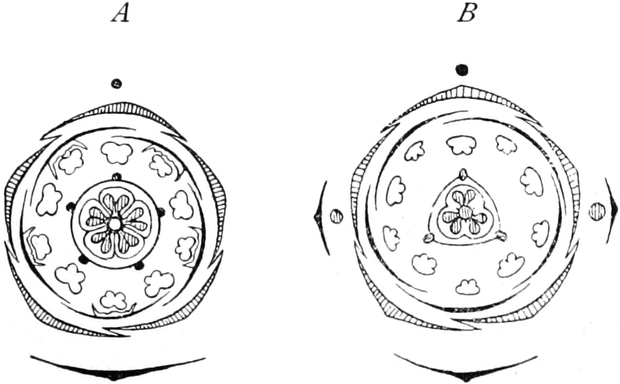
Fig. 671. Diagramme von Caryophyllaceen. A Viscaria,
Scheidewände im unteren Teile des Fruchtknotens vorhanden. B Silene,
Scheidewände fehlen. Nach A. W. EICHLER.
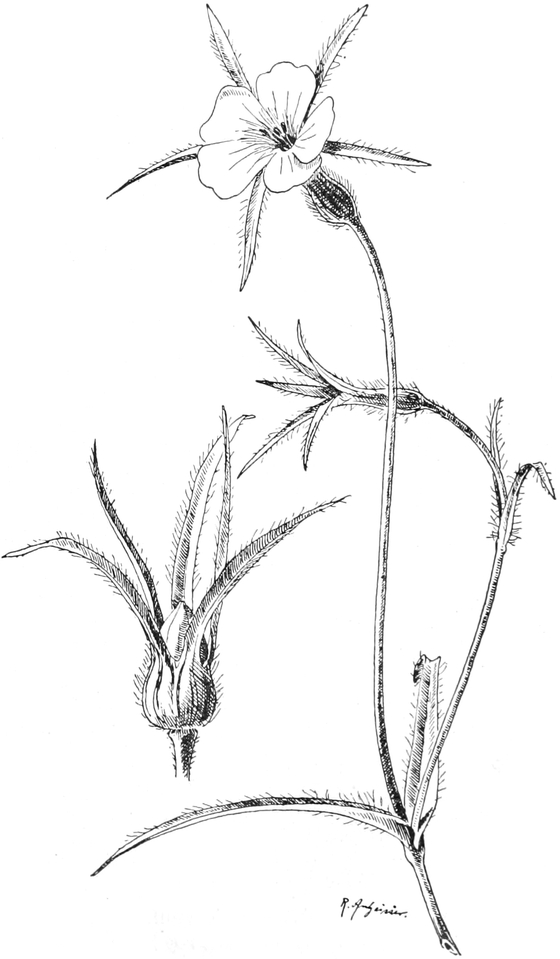
Fig. 672. Agrostemma Githago. Blühender Zweig und
Frucht. 1⁄2 nat. Größe. — Giftig.
Cerastium- und Stellaria-Arten gehören zu dem Frühjahrsschmuck
unserer Fluren und Wälder mit ihren zierlichen weißen Blüten
und gespaltenen Kronblättern. Dianthus-Arten, Nelken, sind an
Wegrändern, sonnigen Hügeln, im Mittel- und Hochgebirge durch ihre
leuchtenden Farben oder ihren Geruch auffallend. Als Ackerunkraut
ist die filzig-behaarte Kornrade, Agrostemma[S. 554] Githago (Fig. 672),
mit blauroten Blüten häufig, ihre Samen sind giftig. Saponaria
officinalis, ein meterhohes Kraut mit gegenständigen breiten
Blättern, ist saponinhaltig und darum giftig (Fig. 670).
Offizinell: Herba Herniariae (Pharm. austr.) von
Herniaria glabra und H. hirsuta.
4. Familie Chenopodiaceae enthält Kräuter mit wechselständigen
Blättern und typisch fünfzähligen Blüten mit einem Perigon- und
Andröceumkreis: P5, A5, G(2–5). Die Staubblätter stehen vor
den Perianthblättern, Reduktionen zu eingeschlechtigen Blüten sind
nicht selten. Eine grundständige, kampylotrope Samenanlage ist im
einfächerigen Fruchtknoten enthalten, der zu einer Nuß wird. Der Samen
birgt einen gekrümmten Embryo, der das mehlige Perisperm rings umlagert.
Fig. 673. Diagramm der Primulaceen (Primula). Nach
F. NOLL.
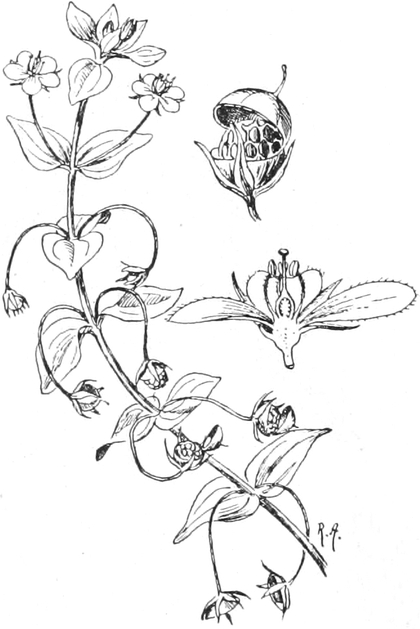
Fig. 674. Anagallis arvensis. 1⁄2 nat. Gr.
Längsdurchschnittene Blüte und aufgesprungene Kapsel. Vergr. —
Giftig.
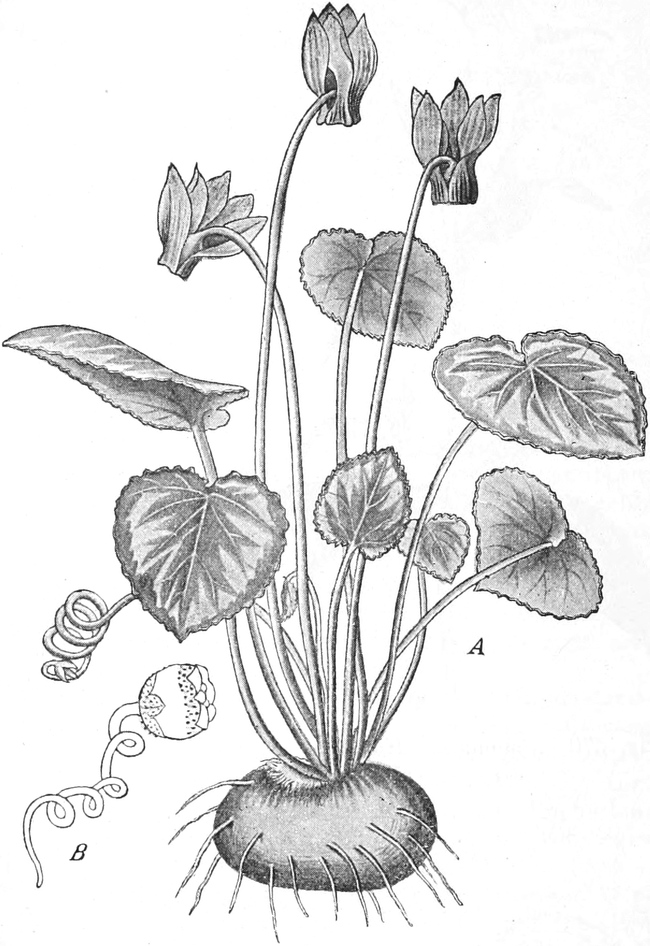
Fig. 675. Cyclamen europaeum. Verkl. A Blühende
Pflanze. B Frucht mit sich schraubig in die Erde einrollendem
Fruchtstiel. Nach H. G. REICHENBACH. — Giftig.
Chenopodiaceen sind vielfach Bewohner des Meeresstrandes und
überhaupt auf salzhaltigem Boden verbreitet, so in den großen
asiatischen Salzsteppen und Wüsten. Salicornia am Meeresstrande
und sonst auf Salzboden (vgl. S. 147). — Neben dem als
Gemüsepflanze zu erwähnenden Spinat (Spinacia oleracea Winter-,
Sp. glabra Sommerspinat) ist vor allem die Zuckerrübe,
Beta vulgaris var. Rapa, von Wichtigkeit. Sie liefert den
offizinellen Zucker (Pharm. germ., austr., helv.). Die Pflanze ist
zweijährig. Im ersten Jahre bildet sie eine dick anschwellende,
fleischige Wurzel, die von der Knospe[S. 555] und einer Anzahl saftiger,
dickstieliger, ganzrandiger, oft etwas gekrauster Blätter gekrönt
wird. Aus dieser Blattrosette treibt im zweiten Jahre ein reich
verzweigter rispenförmiger Blütenstand hervor, mit unscheinbaren,
grünlichen Einzelblüten reich besetzt, deren Fruchtknoten
dreikarpellig ist. Am Schluß des ersten Jahres geerntet, liefert
die Wurzel Rohrzucker, welcher die gespeicherte Reservenahrung
darstellt. Durch stete Zuchtwahl ist der Zuckergehalt von 7–8% auf
etwa 15% durchschnittlich gebracht, steigt jedoch bis 21%, ja 26%.
Die Stammpflanze der Zuckerrübe ist Beta patula. Chenopodium
und Atriplex sind häufige Unkräuter in der Nähe menschlicher
Wohnungen. Von Chenopodium ambrosioides stammt die offizinelle
Herba Chenopodii (Pharm. austr.).
8. Ordnung.
Primulinae.
Familie der Primulaceen. Das Diagramm (Fig. 673) zeigt
nur einen Staubblattkreis, der aber nicht alterniert, sondern
vor den sympetalen Kronblättern steht. Die zentrale Placenta ist
charakteristisch. Die Gattung Primula ist weit verbreitet;
einheimisch sind zahlreiche Arten, die mit dem der Familie
eigenen einfächerigen oberständigen Fruchtknoten und einfachem
Griffel ausgerüstet sind. Primula besitzt sehr ausgeprägte
dimorphe Heterostylie (Fig. 545). Anagallis mit seiner
Deckelkapsel (Fig. 674) und Cyclamen (Fig. 675) sind bekannte
Primulaceen-Gattungen. Die frischen Knollen von Cyclamen,
das Kraut von Anagallis und die Drüsenhaare verschiedener
Primula-Arten [Pr. obconica, Corthusa Mathioli[482]] sind
giftig.
9. Ordnung.
Polygoninae.
Die einzige Familie, Polygonaceae, enthält meist
perennierende Kräuter mit hohlen, knotig gegliederten Stengeln und
einfachen wechselständigen Blättern. Ihre häutigen Nebenblätter
sind zu einer die Stammknospe überziehenden Tüte, Ochrea,
verwachsen die vom Stamme durchbrochen wird und als röhrenartiges
Organ an der Internodiumbasis erhalten bleibt (Fig. 676). Die
Familie ist vorwiegend in der nördlichen gemäßigten Zone heimisch.
Fig. 676. Blatt von Polygonum amplexicaule mit der
Ochrea st. 1⁄3 nat. Gr.
Fig. 677. Rheum officinale. A Blüte. B
Blütenlängsschnitt. C Gynäceum mit Diskus. D Rheum compactum,
Frucht. Vergr. Nach LÜRSSEN.
Gattungen: Die Gattung Rheum, Rhabarber, stammt
aus Ostasien und trägt mächtige, grundständige, einfache,
handförmig geaderte Blätter und aufstrebende, sehr große, rispige
Blütenstände. Die Einzelblüten besitzen zwei Perigonkreise von
gleichartiger Ausbildung, zwei Staubblattkreise, deren äußerer
durch Spaltung verdoppelt[S. 556] ist, also P3 + 3, A6 + 3, G(̲3). —
Große drüsige Diskusschuppen deuten auf Insektenbesuch hin. Der
dreikantige Fruchtknoten ist in der Reife dreiflügelig (Fig. 677
D). Rheum-Arten werden vielfach als Zierpflanzen, mehr noch als
Gemüse gebaut. Rumex acetosa, Sauerampfer, einheimische
Gemüsepflanze mit spießförmigen Blättern. Ihr Blütenbau gleicht
demjenigen von Rheum, doch fehlt der innere Staubblattkreis.
Polygonum-Arten mit gefärbtem, fünfblätterigem Perigon und
wechselnder Zahl im Andröceum; Fagopyrum esculentum liefert in
seinen dreikantigen Früchten Buchweizen (Fig. 585 B).
Offizinell: Rhizoma Rhei von Rheum officinale,
Rh. palmatum und wohl noch anderen Arten. (Pharm. germ., austr.,
helv.) Herba Polygoni von Polygonum aviculare (Pharm. austr.).
10. Ordnung.
Loranthiflorae.
1. Familie Santalaceae. Grüne, im Boden wurzelnde
Halbparasiten, die den Wurzeln anderer Pflanzen durch
Haustorien Nährstoffe entziehen. Thesium-Arten einheimisch.
Offizinell: Oleum Santali (Pharm. germ., austr.,
helv.) aus dem Holze von Santalum album, das für Kunsttischlerei
geschätzt ist. 2. Familie Loranthaceae. Auf Baumästen
lebende, belaubte, halbparasitische Sträucher, die besonders in
den Tropen häufig sind und z. B. im tropischen Amerika durch die
Farbenpracht ihrer Blüten zu Zeiten auffallen. Einheimisch:
Der seltene Loranthus europaeus lebt auf Eichen; Viscum album
(Fig. 678), die Mistel, ein häufiger immergrüner Halbparasit
auf den verschiedensten Bäumen, trägt gegenständige, am Ende
verbreiterte, ganzrandige Blätter am knotig gegliederten Stamm. Die
weißen Beerenfrüchte werden von Vögeln verbreitet. Aus den Samen
treibt ein der Wurzelhaube entbehrender „Senker“ durch die Rinde
bis ans Holz, in das er nicht eindringen kann. Sein Scheitel wird
vom Jahreszuwachs des Holzes umwallt; die dem jeweiligen Kambium
des Wirtes entsprechende Region vermittelt weiteren Zuwachs.

Fig. 678. Viscum album. 1⁄2 nat. Gr. Mit Blüten und
Früchten.
11. Ordnung.
Juglandiflorae.
Die Familie Juglandaceae enthält stattliche, monözische
Bäume der nördlichen Hemisphäre mit unpaarig gefiederten,
aromatischen Blättern in wechselständiger Anordnung ohne
Nebenblätter und mit stark reduzierten Blüten.
Der Walnußbaum, Juglans regia (Fig. 679), ist in Westasien
und im östlichen Mittelmeergebiet einheimisch und wird jetzt
in ganz Europa viel kultiviert. Im Frühjahr strecken sich aus
vorjährigen Achselknospen lang herabhängende, dicke Kätzchen
hervor, deren Achse mit zahlreichen Einzelblüten bedeckt ist. Jede
besteht aus 3–5 Perianthblättern, welche mit zwei Vorblättern
zusammen dem Deckblatt angewachsen sind und zahlreiche Staubblätter
umhüllen. Sie sind gegen die Infloreszenzspitze gekehrt.
Weibliche Blüten stehen in geringer Zahl am Gipfel der jungen
Sprosse. Ihre zwei Fruchtblätter[S. 557] enden in großen, federigen,
auseinanderspreizenden Narben, und das Perigon ist ebenfalls
mit Deckblatt und Vorblättern verwachsen. Der unterständige,
einfächerige Fruchtknoten entwickelt seine grundständige atrope
Samenanlage zur Steinfrucht. In der Fruchtschale liegt ein von
dünner Samenschale umschlossener Embryo mit großen, ölreichen
Kotyledonen, die durch unvollkommene, falsche Scheidewände
mannigfach gelappt sind; ein Endosperm fehlt. Auch andere
Juglans-, und Carya-Arten liefern eßbare Samen und geschätztes
Nutzholz.
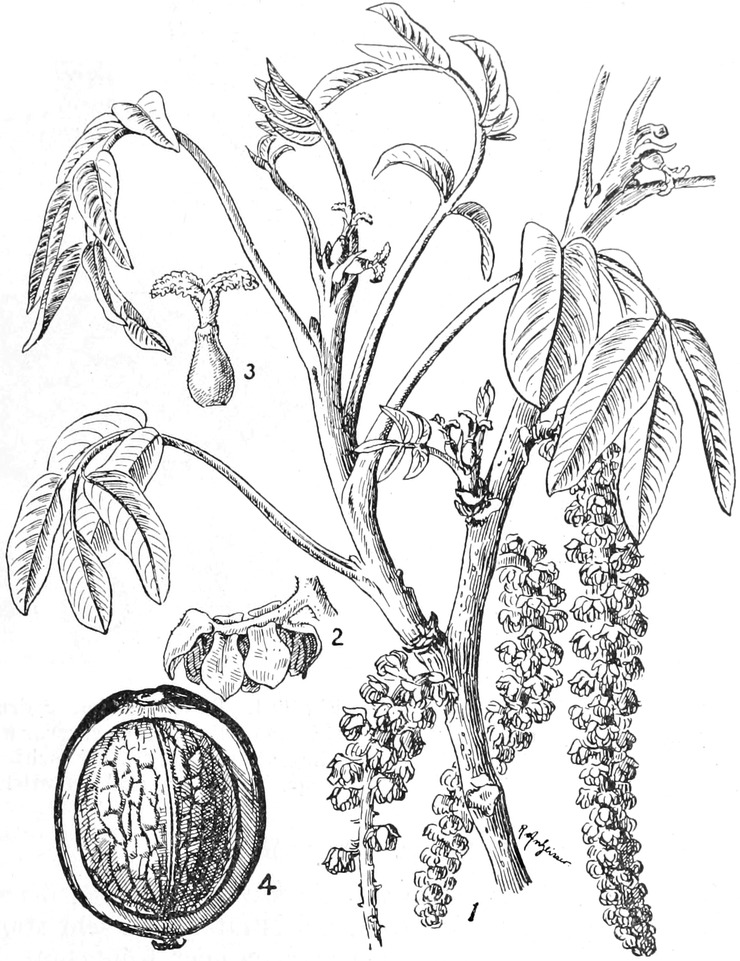
Fig. 679. Juglans regia. 1⁄2 nat. Gr. 1 Zweig mit
jungem Laub, männlichen Kätzchen und weiblichen Blüten am Gipfel. 2
Männliche, 3 weibliche Einzelblüte. 4 Frucht, deren äußere Schale
zum Teil entfernt ist. — Offizinell.
Offizinell: Folia Juglandis (Pharm. germ., austr., helv.)
von Juglans regia.
12. Ordnung.
Piperinae.
Einzige Familie Piperaceae. Wichtig ist die Gattung Piper
mit dreizähligen, meist aber reduzierten Blüten, einfächerigem
Fruchtknoten, der eine Steinfrucht mit Perisperm ergibt, welches
den von spärlichem Endosperm umhüllten Embryo einschließt.
Zerstreute Leitbündel auf dem Stammquerschnitt erinnern an die
Monokotylen, doch ist Dickenwachstum vorhanden. Die Stammpflanze
des Pfeffers Piper nigrum, ist ein im malayischen Gebiete
heimischer, jetzt überall in den Tropen angebauter Wurzelkletterer
(Fig. 680). Seine unreifen Früchte geben den schwarzen
Pfeffer, nach der Reife und Entfernung der fleischigen äußeren
Fruchtschale den weißen Pfeffer.
[S. 558]
Offizinell: Cubebae (Pharm. germ., austr., helv.).
Früchte von Piper Cubeba (Fig. 681), durch stielartigen Fortsatz
der Fruchtbasis vom schwarzen Pfeffer unterschieden. Fructus
piperis nigri (Pharm. austr.), Früchte von P. nigrum.
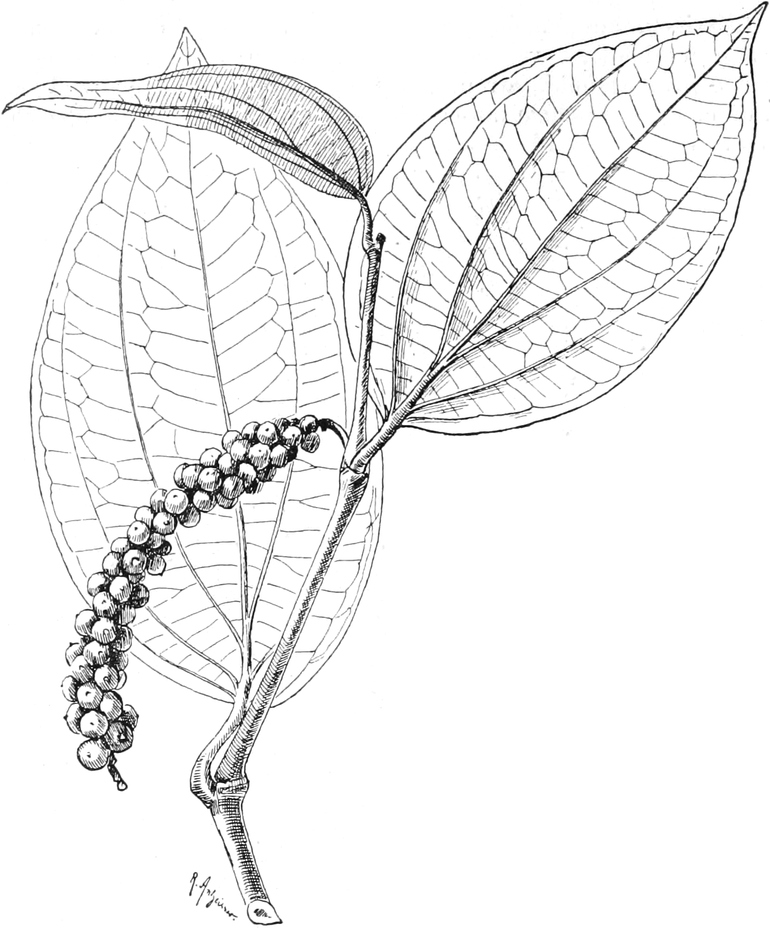
Fig. 680. Piper nigrum. 1⁄2 nat. Gr.
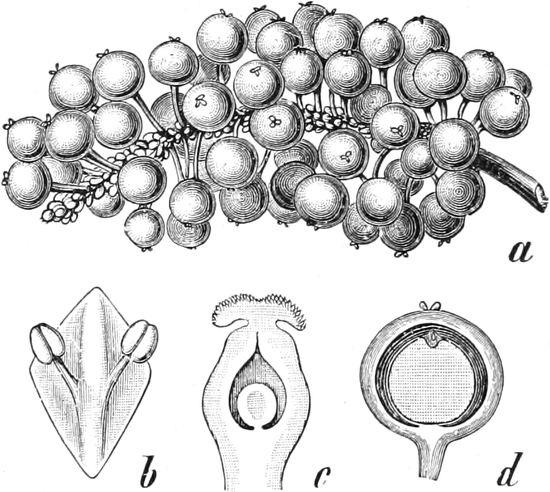
Fig. 681. Piper Cubeba. a Fruchtstand, nat. Gr., b
männliche Blüte, vergr., c weibliche Blüte, Längsschnitt, vergr., d
Frucht. Längsschnitt, vergr. Nach BERG und SCHMIDT.
Offizinell.
13. Ordnung.
Querciflorae[483].
Diese Ordnung umfaßt unsere wichtigsten Laubbäume, deren ungeteilte
Blätter hinfällige Nebenblätter besitzen. Die Blüten sind sehr stark
reduziert, stets monözisch verteilt und stehen in Kätzchen oder
Köpfchen. Der Fruchtknoten ist unterständig mit hängenden Samenanlagen,
welche einsamige, endospermlose Nüsse bringen.
1. Familie Betulaceae. Die männlichen Blüten sind der
Deckschuppe angewachsen. Der zweifächerige Fruchtknoten hat zwei
lange Narben und eine hängende anatrope Samenanlage in jedem Fache.
Die Pflanzen sind vorzugsweise auf der Nordhemisphäre verbreitet.
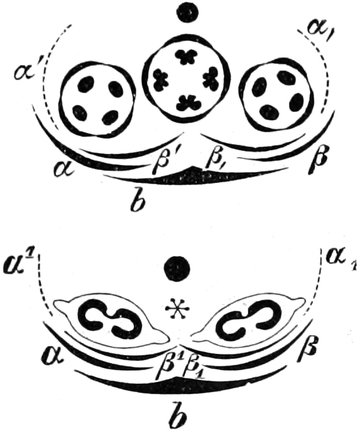
Fig. 682. Alnus glutinosa. Diagramme der männlichen
und weiblichen Blüten (nach A. W. EICHLER). Deckblatt b,
die zugehörigen Vorblätter α β, α′ β′.
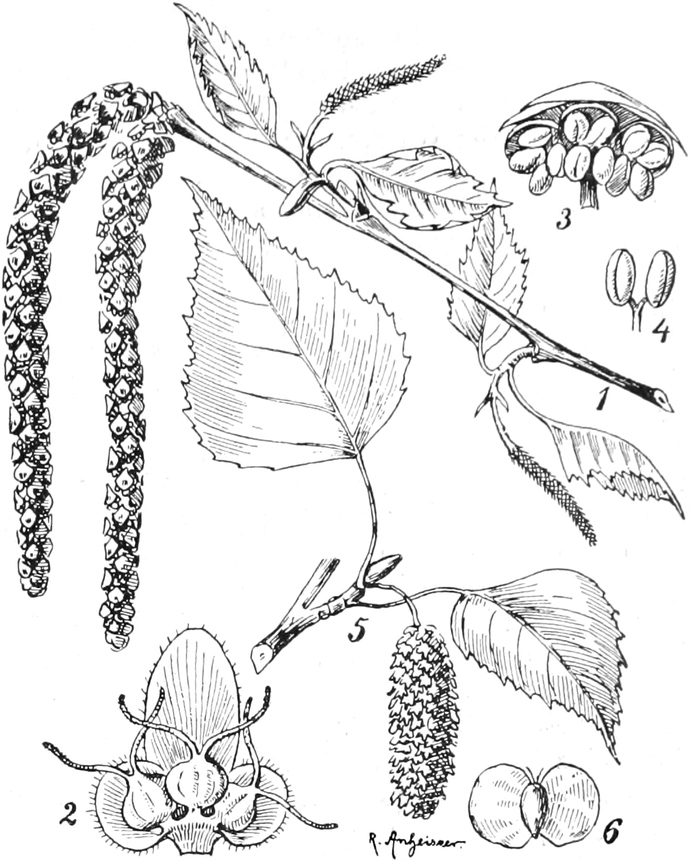
Fig. 683. Betula verrucosa. 1 Blütenzweig,
männliche Infloreszenzen gipfelständig, weibliche an kleineren
Seitenzweigen. 2 Dichasium aus drei weiblichen Blüten. 3
Männliche Einzelblüte. 4 Ein Staubblatt. 5 Ein Fruchtsproß.
6 Frucht. 1 und 5 2⁄3 nat. Gr., 2–4 und 6 vergr. —
Offizinell.
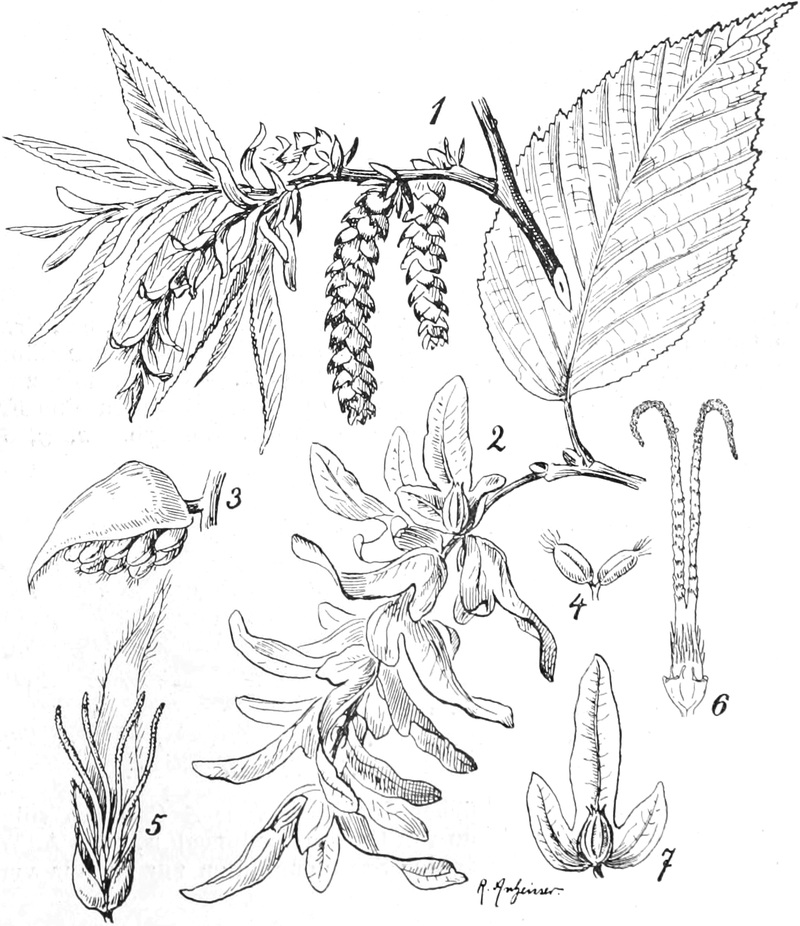
Fig. 684. Carpinus Betulus. 1 Blühender Zweig,
männliche Kätzchen aus vorjährigen Knospen, weibliche am jungen
neuen Jahrestriebe. 2 Fruchtstand. 3 Männliche Einzelblüte.
4 Ein Staubblatt. 5 Deckblatt mit zwei weiblichen Blüten. 6
Weibliche Einzelblüte (nach BÜSGEN). 7 Frucht. 1, 2,
7 2⁄3 nat. Gr., 3–6 vergr.
Wichtige deutsche Gattungen: Alnus glutinosa, unsere
Schwarzerle, ist ein stattlicher Baum feuchter Wälder, er
wächst auch an Sümpfen und Flußufern der Ebene. Seine Blütenstände
erscheinen bereits im Herbste des Vorjahres als gestielte Kätzchen;
die männlichen hängen lang herab, die kurzen weiblichen sind
aufgerichtet. Männliche Blüten P 4, A 4 stehen in dreizähligen
Dichasien (Fig. 538, 682). Weibliche sind in Zweizahl vorhanden;
ihr Deckblättchen verwächst mit den vier Vorblättern zu einer
dauernden, holzigen, fünflappigen Schuppe des Zäpfchens. Alnus
incana ist durch unterseits graufilzige Blätter verschieden.
Betula verrucosa (Fig. 683), unsere weißrindige Birke,
trägt langgestielte dreieckige Blätter; auf allen jugendlichen
Teilen stehen zahlreiche Drüsenhaare, denen der harzig-aromatische
Geruch entströmt. Männliche Blütenstände werden einzeln oder
zu wenigen im Herbst des Vorjahres am Gipfel von Langtrieben
gebildet. Die Blüten (P 2, A 2 in dreizähligen Dichasien) haben
tief zweispaltige Antheren (Fig. 683 3, 4). Weibliche
Infloreszenzen stehen einzeln am Gipfel kleiner dies[S. 559]jähriger
Kurztriebe, ihre Blüten zeigen dreizählige Dichasien unter jeder
der dreilappigen, aus Deckblättchen und den beiden Vorblättern
verwachsenen Schuppen. Der Fruchtstand hängt; nach dem Ausfallen
der geflügelten Früchte zerfällt der Zapfen durch Ablösung der
Schuppen. — Carpinus Betulus. Hainbuche, Weißbuche
(Fig. 684). Ein wichtiger Waldbaum. Seine Infloreszenzen erscheinen
erst im Frühjahre, die männlichen aus vorjährigen Achselknospen
ohne Blätter oder von nur wenigen begleitet, weibliche aber
meist gipfelständig. Die Deckblättchen der männlichen Kätzchen
tragen 4–10 bis zum Grunde gespaltene Staubblätter, aber ohne
Vorblätter oder[S. 560] Perigon. Weibliche Blüten stehen in Zweizahl
unter jedem Deckblatt, jede von besonderem Deckblättchen und zwei
Vorblättern begleitet. Diese drei verwachsen zu der dreilappigen
Hülle, welche als Flugorgan der Verbreitung dient. — Corylus
Avellana, der stattliche Haselstrauch unserer Wälder,
entwickelt seine Infloreszenzen bereits im Vorjahre; männliche
Kätzchen hängen während des Winters nackt herab; die weiblichen
bleiben von den Knospenschuppen umhüllt und strecken nur zur
Zeit der Blüte ihre langen roten Narben zwischen jenen hindurch
ins Freie. Den männlichen Deckblättchen ist nur eine
perianthlose, doch mit zwei Vorblättern versehene Blüte mit vier
bis zum Grunde gespaltenen Antheren angewachsen. In den weiblichen,
sehr gestauchten Kätzchen findet sich ebenso wie bei Carpinus ein
zweiblütiges Dichasium unter jedem Deckblatte vor; die geschlitzte
Fruchthülle entsteht auch hier aus den Vorblättern und dem
besonderen Deckblatt jeder Einzelblüte. Corylus tubulosa, die
Lambertsnuß, aus Südeuropa.
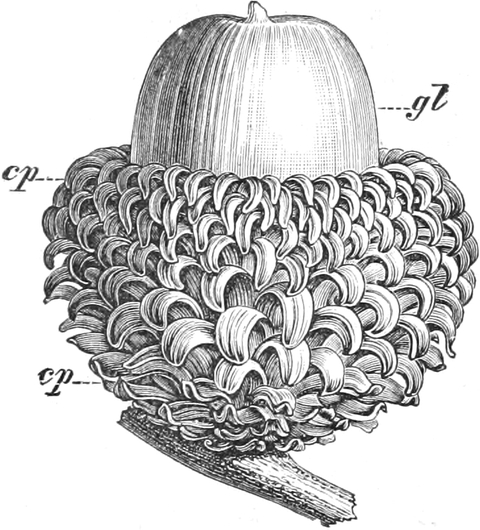
Fig. 685. Frucht mit Cupula von Quercus Aegilops, nach
DUCHARTRE. cp Cupula, gl Frucht.
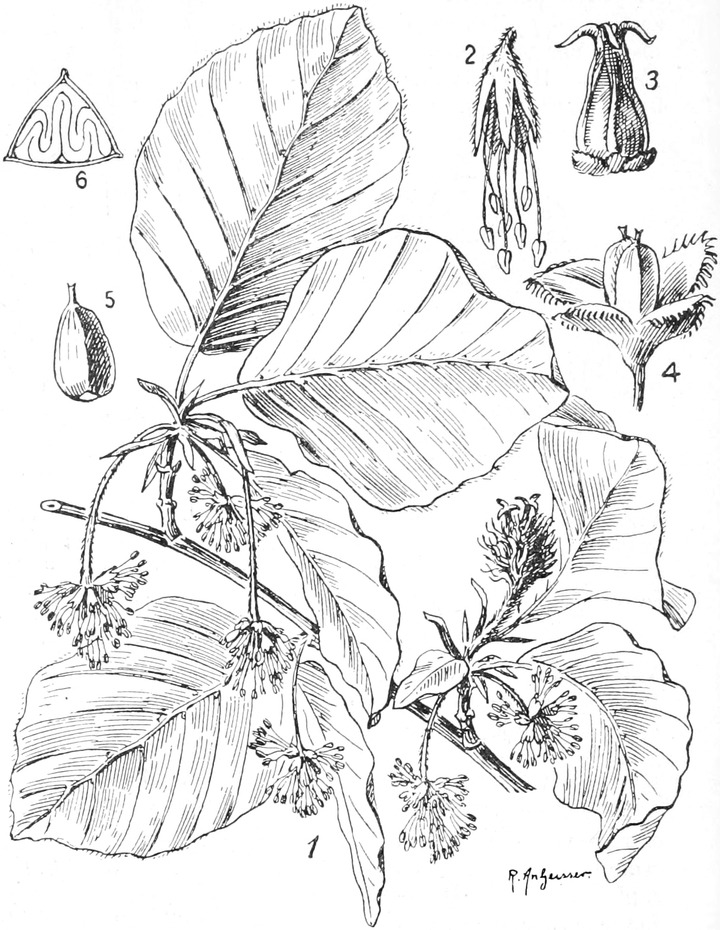
Fig. 686. Fagus silvatica. 2⁄3 nat. Gr. 1 Zweig mit
männlichen und weiblichen Blütenständen. 2 Männliche, 3 weibliche
Einzelblüte. 4 Cupula, geöffnet, mit zwei Früchten. 5 Frucht,
isoliert. 6 Querschnitt durch eine Frucht, zeigt die gefalteten
Kotyledonen des Embryo. 2, 3, 6 vergr. — Offizinell.
Fig. 687. Diagramme weiblicher Dichasien von: A
Castanea vulgaris, Fig. 688, B Fagus silvatica und C der
Einzelblüte von Quercus pedunculata, nach A. W. EICHLER.
Deckblatt b. Vorblätter α, β. Vorblätter der Sekundanblüten zur Cupula
verwachsen α, β, α′, β′.
[S. 561]
Offizinell: Oleum Betulae empyreumaticum, Birkenteer
(Pharm. austr., helv.), von Betula verrucosa.
2. Familie Cupuliferae. Ihre blattachselständigen
Infloreszenzen tragen teils männliche, mit Perianth versehene Blüten,
teils weibliche, die einzeln oder zu mehreren von einer Cupula (Fig.
685 cp) umhüllt werden, d. h. einem aus verwachsenen Vorblättern
entstandenen Gebilde. Der dreifächerige Fruchtknoten trägt je zwei
hängende anatrope Samenanlagen im Fache und endet in drei Narben.
Fig. 689. Quercus pedunculata. Längsschnitt durch den
jungen Fruchtknoten. b Becher, e Samenanlagen, d Fruchtknoten,
c Perigon, f Griffel, g Narbe. Vergr. Nach BERG u.
SCHMIDT.
Fig. 690. Quercus pedunculata. A Blühender Zweig. B
Männliche Blüte. C Staubblätter. D Weibliche Blüte. B–D vergr.
E Fruchtstand. F Cupula. G–H Samen. — Offizinell. Nach
A. F. W. SCHIMPER.
Die Cupuliferen sind hauptsächlich in der gemäßigten Zone
der nördlichen Hemisphäre verbreitet, außerdem im tropischen
Asien. Wichtige einheimische Arten: Fagus silvatica,
Buche, Rotbuche (Fig. 686), einer der wichtigsten
Laubbäume Deutschlands, in schönen Waldungen besonders an der
Ostseeküste vertreten, wo seine Ostgrenze jedoch Königsberg nicht
erreicht. Die zweizeilig stehenden Blätter sind ganzrandig,
elliptisch, kurzgestielt und besonders in der Jugend ringsum fein
bewimpert. Blütenstände finden sich durchweg an diesjährigen
Trieben. Die männlichen sind seitenständig, kopfförmig und hängend,
ihre Einzelblüten mit schief glockenförmigem Perianth haben meist
8–12 Staubblätter. Weibliche Infloreszenzen sind gipfelständige,
zweiblütige Dichasien, welche die Cupula (Fig. 688 B) bis zur
Reife der zu dreieckigen Nüssen heranwachsenden Früchte völlig
einschließt, um alsdann in vier Nähten aufzuspringen. Castanea
vulgaris, Edelkastanie, im Mittelmeergebiet zu Hause,
reift ihre Früchte noch in Süddeutschland. Ihre Infloreszenzen
stehen an diesjährigen Trieben aufrecht; sie führen teils nur
männliche Blüten, teils unten weibliche, oben männliche, in
dichasialer Anordnung. Die weiblichen dreiblütigen Dichasien (Fig.
687 A) liefern drei von der scharf stachlichten Cupula völlig
umschlossene Nüsse. Die Cupula öffnet sich mit vier Klappen.
Quercus pedunculata, Stieleiche (Fig. 689 und 690) und
Quercus sessiliflora, Traubeneiche, die mächtigsten
Laubbäume der europäischen Wälder, haben ovale, rings mehrfach
stumpf ausgebuchtete Blätter. Die männlichen Infloreszenzen brechen
zugleich mit dem frischen[S. 562] Laube aus vorjährigen Blattachselknospen
hervor, ihre Blüten stehen einzeln mit 5–7 Perianth- und 6–12
Staubblättern an der hängenden Achse. Weibliche Infloreszenzen
finden sich am Gipfel diesjähriger Sprosse blattachselständig,
aufrecht, wenigblütig mit einzelnen Blüten, die bei der Stieleiche
langgestielt, bei der Traubeneiche ungestielt sitzend sind. Jede
Blüte wird von ihrer (Fig. 688 C) erst an der reifen Frucht voll
entwickelten Cupula umhüllt.
Die Buche liefert Brennholz, Teer, Holzessig; Quercus technisch
verwertetes Holz, gerbstoffreiche Rinde für die Gerberei. Kork von
der Korkeiche.
Offizinell: Cortex Quercus (Pharm. germ., austr.,
helv.) und Semen Quercus (Pharm. austr.). Die im Orient
heimische Q. infectoria erzeugt an jungen Zweigen, wenn von der
Gallwespe, Cynips tinctoria Hart., gestochen, die offizinellen
Galläpfel, Gallae (Pharm. germ., austr., helv.). Fagus
silvatica gibt Pix liquida (Pharm. austr.).
14. Ordnung.
Saliciflorae.
Familie Salicaceae. Diözische Bäume und Sträucher mit
einfachen wechselständigen Blättern und Nebenblättern, die vor der
Beblätterung blühen. Ihr einfächeriger zweikarpelliger Fruchtknoten
ergibt eine Kapselfrucht mit zahlreichen wandständigen, beschopften
und endospermfreien Samen.
Fig. 691. Salix viminalis. A Blühender, männlicher
Zweig. Nat. Gr. B Männliche Blüte, mit Deckblatt, vergr. C
Weibliches Kätzchen. D und E Weibliche Blüten. vergr. F Frucht.
Nat. Gr. G Dieselbe, vergr. H Samen, vergr. Nach A. F. W.
SCHIMPER.
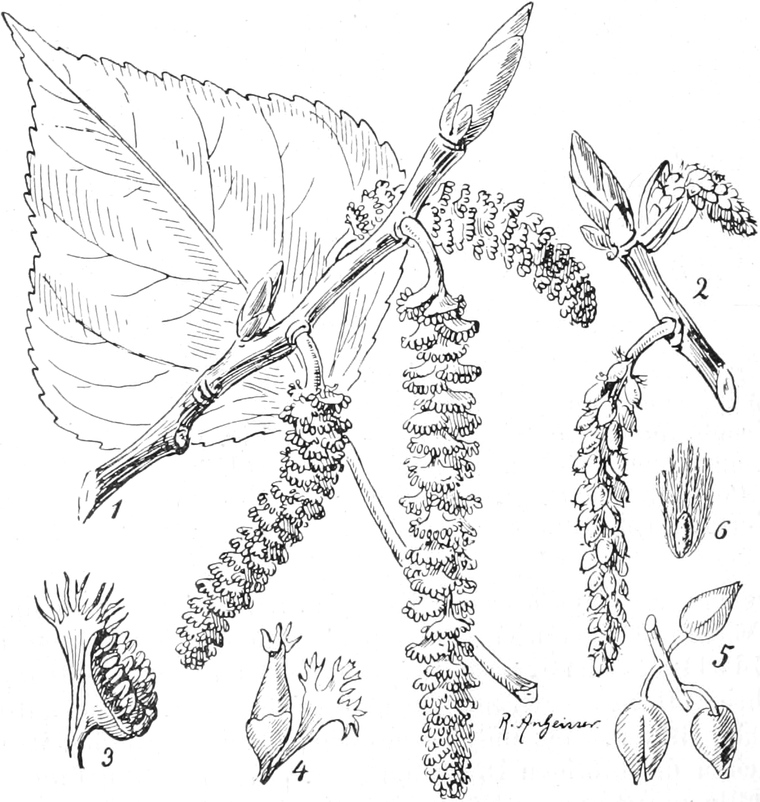
Fig. 692. Populus nigra. 1 Männliche Infloreszenz. 2
Weiblicher Blütenstand. 3 Männliche, 4 weibliche Einzelblüte. 5
Früchte. 6 Samen. 1 und 2 3⁄4 nat. Gr. 3–6 vergr.
Salix, Weide, und Populus, Pappel, sind die
einzigen Gattungen der Familie. Salix, mit aufrechten Kätzchen
versehen, ist an Insektenbestäubung angepaßt und besitzt daher
männliche wohlriechende Blüten, klebrigen Pollen und Nektarien
in Gestalt kleiner Diskusschuppen. Die Zahl der Staubblätter
wechselt in den verschiedenen Arten von 2–5. Deckschuppen sind
ganzrandig (Fig. 691). Bei uns an Flußufern weit verbreitet.
Als unterirdische niedrige Stämmchen, die nur ihre jedesmaligen
jungen Triebe über den Boden emporsenden, gehören Salix-Arten im
hohen Norden und Hochgebirge zu den häufigeren[S. 563] Pflanzenformen.
Populus mit anemophilen Blüten. Der Diskus ist hier mehr krug-
bis schüsselförmig ausgebildet und Honigabsonderung fehlt. Durch
langgestielte, pfeilförmige Blätter unterscheiden sich die
einheimischen Pappelarten habituell erheblich von den Salices. Die
Blüten stimmen bis auf die zerschlitzten Deckblättchen mit denen
von Salix überein. Kätzchen hängend (Fig. 692).
Offizinell: Cortex Salicis von Salix alba, Salix
fragilis usw. (Pharm. helv.).
15. Ordnung.
Urticinae.
Die Ordnung enthält Pflanzen mit unscheinbaren, kleinen Blüten
in dichten Blütenständen. Staubblätter sind den Perigonblättern
gleichzählig und stehen vor ihnen. Der oberständige, ein- oder
zweikarpellige Fruchtknoten ist meist einfächerig mit einer hängenden
Samenanlage, die Nüsse oder Steinfrüchte ergibt. Samen in der Regel
endospermhaltig.
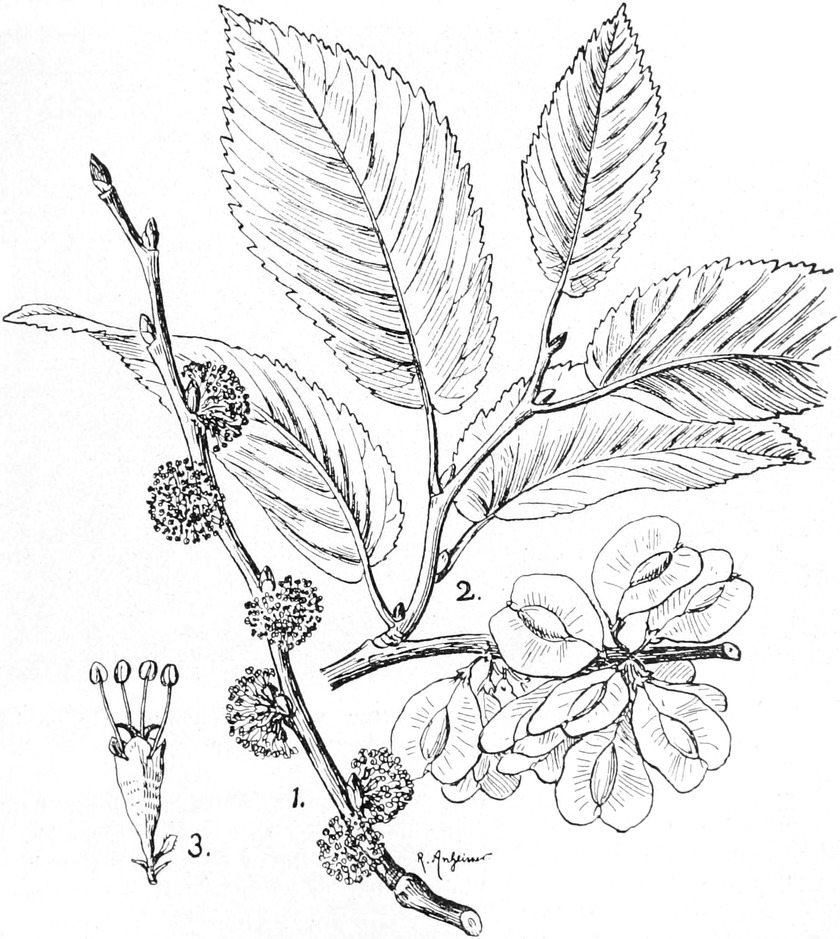
Fig. 693. Ulmus campestris. 2⁄3 nat. Gr. 1
Blütenzweig. 2 Zweig mit Früchten. 3 Einzelblüte, vergr.
1. Familie Ulmaceae. Ulmus campestris (Fig. 693),
Rüster, ein häufiger Baum Mitteleuropas. Bei ausgesprochen
zweizeiliger Beblätterung und dorsiventraler Verzweigung aller
Triebe kommt die regelmäßige Rundung der Krone älterer Exemplare
nur dadurch zustande, daß die Blattflächen eines jeden Nebenzweiges
um einen gewissen Winkel von denen des Hauptzweiges abweichen.
Die Blätter sind stets asymmetrisch. Blüten stehen geknäuelt in
den Achseln vorjähriger Blätter, sie sind zwitterig oder durch
Fehlschlagen eingeschlechtig, ihre Staubblätter in der Knospe
gerade. Die Früchte reifen meist vor der Beblätterung des Baumes,
der bereits im Februar oder März blüht. Sie sind breit[S. 564] geflügelt
und werden durch den Wind verbreitet. U. montana, U. effusa
nahe verwandte Formen. Celtis, mit Steinfrüchten, wird in
verschiedenen Arten häufig angepflanzt.
2. Familie Moraceae. Hierher gehören meist Bäume oder
Sträucher mit reichem Milchsaftgehalt, deren Blätter wechselständig
stehen und hinfällige Nebenblätter haben. Ihre Blüten sind
eingeschlechtig, in köpfchenähnlichen oder schüsselförmigen
Infloreszenzen vereinigt und meist vierzählig.

Fig. 694. Ficus bengalensis im botanischen Garten von
Buitenzorg. Einige Epiphyten auf den wagerecht abspreizenden Ästen.
Wichtige Vertreter: Außer den Maulbeerbäumen, die
zur Seidenraupenzucht (Morus alba) oder als Fruchtbäume (Morus
nigra) (Fig. 588 B) vielfach gezogen werden, ist vor allem
die Gattung Ficus zu nennen. Der nördlichste Vertreter ist
der gewöhnliche Feigenbaum, Ficus carica[484] (Fig. 541), im
Mittelmeergebiet einheimisch und seit langer Zeit kultiviert;
ein niedriger Baum mit großen, fingerförmig eingeschnittenen
Blättern, der seine Nebenblätter als Schutzkappe über der Knospe
geschlossen behält. Blütenstände in Form krugförmiger Gebilde mit
enger Mündung tragen ihre Einzelblüten auf der inneren Oberfläche
dicht beisammen. Die flach scheibenförmigen Infloreszenzen von
Dorstenia-Arten, welche auf der Oberseite mit Einzelblüten
besetzt sind, stellen in mancher Beziehung ähnliche Bildungen
dar, über das Fortschleudern ihrer Fruchtsteinkerne vgl.
GOEBEL[485]. Über die Bestäubung der Feigen vgl. S. 481,
Fig. 541. Die verbreiterte Blütenstandsachse mit dem Perigon der
Einzelblüten bildet den fleischigen, süßschmeckenden Teil der
Eßfeigen. Die kleinen harten Kernchen sind aus dem Fruchtknoten
hervorgegangene Einzelfrüchtchen, Nüsse. — Ficus-Arten gehören
mit zu den gewaltigsten Baumgestalten tropischer Wälder. Vor allem
merkwürdig ist der Banyan, Ficus bengalensis in Ostindien. Auf
Baumästen keimend aus Samen, der von fruchtfressenden Vögeln
dorthin gebracht war, entwickelt er sich zu einem stattlichen
Epiphyten. Aber erst wenn seine Wurzeln den Boden erreicht haben
und die Pflanze nicht mehr auf die karge Epiphytenernährung
angewiesen ist, zeigt sie ihre Eigenart. Der Wirtsbaum, auf dem sie
sitzt, wird nach und nach erdrosselt, immer neue Wurzeln erreichen
den Boden und bilden säulengleiche Stämme, so daß schließlich
ein ganzer Wald aus dem kleinen Keimling hervorgeht, und in dem
Schatten der Krone ein Dorf Raum genug findet (Fig. 694). Milchsaft
von Ficus elastica wird zur Gewinnung von Kautschuk den Bäumen
durch Einschnitte entzogen. Castilloa elastica ist ein wichtiger
Kautschuk liefernder Baum Zentralamerikas. Artocarpus-Arten,
Brotbäume, sind bekannte Fruchtbäume der Tropen, deren riesige
Fruchtstände roh oder geröstet genossen werden.
[S. 565]
Offizinell: Morus nigra liefert Syrupus mororum
(Pharm. helv.); Ficus elastica (Ostindien), Castilloa elastica
(Mexiko) und andere tropische Moraceen liefern in ihrem Milchsafte
Kautschuk[486] (Pharm. germ.).
3. Familie Cannabinaceae. Humulus Lupulus,
Hopfen, ist in Mitteleuropa einheimisch, sein ausdauerndes
Rhizom bringt jedes Jahr neue schlingende Triebe (Fig. 695).
Der Stamm und seine gegenständigen handnervigen Blätter sind
rauhhaarig; ersterer trägt eigenartige Widerhaken, die ein
Herabgleiten von einer einmal gefaßten Stütze hindern. Die
männlichen Blüten der diözischen Pflanze sind fünfzählig, mit
geraden Staubblättern versehen und in Dichasien angeordnet mit
mittelständigem Bereicherungssproß. Die weiblichen Teilblütenstände
sind kätzchenförmig; an ihrer Achse sitzen spreitenlose Hochblätter
die auf ihre Nebenblattpaare reduziert sind. Der Achselsproß dieses
Hochblattes ist unterdrückt; jedes Nebenblatt trägt zwei Blüten,
je von einem eigenen Deckblatte umhüllt, in der Achsel. Diese
Deckblätter wachsen bei der Reife über die Nebenblätter hinaus
und bedingen das zäpfchenähnliche Aussehen des Fruchtstandes. Sie
tragen die Hopfendrüsen, derenwegen die Pflanze kultiviert wird.
Cannabis sativa, Hanf. Einjähriges, in Indien heimisches
Kraut, mit fingerförmig zerteilten rauhen Blättern, die unten
gegenständig, in der Blütenregion wechselständig angeordnet
sind. Blütenstände wie beim Hopfen, doch ist der bei den
weiblichen unterdrückte Mitteltrieb hier zu einem laubigen Sproß
ausgewachsen. Die Deckblättchen tragen nur je eine Blüte in
der Achsel. In den Laubblattachseln des Mitteltriebes wiederholt
sich dasselbe, und so kommt die dichtbuschige Infloreszenz des
weiblichen Hanfes zustande. Die Pflanze wird in Europa ihrer ein
bis einige Zentimeter langen Bastfasern wegen gebaut. Weibliche
Blütenstände indischer Herkunft sind auf allen Teilen mit einer
Menge von Drüsenhaaren bedeckt, die eine harzige, klebrige Masse
absondern; sie werden für medizinische Zwecke benutzt und dienen
im Orient zur Bereitung eines narkotisch wirkenden Genußmittels
„Haschisch“.
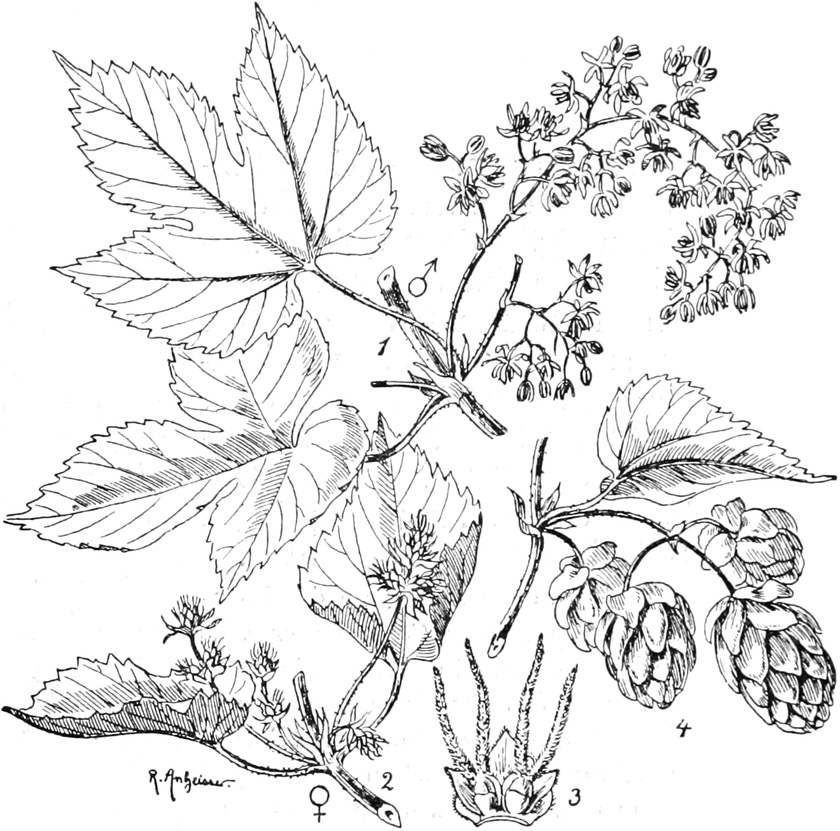
Fig. 695. Humulus Lupulus. 1 Männlicher Blütenstand.
2 Weibliche Infloreszenz. 3 Zwei weibliche Blüten in der
Deckblattachsel. 4 Fruchtzäpfchen. 1⁄2 nat. Gr.
Offizinell: Cannabis sativa liefert Fructus
Cannabis (Pharm. helv.); Cannabis sativa var. indica
liefert Herba Cannabis indicae (Pharm. austr., helv.).
— Die Drüsen der Zapfenschuppen von Humulus Lupulus sind als
Lupulinum, s. Glandulae Lupuli (Pharm. austr.,
helv.), gebräuchlich, der ganze Zapfen als Strobilus Lupuli
(Pharm. helv.).
4. Familie Urticaceae. Hierher gehören meist perennierende
Kräuter mit einfachen Blättern und Nebenblättern, deren zweizählige
anemophile Blüten durch Fehlschlag eingeschlechtig geworden
sind, also Blütenformel P 2 + 2, A 2 + 2. Die Staubblätter in
der Knospenlage durch Einwärtskrümmung gespannt. schlagen beim
Aufblühen elastisch zurück und verstäuben dabei ihren Pollen.
Weibliche Blüten besitzen ein verwachsenes Perianth und nur ein
Fruchtblatt, mit einer grundständigen atropen Samenanlage. Die
reifen Früchte von Pilea- und Elatostema-Arten werden durch
aus Staminodien entwickelte Schleuderorgane fortgeschnellt. In den
Tropen reicher als bei uns verbreitete Pflanzenfamilie.
[S. 566]
Manche Urticaceen sind durch den Besitz von Brennhaaren (vgl. Fig.
52) ausgezeichnet, wie unsere Brennesseln, Urtica dioica und
Urtica urens, und die gefährlichen tropischen Laportea-Arten.
Einige Urticaceen sind wichtige Faserpflanzen, wie Boehmeria
nivea, Ramie. Unsere einheimischen Nesselarten lieferten vor
Bekanntwerden der Baumwolle Gespinstfasern und kamen infolge des
Krieges und seiner Folgen wieder zu Ehren.
Greifen wir hier vor Einschiebung des Columniferenastes auf den
Stammbaum der Hauptreihe (S. 524) zurück, so würden sich zunächst die
Rhoeadinae, denen morphologische Beziehungen zu den Polycarpicae nicht
wohl abgesprochen werden können, anreihen, denen sich die Cistiflorae
anschließen, worauf dann auf die Resedaceae die Columniferae folgen. Da
die Resedaceae von jeher in Beziehung zu den Capparidaceen einerseits,
den Cistiflorae (Parietales pro parte) andererseits gebracht sind,
so wird diese Anordnung, die sich nicht anders als durch Anreihung
ausführen läßt, den morphologischen wie den serodiagnostischen
Beziehungen am besten entsprechen.
Die 16. Ordnung Rhoeadinae enthält meist Kräuter mit
wechselständigen Blättern. Ihre zwittrigen Blüten sind aus meist
zweigliedrigen Quirlen aufgebaut; sie besitzen einen oberständigen
einfächerigen Fruchtknoten, der die Samenanlagen an den verwachsenen
Rändern der Karpelle, den Plazenten, trägt (Fig. 696). Die Narben
stehen über den Nähten der Fruchtblätter, und die Öffnung der Frucht
erfolgt durch Abspringen der Fruchtblätter von den Plazenten.
Den Anschluß der Ordnung an die Polycarpicae stellt die 1. Familie
der Papaveraceen[487] her durch gewisse Merkmale,
wie Gehalt an Milchsaftschläuchen (Nymphaeaceen), Vorkommen
dreizähliger Blüten bei Bocconia (Berberidaceen), Anordnung der
Narben direkt auf den Fruchtblättern und das, wenn auch seltene
Vorkommen apokarper Gynäceen wie bei typischen Vertretern der
Polycarpicae (z. B. Platystemon). Die große Vermehrung der
Staubblätter, die zyklisch stehen, wird auf Spaltung zurückgeführt.
Die Samen haben reichliches Endosperm.
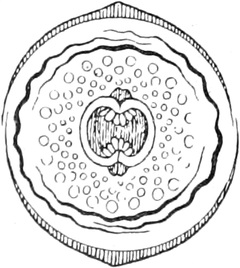
Fig. 696. Blütendiagramm der Papaveraceen. Glaucium.
Nach A. W. EICHLER.

