
Title: The Evolution Theory, Vol. 1 of 2
Author: August Weismann
Translator: J. Arthur Thomson
Margaret R. Thomson
Release date: January 6, 2021 [eBook #64227]
Most recently updated: October 18, 2024
Language: English
Other information and formats: www.gutenberg.org/ebooks/64227
Credits: Constanze Hofmann, Alan, Marilynda Fraser-Cunliffe and the Online Distributed Proofreading Team at https://www.pgdp.net (This book was produced from images made available by the HathiTrust Digital Library and The Internet Archive.)
VOLUME I
THE
EVOLUTION THEORY
BY
Dr. AUGUST WEISMANN
PROFESSOR OF ZOOLOGY IN THE UNIVERSITY OF FREIBURG IN BREISGAU
TRANSLATED WITH THE AUTHOR'S CO-OPERATION
BY
J. ARTHUR THOMSON
REGIUS PROFESSOR OF NATURAL HISTORY IN THE UNIVERSITY OF ABERDEEN
AND
MARGARET R. THOMSON
ILLUSTRATED
IN TWO VOLUMES
VOL. I
LONDON
EDWARD ARNOLD
41 & 43 MADDOX STREET, BOND STREET, W.
1904
All rights reserved
AUTHOR'S PREFACE
When a life of pleasant labour is drawing towards a close, the wish naturally asserts itself to gather together the main results, and to combine them in a well-defined and harmonious picture which may be left as a legacy to succeeding generations.
This wish has been my main motive in the publication of these lectures, which I delivered in the University of Freiburg in Breisgau. But there has been an additional motive in the fact that the theory of heredity published by me a decade ago has given rise not only to many investigations prompted by it, but also to a whole literature of 'refutations,' and, what is much better, has brought to light a mass of new facts which, at first sight at least, seem to contradict my main theory. As I remain as convinced that the essential part of my theory is well grounded as I was when I first sketched it, I naturally wish to show how the new facts may be brought into harmony with it.
It is by no means only with the theory of heredity by itself that I am concerned, for that has served, so to speak, as a means to a higher end, as a groundwork on which to base an interpretation of the transformations of life through the course of the ages. For the phenomena of heredity, like all the functions of individual life, stand in the closest association with the whole evolution of life upon our earth; indeed, they form its roots, the nutritive basis from which all its innumerable branches and twigs are, in the long run, derived. Thus the phenomena of the individual life, and especially those of reproduction and inheritance, must be considered in connexion with the Theory of Descent, that the latter may be illumined by them, and so brought nearer our understanding.
I make this attempt to sum up and present as a harmonious[Pg vi] whole the theories which for forty years I have been gradually building up on the basis of the legacy of the great workers of the past, and on the results of my own investigations and those of many fellow workers, not because I regard the picture as complete or incapable of improvement, but because I believe its essential features to be correct, and because an eye-trouble which has hindered my work for many years makes it uncertain whether I shall have much more time and strength granted to me for its further elaboration. We are standing in the midst of a flood-tide of investigation, which is ceaselessly heaping up new facts bearing upon the problem of evolution. Every theory formulated at this time must be prepared shortly to find itself face to face with a mass of new facts which may necessitate its more or less complete reconstruction. How much or how little of it may remain, in face of the facts of the future, it is impossible to predict. But this will be so for a long time, and it seems to me we must not on that account refrain from following out our convictions to the best of our ability and presenting them sharply and definitely, for it is only well-defined arguments which can be satisfactorily criticized, and can be improved if they are imperfect, or rejected if they are erroneous. In both these processes progress lies.
This book consists of 'Lectures' which were given publicly at the university here. In my introductory lecture in 1867 I championed the Theory of Descent, which was then the subject of lively controversy, but it was not till seven years later that I gave, by way of experiment, a short summer course with a view to aiding in the dissemination of Darwin's views. Then very gradually my own studies and researches and those of others led me to add to the Darwinian edifice, and to attempt a further elaboration of it, and accordingly these 'Lectures,' which were delivered almost regularly every year from 1880 onwards, were gradually modified in accordance with the state of my knowledge at the time, so that they have been, I may say, a mirror of the course of my own intellectual evolution.
In the last two decades of the nineteenth century much that is new has been introduced into biological science; Nägeli's idea of 'idioplasm'—the substance which determines form; Roux's Struggle of the Parts, the recognition of a special hereditary substance, 'the germ-plasm,' its analysis into chromosomes, and its continuity from generation to generation; the potential immortality of unicellular organisms and of the germ-cells in contrast to the natural death of higher forms and 'bodies'; a deeper interpretation of mitotic nuclear division, the discovery of the centrosphere—the marvellous dividing apparatus of the cell—which at once allowed us to penetrate a whole stratum deeper into the unfathomable mine of microscopic vital structure; then the clearing up of our ideas in regard to fertilization, and the analysis of this into the two processes combined in it, reproduction and the mingling of the germ-plasms (Amphimixis); in connexion with this, the phenomena of maturation, first in the female and then in the male cell, and their significance as a reduction of the hereditary units:—all this and much more we have gained during this period. Finally, there is the refutation of the Lamarckian principle, and the consequent elaboration of the principle of selection by applying it to the hitherto closed region of the ultimate vital elements of the germ-plasm.
The actual form of these lectures has developed as they were transcribed. But although the form is thus to some extent new, I have followed in the main the same train of thought as in the lectures of recent years. The lecture-form has been adhered to in the book, not merely because of the greater vividness of presentation which it implies, but for many other reasons, of which the greater freedom in the choice of material and the limiting of quotation to a minimum are not the least. That all polemics of a personal kind have thus been excluded will not injure the book, but it is by no means lacking in discussions of opinion, and will, therefore, I trust, contribute something towards the clearing up of disputed points.
I have endeavoured to introduce as much of the researches[Pg viii] and writings of others as possible without making the book heavy; but my aim has been to write a book to be read, not merely one to be referred to.
If it be asked, finally, for whom the book is intended, I can hardly answer otherwise than 'For him whom it interests.' The lectures were delivered to an audience consisting for the most part of students of medicine and natural science, but including some from other faculties, and sometimes even some of my colleagues in other departments. In writing the book I have presupposed as little special knowledge as possible, and I venture to hope that any one who reads the book and does not merely skim it, will be able without difficulty to enter into the abstruse questions treated of in the later lectures.
It would be a great satisfaction to me if this book were to be the means of introducing my theoretical views more freely among investigators, and to this end I have elaborated special sections more fully than in the lectures. Notwithstanding much controversy, I still regard its fundamental features as correct, especially the assumption of 'controlling' vital units, the determinants, and their aggregation into 'ids'; but the determinant theory also implies germinal selection, and without it the whole idea of the guiding of the course of transformation of the forms of life, through selection which rejects the unfit and favours the more fit, is, to my mind, a mere torso, or a tree without roots.
I only know of two prominent workers of our day who have given thorough-going adherence to my views: Emery in Bologna and J. Arthur Thomson in Aberdeen. But I still hope to be able to convince many others when the consistency and the far-reachingness of these ideas are better understood. In many details I may have made mistakes which the investigations of the future will correct, but as far as the basis of my theory is concerned I am confident: the principle of selection does rule over all the categories of vital units. It does not, indeed, create primary variations, but it determines the paths of evolution which these are to[Pg ix] follow, and thus controls all differentiation, all ascent of organization, and ultimately the whole course of organic evolution on the earth, for everything about living beings depends upon adaptation, though not on adaptation in the sense in which Darwin used the word.
The great prominence thus given to the idea of selection has been condemned as one-sided and exaggerated, but the physicist is quite as open to the same reproach when he thinks of gravity as operative not on our earth alone, but as dominating the whole cosmos, whether visible to us or not. If there is gravity at all it must prevail everywhere, that is, wherever material masses exist; and in the same way the co-operation of certain conditions with certain primary vital forces must call forth the same process of selection wherever living beings exist; thus not only are the vital units which we can perceive, such as individuals and cells, subject to selection, but those units the existence of which we can only deduce theoretically, because they are too minute for our microscopes, are subject to it likewise.
This extension of the principle of selection to all grades of vital units is the characteristic feature of my theories; it is to this idea that these lectures lead, and it is this—in my own opinion—which gives this book its importance. This idea will endure even if everything else in the book should prove transient.
Many may wonder, perhaps, why in the earlier lectures much that has long been known should be presented afresh, but I regard it as indispensable that the student who wishes to make up his own mind in regard to the selection-idea should not only be clear as to what it means theoretically, but should also form for himself a conception of its sphere of influence. Many prejudiced utterances in regard to 'Natural Selection' would never have been published if those responsible for them had known more of the facts; if they had had any idea of the inexhaustible wealth of phenomena which can only be interpreted in the light of this principle, in as far, that is, as we are able to give explanations[Pg x] of life at all. For this reason I have gone into the subject of colour-adaptations, and especially into that of mimicry, in great detail; I wished to give the reader a firm foundation of fact from which he could select what suited him when he wished to test by the light of facts the more difficult problems discussed in the book.
In conclusion, I wish to thank all those who have given me assistance in one way or other in this work: my former assistant and friend Professor V. Häcker in Stuttgart, my pupils and fellow workers Dr. Gunther and Dr. Petrunkewitsch, and the publisher, who has met my wishes in the most amiable manner.
Freiburg-I-Br.,
February 20, 1902.
PREFATORY NOTE TO ENGLISH EDITION
Professor Weismann's Evolution Theory, here translated from the second German edition (1904), is a work of compelling interest, the fruit of a lifetime of observation and reflection, a veteran's judicial summing up of his results, and certainly one of the most important contributions to Evolution literature since Darwin's day.
As the author's preface indicates, the salient features of his crowning work are (1) the illumination of the Evolution process with a wealth of fresh illustrations; (2) the vindication of the 'Germ-plasm' concept as a valuable working hypothesis; (3) the final abandonment of any assumption of transmissible acquired characters; (4) a further analysis of the nature and origin of variations; and (5), above all, an extension of the Selection principle of Darwin and Wallace, which finds its logical outcome in the suggestive theory of Germinal Selection.
The translation will be welcomed, we believe, not only by biological experts who have followed the development of 'Weismannism' during the last twenty years, and will here find its full expression for the time being, but also by those who, while acquainted with individual essays, have not hitherto realized the author's complete system. Apart from the theoretical conceptions which unify the book and mark it as an original contribution of great value, there is a lucid exposition of recent biological advances which will appeal to those who care more for facts than theories. To critics of evolutionism, who are still happily with us, the book ought to be indispensable; it will afford them much material for argumentation, and should save them many tilts against[Pg xii] windmills. But, above all, the book will be valued by workers in many departments of Biology, who are trying to help in the evolution of Evolution Theory, for it is characteristic of the author, as the history of recent research shows, to be suggestive and stimulating, claiming no finality for his conclusions, but urging us to test them in a mood of 'thätige Skepsis.'
The translation of this book—the burden of which has been borne by my wife—has been a pleasure, but it has also been a serious responsibility. We have had fine examples set us by previous translators of some of Weismann's works, Meldola, Poulton, Shipley, Parker, and others; and if we have fallen short of their achievements, it has not been for lack of endeavour to follow the original with fidelity, nor for lack of encouragement on the part of the author, who revised every page and suggested many emendations.
J. ARTHUR THOMSON.
University of Aberdeen,
October, 1904.
CONTENTS
| LECTURE | PAGE | |
| I. | Introductory | 1 |
| II. | The Darwinian Theory | 25 |
| III. | The Darwinian Theory (continued) | 42 |
| IV. | The Coloration of Animals and its relation to the Processes of Selection |
57 |
| V. | True Mimicry | 91 |
| VI. | Protective Adaptations in Plants | 119 |
| VII. | Carnivorous Plants | 132 |
| VIII. | The Instincts of Animals | 141 |
| IX. | Organic Partnerships or Symbiosis | 161 |
| X. | The Origin of Flowers | 179 |
| XI. | Sexual Selection | 210 |
| XII. | Intra-Selection or Selection among Tissues | 240 |
| XIII. | Reproduction in Unicellular Organisms | 253 |
| XIV. | Reproduction by Germ-cells | 266 |
| XV. | The Process of Fertilization | 286 |
| XVI. | Fertilization in Plants and Unicellular Organisms and its immediate significance |
312 |
| XVII. | The Germ-plasm Theory | 345 |
| XVIII. | The Germ-plasm Theory (continued) | 373 |
| XIX. | The Germ-plasm Theory (continued) | 392 |
LIST OF ILLUSTRATIONS
| FIGURE | PAGE | |
| 1. | Group of various races of domestic pigeons | 35 |
| 2. | Longitudinally striped caterpillar of a Satyrid | 67 |
| 3. | Full-grown caterpillar of the Eyed Hawk-moth (Smerinthus ocellatus) | 67 |
| 4. | Full-grown caterpillar of the Elephant Hawk-moth (Chærocampa elpenor) | 68 |
| 5. | The Eyed Hawk-moth in its 'terrifying attitude' | 69 |
| 6. | Under surface of the wings of Caligo | 70 |
| 7. | Caterpillar of a North American Darapsa | 71 |
| 8. | Caterpillar of the Buckthorn Hawk-moth (Deilephila hippophaës) | 73 |
| 9. | Hebomoja glaucippe, from India; under surface | 76 |
| 10. | Xylina vetusta, in flight and at rest | 77 |
| 11. | Tropidoderus childreni, in flying pose | 79 |
| 12. | Notodonta camelina, in flight and at rest | 80 |
| 13. | Kallima paralecta, from India, right under side of the butterfly at rest | 83, 357 |
| 14. | Cœnophlebia archidona, from Bolivia, in its resting attitude | 85 |
| 15. | Cærois chorinæus, from the lower Amazon, in its resting attitude | 86 |
| 16. | Phyllodes ornata, from Assam | 87 |
| 17. | Caterpillar of Selenia tetralunaria, seated on a birch twig | 90, 360 |
| 18. | Upper surfaces of Acræa egina, Papilio ridleyanus, and Pseudacræa boisduvalii | 102 |
| 19. | Barbed bristles of Opuntia rafinesquii | 123 |
| 20. | Vertical section through a piece of a leaf of the Stinging-nettle (Urtica dioica) | 123 |
| 21. | A piece of a twig of Barberry (Berberis vulgaris) | 124 |
| 22. | Tragacanth (Astragalus tragacantha) | 125 |
| 23. | Bladderwort (Utricularia grafiana) | 133 |
| 24. | Pitcher of Nepenthes villosa | 134 |
| 25. | Butterwort (Pinguicula vulgaris) | 136 |
| 26. | The Sundew (Drosera rotundifolia) | 137 |
| 27. | A leaf of the Sundew | 137 |
| 28. | Leaf of Venus Fly-trap | 138 |
| 29. | Aldrovandia vesiculosa | 138 |
| 30. | Aldrovandia, its trap apparatus | 139 |
| 31. | Sea-cucumber (Cucumaria) | 148 |
| 32. | Metamorphosis of Sitaris humeralis, an oil-beetle | 150 |
| 33. | Cocoon of the Emperor Moth (Saturnia carpini) | 158 |
| 34. | Hermit-crab | 163 |
| 35. | Hydra viridis, the Green Freshwater Polyp | 169 |
| 36. | Amœba viridis | 170 |
| 37. | Twig of an Imbauba-tree, showing hair cushions | 172 |
| 38.[Pg xv] | A fragment of a Lichen | 173 |
| 39. | A fragment of a Silver Poplar root | 176 |
| 40. | Potentilla verna | 181 |
| 41. | Flower of Meadow Sage | 183 |
| 42. | Alpine Lousewort (Pedicularis asplenifolia) | 184 |
| 43. | Flower of Birthwort (Aristolochia clematitis) | 185 |
| 44. | Alpine Butterwort (Pinguicula alpina) | 185 |
| 45. | Daphne mezereum and Daphne striata | 187 |
| 46. | Common Orchis (Orchis mascula) | 188 |
| 47. | Head of a Butterfly | 190 |
| 48. | Mouth-parts of the Cockroach | 191 |
| 49. | Head of the Bee | 192 |
| 50. | Flowers of the Willow | 194 |
| 51. | The Yucca-moth (Pronuba yuccasella) | 201 |
| 52. | The fertilization of the Yucca | 202 |
| 53. | Scent-scales of diurnal Butterflies | 217 |
| 54. | A portion of the upper surface of the wing of a male 'blue' (Lycæna menalcas) | 218 |
| 55. | Zeuxidia wallacei, male | 218 |
| 56. | Leptodora hyalina | 224 |
| 57. | Moina paradoxa, male | 225 |
| 58. | Moina paradoxa, female | 226 |
| 59. | An Amœba: the process of division | 253 |
| 60. | Stentor rœselii, trumpet-animalcule | 254 |
| 61. | Holophrya multifiliis | 256 |
| 62. | Pandorina morum | 257 |
| 63. | Volvox aureus | 270 |
| 64. | Fucus platycarpus, brown sea-wrack | 272 |
| 65. | Copulation in a Daphnid (Lyncæid) | 276 |
| 66. | Spermatozoa of various Daphnids | 277 |
| 67. | Spermatozoa of various animals | 278 |
| 68. | Diagram of a spermatozoon | 279, 338 |
| 69. | Ovum of the Sea-urchin | 281, 338 |
| 70. | Daphnella | 283 |
| 71. | Bythotrephes longimanus | 283 |
| 72. | Sida crystallina, a Daphnid | 284 |
| 73. | Diagrammatic longitudinal section of a hen's egg before incubation | 285 |
| 74. | Diagram of nuclear division | 288 |
| 75. | Process of fertilization in Ascaris megalocephala | 296 |
| 76. | Diagram of the maturation divisions of the ovum | 299 |
| 77. | Diagram of the maturation divisions of the sperm-cell | 301 |
| 78. | Diagram of the maturation of a parthenogenetic ovum | 305 |
| 79. | The two maturation divisions of the 'drone eggs' of the Bee | 307, 337 |
| 80. | Fertilization of the ovum of a Gasteropod | 310 |
| 81. | Formation of polar bodies in a Lichen | 313 |
| 82. | Fertilization in the Lily | 314 |
| 83. | Conjugation of Noctiluca | 317 |
| 84. | Conjugation and polar body formation in the Sun-animalcule | 319 |
| 85.[Pg xvi] | Diagram of the conjugation of an Infusorian | 321 |
| 86. | Conjugation of an Infusorian | 323 |
| 87. | Diagram to illustrate the operation of amphimixis | 348 |
| 88. | Sperm-mother-cells (spermatocytes) of the Salamander | 350 |
| 89. | Anterior region of the larva of a Midge | 364, 393 |
| 90. | The Common Shore-Crab, seen from below | 367 |
| 91. | Hind leg of a Locustid | 371 |
| 92. | Echinoderm-larvæ | 387 |
| 93. | Development of a limb in the pupa of a Fly | 395 |
| 94. | Diagram to illustrate the phylogenetic shifting back of the origins of the germ-cells in medusoids and hydroids | 412 |
| 95. | Diagram to illustrate the migration of the germ-cells in Hydromedusæ | 414 |
COLOURED PLATES
SOME MIMETIC BUTTERFLIES AND THEIR IMMUNE MODELS
| Plate I | to face page | 112 | |
| Plate II | " " | 114 | |
| Plate III | " " | 116 |
INTRODUCTORY
Every one knows in a general way what is meant by the doctrine of descent—that it is the theory which maintains that the forms of life, animals and plants, which we see on our earth to-day, have not been the same from all time, but have been developed, by a process of transformation, from others of an earlier age, and are in fact descended from ancestors specifically different. According to this doctrine of descent, the whole diversity of animals and plants owes its origin to a transformation process, in the course of which the earliest inhabitants of our earth, extremely simple forms of life, were in part evolved in the course of time into forms of continually increasing complexity of structure and efficiency of function, somewhat in the same way as we can see every day, when any higher animal is developed from a single cell, the egg-cell, not suddenly or directly, but connected with its origin by a long series of ever more complex transformation stages, each of which is the preparation for, and leads on to the succeeding one. The theory of descent is thus a theory of development or evolution. It does not merely, as earlier science did, take for granted and describe existing forms of life, but regards them as having become what they are through a process of evolution, and it seeks to investigate the stages of this process, and to discover the impelling forces that lie behind it. Briefly, the theory of descent is an attempt at a scientific interpretation of the origin and diversity of the animate world.
In these lectures, therefore, we have not merely to show on what grounds we make this postulate of an evolution process, and to marshall the facts which necessitate it; we must also try to penetrate as far as possible towards the causes which bring about such transformations. For this reason we are forced to go beyond the limits of the theory of descent in the narrow sense, and to deal with the general processes of life itself, especially with reproduction and the closely associated problem of heredity. The transformation of species can only be interpreted in one of two ways; either it depends on a peculiar internal force, which is usually only latent in the organism, but from time to time becomes active, and then, to a[Pg 2] certain extent, moulds it into new forms; or it depends on the continually operating forces which make up life, and on the way in which these are influenced by changing external conditions. Which of these alternatives is correct we can only undertake to determine when we know the phenomena of life, and as far as possible their causes, so that it is indispensable to make ourselves acquainted with these as far as we can.
When we look at one of the lowest forms of life, such as an Amœba or a single-celled Alga, and reflect that, according to the theory of evolution, the whole realm of creation as we see it now, with Man at its head, has evolved from similar or perhaps even smaller and simpler organisms, it seems at first sight a monstrous assumption, and one which quite contradicts our simplest and most certain observations. For what is more certain than that the animals and plants around us remain the same, as long as we can observe them, not through the lifetime of an individual only, but through centuries, and in the case of many species, for several thousand years?
This being so, it is intelligible enough that the doctrine of evolution, on its first emergence at the end of the eighteenth century, was received with violent opposition, not on the part of the laity only, but by the majority of scientific minds, and instead of being followed up, was at first opposed, then neglected, and finally totally forgotten, to spring up anew in our own day. But even then a host of antagonists ranged themselves against the doctrine, and, not content with loftily ignoring it, made it the subject of the most violent and varied attacks.
This was the state of affairs when, in 1858, Darwin's book on The Origin of Species appeared, and hoisted the flag of evolution afresh. The struggle that ensued may now be regarded as at an end, at least as far as we are concerned—that is, in the domain of science. The doctrine of descent has gained the day, and we can confidently say that the Evolution theory has become a permanent possession of science that can never again be taken away. It forms the foundation of all our theories of the organic world, and all further progress must start from this basis.
In the course of these lectures, we shall find at every step fresh evidence of the truth of this assertion, which may at first seem all too bold. It is not by any means to be supposed that the whole question in regard to the transformation of organisms and the succession of new forms of life has been answered in full, or that we have now been fortunate enough to solve the riddle of life itself. No! whether we ever reach that goal or not, we are a long way from it as yet, and[Pg 3] even the much easier problem, how and by what forces the evolution of the living world has proceeded from a given beginning, is far from being finally settled; antagonistic views are still in conflict, and there is no arbitrator whose authoritative word can decide which is right. The How? of evolution is still doubtful, but not the fact, and this is the secure foundation on which we stand to-day: The world of life, as we know it, has been evolved, and did not originate all at once.
Were I to try to give, in advance, even an approximate idea of the confidence with which we can take our stand on this foundation, I should be almost embarrassed by the wealth of facts on which I might draw. It is hardly possible nowadays to open a book on the minute or general structural relations, or on the development of any animal whatever, without finding in it evidences in favour of the Evolution theory, that is to say, facts which can only be understood on the assumption of the evolution of the organic world. This, too, without taking into account at all the continually increasing number of facts Palæontology is bringing to light, placing before our eyes the forms which the Evolution theory postulates as the ancestors of the organic world of to-day: birds with teeth in their bills, reptile-like forms clothed with feathers, and numerous other long-extinct forms of life, which, covered up by the mud of earlier waters, and preserved as 'fossils' in the later sedimentary rocks, tell us plainly how the earlier world of animals and plants was constituted. Later, we shall see that the geographical distribution of plant and animal species of the present day can only be understood in the light of the Evolution theory. But meantime, before we go into details, what may justify my assumption is the fact that the Evolution theory enables us to predict.
Let us take only a few examples. The skeleton of the wrist in all vertebrate animals above Fishes consists of two rows of small bones, on the outer of which are placed the five bones of the palm, corresponding to the five fingers. The outer row is curved, and there is thus a space between the two rows, which, in Amphibians and Reptiles, is filled by a special small bone. This 'os centrale' is absent in many Mammals, notably, for instance, in Man, and the space between the two rows is filled up by an enlargement of one of the other bones. Now if Mammals be descended from the lower vertebrates, as the theory of descent assumes, we should expect to find the 'os centrale' even in Man in young stages, and, after many unsuccessful attempts, Rosenberg has at last been able to demonstrate it at a very early stage of embryonic development.
This prediction, with another to be explained later, is based upon the experience that the development of an individual animal follows, in a general way, the same course as the racial evolution of the species, so that structures of the ancestors of a species, even if they are not found in the fully developed animal, may occur in one of its earlier embryonic stages. Further on, we shall come to know this fact more intimately as a 'biogenetic law,' and it alone would be almost enough to justify the theory of evolution. Thus, for instance, the lowest vertebrates, the Fishes, breathe by means of gills, and these breathing organs are supported by four or more gill-arches, between which spaces, the gill-slits, remain open for the passage of water. Although Reptiles, Birds, and Mammals breathe by lungs, and at no time of their life by gills, yet, in their earliest youth, that is, during their early development in the egg, they possess these gill-arches and gill-slits, which subsequently disappear, or are transformed into other structures.
On the strength of this 'biogenetic law' it could also be predicted that Man, in whom, as is well known, there are twelve pairs of ribs, would, in his earliest youth, possess a thirteenth pair, for the lower Mammals have more numerous ribs, and even our nearest relatives, the anthropoid Apes, the gorilla and chimpanzee, have a thirteenth rib, though a very small one, and the siamang has even a fourteenth. This prediction also has been verified by the examination of young human embryos, in which a small thirteenth rib is present, though it rapidly disappears.
During the seventies I was engaged in investigating the development of the curious marking which adorns the long body of many of our caterpillars. I studied in particular the caterpillars of our Sphingidæ or hawk-moths, and found, by a comparison of the various stages of development from the emergence of the caterpillar from the egg on to its full growth, that there is a definite succession of different kinds of markings following each other, in a whole range of species, in a similar manner. From the standpoint of the Evolution theory, I concluded that the markings of the youngest caterpillars, simple longitudinal stripes, must have been those of the most remote ancestors of our present species, while those of the later stages, oblique stripes, were those of ancestors of a later date.
If this were the case, then all the species of caterpillar which now exhibit oblique stripes in their full-grown stage must have had longitudinal stripes in their youngest stages, and because of this succession of markings in the individual development, I was able to predict that the then unknown young form of the caterpillar of our[Pg 5] privet hawk-moth (Sphinx ligustri) must have a white line along each side of the back. Ten years later, the English zoologist, Poulton, succeeded in rearing the eggs of Sphinx ligustri, and it was then demonstrated that the young caterpillar actually possessed the postulated white lines.
Such predictions undoubtedly give the hypothesis on which they are based, the Evolution theory, a high degree of certainty, and are almost comparable to the prediction of the discovery of the planet Neptune by Leverrier. As is well known, this, the most distant of all the planets, whose period of revolution round the sun is almost 165 of our years, would probably never have been recognized as a planet, had not Adams, an astronomer at the Greenwich Observatory, and afterwards Leverrier, deduced its presence from slight disturbances in the path of Jupiter's moons, and indicated the spot where an unknown planet must be looked for. Immediately all telescopes were directed towards the spot indicated, and Galle, at the Berlin Observatory, found the sought-for planet.
We might with justice regard as lacking in discernment those who, in the face of such experiences, still doubt that the earth revolves round the sun, and we might fairly say the same of any one who, in the face of the known facts, would dispute the truth of the Evolution theory. It is the only basis on which an understanding of these facts is possible, just as the Kant-Laplace theory of the solar system is the only basis on which an adequate interpretation of the facts of the heavens can be arrived at.
To this comparison of the two theories it has been objected that the Evolution theory has far less validity than the other, first, because it can never be mathematically demonstrated, and secondly, because at the best it can only interpret the transformations of the animate world, and not its origin. Both objections are just: the phenomena of life are in their nature much too intricate for mathematics to deal with, except with extreme diffidence; and the question of the origin of life is a problem which will probably have to wait long for solution. So, if it gives pleasure to any one to regard the one theory as having more validity than the other, no one can object; but there is no particular advantage to be gained by doing so. In any case, the Evolution theory shares the disadvantage of not being able to explain everything in its own province with the Kant-Laplace cosmogony, for that, too, must presuppose the first beginning, the rotating nebula.
Although I regard the doctrine of descent as proved, and hold it to be one of the greatest acquisitions of human knowledge,[Pg 6] I must repeat that I do not mean to say that everything is clear in regard to the evolution of the living world. On the contrary, I believe that we still stand merely on the threshold of investigation, and that our insight into the mighty process of evolution, which has brought about the endless diversity of life upon our earth, is still very incomplete in relation to what may yet be found out, and that, instead of being vainglorious, our attitude should be one of modesty. We may well rejoice over the great step forward which the dominant recognition of the Evolution theory implies, but we must confess that the beginnings of life are as little clear to us as those of the solar system. But we can do this at least: we can refer the innumerable and wonderful inter-relations of the organic cosmos to their causes—common descent and adaptation—and we can try to discover the ways and means which have co-operated to bring the organic world to the state in which we know it.
When I say that the theory of descent is the most progressive step that has yet been taken in the development of human knowledge, I am bound to give my reasons for this opinion. It is justified, it seems to me, even by this fact alone, that the Evolution idea is not merely a new light on the special region of biological science, zoology and botany, but is of quite general importance. The conception of an evolution of the world of life upon the earth reaches far beyond the bounds of any single science, and influences our whole realm of thought. It means nothing less than the elimination of the miraculous from our knowledge of nature, and the placing of the phenomena of life on the same plane as the other natural processes, that is, as having been brought about by the same forces, and being subject to the same laws. In the domain of the inorganic, no one now doubts that out of nothing nothing can come: energy and matter are from everlasting to everlasting, they can neither be increased or decreased, they can only be transformed—heat into mechanical energy, into light, into electricity, and so on. For us moderns, the lightning is no longer hurled by the Thunderer Zeus on the head of the wicked, but, careless alike of merit or guilt, it strikes where the electric tension finds the easiest and shortest line of discharge. Thus to our mode of thought it now seems clear that no event in the world of the living depends upon caprice, that at no time have organisms been called forth out of nothing by the mighty word of a Creator, but they have been produced at all times by the co-operation of the existing forces of nature, and every species must have arisen just where, and when, and in the form in which it actually did arise, as the necessary outcome of the existing conditions of energy and[Pg 7] matter, and of their interactions upon each other. It is this correlation of animate nature with natural forces and natural laws which gives to the doctrine of evolution its most general importance. For it thus supplies the keystone in the arch of our interpretation of nature and gives it unity; for the first time it makes it possible to form a conception of a world-mechanism, in which each stage is the result of the one before it, and the cause of the succeeding one.
How deeply all our earlier opinions are affected by this doctrine will become clear if we fix our attention on a single point, the derivation of the human understanding from that of animal ancestors. What of the reason of Man, of his morals, of his freedom of will? may be asked, as it has been, and still is often asked. What has been regarded as absolutely distinct from the nature of animals is said to differ from their mental activities only in degree, and to have evolved from them. The mind of a Kant, of a Laplace, of a Darwin—or to ascend into the plane of the highest and finest emotional life, the genius of a Raphael or a Mozart—to have any real connexion, however far back, with the lowly psychical life of an animal! That is contrary to all our traditionary, we might say our inborn, ideas, and it is not to be wondered at that the laity, and especially the more cultured among them, should have opposed such a doctrine whose dominating power was unintelligible to them, because they were ignorant of the facts on which it rests. That a man should feel his dignity lowered by the idea of descent from animals is almost comical to the naturalist, for he knows that every one of us, in his first beginning, occupied a much lowlier position than that of our mammalian ancestors—was, in fact, as regards visible structure, on a level with the Amœba, that microscopically minute unicellular animal, which can hardly be said to possess organs, and whose psychical activities are limited to recognizing and engulfing its food. Very gradually at first, and step by step, there develop from this single cell, the ovum, more and more numerous cells; this mass of cells segregates into different groups, which differentiate further and further, until at last they form the perfect man. This occurs in the development of every human being, and we are merely unaccustomed to the thought that it means nothing else than an incredibly rapid ascent of the organism from a very low level of life to the highest.
Still less is it to be wondered at that the Evolution doctrine met with violent opposition on the part of the representatives of religion, for it stood in open contradiction to that remarkable and venerable cosmogony, the Mosaic story of Creation, which people had been accustomed to regard, not as what it is—a conception of nature[Pg 8] at an early stage of human culture—but as an inalienable part of our own religion. But investigation shows us that the doctrine of evolution is true, and it is only a weak religion which is incapable of adapting itself to the truth, retaining what is essential, and letting go what is unessential and subject to change with the development of the human mind. Even the heliocentric hypothesis was in its day declared false by the Church, and Galilei was forced to retract; but the earth continued to revolve round the sun, and nowadays any one who doubted it would be considered mentally weak or warped. So in all likelihood the time is not far distant when the champions of religion will abandon their profitless struggle against the new truth, and will see that the recognition of a law-governed evolution of the organic world is no more prejudicial to true religion than is the revolution of the earth round the sun.
Having given this very general orientation of the Evolution problem, which is to engage our attention in detail, I shall approach the problem itself by the historical method, for I do not wish to bring the views of present-day science quite suddenly and directly into prominence. I would rather seek first to illustrate how earlier generations have tried to solve the question of the origin of the living world. We shall see that few attempts at solution were made until quite recently, that is, until the end of the eighteenth and the beginning of the nineteenth century. Only then there appeared a few gifted naturalists with evolutionist ideas, but these ideas did not penetrate far; and it was not till after the middle of the nineteenth century that they found a new champion, who was to make them common property and a permanent possession of science. It was the teaching of Charles Darwin that brought about this thorough awakening, and laid the foundations of our present interpretations, and his work will therefore engross our attention for a number of lectures. Only after we have made ourselves acquainted with his teaching shall we try to test its foundations, and to see how far this splendid structure stands on a secure basis of fact, and how deeply its power of interpretation penetrates towards the roots of phenomena. We shall examine the forces by which organisms are dominated, and the phenomena produced, and thereby test Darwin's principles of interpretation, in part rejecting them, in part accepting them, though in a much extended form, and thus try to give the whole theoretic structure a more secure foundation. I hope to be able to show that we have made some real progress since Darwin's day, that[Pg 9] deductions have been drawn from his theory which even he did not dream of, which have thrown fresh light on a vast range of phenomena, and, finally, that through the more extended use of his own principles, the Evolution theory has gained a completeness, and an intrinsic harmony which it previously lacked.
This at least is my own opinion, but I cannot ignore the fact that it is by no means shared by all living naturalists. The obvious gaps and insufficiencies of the Darwinian theory have in the last few decennia prompted all sorts of attempts at improving it. Some of these were lost sight of almost as soon as they were suggested, but others have held their own, and can still claim numerous supporters. It would only tend to bewilder if I gave an account of those of the former class, but those which still hold their own must be noticed in these lectures, though it is by no means my intention to expound the confused mass of opinions which has gathered round the doctrine of evolution, but rather to give a presentation of the theory as it has gradually grown up in my own mind in the course of the last four decades. Even this will not be the last of which science will take knowledge, but it will, I hope, at least be one which can be further built upon.
Let us, then, begin at once with that earliest forerunner of the modern theory of descent, the gifted Greek philosopher Empedocles, who, equally important as a leader of the state of Agrigentum, and as a thinker in purely theoretical regions of thought, advanced very notable views regarding the origin of organisms. We must, however, be prepared to hear something that is hardly a theory in the modern scientific acceptation of that term; and we must not be repelled by the unbridled poetical fancy of the speculative philosopher; we have to recognize that there is a sound kernel contained in his amusing pictures—a thought which we meet with later, in much more concrete form, in the Darwinian theory, and which, if I mistake not, we shall keep firm hold of in all time to come.
According to Empedocles the world was formed by the four elements of the ancients, Earth, Water, Fire, and Air, moved and guided by two fundamental forces, Hate and Love, or, as we should now say, Repulsion and Attraction. Through the chance play of these two forces with the elements, there arose first the plants, then the animals, in such a manner that at first only parts and organs of animals were formed: single eyes without faces, arms without bodies, and so on. Then, in wild play, Nature attempted to put together these separate parts, and so created all manner of combinations, for the most part inept monsters unfit for life, but in a few[Pg 10] cases, where the parts fitted, there resulted a creature capable not only of life, but, if the juxtaposition was perfect, even of reproduction.
This phantastic picture of creation seems to us mad enough, but there slumbers in it, all unsuspected though it may have been by the author, the true idea of selection, the idea that much that is unfit certainly arises, but that only the fit endures. The mechanical coming-to-be of the fit is the sound kernel in this wondersome doctrine.
The natural science of the ancients, in regard to life and its forms, reached its climax in Aristotle (died 322 B. C.). A true polyhistorian, his writings comprehended all the knowledge of his time, but he also added much to it from his own observation. In his writings we find many good observations on the structure and habits of a number of organisms, and he also had the merit of being the first to attempt a systematic grouping of animals. With true insight, he grouped all the vertebrates together as Enaimata or animals with blood, and classed all the rest together as Anaimata or bloodless animals. That he denied to the latter group the possession of blood is not to be wondered at, when we take into account the extremely imperfect means of investigation available in his time, nor is it surprising that he should have ranked this motley company, in antithesis to the blood-possessing animals, as a unified and equivalent group. Two thousand years later, Lamarck did exactly the same thing, when he divided the animals into backboned and backboneless, and we reckon this nowadays as a merit only in so far that he was the first, after Aristotle, to re-express the solidarity of the classes of animals which we now call vertebrates.
Aristotle was, however, not a systematic zoologist in our sense of the term, as indeed was hardly possible, considering the very small number of animal forms that were known in his time. In our day we have before us descriptions of nearly 300,000 named species wherefrom to construct our classification, while Aristotle knew hardly more than 200. Of the whole world of microscopic animals he could, of course, have no idea, any more than of the remains of prehistoric animals, of which we now know about 40,000 named and adequately described species. One would have thought that it would have occurred to a quick-witted people like the Greeks to pause and ponder when they found mussel-shells and marine snail-shells on the hills far above the sea; but they explained these by the great flood in the time of Deucalion and Pyrrha, and they did not observe that the fossil molluscs were of different species from the similar animals living in the sea in their own day.
Thus there was nothing to suggest to Aristotle and others of his time the idea that a transformation of species had been going on through the ages, and even the centuries after him evoked no such idea, nor did there arise new speculations, after the manner of Empedocles, in regard to the origin of the organic world. On the whole, the knowledge of the living world retrograded rather than advanced until the beginning of the Roman Empire. What Aristotle had known was forgotten, and Pliny's work on animals is a catalogue embellished with numerous fables, arranged according to a purely external principle of division. Pliny divided animals into those belonging to earth, water, and air, which is not very much more scientific than if he had arranged them according to the letters of the alphabet.
During the time of the Roman Empire, as is well known, the knowledge of natural history sank lower and lower; there was no more investigation of nature, and even the physicians lost all scientific basis, and practised only in accordance with their traditional esoteric secrets. As the whole culture of the West gradually disappeared, the knowledge of nature possessed by earlier centuries was also completely lost, and in the first half of the Middle Ages Europeans revealed a depth of ignorance of the natural objects lying about them, which it is difficult for us now to form any conception of.
Christianity was in part responsible for this, because it regarded natural science as a product of heathendom, and therefore felt bound to look coldly on it, if not even to oppose it. Later, however, even the Christian Church felt itself forced to give the people some mental nourishment in the form of natural history, and under its influence, perhaps actually composed by teachers of the Church, there appeared a little book, the so-called Physiologus, which was meant to instruct the people in regard to the animal world. This remarkable work, which has been preserved, must have had a very wide distribution in the earlier Middle Ages, for it was translated into no fewer than twelve languages, Greek, Armenian, Syriac, Arabic, Ethiopic, and so on. The contents are very remarkable, and come from the most diverse sources, that is, from the most different writers of antiquity, from Herodotus, from the Bible, and so forth, but never from original observation. The compilation does not really give descriptions of animals or of their habits, but, of each of the forty-one animals which the Physiologus recognizes, something remarkable is briefly related in true lapidary style, sometimes a mere curiosity without further import, or sometimes a symbolical interpretation. Thus the book says of the panther: 'he is gaily coloured; after satiating himself he[Pg 12] sleeps three days, and awakes roaring, giving forth such an agreeable odour that all animals come to him.' Of the pelican the well-known legend is related, that it tears open its own breast to feed its young with its blood, thus standing as a symbol of mother-love. Fabulous creatures, too, appear in these pages. Of the Phœnix, that bird whose plumage glitters with gold and precious stones, which was known even to Herodotus, and which has survived through Eastern fairy-tales on to the time of our own romanticists (Tieck), we read: 'it lives a thousand years, because it has not eaten of the tree of knowledge'; then 'it sets fire to itself and arises anew from its own ashes,' a symbol of nature's infinite power of renewing its youth.
But while among the peoples of Europe all the science of the ancients was lost, except a few barely recognizable fragments, the old lore was preserved, both as regards organic nature and other orders of facts, among the Arabs, through whom so many treasures of antiquity have eventually been handed down to us, coming in the track of the Arabian conquests across North Africa and Spain to the nations of Europe.
It was in this way, too, that the writings of Aristotle again found recognition, after having been translated into Latin at Palermo at the order of that enthusiast for Science and Art, the Hohenstaufen Emperor, Frederick the Second. Our Emperor presented one copy of Aristotle's writings to the University of Bologna, and thus the wisdom of the ancient Greeks again became the common property of European culture. From the thirteenth century to the eighteenth, the study of natural science was limited to repeating and extending the work of Aristotle. Nothing new, depending upon personal observation, was added, and it does not even seem to have occurred to any one to subject the statements of the Stagirite to any test, even when they concerned the most familiar objects. No one noticed the error which ascribed to the fly eight legs instead of six; there was in fact as yet no investigation, and all knowledge of natural history was purely scholastic, and gave absolute credence to the authority of the ancients.
A revulsion, however, occurred in the century of the Reformation, with the breaking down of the blind belief in authority which had till then prevailed in all provinces of human knowledge and thought. After a long and severe struggle, dry scholasticism was finally overcome, and natural science, with the rest, turned from a mere reliance on books to original thinking and personal observation. Thenceforward interpretations of natural processes were sought for no longer in the writings of the ancients, but in Nature herself.[Pg 13] Of the magnitude of this emancipation, and of the severity of the struggle against deep-rooted authority, one could form a faint idea from experience even in my own youth. Our young minds were so deeply imbued with the involuntary feeling that the ancients were superior to us moderns in each and every respect, that not only the hardly re-attainable plastic art of the Greeks and the immortal songs of Homer, but all the mental products of antiquity seemed to us models which could never be equalled; the tragedies of Sophocles were for us the greatest tragedies that the world had ever seen, the odes of Horace the most beautiful poems of all time!
In the domain of natural science the new era began with the overthrow of the Ptolemaic cosmogony, which, for more than a thousand years, had served as a basis for astronomy. When the German canon, Nicolas Copernicus (born at Thorn, 1473, died 1543), reversed the old theory, and showed that the sun did not revolve round the earth, but the earth round the sun, the ice was broken and the way paved for further progress. Galilei uttered his famous 'e pur si muove,' Kepler established his three laws of the movements of the planets, and Newton, a century later, interpreted their courses in terms of the law of gravitation.
But we have not here to do with a history of physics or astronomy, and I only wish to recall these well-known facts, in order that we may see how increased knowledge in this domain was always accompanied by advances in that of biology.
Here, however, we cannot yet chronicle any such thoroughgoing revolution of general conceptions; the basis of detailed empirical knowledge was not nearly broad enough for that, and it was in the acquiring of such a foundation that the next three centuries, from the sixteenth to the end of the eighteenth, were eagerly occupied.
The first step necessary was to collate the items of individual knowledge in regard to the various forms of life, and to bring the whole in unified form into general notice. This need was met for the first time by Conrad Gessner's Thierbuch, a handsome folio volume, printed at Zurich in 1551, and embellished with numerous woodcuts, some of them very good. This was followed, in 1600, by a great work in many volumes, written in Latin, by a professor of Bologna, Aldrovandi. Not native animals alone but foreign ones also were described in these works, for, after the discovery of America and the opening up of communication with the East Indies, many new animal and plant forms came to the knowledge of European nations by way of the sea. Thus Francesco Hernandez (died 1600), physician in ordinary to Philip II, described no fewer than forty new[Pg 14] Mammals, more than two hundred Birds, and many other American animals.
Again, in a quite different way, the naturalist's field of vision was widened, namely, by the invention of the simple microscope, with which Leeuwenhoek first discovered the new world of Infusorians, and Swammerdam made his notable observations on the structure and development of the very varied minute animal inhabitants of fresh water. In the same century, the seventeenth, anatomists like Tulpius, Malpighi, and many others extended the knowledge of the internal structure of the higher animals and of Man, and a foundation was laid for a deeper insight into the nature of vital functions by the discovery of the circulation of the blood in Man and the higher animals. In the following century, the eighteenth, this path of active research was eagerly followed, and we need only mention such names as Réaumur, Rösel von Rosenhof, De Geer, Bonnet, J. Chr. Schäfer, and Ledermüller, to be immediately reminded of the wealth of facts about the structure, life, and especially the development of our indigenous animals, which we owe to the labours of these men.
All these advances, great and many-sided as they were, did not at once lead to a renewal of the attempt of Empedocles to explain the origin of the organic world. This was as yet not even recognized as a problem requiring investigation, for men were content to take the world of life simply as a fact. The idea of getting beyond the naïve, poetic standpoint of the Mosaic story of Creation was as yet remote from the minds of naturalists, partly because they were wholly fascinated by the observation of masses of details, but chiefly because, first by the English physician, John Ray (died 1678), then by the great Swede, Carl Linné, the conception of organic 'species' had been formulated and sharply defined. It is true enough that before the works of these two men 'species' had been spoken of, but without being connected with any definite idea; the word was used rather in the same vague sense as the word 'genus,' to designate one of the smaller groups of organic forms, but without implying any clear idea of its scope or of its limitations. Now, however, for the first time, the term 'species' came to be used strictly to mean the smallest homogeneous group of individual forms of life upon the earth. John Ray held that the surest indication of a 'species' was that its members had been produced from the same seed; that is, 'forms which are of different species maintain this specific nature constantly, and one species does not arise from the seed of another.' Here we have the germ of the doctrine of the absolute nature and the[Pg 15] immutability of species which Linné briefly characterized in these words: 'Species tot sunt, quot formæ ab initio creatæ sunt,' 'there are just so many species as there were forms created in the beginning.' It is here clearly implied, that species as we know them have been as they are from all time, that, therefore, they exist in nature as such and unchangeably, and have not been merely read into nature by man.
This view, though we cannot now regard it as correct, was undoubtedly reasonable, and thoroughly in accordance with the spirit of the time; it was congruent with the knowledge, and above all with the scientific endeavours of the age. In the eighteenth century there was danger that all outlook on nature as a whole would be lost—smothered under the enormous mass of isolated facts, and especially under the inundation of diverse animal and plant forms which were continually being recognized. It must therefore have been regarded as a real deliverance, when Linné reduced this chaos of forms to a clearly ordered system, and relegated each form to its proper place and value in relation to the whole. How, indeed, could the great systematist have performed his task at all, if he had not been able to work with definite and sharply circumscribed groups of forms, if he had not been able to regard at least the lowest elements of his system, the species, as fixed and definite types? On the other hand, Linné was much too shrewd an observer not to entertain, in the course of his long life, and under the influence of the continually accumulating material, doubts as to the correctness of his assumption of the fixity and absoluteness of his species. He discovered from his own experience, what is fully borne out by ours, that it is easy enough to define a species when there are only a few specimens of a form to deal with, but that the difficulty increases in proportion to the number and to the diversity of habitat of those that are to be brought under one category. In the last edition of the Systema Naturæ we find very noteworthy passages in which Linné wonders whether, after all, a species may not change, and in the course of time diverge into varieties, and so forth. Of these doubts no notice was taken at the time; the accepted doctrine of the fixity of species was held to and even raised to the rank of a scientific dogma. Georges Cuvier, the great disciple of the Stuttgart 'Karlschule,' accentuated the doctrine still further by his establishment of animal-types, the largest groups of forms in the animal kingdom within which a definite and fundamentally distinct plan of architecture prevails. His four types, Vertebrates, Molluscs, Articulate and Radiate animals, furnished a further corroboration of the absolute nature of species, since they[Pg 16] seemed to show that even the highest and most comprehensive groups are sharply defined off from one another.
Let me add that this doctrine of the absolute nature of species was not fully elaborated till our own day, when the Swiss (afterwards American) naturalist, Louis Agassiz, went so far as to maintain that not only the highest and the lowest categories, but all those coming between them, were categories established and sharply separated by Nature herself. But in spite of much ingenuity and his wide and comprehensive outlook he exerted himself in vain to find satisfactory and really characteristic definitions of what was to be considered a class, an order, a family, or a genus. He did not succeed in finding a rational definition of these systematic concepts, and his endeavour may be regarded as the last important attempt to prop up an interpretation of nature already doomed to fall. But in referring to Louis Agassiz I have anticipated the historical course of scientific development, and must therefore go back to the last quarter of the eighteenth century.
The first unmistakable pioneer of the theory of descent, which now emerged for the first time as a scientific doctrine, was our great poet Goethe. He has indeed been often named as the founder of the theory, but that seems to me saying too much. It is true, however, that the inquiring mind of the poet certainly recognized in the structure of 'related' animals the marvellous general resemblances amid all the differences in detail, and he probed for the reason of these form-relations. Through the science of 'comparative anatomy,' as it was taught at the close of the century by Kielmeyer, Cuvier's teacher, and later by Cuvier himself, Blumenbach, and others, numerous facts had become known, which paved the way for such questions. It had, for instance, been recognized that the arm of man, the wing of the bird, the paddle of the seal, and even the foreleg of the horse, contain essentially the same chain of bones, and Goethe had already expressed these relations in his well-known verse,
As to what this law was he did not at that time pronounce an opinion, though he may even then have thought of the transformation of species. At first he contented himself with seeking for an ideal archetype or 'Urtypus' which was supposed to lie at the foundation of a larger or smaller group. He discovered the archetypal plant or 'Urpflanze,' when he rightly recognized that the parts of the flower are nothing more than modified leaves. He spoke plainly of the 'metamorphosis of plants,' meaning by that the transformation of his 'archetype' into the[Pg 17] endless diversity of actual plant forms. But at first he certainly thought of this transformation only in the ideal sense, and not as a factual evolutionary process.
The first who definitely maintained the latter view was, remarkably enough, the grandfather of the man who, in our own day, made the theory of descent finally triumphant, the English physician Erasmus Darwin, born 1731. This quiet thinker published, in 1794, a book entitled Zoonomia, and in it he takes the important step of substituting for Goethe's 'secret law' a real relationship of species. He proclaims the gradual establishment and ennobling of the animal world, and bases his view mainly on the numerous obvious adaptations of the structure of an organ to its use. I have not been able to find any passage in the book in which he has expressly indicated that, because many of the conditions of life could not have existed from the beginning, these adaptations are therefore, as such, an argument for the gradual transformation of species. But he assumed that such exact adaptations to the functions of an organ could only arise through the exercise of that function, and in this he saw a proof of transformation. Goethe had expressed the same idea when he said, 'Thus the eagle has conformed itself through the air to the air, the mole through the earth to the earth, and the seal through the water to the water,' and this shows that he too at one time thought of an actual transformation. But neither he nor Erasmus Darwin were at all clear as to how the use of an organ could bring about its variation and transformation. The latter only says that, for instance, the snout of the pig has become hard through its constant grubbing in the ground; the trunk of the elephant has acquired its great mobility through the perpetual use of it for all sorts of purposes; the tongue of the herbivore owes its hard, grater-like condition to the rubbing to and fro of the hard grass in the mouth, and so on. How acute and thoughtful an observer Erasmus Darwin was, is shown by the fact that he had correctly appreciated the biological significance of many of the colour-adaptations of animals to their surroundings, though it was reserved for his grandson to make this fully clear at a much later date. Thus he regarded the varied colouring of the python, of the leopard, and of the wild cat as the best adapted for concealing them from their prey amid the play of light and shadow in a leafy thicket. The black spot in front of the eye of the swan he considered an arrangement to prevent the bird from being dazzled, as would happen if that spot were as snow-white as the rest of the plumage.
At the end of the book he sums up his views in the following[Pg 18] sentences: 'The world has been evolved, not created; it has arisen little by little from a small beginning, and has increased through the activity of the elemental forces embodied in itself, and so has rather grown than suddenly come into being at an almighty word.' 'What a sublime idea of the infinite might of the great Architect! the Cause of all causes, the Father of all fathers, the Ens entium! For if we could compare the Infinite it would surely require a greater Infinite to cause the causes of effects than to produce the effects themselves.'
In these words he sets forth his position in regard to religion, and does so in precisely the same terms as we may use to-day when we say: 'All that happens in the world depends on the forces that prevail in it, and results according to law; but where these forces and their substratum, Matter, come from, we know not, and here we have room for faith.'
I have not been able to discover whether the Zoonomia, with its revolutionary ideas, attracted much attention at the time when it appeared, but it would seem not. In any case, it was afterwards so absolutely forgotten, that in an otherwise very complete History of Zoology, published in 1872 by Victor Carus, it was not even mentioned. About a year after the appearance of Zoonomia, Isidore Geoffrey St.-Hilaire in Paris expounded the view that what are called species are really only 'degenerations,' deteriorations from one and the same type, which shows that he too had begun to have doubts as to the fixity of species. Yet it was not till the third decade of the nineteenth century that he clearly and definitely took up the position of the doctrine of transformation, and to this we shall have to return later on.
But as early as the first decade of the century this position was taken up by two noteworthy naturalists, a German and a Frenchman, Treviranus and Lamarck.
Gottfried Reinhold Treviranus, born at Bremen in 1776, an excellent observer and an ingenious investigator, published, in 1802, a book entitled Biologie, oder Philosophie der lebenden Natur [Biology, or Philosophy of Animate Nature], in which he expresses and elaborates the idea of the Evolution theory with perfect clearness. We read there, for instance: 'In every living being there exists a capacity for endless diversity of form; each possesses the power of adapting its organization to the variations of the external world, and it is this power, called into activity by cosmic changes, which has enabled the simple zoophytes of the primitive world to climb to higher and higher stages of organization, and has brought endless[Pg 19] variety into nature.' But where the motive power lies, which brings about these transformations from the lowliest to ever higher forms of life, was a question which Treviranus apparently did not venture to discuss. To do this, and thus to take the first step towards a causal explanation of the assumed transformations, was left for his successor.
Jean Baptiste de Lamarck, born in 1744 in a village of Picardy, was first a soldier, then a botanist, and finally a zoologist. He won his scientific spurs first by his Flora of France, and zoology holds him in honour as the founder of the category of 'vertebrates.' Not that he occupied himself in particular detail with these, but he recognized the close alliance of the classes of animals in question—an alliance which was subsequently expressed by Cuvier by the systematic term 'type' or 'embranchement.'
In his Philosophie zoologique, published in 1809, Lamarck set forth a theory of evolution whose truth he attempted to vindicate by showing—as Treviranus had done before him—that the conception of species, on the immutability of which the whole hypothesis of creation had been based, was an artificial one, read into nature by us; that sharply circumscribed groups do not exist in nature at all; and that it is often very difficult, and not infrequently quite impossible, to define one species precisely from allied forms, because it is connected with these on all sides by transition stages. Groups of forms which thus melted into one another indicated that the doctrine of the fixity of species could not be correct, any more than that of their absolute nature. Species, he maintained, are not immutable, and are not so old as nature; they are fixed only for a certain time. The shortness of our life prevents our directly recognizing this. 'If we lived a much shorter time, say about a second, the hour-hand of the clock would appear to us to stand still, and even the combined observations of thirty generations would afford no decisive evidence as to the hand's movement, and yet it had been moving.'
The causes on which, according to Lamarck, the transformation of species, their modification into new species, depends, lie in the changes in the conditions of life which must have occurred ceaselessly from the earliest period of the earth's history till our own day, now here, now there, due in part to changes in climate and in food-supply, in part to changes in the earth's crust by the rising or sinking of land-masses, and so forth. These external changes have sometimes been the direct cause of changes in bodily structure, as in the case of heat or cold; but they have sometimes and much more effectively operated indirectly. Thus changed conditions may have[Pg 20] prompted an animal of a given species to use certain parts of its body in a new way, more vigorously, or less actively, or even not at all, and the more vigorous use, or, conversely, the disuse, has brought about variations in the organ in question.
Thus the whales lost their teeth when they abandoned their fish diet, and acquired the habit of feeding on minute and delicate molluscs, which they swallowed whole without seizure or mastication. Thus, too, the eyes of the mole degenerated through its life in the dark, and a still greater degeneration of the eyes has taken place in animals, like the proteus-salamander, which always inhabit lightless caves. In mussels both head and eyes degenerated because the animals could no longer use them after they became enclosed in opaque mantles and shells. In the same way snakes lost their legs pari passu with the acquisition of the habit of moving along by wriggling their long bodies, and of creeping through narrow fissures and holes. On the other hand, Lamarck interpreted the evolution of the web-feet of swimming birds by supposing that some land-bird or other had formed the habit of going into the water to seek for food, and consequently of spreading out its toes as widely as possible so as to strike the water more vigorously. In this way the fold of skin between the toes was stretched, and as the extension of the toes was very frequent and was continued through many generations, the web expanded and grew larger, and thus formed the web-foot.
In the same way the long legs of the wading birds have been, according to Lamarck, gradually evolved by the continual stretching of the limbs by wading in deeper and deeper water, and similarly for the long necks and bills of the waders, the herons and the storks. Finally we may mention the case of the giraffe, whose enormously long neck and tall forelegs are interpreted as due to the fact that the animal feeds on the foliage of trees, and was always stretching as far as possible, in order to reach the higher leaves.
We shall see later in what a different way Charles Darwin explained this case of the giraffe. Lamarck's idea is at once clear; it is true that exercising an organ strengthens it, that disuse makes it weaker. Through much gymnastic exercise the muscles of the arm become thicker and more capable, and memory too may be improved, that is to say, even a definite part of the brain may be considerably strengthened by use. Indeed, we may now go so far as to admit that every organ is strengthened by use and weakened by disuse, and so far the foundations of Lamarck's interpretations are sound. But he presupposes something that cannot be admitted so readily, namely, that such 'functional' improvement or diminution in the strength of[Pg 21] an organ can be transmitted by inheritance to the succeeding generation. We shall have to discuss this question in detail at a later stage, and I shall only say now that opinions as to whether this is possible or not are very much divided. I myself doubt this possibility, and therefore cannot admit the validity of the Lamarckian evolutionary principle in so far as it implies the directly transforming effect of the functioning of an organ. But even if we recognize the Lamarckian factor as a vera causa, it is easy to show that there are a great many characters which it is not in a position to interpret. Many insects which live upon green leaves are green, and not a few of them possess exactly the shade of green which marks the plant on which they feed; they are thus protected in a certain measure from injuries. But how could this green colour of the skin have been brought about by the activity of the skin, since the colour of the surroundings does not usually stimulate the skin to activity at all? Or how should a grasshopper, which is in the habit of sitting on dry branches of herbs, have thereby been incited to an activity which imparts to it the colour and shape of a dry twig? Just as little, or perhaps still less, can the protective green colour of a bird's or insect's eggs be explained through the direct influence of their usually green surroundings, even if we disregard the fact that the eggs are green when they are laid—that is, before the environment can have had any influence on them.
The Lamarckian principle of modification through use does not, in any case, nearly suffice as an interpretation of the transformations of the organic world. It must be allowed that Lamarck's theory of transformation was well founded at the time when it was advanced; it not only attacked the doctrine of the immutability of species, but sought for the first time to indicate the forces and influences which must be operative in the transformations of species; it was therefore well worth careful testing. Nevertheless it did not divert science from its chosen path; very little notice was taken of it, and in the great Cuvier's chronicle of scientific publications for 1809, not a syllable is devoted to Lamarck's book, so strong was the power of prejudice.
But, although the new doctrine was thus ignored, it did not altogether fall to the ground; it glimmered for a while in Germany, where it found its champions in the 'Naturphilosophie' of the time, and especially in Lorenz Oken, a peasant's son, born at Ortenau, near Offenburg, in 1783.
Oken professed views similar to those of Erasmus Darwin, Treviranus, and Lamarck, though they were not clothed in such[Pg 22] purely scientific garb, being, in fact, bound up with the general philosophical speculations which came increasingly into favour at that time, chiefly through the writings of Schelling. In the same year, 1809, in which Lamarck published his Philosophie zoologique, Oken's Lehrbuch der Naturphilosophie appeared.
This book is by no means simply a theory of descent; its scope is much wider, including the phenomena of the whole cosmos; on the other hand, it goes too little into details and is too indefinite to deserve its title. Its way of playing with ideas, its conjectures and inferences from a fanciful basis, make it difficult for us now to think ourselves into its mode of speculation, but I should like to give some indication of it, for it was just these speculative encroachments of the 'categories' of the so-called 'Naturphilosophie' which played a fatal part in causing the temporary disappearance of the Evolution-theory from science, so that, later on, it had to be established anew.
Oken defines natural science as 'the science of the everlasting transmutations of God (the Spirit) in the world': Every thing, considered in the light of the genetic process of the whole, includes, besides the idea of being, that of not-being, in that it is involved in a higher form. 'In these antitheses the category of polarity is included. The simpler elementary bodies unite into higher forms, which are thus merely repetitions at a potential higher than that of their causes. Thus the different genera of bodies form parallel and corresponding series, the reasonable arrangement of which results as an intrinsic necessity from their genetic connexion. In individuals these lowlier series make their appearance again during development. The contrasts in the solar system between planets and sun are repeated in plants and animals, and, as light is the principle of movement, animals have the power of independent movement in advance of the plants which belong to the earth.'
Obviously enough, this is no longer the study of nature; it is nature-construction from a basis of guesses and analogies rather than of knowledge and facts. Light is the principle of motion, and as animals move, they correspond to the sun, and plants to the planets! Here there is not even a hint of a deepening of knowledge, and all these deductions now seem to us quite worthless.
On the other hand, it must be allowed that good ideas are by no means absent from this 'philosophy,' nor can we deny to this restlessly industrious man a great mind always bent on discovering what was general and essential. Much of what we now know he even then guessed at and taught, as, for instance, that the basis of all forms of life in this infinitely diverse world of organisms was one and the[Pg 23] same substance—'primitive slime,' 'Urschleim' as he called it, or, as we should now say, 'protoplasm.' We can therefore, mutatis mutandis, agree with Oken when he says,'Everything organic has come from slime, and is nothing but diversely organized slime.' Many naturalists of the present day would go further, and agree with Oken when he suggests that 'this primitive slime has arisen in the sea, in the course of the planet's (the earth's) evolution out of inorganic material.'
Thus Oken postulated, as the specific vehicle of life, a primitive substance, in essence at least homogeneous. But he went further, and maintained that his 'Urschleim' assumed the form of vesicles, of which the various organisms were composed. 'The organic world has as its basis an infinitude of such vesicles.' Who is not at once reminded of the now dominant Cell-theory? And, in fact, thirty years later, when the cell was discovered, Oken did claim priority for himself. In so doing, he obviously confused the formulating of a problem with the solving of it; he had, quite rightly, divined that organisms must be built up of very minute concentrations of the primitive substance, but he had never seen a cell, or proved the necessity for its existence, or even attempted to prove it. His vesicle-theory was a pure divination, a prevision of genius, but one which could not directly deepen knowledge; it did not prompt, or even hasten, the discovery of the cell. Here, as throughout in his natural philosophy, Oken built, not from beneath upwards, by first establishing facts and then drawing conclusions from them, but, inversely, he invented ideas and principles, and out of them reconstructed the world. In this he differs essentially from his predecessors Erasmus Darwin, Treviranus, and Lamarck, who all reasoned inductively, that is, from observed data.
Thus the whole evolutionary movement was lost in indefiniteness; because men wanted to find a reason for everything, they missed even what might then have been explained. Moreover, the theory of evolution still lacked a sufficiently broad basis of facts; the 'Naturphilosophie,' by its want of moderation, robbed it of all credit; and it is not to be wondered at that men soon ceased to occupy themselves with the problem of the evolution of the living world. A few indeed held fast to the doctrine of evolution during the first third of the century, but then it disappeared completely from the realm of science.
Its last flicker of life was seen in France, in 1830, at the time of the July revolution, when the legitimate sovereignty of Charles X was overthrown. It is interesting to note the lively interest that[Pg 24] Goethe, the first forerunner of the theory, and then aged eighty-one, had in the intellectual combat that took place in the French Academy between Cuvier and Isidore Geoffroy St.-Hilaire. A friend of Goethe's, Soret, relates that on August 2, 1830, he went into the poet's room, and was greeted with the words: 'Well, what do you think of this great event? The volcano is in eruption, and all is in flames. There can no longer be discussion with closed doors.' Soret replied: 'It is a terrible business! But what else was to be expected with things as they are, and with such a ministry, than that it should end in the expulsion of the reigning family?' To which Goethe answered: 'We don't seem to understand each other, my dear friend. I am not talking of these people at all; I am thinking of quite different affairs. I refer to the open rupture in the Academy between Cuvier and Geoffroy St.-Hilaire; it is of the utmost importance to science.'
In this conflict of opinions, Cuvier opposed Geoffroy's conception of the unity of the plan of structure in all animals, confronting him with the four Cuvierian types, in each of which the plan of structure was altogether different, and strongly insisting on the doctrine of the fixity of species, which he maintained to be the necessary postulate of a scientific natural history.
The victory fell to Cuvier, and it cannot be denied that there was much justification for his opinions at the time, for the knowledge of facts at that stage was not nearly comprehensive enough to give security to the Evolution theory, and moreover the quiet progress of science might have been hindered rather than furthered by premature generalization and theorizing. It had now been seen how far the interpretation of general biological problems could be carried with the available material; the 'Naturphilosophie' had not merely exploited it as far as possible, but had burdened it much beyond its carrying power, and the world was weary of insecure speculations. The 'Naturphilosophie' was for the time quite worked out, and a long period set in, during which all energies were devoted to detailed research.
THE DARWINIAN THEORY
Period of detailed research—Appearance of Darwin's Origin of Species—Darwin's life—Voyage round the world—His teaching—Domesticated animals, dog, horse—Pigeons—Artificial selection—Unconscious selection—Correlated variations.
The period of wholly unphilosophical, purely detailed research may be reckoned as from about 1830 to 1860, though, of course, many of the labours of the earlier part of the century must be counted among the investigations which were carried out without any reference to general questions, and even after 1860 numerous such works appeared. Nor could it be otherwise, for the basis of all science must be found in facts, and the thorough working up of the fact-material will always remain the first and most indispensable condition of our scientific progress. During the period referred to, however, it had become the sole end to be striven for; and all energies were concentrated exclusively on the accumulation of facts.
The previous century had added much to the knowledge of the inner structure of animals, the so-called 'comparative anatomy,' and in the nineteenth century this line of investigation was pursued even more extensively and energetically, so that the knowledge increased enormously. Up till this time it was chiefly the structure of the backboned animals and of a few 'backboneless' animals, so called, that had been studied, but now all the lower groups of the animal kingdom were also investigated, and became known better and in more detail as the methods of research improved.
Not content, however, with a knowledge of the adult animal, naturalists began to investigate its development. In the year 1814 the first great work on development appeared, on the development of the chick, by Pander and Von Baer. It was there shown for the first time, how the chick begins as a little disk-shaped membrane on the surface of the yolk of the egg, at first simply as a pale streak, the 'primitive streak,' then as a groove, the 'primitive groove,' by the side of which arise two folds, the 'medullary folds,' and further how a system of blood-vessels is developed around this primitive rudiment on the upper surface of the yolk, how a heart[Pg 26] arises before the rest of the body is complete, and how the blood begins to circulate; in short, there was disclosed all the marvel of development to which we are now so much accustomed, that we can hardly understand the sensation it made at that time.
Later on, attention was turned to the development of Fishes and Amphibians (Agassiz and Vogt, later Remak), then to that of the Worms (Bagge), of Insects (Kölliker), and gradually the development of all the groups of the animal-kingdom—from Sponges to Man—was so thoroughly investigated that it almost seems to-day as if there could not be much that is new to discover in this department. This impression may indeed be true as far as the less complex processes and the more obvious questions are concerned, but it is impossible to predict what new problems may confront us, whose solution will depend on a still more detailed study of development.
As embryology is a science of the nineteenth century, so also is histology, the science of tissues. Its pioneer was Bichat, but its real foundations were not laid till Schwann and Schleiden formulated the conception of the 'cell,' and proved that all animals and plants were composed of cells. What Oken had only guessed at they now proved, that there are very minute form-elements of life which build up all the parts of animals and plants or produce them by processes of secretion. New light was thus shed on embryonic development, and this gradually led to the recognition of the fact that the egg, too, is a cell, and that development depends on a cell-division process in this egg-cell. This led further to the conception of many-celled and single-celled organisms, and so on to many items of knowledge to speak of which here would carry us too far.
For it is not my intention to attempt a complete review of the development of biology in the nineteenth century, or even in the period which we have mentioned as devoted to detailed research; it is rather my desire to convey a general impression of the enormous extent and many-sidedness of the progress that was made in this time. Let us therefore briefly recall the entirely new facts which were brought to light in this period with regard to the reproduction of animals. Asexual reproduction by budding and division was already known, but parthenogenesis is a discovery of this period, and so also is alternation of generations, so far-reaching in its bearing on general problems. It was first observed (1819) by Chamisso in Salpa, then by Steenstrup in Medusæ and trematodes, and was later made fully clear in its most diverse forms and relations by the researches of Leuckart, Vogt, Kölliker, Gegenbaur, Agassiz, and other illustrious investigators. Reproduction by heterogony, too,[Pg 27] which occurs in many crustaceans, and in aphides and certain worms, was recognized at that time, and in the sixties Carl Ernst von Baer added to the list precocious reproduction, or pædogenesis, which is illustrated in certain insects which reproduce in the larval state.
This may suffice to convey some idea of the great mass of new, and in some cases startling facts previously unguessed at, which were then brought to light in the department of animal biology alone. To this must be added the vast increase in the number of known species and varieties, their distribution on the earth, and all this, mutatis mutandis, for plants also. Nor can we omit to mention the rapidly growing number of fossil species of animals and plants.
Thus there gradually accumulated a new mass of material; investigation became more and more specialized, and the danger became imminent that workers in the various departments would be unable to understand each other, so completely were they independent of one another in their specialist researches. There was lack of any unifying bond, for workers had lost sight of the general problem in which all branches of the science meet, and through which alone they can be united into a general science of biology. The time had come for again combining and correlating the details, lest they should grow into an unconnected chaos, through which it would be impossible to find one's way, because no one could overlook it and grasp it as a whole. In a word, it was high time to return to general questions.
Though I have called the period from 1830 to 1860 that of purely detailed research, I do not mean to ignore the fact that, during that time, there were a few feeble attempts to return to the great questions which had been raised at the beginning of the century. But the point is, that all such attempts remained unnoticed. Thus there appeared, in 1844, a book entitled Vestiges of the Natural History of Creation, the anonymous author of which revealed himself much later as Robert Chambers, an Edinburgh publisher. In this book the evolution of species was ascribed to two powers, a power of transformation and a power of adaptation. Two Frenchmen, Naudin and Lecoq, also published a work in which the theory of evolution was set forth, and from 1852 to 1854 the well-known German anthropologist Schaafhausen was writing on similar lines. But all these calls sounded unheard, so deeply were naturalists plunged in detailed investigations, and it required a much mightier voice to command the ear of the scientific world.
It is impossible to estimate the effect of Darwin's book on The[Pg 28] Origin of Species, published in English in 1858, in German in 1859 unless we fully realize how completely the biologists of that time had turned away from general problems. I can only say that we, who were then the younger men, studying in the fifties, had no idea that a theory of evolution had ever been put forward, for no one spoke of it to us, and it was never mentioned in a lecture. It seemed as if all the teachers in our universities had drunk of the waters of Lethe, and had utterly forgotten that such a theory had ever been discussed, or as if they were ashamed of these philosophical flights on the part of natural science, and wished to guard their students from similar deviations. The over-speculation of the 'Naturphilosophie' had left in their minds a deep antipathy to all far-reaching deductions, and, in their legitimate striving after purely inductive investigation, they forgot that the mere gathering of facts is not enough, that the drawing of conclusions is an essential part of the induction, and that a mass of bare facts, however enormous, does not constitute a science.
One of my most stimulating teachers at that time, the gifted anatomist, Jacob Henle, had written as a motto under his picture, 'There is a virtue of renunciation, not in the province of morality alone, but in that of intellect as well,' a sentence which expressly indicated the desirability of refraining from all attempts to probe into the more general problems of life. Thus the young students of that time were nourished only on the results of detailed research, in part indeed interesting enough, but in part dry and, because uncorrelated, unintelligible in the higher sense, and only here and there awakening a deeper interest, when, as in physiology and in embryology, they formed a connected system in themselves. Without being fully clear as to what was lacking, we certainly missed the deeper correlation of the many separate disciplines.
It is therefore not to be wondered that Darwin's book fell like a bolt from the blue; it was eagerly devoured, and while it excited in the minds of the younger students delight and enthusiasm, it aroused among the older naturalists anything from cool aversion to violent opposition. The world was as though thunderstruck, as we can readily see from the preface with which the excellent zoologist of Heidelberg, Bronn, introduced his translation of Darwin's book, where he asks this question among others, 'How will it be with you, dear reader, after you have read this book?' and so forth.
But before I enter on a detailed examination of the contents of this epoch-making book, I should like to say a few words about the man himself, who thus revolutionized our thinking.
Charles Darwin was born in 1809, the year of the publication of Lamarck's Philosophie zoologique, and of Oken's Lehrbuch der Naturphilosophie. There was thus a whole generation between the first emergence of the Evolution theory and its later revival. Darwin's father was a physician, and his education was not a regular one. In his youth he seems to have devoted much time and enthusiasm to hunting, and only very slowly to have taken up regular studies towards a definite end. In accordance with his father's wishes, he studied medicine for a time, but soon abandoned it to devote himself to botany and zoology. Before he had had time to distinguish himself in any special way in these subjects, he was offered, in his twenty-first year, the post of naturalist on an English war-ship which was to make a voyage round the world, and that at a leisurely rate.
This was decisive not only for Darwin's immediate studies, but for the work of his life, for, as he tells us himself, it was during this voyage on the Beagle that the idea of the Evolution theory first came to him. While the vessel made a stay at the Galapagos Islands, west of South America, he noticed that quite a number of little land-birds occurred there which closely resembled those of the neighbouring mainland, but yet were different from them. Almost every little island had its own species, and so he concluded that all these might be descended from representatives of a few species which had long before drifted over from the mainland to these volcanic islands, become established there, and in the course of time taken on the character of new species. The problem of the transformation of species opened up before him, and he made up his mind to follow up the idea after his return, in the hope that by a patient collecting of facts, he would by and by arrive at some security with regard to this great question.
I need not linger over any detailed account of his travels; one can readily understand how a voyage round the world, lasting for five years, would offer to the inquiring mind of a Darwin rich opportunities for the most varied observations. That he did not fail to make use of these is evidenced not only by his book on The Origin of Species, but by several more special works, published shortly after his return—his natural history of those remarkable sessile crustaceans, the barnacles or Cirripedia, and his studies on the origin of coral reefs. The first-named book still holds its own as a classic monograph on this animal group, with its wealth of forms; and the theory of the origin of coral reefs which Darwin elaborated has still many adherents, in spite of various rival interpretations.
But Darwin would hardly have achieved what he did if he had been compelled to secure for himself a professional position in order to obtain bread and butter. Such great problems demand not only the whole of a man's mental energy, they monopolize his time. Studies of detail may well be taken up in leisure hours, but big problems absorb all the thoughts and must always be present to the mind, lest the connexion between the many individual inquiries, which make up the whole task, be lost sight of. Darwin had the good fortune to be a free investigator, and to be able to retire, on his return from his travels, to a small property at Down in Kent, there to live for his family and his work. Here he followed up the idea of evolution which he had already formulated, and it has always seemed to me the most remarkable thing about him, that he was able to keep in mind and work up the hundreds of isolated inquiries that were eventually to be brought together to form the main fabric of his theory. When one studies his many later writings, one cannot but be surprised afresh by the number of different sets of facts he collected at the same time, partly from others, partly from personal observation, and continually also from his own experiments. He made experiments on plants and on animals, and the number of people with whom he carried on a scientific correspondence is simply astounding. In this way he brought together, in the course of twenty years, an extraordinarily rich material of facts, from the fullness of which he was able later to write his book on The Origin of Species. Never before had a theory of evolution been so thoroughly prepared for, and it is undoubtedly to this that it owed a great part of its success; not to this alone, however, but still more, if not mainly, to the fact that it presented a principle of interpretation that had never before been thought of, but whose importance was apparent as soon as attention was called to it—the principle of selection.
Charles Darwin championed, in the main, the same fundamental ideas as had been promulgated by his grandfather, Erasmus Darwin, by Treviranus, and by Lamarck: species only seem to us immutable; in reality they can vary, and become transformed into other species, and the living world of our day has arisen through such transformations, through a sublime process of evolution which began with the lowest forms of life, but by degrees, in the course of unthinkably long ages, progressed to organisms more and more complex in structure, more and more effective in function.
It is interesting to note at what point Darwin first put in his lever to attempt the solution of the problem of evolution. He started from quite a different point from the investigators of the early part[Pg 31] of the century, for he began with forms of life which had previously been markedly neglected by science, the varieties of our domesticated animals and cultivated plants.
Previously these had been in a sense mere step-children of biology, inconvenient existences which would not fit properly into the system, which were therefore as far as possible ignored or dismissed as outside the scope of 'the natural,' because it was difficult to know what else to do with them. I can quite well remember that, even as a boy, I was struck by the fact that one could find nothing in the systematic books about the many well-established garden forms of plants, or about our domestic animals, which seemed to be regarded as in a sense artificial products, and as such not worthy of scientific consideration. But it was in these that Darwin particularly interested himself, making them virtually the basis of his theory, for he led up from them to the very principle of transformation, which was his most important addition to the earlier presentations of the Evolution theory.
He started from the existence of varieties which may be observed in so many wild species. His line of thought was somewhat as follows: If species have really arisen through a gradual process of transformation, then varieties must be regarded as possible first steps towards new species; if, therefore, we can only succeed in finding out the causes which underlie the formation of any varieties whatever, we shall have discovered the causes of the transformation of species. Now we find by far the greatest number of varieties, and the most marked ones, among our domesticated animals and plants, and unless we are to assume that each of these is descended from a special wild species, the reason why there has been such a wealth of variety-formation among them must lie in the conditions which influence the relevant species in the course of domestication; and it remains for us to analyse these conditions till we come upon the track of the operative factors. With this conviction, Darwin devoted himself to the study of domesticated animals and plants.
The first essential was to prove that every variety had not a separate wild species as ancestor, but that the whole wealth of our domesticated breeds originated, in each case, from one, or at least from a few wild species. Of course I cannot here recapitulate the multitudinous facts which were marshalled by Darwin, especially in his later works, notably his Animals and Plants under Domestication, but this is not necessary to an understanding of his conclusions, and I shall therefore restrict myself to a few examples.
Let us take first the domestic dog, Canis familiaris, Linné. We[Pg 32] have at the present day no fewer than seven main breeds, each of which has its sub-breeds, often numerous. Thus there are forty-eight sub-breeds which are used as guardians of our houses, 'house-dogs' in the restricted sense, thirty sub-breeds of dogs with silk-like hair (King Charles dogs, Newfoundland dogs, &c.), twelve of terriers, and thirty-five of sporting dogs, among them such different forms as the deerhound and the pointer. We have further nineteen sub-breeds of bulldogs, thirty-five of greyhounds, and six of naked or hairless dogs. Not only the main breeds, but even the sub-breeds often differ as markedly from one another as wild species do, and the question must first be decided whether each of the very distinct breeds has not a special wild species as ancestor.
Obviously, however, this cannot be maintained, for so many species of wild dog have never existed on the earth at any time. We know, too, that 4,000 or 5,000 years ago a large number of breeds of dogs were in existence in India and Egypt. There were Pariah dogs, coursers, greyhounds, mastiffs, house-dogs, lapdogs and terriers. It is not possible that the products of all lands could, at that time, have been gathered into one, and it is inconceivable that so many wild species could have existed in the one country of India.
On the other hand, however, it cannot be maintained that all our present breeds have descended from a single wild species; it is much more probable that several wild species were domesticated in different countries.
It has often been supposed that the manifold diversity of our present breeds has been brought about by crossing the various tamed species. That cannot be the case, however, because crossing gives rise only to hybrid mongrel forms, not to distinct breeds with quite new characters. It is true that all breeds of dogs can be very readily crossed with each other, but the result is not new breeds, but those numberless and transient intermediate forms which the dog-breeder despises as worthless for his purpose. It must therefore have been through the influence of domestication, combined with crossing, that a few wild species gave rise to the various breeds of dogs.
The pedigree of the horse is rather more clear than that of the dog. Even in this case, indeed, one cannot definitely name the ancestral wild form, but it is very probable that it was of a grey-brown colour, and similar to the wild horses of our own day. Darwin supposes that it must also have had the black stripe on the back which is exhibited by the domestic ass, and by several wild species of ass, basing his opinion on the fact that the spinal stripe often occurs in foals, especially in those of a grey-brown colour.
But though there can be no doubt that this is to be interpreted as a reversion to a character of a remote ancestor, it by no means follows that the direct ancestral form must have had this stripe. I am more inclined to believe that the ancestor which bore this mark was considerably more remote, and lived before the differentiation of the horse from the ass. Darwin himself noted the remarkable fact that in rare cases, especially in foals, not only may the stripe on the back be present, but there may be more or less distinct zebra-striping on the legs and withers: this, however, must be interpreted as a reversion to the character of a very much more remote ancestor, to a common ancestor of all our present-day horses and asses, which must have been striped over its whole body, like the zebra living in Africa now.
It cannot be proved of any of the wild horses of to-day that they are not descended from domesticated ancestors; indeed, we can say with certainty that the thousands of wild horses which roam the plains of North and South America are descended from domestic horses, for there was no horse in America at the time it was discovered by the Europeans. In all probability our horse originated in Middle Asia, was there first domesticated, and has thence been gradually introduced into other countries. In Egypt it appears first on the monuments in the seventeenth century B.C., and it seems to have been introduced by the conquering Hyksos. On the ancient Assyrian monuments the chase after wild horses is depicted, and they were not caught, but killed with arrow and lance, like the lion and the gazelle.
But even if two wild species of horse had been tamed in different parts of the great continent of Asia, these two domesticated animals would have varied much and in the most diverse manner, as we may infer from our different breeds of horses at the present day. There are a great many of these, and many of them differ very considerably from each other. If we think of the lightly built Arab horse, and place beside it the little pony, or the enormous Percheron, the powerful cart-horse from the old French province of La Perche, which easily draws a load of fifty kilograms, we are face to face with differences as great as those between natural species. And we may realize how many breeds of horses there are now upon the earth if we remember that nearly every oceanic island has its special breed of ponies. Not only in the cold Shetland Islands, England, Sardinia and Corsica, but in almost every one of the larger islands of the extensive Indian Archipelago there is one, and Borneo and Sumatra have several.
But the most conclusive proof of descent from a single wild species is afforded by the pigeons, and as the production of new breeds among them has been, and will continue to be, carried on with particular enthusiasm and deliberateness, I propose to deal with them somewhat more in detail.
Darwin's work proves beyond a doubt that all our present-day breeds of pigeons are descended from a single wild species, the rock-dove, Columba livia. In appearance, this form, which still lives in a wild state, differs little from our half-wild blue-grey field-pigeon. It has the same metallic shimmer on the feathers of the neck, the same two black cross-bars on the wings as well as the band over the tail, and it has also the same slate-blue general colour. Now, all breeds of pigeons are without restriction fertile inter se, so that any breed can be crossed with any other, and it often happens that, in the products of such crossing, characters appear which the parents, that is, the two or more crossed breeds, did not possess, but which are among the characters of the rock-dove. Thus Darwin obtained, by crossing a pure white fantail with a black barb, hybrids which were partly blackish brown, partly mixed with white, but when he crossed these hybrids with others from two breeds which were likewise not blue, and had no bars, he obtained a slate-blue rock-pigeon, with bars on the wings and tail. We shall inquire later on how far it is correct to regard such cases as reversions to remote ancestors, but if we take it for granted in the meantime, we have here a proof of the descent of our breeds from a single wild species. This is corroborated, too, by everything that we know about the distribution of the rock-pigeon and the place and time of its domestication. It still lives on the cliff-guarded shores of England, Brittany, Portugal, and Spain, and both in India and in Egypt there were tame pigeons at a very early period. Pigeons appear on the menu of a Pharaoh of the fourth dynasty (3000 B.C.), and of India we know at least that in 1600 A.D. there were 20,000 pigeons belonging to the court of one of the princes.
The beauty of this bird, and the ease with which it can be tamed, obviously called man's attention to it at a very early date, and it has been one of man's domestic companions for several thousands of years. Now we can distinguish at least twenty main races (Fig. 1), which differ from each other as markedly as, if not more markedly than, the most nearly allied of the 288 wild species of pigeons which inhabit the earth. We have carriers and tumblers, runts and barbs, pouters, turbits and Jacobins, trumpeters and laughers, fantails, swallows, Indian pigeons, &c.

Fig. 1. Group of various races of domestic pigeons (after Prütz). 1. Pouter. 2. Indian barb. 3. Bucharest trumpeter with a whorl of feathers (Nelke) on its forehead. 4. Nürnberger swallow. 5. Nürnberger bagadotte. 6. English carrier. 7. Fantail. 8. Eastern turbit. 9. Schmalkaldener Jacobin. 10. Chinese owl. 11. German turbit.
Each of these races falls into sub-races; thus there is a German, an English, and a Dutch pouter-pigeon. The books on pigeons mention over 150 kinds which are quite distinct from one another, and breed true, that is, always produce young similar to themselves.
Without entering upon a detailed description of any of these, I should like to call attention to the way in which certain characters have varied among them. Colour is a subordinate race-character, in so far that colour alone does not constitute a race, yet the colouring within a particular sub-race is usually very sharply defined, and in every breed there are sub-races of different colours. Thus there are white, black, and blue fantails, there are white turbits with red-brown wings, but also red ones with white heads, and white tumblers with black heads, &c. Very unusual colours and colour-markings sometimes occur. Thus one sub-race of tumblers exhibits a peculiar clayey-yellow colour splashed with black markings, otherwise rare among pigeons, and almost suggestive of a prairie-hen; there is also a copper-red spot-pigeon, a cherry-red 'Gimpel'-pigeon, lark-coloured pigeons, &c. Then we find all possible juxtapositions of colours, limited to quite definite regions of the body; thus we have white tumblers with a red head, red tail, and red wing-tips, or white tumblers with a black head, red turbits with white head, Indian pigeons quite black except for white wing-tips, and so on. The distribution of colour is often very complicated, but nevertheless, all the individuals of the breed show it in exactly the same manner. Thus there are the so-called blondinettes in which almost the whole body is copper-red, but the wings white, save that each quill bears at the rounded end of its vane a black and red fringe. I should never come to an end, if I were to try to give anything like a complete idea of the diversity of colouring among the various breeds of pigeons.
Even such an important and, among wild species, unusually constant organ as the bill has varied among pigeons to an astonishing degree. Carrier-pigeons (Fig. 1, No. 6) have an enormously long and strong bill, which is moreover covered with a thick red growth of the cere, while in the turbits and owls (Fig. 1, Nos. 8 and 10) the bill is shorter than any we find among wild birds. In many breeds even the form of the bill deviates far from the normal, as in the bagadottes (No. 5) with crooked bill.
Like the bill, the legs vary in regard to their length. The pouters (No. 1) stand on their long legs as on stilts, while the legs of the 'Nürnberger swallow' are strikingly small. Remarkable, too, and very different from the wild species, is the thick growth of feathers on the feet and toes of the pouters and trumpeters (Fig. 1, No. 1),[Pg 37] as well as of some other breeds, which suggests the arrangement of feathers on a wing.
Furthermore, the number and size of wing and tail-feathers in the different breeds often deviate considerably from the normal. The fantail (No. 7) in its most perfect form possesses forty tail-feathers, instead of the twelve usual in the wild rock-pigeon, and they are carried upright like a fan, while the head and neck of the bird are bent sharply backwards. In the hen-like pigeons the tail-feathers are few and short, so that they show an upright tail like that of a hen. I have already referred to the extraordinary carunculated skin-growth on the bill of many breeds; such folds also often surround the eye, and, as in the Indian barb (No. 3), are developed into well-formed thick circular ridges, while in the English carrier (No. 6) they lie about the bill as a formless mass of flesh.
Even the skull has undergone many variations, as can be observed even in the living bird in many of the breeds with short forehead. Differences are to be found, too, in the number and breadth of the ribs, the length of the breast-bone, the number and size of the tail-vertebræ in different breeds. Of the internal organs, the crop in many breeds, but particularly in the pouters (No. 1), has attained an enormous size, and with this size is usually associated the habit of blowing it out with air, and assuming the characteristically upright position.
That variations have taken place, too, in the most delicate structure of the brain, is shown by certain new instincts, such as the trumpeting of the trumpeters, the cooing of others, and the silence of yet other breeds, as well as by the curious habit of the tumblers of ascending quickly and vertically to a considerable height, and then turning over once, or even several times, in the course of their descent. In contrast to this, other breeds like the fantails have altogether given up the habit of flying high, and usually remain close to the dove-cot.
Lastly, let me mention that the unusual development of individual feathers, or of groups of feathers, has become a race-character, upon which depend such remarkable structures as the feather-mantle turned over the head in the Jacobins (No. 9), the cap or plume on the head of various breeds, the white beard in the bearded tumbler, the collars which lie like a shirt-collar on the breast, or run down the sides of the neck (Nos. 8 and 10), and the circle of feathers which marks the root of the bill in the Bucharest trumpeter (No. 3).
After what has been said, it is hardly necessary to add that the size of the whole body differs in different races. But the differences[Pg 38] are very considerable, for, according to Darwin, one of the largest runt-pigeons weighed exactly five times as much as one of the smallest tumblers with short forehead, and in the illustration (Fig. 1) the pouter looks a giant beside the little barb to its left.
Thus we see that nearly every part of the body of the pigeon has varied under domestication in the most diverse ways, and to a high degree; and the same is true of several other domesticated animals, poultry, horses, sheep, cattle, pigs, and so on, though the matter is not altogether so clear in their case, since descent from a single wild species cannot be proved, and is in many cases improbable. But in the case of pigeons this common descent is certain, and we have now to inquire in what manner all these variations from the parent form have been brought about.
The answering of this question is rendered easier by the fact that new breeds arise even now, and that, to some extent at least, they can be caused to arise, consciously and intentionally. In England, as well as in Germany and France, there are associations for the breeding of birds, and in England especially pigeon and poultry clubs are numerous and highly developed. These by no means confine themselves to simply preserving the purity of existing breeds, they are continually striving to improve them, by increasing and accentuating their characters, or even by introducing quite new qualities, and in many cases they succeed even in this last. Prizes are offered for particular new variations, and thus a spirit of rivalry is fostered among the breeders, and each strives to produce the desired character as quickly as possible. Darwin says: 'The English judges decided that the comb of the Spanish cock, which had previously hung limply down, should stand erect, and in five years this end was achieved; they ordained that hens should have beards, and six years later fifty-seven of the groups of hens exhibited at the Crystal Palace in London were bearded.' The transformation does not always come about so quickly, however; thus, for instance, it required thirteen years before a certain breed of tumblers was furnished with a white head. But the breeders cause every visible part of the body to vary as seems good to them, and within the last fifty years they have really brought about very considerable changes in many breeds. Their method of procedure is carefully to select for breeding those birds which already possess a faint beginning of the desired character. Domesticated animals have on the whole a higher degree of variability than wild species, and the breeder takes advantage of this. Suppose it is a question of adding a crown of feathers to a smooth-headed breed, a bird is chosen which has the feathers on the back of the[Pg 39] head a little longer than usual, and mated for breeding. Among its descendants there will probably be some which also exhibit these slightly prominent feathers, and possibly there may be one or other of them which has these feathers considerably lengthened. This one is then used for breeding, and by continually proceeding thus, and selecting for breeding, from generation to generation, only the individuals which approach most nearly to the desired end, the wished-for character is at last secured.
Thus it is not by crossing of different breeds, but by a patient accumulating of insignificant little variations through many generations, that the desired transformations are brought about. That is the magic wand by means of which the expert breeder produces his different breeds, we might almost say, as the sculptor moulds and remoulds his clay model according to his fancy. Quite according to his fancy the breeder has brought about all the fantastic forms we are familiar with among pigeons, mere variations which are of no use either to the bird itself or to man, which simply gratify man's whim without in many cases even satisfying his sense of beauty. For many of the existing breeds of pigeons, hens, and other domesticated animals, are anything but beautiful, the body being often unharmonious in structure and sometimes actually monstrous.
Among pigeons, as well as among other domesticated animals, some changes have been brought about, which are not only of no use to their possessors, but would be actually disadvantageous if they were living under natural conditions. Some of the very short-billed breeds of pigeons have the bill so short and soft that the young can no longer use it to scratch and break the egg-shell, and would perish miserably if human aid were not at hand. The Yorkshire pig has become such a colossus of fat on weak, short legs, that if it were dependent on its own resources, it could not secure its food, much less escape from a beast of prey; and among horses the heavy cart-horse and the racer are alike unfit to cope with the dangers of a wild life, or the vicissitudes of weather.
Breeding has done much to bring about variations useful to man. Thus we have breeds of cattle which excel in flesh, or in milk, or as draught animals, and sheep which excel in flesh or in wool, and to what a height the perfecting of a useful quality can be brought is shown, in regard to fineness of wool, by that finest breed of sheep, the merino, which instead of the 5,500 hairs borne by the old German sheep on a square inch, possesses 48,000.
Not infrequently it is a particular stage of a species that has been bred by man, and the other stages have remained more or less[Pg 40] unaltered. Thus it is with one of the few domesticated insects, the silk-moth. Only the cocoon is of use to man, and according to the cocoon different breeds are distinguished, differing in fineness, colour, &c.; but no breeds can be distinguished in reference to the larvæ, or the perfect insects. Among gooseberries there are about a hundred varieties distinguished according to the form, colour, size, thickness of skin, hairiness, &c., of the fruits, but the little, inconspicuous, green blossoms, of which the breeders take no account, are alike in them all. In the pansies (Viola tricolor), on the other hand, it is only by the flowers that the varieties are distinguished, while the seeds have remained alike in all.
It may be asked how it could have occurred to any one, when pigeons, for instance, first began to be domesticated, to wish to produce fantails or pouters, since he could have no mental picture of them in advance. Darwin replies to this objection, that it was not always conscious and methodical artificial selection, such as is now practised, that brought about the origin of breeds, but that they have very often resulted, and at first perhaps always, from unconscious selection. When savages tamed a dog, they used the 'best' of their dogs for breeding, that is, they chose those which had in the highest degree the qualities they valued, watchfulness, for instance, or if the dog were intended for the chase, keen scent and swiftness. In this way the body of the animal would be changed in a definite direction, especially if rivalry helped, and if it was the ambition of each to possess a dog as good as, or better than those of his tribal companions. That perfectly definite changes in bodily form can thus be brought about unconsciously is well illustrated by the case of a racehorse. This has arisen within the last two hundred years simply because the fleetest of the products of crossing between the Arab and the English horse were always chosen for breeding. It could not have been predicted that horses with thin neck, small head, long rump, and slender legs would necessarily be the swiftest runners; but this is the form which has resulted from the selection,—a very ugly, but very swift horse. This unconscious selection must undoubtedly have played a large part in the early stages of the evolution of the breeds of our domestic animals.
But even in the fully conscious and methodical selective breeding of particular characters, the breeder rarely alters only the one his attention is fixed on; generally quite a number of other characters alter apart from his intention as an inevitable accompaniment of the desired variation on which attention was riveted. There are breeds of rabbits whose ears hang limply down instead of standing erect,[Pg 41] and in these so-called lop-eared rabbits the ear-muscles are partly degenerated, and as a consequence of this lack of muscular strain the skull has assumed another form. Thus the variation of one part may influence the development of a second and a third organ, and may even not stop there, for very often the influence has penetrated much deeper and affected quite remote parts of the body.
If any one were to succeed in adding a heavy pair of horns to a breed of hornless sheep, there would run parallel with the course of this variation, which was directly aimed at, a long series of secondary changes which would affect at least the whole of the anterior half of the body; the skull would become thicker and stronger to support the weight of the heavy horns; the neck-tendon (ligamentum nuchæ) would have to become thicker to hold up the heavy head, and so also with the muscles of the neck; the spinous processes of the cervical and dorsal vertebrae would become longer and stronger, and the forelegs, too, would need to adapt themselves to the heavier burden. Every organism thus resembles, as it were, a mosaic, out of which no individual group of pieces can be taken and replaced by another without in some measure disturbing the correlation and harmony of the whole: in order to restore this, the pieces all round about the changed part must be moved or replaced by others.
According to Darwin, it is to this correlation of parts that we must refer the variation of other parts besides the one intentionally altered in the course of breeding. It must be admitted that the mutual dependence of the parts plays a very important rôle in the economy and development of the animal body, as we shall see later, and these connexions still remain very mysterious to us. Especially is this the case with the connexion between the reproductive organs and the so-called secondary sexual characters. Removal of the reproductive organs or gonads induces, in Man, for instance, if it be effected in youth, the persistence of the childish voice and the non-development of the beard; in the stag the antlers do not appear, and in the cock the comb does not develop perfectly, &c., but we are not yet able to understand clearly why this should be so.
THE DARWINIAN THEORY (continued)
Natural selection—Variation—Struggle for existence—Geometric ratio of rate of increase—Normal number and ratio of elimination in a species—Accidental causes of extinction—Dependence of the strength of a species on enemies—Struggle for existence between individuals of the same species—Natural selection affects all organs and stages—Summary.
In artificial selection, through which, with or without conscious intention, our domesticated animals and cultivated plants have arisen, there must obviously be three kinds of co-operative factors: first, the variability of the species; second, the capacity of the organism for transmitting its particular characters to its progeny; and third, the breeder who selects particular qualities for breeding. No one of the factors can be dispensed with; the breeder could effect nothing, were there not presented to him the variations of parts in the particular direction in which he wishes them to vary; an indefinite variation, that is, a variation not guided by selection, would never lead to the formation of new breeds; the species would probably become in time a motley mixture of all sorts of variations, but a breed with definite characters, transmissible in their purity to its descendants, could never be formed. Finally, every process of selective breeding would be futile, if the variations which appeared could not be transmitted.
Darwin assumes that processes of transformation quite similar to those which take place under the guidance of Man occur also in nature, and that it is mainly these which have brought about and guided the transformations of species which have taken place in the course of the earth's history. This process he calls natural selection.
It will readily be admitted that two out of the three factors necessary to a process of selective breeding are present also in the natural conditions of the life of species. Variability in some degree or other is absent from no species of animal or plant, though it may be greater in one than in another, and it cannot be doubted that the inborn differences which distinguish one individual from another are capable of transmission. It is only to untrained observers that all the individuals of a species appear alike; for instance, all garden whites, or all the individuals of the small tortoiseshell butterfly (Vanessa[Pg 43] urticæ), or all the chaffinches. If the individuals are carefully compared it will be recognized that, even in these relatively constant species, no individual exactly resembles another; that even among butterflies twenty black scales may go to form a particular spot on the wings in one individual and thirty or twenty-five in others; that the length of the body, the legs, the antennæ, the proboscis exhibit minute differences; and it is probable that the same combination of quite similar parts never occurs twice. In many animals this cannot, of course, be proved, because our power of diagnosis is not fine enough to be able to estimate the differences directly, and because a comparison of measurements of all the parts in detail is not practicable. So we may here confine ourselves to the differences in the human race, which we can recognize with ease and certainty. Even as regards the face alone, all men differ from one another, and, numerous and complete as likenesses may be, it is impossible to find two human beings in which even the characters of the face are exactly similar. Even so-called 'identical twins' can always be distinguished if they are directly compared either in person or in a photograph, and if the rest of the body be also taken into consideration we find numerous small, sometimes even measurable differences.
The same is true of animals, and it is only our lack of practice that is at fault if we frequently fail to detect their individual differences. The Bohemian shepherds are said to know personally, and be able to distinguish from all the rest, every sheep in their herds of many thousands. Thus the factors of variability and transmissibility must be granted, and it remains only to ask: Who plays the part of selecting breeder in wild nature? The answer to this question forms the kernel to the whole Darwinian theory, which ascribes this rôle to the conditions of life, to definite relations of individuals to the external influences which they meet with during the course of their lives, and which together make up their 'struggle for existence.'
To make this idea clear I must to some extent diverge.
It is a generally observed fact that, in every species of animals or of plants, more germs and more individuals are produced than grow to maturity, or become capable of reproduction. Numerous young individuals perish at an early stage, often because of unfavourable circumstances—cold, drought, damp, or through hunger, or at the hands of their enemies. When we ask which of the progeny perish early, and which survive to carry on the species, we are at first sight inclined to suppose that this is entirely a matter of chance; but this is just what Darwin disputed. It is not chance alone, it is, above all,[Pg 44] the differences between individuals, which enable them to withstand adverse circumstances better or worse, and thus decide, according to his view, which shall perish and which shall survive. If this be so, then we have a veritable process of selection, and one which secures that the 'best,' that is, the most capable of resistance, survive to breed, being thus, so to speak, 'selected.'
It may be asked, however, why so many individuals must perish in youth, and whether it could not have been arranged that all, or at least most, should survive till they had reproduced. But this is an impossibility, unrealizable for this among other reasons, that organisms multiply in geometrical progression, and that their progeny would very soon exceed the limits of computability. This does not occur, for there is a limit set which they can in no case overstep,—which, indeed, as we shall see, they never reach—I mean the limits of space and food-supply. Every species, by the natural requirements of its life, is restricted to a particular habitat, to land or to water, but most are still more strictly limited to a definite area of the earth's surface, which alone affords the climate suited to them, or where alone the still more specialized conditions of their existence can be realized. Thus, for instance, the occurrence of a particular species of plant determines that of the animal which is dependent on it for its food-supply. If they could multiply unchecked, that is, without the loss of many of their progeny, every species would fill up its area of occurrence and exhaust the whole of its food-supply, and thus bring about its own extermination. This seems to be prevented in some way, for as a matter of fact it does not happen.
It may, perhaps, be imagined that this might be prevented by a regulation of the productivity of the species, and that those which have not a large area of distribution, or can only count on a relatively limited food-supply, have also a low rate of multiplication, but this is not the case; even the lowest rate of multiplication would very soon suffice to make any species fill up its whole available space and completely exhaust its food-supply. Darwin takes as an example the elephant, which only begins to breed at thirty years of age, and continues to do so till about ninety, but so slowly that in these sixty years only three pairs of young are produced. Nevertheless, in 500 years an elephant pair would be represented by fifteen millions of descendants, if all the young survived till they were capable of reproduction. A species of bird with a duration of life of five years, during which it breeds four times, producing and rearing four young each time, would in the course of fifteen years have 2,000 millions of descendants.
Thus, although the fertility of each species is, as a matter of fact, precisely regulated, a low rate of multiplication is not in itself sufficient to prevent the excessive increase of any species, nor is the quantity of the relevant food-supply. Whether this be very large or very small, we see that in reality it is never entirely used up, that, as a matter of fact, a much greater quantity is always left over than has been consumed. If increase depended only on food-supply, there would, for instance, be food enough in their tropical home for many thousand times more elephants than actually occur; and among ourselves the cockchafers might appear in much greater numbers than they do even in the worst cockchafer year, for all the leaves of all the trees are never eaten up; a great many leaves and a great many trees are left untouched even in the years when the voracious insects are the most numerous. Nor do the rose-aphides, notwithstanding their enormously rapid multiplication, ever destroy all the young shoots of a rose-bush, or all the rose-bushes of a garden, or of the whole area in which roses grow.
At the same time it must be noted, that the number of individuals in a species undoubtedly does bear some relation to the amount of the food-supply available; for instance, it is very low among the large carnivores, the lion, the eagle, and the like. In our Alps the eagles have become rarer with the decrease of game, and where one eagle pair make their eyrie they rule alone over a hunting territory of more than sixty miles, a preserve on which no others of the same species are allowed to intrude. If there were several pairs of eagles in such a preserve, they would soon have so decimated the food-supply that they would starve. On the other hand, numerous herbivores, e.g. chamois and marmots, live within the bounds of the pair of eagles' hunting grounds, since the food they require is present in enormously greater quantity.
