The Project Gutenberg eBook of Some possible bearings of genetics on pathology
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms
of the Project Gutenberg License included with this eBook or online
at www.gutenberg.org. If you are not located in the United States,
you will have to check the laws of the country where you are located
before using this eBook.
Title: Some possible bearings of genetics on pathology
Author: Thomas Hunt Morgan
Release date: July 20, 2023 [eBook #71234]
Language: English
Original publication: United States: Press of the New Era Print. Co, 1922
Credits: Bryan Ness and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive/Canadian Libraries)
*** START OF THE PROJECT GUTENBERG EBOOK SOME POSSIBLE BEARINGS OF GENETICS ON PATHOLOGY ***
[1]
SOME POSSIBLE BEARINGS OF
GENETICS ON PATHOLOGY
THOMAS HUNT MORGAN Professor of Experimental Zoology, Columbia University, New York.
Middleton Goldsmith Lecture delivered before the New
York Pathological Society on February 3, 1922.
PRESS OF
THE NEW ERA PRINTING COMPANY
LANCASTER, PA.
1922
SOME POSSIBLE BEARINGS OF
GENETICS ON PATHOLOGY
Thomas Hunt Morgan,
Professor Experimental Zoology, Columbia University.
It has been pointed out in derision that modern genetics deals,
for the most part, with the inheritance of abnormalities and disorders
of various kinds—albinos, brachydactyls, cretins, dwarfs,
freaks, giants, hermaphrodites, imbeciles, Jukes, Kallikaks, lunatics,
morons, polydactyls, runts, simpletons, twins, and Zeros: in
a word, with pathological phenomena in a very broad sense. This
statement, intended as a reflection on genetics, carries with it an
implication that a study dealing with such material cannot be of
first rate importance. Such condemnation will probably be received
by pathologists with the kind of smile it deserves, and I feel
that I am not likely to be called upon here to answer such an indictment.
Nevertheless, I am going to ask your indulgence, for a
moment, since this slightly malicious statement should not be allowed
to pass unchallenged, both because it is inaccurate, and because,
even were it true, the result of such work might still be of
more importance than its critics seem to realize. The source of
this criticism is not without significance. It comes almost always
from those whose interests lie in the field of evolution—in the old-fashioned
use of that word. Now the articles of all evolutionary
platforms include a plank about heredity. This plank is for the
most part an ancient article that has been worn pretty thin. It is
difficult to replace it (or at least it is supposed to be difficult to replace
it) with the new wood of Mendelian genetics. Hence, I
think, originates the criticism referred to.
It is true that the student of Mendelian heredity does not often
trouble himself about the nature of the character that he studies.
He is concerned rather with its mode of inheritance. But the
geneticist knows that opposed to each defect-producing element[2]
in the germ-plasm there is a normal partner of that
element which we call its allelomorph. We can not study the inheritance
of one member of such a pair of genes without at the
same time studying the other. Hence whatever we learn about
those hereditary elements that stand for defects, we learn just as
much about the behavior of the normal partners of those elements.
In a word, heredity is not confined to a study of the shuffling of
those genes that produce abnormal forms, but is equally concerned
with what is going on when normal genes are redistributed. This
method of pitting one gene against the other furnishes the
only kind of information relating to heredity about which we have
precise knowledge.
In man and in domesticated animals we find that individuals
appear occasionally that are defective in one or another respect.
Some of the defects are inherited. Rarely a new one appears that
has not been seen before. But the majority of them are reappearances
of characters that have been carried under the surface
as recessive genes in the germ-plasm. Today we recognize that
each of these modifications, if recessive, has first arisen as a mutational
change in a single gene before it appeared on the surface
as a character by the coming together of two such genes. Mendelism
has furnished some information as to the way in which
these hidden genes may get dispersed in the race. An example
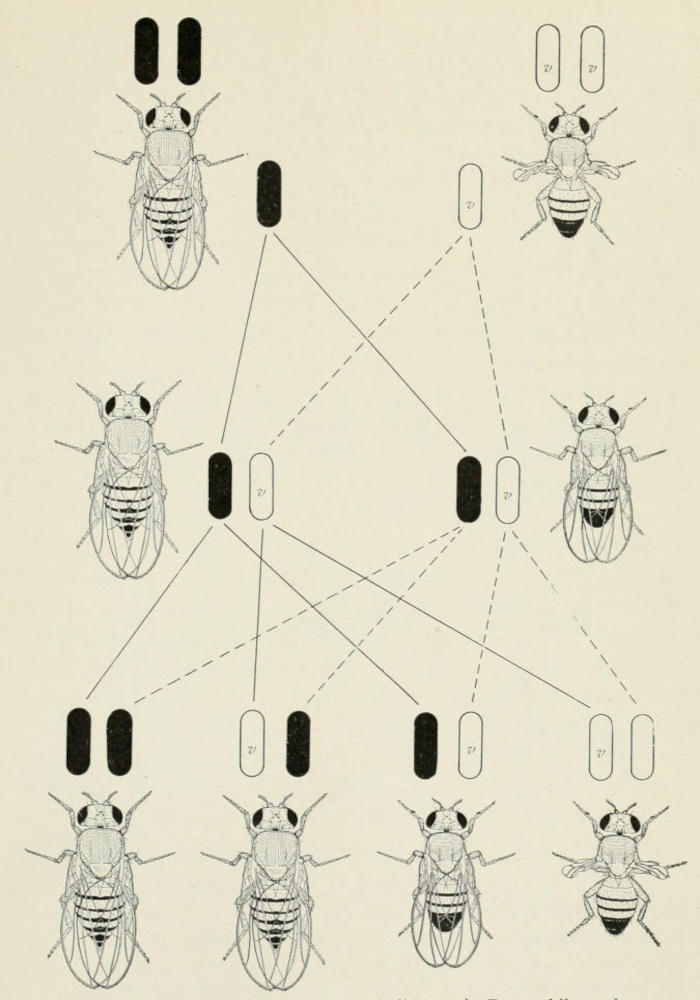
will serve to make this clear, Fig. 1.
If a fly with vestigial wings, a recessive character, is crossed
to a wild fly with long wings, all the offspring (F₁’s) will have
long wings. If these are bred to each other the offspring will be
of two kinds, like their grandparents, in the ratio of three long
winged to one vestigial fly. The extracted vestigials will breed
true to vestigial. The fact that the gene for vestigial has been
carried by long winged F₁ parents has not affected the gene in any
way, for the second generation of vestigials has wings as short
as those of their grandparents.
I have brought forward this case not so much to illustrate
Mendel’s law of segregation as to use the facts for another purpose.
[3]
Fig. 1. Cross between long-winged (wild type) Drosophila melanogaster
and vestigial-winged fly, producing long-winged offspring (F₁), which if bred
to each other give in the next generation 3 long to 1 vestigial. In the middle
of the diagram, the pair of chromosomes that are involved in this cross are
represented. The chromosome with the factor (gene) for long wings is here
black; that for vestigial is open (v).