Fig. 697. Papaver Rhoeas. 1⁄2 nat. Gr. —
Offizinell.
Chelidonium majus, Schöllkraut, hat gelben Milchsaft und
zweikarpelligen Fruchtknoten. Als Zierpflanzen dienen verschiedene
Arten von Escholtzia, Argemone und Papaver. Papaver
rhoeas, Mohn, als Unkraut in Kornfeldern oder auf trockenen
Wiesen verbreitet (Fig. 697).[S. 567] Charakteristisch ist die scharf
abwärts gekrümmte Lage der Blütenknospen. Papaver somniferum
stammt aus dem Orient und ist in allen Teilen reich an weißem
Milchsafte. Die mit Wachs hellblau bereifte, völlig glatte
Pflanze, deren Blütenstiel allein vereinzelte grobe Borsten
trägt, hat sitzende, am Rande unregelmäßig gesägte bis gekerbte
Blätter und eine violette oder weiße Blumenkrone mit dunklen
Flecken am Grunde. Der einfächerige Fruchtknoten springt bei der
Reife nur an den Fruchtblattspitzen von den zahlreichen, tief
einschneidenden Plazenten ab, die Fruchtblätter biegen sich unter
dem überstehenden, flachen narbentragenden Deckel weg nach außen,
und ihre nierenförmigen Samen werden aus den Löchern vom Winde
herausgeschleudert.
Offizinell: Semen Papaveris (Pharm. germ., helv.),
Fructus Papaveris immaturi (Pharm. germ., austr., helv.),
Opium (aus dem Milchsafte) (Pharm. germ., austr., helv.),
Morphium (ibid.) alles von Papaver somniferum. Flores
Rhoeados (Pharm. austr., helv.) von Papaver Rhoeas.
Die 2. kleine Familie der Fumariaceen ist interessant
durch das Vorkommen transversal zygomorpher Blüten bei Corydalis
(Fig. 698) und zweisporniger bisymmetrischer Krone bei Dicentra
spectabilis. Die Früchte sind bei Fumaria Nüßchen, bei
Corydalis und Dicentra Kapseln. Die Samen haben Endosperm.

Fig. 698. Diagramm von Corydalis cava, nach A.
W. EICHLER. Am Grunde des Staubblattes, über dem Sporn, eine
Nektardrüse.
Fig. 699. Cruciferae. Diagramm (Brassica). Nach F.
NOLL.
Fig. 700. Cardamine pratensis. Blüte ohne Perianth.
Vergr. 4. Nach H. BAILLON.
Fig. 701. Früchte der Cruciferae. A Cheiranthus
Cheiri. B Lepidium sativum. C Capsella Bursa pastoris. D Lunaria
biennis. E Crambe maritima. Nach H. BAILLON.
3. Familie Cruciferae[488]. Die Familie der Kreuzblütler
ist besonders auf der Nordhemisphäre zu Hause und zählt bei uns und
im Mittelmeergebiet zu den arten- und individuenreichsten, der man
überall begegnet. Es sind meist ein- oder zweijährige Kräuter oder
Stauden mit wechselständigen Blättern, traubigen, meist deck- und
vorblattlosen Blütenständen und radiären, stets seitlich stehenden
Einzelblüten. K2 + 2, C4, A2 + 4, G(̲2) (Fig. 699). Der Kelch beginnt
mit einem median stehenden Wirtel, vier Kronblätter alternieren mit dem
Kelche. Zwei äußere Staubblätter sind kürzer als die vier (bzw. zwei
bis auf den Grund gespaltenen) medianen (S. 107) inneren (Fig. 700).
Die Fruchtblätter bilden einen oberständigen, meist schotenförmigen
Fruchtknoten, der durch eine falsche (S. 471), zwischen den parietalen
Plazenten ausgespannte Scheidewand (Fig. 701 A, C, D)
zweifächerig wird und sich durch Abheben der Frucht[S. 568]blattmittelstücke
klappig öffnet. Die Samen bleiben dabei mit der falschen Scheidewand
und den Plazenten am Tragstiele erhalten. Bisweilen, z. B. bei Isatis,
finden sich Schließfrüchte. Der gekrümmte Keimling liegt von einer
einzigen Zellschicht Endosperm umgeben in der Samenschale (Fig. 702,
703).
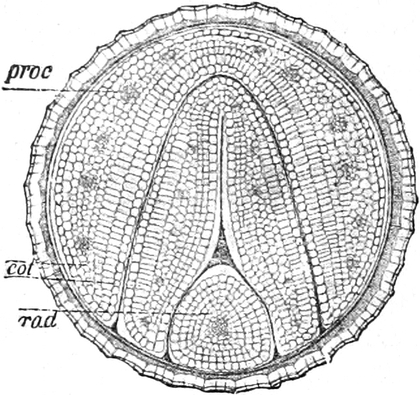
Fig. 702. Querschnitt durch den Samen des schwarzen
Senfs, Brassica nigra. rad Radicula, cot Kotyledonen, proc
Leitbündelanlagen. Nach A. MÖLLER.
Fig. 703. Samen, quer durchschnitten; Würzelchen und
Keimblätter in verschiedener Lagerung sichtbar. A Cheiranthus Cheiri.
Vergr. 8. B Sisymbrium Alliaria. Vergr. 7. Nach H. BAILLON.

Fig. 704. Brassica nigra. 1⁄2 nat. Gr. —
Offizinell.
Fig. 705. Capparis spinosa, blühender Zweig und junge
Frucht auf ihrem Gynophor. 1⁄2 nat. Gr.
Nicht nur ihrer Häufigkeit und Artenzahl nach zählen die Cruciferen
zu den wichtigeren heimischen Familien, sondern auch die große
Zahl der ihnen entstammenden Nutz- und Gartenpflanzen verleiht
ihnen größere Bedeutung; auch zählen manche Cruciferen zu den
ertragreicheren Honigblumen vermöge der am Grunde der[S. 569] Staubblätter
aus der Blütenachse hervorgehenden Honigdrüsen. Cheiranthus
Cheiri (Fig. 701 A). Goldlack, Matthiola, Levkoje, beliebte
Zierpflanzen. Brassica oleracea liefert den Kohl in seinen
verschiedenen Formen: a) silvestris, an den nordeuropäischen
Küsten ist als wilde Form anzusehen, b) acephala, Blätterkohl, c)
gongylodes, Kohlrübe, d) gemmifera, Rosenkohl, e) sabauda,
Wirsing, f) capitata, Kopfkohl, g) botrytis, Blumenkohl,
Brassica campestris, Rübsen mit den Kulturformen: a) annua,
Sommerrübsen, b) oleifera, Winterrübsen, c) rapifera, Teltower
Rübchen. Br. napus, Raps: a) annua, Sommerraps, b) oleifera,
Winterraps c) Napobrassica, Wruke. Brassica nigra, Senf (Fig.
704), eine einjährige, bereits im Altertume vielfach angebaute
Pflanze. Ihre grundständigen Blätter sind fiederteilig mit
stumpfen Endlappen, völlig unbehaart bis auf einige grobe Borsten
der Blattoberseite. Die dottergelben Einzelblüten stehen von der
Spindel ab, die abgeblühten, glatten Fruchtknoten und Früchte
dagegen sind ihr angedrückt und ragen gerade aufwärts. Sinapis
alba, Weißer Senf, ist eine rauhbehaarte Pflanze und durch
weit von der Spindel abspreizende, lang und flach geschnäbelte
Früchte, deren Klappen grob borstenhaarig sind, wie durch
doppelt so große, weiß-gelbe Samen, leicht vom schwarzen Senf
zu unterscheiden. Anastatica hierochuntica, die Jerichorose,
eine durch hygroskopische Bewegung (S. 295) ihrer Zweige bekannte
einjährige Wüstenpflanze Nordafrikas. Crambe (Fig. 701 E),
mit im unteren Teil unfruchtbaren Schoten, und Cakile sind
dickblättrige Strandpflanzen, Raphanus sativus, ist der Rettich
und das Radieschen. Cochlearia, Löffelkraut. Vesicaria,
Aubrietia, Draba, Lunaria (Fig. 701 D), Erophila-Arten,
das Hungerblümchen, Iberis mit etwas dorsiventralen Blüten,
Capsella bursa pastoris, das Hirtentäschl (Fig. 701 C) sind
bekannte Cruciferen. Isatis tinctoria, der Waid, früher als
Farbstoff liefernde Pflanze benutzt.
Offizinell: Semen Erucae (Pharm. helv.) von Sinapis
alba. Semen Sinapis (Pharm. germ., austr., helv.) von
Brassica nigra. Oleum Sinapis (ibid.) von beiden Pflanzen.
Zur 4. Familie Capparidaceae gehört Capparis spinosa,
ein kleiner Felsenstrauch der Mittelmeerländer, mit einfachen
Blättern, kurzdornigen Nebenblättern und blattwinkelständigen,
radiären Einzelblüten, die sich von denen der Cruciferen durch
Spaltung der Staubblätter in unbestimmt viele, und durch
Einschiebung eines Gynophors, unterscheiden, welches den
Fruchtknoten hoch über die Blüte emporhebt (Fig. 705). Die Frucht
ist eine Beere von etwa Pflaumengröße mit zahlreichen Samen. Junge
Blütenknospen liefern die „Kappern“.
17. Ordnung.
Cistiflorae.
Regelmäßig fünfzählige Blüten mit vermehrten, gespaltenen oder
bündelig verwachsenen Staubblättern und einem dreizähligen
oberständigen Fruchtknoten kennzeichnen die Mehrzahl der
Cistifloren.
Fig. 706. Blütendiagramm von Helianthemum vulgare
(Cistaceae). Nach A. W. EICHLER.
Fig. 707. Diagramm von Viola. Nach F. NOLL.
Die 1. Familie der Cistaceen ist charakterisiert durch
strahlige, fünfzählige Blüten mit zahlreichen Staubblättern und
drei oder fünf zu einem Fruchtknoten verwachsenen Fruchtblättern
mit parietalen Plazenten. Das einheimische kleine Sonnenröschen
Helianthemum vulgare (Fig. 706) und die Cistus-Arten des
Mittelmeergebietes entsprechen normalen Cistaceen. Angehörige der
2. Familie der Violaceen unterscheiden sich durch ihre
meist dorsiventralen Blüten, die nur fünf Staubgefäße und einen
einfächerigen Fruchtknoten besitzen (Fig. 707). In das gespornte
vordere Kronblatt ragen die beiden vorderen Staubblätter mit Nektar
absondernden Fortsätzen hinein. Offizinell ist Herba
violae tricoloris (Pharm. germ., austr., helv.).
[S. 570]
In der 3. Familie der Ternstroemiaceen, zu der die
Teepflanze und die Kamelie gehören, fällt der allmähliche Übergang
von Kelch- in Kronblätter auf, wie ihn die Magnoliaceen zeigen,
zahlreiche Staubblätter und ein dreifächeriger Fruchtknoten mit
zentralwinkelständigen Plazenten vervollständigen das Bild.
Offizinell: Folia Theae (Pharm. austr.) von Thea
chinensis (Fig. 708). Durch bündelweise Vereinigung der
Staubblätter und schizogene Sekretbehälter ist die 4. Familie
der Guttiferae unterschieden. Die Gattung Hypericum
ist ein einheimischer Vertreter. Offizinell: Gummigutt
oder Gutti, der eingetrocknete Sekretsaft von Garcinia
Hanburyi (Pharm. germ., austr., helv.). Als 5. Familie
schließen sich die Dipterocarpaceen an; sie sind durch
starke Vergrößerung aller oder einzelner Kelchblätter nach der
Befruchtung charakterisiert. Dryobalanops Camphora liefert
den Borneokampfer. Offizinell: Dammar (Pharm. germ.,
austr.) von Shorea Wiesneri.
18. Ordnung.
Columniferae.
Die Angehörigen dieser Ordnung besitzen fünfzählige, strahlige
Zwitterblüten, in denen einer, meist der äußere, der beiden
Staubblattwirtel unterdrückt oder nur staminodial vorhanden ist,
während der andere eine mehr oder minder große Vermehrung seiner
Glieder durch Spaltung erfährt. Vielfach geht eine Verwachsung der
Filamente daneben her. Auch die Fruchtblätter sind bisweilen durch
Verzweigung vermehrt. Der oberständige Fruchtknoten ist entsprechend
gefächert.
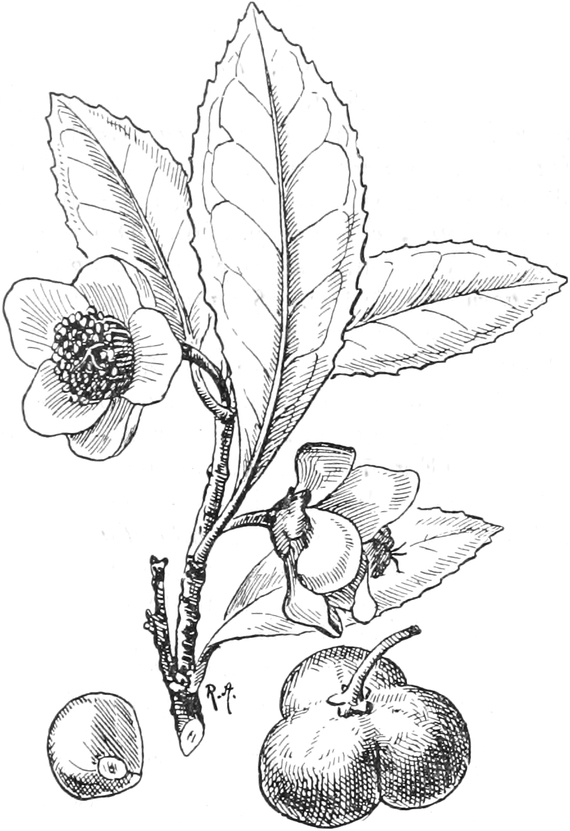
Fig. 708. Thea chinensis. Blühender Zweig. 2⁄3 nat. Gr.
Frucht und Samen. — Offizinell.
1. Familie Malvaceae. Charakteristisch für die Familie sind
in der Knospe gedrehte protandrische Blüten, deren Staubblätter zu
einer der Krone angewachsenen Röhre verbunden sind, welche die Griffel
umhüllt (Fig. 709, 710) und nur oben in zahlreiche freie Enden mit je
einer nierenförmigen Theca gespalten ist. K5, C5, A(∞), G(3) oder ∞.
Die Pollenkörner sind mit stachliger Exine versehen, so daß sie leicht
am Haarkleid der bestäubenden Insekten haften (Fig. 516).
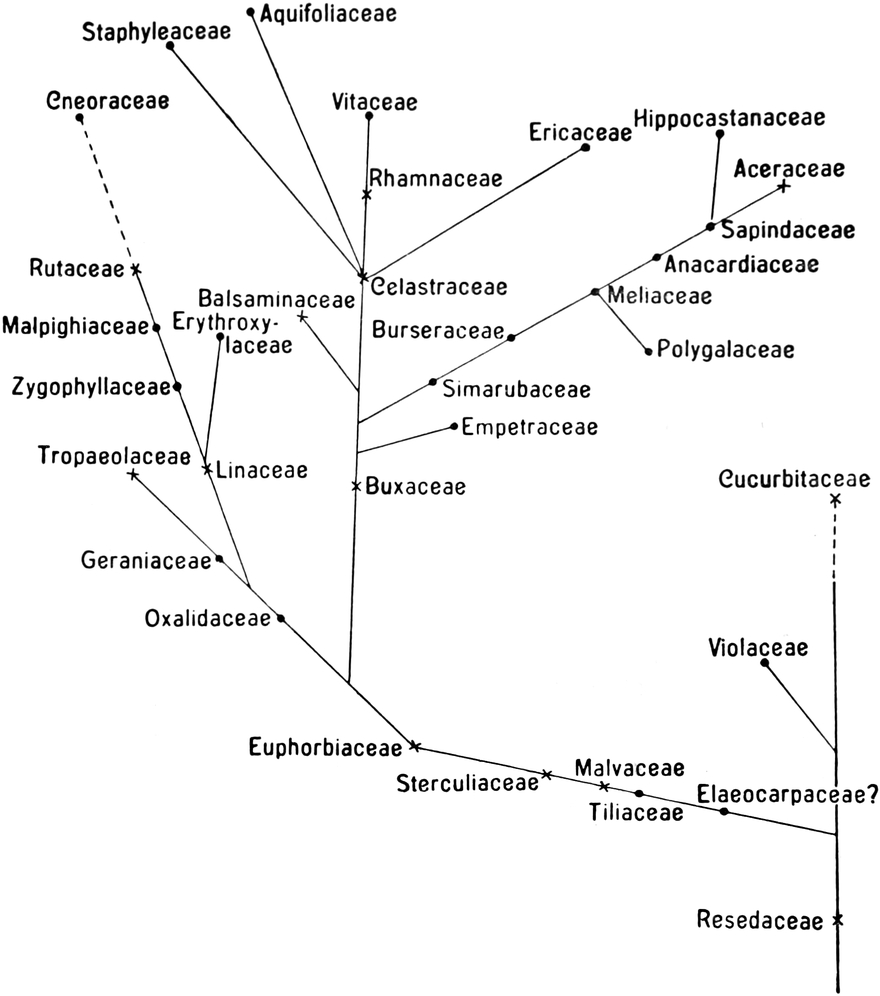
Stammbaum des Columniferenastes und der anschließenden Familien nach
serodiagnostischen Untersuchungen von F. HOEFFGEN.
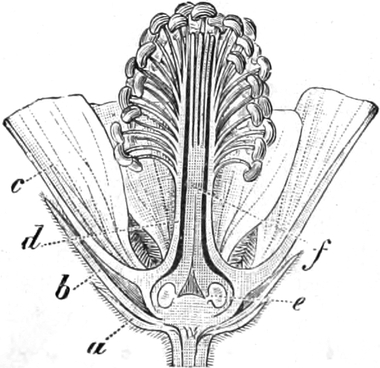
Fig. 709. Althaea officinalis. Längsdurchschnittene
Blüte mit teilweise entfernten Kronblättern. a Der Außenkelch,
b der Innenkelch, c die Kronblätter, d das Andröceum, f der
Griffel, e die Samenanlagen. Nach BERG und SCHMIDT.
— Offizinell.
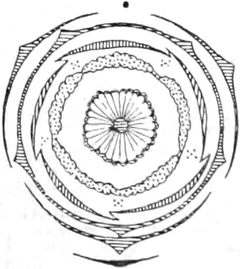
Fig. 710. Malvaceae. Diagramm (Malva).
Fig. 711. Malva silvestris. a Blüte. b Blütenknospe,
c Frucht. Nat. Gr. Nach H. SCHENCK. — Offizinell.
Die einheimische Gattung Malva umfaßt ausdauernde Kräuter mit
langgestielten, handnervigen Blättern. Die Blüten sind einzeln oder
in kleinen Wickeln blattachselständig verteilt; sie besitzen drei
freie Außenkelchblätter und tief ausgerandete, meist rosa gefärbte
Kronblätter (Fig. 711). Die Vegetationsorgane der nahe verwandten
Gattung Althaea sind dicht mit Büschelhaaren bekleidet, sammetig
weich. Ihr Außenkelch zählt sechs bis neun am Grunde verwachsene
Blätter. Die Früchte sind Spaltfrüchte, den zahlreichen,
kreisförmig angeordneten Fruchtblättern entsprechend. Hibiscus
und Gossypium-Arten sind Sträucher oder Bäume mit drei- bis
fünfzipfligen, langgestielten Blättern, deren Blüten mit drei
großen, die Knospen völlig verdeckenden Außenkelchblättern versehen
sind; ihre Früchte sind drei- bis fünfkarpellige, fachspaltige
Kapseln. Der dichte Filz langer Flughaare auf den Gossypiumsamen
dient ihrer Verbreitung und liefert, durch Kultur und Auslese
verfeinert, die Baumwolle (Fig. 712).
Offizinell: Folia Malvae (Pharm. germ., austr.,
helv.) von Malva neglecta und M. silvestris. Flores
Malvae (ibidem) von M. silvestris. Folia Althaeae
und Radix[S. 571]
Althaeae (ibidem) von Althaea officinalis;
Gossypium (ibidem) von Gossypium arboreum, G.
barbadense, G. herbaceum.

Fig. 712. Blütenzweig und aufgesprungene Frucht von
Gossypium herbaceum. 1⁄2
nat. Gr. — Offizinell.
Fig. 713. Tiliaceae, Diagramm (Tilia). Nach A. W.
EICHLER.
Fig. 714. Tilia ulmifolia. A Blütenstand,
nat. Gr. a Infloreszenzstiel, b das Hochblatt. B Frucht,
längsdurchschnitten und vergrößert, o Perikarp, p verkümmerte
Scheidewände und Samenanlagen, q Samen, r Endosperm, s Keim, t
Würzelchen des letzteren. Nach BERG und SCHMIDT. —
Offizinell.

Fig. 715. Sterculiaceae. Diagramm (Theobroma). Nach
A. W. EICHLER.
[S. 572]
Die 2. Familie, Tiliaceae, enthält Pflanzen mit einfachen
Blättern und hinfälligen Nebenblättern. Der freiblättrige Kelch und
die Krone haben klappige Knospenlage. Die Staubblätter besitzen
introrse Antheren; dem Charakter der Ordnung gemäß ist meist
nur der innere Kreis vorhanden und gespalten (Fig. 713). Der
Griffel ist einfach. Hierher gehören meist tropische Gattungen,
darunter die krautigen, Jute liefernden Corchorus-Arten.
Einheimisch ist nur Tilia, die Linde, in zwei Arten. Die Blätter
sind asymmetrisch und ihr Rand ist gesägt. Die Blütenstände (Fig.
714 A) sind mit einem als Flugblatt ausgebildeten Vorblatte bis
zur halben Länge verwachsen und aus Dichasien zusammengesetzt.[S. 573]
Die breitblättrige Linde hat 3–7, die ulmenblättrige dagegen 5–15
Einzelblüten im Blütenstande. Der behaarte Fruchtknoten enthält
zwei Samenanlagen in jedem seiner fünf Fächer, eine davon verdrängt
alle übrigen (Fig. 714 B).
Offizinell: Flores Tiliae (Pharm. germ., austr.,
helv.) von T. platyphyllos und T. ulmifolia.
3. Familie Sterculiaceae. Den Tiliaceen ähnlich
weicht diese in den Tropen verbreitete Familie ab durch
verwachsenblättrigen Kelch, gedrehte Knospenlage und zu einer Röhre
verwachsene Staubblätter, von denen die kelchständigen staminodial
bleiben, während die Kronstaubblätter oft durch Spaltung vermehrt
werden (Fig. 715). Die Antheren sind extrors.
Fig. 716. Theobroma Cacao. 1 Fruchttragender Stamm.
2 Blühender Ast. 3 Einzelblüte. 4 Staubblattring. 5 Staubblatt
von vorn. 3, 4 Etwa nat. Gr. 5 Vergr. 1, 2 Stark verkleinert.
— Offizinell. Stamm nach einer Photographie.
Theobroma Cacao, der Kakaobaum (Fig. 716), ist im tropischen
Zentral- und Südamerika einheimisch, dort auch seit langer
Zeit in Kultur. Der niedrige Baum trägt kurzgestielte, steife,
brüchig-harte, einfache Blätter von erheblicher Größe, ovaler Form
und dunkelgrüner Farbe, die in der Jugend mehr hellrötlich sind
und wie bei zahlreichen tropischen Bäumen schlaff herabhängen.
Die Blüten brechen am Stamme oder an älteren Ästen aus früher
blattachselständigen, erhaltenen Knospen hervor; der Baum ist
cauliflor. Jedes am Grunde bauchige Kronblatt verschmälert
sich nach außen erheblich[S. 574] und endet mit verbreitertem Zipfel. So
ist die Gesamtform der rötlichen Blüten etwa urnenförmig, mit fünf
auseinander strahlenden Zipfeln. Der fünffächerige Fruchtknoten
enthält zahlreiche Samenanlagen in jedem Fache; bei dem
Heranwachsen der Frucht drängen sich die erweichenden Gewebemassen
der Scheidewände zwischen die einzelnen Samen ein und so wird die
reife Frucht einfächerig, vielsamig. Zwei mächtige gefaltete,
brüchige Kotyledonen des kleinen Embryos füllen die Samenschale
aus. — Cola acuminata und C. vera im tropischen Afrika liefern
die Kolanüsse. Offizinell: Oleum Cacao (Pharm. germ.,
austr., helv.) und Theobromin (Pharm. germ., helv.) von
Theobroma Cacao Semen Colae von Cola acuminata und C.
vera (Pharm. austr.).
19. Ordnung.
Tricoccae[489].
Euphorbiaceae. Gewächse von außergewöhnlich verschiedenartigem
Habitus wie Kräuter, Sträucher, blattlose Stammsukkulenten, Bäume mit
normalen Blättern oder auch mit Blattschuppen und assimilierenden
Phyllokladien, stimmen die Euphorbiaceen darin überein, daß sie
eingeschlechtige, radiäre Blüten mit meist einfacher Blütenhülle
oder ganz ohne solche besitzen. Das Andröceum ist diplostemon oder
vielzählig. Die weiblichen Blüten sind durch einen oberständigen,
dreikarpelligen und dreifächerigen Fruchtknoten charakterisiert, der
in jedem Fache 1–2 hängende Samenanlagen enthält mit ventraler Raphe,
Mikropyle also aufwärts und auswärts gekehrt. Sie wird von einem
plazentaren Auswuchs, dem Obturator, gedeckt (Fig. 717); dieser
vermittelt Leitung und Ernährung der Pollenschläuche und schwindet
nach der Befruchtung (vgl. S. 495). Die von dem äußeren Integument
ausgebildete Caruncula (Fig. 719 D) bleibt dagegen an dem reifen
Samen noch vorhanden, dessen Ablösung von der Plazenta ihrer Mitwirkung
zu danken ist. Früchte sind Kapseln, deren Außenwände elastisch von
einer Mittelsäule zurückschnellen und so die Fächer öffnen. Die Familie
ist über die ganze Erde verbreitet.
Fig. 717. Obturator an der Samenanlage von Euphorbia
dioica. Nach PAX in ENGLER-PRANTL.
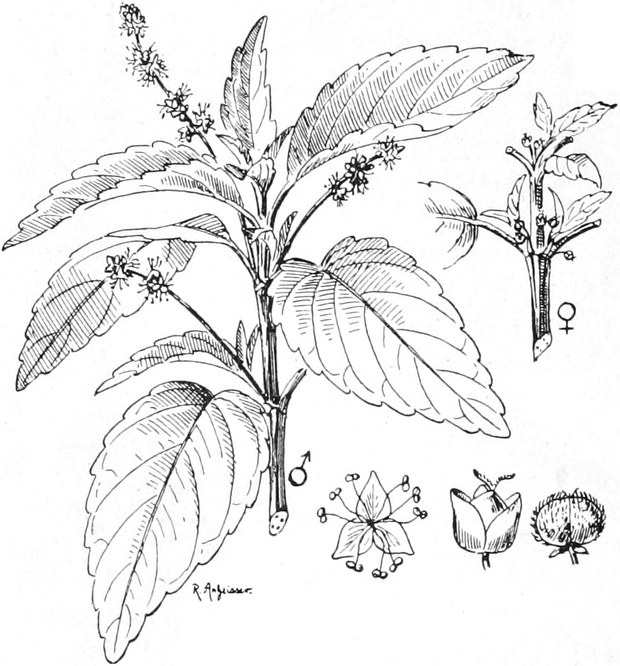
Fig. 718. Mercurialis annua. 1⁄2 nat. Gr. Männliche
blühende Pflanze und Einzelblüte. Stück einer weiblichen Pflanze,
Einzelblüten und Frucht. — Giftig.
Wichtige Gattungen: Zahlreiche Euphorbiaceen zeigen bei
diözischer oder monözischer Blütenverteilung einen sehr einfachen
Blütenbau, so Mercurialis (Fig. 718), durch zweikarpelligen
Fruchtknoten abweichend; die Gattung Croton, männliche Blüten mit
doppeltem, weibliche mit einfachem Perianth, enthält die wichtigen
offizinellen Arten: C. Eluteria und C. Tiglium. Dagegen ist
die in vielen einheimischen Arten vertretene Gattung Euphorbia,
Wolfsmilch, neben anderen ausgezeichnet durch Vereinigung
zahlreicher, einfachst gebauter Blüten in sehr komplizierten
Blütenständen, Cyathium genannt (Fig. 719, 720, 721),
die besonders in zygomorpher Ausbildung wie bei Pedilanthus,
ganz den Eindruck einer Einzelblüte machen. Das Cyathium besteht
aus einer nackten,[S. 575] langgestielten und nach unten umgewendeten
weiblichen Gipfelblüte, die von mehreren Gruppen ebenfalls
gestielter, auf je ein vom Stiel abgegliedertes Staubblatt
beschränkter, männlicher Blüten umgeben wird. In einigen Fällen ist
die weibliche Einzelblüte und jede männliche Blüte mit eigenem,
kleinem Perianth versehen. Stets aber wird der ganze Blütenstand,
das Cyathium, durch fünf Hüllblätter umschlossen, mit denen vier
große elliptische oder zweihörnige Nektardrüsen alternieren, welche
den Eindruck der Einzelblüte erhöhen. Dort, wo die übergebogene
weibliche Blüte herabhängt, fehlt die fünfte Drüse. Zwischen den
vor je einem Hüllblatte (Fig. 720) stehenden Gruppen männlicher
Blüten sind zerschlitzte Haarbildungen auf der Infloreszenzachse
vorhanden, die im Längsschnitte (Fig. 719 B) sichtbar werden.
Solche Blütenstände pflegen nun, in dichasialen Zweigen zu
mehreren angeordnet, sich zu drei- bis vielstrahlig-trugdoldigen
Gesamtinfloreszenzen zu vereinigen. Häufig ist die weibliche Blüte
nur in einzelnen von ihnen entwickelt, in den anderen rudimentär
geblieben. Zahlreiche, besonders afrikanische Euphorbia-Arten
haben die Gestalt von kaktusähnlichen Stammsukkulenten (Fig. 721).
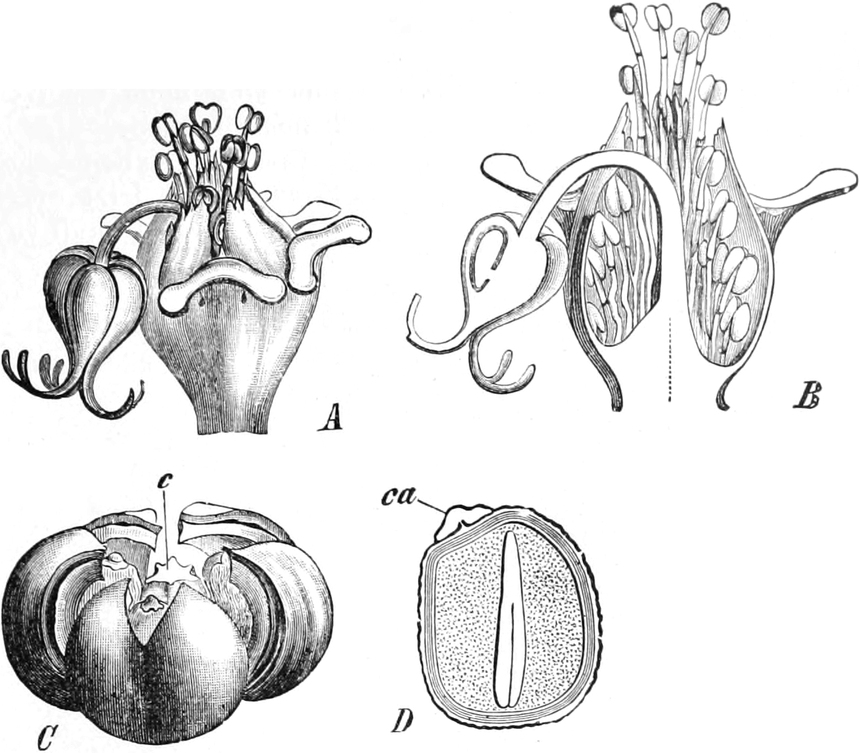
Fig. 719. Euphorbia Lathyris. A Cyathium. Vergr. 4.
B Cyathium längsdurchschnitten. Vergr. 7. C Frucht aufgesprungen
zeigt das Mittelsäulchen (c). D Samenlängsschnitt, Keimling im
Endosperm, ca Caruncula, Vergr. 4. (A–D nach H. BAILLON.
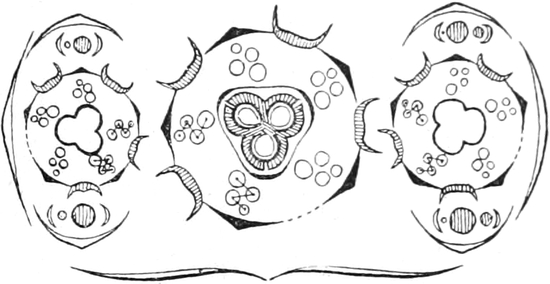
Fig. 720. Diagramm eines Dichasialzweiges von Euphorbia,
fertile weibliche Blüte nur im Mittelcyathium. Nach A. W.
EICHLER.
Fig. 721. Euphorbia resinifera. Nat. Gr. Nach
BERG und SCHMIDT. — Offizinell.
Euphorbia und andere, aber durchaus nicht alle Angehörigen
der Familie, enthalten einen bei jeder Verletzung aus den
ungegliederten (bisweilen auch gegliederten wie bei Hevea)
Schläuchen reichlich ausfließenden Milchsaft, der in vielen Fällen
giftig ist.
Ein wichtiger Bestandteil des Milchsaftes von Hevea-Arten
(H. Sieberi, discolor, rigidifolia, paucifolia,
lutea, guyanensis, Spruceana) ist der Kautschuk
(vgl. [486]), welcher als Parakautschuk in den Tropen
Südamerikas, speziell im Gebiet des Amazonas, gewonnen, lange
Zeit etwa 50 % der Gesamtkautschukproduktion der Erde deckte,
jetzt aber durch den in Kolonien gewonnenen Hevea-, Ficus- und
Castilloa-Kautschuk weit überholt ist. Daneben ist der von Manihot
Glaziovii, einer ebenfalls südamerikanischen[S. 576] Euphorbiacee,
erhaltene Cearakautschuk zu erwähnen. Eine nahe verwandte
Pflanze, Manihot utilissima, gibt in ihren Wurzelknollen
ein außerordentlich wichtiges Nahrungsmittel der Tropen ab:
Maniok oder Cassave, ein Stärkemehl, dessen feinste
Sorten als Tapioka oder brasilianisches Arrowroot eine Rolle
im Welthandel spielen. Der in Brasilien einheimische Strauch wird
jetzt überall in den Tropen kultiviert.
Fig. 722. Ricinus communis. Stark verkl. Nach
BAILLON. — Giftig und offizinell.
Ricinus communis (Fig. 722) ist ein hoher Strauch des tropischen
Afrika, der als Zierpflanze gezogen, in unserem Klima alljährlich
erfriert. Der hohle Stamm ist an den massiven Knotenstellen mit
langgestielten schildförmigen, handförmig eingeschnittenen Blättern
besetzt. Infloreszenzen (Fig. 723) stehen terminal, werden jedoch
durch vegetative Seitensprosse überholt. Sie tragen unten männliche
Blüten mit häutigem, 4- bis 5blättrigem Kelch und bäumchenartig
verzweigten Staubblättern; jede letzte Endigung trägt eine Theca.
Darüber weibliche Blüten mit 3–5 Kelchblättern, einem großen
dreiteiligen Fruchtknoten, der außen mit warzigen Stacheln übersät
ist und in drei große gegabelte, rote Narben ausläuft. Bei der
Reife liegt ein gefleckter Samen mit weißer Caruncula in jedem
Fache.

Fig. 723. Ricinus communis. Blütenstand. 1⁄2 nat. Gr.
Junge Frucht längsdurchschnitten. — Offizinell.
Offizinell: Euphorbium (Pharm. germ., austr.,
helv.) von Euphorbia resinifera (Marokko). Cortex
Cascarillae (Pharm. germ., austr., helv.) von Croton Eluteria
(Bahama-Inseln). Oleum Crotonis (Pharm. germ., austr.,
helv.) von Croton Tiglium (Ostindien). Kamala (Pharm.
germ., austr., helv.), Drüsenhaare der Fruchtkapsel von
[S. 577]
Mallotus philippinensis (ostasiatische Tropen), Oleum Ricini
(Pharm. germ., austr., helv.) von Ricinus communis. Kautschuk
(Pharm. germ.) von Hevea-Arten und Manihot Glaziovii.
20. Ordnung.
Gruinales.
Zwittrige, durchaus fünfgliedrige, strahlige Blüten mit oberständigem,
gefächertem Fruchtknoten [also K5, C5, A5 + 5, G(̱5)] finden sich
bei den meisten Angehörigen der Ordnung. Sind die Blüten dorsiventral,
so tritt häufig Reduktion ein (Polygalaceen). Die Staubblätter sind am
Grunde verwachsen, obdiplostemon oder haplostemon und führen an ihrer
Außenseite Nektarien oder solche sind als ringförmiger intrastaminaler
Diskus vorhanden (Rutaceen). Die Samenanlagen sind meist hängend
orientiert, mit aufwärts gerichteter Mikropyle und ventraler Raphe oder
haben bei abwärts gerichteter Mikropyle eine dorsale Raphe.
Fig. 724. Blütendiagramme von Geraniaceen. A Geranium
pratense. B Pelargonium zonale. Nach A. W. EICHLER.
Fig. 725. Frucht von Pelargonium inquinans. Vergr. 3.
Nach H. BAILLON.
Fig. 726. Linum usitatissimum. A Nat. Gr. B und C
Vergr. 3. — Offizinell. Gez. H. SCHENCK.

Fig.727. Erythroxylon Coca. 2⁄3 nat. Gr. —
Offizinell.
Zu der 1. Familie Geraniaceae gehören die Gattungen
Geranium mit radiären und Pelargonium mit dorsiventralen
Blüten (Fig. 724); beide haben handnervige[S. 578] Blätter. Die fünf
geschnäbelten Fruchtblätter führen zwei Samenanlagen in jedem
Fache; sie lösen sich bei der Reife von einem Mittelsäulchen ab,
öffnen sich dabei und entlassen den Samen oder bleiben geschlossen
und können sich durch hygroskopische Krümmungen der Grannen in die
Erde einbohren (Fig. 725, vgl. Fig. 295, S. 276).
2. Familie Linaceae. Linum usitatissimum (Fig. 726).
Der Lein ist eine alte, einjährige Kulturpflanze. Ihre
zahlreichen radiären, blauen, kurzlebigen Blüten mit am Grunde
verwachsenen Staubblättern und fünf freien Griffeln beschließen, in
traubigen Wickeln stehend, den mit kleinen schmalen Blättern reich
besetzten Stengel, dessen Bastfasern nach geeigneter Zubereitung zu
Leinwand verwebt werden. Die Samen der fünffächerigen Kapsel dienen
zur Ölgewinnung Oleum Lini und sind auch ihrer Schleimepidermis
wegen offizinell: Semen Lini (Pharm. germ., austr.,
helv.) von Linum usitatissimum.
Zu der kleinen 3. Familie der Erythroxylaceae gehört
der offizinelle kleine Strauch Erythroxylon Coca aus
Peru mit ungeteilten ganzrandigen lebhaft grünen Blättern und
blattachselständigen Gruppen kleiner weißer Blüten (Fig. 727). Er
liefert: Cocaïnum (Pharm. germ., austr., helv.) und Folia
Coca (Pharm. germ., helv.).
Guajacum officinale, ein Bäumchen Westindiens mit gegenständigen,
paarig gefiederten Blättern gehört zu der 4. Familie
Zygophyllaceae und liefert das offizinelle
Guajakholz und Guajakharz: Lignum Guajaci (Pharm. germ.,
austr., helv.). Resina Guajaci (Pharm. austr., helv.).
Fig. 728. Ruta graveolens. 1⁄3 nat. Gr. —
Offizinell.
Fig. 729. Blütendiagramm von Citrus vulgaris. Nach
A. W. EICHLER.
Fig. 730. Citrus vulgaris. 1⁄2 nat. Gr. —
Offizinell.
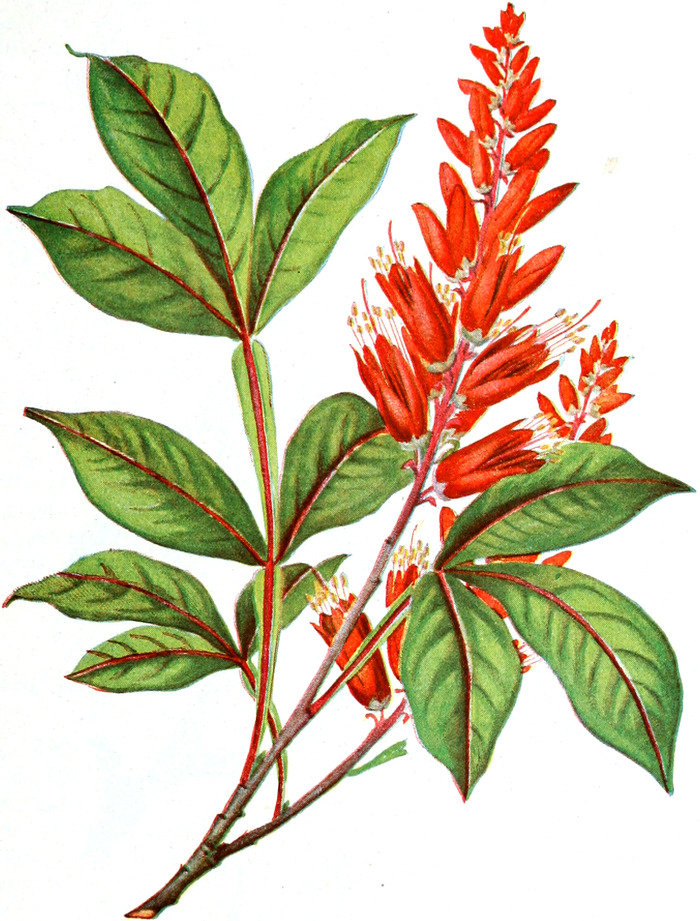
Fig. 731. Quassia amara. 1⁄2 nat. Gr. Nach BERG
und SCHMIDT. — Offizinell.
Fig. 732. Blütendiagramm von Polygala myrtifolia. Nach
A. W. EICHLER.
Fig. 733. Polygala Senega. A Blüte, a kleine
Kelchblätter, b große Kelchblätter, c Kahn, e seitliche
Kronblätter, d Anhängsel des medianen Kronblattes. B Andröceum,
h Antheren, vergr. — Nach BERG und SCHMIDT. —
Offizinell.
Fig. 734. Polygala Senega. 1⁄2 nat. Gr. —
Offizinell.
Wichtige Gattungen enthält die 5. Familie Rutaceae.
Ruta graveolens (Fig. 728), die Raute, eine ausdauernde,
halbstrauchige Pflanze mit doppelt gefiederten bis fiederteiligen
Blättern. Die Endblüten ihrer dichasialen Blütenstände sind bei
kräftigen Exemplaren fünfzählig, alle anderen vierzählig, strahlig,
mit großem intrastaminalem Diskus. Dictamnus fraxinella, der
einheimische Diptam, besitzt ansehnliche, dorsiventrale Blüten
in rispigen Infloreszenzen mit oben freien Fruchtblättern. Die
wichtigste Nutzpflanzen-Gattung ist Citrus[490]. Abweichend vom
Typus zeigen ihre Blüten (Fig. 729, 730) zahlreiche Staubblätter
in einem Kreise bündelweise vereinigt und eine Vermehrung der
Beerenfrüchte liefernden Fruchtblätter. Das Fruchtfleisch besteht
aus Zotten von saftreichen Zellen, welche in die Fächer einwachsen;
ihre Samen sind durch Adventivembryonen[S. 579] oft mehrkeimig (vgl. S.
500). Viele Arten haben einfache Blätter mit mehr oder minder
geflügeltem Blattstiel; diejenigen anderer Arten jedoch sind
dreizählig, und das Gelenk an der Einfügungsstelle der Spreite
zeigt, daß jene scheinbar einfachen Blätter unpaarig gefiederten
entsprechen, deren Endfiederchen allein erhalten blieb. Dornen
an der Blattansatzstelle entsprechen den ersten Blättchen der
Achselknospe. Citrus ist ostasiatischen Ursprungs; mehrere
Arten haben in den wärmeren Himalayatälern ihre Heimat,[S. 580] und
wohl alle wichtigen Kulturformen sind von den Chinesen zuerst
gezogen worden. Citrus decumana, Pompelmus, tropisch; Citrus
medica, diejenige Form, welche den Griechen beim Zuge Alexanders
als „medischer Apfel“ bekannt, wurde, ist jetzt in verschiedenen
Varietäten verbreitet, von denen Citrus (medica) Limonum
unserer Zitrone entspricht. Dieser Baum dürfte vom 3.
oder 4. Jahrhundert unserer Zeitrechnung ab im Mittelmeergebiet
gezogen worden sein. Citrus (medica) Bajoura (Cedro ital.,
Cedratier franz.), die Cedrate liefert aus ihren dickschaligen
Früchten das Zitronat. Citrus Aurantium kommt in zwei
verschiedenen Formen vor, als Citrus (Aurantium) vulgaris,
Fig. 730, Pomeranze, und C. (Aurantium) sinensis
Apfelsine; Citrus nobilis, Mandarine, ebenfalls
chinesischer Abstammung. Durch Pfropfung erhaltene Chimären von
Citrus aurantium und C. limonum heißen Bizzaria.
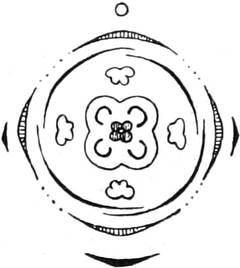
Fig. 735. Blütendiagramm von Ilex Aquifolium. Nach
A. W. EICHLER.
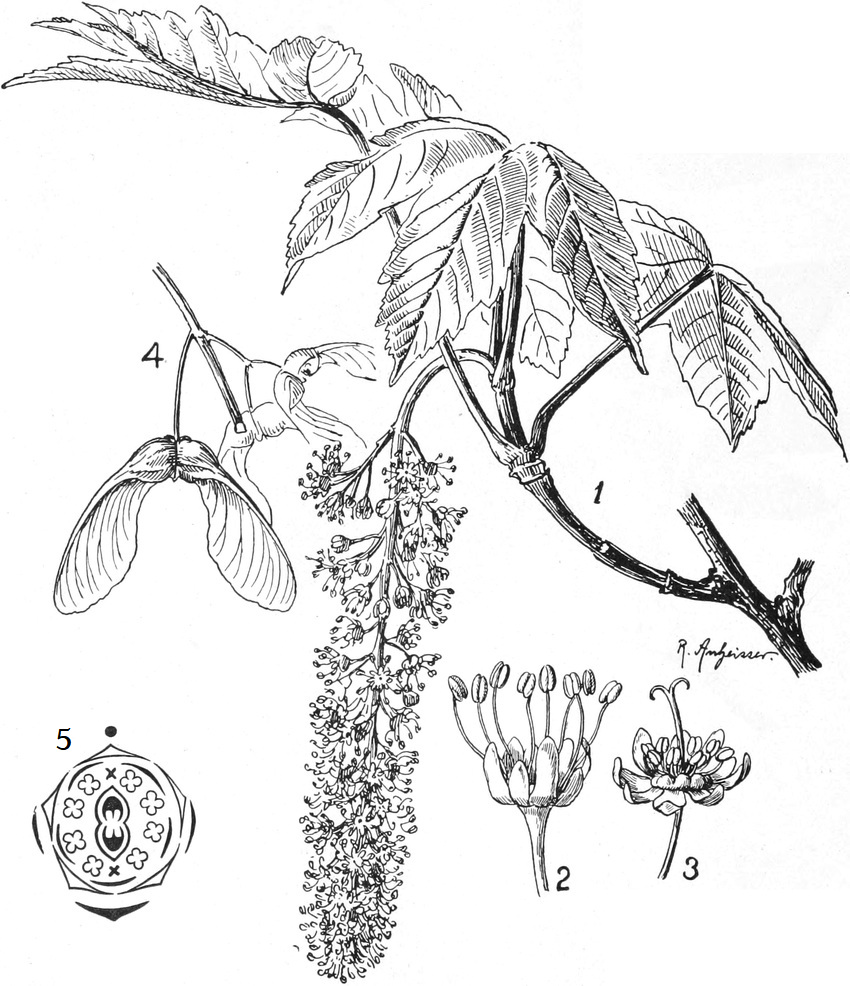
Fig. 736. Acer Pseudoplatanus. 1⁄2 nat. Gr. 1 Zweig
mit terminaler, hängender Blütenrispe. 2 Männliche, 3 weibliche
Einzelblüte. 4 Früchte. 5 Diagramm nach A. W. EICHLER.
Offizinell: Citrus vulgaris liefert: Fructus Aurantii
immaturi, Cortex Aurantii fruct. (Pharm. germ., austr.,
helv.), Oleum Aurantii florum und Folia Aurantii
(Pharm. austr., helv.), Oleum Aurantii Pericarpii (Pharm.
austr.). — Citrus limonum: Cortex Citri fructus und
Oleum Citri (Pharm. germ., austr.,[S. 581] helv.). — Citrus
Bergamia: Oleum Bergamottae (Pharm. helv.). — Ruta
graveolens: Herba Rutae (Pharm. helv.). — Pilocarpus
pennatifolius und P. Jaborandi, baumförmige Sträucher des
östlichen Brasiliens mit großen, unpaarig gefiederten Blättern:
Folia Jaborandi und Pilocarpinum (Pharm. germ.,
austr., helv.).
Wichtige offizinelle Pflanzen enthalten die 6. Familie
der Simarubaceae, nämlich Quassia amara (Fig. 731), Surinam,
und Picrasma excelsa, westindische Inseln, die Bitterholz
liefern: Lignum Quassiae (Pharm. germ., austr., helv.),
außerdem Cortex Simarubae Wurzelrinde von Simaruba amara
(Pharm. germ.), und die 7. Familie Burseraceae, die
offizinelle Harze ausscheidende Pflanzen umfaßt: Commiphora
abyssinica und C. Schimperi, arabisch-ostafrikanische Bäumchen,
geben Myrrha (Pharm. germ., austr., helv.), Boswellia
Carteri und B. Bhau dajianae, Bäumchen derselben Heimat wie
die Commiphora-Arten, liefern Olibanum, Weihrauch
(Pharm. austr.). Canarium-Arten der Philippinen endlich
Elemi (Pharm. austr., helv.).
Zwei seitliche kronblattähnliche Kelchblätter sind das beste
Kennzeichen der 8. Familie Polygalaceae, daneben
das kahnförmige untere Kronblatt und die röhrig verwachsenen
acht Staubblätter. (Fig. 732, 733, 734) K5, C3, A(8), G(̲2).
Einheimische Arten sind: Polygala vulgaris und P. amara,
auf Wiesen häufig; P. Chamaebuxus, kleiner Halbstrauch der Alpen.
Offizinell: Polygala Senega, Nordamerika, liefert Rad.
Senegae (Pharm. germ., austr., helv.).
9. Familie der Sapindaceae. Meist in tropischen Gebieten
heimische Pflanzen. Offizinell: Guarana (Pharm.
austr., helv.) aus dem zerquetschten Samen von Paullinia cupana,
einer Liana Brasiliens. Ebenso sind die Vertreter der 10. Familie
Anacardiaceae meist Tropenbewohner wie Mangifera
indica. Rhus toxicodendron, der Giftstrauch Nordamerikas
und die Pistacia-Arten gehören hierher. Offizinell:
Mastix (Pharm. austr.) von Pistacia Lentiscus. Zur 11.
Familie, den Aquifoliaceae, gehören die Ilex-Arten:
(Fig. 735) Ilex aquifolium die einheimische sog. Stechpalme,
I. paraguariensis, die Matepflanze, die den Paraguaytee
liefert. Endlich als 12. Familie sind die Aceraceae zu
nennen, denen die Ahorn-Arten angehören mit den charakteristischen
geflügelten Spaltfrüchten (Fig. 736), und 13. Familie die
Hippocastanaceae mit der Roßkastanie Aesculus
hippocastanum.
21. Ordnung.
Frangulinae.
Die Ordnung wird charakterisiert durch nur einen Staubblattkreis
vor den Kronblättern und einen intrastaminalen Diskus.
Die einzige einheimische Gattung der sonst in den Tropen verbreiteten
1. Familie Rhamnaceae ist Rhamnus. Rh. Frangula,
Faulbaum (Fig. 737 B, 738, 739), ist ein in Wäldern und an
Wegen häufiger Strauch mit wechselständigen ganzrandigen Blättern
und kleinen Nebenblättchen, dessen Blüten in Gruppen oder einzeln
blattachselständig stehen und bis auf das Gynäceum fünfzählig sind;
die schüsselförmige Blütenachse wird als Diskus ausgebildet. Zwei bis
drei Fruchtblätter mit ungeteilter Narbe liefern Steinfrüchte mit
ebensoviel Samen. Rh. cathartica, Kreuzdorn, ist die zweite
einheimische Art. Die Zweige sind meist dornig mit gegenständigen
gesägten Blättern. Die durchweg vierzähligen Blüten (Fig. 737 A)
werden durch Fehlschlagen diözisch; weibliche enthalten vier freie
Griffel und bringen viersamige Steinfrüchte, die Samen mit dorsaler
Raphe. Colletia spinosa und C. cruciata sind xerophile, blattlose
Sträucher Südamerikas. Offizinell: Cortex Frangulae
(Pharm. germ., austr., helv.) von Rh. Frangula. Cortex Rhamni
Purshianae (Pharm. austr., helv.) = Cascara sagrada von Rh.
Purshiana aus Nordamerika. Syrupus Rhamni catharticae (Pharm.
germ.) von Rh. cathartica.
Die nordhemisphärischen Gattungen Vitis, Ampelopsis,
Parthenocissus und die tropische Gattung Cissus gehören zur
2. Familie Vitaceae (Fig. 740, 741). Vitis vinifera,
die Weinrebe, ist als alte Kulturpflanze in zahlreichen Rassen und
Varietäten verbreitet. Die Blätter sind handförmig gelappt, die
blattgegenständigen Rankensprosse sind ursprünglich endständig,
aber durch einen Achselsproß zur Seite gedrängt. Oft stehen die
rispigen Blütenstände an Stelle von Ranken, und Zwischenbildungen
beider sind häufig. Der Kelch ist nur in Form eines kurzen Randes
vorhanden, und die am Scheitel verwachsene fünfzählige Krone
wird beim Öffnen der Blüten abgeworfen. Korinthen sind
die samenlosen Früchte der Vitis vinifera var. apyrena. Die
in Nordamerika und Asien verbreiteten Parthenocissus-Arten,
wilder Wein, haben geteilte Blätter und[S. 582] klettern zum Teil
mit Haftscheiben tragenden Ranken (Fig. 208). Offizinell:
Vinum (Pharm. germ., austr., helv.).
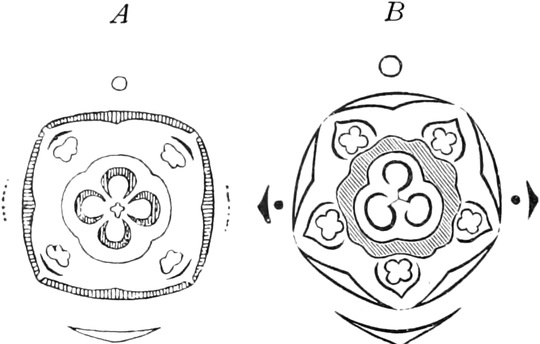
Fig. 737. Blütendiagramme. A Von Rhamnus cathartica
(hermaphrodit gedacht). B Von Rh. Frangula. Nach A. W.
EICHLER.
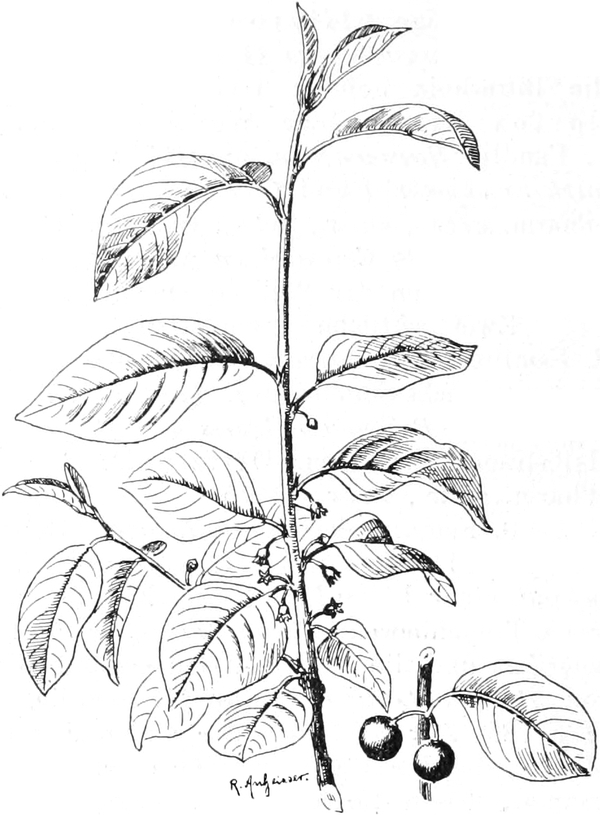
Fig. 738. Rhamnus Frangula. 1⁄2 nat Gr. Blühender Zweig
und Zweigstückchen mit Früchten.
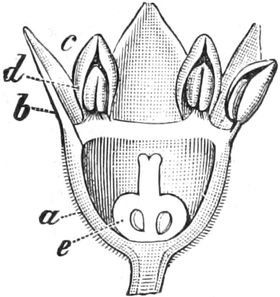
Fig. 739. Rhamnus Frangula. Blüte längsdurchschnitten.
a Blütenbecher, b Kelch, c Kronblatt, d Staubblätter, e
Fruchtknoten, Vergr. Nach BERG und SCHMIDT.
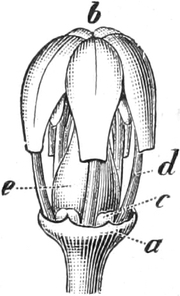
Fig. 740. Vitis vinifera. Öffnung der Blüte. a Kelch,
b Krone, c Diskus, d Staubblätter, e Fruchtknoten. Vergr. Nach
BERG und SCHMIDT.
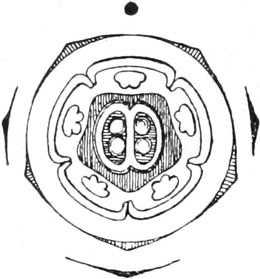
Fig. 741. Blütendiagramm von Ampelopsis hederacea. Nach
A. W. EICHLER.
[Die im Vorhergehenden gegebene Reihenfolge ist im wesentlichen
derjenigen des sero-diagnostischen Stammbaumes gefolgt; daß die
Aquifoliaceen zu den Sapindaceen gestellt sind, ist wohl eine kleine
Vereinfachung für die Aufzählung der Familien. Nun sollen auch die
Ericaceen in diesen Zweig eingegliedert werden. Wenn es auch seit
langem klar war, daß sie nicht zu den typischen Sympetalen gehören, so
hätte man nach morphologischen Gesichtspunkten eine Anreihung an die
Guttiferen oder die Ternstroemiaceen erwarten können, wohin teilweise
Sympetalie, apikale Poren der Antheren und Eigentümlichkeiten im Bau
der Fruchtknoten zu weisen scheinen. Somit reihe ich die Ericaceen nur
mit allem Vorbehalt vorläufig als letztes Glied dem Columniferenaste
ein.]
[S. 583]
22. Ordnung.
Ericinae.
mit der Familie der Ericaceae. Es sind hier immergrüne,
kleinblättrige, häufig nadelförmig belaubte Sträucher oder
Halbsträucher vereinigt, deren Blüten Antheren besitzen, die einmal
durch ein „Exothecium“ (S. 470) ausgezeichnet sind, die sich
andererseits in Poren oder Spalten öffnen und hornförmig abstehende
Anhänge tragen, daher auch wohl Bicornes genannt werden.
Fünfzählige Blüten in allen fünf Kreisen tragen die
Rhododendron-Arten, die Alpenrosen, deren wir drei einheimische
Arten besitzen. Ebenso sind die Blüten des Porst, Ledum
palustre, und der Gattung Andromeda fünfblättrig, und alle
genannten besitzen einen oberständigen Fruchtknoten, der zu einer
Kapselfrucht wird. Ebenso verhält sich Arctostaphylos uva ursi
(Fig. 742), mit dem einzigen Unterschiede, daß hier Steinfrüchte
entstehen. Fünfzählige Blüten mit unterständigem Fruchtknoten
finden wir bei mehreren Beerensträuchern aus der Gattung
Vaccinium (Fig. 743): der Heidelbeere, Vaccinium Myrtillus,
und Preißelbeere, V. Vitis idaea, welche dementsprechend die
Reste des Kelches auf jeder Beere tragen. — Eine Reduktion der
Gliederzahlen auf vier findet sich in der Gattung Erica, die
besonders im trockenen Mittelmeer- und Kapgebiet verbreitet ist;
dabei hat Erica oberständigen Fruchtknoten. Verbreitet ist bei
uns außer Erica Tetralix besonders das nahe verwandte Heidekraut,
Calluna vulgaris[491], von Erica durch den die Krone
überragenden Kelch unterschieden.
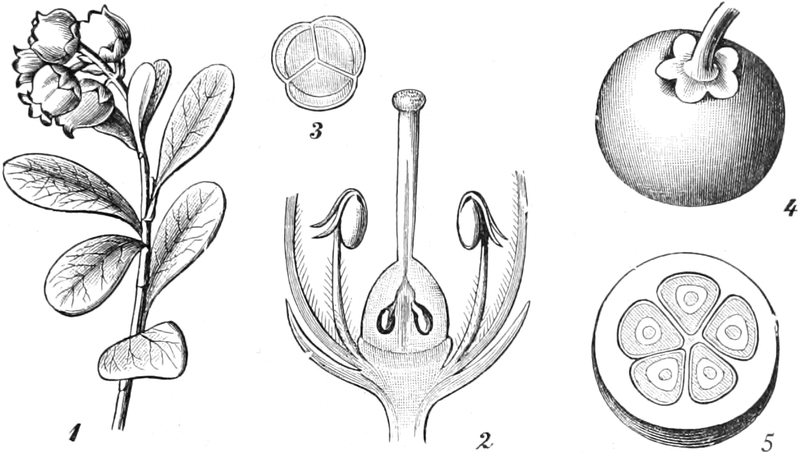
Fig. 742. Arctostaphylos uva ursi. 1 Blühender
Zweig. 2 Blüte im Längsschnitt. 3 Pollentetrade. 4 Frucht.
5 Fruchtquerschnitt. 2–5 vergr. Nach BERG und
SCHMIDT. — Offizinell.
Fig. 743. Diagramm von Vaccinium (Ericaceae). Nach
F. NOLL.
Offizinell sind Folia uvae ursi (Pharm. germ., austr.,
helv.) von Arctostaphylos uva ursi und Fructus Myrtilli
(Pharm. austr., helv.) von Vaccinium Myrtillus.
[S. 584]
B. Sympetalae.
Das einzige den Sympetalen gemeinsame Merkmal ist die
verwachsenblättrige Krone. Die Mehrzahl der nach Ausschluß der
Primulinae und Ericinae verbliebenen Sympetalen haben nur einen
Staubblattkreis, da der innere Kreis ausgefallen ist. Sie werden daher
als Tetracyclicae bezeichnet. Nur die einzige Ordnung der Diospyrinae
besitzt auch diesen inneren Staubblattkreis, sie werden daher als
Pentacyclicae jenen Vierkreisigen gegenübergestellt.
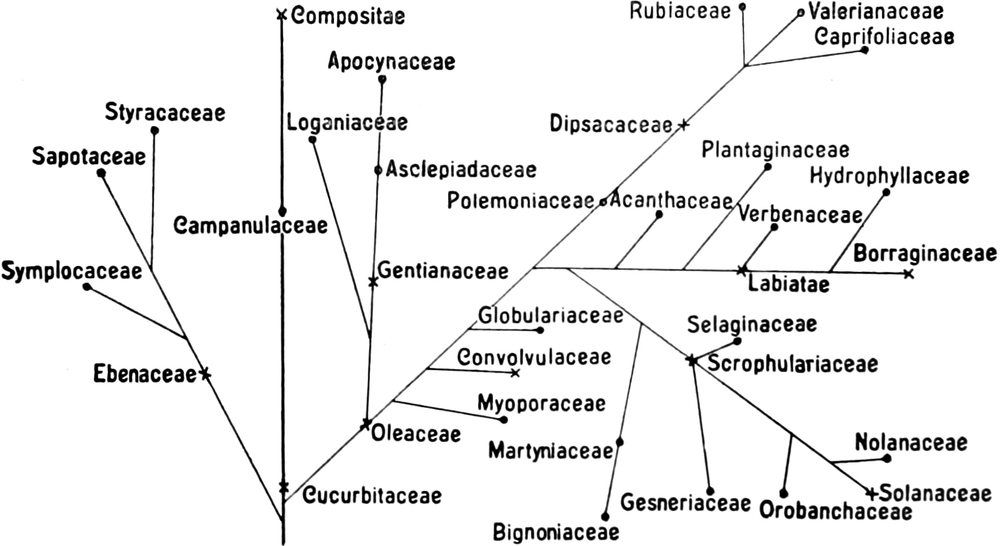
Stammbaum der Sympetalen nach den sero-diagnostischen Untersuchungen
von W. ALEXNAT.
Die 1. Ordnung der
Diospyrinae,
zu der die tropischen Familien der Ebenaceae, Sapotaceae und
Styracaceae zu zählen sind, ist von Bedeutung wegen der den
Sapotaceen angehörigen Palaquium- (Fig. 744) und Payena-Arten
des Malayischen Archipels, welche Guttapercha (Pharm. germ.)
liefern; Mimusops-Arten (Sapotaceae) geben einen ähnlichen
Körper, Balata, und von den Styracaceen leitet man das
Benzoëharz (Pharm. germ., austr., helv.) ab, doch ist seine
Herkunft von Styrax Benzoin zweifelhaft geworden.