While it is true that the number of individuals of a given species which live in a particular area is not exactly the same year in year out, being subject to small, and sometimes, as in the case of the aphides and cockchafers, to very great fluctuations, nevertheless we may assume that the average number remains the same, that in the course of a century, or, let us say, of a thousand years, the number of mature individuals inhabiting the particular area remains the same. This, of course, only holds true on the supposition that there has been no great change in the external conditions of life during this period. But before Man began to interfere with nature, these external conditions would remain uniform for much longer periods than we have assumed. Let us call the average number of individuals[Pg 46] occurring on such a uniform area, the normal number of the species; this number will be determined in the first instance by the number of offspring that are annually brought forth, and secondly by the number that annually perish before reaching maturity. As the fertility of a species is a definite quantity, so also will its elimination be definite, or, as we may say, when the normal number under uniform conditions of life remains constant, the ratio of elimination will also remain constant. Each species is therefore subject to a perfectly definite ratio of elimination which remains on the average constant, and this is the reason why a species does not multiply beyond its normal number notwithstanding the great excess of the food-supply, and notwithstanding the fertility which, in all species, is sufficient to lead to boundless multiplication.
It is not difficult to calculate the ratio of elimination for a particular species, if one knows its rate of multiplication; for if the normal number remains constant, it follows that only two of all the offspring which a pair brings forth in the course of its life can attain to reproductive maturity, and that all the rest must perish.
Suppose, for instance, a pair of storks produced four young ones annually for twenty years, of these eighty young ones which are born within this period, on an average seventy-eight must perish, and only two can become mature animals. If more than two attained maturity the total number of storks would increase, and this is against the presupposition of constancy in the normal number. It is important, in reference to the fact on which we are now focusing our attention, that we should consider some other illustrations from the same point of view. The female trout yearly produces about 600 eggs; let us assume that it remains capable of reproduction for only ten years, then the elimination-number of the species will be 6,000 less two, that is, 5,998, for of the 6,000 eggs only two can become mature animals. But in the majority of fishes the ratio of extermination is enormously greater than this. Thus a female herring brings forth 40,000 eggs annually, the duration of life is estimated at ten years, and this means an elimination number of 400,000 less two, that is, 399,998. The carp produces 200,000 eggs a year, and the sturgeon two millions, and both species live long, and remain capable of reproduction for at least fifty years. But of all the 100 million eggs which are produced by the sturgeon, only two reach their full development and reproduce; all others perish prematurely.
But even with these examples we have not reached the highest elimination number, for many of the lower animals—not to speak of many plants—produce an even greater number of offspring.[Pg 47] Leuwenhoek calculated the fertility of a thread-worm at sixty million eggs, and a tape-worm produces hardly less than 100 millions.
There exists, therefore, a constant relation between fertility and the ratio of elimination; the higher the latter is, the greater must the former be, if the species is to survive at all. The example of the tape-worm makes this very obvious, for here we can readily understand why the fertility must be so enormous, as we are aware of the long chain of chances on which the successful development of this animal depends. The common tape-worm of Man, Tænia solium, does not lay its eggs, they remain enclosed within one of the liberated joints or 'proglottides.' Only if this liberated joint or one of the embryos within it happens to be fortuitously eaten by a pig or other mammal can there be successful development, and even then under difficulties and possible failures, and not right away into adult animals, but first into microscopically minute larvæ which may bore their way into the walls of the intestine, or, if they are fortunate enough, may get into the blood-stream and be carried by it to a remote part of the body. There they develop into 'measles,' the so-called bladder-worms, within which the head of the tape-worm arises. But in order that this may become a complete and reproductive adult worm the pig must die, and the next step necessary is that a piece of the flesh of the infected first host must happen to be swallowed raw by a man or other mammal! Only then does the fortunate bladder-worm—swallowed with the flesh—attain the goal of its life, that is, a suitable place to mature in, the food-canal of a human being. It is obvious that countless eggs must be lost for one that succeeds in getting through the whole course of a development depending so greatly on chance. Hence the necessity for such enormous productivity of eggs.
In many cases the causes of elimination, which keep a species within due bounds, are very difficult to determine. Enemies, that is to say, other species which use the species in question as food, play an important rôle; often, however, the cause lies in the unfavourableness of external conditions, in chance, which is favourable only to one of a thousand. The oak would only require to produce one seed in the 500 years of its life, if it were certain that that one would grow into an oak-tree; but most of the little acorns are eaten up by pigs, squirrels, insects, &c., before they have had time to sprout, thousands fall on ground already thickly covered with growth where they cannot take root, and even if they do succeed in finding an unoccupied space in which to germinate, the young plants are still surrounded by a thousand dangers—the possibility of being devoured by many animals large and[Pg 48] small, of being suffocated by the surrounding vegetation, and so on. We can thus understand, to some extent, though only approximately, why it is that the oak must year by year produce thousands of seeds in order that the species may maintain its normal number, and not be exterminated; for it is obvious that a constant, even though slow diminution of the normal number, a regular deficit, so to speak, can end in nothing else than the gradual extinction of the species.
But even this prodigality of seeds is not the greatest reach of fertility that we meet with in nature; it is, perhaps, amongst the simpler flowerless plants that we find the climax. It has been calculated that a single frond of the beautiful fern so common in our woods, Aspidium filix mas, produces about fourteen million spores. They serve to distribute the species, and are carried as motes by the wind, but comparatively few of the millions ever get the length of germinating at all, much less of attaining to full development into adult plants. Thus we see that the apparent prodigality of nature is a real necessity, an indispensable condition of the maintenance of the species; the fertility of each species is related to the actualities of elimination to which it is exposed. This is clearly seen when a species is placed under new and more favourable conditions of life, in which it has an abundant food-supply and few enemies. This was the case, for instance, with the horses introduced from Europe into South America, where they reverted to a feral state, and are now represented by herds of many thousands roaming the great grassy plains. If the small singing-birds of a region diminish in number, there is a great increase of caterpillars and other injurious insects which form part of their food-supply. The colossal destruction which the much-dreaded nun-moth from time to time brings about in our woods probably depends in part on the diminution of one or another of the many animals inimical to insects; but the occurrence of several years of weather-conditions favourable to the larvæ must also be taken into account. How enormously, indeed almost inconceivably, the number of larvæ may increase under favourable conditions is shown by such devastations as that in Prussia in 1856, when many square miles of forest were absolutely eaten up. The caterpillars were so numerous that even from some distance the falling excrement could be heard rustling like rain, and ten hundredweights of the eggs were collected, with an average of 20,000 eggs to the half-ounce!
But it would be a great mistake to conclude, from this enormous and sudden increase in the number of individuals, that the normal number of individuals is determined by the number of enemies alone.[Pg 49] The average number of individuals in a species depends on many other conditions, especially on the extent of the available area, and on the amount of the food-supply in relation to the size of body in the species. I cannot dwell on this now, but I wish to point out that, for the continuance of a species, it is indifferent whether it is 'frequent' or 'rare,' if we presuppose that its normal number remains on an average constant for centuries, that is, that its fertility suffices to make good the continual losses through enemies and other causes of elimination. One would be inclined to conclude from such cases of sudden and enormous increase in the number of individuals as these caterpillar-blights, that enemies and other causes of destruction played the major part in the regulation of the normal number of the species. But this is only apparently the case. Enemies necessitate a certain fertility in the species on which they prey, so that the elimination in each generation may be made good; but the number of pairs capable of reproduction is not thereby decisively determined. We must not forget that the number of enemies is also, on the other hand, dependent on the number of victims, and that the normal number of enemies must rise and fall with that of the species preyed upon.
For this reason, such an enormous increase as that of the caterpillars cannot last long; it carries its corrective in itself. The appearance of the caterpillars in such enormous numbers in itself increases the host of their enemies; singing-birds, ichneumon-flies, beetle-grubs, and predaceous beetles find abundant and available food, and therefore reproduce and multiply so rapidly, that, with the help of the caterpillar's plant-enemies, especially the insect-destroying fungi, they soon reduce the caterpillars to their normal number, or even below it. But then the reverse process begins; the enemies of the caterpillars diminish because their food has become scarce, and their normal number is lowered, while that of the caterpillars gradually rises again.
When the number of foxes in a hunting district increases, the number of the hares that they prey upon diminishes, and, on the other hand, the decimating of the foxes by Man brings about an increase in the number of hares in the district. Under natural conditions, that is, without the intervention of Man, there would be a constant balancing of the numbers of hares and foxes, for every noteworthy increase of the hares would be followed by a similar increase of foxes, and this, in its turn, would diminish the number of hares, so that they would no longer suffice for the support of so many foxes, and these would decrease in number again, until the number of hares had[Pg 50] again increased because of the lessened persecution and elimination. In nature the case is not quite so simple, because the fox does not live on hares alone, and the hare is not preyed upon only by the fox; but the illustration may serve to elucidate the point that a moving equilibrium is maintained between the species of a district, between persecutors and persecuted, in such a way that the number of individuals in the two species is always varying a little up and down, and that each influences the other so that a regulative process results. Throughout periods of considerable length the average remains the same; that is to say, a normal number is established. This normal strength of population is the mean above and below which the number of individuals is constantly varying. It is, of course, seldom that the mutual influences and regulations are so simple as in the example given; usually several or even many species interact upon each other, and not beasts of prey and their victims alone, but the most diverse species of animals and plants, which do not stand in any obvious relation to one another at all. Moreover, the physical, and especially the climatic conditions, also cause the normal number of the species to rise and fall.
The inter-relations between species living together on the same area are so intricate that I should like to give two other illustrations. Let us first take Darwin's famous instance of the fertility of clover, which depends on the number of cats. It is of course only an imaginary one, but the facts it is based upon are quite correct. The number of cats living in a village to a certain extent determines the number of field-mice in the neighbourhood. These again destroy the nests of the humble-bees, which live in holes in the ground, and thus the number of humble-bees depends on that of the field-mice and cats. But the clover must be pollinated by insects if it is to produce fertile seed, and only the humble-bee has a proboscis long enough to effect the pollination. Therefore the quantity of clover-seed annually produced depends on the number of humble-bees, and ultimately upon the number of cats. And, as a matter of fact, humble-bees were introduced into New Zealand from England, because without them the clover would produce no fertile seeds.
On the grassy plains of Paraguay there are no wild cattle and horses, because of the presence of a fly which has a predilection for laying its eggs in the navel of the newly-born calves and foals, with the result that the calves or foals are killed by the emerging maggots. We may reasonably assume that the numerical strength of this fly-species depends on the distribution of insect-eating birds, whose numbers in turn are determined by certain beasts of prey. These[Pg 51] again vary in number in relation to the extent of the forest-land, and this is determined by the number of ruminants which browse on the young growth of the woods (Darwin).
That forests can actually be totally destroyed by ruminants is proved by the case of the island of St. Helena among others. On its discovery the island was covered with thick wood, but in the course of 200 years it was transformed into a bare rock by goats and pigs, which devoured the young growth so completely that trees which were felled or which died were not replaced.
This point is vividly illustrated by Darwin's observation of a wide heath on which stood only a few groups of old pine-trees. The mere fencing in of a portion of the heath sufficed to call forth a thick growth of young seedling pines within the enclosure, and an examination of the open part of the heath revealed that the grazing cattle had eaten up all the young pine-trees which sprang from seed, and that again and again. In one small space thirty-two little trees stood concealed in the grass, and several of these showed as many as twenty-six yearly rings.
How definitely the number of individuals in different species living on the same area mutually limit and thereby regulate each other, Darwin sought to illustrate also by the case of the primitive forest, where the numerous species of plants occur, not mixed together irregularly, but in a definite proportion. We can find examples of the same kind wherever the plant-growth of a district has been left to itself. If we walk along the banks of our little river, the Dreisam, we see a wild confusion of the most diverse trees, shrubs and herbaceous plants. But, even though it cannot be demonstrated, we may be certain that these are represented in definite numerical proportions, dependent on the natural qualities and requirements of each species, on the number of their seeds and the facilities for their distribution, on the favourable or unfavourable season at which they ripen, and on their varying capacity for taking root in the worst ground, and springing quickly up, &c. They limit each other mutually, so that the whole flora of the river-bank will be made up of one per cent. of this species, one per cent. of that, and, it may be, five per cent. of a third, and the same combination will repeat itself in the same proportions on the banks of other rivers of our country in as far as the external conditions are the same. The same must be true of the fauna of such a plant-thicket; the animal species also limit one another mutually, and thereby regulate the number of individuals, which becomes relatively stable over any area on which the conditions remain the same. That is to say, a 'normal number' is attained and persists.
Thus we see that the capacity for boundless multiplication inherent in every species is limited by the co-existence of other species; there is, metaphorically speaking, a continuous struggle going on between species, plant and animal alike; each seeks as far as possible to multiply, and each is hemmed in by the others and as far as possible prevented from doing so. The 'struggle' is by no means only the direct limitation of the number of individuals, which consists in the use of one species by another as food, as in beasts of prey and their victims, or locusts and plants; it is much more the indirect limitation—figuratively speaking, the struggle for space, for light, for moisture among plants, for food among animals. But all this, important as it is, does not yet exhaust the content of that 'struggle for existence' to which Darwin and Wallace ascribe the rôle of the breeder in the process of natural selection. The struggle, that is, the mutual limiting of species, may indeed restrict a species in its distribution, and may reduce its normal number possibly to nil. In other words, it may bring about extinction, but it cannot make a species other than it is. This can only be done by a struggle within the limits of the species itself, and this struggle is due to the fact that of the numerous offspring, on an average those survive—that is, attain to reproduction—which are the most fit, whose constitution makes it most possible for them to overcome the difficulties and dangers of life, and so to reach maturity. We see, in fact, that a large percentage of each generation in all species always perishes before attaining maturity. If, then, the decision as to which is to perish and which is to reach maturity is not a matter of chance alone, but is in part due to the constitution of the growing individual; if the 'fittest' do on the average survive, and the 'least fit' are on the average eliminated, we have here a process of selection entirely comparable to that of artificial selection, and one whose result must be the 'improvement' of the species, whether that depends on one set of characters or on another. The victorious qualities, which earlier were peculiar to certain individuals, must gradually become the common property of the species, if in each generation the individuals which attained to reproduction all possessed them, and thus could transmit them to their progeny. But those of the descendants which did not inherit them would again be at a disadvantage in the struggle for existence, or rather for reaching maturity, if in each generation a higher percentage of individuals which possess these characters reach maturity than of those which do not possess them. This percentage must increase in each generation, because, in each, natural selection again chooses out the fittest, and it must finally rise to 100 per[Pg 53] cent., that is to say, none but individuals of this fittest type will be left surviving.
This does not yet exhaust the process, however, for we can infer from the results of artificial breed-forming that the selected characters may intensify from generation to generation, and that they will continue to do so as long as it gives them any advantage in the struggle for existence, for so long will it lead to the more frequent survival of its possessors. The increase will only stop when it has reached the highest degree of usefulness, and in this way new characters may be formed, just as, in artificial selection, the short upward-turning feathers of the Jacobin pigeon have been intensified into the peruke, a feather canopy covering the head.
A few examples of natural selection will make the process clearer. Our hare is well secured from discovery by his fur of mixed brown, yellow, white, and black, when he cowers in his form among the dry leaves of the underwood. It is easy to pass close to him without seeing him. But if the ground and the bushes are covered with snow, he contrasts conspicuously with them. Suppose, now, that our climate became colder, and that the winter brought lasting snow, the hares which had the largest mixture of white in their fur would have an advantage in their 'struggle for existence' over their darker fellows; they would be less easily discovered by their enemies—the fox, the badger, the horned owl, and the wild cat. Of the numerous hares which would annually become the prey of these enemies, there would be, on an average, more dark than light individuals. The percentage of light-coloured hares would, therefore, increase from generation to generation, and the longer the winter the keener would be the selection between dark and light hares, until finally none but light ones would remain. At the same time, the colour of the hares would become increasingly light, first, because it would happen more and more frequently that two light hares would pair, and secondly, because, after a time, the struggle for existence would no longer be between light and dark hares, but between light hares and still lighter ones. Thus ultimately a race of white hares would arise, as has actually happened in the Arctic regions and on the Alps.
Or let us think of a herbaceous plant, in appearance something like a belladonna, rich in leaves and very juicy, but not poisonous. It would doubtless be a favourite food with the animals of the forest, and it would not, therefore, attain to more than a sparse occurrence, since few of the individuals would be able to form seeds. But now let us assume that a stuff of very unpleasant taste develops in[Pg 54] the stem and leaves of some of the individuals, as may easily happen through very slight changes in the chemical metabolism of the plant, what, then, could result but that such individuals would be less readily eaten than the others? A process of selection must, therefore, ensue, and the unpleasant-tasting specimens of the plant would be much more frequently spared, and consequently would bear seed much oftener than the palatable ones. Thus the number of unpalatable plants would increase from year to year. If the stuff in question were not only unpalatable but poisonous, or gradually became so, a plant would in time be evolved which would be absolutely safe from being devoured by animals, just as the deadly nightshade (Atropa belladonna) actually is.
Or let us suppose that a stretch of water is inhabited by a species of carp, which have hitherto had no large enemy, and so have become lazy and slow, and that there migrates from the sea into this stretch of water a large species of pike. At first numerous carp will fall victims to the pike, and the pike will rapidly increase in number. But if all the carp were not equally lazy and dull-witted, if some of them were quicker and more intelligent, these would, on an average, become more rarely the victims of the pike, and numerous individuals with these better qualities would survive in each generation, till ultimately there were no others, and the useful characters would gradually become intensified, and so a more active and wary race of carp would arise.
Let us suppose, however, that the increased activity and wariness would not alone suffice to preserve the colony from extinction; it might require also an increased fertility to prevent the normal number from being permanently lowered; but even this could eventually be brought about by natural selection, if the nature of the species and the general conditions of its life permitted. For there are variations of fertility in every species, and if the chance of seeing some of its eggs become mature animals were greater for the more fertile female than for the less fertile, ceteris paribus, a process of selection must take place, which would result in an increase of fertility as far as that was possible.
Obviously, such processes of natural selection can affect all parts and characters—size and form of the body, as well as isolated parts, the external skin and its colour, every internal organ—and not bodily characters alone, but psychical ones as well, such as intelligence and instincts. According to this principle, it is only characters which are biologically indifferent that cannot be altered through natural selection.
Natural selection can also bring about changes at every age, for the elimination of individuals begins from the egg, and any kind of egg which is in some way better able to escape elimination will transmit its useful characters to its descendants, because the resulting young animals will thus more frequently reach full development than the young from other eggs. In the same way, at every succeeding stage of development, every character favourable to the preservation of the individual will be maintained and intensified.
We see from all this that natural selection is vastly more powerful than artificial selection by Man. In the latter, only one character at a time can be caused to change, while natural selection may influence a whole group of characters at the same time, as well as all the stages of development. Through the weeding out of the individuals which are annually exterminated, it is always on an average the 'fittest' which survive, that is to say, those which have the greatest number of bodily parts and rudiments of parts in the fittest possible condition of development at every stage. The longer this process of selection continues, the smaller will be the deviations of the individual from this standard, and the more minute will be the differences of fitness determining which is to be eliminated and which is to survive to reproduce its characteristics. In the immeasurable periods of time which are at the disposal of natural selection, and in the inestimable numbers of individuals on which it may operate, lie the essential causes of superiority of natural selection over the artificial selection of Man.
To sum up briefly: Natural selection depends essentially on the cumulative augmentation of the most minute useful variations in the direction of their utility; only the useful is developed and increased, and great effects are brought about slowly through the summing up of many very minute steps. Natural selection is a self-regulation of the species which secures its preservation; its result is the ceaseless adaptation of the species to its life-conditions. As soon as these vary, natural selection changes its mode of action, for what was previously the best is now no longer so; parts that before had to be large must now perhaps be small, or vice versa; muscle-groups which were weak must now become strong, and so on. The conditions of life are, so to speak, the mould into which natural selection is continually pouring the species anew.
But the philosophical significance of natural selection lies in the fact, that it shows us how to explain the origin of useful, well-adapted structures purely by mechanical forces and without having to fall back on a directive force. We are thus for the first time in[Pg 56] a position to understand, in some degree, the marvellous adaptation of the organism to an end, without having to call to our aid any supernaturally intrusive force on the part of the Creator. We understand now how, in a purely mechanical way, through the forces always at work in nature, all forms of life must conform to, and adapt themselves precisely to the conditions of their life, since only the best possible is preserved, and everything less good is continually being rejected.
Before I go on to expound in detail the phenomena which we refer to natural selection, I must briefly state that Darwin did not ascribe to natural selection by any means all the changes which have taken place in organisms in the course of time. On the one hand, he ascribed a not inconsiderable importance to the correlated variations we have already mentioned; still more, however, he relied on the direct influence of altered conditions of life, whether these consist in climatic and other changes in the environment, or in the assumption of new habits, and the increased or diminished use of individual parts and organs thereby induced. He recognized the principle so strongly emphasized by Lamarck, of use and disuse as a cause of heritable increase or decrease of the exercised or neglected part, though he did so with a certain reserve. I shall return later to these factors of modification, and shall then attempt to show that these too are to be referred to processes of selection, which are, however, of a different order from the phenomena which the Darwin-Wallace principle of natural selection serves to interpret. But, in the first instance, it appears to me to be necessary to show how far the Darwin-Wallace interpretation will suffice, and in the next lectures we shall occupy ourselves with this question exclusively.
THE COLORATION OF ANIMALS AND ITS RELATION TO THE PROCESSES OF SELECTION
Biological significance of colours—Protective colours of eggs—Animals of the snow-region—Animals of the desert—Transparent animals—Green animals—Nocturnal animals—Double colour-adaptation—Protective marking of caterpillars—Warning markings—Dimorphism of colouring in caterpillars—Shunting back of colouring in ontogeny—'Sympathetic' colouring in diurnal Lepidoptera—In nocturnal Lepidoptera—Theoretical considerations—The influence of illumination in the production of protective colouring, Tropidoderus—Harmony of protective colouring in minute details—Notodonta—Objections—Imitation of Strange objects, Xylina—Leaf-butterflies, Kallima—Hebomoja—Nocturnal Lepidoptera with leaf-markings—Orthoptera resembling leaves—Caterpillars of the Geometridæ.
We have seen what Darwin meant by natural selection, and we understand that this process really implies a transformation of organisms by slow degrees, in the direction of adaptive fitness—a transformation which must ensue as necessarily as when a human selector, prompted by conscious intention, tries to improve an animal in a particular direction, by always selecting the 'fittest' animals for breeding. In nature, too, there is selection, because in every generation the majority succumb in the struggle for life, while on an average those which survive, attain to reproductive maturity, and transmit their characters to their descendants, are those which are best adapted to the conditions of their life—that is, which possess those variations of most advantage in overcoming the dangers of life. Since individuals are always variable in some degree, since their variations can be inherited by their progeny, and since the continually repeated elimination of the majority of those descendants is a fact, the inference from these premisses must be correct; there must be a 'natural selection' in the direction of a gradually increasing fitness and effectiveness of the forms of life.
We cannot, however, directly observe this process of natural selection; it goes on too slowly, and our powers of observation are neither comprehensive nor fine enough. How could we set about investigating the millions of individuals which constitute the numerical strength of a species on a given area, to find out whether they possess some variable character in a definite percentage, and whether this percentage increases in the course of decades or centuries? And[Pg 58] there is, furthermore, the difficulty of estimating the biological importance of any variation that may occur. Even in cases where we know its significance quite well in a general way, we cannot estimate its relative value in reference to the variation of some other character, though that other may also be quite intelligible. Later on, we shall speak of protective colouring, and in so doing we shall discuss the caterpillars of one of the Sphingidæ, which occur in two protective colours, some being brown, others green. From the greater frequency of the brown form we may conclude that brown is here a better adaptation than green, but how could we infer this from the character itself, or from our merely approximate knowledge of the mode of life of the species, its habits, and the dangers which threaten it? A direct estimation of the relative protective value of the two colours is altogether out of the question. The survival of the fittest cannot be proved in nature, simply because we are not in a position to decide, a priori, what the fittest is. For this reason I was forced to try to make the process of natural selection clear by means of imagined examples, rather than observed ones.
But though we cannot directly follow the uninterrupted process of natural selection which is going on under natural conditions, there is another kind of proof for this hypothesis, besides that which consists in logically deducing a process from correct premisses; I should like to call this the practical proof. If a hypothesis can be made to explain a great number of otherwise unintelligible facts, it thereby gains a high degree of probability, and this is increased when there are no facts to be found which are in contradiction to it.
Both of these criteria are fulfilled by the selection-hypothesis, and indeed the phenomena which may be explained by it, and are intelligible in no other way, present themselves to us in such enormous numbers, that there can be no doubt whatever as to the correctness of the principle; all that can be still disputed is, how far it reaches.
Let us now turn our attention to this practical way of proving the theory by the facts which it serves to interpret, beginning with a consideration of the external appearance of organisms, their colour and form.
The Colour and Form of Organisms.
Erasmus Darwin had in many cases already rightly recognized the biological significance of the colouring of an animal species, and we may be sure that many of the numerous good observers of earlier times had similar ideas. I can even state definitely that Rösel von Rosenhof, the famous miniature-painter and naturalist of Nürnberg[Pg 59] in the middle of the eighteenth century, recognized clearly, and gave beautiful descriptions of what we now call colour-adaptation. It is true that he gave them only as isolated instances, and was far from recognizing the phenomenon of colour-adaptation in general, or even from inquiring into its causes. From the time of Linné, the endeavour to establish new species overshadowed all the finer observation of life-habits and inter-relations, and, later on, after Blumenbach, Kielmeyer, Cuvier, and others, the eager investigation of the internal structure of animals also tended to divert attention from these œcological relations. In systematic zoology, colour ranked only as a diagnostic character of subordinate value, because it is often not very stable, and indeed is sometimes very variable; it was therefore found preferable to keep to such relatively stable differences as are to be found in the form, size, and number of parts.
Charles Darwin was the first to redirect attention to the fact that the colouring of animals is anything but an unimportant matter; that, on the contrary, in many cases it is of use to the animal, e.g. in making it inconspicuous; a green insect is not readily seen on green leaves, nor a grey-brown one on the bark of a tree.
It is plain that the origin of such a so-called 'sympathetic' coloration, harmonizing with the usual environment of the animal, can be easily interpreted in terms of the principle of selection; and it is equally evident that it cannot be explained by the Lamarckian principle of transformation. Through the accumulation of slight useful variations in colour, it is quite possible for a green or a brown insect to arise from a previous colour, but a grey or a brown insect could not possibly have become a green one simply by getting into the habit of sitting on a green leaf; and still less can the will of the animal or any kind of activity have brought the change about. Even if the animal had any idea that it would be very useful to it to be coloured green, now that it had got into the habit of sitting on a leaf, it could not have done anything towards attaining the desirable green colour. Quite recently the possibility of a kind of colour-photography on the skin of the animal has been suggested, but there are many species whose colouring is in contrast to their environment, so that the skin in these cases does not act as a photographic plate, and it would, therefore, have to be explained how it comes to pass that it functions as such in the sympathetically coloured animals. I do not ask for proof of the chemical composition of the stuff which is supposed to be sensitive to light. Whether this be iodide of silver or something quite different, the question remains the same: how comes it that it has only appeared in animals to which a sympathetic[Pg 60] colouring is advantageous in the struggle for life? And the answer, from our point of view, must read: it has arisen through natural selection in those species to which a sympathetic colouring is useful. Thus even if the supposition that sympathetic colouring is due to automatic photography on the part of the skin were correct, we should still have to regard it as an outcome of natural selection; but it is not correct—at least in general—as the above objection shows, and as will be further apparent from many of the phenomena of colour-adaptation which I shall now adduce.
To explain sympathetic coloration, then, we must assume, with Darwin and Wallace, a process of selection due to the fact that, as changes took place in the course of time in the colouring of the surroundings, those individuals on an average most easily escaped the persecution of their enemies which diverged least in colour from their surroundings, and so, in the course of generations, an ever greater harmony with this colouring was established. Variations in colouring crop up everywhere, and as soon as these reached such a degree as to afford their possessors a more effective protection than the colouring of their fellows, then natural selection of necessity stepped in, and would only cease to act when the harmony with the environment had become complete, or, at least, so nearly so that any increase of it could not heighten the deception.
Of course, it is presupposed in the working out this selective process that the species has enemies which see. This is the case, however, with most animals living on the earth or in the water, unless they are of microscopic minuteness. Many animals, too, are subject to persecution not only in their adult state, but at almost every period of their life, and so, in general, we should expect that many of them would have attained at each stage that coloration of body that would render them least liable to discovery by their enemies.
And this is in reality the case: numerous animals are protected in some measure by so-called sympathetic colouring, from the egg to the adult state.
Let us begin with the egg, and of course there is no need to speak of any eggs except those which are laid. Of these many are simply white in colour, e.g. the eggs of many birds, snakes, and lizards, and this seems to contradict our prediction; but these eggs are either hidden in earth, compost, or sand, as in the case of the reptiles, or they are laid in dome-shaped nests, or concealed in holes in trees, as in many birds; thus they require no protective colouring.
In other cases, however, numerous eggs, especially of insects and[Pg 61] birds, possess a colouring which makes it very difficult to distinguish them from their usual surroundings. Our large green grasshopper (Locusta viridissima) lays its eggs in the earth, and they are brown, exactly like the earth which surrounds them. They are enough in themselves to refute the hypothesis that sympathetic colouring has arisen through self-photography, for these eggs lie in total darkness in the ground. Insect-eggs which are laid on the bark of trees are often grey-brown or whitish like it, and the eggs of the humming-bird hawk-moth (Macroglossa stellatarum), which are attached singly to the leaves of the bedstraw, have the same beautiful light-green colour as these leaves, and, in point of fact, green is a predominant colour of the eggs in a very large number of insects.
But the eggs of many birds, too, exhibit 'sympathetic' colouring; thus the curlew (Numenius arquata) has green eggs, which are laid in the grass; but the red grouse (Lagopus scoticus) lays blackish-brown eggs, exactly of the colour of the surrounding moor-soil; and it has been observed that they remain uncovered for twelve days, for the hen lays only one egg daily, and does not begin to brood until the whole number of twelve is complete. Herein lies the reason of the colour-adaptation, which the eggs would not have required, if they had always been covered by the brooding bird.
The eggs of birds are frequently not of one colour only; those of the Alpine ptarmigan (Lagopus alpinus), for instance, are ochre-yellow with brown and red-brown dots, resembling the nest, which is carelessly constructed of dry parts of plants. Sometimes this mingling of colours reaches an astonishing degree of resemblance to surroundings, as in the golden plover (Charadrius pluvialis), whose eggs, like those of the peewit (Vanellus cristatus), are laid among stones and grasses, not in a true nest, but in a flat depression in the sand, and, protected by a motley speckling with streaking of white, yellow, grey and brown, are excellently concealed. Perhaps the eggs of the sandpipers and gulls are even better protected, for their colouring is a mingling of yellow, brown, and grey, which imitates the sand in which they are laid so perfectly, that one may easily tread on them before becoming aware of them.
But let us now turn from eggs to adult animals. Darwin first pointed out that the fauna of great regions may exhibit one and the same ground-colouring, as is the case in the Arctic zone and in the deserts. The most diverse inhabitants of these regions show quite similar coloration, namely, that which harmonizes with the dominant colour of the region itself. It is not only the persecuted animals, which need protection, that are sympathetically coloured in these[Pg 62] cases, the persecutors themselves are likewise adapted, and this need not surprise us, when we remember that the very existence of a beast of prey depends on its being able to gain possession of its victims, and that therefore it must be of the greatest use to it to contrast as little as possible with its surroundings, and thus be able to steal on its quarry unperceived. Those that are best adapted in colour will secure the most abundant food, and will reproduce most prolifically; and they will thus have a better prospect of transmitting their usual colouring to their offspring. The Polar bear would starve if he were brown or grey, like his relatives; among the ice and snow of the Polar regions his victims, the seals, would see him coming from afar.
In the Arctic zone the adaptation of the colouring of the animals to the white of the surroundings is particularly striking. Most of the mammals there are pure white, or approximately white, at least during the long winter; and it is easily understood that they must be so if they are to survive in the midst of the snow and ice,—both beasts of prey and their victims. For the latter the sympathetic colouring is of 'protective' value; for the former, of 'aggressive' value (Poulton). Thus we find not only the Polar hare and the snow-bunting white, but also the Arctic fox, the Polar bear, and the great snowy owl; and though the brown sable is an exception, that is intelligible enough, for he lives on trees, and is best concealed when he cowers close to the dark trunk and branches. For him there would be no advantage in being white, and therefore he has not become so.
Desert animals are also almost all sympathetically coloured, that is, they are of a peculiarly sandy yellow, or yellowish-brown, or clayey-yellow, or a mixture of all these colours; and here again the beasts of prey and their victims are similarly coloured. The lion must be almost invisible from a short distance, when he steals along towards his prey, crouching close to the ground; but the camel too, the various species of antelope, the giraffe, all the smaller mammals, and also the horned viper (Vipera cerastes), the Egyptian spectacled snake (Naja haje), many lizards, geckos, and the great Varanus, numerous birds, not a few insects, especially locusts, show the colours of the desert. It is true that the birds often have very conspicuous colours, such as white on breast and under parts, but the upper surface is coloured like the desert, and conceals them from pursuers whenever they cower close to the ground. It has even been observed that a locust of the genus Tryxalis is of a light sand-colour in the sandy part of the Libyan desert, but dark brown in its rocky parts, thus illustrating a double adaptation in the same species.
Another group, which agrees in colour with the general surroundings, is that of the 'glass-animals,' as they have been called, though perhaps 'crystal animals' is a better term. A great number of simple free-swimming marine forms, and a few fresh-water ones, are quite colourless, and perfectly transparent, or have at most a bluish or greenish tinge, and on this account they are quite invisible as long as they remain in the water. In our lakes there lives a little crustacean about a centimetre in length, of the order of water-fleas (Leptodora hyalina), a mighty hunter among the smallest animals, which swims forward jerkily with its long swimming-appendages, and widely spreads its six pairs of claws, armed with thorny bristles, like a weir basket, to seize its prey. We may have dozens of these in a glass of water without being able to see a single one, even when we hold the glass against the light, for the creatures are crystal-clear and transparent, and have exactly the same refractive power as the water. It requires a very sharp scrutiny and a knowledge of the animals to be able to detect in the water little yellowish stripes, which are the stomachs of the animals filled with food in process of digestion, for which, as we can readily understand, invisibility cannot very well be arranged. If the water be then strained through a fine cloth, a little gelatine-like mass of the bodies of the Leptodora will remain on the sieve.
A great many of the lower marine animals are equally transparent, and as clear as water; most of the lower Medusæ, the ctenophores, various molluscs, the barrel-shaped Salpæ, worms, many crustaceans of quite different orders, and above all an enormous number of larvæ of the most diverse animal groups. I can remember seeing the sea at the shore at Mentone so full of Salpæ, that in every glass of sea-water drawn at random there were many of them, and sometimes a glass held a positive animal soup. But one did not see them in the glass of water, and only those who knew what to look for recognized them by the bluish intestinal sac that lies posteriorly in the invisible body. But when the water was poured off through a fine net, there remained on the filter a large mass of a crystalline gelatinous substance.
It is obvious that this must serve as a protective arrangement, for the animals are not seen by their pursuers; but it is not an absolute protection, for they have many pursuers who do not wait till they see their prey, but are almost constantly snapping the mouth open and shut, leaving it to chance to bring them their prey. No protective arrangement, however, affords absolute security; it protects against some enemies, perhaps against many, but never against all.
But now let us turn to a group of a different colouring, the green[Pg 64] animals. We are familiar with our big grass-green grasshopper, and we know how easily it is overlooked when it sits quietly on a high grass-stem, surrounded by grasses and herbage; the light grass-green of its whole body protects it most effectively from discovery: for myself, at least, I must confess that in a flowery meadow I have stood right in front of one, and have looked close to it for a long time without detecting it. In the same way countless insects of the most diverse groups—bugs, dipterous flies, sawflies, butterflies—and especially the larvæ (caterpillars) of the last, are of the same green as the plants on which they live, and this again applies to the predaceous species, as well as the species preyed upon. Thus the rapacious praying-mantis (Mantis religiosa) is as green as the grass in which it lurks motionless for its victim—a dragonfly, a fly, or a butterfly.
There are also green spiders, green amphibians like the edible frog, and especially the tree-frog, green reptiles like lizards and the tree-snakes of tropical forests. It is always animals which live among green that are green in colour.
We may wonder, for a moment, why there are so few green birds, since they spend so much of their time among the green leaves. But this paucity of green birds is only true of temperate climates. In Germany we have only the green woodpecker, the siskin, and a few other little birds, and even these are not of a bright green, but are rather greyish-green. The explanation lies in the long winter, when the trees are leafless. In the evergreen forests of the tropics there are numerous green birds belonging to very diverse families.
Yet another group with a common colour-adaptation deserves mention—the beasts of the night. They are all more or less grey, brown, yellowish, or a mixture of these colours, and it is obvious that, in the duskiness of night, they must blend better with their environment on this account. White mice and white rats cannot exist under natural conditions, since they are conspicuous in the night, and the same would be true of white bats, nightjars, and owls; but all of these have a coloration suited to nocturnal habits.
A very remarkable fact is that in many animals the colour-adaptation is a double one. Thus the Arctic fox is white only in winter, while in summer he is greyish-brown; the ermine changes in the same way, and the great white snowy owl of the Arctic regions has in summer a grey-brown variegated plumage. Many animals which are subject to persecution also change colour with the seasons, like the mountain hare (Lepus variabilis), which is brown in summer and pure white in winter, the Lapland lemming, and the ptarmigan (Lagopus alpinus), which do the same. It has been[Pg 65] doubted whether natural selection can explain this double coloration, but I do not know where the difficulty lies, and there is certainly no other principle whose aid we can evoke. The mountain hare must have had some sort of colour before it attained to seasonal dimorphism. Let us assume that it was brown, that the climate became colder and the winter longer, then those hares would have most chance of surviving which became lighter in winter, and so a white race was formed. Poulton has shown that the whiteness is due to the fact that the dark hairs of the summer coat grow white as they lengthen at the beginning of winter, and the abundance of new hairs which complete the winter coat are from the first white throughout. If the white hairs were to persist throughout the summer it would be very disadvantageous to their wearer; so a double selection must take place, in summer the individuals which remain white, in winter those which remain brown, being most frequently eliminated, so that only those would be left which were brown in summer and white in winter. This double selection would be favoured by the fact that there would be, in any case, a change of fur at the beginning of summer; the winter hairs fall out and the fur becomes thinner. The process does not differ essentially from that which takes place in any species when two or more parts or characters, which are not directly connected, have to be changed, such as, for instance, colour and fertility. The struggle for existence will in this case be favourable, on the one hand, to the advantageously coloured, and on the other to the most fertile, and though the two characters may at first only occur separately, they will soon be united by free crossing, until ultimately only those individuals will occur which are at once the most favourably coloured and the most fertile. So in this case there remain only those which are brown in summer and white in winter.
We must ascribe to the influence of the processes of selection the exact regulation of the duration of the winter and summer dress, which has been carefully studied in the case of the variable hare. In the high Alps it remains white for six or seven months, in the south of Norway for eight months, in Northern Norway for nine months, and in Northern Greenland it never loses its white coat at all, as there the snow, even in summer, melts only in some places and for a short time. But apart from concealment there is certainly another adaptation involved here—namely, the growth of the hair as a protection against the cold. From an old experiment made in 1835 by Captain J. Ross, and recently brought to light again by Poulton, we learn that a captive lemming kept in a room in winter did not change colour until it was exposed to the cold. The constitution of[Pg 66] animals which become white in winter is thus so organized that the setting in of cold weather acts as a stimulus which incites the skin to the production of white hairs. This predisposition also we must refer to the influence of natural selection, since it must have been very useful to the species that the winter coat should grow just when it was necessary as a protection against cold. This explains at the same time why the predisposition to respond to the stimulus of cold by a growth of winter fur finds expression earlier in those colonies of Arctic animals, such as the hare, which live in Lapland, than in those which live in the south of Norway.
But that it is not the direct influence of cold which colours the hair of a furred animal white we can see from our common hare (Lepus timidus), which, in spite of the winter's cold, does not become white, but retains its brown coat, and not less so from the mountain hare (Lepus variabilis), which in the south of Sweden also remains brown, although the winter there may be exceedingly cold. But as the covering of the ground with snow is not so uninterrupted there as in the higher North, a white coat would be not a better protection than a brown one, but a worse. The white colouring of Arctic animals is therefore not directly due to the influence of the climate, as has often been maintained, but is due to it indirectly, that is, through the operation of natural selection. I have tried to make this clear by means of this example, so that we may not have to repeat it in considering those which are to follow.
But all attempts at any other explanation are even more decidedly excluded when we turn our attention to more complicated cases of colour-adaptation, which are not confined to the simple, general coloration, but are helped by markings and colour-patterns, that is, by schemes of colour.
Thus numerous caterpillars exhibit definite lines and spots on their ground-colouring, which, in one way or another, aid in protecting them from their enemies.
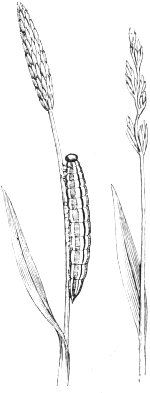
Fig. 2. Longitudinally
striped
caterpillar of a Satyrid.
After Rösel.
The green grass-eating caterpillar of many of our Satyridæ has two or more darker or lighter lines running down the sides of its body, which make it much less conspicuous among the grasses on which it feeds than if it were a uniform green mass (Fig. 2). Not infrequently the colour and form present a remarkably close resemblance to the inflorescences or fruit-ears of the grasses. Caterpillars marked thus are never found on the leaves of trees, where they would immediately catch the eye. It is true that longitudinal striping often occurs on caterpillars which live on other plants besides grass, but as these other plants grow among the grasses the[Pg 67] protective efficacy is just the same. This is the case with the Pieridæ (Garden Whites).
All the caterpillars of our Sphingidæ, on the other hand, which live on bushes and trees, have on the sides of the segments light oblique stripes, seven in number, which are disposed to the longitudinal axis of the body at the same angle as the lateral veins of a leaf of their food-plant have to the mid-rib. It cannot of course be said that the caterpillar thereby gains the appearance of a leaf, indeed, if one sees it apart from its food-plant it does not look in the least like a leaf, but among the leaves of a bush or tree this marking secures it in a high degree from discovery. Thus the caterpillar of the eyed hawk-moth (Smerinthus ocellatus), when it is sitting among the crowded foliage of a willow, is often very difficult to find, because its large green body does not appear as a single green spot, but is divided by the oblique lateral stripes into sections like the half of a willow leaf, so that even a searching glance is led astray, there being nothing to focus attention on the animal as distinguished from its surroundings (Fig. 3). As a boy I often had the interesting experience of overlooking a caterpillar which was sitting just before me, until after a time I chanced to hit upon the exact spot in the field of vision.

Fig. 3. Full-grown caterpillar of the Eyed
Hawk-moth, Smerinthus ocellatus. sb, the subdorsal
stripe.
In the majority of these caterpillars with oblique stripes, the likeness to the half of a leaf is heightened by the fact that the light oblique row is accompanied by a broader coloured band, suggesting the shade of the leaf's mid-rib. The caterpillar of Sphinx ligustri has a lilac band, and that of Sphinx atropos a blue one. In both cases it is difficult to believe that such striking colours can secure the animals from discovery, yet among the blending shadows of the leaf-complex of their food-plant they greatly increase their resemblance to a leaf-surface. Of the death's-head caterpillar (Sphinx[Pg 68] atropos) this sounds almost incredible, for this form is chiefly a bright golden yellow, and the narrow white oblique stripes have sky-blue borders becoming darker towards the under side; but it must not be forgotten that the potato is not the true food-plant of the species, for it lives, in its true home in Africa, and also in the south of Spain, on wild solanaceous plants, which, we are informed by Noll, have precisely these colours—golden-yellow and blue in the blossom, the fruit, and in part also in the leaves and stem. There the caterpillars sit the whole day long on the plants, while with us they have formed the habit of feeding only in the twilight and at night, and concealing themselves in the earth by day, a habit that is found in other caterpillars also, and which we must again ascribe to a process of natural selection.

Fig. 4. Full-grown caterpillar of the Elephant Hawk-moth (Chærocampa elpenor) in its
"terrifying attitude."
Some caterpillars exhibit other, more complex markings, which do not protect them by rendering them difficult to detect, but by terrifying the enemy who has discovered them, and warning him away. Such terrifying or aggressive colours are to be found, for instance, in the caterpillars of the Sphingid genus Chærocampa in the form of large eye-like spots, which occur in pairs close together on the fourth and fifth segments of the animal. Children and those unfamiliar with animals take these for true eyes; and as the caterpillar, when it is threatened by an enemy, draws in the head and anterior segments, so that the fourth one is greatly distended, the eye-spots seem to stand on a thick head (Fig. 4), and it cannot be wondered at that the smaller birds, lizards, and other enemies are so terrified that they refrain from attacking. Even hens hesitate to seize such a caterpillar in its defiant attitude, and I once looked on for a long time in a hen-coop while one hen after another rushed to pick up a caterpillar I had placed there, but, when close to it, hastily drew back the head already prepared to strike. Even a gallant cock was a long time in making up his mind to attack the terrible beast, and drew back repeatedly before he at length ventured to strike a deadly blow with his bill. After the first stroke the caterpillar, of course, was lost. Thus even this disguise is only a relative protection, effective only against smaller enemies. But that these are really frightened away, I had once an opportunity of observing, when I put[Pg 69] a caterpillar of the common elephant hawk-moth (Chærocampa elpenor) in the feeding-trough of a hencoop, and a sparrow flew down to feed from the trough. It descended at first with its back to the caterpillar and fed cheerily. But when by chance it turned round, and spied the caterpillar, it scurried hastily away.
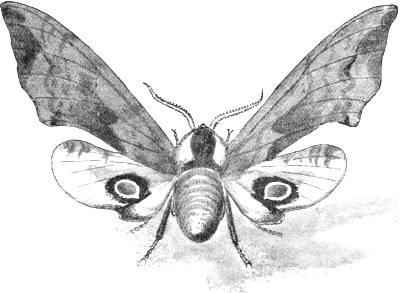
Fig. 5. The Eyed Hawk-moth in its 'terrifying attitude.'
Among Lepidoptera, too, eye-spots often occur on the wings, and to some extent, at least, they have in this case also the significance of warning marks. Take, for instance, the large blue and black eye-spots on the posterior wings of the eyed hawk-moth (Smerinthus ocellatus). When the insect is sitting quietly the two spots are not visible, as they are covered by the anterior wings, but as soon as the creature is alarmed it spreads all four wings, and now both eyes stand boldly out on the red posterior wings and alarm the assailant, as they give the impression of the head of a much larger animal (see Fig. 5). There are also eye-like spots which have not this significance and effect, as, for instance, the 'eye-spots' on the train-feathers of the peacock and the Argus pheasant, or the little eye-like spots on the under surface of many diurnal butterflies. In the first case, it is a matter of decoration; in the second, perhaps of the mimicry of dewdrops, which increases still further the resemblance to a withered leaf; but there are undoubtedly many cases in which the eye-spots serve as means of frightening off enemies, and these cases are especially common among butterflies.
Such warning marks are in no way contradictory to the sympathetic colouring of the rest of the body, and indeed we usually find them in combination with it. In some cases the eye-spot, though very conspicuous, is covered, as in the eyed hawk-moth, when at rest,[Pg 70] by the sympathetically coloured parts—in this instance the anterior wings. In other cases eye-spots of considerable size lie clearly exposed, but exhibit the same sympathetic colours as the whole of the rest of the wing-surface. In this case they do not interfere with the protective influence of general colouring, because they are only visible from a very short distance. This is the case in the large Caligo species of South America, which only fly for a short time in the early morning and in the evening, remaining concealed throughout the day in dark shadowy places, where the mingled colouring of brown, grey, yellow, and black on the under surfaces of the wings prevents their being recognized from a distance as butterflies at all. But even the best sympathetic colouring is not an absolute protection, and when the insect is discovered by an enemy near at hand, the terrifying mark, a large deep-black spot on the posterior wing, comes into effect, and scares the assailant away.
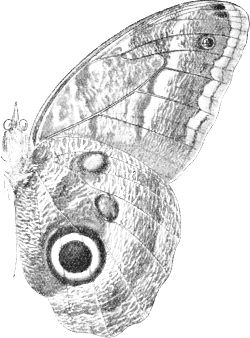
Fig. 6. Under surface of the wings of Caligo.
In such cases the sympathetic colouring was probably the first to arise, and the eye-spot was developed later by a new process of selection, brought about by the necessity of protecting the species more effectively than by mere inconspicuousness alone. In many cases it can be proved that the power of scaring off an enemy did not begin with the formation of the eye-spot, but with the development of a new instinct. When the caterpillar of Chærocampa elpenor is attacked it immediately assumes the defiant attitude described above, but the same striking attitude is assumed by the caterpillars of the allied American genus Darapsa, as I learn from an old illustration by Abbot and Smith, although this form possesses no eye-spots (Fig. 7). Thus, then, metaphorically speaking, the caterpillar at first attempted to scare off its enemy by a terrifying attitude alone, and it was only subsequently, in the course of the phyletic evolution, that the eye-spots were added, in the elephant hawk-moths and other species, to heighten the terrifying effect. But that the eye-spot did not make its appearance suddenly is proved by several[Pg 71] American species of Smerinthus, in which they are much less perfectly developed than in the European species. In these Sphingidæ, too, the defiant attitude was evolved earlier than the eye-spots, as we may see from our poplar hawk-moth (Smerinthus populi), which, when alarmed, spreads out all four wings in the same peculiar manner which in the eyed hawk-moth (Smerinthus ocellatus) displays the eye-spots; it strikes about with its wings as if to scare off the enemy, an effect which will certainly be more surely achieved if, at the same time, a pair of eyes becomes suddenly visible.
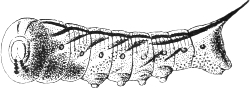
Fig. 7. Caterpillar of a North American
Darapsa in its "terrifying attitude" (after
Abbot and Smith).
Sympathetically coloured caterpillars are, however, by no means the only ones; there are some with such striking, glaring colours that, far from rendering their possessors inconspicuous, they make them visible from a long way off; but this apparent contradiction of the theory of the colour-adaptation of animals that require protection has been explained by the acuteness of Alfred Russel Wallace. We know that among insects, and also among caterpillars, there are many which have a repulsive taste. In any case, certain caterpillars are rejected by many birds and lizards. Such species are, therefore, relatively safe from being devoured. If they were protectively coloured, or if, moreover, they resembled caterpillars with an agreeable taste, they would gain little advantage from their unpalatability; for the birds would at first take them for eatable, and would only discover their repulsiveness on attempting to eat them. But a caterpillar which has received a single stroke from a bird's bill is doomed to death. It must therefore be of the greatest advantage for unpalatable caterpillars, and unpalatable animals generally, to be in their colouring as conspicuously distinguishable as possible from the edible species. Hence, then, the glaring colours, which we can now refer without any further difficulty to the process of natural selection, for every individual of an ill-tasting species that is more conspicuously coloured than its fellows must have an advantage over them, and must have a better chance of surviving, because it will be less easily mistaken for a member of an edible species.
I should like to discuss one other phenomenon, which is well calculated to give us a deeper insight into the transformation processes of organisms—I refer to the remarkable dimorphism of colour which occurs in many of the species of caterpillar just described.
The caterpillar of the convolvulus hawk-moth (Sphinx convolvuli)[Pg 72] is in its full-grown stage green, like the wild convolvulus on which it lives, or brown like the ground on which its food-plant grows. It thus shows a double adaptation, each of which is capable of protecting it to a certain extent, and we might think to the same extent. But that is not so, the brown colouring is a more effective protection than the green, as we may learn from two facts. In the first place, the four young stages of the caterpillar are green, and it only becomes brown in the last stage, though sometimes even then it remains green. This shows that the brown is a relatively modern adaptation, and it could not have arisen had it not been better than the original green. In the second place, the green-coloured caterpillars of the convolvulus hawk-moth are nowadays much less numerous than the brown ones, and this implies that the latter survive oftener in the struggle for existence. We have here an interesting case of an easily recognizable process of selection still going on between the old green and the newer brown variety.
It is hardly necessary to ask why the brown colour should in this case be a better protection than the green, for it is obvious that such a large green body as that of the full-grown convolvulus-caterpillar would be but badly concealed among the little leaves of the convolvulus plant in spite of its green colour; while the brown caterpillar, on the brown soil, with its pebbles, hollows, and irregular shadows, is excellently protected, especially if it passes the day concealed in the ground, as is actually the case.
Our view is materially strengthened by the fact that the same phenomenon of double colouring occurs in several allied species of Sphingidæ, but in a manner which shows us that we have to do with a similar process of transformation, only at a more advanced stage. The caterpillar of Chærocampa elpenor (Fig. 4) shows the same state of things as that of the convolvulus hawk-moth; it is brown or green, and the green form is the less common. But in the two other European species of Chærocampa the full-grown caterpillar is always brown, and indeed it becomes brown in the fourth stage, instead of, like Chærocampa elpenor, only in the fifth and last. Another indigenous sphingid species, Deilephila vespertilio, only remains green during the first two stages, and assumes in the third stage the grey-brown colour which it afterwards retains. The dark colour has obviously prevailed among the full-grown caterpillars for a considerable length of time, for it is in this, the largest and most conspicuous stage, that the change of colour must have been most necessary, and consequently the process of selection must have begun in it, and only after the more protective brown became general would it have extended to the next[Pg 73] stage below, if it were of use there too, and, later on, to still earlier stages in the life-history.
One might be inclined to ascribe this shunting back of a new character from the later to the earlier stages of development to purely internal forces, which brought it about of necessity, and quite independently of whether the extension of the character was useful or injurious. We shall come back to this later, and try to find out how far this is the case, but in the meantime we may regard at least so much as established, that this shunting back does not take place everywhere and without limits, but that natural selection calls a halt as soon as its effect would be injurious.
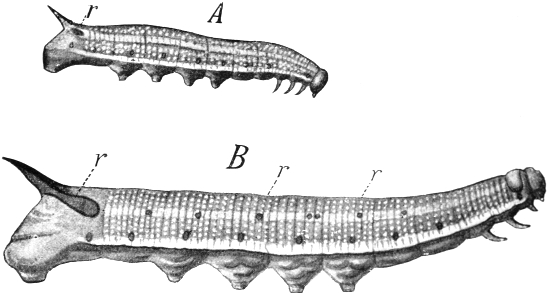
Fig. 8. Caterpillar of the Buckthorn Hawk-moth, Deilephila hippophaës. A, Stage III. B, Stage V. r, ring-spots.
There could be no continuance of insect-metamorphosis if every character of the final stage had to be shunted back to the one next below, for then, for instance, the characters of the butterfly must, in the course of the phyletic evolution, be carried back to the pupa and larva. But even in the larval stage alone it can be seen that this carrying back is kept within well-defined limits. Thus, for instance, in the dimorphic caterpillars of the Sphingidæ the brown of the full-grown stage never comes so far down as the earliest stages, for the little caterpillars are all green, like the leaves and stems on which they sit. On the other hand, there are species in which the green persists, as apparently the most advantageous colour. Thus in the buckthorn hawk-moth (Deilephila hippophaës) (Fig. 8), which lives in the warm valleys of the Alps, and especially in Valais, the caterpillars are grey-green in all stages, and are exactly of the shade of the lower surface of the buckthorn leaves; they possess no oblique lines, for these would not make them more like the leaves, as the full-grown caterpillars are much bigger than an individual leaf of buckthorn,[Pg 74] on which, moreover, the lateral veins are not very conspicuous. Nevertheless the caterpillar enjoys very fair security, as it does not feed through the day, but only in twilight and at night; it passes the daytime concealed in the dry leaves and earth about the base of the bush. Its resemblance to the leaves is very great, and is increased by the fact that it bears on the last segment a comparatively large orange-coloured spot (r), exactly the colour of the buckthorn berry, which ripens just at the time that the caterpillar attains its full growth.
But butterflies are as much persecuted, and have as much need of protection, as caterpillars, and among them, too, we find many instances of protective colouring, which are the more interesting in that they occur, as a rule, only on such parts of the body as remain visible when the insect is at rest, which is exactly what we should expect if the coloration has been wrought out in the course of natural selection. But it is well known that the resting position of diurnal Lepidoptera is quite different from that of the nocturnal forms, and is not even the same among all families, and in accordance with this we find the sympathetic colouring occurs on quite different areas in the different families.
The reason why the butterflies only require to be protected by their colour in the sleeping or resting position is that no colour whatever could make a flying butterfly invisible to its enemies, because the background against which its body shows is continually changing during its flight, and, moreover, the movement alone is enough to betray it, even if it is of a dull colour.
Thus, in general, only those parts of a butterfly's wing that are invisible at rest could safely bear bright or conspicuous colour, while the visible portions had to acquire sympathetic coloration through natural selection.
As the diurnal butterflies, when at rest, turn their wings upward and bring them together, it is only the under side which is sympathetically coloured, and that only as far as it is visible, that is, the whole of the posterior wing, and as much of the anterior one as is not covered by it. Many diurnal butterflies, when at rest, fold the anterior wing so far back that only its tip remains visible, and in such cases only this tip is protectively coloured, while in other forms, which have not this habit, almost the whole surface of the wing is sympathetically coloured.
A very simple protective colouring is exhibited by our 'lemon butterfly' (Rhodocera rhamni), in which the under surface is a whitish yellow, which protects the insect well when it settles on[Pg 75] the dry leaves on the ground in the light woods which it is fond of frequenting.
Our gayest diurnal butterflies, the species of Vanessa, all have the under surface of a dusky colour, sometimes passing into a blackish brown, as in the peacock-butterfly, Vanessa (v. io), sometimes more into greyish brown, or brown-yellow, or reddish brown. They are never simple colours, but always consist of mixtures of different colour-tones—indeed, there is often a complex mingling of many colours, as grey, brown, black, white, green, blue, yellow, and red, made up of dots, strokes, spots, and rings, into a wonderful and very constant pattern, which, taken as a whole, has the effect of being uniform, and harmonizes with the soil, or with the highway, on which the species loves to settle, with much greater accuracy than a monochrome grey or brown would do. When the 'painted lady' (Vanessa cardui) settles on the ground it is hardly distinguishable from it, and this species in particular has a preference for settling on the ground. Other species of Vanessa, such as the peacock and the Camberwell beauty (Vanessa antiopa), are underneath of a dark blackish grey, or even black; when resting they press themselves into the darkest corners and crevices, and are thus most effectively secured from discovery.
Many diurnal Lepidoptera, on the other hand, especially the wood-butterflies of the family Satyridæ, have the habit of resting on the trunks of trees, as Satyrus proserpina does on the great beech-trunks of the forest clearings. These large butterflies, coloured conspicuously on the upper surface in deep velvety black and white, are marked on the under surface exactly to match the whitish bark of the great beech, covered over with white, grey, blackish-brown, and yellow spots, and the butterfly whose flight one has just been carefully following disappears as it suddenly alights on such a tree-trunk. As I have already stated, the protective colour only extends over as much of the insect as is seen when it is at rest. As the anterior wings are folded far back between the posterior ones, the protective colouring is limited to the whole surface of the posterior wing, and the tip of the anterior one, as far as that is visible in the resting attitude; the protectively coloured area is somewhat sharply bounded, and it is often of very different extent in quite nearly allied species, according to whether the species folds the anterior wing far back or not. Thus in our common small tortoiseshell-butterfly (Vanessa urticæ) the protective area is considerably wider than in the large tortoiseshell (Vanessa polychloros), much as the two resemble each other in other details.
This harmony between the wing tips and the posterior wings is[Pg 76] nowhere wanting, where the under side is protectively coloured at all, but in many cases the protective colouring spreads over almost the whole of the anterior wings, and these are then not folded far back when at rest, as will be seen later in the so-called leaf-butterflies.
There is one genus of diurnal butterflies which seems to contradict the law that all the surface that is visible in the resting position exhibits the protective coloration—the South American wood-butterflies of the genus Ageronia. They have on the upper surface a very complicated bark-like pattern of mingled grey on grey, and this confirms the usual rule, for we know that these butterflies—a striking exception among all the other diurnal forms—settle with outspread wings on the trunk of a tree in exactly the same attitude as many of the nocturnal Lepidoptera of the family of the Loopers or Geometridæ, in which the upper surface is also deceptively like the bark of the tree on which they rest.
Fig. 9. Hebomoja glaucippe, from India; under surface. A, in flight. B, in resting attitude.
In all the nocturnal Lepidoptera it is the upper side of the wing which is sympathetically coloured, if protective coloration has been developed at all. In all the Sphingidæ, many 'Owls' and Bombycidæ, the anterior wings are grey banded with darker zigzag lines, and mottled with many shades of black, grey, yellow, red, and even violet. As the anterior wings cover the body and the posterior wings like a roof, they make the resting insect very inconspicuous when it has settled on wooden fences, trunks of trees, or even old timber. When bright colours—red, yellow, or blue—occur in these moths[Pg 77] it is always on the posterior wings, which are covered when at rest. This can best be observed in the species of the genus Catocala.
Let us now, however, interrupt our survey of the facts for a moment, and let us inquire whether all the cases of protective colouring in Lepidoptera we have considered can be referred to natural selection, or whether it is not conceivable that other causes may have evoked them.
Fig. 10. Xylina vetusta, after Rösel. A, in flight. B, at rest.
The first thing to be said is that the Lamarckian principle of the inherited effects of use and disuse cannot here be taken into account, as the colours of the surface of the body do not exercise any active function at all; their effect is due simply to their presence, and it is for them quite indifferent whether and how often they have opportunity to protect their bearers from enemies, or whether no enemies ever chance to appear. It has frequently been suggested, too, that these colorations are associated with the differences in the strength of the illumination to which the different parts and surfaces are exposed. But this again is untenable, as is proved even by the dimorphism frequently occurring in caterpillars, for the green and the brown individuals are exposed to precisely the same light; and still more clearly by the sympathetic colouring, which is so exactly defined and yet so different on the under surface of the diurnal butterflies. Yet there are isolated cases in which it seems as if the direct influence of the light had brought about certain striking differences in the colouring of the parts of an insect, and I shall describe perhaps the prettiest of these cases, to which Brunner von Wattenwyl directed attention. It concerns one of the Orthoptera of Australia, a Phasmid, Tropidoderus childreni, Grey, which has a general colouring of leaf-green, but with singular deviations from it on certain areas of the body. In this insect the anterior wings which form the wing covers or elytra (Fig. 11, V) are so short that they scarcely cover the half of the long abdomen. Their place is taken by the anterior margin of the posterior wing (H. horn), which is hard and horny like the elytra, and in the resting position protects the whole abdomen. All these covering parts are grass-green, except at the places where they overlap; on these areas they have a faded look, and[Pg 78] are yellowish instead of green. Brunner says of this: 'The phenomenon gives the impression that the more brilliant colour is a character due to daylight. If several sheets of white paper of unequal dimensions be placed one above the other, ... and exposed to the sun, after a short time silhouettes of the smaller sheets will appear on the larger ones, either in a lighter or in a darker colour. Probably this "fading" of the covered parts in the Phasmid also belongs to this "category of photographs."' This seems convincing, but analogous phenomena in other insects prevent our regarding the pretty comparison with the photograph as a sufficient explanation. If it were a question of a diurnal butterfly, such an assumption would have to be rejected on this ground alone, that the wing colouring is developed in the pupa, and appears perfect and unalterable as soon as the perfect insect emerges. But in the pupa the position of the wings is exactly the reverse of that seen in the resting attitude of a butterfly, that is, the protectively coloured under side of the wing is not turned towards the light but away from it. Moreover, in the pupa the anterior wings cover the posterior ones completely, no matter what the wing position may be later in the perfect insect. Furthermore, the thick and often darkly coloured sheath of the pupa prevents the light having any effect, and not a few species pass their pupal stage in such dark places—for instance, under stones, as in the case of many 'Blues'—that the light can hardly reach them. And if the light did exercise an influence, how could it produce such diverse coloration as the protective colours of diurnal butterflies, on the one side dark, even to blackness, on the other side, yellow, reddish, and even white and pure green; and how should the same rays of light call forth complicated colour patterns on one and the same surface, for instance, the white, sprinkled with green, of the Aurora butterfly (Anthocharis cardaminis)? Finally, we have only to remember that numerous nocturnal Lepidoptera pass through their pupa stage underground, although they exhibit brilliant as well as protective colours in the most appropriate distribution, to reject once for all the hypothesis that the influence of light plays any decisive rôle in determining the distribution of the colours on the wings of Lepidoptera.
But it is otherwise with Tropidoderus. In this case the wings grow gradually during the slow growth of the animal, which takes place in full light, and the wings of the young insect probably lie one above the other, in exactly the same position, and cover the same places as in the full-grown form; we might, therefore, from the facts of the case, admit the possibility that the yellow of the covered portions is due to the exclusion of light.
Fig. 11. Tropidoderus childreni, after Brunner von Wattenwyl, in flying pose. V anterior wing. H. häut, membranous part of posterior wing. H. horn, horny portion.
But as soon as the conditions that obtain among Lepidoptera are also taken into consideration we recognize the insufficiency of the interpretation suggested, for among butterflies we have precisely the same phenomenon—sharp limitation of the protective colouring to the parts visible in the resting position, a fact which, in the case of the said butterflies, admits of no other interpretation than that of natural selection. Let us therefore see if we cannot, in the case of Tropidoderus, arrive at some better understanding of the phenomenon than that implied in the theory of direct light-influence. Obviously, the yellow parts of the animal do not require to be green, since they are not visible in the sitting position, and the locust in flight could not by any device be made invisible. It therefore only remains to be explained why the yellow parts are not colourless, and why they are not also green. We cannot at present answer with any confidence; it is possible that the colouring matter which causes the green only becomes green under the influence of direct sunlight, and otherwise remains yellow; it is possible, too, that, as in Lepidoptera (see Fig. 9), the full protective colour is only developed by natural selection in the places which are visible in the sitting position, and[Pg 80] that the covered parts take on any indifferent colour, which might be readily afforded by the metabolism of the insect. But this much is certain, that the covered parts would be green, if that were advantageous to the survival of the species, just as the under surface of the wings of some diurnal butterflies is green. Had it been required, the green colour would have resulted in the course of natural selection, just as it has resulted in the most different parts of the most diverse insects, even in those whose development takes place entirely removed from the influence of light. Therein lies the difference between our interpretation and that of Brunner von Wattenwyl: without natural selection no explanation of this case is possible.
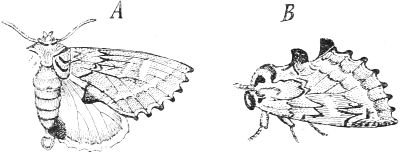
Fig. 12. Notodonta camelina, after Rösel. A, in flight. B, at rest.
Hitherto I have spoken only of the diurnal butterflies in which the anterior wings show an extension of the protective colouring which marks the whole surface of the posterior wings, and it was always the tips of the anterior wings that were thus coloured. But among the nocturnal Lepidoptera there are corresponding cases, in which a little tip of the posterior wing forms the continuation of the protective surface of the anterior wing. Some species of Notodonta and allied genera show in the posterior corner of the otherwise whitish posterior wings a little grey spot, and a hair tuft which in colour, and—when it is big enough—in marking, exactly resembles the protectively coloured anterior wings (Fig. 12). The 'why' is at once clear, when one looks at the insect in the resting position, for only this little corner of the wing projects beyond the covering anterior wing. This has been regarded as telling against natural selection, for such a little spot could not possibly, by its colour, turn the scale as to the life or death of the individual, and so could not be selected. But one might say the same of the tip of the anterior wing in the diurnal forms, although there the protective surface is larger, often much larger. But who is to decide how large an exposed, unprotected spot must be in order to attract the attention of an enemy on the look out for food? Or[Pg 81] who can prove that the best and most familiar protective colouring really protects its possessors? What if, after all, it is all a game, a joke, which the Creator is playing with us poor mortals? Did not a trustworthy observer recently watch carefully, and see how a pair of sparrows daily cleared a wooden fence on which moths of the genus Catocala and other species of nocturnal Lepidoptera, excellently furnished with protective colours, were wont to settle by day? They did their work thoroughly, and hardly overlooked a single individual. But who has a right to see anything more in this than—what surely goes without saying—that the best protective colouring is not an absolute protection, and never preserves all from destruction, but always only some, and it may be very few.
How else could there be such a high ratio of elimination, and such a constancy in the number of individuals of a species on any unchanging area? These sparrows had simply made full use of an experience, probably acquired by chance to begin with, and their vision had become sharpened for this particular species on the almost similarly coloured wooden fence, just as that of the expert butterfly collector does. It certainly does not follow from this that the protective colouring was useless, nor can we regard the harmony between the protruding tip of the anterior or posterior wing and the large protectively coloured surface of the covering wing as of no importance. On the contrary, if the tips were white or conspicuously coloured like the rest of the posterior wing, they would assuredly attract the sharp eye of hungry enemies to the spot, and so betray the victim. Instead of this, the spot in question is not only dark, but, in the case of Notodonta, is furnished with a tuft of hairs, which, in the insect's resting position (Fig. 12, B), lies on the back, and looks like a dark, somewhat curved projecting tooth, in front of which there stands another, quite similar, which arises from the anterior wing, and behind there are other seven, rather smaller, dark teeth of the same kind, springing from the outer edge of the anterior wing. Taken altogether, they mimic the dentated edge of a withered leaf, and thus, in spite of their diverse origins, form a unified picture, and one with a considerable protective value. How is it possible to doubt that each of these hair-tufts has arisen under the influence of natural selection, and that its absence or imperfect development might result in the discovery and elimination of the insect concerned?
These cases seem to me particularly beautiful proofs of the productive efficiency of selection. The wing is protected just as far as it protrudes from beneath the other—not a millimetre[Pg 82] further! How should it be otherwise, when the colouring of the parts just beside these is indifferent for the species, so that any variations in these parts in the direction of protective colouring never survive to be transmitted and accumulated?
It is precisely this restriction to what is absolutely necessary that is the surest sign, here and elsewhere, that the character in question has been brought about by natural selection. And if this is the only possible, and at the same time quite sufficient explanation of the remarkably well-defined colour deliminations in all Lepidoptera, there can be no reason why we should try to drag in any other factor to explain the case of Tropidoderus, the less so as here again selection alone can account for the green of the exposed surfaces; and furthermore, the modification, common in other Phasmidæ, of the most anterior green stripe of the posterior wing into a firm cover protecting the soft abdomen, also points to natural selection; the cover-wings proper have here become too short, and so the edge of the posterior wing has been modified into a hard rib, which protects the soft body of the insect (Fig. 11, H. horn). No differences in illumination, and no direct effect of any external influence whatever could have brought that about.
How much more I might adduce in this connexion! The manifold diversity of colour and form adaptation is so great among insects, to which protection from their enemies is so necessary, and especially among butterflies, that I should never come to an end if I were to try to give even an approximate idea of it. Let us, therefore, turn now from such cases to a higher—the highest—grade of adaptation, that in which there is not only a mimicry of special and complex coloration, but in which the whole animal has become like some external object, and is thereby secured from discovery.
We must first consider the case of our lappet moth (Gastropacha quercifolia), which in its copper-red colour and in the remarkable shape and dentated edges of its wings, and finally in the quite extraordinary clucking-hen-like attitude of the wings when at rest, greatly resembles some dry oak-leaves lying one above the other.
Not unlike this is a 'shark' moth found in this country, Xylina obsoleta, which, as the name indicates, looks when at rest like a broken bit of half-rotten wood (Fig. 10, p. 77). It 'feigns death,' as we commonly say, that is, it draws the legs and antennæ close to the body, and does not move; indeed, one may lift it up and throw it on the ground without its betraying by a single twitch that it lives. Only after it has been left undisturbed for some time does it show[Pg 83] signs of life again, and makes off hastily, to find a better hiding-place. The colouring of this moth is so curiously mingled—brown, whitish, black, and yellow—and traced with acute-angled lines and curves, that one cannot distinguish it at sight from a bit of rotten wood. I experienced that myself once when, passing a hedge, I thought I saw a Xylina sitting on the ground, and picked it up to examine it. I thought it was a bit of wood, and, disappointed, I threw it down again on the grass, but then I felt uncertain, and picked it up once more—to find that it was a moth after all[1]!
[1] Rösel says in this connexion: 'The marvellous form of this Papilio preserves it from injuries, for, when he hangs freely on a trunk of a tree, he would be taken ten times sooner for a piece of bark than for a living creature. By day, too, he is so little sensitive, that if he be thrown down from his resting-place he falls to the ground as if lifeless, and remains lying motionless. One may also throw him into the air, or turn him about, and he will rarely give a sign of life. I have impaled many of them on needles, without seeing any sign of sensitiveness on their part. This is the more remarkable that these birds (sic), after they have submitted to all the torment and misery one can inflict on them, without showing any sign of feeling, will, whenever they are left in peace and have no further disturbances to fear, quickly creep off to a dark corner and attempt to conceal themselves from future attacks.'—Insektenbelustigungen, Nürnberg, 1746, vol. i. p. 52.
This case of Xylina is hardly less remarkable, and its likeness to the mimicked object is scarcely less wonderful than that of the often discussed mimicry of a leaf, with stalk, midrib, and lateral veins, by many of the forest butterflies of South America and India.
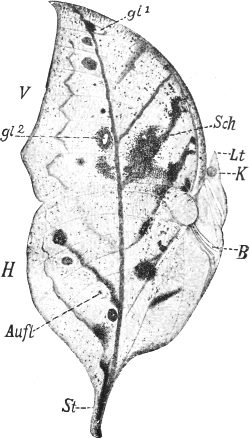
Fig. 13. Kallima paralecta, from India,
right under side of the butterfly at rest.
K, head. Lt, maxillary palps. B, limbs. V,
anterior wing. H, posterior wing. St, 'tail'
of the latter, corresponding to the stalk of
the leaf. gl1 and gl2, transparent spots. Aufl,
eye-spots. Sch, mould-spots.
The best known of these is the Indian Kallima paralecta, which, when it settles, is deceptively like a dead leaf, or rather like a dry or a half-withered one, on which brown alternates with red, and on which there are one or two translucent spots, without scales, presumably representing dewdrops. The upper surface of this butterfly is simply marked, but gorgeously coloured—blue-black[Pg 84] with a reddish yellow, or bluish white band—and quite constant. The under surface, on the other hand, although it always resembles a dead leaf, shows very varied ground colours, being sometimes greyish, sometimes yellowish, or reddish yellow, or even greenish. Often it shows the lateral veining of the leaf quite as distinctly as in Fig. 13, but often quite indistinctly, and the black, mouldy spots (Sch) of our figure may be more strongly marked, or they may be absent. It would seem as if the mimicry of different kinds of leaves was here aimed at—so to speak—just as in the case of the varied and numerous species of the South American genus Anæa, which usually live in the woods, and are all more or less leaf-like, but each species is like a different leaf, or like a leaf in a different condition, dry, moist, or decomposing. It is simply astounding to see this diversity of leaf mimicry, and the extraordinary faithfulness with which the impression of the leaf is reproduced. But it is by no means always the venation which causes the resemblance, for this is often inconspicuous; the high degree of deceptiveness is due to the silvery-clear yellow, dark yellow, red-brown to dark black-brown ground-colouring, which is never quite uniform, and over which there usually spreads a whitish ripple, combined with the remarkable imitation of the sheen of many leaves. The upper side of this butterfly is almost always conspicuously decorated with violet, dark blue or red, but always without any relation to the under surface. Not in all, but in many of the species of this genus, we find the round, translucent mirrors on the wing already mentioned in the case of Kallima, and in some species quite remarkable means are made use of to make the resemblance to a leaf thoroughly deceptive. Thus Anæa polyxo, when sitting, looks like a leaf out of the edge of which a caterpillar has eaten a little piece; in reality there is nothing missing from the wing, but on the front margin of the anterior wing a semicircular spot of a bright, soft, yellow colour stands out so sharply from the rest of the chestnut-brown wing surface, that it has the effect of a hole in the leaf.
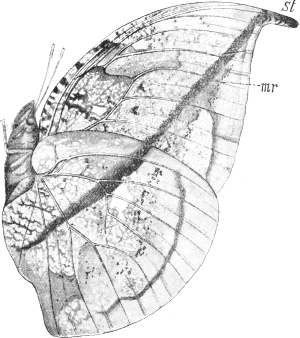
Fig. 14. Cœnophlebia archidona, from Bolivia, in its
resting attitude. mr, midrib of the apparent leaf.
st, the apparent stalk.
A modern opponent of the selection theory (Eimer) has suggested that the marking of the lateral veins, and other resemblances to a leaf in Kallima, represent nothing more than the pattern which was present in any case, inherited from ancestors, and which in the course of time arranged itself in a particular manner according to internal developmental laws. Not selection—that is, adaptation to surroundings—but the internal developmental impulse has brought about the resemblance to the leaf. It is astonishing how a preconceived idea can blind a man and weaken his judgment! It goes [Pg 85]without saying that the adaptations do not start from a tabula rasa, but from what is already present; of course, natural selection makes use of the markings inherited from ancestors; it takes what already exists, and alters or extends it as suits best. Thus it is easy to prove that the clear mirrors (Fig. 13, gl1 and gl2) on the wings of Kallima have arisen from a modification of the nuclei of eye-spots, just as the dark mould-spots which often occur, frequently develop in association with the inherited eye-spots; not always however, for many such accumulations of black scales occur in spots on which there has never been an eye-spot. Thus, too, the 'midribs' of the butterfly have in part arisen from a gradual displacing, extending, and altering of the direction of inherited stripes as, for instance, is clearly recognizable in the posterior wing of Fig. 13, but sometimes they are new formations. But the veining of a leaf is never found on the wing of any butterfly of a species which has not the habit of resting among leaves, or which has not had it at one time, and it never corresponds to the natural marking of any genus which does not live in forests. This impression of leaf-venation has obviously arisen from quite different patterns of markings, and it has been reached now by one way, now by another. We can see this from the fact that, in different butterflies, it lies in quite different positions on the wing. In the Kallima species the stalk of the leaf lies in the tail of the posterior wing, the tip of the midrib lies near the tip of the wing; in Cœnophlebia archidona it is exactly reversed, the tip of the anterior wing (Fig. 14) is prolonged, and forms the stalk, while a broad, dark, stripe, the midrib (mr), runs from there across the middle of both wings, and seems to give off two or three lateral ribs running outwards. If it be asked whether this butterfly always sits[Pg 86] down so artistically that the 'upward turning leaf-stalk is in juxtaposition to a twig,' we may answer that a bird flying fast is not likely to look to see whether every leaf in the profusion of foliage in the primitive forests is properly fastened to its stalk or not, any more than we should do in the case of a painted bush, on which many a leaf has the appearance of floating in the air, just as in nature, or in its faithful copy, the photograph.
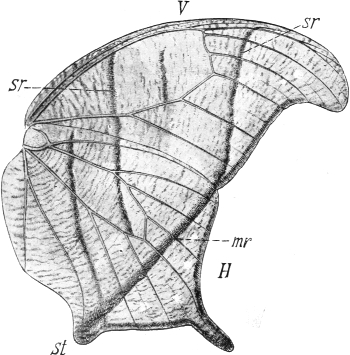
Fig. 15. Cærois chorinæus, from the lower Amazon, in its
resting attitude. V, anterior wing. H, posterior wing. mr,
midrib of the apparent leaf. sr, lateral veins. st, hint of
a leaf-stalk.
Quite different from the leaf-marking either of Cœnophlebia or Kallima is that of one of the Satyrides of the lower Amazon valley, Cærois chorinæus (Fig. 15). If one spreads this butterfly out in the usual way it does not look in the least like a leaf, and one only sees a number of curiously placed disconnected stripes on the under surface of the wing. But if the wings be folded together to correspond with the sitting position of the butterfly, there appears the figure of a leaf, of which, however, only half is present, and whose midrib (mr) runs obliquely forward from the inner angle of the posterior wing. Here, again, it is not difficult to guess that this straight stripe has arisen, by displacement and straightening, from a curved line inherited from some remote ancestor, and it is these precise changes which are the work of the adaptive processes of natural selection. The same applies to the lateral ribs (sr), which are here four in number.
But even the division of the wing surface by a single dark line, such as that which crosses the middle of the posterior wing of Hebomoja (Fig. 9), an Indian butterfly, heightens not inconsiderably the resemblance of the resting butterfly to a leaf, a resemblance which[Pg 87] has already been shown in the form and colour. Indeed, even the sharp division of the wing surface into a darker inner and a lighter outer portion, which occurs in many species of Anæa, gives a very vivid impression of a leaf crossed by a midrib.
It is not without a purpose that I have lingered so long over the leaf-butterflies. I wished to make it clear that we have by no means to do with a few exceptional cases, but with a great number, in all of which resemblance to a leaf has been aimed at, although it has been attained in varying degrees, and by very diverse ways. Whoever surveys this wealth of fact must certainly receive the impression, that, wherever it was advantageous to the existence of the species, the evolution of such a deceptive resemblance has also been possible. In any case one cannot but be convinced that it is not a case of chance resemblance, as some naturalists have recently tried to maintain.
But I have not yet quite finished my outline-survey of the facts, for I must not omit to mention that, in the evergreen tropical forests, there are also large nocturnal Lepidoptera, which mimic leaves, sometimes green ones, sometimes brown, dead ones.
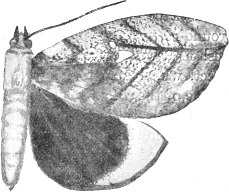
Fig. 16. Phyllodes ornata, from Assam.
Upper surface with leaf-like marking only
on the anterior wing, which is the only
part visible when at rest; 2/3 nat. size.
Fig. 16 gives a good picture, reduced to two-thirds, of such a species, Phyllodes ornata, from Assam. The posterior wings are conspicuously coloured in deep black and yellow; in the resting position they are covered by the anterior wings, and these are red-brown with black markings which precisely and clearly mimic the ribs of a leaf. The midrib begins near the tip of the anterior wing, but breaks off half-way across the wing at two silvery white spots, similar to those in many of the diurnal forms, which also mimic decaying leaves. Three pairs of side veins go off backwards and forwards with remarkable regularity from the midrib, almost at the same angle, and parallel to one another, and three more are indicated by vague shading. Then the midrib begins again in the internal half of the wing, though only represented by a broad shading. The whole suggests two torn, rotten leaves, one partly covering the other; and the deception will certainly be perfect when the moth rests on the ground or among decaying leaves.
That all these extremely favourable protective colorations find[Pg 88] their explanation in the slow and gradually cumulative effects of natural selection cannot be disputed; it is beyond doubt that they cannot be explained, so far as we know, in any other way.
If, however, it were possible for a species of butterfly living in the forest and among leaves to become, through natural selection, in any degree, and in a continually increasing degree, like a leaf, surely many insects living in the woods, and especially in the tropical woods, would also have followed such an advantageous path of variation—at least, so we should be inclined to think. And this is indeed the case; numerous insects, of different orders, if they are as large as a leaf, have taken on the colour, form, and usually also the markings, of a leaf. Thus green and also decaying and dead leaves are most realistically imitated by many tropical Locustidæ. Besides Tropidoderus, figured on p. 79, a Pterochroa of South Brazil affords a particularly fine illustration of this, for not only does the ground-colour, brown or green, harmonize with that of a dead or fresh leaf, but, at the same time, all sorts of details are marked on the insect, which help to heighten the deceptive impression. Even the outline of the wings is leaf-like, and leaf-veins are marked on the wing-covers with the most beautiful distinctness, and finally there is, especially in the light-green individuals, a spot at the wing tip which, by means of a mixture of brown, yellow, reddish, and violet colour-tones, mimics a decaying spot with astonishing fidelity. Here, again, the origin of this special adaptation can be clearly recognized, for the vaguely concentric arrangement of the colours indicates that, in the ancestors of the species, an eye-spot had occurred on this area, of the same kind as we still see on the posterior wing, which is covered in the resting position. Thus we can again look back on the history of the species and conclude that the dissolution and degeneration of the eye-spot began at the time when the leaf resemblance was evolved, and this was probably caused by some change of habitat, which we can now no longer guess at.
Many species of leaf-like Orthoptera, both in the Old and New World, have tough, green, parchment-like wing-covers which bear a remarkable resemblance to the thick Magnolia-like leaves of tropical plants. Along with these we must also mention the 'walking leaf,' which has been well known for centuries. In its case, not the wing-covers alone, but the head and thorax, and even the legs, are of the colour and shape of a leaf.
The stick-insects, too, must not remain unnoticed; those quaint inhabitants of warm countries, whose elongated brown body looks like a knotted twig, and whose long legs, likewise stick-like, are stretched[Pg 89] out irregularly at different angles to the body, and usually remain motionless when the insect is resting. These creatures are vegetarian, and generally keep so still, that even the naturalist who is on the look-out for them may easily overlook them. Even such an experienced student of insects as Alfred Russel Wallace was deceived, for a native of the Phillipines once brought him a specimen as a 'walking-stick' insect, which he rejected, saying that this time it was no animal but really a twig, until the native showed him that it was an insect whose likeness to a twig was increased by the fact that it bore on its back a ragged green growth, which looked exactly like a liverwort (Jungermannia), which occurs on the twigs of the trees in that region.
We must also notice here the thorn-bugs, which are numerous on the prickly shrubs of tropical deserts and plateaux, especially in Mexico. These bear on the relatively very small body two or three large spines, which make them look like a part of the thorny bush on which they sit. But this masking by mimicry of thorns is not confined to insects, it is seen in lizards as well, notably in Moloch horridus, a lizard that lives in the Australian bush, and is covered all over with thorn-like scales.
These examples should be enough to show that mimicry of the usual surroundings on the part of animals which are in need of protection, or are wont to lurk on the watch for their prey, are not isolated exceptions, chance resemblances, or, as they used to be called, 'freaks of nature,' but that, on the contrary, they are the rule, depending on natural causes, and always occurring when these causes are operative. That such protective resemblances seem to be much more frequent in warmer climates than with us is probably a fallacy due to the fact that the number of species (especially of insects) is very much greater there, and that many insect types have their representatives of considerable size of body, which not only makes them more conspicuous to us, but makes some protective device in relation to their enemies or victims much more necessary.
But we must here take account of one more example which occurs in our fauna in many modifications: the caterpillars of Geometridæ. Many of these soft and easily injured caterpillars resemble closely, in colour and shade, the bark of the tree or shrub on which they live (Fig. 17). At the same time they have the habit, when at rest, of stretching themselves out straight and stiff, so that they stand out free, at an acute angle from the branch, thus seeming like one of its lateral twigs. In many species the resemblance is heightened by the extraordinary pose of the head (K) and of the[Pg 90] claw-like feet (F), which, partly pressed close to the head, partly standing out from it, give the anterior end of the caterpillar the appearance of two terminal buds, while various little pointed, knotlike warts, scattered over the body, represent the sleeping buds of the little twig. Who has not at one time or other taken such a caterpillar for a little branch, and not inexpert observers only, but even trained naturalists? Many a time I have not been able to make quite sure of what I had before me until I touched it!
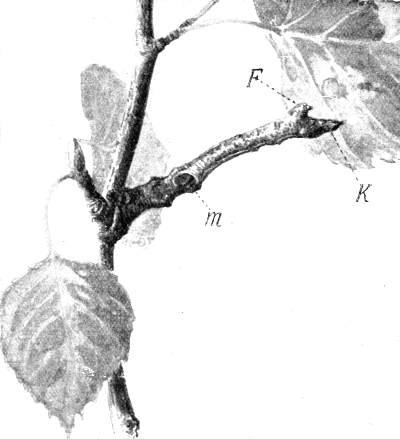
Fig. 17. Caterpillar of Selenia tetralunaria, seated on a birch twig. K, head. F, feet. m, tubercle, resembling a 'sleeping bud'; nat. size.
TRUE MIMICRY
Mimicry: its discovery by Bates—Heliconiidæ and Pieridæ—Danaides—Papilio merope and its five females—The females lead the way—Species with mimicry in both sexes—Objections—Enemies of butterflies—The immunity of the models—Poisonousness of the food-plants of immune species—Several mimics of the same immune species—Persecuted species of the same genus resemble quite different models—Elymnias—Degree of resemblance—Differences between the caterpillars of the model and the copy—The same resemblance arrived at by different ways—Transparent-winged butterflies—The gradually increasing resemblance points to causes operating mechanically—Rarity of the mimetic species—Danger to the existence of the species not a necessary condition of mimetic transformation—Papilio meriones and Papilio merope—Comparison with the dimorphic caterpillars—Papilio turnus—'Mimicry rings' of immune species—Danais erippus and Limenitis archippus—Marked divergence of mimetic species from their nearest relatives—Mimicry in other insects—Imitators of ants and bees.
Let us now turn to the most remarkable of all protective form- and colour-adaptations, the so-called Mimicry, including all cases of the imitation of one animal by another, which we came to know first through Bates, and to a fuller understanding of which A. R. Wallace and Fritz Müller have especially contributed.
While the English naturalist, Bates[2], was collecting and observing on the banks of the Amazons—as he did for twelve years—it sometimes occurred that, among a swarm of those gaily coloured, quaintly shaped butterflies, the Heliconiidæ (Pl. II, Fig. 13), he caught one which, on closer examination, proved to be essentially different from its numerous companions. It was certainly like them both in colour and form, but it belonged to quite a different family of butterflies, that of the Pieridæ or Whites (Pl. II, Fig. 19). These whites with the colours of the Heliconiidæ always occurred singly in swarms of the latter form, and Bates found that, in the different districts of the Amazon, they always resembled in a striking manner the species of Heliconiidæ there prevalent. Many of them had been previously known to entomologists, and because they diverged so far from the usual type of the Pieridæ, especially in the form of the wing, the name Dysmorphia, the 'mis-shapen,' had been given to them, although the meaning of this 'mis-shapenness' long remained[Pg 92] a mystery. The French Lepidopterist, Boisduval, went a step further when he pointed out as something remarkable that nature sometimes makes several species of quite different families exactly alike, and called attention to three African butterflies, of which we shall have to speak later in detail. But even he was too much fettered by the old views of the immutability of species to arrive at a correct interpretation. Thus it was reserved for Bates to take the decisive step. Observing that the Heliconiidæ occurred frequently, and usually in large swarms, he concluded that they must have few enemies, and as he never saw the numerous insectivorous birds and insects hunting them, he further concluded that they must have something disagreeable which secured them from the attacks of these predaceous forms. On the other hand, he found that the heliconid-like Whites were always rare, and he took this as a sign that they were much persecuted, and that they must, therefore, be palatable tit-bits for the insectivores. If it were possible, then, that a species of Whites with the usual white colour of the family should give rise to variations, which would make them in any degree resemble the Heliconiidæ, which are secure from persecution, and if, in addition, those that exhibited the profitable variation attached themselves to swarms of the mimicked form, then these variants would be to a certain extent secured from attack, and more and more so in proportion as the resemblance to the protected model increased. The great likeness of these Whites to the Heliconiidæ, Bates further argued, would depend on a process of selection, based on the fact that, in each generation, those individuals would on the average survive for reproduction which were a little more like the model than the rest, and thus the resemblance, doubtless slight to begin with, would gradually reach its present degree of perfection.
[2] Contributions to an Insect Fauna of the Amazon Valley, Trans. Linn. Soc., Vol. XXIII, 1862.
Bates's hypotheses have been subsequently confirmed in the most striking way. The Heliconiidæ do possess a disagreeable taste and odour, and are utterly rejected by birds, lizards, and other animals. It has been directly observed that puff-birds, species of Trogon, and other insectivorous birds, looking down from the tops of trees in search of food, allowed to pass unheeded the swarms of gaily coloured Heliconiidæ which were fluttering among the leaves, and experiments with various insectivorous animals yielded the same result: the Heliconiidæ are immune. We can, therefore, not only understand that it must be advantageous to resemble them, we can also appreciate many of their peculiar characters, such as their gay coloration, which must serve as a sign of their disagreeable taste, and their slow, fluttering flight, as well as their habit of flocking together,[Pg 93] which must make it easier for the birds to recognize them as uneatable. Everything which marks out these unpalatable morsels, and makes them more readily recognizable, must be to their advantage, and therefore must have been favoured by natural selection (Pl. II, Fig. 13).
In the same way, every increase of resemblance on the part of the mimics would increase their chances of escaping notice, and any one who is accustomed to observe butterflies in nature can well understand that even very slight resemblances may have formed the beginning of the selection process; perhaps even a mere variation in the manner of flight, combined with the habit of associating with the swarms of Heliconiidæ. I myself have many times been momentarily deceived in our own woods by a White of unusually majestic flight, so that I took it for an Apatura or a Limenitis. If, therefore, individual Whites occurred here and there in the Amazon valley, which flew somewhat after the manner of the Heliconiidæ, and associated with them, they might possibly have attained a certain degree of security through that alone, and it would be greatly increased if at the same time they varied somewhat in colour in the direction of their companions.
In any case there can be no doubt whatever that in these cases a real transformation of the species in colour and marking, and perhaps often, too, in form of wing, has taken place, and that within comparatively modern times—let us say during the distribution of a species which required protection over a large continent, or since the last breaking up of an immune species into local species. Various facts prove this; above all, the circumstance that it is often only the females which exhibit this protective mimicry; and that one and the same species may mimic a different immune species in different areas, but always the one occurring abundantly in that area, and so on.
Definite examples will make this clearer, and I will only say in advance that, since the discovery of Bates, numerous cases of mimicry in butterflies have been found, not only in South America, but in all tropical countries which have a rich Lepidopteran fauna. And it is not only between the Heliconiidæ and the Pieridæ that such relations have been evolved; many much-persecuted, unprotected species of different families everywhere mimic species which are rejected on account of their nauseous taste, and these, too, belonging to different families. The Heliconiidæ are a purely American group, but in the Old World and in Australia their place is taken by the three great families of Danaides, Euplœides, and Acræides, since, as it seems, they all taste unpleasantly, and are rejected by all, or at least by most, of[Pg 94] the insectivorous birds. Numerous species of the genus Danais (Pl. I, Fig. 8), Amauris (Pl. I, Fig. 5), Euplœa (Pl. III, Fig. 25, 27), and Acræa (Pl. II, Fig. 2), and also many species of Papilio and other genera, enjoy the advantage of unpleasant taste, if not even of poisonousness; they are, therefore, secure from pursuit, and are, in consequence, much mimicked by palatable butterflies.
As a further example, I now select a diurnal butterfly from Africa, Papilio merope Cramer[3], which was shown by Trimen in 1868 to be mimetic. The species has a wide distribution, for, if we except slight local differences in the marking of the male, its range extends over the greater part of Africa, from Abyssinia to the Cape, and from East Africa to the Gold Coast.
[3] The West African form of Papilio merope has been quite recently distinguished from the southern form and regarded as a distinct species, the latter being now called Papilio cenea. The differences in the males are very slight—somewhat shorter wings, shorter wing-tail, and so on—differences which seem relatively unimportant in comparison with the differences between the males and the females.
The male is a beautiful large butterfly, yellowish white, with a touch of black, and with little tails to the posterior wings (Pl. I, Fig. 1), like our own swallowtail. A very nearly related species occurs in Madagascar, and there the female is similarly coloured, though it may be distinguished by having a little more black on the wing. On the mainland of Africa, however, the females of Papilio merope are so different in colour and form of wing that it would be difficult to believe them of the same species as the male had not both sexes more than once been reared from the eggs of one mother. The females (Pl. I, Fig. 6) in South Africa imitate a species of Amauris, A. echeria (Pl. I, Fig. 7), of a dark ground-colour with white, or brownish-white, mirrors and spots, and they resemble it most deceptively. But what makes the case more interesting in its theoretical aspect is that Danais echeria of Cape Colony is markedly different from Danais echeria of Natal, and the female of Papilio merope has followed those two local varieties, and has likewise a Cape and a Natal local form. Even this is not all, for in Cape Colony there are two other females of Papilio merope. One of them has a yellow ground-colour, and resembles Danais chrysippus, which is extremely abundant there (Pl. I, Fig. 3); the other is entirely different (Pl. I, Fig. 4), for it closely mimics another Danaid occurring in the same districts of Africa, and also immune, Amauris niavius (Pl. I, Fig. 5), not only in the beautiful pure white and deep black of the wing surface, but also in the distribution of these colours to form a pattern.
We have thus in Africa four different females of Papilio merope, each of which mimics a protected species of Danaid. They are not always[Pg 95] locally separate, so that each is exclusively restricted to a particular region, for their areas of distribution often overlap, and, at the Cape for instance, one male form and three different forms of female have been reared from one set of eggs. In addition, we have the fact that between the two local forms of Danais echeria transition forms occur, and that the mimetic females of Papilio merope show the same transition forms locally, and we must admit that all these facts harmonize most beautifully with the selection interpretation, but defy any other. And that the last doubt may be dispelled, nature has preserved the primitive female form on the continent of Africa—namely, in Abyssinia, where, along with the mimetic females, there are others which are tailed like the males (Pl. I, Fig. 1), and are like them in form and colour, a few minor differences excepted.
Thus we have in Papilio merope a species which, in the course of its distribution through Africa, has scarcely varied at all in the male sex, but in the female has almost everywhere lost the outward appearance of a Papilio, and has assumed that of a Danaid, which is protected by being unpalatable, and not even everywhere the appearance of the same species, but in each place that of the prevailing one, and sometimes of several in one region. These females thus show at the present day a polymorphism which consists of four chief mimetic forms, to which has to be added the primitive form—that resembling the male. This has survived in Abyssinia alone, and even there it is not the only one, but occurs along with some of the mimetic forms.
To the question why only the females are mimetic in this and other cases, Darwin and Wallace have answered that the females are more in need of protection. In the first place, the males among butterflies are considerably in the majority, and, secondly, the females must live longer in order to be able to lay their eggs. Moreover, the females, which are loaded with numerous eggs, are heavier in flight, and during the whole period of egg-laying—that is, for a considerable time—they are exposed to the attacks of numerous enemies. Whether one of the abundant males is devoured sooner or later is immaterial to the persistence of the species, since one male is sufficient to fertilize several females. The death of a single female, on the other hand, implies a loss of several hundred descendants to the species. It is, therefore, intelligible that, in species already somewhat rare, the female must first of all be protected; that is to say, that all variations tending in the direction of her protection would give rise to a process of selection resulting in an augmentation of the protective characters.
But there are also butterflies in which both sexes mimic a protected model. Thus many imitators of the unpalatable Acræides (Pl. II, Fig. 21) resemble the model in both sexes, and of the South American Whites which mimic the Heliconiidæ there are some which have the appearance of the Heliconiidæ even in the male sex (Pl. II, Fig. 18, 19), while others look like ordinary Whites (for instance, Archonias potamea). But in many of these species, which are mimetic in the female sex, we find also in the male some indications of the mimetic colouring, but in the first instance only on the under surface. Thus the females of Perhybris pyrrha (Pl. II, Fig. 17) resemble in their black, yellow, and orange-red colour-pattern the immune American Danaid, Lycorea halia (Pl. II, Fig. 12), but their mates are, on the upper surface, like our common Whites, though they already show on the under surface the orange-red transverse stripes of the Lycorea (Pl. II, Fig. 16). In other mimetic species of Whites a similar beginning is even more faintly hinted at, and in others, again, the upper surface of the male is also provided with protective colours, and only a single white spot on the posterior, or sometimes even on the anterior wing as well, shows the original white of the Pieridæ (Fig. 18).
I do not know how any one can put any other construction on these facts than that the females first assumed the protective colouring, and that the males followed later, and more slowly. Whether this is due to inheritance on the female side, and thus ensues as a mechanical necessity, in virtue of laws of inheritance still unknown to us, or whether it arose because there was a certain advantage in protection to the males—though not such a marked one—and that these, therefore, followed independently along the same path of evolution as the females, has yet to be investigated. Personally, I incline to the latter view, because there are protected mimetic species, in which the female mimics one immune model, and the male another, quite different from the female's. A case in point is that of an Indian butterfly, Euripus haliterses, and also Hypolimnas scopas, in the latter of which the male resembles the male of Euplœa pyrgion, and the female is like the somewhat different female of the same protected species. The Indian Papilio paradoxus, too, seems to show the independence of the processes of mimetic adaptation, for the male is like the blue male of the immune Euplœa binotata (Pl. III, Fig. 25), while the female resembles the radially-striped female of Euplœa midamus (Pl. III, Fig. 27), and this double adaptation is repeated in another of the persecuted butterflies, Elymnias leucocyma (Pl. III, Fig. 26, 28).
Many objections have been made to the interpretation of mimicry[Pg 97] by selection. It has been asserted that butterflies are exposed to injury from birds only to an inconsiderable extent, not sufficient to account for such an intense and persistent process of selection, because they are not very welcome morsels, on account of the large and uneatable wings and the relatively small body. Doubt has also been raised as to the immunity of the models, which has not been proved in many of the species in regard to which it is assumed. Finally, it is maintained that the advantage which resemblance to an immune model brings is not proved, but is purely hypothetical; and that it is probable that the birds do not distinguish the colours and markings of the flying butterflies at all, but are at the most only deceived by resemblances in their manner of flight.
The last objection contains a certain amount of truth, inasmuch as the manner of flight always plays a part in the mimicry of a strange species. We shall see later how much the instincts of a species contribute to the deception in all cases of protective colouring. It is, therefore, not improbable that, in many cases, the imitation of the flight of an immune species, and a gradually increasing familiarity with the habitats of the same immune species, preceded the modification of the colour. Indeed, the slow flight of immune species (Heliconiidæ) has been unanimously emphasized by observers, as a factor in facilitating the recognition of the butterflies by the sharp-sighted birds.
That it was not only in earlier ages of the world's history that butterflies were much persecuted, as some have supposed, but that they are so still, seems to me indisputable in view of the observations of the last quarter of a century. Even in this country, where both butterflies and insect-eating birds are being more and more crowded out through cultivation, a considerable number of butterflies in flight fall victims to the birds. Kennel gives observations on this point in regard to the white-throat; Caspari for the swallows. The latter let about a hundred little tortoiseshell butterflies (Vanessa antiopa) fly from his window, 'but not ten of them reached the neighbouring wood,' all the rest being eaten by swallows, 'which congregated in numbers in front of his window.' Kathariner observed, in the highlands of Asia Minor, a flock of bee-eaters (Merops) which caught in flight and swallowed a great many individuals of a very beautiful diurnal butterfly (Thais cerisyi).
Finally, Pastor Slevogt has collected much evidence to show that our indigenous butterflies have a great deal to suffer in the way of persecution from birds. And in regard to tropical countries, the chase of butterflies by insectivorous birds has long been known.[Pg 98] Thus Pöppig says that in the primitive forests one can easily recognize the place which has been selected by one of the Jacamars (Galbulidæ) as its favourite resting-place, for the wings of the largest and most beautiful butterflies, whose bodies alone are eaten, lie on the ground in a circle for a distance of several paces. We owe direct observations on the hunting of insects by birds of the primitive forest especially to Dr. Hahnel, who found many opportunities for observation in the course of his enthusiastic collecting journeys in Central and South America. He writes: 'No other family of butterflies suffered so much from birds as the Pieridæ (Whites), and these freebooters often snapped away the prettiest and freshest specimens from quite close to me. Every time I was amazed anew at the unfailing security of their flight, and I gladly paid for the spectacle by the loss of a few specimens.' Of the pursuit of one of the large Caligo species, whose leaf-like under surface, marked with eye-spots, I have already described, (Fig. 6, p. 70), he says: 'With incredible skill this fairly large insect avoided every blow of the bill of the bird which followed it in close chase, and saved itself by flying from one shrub to another, till at last it was lost to sight in the thickest tangle of branches, and the exhausted bird gave up further attempts at pursuit.'
But, in addition to the birds, the butterflies of the primitive forest have to dread the persecution of other insects, especially of the large predaceous dragon-flies, which throw themselves upon them in the midst of their flight. Hahnel often saw a specimen of the large, beautiful, blue Morpho cisseis, which was fluttering peacefully about the crown of a tree, suddenly shoot head downwards, 'like an ox with horns lowered, and then reascended apparently with difficulty, after it had torn itself free from its sudden assailant, whose jaws left distinct short scars.'
In addition to birds and predatory insects the butterflies are persecuted by the whole army of lizards. In order to entice the butterflies, Hahnel laid bait in the wood, 'sugar-cane, little sweet bananas, and such like.' Various kinds of butterfly settled on it, 'Satyrides, Ageroniæ, Adelpha and other Nymphalidæ.' He saw that they 'were persistently stalked and attacked by greedy lizards, which, in spite of their plump figure and uncouth gait, showed themselves able to spring suddenly out and snatch their prey with great adroitness. It is, however, very wonderful to see the agility such a persecuted insect displays in evading the repeated attacks of these marauders.' Thus on one occasion an Adelpha was driven off a dozen times from the exposed bait by a lizard, which pounced upon[Pg 99] it, but it always settled down for a short time on a leaf, and soon returned to its repast, whereupon the enemy 'instantaneously rushed upon it in a fury, until at last he was obliged to give in,' abandoning the attempt to catch a creature so adept in retreat.
Many butterflies assemble at midday on sandbanks in the middle of the river, in order to drink, and there, too, the lizards are always lurking about. Hahnel gives a pretty and undoubtedly accurate description of the protective value of the long tail borne by many of the sail-like Papilios at the end of the posterior wing; they 'quite obviously' afford protection against the lizards, 'which, after snapping, often find themselves obliged to be content with the tail alone, while the rest of the animal flies away practically uninjured.'
Not only is the great persecution of the butterflies a fact, the immunity of the known species, which are models for mimicry, is also certain. For numerous species, at any rate, this has now been established. First of all—as has already been said—this is true of the Heliconiidæ, in regard to which Wallace long ago showed that, if the thorax be pressed, they exude a yellowish juice of unpleasant smell. This is probably the blood of the insect, but that does not hinder the repulsive odour of the living butterfly being perceptible at a distance of 'several paces,' as Seitz observed in Heliconius besei.