[4]
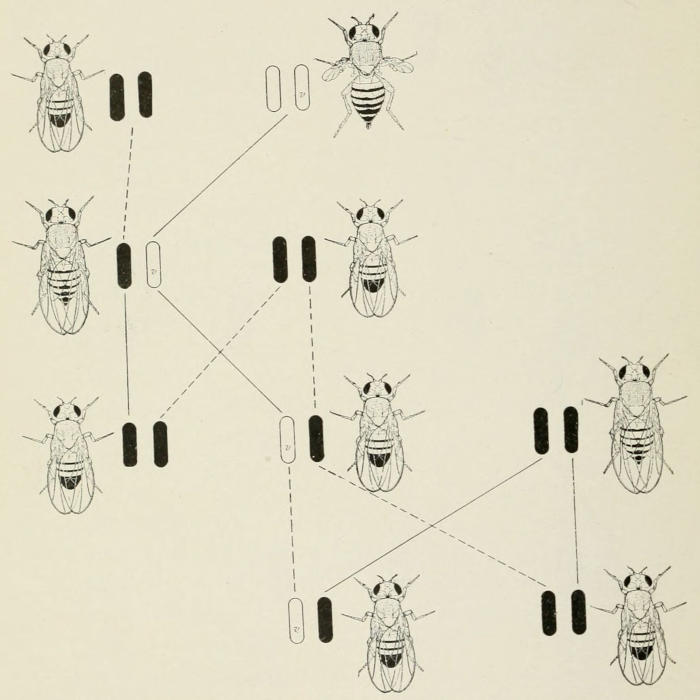
Fig. 2. Cross between long and vestigial wings, giving long in F₁. The
long-winged hybrid F₁ female is then represented as out-bred to a wild-type
male, giving long-winged offspring again—half pure-long and half hybrid-long.
The last are represented as again out-bred to wild type, giving long-winged
offspring again and of the same two genetic kinds as above.
When the vestigial fly was crossed to normal the mutant character
disappeared in the hybrid. If such a hybrid is out-bred to
normal all the offspring are again normal, but half of them carry
the vestigial gene. If these are out-crossed again still only normal
flies appear, Fig. 2. If such out-breeding is continued the vestigial
gene will become widely distributed without ever showing itself
at the surface, so to speak. If, however, at any time two[5]
hybrid flies mate, then a quarter of the offspring will have vestigial
wings. It might seem then that the character had appeared
for the first time in the race, if one did not know its past. In reality
its gene may have been there for some time. Probably many
of the recessive defects and malformations that appear in the
human race—at least those due to hereditary factors—have had
representative genes in the germ-plasm for several generations before
they have appeared on the surface.
We do not know how widespread recessive genes are in the
human germ-plasm. The fact that defective individuals appear
in certain communities may be safely interpreted to mean that individuals
bearing the same gene have at last come together. On
the other hand, the absence of such individuals from the community,
at large, may only mean that the chance of suitable combinations
is small, and does not mean necessarily that the gene in
question is confined to the community within which the defects
have been recorded.
My illustration may give, however, an entirely erroneous idea
as to the chance of a recessive character contaminating the race.
If one can control the matings, so that out-breeding takes place
each time, the result would undoubtedly be like that in our diagram;
but what chance is there for a recessive character, that is
neither beneficial nor injurious, if left to itself, to contaminate
widely the race with its gene? The answer is that for any one
defect there is hardly any chance at all. On the other hand, there
is always a possibility that a defect may become widespread despite
the chances against each in turn. If a recessive character is
selected against each time it appears on the surface, the chance is
extraordinarily small that the gene for such a character could
ever become widespread in a race. If the recessive character is
advantageous, its chance is somewhat better, but still the chance
that it may be lost is very great.
Let us turn for a moment to the inheritance of a Mendelian
dominant character, and to simplify the situation let us first assume
that the character itself is neither advantageous nor disadvantageous.
[6]
It is popularly supposed that if a trait is dominant it will be
expected to spread more widely in the race than will a recessive
character. This is owing largely to a verbal confusion. Colloquially
we think of dominance as meaning spreading. A dominant
nation, for example, is one that is spread widely over the face
of the earth. But a Mendelian dominant should carry no such
implications. A dominant gene, if crossed into a race, will stand
the same chances of being lost as a recessive gene, Fig. 3.
The situation is similar in many ways to the inheritance of
surnames in any human population. A new surname introduced
is likely to disappear after a few generations. There is a bare
chance, however, that it may spread.
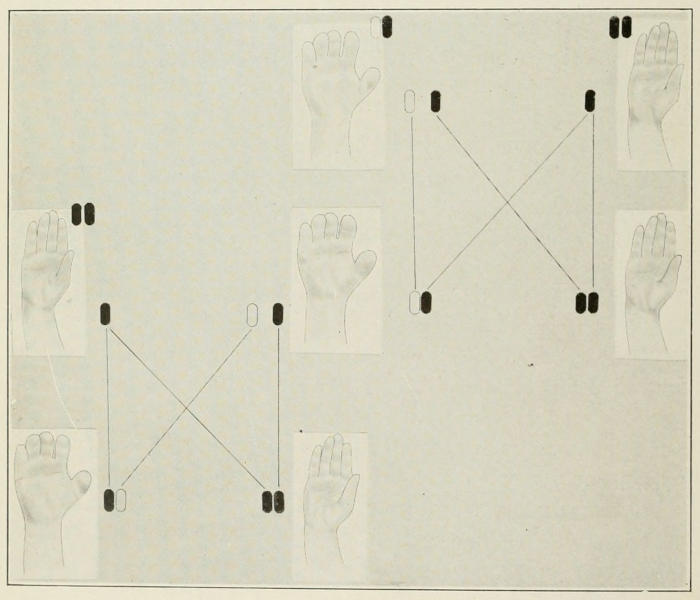
Fig. 3. Mating of short-fingered and normal individual (the short-fingered
character is dominant), giving in F₁ normal and short-fingered individuals
in equal numbers. If the latter is out-bred to normal again, half the
next generation is normal and half short-fingered.
[7]
Of course if a dominant character is advantageous in itself, it
will have a better chance of spreading through the race, than will
an advantageous recessive character, because every hybrid that
carries one dominant gene shows also the character, which increases
the chance that it will propagate and spread the genes.
But, on the other hand, if a dominant character is injurious it will
have a smaller chance of spreading than will an injurious recessive
character; for, the recessive may be carried by the hybrid without
showing itself, and therefore will not place the hybrid individual
at a disadvantage.
An excellent illustration of dominance is that recently published
by Mohr. He has traced, through five generations of a
Norwegian family, the inheritance of a shortened first digit. In
the history of this case there is one record that is extraordinarily
interesting. A child was born that was so completely crippled
that it died in infancy. One parent was short fingered; the other,
a cousin, was probably also short fingered. It is possible that the
child had a double inheritance of this character; it was a pure
dominant. If this is true, then it appears that this character can
survive to maturity only in the hybrid condition. As a matter of
fact, in other animals there are some well-recognized cases of this
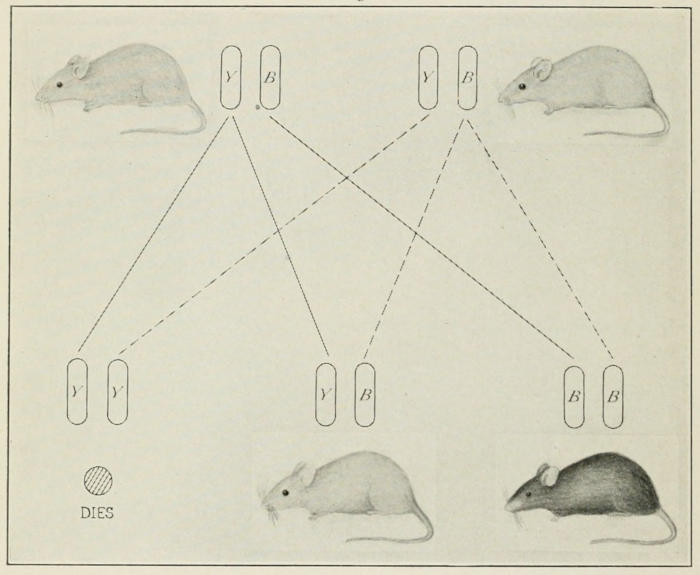
sort. That of the yellow mouse is the best known. Yellow is a
dominant and in double dose it kills; therefore when yellow is
bred to yellow all the pure yellows die. The hybrid yellows and
the pure blacks (in Fig. 4) survive. Here yellow is discriminated
against in the embryo; but, being dominant, it still appears
twice as frequently in each generation as does the alternate
character (here black). In the fly, Drosophila, we have at least
25 dominant lethal characters, but as yet we have no knowledge
as to why such a high percentage of dominant characters should
be lethal when homozygous.
