UNITED STATES
DEPARTMENT OF AGRICULTURE
CIRCULAR No. 363
Washington, D. C. October 1935
THE MIGRATION
OF NORTH AMERICAN BIRDS
By
FREDERICK C. LINCOLN
Senior Biologist in Charge, Section of Distribution
and Migration of Birds Division of Wildlife Research,
Bureau of Biological Survey
For sale by the Superintendent of Documents,
Washington, D. C. Price 10 cents
- 1 -
|
CIRCULAR No. 363 OCTOBER 1935
UNITED STATES DEPARTMENT OF AGRICULTURE
WASHINGTON, D.C.
|
|
|
THE MIGRATION OF
NORTH AMERICAN BIRDS[1]
By Frederick C. Lincoln, senior biologist, in charge Section of Distribution and
Migration of Birds, Division of Wildlife Research, Bureau of Biological Survey
CONTENTS
|
Page |
| Introduction |
1 |
| Mystery of migration |
3 |
| Historical accounts |
3 |
| Advantages of migration |
4 |
| Theories of causes of migration |
5 |
| When birds migrate |
7 |
| Movements of species and groups |
8 |
| Nocturnal and diurnal migration |
11 |
| How birds migrate |
13 |
| Speed of flight and speed of migration |
13 |
| Altitudes at which birds travel |
22 |
| Orientation |
23 |
| Segregation during migration |
25 |
| Where birds migrate |
27 |
| Distances of migration vary |
27 |
| Short and undetermined migration |
27 |
| Variable migrations within species |
28 |
| Fall flights not far south of breeding ranges |
29 |
| Long-distance migrations |
30 |
| Routes of migration |
33 |
| Wide and narrow migration lanes |
34 |
| Atlantic oceanic route |
39 |
| Atlantic coast route and tributaries |
42 |
Mackenzie Valley-Great Lakes-Mississippi Valley
route and tributaries |
45 |
| Pacific coast route |
47 |
| Pacific oceanic route |
49 |
| Arctic routes |
52 |
| Evolution of migration routes |
52 |
| Vertical migration |
55 |
| Vagrant migration |
55 |
| Perils of migration |
56 |
| Storms |
56 |
| Aerial obstructions |
57 |
| Exhaustion |
59 |
| Influence of the weather on migration |
59 |
| Problems of migration |
61 |
| Banding studies |
61 |
| Movements of residents |
62 |
| Migration of the white-throated sparrow |
63 |
| Migration of the yellow-billed loon |
63 |
| Conclusions |
65 |
| Bibliography |
66 |
| Index |
69 |
INTRODUCTION
Where do the birds go each fall that have nested in our dooryards
and frequented the neighboring woods, hills, and marshes? Will the
same ones return again to their former haunts next spring? What
dangers do they face on their round-trip flight and in their winter
homes? These and other questions on the migratory habits of birds
puzzle the minds of many who are interested in the feathered species,
whether it be the farmer who profits by their tireless warfare against
the weed and insect pests of his crops, the bird student who enjoys an
abundance and variety of feathered inhabitants about him, or the
hunter who wants a continuation from year to year of the sport of
wild-fowling. Lack of information on the subject may mean the loss
of an important resource by unconsciously letting it slip from us.
Ignorance of the facts may be responsible for inadequate legal protection
for such species as may urgently need it. More general knowledge
on the subject will aid in the perpetuation of the various migrants,
the seasonal habitats of some of which are in grave danger from man's
- 2 -
utilization, sometimes unwisely, of the marsh, water, and other areas
they formerly frequented.
The migrations of birds were probably among the first natural
phenomena to attract the attention and intrigue the imagination of
man. Recorded observations on the subject date back nearly 3,000
years, to the times of Hesiod, Homer, Herodotus, Aristotle, and others.
In the Bible are several references to the periodic movements of birds,
as in the book of Job (39:26), where the inquiry is made: "Doth the
hawk fly by Thy wisdom and stretch her wings toward the south?"
Jeremiah (8:7), wrote: "The stork in the heavens knoweth her
appointed time; and the turtle [dove], and the crane, and the swallow,
observe the time of their coming." And the flight of quail that saved
the Israelites from starvation in their wanderings in the wilderness of
Sinai is now recognized as a vast movement of migratory quail
(Coturnix coturnix) between their breeding grounds and their winter
home in Africa.
Throughout the ages the return flights of migratory birds have
been important (1) as a source of food after a lean winter, and (2) as
the harbinger of a change in season. The arrival of certain species
has been heralded with appropriate ceremonies in many lands, and
among Eskimo and other tribes the phenomenon to this day is the
accepted sign of the imminence of spring and of warmer weather.
The pioneer fur traders in Alaska and Canada offered rewards to the
Indian or Eskimo who saw the first goose of the spring, and all joined
in jubilant welcome to the newcomer.
Always hunted for food, the large flocks of ducks and geese became
objects of the enthusiastic attention of an increasing army of sportsmen
as the North American Continent became ever more thickly
settled. Most of the nongame species were found to be valuable also
as allies of the farmer in his never-ending warfare against weed and
insect pests. The need for laws protecting the valuable game and
nongame birds and for regulated hunting of the diminishing game
species followed as a natural course. In the management of this wildlife
resource it has become obvious that continuous studies must be
made of the food habits of the various species, their environmental
needs, and their travels. Hence bird investigations are made by the
Biological Survey, the Bureau charged by Congress under the Migratory
Bird Treaty Act with the duty of protecting those species that
in their yearly journeys pass back and forth between the United States
and Canada.
For half a century the Biological Survey has been collecting data
on the interesting and important phenomenon of the migration of
North American birds. The field men of the Bureau have gathered
information concerning the distribution and seasonal movements of
the different species in many extended areas, from the Arctic coast
south to the pampas of Argentina. Supplementing these investigations
is the work of hundreds of volunteer ornithologists and bird
students throughout the United States and Canada, who each year,
spring and fall, forward to the Bureau reports on migrations observed
in their respective localities. Added, to the mass of data thus assembled
is a rapidly growing file of records of birds that have been banded
and of the subsequent recovery of the marked individuals.
These data, together with other carded records gleaned by the
Biological Survey from a vast literature, constitute a series of files
- 3 -
that now contain well over 2,500,000 entries, easily the greatest
existing accumulation of information pertaining to the distribution
and movements of North American birds. Not only do the facts
thus assembled form the basis of regulatory action for the protection
of the birds, but they also make it possible to publish scientific
accounts of the ranges and migrations of the different species. They
furnish the basis of this publication.
The several important bird-protective measures adopted by State
and Federal Governments, particularly those having as their objective
the conservation of the migratory song, insectivorous, and game
species, can be effective only if they have intelligent public support.
To increase such support, information must be more generally available
on that little understood but universally fascinating subject
of bird migration. A brief presentation of facts on the migratory
habits of the birds, scientifically gathered by the Bureau of Biological
Survey over many years, will be helpful to bird-study classes, to
conservation organizations, and to farmers and others individually
interested in the welfare of the birds.
THE MYSTERY OF MIGRATION
HISTORICAL ACCOUNTS
Of observers whose writings are extant, Aristotle, naturalist and
philosopher of ancient Greece, was one of the first to discuss the
subject of bird migration. He noted that cranes traveled from the
steppes of Scythia to the marshes at the headwaters of the Nile,
and that pelicans, geese, swans, rails, doves, and many other birds
likewise passed to warmer regions to spend the winter. In the
earliest years of the Christian era, the elder Pliny, Roman naturalist,
in his Historia Naturalis, repeated much of what Aristotle had said
on migration, and added comments of his own concerning the movements
of the European blackbird, the starling, and the thrushes.
In spite of the keen perception shown in some of his statements,
Aristotle also sponsored some superstitions on bird migration that
persisted for several centuries. One of these, that of hibernation,
became so firmly rooted that in 1878, the American ornithologist
Coues (20)[2] listed the titles of no less than 182 papers dealing with
the hibernation of swallows (Hirundinidae). The hibernation theory
accounted for the autumnal disappearance of certain species of birds
as their passing the cold season in a torpid state, hidden in hollow
trees, caves, or the mud of marshes. Aristotle ascribed hibernation
not only to swallows but also to storks, kites, doves, and others.
Some early naturalists wrote fantastic accounts of flocks of swallows
seen congregating in the marshes until their accumulated weight
bent into the water the reeds on which they clung and thus submerged
the birds. It was even recorded that when fishermen in
northern waters drew up their nets they sometimes had a mixed
"catch" purported to consist of fish and hibernating swallows.
Clarke (4) quotes Olaus Magnus, Archbishop of Upsala, who in 1555
published a work entitled "Historia de Gentibus Septentrionalibus
et Natura", wherein he observed that if swallows so caught were
taken into a warm room they soon begin to fly about but would live
only a short time.
- 4 -
The hibernation theory survived for more than 2,000 years and is
still occasionally repeated by credulous persons to account for failure
to locate definitely the winter home of the chimney swifts (Chaetura
pelagica), which each autumn gather in immense flocks in southern
Georgia and northern Florida and then suddenly disappear. There are,
however, records of occurrence during migration for a few points
in the West Indies, Mexico, and Central America, and it is probable
that these birds spend the winter season in the great rain-forest area
of the Amazon Valley in Brazil, passing most of the daytime high
in the air with other swifts that are local residents.
Aristotle was also the originator of the theory of transmutation,
basing it upon the fact that frequently one species will arrive from the
north just as another species departs for more southerly latitudes.
From this he reasoned that although it was commonly believed that
such birds were of two different species, there really was only one,
and that this one assumed the different plumages to correspond with
the summer and winter seasons.
Probably the most remarkable theory that has been advanced to
account for migration is contained in a pamphlet mentioned by
Clarke (4, v. 1, pp. 9-11), as published in 1703 under the title: "An
Essay Toward the Probable Solution of this Question: Whence
Come the Stork and the Turtle, the Crane, and the Swallow, when
they Know and Observe the Appointed Time of their Coming."
It was written "By a Person of Learning and Piety", whose "probable
solution" was that migratory birds flew to the moon and there
spent the winter.
Some who easily accepted the disappearance of the larger birds as
migratory travelers were unable to understand how the smaller species,
some of them notoriously poor fliers, could make similar journeys.
They contended that the larger species, as the storks and cranes,
carried their smaller companions as living freight. In some of the
Mediterranean countries it is still believed that these broad-pinioned
birds serve as aerial transports for the hosts of small birds that congregate
on the shores of the Mediterranean Sea, awaiting opportunity
for this kind of passage to their winter homes in Africa. Similar
beliefs are found among some tribes of North American Indians.
ADVANTAGES OF MIGRATION
Before presenting some of the present theories concerning the
origin of bird migration, it seems well to consider briefly the ends
served by this annual round trip between breeding grounds and
winter quarters. It is apparent that the migratory habit enables
a species to enjoy the summers of northern latitudes while avoiding
the severity of the winters. In other words, migration makes it
possible for some species to inhabit two different areas at seasons
when each presents favorable conditions. In the performance of its
reproductive duties every pair of birds requires a certain domain,
the extent of which varies greatly in different species. Generally,
however, this territory must be large enough to provide adequate
food, not only for the parent birds but also for the lusty appetites
that come into being with the hatching of the eggs. Thus, if all
birds were to remain constantly either in tropical or in temperate
regions, there might be intolerable overcrowding during the breeding
season. By the spring withdrawal to regions uninhabitable earlier
- 5 -
in the year, the migrants are assured of adequate space and ample
food upon their arrival in the winter-freed North, and it may be
assumed that nonmigrant species resident in the South are benefited
by the withdrawal of the migrants.
Nevertheless it cannot be said that the winter or summer areas are
entirely unsuited to the requirements of every migrating species at
other seasons, for some individuals pass the winter in areas that are
frequented only in summer by other individuals of their species. The
extensive breeding ranges of such species present wide climatic variations,
so that some individuals may actually be resident in a region
where others of their kind are present only in winter.
The tendency in many species to move southward at the approach
of winter is not always due to the seasonal low temperatures, since
experiments have demonstrated that many summer insect feeders,
when confined in outdoor aviaries, comfortably withstand temperatures
far below zero. The main consideration is the depletion of the
food supply, caused either by disappearance or hibernation of insects,
or by the mantle of snow or ice that prevents access to the seeds and
other forms of food found on or close to the ground or submerged in
water. Possibly also the shortened hours of daylight materially
restrict the ability of the birds to obtain sufficient food at a time when
the cold requires an increased supply to maintain body heat. It is
noteworthy that chickadees (Penthestes atricapillus) and some other
of our smaller birds have no fear of Arctic weather, as their food supplies
are mainly arboreal and so are always available. Also, when
there is a good supply of food in the form of pine seeds in Canadian
woods, nuthatches (Sitta carolinensis and S. canadensis) and crossbills
(Loxia curvirostra and L. leucoptera) will remain through the winter.
When these birds appear abundantly in winter at points in southern
latitudes, it may be concluded that there is a shortage of their food in
the North, or that they have been lured farther south by the greater
abundance of this food there.
THEORIES OF THE CAUSES OF MIGRATION
Migration has long since become a definite hereditary habit that
recurs in annual cycles, probably because of physiological stimuli
associated with the reproductive period. In seeking its origin it is
necessary to study the history of the birds' occupation of their present
ranges, and from the information available to consider what appear
to be reasonable theories. The two now most commonly accepted
are diametrically opposed to each other.
NORTHERN ANCESTRAL HOME THEORY
According to one of these, nonmigratory birds swarmed over the
entire Northern Hemisphere in earlier ages, the conditions of food and
habitat being such as to permit them to remain in their haunts throughout
the year. The entire northern area then afforded the two important
avian requirements—suitable breeding conditions and a year-long
food supply. This is the condition today in the Tropics, and it
is noteworthy that many tropical birds are nonmigratory. Gradually,
however, in the Northern Hemisphere the glacial ice fields advanced
southward, forcing the birds before them, until finally all bird life was
concentrated in southern latitudes. As the ages passed and the ice
- 6 -
cap gradually retreated, each spring the birds endeavored to return
to their ancestral homes in the North, only to be again driven south
at the approach of winter. As the size of the ice-covered area diminished,
the journeys made became ever longer, until eventually the
habits of migration were fixed to accord with the climatic conditions
of the present age.
Thus, this theory supposes that today migratory birds follow the
path of a great racial movement that took place in a distant past,
associated with advances and recessions of the ice. The actions of
the birds themselves lend some support to this theory, as every bird
student has noted the feverish impatience with which certain species
push northward in spring, sometimes advancing so rapidly upon the
heels of winter as to perish in great numbers when overtaken by late
storms. It is probable, however, that at that season the reproductive
impulse urges the birds on to their northern breeding grounds.
SOUTHERN ANCESTRAL HOME THEORY
The opposing theory is simpler in some respects and supposes that
the ancestral home of all birds was in the Tropics and that as all bird
life tends to overpopulation there was a constant effort to seek breeding
grounds where competition would be less keen. Species that
strove for more northern latitudes would be kept in check by the ice
and forced to return southward with the recurrence of winter conditions.
As the ice retreated, vast areas of virgin country became
successively suitable for summer occupancy, but the winter habitat
remained the home to which the birds returned after the nesting
season. It is a fact that some species spend very little time on their
breeding grounds; the orchard oriole (Icterus spurius), for example,
spends only 2½ months in its summer home, arriving in southern
Pennsylvania about the first week in May and leaving by the middle
of July.
Both theories assume that migration is an ingrained habit, but
neither is supported by positive biological data. Both have been
criticized also on geological grounds, and neither can be accepted
without qualification. It is apparent, however, that whether the
ancestral home of any species was at the northern or at the southern
limits of its present range, or even in some intermediate region, the
search for conditions favorable for breeding in summer and for feeding
in winter has been a principal factor underlying the origin of migration.
THEORY OF PHOTOPERIODISM
A modern view, based on studies of living behavior, favors the
theory of photoperiodism, propounded by recent investigators as the
cause of the annually induced movements of the birds. This theory
holds as its major premise that quantity of light and length of day are
the stimulating causes of migration. Its proponents urge that migration
is a phenomenon far too regular to be created anew each season
merely under stress of circumstances, such as need for food; and that
it begins before the necessity for a change in latitude becomes at all
pressing. Swallows, nighthawks (Chordeiles minor), shore birds, and
others may start their southward movement while the summer food
supply in the North is at peak abundance; while robins (Turdus
migratorius), bluebirds (Sialia sialis), and others may leave an abundant
- 7 -
food in the South in spring and press toward northern points
when the food supplies there are almost entirely lacking and when
severe cold and storms are likely to play havoc with the advance
migrants. The regularity of arrival and departure is one of the most
impressive features of migration, and since birds travel in almost
strict accordance with the calendar, the proponents of the theory ask:
What phenomenon to which we may attribute the stimulating impulse
occurs with such precise regularity as the constantly increasing
light in spring?
Experimental work has abundantly demonstrated the effect of
increased light upon the growth, flowering, and fruiting of plants.
Similarly, experiments with the common junco, or snowbird (Junco
hyemalis), reported by Rowan (42, p. 121), resulted in increased
development of the sexual organs by the end of December, although
the birds were confined in outdoor aviaries in Canada and had been
exposed to temperatures as low as -44° F. From the first of November
until early in January, the juncos were subjected to ever-increasing
light, supplied in the aviaries by electric bulbs. As regards illumination,
they were thus artificially provided with conditions approximating
those of spring. At the close of this period, it was found that
the sexual organs of the birds had attained the maximum development
normally associated with spring. With gradual reduction of the
lighting over a period of little more than 1 month, the organs returned
to their normal winter condition.
After a consideration of all evidence, including the fact that no
ultraviolet rays were used, it was concluded that the explanation lay
in the increased exercise taken during the periods of increased light.
A simple test whereby certain birds were forced by mechanical means
to take more exercise, the light being so reduced that there was merely
sufficient glow for them to see the advancing mechanism that forced
them into movement, showed that the rate of development of the
sexual organs exactly paralleled that in the birds that were exposed
to extended periods of illumination in the outdoor aviaries. Other
features in this experiment—such as the behavior of the birds themselves—also
indicated that more activity due to increased light is the
governing cause of the spring development of the sexual organs. If
this development be accepted as a controlling cause of migration, then
this experiment must be recognized as of great importance.
Upon closer analysis, however, it is found that this theory, like
those before discussed, is open to serious objections. First, some of
our summer residents that migrate south for the winter do not stop in
equatorial regions, where they might find the periods of day and
night about equally divided, but push on beyond, some penetrating as
far south as Patagonia. Also it might be asked: If the lengthening
day is the stimulating factor, why should our summer birds wintering
in the Tropics ever start northward, as in their winter quarters the
variation in the length of day from winter to summer is imperceptible.
Like all the other theories advanced, this also, as at present understood,
is subject to unanswered criticism.
WHEN BIRDS MIGRATE
It is known that at any given point many species leave in fall and
return in spring. Since bird banding has had such wide application
as a method of study, it is known also that in some species one of the
- 8 -
parent birds (rarely both) frequently returns and nests in the same
tree, bush, or box that held its nest in the previous season (fig. 1.
One ordinarily thinks of the world of birds as quiescent during two
seasons each year, at nesting time and in winter. For individual
species this is obviously the case, but when the entire avifauna of the
continent is considered
it is found that there
are at almost all periods
some latitudinal
movements.

B12M
Figure 1.—The bluebird may return regularly year after year to nest
in the same hole or box that was occupied in previous seasons.
MOVEMENTS OF SPECIES
AND GROUPS
Some species begin
their fall migrations
early in July, and in
some parts of the country
distinct southward
movements can be
detected from then
until the beginning or
middle of winter. For
example, many shore
birds start south in the
early part of July,
while the goshawks
(Astur atricapillus),
snowy owls (Nyctea
nyctea), redpolls
(Acanthis linaria),
Bohemian waxwings
(Bombycilia garrula),
and many others do
not leave the North
until forced to do so
by the advent of severe
winter weather or by
lack of the customary
food. Thus, an observer in the northern part of the United States
may record an almost unbroken southward procession of birds from
midsummer to winter, and note some of the returning migrants as
early as the middle of February. Purple martins (Progne subis)
have been known to arrive in Florida late in January on their way
north, and the northern movement may continue among late arrivals
into the first week of June. In some species the migration is so
prolonged that the first arrivals in the southern part of the breeding
range will have performed their parental duties while others of that
species are still on their way north.
A study of these facts indicates that sometimes there exists a very
definite relationship between what we may term northern and
southern groups of individuals of the same species. A supposition,
on which additional banding work is expected later to give definite
facts, is that in the case of some species that have an extensive latitudinal
- 9 -
breeding range and a normal migration, the individuals that
nest farthest south migrate first in fall and proceed to the southern
part of the winter range; those that occupy the central parts of the
breeding range migrate next, and travel to regions in the winter range
north of those occupied by the first group; and finally the individuals
breeding farthest north are the last to begin their fall migration and
these remain farthest north during the winter. In other words, this
theory supposes that the southward movement of the species is
normally such that the different groups maintain their relative
latitudinal positions, both spring and fall.
