Tafel X und XI
Bei der Bearbeitung der von den Hrn. Sarasin in Celébes gesammelten Säugethiere beobachtete Hr. A. B. Meyer (s. oben S. 17) im Gesichte der von ihm aufgestellten neuen Fledermausart Nyctinomus sarasinorum Haare von sehr eigenthümlicher, einem Senflöffelchen vergleichbarer Form, die, wie
eine sogleich vorgenommene Prüfung des im Museum vorhandenen Materials ergab, bei
den Arten der Gruppe Molossi ziemlich allgemein verbreitet sind.
Die anfänglich gehegte Vermuthung, dass diese Haare überhaupt noch nicht bekannt geworden
seien, bestätigte sich bei genauerer Durchforschung der Literatur allerdings nicht.
Abgesehen von einigen Angaben älterer Autoren, aus denen hervorgeht, dass sie etwas
von der Form der Haare erkannt haben, und die im Verlaufe der Darstellung einzeln
Berücksichtigung finden werden, erwähnt Dobson1 1876 bei der Beschreibung des Molossus abrasus Temm. diese Haare als „short spoon-shaped hairs“ ohne jeden anderen erläuternden Zusatz. Da er die Beobachtung nicht weiter verfolgt
hat und die kurze Notiz auch im Texte seines umfangreichen Werkes ganz versteckt ist,
so ist es erklärlich, dass sie in die Lehrbücher und andere zusammenfassende Werke,
soweit meine Kenntniss reicht, keinen Eingang gefunden hat2.
[33]
Es wird daher gerechtfertigt sein, diese eigenthümliche Form von Säugethierhaaren
durch eingehendere, von Abbildungen begleitete Schilderung weiteren Kreisen bekannt
zu machen.
Ich bemerke vorweg, dass ich ausserhalb der Gruppe Molossi solche Haare nicht beobachtet habe. Nachdem ich mich hier mit ihnen vertraut gemacht,
wurde das gesammte in Spiritus aufbewahrte Chiropterenmaterial des Dresdener Museums,
worin alle Abtheilungen der Ordnung vertreten sind, ohne Erfolg durchgesehen.
Von den Molossi standen mir zur Verfügung Cheiromeles torquatus Horsf., vier Arten von Molossus und vierzehn von Nyctinomus, die theils dem Bestande des Dresdener Museums angehören, theils von Herrn Prof.
Lampert, dem Director des Königlichen Naturalien-Cabinets in Stuttgart, für die Zwecke dieser
Untersuchung in entgegenkommendster Weise hergeliehen waren.
Die Untersuchung führte ich derart aus, dass jedes einzelne Exemplar zunächst unter
dem Zeiss’schen binocularen Präparirmicroscope, das sich für diesen Zweck als sehr
geeignet erwies, auf das Vorkommen und die topographische Anordnung der fraglichen
Haare genau geprüft wurde. Sodann wurden von den Haaren einer jeden Art eine Anzahl
microscopischer Präparate angefertigt.
Um Wiederholungen möglichst zu vermeiden, werde ich bei der Darstellung der Befunde
dagegen so vorgehen, dass ich zuerst die allgemeinen Eigenschaften dieser Haare schildere,
wie sie sich aus der Vergleichung der Einzelbeobachtungen ergeben, und dann erst das
besondere Verhalten bei den einzelnen Arten beschreibe.
Am Haarkleide der Säugethiere kann man allgemein unterscheiden das eigentliche Körperhaar, das die gleichmässige Bedeckung für den grössten Theil des Leibes bildet, und die
von ihm nach Form und Grösse mehr oder weniger abweichenden, besonderen Zwecken angepassten Haare, die sich nur an bestimmten Stellen, namentlich, als sogenannte Spürhaare, im Gesichte,
vorfinden. Die Haare, die den eigentlichen Gegenstand dieser Mittheilung bilden, gehören
zwar in die zweite Gruppe, doch wird es ihre Charakterisirung erleichtern, wenn wir
vorher einen Blick auch auf das gewöhnliche Körperhaar werfen.
Durch seine Beschaffenheit, die schon öfter die Aufmerksamkeit der Forscher3 auf sich gezogen hat, nehmen die Fledermäuse unter den Säugethieren eine Sonderstellung
ein. „Die Haare der Fledermäuse sind abgeplattet und deutlich charakterisirt durch
den Mangel an Mark im grössten Theile des Schaftes, durch die stark vorspringenden
Cuticularschuppen und besonders durch die eigenthümliche Spiraltour, in welcher die
Schuppen gestellt sind“ (Waldeyer4). Die eigenthümliche Ausbildung der Cuticula ist zwar nicht bei allen Chiropteren
gleich ausgeprägt, namentlich unter den Frugivoren tritt sie zurück (Koch5, Marchi6) und andererseits findet sie sich gelegentlich in ähnlicher Weise auch bei Angehörigen
anderer Säugethiergruppen (vgl. Gegenbaur, Vergl. Anat. I. 1898, p. 147, Fig. 59), aber als Ordnungscharakter bleibt sie doch
für die Chiropteren bezeichnend. Einen guten Überblick über die dadurch bedingten,
oft sehr zierlichen und complicirten Formen der Körperhaare in den verschiedenen Abtheilungen
der Ordnung gewährt die Arbeit von Marchi7.
Von der uns beschäftigenden Unterfamilie Molossi besitzt das Haar der zahlreichen Arten der beiden Gattungen Molossus und Nyctinomus durchweg den typischen Fledermauscharakter, abweichend verhält sich dagegen die dritte
Gattung Cheiromeles mit der einzigen Art torquatus.
Die Arten der beiden erstgenannten Gattungen stimmen wieder unter einander im allgemeinen
soweit überein, dass es für die Zwecke der vorliegenden Abhandlung genügt, das Verhalten
einer Art zu berücksichtigen, und zwar mag als Beispiel die neue Species Nyctinomus astrolabiensis A. B. M. dienen.
[34]
In einer von der Brust genommenen Probe (Fig. 30, Taf. X) erscheinen die einzelnen Haare im ganzen fein, wenn auch in den einzelnen Abschnitten
ihrer Länge verschieden dick, ziemlich kurz (c 4 mm), marklos und jederseits am Rand
einer feinen Säge ähnlich gezähnt. Bei passender Einstellung des Microscops erkennt
man indessen, dass sich von der anscheinend freien Spitze einer jeden Zacke über die
Oberfläche des Haares eine feine, selbst wieder gezackte Linie herüberzieht. Die Deutung
der Beobachtung ist leicht. Die Cuticularschuppen sind in trichter- oder manschettenartigen
Segmenten angeordnet, von denen jedes höher gelegene mit seiner verengerten Basis
in den erweiterten Endabschnitt des vorhergehenden hineingesteckt ist. Stellt man
daher genau auf den Rand des Haares ein, sodass hier der trennende Raum zwischen zwei
Trichterstücken scharf hervortritt, die Oberfläche des Haares aber undeutlich wird,
so muss der Eindruck entstehen, als sei der Schaft mit zwei gegenüberstehenden Zeilen
stark vorspringender isolirter Schuppen besetzt. Die trichterförmigen Segmente sind
zur Längsaxe des Haares meist etwas schräg abgestutzt, so dass die Zacken an der einen
Seite des Haares gegen die an der andern ein wenig verschoben erscheinen. Die Schuppen
sind also hier nicht in einer eigentlichen fortlaufenden Spirallinie, sondern in etwas
schräg am Schafte stehenden Wirteln angeordnet. Nach Dobson8 gilt dies ausser für die Molossi allgemein für die Emballonuridae und Phyllostomidae, während für die Vespertilionidae, Nycteridae und Rhinolophidae ein anderer Typus — einfache Spirale mit dachziegelartiger Deckung der einzelnen
Schuppen — bezeichnend wäre.
Das einzelne Haar zeigt nicht durch seine ganze Länge den gleichen Bau. Innerhalb
der Haut ist der Schaft glatt, auch der freie Theil zeigt am Grund eine verschieden
lange Strecke weit keine oder undeutliche und wenige, darauf im grössten Theile seiner
Länge sehr deutliche und regelmässige Zacken. Im letzten Drittel etwa der Schaftlänge
werden die Segmente der Cuticula viel kürzer, und die freien Enden der Schuppen stehen
weniger stark ab, so dass dieser Theil des Haares viel glatter als der untere erscheint.
Die Dicke des Haares ist im unteren, stark gezähnten Theil am geringsten (circa 0,009
mm) und ziemlich gleichmässig, doch entsteht, da das Haar im ganzen etwas abgeplattet
ist, infolge Drehung des Schaftes öfter der Anschein, als nehme die Dicke wiederholt
zu und ab. Im oberen glatteren Abschnitte wächst sie aber in der That ziemlich rasch
etwa auf das Doppelte an, worauf die Zuspitzung dieses Theils zum freien Ende des
Haares ohne Änderung seiner Structur ganz allmählich erfolgt. Kolenati, der wohl zuerst auf diese Verschiedenheiten der Structur des einzelnen Haares aufmerksam
gemacht und sie als regelmässig wiederkehrend bei vielen Arten nachgewiesen hat, bezeichnet
den untern Abschnitt, der die für die Art charakteristische Ausbildung am deutlichsten
zeigt, als das „charakteristische Haardrittel“9. Bei den hier behandelten Haaren beträgt dieser Abschnitt aber immer weit mehr, über
die Hälfte bis 3–4 Fünftel, und ungefähr dasselbe Verhältniss liegt übrigens auch
in der von Kolenati10 gegebenen Abbildung des Haares von Rhinolophus hippocrepis (Hermann) [gleich Rhin. hipposideros (Bchst.)] vor.
Am einzelnen Haar erscheint unter dem Microscope der untere „charakteristische“ Abschnitt
fast farblos, im oberen findet sich körniges Pigment gewöhnlich in segmentweiser,
durch farbstoffreie Strecken unterbrochener Anhäufung.
Das Haarkleid ist am ganzen Körper gleichartig, Haare vom Rücken bieten die gleichen
Verhältnisse wie die vom Bauch. Ein Unterschied von Stichel- und Wollhaar ist nicht
zu bemerken, ebensowenig eine Anordnung in Gruppen.
Die functionelle Bedeutung der Eigenthümlichkeiten des Fledermaushaares hat Kolenati wohl richtig beurtheilt: „Da alle Haare gleich sind, somit das die Körperwärme zusammenhaltende
Wollhaar nicht vorhanden ist, so muss das Haar an der Basis, hiermit in der Nähe des
Körpers die erwärmte Luftschichte abschliessen und sich vermöge der an jener Stelle
vorragenden Spiralen stauen, in der breiteren Stelle des Haares dagegen dicht aneinander
anschliessen“11.
[35]
Gewissermaassen einen negativen Beweis für die Richtigkeit dieser Anschauung bietet
der oben erwähnte Cheiromeles torquatus Horsf., der sich durch die Beschaffenheit seines Körperhaares wie in vielen anderen Beziehungen
von den Molossus- und Nyctinomus-Arten wesentlich unterscheidet.
Das Thier besitzt eine sehr dicke und derbe Haut, die zunächst völlig nackt erscheint,
erst bei genauerer Prüfung erkennt man, dass sie grössten Theils von einem spärlichen,
kurzen Flaum überzogen ist. Untersucht man eine Probe des letzteren, etwa von der
Brust (Fig. 21, Taf. X), so findet man, dass er aus längeren und kürzeren Haaren besteht, die im übrigen
gleichen Bau zeigen. Es scheinen um ein grösseres Haar von circa 1,2 mm Länge einige
kürzere und einige ganz kleine herumzustehen. Eine genauere Prüfung, ob hier eine
gesetzmässige Gruppirung im Sinne de Meijeres12 vorliegt, wurde nicht vorgenommen. Diese Haare sind spindelförmig, mit der dicksten
Stelle nahe dem Grunde des freien Schafttheiles, ohne wahrnehmbares Mark und besitzen
sehr wenig hervortretende Cuticularschuppen (Fig. 21 e), die das Haar bei geringer
Vergrösserung nur fein quergestreift erscheinen lassen. — Das eigentümliche Verhalten
des Cheiromeles-Haares erklärt sich offenbar daraus, dass bei diesem Thiere dem mächtig entwickelten
Integumente die Aufgabe des Wärmeschutzes zugefallen ist, was zu einer Reduction des
nun überflüssigen Haarkleides geführt hat, die sich nicht nur auf dessen Menge im
ganzen, sondern auch auf die dem Zwecke des Wärmeschutzes angepasste feinere Structur
des einzelnen Haares erstreckt hat.
Bei den Molossus- und Nyctinomus-Arten ändert sich die Beschaffenheit des Haarkleides, wenn wir von der Flughaut und
dem Schwanze, die zum grössten Theile nackt sind, absehen, an zwei Stellen in schon
für die Betrachtung mit freiem Auge merkbarer Weise, am Gesicht und an den Füssen. Eine kurze Schilderung der äusseren Formverhältnisse dieser Theile wird das Verständniss
der folgenden Ausführungen, in denen öfter auf die topographischen Beziehungen Rücksicht
zu nehmen ist, erleichtern und möge daher hier zunächst Platz finden.