Fig. 744. Palaquium Gutta. 1⁄2 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
B. Tetracyclicae.
1. Fruchtknoten oberständig.
Die Tetracyclicae haben also nur vier regelmäßig miteinander
alternierende Blütenwirtel aufzuweisen. Sie lassen sich nach der
Stellung des Fruchtknotens[S. 585] in zwei Gruppen von Ordnungen zerlegen,
entweder ist der Fruchtknoten oberständig, hierher gehören die
Ordnungen der Contortae, Tubiflorae und Personatae. Alle diese
Ordnungen stimmen auch darin überein, daß der Fruchtknoten aus
zwei Fruchtblättern besteht. Die Ordnungen mit unterständigem
Fruchtknoten sind die Rubiinae und die den Rubiinae nahestehenden
Dipsacaceae, die von den Synandrae (Campanulaceae-Compositae) getrennt
werden. Diese bleiben in der Hauptreihe; die mit den Rubiinae endende
Abzweigung setzt ebenso wie die einzige pentazyklische Ordnung der
Diospyrinae an die Cucurbitaceae als Abzweigung an. Die Zahl der
durchweg unterständigen Fruchtblätter ist bei der Mehrzahl der Rubiinae
zwei, Cucurbitaceae, Cuprifoliaceae und Valerianaceae besitzen drei,
die Campanulaceae zwei oder fünf und Dipsacaceae und Compositae nur
ein Fruchtblatt.
Nachdem so einzelne gemeinsame Merkmale herausgefunden sind, sollen die
Ordnungen nach ihren Familien besprochen werden.
Fig. 745. Oleaceae. Diagramm (Syringa). Nach F.
NOLL.

Fig. 746. Olea europaea mit Früchten. 1⁄2 nat. Gr. —
Offizinell.

Fig. 747. Olea europaea. A Ausgebreitete
Krone. B Kelch und Fruchtknoten im Längsschnitt. Vergr. Nach
ENGLER-PRANTL.
Fig. 748. Olea europaea. Steinfrucht.
Die 2. Ordnung,
Contortae,
umfaßt Pflanzen mit dekussierten, meist einfachen Blättern, durchweg
strahligen Blüten, deren Krone in der Knospenlage häufig gedreht und
deren Andröceum der Krone angewachsen ist.
Die 1. Familie der Oleaceen ist an der Zweizahl des
Andröceums leicht zu erkennen; die Krone ist meist vierzipfelig,
wie das Diagramm von Syringa zeigt (Fig. 745). Neben Ligustrum,
Jasminum, Forsythia und Syringa ist Olea europaea,
der Ölbaum oder die Olive, die wichtigste Pflanze der Familie
(Fig. 746). Im Mittelmeergebiete heimisch, wird sie hier auch
vorzugsweise kultiviert. Blüten und Früchte entsprechen dem
Familientypus[S. 586] (Fig. 747). Die Steinfrucht enthält sowohl im
Fruchtfleisch wie im Endosperm des Samens fettes Öl (Fig. 748).
Vom Familientypus weicht durch gefiederte Blätter die Esche,
Fraxinus, ab. Fraxinus excelsior mit anemophilen Blüten blüht
vor der Belaubung, Fr. Ornus, die in Sizilien zur Mannagewinnung
kultivierte Bergesche oder Mannaesche, hat ansehnliche
gipfelständige Infloreszenzen, deren Einzelblüten doppeltes
Perianth mit tiefgespaltener weißer Krone besitzen. Es sind neben
Zwitterblüten (Fig. 749) rein weibliche schwarze Blüten vorhanden;
die Pflanze ist polygam.
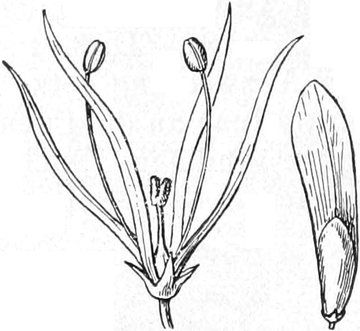
Fig. 749. Fraxinus Ornus. Einzelblüte und Frucht. —
Offizinell.
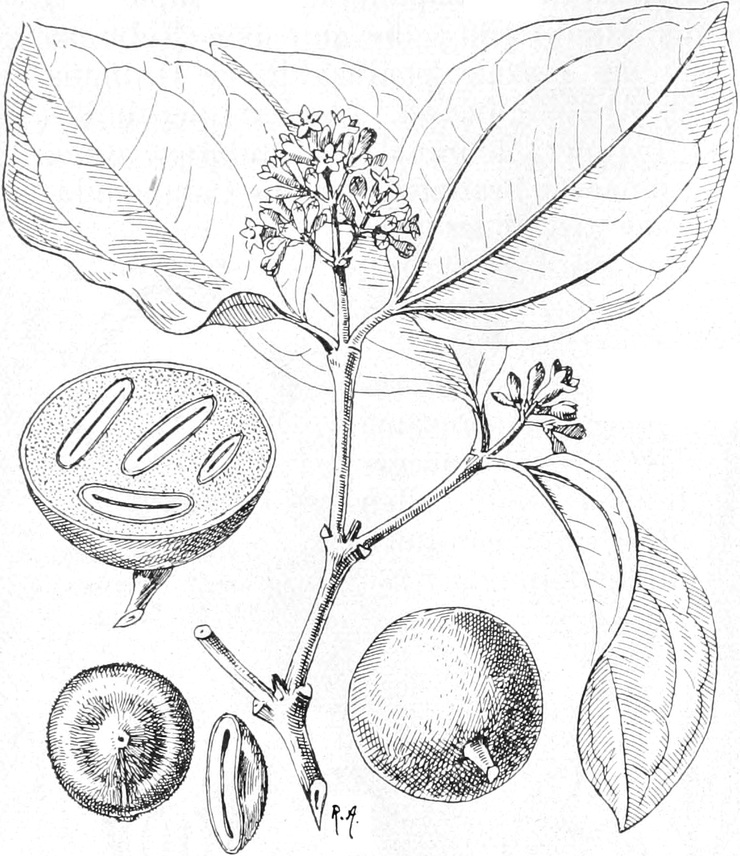
Fig. 750. Strychnos nux vomica. 1⁄2 nat. Gr. Frucht
und Samen, Querschnitte durch beide. — Offizinell und
giftig.
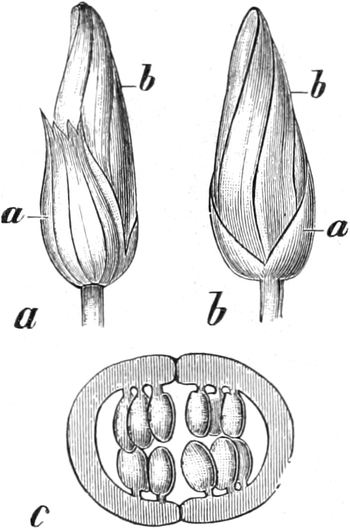
Fig. 751. Gentiana lutea. a und b Blütenknospe mit
Kelch (a) und der gedrehten Krone (b). Nat. Gr. c Fruchtknoten
im Querschnitte, vergr. Nach BERG und SCHMIDT. —
Offizinell.

Fig. 752. Vinca minor. 2⁄3 nat. Gr.
Offizinell ist Oleum Olivarum (Pharm. germ., austr.,
helv.) von der Olive und Manna (ibidem) von der Mannaesche.
[S. 587]
Die 2. kleine Familie der Contortae, die
Loganiaceae[492], ist nur wegen der wichtigen Gift-
und offizinellen Pflanze Strychnos nux vomica (Fig. 750) zu
erwähnen, die auch das Pfeilgift der Malayen liefert; aus anderen
Strychnos-Arten wird das Curare Südamerikas gewonnen; ruminiertes
Endosperm bei Spigelia.
Offizinell: Semen Strychni (Pharm. germ., austr.,
helv.) und Radix Gelsemii (Pharm. germ., austr., helv.) von
Gelsemium nitidum.
Wichtiger ist die 3. Familie der Ordnung, die
Gentianaceae, durch einfächerigen Fruchtknoten kenntlich,
mit deutlich gedrehter Krone in der Knospe (Fig. 751). Einheimisch
ist die Gattung Gentiana mit zahlreichen Arten, deren lebhaft,
meist blau gefärbte große Blüten ein Schmuck der Alpenwiesen sind.
Der umstrittene Saisondimorphismus soll durch Entwicklung nahe
verwandter Formen, deren eine vor dem ersten Wiesenschnitt blüht
und fruchtet, während die andere ihre Blütezeit nach dem Schnitt
hat, an Gentiana-Arten[493] gute Bestätigung finden.
[S. 588]

Fig. 753. Nerium Oleander. 1⁄2 nat. Gr. —
Giftig.
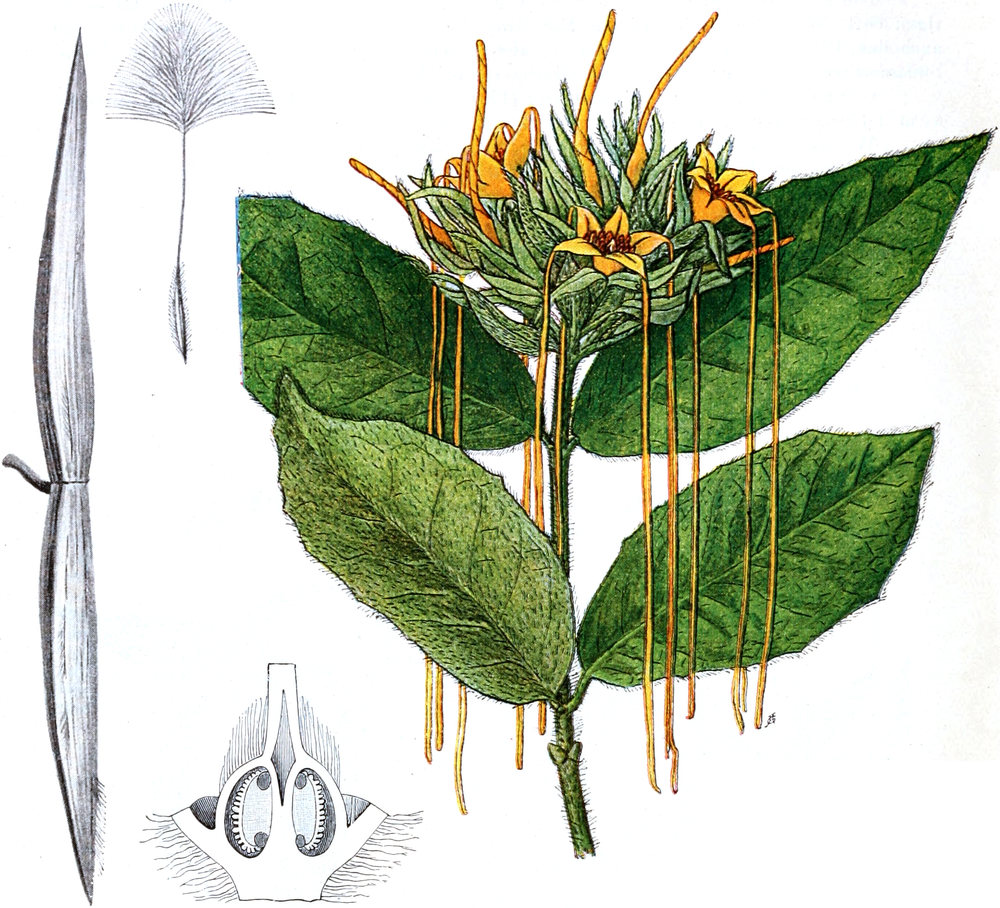
Fig. 754. Strophanthus hispidus. 1⁄2 nat. Gr. Nach
E. GILG kombiniert mit MEYER und SCHUMANN.
Fruchtknoten im Längsschnitt 10⁄1, Frucht ca. 1⁄7, Samen 1⁄3 nat. Gr.
Nach SCHUMANN in ENGLER-PRANTL. — Offizinell.
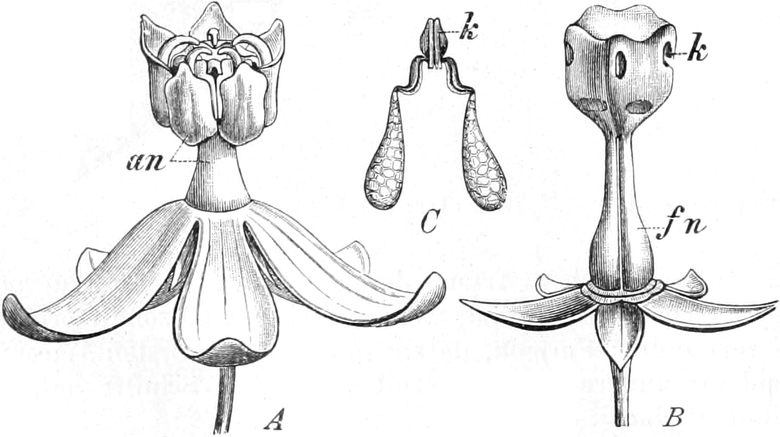
Fig. 755. Asclepias curassavica. A Blüte, an
Andröceum. Vergr. 4. B Kelch und Gynäceum, fn Fruchtknoten, k
Klemmkörperchen. Vergr. 6. C Zwei Pollinien. Stärker vergr. Nach
H. BAILLON.
Fig. 756. Vincetoxicum officinale. 1⁄2 nat. Gr. —
Giftig. Nach H. SCHENCK.
Erythraea, Tausendgüldenkraut, Menyanthes,
Bitterklee, Limnanthemum, eine westdeutsche Wasserpflanze
mit Schwimmblättern, sind bekannte Gentianaceen.
Offizinell: Radix Gentianae (Pharm. germ., austr.,
helv.) von G. lutea, pannonica, punctata, purpurea, den
größten Gentiana-Arten unserer Gebirge, Herba Centaurii[S. 589]
(ibidem) von Erythraea Centaurium und Folia Trifolii
fibrini (ibidem) von Menyanthes trifoliata.
Die 4. Familie der Apocynaceae, hauptsächlich in den Tropen
verbreitet, umfaßt nur immergrüne Pflanzen mit Milchsaft. Die beiden
Fruchtblätter sind unterwärts frei, oben hält der gemeinsame
Griffel sie mit ringförmiger Narbe zusammen; so streben sie nach
der Befruchtung auseinander und bilden zwei große Balgfrüchte mit
zahlreichen, meist durch Besitz eines Haarschopfes ausgezeichneten
Samen (Fig. 754).
Einheimisch ist nur Vinca minor, das kleine Immergrün der Wälder
in West- und Süddeutschland (Fig. 752). Im Mittelmeergebiet
heimisch, bei uns vielfach in Kultur, ist der giftige Oleander,
Nerium Oleander (Fig. 753). Cerbera Odollam (Fig. 590),
Schwimmfrucht der Mangrove, ist eine Apocynacee.
Offizinell ist Semen Strophanti (Fig. 754)[494] von der
afrikanischen Liane Str. Kombé (Pharm. germ., austr., helv.),
Kautschuk[495] (Pharm. germ.) von Kickxia elastica,
Landolphia-Arten, Carpodinus-Arten, alle im tropischen Afrika
beheimatet, ferner von dem brasilianischen Baum Hancornia
speciosa und der malayischen Lianengattung Willoughbeia;
Guttapercha (Pharm. germ.) von Tabernaemontana Donnell
Smithii (Mittelamerika) und endlich Cortex Quebracho von
Aspidosperma Quebracho (Pharm. helv.).
Die 5. Familie der Asclepiadaceae gleicht in allen Punkten der
vorigen, besitzt aber freie, nur durch die prismatische (Fig.
755) Narbe zusammengehaltene Fruchtblätter. Ihre Staubblätter
sind an der Basis vereinigt und tragen dorsale nektarführende
Anhängsel, welche eine Nebenkrone bilden. Die Pollenmassen bleiben
fachweise zu Pollinien verklebt, deren Stiele an drüsigen Schwellungen
des kantigen Narbenkopfes, den Klemmkörperchen, festsitzen.
Diese alternieren mit den Staubblättern, so daß von den an einem
Klemmkörperchen hängenden Pollinienpaaren je eines dem rechten und
dem linken Nachbarstaubblatte entstammen. Besuchende Insekten nehmen,
ähnlich wie bei den Orchidaceen, die ganzen Pollinien mit fort und
übertragen sie auf andere Blüten (Fig. 755).
Einheimisch ist nur Vincetoxicum officinale (Fig. 756), eine
unscheinbare, aber giftige Staude mit kleinen weißen Blüten und
langbehaarten Samen in den Balgfrüchten. Die übrigen Asclepiadaceen
sind meist Bewohner der Tropen und Subtropen. Besondere Erwähnung
verdienen die sukkulenten Stapelia-, Hoodia-, Trichocaulon-
usw. Arten, kaktusähnliche (Konvergenz) Pflanzengestalten der
südafrikanischen Wüsten, und die eigenartige Kannenpflanze des
Malayischen Gebietes, Dischidia Rafflesiana[496] (Fig. 211),
deren Kannen als Wasserkondensatoren dienen. Vielfach kultiviert
wird Hoya carnosa, die Wachsblume, und zahlreiche Ceropegia-Arten
mit eigenartig überdachten Blüten. Offizinell: Cortex
Condurango (Pharm. germ., austr., helv.) von Marsdenia
Cundurango, einer in Ecuador und Columbien heimischen Liane.
Die 3. Ordnung
der Sympetalen sind die Tubiflorae,
denen man Beziehungen zu den Gruinales und Rosifloren zuschreibt.
Fünfzählige, oft dorsiventrale Blüten mit zweifächerigem Fruchtknoten,
der in jedem Fache zwei durch falsche Scheidewände getrennte
Samenanlagen führt. Die normale Zahl der Staubblätter wird bei den
dorsiventralen Blüten auf vier, bisweilen auf zwei reduziert.
An den Anfang sei die strahlblütige 1. Familie der
Convolvulaceae, der Windengewächse, gesetzt, deren
Angehörige vielfach Schlingpflanzen sind mit wechselständigen,
pfeilförmigen Blättern und weiten trichterförmigen, in der
Knospenlage längsgefalteten Blumenkronen. Die Frucht wird zu einer
Kapsel mit aufrechten Samenanlagen. Unsere häufige Ackerwinde,
Convolvulus arvensis, an allen Wegrainen, in Getreidefeldern usw.
verbreitet, trägt blattachselständige langgestielte Einzelblüten.
Die großblütige Zaunwinde, Calystegia sepium, hat zwei große
Vorblätter unter dem Kelche. Auch die auf Weiden, Urticaceen
und sonst parasitierenden Cuscuta-Arten mit ihren bleichen
Stengeln und geknäuelten kleinen Blüten gehören hierher (Fig.
219). Ipomoea, als schönfarbige Sommerblume[S. 590] bekannt, liefert
das pantropische Strandgewächs I. pes caprae. Nahe verwandt
ist Pharbitis (Fig. 303, S. 282). Offizinell sind
Tubera Jalapae (Pharm. germ., austr., helv.) von Exogonium
Purga (Fig. 757), ferner Scammonium (Pharm. helv.) von
Convolvulus Scammonia (Kleinasien).

Fig. 757. Exogonium Purga. 1⁄2 nat. Gr. Nach
BERG und SCHMIDT. — Offizinell.
Fig. 758. Borrago officinalis. a Blüte, b und c
Frucht. Nat. Gr.
Fig. 759. Blütendiagramme. A Verbena officinalis. Nach
A. W. EICHLER. B Lamium (Labiatae). Nach F. NOLL.
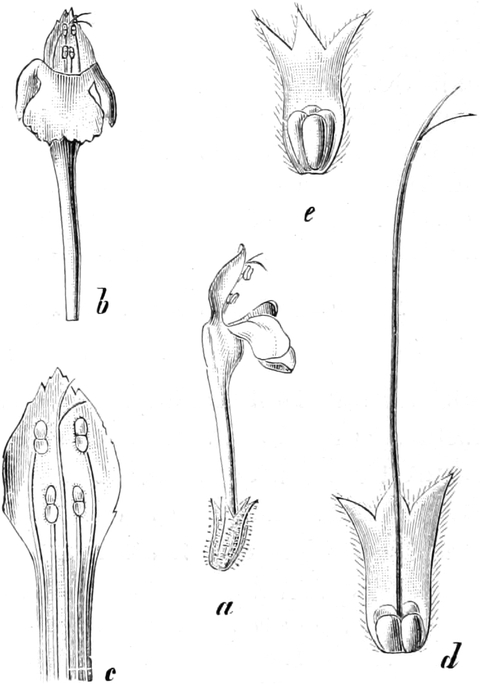
Fig. 760. Galeopsis ochroleuca. a Blüte, nat. Gr.,
b dieselbe ohne Kelch, nat Gr., c Korolle, aufgeschnitten, mit den
Staubblättern und dem Griffel, d Kelch mit dem Gynäceum, e Frucht,
c–e vergr. 2. — Offizinell. Nach H. SCHENCK.
In der 2. Familie der Borraginaceae, der
Rauhblättrigen, sind borstig behaarte Stauden, wie das
Ochsenauge Anchusa, die Natternzunge Echium, die Wallwurz
Symphytum, das Vergißmeinnicht Myosotis vereinigt, deren
radiäre, in einzelnen Fällen bereits ein wenig zygomorphe Blüten
(Echium) in Wickeln oder meist Doppelwickeln vereinigt sind.
Durch die andersfarbigen Schlundschuppen und den mittels der tief
einschneidenden falschen Scheidewand in vier einsamige Klausen[S. 591]
zerteilten Fruchtknoten, in deren Mitte der Griffel aufragt (Fig.
758), ist die Familie scharf charakterisiert. Deutlich dorsiventrale
Blüten mit nur vier Staubblättern und einem auf den ebenfalls vier
Samen beherbergenden Fruchtknoten (Fig. 759) endständig aufgesetzten
Griffel unterscheiden die 3. Familie der Verbenaceae, zu der
der wertvolle Teakholzbaum Tectona grandis gehört; außerdem ist der
vivipare Mangrovebaum Avicennia[496] eine Verbenacee.

Fig. 761. Lavandula vera. 1⁄2 nat. Gr. —
Offizinell.

Fig. 762. Salvia officinalis. Blühender Sproß. 1⁄2 nat.
Gr. Der Länge nach aufgeschnittene Kronröhre mit den Staubblättern.
Vergr. — Offizinell.
[S. 592]
Wichtiger ist die 4. große Familie Labiatae, die typischen
Lippenblütler, die durch vierkantigen Stengel, dekussierte
Blattstellung und aromatische Drüsenhaare schon in ihren vegetativen
Organen scharf hervortreten. Die in achselständigen Dichasien oder
Doppelwickeln vereinigten Blüten sind stets dorsiventral gebaut; sie
bestehen aus verwachsenem Kelch, zweilippiger Krone, deren Oberlippe
zwei, die Unterlippe drei Zipfel besitzt. Ihre vier Staubblätter sind
ungleich lang. Salvia, der Salbei und Rosmarinus haben deren nur
zwei. Der Fruchtknoten (Fig. 759) entspricht genau demjenigen der
Borraginaceen; in seinem Grunde liegt ein ringförmiges Nectarium.
Ein großer Teil unserer Flora besteht aus Lippenblütlern.
Lamium, die Taubnessel, Stachys, der Ziest, Galeopsis (Fig.
760) mit helmförmiger, Ajuga mit sehr kurzer, Teucrium mit
tiefgespaltener Oberlippe zeigen einige Formverschiedenheiten.
Bei Glechoma und Nepeta sind die hinteren Staubblätter die
längeren, umgekehrt wie bei den übrigen Labiaten. Bei Salvia,
dem Salbei, sind die beiden allein vorhandenen Staubblätter für
Bestäubungszwecke eigenartig gebaut (vgl. S. 481, Fig. 540). Ihrer
aromatischen Eigenschaften wegen sind zahlreiche Lippenblütler
für offizinelle Zwecke herbeigezogen. Einen besonders
reichen Beitrag dafür stellen auch die trockenen Gesträuche der
Mittelmeerländer, die Macchien. So liefern Lavandula vera (Fig.
761) Flores Lavandulae und Oleum Lavandulae (Pharm.
germ., austr., helv.), Salvia officinalis (Fig. 762) Fol.
Salviae (ibid.), Melissa officinalis Fol. Melissae
(ibid.), Thymus Serpyllum Herba Serpylli (ibid.),
Thymus vulgaris Herba Thymi, Oleum Thymi und
Thymolum (ibid.), Rosmarinus officinalis Folia und
Oleum Rosmarini (ibid.), Mentha piperita Folia
und Oleum Menthae piperitae wie Mentholum (ibid.),
Galeopsis ochroleuca Herba Galeopsidis (Pharm. austr.),
Origanum vulgare Herba Origani (ibid.), Origanum
Majorana Herba Majoranae (Pharm. helv.).
4. Ordnung.
Personatae.
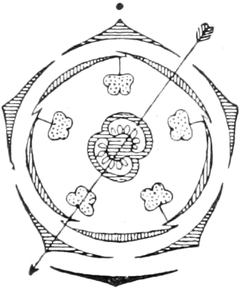
Fig. 763. Solanaceae. Diagramm (Petunia). Nach F.
NOLL.

Fig. 764. Solanum Dulcamara. 1⁄2 nat. Gr. —
Offizinell und giftig.
Gemeinsamen Ursprung mit den Tubifloren scheinen die Personatae zu
haben. Sie umfassen ebenfalls radiäre und dorsiventrale Blütenformen,
doch fehlen die falschen Scheidewände, und die Zahl der Samenanlagen
ist erheblich größer. Die 1. wichtige Fa[S. 593]milie, Solanaceae,
besitzt meist radiäre Blüten, deren Kronblätter in der Knospenlage
gefaltet sind. Der Fruchtknoten wird durch eine schräg zur Mediane
stehende Wand geteilt (Fig. 763). Die verschiedenartigen Früchte
umschließen Samen mit stark gekrümmtem Embryo im Endosperm. Anatomisch
ist der Besitz bikollateraler Leitbündel hervorzuheben.

Fig. 765. Atropa Belladonna. 1⁄2 nat. Gr. —
Offizinell und giftig.
Radiäre Blüten und Beerenfrüchte zeichnen die Gattung Solanum
aus. Solanum tuberosum ist die Kartoffel (Fig. 201), S. nigrum
der Nachtschatten, S. Dulcamara (Fig. 764), der Bittersüß, und
Lycopersicum die Tomate, sind weitere bekannte Solanum-Arten.
Über Pfropfbastarde, Periklinalchimären und Gigasformen der
Solanum-Arten vgl. S. 262 und H. WINKLER[497]. Atropa
Belladonna, die Tollkirsche (Fig. 765), eine sehr giftige Staude
Europas, ist an der radiären, röhrig aufgedunsenen Blüte von trüb
purpurner oder auch gelber Färbung, wie an den schwarzglänzenden
Beerenfrüchten auf stark vergrößertem Kelche zu erkennen. Der
zunächst radiäre Hauptsproß verzweigt sich unter der Endblüte
in meist drei gleich starke, plagiotrope Zweige, die sich
wickelartig weiter verzweigen; durch Hinaufwachsen des Tragblattes
an dem Achselsproß wird der Anschein gepaarter Blätter erweckt
(Fig. 765). Capsicum annuum hat ähnliche Verzweigung; seine
trockenen Beerenfrüchte liefern den spanischen Pfeffer. Datura
Stramonium, der Stechapfel, ist ebenfalls eine Atropa-ähnlich
verzweigte einjährige Pflanze, die mit ihren ausgeschweiften
Blättern, den großen, in der Knospe gefalteten weißen Blüten und
den charakteristischen scharf bewehrten Kapselfrüchten leicht
kenntlich ist (Fig. 766). Nicotiana tabacum, die Tabakpflanze
(Fig. 767), ist in zahlreichen Kulturformen verbreitet; die großen,
wechselständigen, stark drüsig behaarten Blätter liefern nach
Trocknung und Fermentation den Tabak; die Früchte der Gattung sind
kapselförmig. Hyoscyamus niger, das Bilsenkraut, eine einjährige
Giftpflanze[S. 594] der alten Welt; die mit stark drüsig behaarten,
sitzenden, wechselständigen Blättern besetzte Achse endigt in einem
Wickel etwas zygomorpher Einzelblüten von trübgelber Farbe mit
blauer Aderung (Fig. 768); die Frucht ist eine Deckelkapsel.

Fig. 766. Datura Stramonium. 1⁄2 nat. Gr. Reife Frucht
aufgesprungen. — Offizinell und giftig.
Infolge ihres Gehaltes an giftigen Alkaloiden, die in der
Medizin Verwendung finden, zählen viele Solanaceen zu den
offizinellen Gewächsen, so liefert Atropa Belladonna
Folia Belladonnae, Radix Belladonnae und
Atropin (Pharm. germ., austr., helv.), Datura Stramonium
Semen und Folia Stramonii (ibid.), Hyoscyamus niger
Folia Hyoscyami (ibid.), Capsicum annuum Fructus
Capsici (Pharm. germ., helv.), Nicotiana tabacum Folia
Nicotianae (Pharm. helv.), Solanum Dulcamara Caules
Dulcamarae (Pharm. austr., helv.); Scopolia carniolica
Scopolaminum (Pharm. germ.).
[S. 595]
Durch dorsiventrale Blüten, nicht gefaltete Knospenlage der
Krone und Unvollständigkeit des Andröceums ist die 2. Familie
Scrophulariaceae von den Solanaceen unterschieden. Außerdem
hat die zweifächerige Kapselfrucht keine schrägstehende Scheidewand.
Von den bekannten Gattungen hat nur Verbascum (Fig. 769, 770
A), die Königskerze, fünf Staubblätter, doch sind die drei
hinteren mit wollig behaarten Filamenten und quergestellten
Antheren abweichend und nur zwei vordere normal ausgebildet.
Die Pflanzen sind zweijährig und durch stark wollige Behaarung
der mächtigen Blattrosette kenntlich. Scrophularia, Linaria
und Antirrhinum, Löwenmaul, haben nur vier Staubblätter bei
einer zweilippigen Krone, bei Gratiola (Fig. 770 B) und
Veronica, Männertreu, sinkt die Zahl der Staubblätter auf zwei
herab. Mimulus, Torenia mit reizbarer Narbe vgl. S. 321.
Maurandia Blattstielkletterer. Digitalis purpurea (Fig. 771),
der Fingerhut, mit einseits gewendeten Blüten am Schaft des im
zweiten Jahre aus der Blattrosette aufschießenden Blütenstandes,
ist giftig, und seine Blätter sind als Folia Digitalis
offizinell (Pharm. germ., austr., helv.). Ebendort die
Blüten von Verbascum thapsiforme und V. phlomoides als
Flores Verbasci.

Fig. 767. Nicotiana Tabacum. 1⁄2 nat. Gr. —
Offizinell und giftig. a Blüte, nat. Gr., b Krone
aufgeschnitten, nat. Gr., c Fruchtknoten, nat. Gr., d und e junge
Frucht. Vergr. 2.
Einen besonderen Verwandtenkreis innerhalb der Scrophulariaceen
bilden die Parasiten und Hemiparasiten wie die völlig
chlorophyllfreie Lathraea[498] und die grün beblätterten[S. 596] aber
mit Wurzelhaustorien auf anderen Pflanzen parasitierenden Arten von
Tozzia, Bartschia, Pedicularis, Euphrasia, Odontites,
Melampyrum und Alectorolophus.
Ebenso sind die Angehörigen der 3. Familie Orobanchaceae
mit der durch einfächerigen Fruchtknoten ausgezeichneten Gattung
Orobanche, Würger, rein parasitisch (Fig. 772).

Fig. 768. Hyoscyamus niger, blühender Sproß und Frucht.
1⁄2 nat. Gr. — Offizinell und giftig.
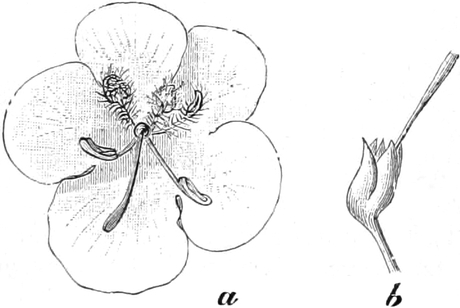
Fig. 769. Verbascum thapsiforme. a Blüte. b Kelch
und Griffel. Nat. Gr. — Offizinell. Nach H. SCHENCK.
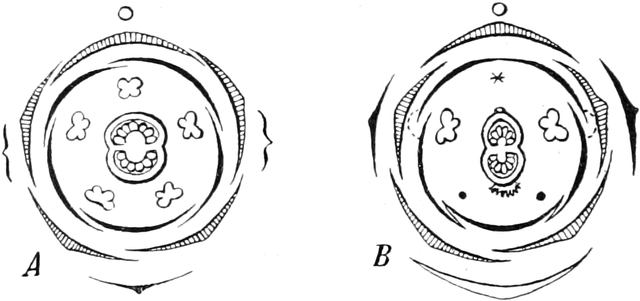
Fig. 770. Scrophulariaceae. Blütendiagramme. Nach A.
W. EICHLER. A Verbascum. B Gratiola.
In der Lebensweise bieten auch die Angehörigen der 4. Familie
Lentibulariaceae Besonderheiten, so sind die Sumpf- und
Wasserpflanzen der Gattungen Utricularia[499] und Pinguicula
insektivor.
In der reduzierten 5. Familie der Plantaginaceae, sind
anemophile Gattungen wie Litorella lacustris und stark dichogame,
wie der protogyne Plantago, Wegerich, vereinigt.
[S. 597]
2. Fruchtknoten unterständig.
Die 5. Ordnung,
Rubiinae,
soll ihre Verwandten in der ebenfalls durch unterständigen Fruchtknoten
ausgezeichneten Ordnung der Umbellifloren besitzen, an die sie also
anschließen würde. Die Blüten sind vier- oder fünfzählig, bei den
verwandten Familien mit zygomorphen (Caprifoliaceen) und asymmetrischen
(Valerianaceen) Blüten ändern sich die Zahlen im Andröceum und Gynäceum.

Fig. 771. Digitalis purpurea. 1⁄2 nat. Gr. a
Corolle, b Kelch und Fruchtknoten, c Frucht aufgesprungen,
d Fruchtquerschnitt. a–d nach H. SCHENCK. —
Offizinell und giftig.
[S. 598]
Die erste Familie der Rubiaceae[500] besitzt radiäre Blüten
und in den vegetativen Teilen einfache Blätter mit Nebenblättern.
Einheimische Rubiaceen gibt es nur wenige, die alle dem Formenkreis
von Asperula, Waldmeister, Galium und Rubia angehören, dadurch
ausgezeichnet, daß die Nebenblätter den Blättern gleichgestaltet sind
und scheinbare Blattquirle darstellen, die normal sechszählig sein
müßten, durch Verwachsung der benachbarten Nebenblätter vierzählig
werden, aber in den Zahlen etwas variieren.
In den Tropen sind Rubiaceen reich vertreten als Bäume, Sträucher,
Kletterpflanzen und Epiphyten.
Eine der wichtigsten Rubiaceen-Gattungen ist Cinchona (Fig. 773);
sie liefert in verschiedenen kultivierten Arten die Chinarinde und
die daraus gewonnenen fieberwidrigen Alkaloide. Stattliche Bäume
der Südamerikanischen Anden, werden die Cinchona-Arten jetzt in
allen tropischen Kolonien angebaut. Ihre Kapselfrüchte beherbergen
zahlreiche geflügelte Samen (Fig. 774). Ebenso wichtig aber als
menschliches Genuß- und Nahrungsmittel ist der Kaffeestrauch,
Coffea arabica (Fig. 775), Gebirgsbewohner Afrikas, und daneben
die im tropischen Tieflande fortkommende Coffea liberica. Die
Früchte sind zweisamige Steinfrüchte. Das Exokarp wird fleischig;
das Endokarp besteht aus einer dünnen Lage Steinzellen, welche die
von ihrer Silberhaut, der Samenschale, eingehüllten zwei Samen,
die Kaffeebohnen, umschließt. Zu den beerenfrüchtigen Formen
gehören die bekannten merkwürdigen Knollenepiphyten Hydnophytum
und Myrmecodia[500], die nach den neuesten Untersuchungen
aus den Exkrementen der sie bewohnenden Ameisenkolonien Nutzen
ziehen. Ebenso sind ernährungsphysiologisch Psychotria und
Pavetta-Arten von Interesse[501], die in ihren Blättern
Stickstoff assimilierende Bakterien beherbergen; sie sind insofern
höher organisiert als die Leguminosen mit ihren Wurzelknöllchen,
als sie die Bakterien auch mit in ihre Samen einschließen und so
gleich Vorsorge für die nächste Generation treffen.
Von offizineller Bedeutung sind außer dem Chininum
und Cortex Chinae (Pharm. germ., austr., helv.) von
Cinchona succirubra und C. Ledgeriana die Radix
Ipecacuanhae (ibid.) von dem kleinen beerenfrüchtigen
Halbstrauch Brasiliens Uragoga Ipecacuanha (Fig. 776) und
Catechu (Gambir), der aus den Blättern der Liane
Ourouparia Gambir gewonnene Extrakt.

Fig. 772. Orobanche minor auf Trifolium repens
schmarotzend. 1⁄2 nat. Gr. Einzelblüte vergr.
[S. 599]
Die 2. Familie der Rubiinae ist die der Caprifoliaceae.
Sie enthält Holzgewächse, deren verschieden gestalteten
Blättern die Nebenblätter meist fehlen. Mit radiären Blüten und
dreifächerigem Fruchtknoten ist Viburnum, der Schneeball,
ausgestattet. Die Früchte enthalten nur einen Samen, die
unfruchtbaren Randblüten dienen als Schauapparat; in der
Zierpflanze sind nur diese unfruchtbaren Blüten in den kugeligen
Trugdoldeninfloreszenzen vorhanden. Sambucus, Holunder, hat
unpaarige Fiederblätter, drüsige Nebenblätter und radiäre Blüten.
Die Frucht enthält drei Samen. Dorsiventrale Blüten finden sich
beim Gaisblatt, Lonicera periclymenum, einem schlingenden Strauch
unserer Haine, dessen langröhrige stark duftende Blüten durch
langrüsselige Sphingiden besucht werden. Diervilla (Weigelia)
beliebter Zierstrauch.
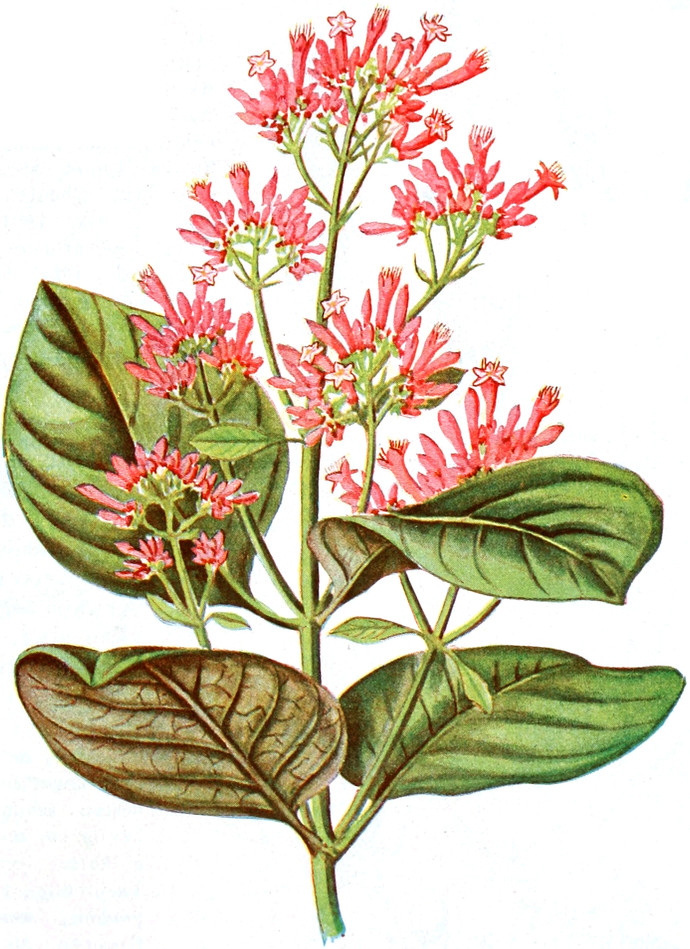
Fig. 773. Cinchona succirubra. 1⁄2 nat. Gr. Nach A.
MEYER und SCHUMANN. — Offizinell.
Offizinell sind Flores Sambuci (Pharm. germ., austr.,
helv.) von Sambucus nigra und Cortex Viburni (Pharm.
austr.) von Viburnum prunifolium.
In der 3. Familie Valerianaceae finden sich Stauden mit
asymmetrischen Blüten, deren Kelch sich erst an den Früchten
als „Pappus“ entwickelt, d. h. zu einer als Flugapparat
dienenden Federkrone. Valeriana, der Baldrian, besitzt eine
gespornte fünfzählige asymmetrische Krone (Fig. 777, 778), drei
Staubblätter und drei Fruchtblätter, von diesen ist aber nur eines
fertil. Andere Valerianaceen haben nur zwei (Fedia) oder ein
(Centranthus) Staubblatt in der Blüte.
Valeriana officinalis liefert das offizinelle Oleum
Valerianae und die Radix Valerianae (Pharm. germ.,
austr., helv.).
[S. 600]
Als 4. Familie sei angefügt die der Dipsacaceae, die als krautige
Pflanzen mit gegenständigen Blättern und vier- bis fünfzähligen,
teils radiären, teils zygomorphen Einzelblüten sich hier gut
einfügen. Die Blüten stehen jede von einem Außenkelch umgeben,
der als Verbreitungsmittel dient, in Köpfchen zusammen, die von
sterilen Hüllblättern eingeschlossen werden.
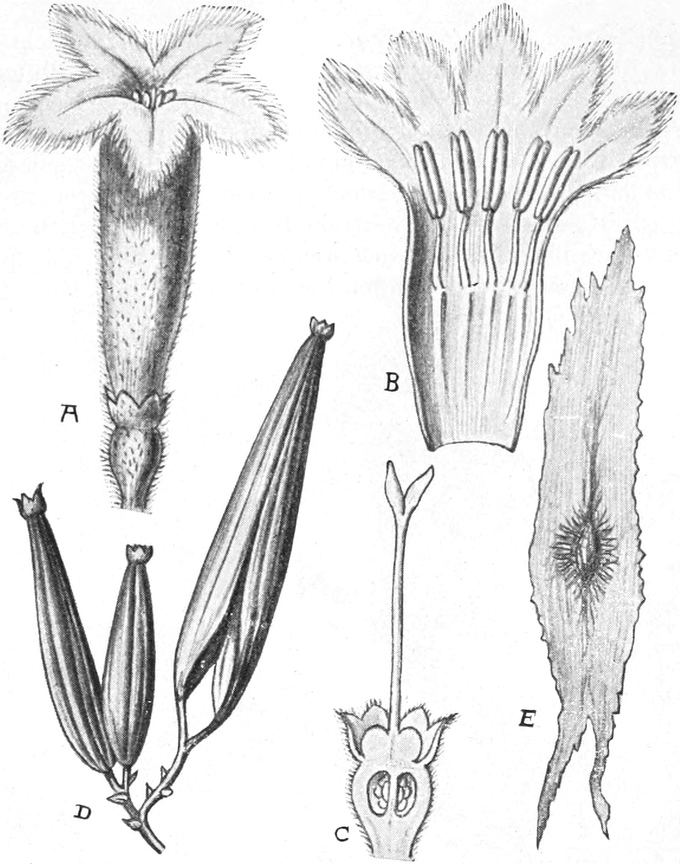
Fig. 774. Cinchona succirubra. A Blüte. B Korolle
aufgeschnitten. C Fruchtknoten-Längsschnitt. D Früchte. E
Samen. A–C und E vergr. D nat. Gr. Nach A. MEYER und
SCHUMANN.
Dipsacus, die Weberkarde, mit stechenden Hüll- und Spreublättern,
hat vierteilige radiäre Krone, vier Staubblätter und ein
Fruchtblatt, das eine hängende anatrope Samenanlage enthält; im
Samen findet sich Endosperm (Fig. 779). Ebenfalls vierteilig ist
Succisa (Fig. 780); fünfteilige Krone und größere dorsiventrale
Randblüten führt Scabiosa; vierteilig und einzige Gattung ohne
Spreublätter ist Knautia.
Der gemeinsame Charakterzug der
Synandrae
als 6. Ordnung ist darin zu finden, daß ihre Antheren, seltener die
ganzen Staubblätter, in irgendeiner Weise miteinander verwachsen oder
verklebt sind. Im übrigen kann die Blüte radiär oder zygomorph sein.
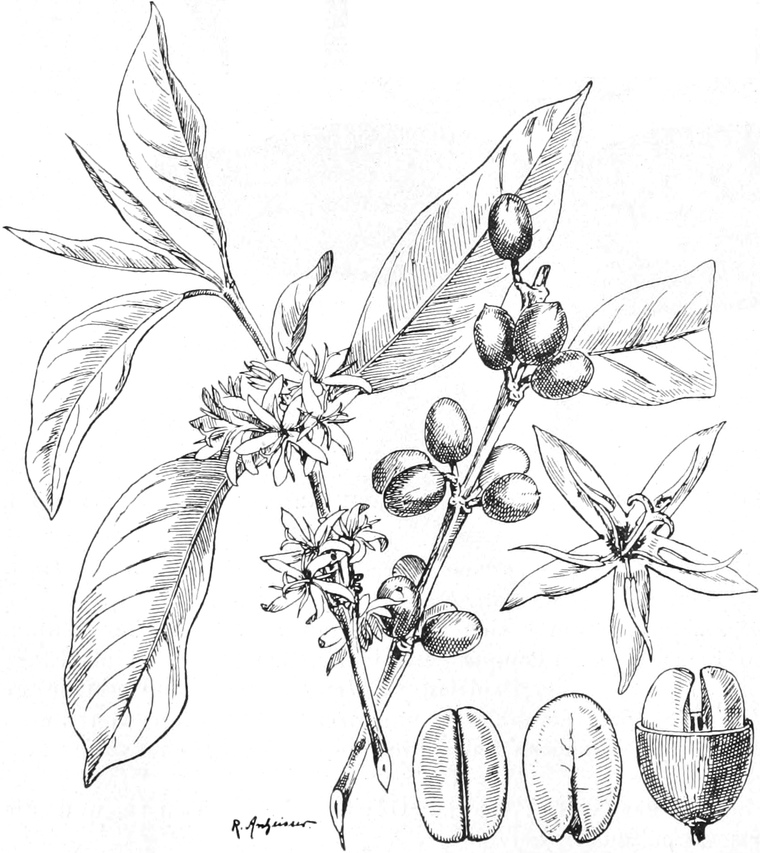
Fig. 775. Coffea arabica. 1⁄2 nat. Gr. Einzelblüte.
Frucht, Samen im Endokarp und daraus befreit. Etwa nat. Gr.
Als 1. Familie sei diejenige der Cucurbitaceae angeführt,
welche am besten schon durch die häufig bei ihr nicht durchgeführte
Sympetalie die Verbindung zu den Choripetalen, wenn auch zu
Gruppen, die in unserem kurzen Auszuge keine Erwähnung finden
konnten, aufrecht erhält. An die Cucurbitaceen sind daher auch
die übrigen Sympetalen angegliedert[S. 601] (vgl. Stammbaum S. 583).
Die Familie umfaßt krautige, rauhhaarige, großblättrige Pflanzen
mit meist monözisch verteilten diklinen Blüten. Kelch und Krone
verwachsen unterwärts, und die Antheren vereinigen sich paarweise,
in anderen Fällen sämtlich miteinander, wobei sie eine ∞förmige
Krümmung annehmen (Fig. 781). Der dreifächerige Fruchtknoten wird
zu einer derbschaligen Beerenfrucht. Die Ranken sind verzweigt
oder unverzweigt und entsprechen in ihrer seitlichen Stellung
einem Vorblatte. Cucumis sativus, die Gurke, und Cucumis
Melo, die Melone, werden vielfach kultiviert. Die Gurkenpflanze
ist parthenokarp[502], d. h. Bestäubung der Narbe ist zum
Fruchtansatz nicht nötig. Cucurbita Pepo, der Kürbis,
Bryonia, die Zaunrübe.
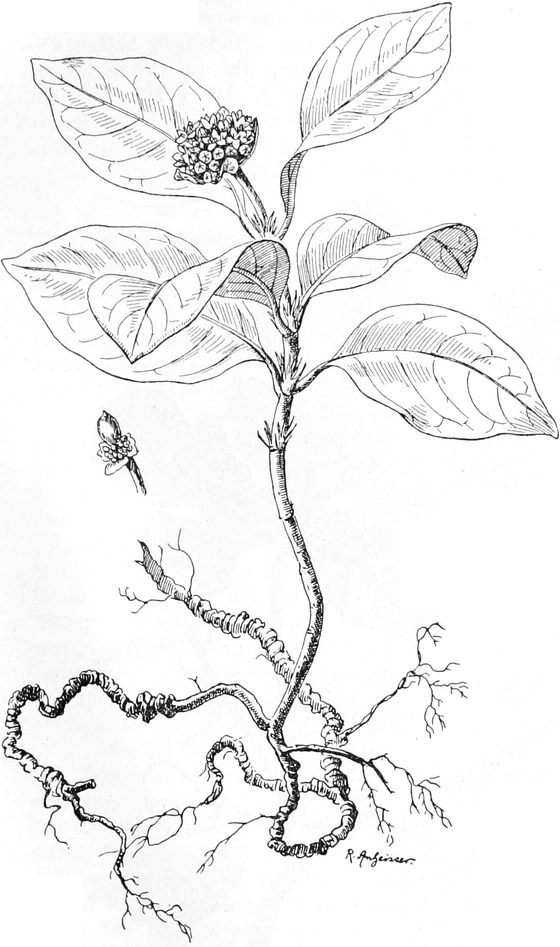
Fig. 776. Uragoga Ipecacuanha. 1⁄2 nat. Gr. Fruchtstand
daneben. — Offizinell.
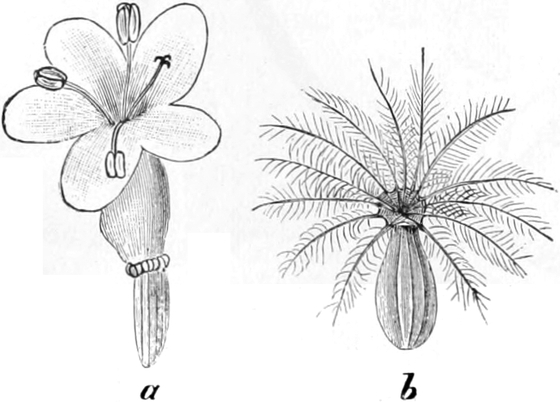
Fig. 777. Valeriana officinalis. a Blüte. Vergr. 8.
b Frucht. Vergr. — Offizinell. Nach H. SCHENCK.
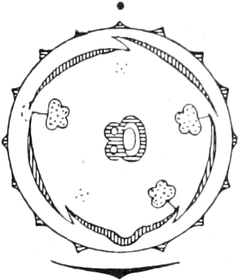
Fig. 778. Valeriana. Diagramm. Nach F. NOLL.
Fig. 779. Frucht von Dipsacus fullonum im Längsschnitt,
hk Hüllkelch, end Endosperm, em Embryo. Nach H. BAILLON.
Fig. 780. Succisa pratensis. a Blüte mit Außenkelch,
b ohne Außenkelch, c Frucht im Längsschnitt, f Fruchtknoten, hk
Außenkelch. Nach H. SCHENCK.
[S. 602]
Offizinell ist Citrullus Colocynthis, die Koloquinte,
eine ausdauernde Pflanze der asiatischen und afrikanischen Wüsten
nördlich des Äquators; in den Achseln ihrer tief dreilappig
fiederschnittigen Blätter stehen einzeln männliche Blüten mit drei
Antheren, von denen zwei paarig verwachsene die doppelte Zahl von
Windungen haben wie die fünfte unpaare, die weiblichen Blüten in
gleicher Stellung. Die Früchte werden zu trockenen Beeren, die
Pflanze liefert Fructus Colocynthidis (Pharm. germ., austr.,
helv.) (Fig. 783).
Fig. 781.
Fig. 782.
Fig. 781 u. 782. Ecballium (Cucurbitaceae). Diagramm. Fig. 781
Männlich. Fig. 782 Weiblich. Nach A. W. EICHLER.

Fig. 783. Citrullus Colocynthis. 1⁄2 nat. Gr.
1 Sproß mit männlicher und weiblicher Blüte. 2 Sproßgipfel
mit männlichen Blütenknospen und Ranken. 3 Männliche Blüte
ausgebreitet. 4 Weibliche Blüte längsdurchschnitten. 5 Junge Frucht
querdurchschnitten. — Offizinell.
[S. 603]
Die Zusammenfügung der folgenden Familien mit den Cucurbitaceen ist
lediglich auf Grund des morphologischen Merkmals der verwachsenen
Antheren möglich. Eine wirkliche Verwandtschaft erscheint, vom
morphologischen Standpunkte aus, unwahrscheinlich, nachdem durch
die Untersuchungen von KRATZER die sehr verschiedenartig
verlaufende Samenentwicklung dargelegt ist. Immerhin soll
Plasmaverwandtschaft nach dem serodiagnostischen Stammbaum
vorhanden sein.
Fig. 784. Blütendiagramm von Campanula medium. Nach
A. W. EICHLER.
Fig. 785. Campanula rotundifolia. Blüte. Nat. Gr. a
Ganz, b im Längsschnitte. H. SCHENCK.