B4503M
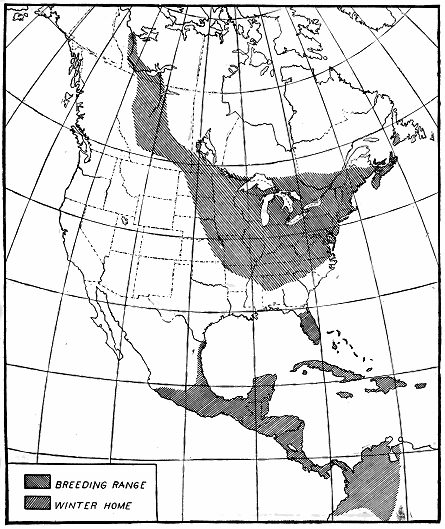
Figure 2.—Summer and winter homes of the black-and-white warbler. A very slow migrant, as the
birds nesting in the northern part of the country take 50 days to cross the breeding range. The rapidity
of their advance is shown in figure 3.
The black-and-white warbler (Mniotilta varia) furnishes an example.
The breeding range of this bird extends west and northwest from
South Carolina and New Brunswick as far as Great Bear Lake in
northwestern Canada (fig. 2). The bird spends the winter in southern
- 10 -
Florida, the West Indies, central Mexico, Central America, and
northwestern South America. In the southern part of its breeding
range it is nesting in April, but the summer residents of New Brunswick
do not reach their breeding grounds before the middle of May.
Therefore, about 50 days are required for these northbound birds to
cross the breeding range, and if 60 days be allowed for nest building,
egg laying, incubation, care of the young, and molt, they would not
be ready to start southward before the middle of July (fig. 3). Then
another 50-day trip south, and the earliest migrants from the northern
areas would reach the Gulf coast in September. But both adults and
young have been observed at Key West, Fla., by the middle of July,
and on the northern coast of South America by August 21. Since
the birds at Key West were fully 500 miles south of the breeding-range,
it is evident that they must have come from the southern part
of the nesting area.
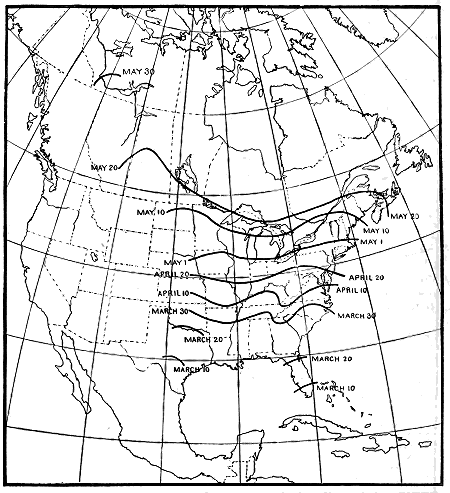
B4504M
Figure 3.—Isochronal migration lines of the black-and-white warbler, showing a slow and uniform migration,
the advance across the United States being apparently only about 20 miles a day.
Many similar cases might be mentioned, such as the black-throated
blue warblers (Dendroica caerulescens), which are still observed in
- 11 -
the mountains of Haiti in the middle of May when others of the
species are en route through North Carolina to breeding territory
in New England or have even reached that region. Redstarts
(Setophaga ruticilla) and yellow warblers (D. aestiva), evidently the
more southern breeders in each case, are seen returning southward
on the northern coast of South America just about the time that
the earliest of those
breeding in the North
reach Florida on their
way to winter quarters.

B2282M
Figure 4.—The Kentucky warbler, a night migrant, in traveling to
its winter quarters in Central America and northwestern South
America, uses route no. 5, figure 20.
NOCTURNAL AND DIURNAL MIGRATION
When one recalls
that most birds appear
to be more or less
helpless in the dark,
it seems remarkable
that many should
select the night hours
for extended travel.
Among those that do,
however, are the great
hosts of shore birds,
rails, flycatchers, orioles,
most of the great
family of sparrows,
the warblers (fig. 4),
vireos, and thrushes,
and, in fact, the majority
of small birds.
That it is common to
find woods and fields
on one day almost
barren of bird life and
on the following day
filled with sparrows,
warblers, and
thrushes, would indicate
the arrival of
migrants during the
night. The passage
of flocks of ducks and
geese is frequently observed
by sportsmen
sitting in their blinds, but great numbers of these birds also pass
through at night, the clarion call of the Canada goose (Branta canadensis)
or the conversational gabbling of a flock of ducks being
common night sounds in spring and fall in many parts of the country.
The sibilant, nocturnal calls of the upland plover, or Bartramian
sandpiper (Bartramia longicauda), and other shore birds during their
spring and fall flights form vivid memories in the minds of many
- 12 -
students of migration. Observations made with telescopes focused
on the full moon have shown processions of birds. The estimate of
one observer that birds passed his point of observation at the rate
of 9,000 an hour gives some indication of the numbers of birds that
are in the air during some of the nights when migration is at its
height. While the passage of migratory birds has thus been recorded
throughout the night, the bulk of the flocks pass during the earlier
hours of the evening and toward daylight in the morning, the periods
from 8 o'clock to midnight and from 4 to 6 a. m. seeming to be
favorite times for nocturnal flight.
It has been claimed, with some reason, that small birds migrate
by night the better to avoid their enemies, and that most of the
nocturnal travelers are those that are naturally timid, sedentary, or
feeble-winged. Included in this group are not only small song and
insectivorous birds, but also such weak fliers as the rails, as well as
the wrens, the small woodland flycatchers, and other species, which,
living habitually more or less in concealment, are probably much
safer making their long flights under the protecting cloak of darkness.
This cannot fully account for the nocturnal habit, however,
since among the night migrants are the snipe, sandpipers, and
plovers, birds that are generally found in the open and are among
the more powerful fliers, some of them making flights of more than
2,000 miles across the ocean. Such exceptionally long flights, of
course, require both day and night flying.
Night travel is probably best for the majority of birds, chiefly
from the standpoint of feeding. Digestion is rapid in birds, and
yet the stomach of a bird killed during the day almost always contains
food. To supply the energy required for long flight, it is
essential that food be obtained at comparatively short intervals, the
longest of which in most species is during the hours of darkness.
If the smaller migrants were to make protracted flights by day they
would be likely to arrive at their destination at nightfall almost
exhausted, but unable to obtain food until the following morning,
since they are entirely daylight feeders. This would delay resumption
of flight and result in great exhaustion or possibly even death
were they so unfortunate as to have their evening arrival coincident
with unusually cold or stormy weather. Traveling at night, they
pause at daybreak and devote the entire period of daylight to alternate
feeding and resting. This permits complete recuperation and
resumption of the journey at nightfall.
Many species of wading and swimming birds migrate indifferently
by day or by night, as they are able to feed at all hours and are
not accustomed to seek safety in concealment. Some diving birds,
including ducks that submerge when in danger, sometimes travel
over water by day and over 1 and at night. The day migrants include,
in addition to some of the ducks and geese, the loons, cranes, gulls,
pelicans, hawks (fig. 7), swallows, nighthawks, and the swifts (fig. 6),
all strong-winged birds. The swifts, swallows, and nighthawks
(sometimes called bullbats) feed entirely on flying insects, and use
their short, weak feet and legs only for grasping a perch during
periods of rest or sleep. Thus they feed as they travel, the circling
flocks being frequently seen late in summer working gradually southward.
Years ago, before birds of prey were so thoughtlessly
slaughtered, great flocks of red-tailed hawks (Buteo borealis), Swainson's
- 13 -
hawks (B. swainsoni), and rough-legged hawks (B. lagopus and
B. regalis) might be seen wheeling majestically across the sky in the
Plains States, and in the East the flights of broad-winged hawks
(B. platypterus), Cooper's hawks (Accipiter cooperi), and sharp-shinned
hawks (A. velox) are still occasionally seen, although these
birds do not actually travel in flocks. To the birds of prey and
possibly to the gulls also, a day's fasting now and then is no hardship,
particularly since they frequently gorge themselves to repletion
when opportunity is afforded.
The spring migrations of the blackpoll warbler (Dendroica striata)
and the cliff swallow (Petrochelidon albifrons) afford an interesting
comparison of the flights of day and night migrants. Both spend the
winter in South America, at which season they are neighbors. But
when the impulse comes to start northward toward their respective
breeding grounds, the warblers strike straight across the Caribbean
Sea to Florida, while the swallows begin their journey by a westward
flight of several hundred miles to Panama (fig. 5). Thence they
move leisurely along the western shore of the Caribbean Sea to
Mexico, and continuing to avoid a long trip over water, they go
completely around the western end of the Gulf of Mexico. This
circuitous route adds more than 2,000 miles to the journey of the
swallows that nest in Nova Scotia. The question may be asked,
Why should the swallow select a route so much longer and more
roundabout than that taken by the blackpoll warbler? The simple
explanation is that the swallow is a day migrant while the warbler
travels at night. The migration of the warbler is made up of a
series of long, nocturnal flights, alternated with days of rest and feeding
in favorable localities. The swallow, on the other hand, starts its
migration several weeks earlier and catches each day's ration of
flying insects during a few hours of aerial evolutions, which at the
same time carry it slowly in the proper direction. Flying along the
insect-teeming shores of the Gulf of Mexico, the 2,000 extra miles
that are added to the migration route are but a fraction of the distance
that these birds cover in pursuit of their food.
Although most of our smaller birds make their longest flights at
night, close observation shows that travel is continued to some
extent by day. This is particularly the case during the latter half of
a migratory season, when the birds manifest a desire to hasten to
their breeding grounds. At this time flocks of birds while feeding
maintain a movement in the general direction of the seasonal journey.
Sometimes they travel hurriedly, and while their flights may be
short, they cover considerable distances in the course of a day.
HOW BIRDS MIGRATE
SPEED OF FLIGHT AND SPEED OF MIGRATION
There is wide-spread misconception concerning the speed at which
birds normally fly, and even regarding the speed they can attain
when occasion demands, as when closely pursued by an enemy. It
is not unusual to hear accounts of birds flying "a mile a minute."
While undoubtedly some birds can and do attain a speed even greater
than this, such cases are exceptional, and it is safe to say that even
when pressed, few can develop an air speed of 60 miles an hour.
They do, however, have two speeds, one being the normal rate for
- 14 -
everyday purposes and also for migration, and an accelerated speed
for escape or pursuit; this in some cases may be nearly double the
normal rate of movement. Nevertheless, the effort required for the
high speeds could not be long sustained, certainly not for the long-distance
migratory journeys that are regularly made by most birds.
B4749M
Figure 5.—Migration of the cliff swallow; a day migrant that, instead of flying across the Caribbean Sea
as does the blackpoll warbler (fig. 11), follows around the coast of Central America, where food is readily
obtained.
The theory that migrating birds attain high speeds received encouragement
from the German ornithologist Gätke, who for
many years, made observations on birds at the island of Heligoland
- 15 -
and published thereon in 1891 (21). He postulated that the blue-throat
(Cyanosylvia suecica), a species of thrush smaller than the
American hermit thrush, would leave African winter quarters at dusk
and reach Heligoland at dawn, which would mean a sustained speed
of 200 miles an hour; and that the American golden plover (Pluvialis
dominion) flew from the coast of Labrador to Brazil in 15 hours, or at
the tremendous speed of 250 miles per hour. These conclusions are
now considered unwarranted by most ornithologists.
Sportsmen also often greatly overestimate the speed at which ducks
and geese fly and sometimes attempt to substantiate their estimates
by mathematical calculations, based upon the known velocity of a
charge of shot, the estimated distance, and the estimated "lead" that
was necessary to hit the bird. If all three elements of the equation
were known with certainty, the speed of the bird could be determined
with a fair degree of accuracy. The majority of the ducks that are
reported as killed at 40, 50, or even 60 yards, however, actually are
shot at distances much less than estimated. To sight along a
gun barrel and estimate correctly the distance of a moving object
against the sky is so nearly impossible for the average gunner as to
make such calculations of little value.
During the past few years reliable data on the speed of birds have
accumulated slowly. It has been found that the common flying speed
of ducks and geese is between 40 and 50 miles an hour, and that it is
much less among smaller birds. Herons, hawks, horned larks, ravens,
and shrikes, timed with the speedometer of an automobile, have been
found to fly 22 to 28 miles an hour, while some of the flycatchers are
such slow fliers that they attain only 1 to 17 miles an hour. Even such
fast-flying birds as the mourning dove rarely exceed 35 miles an hour.
All these birds can fly faster, but it is to be remembered that at training
camps during the World War, airplanes having a maximum speed of
about 80 miles an hour easily overtook flocks of ducks that it may be
supposed were making every effort to escape. Aviators have claimed
that at 65 miles an hour they can overtake the fastest ducks, though
cases are on record of ducks passing airplanes that were making 55
miles an hour.
The greatest bird speeds that have been reliably recorded are of the
swifts (fig. 6) and the duck hawk, or peregrine falcon (Falco peregrinus)
(fig. 7). An observer in an airplane in Mesopotamia reported
that swifts easily circled his ship when it was traveling at 68
miles an hour. To do this, the birds certainly were flying at a speed
as high as 100 miles an hour. Once a hunting duck hawk, timed with
a stop watch, was calculated to have attained a speed between 165
and 180 miles an hour.
The speed of migration, however, is quite different from that attained
in forced flights for short distances. A sustained flight of 10
hours a day would carry herons, hawks, crows, and smaller birds from
100 to 250 miles, while ducks and geese might travel as much as 400
to 500 miles in the same period. Measured as air-line distances, these
journeys are impressive and indicate that birds could cover the ordinary
migration route from the northern United States or even from
northern Canada to winter quarters in the West Indies or in Central
America or South America in a relatively short time. It is probable
that individual birds do make flights of the length indicated and that
barn swallows (Hirundo erythrogaster) seen in May on Beata Island,
- 16 -
off the southern coast of the Dominican Republic, may have reached
that point after a nonstop flight of 350 miles across the Caribbean
Sea from the coast of Venezuela. Nevertheless, whether they continue
such journeys day after day is doubtful.

B988M
Figure 6.—Chimney swift: a speedy day migrant, the flight, of which
on occasions probably exceeds 100 miles an hour.
It seems more likely that migrations are performed in a leisurely
manner, and that after a flight of a few hours the birds pause to feed
and rest for 1 or several days, particularly if they find themselves in
congenial surroundings. Some indication of this is found in the records
of banded birds. Considering only the shortest intervals that have
elapsed between banding in the North and recovery in southern regions,
it is found that
usually a month or
more is taken to cover
an air-line distance
of a thousand miles.
For example, a black
duck (Anas rubripes)
banded at Lake
Scugog, Ontario, was
killed 12 days later at
Vicksburg, Miss. If
the bird was taken
shortly after its arrival,
the record would
indicate an average
daily flight of only 83
miles, a distance that
could have been covered
in about 2 hours'
flying time. Among
the thousands of banding
records obtained
in recent years, evidences
of such rapid
flight are decidedly
scarce, for with few
exceptions all thousand-mile flights have
required 2 to 4 weeks
or more. The greatest
speed thus far recorded
for a banded
bird is that of a mallard (A. platyrhynchos) banded on November 23,
1930, in Green Bay, Wis., and shot 5 days later, 900 miles away, near
Georgetown, S. C. This bird doubtless flew at least 1,000 miles in the
5 days, as its route probably was not in a direct air-line, but, even so,
the average daily distance was only 200 miles, which could easily have
been covered in 5 hours.
It seems certain that migratory journeys are performed at the
normal rate of flight, as this would best conserve the strength of the
birds and eliminate the fatigue that would result from effort required
for great speed. Migrating birds passing lightships and lighthouses,
- 17 -
or crossing the face of the moon, have been observed to fly without
hurry or evidence of straining to attain high speed.

B213M
Figure 7.—Duck hawk, or peregrine falcon (Falco peregrinus): a day
migrant and one of the fastest and most graceful fliers of all North
American birds.
The speed of migration also is demonstrated by the dates of arrival,
particularly during the spring movement. The Canada goose affords
a typical example of regular, but slow, migration. Its advance northward
at this season is at the same rate as the advance of the season
(fig. 8). In fact, the isotherm of 35° F. appears to be a governing
factor in the speed at which these geese move north, and over their
entire trip the vanguard
follows closely
the advance of this
isotherm.
Few species perform
such regular migrations,
many waiting in
their winter homes
until spring is well
advanced and then
moving rapidly to
their breeding-grounds.
Sometimes
this advance is so rapid
that the later migrants
actually catch
up with species that
for a month or more
may have been pressing
slowly but steadily
northward.
One of the best examples
of rapid migration
is found in the
gray-cheeked thrush
(Hylocichla minima
aliciae). This bird
winters in Colombia,
Ecuador, Peru, Venezuela,
and British
Guiana and does not
start its northward
journey until many
other species are well
on their way. It does
not appear in the
United States until the
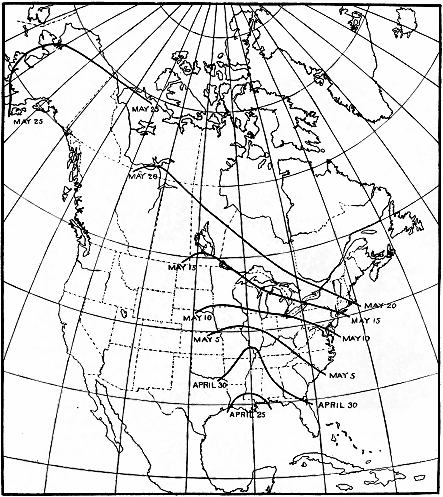
last of April—April 25, near the mouth of the Mississippi, and April 30
in northern Florida (fig. 9). A month later, or by the last week in
May, the bird is seen in northwestern Alaska, the 4,000-mile trip
from Louisiana having been made at an average speed of about 130
miles a day.
Another example of rapid migration is furnished by the yellow, or
summer, warbler. Coming from the Tropics, the birds reach New
- 18 -
Orleans about April 5, when the average temperature is 65° F. Traveling
north much faster than does the season, they reach their
breeding grounds in Manitoba the latter part of May, when the
average temperature is only 47°. Encountering progressively colder
weather over their entire route, they cross a strip of country in the
15 days from May 11 to 25 that spring takes 35 days to cross. This
"catching up" with spring is habitual in species that winter south of
the United States and in most of the northern species that winter in
the Gulf States. To this rule there appear to be only six exceptions—the
Canada goose, the mallard, the pintail (Dafila acuta), the
crow (Corvus brachyrhynchos), the red-winged blackbird (Agelaius
phoeniceus), and the robin.
B4508M
Figure 8.—Migration of the Canada goose. The northward movement keeps pace with the advance of
spring, in this case the advance of the isotherm of 35° F. agreeing with that of the birds.
The blue goose (Chen caerulescens) (fig. 10) presents a striking
example of a late but very rapid spring migration. Practically all
members of the species winter in the great coastal marshes of Louisiana,
where 50,000 or more may be seen grazing in the "pastures"
or flying overhead in flocks of various sizes. Their breeding grounds
are chiefly on Baffin Island and on Southampton Island in the northern
part of Hudson Bay, in a region where conditions of severe cold prevail
except for a few weeks each year. The birds seem to realize that
even though the season in their winter quarters is advancing rapidly,
their nesting grounds are still covered with a heavy blanket of ice
and snow. Accordingly they remain in the coastal marshes until the
last of March or the first of April, when the local birds are already
- 19 -
busily engaged with the duties of reproduction. The flight northward
is rapid, almost nonstop, so far as the United States is concerned,
for although the birds are sometimes recorded in large numbers
in the Mississippi Valley, including eastern South Dakota, and
in southeastern Manitoba, there are few records anywhere along the
route of such great flocks as are known to winter in Louisiana. When
the birds arrive in the James Bay region of Canada they apparently
enjoy a prolonged period of rest, as they are not noted in the vicinity
of their breeding grounds until the first of June. During the first 2
weeks of that month they pour into the tundra country by the
thousands, and each pair immediately sets about the business of
rearing a brood.
B4505M
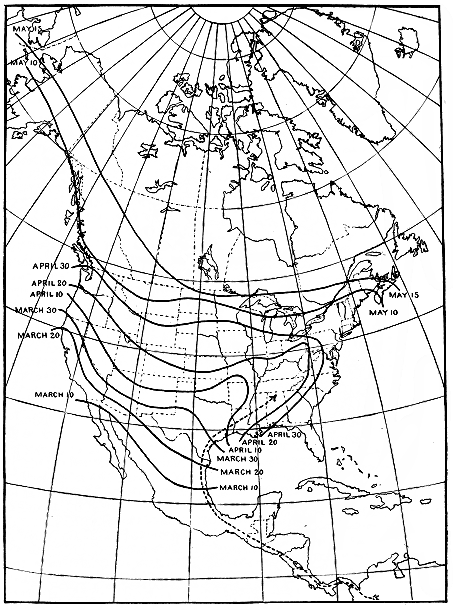
Figure 9.—Isochronal migration lines of the gray-cheeked thrush; an example of rapid migration. The
distance from Louisiana to Alaska is about 4,000 miles and is covered at an average speed of about 130
miles per day. The last part of the journey is covered at a speed that is several times what it is in the
Mississippi Valley.