In man there are no certain cases known of lethal dominants
unless some of the short-fingered types come under this heading.
Dominant and recessive characters have been so much discussed
in modern Mendelian literature that it is popularly supposed
that all Mendelian characters must be either dominant or[8]
recessive when bred to the type. This is not the case. The
hybrid (or heterozygote) is frequently intermediate. In fact,
it might be said, almost without exaggeration, that the heterozygote
nearly always shows some traces of its double origin. Sometimes
the hybrid character is nearly midway between the parent
types, sometimes more like one, or like the other. The important
fact, however, is that in the germ cell of such intermediate
hybrids, there is the same clean separation of the parental genes.
In consequence, we find in the second generation the two grandparental
types in pure form and an array of intermediates connecting
them.
Fig. 4. Yellow mouse (YB) crossed to yellow mouse (YB) produces
here black and yellow offspring, in the ratio of 2:1. These yellow are
again hybrid, and if bred to each other give the same result again. Pure yellow
(YY) offspring die at early stage. They constitute one quarter of all
the offspring.
[9]
In connection with the question of spreading of mutant genes
in the race there is another consideration, seldom referred to, that
may occasionally have some weight in accounting for the dispersal
of genes. In some combinations the hybrid may be more
vigorous and more fertile than either parental race. Hence it
may have a better chance of survival than an individual of either
parent stock. It is a difficult question, that we cannot answer at
present, whether a mixed strain has a better chance of survival
than one or another of the strains of which it is made up. The
possibility that some hybrid strains may be better than either pure
strain is enough to put one on his guard against the popular doctrine
of racial purity so-called. Whatever advantages some kinds
of pure races of mankind may have, from a political, religious or
militaristic viewpoint, this should not blind us to the possibility of
the biological advantages that certain mixtures may bring about.
I emphasize the statement that certain mixtures of races may have
a biological advantage. It is equally possible that other combinations
may have a biological disadvantage. We are far from being
able to state at present what combinations are beneficial and what
are biologically injurious. It is an interesting problem, one of
deep significance I think for the future of the human race, but
mixed up as it is at present with difficult social and political questions
it is a problem that only a light-hearted amateur or a politician
is likely to be dogmatic about.
Before we take up the main questions before us this evening,
I must speak of one other form of heredity. In many instances
we have evidence that a character is the product of more than a
single mutant gene. I say “mutant gene” because in fact every
character is no doubt the product of the combined action of many
genes, but in addition to this general relation there are many cases
now known where there are several specific genes whose chief
effect is on one character. Size differences furnish abundant
data of this sort. One of the clearest cases is that of the size of
the ear of corn. Some races of corn have short ears (and cobs),
some long. If two such races are crossed, the hybrid is intermediate
with a considerable range of variation. If the hybrid is
self-fertilized, the progeny in the next generation shows a still
wider range of variation, extending from that of the shorter
grandparent to that of the longer. Both grandparental cobs have
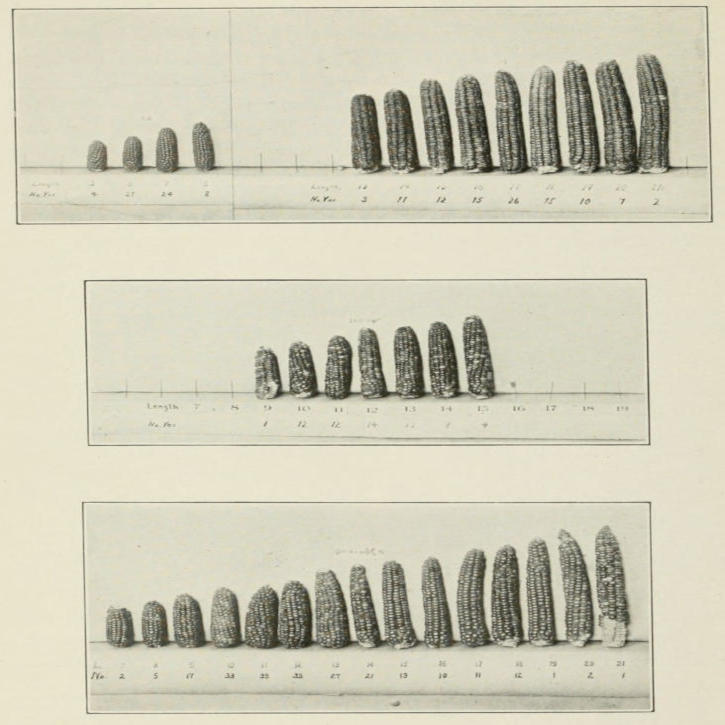
reappeared, but also many intermediate grades, Fig. 5.
[10]
Fig. 5. Cross between long- and short-eared corn. Samples of two original
types shown in upper part of figure, hybrid offspring in the middle of
figure, and samples of 2d generation in the lower part. (After East and
Hays.)
[11]
Such cases were formerly spoken of as blended inheritance.
It was supposed that the materials of the two parents have, as it
were, fused in the offspring and have remained fused. Today we
have a better explanation. It is this. Besides two major factors
that here determine cob length, there are other minor factors,
some of which make the short cob longer, others that make the
long cob shorter. These go over into the first generation hybrids,
and are sorted out in the germ cells of the hybrid. Consequently,
when the F₁’s are inbred, there are all sorts of recombinations of
the minor factors. This explains the greater variability of the
second generation.
It is probable that in most of our domesticated animals, including
man, much of the variability is due to multiple factors, which
makes a study of inheritance in these groups extremely difficult,
especially when, as in the case of man, the number of offspring
from a pair is small, and critical combinations for study can not
be made.
If then it is highly improbable that any particular defective
trait could ever become widely spread in the human germ-plasm,
how does it come about that such defects as feeblemindedness and
insanity are so widespread in the racial inheritance? There are
several possibilities here to keep in mind, but I think we ought not
to pretend that we can give a completely satisfactory account of
the situation.
First. While the chance is heavily against any one defect establishing
itself, there is always the possibility that some one defect
may establish itself. It must be remembered that while many
defective strains may be lost, one would notice only those that had
taken root. It is the presence of these that may give us an exaggerated
idea of the generality of such occurrences.
[12]
Second. If the human germ-plasm is continually mutating to
produce one or another kind of specific defect, this will increase
the chance for any recurrent defect to finally establish itself.
That particular mutations do recur in other animals is now
abundantly established by evidence that comes from several
sources.
Third. There is a growing impression that a good deal of
feeblemindedness and insanity are environmental rather than hereditary
traits; poverty, malnutrition, and especially syphilis are
said to play a considerable rôle in their production. It is unsafe
therefore to conclude that the human germ-plasm is as badly contaminated
as some pessimists seem to think.