Repeated experiments have been made, which have shown that such butterflies are rejected not only by the insectivorous birds of the primitive forest, but also by tame turkeys, pheasants and partridges, usually so greedy. Hahnel has recently repeated these experiments in Brazil with hens, and he obtained the same result. The hens, 'which otherwise devoured all butterflies eagerly,' rejected all Ithomidæ, Heliconiidæ, the white Papilios, as also some of the gaily coloured Heliconiid-like moths which fly by day, such as Esthema bicolor and Pericopis lycorea. Obviously, the gay or conspicuous colour of these Lepidoptera acts as a warning signal of their unpalatability, and protects them from attempts on the part of the birds to investigate their flavour. Hence we find that the under surface of these insects is coloured like the upper. Even the numbers of these species which fly about indicates that they must be little decimated, and, in point of fact, we never find the wings of Heliconiidæ lying on the ground in the forests of South America, while those of the Nymphalidæ and other butterflies are by no means uncommonly seen as the remains of birds' meals.
There is just as little room for doubt, as in the case of the Heliconiidæ and their allies, that the Danaidæ, Acræidæ, and the[Pg 100] Euplœidæ in the tropical regions of the Old World enjoy a certain immunity on account of their repulsive odour and taste. Here, too, observation and experiment have shown that birds, lizards, and predaceous insects leave the butterflies of these families unmolested. I need only mention the observation of Trimen that, under an acacia much visited by butterflies, on which Mantides—the so-called praying-insects—caught and devoured large numbers, the wings of an Acræa or a Danais were never found. These unpalatable butterflies also possess a motley or at least striking dress, recognizable from afar, and alike on both surfaces; and they also have a slow flight, by which they are readily recognized. They, too, usually assemble in large swarms, and both sexes are alike, or resemble each other closely in colouring, or at least they are both equally conspicuous. But even these cases do not complete the list of butterflies which are protected by their unpalatability; among the otherwise much-persecuted and therefore palatable Pieridæ (Whites) there is an Asiatic genus, Delias, which in all probability belongs to the immune butterflies, as their gaily coloured under surface indicates, and among the nocturnal Lepidoptera of different countries and families there are isolated generations which are very gaily and conspicuously coloured, and which are rejected by birds, their unpleasant odour being perceptible at a distance of several feet (Chalcosiidæ and Eusemiidæ). The latter no longer fly under cover of night, like their relatives, but have assumed diurnal habits.
It is to be supposed that the repulsiveness of such 'unpalatable' butterflies is associated with the food-plant on which the caterpillar lives. Acrid, nauseous, astringent, and actually poisonous substances are produced in many plants, and we shall see later that this is to their own advantage; these substances pass into the insect, and they do so probably in part unaltered, in part certainly altered, but still they are protective, perhaps even in an increased degree. This is borne out by the fact that many caterpillars of immune butterflies live on more or less poisonous plants: the Acræidæ and Heliconiidæ on Passiflores, which contain nauseous substances; the Danaidæ on the poisonous Asclepiadæ, which are rich in milky juice or latex; the Euplœæ on the poisonous species of Ficus, the Neotropinæ on the Solanaceæ, and so on. But there are many genera, rich in species, and distributed over the whole earth, the caterpillars of which live on plants of very various families and characters, and of these the majority of species are palatable, though a few are repulsive in taste and odour, and therefore immune. This is the case in the genus Papilio. As far back as the sixties Wallace discovered that there were immune[Pg 101] species of Papilio, and that these were mimicked by other species. Later it was shown that these immune species live chiefly on poisonous plants (in the wide sense), on various Aristolochiæ; and Haase has recently grouped these together as poison-eaters (Aristolochia-butterflies or Pharmacophagæ). They are distinguished by a conspicuous red on the body. In some of them, as in Papilio philoxenus, a repulsive odour as of decomposing urine has been detected in the living animal.
We see, then, that the much-persecuted and easily injured butterflies make use of a poisonous substance (in the widest sense), prepared in the plant for its own protection, and, wherever their own metabolism makes it possible, they use it to protect themselves. We need not wonder, therefore, that so many butterflies are immune, nor that among the numerous palatable species a small proportion have endeavoured to become like the protected species, as far as natural selection was able to bring such a resemblance about.
There is hardly any adaptation phenomenon so widely distributed and diverse in its manifestations, which has been at the same time so much observed and followed out into all its details, as Mimicry; and it must surely be regarded as a justification of the validity of interpreting it in terms of Natural Selection that all the observed phenomena tally so beautifully with the deductions from the theory. I at least know of no facts which contradict the theory, but of many which might have been predicted from it.
For instance, it might have been predicted from the theory alone that an immune species would often have several mimics, as, in point of fact, is frequently the case, and it would be easy to give numerous examples of this. Thus the two Danaids of South and Central Africa, Amauris echeria and Amauris niavius, are mimicked, not only by the two female forms of Papilio merope, as we have already described in detail, but the latter is also mimicked by Nymphalid, which requires protection, Diadema anthedon, and the former by two diurnal butterflies of different families, Diadema nuina and Papilio echerioides.
Similarly, the black-and-red coloured Heliconius melpomene in Brazil is mimicked both by the female of a White (Archonias teuthamis), and by a Papilio, which has received the name of P. euterpinus on account of this resemblance. Thus, too, the immune Methona psidii, Cr. of Brazil, with its half-transparent wings marked with black bands, has five mimics, belonging to five different genera, and one of these is not a true diurnal butterfly at all,[Pg 102] but one of the day-flying species of the genus Castnia, whose systematic position is doubtful.
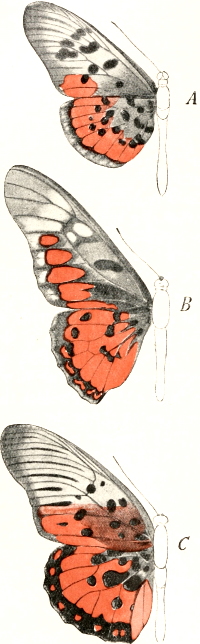
Fig. 18. Upper surfaces of A, Acræa
egina, from the Gold Coast, immune.
B, Papilio ridleyanus, from Gaboon, not
immune. C, Pseudacræa boisduvalii,
from the Gold Coast, not immune.
The West African immune Acræid, Acræa gea (Pl. II, Fig. 21), is deceptively mimicked, both as to the narrow, long shape of the wing and its blackish-brown and white mottled markings, by a Nymphalid, Pseudacræa hirce, by the female of a Papilio (P. cynorta) whose mate is quite different, and by the female of a Satyrid (Elymnias phegea) (Pl. II, Fig. 20). In the Papilio the resemblance extends to the peculiar pitch-black shining spot on the under side of the base of the posterior wing, and all three are like the model on both surfaces, and therefore in flight as well as in the resting attitude.
On the same West African coast occurs the strange greyish-black Acræa egina, with brick-red spots and bands, and coal-black dots (Fig. 18, A). This immune species is deceptively mimicked in its native country by two other butterflies—a Nymphalid, Pseudacræa boisduvalii (Fig. 18, C), and by a female Papilio (P. ridleyanus) (Fig. 18, B), by the latter not so exactly as by the former, but quite sufficiently to be confused with its model in flight.
It would have been less easy to predict with certainty from the theory that, conversely, the different species of a genus which stood in need of protection would be able to mimic quite different immune models, for who would have ventured to prophesy how far the capacity of a species for variation might go, and how many different kinds of coloration it was able to assume? But the facts teach us that there is a wide range of possibility in this respect.
Most interesting in this respect is, perhaps, the Asiatic-African genus Elymnias, a Satyrid whose numerous (over thirty) species all seem to be in need of protection, for many of them mimic immune butterflies, while the rest are inconspicuous and are provided with protective colouring on the under surface. On Plates II and III some of the former are depicted beside their models. The single African species (Elymnias phegea) (Pl. II, Fig. 20) mimics, as has been already mentioned, the prevalent Acræa gea (Pl. II, Fig. 21). Many of the Asiatic Elymniidæ are mimics of the immune Euplœæ, especially the dark-brown species with steel-blue shimmer, such as E. patna in India, E. beza in Borneo, and E. penanga in Borneo. In Amboina there flies an E. vitellia, the female of which mimics accurately the plain, light-brown, inconspicuous Euplœa climena which occurs there. The male of Elymnias leucocyma (Pl. III, Fig. 26) resembles the brown and blue shimmering Euplœa binotata (Pl. III, Fig. 25), while the female mimics the dusky, radially-striped female of Euplœa midamus (Pl. III, Figs. 27 and 28): the male of Elymnias cassiphone resembles the blackish-brown and deep-blue iridescent Euplœa claudia, while the female is like the female of Euplœa midamus. A number of species of Elymnias copy Danaids: thus both sexes of E. lais are like Danais vulgaris (Pl. III, Figs. 29 and 30), and E. ceryx and E. timandra are like another similar Danaid, D. tytia. The female only of E. undularis of Ceylon mimics the brown-yellow D. genutia (Pl. II, Fig. 22) in general appearance, though not minutely, while the male (Pl. II, Fig. 24) seems to attempt an imitation of the blue Euplœæ. A rare form, not often represented in collections, Elymnias künstleri, bears a striking resemblance to the Danaid, Ideopsis daos Boisd., with its white wings spotted with black, while three species mimic the probably immune Pierid genus Delias, especially on the under surface, which is decorated with yellow and red. Perhaps the one which has diverged farthest from the original type is Elymnias agondas Boisd. (Pl. II, Fig. 32) of the Papua region and the island of Waigeu, for it bears two large blue eye-spots on the posterior wings, and thus, especially in the case of the almost white female, closely resembles Tenaris bioculatus (Pl. III, Fig. 31). There are thus seven or eight types of marking and colouring differing from one another, and belonging to six different genera and a much greater number of species, which are mimicked by this one genus Elymnias.
It is most interesting to note how these mimetic species give up, more or less, the original sympathetic colouring of the under surface, and use in establishing their mimicry the marking elements[Pg 104] which were originally directed towards concealment. According to the beautiful observations of Erich Haase on this genus Elymnias, the ground-colouring on the under surface must have been 'a grey, darkly mottled protective one,' as still occurs, for instance, in several mimetic species, such as Elymnias lais (Pl. III, Fig. 30). This leaf-colouring disappears more and more the more perfect the mimicry of the model becomes, until, finally, the model is repeated on the under surface also. Compare, for instance, Figs. 30 and 32. From this we may conclude that a dress which makes Lepidoptera appear unpalatable morsels is a more effective protection than resemblance to a leaf. That might indeed be deduced even from the theory, for resemblance to a leaf never protects absolutely, and does so, in any case, only during rest, while apparent unpalatability repels assailants at all times.
Those unversed in butterfly lore usually ask, when these mimetic relations are expounded to them, how we know that copies which are so like their models really belong to a different genus, or even family. There are certainly cases in which model and copy resemble each other so closely that even a zoologist cannot tell one from the other without close examination, as, for instance, in the case of certain transparent-winged Heliconiidæ of Brazil (Ithomiides) and their mimics belonging to the family of Whites. But even in such cases the likeness only extends as far as is theoretically requisite, that is, only to those characters that make the butterfly appear to the eye of its pursuer like another species, known to it to be unpalatable. The likeness does not extend to details, which can only be seen with a magnifying-glass or a microscope, and above all, it does not extend to the caterpillar, pupa, or egg. Thus, in the case cited, we may be certain that the caterpillar of Ithomia is quite different from that of the mimicking White, since the former will be, in structure, of the type of Ithomia caterpillar, and the other of the usual type of Whites. As yet, indeed, these two species are not known in their caterpillar stages, but other cases are known. A species belonging to the same genus as our indigenous 'kingfishers' (Limenitis populi), a diurnal butterfly of North America, Limenitis archippus (Pl. I, Fig. 9), strongly resembles the brown-yellow, immune Danais erippus (Pl. I, Fig. 8), while the caterpillars of both species are quite different, that of Danais erippus possessing the remarkable, soft and flexible horn-like processes of the Danaid caterpillars (Pl. I, Fig. 10a), while the caterpillar of Limenitis archippus (Pl. I, Fig. 11a) is at once recognizable by its blunt, club-shaped and spinose papillæ as a Limenitis caterpillar. The adaptation of the butterfly to its protected model has[Pg 105] thus exercised no influence upon the caterpillar. Nor has it affected the pupa, which in both cases exhibits the very different and quite characteristic form of the Danais pupa and the Limenitis pupa respectively (Pl. I, Fig. 10b, and 11b).
But even in the butterfly itself nothing is altered, except what increases the resemblance to the model. All else has remained unchanged, above all, the venation of the wings. Since the painstaking and valuable work of Herrich-Schäfer the venation has been made the basis of the whole systematic arrangement of butterflies, and it enables us, in point of fact, to distinguish with precision, not the families alone, but often even the genera, for the course of the veins in the different species of a single genus is the same, and that is true for the mimetic species as well as for others. Thus the Danaid-like Limenitis has the usual Limenitis venation, of the kind seen in our own indigenous species of Limenitis, and the already described Elymnias species of the African and Indian forests and grassy plains have always the venation characteristic of this genus, whether they be protected only by sympathetic colouring or imitate an immune Euplœa, a Danais, an Acræa, or a Tenaris. However much the contour of the wing may vary, the venation is unaffected, and we can distinguish model from copy by this means alone, so that, even when there is the closest resemblance, no doubt is possible. In its theoretical aspect this constancy of venation is obviously important, for as nothing about the organism is incapable of variation, the veining of the wings might have varied, as indeed it has varied from genus to genus in the course of the phylogenetic history; but as changes in venation could not be detected by the butterflies' enemies, however sharp-sighted, there has been no reason in these cases for variation in this respect.
In this connexion Poulton has brought forward interesting facts showing that the mimics of one model, belonging to different genera, often secure the same effect in quite different ways. Thus the glass-like transparency of the wings in the Heliconiidæ of the genus Methona depends on a considerable reduction of the size of the scales, which ordinarily cover both sides of the wing as thickly as the tiles on a roof, and produce the colour. In another quite similar species, also transparent-winged, the Danaid Ituna ilione, the transparency is due to the absence of most of the scales, and in a third mimic, Castnia linus, var. heliconoides, the scales are not altered either in size or number, but have become absolutely unpigmented and transparent. In a fourth mimic, a Pierid, Dismorphia crise, the scales have not decreased in number, but have become quite minute, while[Pg 106] in a fifth case, the nocturnal Hyelosia heliconoides Swains., the same thing has happened as in Castnia, but the scales are also fewer in number. Thus in each of the mimics the changes which have taken place in the scales are quite different, but they bring about the same effect, the glass-like transparency of the wings, on which the resemblance to the model depends: what we have before us is, therefore, not a similarity of variation, but only an appearance of similarity in external features.
In the face of such facts there can be no further question of the often repeated objection, that the resemblance of model and copy depend on the similarity of external influences upon species living in the same latitude, even if that were not already sufficiently refuted by the frequent restriction of the mimicry to the female. And that mimicry should be a mere matter of chance is negatived even by the single fact that model and copy always live in the same area, and that the local varieties of the model are faithfully followed by the mimic. An interesting example of this is furnished by Elymnias undularis, already mentioned, for in this case the female (Pl. II, Fig. 23) mimics the brown-yellow Danais plexippus (Pl. II, Fig. 22), not wherever E. undularis occurs, but only in Ceylon and British India. In Burmah, where another Danaid, D. hegesippus, is common, it mimics that; and in Malacca it does not copy a Danaid at all, but resembles the male of its own species, which in India is very different from it, since there the female mimics one of the blue iridescent Euplœæ (Pl. III, Fig. 24). It cannot therefore be a matter of 'chance,' and we should have to give up all attempt at a scientific interpretation if we were not prepared to accept that of natural selection. Even the interference of a purposeful Power can hardly be seriously considered in this case, even by those who are inclined to such a view, for the gradual approximation to the model, which is a matter of course in a process of evolution, could only appear, if referred to the benevolent intelligence of a Creator, as an unworthy trick, designed to lead humanity astray in its strivings after knowledge. On the other hand, this gradual increase of resemblance, which becomes apparent when we compare several mimetic species—this carrying over, step by step, from the female to the male—and many other facts point to the working of natural forces according to law, and, if there is to be found anywhere in living nature a complicated process of self-regulation, it certainly lies before us here, clearer and less open to objections than almost anywhere else. I do not mean to say, however, that we can verify it statistically in detail, as has been demanded by the fanatical opponents of natural selection. A direct testing of[Pg 107] natural selection is, as has been already shown, nowhere possible: we can never exactly estimate how great the advantage is which a species requiring protection derives from a slight increase in the resemblance to an immune model; and I for one do not know how we could even definitely prove that a certain species needed a greater degree of protection than it had previously enjoyed in order to ensure its persistence in the struggle. It would be necessary to know the total number of individuals living on a certain area for many generations. If it appeared that there was a progressive diminution in the number of individuals, we should be justified in concluding that the species had not an adequate power of persistence, and that it therefore required a more effective protection. But it is impossible for us to collect such exact data for any species living under natural conditions, although we can often say approximately that a species is progressively decreasing in numbers. Even this, however, we can usually do only in cases which are influenced directly or indirectly by the interference of Man in nature, and in which the falling off in the species occurs so rapidly that there is no time for the slow counteractive influence of natural selection. We shall see later that in this way many species have been eliminated even within historic times.
I have just spoken of the 'need of protection,' and I have a few remarks to add on that subject. It is a mistake to believe that every 'rare' species, that is, one represented by few individuals, is already in process of disappearing. It is not the absolute number of individuals that determines the survival of a species, but the fact of the number remaining the same. It is equally mistaken to suppose that an amelioration of the conditions of existence for any species by natural selection is possible only when its persistence is already threatened; that is, when the number of individuals (the 'normal number') is steadily decreasing. On the contrary, it is of the essence of natural selection that every favourable variation which crops up is, ceteris paribus, preserved, and becomes the common possession of the species, quite independently of whether this improvement is absolutely necessary to its preservation or not. In the latter case it will simply become a commoner species instead of a rare one; and every species is, so to speak, striving to become common and widely distributed, since every advantageous variation that can possibly be produced is accumulated and made the common property of the species. But this has its limits, not only in the constitution and the structure of each species, but also in the external conditions of its life. If a species of butterfly be restricted, in the caterpillar stage, to a single, rare species of plant, its normal number will be, and must remain,[Pg 108] a small one. But if there arise within it a variation in the food-instinct whereby a second and it may be a commoner plant becomes available, then the normal number of the species will rise, and perhaps the original number of individuals may be more than doubled. It is, however, by no means necessary to assume that the species was previously in process of decadence; on the contrary its normal number may have remained quite constant.
So, in the case of the mimetic butterflies, we do not need to assume that they all previously required protection in the sense that they would have become extinct had they not assumed a likeness to an immune species. We may indeed conclude, on other grounds, that it was the rarer species which increased their number of individuals by the mimetic protection, and in doing so they certainly enhanced at the same time their chance of survival as a species. In the more abundant species mimetic resemblance to species whose unpalatability rendered them immune could not have been evolved, as it would have been disadvantageous, not only for the model, but for the mimicking species itself, while in species less rich in individuals, such resemblance would necessarily have a protective value, no matter whether the species was in danger of extinction or not. The process of selection must have started simply because the mimetic individuals survived more frequently than the others, and the mimetic resemblance must have gone on increasing as long as the increase brought with it a more effective protection. It is, therefore, a fallacious objection to say that a species, whose existence was threatened, would, considering the slowness of the process of selection, have died out altogether before it could have acquired effective protection by mimicking an immune species. The assumption is false—the widespread, hazy idea that the process of natural selection can only begin when the existence of the species is threatened. On the contrary, every species utilizes every possibility of improvement; and every improvement for which variation supplies the necessary material is possible. The augmentation of the profitable variations follows as a necessity from the more frequent survival of the best-adapted individuals, and this 'more frequent survival' will be not only a relative one, due to the fact that the better adapted individuals will be less decimated, it will also be absolute, because more individuals of the species will survive than before. Of this Papilio merope may serve as an example; in Madagascar it now flies about only slightly varied from the original form, var. meriones. Here, therefore, the species is maintained, without the aid of mimetic protection. We do not know if the reason for this lies in the absence[Pg 109] of an immune model, or in the non-appearance of suitable mimetic variants, or in other conditions; but we know that without mimicry the species holds its own against its enemies. But if, in Abyssinia, a female of this butterfly exhibited variations which would make her resemble, in any degree, the unpalatable Danias chrysippus, these mimetic variants would be less decimated than the original form of female, and would, therefore, gain stability, and gradually increase both in mimetic resemblance and in the number of individuals. But is this any reason why the original form of the female should diminish in numbers? In itself, certainly not; the red mimetic females could increase in number without causing any decrease of the yellow ones, for the red are in no way in conflict with the yellow, and we must not think of the number of individuals as so fixed for each species that it cannot increase. On the contrary, it must increase, as soon as the conditions of existence are permanently improved, and this happens, in this case, through the mimetic protection of the red female. We can thus easily understand how mimetic and non-mimetic females can live side by side in Abyssinia.
In all the rest of Africa, however, there are only mimetic females of Papilio merope, and none of the colour of the male; these last, therefore, have been crowded out by the mimetic form, not actively, but through the more frequent survival of the mimetic form, so that those like the male became gradually rarer, and finally died out—that is, ceased to occur. The matter is not so simple as it seems, and we shall best understand it by thinking of the dimorphism of the caterpillars of our hawk-moths, which we discussed before, in which the green form in the full-grown caterpillar is less well protected than the brown. In many species the brown form has crowded out the green, in others brown and green occur side by side, but the green is less abundant, and in some species very rare. This must be regarded as the simple result of the circumstance that a higher percentage of the green than of the brown caterpillars fall victims to enemies, and thus, in the course of generations, the green form becomes slowly but steadily rarer. This will be the case even if the newer and better adaptation raises the number of individuals (the 'normal number') in the species, for this increase must always be a limited one, even if it be very great, which is hardly likely in this case. For the normal number is not determined by the mortality at one stage, but by that at all the stages of life taken together. Thus a normal number always persists, notwithstanding the improved conditions for the species, and, on this assumption, the form under less favourable conditions cannot permanently hold its own with that under better con[Pg 110]ditions, but must gradually disappear. We can understand, then, that the primitive form of the Papilio merope female may persist even for a long time side by side with the mimetic form in certain habitats. It is, probably, not a mere chance, that this should have happened just in Abyssinia, for, in that region, the mimetic female is still tailed—that is, she has not yet reached the highest degree of resemblance to her immune model. In the whole of the rest of Africa the process of the transformation of the female has already reached its highest point, and on the east and west coasts, as well as in South Africa, the primitive form of the species is now represented only by the male.
The gradual dying out of the less favourably conditioned forms of a species is a law which follows as a logical necessity from the essence of the process of selection, but its reality may be inferred from the phenomena themselves. On it depends, as far at least as adaptations are concerned, the transformation of species.
A beautiful example of the crowding out of a less favoured form of a species by a more favoured one is afforded by a butterfly of North America, of which the two female forms have long been known, although the reason for their dimorphism was not understood. A yellow butterfly, Papilio turnus, not unlike our swallow-tail, has yellow females in the north and east of the United States, but black ones in the south and west. There was much guessing as to what the cause of this striking phenomenon might be, and it was for a time thought that this difference was directly due to the influence of climate, and, later, the black form of female was regarded as protectively coloured, because of the supposed greater persecution by birds in the south, since the female would be less easily recognized if of a dark colour, and would thus be better protected. This last explanation could hardly be looked upon as satisfactory, for a black butterfly in flight would be very easily seen by sharp-sighted birds; indeed, against a light background, it would be even more readily seen than a light one.
Since we have acquired a more exact knowledge of the immune species of Papilio this case has become clear to us. For on those stretches of country on which the black female of Papilio turnus lives there occurs another Papilio which is black in both sexes, Papilio philenor, and this is one of those species which are protected by their unpleasant taste and odour. Here, therefore, we have a case of mimicry, the female of Papilio turnus imitates the immune Papilio philenor, and thereby secures protection for itself; but as the immune model only occurs in the southern half of the distribution of Papilio [Pg 111]turnus a somewhat sharp separation of the two forms of female has been evolved; the black, mimetic form, being the most fit, has completely crowded the primitive yellow form out of the area inhabited by Papilio philenor, while beyond this area, to the north and west, the yellow form alone prevails. The extensive and careful studies of Edwards have shown that the two forms occur together only in a very narrow transition region.
We thus see that the facts, wherever we scrutinize them carefully, harmonize with the theory. Of course we can only penetrate to a certain depth with the theory of selection, and we are still far from having reached the fundamental causes of the phenomena. Indeed, our understanding must in the meantime stop short before the causes of variations and their accumulation, but up to that point the theory gives us clearness, and discloses the causal connexion of phenomena in the most beautiful way. Although we do not yet understand how the southern female Papilio turnus was able to produce the advantageous black, we do see why a black variation, when it did occur, should increase and be strengthened, until it crowded out the yellow form from the area of the immune model, and we are able in a general way to refer the whole complicated phenomena of mimicry to their proximate causes.
This is true also of other phenomena which have had no part in establishing the theory, since attention was only directed to them later, and it is true even of some which, at first sight, seem to contradict the theory altogether. To this class belongs, for instance, the phenomenon that immune species not unfrequently mimic each other, as was first observed among the Heliconiid-like butterflies of South America. In four different families, the Danaidæ, the Neotropidæ, the Heliconiidæ, and the Acræidæ, there are species, distributed over the same area, which resemble each other in their conspicuous colouring and marking, and also in the peculiar shape of the wings. After what has been said one might be inclined to regard one of these species as the unpalatable model and the others as the palatable mimics, but they are all unpalatable, and are not eaten by birds. The puzzle of this apparent contradiction was solved by Fritz Müller[4], who pointed out that the aversion to non-edible butterflies is not innate in birds, but must be acquired. Each young bird has to learn from experience which victim is good to eat, and which bad. If every inedible species had its particular and distinctive colour-dress a considerable number of individuals of each species would fall victims to the experiments of young birds in each generation, for a butterfly[Pg 112] which has once been pecked at, or squeezed by the bill of a bird, is doomed to die. But if two inedible species which resemble each other inhabit the same area they will be regarded by the birds as one and the same, and if five or more inedible species resemble each other all five will present the same appearance to the bird, and it will not require to repeat on the other four the experience of unpalatability it has gained from one. Thus the total of five species will be no more severely decimated by the young birds than each of them would have been if it had occurred alone; the same number of victims of experiment, which are necessary every year in the education of the young birds, will, when all five species look alike, be divided among the whole 'mimicry ring,' as we may say. The advantage of the resemblance is thus obvious, and we can understand why a process of selection should develop among such inedible species which should result in their being readily mistaken for one another; we can understand why, in the neighbourhood of Fritz Müller's home, Blumenau, in the province of Santa Catarina in South Brazil, the Danaidæ, species of Lycorea; the Heliconiidæ, Heliconius eucrate and Eueides isabella; and the Neotropinæ, Mechanitis lysimnia and species of Melinæa, should all exhibit the same colours, brown, black and yellow, in a similar pattern, on similarly shaped wings. The agreement is by no means perfect in detail, but it can be noticed in all parts of South America inhabited by species of these genera, and the same differences which distinguish, for instance, the two species of Heliconius flying in two different regions, also distinguish the two species of Eueides and the two species of Mechanitis. In Honduras we find the same mutually protective company of inedible genera as in Santa Catarina, but represented by other species, which all differ from the species in Santa Catarina in the same characters, as, for instance, that they have two instead of one pale yellow cross-stripe on the anterior wings. The species are: Lycorea atergatis, Heliconius telchinia, Eueides dynastes, Mechanitis doryssus, and Melinæa imitata[5]. In the environs of Bahia this mimicry ring consists of the following species: Heliconius eucrate, Lycorea halia, Mechanitis lysimnia, and Melinæa ethra, as figured on Pl. II, Fig. 12, iv, and such a mutual assurance society has always one or other edible species as mimic. The larger the mimetic assurance company is, the less harm can mimics do to it. In the case figured it is two Pieridæ already known to us that have fairly well assumed the Heliconiid guise, namely, Dismorphia astynome [Pg 113](Pl. II, Figs. 18 and 19) and Perhybris pyrrha (Pl. II, Figs. 16 and 17). In the latter of these the male still has, on the upper surface, just the appearance of one of our common Garden-whites, while the female is coloured quite like the Heliconiidæ, but without having lost the form of wing of the Whites. The larger the mimetic company is the greater will be the protection afforded to its palatable mimics, since they will be the more rarely seized by way of experiment. It is, of course, obvious that in this kind of mimicry—that is, in the imitation of an unpalatable and rejected species for protection—it is presupposed as a general postulate that the edible mimics are considerably in the minority, as Darwin showed; for if it were otherwise their enemies would soon discover that among the apparently unpalatable species there were some which were pleasant to taste. Here, too, the facts bear out the theory, although exceptions can easily be imagined, and do seem to occur.
[4] Kosmos, vol. v, 1881, p. 260 onwards.
[5] According to Poulton's report in Nature, July 6, 1889, of 'Sykes, Natural Selection in the Lepidoptera,' Trans. Manchester Microscop. Soc. 1897, p. 54.

PLATE I

| FIG | |
| 1. | Papilio merope, male, Africa. |
| 2. | The same species, one form of mimetic female. |
| 3. | Danais chrysippus, Africa, immune model of Fig. 2. |
| 4. | Papilio merope, second form of mimetic female, S. Africa. |
| 5. | Amauris niavius, S. Africa, immune model of Fig 4. |
| 6. | Papilio merope, third form of mimetic female, S. Africa. |
| 7. | Amauris echeria, S. Africa, immune model of Fig. 6. |
| 8. | Danais erippus, immune model of Fig. 9, Central N. America. |
| 9. | Limenitis archippus, Central N. America, mimics the foregoing species. |
| 10. | Danais erippus, (a) CATERPILLAR, (b) PUPA. |
| 11. | Limenitis archippus, (a) CATERPILLAR, (b) PUPA. |
To face Plate I
This comparative rarity is true of the imitators of the Heliconiidæ and their great mimicry ring of unpalatable species, and is very general. Thus, for instance, there is a series of palatable mimics of the beautiful blue Euplœæ of the Indo-Malayan region (Pl. III, Figs. 25 and 27), but each of these mimics is rare compared with the hosts of the blue unpalatable company, for these immune butterflies also occur in many species, all similar to Euplœa midamus or binotata (Pl. II, Figs. 1 and 3); and the same applies to the mimics of the Indo-Malayan Danaidæ. There are a great many Danais species, all of them resembling Danais vulgaris (Pl. III, Fig. 20), which, when they occur together, form an inedible ring, and this ring is imitated by a whole series of edible species, each of which is comparatively rare. And there are no fewer than six species of Papilio which resemble these Danaids to the point of being easily mistaken for them, while another rare Papilio effectively copies the iridescence of the blue Euplœæ—a coloration so unusual in the genus that the species has received the name of Papilio paradoxus.
But even in single species of butterflies immune through unpalatability there is usually a great abundance of individuals. Thus Danais chrysippus, which is distributed over the whole of Africa, is a very common butterfly wherever it can live at all; and in North America, in which country there are only two widely distributed species of Danais, these often occur in enormous numbers. The beautiful large Danais erippus Cramer (Pl. I, Fig. 8), is distributed over almost all America, and in many places is not only frequent, but occurs in great swarms. Usually it peoples the broad, open stretches of the western prairies of the United States, but when[Pg 114] violent winds blow, as they do there in September especially, the insects are driven together into the small wooded spots of the prairie, and then they cover the trees in incredibly large crowds, often so thickly that the leaves are entirely hidden, and the trees look brown instead of green. Millions of butterflies go to make up such swarms, which have been observed in many parts of the United States, even quite in the East, in New Jersey, and elsewhere.
Considering this extraordinary abundance of the immune species, it is not surprising that its palatable copy, Limenitis archippus (Pl. I, Fig. 9), should also be widely distributed in North America, and in many places it is not rare, but even abundant. The enormous majority of Danais erippus will protect the species which resembles it so closely, even though it is not rare. Any doubt as to this being a case of mimicry disappears in face of the fact that, in Florida, there flies a second very similar but much darker brown North American Danais, and that it is accompanied there by an equally dark variety of Limenitis archippus (L. eros).
To prove the correctness of the hypothesis of an actual process of selection—which we assume in our interpretation of mimicry—I mean the assumption that the disguise of the species seeking protection really deceives the enemy, and thus actually affords protection, I need only cite the evidence of an acute and experienced entomologist who was himself deceived by it. Seitz[6], to whom we owe many valuable biological observations on butterflies, relates that, while he was collecting in the neighbourhood of the town of Bahia, he was surrounded by swarms of Catopsiliæ, similar to our lemon butterfly, especially the common Catopsilia argante, but he took no notice of these, as he 'had already collected as many of them as he wanted.' It was only when he saw a pair in copula that he caught them in his net. But to his extreme surprise he found that he had not caught a Catopsilia, but a butterfly of the family Nymphalidæ, one of those Anææ whose numerous species are distributed over South America. These Anææ are dark, or beautifully bright on the upper surface, but on the under side are leaf-coloured, and one of them bears the name Anæa opalina, because it is quite clear and pale, and of opal-like brilliance. The captive was nearly related to [Pg 115]this species. Seitz was so much surprised by the discovery that the male, which had quickly detached itself from the female, escaped him, and he could only make out that, 'as it flew away, it unfolded dark wings, which certainly bore little resemblance to those of the lemon butterfly.' In the hope of securing more of this rare booty he then hunted only for Catopsilia argante, without however securing another coveted specimen—he caught no more Anœæ, which shows that in this case, too, the mimetic species was much rarer.
[6] In citing this observation of Seitz, I do not mean to assert that there is true mimicry between Anæa opalina, or its allied species in Bahia, and the Catopsilia, though I regard this as extremely probable, because of the marked dimorphism between the male and the female, in conjunction with the very striking resemblance of the female to the Catopsilia. The example was given only to show how very deceptive such resemblances may be. To assert with confidence that it is a case of mimicry we should require to know that Catopsilia is immune, and on that point we have as yet no information.

PLATE II
| FIG. | |
| 12-15 | REPRESENT A 'MIMICRY-RING' COMPOSED OF FOUR IMMUNE SPECIES BELONGING TO THREE DIFFERENT FAMILIES AND FOUR DIFFERENT GENERA. |
| 12. | Heliconius eucrate, Bahia. |
| 13. | Lycorea halia, Bahia. |
| 14. | Mechanitis lysimnia, Bahia. |
| 15. | Melinæa ethra, Bahia. |
| 16, 17. | Perhybris pyrrha, male and female, S. American 'Whites' (Pieridæ). The female mimics an immune Heliconiid, while the male shows only an indication of the mimetic colouring on the under surface. |
| 18, 19. | Dismorphia Astynome, male and female, also belonging to the family of 'Whites,' and mimicking immune Heliconiids; a white spot on the posterior wing of the male is all that remains of the original 'White' coloration. |
| 20. | Elymnias phegea, W. Africa, of the family Satyrides, mimics the foregoing species. |
| 21. | Acræa gea, an immune W. African species. |
| 22. | Danais genutia, an immune Danaid from Ceylon. |
| 23. | Plymnias undularis, female, one of the mimics of Fig. 22. The male, which is quite different, is figured on Plate III (Fig. 24). |
To face Plate II
We see, then, that the need for protection in butterflies has a great influence on their external appearance, especially as regards their colour and marking. First, because the resting insect frequently has the visible surfaces sympathetically coloured, and also, because there are numerous species, indeed whole families, which contain nauseous, perhaps even actually poisonous, juices, and these have been subject to a double process of selection, directed towards the increase of the nauseousness, and at the same time towards acquiring as conspicuous a dress as possible. Thus the whole surface of these butterflies became gaily coloured, and often—as in many of the tropical nocturnal Lepidoptera which fly by day, the Agaristidæ, Euschemidæ, and Glaucopidæ—quite glaringly bright. We thus understand the striking or at least readily recognizable colours of the Heliconiidæ, the Euplœæ, the Danaidæ, and the Acræidæ. Finally, the unpalatable species influence many others which are edible, since the latter strive to resemble an immune species; and how considerable the variations and colour transformations thus induced can be is shown by the Whites of the genus Perhybris (Pl. II, Figs. 16 and 17) and Archonias, in which the male has wholly or partially retained the primitive dress of the Whites, and in which, side by side with wholly mimetic species, other species occur in which both sexes exhibit the garb of the Whites unaltered. Such cases tell decidedly against the often expressed view that mimetic species must have had from the outset a great resemblance to the model; they show rather that very great deviations in form, but more especially in colour, have been brought about solely by the necessity for mimetic adaptation, and that they have come about only slowly and step by step, as the different grades of resemblance to the model in different species of the same genus clearly show.
Lepidoptera are by no means the only insects which exhibit the phenomenon of mimicry, nor are insects the only animals in which it occurs; and unpleasant taste and odour are not the only protective characters; there are many others, as, for instance, among insects, the hardness of the chitinous cuticle.
One of the most beautiful examples of mimicry was discovered by Gerstäcker, not in free nature, but in the entomological collection at Berlin. There he found beside a green, metallic weevil-beetle, one of the Pachyrhynchidæ from the Philippines, two other insects with the same metallic sheen and very similar form of body. They had been put in beside the weevil as duplicates, but more careful observation showed that they were delicate Gryllidæ, which mimicked the hard beetles so deceptively that even the practised eye of the entomologist was misled by them. Later on it was shown that these Gryllids live in the Philippines beside the weevils, and even on the same leaves with them, and that the beetles are protected from the attacks of birds and other enemies by the extraordinary hardness of their cuticle. The case is especially remarkable because in general the Gryllidæ have no metallic shimmer, and the form of body must have been considerably altered to make them resemble the beetle. The usually broad head of the Gryllids is in this case narrower, the usually flat wing-covers are arched and pear-shaped, and the legs have become quite beetle-like. The security enjoyed by the weevil must be very perfect, for it is mimicked by three other species of beetle in the Philippines.
Animals can also be protected from attack by the possession of dangerous weapons. To this class belong insects with poisonous stings, like the bees, wasps, and ants, and in some degree also the ichneumon-flies. We cannot wonder, therefore, that these dreaded species find imitators. In this case it is not of so much importance that the copy should be rarer than the model, for anything that looks like a dangerous insect will be avoided, since close investigation is in this case attended with danger. So we find that hornets, wasps, and bees are frequently imitated by other insects, by beetles, flies, and butterflies; and these must derive a certain advantage, even when the resemblance is only a general one. Many Longicorns, which visit flowers, are striped black and yellow, like a wasp, and so are many flies, like the species of Syrphus, and so on. The Longicorn Necydalis major bears a strong resemblance to a large ichneumon-fly; it has the same long-drawn-out body, the same swellings on the femur and tibia, the curved antennæ, the glossy brown colour, and its wing-covers are quite short, leaving the wings free, so that the deception is very complete.
Bees, too, are sometimes so well imitated that they are hardly to be distinguished from their mimics, not in flight only, but also when visiting flowers. The best and commonest mimic of our honey-bee is a perfectly harmless fly of the same size and colour, the drone-fly (Eristalis tenax). The two are often to be seen together on the [Pg 117]same flowering shrub, as, for instance, in autumn, on the Japanese buckwheat of our gardens (Polygonum sieboldii), both busily seeking for honey. I once noticed a boy catching the flies with a net in order to imprison them, but a bee stung him severely in the finger. He immediately abandoned the chase, and gave up the flies, perceiving the dangers of confusion. So the animal enemies of Eristalis will often prefer to leave it in peace rather than run the risk of being stung.

PLATE III
| FIG. | |
| 24. | Elymnias undularis, male of the species of which the mimetic female is depicted in Fig. 23. |
| 25. | Euplœa binotata, immune Indian species, mimicked by |
| 26. | Elymnias leucocyma, male, of which |
| 27. | Euplœa midamus. |
| 28. | THE FEMALE MIMICS FAIRLY CLOSELY |
| 29. | Danais vulgaris, immune Indian Danaid. |
| 30. | Elymnias lais, mimetic of the foregoing species, but only on the upper surface. The lower surface retains the original protective colouring representing a decaying leaf. |
| 31. | Tenaris bioculatus, from the Papua region. |
| 32. | Elymnias agondas, mimics the foregoing species from the same locality. |
To face Plate III
There is still another relation between two species which can be induced by mimicry—namely, parasitism, when, for instance, the so-called cuckoo-bees and parasitic humble-bees deceptively resemble in colour, arrangement of hair, and form of body, the species into whose nests they smuggle their eggs, to have them brought up at the expense of the bee or humble-bee in question. In the same way, among the numerous parasites of ant nests, there are some which copy the ants themselves, and so secure themselves from molestation, although they devour the ants' eggs and pupæ. Thus, among the hosts of South American driver-ants (Eciton prædator) there lives a predaceous beetle of the family Staphylinæ, which has received the name Mimeciton because it resembles the ant in form and in the nature of the external surface, though not in colour, which is to be explained by the fact that this ant has no compound eyes, and is therefore almost blind, or at any rate cannot see colours.
I should never come to an end were I to attempt to exhibit the great wealth of observations now available in regard to mimicry. But this at least may be added, that isolated cases of mimicry have been found even among Vertebrates. Thus, according to Wallace, the red-and-black striped poisonous coral snake of South America (Elaps) is most realistically imitated by a non-poisonous snake (Erythrolampus) of the same region. Among birds, Wallace cites a few cases which may be regarded as mimicry, but none are known among mammals, which is not to be wondered at when we consider how very much less numerous in individuals the species are which live together on one area, and how much less likely it is that two species should be, to begin with, so near each other in size, habit, and form that the process of natural selection could bring about a deceptive degree of resemblance. Without doubt it is among insects that the conditions for mimicry are especially favourable, partly because of the enormous number of species which live together and have interrelations on the same area, even in our latitudes and much more so in the tropics, and also because of their usually great fecundity, and their rapid multiplication, both of which are factors[Pg 118] favourable to starting and continuing the processes of natural selection. Furthermore, we have to take into account the hosts of enemies which depend wholly or in great part on insects for food, and destroy them in enormous numbers, eliminating them in inverse proportion to the perfection of their adaptation. Finally, there is the extreme susceptibility of many insects to injury. This makes it very desirable that they should have some disguise sufficient to protect them from even the first attempt at an attack, since that would in many cases prove fatal.
PROTECTIVE ADAPTATIONS IN PLANTS
Protection against large animals—Poisons—Ethereal oils—Spines and thorns—Sharp and stinging-hairs—Felt-hairs—Position of the thorns: buckthorn—Tragacanth shrub—Prigana scrub—Alpine shrubs—Protection against small enemies—Chemical substances—Mechanical protective arrangements—Raphides—Conclusion.
We have seen in how many different ways animals are able to adapt themselves to the conditions of life, both protectively and aggressively; how they approximate in their colour to that of their surroundings so that they harmonize with it; how they copy lifeless objects, or parts of plants, leaves, or twigs, or even mimic, in form and colour, other animals which are in some way protected. When we consider that by far the greater number of species find protection in some degree through their colouring, and often through their form, and when, at the same time, we remember how different this colouring is in nearly related species, and even within the same species (dimorphism), we can scarcely avoid the impression that the forms of life are made of a plastic material, which, like the sculptor's clay, can be kneaded at will into almost any desired form.
This impression is corroborated when we turn our attention to plants, and consider the different ways in which they are able to protect themselves from the attacks of animals.
That plants stand in need of some protection is obvious enough, since their leaves and other green parts contain much nourishment, and an endless army of animals, large and small, depends upon these alone for sustenance. Indeed, the existence of animals depends altogether on the occurrence of plants, for carnivorous and saprophytic animals could only arise after vegetarian forms had been already in existence. But if the green parts of the plants were left defenceless at the mercy of the multitude of herbivorous animals, it would not be long before they were exterminated from the face of the earth, for the animals would devour unsparingly whatever was within their reach, and, as their increase does not depend on their ratio of elimination alone, but also on their fertility, and on their rapidity of multiplication, they would go on increasing in numbers at the expense of the[Pg 120] superabundant nourishment until the plants on which they depended were themselves consumed.
When we inquire into the means whereby plants evade such a fate we are astonished at the endless diversity of the devices employed.
Let us consider first of all the menace to plants from the larger herbivores, from elephants and cattle down to the hare and the roe-deer; we find that many plants are protected by poisons, which develop in the sap of their stems, leaves, roots, and fruits. The juicy and beautifully leaved Belladonna (Atropa belladonna) is never touched by roe-deer, stags, or other herbivores, and the same is true of the thorn-apple (Datura stramonium), the henbane (Hyoscyamus niger), the spotted hemlock (Conium maculatum), the danewort of our woods (Sambucus ebulus), and many others; they all contain a poison. Like the unpalatable butterflies, these unpalatable plants are also furnished with a warning sign of their undesirability, namely, a disagreeable odour, perceptible even by man, which scares off animals from touching them. The development of this through natural selection presents no very serious difficulty.
But, strangely enough, there are not a few poisonous plants in which we, at least, are unable to detect any such warning sign. Among these are the blue aconite (Aconitum), the black hellebore (Helleborus niger), the meadow-saffron (Colchicum autumnale), species of Gentian, of spurge (Euphorbia), and others. Yet these are avoided by deer, roe-deer, chamois, hares, and marmots, and our cattle, horses, and sheep also usually leave them untouched. A case has, however, been reported from the valley of the Aur, on the lower Rhine, which seems to contradict this. On the rocky grass-slopes of the valley the poisonous hellebore (Helleborus viridis) grows in great abundance, and the sheep of that region, which were wont to graze on the slopes, avoided these plants. But some sheep from another part were imported into the valley, and these ate the hellebore, with the result that many died. If these poisonous plants, then, were furnished with a warning sign such as a disagreeable odour, not perceptible to us, we should have to assume that the imported sheep had a less acute sense of smell than the others, which is not impossible in domesticated animals. If there were no such warning sign, then it must have been not an instinct but a continuous tradition which prevented the native sheep from touching the inedible plants.
A more naïve interpretation of nature than that of our day would have regarded the fragrant ethereal oils developed in the seeds of many plants, as in those of fennel, cummin, and other Umbelliferous[Pg 121] plants, as a peculiarity designed for the use and profit of man. But these ethereal substances are obviously a means of protection against the depredations of seed-eating birds, for a sparrow which was allowed to eat three or four seeds of cummin died very soon afterwards.
Many plants produce bitter substances in their green parts, and so secure at least some measure of protection, as is the case with the majority of mosses, the ferns, and species of Plantago and Linaria. Others, again, deposit silicic acid in their cell-walls, or develop in addition a very thick epidermis, so that they afford at the best an unpleasant food, e.g. many grasses, the horse-tails, the rhododendron, and the bilberry. Others, again (Alchemilla vulgaris), have cup-shaped leaves, which retain rain and dew for a long time, and this protects them from grazing animals, which are unwilling to touch wet grass and plants.
Especially widely distributed and diverse is the protection of plants by sharp thorns and spines. It is extremely interesting to note in how many different and advantageous ways this armature is disposed.
Obvious at once is the fact that thorns and spines only occur on those parts which are naturally exposed to attack. Thus we find them particularly strong in young plants, and on the lower parts of older ones. The holly, for instance, has crenate, spinose leaves only to the height to which grazing animals can reach; beyond that the leaves are smooth-edged and spineless, like those of the camelia. It is almost the same with some wild pear-trees, which are quite covered with thorns as long as they are low, but afterwards grow a thornless crown.
Similarly, low bushes, when they are armed with thorns or the like at all, are covered with them all over, like the rose-bush.
When the leaves of a plant are spinose the spines are disposed on the parts usually attacked; and thus we understand why the enormous floating leaves of Victoria regia should have on their under surface long, pointed spines which, especially at the upturned margin, attain a length of several inches; it is from water animals—water snails—that danger threatens them.
Thorns are developed in the most diverse ways. In many of the bushes on the coast of the Mediterranean true leaves are wanting altogether, the green branches and twigs being themselves the assimilating parts, and these are so stiff and rigid, so like some kind of thorn, that they suffice to scare off any greedy herbivore. Among our own bushes the Broom (Spartium scoparium) may be taken as an example of this class.
In other cases the spines are found on the leaves themselves, but there is great diversity in their mode of arrangement. In many tropical plants, such as the Yucca and the Aloe, the point of the long, reed-shaped leaf is transformed into a spine, and this is the case in many of our native grasses. Kerner von Marilaun notes that, in the Southern Alps, two such grasses, Festuca alpestris and Nardus stricta, occur frequently in certain localities, and they prick the muzzles of the cattle so badly that they return bleeding from the pasture. This prevents these Alpine runs from being made full use of, so the grasses are as far as possible extirpated by man, and, curiously enough, also by the cattle themselves, for they seize the grass at the base of the tuft with their teeth, pull it out, and let it fall, so that it withers. Kerner saw thousands of such pieces of turf which had been pulled up by the cattle lying dried and bleached by the sun on some of the Alpine grazing grounds in the Tyrolese Stubaithal.
Again, in many plants the whole leaf-edge is transformed into a spiny wall, which may be enlarged by indentations and lobate projections, as in the holly, and, in a much higher degree, in the thistles (Carduus), in Eryngium, in Acanthus, and in many Solanaceæ. Often, too, there are barbed hooks on the leaf-edge, which work like a saw; or the leaf-edge, though without spines, may be made sharp by deposits of silicic acid, as in the sedges, whose sharp edges are moved to and fro in the mouths of ruminants, and thus injure the mucous membrane. The hook-bristles of the fig-cactus (Opuntia), which, though small, are abundantly provided with barbs, must also be mentioned; for they are to be found in great numbers surrounding the buds of these plants, and most effectively protect them from being eaten away by animals (Fig. 19).
To this category, too, belong the short, prickly bristles of the rough-leaved plants, which cover the whole plant as with an overcoat of sharp needles; of these we may mention the adder's tongue (Echium vulgare), the comfrey (Symphytum officinale), and the borage (Borago officinalis).
Very well known are the stinging-hairs of the Urticaceæ, long hairs (Fig. 20) with an elastic base, but with glass-like, brittle, rounded heads, which break off at the lightest touch, whereupon the sharp point of the broken hair penetrates the skin of the creature which has touched it, and the poisonous contents of the hair are poured into the wound. Even our large stinging-nettle (Urtica dioica) can cause intense irritation, and evoke the 'nettle-rash,' named after it, on the human skin; but there are many tropical[Pg 123] species of nettle, e.g. Urtica stimulata in Java, and others, which have an effect similar to that of snake-poison and produce tetanoid spasms, and so on. In addition to formic acid these hairs contain an undefined ferment, a so-called Enzyme. It need scarcely be said that these stinging-hairs must have much more severe effects on the mucous membrane of the mouth of grazing animals than on the human skin, and that they are therefore an excellent protection for the plants. As a matter of fact we never find our nettle patches eaten away, and even the donkey, which eats thistles freely, turns away from the stinging-nettle. But even these stinging-hairs, like all other protective devices, do not afford an absolute protection. The caterpillars of several of our diurnal butterflies feed exclusively on the stinging-nettle, and they eat up the leaves, stinging-hairs and all. This is the case with five species of the genus Vanessa, namely: Vanessa io, the 'peacock,' Vanessa urticæ, the small tortoiseshell, Vanessa prorsa, Vanessa C. album, the C. butterfly, and Vanessa atalanta, the admiral.
Fig. 19. Barbed bristles
of Opuntia rafinesquii; enlarged.
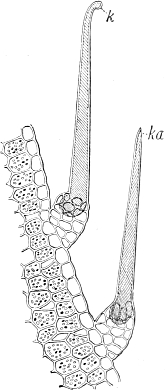
Fig. 20. Vertical section through
a piece of a leaf of the Stinging-nettle
(Urtica dioica), bearing two stinging-hairs;
magnified 85 times; adapted
from Kerner and Haberlandt.
We are all familiar with our mulleins (Verbascum), those[Pg 124] beautiful flower-spikes with the thick, soft felt leaves, which grow on stony or sandy soil. Harmless as they look, they are much disliked by animals as food, for the thick hairy felt which covers them breaks up in the mouth, and sticks in the folds of the mucous membrane, causing burning sensations and other discomforts. They, too, are therefore spared by grazing animals, but they have smaller enemies, like the caterpillars of the genus Cucullia, which, however, never completely destroy them, but only eat large holes in their leaves.
Let us now consider in somewhat greater detail the true thorns, the most conspicuous protection of many plants. It is very remarkable that these are always so placed, and so regulated as to their length and character, as to afford protection to the most important and the most exposed parts of the plant. Thus many bushes, which would otherwise be in danger of being completely devoured by cattle, are stiff with thorns which are nothing else than pointed, hard twigs without, or with very little foliage. Among these are the sloes, the buckthorn (Rhamnus), the sea-buckthorn (Hippophäe), and the barberry (Berberis). In the last-named three thorns arise in a group, and protect the young bud from danger in three directions (Fig. 21).
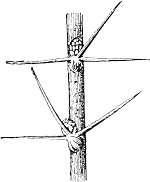
Fig. 21. A piece of a
twig of Barberry (Berberis
vulgaris) in spring; after
Kerner.
The fine-leaved mimosas of the tropics have similar but very long and sharp thorns, and their leaves are movable and sensitive, so that, when they are touched, they shut up and draw back behind the rampart of stiff thorns, which are just of the right length to protect them.
In many thorny bushes only the young shoots of each spring remain green through the summer, and in autumn they become transformed into thorns, under whose protection the shoots of the following spring will develop. Sometimes, too, the leaf-stalks are modified in the course of the summer into thorns, as in Tragacanth (Astragalus tragacantha). In this case the young leaves are protected by a circle of thorns, consisting of the leaf-stalks of the preceding year which have not fallen off (Fig. 22, A, B, C).
I should have to go on for a long time with my exposition, even if I were to confine attention to the essential facts; we shall, therefore, only recall the well-known phenomenon of the Cactuses, in which the leaves are entirely transformed into spines, which may attain a length of eight centimetres, while the fleshy stem alone represents the green—that is, the assimilating parts of the plant.[Pg 125] The species of Cactus are almost the only plants which grow on the stony, hard, and hot plateaux of Mexico, and they are protected from desiccation by the thickness of their epidermis. But, enticing as is the food promised by the juicy stem, animals rarely venture to approach them, and it is only when tortured by thirst that horses and asses occasionally knock off the spines with their hoofs, and so reach the soft tissues rich in water. For this attempt, however, as Alexander von Humboldt pointed out, they often suffer, as the sharp spines are apt to pierce the hoof. In any case, the cactuses are effectively protected from the danger of extermination by grazing animals.
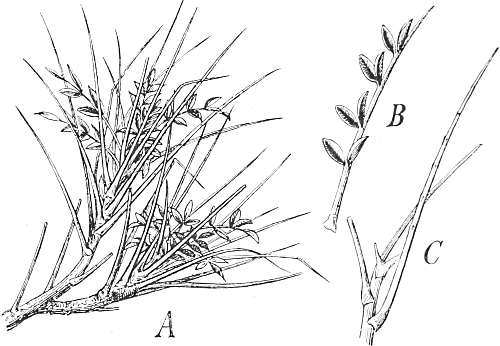
Fig. 22. Tragacanth (Astragalus tragacantha). A, two spring shoots. B, a single leaf, from which the three uppermost leaflets have fallen off. C, leaf midrib, from which all the leaflets have fallen off. After Kerner.
It must certainly strike every one that many districts, especially those which are dry, hot, and stony, are conspicuously rich in thorny plants, and it has often been supposed that the production of thorns must be a direct result of these peculiar conditions of life; indeed, the hard, thorny habit of many of these plants has even been regarded as a protection against desiccation. This, however, is contradicted by all those thorny plants which, like the cactuses, possess tissues extremely rich in sap, and in which desiccation is prevented, not by the thorns, but by the thick epidermis. The only satisfactory explanation is that afforded in terms of natural selection. In such hot, and at the same time dry regions, the plant-growth is often very[Pg 126] scanty, and the food available for the grazing animals is, at least at times, very scarce; on this account, if the plants are to survive there at all, they must be armed with the most perfect means of protection possible against the attacks of hungry and thirsty animals. The struggle for existence in relation to such enemies is much more severe than in more luxuriant regions, and the protection by thorns has been developed to the highest possible pitch of perfection; species which were unable to develop this protection died out altogether. Hence the cactuses of Mexico, and the many thorny bushes and shrubs of the hot, and, in the summer, dried-up stony coast-lands of the Mediterranean in Spain, Corsica, Africa, and other countries. This so-called 'Prigana scrub' embraces a number of species, whose nearest relatives in our climate are not provided with spines, as, for instance, Genista hispanica, Onobrychis cornuta, Sonchus cervicornus, Euphorbia spinosa, Stachys spinosa, and others.
Why do so few thorny plants grow on the rich and well-watered Alpine pastures? Probably because there is to be found there a rich and luxuriant plant-growth which can never be wholly exterminated by the grazing of animals, so that an individual species would not, by developing thorns, have gained any advantage in the way of increased capacity for existence.
But these Alpine grazing grounds serve well to illustrate how great may be the advantage which protective devices give to a species. Much to the annoyance of the herdsmen, who endeavour to extirpate them as far as possible, enormous masses of rhododendrons often cover whole stretches, because their hard silicious leaves cannot be eaten, and many other plants despised of cattle flourish and increase on the grazing runs, like the repulsively bitter, large Gentiana asclepiadea, the malodorous Aposeris fœtida, and various ferns of disagreeable taste.
The advantage derived by plants from the possession of any kind of protective device against grazing animals is perhaps best of all seen in the 'shrubbery,' which on every Alp is to be found in the immediate neighbourhood of the herdsman's hut. There, where the cattle daily assemble, and where the soil is continually being richly manured by them, we always find a large, luxuriantly growing company of the poisonous aconite, the bitter goosefoot (Chenopodium bonus henricus), the stinging-nettle, the thistle (Cirsium spinosissimum), the ill-smelling Atriplex, and some other inedible species, while the palatable herbs are gradually exterminated by the cattle which daily gather round the hut (Kerner).
To sum up. We have seen that there is among plants an[Pg 127] extraordinary diversity of protective adaptations, which secures them from extermination by the larger herbivores.
Since all useful contrivances, or, as we say, all adaptations, are capable of interpretation in terms of the process of selection, we must refer this great array of the most diverse protective devices to natural selection; and again, as among animals, we receive the impression that the organism is, to a certain extent, really capable of producing every variation necessary to its maintenance. Literally speaking, this would not be correct, but at any rate the number of adaptations possible to each form of life must be an enormous one, so great, indeed, that ultimately every species does secure protection for itself in some manner and in some degree, whether it be by the production of a poison or a nauseous substance within itself, or by surrounding itself with thorns or spines. And if it be, in a certain sense, a matter of 'chance' whether a plant has taken to one method of defence or to another, according as its innate constitution favoured the production of one rather than of any other, yet it would not be easy to prove, even in the case of the purely chemical means of protection, that these would have occurred in the same distribution and concentration as a necessary result of the metabolism of the plant, even if they had not been useful and consequently augmented by selection. But in the case of the mechanical means of protection this mode of explanation fails as utterly as that of the direct effect of the conditions of life. Why the holly should have spinose leaves beneath and smooth ones above can never be deduced from the constitution of the species.
While the protective adaptations of plants against the larger herbivores always point to natural selection, our appreciation of the adaptability of plants, and at the same time of the potency of natural selection, will be strengthened still more if we turn our attention for a little to the arrangements which prevent the extermination of plants by the lower and small animals.
It might indeed be supposed that extermination by these could hardly be an imminent danger, but if we think of the cockchafer blight, or of the destruction of whole woods by the caterpillar of the 'white nun,' or even of the destruction of several successive plantings of young salad plants which the snails often cause in our gardens, it cannot be doubted that all plants would be exterminated by insects and snails alone unless they were protected against them in some degree.
We owe our detailed knowledge of the means by which plants protect themselves against the menace of the greedy and prolific[Pg 128] snails to the beautiful investigations of Stahl, Professor of Botany in the University of Jena.
In this case, too, both chemical and mechanical means are made use of. The minute quantity of tannic acid which is contained in the leaves of the clover prevents many snails from eating them, as, for instance, the garden snail (Helix hortensis). If the leaves be soaked so as to wash out the tannin the snail readily accepts them as food. It is true that the small, whitish field-slug (Limax agrestis) does not object to the presence of the tannin, and eats the fresh leaves of the clover; indeed, there is no such thing as absolute protection. In discussing the herbivorous mammals I have already mentioned that many trees and shrubs, mosses and ferns are effectively protected by the large amount of tannin they contain; this protection is effective also against snails, for all these plants are fairly free from their attacks; and the same is true of many other tannin-containing plants, species of saxifrage and sedum, the strawberry, many water-plants, like the pond-weeds (Potamogeton), the horn-nut (Trapa), the mare's tail (Hippuris). All these plants are only eaten by snails in case of necessity, or in the washed-out state.
In other plants protection is gained by means of some acid, especially oxalic acid, like the wood-sorrel (Oxalis acetosella), the sorrel (Rumex), and the species of Begonia. When Stahl smeared slices of carrot, which is a favourite food of snails, with a weak (one per cent.) solution of oxalate of potassium, they were refused by the snails, and this is not surprising when we remember that even the external skin of the snail is very sensitive, and the mucous membrane of the mouth is not likely to be less so.
Similarly, many plants develop ethereal oils in the hairs which cover them, as in the herb-Robert (Geranium robertianum). Even the almost omnivorous field-slug (Limax agrestis) does not touch this plant, and if it be placed upon it, escapes with all dispatch from the ethereal oil, which burns its naked skin, by covering itself with mucus and letting itself down to the ground by a thread. The mints (Mentha) and the dittany (Dictamnus albus) also produce such oils.
Among chemical means of protection must be named the pure bitter stuffs, such as are found in the species of gentian, the milkwort (Polygala amara), and in many other plants, and also the curious 'oil-bodies' of the liverworts.
But some plants also defend themselves against the attacks of snails by mechanical means.
First there are the various kinds of bristle arrangements, which[Pg 129] prevent the snails from creeping up the stalks. We never find the comfrey (Symphytum officinale) of our meadows eaten by snails, for it is thickly covered over with stiff bristles, which are most disagreeable to the snail, and the stinging-nettle (Urtica dioica) is similarly protected by bristle hairs, while, as we have already seen, its stinging-hairs secure immunity from the attacks of larger animals.
And although it is true that the majority of plants do not prevent the snails from creeping up their stalks, yet they do not serve them in any great degree as food, since the green parts often offer resistance to mastication and digestion. Thus the lime encrustations which cover the stoneworts (Chara) prevent snails from eating them. If the lime be dissolved by means of acids, and the plants then offered to the snails, they will eat them greedily. The same is true of the silicifying of the cell-walls, so widely distributed among mosses and grasses, and when this occurs in a high degree it forms an effective protection even against the large herbivores. Our slightly siliceous grasses are secure from snails, and that it is really the presence of the silicic acid which deters them from an otherwise welcome kind of food is proved by Stahl's experiment of growing maize in pure water, and so obtaining plants poor in silica. These were devoured without ceremony by the snails.
Of the many other protective peculiarities which make it difficult for snails to eat plants I shall only recall the so-called 'Raphides,' those microscopic crystal-like needles of oxalate of lime, pointed at both ends, which lie close together in the tissues of many plants. Cuckoo pint (Arum maculatum), the narcissi, the snowdrops (Leucojum), the squill (Scilla), and the asparagus contain them, and all these plants are spared by snails obviously because during mastication they are unpleasantly affected by the raphides. Even the voracious field-slug rejects these.
Of course it cannot be said that these raphides protect against all other enemies. They are effective against rodents and ruminants, and also against locusts, but a number of caterpillars seek out by preference just those plants which contain raphides. Thus certain caterpillars of the Sphingidæ feed on species of Galium and Epilobium, the leaves of the vine, and the wild balsam (Impatiens). The caterpillar of Chærocampa elpenor, which especially prefers Vitis and Epilobium, has transferred its affections to the fuchsias in our gardens, which came from South America; the butterfly not infrequently lays its eggs on these plants, and the caterpillars devour them readily; but the fuchsias may also contain raphides.
We may say, indeed, that almost all wild Phanerogams are protected in some degree against snails, and this almost suggests the question: What then is left for the snails to feed on if everything is thus armed against them? But, in the first place, there remain our cultivated plants, which, like the garden lettuce (Lactuca), are quite without defence; and secondly, the snails often eat the plants only after they have been rooted up and lie rotting on the ground, that is, when the protective ingredient has been dissolved out by the rain; finally, no means of protection, as I have often said already, is absolute or effective against all snails. Many of these are, as Stahl calls them, 'specialists.' Thus, the large slug of our woods eats the poisonous fungi which are rejected by other snails, and in the same way there are many other specialists which, however, are not likely to eliminate unaided the plants to which they have adapted themselves. There are certainly also omnivorous forms, like the field-slug (Limax agrestis), to which we have referred so often, and Arion empiricorum, the red slug, but just because these eat so many kinds of plant they are less dangerous to any one species.
These manifold devices for protecting plants against the depredations of snails afford another proof that innumerable details in the organization of plants, as of animals, must be referred to natural selection, since they are capable of interpretation in no other way. If these protective devices were to be found only in isolated plants, we might perhaps talk of 'chance'; we might refer them to the inborn constitution of the plant, which made the production of bristles, or bitter stuffs, or the deposition of silicic acid a necessity, and which 'happened' to make the plants distasteful to certain snails. But as it appears that all plants are protected against snails, one in this way, another in that, this objection cannot be sustained. Furthermore, some of the beautiful experiments made by Stahl to prove the protective effect of these devices showed, at the same time, that they were not in themselves indispensable to the existence of the plant; maize, for instance, develops a plant perfectly capable of life, even though silicic acid be withheld, and the acid is, therefore, not an element essential to its constitution, but a means of protection against voracious animals. The clearest proof of this is afforded by plants like the lettuce (Lactuca), which formed protective stuffs in the wild state, but have lost them altogether under cultivation, through disuse, as we shall see more precisely later on. As the eyes of animals which live in darkness have degenerated, so the plants which have been taken under the protection of man have lost their natural means of defence, because these were no longer necessary to the maintenance[Pg 131] of the species. Even the protective bitter substances (tannin-compounds) are not essential to the constitution of the genus Lactuca; their formation may be discontinued without the plant being otherwise affected. And in this case it is not a question of the withdrawal of something which has to be taken in from outside, it is the non-development of what is purely a product of the internal metabolism.
The adaptations of plants against snails are instructive in another way, namely, in their extraordinary diversity. Here again we see how great is the plasticity of organic forms, and how precisely, though in many very different ways, they adapt themselves to the conditions of their life, in this case the weaknesses of their greedy enemies, and all to attain the same end, the security of their existence as a species. We see at the same time that innumerable minute details in the structure and character of a species, which may appear unimportant, may yet have their definite uses—hairs, bristles, and raphides, as well as bitter substances, ethereal oils, acids, and tannin-compounds. But we must, of course, have minute and exhaustive investigations, like those of Stahl, in regard to the biological relations of these peculiarities before their utility can become clear to us.
CARNIVOROUS PLANTS
Introduction—The Bladderworts or Utriculariæ—Pitcher-plants, Nepenthes—The Toothwort, Lathræa—The Butterwort, Pinguicula—The Sundew, Drosera—The Flytrap—Aldrovandia—Conclusions.
That the principle of selection dominates, to a large extent at least, all the structural characters of plants, and moulds these in direct relation to the prospects of greater success which may be offered in the vicissitudes of the life-conditions of a single species or group of species, is nowhere more apparent than in the case of the so-called 'insectivorous' or 'carnivorous' plants. Here again it was Charles Darwin who led the way, for while many plants had long been known on the sticky leaves of which insects were often caught and killed, it had occurred to no one to regard this as of any special use for the plant, much less to look on the peculiar dispositions of such leaves as especially determined for this purpose. Darwin was the first to show that there is no small number of plants—we now know about 500—which secure only a portion of their nutritive material by the usual method of assimilation, and gain another and smaller portion by dissolving and utilizing animal protoplasm, especially nitrogenous muscle substance. The correctness of this interpretation was at first disputed, but Darwin showed that pieces of muscle, or any nitrogenous organic substance, were really dissolved by the relevant parts of the plant, and were afterwards absorbed. It can therefore no longer be doubted that the remarkable contrivances by which animals are laid hold of by plants—are in a certain sense caught and killed—have arisen with reference to this particular end; or, to speak less metaphorically, that existing structural and functional peculiarities in a plant which caused animals to be held fast were of advantage to the nutrition of the plant, and were therefore augmented and perfected by natural selection. That this was possible is obvious from the number of insectivorous plants which now live upon the earth, and that these processes of selection ran their courses quite independently of one another, and even that they started from different parts of the plant, is shown by the diversity of the con[Pg 133]trivances which occur in plants of several different families. A few of these I wish to discuss in some detail.
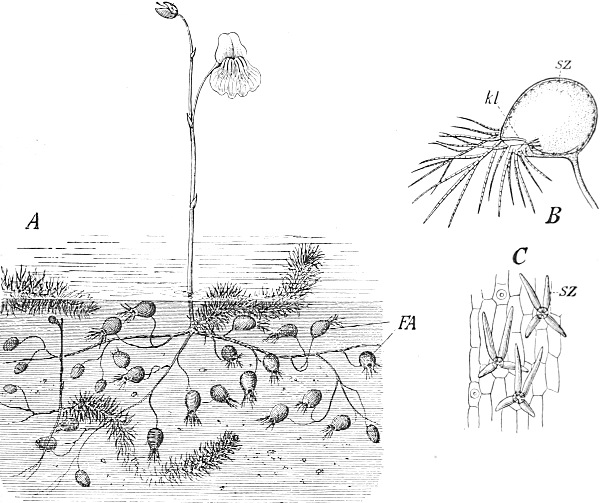
Fig. 23. Utricularia grafiana, after Kerner. A, a plant in its natural position, floating in the water. FA, traps. B, a trap enlarged four times. sz, suctorial cells. kl, valve, which closes the entrance to the trap. C, suctorial cells on the internal wall of the trap, enlarged 250 times.
The marshes of European countries, and also those of warmer lands, often contain bladderworts, or Utriculariæ (Fig. 23)—floating water-plants, without roots, and with horizontally spread, long-drawn-out, tendril-like shoots, in part thickly covered with whorls of delicate, needle-shaped leaves, in part bearing sparse leaves of quite peculiar structure. These are stalked, hollow bladders (Fig. 23 A, FA), with quite a narrow entrance at the apex, which is closed, as far as larger animals are concerned, by projecting bristle-like hairs (B). Small animals, such as water-fleas (Daphnia), species of Cyclops, and Ostracods, can swim in between the bristles, and they then come in contact with a valve which opens easily inwards (B, kl) and allows them to penetrate into the interior of the trap. Once inside they are[Pg 134] captives, for the valve does not open outwards; therefore they soon die and decompose, and are then taken up by special absorptive cells (B, C, sz) and utilized as nourishment for the plants. In this way the Utriculariæ catch numerous little crustaceans and insect larvæ, which slip into their traps, presumably for concealment.
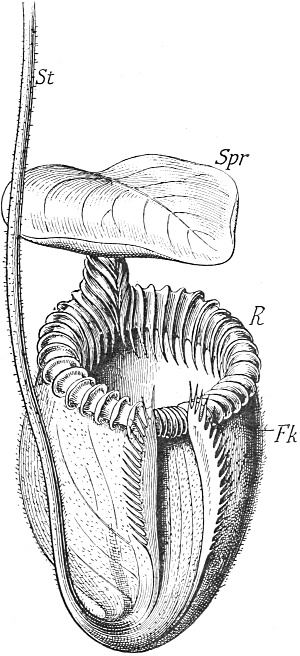
Fig. 24. Pitcher of Nepenthes villosa, after
Kerner. St, stalk of the leaf. Spr, its apex.
Fk, the pitcher. R, the margin beset with
incurved spines.
Another example is found in the marsh plants of the genus Nepenthes, some species of which live as climbers on the outskirts of tropical forests, climbing up the trees and letting their long, thin tendrils hang downwards, often over ponds and stagnant pools, where swarms of small flying insects abound. These plants have developed exceedingly remarkable contrivances for catching insects and using them as food (Fig. 24). The long stalks (St) of their leaves (Spr) are first bent downwards, then they suddenly turn sharply upwards, and the upturned portion is modified into a pitcher-like structure, in the bottom of which a fluid gathers, acid in taste, containing pepsin, and therefore a digestive fluid. Nitrogenous substances, such as flesh, dissolve in this fluid, and insects which fall into the pitcher from the rim are killed and dissolved. There are many species of Nepenthes, but not all of them possess the trap-structure in equal perfection, so that we are able, to some extent, to follow the course of its evolution, from a broad leaf-stalk, somewhat bent over at the edges, to the marvellous closed pitcher shown by Nepenthes villosa (Fig. 24) of Borneo. In this species the pitchers attain a length of fifty centimetres, and are beautifully coloured, resembling in that respect, as well as in their form, the tobacco-pipe-like flowers of the tropical Aristolochiæ. When we come to discuss the origin of flowers,[Pg 135] we shall see that the bright, conspicuous colour possesses a very considerable value in attracting insects; and in the case of the pitcher-plant, too, the gorgeous colour probably allures insects to settle on the rim of the pitcher, and they are tempted to dally the longer since it secretes honey. But the thick, swollen rim of the pitcher is as smooth as if it were made of polished wax, and resembles the petals of those magnificent large orchids, the Stanhopeæ; the inner surface of the pitcher below the margin is also smooth, so that insects which creep about seeking honey are apt to slip and fall to the bottom. Even if many of them are not at once killed by the digestive fluid, but are able to climb up the smooth wall again, they cannot escape, for beneath the swollen rim, which projects inwards, there is a circle of strong bristles or teeth, with the points directed downwards, which, like thorns, prevent the captive's escape. Thus the pitchers of Nepenthes secure and digest a large number of insects, and we can easily understand that the plant acquires a considerable amount of valuable nourishment in this way, for ready-made protoplasm is a convenient food to which the plant has to do but little in order to convert it into its own particular kind of living matter.
The toothwort (Lathræa squamaria) must also be briefly noticed here, because it does not catch insects through the medium either of air or of water, but through the earth. As is well known, this plant is parasitic on the roots of various foliage-trees. It is of a pale yellowish colour, and has no green assimilating parts. For such a plant it must be of particular value to be able to catch animals and to use them as food. To this end the short, pale leaves, which surround the creeping, underground stem in the form of closely appressed scales, have been modified into snares for minute animals. The leaves have their upper parts recurved downwards, and the edges have grown together, so that only a small opening is left at the base, and this leads into a system of tunnels. Aphides, rotifers, bear-animalcules, but especially springtails (Podurids), creep into these hollow leaves, are held fast by a sticky secretion, and are dissolved and absorbed.
Another example, also indigenous, is that graceful marsh plant, the butterwort (Pinguicula vulgaris), whose broad, tongue-shaped leaves, arranged in the form of a rosette, have been modified into an insect trap by the turning up of their edges, while the middle is deepened into a longitudinal groove (Fig. 25). The whole upper surface of the leaf is covered with an enormous number of little mushroom-shaped glands (B, C, Dr), which secrete a viscid slime. Insects which settle on the leaf stick fast, and as the glands continue[Pg 136] to pour out more and more slime, while at the same time the edges of the leaf, stimulated by the struggling of the insect, curl over still farther, the victims are drowned in the slime, and ultimately absorbed; for this secretion is so powerful that even fragments of cartilage are dissolved by it in forty-eight hours. Midges and mayflies in particular fall victims to this plant, which is common in marshy places both in mountain and plain.
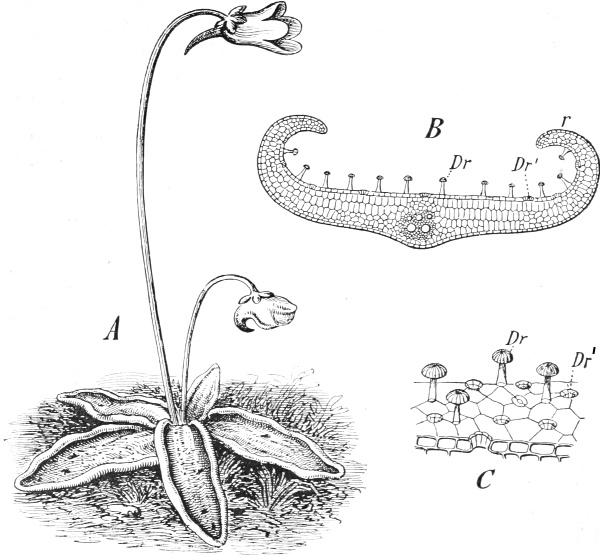
Fig. 25. Butterwort (Pinguicula vulgaris). A, the entire plant, showing the incurved margins of the leaves and some insects caught by the secretion. B, cross-section through a leaf, enlarged 50 times. r, the margin. Dr, Drl, two kinds of glands. C, a portion of the leaf-surface, magnified 180 times.
We must also mention the sundew (Drosera rotundifolia), which takes its name from the seeming dewdrops that sparkle in the sun on the leaves, or rather on the rounded extremities of long and rather thick cilia-like hairs which cover the whole upper surface of the leaf. In reality the apparent dewdrops consist of a sticky, clear, viscid slime, which is secreted by the glandular ends of the pin-shaped hairs or 'tentacles.' Insects which settle on the leaf are caught by the slime, and in this case also an acid, pepsin-containing fluid is secreted, which gradually effects the digestion of the soluble parts of the insect. It is especially noteworthy that it is not only those tentacles which[Pg 137] are in contact with the insect that take part in its digestion and absorption, for all the others gradually alter their position from the moment when any nitrogenous body, be it a fragment of flesh or an insect, touches any of them. All begin to curve slowly towards the stimulating object (Fig. 27), so that, after one to three hours, all the tentacles have their heads towards it, and collectively pour out their digestive juice upon it.
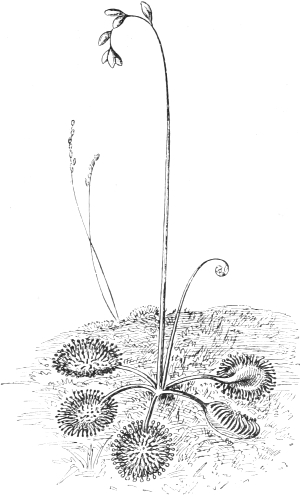
Fig. 26. The Sundew (Drosera rotundifolia), after Kerner.
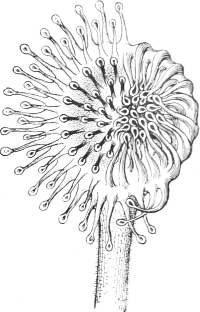
Fig. 27. A leaf of the Sundew, with half of the tentacles curved in upon a captured insect; enlarged 4 times.
The sundew grows in marshes, as, for instance, those of the Black Forest, and also on the moss-covered ridges there, and it is easy to observe that a leaf often shows not merely a single gnat, midge, or little dragon-fly, but several, sometimes as many as a dozen. In this case, again, the value of the arrangement from the point of view of nourishment can be no inconsiderable one.
In the case of the sundew we are obviously face to face with an exceedingly complex adaptation, for not only is there a secretion of the peculiar digestive juices, which occur only in carnivorous plants, but the secreting tentacles are actively motile. That the tentacles more remote from the captive may be excited to curve towards it, it is necessary that the stimulus exerted by it on the heads of the tentacles connected with it be conveyed to the base, and[Pg 138] thence to the tips of the other tentacles, for they curve throughout their whole length. The utility of the contrivance is obvious, but that an arrangement so divergent from the ordinary dispositions of plants could be brought about points to the length of time that the processes of natural selection must have gone on, preserving every new little variation, and adding it to the rest.
Fig. 28. Leaf of Venus Fly-trap
(Dionæa muscipula), after Kerner. A,
leaf-blade (Spr) open. St, leaf-stalk.
Stch, sensitive hairs. B, vertical section
through the closed leaf-blade.
Fig. 29. Aldrovandia vesiculosa, a branch with the
traps FA.
Two plants remain to be noticed in conclusion, both possessing movable, closing traps for catching animals. The so-called Venus fly-trap (Dionæa muscipula) is a marsh plant of North America, the leaves of which, like those of Pinguicula and Drosera, are arranged in a rosette on the ground. The individual leaf has a spatula-like stalk and a blade in two halves (Fig. 28, A), each edged with long and strong spinous processes, directed obliquely inwards. The halves of the blade, when the necessary stimulus is applied to the surface, can close together in a very short time, from 10 to 30 seconds. The two rows of marginal spines then cross, as the interlocking fingers of the hands do, and thus form a cage out of which the imprisoned insect cannot escape. The appropriate stimulus to set the mechanism in motion is a light touch, while a more violent shock, or strong pressure, or a current of air, does not cause the trap to close. But if a fly comes to creep about on the leaf, and in doing so touches one of six short jointed hairs rising erect from a minute cushion of cells, then the leaf closes, quickly indeed, but at the same time so gently and imperceptibly that the fly is unaware of danger and does not try to escape. Then numerous purple mucous glands begin to surround the victim with pepsin-containing, acid, digestive juice which gradually dissolves it.
One of the water-plants of Southern Europe, Aldrovandia vesiculosa, which is also to be found in swamps on the northern ridge of the Alps, possesses, in addition to the capturing and digesting apparatus proper, an active motile apparatus, which is set in motion through sensitive hairs. When I found the plant for the first time in a swamp at Lindau, on the Lake of Constance, I took it at first sight for an Utricularia, for the two plants resemble each other in external appearance (cf. Figs. 22 and 29), but the modification of the leaves into traps is quite different. On both halves of the leaf-blade there are numerous bristles (Fig. 30, A), and the lightest touch on these by a little water animal acts as a releasing stimulus to the motile elements of the leaf (Stch). As in the Venus fly-trap, the two halves of the leaf close together somewhat quickly, but quite quietly, and the animal is caught. Fig. 30 shows a section of one of these traps in its closed state. The captive animals cannot escape, because the margins of the leaf shut quite tightly on one another, and are beset with little teeth. Numerous little glands (Dr) secrete a digestive juice, and after some days, or even weeks, the insoluble remains of the minute animals may be found inside the trap.
Fig. 30. Aldrovandia: its trap apparatus. A, open. St, stalk of the leaf. Spr, blade of the leaf. Stch, sensitive bristles. Dr, glands. B, closed, a cross-section.
Many more cases of animal-catching plants might be adduced, but it is far from my intention to try to describe all the existing contrivances; those already mentioned may suffice to give an idea of the diversity and of the detailed effectiveness of these adaptations. They amplify—so it seems to me—our conception of the scope of natural selection, by showing us that adaptations may arise which[Pg 140] are quite foreign to the original mode of life of the organism in question, and stand, indeed, in apparent contradiction to its fundamental physiological processes. It is hardly necessary to enter into a special argument to show that they can only have been brought about in the course of natural selection, since every other interpretation of their occurrence fails. Neither climatic nor any other external direct influence could have effected these modifications of the parts of plants, which are all so different, yet all so well suited to their purpose; they are different even in plants growing quite close together, like the sundew and the butterwort. The Lamarckian principle of use and disuse hardly enters into the question at all, since plants do not possess a will, and we can hardly speak of 'chance' where we have to do with such complex and diversely combined transformations. A process of selection actually operative in each of these cases can easily be thought out, and I shall leave it to my readers themselves to do this, and shall only indicate that we have to do with increasing elaboration in two different directions: first, improvements in the ability to utilize animal substances which happened to stick to the leaves, and second, an increase in the probability of animals sticking to the leaves, and so becoming available. Thus there arose, on the one hand, dissolving and digestive juices, and arrangements for absorption; and, on the other hand, viscid slime, and traps of various kinds to secure the animals, as well as honey and bright colours to attract them.
But it is not merely transformations in the form of the stems and leaves which have come about; there are also important physiological changes. The sensitiveness to stimulus of various parts of the leaf is greatly increased, to a certain extent in the butterwort, the edges of whose leaves turn inwards in response to stimulus, still more in the sundew, in which the stimulus is conveyed from the tentacles touched to all the others, but most wonderfully of all in the Venus fly-trap and Aldrovandia, whose sensitive hairs so transmit the stimulus that the whole leaf is affected by it, and is set in motion, in a manner quite comparable to the effects of a nerve-stimulus in animals.
Thus the case of carnivorous or insectivorous plants shows us that, in the course of natural selection, quite new organs can be produced in a plant by a thoroughgoing transformation of old ones, as, for instance, the pitchers of Nepenthes, and that, furthermore, even the physiological capacities of the plant may be changed in the most far-reaching manner, increasing and varying until they come to resemble the functions of the animal body.
THE INSTINCTS OF ANIMALS
The robber-wasp—Statement of the problem—Material basis of instincts—Instincts are not 'inherited habits'—Instinct of self-preservation—Fugitive instinct: death-feigning—Masking of crabs—Nutritive instinct—Monophagous caterpillars—Diverse modes of acquiring food: May-flies, sea-cucumbers, fishes that snare—'Aberration' of instinct—Change of instinct during metamorphosis: Eristalis, Sitaris—Imperfection of adaptation points to origin through natural selection—Instinct and will—Instincts and protective coloration—Leisurely flight of Heliconiidæ—Rapid flight of Papilionidæ—Instincts which act only once in a lifetime—Pupation of butterflies—Pupation of the Longicorns—Pupation of the silk-moth—The emperor moth—The cocoons of Atlas—Oviposition of butterflies.
We have hitherto considered animals with especial regard to the variation and re-adaptation of morphological characters, e.g. modifications of form and colour; and we have now to ask whether their behaviour also is to be referred as to its origin, in whole or in part, to the principle of selection. All around us we can see that animals know how to use their parts or organs in a purposeful manner: the duckling swims at once upon the water; the chicken which has just been hatched from the egg pecks at the seeds lying on the ground; the butterfly but newly emerged from the pupa, as soon as its wings have dried and hardened, knows how to use them in flight; and the predatory wasp requires no instruction to recognize her victim, a particular caterpillar, a grasshopper, or some other definite insect; she knows how to attack it, to paralyse it by stings, and then hesitates not a moment as to what she has to do next; she drags it to her nest, deposits it in one of the cells already prepared for her future brood, lays a single egg upon it, and roofs the cell carefully over. It is only because all these complex acts are so precisely performed, as precisely as if the wasp knew why she performed them, that the species is able to maintain its existence, for only thus can the rearing of the next generation be secured. Out of the egg there slips a little larva, which at once makes for the paralysed victim, feeds upon it, and grows thereby, then, within the shelter of the closed cell, passes through the pupa stage and is transformed into a perfect wasp. Many species of these predatory wasps do not lay the egg directly beside or upon their prey, but lest its movements should endanger their offspring,[Pg 142] they hang the egg above it by a silken thread. It is thus in security, and the young larva, too, when it appears, can withdraw to its safely swinging resting-place as soon as danger threatens from the convulsive struggles of the unfortunate victim at whose body it is gnawing.
Every animal has a great many such 'instincts,' which lead it, indeed force it, to act appropriately towards an end, without having any consciousness of that end. For how should the butterfly know what flying is, or that it possessed the power of flight at all, or who could have shown the predatory wasp, when she wakened from the pupa sleep to quite a new kind of life, all that she had to do in order to procure food for herself and to secure shelter and nourishment for the brood which was still enclosed within her ovary? Since species have developed from other species, these regulators of the body, the instincts, cannot have been the same in earlier times; they must have evolved out of the instincts of ancestors, and the questions we have to ask are: By what factors? In what way? Has the principle of selection been operative here too, or can we refer instincts to the inherited effects of use and disuse?
Before I enter upon this question it is necessary to consider for a little the physiological basis of instinct. We can distinguish three kinds of actions: purely reflex, purely instinctive, and purely conscious actions. In the case of the first, we see most clearly that they depend on an existing mechanism, for they follow of necessity on a particular stimulus, and cannot always be suppressed. Bright light striking our eye makes the pupil narrower by a contraction of the iris, and in the same way our eyelids close if a finger be thrust suddenly towards them. We know, too, the principle of these reflex mechanisms; they depend on nerve connexions. Sensory nerves are so connected in the nerve-centres with motor nerves, that a stimulus affecting the former at the periphery of the body, as at the eye, is carried to certain nerve-cells of the brain, and from these it excites to activity certain motor centres, so that definite movements are set up. It is rarely only one muscle that is thus excited to activity, there are usually several, and here we have the transition to instinctive action, which consists in a longer or shorter series of actions, that is, of motor combinations. These, too, are originally, at least, set a-going by a sense impression, an external stimulus which affects a sensory nerve exactly in the same way as in the reflex mechanism, and this stimulus is carried to a particular group of sensory nerve-cells in the central nervous organ, and from these transmitted by very fine inter-connexions to motor centres. There are extraordinarily complex[Pg 143] instinctive actions, and in these the completion of one action is obviously the stimulus to the second, the completion of the second to the third, and so on, until the entire chain of inter-dependent movements which make up the whole performance has been completed.
Instincts have thus a material basis in the cells and fibres of the nervous system, and through variations in the connexions and irritability of these nervous parts they too can be modified, like any of the other characters of the body, such as form and colour.
Conscious actions depend directly on the will, and they have a close connexion with instinctive actions in as far as these also can be controlled by the will, that is, can be set a-going or inhibited, and also, on the other hand, in as far as purely voluntary actions may become instinctive through frequent repetition. The first case is illustrated, for instance, when the suckling of a child at the mother's breast is continued into the second year of life, as not infrequently happens in the southern countries of Europe. Such a child knows exactly why it wants the breast, and its action is a conscious one, while the newborn child seeks about with the mouth instinctively, and when it has found what it sought performs the somewhat complex sucking movements automatically. The second case is illustrated, when, for instance, we have made a habit of winding up a watch on going to bed, and do it when we happen to change our clothes through the day, although it is then purposeless and would have been omitted if the action had required a conscious effort of will. One can often observe on oneself in how short a time a conscious action may become instinctive. I once sent my keyless watch to a watchmaker for repairs, and received from him for the time an ordinary watch, which had to be wound with a key, which key I kept for safety in my purse. At the end of eight days I got back my own watch, and on undressing the first night I found myself 'instinctively' taking my purse from my pocket in order to get the key, which, as I very well knew, I no longer needed. And that a long series of complex movements, originally performed only consciously, may be gone through instinctively, is shown by the fact that pieces of music which have been learnt by heart can often be played without mistake from beginning to end while the player is thinking of quite other things. The complex instinctive actions of animals are performed in quite a similar manner.
There is thus no sharp boundary line between reflex and instinctive actions, nor between instinctive and conscious actions, but one passes over into the other, and the thought suggests itself, that in[Pg 144] the phyletic development also transitions from one kind of action to the other must have taken place. As long as one believes the Lamarckian principle to be really operative one can suppose that actions, which were originally dependent on the will, when they were often repeated, became instinctive, or, in other words, that instincts, many of them at least, are inherited habits.
I shall endeavour to show later on that this assumption, plausible as it seems at first sight, cannot be correct; but in the meantime I must confine myself to saying that there are a great number of instincts which must be referred to the process of selection, and that the rest can be similarly interpreted in their essentials at least.
The instinct of self-preservation is universally distributed, and it is exhibited in many animals by flight from their enemies. The hare flees from the fox and from men; the bird flies away at the approach of the cat; the butterfly flies from even the shadow of the net spread to catch it. These might be regarded as purely conscious actions, and in the case of the hare and the bird experience and will have undoubtedly some part in them, but even in these the basis of the action is an organic impulse; this, and not reflection, causes the animal to flee at sight of an enemy. In the butterfly, indeed, this must be purely instinctive, since it is done with the same precision immediately on leaving the pupa state, before the animal has had any experience. But even in the case of the hare and the bird, taking to flight would in most cases come too late if reflection were necessary first; if it is to be effective it must take place as instantaneously as the shutting of the lids when danger threatens the eye.
The hermit-crab (Fig. 34, p. 163), which conceals its soft abdomen in an empty mollusc shell, and drags that about with it on the floor of the sea, withdraws with lightning-like rapidity into its house as soon as any suspicious movement catches its eye, and it is very difficult to grip one of its legs with the forceps in time to draw it out of its shell. The same is the case with the so-called Serpulids, worms of the genus Serpula, and its allies; it is not easy to seize them, because, however quick one is with the forceps, their instinct of fugitive self-preservation acts more quickly still, and they shoot back into their protecting tubes before one has had time to grasp them. But this impulse to flee from enemies, though it seems almost a matter of course, is by no means common to all animals, for in quite a large number the instinct of self-preservation finds expression in an exactly contrary manner, namely, in the so-called 'death-feigning,' that is, remaining absolutely motionless in a definite position precisely prescribed to the animal by its instinct. In speaking of protective colouring,[Pg 145] I drew attention to the 'wood-moth' (Xylina), which resembles a broken fragment of half-decayed wood so deceptively, and I pointed out that the colour-resemblance to wood would be in itself of but little use to the insect if it were not combined with the instinct to remain motionless in danger, to 'feign death.' The antennæ and legs are drawn close to the body, so that they rather heighten the disguise, and, instead of running away, the insect does not move a muscle until the danger is past. This instinct must have evolved hand in hand with the resemblance to a piece of wood, and, just as we sought to interpret the latter from the fact that the moths which most resembled the wood had always the best chance of surviving, so we maintain that those moths would profit most by their resemblance which drew in their legs and antennæ closely and lay most perfectly still. Thus the brain-mechanism, which effected the keeping still whenever the senses announced danger, would be more and more firmly established and perfected in the course of selection.
Even nearly related animals may have quite different instincts which secure them against danger. Thus in the group of pocket crabs (Notopoda) there are some species which run away when danger threatens, but others which anticipate the risk of discovery by masking themselves to a certain extent. With the last pair of legs they hold over themselves a large piece of sponge, which then grows till it often leaves only the limbs and frontal region uncovered. Of course there can be no question of consciousness in what the crab does, as is proved by the fact that these crabs will, in case of necessity, take a transparent piece of glass instead of the sponge; but the impulse to cover themselves with something is strong in them, and finds expression not only when they see a really protective substance, but even when they see one which is transparent and therefore wholly useless for the purpose. Crabs from which the sponge has been taken away wander about until they find another; the impulse is thus set up not only by the sight of the sponge or of a stone, but also by the feeling that their back is uncovered. The large spider-crab of the Mediterranean (Maja squinado) effects its disguise in a somewhat different manner. It has peculiar hooked bristles on the back, and on these it hooks little bunches of seaweed, often many of them, so that it is entirely covered and looks like a bunch of wrack rather than like an animal. Here again a bodily variation has gone hand in hand with the development of the instinct to cover itself: the bristles of the back have become hooked. Many instincts are accompanied by structural modifications, and in the crabs which cover themselves with sponge or stone this is the case, for the last pair of[Pg 146] thoracic legs is turned towards the back, instead of being set at the side of the body, as is usual among crabs. They are thus enabled to hold the sponge much better and more permanently, and as this is advantageous we may well ascribe the change to natural selection.
Let us now turn our attention to another category of instincts, the most common and most indispensable of all, those which lead to the seeking and devouring of food.
The chicken just emerged from the egg picks up the seeds thrown to it with no experience of what eating is, or what can be made to serve it as food; its instinct for food expresses itself in picking up, and it is awakened or stimulated to action by sight of the seeds. As Lloyd Morgan in his excellent book on Habit and Instinct well says, 'It does not pick at the seeds because instinct says to it that this is something to be picked up and tested, but because it cannot do anything else.'
In the same way the instinct to seek for food wakes in the kitten at the sight of a mouse. I once set before a kitten which had never seen a mouse a living one in a trap. The kitten became greatly excited, and when I opened the trap and let the mouse run away she overtook and caught it in a few bounds. The instinct in this case does not express itself as in the chicken by the rapid lowering of the head and seizing the food, but in quite a different combination of movements, in running after and grasping the fleeing victim. But that is not all that is included in the instinctive action in the case of the cat, for there is also the whole wild and gruesome play, the familiar letting go and catching again, the passionate growling of satisfaction which, in its wildness, reminds us much more of a blood-thirsty tiger than of a tame domestic animal.
As the egg-laying instinct of the female butterfly is excited only by the sight and odour of a particular plant, so also is the food instinct of the caterpillar. If we put a silkworm caterpillar (Bombyx mori) just out of the egg upon a mulberry leaf it will soon begin to gnaw at it; but put it on a beech leaf or on that of any other indigenous tree, shrub, or herb and it will not touch it, but simply die of hunger. And yet it could quite well eat many of these leaves, and thrive on them too, but the smell and perhaps also the sight of them is not the appropriate stimulus to liberate the instinct of eating. There are many species of caterpillar which are 'monophagous,' that is to say, restricted to a single species of plant in a country. One may ask how such a restriction of the liberating stimulus to a single species could have been brought about by natural selection, since it could not possibly be advantageous to be so much restricted in food.[Pg 147] The answer to this will be found in the following facts. On the Belladonna plant there lives a little beetle whose feeding instinct is aroused by this plant alone. But as Atropa belladonna is avoided entirely by other animals on account of its poisonousness, this beetle is, so to speak, sole proprietor of the Belladonna; no other species disputes its food, and in this there must assuredly be a great advantage, as soon as the other instincts, above all that of egg-laying, are so regulated as to secure that the larva shall have access to its food-plant; and this is the case. The monophagy of many caterpillars is to be understood in the same way; it is an adaptation to a plant otherwise little sought after, and it is combined with a more or less complete loss of sensitiveness to the stimulus of other plants. The establishment of such a specialized food-instinct depends on its utility, and has resulted from the preference given, through natural selection, to those individuals in which the food-instinct responded to the stimulus of the smallest possible number of plants, and at the same time to those which showed themselves best adapted to a plant especially favourable to their kind, whose food-instinct was not only most strongly excited by this one plant, but also whose stomach and general metabolism made the best use of it. So we understand why so many caterpillars live on poisonous plants, not only some of our indigenous Sphingidæ, like Deilephila euphorbiæ, but whole groups of tropical Papilionidæ, Danaides, Acræides, and Heliconiidæ. With this again is connected the poisonousness or nauseousness of these butterflies.
How diversely the instinct to procure food may be developed in one and the same group of animals is shown by the fact that not infrequently plant-eating, saphrophytic, and flesh-eating animals occur in a single group of organisms, as, for instance, in the order of water-fleas or Daphnidae, or in the class of Infusorians. Many species find their food by making an eddy in the water, which brings a stream towards the mouth, and with it all sorts of vegetable or dead particles; others live by preying upon other animals like themselves.
But even when the food-instinct in all the species of a group is directed towards living prey, the procuring of it may be achieved by means of quite different instincts. Such finer graduations of the food-instinct are found not infrequently within quite small groups of animals, as in the Ephemeridæ or Day-flies. All their larvæ live by preying on other animals, but those of one family, represented by the genus Chloëon, seek to secure their victims by agility and speed, while the larvæ of the second family, with the typical[Pg 148] genus Baëtis, have the instinct to press their smooth broad bodies, with large-eyed head, close to the brook pebbles on which they sit. They are exactly like these in colour, and thus they lurk almost invisible, until a victim comes within their reach, when they throw themselves upon it with a bound. The third group, with the typical genus Ephemera, follows its instinct to dig deep tubes in the mud at the bottom of the water, and to lurk in these for their prey. We have thus within this small group of Day-flies three distinct modifications of the food-instinct, which differ essentially from one another, are made up of quite different combinations of actions, and, consequently, must have their foundation in essentially different directive brain-mechanisms. All these cases have only one feature in common; the animals all throw themselves upon their prey as soon as they are near enough.
Fig. 31. Sea-cucumber (Cucumaria), with
expanded tentacles (a), and protruded
tube-feet (b); after Ludwig.
But even this common feature is not everywhere part of the food-instinct. The sea-cucumber (Cucumaria) (Fig. 31), according to the observations made on it by Eisig in the Aquarium of the Zoological Station at Naples, gets its food in the following manner. The animal sits half or entirely erect on a projecting piece of rock and unfolds its ten tree-like tentacles which surround the mouth. These are branched, and have quite the effect of little tufts of seaweed. They are probably taken for such by many minute animals; for larvæ of all kinds, Infusorians, Rotifers and worms settle down on them. But the sea-cucumber bends inwards first one tentacle and then another, so slowly as barely to be noticeable, brings the point to its mouth, lets it glide gradually deeper into the gullet, until the whole tentacle is within, and after a time draws it out again equally slowly and unfolds it anew. Obviously it wipes the tentacle inside the gullet, and retains everything living that was upon it. This performance is repeated continually, day and night, and it is usually the only externally visible sign of life which the animal displays.
This remarkable instinct is associated with a structural peculiarity, for without the arborescent tentacles the fishing would not be nearly so successful. Other sea-cucumbers or Holothurians have different tentacles, and use them in quite a different manner, filling the mouth with mud by means of them.
Very frequently, indeed, there are visible structural changes associated with the modified food-instinct. Most predatory fishes chase their prey, like the pike, the perch, and the shark, but there are also lurkers, and these show in addition to the lurking instinct certain definite bodily adaptations, without which this instinct could not have such full play.
Thus in a marine fish known as the 'star-gazer' (Uranoscopus) the eyes are situated not on the sides but on the top of the head, and the mouth is also directed upwards. Its instinct leads it to bury itself in the sand so that only the eyes are uncovered. It lies in wait in this way until a suitable victim comes within reach, and then snaps at it with a sudden movement. But it also possesses a decoying organ, a soft worm-shaped flap, which it protrudes from the mouth as soon as little fishes draw near. They make for this bait, and are thus caught.
Such ingenious fishing, which is quite suggestive of the human method of catching trout with artificial bait, occurs in many predatory fishes; but in every case the fish acts instinctively, without reflection, on becoming aware of approaching prey. The suitability of the action to the end does not depend upon consciousness of the end, or upon reflection, but is a purely mechanical action, performed in response to the stimulus of a sense-impression.
This is best shown by the fact that the instincts may lead their possessors astray, which always happens when an animal is transferred to an unnatural situation, to which its instincts are not adapted, so to speak. The mole-cricket, which is in the habit of escaping pursuit by burrowing in the earth, makes violent motions with the forelegs, even if it be placed upon a plate of glass into which it could not possibly burrow; an ant-lion (Myrmeleo), whose instinct impels it to bore into loose sand by pushing backwards with the abdomen, goes backwards on a plate of glass as soon as danger threatens, and endeavours, with the utmost exertions, to bore into it. It knows no other mode of flight, and its intellect is much too weak to suggest any novel mode. Even the mode of escape most universal among animals, that of simply running away, does not occur to it; it acts as it must in accordance with its inborn instinct; it cannot do otherwise.
The change of instincts in the different stages of development of[Pg 150] one and the same animal have always seemed to me very remarkable; for instance, the change of the food-instinct in the caterpillar and the butterfly, where the food-instinct is liberated in the caterpillar by the leaf of a particular plant, but in the butterfly by the sight and fragrance of a flower, the nectar of which it sucks. In this case everything is different in the two stages of development, the whole apparatus for seeking and taking food, as well as the nerve-mechanism which determines these modes of action. And how far apart often are the stimuli which liberate the instinct! The larva of the flower-visiting, honey-sucking Eristalis tenax is the ugly, white, so-called rat-tailed larva, well described by Réaumur, which lives swimming in liquid manure, and feeds on that! What complete and far-reaching changes, not only in the visible structure, but also in the finer nervous mechanisms, which we cannot yet verify, must have taken place in the vicissitudes of time and circumstance during the life-history of this insect!
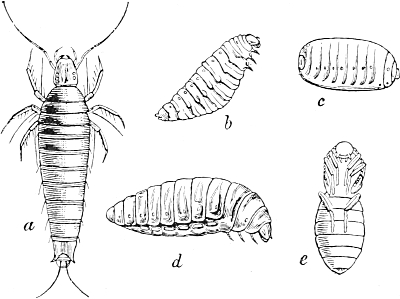
Fig. 32. Metamorphosis of Sitaris humeralis, an oil-beetle, after Fabre. a, first larval form, much enlarged. b, second larval form. c, resting stage of this larva (so-called 'pseudo-pupa'). d, third larval form. e, pupa.
Not the food-instinct alone, but the instinct of self-preservation, of mode of motion, in short, every kind of instinct, may vary in the course of an individual life. Let us follow the somewhat complex life-history of a beetle of the family of the Blister-beetles or Cantharides, as we learnt it first from Fabre. The female of the red-shouldered bee-beetle (Sitaris humeralis) lays its eggs on the ground in the neighbourhood of the underground nest of a honey-gathering burrowing-bee (Anthophora). The larvæ, when they emerge, are agile, six-legged, and furnished with a horny head and biting mouth-parts, as well as with a tail-fork for springing (Fig. 32, a).[Pg 151] The little animals have at first no food-instinct, or at least none manifests itself, but they run about, and as soon as they see a bee of the genus Anthophora they spring upon it and hide themselves in its thick, hairy coat. If they have been fortunate the bee is a female, who founds a new colony and builds cells, in each of which she deposits some honey and lays an egg upon it. As soon as this has been done the Sitaris larva leaves its hiding-place, bites the egg of the bee open, and gradually eats up the contents. Then it moults, and takes the form of a grub with minute feet and imperfect masticating organs; the tail-fork, too, is lost, for all these parts are now useless, since it can obtain liquid nourishment without further change of place, from the honey in the cell, in exactly the quantity necessary to its growth. Then it spends the winter in a hardened, pupa-like skin, and it is not till the next year (the third), after another short larval stage (d) and subsequent true pupahood (e), that the fully-formed beetle emerges. This again possesses biting mouth-parts, and eats leaves, and has legs to run with and wings to fly with.