The robin has been mentioned as a slow migrant, and as a species
it takes 78 days to make the 3,000-mile trip from Iowa to Alaska, a
- 20 -
stretch of country that is crossed by advancing spring in 68 days. In
this case, however, it does not mean that individual robins are necessarily
slow, for probably the northward movement of the species
depends upon the continual advance of birds from the rear, the first
individuals arriving in a suitable locality remaining to nest, while the
responsibility of the northward movement of the species is continued
by those still to come.

B4766M
Figure 10.—Blue goose, a late but rapid spring migrant that winters
on the coast of Louisiana and breeds on Baffin and Southampton
Islands and flies between summer and winter areas practically
without stops. Photograph by Paul W. Hoffman.
Special interest attaches to the great variation in the speed at which
birds travel in different sections of the broad flyway extending from
the Gulf of Mexico to the Arctic Ocean by way of the Mississippi and
Mackenzie Valleys. The blackpoll warbler furnishes an excellent
example (fig. 11). This species winters in north-central South America
and migrates in
April across the West
Indies to Florida.
From this point some
individuals fly northwest
to the Mississippi
Valley, north to Manitoba,
northwest to the
Mackenzie River, and
thence almost due west
to western Alaska. In
tracing the long route
of these birds it is found
that a fairly uniform
average speed of 30 to
35 miles a day is maintained
from the Gulf to
Minnesota. Then
comes a spurt, for a
week later the blackpolls
have reached the
central part of the
Mackenzie Valley, and
by the following week
they are observed in
northwestern Alaska.
During the latter part
of the journey, therefore,
many individuals
must average more than 200 miles a day. They use 30 days
in traveling from Florida to southern Minnesota, a distance of
about 1,000 miles, and scarcely half that time to cover the remaining
2,500 miles to Alaska. It should be noted that the increased
speed is directly associated with the change in direction, the north-and-south
course in the Mississippi Valley being accomplished
slowly, while the northwesterly course across Canada is made at
a much greater speed. Increased speed across western Canada to
Alaska is also shown by many other species. A study of all species
traveling up the Mississippi Valley indicates an average speed of
about 23 miles a day. From southern Minnesota to southern Manitoba
16 species maintain an average speed of about 40 miles a day.
- 21 -
From that point to Lake Athabaska, 12 species travel at an average
speed of 72 miles a day; while 5 others travel to Great Slave Lake
at 116 miles a day; and another 5 species cover 150 miles a day to
reach Alaska. This change is in correlation with a corresponding
variation in the isothermal lines, which turn northwestward west of
the Great Lakes.
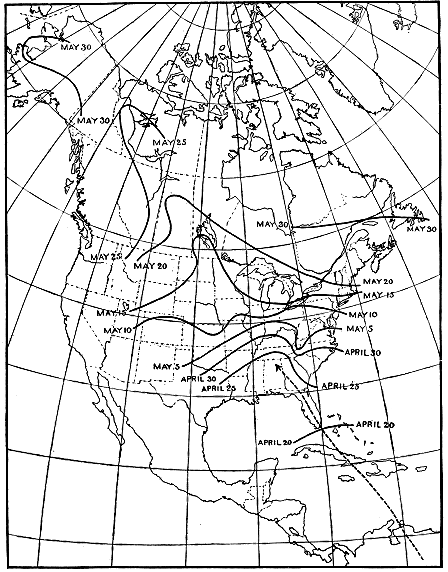
B4495M
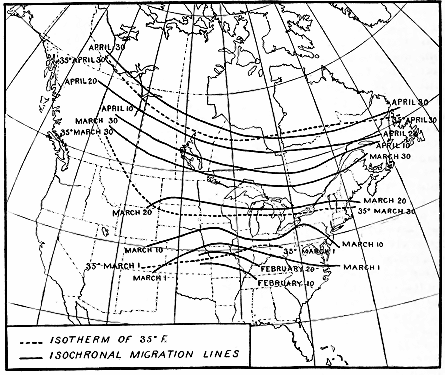
Figure 11.—Migration of the blackpoll warbler. The solid isochronal lines show the places at which
these birds arrive at the same time. As the birds move northward these lines become farther apart,
showing that the warblers move faster with the advance of spring. From April 30 to May 10 the average
speed is about 30 miles a day, while from May 25 to May 30 it is increased to more than 200 miles.
As has been previously indicated, the advance of spring in the
northern interior is much more rapid than in the Mississippi Valley
and on the Gulf coast. In other words, in the North, spring comes
- 22 -
with a rush, and during the height of the migration season in Saskatchewan
the temperature in the southern part of the Mackenzie Valley
just about equals that in the Lake Superior area, which is 700 miles
farther south. Such conditions, coupled with the diagonal course of
the birds across this region of fast-moving spring, exert a great influence
on migration and are the chief factors in the acceleration of speed
of travel.
Variations in speed of migration in different parts of the country
are illustrated also by the movements of the cliff swallow (fig. 5),
which breeds from Mexico to Alaska and winters in Brazil and
Argentina. It would be expected in spring to appear in the United
States first in Florida and Texas, then in the southern Rocky Mountain
region, and finally on the Pacific coast. As a matter of fact,
however, the earliest spring records come from north-central California,
where the bird usually is common before the first arrivals are
observed in Texas or Florida. The route taken, for many years a
migration problem, was solved when it was found that these swallows
went around the Gulf of Mexico rather than across it. The isochronal
lines on the map show the more rapid advance along the Pacific coast.
By March 20, when the vanguard has not quite reached the lower
Rio Grande in Texas, the species is already north of San Francisco
in California.
ALTITUDES AT WHICH BIRDS TRAVEL
At one time students of bird migration held firmly to the theory
that normal migration takes place at heights above 15,000 feet, reasoning
(somewhat uncertainly) that flying becomes easier as altitude
is gained. Since the development of the airplane, however, and with
it man's exploration of the upper regions of the air, it has become
common knowledge that rarefied atmosphere adds greatly to the
difficulties of flight. This is due not only to the reduction in oxygen
(whether for gasoline engine or the lungs of a bird) but also to the
lack of buoyancy of rarefied air. Such birds as vultures, pelicans,
cranes, and some of the hawks feel this the least, since compared
with body weight the supporting surface of their wings is very great,
but for the smaller and shorter winged birds lack of buoyancy at high
altitudes presents a difficult obstacle in flight. Even when flying
close to the earth, small birds have to keep their wings in rapid
motion.
Another postulate favoring the high-altitude flying theory was that
the wonderful vision of birds was their sole guidance during migratory
flights; and to keep landmarks in view the birds were obliged to fly
high, particularly when crossing wide areas of water. This will be
considered in greater detail under Orientation (p. 23), so here it will
be sufficient to say that birds rely only in part upon vision to guide
them on migration. Also, it is to be remembered that there are
definite physical limitations to the range of visibility even under perfect
atmospheric conditions. Chief of these is the curvature of the
earth's surface. Thus, if birds flew over the Gulf of Mexico to
Louisiana and Florida at a height of 5 miles, they would still be unable
to see a third of the way across. And yet this trip is made twice each
year by thousands of thrushes, warblers, and others.
Actual knowledge of the altitude of migratory flight is scanty,
though estimates obtained by means of the telescope, and still more
- 23 -
accurate data resulting from altimeter observation from airplanes,
are slowly accumulating. It is, of course, obvious that some birds
that cross mountain ranges during migration must attain a great altitude.
Observers at an altitude of 14,000 feet in the Himalayas have
recorded storks and cranes flying so high that they could be seen only
through field glasses. Being beyond the range of unaided vision they
must have been at least 6,000 feet above the observers, or at an actual
altitude of 20,000 feet above sea level. Such cases, however, are
exceptional, as aviators have reported that they rarely meet birds
above an altitude of 5,000 feet.
It is now known that migration in general is performed below an
altitude of 3,000 feet. Some proof of this statement is available.
Observations made from lighthouses and other points of vantage
indicate that migrants commonly travel at altitudes of a very few feet
to a few hundred feet above sea or land. Sandpipers, sanderlings
(Crocethia alba), and northern phalaropes (Lobipes lobatus), observed
in migration on the Pacific oceanic route, have been noted to fly so
low that they were visible only as they topped a wave. Observers
stationed at lighthouses and lightships off the English coast have
similarly recorded the passage of land birds, which sometimes flew
just above the surface of the water, and rarely above 200 feet. During
the World War broad areas in the air were under constant close
surveillance, and among the airplane pilots and observers many took
more than a casual interest in birds. Of the several hundred records
resulting from their observations only 36 were of birds flying above
5,000 feet, and only 7 above 8,500 feet. Cranes were once recorded
at an altitude of 15,000 feet, while the lapwing (Vanellus vanellus)
was the bird most frequently seen at high levels, 8,500 feet being its
greatest recorded altitude.
These observations naturally relate only to daytime travelers, but
there is no reason to believe that nocturnal migration is performed at
higher altitudes. The fact that many birds are killed each year by
striking the lanterns at lighthouses, or other man-made obstructions,
does not, however, furnish conclusive proof that low altitudes are
generally used during nocturnal flight, for it should be recalled that
these accidents occur chiefly in foggy or unsettled weather, and also,
that powerful lights have a great attraction for many species of birds.
The altitude at which birds travel is affected by other weather conditions
also. For example, flight at the higher elevations is facilitated
on clear, warm days by the currents of warm air that ascend from
broad areas.
ORIENTATION
There probably is no single aspect of the entire subject of bird
migration that challenges our admiration for birds so much as the
unerring certainty with which they cover thousands of miles of land
and water to come to rest in exactly the same spot where they spent the
previous summer or winter. The records of birds marked with
numbered bands afford abundant proof that the same individuals of
many species will return again and again to their identical nesting
sites. These data show also that many individuals migrate in fall
over the same route, year after year, making the same stops, and
finally arriving at the precise thicket that served them in previous
winters.
- 24 -
The faculty that enables these birds to point their course accurately
over vast expanses of land and water may for want of a better term be
called a "sense of direction." Man recognizes this sense in himself,
though usually it is imperfect and frequently at fault. Nevertheless
the facility with which experienced hunters and woodsmen locate tiny
camps or other points in forested or mountainous country, frequently
cloaked by darkness or fog, with all recognizable landmarks obliterated
seems due to this faculty. Ability to travel with precision over
unmarked trails is not limited either to birds or to man. It is likewise
possessed by many mammals as well as by some insects and
fishes, the well-known migrations of the salmon (Oncorhynchus) and
the eel (Anguilla) being notable examples.
Ability to follow a more or less definite course to a definite goal is
evidently part of an inherited faculty. Both the path and the goal
must have been determined either when the habit originated or in the
course of its subsequent evolution. The theory is sometimes advanced
that the older and more experienced birds lead the way, showing the
route to their younger companions. This explanation may be
acceptable for some species, but not for those in which adults and the
young migrate at different times. The young cowbird that is reared
by foster parents flocks with others of its kind when grown and in many
cases can hardly be said to have adult guidance in migration. An
inherited migratory instinct with a definite sense of the goal to be
reached and the route to be followed must be attributed to these
birds.
It is known, however, that birds possess wonderful vision. If they
also have retentive memories, subsequent trips over the route may well
be steered in part by recognizable landmarks. The arguments against
the theory of vision and memory are chiefly that much migration takes
place at night and that great stretches of the open sea are crossed
without hesitation. Nevertheless, the nights are rarely so dark that
all terrestrial objects are totally obscured, and such features as
coast lines and rivers are just those that are most likely to be seen
in the faintest light, particularly by the acute vision of a bird and
from its aerial points of observation. But some birds fly unerringly
through the densest fog. Members of the Biological Survey, proceeding
by steamer from the island of Unalaska to Bogoslof Island in
Bering Sea through a fog that was so heavy as to make invisible
every object beyond a hundred yards, recorded the fact that flocks of
murres, returning to Bogoslof, after quests for food, broke through the
wall of fog astern, flew by the vessel, and disappeared into the mists
ahead. The ship was heading direct for the island by the use of compass
and chart, but its course was no more sure than that of the
birds.
Some investigators have asserted that the sense of direction has
its seat in the ears or nasal passages and thus that the bird is enabled
to identify air currents and other phenomena. It has been found that
disturbance of the columella, or the semicircular canals of the inner
ear, will destroy the homing instinct of the racing pigeon, but experiments
in the form of delicate operations, or closing the ears with wax,
prove such a serious shock to the sensitive nervous system of the bird
that they cannot be considered as affording conclusive evidence.
Several years ago careful studies were made of the homing instinct
of the sooty and noddy terns (Sterna fuscata and Anoüs stolidus),- 25 -
tropical species that in the Atlantic region reach their most northern
breeding point on the Dry Tortugas Islands, off the southwest coast
of Florida. They are not known to wander regularly any appreciable
distance farther north. It was found that some were able to return
to their nests on the Tortugas after they had been taken on board
ship, confined in cages below deck, and carried northward distances
varying from 400 to 800 miles before being released. Landmarks of
all lands were entirely lacking, and the birds certainly were liberated
in a region in which they had had no previous experience.
Possibly the "homing instinct" as shown by these terns, by the
man-of-war birds (Fregata minor), that are trained and used as message
carriers in the Tuamotu, Gilbert, and Marshall Islands, and by
the homing pigeon, is not identical with the sense of perceptive
orientation that figures in the flights of migratory birds. Nevertheless,
it seems closely akin and is probably caused by the same impulses,
whatever they may be and however they may be received. It
is to be remembered, however, that while homing may involve flight
from a point that the bird has never before visited, the flight is always
to a known point — that is, the bird's nest— while, on the other hand,
the first migratory flight is always from the region of the bird's birth
to a region it has never before visited. The spring migration might,
of course, be more nearly considered as true "homing."
At the present time some students lean strongly toward the possible
existence of a "magnetic sense" as being the important factor in the
power of geographical orientation. No direct evidence in support of
this has been obtained, but it is not impossible that there may exist
some form of physiological sensibility to the phenomena of terrestrial
magnetism. The theory as laid down (chiefly by European investigators)
is highly complex, but briefly stated it is based on a supposed
sensitiveness of birds to the magnetic influences that cause variations
in the declination and dip of magnetic needles. Some experimental
work already done lends a little support to the theory but it is still
far from established.
In concluding this discussion of orientation it is pertinent to point
out that the migratory instinct appears to be more or less transitory,
that it is not persistent over an extended period. Migratory birds
may be arrested en route, either by natural conditions, such as unusual
food supplies, or forcibly by the act of man, and detained until the
end or nearly the end of the migratory season, and then may not
attempt to finish the journey, apparently having lost the migratory
impulse. In the fall and early winter of 1929, abundant food and an
open season caused an unusual number of mallard ducks to arrest
their migration and remain in western Montana and northern Idaho.
Later, however, a heavy snowfall with subzero temperatures suddenly
cut off the food supply, with the result that great numbers of the
birds starved to death, when a flight of a few hours would have
carried them to a region of open water and abundant food.
SEGREGATION DURING MIGRATION
During the height of the northward movement in spring the woods
and thickets may be suddenly filled with several species of wood
warblers, thrushes, sparrows, flycatchers, and others, which it is
natural to conclude have traveled together and arrived simultaneously.
- 26 -
Probably they did, but such combined migration is by no means the
rule for all species.
As a group the wood warblers (Compsothlypidae) probably travel
more in mixed companies than do any other single family of North
American birds. The flocks are likely to be made up of several species,
spring and fall, with both adults and young. Sometimes swallows,
sparrows, blackbirds, and some of the shore birds also migrate in
mixed flocks. In fall, great flocks of blackbirds frequently sweep
south across the Plains States, and occasionally one flock will contain
bronzed grackles (Quiscalus quiscula), red-winged blackbirds, yellow-headed
blackbirds (Xanthocephalus xanthocephalus), and Brewer's
blackbirds (Euphagus cyanocephalus).
On the other hand, many species keep strictly to themselves. It
would be difficult for any other kind of bird to keep in company with
a bird of such rapid movements as the chimney swift, which is rarely
found associated with any other species at any season. Nighthawks,
or bullbats, also fly in separate companies, as usually do crows, waxwings,
crossbills, bobolinks, and kingbirds. Occasionally, a flock of
ducks will be observed to contain several species, but generally when
they are actually on migration the individuals of each species separate
and travel with others of their own kind. The flocks of blue geese,
previously mentioned in connection with speed of flight (p. 18), frequently
have with them a few of the closely related snow geese (Chen
hyperborea), particularly in the eastern part of their winter range.
The proportion here is usually about 10 to 1, but farther west the
numbers of snow geese increase until they outnumber their blue
relatives.
The adults of most perching birds drive the young away when they
are grown, probably to be relieved of the necessity of providing for
them, and also in order that the parents may have opportunity to
rest and renew their plumage before starting for winter quarters.
The young birds are therefore likely to drift together and, having no
further responsibility, may start south ahead of their parents. In
contrast with this indifference on the part of the adults of perching
birds, Canada geese and some others, remain in family groups, the
parent birds undergoing the wing molt that renders them flightless
during the period of growth of their young, so that old and young
acquire their full plumage at the same time and thus are able to start
south together. The large flocks, therefore, are composed of many
families that band together, and when they separate into V-shaped
units it is probably correct to assume that it is an old bird that leads
the group. Where there is segregation of the sexes, the young birds
usually accompany their mothers, as is the case with some of the
ducks. After the females start to incubate their eggs, the males of
most species of ducks flock by themselves and remain together until
fall.
The males and females of some species may migrate either simultaneously
or separately. In the latter case it is usually the males
that arrive first, sometimes great flocks of male birds, as in the red-winged
blackbird, reaching a locality several days before any of the
females. This is particularly the rule in spring; the first robins are
usually found to be males, as also are the first song sparrows (Melospiza
melodia), rose-breasted grosbeaks (Hedymeles ludovicianus), and
scarlet tanagers (Piranga erythromelas). This early arrival of the
- 27 -
males has been explained on the theory of territorial possession, under
which the male selects the area where it elects to breed, each individual
attempting to protect a definite territory from trespass by other males
of his own kind, at the same time singing or otherwise announcing his
presence and inviting the later arriving females to examine the territory
that he has selected for nesting. The long-billed marsh wren
(Telmatodytes palustris) is a noteworthy example, and the males of
this species may enthusiastically build several dummy nests before
the females arrive.
In a few species, the males and females apparently arrive at the
breeding grounds together and proceed at once to nest building. In
fact among the shore birds, ducks, and geese, courtship and mating
may take place, in whole or in part, while the birds are in the South
or on their way north, so that when they arrive at the northern nesting
grounds they are paired and ready to proceed at once with the
raising of their families. Mallards and black ducks may be observed
in pairs as early as January, the female leading and the male following
when they take flight. Naturally these mated pairs migrate north
in company, and it was largely to protect such species that duck
shooting in spring was abolished by Federal law several, years ago.
Many shore birds nest well within the Arctic Circle, and it is the
opinion of ornithologists that most of these birds share, at least in
part, the habits of the phalaropes, a family in which the male assumes
the entire care of the eggs and young. If this be true, it explains why
in southern latitudes so many of the earliest fall arrivals are females
that may have deserted the breeding grounds after the eggs were laid.
Migratory flights are frequently accomplished in close flock formation,
as with the shore birds, blackbirds, and waxwings, and especially
some of the sparrows—the snow buntings (Plectrophenax
nivalis), longspurs, juncos, and tree sparrows (Spizella arborea).
Other species, however, though they travel in flocks, maintain a very
loose formation; examples are the turkey vultures (Cathartes aura),
the hawks, swifts, blue jays, swallows, warblers, and bluebirds.
Still others, the grebes, great horned owls (Bubo virginianus), winter
wrens (Nannus hiemalis), shrikes, and belted kingfishers (Megaceryle
alcyon), for example, ordinarily travel alone, and when several are
found in close proximity it is an indication that they have been drawn
together by unusual conditions, such as abundant food.
WHERE BIRDS MIGRATE
DISTANCES OF MIGRATION VARY
Definite evidence shows that both the length and the duration of
the migratory journey vary greatly. The bobwhite and the western
quails, the cardinal (Richmondena cardinalis), the Carolina wren
(Thryothorus ludovicianus), and probably some of the titmice and
woodpeckers, which are apparently nonmigratory, may round out
their full period of existence without at any time going more than
10 miles from the nest where they were hatched.
SHORT AND UNDETERMINED MIGRATIONS
Song sparrows, meadow larks (Sturnella), blue jays (Cyanocitta
cristata), and some other species make such short migrations that
the movement is difficult to detect, as individuals may be found in
- 28 -
one area throughout the year. Thus, at the southern part of the
range there is merely a concentration in winter, the summer individuals
being entirely sedentary. Speculation is useless on the distances
of individual migration without definite evidence concerning the
precise winter quarters of birds that summer in a particular part of
the breeding range of the species, but from the records of banded
birds important evidence is becoming available. Eventually it may
be possible to say definitely just how far the song sparrows that nest
in northern New England and the Maritime Provinces of Canada
travel to their winter quarters, and whether the blue jays of New
York and the upper Mississippi Valley remain throughout the winter
in their breeding areas, or move farther south and relinquish their
places to individuals from southern Canada.