Dem Kopf ist in den Gattungen Molossus und Nyctinomus durchgehends der Besitz sehr breiter Ohren eigenthümlich, die, seitlich weit herabreichend
und in der Medianlinie sich dicht berührend oder häufig sogar verwachsend, für das
Gesicht nach oben und rückwärts eine scharfe Begrenzung bilden (vgl. die Figuren auf
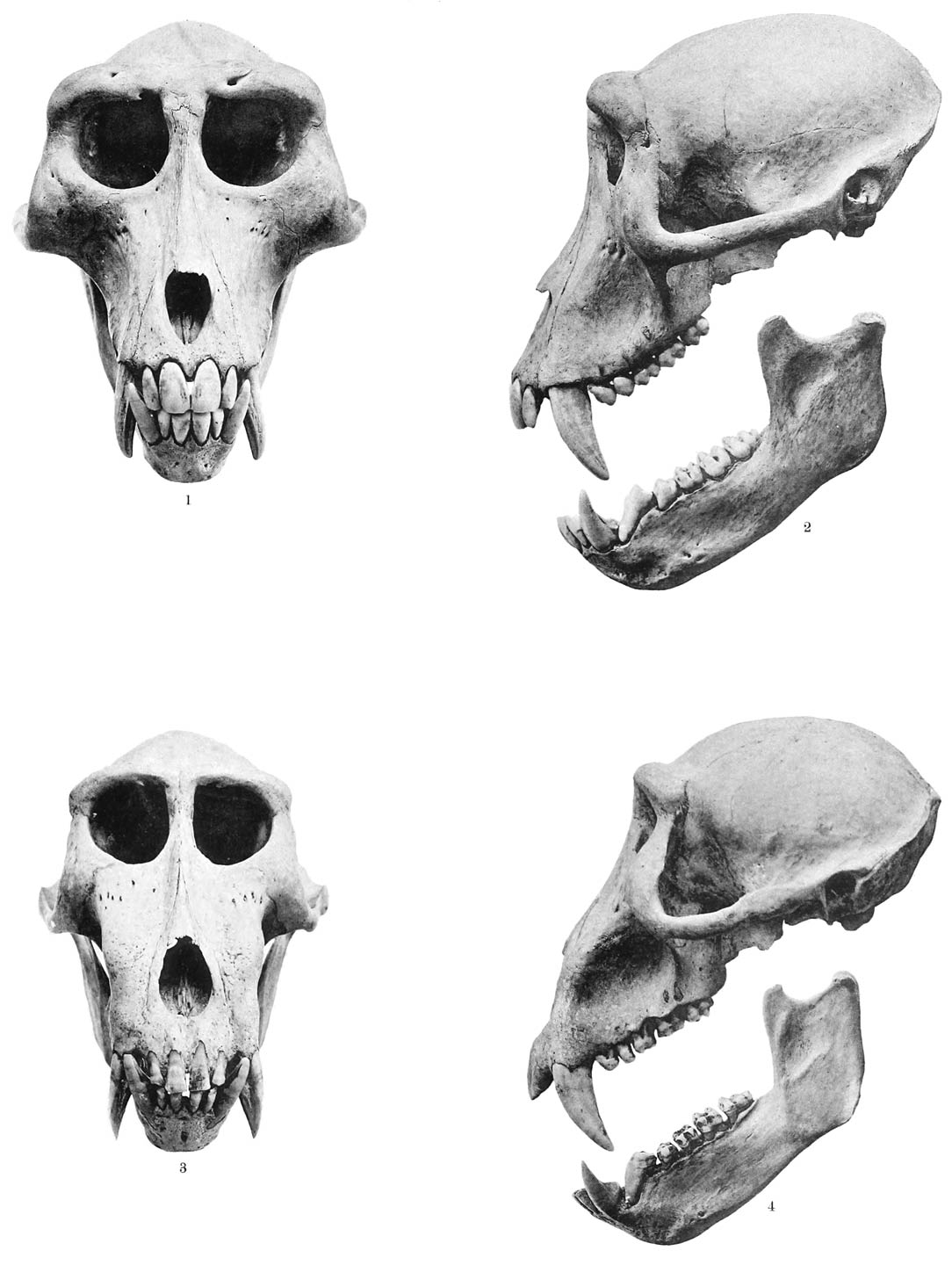
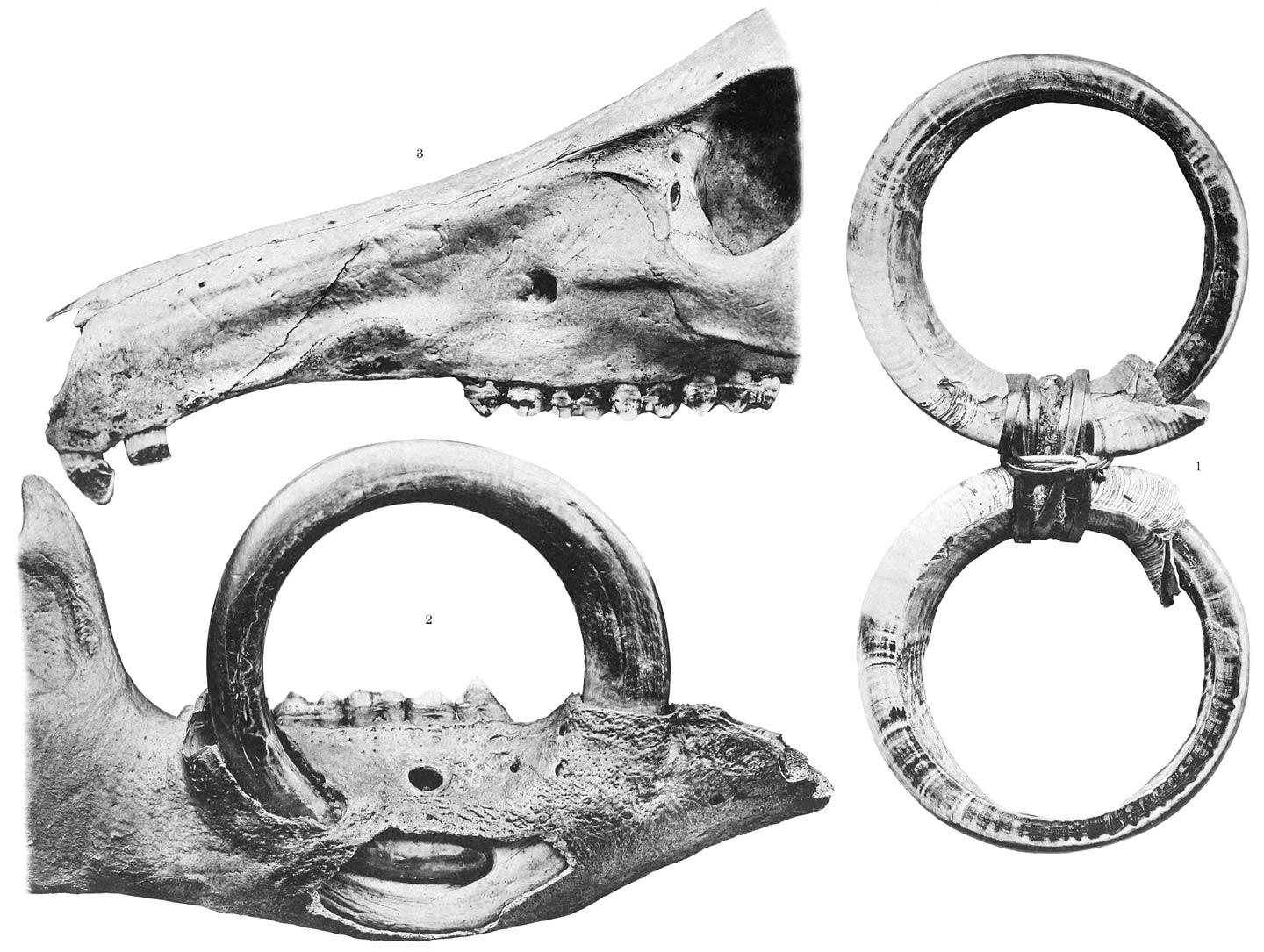
Tafel XI). Die Schnauze ist breit, kurz und frontal von oben vorn nach unten hinten schräg
abgestutzt (Taf. X, Fig. 3), so dass die, häufig von einem verdickten und mit kleinen Vorsprüngen besetzten
Rand eingefassten, Nasenlöcher ihren vordersten Punkt einnehmen. Die Oberlippe ist
sehr dick und bei vielen Arten mit senkrecht zum Mundspalte stehenden tiefen Falten
versehen, über die Unterlippe greift sie etwas über. Was die Behaarung anbelangt,
so nimmt man mit freiem Auge nur wahr, dass manche Stellen nahezu nackt sind, während
an anderen die Haare im Gegensatze zu denen am Körper theilweise eine besondere Anordnung,
wie in Büscheln und Reihen erkennen lassen, bisweilen auch sich durch grössere Länge
auszeichnen.
Die Füsse, die von der Flughaut vollständig freigelassen werden, sind kurz, breit und platt,
die Zehen ziemlich gleich lang, aber die erste und fünfte sind breiter als die anderen
und besitzen je längs der Aussenseite eine umgrenzte verdickte Stelle, eine Art Schwiele.
Auffallend ist die starke Behaarung der Füsse, wodurch die Molossi, wie zuerst Geoffroy Saint-Hilaire13 hervorgehoben hat, unter den Chiropteren ganz vereinzelt dastehen. Die Haare sind
theils sehr lang, gekrümmt und stehen auf der Dorsalseite der Nagelglieder, sodass
sie über die Zehenenden frei herüberragen, theils sind sie sehr kurz und überziehen
dicht die beiden schwielenartigen Felder an den Aussenseiten der ersten und fünften
Zehe. Soviel erkennt man hier mit blossem Auge.
Die genauere Prüfung der an Kopf und Füssen vorkommenden Haare ergiebt, dass man sie
füglich in zwei Gruppen scheiden kann, zwischen denen es allerdings auch Übergangsformen giebt.
Die Haare der ersten Gruppe besitzen die gewohnte spindlig-fadenförmige Gestalt, sie gleichen dem Körperhaare
durch die immer noch ziemlich stark vorspringenden Cuticularschuppen, die auch die
charakteristische Anordnung in trichterförmigen Segmenten besitzen, unterscheiden
sich aber von ihm durch meist bedeutendere Dicke, oft auch Länge, sowie stärkere Pigmentirung.
Im einzelnen treten diese Haare [36]nicht nur bei verschiedenen Arten, sondern z. Th. auch bei derselben je nach dem Standort
in mancherlei Formen auf, die alle anzuführen, ausserhalb des Rahmens dieser Arbeit
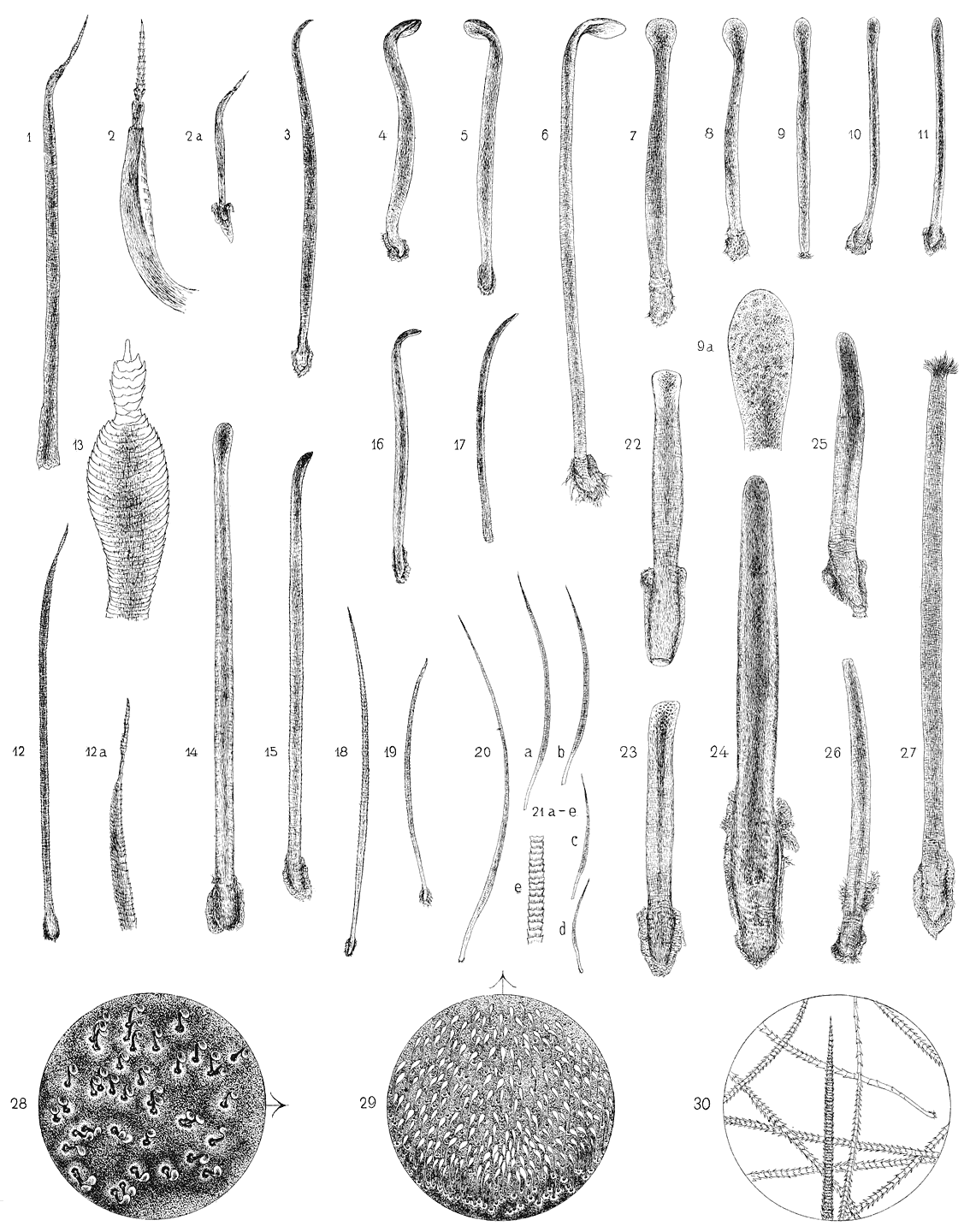
liegt. Es muss auch hier genügen, ein paar Beispiele herauszugreifen. Fig. 12 und
18 auf Taf. X geben Haare vom Gesichte von Nyctinomus bivittatus Hgl. wieder, die unter sich in der Dicke und der Zähnelung der Cuticula ziemlich übereinstimmen.
Von dem eigentümlichen Aufsatz auf der Spitze, den das eine aufweist, wird weiter
unten noch die Rede sein. Ein Haar von der Oberlippe von Nyctinomus astrolabiensis (Fig. 19) ist dagegen schmächtiger und glatter, es gleicht einigermaassen dem Körperhaare
von Cheiromeles (Fig. 21 a).
Die zweite Gruppe umfasst sodann die eigentümlich ausgebildeten, in den ausgeprägten Fällen dem Körperhaare
ganz unähnlichen Haare, die den eigentlichen Gegenstand dieser Mittheilung bilden.