If we turn now more directly to special kinds of human inheritance
we shall find a great deal of evidence showing that the
same laws of inheritance that hold for animals and for plants apply
to man. It would be surprising if this were not the case.
On the other hand, when we scrutinize the pedigrees that have
been published to illustrate heredity in man, we shall find many of
them very unsatisfactory in two main respects. (1) The number
of offspring in a family is usually too small to serve as a
sample of the germ-plasm of the parents. (2) Therefore, since
recourse must be had to many families for sufficient data, it is
essential that the diagnosis of the defects of the parents and of the
children is correct. A single mistake may throw the result into
confusion. In cases where the defect is structural, a correct classification
may be possible, but in other cases, especially where psychological
defects are involved, the diagnosis is difficult and the
results, in consequence, less certain. Often the best that we can
do in the case of man is to try to find the simplest Mendelian formula
to which the evidence will fit. If one factor-difference will
not suffice, then two must be tried; if two will not do, then three
must be tried, etc. Now I need hardly point out that we can explain
almost anything if we are allowed enough factors. It is, at
best, a dangerous practice, one to be used only with great caution
and the conclusion stated as provisional and checked in every possible
way.
[13]
I propose now to pass in review some characters in man known
to be inherited, choosing preferably those that come nearest to
the field of pathology, or belonging to it. I shall begin with
comparatively simple cases, about which there can be little doubt,
and pass to more and more difficult situations. I am taking the
risk of reaching an anticlimax, but nevertheless such a procedure
will, I hope, serve our purpose this evening if I can point out
where the evidence is satisfactory and where it is deficient.
My first illustration of inheritance in man may be said to be
a physiological one, mainly because we do not know at present any
structural or chemical basis for the reaction.
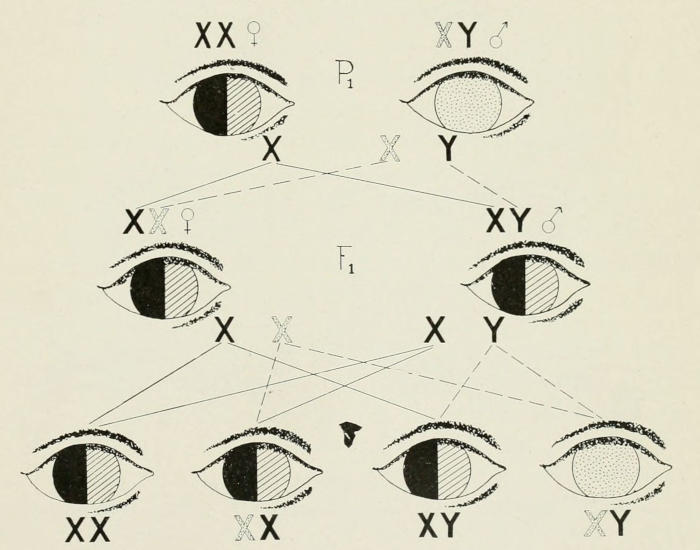
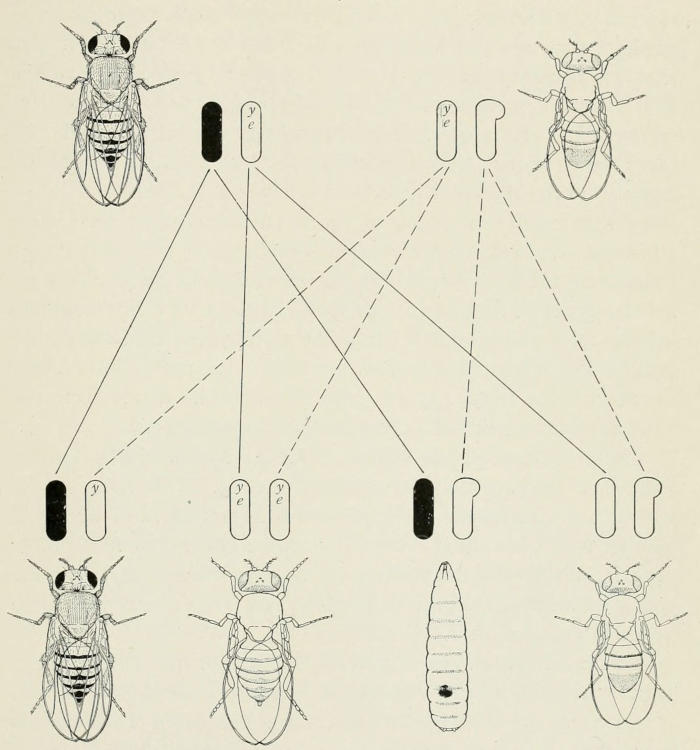
Fig. 6. Inheritance of color blindness of man which is sex-linked (i.e.,
the factor for color blindness is carried in the X-chromosome). This X-chromosome
is stippled in the figure while the X-chromosomes for normal
eyes are represented by black X’s. The color-blind eye is also stippled, and
the normal eye (which distinguishes between red and green) is here represented
by an eye half black and half cross-lined. The 1st generation offspring
(F₁) are normal eyed. In the 2d generation offspring, half the sons
are color blind.
Color-blindness in man is clearly a case of sex-linked inheritance.[14]
It conforms to the general scheme of inheritance in other
animals; in Drosophila, for example, we have about sixty mutant
characters which show this form of inheritance.
A color-blind man married to a normal woman has only normal
daughters and sons; all of the daughters, however, transmit
color-blindness to half of their sons, Fig. 6.
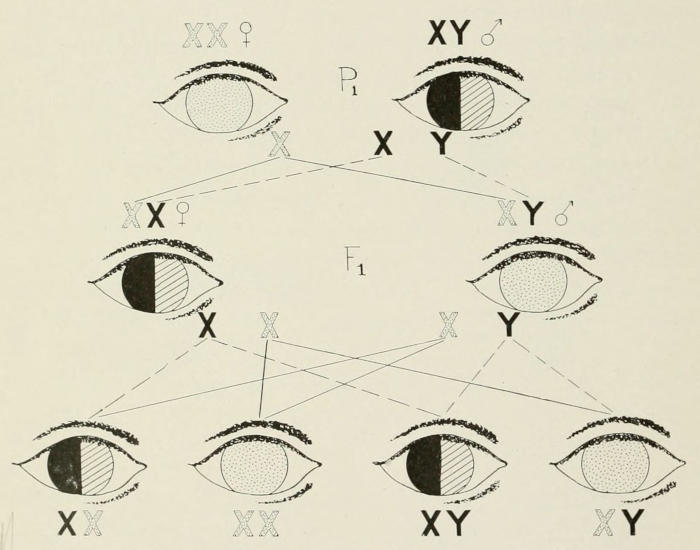
Color-blind women are rare, because they can never arise unless
a color-blind man marries a woman who is color-blind, or
else marries a normal woman who had a color-blind father, or
had a mother heterozygous for color-blindness, Fig. 7.
Fig. 7. Reciprocal of the cross shown in Fig. 6. Here a normal-eyed
male marries a color-blind female, giving all color-blind sons and normal
daughters. When two individuals like these marry, the expectation is for
half of the daughters and half of the sons to be color blind, and half of the
daughters and half of the sons to be normal eyed.
The pedigrees of color-blind families—and they are many—leave
little doubt as to the mode of inheritance of this character.