In this beetle, then, the food-instinct changes three times in the course of its life; first the egg of the bee is the liberating stimulus, then the honey, and finally leaves. The instinct of moving about varies likewise, expressing itself first in running and jumping and in catching on, then in lying still within the cell, and, lastly, in flying and running about on bushes and trees.
We can well understand that, in the course of innumerable generations and species of insects, the various stages of development would, by means of selection, become more and more different from each other, both structurally and in their instincts, as they adapted themselves better to different conditions of life; and thus ultimately instincts frequently and markedly divergent have been developed in the successive stages of life. No other interpretation is possible; through natural selection alone can we understand even the principle of such adaptations. An animal can thus very well be compared to a machine which is so arranged that it works correctly under all ordinary circumstances, that is to say, it performs all the actions necessary to the preservation of the individual and of its kind. The parts of the machine are fitted together in the best possible way, and work on each other so ingeniously that, under normal circumstances, a result suited to the end is achieved. We have seen how precisely the liberating stimulus for an action may be defined, and this secures a far-reaching specialization of instincts. But as every machine can work only with the material for which it was constructed, so the instinct can only call forth an action[Pg 152] effectively adjusted to its end when the animal is under natural conditions. Its specialization has its limits, and in this lies the reason of its limited purposiveness. For instance, if the larva of Sitaris were not impelled by the sight of every bee to spring on it and cling to it, but only by the females, then many of them would be saved from the fate that awaits them if they attach themselves to male bees, which make no nest, or even to other flying insects, in which case also there is no possibility of further development. But both these things happen, although the latter has not yet, to my knowledge, been recorded of Sitaris, but only of its relative, the larva of Melöe.
'Instinct goes astray,' it is often said; but in truth it does not go astray, but is only not so highly specialized in relation to the liberating stimulus of the action as seems to us necessary for perfect purposiveness. But in this very imperfection there lies, as it seems to me, another proof that we have to do with the results of a process of selection, for it is of the very nature of these never to be perfect, but only relatively perfect, that is to say, just as perfect as is necessary to the maintenance of the species. At the moment at which this grade of perfection is reached every possibility of a further increase in the effectiveness of adjustment to the end ceases, because it would then no longer directly further the end. Why, for instance, should the liberating stimulus in this case be more highly specialized, since enough of the Sitaris larvæ already succeed in attaching themselves to female bees? It is not for nothing that the beetles of this family are so prolific; what is lacking in the perfection of the instinct is made up for by the multitude of young larvæ. A single female of the oil-beetle (Melöe) lays several hundred eggs.
In speaking of the animal as a machine, it must be added that it is a machine which can be altered in varying degrees, which can be regulated to work at high or low pressure, slowly or quickly, finely or roughly. This regulating is the work of the intelligence, the limited 'thinking-power,' which must be ascribed to the higher animals in a very considerable degree, but which in the lower animals becomes less and less apparent, until finally it is unrecognizable. That instinctive actions can be modified or inhibited by intelligence and will is proved by any trained beast of prey which masters its hunger and the impulse to snap at the piece of flesh held before it, because it knows that if it does not control itself painful blows will be the consequence. In a later lecture I shall return to the connexion between will and instinct; all that concerns us here is to regard instincts as the outcome of the processes of selection, and as an indirect proof of the reality of these.
From what I have already said at least so much must be clear, that nothing, in principle, stands in the way of referring instincts to selection, since their very essence is their adaptation to an end, and such purposive changes are precisely those that are preserved in the struggle for existence. It might, however, be supposed that in all this the principle of use and disuse also had a share, and that without it no changes in instincts could have come about.
There are, however, numerous instincts in considering which this can be entirely excluded.
At an earlier stage we discussed in detail the protective colourings which secure insects, and especially butterflies, from extermination by their numerous enemies, and it was mentioned that this was always accompanied by corresponding instincts, without which the protective colouring and the deceptive form would have profited nothing, or at any rate not nearly so much. If the caterpillar of the Catocala sponsa, which resembles the bark of an oak so deceptively, did not possess at the same time the instinct to creep away from the leaves and hide in the clefts of the bark on the trunk of the oak-tree, its disguise would be of very little use to it; and if the predatory and grass-coloured praying mantis was not impelled by instinct to lie in wait among the grass for its prey, instead of pursuing it, it would rarely succeed in seizing any of its victims, because of its somewhat leisurely mode of movement. This adaptation of the instincts to the protective colouring is carried into the most minute and apparently trifling details. Thus different observers have established the fact that the nauseous, sometimes even poisonous, butterflies, which are distinguished by their glaring or sharply contrasted colour-pattern, are all slow fliers. This is the case with the Danaides and Euplœides of the Old World and the Heliconiides of the New; many of their mimetic imitators also fly slowly.
If we inquire how this instinct of fluttering, careless flight has come to be, we may leave habit as primum movens out of the question altogether, for there are no external conditions which could have induced the butterfly to take to slower flight than its ancestors exhibited. That it is now advantageous for it—since it acts as a signal of its nauseousness—to be as clearly seen and recognized as possible can exercise no direct influence on its manner of flight, since it knows nothing about it. Even if we assume that individual variations cropped up which had an instinct for slower flight, there would still, without selection, be no reason why this variation in particular should multiply, still less why the originally slight slowing of the flight should become more marked in the course of generations. On[Pg 154] the contrary, the butterflies fly a great deal, just as all other diurnal butterflies do; they exert their power of flight as long as the sun shines, and if the exercise of one generation influences the next, they ought to become gradually more capable of rapid flight. In this case exactly the opposite takes place to what is ascribed to the Lamarckian principle; more constant use must here have brought about a diminution of the activity of the relevant parts. It is quite otherwise when we look at it from the point of view of selection. The variants which cropped up by chance with slower flight survived because they were most easily recognized and avoided; they are the most frequent survivors; they leave descendants which inherit the slower flight-instinct, and this goes on increasing in them as long as the increase carries any advantage with it. As soon as this ceases to be the case the variation comes to a standstill, for it is adapted to the average of the conditions at a given time.
We may picture to ourselves the thousand kinds of regulations of animal movements through instinct as having come about in a similar way; in the majority of cases we must picture it thus. For it is only in the case of those with high intelligence that we can ask whether the animal did not by deliberation help in establishing the purposive variation in its movements. Among insects in any case this could only be taken into account to a very limited extent, although I do not dispute that the more intelligent among them may learn, and may make experiments, and can modify their actions accordingly. But in fleeing from an enemy experience has nothing to do with it, for the first time it is caught it pays the penalty with its life. Without care, and with no idea of the dangers surrounding them on all sides, the butterflies float about, guided only by their instinct, which, however, is so exactly adapted to the conditions of their life that a sufficient number of them to preserve the species always happily escapes all the many dangers. I may remind you of Hahnel's case of the butterfly, already mentioned, which escaped the agile lizard by flying rapidly up from the sweet bait, but settled again upon it without fear immediately afterwards, to fly from the lizard as before, and did so several times in succession. We usually judge such actions far too much from the human standpoint; the butterfly does not wish to escape the death which threatens it; it knows nothing about death; it is not with it as it was with Dr. Hahnel himself, who when he was once in danger from a jaguar in a thicket was so affected by the thought of the death he had happily escaped that he never cared to pass the place again, but made a long circuit to his home. The butterfly does not act according to reflection and imagination; it flies[Pg 155] up with lightning-like rapidity when the lizard rushes at it, because this rapid movement, which it sees, acts as the stimulus which liberates the flight-instinct, and this works so promptly that in most cases the insect is rescued from danger. Its disposition, however, is not otherwise affected by its narrow escape, and it obeys anew the food-instinct which impels it to settle again on the bait, until the flight-instinct is again set a-going by the visual impression of the re-advance of the lizard. It is the plaything of its instincts, a machine which works exactly as it must. That it is only sense-impressions and not conceptions which here liberate the actions can be well seen in the case of shy species of butterfly like our purple emperor (Apatura iris), which flies up like lightning from the moist wood-paths on which it loves to settle as soon as any rapidly moving visual image, even if it be only a shadow, strikes its eyes. For this reason the collector tries to approach it so as not to throw his shadow before him, for then the insect lets the advancing enemy get quite close, and only flies up when the net is quickly thrust towards it. In all probability the eye of this insect is particularly well adapted for perceiving movements, and certainly the flight-instinct reacts very promptly to such visual impressions, and we can understand that it must have been so regulated if, as we assume, the regulation came about through processes of selection, for the enemies of the butterflies, such as birds, dragon-flies, and lizards shoot quickly out on their prey, and therefore those butterflies must always have survived whose instinct impelled them to take to flight most quickly.
In this, then, as in a thousand other cases, the instinct of flight, or indeed any other mode of movement, cannot be interpreted as an 'inherited habit,' because there is no evidence of the possession of that degree of intelligence which could have induced the variation in the previous habit, that is, in manner of movement. The same is true of animals of low intelligence in regard to all the other instincts, which otherwise might seem to be explicable in terms of the Lamarckian principle.
In addition, there is a whole large group of instincts in regard to which the idea of the Lamarckian principle cannot be entertained, as I showed years ago, and it consists of all those instincts which are only exercised once in the course of a lifetime. These cannot possibly depend on practice in an individual lifetime, and transmission of the results of this exercise to the following generation; they can therefore only be interpreted in terms of selection, unless we are to give up all attempts at a scientific interpretation, and simply accept them as 'marvels.'
To this class belong all the diverse instincts by which insects protect themselves against attack during the pupa stage. Even the way in which the caterpillars of many diurnal butterflies hang themselves up in pupation is not by any means a very simple instinctive action. The caterpillar first spins, in a suitable place, a small round disk of silk threads, to which it then attaches the posterior end of its body, so securely that it cannot be easily torn away. More complicated still is the securing of the pupa when it does not hang freely, but is to remain pressed against a wall or a tree, as is the case in the Papilionidæ and the Pieridæ. In this case the caterpillar must, in addition to the usual cradle, spin a thread of silk, in an ingenious way, diagonally across the thorax, so that it may cross about the middle of the wing rudiments, and not be too loose, lest the pupa fall out, yet not too tight, lest the thread cut too deeply into the wing rudiments and hinder their development. When one remembers that it is the caterpillar that does all this, before it has taken the form of the pupa, and that it must all be adapted to the pupa's form, we are amazed at the extraordinary exactness with which instinct prescribes all the individual movements which make the whole of the complex performance effective. And yet, as each caterpillar only accomplishes this performance once in its life, it could at no time in the development of the species have become a habit in the case of any individual caterpillar, and it cannot therefore be an 'inherited habit.'
But however diverse are the methods of securing the safety of the pupæ in the different families of butterflies, they must all be referred back to a single root, if the butterfly pedigree can be traced back to a single ancestral group. The caterpillar of the Sphingidæ does not creep up walls and trees when it is ready to enter on the pupa stage, as so many of the caterpillars of the diurnal butterflies do, but instead its instinct compels it to run about on the ground until it has found a spot which seems to it suited for boring into the earth, or, to speak less metaphorically, until it comes to a place which, from its nature, acts as a liberating stimulus to the instinct to burrow. Then it penetrates more or less deeply, according to the species, and makes a small chamber, which it lines with silken threads to prevent it collapsing; this done, it moults, and enters on the pupa stage. The exactness with which the individual movements are prescribed by instinct is seen in the way in which the size of the chamber is regulated so as to be exactly as large as is necessary to give the pupa room enough without leaving any superfluous free space. This is not so simple as it seems, and is not directly conditioned by the size of the[Pg 157] animal, for the caterpillar is longer and altogether of greater volume than the pupa. The same thing is seen in the stag-beetle (Lucanus cervus), the largest of our indigenous beetles, which gets its name from the powerful antler-like jaws which distinguish the male. It also undergoes its pupal metamorphosis in the earth, and makes a large hard ball of clay, hollow inside, and as smooth as if polished, and its cavity is exactly the size of the future pupa, or to speak more precisely, of the fully-formed beetle. For, as Rösel von Rosenhof in his day 'observed with amazement,' the balls in which the males lie have a much longer cavity than those built by the females, and for this reason, that when the fully-formed beetle emerges from the pupa it must, if it is a male, have room to stretch out its horns, which have till then lain upon the breast. 'For the beetles do not leave their dwelling-place until all their parts are sufficiently strong and properly hardened, and till the season has arrived in which they are wont to fly about.' The male larva thus makes a much longer pupa-house than the female larva, in anticipation, so to speak, of the enormous size of the jaws which will grow out later!
Here the instinct has two modes of expression, according as the bodily parts are male or female. Here we have to do with an action which is performed once in a lifetime, and thus the possibility of any other explanation of the origin of this instinct than through natural selection is excluded.
Not less significant is the case of the silk-cocoons. The cocoons spun by the silkworm are egg-shaped, and consist of a single thread many thousand yards in length, which is wound round the spinning caterpillar so that not a space is left uncovered. The web is firm, tough, and very difficult to tear; therefore we must grant that the pupa resting within will enjoy a very considerable degree of security against injury. But the moth must be able to get out, and that this may be possible the caterpillar is impelled by instinct to make its spinning movements such that the cocoon is eventually looser at the anterior end, so that the insect, when it is ready to emerge, can tear it asunder with its feet and make a way out for itself. For this very reason, because the silk must be torn and spoilt by the emerging insect, silk-breeders kill the pupating insect before it begins to make its way out.
But there are species whose cocoons are provided from the very start with an outlet, for the caterpillar spins the silk round itself in such a way that a round opening is left. But this opening would be not only a convenient door for the butterfly to emerge by, but an equally convenient entrance for all its enemies. It is, therefore, closed[Pg 158] up. In the case of the 'emperor moth' (Saturnia carpini) this is effected by means of a circle of stiff bristles of silk on the inside (Fig. 33), the points of which bend outwards like those of a weir-basket (r); from the inside the emerging moth can easily push aside the bristles, while the threatening enemy from without is scared off by the stiff points of the bristles.
Fig. 33. Cocoon of the Emperor Moth
(Saturnia carpini), after Rösel. A, enclosed
pupa. B, emerging moth. r, hedge of bristles.
fl, wings.
Such a cocoon is comparable to a work of art in which every part harmonizes with the rest, and all together are adapted as well as possible to their purpose. And yet it is all accomplished without the caterpillar having the remotest conception of what it is aiming at when it winds the endless silken thread about itself in the artistic and precisely prescribed coils. Nor has it any time for trying experiments or for learning; it must make all the complex bendings and turnings of the head which spins the thread, and of the anterior part of the body which guides the thread, quite exactly and correctly the first time if a good cocoon is to be produced. Here every possibility of interpreting this instinct as 'an inherited habit' is excluded, for each caterpillar becomes a pupa only once; and it is just as impossible to suppose that it can be directed by intelligence, since it can neither know that it is about to become a pupa, nor that, in the pupa stage, it will be in danger from enemies which will attempt to force their way into the cocoon, nor that the hedge of bristles will protect it from such enemies. Our only clue to an interpretation is in the slow process by which minute useful variations in the primitive instinct of spinning are accumulated through selection; and it is wonderful to see how exactly these spinning powers are adapted to the particular life-conditions of individual species.
Thus there are several of the Saturnides whose enormous caterpillars live on large-leaved trees, and these make use of the large leaves to form a shelter for the pupa stage, spinning them together so that the cocoon is for the most part surrounded by leaf. But as the leaf might easily fall off with the weight of the pupa, they make the leaf-stalk fast to the twig from which it grows by binding the two firmly together with a broad, strong, closely-apposed silken band.[Pg 159] Seitz relates of the largest of all these spinners, the Chinese Attacus atlas, that this silk sheath 'is continued to the nearest strong branch, so that it is impossible with the hand to detach the leaves that conceal an Atlas-pupa from the tree.' To be sure, this pupa weighs about eleven grammes!
Since instincts vary, as well as the visible parts of an animal, a fulcrum is afforded by means of which selection can bring about all these very special adaptations to given conditions, since it always preserves for breeding the best suited variations of an already existing instinct. Any other interpretation is once more excluded.
The same may be said of insects and their egg-laying. This, too, is in many cases only performed once in a lifetime, and the insect dies before it has seen the fruit of its labour. Yet egg-laying is performed in the most effective manner, and with the most perfect security of result. It seems as if the insect knew, so to speak, exactly where, in what numbers, and how it should lay its eggs. Many Mayflies (Ephemeridæ) let their eggs fall all at once into the water in which the larvæ live; many Lepidoptera, such as Macroglossa stellatarum, lay their eggs singly, and on definite plants—the humming-bird hawk-moth, just referred to, on Galium mollugo; others, like Melitæa cinxia, lay their eggs in heaps on the leaves of the way-bread (Plantago media), or, like Aglia tau, on the bark of a large beech-tree. Nothing in these different modes of egg-laying is due to chance or caprice; all is determined and regulated by instinct, and all, as far as we can see, is as well adapted to its purpose as possible. When, for instance, Macroglossa stellatarum lays her eggs singly, or in twos or threes, on the green leaves of the food-plant, it thereby obviates the danger of scarcity of food for the comparatively large caterpillars, since not many of them could subsist together on a single plant of Galium, while Aglia tau can place several hundred eggs on the same beech-tree trunk without having to fear that its caterpillars will not find abundant nourishment. The precision with which the egg-laying instinct works is even greater in other species in which there are more special requirements, e.g. when the eggs have to be laid on the under side of the leaves, as in Vanessa prorsa, or where they have to be cemented together in a little pillar, so that they bear a deceptive resemblance to the green flower-buds of the food-plant (the stinging-nettle).
It is certainly astonishing how exactly the stimulus in these cases is specialized to the liberation of the instinct. In general the smell of the food-plant of the caterpillar is enough for most butterflies, and this attracts the female ready to deposit its eggs, but[Pg 160] complete liberation of the instinct is only effected by the visual impression of the under side of the leaf. We cannot but be astonished that there is room for such finely graded nerve-mechanisms in the little brain of a butterfly, and yet it would be easy enough to adduce still more complex instincts connected with oviposition in insects. The large water-beetle, Hydrophilus piceus, lays its eggs on a floating raft made by itself; the gall-wasps must first pierce with their ovipositor into a particular part of a particular plant to be able to lay the eggs in the proper place, and this in no haphazard way but with great carefulness and in a perfectly definite manner. But there is no necessity to refer here to many or to the most complicated cases of egg-laying; I only wish to show that, even in the simple cases, such as that of the butterflies just referred to, there is a precisely regulated combination of actions which is executed mechanically, and which cannot be interpreted as inherited habit, because it never was a habit in any individual of any generation.
It is thus placed beyond the possibility of doubt that very many instincts, at least, must depend on selection, and it would be useless to go further in this direction by extending our survey to other groups of instincts. I shall, however, return later on to the study of instincts, and, after we have become acquainted with the main features of the laws of inheritance, it will then be seen that, even among higher animals, instincts can never be interpreted in terms of the Lamarckian principle.
ORGANIC PARTNERSHIPS OR SYMBIOSIS
Hermit-crabs and sea-anemones—Hermit-crabs and hydroid polyps—Fishes and sea-anemones—Green fresh-water polyps—Green Amœba—Sea-anemones and yellow Algæ—Cecropia trees and ants—Lichens—Root fungi—Origin of Symbiosis—Nostoc and Azolla apparently contradict the origin through natural selection.
We have already seen, by means of many examples, to what a great degree animals and plants are able to adapt themselves to new conditions of life; how animals imitate their surroundings in colour and form, how instincts have varied in all directions, how plants have made use of the chance of frequent contact with little animals to obtain nourishment from them, and have developed contrivances adapted for bringing as many of these as possible into their power and causing them to yield them the largest possible amount of food. A great many of these could only be interpreted in terms of natural selection, and in others it seemed at least very probable that selection was one of the factors in bringing them about.
Particularly clear proof of the reality of natural selection is afforded by those cases where one form of life associates itself with a very different one so intimately that they are dependent on one another and cannot live without one another—at least in extreme cases—and that new organs, and, indeed, new dual organisms, are sometimes produced by this interdependence of life. This phenomenon—so-called 'Symbiosis'—was discovered by two sharp-sighted botanists, Anton de Bary and Schwendener. But Symbiosis occurs not only between plants; it occurs also between plants and animals and between two species of animal, and we understand by it a life of partnership depending on mutual benefits, so that each of the two species affords some advantage to the other, and makes existence easier for it. In this respect Symbiosis differs from Parasitism, in which one species is simply preyed upon by another without receiving any benefit from it in return, and also from the more innocent Commensalism of Van Beneden, the table-companionship in which one species depends for its existence on the richly-spread table of another. Symbiosis is particularly interesting, because, in addition to extreme cases with marked adaptations, many occur which are of[Pg 162] great simplicity, and which seem to have brought about almost no change in the two associated species.
We shall take our first examples from the Animal Kingdom.
The partnership between certain sea-anemones (Actiniæ) and hermit-crabs (Paguridæ) had been noticed long before any particular attention was devoted to it. Many species of hermit-crab frequently carry a large sea-anemone about with them on the mollusc shell which they use as a protecting-house; indeed, two or three of these beautiful many-tentacled polyps are often attached to them, and this is not at all a matter of chance, but depends upon instinct on the part of both animals; they have the feeling of belonging to each other. If the sea-anemone be taken away from the hermit-crab and put in a distant part of the aquarium, the crab seeks about till it finds it, then seizes it with its claws and sets it on its house again. The instinct to cover itself with Actiniæ is so strong within it that it loads itself with as many of these friends as it can procure, sometimes with more than there is room for on the shell. The sea-anemone on its part calmly submits to the crab's manipulations—a fact very surprising to any one who is aware of the anemone's ordinarily extreme sensitiveness to contact, and knows how it immediately draws itself together on any attempt to detach it from the ground, and will often let itself be torn in pieces rather than give way. The mutual instincts of the two creatures are thus adapted to each other; but it does not at first sight seem as if any structural changes had taken place in favour of the partnership. This is true, indeed, as regards the hermit-crab, but not as regards the sea-anemone, although the nature of the adaptation on the sea-anemone's part only becomes apparent when the two animals are closely observed in their life together.
We owe our understanding of this adaptive change in the sea-anemone, and, indeed, our knowledge of this whole case of Symbiosis, to the beautiful observations of Eisig. Starting from the hypothesis that the mutual relations could only be the outcome of natural selection, Eisig pointed out that this partnership must offer some advantage not to one partner only, but to both; otherwise it could not have arisen through selection. The advantage to the sea-anemone is obvious enough; since of itself it can only move very slowly, and is usually firmly fixed in one place, it is easy to see that it would be useful to it to be carried about on the floor of the sea by the hermit-crab, and to get its share of the hermit-crab's food. But the service yielded to the hermit-crab by the sea-anemone in return is not nearly so apparent. Eisig made an observation in the Zoological Station at Naples which solved this riddle. He saw an octopus attack the[Pg 163] hermit-crab and attempt to draw it out of its shell with the point of one of its eight arms. But before this had succeeded there sprang from the body of the sea-anemone a large number of thin worm-like threads which spread over the arm of the robber, who immediately let go his hold of the crustacean and troubled himself no further with it. These threads, called acontia, are thickly beset with stinging-cells, which must at least cause a violent smarting on the soft skin of the octopus. Thus we see that the Actinia instinctively defends its partner from attacks, and does it so effectively that we need not wonder how the instinct to provide itself with Actiniæ could have arisen in the hermit-crab. But the acontia seem to have been greatly strengthened in the course of the sea-anemone's association with hermit-crabs, for they do not occur in all forms, and they are most highly developed in those which live in Symbiosis with crustaceans.
Fig. 34. Hermit-crab (E), within a Gasteropod shell, on which a colony of Podocoryne carnea has established itself. From the common root-work (which is not clearly shown) there arise numerous nutritive polyps with tentacles (np), among which are smaller 'blastostyle' polyps with a circle of medusoid buds (mk), spine-like personæ (stp), and on the margin of the mollusc shell a row of defensive individuals (wp). F, antennæ. Au, eyes of the hermit-crab; slightly enlarged.
In this case the structural change, the transformation of the mesenteric filaments that occur in all Actiniæ into projectile acontia, is comparatively slight, but in another partnership between hermit-crabs and polyps the latter have undergone a much more marked adaptation. At Naples Eupagurus prideauxii is one of the commonest hermit-crabs. It lives at a depth of about a hundred feet, and is often brought to the Zoological Station by the fishermen in large quantities. Its borrowed mollusc shell often bears a little polyp, Podocoryne carnea (Fig. 34), which forms colonies of often[Pg 164] several hundred individuals, arising from a common root-work of stolons which covers the shell. The polyp colony is composed of different kinds of individuals or personæ, illustrating the principle of division of labour: it includes (1) nutritive persons (np) which possess a proboscis, mouth, and tentacles on their club-shaped bodies; (2) much smaller blastostyles (bl), that is to say, polyps with degenerate mouth and tentacles, which are wholly given over to the production of buds (mk), which then develop into sexual animals, little free-swimming medusoids; and (3) protective personæ in the form of hard spines (stp), beneath the shelter of which the soft polyps withdraw when the mollusc shell is rocked about on the sea-floor by the rolling of the waves. In addition to these three different kinds of individuals or personæ there are also (4) defensive polyps (wp) of long, thread-like shape, thickly set with stinging-cells, but possessing neither mouth nor tentacles. It might at first be thought that these are for the defence of the colony, but this is not so; the fact is that they rather serve for the direct defence of the hermit-crab. This is indicated by the position they occupy in the colony; they are not regularly distributed over the surface, but are ranged round the edge, and, indeed, only on the edge which surrounds the opening of the mollusc shell. Here these defensive polyps stand in close array, sometimes spirally contracted, sometimes hanging loosely down over the hermit-crab like a fringe. Their function, like the acontia of Actiniæ, is to defend the crab when an enemy tries to follow it within the shelter of its domicile. This can easily be demonstrated by drawing out the hermit-crab from the Gasteropod shell, and, when the colony has settled down again, seizing the shell with the forceps and drawing it slowly through the water. The water-stream which then flows upon the shell mimics the attack of an enemy, and immediately all the defensive polyps, as at a given signal, strike from above downwards, and repeat this three or four times; they are scaring off the supposed enemy.
In this species of polyp a special form of individual has developed with a quite definite position in the colony, and furnished with a special instinct or reflex mechanism which is directly useful only to the crab, and has therefore, in a sense, arisen for its advantage. This can quite well be explained through natural selection, for indirectly these polyps are also of use to the colony, inasmuch as they protect their valuable partner, and thus render it possible for the hydroid colony to make the partnership of use to the hermit-crab as well as to itself.
This mutual arrangement thus satisfies the requirement which,[Pg 165] from the selectionist point of view, must be made in regard to all that is new—that it must be useful to its possessor.
If it be asked what service the hermit-crab renders to the polyp colony in return, the answer is that, as in the symbiosis with sea-anemones, the hermit-crab carries the polyps to their food, which is also its own. Hermit-crabs eat all sorts of animal food, living or dead, which they find on the sea-floor, and the remains of their meal fall to the share of the polyps. Once, without special intention, I laid a hermit-crab with its polyp colony in a flat vessel of sea-water beside a bright green living sponge. After some time the majority of the polyps had become bright green; they had filled themselves with the green cells of the sponge.
I do not know how else we should picture to ourselves the origin of symbiotic instincts in such lowly animals except through the transmission and augmentation of variations in the instincts of the two partners—variations which made their possessors more capable of survival. Mollusc shells, ever since there were any, must have served as a foundation and point of attachment for polyp colonies; as a matter of fact, we find to-day on mollusc shells many kinds of polyp colonies which show no special adaptation to a life of partnership with hermit-crabs. From such indifferent associations a symbiotic one must gradually have been evolved in some instances, through the preservation and augmentation of every useful variation, both of instincts and reflex actions, as well as of form and structure. I shall not attempt to trace the course of this evolution in detail, but it is obvious that the development of defensive polyps, and of their instinct to defend the crustacean, can be interpreted neither as due to any direct influence nor as due to the effect of use, but only to the utility of this arrangement, the beginnings of which—polyps with stinging-cells—were already present. Their augmentation and perfecting must be referred entirely to natural selection. It is the same with adaptations which do not refer directly to the crustacean partner, but rather to the disposition of the polyps on the shell. The spinous personæ which protect the softer polyps from being crushed by being rolled about on the pebbles by the waves cannot possibly be regarded as the direct result of this crushing. But it is obvious that some such colonies must have had among their members some with a stronger external skeleton, and therefore less easily crushed than the rest, and this would lead to their more frequent survival.
No adaptation seems to have taken place in the hermit-crab in this case, but that is probably only apparently the case; the probability is that it would not tolerate the presence of the polyp-colony on the[Pg 166] shell unless its instinct compelled it thereto, just as its instinct impels it to cover itself with sea-anemones, and fearlessly to grasp the dangerous animal, which, however, only shows its partner its softer side. Truly, such transformations of instinct are wonderful enough, but that they should have come about through intelligence is here quite inconceivable; there remains nothing but natural selection.
A case in which no apparent corporeal adaptations have occurred, but which depends altogether on slight modifications of the instincts, is afforded by the well-known relations between ants and aphides. These two groups of insects live in a kind of symbiosis, although they are by no means inseparably connected with each other. Wherever strong colonies of aphides cover the young shoots of a plant, such as a stinging-nettle, a rose, or an elder, we almost always find ants which walk cautiously about among the plant-lice, often in great numbers, stopping now and again to stroke them with their antennæ, and then licking up the sweet juice from the intestine which they now give forth. Darwin showed by experiment that the aphides retain this juice if no ants are on the spot, and only give it off when ants are put beside them. Herein lies the proof that we have again to do with a case of modification of instincts. This juice is, of course, not the secretion of special glands, as it was still believed to be in Darwin's time, and it does not come from the so-called 'honey-tubes' situated on the back of the abdomen of the aphides; it is simply their excrement, which is liquid like their food, and the voiding of it has become instinctively connected with the presence of the friendly ants.
That the aphides are not in any way afraid of the ants implies, in itself, a modification of their instinct, for these poisonous insects, prone to biting, are otherwise much dreaded in the insect world. Moreover, the aphides, harmless as they seem, are not quite without means of defence, although these are never used against the ants. Other animals which approach them they bespatter with the sticky, oily secretion prepared in the so-called 'honey-tubes' already noted, squirting it especially into the eyes of an assailant, so that the attack is abandoned.
Of course the aphides have no idea wherein the utility of their friendship for the ants consists, but it is not difficult for us to discover it, since the ants, by their mere presence in the aphid colony, frighten and keep off their enemies. We see, then, that the conditions for a process of natural selection are here afforded: the instinct to be friendly to the ants is thoroughly useful, and the instinct of the ants to seek out the aphides, and, instead of devouring[Pg 167] them, to 'milk' them, is also advantageous; it must be an old acquisition, an instinct early developed, for in several species it has gone so far that the aphides are carried into the ants' nest, and are there (as one might say) kept and tended as domesticated animals.
A pretty case of symbiosis between two animals is reported by Sluiter, and I mention it because it concerns a vertebrate animal, and intelligence has something to do with it. In the neighbourhood of Batavia there are frequently to be found on the coral reefs large yellow sea-anemones, with very numerous and comparatively long tentacles, and a little brightly-coloured fish, of the genus Trachichthys, makes use of these forests, beset with stinging-cells, to find security from its enemies. These appear to be numerous, for in an aquarium, at any rate, the little fish very soon falls a victim to one or other of them, unless he is supplied with the protective sea-anemones. When this is the case it swims blithely about among the tentacles, and the sea-anemone does not sting it; for there has been a modification of instinct on its part as well as on that of the fish. The advantage it gains from the fish is, that the latter brings large morsels of food—in the aquarium, pieces of meat—into the anemone's mouth. In doing so it tears away fibres for itself, and even if the Actinia has swallowed pieces too quickly, the fish pulls them half out of the gullet again, and only relinquishes them to be consumed by its partner when it has satisfied its own appetite. In this case, again, the modification of the instinct is the only adaptation which has been brought about by the symbiosis, and its origin seems difficult to understand. How can the fish have first formed the habit of putting its prey into the mouth of the anemone instead of eating it directly? Although in many cases it is difficult to guess at the beginnings of a process of selection, because they are scarcely discoverable in the subsequently accumulated variations, yet in this instance we may perhaps picture them to ourselves in this way: The fish was in the habit of letting fall pieces of food which could not be swallowed whole, and of diving down upon them repeatedly, to tear off a fragment each time. As the sea-floor in flat places is often covered with sea-anemones, these pieces would often sink down upon one, which would welcome it as a dainty, and set about swallowing it, slowly in its own fashion. The fish must then have found by experience that it could tear off little bits much more easily from a piece that was held firmly by the anemone than from one that was lying loose upon the ground, and this may have caused it to do intentionally what was at first done by chance. But the sea-anemone, suffering no harm from the fish—indeed, its association of[Pg 168] ideas, if I may use the expression, must rather have been little fishes and unexpected food—had no cause to shoot its microscopic arrows at it, and did not do so even when the fish concealed itself among the tentacles. This latter habit on the part of the fish would be developed into an instinct through natural selection, since the individuals that most frequently exhibited it would be the best protected, and therefore, on an average, the most likely to survive. Whether the benevolent attitude of the anemone towards the fish is to be regarded as the expression of an instinct is open to dispute, for it is quite conceivable that each individual sea-anemone is disposed to gentleness by the behaviour of the fish, and so the development of a special hereditary instinct was unnecessary, because without it each anemone reacted in the manner most likely to secure its own advantage[7].
[7] Since the above was written Plate has observed several similar cases in the Red Sea. A little fish lives along with the anemone, Crambactis aurantiaca, a foot in size, and not only conceals itself among its tentacles, but remains among them when the anemone draws them in. These fishes, therefore, must be immune against the stinging-cells of the sea-anemone; and in the same way another species of fish appears to be immune from the strong poison secreted by sea-urchins of the genus Diadema from the points of their spines, among which the fishes live. This relation certainly seems more like a one-sided adaptation on the part of the fishes than a true symbiosis, but in the cases observed by Sluiter the return service of the fishes seems to be regularly rendered. Here, as everywhere else in nature, there are transition stages, and a one-sided protective relation may gradually, under favourable circumstances, be transformed into a symbiosis.
The same may be true of the fish as far as laying its booty in the mouth of the anemone is concerned; there may be no inherited instinct in this; it may be an intelligent action, which is learnt anew in the lifetime of each individual.
It might of course be objected to this interpretation that the beginning of the process, namely, the assumption that chance fragments from the food of the fish falling just on the anemone is very improbable; but I once observed that flat rocks washed over by the sea on the Mediterranean coast (not far from Ajaccio) were so thickly covered with green anemones that at first I took the green growth for some strange sea-grass new to me until I had pulled up a little tuft of the supposed plants and identified them as the soft tentacles of Anthea cereus. Anemones must be equally abundant in the tropical seas of Java, and a sinking fragment must often alight on the mouth of one of them.
Much attention and keen discussion have in the last few decades been focussed on cases of symbiosis between unicellular Algæ and simple animals. A good example is our green fresh-water polyp, Hydra viridis (Fig. 35, A). Its beautiful colour is due to chlorophyll, and it was long a matter of surprise that animals should produce[Pg 169] chlorophyll, which is a characteristic and fundamental important substance of assimilating plants, until Geza Entz and M. Braun demonstrated that the green did not belong to the animal at all, but to unicellular green Algæ, so-called Zoochlorellæ, which are embedded in the endoderm cells of the polyps in great numbers (Fig. 35, zchl). As these algoid cells assimilate, and thus liberate oxygen, their presence is of advantage to the polyp. That—as was at first believed—they also yield nourishment to the polyp I consider very probable, notwithstanding the apparently opposed results of the experiments of so acute an observer as von Graff, for I have seen a large number of these animals thrive for months, and multiply rapidly by budding in pure water which contained no food of any kind. In favour of this view, too, are some observations, to be cited presently, on unicellular animals, in regard to whose nourishment by the zoochlorellæ living within them there can be no doubt at all.
Fig. 35. Hydra viridis, the Green Fresh-water Polyp. A, the entire animal, greatly
enlarged. M, the mouth. t, tentacles. sp, testis. ov, ovary, both in the ectoderm. Ei, a
ripe ovum, already green, in process of being extruded. After Leuckart and Nitsche.
B, section of the body-wall, about the position of the ovary in A. Eiz, the ovum
lying in the ectoderm (ect), in which zoochlorellæ (zchl), belonging to the endoderm
(ent), have already migrated through the supporting middle lamella (st). eik, nucleus
of ovum. After Hamann.
The little algæ on their part find a peaceful and relatively secure abode within the polyp, and they apparently do not occur outside of it, at least they do not now migrate from outside into the animal, but[Pg 170] are carried over as a heritable possession of the polyps from one generation to another, and in a very interesting manner, namely, by means of the eggs, and by these alone. As Hamann has shown, the zoochlorellæ migrate at the time when an egg is formed in the outer layer of the body of the polyp (Fig. 35) from the inner layer outwards, piercing through the supporting layer between them (st) and penetrating into the egg (B, Eiz). They make their way only into the egg, not into the sperm-cells, which in any case are too small to include them. Thus they are absent from no young polyp of this species, and it is easy to understand why earlier experimental attempts to rear colourless polyps from eggs could never succeed even in the purest water.
Fig. 36. A, Amœba viridis. k, the
nucleus. cv, contractile vacuole. zchl,
the zoochlorellæ. B, a single zoochlorella
under high power. After
A. Gruber.
Quite similar green algæ live in symbiosis with unicellular animals, as, for instance, with an amœba (Fig. 36) and with an Infusorian of the genus Bursaria. In the Zoological Institute in Freiburg there is a living colony of a green amœba and a green Bursaria, both of which came from America, sent to us some years ago by Professor Wilder, of Chicago, inside a letter with dried Sphagnum, or bog-moss. The plants came from stagnant water in the Connecticut valley in Massachusetts. That in this case the zoochlorellæ are of use to the animals within which they live, not only by giving off oxygen, but also by yielding food-stuff, has been proved by A. Gruber, who bred the two green species for seven years in pure water which contained no trace of any kind of organic food for them. Nevertheless, they multiplied rapidly, and still form a green scum on the walls of the glass in which they are kept. They only die away when they are kept in the dark, where the algæ are unable to assimilate; then one green cell after another wanes and disappears, and, in consequence, their hosts also die from the double cause of lack of oxygen and lack of food.
Even in this case the symbiotically united organisms have not remained unaltered. The algæ at least differ from others of their kind in their power of resistance to living animal protoplasm. They are not digested by it, and we may infer from this that they possess some sort of protective adaptation against the dissolving power of[Pg 171] animal digestive juices; they must, therefore, have undergone some variation, and adapted themselves to the new situation. Probably their cell-membrane has become impenetrable to the stuffs which would naturally digest them, an adaptation which could not be referred to direct effect or to use, but only to the accumulation of useful variations which cropped up—in other words, to natural selection. That any adaptive variation has taken place on the part of the host, whether polyp, amœba, or Infusorian, cannot be made out. None of these have altered their original mode of life; they do not depend on the nourishment afforded by the algæ, but feed on other animals, if these come in their way, and they live in water rich in oxygen like other species allied to them, and therefore are not altogether dependent on the algæ in this connexion; but they can no more help having their partners than the pig can help having Trichinæ in its muscles.
Similar plant-cells, not green however, but yellow, called zooxanthellæ, live in great numbers in the endoderm of various sea-anemones and in the soft plasmic substance of many Radiolarians. In both these cases we must look for the benefit they confer on their host in the oxygen they give off, for, like the green zoochlorellæ, they break up carbonic acid gas in the light, and give off oxygen; they no longer occur, as far as is known, in a free state, but are always associated with the host, and they must therefore have altered in constitution, and have adapted themselves to the conditions of the symbiosis.
Higher plants, too, sometimes have symbiotic relations with animals; the most remarkable and best-known example is the relation between ants and certain trees, in which the ants protect trees which afford them in return both a dwelling-place and food. We owe our knowledge of these cases to Thomas Belt and Fritz Müller, and more recently it has been materially increased by Schimper's researches.
In the forests of South America there grow 'Imbauba,' or candelabra-trees, species of the genus Cecropia, which well deserve their name, for their bare branches stretch out like candelabra, and bear little bunches of leaves only at their tips. These leaves are menaced by the leaf-cutting ants of the genus Œcodoma, which attack numerous species of plants in these regions, often in tens of thousands, biting off the leaves, cutting them in pieces on the ground, and carrying them on their backs piece by piece to their nests. There they use them to make a kind of compost heap, on which fungi, to which the ants are very partial, readily grow. The candelabra-tree protects itself from these dangerous robbers, inasmuch as it has established an association with another ant (Azteca instabilis), which finds a safe[Pg 172] dwelling-place in its hollow, chambered stem (Fig. 37, A), and feeds on a brown sap which oozes from the inside. On the stem there are even little pits regularly arranged in definite places (E), through which the female of Azteca can easily bore her way into the interior. There she lays her eggs, and soon the whole interior of the trunk teems with ants, which come trooping out whenever the tree is shaken.
Fig. 37. A, a piece of a twig of an
Imbauba-tree (Cecropia adenopus), with
the leaves cut off. At the leaf-bases are
the hair-cushions (P). E, the opening
for the associated ant (Azteca instabilis).
B, a piece of the hair-cushion with the
egg-shaped nutritive corpuscles (nk).
After Schimper.
This alone would not suffice to protect the tree against the leaf-cutting ants, for how should the Aztec ants living inside notice the presence of the lightly climbing leaf-cutters? But that is provided for, for the Aztecs also frequent the outside of the trunk, and just where attack would be most disastrous, namely, at the stalks of the young leaves. At these places there is a peculiar velvet-like cushion of hair (P), from which grow little stalked white papillæ (Fig. 37, B), which are rich in nourishment, and are not only eaten by the ants, but are harvested by them, being carried off into the ants' dwellings, presumably to feed their larvæ. In this case, then, a particular organ, offering special attraction to ants, has been developed by the plant at the places more especially threatened; while, as regards the ants, it is probable that only the instincts of feeding and habitat require to be modified, since courage and thirst for battle are present in all ants, almost any species being ready at any time to throw itself on any other which intrudes into its domain.
It should be noted that not all the candelabra-trees live in symbiosis with ants, and so secure a means of defence against the leaf-cutters. Schimper found in the primitive forests of South America several species of Cecropia which never had ants in the chambers of their hollow stem. But these species did not exhibit the nutritive cushions at the base of the leaf-stalk; these contrivances for attracting and retaining the presence of partner ants were altogether absent. Indeed, only one species, Cecropia peltata, has produced these[Pg 173] peculiar structures, and, as they are of no direct use to the tree, we must say that it has produced them only for the ants. Here, again, natural selection must have gradually brought about the development of these nutritive cushions, though as yet we do not know what the beginnings of the process may have been. In no case can the origin of these cushions be referred to any direct influence of the environmental conditions.
We may now pass to the association of two species of plants, of which the lichens furnish the best-known and probably most complete illustration. Till about twenty years ago the lichens, which in so many diverse forms clothe the bark of trees, the stones, and the rocks, were regarded as simple plants like the flowering plants, the ferns, or the mosses; and many lichenologists occupied themselves with the exact systematic distinction of about a thousand species, each of which could be as well and exactly classified, according to form, colour, habitat, and minute structure, as any other kind of plant. Then De Bary and Schwendener discovered that the lichens were made up of two kinds of plants, fungi and algæ, so intimately associated with and adapted to one another, that on coming together they always assume the same specific form.
Fig. 38. A fragment of a Lichen (Ephebe kerneri), magnified 450 times.
a, the green alga-cells. P, the fungoid filaments. After Kerner.
The framework, and therefore the largest part, and the one which determines the form of a lichen, is due to the fungus (Fig. 38). Colourless threads of fungus ramify in a definite manner according to the species of fungus, and in the network of spaces left by this ramification green alga-cells (a) lie singly, or in rows, or groups. The fungus is propagated by multitudes of minute spores, which it[Pg 174] produces periodically, and these are disseminated in the air by the bursting of the sporangia and are carried away by the wind in the form of fine dust; the alga multiplies simply by continual division into two, but it also, like the whole lichen, can survive desiccation, and, after falling to pieces, is likewise carried through the air as microscopic dust.
The partnership of the two plants rests on a basis of mutual benefit; the fungus, like all fungi, is without chlorophyll, and cannot therefore decompose carbonic acid gas or elaborate its own organic food-stuffs; it receives these from the alga. The alga has in the network of the fungus a safe shelter and basis of attachment, for the fungus is able to bore into the bark of trees and even into stones; besides which it absorbs water and salts, and supplies these to the partner alga. We here see the mutual advantage derived from the partnership, which is really an extremely intimate one. Fungus spores, sown by themselves, spring up and develop some branchings of fungoid hyphæ, a so-called mycelium, but without the requisite partner alga these remain weak and soon die away. The alga, on the other hand, can, in some cases, though not in all, survive without the fungus if the necessary conditions of its life be supplied to it, but it grows differently and more luxuriantly in association with the fungus.
The same species of alga may be found associated with different species of fungi, and then each partnership forms a distinct species of lichen of definite and characteristic appearance; Stahl even succeeded in making new species of lichen artificially by bringing the spores of a lichen-forming fungus into contact with alga-cells, with which they had never been associated in free nature.
The most remarkable feature of this remarkable association seems to me to be the formation of common reproductive bodies—an adaptation in face of which all doubt as to the theory of selection must disappear. Periodically there are developed in the substance of the lichen small corpuscles, the so-called soredia, each of which consists of one or more alga-cells surrounded and kept together by threads of the fungus. When they are developed in large numbers they form a floury dust over the maternal lichen, which 'breaks up' and leaves them, like the spores of the fungus, to be carried away by the wind. If these alight on favourable soil nothing more is needed than the external conditions of development, light, warmth, and water, to enable the lichen to spring up anew. The great advantage to the preservation of 'species' is obvious, for, when multiplication by the ordinary method occurs among lichens, the spores of the fungus,[Pg 175] even if they have fallen on good ground, can only develop into a new lichen if chance bring to them the proper partner alga.
Obviously there must be, in the formation of the soredia, great advantage for the species, or rather 'for the two species,' for the fungi as well as the algæ benefit by the arrangement, which ensures the continuance of the partnership. It was not without reason, however, that the dual organism was so long regarded as a simple species in the natural history sense, for that is what it really is, although it has arisen in a manner quite different from the usual origin of species. As we know species which consist only of single cells, and others which consist of many cells, differentiated in different ways, and forming a cell-community or 'person,' and, finally, others which consist of a community of diversely differentiated personæ, making up a 'stock'; so in the lichens we see that even different species may combine to form a new physiological whole, a vital unit, an individual of the highest order. When, at the outset of these lectures, I said that the theory of evolution was now no longer a mere hypothesis, and that its general truth could no longer be doubted by any one acquainted with the facts available, I had in my mind, among other facts, especially that of symbiosis, and above all the case of the lichens.
There are many other interesting cases of symbiosis between two different kinds of plants, and one side of the partnership is represented by fungi in a relatively large number of instances. The reason is not far to seek: fungi must always be dependent on other plants for their food; they must be parasitic, because they cannot themselves produce the organic substances they require. They must therefore associate themselves in some way with other organisms, living or dead, and as a general rule they simply prey upon their associate, sucking up its juices and killing it. But in not a few cases they can render services in return, and, as we have seen in the case of the lichens, symbiosis may then occur. Fungi in general have the power of discovering and absorbing the least trace of water in the soil, and with it they absorb the salts necessary to the plant, and in this, apparently, consists the service which they are able to render even to large plants fixed deep in the earth, such as shrubs and trees. The roots of many of our forest trees, e.g. beech, oak, fir, silver poplar, and bushes like broom, heaths, and rhododendrons, are thickly wrapped round with a network of fungoid threads, and the mutual relations just indicated exist between these and the plants in question (Fig. 39, A and B). The plants give to the fungi some contribution from the superfluity of their food-stuffs, and receive in return water[Pg 176] and salts, which are of value especially in times of drought. Perhaps there is some connexion between this and the fact that limes wither and lose their leaves so quickly during great summer-heat; these and many other of our trees possess no root-fungi or mycorhizæ.
It is easy to understand, therefore, that genuine 'symbiosis' may have arisen from parasitism. But that this is not the only path that leads to symbiosis is shown by the cases of animal symbiosis we have already discussed.
Fig. 39. A, fragment of a Silver Poplar root, with an envelope of symbiotic fungoid filaments (mycelium); after Kerner. B, apex of a Beech root, with the closely enveloping mantle of mycelium; enlarged 480 times.
The partnership between polyps and hermit-crabs may have arisen from a one-sided commensalism, since polyps establishing themselves on mollusc shells which were often made use of by hermit-crabs would be better fed than those which settled down on stones. There are still species which make use of both modes of settlement. Then followed the adaptation of the crustacean to the polyp, for, first, those hermit-crabs would thrive best which tolerated the presence of the polyp; then those which sought its presence, that is to say, which gave a preference to shells covered with polyps; and, finally, those which would take no others, and even themselves fixed the sea-anemone upon it, if it chanced to be removed. Intelligence need not be taken into account in the matter at all, not even in the hermit-crab's case. We have only to recall the complex instincts, exercised only once in a lifetime, which compel the silkworm and the emperor moth to elaborate their effective cocoons. The elaboration of the spinning-instinct can only be due to natural selection, for the insect can have had no idea of the utility of its performance, and the same is true in the case of the sea-anemones or[Pg 177] the hydroid polyps and the hermit-crab. The sea-anemone is quite unconscious that it is defending its partner, the hermit-crab, when it lashes out its stinging acontia on any disturbance, and the hermit-crab is equally unaware that the sea-anemone is contributing to its safety; both animals act quite unconsciously, purely instinctively, and the origin of these instincts, on which the symbiosis is based, must be due, not to intelligent activities which have become habitual, but only to the survival of the fittest.
According to the principle of natural selection nothing can arise but that which is of use directly or indirectly to its possessor. Nevertheless, there are cases in which it appears as if something had arisen, which was of no use to the species in which the variation appeared, but only to the species protected by it. This is the case in the remarkable symbiosis between algæ of the family Nostocaceæ and the floating, moss-like water-fern Azolla. This plant, in external appearance almost like duckweed, has on the under surface of its leaves a minute opening, leading into a relatively roomy hair-lined cavity, and in this cavity there is always, enclosed in jelly, a bluish green unicellular alga, Anabæna. The cavity is present in every leaf, and the alga is present in every cavity, making its way in from a deposit of alga-cells which is found on the incurved tip of every young shoot. As soon as a young leaf of Azolla unfolds from the bud it receives its Anabæna cells from this deposit, and no one has yet found either twigs or leaves which were free from the algæ. But no one has succeeded in discovering any benefit derived by the Azolla from this partnership.
This looks like a contradiction of the theory of selection, but there remains the possibility that there is some benefit rendered to the Azolla by the alga, though we cannot see it as yet. There is also the possibility that the cavity is an organ which was of use to the plant at an earlier time, perhaps as an insect-trap, but has now lost its significance, and is utilized by the alga as a dwelling-place. This, however, is contradicted by the remarkable distribution of the four known species of Azolla. Two of these are widely distributed in America; the third lives in Australia, Asia, and Africa; the fourth in the region of the Nile: all four have cavities in their leaves, and in all these forms the cavity is inhabited by the same species of Anabæna. This indicates that the leaf-cavity and the partnership with the alga must have originated in remote antiquity; the symbiosis must date from a time before the four modern species of Azolla had split off from a single parent-species. But no rudimentary organ, that is to say, no organ not of use to the plant itself,[Pg 178] would have been preserved through such a vast period of time, as we shall see later, for useless organs disappear in the course of ages. As the cavity has not yet disappeared, we may assume with some probability that it is useful to the plant, whether by means of the Anabæna, or in some other unknown way. To draw an argument against the reality of the processes of selection from our lack of knowledge of what this advantage may be would be as unreasonable as if, notwithstanding our experience that stones sink in the water, we were to assume of a particular stone which we did not see sink, because it was hidden from our sight by bushes, that perhaps it had not sunk, but was capable of floating.
THE ORIGIN OF FLOWERS
Introduction—Precursors of Darwin—Pollination by wind—Arrangements in flowers for securing cross-fertilization—Salvia, Pedicularis—Flowers visited by flies—Aristolochia—Pinguicula—Daphne—Orchids—Flowers are built up of adaptations—Mouth-parts of insects—Proboscis of butterflies—Mouth-parts of the cockroach—Of the bee—Pollen baskets of bees—Origin of flowers—Attraction of insects by colour—Limitation of the area visited—Nägeli's objection to the theory of selection—Other interpretations excluded—Viola calcarata—Only those changes which are useful to their possessors have persisted—Deceptive flowers—Cypripedium—Pollinia of Orchis—The case of the Yucca-moth—The relative imperfection of the adaptations tells in favour of their origin through natural selection—Honey thieves.
When one species is so intimately bound up with another that neither can live for any length of time except in partnership, that is certainly an example of far-reaching mutual adaptation, but there are innumerable cases of mutual adaptation, in which, although there is no common life in the same place, yet the first form of life is adjusted in relation to the peculiarities of the second, and the second to those of the first. One of the most beautiful, and, in regard to natural selection, the most instructive of these cases is illustrated by the relations between insects and the higher plants, relations which have grown out of the fact that many insects have formed the habit of visiting the flowers of the plants for the sake of the pollen. In this connexion the theory of selection has made the most unexpected and highly interesting disclosures, for it has informed us how the flowers have arisen.
In earlier times the beauty, the splendour of colour, and the fragrance of flowers were regarded as phenomena created for the delight of mankind, or as an outcome of the infinite creative power of Mother Nature, who loves to run riot in form and colour. Without allowing our pleasure in all this manifold beauty to be spoilt, we must nowadays form quite a different conception of the way in which the flowers have been called into being. Although here, as everywhere else in Nature, we cannot go back to ultimate causes, yet we can show, on very satisfactory evidence, that the flowers illustrate the reaction of the plants to the visits of insects, and that they have been in large measure evoked by these visits. There might, indeed,[Pg 180] have been blossoms, but there would have been no flowers—that is to say, blossoms with large, coloured, outer parts, with fragrance, and with nectar inside, unless the blossoms had been sought out by insects during the long ages. Flowers are adaptations of the higher flowering plants to the visits of insects. There can be no doubt about that now, for—thanks to the numerous and very detailed studies of a small number of prominent workers—we need not only suppose it, we can prove it with all the certainty that can be desired. The mutual adaptations of insects and flowers afford one of the clearest examples of the mode of operation and the power of natural selection, and the case cannot therefore be omitted from lectures on the theory of descent.
That bees and many other insects visit flowers for the sake of the nectar and pollen has been known to men from very early times. But this fact by itself would only explain why adaptations to flower-visiting have taken place in these insects to enable them, for instance, to reach the nectar out of deep corolla-tubes, or to load themselves with a great quantity of pollen, and to carry it to their hives, as happens in the case of the bees. But what causes the plants to produce nectar, and offer it to the insects, since it is of no use to themselves? And further, what induces them to make the pillage easier to the insects, by making their blossoms visible from afar through their brilliant colours, or by sending forth a stream of fragrance that, even during the night, guides their visitors towards them?
As far back as the end of the eighteenth century a thoughtful and clear-sighted Berlin naturalist, Christian Konrad Sprengel, took a great step towards answering this question. In the year 1793 he published a paper entitled 'The Newly Discovered Secret of Nature in the Structure and Fertilization of Flowers[8],' in which he quite correctly recognized and interpreted a great many of the remarkable adaptations of flowers to the visits of insects. Unfortunately, the value of these discoveries was not appreciated in Sprengel's own time, and his work had to wait more than half a century for recognition.
[8] Das neu-entdeckte Geheimniss der Natur im Bau u. der Befruchtung der Blumen, Berlin, 1793.
Sprengel was completely dominated by the idea of an all-wise Creator, who 'has not created even a single hair without intention,' and, guided by this idea, he endeavoured to penetrate into the significance of many little details in the structure of flowers. Thus he recognized that the hairs which cover the lower surface of the[Pg 181] petals of the wood-cranesbill (Geranium sylvaticum) protect the nectar of the flower from being diluted with rain, and he drew the conclusion, correct enough, though far removed from our modern ideas as regards the directly efficient cause, that the nectar was there for the insects.
He was also impressed by the fact that the sky-blue corolla of the forget-me-not (Myosotis palustris) has a beautiful yellow ring round the entrance to the corolla-tube, and he interpreted this as a means by which insects were shown the way to the nectar which is concealed in the depths of the tube.
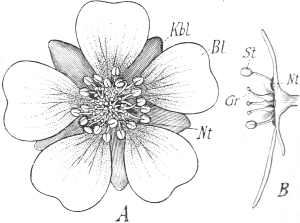
Fig. 40. Potentilla verna, after Hermann Müller. A,
seen from above. Kbl, sepals. Bl, petals. Nt, nectaries
near the base of the stamens. B, section through the
flower. Gr, stigma. St, stamen. Nt, nectary.
We now know that such 'honey-guides' are present in most of the flowers visited by insects, in the form of spots, lines, or other marking, usually of conspicuous colour, that is, of a colour contrasting with the ground colour of the flower. Thus, in species of Iris, regular paths of short hairs lead the way to the place where the nectar lies. In the spring potentilla (Potentilla verna) (Fig. 40) the yellow petals (A, Bl) become bright orange-red towards their bases, and this shows the way to the nectaries, which lie at the bases of the stamens (st), and are protected by hairs, the so-called 'nectar-covers' (Saftdecke) of Sprengel, from being washed by rain.
The recognition of the honey-guides led Sprengel on to the idea that the general colouring of the flower effects on a large scale what the honey-guides do in a more detailed way—it attracts the attention of passing insects to where nectar is to be found; indeed, he went an important step further by recognizing that there are flowers which cannot fertilize themselves, in which the insect, in its search for honey, covers itself with pollen, which is then rubbed off on the stigma of the next flower visited, fertilization being thus effected. He demonstrated this not only for the Iris, but for many other flowers, and he drew the conclusion that 'Nature does not seem to have wished that any flower should be fertilized by its own pollen.'[Pg 182] How near Sprengel was to reaching a complete solution of the problem is now plain to us, for he even discovered that many flowers, such as Hemerocallis fulva, remained infertile if they were dusted with their own pollen.
Even the numerous experiments of that admirable German botanist, C. F. Gärtner, although they advanced matters further, did not suffice to make the relations between insects and flowers thoroughly clear; for this the basis of the theory of Descent and Selection was necessary. Here, again, it was reserved for Charles Darwin to lead the way where both contemporaries and predecessors had been blindly groping. He recognized that, in general, self-fertilization is disadvantageous to plants; that they produce fewer seeds, and that these produce feebler plants, than when they are cross-fertilized; that, therefore, those flowers which are arranged to secure cross-fertilization have an advantage over those which are self-fertilized. In many species, as Sprengel had already pointed out, self-fertilization leads to actual infertility; only a few plants are as fertile with their own pollen as with that of another plant; and Darwin believed that, in all flowering plants, crossing with others of the same kind, at least from time to time, is necessary if they are not to degenerate.
Thus the advantage which the flowers derive from the visits of insects lies in the fact that insects are instrumental in the cross-fertilization of the flowers, and we can now understand how the plant was able to vary in a manner favourable to the insect-visits, and to exhibit adaptations which serve exclusively to make these visits easier; we understand how it was possible that there should develop among flowers an endless number of contrivances which served solely to attract insects, and even how, for the same end, the insignificant blossoms of the oldest Phanerogams must have been transformed into real flowers.
We must not imagine, however, that the obviously important crossing of plant-individuals, usually called 'cross-pollination,' can be effected only by means of insects. There were numerous plants in earlier times, and there is still a whole series in which cross-fertilization is effected through the air by the wind; these are the anemophilous or wind-pollinated Angiosperms.
To these belong most of the catkin-bearers, such as hazel and birch, and also the grasses and sedges, the hemp and the hop, and so forth. In these plants there is no real flower, but only an inconspicuous blossom, without brightly-coloured outer envelopes, without fragrance or nectar; all of them have smooth pollen grains, which[Pg 183] easily separate into fine dust and are carried away by the wind until they fall, by chance, far from their place of origin, on the stigma of a female blossom.
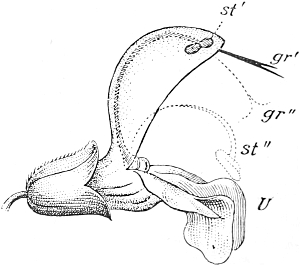
Fig. 41. Flower of Meadow Sage (Salvia pratensis),
after H. Müller. st´, immature anthers concealed
in the 'helmet' of the flower. st´´, mature anther
lowered. gr´, immature stigma. gr´´, mature
stigma. U, the lower lip of the corolla, the
landing-stage for the bee.
By far the greater number of the phanerogams, however, especially all our indigenous 'flowers,' are, as a rule, fertilized by means of insects, and it is amazing to see in what diverse ways, often highly specialized, they have adapted themselves to the visits of insects. Thus there are flowers in which the nectar lies open to view, and these can be feasted on by all manner of insects; there are others in which the nectar is rather more concealed, but still easily found, and reached by insects with short mouth-parts, e.g. large flowers blooming by day and bearing much pollen, like the Magnolias. These have been called beetle-flowers, because they are visited especially by the honey-loving Longicorns.
Other flowers blooming by day are especially adapted to fertilization by means of bees; they are always beautifully coloured, often blue; they are fragrant, and contain nectar deep down in the flower, where it can only be reached by the comparatively long proboscis of the bee. Different arrangements in the different flowers secure that the bee cannot enjoy the nectar without at the same time effecting the cross-pollination. Thus the stamens of the meadow sage (Salvia pratensis) are at first hidden within the helmet-shaped upper lip of the flower (Fig. 41, st´), but bear lower down on their stalk a short handle-like process, which turns the pollen-bearing anther downwards (st´´) as soon as it is pressed back by an intruding insect. The pollen-sacs then strike downwards on the back of the bee, and cover it with pollen. When the bee visits another more mature flower, the long style, which was at first hidden within the helmet, has bent downwards (gr´´), and now stands just in front of the entrance to the flower, so that the bee must rub off a part of the pollen covering its back on to the stigma, and fertilization is thus effected.
There are other flowers which are specially disposed to suit the[Pg 184] visits of the humble-bees, as, for instance, Pedicularis asplenifolia, the fern-leaved louse-wort, a plant of the high Alps (Fig. 42). The first thing that strikes us about this plant is the thickly tufted hair covering on the calyx (k), which serves to keep off little wingless insects from the flower; then there is the strange left-sided twisting of the individual flowers, whose under lip allows only a strong insect like the humble-bee to gain access, towards the left, to the corolla-tube (kr), in the depths of which the nectar is concealed. While the humble-bee is sucking up the nectar it becomes dusted over with pollen from the anthers, which falls to dust at a touch, and when it insinuates itself into a second flower its powdered back comes first into contact with the stigma of the pistil (gr) which projects from the elongated bill-shaped under lip, dusting it over with the pollen of the first visited flower. Butterflies and smaller bees cannot rob this flower; it is strictly a humble-bee's flower.
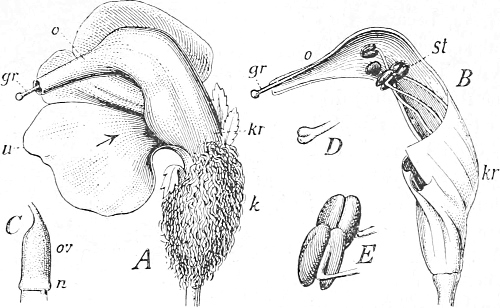
Fig. 42. Alpine Lousewort (Pedicularis asplenifolia). A, flower seen from the left side, enlarged three times; the arrows show the path by which the humble-bee enters. B, the same flower, seen from the left, after removal of the calyx, the lower lip and the left half of the upper lip. C, ovary (ov), nectary (n), and base of style. D, tip of style, bearing the stigma. E, two anthers turned towards one another. o, upper lip. u, lower lip. gr, style. st, anthers. kr, corolla-tube. k, calyx.
There are not a few of such flowers adapted to a very restricted circle of visitors, and in all of them we find contrivances which close the entrance to all except what we may call the welcome insects; sometimes there are cushions of bristles which prevent little insects from creeping up from below, or it is the oblique position of the flower which prevents their getting in from the stem; sometimes it is the length and narrowness of the corolla-tube, or the deep and hidden situation of the nectar, which only allows intelligent insects to find the treasure.
Very remarkable are those flowers which are adapted to the visits of flies, for they correspond in several respects to the peculiarities of these insects. In the first place, flies are fond of decaying substances and the odours given off by these, and so the flowers which depend for their cross-fertilization on flies have taken on the dull and ugly colours of decay, and give out a disagreeable smell. But flies are also shy and restless, turning now hither, now thither, and cannot be reckoned among the 'constant' insect visitors, that is to say, they do not persistently visit the same species; it is, therefore, evident that they might easily carry away the pollen without any useful result ensuing. Moreover, their intelligence is of a low order, and they do not seek nectar with the perseverance shown by bees and humble-bees. It is not surprising, therefore, to find that many of the flowers adapted for the visits of flies are so constructed that they detain their visitors until they have done their duty, that is to say, until they have effected, or at least begun, the process of cross-pollination.
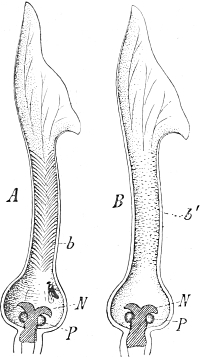
Fig. 43. Flower of Birthwort (Aristolochia
clematitis) cut in half. A, before
pollination by small flies. b, the
bristles. B, after pollination. P, pollen
mass. N, stigma, b, the bristles.
b´, their remains. After H. Müller.
Fig. 44. Alpine Butterwort
(Pinguicula alpina).
A, section through the flower.
K, calyx. bh, bristly prominences.
sp, spur. st, stamen.
n, stigma. B, stigma and
stamen more magnified.
After H. Müller.
Our birthwort (Aristolochia clematitis) and the Cuckoo-pint (Arum maculatum) are pit-fall flowers, whose long corolla-tubes have[Pg 186] an enlargement at the base, in which both pistil and stamens are contained. In the birthwort (Fig. 43) the narrow entrance-tube is thickly beset with stiff hairs (A, b), whose points are all directed towards the base. Little flies can creep down quite comfortably into the basal expansion, but once there they are kept imprisoned until the flower, in consequence of the pollination of the stigma, begins to wither, the first parts to go being these very bristles (B, b´), whose points, like a fish-weir, prevented the flies from creeping out. Other 'fly-flowers,' as for instance the Alpine butterwort (Pinguicula alpina) (Fig. 44), securely imprison the plump fly as soon as it has succeeded in forcing itself in far enough to reach, with its short proboscis, the nectar contained in the spur (sp) of the corolla. The backward-directed bristles hold it fast for some time, and it is only by hard pressing with the back against the anthers (st) lying above it, and against the stigma (n), that it ultimately succeeds in getting free, but it never does so without having either loaded itself with pollen, or rubbed off on the stigma the pollen it brought with it from another similar flower. The Alpine butterwort is protogynous, that is to say, the pistil ripens first, the pollen later, so that the possibility of self-fertilization is altogether excluded.
It would be impossible to give even an approximate idea of the diversity of the contrivances for securing fertilization in flowers without spending many hours over them, for they are different in almost every flower, often widely so, and even in species of the same genus they are by no means always alike; for not infrequently one species is adapted to one circle of visitors, and its near relative to another. Thus the flower of the common Daphne (Daphne mezereum) (Fig. 45, A and C) is adapted to the visits of butterflies, bees, and hover-flies, while its nearest relative (Daphne striata) (Fig. 45, B and D) has a somewhat narrower and longer corolla-tube, so that only butterflies can feast upon it. This example shows that there are exclusively 'butterfly flowers,' but specialization goes further, for there are flowers adapted to diurnal and others to nocturnal Lepidoptera. The former have usually bright, often red colours, and a pleasant aromatic fragrance, and in all of them the nectar lies at the bottom of a very narrow corolla-tube. To this class belong, for instance, the species of pink, many orchids, such as Orchis ustulata, and Nigritella angustifolia of the Alps, which smells strongly of vanilla; also the beautiful campion (Lychnis diurna) and the Alpine primrose (Primula farinosa). The flowers adapted to nocturnal Lepidoptera are characterized by pale, often white colour, and a strong and pleasant smell, which only begins to stream out after sunset, and indeed many of these flowers[Pg 187] are quite closed by day. This is the case with the large, white, scentless bindweed (Convolvulus sepium), which is chiefly visited and fertilized by the largest of our hawk-moths (Sphinx convolvuli). The pale soapwort (Saponaria officinalis) exhales a delicate fragrance which attracts the Sphingidæ from afar, and the sweet smell of the honeysuckle (Lonicera periclymenum) is well known, and has the same effect; an arbour of honeysuckle often attracts whole companies of our most beautiful Sphingidæ and Noctuidæ on warm June nights, to the great delight of the moth-collecting youth.
Fig. 45. Daphne mezereum (A and C) and Daphne striata (B and D). The former visited by butterflies, bees, and flies, the latter by butterflies only. A and B, vertical sections of the flowers. St, stamens. Gr, style. n, nectary. C and D, flowers seen from above. After H. Müller.
I cannot conclude this account of flower-adaptations without considering the orchids somewhat more in detail, for it is among them that we find the most far-reaching adaptations to the visits of insects. Among them, too, great diversity prevails, as is evident from the fact that Darwin devoted a whole book to the arrangements for fertilization in orchids, but the main features are very much the same in the majority. Figure 46 gives a representation of one of our commonest species (Orchis mascula), A shows the flower in side view, B as it appears from in front. The flower seems as it were to float on the end of the stalk (st), stretching out horizontally the spur (sp) which contains the nectar. Between the large, broad under lip (U), marked with a honey-guide (sm), and offering a convenient alighting surface, and the broad, cushion-like stigma (n) lies the entrance to the spur. Fertilization occurs in the following way:—The fly or bee, when it is in the act of pushing its proboscis into the nectar-containing spur,[Pg 188] knocks with its head against the so-called rostellum (r), a little beak-like process at the base of the stamens (p). The pollen masses are of very peculiar construction, not falling to dust, but forming little stalked clubs, with the pollen grains glued together, and so arranged that they spring off when the rostellum is touched and attach themselves to the head of the insect, as at D on the pencil (Fig. 46). When the bee has sucked up the nectar out of the spur, and then proceeds to penetrate into another flower of the same species, the pollinia have bent downwards on its forehead (E), and must unfailingly come in contact with the stigma of the second flower, to which they now remain attached, and effect its fertilization. What a long chain of purposeful arrangements in a single flower, and no interpretation of them is available except through natural selection!
Fig. 46. Common Orchis (Orchis mascula). A, flower in side view. st, stalk. sp, spur with the nectary (n). ei, entrance to the spur. U, lower lip. B, flower from in front. p, pollinia. Sm, honey-guide. ei, entrance to the nectar. na, stigma. r, rostellum. U, lower lip. C, vertical section through the rostellum (r), pollinium (p). ei, entrance. D, the pollinia removed and standing erect on the tip of a lead-pencil. E, the same, somewhat later, curved downwards.
And how diversely are these again modified in the different genera and species of orchids, of which one is adapted to the visits of butterflies exclusively, as Orchis ustulata, another to those of bees, as Orchis morio, and a third to those of flies, as Ophrys muscifera. These flowers are adapted to insect visits in the minutest details of the form of the petals, which are smooth, as if polished with wax, where insects are not intended to creep, but velvety or hairy where the path leads[Pg 189] to the nectar, and at the same time to the pollen and the stigma. And then there is the diversity in the form and colour of the 'honey-guides' on the 'alighting surface,' that is, the under lip of the flower, upon which the insect sits and holds fast, while it pushes its head as far as possible into the spur, so that its proboscis may reach the nectar lying deep within it! Even though we cannot pretend to guess at the significance of every curve and colour-spot in one of the great tropical orchids, such as Stanhopea tigrina, yet we may believe, with Sprengel, that all this has its significance, or has had it for the ancestors of the plant in question, and in fact that the flower is made up of nothing but adaptations, either actual or inherited from its ancestors, although sometimes perhaps no longer of functional importance.
So far, then, we have illustrated the fact that there are hundreds and thousands of contrivances in flowers adapted solely to the visits of insects and to securing cross-fertilization, and these adaptations go so far that we might almost believe them to be the outcome of the most exact calculation and the most ingenious reflection. But they all admit of interpretation through natural selection, for all these details, which used to be looked upon as merely ornamental, are directly or indirectly of use to the species; directly, when, for instance, they concern the dusting of the insect with the pollen; indirectly, when they are a means of attracting visits.
Moreover, the evidence of the operation of the processes of selection becomes absolutely convincing when we consider that, as in symbiosis, there are always two sets of adaptations taking place independently of one another—those of the flowers to the visits of the insects, and those of the insects to the habit of visiting the flowers. To understand this clearly we must turn our attention to the insects, and try to see in what way they have been changed by adapting themselves to the diet which the flowers afford.
As is well known, several orders of insects possess mouth-parts which are suited for sucking up fluids, and these have evolved, through adaptation to a fluid diet, from the biting mouth-parts of the primitive insects which we see still surviving in several orders. Thus the Diptera may have gradually acquired the sucking proboscis which occurs in many of them by licking up decaying vegetable and animal matter, and by piercing into and sucking living animals. But even among the Diptera several families have more recently adapted themselves quite specially to a flower diet, to honey-sucking, like the hover-flies, the Syrphidæ,and the Bombyliidæ, whose long thin proboscis penetrates deep into narrow corolla-tubes, and is able to suck up the nectar from the very bottom. The transformation was not so impor[Pg 190]tant in this case, since the already existing sucking apparatus only required to be a little altered.
Again, in the order Hemiptera (Bugs) the suctorial proboscis does not owe its origin to a diet of flowers, for no member of the group is now adapted to that mode of obtaining food.
Fig. 47. Head of a Butterfly. A, seen from
in front. au, eyes. la, upper lip. md, rudiments
of the mandibles. pm, rudimentary
maxillary palps. mx´, the first maxillæ
modified into the suctorial proboscis. pl,
palps of labium or second maxillæ, cut off
at the root, remaining in B—which is a side
view. at, antennæ. Adapted from Savigny.
The proboscis of the Lepidoptera, on the other hand, depends entirely on adaptation to honey-sucking, and we may go the length of saying that the order of Lepidoptera would not exist if there were no flowers. This large and diverse insect-group is probably descended from the ancestors of the modern caddis-flies or Phryganidæ, whose weakly developed jaws were chiefly used for licking up the sugary juices of plants. But as flowering plants evolved the licking apparatus of the primitive butterflies developed more and more into a sucking organ, and was ultimately transformed into the long, spirally coiled suctorial proboscis as we see it in the modern butterflies (Fig. 47). It has taken some pains to trace this organ back to the biting mouth-parts of the primitive insects, for nearly everything about it has degenerated and become stunted except the maxillæ (mx´). Even the palps (pm) of these have become so small and inconspicuous in most of the Lepidoptera that it is only quite recently that remains of them have been recognized in a minute protuberance among the hairs. The mandibles (md) have quite degenerated, and even the under lip has disappeared, and only its palps are well developed (B, pl). But the first maxillæ (mx´), although very strong and long, are so extraordinarily altered in shape and structure that they diverge from the maxillæ of all other insects. They have become hollow, probe-like half-tubes, which fit together exactly, and thus form a closed sucking-tube of most complex construction, composed of many very small joints, after the fashion of a chain-saw, which are all moved by little muscles, and are subject to the will through nerves, and are also furnished with tactile and taste papillæ. Except this remarkable sucking proboscis there are no peculiarities in the body of the butterfly which might be[Pg 191] regarded as adaptations to flower-visiting, with a few isolated exceptions, of which one will be mentioned later. This is intelligible enough, for the butterfly has nothing more to seek from the flower beyond food for itself; it does not carry stores for offspring.
The bees, however, do this, and accordingly we find that in them the adaptations to flower-visiting are not confined to the mouth-parts.
As far as we can judge now, the flower-visiting bees are descended from insects which resembled the modern burrowing-wasps. Among these the females themselves live on nectar and pollen, and build cells in holes in the ground, and feed their brood. They do not feed them on honey, however, but on animals—on caterpillars, grasshoppers, and other insects, which they kill by a sting in the abdomen, or often only paralyse, so that the victim is brought into the cells of the nest alive but defenceless, and remains alive until the young larva of the wasp, which emerges from the egg, sets to work to devour it.
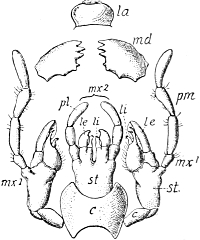
Fig. 48. Mouth-parts of the
Cockroach (Periplaneta orientalis),
after R. Hertwig. la, upper lip
or labrum. md, mandibles. mx1,
first maxillæ, with c, cardo, st,
stipes, li, internal lobe or lacinia,
le, external lobe or galea, and pm,
the maxillary palp. mx2, the
labium or second maxillæ, with
similar detailed parts.
Before I go on to explain the origin of the sucking proboscis of the bee from the biting mouth-parts of the primitive insects I must first briefly consider the latter.
The biting mouth-parts of beetles, Neuroptera, and Orthoptera (Fig. 48), consist of three pairs of jaws, of which the first, the mandibles (md), are simply powerful pincers for seizing and tearing or chewing the food. They have no part in the development of the suctorial apparatus either in bees or in butterflies, so they may be left out of account. The two other pairs of jaws, the first and second maxillæ (mx1 and mx2), are constructed exactly on the same type, having a jointed basal portion (st) bearing two lobes, an external (le) and an internal (li), and a feeler or palp, usually with several joints, directed outwards from the lobes (pm and pl). The second pair of maxillæ (mx2) differs from the first chiefly in this, that the components of the pair meet in the median line of the body, and fuse more or less to form the so-called 'under lip' or labium. In the example given, the cockroach (Periplaneta orientalis), this fusion[Pg 192] is only partial, the lobes having remained separate (le and li); and the same is true of the bee, but in this case the inner lobes have grown into a long worm-like process which is thrust into the nectar in the act of sucking.
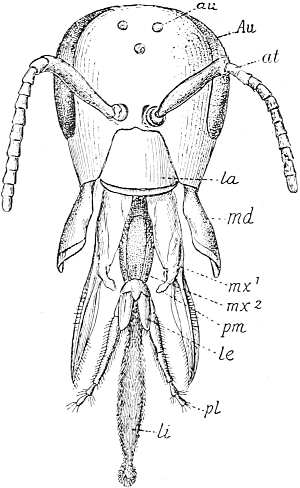
Fig. 49. Head of the Bee. Au, compound
eyes. au, ocelli. at, antennæ. la, upper lip.
md, mandibles. mx1, first maxillæ, with pm,
the rudimentary maxillary palp. mx2, second
maxillæ with the internal lobes (li) fused to
form the 'tongue.' le, the external lobes of
the second maxillæ, known as 'paraglossæ.'
pl, labial palp.
Even the burrowing-wasps exhibit the beginnings of variation in this direction, for the under lip is somewhat lengthened and modified into a licking organ. The adaptation has not gone much further than this, even in one of the true flower-bees, Prosopis, which feeds its larvæ with pollen and honey, and it is only in the true honey-bee that the adaptation is complete (Fig. 49). Here the so-called 'inner lobe' of the under lip (li) has elongated into the worm-shaped process already mentioned; it is thickly covered with short bristles, and is called the 'tongue' of the bee (li). The outer lobes of the under lip have degenerated into little leaf-like organs, the so-called accessory tongue or paraglossa (le), while the palps of the under lip (pl) have elongated to correspond with the tongue, and serve as a sensitive and probably also as a smelling organ, in contrast to the palps of the first maxillæ, which have shrunk to minute stumps (pm). The whole of the under lip, which has elongated even in its basal portions, forms, with the equally long first maxillæ, the proboscis of the bee. The first maxillæ are sheath-like half-tubes, closely apposed around the tongue, and form along with it the suctorial tube, through which the nectar is sucked up. Thus, of the three pairs of jaws in insects, only the first pair, the mandibles, have remained unaltered, obviously because the bee requires a biting-organ for eating pollen, for kneading wax, and for building cells.
But bees do not only feast on nectar and pollen themselves, they carry these home as food for their larvæ. The form already[Pg 193] mentioned, Prosopis, takes up pollen and nectar in its mouth, and afterwards disgorges the pulp as food for its larvæ, but the rest of the true bees have special and much more effective collecting-organs, either a thick covering of hair on the abdomen, or along the whole length of the posterior legs, or finally, a highly developed collecting apparatus, such as that possessed by the honey-bee—the basket and brush on the hind leg. The former is a hollow on the outer surface of the tibia, the latter a considerable enlargement of the basal tarsal joint, which, at the same time, is covered on the inner surface with short bristles, arranged in transverse rows like a brush. The bee kneads the pollen into the basket, and one can often see bees flying back to the hive with a thick yellow ball of pollen on the hind leg. In those bees which collect on the abdomen, like Osmia and Megachile, the pollen mass forms a thick clump on the belly, and in the case of Andrena Sprengel observed long ago that it sometimes flew with a packet of pollen bigger than its own body on the hind leg.
All these are contrivances which have gradually originated through the habit of carrying home pollen for the helpless larvæ shut up in the cells. They have developed differently in the various groups of bees, probably because the primary variations with which the process of selection began were different in the various ancestral forms.
In the ancestors of those which carry pollen on the abdomen there was probably a thick covering of hair on the ventral surface of the body, which served as a starting-point for the selection, and, in consequence, the further course of the adaptation would be concerned solely with this hair-covered surface, while variations in other less hairy spots would remain un-utilized.
After all this it will no longer seem a paradoxical statement that the existence of gaily coloured, diversely formed, and fragrant flowers is due to the visits of insects, and that, on the other hand, many insects have undergone essential transformations in their mouth-parts and otherwise as an adaptation to a flower diet, and that an entire order of insects with thousands of species—the Lepidoptera—would not be in existence at all if there had been no flowers. We must now attempt to show, in a more detailed way, how, by what steps, and under what conditions, our modern flowers have arisen from the earlier flowering plants. In this I follow closely the classic exposition which we owe to Hermann Müller.
The ancestral forms of the modern higher plants, the so-called 'primitive seed plants' or 'Archisperms,' were all anemophilous, as the Conifers and Cycads are still. Their smooth pollen-grains,[Pg 194] produced in enormous quantities, fell like clouds of dust into the air, were carried by the wind hither and thither, and some occasionally alighted on the stigma of a female flower. In these plants the sexes often occur separately on different trees or individuals, and there must be a certain advantage in this when the pollination is effected by the wind.
The male flowers of the Archisperms would be visited by insects in remote ages, just as they are now; but the visitors came to feed upon the pollen, and did not render any service to the plant in return; they rather did it harm by reducing its store of pollen. If it was possible to cause the insect to benefit the plant at the same time as it was pillaging the pollen, by carrying some of it to female blossoms and thereby securing cross-fertilization, it would be of great advantage, for the plant would no longer require to produce such enormous quantities of pollen, and the fertilization would be much more certain than when it depended on the wind. It is obvious that the successful pollination of anemophilous plants implies good weather and a favourable wind.
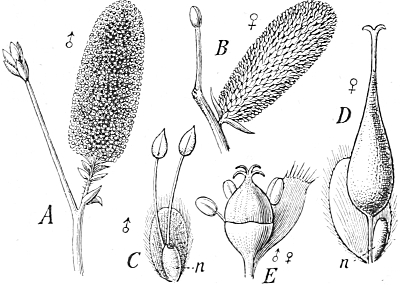
Fig. 50. Flowers of the Willow (Salix cinerea); after H. Müller. A, the male. B, the female catkin. C, individual male flower; n, nectary. D, individual female flower; n, nectary. E, Poplar, an exceptional hermaphrodite flower.
It is plain that the utilization of the insect-visitors in fertilization might be secured in either of two ways; the female blossoms might also offer something attractive to the insects, or hermaphrodite flowers might be formed. As a matter of fact, both ways have been followed by Nature. An example of the former is the willow, the cross-fertilization of which was forced upon the insects by the development in both female and male blossoms of a nectary (Fig. 50, C and D), a little pit or basin in which nectar was secreted. The insects flew now to male and now to female willow-catkins, and in doing so they[Pg 195] carried to the stigma of the female blossom the pollen, which in this case was not dusty but sticky, so that it readily adhered to their bodies.
The securing of cross-fertilization by the development of hermaphrodite flowers has, however, occurred much more frequently, and we can understand that this method secured the advantageous crossing much more perfectly, for the pollen had necessarily to be carried from blossom to blossom, while, in cases like that of the willow, countless male blossoms might be visited for nectar one after the other before the insect made up its mind to fly to a female blossom of the same species. The beginnings of the modification of the unisexual flowers in this direction may be seen in variations which occur even now, for we not infrequently find, in a male catkin, individual blossoms, which, in addition to the stamens, possess also a pistil with a stigma. (Fig. 50 E shows such an abnormal hermaphrodite flower from a poplar.)
As soon as hermaphrodite flowers came into existence the struggle to attract insects began in a more intense degree. Every little improvement in this direction would form the starting-point of a process of selection, and would be carried on and increased to the highest possible pitch of perfection.
It was probably the outer envelopes of the blossoms which first changed their original green into other colours, usually those which contrasted strongly with the green, and thus directed the attention of the insects to the flowers. Variations in the colour of ordinary leaves are always cropping up from time to time, whether it be that the green is transformed into yellow or that the chlorophyll disappears more or less completely and red or blue coloured juices take its place. Many insects can undoubtedly see colour, and are attracted by the size of coloured flowers, as Hermann Müller found by counting the visits of insects to two nearly related species of mallow, one of which, Malva silvestris, has very large bright rose-red flowers visible from afar, while the other, Malva rotundifolia, has very inconspicuous small pale-red flowers. To the former there were thirty-one different visitors, to the latter he could only make sure of four. The second species, as is to be expected, depends chiefly on self-fertilization.
It has recently been disputed from various quarters that insects are attracted by the colours of the flowers, and these objections are based chiefly on experiments with artificial flowers. But when, for instance, Plateau, in the course of such experiments saw bees and butterflies first fly towards the artificial flowers, and then turn away and concern themselves no more about them, that only proves that[Pg 196] their sight is sharper than we have given them credit for; for though they may be deceived at a distance, they are not so when they are near; it is possible, too, that the sense of smell turns the scale[9]. I have myself made similar experiments with diurnal butterflies, before which I placed a single artificial chrysanthemum midst a mass of natural flowers. It rarely happened indeed that a butterfly settled on the artificial flower; they usually flew first above it, but did not alight. Twice, however, I saw them alight on the artificial flower, and eagerly grope about with the proboscis for a few moments, then fly quickly away. They had visited the real chrysanthemums or horse-daisies with evident delight, and eagerly sucked up the honey from the many individual florets of every flower, and they now endeavoured to do the same in the artificial flower, and only desisted when the attempt proved unsuccessful. In this experiment the colours were of course only white and yellow; with red and blue it is probably more difficult to give the exact impression of the natural flower-colours; and in addition there is the absence of the delicate fragrance exhaled by the flower.
[9] The experiments of Plateau have since been criticized by Kienitz-Gerloff, who altogether denies their value (1903).
It must be allowed that the colour is certainly not the sole attraction to the flower; the fragrance helps in most cases, and even this is not the object of the insect's visits. The real object is the nectar, to which colour and fragrance only show the way. The development of fragrance and nectar must, like that of the colour, have been carried on and increased by processes of selection, which had their basis in the necessity for securing insect-visits, and as soon as these main qualities of the flower were established greater refinements would begin, and flower-forms would be evolved, which would diverge farther and farther, especially in shape, from the originally simple and regular form of the blossom.
The reason for this must have lain chiefly in the fact that, after insect-visits in general were secured by a flower, it would be advantageous to exclude all insects which would pillage the nectar without rendering in return the service of cross-fertilization—all those, therefore, which were unsuited either because of their minute size or because of the inconstancy of their visits. Before the butterflies and the bees existed, the regularly formed flat flower with unconcealed nectar would be visited by a mixed company of caddis-flies, saw-flies, and ichneumon-flies. But as the nectar changed its place to the deeper recesses of the flower it was withdrawn from all but the more intelligent insects, and thus the circle of visitors was already narrowed[Pg 197] to some extent. But when in a particular species the petals fused into a short tube, all visitors were excluded whose mouth-parts were too short to reach the nectar; while among those which could reach it the process of proboscis-formation began; the under lip, or the first maxillæ, or both parts together, lengthened step for step with the corolla-tube of the flower, and thus from the caddis-flies came the butterflies, and from the ichneumon-flies the burrowing-wasps (Sphegidæ) and the bees.
At first sight one might perhaps imagine that it would have been more advantageous to the flowers to attract a great many visitors, but this is obviously not the case. On the contrary, specialized flowers, accessible only to a few visitors, have a much greater certainty of being pollinated by them, because insects which only fly to a few species are more certain to visit these, and above all to visit many flowers of the same species one after another. Hermann Müller observed that, in four minutes, one of the humming-bird hawk-moths (Macroglossa stellatarum) visited 108 different flowers of the same species, the beautiful Alpine violet (Viola calcarata), one after the other, and it may have effected an equal number of pollinations in that short time.
It was, therefore, a real advantage to the flowers to narrow their circle of visitors more and more by varying so that only the useful visitors could gain access to their nectar, and that the rest should be excluded. Thus there arose 'bee-flowers,' 'butterfly-flowers,' 'hawk-moth flowers,' and, indeed, in many cases, a species of flower has become so highly specialized that its fertilization can only be brought about by a single species of insect. This explains the remarkable adaptations of the orchids and the enormous length of the proboscis in certain butterflies. Even our own hawk-moths Macroglossa stellatarum and Sphinx convolvuli show an astonishing length of proboscis, which measures 8 cm. in the latter species. In Macrosilia cluentius, in Brazil, the proboscis is 20 cm. in length; and in Madagascar there grows an orchid with nectaries 30 cm. in length, filled with nectar to a depth of 2 cm., but the fertilizing hawk-moth is not yet known.
Thus we may say that the flowers, by varying in one direction or another, have selected a definite circle of visitors, and, conversely, that particular insect-groups have selected particular flowers for themselves, for those transformations of the flowers were always most advantageous which secured to them the exclusive visits of their best crossing agents, and these transformations were, on the one hand, such as kept off unwelcome visitors, and, on the other hand, such as attracted the most suitable ones.
From the botanical point of view the assumption that flowers and flower-visiting insects have been adapted to each other by means of processes of selection has been regarded as untenable, because every variation in the flower presupposes a corresponding one in the insect. I should not have mentioned this objection had it not come from such a famous naturalist as Nägeli, and if it were not both interesting and useful in our present discussion. Nägeli maintained that selection could not, for instance, have effected a lengthening of the corolla-tube of a flower, because the proboscis of the insects must have lengthened simultaneously with it. If the corolla-tube had lengthened alone, without the proboscis of the butterfly being at the same time elongated, the flower would no longer be fertilized at all, and if the lengthening of the proboscis preceded that of the corolla-tube it would have no value for the butterfly, and could not therefore have been the object of a process of selection.
This objection overlooks the facts that a species of plant and of butterfly consists not of one individual but of thousands or millions, and that these are not absolutely uniform, but in fact heterogeneous. It is precisely in this that the struggle for existence consists—that the individuals of every species differ from one another, and that some are better, others less well constituted. The elimination of the latter and the preferring of the former constitutes the process of selection, which always secures the fitter by continually rejecting the less fit. In the case we are considering, then, there would be, among the individuals of the plant-species concerned, flowers with a longer and flowers with a shorter corolla-tube, and among the butterflies some with a longer and some with a shorter proboscis. If among the flowers the longer ones were more certain to be cross-fertilized than the shorter ones, because hurtful visitors were better excluded, the longer ones would produce more and better seeds, and would transmit their character to more descendants; and if, among the butterflies, those with the longer proboscis had an advantage, because the nectar in the longer tubes would, so to speak, be reserved for them, and they would thus be better nourished than those with the shorter proboscis, the number of individuals with long proboscis must have increased from generation to generation. Thus the length of the corolla-tube and the length of the proboscis would go on increasing as long as there was any advantage in it for the flower, and both parties must of necessity have varied pari passu, since every lengthening of the corolla was accompanied by a preferring of the longest proboscis variation. The augmentation of the characters depended on, and could only have depended on, a guiding of the variations in the[Pg 199] direction of utility. But this is exactly what we call, after Darwin and Wallace, Natural Selection.
We have, however, in the history of flowers, a means of demonstrating the reality of the processes of selection in two other ways. In the first place, it is obvious that no other interpretation can be given of such simultaneous mutual adaptations of two different kinds of organisms. If we were to postulate, as Nägeli, for instance, did, an intrinsic Power of Development in organisms, which produces and guides their variations, we should, as I have already said, be compelled also to take for granted a kind of pre-established harmony, such as Leibnitz assumed to account for the correlation of body and mind: plant and insect must always have been correspondingly altered so that they bore the same relation to each other as two clocks which were so exactly fashioned that they always kept time, though they did not influence each other. But the case would be more complicated than that of the clocks, because the changes which must have taken place on both sides were quite different, and yet at the same time such that they corresponded as exactly as Will and Action. The whole history of the earth and of the forms of life must, therefore, have been foreseen down to the smallest details, and embodied in the postulated Power of Development.
But such an assumption could hardly lay claim to the rank of a scientific hypothesis. Although every grain of sand blown about by the wind on this earth could certainly only have fallen where it actually did fall, yet it is in the power of any of us to throw a handful of sand wherever it pleases us, and although even this act of throwing must have had its sufficient reason in us, yet no one could maintain that its direction and the places where the grains fell were predestined in the history of the earth. In other words: That which we call chance plays a part also in the evolution of organisms, and the assumption of a Power of Development, predestinating even in detail, is contradicted by the fact that species are transformed in accordance with the chance conditions of their life.
This can be clearly demonstrated in the case of flowers. That the wild pansy (Viola tricolor), which lives in the plains and on mountains of moderate elevation, is fertilized by bees, and the nearly allied Viola calcarata of the High Alps by Lepidoptera, is readily intelligible, since bees are very abundant in the lower region, and make the fertilization of the species a certainty, while this is not so in the High Alps. There the Lepidoptera are greatly in the majority, as every one knows who has traversed the flower-decked meads of the High Alps in July, and has seen the hundreds and thousands[Pg 200] of butterflies and moths which fly from flower to flower. Thus the viola of the High Alps has become a 'butterfly-flower' by the development of its nectaries into a long spur, accessible only to the proboscis of a moth or butterfly. The chance which led certain individuals of the ancestral species to climb the Alps must also have supplied the incentive to the production of the changes adapted to the visits of the prevalent insect. The hypothesis of a predestinating Power of Development suffers utter shipwreck in face of facts like these.
We have, furthermore, an excellent touchstone for the reality of the processes of selection in the quality of the variations in flowers and insects. Natural selection can only bring about those changes which are of use to the possessors themselves; we should therefore expect to find among flowers only such arrangements as are, directly or indirectly, of use to them, and, conversely, among insects only such as are useful to the insect.
And this is what we actually do find. All the arrangements of the flowers—their colour, their form, their honey-guides, their hairy honey-paths (Iris), their fragrance, and their honey itself—are all indirectly useful to the plant itself, because they all co-operate in compelling the honey-seeking insect to effect the fertilization of the flower. This is most clearly seen in the case of the so-called 'Deceptive' flowers, which attract insects by their size and beauty, their fragrance, and their resemblance to other flowers, and force their visitors to be the means of their cross-fertilization, although they contain no nectar at all. This is the case, according to Hermann Müller, with the most beautiful of our indigenous orchids, the lady's slipper (Cypripedium calceolaris). This flower is visited by bees of the genus Andrena, which creep into the large wooden-shoe-shaped under lip in the search for honey, only to find themselves prisoners, for they cannot get out, at least by the way they came in, because of the steep and smoothly polished walls of the flower. There is only one way for the bee; it must force itself under the stigma, which it can only do with great exertion, and not without being smeared with pollen, which it carries to the next flower into which it creeps. It can only leave this one in the same way, and thus the pollen is transferred to the stigma by a mechanical necessity.
Such remarkable cases remind us in some ways of those cases of mimicry in which the deceptions have to be used with caution or they lose their effect. One might be disposed to imagine that such an intelligent insect as a bee would not be deceived by the lady's slipper more than once, and would not creep into a second flower after discovering that there was no nectar in the first. But this[Pg 201] conclusion is not correct, for the bees are well accustomed in many flowers to find that the nectar has already been taken by other bees; they could therefore not conclude from one unsuccessful visit that the Cypripedium did not produce nectar at all, but would try again in a second, a third, and a fourth flower. If these orchids had abundantly covered flower-spikes like many species of Orchis, and if the species were common, the bees would probably soon learn not to visit them, but the reverse is the case. There is usually only one or, at most, two open flowers on the lady's slipper, and the plant is rare, and probably occurs nowhere in large numbers.
If we could find a flower in which the nectar lay open and accessible to all insects, and which did not require any service from them in return, the case could not be interpreted in terms of natural selection; but we do not know of any such case.
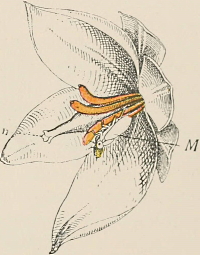
Fig. 51. The Yucca-moth (Pronuba
yuccasella). M, laying eggs in
the ovary of the Yucca flower.
n, the stigma. After Riley.
Conversely, too, there are no adaptations in the insects which are useful only to the flowers, and which are not of some use, directly or indirectly, to the insect itself. Bees and butterflies certainly carry the pollen from one flower to the stigma of another, but they are not impelled to do this by a special instinct; they are forced to do it by the structure of the flower, which has its stamens so placed and arranged that they must shake their pollen over the visitor, or it may be that the anthers are modified into stalked, viscid pollinia which spring off at a touch, and fix themselves, so to speak, on the insect's head. And even this is not all in the case of the orchis, for the insect would never of its own accord transfer these pollinia on to the stigma of the next flower; this is effected by the physical peculiarity which causes the pollinia, after a short time, to bend forwards on the insect's head.
All this fits in as well as possible with the hypothesis: how could an instinct to carry pollen from one flower to the stigma of another have been developed in an insect through natural selection, since the insect itself has nothing to gain from this proceeding? Accordingly, we never find in the insect any pincers or any kind of grasping organ adapted for seizing and transmitting the pollen.
There is, however, one very remarkable case in which this appears to be so, indeed really is so, and nevertheless it is not[Pg 202] contradictory to, but is corroborative of, the theory of selection. The excellent American entomologist, Riley, established by means of careful observations that the large white flowers of the Yucca are fertilized by a little moth which behaves in a manner otherwise unheard of among insects. Only the females visit the flowers, and they at once busy themselves collecting a large ball of pollen. To this end they have on the maxillary palps (Fig. 52, C, mxp) a long process (si), curved in the form of a sickle, and covered with hairs, which probably no other Lepidopteron possesses, with the help of which the moth very quickly sweeps together a ball of pollen, it may be three times the size of her own head. With this ball the insect flies to the next flower, and there she lays her egg, by means of an ovipositor otherwise unknown among Lepidoptera (Fig. 52, A, op), in the pods of the flower. Finally, she pushes the ball of pollen deep into the funnel-shaped stigmatic opening on the pistil (Fig. 51, n), and so effects the cross-fertilization. The ovules develop, and when the caterpillars emerge from the egg four to five days later they feed on these until they are ready to enter on the pupa stage. Each little caterpillar requires about eighteen or twenty seeds for its nourishment (Fig. 52, B, r).
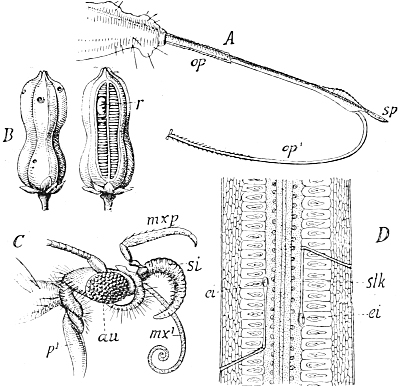
Fig. 52. The fertilization of the Yucca. A, ovipositor of the Yucca-moth. op, its sheath. sp, its apex. op1, the protruded oviduct. B, two ovaries of the Yucca, showing the holes by which the young moths escape, and (r) a caterpillar in the interior. C, head of the female moth, with the sickle-shaped process (si) on the maxillary palps for sweeping off the pollen and rolling it into a ball. mx1, the proboscis. au, eye. p1 base of first leg. D, longitudinal section through an ovary of the Yucca, soon after the laying of two eggs (ei). stk, the canal made by the ovipositor.
Here, then, we find an adaptation of certain parts of the moth's body in relation to the fertilization of the flower, but in this case it is as much in the interest of the moth as of the plant. By carrying the pollen to the stigma the moths secure the development of the ovules, which serve their offspring as food, so that we have here to do with a peculiar form of care for offspring, which is not more remarkable than many other kinds of brood-care in insects, such as ants, bees, Sphex-wasps, ichneumon-flies, and gall-flies.
It might be objected that this case of the Yucca is not so much one of effecting fertilization as of parasitism; but the eggs, which are laid in the seed-pods, are very few, and the caterpillars which emerge from them only devour a very small proportion of the seeds, of which there may be about 200 (Fig. 52, B). Thus the plants also derive an advantage from the moth's procedure, for quite enough seeds are left. The form and position of the stamens and of the stigma seem to be as exactly adapted to the visits of the moth as the moth is to the transference of the pollen, for the Yucca can only be fertilized by this one moth, and sets no seed if the moth be absent. For this reason the species of Yucca cultivated in Europe remain sterile.
Thus the apparent contradiction is explained, and the facts everywhere support the hypothesis that the adaptations between flowers and insects depend upon processes of selection.
This origin is incontrovertibly proved, it seems to me, in another way, namely, by the merely relative perfection of the adaptations, or rather, by their relative imperfection.
I have already pointed out that all adaptations which depend upon natural selection can only be relatively perfect, as follows from the nature of their efficient causes, for natural selection only operates as long as a further increase of the character concerned would be of advantage to the existence of the species. It cannot be operative beyond this point, because the existence of the species cannot be more perfectly secured in this direction, or, to speak more precisely, because further variations in the direction hitherto followed would no longer be improvements, even though they might appear so to us.
Thus the corolla of many flowers is suited to the thick, hairy head and thorax of the bee, for to these only does the pollen adhere in sufficient quantity to fertilize the next flower; yet the same flowers are frequently visited by butterflies, and in many of them there has been no adaptation to prevent these useless visits. Obviously this is because preventive arrangements could only begin, according to our theory, when they were necessary to the preservation of the species; in this case, therefore, only when the pillaging visits of the butterflies[Pg 204] withdrew so many flowers from the influence of the effective pollinating visitor, the bee, that too few seeds were formed, and the survival of the species was threatened by the continual dwindling of the normal number. As long as the bees visit the flowers frequently enough to ensure the formation of the necessary number of seeds a process of selection could not set in; but should the bees find, for instance, that nearly all the flowers had been robbed of their nectar, and should therefore visit them less diligently, then every variation of the flower which made honey less accessible to the butterflies would become the objective of a process of selection.
Everywhere we find similar imperfections of adaptation which indicate that they must depend on processes of selection. Thus numerous flowers are visited by insects other than those which pollinate them, and these bring them no advantage, but merely rob them of nectar and pollen; the most beautiful contrivances of many flowers, such as Glycinia, which are directed towards cross-fertilization by bees, are rendered of no effect because wood-bees and humble-bees bite holes into the nectaries from the outside, and so reach the nectar by the shortest way. I do not know whether bees in the native land of the Glycinia do the same thing, but in any case they can do no sensible injury to the species, since otherwise processes of selection would have set in which would have prevented the damage in some way or other, whether by the production of stinging-hairs, or hairs with a burning secretion, or in some other way. If the actual constitution of the plant made this impossible, the species would become less abundant and would gradually die out.
Thus the relative imperfection of the flower-adaptations, which in general are so worthy of admiration, affords a further indication that their origin is due to processes of selection.
ADDITIONAL NOTE TO CHAPTER X.
It has been remarked that the chapter on the Origin of Flowers in the German Edition contains no discussion and refutation of the objections which have up till recently been urged against the theory of flowers propounded by Darwin and Hermann Müller. I admit that this chapter seemed to be so harmonious and so well rounded, and at the same time so convincing as to the reality of the processes of selection, that the feeble objections to it, and the attempts of opponents to find another explanation of the phenomena, might well be disregarded in this book.
However, the most important of these objections and counter-theories may here be briefly mentioned.