Sie finden sich sowohl am Gesichte wie an den Füssen, und, mit einer geringfügigen
Ausnahme bei Cheiromeles, ist ihr Vorkommen auch auf diese Stellen beschränkt.
Ihre Anordnung im Gesicht ist bei den einzelnen Arten sehr verschieden und wird weiter
unten für jede Species besonders geschildert werden. Vorläufig sei bemerkt, dass sie
sich allgemein finden auf dem Felde zwischen der Nase und dem äusseren Rande der Oberlippe
sowie auf den seitlichen Theilen der letzteren, besonders auf den Kämmen der senkrechten
Falten, wo solche vorhanden sind. Nur bei manchen Arten kommen sie vor in dem Theile
des Gesichts oberhalb der Nase bis gegen den Grund der Ohrmuscheln hin und ferner
auf dem Unterkiefer.
Weit einförmiger ist das Verhalten an den Füssen. Hier bedecken sie die oben erwähnten
Schwielen an den freien Seiten der beiden äusseren Zehen, und die Ausdehnung dieser
Felder ist bei den einzelnen Arten von Nyctinomus und Molossus nur sehr wenig verschieden (Taf. XI Fig. 1 a u. 2 a). Abweichend verhält sich wiederum Cheiromeles, wie weiter unten geschildert werden wird.
Eine allgemeine Anschauung von der Beschaffenheit dieser Haare gewinnt man am leichtesten
bei ihrer Musterung in situ unter dem binocularen Microscope; die Figuren 28 und 29
auf Tafel X versuchen das Bild wiederzugeben, das die charakteristischen Felder an der Schnauzenspitze
von Nyctinomus sarasinorum und Molossus rufus obscurus bei dieser Art der Betrachtung gewähren.
Bei Nyctinomus sarasinorum erscheinen die Haare als kurze, circa 1 mm hohe, derbe, dunkle Borsten, die an der
Spitze nach einer Seite hin ein annähernd halbkugliges Knöpfchen tragen, das gegenüber
dem dunklen Schafte fast weiss erscheint, nur in seiner mittleren Parthie etwas dunkler
gefärbt ist. Bewegt man das Object hin und her, so vermag man schon jetzt mit völliger
Sicherheit zu erkennen, dass die Knöpfchen an der Unterseite ausgehöhlt sind, sodass
sich der Vergleich des ganzen Haares mit der Form eines Senflöffelchens unwillkürlich
aufdrängt. Jedes Haar steht in einer seichten, muldenartigen Vertiefung der Haut,
die etwas heller gefärbt ist als ihre Nachbarschaft und sich gegen diese durch eine
geringe wallartige Erhebung absetzt. Nicht alle Haare sind gleichmässig ausgebildet,
man beobachtet, namentlich in den seitlichen Parthien des Gesichts, auch solche, bei
denen das Knöpfchen viel weniger ausgeprägt ist. So typische Formen wie an der Schnauze
von Nyctinomus sarasinorum findet man überhaupt nur noch bei wenigen Arten.
Über die Zahl und die gegenseitige Anordnung dieser Haare lässt sich etwas allgemein Zutreffendes
nicht sagen. Bei Nyctinomus sarasinorum stehen sie nicht besonders dicht, zeigen aber eine gewisse Neigung, Gruppen von zwei
bis drei zu bilden. Bei anderen Arten habe ich indessen nichts dergleichen bemerkt.
Dagegen ist ganz allgemein eine gewisse Regelmässigkeit in der Orientirung dieser
Haare nicht zu verkennen. Überall im Gesichte sind sie nämlich vorwiegend so gestellt,
dass die verbreiterte Endplatte nach einem Punkt hingerichtet ist, der median zwischen
den Nasenlöchern liegt. In der Figur 28 auf Taf. X, wo der Pfeil am Rande die Medianlinie und die Richtung nach der Nase zu anzeigt,
tritt dies auch deutlich hervor.
Auf dem Feld an der Schnauzenspitze des Molossus rufus obscurus (Fig. 29, Taf. X) stehen die Haare sehr viel dichter als bei Nyctinomus sarasinorum, doch ist ihre Form im ganzen weniger typisch und bei manchen nur schwach angedeutet.
Dagegen gilt in Betreff der Orientirung der einzelnen Haare wieder das für Nyctinomus sarasinorum Bemerkte.
[37]
Die Haare an den Füssen besitzen im allgemeinen noch weniger ausgeprägte Form, orientirt
sind sie in der Regel derart, dass die Enden nach der proximalen Seite des Fusses
und zugleich etwas nach dem zunächst gelegenen freien Zehenrand hin gerichtet sind.
Wir gehen nun zur genaueren Betrachtung des einzelnen Haares über. Die Haare, deren
Gestalt am ausgeprägtesten ist, sind sehr kurz (Taf. X, Fig. 4, 5, 7, 8). Ausnahmen machen einerseits die an der ersten Zehe von Cheiromeles (Fig. 6), die die doppelte Länge und darüber erreichen; andererseits finden sich
bei Nyctinomus pumilus (Crtschm.) solche von kaum der Hälfte der gewöhnlichen Länge. Im Vergleiche mit dem Körperhaar
ist der Schaft meist von beträchtlicher Dicke (0,08 mm und mehr), gewöhnlich ist er
ziemlich gerade, oft aber besitzt er auch eine leichte Krümmung, und bei Nyctinomus sarasinorum bildet eine solche von s-Form sogar die Regel. Sie ist besonders deutlich an den
kürzeren Haaren von wohl ausgeprägter Form, wodurch deren Ähnlichkeit mit einem Senflöffelchen
noch auffallender wird (Fig. 4 auf Taf. X, auch in dem Habitusbilde Fig. 28 zu bemerken), findet sich aber auch an den längeren
weniger typischen (Fig. 3, Taf. X).
Das eigenthümlich gestaltete Endknöpfchen, das den Charakter des Haares bestimmt,
ist gegen den Schaft winklig abgesetzt, und zwar beträgt der Winkel bei den typischen
Formen ungefähr einen Rechten, während er bei weniger typischen im allgemeinen um
so stumpfer wird, je weniger die Endplatte die charakteristische Form zeigt (Taf. X, Fig. 3, 15, 16). Bei den Anfangsformen mit nur wenig verbreitertem Ende (Fig. 10),
wie wir sie neben den ausgeprägten im Gesicht und, bei den meisten Arten vorwiegend,
an den Füssen finden, ist es nur wenig gegen den Schaft gekrümmt.
Meist ist mit der geringeren Ausbildung der Endplatte eine Zunahme der Länge und Verminderung
der Dicke des Schaftes verbunden (Fig. 3, Taf. X), doch kommen solche Formen auch mit kurzem dünnem (Fig. 11) und mit langem dickem
Schafte (Fig. 14, 15) vor.
Für die topographische Vertheilung der verschiedenen Formen gilt allgemein die Regel,
dass sich die am besten entwickelten auf dem Felde zwischen Nase und oberem Mundrande
finden, während mit der Entfernung von dieser Region die minder typischen an Zahl
zunehmen. Die sehr langen und dicken Borsten (Fig. 14, 15) stehen besonders oberhalb
der Nase gegen die Ohrmuscheln hin. Die Haare auf den senkrechten Wülsten der Oberlippe
besitzen meist ziemlich typische Form, während auch auf dem Feld unterhalb der Nase,
wenn es, wie bei Molossus rufus (Fig. 29 Taf. X) sehr dicht bestanden ist, die Haare an seiner Peripherie z. Th. nur Anfangsformen
von Spatelhaaren sind.
Das umgebogene Ende des Haares selbst stellt bei bester Entwicklung nahezu ein Kugelsegment
dar, das so ausgehöhlt ist, dass seine Wand im Centrum am stärksten ist und sich nach
der Peripherie zu allmählich verdünnt, um in scharfem Rande zu enden. Bei einem etwas
geringeren Grade der Ausbildung ist die Endplatte mehr verkehrt eiförmig und flacher
ausgehöhlt.
Hier ist die einfache Bezeichnung „löffelförmig“, wie sie Dobson14 für die Haare an der Schnauze des Molossus abrasus Temm. gebraucht, am Platze. Schliesslich schwindet auch die geringe Ausbuchtung, und es
bleibt nur die mehr oder weniger verbreiterte und gegen den Schaft gekrümmte Endplatte,
wodurch das Haar eine Gestalt gewinnt, die man am passendsten vielleicht mit den bei
microscopischer Präparation gebräuchlichen Spateln vergleichen könnte. Bei manchen
Haaren ist die Verbreiterung und Krümmung des Endes so wenig ausgeprägt (Taf. X, Fig. 11 u. 17), dass man erst nach Kenntniss der typischen Formen durch vergleichende
Beobachtung auf sie als erste Andeutung solcher Bildung aufmerksam wird.
Der Schaft der löffel- oder spatelförmigen Haare ist so glatt, wie etwa der eines
Menschenhaares. Bei stärkerer Vergrösserung treten nur feine Querlinien auf der Oberfläche
als Ausdruck der Cuticularstructur hervor. Er ist an den Haaren vom Gesicht in der
Regel dunkel, öfter, z. B. bei Nyctinomus plicatus und sarasinorum fast schwarz. Gewöhnlich ist in der Axe ein besonders dunkler ziemlich scharf begrenzter
Strang zu bemerken, den man für einen Markcylinder halten könnte. Doch zeigt die genauere
Untersuchung, besonders von Querschnitten, dass auch den Spatelhaaren ein gesondertes
Mark mangelt, jener Eindruck rührt daher, dass das Pigment ganz überwiegend eine ziemlich
genau kreisförmige centrale Parthie des Schaftes erfüllt.
[38]
Die Endplatte muss schon infolge ihrer grösseren Dünne heller als der Schaft erscheinen,
sie ist aber auch absolut ärmer an Pigment, da sie vorwiegend aus einer Fortsetzung
der peripherischen pigmentlosen Parthie des Schaftes besteht. Der dunkle centrale
Strang des Schaftes setzt sich gewöhnlich eine Strecke weit in die Endplatte hinein
fort, bisweilen entspricht dieser Stelle in der Concavität der Endplatte ein kielartiger,
distalwärts allmählich verstreichender Vorsprung. Die Figur 5, Taf. X (von Nyctinomus plicatus), die den Haarlöffel in der Ansicht schräg von unten darstellt, zeigt ihn sehr deutlich.
Von diesem dunklen Mittelstrang aus strahlt dann das Pigment nach der Peripherie hin
in immer feinerer Vertheilung aus. Je allmählicher letztere vor sich geht, um so zierlicher
ist das Bild, das die Endplatte unter stärkerer Vergrösserung gewährt (Fig. 9 a, Taf. X). In dem mittleren Strange bildet das Pigment grobe dichtgedrängte Schollen, die
weiter nach der Peripherie feiner werden und durch weitere Zwischenräume getrennt
sind, dann als Häufchen feiner brauner Körnchen erscheinen, bis endlich in den Randparthieen
der Platte solche Körnchen nur noch ganz vereinzelt zu bemerken sind.
Oft hört die Pigmentirung aber auch ziemlich plötzlich und nahe dem Grunde der Endplatte
auf, wie an dem Haare von Nyctinomus limbatus (Fig. 8) und dem von N. bivittatus (Fig. 7) zu sehen ist.
An Haaren mit nur wenig verbreitertem Ende ist dies in der Regel durchweg ziemlich
dunkel, nur die äusserste Randzone erscheint etwas heller (Fig. 10, 11 auf Taf. X).
Die Spatelhaare an den Füssen sind abweichend von denen im Gesichte meist fast ohne
Pigment und lassen daher auch keinen Unterschied in der Färbung von Schaft und Endplatte
wahrnehmen.
Manche von den Borsten (Fig. 11, 17), die wir als erste Andeutungen von Spatelhaaren
erkannten, bilden offenbar eine Vermittlung zwischen den auf den ersten Blick ganz
isolirt stehenden Haaren von ausgeprägter Löffelform und denen, die oben als erste
Gruppe der vom Körperhaar abweichenden beschrieben wurden. Für diese Auffassung ist
auch bemerkenswerth, dass bei solchen gewissermaassen rudimentären Formen die Cuticularschuppung
stärker als an den echten Spatelhaaren hervortritt (Fig. 17).
In anderer Richtung deuten auf eine innere Verwandtschaft der Spatelhaare mit den
gewöhnlichen des Körpers die eigenthümlichen Formen, zu deren Beschreibung ich nun
übergehe.
Bei einigen Arten von Nyctinomus bemerkte ich unter dem Binoculare neben Spatelhaaren von bekannter Form solche, deren
Endplatte in einen feinen fadenförmigen Fortsatz auszulaufen schien. Prüfung bei stärkerer
Vergrösserung ergab, dass dieser fadenförmige Anhang, der sich an die Endplatte in
der Mitte ihres distalen freien Randes ansetzt, in seiner Structur dem Körperhaare
sehr ähnlich ist (Taf. X, Fig. 1, 2, 2 a, 13). Er ist pigmentlos und besitzt stark vorspringende, in Trichtersegmenten geordnete
Cuticularschuppen. Der Anhang zeigt ziemlich regelmässig eine bestimmte Gliederung.
An der Übergangstelle in die Endplatte (Fig. 13) verjüngt er sich etwas, darauf folgt
ein kurzes cylindrisches Stück, dann wieder eine meist tiefe Einschnürung und hierauf
das Endglied, das etwa viermal so lang ist wie das erste und vom dickeren Grunde ganz
allmählich in eine feine Spitze ausläuft (Fig. 2).
Diese eigentümliche Form hat ein Seitenstück unter den modificirten Haaren unserer
ersten Gruppe. Figur 12, 12 a giebt ein solches Präparat wieder. Das Haar, das sonst
ganz den früher beschriebenen (Fig. 18) gleicht, trägt an der Spitze einen Aufsatz
von ganz derselben Form und Beschaffenheit wie an den eben geschilderten Spatelhaaren.
