
Title: Der Ameisenlöwe
eine biologische, tierpsychologische und reflexbiologische Untersuchung
Author: Franz Doflein
Release date: January 23, 2025 [eBook #75185]
Language: German
Original publication: Jena: Gustav Fischer, 1916
Other information and formats: www.gutenberg.org/ebooks/75185
Credits: Peter Becker and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
Anmerkungen zur Transkription
Der vorliegende Text wurde anhand der Buchausgabe von 1916 so weit wie möglich originalgetreu wiedergegeben. Typographische Fehler wurden stillschweigend korrigiert. Ungewöhnliche und heute nicht mehr verwendete Schreibweisen bleiben gegenüber dem Original unverändert; fremdsprachliche Ausdrücke wurden nicht korrigiert.
EINE BIOLOGISCHE, TIERPSYCHOLOGISCHE
UND REFLEXBIOLOGISCHE UNTERSUCHUNG
VON
DR. FRANZ DOFLEIN,
O. PROFESSOR DER ZOOLOGIE AN DER UNIVERSITÄT FREIBURG I./BR.
MIT 10 TAFELN UND 43 ABBILDUNGEN IM TEXT
JENA
VERLAG VON GUSTAV FISCHER
1916
Alle Rechte vorbehalten.
Die vorliegende Arbeit wurde schon vor einer Reihe von Jahren begonnen. Erst während der Kriegszeit kam ich dazu, sie zum vorläufigen Abschluß zu bringen. Es war nicht immer leicht, beim Donner der Geschütze der nahen Front die Beobachtungen im Freien und im Laboratorium in Ruhe durchzuführen. Um so unvergeßlicher werden mir die Zeit dieser Arbeit und die Eindrücke bleiben, die sie brachte. Meinem Verleger bin ich sehr zu Dank verpflichtet für die Bereitwilligkeit, die Arbeit in der jetzigen Zeit herauszugeben und so liberal auszustatten.
Die Durchführung der experimentellen Untersuchungen wurde mir sehr erleichtert durch neue Einrichtungen meines Instituts, welche ursprünglich zu anderen Zwecken aus einer Zuwendung der Freiburger Wissenschaftlichen Gesellschaft beschafft worden waren. Ihr sei hiermit auch öffentlich gedankt.
Freiburg i./Br. im März 1916.
Franz Doflein.
|
Seite
|
||
|
Einleitung
|
||
|
I.
|
Vorkommen des Ameisenlöwen
|
|
|
II.
|
Bau des Ameisenlöwen
|
|
|
a) Aeußere Morphologie
|
||
|
b) Färbung und Zeichnung
|
||
|
III.
|
Das Verhalten des Ameisenlöwen in
freier Natur
|
|
|
IV.
|
Das Verhalten des Ameisenlöwen
unter experimentellen Bedingungen
|
|
|
1. Das Totstellen
|
||
|
2. Die Bereitschaftsstellung
|
||
|
3. Die Umdrehreaktion
|
||
|
4. Die Wanderbewegungen
|
||
|
5. Das Einbohren in den Sand
|
||
|
6. Der Bau der Sandtrichter
|
||
|
7. Der Ameisenfang
|
||
|
V.
|
Sinnesorgane und Sinnesreaktionen
des Ameisenlöwen
|
|
|
1. Bau und Funktion der Augen
|
||
|
2. Die Lichtsinnesreaktionen
|
||
|
3. Temperatursinn und Thermotaxis
|
||
|
4. Interferenz von Phototaxis und Thermotaxis
|
||
|
5. Der Tastsinn und seine Organe
|
||
|
a) Die Sinneshaare der Körperoberfläche
|
||
|
b) Die Erscheinungen der Tangorezeption
|
||
|
6. Thigmotaxis
|
||
|
7. Chemische Sinne
|
||
|
VI.
|
Die wichtigsten Reflexe des
Ameisenlöwen
|
|
|
VII.
|
Die Reizbarkeit des Ameisenlöwen
|
|
|
VIII.
|
Abriß der Lebensgeschichte des
Ameisenlöwen
|
|
|
IX.
|
Abschluß und Ergebnisse
|
|
|
Literaturverzeichnis
|
||
|
Erklärung der Tafeln
|
||
[S. 1]
Die Ameisenlöwen, die Larven einer Gruppe der planipennen Neuropteren, haben schon seit langer Zeit die Aufmerksamkeit der Naturbeobachter auf sich gelenkt. Die alten Naturforscher haben zahlreiche Schilderungen ihrer eigenartigen Lebensweise gegeben, welche fast stets als Musterbeispiel planmäßigen, intelligenten Handelns beschrieben wurde.
Ameisenlöwen im eigentlichen Sinn des Wortes sind nur die Larven gewisser Arten und Gattungen der Gruppe — bei uns in Deutschland die Larven von Arten der Gattung Myrmeleo, während z. B. die nahe verwandten Arten der in Süddeutschland an vielen Orten vorkommenden Gattung Ascalaphus frei umherschweifend sich meist von Blattläusen ernähren, wie das ja auch viele andere Neuropterenlarven tun.
Die echten Ameisenlöwen trifft man aber nur selten frei umherwandernd. In der Regel findet man sie in der Tiefe eines in feiner Erde oder Sand eingesenkten Trichters bis zu den Mandibeln eingewühlt. Die Mandibel strecken sie aus dem Sand weit klaffend hervor, bereit, jedes Insekt zwischen ihnen zu erfassen, welches den steilen Hang des Trichters mit seinem weichenden Boden herabgeglitten ist. Der Sturz der Ameisen wird häufig herbeigeführt oder befördert dadurch, daß der Ameisenlöwe Sand und Steinchen in die Höhe schleudert, welche das Opfer treffen oder es beim Herabrollen mitreißen, oft auch Partien der Trichterwand zum Herabgleiten bringen.
Die herabstürzende Ameise wird sofort von den zusammenklappenden Mandibeln der räuberischen Larve erfaßt und in einer eigenartigen Weise ausgesaugt, auf die wir später noch eingehen werden.
Der Bau der seltsamen Tierfalle, das Schießen nach den Opfern — das ganze Benehmen des Ameisenlöwen — macht den Eindruck[S. 2] einer ganz besonderen tierischen Leistung, und man glaubt, einem raffinierten Räuber gegenüberzustehen.
So ist das Tier auch meist beurteilt worden, und die zahlreichen Schilderungen in der Literatur laufen fast stets auf eine Hervorhebung der Tierintelligenz hinaus. Es ist aber sehr verwunderlich, daß ein so leicht erreichbares Objekt niemals einer genauen analytischen Untersuchung und gewissenhafter Beobachtung unterzogen wurde. Ja, unter den zahlreichen Schilderungen der Lebensweise des Ameisenlöwen scheint immer eine sich auf die andere zu stützen, und es wurde im Laufe der Zeit nicht allzuviel an kritisch beobachteten Tatsachen den alten Beschreibungen hinzugefügt.
Wie bei so vielen einheimischen Tieren, gehen die Schilderungen der Lebensweise des Ameisenlöwen meist auf die Beschreibung Rösels von Rosenhof zurück. Diese Beschreibung enthält aber neben sehr richtigen Beobachtungen einige auffällige Irrtümer; es berührt einen ganz seltsam, daß man den gewissenhaften Rösel von Rosenhof auch einmal in einigen Dingen richtigstellen muß, während er sonst so oft späteren Beobachtern gegenüber recht behalten hat.
Rösels Schilderung ist ohne weitere Kritik in die meisten Lehrbücher und in Schilderungen des Insektenlebens übergegangen; so steht sie noch in der letzten (3.) Auflage von Brehms Tierleben, wo sie ohne weiteres als richtig hingenommen wird. „Ausdauer und Schlauheit“ werden da als die wesentlichen Eigenschaften hervorgehoben, welche den Ameisenlöwen zu seinen eigenartigen Leistungen befähigen; sie sollen ihm ersetzen, was ihm „durch den Mangel anderer Naturanlagen versagt worden ist“ (Brehm).