Plateau in Ghent was the first to collect facts which appeared to contradict the Darwinian theory of flowers; he observed that insects avoided artificial flowers, even when they were indistinguishable in colour from natural ones as far as our eyes could perceive, and he concluded from this that it is not the colour which guides the insects to the flowers, that they find the blossoms less by their sense of sight than by their sense of smell. But great caution is required in drawing conclusions from experiments of this kind. I once placed artificial marguerites (Chrysanthemum leucanthemum) among natural ones in a roomy frame in the open air, and for a considerable time I was unable to see any of the numerous butterflies (Vanessa urticæ), which were flying about the real chrysanthemums, settle on one of the artificial flowers. The insects often flew quite close to them without paying them the least attention, and I was inclined to conclude that they either perceived the difference at sight, or that they missed the odour of the natural flowers in the artificial ones. But in the course of a few days it happened twice in my presence that a butterfly settled on one of the artificial blooms and persistently groped about with fully outstretched tube to find the entrance to the honey. It was only after prolonged futile attempts that it desisted and flew away. That bees are guided by the eye in their visits to flowers has been shown by A. Forel, who cut off the whole proboscis, together with the antennæ, from humble-bees which were swarming eagerly about the flowers. He thus robbed them of the whole apparatus of smell, and nevertheless they flew down from a considerable height direct to the same flowers. An English observer, Mr. G. N. Bulman, has been led to believe, with Plateau, that it is a matter of entire indifference to the bees whether the flowers are blue, or red, or simply green in colour, if only they contain honey, and that therefore the bees could have played no part in the development of blue flowers, as Hermann Müller assumed they had, and that they could have no preference for blue or any other colour, as Sir John Lubbock and others had concluded from their experiments. This is correct in so far that bees feed as eagerly on the greenish blossoms of the lime-tree as they do on the deep-blue gentian of the Alpine meadows or the red blossoms of the Weigelia, the dog-roses of our gardens or the yellow buttercups (Ranunculus) of our meadows; they despise nothing that yields them honey. But it certainly does not follow from this that the bees may not, under certain circumstances, have exercised a selecting influence upon the fixation and intensification of a new colour-variety of a flower. This is less a question of a colour-preference, in the human sense, on the part of the[Pg 206] bees than of the greater visibility of the colour in question in the environment peculiar to the flower, and of the amount of rivalry the bees meet with from other insects in regard to the same flower. In individual cases this would be difficult to demonstrate, especially since we can form only an approximate idea of the insect's power of seeing colour, and cannot judge what the colours of the individual blossoms count for in the mosaic picture of a flowery meadow. Yet this is the important point, for, as soon as the bees perceive one colour more readily than another, the preponderance of this colour-variety over other variations is assured, since it will be more frequently visited. In the same way we cannot guess in individual cases why one species of flower should exhale perfume while a nearly related species does not. But when we remember that many flowers adapted for the visits of dipterous insects possess a nauseous carrion-like smell, by means of which they not only attract flies but scare off other insects, we can readily imagine cases in which it was of importance to a flower to be able to be easily found by bees without betraying itself by its pleasant fragrance to other less desirable visitors.
Thus, therefore, we can understand the odourless but intensely blue species of gentian, if we may assume that its blue colour is more visible to bees than to other insects. If I were to elaborate in detail all the principles which here suggest themselves to me I should require to write a complete section, and I am unwilling to do this until I can bring forward a much larger number of new observations than I am at present in a position to do. All I wish to do here is to exhort doubters to modesty, and to remind them that these matters are exceedingly complex, and that we should be glad and grateful that expert observers like Darwin and Hermann Müller have given us some insight into the principles interconnecting the facts, instead of imagining whenever we meet with some little apparently contradictory fact, which may indeed be quite correct in itself, that the whole theory of the development of flowers through insects has been overthrown. Let us rather endeavour to understand such facts, and to arrange them in their places as stones of the new building.
Often the contradiction is merely the result of the imperfect theoretical conceptions of its discoverer, as we have already shown in regard to Nägeli. Bulman, too, fancies he has proved that bees do not distinguish between the different varieties of a flower, but visit them indiscriminately with the same eagerness, thus causing intercrossing of all the varieties, and preventing any one from becoming[Pg 207] dominant. But are the varieties which we plant side by side in our gardens of the kind that are evolved by bees? That is to say, are their differences such as will turn the scale for or against the visits of the bees? If one were less, another more easily seen by the bees; or if one were more fragrant, or had a fragrance more agreeable to bees than the other, the result of the experiment would probably have been very different.
One more objection has been made. It is said that the bees, although exclusively restricted, both themselves and their descendants, to a diet of flowers, are not so constant to a particular flower as the theory requires. They do indeed exhibit a 'considerable amount of constancy,' and often visit a large number of flowers of the same species in succession, but the theory requires that they should not only confine themselves to this one species, but to a single variety of this species. These views show that their authors have not penetrated far towards an understanding of the nature of selection. Nature does not operate with individual flowers, but with millions and myriads of them, and not with the flowers of a single spring, but with those of hundreds and thousands of years. How often a particular bee may carry pollen uselessly to a strange flower without thereby lowering the aggregate of seeds so far that the existence of the species seems imperilled, or how often she may fertilize the pistil of a useful variation with the pollen of the parent species, without interrupting or hindering the process of the evolution of the variety, no mortal can calculate, and what the theory requires can only be formulated in this way: The constancy of the bees in their visits to the flowers must be so great that, on an average, the quantity of seeds will be formed which suffices for the preservation of the species. And in regard to the transformation of a species, the attraction which the useful variety has for the bees must, on an average, be somewhat stronger than that of the parent species. As soon as this is the case the seeds of the variety will be formed in preponderant numbers, although they may not all be quite pure from the first, and by degrees, in the course of generations, the plants of the new variety will preponderate more and more over those of the parent form, and finally will alone remain. In the first case we have before our eyes the proof that, in spite of the imperfect constancy of the bees, a sufficient number of seeds is produced to secure the existence of the species. Or does Mr. Bulman conclude from the fact that the bees are not absolutely constant that flowers are not fertilized by bees at all?
I cannot conclude this note without touching briefly upon what the opponents of the flower theory have contributed, and what explanation of the facts they are prepared to offer.
In his important work, Mechanische-physiologische Theorie der Abstammungslehre, published in 1884, Nägeli, as a convinced opponent of the theory of selection, attempted an explanation. He was quite aware that his assumption of an inward 'perfecting principle' would not suffice to explain the mutual adaptations of flowers and insects, and he refers the transformation of the first inconspicuous blossoms into flowers to the mechanical stimulus which the visiting insects exerted upon the parts of the blossom. By the pressure of their footsteps, the pushing and probing with their proboscis, they have, he says, transformed gradually, for instance, the little covering leaves at the base of a pollen vessel into large flower petals, caused the conversion of short flower-tubes into long ones, and of the pollen, once dry and dusty, into the firmly adhesive mass formed in the anther lobes of our modern flowers. The colour of the flowers depends, according to him, upon the influence of light, which certainly no more explains the yellow ring on a blue ground in the forget-me-not than it does the many other nectar-guides which show the insect the way to the honey. Nägeli works with the Lamarckian principle in the most daring way, and with the same naïveté as Lamarck himself in his time, that is, without offering any sort of explanation as to how the minute impression made, say by the foot or by the proboscis of an insect, upon a flower, is to be handed on to the flowers of succeeding generations. He treats the unending chain of generations as if it were a single individual, and operates with his 'secular' stimulus, and with 'weak stimuli, lasting through countless generations,' as though they were a proved fact. But I have not even touched upon the question as to whether these 'stimuli' could produce the changes he ascribes to them, even if they were continually affecting the flower. How the scale-like covering leaves of the pollen vessels could become larger and petal-like through the treading of an insect's foot is as difficult to see as why a honey-tube should become longer because of the butterfly's honey-sucking: might it not just as well become wider, narrower, or even shorter? I see no convincing reason why it should become longer! And even if it did so, it would necessarily continue to lengthen as time went on, and this is not the case, for we find corolla-tubes of all possible lengths, but, it is to be noted, always in harmony with the length of the proboscis of the visiting insect. In a similar way Henslow has recently attempted to refer the origin of flowers to the mechanical stimulus exercised upon it by the visiting insects. 'An insect hanging to the lower petal of a flower elongates the same by its weight, and the lengthened petal is transmitted by heredity.'...'The irritation[Pg 209] caused by its feet in walking along the flower causes the appearance of colouring matter, and the colour is likewise transmitted.'...'As it probes for honey it causes a flow of sweet sap to that part, and this also becomes hereditary!'
In this case, also, it is simply taken for granted that every little passing irritation not only produces a perceptible effect, but that this effect is transmissible. In a later lecture we shall have to discuss in detail the question of the inheritance of functional modifications. It is enough to say here that, if this kind of transmission really took place even in the case of such minute and transitory changes, there could be no dispute as to the correctness of the 'Lamarckian principle,' since every fairly strong and lasting irritation could be demonstrated with certainty to produce an effect. When a butterfly, floating freely in the air, sucks honey from a tube, the irritation must be almost analogous to that caused by a comb lightly drawn by some one through our hair, and this is supposed to effect the gradual lengthening of the corolla-tube of the flower!
The secretion of honey, too, depends upon the persistent irritation of the proboscis! Then 'deceptive flowers,' like the Cypripedium we have mentioned, could not exist at all, for they contain no honey, although the proboscis of the bee must cause the same irritation in them as in other orchids which do contain honey. This whole 'theory' of direct effect is, moreover, only a crude and apparent interpretation, which explains the conditions only in so far as they can be seen from a distance; it fails as soon as they are more exactly examined; all the great differences in the position of the honey, its concealment from intelligent insects, its protection from rain by means of hairs, and against unwelcome guests by a sticky secretion, the development of a corolla-tube which corresponds in length to the length of the visiting insect's proboscis, the development of spurs on the flower, in short, all the numerous contrivances which have reference to cross-fertilization by insects remain quite unintelligible in the light of this theory—it is a mere pis aller explanation for those who continue to struggle against accepting the theory of selection.
SEXUAL SELECTION
Decorative colouring of male butterflies and birds—Wallace's interpretation—Preponderance of males—Choice of the females—Sense by sight in butterflies—Attractive odours—Scent-scales—Fragrance of the females—The limits of natural and sexual selection not clearly defined—Odours of particular species—Odours of other animals at the breeding season—Song of the Cicadas, and of birds—Diversity of decoration successively acquired—Humming-birds—Substitution of other aids to wooing in place of personal decoration—Smelling organs of male insects and crabs—Contrivances for seizing and holding the female—Small size of certain males—Weapons of males used in struggle for the females—Turban eyes of Ephemerids—Hoods that can be inflated on the head of birds—Absence of secondary sexual characters in lower animals—Transference of male characters to the females—Lycæna—Parrots—Fashion operative in the phyletic modifications of colour—Pattern of markings on the upper surface of a butterfly's wing simpler than on the under side—Conclusion.
We found in the process of Natural Selection an explanation of numerous effective adaptations in plants and animals, as regards form, colouring, and metabolism, of the most diverse weapons and protective devices, of the existence of those forms of blossoms which we call flowers, of instincts, and so on. The origin of the most characteristic parts of whole orders of insects can only be understood as adaptations to the environment brought about by means of natural selection. Impressed by this, we have now to ask whether all the transformations of organisms may not be referred to adaptation to the continually changing conditions of life? We shall return to this question later, but in the meantime we are far from being able to answer it in the affirmative, for there are undoubtedly a great many characters, at least in animals, which cannot have owed their origin to natural selection in the form in which we have studied it so far.
How could the splendid plumage of the humming-birds, of the pheasants, of the parrots, the wonderful colour-patterns of so many diurnal butterflies, be referred to the process of natural selection, since all these characters can have no significance for their possessors in the struggle for existence? Or of what use in the struggle for existence could the possession of its gorgeous dress of feathers be to the bird of Paradise; or of what service is the azure blue iridescence of the Morpho of Brazil, which makes it conspicuous from a distance[Pg 211] when it plays about the crowns of the palm-trees? We might indeed suppose that they are warning signs of unpalatableness, like those of the Heliconiides or of the gaily coloured caterpillars, but, in the first place, these gay creatures are by no means inedible, and are indeed much persecuted; and, secondly, the females have quite different and very much darker and simpler colours. The gleaming splendour of all these birds of Paradise and humming-birds, as well as that of many butterflies, is found in the male sex only. The females of the birds just mentioned are dark in colour and without the sparkling decorative feathers of the males; they are plain—just like the females of many butterflies. Alfred Russel Wallace has suggested that the explanation of this lies in the greater need of the females for protection, since, as is well known, they usually perform the labours of brooding, and are thus frequently exposed to the attacks of enemies. It is undoubtedly true that the dark and inconspicuous colouring of many birds and butterflies depends on this need for protection, but this does not explain the brilliant colours of the males of these species. Or can it be that these require no explanation further than that they are, so to speak, a chance secondary outcome of the structural relations of the feathers and wing-scales respectively, which brought with it some other advantage not known to us? Perhaps something in the same way as the red colour of the blood in all vertebrates, from fishes upwards, cannot be useful on the ground that it appears red to us, but because it is the expression of the chemical constitution of the hæmoglobin, a body which is indispensable to the metabolism, which here has the secondary and intrinsically quite unimportant peculiarity of reflecting the red rays of light.
No one can seriously believe this in regard to butterflies who knows that their colours are dependent on the scales which thickly cover the wings, and the significance of which, in part at least, is just to give this or that colour to the wing. They are degenerate or colourless among the transparent-winged butterflies, and their colour depends partly on pigment, partly on fluorescence and interference conditioned by the fine microscopical structure of a system of intercrossing lines on faintly coloured scales. The scales of our male 'blue' butterflies (Lycæna) only appear blue because of their structure, while the brown scales of their mates are due to a brown pigment. If the pigment be removed from the scales of the female by boiling with caustic potash, and they be then dried, they do not look blue like those of the male; the scales of the male, therefore, must possess something which those of the female do not.
Still less will any one be disposed to regard the marvellous splendour of the plumage of the male bird of Paradise, with its erectile collars—glistening like burnished metal—on the neck, breast or shoulders, with its tufts, with its specially decorative feathers standing singly out from the rest of the plumage, on head, wings, or tail, with its mane-like bunch of loose, pendulous feathers on the belly and on the sides, in short, with its extraordinary, diverse, and unique equipment of feathers, as a mere unintentional accessory effect of a feather dress designed for flight and protective warmth. Such conspicuous, diverse, and unusual specializations of plumage must have some other significance than that just indicated.
Alfred Russel Wallace regards these distinctive features of the male as an expression of the greater vigour, and the more active metabolism of the males, but it is unproved that the vigour of the male birds is greater than that of the females, and it is not easy to see why a more active metabolism should be necessary for the production of strikingly bright colours than for that of a dark or protective colour. Moreover, there are brilliantly coloured females, both among birds and butterflies, and in nearly allied species the males may be either gorgeous or quite plain like the females.
Darwin refers the origin of these secondary sexual characters to processes of selection quite analogous to those of ordinary natural selection, only that in this case it is not the maintenance of the species which is aimed at, but the attainment of reproduction by the single individual. The males are to some extent obliged to struggle for the possession of the females, and every little variation which enables a male to gain possession of a female more readily than his neighbour has for this reason a greater likelihood of being transmitted to descendants. Thus, attractive variations which once crop up will be transmitted to more and more numerous males of the species, and among these it will always be those possessing the character in question in the highest degree which will have the best chance of securing a mate, and so the character will continue to be augmented as long as variations in this direction appear.
Two kinds of preliminary conditions, however, must be assumed. As the ordinary natural selection could never have operated but for the fact that in every generation a great many individuals, indeed the majority of them, perish before they have had time to reproduce, so the process of sexual selection could never have come into operation if every male were able ultimately to secure a mate, no matter what degree of attractiveness to the latter he possessed. If the numbers of males and females were equal, so that there was[Pg 213] always one female to one male, there could be no choice exercised either by male or female, for there would always remain individuals enough of both sexes, so that no male need remain unmated.
But this is not the case: the proportions of the sexes are very rarely as 1 : 1; there is usually a preponderating number of males, more rarely of females. Among birds the males are usually in the majority, still more so among fishes; and among diurnal butterflies there are often a hundred males to one female (Bates), although there seem to be a few tropical Papilionidæ among which the females have rather the preponderance. Darwin called attention to the fact that one could infer the greater rarity of the females even from the pricelists of butterflies issued by the late Dr. Staudinger in connexion with his business, for the females in most species, except the very common ones, are priced much higher than the males, often twice as high. In the whole list of many thousands of species there are only eleven species of nocturnal Lepidoptera in which the males are dearer than the females.
Among the Mayflies or Ephemerides, too, the males are in the majority; in many of them there are sixty males to one female: but there are other kinds of insects, such as the dragon-flies (Libellulidæ), in which the females are three or four times as numerous. There are also, it may be remembered, some kinds of insects, such as Aphides, which have become capable of parthenogenetic reproduction, and in which the males are becoming extinct, e.g. in the case of Cerataphis in British orchid-houses.
The first postulate implied in 'sexual selection,' namely, that there be an unequal number of individuals in the two sexes, is therefore fulfilled in Nature; we have now to inquire whether the second condition postulated—the power of choice—may also be regarded as a reality.
This point has been disputed from many sides, and even by one of the founders of the whole selection theory, Alfred Russel Wallace. This naturalist doubts whether a choice is exercised among birds by either sex in regard to pairing, and maintains that, even if there could be a choice, this could not have produced such differences in colour and character of the plumage, since that would presuppose the existence of similar taste in the females through many generations. In a similar way it has been doubted whether butterflies can be said to exercise any real power of sexual choice, whether a more beautiful male is as such preferred to a less beautiful suitor.
It must be admitted that direct observation of choosing is[Pg 214] difficult, and that as yet there is very little that can be said with certainty on this point. But there are, after all, some precise observations on mammals and birds which prove that the female shows active inclination to, or disinclination for, a particular male. If we hold fast to this fact, and add to it that the distinctive markings of the males are wonderfully developed during the period of courtship, and are displayed before the females, and that they only appear in mammals, birds, amphibians, and fishes at the time of sexual maturity, it seems to me that there can be no doubt that they are intended to fascinate the females, and to induce them to yield themselves to the males. The opponents of the theory of sexual selection attach too much importance to isolated cases; they imagine that each female must make a choice between several males. But the theory of sexual selection does not demand this, any more than the theory of natural selection requires the assumption that every individual of a species which is better equipped for the struggle for existence must necessarily survive and attain to reproduction, or, conversely, that the less well equipped must necessarily perish.
All that the theory requires is, that the selective and eliminative processes do, on an average, secure their ends, and in the same way the theory of sexual selection does not need the assumption that every female is in a position to exercise a scrupulous choice from among a troop of males, but only that, on an average, the males more agreeable to the females are selected, and those less agreeable rejected. If this is the case, it must result in the male characters most attractive to the females gaining preponderance, and becoming more and more firmly established in the species, increasing in intensity, and finally becoming a stable possession of all the males.
When we go more into details we shall see that the particular qualities of the distinctive masculine characters are exactly such as they would be if they owed their existence to processes of selection; in other words, from this point of view the phenomena of the decorative sexual characters can be understood up to a certain point. It seems to me that we are bound to accept the process of sexual selection as really operative, and instead of throwing doubt upon it, because the choice of the females can rarely be directly established, we should rather deduce from the numerous sexual characters of the males, which have a significance only in relation to courtship, that the females of the species are sensitive to these distinguishing characters, and are really capable of exercising a choice.
In my mind at least there remains no doubt that the 'sexual[Pg 215] selection' of Darwin is an important factor in the transformation of species, even if I only take into consideration those secondary sexual characters which are related to wooing. We shall see, however, that there are others in regard to whose origin through processes of selection doubt is still less legitimate, and from which, on this account, we can argue back to the courtship characters.
The first beginning of transformation is not, even in ordinary natural selection, to be understood as due to selection, but is to be regarded as a given variation (the causes of which we shall discuss later on); it is only the increase of such incipient variations in a definite direction that can depend on natural selection, and they must depend on it in so far as the transformations are purposeful. Now, all secondary sexual characters can be recognized as useful, save only the decorative distinctions, although these also undoubtedly represent intensifications of originally unimportant variations. Are we then to regard these alone as the mere outcome of the internal impulsive forces of the organism, while in the case of the analogous sexual characters for tracking, catching, and holding the female, and so forth, the augmentation and the directing must be referred to processes of selection? But if there be any utility at all in the decorative sexual characters it can only lie in their greater attractiveness to the females, and it can only be of any account if the females have, in a certain sense, the power of choice. Independently, therefore, of direct observations as to the actual occurrence of choosing, we should be compelled by our chain of reasoning to assume that there was such a power of choice—and I shall immediately discuss it more precisely.
If we consider the decorative, distinctive characters of the males more closely, we find that they are of very diverse kinds. The males of many animals are distinguished from the females chiefly by greater beauty of form, and especially of colour. This is the case in many birds, some amphibians, like the water-salamander, many fishes, many insects, and above all, in diurnal Lepidoptera. Especially among birds the dimorphism between the sexes is in obvious relation to the excess in the number of male individuals, or—what practically comes to the same thing—to polygamy. For when a male attaches to himself four or ten females the result is the same as if the number of female individuals were divided by four or by ten. Thus the fowls and pheasants, which are polygamous, are adorned by magnificent colours in the male sex, while the monogamous partridges and quails exhibit the same colouring in both sexes. Of course 'beautiful' is a relative term, and we must not simply assume that what seems beautiful to[Pg 216] us appears so to all animals; yet when we see that all the male birds which are beautifully decorative according to our taste—whether humming-birds, pheasants, birds of Paradise, or rock-cocks (Rupicola crocea)—unfold their 'feather-wheels, 'fans,' 'collars,' and so forth, before the eyes of the females in the breeding season, and display them in all their brilliance, we must conclude that, in these instances at least, human taste accords with that of the animals. That birds have sharp vision and distinguish colours is well known; it is not for nothing that the service berries and many other berries suitable for birds are red, the mistletoe berries white, in contrast to the evergreen foliage of this plant, the juniper berries black so that they stand out amid the snows of winter; in this direction, then, there is no difficulty in the way of sexual selection.
Even among much lower animals, like the butterflies, there seems to me no reason for the assumption that they do not see the gorgeous colours and often very complicated markings, the bars and eye-spots, on the wings of their fellows of the same species. Of course if each facet of the insect eye contributed only a single visual impression, as Johannes Müller supposed, then even an eye with 12,000 facets would give but a rough and ill-defined picture of objects more than a few feet away, and I confess that for a long time I regarded this as an obstacle in the way of referring the sexual dimorphism of butterflies to processes of selection. But we now know, through Exner, that this is not the case; we know that each facet gives a little picture, and not an 'inverted' but an 'upright' one, and experiment with the excised insect eye has directly shown that it throws on a photographic plate a tolerably clear image of even distant objects, such as the frame of a window, a large letter painted on the window, or even a church tower visible through it.
Furthermore, the structure of the eye allows of incomparably clearer vision of near objects, for in that case the eyes act like lenses, and reveal much more minute details than we ourselves are able to make out. Here again, therefore, there is no obstacle to the Darwinian hypothesis of a choice on the part of the females, for although it cannot be demonstrated from the structure of the eye itself that insects see colour, and that colours have a specially exciting influence on them, yet we can deduce this with certainty from the phenomena of their life. The butterflies fly to gaily coloured flowers, and as they find in them their food, the nectar of the flowers, we may take for granted that the sight of the colour of their food-providing plants is associated with an agreeable sensation, and this[Pg 217] is an indication that similar colours in their fellows may awaken similar agreeable sensations.
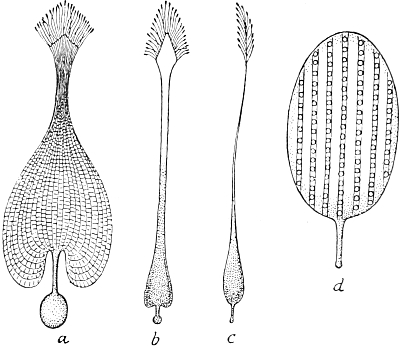
Fig. 53. Scent-scales of diurnal butterflies. a, of Pieris. b, of Argynnis paphia. c, of a Satyrid. d, of Lycæna. All highly magnified.
This conclusion is furthermore confirmed by the fact that, in the male sex, numerous species of butterfly possess another means of exciting the females, namely, by pleasant odours. Volatile ethereal oils are secreted by certain cells of the skin, and exhale into the air through specially constructed scales. Usually the apparatus for dispersing fragrance occurs on the wing in the form of the so-called scent-scales (Duftschuppen), peculiar modifications of the ordinary colour-scales of the wing, but sometimes they take the form of brush-like hair-tufts on the abdomen, and they are in all cases so arranged that the volatile perfume from the cells of the skin penetrates into them, and then evaporates through very thin spots on the surface of the scale, or through brush-like, expanded fringes on their tips. Many of these have long been known to entomologists, because their divergence in form from the ordinary scales attracted attention; and it was also observed that they never occurred on the females, but only on the males. Their significance, however, remained obscure until, by a happy chance, Fritz Müller, in his Brazilian garden, discovered the fact that there are butterflies which give off fragrance like a flower, and then close investigation revealed to him the connexion between this delicate odour and the so-called 'male scales.' One can convince oneself of the correctness of the observation even in some of our own butterflies by brushing the finger over the wing of a newly caught male Garden White (Pieris napi). The finger will be found covered with a white dust, the rubbed-off wing-scales, [Pg 218]and it will have a delicate perfume of lemon or balsam, thus proving that the fragrance adheres to the scales.
Fig. 54. A portion of the upper surface
of the wing of a male 'blue' (Lycæna
menalcas); after Dr. F. Köhler. bl, ordinary
blue scales. d, scent-scales. Highly
magnified.
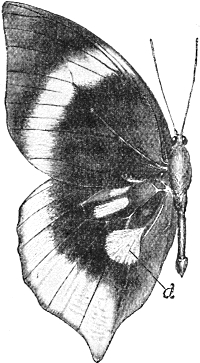
Fig. 55. Zeuxidia wallacei,
male, showing four tufts
of long, bristle-like, bright
yellow scent-scales (d) on
the upper surface of the
posterior wing.
In the last case, that is, among the Whites (Pieridæ) (Fig. 53, a), the scent-scales are distributed fairly regularly over the upper surface of the wing, and the same is true of our blue butterflies, the Lycænnidæ whose minute lute-shaped scales are shown singly in Fig. 53, d, but in their natural position among the ordinary scales in Fig. 54. In many other diurnal, and also in nocturnal Lepidoptera, the fragrant scales are united into tufts and localized in definite areas. They then often form fairly large spots, stripes, or brushes, which are easily visible to the naked eye. Thus the males of our various species of grass-butterflies (Satyridæ) have velvet-like black spots on the anterior wings, while the fritillary, Argynnis paphia, has coal-black stripes on four longitudinal ribs of the anterior wing which are absent in the females, and which are composed of hundreds of odoriferous scales. Certain large forest butterflies of South America, resembling our Apatura, bear in the middle of the gorgeous green shimmering posterior wing a thick expansible brush of long, bright yellow scent-scales, and a similar arrangement obtains in the beautiful violet butterfly of the Malay Islands, the Zeuxidia wallacei depicted in Fig. 55. In many of the Danaides, which we have already considered in relation to mimicry, the scent apparatus is even more perfect, for it is sunk in a fairly deep pocket on the posterior wings, and in this the scent-producing, hair-like scales lie concealed until the butterfly wishes to allow the fragrance to stream forth. In many South American and Indian species of Papilio the fragrant hairs are disposed in a sort of mane on a fold of the edge of the posterior wing, and so on. The diversity of these arrangements is extreme, and they are widely distributed among both diurnal and nocturnal Lepidoptera, in the latter sometimes in the form of a thick, glistening,[Pg 219] white felt which fills a folded-over portion of the edge of the posterior wing. In many cases the perfume can be retained, and then, by a sudden turning out of the wing-fold, be allowed to stream forth. But there are a great many species of butterfly which do not possess odoriferous scales, and they are often wanting in near relatives of fragrant species; they are obviously of very late origin, and arose only after the majority of our modern species were already differentiated. It often seems as if they bore a compensatory relation to beauty of colour, somewhat in the same way as many modestly coloured flowers develop a strong perfume, while, conversely, many magnificently coloured flowers have no scent at all. Although among butterflies, as among flowers, there are species which possess both beauty and fragrance, yet our most beautiful diurnal butterflies, the Vanessas, the Apaturas, and Limenitis, possess no scent-scales; and many inconspicuous, that is, protectively coloured nocturnal Lepidoptera, are strongly fragrant, like most night-flowers: I need only mention the convolvulus hawk-moth (Sphinx convolvuli), whose musk-like odour was known to entomologists long before the discovery of scent-scales.
It is, however, always only in the males that this odoriferous apparatus is present. It must not be believed on this account that this fragrance has the significance of a means of attraction comparable to the perfume of the flowers which induces butterflies to visit them; indeed, we cannot assume that the odour carries to a distance, for, as far as we can make out, it is perceptible only within a very short radius, and this is indicated also by the manifold arrangements of the odoriferous organs, which are all calculated to retain the fragrance, and then—in the immediate neighbourhood of the female—to let it suddenly stream forth. Obviously, this arrangement can have no other significance than that of a sexual excitant; its use is to incline the female to the male, to fascinate her, just as do the beautiful colours, in regard to which we must draw the same inference. It is in this direction that the already mentioned relation of compensation between beautiful colours and pleasant odours is particularly interesting, for it confirms our interpretation of the decorative colours as a means of sexual excitement. The most delicately fragrant or the most beautifully coloured males were those which most excited the females, and thus most easily attained to reproduction. The expression used by Darwin, that the females 'choose,' must be taken metaphorically; they do not exercise a conscious choice, but they follow the male which excites them most strongly. Thus there arises a process of selection among these distinctively male characteristics.
If the odoriferous organs we have been discussing had merely been a means of attraction, serving to announce the proximity of a member of the species, then they should have occurred, not in the males but in the females, for these are sought out by the males, not conversely. The males are able to track their desired mates from great distances, and many remarkable examples of this are known, some of them indeed sounding almost fabulous. The females must therefore also exhale a fragrance, and perhaps continually, but it is much more delicate, carries extraordinarily far, and is quite imperceptible to our weak sense of smell. It is possible that it streams out from all the scales covering the wings and body, for, as I long ago pointed out, all the scales retain a connexion with the living cells of the skin, however minute these may be, and it is therefore quite possible that the cells produce scent imperceptible by us, and let it exhale through the ordinary scales, since the male scent-scales owe their ethereal oil to the large gland-like cells of the hypodermis on which they are placed.
Here we see very clearly the difference between ordinary natural selection and sexual selection. The male odoriferous organs depend on the latter, for they do not serve for the maintenance of the species, but are of advantage in the courting competitions among the males for the possession of the females, while the assumed fragrant cells of the females must depend on natural selection, since they are of general importance for the mutual discovery of the sexes, which would otherwise be in most cases impossible. This hypothetical 'species scent,' as we may call it, is first of all useful in securing the existence of the species, and must therefore be referred to natural selection. The other, the 'male scent,' might be, and actually is, wanting in many species, although it may be necessary to reproduction in cases where it has become a male specific character, and could not be absent from any male without dooming him to sterility.
That the 'species scent' really exists admits of no doubt, although we may be unable to perceive it. Entomologists have long been in the habit of catching the males of the rarer Lepidoptera, especially of the nocturnal forms, by freely exposing a captive female. Some years ago I kept for some time in my study, with a view to certain experiments, females of the eyed hawk-moth (Smerinthus ocellatus), and placed them at first, without any special intention, in a gauze-covered vessel near the open window. The very next morning several males had gathered and were sitting on the window-sill, or on the wall of the room close to the vessel, and by continuing the experiment I caught, in the course of nine nights, no fewer than forty-two males[Pg 221] of this species, which I had never believed to be so numerous in the gardens of the town. The males of the nocturnal Lepidoptera obviously possess an incredibly delicate organ of smell, and its bearers, the antennæ, are usually larger and more complex in structure in the male sex than in the female.
Butterflies are by no means the only creatures that produce a peculiar odour at the breeding season; many other animals do the same, though in their case it does not seem so pleasant to our sense of smell. It is true that the scent of the musk-deer and that of the beaver (Castoreum), when much diluted, are agreeable to man, but others, like the odours exhaled by stags or by beasts of prey, are very disagreeable to us, though they have for the species that produce them the same significance as the others, and are therefore to be referred to sexual selection.
Darwin referred all the different mechanisms for the production of sounds, up to the song of birds, to sexual selection, but it is probable that natural selection has also to do with this in many ways. It is certainly only the males which produce the well-known song of the Cicadas, crickets, grasshoppers and birds, and I do not see any reason to doubt that this 'music' affects the females by arousing sexual excitement. To some extent, then, the rivalry among the males for the possession of the females—that is to say, sexual selection—must have produced these mechanisms of song; and how long-continued and gradual the accumulations must have been which produced the song of the thrush or of the nightingale from the chirping of the sparrow we may learn from the innumerable species which, as regards beauty of song, may be ranged between these two extremes.
My assumption that natural selection has also been operative in the case of the song of insects and birds is based on the fact that many of our songsters live widely scattered, and that the characteristic note must be a means by which the two sexes find each other. That they should find each other is an indispensable condition for the maintenance of the species. Thus it is well known that each species has a characteristic 'note' or love-call, which the male utters during the breeding season, and which is answered by the female. From this simple love-call the modern song of many species must have developed by means of sexual selection.
It is remarkable that here again the various distinguishing characters of the male seem to be often mutually restrictive or mutually exclusive. The best singers among our birds are inconspicuously coloured, grey or brown-grey, and this can hardly be regarded as due to chance, but as the outcome of a greater sensitiveness[Pg 222] on the part of the females either to the song or to the beauty of their mates. And since, according to the theory, only those characters of the males could be increased which decided the choice, it therefore seems to me that this mutual exclusiveness of the two kinds of distinguishing characters is another indication of the reality of sexual selection. It proves—so at least I am inclined to believe—that the excitement of the female has been essentially affected by only one of the characters of the male, that in the bird of Paradise it was mainly the brilliance of the plumage which roused excitement, while in the nightingale it was mainly the song.
It might be objected to this that there are brilliant butterflies which also possess scent-scales. This is really the case; thus a magnificent blue iridescent Apatura from Brazil has on the posterior wings a large yellow brush of scent-hairs, and even the beautiful blue males of our Lycænids have scent-scales in addition to their beautiful colour. But this can hardly be considered as a contradiction, but is rather an exception, which is the easier to explain since the odoriferous apparatus is a relatively simple arrangement, which did not require such a long series of generations for its evolution as the complicated song-box and brain-mechanism of the singing-birds.
Moreover, it may also be that the scent-scales have arisen later than the decorative colouring, and they would do so the more easily since the brilliant blue, when once it was perfectly developed, and was common to all the males of the species in an equal degree, was no longer distinctive, and would have no specially exciting effect, while a novel preferential character in the male would have a much stronger effect. In the same way, the different parts of the body would be furnished in succession with decorative and, therefore, exciting distinctive characters. To understand this effect on the opposite sex we need only think of analogous phenomena in human kind, and of the strongly exciting effect that the sight of the secondary sexual characters of the woman has upon the man.
By the successive additions of new decorative characters after the older ones became general and reached a climax, the origin of the extraordinary diversity of the decorative plumage in one and the same species of bird, can be readily understood, and the same is true of the complicated decorative coloration of the butterflies in so far as it depends on sexual selection, and not on other factors. The details did not arise all at once, but one after the other, and every character went on increasing till it had reached its limit of increase, but whenever it was common in its highest development to all the males it[Pg 223] was no longer an object of preference or the cause of specially violent excitement, so that a new process of selection would begin in reference to some other part of the body. We thus understand how, among male birds of Paradise and humming-birds, such a marvellous diversity of colours and of decorative feathers is found combined in one and the same species.
Whoever has seen the Gould Collection of humming-birds in London must have observed with amazement that among the 130 or so species of these beautiful little birds nearly every group of feathers in the body has been affected by the decorative colouring. In one species the little feathers on the region of the throat are emerald green, metallic blue, or rose; in another the feathers of the neck have been transformed into an erectile collar of rose-coloured feathers with a metallic sheen; or, again, it is the little feathers round the ear that stand erect and are brilliantly coloured. Sometimes we find that the feathers of the tail are lengthened, it may be only two of them, or the various lengths may be graduated like steps; sometimes the tail has assumed the form of a wedge, or is fan-like, or is shaped like the tail of a swallow, and all this in combination with the most diverse colours and patterns, black and white, ultramarine blue, and so forth. Or it may be the outermost tail-feathers which are the longest, the inner ones the shortest, or the four outer feathers are broad, pointed, directed outwards, and only half as long as the other two, which are very long and straight. Some species exhibit a sort of fine swan's down on the legs, others have a gorgeous metallic red cap on the head—in short, the variety is beyond description, just as we should expect it to be if now this and now that chance variation attracted the favourable regard of the selecting sex, and thus attained to its highest pitch of development.
The decorative colouring of male birds may be replaced, not only by the power of song, but in other ways also. Not all the male birds of Paradise possess the familiar feather ornaments. The Italian traveller Beccari has called attention to a species, the males of which are simply coloured brown, like the females of other species. This Amblyornis inornata entices its mate to itself in the pairing time in a very peculiar manner, for it arranges in the midst of the primitive forests of New Guinea a little 'love garden' or bower, a spot several feet in extent, strewn with white sand, on which it places shining stones and shells, and brightly coloured berries. In this case a special instinct has developed, which has replaced the personal charm of the bird in the eyes of the female. For this very reason the case seems to me to have some theoretical importance, for it serves indirectly to[Pg 224] show that the personal excellences do actually function as a means of exciting and attracting, if any one should still doubt it.
All the distinguishing characters of the male which we have hitherto considered have had reference to gaining the favour of the female, but there are many other secondary sexual characters which are employed in quite a different manner to secure possession of the female. I have already mentioned that in many butterflies the males possess a much larger organ of smell. The antennæ of the males of numerous beetles, such as the cockchafer and its relatives, are also much larger, and furnished with much broader accessory branches, than those of the female, and the same is the case in many of the lower crustaceans, like the large transparent Daphnid of our lakes, Leptodora hyalina. Here the anterior antenna bears (Fig. 56, A and B, at´) olfactory filaments; in the female this appendage is small and stump-like, while in the male (A) it grows to a long, somewhat curved rod, which is extended obliquely into the water, and in addition to the nine olfactory filaments of the female (ri) bears from sixty to ninety more (ri´).
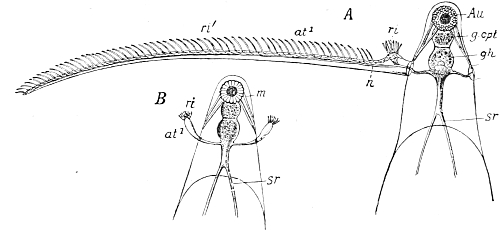
Fig. 56. Leptodora hyalina. A, head of the male. B, head of the female. Au, eye. g. opt, optic ganglion. gh, brain. at´, first antenna with olfactory filaments ri and ri´. sr, œsophageal nerve-ring. n, nerve. m, muscles.
In this and many other such cases it is not the struggle of the species for existence which has so markedly augmented this distinctive characteristic of the male; it is undoubtedly the struggle of the males among themselves, their competition for the possession of the females. In regard to decorative distinctions, the reality of a rivalry in wooing and the ultimate victory of the most decorative may perhaps be still doubted; but it is quite certain that, on an average, the male which can smell and track best will also gain possession of the females more easily than one less well equipped. Exactly the same is also true of those cases in which the male dis[Pg 225]tinguishing character does not refer merely to finding the female, but to holding her fast, or, as we may say, to capturing her.
Thus the males of the Copepods possess on their anterior antennæ an arrangement which enables them to throw a long whiplike structure like a lasso round the head of the female as she rapidly swims away. The antennæ of the male Daphnids, too, are in one genus (Moina) developed into a grasping apparatus, instead of into smelling organs as in Leptodora. Fig. 57 shows the male, Fig. 58 the female of Moina paradoxa; the first antennæ of the male are not only much longer and stronger than those of the female (at1), but they are also armed with claws at the end, so that the males can catch their mates as with a fork, and hold them fast. And even that was not enough, for, in addition, the males of most Daphnids possess a large sickle-shaped but blunt claw on the first pair of legs (Fig. 57, fkr), which enables them to cling to the smooth shell of the female, and to clamber up on it to get into the proper position for copulation.
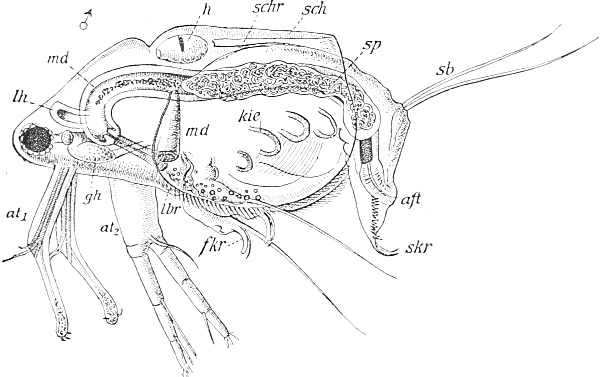
Fig. 57. Moina paradoxa, male. at1, first antennæ, with claws at the tip for capturing the female. at2, second antennæ. fkr, claws on the first pair of legs for clambering. gh, brain. lbr, upper lip. md, mandible. md, mid-gut, with the liver lobes (lh). h, heart. sp, testis. aft, anus. sb, caudal setæ. skr, caudal claws. sch, shell. schr, cavity of the shell. kie, gill-plates. Magnified 100 times.
If we inquire into the manner of the origin of secondary sexual characters of this kind, we shall find that both may have been increased by sexual selection, for a male with a better sickle will succeed more quickly in getting into the proper position for copulation than one with a less perfect mechanism. This assumption does not rest[Pg 226] on mere theory, for I was once able, by a happy chance, to observe for a considerable time, under the microscope, a female to whose shell two males were clinging, each trying to push the other off. Nevertheless it seems to me very questionable whether the origin of this sickle-claw can be referred to sexual selection, for without this clamping-organ copulation in most Daphnids would not be possible. It was thus not as an advantage which one male had over another that the clamping-sickle evolved, but rather as a necessary acquisition of the whole family, which must have developed in all the species at the same time as the other peculiarities, and notably those of the shell. The competition of the males among themselves is thus in this case simply an expression of the struggle for existence on the part of the species as such, and it is not a question merely of a character which makes it easier for the males to gain possession of the females, but of one which had necessarily to arise lest the species should become extinct. In other words, in this case natural selection and sexual selection coincide.
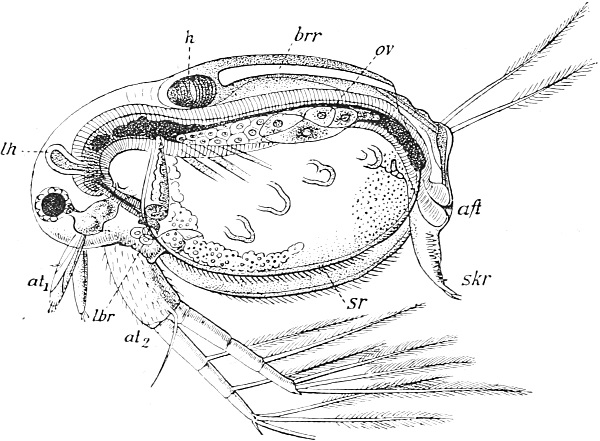
Fig. 58. Moina paradoxa, female. The letters of Fig. 57 apply mutatis mutandis. brr, brood-pouch. ov, ovary. sr, margin of shell.
The case of the antennæ of Moina, which have been modified into grasping organs, is quite different; these owe their origin not to natural selection, but to sexual selection, for antennæ of that kind are not indispensable to the existence of the species, as we can see from the closely related genera, Daphnia and Simocephalus, where the males have quite short stump-like antennæ, furnished with olfactory filaments not much more numerous than the females possess. Just as these supernumerary olfactory filaments were produced by[Pg 227] sexual selection, and not by the ordinary natural selection, because those males with the more acute sense of smell had an advantage over those in which it was blunter, so the males of the genus Moina which could grasp most securely had an advantage over those that gripped less firmly, and thus arose these two different kinds of male characteristics. Neither of them is of advantage to the species as such, but only to the males in their competition for the possession of the females.
But, where the production of a novel character in the male is concerned, natural selection cannot proceed in a different manner from sexual selection; the process of selection is exactly the same: the better equipped males survive, the less well-equipped die without begetting offspring; the difference lies only in the fact that in the one case the improvement is in the species as such, in the other case only in one sex without the existence of the species being thereby made more secure. Such cases are instructive, because they make a denial of the process of sexual selection quite impossible if that of species-selection is admitted. If processes of selection are operative at all as factors in transformation, they must act even where the advantage is not to the species but only 'intra-sexual,' and the one process must often run into the other, so that it is often quite impossible to draw an exact line of demarcation between them.
Numerous secondary sexual differences probably depend purely on species selection, that is to say, they include an improvement of the species in relation to the struggle for existence. We may find a case in point in the dwarf-like smallness of the males in many parasitic crustaceans, in some worms, in many Rotifers, and in the Cirripedes. It can hardly have been of advantage for the individual male to be smaller than his fellows, but it was of advantage for the species to produce as many males as possible in order to ensure a meeting with the females, and, as the enormous production of males made it advantageous for the species that as little material as possible should be used in their individual production, we can readily understand the minuteness of the males, and in some cases, as in the Rotifers and Bonellia, their poor equipment, lack of nutritive organs, and ephemeral existence. The marine worm, Bonellia viridis, whose female may be a foot in length, is not the only case in which a microscopically small male lives like a parasite inside the female. Among the round-worms, too, there is a species called Trichosomum crassicauda, discovered by Leuckart in the rat, the dwarf males of which live in the reproductive organs of the female. All these are arrangements for securing the propagation of the species, which[Pg 228] might have been endangered if the males had had to seek out the females, which, in the case of Bonellia, live in holes in the rocks on the sea-floor, and, in the case of Trichosomum, are concealed in the urinary bladder of the rat. Obviously, this is the reason which, in addition to the one already mentioned, has conditioned and produced, or helped to produce, the remarkable minuteness of certain males.
From another category of sexual differences we see in how many ways species-selection and sexual selection play into each other's hands. In many species of animals the males are eager for combat, and they are equipped with special weapons, or excel the females in general strength of body. As these males struggle, in the literal sense of the word, for the possession of the females, Darwin referred to sexual selection those distinguishing characters which gave the stronger male the victory over the weaker, and thus raised the victorious characters to the rank of general characters of the species. And it certainly cannot be doubted that, for instance, the strength and the antlers of the stag must have been increased through the combats which recurred every year at the breeding season, for the stronger always win in these battles. The case is the same with the strength and the weapons of many other male animals. The lion is effectively protected by his mane from the bite of a rival, and the same protective arrangement occurs in quite a different family of mammals—in an eared seal, which is called the 'sea-lion' for this very reason. Among the seals the secondary sexual characters are often very strongly developed, at least in all the polygamous species, for in these the struggle for the females is very keen. In the 'sea-lions' and 'sea-elephants' there are often fifty females to one male, and the latter are 'enormously larger' than the females, while in monogamous species of seal the two sexes are alike in size.
Darwin has shown that actual combat for the females takes place among most mammals, not only among stags, lions, and seals, but even among the moles and the timid hares. Even among birds such combats occur, and this is sometimes particularly noteworthy in those species in which the males possess the most decorative colouring, like the humming-birds. In some cases among birds there has also been a development of weapons. Witness the spur of the cock, whose merciless combats with his rivals Man has, as is well known, made positively atrocious for his own amusement, by preventing the flight of the vanquished.
In Darwin's great work on sexual selection a considerable number of cases are cited from among lower vertebrates, such as crocodiles and fishes, and even from insects, in which the males fight[Pg 229] for the possession of the females, and exhibit distinctive masculine characters adapted to such combats. But I do not propose to enter upon a discussion of such cases, since my aim is rather to elucidate the relation between sexual selection and species-selection than to discuss all the phenomena of the former in detail. But the combats of males illustrate with particular clearness the relation of sexual selection and species-selection, since many of the weapons or protective arrangements which may have arisen through sexual selection imply at the same time an improvement to the species in relation to the struggle for existence. Thus greater strength or sharper and larger teeth in the males mean a gain to the species, and it is indifferent to the species whether the weaker males succumb to a strange enemy (species-selection) or to their stronger rivals (sexual selection), provided only that the better equipped survive and leave descendants similarly endowed.
I have intentionally begun the consideration of sexual selection with the cases most difficult to interpret on this theory, with those which have called forth the greatest divergence of opinion—the decorative colours and forms, the song of birds and of insects, the alluring odours—in short, all the courtship-adaptations of the males; these are the most difficult to deal with, because it is not easy to demonstrate directly that the females do choose. But if we revise them briefly in reverse order, I believe that all doubt as to the reality of choice on the part of the females will disappear. Thus the last-mentioned sexual characters of greater strength and greater perfection of weapons and defence in the males have been evolved by sexual selection in close co-operation with species-selection. We should have to deny species-selection altogether if we were to dispute this form of sexual selection, which is closely connected with pure species-selection, such, for instance, as is revealed in the production of dwarf males, where there does not seem to be any aid from sexual selection at all.
Then came the cases in which the tracking and grasping organs of the males were strengthened or were increased in number, and here too species-selection may have had its share, for instance, in evolving the sickle-claws of the Daphnids, which were inevitably advanced and perfected through sexual selection, which must in this case have operated independently of any choice on the part of the female. In other cases the result may be referred to pure sexual selection, as in the grasping antennæ of the male Moina, or in the highly developed olfactory antennæ of the male Leptodora. That new organs, too, can arise in this way is shown by the 'turban eyes'—to which little attention has hitherto been paid—of some Ephemerids of the genera[Pg 230] Cloë and Potamanthus, which were long ago described by Pictet, the monographer of this family. These are large turban-shaped compound eyes, occurring beside the ordinary eyes in the males alone, which in these genera are in a majority of sixty to one. Whole swarms of these males fly about over the water on the search for females, and their highly developed organ of vision seems to decide matters for them just as the organ of smell does for Leptodora. Neither of these sense-organs can have any other advantage than that of making their possessors aware of the female, for the whole activity of the short-lived adult Ephemerides is limited to reproduction; they take no food, and have nothing whatever to do except to reproduce.
Finally, when in an enormous number of cases we find in addition to one or the other of the already mentioned male distinguishing characters some which do not directly lead to gaining possession of the female, but do so only by sexually exciting her, can we doubt that the same principle has been operative, that here too processes of selection are fundamental, depending on the fact that in the wooing of the female the successful male is the one who most strongly excites her? There is no question of æsthetic pleasure in this, as the opponents of the theory of sexual selection have often urged, but only of sexual excitement, which may be aroused by very different means, by colours and shapes, but also by love-calls, songs, or odours. There are a few tropical birds (Chasmorhynchus) which have as the only distinguishing character of the male sex a hollow and soft appendage several inches long borne on the head. Usually it hangs down limply at the side of the head, but during the breeding season it is inflated from the mouth-cavity, and then stands erect like a spur. One species of this genus has as many as three of these horns, one of which is upright, while the other two stand out laterally from the head. Can it be supposed that these remarkable horns satisfy the female's 'sense of beauty'? To human beings they appear rather ugly than beautiful, both when limp and when inflated, but at any rate they are striking, and will be regarded by the female bird as something out of the common, and, since they are only fully displayed during the breeding season, that is, when the male is sexually excited, they will have an exciting effect on the female too. These inflated horns are symptoms of excitement, and they arouse it in the female. In exactly the same way the decorative feathers, the ruby-red and emerald-green feather collars of the humming-birds and birds of Paradise, are only erected and displayed when the males are wooing, and they, too, act as signs of excitement. This is not to say[Pg 231] that the gorgeousness of colour, the eye-spots on the train of the peacock and the Argus pheasant, and the hundreds of different kinds of beautiful feathers, do not also exercise a fascinating influence; on the contrary, we cannot avoid assuming this, since otherwise we could find no sufficient reason for their origin. But the primary effect in wooing is not due to the mere pleasure in the sight, or in the odour, or in the song, but to the contagious excitement which these express. The females do not behave as dispassionate judges, but as excitable persons which fall to the lot of the male who is able to excite them most strongly. It may be, however, that a sense of æsthetic satisfaction in perceiving such symptoms of excitement may also have been evolved as an accessory effect, at least in the higher and more intelligent animals.
In the lower animals, which are lacking not only in intelligence but also in the higher and more complex differentiation of the sensory system, the development of such secondary sex characters is rare or altogether absent. Animals which have no sense of hearing can develop no song, and animals which do not see cannot acquire gorgeous colours as a means of exciting one sex through the other. But distinctive sex coloration may arise even in lowly animals, though there can be no question of æsthetic pleasure associated therewith; if the animals are able to see the colours at all, sexual excitement may be associated with these.
We need not wonder, therefore, that in the somewhat stupid fishes, in the butterflies, and in the lower crustaceans, like the Daphnids, we still find brilliant colours, which we can hardly interpret otherwise than as the results of sexual selection. On the other hand, the absence of such characters in animals of a still lower order, with still simpler sense-organs, like the Polyps, Medusæ, Echinoderms, most Worms, and the Sponges, affords an indirect confirmation of the correctness of our view as to the reality of a sexual selection in the more highly organized animals.
We see, then, that numerous peculiarities which distinguish the males of a species from the females depend on the process of sexual selection. This may be said of ornamental outgrowths, colours, remarkable feathers and feather-groups, peculiar odoriferous organs, vocal organs, artistic instincts, and also weapons, like antlers, tusks, and spurs, notable size and strength of body, and protective devices like manes; and again, the various organs for catching and holding the females, or for finding them out by sight or smell, must also be referred, at least in part, to sexual selection. The diversity of the male sexual characters is so great that I cannot give more than[Pg 232] a faint idea of them without entering on a long catalogue; whoever wishes a complete survey has only to consult Darwin's Descent of Man.
But the significance of sexual selection is by no means exhausted with the production of the male sexual characters, for these characters are often more or less completely transferred to the females, and thus give rise to a transformation of the whole species, and not only of the male section of it. This is obviously a very important consequence of sexual selection, one which, as we shall see, materially deepens our insight into the mode of origin of new species.
First let us try to determine the facts. Many male characters are not represented in the female in any degree, and therefore have never been transmitted to them at all. Such are the mane of the lion, the grasping antennæ of Moina, the turban eyes of the Ephemerides, the intensification of the sense of smell in Leptodora, the lasso-like antennæ of the Copepods, the scent-scales of the butterflies, and the musk glands of the alligators and stags. But in other cases there has been transmission, though only to a slight extent. Thus many female humming-birds have a faint indication of the magnificent metallic colouring of the males; many female blue butterflies have a tinge of the beautiful blue of their mates; the females of the stag-beetle (Lucanus cervus) possess a diminutive suggestion of the antler-like jaws of the male, and the female crickets, although they do not chirp, have a slight indication of the 'musical' mechanism of the male on the wing-coverts, and some of them even produce feeble notes at certain times.
It can be proved, however, that such transmissions may, in the course of many successive generations, become intensified until the characters are exhibited by the females in the same degree as in the males. I know no better example of this than that afforded by the beautiful butterflies of the genus Lycæna. In this genus, which is rich in species and widely distributed over the whole earth, and must therefore be an old one, the upper surface of the wing is blue in by far the greater number of species, at least in the male sex. But there are three or four species which are dark-brown, and quite or nearly alike in the two sexes; such are the species Lycæna agestis, L. eumedon, L. admetus, and others. Everything indicates that this is the primitive colour of the genus. Moreover, there are some species with brown females, in which the males are not completely blue, but which have a slight bluish tinge, like L. alsus, the smallest of our indigenous Blues. Then follows a host of beautiful species, like L. alexis, L. adonis, L. damon, L. corydon, and many others, with[Pg 233] brown females, and among these there occasionally occur females more or less tinged with blue. These lead on to L. meleager, which has two forms of female, a common brown and a rarer blue; and thus we reach L. tiresias, L. optilete, and L. argiolus, in which all the females are blue, although less intensely and completely so than their mates. The climax of this evolutionary series is reached by some species like L. beatica, belonging to tropical or at least warm countries, in which both sexes are of an equally intense blue. As we know that, in species with an excess of males, sexual characters always begin in the males, there can be no doubt as to the direction of evolution—from brown to blue—in this series. Furthermore, the entire absence of scent-scales in most of the species with brown males indicates the great age of these species, for, as far as I have been able to investigate, all the males of the blue species possess them.
Darwin regarded this transferring of the male characters to the females as due to inheritance, and it really seems as if it were simply a case of transmission by inheritance to one sex of what has been acquired by the other. Yet we have to ask whether we can continue to regard the facts in this light. In any case this 'transmission' is not an inevitable physiological process, necessarily resulting from the intrinsic conditions of inheritance, for we see that it often does not occur, even in many cases in which we can see no external reasons why it should not do so, though in other cases the failure may be presumably correlated with the external conditions of life. Thus, for instance, the persistent retention of the brown colour in the majority of our female Lycænidæ has probably its reason in the greater need of protection on the part of the much rarer females, and this must be so also in the case of many birds in which the brilliant colours of the males have not been transferred to the females. Wallace first pointed out that all birds whose females brood in exposed nests are inconspicuously coloured in the female sex, even if the males are brightly coloured, while those whose nests are concealed in holes of trees or the like, or which build domes over them, not rarely exhibit brilliant colouring in both sexes. This is the case in woodpeckers and parrots, while the gallinaceous birds, which brood in the open, have usually inconspicuously coloured females, for the most part very well adapted to their surroundings.
If we grasp the fact that a transference of the characters which have arisen through sexual selection can take place, we have a valuable aid in the interpretation of many phenomena which would otherwise remain quite inexplicable. What is the meaning of the gay colours of the parrots, which occur in such incredibly diverse com[Pg 234]binations in this large and widely distributed family? Or of the marvellously complex markings and colour-patterns of the butterflies? In some cases they may be protective, as is the green of many parrots; in others, warning signs of unpalatability, like the bright colours and contrasted markings of many Heliconiidæ and Eusemiidæ and other butterflies with a nauseous taste; but there remain a great many cases to which neither of these explanations applies, which could only be regarded as pure freaks of nature if we did not know that male sexual characters can be transferred to the females, and that thus all the individuals of a species can be totally altered in their colouring.
Thus the occurrence not only of conspicuous, but of complicated, coloration is explained.
Darwin has shown that, in the equipment developed by the males in their competition for the possession of the females, it is by no means only those characters which may be considered 'beautiful' in themselves that have to be considered; it is rather the striking characteristics which mark their possessor and distinguish it from others that are primarily important. In fact, it is the principle of 'mode' or 'fashion' which is operative; something new is demanded, and as far as possible something quite different from that which was previously considered beautiful. Thus the starting-point for such processes of selection may have been afforded by white spots on a black ground, or, indeed, by any light spots on a dark ground, which may have been the primitive colour in most cases. If in the course of a long series of generations these spots became the common property of all the males, a possibility of further change was opened up as soon as a new contrast cropped up as a chance variation, which would then, under favourable conditions, be the starting-point of a new process of selection. Darwin has cited some cases in which, from a comparison of the dress of the young bird with that of the adult, we may conclude that a transformation of the colouring of the whole plumage must have taken place in the course of the phylogenetic history.
In other cases the course of the process of selection has been such that, though the general colouring has not been changed, variations have appeared in particular regions of the body—spots or stripes which accumulated through the ages and co-operated to form an increasingly diverse and complex colour-scheme, such a 'marking' of the animal as we may observe to-day, especially in butterflies, but also in birds.
It is a fine corroboration of the origin of bright colours through[Pg 235] sexual selection that, even in those groups of the animal kingdom which are in general sexually monomorphic, there always occur some species in which male and female are quite different, and a host of species in which both sexes are alike in the main, yet with differences in certain minor points. Among the parrots similarity of colouring prevails as a general rule, but in New Guinea there lives a parrot the female of which is a gorgeous blood-red and the male a beautiful light-green; minor differences occur in many species, for instance, the female of the horned parrot (Cyanorhamphus cornutus Gm.) lacks the two long black and red feathers on the head, that of the grass-parakeet (Melopsittacus undulatus) is a slightly paler green and has not the beautiful blue spots on the cheeks which the male possesses. Innumerable similar instances might be cited, serving to show that all these distinguishing characters of the males have been acquired step by step and piece by piece, and are slowly and independently transferred to the females—if, indeed, at all.
In yet another way the correctness of the Darwinian theory of sexual selection may be deduced from the markings and coloration of birds and butterflies.
It has frequently struck me, during the long period in which I have been studying brightly coloured birds and butterflies, that those colour-patterns which are referable to sexual selection are much simpler than those which must be referred to species-selection, especially in the case of what we call 'sympathetic coloration.' How crude is the decorative pattern of most parrots, notwithstanding all the brilliance of their colour. Large tracts of the body are red, others green, yellow, blue, and occasionally one finds a red and blue striped feather collar, a head which is red above and yellow underneath, but it is seldom that the colours vary enough in a small space to give rise to a delicate decorative pattern. The gayest of parrots are the Brush Tongues (Trichoglossus), and even among them subtlety of coloration does not go further than the combination of three colours on one of the long tail-feathers, or the production of a double band round the neck, and so forth. If we compare with this the complex markings of the inconspicuously coloured females of the pheasants, of the partridges, or that of the upper surface of the many birds in mingled grey, blackish-brown and white, which resemble the ground or the dried leaves when they crouch, we find that the colour-pattern in these cases is infinitely finer and more complex.
This seems to me quite intelligible when we remember, on the one hand, that species-selection must operate far more intensively than[Pg 236] sexual selection, and that in the production of a protective colouring it is a question of deceiving the eye of a sharp-sighted enemy, while the aim of sexual selection is to secure the approval of others of the same species. As long as the enemy on the search for prey perceives the difference between the markings of its victim and those of the surroundings, so long will the gradual and steady improvement of the protective coloration continue, so long will new shades and new lines be added. We can thus understand how there would be gradually reached a complexity of marking to which sexual selection can never attain, or at least only in regard to a few specially favourable points. The eye-spots on the train feathers of the Argus pheasant and the peacock are such points, and these occur among polygamous birds in which sexual selection must be very intense; they are placed, too, on a part of the body, the wheel-shaped train, which is peculiarly suited for communicating the excitement of the male to the female, and must therefore be especially influenced by the latter. In general, however, we may say on a priori grounds that the intensity of species-selection is greater than that of sexual selection, because the former ceaselessly and pitilessly eliminates the less perfect, while the claims of the latter are in any case less imperative, and are also often mollified by a variety of chance circumstances.
But in the case of insects, in particular, we have to add that the protective colours and the decorative colours have been, so to speak, painted by different artists—the former by birds, lizards, and other persecutors endowed with well-developed eyes, the latter by the insects themselves, whose eyes can hardly possess, for objects not quite near, that acuteness of vision which the bird's eye has. Thus we find that the protective coloration of butterflies has often a very complex marking, while the same butterfly may exhibit only a rather crude though brilliant pattern on its upper surface, where the coloration is due to sexual selection. Thus the famous Kallima has on its under surface the likeness of a dry or decayed leaf composed of a number of colour-tones—quite a complex painting. But if we look at the upper surface we see a deep brown with a shimmer of steel blue as the ground-colour of the wings, and on it a broad yellow band and a white spot: that is the whole pattern. We find a similar state of things among many of the forest butterflies of Brazil, and also among our indigenous butterflies. The pattern of our gayest diurnal butterflies, the red Admiral and the tortoiseshell butterfly (Vanessa atalanta and Vanessa cardui), is somewhat crude on the upper surface, and very simple compared with the protective colouring of the under surface, which is made up of hundreds of points, spots, strokes, and[Pg 237] lines of every shape and colour. On the other hand, the upper surface of the anterior wings in the hawk-moths and the Noctuidæ exhibits protective coloration, and is made up of curious zigzag complex lines, strokes, and spots, so that it resembles the bark of a tree or a bit of an old wooden fence—a painting, like the modern impressionist work, which, with an apparently meaningless confusion of colour splashes, conveys a perfect impression even of the details of a landscape. In the owl-moths (Noctuidæ) the wing surfaces, which are brightly coloured, are simple, almost crude, in pattern, as in the moths of the genus Catocala, with their red, blue, or yellow posterior wings, traversed by a large black band; while in the Geometer-moths, whose wings are spread out flat when at rest, the protective upper surface of all four wings is covered with a complex pattern of lines, spots, and streaks in different shades of grey, yellow, white, and black, so that it bears a deceptive resemblance to the bark of a tree or the side of a wall. For a long time I could not understand how such a definite and constant pattern could arise through natural selection if it was a case of mimicking the impression of bark or of any other irregularly covered surface, the colours of which are not mingled in exactly the same way everywhere. But now I think I understand it; for in the apparently meaningless colour-splashes of an 'impressionist' landscape the different splashes must be exactly where they are, otherwise on stepping back from the picture one would see, not a Haarlem hyacinth-field, or an avenue of poplars with their golden autumn leaves, but a mere unintelligible daub. It is the type of the colour-pattern that must be attained, and in nature this is attained very slowly, step by step, spot after spot, and therefore, obviously, no correct stroke once attained will be given up again, since, in combination with the rest, it secures the proper type of colour-pattern. Only thus, it seems to me, can we understand how apparently meaningless lines, like the figures 1840 on the under surface of Vanessa atalanta, could have become a constant characteristic of the species.
To sum up briefly, we may say that sexual selection is a much more powerful factor in transformation than we should at first be inclined to believe. It cannot, of course, have been operative in the case of plants, nor can it be taken into consideration in regard to the lower animals, for these, like the plants, do not pair, or, at any rate, do so without any possibility of choice. Animals which live on the sea-floor, or which are attached there, must simply liberate their reproductive cells into the water, and cannot secure that they unite with those of this or that individual. This is the case among sponges,[Pg 238] corals, and Hydroid polyps. In some other classes the sense organs are too poorly developed, and the eyes in particular too imperfect to be excited in different degrees by any peculiarities in the appearance or behaviour of the males. This is what Darwin meant when he ascribed to these animals 'too imperfect senses and much too low intelligence' 'to estimate the beauty or other attractive points of the opposite sex, or to feel anything like rivalry.' Accordingly, in the Protozoa, Echinoderms, Medusæ, and Ctenophores, secondary sexual characters are entirely absent, as pairing also is.
In those worms that pair we first meet with secondary sexual characters, and from this level upwards they are never quite absent from any large group, and gradually play an increasingly important rôle.
But the significance of sexual selection lies, as we have seen, not only in the fact that one sex of a species, usually the male, is modified, but in the possibility of the transference of this modification to the females, and further, in the fact that the process of variation may start afresh at any time, and thus one variation may be developed upon or alongside of another. In this way we can explain certain complex and often fantastic forms and colourings which we could not otherwise understand; thus the extraordinary number of nearly related species in some animal groups, such as butterflies and birds, in which the differences mainly concern the colour-patterns.
Darwin has shown convincingly that a surprising number of characters in animals, from worms upwards, have their roots in sexual selection, and has pointed out the probability that this process has played an important part in the evolution of the human race also, though, in this case, all is not yet so clearly and certainly known as among animals.
To conclude this section, I should like once again to call attention to the deficiency which is necessarily involved in the assumption of any selection, sexual selection included, namely, that the first beginning of the character which has been intensified by selection remains obscure. Darwin attached importance to the occurrence of ordinary individual variation, but it is open to question whether the insignificant variations thus produced could give an adequate advantage in the competition for the possession of the females; and, further, whether we have not grounds for the assumption that larger variations also occur. This question may also be asked in regard to ordinary natural selection, although in that case we can imagine the beginnings to be smaller, since here the advantage of a variation lies only in the fact that it is useful, not in its being[Pg 239] appreciated by others. As a matter of fact, this very difficulty as to the first beginnings of variations has been frequently urged against both hypotheses of selection, and rightly so, inasmuch as this must be above all else the point of attack for further investigations. But it is a mistake to deny the whole processes of selection simply because this point is not yet clear. Later on we shall attempt to gain some insight into the causes of variation, and then we shall return to this question of the beginnings of the selective processes. In the meantime let it suffice to say that Darwin was very well aware that, in addition to the ordinary individual variations, there were also larger deviations which occurred discontinuously in single forms. He believed, however, that such occurrences were very rare, and, on the whole, he was not inclined to ascribe to them any particular importance in the transformation of species. He rather referred the organic transformations which have taken place in the course of the earth's history, in the main, to the intensification of the ordinary individual variations, and I believe that he was right in so doing, since adaptations from their very nature cannot have been brought about by sudden chance leaps in organization, but can only have become exactly suited to chance conditions of life through a gradual accumulation of minute variations in the direction of utility. Whether, however, purely sexual distinctions may not have had their primary roots in discontinuous variations must be inquired into later. Theoretically, there is nothing against this assumption, when such characters are not adaptations like the lasso antennæ of the Copepods, or the turban eyes of the Ephemerids; mere distinctive markings, decorative coloration, peculiar outgrowths, and the like, may, if they arose discontinuously, very well have formed the basis for further sexual selection, as long as they were not prejudicial to the existence of the species.
INTRA-SELECTION OR SELECTION AMONG TISSUES
Does the Lamarckian principle really play a part in the transformations of species?—Darwin's position in regard to this question—Doubts expressed by Galton and others—Neo-Lamarckians and Neo-Darwinians—Results of exercise and practice: functional adaptation—Wilhelm Roux, Kampf der Theile.
We have devoted a whole series of lectures to studying the Darwin-Wallace principle of Natural Selection and the range of its operation. It seemed to us to make innumerable adaptations intelligible up to a certain point. We now understand how the purposefulness, which we meet with everywhere among organisms, can have arisen without the direct interference of a Power working intentionally towards an end—simply as the outcome and result of the survival of the fittest. The two forms of the processes of selection, 'natural selection' in the narrower sense, and 'sexual selection,' dominate, so to speak, all parts and all functions of the organism, and are striving to adapt these as well as possible to the conditions of their life. And although the range of operation of Natural Selection is incomparably greater, because it actually affects every part, yet we must attribute to sexual selection also, at least among animals, a range of influence by no means unimportant, since through it, as far as we can see at present, not only do the secondary sexual characters in all their diversity arise, but by the transference of these to the other sex that too is modified, and thus the whole species may be influenced, and may indeed be started afresh on an unlimited series of further transformations.
But although the processes of selection play such an important part in the transformations of the forms of life, we have to inquire whether they are the sole factors in these transformations, whether the accumulation of chance variations in the direction of utility has been the sole factor in bringing about the evolution of the animate world, or whether other factors have not also co-operated with it.
We are all aware that Lamarck regarded the direct influence of use and disuse as the most essential factor in transformation, and that Darwin, though hesitatingly and cautiously, recognized and accepted this factor, which he believed to be indispensable. Indeed, it seems[Pg 241] at first sight to be so. There is a whole range of facts which seem to be intelligible only in terms of the Lamarckian theory; in particular, the existence of numberless vestigial or rudimentary organs which have degenerated through disuse, the remains of eyes in animals which live in darkness, of wings in running birds, of hind legs in swimming mammals (whales), and of ear muscles in Man, who no longer points his ears, and so forth through a long list.
According to Wiedersheim, there are in Man alone about two hundred of these vestigial or rudimentary organs, and there is no higher animal which does not possess some. In all, therefore, a piece of the past history of the species is embodied in the actually existing organism, and bears witness to the fact that much of what the ancestors possessed is now superfluous, and is either transformed, or is gradually set aside, or is still in process of being set aside. It seems obvious that this gradual dwindling and degeneration of an organ no longer needed cannot be explained through natural selection in the Darwin-Wallace sense, for the process goes on so exceedingly slowly that the minute differences in the size of an organ, which may occur among individuals of the species at any given time during the retrogressive process, cannot possibly have a selection value. Whether the degenerate and now functionless hind leg of the whale is a little larger or a little smaller can have no importance in the struggle for existence; the smaller organ cannot be considered either as a lesser hindrance in swimming or as a greater economy of material, and the case is the same in regard to most other instances of degeneration through disuse. We therefore require another interpretation, and at first sight this seems to be supplied by the Lamarckian principle.
But the reverse process, the strengthening, the enlarging, and the more perfect development of a part, very often goes on proportionately to its more frequent use, and here again the Lamarckian principle seems to afford a simple explanation. For we know that exercise strengthens a part, as disuse weakens it, and if we could assume that these results of use and disuse were transmitted from the individual who brought them about or 'acquired' them in the course of his life to his offspring, then there would be nothing to object to in the Lamarckian principle. But it is precisely here that the difficulty lies. Can we assume such a transmission of 'acquired' characters? Does it exist? Can it be demonstrated?
That Lamarck did not even put this question to himself, but assumed such transmission as a matter of course, is readily intelligible when we consider the time at which he lived. He was himself one of the first to grasp the idea of the transmutation-hypothesis, and he[Pg 242] was only too glad to have any sort of principle of interpretation ready to work with. But Charles Darwin, too, attributed a not inconsiderable influence to this principle, although the transmission of 'acquired' characters which it took for granted was not accepted without reflective hesitation. He even directed his own particular theory of heredity, as we shall see, especially to the explanation of this supposed form of inheritance, and we can very well understand this, after what I have said as to the impossibility of explaining the disappearance of organs which have become superfluous by the Darwin-Wallace theory of Natural Selection. Darwin needed the Lamarckian principle for the explanation of these phenomena, and it was this that decided him to assume the transmission of 'acquired' characters, although the proofs of it can hardly have satisfied him. For when we are confronted with facts which we see no possibility of understanding save on a single hypothesis, even though it be an undemonstrable one, we are naturally led to accept the hypothesis, at least until a better one can be found. It is in this way, obviously, that we are to understand Darwin's attitude to the Lamarckian principle; he did not reject it, because it seemed to him to offer the only possible explanation of the disappearance of characters which have become useless; he adhered to it, although the transmission of acquired characters which it assumed must have seemed, and, in point of fact, did seem to him doubtful, or at least not definitely proved. Doubts, some faint, some stronger, as to this assumed form of inheritance were hardly expressed till somewhat late in the day—almost twenty years after the appearance of the Origin of Species—first by Francis Galton (1875), then by His, who definitely declared himself at least against any inheritance of mutilations, and by Du Bois-Reymond, who, in his address Ueber die Uebung in 1881, said: 'If we are to be honest, we must admit that the inheritance of acquired characters is a hypothesis inferred solely from the facts which have to be explained, and that it is in itself quite obscure.'
This is how it must appear to every one who examines it simply in respect of its theoretical possibility, its conceivability. This is how it appeared to me when I attempted, in 1883, to arrive at clearness on the subject, and I then expressed my conviction that such a form of inheritance was not only unproved, but that it was even theoretically unthinkable, and that we ought to try to explain the fact of the disappearance of disused parts in some other way, and I attempted to give an explanation, as will be seen later.
Thus war was declared against the Lamarckian principle of the direct effect of use and disuse, and there arose a strife which[Pg 243] has continued down to the present day, the strife between the Neo-Lamarckians and the Neo-Darwinians, as the two disputing parties have been called.
In order to form an independent opinion in regard to this famous dispute, it is, first of all, necessary to examine what actually takes place when an organ is exercised or is left inactive, and further, whether we can assume that the results of this exercise or inaction can be transmitted to descendants.
That exercise in general has a strengthening, and neglect of it a weakening influence on the relevant organ has long been known and is familiar to all; gymnastics make the muscles stronger, the thickness of the exercised muscle and the number of its fibres increases; the right arm, which is much more used than the left, is capable of performing twenty per cent. more work. Similarly, the activity of glands is increased by exercise, and the glands themselves are increased in size, as are the milk-glands of the cow through frequent milking; and that even the nerve-elements can be favourably influenced by exercise is proved by actors and professors of mnemonics, who have by practice increased their powers of memory to an almost incredible degree. I have heard of a singer who had learned by heart 160 operas; and which of us has not experienced how quickly the capacity for learning by rote can be again increased by practice, even after it has been neglected or left unexercised for a long time?
I have always been particularly struck with the practising of a piece of music, with its long succession of periods of different phrase, with its changes in melody, rhythm, and harmony, which nevertheless becomes so firmly stamped on the memory that it can be played, not only consciously, but quite unconsciously, when the player is thinking intensely of other things. It is in this case not the memory alone, but the whole complicated mechanism of successive muscle-impulses, with all the details of fast and slow, loud and soft, that is engraved on the brain elements, just like a long series of reflex movements which set one another a-going. Though in this case we cannot demonstrate the material changes which have taken place in the nervous elements, there can be no doubt that changes have taken place, and that these consist in a strengthening of definite elements and parts of elements. The strengthening causes certain ganglion-cells to give a stronger impulse in a particular direction, and this impulse acquires increasing transmissive power, and so on.
Our first theoretical insight into these relations came through Wilhelm Roux, who, in 1881, gave expression to what had previously[Pg 244] been an open, if not quite conscious, secret, that 'functional stimulus strengthens the organ,' that is to say, that an organ increases through its own specific activity. Up till that time it had been believed that it was merely the increased flow of blood that caused the increase in the size of a much-used part. Roux showed that there is a 'quantitative self-regulation of the organ according to the strength of the stimulus supplied to it'; that the stimulated organ, that is, the organ which is performing its normal function, may, in spite of the increased breaking down or combustion (dissimilation), assimilate all the more rapidly; that its used-up material is 'over-compensated,' and that therefore it grows. He called this the 'trophic' or nutritive effect of the stimulus, and in terms of this he explained the increase and the heightened functional capacity of the much-used organ. Conversely, he referred the decrease of a disused organ to 'functional atrophy,' which sets in when there is a deficient compensation for the substance used up in the metabolism.
But if we press for deeper analysis, we must ask: 'On what does this trophic effect of functional stimulus depend?' Roux could not answer this question when he wrote, nor can we do so now, as Plate has justly emphasized. We are here face to face with the fundamental phenomenon of life, metabolism; and, since we do not understand the causes of this, we are not in a position to say why it varies in this way or in that according to the 'stimulus.' But the fact itself is certain that the organs respond up to a certain point to the claims made upon them; they increase in proportion as they function more frequently or more vigorously, they are able to respond to increased functional demands, and this Roux has called 'functional adaptation.' As an animal adapts itself to the claims of the conditions of its life, for instance, by taking on a green or a brown protective colour according as it lives on green or brown parts of plants, so the individual organ adapts itself to the strength of the stimulus which impels it to function, and increases or decreases in proportion to it. If one kidney in Man degenerate, or be surgically removed, the other begins to grow, and goes on increasing until it has reached nearly twice its former size. The specific stimulus which is brought to bear upon it by the urea contained in the blood, and which forces it to grow, is twice as great in the absence of the other kidney, and therefore the remaining kidney grows in response to the increased stimulus and its 'trophic effect' until its increase in size has reduced the functional intensity to the normal proportion.
Adaptation of an organ in the opposite direction takes place when the function diminishes or ceases. If a nerve supplying a[Pg 245] muscle or a gland be cut through, the organ concerned begins to degenerate and to lose its normal structure to a greater or less degree. Sensory nerves also degenerate in their peripheral part when they are cut through. In such cases there may be no alteration either in the nutritive mechanism or in the blood-vessels, &c., but the functional stimulus—in the case of the muscle, the stimulus from the will—no longer affects the organ, and its metabolism is so much lowered in consequence that it begins to degenerate.
When we admit that the fit adaptation of the organism, as far as we understand it, must depend upon processes of selection, we may refer this 'functional adaptation' also to primitive processes of selection, which prevailed at the very beginning of life upon our earth, and represented, so to speak, the first adaptation that was established, but we can say nothing with certainty in regard to this matter as long as we do not understand the essence of assimilation. It is conceivable, however, that a primary adaptiveness may have arisen, so to speak, abruptly, through a concurrence of favourable circumstances, as we shall endeavour to show later on when we discuss the beginnings of life.
Even although we cannot lay bare the primary roots of 'functional adaptation' we can gain from the fact itself very valuable insight into phenomena which would otherwise be unintelligible and mysterious: the perfectly adapted structure of many tissues and their power of adaptation to changed conditions. In this lies, in the main, the advance in our knowledge which is due to Roux's Kampf der Theile.
If a number of embryonic cells of different capacity, say A, B, and C, be affected by different kinds of functional stimuli, a, b, and c, those cells will grow most rapidly which are most frequently affected by the stimulus appropriate to them. The proportion in which the cells A, B, and C will ultimately be present in the tissues will depend upon the frequency with which the stimuli a, b, and c act upon the tissue. But the tissue will be still more precisely determined as to its structure if the three kinds of stimuli affect the cell-mass, not uniformly all over, but only at certain spots, or along particular paths, one in this, the other in that. Thus the cells A will predominate over the cells B and C at all the places which are most frequently affected by the stimulus a, the cells B in the sphere of the stimulus b, and the cells C in that of the stimulus c; there they will increase most rapidly and so crowd out the other kinds of cells, and thus a spatial arrangement will be established within the tissue, a 'structure' which corresponds and is well adapted to its end. This is what Roux deduced from his[Pg 246] Struggle of the Parts, and I subsequently defined the process as histonal or tissue selection.
Let us first take an example. The anatomist Hermann Meyer showed in 1869 that the so-called 'spongiosa,' that is, the bony tissue of spongy structure within the terminal portions of the long bones in Man and Mammals, has a minute structure conspicuously well adapted to its office. The thin bone lamellæ of this 'spongiosa' lie precisely in the direction of the strongest strain or pressure which is exerted upon the bone at the particular area. Arch-like in form, they are kept apart by means of buttresses, and no architect could have done better if he had been entrusted with the task of making a complicated system of arches with the greatest possible carrying and resisting power combined with the greatest possible economy of material.
This well-adapted structure is now interpreted through the Struggle of the Parts as a self-differentiation, for if there be in the rudiments or primordia of the bone differently endowed elements[10], that is, cells which respond in diverse ways to different stimuli, these must arrange themselves locally, owing to the struggle for space and food, in a manner corresponding to the distribution of the different stimuli in the bone. The largest amount of bone substance will be formed in the directions of the strongest strain and the greatest pressure, because the bone-forming cells are excited by this, their functional stimulus, to growth and multiplication. Thus the buttress and arch structure comes about, and between the delicate bone lamellæ spaces will remain free, and these, being relieved from the burden of strain and pressure by the aforesaid bony lamellæ, will offer suitable conditions of life to cells with other functional properties, such as connective tissue cells or vascular cells.
[10] I do not here enter into the question whether we have not in this case to do with similar elements, which have the power of differentiating into one or another kind of cell according to the nature of the external stimuli by which they are influenced.
The structure of the bone spongiosa is not everywhere the same, and it is demonstrably related with precision to the conditions of strain and pressure at each particular region. Thus, just below the soft cartilaginous covering of the joints there are no long pillars with short arches, but only rounded meshes, because the pressure is here almost equally strong from all sides. The long parallel pillars only occur further down in the bone, and they lie in two directions which intersect each other obliquely, corresponding to the two main directions of pressure. But it is only under the functional stimulus of pressure that the bone-forming cells have an advantage over the[Pg 247] others, and multiply more quickly, thus crowding out those that are not attuned to the appropriate functional stimulus.
In a similar manner Roux interprets, in the light of the struggle of the parts, the striking adaptations in the course, the branching, and the lumen-formation of the blood-vessels, in the direction of the intersecting connective tissue strands in the tail-fin of the dolphin, in the direction of the fibres in the tympanum, and in many other adaptations in the histological structure of complex tissues.
In this there is manifestly an important step of progress, for it is obvious that the direction of the bone-lamellæ and such like could not have been determined by individual selection, and the same is true in regard to many other histological details. It cannot be disputed, however, that there is a kind of selection-process here also, similar to that which we think of, with Darwin and Wallace, as occurring between individual organisms. Just as in the latter, which we shall henceforward call personal selection, variability and inheritance lead, in the struggle for existence, to the survival of the fittest, so, in histonal differentiation, the same three factors lead to the victory of what is best suited to the parts of the body in question. The tissues and the parts of the tissues have to distribute and arrange themselves so that each comes to fill the place in which it is most effectively and frequently affected by its specific stimulus, that is, the stimulus in regard to which it is superior to other parts; but these places are also those the occupation of which by the best re-acting parts makes the whole tissue capable of more effective function, and therefore makes its structure the fittest. Variability—in this case that of embryonic cells, with different primary constituents—must be assumed; inheritance is implied in the multiplication of the cells by division; and the 'struggle for existence' here assumes its frequent form of a competition for food and space; the cells which assimilate more rapidly because of the more frequent functional stimulus increase more rapidly, draw away nourishment from the more slowly-multiplying cells around them, and thus crowd these out to a greater or less extent.
We might even speak of histonal selection among unicellulars, for it is conceivable that in primitive living substance, such as that of a moneron, there may be minute differences among the vital particles, involving also functional distinctions, which, under the influence of diverse stimuli, may gradually give rise to an increasingly complex differentiation. For the variations in the primary living substance most strongly affected by a particular stimulus would tend to accumulate at the places most frequently reached by that stimulus, and[Pg 248] would crowd out other variations at that spot, just as the body and its individual parts may be said to have taken their architectural form in exact response to the demands made upon them by function. In this case, of course, personal selection and histonal selection co-operate, for every improvement in the organization of the fundamental living substance means at the same time a lasting improvement in the whole individual.
In many-celled organisms, however, we must admit that there is an essential difference between personal and histonal selection, inasmuch as the latter can give rise to adaptive structural modifications corresponding to the needs of the tissue at the moment, but not to permanent and cumulative changes in the individual elements of the tissue. If a broken bone heals crookedly, the spongy substance within the healed portion does not remain as it was before, for the pillars and arches, which now no longer run in the direction best suited to their function, break up, and a new system of arches is formed, not in line with the earlier one, but adapted to the new conditions of pressure. This is certainly an adaptation through selection, but the elements, that is the cells which form the bone substance in response to strain and pressure, or those which in response to the stimulus of the blood flowing into the spaces form the blood-vessels, or those which being quite freed from one-sided pressure develop into connective tissue, must be presupposed. These kinds of cells must be virtually implied in the germ-rudiment; they are themselves adaptations of the organism, and can therefore only be referred to personal selection. And this is true of all adaptations of the elements of multicellular organisms, and thus of the cells. Their adaptation according to the principle of division of labour, their differentiation into muscle, nerve, and gland cells can only be referred to natural selection in the Darwin-Wallace sense, and cannot depend upon histonal selection. In the spongy substance of the bone a better bone-cell does not struggle with an inferior one and leave behind it by its survival a host of descendants which are, if possible, better than itself; the struggle for existence and for descendants, in this case, is between two kinds of cell which were different from the beginning, and of which one has the advantage at one spot, another at another. The case may be compared to that of a flock of nearly allied species of bird, of which one species thrives best in the plains, another among the hills, and a third among the mountain forests, all mingled together in a vast new territory to which they had migrated, and in which all three kinds of conditions were represented. A struggle would arise among the different species, in which in every[Pg 249] case the particular species would be victorious which was best adapted to the local conditions. But each would thrive best in the region in which it was superior to the others, and very soon the three species would be distributed as they were in the land from which they came—in the plains, the high lands, and the mountain forests. This would be the result of a struggle between the three species, not between individuals within each species, and it could not therefore bring about an improvement of a single species, but only the local prevalence of one or another. The characters which made one species adapted for the plain, another for the mountain forest were already there; they can only be referred to personal selection, which brought about the adaptation of their ancestors in the course of ages to the conditions of their life. Something similar is true of the adaptations of the tissues; the differentiation of the individual kinds of cells is an ancient inheritance, and depends upon personal selection, but their distribution and arrangement into specially adapted tissues, so far as there is any plasticity at all, depends upon histonal selection. Obviously, however, only as far as the tissue is plastic, that is, with the power of adjusting itself to particular local conditions. Only adaptations of this kind can be referred to histonal selection; the ground-plan, even of the most complicated tissue, such as the large glands of mammals, the kidneys, the liver, and so on, must have been implicit in the germ, and must therefore be referred to personal selection. A precise limitation of the respective spheres of action of personal selection and histonal selection is not possible as yet, since hardly any investigations on the subject are available.
Roux undoubtedly over-estimated the influence of his 'struggle of parts' when he believed that the most delicate adaptations of the different kinds of cells depended on it. I admit that, for a considerable time, I made the same mistake, until it became clear to me, as it did first in regard to the sex-cells, that this is not, and cannot be the case. How, for instance, could the diverse and minutely detailed adaptations of the sex-cells—which we are to discuss in a subsequent lecture—have arisen in this way? As far as the individual sperm-cell is concerned, it is a matter of indifference whether its head is a little thinner or thicker, its point a little sharper or blunter, its tail a little stronger or weaker. This does not decide whether the cell is to thrive better, or to occur in greater numbers than some other variety. But it does decide whether it is to be able to penetrate through the minute micropyle, or through the firm egg-envelope, into the egg, there to effect fertilization. An individual with less well formed sperm-cells will be able to fertilize fewer eggs, and therefore to leave[Pg 250] fewer descendants which might inherit its tendency to produce inferior sperm-cells, and conversely. Thus it is not the sperm-cells of any one individual which are selected according to their fitness, it is the individuals themselves which compete with one another in the production of germ-cells which shall fertilize best, that is, most certainly. The struggle is thus not intercellular, but a struggle between persons.
The same is true of all cells differentiated for particular functions; every new kind of glandular, muscular, or nerve cell, such as have arisen a thousandfold in the course of phylogeny, can only have resulted from a struggle between individuals which turned on the possession of the best cells of a particular kind, not from a struggle between the cells themselves, since these would gain no advantage from serving the organism, as a whole, better than others of their kind. In regard to the sex-cells we might admit, in addition to personal selection, the possibility of an internal struggle between the sperm-cells or egg-cells of the same individual, inasmuch as each of these cells is the primordium of a new individual, and as those better adapted for reproduction might transmit their better quality to these new individuals. I will not here enter into my reasons for regarding this idea as erroneous, for in any case this interpretation would not apply to any other kind of cells. If, for instance, it were a question of the transformation of an ordinary mucus or salivary gland into a poison gland, it would not matter in the least to the individual cell whether it yielded a harmless or a poisonous secretion; but individuals with many poisonous cells would have an advantage in the struggle for existence.
I agree so far with Plate when he refers the differentiation of the tissues entirely to personal selection, but not in his further conclusion that histonal selection does not exist. The ground-plan of the architectural structure of the organ depends upon personal selection, but the realization of the plan in particular cases is not predetermined down to the minutest details, but is regulated by histonal selection, and is thus to a certain extent an adaptation to local conditions of stimulus. The direction, strength, and size of every single bone lamella is not predetermined from the germ, but only the occurrence and nature of bone-cells and bone lamellæ in general. The direction and the strength which these bone lamellæ may assume depends on the local conditions of strain and pressure which affect the cell-mass, as is shown very clearly by the spongiosa of an obliquely healed bone, which we have already described.
But let us now turn to the question which is here most important for us: whether functional adaptations can be transmitted. We must[Pg 251] admit that the insight we have so far gained into the causes of these adaptations does not make it much easier to answer the question. Histonal selection is a purely local process of adaptation to the conditions of stimuli prevailing at the moment, and no one will be likely to suppose that the distorted position of the spongiosa of a badly healed fracture could reappear in the straight bone of a descendant; this would be quite contrary to the principle, for the crooked lamellæ would in that case no longer be the best adapted. Even the question whether the strengthening of a muscle through use can be transmitted cannot be answered in the light of the knowledge we have hitherto gained. The 'trophic effect of the functional stimulus' is brought into activity through entirely local influences, through which only the parts most strongly affected by the stimulus can be caused to vary. Thus the problem remains unaltered, How can purely local changes, not based in the germ, but called forth by the chances of life, be transmitted to descendants?
If all species, even in the highest groups, reproduced by dividing into two, we might imagine that a direct transmission of the changes acquired in the course of the individual life through use or disuse took place, though this would presuppose a much more complicated mechanism than is apparent at first sight. But it is well known that multiplication by fission is for the most part restricted to simple organisms, and that the great majority of modern plants and animals reproduce by means of germ-cells, which develop within the organism in regions often very remote from the parts, the results of the exercise of which are said to be transmitted. Moreover, the germ-cells are of very simple structure, at least as far as our eyes can discern; for we see in a germ-cell neither muscles nor bones nor ligaments, glands nor nerves, but only a cell-body consisting of that semifluid living matter to which the general name of protoplasm has been given, and of a nucleus, in regard to which we cannot say that it differs in any essential or definite way from the nucleus of any other cell. How then could the changes which take place in a muscle through exercise, or in the degeneration of a joint in consequence of disuse, communicate themselves to a germ-cell lying inside the body, and do so in such a fashion that this germ-cell is able, when it grows into a new organism, to produce of itself, in the relevant muscle and joint, a change the same as that which had arisen in the parent through use and disuse? That is the question which forced itself upon me very early, and in following it up I have been led to an absolute denial of the transmission of this kind of 'acquired characters.'
In order to explain how I reached this result, and what it is[Pg 252] based upon, it is indispensable that we should first make ourselves acquainted with the phenomena of heredity in general, and the inseparably associated phenomena of reproduction, so that we may form some sort of theoretic conception of the process of inheritance—a picture, necessarily provisional and imperfect, of the mechanism which enables the germ-cell to reproduce the whole organism, and not merely, like other cells, others like itself. We are thus led to an investigation of reproduction and heredity, at the conclusion of which we shall feel justified in returning to the question of the inheritance of acquired characters, in order to give a verdict as to the retention or dismissal of the Lamarckian principle.
REPRODUCTION IN UNICELLULAR ORGANISMS
Reproduction by division—In Amœbæ—In Infusorians—Divisions following one another in immediate succession—Formation of germ-cells in the Metazoa—Contrast between germ-cells and body-cells—Potential immortality of unicellular organisms—Beginning of natural death—Budding and division in the Metazoa.
We wish to consider the reproduction of organisms with special reference to the problem of heredity, and it is most instructive to begin with the lowest forms of life—the unicellulars—because their structure, as far as we can see with the instruments at our command, is very simple, and, what is even more important, is relatively homogeneous.
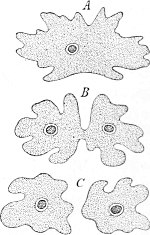
Fig. 59. An Amœba: the process
of division. A, before the
beginning of the division. B,
the nucleus divided into two.
C, the two daughter-Amœbæ.
Magnified about 400 times.
Suppose that there are bacteria-like organisms of quite homogeneous structure, and that these multiply by simply dividing into two, each rod-like creature dividing transversely in the middle of its length, the two halves would represent independent daughter-organisms, whose structure would correspond exactly with that of the mother-organism, could not indeed in any way deviate from it, and consequently would take over all its characters, that is, would inherit them. The size of body is the only feature which is not obviously inherited, but in reality it is potentially heritable, since the structure of the divided portions involves the capacity and the limits of their possible growth. Moreover, the size of body is not invariable in any species; a particular size is only reproduced under similar conditions of development. Inheritance here consists simply in a continuation of the mother-organism into its two daughter-cells.
Even in an Amœba (Fig. 59) we might picture the process of inheritance as equally simple, though in so doing we should probably be making a fallacious inference, for the structure of these lowest [Pg 254]unicellular animals probably seems to us simpler and more homogeneous than it really is. Among Infusorians it is quite obvious that inheritance implies more than the mere halving of the mother-animal into the two daughter-cells; something more must be involved. For among these unicellular animals the differentiation of the body is not only great, but it is unsymmetrical. The posterior and the anterior ends are different, and the transverse division of the animal, in which the process of reproduction here consists, does not produce two halves, but two very unequal portions. In the division of Stentor, the so-called trumpet-animalcule (Fig. 60), the anterior portion contains the funnel-shaped mouth and gullet with its complicated nutritive apparatus, the circular peristome with its spirally curved rows of composite ciliated plates, the so-called membranellæ, and so forth; the posterior half contains nothing of all this, but possesses the foot of the mother-Stentor with its attaching organ, which the anterior half lacks. But each of the two portions possesses the power of 'regeneration,' that is, it is able to develop anew the missing parts, mouth or foot, and so on. So that here there is no longer merely a simple continuance of the maternal organization in the daughter-animals, there is[Pg 255] something new added, something which requires explanation; we are confronted with the first riddle of heredity. Simple growth does not explain the phenomenon, for what has to be added to complete the halved portions has a different structure, a different form, different accessory apparatus from any that the halves themselves possess. It in no way affects this state of matters that in the normal process of division in Infusorians the formation of the new mouth and peristome-region begins before the halves have actually separated, for even if a Stentor be cut in two artificially the cut halves form complete animals. And, indeed, a Stentor may be cut into three or four pieces, and in certain conditions each piece will develop into a complete animal. These pieces must therefore possess something more than the mere power of growth. We shall try later on to discover whether this marvellous invisible transmission of characters, this implication of the whole in each of the parts, can be in any way theoretically expressed and included in our scheme of conceptual formulation.
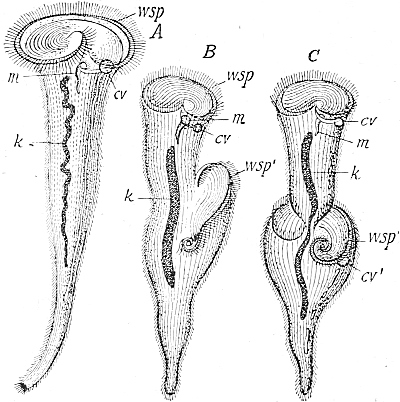
Fig. 60. Stentor rœselii, trumpet-animalcule. Process of division. wsp, ciliated spiral leading to the mouth (m); cv, contractile vacuole. A, in preparation for division, the nucleus (k) has coalesced into a long twisted band. B, a second ciliated spiral (wsp´) has begun to be formed; the nucleus (k) is contracted. C, just before the constricting off of the two daughter-Stentors. Magnified about 400 times. After Stein.
Now that we have become familiar with these facts it will no longer surprise us to learn that the reproduction of unicellular animals does not always depend on equal division, but that unequal spontaneous divisions are also possible, so that one or several smaller portions of the cell-body, containing a portion of the cell-nucleus, can separate off from the mother-animal. This occurs especially among the suctorial Infusorians or Acinetæ. In relation to the phenomena of inheritance the problem raised by the equal division of the Infusorians repeats itself, and it is in no way affected by the fact that equal division can take place several times, or many times in succession, so that from one animal a large number of small pieces of the same size may be produced in rapid succession. The characteristic marks of the mother-animal are not infrequently lost sight of, wholly or partially, when this occurs, and the divided portions seem to consist of a homogeneous cell-body and nucleus; but they possess the power of regenerating themselves, or of developing, if the expression be preferred, into animals similar to the maternal-organism. Such divided portions might very well be called germs, only it must not be forgotten that the relation of the mother-animal to these germs is a different one from that of a higher animal or plant to its germ-cells; the unicellular animal breaks up by continued division into these 'germs,' while the Metazoon continues its individual existence unimpaired by the production of its germ-cells.
The beginning of a so-called 'spore-formation' is to be found in many Infusorians. Thus the holotrichous species, Holophrya multifiliis (Fig. 61), reproduces by first becoming enclosed in a cyst or capsule,[Pg 256] and then dividing many times in rapid succession, so that 2, 4, 8, 16, &c. individuals arise consecutively, and subsequently burst forth from the cyst (Fig. 61, B). In the Gregarines and other Sporozoa the period of division lasts much longer, and the encysted animal divides into 128, 256, or even more portions; but in this case also each part or 'spore' receives a piece of the maternal cell-body and cell-nucleus, so that there is no difference in principle between this and the simple division into two exhibited by Stentor; as in that case, so here, it is not the fully differentiated structure of the animal which is handed on to the divided parts; it is only the power to redevelop this anew on their own account. Thus here again we are face to face with the fundamental problem of heredity: How is it possible that the power of reproducing the complex whole can be inherent in the simple parts?
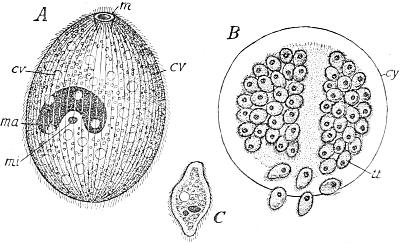
Fig. 61. Holophrya multifiliis, an Infusorian parasitic on the skin of fishes. A, in its usual condition; ma, macronucleus; mi, micronucleus; cv, contractile vacuole; m, mouth. B, after binary fission has been several times repeated within the cyst (cy); tt, results of the division. C, one of these units much enlarged.
In contrast to the unicellular organisms, the great majority of the multicellulars, the Metazoa and Metaphyta, many-celled animals and plants, differ not only in the multitude of their cells, but even more in the manifold differentiation of these cells according to the principle of division of labour, so that the various functions of the animal are not performed by all the cells uniformly, but each function is relegated to a particular set of cells specially organized with reference to it. Thus there is differentiation between motile, nutritive, and reproductive cells, and there may also be glandular, nerve, muscle, and skin cells, and we know how this differentiation into a great number of different kinds of cells with highly specialized functions has arisen, especially among the higher animals, in a multiplicity which cannot easily [Pg 257]be overlooked. Thus we find a large number of the most diverse kinds of cells, all of which serve for the maintenance of the body, in contrast to the simply reproductive cells or germ-cells. These alone possess the power of reproducing, under certain conditions, a new individual of the same species. We can contrast with these germ-cells, which serve, not for the maintenance of the individual, but only for that of the species, all the other kinds of cells under the name of somatic or body-cells. The problem which we have to solve now lies before us in the question, How comes it that the germ-cell is able to bring forth from itself all the other cells in definite sequence and arrangement, and is thus able to build up the body of a new individual?
The similarity of this problem to that formulated in regard to unicellular organisms is at once obvious, but it becomes still more emphatic when we remember that the gulf between unicellular organisms and the higher animals and plants is bridged over by certain transition forms which are of the greatest interest, especially in relation to the problems of inheritance.
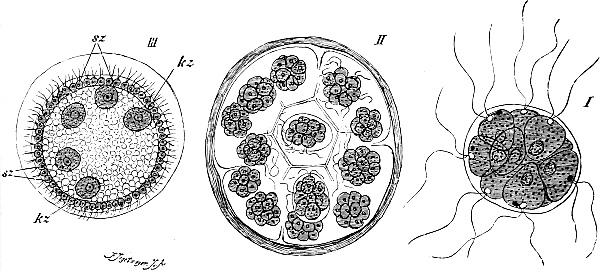
Fig. 62. Pandorina morum; after Pringsheim. I, A young colony, consisting of 16 cells. II, Another colony, whose cells have reproduced daughter-colonies; all the cells uniformly alike. III, A young Volvox-colony; sz, somatic cells; kz, germ-cells.
Among the lower Algæ there is a family, the Volvocineæ, in which the differentiation of the many-celled body on the principle of division of labour has just set in; in some genera it has been actually effected, though in the simplest way imaginable, and in others it has not yet begun. Thus in the genus Pandorina the individual consists of sixteen green cells, united into a ball (Fig. 62, I), each one exactly like the other, and all functioning alike. They are all united into a[Pg 258] spherical body, a whole, by a gelatinous matrix which they all secrete, and thus they form a cell-colony, a cell-stock, a many-celled individual; but each of these cells has not only all the typical parts—cell-body, nucleus, and contractile vacuole—but each possesses a pair of flagella or motor organs, an eye-spot, and a chlorophyll body which enables them to assimilate nourishment from the water and the air. Each one of these cells thus performs all the somatic functions, that is, all that are necessary to the maintenance of the individual life. But each also possesses the power of reproducing the whole colony from itself, that is, it also performs the function of reproduction necessary to the maintenance of the species. When such a colony, whose sixteen cells are continually growing, has led for some time a free-swimming life in the water, the cells retract their flagella, and each begins to multiply by dividing into 2, 4, 8, finally into 16 cells of the same kind, which remain together, forming a spherical mass enclosed in a gelatinous secretion (Fig. 62, II). Thus there are now, instead of sixteen cells in the mother-colony, sixteen daughter-colonies, each with sixteen cells which soon acquire flagella and eye-spots, and are then ready to burst forth from the dissolving jelly of the maternal stock as independent individuals. This Pandorina shows no trace of a differentiation of its component cells to particular and different functions, but a nearly allied genus of the same family, the genus Volvox (Fig. 62, III), consists of two kinds of cells—on the one hand of small cells (sz) which occur in large numbers and compose the wall of the hollow gelatinous mass, forming, so to speak, the skeleton of the Volvox; and, on the other hand, of a much smaller number of cells which are very much larger (kz). The former, the 'body' or 'somatic' cells, are green, and have a red 'eye-spot' and two flagella; they are connected with each other by processes from their cell-bodies, and are able, by means of the co-ordinated action of their flagella, to propel the whole colony with a slow rotatory movement through the water. Many of my readers are doubtless familiar with these light green spheres, which are quite recognizable with the naked eye, and people our marsh pools and ponds in Spring in such abundance that it is only necessary to draw a glass of water to procure a large number of them.
The little flagellated cells just described serve not only for the locomotion of the colony, but also for nutrition, for the secretion of the jelly, and for the excretion of waste products; in short, they perform all the functions necessary to the maintenance of life, but not that of reproduction. They can, indeed, multiply by dividing when the colony is young, like the cells of Pandorina, but they cannot[Pg 259] reproduce the whole colony but only cells like themselves, that is, other somatic cells. In Volvox the maintenance of the species, the production of a daughter-colony, is the function of the second and larger kind of cells, the reproductive cells, which are contained in the interior (filled with a watery fluid) of the gelatinous sphere. They possess no flagella (kz), and so take no share in the swimming movements of the somatic cells. For the present we need not allude to the fact that there are several kinds of these cells, and need only state that the simplest among them, the so-called 'Parthenogonidia,' after they have reached a considerable size, begin a process of division which results in the formation of a daughter-colony. Usually there are several of these large reproductive cells in a Volvox colony, and as soon as these have developed into a similar number of daughter-colonies they burst out through a rupture in the now flaccid jelly of the maternal sphere and begin to lead an independent life. The mother-sphere, which now consists only of somatic cells, is unable to produce new reproductive cells; it gradually loses its spherical form, sinks to the ground, and dies.