Als ich, noch im Anfange der Untersuchung, den Anhang an den Spatelhaaren bemerkte,
lag die Frage nahe, ob er nicht regelmässig vorkomme und, wo er fehle, erst nachträglich
verloren gegangen sei. Indessen musste diese Frage nach eingehender Prüfung verneint
werden. Der Ansatz haftet an der Platte ziemlich fest und ist auch in sich gegen Zerrung
und Biegung widerstandsfähig, am leichtesten erfolgt die Zerreissung an der eingeschnürten
Stelle, aber auch erst bei einer bestimmten Gewalteinwirkung (wie am Präparate der
Fig. 13, Taf. X geschehen). Es wäre also schwer denkbar, dass ihn bei der Mehrzahl der Exemplare
alle und bei den übrigen die meisten Haare verloren haben sollten. Auch ergab die
genaue Untersuchung der isolirten Spatelhaare, dass die Endplatten immer einen vollkommen
unversehrten Rand besassen. Man muss daher wohl annehmen, dass die Bildung nur manchen
Spatelhaaren und vielleicht nur bei bestimmten Species zukommt.
Vergegenwärtigen wir uns die Entwicklung dieser Haare, so ist klar, dass die die Papille
überkleidende epitheliale Matrix hier zuerst eine Zeit lang ein Gebilde producirt,
das einem Körperhaare gleicht, [39]und dass dann mit einem Mal in der Production eine Änderung eintritt, sodass das Erzeugniss
ein Spatelhaar wird.
Einen analogen Vorgang können wir vielleicht aus der Entwicklungsgeschichte der Feder
heranziehen. Dieselbe Papille und Matrix, die eine Dune des jungen Vogels erzeugt
hat, producirt später auch eine bleibende Feder des ersten Gefieders, die daher die
Dune auf ihrer Spitze trägt (vergl. Gegenbaur, Vergl. Anat. Bd. 1, p. 139; Gadow in Bronns Klassen und Ordnungen des Thierreichs, Vögel, I. Anatomischer Theil. Leipzig
1891, p. 524 u. 525). So spielt sich der Vorgang aber nur bei der Bildung des ersten
Gefieders ab, später entwickeln sich nach der Mauser die Ersatzfedern direct ohne
vorhergehende Erzeugung einer Dune.
Auch in unserem Falle scheint mir der Gedanke nicht ohne weiteres abzuweisen, ob es
vielleicht nur die zum ersten Male gebildeten Haare sind, die einen solchen Anhang
besitzen. Zur Entscheidung dieser Frage wäre es nöthig, von den einzelnen Arten Reihen
verschiedener Alterstufen zu untersuchen, die mir nicht zur Verfügung standen.
Bei einigen Arten der Gattung Nyctinomus kommen im Gesicht Haare, die in der Form irgendwelche Ähnlichkeit mit Spatelhaaren
besitzen, überhaupt nicht vor. Immer aber finden sich dann an den Stellen, die bei
anderen Arten welche tragen, Borsten oder Stacheln, die sich durch Dicke, Länge und
histiologische Structur von den anderen im Gesichte vorkommenden längeren und steiferen
Haaren deutlich unterscheiden. Ihr Schaft ist glatt wie bei den Spatelhaaren, dunkel
pigmentirt, meist mit deutlichem axialem Strange (Fig. 24). Ich glaube daher, dass
diese Borsten auch im morphologischen Sinn als Vertreter der bei den anderen Arten
vorkommenden Spatelhaare zu betrachten sind.
Besonderes Interesse bietet in dieser Hinsicht Cheiromeles torquatus. Typische Spatelhaare finden sich hier nur an den Füssen und zwar abweichend von
Molossus und Nyctinomus ausschliesslich auf einem am Aussenrande der ersten, nach Art eines Daumens frei
beweglichen Zehe, gelegenen Felde, das die Figuren 11 b–d, Taf. XI in verschiedenen Ansichten wiedergeben. Die hier vorhandenen Spatelhaare sind, wie
schon erwähnt, die längsten von ziemlich typischer Form, die ich überhaupt beobachtet
habe. Von den Spatelhaaren an den Füssen der Molossus- und Nyctinomus-Arten unterscheiden sie sich ausser durch die viel ausgeprägtere Form und die grössere
Länge auch durch bedeutend dunklere Färbung, die freilich immer noch heller ist als
bei den Spatelhaaren im Gesicht.
Im Gesichte besitzt Cheiromeles unterhalb der Nasenlöcher eine dichte Gruppe kurzer, steifer und dicker Borsten,
die im ganzen etwas abgeplattet sind (Taf. X, Fig. 22, 23). Kurz vor dem Ende verjüngt sich der Schaft ein wenig, während das
Ende selbst wieder eine breitere, quer abgeschnittene Platte vorstellt, die etwas
gegen den Schaft gekrümmt ist. Offenbar sind diese Haare als modificirte Spatelhaare
aufzufassen.
An den seitlichen Theilen des Gesichts, wo die Molossus- und Nyctinomus-Arten Spatelhaare besitzen, finden sich bei Cheiromeles nur dicke Borsten verschiedener Länge. An manchen der kürzeren von ihnen ist das
Ende gegen den Schaft etwas abgesetzt und gekrümmt, wodurch sie sich den Borsten unterhalb
der Nase anschliessen (Fig. 25). So zeigt uns denn Cheiromeles nebeneinander eine Reihe von Übergängen von typischen Spatelhaaren zu den Borsten
und Stacheln gewöhnlicher Form, die wir im Gesicht einiger Arten ausschliesslich antreffen.
Eine Eigenthümlichkeit dieser Borsten bei N. brasiliensis und africanus mag noch Erwähnung finden. Sie scheinen vielfach, unter Lupenvergrösserung betrachtet,
weissliche Knöpfchen zu tragen, sodass man sie für Spatelhaare halten könnte. Thatsächlich
rührt die Erscheinung daher, dass die Borsten am Ende eine kurze Strecke pinselartig
sehr fein aufgefasert sind (Taf. X, Fig. 27) und die zwischen den Fasern festsitzende Luft das Licht diffus reflectirt.
Zwischen Borsten mit intactem und aufgefasertem Ende findet man Übergänge; in Fig.
26, Taf. X ist z. B. eine Borste abgebildet, bei der eine geringe Verbreiterung und dellenartige
Einziehung des Endes die bevorstehende Aufsplitterung anzeigt. Es handelt sich hier
also jedenfalls nicht etwa um ein durch Abbrechen des Schaftes verursachtes Kunstprodukt,
und auch für die Annahme, dass die Zerfaserung erst post mortem durch Maceration im
Spiritus aufgetreten sei, scheint mir kein Grund vorzuliegen. Ähnliche, wenn auch
nicht so regelmässige, Zersplitterungen der Spitze beobachtet man auch sonst gelegentlich
an menschlichen und thierischen Haaren (vergl. Waldeyer, Atlas 112 u. 175, 1884; Fig. 38 u. 139).
[40]
Was die Frage nach der functionellen Bedeutung der löffel- und spatelförmigen Haare und der ihnen entsprechenden Borsten und Stacheln
anlangt, so würde es sich in erster Linie darum handeln, ob sie nur als eine besondere
Form der gewöhnlichen Haare oder als „Tasthaare“ im engeren Sinne betrachtet werden
müssen. Da eine Untersuchung auf nervöse Endapparate an unserem Materiale von vornherein
ausgeschlossen war, musste ich mich darauf beschränken, aus der Structur des Haarbalges
vielleicht einigen Anhalt zur Beurtheilung zu gewinnen. Am geeignetsten wäre zu dem
Zwecke das Exemplar von Nyctinomus sarasinorum gewesen, das sich, wie alles von den Hrn. Sarasin gesammelte Material, durch vortrefflichen Erhaltungszustand auszeichnete. Da es aber
der Typus und bis jetzt das einzige vorhandene Exemplar der Art ist, so mochte ich
es nicht beschädigen, und entnahm daher Hautstücke von der Oberlippe und den Feldern
an den Zehen eines Exemplars von Nyctinomus plicatus, wo allerdings die Conservirung sehr viel zu wünschen liess.
Die Haarbälge sind recht derb und massig, besonders mit Rücksicht auf die geringe
Grösse der Haare, und ihre dichte Anhäufung ist es wesentlich, wodurch die schwielige
Verdickung an den Zehen bedingt wird. Doch konnte ich von einer cavernösen Structur
der Balgwandung, wie sie für „Tasthaare“ als charakteristisch gilt und letztere auch
als „Sinushaare“ bezeichnen lässt, an Durchschnitten hier nichts wahrnehmen. Da ich
indessen bei der Kürze der verfügbaren Zeit erst wenige Präparate anfertigen konnte
und da die Gewebe durch jahrelanges Liegen in dünnem Spiritus sehr gelitten hatten,
will ich ein abschliessendes Urtheil hiermit keineswegs ausgesprochen haben.
Auch die Angaben über die Lebensgewohnheiten der Molossiden, die bisher vorliegen,
sind zu dürftig, um über die besondere Leistung dieser Haare mehr als Vermuthungen
zu gestatten.
Die Haare im Gesichte mögen, wenn sie doch als Tasthaare zu betrachten sein sollten,
im Dienste des allgemeinen, bei den Fledermäusen so hoch entwickelten Hautsinnes stehen,
man könnte aber vielleicht auch daran denken, dass ihnen eine Funktion bei der Nahrungsaufnahme
zufällt. Nach Dobson (Catal. 1878, 403) wäre die grosse Dehnbarkeit der oft mit tiefen Falten versehenen
Lippen der Molossiden von günstiger Wirkung beim Verschlingen der vorzüglich aus „grossen
rundleibigen Käfern“ bestehenden Beute. Dabei könnten die vornehmlich auf der Oberlippe
befindlichen Haare wohl eine Rolle spielen.
Die Zehen benutzen die Fledermäuse allgemein, um den Pelz zu ordnen und von Parasiten
zu säubern, wobei die Spatelhaare an den Füssen, ob ihnen nun eine specielle nervöse
Funktion zukommt oder nicht, ganz dienlich sein könnten. Von Interesse ist in der
Beziehung folgende Angabe Osburns15 über Nyctinomus brasiliensis: „First one and then another wakes up, and withdrawing one leg and leaving himself suspended
by the other alone, adroitly uses the foot at liberty as a comb16, with a rapid effective movement dressing the fur of the under part and head16. The foot is then cleaned quickly with the teeth or tongue, and restored to its first
use. Then the other leg does duty. Perhaps the hairs with which the foot is set may aid to this end16. I often have seen them do this in confinement; and probably the numerous Bat-flies
with which they are infested may be the cause of extra dressing. It is impossible
to imagine a more perfect or effective comb than the little foot thus used makes ..“. Aus den Angaben Osburns ist allerdings nicht zu entnehmen, ob er die kurzen Haare an den Aussenseiten der
Zehen bemerkt hat oder ob nur die langen gekrümmten Haare auf den Nagelgliedern aller
Zehen gemeint sind, die Dobson (Catal. 1878, 403) „prehensile hairs“ nennt, ohne aber diese sonderbare Bezeichnung näher zu begründen17. — Merkwürdig bliebe dann freilich die besonders mächtige [41]Entwicklung der Spatelhaare an der grossen Zehe des Cheiromeles, der doch nur ein sehr rudimentäres Haarkleid besitzt. — Hier sei auch darauf hingewiesen,
dass die Molossiden unter den Fledermäusen am meisten geschickt sind, sich auf ebener
Erde laufend fortzubewegen, wobei natürlich die Hintergliedmaassen die Hauptarbeit
zu leisten haben (vgl. Dobson, Catal. 1878, 403; Schneider, Nouv. Mém. Soc. Helv. XXIV, 1871, S.A. 8–9).
Bei alle dem wird man sich bezüglich der Function der Spatelhaare auch gegenwärtig
bei der Ansicht bescheiden müssen, die Horsfield (Zool. Res. 1824, VIII. Cheiromeles, 6. S.) aussprach, als er zum ersten Male die
Felder an den Füssen von Cheiromeles und Nyctinomus plicatus beschrieb: „It is doubtless of importance in the economy of the animal, but its use remains to
be determined.“
Für eine solche Untersuchung käme zunächst der europäische Vertreter der Molossiden,
Nyctinomus cestonii, in Betracht, der u. a. in Italien und Griechenland heimisch ist. Doch ist das Thier
dort selten und gehört überdies zu den Arten, denen im Gesichte Spatelhaare fehlen.
Dagegen finden sich Arten mit typischen Haaren gemein und in Menge in manchen tropischen
Gegenden (vgl. die Bemerkung A. B. Meyers oben S. 18), und da ja neuerdings immer häufiger auch in feineren anatomischen Untersuchungen
geübte und entsprechend ausgerüstete Forscher dorthin kommen, so bietet sich wohl
einmal Gelegenheit, die wahre Bedeutung der eigenthümlichen Gebilde aufzuklären.
Im Anschluss an die Schilderung der löffelförmigen Haare der Molossiden ist es vielleicht
angezeigt, einen Blick auf die bisher überhaupt beschriebenen Formen von Säugethierhaaren
zu werfen. Eine daraufhin vorgenommene Durchsicht der Literatur lieferte mir ein ziemlich
dürftiges Ergebniss. Zwar darin stimmen alle Autoren überein, dass die Mannigfaltigkeit
ausserordentlich gross ist, was die Massenentwicklung und die feinere Structur des
einzelnen Haares anlangt, aber die Grundform, die mit ganz verschwindenden Ausnahmen
überall wiederkehrt, ist immer dieselbe spindlig-fadenförmige.
Als merklich abweichend verdienen vor allem die Grannenhaare von Ornithorhynchus Erwähnung. Maurer18 beschreibt sie folgendermaassen: „Der Schaft beginnt in der Tiefe zugespitzt, er
verbreitert sich dann rasch, so dass sein Querschnitt die Form eines langen Ovals
mit leicht bogenförmig gekrümmter Längsachse zeigt.