An illustration of what is now known on this subject is found in
the case of the robin. This bird occurs in the Middle Atlantic States
throughout the year, in Canada only in summer, and along the Gulf
coast only as a winter resident. On the Atlantic coast its movements
are readily ascertained, since, for example, in the section about
Washington, D. C., the breeding robin is the southern variety (Turdus
migratorius achrusterus), which is found there from the first of April
to the last of October, when its place is taken (in smaller numbers)
by the northern robin (T. m. migratorius), which arrives about the
middle of October and remains until the following April. It is
probable that a similar interchange of individual robins occurs
throughout a large part of the rest of its range, the hardy birds from
the north being the winter tenants in the abandoned summer homes
of the southern birds.
The red-winged blackbirds that nest in northern Texas are almost
sedentary, but in winter they are joined by representatives of other
subspecies that nest as far north as the Mackenzie Valley.
VARIABLE MIGRATIONS WITHIN SPECIES
The difference in characters between subspecies has been used by
students of migration to discover other interesting facts concerning
variations of the migratory flight between closely related birds that
breed in different latitudes. The familiar eastern fox sparrow
(Passerella iliaca iliaca), for example, breeds from northwestern
Alaska to Labrador, and in winter is found concentrated in the southeastern
part of the United States. It thus travels a long distance
each year. On the west coast of the continent, however, six subspecies
of this bird breed in rather sharply delimited ranges, extending
from the region of Puget Sound and Vancouver Island to Unimak
Island, at the end of the Alaska Peninsula. One of these, known as
the sooty fox sparrow (P. i. fuliginosa), breeds in the Puget Sound
area and makes practically no migration at all, while the other races,
nesting on the coast of British Columbia and Alaska, are found in
winter chiefly in California. The races that breed farthest north are
in winter found farthest south, illustrating a tendency for those birds
that are forced to migrate to pass over those so favorably located that
they have no need to leave their breeding areas, while the northern
birds settle for the winter in the unoccupied areas farther south
(fig. 12).
Another example of the same kind is found in the case of the
Maryland yellowthroat (Geothlypis trichas) of the Atlantic coast.
- 29 -
Birds occupying the most southern part of the general range are
almost nonmigratory, residing throughout the year in Florida, while
those breeding as far north as Newfoundland go to the West Indies
for the winter, thus passing directly over the home of their southern
relatives.
B4774AM
Figure 12.—Migration of Pacific-coast forms of the fox sparrow. The
breeding ranges of the different races are encircled by solid lines,
while the winter ranges are dotted. The numbers indicate the areas
used by the different subspecies, as follows: 1, Shumagin fox sparrow;
2, Kodiak fox sparrow; 3, Valdez fox sparrow; 4, Yakutat fox sparrow;
5, Townsend fox sparrow; 6, sooty fox sparrow. (After Swarth; courtesy
of the Museum of Vertebrate Zoology, University of California.)
The palm warbler (Dendroica, palmarum), which breeds from Nova
Scotia and Maine west and northwest to southern Mackenzie, has
been separated into two subspecies. Those breeding in the interior
of Canada (D. p. palmarum) make a 3,000-mile journey from Great
Slave Lake to Cuba,
passing through the
Gulf States early in
October. After the
bulk have passed, the
palm warblers from
the Northeastern
States and Provinces
(D. p. hypochrysea)
drift slowly into the
Gulf coast region,
where they remain for
the winter. Their
migratory journey is
about half as long as
that of the northwestern
subspecies.
There is no invariable
law governing the
distance of migration,
although in general it
is found that where a
species has an extensive
range, the subspecies
that breed farthest
north go farthest
south to spend
the winter
FALL FLIGHTS NOT FAR
SOUTH OF BREEDING
RANGES
Some other species
that have extensive
summer ranges, for instance,
the pine warbler (Dendroica pinus), rock wren (Salpinctes obsoletus),
field sparrow (Spizella pusilla), loggerhead shrike (Lanius ludovicianus),
and black-headed grosbeak (Hedymeles melanocephalus), are
found to concentrate during the winter season in the southern part
of the breeding range, or to occupy additional territory that is only
a short distance farther south. The entire species may thus be confined
within a restricted area for the period of winter, and then, with
the return of warmer weather, spreads out to reoccupy the full range.
- 30 -
There are many species, including the tree sparrow, slate-colored
junco, and Lapland longspur (Calcarius lapponicus), that nest in
Canada and winter in the United States; while others, including the
vesper sparrow (Pooecetes gramineus), chipping sparrow (Spizella
passerina), grackles, red-winged blackbird, bluebird, the woodcock
(Philohela minor), and several species of ducks, nest in the northern
United States and move south for the winter to areas along the Gulf
of Mexico. This list includes the more hardy species, some individuals
of which may linger in protected places well within the reach of
severe cold, as, for example, Wilson's snipe, or jacksnipe (Capella
delicata), which frequently
is found
during subzero weather
in parts of the
Rocky Mountain
region where warm
springs assure a food
supply. More than
100 of our summer
birds leave the United
States entirely and
spend the winter id
the West Indies or
in Central America
or South America.
For example, the
Cape May warbler
(Dendroica tigrina),
which breeds from
northern New England,
northern Michigan,
and northern
Minnesota, north to
New Brunswick,
Nova Scotia, and
nearly to Great Slave
Lake, is concentrated
in winter chiefly
in the West Indies,
its metropolis at
this season being the
island of Hispaniola.

B3763M
Figure 13.—Barn Swallow, a bird that has so long a migration route
that some individuals breed north to Yukon and Alaska, while
the winter range extends south to Argentina, 7,000 miles away.
LONG-DISTANCE MIGRATIONS
Some of the common summer residents are not content with a trip
to northern South America, but push on across the Equator and
finally come to rest for the winter in the pampas of Argentina, or even
in Patagonia. Thus some species that are more or less associated
with, each other in summer, as nighthawks, barn swallows, cliff swallows,
and some of the thrushes may also occupy the same general
winter quarters in Brazil. Some individual nighthawks and barn
swallows (fig. 13) travel still farther, and of all North American land
birds these species probably have the longest migration route, as they
occur north to Yukon and Alaska, and south to Argentina, 7,000
- 31 -
miles away. Such seasonal flights are exceeded in length, however,
by the journeys of several species of water birds, chiefly members of
the suborder of shore birds. In this group are 19 species that breed
north of the Arctic Circle and winter in South America, 6 of them
going as far south as Patagonia, and thus having a migration route
more than 8,000 miles in length.

B1035M
Figure 14.—Arctic tern. The longest flight known for an individual
bird was accomplished by an arctic tern that in 3 months
flew from the coast of Labrador to the coast of southeastern
Africa.
The arctic tern (Sterna paradisaea) is the champion "globe trotter"
and long-distance flier (figs. 14 and 15). Its name "arctic" is well
earned, as its breeding range is circumpolar and it nests as far north
as it can find a suitable place. The first nest to be found in this
region was only 7½° from the North Pole, and it contained a downy
chick surrounded by a wall of newly-fallen snow that had been scooped
out by the parent.
In North America it
breeds south in the interior to Great Slave
Lake, and on the Atlantic
coast to Massachusetts. After the
young are grown, the
arctic terns disappear
from their North
American breeding
grounds, and a few
months later they
may be found in the
Antarctic region,
11,000 miles away.
Until very recently
the route followed by
these hardy fliers was
a complete mystery,
for although a few
scattered individuals
have been noted south
as far as Long Island,
the species is otherwise
practically unknown
along the Atlantic
coasts of North
America and South
America. It is, however,
known as a migrant on the west coast of Europe and Africa.
By means of numbered bands the picture is now developing of what
is apparently not only the longest but also one of the most remarkable
of all migratory journeys.
Judging by the evidence at present available, it seems likely that
the arctic terns of eastern North America originally found their way
here from the Old World, probably by way of Iceland and Greenland.
Consequently when the time comes for them to migrate to winter
quarters they do not go directly south as do the common (Sterna
hirundo) and Forster's terns (S. forsteri), but instead, they fly back
eastward along their ancestral route across the Atlantic to the shores
of Europe and then go south along the African coast to their winter
home; those that breed in the northwestern part of the continent, as
- 32 -
in Alaska, probably migrate chiefly down the western coast, as the
species is not infrequently reported on the coast of California and
also on the western coast of South America.
B4750M
Figure 15.— Distribution and the migration of the arctic terns of eastern North America. The route
indicated for this bird is unique, as no other species is known to breed abundantly in North America and
to cross the Atlantic Ocean to and from the Old World. The extreme summer and winter homes are
11,000 miles apart, and as the route taken is circuitous, these terns probably fly at least 25,000 miles each
year.
The evidence yielded by banding consists of only three definite
cases, but their interpretation seems to permit but one conclusion:
All three birds were banded as downy chicks, one on July 3, 1913, at
- 33 -
Eastern Egg Rock, Maine,[3] and the other two at the Red Islands,
Turnevik Bay, Labrador, on July 22, 1927, and July 23, 1928. The
first was found dead in the Niger River delta, West Africa, in August
1917, while the Labrador birds were recovered near La Rochelle,
France, on October 1, 1927, and at Margate, near Port Shepstone,
Natal, South Africa, on November 14, 1928. The flight shown by
this last record is the longest known, the trip, between 8,000 and
9,000 miles, being accomplished in less than 3 months.
Probably no other animal in the world enjoys as many hours of
daylight as does the arctic tern, since for these birds the sun never
sets during their nesting season in the northern part of the range,
while during their sojourn in the south, daylight is continuous.
During several months of the year they have 24 hours of daylight
and during the other months considerably more daylight than
darkness.
ROUTES OF MIGRATION
While it is beyond question that certain general directions of
flight are constantly followed by migratory birds, it is well to remember
that the term "migration route" is to some extent a theoretical
concept, concerned entirely with the lines of general advance or
retreat of a species, rather than the exact course followed by individual
birds. Even the records of banded birds usually show no more than
the places of banding and recovery, and one must have recourse to
intermediate records and to reasoning based on probabilities to fill
in details of the flyway actually traversed between the two points.
There is also infinite variety in the routes covered during migration
by different species. In fact, the choice of migration highways is so
wide that it seems as if the routes of no two species coincide. Differences
in distance traveled, in time of starting, in speed of flight,
in geographical position, in latitudes of breeding and of wintering
grounds, and in other factors, all contribute to this great variation
of migration routes. Nevertheless, there are certain factors that
serve to guide the avian travelers along more or less definite lines, and
it is possible to define general lines of migration for the majority of
species.
It has frequently been observed that migrating birds have a
tendency to follow major topographic lines on the earth's surface
when their trend is in the general direction of the birds' journey.
Bird migration is generally thought of as a north-and-south movement,
with the lanes of heavier concentration following the coasts,
mountain ranges, and principal river valleys. To a considerable
extent this is the case, particularly in North America, where the coast
lines, mountain chains, and the larger rivers in general run north and
south. Students of American birds thus have exceptionally good
opportunities to study migratory movements. In cases where the
migration is a long one, however, the notion must be abandoned
that the birds' flight is restricted to particular narrow routes that
follow river valleys and the like, as many species seem to disregard
utterly such apparently good natural flyways as river valleys. For
example, the Arkansas River has a general east and west course for
a great part of its length, and while it does constitute a highway for
- 34 -
many perching birds en route from the Mississippi Valley to the
Rocky Mountain region, some of the hawks and many ducks and
shore birds pay the valley scant attention. They may arrest their
fall journey to feed among cottonwoods or along sand bars, but when
ready to resume their flight they leave the river and fly directly
south over the more or less arid region that lies between the Arkansas
and the Rio Grande.
WIDE AND NARROW MIGRATION LANES
When birds start their southward migration, the movement
necessarily involves the full width of the breeding range. Later,
there is a convergence of the lines of flight taken by individual
birds, owing to the conformation of the land mass, and as the species
proceeds southward the width of the occupied region becomes less
and less. An example of this is provided by the common kingbird
(Tyrannus tyrannus), which breeds from Newfoundland to British
Columbia, a summer range 2,800 miles wide. On migration, however,
its paths converge, until in the southern part of the United States the
occupied area extends from Florida to the mouth of the Rio Grande,
a distance of only 900 miles, and still farther south the migration
path is further restricted. In the latitude of Yucatan it is not more
than 400 miles wide, and it is probable that the great bulk of the
species moves in a belt that is less than half that width.
A migration route, therefore, may be anything from a narrow path
that adheres closely to some definite geographical feature, such as a
river valley or a coast line, to a broad boulevard that leads in the
desired direction and follows only the general trend of the land mass.
Also it is to be remembered that whatever main routes are described,
there remain a multitude of tributary and separate minor routes.
In fact, with the entire continent of North America crossed by migratory
birds, the different groups or species frequently follow lines that
may repeatedly intersect those taken by others of their own kind or
by other species. The arterial routes, therefore, must be considered
merely as indicating paths of migration on which the tendency to
concentrate is particularly noticeable.
In considering the width of migration lanes it will be obvious that
certain species, as the knot (Calidris canutus) and the purple sandpiper
(Arquatella maritima), which are normally found only along the
coasts, must have extremely narrow routes of travel. They are limited
on one side by the broad waters of the ocean and on the other by
land and fresh water, both of which are unsuited to furnish the food
that is desired and necessary to the well being of these species.
Among land birds that have a definite migration, the Ipswich sparrow
(Passerculus princeps) has what is probably the most restricted
migration range of any species. It is known to breed only on Sable
Island, Nova Scotia, and it winters along the Atlantic coast south to
Georgia. Living constantly within sound of the surf, it is rarely
more than a quarter of a mile from the outer beach, and is entirely at
home among the sand dunes and their sparse covering of coarse grass.
- 35 -
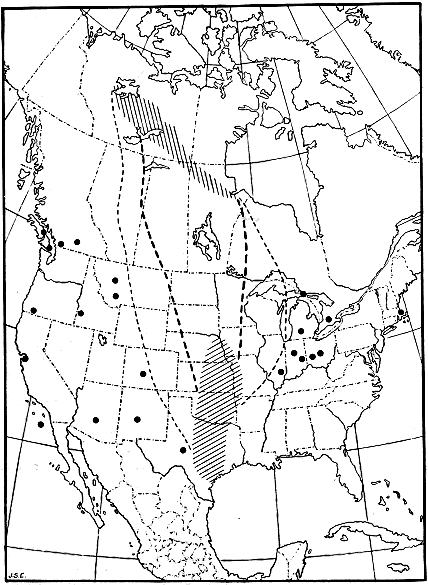
B4630M
Figure 16.—Breeding and wintering ranges and migration of Harris's sparrow, an example of a narrow
migration route through the interior of the country. The heavy broken lines enclose the region traversed
by the majority of these finches; the light broken line encloses the country where they occur with more or
less regularity; while the spots indicate records of accidental or sporadic occurrence.
Harris's sparrow (Zonotrichia querula) supplies an interesting example
of a narrow migration route in the interior of the country (fig. 16).
This fine, large finch is known to breed only in the region from Fort
Churchill, on the west shore of Hudson Bay, northwest to the shores
of Great Bear Lake. Very few actual breeding records of the species
are available, but these are sufficient to indicate that the breeding
range is in the strip of country characterized by more or less stunted
timber just south of the limit of trees. When it begins its fall migration,
this bird necessarily covers the full width of its breeding area.
Then it proceeds almost directly south, or slightly southeasterly, the
area covered by the majority of the species becoming gradually constricted,
so that by the time it reaches the United States it is most numerous
in a belt about 500 miles wide, extending across North Dakota
to central Minnesota. Harris's sparrows are noted on migration with
- 36 -
fair regularity east to the western shore of Lake Michigan, and west
to the foothills of the Rocky Mountains, but the great bulk of the
species moves north and south through a relatively narrow path in
the central part of the continent. Present knowledge suggests that
the reason for this narrow migration range is the close association that
Harris's sparrow maintains with a certain type of habitat, including
brushy places, thickets, edges of groves, and weed patches. While
these environmental conditions are found in other parts of the country,
the region crossed by this sparrow presents almost a continuous succession
of habitat of this type. Its winter range extends from southeastern
Nebraska and northwestern Missouri, across eastern Kansas
and Oklahoma and through a narrow section of central Texas, at
places hardly more than 150 miles wide.
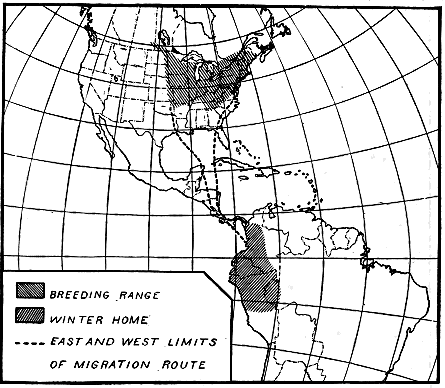
B4502M
Figure 17.—Distribution and migration of the scarlet tanager. During the breeding season individual
scarlet tanagers may be 1,900 miles apart in an east-and-west line across the breeding range. In migration,
however, the lines converge until in southern Central America they are not more than 100 miles
apart. For migration paths of other widths see
figures 16, 18, and 19.
The scarlet tanager presents another extreme case of narrowness of
migration route (fig. 17), its breeding range extending in greatest
width from New Brunswick to Saskatchewan, a distance of about
1,900 miles. As the birds move southward in fall their path of migration
becomes more and more constricted, until at the time they leave
the United States all are included in the 600-mile belt from eastern
Texas to the Florida peninsula. Continuing to converge through
Honduras and Costa Rica, the boundaries there are not more than
- 37 -
100 miles apart. The species winters in northwestern South America,
where it spreads out over most of Colombia, Ecuador, and Peru.
The rose-breasted grosbeak also leaves the United States through
the 700-mile stretch from eastern Texas to Appalachicola Bay, but
thereafter the lines do dot further converge, as this grosbeak enters the
northern part of its winter quarters in Central America and South
America through a door of about the same width (fig. 18).
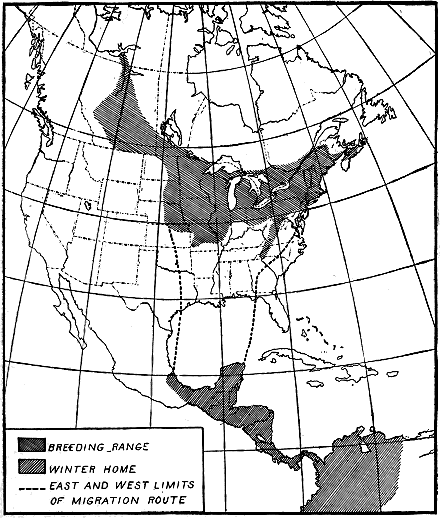
B4501M
Figure 18.—Distribution and migration of the rose-breasted grosbeak. Though the width of the breeding
range is about 2,500 miles, the migratory lines converge until the boundaries are only about 700 miles apart
when the birds leave the United States. For migration paths of other widths see
figures 16, 17, and 18.
While the cases cited represent extremes of convergence, a narrowing
of the migratory path is the rule to a greater or less degree for the
majority of North American birds. The shape of the continent tends
to effect this, and so the width of the migration route in the latitude
of the Gulf of Mexico is usually much less than in the breeding
territory.
- 38 -
The redstart (Setophaga ruticilla) represents a notable case of a
wide migration route, although even in the southern United States
this is much narrower than the breeding range (fig. 19). These birds,
however, cross all parts of the Gulf of Mexico and pass from Florida
to Cuba and Haiti by way of the Bahamas, so that here their route
has a width of about 2,500 miles.
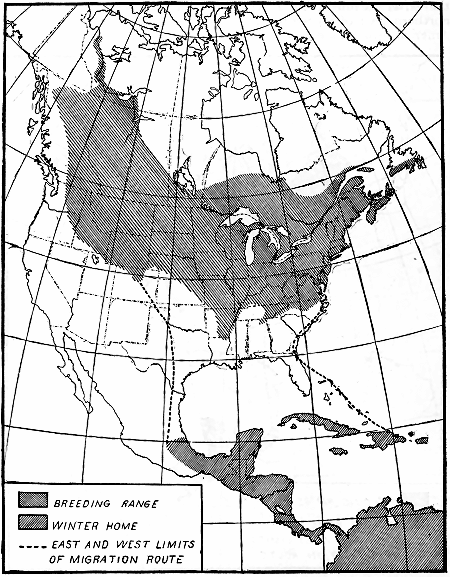
B4500M
Figure 19.—Distribution and migration of the redstart. An example of a wide migration route, since birds
of this species cross all parts of the Gulf of Mexico, or may travel from Florida to Cuba and through the
Bahamas. Their fly way thus has an east and west width of more than 2,000 miles. For migration paths
of greater or less extent see
figures 16, 17, and 18.