Der Bau des Trichters wird im Anschluß an Rösel folgendermaßen geschildert: Das Tier baut unter ruckweisen, rückwärts gerichteten Bewegungen. „Es beginnt den Bau mit einem kreisförmigen Graben, dessen Größe durch seine eigene bedingt wird, und dessen Außenrand gleichzeitig den der zukünftigen Wohnung absteckt. In der Mitte steht demnach ein stumpfer Sandkegel, welchen er auf eine ebenso fördernde, wie sinnreiche Weise zu beseitigen versteht. Er wühlt sich da, wo er den ersten Kreis eben vollendete, mit dem Hinterleib in den Sand, und in einer immer enger werdenden Schraubenlinie zurückweichend, bringt er mit dem nach innen liegenden Vorderfuß den Sand auf seinen breiten, schaufelartigen Kopf und wirft ihn mit demselben so gewandt und mit[S. 3] solcher Gewalt über den Außenrand des ersten Grabens, daß er mindestens 5 cm weit wegfliegt. Dann und wann ruht er aus; ist er aber bei der Arbeit, so erzeugen die flinken Bewegungen einen ununterbrochenen Sandregen. Der innere Kegel nimmt mit jedem Umgang immer mehr ab, wie sich von selbst versteht, und schwindet vollständig mit der Ankunft des kleinen Minengräbers im Mittelpunkt, wo er sich mit Ausschluß der Zangen einwühlt und Platz greift. Um sich die Arbeit, welche eine bedeutende Muskelkraft in Anspruch nimmt, zu erleichtern, geht er nicht von Anfang bis zu Ende in derselben Richtung, sondern dreht sich von Zeit zu Zeit um, damit einmal das linke Bein Handlangerdienste verrichte, wenn es bisher das rechte getan hatte. Kommen gröbere Sandkörner in den Weg, was nicht ausbleibt, so werden sie einzeln aufgeladen, noch größere, welche sich nicht werfen lassen, wohl gar auf dem Rücken hinausgetragen. Man hat beobachtet, daß in dieser Hinsicht mißlungene Versuche öfter wiederholt werden, und daß erst dann, wenn sich alle Bemühungen erfolglos zeigten, ein anderer Platz in der Nachbarschaft ausgesucht wurde, um hier die Arbeit in Erwartung eines glücklicheren Erfolgs von vorn zu beginnen.“
In fast allen zusammenfassenden Darstellungen findet sich diese Schilderung nach Rösel wiederholt; neue Originaluntersuchungen des interessanten Vorgangs finden sich kaum. Nur eine Beschreibung von Redtenbacher fand ich, welche auf eigenen Beobachtungen fußt.
Redtenbachers Beobachtungen stimmen in vielen Punkten mit den meinigen überein, doch weichen sie auch in einigen nicht unwesentlichen Einzelheiten von ihnen ab. Dies ist vielleicht darauf zurückzuführen, daß er die nahe verwandte Art Myrmeleo europaeus M’L. untersuchte, während mir M. formicarius L. wohl in all meinen Versuchen vorlag. In späteren Abschnitten, so besonders in demjenigen über Trichterbau, wird auf die verschiedenen Abweichungen unserer Beobachtungen im einzelnen hingewiesen werden.
Welch seltsam durcheinandergehende Anschauungen über die Handlungen des Ameisenlöwen von den verschiedenen Autoren geäußert werden, lehrt jeder Blick in die Literatur, welche allerdings mehr von Dilettanten als von sorgfältig und kritisch beobachtenden Fachleuten herrührt. Man beachte nur, wie schwer den Beobachtern einzelner Vorgänge, wenn sie ohne größeren Zusammenhang, ohne planmäßige Arbeit registriert werden, ihre Beurteilung fällt. So als Beispiel die kurzen Notizen von Chapman und Lucas in des letzteren Arbeit (1906).
[S. 4]
Meine Beobachtungen am Ameisenlöwen weichen von der Mehrzahl der vorhandenen Beschreibungen so sehr ab, ihre Analyse ergibt zudem eine so von der herrschenden sich unterscheidende theoretische Beurteilung der physiologischen Grundlagen der Handlungen des Tieres, daß eine ausführliche Darstellung gerechtfertigt erscheint.
Seit einer Reihe von Jahren (seit 1910) beobachte ich Ameisenlöwen, experimentiere mit ihnen und beachte ihr Vorkommen. Ich habe sie auf der oberbayrischen Hochebene, in der weiteren Umgebung von München, beobachtet, erhielt sie aus Landshut a. d. Isar zugeschickt, fand sie in den bayrischen Alpen bei Garmisch. In großer Menge kommen sie an der Salzach in der Nachbarschaft von Burghausen vor. Mein wichtigstes Beobachtungs- und Versuchsmaterial stammt aber aus der Umgebung von Freiburg i. Br., wo Ameisenlöwen sowohl auf den Schwarzwaldausläufern in der Nähe der Stadt, als auch im eigentlichen Schwarzwald, wie in Kaiserstuhl und Vogesen sehr häufig sind.
Meist fand ich die Trichter der Larven an Waldrändern, auf Lichtungen, an Wegböschungen, am Rand von Hecken und Buschwerk, auch an heidekrautbewachsenen offenen Stellen. Wo Kiefernwälder gedeihen, pflegen auch für Ameisenlöwen die geeigneten Existenzbedingungen vorzuliegen.
Nie fand ich sie im tiefen Schatten des Waldes, immer ist Sonnenschein eine der notwendigen Bedingungen für ihr Vorkommen. Doch beobachtete ich sie bei uns in Deutschland nie in ganz offenem Gelände, wo sie wohl im Mittelmeergebiet und in südlichen, regenarmen Gebieten leben. Schutz vor Regen, der die Trichter zusammenschwemmen, die Erd- und Sandkörner zusammenbacken würde, ist eine notwendige Vorbedingung für ihr Vorkommen.
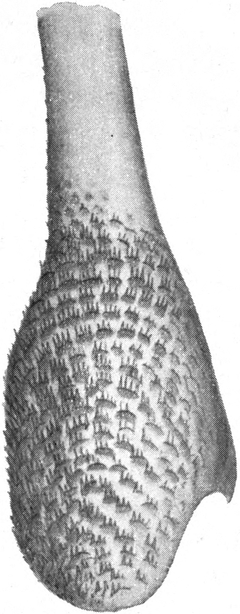
So finden wir denn die Trichter meist an überdachten Stellen, wo Wurzeln eines Baumes, Rasenpolster, Felsen und Steine etwas unterwühlt und beständigen atmosphärischen Einflüssen unterworfen sind. Unter solchen vorstehenden Böschungen, also vor allem an Wald- und Straßenrändern gibt es immer Stellen, welche in der Regel dem Regen unzugänglich sind und nur bei ganz schweren Regengüssen naß werden. Der Wind und das langsam durchrieselnde Wasser haben aber eine verarbeitende Wirkung auf deren Material.[S. 5] Es wird in feine Körner und Staub zerlegt, welche an der Böschung herabrieseln und sich da der Schwere nach anordnen, so daß oben das feinste Material liegen bleibt, während die größeren und schwereren Stücke die Böschung hinabrollen. Besonders da, wo unter dem schützenden Dach eine der Wagrechten sich annähernde Ebene feinsten Materials sich bildet und wo dessen Schicht eine gewisse Dicke erreicht, siedeln sich die Ameisenlöwen mit Vorliebe an (Abb. 1).

Dabei machen sie keinen großen Unterschied in dem Material, aus welchem ihr „Sand“ besteht. Ich fand sie ebenso im Quarzsand, wie in dem aus Tuff und Löß stammenden Staubsand des Kaiserstuhls. Auch in Humus bauen sie ihre Trichter. Es muß das Material nur hinreichend trocken sein, und aus Gründen, die wir später kennen lernen werden, müssen die einzelnen Partikel ein gewisses Gewicht besitzen. So kommen sie nicht leicht in dem feinen Kalkstaub der bayrischen Hochebene vor, wohl aber in dem grobkörnigen Kalksand[S. 6] des Gebirgsrandes, bevorzugen aber den aus Buntsandstein, Granit, Gneis usw. sich bildenden schweren, feinkörnigen Sand.
Nach heftigen Regengüssen findet man die Ameisenlöwen bewegungslos unter den eingestürzten Trichtern im nassen Sand. Erst wenn dieser wieder ziemlich trocken geworden ist, wird der Trichter neu gebaut.
Das Vorkommen an den charakteristischen Fundorten ist wohl zum Teil dadurch bedingt, daß die Muttertiere die Eier gleich an geeigneten Stellen ablegen werden. Meine Experimente weisen aber auch darauf hin, daß die Larven zum Trichterbau geeignete Stellen durch aktive Wanderung aufsuchen können.
Zum Verständnis der Bewegungen und der Reizreaktionen des Ameisenlöwen ist — wie aus den späteren Abschnitten dieser Untersuchung klar werden wird — eine genaue Kenntnis des Baues dieses merkwürdigen Tieres unbedingt notwendig. Die bisher vorliegenden Beschreibungen berücksichtigen die für uns wichtigen Einzelheiten kaum. Daher habe ich im Verlauf meiner Forschungen — wie es jeweils die Experimente und Beobachtungen am lebenden Tier nötig machten — viele Einzelheiten seines Baues untersucht, welche ich im nachfolgenden gemeinsam mit dem früher schon von dem Tier Bekannten darstellen will.