„Nach oben gegen die Talgdrüse zu wird der Schaft etwas dünner und rundlich, und so
tritt er aus der Balgöffnung hervor. Er setzt sich in einen langen drehrunden Abschnitt
fort, der sich endlich zu einem lanzettförmigen Plättchen verbreitert. Dasselbe endet
in einer abgestutzten Spitze.“
Da mit dieser Schilderung die Abbildung in Waldeyers Atlas (Tat. VIII, Fig. 100) nicht übereinstimmt, untersuchte ich selbst diese Haare
und fand Maurers Darstellung bestätigt. Die Abbildung bei Waldeyer giebt, wie mir scheint, nicht ein „ganzes Grannenhaar“ wieder, wie es in der Figurenerklärung
heisst (Waldeyer, Atlas, 189), sondern nur den oberen Theil eines solchen. Man müsste sich das untere
dünne Stück reichlich doppelt so lang denken wie die breite Endplatte, um eine zutreffende
Vorstellung von der Gesammtform dieses merkwürdigen Haares zu erhalten. Die feinere
Structur der einzelnen Theile finde ich vollkommen in Übereinstimmung mit den Beschreibungen
und Abbildungen Waldeyers (Atlas, 190; Taf. VIII, Fig. 101–103).
Ähnliche Haare wie die eben beschriebenen des Schnabelthiers finden sich nach Maurer19 auch bei Perameles gunni Gr.
[42]
Ausser diesen habe ich keine von dem allgemeinen „faden-spindligen“20 Grundtypus wesentlich abweichenden Haarformen erwähnt gefunden, und die löffelförmigen Haare der Molossi sind ihnen als ein weiterer interessanter und bis jetzt isolirt dastehender Befund
dieser Art anzureihen.
Die Liste auffallend gestalteter Haare wird sich möglicherweise am ehesten bereichern
lassen, wenn man mehr, als es bisher im allgemeinen geschehen zu sein scheint, sich
nicht auf die Untersuchung des gewöhnlichen Körperhaares beschränken, sondern grundsätzlich
bei jeder Thierart alle Stellen des Körpers einer genauen Prüfung unterwerfen wird,
an denen der Charakter der Behaarung modificirt erscheint. Hätte Marchi dies beachtet, so würden ihm, der eine ganze Reihe von Molossiden-Arten sehr gründlich
auf die Beschaffenheit des Körperhaares untersucht hat21, die löffelförmigen Haare gewiss nicht entgangen sein.
Es soll nunmehr das Verhalten der löffelförmigen und der ihnen gleichwertigen Haare
bei jeder der untersuchten Arten, besonders mit Rücksicht auf die topographische Vertheilung, kurz beschrieben werden.
Wo von derselben Art mehrere Exemplare vorlagen, liessen sich öfter gewisse individuelle
Schwankungen in der Zahl und wohl auch in der Formentwicklung dieser Haare wahrnehmen,
dagegen fand ich die Verbreitung und Anordnung immer durchaus übereinstimmend, so
dass sich für jede einzelne Art eine für alle Exemplare zutreffende Schilderung geben
lässt. Auch das Geschlecht ist in dieser Hinsicht ohne Einfluss.
Die Anordnung der Arten folgt im allgemeinen der in Dobsons Catalogue 1878, mit einigen im Interesse der bequemeren Darstellung gebotenen Abweichungen.
Es empfahl sich mit dem Genus Nyctinomus zu beginnen und mit Cheiromeles zu schliessen. Von den Nyctinomus-Arten sind zunächst die behandelt, die im Gesicht echte Spatelhaare besitzen, dann
erst jene, denen sie dort fehlen. Die drei Dobson noch nicht bekannten Arten (sarasinorum, astrolabiensis und loriae) habe ich den nächstverwandten angereiht.
Für das Verständniss der Beschreibungen seien folgende Bemerkungen vorausgeschickt.
Die Haare sind gemeinhin als „Spatelhaare“ bezeichnet, und es werden im allgemeinen
drei Grade der Ausbildung unterschieden: typische Formen, entsprechend den Figuren 4, 5, 7, 8 auf Tafel X, mittlere (Fig. 9, 16, 3) und wenig ausgeprägte (Fig. 10, auch 14 und 15). Die absolute Länge der Haare ist, wie früher schon bemerkt,
immer sehr gering. Als gewöhnliches Durchschnittsmaass ist circa 1 mm anzusehen, unter
„sehr langen“ Haaren sind solche von etwa 2–2,5 mm Länge (Fig. 1, 6, 14, 15) verstanden,
unter „kurzen“ solche unter 1 mm (Fig. 2 a).
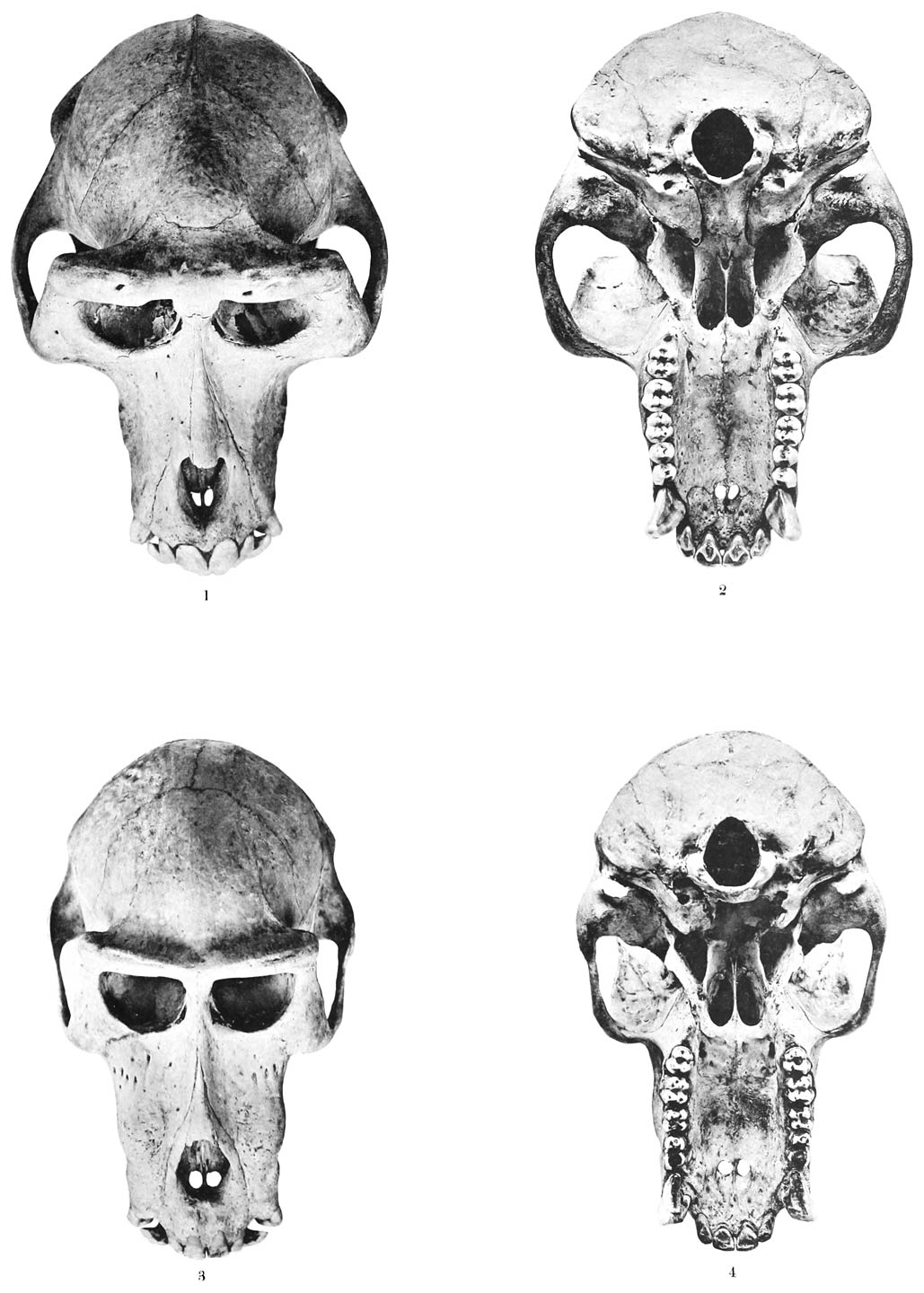
Die Figuren auf Tafel XI sollen in der Mehrzahl dazu dienen, eine Anschauung von einigen typischen Anordnungen
der Spatelhaare im Gesichte verschiedener Molossiden-Arten zu geben. Es ist zu dem
Zwecke der Kopf fast durchweg in der Ansicht von vorn und etwas von unten gezeichnet,
sodass das Gebiet der Schnauze, der Ober- und Unterlippe möglichst vollständig vor
Augen liegt. In die Umrisszeichnung aller dieser Theile sind dann unter Controlle
mittelst des Binoculars die Spatelhaare oder die ihnen entsprechenden Borsten nach
Zahl und Anordnung möglichst genau eingetragen und durch Punkte oder durch Striche
mit verdickten Enden angedeutet. Andere als Spatelhaare oder ihre Vertreter sind dabei
nicht berücksichtigt.
Bei jeder Art werde ich, soweit sie mir bekannt geworden sind, auch die Angaben früherer
Autoren, die sich auf das Vorkommen dieser Haare beziehen, anführen, in der Synonymie
folge ich dabei der Autorität von Dobsons Catalogue 1878.
Exemplare, die aus der Sammlung des Königlichen Naturalien-Cabinets in Stuttgart hergeliehen
waren, sind als solche gekennzeichnet, die anderen, bei denen nichts bemerkt ist,
gehören dem Dresdener Museum.
[43]
[Inhalt]
Molossi
Als erster und wohl auch einziger Autor, der auf den Besitz von Spatelhaaren als einen
allgemeinen Charakter der Gruppe aufmerksam gemacht hat, ist Burmeister zu nennen. In der „System. Übers. d. Thiere Brasil. I. Säugethiere. Berl. 1854“ sagt
er (S. 66) bei der allgemeinen Charakteristik der Gattung Dysopes (= Molossus und Nyctinomus): „Die breiten Lippen sind … mit einem dichten Wimpernsaume besetzt; Schnurrhaare
fehlen oder stehen sehr vereinzelt, dagegen sieht man kurze, hakenförmig aufwärts gebogene Borsten in den Lippen.“22 Ferner S. 67: „die Zehen sind kurz, dick, klein, ausserhalb mit langen, steifen,
gebogenen, abstehenden Wimperhaaren besetzt; die erste und letzte Zehe etwas erweitert
und unterhalb mit steifen Häkchen bekleidet.“22 Offenbar sind hier die Spatelhaare gemeint, die ja in der That bei geringer Lupenvergrösserung
von ihrer eigenthümlichen Form kaum mehr als die Krümmung des oberen Endes erkennen
lassen.
Aus Burmeisters weiterer Darstellung geht übrigens hervor, dass er die Haare mit Sicherheit nur bei
zwei Molossus-Arten (temmincki und perotis) und möglicherweise noch bei Molossus rufus obscurus und Nyctinomus macrotis beobachtet hat. Die älteren Angaben von Horsfield über Nyctinomus plicatus (vergl. unter diesem) erwähnt er nicht. Aber wenn auch Burmeisters Verallgemeinerung demnach für jene Zeit wohl nicht ganz begründet war, so ist sie
doch jedenfalls richtig gewesen.
Daneben wäre noch Allen zu nennen, der in der allgemeinen Einleitung zu seinem „Monograph of the Bats of
North America“ (Bull. U. S. Nat. Mus. 43, 12–13; 1893) unter der Rubrik „Haar“ folgende,
allerdings viel weniger bestimmte, Bemerkung macht: „Bristles (setae) usually surmount warts (verrucae) [sc. in der ganzen Ordnung]. They are best developed on the face of Molossi, though they may be found in the
group last named on the upper surface of the interfemoral membrane.“ „Fringes of bristles adorn the margins of the toes in Molossi.“ Ich bemerke dazu, dass ich solche Borsten auf der Rückenseite der Interfemoralmembran
nur bei Cheiromeles torquatus beobachtet habe, wo sie schon Horsfield bekannt waren (vgl. später unter Cheiromeles). Allen giebt, soviel ich sehe, für seine allgemein gehaltene Bemerkung keine speciellen
Belege.
Nyctinomus Geoffr.
Arten mit Spatelhaaren im Gesicht
1. Nyctinomus plicatus (Buch. Ham.)
Tafel XI, Fig. 1 u. 1 a, Tafel X, Fig. 5, 9, 9 a, 10, 11, 16 u. 17
Von dieser weit — von Vorderindien bis Tasmanien — verbreiteten Art standen mir zur
Verfügung Exemplare von Port Darwin (N Australien) 5 (Stuttgart), Jobi (Insel im Norden
von NW Neu Guinea) 3, Java (SW) 1, Sumátra (NO) 1.
Kopf: Der obere Nasenlochrand tritt wulstig hervor und ist mit Höckerchen besetzt, nach
abwärts fliesst er in der Medianlinie mit dem der anderen Seite zu einer kurzen Leiste
zusammen. Letztere besonders deutlich an dem Exemplare von Sumátra, demnächst an denen
von Australien. Oberlippe sehr dick, mit circa 10 tiefen senkrecht zum Mundrande stehenden
Falten. — Spatelhaare sehr typischer Form mässig zahlreich auf dem Felde zwischen
Nasenlöchern und Mundrand, doch erst unterhalb der medianen Leiste. Am seitlichen
Mundrande weniger zahlreich, zu je 2–8 auf den Kämmen der Falten bis nahe an den Mundwinkel, in den Furchen nicht. Form hier, je weiter seitwärts, um so weniger typisch, doch vorwiegend noch
mittel, aber auch Übergänge zu Borsten einfacher Art (Fig. 11, 17 auf Tafel X). In den seitlichen Parthieen des Gesichts entfernt vom Mundrande keine Spatelhaare,
wohl aber in den medianwärts gelegenen Theilen oberhalb der Nase bis gegen die Ohren
hin und zwar hier vorwiegend längere dicke Borsten wenig ausgeprägter Form (wie in
Fig. 14, 15, Tafel X). — Auf dem Unterkiefer bei den Exemplaren von Port Darwin zerstreut in zwei Gruppen
nahe dem Mundwinkel, bei denen von Neu Guinea und Java vereinzelt hier und da, bei
dem von Sumátra fehlend. Diese Borsten dick und wenig typisch, ähnlich Fig. 25, Tafel X.