Fig. 786. Blütendiagramm von Lobelia fulgens. Nach
A. W. EICHLER.
Die 2. Familie, die der Campanulaceae, enthält
milchsaftführende Kräuter mit radiären Blüten und drei- oder
fünfzähligen Fruchtknoten. Die Staubblätter sind der Blütenachse
eingefügt und mit ihren Antheren verklebt oder verwachsen. Die
Gattung Campanula (Fig. 784, 785) ist bei uns vielfach vertreten
und bildet mit ihren blauen Glocken einen Schmuck der sommerlichen
Vegetation. Phyteuma hat ährenförmige Blütenstände, deren Blüten
ihre Kronblätter nur an der Basis öffnen. Erst wenn der in der
Knospe entleerte Pollen von den Griffelfeghaaren[503] hinausgefegt
ist, öffnen sich die Kronblätter und spreizen die Narbenschenkel
auseinander. Jasione hat kopfige, an Kompositen erinnernde
Infloreszenzen.
Die 3. Familie der Lobeliaceen unterscheidet sich
nur durch dorsiventrale Blüten und zwei Fruchtblätter von den
Campanulaceen. Das mediane Kelchblatt steht auf der Vorderseite
vor einem tiefen Schlitz der Krone; durch Drehung oder Übernicken
der ganzen Blüte wird die normale Stellung hergestellt (Fig. 786).
Lobelia Dortmanna hat als nordische Wasserpflanze Interesse, da
sie völlig den Habitus einer Litorella angenommen hat.
Offizinell ist Herba Lobeliae von L. inflata,
Nordamerika (Fig. 787).
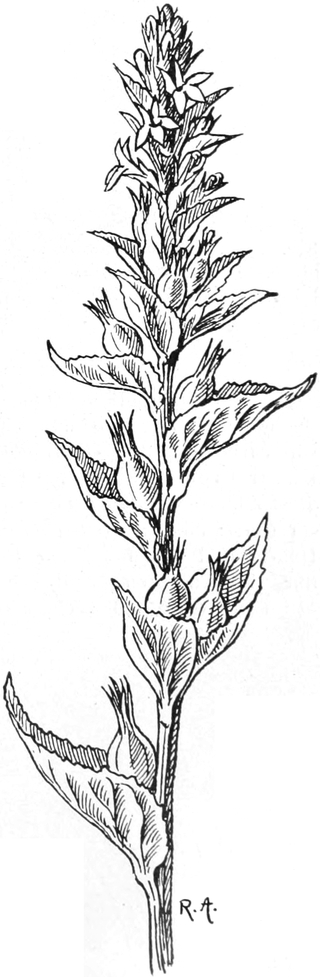
Fig. 787. Lobelia inflata. Spitze einer blühenden und
fruchtenden Pflanze.
3. Familie Compositae[504]. Über die ganze Erdoberfläche
verbreitete große Familie mit meist krautigen Angehörigen — baumförmig
ist z. B. Senecio Johnstoni. Die Blüten stehen in Köpfchen beisammen.
Die Einzelblüten sind radiär oder dorsiventral, und es finden sich
entweder nur gleiche oder auch verschiedene im Köpfchen vereinigt.
Staubblätter sind fünf vorhanden, der Kronröhre angewachsen; ihre
Antheren sind intrors und bilden durch Verwachsung ihrer Kutikula
eine Röhre (Fig. 788), welche vom noch unentwickelten Griffel unten
geschlossen wird. Die Blüten sind[S. 604] protandrisch, so daß der frühzeitig
entleerte Pollen bei der Streckung des Griffels von den Feghaaren nach
oben hinausgedrängt wird. Der fertig entwickelte Griffel ist oben stets
in zwei Narben gegabelt. Die einzige Samenanlage des einfächerigen
Fruchtknotens ist stets anatrop und aufrecht (Fig. 791), die Samen
sind endospermlos. Die Früchte sind vielfach von einem als Kelch
anzusehenden Haarkranze, dem Pappus (Fig. 789), gekrönt, der ihrer
Verbreitung durch den Wind dient. Als Reservestoff findet sich in
Wurzeln und Knollen (Fig. 203) meist Inulin, in den Samen Aleuron und
fettes Öl.
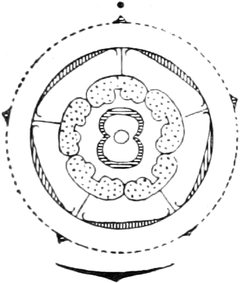
Fig. 788. Compositae. Diagramm (Carduus). Nach F.
NOLL.
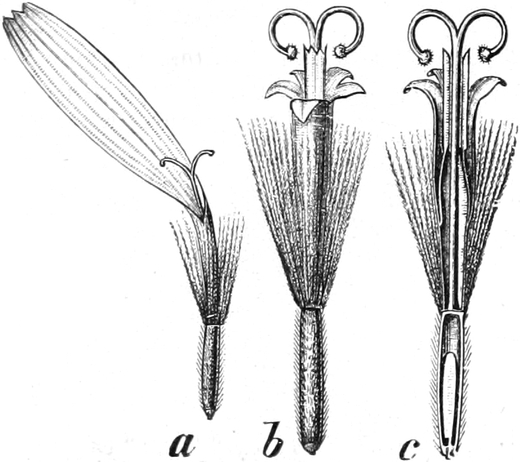
Fig. 789. Arnica montana. a Randblüte, b
Scheibenblüte, c diese im Längsschnitt. Vergr. Nach BERG und
SCHMIDT.
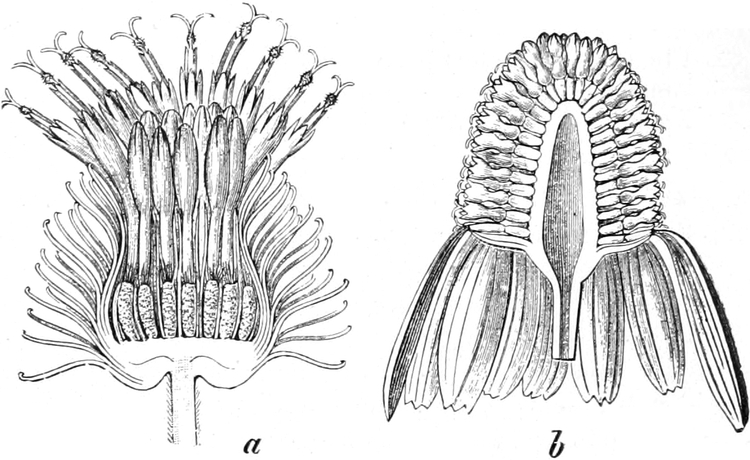
Fig. 790. a Lappa major. Köpfchen im Längsschnitt mit
hakenförmigem Hüllkelch und mit Spreublättchen auf dem Blütenboden. b
Matricaria Chamomilla, ohne Spreublättchen. Vergr. Nach BERG
und SCHMIDT. — Offizinell.
Fig. 791. Arnica montana. a Köpfchenachse, nach
Entfernung der Früchte. Vergr. b Frucht im Längsschnitt, vom
Pappus ist nur der untere Teil gezeichnet. Nach BERG und
SCHMIDT. Offizinell.
Fig. 792. Andröceum von Carduus crispus. Vergr. 10. Nach
H. BAILLON.
Die Einzelblüten sind entweder radiär mit fünflappiger Krone,
sie heißen Röhrenblüten (Fig. 789, b, c), oder es
kommen durch Unterdrückung der Oberlippe einlippige Blüten
zustande, deren Unterlippe drei Zipfel zeigt (Fig. 789
a). Sehr ähnlich sind diesen letzteren die zungenförmigen
Blüten, wie Taraxacum sie führt: einseitig tief gespalten,
am Rande mit fünf Zipfeln. Häufig sind neben solchen
Kompositen, die nur Röhrenblüten oder nur Zungenblüten in
ihren Köpfchen besitzen, andere, die in der Mitte Röhrenblüten
(Scheibenblüten), am Rande einlippige Blüten[S. 605] (Randblüten) tragen.
Meist tritt dann zu einer etwa vorhandenen Differenz der Färbung
eine solche des Geschlechts hinzu, indem die röhrenförmigen
Scheibenblüten zwittrig, die einlippigen Randblüten rein weiblich
sind, d. h. die Köpfchen sind heterogam (Matricaria,
Arnica). Endlich finden sich bisweilen am Rande völlig steril
gewordene Blüten (Centaurea Cyanus), die nur als Schauapparat zur
Anlockung von Insekten dienen können.
Fig. 793. Kompositenfrüchte. A Helianthus annuus. B
Hieracium virosum. C Cichorium Intybus. Nach H. BAILLON.

Fig. 794. Fruchtköpfchen von Taraxacum officinale.
Haarkelch auf den erhaltenen Früchtchen durch einen stielförmigen
Schnabel emporgehoben; Infloreszenzboden umgeschlagen. Etwa nat. Gr. —
Nach K. GOEBEL.
Eine Reihe von Gattungen führt nur Röhrenblüten im
Köpfchen, so Carduus (Fig. 792), die Distel, durch
haarförmige Pappusborsten ausgezeichnet, Cirsium an federigen
Pappushaaren kenntlich, Echinops, mit einblütigen Köpfchen,
die zu vielen vereinigt sind. Lappa, die Klette (Fig. 790
a), hat Hüllkelchblätter mit hakenförmig gekrümmter Spitze.
Cynara Scolymus, die Artischocke. Cnicus benedictus, das
Benediktenkraut (Fig. 795), besitzt einzelne, endständige Köpfchen,
deren Hüllkelchblätter mit großem, zum Teil gefiedertem Endstachel
versehen und spinnwebig behaart sind. Centaurea, die Kornblume,
hat einen trockenhäutigen Hüllkelch und größere, aber sterile
Randblüten. — Andere Gattungen haben nur zwittrige Zungenblüten
im Köpfchen und führen gleichzeitig Milchsaftgefäße in allen
Teilen. Taraxacum officinale, der überall verbreitete Löwenzahn;
aus einer Rosette grob schrotsägeförmiger Blätter erheben sich
die Blütenköpfchen, jedes einzeln auf hohlem Stiele, der nach
der Blüte eine zweite Wachstumsperiode durchläuft (S. 244, Fig.
261). Die Früchte sind schnabelförmig verlängert, so daß der
haarige Pappus zu einer fallschirmartigen, gestielten Krone wird
(Fig. 794). Lactuca sativa liefert den Kopfsalat. L. virosa
ist der Giftlattich. L. Scariola, als Kompaßpflanze durch ihre
vertikal und meridional stehenden Blattflächen bekannt (vgl.
S. 310). Cichorium Intybus, die Zichorie (Fig. 793 C) hat
blaublütige Köpfchen und einen Pappus in Form kurzer aufgerichteter
Schüppchen. C. Endivia, Endivie, Tragopogon und Scorzonera
mit federigem Pappus. Sc. hispanica, die als Gemüse dienende
Schwarzwurzel. Crepis-Arten haben einen haarförmigen Pappus, der
weich, biegsam und bräunlich ist.[505] Sonchus, mit vielreihigen
Pappusborsten. Hieracium, eine ungeheuer formenreiche europäische
Gattung, hat einen weißen, steifen, zerbrechlichen Pappus (Fig.
793 B). — Meist aber sind zweierlei Blütenformen im Köpfchen
vereinigt. Aster, Solidago, Erigeron sind drei in
zahlreichen Arten in Europa, Amerika, Asien weitverbreitete
Gattungen, von denen besonders die Aster-Arten als Zierpflanzen
beliebt sind. Haastia- und Raoulia-Arten Neuseelands (Fig.
189), polsterförmige, wollig behaarte Pflanzen, täuschen durch
ihre weiße Farbe und ihre Form das wichtigste Heerdenvieh des
Landes vor, daher „vegetable sheep“ genannt. Inula, eine
Gattung einheimischer Kräuter mit trockenhäutigen Hüllblättern.
Bei Gnaphalium, Antennaria, Helichrysum (Immortellen),
Leontopodium (Edelweiß) u. a. ist der trockene Hüllkelch
blumenkronartig gefärbt. Helianthus annuus (die Sonnenblume)
(Fig. 793 A)[S. 606] wird wegen ihrer ölreichen Früchte vielfach
angebaut, ebenso H. tuberosus, der Topinambur, wegen seiner
eßbaren Knollen. Silphium laciniatum, nordamerikanische
Kompaßpflanze. Dahlia, die Georgine, stammt aus Amerika, ebenso
Bidens; dieser ist auch in einheimischen Arten verbreitet, die
durch ihre gegenständigen, bei Wasserformen zum Teil heteromorphen
Blätter auffallen. Achillea, die Schafgarbe, ist eine überall
verbreitete Staude. A. moschata und A. atrata, hochalpine,
vikariierende Arten, diese auf Kalk-, jene auf Schieferboden.
Anthemis nobilis, die römische Kamille, hat entweder nur
Scheibenblüten im Köpfchen, oder diese können mehr oder minder
durch Lippenblüten ersetzt sein. Matricaria Chamomilla (Fig.
790 b und 796), die echte Kamille, ist ein einjähriges, vielfach
verästeltes Kraut mit kegelförmig emporwachsendem, hohlem
Blütenboden, gelben Scheibenblüten und zurückgeschlagenen weißen
weiblichen Randblüten in den endständigen einzelnen Köpfchen.
Tanacetum hat nur röhrige Blüten, seine Randblüten sind rein
weiblich. Auch bei Artemisia sind die Blüten alle röhrenförmig
und die Randblüten meist weiblich, so A. Absinthium (Wermut);
dagegen sind in dem armblütigen Köpfchen von A. Cina (Fig. 797)
alle Blüten zwittrig. Tussilago Farfara, der Huflattich blüht
vor Erscheinen der Blätter. Der einköpfige Blütenstiel ist nur
mit Schuppenblättern besetzt, die Einzelblüten stehen auf kahlem
Blütenboden und haben einen haarförmigen, feinen weißen Pappus;
die Randblüten sind weiblich. Das Köpfchen wird von ein- bis
zweireihigen Hüllblättern umgeben (Fig. 798). Die großen, später
erscheinenden Blätter sind herzförmig, unten weißfilzig. Petasites
officinalis ist eine als Pestwurz bekannte großblättrige Staude.
Senecio vulgaris führt nur zwittrige Röhrenblüten und keine
Randblüten. Doronicum, Cineraria sind verbreitete Zierpflanzen.
Arnica montana (Wohlverleih) (Fig. 789, 791, 799) hat eine
grundständige Rosette aus zwei bis vier Paaren gegenständiger
Blätter und eine endständige, einköpfige Blütenachse, deren
gegenständige Vorblätter meist noch je einen einköpfigen
Blütenstand aus den Achselknospen[S. 607] entwickeln. Calendula,
die Ringelblume, und Dimorphotheca zeigen unregelmäßige und
verschieden gestaltete Früchte.
Fig. 795. Cnicus benedictus. Nach H. BAILLON.
Offizinell.
Fig. 796. Matricaria Chamomilla. 2⁄3 nat. Gr. —
Offizinell.

Fig. 797. Artemisia Cina. Nach A. MEYER und
SCHUMANN — Offizinell.
Offizinell: Arnica montana liefert: Rad. Arnicae
(Pharm. austr.), Flores Arnicae (Pharm. germ., austr.,
helv.). — Artemisia Absinthium: Herba Absinthii
(ibid.). — Artemisia Cina, Turkestan: Flores Cinae und
Santoninum (ibid.). — Matricaria Chamomilla: Flores
Chamomillae (ibid.) und Oleum Chamomillae (Pharm.
helv.). — Cnicus benidictus (Südeuropa): Herba Cardui
benedicti (Pharm. germ., helv.). — Tussilago Farfara:
Folia Farfarae (Pharm. germ., austr.). — Achillea[S. 608]
Millefolium: Herba Millefolii (Pharm. austr.). — Anthemis
nobilis: Flores Chamomillae romanae (Pharm. austr., helv.).
— Lappa vulgaris: Rad. Bardanae (ibid.). Anacyclus
Pyrethrum (Südeuropa): Rad. Pyrethri (ibid.). — Taraxacum
officinale: Rad. et herba Taraxaci (Pharm. germ., austr.,
helv.) und Folia Taraxaci (Pharm. austr). — Vorderasien und
Kaukasus liefern das persische Insektenpulver von Pyrethrum
roseum. Das dalmatinische stammt ab von dem dort heimischen
Chrysanthemum cinerariaefolium.

Fig. 798. Tussilago Farfara. Nach H. BAILLON.
— Offizinell.

Fig. 799. Arnica montana. 1⁄2 nat. Gr. —
Offizinell.
[S. 609]
2. Unterklasse. Monokotylae.
Die mit einem Keimblatt versehenen Angiospermen, die Monokotylen, sind
ihrem Gesamtaufbau nach meist Kräuter oder Stauden, seltener Sträucher
oder Bäume.
Ihr kleiner Embryo streckt bei der Keimung sein Würzelchen und
Hypokotyl aus der Samenschale, während der scheidenförmige Kotyledon
häufig mit seinem oberen Ende darin stecken bleibt und die Aufsaugung
des meist reichlich vorhandenen Nährgewebes besorgt. Die Hauptwurzel
stellt ihr Wachstum früher oder später ein und wird durch zahlreiche
Adventivwurzeln, die aus dem Stamm entspringen, ersetzt. Bei den
Gräsern sind solche bereits am Embryo angelegt. So fehlt den
Monokotylen durchweg ein einheitliches, auf eine Hauptwurzel und ihre
Verzweigung zurückführbares Wurzelsystem, wie die Gymnospermen und
Dikotylen es meist besitzen.
Stammbaum der Monokotylen nach den serodiagnostischen Untersuchungen
von E. WORSECK.
Der Stammvegetationspunkt bleibt mehr oder minder lange von dem
Scheidenteil des Kotyledons umschlossen. Er bringt auch weiterhin
langscheidige, am Grunde längere Zeit fortwachsende Blätter in
zweizeiliger oder wechselständiger Anordnung hervor. Das Stammwachstum
ist häufig beschränkt, Verzweigung fehlt in vielen Fällen ganz oder
führt doch nur selten zur Bildung einer reichverzweigten Krone.
Die Blätter pflegen ungestielt und parallelnervig, von schmaler
langgestreckter, linearer oder elliptischer Form zu sein (Fig. 800).
Durch frühzeitiges Absterben bestimmter Teile der[S. 610] Blattspreite kommen
die gefiederten oder gefächerten Palmblätter wie die durchlöcherten
Blätter einiger Araceen zustande.
In anatomischer Hinsicht sind die Monokotylen durch
geschlossene, über den ganzen Stammquerschnitt verteilte
Leitbündel (vgl. Fig. 164, S. 123) ausgezeichnet, welche kein
Kambium zu entwickeln vermögen. Infolgedessen fehlt den Monokotylen
das Dickenwachstum entweder gänzlich, oder es tritt in den seltenen
Fällen, wo es sich findet, in der Weise auf, daß am Außenrande des
Zentralzylinders vollkommen neue, geschlossene Bündel und zwischen
ihnen Grundgewebe gebildet werden.
Fig. 800. Blatt mit streifiger Nervatur (Polygonatum
multiflorum). 3⁄4 nat. Gr.
Fig. 801. Diagramm einer typischen Monokotylenblüte.
Monokotylenblüten sind in der Regel pentazyklisch gebaut, besitzen also
zwei Perianthkreise, zwei Andröceum- und einen Gynäceumwirtel. Die
typische Zahl der Glieder eines jeden Wirtels ist drei. Beide
Perianthkreise sind meist gleichartig ausgebildet, somit als Perigon zu
bezeichnen (Fig. 801). Demnach entspricht die Blütenformel P 3 + 3, A 3
+ 3, G(3) der typischen Monokotylenblüte.
a)
Blüten radiär.
1. Ordnung.
Helobiae.
Die Ordnung begreift in sich nur Wasser- oder Sumpfpflanzen. Ihre
radiären Blüten haben ein in zwei Kreisen angeordnetes, häufig
apokarpes Gynäceum, das Schließ- oder Balgfrüchtchen entwickelt, deren
Samen kein Nährgewebe um den großen Embryo ausbilden. Die Ordnung
vermittelt durch ihren Blütenbau den Anschluß der Monokotylen an die
Polycarpicae, vgl. S. 523[474] und den Stammbaum.
Die 1. Familie Alismaceae ist in der warmen und
gemäßigten Zone weit verbreitet. Einheimische Arten: Alisma
Plantago, Sagittaria sagittifolia und Butomus umbellatus sind
häufige deutsche Sumpfpflanzen mit langgestielten, rispigen oder
doldenartigen Blütenständen. Ihre Einzelblüten sind durch Besitz
von Kelch und weißer, bei Butomus rötlicher Krone ausgezeichnet.
Das Andröceum ist sechs- oder mehrzählig, das Gynäceum ist apokarp,
sechs- oder vielzählig, wirtelige und spiralige Stellung kommt
vor (Fig. 802). Sagittaria dagegen ist monözisch, ihre Blüten
werden eingeschlechtig durch Fehlschlagen des anderen Geschlechtes;
die männlichen enthalten zahlreiche Staubblätter und unfruchtbare
Fruchtblätter, die weiblichen nur Staminodien und zahlreiche freie
Fruchtblätter auf stark gewölbter Blütenachse (Fig. 803). Die
Blätter sind bei Butomus lang lineal, gerinnt dreikantig; bei
Alisma und Sagittaria langgestielt mit löffelförmiger bzw.
pfeilförmiger Spreite. Individuen beider Gattungen, die tief in
flutendem Wasser stehen, zeigen lang[S. 611] bandförmige Blätter, wie sie
bei der Keimung als Übergangsformen auftreten; solche Pflanzen
kommen nicht zur Blüte.
Fig. 802. Blütendiagramm von Echinodorus parvulus, einer
Alismacee. Nach A. W. EICHLER.
Fig. 803. Sagittaria sagittifolia. a Blüten. b
Frucht, nach Entfernung eines Teiles der Karpelle. Vergr. b nach
ENGLER und PRANTL.
2. Familie Potamogetonaceae. Potamogeton ist in vielen
verschiedenen Arten in stehenden wie fließenden Gewässern auf der
ganzen Erde verbreitet. Seine Blätter sind meist untergetaucht mit
langen, einseitig geschlitzten Axillarstipeln. Die zwittrigen,
vierzähligen Blüten mit apokarpem Gynäceum sind in einem über
das Wasser emporragenden ährenförmigen Blütenstand vereinigt.
P. natans unserer Tümpel führt zur Blütezeit meist nur
Schwimmblätter, da die untergetauchten stielrunden Wasserblätter
bereits vergangen sind (Fig. 804). Ruppia maritima, Zanichellia
palustris sind Brackwasserformen; Zostera marina ist das an
allen nördlich temperierten Meeresküsten häufige Seegras,
vielfach als Polstermaterial verwendet.
Fig. 804. Potamogeton natans. Blühender Sproß. 1⁄2 nat.
Gr.
3. Familie Najadaceae. Najas marina diözisch. Blüte
mit einem Staubblatt. mit einem Fruchtblatt in becherförmigen
Hüllen.
[S. 612]
4. Familie Hydrocharitaceae. Hydrocharis morsus
ranae und Stratiotes aloides sind schwimmende einheimische
Wasserpflanzen, die sich besonders durch Ausläuferbildung vermehren
und ganz oder in Form von Winterknospen überwintern. Ihre Blüten
sind diözisch und entomophil. In den männlichen Blüten finden
sich mehrere dreizählige Wirtel von Staubblättern; die weiblichen
haben Staminodien und zwei dreizählige Kreise von Fruchtblättern.
Vallisneria spiralis, eine Süßwasserpflanze der Tropen, ist bis
in die oberitalienischen Seen verbreitet; Helodea canadensis,
die Wasserpest aus Nordamerika, ist in jedem Wasserloch zu finden;
beide sind hydrophil (vgl. S. 479).
2. Ordnung.
Spadiciflorae.
Der gemeinsame Charakter dieser Ordnung wird bedingt durch den
eigenartigen Blütenstand: er ist ährenartig, besitzt aber eine
angeschwollene dicke, oft fleischige Achse, stellt also einen
Kolben, Spadix, dar. Die Einzelblüten sind meist diklin,
monözisch oder seltener diözisch.
Die 1. Familie Typhaceae umfaßt einheimische Sumpfpflanzen
mit langen linealen Blättern und langgestielten Blütenkolben,
die oben die männlichen, darunter die weiblichen Blüten tragen,
beide sind in großer Zahl ohne Blütenhülle zusammengedrängt. Daran
schließt unmittelbar die zweite Familie Sparganiaceae
an, deren Kolben kugelig sind und deren Einzelblüten ein Perigon
besitzen.
3. Familie Pandanaceae. Eigenartige, auf Stützwurzeln
stehende Bäume oder kletternde Sträucher, die in allen den
Indischen Ozean umlagernden Tropenländern und pazifischen Inseln
heimisch sind und auch wohl als Schraubenpalmen bezeichnet werden,
weil ihre langen, scharf bewehrten, rinnenförmigen Blätter in
dreizeiliger Schraube lückenlos den Stamm umlaufen. Ihre männlichen
und weiblichen diklinen Blütenstände sind endständige Kolben, die
perianthlose Einzelblüten tragen und in der Achsel scheidiger
Deckblätter stehen. Pandanus (vgl. Fig. 807 vor den Palmen),
Freycinetia (vgl. S. 482).
4. Familie Palmae[506]. Die Palmen sind eine ausschließlich
tropische und subtropische Familie von meist baumförmigen Angehörigen.
Ihr schlanker, fast immer einfacher, nur bei den afrikanischen
Hyphaene-Arten gabelig verzweigter Stamm besitzt meist auf seiner
ganzen Länge denselben Durchmesser. Einzelne Formen weisen jedoch
eine deutliche Dickenzunahme gegen die Basis hin, bisweilen auch in
der halben Stammhöhe, auf, die nur auf Vergrößerung der vorhandenen
Elemente und eventuell örtlich beschränkten Neubildungen beruht. Genau
so verhalten sich übrigens auch die Pandanaceen. Die Blätter bilden
eine gipfelständige Krone und erreichen oft gewaltige Dimensionen.
Es sind entweder Fieder- oder Fächerblätter, meist langgestielt,
deren Zerteilung durch Einreißen der ursprünglich einfachen, in der
Knospenlage gefalteten Spreiten an den absterbenden Faltungsstellen
zustande kommt. Der Blütenstand ist in einigen Fällen terminal, wie
bei der Sagopalme, Metroxylon; das Individuum stirbt dann mit
der Fruchtbildung ab. Häufiger jedoch stehen die Infloreszenzen
blattachselständig. Sie sind während der Entwicklung von einer
mächtigen, sehr widerstandsfähigen Scheide, der Spatha,
umhüllt, die zur Zeit der Blüte aufplatzt und den einfachen oder meist
verzweigten Blütenstand ins Freie treten läßt (Fig. 805).
Die Einzelblüten sind in der Regel eingeschlechtig und nach
dem regelmäßigen Monokotylentypus gebaut, also P3 + 3, A3 + 3
für die männlichen, P3 + 3, G(̲3) für die weiblichen. Fig. 805
zeigt den Blütenstand von Cocos nucifera mit der Spatha, die
ihn am Grunde noch umgibt. Man bemerkt neben zahlreichen, nach
den Infloreszenzenden hin in dicht gedrängten Ähren stehenden
männlichen Blüten einzelne am Grunde befindliche, noch ungeöffnete
weibliche von erheblich größeren Dimensionen; Cocos ist also
monözisch. Der einfächerige Fruchtknoten besteht aus drei
miteinander verwachsenen Fruchtblättern, in denen sich jedoch nur
ein Fruchtfach entwickelt. Die reifen Früchte hängen zu mehreren
an einem Fruchtstande. Jede Frucht ist von einem glatten Exokarp,
einem grobfaserigen Mesokarp und einem steinharten Endokarp
umhüllt. Das lufthaltige Mesokarp (Kokosfaser) bedingt die
Schwimmfähigkeit und damit die große Verbreitung[S. 613] der Palme über
alle Tropenküsten. An der Basis zeigt jedes Karpell „ein Keimloch“
(Fig. 806) im Endokarp, das zuwächst; am wenigsten Widerstand
bietet dasjenige, dem der Keimling anliegt. Endosperm wird in
dicker Lage rings an der Wandung abgelagert; es ist sehr fettreich
und bildet die Kopra des Handels. Der Innenraum wird zum
Teil von einer Flüssigkeit, der sog. Kokosmilch, ausgefüllt, die
für die Keimung von Bedeutung sein dürfte. Der keimende Embryo
entwickelt in den Hohlraum hinein ein mächtiges Saugorgan, welches
dem Keimling Reservestoffe zuführt und ihn schließlich ganz
ausfüllt. Fig. 807 gibt den Habitus eines Kokoswäldchens wieder.
Fig. 805. Cocos nucifera. Blütenstand der Kokospalme.
Stark verkleinert.
[S. 614]
Doch nicht überall sind die Verhältnisse die gleichen. Bei Areca
Catechu (Fig. 833) ergibt die aus ähnlichem Fruchtknoten entwickelte
Frucht eine Beere, indem das Mesokarp grobfaserig-fleischige Konsistenz
annimmt. Das weiße, durch einwachsende dunkle Zellstreifen der
Samenschale ruminierte Endosperm wird hier steinhart, weil
Zellulose als Reservestoff abgelagert ist. Ebenso ist die Frucht der
diözischen Dattelpalme, Phoenix dactylifera, eine Beere, die aber aus
einem apokarpen Gynäceum hervorgeht; von den drei freien Fruchtblättern
wird nur eines völlig ausgebildet. Sonstige wichtige Nutzpflanzen unter
den Palmen sind: Elaëis guineensis, die afrikanische Ölpalme.
Calamus-Arten liefern das Stuhlrohr, Metroxylon-Arten
Sago, beide im asiatisch-australischen Tropengebiet zu Hause:
Phytelephas macrocarpa, eine amerikanische stammlose Palme, liefert
vegetabilisches Elfenbein im harten Endosperm ihrer Samen.
Verschiedene Arten lassen nach Abschneiden der Infloreszenzanlagen eine
Menge zuckerhaltigen Saftes ausfließen, der bald zu Palmwein vergoren,
bald (Arenga saccharifera) zur Gewinnung von Rohrzucker eingedickt
wird.
Offizinell: Areca Catechu (trop. Asien) liefert Semen
Arecae (Pharm. germ., helv.).
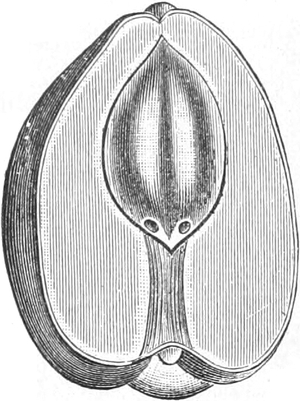
Fig. 806. Kokosnuß, nach teilweiser Entfernung des
faserigen Mesokarps, verkleinert. Nach E. WARMING.

Fig. 807. Kokosinseln bei Hilo, Hawaii. Pandanus
odoratissimus vor den Palmen.
[S. 615]
5. Familie Araceae. Die Araceen sind meist Kräuter oder
Stauden; im feuchten Walde der Tropen treten sie als Wurzelkletterer
auf (Fig. 808) und spielen dort eine hervorragende Rolle. Einige Arten,
wie Monstera, zeigen Zerteilung und Durchlöcherung ihrer mächtigen
Blattspreiten durch Absterben genau umschriebener Stellen (vergl.
Palmae). Die Araceenblüten sind sehr reduziert und meist diklin an
einer unverzweigten, fleischig-kolbigen Achse, Spadix, angeordnet, die
an der Basis eine Spatha von oft lebhafter Färbung als Schauapparat
besitzt, z. B. bei Anthurium Scherzerianum „Krebsschere“ und
Richardia aethiopica „Kalla“ genannt, zwei in unseren Gewächshäusern
häufig kultivierten Araceen. Die Früchte sind meist Beeren, die
häufig lebhaft rote, bläuliche oder weiße Farben zeigen. Colocasia
(s. S. 200) und Caladium vielfach ihrer riesigen und zum Teil
Blumenblatt-ähnlich schön gefärbten Blätter wegen in Kultur. Ariopsis
und Spathicarpa mit charakteristischen Infloreszenzen.

Fig. 808. Wurzelkletternde Araceen im feucht-tropischen
Walde von Chiapas. (Cafetal Trionfo.) Von den hoch oben befindlichen
Exemplaren werden Wurzeltaue zum Boden entsandt, die ihnen nach
Absterben des Stammes zur Ernährung dienen.
Deutsche Vertreter: Acorus Calamus, der Kalmus, ist
erst im Laufe der letzten 2 bis 3 Jahrhunderte aus dem wärmeren
Asien zu uns gewandert. Seine Blüten sind vollständige, zwittrige
Monokotylenblüten, die an einem kurzen Kolben sitzen, der
endständig ist, aber von seiner blattartigen Spatha zur Seite
gedrängt wird (Fig. 809). Die in unseren Torfsümpfen verbreitete
Calla palustris und Arum maculatum (Fig. 810), eine[S. 616] mit
knolligem Rhizom ausdauernde Staude unserer Laubwälder, sind
wie viele andere Araceen giftig. Arum entwickelt eine Anzahl
langgestielter, pfeilförmiger Blätter, deren braune Flecken der
Pflanze den Beinamen gegeben haben. Die monözischen, perianthlosen
Blüten sind an einem endständigen Kolben, dem Aronstab, angeordnet
und werden von einer oben weit geöffneten grünlichen Spatha völlig
umhüllt. An der Basis des Kolbens tief im Grunde der Spatha sitzen
die weiblichen Blüten, in geringem Abstand darüber die männlichen
und weiter oben, gerade der Einschnürung und Verengung der Spatha
entsprechend, einige steril gewordene Blüten von haarförmiger
Gestalt. Sie sind wie Reusenhaare abwärts gebogen und gestatten
kleinen, durch den eigenartigen Geruch und die angenehme Wärme
in den kalten Frühjahrsnächten angelockten Insekten wohl den
Eintritt in den unteren erweiterten Kessel, hindern aber ihren
Wiederaustritt, bis die weiblichen Blüten durch von anderen Blüten
mitgebrachten Pollen bestäubt sind, worauf die Haare schrumpfen
und den Ausgang freigeben. Beim Verlassen des Gefängnisses müssen
die Insekten an den inzwischen geöffneten männlichen Blüten
vorbeikriechen, mit deren Pollen bedeckt sie andere Blüten
aufsuchen.
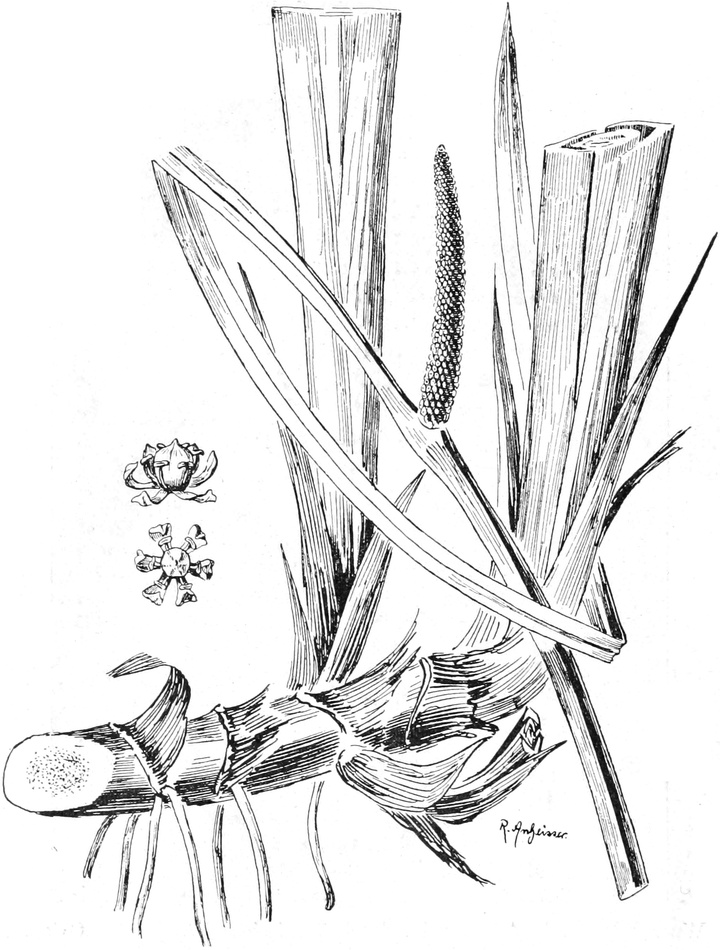
Fig. 809. Acorus Calamus, blühende Pflanze. Einzelblüte
von oben und im Profil. 1⁄2 nat. Gr. — Offizinell.
[S. 617]

Fig. 810. Arum maculatum, 1⁄2 nat. Gr. Blütenstand und
Fruchtstand, 2⁄3 nat. Gr. — Giftig.
Offizinell: Acorus Calamus: Rhizoma Calami (Pharm.
germ., austr., helv.) und Oleum Calami (Pharm. germ.).
3. Ordnung.
Liliiflorae
Strahlige fünfwirtelige Monokotylenblüten mit oberständigem oder
unterständigem Fruchtknoten sind das Charakteristikum der Ordnung
(Fig. 801). Das Perianth ist in beiden Kreisen gleichartig als Perigon
ausgebildet. Im Andröceum wird nur bei den Iridaceen ein Wirtel
unterdrückt.[S. 618] Das Gynäceum wechselt in der Stellung, doch ist stets ein
aus drei Karpellen zusammengesetzter, meist dreifächeriger Fruchtknoten
vorhanden.
In der 1. Familie Juncaceae sind grasähnliche Gewächse
mit vollständiger Liliiflorenblüte vereinigt, deren Perigon
spelzenähnlich ist, so daß auf Windbestäubung zu schließen ist. Der
Pollen bleibt in Tetraden vereinigt. Der oberständige Fruchtknoten
ist ein- oder dreifächerig und wird von drei langen papillösen
Narben gekrönt. Ein mehliges Endosperm umgibt den in Kapselfrüchten
entwickelten Samen. Die Familie ist in der gemäßigten Zone beider
Hemisphären verbreitet.
Die Gattung Juncus, Binse, ist in zahlreichen Arten bei
uns vertreten. Ihre stielrunden, mit Luftkammern versehenen Halme
und Blätter finden sich überall an Wassertümpeln und Flußläufen.
Die auf Windbestäubung eingerichteten Blüten (Fig. 811) sitzen
knäuelig am Gipfel der Sprosse, oft durch das in der Richtung der
Achse gestellte Stützblatt zur Seite gedrängt. Ihre Früchte sind
vielsamig. Luzula mit flachen Blättern und dreisamigen Früchtchen
ist in mehreren im Frühling blühenden Arten verbreitet.
Die Blüte der 2. Familie Liliaceae entspricht vollkommen
einer typischen Liliiflorenblüte mit gefärbtem Perigon und zumeist
entomophiler Bestäubung. Der Fruchtknoten ist oberständig und enthält
zahlreiche Samen, deren Endosperm hornig oder fleischig sein kann.
Die Früchte sind septicide oder loculicide Kapseln, in anderen Fällen
Beeren.
Die Mehrzahl der Liliaceen sind mit Zwiebeln, Knollen oder
sonstwie gestalteten Rhizomen perennierende krautartige Gewächse,
die vorzugsweise in den warm temperierten Gebieten beheimatet
sind. Colchicum autumnale, Herbstzeitlose (Fig. 812), ist
eine häufige, sehr giftige, ausdauernde Staude unserer Wiesen;
untersucht man die Pflanze im Herbst, wenn sie ihre Blüten
öffnet, so findet sich eine braune feste Hülle, die eine Knolle
und den ihr an der Basis angewachsenen, blühenden Seitentrieb
umgibt. Dieser bringt drei kurze ringförmige Scheidenblätter,
und in der Achsel des dritten befindet sich eine Knospe, die
Anlage des nächstjährigen Blütensprosses, dessen basales Ende zur
Tochterknolle wird. Im Frühjahr ist die Knolle ausgesogen und
von der Tochterknolle verdrängt. Die drei Laubblätter erheben
sich mit ihrer dunkelgrünen, langen, rinnenförmigen Spreite über
den Boden und umscheiden die gestreckte Achse, die am Gipfel die
septiciden Kapselfrüchte trägt (Fig. 812 f), deren Samen als
Semen Colchici offizinell sind. — Der giftige Germer, Veratrum
album, ist eine ansehnliche Staude unserer Bergwiesen; die
großen, elliptischen, längsgefalteten Blätter bilden eine buschige
Rosette. Eine endständige, stattliche, pyramidale Rispe trägt die
grünlich-weißen, polygamen Blüten. Ebenfalls septicide Kapseln hat
Schoenocaulon (Sabadilla) officinale, ein grasblättriges
Zwiebelgewächs der mittelamerikanischen und venezolanischen Anden,
das offizinelle Bedeutung besitzt. Hierher gehört auch die windende
Bowiea (Südafrika), die schön blühende Gloriosa und Littonia,
beides Blattspitzenranker.
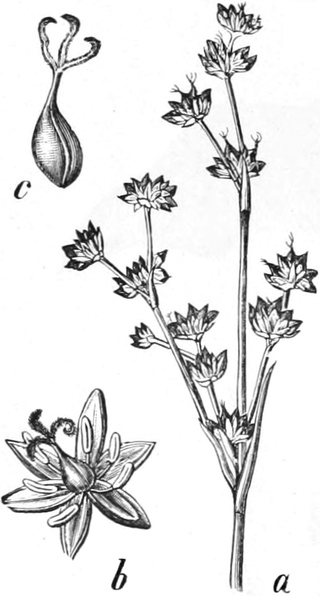
Fig. 811. Juncus lamprocarpus. a Teil der
Infloreszenz. b Blüte, vergr., c Gynäceum, vergr. Nach A. F. W.
SCHIMPER.
Dagegen haben unsere beliebten Ziergewächse wie Tulipa (Fig.
202), Hyacinthus Lilium (Fig. 205) Muscari, Scilla, die
Küchengewächse liefernde Gattung Allium, Lauch, ferner Urginea
(Fig. 813), die Meerzwiebel der Mittelmeerküsten und Galtonia,
Südafrika, ausnahmslos loculicide Kapseln. Ornithogalum
umbellatum (Fig. 814) mag als Beispiel der Lebensweise dienen. Im
Herbst untersucht, zeigt die Pflanze eine Zwiebel aus fleischigen
Schuppen, deren Narben den vergangenen Blattspreiten entsprechen.
In der Achsel der innersten Zwiebelschuppe neben dem abgeblühten
Infloreszenzstiel steht ein junges, aus einer Anzahl von Blättern
gebildetes Knöspchen, seinen Schluß bildet die Blütenstandsanlage.
Im Frühjahr wachsen die Blätter zu linealen langgestreckten
Gebilden heran und erheben sich mit der Infloreszenz über den
Boden. Ihre weißen Einzelblüten enden in einem dreifächerigen
Fruchtknoten, den ein gemeinsamer Griffel krönt. Die Blattbasen,
die inzwischen fleischig angeschwollen und mit Rerservestoffen
gefüllt[S. 619] sind, bilden die Zwiebelschuppen, während ihre
oberirdischen Teile zugrunde gehen. In ähnlicher Weise verläuft bei
allen genannten Zwiebelpflanzen der jährliche Entwicklungsgang.
Sie können nach der kurzen Vegetationszeit allen Unbilden der
Kälte oder der Trockenheit trotzen, indem sie sich unter den
Erdboden zurückziehen. Von baumförmigen Liliaceen ist Aloë mit
fleischigen, häufig am Rande bewehrten Blättern (Fig. 815, 816)
zu nennen, artenreiche Gattung Afrikas. Hierher gehört auch der
„Neuseeländische Flachs“ Phormium tenax.
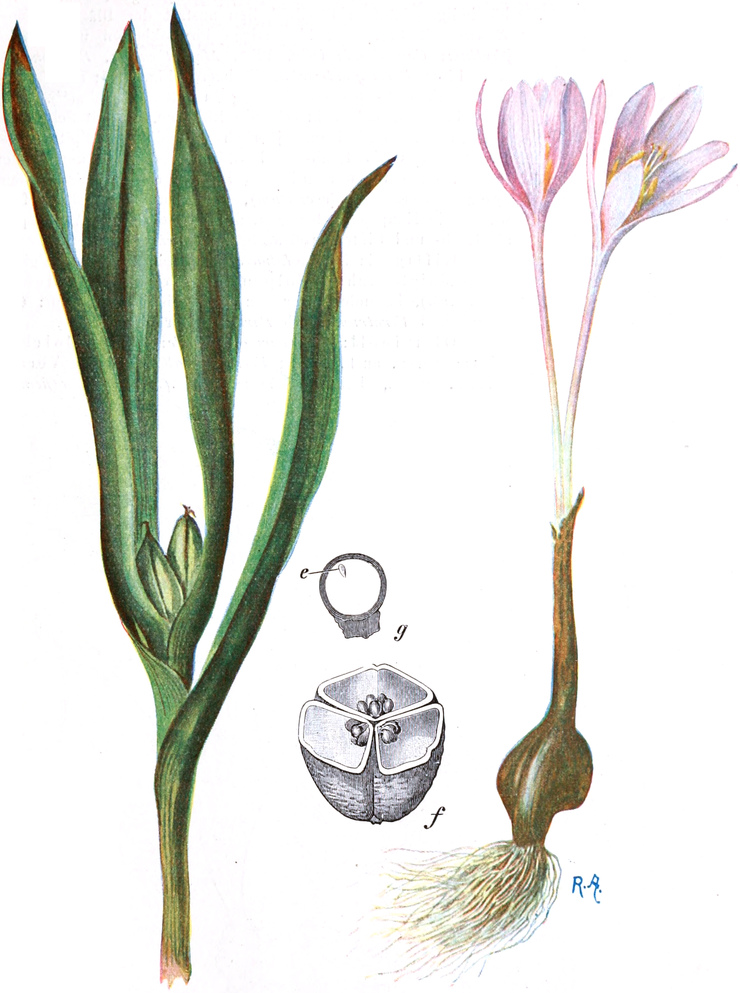
Fig. 812. Colchicum autumnale. 1⁄2 nat. Gr. f Frucht
im Querschnitt, g Samen mit Embryo, vergrößert. — Offizinell
und giftig.
[S. 620]
Die durch hohes Alter ausgezeichnete und durch eigenartigen
Habitus auffallende Dracaena (Fig. 817) trägt, wie die ähnlichen
Gattungen Cordyline und Yucca, Beerenfrüchte. Ebenso Smilax,
Sarsaparille, mit Hilfe rankenartiger Emergenzen ihrer Blattstiele
kletternde Sträucher wärmerer Länder. Hierher ferner Asparagus,
Spargel, mit büschelig gehäuften Phyllokladien anstatt der
Blätter, ebenso Ruscus mit breiteren, blattartigen Phyllokladien
und Myrsiphyllum; Convallaria (Fig. 125), Majanthemum,
Polygonatum (Fig. 143); Paris quadrifolia, Einbeere (Fig.
818); meist vier-, doch auch drei- bis sechsblättrig in allen
Wirteln[507]. Alle diese Pflanzen haben kriechende Rhizome, die
mit Schuppenblättern besetzt sind und jährlich entweder die Spitze
ihres Hauptsprosses als Laub- und Blütensproß über den Boden
senden, dann ihr unterirdisches Rhizom durch einen Seitenzweig
fortsetzen (Polygonatum), oder eine unterirdisch fortwachsende
Hauptachse besitzen, die jährlich einen Achselsproß als Laub- und
Blütensproß ausbildet (Paris).
Giftig: Zahlreiche Liliaceen sind mehr oder minder
giftig, so das Maiglöckchen, Tulpen- und
Kaiserkronzwiebeln (Fritillaria), besonders aber von
einheimischen Pflanzen: Colchicum und Veratrum; auch Paris
gilt für giftig.

Fig. 813. Urginea maritima, ca. 1⁄10 nat. Gr. —
Offizinell. Nach BERG und SCHMIDT.
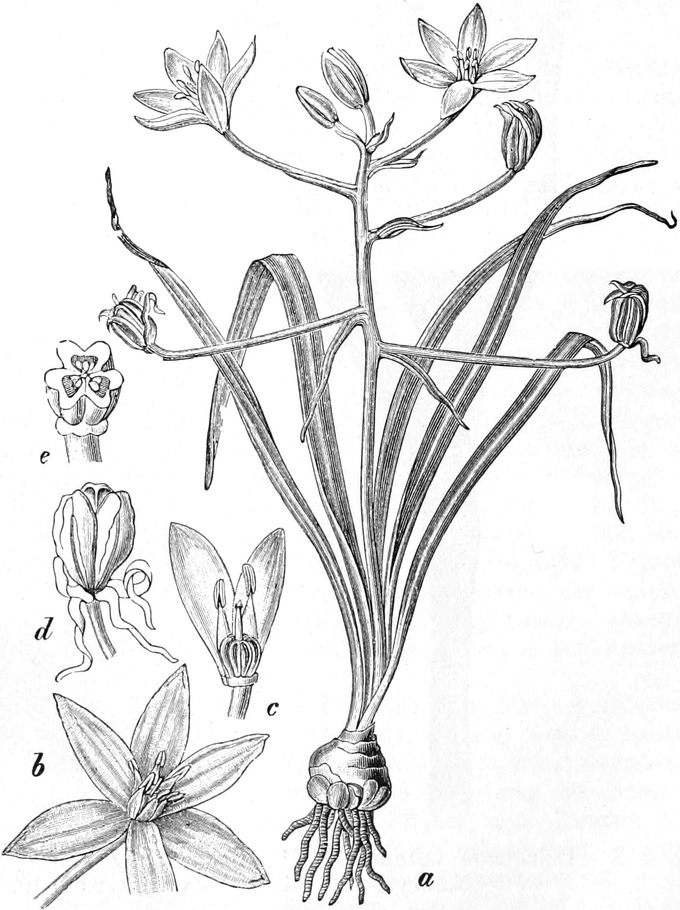
Fig. 814. a–e Ornithogalum umbellatum. a Ganze Pflanze,
verkleinert, b Blüte in nat. Gr., c Blüte im Längsschnitt,
d Frucht, e Querschnitt durch die Frucht. Nach A. F. W.
SCHIMPER.
Offizinell: Colchicum autumnale: Semen Colchici
(Pharm. germ., austr., helv.); Veratrum album: Rhiz.
Veratri (Pharm. germ., helv.); Schoenocaulon (Sabadilla)
officinale:[S. 621] Semen Sabadillae (Pharm. germ.,
austr., helv.) und Veratrinum (ibid.), Aloë ferox ist
die Hauptlieferantin der Kap-Aloë, wie A. vera für
Barbados-Aloë (ibid.), Urginea maritima: Bulbus
Scillae (ibid.), Smilax-Arten: Rad. Sarsaparillae
(ibid.), Convallaria majalis: Herba Convallariae (Pharm.
austr., helv.).

Fig. 815. Aloë speciosa und Aloë ferox, diese mit
verzweigten Blütenständen nach R. MARLOTH. A. ferox
offizinell.
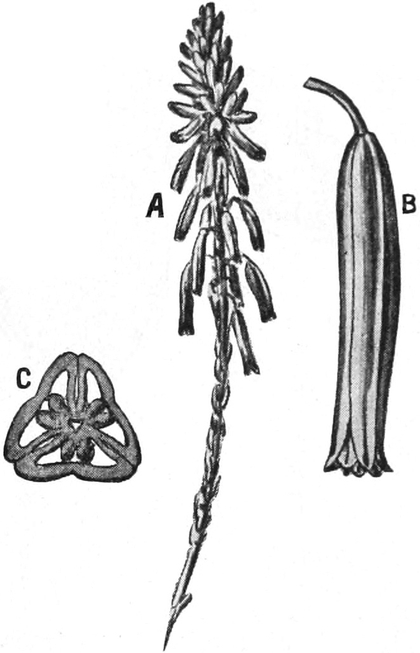
Fig. 816. Aloë succotrina. A Infloreszenz. B
Einzelblüte. C Fruchtknoten-Querschnitt.

Fig. 817. Dracaena draco. Drachenbaum von Laguna,
Kanarische Inseln. Nach C. CHUN.
[S. 622]
Die 3. Familie Amaryllidaceae unterscheidet sich von den
Liliaceen nur durch unterständigen Fruchtknoten. Die einheimischen
Amaryllidaceen Leucojum (Fig. 819), Galanthus (Schneeglöckchen)
und Narcissus sind Zwiebelgewächse und im Habitus den Zwiebeln
besitzenden Liliaceen ähnlich. Die Mehrzahl der Gattungen
gehört aber den Tropen und Subtropen an, wie z. B. die häufig
in Warmhäusern kultivierten Alstroemeria-, Haemanthus-,
Clivia- und Crinum-Arten. Wichtiger ist die Gattung Agave.
Diese mächtigste aller Blattsukkulenten ist in zahlreichen Arten
im wärmeren Amerika zu Hause. Zur Zeit ist Agave Sisalana aus
Yucatan eine der wichtigsten Faserpflanzen, die in großem Maßstabe
z. B. in Ostafrika und anderen Kolonien mit trockenem und doch
warmem Klima angebaut wird. A. Salmiana liefert in ihrem nach
Abschneiden der Infloreszenzknospe überreichlich ausfließenden,
alsdann vergorenen Safte: Pulque, das Nationalgetränk der
Mexikaner. Anspruchslosere Agave-Arten sind vielfach im
Mittelmeergebiet akklimatisiert.

Fig. 818. Paris quadrifolia. 1⁄2 nat. Gr. —
Giftig.
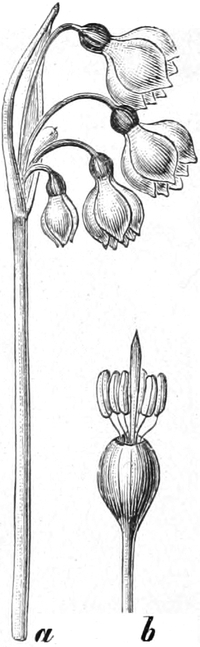
Fig. 819. Leucojum aestivum. a Blütenschaft
(verkleinert), b Gynäceum und Andröceum (nat. Gr.). Nach F. A. W.
SCHIMPER.
Die 4. Familie Iridaceae gleicht den Amaryllidaceen im Besitze
eines unterständigen Fruchtknotens, unterscheidet sich aber von ihnen
und dem Liliaceentypus durch das Fehlen des inneren Andröceumwirtels
(Fig. 820) (vergl. die atavistische Form Iris pallida, Lam. forma
abavia Heinricher S. 474, Fig. 528). Die beiden Perigonkreise sind
nicht immer gleichförmig. Die Iridaceen[S. 623] zeigen stets ungestielte
Blätter und überwiegend knollenförmige oder gestreckte Rhizome, während
Zwiebeln minder häufig sind. Die Früchte werden zu loculiciden Kapseln.
Die Familie ist vorzugsweise im Kapland und den wärmeren Teilen
Amerikas heimisch.
Crocus sativus, der Safran, ist eine alte Kulturpflanze des
Orients mit schmal-grasartigen Blättern und knollenförmigem
Rhizom. Die Blüten sind steril, wenn sie nicht mit Pollen wilder
Formen bestäubt werden. Ihre großen Narben liefern den „Crocus“
oder Safran (Fig. 821). Andere Arten werden häufig als Zierpflanzen
kultiviert. Iris, eine auch in Deutschland mit der Sumpfpflanze
I. Pseudacorus einheimische Gattung, ist durch zweizeilige
reitende Blätter ausgezeichnet, d. h. die Blätter umfassen
das dickfleischige Rhizom mit ihrer Scheide, steigen vertikal empor
und zeigen zwei gleiche Flanken bei schwertförmigem Umriß (Fig.
822). Die ansehnliche Blüte schlägt ihren äußeren Perigonkreis
abwärts, wölbt dagegen den inneren empor, ihre drei Antheren
werden von den drei großen, kronartig entwickelten Griffelästen
völlig überdeckt, die auf ihrer Außenseite einen kleinen
dreieckigen Narbenlappen tragen. In der Gattung Gladiolus ist die
Gleichartigkeit der Perigonblätter noch weiter gestört, die Blüten
werden dorsiventral.
Offizinell: Crocus sativus: Crocus (Pharm. germ.,
austr., helv.). Iris florentina, pallida, germanica des
Mittelmeergebietes: Rhizoma Iridis (ibid.).
Fig. 820. Diagramm der Iridaceae (Iris).
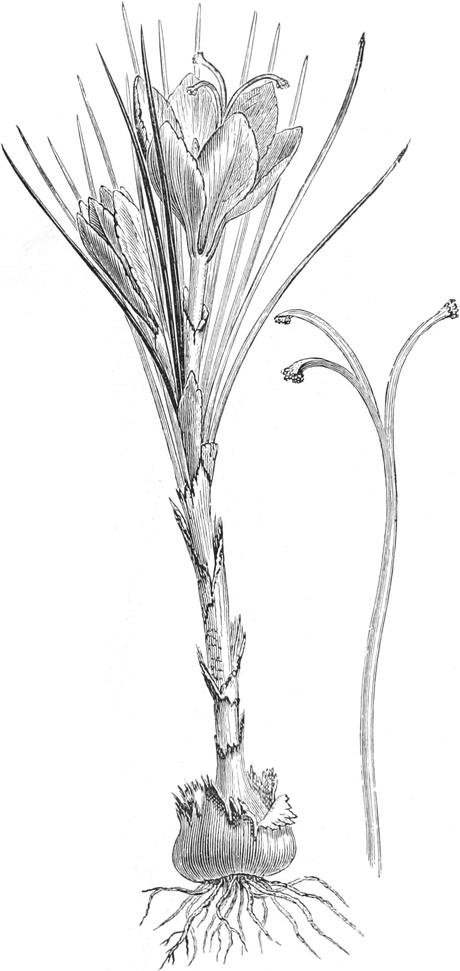
Fig. 821. Crocus sativus. Griffel mit dreiteiliger
Narbe. Nach H. BAILLON. — Offizinell.
5. Familie Bromeliaceae. Diese große, fast ausschließlich
amerikanisch-tropische Familie mit typisch xerophilen Blättern von
rosettenförmiger Anordnung enthält zahlreiche meist epiphytisch
lebende Pflanzen (Gattung Tillandsia) mit zwittrigen Blüten. Bei
erdbewohnenden Formen sind alle Blätter scharf bewehrt. Ananassa
sativa liefert in ihrem Fruchtstande die Ananas.
Die 4. Ordnung der Enantioblastae ist durch
atrope Samenanlagen ausgezeichnet, die sich sonst nur selten
finden. Es liegt also der Keimling dem Nabel[S. 624] gegenüber an der
Spitze des Endosperms. Familie Commelinaceae. Eine nur in den
Tropen und Subtropen verbreitete Familie, deren Perianth in
Kelch und Krone differenziert ist. Commelina, die Haare der
Staubblätter von Tradescantia bilden ein für Plasmaströmung
und Kernteilungsfiguren bekanntes und geeignetes Objekt. Rhoeo
discolor, Mexiko, vielfach in Kultur.