In Volvox we have, for the first time, a cell-colony in which a distinction has been established between body or somatic cells and reproductive or germ-cells. In contrast to Pandorina, a large number, indeed the majority of the cells of the colony, have lost the power of reproducing the whole by division, and only the few reproductive cells possess this, while they, in turn, have lost other functions, notably that of locomotion. Their power of reproducing the whole, that is to say, their hereditary capacity, gives them a greater theoretical interest than the cells of Pandorina, for the latter require only to produce others like themselves, because there is only one kind of cell in the colony, while in Volvox the reproductive cell can not only produce others like itself, by division, but can produce the body-cells as well. The problem is quite analogous to the one which we have had to face in regard to the unicellular animals of complex structure, the Infusorians. The question, How can the part of the trumpet-animalcule which is mouthless develop from itself a new mouth and ciliated apparatus? here transforms itself into the question, How can a cell by division give rise not only to others like itself, but also to the body-cells, which are of quite different structure? This is, in its simplest form, the fundamental problem of all reproduction through germ-cells, to which we must now pass on. But first a short digression.
We have already noted that unicellular organisms multiply by division, and originally, as well as in the great majority of cases[Pg 260] to-day, by division into two. It follows, therefore, that there is no natural death among them, for, if there were, the species would die out as the individuals grew old; but this does not happen. The two daughter organisms which arise from the binary fission of an Infusorian are in no way different in regard to their power of life; each of them possesses an equal power of doubling itself again by division, and so it goes on, as far as we can see, for an unlimited time. Thus the unicellular organisms are not subject to natural death; their body is indeed used up in the course of ordinary life so that the formation of new cilia and so on is necessary, but it is not worn away in the same sense in which our body is and that of all Metazoa and Metaphytes, where, through functioning, the organs are gradually worn away until they become incapable of function. Our body grows old, and can at last no longer continue to live; but among unicellular organisms there is no growing old, and no death in the normal course of the development of the individual. The unicellulars are, as we may say, immortal; that is, while individuals may be annihilated, by external agencies, boiling heat, poisons, being crushed, or eaten, and so on, at every period some individuals escape such a fate, and perpetuate themselves through succeeding ages. For, strictly speaking, the daughter-individual is only a continuation of the mother-individual; it contains not only half of the substance, but also the organization, and life is continued directly from mother to daughter. The daughter is simply half of the mother, which is subsequently regenerated; and the other half of the mother lives on in the other daughter, so that nothing dies in this multiplication. It may be said that the daughter has to develop the other half of its body anew, and that therefore it is a new individuality, and not merely a continuation of the old, and that therefore the unicellular animals are not immortal. The 'immortality' of the Protozoa may be scoffed at; the idea may seem absurd that the 'immortal' Protozoa are still the same individuals which lived upon the earth millions of years ago, but all such objections mean no more than doctrinaire quibbling with the concepts of 'individual' and 'immortality,' which do not exist in nature at all, but are mere human abstractions, and therefore only of relative value. My thesis as to the potential immortality of the Unicellulars aims at nothing more than impressing on Science the fact that the occurrence of physiological, that is, natural, death is causally associated with the transition from single-celled to many-celled organisms; and this is a truth which will not be overthrown by any sophisms. It is the Volvocineæ which show us, so to speak, the exact point at which natural death set in, at which it was introduced into the world of life.[Pg 261] In Pandorina the state of things is still the same as in single-celled organisms, for each cell is still all in all, each can bring forth the whole, none dies from physiological causes involved in the course of development, and they are therefore 'immortal' in the sense stated. But in Volvox the 'individual' dies when it has given off its reproductive cells, because here the contrast between germ-cells and body has developed. Only the body is mortal in the sense of being subject to natural death; the germ-cells possess the potential immortality of the single-celled animals, and it is necessary that they should possess it if the species is to continue to exist.
From this alone it does not seem quite clear why the body or soma should be subject to death, and when I first endeavoured to arrive at clearness in regard to these matters I tried to find out why a natural death of the body was necessitated by the course of evolution. I did not at once discover the true explanation, but without delaying to discuss my mistakes I shall proceed to expound what I believe to be the true reason. It lies simply in the fact, which we shall inquire into later on in more detail, that every function and every organ disappears as soon as it becomes superfluous for the maintenance of the particular form of life in question. The power of being able to live on without limit is useless for the somatic cells, and thus also for the body, since these cannot produce new reproductive cells after those that had been present are liberated; and with this the individual ceases to be of any value for the preservation of the species. What advantage would it be to the species if the Volvox balls were to continue living for an unlimited time after the reproductive cells were developed and had been liberated? Obviously their further fate can have no influence whatever in determining or preserving the characters of the species, and it is quite indifferent to the continuance of the species whether and how long they go on living. Therefore the soma has lost the capacity which conditions endless continuance of life and continued renewal of body-cells.
In regard to these views it has been asked jeeringly, how 'immortality,' if it were really a property of the Unicellulars and of undifferentiated cell-colonies, could be lost, as if the world, which we believe to be everlasting, should give up its everlastingness. But the jeer recoils on the superficial outlook which is unable to distinguish between the immortality dreamed of by the poets, religious and secular, and the real power that certain forms of life have to resist being permanently exhausted by their own metabolism. That we should call this 'immortality' does not seem to me to require any apology, for the right has always been conceded to science to transfer[Pg 262] popular words and ideas in a restricted and somewhat altered sense to scientific conceptions when it seems necessary. That the word 'immortality' in this case expresses the state of matters more precisely and better than any other cannot be doubted, any more than we can doubt that there exists in regard to natural death a real difference, which we must take account of, between the Unicellulars and the higher organisms. What enables the species in the case of the higher organisms, like ourselves for instance, to last through ages is not the immortality of the individual, of the person, but only that of the germ-cells; these alone, among the cells of the whole body, have retained the primæval power. A small piece of the individual is still immortal, but only a minute part, which cannot be considered as equivalent to the whole, either morphologically or from the point of view of the conception of individuality. Can anyone consider himself identical with his children? If any one should imagine this, it would still not be the case, for he himself would in the course of time suffer natural death, and his children would continue to live on until they too had brought forth children, and in their turn also came to die. It is quite different with an Infusorian, which never lies down to die, but simply splits itself afresh into two halves which continue to live.
It is hardly credible that such a simple and clear truth should have remained so long undiscovered, and it is even more incredible that since it was enunciated it should have been until quite recently laughed at as false, as a piece of pseudo-science, and as valueless. But it is the fate of all knowledge which rests on an intelligent and comprehensive working up of facts to be attacked, until it gradually bears down antagonism by the weight of its truth, and compels at least a silent recognition.
The fact that natural death made its appearance with the appearance of a 'body,' a soma, as distinguished from the germ-cells, will sooner or later compel recognition. When I pointed out above that the explanation of natural death lay in the fact that it would be superfluous for the soma to continue to live on unlimitedly, after it had discharged its germ-cells, and so fulfilled its duty to the species, I only intended to say that this was the general reason for the introduction of natural death. I have no doubt that the actual beginning of this phenomenon could have, and probably did come about in other ways. Many kinds of cells in higher animals perish as a result of their function; it is, so to speak, their business to perish, to break up; this is the case with many glandular and epithelial cells. It may very well be that, in many of the highly differentiated tissue-cells, such as nerve, muscle, and glandular cells, the high[Pg 263] differentiation in itself excludes the possibility of unlimited length of life and multiplication. Through this alone, therefore, the exhaustion of the body and an ultimate death may be explicable from internal causes. But the deeper cause remains what I have already indicated, for it is obvious that if the continued life, that is, the immortality of the soma, were necessary to the preservation of the species it would have survived through natural selection; that is to say, had it been so, then histological differentiations incompatible with immortality would not have made their appearance; they would always have been eliminated on their way to development, since only that which is adapted to its end survives. Only if the immortality of the soma were indifferent for the species could the soma have become so highly organized that it became subject to death.
Thus the old song of the transitoriness of life does not apply to all the forms of life: natural death is a phenomenon which made its appearance comparatively late in the development of the organic world, a phenomenon which, up to a certain point, we can quite well understand from the standpoint of purposefulness.
It would take me too far from the goal towards which we are at present making if I were now to attempt to show, in connexion with natural death, that the durability of the soma, or what we usually call the normal duration of life, is also exactly regulated by natural selection, so that each species possesses exactly that duration of life which is most favourable to it, according to its physical constitution, its physiological capacity, and the conditions of life to which it has to adapt itself[11]. But, interesting as this subject is, I must not digress further, but return to our proper subject of study, namely, reproduction in its relation to inheritance.
[11] See Weismann, Ueber die Dauer des Lebens, Jena, 1882. Translated in Essays on Heredity.
We digressed from this study after having seen that all, even the most complex, multicellular plants and animals, in which the differentiation of the cells into a number of cell-groups with the most diverse functions has attained the highest degree of complexity, are able to produce special cells, the germ-cells, which have the power of reproducing from themselves another organism of the same species, and with the same complex structure. It might be thought that such cells must necessarily be very complex in their own structure, but in most cases nothing of the kind is to be seen, and the germ-cells often appear simpler in organization than many of the tissue-cells, such as the glandular-cells; and where there is an unusual size or complexity of structure in the germ-cell it usually[Pg 264] bears no relation to the grade of organization of the young creature that is to arise from it, but is due solely to the special conditions imposed on the particular germ-cell, if a young organism is to be evolved from it. We shall soon see what is meant by this.
I must note here that plants and animals do not multiply by means of germ-cells alone, but that many species—the majority of plants and the simpler forms of animals—also exhibit multiplication by budding or division. All animals and plants which do not stop short at the stage of the individual, the 'person,' but rise to the higher stage of the 'stock' (or corm), illustrate this. The first person from which the formation of the stock proceeds gives rise by budding or division to new persons which remain attached to it, and in turn by repeated production of buds give rise to a third, fourth, or nth generation of persons, all remaining in connexion with the first, and together forming the composite individuality of the animal-colony or plant-stock. Such colonies or stocks are seen in polyps and corals, Siphonophoræ and Bryozoa, and among plants, according to Alexander Braun, in all phanerogams which do not consist only of a single shoot. In these cases we find that definite, or perhaps indefinite groups of cells in the stock may give rise to a new person, and we have to inquire how this power may be theoretically interpreted.
New stocks may also have their origin from such buds, or from single persons of the stock. The fresh-water polyp (Hydra) gives rise by budding to a small stock of at most three or four persons; but the young animals budded off only remain attached to the parent hydra until they have attained their full development; then they detach themselves and settle down independently, and begin to bud off in turn a similar and transitory stock. Among plants there are many which, like Dentaria bulbifera and Marchantia polymorpha, multiply by so-called 'brood-buds,' that is, buds which fall from the stock and grow into new plants. The whole horticultural propagation of plants by cuttings also depends on the process of budding, for what is cut off from the parent plant and stuck into the earth is a single shoot, that is, a 'person' which possesses the power of sending down roots into the earth, and by continual budding giving rise to new shoots or persons which together make up a new plant-stock.
I must not, however, spend much time over this so-called 'asexual' reproduction by budding and division, because it does not suggest any way by which we may penetrate more deeply into the processes of inheritance, and we may be content if we can bring them into harmony with other theoretical views which we deduce from[Pg 265] other phenomena. These forms of reproduction were long regarded as the oldest and the simplest, and it is only since the time of Francis Balfour that the conviction has gradually gained ground that this cannot be so, but that they are rather secondary methods of multiplication in the Metazoa and Metaphyta, which therefore rest on a very complex basis. We have seen that the germ-cells made their appearance along with the multicellular body, and the step from Pandorina to Volvox is as small a step as can be well imagined. It is thus proved that the oldest mode of multiplication among multicellular organisms was that through germ-cells, at least along this line of evolution. Volvox does not reproduce by dividing, or by the development of buds from any part of the spherical colony of cells. What is known as budding among single-celled organisms is only an unequal cell-division, and has nothing but its external appearance in common with the budding of higher plants and animals. The latter, therefore, is something new, of later and independent origin; the primitive mode is reproduction by unicellular germs.
REPRODUCTION BY GERM-CELLS.
Historical—Differentiation of germ-cells into male and female—Pandorina—Volvox—Sperm-cells and ova in Algæ—Zoosperm form of the male germ-cells—Zoosperms of the Barnacles—Adaptation of the sperm-cells to the conditions of fertilization—Daphnids—Spermatozoa in different animal groups—Their minute structure—Form and structure of the egg-cell—Adaptation of the ovum to given conditions—Dimorphic ova in the same species—Nutritive cells associated with egg-cells—Complex structure of the bird's egg.
If we now turn to the reproduction of the Metazoa and Metaphyta by means of germ-cells we find that a great number of lowly plants produce germ-cells which require nothing more for the development of a new plant beyond certain favourable external conditions, above all, moisture and warmth. Such, for instance, are the 'spores' of the ferns, which are formed on the under surface of the fronds in little clusters of a brown or yellow colour, easily visible to the naked eye. These spores are individually very small, so that thousands go to form one spore-cluster or sporangium, and millions of spores are given off annually by a single fern. Each spore is a germ-cell enclosed in a protective capsule, and may, if carried by the wind to a spot favourable to germination, become a young plant, the so-called prothallium, from which the fern-plant proper subsequently develops.
This reproduction by spores has been regarded as a form of 'asexual reproduction' so-called, and has been classed along with budding and fission under this head. But it has nothing in common with these forms of multiplication except the negative character that the act of fertilization, which we shall inquire into later on, does not in this case occur. This mode of classification has no longer any more justification than the division of the animal kingdom into backboned and backboneless animals, in which the negative character of the absence of vertebræ has led to the slumping of quite heterogeneous forms in one group. I do not mean to dispute that both these classifications were fully justified in their own time; indeed they expressed a step of progress. Nowadays, however, the division 'Invertebrata' or 'backboneless animals' as a scientific conception has been abandoned, and the same should be done with the category[Pg 267] 'asexual reproduction,' since it groups together quite different things, such as multiplication by single-celled and many-celled 'germs,' and is moreover based on a quite erroneous idea of what 'fertilization' really is. Both terms may very well be retained as a mere matter of convenience, but it is much to be desired that the two apt designations proposed by Haeckel—Monogony for asexual, and Amphigony for sexual reproduction—should come into general use.
Meanwhile it is enough to say that reproduction by 'spores' occurs normally in Algæ, fungi, mosses, and fern-like plants, and that there are also animals in which the germ-cells possess the power of giving rise of themselves to a new individual. But the cases which I am chiefly thinking of are those of so-called virgin birth or parthenogenesis, which are not to be compared with multiplication by spores in regard to their mode of origin; there is a peculiarity in the origin of this mode of multiplication which I can only make clear after we have studied the normal forms of what is called 'sexual reproduction.'
We shall therefore pass on to this mode of reproduction. It is well known that, in all higher animals, just as in Man, an individual cannot reproduce by itself; the co-operation of two individuals is necessary, and these—the male and the female—differ essentially from each other in many particulars. Their union in the act of procreation induces the development of a new individual, whether this matures within the mother in a special receptacle, or whether it is deposited as a 'fertilized egg,' as in birds, the lower vertebrates, and most 'invertebrates.'
As long as Man has lived he has regarded this process of procreation as the essential factor in the origin of new individuals, and as he had no insight into the essence of the process he had necessarily to regard reproduction as something entirely mysterious, and the co-operation of the two sexes as a conditio sine qua non of reproduction in general; thus copulation and reproduction seemed identical.
This was in the main the state of opinion at the time of the discovery of innumerable minute filaments, the so-called 'spermatozoa' in the 'fertilizing' spermatic fluid of the male. The discovery was made in 1677 by Leeuwenhoek in the case of birds, mammals, and many other animals. Albrecht von Haller (1708-77) was at first inclined to regard these spermatozoa as the rudiments of the embryo, but later on, in the course of his long life, he withdrew this theory, and declared them to be a kind of parasite in the spermatic fluid without anything to do with fertilization. The same opinion was expressed in 1835 by K. E. von Baer, in opposition to the opinion of Prevost and Dumas, who had rightly interpreted the spermatozoa[Pg 268] as the essential elements of the spermatic fluid. When one follows the matter out in detail, one finds it almost incredible that such a number of mistakes should have been made, and so many circuitous paths traversed, before even the limited knowledge current in the middle of the nineteenth century was attained—that is to say, enough to give ground for the assertion that fertilization depends upon the contact of the spermatozoon with the body of the egg. In 1843 Martin Barry had found the spermatozoa within the egg-envelope of the rabbit ovum, but it was some time later (1852) that the investigations of Meissner, Bischoff, and Newport established the fact that the zoosperm penetrates through the egg-envelope. All else remained quite obscure, and could not be cleared up as long as it was believed, on the strength of observations which were in themselves correct enough, that several zoosperms were always necessary to fertilize one ovum.
To an understanding of the process even in its most general outlines there was lacking, apart from technical methods, an appreciation of the morphological value of the ovum and the spermatozoon. It was necessary to recognize both ovum and spermatozoon as cells before their union in fertilization could be regarded as the fusion of two cells, as a copulation or conjugation of two minute elementary organisms. But this knowledge only gained ground very gradually, and even in the sixties opinions on the subject were very much divided. Moreover, there was an entire absence of knowledge in regard to 'sexual' reproduction among the lower plants, the Algæ, Fungi, Mosses, and Ferns, as well as of any detailed acquaintance with the processes of fertilization among flowering plants. All this had to be elucidated by the labours of many distinguished observers before it was possible to say so much even as this, that the process of fertilization depends in general on the union of two cells.
I need not discuss the whole of this long process of scientific development, and have only touched upon it because I wished to emphasize that the conception of the process of fertilization was for a long time quite erroneous, and has only attained to clearness in recent times. Pairing as it is seen in the higher animals was for long regarded as the essential part of the process, and a mysterious life-awakening influence was assumed in regard to it; and even when it was understood that not the copulation, but the union of two living units which was always brought about thereby—the union of the male and the female germ-cells—was the essence of 'fertilization,' this was still regarded as a life-awakening process, and the way to a true understanding of the facts was thus once more blocked.
The simplest form of sexual reproduction in many-celled animals[Pg 269] is found, among others, in the Volvocineæ, those green, spherical, freshwater cell-colonies which we have already studied in relation to reproduction by asexual germ-cells. Among them it is the rule that, after a long series of generations producing only 'asexual' germ-cells, colonies occur in which each germ-cell is no longer able to develop a new colony alone, but can do so only after it has united with another germ-cell.
Now, as we have seen, there are Volvocineæ in which the differentiation of cells into those of the body (soma) and those concerned with reproduction has not been established, and all the cells are therefore alike. In these, as for instance in the genus Pandorina (Fig. 62, p. 257), when sexual reproduction is to occur the whole colony breaks up into sixteen cells; these burst forth from the gelatinous matrix in which they have been hitherto enclosed, swim about in the water with the help of their two flagella, meet other similar free-swimming cells and conjugate with these. The two swimming cells come close to each other, draw in their flagella, sink to the ground in consequence, and fuse completely both as to the cell-body and the nucleus. They assume a spherical form, lose the eye-spot, become surrounded with a tough cell-skin or cyst, and so remain for a longer or shorter time as so-called 'zygotes' or lasting spores. Then they develop by repeated cell-division into one of the sixteen-celled Pandorina colonies with which we are already familiar; this bursts forth from the capsule and swims freely about in the water again.
Here, therefore, the so-called sexual reproduction depends on the fusion of two cells similar in appearance, and when this phenomenon was first known it was regarded as something quite different from the corresponding reproduction in other multicellular organisms. But we now know that quite nearly related Volvocineæ belonging to the genus Volvox and to other genera, which exhibit a differentiation into body-cells and reproductive cells, may reproduce sexually by means of two different kinds of germ-cells; and we have also learned through Goebel and others that even genera like Pandorina, which consist of only one kind of cells, may yet produce male and female reproductive cells differing essentially in form from one another. In Eudorina, for instance, a gelatinous sphere containing sixteen or thirty-two individual cells, asexual reproduction occurs in exactly the same way as in Pandorina, that is, each of these cells divides four or five times in rapid succession, and thus forms a new colony, which then bursts forth; but when the time for sexual reproduction comes the colonies behave differently, for some become female and some [Pg 270]male. In the former the cells remain as they were before, but in the male colonies the sixteen or thirty-two cells undergo a peculiar process of division, which ends in each becoming a mass (16-32) of so-called 'zoosperms,' that is, minute, narrow, longitudinally elongated cells with two flagella (Fig. 63 at D shows those of Volvox). In Eudorina they differ from the female germ-cells or ova externally in form and size, as well as by being much more actively motile, and they contain green and subsequently yellow colouring matter, and a red eye-spot. We here find, for the first time among multicellular organisms, the differentiation of male and female germ-cells; and we learn from this that the essence of fertilization does not lie in this[Pg 271] differentiation, since it may be absent, but that this distinction of female and male cells is only of secondary moment. From the fact that the egg-cells are larger and less active, the 'sperm-cells' or zoosperms smaller and livelier, we can already anticipate what will be more definitely established as our knowledge of the facts increases—that a differentiation according to the principle of division of labour has taken place even in the germ-cells, and that the first effect of this is to render the meeting of the cells destined for conjugation easier and more certain. The much smaller and more slender zoosperms swim about in the water in clusters until they come in contact with a female colony; then they separate from each other, bore their way into the soft jelly of the female colony, and 'fertilize' the egg-cell, that is to say, each male cell fuses with a female cell and forms a 'lasting spore,' exactly as in Pandorina.
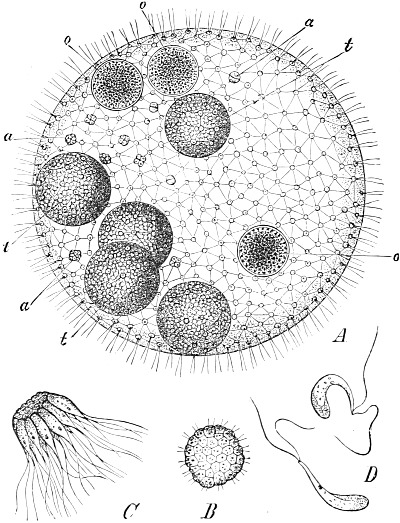
Fig. 63. Volvox aureus, after Klein and Schenck. A, besides the small flagellate somatic cells of the colony there are five large egg-cells (t) which are capable of parthenogenetic development, three recently fertilized egg-cells (o) and a number of male germ-cells (a) in process of multiplication. From each of these, by continued division, a bundle of spermatozoa arises. B, a bundle of thirty-two sperm-cells in process of development, seen from above. C, the same seen from the side. Magnified 687 times. D individual spermatozoa, magnified 824 times.
In Volvox the state of matters is similar to that in Eudorina; here again, in addition to the 'asexual' reproduction through the 'Parthenogonidia' which are like egg-cells in appearance (Fig. 63, A, t), there are also male and female germ-cells which are usually produced alternately with the former, but sometimes at the same time, as in Fig. 63. The egg-cells are large and have no flagella, the sperm-cells lie together in clusters, and after they are mature (D) they swim freely in the water and then bore into another colony, where each unites with an egg-cell. The difference between the two kinds of cells consists therefore in the much greater number, the much smaller size, and the greater activity of the male cells, and in the smaller number but much larger size of the female cells—a differentiation in accordance with the principle of division of labour, depending on the fact that the two kinds of cells must reach each other, and yet must contain a certain mass of living protoplasm. While the small size but large number of male cells, combined with their motility, gives them an advantage in seeking out and boring into the female cells, the large size of the latter, on the other hand, makes up for the loss in mass which the fertilized egg would otherwise suffer from the diminution in size of the male cell. This difference in size may be greatly accentuated; thus in one of the brown sea-wracks, for instance, the spermatozoa are only 5 micro-millimetres in length, while the ova are spherical and have a diameter of 80-100 micro-millimetres, thus containing a mass 30-60,000 times greater (Möbius). Fig. 64 shows an ovum of this species surrounded by spermatozoa
In the course of the evolution of species this contrast between female and male germ-cells became more and more marked, not always in the same direction, however, but in one or another according[Pg 272] to the conditions of fertilization. It would be erroneous to suppose that, with the higher differentiation of the organism as a whole, the differentiation of the germ-cells became increasingly complex. On the contrary we find even among Algæ, as the case of Fucus shows, a marked difference between the sex-cells, which rather decreases than increases among many of the higher plants. It is not on the more or less complex structure of the organism itself that the nature and degree of the dimorphism of the germ-cells depends, but on the special conditions which are involved in each case, both in the union of the two kinds of sex-cells and in the subsequent development of the product of this union, the 'fertilized ovum.'

Fig. 64. Fucus platycarpus, brown sea-wrack.
Ei, ovum, surrounded by swarming
sperm-cells (sp). After Schenck.
Thus it comes about that the male or 'sperm-cells' of the lower plants, of the lower animals, and, again, of the highest animals are similar in structure. In all these organisms the male germ-cells exhibit the minuteness, the form, and the activity of the so-called 'zoosperms' or 'spermatozoa,' that is to say, they are thread-like, very minute corpuscles, which move rapidly forwards in water or other fluid with undulatory movements, and penetrate into the ovum with similar boring movements when they have been fortunate enough to reach their goal. At the anterior end they possess a more or less conspicuous thickening, the so-called 'head' in which the nucleus lies, and this is followed by the 'tail,' a thread-like structure consisting of cytoplasm which effects undulatory movements comparable to those of the flagella of Infusorians and Volvocineæ. The whole spermatozoon is thus a specialized 'flagellate cell.'
When these 'zoosperms' were recognized as the 'fertilizing elements' in higher animals, and when 'sperm-threads' had been found, not only in all mammals and birds, reptiles, amphibians, and fishes, but even in many 'invertebrates,' the conclusion was suggested that the function of fertilization might be discharged by this lively motile substance; for until the eighth decade of the nineteenth century fertilization was still regarded by many as an 'awakening of life' in the egg. Since life depends on movement, in truth on infinitely fine molecular movements, of which the movement of the whole germ-cell[Pg 273] from place to place is only a visible outcome, fertilization was pictured, by a not very luminous process of reasoning, as the awakening of life in the ovum—in itself incapable of further life—through the transference to it of movement through the agency of the zoosperm. Some investigators even went the length of regarding the ovum as 'dead organic material.'
I mention this at this point, though I do not propose in the meantime to inquire further into the significance of the conjugation of the sex-cells. But the view just referred to is so completely refuted even by the external form of the male germ-cells in many groups of plants and animals, that I cannot discuss these differences in form without at the same time indicating the conclusions which they directly suggest.
The great majority of plants and animals exhibit the zoosperm form of male germ-cells, and this must obviously be interpreted in the light of the fact that the ova to be fertilized are not generally to be found in direct proximity to the sperms shed by the male organism, but are at some distance from them. Among Medusæ and Polyps both male and female germ-cells are liberated into the water, simultaneously it may be, but separated from each other by distances of some feet or yards. The spermatozoa then swim about seeking the ova, which are also floating freely in the sea, guided by a power of attraction on the part of the latter—an attraction of whose nature we know nothing, though in the case of certain fern-ova it has been traced to the secretion of malic acid (Pfeffer).
The same conditions obtain among Sponges. Here, again, the persons or stocks are either male or female; the latter produce large delicate ova, which lie in the interior of the sponge and there await the fertilizing sperms; the former give off the ripe sperms into the water in such abundance that thousands and millions of zoosperms burst forth simultaneously in all directions; these seek about for a female sponge, penetrate into its canal system, and so ultimately reach the ova. Of course only a very few of them reach their goal; the greater number are lost in the water and become the prey of Infusorians, Radiolarians, or other lowly animals. The fact that enormous numbers thus miss their true aim shows us why these zoosperms must be produced in such quantities; it is simply an adaptation to the extraordinarily high ratio of elimination in these cells, just as the number of young annually produced by an animal, or of seeds by a plant, is regulated by natural selection according to the ratio of elimination of the particular species. The more numerous the descendants which succumb each time to unfavourable circum[Pg 274]stances, to enemies, or to lack of food, the more prolific must the species be. The same holds true of the regulation of the number of male germ-cells to be produced by an individual; there must be so many developed that, in spite of the unavoidable enormous loss, on an average the number of mature ova necessary to the maintenance of the species always receive spermatozoa.
Also associated with the prodigal production of zoosperms is their minuteness, for the more zoosperms that can be developed out of a given mass of organic substance the smaller they are. Each species is restricted within definite limits of production by its size and the volume of its body, and there is thus an advantage in having the zoosperms of the smallest possible size whenever the chance of the individual sperm successfully reaching an ovum is very small. In all such cases nature has abstained from burdening the male germ-cell with an appreciable contribution of material to the result of conjugation, that is, to the foundation of the new organism; the passive ovum contains in itself alone almost all that is necessary to the building up of the embryo. Fertilization of the ovum by the liberation of the sperm-cells into the water occurs not only in animals of low degree, such as Sponges, Medusæ, Star-fishes, Sea-urchins and their relatives, but also in much higher animals, such as many Fishes and Amphibians, and in these the male cells have the form of motile threads. But the spermatozoon-form of male cell does not occur only in animals and plants which live in the water, or in those which, like mosses and many vascular plants, are at least occasionally covered by a thin layer of rain or dew, in which the zoosperms can swim to the ova, it occurs also in a very large number of animals in which the sperms pass directly into the body of the female, in those, therefore, in which copulation takes place.
But even where copulation occurs we find that in most cases, as, for instance, in Vertebrates, Molluscs, and Insects, the zoosperm-form is retained. The reason for this is obviously twofold: in the first place, in many cases the sperms do not directly reach the ovum as a consequence of copulation, but may have to go a long way within the body of the female, as in mammals; or even when the way is short and certain, the ovum may be encased in a firm covering or shell difficult to penetrate, and the thread-like zoosperm has to face the task of boring its way through this shell, or slipping in through a minute opening, the so-called micropyle. In either case it would be difficult to imagine a form of sperm-cell better adapted to the fulfilment of this task than that of a thread with a thin, pointed head-portion and a long motile tail, which enables the zoosperm to twist[Pg 275] itself like a screw through a narrow opening in the egg-envelope, whether the opening was previously present or not.
We can thus understand why, among insects for instance, the male cells should always occur in the form of zoosperms, although in this case they reach a special receptacle in the female reproductive organs, the 'receptaculum seminis,' and are stored up in this. When a mature ovum gliding downwards through the oviduct comes to the place where this receptacle opens into it, the liberation of a few sperm-cells suffices to fertilize it with certainty, provided that they possess the thread-like form, which allows them to slip in through the very minute opening in the egg-envelope. It might be inferred from the certainty with which the ovum must in this case be found by the spermatozoon that only a small number of the latter would require to be produced, and yet even here the number is very large, though not so enormous as in the sea-urchins and other marine animals, which simply allow the sperm-cells to escape into the water. The large number in insects is due to the fact that many of the sperms may miss the micropyle; and also that in many insects a very large number of eggs have to be fertilized in succession. In the course of a life lasting three or four years the queen bee lays many thousand of eggs, most of which are fertilized, and that from a seminal receptacle which has been filled only once.
There are, however, other sperm-cells of thread-like form which are not produced in such enormous multitudes, but in a much more moderate number, perhaps a few hundreds in the testicle. This is so in the little Crustaceans, known as Ostracods, all the freshwater species of which possess zoosperms only moderately numerous and of quite unusual size.
The comparatively small number is explained by the certainty with which each of them reaches the ovum, and the large size may be accounted for in part by the small number which suffices, and which, therefore, admits of the male cell also carrying a considerable portion of the material for the building up of the embryo. Probably, however, the thickness and firmness of the covering of the ovum has something to do with it, for it has no opening for the entrance of the male cell, and it is fully hardened by the time fertilization takes place. Perhaps nowhere can we see more clearly how every little detail of the structure of the organism is dominated by the principle of adaptation than in the arrangements for fertilization, and notably in those which obtain in the Ostracods. I pass by the complicated apparatus for copulation, since we do not yet understand it fully in all particulars. According to my own investigations and those of my[Pg 276] former students, Dr. Stuhlmann and Dr. Schwarz, the essential point seems to be that the colossally large zoosperms, which show no activity within the body of the male, leave it one at a time, so to speak, in single file. In copulation they are pressed out singly, one after the other, through a very fine tube, and then they enter, still singly, through the reproductive aperture of the female into an equally fine passage with spiral windings, through which they ultimately reach a roomy pear-shaped receptacle, the 'receptaculum seminis' of the female. There they lie in a long band composed of several hundreds, and only now attain their full maturity by throwing off an outer cuticle—moulting, so to speak. It is only when they get into a fluid medium that they show the power of undulatory movement, feeble at first, but gradually more energetic and more violent. And these movements enable them to penetrate like gimlets into the calcareous egg-shell. In the normal course it happens that when a mature ovum is deposited from the opening of the oviduct, one of the giant zoosperms at the same time, or shortly afterwards, leaves the 'receptaculum seminis' of the female by way of the spiral passage, and reaches the exterior just behind the ovum. The actual process of penetration has not been observed as yet, but the zoosperm has been seen at a slightly later stage spirally coiled inside the ovum.
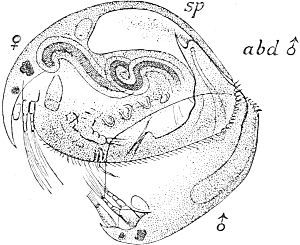
Fig. 65. Copulation in a Daphnid (Lyncæid).
Emptying of the sperm (sp) into the brood-chamber
of the female (♀). abd ♂, the abdomen of the male.
Magnified 100 times.
In these Ostracods the sperms are often visible with the naked eye, and in some species they are twice the length of the animal; they are thus emphatically giant cells, which can develop a very considerable boring power.
In respect to the various adaptations of the sperm-cells to the conditions of fertilization there is hardly any group more interesting than the water-fleas or Daphnids.
It is amazing how greatly the size of the sperms varies among the Daphnids, and how it stands in inverse proportion to their [Pg 277]number, and how both are obviously regulated in relation to the difficulties which stand in the way of each sperm-cell before it can reach the ovum. In some species the sperm-cells are very large, in others extremely small. In the genera Daphnia, Lynceus, and others, copulation occurs as shown in Fig. 65; the sperm-cells (sp) are liberated by the male into the capacious brood-cavity of the female, which at the moment is closed to some extent by the abdomen of the male, in reality closed only partially at the posterior end and at the sides. It seems inevitable that a large proportion of the male elements should stream out again and be lost because of the violent movements of both animals. Accordingly, we find that the sperm-cells are only about the hundredth part of a millimetre in length and of round or rod-like form, and are voided in multitudes into the brood-cavity. Fig. 66, f, g, and h, show such cells in different species, as they occur in the testes to the number of many thousands. But in all the species in which the brood-cavity is closed, and in which therefore there is not such a serious loss of sperm-cells, the elements are much larger, and at the same time less numerous. They are largest and least numerous in species of genera like Daphnella, Polyphemus, and Bythotrephes, in which the males have a copulatory organ, so that the possibility of loss of the male cells is excluded. Thus the round, delicate, and viscid sperm-cells of Bythotrephes (Fig. 66, b) are more than a tenth of a millimetre in length, but they are developed in proportionately smaller numbers, so that more than twenty are never found in the testis, and often only six or eight, while in copulation only from three to five are ejected. But as there are only two eggs to be fertilized at a time, and as the male cells are expressed into the brood-cavity directly upon the eggs, so that they immediately adhere to them, this small number is amply sufficient.
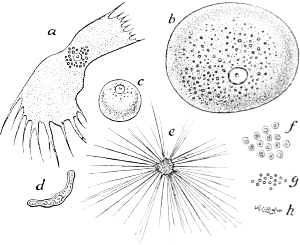
Fig. 66. Spermatozoa of various Daphnids. a, Sida.
b, Bythotrephes. c, Daphnella. d, Moina paradoxa. e,
Moina rectirostris. f, Eurycercus lamellatus. g, Alonella
pygmæa. h, Peracantha truncata. All magnified 300
times.
It is remarkable how different the sperm-cells sometimes are in quite nearly related species of Daphnids, as a glance at Fig. 66 will show; and, on the other hand, how similar they may be in two species which belong to different families, like Bythotrephes longimanus (b), and Daphnella hyalina (c). The last fact may be explained as an adaptation to similar conditions of fertilization. Both species have effective copulatory organs, and their large delicate sperm-cells must immediately adhere when they come into contact with the shell-less ovum, and penetrate into it by means of amœboid processes. Conversely, the difference between sperm-cells of allied species like Sida crystallina (a), Moina rectirostris (e) and M. paradoxa (d) is related to different adaptations to nearly the same conditions of fertilization. In Sida (Fig. 66 a) the large flat sperm-cells, with their fringed ends and their large soft surface, adhere easily to the ova, and the same end is attained in Moina rectirostris by means of stiff radiating processes, while in the nearly related species, Moina paradoxa, the male cell (d) resembles an Australian boomerang and presses in like a wedge between the ova and the wall of the brood-sac.
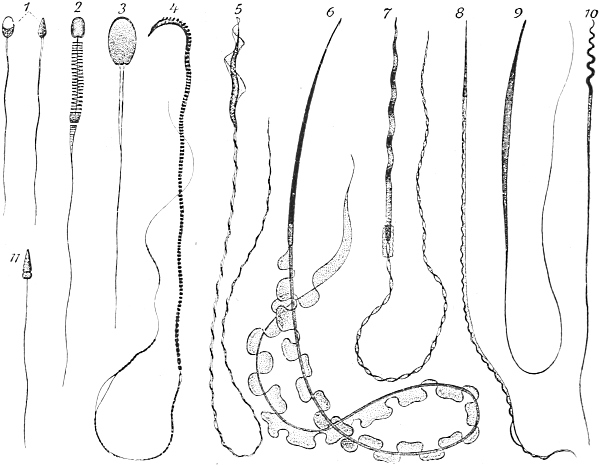
Fig. 67. Spermatozoa of various animals, after Ballowitz, Kölliker, and vom Rath. 1, man. 2, bat (Vesperugo). 3, pig. 4, rat. 5, bullfinch. 6, newt. 7, skate (Raja). 8, beetle. 9, mole-cricket (Gryllotalpa). 10, freshwater snail (Paludina). 11, sea-urchin. Much magnified.
In Fig. 67 a small selection of animal male cells is figured, all of[Pg 279] which take the form of sperm-threads or spermatozoa, and yet they differ very much from one another in detail. It would undoubtedly be of great interest to follow out these minute adaptations of the sperm-cells to the conditions of fertilization, and to demonstrate that their size, and especially their form, in the different species of animals are adjusted to the special constitution of the ovum, its envelope, and its micropyles, and to the ease or difficulty with which it can be reached; but much information must be forthcoming before we can even suggest, for instance, why the sperm-cell of the salamander is so enormously long, large, and pointed at the head, while that of Man (Fig. 67, 1) is comparatively short, with broad, flat head and a recently discovered minute apex; or why those of Man and many fishes (such as Cobitis) should be so much alike, and so on. From many sides, however, we are led to conclude that even down to the minutest details nothing is in vain, and that everything depends on adaptation.
In general, even the peculiarities of form already indicate this; thus the spirally coiled structure of the head, which is especially well developed in the spermatozoa of birds (Fig. 67, 5), in those of the skate (7), and of the freshwater snail (Paludina) (10), works like a corkscrew, and makes it possible for the spermatozoon to pierce through the resistant envelope of the ovum. Similarly, the sharply pointed head of the insect spermatozoon (Fig. 67, 8 & 9) seems adapted for slipping through the minute pre-formed micropyle in the hard egg-shell.
Of the detailed and complicated structure of spermatozoa we have only recently been made aware through the increasing perfection of the microscope and of technical methods of investigation. Fig. 68 shows one after a diagrammatic figure by Wilson. We see the apical point (sp) for boring into the ovum, the nucleus (n) surrounded by a thin layer of protoplasm, which together form the head, then the middle portion (m) which contains the 'centrosome' (c), and the 'tail' or 'flagellum' which effects the movement of the whole and which itself possesses a complex structure with an 'axial filament' (ax) and an enveloping layer, the latter often drawn out into a spirally twisted, undulating membrane of the most extreme delicacy, as is most clearly seen in the newt (Fig. 67, 6).

Fig. 68. Diagram of a
spermatozoon, after Wilson.
sp, apical point.
n, nucleus.
c, centrospere.
m, middle piece.
ax, axial filament.
e, terminal filament.
Not in the Daphnids alone, but in other groups of Crustaceans as[Pg 280] well, sperm-cells of quite peculiar form occur, as, for instance, in the crayfish and its marine relatives, the crabs and the long-tailed Decapods. In these cases the spermatozoa bear long and stiff thorn-like processes, which, as in the sperm-cells of Moina, make them adhesive, and, according to Brandes, render it possible for them to cling among the bristles on the abdomen of the female until one of the many eggs leaving the oviduct comes within reach. For among these Crustacea there is no true copulation, but the masses of sperm-cells are packed together into sperm-packets or 'spermatophores,' and are affixed by the male near the opening of the oviduct, where they burst and pour forth their contents between the appendages of the female.
All these remarkable and widely divergent structures and arrangements depend not upon chance or on the fantastic expression of a 'formative power,' as an earlier generation was wont to phrase it; they are undoubtedly without exception adaptations to the most intimate conditions of fertilization in each individual case. I lay particular stress upon a recognition of this, because it permits us to infer with certainty that even the variations of the single cell, if they are sufficiently important for the species, may be controlled by natural selection. It is obvious that the adaptations of the sex-cells must depend not on histonal selection, but only upon personal selection, since it is indifferent for the individual sperm-cells whether fertilization is accomplished successfully or not, while it is by no means indifferent for the species. The organism dies without descendants if its sperm-cells do not fertilize, and the carrying on of the species must be left to those of its fellows which produced sperm-cells which fertilize with more certainty; thus it is not the sperm-cells themselves, but the individual organisms which are selected, and that in relation to the quality of the sex-cells they produce.
In contrast with the great diversity of form exhibited by the spermatozoa, the differentiation of the ovum appears very uniform, at least in regard to form and activity. The main form is spherical, but it is subject to many variations in the way of elongation or flattening. In the lower forms of life, as, for instance, among the sponges, and also in the polyps and Medusæ the egg-cells possess, until they are mature, the locomotor capacity of unicellular organisms; they creep about after the manner of amœbæ, and indeed, as I showed years ago, this movement from place to place in many polyps is exactly regulated; thus at a definite time they may leave the place where they originated and may, for instance, creep from the outer layer of cells (ectoderm) of the animal into the inner layer (endoderm) by boring through the so-called 'supporting lamella,' then they may creep further in the[Pg 281] endoderm, and finally return to quite definite and often remote spots in the ectoderm (Eudendrium, Fig. 95). In another hydroid polyp (Corydendrium parasiticum) the mature egg-cells leave their former position within the endoderm and creep entirely outside of the animal which produced them, establishing themselves in a definite spot on its external surface, where they await the fertilizing zoosperms. Many ova can accomplish slight amœboid movements, but in most animals these do not suffice for movement from place to place, and the ova remain quietly in the spot where they were developed, or are passively pushed to another. Cases such as that of the polyp I have cited, in which the ovum actually comes to meet the male element, are quite exceptional, for in general the ovum is the passive and the spermatozoon the active or exploring element in fertilization. The female cell is entrusted with procuring and storing the material necessary to the building up of the embryo; and its peculiarities depend chiefly on this.
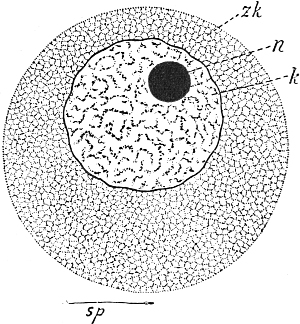
Fig. 69. Ovum of the Sea-urchin, Toxopneustes lividus,
after Wilson. zk, cell-body. k, nucleus or so-called
'germinal vesicle,' n, nucleolus or so-called 'germinal
spot.' Below there is a spermatozoon of the
same animal, with the same magnification (750
times).
It is true that in plants this stored material is seldom considerable, and that is because the ovum so frequently remains even after fertilization within the living tissues of the plant, and is thence supplied, often very abundantly, with food-stuffs; and, moreover, because the young plant that springs from the fertilized ovum maybe very small and simple, and yet capable of immediately procuring its own nourishment. But there are exceptions to this; thus the ova of the brown sea-wracks, or Fucaceæ, for instance, are quite twenty times as large as the ordinary cells of the algæ (Fig. 64), and contain a quantity of food-stuff within themselves. In this case the ova are liberated into the water even before fertilization, and the nutrition of the embryo from the mother-plant is excluded.
In these Algæ we meet, for the first time, with a special organ in which the ova arise. In animals this is much more generally the case, and from sponges upwards there are always quite definite parts and tissues of the body which are alone able to develop eggs, and these are usually well-defined organs of special structure, the ovaries. Similarly, in male animals the spermatozoa arise in special places, and usually in special organs, the spermaries or testes.
Animal ova often consist of more than the simple cell-body, the protoplasm and its nucleus; they almost always contain in the cell-body a so-called 'Deutoplasm,' as Van Beneden has fittingly named the yolk-substance. This consists of fats, carbohydrates, or albuminoids, which often lie in the cell-body in the form of spherules, flakes, or grains—a nutritive material that is often surrounded and enclosed by a small quantity of living matter or formative protoplasm. Apart from these stores of yolk it would be impossible for a young animal to develop from the ovum of a snake or a bird, for such highly differentiated animals could not be formed from an egg of microscopic dimensions if this remained without some supply of food from outside of itself during the period of development. There is obviously need for a considerable amount of building material, so that all the organs and parts, which are composed of thousands and millions of cells, may be developed.
Thus the size of the animal-ovum depends essentially on the quantity of yolk that has to be supplied to the egg, and this depends in the main on whether the egg is still drawing nourishment from the mother during the development of the young animal. Therefore, as a general rule, eggs which are laid, and are surrounded and protected by a shell, are usually much larger than the eggs of animals which go through their development within the body of the mother. The best known illustration of this proposition is offered by mammals and birds, animals of equally high organization and comparable in bodily size. While the eggs of birds may be as much as 15 centimetres in length, and may weigh 1½ kilogrammes, those of most mammals remain microscopically minute, and scarcely exceed a length of 0.3 millimetres. The same principle is often illustrated within one and the same small group of animals, and even in the same species. Here, again, the Daphnids or water-fleas may serve as an example.
Among these there are two kinds of eggs, summer and winter eggs, of which the former go through their development into a young animal within a brood-cavity on the back of the female, while the others are liberated into the water, and are surrounded by a hard shell. The summer eggs receive more or less nourishment from the mother by[Pg 283] the extravasation of the nutritive constituents of the blood into the brood-cavity, and they thus require a smaller provision of yolk than the winter eggs, which are thrown entirely upon their own resources. Accordingly we find that in all Daphnids the summer eggs are at least a little smaller and have less yolk than the winter eggs, as in the genus Daphnella (Fig. 70, A and B), while in some species, e.g. of Bythotrephes, this difference increases so much that the summer eggs are almost without yolk, and therefore very minute (Fig. 71, B). The reason of this lies in the fact that in this case the brood-sac is filled with a nutritive fluid rich in albuminoid substances, so that the embryo during its development is continually supplied with concentrated nourishment. This is not the case with the winter eggs, because these are liberated into the water, and we therefore find that they are of enormous size and quite filled with yolk (Fig. 71, A).
Fig. 70. Daphnella. A, summer
egg. B, winter egg. Oe,
'oil-globules' of the summer
egg.
Fig. 71. Bythotrephes longimanus. A, the brood-sac
(Br) of the female containing two winter-ova
(Wei), on which five large sperm-cells (sp) are lying.
R, dorsal surface of the animal. Dr, glandular layer
which secretes the shell-substance. BK, copulatory
canal. B, the brood-sac (Br) containing two summer-ova
(Sei). Both figures under the same magnification
(100).
In this instance, as in all the simpler eggs, the yolk constituents are secretions of the cell-body of the ovum; but nature employs many devices, if I may so speak, to bring up the mass of the egg, and especially of the yolk, to the highest attainable point. Thus in many orders of Crustaceans, for instance in the water-fleas just mentioned, there are special egg-nourishing cells, that is, young ovum-cells which[Pg 284] do not differ from the rest either in origin or in appearance, only they do not become mature eggs, but at a definite time cease to make progress, and then slowly break up, so that their substance may be absorbed as food by the true ova. Thus there is a much greater and at the same time more rapid growth than could be attained through nourishment from the blood alone. In the Daphnids the ovaries consist of groups of four cells each, only one of which becomes an ovum (Fig. 72, Ei), while the other three (1, 2, and 4) form nutritive cells which break up. This is so in all summer eggs; but in the winter eggs a much larger number of nutritive cells may take part in equipping a single ovum, and in the genus Moina over forty do so. But here the difference in size between the two kinds of eggs is very marked, the winter eggs being twice the diameter of the summer eggs.
Fig. 72. Sida crystallina, a Daphnid: a fragment of the ovary showing one of the groups of four cells, of which 1, 2, and 4 are nutritive cells, and only 3 becomes an ovum. Magnified 300 times.
In many insects also, e.g. in beetles and bees, similar nutritive cells occur, but there is in these forms a different arrangement which serves at the same time for the formation of the shell, and the supplying to the ovum of the necessary yolk-stuffs—the ovum is surrounded with a dense layer of epithelial cells, a so-called 'follicle.' In mammals and birds also these 'follicle cells' certainly play an important part in the nutrition of the ovum, though it is not yet quite clear how they act—whether they produce within themselves grains of yolk and other nutritive substances and convey these to the ovum by means of fine radiating processes, or whether they themselves ultimately migrate into the ovum and there break up. In any case it is worthy of note that all these follicular cells in insects and vertebrates have the same origin as the egg-cells, that is, they are modified germ-cells. The case is therefore essentially the same as in the nutritive cells of the Daphnids; nature sacrifices the greater number of the germ-cells in order to be able to provide more abundantly for the minority. She thus succeeds in raising the egg[Pg 285] beyond itself, so to speak, and provides the means for a growth which could obviously not be attained by means of the ordinary nourishment supplied by the blood.
Fig. 73. Diagrammatic longitudinal section of a
hen's egg before incubation, after Allen Thomson.
Bl, germinal disk. GD, yellow yolk. WD, white yolk.
DM, vitelline membrane. EW, albumen. Ch, chalaza.
S, shell membrane. KS, shell. LR, air chamber.
We now understand why the eggs of many animals should be of such enormous size and often of such complex structure. The eggs of birds are especially remarkable in this respect, and it has till recently been disputed whether they are really morphologically equivalent to a single cell. But this is undoubtedly the case, and though only the small thin germinal disk (Fig. 73, Bl) with its nucleus is the active part of this cell—the cell-body proper—yet all the rest—the enormous sphere of yolk with its regular layers of yellow (GD) and white (WD) yolk, the concentric layers of fluid albumen (EW) round about this, the chalazæ (Ch), and finally, the delicate shell membrane (S) and the limy shell (KS)—belong to this cell, and have arisen in connexion with it (Fig. 73).
THE PROCESS OF FERTILIZATION
Cell-division and nuclear division—The chromatin as the material basis of inheritance—The rôle of the centrosphere in the mechanism of division—The Chromosomes—Fertilization of the egg of the sea-urchin according to Hertwig—Of the egg of Ascaris according to Van Beneden—The directive divisions, or the extrusion of the polar bodies—Halving of the number of chromosomes—The same in the sperm-cell—Reducing division in parthenogenetic eggs—In the bee—Exceptional and artificial parthenogenesis—Rôle of the centrosphere in fertilization and in parthenogenesis.
Now that we have made ourselves acquainted with the two kinds of germ-cells on the union of which 'sexual reproduction' depends, we may proceed to a more detailed discussion of the process of fertilization itself. But it is indispensable that we should take account of the processes of nuclear and cell-division, as these have been gradually recognized and understood in the course of the last decade. It may appear strange that the processes of division should throw light on the apparently opposite processes of cell-union, but it is the case, and no understanding of the latter is possible without a knowledge of the former.
From the time of the discovery of the cell until well on in the sixties the process of cell-division was looked on as a perfectly simple process, as a mere constriction in the middle of the cell. It was observed that a cell in the act of dividing (Fig. 59, A) stretched itself out, that its nucleus also became longer, became thinner in the middle, assumed a dumb-bell form, and was then gradually constricted, giving rise to two nuclei (B), whereupon the body of the cell also constricted and the two daughter-cells were formed (C). In certain worn-out or highly differentiated cells a cell-division of this kind really seems to occur—the so-called 'direct' division—but in young cells, and indeed in all vigorous cells, the process, which looks simple, is, in reality, exceedingly complex. Not only is the structure of the nucleus incomparably more complex than was recognized a quarter of a century ago, but nature has placed within the cell a special and marvellously intricate apparatus, by means of which the component parts of the nucleus are divided between the two daughter-nuclei.
For a long time all that was distinguished in the cell-nucleus was[Pg 287] the nuclear membrane and a fluid content in which one or more nuclear bodies or nucleoli float. But this does not by any means exhaust what can now be recognized in the structure of the nucleus, and the most important constituents are not even among these, for recent researches, especially those of Häcker, have shown that the nucleolus or the nucleoli, to which there was formerly an inclination to attach a very high importance, must be regarded as only transient formations and not living elements—in fact, as mere collections of organic substance—'bye-products of the metabolism,' which at a definite time, that is just before the division of the nucleus, disappear from the nuclear space and are used up. We now know that in the resting cell, that is, in the cell which is not in the act of dividing (Fig. 74, A), a very fine network of pale threads, often very difficult to make visible, fills the whole nuclear cavity, like a spider's web or the finest soap bubbles, and that in this so-called nuclear framework there are embedded granules of rounded or angular form (A, chr) which consist of a substance which stains deeply with such pigments as carmine, hæmatoxylin, all aniline dyes, &c., and which has therefore received the name of chromatin. Often, indeed generally, these granules are exceedingly small, but sometimes they are bigger, and in that case they are less numerous and more easily made visible; in all cases, however, they are in a certain sense the most important part of the nucleus, for we must assume that it is their influence which determines the nature of the cell, which, so to speak, impresses it with the specific stamp, and makes the young cell a muscle-cell or a nerve-cell, which even gives the germ-cell the power of producing, by continued multiplication through division, a whole multicellular organism of a particular structure and definite differentiation, in short, a new individual of the particular species to which the parents belong. We call the substance of which these chromatin granules consist by the name first introduced into science by Nägeli, though only to designate a postulated substance which had not at that time been observed, but which he imagined to be contained within the cell-body—by the name Idioplasm, that is to say, a living substance determining the individual nature (εἶδος = form). I am anticipating here, and I reserve a more detailed explanation until I can gradually bring together all the facts which justify the conception I have just indicated of the 'chromatin grains' as an 'idioplasm,' or, as we may also call it, a 'hereditary substance.'
That this chromatin must be something quite special we see from the processes of cell and nuclear division, which I shall now briefly describe.
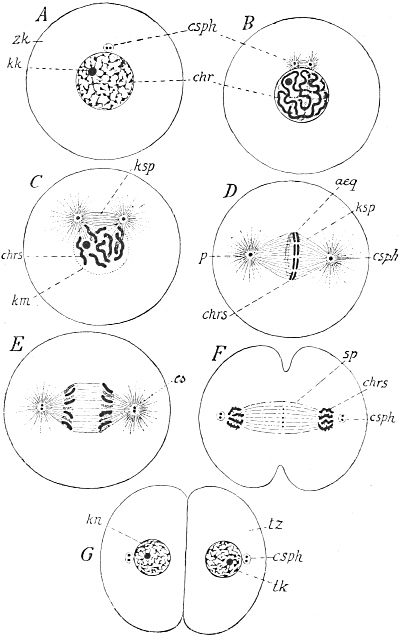
Fig. 74. Diagram of nuclear division, adapted from E. B. Wilson. A, resting cell with cell-substance (zk), centrosphere (csph) which contains two centrosomes, nucleolus (kk); and chromosomes (chr), the last distributed in the nuclear reticulum. B, the chromatin united in a coiled thread; the centrosphere divided into two and giving off rays which unite the halves. C, the nuclear spindle (ksp) formed, the rays more strongly developed, the nuclear membrane (km) in process of dissolution, the chromatin thread divided into eight similar pieces (chrs), the rays are attaching themselves to the chromosomes. D, perfected nuclear spindle with the two centrospheres at the poles (csph) and the eight chromosomes (chrs) in the equator of the spindle, all now longitudinally split. E, daughter-chromosomes diverging from one another, but still united by filaments, the centrosomes (cs) are already doubled for the next division. F, daughter-chromosomes, quite separated from one another, are already beginning to give off processes; the cell-substance is beginning to be constricted. G, end of the process of division: two daughter-cells (tz) with similar nuclear reticulum (tk) and centrospheres (csph), as in A.
When a cell is on the eve of dividing we observe first that the chromatin grains, which have till then been scattered throughout the network of the nucleus, approach each other and arrange themselves[Pg 289] into a long thin thread which, irregularly intertwined, forms a loose skein, the so-called coil-stage (Fig. 74, B). The thread then begins to thicken, and somewhat later it can be seen to have broken up into a number of pieces of equal length, as if it had been cut into equal pieces with scissors (C).
These pieces or chromosomes become shorter by slowly contracting, and thus each takes the form of an angular loop, a straight rod, or a roundish, oval, or spherical body (Fig. 74, C, chrs). While this is happening, we can see at the side of the nucleus, and closely apposed to it, a pale longitudinally striped figure with a swelling, similar to a handle, at both ends—the so-called nuclear spindle or central spindle (ksp). This is the apparatus for the division of the nucleus, and it was previously represented by a small body susceptible to certain stains—the centrosome, which was surrounded by a halo-like zone, the centrosphere or 'sphere.' This body was long overlooked, but now the majority of investigators assume that, though it is often inconspicuous and very difficult to make visible, it is nevertheless present in every cell which is capable of division, and that it is therefore a permanent and indispensable constituent of the cell (Fig. 74, A and B, csph).
When a cell is on the point of dividing, this remarkable cell-organ, which has hitherto seemed no more than an insignificant, pale, little sphere, now becomes active. First of all, often before the formation of the chromatin coil, it doubles by division (A and B csph), at first only as regards the centrosome, and then as regards the sphere also (B); and while division is going on fine protoplasmic filaments issue from the dividing sphere and radiate like rays from a sun into the cell-substance. As they only retain their connexion with each other at the surfaces of the dividing halves of the sphere which are turned towards each other, we might almost say that fine threads are drawn out between the two halves as they separate, and these become longer the further apart the halves diverge. In this manner the much-talked-of 'spindle figure' arises, which was first described in the seventies through the researches of A. Schneider, Auerbach, and Bütschli, but the significance and origin of which have claimed the labours of many later investigators down to our own day.
The processes now to be described do not always take place in exactly the same manner, but the gist of the business is everywhere the same, and it consists in this, that the two ends or 'poles' of the spindle diverge further and further apart, and between them lies the nucleus whose membrane now disappears (C, km) while the spindle threads traverse its interior. Sometimes the membrane is retained,[Pg 290] but nevertheless the spindle threads penetrate into the interior of the nucleus. But the chromosomes always range themselves quite regularly in the 'equatorial plane' of the spindle (D, aeq)—a process the precise mechanism of which is by no means clearly understood, and indeed the play of the forces in the whole process of nuclear division is still very imperfectly revealed to our intelligence.
Thus we have now before us a pale, spindle-shaped figure, which takes only a faint stain, with the 'suns' (cs) at its 'poles,' and in its equatorial plane the loop- or rod-shaped, or spherical chromosomes (chrs). The whole is designated the 'karyokinetic,' the 'mitotic,' or the 'nuclear division figure.'
The meaning and importance of this, at first sight, puzzling figure will at once become clear from what follows. It may be observed at this stage, if not even long before, that each of the chromatin rods or loops has split along its whole length like a log of wood, and that the split halves are beginning slowly and hardly noticeably to move away from each other, one half towards one, the other towards the other pole of the spindle (Fig. D and F). Directly in front of the centrosome they make a halt, and now the material for the two daughter-nuclei is in its proper place (F, chrs). These develop quickly, each chromosome group surrounding itself with a nuclear membrane (Fig. G) within which the chromosomes gradually become transformed again into a nuclear network. Within the chromatin substance proper this is scattered about in small roundish or angular granules, lying especially at the intersecting points of the network. It may be stated at once, though the full significance of the statement can only be appreciated later, that we may assume with probability that this breaking up of the chromosomes is only apparent, and that these rods or spheres really continue to exist in the nuclear network, only in a different form, greatly spread out, somewhat after the manner of a Rhizopod which stretches out fine processes in all directions. These processes branch and anastomose, so that the body, which previously seemed compact, now appears as a fine network. In point of fact, it can be directly observed that the chromosomes, after the nucleus is completely divided into two daughter-nuclei, send out pointed processes (F and G) which gradually increase in length and branch, while the body of the chromosome itself becomes gradually smaller. It is thus probable that, when such a daughter-nucleus is on the point of dividing anew, it may, by a drawing together of the processes or pseudopodia of the chromosomes, produce the same rods or spheres as those which previously gave rise to the network. More definite reasons for this interpretation will be adduced later on. In[Pg 291] any case, the chromosomes, even in their compact rod-like state, consist of two kinds of substance, the chromatin proper, which stains deeply, and the linin, which is difficult to stain; and it is the latter which, by breaking up, forms the pale part of the nuclear network.
Thus we can understand that the number of chromosomes remains the same in every cell-generation throughout development, as it is the same in all the individuals of a species. The numbers are known for many species: in some worms there are only two or four chromosomes, while in other related worms there are eight; in the grasshopper there are twelve, and in a marine worm, Sagitta, eighteen; in the mouse, the trout, and the lily there are twenty-four; in some snails thirty-two; in the sharks thirty-six, and in Artemia, a little salt-water crustacean, 168 chromosomes. In Man the chromosomes are so small that their normal number is not certain—sixteen have been counted. This counting can only be done during the process of nuclear division, for afterwards the chromosomes flow indistinguishably together, or rather apart, only to reappear, however, in the old form and number whenever the nucleus again begins to divide.
It remains to be told what becomes of the centrosphere in cell-division. As soon as the formation of the daughter-nuclei has been brought about by the divergence of the split halves of the loops, the spindle figure begins to retrograde, its threads become pale and gradually disappear, as does the whole radiate halo of the centrosphere (Fig. F and G). The cell-body has by this time also divided in the equatorial plane of the nuclear spindle, and the centrosome remains usually as a very inconspicuous pale body lying in the cytoplasm close to the nucleus, reawakening to renewed activity when cell-division is about to recommence (G, csph).
These, briefly, are the remarkable processes of nuclear division. Their net result is obvious; the chromatin substance is divided between the daughter-nuclei with the greatest conceivable accuracy.
It is not so easy to understand the mechanism of this partition, and there are various divergent theories on this point. According to the older idea of Van Beneden, the spindle fibres work like muscles, and by contracting draw the halves of the chromosomes which adhere to them towards the pole, while the rest of the fibres radiating out from the polar corpuscles act as resisting and supporting elements. This view, with many modifications however, has still its champions, and M. Heidenhain in particular has made a notable attempt to establish it and to work it out in detail. Opposed to it stand the views of those who, like O. Hertwig, Bütschli, Häcker, and others regard the rays not as specific elements which were pre-formed in the[Pg 292] cell, but as the expression of the orientation of certain protoplasmic particles—an orientation evoked by forces which have their seat within the central corpuscles, and act in the manner of magnetic or electric forces. That the central corpuscles are centres of attraction seems to me hardly open to doubt, and I cannot regard the regular arrangement of the chromosomes in the equatorial plane of the spindle as due to a mere adhesion to contractile threads. Some still unknown forces—chemotactic or otherwise—must be at work here. Later on we shall study the phenomenon of the migration of the sperm-nucleus into the ovum, when it is accompanied by its central body and its halo of rays. Häcker seems to me justified in inferring from this phenomenon alone that the sudden origin of the rays is due to forces resident in the central corpuscle. But undoubtedly even this 'dynamic' explanation of karyokinesis is still only at the stage of hypothesis and reasoning from analogy, and is far removed from a definite knowledge of the forces at work.
For the problems with which we are here chiefly concerned, the problems of heredity, it is enough to know that the cells of multicellular organisms possess an extremely complex apparatus for division, whose chief importance lies in the fact that through it the chromatin units of the nucleus are divided into precisely equal parts, and so separated from each other that one half forms one daughter-nucleus, the other half the other. It is not merely that there is an exact division of the whole chromatin in the mass, which could have been effected much more simply, but that there is a regulated distribution of the different qualities of the chromatin, as we shall see later.
It must here be emphasized that the splitting of the chromosomes does not depend on external forces, but on internal ones involved in their organization, and in the definite attractions and repulsions of their component particles which come about in the course of growth. The chromosomes do not split like a trunk that has been broken open with an axe, but rather like a tree burst apart by the frost, that is, by the freezing of the water within itself. I consider it very important that we should recognize this, even though we do not yet know what the forces are that have control in this case, because it leads us to conclude that the structure of the chromosomes is extremely complex, that they are, so to speak, a world in themselves, that they possess an infinitely complex and delicate though invisible organization, in which intrinsic chemico-physical forces produce the regulated succession of changes which we observe. We shall afterwards see that we are led to the same conclusion from another direction—that is, from[Pg 293] the phenomena of inheritance. We shall then recognize that the rod- or loop-shaped chromosomes cannot be simple elements, but are composed of linear series of 10, 20, or more globular single-chromosomes, each of which represents a particular kind of chromatin or hereditary substance. If we consider this carefully, we shall see that it would hardly be possible to think out a mode of nuclear division which would so exactly and securely fulfil the purpose of conveying these many kinds of chromatin to the two daughter-nuclei in like proportions as does the mechanism of distribution actually brought about by nature. The longitudinal splitting of the rods halves the chromosomes, and the spindle apparatus secures the proper distribution of the halves between the two daughter-nuclei.
So much, at least, is certain, that no such complicated mechanism for 'mitotic' division would have arisen if the very precise division of a substance of the highest importance had not been concerned, and in this conclusion lies the first hint of the interpretation of the chromatin substance as the bearer of the hereditary qualities.
We are now familiar with the cell-nucleus and the apparatus for its division, and we are thus fully prepared to begin the study of the phenomena of 'fertilization.' Here also the processes depend essentially on the behaviour of the cell-nuclei, for even the first observations made by O. Hertwig on the behaviour of the spermatozoon after it has penetrated into the ovum led to the suggestion that the essential fact is the union of two nuclei; and numerous later, more and more deeply penetrating researches have furnished abundant evidence that the so-called 'fertilization' is essentially a nuclear fusion.
Let us begin with O. Hertwig's observations on the ovum of the sea-urchin. Eggs of this animal, which have been taken out of the ovary of the female, may easily be fertilized artificially by pouring over them spermatic fluid taken from a male, and diluted with sea-water. Before this is done only one nucleus can be observed in the ovum, but shortly afterwards two nucleus-like structures of unequal size can be seen within the ovum, and the smaller is surrounded by a circle of rays. Hertwig rightly interpreted this smaller nucleus as the modified remains of the penetrating spermatozoon, which then slowly approaches the nucleus of the egg, and ultimately fuses with it to form a 'segmentation nucleus.' From this starts the so-called 'segmentation' of the ovum, that is, the series of repeated divisions resulting in the formation of an ordered mass of cells, which by continued division of cells builds up the embryo.
Simple as this process of nuclear conjugation may seem, it was by no means so easy to recognize, and several investigators, especially[Pg 294] Auerbach, Schneider, and Bütschli, had seen stages of the process at an earlier date without arriving at the true interpretation of the phenomena. This was chiefly due to the fact that, in addition to the phenomena of fertilization proper, which we have briefly sketched, other nuclear changes take place in the maturing ovum, and these are not very easy to distinguish from the former; we refer to the phenomena of the so-called 'maturation of the ovum.' When the ovum-cell has attained its full size within the ovary it is not yet capable of being fertilized, but must first undergo two processes of division, to the right understanding of which Hertwig's investigations, and afterwards those of Fol, have contributed much.
For a long time it had been a familiar observation that small refractive corpuscles were extruded from one pole of the ovum shortly before the beginning of embryonic development. These were called 'polar bodies,' because it was believed that they marked the place which would afterwards be intersected by the first plane of division; it was only known at that time that they had to be extruded from the egg, but no one had the remotest idea of their real nature.
We now know that they are cells, and that their origin depends on a twice repeated division of the egg-cell; but it is a very unequal division, for these 'directive cells' or 'polar bodies' are always much smaller than the ovum, and indeed are usually so small that it is easy to understand why their cellular nature was for so long overlooked. Yet they have always a cell-body, and in many ova, for instance those of certain marine Nudibranchs, this is quite considerable; and they have likewise always a nucleus, which, notwithstanding the smallness of the cell-body, is in all cases exactly of the same size as the sister nucleus which remains behind in the ovum after division—a fact which is in itself enough to indicate that we have here to do essentially with readjustments and changes in the nucleus of the ovum.
Long before the polar or directive divisions were recognized as divisions of the egg-cell it was known that the nucleus of the ovum disappeared as soon as the latter attained to its full size within the ovary. It was also known that this nucleus—the large so-called 'germinal vesicle' lying in the middle of the ovum—left its central position and moved to the upper surface of the ovum, there to become paler and paler, and ultimately to disappear altogether from the sight of the observer. By many it was believed that it broke up, and that the 'segmentation nucleus,' which is afterwards obvious, is a new formation. The truth is that the germinal vesicle, at the time of its disappearance, is transformed into a division figure which is invisible[Pg 295] without the aid of artificial staining. The nuclear membrane breaks up; the centrosome of the ovum, which, although hardly visible, had previously lain beside the germinal vesicle, divides into two centrosomes and their centrospheres, and these now form the 'mitotic figure' by moving away from each other and sending out their protoplasmic rays. This nuclear spindle soon ranges itself at right angles to the surface of the egg, which at the same time arches itself into a protuberance, and soon two daughter-nuclei are formed, one of them lying within the protuberance (Fig. 75, A, Rk1). This soon separates itself off from the ovum, surrounded by a small quantity of cell-substance. The other daughter-nucleus remains within the ovum, but neither of them remains in a state of rest; both are again transformed into a spindle and divide once more; the minute first 'polar body' dividing into two 'secondary polar bodies' of half the size (B, Rk1), while the nuclear spindle within the egg brings about a second division of the ovum (B, Rk2) whose unequal products are the second polar cell and the mature ovum—that is, the ovum ready for fertilization. The process is now complete; the egg-cell, which has lost very little plasmic material through the 'polar bodies' and has not become visibly smaller, has now a nucleus (B, Eik) which has become considerably smaller through the two rapidly successive divisions, and, as we shall see later, has also undergone internal changes. In this state it is 'ripe,' that is, it is ready to enter into conjugation with the nucleus of a male cell, and this we have already recognized as the essential element in the process of fertilization.
These processes of 'maturation of the ovum' are common to all animal ova which require fertilization, and they follow almost the same course, only that in many cases the second division of the first polar body does not take place, so that only two polar bodies in all are formed. All these processes have nothing directly to do with fertilization, but it is only through them that the ovum becomes capable of fertilization. This does not prevent the spermatozoon from previously making its way into the ovum, for this is usually the case (Fig. 75, A, sp); there it waits until the second 'directive division' of the ovum has been accomplished, utilizing the time to become transformed in the manner necessary for the conjugation of the two nuclei. Only in a few species, for example in the sea-urchin, does the egg complete its polar divisions within the ovary, therefore before it has come into contact with the sperm at all.
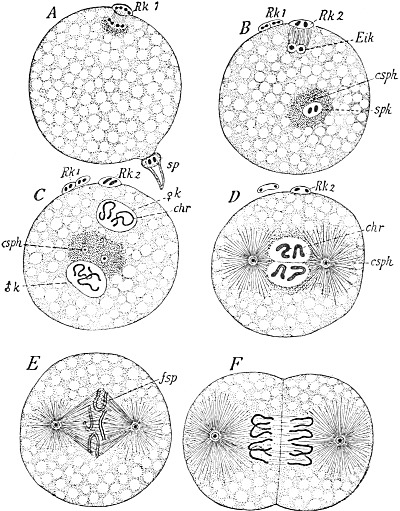
Fig. 75. Process of fertilization in Ascaris megalocephala, the thread-worm of the horse, adapted from Boveri and Van Beneden. A, ovum in process of the first directive division; Rk 1, first polar body; sp, spermatozoon with two chromosomes in its nucleus, attaching itself to the ovum, and about to penetrate into it; a protrusion of the egg-protoplasm is meeting it. B, the second directive division has been completed; Rk2, the second polar body; Eik, the reduced nucleus of the ovum. The first polar body (Rk 1) has divided into two daughter-cells, spk; the nucleus of the spermatozoon remains visible with its two centrospheres (csph). C, the sperm nucleus (♂k) and the ovum nucleus (♀k) have grown, each has two loop-like chromosomes; only the male nucleus has a centrosphere, which has already divided into two (csph). D, the two nuclei lie apposed between the poles of the nuclear spindle. E, the four chromosomes have split longitudinally; the spindle for the first division of the ovum (the segmentation spindle, fsp) has been formed. F, divergence of the daughter-chromosomes towards the two poles; division of the ovum into the first two cleavage cells or embryonic cells.
That we may be able to penetrate still more deeply into the processes of fertilization, the best illustration to take seems to me to be, as yet, the ovum of the thread-worm of the horse (Ascaris[Pg 296] megalocephala), which has become famous through the classical observations of Ed. van Beneden. Many favourable circumstances unite in this case to make the essentials of the process clearly recognizable. Fertilization takes place within the body of the female, in an[Pg 297] enlarged portion of the oviduct, within which a number of the remarkable sperm-cells are always found in a mature female. They are remarkable in being not thread-like, but rather spheroidal cells, bearing, however, a small protuberance something like a pointed horn (Fig. 75, A, sp). When such a sperm-cell comes in contact with the upper surface of an ovum a swelling forms at the place touched, and the sperm-cell attaches itself firmly to this, and is drawn by it into the ovum. Without doubt, amœboid movements on the part of the sperm-cell itself play some part in this, as can be most plainly seen in the large sperm-cells of many Daphnids which we have already discussed. In the egg of the thread-worm the whole sperm-cell with its nucleus can soon be detected within the substance of the ovum, and it then changes rapidly. Its main body fades more and more completely, until at last it disappears altogether, while the nucleus becomes vesicle-like and soon attains a considerable size (Fig. 75, B, spk). Meanwhile the residue of the germinal vesicle which remained behind in the ovum after the second directive division (B, Eik) has changed into a large vesicle-like nucleus (C, ♀ k), which in the ovum of Ascaris, as well as in the spermatozoon, at first contains a nuclear reticulum with irregular fragments of chromatin. Later on, these form a spiral coil in the manner we have already described, and finally this breaks up into two large and relatively thick angular loops or chromosomes (Fig. 75, C and D, chr).
At the same time a nuclear division apparatus has formed in the space between the two nuclei—the so-called male and female 'pronuclei' (♂ k, ♀ k)—two centrospheres (csph) become visible, at first lying close together, but afterwards moving apart (D) to form the poles of a nuclear spindle, in the equatorial plane of which the four chromosomes of the male and female pronuclei are now arranged. The nuclear membranes disappear, and the two nuclei now unite to form one, the segmentation nucleus (D). A dividing spindle then develops and brings about the first embryonic cell-division (E), and thus the beginning of the 'segmentation' of the ovum; each of the four chromatin loops splits longitudinally, and each of the split halves migrates, one to one, the other to the other daughter-nucleus (F). As this same method of distribution of the chromatin substance is repeated at every successive cell-division throughout embryogenesis, and indeed through the whole of development, it follows that the result of fertilization is, that all the cells of the body of the new animal which develops from the ovum contain an equal quantity of paternal and of maternal chromatin. If we are right in regarding the chromatin substance as the hereditary substance, it becomes immediately apparent[Pg 298] that this equal division is of the most far-reaching importance, for it shows us that the so-called process of fertilization is the union of equal quantities of hereditary substance of paternal and maternal origin.
The process of fertilization is now known in all its details in a great number of animals in the most diverse groups; it is everywhere the same in its essential features; there is always only one sperm-cell which normally enters into conjugation with the ovum-nucleus, and in every case the sperm-cell, however minute it may be to begin with, forms a nucleus nearly or exactly as large as the nucleus of the ovum, and in all cases it contains the same number of chromosomes as the nucleus of the ovum. Of special interest, however, is the fact that this number is always half the number of the chromosomes exhibited by the somatic cells of the particular animal in question, and that the reduction of the number of chromosomes to half the normal is effected in both male and female germ-cells by the last divisions of these cells, which take place before they have attained to a state of maturity. In the ovum the reduction occurs in the directive divisions, to which we must therefore turn our attention once more, with special reference to the number of chromosomes.
We saw that, in the full-grown ovarian egg, the germinal vesicle rises to the surface and there becomes transformed into the first polar spindle. Now this shows, in its equatorial plane, double the number of chromosomes normal to the species. This duplication comes about, not directly before the nuclear division, but much earlier in the young mother-egg-cell; it is only the change in the time of the splitting of the chromosomes that is unusual. The first maturation division takes place nevertheless in accordance with the usual plan of nuclear division; it is, as I have called it, an 'equation division,' that is, both daughter-nuclei again receive the same number of chromosomes as the young mother-egg-cell had to start with, namely, the normal number of the species. Thus, if the young mother-egg-cell had four chromosomes (Fig. 76, A), this number would double to eight at an early stage (B), but the first maturing division would give each daughter-nucleus four (C and D). In the second maturation division the case is different, for here no splitting and duplicating of the number of chromosomes takes place, but the existing number, by being distributed between the two daughter-nuclei, is reduced to half in each (E and F). For this reason I have called it a 'reducing division.' In our example, therefore, the ovum, as well as the second polar body, would contain only two chromosomes (Fig. 76, F).
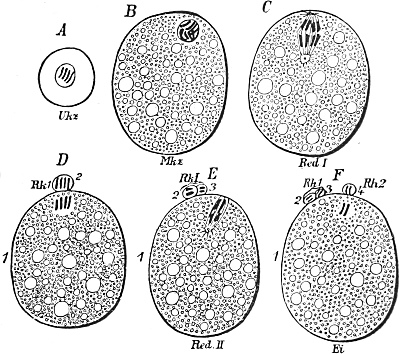
Fig. 76. Diagram of the maturation divisions of the ovum. A, primitive germ-cell. B, mother-egg-cell, which has grown and has doubled the number of its chromosomes. C, first maturation division. D, immediately thereafter; Rk1, the first directive cell or polar body. E, the second maturation spindle has been formed; the first polar body has divided into two (2 and 3); the four chromosomes remaining in the ovum lie in the second directive spindle. F, immediately after the second maturation division; 1, the mature ovum; 2, 3, and 4, the three polar cells, each of these four cells containing two chromosomes.
I cannot enter into the details of the process here, for we are[Pg 299] dealing with essentials and not with isolated and, so to speak, chance details, but I must emphasize the fact that the same process of reduction of the number of chromosomes takes place in this or an analogous manner in all animal ova, and can be demonstrated also in most of the chief groups of plants. Whether it be, as many have maintained, that the reduction is not always first effected by the 'maturation divisions,' but in some cases takes place earlier in the primitive egg-cell[12], so much is certain, that the nuclei which come together for 'fertilization' only contain half the normal number of chromosomes, and this is true not only of the ovum but also of the sperm-nucleus.
[12] See the discussion of this point in chapter xxii.
Arguing from general considerations, but especially from the theory which regards the chromosomes as the bearers of the hereditary substance, I had come to the conclusion, before there was any full[Pg 300] knowledge of the phenomena of the maturation of the ovum, that a reduction of the chromosomes by half must take place, and had postulated a similar 'reducing division' for the sperm-cell, and further, for plants as well as animals—indeed, for all sexually reproducing forms of life. The two divisions in the sperm-cell corresponding to the polar divisions of the ovum with their reduction of chromosomes were demonstrated by Oscar Hertwig in the case of the thread-worm of the horse (Ascaris megalocephala)—a form which has proved so very important in relation to the whole theory of fertilization. It is true that in this case the course of the phenomena of reduction is less convincing than in some other forms which have been investigated more recently, as, for instance, the mole-cricket and the bugs. In these instances, at any rate, a 'reducing division' in spermatogenesis, quite corresponding to that of the egg-cell, has been demonstrated, and this demonstration is of particular value owing to the fact that the development of the sperm-cell, as we shall presently see, throws an entirely new light on that of the ovum, and especially on the phyletic significance of the polar bodies.
We began our consideration of the processes of reduction with the full-grown egg-cell, but now let us go back to the earliest rudiments of the ovary of the embryo, and we find that it consists of a single primitive egg-cell, from which, by division, all the other egg-cells arise. In the same way the first rudiment in the testis or spermary is formed by a primitive sperm-cell, which does not differ visibly from the primitive egg-cell. Both now multiply by division for a considerable time, and in the ovary this is followed by a period of growth, during which multiplication ceases, and each cell increases considerably in size and lays in a store of yolk. Each cell thus ultimately reaches the condition with which we started previously, that of the full-grown mother-egg-cell.
Although the primitive sperm-cells do not exhibit such pronounced growth as the ova, they have likewise their period of growth, during which multiplication by division ceases, and the cells increase only in size (Fig. 77, A). When they have attained their maximum of growth the number of chromosomes is seen to have been doubled by longitudinal splitting (as in the diagram, Fig. 77, B, from four to eight). From this mother-sperm-cell there now arise by two divisions in rapid succession (C-F) four sperm-cells, and the same reduction of the number of chromosomes to half is effected as in the polar divisions of the egg-cell. In the first division, four chromosomes go to each daughter-cell (D), in the second, two (F). The only essential difference between the corresponding processes in the egg-cell and the sperm-[Pg 301]cell lies in the fact that the divisions of the so-called 'spermatocyte' or mother-sperm-cell are equal, so that four granddaughter-cells of equal size arise, while in the mother-egg-cell or 'ovocyte' the divisions are very unequal. In the former the result of the divisions is four cells capable of fertilizing, in the latter one cell capable of being fertilized and three minute 'polar cells' which are incapable of conjugating with a sperm-cell and giving rise to a new individual.
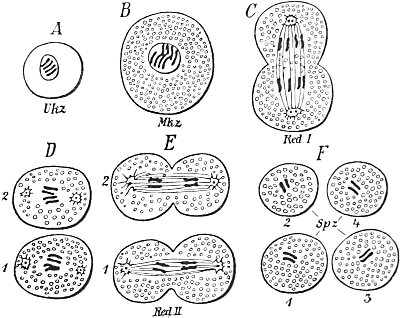
Fig. 77. Diagram of the maturation-divisions of the sperm-cell, adapted from O. Hertwig. A, primitive sperm-cell. B, mother-sperm-cell. C, first maturation division. D 1 and 2, the two daughter-cells. E, the second maturation division, by which the four cells of F arise, each with half the number of chromosomes.
There can thus be no doubt that the polar cells, as Mark and Bütschli long ago suggested, are abortive ova, that is, that, at a remote period in the evolution of animal life, each of these four descendants of a mother-egg-cell became a germ-cell capable of development. It is not difficult to infer that the unequal division, which now leads to an insufficient size in three of these descendants, has gone on pari passu with the continually increasing size of the mature ovum, and had its reason in the fact that it was important above all things to store in the ovum as much protoplasm and yolk as possible. We have already seen that even the dissolution of a number of the sister-cells of the ovum is sometimes demanded, so that the ovum may be surrounded by nutritive follicular cells. In short, the greatest possible quantity of nourishment is conveyed to the[Pg 302] ovum in every conceivable way, and it is thus stimulated to a growth which no single cell could attain to if it were dependent on the ordinary nutrition supplied by the blood. And we can understand that nature—to speak metaphorically—did not wish to destroy her own work by finally distributing among four ova all the nourishment she had succeeded in heaping up in all sorts of ways within the mother-egg-cell.
But it may be asked, Why have all these unnecessary divisions been maintained up till the present day? Why have they not long ago been given up, since they can and do only lead to the production of three abortive ova, which are foredoomed to perish? Are they mere vestiges, processes which are in themselves meaningless, but have, so to speak, been maintained by the principle of inertia? This principle is certainly operative in some sense and to some extent even in living nature; a process which has been regularly repeated through a long series of generations does not at once cease to be performed when it is no longer of use to the organism concerned. The eyes of animals which have migrated to lightless depths do not disappear all at once and leave no trace; they degenerate very gradually and only in the course of many generations; and it would thus be quite possible to defend the position that these polar or 'maturation divisions' of the ovum are purely phyletic reminiscences without actual significance.
But I cannot agree with this opinion. If it were actually so we should expect that the formation of the polar bodies would not still take place in all cases in almost the same manner, for all rudimentary parts and processes vary greatly; we should expect that in many animal groups the polar divisions would not occur, or perhaps that only half the number would occur. But this is not so; in all multicellular organisms, from the lowest to the highest, two reducing divisions take place, and always in almost the same manner, with the exception of a single category of ova, of which I shall presently have to speak. We shall see later that even in unicellular organisms analogous processes may be observed.
But it is also intelligible that this twice repeated division of the mother-egg-cell is necessary if the reduction in the number of chromosomes to half is only possible in this way, since this reduction is indispensable. If each of the two conjugating germ-cells contained the full normal number of chromosomes, the segmentation-nucleus would contain a double number, and if that went on, the number of chromosomes would increase in arithmetical proportion from generation to generation, and would soon become enormous. Even though we were not otherwise certain that these chromosomes are units of a[Pg 303] permanent nature, which only apparently break up in the nuclear reticulum, but in reality persist, the fact of reduction would point in this direction. For if they were not permanent structures and distinct from one another, and if their number depended solely on the quantity of chromatin which the nucleus contains, the reduction in number might be secured if the chromosomes in the growing egg and sperm-cells increased in size more slowly than the cell-body and the other parts of the cell. But from the fact that the reduction takes place not in this simple way, but, in sperm-cells and in ova which require to be fertilized, only through cell-division and a specific mode of nuclear division, we may conclude that it cannot happen otherwise, that chromosomes are not mere aggregates of organic substance, but organs whose number can only be reduced by the extrusion of some of them from the cell.
It is true that there are ova in which the process of reduction does not follow the course we have described, but the exceptions only serve to confirm our view of the reducing significance of the polar divisions, and of their persistence because of the necessity for reduction.
As far back as the middle of the nineteenth century it was known that in various animals the eggs develop without fertilization. This reproduction by 'parthenogenesis' was first established with certainty by the German bee-keeper Dzierzon in 1845, and then scientifically corroborated by Rudolph Leuckart and C. Th. von Siebold. Although parthenogenesis was at first observed only in a few groups of the animal kingdom, in bees and some nocturnal Lepidoptera (Psychidæ and Tineidæ), it has become more and more apparent in the course of years that this 'virgin reproduction' is by no means a rare form of reproduction, and that it occurs regularly and normally in many cases, especially in the very diverse groups of the great series of Arthropoda. Thus among insects it is found in certain saw-flies, gall-flies, ichneumon-flies, in the honey bee, and in common wasps, and it is particularly widespread among plant-lice (Aphides) such as the vine-aphis (Phylloxera), whose prodigious multiplication in a short time depends partly on the fact that all the generations, with the exception of one, consist only of females with a parthenogenetic mode of reproduction.
Among the lower Crustaceans also parthenogenesis plays a large rôle, and in many species it even occurs as the sole mode of reproduction, but more often—as is also the case among insects—it occurs alternately with bi-sexual reproduction. For parthenogenesis must not be regarded as asexual reproduction, but rather as unisexual, that is,[Pg 304] as arising from sexually differentiated individuals (females), and from germ-cells (true ova), but brought about by the agency of individuals of only one sex, the female. These parthenogenetic eggs emancipate themselves, so to speak, from the law that was previously regarded as without exception, that all ova require fertilization to enable them to develop. That this law admits of many exceptions is now universally admitted; thus in the small family of water-fleas (Daphnids) there are even two kinds of eggs, the summer and winter eggs we have already mentioned, which are produced by the same female, and yet the former kind develop without fertilization, while the latter require to be fertilized before they can develop.
It was obviously important to learn the state of affairs in regard to reducing divisions in parthenogenetic ova, to find out whether here also, three, or, in some circumstances, two polar bodies were formed, and whether the second polar division reduced the number of chromosomes to half. If the theory previously advanced as to the importance of the chromatin, and especially of the reducing effect of the second maturing division be correct, we should expect the second division to be wanting in parthenogenetic eggs, since otherwise the number of chromosomes would be reduced to half in each generation, and would thus gradually disappear or sink to one.
Having directed my attention to this problem, I succeeded in establishing for a Daphnid, Polyphemus, that the second polar division does not occur, and that only one polar body is formed. Blochmann found the same in the parthenogenetic eggs of plant-lice or Aphides, in which, moreover, the eggs requiring fertilization exhibit, like the winter eggs of Daphnids, two polar divisions. It was thus established that at least those eggs of Aphides and Daphnids which are wholly parthenogenetic retain the full number of chromosomes of their species, as is represented in the diagram, Fig. 78. When parthenogenesis set in the polar divisions were limited to one, and that this could happen justifies us in concluding a posteriori that it could have happened also in the case of ova which required fertilization if that had been necessary or even merely indifferent. The polar divisions are thus not mere 'vestigial' processes; they have an immediate significance, and it lies in the reduction of the number of chromosomes.
But I must make a reservation here; it is not universally true of parthenogenetic eggs that maturation takes place without the second polar division. The first exception was observed in the salt-water crustacean, Artemia salina. In this case only one polar body is actually extruded and the number of chromosomes remains normal,[Pg 305] as I was able to demonstrate with the small number of ova at my disposal; but according to the investigations of Brauer on more abundant material it appears that, while the second polar division is suppressed in the majority of the ova, and the external extrusion of a second polar body never occurs, the second polar division does nevertheless sometimes take place. The two daughter-nuclei arising from this division unite again immediately afterwards to form a single nucleus, and this now functions as a segmentation nucleus. Of course it again contains the full number of chromosomes, namely, twice 84=168.
In Artemia, therefore, the adaptation of the ova to parthenogenetic development is not yet fully established, and the complete abandonment of the second polar division seems to be phyletically striven for, since, although the division still takes place, its effect is neutralized immediately afterwards.
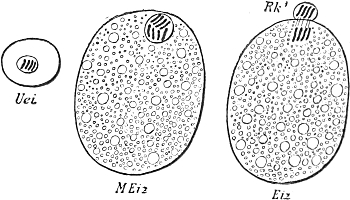
Fig. 78. Diagram of the maturation of a parthenogenetic ovum. The number of chromosomes normal to the species has been assumed to be four. Uei, a primitive germ-cell. M Eiz, a mother-egg-cell, with twice the normal number of chromosomes. Eiz, mature ovum after the separation of the first and only polar body. Rk1.
Among bees the state of affairs is again exceptional. Here the female, the so-called queen bee, possesses a capacious sperm-sac, in which the spermatozoa received in copulation remain living for years, and the fertilization of an ovum is effected in the usual way from this sac while the egg from the ovary is passing down the oviduct. The queen bee has the power of releasing some spermatozoa from the receptacle, or of not doing so, and thus of fertilizing the egg, or of not fertilizing it. Since the notable observations of Dzierzon and the investigations of von Siebold and Leuckart which followed them, it has been assumed that only those eggs were fertilized which were laid in the cells destined for rearing females (workers or queens), while those which were to give rise to 'drones' or males remained[Pg 306] normally unfertilized. Only in the last decade of the past century did the bee-keepers begin to cast doubt on this so-called 'Dzierzon theory'; various violent and obstinate attacks were made upon it, and these were supported by new and apparently convincing experiments. Dickel, a teacher in Darmstadt, has been particularly strenuous in attempting to overthrow the old theory, by emphasizing the fact that von Siebold's old investigations on bee eggs afforded no convincing proof. Von Siebold made his investigations on eggs freshly taken from the hive, and was never able to find spermatozoa in 'drone eggs' (that is, eggs laid in drone cells and therefore destined to develop into drones), while he was often able to demonstrate the presence of from one to four spermatozoa in 'worker eggs.' But he only examined 'drone eggs' which were already twelve hours old, and in these, as we now know, he would not have found spermatozoa in any case, even if they had been fertilized, because in ova at that stage the development of the embryo has already fully begun, and nothing remains of the spermatozoa. In the bee, according to Buttel-Reepen, the fertilizing spermatozoon is transformed in twenty minutes after it has penetrated into the egg into a minute 'sperm-nucleus' which is almost invisible even in sections, and certainly nothing whatever could be seen of it by the old method of squeezing the fresh ovum.
It had therefore to be admitted that Dzierzon's theory rested on an insecure foundation, and I accordingly set two of my students at that time, Dr. Paulcke and Dr. Petrunkewitsch, to examine the eggs of the bee anew with regard to the point in question, using the greatly improved methods at their disposal. These investigations have been carried out in the Freiburg Zoological Institute during the last three years, and have resulted in establishing the absolute correctness of Dzierzon's theory: the 'drone eggs' do remain unfertilized, while the eggs from which females are to develop are fertilized without exception.
In this case, therefore, we have, in the same animal, eggs which can be fertilized and eggs which, without fertilization, develop parthenogenetically, and it is therefore of the greatest possible interest to know the state of matters in them in regard to the directive divisions and the reduction of the chromosomes.
Dr. Petrunkewitsch's investigations have shown that in both cases, that is, whether a spermatozoon penetrates into the ovum or does not, a twice-repeated division of the nuclear material in the ovum takes place. Moreover, the two daughter-nuclei which result from the second division do not, as Brauer showed was sometimes the[Pg 307] case in Artemia, unite again afterwards; they remain separate, and the number of chromosomes—there are sixteen of them—is thereby reduced to half in the segmentation nucleus. But this is not all, for before embryonic development has begun the normal number can be again seen in the segmentation nucleus; the chromosomes must therefore have doubled their number by division within the nucleus.
It is probable that something similar takes place in the cases of exceptional parthenogenesis which have long been known, but this point has not yet been sufficiently investigated. Nevertheless I cannot pass them over, as they are instructive from another point of view.
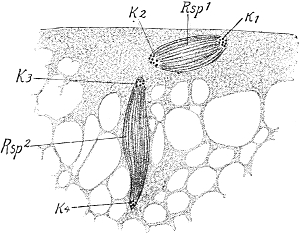
Fig. 79. The two maturation divisions of the
'drone eggs' (unfertilized eggs) of the Bee, after
Petrunkewitsch. Rsp 1, the first directive spindle.
k 1 and k 2, the two daughter-nuclei of the same.
Rsp 2, the second directive spindle. k 3 and k 4, the
two daughter-nuclei. In the next stage k 2 and k 3
unite to form the primitive sex-nucleus. Highly
magnified.
In some silk-moths (Bombycidæ) and hawk-moths (Sphingidæ), especially in the silk-moth proper (Bombyx mori), in Liparis dispar, and in quite a number of other Lepidoptera, it sometimes happens that, out of a large number of unfertilized eggs, a few will develop and produce caterpillars. This is interesting enough, but it gains increased importance through the investigations of the Russian naturalist, Tichomiroff, who succeeded in considerably increasing the number of unfertilized eggs that developed by gently rubbing them with a paint-brush, or by dipping them for a little in dilute sulphuric acid. It is thus possible to make eggs, which would not ordinarily develop without being fertilized, capable of parthenogenetic development by means of mechanical or chemical stimulus. This sounds almost incredible, but it is beyond a doubt, and it is still further corroborated by the fact that Prof. Jacques Loeb has succeeded in inciting the eggs of a sea-urchin to parthenogenetic development by means of a chemical stimulus. When he added to the sea-water in which the eggs were laid a certain quantity of chloride of magnesium the ova developed, and not only went through the process of segmentation, but even reached the stage of the quaint easel-like Pluteus larva. Quite recently Hans Winkler has made the interesting observation[Pg 308] that from sea-urchin sperms which have been killed by heat it is possible to extract in aqueous solution a substance capable of exciting unfertilized sea-urchin eggs to development, although they only go as far as to the sixteen-cell stage.
From all these results we can at least infer so much, that chemical changes and influences may determine whether the ripe ovum shall go on to embryonic development or not, and that these influences, may be very diverse in nature in different cases. I shall return later to these important facts.
When we sum up the facts we have cited with reference to the reduction of the number of chromosomes, it appears that nature is, as it were, striving to keep the number constant for each species; that in germ-cells which are destined for amphimixis they are reduced to half the normal number, but that this halving of the number is suppressed where fertilization is always absent, or that the reduction to half is compensated for again in various ways, whether by subsequent fusion of the two daughter-nuclei, which have arisen from the process of reduction, or by an independent duplicating of the chromosomes in the segmentation nucleus.
We might perhaps be inclined to conclude from all this that the occurrence of development depended on the presence of the normal number of chromosomes; and I used to regard this as possible. But facts which have been more recently brought to light have excluded this view. Above all, we now know that every nuclear division depends on the presence of a dividing apparatus, a centrosphere, but that this organ degenerates in the ova of most animals and is completely lost after the second polar division has been effected. The mature ovum is therefore in itself incapable of entering on its embryonic development, no matter how many chromosomes its nucleus contains; it is only capable of further division when the fertilizing sperm-cell brings with it its dividing apparatus or centrosphere. In thread-like sperms this lies in the median portion (Fig. 68C), and after the tail-piece has been dissolved, which happens soon after the sperm enters the egg, the central corpuscle, at first very small, can be recognized in front of the sperm-nucleus, where it is soon transformed into an 'aster' and divides into two. Then both spheres move apart (Fig. 75D, p. 296) and form the nuclear spindle between them by the confluence of their rays.
From this the division of the ovum into the two first embryonic cells proceeds. The two pronuclei in the ovum, the male and the female, are thus exactly alike as to number of the chromosomes, and frequently at least as to size and appearance (Fig. 75C). But they[Pg 309] differ in the possession or absence of a dividing apparatus, and in the great majority of cases it is the male nucleus that brings with it the central corpuscle which seems to be indispensable to embryonic development (B, cspt). Hitherto, at least, only two exceptions to this are known. In the little segmented worm, Myzostoma, which is parasitic on sea-lilies or Crinoids, Wheeler observed that the ovum retained its central corpuscle even after the polar divisions, while the sperm-cell which penetrated into the egg had none. More recently Conklin made the interesting discovery that in the egg of a marine Gasteropod (Crepidula) both the egg-nucleus and the sperm-nucleus retain their centrosphere and together form the segmentation spindle, one lying at one pole and the other at the opposite.
All these observations confirm the view that the sperm and the egg-cell are alike in this respect also. Each of them can, in certain circumstances, bring with it the dividing apparatus indispensable to development, though it is usually the sperm-cell that does so.
I should indeed assume that the sperm-cell and the egg-cell were essentially alike, even although there were no exception to this rule, that is, although the centrosome of the ovum perished in all eggs which were fertilized. For this is obviously a secondary arrangement, an adaptation to fertilization, that the ovum should be incapable of development without fertilization, and it is made so by the disappearance of its centrosome. In all other cells, as far as is known, the central corpuscle persists after division, so that this remarkable cell-organ is transmitted from cell to cell just like the nucleus, and like it, never rises de novo. It is only in the egg-cell that it disappears, though even there often very late, for it may be present, as an aster, even after the sperm has penetrated into the ovum and disclosed its own central body, or even brought it the length of dividing into two (Fig. 80, A and B). But the ovum-centrosome disappears as soon as the second polar division is accomplished.
That this disappearance is really a secondary arrangement, which may be again departed from, is proved by the case of those eggs which are able to develop parthenogenetically, for in them the central body does not disappear, but persists in the ovum after the first polar division, as Brauer showed in Artemia. It then behaves exactly like the sphere of the sperm-nucleus in the fertilized ovum, that is, it duplicates itself and forms the segmentation spindle.
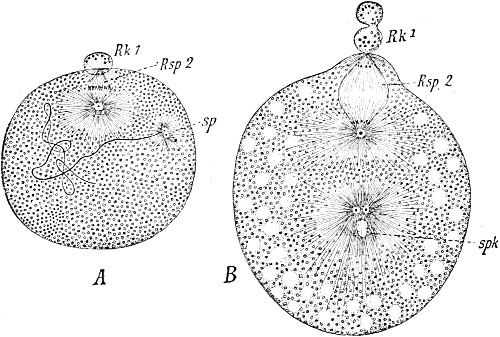
Fig. 80. Fertilization of the ovum of a Gasteropod (Physa), after Kostanecki and Wierzejski. A, the whole spermatozoon lies in the ovum. sp, its already divided centrosphere. Rk 1, the first polar body. Rsp 2, the second directive spindle. B, spk, the sperm-nucleus, the second directive spindle still has its centrosphere, which afterwards disappears. The first polar body (Rk 1) has divided into two. Highly magnified.
Thus the beginning of embryonic development in the ovum depends not on a definite number of chromosomes, but on the presence of an apparatus for division. Upon what the awakening of this to activity just at that time depends cannot as yet be exactly stated; we [Pg 310]can only indicate that all parts of the cell have interrelations with each other, and that, therefore, the division mechanism is dependent on the condition of the rest of the cell-parts at the moment, and on the substances which they contain or produce. From what we know experimentally in regard to artificial parthenogenesis it is not difficult to imagine that some sort of chemical substances are necessary to stimulate the central corpuscle to activity. In any case, the whole nutrition of the central corpuscle depends on the cell in which it lies, as is shown by the fact that the sperm-nucleus, whose centrosome before the entrance of the sperm into the ovum was inactive and scarcely recognizable, grows rapidly after entrance and forms a large aster round itself—is, in short, in the highest degree active (Fig. 80). As the chromosomes certainly play an important part in the life of the cell, and materially help to determine its various phases, it cannot be disputed that they also may share in awakening the activity of the central corpuscle. But this influence is only indirect; it is not the mere number of chromosomes that decides whether the central corpuscle is to become active or remain inactive. This cannot be assumed, because we have in the maturation divisions a proof that[Pg 311] division may take place with a double number of chromosomes as well as with the undoubled number; while in the divisions of the mother-egg-cells and the mother-sperm-cells we have proof that a doubled number of chromosomes does not in itself compel to division.
The exceptional and artificially produced cases of parthenogenesis which we have discussed above are probably to be interpreted thus: through slight differences in the constitution of the ovum, or through certain mechanical or chemical stimuli, the metabolic processes in the ovum are so altered that the centrosome of the ovum, instead of breaking up, is stimulated to growth, and thus produces the active dividing apparatus which is otherwise only brought into it by the sperm. This is a more exact definition of the interpretation I gave earlier (1891) of the 'chance' parthenogenesis of the silk-moth, which was then the only case known, when I said 'the nucleoplasm of some ova must possess the power of growth in a greater degree than the majority.'
But we are not yet in a position to go further, or to define more exactly the nature of the processes of metabolism which are involved.
FERTILIZATION IN PLANTS AND UNICELLULAR
ORGANISMS, AND ITS IMMEDIATE
SIGNIFICANCE
Fertilization in a lichen, Basidiobolus—In Phanerogams—Here too there is reduction of the number of chromosomes by a half—'Polar cells' in lower and higher plants—Conjugation among unicellular organisms—Noctiluca—The maternal and paternal chromosomes remain apart—Actinophrys—Infusoria—Sexual differentiation of the two conjugates in Vorticella—Importance of the process of Amphimixis—Not a 'life-awakening' process—May occur independently of multiplication—The Rejuvenescence hypothesis—Pure parthenogenesis—The cycle idea—Does Amphimixis prevent natural death?—Maupas' experiments with Infusorians—Bütschli's view—Potential immortality of unicellular organisms—The immortality of unicellular organisms and of the germ-cells depends on the fact that there is no time-limit to the multiplication of the smallest living particles—Parthenogenesis is not self-fertilization—Petrunkewitsch's observations on the ova of bees—Is the chromatin really the 'hereditary substance'?—Nägeli's conclusion from the difference in size between ovum and spermatozoon—Artificial division of Infusorians—Boveri's experiments with the fertilization of pieces of ova not containing a nucleus—Fertilization gives an impulse to development even to non-nucleated pieces of ova—Merogony—The female and male nuclear substances are essentially alike—Summary.
I now turn to the consideration of the process of fertilization in plants and unicellular organisms.
With regard to plants, it can now be definitely asserted that in them, too, fertilization is essentially a conjugation of nuclei; it depends on the union of the nuclei of the two 'sex-cells.' These sex-cells are usually very small among lower plants, indeed up to the phanerogams; this is especially true of the zoosperm-like male germ-cells, but it usually holds also true of the ovum, which is but seldom burdened with an abundant supply of yolk. In spite of the many difficulties which this smallness of size puts in the way of observation, the untiring exertions of a host of excellent investigators have succeeded in following the process of fertilization in all the larger groups of plants—in algæ, fungi, mosses, ferns, and horse-tails among cryptogams, and in phanerogams.
I shall first give an example from among the lower plants (Fig. 81). In one of the lichens, Basidiobolus ranarum, each of two adjacent cells in the fungus-thread gives off a bill-like process, and the[Pg 313] two processes become closely apposed (Fig. 81, a). The nucleus of each cell moves into the bill-shaped process, is there transformed into a nuclear spindle (B, ksp) and divides, so that one daughter-nucleus comes to lie in the apex point of the bill, the other at the base. The cell-body also divides, though very unequally, and the final outcome of the process is two cells in each, of which one is small and occupies the apex of the bill, while the other is large and fills all the rest of the cell-space. The former do not play any further part of importance, but break up, the latter are the sex-cells, the cytoplasm of which now coalesces through a gap in the cell-walls, while their nuclei become closely apposed and ultimately unite (C, ♂ and ♀ k). From this union arises the fertilized spore, the so-called 'zygote' (D). The two small abortive cells so greatly resemble in their origin the polar cells of the animal ovum that it is difficult to resist the supposition that they bring about a reduction in the number of chromosomes. But the number of the chromosomes has not yet been determined either in them or in the sex-nuclei.