[44]
Füsse: Die Felder am äusseren Rande der ersten und fünften Zehe dicht bestanden mit Spatelhaaren
meist wenig ausgeprägter, seltener der mittleren sich nähernder Form. Vom proximalen
zum distalen Ende des Feldes nehmen die Haare an Länge zu, so dass sie z. Th. denen
von Cheiromeles (Tafel X, Fig. 6) gleich kommen, doch sind diese langen Formen am Ende nur ganz wenig verbreitert.
— Die langen (bis gegen 1 cm) gekrümmten Haare, die sich auf der Dorsalseite der Nagelglieder
aller Zehen finden, endigen zugespitzt, sind aber wie die Spatelhaare hell und glatt. Vollständige
Übergänge zwischen beiden Haarformen habe ich indessen nicht beobachtet. Dies gilt
auch für die folgenden Arten.
Horsfield in den Zoolog. Researches, London 1824 bemerkt über das Gesicht seines N. tenuis (= plicatus) unter „Character naturalis“: „Labrum laxum plicatum … verrucis pluribus rugosum, setisque obtusis obsitum23.“ Und später: „The lips and lateral parts of the face are extremely rough, being covered with numerous minute warty points, which are individually terminated by a short stiff bristle23.“ Vermuthlich sind hier die Spatelhaare nebst der muldenartigen Vertiefung der Haut
an ihrem Grunde gemeint.
Ganz klar und zutreffend sind dagegen die Felder an den Füssen beschrieben (Horsfield, Zool. Research. 1824, Nyctinomus tenuis): „A series of delicate hairs, about one line in length, extends along the whole of the
exterior side both of the thumb and of the little finger; a few hairs of a greater
length are scattered through these and likewise stretch forward, and spread over the
claw. These hairs rise nearly erect or vertically from the finger, and are not directed
horizontally outward, as in Cheiromeles. The separate hairs are bent or hooked at the extremity; their colour is silvery
gray. This regularly defined series of hooked hairs must not be confounded with the
long lax hairs which are observed in all the fingers of the Nyctinomi, and which, according to M. Geoffroy, must also be placed among the generic characters.“
2. Nyctinomus sarasinorum A. B. M.
Tafel XI, Fig. 2 u. 2 a, Tafel X, Fig. 3, 4 u. 28
1 Exemplar von Central Celébes. Verhält sich dem vorhergehenden sehr ähnlich. Spatelhaare
auf dem Felde zwischen Nasenlöchern und Mundrand sehr typisch, vielfach mit s-förmig
gebogenem Schafte. Seitwärts auf den senkrechten Wülsten verschwinden die Haare in
grösserer Entfernung vom Mundwinkel als bei plicatus. Oben und medianwärts von der Nase lange Spatelhaare (wie Fig. 14, 15, Tafel X), aber sparsamer als bei plicatus. Unterkiefer ohne Spatelhaare. — Die Felder an den Füssen etwas länger und schmäler
als bei plicatus, dicht mit Haaren wenig ausgeprägter Form bestellt.
3. Nyctinomus bivittatus Hgl.
Tafel XI, Fig. 3, Tafel X, Fig. 7, 12, 12 a, 14, 15 u. 18
3 Exemplare von Keren, Bogos, NO Afrika (Stuttgart). Zwischen den Nasenlöchern eine
kurze senkrechte Leiste; Oberlippe mit tiefen Falten. Unterhalb der Nasenlöcher und
der medianen Leiste typische Spatelhaare nicht sehr dicht gestellt, gegen den Mund
hin in scharfer gerader Linie abschneidend. Seitwärts unter Abnahme der typischen
Form zu 3–5 auf den Kämmen der Falten. Oberhalb der Nase lange, wenig typische dicke
Haare. Um von dieser Region eine Anschauung zu geben, ist der Kopf dieser Art von
der Seite dargestellt. Auf dem Unterkiefer vereinzelt helle kurze, aber dicke, wenig
typische Spatelhaare. Unter den Haaren der „ersten Gruppe“ fand ich am Kopfe dieser
Art solche mit fadenförmigem Anhang an der Spitze (Fig. 12). — Form der Haare an den
Füssen wenig ausgeprägt.
4. Nyctinomus brachypterus (Ptrs.)
Tafel X, Fig. 13
1 Exemplar von Lagos, W Afrika (Stuttgart). Auf dem Felde zwischen Nase und Mund ziemlich
weitläufig kurze Spatelhaare, deren Endplatte breit und ziemlich flach ist. Auf den
Falten des seitlichen Theiles der Oberlippe ebensolche, nur noch kürzer, so dass die
Endplatte fast unmittelbar über der Hautoberfläche steht. Die mittlere Parthie des
Gesichts oberhalb der Nase ganz ohne Spatelhaare. Unter den am weitesten nach oben
und medianwärts stehenden sind einzelne von wenig ausgeprägter Form, deren [45]Endplatte einen fadenförmigen Anhang trägt. — Unterkiefer frei. — Haare der Felder
an den Füssen ziemlich dicht und mittlerer Form sich nähernd.
5. Nyctinomus pumilus (Crtschm.)
Tafel X, Fig. 2 u. 2 a
2 Exemplare von Ägypten (Stuttgart), 1 von Massaua, O Afrika, 3 von Akusi, W Afrika (Stuttgart). Spatelhaare
ähnlich wie bei plicatus zwischen Nase und Mund, auf den Falten der Oberlippe und median oberhalb der Nase,
aber überall spärlich und vorwiegend nur mittlere Formen. Einzelne, z. Th. sehr kurze,
Haare (siehe die Figur) auf der Spitze mit fadenförmigem Anhange. Die Haare auf dem
Felde vorn an der Schnauze zeigen, namentlich an einigen Exemplaren, eine Sonderung
in eine obere und untere Gruppe angedeutet, wie sie schärfer bei der folgenden Art
besteht. — Am Unterkiefer wenig ausgeprägte Spatelhaare sehr spärlich jederseits nahe
dem Mundwinkel. — Felder an den Füssen ziemlich dicht behaart, Haare am Ende meist
nur wenig verbreitert.
6. Nyctinomus limbatus (Ptrs.)
Tafel XI, Fig. 4, Tafel X, Fig. 1 u. 8
Je 1 Exemplar von Quelimane, O Afrika (Stuttgart) und von Kama, W Afrika. Unterhalb
der Nasenlöcher und der zwischen ihnen befindlichen medianen Leiste ziemlich typische
Spatelhaare in zwei zwar nahen, aber doch deutlich unterscheidbaren Gruppen, die sich
hauptsächlich in querer Richtung ausdehnen. Die Haare der oberen Gruppe sind kürzer
als die der unteren und stehen in 2–3 Reihen, die der unteren nur in 1–2. Seitwärts
auf den Falten der Oberlippe minder typische Formen. Sehr lange dicke wenig ausgeprägte
Spatelhaare oberhalb der Nase bis gegen die Ohren hin. Unter letzteren Haaren auch
solche mit fadenförmigem Anhang an der Endplatte (Fig. 1). — Am Unterkiefer wenig
typische Haare ziemlich spärlich nach den Mundwinkeln hin. — Auf den Feldern der Füsse
die Haare locker gestellt, am Ende nur wenig verbreitert.
Über diese Art bemerkt Peters (Reise n. Mossambique. Zool. I. Säugeth. 56, Berl. 1852): „Die Oberlippe ist dick,
faltig, am Rande gekerbt und mit kurzen steifen Haaren besetzt“24. Dass hier die Spatelhaare gemeint sind, ergiebt ganz klar die Betrachtung der Fig.
1 a auf Tafel XIV (daselbst). Die Spatelhaare sind da vollkommen richtig in der Seitenansicht
des Kopfes (ähnlich wie in unserer Fig. 3 auf Tafel XI) durch Punkte und Striche vom Zeichner angedeutet.
7. Nyctinomus angolensis Ptrs.
Tafel XI, Fig. 5
2 Exemplare von Madagascar. Das Feld zwischen Nase und Mundrand mit Spatelhaaren mässig
dicht bestanden; die einzelnen Haare im oberen Theile des Feldes kürzer, nach unten
hin länger, in ziemlich scharfer Linie aufhörend; Endplatten ziemlich typisch, aber
nur mässig gegen den Schaft gebogen. Haare geringerer Ausbildung auf den Wülsten der
Oberlippe und oberhalb der Nase, hier wieder sehr lange und wenig ausgeprägte Formen.
— Auf dem Unterkiefer kurze Spatelhaare von nur angedeuteter Form spärlich in zwei
Gruppen nach den Mundwinkeln hin. — An den Füssen die erste Zehe sehr stark verdickt,
die Haare auf ihr und der fünften Zehe sehr locker gestellt und kaum am Ende verbreitert.
8. Nyctinomus astrolabiensis A. B. M.
Tafel XI, Fig. 6, Tafel X, Fig. 19 u. 30
1 Exemplar von Deutsch Neu Guinea. Spatelhaare mittlerer und geringerer Ausbildung
in einfacher Reihe längs des medianen Theils des oberen Mundrandes. Nach letzterem
zu schliesst sich dann, wie auch bei norfolcensis, noch eine Reihe von Borsten anderer Art (Fig. 19) an. Auf den seitlichen Parthieen
der schwach gerunzelten Oberlippe spärlich meist wenig ausgeprägte Spatelhaare. Sonst
im Gesichte fehlend. — Felder an den Füssen nach Form und Ausdehnung ähnlich wie bei
sarasinorum (vgl. Fig. 2 a auf Tafel XI), sehr dicht mit am Ende wenig verbreiterten Haaren besetzt.
[46]
9. Nyctinomus norfolcensis (Gr.)
1 Exemplar von N. S. Wales. Spatelhaare mittlerer Form in einfacher Reihe im medianen
Theile der Schnauze längs des äusseren Randes der Oberlippe; spärlich an ihrem seitlichen
ziemlich glatten Abschnitt. Oberhalb der Nase fehlend. — Auf dem Unterkiefer sehr
vereinzelt helle Haare von kaum angedeuteter Form. — Haare an den Füssen mässig dicht,
wenig ausgeprägt.
10. Nyctinomus loriae Thos.
2 Exemplare von Brit. Neu Guinea. Am Kopf helle, wenig ausgeprägte Spatelhaare spärlich
in einer Reihe nahe dem Mundrand auf dem medianen Theile der Oberlippe, sonst fehlend.
— Haare an den Füssen mit kaum verbreitertem Ende.
Arten ohne Spatelhaare im Gesicht
11. Nyctinomus brasiliensis Is. Geoffr.
Tafel XI, Fig. 7, Tafel X, Fig. 26 u. 27
Da bei dieser Art die den Spatelhaaren homologen Borsten am besten entwickelt sind,
so möge sie hier ohne Rücksicht auf die systematische Ordnung zuerst besprochen werden.
2 Exemplare von Brasilien, 1 von Guatemala (Stuttgart). Unterhalb der Nase zahlreiche
dünne in unsere „erste Gruppe“ zu rechnende Borsten. Nur bei éinem Exemplare von Brasilien
unter ihnen spärlich auch dicke der anderen Art. Letztere bei allen Exemplaren zahlreich
weiter seitwärts auf den Wülsten der Oberlippe sowie oberhalb der Nase, hier z. Th.
von beträchtlicher Länge (Fig. 27). — Am Unterkiefer solche Borsten zerstreut jederseits
von der Mittellinie. Die Enden der Borsten, besonders der längeren, vielfach pinselartig
aufgefasert. — Felder an den Füssen ziemlich dicht mit Haaren besetzt, die im ganzen
etwas abgeplattet, am oberen Ende aber nicht verbreitert sind, im übrigen den Haaren
an den Füssen der bisher besprochenen Arten gleichen.
Auf die Borsten im Gesichte von brasiliensis hat Allen aufmerksam gemacht (Monogr. Bats N. Am. Bull. U. St. Nat. Mus. 43, 1893). Er sagt
(S. 165): „The face is naked over the dorsal surface except in the space between the median border
of the auricle and the nostril where a number of long 3 mm to 4 mm stout bristles
are found“, und er macht dazu die Anmerkung: „The bristles about the nostrils have been neglected in describing species of Nyctinomus. They are conspicuous in N. brasiliensis, while almost absent in N. plicatus“. Die Borsten auf den Wülsten der Oberlippe werden im Texte nicht erwähnt, in den
Abbildungen (Fig. 1 u. 2, Taf. XXXII) sind sie dagegen richtig angedeutet. Das gleiche
gilt für die Felder an den Füssen (daselbst Fig. 4, 5 u. 6).
12. Nyctinomus africanus Dobs.
1 Exemplar von Keren, Bogos, NO Afrika (Stuttgart). Neben zahlreichen Borsten anderer
Art ähnliche wie bei brasiliensis spärlich unterhalb der Nasenlöcher sowie etwas seitlich von ihnen, ebensolche, z.
Th. sehr lange, Borsten oberhalb der Nase. — Am Unterkiefer vereinzelt. Manche von
diesen Borsten an der Spitze ähnlich denen von brasiliensis aufgefasert. — Haare an den Füssen locker stehend, am oberen Ende wenig, aber deutlich,
verbreitert.
13. Nyctinomus cestonii (Savi)
1 Exemplar aus Italien (Stuttgart), sehr ausgeblasst, was die genauere Untersuchung
erschwert. Borsten, ähnlich denen (Fig. 26, Taf. X) von brasiliensis, bemerkte ich nur auf den seitlichen Abschnitten der etwas gewulsteten Oberlippe.
— Haare an den Füssen stehen ziemlich dicht, Enden wenig verbreitert.
[47]
14. Nyctinomus gracilis (Natt.)
1 Exemplar von Panama (Stuttgart). Am Kopfe sind nur einige helle, ziemlich kurze,
etwas abgeplattete Borsten auf den Falten der Oberlippe möglicherweise als Vertreter
der Spatelhaare zu betrachten.—An den Füssen die gewöhnliche Anordnung, aber die einzelnen
Härchen sind wie bei brasiliensis nur im ganzen etwas abgeplattet und nicht am Ende
verbreitert.
Molossus Geoffr.
1. Molossus rufus Geoffr. und M. rufus obscurus (Geoffr.)
Tafel XI, Fig. 8, Tafel X, Fig. 29
Art und Unterart verhalten sich in Bezug auf das Vorkommen von Spatelhaaren ganz übereinstimmend.