Bei dem ruhig auf einer Unterlage mit der Bauchseite aufliegenden Ameisenlöwen — es sei hier zunächst nur von den vollkommen oder annähernd erwachsenen Individuen die Rede — sehen wir von oben auf einen ungefähr wappenschildförmigen Körper, dessen Vorderrand einen bogenförmigen Ausschnitt zeigt; von den „Schultern“ aus erweitert sich der Körperumriß noch etwas, um dann nach hinten bis zur abgerundeten Hinterleibsspitze allmählich zuzulaufen. Der Querschnitt des Körpers ist hochoval, der Leib beim gut genährten Tier hochgewölbt und prall. Den vorderen Teil des Leibes bilden die beiden großen und breiten hinteren Thorakalsegmente; der Metathorax sieht fast genau so aus, wie die an ihn sich anschließenden Segmente des Abdomens; er grenzt an den Mesothorax mit einem schwach konkaven Bogen, während seine hintere Begrenzung schwach konvex nach vorn ist. Auch der Mesothorax hat an der Rückenseite[S. 7] eine konkave Vorderkontur, hinter welcher eine tiefe Einbuchtung nach hinten verläuft, die von einem wulstigen Rand eingefaßt ist. Der vordere Rand des Mesothorax überwölbt das hintere Ende des Prothorax. Vor dem Hinterrand des Mesothorax bildet sich bei Längskontraktion des Körpers, wie auch auf dem Metathorax und[S. 10] den Abdominalgliedern ein dem Segmentrand paralleler Wulst. Ueber den eigentlichen Körperumriß ragt wie ein Hals der bedeutend schmälere Prothorax hervor (Abb. 2). Er ist nur etwa so breit wie der Kopf, hat bogenförmige Seitenränder und auf der Mitte der Rückenfläche einen Längswulst. Der hintere Teil des Prothorax kann unter die oben erwähnte Vorwölbung des Mesothorax zurückgezogen werden. Von unten sind die drei Thorakalglieder relativ glatt, abgesehen von einem Wulst in der Mittellinie und den durch die Einlenkung der Beinpaare bedingten Vertiefungen. Dazu kommen an den Seiten des Meso- und Metathorax jene „Schultern“ mit ihren Borstenbüscheln, die nachher im Zusammenhang mit dem Borstenkleid des ganzen Tieres besprochen werden sollen.
Der Hinterleib (Abb. 2) besteht aus 9 deutlich sichtbaren Ringen, welche bis zum sechsten allmählich, dann plötzlich kleiner werden. In der Richtung von vorn nach hinten sind die Hinterleibsringe untereinander etwa gleich breit. Doch sind infolge der Verlängerung der Zwischensegmenthäute zwischen je zwei der Chitinringe sogenannte Zwischensegmente eingeschoben, deren es also bei 3 Brust- und 9 Hinterleibsringen im ganzen auch 12 gibt. Jeder Ring stellt infolge seiner Wölbung einen den Körper umfassenden, wulstartigen Gürtel dar; über das ganze Abdomen verlaufen sowohl auf der Ober- wie auf der Unterseite parallel dem hochgewölbten Mittelteil je zwei Furchen von vorn nach hinten, welche meist ziemlich seicht sind und nur bei ausgehungerten Individuen sich stärker vertiefen. Sie liegen etwas näher der Rückenmitte als dem Seitenrand.
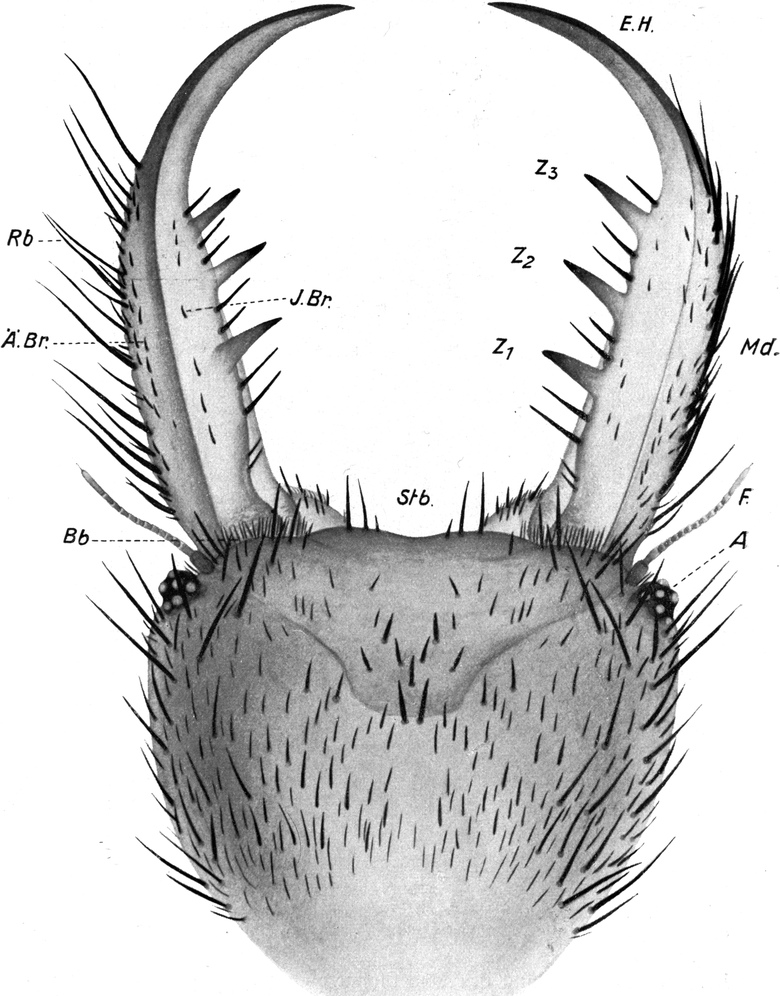
Der Kopf ist, von oben (Abb. 3) betrachtet, etwa herzförmig, da die Stirn konkav ausgeschnitten ist und sein Umriß gegen den „Hals“ zu etwas zuläuft. Gegen die Stirnbucht vertieft sich auch die Stirnfläche, während die beiden Vorderecken wulstig verdickt sind. Letztere beiden tragen die Augen und Fühler. Von unten ist der Kopf stark gewölbt, sehr glatt und glänzend (Abb. 4). In der Mundregion ist eine tiefe Bucht. Gegen den Hals hin ist die Unterseite des Kopfes schwach kinnartig vorgewölbt.
Von den Mundteilen fallen äußerlich zunächst nur die mächtigen Mandibel, die Oberkiefer, auf. Sie sind große, hakenförmige Bildungen, etwa von gleicher Länge wie der ganze Kopf (Abb. 3).
Die Mandibel verlaufen in ihrem basalen Teil ziemlich gerade, um gegen das Ende in einer säbelförmigen Krümmung nach innen[S. 11] umzubiegen. Außer einer Einbuchtung an der Basis ist der Außenrand glatt, allerdings sehr dicht mit Borsten bestanden. Der Innenrand zeigt an entsprechender Stelle wie der Außenrand eine Ausbuchtung, so daß an der Basis der Mandibel auf diese Weise eine Einbuchtung entsteht (Abb. 3 und 4). Die gerade Strecke des Innenrandes trägt in gleichmäßigem Abstand drei mächtige, feste, spitze Zähne, welche schief nach vorn gerichtet sind. Sie sind wie die Mandibel selbst von oben nach unten abgeflacht. Der Endhaken der Mandibel ist auch flach und an der Außenseite ganzrandig. Auch ist er borstenlos (Abb. 3). Die Innenseite der Spitze ist jedoch gezähnelt; und zwar sind es 8–10 Sägezähne, welche mit ihrer Spitze nach hinten gerichtet sind. Die Reihe dieser Sägezähne geht dann auf die Unterseite der Mandibel über und bildet da eine Reihe schiefer Rillen und Höcker (vgl. Abb. 5). Das Ganze ist ein vorzügliches Hilfsmittel, um Beutetiere während des Aussaugens festzuhalten. An der Außenseite sind nach vorn gerichtete Zähne vorhanden, welche wohl beim Bohren der Wunde Dienste leisten.