In the following, the discussion of the principal routes of North
American birds relates chiefly to the fall migration, for, except as
otherwise noted, the spring flight generally retraces the same course.
- 39 -
The routes indicated on the maps (figs. 20 and 21) must not be considered
as representing paths with clearly defined borders, but rather
as convenient subdivisions of the one great flyway that covers practically
the entire width of the North American Continent and extends
from the Arctic coast to South America.
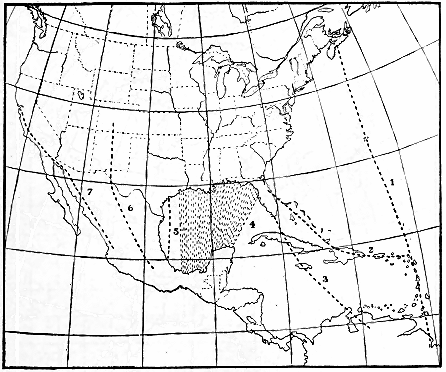
B1032M
Figure 20.—Principal migration routes used by birds in passing from North America to winter quarters in
the West Indies, Central America, and South America. Route no. 4 is the one used most extensively; only
a few species make the 2,400-mile flight from Nova Scotia to South America.
ATLANTIC OCEANIC ROUTE
By reference to figure 20 it will be noted that route no. 1 is almost
entirely oceanic, passing directly over the Atlantic Ocean from
Labrador and Nova Scotia to the Lesser Antilles, and then through this
group of small islands to the mainland of South America. It is not
used by any of the smaller land birds, but is followed chiefly by thousands
of water birds and by shore birds of several species, the adult
golden plover being a notable example. Since it lies entirely over the
sea, this route is definitely known only at its terminals and from
occasional observations made on Bermuda and other islands in its
course. Some of the shore birds that breed on the Arctic tundras of
Mackenzie and in Alaska fly southeastward across Canada to the
Atlantic coast and finally follow the oceanic route to the mainland of
South America. The golden plover may accomplish the whole 2,400
miles without pause or rest, in fair weather the flocks passing Bermuda
and sometimes even the islands of the Antilles without stopping.
Although most birds make their migratory flights either by day or by
- 40 -
night, the golden plover in this remarkable journey flies both day and
night. As it swims lightly and easily it may make a few short stops
along the way, and it has been actually seen resting on the ocean.
Other shore birds have been observed busily feeding in that great area
of ocean known as the Sargasso Sea, where thousands of square miles
of floating seaweed teem with marine life.
B4767M
Figure 21.—Migration routes of North American birds. Though this map was prepared chiefly to show
the flyways used by waterfowl, most of these routes also are utilized by innumerable land birds. For
example, the important Mackenzie Valley-Great Lakes-Mississippi Valley route is shown (with its
tributaries) from the Arctic coast to the delta of the Mississippi River.
- 41 -
B4768M
Figure 22.—Distribution and migration of the golden plover, Pluvialis dominica. Adults of the eastern
form (P. d. dominica) migrate across northeastern Canada and then by a nonstop flight reach South
America. In spring they return by way of the Mississippi Valley. Their entire route is therefore in
the form of a great ellipse with a major axis of 8,000 miles and a minor axis of about 2,000 miles. The
Pacific golden plovers (P. d. fulva), which breed in Alaska, apparently make a nonstop flight across the
ocean to Hawaii, the Marquesas Islands, and the Low Archipelago, returning in spring over the same
route.
The annual flight of the adult golden plover is so wonderful that
it may be given in some detail, particularly since it is one of the
exceptions to the general rule that spring and fall movements are
over the same routes (fig. 22). After reaching the South American
coast the birds make a short stop and then continue overland to the
pampas of Argentina, where they remain from September to March.
Leaving their winter quarters, they cross northwestern South America
and the Gulf of Mexico, reaching the North American mainland
on the coasts of Texas and Louisiana. Thence they proceed slowly
- 42 -
up the Mississippi Valley, and by the early part of June are again on
their breeding grounds, having performed a round-trip journey in the
form of an enormous ellipse with the minor axis about 2,000 miles
and the major axis 8,000 miles, reaching from the Arctic tundras to
the pampas of Argentina. The older birds are probably accompanied
by some of the young, perhaps those from early nestings, but most
of the immatures leave their natal grounds late in summer and move
southward through the interior of the country, re turning in spring
over essentially the same course. The elliptical route is therefore
used chiefly by fully adult birds.
ATLANTIC COAST ROUTE AND TRIBUTARIES
The Atlantic coast is a regular avenue of travel, and along it are
many famous points for observing both land and water birds. About
50 different kinds of land birds that breed in New England follow
the coast southward to Florida and travel thence by island and mainland
to South America (fig. 20, route 2). As will be seen from the
map, a seemingly natural and convenient highway extends through
the Bahamas, Cuba, Hispaniola, Puerto Rico, and the Lesser
Antilles to the South American coast. Resting places are afforded at
convenient intervals, and at no time need the aerial travelers be out
of sight of land. It is not, however, the favored highway, and only
about 25 species of birds go beyond Cuba to Puerto Rico along this
route to their winter quarters, while only 6 species are known to
reach South America by way of the Lesser Antilles. The obvious
drawback is lack of adequate food. The total area of all the West
Indies east of Puerto Rico is less than that of Rhode Island, so that
if only a small part of the birds of the eastern United States were to
travel this way, it is doubtful whether even the luxuriant flora and
fauna of tropical habitats would provide food sufficient for their
needs.
In the northern part of the Atlantic coast route is a tributary
route used by the brant (Branta bernicla) that is of special interest.
The southward movement of these birds is chiefly along the western
shores of Hudson Bay and thence southeastward to the Atlantic
coast. Returning in spring, they follow the coast line north to the
Gulf of St. Lawrence and then fly almost due north to their breeding
grounds on the west coast of Greenland and the islands of the Arctic
archipelago. The round trip is therefore in the form of a great ellipse,
probably 3,000 miles long by 1,000 miles wide.
The Atlantic flyway receives accretions of waterfowl from three or
four interior migration paths, one of which is of first importance, as it
includes great flocks of canvasbacks (Nyroca valisineria), redheads (N.
americana), scaup ducks (N. marila and N. affinis), Canada geese,
and many of the black ducks that winter in the waters and marshes
of the coastal region south of Delaware Bay. The canvasbacks, redheads,
and scaups come from their breeding grounds on the great
northern plains of central Canada, follow the general southeasterly
trend of the Great Lakes (fig. 21), cross Pennsylvania over the mountains,
and reach the Atlantic coast in the vicinity of Delaware and
Chesapeake Bays. Black ducks, mallards, and blue-winged teals
(Querquedula discors) that have gathered in southern Ontario during
the fall leave these feeding grounds and proceed southwest over a
- 43 -
course that is apparently headed for the Mississippi Valley. Many
do continue this route down the Ohio Valley, but others, upon reaching
the vicinity of Lake St. Clair, between Michigan and Ontario,
swing abruptly southeast and, crossing the mountains in a single
flight, reach the Atlantic coast south of New Jersey. This route,
with its Mississippi Valley branch, has been fully demonstrated by
the recovery records of ducks banded by a cooperator of the Biological
Survey at Lake Scugog, Ontario.
The white-winged scoter (Melanitta deglandi), which also breeds in
the interior country from northern North Dakota north to the Arctic
coast, is another bird having an elliptical migration route, so far as
those wintering on the Atlantic coast are concerned. This duck
breeds only near fresh water and winters entirely on the ocean along
both the Atlantic and Pacific coasts of the United States. Those
wintering on the Atlantic side leave their breeding grounds west of
Hudson Bay and fly 1,500 miles almost due east to the most eastern
part of Labrador, whence they proceed southward across the Gulf of
St. Lawrence to their winter home, which extends from southwestern
Maine to Chesapeake Bay. The spring flight is made by an interior
route that traverses the valleys of the Connecticut, Hudson, and
Ottawa Rivers, and thence passes west and north to the breeding
grounds.
A study of the Canada geese that winter abundantly in the waters
of Back Bay, Va., and Currituck Sound, N. C., reveals another important
tributary to the Atlantic coast route. Banding has shown that
the principal breeding grounds of these birds are among the islands
and on the eastern shores of Hudson Bay (fig. 21). From this region
they move south in fall to the point of lower Ontario between Lakes
Erie and Huron. Some of the banded geese are recovered in the
Mississippi Valley, but the great majority are retaken either on their
breeding grounds or on the Atlantic coast south of Delaware Bay,
showing another instance of a long cross-country flight by waterfowl.
Although Canada geese are abundant in migration on the coast of
New England, the birds taken there do not include any that were
banded in southern Ontario. Again, banding has shown that the
New England visitants come from other breeding areas, chiefly Newfoundland
and the desolate coast of Labrador, and that their migration
is entirely coastwise.
Still another cross-country route between the Mississippi Valley
and the Atlantic coast may be briefly described. While not yet well
understood, a hitherto unsuspected migration route across the Alleghenies
to the Mississippi Valley has been revealed by the banding of
blue-winged teal, on the coastal sawgrass marshes of South Carolina.
Birds marked in these marshes have been retaken in Tennessee and
Kentucky as well as in States farther north in the Mississippi Valley.
Several species of shoal-water ducks, including this dainty little teal
and the shoveler (Spatula clypeata), are more or less common winter
residents in the South Carolina marshes, but are less common or
even decidedly rare in most of the coastal marshes farther north, so
this cross-country route connecting two main arteries of migration
seems to be of considerable importance.
- 44 -
B4762M
Figure 23.—Distribution and migration of the bobolink. In crossing to South America most of the bobolinks
use route no. 3 (fig. 20), directly from Jamaica across an unbroken stretch of ocean. Colonies of these
birds have established themselves in several areas in the western United States, but in migration they adhere
to the ancestral flyways and show no tendency to take the short cut across Arizona, New Mexico,
and Texas.
Referring again to figure 20, it is noted that route no. 3 presents a
much more direct line of flight for the Atlantic coast migrants to
South America than the others, although it involves much longer
flights. It is used almost entirely by land birds. After taking off
from the coast of Florida the migrants find only two land masses on
the way where they can pause for rest and food. Nevertheless, tens
of thousands of birds of some 60 species cross the 150 miles from Florida
to Cuba, where about half of them elect to remain for the winter.
The others fly the 90 miles between Cuba and Jamaica. From that
point to the South American coast, however, there is a stretch of unbroken
ocean fully 500 miles across, and scarcely a third of the North
American migrants leave the forested mountains of Jamaica to risk
the perils of this ocean trip. Chief among those that do is the bobolink
(Dolichonyx oryzivorus), which so far outnumbers all other birds
using this fly way that route no. 3 may well be called "the bobolink
route" (fig. 23). As traveling companions along this route the bobolinks
may meet vireos, kingbirds, and nighthawks from Florida;
the chuck-will's-widow (Antrostomus carolinensis) of the Southeastern
States; black-billed and yellow-billed cuckoos (Coccyzus erythropthalmus
and C. americanus) from New England; gray-cheeked thrushes
from Quebec, bank swallows (Riparia riparia) from Labrador; and
- 45 -
blackpoll warblers from Alaska. Sometimes this scattered assemblage
will be joined by a tanager or a wood thrush but "the bobolink
route" is not popular with the greater number of migrants, and although
many individuals traverse it, they are only a small fraction
of the multitudes of North American birds that spend the winter in
South America.
MACKENZIE VALLEY-GREAT LAKES-MISSISSIPPI VALLEY ROUTE AND TRIBUTARIES
Easily the longest flyway of any in the Western Hemisphere is that
extending from the Mackenzie Valley past the Great Lakes and down
the Mississippi River, including its tributaries. Its northern terminus
is on the Arctic coast in the regions of Kotzebue Sound, Alaska,
and the mouth of the Mackenzie River, while its southern end lies
in Patagonia (fig. 21). During the spring migration some of the
shore birds traverse the full extent of this great path, and it seems
likely that the nighthawk, the barn swallow, the blackpoll warbler,
and individuals of several other species that breed north to Yukon
and Alaska must twice each year cover the larger part of it.
For more than 3,000 miles—from the mouth of the Mackenzie to
the Delta of the Mississippi—this flyway is uninterrupted by mountains.
In fact, there is not even a ridge of hills on the route high
enough to interfere with the movements of the feathered travelers,
and the greatest elevation above sea level is less than 2,000 feet. Well
timbered and watered, the entire region affords ideal conditions for the
support of its great hosts of migrating birds. This route is followed
by such vast numbers of ducks, geese, shore birds, blackbirds, sparrows,
warblers, and thrushes, that observers stationed at favorable
points in the Mississippi Valley during the height of migration can
see a greater number of species and individuals than can be noted anywhere
else in the world.
Starting in the region of Kotzebue Sound, Alaska, the route extends
eastward across northern Alaska and joins another that has its origin
at the mouth of the Mackenzie River (fig. 21). The line of flight
then trends a little east of south through the great lake system of
central Canada, where it is joined by 2 or 3 other routes from the
northeast that have their origin on the central Arctic coast. Continuing
southward the migrating flocks are constantly augmented by
additions to their numbers as they pass over the great breeding
grounds of central and southern Canada. Upon reaching the headwaters
of the Missouri and Mississippi Rivers the route follows these
streams to the Gulf coast. Arriving in this latitude many species,
including ducks and geese, the robin, the myrtle warbler, and some
others spread out east and west for their winter sojourn. Others,
despite the perils of a trip involving a flight of several hundred miles
across the Gulf of Mexico, strike out boldly for Central America and
South America. This part of the route is a broad "boulevard"
extending from northwestern Florida to eastern Texas and reaching
southward across the Gulf of Mexico to Yucatan and the Isthmus of
Tehuantepec (fig. 20, route 4).
Many of the birds that breed east of the Allegheny Mountains
parallel the sea coast as they move southwestward in fall and, apparently
maintaining the same direction from northwestern Florida,
cross the Gulf to the coastal regions of eastern Mexico. On the other
- 46 -
hand, the birds that have come south directly through the Mississippi
Valley and the region west to the Rocky Mountains reach the coastal
plains of Mississippi, Louisiana, and Texas and continue directly
across the Gulf. The great majority of North American birds seeking
winter homes in the Tropics thus elect the short cut across the Gulf
of Mexico in preference to the longer although safer land or island
journey by way of Texas or Florida. During the height of the migration
some of the islands off the coasts of Louisiana and Texas are
wonderful observation places for the student of birds, as the feathered
travelers literally swarm over them.
One of the short cuts (fig. 20, route 5) that may be considered a part
of this great water artery of migration extends a few hundred miles
from the coast of Texas to the northern part of the State of Vera
Cruz. As the neighboring coast is arid and thus entirely unsuited to
the needs of birds that are frequenters of moist woodlands, it is not
surprising to find that this Gulf route is used by such woodland species
as the golden-winged warbler (Vermivora chrysoptera), the worm-eating
warbler (Helmitheros vermivorus), and the Kentucky warbler
(Oporornis formosus).
Formerly it was thought that most of the North American birds
that migrate to Central America made a leisurely trip along the
Florida coast, crossed to Cuba, and thence made the short flight from
the western tip of Cuba to Yucatan. A glance at the map would
suggest this as a most natural route, but as a matter of fact it is practically
deserted except for a few swallows and shore birds, or an occasional
land bird storm-driven from its accustomed course.
Present knowledge of the chief tributaries to the Mackenzie Valley-Great
Lakes-Mississippi Valley highway relates chiefly to waterfowl.
Reference already has been made to the flight of the black ducks
(p. 42) that reach the Mississippi Valley from southern Ontario.
Some individuals of this species banded at Lake Scugog, Ontario,
have been recaptured in succeeding seasons in Wisconsin and Manitoba,
but the majority have been retaken at points south of the junction
of the Ohio River with the Mississippi, definitely indicating their
route of travel from southern Ontario.
A second route that joins the main artery on its eastern side is the
one used by the blue goose (fig. 10), the migration route of which is
probably more nearly due north and south than that of any other
North American bird. The breeding grounds, which only recently
have been discovered, are in the Fox Basin region of Baffin Island
and on Southampton Island. In fall these geese work southward,
chiefly along the eastern shore of Hudson Bay, and upon reaching
the southern extremity of James Bay they take off for what is practically
a nonstop flight to the great coastal marshes of Louisiana west
of the delta of the Mississippi River (fig. 21). In some seasons the
flocks make intermediate stops among the islands and sand bars of
the Mississippi, as they are occasionally common in the general vicinity
of Memphis, Tenn. Most of the birds push on, however, and
during the period from the first of November to the last of March
fully 90 percent of the species are concentrated in the area between
the Sabine and the Mississippi Rivers. On the return trip northward
there is sometimes a tendency for some of the blue geese to veer off
toward the Northwest, as they are occasionally abundant in eastern
South Dakota and southeastern Manitoba. It is of particular interest
- 47 -
to note that while some other geese and many ducks start their
northward journey at the first sign of awakening spring, the blue
goose remains in its winter quarters until the season there is far
advanced, seemingly aware that its own breeding grounds in the
Arctic are still in the grip of winter.
As shown by the route map (fig. 21) a great western highway also
has its origin in the Mackenzie River delta area and in Alaska. This
is used chiefly by the pintail and the baldpate (Mareca americana),
which fly southward through eastern Alberta to western Montana.
Some localities in this area, as for example, the National Bison Range,
at Moiese, Mont., normally furnish food in such abundance as to induce
these birds to pause in their migratory movement. Upon resuming
travel, some flocks move almost directly west across Idaho to
the valley of the Columbia River, from which they turn abruptly
south to the interior valleys of California. Others leave the Montana
feeding and resting areas and turn southeastward across Wyoming and
Nebraska to join the flocks that either are moving southward through
the Great Plains, or are to continue across Arkansas to the main
Mississippi Valley fly way.
Many redheads that breed in the Bear River marshes in Utah,
take a westerly route across Nevada to California, but some leave
these breeding grounds and fly northeastward across North Dakota
and Minnesota to join the flocks of these ducks that come out of the
prairie regions of Canada, and travel southeastward to the Atlantic
coast. This route can be well traced by the records of ducks banded
in summer in the Bear River marshes and retaken the following fall
at points in eastern Montana, Wyoming, South Dakota, North Dakota,
Minnesota, Wisconsin, Michigan, and Maryland.
Another route from these great marshes crosses the mountains in
an easterly direction, but almost immediately turns southward
through Colorado and New Mexico, and continues to winter quarters
in the Valley of Mexico (fig. 21 and fig. 20, route 6). This route also
represents the travels of many of the land birds of the Rocky Mountain
region. Such birds perform comparatively short migrations,
most of them being content to stop when they reach the middle districts
of Mexico, only a few passing east beyond the southern part of
that country.
This account of the Mackenzie Valley-Great Lakes-Mississippi
Valley route shows the great importance of this highway and also its
complicated nature. It receives accretions from both the northeast
and the northwest, while branch routes make wide detours toward
the Pacific coast before turning back to the parent fly way. This
flyway is notable for its great length, as it extends from the Arctic-coast
to Patagonia; and for its width east and west, as in North
America it reaches from the Allegheny Mountains westward to the
Great Basin.
PACIFIC COAST ROUTE
Although it does present features of unusual interest, the Pacific-coast
route is not of so great importance as some of the others described.
Because of the equable conditions that prevail, many species
of birds along the coast from the Northwestern States to Southern
Alaska either do not migrate at all or else make relatively short
journeys. This route has its origin chiefly in Alaska, the general
region of the delta of the Yukon River marking its northern terminus,
- 48 -
although a few species join it after a flight westward along the Alaskan
Arctic coast (fig. 21). Some of the scoters (Melanitta and Oidemia)
and other sea ducks of the north Pacific region, and the diminutive
cackling goose (Branta canadensis minima), which breeds in the delta
of the Yukon River, use the coastal sea route for all or most of their
southward flight. The journey of the cackling geese, as shown by
return records from birds banded at Hooper Bay, Alaska, has been
traced southward across the Alaska Peninsula and apparently across
the Gulf of Alaska to the Queen Charlotte Islands, the birds following
the coast line south to near the mouth of the Columbia River. There
the route swings toward the interior for a short distance before continuing
south by way of the Willamette River Valley. The winter
quarters of the cackling geese are chiefly in the vicinity of Tule Lake,
on the Oregon-California line, and in the Sacramento Valley of California,
though a few push on to the San Joaquin Valley.
B4509M
Figure 24.—Probable breeding range, the winter range, and the migration route of Ross's goose.
This is the only species of which all members apparently breed in the Arctic regions, migrate south
through the Mackenzie Valley, and upon reaching the United States turn to the southwest rather than
the southeast. The southern part of this route, however, is followed by some mallards, pintails, baldpates,
and possibly by other ducks.