Fig. 822. Iris germanica, 1⁄2 nat. Gr. —
Offizinell.
[S. 625]
b)
Blüten mehr oder minder reduziert.
5. Ordnung.
Glumiflorae.
Die Ordnung der Spelzenblüher umfaßt ausschließlich Gewächse von
grasartigem Habitus und ein- bis mehrjähriger Dauer. Sie ist in
ihren beiden Familien über die ganze Erdoberfläche verbreitet.
Holziger Schaft eignet nur der Gattung Bambusa. Das allen gemeinsame
Merkmal liegt in der Vereinigung zahlreicher Einzelblüten, die eines
ausgebildeten Perianthes entbehren, dagegen von trockenhäutigen
Hochblättern, den Spelzen, gestützt werden, zu mehr oder minder
reich zusammengesetzten Blütenständen. Ebenso wie das Perianth,
das entweder vollständig ausfällt oder zu Borsten oder Schüppchen
verkümmert, fehlt häufig der innere Andröceumwirtel. Der oberständige
Fruchtknoten ist stets einfächerig und enthält nur eine Samenanlage; er
entspricht bald drei Fruchtblättern (Cyperaceen), bald zweien (einige
Carices), bald nur einem (Gramineen). Die Narben sind von erheblicher
Größe, papillös fadenförmig oder federig, wie es die Windbestäubung
verlangt. Die Früchte sind Schließfrüchte.
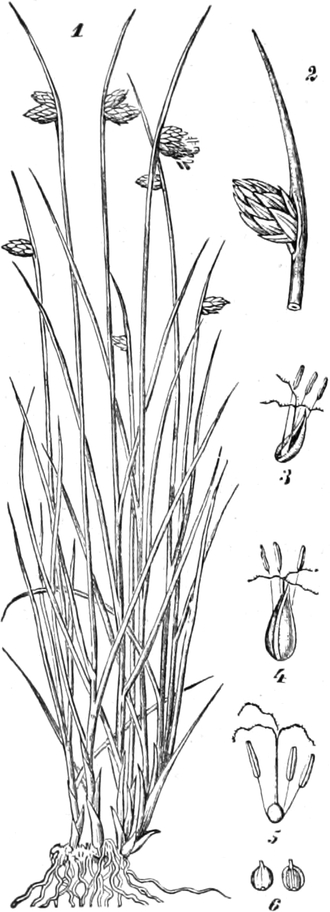
Fig. 823. Scirpus setaceus. Nat. Gr. 1 Blühende
Pflanze. 2 Gipfel eines fertilen Halmes. 3 Einzelblüte. 4
Dieselbe vom Rücken. 5 Dieselbe ohne Deckspelze. 6 Früchte. 2–6
vergr. Nach G. F. HOFFMANN.
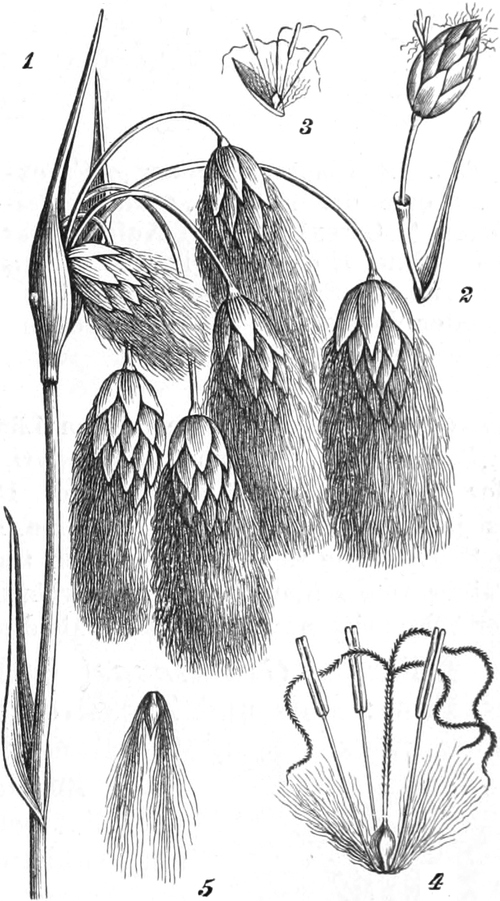
Fig. 824. Eriophorum augustifolium. Etwa nat. Gr. 1
Fruchtender Halm. 2 Ein blühendes Ährchen. 3 Einzelblüte. 4
Dieselbe ohne Spelze. 5 Früchtchen. 3–5 vergr. Nach G. F.
HOFFMANN.
1. Familie Cyperaceae. Die Riedgräser sind durch ihren meist
dreikantigen, in der Regel weder knotig gegliederten noch hohlen Halm
und die geschlossenen Scheiden ihrer Blätter kenntlich. Ihre Blüten
sind entweder eingeschlechtig, und dann meist monözisch (Carex), oder
zwittrig, wie bei der Mehrzahl der Gattungen. Der Fruchtknoten ist
zwei- oder dreikarpellig[S. 626] mit grundständiger, aufrechter, anatroper
Samenanlage. Die Fruchtschale ist nicht mit der Samenschale verwachsen,
die einen kleinen, rings von Endosperm umschlossenen Embryo enthält.
Wichtige Gattungen: Cyperus, Scirpus und Eriophorum
haben zwittrige Blüten, Fig. 823 zeigt eine blühende Pflanze des
einjährigen Scirpus setaceus, mit steifen, oberseits gerinnten
Blättern. Fertile Halme mit langem oberstem Internodium tragen die
1–3 Ährchen endständig; sie werden durch das in der Richtung
des Halmes aufstrebende Hüllblättchen zur Seite gedrückt und sind
mit zahlreichen dachziegeligen Spelzen bedeckt. Nur die untersten
größeren bleiben steril, alle anderen decken je eine nackte
Zwitterblüte. Eriophorum angustifolium, das zur Blütezeit wenig
auffallende Wollgras, bringt am Gipfel des fertilen Halmes
drei bis sieben langgestielte Ährchen mit zahlreichen dachziegelig
deckenden Spelzen. Die Einzelblüten sind am Grunde von vielen
Haaren umgeben, die von Staubblättern und Griffeln überragt werden.
Zur Fruchtzeit dagegen sind die Haare bis etwa 3 cm lang geworden
und ragen weit über die Spelzen hervor. Sie bilden ein für die
Verbreitung der Früchtchen wichtiges Flugorgan. Durch ihre weiße
Farbe machen sie die Pflanze und ihre dann herabhängenden Ähren
zu einem auffälligen Bestandteil unserer torfigen Wiesen (Fig.
824). Cyperus papyrus in Ägypten lieferte in den Längsscheiben
seiner schenkeldick werdenden Halme das „Papier“ des Altertums,
die Papyri. Carex hat nackte eingeschlechtige Blüten, welche
in der Regel monözisch verteilt sind. Die männlichen Ähren sind
einfach; in der Achsel eines jeden Deckblättchens sitzt eine
männliche Blüte, aus drei Staubblättern gebildet (Fig. 825
A). Die weiblichen Ährchen tragen in der Deckblattachsel je
ein Seitensprößchen, aus einer vom schlauchförmigen Vorblatt,
dem Utriculus, umgebenen Spindel a und dem in seiner Achsel
sitzenden, bald zwei- bald dreikarpelligen Fruchtknoten bestehend
(Fig. 825 B–E).
Fig. 826. Schema des Grasährchens. g Die Hüllspelzen,
p1 und p2 palea inferior und superior. B Die Blüte, e Die
Lodiculae. Sämtliche Achsenteile verlängert gedacht.
2. Familie Gramineae[508]. Die echten Gräser besitzen
stielrunde hohle (Ausnahme: Mais und Zuckerrohr), durch massive
Knotenstellen gegliederte Halme, zweizeilige Blattstellung und eine
meist offene Scheide, die an der Basis knotig verdickt zu sein pflegt.
An der Grenze der Blattscheide und -spreite ragt fast ausnahmslos
ein erhabener häutiger Rand über das Blatt hervor: die Ligula
(vgl. Fig. 138). Gramineenblüten finden sich in ähren-, trauben-
oder rispenartigen Gesamtblütenständen vereinigt, die jedesmal aus
ährenartigen Teilinfloreszenzen, den „Ährchen“, zusammengesetzt
sind. Meist ist das Ährchen mehrblütig. Es beginnt in der Regel (Fig.
826, 827) mit zwei (in einzelnen Fällen einer, oder 3–4) sterilen
Hüllspelzen (gluma); in zweizeiliger Anordnung wie diese
folgen die fertilen Deckspelzen (palea inferior) mit je
einer Blüte in ihren Achseln. Die Deckspelzen sind oft begrannt,
d. h. sie tragen eine steife widerhaarige Borste auf dem Rücken oder
an ihrer Spitze, die Granne. Jedem Einzelblütchen geht eine
Vorspelze (palea superior) vorauf. Es folgen zwei kleine
Schüppchen, die als Schwellkörper zur Öffnung der Blüte beitragen (Fig.
828 B, C) und Lodiculae heißen; endlich das meist[S. 627] aus einem
dreigliedrigen Wirtel bestehende Andröceum und der mit zwei federartig
verzweigten papillösen Narben gekrönte Fruchtknoten. Er umschließt eine
anatrope oder schwach kampylotrope Samenanlage.
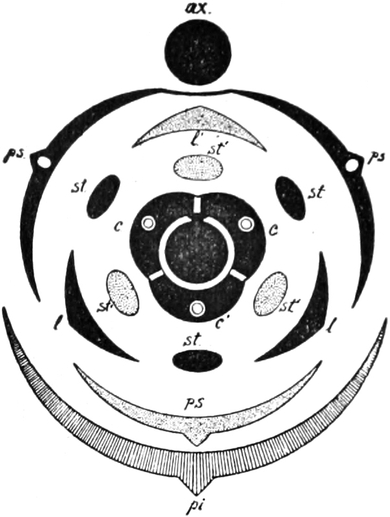
Fig. 827. Diagramm der Grasblüte. Die fehlenden Teile
matt punktiert. ax Achsenende der Ährchenachse, pi palea inferior,
ps palea superior (äußeres Perigon), l Lodiculae (inneres Perigon),
st äußerer, st′ innerer Staubblattkreis, c laterale Fruchtblätter
c′ dorsales Fruchtblatt. Nach J. SCHUSTER.
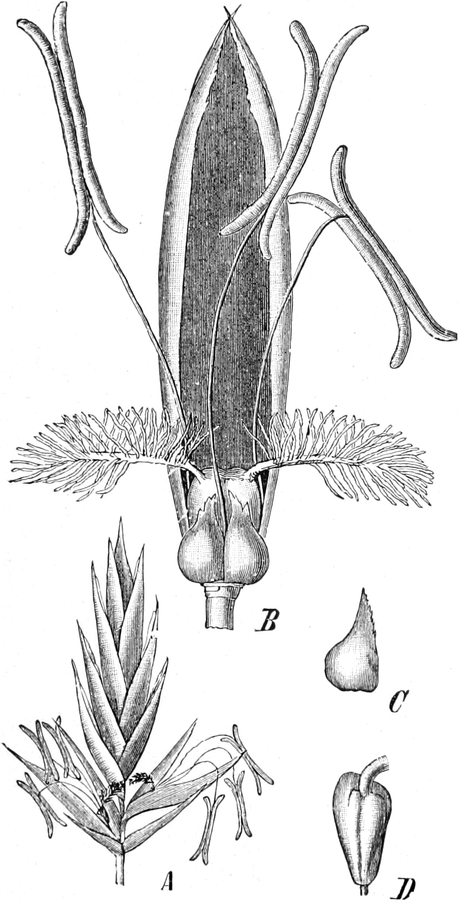
Fig. 828. Festuca elatior. A Ährchen (vgl. Fig.
826) mit zwei offenen Blüten, unten die beiden sterilen Hüllspelzen.
Vergr. 3. B Die Blüte; vorn die beiden Lodiculae, hinten die
Vorspelze (palea superior), Fruchtknoten mit federartigen Narben. C
Eine Lodicula. D Fruchtknoten, von der Seite, mit dem Stiel einer
abgeschnittenen Narbe. B-D Vergr. 12. Nach H. SCHENCK.
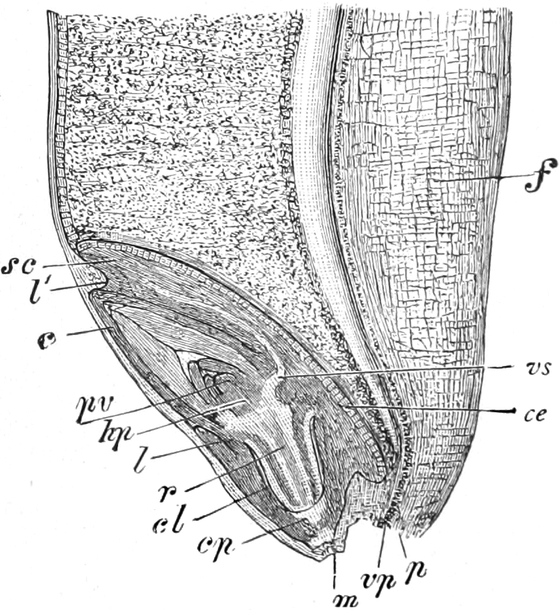
Fig. 829. Medianer Längsschnitt durch den unteren Teil
eines Weizenkorns. Links unten der Keim mit dem Scutellum sc, l′
Ligula, vs Leitbündel des Scutellum, ce sein Zylinderepithel,
c Scheidenteil des Kotyledons, pv Stammvegetationskegel, hp
Hypokotyl, l Epiblast, r Radicula, cl Wurzelscheide, m
Austrittsstelle der Radicula, p Fruchtstiel, vp sein Leitbündel,
f Seitenwandung der Furche. Vergr. 14. Nach E. STRASBURGER.
Nicht immer ist der Bau so stark reduziert; so hat die Reisblüte
(Fig. 832) ein vollzähliges Andröceum, ebenso die Bambuseen,
welche daneben drei Griffel besitzen und auch drei Lodiculae
aufweisen. Streptochaeta endlich hat eine normale pentazyklische
Monokotylenblüte, deren Gynäceum der Anlage nach dreizählig
ist. Man ist daher berechtigt, die Lodiculae als dem inneren
Perianthkreis entsprechend anzusehen, während die Vorspelze zwei
verwachsene Blätter des äußeren Perianthkreises, dessen drittes
fehlt, darstellen könnte. Im Gynäceum ist von[S. 628] den ursprünglichen
drei Karpellen meist nur ein, aus den zwei lateralen Fruchtblättern
gebildetes, Doppelblatt übrig geblieben. Nach dieser Auffassung,
die z. B. von GOEBEL vertreten wird, gelangt man zu dem
umstehenden Diagramm (Fig. 827).
Über die Windblütigkeit der Gräser vgl. S. 479. Die Frucht zeigt
eine innige Verwachsung von Frucht- und Samenschale, sie wird
Karyopse genannt. Der Embryo liegt dem stärkereichen Endosperm
seitlich mit seinem Kotyledon, dem Scutellum, an, welches
bei der Keimung als Saugorgan die Aufnahme der gespeicherten
Reservestoffe bewirkt (Fig. 829).

Fig. 830. Getreidearten. A Roggen. Secale cereale. B
Spelt, Triticum Spelta. C Zweizeilige Gerste. Hordeum distichum. D
Weizen, Triticum vulgare. — D Offizinell.
Zu den Gräsern zählen als wichtige Nutzpflanzen vor allem die
eigentlichen Brotpflanzen: Der Weizen, Triticum (Fig.
830 B und D), mit einzelstehenden zwei- bis vielblütigen
Ährchen, deren Hüllspelzen breiteiförmig sind (Fig. 831 B). Von
Weizenarten unterscheidet F. KOERNICKE 1. Tr. vulgare,
Saatweizen mit verschiedenen Unterarten; 2. Tr. polonicum,
Polnischer Weizen; 3. Tr. monococcum, Einkorn.
Der Roggen, Secale cereale (Fig. 830 A). Die Ährchen
stehen einzeln und sind zweiblütig, ihre Hüllspelzen pfriemlich
(Fig. 831 A). Die Gerste, Hordeum vulgare (Fig. 830
C). Die einblütigen Ährchen stehen zu dreien, bei den Unterarten
H. hexastichum und H. tetra[S. 629]stichum sind alle Reihen,
bei H. distichum ist nur die Mittelreihe fruchtbar. Der
Hafer, Avena sativa, und der Mais, Zea Mays. Alle
diese sind der Kultur in gemäßigtem Klima zugänglich. Ihre Heimat
ist, bis auf die des amerikanischen Mais, voraussichtlich Westasien
oder Südosteuropa; in wildem Zustand bekannt sind nur Triticum
monococcum var. aegilopodioides als Stammform des Einkorns, Tr.
dicoccoides als wahrscheinliche Stammform des Weizens, Secale
montanum als Stammform des Roggens, Hordeum spontaneum, dem H.
distichum nahestehend, Stammform der Gerste. Diese wilden Formen
sind durch Auseinanderfallen ihrer Spindel bei der Fruchtreife
gekennzeichnet, eine Eigenschaft, die für Kulturformen höchst
unvorteilhaft wirken müßte.
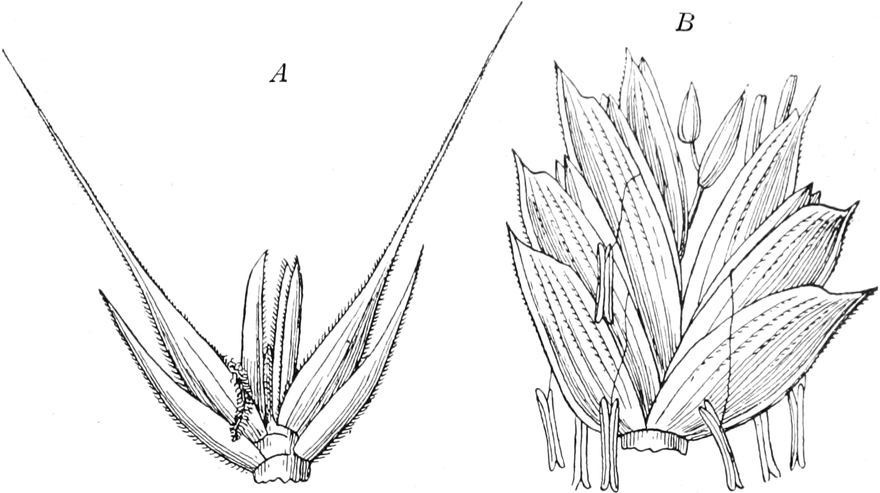
Fig. 831. A Ährchen des Roggens, zweiblütig. B
Ährchen des Weizens, mehrblütig.

Fig. 832. Oryza sativa. Blütenrispe, 1⁄2 nat. Gr.
Einzelnes Ährchen, vergr. — Offizinell.
Das wichtigste tropische Getreide ist der Reis, Oryza
sativa (Fig. 832), bis in die warm temperierten Länder hinein
in größtem Maßstabe kultiviert (Fig. 833) und bei hinreichender
Feuchtigkeit von unerreichter Fruchtbarkeit. Speziell in Afrika
ist die Mohrhirse, Andropogon Sorghum, in mehreren
Varietäten zu Hause. Sie bildet als Durrha die wichtigste
Brotpflanze für diesen ganzen Kontinent; schließlich bleiben die
im Mittelmeergebiet und in Asien kultivierten Panicum miliaceum,
echte Hirse, und P. italicum, Kolbenhirse, zu
erwähnen, beide asiatischen Ursprungs. Als Nahrungsmittel nimmt
ferner das Zuckerrohr, Saccharum officinarum, ein
übermannshohes, im tropischen Asien, Vorder- und Hinterindien
beheimatetes perennierendes Gras, eine wichtige Stelle ein. Es wird
zur Zeit überall in den Tropen kultiviert, um aus dem fleischigen
Mark des hier nicht hohlen Stengels durch Auspressen und
Eindicken des Saftes Rohrzucker zu gewinnen.
Als wichtige heimische Futtergräser unserer Wiesen mögen
genannt sein:
Agrostis alba, Alopecurus pratensis, Anthoxanthum odoratum,
Arrhenatherum elatius, Avena flavescens und pubescens,
Briza[S. 630] media, Dactylis glomerata, Holcus lanatus, Lolium
perenne, Phleum pratense, Poa pratensis; außerdem sind
zu beachten Aira-, Bromus-, Calamagrostis-, Festuca-,
Melica- usw. Arten. Eine außerordentlich mannigfaltige Anwendung
finden endlich die baumförmigen tropischen Bambus-Arten in
ihren stattlicheren Vertretern: Häuser, Wände, Fußböden, Leitern,
Brücken, Stricke, Wasserkrüge, Kochgefäße, Wasserleitungsröhren
usw. werden aus den Stämmen angefertigt, so daß diese Pflanzen für
die dortigen Verhältnisse geradezu unentbehrlich genannt werden
müssen.
Giftig: Der einjährige Taumellolch, Lolium
temulentum (Fig. 834), hat in der Regel von Pilzhyphen umsponnene
und dann durch Alkaloidgehalt giftige Früchte; pilzfreie Früchte
der Pflanze sind unschädlich[509]. Die Pflanze ist einjährig,
entbehrt der sterilen Triebe und kann dadurch leicht von den
häufigen Lolium-Arten, perenne und multiflorum, unterschieden
werden. Der Taumellolch ist das einzige giftige Gras.

Fig. 833. Für Reiskulturen hergerichtetes Terrassenland
in Ceylon. Das für den jungen, einzeln zu pflanzenden Reis notwendige
Wasser läuft von Terrasse zu Terrasse, deren erhöhte Ränder eine
Abflußstelle besitzen. Im Vordergrunde Bananen (Musa), in der
Mitte eine Arecapalme, vorn rechts eine Kaffeeplantage. Nach einer
Photographie.
Offizinell: Saccharum officinarum: Saccharum
(Pharm. germ., austr., helv.). Agropyrum repens: Rhizoma
Graminis (Pharm. austr., helv.). Triticum vulgare: Amylum
Tritici (Pharm. germ., austr., helv.). Oryza sativa:
Amylum Oryzae (ibid.).
c)
Blüten zygomorph.
6 Ordnung.
Scitamineae.
Tropische Stauden von zum Teil gewaltigen Dimensionen, in einzelnen
Fällen baumartig mit dorsiventralen oder asymmetrischen Blüten
entsprechen dem Typus dieser Ordnung. Das Perianth ist in Kelch
und Krone gesondert und das Andröceum stark reduziert, zum Teil
staminodial, kronblattähnlich. Der unterständige, dreifächerige
Fruchtknoten enthält Perisperm führende Samen.
[S. 631]
1. Familie Musaceae. Die Banane, Musa (Fig. 835), ist
eine der wichtigsten Fruchtpflanzen aller tropischen Gegenden.
Einander dicht umschließende Scheiden der mächtigen Blätter bilden
ein stammartig aussehendes Gebilde, aus dem die endständige
Infloreszenz ihre großenteils parthenokarpen[510] dichtgedrängten
Beerenfrüchte herabhängen läßt. Musa textilis liefert Manilahanf;
Ravenala besitzt einen Holzstamm, Strelitzia reginae (Fig. 542)
vom Kap wird ihrer prächtigen, ornithophilen Blüten halber häufig
kultiviert.
2. Familie Zingiberaceae. Die dorsiventralen Einzelblüten
stehen in Ähren von bisweilen köpfchenartigem Habitus. Eine
dreizipfelige Krone ragt aus dem unscheinbaren röhrigen Kelch
hervor. Wenn der äußere Staubblattkreis nicht gänzlich fehlt, ist
er in zwei seitlichen Staminodien vertreten (Fig. 836 sst1,
sst2). Im inneren Staubblattkreis ist das hintere Staubblatt
allein fertil (st), die beiden übrigen sind vereinigt und in Form
petaloider Staminodien ausgebildet; sie stellen als Labellum
(l) durch ihre Größe und hervortretende Lage, wie durch lebhafte
Färbung den Schauapparat der Zingiberaceenblüte dar. Der Griffel
verläuft in dem röhrenartigen Einschnitt zwischen den beiden
Thecae des Staubblattes. Aus dem Fruchtknoten wird eine
Kapsel. Die Zingiberaceen gehören meist dem tropischen Asien an.
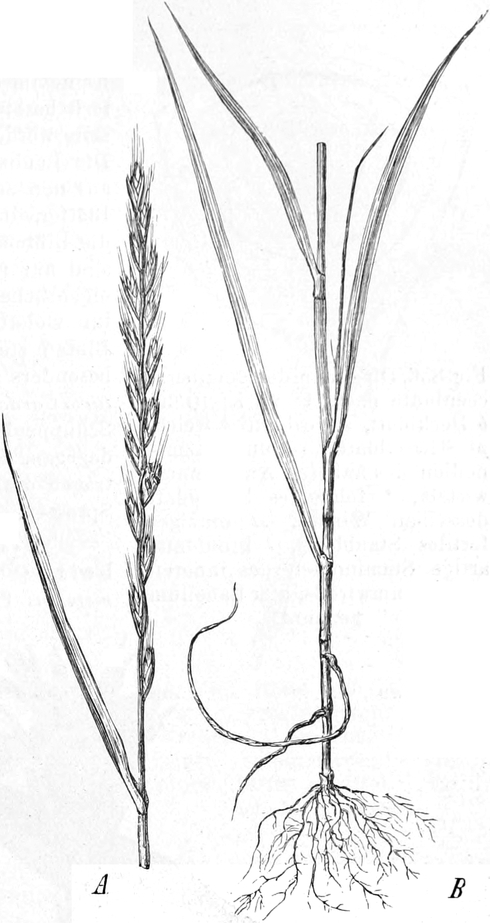
Fig. 834. Lolium temulentum. — Giftig. Nach
H. SCHENCK.

Fig. 835. Gruppe von Musa sapientum mit Manihot
utilissima. Ceylon, nach einer Photographie.
Zingiber officinale, der Ingwer, eine alte Kulturpflanze
Südostasiens, wird jetzt[S. 632] überall in den Tropen kultiviert (Fig.
837). Das flache, geweihartig verzweigte Rhizom steht auf seiner
hohen Kante im Boden. Es ist mit zweizeilig stehenden Blättern
besetzt, nur die Achselknospen der Rhizomunterseite werden
gefördert und setzen den Hauptsproß fort. Die Laubzweige bestehen
trotz ihrer Länge fast nur aus den Scheiden der großen, ungeteilten
ganzrandigen Blätter, ihre Achse bleibt außerordentlich kurz. Nur
die Blütensprosse sind massiv, sie bleiben niedriger und sind
nur mit den langscheidigen Schuppenblättern ohne eigentliche
Spreite bekleidet. Die hellgelben, durch ihr violettes, heller
geflecktes Labellum auffallenden Blüten stehen in der Achsel
großer Hochblätter, die besonders an ihrem Rande lebhaft gefärbt
sind. Elettaria Cardamomum und Curcuma haben ebenfalls nur
mit Schuppenblättern bestandene Infloreszenzstiele, Alpinia
dagegen, wie die häufig kultivierten Hedychium-Arten tragen die
Blütenstände terminal an normal belaubten Sprossen.
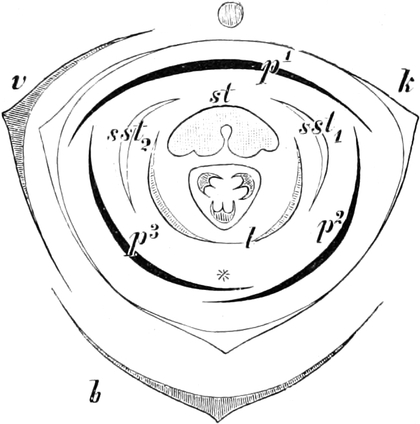
Fig. 836. Diagramm der Zingiberaceenblüte nach
A. W. EICHLER. b Deckblatt, v Vorblatt, k Kelch,
p1–3 Kronblätter, sst1 und 2 Staminodien des äußeren
Andröceumwirtels, * fehlendes Staubblatt desselben Wirtels, st
einziges fertiles Staubblatt, l kronblattartige Staminodien des
inneren Andröceumwirtels, hier Labellum genannt.
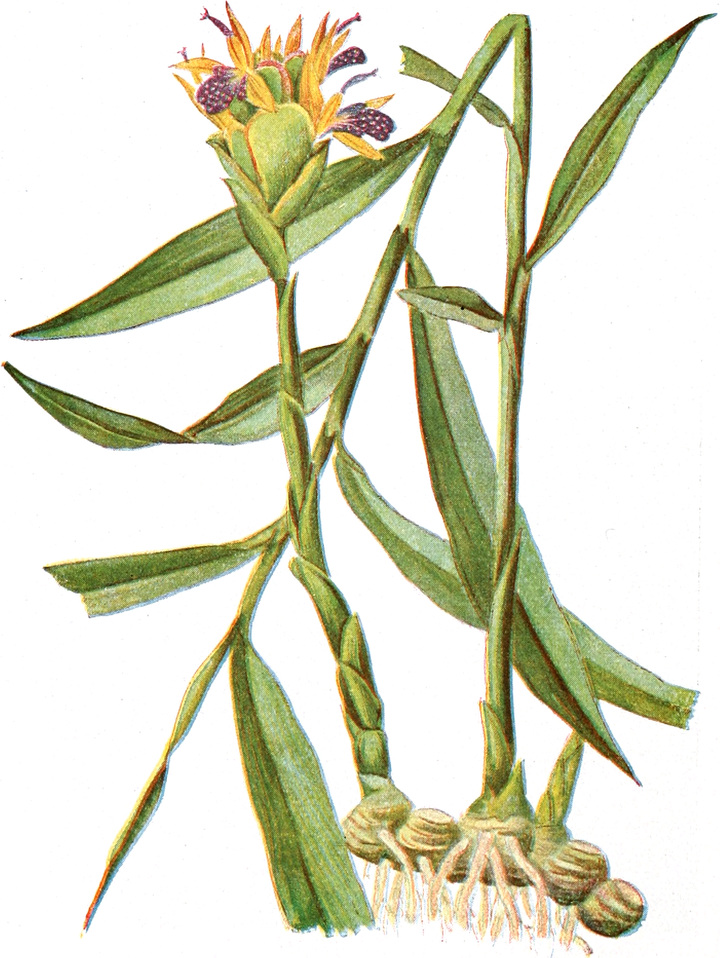
Fig. 837. Zingiber officinale. 1⁄2 nat. Gr. Nach
BERG und SCHMIDT. — Offizinell.
Offizinell: Zingiber officinale: Rhiz. Zingiberis
(Pharm. germ., austr., helv.). Elettaria Cardamomum: Fructus
Cardamomi (ibid.), Cardamomen,[S. 633] Curcuma Zedoaria, Zittwer:
Rhiz. Zedoariae (ibid.), Alpinia officinarum, Galgant:
Rhiz. Galangae (Pharm. germ., helv.).
Großblättrige häufig kultivierte Stauden mit asymmetrischen
Blüten (Fig. 838) gehören zu den Cannaceae, die nur eine
halbe, also monothezische, Anthere haben, die andere Hälfte ist
blumenblattartig.
Den gleichen Habitus und ebenso gebaute, aber minder große Blüten
besitzen die Marantaceae, deren Blätter am Ansatz der
Spreite mit Gelenkpolstern versehen sind.
Das Rhizom von Maranta arundinacea liefert westindisches
Arrowroot.
7. Ordnung.
Gynandrae.
Die Familie Orchidaceae enthält perennierende, krautige,
erdbewohnende oder epiphytische Gewächse mit zwittrigen, stark
dorsiventralen Blüten, deren Perianth kronartig ist; das hintere Blatt
des inneren Kreises wird als Lippe, Labellum ausgebildet
und läuft häufig in einen Sporn aus.
Das bei den Scitamineen erwähnte „Labellum“ ist morphologisch
ganz anderer Natur, da es staminodialen und petaloid gewordenen
Staubblättern entspricht.
Das Andröceum wird auf die drei vorderen Glieder beschränkt, von
denen meist das mittlere, dem äußeren Kreise angehörige allein fertil
ist, während die anderen fehlen oder Staminodien darstellen. Das
einfächerige, unterständige Gynäceum ist aus drei Fruchtblättern
verwachsen, die eine Kapselfrucht liefern mit äußerst zahlreichen an
den randständigen, d. h. parietalen Plazenten sitzenden Samen (Fig.
839 u. 842). Das fertile Staubblatt verwächst mit dem Griffel zu einem
Säulchen, Gynostemium, welches in der Mitte der Blüte
mehr oder minder emporragt. Durch Drehung der ganzen Blüte um 180°
(vgl. Fig. 839 u. 842) oder Übernicken gelangt die als Anflugstelle für
Insekten dienende Lippe auf die Vorderseite.
Die Orchidaceen erreichen ihre reichste Entwicklung in den Tropen
aller Erdteile, wo sie meist unter den Epiphyten eine wichtige
Rolle spielen.
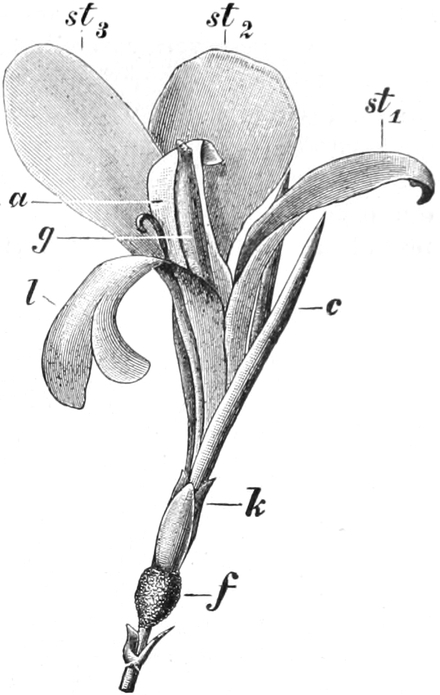
Fig. 838. Blüte von Canna iridiflora. f Fruchtknoten,
k Kelch, c Krone, l Labellum, st 1–3 die übrigen Staminodien,
a fertile Antherenhälfte, g Griffel. 1⁄2 nat. Gr. Nach H.
SCHENCK.

Fig. 839. Orchidaceen-Diagramm (Orchis). Nach F.
NOLL verändert.
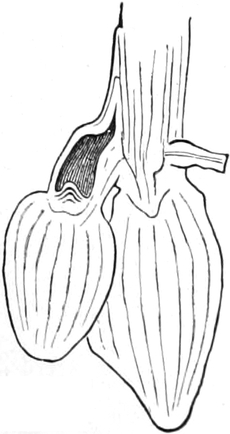
Fig. 840. Orchis militaris. Längsschnitt durch Mutter-
und Tochterknolle. Nach LÜRSSEN. — Offizinell.
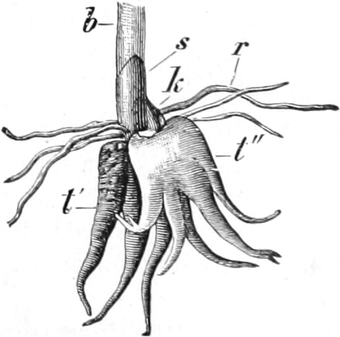
Fig. 841. Wurzelsystem von Orchis latifolia. b Basis
des Stengels, s Niederblatt, t′ alte, t″ junge Knolle, k
Knospe, r Wurzeln. Nach H. SCHENCK.
[S. 634]
Orchis, Knabenkraut, Ophrys, Gymnadenia, Platanthera
sind alle mit Knollen versehen. Epipactis, Cephalanthera,
Listera haben ein verzweigtes Rhizom. Neottia, Nestwurz,
Coralliorrhiza, Epipogon, Limodorum leben saprophytisch
oder richtiger parasitisch auf Kosten ihrer Mykorrhizen[511] und
sind daher fast oder ganz chlorophyllfrei. Cypripedium, der
Frauenschuh, hat zwei fertile seitliche Staubblätter des inneren
Wirtels.
Zu genauerer Darstellung mag eine unserer häufigeren einheimischen
Orchis-Arten, Orchis militaris, dienen, die in Fig. 842,
844 wiedergegeben ist. Untersucht man eine solche Pflanze
zur Zeit ihrer Blüte, so findet sich ein Paar fleischiger
Knollen als Ausgangspunkt. Beide sind mit einem Flaum von
Wurzelhaaren überdeckt. Die größere braune Knolle, von mehr
schwammiger Beschaffenheit, setzt sich nach oben in den von
ein paar Niederblättern und den Scheiden der 2–4 Laubblätter
länglich-elliptischer Form umhüllten Infloreszenzstiel fort,
welcher mit einer pyramidalen Blütentraube abschließt. Die
kleinere Knolle ist von weißer Farbe und fester Konsistenz; sie
trägt, wie der Längsschnitt (Fig. 840) zeigt, eine Knospe auf dem
Scheitel, die bereits ein paar Scheidenblätter entwickelt hat. Es
handelt sich um den in der Achsel eines der ersten Scheidenblätter
der Pflanze, dicht über der älteren braunen Knolle gebildeten
Achselsproß, welcher mit seiner als Reservestoffbehälter knollig
anschwellenden Wurzelanlage das Scheidenblatt durchbrochen hat
(Fig. 844) und die Mutterpflanze im nächsten Jahre ersetzen wird.
Fig. 842. Orchis militaris. A Eine von der kleinen
Braktee (a) gestützte Blüte, b Fruchtknoten, c die äußeren, d
die beiden oberen inneren Perigonblätter, e Labellum mit dem Sporn,
f, g Gynostemium. — B Dieselbe nach Entfernung des Perigons
mit Ausnahme des oberen Teils des Labellum, h Narbe, l Rostellum,
k zahnartiger Fortsatz des Rostellum, m Fach der Anthere, n
Konnektiv, o Pollinium, q Klebmasse, p Staminodium, vergr. —
C Einzelnes Pollinium, r Kaudikula, s Pollen, stärker vergr.
— D Frucht im Querschnitt, schwach vergr. Nach BERG und
SCHMIDT.
Fig. 843. Vanilla planifolia (nach BERG und
SCHMIDT aus ENGLER-PRANTL), verkleinert. A Lippe
und Gynostemium. B Gynostemium von der Seite. C Gynostemiumspitze
von vorn. D Anthere. E Samen, vergr. — Offizinell.
Bei Betrachtung der Blüten fällt sogleich die schraubige Drehung
des Fruchtknotens auf, der hier die Abwärtskehrung der „Unterlippe“
zu danken ist. Diese ist dreizipflig, und ihr Mittellappen teilt
sich am Ende abermals in zwei gabelig auseinanderstehende Läppchen.
Ganz an der Basis dieses Labellums ist ein Sporn als Aussackung zu
erkennen. Er dient als Nektarium, und seine Öffnung findet sich
direkt unter dem Gynostemium (Fig. 842 A, B). Dieses trägt
auf seiner der Unterlippe und den sich[S. 635] darauf niederlassenden
Insekten zugekehrten Seite eine große Narbenfläche (h),
welche zwei vereinigten Narben entspricht. Die dritte Narbe ist
zu einem als Rostellum bezeichneten Gebilde (l, k)
umgeformt und dient der Ausrüstung der männlichen Organe. Die
eine fruchtbare Anthere besitzt zwei Thecae, durch das als
Abschluß des Gynostemiums sichtbare Konnektiv (n) verbunden.
Die ganze Pollenmasse jedes der beiden Fächer wird durch eine
Bindesubstanz zusammengehalten, welche gleichzeitig nach unten in
einen Stiel ausläuft. Dieser heißt Kaudikula; das gesamte,
Pollinium genannte Gebilde ist von wachsartiger Konsistenz.
Die Kaudiculae enden nun an jenem Rostellum, welches eine
Klebmasse von zäher Beschaffenheit enthält, die geeignet ist,
einmal die Pollinien, welche frei im Fache liegen, an Ort und
Stelle festzuhalten, andererseits aber auch sie an andere damit in
Berührung gelangende Körper anzukleben. Versucht nun ein auf der
Unterlippe sitzendes Insekt den im Sporn ausgeschiedenen Nektar zu
erreichen, so muß es mit Kopf oder Rüssel das Rostellum berühren
und die Pollinien mitnehmen. Beim Eintrocknen der Kaudiculae biegen
die Pollinien sich nach vorn und werden daher bei einer nächst
besuchten Blüte genau auf die Narbenfläche gelangen müssen.
In ähnlicher, vielfach aber noch weit komplizierterer Art sind alle
Orchidaceen auf Insektenbesuch angepaßt, dessen sie zur Bestäubung
ihrer Blüten nicht entbehren können[512]. In vielen Fällen ist
die Anpassung so speziell auf den Bau eines bestimmten Insektes
gerichtet, daß kein anderes dieselbe Leistung zu vollziehen vermag;
so blieb z. B. die aus ihrer amerikanischen Heimat in andere
tropische Länder gebrachte Vanilla (Fig. 843) stets unfruchtbar,
da das bestäubende Insekt fehlte. Nachdem dies erkannt war, wird
sie jetzt durch Menschenhand einzeln bestäubt und setzt daraufhin
regelmäßig Früchte an. Zur Vervollständigung ist noch hinzuzufügen,
daß bei manchen Formen, so auch bei Vanilla, der Pollen körnig
bleibt. Zahlreiche tropische Orchidaceen werden ihrer herrlich
duftenden und schön geformten, farbenprächtigen Blüten wegen bei
uns in Gewächshäusern kultiviert, so Cattleya, Laelia, Vanda,
Dendrobium, Stanhopea u. v. a.