Der Innenrand der Mandibel trägt eine Reihe mittellanger, sehr kräftiger Borsten, welche stumpf, ungezähnt und ungefiedert sind. Eine von ihnen steht vor dem vordersten Zahn, zwischen den Zähnen meist zwei, manchmal auch drei, hinter dem hintersten Zahn auch zwei bis drei, in der inneren Einbuchtung noch zwei kleinere. Der Außenrand trägt eine größere Anzahl sehr kräftiger und besonders langer Borsten, deren erste etwa gegenüber dem ersten Zahn, meist etwas weiter vorn steht. Zwischen ihnen stehen kleinere Borsten. Sie alle sind am vorderen Teil der Mandibel glattrandig, an deren Basis eigenartig gezähnelt. Auf der Oberseite der Mandibel finden sich ferner zwei Reihen von kurzen, kräftigen Borsten, welche schwach gekrümmt sind und in zwei Hauptgruppen stehen. Eine meist einfache Reihe (Abb. 3 J.Br) verläuft parallel dem Innenrand der Mandibel, eine im proximalen Teil doppelte Reihe parallel dem Außenrand (Abb. 3 Ä.Br). Alle diese Borsten sind schief nach vorn geneigt (Abb. 3 und 4).
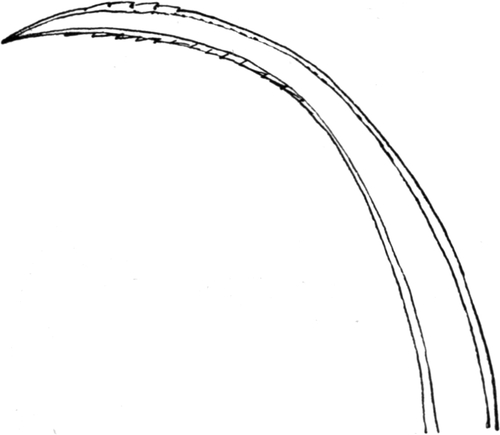
[S. 12]
An der Unterseite findet sich nur am Außenrand in der Nähe der Basis eine Gruppe von Borsten, die einigermaßen dicht stehen, glatt, gekrümmt und nach vorn gerichtet sind (Abb. 4). An der Unterseite zieht sich ferner bis zur Spitze der Mandibel eine Rinne hin, welche an der Basis breiter, an der Spitze schmäler ist, und deren äußerer Rand dem Außenrand der Mandibel parallel verläuft (Abb. 4 Ri). Sie wird von Wülsten begrenzt, von denen der äußere stärker entwickelt ist.
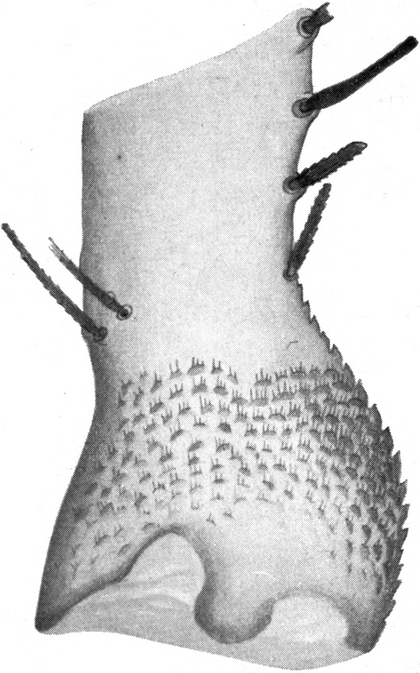
Der untere Teil der Mandibel steckt im Kopfe drin; er endet mit verschiedenen zipfelförmig ausgezogenen Ansatzstellen für die Beißmuskeln, nach einwärts ziehenden Abductores und nach abwärts ziehenden Adductores. Oberhalb derselben ist die Mandibel nach innen halbkugelig angeschwollen. Die Wölbung dieses Gebildes ist durch zahlreiche schuppenförmige, nach vorn gerichtete, gezähnelte Granulationen rauh. Offenbar dienen diese Granulationen dazu, um die Mandibel in den verschiedensten Stellungen zu sperren. Auch werden sie ermöglichen, daß sie ohne Muskelwirkung zuhalten, wenn sie über einer Ameise zusammengebissen wurden (Abb. 6).
[S. 13]
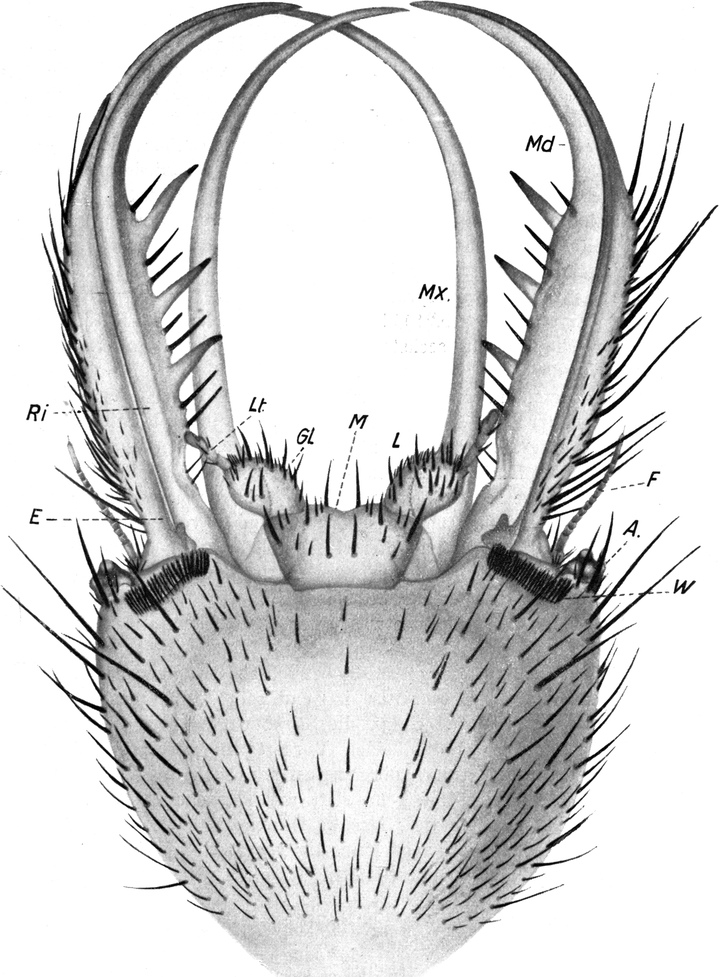
In derselben Region zeigt die in der Rille der Mandibel verborgene Maxille eine entsprechende Verbreiterung, die ebenfalls mit solchen Schuppen bedeckt ist (Abb. 7). Sonst zeigt die Maxille auf ihrer Oberfläche weder Haare noch Zähne. Nur an der Spitze ist sie mit Zähnchen versehen, welche umgekehrt wie beim Oberkiefer an der Innenseite nach vorn, an der Außenseite nach hinten gerichtet sind (Abb. 5). Sie ist sehr schlank, säbelförmig und hat auf beiden Seiten ganz glatte Ränder (Abb. 4 Mx). Sie ist abgeplattet und hat selbst auf der Oberseite eine Rinne und bildet so mit der Rinne der Mandibel, in welche sie ganz genau hineinpaßt, eine Röhre, durch welche der Ameisenlöwe seine Nahrung einsaugt und der Mundhöhle zuleitet. Nach Dewitz sind die am Ober- und Unterkiefer befindlichen beiden Längsleisten eine „Führung“, dazu bestimmt, den beiden eng vereinigten Mundgliedmaßen eine Verschiebung in der Längsrichtung zu gestatten, ohne daß die beiden Teile auseinanderklappen.
Ventral schließen sich an die Maxillen die Bestandteile der Unterlippe an. Ich unterscheide ein in der Mitte leicht eingekerbtes einheitliches Mittelstück (Abb. 4 M), aus welchem jederseits eine schwach bewegliche Glossa (Abb. 4 Gl) hervorragt, welche den viergliedrigen Labialtaster (Abb. 4 Lt) trägt. Alle Teile tragen kurze starre und einige längere Borsten. Auffallend ist die sehr regelmäßige Verteilung der letzteren auf dem Mittelstück. Zwischen den Glossen stehen 2 große Borsten, 2 ebensolche weiter hinten auf der Kinnfläche, je eine in den äußeren Winkeln, und auch die kleineren Borsten sind symmetrisch verteilt (vgl. Abb. 3).