Accepting this evidence as on the whole satisfactory, there is[15]
still something more to be said. As is well-known there are many
grades of color-blindness. We do not know whether these grades
are due to fluctuating (individual) variations—assuming it to be
due to one gene: or whether there are several genes that differ in
the degree to which they produce the defect. In fact we know
now of a good many cases in other animals where there are several
mutations of the same gene. For instance, in Drosophila
there is a series of ten such multiple allelomorphs for eye colors
that range from pure white to deep wine-red. There is still another
possible interpretation of the different kinds of color-blindness—one
which a priori would seem to be the most probable—namely,
that the differences are due to other modifying genes that
affect the extent to which the character develops.
While in the great majority of cases, the scheme of color-blindness
is that shown by the diagram, we know that occasionally the
machinery may be changed to give a somewhat different result.
It is possible, for example, that a color-blind man married to a
perfectly normal woman may rarely produce a color-blind son. A
few years ago such a result would have appeared to upset the entire
scheme of sex-linked inheritance, today we understand how
such cases may arise through a process that is called non-disjunction,
which is best illustrated by numerous cases well worked out
in Drosophila.
My second illustration has a more obvious chemical basis.
Hemophilia is also sex-linked in inheritance. It is known to be
much more common in men than in women, the explanation for
this is the same as in the other case. In affected individuals the
blood fails to coagulate quickly and the difference in chemical
composition of the blood is, in contrast to normal, the inherited
character.
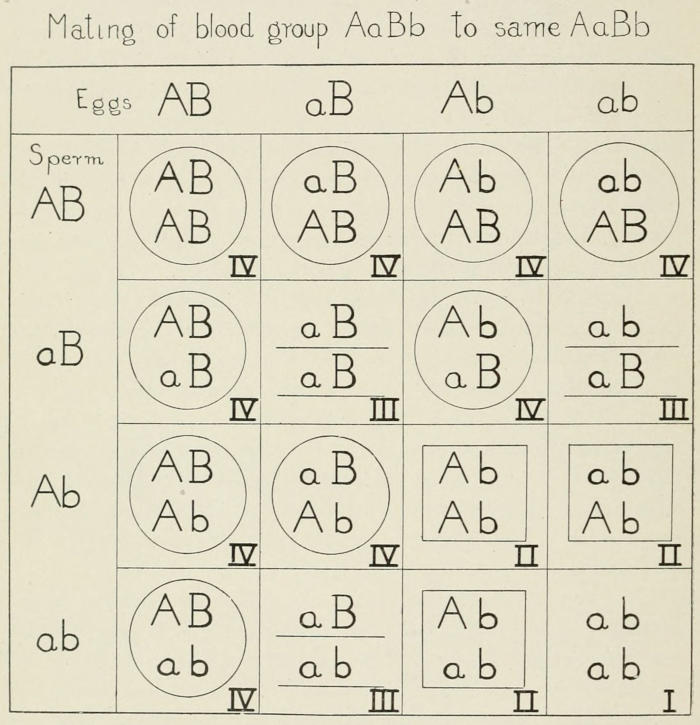
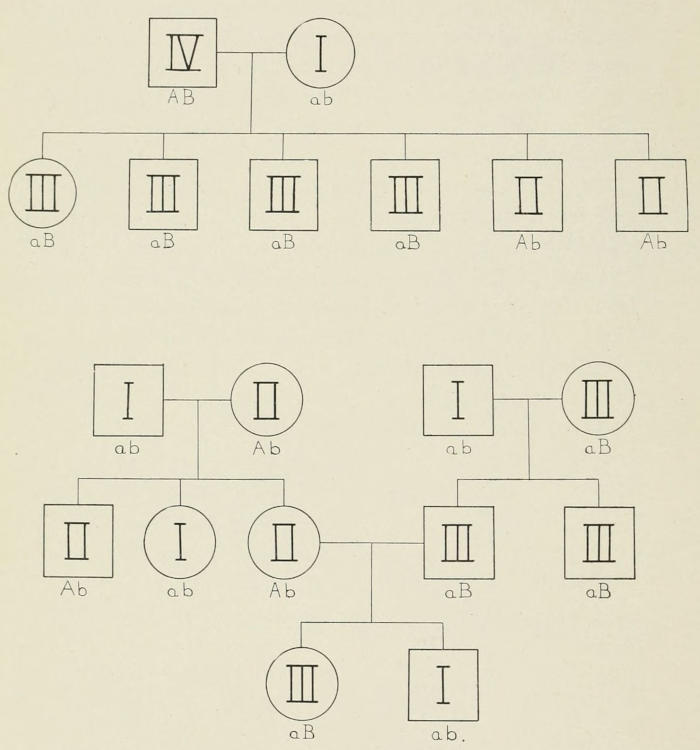
Fig. 8. Representing the kinds of individuals expected when an individual
of the blood group type AaBb marries individual of the same blood
type, namely AaBb. Sixteen kinds of individuals are possible in the ratio of
9:3:3:1. These belong to four blood types, namely, class IV that contains
at least one A and one B; class II that contains at least one A but no B;
class III that contains at least one B but no A; and class I that contains
neither A nor B.
[16]
One of the most remarkable cases of heredity in man is found
in the so-called blood groups. As first definitely shown by Von
Dungern and Hirschfeld in 1910, the inheritance of the four blood
groups conforms to Mendel’s laws. So consistent is this relation
that, as Ottenberg pointed out in 1921, the evidence might be
used in certain cases to determine the parentage of the child.
Since this statement has recently been disputed by Buchanan, from
an entirely wrong interpretation of Mendel’s principles, I should
like to point out that on the Mendelian assumption of two pairs
of factors, all the known results are fully accounted for. If we
represent one pair of genes by A and a and the other pair by B[17]
and b, and if we represent an individual with the genetic constitution
AaBb mating with another individual of like constitution
(AaBb), then each will contain four kinds of germ cells, viz., AB,
Ab, Ba, and ab. The sixteen possible combinations formed if
any sperm may fertilize any egg are shown in Fig. 8.
These sixteen individuals fall into four groups according to
whether they have both A and B, or only A, or only B, or neither
A nor B (i.e., ab) in the proportion of 9AB:3A:3B:1ab. These
four genetic classes correspond to the four recognized blood types
IV, II, III, I, as indicated in the diagram. Now these sixteen
kinds of individuals are found in all populations, so far studied,
although in somewhat different proportions in different “races.”
It is very simple to tell what the kinds of genetic offspring
will be where any one of these sixteen individuals marries any
other one. These possibilities are summarized in the following
statement taken from Ottenberg:
Unions of
I
and
I
give
I
I
II
}
I, II
II
II
}
I
III
}
I, III
III
III
}
Unions of
II
and
III
give
I, II, III, IV.
IV
I
I, II, III, IV.
IV
II
I, II, III, IV.
IV
III
I, II, III, IV.
IV
IV
I, II, III, IV.
Two actual pedigrees, one of them carried through three generations,
will serve to illustrate particular cases, Fig. 9.
From a knowledge of the blood group to which the child belongs
it is possible to predict to what groups its parents may have
belonged, and in certain cases it is possible to state that an individual
of a certain group could not have been the parent of a particular
child.
[18]
Fig. 9. The upper pedigree gives the children from the family in which
types I and IV were the parents. The offspring belong to types II and III
(two of the four possible kinds of offspring).
The lower pedigree represents three generations. The grandparents are
I and II and I and III, respectively, while the parents are II and III.