- 49 -
A tributary of this flyway is followed by Ross's goose (Chen
rossi), which is believed to breed on the Arctic islands north of
Mackenzie (fig. 24). Its fall migration is across the barren grounds
to Great Slave and Athabaska Lakes, where it joins thousands of other
waterfowl bound for their winter homes along the eastern coast of the
United States and the Gulf of Mexico. But when Ross's geese have
traveled south approximately to the northern boundary of Montana,
they separate from their companions, and turning to the southwest
cross the Rocky Mountains and settle for the winter in California.
The route taken by the white-winged scoters that winter on the
Atlantic coast already has been indicated (p. 43). Some birds of
this species, however, winter on the Pacific coast from Puget Sound
south to southern California. Their passage by thousands up and
down the coast has been noted as far north as northwestern British
Columbia. The species is known to nest in Alaska, which may be
the home of some at least of the scoters that winter on the Pacific
coast. If such be the case, however, it must be admitted that a part
of the route taken by the birds when on migration is unknown,
though very few observations are available from the interior of
northern British Columbia, across which the route may lie.
The southward route of those migratory land birds of the Pacific
coast that in winter leave the United States extends chiefly through
the interior of California to the mouth of the Colorado River and on
to winter quarters in western Mexico.
The movements of the western tanager (Piranga ludoviciana) show
a migration route that is in some ways remarkable. The species
breeds in the mountains from the northern part of Baja California
and western Texas north to northeastern British Columbia and
southwestern Mackenzie. Its winter range is in two discontinuous
areas—southern Baja California and eastern Mexico south to
Guatemala (fig. 25). On the spring migration the birds enter the
United States about April 20, appearing first in western Texas and
the southern parts of New Mexico and Arizona (fig. 26). By April
30 the van has advanced evenly to an approximate east-and-west line
across central New Mexico, Arizona, and southern California. But
by May 10 the easternmost birds have advanced only to southern
Colorado, while those in the far West have reached northern Washington.
Ten days later the northward advance of the species is
shown as a great curve, extending northeastward from Vancouver
Island to central Alberta and thence southeastward to northern
Colorado. Since these tanagers do not reach northern Colorado until
May 20, it is evident that those present in Alberta on that date,
instead of traveling northward through the Rocky Mountains, which
from the location of their summer and winter homes would seem to
be the natural route, reached there by the Pacific coast route to southern
British Columbia and thence across the mountains, despite the
fact that these are still partly covered with snow at that time.
PACIFIC OCEANIC ROUTE
The route of the Pacific golden plover (Pluvialis dominica fulva) is
fully as interesting and as remarkable as the elliptical course followed
by its eastern cousin (P. d. dominica) (fig. 22). The breeding range
- 50 -
of the eastern golden plover extends through Arctic America west to
the northern coast of Alaska, where in the vicinity of Point Barrow it
meets the nesting grounds of the Pacific form, which is really an Asiatic
subspecies. It breeds chiefly in the Arctic coast region of Siberia
and merely overflows onto the Alaskan coast, some of the birds probably
migrating south along the coast of Asia to winter quarters in
Japan, China, India, Australia, New Zealand, and Oceania, including
the Hawaiian Islands, the Marquesas Islands, and the Low Archipelago.
Golden plovers in migration have been observed at sea on a
line that apparently extends from these islands to the Aleutians, and
it therefore appears certain that at least some of the Alaskan birds
make a nonstop flight across a landless sea from Alaska to Hawaii.
While it would seem incredible that any birds could lay a course so
straight as to attain these small oceanic islands, 2,000 miles south
of the Aleutians, 2,000 miles west of Baja California, and nearly
4,000 miles east of Japan, the evidence admits only the conclusion
that year after year this transoceanic round-trip journey between
Alaska and Hawaii is made by considerable numbers of golden plovers.
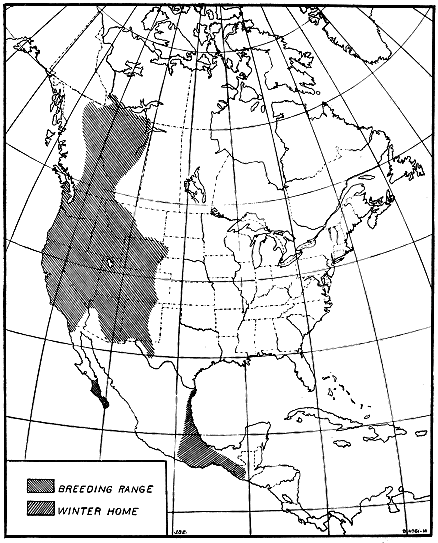
B4761M
Figure 25.—Breeding and wintering ranges of the western tanager.
See figure 26 for the spring route
taken by the birds breeding in the northern part of the range.
- 51 -
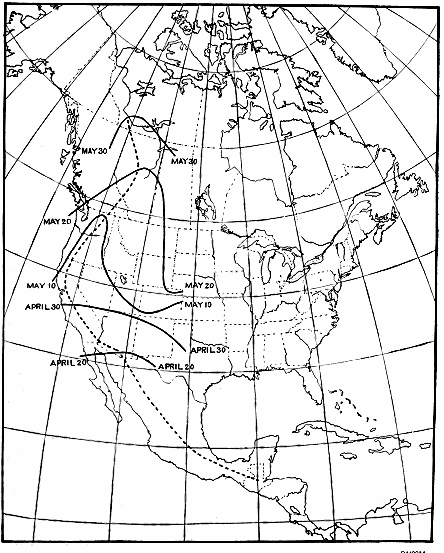
B4499M
Figure 26.—Migration of the western tanager. The birds that arrive in eastern Alberta by May 20 do not
travel northward along the eastern base of the Rocky Mountains, as in that region the van has then only
reached northern Colorado. Instead the isochronal lines indicate that they migrate north through
California, Oregon, and Washington, and then cross the mountains of British Columbia.
The Pacific oceanic route probably is used also by the arctic
terns that breed in Alaska, and possibly by those from the more western
tern colonies of Canada. This species is of regular occurrence on
the western coasts of both the United States and South America,
indicating that the western representatives travel southward to the
Antarctic winter quarters without the spectacular migration features
- 52 -
that appear to characterize the flight of those from the eastern part
of the continent (fig. 15).
ARCTIC ROUTES
In the discussion of the migration of the Arctic tern (p. 31) it was
noted that this species makes a very distinct west-to-east movement
across northern Canada, continuing the flight eastward across the
Atlantic Ocean toward the western coast of Europe. It seems likely
that there are other species, including the parasitic jaeger (Stercorarius
parasiticus) that regularly breed in the northern part of the Western
Hemisphere but migrate back to the Old World for their winter
sojourn. Some others, as the red-legged kittiwake (Rissa brevirostris)
and Ross's gull (Rhodostethia rosea), remain near the Arctic region
throughout the year, retreating southward in winter only a few
hundred miles. The emperor goose (Philacte canagica) in winter is
found only a relatively short distance south of its breeding grounds,
and eider ducks (Somateria and Arctonetta), although wintering in
latitudes well south of the breeding grounds, nevertheless remain
farther north than do the majority of other species of ducks (fig. 21).
The routes followed by these birds are chiefly coastwise, and in the
final analysis may be considered as being tributary either to the
Atlantic or to the Pacific coast routes. The passage of gulls, ducks,
and other water birds at Point Barrow, Alaska, and at other points
on the Arctic coast, has been noted by several observers, and from
present knowledge it may be said that the best defined Arctic route
in North America is the one that follows around the coast of Alaska.
EVOLUTION OF MIGRATION ROUTES
From the foregoing descriptions of migration routes it will be
observed that the general trend of migration in most species of North
American birds is northwest and southeast. It is comparatively
easy to trace the probable steps in the evolution of the migrations
of some species, and some routes have developed so recently that they
still plainly show their origin.
The tendency is for eastern species to extend their ranges by pushing
westward, particularly in the north. For example in the Stikine
River Valley of northern British Columbia and southwestern Alaska
the eastern nighthawk (Chordeiles minor minor), eastern chipping
sparrow (Spizella passerina passerina), rusty blackbird (Euphagus
carolinus), eastern yellow warbler (Dendroica aestiva aestiva), redstart,
and others have established breeding stations at points 20 to 100 miles
from the Pacific Ocean. The robin, flicker (Colaptes auratus),
slate-colored junco (Junco hyemalis hyemalis), blackpoll warbler,
myrtle warbler (Dendroica coronata), and ovenbird (Seiurus aurocapillus),
all common eastern species, also are established as breeding
birds in western Alaska, the ovenbird having been detected on
the lower Yukon River. These birds, however, do not migrate in
fall by any of the Pacific routes, but instead retrace their journey
across the mountains and move southward along the broad flyways
of the interior.
The red-eyed vireo (Vireo olivaceus), a striking example of an
abundant woodland bird, is essentially an inhabitant of States east
of the Great Plains, but an arm of its breeding range extends north
- 53 -west
to the Pacific coast in British Columbia (fig. 27). It seems
evident that this is a range extension that has taken place comparatively
recently by a westward movement from the upper Missouri
Valley, and that the invaders retrace in spring and fall the general
route by which they originally entered the country.
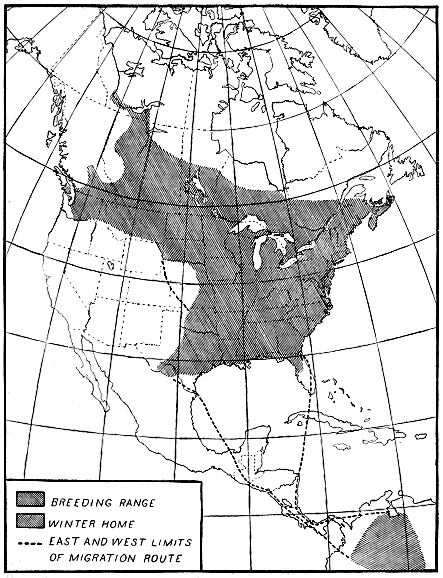
B4506M
Figure 27.—Distribution and migration of the red-eyed vireo. It is evident that the red-eyed vireo has
only recently invaded Washington by an extension of its breeding range almost due west from the upper
Missouri Valley. Like the bobolink, however (fig. 23), the western breeders do not take the short cut
south or southeast from their nesting grounds, but migrate spring and fall along the route traversed in
making this extension.
In the case of the bobolink, a new extension of the breeding range,
and a consequent change in the migration of the species, has taken
place since the spread of settlement in this country (fig. 23). A bird
- 54 -
of damp meadows, it was originally cut off from the Western States
by the intervening arid regions. But with the advent of irrigation and
the bringing of large areas under cultivation, small colonies of nesting
bobolinks have appeared at various western points, and now the
species is established as a regular breeder in the great mountain parks
and irrigated valleys of Colorado and elsewhere almost to the Pacific
coast. In retracing their course to reach the western edge of the
route followed by the bulk of the bobolinks that breed in the northern
United States and southern Canada, these western pioneers must fly
long distances along a line that runs almost due east and west.
Similarly it is possible to sketch what seems to be the logical
evolution of the remarkable routes of the golden plover (fig. 22).
It may be assumed that the eastern birds of this species first followed
an all-land route from the South American winter quarters through
Central America, Mexico, and Texas to the western parts of the
Mississippi Valley. As the migration route lengthened northward
with the retreat of the ice and the bird's powers of flight developed,
it would have a tendency to straighten the line and to shorten it by
cutting off some of the great curve through Mexico and Texas.
First a short flight across the western part of the Gulf of Mexico was
probably essayed. Proving successful, this was followed by flight
lines that moved farther east, until finally the roundabout curve
through Texas was entirely discarded and the flight made directly
across the Gulf to southern Louisiana.
As the great areas in Canada were gradually added to the bird's
domain, other important factors arose, the chief being the attractiveness
of the vast stretches of coast and plain of the Labrador Peninsula,
which in fall offered a bountiful store of berries. The fall route therefore
worked eastward to the Gulf of St. Lawrence thence southwest
through the interior to the coast of Florida and across the Gulf of
Mexico to the Central American mainland. A series of shortening
flights followed to take out the great curve of the New England coast.
A relatively short ocean flight was probably attempted, say from
Cape Cod to the Bahama Islands, Cuba, and Jamaica, followed
eventually by the long direct oceanic route as it is now known.
As the Labrador Peninsula in spring is bound by frost and shrouded
by fog while the season advances rapidly through the interior, the
oceanic route proved useful only in fall, and the spring flight continued
through the Mississippi Valley. The above outline gives a probable
and fairly plausible explanation of the origin of this wonderful route,
particularly when it is remembered that migration routes as now
known are evolutions—age-long modifications of other routes.
The evolution of the migration of the Pacific golden plover may be
explained in a similar fashion. At first the route probably followed
the Asiatic coast, through the Malay Peninsula and Oceania, thence
east in a great curve to the Low Archipelago, with individuals and
flocks dropping out to winter at many points along the way. The
Siberian birds probably continue to follow this ancient flyway, but
those nesting in Alaska began a long evolutionary series of flights
that cut down the length of their journey by shortening the curve,
until finally the transoceanic route of the present day was developed.
- 55 -
VERTICAL MIGRATION
In the effort to find winter quarters furnishing satisfactory living
conditions, many North American birds fly hundreds of miles across
land and sea. Others, however, are able to attain their objective
merely by moving down the sides of a mountain. In such cases a
few hundred feet of altitude corresponds to hundreds of miles of
latitude. Movements of this kind, known as "vertical migrations",
are found wherever there are large mountain ranges. In the Rocky
Mountain region they are particularly notable, as chickadees, rosy
finches (Leucosticte), juncos, pine grosbeaks (Pinicola), and some
other species that nest in the Alpine Zone move down to the lower
levels to spend the winter. It has been noted that such species as
Williamson's sapsucker (Sphyrapicus thyroideus), and the western
wood pewee (Myiochanes richardsoni), which nest in the higher mountains,
move down to the lower regions in August following the breeding
season. At this time there is a distinct tendency also among the
young of mountain-breeding birds to work down to the lower levels
as soon as the nesting season is over. The sudden increases among
birds in the edges of the foothills are particularly noticeable when
cold spells with snow or frost occur at the higher altitudes.
Some species that normally breed in the Hudsonian or Arctic
Zones find suitable breeding areas on the higher levels of the mountains,
as for example the pipit, or titlark (Anthus spinoletta rubescens),
which breeds on the tundras of Alaska and northern Canada and
also south as far as Colorado on the summits of many peaks in the
Rocky Mountains. On the other hand a few species, as the Clark's
crow, or nutcracker (Nucifraga columbiana), nest at relatively low
altitudes in the mountains and as the summer advances move higher
up, thus performing a vertical migration that in a sense is comparable
with the post-breeding movements of herons on the Atlantic coast.
These illustrations show that the length of a migration route may
depend upon factors other than latitude.
VAGRANT MIGRATION
The most striking feature of the migrations of some of the herons
is a northward movement after the nesting season. The young of
some species commonly wander late in summer and in fall, sometimes
traveling several hundred miles north of the district in which they
were hatched. The little blue heron (Florida caerulea caerulea)
breeds commonly north to South Carolina, and by the last of July
the young birds begin to appear along the Potomac, Patuxent, and
Susquehanna Rivers, tributary to Chesapeake Bay. Although
almost all are immature individuals, as shown by their white plumage,
an occasional adult may be noted. With them come snowy herons
(Egretta thula thula) and egrets (Casmerodius albus egretta), and on
occasion all three species will travel in the East as far north as New
England, and in the Mississippi Valley to southeastern Kansas and
Illinois. In September most of them disappear, probably returning
south by the same route.
The black-crowned night heron (Nycticorax nycticorax hoactli) has
similar wandering habits, and young birds banded in a large colony
at Barnstable, Mass., have been recaptured the same season north to
Maine and Quebec and west to New York. This habit seems to be
- 56 -
shared by some of the gulls also, although here the evidence is not so
conclusive. Herring gulls (Larus argentatus smithsonianus) banded
as chicks at colonies in the Great Lakes have scattered in all directions
after the breeding season, some having been recovered well north in
Canada.
These movements may be considered as migration governed only by
the availability of food, and they are counteracted in fall by a directive
migratory impulse that carries back to their normal winter homes in
the south such birds as after the mating period have attained more
northern latitudes. They are not to be compared with the great
invasions of certain birds from the North. Classic examples of the
latter in the eastern part of the country are the periodic flights of
crossbills. Sometimes these migrations will extend well south into the
Carolinian Zone.
Snowy owls are noted for occasional invasions that probably are
caused by a shortage of the lemmings and rabbits that constitute their
normal food in the North. At least nine notable flights of these birds
occurred during the period 1876 to 1927. In the great flight of 1926-27
they were noted as far south as Iowa, Ohio, West Virginia, and
North Carolina.
In the Rocky Mountain region great flights of the beautiful Bohemian
waxwing (Bombycilla garrula pallidiceps), are occasionally
recorded. The greatest invasion in the history of Colorado ornithology
occurred in February 1917, at which time the writer estimated
that at least 10,000 were within the corporate limits of the city of
Denver. The last previous occurrence of the species in large numbers
in that section was in 1908.
Evening grosbeaks (Hesperiphona vespertina) likewise are given to
performing more or less wandering journeys, and curiously enough, in
addition to occasional trips south of their regular range, they travel
east and west, sometimes covering long distances. For example,
grosbeaks banded at Sault Ste. Marie, Mich., have been recaptured on
Cape Cod, Mass., and in the following season have been re trapped at
the banding station. Banding records demonstrate that this east-and-west
trip across the northeastern part of the country is sometimes
made also by purple finches (Carpodacus purpureus).
PERILS OF MIGRATION
The period of migration is a season full of peril for birds. Untold
thousands of the smaller migrants are destroyed each year by storms,
in unfamiliar habitats, and through attacks of predatory birds,
mammals, and reptiles. If each pair of adult birds should succeed in
raising two fledglings to maturity, the population of migratory birds
would have a potential annual increase of 100 percent and the world
would soon be heavily overpopulated with them. It is evident,
therefore, that there is no such increase, and that the annual mortality
from natural causes is heavy enough to keep it in check.
STORMS
Of the various factors limiting the abundance of birds, particularly
the smaller species, storms are the most potent. Special sufferers
are those birds that in crossing broad stretches of water are forced
by a storm down within reach of the waves. Such a catastrophe was
- 57 -
once seen from the deck of a vessel in the Gulf of Mexico, 30 miles
off the mouth of the Mississippi River. Great numbers of migrating
birds, chiefly warblers, had accomplished nearly 95 percent of their
long flight and were nearing land when, caught by a norther, against
which they were unable to contend, hundreds were forced into the
waters of the Gulf and drowned. On another occasion, on Lake
Michigan, a severe storm, coming up at a time when large numbers of
migratory birds were crossing, forced numerous victims into the waves.
During the fall migration of 1906, when thousands of birds were crossing
Lake Huron, a sudden drop in temperature, accompanied by a
heavy snowfall, resulted in the death of incredible numbers.
Literally thousands were forced into the water and subsequently cast
up along the beaches, where in places their bodies were piled in windrows.
On one section of the beach the dead birds were estimated at
1,000 per mile, and at another point at five times that number. Most
of them were species that rank among our most desirable birds as
destroyers of insects and weed seeds, including slate-colored juncos,
tree sparrows, white-throated sparrows, swamp sparrows, winter
wrens, and golden-crowned kinglets, together with many brown
creepers, hermit thrushes, warblers, vireos, and others.
Of all species of North American birds, the Lapland longspur
(Calcarius lapponicus) seems to be the most frequent victim of mass
destruction from storms. These birds sometimes congregate in
enormous numbers where grass or weed seed is abundant. Almost
every winter brings in reports of their death by thousands somewhere
in the Middle West. While migrating northward at night they have
encountered blinding storms of wet, clinging snow, which have so
bewildered them that they have flown into various obstructions, or
have sunk to the ground and perished of exposure and exhaustion.
In 1907 an experienced ornithologist estimated that 750,000 longspurs
were lying dead on the ice of two lakes in Minnesota, each about 1
square mile in extent, and dead birds were reported in greater or less
abundance on this occasion over an area of more than 1,500 square
miles. The heaviest mortality occurred in towns, where, bewildered
by the darkness and the heavy falling snow, some of the birds congregating
in great numbers, flew against various obstacles and were
killed or stunned, while many others fell to the ground exhausted.
Similar catastrophes have been reported from eastern Colorado,
Nebraska, and North Dakota.
During the early part of June 1927, a hailstorm of exceptional
severity in and around Denver, Colo., killed large numbers of robins,
meadow larks, sparrows, and others. The lawns of parks were strewn
with the bodies of these birds, and many lay dead in their nests where
they were covering their eggs or young when the storm broke.