Der Zugang in die Mundhöhle befindet sich, wie besonders Lozinski nachgewiesen hat, in den äußersten Winkeln der eigenartig zusammengepreßten Mundspalte. Dort, wo die Kiefer ihren Gelenken zustreben, steht die Mundspalte beiderseits etwas offen, und hier findet die Verbindung zwischen der Mundhöhle und den Längskanälen der Mandibel statt. Die Mundspalte ist nicht etwa zugewachsen, sondern zusammengepreßt und wird durch Einfügung von Fortsätzen in Rinnen zusammengehalten. Dabei spielt ein als Labrum gedeutetes Plättchen am Oberrand des Kopfes eine besondere Rolle.
Zu beiden Seiten der Unterlippe finden sich zwei bewegliche Stücke im Skelett des Kopfes. Sie werden von verschiedenen Untersuchern als Cardo und Stipes der Mandibel gedeutet.
Im untersten Teil der Maxille findet sich ferner nach Lozinski eine Drüse, welche ihr Sekret in den Kanal entleert und wohl als Giftdrüse zu deuten ist.
[S. 14]
Oberhalb des Ansatzes der Mandibel ragt der Kopf bogenförmig vor. An der äußersten Kante sitzen die Augen. Diese stellen kegelförmige Höcker dar, welche schief nach außen und etwas nach oben gerichtet sind (Abb. 3 und 4 A). Die exponierte Lage der Augen bringt es mit sich, daß sie leicht aus dem Sande hervorragen, wenn der übrige Tierkörper in ihm verborgen ist. Das Auge besteht aus einem Chitinzapfen, dessen Wand dunkelbraun pigmentiert ist. Nur an einigen Stellen sind kreisförmige Unterbrechungen des pigmentierten Chitins; hier befinden sich die Linsen, welche sehr stark kuppelförmig über den äußeren Umriß des Augenkegels vorragen (Abb. 4). Ich zähle auf der unteren Fläche 1 Linse und deren 6 auf der oberen Fläche des Augenstiels, also insgesamt 7 Linsen. Der feinere Bau der Augen wird weiter unten bei Besprechung des Lichtsinnes des Ameisenlöwen beschrieben. Unterhalb des Augenkegels zieht sich im Bogen bis zur Stirn, fast wie eine Wimperreihe aussehend, eine Reihe feiner, flacher und gefiederter Borsten hin (Abb. 3 Bb). Etwas vor dem Auge, unterhalb dieser Wimperreihe und oberhalb des Mandibelursprungs entspringt jederseits die Antenne (Abb. 3 und 4 F). Sie beginnt mit einem starken Basalglied, auf welches eine aus kleineren Gliedern zusammengesetzte Antennengeißel folgt. Das erste Geißelglied ist doppelt so lang und etwas dünner als die folgenden Glieder; diese, 11–13 an der Zahl, sind untereinander gleich groß, jeweils am inneren Rande etwas schmäler als am äußeren. Sie stecken infolgedessen dütenförmig eins in dem anderen. Das letzte Glied der Antenne ist etwa dreimal so lang wie diese Geißelglieder, ebenso breit wie diese und am äußersten Ende mit einem feinen, stiftchenförmigen, wahrscheinlich hohlen Fortsatz versehen. Auf den Gliedern der Antennen finden sich keine Haare. Der erwähnte Unterschied in der Zahl der Antennenglieder bei den verschiedenen Individuen ist wahrscheinlich darauf zurückzuführen, daß die beiden Geschlechter schon im Larvenzustand zu unterscheiden sind. Wir werden später noch weitere Angaben hierüber machen.
Auch an der Unterseite des Kopfes, zwischen dem Auge und der Antenne einerseits und der Mandibelbasis andererseits verläuft eine ähnliche Wimperreihe, wie wir sie oberhalb der Mandibel antrafen (Abb. 4 W). Doch besteht diese untere Wimperreihe aus sehr groben, kurzen, stumpfen und an der Längskante feingesägten Borsten. Zwischen ihnen stehen einige feine, dünne Borsten, wie wir sie in der oberen Wimperreihe antrafen. Da einzelne der Borsten dieser[S. 15] unteren Reihe Spitzen besitzen, so nehme ich an, daß der Mangel von Spitzen auf der Abnützung derselben beruht. Die nach vorn gerichteten Spitzen erfahren sicher bei den Bewegungen des Tieres durch den Widerstand des Sandes eine starke Abnützung. Auch andere Borsten der Kopfregion zeigen abgebrochene oder abgenützte Spitzen. Jene untere Borstenreihe vermag wohl auch infolge ihrer Anordnung das Eindringen von feinen Sandteilchen in das Gelenk der Maxille zu verhindern.
Die Oberseite des Kopfes ist ähnlich wie die Mandibel mit zahlreichen schwarzen, starken Borsten besetzt (Abb. 3). Dieselben stecken alle mit Kugelgelenken in wulstigen Pfannen, welche von der Chitinbedeckung des Kopfes gebildet werden. Vorn und am Seitenrand sind die Borsten lang und dick, vor allem auch in der Umgebung der Augen. Am Vorderrande des Kopfes sind sie gerade nach vorn gestreckt; an den Seitenrändern des Kopfes sind sie etwas nach innen gebogen und ragen mit ihren Spitzen gerade nach vorn. Auf der Mitte der Stirnregion steht eine regelmäßig angeordnete Gruppe von 9 starken Borsten (vgl. Abb. 3). Weiter nach hinten auf der Oberseite des Kopfes sind die Haare etwas kürzer und dünner; alle stehen nach vorn und sind etwas nach unten gekrümmt. Ganz ähnlich verhält sich die Behaarung auf der Unterseite des Kopfes, doch ist sie häufig sehr viel schwächer als auf der Oberseite. Auch besteht sie in der Hauptsache aus untereinander gleich großen, relativ kleinen Haaren; nur am Rand und vor allem vorn in der Augenregion steht eine Anzahl langer und starker Borsten. Die Region hinter der Unterlippe und das Kinn sind fast borstenfrei; doch lassen die wenigen hier vorhandenen Haare eine auffallend symmetrische Anordnung erkennen (Abb. 3).
Der Kopf ist an seinem Uebergang zum Hals stark verschmälert und steckt mit seinem Ansatz in dem letzteren wie in einem Futteral. Der Hals oder das erste Thorakalglied ist lang und schmal. Auf reinen Chitinpräparaten macht er den Eindruck, als sei er aus zwei Segmenten aufgebaut. Der hintere Teil kann unter den Vorderrand des zweiten Brustringes zurückgezogen werden. Der vordere Teil wird als ein sogenanntes Zwischensegment angesehen. In der Mitte befindet sich eine Gelenkstelle. Der Vorderrand des ganzen Halsgliedes sowie derjenige des mittleren Gelenkes ist mit einer Reihe nach vorn stehender Borsten bestanden. Ueberhaupt sind Ränder und Oberfläche des Halses mit Borsten vollkommen bedeckt. Seitlich vorn findet sich ein zitzenförmiger Höcker, der einige Borsten[S. 16] trägt (Abb. 2). Im hinteren Teil der zweiten Hälfte dieses Gliedes befindet sich der Ursprung des ersten Beinpaares. Die Beinpaare werden wir nachher im Zusammenhang beschreiben.