In the transfusion of blood from one individual to another,
that is sometimes necessary, it is essential that the blood corpuscles
of the donor are not agglutinated by the serum of the recipient.
Thus it is a matter of great importance to select a donor that
does not bring about such a catastrophe. The simple rules are
that individuals belonging to the same blood group (I, II, III,[19]
or IV) do not agglutinate each other’s blood, but the blood corpuscles
of an individual represented by AA or Aa will be precipitated
if the donor contains the agglutinin represented by aa, and
conversely the blood corpuscles of an individual represented by
BB or Bb will be precipitated if the donor contains the agglutinin
represented by bb. Inspection of the diagram will show that
group II (with serum bb) precipitates III and IV, and group III
(with serum aa) precipitates II and IV. Further the serum of
group I (aa bb) precipitates all of the other groups; while the
serum of group IV precipitates none of the others.
My fourth illustration has probably in some cases a glandular
basis, and in this sense has probably also a quantitative chemical
background. Height or stature in man is, in part, an hereditary
trait. It is sometimes said that short is dominant to tall, because
short parents may have both tall and short children, but tall
parents produce only tall children. This is probably an overstatement,
or at least a rather loose generalization. Height may
be due to long legs, or to a long body, or to a long neck or to time
of reaching maturity or to any combination of these; and these
differences may themselves be due to independent factors in inheritance.
The best that we can do with height at present is to
refer it to a multiple factor basis, the actual factors being little
understood.
In addition to these differences in stature, all of which we call
normal differences, there are certain extreme conditions superimposed
on these as a background, in which the endocrine glands
probably play an important rôle. While it may well be that many
of these cases are caused by tumors of one of the glands, more
especially of the pituitary, thyroid, or testis, it is quite possible
that there may be actual inherited differences in the size and activity
of these glands.
So far as I know there are no thoroughly worked out cases
of the inheritance of such differences in man or in mammals, but
in the case of certain races of birds I have been able to show both
by breeding tests and by castration experiments that glandular
differences are inherited according to the Mendelian scheme.
[20]
Fig. 10. Above (A) normal adult hen-feathered Campine cock. Below
(B) castrated cock about one year after operation. The castrated bird has
developed the secondary sexual characters of cock-feathering.
[21]
There is a race of fowls known as Campines in which there
are two kinds of males, hen-feathered males and cock-feathered
males. If the hen-feathered male is castrated, the new feathers
that develop are the long feathers of the cock-feathered male, Fig.
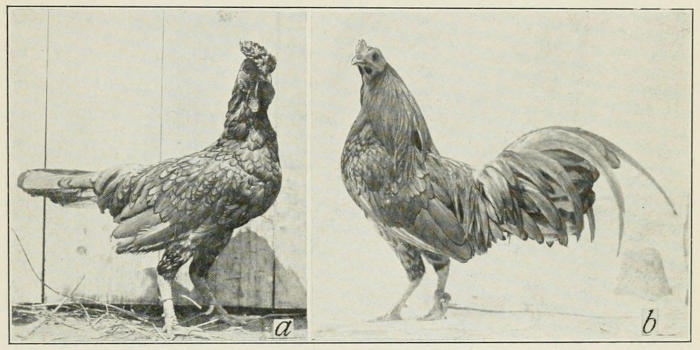
10. In another race of fowls, Sebright bantams, only the hen-feathered
males are known. If these are castrated, the new feathers
that develop are the long feathers characteristic of all other
races of poultry, Fig. 11.
Fig. 11. To left (a) hen-feathered Sebright cock. To right (b) castrated
Sebright cock that has developed characteristic cock-feathering.
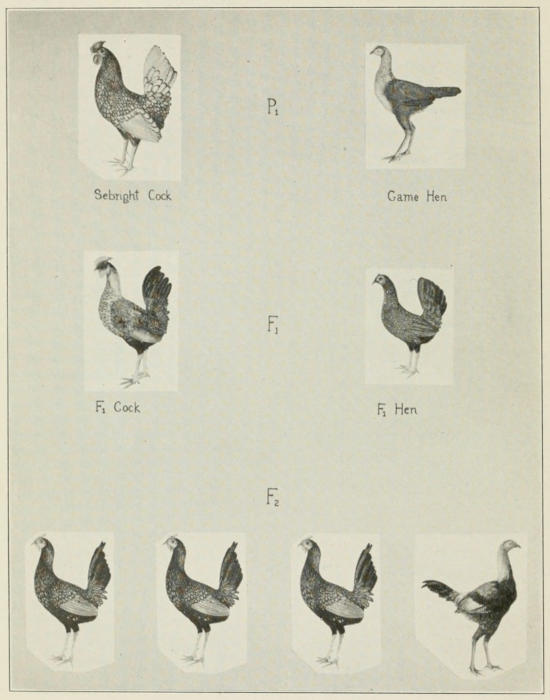
If the Sebright male is out-crossed to a hen of another breed
in which only cock-feathered males occur, it will be found that
all the first generation males are hen-feathered. If these are now
bred to their sisters there are produced, in the second generation,
three hen-feathered males to one cock-feathered male, showing
that the difference between the two races is inherited, Fig. 12.
[22]
Fig. 12. Cross between hen-feathered Sebright cock and black-breasted
game female belonging to a race with cock-feathered males. The offspring
(F₁) are hen-feathered males and normal hens. These inbred give 3 hen-feathered
to 1 cock-feathered son.
[23]
Now in this case we can perhaps go further. An examination
of sections of the testes has shown that in the hen-feathered Sebright
male there are certain kinds of cells, called luteal cells, while
these are absent in the sections of the testes of normal cocks.
These same luteal cells are like those present in the stroma of the
ovary of all female birds. If we assume that they make an internal
secretion that prevents the development of cock-feathering,
both in the normal hen and in hen-feathered cocks, we have a complete
explanation of all the facts. This explanation is made more
probable by the results of removing the ovary of the hen, when, as
Goodale has shown, the spayed hen develops the full male plumage
of her breed. Since the luteal cells are present in the hen and
in the hen-feathered cock, and are absent in the adult cock-feathered
male, it seems not a far-fetched hypothesis to assume that
these cells (or their secretions) are those involved.
The next illustration carries its into a more debatable field.
Many human defects are connected with the nervous system, and
it is interesting to find that many of them are believed to be inherited;
even when no corresponding structural basis in the brain
can be made responsible for the defect.
Feeblemindedness, insanity, and even some types of criminality
have been said to be inherited according to a simple one-factor
Mendelian difference. Owing to the difficulty of diagnosis
it is obvious that the student of genetics would be expected to
approach these problems with the utmost caution. The data, on
which some rather sweeping conclusions have been based, sometimes
show, on closer scrutiny, obvious contradictions. Take,
for example, the case of feeblemindedness which has been represented
as though it differed from the normal (whatever that may
be) by a single Mendelian factor difference. The evidence for
this is far from convincing, and all that can be safely said, I think,
is that there are types of imbecility that may possibly be due to
multiple factors, but until the relation of imbecility to various
disorders of the glands and to syphilis has been thoroughly
studied, even my cautious statement may seem to go too far.
Curiously enough no one has as yet had the temerity to suggest
that some of the high-grade imbecile types—the moron, for example—might
represent an ancestral stage of the human race. If
this were true, intelligence would then be looked upon as an innovation
in the race, that has not yet spread to all of its members.