AERIAL OBSTRUCTIONS
The destruction of migratory birds by their striking lighthouses,
light ships, tall bridge piers, monuments, and other obstructions has
been tremendous. Beams of the lanterns at light stations have a
powerful attraction for nocturnal travelers of the air. It may be
likened to the fascination for lights that is shown by many insects,
particularly night-flying moths. The attraction is not so potent in
clear weather, but when the atmosphere is moisture laden, as in a
heavy fog, the rays have a dazzling effect that lures the birds to their
- 58 -
death. They may fly straight up the beam and dash themselves
headlong against the glass, or they may keep fluttering around the
source of the light until exhausted, and then drop to the rocks or
waves below. The fixed, white, stationary light located 180 feet
above sea level at Ponce de Leon Inlet (formerly Mosquito Inlet),
Fla., has caused great destruction of bird life even though the lens
is shielded by wire netting. On one occasion an observer gathered up
a bushel-basketful of warblers, sparrows, and other small passerine
birds that had struck during the night. The birds apparently beat
themselves to death against the wire or fell exhausted to the concrete
pavement below, frequently to be destroyed there by cats or skunks.
Two other lighthouses at the southern end of Florida, Sombrero
Key and Fowey Rocks, have been the cause of a great number of
bird tragedies, while heavy mortality has been noted also At some of
the lights on the Great Lakes and on the coast of Quebec. It is
the fixed white lights that cause such disasters to birds, as the stations
equipped with flashing or red lights do not present such strong attractions.
That it is not a mere case of geographical location has
been demonstrated, for it is observed that when fixed white lights have
been changed to red or flashing lights, the migrating birds are no
longer endangered. At some of the light stations in England and
elsewhere, shelves and perches have been placed below the lanterns
to afford places where birds can rest until they have overcome their
bewilderment.
For many years at the National Capital, the Washington Monument,
although unilluminated, caused the destruction of large numbers
of small birds, due apparently to their inability to see this obstacle
in their path, towering more than 555 feet into the air. One morning
in the spring of 1902 the bodies of nearly 150 warblers, sparrows, and
other birds were found about its base. Then, as the illumination of
the city was improved and the Monument became more visible at
night, the loss became steadily less, until by 1920 only a few birds
would be killed during an entire migration. On November 11, 1931,
however, as part of the Armistice Day celebration, batteries of
brilliant floodlights grouped on all four sides about the base of the
Monument, were added to the two searchlights already trained on
the apex, so that the lighted shaft probably corresponded in brilliancy
to a very low magnitude lighthouse lantern. Airplane pilots
have ventured opinions that on a clear night it could be seen for 40
miles. It is certain that there is an extensive area of illumination,
and on clear, dark nights when the nocturnal travelers seem to fly at
lower altitudes, many of them are attracted to the Monument as to
a lighthouse beacon, and wind currents prevent a last-minute avoidance.
During the fall migration of 1932 more than 500 warblers,
vireos, thrushes, kinglets, sparrows, and others were killed. In 1933
the mortality was less, but the Monument at times still remains a
serious menace to birds during migration.
When the torch on the Statue of Liberty in New York Harbor was
kept lighted, it caused an enormous destruction of bird life, tabulations
showing as many as 700 birds in a single month.
- 59 -
EXHAUSTION
Although it would seem that the exertion incident to the long
flights of many species of migratory birds would result in their arrival
at their destination in a state bordering on exhaustion, this is contrary
to the truth. Both the soaring and the sailing of birds show them to
be proficient in the use of factors employed in aerial transportation
that only recently have become understood and imitated by aeronautical
engineers. The use of ascending currents of air, employed
by all soaring birds, and easily demonstrated by observing the gulls
that glide hour after hour along the windward side of a ship, are now
utilized by man in his operation of gliders. Moreover, the whole
structure of a bird renders it the most perfect machine for extensive
flight that the world has ever known. Hollow, air-filled bones, making
an ideal combination of strength and lightness, and the lightest
and toughest material possible for flight in the form of feathers, combine
to produce a perfect flying machine. Mere consideration of a
bird's economy of fuel or energy also is enlightening. The golden
plover, traveling over the oceanic route, makes the entire distance
of 2,400 miles from Nova Scotia to South America without stop,
probably requiring about 48 hours of continuous flight. This is
accomplished with the consumption of less than 2 ounces of fuel in the
form of body fat. To be as economical in operation, a 1,000-pound
airplane would consume in a 20-mile flight not the gallon of fuel
usually required, but only a single pint.
The sora, or Carolina rail (Porzana Carolina), which is such a
notoriously weak flyer that at least one writer was led to infer that
most of its migration was made on foot, has one of the longest migration
routes of any member of its family, and easily crosses the wide
reaches of the Caribbean Sea. The tiny ruby-throated hummingbird
(Archilochus colubris) crosses the Gulf of Mexico in a single flight
of more than 500 miles.
While birds that have recently arrived from a protracted flight over
land or sea sometimes show evidences of being tired, their condition
is far from a state of exhaustion, unless unusual conditions have been
encountered, and with a few hours' rest and a crop well filled with
proper food they exhibit eagerness to resume their journey. The
popular notion that birds find the long ocean flights excessively
wearisome and that they sink exhausted when terra firma is reached
does not agree with the facts. The truth lies in the opposite direction,
as even small land birds are so little averse to ocean voyages
that they not only cross the Gulf of Mexico at its widest point, but
may even pass without pause over the low, swampy coastal plain
to the higher regions beyond. Under favorable conditions birds can
fly when, where, and how they please. Consequently the distance 4
covered in a single flight is governed chiefly by the food supply.
Exhaustion, except as the result of unusual factors, cannot be said to
be an important peril of migration.
INFLUENCE OF THE WEATHER ON MIGRATION
The state of the weather at any point has little if anything to do
with the time of arrival of migratory birds. This is contrary to the
belief of observers who have thought that they could foretell the
appearance of various species by a study of the weather conditions.
- 60 -
Though the insistent crescendo note of the ovenbird is ordinarily
associated with the full verdure of May woods, this bird has been
known to reach its breeding grounds in a snowstorm and the records
of its arrival in southern Minnesota show a temperature variation
from near freezing to full summer warmth. Temperatures recorded
at the time of arrival of several other common birds show variations
of 14° to 37° F., the average variation being about 24°.
It should be remembered that North American species spending
the winter months in tropical latitudes experience no marked changes
in climatic conditions from November to March or April, yet frequently
they will start the northward movement in January or
February. This is in obedience to physiological promptings and has
no relation to the prevailing weather conditions. For migratory
birds the winter season is a period of rest, a time when they have no
cares other than those associated with the daily search for food or
escape from their natural enemies. Their migrations, however, are
a vital part of their life cycles, which have become so well adjusted
that the seasons of travel correspond in general with the major
seasonal changes on their breeding grounds. With the approach of
spring, therefore, the reproductive impulse awakens, and each
individual bird is irresistibly impelled to start the journey that ends
in its summer home.
Through their influence upon the food supply, weather conditions
in the breeding areas do, of course, figure prominently in the picture.
In fact, they are the major factors in determining the average dates
of arrival of the different species. The word "average" must be
emphasized, for the migrations of birds have so evolved that in
general they synchronize with average climatic conditions. In other
words, migration is so ingrained that each species moves north in
spring when the average weather that will be encountered is not
unendurable. The hardy birds travel early, fearless of the blasts of
retreating winter. The more delicate species come later, when there
is less danger of encountering prolonged periods of inclement weather.
Some of these hardy birds pause and allow the spring season to
advance, and then by rapid travel again overtake it, or in some cases
actually outstrip it. At times this results in some hardship and
occasionally in the destruction of large numbers of individuals, as
has happened when early migrating bluebirds have been overwhelmed
by a late winter storm. Unless such conditions are prolonged, however,
no serious effect on the species is noted. Nevertheless, the
soundness of the bird's instincts is evidenced by the fact that natural
catastrophes, great though they may be, do not permanently diminish
the avian population.
As has been pointed out, the advance of average temperature lines,
known as isotherms, is found to correspond closely with the northward
movements of certain species. For example, the northward
travels of the Canada goose are found to coincide with the advance
of the isotherm of 35° F. (fig. 8).
The spring flight of migrants, if interrupted for any reason, is
resumed when weather conditions again become favorable, and it is
probable that all instances of arrival of birds in stormy weather can
be explained on the theory that the flight was begun while the weather
was auspicious. The state of the weather when a flight starts at any
southern point, the relation of that place to the average position of
- 61 -
the bird under normal weather conditions on that date, and the
average rate of migratory flight, are data basic to any reasonably
accurate prediction of the time arrival may be expected in northern
areas.
Head winds are as unfavorable to migration as is rain or snow, as
they greatly increase the labor of flight and cut down the speed of
cross-country travel. If such winds have a particularly high velocity
they may force down the weaker travelers, and should this happen
over water areas, large numbers of birds would be lost. Even strong
winds that blow in the direction of aerial travel are unfavorable for
the birds, as they interfere with their balance and disarrange their
feathers. Moderate tail winds and cross or quartering breezes appear
to offer the best conditions for the passage of the migrants.
B3712M
Figure 28.—A banded belted kingfisher, one of about 2,000,000 birds that have been marked with Biological
Survey bands.
PROBLEMS OF MIGRATION
BANDING STUDIES
The study of living birds by the banding method, whereby great
numbers of individuals are marked (fig. 28) with numbered aluminum
leg rings, has come to be recognized as a most accurate means of
ornithological research. Since 1920, banding work in North America
has been under the direction of the Bureau of Biological Survey, in
cooperation with the National Parks Branch of Canada. Every year
volunteer cooperators, working under permit, place bands on thousands
of birds, game and nongame, large and small, migratory and
nonmigratory, each band carrying a serial number and the legend,
NOTIFY BIOLOGICAL SURVEY, WASHINGTON, D. C., or on the smaller
sizes, NOTIFY BIOL. SURV., WASH., D. C. When a banded bird is
- 62 -
reported from a second locality, a definite fact relative to its movements
becomes known, and a study of many cases of this nature
develops more and more complete knowledge of the details of
migration.
The records of banded birds are also yielding other pertinent information
relative to their migrations, such as the exact dates of arrival
and departure of individuals, the length of time that different birds
pause on their migratory journeys to feed and rest, the relation
between weather conditions and the starting times for migration, the
rates of travel of individual birds, the degree of regularity with which
birds return to the exact summer or winter quarters used in former
years, and many other details that could be learned in no other
manner. Banding stations that are operated systematically throughout
the year, therefore, are supplying much information concerning
the movements of migratory birds that heretofore could only be
surmised.
MOVEMENTS OF RESIDENTS
Typical migration consists of definite movements that are repeated
regularly year after year, and it is to these that the term is generally
restricted. It is desirable, however, if only for purposes of comparison,
that some account be taken of the movements of some other
birds, which, while not typical, do possess some of the characteristics
of true migration. Data on this subject are being collected through
bird banding.
There are several species that are customarily grouped under the
heading "permanent residents", the term implying that these birds
do not travel but remain throughout the year in one locality. Among
these are the cardinal, the tufted titmouse (Baeolophus bicolor), the
wren tit (Chamaea fasciata), the Carolina wren, the house finch
(Carpodacus mexicanus frontalis), the bobwhite (Colinus virginianus),
the California quail (Lophortyx californica), and the ruffed grouse
(Bonasa umbellus). Each species may be present constantly throughout
the year, although in the northern part of the range there is
probably a slight withdrawal of the breeding birds in winter. The
individuals to be seen at that season, therefore, may not always be
the same as those observed during the summer. It is certain, however,
that these species do not regularly perform extensive journeys.
While the blue jay is disposed to be secretive, it is such a showy
and noisy bird that is not likely to escape notice. In the vicinity of
Washington, D. C., as in many other places, it is present the year
round, but at the end of September or early in October when the
weather is becoming cooler, troops of jays are sometimes seen working
southward through the trees. A corresponding northward movement
occurs again in May. This is unquestionably a migration to
and from some winter range, but its extent or significance is not now
known. Some light is being shed on the matter, however, through
the records of banded birds, and these eventually will fill in a more
perfect picture of the movements of this species. One jay, banded
on September 14, 1923, at Waukegan, Ill., was killed at Peruque,
Mo., on November 15 of the same year; another, banded at Winnetka,
Ill., on June 16, 1925, was retaken at Sulphur Rock, Ark., the following
December 10; a third, banded on May 6, 1925, at Whitten, Iowa,
was recaptured at Decatur, Ark., on January 22, 1926. These three
- 63 -
birds unquestionably had made a flight that had every appearance
of being a true migration to winter quarters in Missouri and Arkansas.
The black-capped chickadee is apparently resident in many places,
but occasionally in winter it invades the range of the southern Carolina
chickadee (Penthestes carolinensis) and in northern Canada it is
regularly a migrant.
In the coastal plain between Washington, D. C., and the Atlantic
Ocean, the white-breasted nuthatch is usually absent during the
summer, nesting at that season in the higher, or piedmont, country.
Late in fall, however, it appears in fair abundance in the wooded
bottoms, remaining at the lower levels until the following March
or April.
Some birds, including the screech owl (Otus asio), bobwhite,
Carolina wren, and mockingbird (Mimus polyglottos), seem to be
actually sedentary, but even these are sometimes given to post-breeding
wanderings. Ordinarily bob whites that are marked with
numbered bands are seldom retaken far from the area where banded,
but sometimes they will travel 10 miles or more. A screech owl
banded at Glenwood, Minn., in March, was recovered the following
December at Emmetsburg, Iowa, 180 miles south. Such flights,
however, are probably more in the nature of a search for new feeding
areas, or to escape from a winged enemy, than a true migratory
journey.
MIGRATION OF THE WHITE-THROATED SPARROW
The white-throated sparrow (Zonotrichia albicollis) (fig. 29), one of
the most abundant members of its family, breeds from northern
Mackenzie and the southern part of the Ungava Peninsula south to
southern Montana, northern Pennsylvania, and Massachusetts. The
winter range extends from the southern part of the breeding range
south to the Gulf coast and northeastern Mexico. It is therefore
a common migrant in many sections. Since it is a ground-feeding
bird and is readily attracted to the vicinity of dwellings, it has been
banded in large numbers, the total to February 1, 1935, being nearly
100,000. It would be expected that these would yield a comparable
number of return records, and that the facts would furnish basic
data relative to the migrations of the species. Such, however, is
not the case. Banded white-throated sparrows are rarely recaptured
at stations between the breeding and the wintering grounds. Operators
of stations in the winter area, as Thomasville, Ga., and Summerville,
S. C., have obtained return records showing that these birds
do come back to the exact winter quarters occupied in previous
seasons. The fact that they do not again visit banding stations on
their migration routes indicates some unusual aspects of their travels,
which it is hoped will eventually be discovered by banding studies.
Problems of this type constitute definite challenges to the student of
bird migration.
MIGRATION OF THE YELLOW-BILLED LOON
The semiannual movements of the yellow-billed loon present an
unusual problem in migration. It breeds along the Arctic coast,
probably from Cape Prince of Wales eastward to Franklin Bay, and
also in the interior of northern Canada south to Clinton-Colden,
Aylmer, and Artillery Lakes, where it is rather common. It has been
reported as already present by May 25 at the mouth of the Liard
- 64 -
River, in southwestern Mackenzie. This coincides with the time that
first arrivals are noted fully 700 miles north, at Point Barrow, Alaska.
The problem has been to ascertain the route used by these birds
to then principal nesting grounds in the interior.
For a long time it was believed that this big diver did not winter in
large numbers anywhere on the Pacific coast, and it had been supposed
that the spring route extended 2,000 miles northeastward from a
wintering ground somewhere in eastern Asia to Bering Strait, then
500 miles still northeast to round Point Barrow, then 500 miles east
to the coast of Mackenzie,
and finally
700 miles south—in
spring—to the region
near the eastern end of
Great Slave Lake.
The yellow-billed
loon is a powerful flier,
and it is probable that
this suggested route
is correct for those
birds that breed in the
northern coastal regions.
A reasonable
doubt may be entertained,
however,
whether the breeding
birds of Great Slave
Lake and contiguous
areas reach their
breeding grounds by
the 700-mile flight
south from the Arctic
coast. Within recent
years it has been
found that these
birds are fairly common
in the maze of
channels and islands
off the coast of
southeastern Alaska
as late as the last
of October and in
February. Possibly
they are present there
during the period from
November through January also, or they may at that time move
farther offshore and so escape detection. If this region is an important
wintering ground, as seems probable, then it is likely that the
breeding birds of the interior reach their nesting grounds by a flight
eastward across the mountains, a trip that is well within their flying
ability, rather than by a circuitous route around the northern coast.
The air-line distance from southeastern Alaska to the mouth of the
Liard River is in fact less than the distance to that point from the
mouth of the Mackenzie.

B2216M
Figure 29.—White-throated sparrow, a bird that apparently breeds
and winters each year in the same areas, but either travels by
different routes or, at least, does not make the same stops, while
on migration.
- 65 -
Differing routes to various parts of a large breeding or wintering
ground, and used by large groups of individuals of other species, are
not unknown. For example, the redhead duck is one of the common
breeding ducks of the Bear River marshes of Utah, where a great
many have been banded each summer. The recovery records of
banded redheads show that while many travel westward to California,
others start their fall migration in the opposite direction and, flying
eastward across the Rocky Mountains, either turn southeast across
the plains to the Gulf of Mexico, or deliberately proceed in a northeasterly
direction to join the flocks of this species moving toward the
Atlantic coast from the prairie regions of southern Canada.
CONCLUSIONS
The migration of birds as it is known today had its beginning in
times so remote that its origins have been entirely obscured, and it
can be interpreted now only in terms of present conditions. The
causes underlying migration are exceedingly complex. The mystery
that formerly cloaked the periodic travels of birds, however, has been
largely dispelled through the fairly complete information that is now
available concerning the extent and times of the seasonal journeys
of most of the species. Many gaps, however, still remain in our
knowledge of the subject. Much has been learned, and present
knowledge is being placed on record, but it must be left to future
study to clear away many of the uncertainties that continue to make
bird migration one of the most fascinating subjects in the science of
ornithology.
Each kind of bird seems to have its own reaction to its environment,
so that the character of movement differs widely in the various species,
and seldom do any two present the same picture. In fact, bird migration
has been described as a phase of geographic distribution
wherein there is a more or less regular seasonal shifting of the avian
population caused by the same factors that determine the ranges of
the sedentary species. If this view is correct, then it must be recognized
that the far-reaching works of man in altering the natural condition
of the earth's surface can so change the environment necessary
for the well being of the birds as to bring about changes in their yearly
travels. The nature and extent of the changes wrought by man on
the North American Continent are easily apparent. Forests have
been extensively cut away and their places have been taken by second
growth or cultivated land, and wide stretches of prairie and plain
have been broken up, irrigated, and devoted to agriculture. These
great changes are exerting a profound effect upon the native bird
populations, and the various species may be either benefited or
adversely affected thereby.
The Federal Government, has recognized its responsibility to the
migratory birds under changing conditions brought about by man,
and by enabling acts for carrying out treaty obligations, it is now
giving many important species legal protection under regulations
administered by the Bureau of Biological Survey. Much is being
done by legislation for the welfare of the birds. The effectiveness of
these conservation laws, however, is increased in the same measure
that the people of the country become acquainted with the facts in the
life histories of the migrants and interest themselves personally in
- 66 -
the well being of the various species. Long before the white man
came to America the birds had established their seasonal lanes of migration
throughout the Western Hemisphere, as here outlined. The
economic, inspirational, and esthetic values of these migratory species
dictate that they be permitted to continue their long-accustomed and
still mysterious habits of migration from clime to clime.
(1) Allard, H. A.
1928. BIRD MIGRATION FROM THE POINT OF VIEW OF LIGHT AND LENGTH OF
DAY CHANGES. Amer. Nat. 62: 385-408.
(2) Austin, O. L., Jr.
1928. MIGRATION-ROUTES OF THE ARCTIC TERN (STERNA PARADISAEA
BRUNNICH). Northeastern Bird Banding Assoc. Bull. 4: 121-125.
(3) Baird, S. F.
1866. THE DISTRIBUTION AND MIGRATIONS OF NORTH AMERICAN BIRDS.
Amer. Jour. Sci. (2) 41: 78-90, 184-192, 337-347.
(4) Clarke, W. E.
1912. STUDIES IN BIRD MIGRATION. 2 V., illus. London.
(5) Cooke, W. W.
1888. REPORT ON BIRD MIGRATION IN THE MISSISSIPPI VALLEY IN THE
YEARS 1884 AND 1885. U. S. Dept. Agr., Div. Econ. Ornith.
Bull. 2, 313 pp., illus.
(6) ——————
1904. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN WARBLERS.
U. S. Dept. Agr., Div. Biol. Survey Bull. 18, 142 pp.
(7) ——————
1904. THE EFFECT OF ALTITUDE ON BIRD MIGRATION. Auk 21: 338-341
(8) ——————
1905. ROUTES OF BIRD MIGRATION. Auk 22: 1-11.
(9) ——————
1905. THE WINTER RANGES OF THE WARBLERS (MNIOTILTIDAE). Auk
22: 296-299.
(10) ——————
1906. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN DUCKS, GEESE,
AND SWANS. U. S. Dept. Agr., Bur. Biol. Survey Bull. 26,
90 pp.