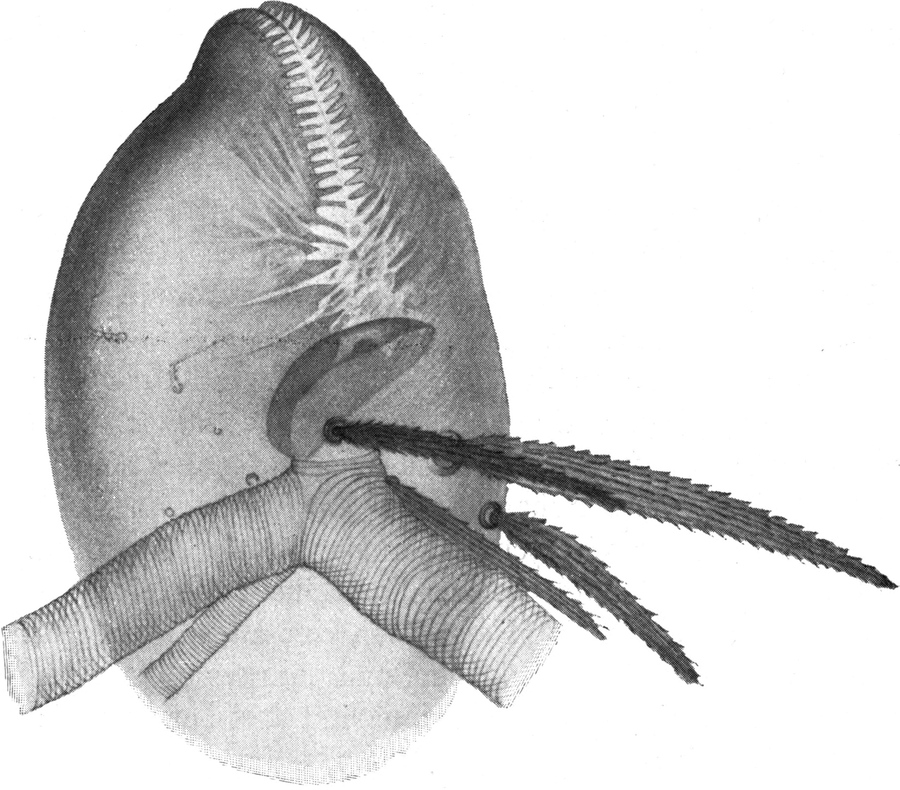
Es folgt nun das zweite Thorakalglied, welches schon um ein gutes Stück breiter ist als der Hals. Es zeigt an den Seiten des Körpers wulstige Vorragungen, welche mit Gruppen langer, dicker Borsten besetzt sind. Man unterscheidet beiderseits zwei solche Wülste, von denen der eine kleinere dem Hals näher liegt, während der größere mehr nach außen gewendet ist. Die Region dieses äußeren Wulstes habe ich oben als die „Schulter“ bezeichnet (vgl. Abb. 2). Der Vorderrand des zweiten Thorakalgliedes zeigt jene muldenförmige Eindellung, welche wir schon früher geschildert haben. An deren Vorder- und Hinterrand stehen mit den Spitzen nach vorn gekehrte Reihen starker Borsten. Auch auf der Oberfläche des Gliedes finden sich zahlreiche Borstengruppen. Die Spitzen aller sind nach vorn gekehrt. Die Eindellung hinter dem wulstigen Vorderrand verbreitert sich nach den Seiten hin; an ihrer tiefsten Stelle ist eine scheibenförmige Region dicken, glänzenden und harten Chitins erkennbar.[S. 17] Diese Stellen entsprechen den Bildungsstellen der Flügel. Sie wiederholen sich auf dem dritten Thorakalglied. An der Unterseite des zweiten Thorakalgliedes sitzen an den vorderen Ecken die zwei Oeffnungen des ersten Stigmenpaares; diese befinden sich auf vorragenden kapselförmigen Ausbauchungen der Körperwand, welche ungefähr die Form einer Taschenuhr besitzen. Sie stehen mit ihrer vorderen Schmalkante nach vorn und unten vom Körper ab. Sie sind an ihrem Vorderrand mit einem schmalen, längsverlaufenden Schlitz versehen. Die Ränder dieses Schlitzes sind mit Zähnen besetzt, welche die Enden von rippenartigen Verdickungen der Chitinwand des ganzen Gebildes darstellen. Die Zähne greifen eng zusammen und bilden so einen sehr schmalen Durchgang, durch welchen Staub und Sand nicht in die Atemröhre eindringen können. Letztere beginnt mit einer weiten Tracheenblase, von der große, bald sich verästelnde Tracheen von typischem Bau in den Körper eindringen. Der Eingang in die Haupttrachee ist von einer deckelartigen Bildung überdacht (Abb. 8).
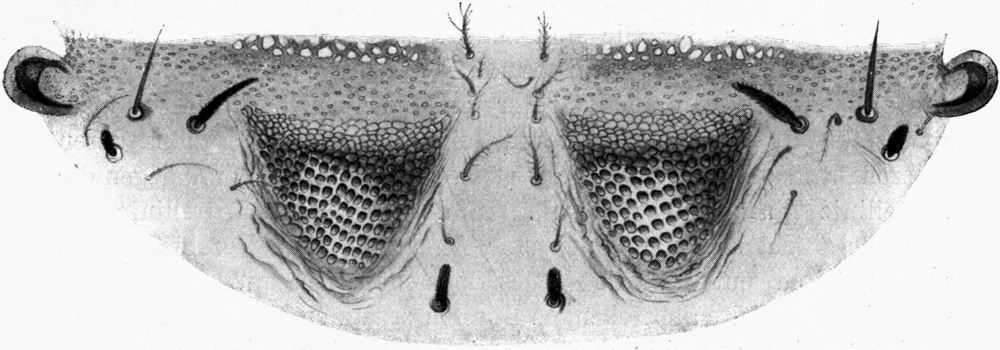
Nicht weniger eigenartig als diese Stigmenkapseln ist die Fläche, welche die Gelenkverbindung zwischen dem ersten und zweiten Thorakalsegment bildet. Rechts und links von der Mittellinie, welche durch dünneres Chitin und zwei parallele Reihen von Borsten gekennzeichnet ist, befinden sich je zwei Höcker, welche seitlich fast bis zu den Stigmenkapseln reichen (Abb. 9). Die Oberfläche dieser Höcker ist vollständig mit kleinen Granulationen bedeckt, wie wir sie schon an der Basis der Mandibel und Maxillen kennen gelernt haben. Sie sind aber mehr kugelig abgerundet als jene. Wie dort, so dienen sie offenbar auch hier zur Bremsung der Bewegung der beiden Thorakalglieder gegeneinander. Wir werden später sehen,[S. 18] daß gerade in diesem Gelenk die für den Schnapp- und Umdrehreflex wichtigste Bewegung ausgeführt wird. Ich nehme daher an, daß wie bei den Mundgliedmaßen das rauhe Höckerfeld insofern als Bremse wirkt, als es bei kontrahiertem Muskel die Gelenkbewegung bis zu einem gewissen Moment hindert. Wird die Bremsung aufgehoben, so erfolgt ganz plötzlich die schnellende Bewegung des betreffenden Gliedes. Auch kann mit Hilfe der Bremsung die Bewegung in jeder Phase gestoppt werden. Aehnliche Höckerfelder finden sich an den ventralen Gelenkflächen der hinteren Abdominalsegmente.
Auf dem hinteren Teil der Unterfläche des zweiten Thorakalgliedes, ziemlich weit gegen den Außenrand hin, liegt die Einlenkungsstelle des zweiten Beinpaares.
Das dritte Thorakalglied fügt sich direkt in den Verband der Hinterleibsglieder ein. Sie sind alle durch tiefe Einkerbungen voneinander getrennt. Am Hinterrande jedes Gliedes verläuft ähnlich wie am Vorderrand ein verstärkter Wulst des Chitins. Diese Wülste sind stets mit einer oder mehreren Reihen von Borsten besetzt, deren Spitzen nach vorn gekehrt sind, meist auch eine Wendung gegen die Mittellinie hin zeigen (Abb. 2). In der Nähe der Mittellinie des Rückens findet sich überhaupt eine Anhäufung von Borsten. Auch an den Seiten entlang zeigt jedes Segment einen mit langen, nach verschiedenen Seiten abstehenden Borsten besetzten, fast halbkugeligen Wulst, ähnlich wie wir deren zwei am Vorderrande des zweiten Thorakalgliedes an der „Schulter“ kennen lernten. Derjenige auf dem dritten Thorakalglied ist der größte von allen und zeigt auch die stärkste Borstenbedeckung (Abb. 2). Vom dritten Thorakalglied an bilden diese Wülste an den beiden Seiten des Hinterleibes zwei gegen die Schwanzspitze konvergierende Reihen.
Der Metathorax ist auch ähnlich dem Mesothorax hinter dem Wulst, welcher seinen Vorderrand einfaßt, muldenförmig eingedellt. Dahinter befindet sich eine Rinne, welche in der Mittellinie nach vorn ausgebogen, seitlich um zwei flache Mulden nach hinten gebogen ist. Diese Mulden sind mit hartem glänzendem Chitin ausgekleidet, wie jene auf dem Mesothorax, und sie entsprechen auch den Stellen, an denen sich ein Flügelpaar entwickeln wird.