Untersucht wurden von M. rufus: 3 Exemplare von Peru (2 Stuttgart), 1 von Surinam (dsgl.) und 1 von Jamaica (dsgl.);
von M. rufus obscurus: 1 von Central Peru (Stuttgart), 3 von Surinam (dsgl.), 1 von Cuba und 1 von Tobago
(Antillen).
Etwas unterhalb der Nasenlöcher bis nahe zum Mundrand ein meist scharf begrenztes,
annähernd dreieckiges oder trapezoides Feld, sehr dicht bestanden mit Spatelhaaren
vorwiegend mittlerer, aber auch geringerer Ausbildung, letztere Formen besonders an
den seitlichen Rändern des Feldes und in seinem unteren Theile, wo die Länge der einzelnen
Haare grösser wird. Sonst am Kopfe keine Spatelhaare. — Felder an den Füssen locker
bestanden mit Haaren von wenig ausgeprägter bis annähernd mittlerer Form.
Gervais (Expéd. de Castelnau, Zool. Mammif. 57, Paris 1855) hat das Feld an der Schnauze
bei M. rufus und rufus obscurus bemerkt und kennzeichnet seine gröberen Verhältnisse ganz zutreffend: „… la supérieure [sc. lèvre] est garnie au-dessous du nez de poils sétiformes très
courts et en brosse.“
Dobson (PZS. 1876, 709; auch Catal. 1878, 410) giebt für M. rufus Folgendes an: „… the nasal apertures … separated by a considerable interval (which is covered with short erect hairs dilated at their extremities25) from the margin of the lip …“ Auffallend bleibt es, dass diese und ähnliche Beobachtungen an noch einigen Molossus-Arten im Vereine mit den Angaben der früheren Autoren, die ihm sicher bekannt waren,
Dobson nicht anregten, den Gegenstand weiter zu verfolgen, und dass er diese eigentümliche
Haarform nicht wenigstens einer Erwähnung in der vortrefflichen allgemeinen Schilderung
der Fledermausorganisation, die er seinem Catal. 1878 vorausschickt, werth gefunden
hat.
2. Molossus nasutus Spix
1 Exemplar von Guatemala (Stuttgart). Am Kopfe Spatelhaare, meist wenig ausgeprägter
Form, nur auf einem Felde zwischen Nase und Mundrand ähnlich wie bei vorigen, aber
in weiterer Ausdehnung, und ebenfalls sehr dicht gedrängt. — Haare an den Füssen am
Ende nur wenig verbreitert.
Dobson (PZS. 1876, 711, auch Catal. 1878, 414) erwähnt das Feld an der Schnauze: „… the wide space between the nostrils and the margin of the upper lip occupied by
a quadrilateral patch of thickly spread short hairs …“
3. Molossus abrasus (Temm.)
Tafel XI, Fig. 9
4 Exemplare von Surinam (3 Stuttgart). Obere Ränder der Nasenlöcher wulstig und mit
warzigen Vorsprüngen, vereinigen sich nach abwärts zu einer medianen Leiste. Letztere
dicht besetzt mit wenig ausgeprägten, nur bei éinem Exemplare mittlerer Form sich
nähernden, Spatelhaaren, die vereinzelt auch noch auf die medialsten Theile des oberen
Nasenlochrandes übergreifen. Sonst am Kopfe keine Spatelhaare. — Felder an den Füssen
locker behaart, Haare verhältnissmässig dunkel, denen am Gesichte sehr ähnlich, z.
Th. ziemlich lang, namentlich am äusseren Rande des Feldes der fünften Zehe.
[48]
Dobson (PZS. 1876, 712; Catal. 1878, 415) sagt über die Haare an der Schnauze: „… the upper margin of the nasal disk thus formed on each side is finely and evenly
toothed, and the internasal ridge covered with short spoon-shaped hairs, similar to those forming a broad patch between the nostrils
and upper lip in M. rufus, but strictly limited to this ridge ..“26. Ich bemerke dazu, dass für die mir vorliegenden Exemplare gerade dieser Art der
Ausdruck „löffelförmige Haare“ kaum gerechtfertigt erscheint, doch kann ja an Dobsons Exemplaren die Form besser ausgebildet gewesen sein.
4. Molossus perotis (Wied)
Tafel XI, Fig. 10
1 Exemplar von Surinam (Stuttgart). Am Kopf ähnlich wie bei vorigem wenig ausgeprägte
Spatelhaare in geringer Zahl ausschliesslich auf einem senkrechten leistenartigen
Felde zwischen den Nasenlöchern, auch jederseits oben auf ihren Rand übergreifend.
— Haare an den Füssen am Ende wenig verbreitert.
Burmeister (Syst. Übers. Thiere Brasil. I. Säugeth. 66, Berl. 1854) sagt über diese Art: „…
die Hinterzehen sind kräftig, besonders die erste und letzte, welche die steifen Häkchen26 tragen …“ Auch erwähnt er (ebendort) die „mittlere stark behaarte Längsschwiele“
an der Schnauze, ohne jedoch etwas über die Form der Haare zu sagen. Vielleicht aber
ist die früher citirte Angabe über das Vorkommen von „hakenförmig aufwärts gebogenen
Borsten“ auf den Lippen des Genus Dysopes (= Molossus und Nyctinomus) überhaupt (ebendort S. 66) auch speciell auf diese Art zu beziehen.
Cheiromeles Horsf.
Cheiromeles torquatus Horsf.
Tafel XI, Fig. 11 u. 11 a–d, Tafel X, Fig. 6, 20, 21 a–e, 22, 23, 24 u. 25
1 Exemplar in Spiritus von Java, ausserdem 1 ausgestopftes und 1 trockener Balg von
Bórneo (SO u. W). Wie schon früher bemerkt, macht der Körper dieses interessanten
Thieres auf den ersten Anblick den Eindruck völliger Nacktheit, thatsächlich aber
ist die derbe schwartenartige Haut, wenn wir von der Flugmembran absehen, überall
in verschiedenem Maasse, doch immer spärlich, behaart. Am dichtesten stehen die Haare
auf Brust und Bauch, aber auch der Rücken ist nicht ganz nackt, wie Dobson (Catal. 1876, 178; Catal. 1878, 406) angiebt, sondern trägt auf seiner chagrinartig
rauhen Oberfläche in regelmässiger Vertheilung einzeln und in sehr weiten Zwischenräumen
stehende kurze Haare. Alle diese Haare besitzen die früher geschilderte Structur (siehe
Tafel X, Fig. 21). Das Gleiche gilt auch für die meisten längeren Haare, die sich an bestimmten
Körperstellen finden, besonders neben den Spatelhaaren und den ihnen verwandten Stacheln
im Gesichte (Fig. 20, Taf. X) und, eine Art Krause bildend, um den Hals. Hier erreichen diese Haare die grösste
Länge, 1–2 cm.
Die Form des Kopfes, der abgeplattet und langgestreckt, dabei zugleich sehr breit
ist (wie man aus einem Vergleiche der Seiten- mit der Vorderansicht auf Tafel XI, Fig. 11 u. 11 a ersehen kann), finde ich unter den mir vorliegenden Abbildungen
nur in der bei Temminck (Monogr. Mammal. II, Pl. 66, Leiden 1835–41) ziemlich naturgetreu wiedergegeben,
doch ist auch hier im Vergleiche zu dem Dresdener Spiritus-Exemplare die Breite zu
gering. In den ersten, nach einem getrockneten Balg entworfenen, Abbildungen Horsfields (Zool. Res. Lond. 1824, No. VIII, 2 Taf., Kopie der Oberansicht bei Temminck: Monogr. Mammal. I, Pl. 17. Paris 1827), sind die Formen verschiedentlich verzerrt,
und bei Dobson (Catal. 1878, Pl. XXI, Fig. 1 u. 1 a) erscheint der Kopf in unnatürlicher Verkürzung
und übertrieben hoch. Abweichend von dem Verhalten der übrigen Molossiden sind bei
Cheiromeles die Ohren verhältnissmässig klein und durch einen weiten Zwischenraum getrennt.
Die Schnauze trägt am äussersten Ende in der Mitte unterhalb der Nasenlöcher ein scharf
begrenztes Feld von der Gestalt eines Dreiecks, dessen Basis dem Mundrande genähert
und parallel ist (Fig. 11 u. 11 a, Taf. XI). Es ist dicht mit Spatelhaaren der in Fig. 22 u. 23 auf Tafel X dargestellten [49]modificirten Form besetzt, und trotz der Kürze der einzelnen Borsten, die noch nicht
1 mm frei hervorragen, markirt es sich doch auf der sonst fast nackten Haut am Spiritus-
wie an den trockenen Exemplaren schon für die Betrachtung mit blossem Auge sehr deutlich.
Indessen ist weder im Texte noch in den Abbildungen Horsfields (Zool. Res. 1824), Temmincks (Monogr. Mammal. II, 1835–41) und Dobsons (Catal. 1876 u. Cat. 1878) dieses Feld berücksichtigt.
Ausserdem finden sich am Kopfe neben spärlichen Haaren, die denen des Körpers gleichen
(Fig. 20, Taf. X), noch in geringer Zahl die derben bald kürzeren, bald längeren Borsten oder Stacheln,
die in Fig. 25 und 24 auf Tafel X abgebildet sind. Ein paar stehen einzeln seitwärts von dem medianen Feld an der Vorderseite
der Schnauze (Fig. 11 a, Tafel XI), und weiter kann man längs des obern Randes ihrer Seitenfläche vom Nasenloche bis
gegen das Ohr hin etwa drei bis vier kleine Gruppen von je zwei bis fünf Borsten unterscheiden.
Die Anordnung, wie sie die Figur 11 (Taf. XI) von dem Spiritusexemplare von Java zeigt, ist an den beiden trockenen von Bórneo
fast genau dieselbe.
Diese Gruppen hat schon Horsfield gesehen. Er unterscheidet (Zool. Res. 1824, No. VIII. Cheiromeles, 3. S.) genau „three small fascicles of short, stiff bristles, conical and glandular at the base,
rigid and spinous at the point“. Auch in der Artdiagnose berücksichtigt er diesen Charakter: „Rostrum conicum, sulcatum, glandulis confertis setiferis in paribus tribus oppositis coronatum“. Der Ausdruck „glandular at the base“ bezieht sich vermuthlich auf das Grübchen, das die Haut, ähnlich wie an den Spatelhaaren,
auch am Grunde dieser Stacheln öfter bildet. Auch Temminck (Monogr. Mammal. II, 349; Pl. 66, Fig. 3, 1835–41) erwähnt „de très petits mammelons qui donnent naissance à quelques poils rares et très courts“, Dobson (Catal. 1876 u. Cat. 1878) dagegen sagt nichts darüber.
Vereinzelt stehen solche Stacheln auch auf dem Unterkiefer in einiger Entfernung von
der Medianlinie, und, merkwürdigerweise, auch auf der Rückseite der Interfemoralmembran,
während ich hier bei den übrigen Molossiden vergeblich nach Spatelhaaren oder gleichwerthigen
Borsten suchte. Beide Standorte sind auch bei Horsfield angegeben.
Die Füsse, deren Beschaffenheit nur am Spiritusexemplare klar zu erkennen ist, sind
derb fleischig, mit dicken Zehen, deren Haut regelmässig quer gerunzelt ist (Taf. XI, Fig. 11 b). Die grosse Zehe ist doppelt so breit wie die anderen und wie ein Daumen
frei beweglich. Auf der ganzen äusseren Fläche, nahe vom Grunde bis zum Nagel, trägt
sie einen Besatz von langen, weit vorragenden Spatelhaaren, der bei der Betrachtung
des Thieres sogleich ins Auge fällt (Tafel XI, Fig. 11 b–d). Horsfield hat davon schon in der ersten Beschreibung des Cheiromeles (Zool. Res. 1824. VIII, 6. S.) eine ganz zutreffende Schilderung gegeben, der ich
mich nur anschliessen kann: „Together with the structure and position of the thumb, a very peculiar character is
afforded to the Cheiromeles by an appendage or brush, consisting of bristly hairs,
directed to one side, and forming a regular series along the outer margin of the thumb.
At the extremity the hairs are long, and spread as a fan over the nail; they gradually
decrease in length as they approach the base, preserving throughout the same direction.
The separate hairs are rigid, thicker at the base, then gradually attenuated, and
terminated by a hook.“ Ergänzend ist hierzu noch zu bemerken, dass die einzelnen Haare an ihrem Ende nicht
nur hakig gekrümmt, sondern zugleich auch spatelförmig verbreitert sind. Die Form
der Endplatte ist meist von mittlerer Ausprägung, am besten bei den kürzeren Haaren
entwickelt. Mit der wachsenden Länge der Haare wird die Abplattung und Erweiterung
am Ende immer geringer bis zu dem Grade der Fig. 11 auf Tafel X, und manchmal erscheint das Ende auch gleichmässig zugespitzt. Die längsten Haare
erreichen bis 8–10 mm, aber auch die von mittlerer Ausbildung werden 2 mm und darüber
lang (Fig. 6, Taf. X). Die Richtung der Krümmung ist durchweg plantar und proximal. Das einzelne Haar
ist röthlich braun gefärbt.
Der fünften Zehe fehlen Spatelhaare durchaus, im Gegensatze zu dem Verhalten aller
anderen Molossiden, dagegen finden sich wie dort so auch bei Cheiromeles an den Nagelgliedern der zweiten bis fünften Zehe einige längere Haare, die aber
immer kürzer sind als die bei Molossus und Nyctinomus und, anders wie bei diesen Gattungen, vorwiegend an der ventralen Seite des Klauengliedes
entspringen. In der Structur gleichen sie sehr den Spatelhaaren der ersten Zehe, sind
aber heller. An der Spitze sind sie etwas abgestumpft. Horsfield (Zool. Res. VIII, 6. S. 1824) bemerkt über diese Haare wieder ganz richtig: „This character [d. i. der Haarbesatz der grossen Zehe] must not be confounded with the long, solitary, bristly hairs which, in Cheiromeles
as well as in Nyctinomus, are loosely scattered over the [50]fingers, and particularly over the extremity, near the insertion of the claws.“ Die späteren Autoren Temminck (Monogr. Mammal. II, 348, 1835–41) und Dobson (Catal. 1876, 178; Catal. 1878, 406) schildern die Verhältnisse der Behaarung an
den Füssen lange nicht so eingehend und klar wie Horsfield.