Fig. 844. Orchis militaris. 1⁄2 nat. Gr. —
Offizinell.
Offizinell: Orchis-Arten und verwandte Formen mit
eiförmigen, nicht handförmig zerteilten (vgl. Fig. 840 u. 844)
Knollen: Tubera Salep (Pharm. germ., austr., helv.). —
Vanilla planifolia, ein in Mexiko einheimischer, vielfach in
den Tropen kultivierter Wurzelkletterer (Fig. 843): Fructus
Vanillae (Pharm. austr., helv.).
[S. 636]
Die fossilen Angiospermen[471].
Die ersten zweifellosen Angiospermen zeigen sich in der oberen
Kreide, und zwar gleich in mannigfachen Formen, die ungefähr in
gleichem Verhältnis wie jetzt zu Monokotylen und Dikotylen gehören.
Gefunden sind zunächst nur Blätter, die große Ähnlichkeit mit denen
jetzt lebender Angiospermen zeigen, hingegen gar keine mit solchen
von Gymnospermen oder gar Pteridophyten. Die hier vorhandene Kluft
kann also durch die paläontologischen Funde nicht überbrückt werden.
Im Eozän und Oligozän werden die Angiospermen sicher bestimmbar;
es sind Angehörige noch existierender Familien, und zwar, sogar
im nördlichen Europa, teilweise von tropischem Charakter, nämlich
Palmen, Dracaena, Smilax usw. unter den Monokotylen,
zahlreiche Quercifloren (namentlich Quercus), Lauraceae
(Cinnamomum u. dgl.), Leguminosae usw. von Dikotylen. Vom
Miozän an werden die Arten teilweise mit lebenden identisch;
im Quarternär fehlen eigene, von den jetzt lebenden erheblich
abweichende Formen. Der Florencharakter war zur Tertiärzeit in
Europa ganz wesentlich von dem gegenwärtigen verschieden; er trug
das Gepräge eines viel wärmeren Klimas und wies, wie für die
Gymnospermen, Typen auf, die gegenwärtig nur noch in fernen Ländern
existieren.
Literatur zur Einleitung und Morphologie von H. Fitting.
Einleitung.
Erster Abschnitt. Zytologie.
Der lebende Inhalt der Zellen.
Gröbere leblose Einschlüsse der Protoplasten.
Die Zellmembranen.
Zweiter Abschnitt. Histologie.
Dritter Abschnitt. Organographie.
Bau des Thallus.
Bau des typischen Kormus.
Anpassungen des Kormus an die Lebensweise
und an die Umwelt.
Fortpflanzungsorgane.
Vierter Abschnitt.
Die Deszendenzlehre und die Entstehung der Anpassungen.
Literatur zur Physiologie von L. Jost.
Einleitung.
Chemische Zusammensetzung, Stoffaufnahme.
Assimilation und Stoffwanderung.
Atmung und Gärung.
Entwicklung.
Vorbemerkungen.
Faktoren der Entwicklung.
Der Entwicklungsgang.
Bewegungen.
Tropismen.
Nastien.
Literatur zu Thallophyten, Bryophyten, Pteridophyten von H. Schenck.
Niedere Thallophyten.
.
Algen.
Pilze.
.
Lichenes.
Bryophyta.
Pteridophyta.
Fossile Kryptogamen.
Literatur von Spermatophyta von G. Karsten.
Übergang von den Farnpflanzen zu den
Samenpflanzen.
Morphologie und Ökologie der Blüte.
Entwicklung der Geschlechtsgeneration.
Gymnospermae.
Angiospermae Dikotylae.
Monokotylae.
Systematisches
Verzeichnis der offizinellen und wichtigsten giftigen Gewächse.
 bedeutet offizinell in Deutschland, Österreich oder Schweiz;
bedeutet offizinell in Deutschland, Österreich oder Schweiz;
+ bedeutet giftig;
bedeutet offizinell und giftig;
* vor den Seitenzahlen bedeutet Abbildung.
Thallophyta.
- Laminaria digitata, Cloustoni 361. *362. *366. 368.
- Chondrus crispus, Carrageen *372. 376.
- Gigartina mamillosa, Carrageen *372. 376.
- Elaphomyces granulatus, Boletus cervinus, Hirschtrüffel 388.
- Claviceps purpurea, Secale cornutum, Mutterkorn 389. *390.
- Fomes fomentarius, Feuerschwamm 407. 409.
- Polyporus officinalis, Agaricus albus, Lärchenschwamm 407. 409.
- + Boletus Satanas, Satanspilz *406.
- + Amanita muscaria, Fliegenschwamm *408.
- + — phalloides, Grüner Knollenblätterschwamm *408.
- + — verna, Weißer Knollenblätterschwamm 408.
- + — mappa, Gelber Knollenblätterschwamm *408.
- + Lactaria torminosa, Giftreizker 408.
- + Russula emetica, Speiteufel 408.
- + Scleroderma vulgare, Hartbovist 409.
- Cetraria islandica, Isländisches Moos *412. 415.
Pteridophyta.
Gymnospermae.
- + Taxus baccata, Eibe *512. 513.
- Juniperus communis. Wacholder *514. 519.
- — Sabina, Sadebaum *515. 519.
- — Oxycedrus 519.
- Picea excelsa, Fichte *517.
- Abies pectinata, Weißtanne *516.
- Larix europaea, Lärche *519.
- — sibirica 519.
- Pinus silvestris, Kiefer *518. 519.
- — montana, Zwergkiefer 519.
- — Laricio, Schwarzkiefer 519.
- — Pinaster, Seestrandskiefer 519.
- — Taeda, Weihrauchkiefer 519.
Polycarpicae.
- + Ranunculus sceleratus, Hahnenfuß *530.
- + — arvensis und andere Arten *530.
- + Caltha palustris, Butterblume *532.
- + Anemone pulsatilla, Küchenschelle *531.
- + — nemorosa, Windröschen 532.
- + Clematis-Arten 532.
- + Delphinium-Arten 532.
- + Adonis autumnalis 532.
- Helleborus-Arten, Nießwurz 532.
- Aconitum Napellus, blauer Eisenhut 532. 533.
- + — Lycoctonum und andere Arten 532.
- Hydrastis canadensis 532. *534.
- Illicium anisatum, Sternanis 525.
- + — religiosum 525.
- Myristica fragrans, Muskatnuß *526.
- Podophyllum peltatum 526. *527.
- Jatrorrhiza palmata 526.
- Cinnamomum Camphora, Kampfer 526.
- — zeylanicum, Ceylonzimt 526.
- Laurus nobilis, Lorbeer 526. *528.
- Sassafras officinale 526. *527.
[S. 655]
Hamamelidinae.
- Liquidambar orientalis, Styraxbaum 533.
- Hamamelis virginiana 533.
Rosiflorae.
- Ribes rubrum, Johannisbeere 534. *535.
- Pirus malus, Apfel 534.
- Cydonia vulgaris, Quitte 536.
- Quillaja Saponaria *536. 537.
- Potentilla Tormentilla 537.
- Spiraea Ulmaria 537.
- Rubus Idaeus, Himbeere 537.
- — fruticosus 537.
- Hagenia abyssinica *536. 537.
- Rosa centifolia 537.
- Prunus Laurocerasus, Kirschlorbeer 537.
- — Cerasus, Kirsche *537.
- — Amygdalus, Mandel 537.
- — domestica, Pflaume 537.
Leguminosae.
- Acacia Catechu *539.
- — Suma 539.
- — Senegal 539.
- Cassia angustifolia, Sennesstrauch *540.
- — acutifolia 540.
- — obovota 540.
- — Fistula 540.
- Copaïfera Langsdorffii und andere Arten 540.
- Krameria triandra 540. *541.
- Tamarindus indica, Tamarinde 540. *541.
- Haematoxylon campechianum, Blauholz 540.
- Myroxylon balsamum var. Pereirae, Perubalsambaum *542. 544.
- — balsamum var. genuinum 544.
- + Laburnum vulgare, Goldregen 544.
- + Cytisus-Arten 544.
- Astragalus gummifer Tranganthsträucher *543 und andere Arten 544.
- Glycyrrhiza glabra, Süßholz 546.
- Melilotus officinalis, Steinklee 544.
- Trigonella foenum graecum, Bockshornsamen 544.
- Spartium scoparium 544.
- Ononis spinosa, Hauhechel 544.
- + Wistaria sinensis, Glycine 544.
- Physostigma venenosum, Kalabarbohne 544.
- Arachis hypogaea 544.
- Andira Araroba 544.
- Pterocarpus santalinus 544.
- — Marsupium 544.
- + Coronilla varia, bunte Kronwicke 544.
Myrtiflorae.
- Daphne Mezereum, Seidelbast *544. 545.
- Jambosa caryophyllus. Nelkenbaum *546. 547
- Eucalyptus globulus 547.
- Melaleuca Leucadendron 547.
- Punica Granatum, Granatbaum *546. 547.
Umbelliflorae.
- Carum Carvi, Kümmel *548. 550.
- Pimpinella Anisum, Anis *548. 550.
- — Saxifraga, Bibernell 550.
- — magna, Bibernell 550.
- Foeniculum capillaceum, Fenchel *548. 550.
- Levisticum officinale, Liebstöckel 550.
- Petroselinum sativum, Petersilie 550.
- + Cicuta virosa, Wasserschierling *549.
- + Sium latifolium, Merk 549.
- + Berula angustifolia, Berle 549.
- + Aethusa Cynapium, Hundspetersilie *550.
- + Oenanthe-Arten, Pferdesaat 549.
- Archangelica officin., Engelwurz 550.
- Imperatoria Ostruthium, Meisterwurz 550.
- Conium maculatum, Schierling 550. *551.
- Coriandrum sativ., Koriander *548. 550.
- Dorema Ammoniacum 550.
- Ferula galbaniflua 550.
- — Narthex 550.
- — Asa foetida 550.
Centrospermae.
- Beta vulgaris, Zuckerrübe 554.
- Chenopodium ambrosioides 554.
- + Agrostemma Githago, Kornrade *553.
- + Saponaria officinalis, Seifenkraut *553.
- Herniaria glabra 554.
- — hirsuta 554.
- + Anhalonium-Arten 552.
Primulinae.
- + Cyclamen europaeum, Alpenveilchen *554. 555.
- + Anagallis arvensis, Gauchheil *554. 555.
- + Primula obconica 555.
- + — sinensis 555.
- + Corthusa Matthioli 555.
Polygoninae.
Loranthiflorae.
- Santalum album, Santelholz 556.
- + Viscum album, Mistel *556.
Juglandiflorae.
- Juglans regia, Walnuß *557.
Piperinae.
- Piper Cubeba, Cubebenpfeffer *558.
- — nigrum *558.
Querciflorae.
- Betula verrucosa, Birke *559. 561.
- Quercus pedunculata, Stieleiche *561. 562.
- — sessiliflora, Traubeneiche 562.
- — infectoria 562.
- Fagus silvatica, Buche *560. 562.
[S. 656]
Saliciflorae.
Urticinae.
- Morus nigra, Maulbeere 565.
- Ficus elastica 565.
- Castilloa elastica 565.
- Cannabis sativa, Hanf 565.
- — sativa var. indica 565.
- Humulus Lupulus, Hopfen *565.
Rhoeadinae.
- Papaver somniferum, Mohn 567.
- — Rhoeas *566. 567.
- Brassica nigra, schwarzer Senf *568.
- Sinapis alba, weißer Senf 569.
Cistiflorae.
- Thea chinensis, Teestrauch *570.
- Garcinia Hanburyi, Guttibaum 570.
- Shorea Wiesneri, Dammarbaum 570.
- Viola tricolor, Stiefmütterchen 569.
Columniferae.
- Malva silvestris, Malve *571.
- — neglecta 570.
- Althaea officinalis, Eibisch *571.
- Gossypium herbaceum, Baumwolle *572. und andere Arten 572.
- Tilia ulmifolia, Winterlinde *572. 573.
- — platyphyllos, Sommerlinde 573.
- Theobroma Cacao, Kakaobaum *573. 574.
- Cola acuminata 574.
- Cola vera 574.
Tricoccae.
- + Mercurialis annua, Bingelkraut *574.
- Croton Tiglium 576.
- — Eluteria 576.
- Euphorbia resinifera *574. 576.
- Hevea-Arten, Parakautschukbäume 577.
- Ricinus communis, Wunderbaum *576. 577.
- Mallotus philippinensis 576.
- Manihot Glaziovii, Ceara Kautschukbaum 577.
Gruinales.
- Linum usitatissimum, Lein *577. 578.
- Erythroxylon Coca. Cocastrauch *577. 578.
- Guajacum officinale, Pockholz 578.
- Ruta graveolens, Raute *578.
- Citrus vulgaris, Pomeranze *578.
- — Limonum. Zitrone 580.
- — Bergamia, Bergamotte 580.
- Pilocarpus pennatifolius 581.
- — Jaborandi 581.
- Picrasma excelsa, Jamaïsches Bitterholz 581.
- Quassia amara, Surinam Bitterholz *579. 581.
- Simaruba amara 581.
- Commiphora abyssinica 581.
- — Schimperi 581.
- Boswellia Carteri 581.
- — Bhau Dajianae 581.
- Canarium-Arten 581.
- Polygala Senega, Senegastaude *579. 581.
Sapindinae.
- Pistacia Lentiscus, Mastixstrauch 581.
- + Rhus toxicodendron und andere Arten 581.
- Paullinia cupana 581.
Frangulinae.
- Rhamnus Frangula, Faulbaum 581. *582.
- — cathartica, Kreuzdorn 581. *582.
- — Purshiana 581.
- Vitis vinifera, Weinstock *582.
Ericinae.
- Arctostaphylos uva ursi, Bärentraube *583.
- Vaccinium Myrtillus, Heidelbeere 583.
- + Rhododendron-Arten 583.
- + Azalea-Arten 583.
- + Ledum palustre, Porst 583.
Diospyrinae.
- Palaquium-Arten *584.
- Payena-Arten 584.
- Styrax Benzoin 584.
- Mimusops 584.
Contortae.
- Fraxinus Ornus, Manna-Esche *586.
- Olea europaea, Ölbaum *585. 586.
- Strychnos nux vomica, Brechnuß *586. 587.
- Gelsemium nitidum 587.
- Gentiana lutea, gelber Enzian *586. 587.
- — pannonica 587.
- — punctata 587.
- — purpurea 587.
- Erythraea centaurium, Tausendgüldenkraut 589.
- Menyanthes trifoliata, Bitterklee 589.
- + Nerium Oleander, Oleander *587. 589.
- Strophanthus Kombé und Str. hispidus *588. 589.
- Kickxia elastica und andere Arten 589.
- Landolphia-Arten 589.
- Hancornia-Arten 589.
- Willoughbeia-Arten 589.
- Tabernaemontana Donnell Smithii 589.
- Aspidosperma Quebracho 589.
- + Vincetoxicum officinale *588. 589.
- Marsdenia Condurango 589.
Tubiflorae.
- Exogonium Purga *590.
- Convolvulus Scammonia 590.
- Galeopsis ochroleuca *590. 592.
- [S. 657]
Lavandula vera, Lavendel *591. 592.
- + Salvia officinalis, Salbei *591. 592.
- + Mentha piperita, Pfefferminze 592.
- + — crispa, Krauseminze 592.
- + Melissa officinalis, Melissa 592.
- + Thymus Serpyllum, Quendel 592.
- + — vulgaris, Thymian 592.
- + Rosmarinus officinalis, Rosmarin 592.
- + Origanum vulgare, Dost 592.
- + — Majorana, Majoran 592.
Personatae.
- Nicotiana Tabacum, Tabak 594. *595.
- + Lycopersicum esculentum, Tomate 593.
- Solanum Dulcamara, Bittersüß *592. 594.
- + — tuberosum, Kartoffel 593.
- + — nigrum, Nachtschatten 593.
- Hyoscyamus niger, Bilsenkraut 594. *596.
- Datura Stramonium, Stechapfel *594.
- Scopolia carniolica 594.
- Atropa Belladonna, Tollkirsche *593. 594.
- Capsicum annuum, Spanischer Pfeffer 594.
- Digitalis purpurea, Fingerhut 595. *597.
- Verbascum phlomoides, Wollkraut 595.
- — thapsiforme 595. *596.
Rubiinae.
- Uragoga Ipecacuanha, Brechwurzel 598. *601.
- Cinchona succirubra, Chinabaum 598. *599.
- — Ledgeriana 598.
- Ououparia Gambir 598.
- Sambucus nigra, Holunder 599.
- Valeriana officinalis, Baldrian 599. *601.
Synandrae.
- Lobelia inflata *603.
- Citrullus Colocynthis, Bittergurke *602.
- + Bryonia dioica, Zaunrübe 601.
- Arnica montana 606. *608.
- Artemisia Cina *607.
- Artemisia Absinthium, Wermut 607.
- Matricaria Chamomilla, Kamille *607. 608.
- Cnicus benedictus, Kardobenediktenkraut *606. 608.
- Tussilago Farfara, Huflattich *608.
- Achillea millefolium, Schafgarbe 608.
- Anthemis nobilis, röm. Kamille 608.
- Lappa vulgaris, Klette 608.
- Anacyclus Pyrethrum 608.
- Taraxacum officinale, Löwenzahn *605. 608.
- + Lactuca virosa, Giftlattich 605.
Spadiciflorae.
- Areca Catechu 614.
- + Arum maculatum, Aronstab *617.
- + Calla palustris 615.
- Acorus Calamus, Kalmus *616.
Liliiflorae.
- Veratrum album, Nießwurz 620.
- Schoenocaulon officinale 620.
- Colchicum autumnale, Herbstzeitlose *619. 620.
- Aloë-Arten *621.
- Urginea maritima, Meerzwiebel *620. 621.
- Smilax-Arten 621.
- + Paris quadrifolia, Einbeere 620. *622.
- Convallaria majalis, Maiglöckchen 621.
- Crocus sativus, Safran *623.
- Iris florentina, Schwertlilie 623.
- — germanica *624.
- — pallida 623.
Glumiflorae.
- + Lolium temulentum, Taumellolch 630. *631.
- Saccharum officinarum, Zuckerrohr 630.
- Agropyrum repens, Quecke 630.
- Triticum vulgare, Weizen *628. 630.
- Oryza sativa, Reis *629. 630.
Scitamineae.
- Zingiber officinale, Ingwer *632.
- Elettaria Cardamomun, Kardamome 632.
- Curcuma Zedoaria, Zittwer 633.
- Alpinia officinarum, Galgant 633.
- Maranta arundinacea, Arrowroot 633.
Gynandrae.
- Orchis-Arten, Knabenkraut *634. *635.
- Vanilla planifolia, Vanille *634. 635.
* vor den Seitenzahlen bedeutet Abbildung.
- Abies *516. 519.
- Abietineae 513. 515.
- Abkürzung der Ruhe 267.
- Abschlußgewebe 42 ff.
- Absorption im Boden 208.
- Abstoßung 292.
- Abstammungslehre 1. 176 ff.
- Acacia *102. *146. *538. *539.
- Acer, Aceraceae *92. *580.
- Acetabularia *359.
- Acetaldehyd 239.
- Achillea 606. 607.
- Achnanthes 347.
- Achlya *379.
- Achse 73.
- Achselknospen, -sprosse 106 ff.
- — Anschluß ihrer Blattstellung 108.
- Acidum agaricinum 409.
- Ackerwinde 589.
- Aconitum 532. *533.
- Acontae 348.
- Acorus *116. 276. *616.
- Acrocarpi 432.
- Acrocomia, Austrittsstelle d. Keimlings *508.
- Acrogynae 426.
- Acrostichum 440.
- Actaea 532.
- Actinomyces 395.
- Adern 95.
- Adiantum 268. *434. 444.
- Adlerfarn *57. 443.
- Adonis 316.
- Adossiertes Vorblatt 108.
- Adoxa 304.
- Adventiv-Keime 171.
- — -Knospen 166.
- — -Sprosse 107. 265.
- — -Wurzeln 199.
- Äcidiosporen 400.
- Äcidium 258. 400.
- Ährchen (Gramineae) *627.
- Ähre 477.
- Äpfelsäure 23. 240. 293.
- Äquationsteilung *174.
- Äquatorialplatte 18.
- Äquifazial 96.
- äquimolekular 191.
- Aërenchym 42. 142 ff.
- Aërobe, Aërobionten 238. 290. 293. 331.
- Aërotaxis 293.
- Aërotropismus 311.
- Aesculus 581.
- Ästivation 75.
- Aethalium 341.
- Äther 206. 267. 318.
- Ätherische Öle 25. 58. 61. 232.
- Aethusa *550.
- Ätzfiguren der Wurzeln 208.
- Äußere Bedingungen 185.
- — Faktoren 250.
- — Ursachen 185.
- Agar-Agar 220. 376.
- Agaricaceae 407.
- Agaricinum 409.
- Agaricus 409.
- — albus 409.
- Agathis 515.
- Agave 201. 242. 622.
- Aglaozonia 365.
- Agrimonia 537.
- Agropyrum 630.
- Agrostemma *553.
- Agrostis 629.
- Ahnenmerkmale 284.
- Ahorn *580.
- Aira 630.
- Aïzoaceae 551.
- Ajuga 592.
- Akropetal 66. 73.
- Aktinomorph 61. 186. *475.
- Aktives Leben 186. 266.
- Albizzia 186.
- Albugo 380. *381.
- Albumosen 231.
- Alchimilla 200. 277. *473. 537.
- Aldrovanda, Tierfang 160.
- Alectorolophus 596.
- Aleurodiscus 405.
- Aleuron *26. 230.
- Algen (Algae) 221. 257. 276. 309. 327.
- — Bauvariationen des Thallus 63 ff.
- — Gewebedifferenzierung 68 ff.
- Algenpilze 376.
- Alisma, Alismaceae 610.
- Alkalisalze 207.
- Alkaloide 12. 24. 58. 222. 232.
- Alkohol 206. 236. 238 ff.
- Allium *116 ff. 303. 618.
- Allogamie 170. 483.
- Allseitsempfindlich 313.
- Alnus 226. *478. *559.
- Aloë 122. *145. *621.
- Alopecurus 629.
- Alpenpflanzen 251. 253. 285.
- Alpenrose 583.
- Alpenveilchen *554.
- Alpinia 632.
- Alsophila 439. *440. *442.
- Alstroemeria 303. 622.
- Alter 272. 273.
- Alternieren 77.
- Althaea *571.
- Aluminium 206.
- Amanita 407. *408.
- Amarantus *119. 222. 267.
- Amaryllidaceae 622.
- Ameisensäure 220.
- Amicia *317.
- Amide 12. 24. 231.
- Amine 221.
- Aminosäuren 221. 231.
- Amitose 20.
- Ammoniacum 550.
- Ammoniak 187. 231. 240.
- — -Assimilation 222 ff.
- — -Bildung 187. 231.
- — -Düngung 209. 221.
- — -Oxydation 238.
- Ammonium bicarb. 210.
- Ammoniumphosphat 318.
- Ammoniumsulfat 209. 221.
- Amöben 337. 340.
- Amöboid 289.
- Ampelopsis 582.
- Amphibisch 180. 257. 285.
- Amphigastrien 426.
- Amphithecium 421.
- Amygdaleae 261. 537.
- Amylodextrin 28.
- Amylose 28.
- Amylum Tritici 630.
- — Oryzae 630.
- Anabaena 336. 445.
- Anacardiaceae 581.
- Anaërobe, Anaërobionten 238. 257. 290. 293. 331.
- Anagallis 555.
- Anacrogynae 426.
- Analog 7.
- Analogien im Bau der Kormuspflanzen 141.
- Ananassa 623.
- Anaphase 19.
- Anaptychia *414.
- Anastatica 295. 569.
- Anastomosen 96. 202.
- Anatomie 7.
- Anatrope Samenanlagen *464.
- Anchusa 590.
- Andira 544.
- Andreaea 420. *429.
- Andröceum 470.
- Androdiözie 470.
- Andromeda 583.
- Andromonözie 470.
- Andropogon 629.
- Aneimia *442.
- Anemone *531. 532.
- Anemophile Pflanzen 170. 478.
- Anethum 549.
- Aneura 426.
- Angiopteris 437.
- Angiospermen 523.
- — fossile 636.
- — sexuelle Generation 493 f.
- — Befruchtung 171. 497.
- Anhalonium 552.
- Anhomogen 323.
- Anis 550.
- Anisophyllie 101 ff.
- Anlagen 259. 277 ff.
- Anlockende Stoffe 292.
- Annuelle Pflanzen 121. 154 ff.
- Annularia *451.
- Annulus 296. 407. 440.
- Anomalien des Dickenwachstums 127 ff.
- — bei Lianen *156.
- Anonaceen 525.
- Anpassungen 3. 5. 141 ff. 257.
- — Entstehung 179 ff.
- Anpassungsmerkmale 6.
- Anschluß der Achselknospen 107 ff.
- Antennaria 605.
- Anthemis 606.
- Anthere 470.
- Antheridien *169. 329. 417. 433.
- Antheridium-Mutterzelle *486. *493.
- Anthoceros 417. 420. *422.
- Anthochlore 24.
- Anthophaein 24.
- Anthoxanthum 629.
- Anthozyane 24.
- Anthriscus 549.
- Anthurium 615.
- Anthyllis 543.
- Antidrome Grundspirale 108.
- Anti-Enzyme 229.
- Antigene 524.
- Antiklinen 40. 75. 269.
- Antikörper 258.
- Antipoden 495.
- Antirrhinum 287. 595.
- Apfelbaum 261. 266. 275. 535.
- Apfelsine 276. 580.
- Apikaler Vegetationspunkt 73.
- Apikales Wachstum 39. 64.
- Apikalöffnungen 99. 200.
- Apium 549.
- Aplanogameten 328.
- Aplanosporen 328.
- Apocynaceae 589.
- Apogamie 172, 443. *499.
- Apokarp 470.
- Apophyse 430.
- Aposporie 175. 443. *500.
- Apothecium 386. 412.
- Appositionswachstum 30. 248.
- Aprikose 261. 537.
- Aqua Laurocerasi 537.
- Aquifoliaceae 581.
- Aquilegia 532.
- Araceen 200. 235. 241. 246. *615.
- Arachis 244. 304. 544.
- Araliaceae 547.
- Araroba 544.
- Araucaria *489. 515.
- Archangelica 549. 550.
- Archegoniaten 327.
- Archegonium *169. 418. 433. 487.
- Archegoniumkammer 487.
- Archespor 420.
- Archidium 432.
- Archimycetes 377.
- Arctostaphylos 583.
- Arcyria *340.
- Areca 614.
- Arenga 614.
- Argemone 566.
- Arginin 231.
- Arillus 502.
- Ariopsis *218. 615.
- Aristolochia *82. *123 ff. *124. 528.
- — Clematitis, Bestäubungseinrichtung 484.
- — Stengelquerschnitt *82.
- Armilla 407.
- Armillaria 241. *396. *404. 408.
- Armleuchteralgen 369.
- Arnica 603. 604. 608.
- Arrhenatherum 629.
- Arrowroot 633.
- Arsenpilz 388.
- Artbildung 189. 288.
- Artemisia *607.
- Arten, elementare 285.
- Arten, Wesen derselben 176 ff.
- Artischocke 605.
- Artocarpus 564.
- Arum 616. *617.
- Arve 519.
- Asa foetida 550.
- Aschenbestandteile 187 ff. 203 ff. 220.
- Asclepias 588.
- Ascodesmis 385.
- Ascolichenes 412.
- Ascomycetes 384.
- Askogon 384.
- Askolichenen 412.
- Askomyceten 384.
- Askosporen *22. 385.
- Askus 386.
- Asparagin 201. 222. 231. 311.
- Asparagus 620.
- Aspergillaceae 388.
- Aspergillus 220. 385. *388.
- Asperula 598.
- Aspidistra 234.
- Aspidium 432. 439. 440. 441. 442. 444.
- Aspidosperma 589.
- Asplenium *434. *435. 440.
- — nidus 158.
- Assimilate 218 ff. 227 ff.
- Assimilation 212.
- — des Kohlenstoffs 212. *213 ff. 219 ff.
- — der Nährsalze 226.
- — des Stickstoffs 221 ff.
- Assimilationsparenchym 41. 98.
- Assimilationsstärke 15. 27. 217.
- Assimilatoren 68.
- Aster 605.
- Asterfigur 18.
- Asterocalamites *451.
- Astragalus *543.
- Asymmetrisch 63. *475.
- Atavismus 178. 284.
- Atemhöhle 44.
- Atemöffnungen *70.
- Atemwurzeln *143. 211. 301.
- Atmung 233 ff. *234. 236.
- — Chemismus 236.
- — intramolekulare 236.
- Atmungsenzyme 237.
- Atmungspigmente 237.
- Atmungsquotient 234.
- — fettreicher Samen 235.
- — von Sukkulenten 235.
- Atmungsversuch 234.
- Atriplex 555.
- Atropa *593.
- Atrope Samenanlagen *464.
- Atropin 594.
- Aubrietia 569.
- Auferstehungssterne 295.
- Aufnahme von Gasen 211.
- — — Nährsalzen 206.
- — — Wasser 189. 194.
- Auge s. Knospe.
- Augenfleck 17.
- Augen, schlafende 109.
- Augentrost (Euphrasia) 596.
- Auricularia 403.
- Ausbildung, innere 250.
- Ausdauernde Pflanzen 121. 151 ff.
- Ausläufer 165.
- Auslese 181.
- Auslösungen 184. 305. 322.
- Aussäung 171.
- Ausschleudern 295. *296. 564.
- Außenwelt 185.
- Austern, grüne 347.
- Austrocknung 186. 189.
- Autogam, Autogamie 170. 483.
- Autonome Bewegungen 296.
- Autonomie d. Merkmale 282.
- Autotroph 220. 223.
- Autotrophe Kormophyten, Bau 141 ff.
- Autotropismus 301. 315.
- Auxanometer *242.
- Auxiliarzellen 374.
- Auxosporen 345.
- Avena *27. 251. *254. 295. 629.
- Avicennia 591.
- Axillarstipeln 101.
- Azetylen 267.
- Azolla 445. 446.
- Azotobacter 224. 332.
- Azygosporen 383.
- Bacca 504.
- Bacillus *330, *331. *332. *333.
- — calfactor 241.
- — coli 241.
- — radicicola *224.
- Bacillariaceae 343.
- Bacteria 329.
- Bacterium 330. 332.
- — phosphoreum 241.
- Bärlapp 452.
- Bärlappgewächse 452.
- Bäume 121. 271.
- Bakterien *63. 208. 213. 215. 217. 219 ff. 241. 288. 290. 292. 329.
- — autotrophe 219.
- — stickstoffbindende 224.
- Bakterienknöllchen *224. 543.
- Bakteriochlorin 332.
- Bakteriopurpurin 332.
- Bakteriosen 332.
- Bakteroiden *224.
- Balata 584.
- Baldrian 599. *601.
- Balgfrucht 504.
- Balsamum Copaïvae 540.
- — peruvianum 544.
- — tolutanum 544.
- Bambus 242. 243. 244. 630.
- — -blüte 627.
- Banane 97. 243. 275. *631.
- Banyan 564.
- Barringtonia 506.
- Bartschia 163. 596.
- Bartflechte 413.
- Basen, organische 222.
- Basidien 396.
- Basidiobolus 383.
- Basidiolichenes 415.
- Basidiomyceten 396.
- Basidiosporen 396.
- — Entstehung 22.
- Basis 61.
- Bast 58. 125. 136 ff.
- Bastarde 278 ff. *279. *280. *281
- Bastardendosperm 278.
- Bastfasern 51. 136.
- Bastparenchym 136.
- Baststränge 136.
- Bastteil 58.
- Batrachium *102. 531.
- Batrachospermum 371. *373. 374. 376.
- Bauchpilze 409.
- Baumfarne 439. 442.
- Baumwolle 46. *47. 204. *572.
- Bazillus 330.
- Becherflechte 413.
- Becherrost 402.
- Beere 504.
- Befruchtung 165. 168 ff. 171 ff. 267. 275. *491. *497.
- Beggiatoa *334.
- Begonia 62. *246. *247. 248. 267. 308.
- Beiknospen, Beisprosse 106.
- Bennettitaceae 521.
- Bennettites 522.
- Benthos 64.
- Benzoë 584.
- Berberis 321. 526.
- Bereicherungssprosse 109.
- Bergmehl 348.
- Berle, Berula 549.
- Berührungsreize 256. 312.
- Bestäubung 170. 477.
- Beta 554.
- Betriebskraft 237.
- Betriebsstoffe 227.
- Betula *559.
- Betulin 140.
- Bewegung der Gase 212.
- — lokomotorische 289.
- — des Protoplasmas 10.
- — des Wassers 195.
- Bewegungserscheinungen 183. 288.
- Bibernell 549.
- Bicornes 583.
- Bidens 606.
- Biddulphia *346.
- Biegungsfeste Konstruktion *80
- Biegungsfestigkeit 52. 80 ff.
- Bienensprache 480.
- Bienne 121. 151.
- Bier 238.
- Bierhefe *22. 238. 394.
- Bifazial 96.
- Bignonia, Stammquerschnitt *156.
- Bikarbonate 217.
- Bikollaterale Leitbündel 87.
- Bilateral 62. 254. 268.
- Bildungsgewebe 39 ff.
- Bilsenkraut *596.
- Binse 618.
- Biogenetisches Grundgesetz 178.
- Biologie 1.
- Birke 198. 201. *559.
- Birkenharz 140.
- Birkenteer 561.
- Birkenwein 201.
- Birnbaum 261. 266. 275. 535.
- Bitterklee 587.
- Bitterstoffe 24.
- Bittersüß 592.
- Blätterschwämme 407.
- Blasenausscheidung *213.
- Blasentang 363.
- Blasia *69. 425. *426.
- Blatt 92 ff.
- — Assimilationsorgan 215.
- — Lebensdauer 104 ff.
- — Lichtstellung 310.
- — Transpirationsorgan 198.
- — Wachstum 244.
- Blattachsel 105.
- Blattaderung *95 ff. 524. *610.
- Blattanlage *73 ff. *92.
- Blattausgestaltung 93 ff.
- Blattdornen 148.
- Blattentwicklung 92 ff. 94.
- Blattfall 104. 150.
- Blattfolge 93.
- Blattgelenke *100. *298.
- Blattgrund 94. 100 ff.
- Blatthälftenmethode 219.
- Blattkissen 94. *100.
- Blattläuse 258.
- Blattmosaïk 100.
- Blattnarbe 104.
- Blattnervatur 95 ff. 434. *610.
- Blattpolster 94. *100. *298. 310. 317. 319.
- Blattprimordien 92.
- Blattquirle 76.
- Blattrand 94.
- Blattranken *155.
- Blattrippen 95 ff.
- Blattscheide 94. *102.
- Blattspitze 92.
- Blattspreite 94 ff.
- — Funktionen 99 ff.
- Blattspuren 83. *84 ff.
- Blattstellung *76 ff.
- Blattstiel 94. 100.
- — -ranker 289.
- Blattstruktur 96 ff.
- Blattsukkulenten 149.
- Blaugrüne Algen 335.
- Blauholz 136.
- Blausäure 537.
- Blechnum 440.
- Blendlinge 278.
- Blepharoplast 447. 451. 486.
- Blühreife 274.
- Blüte 167. *168.
- — Pteridophyten 449. 453.
- — Angiospermen 469.
- — Gymnospermen 468.
- Blütenachse 469.
- Blütenbecher 473.
- Blütenbestäubung 477.
- Blütenbildung 273.
- Blütenblätter *168.
- Blütenboden 473.
- Blütendiagramme *168. *474.
- Blütendüfte 480.
- Blütenfarbstoffe 24.
- Blütenformel 474.
- Blütenhülle 469.
- Blütennektarien 60. 473.
- Blüten-Ökologie 477.
- Blütensproß 167. *168. 262.
- Blütenstand 110.
- Blütensymmetrie *475.
- Bluten 201.
- Blutfarbige Laubblätter 24.
- Blutsverwandtschaft 2.
- Blutungsdruck *201. 202.
- Blutungssaft 201.
- Bocconia 566.
- Bockshornklee, Bockshornsamen 543.
- Boden, Aufschließung 208.
- — und Pflanzenverteilung 210.
- Bodenabsorption 208.
- Bodenbakterien 208. 224. 238. 340.
- Boden-Impfung 226.
- Bodenwurzel 112 ff.
- Boehmeria 566.
- Bohne *286. *298. 544.
- Boletus 384. *406.
- — cervinus 388.
- Borke 50. *139 ff.
- Borrago, Borretsch 272. 275. 590.
- Borstenhaare 46.
- Boswellia 581.
- Botanik, allgemeine 4.
- — angewandte 4.
- — reine 4.
- — spezielle 4.
- — theoretische 4.
- Botrychium *438.
- Botrydium *352.
- Botrys = Traube.
- Botrytis 243 ff.
- Boudiera 385. *392.
- Boussingaultia 271.
- Bovista 409.
- Bowenia 509.
- Bowiea 618.
- Brachsenkraut 458.
- Brätling 407.
- Brakteen 103.
- Brandpilze 397.
- Brandsporen 397.
- Branntwein 238.
- Brassica 187. *568. 569.
- Braunalgen 67 ff. 361.
- Braunkohle 215.
- Brennessel 566.
- Brennhaare *46 ff.
- Brenztraubensäure 239.
- Brettwurzeln 120.
- Briza 630.
- Brombeere 285. 537.
- Bromeliaceen 623.
- — als Epiphyten 158.
- Bromus 630.
- Bronveauxhybriden 262.
- Brotbaum 564.
- Brotschimmel 388.
- Bruguiera 545.
- Brunnenfaden 334.
- Brutbecher 424.
- Brutknospen 165. 273. 424.
- Brutkörper 165 ff. 424.
- Brutzwiebeln 165.
- Bryales 420. 429.
- Bryonia 284. 312. 601.
- Bryophyllum 248. 267. 534.
- — Adventivknospen auf Blättern 166.
- Bryophyten 70 ff. 415.
- Bryopsis 357. 360.
- Buche *97. *150. 198. 266. *560.
- Buchweizen 205. 556.
- Bündelscheide 89.
- Bulbillen *166.
- Bulbochaete 357. *358.
- Bulbus Scillae 621.
- Burseraceae 581.
- Butomus 610.
- Butterblume 532.
- Butterpilz 406.
- Buttersäurebakterien 239. 331.
-
- Cabomba 529.
- Cactaceae *149. 316. 552.
- Caeoma 403.
- Caesalpiniaceae 539.
- Cakile 569.
- Calabarbohne 544.
- Caladium 615.
- Calamagrostis 630.
- Calamariaceen 451.
- Calamites 451.
- Calamostachys *452.
- Calamus 614.
- Calendula 607.
- Calla 615.
- Callithamnion 371. *373.
- Callitris 519.
- Calluna 583.
- Calobryum 427.
- Caltha 532.
- Calycanthaceae 525.
- Calyptra 419.
- Calyptrosphaera *338.
- Calystegia 589.
- Campanula 230. 263. 270. *603.
- Campecheholz 136.
- Camphora 526. 570.
- Campylotrope Samenanlage *464.
- Canarium 581.
- Canna, Cannaceae *633.
- Cannabis 565.
- Cantharellus 407.
- Capillitium 340. 409.
- Capparis 568.
- Caprifoliaceae 599.
- Capsella 567.
- — Keim 497.
- Capsicum 594.
- Capsula 504.
- Cardamine 567.
- — Adventivknospen 166.
- Cardamome 632.
- Carduus 604.
- Carex 626.
- Carnivore Pflanzen s. Insektivoren.
- Carpinus 559.
- — Keimpflanze *115.
- Carpodinus 589.
- Carrageen 376.
- Carum *548.
- Caruncula 575.
- Caryophyllaceae 278. 317. 552.
- Caryophylli 547.
- Cascara sagrada 581.
- Cassave 576.
- Casparyscher Punkt (Streifen) *49. 116.
- Cassia 540.
- Cassytha 526.
- Castanea 566.
- Castilloa 565.
- Casuarina 226.
- Catechu 539. 598.
- Caulerpa 359. *360.
- Caules Dulcamarae 594.
- Cauliflor 573.
- Cecidien 258.
- Cecidomyia rosaria 258.
- Cedrate 580.
- Cedrus 517.
- Celtis 564.
- Centaurea *321. 605.
- Centranthus 599.
- Centricae 345.
- Centrospermae 551.
- Cephalanthera 634.
- Cephalotaceen 528.
- Cephalotus, Tierfang 160.
- Cephaëlis s. Uragoga *601.
- Ceramiaceae 373.
- Cerastium 553.
- Ceratiomyxa 340. 341.
- Ceratium *343.
- Ceratocorys *343.
- Ceratonia 540.
- Ceratozamia *511.
- Cerbera *506.
- Cercis *539. 540.
- Cereus *149. *552.
- Ceriops 545.
- Ceropegia, Milchröhre *58. 589.
- Cetraria *411. *412. 413. 415.
- Ceylonzimt 526.
- Chaerophyllum 550.
- Chaetocladium 383.
- Chalaza 464.
- Chalazogamie *496.
- Champignon 407.
- Chara 277. *369. *370. 371.
- Characeen 238, 290, 369.
- Charophyten 369.
- Chasmogam 485.
- Cheiranthus *567.
- Cheirostrobus 452.
- Chelidonium *287. 566.
- Chemische Reize 267. 311. 318.
- Chemonastie 318.
- Chemosynthese 219.
- Chemotaxis 292.
- Chemotropismus 311.
- Chenopodiaceae, Chenopodium 222. 317. 554.
- Chilisalpeter 209.
- Chimaere 262 ff. *263. *264.
- Chinarindenbaum 598. *599. *600.
- Chininum 598.
- Chiropterophil 482.
- Chitin 31 ff.
- Chlamydomonas 291. *353. 354.
- Chlamydosporen 396.
- Chlamydothrix 334.
- Chlor 187. 206.
- Chloralhydrat 206.
- Chloramoeba *351.
- Chlorella *355.
- Chlorococcum *355.
- Chlorophyceae 352.
- Chlorophyll 14 ff. *214. 215. 227. 252.
- Chlorophyllkörner, -körper *14. *20. 214. 291. *292.
- — mit Assimilationsstärke, in Teilung *20.
- Chloroplasten *14. 216.
- Chlorose 205. 227.
- Chlorosphaera *355.
- Chlorotisch 215.
- Choanephora 383.
- Choiromyces 393.
- Choleravibrio *333.
- Cholesterin 207.
- Cholin 222.
- Chondrioderma *10. 340. *341.
- Chondriosomen 13.
- Chondromyces *336.
- Chondrus 371. *372. 376.
- Chorda *364.
- Choripetalae 525.
- Chromatin 14. 17.
- Chromatophoren 9. 14.
- — Einschlüsse 27 ff.
- — Teilung *20.
- Chromoplasten *16.
- Chromosomen 17. *18. 276. 277. 285. 288.
- — Paare *19.
- Chromosome als Vererbungsträger 19. 285.
- — Reduktion 172 ff.
- — Verdoppelung 172.
- — Zahl 19. 172 ff.
- Chroococcus 335.
- Chroolepus 356.
- Chrysamoeba *337.
- Chrysarobinum 544.
- Chrysidella 338.
- Chrysomonadinen 337.
- Chrysophyta 338.
- Chrysophlyctis 378.
- Chrysosplenium 482.
- Chytridiaceae 377.
- Cibotium 444.
- Cichorie, Cichorium *605.
- Cicuta *549.
- Cimicifuga *529.
- Cinchona *599. *600.
- Cincinnus = Wickel.
- Cineraria 606.
- Cinnamomum 526.
- Circaea 546.
- Cirsium 278. 605.
- Cissus 313.
- Cistus, Cistiflorae 569.
- Citronat 580.
- Citrullus 602.
- Citrus *578. 580.
- Cladonia *413. 415.
- Cladophora *13. *14. *21. *66. 247. *359.
- Cladostephus *69. 361.
- Cladothrix *330. 334.
- Clavaria 405. *406.
- Claviceps *34. 389. *390.
- Clematis *35. *85. 532.
- Closterium *349.
- Clostridium 224. *332.
- Cnicus *607.
- Cobaea 187. 313.
- Cocaïn 578.
- Coccolithophoriden 338.
- Cocconeïs *347.
- Cochenillekultur 552.
- Cochlearia 186. 569.
- Cocos *613. *614.
- Coenobien 354.
- Coenogameten 377. 385.
- Coffea 598. *600.
- Cola 574.
- Colchicum 316. 618. *619.
- Coleochaete *358.
- Colletia 581.
- Collema 411. *414.
- Colocasia 200. 615.
- Columella 382. 420. 428.
- Columniferae 570.
- Commelinaceae 624.
- Commiphora 581.
- Compositae 230. 277. *316. *321. 604.
- Conferva 351. *352.
- Coniferae 278. 511.
- Conium *551.
- Conjugatae 348.
- Contortae 585.
- Convallaria *93. 620.
- Convolvulus 278. 589.
- Copaïfera 540.
- Copra 613.
- Coprinus 404. 405.
- Cora 412. *415.
- Corallinaceen 372.
- Corallorrhiza *163. 226. 634.
- Corchorus 572.
- Cordaites *521.
- Cordyline *123. 620.
- Corethron *345.
- Coriandrum *548. 550.
- Cornaceae, Cornus *547.
- Cornelkirsche 547.
- Coronilla 544.
- Corsinia *420.
- Cortex Cascarillae 576.
- — Chinae 598.
- — Cinnamomi zeylanici 526.
- — Condurango 589.
- — Frangulae 581.
- — Fruct. Aurant. 580.
- — Fruct. Citri 580.
- — Granati 547.
- — Mezereï 545.
- — Quebracho 589.
- — Quercus 562.
- — Quillajae 537.
- — Rhamni Purshianae 581.
- — Salicis 563.
- — Sassafras 526.
- — Simarubae 581.
- — Viburni 599.
- Corthusa 555.
- Corydalis 314. *567.
- Corylus 560.
- Coscinodiscus *346.
- Cosmarium *349.
- Crambe *567.
- Crassula 235. 534.
- Crataegomespilus 262.
- Crataegus *95. 262. 536.
- Craterellus 405.
- Crenothrix 334.
- Crepis 605.
- Cribraria *340. 342.
- Crocus 152 ff. 316. 623.
- Crossotheca 463.
- Croton 574.
- Cruciferae 567.
- Cryptomonas *338.
- Cryptospora *389.
- Cubebae *558.
- Cucumis 601.
- Cucurbita *30. *53. *54. *55. 313.
- — Ranken 601.
- Cucurbitaceae 600.
- Cuphea, Verschiebung der Achselknospe *106.
- Cupressus 513.
- Cupula *561.
- Cupuliferae 561.
- Curare 587.
- Curcuma 632.
- Cuscuta *162. 256. 315. 589.
- Cutleria *364. 365.
- Cyanophyceen 185. 221. 335.
- Cyathea 439. 442.
- Cyathium 575.
- Cycadaceae, Cycas 297. *509. *510.
- Cycadeen, Entwicklung 485.
- Cycadeoidea 522.
- Cycadinae 509.
- Cycadofilices 463.
- Cycadophyta 522.
- Cyclamen 554.
- Cydonia 535.
- Cylindrocystis *348.
- Cymöse Verzweigung 110 ff.
- — Blütenstände *112. 488.
- Cynara 605.
- Cynips 258.
- Cyperaceae, Cyperus 625.
- Cypresse 514.
- Cypripedium 634.
- Cystococcus *411.
- Cystodinium *342. 343.
- Cystopus 380.
- Cytisus 262. *263. 544.
- Dactylis 630.
- Dahlia *153. 606.
- Dammar 570.
- Daphne 544.
- Darlingtonia, Tierfang 160.
- Darwin, Ch. 1.
- Darwinismus 1. 181.
- Dattelpalme 614.
- Datura 594.
- Daucus *16. 549.
- Dauer des Wachstums 245.
- Dauergewebe 39. 40 ff. 250.
- — primäre, sekundäre 40.
- Dauerzellen 9.
- Dauerzustände 186.
- Dawsonia 421.
- Deckblatt 106. *107.
- Deckelkapsel *503.
- Deckschuppen in Winterknospen 151.
- Deckspelzen 626.
- Deformation 257.
- Degeneration 276.
- Dehnbarkeit der Sklerenchymfasern 52.
- Dehydrierung 237.
- Dekussierte Blattstellung *77.
- Delesseria *67. 371. 376.
- Delphinium 532.
- Dendrobium 505. 634.
- Denitrifikation 240. 332.
- Denken 323.
- Dentaria, Brutzwiebeln *166.
- Dermatogen 42. 74. 114.
- Desamidasen 231.
- Desmidiaceen 290. 348.
- Desmodium 298.
- Deszendenztheorie 1. 176 ff. 288.
- Determination 269.
- Deutzia 534.
- Dewargefäße 241.
- Dextrin, technisches 28.
- Dextrose 32. 230.
- Diageotropismus 302.
- Diagonal 108.
- Diagramm *76 ff. *107. *168. 474.
- Diakinese 173.
- Dianthus 278. 553.
- Diarch 117.
- Diastase 28. 229.
- Diasterfigur 18.
- Diatomeen *63. 186. 206. 290. 343.
- Dicentra 567.
- Dichasium *111. *112.
- Dichogamie 483.
- Dichotome Verzweigung 65.
- — bei Sproß *105.
- — bei Wurzeln 119.
- Dichotomie *65.
- — falsche 109.
- Dichotomosiphon 360.
- Dickenwachstum 120 ff. 248.
- — Anomalien *127. *156.
- — der Dikotylen 123 ff.
- — der Farnstämme 122 ff.
- — der Monokotylen 122 ff. *123. *125.
- — primäres, sekundäres 121 ff.
- — des Stammes 123 ff.
- — der Wurzel 123. 127. 156.
- Dickenzunahme, primäre, des Urmeristems 121.
- Dicksonia 439.
- Dicotylae 524.
- Dictamnus 578.
- Dictyonema 415.
- Dictyophora 243.
- Dictyota *65. *69. 361. 365. *366.
- Diervilla 599.
- Differenzierung 269.
- Diffuse Reize 315. 323.
- Diffusion 190. 208. 211. 232. 293.
- Digene Fortpflanzung 165. 276.
- Digestionsdrüsen *60. *159.
- Digitalis 274. *597.
- Dihybriden 283.
- Diklin 468.
- Dilatation 137.
- Dill 549.
- Dimorpha *339.
- Dimorphe Heterostylie 484.
- Dimorphotheca 607.
- Dinobryon *337.
- Dinoflagellatae 342.
- Diözisch 468.
- Dionaea *160. 318. 326.
- Dioon *487. 509.
- Dioscoreaceae 276.
- Diospyrinae 584.
- Diphtheriebazillus 333.
- Diploide Phase 172. 283. 329.
- Diplokaulisch 109.
- Diplococcus 333.
- Diplostemon 473.
- Dipsacaceae, Dipsacus 600. *601.
- Diptam 578.
- Dipterocarpaceae 570.
- Disaccharide 23.
- Dischidia Rafflesiana *157. 589.
- Discomycetes 390.
- Diskus 578.
- Dislokator 485.
- Dispirem 19.
- Dissimilation 233.
- Distel 605.
- Distephanus *338.
- Distomatinen 339.
- Divergenzwinkel 77.
- Doassansia 398.
- Dolde 476.
- Dominieren 280. 281.
- Doppelchromosomen 19. 173.
- Dorema 550.
- Dornen, Dornsproß *148.
- Doronicum 606.
- Dorsal 472.
- Dorsiventral 62. 254. 268. 302. 476.
- Dorsiventralität, ihre Ausbildung 65.
- Dorstenia 564.
- Dotterblume 532.
- Draba 285. 569.
- Dracaena *25. *123. 621.
- Drachenbäume 621.
- Dreiachsig 109.
- Drepanium = Sichel.
- Drimys, Holzbau 129. 525.
- Drosera, Tierfang 158 ff. *159. 528.
- — Chemonastie 319.
- Druck, negativer 203; osmotischer 190 ff.
- Druckfestigung 52. *117.
- Drüsen *60 ff. 61.
- Drüsenepithel 60.
- Drüsenflächen 60.
- Drüsengewebe 60.
- Drüsenhaare 48. *60.
- Drüsenschuppen *48. *60.
- Drüsenzellen 60.
- Drüsenzotten *48.
- Drupa 504.
- Drusen 25.
- Dryas 537.
- Dryobalanops 570.
- Dryopteris 432. 439. *441. *442. 444.
- Dudresnaya 374. *375.
- Düngung 209. 217.
- Dunkelheit *252. *254. 267. *292. *316. *317.
- Dunkelkeimer 267.
- Dunkelstarre 297. 320.
- Durchlässigkeit 191. 207.
- Durchlaßstreifen 89.
- Durchlaßzellen 49. 116. *118.
- Durchlüftung 212.
- Durchlüftungsgewebe 42.
- Durrha 629.
- Ebenaceae 584.
- Ebenenform 285.
- Ebenholz 135.
- Ecballium 602.
- Eccremocarpus 313.
- Echinocactus 552.
- Echinodermen 277.
- Echinodorus 611.
- Echinops 605.
- Echium 590.
- Ectocarpus 292. 361. *364. 365.
- Edelreis 260. *261.
- Edeltanne *516.
- Edelweiß 605.
- Efeu 102. 254. 274. 547.
- Egerling 407.
- Ei 169. 266. 292. 337.
- Eiapparat 494.
- Eibe *512. *513.
- Eibefruchtung 169.
- Eiche *139. 261. 266. 272. *561.
- Eichenmehltau 387.
- Eierschwamm 407.
- Eikern 171.
- Einachsig 108. 488. 491.
- Einbeere *622.
- Eingeschlechtige Blüten 468.
- Eingraben der Früchte und Samen 507.
- Einhäusig 468.
- Einheiten, systematische 176 ff.
- Einjährige Pflanzen 121. 154. 271.
- Einkorn 628.
- Einschlüsse der Chromatophoren 27 ff.; des Protoplasmas 23.
- Eingipsen 260.
- Einseitig empfindlich 313.
- Einseitiger Reiz 323.
- Einwirkungen, äußere 185. 258. 299.
- Eisbildung 185.
- Eisen 187. 205 ff. 227.
- Eisenbakterien 219. 236. 334.
- Eisenhut *533.
- Eisenmangel 205.
- Eiterkokken *333.
- Eiweiß, Abbau 220. 221. 230. 240.
- — Synthese 221.
- — Vergärung 240.
- Eiweißkörper 11. 26.
- Eiweißkristalle 16. *26. 230.
- Eiweißstoffe 11. 26. 201. 220. 227. 293. 311.
- Eizelle *169. 254. 266. 267. 277. 292. 329. 495.
- — diploide 172.
- Ektotroph 225.
- Elaeagnaceae 226. 545.
- Elaeïs 614.
- Elaphomyces 388.
- Elateren 420. 425. 449.
- Elektion d. Nährstoffe 220.
- Elektrischer Schlag 320.
- Elementarart 285.
- Elemente 188.
- Elemi 581.
- Elettaria, Samen *502. 632.
- Elfenbeinpalme *37. 230. 614.
- Elyna *626.
- Embryo 170. 171. 276. 492. *497.
- Embryonal 9. 245. 250.
- — Schläuche 492.
- Embryonale Zelle 9.
- Embryosack *22. *487. *490. *493. *495.
- — Entwicklungsabweichungen 495. 499.
- Embryosackkern, primärer 494.
- — sekundärer *495.
- Embryoträger 433. *488. *492. *497.
- Emergenzen *48. *159.
- Empfindlichkeit 293.
- Empfindung 323.
- Empusa *383.
- Emulsoid 12.
- Enantioblastae 623.
- Encephalartos 509.
- Endarch 90.
- Endemismen, progressive, Relikt- 178 ff.
- Endivie 605.
- Endocarpon 412.
- Endodermis *49.
- — des Stengels 81.
- Endospermie der Wurzel 116.
- Endogene Entstehung von Adventivknospen 107.
- Endogene Wurzelentstehung 119.
- Endokarp 503.
- Endophyllum *403.
- Endosperm 228. 230. 488. *502.
- Endospermkern 496.
- Endospor 435.
- Endosporen 166. 331.
- Endothecium 421. 470.
- Endotroph 226.
- Endprodukte des Stoffwechsels 23.
- Energie 233. 237.
- Engelsüß 439.
- Engelswurz 549.
- Engholz 131.
- Enteromorpha *356.
- Entfaltung 297.
- Entomophilie 480.
- Entomophthoraceae 383.
- Entwicklung 183. 242. 277.
- Entwicklungsbeginn 266.
- — -gang 265 ff.
- — -physiologie 7. 183. 242.
- Enzian s. Gentiana.
- Enzyme 12. 28. 32. 216. 220. 229 ff. 237.
- Ephebe 411.
- Ephedra 519. *520.
- Ephemerum 432.
- Epidermis *42 ff. 115. *246. *313.
- — des Blattes 97.
- — des Stengels 79.
- — mehrschichtige 97.
- — mehrschichtige als Wasserspeicher 149.
- — bei Xerophyten 144 ff.
- Epigäische Keimung 103. 508.
- Epigyn *473.
- Epikotyl *115.
- Epilobium 280. 545.
- Epinastie 297.
- Epipactis *20. 634.
- Epiphyllum 552.
- Epiphyten 157 ff. 195.
- Epipogon 163. 226. 634.
- Epithel 60.
- Epitheme 98 ff.
- Equisetinae 448.
- Equisetum *74. 254. 295. 448. *449. *450. *451.
- Eranthis 532.
- Erbse *155. 204. 257. 544.
- Erdbeere 166. 276. 278. 537.
- Erdbrot 415.
- Erdnuß 544.
- Erdsproß 73. 151 ff.
- Erdstern 409. 410.
- Erdwurzel 112 ff.
- Erepsin 231.
- Erfrieren 185.
- Erhaltungsfähigkeit 181.
- Ericaceae, Erica 583.
- Erigeron 605.
- Eriobotrya 536.
- Eriophorum *625.
- Erle *478. *559.
- Erneuerungsknospen 151.
- Erneuerungssproß 109.
- Ernolith 394.
- Erodium *295. 577.
- Erophila 569.
- Ersatzbildung 246.
- Ersatzfasern *128. 129.
- Erschütterung 319.
- Erstarkung 121. 271.
- Erstlinge 90.
- Erysibe (Erysiphe) *22. 386. 387.
- Erysipelkokken 333.
- Erythraea 587.
- Erythroxylum *577.
- Esche *586.
- Escholtzia 566.
- Essigbakterien 240. 331.
- Etagenkork 138.
- Etiolement 215. *252. 259.
- Eucalyptus 202. 546.
- Eucheuma 376.
- Eudorina 354.
- Euglena, Eugleninae 338. *339.
- Eumycetes 383.
- Euphorbia 258. 295. 575.
- Euphorbiaceae als Sukkulenten *149. 575.
- Euphorbium 576.
- Euphrasia als Halbschmarotzer 163. 596.
- Eurotium 388.
- Euryale 529.
- Eusporangiatae 437.
- Evonymus, Vegetationsscheitel *73.
- Exarch 90.
- Exine 416.
- Exkrete 227.
- Exkretion 200.
- Exoascus 393.
- Exobasidium *403.
- Exodermis 116.
- Exogen 73.
- Exogene Entstehung von Seitenknospen 105 ff.
- Exogonium *590.
- Exokarp 503.
- Exospor 435.
- Exosporen 166.
- Exothecium 470.
- Experiment 182.
- Extraaxilläre Seitenknospen 105.
- Extractum ferri pomati 537.
- Extranuptiale Nektarien 60.
- Extrors 470.
- Fadenbakterien 334.
- Fadenpilze 383.
- Fächel 111. *112.
- Färbungsverfahren 12.
- Fäulnis 240.
- Fäulnisbakterien 331.
- Fagopyrum *204. *504. 556.
- Fagus *97. *150. *560.
- Faktoren, äußere 250.
- — der Entwicklung 250.
- — innere 259.
- Fallverzögerung der Samen 505.
- Faltenparenchym *98.
- Farbhölzer 136.
- Farbkörper *14. *16.
- Farbstoffe in. Pflanzen 14 ff. 24. 232.
- — der Farbhölzer 33.
- Farne 252. 254. 276. 293. *296. 297. 309. 437.
- Farnpflanzen 432.
- Fasertracheïden *128.
- Faserschicht *468.
- Faszikularkambium 124.
- Faulbaum 581.
- Federharze 232.
- Fedia 599.
- Fegatella *417.
- Feige 275. *481. 564.
- Feigengallwespe 481.
- Feldspat 208.
- Fenchel *548. 550.
- Ferrozyankupfer 190.
- Ferula 550.
- Festigung des Blattes *97.
- — im Holz 128.
- Festigung der Pflanze 29. 51 ff. 121.
- Festigungsgewebe 51 ff.
- Festuca *627. 629.
- Fette 12. 25. 218. 220. 230. 235.
- Fettpflanzen 149.
- Fettsäure 222. 230.
- Feuerschwamm 407.
- Fibröse Schicht 470.
- Fibrovasalbündel 58.
- Fichte 204. *517.
- Fichtenzapfen. Blattstellung *78.
- Ficus *30. 277. 308. *481. 564.
- Filament *470.
- Filices 439.
- Filicinae 437.
- Fingerhut *597.
- Fixe Lichtlage 310.
- Fixierung 11. 12.
- Flachs vgl. Lein.
- Flachsprosse *147.
- Flächen minimae areae 269.
- Flächenstellung *292.
- Flächenwachstum 248.
- Flagellaria 314.
- Flagellata 337.
- Flaschenkork *8. *49. 139.
- Flavone 24.
- Flechten 208. 221. 226. 258. 327. 410.
- Flechtgewebe *34. 384.
- Fleischextrakt 292.
- Fleischfressende Pflanzen 158 ff. 222. 318.
- Fliegenschimmel 383.
- Fliegenschwamm *408.
- Flores Arnicae 607.
- — Chamomillae 607.
- — Ch. romanae 608.
- — Cinae 607.
- — Koso 537.
- — Lavandulae 592.
- — Malvae 570.
- — Rhoeados 567.
- — Rosae 537.
- — Sambuci 599.
- — Spiraeae 537.
- — Tiliae 573.
- — Verbasci 595.
- Florideen 314. 371.
- — „Stärke“ 29. 218. 372.
- Flügelsamen und -früchte *505.
- Flugblasen 468.
- Flughaare 505.
- Fluoreszenz des Chlorophylls 15
- Foeniculum *548. 550.
- Folgeblätter 102. 253. 270.
- Folgemeristeme 40.
- Folia Aconiti 532.
- — Adianti 404.
- — Althaeae 570.
- — Aurantii 580.
- — Belladonnae 594.
- — Coca 578.
- — Digitalis 595.
- — Eucalypti 547.
- — Farfarae 607.
- — Hamamelidis 533.
- — Hyoscyami 594.
- — Jaborandi 581.
- — Juglandis 557.
- — Malvae 570.
- — Melissae 592.
- — Menthae crispae 598.
- — M. piperitae 592.
- — Nicotianae 594.
- — Rosmarini 592.
- — Rubi fruticosi 537.
- — Salviae 592.
- — Sennae 540.
- — Stramonii 594.
- — Taraxaci 608.
- — Theae 570.
- — Trifolii fibrini 589.
- — Uvae ursi 583.
- Fomes *407. 409.
- Fontinalis 432.
- Formaldehyd 213.
- Formative Wirkung 250. 252.
- Formwechsel 183.
- Forsythia 585.
- Fortpflanzung 62. 163 ff. 183. 272 ff. 282.
- Fortpflanzungsorgane 62. 163.
- Fortschleudern der Samen u. Sporen 295. 564.
- Fovea 459.
- Fragaria 165. 537.
- — monophylla 287.
- Fragmentation 20.
- Fraktionierte Endospermbildung 498.
- Frangulinae 581.
- Frauenhaar 444.
- Frauenschuh 634.
- Fraxinus *586.
- Freie Kernteilung 21.
- Freie Zellbildung *22.
- Fremdbestäubung 170. 286. 483.
- Freycinetia 482. 612.
- Fritillaria 620.
- Froschlaichpilz 332.
- Frost 267.
- Frucht 171. 187. 232. 275. *502.
- Fruchtbarkeit 280.
- Fruchtblätter 167. 464.
- Fruchtfarben 24.
- Fruchtknoten 167. 470.
- Fruchtkörper von Pilzen 170.
- Fruchtstand 504.
- Fructus Anisi 550.
- — Anisi stellati 525.
- — Aurantii immaturi 580.
- — Cannabis 565.
- — Capsici 594.
- — Cardamomi 632.
- — Carvi 550.
- — Cassiae fistulae 540.
- — Colocynthidis 602.
- — Conii 550.
- — Coriandri 550.
- — Foeniculi 550.
- — Juniperi 519.
- — Lauri 526.
- — Myrtilli 583.
- — Papaveris 567.
- — Petroselini 550.
- — Piperis nigri 558.
- — Rhamni cathart. 581.
- — Sennae 540.
- — Vanillae 635.
- Frühholz 131.
- Frühjahrspflanzen 251.
- Frühlingsholz 131.
- Frühtreiberei 267.
- Frullania *426.
- Fucaceae 367.
- Fuchsia 247. 545.
- Fucosan 363.
- Fucoxanthin 362.
- Fucus *362. 363. *367. *368.
- — amylaceus 376.
- Fühl-Borsten, -Haare, -Papillen, -Tüpfel 48.
- Füllungen *80.
- Füllzellen 51.
- Fuligo 289. 341.
- Fumaria 314. 567.
- Funaria *14. *20. *416. *418. *420. *421. 430. 432.
- Fungi 327.
- — imperfecti 386.
- Fungus Chirurgorum 409.
- — Laricis 409.
- Funiculus 464.
- Funkia, Adventivkeime *499.
- Fusionen 53.
- Fußstück der Haare 46.
- Futtergräser 629.
- Gabelung *65.
- — beim Kormus *105.
- Gärung 233. 238. 241. 394.
- Gärungsbakterien 331.
- Gaisblatt 303. 599.
- Galanthus 612.
- Galbanum 550.
- Galeopsis *592.
- Galgant 633.
- Galium 101. 598.
- Gallae 562.
- Gallen 257.
- Gallenröhrling 407.
- Gallmücken 258.
- Gallwespen 258.
- Galtonia 618.
- — Kernplatte *19.
- Galvanotaxis 299.
- Galvanotropismus 312.
- Gambir 598.
- Gametangien 168. 328.
- Gameten 168 ff. 328.
- Gametophyt 72. 175. 329.
- Garcinia 570.
- Gasblasenmethode *213.
- Gasdiffusionsmethode 199.
- Gase 188. 210 ff. 311.
- Gasteromyceten 409.
- Gasvacuolen 336.
- Geaster *409. 410.
- Gefäßbündel 58.
- Gefäße *55 ff. *88. 201.
- — im Holz *128.
- Gefäßglieder 55.
- Gefäßpflanzen 326
- Gefäßstränge, Bau 58 ff. 87.
- — im Dikotylenholz 133.
- Gefäßteil 58. 84.
- Gegenfüßlerinnen 494.
- Gehilfinnen 494.
- Geißelbewegung 11. 64.
- Geißeln 11. 64. 166. *169. 289.
- Geitonogamie 170.
- Geleitzellen *54. 87 ff. 136.
- Gelenke *100. 297. *298. 310. *317. *319.
- Gel 189.
- Gelatine 220.
- Gelidium 376.
- Gelsemium 587.
- Gemini 173.
- Gene 277 ff. 285.
- Generatio spontanea 3.
- Generation 175.
- Generationswechsel 175. 329.
- — Übersicht 467.
- — und Kernphasenwechsel 175.
- Generative Kerne 493. *489.
- — Zellen *485.
- Genista 543.
- Gentiana, Gentianaceae 210. 587.
- Geograph. Verbreitung 210.
- Geophil 151.
- Geophyten 151 ff.
- Georgine *153. 606.
- Geotropismus 299 ff. *301. 302.
- Geradzeilen 77.
- Geranium 295. *577.
- Gerbstoffe 24. 32. 58. 207. 232.
- — in Borke 140.
- — im Kernholz 135.
- Gerinnungsmittel 11.
- Gerste *114. *628.
- Geschlechtsbestimmung 275. 284.
- — -organe 168.
- — -verlust 165.
- — -zellen 165.
- männliche () 168.
- weibliche () 168.
- Geschwindigkeit des Wachstums 244.
- Gesteinlösung 208.
- Gesetz des Minimums 217.
- Getreidebrand 397.
- Getreiderost 402.
- Geum 278. 537.
- Gewebe 33 ff.
- — -Arten 38 ff.
- — -Differenzierung 39.
- — -Entstehung 34.
- — primäre 40.
- — sekundäre 40. 122. 128 ff.
- Gewebelehre 7. 33.
- Gewebemutterzellen 40.
- — im Kambium 125.
- Gewebespannung 248. *249.
- Gewebesysteme 38.
- Gewürznelken 546.
- Gießkannenschimmel *166. 388.
- Gifte 185. 232. 254. 257. 288. 297. 318.
- Giftlattich s. Lactuca virosa 605.
- Giftreizker 408.
- Gigartina 371. *372. 376.
- Gigasformen 593.
- Ginkgo, Ginkgoinae *511.
- Gladiolus 623.
- Glandulae Lupuli 565.
- Glanzlichter 155.
- Glaucium *566.
- Gleba 409.
- Glechoma 275. 592.
- Gleditschia *148. 540.
- Gleicheniaceae 442.
- Gleitendes Wachstum 40. 125. *128.
- Glieder der Gefäße 55.
- — eines Wirtels 76.
- Glimmer 208.
- Globoide *26. 230.
- Globuline 26.
- Glochiden 447.
- Glockenblume *603.
- Gloeocapsa *34. 335.
- Gloriosa 314. 618.
- Gluma 626.
- Glumiflorae 625.
- Glutamin 231.
- Glycine s. Wistaria 544.
- Glycyrrhiza 544.
- Glykogen 23 ff. 384.
- Glykokoll 221.
- Glykose 23. 237. 248.
- Glykoside 12. 24. 232.
- Glyzerin 220. 222. 230.
- Gnadenkraut s. Gratiola 595.
- Gnaphalium 605.
- Gnetinae, Gnetum 519. *520.
- Goldfussia 321.
- Goldlack *567.
- Goldregen 543.
- Gonimoblast 374.
- Gossypium *572.
- Gracilaria 376.
- Gräser 200. 206. 243. 249. 275.
- Gramineae *102. 626.
- Grana 17.
- Granatbaum *546.
- Granne 626.
- Graphis 413.
- Grasknoten *102.
- — geotropisch *302.
- Gratiola *595.
- Grenzhäutchen d. Zellhaut 35.
- Grenzwert, plasmolytischer 193.
- Griffel *472.
- Griffithia 376.
- Größe von Pflanzen 202. 245. 254. 268. 275.
- Grünalgen 352.
- Grünling 407.
- Gruinales 577.
- Grundform 6. 72.
- Grundgesetz, biogenetisches 178.
- Grundgewebesystem 41.
- Grundorgane des Kormus 72.
- Grundspirale 78.
- — antidrome 108.
- — homodrome 108.
- Grundstoffe 187.
- Guajacum 578.
- Guano 209.
- Guarana 581.
- Guajava 546.
- Gummi 33. 58.
- — in Kernholz 135.
- — in Winterknospen 151.
- — in Wundholz 140.
- Gummi arabicum 539.
- Gummigänge 61.
- Gummigutt 570.
- Gummiharze 58. 232.
- Gummosis 33. 140.
- Gurke 601.
- — parthenokarp 275. 601.
- Gurtungen *80.
- Guttapercha 58 ff. 232. 577. 584. 589.
- Guttation 144. *200.
- Gutti 570.
- Guttiferae 570.
- Gymnadenia 634.
- Gymnodiniaceae 343.
- Gymnospermae 509.
- — fossile 519.
- Gymnospermenblüte *468. 509 ff.
- Gymnosporangium 400.
- Gymnostomum 272.
- Gynäceum 470.
- Gynandrae 633.
- Gynodiözisch 470.
- Gynomonözisch 470.
- Gynophor 569.
- Gynostemium 276. 528. *634.
- Gyromitra 392.
- Gyrophora 415.
- Haare *46 ff. 145.
- Haastia 605.
- Habichtsschwamm 406.
- Habitus 108.
- Hadrom 58.
- Haematococcus *353.
- Haematochrom 17. 353.
- Haematoxylin 136.
- Haematoxylon 540.
- Haemoglobin 273.
- Hafer *27. 629.
- Haftdruck 207.
- Haftorgane 64.
- Haftscheiben 64.
- — an Ranken *156. 314.
- Haftwurzeln bei Epiphyten 158.
- Hagenia *536.
- Hahnenkamm 405.
- Hainbuche *559.
- Halbschmarotzer 163.
- Halimeda 359.
- Hallimasch 408.
- Halophyten 144. *147. 200. 206.
- Halosphaera 351.
- Hamamelidinae 532.
- Hancornia 589.
- Hanf 565.
- Haplobacteria 331.
- Haploide Phase 172. 337.
- Haplokaulisch 108.
- Haplomitrium *426. 427.
- Haplostemon 473.
- Hapteren 64.
- Haptotropismus 312.
- Harnstoff 210. 231.
- Hartbovist *409.
- Hartlaubgewächse 145.
- Hartschaligkeit 267.
- Harveyella 376.
- Harz 25. 58. 61. 129. 151. 232.
- Harzdrüsen 61. 129. 151.
- Harzgänge 61. *98. *130.
- Harzkanäle *130.
- Haschisch 565.
- Haselstrauch 560.
- Hauhechel 543.
- Hauptachse *65. 66. 110 ff.
- Hauptreihe b. wechselständ. Blattstellungen 79.
- Hauptwurzel 120.
- Hausschwamm 407.
- Haustorien *67. *162. 163. 223.
- Hautgelenke 45.
- Hautgewebesystem 42.
- Hautschicht 12. 207.
- Hedera 102. 156. 547.
- Hedychium 632.
- Hefepilze 238. 394.
- Hefesprossung 394.
- Hefezellen *22.
- Heidekraut 583.
- Heidelbeere 583.
- Heilung 256.
- Helianthemum *569.
- Helianthus 218. 249. 308. *497. *605.
- Helichrysum 605.
- Heliotropismus s. Phototropismus.
- Heliotropium *477.
- Helleborus *44. *103. *197. 532.
- Helobiae 611.
- Helodea *213. 612.
- Helvellaceen 392.
- Helwingia, Blattständige Blüten *106.
- Hemicellulosen 31 ff.
- Hepaticae 422.
- Herba Absinthii 607.
- — Adonidis 532.
- — Cannabis 565.
- — Capilli Veneris 444.
- — Cardui benedicti 607.
- — Centaurii 587.
- — Chenopodii 555.
- — Conii 550.
- — Convallariae 621.
- — Equiseti 451.
- — Galeopsidis 592.
- — Herniariae 554.
- — Lobeliae 604.
- — Majoranae 592.
- — Meliloti 544.
- — Millefolii 608.
- — Origani 592.
- — Polygoni 556.
- — Rutae 581.
- — Sabinae 519.
- — Serpylli 592.
- — Thymi 592.
- — Violae tricoloris 569.
- Herbstfärbung der Laubblätter 16. 24. 232.
- Herbstzeitlose *618. Knolle 152 ff.
- Herkogamie 484.
- Hermaphrodit 469.
- Herniaria 554.
- Heterocontae 351.
- Heterogamie 168. 328. 605.
- Heteromer 411.
- Heterophyllie 101 ff. *102. 270.
- Heterospor 435.
- Heterostylie 484.
- Heterothallisch 382. 405.
- Heterotroph 219.
- Heterotrophe Kormophyten, Bau 161 ff.
- Heterozygoten 278.
- Heterozysten 336.
- Heu, Selbsterhitzung 241.
- Heubazillus 331.
- Hevea 577.
- Hexenbesen 393.
- Hexenei 410.
- Hexenkraut 545.
- Hexenmehl 456.
- Hexenringe 405.
- Hibernakeln 166.
- Hibiscus 570.
- Hieracium 278. 280. *500. 605.
- Hilum 500.
- Himbeere 537.
- Hinterhof 44.
- Hippocastanaceae 581.
- Hippophaë 545.
- Hippuris *74. 257.
- Hirschtrüffel 388.
- Hirschzunge 439.
- Hirse 629.
- Histoid 258.
- Histologie 7. 33.
- Hitzetod 185.
- Hochblätter 93. *103. 270.
- Hochgebirgspflanzen 144. 285.
- Hoftüpfel 56. *57.
- Holcus 630.
- Holunder 599.
- Holunderschwamm 403.
- Holz 58. 125. 128 ff. 217.
- Holzbau bei Dikotylen 132 ff.
- — bei Gymnospermen 129.
- Holzelemente *128.
- Holzfarbstoffe 136.
- Holzfaser *128.
- Holzfaserstränge im Dikotylenholz 134.
- Holzgewächse 121.
- — immergrüne 104. 150.
- — sommergrüne 104. 150.
- — als Tropophyten 150.
- Holzparenchym *128.
- — Anordnung im Dikotylenholz 133 ff.
- — im Gymnospermenholz 129.
- Holzstoff 32.
- Holzteil 58.
- Homodrome Grundspirale 108.
- Homoeomer 411.
- Homogener Reiz 323.
- Homolog 7.
- Homologien im Bau der Kormusorgane 141.
- Homospor 435.
- Homothallisch 383. 405.
- Homozygoten 278.
- Honigtau 389.
- Hoodia 589.
- Hopfen *60. 303. 565.
- Hordeum *114. 628.
- Hormogonien 336.
- Hornea 458.
- Hornklee 543.
- Hortensie 534.
- Hoya 589.
- Huflattich 606. *608.
- Hüllspelzen 626.
- Hülse 504.
- Humulus *60. *565.
- Humussammelnde Epiphyten 158.
- Hundspetersilie *550.
- Hungerzustand 241.
- Hyacinthus 618.
- Hyaloplasma 12.
- Hybride 278.
- Hydathoden 60. 144. 200.
- Hydnophytum 598.
- Hydnum *406.
- Hydrangea 534.
- Hydrastin 532.
- Hydrastis 534.
- Hydrierung 237.
- Hydrocharis 612.
- Hydrodictyon 356.
- Hydrolapathum 371.
- Hydrolyse 228 ff.
- Hydrophil 170. 479.
- Hydrophyten 141 ff.
- Hydropteriden 444.
- Hydrotaxis 293.
- Hydrotropismus 311.
- Hydrurus 337. *338.
- Hygrochasie 295.
- Hygrometer 296.
- Hygromorphie 143.
- Hygrophil 143.
- Hygrophyten 143 ff. 159.
- Hygroskopische Bewegungen 294. 295 ff.
- Hymenium 385. 403.
- Hymenogastreen 409.
- Hymenolichenen 415.
- Hymenomyceten 403.
- Hymenophyllum 442.
- — Wasseraufnahme durch den Sproß 143.
- Hyoscyamus *596.
- Hypanthium 473.
- Hypericum 570.
- Hyphaene 612.
- Hyphen *13. *67. 376. 384.
- Hypholoma *396.
- Hypnaceae 432.
- Hypnodinium 343.
- Hypochnus 405.
- Hypogäische Keimung 103. 508.
- Hypogyn *473.
- Hypokotyl 93. *115.
- Hyponastie 297.
- Hypophyse *498.
- Iberis 569.
- Idioblasten 38.
- Ilex *580.
- Illicium 525.
- Immergrün 586.
- Immortellen 605.
- Immunität 258.
- Impatiens *96. 198.
- Imperatoria 550.
- Indigkarmin 240.
- Individualität der Chromosomen 19.
- Indol 240.
- Indusium 440.
- induziert 298 ff.
- Infloreszenz 110.
- Influenzabazillus 333.
- Infiltrationsmethode 198.
- Ingwer 632.
- Initialschichten im Kambium 123.
- — zellen der Haare 46, im Kambium 39. 40.
- Innere Ursachen 185. 250. 259. 266.
- Innovationssproß 109.
- Insekten-Bestäubung 480.
- — als Gallbildner 258.
- Insektenblütige 480.
- Insektenpulver 608.
- Insektivoren *159 ff. 200. 222. 318. 528.
- Integumente 464.
- Interfaszikularkambium 124.
- Interkalares Wachstum 40. 64. 76. 244.
- Intermediär 280.
- Internodien 76.
- Interpetiolarstipeln 102.
- Interzellularräume *35. *36. 37. 211.
- Intine 416. 465.
- Intramolekular 236.
- Intrors 470.
- Intumeszenz 256.
- Intussuszeptionswachstum 29. 31. 248.
- Inula 605.
- Inulin 23. 228. 230.
- Inversstellung 255.
- Invertin 230.
- Ionen 205.
- Ipomoea 589.
- Iridaceae, Iris 278. 622. *624.
- — Diagramm 474.
- Irländisches Moos 376.
- Isatis 569.
- Isländisches Moos 413.
- Isocontae 353.
- Isoëtes 189. 458. *459.
- Isogamie 168. 328.
- Isolateral 96.
- Isosmotisch 191.
- Isospor 435.
- Ithyphallus *410.
- Jahresperiode 149.
- Jahresringe 130. *132. 202.
- Jambosa *546.
- Jasione 604.
- Jasminum 585.
- Jatrorrhiza 526.
- Jerichorose 295.
- Jod, in Meeresalgen 206. 207. 368.
- Jodprobe *218.
- Johannisbeere *535.
- Johannisbrotbaum 540.
- Judasohr 403.
- Jugendblätter 102. 270.
- Juglandiflorae, Juglans 556. *557.
- Juncaceae, Juncus *618.
- Jungermannia 426.
- Jungermanniales 417. 420. 425.
- Juniperus *84. *514.
- Jute 572.
-
- Kälberkropf 550.
- Kälte-Starre 290. 297. 320.
- — -Perioden 266.
- — Widerstandsfähigkeit 186.
- Käse 239.
- Kaffee 186. *600.
- Kaffernhirse s. Mohrhirse 629.
- Kahmhaut 331.
- Kaiserkrone 620.
- Kaiserling 407.
- Kakao 186. *573.
- Kalabarbohne 544.
- Kalium 187. 205. 209. 223. 227. 232.
- Kalkalgen 359. 372.
- Kalk, Giftwirkung 207. 210.
- Kalkschüppchen 200.
- Kallose 55.
- Kallus 53. 246. 261.
- — im Bast 136.
- — Wundgewebe 140.
- Kallusplatten *54.
- Kalmus 276. *616.
- Kalyptra *113 ff. 429.
- Kalyptrogen 114.
- Kalzium 187. 205 ff. 227.
- Kalziumcyanamid 210.
- Kalziumkarbonat 32.
- — im Kernholz 135.
- — in Membranen 33.
- — im Sekret von Wasserspalten 99.
- Kalziummalat 59.
- Kalziumoxalat *25. 32. 238.
- Kalziumoxalatkristalle 25. 26.
- — in Membranen 33.
- Kamala 576.
- Kambium 40. 122 ff. 302. 309.
- — faszikulares 124.
- — im Gymnosperm. u. Dikotylenstamm 123 ff.
- — interfaszikulares 123.
- — in offenen Leitbündeln 87.
- — in Monokotylenwurzeln 123.
- — Periodizität 130.
- — beim sekundären Zuwachs 122.
- Kambiumbildung in Gymnospermen- und Dikotylenstengeln 123 ff.
- — in Monokotylenstengeln 122.
- — wiederholte in Stämmen u. Wurzeln 127.
- Kambiummantel bei Dikotylen und Gymnospermen im Stamm 123 ff.
- — bei Monokotylen 122.
- — desgl. Wurzel 127.
- — Typen seiner Entstehung 126.
- Kambiumzellen *125.
- Kamelie 570.
- Kamille 606.
- Kampf ums Dasein 181.
- Kampfer 526. 570.
- Kampylotrop *464.
- Kandelia 545.
- Kannenpflanzen, Tierfang 160.
- Kantenkollenchym 53.
- Kappern 569.
- Kapsel 504.
- Kapuzinerkresse *16. *99.
- Karde 600.
- Kardinalpunkte 186. 217. 251.
- Karnallit 210.
- Karnivore 158 ff. 222.
- Karotine 15.
- Karotte 549.
- Karpell 470.
- Karpogon 374. 384. 414.
- Karpophor 548.
- Karposporen 373. 374.
- Kartoffel *27. *152. 204. 228. 270. 593.
- — Krebs 378.
- — Schwarzbeinigkeit 332.
- — Pilz 380.
- — Schorf 395.
- Karyokinese 17. *18.
- Karyosomkerne 14.
- Kastanie 561.
- Katalysator 229.
- Kätzchen 558.
- Kaudikula 634.
- Kausal 182.
- Kautschuk 59. 232. 565. 577. 589.
- Kefir 239.
- Keilblattgewächse 452.
- Keim 164. 277.
- Keimblätter 93. 102 ff. 498.
- Keimkern *488.
- Keimlinge 164.
- Keimpflanze *115. 507.
- Keimung 103. 164. 171. 267.
- Keimzelle 164.
- Kelch 470.
- Kelchblätter 167. 470.
- Kelp 368.
- Kerbel 549.
- Kern *9.
- Kerngerüst 14.
- Kerngröße 13.
- Kerngummi 135.
- Kernhöhle 14.
- Kernholz 135 ff. 202. 272.
- Kernkörperchen 14.
- Kernphasenwechsel 172.
- — und Generationswechsel 175.
- Kernpilze 388.
- Kernplatte 18. *19.
- Kernsaft 14.
- Kernspindel 18.
- Kernteilung 17 ff. *18. 268.
- — Äquationsteilung 174.
- — direkte, amitotische 20.
- — freie 21.
- — heterotypische 172.
- — homöotypische 174.
- — indirekte, mitotische 17. *18.
- — typische 17. *18. 172.
- Kernwand 14.
- Kickxia 589.
- Kiefer *57. *98. *130 ff. *518. 519.
- Kieselalgen 343.
- Kieselgur 206. 348.
- Kieselkörper 26.
- Kieselsäure 32. 207.
- — Einlagerung in Epidermis 43.
- — im Kernholz 136.
- — in Membranen 33.
- Kino 544.
- Kirsche 274. *537.
- Kirschgummi 33. 140.
- Kirschlorbeer 537.
- Kladodien *147.
- Klappertopf s. Alectorolophus.
- Klausen 590.
- Kleber *26.
- Klee 543.
- Kleeseide s. Cuscuta 589.
- Kleie 26.
- Kleinarten 176.
- Kleinkörner 28.
- Kleistogame Blüten 274. 485.
- Klemmkörperchen (Asclepiadaceen) *588.
- Kletterhaken 314.
- Kletterpflanzen 155 ff. 312 ff.
- Kletterwurzeln 156. 254. 308.
- Klimaperiodizität 149 ff.
- Klimate, trockene 149 ff.
- — wechselfeuchte 150.
- Klimmhaare 156.
- Klinostat 301. 309.
- Knautia 600.
- KNIGHTS geotropische Versuche 300.
- Knoblauch 276. Vgl. Lauch.
- Knöterich (Polygonum) 555.
- Knollen 151 ff. 228. 270.
- Knollenblätterschwamm *408.
- KNOPsche Nährlösung 205.
- Knospen 75. 266.
- — schlafende 109.
- Knospendeckung 75.
- Knospenentfaltung 297.
- Knospenlage *75.
- Knospenschuppen *93. *150.
- — in Winterknospen *150 ff.
- Knoten 76. *302.
- Kobaltprobe 195.
- Kochsalz 207. 210.
- Königsfarn 442.
- Köpfchen 477.
- Köpfchenhaare 60.
- Körnerplasma 12.
- Kohäsionsmechanismen *296. 426. 428. 440. 449. 456.
- Kohäsionstheorie 203.
- Kohl 198. 569.
- Kohle 204. 215.
- Kohlehydrate 12. 23. 201. 218. 220. 228. 240.
- Kohlenoxyd 216.
- Kohlensäure 187. 240.
- — Assimilation 212. 219.
- — Aufnahme 210.
- — Bildung 233 ff. 238 ff.
- — in der Luft 216.
- — im Wasser 217.
- Kohlenstoff 187. 188.
- Kohlenstoffassimilation der autotrophen Pflanze 212.
- — gewisser Bakterien 219.
- — und Blütenbildung 273.
- — der Heterotrophen 219.
- Kohlenwasserstoff 220.
- Kohlhernie 342.
- Kohlrabi, Knolle 153.
- Kokken 330.
- Kokos *613. *614.
- Kolanuß 574.
- Kolben 512.
- Kollaterale Beiknospen *106.
- — Leitbündel *88 ff. *90. *124.
- Kollenchym 51. *53.
- Kolleteren 151.
- Kolloidale Lösung 12. 189.
- Kolonbazillus *333.
- Kombinationen 285 ff.
- Kompaßpflanzen 145. 310.
- Kompensationen 260.
- Konidien *166. 383. 386. 396.
- — Entstehung 22.
- Koniferen 511.
- Konjugaten 348.
- Konnektiv 470.
- Konsortium 410.
- Konstanz der Arten 176.
- Kontaktreize 256. 313.
- Kontraktion der Wurzel 115. *154. 249.
- Konvergenz 149. 555.
- Konvergentes Licht 291.
- Konzentrische Leitbündel *86. 123.
- Kopfschimmel 381.
- Kopra 613.
- Koppelung 285.
- Kopulation 168.
- Kopulieren 168. *261.
- Korallenschwamm 405.
- Koriander *548. 550.
- Korinthe 581.
- Kork 49 ff. *50. 138 ff. 246.
- Korkeiche 50. 139. 562.
- Korkhaut 49 ff. 138.
- Korkkambium 49. 138.
- Korkkrusten 49 ff. 138.
- Korkrinde 138.
- Korkstoff 33.
- Kormophyten 63.
- Kormus 62. 72 ff.
- — Anpassungen an Umwelt 141 ff.
- Kornelkirsche *547.
- Kornrade *553.
- Korolle 470.
- Korrelationen 259. 269. 272. 275.
- Korrosion der Stärke *229.
- Kotyledonen 102 ff. *115. 228. 497.
- Kräuter 121.
- Krameria 541.
- Krausblättrigkeit 287.
- Kreisende Bewegung 297.
- Kreislauf der Stoffe 240.
- Kreuzdorn 581.
- Kreuzung 285.
- Kribralteil 58.
- Kriechen 289.
- Kristalle 182.
- Kristalldrusen 25.
- Kristallsand 25.
- Kristallschläuche im Bast 136.
- Kristallzellen 25 ff. 58.
- Krone 470.
- Kronblätter 168.
- Krümmungsarten *294.
- Krümmungsbewegungen *294. 296.
- Krummholzkiefer s. Zwergkiefer 519.
- Kryptogamen 325.
- Küchenschelle *531.
- Küchenzwiebel vgl. Allium 618.
- Kümmel *549.
- Künstliche Pflanze 183.
- Kürbis *53. *54. *56. 196. 245. 600.
- Kumys 239.
- Kupfervitriol 257.
- Kurztriebe 66. 109. 110. 255.
- Kutikula 42. 194. 211.
- Kutikulär 196. 197.
- Kutin 33.
- Kutinisierung 33. 42.
- Kutisgewebe 48.
- Kutiszellen 48.
- Labellum 631. 633.
- Labiatae 592.
- Laboulbenieae 395.
- Laburnum 262. *263. 543.
- Lachnea 385. *390.
- Lackmus 415.
- Lactaria 407. 408.
- Lactuca 146. 310. 605.
- Längenwachstum 242 ff.
- Längsachse 61.
- Längsschnitt, radialer 62.
- — tangentialer 62.
- Lärche 110. *519.
- Lärchenschwamm 407. 409.
- Lävulose 230.
- Lagenostoma *463.
- Lamarck 1.
- Lamarckismus 180 ff.
- Lambertsnuß 560.
- Lamina 94.
- Laminaria, Laminariaceen 69. 361. *362. *366. 367. 368.
- Laminarin 363.
- Lamium *592.
- Landform 285.
- Landolphia 589.
- Landpflanzen 143 ff. 194.
- Langtriebe 66.
- — beim Kormus 109.
- Laportea 566.
- Lappa *604. 605.
- Larix *519.
- Latentes Leben 186. 189. 266.
- Lathraea 257.
- Lathyrus *155. 544.
- Laubabwerfende Holzgewächse 150.
- Laubblatt *93 ff. 270.
- — -funktion 198. 215.
- — Innerer Bau 96 ff.
- — Reduktion bei Parasiten 161; bei Xerophyten 146 ff.
- — Stoffauswanderung 232.
- Laubfall 104. 271.
- Laubknospe 75.
- Laubmoose 254. 293. 427.
- Laubsproß 73. 270.
- Lauch 618.
- Laudatea 415.
- Lauraceae 526.
- Laurus *528.
- Lavandula, Lavendel *591.
- Lebensbaum 514.
- Lebensbedingungen 184, 185. 260. 290.
- Lebensdauer 121. 261. 271.
- Lebenserscheinungen 182.
- Lebenstätigkeit 184. 186.
- Lebenszustände 186. 266.
- Lebermoose 254. 309. 422.
- — Thallusbau 69 ff.
- Lecanora 415.
- Ledum 583.
- Legumen 538.
- Leguminosae 224. 297. 317. 538.
- Leguminosenknöllchen *224. 332. 543.
- Leimzotten in Winterknospen 151.
- Leitbündel *58. *86 ff. *88. 89. 96 ff. *97. *116 ff. *124.
- — bikollaterale 87.
- — blatteigene 83.
- — endarche 90.
- — exarche 90.
- — gemeinsame 83.
- — geschlossene 87. *88.
- — im Blatt 96 ff.
- — im Stengel 81 ff. *84 ff.
- — kollaterale 87. *88. *90.
- — kollaterale, offene *124.
- — konzentrische *86. 87. 123.
- — der Laubmoose *71.
- — mesarche 90.
- — offene 87. *89.
- — Ontogenie 89 ff.
- — Phylogenie *91.
- — primanen 90.
- — primäre 58. 84 ff.
- — radiale *86. *116 ff.
- — Reduktion bei Parasiten 161.
- — sekundäre 58 ff.
- — stammeigene 83.
- — Übergang der Wurzel in die Stengelbündel *117 ff.
- — unvollständige 58.
- — vollständige 58.
- — der Wurzeln *116 ff.
- Leitbündelanordnung im Blatt 96 ff.
- — in Sproßachsen 82 ff.
- Leitbündelendigung *96.
- Leitbündelscheiden 89.
- Leitbündelsystem 57. 82 ff.
- Leitbündeltypen der Farnpflanzen *91.
- Leitbündelverlauf 82 ff. *84 ff.
- Leitergefäß *56.
- Leitgewebe 53 ff.
- Leitparenchym 41.
- Leitung des Wassers 201.
- — von Reizen 306. 310. 320.
- Lemna *292.
- Lens 544.
- Lentibulariaceae 596.
- Lentizellen *50 ff. 139. 212.
- Leocarpus *340.
- Leontodon *316.
- Leontopodium 605.
- Lepidium *567.
- Lepidocarpon 461.
- Lepidodendron *460. 461.
- Lepidospermeae 461.
- Lepidostrobus *461.
- Lepiota 407.
- Leptom 58.
- Leptomitus 380.
- Leptosporangiate Farne 439.
- Leptothrix *63. 238. 334.
- Lessonia 362.
- Leuchtbakterien 241. 332.
- Leuchten der Pflanzen 241.
- Leuchtende Peridineen 343.
- Leuchtmoos 432.
- Leuchtpilze 241. 408.
- Leucin 222. 273.
- Leucobryum 427.
- Leucojum *622.
- Leuconostoe 332.
- Leucosin 337.
- Leukoplasten *16. *28. 216.
- Levisticum 549.
- Levkoje 569.
- Lezithin 12. 207. 222. 227. 230.
- Lianen 155 ff.
- — Gefäße 55.
- Lichen islandicus 415.
- Lichenes 327. 410.
- Licht, Lebensbedingung 186.
- — Absorption 214.
- — Bedingung der Assimilation 214. *218.
- — — — Bildung von Chlorophyll 216.
- — — — Blütenbildung 274.
- — — — Entwicklung 251.
- — — des Wachstums 251.
- — — von Phototaxis 291.
- — — — Phototropismus 307.
- — Menge 310.
- — Richtung 254. 309.
- — Wellenlänge *214. 254. 273.
- Lichtbedürfnis 186.
- Lichtblätter 98. 253.
- Lichtenergie 215.
- Lichtfalle 291.
- Lichtkeimer 267.
- Lichtlage, fixe 310.
- Lichtperzeption 310.
- Lichtwachstumsreaktion 252. 309.
- Lichtwechsel 252. *316. *317.
- Licmophora *344.
- Liebstöckel 549.
- Lignine 32.
- Lignum Guajaci 578.
- — Haematoxyli 540.
- — Juniperi 519.
- — Quassiae 581.
- — Santali rubrum 544.
- — Sassafras 526.
- Ligula der Gräser *102. 626.
- — der Selaginellen 457.
- — von Isoëtes 459.
- Ligulatae 459.
- Ligustrum 585.
- Liliaceae 302. 618.
- — Diagramm *168.
- Liliiflorae 301. 617.
- Lilium *154. *173. 276. 617.
- Limnanthemum 587.
- Limodorum 634.
- Limone vgl. Citrus 579.
- Linaceae 578.
- Linaria 278. 595.
- — cymbalaria 244. 310.
- Linde *133 ff. 272. 572.
- Linie, reine 285. 286.
- Linin 14. 17.
- Linkswinder *303.
- Linse 544.
- Linum *578.
- Lipase 230.
- Lipoide 12. 207.
- Liquidambar 533.
- Liriodendron 525.
- Listera 634.
- Lithium 205.
- Littonia 314. 618.
- Loasa 303.
- Lobaria 415.
- Lobelia *603.
- Loculicid 503.
- Lodiculae *626.
- Löcherschwämme 406.
- Löffelkraut vgl. Cochlearia 569.
- Lösungen 189.
- Lösung von Gestein 208.
- Lodoicea 506.
- Löwenmaul s. Antirrhinum 595.
- Löwenzahn, Milchsaftgefäße 59.
- Loganiaceae 587.
- Lohblüte 341.
- Lokalisierte Empfindung 310.
- Lokomotion 289 ff.
- Lolium 630. *631.
- Lonicera 303. 599.
- Loranthiflorae, Loranthus 163. 223. 556.
- Lorbeer 526.
- Lorchel 392.
- Lotus *542. 543.
- Lotosblume vgl. Nelumbium 529.
- Lücken (Sekret-) 61.
- Luftgewebe 42. 142 ff.
- Luftspalten *44 ff.
- Luftsproß 73.
- Lufttrocken 187. 189.
- Luftstickstoff 221. 224 ff.
- Luftverdünnung im Holzkörper 203.
- Luftwurzeln 112. 120. 158. 249.
- Lumen 8.
- Lunaria *569.
- Lungenflechte 415.
- Lupine, Lupinus 225. *256. 543.
- Lupulinum 565.
- Luxurierendes Wachstum 280.
- Luzula 618.
- Lycoperdon *409.
- Lycopersicum *264. 593.
- Lycopodiaceen 453.
- — Verzweigung *105.
- Lycopodinen 452.
- Lycopodium 206. 293. *434. 453. *454. *455. 456.
- Lyginodendron *462. 463.
- Lysigene Interzellularen 38.
- Lythraceae, Lythrum 267. 545.
- Macchia 592.
- Macis 525.
- Macrocystis 361. *362.
- Macrozamia 509.
- Magnesium 187. 206. 227.
- Magnolia, Magnoliaceae 266. 525.
- Maiblume *93.
- Maiglöckchen *93. 620.
- Mairan s. Origanum 592.
- Mais *80. *88. 283. 629.
- Majanthemum 620.
- Makrosporangium 435. 464.
- Makrospore 435. 464.
- Makrosporophyll 464.
- Malakophil 482.
- Mallotus 577.
- Maltose (Malzzucker) 23. 229.
- Malva, Malvaceae *570.
- Mamillaria 552.
- Mandarine 275. 580.
- Mandel 266. 537.
- Mangifera 581.
- Mangrove *143. *545.
- — Atemwurzeln *143. 211.
- — Aussaat 508.
- — Xerophytenstruktur 144.
- Manihot, Maniok 576.
- Manna, Mannaesche *586.
- Mannaflechte 415,
- Mannit 363. 368.
- Mantelblätter bei Epiphyten 158.
- Mantelchimäre 263.
- Maranta, Marantaceae 633.
- Marattiaceen 437.
- Marcgravia 482.
- Marchantia *70. 254. 293. *417. *418. *423. *424.
- Marchantiales 417. 420.
- Marennin 347.
- Mark des Stengels 82.
- Markbündel 82.
- Markkrone 127.
- Markstrahlen 82. 125. 129 ff. *130 ff. 136 ff.
- — primäre *125.
- — sekundäre 126.
- Markstrahlparenchym im Bast 136.
- Markstrahltracheïden *131.
- Markstrahlzellen 134.
- — liegende 134.
- — stehende 134.
- Maronenpilz 406.
- Marsdenia 589.
- Marmor 208.
- Marsilia 277. 297. *444. *447. *448.
- Marsupium 419.
- Martensia 373.
- Martynia 321.
- Maschine 184.
- Maser, Maserbildung im Holz 135.
- Massenströmung 208.
- Massula 447.
- Mastigamoeba *339.
- Mastix 581.
- Mate 580.
- Matricaria *607.
- Matthiola 569.
- Maulbeerbaum 564.
- Maurandia *314.
- Maximum 185. 186. 243. 250. 290.
- Mazeration 35. 128.
- Mechanismus 184.
- Mechanische Einflüsse 255.
- Mechanische Elemente *52 ff.
- — Gewebe *52 ff.
- — im Blatt *97.
- — im Holz 128.
- — im Stengel *80 ff.
- — in Wurzeln *117.
- Mechanische Reize 255. 313. 318.
- Mediane 62. 106. *107.
- Median, vorn, hinten an Seitenknospen 107.
- Medicago 186. 543.
- Meeresleuchten 241. 343.
- Meerespflanzen 194.
- Meersalat 356.
- Meerstrandpflanzen 144.
- Meerzwiebel *620. 621.
- Mehl 27.
- Mehltau 386.
- — falscher 380.
- Meiosis 172.
- Meiotische Teilung 20. 172.
- Melaleuca 547.
- Melampyrum 163. 596.
- Melandrium 284.
- Melica 630.
- Melilotus 543.
- Melissa 592.
- Melone 601.
- Melosira 346.
- Membranstoffe 31 ff.
- MENDELsche Regeln 280 ff.
- Menispermaceae 526.
- Mentha 592.
- Mentholum 592.
- Menyanthes 589.
- Mercurialis *574.
- Meringosphaera 351.
- Meristeme 39 ff.
- — sekundäre 40.
- Meristemzelle *9.
- Merk 549.
- Merkmale, Autonomie 282.
- Merkmalspaare 282.
- Merulius 407.
- Mesarch 90.
- Mesembryanthemum 149. 295. 551.
- Mesocarpus 292. 310. 350.
- Mesokarp 503.
- Mesophyll 97.
- Mesophyllscheiden 96.
- Mesotaenium 348.
- Mespilus 262. 536.
- Mestom 58.
- Metalle 227.
- Metamorphose 6.
- Metaphase 19.
- Metasyndese 174.
- Methan 240.
- Methanbakterien 219. 238. 331.
- Methylenblau 207. 240.
- Methylglyoxal 239.
- Metroxylon 612.
- Metzgeria *70. 425. 426.
- Miadesmia 461.
- Micrasterias *349.
- Micrococcus *63. 240. *333.
- Microcycas 509.
- Microphasen 387.
- Mikropyle 464.
- Mikroskop 242.
- Mikrosomen 12.
- Mikrosporangium 435.
- Mikrosporen 435. 465.
- Mikrosporophyll 465.
- Mikrozyste 341.
- Milchröhren *58 ff.
- — im Bast 136.
- — gegliederte *59.
- — ungegliederte *58.
- Milchgefäße *59.
- Milchsaft 58 ff.
- Milchsäurebazillus 331.
- — -gärung 239.
- Milzbrandbazillus *333.
- Mimosa *538.
- — Bewegungen *319.
- Mimulus 274. 321. 595.
- Mimusops 584.
- Mineralisierung 240.
- Mineralstoffe im Plasma 12.
- — in Pflanzen 188. 203. 226.
- Minimalflächen 269.
- Minimum 185. 186. 250. 290.
- — Gesetz 217.
- Minze s. Mentha 592.
- Mirabelle 261.
- Mirabilis *280.
- Mischlinge 277.
- Mispel 263. 536.
- Mißbildungen 259.
- Mist 209. 241.
- Mistel *37. 163. 223. 258. 308. 556.
- Mitochondrien 13.
- Mitose 17.
- Mittelbildung 262. 280.
- Mittellamelle 35.
- Mittelständiger Fruchtknoten 483.
- Mnium *71. 427. *430. *431. 432.
- Mobilisierung der Reservestoffe 228.
- Modifikation 258. 285.
- Modifikationsfähigkeit 180.
- Möhre *16. 549.
- Mohn 566.
- Mohrhirse 629.
- Monascus 385. 386.
- Mondraute 438.
- Monoblepharis *378.
- Monochasium *111. 477.
- Monogene Fortpflanzung 165. 276.
- Monohybriden 282.
- Monokarpisch 271.
- Monokotylen 218. 609.
- — Dickenwachstum *122 ff.
- Monopodium 66. 105. 110.
- Monosaccharide 23.
- Monosporen 373.
- Monostelie 92.
- Monotropa 163. 225.
- monözisch 470.
- Monstera 246. 615.
- Mooskapsel 295.
- — als Sporophyt 175.
- Moospflanzen 292. 415.
- — Bauvariationen des Thallus 69 ff.
- — Gewebesonderung 70 ff.
- Moraceae 564.
- Morchella *386. *392.
- Morchelpilze 392.
- Morphium 567.
- Morphologie 3. 5.
- — äußere 7. innere 7.
- Morphologischer Wert 7.
- Morus *504. 564.
- Mosaikbastarde 278.
- Mucor 312. 381. *382. 383. 422.
- Mucorineae 312. 381.
- Mucuna, Stammquerschnitt *127.
- Musa, Musaceae 186. *631.
- Muscari 618.
- Musci 427.
- Muscus pulmonarius 415.
- Muskatnuß *526.
- Mutation 285 ff.
- Mutterachse 66.
- Mutterkorn 389.
- — Sklerotium *390.
- Mycetozoa 339.
- Mycobacterium *333.
- Mykorrhiza 163. *228. 229. 384. 433.
- Myosotis 590.
- Myosurus 529.
- Myristica *525. *526.
- Myrmecodia 148. 598.
- Myroxylon *542.
- Myrrha 581.
- Myrsinaceae 226.
- Myrsiphyllum *303. 620.
- Myrte, Myrtus 546.
- Myrtiflorae 544.
- Myxamöben 340.
- Myxobacteriaceae 336.
- Myxococcus *336.
- Myxogasteres 340.
- Myxomyceten *10. 289. 293. 339.
- Myzelium *67. 377. 384.
- Myzelschnallen 404.
- Myzetozoa 340.
- Nabel 500.
- Nachreife 267.
- Nachruhe 267.
- Nachtblüher 316.
- Nachtstellung *292. *317.
- Nadelhölzer 511.
- Nägelchen 547.
- Nährgewebe *501.
- Nährlösung 204. 205.
- Nährsalze 203 ff. 226. 274.
- Nährstoffe 188.
- Nährwurzeln bei Epiphyten 158.
- Najas 611.
- Narbe 170. 200. 276. 321.
- Narcissus 622.
- Narkotika 322.
- Nastie 299. 315.
- Natrium 187. 205. 206. 227.
- — -Selenit 240.
- — -Thiosulfat 240.
- Natterzunge 438.
- Navicula 346. *347.
- Nebenblätter 94. 100 ff.
- Nebenprodukte 232.
- Nebenwurzeln 119.
- — an Konvexseite *256.
- Nebenzellen *44. 45.
- Neckeraceae 432.
- Nectria 388. 389.
- Negative Reaktion 291. 299. 301. 308. 310. 311. 312.
- Nektarien 60. 99. 200. 473.
- Nektarspalten 99.
- Nelke 553.
- Nelumbium 529.
- Nemalion 373. 374. 376.
- Neottia 163. *225. 268. 634.
- Nepenthaceen 528.
- Nepenthes 314.
- — Tierfang *160. 223.
- Nepeta 592.
- Nephromium 414.
- Nerium *587.
- Nervatur 95 ff. 408. 423. 524. *610.
- Nerven 320.
- Nessel 566.
- Nestwurz 634.
- Netzgefäß *55.
- Neubildung 245.
- Neuropteris 463.
- Nicotiana 278. 287. 316. 595.
- Niederblätter 93, 103. 152. 270.
- Niederschlagsmembran 190.
- Nischenblätter bei Epiphyten 158.
- Nitella 290. 369. 370.
- — Turgordehnung *192.
- Nitophyllum 373. 376.
- Nitratbakterien 219. 238. *334.
- Nitrate 23. 221.
- Nitrifikation 219. 236. 332.
- Nitritbakterien 219. 238. 334.
- Nitrobacter *334.
- Nitrosomonas *334.
- Nitzschia 347.
- Nodi 76.
- Nopalea 552.
- Nostoc *335. 336.
- Nucellus 464.
- Nützlichkeit, Zustandekommen 181.
- Nukleoalbumin 227.
- Nukleolen 13. 18.
- Nukleoproteïde 12. 227.
- Nukleus (Zellkern) 9. 13.
- Nuphar 528.
- Nuptiale Nektarien 60.
- Nuß 504.
- Nutationen 294. 297.
- Nux 504.
- Nyktinastische Bewegungen *316. *317.
- Nymphaea 257. 316. *529.
- Obdiplostemon 473.
- Oberblatt 94.
- Oberfläche, freie 63.
- Oberflächenspannung 289.
- Oberhaut (Epidermis) 42 ff.
- Oberständiger Fruchtknoten *472.
- Obst 266.
- Obstbau 255.
- Obturator *574.
- Ochrea 101. *555.
- Ochrolechia 415.
- Ochsenauge 590.
- Octarch 117.
- Odontites 596.
- Odontospermum 295.
- Oedogonium *357.
- Ökologie 3.
- — der Bestäubung 170. 477.
- Ölbaum 230. *585.
- Öldrüsen *60.
- Öle, ätherische 25. 58 ff. 232.
- — fette 25. 218. 230.
- Ölgänge 61.
- — -körper bei Lebermoosen *70.
- Ölpalme 230. 614.
- Ölräume 60 ff.
- Ölweide 545.
- Ölstriemen 549.
- Oenanthe 549.
- Oenothera 288. *545.
- Oidium 387.
- Okenia 507.
- Okulieren *261.
- Olea, Oleaceae *585.
- Oleander 145. 247. 587.
- Oleum Anisi 550.
- — Amygdalarum 537.
- — Arachidis 544.
- — Aurantii corticis 580.
- — Aurantii florum 580.
- — Bergamottae 581.
- — Betulae, empyreumaticum 561.
- — Cacao 574.
- — cadinum 519.
- — Cajuputi 547.
- — Calami 617.
- — Carvi 550.
- — Caryophyllorum 547.
- — Chamomillae 607.
- — Cinnamomi 526.
- — Citri 580.
- — Crotonis 576.
- — Foeniculi 550.
- — Juniperi 519.
- — Lauri 526.
- — Lavandulae 592.
- — Lini 578.
- — Macidis 525.
- — Menthae 592.
- — Nucistae 525.
- — Olivarum 586.
- — Pini Pumilionis 519.
- — Ricini 577.
- — Rosae 537.
- — Rosmarini 592.
- — Santali 556.
- — Sinapis 561.
- — Terebinthinae 519.
- — Thymi 592.
- — Valerianae 599.
- Olibanum 581.
- Olive s. Ölbaum 585.
- Olpidiopsis 378.
- Olpidium *377. 378.
- Omnivor 220.
- Onagraceae 545.
- Onoclea *434.
- Ononis 543.
- Ontogenie 2. 6.
- Oogamie 169. 329.
- Oogonien *169. 327.
- Oomyceten 378.
- Oosphäre 329.
- Oospore 170. 275. 329.
- Ophiocytium 350.
- Ophioglossum 268. *438. *439.
- Ophrys 634.
- Opium 567.
- Opponierte Stipel 101.
- Optimum 185. 217. 250. 291.
- Opuntia *147. 552.
- Orchidaceae 226. 276. 303. 633.
- Orchis *153. *634. *635.
- Organe 5.
- Organbildung 245.
- Organische Säuren 23. 204. 220. 232. 235.
- Organische Substanz 187. 208.
- Organismus 5.
- Organographie 7. 61 ff.
- Organoid 258.
- Origanum 592.
- Ornithogalum *36. 316. 618. *620.
- Ornithophilie *482.
- Ornithopus 544.
- Orobanche 163. 257. 267. *598.
- Orseille 415.
- Orthostichen 77.
- Orthotrop 62. 299. 307.
- Ortsveränderung 288.
- Oryza *629. *630.
- Oscillaria *335. 336.
- Osmometer *190.
- Osmose 190.
- Osmotaxis 293.
- Osmotischer Druck 190 ff.
- — — Wert 193. 198.
- Osmunda *442.
- Osterluzei, Bestäubungseinrichtung *484.
- Ourouparia 598.
- Ovium 168. 329.
- Oxalis 297. 298. 317.
- Oxalsäure 220. 235. 240. 248.
- Oxydase 237.
- Oxydation 236. 238.
- Oxydationsgärung 240.
- Padina 365.
- Paeonia 529.
- Pahudia 592.
- Pakoe-Kidang 444.
- Palaeostachya *452.
- Paläontologie 2.
- Paläophytologie 3.
- Palaquium *584.
- Palea 440.
- Paleae haemostaticae 444.
- Palisadenzellen 98. 253.
- Palmae, Palmen 211. 235. 301. 612.
- Palmella 354.
- Palmentypus des Leitbündelverlaufes *85.
- — des Stammes 121.
- Palmöl 614.
- Palmwein 201. 614.
- Panaschierung 15. 215. 218.
- Pandanaceae, Pandanus *612.
- Pandorina 354.
- Paniceen 306. 311.
- Panicula = Rispe.
- Panicum 629.
- Pantostomatinen 339.
- Papaver *566.
- Papilionaceae 586. 278.
- Papillenhaare *46.
- Pappel *562.
- Pappus *601. *603.
- Papyrus 626.
- Paraffinöl 256.
- Paraguaytee 581.
- Parallelotrop 299.
- Paraphysen 367. 403. 385. 428.
- Parasiten 67. *161 ff. 220 ff. 223. 257. 271.
- — Haustorien *67. *162.
- Parasolschwamm 407.
- Parastichen *78.
- Parasyndese 174.
- Paratonisch 297. 298.
- Parenchym 38. 40 ff.
- Parenchymsystem 41.
- Parenchymzelle 41.
- Parietal 479.
- Paris 620. *622.
- Parmelia *411. 412. *413. 414.
- Parnassia 534.
- Parthenocissus *156. 256. 297. 314. 581.
- Parthenogenesis 165. 172. 277. 499.
- Parthenokarpe Früchte 275. 601. 631.
- Passiflora 187. 312.
- Pastinaca, Pastinak 549.
- Pathologie 4.
- Paullinia 581.
- Pavetta 598.
- Payena 584.
- Pediastrum *355.
- Pedicularis 163. 596.
- Peireskia 552.
- Pektinase 35.
- Pektinstoffe 31 ff.
- — in Mittellamellen 36.
- Pelargonium 246. 247. *577.
- Pellia 425. 426.
- Pelorie 476.
- Peltigera 414.
- Penicillium *67. 220. *388.
- Pennatae 346.
- Pennawar Djambi 444.
- Pentazyklisch 473.
- Pentacyclicae 584.
- Pentarch 117.
- Pentosane 31 ff.
- Pepsine 231.
- Pepton 231. 292. 311.
- Peptonisierende Enzyme 59. 231.
- Perennierend 121. 151 ff.
- Perianth 425. 469.
- Periblem 74. 114.
- Perichaetium 428.
- Periderm 138 ff.
- Peridineae, Peridinium *342. 343.
- Peridium 340. 409.
- Perigon 470.
- Perigyn 473.
- Perikarp 503.
- Periklinalchimäre 262.
- Periklinen 40. 75. 269.
- Periode, große Wachstums- 243.
- Periodische Bewegung 317.
- Periodizität 149. 201. 266. 270.
- — der Kambiumtätigkeit 130 ff.
- Periphysen 388.
- Periplasma 22. 31. 381. 385.
- Periplasmodium 31. 435.
- Perisperm 501.
- Perispor 435.
- Peristom 295. 430.
- Perithecium 386.
- Perizykel im Stengel 82.
- — in Wurzeln 116.
- Permeabilität 190. 207.
- Peronospora, Peronosporeen *67. *380. *381.
- Persea *527.
- Persio 415.
- Personatae 592.
- Perzeption 306. 311. 323.
- — Organe 311.
- Pestbazillus 333.
- Petasites, Pestwurz 606.
- Petiolus 94.
- Petroselinum, Petersilie 549.
- Petunia *592.
- Peziza *390.
- Pfahlwurzel 120.
- Pfeffer *558.
- Pferdekümmel 549.
- Pfifferling 407.
- Pfirsich 261. 537.
- Pflanze und Tier 1. 2 ff. 182. 321 ff.
- Pflanzendrogen 4.
- Pflanzengeographie 3. 210.
- Pflanzenpathologie 4.
- Pflanzenverteilung 210.
- Pflanzenwachs *43.
- Pflaume 261. 537.
- Pfropfbastarde 262 ff.
- Pfropfen *261.
- Pfropfhybride 262 ff.
- Pfropfreis 260.
- Phaeocystis 338.
- Phaeophyceen 361.
- Phaeosporeen 364.
- Phaeothamnion 338.
- Phallaceae 410.
- Phanerogamen 325.
- — Blüte 167 ff. 468.
- Pharbitis *303. 590.
- Pharmakognosie 4.
- Phascum 432.
- Phasen des Wachstums 245.
- Phasenwechsel 172.
- Phaseolus *27. *298. 318. 544.
- Phelloderm 138.
- Phellogen 138.
- Phelloid 50. 140.
- Phellonsäure 33.
- Philadelphus 534.
- Phleum 630.
- Phloëm 58. 85.
- Phloëmparenchym 87.
- — primanen 90.
- Phoenix 614.
- Phobisch 291 ff.
- Phormium, Blattquerschnitt *97. 619.
- Phosphate 23. 205. 227. 275. 292. 311. 318.
- Phosphor 187. 205. 223. 227. 232. 275.
- Phosphorsäure 209. 227.
- Photogene Bakterien 241. 332.
- — Pflanzen 241.
- — Pilze 408.
- Photonastie 316.
- Photosynthese 219.
- Phototaxis 291.
- Phototropismus 254. *307. *308.
- Phragmidium 400. *401.
- Phycomyces 382. 383.
- Phycomycetes 376.
- Phykoërythrin 15. 335. 372.
- Phykoxanthin 16. 344. 362.
- Phykozyan 16. 335. 372.
- Phyllactinia 387.
- Phyllocactus 552.
- Phyllodium 145. *146.
- Phyllokladien *147.
- Phylogenie 2. 6.
- Physalis *504.
- Physarum *341.
- Physcia 414.
- Physiologie 3. 182.
- Physiognomie 141.
- Physostigma 544.
- Phytelephas *37. 614.
- Phyteuma 603.
- Phytin 26. 227.
- Phytol 15.
- Phytophthora *380.
- Phytosterine 12.
- Picea *490. *491. *518. 519.
- Picrasma 581.
- Pigmentbakterien 332.
- Pilobolus *308. 382.
- Pilocarpus, Pilokarpin 581.
- Pilostyles, Schmarotzertum *161.
- Pilularia *444. 447. 448.
- Pilze *67. 166. 194. 208. 220 ff. 226. 235. 238. 240. 245. 252.
258. 268. 276. 311. 327.
- Pimpinella *545. 549. 550.
- Pinaceae 512.
- Pinguicula 158. 318. 614.
- Pinie 519.
- Pinnularia *63.
- Pinselschimmel 388.
- Pinus *57. *98. *130 ff. *225. 230. *468. *489. *491. *492.
*518. 519.
- Piper, Piperaceae *558.
- Pirus *138. 535.
- Pistacia 581.
- Pisum *155. 544.
- Pithecoctenium, Flügelsame *505.
- Pix liquida 519. 562.
- Placenta 475.
- Plagiochila *70. *426.
- Plagiogeotropismus 302.
- Plagiotrop 62. 299. 302.
- Plankton 64. 342. 345.
- Planktoniella *344.
- Planogameten 328.
- Plantago *475. 596.
- Plasma 9. 12.
- Plasmabelag 10.
- Plasmabewegung 10 ff.
- Plasmahaut 12.
- Plasmaverbindungen 36.
- Plasmodesmen *36. 209,
- Plasmodiophora 342.
- Plasmodium *10. 289. 293. 340.
- Plasmolyse 10. 192. *193. 206. 298.
- Plasmolytische Grenzkonzentration 193.
- Plasmopara 380.
- Plastiden 9.
- Plastische Dehnung 248.
- Platanaceae, Platanus 533.
- Platanthera 634.
- Platin 204.
- Plattenkollenchym 53.
- Platterbse 544.
- Platycerium 158. 268.
- Platystemon 566.
- Plectascineae 388.
- Plectridium *332.
- Pleiochasium 111. 477.
- Plektenchym 34. 384.
- Plerom 74. 114.
- Pleurocarpi 432.
- Pleurocladia *364.
- Pleurosigma 346. 348.
- Plumula 171. *497.
- Pneumathoden 143.
- Pneumatophoren *143. 211.
- Poa 630.
- Poaeoideen 306.
- Podetien 413.
- Podocarpus 225. 512.
- Podophyllum *527.
- Podospora *389.
- Polarität 61. 64. 247. 254. 255. 261. 267.
- Polarpflanze 251.
- Polioplasma 12.
- Polkappen 18.
- Pollenkammer *487.
- Pollenkörner *30. 167. 257. 276. *468. *470. *485. *489.
- Pollenmutterzelle *466.
- Pollensack 167. *466. *468.
- Pollenschlauch 171. 245. 311. *487. *489. *493. *494.
- Pollenübertragung 170. 478.
- Pollinium *588. *634.
- Polstergewächse *145.
- Polyangideae 336.
- Polyangium *336.
- Polyarch 117.
- Polycarpicae 525.
- Polyembryonie 171. *504.
- Polygala *579. 581.
- Polygam 470.
- Polygonatum *103. *305. *610. 620.
- Polygonum 303. 555.
- — Embryosackentwicklung *495. *496.
- Polypeptide 221.
- Polypodiaceae 440.
- Polypodium 296. *434. 439. 442. *443.
- Polyporaceae 406.
- Polyporus 407. 409.
- Polysaccharide 23. 31.
- Polysiphonia 376.
- Polysporangium 373.
- Polystelie 92.
- Polytoma *353. 354.
- Polytrichum *419. 427. 428. *430. 432.
- Pomeranze 580.
- Pompelmus 580.
- Populus *75. *562.
- Porenkork 50.
- Poricid 504.
- Porogamie 495.
- Porometer 199.
- Porst 583.
- Porus 43.
- Positive Reaktion 291. 299. 302.
- Potamogeton 213. *611.
- Potentilla 278. 537.
- Prähaustorien von Cuscuta 162.
- Präsentationszeit 307. 310.
- Prävalieren 281.
- Preißelbeere 583.
- Primärblatt 102.
- Primäre Verdickungsschichten 35.
- Primanen 90.
- Primordialblatt 92. 94.
- Primordium 92.
- Primula *60. *484. *554. 555.
- Proëmbryo *488.
- Profilstellung *292. 310.
- Promycel 397. 400.
- Propeller 289.
- Prophase 19.
- Prosenchyme 40.
- Protandrie 483.
- Proteasen 231.
- Proteïde 12.
- Proteïne 11.
- Proteïnkörner *26. 59.
- Proteïnsubstanzen 311. s. auch Eiweiß.
- Proteolytische Enzyme 231.
- Prothallienzellen, vegetative *485. *489.
- Prothallium *72. 175. 252. 309. 432.
- — von Samenpflanzen 175.
- Protococcales 354.
- Protogynie 483.
- Protomastiginen 339.
- Protonema 416. 427.
- Protophloëm 90.
- Protoplasma 1. 9. 183. 184. 189. 191 ff. 206. 248.
- — Aggregatzustand 10.
- — Chemische Eigenschaften 11.
- — Färbung 12.
- — Fixierung 12.
- — physikalische Eigenschaften 10.
- — Reaktionen 12.
- Protoplasmabewegung 10. 209. 290.
- Protoplasmaeinschlüsse 23.
- Protoplasmaströmung 10 ff. 209. 290.
- Protoplasmaverbindungen 36.
- Protoplast 8. 9.
- Protosiphon 277. 356.
- Protoxylem 90.
- Prunus *93. *535. 537.
- Psalliota *407.
- Pseudomonas 241.
- Pseudoparenchym 384.
- Pseudopodium 337.
- Pseudotsuga 516.
- Psidium 546.
- Psilotum 458.
- Psyche 323.
- Psychotria 598.
- Pteridium *57. *86. *113. *433. *434. 442. 443.
- Pteridophyten 432.
- Pteridospermen 461.
- Pteris *433.
- Pterocarpus 544.
- Puccinia *399. *400. 401. 402.
- Pulpa prunorum 537.
- — Tamarindorum 540.
- Pulque 201. 622.
- Pulsierende Vakuole 11.
- Pulu 444.
- Punica, Punicaceae *546.
- Purpurbakterien 215. 332.
- Pykniden *389.
- Pyknosporen 389.
- Pyknokonidien 389.
- Pyramidenwuchs 287.
- Pyrenoide *14.
- Pyrenomyceten 388.
- Pyronema 385. 386. *391. *392.
- Pythium 380.
- Quarz 204.
- Quassia *581.
- Quellstifte 368.
- Quellung 189. 295.
- Quellungsmechanismus 295.
- Quercus *139. *560. *561.
- Querschnitt 61.
- Quillaja *536.
- Quirl 76.
- Quitte 261. 536.
- Radiär 61. 268. *475.
- Radiale Leitbündel *86. 117.
- Radieschen 153. 569.
- Radikula 171. 498.
- Radiumstrahlen 254.
- Radix Althaeae 570.
- — Angelicae 550.
- — Arnicae 607.
- — Bardanae 608.
- — Belladonnae 594.
- — Colombo 526.
- — Gentianae 587.
- — Gelsemii 587.
- — Ipecacuanhae 598.
- — Levistici 550.
- — Liquiritiae 544.
- — Ononidis 544.
- — Petroselini 550.
- — Pimpinellae 550.
- — Pyrethri 608.
- — Ratanhiae 540.
- — Rheï s. Rhizoma Rheï 556.
- — Sarsaparillae 621.
- — Senegae 581.
- — Taraxaci 608.
- — Valerianae 599.
- Rafflesia, Rafflesiaceae 162. 528.
- Ramalina 413.
- Ramie 566.
- Randwachstum 74.
- Ranken *155 ff. 256. 297. 303. 308. 312. *313. 318.
- Rankenpflanzen 156.
- Rankenplatterbse *155. 544.
- Ranunculus *89. 276. *530.
- Raoulia *145. 605.
- Raphanus 569.
- Raphe 465.
- Raps 569.
- Rasse 277. 286.
- Raute *578.
- Ravenala 631.
- Razemös 66. 110.
- Reaktion 306.
- Reaktionszeit 307.
- Rebe 201. 276.
- Rebenzucht 255.
- Rechtswinder *303.
- Reduktion der Oberfläche bei Xerophyten *146 ff.
- Reduktionsgärung 240.
- Reduktionsteilung 20. 172 ff. *174. 276. 285.
- Reduzierte Organe 7.
- — u. Abstammungslehre 178.
- Regenwald 155. 157. 186.
- Regulationen 183. 184. 194. 197. 248. 260.
- Reif (aus Wachs) 43.
- Reifen der Früchte 232.
- Reis *629. *630.
- Reiz 184. 257. 267. 275. 290. 291. 297. 298 ff. 321.
- Reizanlaß 323.
- — -mittel 323.
- — -schwelle 293. 322.
- Reizbarkeit 183. 321.
- Reizbewegungen 298 ff.
- Reizempfänglichkeit 322.
- Reizerscheinungen 184.
- Reizker 407.
- Reizleitung 306. 310. 323.
- Reizmengengesetz 306. 310. 324.
- Reizperzeption 306. 310.
- Reizreaktion 306. 310. 323.
- Reizschwelle 293. 322.
- Reliktendemismen 179.
- Renntierflechte 413. 415.
- Reservestärke 27.
- Reservestoffe 23. 24. 227 ff. 231.
- Reservezellulose 32. 36. 230. *501.
- Resina Guajaci 578.
- Restitution 107. 245. 250. 261. 267. 278.
- Resultante 291. 305. 306. 308. 315. 324.
- Rettich, Knolle 153. 569.
- Rezessiv 281.
- Rhabarber 555.
- Rhabdonema 347.
- Rhamnaceae, Rhamnus *581.
- Rhaphiden *25.
- Rheotaxis 294.
- Rheotropismus 312.
- Rheum *555.
- Rhexigene Interzellularen 38.
- Rhinantheae 163. 223.
- Rhipidium = Fächel.
- Rhipsalis 552.
- Rhizinen 411.
- Rhizoiden 70. 427.
- Rhizom 73. *103. 151 ff. 270. 302. 304.
- Rhizoma Calami 617.
- — Filicis 444.
- — Galangae 633.
- — Graminis 630.
- — Hydrastis 532.
- — Imperatoriae 550.
- — Iridis 623.
- — Rheï 556.
- — Tormentillae 537.
- — Veratri 620.
- — Zedoariae 633.
- — Zingiberis 632.
- Rhizomorpha 241. 408.
- Rhizophora *143. *545.
- Rhizopodien 337.
- Rhizopus *381. 383.
- Rhododendron 280. 583.
- Rhodomela 376.
- Rhodophyceen *67. 371.
- Rhoeadinae 566.
- Rhoeo 624.
- Rhus 581.
- Rhyniaceae 458.
- Ribes *535.
- Riccia *69. 420. *425.
- Richardia 615.
- Richtende Reize 291. 299.
- Richtung der Zellwand 269.
- Richtungsbewegungen 299.
- Ricinus *26. 230. *576.
- Riedgras 625.
- Rinde der Luftwurzeln 116.
- — sekundäre 125.
- — des Stengels 79 ff.
- — der Erdwurzeln 116.
- Rindenbündel 82.
- Rindengrenze in Stengeln 81.
- — in Wurzeln 116.
- Ringelborke 140.
- Ringelblume 607.
- Ringelung 202.
- Ringgefäß *55.
- Ringtracheïde *55.
- Rippen 95.
- Rispe *476.
- Rizinusöl 25. 577.
- Robinia *135. *148. 317. 320. 543.
- Roccella *413. 415.
- Röhrling 406.
- Röntgenstrahlen 254.
- Roggen 204. *628. *629.
- Rohr (Stuhl-) 614.
- Rohrzucker 23. 230. 293. 554. 614. 630.
- Rosa 285. *535. 537.
- Rosiflorae 533.
- Rose von Jericho 295. 569.
- Rosettenpflanzen 151. 249. 270.
- Rosine 275.
- Rosmarinus 592.
- Roßkastanie 251. 581.
- Rostellum 634.
- Rostpilze 399.
- Rotation des Protoplasmas 11. 290.
- Rotalgen 215. 371.
- Rotbuche *560.
- Rotdorn 536.
- Rotierende Nutation 297.
- Rottanne 517.
- Rozites 408.
- Rubia 598.
- Rubidium 205.
- Rubiinae 226. 597.
- Rubus 278. *504. 537.
- Rübe 153. 193. 228. 274.
- — Dickenwachstum 127.
- — Teltower 569.
- Rübsen 569.
- Rückkreuzung 283.
- Rückschlag 262. *263.
- Rüster *563.
- Ruhelage 302.
- Ruheperioden 149. 266.
- — der Keime 164.
- — der Samen 465.
- Ruhezustände 266.
- Rumex 556.
- Ruminiertes Endosperm *526. 614.
- Runkelrübe 127. 554.
- Ruppia 611.
- Ruscus *147. 620.
- Russula *404.
- Ruta, Rutaceae 578.
- Rutengewächse 146.
- Sabadilla 618.
- Saccharomyces *22. *394.
- Saccharum *43. 554. 614. 630.
- SACHSsche Jodprobe *218.
- Sadebaum *515.
- Säulenwurzeln *120.
- Säureausscheidung 208.
- Säuren 201. 208. 220. 232. 267. 292. 311.
- Safran *623.
- Saftfäden 367. 403. 428. 385.
- Saftraum *9. 10. 248.
- Sagittaria *611.
- Sago 614.
- Saison-Dimorphismus 587.
- Salat 605.
- Salbei *591.
- Salep 635.
- Salicornia *147. 554.
- Salix *255. 278. *562.
- Salpeterbakterien 219. 238. 334.
- Salpetersäure 222. 227. 238.
- Salpetersaurer Kalk 209.
- Salpeterpflanzen 222.
- Salpeterwert 193.
- Salpetrige Säure 221. 238.
- Salvia *592.
- — Bestäubung *481.
- Salvinia 444. *445. *446.
- Salzdrüsen 60.
- Salze 188. 204. 206.
- Salzpflanzen 144.
- Salzsäure 227.
- Sambucus *50. 599.
- Samen 165. 171. 228. 266. 267. 326. 465.
- — Ausschleuderung 295. 564.
- — Keimung 171. 235. 507.
- — Ruhe 266. 465.
- — Verbreitung 171. 295. 505.
- Samenanlage 167. 464.
- — dorsal, — ventral 472.
- Samenfarne 461.
- Samenlose Früchte 275.
- Samenmantel 502.
- Samennaht 465.
- Samenpflanzen 334. 464.
- Samenschale 171. 500.
- Sammelart 285.
- Sammelfrucht 504.
- Samolus *106.
- Sandaraca 519.
- Sandelholz 556. 544.
- Sanguisorba *535.
- Santalaceae, Santalum 223. 556.
- Santoninum 607.
- Sapindaceae 156 ff. 581.
- Saponaria *553.
- Sapotaceae 584.
- Saprolegnia *166. 273. *379.
- Saprophyten 220 ff.
- Sarcina 334.
- Sargassum 363.
- Sarothamnus 543.
- Sarraceniaceen 160. 528.
- Sassafras 526. *527.
- Satanspilz 406.
- Saubohne 544.
- Sauerampfer 556.
- Sauerkraut 240.
- Sauermilch 239.
- Sauerstoff 187. 188. 208. 210. 211. 212. 233 ff. 235. 257. 267.
283. 290. 320.
- — als Nebenprodukt der Assimilation *213. 236.
- — bei Atmung verbraucht 233 ff.
- — Bedeutung für Wachstum 257.
- — Bindung 236. 240.
- — Gewinn 240.
- Saugfortsätze *67. 163.
- Saugkraft 192 ff. 196.
- — transpirierender Sprosse 202.
- Saugnäpfe 256.
- Saugung, osmotische 203.
- Saugvermögen 192. 195. 196. 203.
- Saugwürzelchen 120.
- Savanne 186.
- Saxifraga, Saxifragaceae 534.
- Scabiosa 600.
- Scammonium 590.
- Scandix 549.
- Scenedesmus *355.
- Schachtelhalme 206. 295. 448.
- Schafgarbe 606.
- Schattenblätter 98. 253. 257. 270.
- Schattenpflanzen 155. 185. 196. 234. 251.
- Schauapparate 480.
- Scheibenpilze 390.
- Scheide *102.
- — des Blattes 101.
- — Gewebe- 42.
- — Leitbündel- 89.
- Scheinachse *105. 110. *111.
- Scheinfrucht 504.
- Scheitel 64. 73.
- Scheitelwachstum 64. 73.
- Scheitelzellen 39. *69. *70. 73. *74. *113.
- — dreischneidige 70.
- — zweischneidige 70.
- Schichtung d. Zellmembranen im Gewebe 34 ff.
- Schichtzellen 465.
- Schierling 550.
- Schistostega *430. 432.
- Schizaeaceen 442.
- Schizocarpium 504.
- Schizogene Interzellularen 37.
- Schizomycetes 329.
- Schizonema 344.
- Schizophyceen 335.
- Schizophyta 335.
- Schizosaccharomyces 394.
- Schlafstellung *317.
- Schläuche 58.
- Schlauchalgen 33. 67 ff. 359.
- Schlauchpilze 384.
- Schleim 24. 68. 60 ff.
- Schleimdrüsen 60.
- Schlehe 261.
- Schleimgänge 61.
- Schleimpilze 291. 339.
- Schleimröhren 59.
- Schleimsaft 61.
- Schleudern 420. 449.
- Schließfrucht 504.
- Schließhaut von Tüpfeln 30. 36.
- Schließzellen *44 ff. *197. 316.
- Schlingpflanzen 156. 187. *303. 315.
- Schmarotzer *161.
- Schmarotzerpilze *67.
- Schmetterlingsblüte *542.
- Schnecken als Befruchtungsvermittler 482.
- Schneckenklee 543.
- Schneeball 599.
- Schneeglöckchen 622.
- Schnittlauch vgl. Lauch 618.
- Schöllkraut 566.
- Schöpfungstheorie 176.
- Schoenocaulon 618.
- Schote 504.
- Schrägzeilen 78.
- Schraubel 111. *112.
- Schraubengefäß *55.
- Schraubenstellung 78.
- Schraubentracheïde *55.
- Schreckfarben 232.
- Schriftflechte 413.
- Schrumpfung 295.
- Schubfestigkeit *97.
- Schuppen 103.
- — in Winterknospen 150 ff.
- Schuppenbäume 461.
- Schuppenborke 140.
- Schuppenhaare 46. *47.
- Schuppennarben 151.
- Schuppenwurz 595.
- Schutzgummi 140.
- Schutzmittel gegen Bestrahlung 155.
- — gegen Erwärmung 144 ff.
- — gegen Kälte 150 ff.
- — gegen Licht 145 ff.
- — gegen Transpiration 144 ff. 196.
- — gegen Vertrocknung 150 ff.
- Schutzwirkung 227.
- Schwämme 405.
- Schwammparenchym 98.
- Schwärmsporen 166. *169. 290. 328.
- Schwarzkiefer 519.
- Schwarzwurzel 605.
- Schwebeeinrichtungen 64.
- Schweber 64.
- Schwefel 187. 205. 226.
- Schwefelbakterien 219. 238. 334.
- Schwefelregen 478.
- Schwefelsäure 226. 227.
- Schwefelwasserstoff 238. 240.
- Schwellkörper 626.
- Schwellwasser 248.
- Schwerkraft 255. 300. 305.
- Schwertlilie s. Iris *624.
- Schwimmblätter *102. 270.
- Schwimmen 289.
- Schwimmer 64.
- Scilla 618.
- Sciadium 352.
- Sciadopitys 515.
- Scinaia 376.
- Scirpus *625.
- Scitamineae 630.
- Scleroderma *409.
- Scleropodium *430.
- Sclerospora 380.
- Scolopendrium 439. *441.
- Scopolaminum 594.
- Scorzonera *508. 605.
- Scrophularia 595.
- Scutellum *627.
- Scytonema 415.
- Secale *628. *629.
- Secale cornutum 389.
- Sedum *534.
- Seegras 611.
- Seele 323.
- Seerose *529.
- Segmente 68.
- Seidelbast s. Daphne *544.
- Seifenkraut s. Saponaria *553.
- Seismonastie 318.
- Seitenachse 65 ff.
- Seitenwurzeln 65. *115. *119. 194.
- Seitliche Verzweigung 65.
- — bei Wurzeln 119.
- Sekret 58 ff. 227.
- Sekretbehälter 58 ff.
- Sekretgewebe 58.
- Sekretlücken 61.
- Sekretschläuche 58.
- Sekretzellen 58.
- — im Bast 136.
- Sektorialchimäre 262.
- Sekundäres Dickenwachstum des Kormus 120 ff.
- — bei Laminaria 69.
- — bei Wurzeln *126 ff.
- Sekundäre Gewebe 122 ff.
- — Rinde 125.
- — Verdickungsschichten 35.
- Sekundärzuwachs 122 ff.
- Selaginella *103. 189. *456. *457.
- Selbstbestäubung 170. 286. 483.
- Selbsterhitzung 241.
- Selbststerile Blüten 483.
- Selbststeuerung 184.
- Selektion 181. 286.
- Sellerie 549.
- Semen Arecae 614.
- — Calabar 544.
- — Colae 574.
- — Colchici 620.
- — Cydoniae 537.
- — Erucae 569.
- — Foenu graeci 544.
- — Lini 578.
- — Myristicae 526.
- — Papaveris 567.
- — Quercus 562.
- — Sabadillae 621.
- — Sinapis 569.
- — Stramonii 594.
- — Strophanthi 589.
- — Strychni 587.
- Semipermeabel 12. 190. 206.
- Sempervivum 273. 274. 275. 534.
- Senecio 266. 271.
- Senf *307. 569.
- Septicid 504.
- Sequoia 201. 272. 515.
- Seriale Beiknospen 106.
- Serjania, Dickenwachstum *156.
- Serodiagnostik 524.
- Serodiagnostische Stammbäume:
- — bei Centrospermae 552.
- — bei Columniferae 571.
- — bei Dikotylen 524.
- — bei Monokotylen 609.
- — bei Polycarpicae 524.
- — bei Sympetalae 583.
- Serradella 226. 544.
- Seta 428.
- Sexualität 165. 168 ff. 171. 273 ff.
- Sexualorgane 168 ff.
- Sexualsystem 333.
- Sexualzellen, Vereinigung *169.
- Sexuelle Fortpflanzung 165. 168 ff. 243 ff.
- Shepherdia *47. 545.
- Shorea 570.
- Sichel 111. *112.
- Sicyos, Ranke 312. *313.
- Siebparenchym 87.
- Siebplatten 53. *54.
- Siebröhren 54 ff. 232.
- — im Bast 136.
- Siebstränge 57 ff. 87.
- — im Bast 136.
- Siebteil 58. 85.
- Siebtüpfel *54.
- Siegelbäume 460.
- Sigillariaceen 460.
- Silene 278. 316. *553.
- Siler 549.
- Silicium 187. 206.
- Silicoflagellatae 338.
- Silphium 310. 605.
- Simarubaceae 581.
- Sinapis *307. 308. 569.
- Sinne 324.
- Sinnpflanze *319.
- Sinus 305. 306. 324.
- Siphonales 34. 67 ff. 290. 359.
- Siphonocladus 359.
- Siphonogamen 466.
- Sisymbrium *568.
- Sium 549.
- Skatol 240.
- Sklereïden *29. 30.
- Sklerenchym 51 ff. 148.
- Sklerenchymfasern *52. 128.
- Sklerenchymstränge 52. 80 ff. 97. 117 ff.
- Sklerenchymzellen 52.
- Sklerokaulen 146.
- Sklerophyllen 145.
- Sklerotien *34. 341. 380. 389.
- Smilax 312. 620.
- Solanum *27. *152. 262. 278. 314. *592.
- — Chimären 262. *264. 609.
- Sole 12. 189.
- Solidago 605.
- Solorina 414.
- Somatisch 250.
- Sommergrüne 150. 186.
- Sommerholz 131.
- Sonchus 605.
- Sonnenblätter 98. 253.
- Sonnenblume 197. *249. 605.
- Sonnenröschen *569.
- Sonnentau, Tentakeln *159.
- Soral 412.
- Sonneratia *143. 301. 547.
- Sorbus 278. *279. 536.
- Soredien 411.
- Sorghum 629.
- Sorus 365. 440.
- Spadiciflorae, Spadix 612.
- Spätholz 131.
- Spaltalgen 335.
- Spaltfrucht 504.
- Spaltöffnungen 43 ff. *44. 71. 196. *197. 316.
- — Funktion 43 ff. 196. *197. 211 ff.
- — bei Moosen 71. 411. 417. 419.
- Spaltöffnungsapparate 43 ff. *45
- Spaltöffnungszahl in Blättern 97. 197.
- Spaltpflanzen 335.
- Spaltpilze 329.
- Spaltung der Bastarde 281.
- Spanischer Pfeffer 593.
- Spannungen 192. *249. *256.
- Sparassis 405.
- Sparganiaceae 612.
- Spargel 620.
- Sparteïnum 544.
- Spartium 544.
- Spatha 612.
- Spathicarpa 615.
- Speichergewebe 41.
- Speichern 207.
- Speicherorgane 151 ff. *152 ff.
- Speichertracheïden 55. 58.
- Speicherung von Stoffen 151 ff. 207.
- Speiteufel 408.
- Spektrum *214.
- Spelt *628.
- Spelzen 626.
- Spermakern 171. *489.
- Spermatangium 373.
- Spermatien 373. 384. 400. 414.
- Spermatochnus 369.
- Spermatogene Zelle 489.
- Spermatogenes Gewebe 169.
- Spermatophyten 326. 464.
- Spermatozoen 329.
- Spermatozoid 329.
- Spermazelle *466.
- Spermien 168. 290. 292 ff. 329. *486.
- Spermogonien 400. 414.
- Spezielle Botanik 4.
- Sphacelaria *364.
- Sphaeria 388.
- Sphärite 23. 28.
- Sphaerococcus 376.
- Sphärokristalle 23. 28.
- Sphaeroplea 359.
- Sphaerotheca *387.
- Sphagnum 71. 272. 428. *429.
- Sphagnales 420. 427.
- Sphenophyllum *452.
- Sphenopteris *462. 463.
- Spezifische Eigenschaften 262.
- Spica = Ähre.
- Spielarten 287.
- Spinacia, Spinat 554.
- Spindelfasern 18.
- Spiraea, Spiraeaceae *535.
- Spiralstellung 78.
- Spiremstadium 17.
- Spirillum *63. 289. *330.
- Spirochäte *63.
- Spirodinium 343.
- Spirogyra *21. 257. *350.
- Spirophyllum 334.
- Spirotaenia 348.
- Spitze 61.
- Spitzenwachstum 64.
- Splachnum 430.
- Splint, Splinthölzer 135.
- Spontane Bewegungen s. autonome B.
- Sporangien 166. 296. 328.
- Sporangiensporen 166.
- Sporen 164. 166 ff. 186. 254. 266. 276. 326. 328.
- Sporenpflanzen 326.
- Sporenschlauch 385.
- Sporenverbreitung 166. 295.
- Sporidien 397. 400.
- Sporodinia *382.
- Sporogenes Gewebe 166.
- Sporogon 419.
- Sporokarpien 444.
- Sporophyll 167. 435. 475.
- Sporophyt 72. 175. 329. 464.
- Sporotheca 436.
- Spreizklimmer 156.
- Spreuschuppen 46. 440.
- Sproß 62. 73 ff.
- Sproßachse 73.
- — äußerer Bau 75 ff.
- — innerer Bau 79 ff.
- Sproßanlagen *73. *105.
- Sproßentwicklung 73 ff.
- Sproßdornen *148.
- Sproßfolge 108 ff.
- Sproßknollen *152.
- — bei epiphytischen Orchideen 157.
- — bei Sukkulenten 149.
- Sproßpilze 394.
- Sproßranken *156.
- Sproßreduktion bei Parasiten 161.
- — bei Xerophyten 147.
- Sproßscheitel *73 ff.
- Sproßvegetationspunkt *73 ff.
- Stachelbeere 534.
- Stacheln 48.
- Stachelschwämme 406.
- Stachys 592.
- Stärke *27 ff. 217. 220. 228. 232.
- — korrodierte *229.
- Stärkebildner 16. 27.
- Stärkeherde *14.
- Stärkekörner *27. 305.
- Stärkemehl 27.
- Stärkescheide des Stengels 81. 89.
- Stäubling 409.
- Stamen, Stamina 470.
- Staminodien 470.
- Stamm, Dickenwachstum 120 ff.
- — Palmentypus 122.
- Stammbaum 177.
- Stammbäume der Centrospermae 552.
- — der Columniferae 571.
- — der Dikotylen 524.
- — der Monokotylen 609.
- — der Polycarpicae 524.
- — der Sympetalae 583.
- Stammesgeschichte 2.
- Stammglieder 76.
- Stammsukkulenten 149.
- Stangeria 509.
- Stanhopea 505.
- Stapelia *149. 589.
- Staphylococcus *333.
- Starrezustände 290. 297. 320.
- Starrkrampfbazillus 333.
- Stationäre Reize 322.
- Statolithen 305.
- Staubbeutel 296.
- Staubblatt *168. 243. *321. 470.
- Stauden 121. 151 ff.
- Stechapfel *594.
- Stechpalme 581.
- Stecklinge 107. 247. 277.
- Steinbrand 398.
- Steinbrech 200. 534.
- Steineiche, Borke *139.
- Steinfrucht 504.
- Steinklee 544.
- Steinkohle 216.
- Steinkork 50.
- Steinpilz *34. 406.
- Steinzellen *29. 30. 51.
- Stelärtheorie 91.
- Stele 91.
- Stellaria 553.
- Stelzwurzeln *143.
- Stemonitis *340. 342.
- Stempel s. Fruchtknoten.
- Stengel 73.
- — innerer Bau 79 ff.
- — -wachstum 244.
- Stengelglieder 76.
- Stengelstruktur 79 ff.
- Steppengräser 146.
- Sterculiaceae 573.
- Stereome 51.
- Stereum 405.
- Sterigmen 396.
- Stern-Anis 525.
- Sternhaare 46. *47.
- Stickstoff 187. 205. 209. 221 ff. 232. 240. 275.
- Stickstoffbinder *224. 332.
- Stiefmütterchen *46. *48. *569.
- Sticta 414.
- Stigma 472.
- Stigmatomyces *395.
- Stilophora 370.
- Stimmung 299. 304. 310. 324.
- Stimmungswechsel 310.
- Stinkbrand 398.
- Stinkmorchel 410.
- Stipa pennata 295. 520.
- Stipeln 101.
- Stipites Laminariae 368.
- Stipulae 94. 100 ff.
- Stockausschlag 107.
- Stoffaufnahme 188 ff.
- Stoffe, Herkunft 188 ff.
- — Speicherung 207.
- Stoffliche Einflüsse 186. 256. 258.
- Stoffumwandlung 227 ff.
- Stoffwanderung 227 ff.
- Stoffwechsel 182. 187. 188.
- Stoffwechselprodukte 232.
- Stolonen 165.
- Stomata *44 ff.
- Stomatär 197.
- Stoppelschwamm 406.
- Stoßreize 319 ff.
- Sträucher 121. 150.
- Strahlenarten 254.
- Strahlenpilze 395.
- Strahlige Blüten *475.
- Strandpflanzen 194.
- Stratiotes 612.
- Straußfarn 440.
- Streckung der Organe 248.
- Streifung der Zellhaut *31.
- Strelitzia *482. 631.
- Streptochaeta *627.
- Streptococcus 332. *333.
- Strickeria, Pyknide *389.
- Strobilus Lupuli *565.
- Strömung des Plasmas 10.
- Strohblume 605.
- Stroma 17. 398.
- Strophanthus *588. 589.
- Struthiopteris 440. *443.
- Strychnos *586. 587.
- Stützblatt 106.
- Stützfasern 20.
- Stützwurzeln 120.
- Stylus *472.
- Styracaceae, Styrax 589.
- Styrax liquidus 532.
- Suberin 32.
- — -lamellen 48.
- Suberinsäure 33.
- Succisa 600.
- Sukkulente *149. 187. 189. 235. 534. 551. 552. 589. 621. 622.
- Sulfate 23. 205. 226. 240.
- Sulfatreduktion 240.
- Sultanine 275.
- Summierung unterschwelliger Reize 307. 310.
- Sumpfpflanzen, Luftkanäle 142 ff.
- Superphosphat 210.
- Surirella 347.
- Suspensor 433. *488. *492. *497.
- Symbiose 225. 258. 336. 410. 422. 423. 437. 438. 445.
- Symmetrisch 61.
- Symmetrieverhältnisse 61. 245. 254. 268.
- Sympetalae 584.
- Symphytum 590.
- Sympodial 110. *111 ff.
- Sympodium 105. 110. *111 ff.
- Synandrae 600.
- Synapsis 172.
- Synchytrium 378.
- Synkarp 470.
- Synedra 347.
- Synergiden *496. *497.
- Syringa *585.
- Syrupus Cerasorum 537.
- — Mororum 565.
- — Ribium 534.
- — Rhamni catharticae 581.
- — Rubi idaei 537.
- Systematik 3.
- Systeme 325.
- Tabak 204. 241. 260. *595.
- Tabakfermentation 241.
- Tabakrauch 267.
- Tabernaemontana 589.
- Taeniophyllum Zollingeri *148
- Tagesperiode 318.
- Tagesschlaf 317.
- Tagesstellung *317.
- TALBOTsches Gesetz 310.
- Tamarindus 540. *541.
- Tanacetum 606.
- Tanne 110. 260.
- Tannin 24.
- Tapetenzellen 31. 435. 465.
- Taphrina 393. *394.
- Tapioka 576.
- Taraxacum *59. 244. 248. *249. *253. 605. 608.
- Taubnessel s. Lamium 592.
- Taumellolch 631.
- Tausendgüldenkraut 587.
- Taxis 291 ff.
- Taxodium 272. *515.
- Taxus *84. 272. *512. *513.
- Tectona, Teakholz 591.
- Tee *570.
- Teilung der Zelle 17 ff. 268.
- Teleutosporen 400.
- Telophase 19.
- Temperatur, Lebensbedingung 185.
- — Einfluß auf Assimilation 217.
- — Einfluß auf Atmung 235.
- — Einfluß auf Wachstum 250. *251. 267.
- — Einfluß auf Blütenbildung 274.
- Temperaturwechsel als Bewegungsreiz 316.
- Tentakeln *159. 318.
- Terebinthina 519.
- Terfezia 388.
- Terminalia 506.
- Ternstroemiaceae 570.
- Tertiäre Verdickungsschichten 35.
- Testobjekte 348.
- Tetanusbazillus 333.
- Tetraden 419. 435.
- Tetrarch 117.
- Tetracyclicae 584.
- Tetragonolobus 544.
- Tetraploidie bei Moosen 176.
- Tetrasporangien, Tetrasporen 365. 379.
- Teucrium 592.
- Teufelsei 410.
- Teufelszwirn s. Cuscuta 589.
- Thalictrum 277.
- Thallöse Pflanzen 63.
- Thallophyta 327.
- Thallus 63 ff.
- Thamnidium 383.
- Thea *570.
- Theca 470.
- Thecotheus 386.
- Thelephora 405. 412. 415.
- Theobroma *573.
- Thermonastie 316. 318.
- Thermotropismus 312.
- Thermotaxis 294.
- Thermophile 241.
- Thesium 163. 223. 556.
- Thigmonastie 318.
- Thigmotropismus 312.
- Thomasmehl 210.
- Thomasschlacke 210.
- Thuja 514.
- Thyllen *135.
- Thymelaeaceae 544.
- Thymolum 392.
- Thymus, Thymian 592.
- Tiefenlage 153 ff. 304. *305.
- Tiere und Pflanzen 1. 2. 3. 182.
- — als Bestäubungsvermittler 170. 491 ff.
- — Samenverbreiter 171. 520.
- Tier 223. 232. 235. 248. 322 ff.
- — Ernährung 220.
- — Stoffaufnahme 188.
- Tierfangende Pflanzen 158 ff. 528.
- Tilia *133 ff. 572.
- Tilletia *398.
- Tilopteris 365.
- Tmesipteris 458.
- Tochterachse *65. 66.
- Tochterchromosomen 18.
- Tod 186. 272.
- Tollkirsche *593.
- Toluifera s. Myroxylon 544.
- Tolypellopsis 371.
- Tomate *264. 593.
- Tonsille 190.
- Topinambur 605.
- Topisch 291.
- Torenia 247. 321. 595.
- Torreya *489. *491.
- Torf 215.
- Torfmoose 71. 428.
- Torsion 294. 295. 303. 309.
- — hygroskopische *295.
- — der Schlingpflanzen 304.
- Torus *57.
- Totentrompete 405.
- Tozzia 163. 223. 267. 596.
- Trabeculae 460.
- Tracheale Gewebeart *55 ff.
- Tracheen *55 ff. *128. 209.
- Tracheïden 55. *57. *128. 129. 209.
- Tradescantia *11. *16. *44. 247. 624.
- Träger 81.
- Tränen s. Bluten.
- Träufelspitze 144.
- Tragacantha *543. 544.
- Tragblatt 106.
- Tragmodul d. Sklerenchymfasern 52.
- Tragopogon 605.
- Transfusionsgewebe in Coniferennadeln 96.
- Transpiration 143 ff. 195 ff. 256.
- Transpirationsschutz 259.
- Transpirationsstrom 196 ff. 199 ff. 202. 209.
- Transplantation 260.
- Transversal, rechts, links 108.
- Transversale 108.
- Transversalgeotropismus 302.
- Transversalphototropismus 309.
- Trapa 545.
- Traube 476.
- Traubenzucker 23. 217. 228.
- Trauerwuchs 287.
- Traumatonastie 318 ff.
- Traumatotropismus 312.
- Tremella *396. 403.
- Trennungsphelloide 140.
- Trennungsschicht bei Blattfall 104.
- Trentepohlia 356. 411.
- Treppentracheïde *57.
- Triarch 117.
- Tribonema 351.
- Trichia *340.
- Trichobakterien 334.
- Trichocaulon 589.
- Trichogyn 374. 414.
- Tricholoma 407.
- Trichomanes *442.
- Trichome 46 ff.
- Trichterzellen 98.
- Tricoccae 574.
- Trifolium 298. 304. 589.
- Trigonella 544.
- Triplokaulisch 109.
- Tripperkokken 333.
- Triticum *26. 243. *627. *628. *629. 630.
- Trockenperioden 266.
- Trockenstarre 297.
- Trockensubstanz 187. 212.
- Tropaeolaceae, Tropaeolum *16. *99. 200. 244. 314.
- — Tropfenausscheidung *200.
- Tropen 266.
- Tropismus 299 ff.
- Tropophyten 149 ff.
- Trüffeln 393.
- Trypanosoma *339.
- Trypsin 231.
- Tsetse 339.
- Tsuga 517.
- Tuber *393.
- Tubera = Wurzelknollen.
- Tubera Aconiti 532.
- — Jalapae 590.
- — Salep 635.
- Tuberkelbazillus *333.
- Tubiflorae 589.
- Tüpfel 30. 36. 209. *313.
- — Schließhaut 30.
- — verzweigte 30.
- Tüpfelgefäß *56 ff.
- Tüpfelkanäle 30.
- Tüte beim Ficusblatt 101.
- Tulipa, Tulpe 316. 618.
- Tulpenbaum 525.
- Turgor 29. 191 ff. 248. 294.
- — Aufhebung *192.
- — -schwankungen als Bewegungsursache 197. 294. 298. 317. 319.
- Turio Pini 519.
- Tussilago 606. *608.
- Typhaceae 612.
- Typhusbazillus *333.
- Tyrosin 222.
- Übergang der Wurzel- in die Stengelbündel 117 ff.
- Übergangszellen 96.
- Überwallung 140.
- Ulex 543.
- Ulmus, Ulme *563.
- Ulothrix *169. 356. *357.
- Ulotrichales 356.
- Ultrafilter 207.
- Ultraviolettes Licht 254.
- Ulva *64. 289. 356.
- Umbella = Dolde 475.
- Umbelliflorae 547.
- Umbildungen 6.
- Umstimmung 291. 304. 310.
- Uncaria s. Ourouparia 598.
- Uncinula *387.
- Ungeschlechtliche Fortpflanzung 165 ff.
- Uniformität 280.
- Unterschwellig 310. 322.
- Unterlage 260.
- Unterständiger Fruchtknoten *473.
- Uragoga 598. *601.
- Uredineen 399.
- Uredosporen 401.
- Urginea 620. 621.
- Urmeristeme 39.
- Urnenblätter *157. 589.
- Ursachen des Lebens 182. 185.
- Urtica *46. 222. 565.
- — Bastarde *281.
- Urticinae 563.
- Urwälder, tropische 155.
- Urzeugung 3.
- Usnea *412. 413.
- Ustilago, Ustilagineen. *397. 398. *399.
- Utricularia *159. 596.
- Vaccinium *583.
- Vagina 94.
- Vaginalstipeln 101.
- Vakuolen *9. 10. 189. 248.
- — pulsierende 11.
- Vakuolenwand 12. 207.
- Valeriana, Valerianaceae 599. *601.
- Vallisneria 612.
- Vanilla *634. 635.
- Varec 368.
- Variabilität 277. 280. 285.
- Variation 280.
- Variationsbewegungen 294. 297. 303. 310. 317.
- Variationskurve *286.
- Varietäten 176.
- Vasalteil 58.
- Vaucheria 268. *360. *361.
- Vegetabilisches Elfenbein 36. 614.
- Vegetationskegel *73 ff. *76. *85. *113 ff.
- Vegetationsorgane 62 ff.
- Vegetationsperiode 149 ff.
- Vegetationspunkt 64. 68. *73 ff. *85. 245. 260. 269.
- — apikaler 64.
- — interkalarer 64.
- — der Wurzel *113 ff.
- Vegetationsruhe 149 ff.
- Vegetationsscheitel 64. 73.
- Vegetative Fortpflanzung 165 ff. 276 ff.
- Veilchen 569.
- Veilchenstein 356.
- Velamen radicum 158.
- Velum 407.
- Venation bei Farnen 435.
- Ventral 472.
- Venusfliegenfalle *160.
- Veratrinum 621.
- Veratrum 618. 620.
- Verbascum *595. *596.
- Verbena *590. 591.
- Verbindungsfäden 20.
- Verbreitung, Samen 171. 505.
- Verdauungsdrüsen *60. *159. 200.
- Verdickungsring 122.
- Verdickungsschichten der Zellmembran 35.
- Verdunstung = Transpiration.
- Veredelung *260. *261.
- Vererbung 277 ff.
- Vergeilung *252.
- Vergiftung 185.
- Vergilbung 232.
- Vergißmeinnicht 590.
- Vergrünung 258.
- Verholzung 32. 121.
- Verjüngung 17.
- Verkalkung 43.
- Verkieselung 32. 43.
- Verkorkte Zellen 42.
- Verkorkte Zellgewebe 48 ff.
- Verkorkung 32 ff.
- Verkürzung der Ruheperiode 267.
- Verlängerung 242.
- Vermehrung 169.
- — künstliche 247.
- Vernatio 75.
- Veronica 274. 595.
- Verrucaria 412.
- Verschiebung der Achselknospen 106 ff.
- Verteilung des Wachstums 243.
- Vertrocknen 185.
- Verwachsung, künstliche 260 ff.
- Verwandtschaft 2.
- Verwesung 240.
- Verwundung 241. 256. 267. 290. 320.
- Verzweigung 65. 104 ff. 119.
- — dichotome 66.
- — echte 66.
- — gabelige 65.
- — bei Moosen 70 ff.
- — razemöse 66.
- — seitliche 66.
- — der Sprosse 104 ff.
- — bei Thallophyten 65 ff.
- — unechte *66.
- — der Wurzel 119.
- Verzweigungssysteme *65. 108 ff.
- Vesicaria 569.
- Vibrio 289. *330. *333.
- Viburnum 599.
- Vicia *244. *302. *541. 544.
- Victoria 241. 316. 529.
- Vielkernige Zellen *13.
- Vielzellbildung 21. *22.
- Vinca *586. 589.
- Vincetoxicum *588. 589.
- Vinum 582.
- Viola *46. *48. *503. 569.
- Viscaria *553.
- Viscum *37. 163. 223. 258. *558.
- Vitis 312. 581.
- Viviparie 508. *545.
- Vogelbeerbaum 536.
- Vollruhe 267.
- Volva 407.
- Volvocineen 291. 353.
- Volvox *354.
- VON DER CRONE-Lösung 205.
- Vorblatt *107. 108.
- Vorhof 44.
- Vorläuferspitze 93.
- Vorruhe 267.
- Vorspelze 626.
- Wacholder *514.
- Wachs *43.
- Wachsblume 589.
- Wachstum 182. 242 ff. 297.
- — apikales 39.
- — der Erdwurzeln 115.
- — gleitendes 40.
- — interkalares 40.
- — der Luftwurzeln 115.
- Wachstumsänderungen, stoßweise 243.
- — -bewegungen 289. 297 ff.
- — -dauer 245.
- — -geschwindigkeit 244.
- — -korrelationen s. Korrelation.
- — -krümmungen 294 ff.
- — -messung 242. 243.
- — -perioden 243.
- — -verteilung 243.
- — -im Blatt 92.
- — -in Sproßachsen 75 ff.
- — -in Wurzeln 115.
- Wahlvermögen 207.
- Wald, Regen- 186.
- — sommergrüner 186.
- Waldmeister 598.
- Wallwurz s. Symphytum 590.
- Walnuß *557.
- Wandbelag des Plasmas 10.
- Wanderstoffe 228.
- Wanderung der Assimilate 231.
- — der Nährstoffe 208.
- Wärme, Bedingung zum Leben 185.
- — bei Atmung 241.
- — bei Gärung 241.
- Wärmestarre 290. 297. 320.
- Wärmeverlust 241.
- Wasser 186 ff. 240. 267.
- — Assimilation 227.
- — Atmungsprodukt 234.
- — Aufnahme durch Pflanzen 115. 186. 189 ff.
- — Aufnahme bei Epiphyten 158.
- — Aufnahme durch Sprosse 143.
- — Ausscheidung 60. 144. 200.
- — Funktion in der Pflanze 188.
- — Lebensbedingung 186.
- — Leitung 201 ff.
- — Menge 187.
- — Verbrauch bei Assimilation 212.
- — Schwell- 248.
- — bei Pollenübertragung 170. 479.
- — bei Samenverbreitung 171. 506.
- — bei Wachstum 256.
- Wasserbahnen 55 ff. 202.
- Wasserbehälter 43.
- — bei Epiphyten 157 ff.
- — bei Xerophyten 149.
- Wasserbewegung 201 ff. 209.
- Wasserblätter 270.
- Wasserblüte 336. 338.
- Wasserblütige Pflanzen 170. 479.
- Wasserdampf 311.
- Wasserdrüsen 60.
- Wasserfarne 444.
- Wassergehalt (Boden u. Luft) 256.
- Wassergewebe 42. 149.
- — epidermales 43.
- — im Mesophyll 98.
- — bei Xerophyten 149.
- Wasserhahnenfuß *102. 531.
- Wasserkultur *204 ff.
- Wassernetz 356.
- Wassernuß 545.
- Wasserpest s. Helodea 612.
- Wasserpflanzen 141 ff. 187. 208. 213. 253. 259. 267. 276.
- — amphibische 180. 259.
- — Sauerstoffausscheidung im Licht *213.
- — Luftkanäle 142 ff.
- Wasserreservoir *149.
- Wassersättigung 190.
- Wasserschierling *549.
- Wasserschwankung 294.
- Wasserspalten 98 ff. *100. 144. 200.
- Wasserspeicher 97. *149.
- Wasserstoff 187. 205. 238.
- — -akzeptor 237. 240.
- — -bakterien 219. 238.
- — -entzug 237.
- Wasserversorgung bei Epiphyten 157 ff.
- Watte 46. *47.
- WEBERsches Gesetz 293. 315. 324.
- Wechselfeuchte Klimate 150.
- Wegerich s. Plantago 596.
- Weide 247. *255. *562.
- Weidenröschen 545.
- Weidenrosen 258.
- Weiderich 545.
- Weihrauch 581.
- Wein 238. 581.
- Weinranken 581.
- Weinsäure 23. 220.
- Weinstock *54. 581.
- — Falscher Mehltau 380.
- — Mehltau 387.
- Weißbuche *559.
- Weißbunte Pflanzen 15.
- Weißdorn 261. 536.
- Weißtanne *516.
- Weitholz 131.
- Weizen *26. 228. *627. *628. *629.
- Welken 185. 195.
- Welwitschia 519. *520.
- Wermut 605.
- Weymouthskiefer 519.
- Wicke 544.
- Wickel *112. 477.
- Wickstroemia 277.
- Widerstände, Wachstum gegen 255.
- Widerstand gegen Hitze 185.
- — Kälte 185.
- Widerton 428.
- Wilder Wein 581.
- Willoughbeia 589.
- Windblütig 170. 478.
- Winde 589.
- Windepflanzen 156. 270. *303.
- Wind als Pollen- und Samen- Transportmittel 170 ff. 478. 505.
- Windung *294.
- Winterknospen *150.
- Winterruhe 149. 266.
- Wirt 162. 221.
- Wirtel 76.
- Wistaria 544.
- Wohlverleih s. Arnica 606.
- Wolfsmilch 574.
- Wollen 323.
- Wollgras 626.
- Wollhaare *47.
- Wollkraut = Verbascum 595.
- Woodsia *443.
- Würger 596. *598.
- Würmer 258.
- Wüste 186.
- Wüstenpflanzen 144 ff. 195.
- Wundenheilung 140.
- Wundgummi 140.
- Wundholz 140.
- Wundklee 543.
- Wundkork 140.
- Wundreiz 319.
- Wundstoffe 269.
- Wundverschluß 140 ff. 232.
- Wurfmaschine 296.
- Wurmfarn 439.
- Wurzel 112 ff.
- — Äußerer Bau 114 ff.
- — blattbürtige 119.
- — Dickenwachstum *126.
- — fleischige, ihr Bau 127.
- — Geotropismus *302.
- — Gesamtlänge 120.
- — Innerer Bau 115.
- — kontraktile *154. 249.
- — Restitution 245.
- — Säureausscheidung 208.
- — sproßbürtige 119.
- — stengelbürtige 119.
- — Struktur 115 ff.
- — Vegetationsscheitel *113 ff.
- — Verkürzung 249.
- — Wachstumsverteilung 243. *244.
- Wurzelanlagen *119.
- Wurzeldornen 148.
- Wurzeldruck *201. 202.
- Wurzelhaare *46. 47. *115. *194. 208.
- Wurzelhaube *113 ff. 208.
- Wurzelhülle, Epiphyten 158.
- Wurzelkletterer 156. 254.
- Wurzelknöllchen *224. 332.
- Wurzelknollen *153.
- Wurzelkontraktion 115. *154. 249.
- Wurzelmetamorphosen 153.
- Wurzelreduktion bei Parasiten 161 ff.
- Wurzelstock 73. *103. *152.
- Wurzelsystem 120.
- Wurzeltasche 113.
- Wurzelträger 456.
- Wurzelverkürzung *154.
- Wurzelverzweigung *119.
- Xanthophylle 15.
- Xenien 278.
- Xenogamie 170.
- Xerochasie 295.
- Xeromorphie 144.
- Xerophile 144.
- Xerophyten 144 ff. 199.
- Xylem 58. 84.
- Xylemparenchym 87.
- Xylemprimanen 90.
- Yucca 122. 620.
- — Motte 481.
-
- Zahnschleimbakterien *63.
- Zahnwurz, Bulbillen *166.
- Zamia *486. 509.
- Zanardinia 365.
- Zanichellia 611.
- Zaunrübe 601.
- Zaunwinde 589.
- Zea *80. *88. 629.
- Zeiger am Bogen *242.
- Zellarten 38.
- Zellbildung, freie *22.
- Zelle 8. *9 ff.
- — in Befruchtung 171 ff.
- — Dauer- 9.
- — embryonale *9.
- — künstliche 183.
- — nackte 8.
- — Ontogenie 17 ff.
- — somatische 250.
- — vielkernig *13.
- Zellengestalt 8.
- Zellenlehre 8.
- Zellenleib 8.
- Zellenpflanzen 326.
- Zellfäden 68.
- Zellfamilien 34.
- Zellflächen 68.
- Zellfusionen 37. 53. 59.
- Zellgewebe 33.
- Zellgröße 8. 268.
- Zellhaut 8. 29 ff. 189. 209. 248.
- Zellkern 9. 13. 292.
- — Funktionen 14.
- Zellkolonien 34.
- Zellumen 8.
- Zellmembran 29 ff.
- — Chemie 31.
- — Dickenwachstum 29. 35.
- — Flächenwachstum 29.
- — Schichtung 30.
- — Streifung *31.
- — im Zellgewebe 34 ff.
- Zellobiose 32.
- Zellplasma 9. 12.
- Zellplatte 20.
- Zellräume 8.
- Zellsaft 10. 23. 190.
- Zellsprossung *22.
- Zellteilung *18 ff. *21. 252. 268.
- Zellulase 32. 230.
- Zellulose 31 ff. 220. 230. 239.
- Zellverjüngung 17.
- Zellverschmelzungen 37.
- Zellwand 29 ff. 189. 269.
- Zellwandbildung, simultane 20.
- — succedane 20.
- Zellwandverdickungen 29 ff.
- Zentralkörper 343.
- Zentralplacenta *471.
- Zentralspalte 45.
- Zentralwinkelständig 471.
- Zentralzylinder im Stengel 79. 82 ff.
- — der Wurzeln 116.
- Zentrifugale Wandverdickungen 31.
- Zentripetale Wandverdickungen 30 ff.
- Zentrifugalkraft, Wirkung auf Pflanzen 300.
- Zentriolen 9.
- Zentrische Blätter 96.
- Zerteilung 67.
- Ziegenbart 405.
- Zilien 11. 64. 166. *169. 289.
- Zimt 526.
- Zingiber *632.
- Zirbelkiefer s. Arve 519.
- Zirkulation des Protoplasmas *11. 290.
- Zirkumnutation 297.
- Zisternenepiphyten 158.
- Zitrone 580.
- Zitronensäure 293.
- Zitterpilze 403.
- Zittwer 632.
- Zoidiophil 170.
- Zoogloea 330.
- Zoosporen 166. 328.
- Zooxanthella 338.
- Zostera 611.
- Zotten *48.
- Zuchtwahl 181.
- Zucker 23. 201. 212. 217. 218. 232. 234. 236. 293. 311.
- Zuckerahorn 201.
- Zuckerkiefer 519.
- Zuckerrohr *43. 629.
- Zuckerrübe 554.
- Zuckertang 361.
- Zuckerwert 193.
- Zufallkurve 286.
- Zugfasern 20.
- Zugfestigung 62. *117.
- Zugwurzeln *154.
- Zunderschwamm 407.
- Zusammensetzung, chemische 187.
- Zuwachsgröße 242.
- Zweiachsig 109.
- Zweihäusig 485.
- Zweijährige Pflanzen 121. 151.
- Zweizeilige Blattstellung *77. 78.
- Zwerge 256.
- Zwergmännchen 358.
- Zwergkiefer 519.
- Zwetsche 543.
- Zwickel 35.
- Zwiebel *152. 297. 618.
- Zwischenformen 278.
- Zwischenzellräume *35. *36. 37.
- Zwittrig 170. 469.
- Zyanidine 24.
- Zygnema 350.
- Zygogynum 525.
- Zygomorph 62. *475.
- Zygomycetes 381.
- Zygophyceae 348.
- Zygophyllaceae 578.
- Zygosaccharomyces 394.
- Zygospore 168. 276. 328.
- Zygote 168. 328.
- Zyklus 78. 168.
- Zymase 239.
- Zymöse Verzweigung 110 ff.
- — Blütenstände *112. 477.
- Zymogene Bakterien 331.
- Zypresse 514.
- Zytase 36. 230.
- Zysten 343.
- Zystiden 403.
- Zystokarp 374.
- Zystolithen *30.
- Zytologie 7. 8.
- Zytoplasma 9. 12.
- — Bewegung 10 ff.
Druckfehler:
S. 32 Zeile 1 von unten lies statt teilweise: technisch.
S. 67 stelle Zeile 9 von oben hinter Zeile 5.
S. 102 bei Fig. 139 lies Batrachium aquatile statt B. aquatiles.