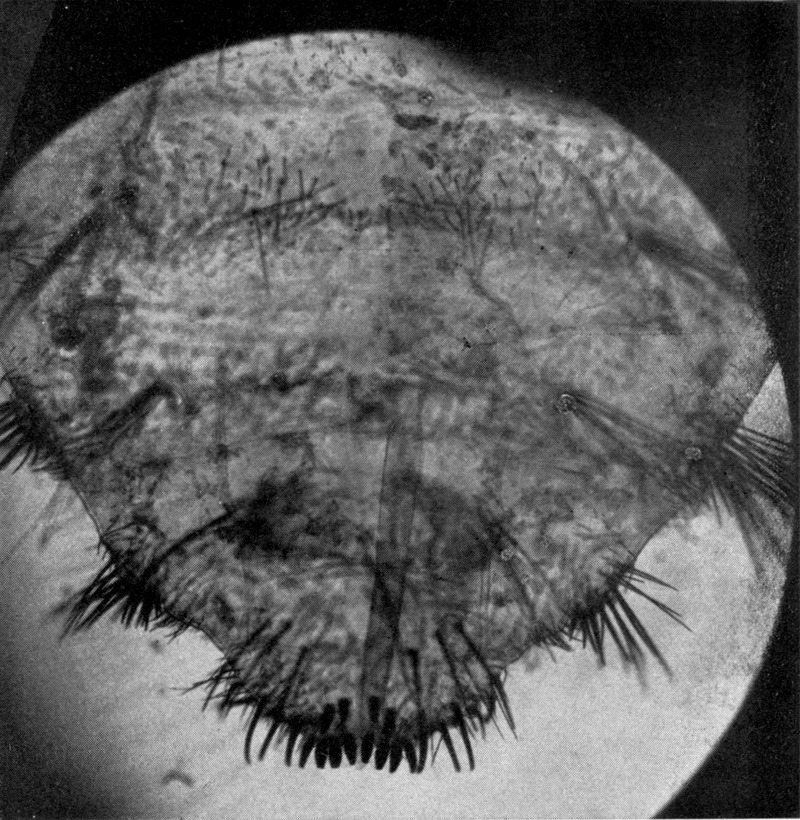
Auch die Bauchseite des ganzen Hinterleibes ist mit Borstenreihen besetzt, deren Spitzen im allgemeinen nach vorn gekehrt sind. Die Borsten der Unterseite sind meist viel länger und zarter als diejenigen der Oberseite (Abb. 10). Auf der Mitte jedes Abdominalsegments zieht sich quer über die Breite des ganzen Tieres ein breites Band[S. 19] von mittelgroßen, nach vorn gerichteten Borsten. Zwischen diesen stehen in regelmäßigen Abständen, symmetrisch zur Mittellinie angeordnet, sechs ähnliche, nur kleinere Wülste, wie wir sie am Seitenrand des zweiten und dritten Thorakalsegments kennen lernten. Dieselben sind mit einer Lage stärkeren Chitins bedeckt als ihre Umgebung und tragen auf stark entwickelten Gelenkknöpfen 6–14, ja oft noch mehr sehr lange Borsten. Diese Wülste samt ihrem Borstenbesatz werden auf den hinteren Abdomensegmenten immer kräftiger. Es sind schließlich nur mehr 4, zuletzt 2. Ihre starren Borsten stehen nach allen Seiten ab. Die letzten Segmente des Abdomens sind mit sehr zahlreichen, sehr starken Borsten bedeckt, welche fast alle nach hinten stehen; nur wenige sind nach der Seite gerichtet. Auf dem letzten Abdomensegment kann man jederseits eine seitliche Gruppe von langen, starken Borsten unterscheiden. Zwischen ihnen finden sich kurz vor dem Hinterende, nahe der Mittellinie 4 sehr starke, kurze, stumpfe, nach hinten gerichtete[S. 20] Borsten. An der Hinterleibsspitze selbst stehen etwa 8 ebensolche Borsten, umgeben von einem Kranz etwas kleinerer, um welche dann die vorhin erwähnten langen Borsten sich anordnen (Abb. 10). Jene kurzen Borsten der Hinterleibsspitze zeigen fast stets deutliche Spuren starker Abnützung. Sie spielen eine wesentliche Rolle bei der Arbeit, welche die Hinterleibsspitze beim Einbohren in den Sand zu leisten hat.

Es bleibt noch nachzutragen, daß sich auf der Unterseite des dritten Thorakalsegments, ziemlich weit gegen die Außenkante gelegen,[S. 21] jederseits eine zweite Stigmenöffnung findet, welche ähnlich wie die ersten eine senkrecht auf die Längsachse des Tieres verlaufende, spaltförmige Oeffnung besitzt. Auch sie ist mit dem fein gezähnelten Verschluß versehen wie das vorige Stigmenpaar, hinter welchem sie aber bei weitem an Größe zurückbleibt. Diese Stigmenöffnung hat nur etwa ein Viertel des Durchmessers der ersten, ist aber etwas größer als die abdominalen Stigmen, deren jedes Segment außer dem letzten ein Paar an der Bauchseite nahe dem Seitenrand trägt (Abb. 11). Nur das Stigma des dritten Thorakalsegments ist stark nach oben gerückt.
Auch das dritte Beinpaar ist auf seinem Segment weit gegen den Außenrand der Bauchseite eingelenkt.
Alle Beinpaare sitzen so am Körper, daß in der Regel ihre Endglieder nach vorn oder höchstens nach den Seiten gerichtet sind, was für die Rückwärtsbewegung des Tieres von Bedeutung ist. Sie sitzen alle auffallend weit außen am Körper.
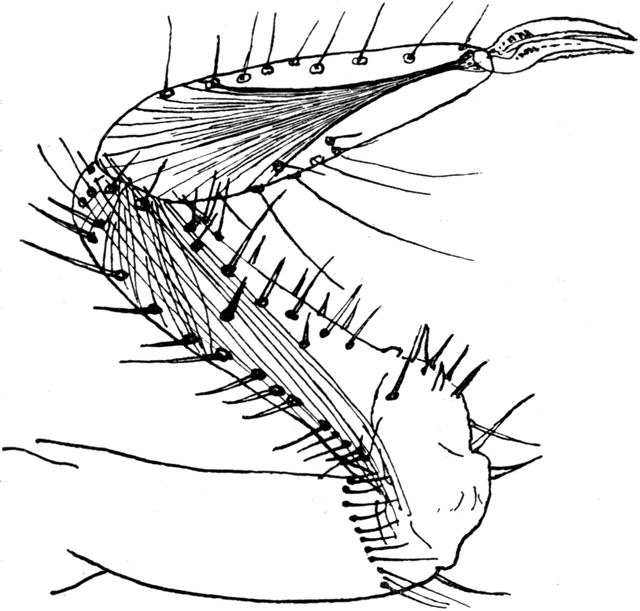
Die Beine selbst sind alle stark behaart. Auf den beiden vorderen Beinpaaren finden sich fast ausschließlich lange und dünne Haare, deren Spitzen nach vorn gerichtet sind. Am ersten und zweiten Beinpaar sind besonders an Femur und Tibia die Haare sehr regelmäßig in geraden Reihen den Kanten der Glieder entlang eingepflanzt. An den Gelenken, besonders der proximalen Beinglieder, finden sich Borstenreihen, wie ich sie seinerzeit bei anderen Arthropoden als „Stellungshaare“ bezeichnet habe. Ich gab ihnen diesen Namen, weil sie als Tasthaare infolge ihrer Stellung an den Gelenken durchaus geeignet sind, um das Tier über die gegenseitige Haltung seiner Glieder zu unterrichten. Bekanntlich fehlt bei den Arthropoden eine sensible Innervation der Muskeln. Es kann also bei diesen Tieren keinen dem Muskelsinn der Wirbeltiere vollkommen entsprechenden Sinn geben. Bei den äußerst präzisen Bewegungen, welche sehr viele Arthropoden haben, müssen wir aber doch annehmen, daß der Muskelsinn irgendwie ersetzt ist. Es ist durch neue Untersuchungen bekannt geworden, daß tatsächlich sensible Nerven zu den Gelenken der Arthropoden ziehen. Diese Nerven werden wohl in der Regel die von mir beschriebenen Stellungshaare innervieren. Auch sonst zeigen die Beine des Ameisenlöwen einige bemerkenswerte Eigentümlichkeiten. Zunächst ist schon ihre Stellung am Körper eine sehr eigenartige. Das erste Beinpaar ist meist geradeaus zu beiden Seiten des Kopfes nach vorn gestreckt, dabei liegen Coxa, Trochanter und Femur dem Kopf und Hals ziemlich[S. 22] parallel, während Tibia und Endglieder etwas abgespreizt sein können. Das zweite Beinpaar ist meist vom Körper sehr stark abgestreckt, wobei alle Glieder beinahe eine gerade Linie bilden können. Das dritte Beinpaar dagegen ist stets stark eingebogen. Die Tibia ist im Winkel gegen das Femur stark eingeknickt. Der nach vorn gerichtete Rand dieses Beinpaares, also speziell der Vorderrand der Tibia, muß beim Graben im Sand stark mithelfen. So sehen wir ihn denn nicht, wie die übrigen Beinglieder, mit relativ langen und dünnen Haaren, sondern mit starken und kurzen Borsten bedeckt (Abb. 12). An dem Endglied jedes der 6 Beine sitzt ein Paar starke, gekrümmte Klauen, welche gegeneinander beweglich sind. Diese Beweglichkeit wird durch einen eigenartigen Mechanismus bewirkt. Die beiden Klauen, deren konvexer Rand abgerundet, deren konkaver Rand jedoch scharf ist, bestehen ausschließlich aus Chitin. Sie sind miteinander durch eine starke Chitinspange verbunden, welche im Endglied des Beines sich befindet. Diese elastische Spange hat die Gestalt eines Halbringes; die Enden dieses Halbringes sind mit den Klauen in fester Verbindung. In der Ruhestellung werden durch die Elastizität dieses Ringes die Klauen auseinandergespreizt. Nun sitzt an der höchsten Stelle der Wölbung des Ringes die Sehne eines Muskels an. Es ist dies ein kräftiger, aus mehreren Bündeln zusammengesetzter Muskel, welcher das ganze letzte Fußglied (die Tibia) durchzieht, in dessen oberen Teil er fächerförmig sich ausbreitet. Die Klauen mit ihrem Ring und ihrer basalen Umhüllung durch das Chitin des sehr kurzen eigentlichen Tarsalgliedes sind in der kreisförmigen unteren Oeffnung des Chitinpanzers des letzten Fußgliedes fest verankert. Wenn nun der Muskel, welchen ich den Klauenmuskel nennen will, sich kontrahiert, so muß, da eine[S. 23] Bewegung sonst nicht möglich wäre, der Chitinring seine Form verändern; er wird in die Länge gezogen, und dadurch werden automatisch die beiden Klauen, besonders mit ihrem Spitzenteil, einander genähert. Sie greifen zusammen, und ich habe öfter gesehen, daß dabei Sandkörnchen und andere Gegenstände zwischen den Klauen erfaßt werden. Die Sehne des Klauenmuskels ist aus zwei parallel laufenden dünnen Fasern zusammengesetzt. Beim ersten und zweiten Beinpaar ist das kurze Tarsalglied frei, beim dritten Beinpaar jedoch mit der Schiene verwachsen.