(11) ——————
1908. AVERAGING MIGRATION DATES. Auk 25: 485-486.
(12) ——————
1910. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN SHOREBIRDS.
U. S. Dept. Agr., Bur. Biol. Survey Bull. 35, 100 pp., illus.
(13) ——————
1913. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN HERONS AND
THEIR ALLIES. U. S. Dept. Agr., Bur. Biol. Survey Bull. 45,
70 pp., illus.
(14) ——————
1913. THE RELATION OF BIRD MIGRATION TO THE WEATHER. Auk 30:
205-221, illus.
(15) ——————
1914. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN RAILS AND
THEIR ALLIES. U. S. Dept. Agr. Bull. 128, 50 pp., illus.
(16) ——————
1915. BIRD MIGRATION. U. S. Dept. Agr. Bull. 185, 47 pp., illus.
(17) ——————
1915. BIRD MIGRATIONS IN THE MACKENZIE VALLEY. Auk 32: 442-459,
illus.
(18) ——————
1915. DISTRIBUTION AND MIGRATION OF NORTH AMERICAN GULLS AND
THEIR ALLIES. U. S. Dept. Agr. Bull. 292, 70 pp., illus.
- 67 -
(19) Cooke, W. W.
1915. THE YELLOW-BILLED LOON: A PROBLEM TN MIGRATION. Condor
17: 213 214.
(20) Coues, E.
1878. BIRDS OF THE COLORADO VALLEY, A REPOSITORY OF SCIENTIFIC
AND POPULAR INFORMATION CONCERNING NORTH AMERICAN
ORNITHOLOGY. U. S. Dept. Int. Misc. Pub. 11, 807 pp., illus.
(21) Gätke, H.
1895. HELIGOLAND AS AN ORNITHOLOGICAL OBSERVATORY, THE RESULTS
OF FIFTY YEARS' EXPERIENCE. (Transl. from the German by
R. Rosenstock.) 599 pp., illus. Edinburgh.
(22) Grinnell, J.
1931. SOME ANGLES IN THE PROBLEM OF BIRD MIGRATION. Auk 48:
22—32.
(23) Gross, A. O.
1927. THE SNOWY OWL MIGRATION OF 1926-27. Auk 44: 479-493, illus.
(24) Harrison, T. H.
1931. ON THE NORMAL FLIGHT SPEEDS OF BIRDS. Brit. Birds 25: 86-96.
(25) Lincoln, F. C.
1917. BOHEMIAN WAXWING (BOMBYCILLA GARRULA) IN COLORADO. Auk
34: 341.
(26) ——————
1922. TRAPPING DUCKS FOR BANDING PURPOSES: WITH AN ACCOUNT OF THE
RESULTS OBTAINED FROM ONE WATERFOWL STATION. Auk 39:
322-334, illus.
(27) ——————
1924. BANDING NOTES ON THE MIGRATION OF THE PINTAIL. Condor 26:
88-90.
(28) ——————
1924. RETURNS FROM BANDED BIRDS, 1920 TO 1923. U. S. Dept. Agr. Bull.
1268, 56 pp., illus.
(29) ——————
1926. THE MIGRATION OF THE CACKLING GOOSE. Condor 28: 153-157,
illus.
(30) ——————
1927. NOTES ON THE MIGRATION OF YOUNG COMMON TERNS. North
eastern Bird Banding Assoc. Bull. 3: 23-28, illus.
(31) ——————
1927. RETURNS FROM BANDED BIRDS, 1923 TO 1926. U. S. Dept. Agr. Tech
Bull. 32, 95 pp., illus.
(32) ——————
1928. THE MIGRATION OF YOUNG NORTH AMERICAN HERRING GULLS. Auk
45: 49-59.
(33) ——————
1934. THE WATERFOWL FLYWAYS OF NORTH AMERICA. U. S. Dept. Agr.
Circ. 342, 12 pp., illus.
(34) Magee, M. J.
1928. EVENING GROSBEAK RECOVERIES. Northeastern Bird Banding
Assoc. Bull. 4: 56-59.
(35) May, J. B.
1929. RECOVERIES OF BLACK-CROWNED NIGHT HERONS BANDED IN MASSACHUSETTS.
Northeastern Bird Banding Assoc. Bull. 5: 7-16,
illus.
(36) Palmén, J. A.
1893. REPORT ON THE MIGRATION OF BIRDS. Transl. from the German by
C. W. Shoemaker. Smithsn. Inst. Ann. Rept. 1892: 375-396,
illus.
(37) Phillips, J. C, and Lincoln, F. C.
1930. AMERICAN WATERFOWL: THEIR PRESENT SITUATION AND THE OUTLOOK
FOR THEIR FUTURE. 312 pp., illus. Boston and New York.
(38) Rowan, W.
1925. RELATION OF LIGHT TO BIRD MIGRATION AND DEVELOPMENTAL
CHANGES. Nature [London] 115: 494-495.
(39) ——————
1926. ON PHOTOPERIODISM, REPRODUCTIVE PERIODICITY, AND THE ANNUAL
MIGRATIONS OF BIRDS AND CERTAIN FISHES. Boston Soc. Nat.
Hist. Proc. 38: [147]-189.
- 68 -
(40) Rowan, W.
1930. EXPERIMENTS IN BIRD MIGRATION. II. REVERSED MIGRATION.
Natl. Acad. Sci. Proc. 16: 520-525.
(41) ——————
1930. THE MECHANISM OF BIRD MIGRATION. Sci. Prog. 25: 70-78.
(42) ——————
1931. THE RIDDLE OF MIGRATION. 151 pp., illus. Baltimore.
(43) Watson, J. B., and Lashley, K. S.
1915. AN HISTORICAL AND EXPERIMENTAL STUDY OF HOMING. Carnegie
Inst. Washington, Dept. Marine Biol. Papers 7: 1-60, illus.
(44) Wetmore, A.
1923. MIGRATION RECORDS FROM WILD DUCKS AND OTHER BIRDS BANDED
IN THE SALT LAKE VALLEY, UTAH. U. S. Dept. Agr. Bull. 1145,
16 pp., illus.
(45) ——————
1926. THE MIGRATIONS OF BIRDS. 217 pp., illus. Cambridge, Mass.
(46) Winkenwerder, H. A.
1902. THE MIGRATION OF BIRDS WITH SPECIAL REFERENCE TO NOCTURNAL
FLIGHT. Wis. Nat. Hist. Soc. (n. s.) 2: [177]-263, illus.
[
A ] [
B ] [
C ] [
D ] [
E ] [
F ] [
G ]
[
H ] [
I ] [
J ] [
K ] [
L ] [
M ]
[
N ]
[
O ] [
P ] [
Q ] [
R ] [
S ] [
T ]
[
V ] [
W ] [
X ] [
Y ] [
Z ]
Acanthis linaria, 8.
Accipiter cooperi, 13.
velox, 13.
Aerial obstructions, 57.
Agelaius phoeniceus, 18.
Altitudes of travel, 22.
Anas platyrhynchos, 10.
rubripes, 16.
Ancestral home theory, 5, 6.
Anguilla, 24.
Anoüs stolidus, 24.
An thus spinoletta rubescens, 55.
Antrostomus carolinensis, 44.
Archilochus colubris, 59.
Arctic route, 52.
Arctonetta, 52.
Aristotle, 2, 3.
Arquatella maritima, 34.
Astur atricapillus, 8.
Atlantic coast route, 42.
Atlantic oceanic route, 39.
Baeolophus bicolor, 62.
Baldpate, 47.
Banding studies, 61.
Bartramia longicauda, 11.
Blackbird, 26, 27, 45.
Brewer's, 26.
European, 3.
red-winged, 18, 26, 28, 30.
rusty, 52.
yellow-headed, 26.
Bluebird, 6, 27, 30, 60.
Bluethroat, 15.
Bobolink, 26, 44, 53, 54
Bobwhite, 27, 62, 63.
Bombycilla garrula, 8.
garrula pallidiceps, 56.
Bonasa umbellus, 62.
Brant, 42.
Branta bernicola, 42.
canadensis, 11.
canadensis minima, 48.
Bubo virginianus, 27.
Bullbat, 26.
Bunting, snow, 27.
Buteo borealis, 12.
lagopus, 13.
platypterus, 13.
regalis, 13.
swainsoni, 13.
Calcarius lapponicus, 30, 57.
Calidris canutus, 34.
Canvasback, 42.
Capella delicata, 30.
Cardinal, 27, 62.
Carpodacus mexicanus frontalis, 62.
purpureus, 56.
Casmerodius albus egretta, 55.
Cathartes aura, 27.
Causes of migration, 5.
Chaetura pelagica, 4.
Chamaea fasciata, 62.
Chen caerulescens, 18.
hyperborea, 26.
rossi, 49.
Chickadee, 5, 55.
black-capped, 63.
Carolina, 63.
Chordeiles minor, 6.
minor minor, 52.
Chuck-will's-widow, 44.
Clarke, W. E., 3.
Coccyzus americanus, 44.
erythropthalmus, 44.
Colaptes auratus, 52.
Colinus virginianus, 62.
Compsothlypidae, 26.
Corvus brachyrhynchos, 18.
Coturnix coturnix, 2.
Coues, Elliott, 3.
Cowbird, 24.
Crane, 2, 3, 4, 12, 22, 23.
Creeper, brown, 57.
Crocethia alba, 23.
Crossbill, 5, 26.
Crow, 15, 18, 26.
Clark's, 55.
Cuckoo, black-billed, 44.
yellow-billed, 44.
Cyanocitta cristata, 27.
Cyanosylvia suecica, 15.
Dafila acuta, 18.
Dendroica aestiva, 11.
aestiva aestiva, 52.
caerulescens, 10.
coronata, 52.
palmarum, 29.
palmarum hypochrysea, 29.
palmarum palmarum, 29.
pinus, 29.
striata, 13.
tigrina, 30.
Distances, 27.
Diurnal migration, 11.
Diving bird, 12.
Dolichonyx oryzivorus, 44.
Dove, 3.
mourning, 15.
turtle, 2.
Duck- 70 -, 2, 11, 12, 15, 26, 27, 30, 34, 43, 45, 47, 52.
black, 16, 27, 42, 46.
eider, 52.
scaup, 42.
sea, 48.
Eel, 24.
Egret, 55.
Egretta thula thula, 55.
Euphagus carolinus, 52.
Exhaustion, 59.
Falco peregrinus, 15.
Falcon, peregrine, 15.
Finch, 34.
house, 62.
purple, 56.
rosy, 55.
Fishes, 24.
Flicker, 52.
Flight speed, 13.
Florida caerulea caerulea, 55.
Flycatcher, 11, 12, 15, 25.
Fregata minor, 25.
Gätke, Heinrich, 14.
Geothlypis trichas, 28.
Goose, 2, 3, 11, 12, 15, 17, 27, 43, 45, 46, 47.
blue, 18, 26, 46, 47.
cackling, 48.
Canada, 11, 17, 18, 26, 42, 43, 60.
emperor, 52.
Ross's, 49.
snow, 26.
Goshawk, 8.
Grackle, 30.
bronzed, 26.
Great Lakes route, 45.
Grebe, 27.
Grosbeak, 37, 56.
black-headed, 29.
evening, 56.
pine, 55.
rose-breasted, 26, 37.
Groups, movements of, 8.
Grouse, ruffed, 62.
Gull, 12, 13, 52, 56, 59.
herring, 56.
Ross's, 52.
Hawk, 2, 12, 15, 22, 27, 34.
broad-winged, 13.
Cooper's, 13.
duck, 15.
red-tailed, 12.
rough-legged, 13.
sharp-shinned, 13.
Swainson's, 12, 13.
Hedymeles ludovicianus, 26.
melanocephalus, 29.
Helmitheros vermivorus, 46.
Herodotus, 2.
Heron, 15.
black-crowned night, 55.
little blue, 55.
snowy, 55.
Hesiod, 2.
Hesperiphona vespertina, 56.
Hirundinidae, 3.
Hirundo erythrogaster, 15.
Historical accounts, 3.
Homer, 2.
Hummingbird, ruby-throated, 59.
Hylocichla minima aliciae, 17.
Icterus spurius, 6.
Insects, 24.
Jacksnipe, 30.
Jaeger, parasitic, 52.
Jay, 62.
blue, 27, 28, 62.
Junco, 7, 27, 55.
common, 7.
slate-colored, 30, 52, 57.
Junco hyemalis, 7.
hyemalis hyemalis, 52.
Kingbird, 26, 44.
common, 34.
Kingfisher, belted, 27.
Kinglet, 58.
golden-crowned, 57.
Kite, 3.
Kittiwake, red-legged, 52.
Knot, 34.
Lanius ludovicianus, 29.
Lapwing, 23.
Lark, horned, 15.
meadow, 27, 57.
Larus argentatus smithsonianus, 56.
Leucosticte, 55.
Lobipes lobatus, 23.
Longspur, 27, 57.
Lapland, 30, 57.
Loon, 12.
yellow-billed, 63, 64.
Lophortyx californica, 62.
Loxia curvirostra, 5.
leucoptera, 5.
Mackenzie Valley route, 45.
Magnus, Olaus, 3.
Mallard, 16, 18, 25, 27, 42.
Man-o'-war bird, 25.
Mareca americana, 47.
Martin, purple, 8.
Megaceryle alcyon, 27.
Melanitta, 48.
deglandi, 43.
Melospiza melodia, 26.
Migration, advantages of, 4.
altitudes, 22.
distances, 27.
diurnal, 11.
fall, 29.
historical accounts, 3.
long-distance, 30.
mystery of, 3.
nocturnal, 11.
perils, 56.
problems, 61.
routes, 33.
segregation during- 71 -, 25.
short, 27.
species and groups, 8.
speed, 13.
theories of causes, 5.
undetermined, 27.
vagrant, 55.
variable, 28.
vertical, 55.
weather, 59.
Mimus polyglottos, 63.
Mississippi Valley route
Mniotilta varia, 9.
Mockingbird, 63.
Movements of residents
Murre, 24.
Myiochanes richardsoni, 55
Mystery of migration, 3.
Nannus hiemalis, 27.
Nighthawk, 6, 12, 26, 30, 44, 45.
eastern, 52.
Nocturnal migration, 11.
Nucifraga columbiana, 55.
Nutcracker, 55.
Nuthatch, 5.
white-breasted, 63.
Nyctea nyctea, 8.
Nycticorax nycticorax hoactli, 55.
Nyroca affinis, 42.
americana, 42.
marila, 42.
valisineria, 42.
Oidemia, 48.
Oncorhynchus, 24.
Oporornis formosus, 46.
Orientation, 23.
Oriole, 11.
orchard, 6.
Otus asio, 63.
Ovenbird, 52, 60.
Owl, great horned, 27.
screech, 63.
snowy, 8, 56.
Pacific coast route, 47.
Pacific oceanic route, 49.
Passerculus princeps, 34.
Passerella iliaca fuliginosa, 28.
iliaca iliaca, 28.
Pelican, 3, 12, 22.
Penthestes atricapillus, 5.
carolinensis, 63.
Perils, aerial obstructions, 57.
exhaustion, 59.
migration, 56.
storms, 56.
Petrochelidon albifrons, 13.
Pewee, western wood, 55.
Phalarope, 27.
northern, 23.
Philacte canagica, 52.
Philohela minor, 30.
Photoperiodism theory, 6.
Pigeon, homing, 25.
racing, 24.
Pinicola, 55.
Pintail, 18, 47.
Pipit, 55.
Piranga erythromelas, 26.
ludoviciana, 49.
Plectrophenax nivalis, 27.
Pliny, 3.
Plover, 12.
American golden, 15.
eastern golden, 50.
golden, 39, 40, 50, 51, 59.
Pacific golden, 49, 54.
upland, 11.
Pluvialis dominica, 15.
dominica dominica, 49.
dominica fulva, 49.
Pooecetes gramineus, 30.
Porzana Carolina, 59.
Problems, migration, 61.
Progne subis, 8.
Quail, 2.
California, 62.
migratory, 2.
western, 27.
Querquedula discors, 42.
Quiscalus quiscula, 26.
Rail, 3, 11, 12.
Carolina, 59.
Raven, 15.
Redhead, 42, 47, 65.
Redpoll, 8.
Redstart, 11, 38, 52.
Residents, movements of, 62.
Rhodostethia rosea, 52.
Richmondena cardinalis, 27.
Riparia riparia, 44.
Rissa brevirostris, 52.
Robin, 6, 18, 19, 20, 26, 28, 45, 52, 57.
Routes, 33.
Arctic, 52.
Atlantic coast, 42.
Atlantic oceanic, 39.
evolution of, 52.
Great Lakes, 45.
Mackenzie Valley, 45.
Mississippi Valley, 45.
narrow, 34.
Pacific coast, 47.
Pacific oceanic, 49.
wide, 34.
Rowan, William, 7.
Salmon, 24.
Salpinctes obsoletus, 29.
Sanderling, 23.
Sandpiper, 12, 23.
Bartramian, 11.
purple, 34.
Sapsucker, Williamson's, 55.
Scaup, 42.
Scoter, 48, 49.
white-winged, 43, 49
Segregation, 25.
Seiurus aurocapilla, 52.
Setophaga ruticilla, 11, 38.
Shore bird- 72 -, 6, 11, 26, 27, 31, 34, 39, 40, 45, 46.
Shoveler, 43.
Shrike, 15, 27.
loggerhead, 29.
Sialia sialis, 6.
Sitta canadensis, 5.
carolinensis, 5.
Snipe, 12.
Wilson's, 30.
Snowbird, 7.
Somateria, 52.
Sora, 59.
Sparrow, 11, 25, 26, 27, 45, 57, 58.
chipping, 30.
eastern chipping, 52.
eastern fox, 28.
field, 29.
Harris's, 34, 35, 36.
Ipswich, 34.
song, 26, 27, 28.
sooty fox, 28.
swamp, 57.
tree, 27, 30, 57.
vesper, 30.
white-throated, 57, 63.
Spatula clypeata, 43.
Species, movements of, 8.
Speed, flight, 13.
migration, 13.
Sphyrapicus thyroideus, 55.
Spizella arborea, 27.
passerina, 30.
passerina passerina, 52.
pusilla, 29.
Starling, 3.
Stercorarius parasiticus, 52.
Sterna forsteri, 31.
fuscata, 24.
hirundo, 31.
paradisaea, 31.
Stork, 2, 3, 4, 23.
Storms, 56.
Sturnella, 27.
Swallow, 2, 3, 4, 6, 12, 13, 22, 26, 27, 46.
bank, 44.
barn, 15, 30, 45.
cliff, 13, 22, 30.
hibernating, 3.
Swan, 3.
Swift, 4, 12, 15, 27.
chimney, 4, 26.
Tanager, 45, 49.
scarlet, 26, 36.
western, 49.
Teal, 43.
blue-winged, 42, 43.
Telmatodytes palustris, 27.
Tern, 25.
arctic, 31, 33, 51, 52.
common, 31.
Forster's, 31.
noddy, 24.
sooty, 24.
Theories of migration, 5.
Thrush, 3, 11, 15, 25, 30, 45, 58.
American hermit, 15.
gray-cheeked, 17, 44.
hermit, 57.
wood, 45.
Thryothorus ludovicianus, 27.
Titlark, 55.
Titmouse, 27.
tufted, 62.
Turdus migratorius, 6.
migratorius achrusterus, 28.
migratorius migratorius, 28.
Turtle, 4.
Tyrannus tyrannus, 34.
Vagrant migration, 55.
Vanellus vanellus, 23.
Vermivora chrysoptera, 46.
Vertical migration, 55.
Vireo, 11, 44, 57, 58.
red-eyed, 52.
Vireo olivaceus, 52.
Vulture, 22.
turkey, 27.
Warbler, 11, 13, 27, 45, 57, 58.
black-and-white, 9.
blackpoll, 13, 20, 45, 52.
black-throated blue, 10.
Cape May, 30.
eastern yellow, 52.
golden-winged, 46.
Kentucky, 46.
myrtle, 45, 52.
palm, 29.
pine, 29.
summer, 17.
wood, 25, 26.
worm-eating, 46.
yellow, 11, 17.
Waterfowl, 42.
Waxwing, 26, 27.
Bohemian, 8, 56.
Weather, influence of, 59.
Woodcock, 30.
Woodpecker, 27.
Wren, 12.
Carolina, 27, 62, 63.
long-billed marsh, 27.
rock, 29.
winter, 27, 57.
Wren-tit, 62.
Xanthocephalus xanthocephalus, 26.
Yellowthroat, Maryland, 28.
Zonotrichia albicollis, 63.
querula, 34.
U. S. GOVERNMENT PRINTING OFFICE: 1935
Transcriber Note
Illustrations moved to prevent splitting paragraphs. Minor typos
corrected. Hyphenation was standardized to the most prevalent used
except those in the Bibliography which were left as printed. The
Index entry Seiurus noveboracensis (Northern Waterthrush)
points to page 52 where the Seiurus aurocapilla (Ovenbird) is
described. The species name in the Index is likely a typo and has
been changed.