I am aware that a similar suggestion has been made with respect
to the criminal. Lombroso’s “criminal type” is notorious. The[24]
criminal has been painted as the ancestral brute from which the
more docile human animal has arisen through loss of “wild-type”
genes. I need not state, perhaps, that no one takes such speculations
seriously today from a genetic standpoint.
Immunity and resistance to disease are subjects of great interest
to geneticists as well as to pathologists.
Setting aside, of course, cases where the immunity is due to
some temporary physiological state (little understood at present,
I believe), and also setting aside immunity acquired by recovery
from attack or inoculation, there still remain races that have, as
we say, a constitutional resistance.
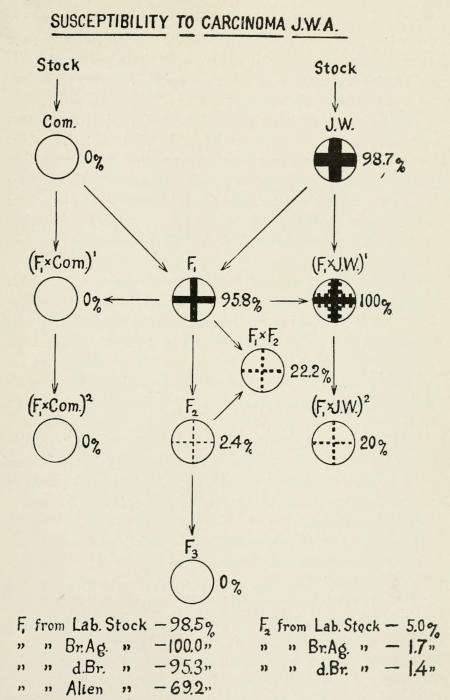
The best ascertained cases in this field are those worked out by
Tyzzer and Tyzzer and Little. A carcinoma that originated in
Japanese waltzing mice grew in practically every individual of
the race when implanted. It failed to grow in “common” mice.
The hybrid mice from these two races were also susceptible in
nearly every case.
When the F₁’s were back-crossed to “common mice” the offspring
were not susceptible. When the F₁’s were back-crossed
to the Japanese waltzer all were susceptible. When the F₁’s were
inbred only about 2.5 per cent. of the offspring were susceptible,
Fig. 13.
These results show at least that there must be more than one,
two or three factor differences between the two races that are
concerned with tumor susceptibility.
Tyzzer and Little suggest in fact that 12 to 14 independently
inherited factors are involved. Larger numbers of tests will be
necessary before it is possible to state how many factors are
needed. A curious feature of the case should not pass unnoticed.
Many or all of the factors for susceptibility must be assumed to
be dominant. It is not generally known, but there is some evidence
that the so-called Japanese waltzer originated from Asiatic
house mice, which according to some writers belong to a distinct
species or at least a distinct variety. The results suggest that
we may be dealing here with species or varietal differences, hence
the large number of factor differences involved. It may be necessary
to work with a simpler situation where fewer factors are
involved; possibly such a case as that of the Jensen tumor will
furnish proper material, but it will be necessary to work with pedigreed
material rather than with “Danish,” “French,” “German,”
or even English breeds of mice.
[25]
Fig. 13. Diagram showing inheritance of immunity to cancer. (From
Tyzzer and Little.)
[26]
In plants also the inheritance of immunity of wheat to rust
has been studied. Biffen’s results with wheat are those best
known. An immune race crossed to a susceptible race gave first
generation plants that were attacked. This means that immunity
is a recessive character. In the next generation there were
64 immune and 194 affected plants (a 1:3 ratio). If the immune
plants are self-fertilized, they yield only immune plants in
later generations.
Nilsson-Ehle and Vavilov think that such simple relations are
rather the exception than the rule. Vavilov found that Persian
wheat, immune to mildew, crossed to different susceptible species
produced offspring that were immune in 13 combinations. In
these cases immunity is dominant.
In the next generation several degrees of resistance were
noted—and a few plants were even more susceptible than their
grandparents.
It is interesting again to note that susceptibility and immunity
are species and variety characters in these cases, but this does
not mean that the differences are not Mendelian. It suggests
however the possibility that several or many factor differences
are often involved.
There is no more interesting field in which genetics and pathology
meet than that of cancer. I realize how careful we on
our side must be in discussing this question with you who are
experts, nevertheless there are certain aspects of the problems
of cancer from the genetic side that I may be allowed briefly to
mention—not, however, without some misgivings.
Suppose all men over seventy-five died of arteriosclerosis.
Could one say that hardening of the arteries is inherited? I
think that it would be proper to use the word heredity to include
such a case, but we would not know how it was inherited unless[27]
there existed another race of men who never died of the malady,
and suitable matings were made between the two races.
Suppose again that all old men died of pneumonia. Could we
say that susceptibility to pneumonia, after eighty, is inherited?
Again, yes! But again we could get no information as to the
way in which this susceptibility is inherited without crossing to
an immune race.
Now suppose there are strains of mice all of which die of cancer
after their first year. Could we say that in them cancer is inherited?
The answer would depend in part on what connotations
the word inherit carries with it, for, either susceptibility
might be meant, or the “spontaneous” development of cancer
might be meant. The latter interpretation is, I think, generally
implied, which carries with it two further implications. First
implication, viz., that when a certain age is reached, a certain inherited
complex leads to the development of cancer in one or
more regions of the body. Here some such process as that of
the hardening of the arteries seems to be vaguely implied.
Second implication, viz., that a change in method of growth (a
release from the ordinary restraining influences) suddenly occurs,
beginning in a single cell of some particular tissue. Stated
in this second way, the appearance of spontaneous cancer suggests
at once a comparison with the mutation process that is known to
occur in somatic cells as well as in germ cells.
Now if the first interpretation is to be placed on the word
heredity, when applied to cancer, there is nothing more to be said,
except that the only way such a situation can be studied as a genetic
problem is to out-cross the strain of cancer mice in question
to another that never develops spontaneous cancer. But if the
second interpretation is implied, then the whole situation is put in
a very different light. Let us examine this a little more closely.
Suppose, as a theoretical possibility, that spontaneous cancer
is due to a recurrent somatic mutation of a specific gene to a dominant
one that leads to cancer. Then the proportion of individuals
that develop spontaneous cancer in such a strain will depend
on the frequency of mutation of this specific gene. Consequently,[28]
if such a strain is out-crossed to another race (that introduces
the allelomorph of the postulated gene), the number of
F₁ offspring that develop the specific cancer would be half as numerous
as in the original cancer strain (since the gene in question
occurs only half as many times as in the original complex). In
the F₂ generation the frequency for the extracted double dominant
will be that of the original strain, that of the F₂ heterozygotes
will be the same as that of the F₁, and the extracted double
recessive class will not develop cancer at all. Now, if it is not
possible to distinguish between these different F₂ classes by inspection,
the difficulty of finding out how cancer is “inherited”
would be very great. In such an imaginary situation, the ratio of
cancer-developing mice may not appear to correspond to any of
the known Mendelian ratios, because superimposed on the genetic
situation there would be added results depending on the frequency
of mutation when a specific gene is present.
Other complicating conditions will also suggest themselves to
any one familiar with genetic and mutation processes; for, the possibility
that the mutation itself is more or less likely to occur in
one or another genetic complex must be reckoned with, as well as
the likelihood of the mutation showing itself or developing in
any tissue or only in cells of specific tissues, etc.
I am far from wishing to suggest that spontaneous cancer is
a mutational process, despite certain rather obvious resemblances
to mutational effects in plants and animals, but I should like to
insist that the appearance of spontaneous cancer is in its nature
so peculiar that one can not afford to ignore such a possibility in
any discussion as to whether spontaneous cancer is or is not “inherited.”