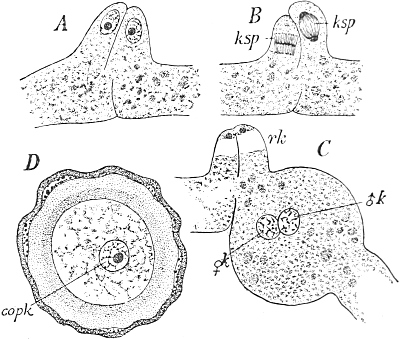
Fig. 81. Formation of polar bodies in a lichen, Basidiobolus ranarum. A, the two conjugating cells with the bill-like processes in which the nuclei lie. B, the nuclei dividing. ksp, the nuclear spindle. C, after the division into a polar body (rk) and a sex-nucleus (♂ k and ♀ k). D, after the union of the nuclei to form a conjugation nucleus (copk); the fertilized ovum is surrounded by envelopes and modified into a lasting spore. After Fairchild.
We have come to know the processes of fertilization among phanerogams chiefly through Strasburger, Guignard, and more recently through the Japanese botanist Hirase. The agreement with the animal process is surprisingly great, notwithstanding the notable differences in the external conditions of fertilization.
As is well known, the male cells in the highest flowering plants are not zoosperms but roundish cells, each of which, enclosed, together with a sister-cell—the so-called 'vegetative' cell—in a thick cellulose capsule constitutes a pollen-grain. The pollen-grains reach the stigma, under which, buried deep within the 'ovule,' the female sex-cell rests, enclosed in a long, sac-like structure called the 'embryo-sac' (Fig. 82, A). Beside it (eiz) there lie several other cells, usually seven in number, two of which, the so-called 'synergidæ' (sy), have their place at one end of the embryo-sac, just in front of the ovum (eiz). Probably these give off a secretion which exercises an attractive (chemotactic) influence on the male fertilizing body ('the pollen-tube'), and thus, so to speak, show it the way to the ovum.
Fig. 82. Fertilization in the Lily, Lilium martagon, after Guignard. A, the embryo-sac before fertilization; sy, synergidæ; eiz, ovum; op and up, upper and lower 'polar nuclei'; ap, antipodal cells. B, the upper part of the embryo-sac, into which the pollen-tube (pschl) has penetrated with the male sex-nucleus (♂k) and its centrosphere; below that is the ovum with its (also doubled) centrosphere (csph). C, remains of the pollen-tube (pschl); the two sex-nuclei are closely apposed. Highly magnified.
When a pollen-grain has reached the stigma it sends out a tube, usually after a few hours, which penetrates into the soft tissue of the style, and grows deep down into the interior of the ovule, ultimately penetrating as far as the embryo-sac through a special little opening in the covering of the ovule, the so-called 'micropyle' (Fig. 82B, pschl). Its blunt end is now closely apposed to this, so that the true sperm-nucleus (B, ♂k), surrounded by some protoplasm, can leave the pollen-tube and wander in among the cells of the embryo-sac. Later on we shall see that two generative nuclei migrate from the pollen-tube, but in the meantime we shall devote our attention only to one[Pg 315] of them, the fertilizing nucleus, which immediately moves towards the ovum-nucleus and apposes itself closely to it. Then follows the fusion or conjugation of the two nuclei, which are alike in size and appearance, just as in the fertilization of the animal ovum (C, ♂ k and ♀ k). Whether in this case, too, the sperm-nucleus brings with it a central corpuscle, or whether, as Guignard believed he observed, the ovum retains its central corpuscle (C, csph), or finally, whether both modes occur, is not yet known with certainty. The fact that, as a rule, seeds capable of reproduction only form in an ovule when the stigma has been previously dusted with pollen, leads us to suppose that, in this case, as among animals, the ovum lacks something that is necessary to induce embryonic development, only retaining this power in very exceptional cases, namely, when adapted for parthenogenesis. And this something may very well be the dividing apparatus of the cell, the centrosome with the centrosphere. But whether this supposition prove correct or not, a nuclear spindle always forms simultaneously with the fusion of the two sex-nuclei into a segmentation nucleus, and this spindle is the starting-point of the young plant, thus exactly corresponding to the first segmentation of the animal ovum. It agrees with it also in the important respect that it again contains the full number of chromosomes—twenty-four in the lily—while the two nuclei, male and female, only exhibit half the number each, that is, twelve.
Thus a reduction in the number of chromosomes to half takes place in plants also, but it is not yet known with certainty whether this is brought about in the same way as among animals, namely, by reducing divisions. Without entering more fully into this still unsolved and very complex problem, I should like to state that I consider this very probable; indeed, I agree with the view of V. Häcker[13], that the reducing divisions of plants are only more difficult to recognize as such, and, furthermore, are often disguised by the fact that they often occur alongside of, or between divisions which are not reducing. If it were possible to reduce the number of chromosomes in a cell to half without the aid of cell-division, if, for instance, only half were to integrate again from the chromatin-network, this must have been quite as possible in the case of animal cells, and then, moreover, the single chromosome would not have had the significance of an individuality, and no special form of nuclear division would have been introduced to reduce their number. That it has been introduced seems to me to prove that it was necessary, and since it[Pg 316] was so among animals, it could not have been dispensed with among plants either.
[13] See V. Häcker, Praxis und Theorie der Zellen- und Befruchtungslehre, Jena, 1899, pp. 144-5.
Moreover, throughout the vegetable kingdom divisions often occur in connexion with the origin of the sex-cells which can be compared, in occurrence and result, with the maturation divisions of animal germ-cells. In the lichen, Basidiobolus, we have already seen that an abortive cell separates itself off from the sex-cell before the latter becomes capable of reproduction (Fig. 81, C). Similar cell-divisions occur in many if not in all groups of plants. In the marine algæ of the genus Fucus it has even been proved that the division of the first primordial cell of the ovary into the so-called 'stalk-cell' and the primitive egg-cell is a reducing division, and brings down the number of chromosomes from thirty-two to sixteen. In vascular plants the reduction is not postponed until the formation of the sex-cells, but occurs earlier in the formation of the spores, as Calkins has demonstrated for ferns; in the Conifers and other Gymnosperms several so-called 'preparatory' divisions precede the formation of the germ-cells, and we know by comparison with the alternation of generations in vascular plants that these are related to the gradual waning of the strictly sexual generation. As the 'polar bodies' or 'directive corpuscles' of the animal ovum are rudimentary egg-cells, so the cells which, in the pollen-grains, separate themselves from the sex-cells proper are rudimentary Prothallium-cells, and, like the animal cells, they perish without playing any further physiological rôle. I will not assert that it is precisely in these divisions that the reducing divisions are concealed, for the analogy with the spore-formation of ferns leads us rather to suppose that it may lie further back; but in any case there is no lack of opportunity in the ontogeny of phanerogamic plants for the interpolation of a reducing division, and as long as it remains unproved that a reduction of the chromosomes can take place directly, that is, without the help of nuclear division, we shall continue to expect with confidence that the reducing divisions of phanerogams will be discovered in the future. Processes of a similar kind are known among unicellular organisms, and there, too, they are associated with nuclear divisions.
In passing to the so-called 'sexual reproduction' of unicellular organisms, I should like first to call attention to the fact that the expression 'reproduction' is not very suitable in this case, for the process in question does not always effect an increase in the number of individuals as reproduction ought to do, but leads, in fact, in many cases, even to a decrease, when two individuals unite to form one. Even if the phenomena of sexual 'reproduction' among higher[Pg 317] organisms, which we have already studied, had not made it clear to us that there are two associated processes, quite different in nature, the conjugation of unicellular organisms would have led us to that conclusion. It has long been known that two unicellular plants or animals occasionally become closely apposed and fuse; and this process of 'conjugation' was many years ago regarded as an analogue to 'fertilization,' although it is only through the laborious investigations of the last two or three decades that this supposition has been proved to be correct. We now know that a process quite analogous to that which we have learnt to know as 'fertilization' takes place among unicellulars, only in this case it is not directly connected with reproduction and multiplication, but occurs independently of them, and, in its most primitive form, it results, not in an increase but—for a short time at least—in a diminution of the number of individuals. This occurrence of the process independently of reproduction appears to me of inestimable value theoretically, for it frees us completely from the old deep-rooted preconceptions in the interpretation of fertilization.
Fig. 83. Conjugation of Noctiluca, after Ischikawa.
A, two Noctilucas beginning to coalesce; pr, the protoplasm
drawn out into processes which traverse the
gelatinous substance of the cell; k, the nucleus.
B, the cells and their gelatinous substance have fused;
the nuclei, in which the chromosomes are visible, are
closely apposed; CK, centrospheres. C, the two nuclei
are united in one nuclear spindle; beginning of
division. D, completion of the division. Highly
magnified.
First let us briefly sketch the process itself in the main forms of its occurrence.
The most primitive form of conjugation is undoubtedly the complete fusion of two unicellular organisms of the same species, as we see it to-day in unicellular plants, and also among the lowest unicellular animals, such as the flagellate Infusorians, Gregarines, and Rhizopods. It is well seen, for instance, in the Noctilucæ, those unicellular flagellate organisms which cause the familiar marine phosphorescence extending uniformly over wide surfaces of water (Fig. 83). In these forms Prof. Ischikawa of Tokio was able to trace the whole process of conjugation. To begin with, two Noctilucas range them[Pg 318]selves side by side (Fig. 83) and coalesce at the surfaces in contact, both as to the spherical gelatinous envelope (A, G) and the protoplasm (pr) itself, which branches in amœboid fashion into the jelly. The union becomes gradually complete, and the two animals form a single sphere (B) with one cell-body. But the two nuclei (K) also place themselves side by side (B), and though they do not actually fuse, they form together, under the guidance of two centrospheres (C), a single nuclear division-figure, which is obviously analogous to the segmentation spindle of the fertilized egg. Then follows a division, by means of which the chromatin substance of the nuclei of both animals is divided between the two daughter-nuclei, and after this has been accomplished the united individual again separates into two independent Noctilucas (D). Although I have spoken here—that is, in referring to the Protozoa—of chromosomes, I must immediately add that these have not yet been seen with full clearness in Noctiluca itself; nothing more has been recognized than deeply staining thickenings of the spindle fibrils, which move from the equator of the nuclear spindle towards the pole. Since, however, in other Protozoa, as, for instance, in the beautiful freshwater Rhizopod (Euglypha alveolata), these thickenings of the nuclear spindle fibrils have been clearly recognized as chromosomes, doubt on this point is hardly justifiable. Apart from this, the assumption that each of the two daughter-nuclei receives half the chromosomes of each of the conjugated nuclei rests on a secure basis, not only because otherwise the whole process would have no meaning, but because the position of the mitotic figure conditions this. Even the fact that the two conjugation-nuclei lying side by side remain apart during nuclear division is not without parallel; Häcker and Rückert observed it also in the segmentation-nucleus of much higher animals, the Copepods, and it has no effect in altering the process of division, but only proves that the chromosomes of maternal and those of paternal origin in the combination-nucleus remain independent—a fact the significance of which I shall discuss later on.
The process of conjugation occurs, in the same manner as in Noctiluca, in a freshwater Rhizopod, the well-known Sun-animalcule, Actinophrys sol (Fig. 84), but in this case complete fusion of the two nuclei takes place (Fig. 84, V) before the formation of the division-spindle (VI, sp), which, with the simultaneous division of the cell-body, gives rise to two new individuals. The process in this case is especially interesting, because Schaudinn has succeeded in observing a maturation division (III, Rsp, directive spindle) as well as in demonstrating polar bodies (IV, Rk). Thus the analogy with the[Pg 319] process of fertilization in the Metazoa and the Metaphyta is almost complete.
But that the conjugation of unicellular organisms, like the fertilization of multicellular organisms, is essentially a matter of nuclear conjugation is shown more distinctly still by the ciliated Infusorians, the most highly organized of the Protozoa.
Fig. 84. Conjugation and polar body formation in the Sun-animalcule, Actinophrys sol, after Schaudinn. I, two free-swimming conjugated individuals, which in II have become surrounded by a transparent gelatinous cyst. III, formation of the directive spindles (RSp). IV, the polar bodies are formed (RK); K, the two sex-nuclei. V, these are fused to form the conjugation-nucleus (K). VI, the conjugation-nucleus is transformed into the division-spindle; the polar bodies (RK) have penetrated the internal cyst-wall, and are in process of degeneration.
Here there is usually no complete union of the cell-bodies of the two animals, but only an adhering of the apposed surfaces. In the relatively large Paramœcium caudatum the process of conjugation is very exactly known through the beautiful investigations of Maupas and R. Hertwig. In this case the mouth-surfaces of the two animals come together and unite over a short area, and then the two animals swim about together in this conjugated state. During this time very remarkable changes take place in their nuclei.
It is well known that these Infusorians have a double nucleus, a large one, the macronucleus (Fig. 85, ma), and one which is usually very small, the micronucleus (mi). We may ascribe to the former of these the guidance and regulation of the everyday processes of life, that is, briefly, of metabolism, and the preservation of the integrity of the whole animal. The small nucleus has often been designated the 'reproductive nucleus,' but as it plays no other part in repro[Pg 320]duction, as far as can be recognized, than that of dividing into two daughter-nuclei, I cannot regard this designation as suitable; it obviously originated in the mistaken interpretation, prevalent till very lately, of conjugation as a 'kind of reproduction,' and this in its turn depends on the conception, transferred from multicellular organisms, of fertilization as a 'sexual reproduction.' We shall immediately see that the micronucleus plays the main part in conjugation, and from this we may suppose that it otherwise fills no rôle in the life of the animal, and therefore it may best be designated the 'supplementary' or reserve nucleus. In every conjugation the macronucleus, which has hitherto been active, breaks up and becomes completely absorbed, very much like a ball of food. This of course takes place slowly; the large nucleus elongates, becomes indented, falls into several pieces, and these are so gradually absorbed that, even after the act of conjugation has been accomplished, irregular fragments of the macronucleus often lie about in the animal (Fig. 85, 9).
But while the macronucleus falls to pieces the previously minute micronucleus grows enormously and forms a distinct longitudinally striated spindle (1, mi). About the same time these divide in both animals, and each of the daughter-nuclei immediately divides again, so that after these two divisions four spindle-shaped descendants of the micronucleus are to be seen in each animal (Fig. 85, 4). We have previously noted that the apparatus for nuclear division in unicellular organisms was similar to that in multicellular organisms, and yet was different from it. In these ciliated infusorians we see an essential difference, for the striated spindle, after the division into daughter-chromosomes has taken place, lengthens out enormously, and becomes so thin in the middle of its length (2) that the two daughter-nuclei at the ends of this long stalk suggest the appearance of a very long and thin dumb-bell, or of a long silk purse. Of asters (centrospheres) there is nothing to be seen, and the mechanism of division is still very obscure; it almost seems as if a rapidly growing substance forced the two groups of chromosomes apart.
Hardly have these four descendants of the micronucleus arisen
when three of them begin to break up and very shortly disappear;
only the fourth is of any further importance, and it divides once
more (5), and so gives rise to the two nuclei which play the chief part
in the process of conjugation—the copulation-nuclei, exactly analogous
to the male and female pronuclei in the fertilized ovum (5, mi4).
But in this case each of the two animals functions doubly, that is,
both as male and female, for each sends one of the two copulation-nuclei
[Pg 321]
[Pg 322]across the bridge formed by the union of the apposed surfaces
into the other animal (6, mi ♂), so that it may form, by union with
the nucleus which has remained there, a double nucleus (7), a structure
which corresponds to the segmentation nucleus of the ovum (copk).
From it there then arises by division a new macronucleus and a new
micronucleus, not usually directly, however, that is, not by a single
division, but through several successive nuclear divisions, into the
meaning of which I cannot here enter. Immediately after the union
of the two sex-nuclei the two animals sever their connexion with each
other; each begins again to feed, and is subject to multiplication by
division just as it was before conjugation took place (8 and 9).
Fig. 85. Diagram of the conjugation of an Infusorian, Paramœcium, after R. Hertwig and Maupas. 1, two animals with the mouth-openings apposed; ma, the macronucleus beginning to degenerate; mi1, the micronucleus has already increased considerably in size and is beginning to divide. 2. each micronucleus has divided into two daughter-nuclei (mi2), which are connected only by the division-strand (ts). 3, to the left each of the daughter-micronuclei (mi2) is beginning to divide; to the right this division is already completed and the grand-daughter-nuclei of the original micronucleus hang together by their division-strands (ts). 4, in each of the animals there are now four grand-daughter-micronuclei (mi3). 5, three of these are in process of dissolution, the fourth is dividing into two great-grand-daughter-nuclei (mi4), which are the two sex-nuclei. 6, one (the male) sex-nucleus (mi ♂) migrates into the other animal, and there unites with the remaining (female) sex-nucleus. 7, the conjugation-nucleus (copk) being formed. 8, the animals have separated; the conjugation-nucleus divides into (9) the new macronucleus (n ma) and the new micronucleus (n mi).
Although the course of this remarkable process exhibits all manner of differences in detail in different species, it is everywhere the same in its essential feature, and this essential feature is undoubtedly the union of an equal quantity of the nuclear substance of two animals to form a new nucleus. It is thus essentially the same process which we have already recognized among higher animals as 'fertilization.' The differences are of minor importance, and they arise partly from the fact that the sex-cells of multicellular animals are not independent self-supporting units, and partly from their differentiation into 'male' and 'female' cells. The minuteness of the sperm-cell, for instance, conditions its penetration of the ovum, which is always much larger and passive, and also the thorough fusion of its cell-body with the cell-body of the ovum. That this difference has very little deep significance is best seen from the fact that, even among Infusorians, there are forms in which the two conjugating individuals are quite different, especially in size, and in which the much smaller 'male' animal fuses completely with the much larger 'female,' and indeed bores its way into it after the manner of a sperm-cell. This is the case among the bell-animalcules (Vorticellinæ) (Fig. 86), the conjugating pairs of which had been observed long before our present insight into these processes had been attained. Indeed, the facts had been interpreted as a kind of 'budding process,' the minute and differently shaped 'male' animal (mi), which at the time of conjugation is attached to the larger 'female' (ma), was regarded as its bud. This supposed bud, however, does not grow out from the animal, but into it!
Thus we see here again that a differentiation of individuals as males and females may occur among unicellular organisms, just as in the sex-cells of higher animals and plants, and this proves to us once more that all these differences of sex, whether in reproductive cells of multicellular organisms, or in the entire multicellular animal or plant,[Pg 323] or finally, in unicellular organisms, are not of essential, but only of secondary significance, however important they may be for securing fertilization or conjugation in each special case. They are always only adaptations to the special conditions, and only occur where they are necessary to ensure the union, and always in such a manner that the union of the two cells is facilitated. In most Infusorians such a differentiation into male and female animals was not necessary, because these organisms are very motile, and are thus readily able to meet and unite; it was therefore sufficient for them to remain hermaphrodite. The bell-animalcules, however, are sedentary, and for them it was obviously an advantage that, at the time of conjugation, smaller, free-swimming, and also more simply organized individuals should arise, which were able to seek out the larger sedentary forms. Here, then, as in many other unicellular animals, these little male individuals only occur when they are necessary, that is, at the time of conjugation. Similarly, in the green alga, Volvox, male and female cells arise only at the time of conjugation, reproduction being at other times effected by means of parthenogonidia, that is, by elements which require no fertilization.
Fig. 86. Conjugation of an Infusorian. Vorticella nebulifera, showing sexual differentiation of the whole organism. After Greef. I, the 'microgonidium' or male individual (mi) attaches itself to the 'macrogonidium' or female individual (ma); cv, contractile vacuole; st, contractile stalk. II, the ciliated circle on the male individual has disappeared. The male has become firmly embedded in the female by means of a sucker-like retraction of its lower end. III, the fusion of the two individuals has been completed; the bristly residue of the male (ct) is about to be thrown off; the stalk (st) is contracted into a spiral. Magnified about 300 times.
As these differences are only adaptations to the necessity that the animals or cells shall find each other and unite, so also are all the other differences of a sexual kind, the thousand-fold differences between the sperm-cell and the egg-cell, and the not less numerous differences between male and female animals, both in 'primary' and especially in the diverse 'secondary' sexual characters which we have[Pg 324] previously discussed; all these are only means for bringing about the process of the union of two germ-cells to form a fertilized 'ovum' which is capable of development. The essential part of this so-called 'sexual reproduction' does not, however, depend on these differences, neither on the sexual differences of the germ-cells nor on those of the whole organism; it lies solely in the actual union of the two germ-cells. Let us remember the idea we have already emphasized, that the essential part of the so-called 'sexual reproduction' does not depend on these differences, and let us hold fast to the idea already indicated, that the chromosomes of the nucleus are the real bearers of the hereditary tendencies; then we see that the mingling, or, better, the union of the hereditary substances of two different individuals, whether single-celled or many-celled, is the result of the process which we have hitherto called fertilization or conjugation, but which we shall henceforward designate by the more general term 'Amphimixis' which means the mingling of substances contributed from two distinct sources.
Having made ourselves acquainted with the phenomena of amphimixis in animals, plants, and unicellular organisms, we have to face the problem of the significance of this remarkable and complicated process. What is it that happens, and what meaning can we attach to it?
The first thing to be done is to show that the old and long-prevailing conception of fertilization as a life-awakening process must be entirely abandoned. That a new individual can arise even among highly organized animals, quite independently of fertilization, is proved by the parthenogenetic eggs of insects and crustaceans; fertilization is not the spark 'which falls into the powder-cask' and causes the explosion; it is only an indispensable condition of development. As we have seen, there are germ-cells which are not sexually differentiated, such as the spores of the lower plants, which are all capable of development without amphimixis; and parthenogenetic ova prove that even differentiated female germ-cells, that is, germ-cells originally adapted for amphimixis, may in certain circumstances develop without it; amphimixis is thus not the fundamental cause of development, but is only, for many germ-cells, one of the conditions which must be fulfilled before development can set in. It is a condition which, under certain circumstances, may be dispensed with.
If, then, the multiplication of individuals by single-celled germs can take place independently of amphimixis, we may conclude that the establishment of amphimixis has nothing to do with the capacity[Pg 325] for multiplication, that it is not a life-awakening process, but is a process of a unique kind, which means something quite different. The whole conception of the awakening of life in the germ is antiquated and out of harmony with the present state of our knowledge. Life never begins anew, as far as we can see, and apart from the possibility that, unknown to us, a spontaneous generation (Urzeugung) of the lowest forms of life is still taking place, life is continuous and consists of an infinite series of living forms between which there is no real interruption. Life, in fact, is like a continuous stream, the larger and smaller waves of which are particular species and individuals. Only a few decennia ago a morphologist, who was rightly held in high esteem, could champion the idea that the mature ovum of animals was lifeless material, which had to be quickened in order to develop, but now such a theory is untenable, since we have become aware of the phenomena of maturation in the ovum, and know that most important vital processes, the reducing divisions, take place at the time of maturation, quite independently of fertilization.
Thus we do not even require to take into account the conjugation of unicellular organisms to make it clear that amphimixis is not the cause of the origin of new individuals, but a process, sui generis, which may indeed be associated with the beginning of embryonic development, but which may also occur independently of it, as we see in the case of unicellular organisms. If, on the one hand, we see development taking place in spores and parthenogenetic ova independently of amphimixis, and on the other hand amphimixis occurring without reproduction in unicellular organisms, we must regard the two phenomena, amphimixis and reproduction, as processes of a distinct kind, which may, however, occur in association with and interdependence upon each other.
It was by chance that human observation brought the latter fact to light first, and therefore led us for so long to accept the idea that fertilization, that is, amphimixis, and development, that is, reproduction, are one and the same; and thus it happens that even now there are many naturalists who cannot rid themselves of the idea that amphimixis, if not a life-awakening, is at least a life-renewing process, a so-called 'process of rejuvenescence.'
More than ten years ago[14] I disputed this view, and since then the facts which make it untenable have become more and more clear. Notwithstanding this I see that it is still adhered to, at least in a[Pg 326] modified form, by many esteemed naturalists, and so it does not seem superfluous to discuss it in more detail.
[14] Die Bedeutung der sexuellen Fortpflanzung für die Selektionstheorie, Jena, 1886.
I have already noted that we see in conjugation an amphimixis without reproduction, and in spores and parthenogenetic ova reproduction without amphimixis, and I do not doubt that every unprejudiced critic will admit this; many among us, however, are not unprejudiced, but are under the spell of earlier ideas, so that they cannot forget that it was long believed that fertilization was an indispensable condition of development; they therefore regard the divisions which recommence sooner or later after conjugation, and which may be repeated hundreds of times, as conditioned by the conjugation which preceded them, and compare them to the series of cells which, in the Metazoa, lead from the fertilized ovum to the fully-formed animal. They regard both series of cell-generations as a developmental cycle, which leads from fertilization to fertilization again, from conjugation to conjugation, and which would be impossible without either fertilization or conjugation.
This play with the idea of a 'cycle' reminds me vividly of similar fantastic play from the time of the much-despised 'Naturphilosophie' of a hundred years ago. As men sought to find the analogues of 'solar' and 'planetary' systems in animal and plant, and believed they had stated something when they compared the motile animals to planets and the sedentary plants to the sun (!), so it is now imagined that a deeper insight has been gained by the recognition of cycles of development. By all means let us regard the development of a multicellular organism as cyclic; it returns again to its starting-point, but this no more explains the forces which produce the cycle, and thus the meaning of fertilization, than a comparison with the circling planets explains the causes of locomotion in animals. With quite as much reason the cycle of development might be made to start from the parthenogenetic ovum, and then the whole conclusion of the fanciful cycle idea in regard to the meaning of fertilization falls to the ground, for in this case the cycle begins without fertilization. Attempts are made to get over this difficulty by showing that in many cases parthenogenesis alternates regularly or irregularly with sexual reproduction, as in the water-fleas (Daphnids), the Aphides, and so on. The mysterious rejuvenating power of amphimixis is supposed to suffice for several generations, a purely gratuitous assumption, which is also in open contradiction to the facts. For there are species which now reproduce exclusively by parthenogenesis, among plants for instance, a number of fungi, among animals a few species of Crustaceans. Of the latter it can be demonstrated that ages ago they[Pg 327] reproduced sexually, for they still possess the sac which serves for receiving spermatozoa, but this sac remains empty, for there are now no males, at least in any habitat of the species known to us. To this set belongs an inhabitant of stagnant water, Limnadia hermanni, a species of Crustacean which was found thirty years ago in hundreds, all of the female sex, near Strassburg, and also many of the little Ostracods (Cypris) which inhabit especially the muddy bottom of our pools and marshes. I bred one of these (Cypris reptans) in numerous aquaria for sixteen years, during which there were about eighty generations, and throughout this time no male ever appeared, nor did the sperm-sac of the female ever contain spermatozoa. The after-effects of the 'rejuvenating' power of an amphimixis supposed to have taken place earlier must in this case have been enduring indeed!
For these reasons it seems to me useless to make comparisons between the developmental cycle of unicellular organisms and the ontogeny of multicellular organisms. Both processes have indeed many points of resemblance—long series of cells, then interruption of the divisions and the occurrence of amphimixis—so that we may quite well speak of cyclic development in the physiological sense, in as far as certain internal conditions periodically recur and compel the organism to conjugation, but we must not suppose that there is more in this than, for instance, in the 'cyclic development' of Man, which consists in the fact that he finds himself periodically impelled to take food. The feeling of hunger which forces him to do so is the signal which warns the organism that it is time to supply fresh combustible material to the metabolism. In the same way, after a long series of generations of Infusorians the necessity for conjugation arises; the whole colony suffers an 'epidemic of conjugation,' and the animals unite in pairs; in the meantime we know not why, and must content ourselves with formulating what is observable, that the nuclear substances of two individuals are thereby mingled in each conjugate.
Obviously the impulse to conjugation is a signal in the same sense as the feeling of hunger is, and we know well from the higher animals what a mighty influence it exerts, an influence hardly less potent than that of hunger. In Schiller's words, 'Durch Hunger und durch Liebe, erhält sich dies Weltgetriebe.'
We can see clearly enough why Nature should have given animals the feeling of hunger, but the reason for the need of conjugation is not so plain; we can only say in the meantime that it must be of some value in maintaining the forms of life, for only that which fulfils a purpose can be permanently established.
I shall return later to the problem of the meaning of 'sexual[Pg 328] reproduction,' and try to probe more deeply into the meaning of its establishment; in the meantime I must restrict myself to having shown its significance in the union of the hereditary substances of two individuals, and at the same time to controverting the theory of the 'rejuvenating power' of amphimixis. I use this expression in its original sense, which indicates that every life is gradually wearing itself away and would become extinct were it not fanned to flame again by amphimixis—by an artifice of Nature, we may say. This conception rests on the fact that the cells of the multicellular body possess for the most part only a limited length of life, for they are used up by the processes of life, and they break up and die, some sooner, some later. As it is observed that all true somatic cells, among higher animals at least, are subject to this law of mortality, but that the germ-cells are not, and that, furthermore, the germ-cells only develop when they are fertilized, the cause of the potential immortality of the germ-cells is believed to lie in amphimixis, and a 'rejuvenating' power in fertilization, or, more generally, in amphimixis, is inferred. Mystical as this sounds, and little as it agrees with our otherwise mechanical conceptions of the economy of life, it was until very recently a widespread view, although perhaps it is now abandoned by many who formerly held it, and has been imperceptibly modified into a quite different conception, for which the word 'rejuvenescence' is retained, but with the altered meaning of a mere 'strengthening of the metabolism' or 'of the constitution.' By many authors, indeed, the two meanings of the word are not clearly kept apart. I shall return later to the modified meaning of the word 'rejuvenescence,' and shall keep in the meantime to the original meaning of the word, which implies a renewal of life which would otherwise die out.
This meaning seemed to gain a firm hold, when, about fifteen years ago, the French investigator Maupas published his remarkable observations on the conjugation of Infusorians. These seemed to show that colonies of Infusorians which were artificially prevented from conjugating gradually died out; not of course at once, but after many, often several hundred, generations; ultimately a degeneration of all the animals in such colonies set in, and ended only with their utter extinction. Maupas himself interpreted this as a senile degeneration which took place because conjugation had been prevented, and he therefore regarded conjugation as a 'rajeunissement karyogamique,' a rejuvenescence, and therefore a means of preventing the ageing and final dying off of the individuals—of obviating, in short, the natural death to which in his opinion they would otherwise be[Pg 329] subject. This conception was greeted with general approval, and there are many people who still regard conjugation as a process by which the capacity for life is renewed—a view which I must still dispute as emphatically as I did some years ago.
In the first place, the observations on which this theory is based admit of another interpretation, quite different from that which has been assumed to be the only possible one. Maupas prevented conjugation, not perhaps because he had isolated individuals and their progeny, but by exposing the whole colony of near relatives to unusual conditions when conjugation was just about to set in, namely, by supplying them with particularly abundant food. The need for conjugation then disappeared, as, conversely, it could be called forth at any time in a colony by hunger. But these are artificial conditions, and indeed the breeding of Infusorians for months in a small quantity of water on the object-glass certainly does not correspond to natural conditions. We must admire the skill of the investigator who was able to keep his colonies alive for months and years under such artificial conditions, but we may venture to doubt whether the fate of extinction which did ultimately overtake them was really due to the absence of conjugation, and not to the unnaturalness of the conditions.
In any case a repetition and modification of Maupas' experiments is very desirable, and would be of lasting value[15].
[15] Since the above was written Calkins has made a series of new experiments, the results of which differed in several respects from those yielded by Maupas' experiments. When his infusorian-cultures began to grow weaker, as happened frequently and at irregular intervals, he was always able to restore them to more vigorous life by a change of diet, and especially by substituting grated meat, liver, and the like for infusions of hay. Certain salts, too, had the same effect: the animals became perfectly vigorous again. Calkins believes that chemical agents, and especially salts, must be supplied to the protoplasm from time to time. He reared 620 generations of Paramœcium without conjugation. But the 620th was weakly and without energy. The addition of an extract of sheep's brains made them perfectly fresh and vigorous again. Further experiments in this direction are to be desired, but, according to those of Calkins, it is probable that Infusorians can continue to live for an unlimited time even without conjugation.
Let us, however, assume for the moment not only that Maupas' observations were correct, which I do not doubt, but also that they were rightly interpreted. Would they in that case afford a proof that amphimixis means a rejuvenescence of the power of life? To my thinking, not in the remotest degree.
It certainly seems as if this were true at the first glance; the colony which is prevented from conjugating goes on multiplying for a considerable time, often indeed for hundreds of generations, but this may be compared with sufferers from hunger, whose life does not cease at once if the feeling of hunger is not appeased.
It was certainly made evident by these experiments that Infusorians which were prevented from conjugating were incapable of unlimited persistence. But even this in no way proves that amphimixis has a power of rejuvenating life, but simply that these animals are adapted for conjugation, and that they degenerate without it, just as the sperm-cell or the ovum dies if it does not attain to amphimixis.
My opponents take it as axiomatic that the life-movement must come to a standstill of itself, and that it therefore requires help. Even so distinguished a specialist on the Protozoa as Bütschli argues that organisms are not perpetua mobilia, and when one remembers the physicist's theory of the impossibility of a perpetuum mobile this looks at first sight like a formidable objection. But does the organism always remain the same as long as it lives, like a pendulum which friction causes to swing more and more slowly till ultimately it comes to a standstill? We know surely that the phenomena of life arise from a continual process of combustion, which is followed by a constant replacement of the used-up particles by new particles; we know that life depends on an unceasing metabolism, which brings about changes in the material basis of the organism every moment, so that it is constantly becoming new again.
I shall attempt to show later on that the cells cannot be the ultimate elements of the organism, but that the life-units visible with the microscope must be made up of smaller invisible units. These, therefore, undergo 'metabolism,' which conditions their multiplication and their destruction, and this 'metabolism' is not to be seen only in the building up and breaking down of 'albuminoid substances,' as the physiologists say, but in the alternation between the multiplication and the dissolution of these smallest vital particles. Therefore, it seems to me that the movement of life, whether in a single-celled or in a many-celled organism, is not to be compared to one pendulum, but to an endless number of pendulums which succeed one another imperceptibly in the course of the metabolism, always producing anew the same result, which therefore may continue ad infinitum. Suppose, then, that we possessed our present conception of life as a process of combustion, and of metabolism as the agency which continually provides new combustible material in the shape of new vital particles, but that we knew nothing about multicellular organisms and their transitory existence, but were acquainted only with unicellular organisms and their unlimited multiplication by division. If we were then to make the observation that all multicellular organisms are mortal, subject to natural and inevitable death, it would at first[Pg 331] appear to us quite unintelligible, since we should be aware that in these also the fire of life is continually being fed by the supply of new combustible material. Not the potential immortality of unicellular organisms would then appear to us remarkable and surprising, but the limitation of the life of multicellular organisms—the occurrence of natural death. Who knows whether, in that case, many of those investigators trained in regard to unicellular organisms alone would not say just the opposite of what Bütschli has said, that there could be no natural death in many-celled organisms, since single-celled organisms prove to us that life is an endless chain of transitory minute vital units?
Furthermore, our physiologists are still far from being able to explain the natural death of many-celled organisms from below—I mean from a knowledge of its necessary causes; on the contrary, they argue from the known occurrence of natural death to the causes which underlie it; and thus they have arrived at the idea, undoubtedly correct, that the somatic cells of the body are gradually so altered by their own activity that they are ultimately unable to function any longer and must die off. Therefore, if we were unacquainted with death, we should not have been able to infer it from our physiological knowledge, and still less from our knowledge of the unicellulars.
As our insight has in point of fact grown by starting from the mortal many-celled organisms, and has only later penetrated down to the unicellular organisms, so we can understand the genesis of the conclusion, deduced from the mortality of the many-celled organisms, that unicellular organisms also are unable to continue without limit the renewal of material and of vital particles, and that consequently they would be subject to natural death if nature had not found in conjugation a 'remedy' for 'the physiological difficulties which ensue automatically and necessarily from the constitution and from the continual functioning' even of unicellular organisms.
But we ask in vain for a shadow of proof of this remarkable conception; it is an axiom deduced from our knowledge of natural death among multicellular organisms, and bolstered up by a mistaken application of the idea of 'perpetual motion.' Or may we regard it as a proof of this if it should be found that all unicellular organisms are adapted for conjugation?
We shall see later on that amphimixis has certainly quite a different and, undoubtedly, a very important effect, namely, that it increases the capacity of the species for adaptation; and a life-renewing effect in Bütschli's sense could only be ascribed to it in addition if the assumption of the necessity of a natural death in[Pg 332] unicellular organisms were not directly contrary to the clear facts of the case; but this is just what it is.
We are acquainted with such contradictory facts, not perhaps among the unicellulars themselves, where it is difficult to procure direct proof, but in regard to the germ-cells of many-celled organisms which correspond to unicellular organisms. We know that under certain circumstances the ovum is capable of persisting by itself—in cases of parthenogenesis—how then can we conclude that amphimixis is in the case of Metazoan germ-cells the cause of their capacity for development? We can only conclude, it seems to me, that their power of developing is usually bound up with the occurrence of amphimixis. So we may conclude in regard to the unicellulars that their unlimited power of multiplication is bound up with the occurrence of amphimixis, but not that amphimixis is the cause of this power, or that it implies a rejuvenescence of life. If unicellular organisms could have been made immortal through amphimixis, then what I maintain would be proved—that they possess potential immortality; but if they did not possess it, no artifice in the world could give it to them; amphimixis could be at most only the condition with the fulfilment of which the realization of their immortality was bound up.
One may ask, How then can amphimixis be a condition of their survival? why should Infusorians which have not conjugated at the proper time be doomed to extinction? And from the standpoint of our present knowledge I am as little able to give a precise answer as my opponents. But I can give one in relation to the amphimixis of multicellular organisms, for in regard to these we know that each of the germ-cells—male and female—uniting in fertilization, is of itself incapable of development and doomed to perish, the sperm-cell because it is too small in mass to be able to develop the whole organism, and the ovum because, in order to become capable of being fertilized, it must undergo certain changes which make it incapable of independent development. We have seen that after the two maturing divisions in the egg-cell have been accomplished the ovum no longer contains a mechanism of division, as the centrosphere breaks up after the second division; embryonic development can therefore only begin when a new centrosphere has been introduced into the ovum, and this is normally brought about by fertilization, that is, by the entrance of the sperm-cell, whose nucleus is accompanied by a centrosphere.
Thus amphimixis is seen to be really a condition of development. But we now know that the ovum can emancipate itself from this condition, [Pg 333]by only going through a part of the processes of maturation which are related to the subsequent amphimixis, and by thus retaining its own centrosome. Nothing is more instructive in this connexion than the cases we have already briefly discussed of facultative or occasional parthenogenesis. We have seen that in some insects, for instance in the silk-moths, there are sometimes, among thousands of unfertilized eggs, a few that develop little caterpillars. If we examine a large number of such unfertilized eggs we not infrequently find among them several which, although they have not gone through the whole course of development, have at least gone through the earlier stages, and others which may have advanced somewhat further and then come to a standstill; in short, we can see that several of these eggs were capable of parthenogenetic development, although in varying degrees.
The cause of this parthenogenetic capacity has not as yet been definitely determined by observation, but we shall hardly go wrong if we seek it in the fact that the centrosphere of the ovum does not always perish immediately and completely during maturation, and may persist, rarely in its integrity, but sometimes in a weakened state. Future observations will probably reveal some differences in the size or aster-forming power of the centrospheres of such eggs; in any case it is of the greatest interest that stimuli of various kinds—mechanical or chemical—can strengthen the disappearing centrosphere of the ovum, although as yet we are far from being able to say how this comes about.
The experiments already mentioned of Tichomiroff, Loeb, and Winkler give us at least an indication how we must picture to ourselves the origin of parthenogenesis, namely, through the fact that the breaking up of the apparatus for division, introduced for the sake of compelling amphimixis, is prevented. Minute changes in the chemistry of the ovum, similar to those caused artificially in the ova of the sea-urchin by the introduction of an infinitesimal quantity of chloride of magnesium (Loeb), in the ovum of the silk-moth by friction or by sulphuric acid (Tichomiroff), or in the sea-urchin ovum by an extract of the sperm of the same animal (H. Winkler), will effect this modification, and normal parthenogenesis is induced.
For the ovum, therefore, amphimixis is certainly not a life-renewing or rejuvenating factor; it only appears as such because the process has in the course of nature been made compulsory by making the two uniting cells each incapable of developing by itself. As we have seen, this is true also of the sperm-cell, for although it contains a centrosphere, and would be capable of division as far as[Pg 334] that is concerned, yet in almost all animals and plants it consists of such a minimal quantity of living matter that it is unable to build up a new multicellular organism by itself. Only in one alga (Ectocarpus siliculosus) has it been observed that not only the female germ-cell can develop parthenogenetically under certain circumstances, but that the male-cell may also do so. In this case, however, the difference in size between the two is not great, and it is noteworthy that the male plant, in correspondence with the smaller size of the zoosperm, tends to be a somewhat poorly developed organism.
If we are forced to the conclusion in regard to multicellular organisms that amphimixis does not supply the power of development to the ovum, but that, on the contrary, the power of development is withdrawn from the ovum, so that amphimixis can, so to speak, be forced, must we not assume something similar for unicellular organisms also? May not amphimixis be made compulsory in their case also, in that the Infusorians in preparation for conjugation go through changes which make their unlimited persistence possible only on condition that they conjugate? In my opinion the division of labour in the nucleus, which is differentiated into a macronucleus and a micronucleus, and the transitory nature of the former, may be regarded as an adaptation in this direction. In any case, it is striking that an organ which otherwise persists without limit among unicellular organisms, the nucleus, is here subject to natural death after the manner of the body of multicellular organisms, that it breaks up and must be reformed from the micronucleus which in this case is alone endowed with potential immortality. I am inclined to regard this as an arrangement for compelling conjugation, since it is only after conjugation that the micronucleus forms a new macronucleus, although the latter is indispensable to life, as we see from experiments in dividing Infusorians artificially.
Suppose we had to create the world of life, and it was said to us that amphimixis must—wherever possible—be secured periodically to all unicellular and multicellular organisms, what better could we do than arrange devices which should exclude individuals which, by chance or constitution, could not attain to amphimixis from the possibility of further life? But would amphimixis then be the cause of persistence or a principle of rejuvenescence?
I do not see that there can be any ground for such an assumption other than the tenacious and probably usually unconscious adherence to the inherited and deep-rooted idea of the dynamic significance of 'fertilization,' no longer, perhaps in its original form, which regarded the sperm as the vital spark which awakened new life in the dead[Pg 335] ovum, but in the modified form of the 'rejuvenating' power of amphimixis.
Quite recently an attempt has been made to modify the idea of the 'rejuvenating' effect of amphimixis so that it should mean only an advantage, not an actual condition of persistence. Hartog, in particular, admits so much, that the occurrence of purely asexual and purely parthenogenetic reproduction excludes the possibility of our regarding the process of amphimixis as a condition of the maintenance of life. But then we must also cease to regard the 'ageing' and dying off of Infusorians which have been prevented from conjugating as an outcome of the primary constitution of the living substance, and should entirely abandon the misleading expression 'rejuvenescence.'
If we fix our attention on the numberless kinds of cells in higher organisms and on multicellular organisms as intact unities, we see that they all die off, that they are subject to a natural death, that is, a cessation of vital movement from internal causes, yet no one is likely to refer their transitoriness to the fact that they do not enter into amphimixis. We find it quite 'intelligible' that the cells of our body should be used up sooner or later as a result of their own function, though we are very far from being able to demonstrate the necessity for this, and so really to 'understand' it.
It is only from the standpoint of utility that we can understand the occurrence of natural death; we see that the germ-cells must be potentially immortal like the unicellular organisms, but that the cells which make up the tissues of the body may be transient, and indeed must be so in the interests of their differentiation—often great and in one direction—which determines the services they render to the body. They required to become so differentiated that they could not continue to live on without limit, and they did become so differentiated because only thus could an ever-increasing functional capacity of the whole organism be rendered possible; but they die not because 'rejuvenescence through amphimixis is denied them, but because their physical constitution is what it is.' And we must explain the death of the whole many-celled individual in a similar way. When we were trying in a previous study to establish the unlimited continuance, the potential immortality, of unicellular organisms, we noted that an eternal continuance of the life of the body of multicellular organisms could certainly not be a necessity, since the continuance of these forms of life is secured by their germ-cells. A continuance of the body cannot even be regarded as useful from any point of view. And what is not useful for a form of life does not arise as a lasting adaptation, which is of course not to say that an immortality of[Pg 336] multicellular organisms, such as they are now, would even have been possible. If these organisms were to attain to such a high degree of functional capacity and of structural complexity as they now exhibit, they obviously could not also have been adapted at the same time to an eternal persistence of life.
This is in perfect harmony with our whole conception of the impelling forces in the development of the organic world; the ever-increasing functional capacity of the structure arose from the advantage which this afforded in the struggle for existence, in comparison with which the apparent advantage of the endless life of the individual was of no account whatever.
I will not here follow out this idea. I have merely touched on it in order to make clear that the death of individuals in all multicellular organisms gives us no ground for thinking of the unlimited life of the germ-cells as dependent on a special artifice of nature, such as amphimixis is often supposed to be. Let us always remember that there is parthenogenesis, and that there are unicellular germs (spores) which are never fertilized, and that the reproduction of many species of animals and plants occurs in this way without the intervention of amphimixis at all.
Attempts have recently been made to prove that parthenogenesis is a kind of self-fertilization, and these have been based on the observations of Blochmann and Brauer, which showed that in the bee and in the salt-water Crustacean, Artemia salina, the reducing second maturation division of the ovum-nucleus is not suppressed, but is regularly accomplished, and that the two daughter-nuclei which result from this division unite with each other subsequently. I have already noted that these statements do not hold true, at least with regard to the bee. In this case the second maturing division takes place without any subsequent fusion of the two daughter-nuclei. According to the observations of Dr. Petrunkewitsch, which I have already mentioned, and for the exactness of which I can vouch, the second maturation-spindle is unusually long, so that the two daughter-nuclei are pushed very far apart (Fig. 79, Rsp 2), and only the inner of the two nuclei (K 4) becomes a segmentation nucleus, while the outer undergoes a remarkable fate; it unites with the inner nucleus which results from the division of the first maturation cell (K 2), and from this union the primitive genital cells of the animal appear to arise—an observation the eventual theoretical importance of which can only be estimated later.
Meantime all we can gain from it is a certain mistrust of the interpretation of the processes of maturation in Artemia which have hitherto been given; at least we are tempted to suppose that the copulation of two nuclei which Brauer observed in Artemia may not have led to the formation of the segmentation nucleus there either, but may have had some other significance.
But, even if we leave this point entirely out of account, there remain all the cases of regular parthenogenesis in which this mode of reproduction occurs alone and not in alternation with the sexual mode. In these only one maturing division is undergone, and only one polar body is formed, and thus there can lie no possibility of supposing a self-fertilization of the ovum.
Fig. 79. The two maturation divisions in the
unfertilized (drone-forming) egg of the bee, after
Petrunkewitsch. Rsp 1, first polar-body in division.
K 1 and K 2, the two daughter-nuclei thereof.
Rsp 2, second directive spindle. K 3 and K 4, the
two daughter-nuclei thereof. In the subsequent
stage K 2 and K 3 unite to form the primordial
sex-cell nucleus. Highly magnified.
It is possible that we may yet discover species among unicellular organisms which multiply without limit in the absence of any amphimixis. R. Hertwig has recently observed phenomena in Infusorians which he is inclined to refer to the suppression of an earlier habit of conjugation, and so to a kind of parthenogenesis. But even if it should be shown that amphimixis plays a part regularly and without exception in the life of all unicellular organisms, the facts in regard to multicellular organisms are not affected; and, finally, the process of amphimixis is one which we have not the slightest ground for assuming to be either an awakener or a maintainer of life, and so I return to the most essential part of the whole problem, the meaning of the chromatin structures, the combination of which is the undoubted result of amphimixis. Do they really represent, as we assumed earlier, the hereditary substance, and what do we mean by this term?
As far as I know the literature and the development of biological theories, the botanist Nägeli was the first to deduce, from the considerable difference in size between the egg-cell and the sperm-cell, the conclusion that the material basis on which the hereditary tendencies[Pg 338] depend must be a minimal quantity of substance. The difference is especially great in animals, even in those species whose eggs may be called small, for instance, those of sea-urchins or of mammals; even in these the mass of spermatozoon is scarcely a thousandth part, often scarcely a hundred-thousandth part of the mass of the ovum. And yet the inheritance from the father and from the mother is equally great. Now as we know that vital powers have always a material basis, a minute quantity, such as is contained, for instance, in the spermatozoon of Man, must have implicitly in it all the hereditary tendencies of the father; and the conclusion is inevitable that in the ovum there can only be an equally minimal quantity of substance which is the bearer of the hereditary powers, for if there were a larger quantity of hereditary substance in the ovum its power of transmission would also be greater[16].
[16] The improbable assumption that the hereditary substance of the father may be in quality altogether different from that of the mother, and so may have the same power of transmission, and yet take up much less room, I leave out of the question altogether.
Fig. 69. Ovum of Sea-urchin (Toxopneustes lividus),
after E. B. Wilson, zk, cell-substance. k, nucleus
(so-called germinal vesicle). n, nucleolus (so-called
germinal spot). Below there is a spermatozoon of
the same animal (sp), magnified in the same proportion,
about 750 times.
Fig. 68. Diagram of a
spermatozoon. After E. B.
Wilson. sp, apex. n, nucleus.
c, centrosphere. m, middle
portion. ax, axial filament.
e, terminal filament.
If we inquire as to the part of the spermatozoon which bears this hereditary substance, we may exclude both the tail-thread and the middle piece (Fig. 68), the former because it obviously fulfils[Pg 339] quite a specialized physiological function and is histologically adapted to this function, the latter because, from observation on the spermatozoon which has made its way into the ovum, we know that it contains the centrosome, the dividing apparatus of the nucleus. Thus there only remains the 'head' of the spermatozoon, which includes the nucleus, as the possible vehicle of the heritable substance. Therefore we are led to seek for the hereditary substance in the nucleus. But the hereditary substance cannot be a perishable substance which may at need be dissolved, in the literal sense of the word, and be formed anew; therefore we cannot look for it in the nuclear membrane, and just as little in the 'nuclear sap' which fills the meshes of the nuclear network, since the material on which heredity depends must necessarily be solid. Nägeli has clearly shown that we must assume a stable, that is, a solid molecular architecture. There thus remains only the nuclear reticulum with its chromatin granules, and when we remember what we have learnt of the behaviour of this chromatin substance during division and amphimixis we can entertain no doubt that the sought-for bearer of the inheritance is contained in the substance of the chromosomes.
The great care with which the chromosomes are halved by means of the complicated division apparatus led us earlier to regard them as a substance of complex and manifold qualities and of great physiological importance; their constant number in any one species, and the reduction of that number to half by means of the reducing divisions, justify us in concluding that they are permanent structures, physiological and morphological units, which undergo no more than an apparent irregular dispersion during the resting state of the nucleus. Finally, the fact that these supposed vehicles of inheritance occur in equal numbers in each of the conjugating germ-cells, and that this number is always, both in animals and in plants, half of the normal number occurring in somatic cells, is decisive. The logical necessity that the hereditary substance of both parents should be transmitted to the offspring in equal quantity could not be more precisely met than it is by the fact that half the normal number of chromosomes occurs in each of the sex-nuclei in the ovum. Personally, I have long been certain, on these grounds, that the chromosomes of the nucleus are the hereditary substance, and I expressed my conviction on this point almost simultaneously with Strasburger and O. Hertwig[17].
[17] More precisely, my conclusions were published several months later than those of the investigators named (1885). I think, however, that no one who is familiar with my writings for the years immediately preceding, which are collected in Aufsätzen über Vererbung und verwandte biologische Fragen (Jena, 1892), will dispute that the idea was reached by me independently. I attach importance to this because all my later work is based upon this idea.
But there is also a physiological proof of the meaning of the nuclear substance; and this we owe, again, to the simultaneous and independent researches of two investigators, M. Nussbaum and A. Gruber, the latter working in the Zoological Institute here (in Freiburg), and at my request. They made experiments on regeneration in unicellular organisms, and found that Infusorians which were artificially divided into two, three, or four pieces were able to build up a whole animal out of each piece, provided that it contained a portion of the nucleus (macronucleus). The large blue trumpet-animalcule, Stentor cœruleus, is well suited for such experiments, not only on account of its size, but because it possesses a very long rosary-like nucleus, which can be easily cut two or three times. When a piece is cut off which does not contain a portion of the nucleus, it may indeed live for some days and swim about and contract, but it is incapable of reconstructing the lost parts, and thus of forming a whole animal, and it perishes. It is in the nucleus, therefore, that we have to look for the substance which stamps the material of the cell-body with a particular form and organization, namely, the form and organization of its ancestors. But that is exactly the conception of a hereditary substance or idioplasm (Nägeli). Some modern biologists deny that there is any hereditary substance per se, and believe that the whole of the germ-cell, cell-body and nucleus together effects transmission. But though it must be admitted that the nucleus without the cell-body cannot express inheritance any more than the cell-body without the nucleus, this is dependent on the fact that the nucleus cannot live without the cell-body; if it be removed from the cell and put, say, into water, it bursts and is dissolved. But the cell-body without the nucleus lives on, though of course only for a few hours or days, and its metabolism ceases only when it is brought to a standstill by the failure to replace by nutrition the used-up material. Thus the argument used by those who deny the existence of a hereditary substance would be paralleled if we denied that Man possesses a thinking substance, and maintained that he thinks with his whole body, and even that the brain cannot think by itself without the body.
I am convinced that it is just as mistaken to maintain that every part of an organism must contain the hereditary tendencies in the same degree, or that in unicellular organisms the cell-body is as important in inheritance as the nucleus (Conklin). If one feels any doubt on this point, one has only to call to mind Nägeli's inference, from the minuteness of the spermatozoon, that the hereditary substance must be minimal in quantity. But even theoretically there[Pg 341] is not the smallest ground for the assumption that the cell-body as well as the nucleus contains the hereditary qualities, since we find in general that functions are distributed among definite substances and parts of the whole organism, and it is just on this division of labour that the whole differentiation of the body depends. And why should this principle not have been employed just here where the most important of all functions is concerned? Why should all living substance be hereditary substance? Although Nägeli thought of his 'idioplasm' otherwise than we now think of hereditary substance, although he wrongly imagined it in the form of strands running a parallel course through the cell-substance and forming a connected reticulum throughout the whole body, he recognized at least so much quite correctly, that there are two great categories of living substance—hereditary substance or idioplasm, and 'nutritive substance' or trophoplasm, and that the former is much smaller in mass than the latter. We now add to this, that the idioplasm must be sought for in the cell-nucleus, and indeed in the chromatin granules of the nuclear network and of the chromosomes.
But incontrovertible proof of the fact that the nuclear substance alone is the hereditary substance was furnished when it was found possible to introduce into a non-nucleated piece of a mature ovum of one species the nucleus of another related species, and when it was seen that the larva that developed from the ovum so treated belonged to the second species. Boveri made this experiment with the ovum and spermatozoon of two species of sea-urchin, and believed that he had succeeded in getting from non-nucleated pieces of the ovum of the first species, fertilized with the sperm of the second, larvæ of this second species; but, unfortunately, later control-experiments made by several investigators, especially by Seeliger, have shown that this result cannot be regarded as quite certain and indubitable.
I must emphasize again that I am far from regarding the cell-protoplasm of the ovum as an indifferent substance. It is certainly not only important but indispensable for the development of the embryo, and it has assuredly its own specific character, as in every other kind of cell. It represents, so to speak, the matrix and nutritive environment in which alone the hereditary substance can unfold its wonderful powers; it has developed historically, like every other kind of cell, but it contains nothing more than the inherited qualities of this one kind of cell-protoplasm, not those of the other cells of the body.
But although the essence of fertilization lies, as we have seen, in the union of the hereditary substance of two individuals, and not[Pg 342] in a 'quickening' of the ovum, we may quite well speak of a quickening by fertilization in another sense, if we mean the impulse to embryonic development, for this is really supplied by the entrance of the sperm-nucleus with its centrosphere into the ovum. But even this impulse can, under certain circumstances, be given in another way, and certainly the awakening of it is not the end of fertilization, but only the condition without which the end, the union of two kinds of nuclear substance, could not be attained. There is no indication whatever that this 'quickening' of the ovum would be necessary for any other reason except that the ovum was previously made incapable of development. There would be no 'fertilization' were not the mingling of hereditary substances of fundamental importance for the organic world.
Moreover, an ovum, or a fragment of an ovum, may also develop of itself, having only one of the sex-nuclei, and the union of the hereditary substance of two cells is therefore not indispensable for the mere production of a new individual.
What has been observed in regard to fragments of ova is particularly interesting in this connexion. Ernst Ziegler first succeeded in halving a newly fertilized sea-urchin ovum, so that one half contained the female and the other the male pronucleus. The latter alone contained a centrosphere, and developed a blastula larva. Delage carried these experiments further, and cut an unfertilized but mature sea-urchin ovum into pieces, and then 'fertilized' the non-nucleated pieces with spermatozoa. These pieces developed and yielded young larvæ of the relevant species; so it is clearly seen that even a piece of mature ovum-protoplasm may undergo embryonic development, provided that a nucleus furnished with a dividing apparatus penetrates into it. Unfortunately it is technically impossible to cut such a non-nucleated and then fertilized fragment of ovum so that one half shall contain the male nucleus the other its centrosphere. Even without this experimentum crucis we may say that the half with the male nucleus would not multiply by division, and that the other probably would, though it would not go through the regular course of segmentation processes, because the hereditary substance absolutely necessary for these was wanting.
But these and similar experiments prove something more, namely, that the nuclei of the sperm-cell and egg-cell do not, as was formerly believed, stand in a primary and essential contrast to each other, which may be described as male and female, but that both are alike in their deeper essence, and may replace each other. They only differ from each other as far as the cells to which they belong differ,[Pg 343] in this, namely, that they are mutually attractive; they find each other and unite, and then go on to develop, which each was previously unable to do by itself. Widely as the sperm-cell and egg-cell differ in size, constitution, and behaviour, in regard to essential character they are alike; they bear the relation—as I expressed it twenty years ago—of 1:1; that is, they both contain an equal quantity of essentially similar hereditary substance, and the quality of this substance is only individually variable. We should, therefore, speak not of a 'male' and 'female,' but of a 'paternal' and a 'maternal' nucleus.
All the more recent experiments on 'merogony,' that is, on the development of fragments of the ovum, confirm this view. Thus Boveri had already observed that even small pieces of sea-urchin ova which did not contain the nucleus of the ovum developed, after the spermatozoon had entered them, into small but otherwise normal larvæ of the species. More recently Hans Winkler proved the same thing for the ova of plants, by dividing the ovum of a marine alga (Cystosira) into two pieces, then fertilizing these with water containing sperms, with the result that he got from both pieces, the nucleated and the non-nucleated, an embryo of normal appearance. In the latter it could only have been a 'paternal' nucleus which directed the development.
To sum up. Our investigation into the meaning of amphimixis has led us to the conclusion that it consists in the union of two equal complements of hereditary substance, contributed by two different individuals, into one unified nucleus, and that the sole immediate result of this is the combination of the hereditary tendencies of two individuals in one. Among multicellular organisms this one individual of dual origin always implies the beginning of a new life, since amphimixis is indissolubly associated with reproduction, and even among unicellular organisms it can hardly be disputed that the two Infusorians which separate after conjugation are no longer the same as they were before. After amphimixis they must contain a different combination of hereditary substance from what they had before, and this must reproduce the parts of the animal in a somewhat modified form. This is theoretically beyond doubt, although it can scarcely be established by observation.
We thus know now what 'fertilization' is. Through the labours of the last decade the veil has been torn from a mystery of nature which for thousands of years confronted humanity as unapproachable; a riddle has been solved for the solution of which a few centuries ago men did not even dare to hope. Not a few have taken part in these labours; some I have already named, but it is impossible that[Pg 344] I should here mention all who have shared in the achievement by observation or reflection. Whoever has helped it on even a single step may say to himself that he has taken an active part in bringing about what must be called essential progress in human knowledge.
But in the science of nature every new solution implies the cropping up of a new riddle, and we are immediately confronted with the problem, Why should nature, in the course of evolution, have interpolated this process of the mingling of different hereditary substances almost everywhere in the organic world? This, however, is a problem which we cannot attack until we have first made ourselves more fully acquainted with the phenomena of inheritance, and have attempted to reason back from these to the nature of the hereditary substance. We must, in short, think out a theory of heredity.
THE GERM-PLASM THEORY
Conception of the 'id' deduced from the process of fertilization—Hereditary substance, 'idioplasm' and 'germ-plasm'—'Idants'—Evolution or Epigenesis—Herbert Spencer's uniform germinal substance—Determinants—Illustrations: Lycæna agestis—The leaf-butterflies—Insect metamorphosis, limbs of segmented animals—Heterotopia—The ultimate living units or biophors—Number of determinants—Stridulating organ of the grasshopper.
In proceeding to expound the theory of heredity which has shaped itself in my mind in the course of my own scientific development, I should like to begin by pointing out that the hereditary substance of the germ-cell of an animal or of a plant contains not only the primary constituents (Anlagen) of a single individual of the species, but rather those of several, often even of many individuals. That this is so can be proved in several ways.
I start from what I hold to be the proved proposition, that the chromatin substance of the nucleus is the hereditary substance. We have seen that this is present in the germ-cells of every species in the form of a definite number of chromosomes, and that in germ-cells destined for fertilization, that is, in sex-cells, this number is first reduced to half, the reduction being effected, as is now proved in regard to a whole series of animals, by the two last cell-divisions, the so-called maturation divisions.
We know that the full number is only reached again through amphimixis, by which process the half number of chromosomes in the male and female germ-cells are united in a single cell, the 'fertilized ovum,' and in a single nucleus, the so-called segmentation nucleus. Thus the hereditary substance of the child is formed half from the paternal, half from the maternal hereditary substance, and we have seen that this remains so during the whole development of the child, since, at every succeeding cell-division each of the paternal and each of the maternal chromosomes doubles by dividing, and the resulting halves are distributed between the two daughter-nuclei.
Now if the complete hereditary substance of a germ-cell before the reducing divisions contains potentially all the primary constituents[Pg 346] of the body, which it does as a matter of course, then it follows that after the reduction each germ-cell must either contain only half the primary constituents of the parents or all the primary constituents must be contained in the half number of chromosomes. The latter seems to me the only possible assumption, as I shall immediately proceed to show, and this is as much as to say that the primary constituents of at least two complete individuals must be contained in the chromosomes of the segmentation nucleus.
That this conclusion is correct is obvious from the fact that a whole, that is, a perfect individual with all its parts, develops from the ovum, and not a defective one. For suppose that each mature germ-cell contained only half the primary constituents of the body, it would be impossible that these halves should always exactly complete themselves to form a whole embryo when they are brought together in fertilization, after having been halved by mere chance during the preceding reducing division; it would be much more likely to happen that they did not complete themselves, and that their union would therefore result in an individual with certain parts wanting. If, for instance, in the sperm-cell only the anterior half of the body was potentially present, and this united with an ovum which likewise contained only the primary constituents of the anterior half, the embryo resulting from their union would lack the posterior half of the body, and so on. Of course so rough a division of the primary constituents is not to be thought of, but however fine we can imagine the halving of the mass of primary constituents to be, there would never be any guarantee that the two cells uniting in amphimixis would complete the mass of primary constituents again; indeed, the chance that the two exactly complementary halves of the mass would meet would rather become less the finer and more complex one imagines the halving by reducing divisions to be. A perfect embryo with all its parts would rarely arise, but now one group of parts, now another would be wanting, while another group might be developed double, or at least would be doubly present in the primary constituents.
But in addition to this the facts of inheritance show us that the resemblance to mother and father may express itself simultaneously in all the parts, or at least in the same parts of the child, as may be seen with especial clearness among plant-hybrids, and thus the conclusion is inevitable that even in the half number of chromosomes all the primary constituents of the whole body are present.
Let us go a generation further. If the species possess four[Pg 347] chromosomes the child will have in its cells two maternal chromosomes (A) and two paternal chromosomes (B); what form will this proportion take in the germ-cells produced by the child? The maturation division can effect the reduction to two chromosomes in different ways; there may, for instance, be two paternal chromosomes (B) left in the one, and two maternal chromosomes (A) in the other daughter-cell, or one paternal (B) and one maternal (A) in the one, and a similar combination in the other cell. Let us follow the latter case further. A sperm-cell which contained the combination A and B might meet in amphimixis with an egg-cell of different origin also containing a similar combination of chromosomes, let us say a chromosome C from the mother, and a chromosome D from the father. We should then have in the segmentation nucleus of the fertilized ovum four different chromosomes, each of which contained the hereditary substance of one grandparent; we should have the four chromosomes, A, B, C, D, as the hereditary substance of the grandchild.
But since, as we have seen, the halved hereditary substance still contains the whole mass of primary constituents, each one of these chromosomes must contain the collective primary constituents of the whole body of the relevant grandparent[18]. The hereditary substance in the fertilized ovum thus consists of several complexes of primary constituents (chromosomes) each of which (an 'id') comprises within itself all the primary constituents of a complete individual.
[18] When I say the 'collective' primary constituents of the whole body of the grandparent this is not expressing it quite precisely, for, as we shall see later, each individual must arise from the co-operation of different chromosomes of different origin, not merely from one of the chromosomes contained in its germ-plasm. In the example given above, the body of each grandparent cannot have arisen only from a single chromosome, which was transmitted to his grandchild, but from the co-operation of this chromosome with three others, which have distributed themselves along other genealogical paths. But this does not affect the above chain of reasoning, for here it is not a question of whether all the primary constituents of the grandparent are present in the child—that can never be the case—but whether the primary constituents transmitted by him represent the whole body of an individual.
It can be made clear in yet another way that, as a consequence of sexual reproduction, the germ-plasm of each species must be composed of several 'ids,' individually different. Let us assume that there was as yet no amphimixis, and that we could look on at its introduction into the organic world; the hereditary substance of the beings which had previously lived and multiplied by division would consist of more or less numerous chromosomes similar to each other, so that, for instance, each individual would contain sixteen identical 'ids.' But if amphimixis were now to take place for the first time, in the same manner as it does to-day—that is, after the reduction of the number of the ids to half—in the first amphimixis [Pg 348]eight paternal ids would unite with eight maternal ids to form the germ-plasm of the new individual, as is indicated in Fig. 87 by a circle of spheres, of which ten are white and ten black as a sign of their difference. We may think of the figure as representing the 'equatorial plate' of a nuclear spindle with its ids arranged in a circle. Now, if two organisms of this generation, with two kinds of ids, unite in amphimixis after previous reduction of the ids, we have figure B, in which the paternal ids (pJ) are seen to the left of the line and the maternal ids (mJ) to the right, while each semicircle is in its turn made up of two kinds of ids, those of the grandparents (p2J and m2J, p2J1 and m2J1). The figures C and D show the two following generations, in which the number of identical ids is each time reduced to half, because eight strange ids are again mingled with them; in C only two ids are still identical, and in D all the ids are individually different, because they have come from different ancestors of the same species. Of course this would only be the case if inbreeding were excluded, because through it the ids of the same forefathers from two or more sides would meet; but prolonged inbreeding is a rare exception in free nature.
Fig. 87. Diagram to illustrate the operation of amphimixis on the composition of the germ-plasm out of diverse ancestral plasms or 'ids.' A-D, the ids of the germ-plasm of four successive generations: A, consisting of only two kinds of ids; B, of four; C, of eight; D, of sixteen kinds. pJ and mJ, paternal and maternal ids. p2J, grandpaternal; p3J, great-grandpaternal; p4J, great-great-grandpaternal ids. The marks in the ids themselves indicate their individually distinct characters.
I shall now call the hereditary substance of a cell its 'idioplasm,' after Nägeli's example, although he sought it in the cell-substance, not in the nucleus, and had a different theoretical conception of its mode of action. It was he, however, who conceived and established the idea of the idioplasm as the bearer of the primary constituents, an Anlagensubstanz, determining the whole structure of the organism in contrast to the general nutritive protoplasm. Every cell contains idioplasm, since every cell-nucleus contains chromatin, but I call the idioplasm of the germ-cells germ-plasm, or the primary-constituent-substance of the whole organism, and the complexes of primary constituents necessary to the production of a complete individual—whose presence we have just shown to be theoretically necessary—I call ids. In many cases these 'ids' might be synonymous with chromosomes, at least in all the cases in which the chromosomes are simple, that is, are not composed of several similarly formed structures. Thus in the salt-water Crustacean, Artemia salina, which possesses 168 minute granular chromosomes, each of these chromosomes must be regarded as an id, for each can in certain circumstances be thrown out from the ovum by the reducing division, or it can be brought into the most various combinations with other chromosomes by fertilization. Each of them must therefore consist of perfect germ-plasm in the sense that all the parts of an individual are virtually contained in it; each is a biological unity, an id. But when we see in many animals larger band-shaped or rod-shaped 'chromosomes,' and when these are composed of a series of granules, as they are, for instance, in the often mentioned Ascaris megalocephala, each of these granules is to be regarded as an id. In point of fact, we find, instead of the two or four large rod-shaped chromosomes of Ascaris megalocephala, a larger number of smaller spherical chromosomes in other species of Ascaris.
Compound chromosomes consisting of several ids, such as all rod or band-like elements of the nuclear substance probably are, I designate 'idants.' That they are composed of several individual ids is not always clearly apparent because of the smallness of the object, and even in larger ones this may only be seen in certain stages. Thus we have in Fig. 88, A and B, two 'mother-sperm-cells' of the salamander; A at an earlier stage, in which the individual ids are not visible; B at a later stage, in which the band has split, and the rosary-like structure has become at once apparent. It is not possible, then, to see at once whether each chromosome corresponds to one or to several ids. A more exact investigation of the processes of reducing division has shown that there are chromosomes of simple[Pg 350] spherical form, that is, composed of several ids whose 'plurivalence' cannot be directly recognized, but can only be inferred from their further development; there are bivalent chromosomes of double value and quadrivalent chromosomes of fourfold value, which we have to think of as made up of two or four ids. It would lead us too far to go into this more precisely, nor does it fall within the scope and intention of these lectures to inquire into these intimate and still disputed details.
The germ-plasm of every species of plant or animal is thus composed of a larger or smaller number of ids or primary constituents of an individual, and it is through the co-operation of these that the individual which develops from the ovum is determined.
Fig. 88. Sperm-mother-cells (spermatocytes) of the salamander. A, cross-section of the cell in the aster-stage; the chromosomes (chr) or idants do not reveal that they are compounded out of many ids, which are, however, quite distinctly seen in B (Jd), where the chromosomes or idants (chr) are already longitudinally split. zk, cell-substance. csp, centrosome. c, centrosome in division. After Hermann and Drüner.
We have further to inquire what conception we can form of the constitution of an id and of its mode of operation. I have already spoken of 'primary constituents' (Anlagen) of which the germ-plasm consists, but what right have we to think of the parts of an animal as already contained in the germ in any form whatever? Is it not equally possible that the germ consists of parts, none of which bear any definite relation in advance to the parts of the finished animal? Might not the germ-cell, along with its nucleus, undergo transformations and regular changes which would successively give rise to new conditions, namely, the different stages of development, until finally the complete animal was attained?
We stand here before an old problem, before the two opposed interpretations—the theory of 'Evolution' and the theory of 'Epigenesis,' which were first ranged against each other long ago, and which are a cause of strife even now, although in somewhat different guise.
The theory of 'Evolution' is especially associated with the name of Bonnet, who elaborated it in detail in the eighteenth century.[Pg 351] It maintains that the development of the ovum to the perfect animal is not really a new creation, but only an unfolding of invisible small parts, which were already present in the ovum. It assumes that the parts of the perfect organism are already preformed in the ovum, and on this account it is called the 'Preformation Theory.' Bonnet often speaks of the preformation of the perfect animal in the germ as a 'miniature model,' although his conception of 'evolution' was not really so crude as has been often alleged. He expressly emphasized that this miniature model was not exactly like the perfect animal, but consisted of 'elementary parts' only, which he thought of as a net whose meshes were filled up during development and by means of nutrition with an infinite number of other parts. But after all, his conceptions, and those of his time generally, were very far removed from the biological thinking of our own day, as may perhaps be most readily understood when I mention that he regarded death and decay as an 'involution,' as a folding back, so to speak, by means of which all the parts gained though nutrition were removed again, so that the net of the miniature model shrank together to the invisible minuteness that it had in the ovum. So it remained, he fancied, till it was reawakened at the resurrection, using the term in the religious sense! He afterwards dropped this fancy, because the objection was made to it that human beings who had lost a leg or an arm in this life would necessarily be maimed at the resurrection!
In Bonnet's time the facts of development were quite unknown, and not even the stages of the development of the chick from the egg had been observed. When this was afterwards done the prevalent theory of 'evolution' necessarily collapsed, for men saw with their own eyes that a miniature model of the chick did not gradually grow into visibility and ultimately into the young chick, but that first of all parts showed themselves in the egg which bore no resemblance at all to the chick, that these first rudiments were then altered, and that through continual new formations and transformations the chick finally appeared. Upon this K. von Wolff based his theory of 'Epigenesis,' or development through new formations and transformations. He maintained that the doctrine of 'Evolutio' was false; that there is no miniature model invisibly contained within the egg; but that from the simple egg-substance there arises, through the agency of the formative powers inherent in it, a long series of stages of development, of which each succeeding one is more complex than the one before, until ultimately the perfect animal is reached.
This certainly marked considerable progress, for it meant the beginning of a science of embryology, that is, the science of the form-development of the animal or plant from the ovum. The result was not so important in its theoretical aspect, for though the knowledge had been gained that the young animal goes through a long series of different stages, it had not been discovered how nature works this wonder and causes an animal of complex structure to arise from the apparently simple substance of the ovum. A solution of the difficulty was found by attributing to the ovum a formative power, afterwards called by Blumenbach the nisus formativus, which possessed the capacity of developing a complex animal from the simple 'slime,' or, as we should say, the simple protoplasm.
If we contrast the strictly theoretical part of the two theories, we find that Bonnet regarded the ovum as something only apparently simple, but in reality almost as complex as the animal which developed from it, and that he thought of the latter, not as being formed anew, but as being unfolded or evolved. That is to say, he thought that rudiments present from the outset in the ovum gradually revealed themselves and became visible. Wolff, on the other hand, regarded the ovum as being what it seemed, something quite simple, out of which only the nisus formativus could, by a series of transformations and new formations, build up a new organism of the relevant species.
Wolff's Epigenesis routed Bonnet's theory so completely from the field that, until quite recently, epigenesis was regarded as the only scientifically justifiable theory, and a return to the 'evolutionist' position would have been looked upon as a retrograde step, as a reversion to a period of fancy which had been happily passed. I myself have been repeatedly told, with regard to my own 'evolutionistic' theory, that the correctness of epigenesis was indisputably established, that is, was a fact, verifiable at any time by actual observation!
But what are the facts? Surely only that there is a succession of numerous developmental stages, which we know very precisely in the case of a great many animals, and that the miniature model which Bonnet assumed to be in the egg does not exist. Both these facts are now no longer called in question. But that does not furnish us with a theory of development, for theory is not the observation of phenomena or of a series of phenomena, it is the interpretation of them. Epigenesis, as formulated first by Aristotle and again by Harvey, Wolff, and Blumenbach, certainly offered an interpretation[Pg 353] of development, not, however, by referring only to what was observable, but by going far beyond it; on the one hand taking the appearance of a homogeneous germ-substance for reality, and, on the other, assuming a special power, which caused a heterogeneous organism to arise from a homogeneous germ.
We cannot now accept either of these assumptions, for we know that the germ-substance is not homogeneous, and indeed is not merely a substance but a living cell of complex structure; and we no longer believe in a special vital force, and therefore not in a special 'power of development,' which could only be a modification of the former. We are thus as little able to accept the old epigenesis as the old evolution, and we must establish a theory of Development and Heredity on a new basis.
What this basis must be is in a general way beyond doubt. Since it is the endeavour of the whole of modern biology to interpret life more and more through the interactions of the physical and chemical forces bound up with matter, development, too, comes within this aim, for development is an expression of life. We seek to understand the mechanism of life, and, as a part of that, the mechanism of development and of heredity which is closely associated with it.
If we wished to attack the problem of heredity at its roots we should first of all have to try to understand the process of life itself as a series of physico-chemical sequences. Perhaps this will be achieved up to a certain point in the future, but if we were to wait for this we should in the meantime have to abandon all attempts at a theoretical interpretation of the phenomena of development and heredity, and might indeed have to postpone them to the Greek Kalends. That would be as though, in the practice and theory of medicine, all investigation into and speculation regarding disease had to wait until the normal, healthy processes of life were thoroughly understood. In that case we should now know nothing of bacteria diseases and the hundred other acquisitions of pathological science: physiology too would have remained far behind its present level if it had lacked the fruitful influence of experience in cases of disease, and the ideas and theories, true and false, which have been based thereon. In the same way we require a theory of development and heredity if we are to penetrate deeper into these phenomena, and must have it in spite of the fact that we are still very far from having a complete causal knowledge of the processes of life. For the raw material of observation, which is to some extent fortuitous, will never bring us any further on; observation must be guided by an idea, and thus directed towards a particular goal.
It is, however, quite possible to leave aside for the present all attempts at an explanation of life, and simply to take the elements of life for granted, and on this basis to build up a theory of heredity. We have already taken a step in that direction by establishing that the whole substance of the fertilized ovum does not take part in heredity in the same degree, but that only a small part, the chromatin of the nucleus, is to be looked upon as the bearer of the hereditary qualities, and by deducing, further, that this chromatin is made up of a varying number of small but still visible units, the ids, each of which virtually represents the whole organism, or, as I have already expressed it, each of which contains within itself, as primary constituents, all the parts of a perfect animal.
It was these 'primary constituents' which led us to the digression in regard to Bonnet's theory of 'Evolutio' and Wolff's 'Epigenesis.'
Let us now inquire what must be the constitution of such a chromatin globule, an id, so that, shut up within the nucleus of a living reproductive cell, it can direct the development of a new organism which resembles its parent. Two fundamental assumptions present themselves, and these can be related to every conception of a 'germ-plasm,' even independently of the assumption of ids. Either we may think of the id as made up of similar or of different kinds of parts, none of which has any constant relation to the parts of the perfect animal, or we think of it as composed of a mass of different kinds of parts, each of which bears a relation to a particular part of the perfect animal, and so to some extent represents its 'primary constituents' (Anlagen), although there may be no resemblance between these 'primary constituents' and the finished parts. The assumption of a germ-plasm composed of similar parts, which has been made, for instance, by Herbert Spencer, may be called the modern form of epigenesis, while the other assumption is the modern form of the 'evolution' theory. As the former theory can no longer call to its aid a 'formative power' as a Deus ex machina, it can only explain development as induced by the influence of external conditions—temperature, air, water, gravity, position of parts—upon the chemical components of the germ-plasm, which are everywhere uniformly mingled; and it makes no difference whether this uniform germ-plasm is thought of as composed of many different kinds of parts, as long as those parts are mingled uniformly to make the germ-plasm and bear no relation to definite parts of the developing animal. Oscar Hertwig has recently outlined such a theory. Although I cannot expound it here I must say at least so much with regard to it, and to[Pg 355] all other theories of development founded on a similar basis, that they could not be accepted even if they were able to offer a workable explanation of the development of the individual, and for this reason, that ontogeny is not an isolated phenomenon which can be interpreted without reference to the whole evolution of the living world, for it is most intimately associated with this, being indeed a piece of it, having, as we shall see, arisen from it, and, furthermore, preparing for its continued progress. Ontogeny must be explained in harmony with phylogeny and on the same principles. The assumption of a germ-plasm without primary constituents, or of a completely homogeneous germ-plasm, as Herbert Spencer maintained, is irreconcilable with this, for, as will be seen, it contradicts certain facts of inheritance and variation. Therefore all theories founded on this assumption must be rejected.
There is another and, I believe, weighty consideration which forbids us to assume a germ-substance without primary constituents. I shall return to this later, but in the meantime I wish to build up more completely my own 'germ-plasm' theory.
I assume that the germ-plasm consists of a large number of different living parts, each of which stands in a definite relation to particular cells or kinds of cells in the organism to be developed, that is, they are 'primary constituents' in the sense that their co-operation in the production of a particular part of the organism is indispensable, the part being determined both as to its existence and its nature by the predestined particles of the germ-plasm. I therefore call these last Determinants (Bestimmungsstücke), and the parts of the complete organism which they determine Determinates, or hereditary parts.
It is easy to show on what basis this assumption rests; the phenomena of inheritance taken in conjunction with those of variation seem to me to compel us to it. We know that all the parts of an organism are variable, and that in one individual the same part may be larger, in another smaller. Not all variations are transmissible, but many of them, and some very minute ones, are. Thus, for instance, in many human families there occurs a small pit, hardly as large as the head of a pin, in the skin of the ear, whose transmission I have observed from the grandmother to the son and to several grandchildren. In such a case there must be a minute something in the germ-plasm, not present in that of other human beings, which causes the origin, in the course of development, of this little abnormality in the skin.
There are human families in which individuals occur repeatedly,[Pg 356] and through several generations, who have a white lock of hair, in a particular spot, on an otherwise dark-haired head. This cannot be referred to external influences, it must depend on a difference in the germ, on one, too, which does not affect the whole body, not even all the hairs of the body, but only those of a particular spot on the surface of the head. It is a matter of indifference whether the white colouring of the hair-tuft is produced by an abnormal constitution of the matrix of the hair, or by other histological elements of the skin, as of the blood-vessels or nerves. It can only depend ultimately on a divergently constituted part of the germ-plasm, which can only affect this one spot on the head, and alter it, if it is itself different from what is usual. On this account I call it the determinant of the relevant skin-spot and hair-group. In Man such minute local variations are usually lost after a number of generations, but in animals there are innumerable phenomena which prove to us that single minute deviations can become permanent. Thus there lives in Central Europe a brown 'blue butterfly,' Lycæna agestis, which has a little black spot in the middle of its wing. The same species also occurs in Scotland, but there, instead of the black spot, it has a milk-white one, and so-called 'eye-spots' on the under surface of the wing have also lost their black centres. The species has thus varied transmissibly, but only in regard to these particular spots on the wing. A slight variation must therefore have taken place in the germ-plasm which only affects these few parts of the body, or, to express it otherwise, the germ-plasms of the ancestral species and of the variety can only be distinguished by a difference which determines exclusively the scale colour of these spots. The two germ-plasms differ, I should say, only as regards the determinants of these wing-scales.
We know from the artificial selection to which Man has subjected and still subjects his domesticated animals and useful plants, that any spots and parts of the body which he chooses can be hereditarily altered, if the desired variations which present themselves are always selected for breeding, and that this does not necessarily cause variation in other parts of the body. When, for instance, in the case cited by Darwin, the comb of a Spanish cock which had previously hung downwards was made to stand upright because a prize had been offered for this character, or when a certain breed of hens was 'furnished with beards,' the results were permanent variations affecting only the parts on which the fancier's attention had been fixed. In the same way, when the tail feathers of the Japanese cock are lengthened to three feet the rest of the plumage does not alter, still less any other part of the body. Of course there are numerous[Pg 357] 'correlated' variations, and in very many cases the breeder causes a second or third character, on which he had not fixed his attention, to vary in addition to the one he was aiming at. But such concomitant variations are not necessary or inevitable in all cases; and indeed we need not refer them all to a true correlation of the parts, but may suppose that they depend not infrequently on the faultiness of our power of observation, which is not sufficiently keen to control several parts of the body at one time, and to notice minimal variations in parts on which we have not specially fixed our attention.
Fig. 13. Kallima paralecta, from India;
showing the right under surface in the
resting pose. K, head. Lt, palps. B, limbs.
V, fore wing. H, hind wing. St, 'tail' of the
latter, representing the stalk of the leaf.
gl1 and gl2, transparent spots, Aufl, remains
of 'eye-spots.' Sch, a 'mould'-spot.
So much, at least, is certain, that in all these cases of the artificial alteration of individual characters the germ-plasm is in some way changed, but always in such a way that it differs from that of the ancestral form through such variations alone, and the effect of these is that only the altered parts are influenced thereby, and not the whole organism. This again is but another way of saying that only the determinants of these parts have altered.
We can see from a thousand cases that exactly the same happens in a state of nature, that there, too, one part changes after another, until the highest possible degree of adaptation to the conditions has been attained. In the mimetic resemblance to leaves exhibited among butterflies this is most clearly seen, for here we are familiar with the model—the leaf—and we see how one species approximates to it in a general way only in the total colour, how others develop a brown stripe crossing the posterior wing obliquely, so that, to a certain extent, it resembles the midrib of a leaf, how in a third species this stripe is continued for some distance forward across the anterior wing, in a fourth it goes a little further, until, finally, in a fifth, it[Pg 358] is continued on to the tip of the anterior wing. This may be seen, for instance, in the genus Anæa, which is rich in species. But even then a still further increase of the resemblance is possible, for, as is well known, there are not infrequently imitations of the lateral veins of the leaf as well, or dark spots which faithfully reproduce the mould-spot on a damp, decaying leaf, or colourless transparent spots which probably simulate dewdrops, and so on. All these are variations relating to individual and distinct groups of wing-scales, which have varied transmissibly and independently, that is, each of them has been produced by a variation in the germ-plasm, which brought about a change in this particular area of the body and in no other.
Let us for a moment assume the impossible, and suppose that we could look on at the evolution of such a leaf-butterfly; the beginning of the leaf-imitation might have its cause in the fact that an ancestral form of Kallima, which had previously lived in the meadows, exhibited on the part of some of its descendants a migration to the woods, and thus divided into two groups, with a different manner of life—a meadow form and a wood form. The latter adapted itself to sitting among leaves, and the midrib of a leaf developed on its wings. In a germ-plasm without 'primary constituents' this variation could only depend on a uniform variation of all the parts, for these parts are either alike among themselves, or at any rate have the same value for every part of the finished organism. But the germ-plasm of the new breed must somehow differ from that of the ancestral form, otherwise it could produce no new variety, but only the ancestral form over again. But how could an animal differing only in one minute part arise from a germ-plasm which has varied in all its parts, and how could such little steps of variation be repeated many times in the course of the phylogeny without the corresponding variations of the germ-plasm becoming so intense that not only the wing-markings but everything about the animal would be altered likewise? And yet these 'leaf-pictures' have not originated suddenly, but by many small steps, so that the germ-plasm must have varied in toto a hundred times in succession if there are no primary constituents.
In the Indian species, Kallima paralecta, there are no fewer than five well-marked varieties, the differences between which depend solely on the manner in which the leaf-picture on the wing is elaborated, for the upper surface of the wing is alike in all. Even a cursory observation of a collection of these butterflies shows that the lateral veins of the leaf-picture are quite different in number, distinctness, and length in the different individuals. On the right half of the wing there may be as many as six of them indicated[Pg 359] (Fig. 13); and it can be observed that the three middle ones are the longest, most sharply defined, and darkest, while those lying near the tip and the base of the mimic leaf are shorter and often even shadowy. On the left side the second lateral vein in particular distinctly shows indentations indicative of the rings, inherited from the ancestral forms, which surrounded the still visible eye-spots (Aufl); the third lateral vein is quite indefinite and shadowy, but nevertheless it runs exactly parallel to the first two, and thus heightens the deceptive effect. We can thus distinguish older and more recent elements in the marking—a proof of the slow and successive origin of the picture.
This is not reconcilable with the conception of a germ-plasm without primary constituents, however complex a mixture it may otherwise be. A substance which had to undergo thousands upon thousands of variations, arising from each other according to law and in the strictest succession, in order that it might become a definite organism, predetermined as to all its thousands of parts down to the most minute, cannot vary over and over again in its whole constitution without the consequences showing themselves in numerous, or indeed in all, the parts of the body. Such variations in the germ-plasm would be comparable to many successive deviations of a ship from her course, which, although the single ones would only cause a minimal deviation from the true course, would, when summed up in a voyage of some length, land the vessel at quite another coast than the one intended. If each individual adaptation of the species depended on a variation of the whole germ-plasm the wood Kallima would soon retain no resemblance to its ancestral form, the meadow species; yet we are acquainted with species of Kallima which do not show the special resemblance to a leaf, but, for instance, still exhibit the perfectly developed eye-spot of the ancestral form, and so forth. It follows, therefore, that the origin of the leaf-picture has not greatly influenced the general character of the species; and the fact that the upper surface of the wings has remained the same in all the varieties is in itself enough to prove this.
Since, then, the resemblance to a leaf cannot have arisen without something in the germ-plasm varying, since the germ-plasm of a forest Kallima and a meadow Kallima must be different in something, and cannot be any more alike than the germ-plasm of a fantail-pigeon and a carrier, there must be 'primary constituents' in the germ-plasm, that is, vital units whose variation occasions the variation of definite parts of the organism, and of these alone.
Fig. 17. Caterpillar of Selenia tetralunaria on a twig of birch. K, head. F, feet. m, protuberances resembling dormant buds. Natural size.
It is on such considerations as these that my assumption, that [Pg 360]the germ-plasm is composed of determinants, depends. There must be as many of these as there are regions in the fully-formed organism capable of independent and transmissible variation, including all the stages of development. Every part, for instance, of the butterfly's wing, which is capable of independent and transmissible variation, must, so I conclude, be represented in the germ-plasm by an element which is likewise variable, the determinant; but the same must be true of every independently and transmissibly variable spot of the caterpillar from which the butterfly developed. We know how markedly caterpillars are adapted in form and colour to their environment. Let us assume that the caterpillar of the butterfly which we chose as an example of wing-marking had the habit of feeding only by night and during the daytime of resting on the trunk of a tree, or, more precisely, in the crevices of the bark. It would then resemble the caterpillar of the moths of the genus Catocala or the Geometers (Geometridæ), and possess the colour of the bark of the tree in question; the determinants of the skin would thus have varied to correspond with this mode of life on the part of the caterpillar, so that the skin would appear grey or brown. But there cannot be only one[Pg 361] determinant of the caterpillar skin in the germ-plasm, for the bark-like colour of, for instance, a Geometer caterpillar is not a uniform grey, but has darker spots at certain places and lighter whitish spots at others, such as are to be seen on the bark of the twig on which the caterpillar is wont to rest, or brown-red spots, like those on the cover-scales of the buds, or little warts and protuberances which exactly correspond to similar roughnesses on the twigs, to cracks in the bark, and so on. All these markings are constant, and are to be found in the same spot in every caterpillar of the species. A large number of regions of the caterpillar skin must therefore be independently determined by the germ-plasm; the germ-plasm must contain parts the variations of which bring about variations only of an independently variable region of the caterpillar skin. In other words, in the germ-plasm of the butterfly ovum there must not only be determinants for many regions of the butterfly's wing, but also for many regions of the caterpillar's skin.
This line of argument, of course, applies to all the bodily parts and organs of the butterfly and of the caterpillar, as well as to all the stages of development of the species as far as these parts are able to vary in such a way that the variation reappears in the following generation, that is to say, as far as it is transmissibly variable.
But all parts must be transmissibly variable which have exhibited independent variation in relation to their ancestors. When, for instance, the eggs of a butterfly (Vanessa levana) bear a deceptive resemblance to the flower-buds of the stinging-nettle on which the caterpillar lives, not only in form and colour, but in their pillar-like arrangement, we may conclude that these eggs have varied transmissibly from those of their ancestors, which had not acquired the habit of living on the stinging-nettle, in these three respects independently, that is, uninfluenced by any other variations the species may have undergone; and that, consequently, the germ-plasm must contain determinants for the egg-shell, egg-colouring, and so on. The manner of laying the eggs in the form of pillars depends on a modification of the egg-laying instinct, which must in its turn depend on the variations of certain nerve-centres, and we learn from this that there must be in the germ-plasm determinants for the individual centres of the nervous system.
It may, perhaps, be suggested that matters could be explained in a simpler way—that it is enough to assume the presence in the egg of determinants for all the parts of the caterpillar, and that those of the butterfly are only formed within the caterpillar.
This suggestion seems justifiable if we confine ourselves to superficial considerations. We read in every handbook of entomology that the wings only arise during the life of the caterpillar, and in a certain sense this is true, for the primary constituents or primordia of wings only develop into the fully formed wing during the larval period. But even if these primordia were only formed during the caterpillar-stage, what could they develop from? Only out of the material parts of the caterpillar, that is, from some of its living cells or cell-groups. The constitution of the wings would therefore be dependent on that of the cells of the caterpillar from which they arose, so that if these varied transmissibly through the variation of their determinants contained in the germ, the determinants of the butterfly which were just developing would vary with them; every transmissible variation of the caterpillar would necessarily cause a similar variation in the butterfly, and this does not happen. If any one hazarded the assumption that the determinants of the butterfly develop only in the caterpillar, but quite independently of its constitution, he would either be making an absurd statement, namely, that the characters of the butterfly were not transmissible at all, or he would be unconsciously admitting that the determinants of the butterfly were already contained in the parts of the caterpillar, and come direct from the germ-plasm.
That the characters of the butterfly do vary independently of those of the caterpillar I demonstrated many years ago, when we were still very far away from the idea of the germ-plasm or of determinants. I demonstrated then that the constancy of the markings of a species can be quite different in the two chief stages; that the caterpillar may be very variable, while the butterfly or the moth may be very constant in all its markings, or conversely. I called attention to the dimorphic caterpillars which are green or brown, and yet become the same moth (for instance, Deilephila elpenor and Sphinx convolvuli); I cited the case of the spurge hawk-moth (Deilephila euphorbiæ), whose dark but at the same time motley caterpillars occur in the Riviera at Nice as a local variety (Nicæa), and there wear quite a different dress—pale clay-yellow, with a double row of large conspicuous dark yellow eye-spots—while the moth does not differ from our variety in a single definite character, except in its larger size. At that time, too, I instituted experiments with the caterpillars of the smallest of our indigenous Vanessa species (Vanessa levana), of which the majority are black with black thorns, while a minority are yellowish-brown with yellow thorns; reared separately, both yielded the same butterfly, though in this case one[Pg 363] would be inclined to suppose that there was some internal connexion between the colour of the caterpillar and that of the butterfly, since the butterfly also occurs in two colours. It was shown, however, that the colour of the butterfly had nothing to do with that of the caterpillar, for it is known to be dependent on the season, and is a seasonal dimorphism, 'while the two forms of caterpillar may occur side by side at all times of the year.'
Subsequently I made a similar experiment with the dimorphic caterpillars of the 'fire'-butterfly (Polyommatus phlæas), and it yielded the same result. The pure green caterpillars became the same butterflies as those marked with broad red longitudinal stripes, and in this case we can definitely describe both colours as protective, for the green form is adapted to the green under surface of the leaf, the red-striped to the green red-edged stalk of the lesser sorrel (Rumex acetosella).
There was really no necessity for special proofs that the caterpillar and butterfly vary transmissibly in complete independence of each other, for the facts of metamorphosis alone are enough to prove it. How would it have been possible otherwise that the jaws adapted for biting should, in the primitive insects, and in the locusts which are nearest to them, remain as a biting apparatus throughout life, while in the caterpillar they are modified during its pupal stage into the suctorial proboscis of the butterfly? The parts of insects, therefore, must be capable of transmissible variation in the stages of life independently of each other. Not only have the jaws of the leaf-eating caterpillars remained unaltered, while in the sexually mature animal they have been gradually modified into a very long and extremely complex suctorial apparatus, but when at a much later time this proboscis became superfluous in a species, because the butterfly or moth, from some cause or another, lost the habit of taking any nourishment at all, its degeneration exercised no effect on the jaws of the caterpillar, as we can observe in many hawk-moths, silk-moths and Geometridæ. How could such a degeneration become transmissible if the caterpillar's jaws, from which those of the adult are developed, remain the same? We are thus forced to assume that there is something in the latter which can vary from the germ, without the jaws themselves being altered thereby. This 'something' it is which I call 'determinants,' vital particles, which—however we may try to picture them—are indeed contained in the cells of the caterpillar's jaws, but are there inactive and do not influence the structure of these, while, on the other hand, it is their constitution which determines the form and structure[Pg 364] of the suctorial proboscis of the butterfly down to the minutest details. It must be these alone which cause the suctorial proboscis to develop, and in some cases to degenerate again, without bringing about any change in the corresponding parts in the caterpillar.
Fig. 89. Anterior region of the larva
of a Midge (Corethra plumicornis). K, head.
Th, thorax. ui, inferior imaginal disks.
oi, superior imaginal disks. ui1, ui2, and
ui3, the primordia of the limbs. oi2
and oi3, the primordia of the wings and
'balancers.' g, brain. bg, chain of ventral
ganglia with nerves which enter the
imaginal disks. trb, tracheal vesicle.
Enlarged about 15 times.
This example seems to me to be preferable to that of the wings of insects in this respect, that there is no organ in the caterpillar with a specific function corresponding to the wing of the butterfly. Yet the two cases are exactly alike, and it would be a mistake to say that the first primordium of the wing within the caterpillar is not a part of the caterpillar at all. At first, certainly, it is only a group of cells on the skin, occurring at a particular spot on the dorsal surface of the second and third segments of the caterpillar, and doubtless arising from a single cell of the embryo, the 'primitive wing-cell,' which, however, has not as yet been demonstrated. But it is nevertheless an integral part of the caterpillar, which could neither be wanting, nor be larger or smaller, and so on; which, in short, does mean something for the caterpillar, although perhaps not more than any other of the skin-cells. For the butterfly, however, this area on the skin means the rudiment of the wing; for from it alone can there arise by multiplication the aggregate of cells which grows out into a hollow protuberance, enlarges by degrees into a disk, the imaginal disk, and eventually develops into the form of wing peculiar to the species. This imaginal disk is connected very early with nerves and with tracheæ, as may be beautifully seen especially in dipterous larvæ (Fig. 89, oi), and these become later the nerves and tracheæ of the wing, while thousands of peculiar scale-like hairs develop on the upper surface; in short, the rudiment becomes a perfect wing with its specific venation, and with the marking and colouring which is often so complicated in Lepidoptera. Almost every little spot and stripe of the latter is handed down with the most tenacious power of transmission from generation to generation, and each can at the same time be transmissibly varied; the same is true of the venation, which is so important systematically just because it is so strictly hereditary, yet it too can vary transmissibly, as can also the hooked[Pg 365] bristles, the odoriferous apparatus, and, in short, the whole complex structure of the wing, with all its specific adaptations to the mode of flight, to the manner of life, and to the colour of the environment. How is it possible that all this can develop from a skin-cell? Is it the influence of position that effects it, and could any other cell of the caterpillar's skin do the same if it were placed in the same position? Could any neighbour-cell of the primitive wing-cell replace it if it were destroyed? It is hardly probable, and I think I can even prove that this is not so. The experiment of killing such a cell in the living animal has not yet been made; if it should succeed, we may venture to say in advance that none of the neighbouring skin-cells will be able to do its work and take its place in developing a wing; the wing in question will simply remain undeveloped. In the summer of 1897 I hatched a specimen of Vanessa antiope from the pupa, which, though otherwise normal and well-developed, lacked the left posterior wing altogether; no trace of it could be recognized. In this case, from some cause which could no longer be discovered, the first formative cell of the wing in the hypodermis, or its descendants, must have been destroyed, and no substitution of another took place, as the defect showed.
The young science of developmental mechanics attributes to the position of a cell in the midst of a group of cells a determining value as regards its further fate, and as far as the cells of the segmenting ovum are concerned this seems to be true in certain cases, but the assumption cannot be generally true except in a very subordinate sense. The formative cell of the wing does not become what it is because of its relative position in the organism. If this were so it could not happen that a wing should develop instead of a leg, as was observed in a Zygæna, nor could there be any of those deformities already referred to, to which the name 'Heterotopia' is applied, and which consist in the development of organs of definite normal structure, or at any rate of apparently normal structure in quite unusual places, e. g. an antenna on the coxa of a leg, or of a leg instead of an antenna (in Sirex), or instead of a wing. It is therefore not some influence from without that makes that particular skin-cell of the caterpillar the rudiment of the wing, but the reason lies within itself, in its own constitution. As the whole mass of determinants for the whole body and for all the stages of its development must be contained within the ovum and the sperm-cell, so the primitive cell of the butterfly's wing must contain all the determinants for the building up of this complicated part;[Pg 366] and if the cell gets into a wrong position in the course of development because of some disturbance or other, a wing may develop from it in that position if the conditions are not too utterly divergent. These heterotopic phenomena afford a further proof of the existence of determinants, because they are quite unintelligible without the assumption of 'primary constituents' or Anlagen.
The hypothesis of determinants in the germ-plasm is so fundamental to my theory of development that I should like to adduce another case in its support and justification. The limbs of the jointed-footed animals, or Arthropods, originally arose as a pair on each segment of the body, and they were at first alike or very similar both in their function and in their form. We find illustration of this in the millipedes, and still more in the species of the interesting genus Peripatus, which resembles them externally, as well as in the swimming and creeping bristle-footed marine worms (Chætopods) belonging to the Annelid phylum. We can quite well picture to ourselves that the whole series of these appendages was represented in the germ-plasm by a single determinant or group of determinants, which only required to be multiplied in development. Without disputing whether this has really been the case in the primitive Arthropods or not, it is certain that it can no longer be the case in the germ-plasm of the Arthropods of to-day. In these each pair of appendages must be represented by a particular determinant. We must infer this from the fact that the several pairs of these appendages have varied transmissibly, independently of each other, for some are jaws, others swimming legs, or merely bearers of the gills or of the eggs; others are walking legs, digging legs, or jumping legs. In Crustaceans a forceps-like claw is often borne by the first of the otherwise similarly constructed appendages, or also by the second or the third, or there may be no forceps, and so on; in short, we see that each individual pair has adapted itself independently to the mode of life of its species. This could only have been possible if each was represented in the germ-plasm by an element, whose variations caused a variation only in that one pair of legs, and in no other.
It may perhaps be objected that the differences in the appendages may quite well have had their origin simply during the development of the animal, while the primary constituents were the same for all, so that a single determinant in the germ-plasm would suffice. But this could only be the case if the differences depended not on internal but on external causes, that is, if the same primary constituents gave rise to a set of appendages which became different because they were[Pg 367] subject in the course of their development to different modifying influences. But this is not the case, at least not to the extent that this supposition would necessitate. Can it be supposed that, for instance, the jumping legs of the water-flea (Gammarus) are a necessary consequence of the somewhat divergent form of the segments from which they grow? A direct proof to the contrary may be found in 'Heterotopia,' for in the place where a posterior limb, modified for holding the eggs, normally occurs in the crab an ordinary walking leg may exceptionally develop (Fig. 90, Bethe), or an appendage resembling an antenna may take the place of an extirpated eye (Herbst). But if there were really only one determinant in the germ-plasm for all the appendages these would of necessity be all alike, apart from the larger or smaller differences which might be stamped upon them by growing from segments different in size and in nutrition. Such differences, however, are far from being sufficient to explain the great deviations seen among the appendages of most kinds of Crustaceans, and still less to explain their adaptation to quite different functions.
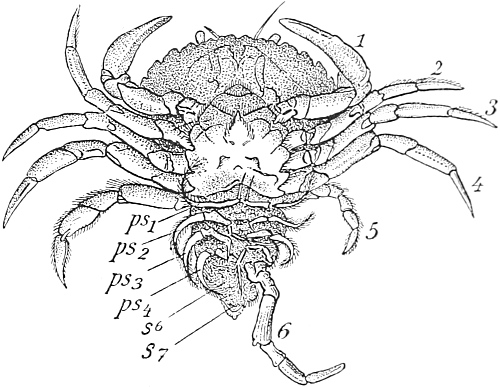
Fig. 90. The Common Shore-Crab (Carcinus mænas), seen from below, with the abdomen forced back. In place of the swimmeret, which ought to be borne by the fifth abdominal swimmeret, a walking leg has grown on the left side, and one which properly should belong to the right side (6). 1-5, thoracic limbs, ps1-4, swimmerets of the right side. s6, s7, posterior segments of the abdomen. After Bethe.
It need not be imagined that my argument can be controverted by saying that one appendage-determinant in the germ may split itself in the course of development into a series of different appendage-determinants. The question would then arise, How is it able to do so? And the answer can be no other than that the single first determinant had within it several different kinds of elements, which subsequently[Pg 368] separated to determine in different ways the various appendages. But that is just another way of saying that this single determinant actually includes within itself several different determinants. For a determinant means nothing more than an element of the germ-substance by whose presence in the germ the specific development of a particular part of the body is conditioned. If we could remove the determinants of a particular appendage from the germ-plasm this appendage would not develop; if we could cause it to vary the appendage also would turn out differently.
In this general sense the determinants of the germ-plasm are not hypothetical, but actual; just as surely as if we had seen them with our eyes, and followed their development. Hypothesis begins when we attempt to make creatures of flesh and blood out of these mere symbols, and to say how they are constituted. But even here there are some things which may be maintained with certainty; for instance, that they are not miniature models, in Bonnet's sense, of the parts which they determine; and, further, that they are not lifeless material, mere substances, but living parts, vital units. If this were not so they would not remain as they are throughout the course of development, but would be displaced and destroyed by the metabolism, instead of dominating it as living matter alone can do—doubtless undergoing oxidation, but at the same time assimilating material from without, and thereby growing. There cannot be lifeless determinants; they must be living units capable of nutrition, growth, and multiplication by division.
And now we have arrived at the point at which a discussion of the organization of the living substance in general can best be interpolated.