Die im Vorstehenden aufgeführten 19 Arten umfassen nur wenig über ein Drittel der
bis jetzt überhaupt bekannt gewordenen Molossiden. Nach dem neuesten Verzeichnisse
der Säugethiere von Trouessart (Catal. Mammal. Nov. Edit. 1898–99. T. I, 142–149, T. II, Append. 1285–1286) und
mit Einschluss der beiden von A. B. Meyer neu aufgestellten Nyctinomus sarasinorum (s. oben S. 16) und astrolabiensis (desgl. S. 19) stehen den 14 von mir untersuchten Nyctinomus-Arten 24, den 4 Species von Molossus 6 gegenüber, von denen ich kein Exemplar zur Verfügung hatte.
Das Verhalten dieser Arten in Bezug auf den Besitz löffelförmiger oder entsprechender
Haare wird sich durch Prüfung des in den Museen vorhandenen Materials mit Leichtigkeit
feststellen lassen, inzwischen mögen hier noch einige Notizen aus der Literatur Platz
finden, die das Vorkommen solcher Haare für mehrere Arten wahrscheinlich machen, z.
Th. sogar mit Sicherheit erweisen.
Dies gilt zunächst für Molossus temmincki (Lund), wie aus der Bemerkung Burmeisters27 „die Lippen mit Hakenborsten28 gleichmässig zerstreut besetzt“ ganz klar hervorgeht.
Sodann ist Nyctinomus macrotis Gr. zu nennen, über den Gray (Ann. Nat. Hist. IV, 5–6, 1840) folgende Angaben macht: „The pads of the great and little toes rather large, covered with white hairs, which are curved and rather dilated at the tip“28. Und ferner: „.. muzzle bald, with a central longitudinal and a transverse marginal ridge of close
set short rigid hairs; upper lip with an oblong tuft of black hairs under the nose
..“. Dass es sich auch in der letzteren Bemerkung um Spatelhaare handelt, ist nicht
unwahrscheinlich, zumal auch in der Abbildung der oberen Parthie des Kopfes dieser
Species bei Dobson (PZS. 1876, 716, Fig. 3) oberhalb der Nasenlöcher vom Zeichner kurze steife Borsten
angegeben sind, die wohl Spatelhaare vorstellen könnten.
Eine Anzahl weiterer, weniger bestimmter Äusserungen stelle ich nach der Reihenfolge
der Arten in Trouessarts Catal. Mammal. 1898–99 hier noch kurz zusammen.
Molossus fluminensis Lat. „Des poils raides et courts occupent l’espace de demi cercle qui sépare le nez de la
lèvre supérieure“. (Lataste, Ann. Mus. Genov. 30, 661; vgl. Fig. 1, 660. 1891). Vermuthlich entsprechen diese
Haare dem Feld unterhalb der Nase bei Molossus rufus, dem diese Art sehr nahe steht (Lataste, l. c. 660).
M. bonariensis (Ptrs.). Die Abbildung bei Peters (Mb. Akad. Berl. 1874, 234. Taf. Fig. 1) zeigt die kurzen Härchen an den Verdickungen
der Zehen sehr deutlich, eine Andeutung ist vielleicht an der Schnauze zu erkennen
(Fig. 2 daselbst). Im Texte wird nichts darüber gesagt.
Nyctinomus orthotis H. Allen. „The face is occupied by a number of stout bristles between the ears and the muzzle“ (Allen, Proc. U. S. Nat. Mus. XII, 638. 1889).
N. kalinowskii Thos. In der Abbildung bei Thomas (PZS. 1893, pl. XXIX, Fig. 10) sind vorn an der Schnauze unterhalb der Nase kurze
steife Borsten angegeben. Im Texte nicht erwähnt.
N. acetabulosus (Comm.). „Face thinly sprinkled with short rigid hairs, or rather bristles“. (Smith, Ill. Zool. S. Afr. Mammal. Dysopes natalensis Smith [pl. 49] 1849).
N. setiger (Ptrs.). „Die breite wulstige, aber nicht quergefaltete Oberlippe ist jederseits mit vier
bis fünf Längsreihen kurzer stachelförmiger dicker Borsten bekleidet, zwischen denen
sparsame feine kurze und längere Härchen hervortreten. Die Unterlippe zeigt noch kürzere
und schwächere sparsame Borsten und Haare“ (Peters, Mb. Akad. Berl. 1878, 196; Taf. 1, Fig. 2–2 c). — Es scheint sich bei N. setiger um ähnliche Borsten wie bei brasiliensis zu handeln.
Hier wäre möglicherweise noch N. jugularis (Ptrs.) anzureihen, der nach der Abbildung bei Peters (M. B. Akad. Berl. 1881, 485, Taf. Fig. 2) auf der Oberlippe und auf der Schnauze
oberhalb der Nase kurze Borsten besitzt. Doch ist die Synonymie der Art zweifelhaft.
Peters hat sie 1865 aufgestellt [51](PZS. 468), Dobson 1876 (PZS. 734; auch Catal. 1878, 440) sie mit N. acetabulosus (Comm.) identificirt, dem sich Trouessart (Catal. Mamm. 1898–99, I, 149) anschliesst. Dagegen hat noch 1881 Peters (M. B. Akad. Berl. 483) dies nicht anerkannt, vielmehr N. jugularis mit dem später von Dobson (PZS. 1876, 733) aufgestellten N. albiventer für identisch erklärt. Ein Vergleich der Figuren bei Peters (M. B. Akad. Berl. 1881, 485, Tafel, 1 a und 2 a) mit denen bei Dobson (Catal. 1878, Pl. XXI, 5 u. 4) spricht mehr zu Gunsten der Auffassung von Peters. Einer vergleichenden Prüfung der vorhandenen Exemplare beider Arten muss es vorbehalten
bleiben, die Sache klarzustellen.
Rechnen wir die zuletzt erwähnten 8–9 Arten den von uns untersuchten hinzu, so würden
wir bis jetzt Spatelhaare oder stellvertretende Borsten bei etwas mehr als der Hälfte
der bekannten Molossiden im Gesicht anzunehmen haben. Für ziemlich sicher können wir
es halten, dass sie allen Arten von Molossus und Nyctinomus an den Aussenseiten der ersten und fünften Zehe zukommen, da die Verdickung dieser
Zehen als allgemeiner Charakter der beiden Gattungen aufgeführt wird (vgl. Dobson, Catal. 1878, 404), die verdickten Stellen aber bei allen untersuchten Arten solche
Haare tragen und durch die Anhäufung ihrer Bälge verursacht sind.
Erst nach genauer Kenntniss des Verhaltens sämmtlicher Arten wird es möglich sein,
die Frage zu beantworten, ob diesen Haaren eine Bedeutung in systematischer Hinsicht beizumessen sei. Einiges lässt sich in dieser Beziehung indessen wohl schon aus den
hier beigebrachten Beobachtungen entnehmen, worauf ich, vorbehaltlich der Controlle
durch eingehendere an reicherem Material anzustellende Prüfung, jetzt im Zusammenhange
kurz aufmerksam machen möchte.
Für jedes der drei Molossiden-Genera ist auch ein besonderes Verhalten der Spatelhaare
charakteristisch. Alle besitzen solche an den Füssen, aber Cheiromeles, der auch im übrigen isolirter steht, nur an der ersten Zehe, Molossus und Nyctinomus an der ersten und fünften. Bei den von mir untersuchten Molossus-Arten sind Spatelhaare am Kopf ausschliesslich auf die Region zwischen den Nasenlöchern
oder zwischen ihnen und dem oberen Mundrande beschränkt29, bei Nyctinomus sind sie oder die entsprechenden Borsten fast immer auch über die seitlichen Parthieen
der Oberlippe verbreitet.
Was die Ausnahmen betrifft, so kann Nyctinomus gracilis, bei dem ich solche Haare oder Borsten am Kopf überhaupt nicht mit Sicherheit nachweisen
konnte, hier ausser Betracht bleiben, da mir von der Art nur éin Exemplar zur Verfügung
stand, dessen Erhaltung nicht besonders günstig war, so dass die Untersuchung eines
reicheren Materiales leicht zu anderen Ergebnissen führen könnte. Von dieser Species
also abgesehen, bilden Nyctinomus norfolcensis, loriae und astrolabiensis eine Gruppe, für die bezeichnend ist, dass die Haare in einfacher Reihe zwischen
Nase und Mundrand angeordnet und fast ausschliesslich auf diese Stelle beschränkt
sind. Hierin ähneln die drei Arten den von mir untersuchten Molossi, bei denen nur nicht die Anordnung in einfacher transversaler Reihe vorkommt.
Nun steht N. loriae dem norfolcensis sehr nahe (Thomas, Ann. Mus. Genov. 38, 609, 1898), und dieser schliesst sich wieder zunächst an die
Untergattung Mormopterus Ptrs. an, die in mehrfacher Hinsicht eine mittlere Stellung zwischen den übrigen Nyctinomus-Arten und der Gattung Molossus einnimmt (Dobson, Catal. 1878, 441–442). Auch N. astrolabiensis steht zu diesen Arten in näherer Beziehung (vgl. das von A. B. Meyer oben S. 19 Bemerkte).
Unter den übrigen Nyctinomus-Arten ist die Anordnung, Menge und Ausbildung der Spatelhaare sehr übereinstimmend
bei plicatus, sarasinorum und bivittatus, von denen letztere beiden auch im übrigen gewissermaassen als celebensischer (A. B.
Meyer, oben S. 17), bezw. afrikanischer (Dobson, Catal. 1878, 426) Vertreter des plicatus zu betrachten sind.
N. pumilus und limbatus, zwischen denen nach Dobson (Catal. 1878, 429) nur geringfügige Unterschiede von kaum specifischem Werthe bestehen,
haben die Sonderung des Feldes unterhalb der Nase in eine obere und untere Abtheilung
gemeinsam.
Die Arten, denen Spatelhaare im Gesichte fehlen (africanus, cestonii, gracilis [?], brasiliensis), lassen sich gegenwärtig wohl nicht unter gemeinsamem Gesichtspunkte betrachten.
[52]
Innerhalb des Genus Molossus bilden die Haare bei rufus, rufus obscurus und nasutus eine scharf begrenzte compacte Gruppe unterhalb der Nasenlöcher, bei abrasus und perotis nehmen sie ein längliches leistenartiges Feld zwischen ihnen ein.
Mit Rücksicht auf die Bezahnung werden Molossus rufus (und rufus obscurus) einerseits, abrasus und perotis andererseits verschiedenen Untergattungen (Molossus Ptrs. und Promops Gerv.) zugetheilt, und nasutus, der danach allerdings zu Promops gehört, ist doch in anderer Beziehung „quite intermediate between M. rufus and M. abrasus“ (Dobson, Catal. 1878, 415), so dass auch die dem Subgenus Molossus entsprechende Anordnung der Spatelhaare bei nasutus nichts Auffälliges hat.
Cheiromeles schliesst sich durch den Besitz des scharf begrenzten dreieckigen Feldes modificirter
Spatelhaare an der Schnauzenspitze näher an Molossus als an Nyctinomus, und zu dieser Auffassung führt auch die Betrachtung der Gesammtorganisation des
merkwürdigen Thieres (Dobson, Catal. 1878, 404).
Kann man, wie wir gesehen haben, mit ziemlicher Sicherheit annehmen, dass jedem Molossiden Spatelhaare (wenigstens an den Füssen, wenn auch nur in rudimentärer Form) zukommen, so bleibt
die Frage, ob man diese merkwürdig geformten Haare als einen ausschliesslich den Molossiden eigenthümlichen Charakter betrachten dürfe. Ich bemerkte schon am Eingange der Abhandlung, dass ich ausserhalb
der genannten Gruppe bei keiner Fledermaus solche gefunden habe. Immerhin hätten bei
der nothgedrungen cursorischen Prüfung des reichen Materiales einzelne löffelförmige
Haare der Beobachtung entgehen können, und die Chiropteren-Sammlung des Dresdener
Museums weist natürlich auch manche Lücke auf, so dass diesem Punkte besondere Aufmerksamkeit
zu schenken sein wird, ehe man ein abschliessendes Urtheil fällen darf.
Hierbei will ich besonders hervorheben, dass in der Dresdener Sammlung die den Molossiden
nächstverwandte Gattung Mystacina Gr. (mit der einzigen Art tuberculata Gr.) nicht vertreten ist, und es auch nicht gelang, sie von anderwärts für diese Untersuchung
zu erhalten. Es spricht aber manches, auch abgesehen von der nahen systematischen
Verwandtschaft, dafür, bei dieser Art am ehesten solche Haare zu vermuthen.
Die Füsse der Mystacina (vgl. die eingehende Schilderung Dobsons, PZS. 1876, 488) sind wie die der Molossi breit, mit verdickter äusserer und innerer Zehe. Die langen gekrümmten Haare werden
als fehlend angegeben, dagegen scheinen nach der Abbildung bei Dobson (PZS. 1876, 487, Fig. b.) die äusseren Seiten der ersten und fünften Zehe mit kurzen
Härchen besetzt zu sein. Was das Gesicht betrifft, so finde ich bei Gray (Zool. Voy. Sulphur, Mammal. II. 23, 1843) eine vielleicht in diesem Zusammenhange
zu deutende Bemerkung: „Nose rather produced, surrounded at the base with a series of short rigid bristles“.
Es wird sich also jedenfalls empfehlen, Mystacina in erster Linie auf das Vorkommen löffelförmiger oder ähnlicher Haare genau zu prüfen.
Sollte es sich durch weitere Untersuchungen endgiltig bestätigen, dass das Vorkommen
löffelförmiger und verwandter Haare auf die Molossiden und etwa noch Mystacina beschränkt ist, so würde diese Thatsache im Vereine mit anderen vielleicht zu erwägen
geben, ob nicht der Gruppe Molossi eine selbständigere Stellung im System, als besondere Familie, anzuweisen sein möchte.
[53]