There are several cases of inheritance of tumors in our Drosophila
material. Here I am on safer ground. One of them,
discovered by Dr. Bridges, worked out by Dr. Stark, I should
like to speak about, because it shows how linkage of characters
can be used in the study of heredity of a character and conversely
in its elimination. In a certain culture one fourth of the maggots
develop one or more black masses of pigment in the body;[29]
such maggots always die. They are always males. Consequently
there are twice as many daughters as sons in such a
strain. The gene is carried by the X-chromosome and its inheritance
is like that of all sex-linked characters as shown in
Fig. 14.
Fig. 14. Diagram showing inheritance of a sex-linked recessive lethal
(“tumor”) factor in Drosophila melanogaster. Here, in the center of the
diagram, the sex-chromosome that carries the lethal factor is represented by
the black rod. A female with the tumor-factor, normal wings and red eyes,
in one of her sex-chromosomes and with the factors for yellow wings and eosin
eyes in the other is bred in each generation to a male with yellow wings and
eosin eyes. In the next generation there are twice as many daughters as sons,
since all the sons that carry the black chromosome die. The half of the
daughters (i.e., those not yellow eosin) that carry the black chromosome repeat
the same history. The linkage of yellow and eosin enables one to pick
out in each generation those daughters that carry the tumor-factor.
[30]
All males that get their single X with this tumor-gene will die;
therefore, since no adult males carry it, normal males must be
used for mating in each generation. They are mated to females
that are heterozygous for the chromosome carrying the tumor
genes. Such matings as I have said always give two daughters
to one son. But since half the daughters are normal and half
carry the gene for tumor it is desirable to be able to pick out the
latter from the stock. Therefore we have made use of a trick we
call “marking the chromosome,” which means that we use a male
whose sex chromosome carries a known gene near the tumor
locus. By using this type of male in successive generations we
get two types of daughters: one type like their surviving brothers
in eye color that do not carry the tumor-gene and the other
daughter with normal eyes that carries it. We use only the latter
to continue the stock, but we could eliminate the tumor from the
stock at once by using the other kind of daughters.
Curiously enough the tumor no longer appears in the inbred
stock but reappears again on out-breeding. Nevertheless the
sex-ratio in the inbred stock continues as before, and since the
missing males are those with red eyes we know that the tumor-gene
is still present and doing its deadly work—only now the
young male larvæ die even before they reach the age at which the
tumor is due to appear.
So far I have spoken of heredity as though that term had become
synonymous with Mendelian heredity. Those of as who
are at work on Mendelian inheritance are often criticized as
too narrow. It is said that we do not recognize that any other
kind of inheritance takes place. I do not think the criticism is
quite fair, because, in the first place, the very great number of
variations studied has been shown to conform to the Mendelian
principles or at least to be capable of such interpretation. There
are, however, a few exceptional cases. In certain albino plants
it has been shown that the inheritance of albinism can be traced
to the behavior of the chlorophyll bodies in the cytoplasm. The[31]
chlorophyll bodies are known to divide and to be distributed to
the two daughter cells at each division independently of the nuclear
division and of the maturation process in the egg.
Why, then, it is asked, may not there be present in the cytoplasm
of the cell other self-perpetuating bodies that are responsible
for certain kinds of inheritance? Why not go further and
ask, why, since the cytoplasm appears to be handed down from
cell to cell, may it not furnish also a different medium for inheritance
of characters? Theoretically such an argument is logical.
No student of Mendelism would I think deny such a possibility.
But, as a matter of fact, it is not going too far to say
that, at present, there is little evidence that such inheritance takes
place, except in a few special cases, like that of the chlorophyll
bodies. It is safe, I think, to say that if cytoplasmic inheritance
played any important rôle in heredity in the higher animals and
plants, we should expect, by now, to have found many cases of it.
None are known to us.
Whether Mendel’s laws of heredity apply to unicellular animals,
to bacteria and to similar types, in which the mechanism
for this type of inheritance has not been shown to exist, can not
be affirmed or denied from the evidence at hand.
There are at present three outstanding cases in the higher animals,
in which an induced variation is said to be inherited afterwards.
These cases are of great interest to pathology. We can
not afford to pass them over. First, there is Brown-Sequard’s
claim that injuries to the nerve cord or to the cervical or sciatic
nerves of guinea pigs produce effects that are transmitted.
Second, there are the cases of the inherited effects caused by
alcohol in guinea pigs discovered by Stockard.
Third, there is Guyer’s evidence that an effect on the eye,
caused by foreign serum, is transmitted.
Brown-Sequard’s experiments have been repeated several
times; almost always with negative results. Today his claims
are practically forgotten.
Stockard’s results with guinea pigs, unlike those of Brown-Sequard,
have been done under carefully controlled conditions.[32]
He has guarded against abnormalities in his stock by using pedigreed
material. The malformations that reappear in successive
generations are general rather than specific. Such organs as the
eye are those hardest hit, but this is supposed to be rather a by-product
of the general debility of the individual. Stockard
points out that the alcohol has affected the germ cells, and it is
through these that the effects are transmitted. Now if one or
more genes had been permanently changed we should expect to
have evidence of Mendelian inheritance. The results do not show
convincingly that the inheritance is not Mendelian, but it does
not appear to be so. There is another possibility. Recent results
have shown that rarely entire blocks of genes—pieces of
the chromosomes—may be duplicated (owing to imperfect separation)
or pieces may be lost. Here the effects on the organism
are more far-reaching than when a single gene is changed. It
remains to be discovered whether, in some such way as this,
Stockard’s remarkable results may be brought into line.
Guyer injected the crushed lens of rabbits into fowls. From
the blood of the fowl he obtained serum that was injected into
pregnant rabbits. The offspring of these rabbits whether male
or female often had defective eyes and lenses. The defect was
even transmitted to later generations. Here also the germ cells
of the embryo may be changed by serum that at the same time
affects the development of the eyes of the embryo in utero.
If this is the case we should expect, as Guyer pointed out,
that the germ cells of the pregnant mother (into which the serum
was injected) would also show effects. It should have been a
simple matter to show this by a proper test. The test that Guyer
made, namely by out-breeding the mother and finding no defective
F₁ young, was quite inadequate if, as appears to be the case,
the character is a recessive.
It is important to keep clearly in mind that there are two distinct
questions involved in these three cases. Genetics has to
deal with only one of them. There is first the question of the
action of environment on the germ cells. Genetics has nothing
to do with this question. There is then to be determined whether,[33]
if variations may be induced in these ways, they fall into one or
another of the Mendelian moulds. This is for the geneticist to
determine, but he finds himself in a curious predicament, for it
can not be claimed that any of these three cases have been shown
to give a direct Mendelian result—but neither can it be denied
that they may possibly come under the scheme, or some modification
of it. There we must leave the matter at present.
If I have appeared at times overcritical concerning the application
of genetics to pathology, it is not because I do not sympathize
with the attempts that have been made to apply genetics to pathology.
I realize, of course, that from the nature of the case
much of this work is pioneer work, where rough and ready
methods have often to be resorted to. So long as this is kept in
view, no harm can be done in attempting to find how far Mendel’s
principles can apply to heredity in man. But I want to
enter a protest against the danger of premature conclusions
drawn from insufficient evidence. In our enthusiasm in applying
Mendel’s laws, we should be careful not to compromise them.