The Viennese physiologist, Ernst Brücke, forty years ago promulgated the theory that living matter could not be a mere mixture of chemical molecules of any kind whatever; it must be 'organized,' that is, it must be composed of small, invisible, vital units. If, as we must certainly assume, the mechanical theory of life is correct, if there is no vital force in the sense of the 'Natur-Philosophie,' Brücke's pronouncement is undoubtedly true; for a fortuitous mixture of molecules could no more produce the phenomena of life than a single molecule could, because, as far as our experience goes, molecules do not live; they neither assimilate, nor grow, nor multiply. Life can therefore arise only through a particular combination of diverse molecules, and all living substance must consist of such definite groups of molecules. Shortly after Brücke, Herbert Spencer likewise assumed the reality of such vital 'units,' and the same assumption[Pg 369] has been made in more recent times by Wiesner, De Vries, and myself. In the meantime we can say nothing more definite about the composition of these bearers of life, or 'biophors,' as I call them, than that albumen-molecules, water, salts, and some other substances play the chief part in their composition. This has been found out by analysis of dead protoplasm; but in what form these substances are contained in the biophors, and how they affect each other in order to produce the phenomena of life by going through a ceaseless cycle of disruptions and reconstructions, is still entirely hidden from us.
We have, however, nothing to do with that here; we content ourselves with recognizing in the biophors the characteristics of life, and picturing to ourselves that all living substance, cell-substance, and nuclear substance, muscle-, nerve-, and gland-substance, in all their diverse forms, consist of biophors, though, of course, of the most varied composition. There must be innumerable kinds of biophors in all the diverse parts of the millions of forms of life which now live upon the earth; but all must be constructed on a certain fundamental plan, which conditions their marvellous capacity for life; all possess the fundamental characters of life—dissimilation, assimilation, growth, and multiplication by division. We must also ascribe to them in some degree the power of movement and sensibility.
As to their size, we can only say that they are far below the limits of visibility, and that even the minutest granules which we can barely perceive by means of our most powerful microscopes cannot be small individual biophors, but must be aggregates of these. On the other hand, the biophors must be larger than any chemical molecule, because they themselves consist of a group of molecules, among which are some of complex composition, and therefore of relatively considerable size.
It may now be asked whether the determinants, whose existence we have already inferred, are not identical with these 'biophors' or smallest living particles; but that is not the case, at least not generally. We called determinants those parts of the germ-substance which determine a 'hereditary character' of the body; that is, whose presence in the germ determines that a particular part of the body, whether it consists of a group of cells, a single cell, or a part of a cell, shall develop in a specific manner, and whose variations cause the variations of these particular parts alone.
Again, it may be asked how large and how numerous such 'hereditary parts' may be, whether they correspond to every distinct part of a cell, or to every cell of the body, or only to the larger cell groups. Obviously the areas which are individually determined from[Pg 370] the germ must differ in size, according as we have to do with an organism which is small or large, simple or more complex. Unicellular organisms, such as Infusorians, probably possess special determinants for a number of cell-organs and cell-parts, although we cannot directly observe the independent and transmissible variation of these organs; lowly multicellular animals, such as the calcareous sponges, will require a relatively small number of determinants, but in the higher multicellular organisms, as, for instance, in most Arthropods, the number must be very high, reaching many thousands if not hundreds of thousands, for in them almost everything in the body is specialized, and must have varied through independent variation from the germ. Thus in many Crustaceans the smelling-hairs occur singly on special joints of the antennæ, and the number of joints furnished with a smelling-hair is different in different species; the size, too, of the smelling-hairs themselves varies greatly, being, for instance, much smaller in our common Asellus than in the blind form from the depths of our lakes, in which the absence of sight is compensated for by an increased acuteness of the sense of smell. Thus the smelling-hairs may vary transmissibly in themselves, while any joint of the antennæ may also produce one independently through variation. In this case accordingly we must assume that there are special determinants for the smelling-hairs, and for the joints of the antennæ. But we cannot always and everywhere refer identical or approximately similar organs, when there are many of them, to a corresponding number of determinants. Certainly the hairs of mammals or the scales of butterflies' wings do not all vary individually and independently, but those of a certain region vary together, and are therefore probably represented in the germ-plasm by a single determinant. These regions often appear to be very small, as is best seen by the fine lines, spots, and bands which compose the marking of a butterfly's wing, and still more in the odoriferous scales occurring in some butterflies, as, for instance, in the blue butterflies (Lycæna). These little lute-shaped scales do not occur in all species, and they occur in very unequal numbers even in those which possess them; there are certain species which exhibit only about a dozen, and these are all on one little spot of the wing. Since these odoriferous scales must have arisen as modifications of the ordinary hair-like scales, as one of my pupils, Dr. Köhler, has demonstrated by comparative studies, these ordinary hair-like scales must have varied transmissibly at certain spots, that is, their determinants have varied while those of the surrounding scales have not.
The case is the same in respect to the sound-producing apparatus[Pg 371] of many insects. Many grasshoppers produce sounds by fiddling with the thigh of the hind leg on the wing, others by rubbing one anterior wing upon the other, and, indeed, always with one particular vein in one upon a particular vein in the other. One of these serves as the bow, the other as the string, of the violin, and the bow is furnished with teeth (Fig. 91), ranged beside each other in a long row, which have the same function as the colophonium of the violin, that is, to grasp and release the strings alternately, and thus to produce resounding vibrations. My pupils, Dr. Petrunkewitsch and Dr. Georg von Guaita, have recently proved that these teeth have arisen as modifications of the hairs which are scattered everywhere over the wing and leg. But only in this one place, on the so-called 'stridulating-vein,' have they been modified to form stridulating teeth (schr). Thus this vein must be capable of transmissible variation by itself alone, that is, there must be parts contained in the germ-plasm, the variation of which causes a variation solely of this individual vein and its hairs, possibly even a variation only on certain hairs on this vein.
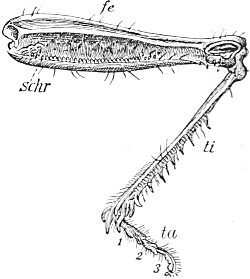
Fig. 91. Hind leg of a Locustid
(Stenobothrus protorma), after Graber. fe,
femur. ti, tibia. ta, tarsal joints. schr,
the stridulating ridge.
On the other hand, there are also large regions, whole cell-masses of the body, which in all probability vary only en bloc, as, for instance, the milliards of blood-cells in Man, the hundreds of thousands or millions of cells in the liver and other glandular organs, the thousands of fibres of a muscle, or of the sinews or fascia, the cells of a cartilage or a bone, and so on. In all these cases a single determinant, or at least a few in the germ-plasm, may be enough. But in numerous cases it is impossible to say how large the region is which is controlled by a single determinant, and it is, of course, of no importance to the theory. In unicellular organisms the determinants will control parts of cells, in multicellular organisms often whole cells and groups of cells.
Perhaps an inference as to the nature of the determinants may be drawn from this with some probability, in as far as mere parts of cells may be supposed to have simpler determinants than whole cells and groups of cells. The determinants in the chromosomes of uni[Pg 372]cellular organisms may therefore often consist of single biophors, so that in this case the conception of biophors would coincide with that of determinants. In multicellular organisms, on the other hand, I should be inclined on the whole to picture the determinant as a group of biophors, which are bound together by internal forces to form a higher vital unity. This determinant must live as a whole, that is, assimilate, grow, and multiply by division, like every vital unit, and its biophors must be individually variable, so that the separate parts of a cell controlled by them may also be capable of transmissible variation. That they are so, every highly differentiated cell of a higher animal teaches us; even the smelling-hairs of a crab exhibit a stalk, a terminal knob, and an internal filament, and many muscle-, nerve-, and gland-cells are much more complex in structure.
THE GERM-PLASM THEORY (continued)
Structure of the germ-plasm—Vital affinities—Division—O. Hertwig's chief objections to this theory—Male and female eggs in the Phylloxera show differential division—Dispersal of the germ-plasm in the course of Ontogeny—Active and passive state of the determinants—Predetermination of cells—There are no determinants of characters—Liberation of the determinants—Accessory idioplasm—Herbst's lithium larvæ—Plant galls—Cells with several facultative determinants—Connective tissue in vertebrates—Mesoderm cells of Echinoderms—Sexual dimorphism—Female and male ids—Polymorphism (Papilio merope)—Ants.
I have endeavoured to prove that the germ-substance proper must be looked for in the chromatin of the nucleus of the germ-cell, and more precisely still in those ids or chromosomes which we conceive of as containing the primary constituents (Anlagen) of a complete organism. Such ids in larger or smaller numbers make up the whole germ-plasm of a germ-cell, and each id in its turn consists of primary constituents or determinants, i.e. of vital units, each of which determines the origin and development of a particular part of the organism. We have now to make an attempt to picture to ourselves how these determinants predetermine those cells or cell-groups to which they correspond. In doing so we have to fall back upon mere hypotheses, and in stating any such hypothesis I wish expressly to emphasize that I am only following up one of the possibilities which our imaginative faculty suggests. Nevertheless, to endeavour to form such a conception is certainly not without use, for it is only by elaborating a theory to the utmost that we are able to use it in application to concrete cases, thus stimulating the search for corroboratory or contradictory facts, and leading gradually to a recognition of the gaps or mistakes in the theory.
The first condition that must be fulfilled in order that a determinant may be able to control a cell or cell-group is that it should succeed in getting into it. It must be guided through the numerous cell-divisions of ontogeny so that it shall ultimately come to lie in the cells which it is to control. This presupposes that each determinant has from the very beginning its definite position in relation to the rest, and that the germ-plasm, therefore, is not a mere loose aggregate of determinants, but that it possesses a structure, an architecture, in[Pg 374] which the individual determinants have each their definite place. The position of the determinants in relation to one another cannot be due to chance, but depends partly on their historical development from earlier ancestral determinants, partly on internal forces, such as we have already assumed for keeping the determinants together. We may best designate these hypothetical forces 'affinities,' and in order to distinguish them from mere chemical affinities we may call them 'vital.' There must be forces interacting among the different determinants which bind them together into a living whole, the id, which can assimilate, grow, and multiply by division, in the same manner as we were forced to assume for the smaller units, the biophors and single determinants. In the ids, however, we can observe the working of these forces quite directly, since each chromosome splits into two halves of equal size at every nuclear division, and not through the agency of external forces, e.g. the attraction which we may assume to be exerted by the fibrils of the nuclear spindle, but through purely internal forces, often long before the nuclear spindle has been formed at all.
But if the determinants must separate from each other in the course of development so as to penetrate singly into the cells they are to control, the id must not only have the power of dividing into daughter-ids of identical composition, it must also possess the power of dividing under certain influences into dissimilar halves, so that the two daughter-ids contain different complexes of determinants. The first mode of division of the id, and with it of the nucleus and of the cell, I call erbgleich, or integral, the second erbungleich, or differential. The first form of multiplication is the usual one, which we observe everywhere when unicellular organisms divide themselves into two equal daughter-units, or when the cells of multicellular bodies produce their like by division into two. The second is not directly observable, because a dissimilarity of the daughter-cells, as long as it lies only in the idioplasm, cannot be actually seen; it can only be inferred from the different rôle which the two daughter-cells play in the building up of the individual. When, for instance, one of two sister-cells of the embryo gives rise to the cells of the alimentary canal and the other to those of the skin and the nervous system, I infer that the mother-cell divided its nuclear substance in a differential way between the two daughter-cells, so that one contained the determinants of the endoderm, the other those of the ectoderm; or when a red and a black spot lie side by side and under exactly the same conditions on the wing of a butterfly, I conclude that the ancestral cells of these two spots have divided differentially, so that[Pg 375] one received the 'red,' the other the 'black' determinants. Our eyes can perceive no difference between the nuclear substance of the two cells, but the same is true of the chromosomes of the paternal and maternal nuclei in the fertilized ovum, although we know in this case that they contain different tendencies. In any case we are not justified in concluding from the apparent similarity of the chromosome-halves in nuclear division that there cannot be differential division. The theoretical possibility that there is such differential division cannot be disputed; indeed, I am inclined to say that it is more easily imagined than the division of the ids into absolutely similar halves. Both are only conceivable on the assumption that there are forces which control the mutual position of the determinants in the ids, that is, on the assumption of 'affinities.' I shall not follow this further, but that there are forces operative within the ids which are still entirely unknown to us is proved at every nuclear division by the spontaneous splitting of the chromosomes.
It has been objected to my theory that such a complex whole as the id could not in any case multiply by division, since there is no apparatus present which can, in the division into two daughter-units, re-establish the architecture disturbed by the growth. But this objection is only valid if we refuse to admit the combining forces, the 'vital affinities' within the ids, and the same is true for the smaller vital units. An ordinary chemical molecule cannot increase by division; if it be forcibly divided it falls into different molecules altogether; it is only the living molecule, that is, the biophor, which possesses this marvellous property of growth and division into two halves similar to itself and to the ancestral molecule, and we may argue from this that in the division of the ids forces of attraction and repulsion must likewise be operative[19].
[19] In my book The Germ-Plasm I have already assumed the existence of 'forces of attraction' in the determinants and biophors, as in the cells. I did not, indeed, enter into details, but I argued on the same basis as now (Germ-Plasm, p. 64, English edition). My critics have overlooked this.
I see no reason why we should not assume the existence of such forces, when we make the assumption that the hundreds of atoms which, according to our modern conceptions, compose the molecule of albumen and determine its nature, are kept by affinities in this definite and exceedingly complex arrangement. Or must we suppose that between the atom-complex of the molecule and the next higher atom-complex of the biophor, determinant, and id there is an absolute line of demarcation, so that we must assume quite different forces in the latter from those we conceive of as operative in the former? The[Pg 376] biophor is ultimately only a group of molecules, the determinants a group of biophors, the id a group of determinants, and all the three inferred stages of vital organization only become real units through the forces operating within them and combining them into a whole. What compels the chromatin granules of the resting nucleus to approach each other at the time of cell-division, to unite into a long, band-like thread, and what is it that subsequently causes this thread to break up again into a definite number of pieces? Obviously only internal forces of which we know nothing further than that they are operative.
We shall see later that this assumption of vital affinities must be made not only in regard to the cells, but also in regard to entire organisms whose parts are united by an internal bond, and whose co-ordination is regulated by forces of which we have as yet no secure knowledge. In the meantime we may designate these forces by the name of 'vital affinities.'
It must be admitted, however, that some objections of a fundamental nature have been urged against the assumption of a differential nuclear division of the hereditary substance. O. Hertwig holds that the assumption of differential division is essentially untenable, because it is contradictory to 'one of the first principles of reproduction,' for 'a physiologically fundamental character of every living being is the power of maintaining its species.'
This certainly seems so, but a closer examination shows that this 'principle,' although correct enough when taken in a very general sense, does not really cover the facts, and is therefore incapable of supporting the inferences drawn from it. If the proposition expressed the whole truth there could have been no evolution from the primitive organisms to higher ones, every living being must have simply reproduced exact copies of itself. Whether the transformations of species have been sudden or gradual, whether they have been brought about by large steps or by very small ones, they could only have come about by breaking through this so-called 'principle' of like begetting like. In fact, we may with more justice maintain the exact converse of the principle, and say that 'no living being is able to produce an exact copy of itself,' and this is true not only of sexual, but of asexual reproduction.
In ontogeny we see exactly the same thing. There are no two daughter-cells of a mother-cell which are exactly alike, and the differences between them, if they increase in the same direction, may lead in later descendants to entire differences of structure. Indeed the whole process of development depends on such an augmentation[Pg 377] of the differences between two daughter-cells—on differences which proceed from within and are definitely pre-established. Here, again, the facts do not justify us in making a dogma of the proposition that it is a 'fundamental power' of every living being to maintain its species by producing replicas of itself. If we look at two directly successive cell-generations, we can hardly, it is true, in most cases, perceive any difference between them, just as in the generations of species; but if we compare the end of a long cell-lineage with the beginning, then the difference is marked, and we recognize that the difference is due to a gradual summing up of minute, invisible deviations. In my opinion these steps of difference cannot possibly depend merely on direct external influences; they proceed rather from the hereditary substance the cell receives from the ovum, which, therefore, in order to attain to such many-sided and far-reaching differentiation, must have undergone a frequently repeated splitting up of its qualities. That this splitting is not merely a variation to which the whole of the hereditary substance of the daughter-cells is uniformly subject, according to the influences dependent on their position in relation to other cells of the embryo, will be made clear from the case of the Ctenophora referred to in the next lecture. A scarcely less striking example is that of those animals in which the ova contain the primary constituents for only one sex, in which, in other words, there are 'male' ova and 'female' ova. This is the case, for instance, among Rotifers, and in plant-lice such as the vine-pest, Phylloxera. Here the eggs from which males develop are smaller than those which produce females. The primary constituents for both male and female are not, as in most animals, contained in the same ovum, to be liberated on one side or the other by influences unknown to us, but in each ovum there is only one of the two sets of primary constituents present, and in this case, therefore, the development of hermaphrodites, which not infrequently occur in other animals, would be impossible. But all these ova have been produced by one primitive reproductive cell, and consequently, at one of the divisions implied in the multiplication of this first cell, a separation of the male from the female primary constituents must have taken place, that is, a differential division of hereditary substance, for which no external and no intercellular influences can possibly account.
If there is, then, a differential division of the ids and with them of the whole idioplasm, the germ-plasm of the fertilized ovum must be broken up in the course of ontogeny into ever smaller groups of determinants. I conceive of this as happening in the following manner.
In many animals the fertilized ovum divides at the first segmentation into two cells, one of which gives rise predominantly to the outer, the other to the inner germinal layer, as in molluscs, for instance. Let us now assume that this is the case altogether, so that one of the first two blastomeres gives rise to the whole of the ectoderm, the other to the whole of the endoderm; we should here have a differential division, for the developmental import (the 'prospective' of Driesch) of the primitive ectoderm-cell is quite different from that of the primitive endoderm-cell, the former giving origin to the skin and the nervous system, with the sense organs, while the second gives rise to the alimentary canal, with the liver, &c. Through this step in segmentation, I conclude, the determinants of all the ectoderm-cells become separated from those of the endoderm-cells; the determinant architecture of the ids must be so constructed in such species that it can be segregated at the first egg-cleavage into ectodermal and endodermal groups of determinants. Such differential divisions will always occur in embryogenesis when it is necessary to divide a cell into two daughter-cells having dissimilar developmental import, and consequently they will continue to occur until the determinant architecture of the ids is completely analysed or segregated out into its different kinds of determinants, so that each cell ultimately contains only one kind of determinant, the one by which its own particular character is determined. This character of course consists not merely in its morphological structure and chemical content, but also in its collective physiological capacity, including its power of division and duration of life[20].
[20] Emery has lately called attention to another direct proof of the existence of differential cell- and nucleus-division. According to observations made by Giardina, in the water-beetle (Dytiscus), one primitive ovum-cell gives rise, through four successive divisions, to fifteen nutritive cells and one well-defined ovum-cell. But only half of the nuclear substance takes part in these divisions, the rest remains inactive in a condensed, cloudy condition. 'The meaning of the whole process is obviously that the germ-plasm mass as a whole is handed over to the ovum-cell, while the nutritive cells receive only the nuclear constituents which belong to them' (Biol. Centralbl., May 15, 1903).
But embryogenesis does not proceed by differential divisions alone, for integral divisions are often interpolated between them, always, for instance, when in a bilateral animal an embryonic cell has to produce by division into two a corresponding organ for the right and left sides of the body; for instance, in the division of the primitive genital cell into the rudiments of the right and left reproductive organs, or in the division of the primitive mesoderm-cell into the right and left initial mesoderm-cell, but also later on in the course of embryogenesis, when, for instance, the right or the left primitive reproductive[Pg 379] cell multiplies into a large number of primitive germ-cells, or in the multiplication of the blood-cells, or of the epithelial cells of a particular region; in short, whenever mother and daughter-cells have the same developmental import, that is, when they are to become nothing more than they already are. In all such cases a similar group of determinants, or a similar single determinant, must in the nuclear division penetrate into each of the two daughter-cells.
It is in this way, it seems to me, that the determinants gain entrance into the cells they are to control, by a regulated splitting up of the ids into ever smaller groups of determinants, by a gradual analysis or segregation of the germ-plasm into the idioplasms of the different ontogenetic stages. When I first developed this idea I assumed that the splitting process would in all cases set in at the same time, namely, at the first division of the ovum. But since then, in the controversies excited by the theory, many facts have been brought to light which prove that the ova of the different animal groups behave differently, and that the splitting up of the aggregate of primary constituents may sometimes begin later—but I shall return to this later on.
If we accept the segregation hypothesis, which is similar in purport to that advanced by Roux as the' mosaic theory,' it must strike us as remarkable that the chromatin mass of the nucleus does not become notably smaller in the course of ontogeny, and even ultimately sink to invisibility. Determinants lie far below the limits of visibility, and if there were really only a single determinant to control each cell there would be no chromatin visible in such a case. This objection has in point of fact been urged against me, although I expressly emphasized in advance the assumption that the determinants are continually multiplying throughout the whole ontogeny, so that in proportion as the number of the kinds of determinants lying within a cell diminishes the number of resting determinants of each kind increases. When, finally, only one kind of determinant is present there is a whole army of determinants of that kind.
It follows from this conception of the gradual segregation of the components of the id in the course of development that we must attribute to the determinants two different states, at least in regard to their effect upon the cell in which they lie: an active state, in which they control the cell, and a passive state, in which they exert no influence upon the cell, although they multiply. From the egg onwards, therefore, a mass of determinants is handed on by the cell-divisions of embryogenesis, which will only later become active.
My conception of the manner in which the determinants become[Pg 380] active is similar to that suggested by De Vries in regard to his 'Pangens,' very minute vital particles which play a determining part in his 'pangen theory,' similar to that filled by the determinants in my germ-plasm theory. It seems to me that the determinants must ultimately break up into the smallest vital elements of which they are composed, the biophors, and that these migrate through the nuclear membrane into the cell-substance. But there a struggle for food and space must take place between the protoplasmic elements already present and the newcomers, and this gives rise to a more or less marked modification of the cell-structure.
It might be supposed that the structure of these biophors corresponded in advance to certain constituent parts of the cell, that there were, for instance, muscle biophors, which make the muscle what it is, or that the plant-cells acquired their chlorophyll-making organs through chlorophyll biophors. De Vries gave expression to this view in his 'pangen theory,' and I confess that at the time there seemed to me much to be said for it, but I am now doubtful whether its general applicability can be admitted. In the first place, it does not seem to me theoretically necessary to assume that the particles which migrate into the cell-bodies should themselves be chlorophyll or muscle particles; they may quite well be only the architects of these, that is to say, particles which by their co-operation with the elements already present in the cell-body give rise to chlorophyll or muscle substance. As we are as yet unacquainted with the forces which dominate these smallest vital particles, as well as the processes which lead to the histological differentiation of the cells, it is useless in the meantime to make any further hypotheses in regard to them. But in any case the biophors which transform the general character of the embryonic cells into the specific character of a particular tissue-cell must themselves possess a specific structure different from that of other biophors, for they must keep up the continuity of the structures handed on from ancestors, chlorophyll and muscle-substance and the like, since we cannot assume that these structures, so peculiar and so complex in their chemical and physical constitution, are formed afresh, so to speak, by spontaneous generation in each new being, as De Vries has very rightly emphasized. A specific biophor, for instance, of muscle substance will produce this substance as soon as it has found its way into the appropriate cell-body, even though it may not be a contractile element itself.
To this must be added that the structure of the body and the distinctive features of an organism do not depend merely on the histological differentiation of the cells, but quite as much on their[Pg 381] number and arrangement, and on the size and on the frequency of repetition of certain parts. These distinctive characters are just as constant and as strictly transmissible, and may be as heritably variable as those which depend on specific cell-differentiation, and they must therefore likewise be determinable by definite elements of the germ-plasm. Obviously enough, however, these elements are not of the same nature as the known specific histological elementary particles; they can be neither nerve-, muscle-, nor gland-biophors. They must rather be vital units of such a kind that they communicate to the cells and lineage of cells, into whose bodies they migrate from within the nucleus, a definite vital power, that is, an organization which regulates the size, form, number of divisions, and so on, of these cells—in short their whole prospective significance. Always, however, they act in co-operation with the cell-body into which they have penetrated.
Throughout we must hold ourselves aloof from the idea that 'characters' are transmissible. It is customary, indeed, to speak as if this were so, and it is also necessary, because we can only recognize the 'characters' of a body, and not the essential 'nature' on which these characters depend; but the determinants are not seed-grains of individual characters, but co-determinants of the nature of the parts which they influence. There are not special determinants of the size of a cell, others of its specific histological differentiation, and still others of its duration of life, power of multiplication, and so on; there are only determinants of the whole physiological nature of a cell, on which all these and many other 'characters' depend. For this reason alone I should object to the assumption that the determinants of the germ are ready-made histological substances. That is as unlikely as that their groups in the germ-plasm are 'miniature models' of the finished parts of the body.
I conceive of the process of cell-differentiation as follows: at every cell-stage in the ontogeny determinants attain to maturity, and break up so that their biophors can migrate into the cell-bodies, so that the quality of each cell is thus kept continually under control, and may be more or less modified, or may remain the same. By the 'maturity' of a determinant I mean its condition when by continual division it has increased in number to such a point that its disintegration into biophors and their migration into the cell-substance can take place.
One more point I must touch upon here, the question of the 'liberation' or 'stimulation' of the determinants. The activity of an organ never depends on itself alone; the contraction of a muscle is induced by a nerve stimulus or by an electric current; the activity[Pg 382] of the nerve-cells of the brain requires the continual stimulus of the blood-stream, and cannot continue to exist without it; the specific sensory-nerves and sense-cells of the eye, ear, olfactory organ, and so on, are all prompted to activity by adequate stimuli. The same is true in regard to the determinants, they must be 'liberated' if they are to distribute themselves and migrate into the cell-body; and we have to ask how that happens, whether it is possibly due only to their own internal condition, which again would, of course, depend on the nutritive conditions of the cell in which they lie, or whether it is perhaps due to some specific stimulus which is necessary in addition to the fact of 'maturity,' just as a muscle is always ready to contract, yet only does so when it is affected by a specific stimulus.
From the very first, therefore, I have considered whether it would not be better to elaborate the determinant theory in such a way that it would not be necessary to assume a disintegration of the id in the course of ontogeny, but simply to conceive of every expression of activity on the part of a determinant as dependent on a specific stimulus, which in many cases can only be supplied by a definite cell, that is, by internal influences, and in other cases may be due to external influences.
Darwin assumed the first of these alternatives in his theory of Pangenesis, which we have still to outline. In it he attributes to his 'gemmules' the power of giving rise to particular cells, which, however, they can only accomplish when they reach the cells which are the genetic antecedents of those which the gemmules are to control. Translated into the language of our theory this view would read as follows: the whole complex of determinants is contained within every cell, as it is contained in the germ-cell, but at every stage of ontogeny, that is, in each of the developing cells, only the determinant which is to control the immediately successive cells is 'liberated,' and that through the stimulus which the specific nature of the cell supplies to the determinant. In that case there would necessarily be in every species of animal as many specific stimuli for determinants as there are determinants. This appeared to me improbable, and I rejected the hypothesis because of the enormous number of specific stimuli which it demands, but also on other grounds which will be touched upon in the course of these lectures.
Although the assumption of an autonomic dissolution of the determinant complexes of the id in the course of ontogeny seems to me imperative, I do not by any means reject the interposition of liberating stimuli, indeed I regard their co-operation as indispensable. Later on we shall discuss cases in which it is definitely[Pg 383] demonstrable that there may be two alternative sets of homologous determinants present in a cell, but that on any occasion only one of these becomes active, a fact which we can only explain on the assumption that only one of these is affected by the specific liberating stimulus. The phenomena of regeneration, of polymorphism, of germ-cell formation, &c., compel us to the assumption that numerous cells, even after the completion of the building up of the body, contain two or more kinds of determinants, as in a sense inactive 'accessory idioplasm,' each of which could control the cell alone, though in reality it only does control it when it is affected by the appropriate liberating stimulus. I stated this view some years ago when I attempted to define more precisely the rôle played by 'external influences as developmental stimuli[21]'. It is not, then, that I underrate the importance of external influences on the organism, but I believe that a still larger part of the determination of what shall happen at a particular point depends on the primary constituents, and that these are not alike at all parts of the body.
[21] Äussere Einflüsse als Entwicklungsreize [External Influences as Stimuli to Development], Jena, 1894.
All living processes, therefore, both those of growing and of differentiation, depend always upon the interaction of external and internal factors, of the environment and the living substance, and the resultants of the interaction, namely, the structure of the body and its parts must necessarily turn out differently, not only when the germ-substance is different, but when the essential conditions of development are changed. But that the constitution of the germ is by far the most potent factor, and that the nature of the results of development depends on it in a much greater degree than on the external conditions, has long been known. The conditions, such as warmth, may vary within certain limits, and yet the frog's egg becomes a frog; though it does not follow that the result of development may not be modified through certain changes in the conditions. The interesting experiments made by Herbst with the eggs of sea-urchins have shown that, in artificially altered sea-water in which sodium-salts are to a slight extent replaced by lithium-salts, these eggs develop into larvæ which only remotely suggest the normal structure, and diverge widely from it both in external shape and in the form of the skeleton.
Such larvæ are not able to survive, but soon perish; they are, however, of great interest from the point of view of our theory, for they show that determinants do not bring forth the same structure under all circumstances, but that, as I have already said, they are vital units of specific composition, which play a part in the course of[Pg 384] development, and give rise under normal external influences to normal parts, while under unusual influences, if these are not such as to prohibit development altogether, they may give rise to an abnormally formed part. It must not be forgotten that most composite parts—indeed, strictly speaking, all the parts—of an animal are not controlled by a single determinant, but by the many which successively determine the character of the cells and define the path of development of the part in question. There are no determinants of 'characters,' but only of parts; the germ-plasm no more contains the determinants of a 'crooked nose' than it does those of a butterfly's tailed wing, but it contains a number of determinants which so control the whole cell-group in all its successive stages, leading on to the development of the nose, that ultimately the crooked nose must result, just as the butterfly's wing with all its veins, membranes, tracheæ, glandular cells, scales, pigment deposits, and pointed tail arises through the successive interposition of numerous determinants in the course of cell-multiplication.
But in both processes we must presuppose normal conditions of development. In regard to the butterfly we know that abnormal conditions, such as cold during the pupal period, can cause considerable variation in the colour and marking of the wing, and in regard to the nose it can scarcely be doubted that, for instance, persistent pressure on the nasal region would result in a considerable deviation from the hereditary form.
The case of the lithium-larvæ is similar. Here the chemical conditions of the first segmentation-cells are modified by the presence of the lithium-salts, and the determinants which make their way out of the nucleus in the first and in subsequent cell-generations find an unusual soil for their activity, which diverges further and further from the normal with each successive cell-generation. Thus the whole animal is abnormally formed. The process may perhaps be compared to a plant which is negatively geotropic and positively heliotropic, that is, the stem of which tends to grow straight upwards, while all its green parts grow towards the light. If a plant of this kind have light shed on it from one side only, the stem with its leaves will grow obliquely towards that side. If the plant be then turned round so that it receives light from the other side, the stem in its further growth will curve in a direction opposite to that which it took before, and so by continually changing the position of the plant in relation to the light one could—theoretically at least—produce a plant with a zigzag stem. But this would not furnish any evidence against the presence of determinants; there are no 'upright deter[Pg 385]minants' any more than there are 'zigzag determinants' or 'crooked nose determinants,' but there are determinants controlling the nature of the cells which give rise, under normal conditions of development, to the straight stem, under abnormal conditions to the zigzag stem, or to a flat nose instead of a crooked one, and so on.
This consideration should make it clear that plant-galls are not in the remotest degree a stone of stumbling for the determinant theory, as some have supposed. Of course there can be no 'gall-determinants,' for galls are not transmissible adaptations of the plants on which they occur; they arise solely through the larvæ of the gall-insect which has laid its eggs within the tissues of the plant. But the specific nature of the different kinds of plant-cells, predetermined by their determinants, is such that, through the abnormal influences exercised upon them by the larvæ, they are compelled to a special reaction which results in the formation of galls. It is marvellous enough that these abnormal stimuli should be so precisely graded and adjusted that such a specifically definite structure should result, and in this case there is obviously a very different state of matters from that obtaining in most other processes of development, in which the chief determining factor is rather implied in the nature of the idioplasm, that is, of the determinants, than in the nature of the external influences. Here, however, the specific structure of the gall depends mainly on the quality, variety, and successive effects of the external influences or stimuli. In discussing the influences of surroundings I shall return once more to the galls.
My determinants have generally been regarded as if they were like grains of seed, from which either nothing may arise, under unfavourable conditions, or just the particular kind of plant from which the seed itself originated.
This simile is, however, to be taken cum grano salis. The whole ovum is certainly comparable to a grain of seed, but single determinants or groups of determinants will always be able to adapt themselves to different influences, and to remain active even under slightly abnormal conditions, though in that case the resulting structures may be somewhat divergent. This relative plasticity is indispensable even in relation to the ceaseless mutual adaptations of the growing parts of the organism. Not only do the cells which live beside each other at the same time influence each other mutually, but the influence extends to the whole cell-lineage. No cell or group of cells develops independently of all the others in the body, but each has its ancestral series of cells on whose determinants it is so far dependent, since these have taken part in determining its own nature,[Pg 386] in, so to speak, supplying the soil in which ultimately its own determinants will be sown from the nucleus, and whose influence modifies these last according to its quality. We might therefore say that every part is determined by all the determinants of its cell-ancestors.
If there be urged against the doctrine of determinants the undoubted fact of the dependence of individual development on external conditions, or the capacity that organisms have of functional adaptation, or especially the power that some parts of the organism have of taking a different form in response to different stimuli, I can only say that I see no reason why certain cells and masses of cells should not be adapted from the first for responding differently to different stimuli.
Therefore I see no contradiction of the determinant theory when, for instance, among the higher vertebrates, the cells of the connective tissue exhibit a great diversity of form, becoming a loose 'filling' connective tissue in one place, a tense fascia, ligament, or tendon tissue in another, according as they are subjected to slight pressure on all sides or to stronger pressure on one side. I see no difficulty in the fact that this connective tissue forms in one case bone-tissue with the most accurate adaptation of its microscopic structure to the conditions of stress and pressure which affect the relevant spot, or in another case cartilaginous tissue, when the cells are exposed to varying pressure (as on the surface of joints), or even that it gives rise to blood-vessels when the pressure of the circulating blood and the tension of the surrounding tissues supply the necessary stimulus. It is easy to see how important, indeed how necessary, the many-sidedness of these cells is for the organism, even leaving out of account such violent interference as the breaking of a bone, the irregular healing of broken ends of bones, new joint formation, and so on, and thinking only of the normal phenomena of growth. While the bone grows it is continually breaking up in the inside and forming anew on the surface, and this occurs through the power of the connective tissue-cells to form different tissues under different influences or stimuli.
We must therefore assume that there are side by side in the connective cells of higher vertebrates determinants of bone, of cartilage, of connective tissue in the narrower sense, and of blood-vessels, and that one or other of these is liberated to activity according to the stimulus affecting it. Phenomena occur also in the development of lower animals which lead us to the same assumption.
Among these is the remarkable behaviour of the primary mesoderm-cells in the young embryo (gastrula) of the Echinoderms (Fig. 92). At the point where the primitive gut or archenteron[Pg 387] invaginates into the interior of the hitherto single-layered blastula (Fig. 92, A), some cells are separated off (M), and move independently, constantly multiplying the while, into the clear gelatinous fluid (G) which fills the cavity of the larva, and there they fix themselves, some on the outer ectodermic layer, others to the various regions and outgrowths of the archenteron (Ms). According as these cells have established themselves at one or another point, they become connective tissue, muscle, or skeleton cells of the dermis, or contribute to the muscular layer of the food-canal and water-vascular system, or, finally, become skeleton-forming cells of the calcareous ring which surrounds the gullet of the sea-cucumber. In all this there is nothing to indicate a determination of the cells in one direction; on the contrary it seems as if the fate of the individual cells depended on the chance conditions which may lead them to one place or to another.
Fig. 92. Echinoderm-larvæ. A, blastula-stage; the primary mesoderm-cells (M) are being formed at the subsequent invagination-area of the endoderm (Ent). Ekt, the ectoderm. B, gastrula-stage; the archenteron (UD) has been invaginated (Ent), and between it and the ectoderm (Ekt) the mesoderm-cells (Ms) migrate into the gelatinous fluid which fills this cavity. There they attach themselves partly to the ectoderm, and partly to the endoderm. After Selenka.
There are thus three possibilities of development, three kinds of reaction, implied in these cells, which are all outwardly alike, and we can only understand their rôle in the building up of this very symmetrical animal if we assume that of these three only one is in each case liberated, by the specific stimulus exerted by the immediate surroundings of the cell, so that it may become, according to the chance position it takes up after its migration, either a skin-cell, a muscle-cell, or a skeleton-forming cell.
This case may be compared in some respects with the permanent colour-adaptation of those caterpillars, in regard to which Poulton demonstrated that they become almost black if they are reared on blackish-brown bark, light brown on light bark, and green if they are kept among leaves, and in all cases permanently so. In this case also the implicated pigment-cells of the skin may develop in three ways, according to whether this or that quality of the light releases this or that determinant.
But in many cases we do not know the quality of the liberating stimulus, and must content ourselves with imagining it. This is so in the case of dimorphism of the sexes. It is clear that in the males of a species the germ-cells develop quite otherwise than they do in the females, that different determining elements attain to activity in each sex, and since the primary constituents of both sexes must be contained in most animals in the ovum and in the spermatozoon, we must assume that in both there are at once 'ovogenic' and 'spermogenic' determinants, of which, however, only one kind becomes active in a given individual. There are, however, both among plants and animals hermaphrodite individuals, in which both kinds of sexual products are developed simultaneously or successively.
It is not only the primary sexual characters, however, that compel us to the assumption of double determinants in the germ-plasm, the secondary sexual characters do so too. We know very well in relation to ourselves that 'the beautiful soprano voice of the mother may be transmitted through the son to the grand-daughter, and that the black beard of the father may pass through the daughter to the grandson.' Thus both kinds of sexual characters must be present in every sexually differentiated being, some visible, others latent. In animals the determinants are sometimes handed on from germ-plasm to germ-plasm through several generations in a latent state, and only make their appearance again in a subsequent generation. This is the case in the water-fleas (Daphnids) and the plant-lice (Aphides), in which several exclusively female generations succeed one another, and only in the last of them do males occur again side by side with the females.
The germ-plasm of the ovum which is ripe for development must thus contain not only the determinants of the specific ova and sperms of the species, but also those of all the male and female sexual characters, which we discussed at length in the section on sexual selection. I then showed that these secondary sexual characters differ greatly in range and in strength, that among lower animals they are almost entirely absent, and that among higher forms, such[Pg 389] Crustaceans, Insects, and Birds, they attain to very different grades of development even among the same species. Thus the birds of Paradise are in most species brilliantly coloured and adorned with decorative feathers only in the male sex, while the females are simply blackish-grey, but there is a single species in which the males are almost as soberly coloured as the females. Conversely, too, we find that in parrots both sexes are usually coloured alike, but a few species exhibit a totally different colouring in the two sexes. In the same way the secondary sex differences may affect only a few parts of the animal or many, while in a few species the sexes are so divergent in structure that almost everything about them may be called different. Examples of this are the dwarf males of most Rotifers, and the males, more minute still in proportion to the females, of the marine worm Bonellia viridis (p. 227).
We have now to inquire what theoretical explanation of these facts we can arrive at in accordance with the germ-plasm theory. That double determinants, male and female, for the differently formed parts of the two sexes must be assumed to exist in the germ-plasm has been already said, and we have to suppose that the same stimulus—usually unknown to us—which incites the determinants of the primary sexual characters to activity also liberates those of the secondary characters. But we may safely go a step further and conclude that there are male and female ids, that is, that the male and female determinants belong to different ids. I infer this from the fact that in some groups, such as the Rotifers and certain plant-lice, the ova are sexually differentiated even at the time of their origin. Males and females of these animals arise from different kinds of eggs, which are even externally recognizable. Both develop parthenogenetically, so that fertilization has nothing to do with it; from the first, therefore, they must contain ids which consist of determinants of one sex alone.
If this conclusion be correct, then the sexual equipment of the determinants of the sexual characters must have taken place in the course of phylogeny in such a way that each id was affected in one direction only, and we should thus have to assume male and female ids, even before the separation of the sexes as males and females, and the same conclusion must be extended to the primary sexual characters. Only in this way can we understand the fact that differences between the sexes, at first small, have increased in the course of phylogeny to such complete divergence of structure as is now exhibited in the forms we have named, Bonellia, the Rotifers, and some parasitic worms.
But there is not only sexual dimorphism, there is also dimorphism of larvæ, e.g. green and brown caterpillars in certain species of hawk-moth (Sphinx), and there are sometimes not only two but three or more forms of a species; and in all these cases determinants of the differential parts must be represented twice, thrice, or several times in each germ-plasm, in each fertilized ovum, at least in all cases in which the different forms live together on the same area. In discussing mimicry we spoke of species of butterfly which were everywhere alike or nearly so in the male sex, while the females were not only quite different from the males, but differed greatly in many respects among themselves. Three different forms of females of Papilio merope occur in the same region of Cape Colony, each of these resembling a protected model. All three forms have been obtained from the eggs of one female. In this case the female ids of the germ-plasm must be represented by three different sets, one of which, when it is in the majority in the fertilized ovum, gives rise to the Danais-form, the second to the Niavius-form, and the third to the Echeria-form of the species. Phylogenetically considered, it is probable that each of these three kinds of ids originated by itself, on a more limited area on which the protected model lived in abundance; but with a wider distribution the different female ids mingled together, were united through the males into a single germ-plasm, and now occasionally exhibit all three forms on the same area. I doubt whether there is any other theory that can offer an interpretation of these facts, and I regard them, therefore, as affording further evidence of the real existence of ids.
The polymorphism of social insects must be thought of as similarly based in the germ-plasm.
In bees there are in addition to the males and females the so-called workers, and this can only depend on the existence of special kinds of ids. Those of the workers were originally truly female, but as many of their determinants underwent variations advantageous for the maintenance of the species, they were modified into special 'worker-ids.' I postpone for the present any inquiry into the causes by which these ids come to dominate the ontogeny; obviously it cannot be by the mere fact of being in a majority over the rest of the ids, as I indicated in the case of the butterflies with polymorphic females.
In many ants the division of labour goes further still; there are two kinds of workers in the colony, the ordinary workers and the so-called 'soldiers,' and in this case the worker-id must have developed in two different directions in the course of phylogeny, and have[Pg 391] separated into two kinds of ids, so that the germ-plasm of these species must contain four kinds of ids.
I might cite many more cases in regard to which the assumption of two or more kinds of determinants seems imperative, but I believe that what has been said is enough to enable any one to think out other cases for himself.
THE GERM-PLASM THEORY (continued)
Co-operation of the determinants to form an organ: insect appendages—Venation of the insect-wing—Deformities in Man—Apex of the fly's leg—Proofs of the existence of determinants—Claws and adhesive lobes—Difference between a theory of development and a theory of heredity—Metamorphosis of the food-canal in insects—Delage's theory—Reinke's theory of the organism-machine—Fechner's views—Apparent contradiction by the facts of developmental mechanics—Formation of the germ-cells—Displacement of the germinal areas in the hydro-medusoid polyps, a proof of the existence of germ-tracks.
It would be futile to attempt to guess at the arrangement of the determinants in the germ-plasm, but so much at least we may say, that the determinants do not lie beside each other in the same disposition as their determinates exhibit in the fully-formed organism. This may be inferred from the complex formative processes of embryogenesis in which many groups of cells, which in their origin were far apart, combine together to form an organ. Thus the arrangement of the determinants in the germ-plasm does not correspond to the subsequent arrangement of the whole animal, nor are primary constituents of the complete organs contained within the germ-plasm. The organ is undoubtedly predetermined in the germ-plasm, but it is not preformed as such.
Here, again, the history of development gives us a certain basis of fact from which to work. Let us consider, for instance, the origin of the appendages in those insects which in the larval state possess neither legs nor wings, but exhibit a gradual emergence of these structures from concealment underneath the integumentary skeleton. In these cases, as I have already shown in regard to the wings, the development of the limbs arises from definite groups of cells in the skin. These must therefore be regarded as the formative, and therefore as the most important and indispensable, parts of the rudiments, and may be designated the imaginal disks, as I many years ago proposed[22] (Fig. 89, ui and oi).
[22] Die Entwicklung der Dipteren, Leipzig, 1864.
But these disks of cells do not contain the whole leg, but only the [Pg 393]skin-layer of it, the 'hypodermis,' which, however, in this case undoubtedly determines the form. But the internal parts of the leg, especially the nerves, tracheæ, and probably also the muscles, are formed from other cell-groups and grow into the imaginal disk from outside. Something similar probably takes place in the case of all organs which are made up of many parts; they are, so to speak, shot together from several points of origin, from various primordia; and determinants are brought into co-operation whose relative value in determining the form and function of the organ may be very diverse.
Fig. 89. Anterior region of the larva
of a Midge (Corethra plumicornis). K, head.
Th, thorax. ui, inferior imaginal disks.
oi, superior imaginal disks. ui1, ui2, and
ui3, the primordia of the limbs. oi2
and oi3, the primordia of the wings and
'balancers.' g, brain. bg, chain of ventral
ganglia with nerves which enter the
imaginal disks. trb, tracheal vesicle.
Enlarged about 15 times.
For it is undoubtedly a very different matter whether a cell bears within it the elements which compel it in the course of growth to develop an organ, for instance a leg, of quite definite size, sculpture, number of joints, and so on, or whether it only bears the somewhat vague power of determining that connective tissue or fatty tissue is to be produced. In the first case it controls the whole formation of the part, in the second it only fills up gaps or lays down fat or other substances within itself if these be presented to it. Between these two extremes of determining power there are many intermediate stages. Cells which contain the determinants of blood-vessels, tracheæ, or nerves need not be so definitely determined that they always give rise to precisely the same blood-vessels, the same branching of the tracheæ, or the same bifurcation of nerves; they may probably possess no more than the general tendency to the formation of such parts, and the special form taken by the nerves, tracheæ, or blood-vessels may be essentially determined by their environment. Thus in the morbid tumours of Man, nerves, and especially blood-vessels, may develop in a quite characteristic manner, which was certainly not determined in advance, but has been called forth by the stimulus, the pressure, and other influences of the cellular basis of the tumour. In short, the cells were only determined to this extent, that they contained the tendency to give rise to blood-vessels under particular influences.
It would be a mistake, however, to think of the primary con[Pg 394]stituents of all cell-groups as so indefinite. Let us call to mind, for instance, the venation of the insect wing. It is well known that this is not only quite different in beetles, bugs, and Diptera from that in the Hymenoptera, and different again in the butterflies, but that it is quite characteristic in every individual family of butterflies, and indeed in every genus. We cannot conceive of the absolute certainty of development of these very characteristic and constant branchings as having its roots elsewhere than in the determinants of the germ-plasm, which, lying within certain series of cells, ultimately cause particular cell-series of the wing-rudiment to become the wing-veins. If this were not so, how would it be possible to understand the fact that every minute deviation in the course of these veins is repeated in exactly the same way in all the individuals of a genus, while in all the individuals of an allied genus the venation turns out slightly different with equal constancy.
But it is quite certain that all determinations are in some degree susceptible to modifying influences, that they are in very different degrees capable of variation.
Many deformities of particular parts in Man and the higher animals may be referred to imperfect or inhibited nutrition of the part in question during embryonic development; the determinants alone cannot make the part, they must have a supply of formative material, and according as this material is afforded more abundantly or more scantily the part will turn out larger or smaller. In the same way the pressure conditions of the surrounding parts must in many cases have a furthering or inhibiting influence, or may even determine the shape. But it is quite possible, indeed even probable, that other specific influences are exerted by the cells or cell-aggregates surrounding an organ which is in process of being formed, just as the stake on which a twining plant is growing may prompt it to coil. If the stake be absent, the predetermined twining of the plant cannot attain to more than very imperfect expression, if indeed it finds any. The spirally coiled sheath of muscle-cells which occurs so often around blood-vessels in worms, Echinoderms, and Vertebrates is probably due to similar processes, that is, on the one hand, to a specific mode of reaction characteristic of these cells, and predetermined from the germ; on the other hand, to the external influence of the cell-surroundings without which the determination of the muscle-cell is not liberated, that is, is not excited to activity.
Fig. 93. The development of a limb in the pupa of a Fly (Sarcophaga carnaria). A, apex of the limb from a pupa four days old; the jointing is hinted at; hy, hypodermis; ps, pupal sheath; ph, phagocytes; tr, tracheal branch. B, the same on the fifth day; the lumen of the limb is quite filled with phagocytes (ph); the last tarsal joint (t5) is beginning to show a bifid apex. C, the same on the seventh day; the claws (Kr) and the adhesive lobes (hl) are formed.
But even if every determinant requires a stimulus to liberate it, whether this stimulus consists in currents of particular nutritive fluids, in contact with other cells, or, conversely, on the removal of some [Pg 395]pressure previously exerted on the cell by its surroundings, the material cause of a structure is to be sought for not in these conditions of its appearance, but in the primary constituents which have been handed on to the relevant cell or cell-group from the germ, in other words, through its determinants. How, for instance, could the blunt rounded knob of the rough and clumsily jointed sac of cells which represents the insect's leg at the beginning of the pupal period (Fig. 93, A) be incited to thicken, to constrict at the root (B), and to form a joint-surface, to broaden out at the end, and produce two sharply cut points (C), which become incurved and form claws (kr), while beneath these a broad flat lobe (hl) grows forward, and with its regularly disposed cells gradually forms the characteristic adhesive organ of the fly—how could all this happen if there were not contained within these cells special formative forces which determine them not only in their form and the rest of their constitution, but above all in their power of multiplication? No special external stimulus affects the still unfinished knob of the fly's leg unless it be the removal of pressure; but this operates regularly, and cannot be[Pg 396] the cause of the growth, at definite places, of claws and adhesive lobes with all their characteristically placed hairs.
We require to assume that each of the cells composing the primary rudiment of the limb possessed a determining power which made it grow and multiply under the given conditions of nutrition and pressure in a prescribed manner and at a prescribed rate; and we must make the same assumption in regard to all the daughter and grand-daughter-cells, and so on. The strictest regulation of the power of multiplication of each of the implicated cells is a necessary condition of the constant production of the same two claws and adhesive lobes, the same form of tarsal joint, the same regular covering of hair, and so on. This exact determination of the cells can only take place through material vital particles, and it is these which I call determinants.
I have already said so much about the assumed 'determinants' of the germ-plasm that it might perhaps be supposed that we have now exhausted the topic; but the assumption of such 'primary constituents' is so fundamental, not only for my own germ-plasm theory of to-day and to-morrow, but also—unless I am much mistaken—for all future theories of development and inheritance. In point of fact, the conception of determinants has as yet penetrated so little into the consciousness of biologists, that I cannot remain content with what I have already said, but must endeavour to test and to corroborate my thesis by additional illustrations.
As far as I am aware, only a few zoologists have expressly and unconditionally agreed with the assumption of determinants; on the other hand, several biologists have rejected it as fanciful and untenable, while others have set it aside as a useless playing with ideas. The last, I am inclined to believe, have not taken the trouble to think out what the idea is. It has even been objected that there can be no determinants because we can see nothing of them, and that they must therefore be pure figments of the imagination, invented to explain facts which could be explained much more easily and simply in some other way. From the very first I have stated emphatically that they have not been, and never will be seen, because they lie far below the limit of visibility, and thus can at best only become visible when they are collected in large aggregates like chromatin granules. Nor have I any objections to make if any one chooses to describe all the details of their activity as mere hypotheses, such, for instance, as their distribution during development, their 'maturation,' their migration from the nucleus, and the manner in which they control the cell. All this is really an imaginative picture which may be[Pg 397] correct to a certain degree, but may also be erroneous; no formal proof of it can be obtained at present; and I am content if it be simply admitted to be possible. On the other hand, the existence of determinants seems to me to be, in the sense indicated, indubitable and demonstrable.
Let us return for a moment to the claws and adhesive lobes which are developed on the foot of the fly. It may perhaps be thought that it is possible to do without the assumption of determinants for these parts, by assuming that although 'external' influences in the ordinary sense could not possibly have determined that certain cells of the apex of the leg should form claws and others adhesive lobes, the result might be due to the differences of intercellular pressure within the apical knob; these may have been stronger in one direction, weaker in another, thus prompting the cells to grow here into claws and there into adhesive lobes. If we had merely to explain from the constitution of the germ-plasm the ontogeny or development of these parts in an individual fly there might perhaps be no radical objection to this view, though it would hardly be possible to explain the assumed differences in pressure otherwise than as due to a different intensity of growth in the cells in the various regions of the limb-apex, which again would have to be referred to differences in the germ-plasm. But when we reflect that these parts vary hereditarily and independently of other parts, and owe their present form to their power of doing so, and that they are differently formed in every genus and species, we see at once that they must be represented in the germ-plasm by particular vital particles, which are the roots of their transmissible variability, that is, which must have previously undergone a corresponding variation if the relevant parts themselves are to vary. Without previous variation of the determinants of the germ no transmissible independent deviation on the part of the claws or adhesive lobes of the animal is conceivable.
All the opponents of my theory have overlooked this fact; both Oscar Hertwig and Kassowitz have forgotten that a theory of development is not a theory of heredity; they only aim at the former, and they therefore dispute the logical necessity for an assumption of determinants.
But as this is the very foundation of the theory, let me further submit the following considerations in its favour.
In insects which undergo metamorphosis, not only the external but the internal parts of the caterpillar or larva go through a more or less complete transformation. In the flies (Muscidæ), for instance, the whole intestinal tract of the larva is reconstructed in the pupa;[Pg 398] in fact it breaks up into a loose, flocculent, dead, but still coherent mass of tissue. Within this there arises a new intestine, as I have shown in an early work (1864); and Kowalewsky and Van Rees have since made us aware of the interesting details of this reconstruction, showing that the new intestine arises from definite cells of the old one, which are present in the larval gut at certain fairly wide distances, and which do not share in the general destruction, but remain alive, grow, and multiply, and form islands of cells in the dead mass. These living islands, continually extending, ultimately come into contact and again form a closed intestinal canal which differs entirely from that of the larva in its form, in its various areas, and in its differentiation. In this case those formative cells of the imago-intestine must have contained the elements which determined their descendants in number, power of multiplication, arrangement, and histological differentiation. In other words, each of these cells must contain the determinants of a particular limited section of the intestine of the imago. The other cells of the intestinal epithelium could not do this, even though they were under exactly the same conditions, were included in the same intimate cell-aggregate, and had the same nutritional opportunities. They break up when the formative cells begin to be active, for till then the latter had remained inactive, and had not multiplied, although they lay regularly distributed among the other cells. Whence, then, could the entire difference in the behaviour of these two sets of cells arise, if it does not depend on the nature of the cells themselves, and how could this difference of nature have developed during the racial history of insect-metamorphosis if determinants did not reach the cell from the germ-plasm—determinants which conditioned that some cells should be hereditarily modified into the cells of the imago-intestine and others into the larval intestine? Quite similar processes have been recently demonstrated in regard to the reconstruction of the larval intestine in other insect-groups. Deegener has done this, for instance, for the water-beetle (Hydrophilus piceus); and it is certain that all these reconstructions start from particular cells, which lie indifferently between the active cells during the larval period, and contain the primary constituents for the formation of a section of the intestine, but which only become active when their hitherto living neighbours die and break up.
The whole of the reconstruction of the external form of the fly takes place in a similar manner. Not only the limb, the head, the stigmata, but the skin itself is formed anew from imaginal disks. In each of the abdominal segments three pairs of little cell-islands[Pg 399] are formed during larval life, and these only enter on the stage of formative activity after pupation, when they multiply rapidly and grow together to form a segment, whose size, form, and external nature is determined by them. But it is well known that the abdominal segments of the fly differ from those of the larva very markedly and in every respect, so that each cell-island must contain determinants which are quite different from those in the skin-cells of the corresponding larval segments. These last break up at the beginning of pupahood, while the former begin to grow vigorously, and to spread themselves out. The most remarkable fact about the whole business, and it seems to me also the most instructive, is that these imaginal disks frequently appear for the first time during larval life, as I found in the case of a midge, Coretha plumicornis, in regard to the disks of the thorax, and as Bruno Wahl[23] has recently demonstrated in the case of the abdominal cell-islands. Since in the young larva the position of the subsequent imaginal disks is occupied by cells which apparently in no way differ from the rest of the skin-cells, and are also exposed to precisely the same external and internal influences, the origination of the imaginal cells from these can only depend on differential cell-division; the primordial cell of each imaginal disk must have separated at the beginning of disk-formation into a larval and an imaginal skin-cell.
[23] Bruno Wahl, Ueber die Entwickelung der hypodermalen Imaginalscheiben im Thorax und Abdomen der Larve von 'Eristalis' L., Zeitschr. f. wiss. Zool., Bd. lxx. 1901.
In insects in which the larva and the imago differ widely, the perfect insect, as regards all its principal parts, is already represented in the larva, namely, in particular cells which lie among those of the corresponding larval parts, and do not visibly differ from these, although they are equipped with quite different determinants, and consequently enter on their formative activity much later, and give rise to quite different structures. As the determinants of the whole animal with all its parts are contained in the ovum, so those of the parts of its imaginal phase are contained in these cells of the imaginal disks.
In addition to all this, we have incontrovertible evidence in favour of the theory of determinants in the independent phyletic variations of the individual stages of development, on which depends the whole phenomenon of 'metamorphosis' which we have just been considering. How could the larval stage have become so different from the imago-stage, if the one were not alterable by variation arising in the germ without the other being affected? If this absolute independence of the transmissible variability of the individual stages[Pg 400] were not an indispensable assumption in the explanation of metamorphosis and other phenomena of development, I should regard an attempt at a theory of development without determinants as justifiable. But I am forced to see in this fact alone an invalidation of all epigenetic theories of development, that is, of all theories which assume a germ-substance without primary constituents, which can produce the complicated body solely by varying step by step under the influence of external influences, both extra- and intra-somatic. It is possible to conceive of an ovum in which the living substance is of such a kind that it must vary in a definite manner under the influence of warmth, air, pressure, and so on, that it must divide into similar, and subsequently also into dissimilar parts, which then interact upon each other in diverse ways and give rise to further variations, which in their turn result in differentiations and variations, till ultimately we have the whole complicated organic machine complete and 'finished' in every detail. Certainly no mortal could make any pronouncement as to the constitution of such a substance, but even if we assume it, for the nonce, as possible, how can we account for the transmissible variation of the individual parts and developmental stages, on which the whole phylogenetic evolution depends?
As the development of the butterfly exhibits the three main stages of caterpillar, pupa, and perfect insect, each of which is independently and hereditarily variable, and therefore implies a something in the germ, whose variation brings about a change in the one stage only, so the ontogeny of every higher animal is made up of numerous stages, which are all capable of independent and transmissible variation. How else should we human beings, in our embryonic phase, still possess the gill-arches of our fish-like ancestors, although much modified and without the gills? Truly, he who would seek to deny that the stages of individual development are capable of independent and transmissible variation must know very little about embryology. But if the facts are as stated, how can they be reconciled with the conception of a germinal substance developing in epigenetic fashion? Every variation in this substance would affect not only the whole succession of stages, but the whole organism with all its parts. In this way too, then, we are driven to the conclusion that there must be something in the germ whose variation causes variation only in a particular part of a particular stage. This something we define in our conception of the 'primary constituents' (Anlagen)—the determinants. These are not to be thought of either as 'miniature models,' or even as the 'seeds' of the parts; they alone cannot produce the part which they determine, but they effect changes in the[Pg 401] cell in which they become active, causing it to vary in such a manner that the formation of the relevant part results. While I conceive of development as a continuous process, I supplement this with the idea that from within, namely, from the nuclear substance, new, directive, 'determining' influences are continually being exerted on the developing cells.
I can hardly think of a better proof of the necessity of this assumption than that furnished by Delage, one of the most acute biologists of France, who, in his comprehensive book on Heredity, has striven to replace the theory of determinants by something simpler. Delage rejects all 'primary constituents' (Anlagen) in the germ, all 'particules représentatives,' as much too complicated an assumption, and thinks it possible to work with the conception of a germ-plasm which is about as simple as the cell-substance of a Rhizopod, that is to say, a protoplasm of definite chemico-physical constitution and composition. Leaving out of account the consideration that the protoplasm of an amœba is scarcely of such extreme simplicity, but is certainly made up of numerous differentiated and definitely arranged biophors, how could such an extremely simple ('éminemment simple') constitution of the ovum as is here assumed give rise to such a complicated organism, the individual parts of which are capable of independent and transmissible variation? According to Delage it does so because the ovum, though not containing 'all the factors requisite for its ultimate resultant,' does contain 'un certain nombre des facteurs nécessaires à la détermination de chaque partie et de chaque caractère de l'organisme futur'! Determinants after all, it may be said, but that is far from the truth! It is not primary constituents that the germ contains, according to Delage, it is chemical substances, for instance muscle substances, probably 'les substances caractéristiques des principales catégories de cellules, c'est-à-dire, celles qui, dans ces cellules, sont la condition principale de leur fonctionnement.' All these must be contained in the ovum. How they are to reach their proper place in the organism, how the 'characteristic chemical substance' of a mole is to land just behind the right or left ear of the fully formed man, is not stated. But apart from this, there is a much deeper error in this assumption of specific chemical substances in the ovum as an explanation of the phenomena of local hereditary variation, and I have already touched upon it: chemical substances are not vital units, which feed and reproduce, which assimilate and which bear a charm against the assimilating power of the surrounding protoplasm. They would necessarily be modified and displaced in the course of ontogeny, and would therefore—no matter [Pg 402]where they had been placed at first—be incapable of performing all that Delage ascribes to them. Either the germ contains 'living' primary constituents, or it is, as Delage maintains, determined chemico-physically; but in the latter case there is no scope for hereditary local variation. Delage must either renounce the attempt to explain this, or he must transform his 'substances chimiques' into real and actually living determinants.
Thus from all sides we are forced to the conclusion that the germ-substance on the whole owes its marvellous power of development not only to its chemico-physical constitution, whether that be eminently simple or marvellously complex, but to the fact that it consists of many and different kinds of 'primary constituents' (Anlagen), that is, of groups of vital units equipped with the forces of life, and capable of interposing actively and in a specific manner, but also capable of remaining latent in a passive state, until they are affected by a liberating stimulus, and on this account able to interpose successively in development. The germ-cell cannot be merely a simple organism, it must be a fabric made up of many different organisms or units, a microcosm.
Yet another train of thought leads us to the same idea, and this has its roots in the extraordinary complexity of the machine which we call the organism.
The botanist Reinke has recently called attention once again to the fact that machines cannot be directly made up of primary physico-chemical forces or energies, but that, as Lotze said, forces of a superior order are indispensable, which so dispose the fundamental chemico-physical forces that they must act in the way aimed at by the purpose of the machine. To produce a watch it is not enough to bring together brass, steel, gold, and stones; to produce a piano it is not enough to lay wood, iron, leather, ivory, steel, &c., side by side, but these stuffs must be brought together in a definite form and combination. In the same way, the mere juxtaposition of carbon and water does not result in a carbohydrate like sugar or illuminating gas; the component elements only yield what is desired when they are placed in a particular and absolutely definite relation to each other, in which they so act upon and with one another that sugar or illuminating gas results, and the same is true of the component elements of a watch or of a piano. In the watch and in the piano this relation is arranged by human intelligence, by the workmen who form the different materials and put them together in the proper manner. In this case, then, human intelligence is, as Reinke says, the 'superior force' which compels the energies to work together in a particular way.
But organisms also are machines which perform a particular and purposeful kind of work, and they are only capable of doing so because the energies which perform the work are forced into definite paths by superior forces; these superior forces are thus 'the steersmen of the energies.' There is undoubtedly a kernel of truth in this view, and I shall return to it. Reinke, however, uses it in a way which I cannot follow; that is, he infers from it a 'cosmic intelligence' which puts these superior forces into the organisms, and thus controls these machines to purposeful work, as the watchmaker puts 'superior forces' into the watch by means of wheels, cylinders, and levers. In one case it is human intelligence which controls the 'superior forces,' in the other 'cosmic' intelligence. I cannot regard this reasoning from analogy as convincing, because, in the first place, these 'superior forces' are not 'forces' at all. They are constellations of energy, co-ordinations of matter and the energies immanent therein under complex and precisely defined conditions, and it is a matter of indifference whether chance or human intelligence has brought them together. If we take Reinke's own example of carbohydrates it is certain that our coal-gas is due to the intelligence of man, which brings together the carbon and the water in such a way that coal-gas must arise. The 'superior forces' must here be looked for in the arrangements of the coke-stove, and, in the second place, in the intelligence of man. But when decaying plants in the marsh form another carbon-compound, marsh-gas, where do the directing 'superior forces' come in? Surely only in the fortuitous concomitance of the necessary materials and the necessary conditions. Or may 'cosmic' intelligence have established this laboratory in the marsh? If not, what can compel us to refer the formation of dextrin or starch in the cells of the green leaves of plants to 'superior forces' which are placed in them by 'cosmic' intelligence? I am far from believing that the great and deep problem here touched upon can be put aside in any off-hand manner, but I feel sure that it will never be solved by word-play about energies and 'superior forces.'
Let us return to the kernel of truth in Reinke's thesis; it lies in this, that, while the working of a machine does really depend on the forces or energies which are bound up with the stuffs of which it consists, it also depends on a particular combination of these stuffs and forces, on a particular 'constellation' of them, as Fechner expressed it. In the watch these 'constellations' are the springs, the wheels, &c., and their position in relation to each other; but in the organism they are the organs, down to the cells and cell-parts; for the cell too is a machine, indeed a very complex one, as its functions[Pg 404] prove. There are thousands of kinds of 'constellations' of elementary substances and forces which condition the activity of the living machine, and only when all these constellations are present in the proper manner and in the proper relations to each other can the functions of the organism be properly discharged.
But the living machine differs essentially from other machines in the fact that it constructs itself; it arises by development from a cell, by going through numerous 'stages of development.' But none of these stages is a dead thing, each is itself a living organism whose chief function is to give rise to the next stage. Thus each stage of the development may be compared to a machine whose function consists in producing a similar but more complex machine. Each stage is thus composed, just like the complete organism, of a number of such 'constellations' of elementary substances and elementary forces, whose number in the beginning is relatively small, but increases rapidly with each new stage.
But whence come these 'constellations' or, to keep to our metaphor, the levers, wheels, and cranks of each successive stage in the making of the organic machine? The epigenetic theory of a germ-plasm without primary constituents answers by pointing to internal and external influences which cause the germ-plasm, originally homogeneous, to differentiate gradually more and more, bringing it into the most diverse 'constellations.' But how can such influences introduce new springs, levers, and wheels of a quite specific kind, as must be the case if apparently similar germinal substances are to give rise to two such different animals as a domestic duck and a teal? The cause must lie in the invisible differences in the protoplasm, opponents will answer, and we with them. But our studies up to this point have shown us that the differences cannot be merely elementary differences, cannot be merely of a physico-chemical nature depending on the composition of the raw material and the implicated energies; they must depend on the definite co-ordination of substances and energies, in other words, on the occurrence of 'constellations' of these. Thus the germ-plasm must be composed of definite and very diverse combinations of living units, which are themselves bound up in a higher 'constellation,' so that they act as a living machine at the first stage of development, and liberate into activity the already existing constellations of the second stage. The second stage in the series of living machines which arise successively from each other liberates the sleeping 'constellations' for the third, and so on.
These 'constellations' of matter and energy are the biophors, the determinants, and the 'groups of determinants' which we may think[Pg 405] of as disposed in a manifold overlapping series. That they do not enter into activity all at once, but successively take their part in development, seems to me a necessary consequence of their successive origin in the phylogeny; and the ontogeny, as we shall see later, arises through a modified condensation of the phylogeny. Now since every new determinant that arises in the course of phylogeny can only develop by division and subsequent variation from the determinants which were previously active at the same place in the organism, it is quite intelligible that later on, when the phylogeny has been condensed in the ontogeny, they should not enter upon their active stage at the same time as their phyletic predecessors, but after them. The theory of Oscar Hertwig, who starts from a germ-plasm without primary constituents, that all parts of the germ-plasm become active at the same time, seems to me quite untenable. How could the wheels, levers, and springs of the complete vital machine, which arose so very slowly in the course of phylogeny, arise to-day in the ontogeny in such rapid succession unless they were already present in the germ-plasm and only required to be incited to activity, that is, liberated by the stage preceding them? Even Fechner supported this view when he supposed that the interaction and mutual influences of the parts in the organism, that is, of the 'constellations,' gave rise of themselves to the succeeding stage, that is to say, to the new constellations peculiar to the succeeding stage. To this Reinke reasonably objected that it was like expecting the window frames of a house in process of building to produce the panes of glass. The panes in the organism only develop in the window frames if their determinants have been present in the germ-plasm from the beginning, and are liberated by the development of the frames, just as the activity of the glazier is liberated by the sight of the completed frames. Neither new panes nor new determinants could be produced rapidly; the former must be manufactured in the glass factory, the latter in the developmental workshop of the form of life in question, which workshop we call its phylogeny. But just as it is unnecessary to erect a new glass factory for each new house that is built, so the development of each individual does not require the establishment each time of those numberless life-factories—the constellations—whose business it is to produce anew the wheels, levers, springs, and cylinders of the developmental machinery at each stage, for they are all provided for in the germ-plasm, and it is only on this account that they are capable of hereditary variation.
I have already directed attention to some embryological facts which seem to be contradictory, if not to the germ-plasm theory itself,[Pg 406] at least to the assumption it makes that the germ-plasm is analysed out during the ontogeny; and something more must be said on this head. I refer to the numerous facts brought to light through the science of developmental mechanics founded by Wilhelm Roux, and particularly to the investigations as to the prospective significance of the segmentation-cells of the animal ovum.
Among these investigations we find experiments in compressing certain eggs (sea-urchin's) in the early stages of segmentation. The blastomeres are prevented by artificial pressure from grouping themselves in the normal manner; they are compelled to spread out side by side in the same plane. If the pressure is removed, they change their grouping, and yield a normal embryo. I will not here discuss whether these results can only be interpreted as showing that each segmentation-cell has the same prospective significance, and that it is only its relative position which decides what part of the embryo is to be formed from it; this could not be done without going into great detail; I therefore assume it to be true, and confine my survey to the second group of experiments, those on isolated segmentation-cells.
It has been shown that in the eggs of the most diverse animals, for instance in the sea-urchin once more, each of the two first blastomeres, if separated from one another, can develop into a complete larva. Indeed, in the eggs of sea-urchin and some other animals each of the first four, or any of the first eight, blastomeres, and indeed any segmentation-cell during the earlier stages, possesses the power of developing to a certain point, namely, as far as the so-called 'blastula-larva.' This seems to contradict a theory which assumes that the primary constituents become separated in the successive stages of ontogeny. But in the first place the blastomeres of all animals do not behave in this way, and, moreover, the facts can be quite well explained without entirely renouncing the assumption of the segregation of the determinant-complexes. It is only necessary to assume that the segmentation-cells, which develop in the isolated condition as if they were intact eggs, still contain the complete germ-plasm, and that the differential segregation into groups of determinants with dissimilar hereditary tendencies takes place later. This would certainly load the theory with further complications, and I shall not enter into the question here, since the facts which we should have to consider are as yet by no means undisputed.
But in any case the facts of developmental mechanics referred to, which we owe to numerous excellent observers of the last decade,—I need only name W. Roux, O. Hertwig, Chun, Driesch, Barfurth,[Pg 407] Morgan, Conklin, Wilson, Crampton, and Fischel—not only leave the essential part of the germ-plasm theory untouched, but rather strengthen than endanger its more subordinate points, such as the assumption of a segregation of the components of the germ-plasm in the course of ontogenesis.
As to the fundamental ideas expressed in the theory, I have already shown that these remain unaltered, even if we do not assume a disintegration or segregation of the germ-plasm, but think of all the developing cells as equipped with the complete germ-plasm. In that case the determinants would be liberated to activity solely by specific stimuli. But in regard to the assumption of disintegration, it must be noted that the facts cited relative to the sea-urchin's ova do not by any means hold true of the eggs of all animals.
In various animal types each of the first two segmentation-cells, when separated from its neighbour, produces only a half-embryo, and any one of the first four cells a quarter-embryo. This 'fractional embryo' is, however, in some cases able later to develop into a whole embryo (to 'postgenerate' itself, as W. Roux says). The isolated blastomere shows, to begin with, an activity of only a half of the primary constituents of the animal, as was first established by W. Roux and maintained conclusively, in spite of many attacks, until it was established beyond doubt by the detailed corroboratory investigations of Endres. The secondary completion of the embryo, which, however, is still disputed, must be regarded as a regeneration, and, to explain it, a co-operation of the complete but not yet wholly active germ-plasm in both segmentation-cells must therefore be assumed.
It would carry us too far if I were to deal in detail even with the most important of the numerous facts that the last decade has brought to light; I shall restrict myself to the most essential.
That isolated segmentation-cells have the capacity of developing into embryos which are complete but correspondingly smaller in size has been demonstrated in animals of various groups, though it does not seem to go to the same length in all. In the Medusæ we find that not only one of the first two, but one of the first four, eight, and even sixteen segmentation-cells may develop a whole larva when isolated (Zoja). In the sea-urchin at least any one of the first eight blastomeres may do so. And Driesch's experiments in cutting up the young larvæ at the blastula-stage (a single-layered ball of cells) leads us to assume that each of these cells still possesses the complete germ-plasm. Beyond that stage, however, the primary constituents obviously divide into those of the ectoderm and those of the endo[Pg 408]derm, for the subsequent two-layered stage in the sea-urchin's development, the gastrula, does not complete itself if it be artificially divided into fragments which consist only of cells from the outer, or only of cells from the inner layer. In corroboration of this experiment made by Barfurth, Samassa was able to demonstrate in regard to the egg of the frog that, even after the third division of the ovum, the segmentation-cells are so different from each other in respect of their primary constituents that they were not able to replace each other mutually. When this investigator killed the ectoderm-cells alone by means of an induction current, or the endoderm-cells alone, the dead half could not be replaced by the half which remained alive, and the whole ovum perished.
If these facts may be adduced in favour of a separation of the primary constituents at an earlier or later stage, we find even stronger proofs among the Ctenophores, Gastropods, Bivalves, and Annelids. In the last-named group Wilson has shown it to be probable that development is really a 'mosaic work,' as Roux and I had assumed. The older observations made by Chun at an earlier date on the Ctenophora, and the more recent experiments of Fischel on the same animals, prove the same thing for this group. In this case complete larvæ are easily distinguished from mere 'partial developments' by the number of the characteristic 'ciliated meridional rows' or ribs, which extend from one pole of the larva to another. In the complete larva there are eight of these, but in larvæ from one of the first two blastomeres (isolated) there are only four, and in those which have arisen from one of the first four blastomeres there are only two. If an ovum at the eight-cell stage can be successfully divided into separate blastomeres, each of these will form an 'eighth larva,' always with only one ciliated rib. Even in the succeeding sixteen-cell stage it could still be demonstrated that the substance responsible for the formation of the ribs only lies in particular places and always suffices only for eight ribs. The sixteen-cell stage consists of eight large cells and eight small ones, the 'macromeres' and the 'micromeres'; if an ovum at this stage be cut so that one piece contains five macromeres and five micromeres, a partial larva will develop which possesses only five ribs, while the larva from the other portion will have only three. But the localizing of the rib-determinants can be followed still further, for in larvæ in which individual micromeres have been displaced from their normal position there is a correlated displacement of the corresponding ribs, and a dislocation of their ciliated comb-plates. The determinants of the ribs must therefore lie in the micromeres, and we must conclude that at the antecedent division they were only imparted[Pg 409] to one daughter-nucleus, while the other, that of the macromere, did not receive this kind of determinant. Here then we have an example of dissimilar or differential division. Those who oppose this theory of qualitative division will hardly be likely to admit this, but will rather seek to maintain that 'external influences,' such as relative position, determine which cells are to give rise to the ciliated ribs and which are not. But the fact that artificial displacement of the micromeres leads to a disarrangement of the ciliated comb-plates, of which the ribs are made up, invalidates this suggestion, and at the same time overthrows the interpretation that it may be the cells which lie on particular meridians that are determined by this position to the production of ciliated plates. Obviously, the converse of this is true; those cells which contain the rib-determinants come to lie in the regular course of development in these eight meridians, and the cells lying between them, though of the same descent (from micromeres), contain no such determinants and therefore form no ribs. But if those cells which are equipped with rib-determinants be artificially displaced, then they give rise to swimming-plates elsewhere than on the aforesaid meridians.
The experiments made by Crampton on a marine Gastropod, Ilyanassa, likewise go to prove that a disintegration or segregation of the primary constituents does occur in the course of development. In this case, when the first two or first four segmentation-cells were artificially separated from each other, they developed exactly as if they still belonged to the complete ovum, that is, each isolated segmentation-cell yielded, respectively, a half or a quarter-embryo. And these 'partial embryos' were not able in this case to give rise subsequently to the missing parts or to form complete embryos.
There are thus two contrasted groups of animals, in one of which a segregation of the mass of primary constituents apparently takes place at the very beginning, while in the other it does not take place in the first stages of development, but apparently occurs later on. We may distinguish these two groups, with Heider, as those having 'regulation ova' and those having 'mosaic ova.' But I do not see that this affords any reason why we should give up our conception of the successive segregation of the germ-plasm into its determinants, even although, as I said before, I may modify it so far as to say that the segregation does not necessarily take place in all groups and species of animals at the same time, but occurs earlier in some and later in others.
Now that I have shown how the germ-plasm theory may be brought into harmony with the phenomena of ontogeny, I wish to go[Pg 410] on to show what the theory can accomplish in clarifying our understanding of the phenomena of reproduction and heredity. I shall at the same time give a brief exposition of some of the most important of these phenomena.
First, a few words in regard to the development of the reproductive cells. We may leave aside in the meantime the question whether they are sexually differentiated or not; we are only concerned just now with the main problem: How is it possible for the organism to produce germ-cells, that is, cells which contain the complete germ-plasm with all its determinants, when the building up of the body in ontogeny, according to our theory, involves a disintegration or segregation of the determinant-architecture into smaller and smaller groups? It is impossible that specific determinants should arise de novo, just as an animal cannot arise otherwise than from its germ, nor a cell otherwise than from a cell, nor a nucleus otherwise than from an already existing nucleus. If vital units ever originate de novo at all, it is only conceivable in the case of the very simplest biophors, as we shall see later when we come to speak of 'Spontaneous Generation.' Specific biophors and the determinants composed of them have behind them a phylogeny, a history, which conditions that they shall arise only from their like.
Thus we see that germ-cells can only arise where all the determinants of the relevant species arranged as ids are already present. If we could assume that the ovum, just beginning to develop, divides at its first cleavage into two cells, one of which gives rise to the whole body (soma) and the other only to the germ-cells lying in this body, the matter would be theoretically simple. We should say, the germ-plasm of the ovum first doubles itself by growth, as the nuclear substance does at every nuclear division, and then divides into two similar halves, one of which, lying in the primordial somatic cell, becomes at once active and breaks up into smaller and smaller groups of determinants corresponding to the building up of the body, while the germ-plasm in the other remains in a more or less 'bound' or 'set' condition, and is only active to the extent of gradually stamping as germ-cells the cells which arise from the primordial germ-cell.
As yet, however, only one group of animals is known to behave demonstrably in this manner, the Diptera among insects; in all others the cell from which the germ-cells exclusively arise, the 'primordial germ-cell,' makes its appearance later in development, usually during embryogenesis and often very early in it, after the first few divisions of the ovum, but sometimes not till long after the end of embryogenesis, and not even in the individual which arises from the ovum,[Pg 411] but in descendants which arise from it by budding. This last case occurs especially in the colonial hydroid polyps, which multiply by budding. Here the primordial germ-cell is separated from the ovum by a long series of cell-generations, and the sole possibility of explaining the presence of germ-plasm in this primordial cell is to be found in the assumption that in the divisions of the ovum the whole of the germ-plasm originally contained in it was not broken up into determinant groups, but that a part, perhaps the greater part, was handed on in a latent state from cell to cell, till sooner or later it reached a cell which it stamped as the primordial germ-cell. Theoretically it makes no difference whether these 'germ-tracks,' that is, the cell-generations which lead from the ovum to the primordial germ-cell, are short or very long, whether they consist of three or six or sixteen cells, or of hundreds and thousands of cells. That all the cells of the germ-track do not take on the character of germ-cells must, in accordance with our conception of the 'maturing' of determinants, be referred to the internal conditions of the cells and of the germ-plasm, perhaps in part also to an associated quantum of somatic idioplasm which is only overpowered in the course of the cell-divisions.
This splitting up of the substance of the ovum into a somatic half, which directs the development of the individual, and a propagative half, which reaches the germ-cells and there remains inactive, and later gives rise to the succeeding generation, constitutes the theory of the continuity of the germ-plasm, which I first stated in a work which appeared in the year 1885. Its fundamental idea had already been expressed much earlier by Francis Galton (1872), without however being fully appreciated at the time or having any influence on the course of science, and the same is true with the later theoretical views of Jäger, Rauber, and Nussbaum, all of whom reached the same idea quite independently of each other, and sought to elaborate it more or less fully.
The hypothesis does not depend for support merely on a recognition of its theoretical necessity; on the contrary, there is a whole series of facts which may be adduced as strongly in its favour.
Thus, even the familiar fact that the excision of the reproductive organs in all animals produces sterility proves that no other cells of the body are able to give rise to germ-cells; germ-plasm cannot be produced de novo. An unmistakable corroboration of this, it seems to me, is to be found in the conditions of germ-cell formation in the medusoids and hydroid polyps, for here it is apparent that the birthplace of the germs, that is, the place at which the germ-cells of the [Pg 412]animal are formed, has been shifted backwards in the course of phylogenetic evolution, that is, has been moved nearer to the starting-point of development. This shifting has exactly followed the 'germ-tracks.' as we shall see, although in some cases it would have been more advantageous if the birthplace of the germ-cells could have lain outside of these. Obviously, then, it is only the existing cell-generations of the germ-track which were able to give rise to germ-cells, or,[Pg 413] in other words, they alone contained the indispensable germ-plasm. With the help of Figs. 94 and 95 I hope to be able to make this matter clear.
Fig. 94. Diagram to illustrate the phylogenetic shifting back of the origins of the germ-cells in medusoids and hydroids. A composite picture. A, branch of a polyp colony. P, polyp-head with mouth (m) and tentacles. St, stalk of the polyp. M, medusoid-bud with the bell (Gl). T, marginal tentacle. m, mouth. Mst, manubrium. GphK, a gonophore-bud. GH, gastric cavity. ekt, ectoderm. ent, endoderm. st, supporting lamella. The germ-cells (kz) arise in the medusoid in the ectoderm of the manubrium—first phyletic stage—where they also attain maturity. In the gonophore-bud (GphK) they arise in the ectoderm (kz´), or further down in the stalk of the polyp at kz´´—third phyletic stage, or in the ectoderm of the branch from which the polyp has arisen, at kz´´´—fourth phyletic stage of the shunting of the originative area of the germ-cells. In the two last cases the germ-cells migrate until they reach their primitive place of origination in the medusoid, or in the corresponding layer of the medusoid gonophore, as may be more clearly seen in Fig. 95. Drawn from my sketch by Dr. Petrunkewitsch.
In the hydroid polyps and their medusoids the germ-cells always arise in the ectoderm; in species which produce sexual medusoids by budding, the germ-cells arise in the ectoderm of the manubrium of these medusoids (Fig. 94, M, kz). But in many species these sexual stages have degenerated in the course of phylogeny into so-called gonophores, that is, to medusoids which still exhibit more or less complete bells, but neither mouth (m) nor marginal tentacles (T), and which no longer break away from the colony to swim freely about, to feed independently, and to produce and ripen germ-cells. The degeneration of the 'gonophores' often goes even further; in many the medusoid bell is represented only by a thin layer of cells, and in some even this token of descent from medusoid ancestry is absent, and they are mere single-layered closed brood-sacs (Fig. 95, Gph).
The adherence of the sexual animal to the hydroid colony has, however, made a more rapid ripening of the germ-cells possible, and nature has taken advantage of this possibility in all the cases known to me, for the germ-cells no longer arise in the manubrium of the mature degenerate medusoid, that is, of the gonophore, but earlier, before the bud which becomes a gonophore possesses a manubrium. The birthplace of the germ-cells is thus shifted back from the manubrium of the medusoid to the young gonophore-bud (Fig. 94, M, kz). The same thing occurs in species in which the medusoids are liberated, but live only for a short time, for instance, in the genus Podocoryne. Although perfect medusoids are formed, these have their germ-cells fully developed at the time of their liberation from the hydroid colony. But in species in which the medusoid-buds have really degenerated and are no longer liberated, the birthplace of the germ-cells is shifted even further back, and in the first place into the stalk (St, kz´´) of the polyp from the gonophore-buds. This is the case in the genus Hydractinia. In the further course of the process the birthplace of the germ-cells has shifted as far back as to the branch from which the polyp has grown out (Fig. 94, A, kz´´´); and finally, in the cases in which the medusoid has degenerated to a mere brood-sac (Fig. 95, Gph), even to the generation of polyps immediately before, that is, into the polyp-stem from which the branch arises that bears the polyps producing the gonophore-bud (Fig. 95, kz´´´). Then we find the birthplace of the germ-cells still further back (Fig. 95, kz´´´´), for the egg and sperm-cells arise in the stem of the principal polyps (the main stem of the colony). The advantage of this arrangement[Pg 414] is easily seen, for the principal polyp is present earlier than those of the secondary branches, and these again earlier than the polyp which bears the sexual buds, and this, finally, earlier than the sexual bud which it bears. Thus this shunting backwards of the birthplace of the germ-cells means an earlier origin of the primordium (Anlage) of the germ-cells, and consequently an earlier maturing of these.
Fig. 95. Diagram to illustrate the migration of the germ-cells in hydro-medusæ from their remotely shunted place of origin to their primitive place of origin in the gonophore, in which they attain to maturity. The state of affairs in Eudendrium is taken as the basis of the diagram. HP, one of the principal polyps. mu, mouth. ma, gut-cavity. t, tentacle. Sta, its stem. A, a branch of the polyp colony. SP, lateral polyp. Gph, a medusoid-bud completely degenerated into a mere gonophore. Ei, ovum. GH, gastric cavity. st, supporting lamella. The originative area of the germ-cells lies in the stem of the principal polyp at kz´´´´, whence the germ-cells first migrate into the endoderm of the branch (A) at kz´´´, creeping within which they reach kz´´ in the lateral polyp (blastostyle), finally reaching the gonophore (kz) and passing again into the ectoderm. Drawn from my sketch by Dr. Petrunkewitsch.
But none of all these germ-cells come to maturity in the birthplace to which they have been shifted, for they migrate independently from it to the place at which they primitively arose, namely, into the[Pg 415] manubrium of the medusoid, which is still present even when great degeneration has occurred, or even—in the most extreme cases of degeneration—into the ectoderm of the brood-sac. This is the case in the genus Eudendrium, of which Fig. 95 gives a diagrammatic representation.
The most interesting feature of this migration of the germ-cells is that the cells invariably arise in the ectoderm (kz´´´´), then pierce through the supporting lamella (st) into the endoderm (kz´´´), and then creep along it to their maturing-place. Once there they break through again to the outer layer of cells, the ectoderm (kz), and come to maturity (Ei). That they make their way through the endoderm is probably to be explained by the fact that they are there in direct proximity to the food-stream which flows through the colony (GH = gastric cavity), and they are thus more richly nourished there than in the ectoderm. But although this is the case, they never arise in the endoderm; in no single case is the birthplace of the germ-cells to be found in the endoderm, but always in the ectoderm, no matter how far back it may have been shunted. Even when the germ-cells migrate through the endoderm, their first recognizable appearance is invariably in the ectoderm, as, for instance, in Podocoryne and Hydractinia. The course of affairs is thus exactly what it would necessarily be if our supposition were correct, that only definite cell-generations—in this case the ectoderm-cells—contain the complete germ-plasm. If the endoderm-cells also contained germ-plasm it would be hard to understand why the germ-cells never arise from them, since their situation offers much better conditions for their further development than that of the ectoderm-cells. It would also be hard to understand why such a circuitous route was chosen as that exhibited by the migration of the young germ-cells into the endoderm. Something must be lacking in the endoderm that is necessary to make a cell into a germ-cell: that something is the germ-plasm.
If we accept the theory of the continuity of the germ-plasm as in the main correct, it appears that higher animals and plants are constructed of two kinds of elements, the somatic cells and the germ-cells; both owe their being to the germ-plasm of the ovum, but the former do not contain it complete but only in individual determinants[24], [Pg 416]and therefore can never give rise again to the rank of germ-cells; the others contain the latent germ-plasm intact, and can therefore produce not only cells like themselves for a certain time by division, but have also the power, when they are mature and the necessary conditions have been fulfilled, of bringing forth a new individual of the same species. The former have only a limited length of life, they die—they must necessarily die—when the life of the individual to which they belong is at an end; the latter are potentially immortal, like the unicellular organisms, that is, they can in favourable circumstances give rise to the germ-cells of a new individual, and so on for all time, as far as we can see. The germ-plasm of a species is thus never formed de novo, but it grows and increases ceaselessly; it is handed on from one generation to another like a long root creeping through the earth, from which at regular distances shoots grow up and become plants, the individuals of the successive generations. If these conditions be considered from the point of view of reproduction, the germ-cells appear the most important part of the individual, for they alone maintain the species, and the body sinks down almost to the level of a mere cradle for the germ-cells, a place in which they are formed, and under favourable conditions are nourished, multiply, and attain to maturity. But the matter can also be looked at in an opposite light, and then the endless root of the germ-plasm, with its germ-cells ever forming new individuals, may be regarded as the means by which alone nature was able to create multicellular organisms, individuals of higher and higher differentiation and capacity, able to adapt themselves to all possible conditions, and to make the fullest use of all the possibilities of life.
[24] Boveri has recently made an observation upon the thread-worm of the horse, which points to the correctness of the conception of the germ-plasm. The two first segmentation-cells both receive the four chromosomes of the species, but, in one of the two, a portion of the chromatin breaks off and degenerates, or dissolves, at least as far as can be seen. The other cell retains the whole mass of chromatin, and from this there arise later the primitive genital-cells. In the germ-track, therefore—so we must interpret it—the whole of the germ-plasm is retained, while a part of it is withdrawn from the soma. I have only partly described the process, and I do not wish to enter in detail on an interpretation of it, since it seems to me obscure and to require further observations before an interpretation can be attempted with any confidence.
MR. EDWARD ARNOLD'S
LIST OF
Scientific and Technical Books.
FOOD AND THE PRINCIPLES OF DIETETICS. By Robert Hutchison, M.D. Edin., F.R.C.P., Assistant Physician to the London Hospital. Seventh and Revised Edition. Illustrated. Demy 8vo., 16s. net.
LECTURES ON DISEASES OF CHILDREN. By Robert Hutchison, M.D. (Edin.), F.R.C.P., Assistant Physician to the London Hospital and to the Hospital for Sick Children, Great Ormond Street, London. With numerous Illustrations. Second Impression. Crown 8vo., 8s. 6d. net.
PRACTICAL PHYSIOLOGY. By M. S. Pembrey, M.A., M.D., Lecturer on Physiology at Guy's Hospital; A. P. Beddard, M.A., M.D., Demonstrator of Physiology, Guy's Hospital; J. S. Edkins, M.A., M.B., Lecturer in Physiology and Demonstrator of Physiology, St. Bartholomew's Hospital; Leonard Hill, M.B., F.R.S., Lecturer on Physiology, London Hospital Medical School; and J. J. R. Macleod, M.B. Copiously Illustrated. New and Revised Edition. Demy 8vo., 12s. 6d. net.
RECENT ADVANCES IN PHYSIOLOGY AND BIO-CHEMISTRY. By Leonard Hill, M.B., F.R.S.; A. P. Beddard, M.A., M.B.; J. J. R. Macleod, M.B.; Benjamin Moore, M.A., D.Sc.; and M. S. Pembrey, M.A., M.D. Demy 8vo., 18s. net.
HUMAN EMBRYOLOGY AND MORPHOLOGY. By A. Keith, M.D., F.R.C.S. Eng., Lecturer on Anatomy at the London Hospital Medical College. With 316 Illustrations. New, Revised, and Enlarged Edition. Demy 8vo., 12s. 6d. net.
SURGICAL NURSING AND THE PRINCIPLES OF SURGERY FOR NURSES. By Russell Howard, M.B., M.S., F.R.C.S., Lecturer on Surgical Nursing at the London Hospital. Crown 8vo., with Illustrations, 6s.
THE PHYSIOLOGICAL ACTION OF DRUGS. An Introduction to Practical Pharmacology. By M. S. Pembrey, M.A., M.D., Lecturer on Physiology in Guy's Hospital Medical School; and C. D. F. Phillips, M.D., LL.D. Fully Illustrated. Demy 8vo., 4s. 6d. net.
PHOTOTHERAPY. By N. R. Finsen. Translated by J. H. Sequeira, M.D. With Illustrations. Demy 8vo., 4s. 6d. net.
A MANUAL OF HUMAN PHYSIOLOGY. By Leonard Hill, M.B., F.R.S. With 173 Illustrations. xii + 484 pages. Crown 8vo., cloth, 6s.
A PRIMER OF PHYSIOLOGY. By Leonard Hill, M.B. 1s.
A MANUAL OF PHARMACOLOGY FOR STUDENTS. By W. E. Dixon, M.A., M.D., B.S., B.Sc. Lond., etc., Assistant to the Downing Professor of Medicine in the University of Cambridge. With Numerous Diagrams. Demy 8vo., 15s. net.
THE CHEMICAL SYNTHESIS OF VITAL PRODUCTS AND THE INTER-RELATIONS BETWEEN ORGANIC COMPOUNDS. By Professor Raphael Meldola, F.R.S., of the City and Guilds of London Technical College, Finsbury. Two vols. Vol. I. now ready. Super Royal 8vo., 21s. net.
LECTURES ON THEORETICAL AND PHYSICAL CHEMISTRY. By Dr. J. H. Van 'T Hoff, Professor of Chemistry at the University of Berlin. Translated by Dr. R. A. Lehfeldt. In three volumes. Illustrated. Demy 8vo., 28s. net; or obtainable separately, as follows:
Vol. I.—Chemical Dynamics. 12s. net.
Vol. II.—Chemical Statics. 8s. 6d. net.
Vol. III.—Relations between Properties and Composition. 7s. 6d. net.
THE ELEMENTS OF INORGANIC CHEMISTRY. For use in Schools and Colleges. By W. A. Shenstone, Lecturer in Chemistry at Clifton College. 554 pages. New Edition, Revised and Enlarged. 4s. 6d.
A COURSE OF PRACTICAL CHEMISTRY. Being a Revised Edition of 'A Laboratory Companion for Use with Shenstone's Inorganic Chemistry.' By W. A. Shenstone. 144 pages. Crown 8vo., 1s. 6d.
A TEXT-BOOK OF PHYSICAL CHEMISTRY. By Dr. R. A. Lehfeldt, Professor of Physics at the East London Technical College. With 40 Illustrations. Crown 8vo., cloth, 7s. 6d.
A TEXT-BOOK OF PHYSICS. With Sections on the Applications of Physics to Physiology and Medicine. By Dr. R. A. Lehfeldt. Fully Illustrated. Crown 8vo., cloth, 6s.
PHYSICAL CHEMISTRY FOR BEGINNERS. By Dr. Ch. M. Van Deventer. Translated by Dr. R. A. Lehfeldt. 2s. 6d.
A FIRST YEAR'S COURSE OF EXPERIMENTAL WORK IN CHEMISTRY. By Ernest H. Cook, D.Sc., F.I.C., Principal of the Clifton Laboratory, Bristol. With 26 Illustrations. Crown 8vo., cloth, 1s. 6d.
AN EXPERIMENTAL COURSE OF CHEMISTRY FOR AGRICULTURAL STUDENTS. By T. S. Dymond, F.I.C., Lecturer on Agricultural Chemistry in the County Technical Laboratories, Chelmsford. With 50 Illustrations. 192 pages. Crown 8vo., cloth, 2s. 6d.
THE STANDARD COURSE OF ELEMENTARY CHEMISTRY. By E. J. Cox, F.C.S. With 90 Illustrations. 350 pages. Crown 8vo., cloth, 3s. Also obtainable in five parts, limp cloth. Parts I.-IV., 7d. each; Part V., 1s.
A PRELIMINARY COURSE OF PRACTICAL PHYSICS. By C. E. Ashford, M.A., Headmaster of the Royal Naval College, Dartmouth. Fcap. 4to., 1s. 6d.
ADVANCED EXAMPLES IN PHYSICS. By A. O. Allen, B.A., B.Sc., A.R.C.Sc., Lond., Assistant Lecturer in Physics at Leeds University. 1s. 6d.
PHYSICAL DETERMINATIONS. A Manual of Laboratory Instructions for the Determination of Physical Quantities. By W. R. Kelsey, B.Sc., A.I.E.E., Principal of the Taunton Technical College. 4s. 6d.
ELECTROLYTIC PREPARATIONS. Exercises for use in the laboratory by chemists and electro-chemists. By Dr. Karl Elbs, Professor of Chemistry at the University of Giessen. Translated by R. S. Hutton, M.Sc. Demy 8vo., 4s. 6d. net.
THE ELECTRIC FURNACE. By Henri Moissan, Professor of Chemistry at the Sorbonne. Authorized English Edition. Translated by A. T. de Mouilpied, M.Sc., Ph.D. With Illustrations. Demy 8vo., 10s. 6d. net.
ELECTRICITY AND MAGNETISM. By C. E. Ashford, M.A., Headmaster of the Royal Naval College, Dartmouth, late Senior Science Master at Harrow School. With over 200 Diagrams. Crown 8vo., 3s. 6d.
MECHANICS. By W. D. Eggar, Science Master at Eton College. Crown 8vo., 3s. 6d.
MAGNETISM AND ELECTRICITY. An Elementary Treatise for Junior Students, Descriptive and Experimental. By J. Paley Yorke, of the Northern Polytechnic Institute, London. With nearly 150 Illustrations. 3s. 6d.
VALVES AND VALVE-GEAR MECHANISMS. By W. E. Dalby, M.Inst.C.E., M.I.M.E., Professor of Engineering, City and Guilds of London Central Technical College. Royal 8vo. Fully Illustrated. 21s. net.
THE BALANCING OF ENGINES. By W. E. Dalby, M.A., B.Sc., M.Inst.C.E., M.I.M.E. With 173 Illustrations. Demy 8vo., 10s. 6d. net.
THE STRENGTH AND ELASTICITY OF STRUCTURAL MEMBERS. By R. J. Woods, Master of Engineering, Royal University of Ireland, Fellow of the Royal Indian Engineering College, and Assistant Professor of Engineering, Cooper's Hill College. Demy 8vo. 10s. 6d. net.
TRAVERSE TABLES. With an Introductory Chapter on Co-ordinate Surveying. By Henry Louis, M.A., A.R.S.M., F.I.C., F.G.S., etc., Professor of Mining and Lecturer on Surveying, Armstrong College, Newcastle-on-Tyne; and G. W. Caunt, M.A. Demy 8vo., 4s. 6d. net.
THE CALCULUS FOR ENGINEERS. By John Perry, M.E., D.Sc., F.R.S., Professor of Mechanics and Mathematics in the Royal College of Science, etc. Crown 8vo., cloth, 7s. 6d.
ELECTRIC AND MAGNETIC CIRCUITS. By Ellis H. Crapper, M.I.E.E., Head of the Electrical Engineering Department in the University College, Sheffield, viii + 380 pages. Demy 8vo., 10s. 6d. net.
AN INTRODUCTION TO THE THEORY OF OPTICS. By Professor Arthur Schuster, Ph.D., F.R.S., Professor of Physics at the University of Manchester. With numerous Diagrams. Demy 8vo., 15s. net.
ASTRONOMICAL DISCOVERY. By H. H. Turner, Savilian Professor of Astronomy in the University of Oxford. Demy 8vo., 10s. 6d. net.
VECTORS AND ROTORS. With Applications. Being Lectures delivered at the Central Technical College By Professor O. Henrici, F.R.S. Edited by G. C. Turner, Goldsmith Institute. Crown 8vo., cloth. 4s. 6d.
THE PRINCIPLES OF MECHANISM. By H. A. Garratt, A.M.I.C.E., Head of the Engineering Department of the Northern Polytechnic Institute, Holloway. Crown 8vo., cloth, 3s. 6d.
ELEMENTARY PLANE AND SOLID MENSURATION. By R. W. K. Edwards, M.A., Lecturer on Mathematics at King's College, London. For use in Schools, Colleges, and Technical Classes. 304 pages. Crown 8vo., 3s. 6d.
FIRST STEPS IN QUANTITATIVE ANALYSIS. By J. C. Gregory, B.Sc., A.I.C. Crown 8vo., cloth, 2s. 6d.
GEOMETRICAL CONICS. By G. W. Caunt, M.A., Lecturer in Mathematics, Armstrong College, Newcastle-on-Tyne, and C. M. Jessop, M.A., Professor of Mathematics, Armstrong College. Crown 8vo., 2s. 6d.
FIVE-FIGURE TABLES OF MATHEMATICAL FUNCTIONS. By J. B. Dale, M.A. Camb., B.A. Lond., Assistant Professor of Mathematics, King's College, University of London. Demy 8vo., 3s. 6d. net.
LOGARITHMIC AND TRIGONOMETRIC TABLES. (To Five Places of Decimals.) By J. B. Dale, M.A. Demy 8vo., 2s. net.
AN ELEMENTARY TREATISE ON PRACTICAL MATHEMATICS. By John Graham, B.A., Demonstrator of Mechanical Engineering and Applied Mathematics in the Technical College, Finsbury. Crown 8vo., cloth, 3s. 6d.
PRELIMINARY PRACTICAL MATHEMATICS. By S. G. Starling, A.R.C.Sc., B.Sc., Head of the Mathematics and Physics Department of the West Ham Municipal Technical Institute; and F. C. Clarke, A.R.C.Sc., B.Sc. 1s. 6d.
A NOTE-BOOK OF EXPERIMENTAL MATHEMATICS. By C. Godfrey, M.A., Headmaster of Royal Naval College, Osborne, and G. M. Bell, B.A., Senior Mathematical Master, Winchester College. Fcap. 4to., paper boards, 2s.
HOUSE, GARDEN, AND FIELD. A Collection of Short Nature Studies. By L. C. Miall, F.R.S., Professor of Biology, University of Leeds. With numerous Diagrams. Crown 8vo., 6s.
THE EVOLUTION THEORY. By August Weismann, Professor of Zoology in the University of Freiburg-im-Breisgau. Translated by Professor J. Arthur Thomson and Margaret Thomson. With numerous Illustrations and Coloured Plates. Two Vols. Royal 8vo., 32s. net.
ANIMAL BEHAVIOUR. By C. Lloyd Morgan, LL.D., F.R.S., Principal of University College Bristol, author of 'Animal Life and Intelligence,' etc. With numerous Illustrations. Large crown 8vo., 10s. 6d.
HABIT AND INSTINCT. By C. Lloyd Morgan, LL.D., F.R.S. With Photogravure Frontispiece, viii + 352 pages. Demy 8vo., cloth, 16s.
A TEXT-BOOK OF ZOOLOGY. By G. P. Mudge, A.R.C.Sc. Lond., Lecturer on Biology at the London Hospital Medical College. With about 200 original Illustrations. Crown 8vo., cloth, 7s. 6d.
ELEMENTARY NATURAL PHILOSOPHY. By Alfred Earl, M.A., Assistant Master at Tonbridge School. Crown 8vo., cloth, 4s. 6d.
A CLASS-BOOK OF BOTANY. By G. P. Mudge, A.R.C.Sc. Lond., F.Z.S., and A. J. Maslen, F.L.S., Lecturer on Botany at the Woolwich Polytechnic. With over 200 Illustrations. Crown 8vo., 7s. 6d.
THE BECQUEREL RAYS AND THE PROPERTIES OF RADIUM. By the Hon. R. J. Strutt, Fellow of Trinity College, Cambridge. With Diagrams. Demy 8vo. 8s. 6d. net.
A MANUAL OF ALCOHOLIC FERMENTATION AND THE ALLIED INDUSTRIES. By Charles G. Matthews, F.I.C., F.C.S., etc. Fully Illustrated. Crown 8vo., cloth, 7s. 6d. net.
WOOD. A Manual of the Natural History and Industrial Applications of the Timbers of Commerce. By G. S. Boulger, F.L.S., F.G.S. Fully Illustrated. Crown 8vo., 7s. 6d. net.
PSYCHOLOGY FOR TEACHERS. By C. Lloyd Morgan, LL.D., F.R.S. xii + 251 pages. Crown 8vo., 3s. 6d.
LONDON: EDWARD ARNOLD, 41 & 43 MADDOX STREET, W.
Transcriber's Notes:
On page 104, the link for 'Elymnias lais' has been corrected from PlII to PlIII.
Illustrations have been moved out of mid-paragraph.
Variations in spelling and hyphenation are retained.
Punctuation has been retained as published.