Verlag von Gustav Fischer in Jena.
Der Preis für die angezeigten Bücher ergibt sich durch
Vervielfältigung der hinter dem Titel stehenden Grundzahl (Gz) mit
der vom Börsenverein der Deutschen Buchhändler jeweils festgesetzten
Schlüsselzahl. Die für gebundene Bücher sich ergebenden Preise
sind nicht verbindlich. — Bei Lieferung nach dem Ausland erfolgt
Berechnung in der Währung des betr. Landes.
Eduard Strasburger
Das botanische Praktikum. Anleitung zum Selbststudium der
mikroskopischen Botanik für Anfänger und Geübtere, zugleich ein
Handbuch der mikroskopischen Technik. Siebente Auflage,
bearbeitet von Dr. Max Koernicke, Prof. d. Botanik a. d.
landwirtschaftl. Hochschule Bonn-Poppelsdorf und d. Univers. Bonn.
Mit 260 Abbild. im Text. XXIV, 883 S. gr. 8o 1923
Gz. 15.—, geb. 17.—
Die Bearbeitung des „Botanischen Praktikums“ durch Professor Koernicke
hat allgemeinen Beifall gefunden, wie der Erfolg der letzten Auflagen
beweist. Die neue Auflage wird den jüngsten wissenschaftlichen
Errungenschaften ebenfalls in hohem Maße gerecht, so daß auch sie ein
unentbehrlicher Begleiter beim botanischen Studium sein wird.
Süddeutsche Apotheker Zeitung 1922, Nr. 14: Das wohl jedem
Mikroskopiker längst bekannte große botanische Praktikum... stellt
sich die Aufgabe, den Anfänger in die mikroskopische Botanik
einzuführen und den Geübteren im Studium zu fördern. Beiden wird
Gelegenheit geboten, nicht nur beobachten zu lernen, sondern sich
auch mit der ganzen modernen mikroskopischen Technik bekannt zu
machen.
Auch Anfänger, die mit dem Gebrauch moderner optischer Instrumente
nicht vertraut sind, die sich also ohne fremde Hilfe in die
mikroskopische Technik einarbeiten wollen, erreichen ihr Ziel
sicher, da das Buch zunächst möglichst wenig voraussetzt und
ganz allmählich zum Schwierigeren übergeht... Am Schluß finden
wir einige sehr ausführliche und mit großer Sorgfalt bearbeitete
Register, die das Buch zu einem idealen Nachschlagewerk und
Ratgeber in allen Fragen der mikroskopischen Technik machen....
Das Praktikum wird in der neuen Auflage noch mehr als früher ein
unentbehrliches Nachschlagewerk bei mikroskopischen Studien
sein und sollte in keinem Laboratorium und in keiner Apotheke
fehlen.
Das kleine botanische Praktikum für Anfänger.
Anleitung zum Selbststudium der mikroskopischen Botanik und
Einführung in die mikroskopische Technik. Zehnte, verbesserte
Auflage, bearbeitet von Dr. Max Koernicke, Prof. d.
Botanik, Bonn. Mit etwa 140 Abbild. im Text. 1923. Im Druck.
Naturwissenschaftl. Wochenschrift 1922, Nr. 14: Das kleine
botanische Praktikum ist im wesentlichen ein Auszug des großen
Praktikums. Es führt deshalb nicht nur wie manche ähnliche Werke
in die Anatomie der höheren Pflanzen, sondern auch in den Bau
und die Fortpflanzungsverhältnisse der Algen und Pilze ein.
Auch die Grundlagen der Fixierungs-, Mikrotom- und Färbetechnik
werden dargestellt. Seine Benutzung empfiehlt sich für jeden, der
eine möglichst umfassende praktische Einführung in die Botanik
erfahren will, also nicht nur für den diese als Hauptfach wählenden
Studierenden, sondern vor allem auch für den zukünftigen Lehrer an
höheren Schulen. Daß sich das Buch in dieser Beziehung bewährt hat,
beweist die rasche Folge der Auflagen.
Nienburg.
Streifzüge an der Riviera. Dritte, gänzlich umgearbeitete
Auflage. Illustriert von Louis Reusch. Mit 85 farbigen
Abbild. im Text. XXVI, 582 S. 1913
Eleg. brosch. 10.—, geb. 13.—
Frankfurter Zeitung, Nr. 130, v. 11. Mai 1913: Strasburger
tritt hier einem größeren Leserkreise nicht nur als Gelehrter,
sondern auch als glänzender Naturbeobachter und feinsinniger
Landschaftsschilderer entgegen. Das Buch enthält weit mehr, als der
Titel vermuten läßt. Sind es auch zunächst botanische Streifzüge,
die der Verfasser mit uns durch die zauberische Frühlingsvegetation
der Riviera unternimmt, so können wir doch auch eine Fülle von
allgemeiner Belehrung daraus schöpfen. Wir verfolgen die Geschichte
der einzelnen Pflanzen, ihre wirtschaftliche Bedeutung, lernen
z. B. die Parfümbereitung in Grasse kennen und lassen uns über
pflanzliche Genuß- und Heilmittel im allgemeinen belehren. Daneben
macht uns Strasburger in angenehmem Plauderton mit dem Klima,
dem Boden, der Geschichte und den Bewohnern der Riviera bekannt,
und einzelne seiner Landschaftsschilderungen (z. B. Mondschein
am Cap Martin) sind in ihrer anschaulichen Naturtreue reine
Kabinettstücke. Einen herrlichen Schmuck des Buches bilden die 85
farbigen Pflanzen- und Landschaftsabbildungen.
Botanik/Zoologie
Lehr- und Hilfsbücher aus dem Verlag von Gustav
Fischer in Jena.
Der Preis für die angezeigten Bücher ergibt sich durch
Vervielfältigung der hinter dem Titel stehenden Grundzahl (Gz.) mit
der vom Börsenverein der deutschen Buchhändler jeweils festgesetzten
Schlüsselzahl. Die für gebundene Bücher sich ergebenden Preise
sind nicht verbindlich. — Bei Lieferung nach dem Ausland erfolgt
Berechnung in der Währung des betr. Landes.
Einführung in die botanische Mikrotechnik. Von Hubert
Sieben, Techniker am Botan. Institut der Univers. Bonn.
Zweite, verm. und verbesserte Auflage. Mit 22 Abbild.
im Text. IX, 114 S. kl. 8o 1920
Gz. 1,75, geb. 3,75
Die botanische Mikrotechnik. Ein Handbuch der
mikroskopischen Arbeitsverfahren. Von Dr. Hans Schneider.
Zweite Auflage des gleichnamigen Werkes von Prof. Dr. A.
Zimmermann. Mit 220 Abbild. im Text. XII, 458 S. gr 8o 1922
Gz. 7.50, geb. 10.—
Inhalt: 1. Das Mikroskop und sein Gebrauch. Allgemeine
Mikrotechnik. Die Freihandtechnik. Das Töten und Aufbewahren
pflanzlicher Objekte. Die Mikrotomarbeit. Das Färben der Präparate.
Das Einschließen der Präparate. Allgemeine Methoden der Verwertung von
Präparaten. — 2. Die wichtigsten qualitativ mikrochemischen Verfahren
zum Nachweis von Pflanzenstoffen. — 3. Die Zellwand: Allgemeines. Die
einzelnen Zellwandstoffe. — 4. Der Protoplast und seine Einschlüsse.
Allgemeines. Der Zellkern und seine Einschlüsse. Zentriolen. Das
Plasma. Die Chromatophoren und ihre Einschlüsse. Andere eiweißartige
Plasmaeinschlüsse. Ölige und gerbstoffhaltige Plasmaeinschlüsse.
Einige andere Plasmaeinschlüsse bei niederen Pflanzen. — 5.
Besondere Methoden zur Untersuchung von Vertretern der verschiedenen
Pflanzengruppen; die wichtigsten Kulturverfahren. — Allgemeines
Register. Register der Objekte.
Erstes mikroskopisches Praktikum. Eine Einführung in den
Gebrauch des Mikroskopes und in die Anatomie der höheren
Pflanzen. Zum Gebrauch in den botanischen Laboratorien und zum
Selbstunterrichte. Für Botaniker, Zoologen, Studierende des höheren
Lehramtes, Pharmazeuten und Chemiker. Von Dr. Arthur Meyer,
o. ö. Prof. d. Botanik u. Direktor d. botan. Gartens a. d. Univers.
Marburg. Dritte, vervollständigte Auflage. Mit 110
Abbild. im Text. V, 255 S. gr. 8o 1915
Gz. 6.50, geb. 8.50
Praktikum für morphologische und systematische Botanik.
Hilfsbuch bei praktischen Übungen und Anleitung zu selbständigen
Studien in der Morphologie und Systematik der Pflanzenwelt. Von
Prof. Dr. Karl Schumann, weil. Kustos am botan. Museum und
Privatdoz. a. d. Univers. zu Berlin. Mit 154 Abbild. im Text. VIII,
610 S. gr. 8o 1904
Gz. 13.—
Anatomie der Pflanze. Von Dr. Hans Molisch, o. ö.
Prof. und Direktor des pflanzenphysiologischen Institutes an der
Univers. Wien. Zweite, neubearbeitete Auflage. Mit
139 Abbild. im Text. VI, 153 S. gr. 8o 1922
Gz. 2.70, geb. 4.20.
Neue Weltanschauung, 1920, Heft 8: .... Das Buch faßt in drei
Abschnitten die wichtigsten Tatsachen übersichtlich und in leicht
verständlicher Sprache zusammen, die die Wissenschaft über die
Anatomie der Pflanzen angesammelt hat. ... Wer sich über den Bau
der Pflanzen schnell unterrichten will, ohne zu den größeren Lehr-
und Handbüchern zu greifen, findet in dem vorliegenden kleinen
Buche das Wissenswerteste zusammengefaßt. Naturgeschichtslehrern an
höheren Schulen wird es im Unterricht gute Dienste leisten können.
Die beigegebenen Abbildungen sind fast sämtlich neu gezeichnet
und durchweg vortrefflich. Freunden der Botanik sei das Buch
angelegentlich empfohlen.
Dr. W. B.
Pathologische Pflanzenanatomie. In ihren Grundzügen
dargestellt von Dr. Ernst Küster, Prof. der Botanik a. d.
Univers. zu Bonn a. Rh. Mit 209 Abbild. im Text. Zweite,
völlig umgearbeitete Auflage. XI, 447 S. gr. 8o 1916
Gz. 12.—, geb. 15.—
Inhalt: Einleitung. — Spezieller Teil: 1. Panaschierung. 2.
Etiolement und verwandte Erscheinungen. 3. Hyperhydrische Gewebe.
4. Wundgewebe und Regeneration. 5. Gallen. — Allgemeiner Teil: 1.
Histogenese der pathologischen Gewebe. 2. Entwicklungsmechanik der
pathologischen Gewebe. 3. Ökologie der pathologischen Gewebe. —
Sachregister.
Naturwissensch. Zeitschr. für Forst- und Landwirtschaft
1916, Heft 5: .... Es befriedigt besonders, nicht nur die sichtende
Hand, sondern auch den klaren, kritischen Geist walten zu sehen
und überall die reiche eigene Erfahrung und Anschauung des Verf.
zu fühlen. Ref. kann getrost sagen, daß es für den Pathologen und
Anatomen das Nachschlagebuch der pathologischen Anatomie ist und
für lange bleiben wird, was allein ausführliche und zuverlässige
Auskunft gibt....
Tubeuf.
Vorlesungen über Pflanzenphysiologie. Von Dr. Ludwig
Jost, o. ö. Prof. an der Univers. Straßburg. Dritte
Auflage. Mit 194 Abbild. im Text. XVI, 760 S. gr. 8o 1913
Vierte Auflage in Vorbereitung.
Pflanzenphysiologie als Theorie der Gärtnerei. Von
Dr. Hans Molisch, o. ö. Prof. und Direktor des
pflanzenphysiolog. Instituts an der Universität Wien. Für
Botaniker, Gärtner, Landwirte, Forstleute und Pflanzenfreunde.
Fünfte, neubearb. Auflage. Mit 151 Abb. i. Text. X,
337 S. gr. 8o 1922 Gz. 6.—, geb. 8.—
Inhalt: I. Ernährung. 1. Die Wasserkultur. 2./3.
Die unentbehrlichen und die entbehrlichen Aschenbestandteile.
4. Stickstoff. 5. Der Boden. 6. Die Düngung. 7. Die
Kohlensäureassimilation. 8. Das Wasser und seine Bewegung. 9. Die
Transpiration und der Transpirationsstrom in Beziehung zu gärtnerischen
Arbeiten. 10. Die Wanderung der Assimilate. 11. Die Ernährung der
Pilze. 12. Ernährungsweisen besonderer Art. — II. Atmung. —
III. Wachstum. 1. Allgemeines. 2. Wachstum und Außenbedingungen.
3. Wachstumsbewegungen. 4. Organbildung. 5. Ruheperiode, Treiberei und
Laubfall. — IV. Vom Erfrieren und Gefrieren der Pflanzen. — V.
Die ungeschlechtliche und die geschlechtliche Fortpflanzung. —
VI. Die Keimung der Samen. — VII. Variabilität, Vererbung
und Pflanzenzüchtung. — Sachregister.
Das Erscheinen von fünf Auflagen innerhalb 7 Jahren (die 1. Aufl.
erschien 1916) ist wohl die beste Empfehlung für dieses Buch; es nimmt
bereits einen ehrenvollen Platz in der gärtnerischen- und in der
botanisch-fachwissenschaftlichen Literatur ein.
Pflanzenphysiologie. Von Prof. Dr. R. Kolkwitz,
Dahlem-Steglitz. Versuche und Beobachtungen an höheren und
niederen Pflanzen einschließlich Bakteriologie und Hydrobiologie
mit Planktonkunde. Zweite, verbesserte und vermehrte
Auflage. Mit 153 Abbild. im Text und 12 zum Teil farbigen
Tafeln. VI, 304 S. gr. 8o 1922
Gz. 5.50, geb. 8.50
Aufgaben und Ziele einer vergleichenden Physiologie auf
geographischer Grundlage. Von Dr. Hans Fitting, o. ö.
Professor der Botanik. 42 S. gr. 8o 1922
Gz. —.90
Die Pflanze als lebender Organismus. Von Dr. Hans
Fitting, o. ö. Prof. der Botanik an der Universität Bonn. 44 S.
gr. 8o 1917
Gz. —.90
Lehrbuch der Pharmakognosie. Von Dr. George
Karsten, o. ö. Prof. a. d. Univers. Halle a. S., und Dr.
Wilhelm Benecke, o. ö. Prof. a. d. Univers. Münster i. W.
Dritte, vollständig umgearbeitete Auflage von G.
Karstens Lehrbuch der Pharmakognosie. Mit 544 zum Teil farbigen
Abbild. im Text. VI, 398 S. gr. 8o 1920
Gz. 7.—, geb. 9.—
Pharmazeut. Zeitung, 1921, Nr. 16: ... Das Werk ist schon
längst zu einem unentbehrlichen Handbuch geworden, so daß es
Eulen nach Athen tragen hieße, darüber noch ein Wort des Lobes
zu verlieren. Es ist für diesen Teil der Wissenschaft eben das
grundlegende Werk.
Dr. R. M.
Mikroskopisches Drogenpraktikum. In Anlehnung an die 5.
Ausgabe des deutschen Arzneibuches. Von Wilhelm Benecke,
a. o. Prof. a. d. Univers. Berlin. Mit 102 vom Verf. gezeichneten
Abbild. im Text. VI, 95 S. gr. 8o 1912
Gz. 3.—, geb. 5.—
Aus pharmazeutischer Unterrichtstätigkeit entstanden, verfolgt das
vorliegende neue Praktikum ein durchaus praktisches Ziel: es gibt eine
kurze und übersichtliche Darstellung der mikroskopischen Charaktere der
wichtigsten Drogen in Wort und Bild, welche den Studenten orientieren
soll über die mikroskopischen Merkmale der Drogen, zu deren genauerer
Durcharbeitung die Zeit im Kolleg nicht reichte. Darüber hinaus wird
es aber auch von Apothekern gewiß gern als ein Atlas zum deutschen
Arzneibuch benutzt werden.
Mikrochemie der Pflanze. Von Dr. Hans Molisch, o. ö.
Prof. und Direktor des pflanzenphysiologischen Instituts a. d.
Univers. Wien. Dritte, neubearbeitete Auflage. Mit
135 Abbild. im Text. XII, 438 S. gr. 8o 1923
Gz. 8.—, geb. 11.—
Mikrokosmos, 1921/22, Heft 5: Ein glänzendes Zeugnis
deutscher Forschertätigkeit. Zwar sagt der Name Molisch schon
alles, aber die peinliche Arbeit, die zahlreichen vorzüglichen
Abbildungen, die erschöpfende Behandlung des Riesenstoffes, die
mehr als ausführlichen Literaturnachweise nach jedem Abschnitt,
die große Übersicht und der ganz wundervolle Stil, der hier ein
so schwieriges Kapitel des Wissens in geradezu genußreicher,
lesenswerter Form bringt, das muß alles betont werden, wenn man
dieser Neuauflage gerecht werden will. Ein Meisterwerk. Wir
können das Buch, das für den ernsten mikroskopierenden Botaniker
unentbehrlich ist, nur angelegentlich empfehlen.
Dr. Stehli.
Allgemeine Biologie. von Oscar Hertwig.
Sechste und siebente, verbesserte und erweiterte
Auflage, bearbeitet von Oscar Hertwig †, o. Prof.
der Anatomie in Berlin und Günther Hertwig, a. o. Prof. der
Anatomie in Rostock i. M. Mit 496 teils farbigen Abbild. im Text.
XVII, 822 S. gr. 8o 1923
Gz. 10.—, geb. 13.—
Naturwissenschaftl. Wochenschrift 1920, Nr. 30: Hertwigs
„Allgemeine Biologie“ bedarf einer besonderen Empfehlung
nicht mehr. Es wird nicht viele Biologen geben, seien es nun
Naturwissenschaftler im engeren Sinne, oder seien es über ihr
Fachgebiet hinaus interessierte Mediziner, denen das Buch unbekannt
geblieben ist. Wer sich über Morphologie und Biologie der Zelle,
dieses Thema im weitesten Sinne gefaßt, unterrichten will, der
findet in der „Allgemeinen Biologie“ ein außerordentlich reiches
Tatsachenmaterial zusammengetragen und wohlverarbeitet, und auch
der Spezialist auf dem Gebiete kann manche Anregung aus dem Buche
schöpfen....
Nachtsheim.
Das Werden der Organismen. Zur Widerlegung von Darwins
Zufallstheorie durch das Gesetz in der Entwicklung. Von Prof. Dr.
Oscar Hertwig, Berlin. Dritte, verbesserte Auflage.
Mit 115 Abbild. im Text. XX, 686 S. gr. 8o 1922
Gz. 10.50, geb. 14.50
Inhalt: 1. Die älteren Zeugungstheorien. 2. Die Stellung
der Biologie zur vitalistischen und mechanistischen Lehre vom
Leben. 3. Die Lehre von der Artzelle als Grundlage für das Werden
der Organismen. 4. Die allgemeinen Prinzipien, nach denen aus den
Artzellen die vielzelligen Organismen entstehen. 5. Die Umwertung des
biogenetischen Grundgesetzes. 6. Die Erhaltung des Lebensprozesses
durch die Generationsfolge. 7. Das System der Organismen. 8. und 9.
Die Frage nach der Konstanz der Arten. 10., 11., 12. Die Stellung der
Organismen im Mechanismus der Natur. 13. Das Problem der Vererbung.
14. Der gegenwärtige Stand des Vererbungsproblems. 15. Lamarckismus
und Darwinismus. 16. Kritik der Selektions- und Zufallstheorie. 17.
Zusammenfassung. Nachwort zur ersten bis dritten Auflage. — Register.
Lehrbuch der Zoologie. Von Dr. Richard Hertwig,
o. ö. Prof. der Zoologie und vergleichenden Anatomie an der
Universität München. Dreizehnte, vermehrte und verbesserte
Auflage. Mit 588 Abbild. im Text. XVI, 682 S. gr. 8o 1922
Gz. 10.—, geb. 12.—
Lehrbuch der Zoologie für Studierende. Von Dr. J. B.
V. Boas, Prof. der Zoologie an der Kgl. landwirtschaftlichen
Hochschule in Kopenhagen. Neunte Auflage. Mit 683 Abbild. im
Text. XI, 735 S. gr. 8o 1922
Gz. 14.—, geb. 17.—
Vorlesungen über allgemeine Histologie. Gehalten an der
Hochschule für Frauen in St. Petersburg. Von Prof. Dr. Alexander
Gurwitsch, St. Petersburg. Mit 204 Abbild. im Text. VI, 345 S.
gr. 8o 1913
Gz. 11.—, geb. 13.—
Anleitung zu makroskopisch-zoologischen Übungen. Von Dr.
H. F. Nierstrasz, Prof. an der Reichsuniversität zu Utrecht,
und Dr. G. Chr. Hirsch, Privatdozent an der Universität zu
Utrecht.
Heft 1: Wirbellose Tiere. VII, 103 S. gr. 8o 1922
Gz. 1.50, geb. 3.50
Leitfaden für das zoologische Praktikum. Von Dr. Willy
Kükenthal, o. ö. Prof. der Zoologie und vergleich. Anatomie
an der Universität Breslau. Achte, umgearbeitete
Auflage. Mit 174 Abbild. im Text. VIII, 322 S. gr. 8o 1920
Z. Zt. vergriffen. 9. Auflage in Vorbereitung.
Leitfaden für das mikroskopisch-zoologische Praktikum.
Von Dr. Walter Stempell, Prof. d. Zoologie u. vergleich.
Anatomie an der Westfäl. Wilhelms-Universität zu Münster i. W.
Zweite, vermehrte und verbesserte Auflage. Mit 86
Abbild. im Text. VI, 105 S. gr. 8o 1919
Gz. 2.75. geb. 4.75
Elemente der Tierphysiologie. Ein Hilfsbuch für Vorlesungen
und praktische Übungen an Universitäten und höheren Schulen, sowie
zum Selbststudium für Zoologen und Mediziner. Von Dr. Walter
Stempell, o. ö. Prof. der Zoologie, vergleich. Anatomie u.
Physiologie, Direktor d. zoolog. Instituts d. Universität Münster
i. W., und Dr. Albert Koch, Privatdoz. d. Zool. a. d.
Univers., Vorst. d. zool. Abtlg. d. Anstalt f. Pflanzenschutz d.
Landwirtschaftskammer Westfalen zu Münster i. W. Zweite,
neubearbeitete und erweiterte Auflage. Mit 373 Abbild. im
Text. XXIX, 758 S. gr. 8o 1923
Im Druck.
Praktikum der Insektenkunde nach biologisch-ökologischen
Gesichtspunkten. Von Prof. Dr. Walter Schoenichen.
Zweite, vermehrte und verbesserte Auflage. Mit 261
Abbild. im Text. X, 227 S. gr. 8o 1921
Gz. 4.—, geb. 6.—
Grundzüge der Hydrobiologie. Von Prof. Dr. Ernst
Hentschel, Leiter der hydrobiolog. Abteil. d. Zool.
Staatsinstituts zu Hamburg, Privatdozent an der Hamburg.
Universität. Mit 100 Abbild. im Text. VII, 221 S. gr. 8o 1923
Gz. 4.—, geb. 6.—
Die Vererbungslehre in gemeinverständlicher Darstellung ihres
Inhalts. Von Dr. Johannes Meisenheimer, ord. Prof. der
Zoologie an d. Universität Leipzig. Mit 49 Abbild. im Text. V, 131
S, gr. 8o 1923
Im Druck.
Erblichkeit und Chromosomen. Eine gemeinverständliche
Darstellung. Von Dr. Theo. J. Stomps, Prof. d. Bot. an d.
Univers. Amsterdam. Aus dem Holländischen ins Deutsche übersetzt
von Dr. Paul von Dall’Armi. Mit 24 Abbild. im Text (nach
Zeichnungen des Verf.). VIII, 158 S. gr. 8o 1923
Gz. 3.50
Inhalt: Einleitung. — I. Chromosomen. 1. Der Bau des
Protoplasten. 2. Die Zell- und Kernteilung. 3. Die Reduktionsteilung.
— II. Erblichkeit. 1. Die stofflichen Träger der erblichen
Eigenschaften. 2. Die Lokalisation der erblichen Eigenschaften. — III.
Die Chromosomen, die stoffliche Basis der Erblichkeit. 1. Beobachtungen
über die Bedeutung des Kernes im Leben der Zelle. 2. Die Untersuchungen
von Th. Boveri über Bastardierung und Merogonie bei Seeigeln.
3. Die Versuche von C. Herbst über künstliche Parthenogenese,
gepaart mit Bastardierung. 4. Die Untersuchungen von Th. Boveri
über doppelte Befruchtung bei Seeigeln. 5. Chromosomen und Geschlecht.
6. Chromosomen und Mendels Regeln. — Schlußwort. Register.
Vorträge über Deszendenztheorie. Gehalten an der Universität
Freiburg i. Br. Von Prof. August Weismann. Dritte,
verbesserte Auflage. Mit 141 Abbild. im Text und 3 farb.
Taf. XXIV, 697 S. 1913
Gz. 11.—, geb. 14.50
Zentralblatt f. Zoologie, allg. u. exper. Biologie, Bd.
6:... Verf. ist wie kein anderer vor ihm tief und erfolgreich in
die schwierigsten Probleme der Abstammungslehre eingedrungen,
mit genialem Blick das Ganze überschauend, und hat uns dabei
nicht nur mit neuen grundlegenden Einsichten beschenkt, sondern
auch der biologischen Forschung Richtung gebend neue Wege
gewiesen... Was auch immer das Schicksal dieses, die Ergebnisse
eines arbeitsreichen und arbeitsfreudigen Forscherlebens
zusammenfassenden Werkes sein möge, eines steht schon heute fest:
Weismanns „Vorträge über Deszendenztheorie“ sind ein
klassisches Werk, dessen ebenso weitspannende, wie tiefgründige
Gedankenwelt noch auf lange Jahre hinaus in der wissenschaftlichen
Biologie befruchtend nachwirken wird.
F. v. Wagner.
Einführung in die Deszendenztheorie. Sechs Vorträge,
gehalten von Karl Camillo Schneider, a. o. Prof. der
Zoologie an der Universität Wien. Zweite, erweiterte
Auflage. Mit einer Karte, 182 teils farbigen Abbild. im Text u.
3 Tafeln. XII, 386 S. gr. 8o 1911
Gz. 9.50, geb. 12.50
Der biologische Lehrausflug. Ein Handbuch für Studierende
und Lehrer aller Schulgattungen. Unter Mitwirkung von
hervorragenden Fachmännern herausgegeben von Prof. Dr. Walther
Schoenichen. Mit 37 Abbild. im Text. XI, 269 S. gr. 8o 1922
Gz. 6.—, geb. 8.—
Inhalt: I. Botanik. 1. Botanische Lehrausflüge. Von
Dr. Eberh. Ulbrich, Kustos am Botan. Museum Berlin-Dahlem. 2.
Führungen im botanischen Garten. Von Prof. Dr. Ludw. Diels,
Direktor d. Botan. Gartens zu Berlin-Dahlem. — II. Zoologie.
3. Der zoologische Lehrausflug. Von Dr. Paul Deegener, Prof.
a. d. Univers. Berlin. 4. Der ornithologische Lehrausflug. Von Prof.
Dr. Bernh. Hoffmann-Dresden. 5. Der entomologische Lehrausflug.
Von Prof. Dr. Rich. Vogel, Privatdoz. d. Zoologie a. d.
Univers. Tübingen. 6. Führungen im zoologischen Garten. Von Prof. Dr.
Walther Schoenichen-Berlin. — III. Allgemeine Biologie.
7. Der hydrobiologische Lehrausflug: I. Binnengewässer. Von Prof. Dr.
August Thienemann-Plön. (Mit 37 Abb.) II. Die Meeresküste.
Von Dr. Arthur Hagmeier, Kustos a. d. Staatl. Biolog. Anstalt
auf Helgoland. 9. Die Untersuchung von Lebensgemeinschaften. Von
Oberstudiendir. Prof. Dr. Karl Matzdorff-Berlin. 10. Botanische
und zoologische Naturdenkmäler. Von Prof. Carl Schulz-Berlin.
— IV. Angewandte Biologie. 11. Der landwirtschaftliche
Lehrausflug. Von Prof. Dr. Wilh. Seedorf-Göttingen.
12. Ausflüge in Baumschulen und Gärtnereien. Von Prof. Dr.
Paul Graeber-Berlin. 13. Volkstümliche und künstlerische
Gartengestaltung. Von Prof. Dr. Ernst Küster-Gießen. 14. Der
forstwirtschaftlich-biologische Lehrausflug. Von Geh. Reg.-Rat Dr.
Karl Eckstein, Prof. a. d. forstl. Hochschule Eberswalde.
15. Der fischereiwirtschaftliche Lehrausflug. Von Geh. Reg.-Rat Dr.
Karl Eckstein, Prof. a. d. forstl. Hochschule Eberswalde. —
Sachregister.
Ein Lehrjahr in der Natur. Anregungen zu biologischen
Spaziergängen für Wanderer und Naturfreunde. Von Prof. Dr. Paul
Deegener. Zwei Teile. VIII, 204 und 298 S. gr. 8o 1922
Gz. 7.50, geb. 11.50

 einjährige „annuelle“,
einjährige „annuelle“,
 zweijährige „bienne“ Pflanzen,
zweijährige „bienne“ Pflanzen,
 ausdauernde „perennierende“ Stauden; Holzgewächse:
ausdauernde „perennierende“ Stauden; Holzgewächse:
 Sträucher
und
Sträucher
und  Bäume. Einen besonderen Typus des Baumes finden wir in
den säulenförmigen, meist unverzweigten Stämmen der Palmen und
Farnbäume, denen mit dem sekundären Dickenwachstum ein echtes
Holzgewebe fehlt.
Bäume. Einen besonderen Typus des Baumes finden wir in
den säulenförmigen, meist unverzweigten Stämmen der Palmen und
Farnbäume, denen mit dem sekundären Dickenwachstum ein echtes
Holzgewebe fehlt.
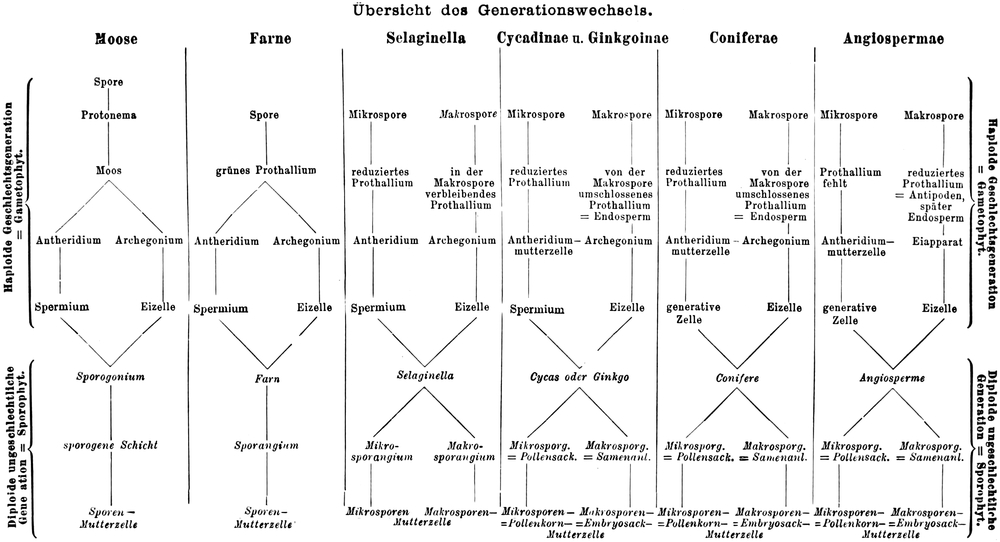
{kind=link}
{kind=link}
{kind=link}
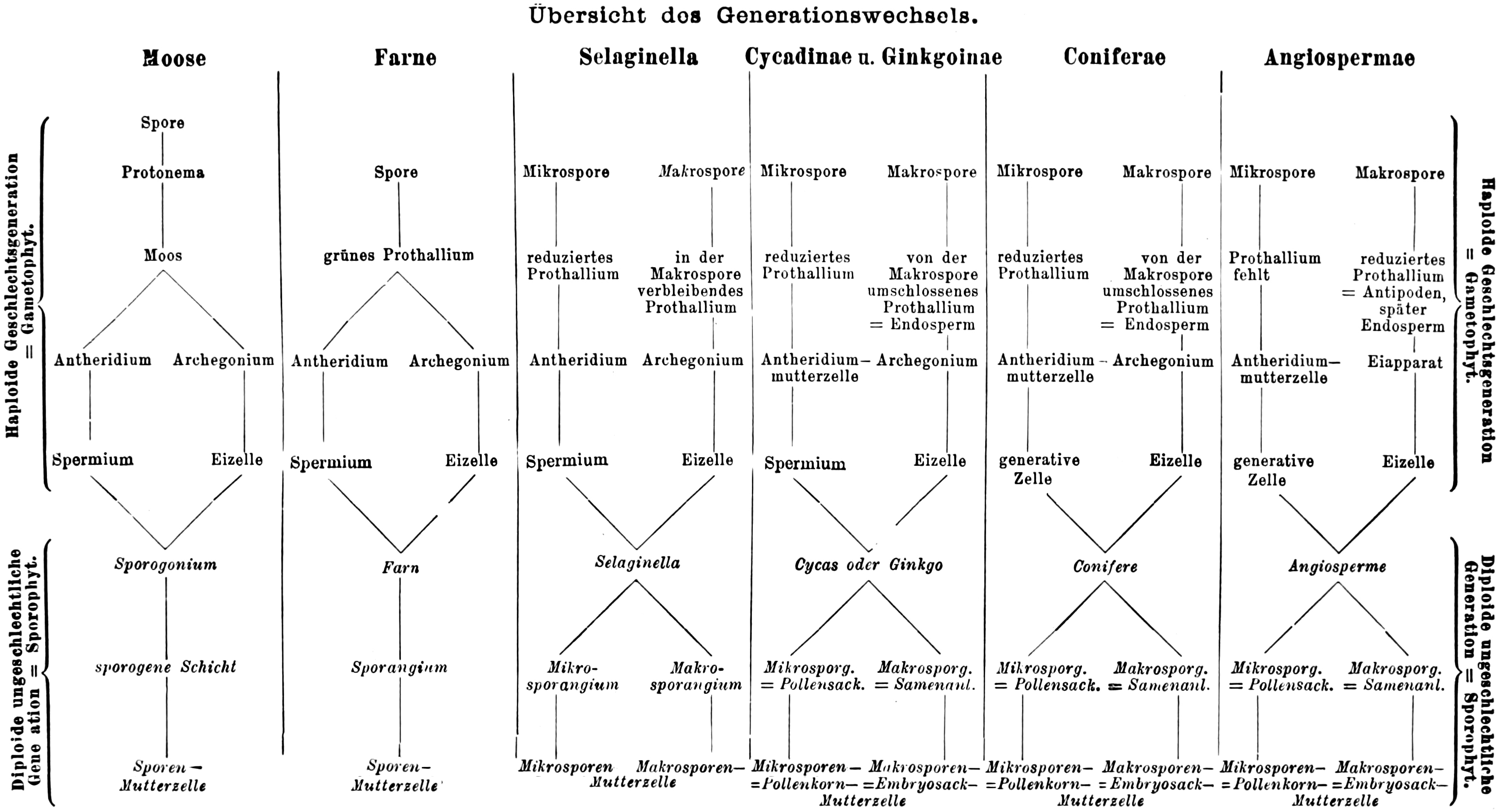{kind=link}