Nimmt man einen Ameisenlöwen aus dem Sand, so ist er infolge des anhaftenden Staubes grau und unscheinbar. Benetzt man ihn jedoch mit Wasser, so tritt seine charakteristische Färbung deutlich hervor.
Die Grundfarbe des ganzen Körpers ist ein fahles, erdiges, helles Gelb. Manche Individuen sind etwas dunkler, gelbbraun, manchmal sogar rotbraun oder gar dunkelbraun gefärbt. Von dem hellen Grundton heben sich die zahlreichen Haare und Borsten, welche dunkelbraun oder schwarz gefärbt sind, scharf ab. Am hellsten ist die Oberseite der Beine.
Ueber die Mitte der ganzen Rückenseite des Tieres zieht sich ein breites, rötliches Band. Diese Färbung, welche bei jüngeren Tieren mehr fleischfarbig, bei älteren mehr braun zu sein pflegt, dehnt sich auf dem dritten Thorakalsegment und dem ersten Abdominalsegment weit nach den Seiten aus (vgl. Abb. 2).
Die Mandibel sind in der Hauptsache bleich gelblich gefärbt; doch zieht sich über die Mitte ihrer Oberfläche ein brauner Streifen; das hakenförmige Ende sowie die Spitzen der großen Stacheln sind dunkelbraun gefärbt. Auch die Taster und Fühler und die Umgebung der Augen sind dunkelbraun.
Ueber die ganze Rückenseite sind nun ferner Flecken dunkelbraunen und schwarzen Pigments verteilt. Auf dem Kopf und den ersten Thorakalsegmenten bilden sie ein Doppelband, welches eine hellere schmale Mittelzone freiläßt. Auf dem Rücken, vor allem des Abdomens sind diese dunkeln Flecken regelmäßig angeordnet. Sie finden sich in Querreihen am Vorderrand besonders der vorderen Segmente; am auffallendsten sind aber die Längsreihen. Von ihnen kann man 9 unterscheiden; am deutlichsten ausgeprägt ist die Mittellinie und zu ihren beiden Seiten je eine parallele Linie[S. 24] großer, in die Quere gezogener Flecken. Zwischen der Mittellinie und den beiden Hauptseitenlinien ziehen je eine, außerhalb der Hauptseitenlinien je zwei Linien kleinerer, rundlicher Pigmentflecke hin. Je ein Flecken ist auf je einem Segment angeordnet; in der Mittellinie liegen vielfach auf jedem Segment zwei in der Quere ausgedehnte strichförmige Flecken; in den Hauptseitenlinien sind solche meist zu einem gelappten Flecken zusammengeflossen. Zwischen diesen 9 Hauptreihen können noch zerstreute oder andeutungsweise in Reihen angeordnete kleinere Flecken vorkommen.
Bemerkenswert ist, daß auf den Pigmentflecken fast stets entweder Gruppen und Büschel von Haaren oder einzelne, besonders starke Borsten stehen.
Noch auffälliger ist die Fleckung der Unterseite. Die Grundfarbe ist da mehr wachsgelb; fast schwarze, hier und da dunkelbraune Pigmentflecken bilden in der Hauptsache 7 Längslinien. Relativ klein und schwach sind die Flecken der Mittellinie, deren Ränder oft verwischt sind. Unregelmäßig gelappt und oft in mehrere Teilflecke zerlegt sind die Flecken der ersten seitlichen Linien. In einigem Abstand von ihnen folgen Doppellinien, welche die Basis des zweiten und dritten Beinpaares umfassen. In diesen 6 seitlichen Längslinien ist im allgemeinen eine Verkleinerung der Flecken von vorn nach hinten festzustellen.
Nahe dem Seitenrand zieht sich schließlich jederseits eine Reihe heller brauner Flecken hin, welche etwa die Spitze der die Borstenbüschel tragenden Warzen einnehmen.
Auch auf der Unterseite des zweiten Beinpaares finden sich einige hellrotbraune, auf der des dritten Beinpaares mehrere (3–5) hell- und dunkelbraune Flecken. Die Klauen aller Beinpaare sind dunkelbraun.
Die Unterseite des Kopfes ist mit kleinen, ziemlich symmetrisch angeordneten dunkelbraunen Fleckchen bedeckt.
Zum Unterschied von seinen Verwandten sieht man unsere gewöhnlichen einheimischen Ameisenlöwen in der Natur selten frei herumlaufen. Man findet sie fast stets in ihrem Trichter eingegraben. Je nach der Größe der Tiere können diese Trichter verschieden[S. 25] groß sein. Die kleinsten, die ich gesehen habe, hatten kaum einige Millimeter im Durchmesser, während die größten 10–12 cm breit waren. Uebrigens hängt die Größe der Trichter außer von der Größe ihrer Erbauer, wie wir später sehen werden, auch von deren physiologischem Zustand und von der physikalischen Beschaffenheit des Sandes oder der Erde, in dem sie gebaut sind, ab.
Wir haben früher schon geschildert, daß sich die Trichter der Ameisenlöwen an solchen Stellen finden, wo unter einer überhängenden Böschung gleichmäßig feine Sand- oder Erdmassen von trockener Beschaffenheit sich angehäuft haben (vgl. Abb. 1). Die Oertlichkeit ist vor Wind geschützt, so daß dort der feine Sand sich ansammeln und die Wärme an schönen Tagen gespeichert werden und nachwirken kann. An solchen Stellen fällt das Licht meist hauptsächlich von einer Seite ein; fast stets kann relativ helles Licht von der Seite, welche der Böschung abgewandt ist, bis in die Tiefe des Trichters dringen. Vielfach kann man nun beobachten, daß die eingegrabenen Ameisenlöwen insofern eine bestimmte Stellung zum einfallenden Licht einnehmen, als das Vorderende ihres Kopfes vom Einfall der Lichtstrahlen abgekehrt ist (Abb. 13). Ich habe so gut wie immer die Tiere in der angegebenen Stellung gefunden, oft 10–20 Trichter hintereinander daraufhin kontrolliert. Nachdem ich diese Feststellung gemacht hatte und vorher schon ihre physiologischen Grundlagen experimentell erforscht hatte, konnte ich ohne Suchen aus jedem Trichter den Ameisenlöwen mit einem Griff der Pinzette herausfischen.
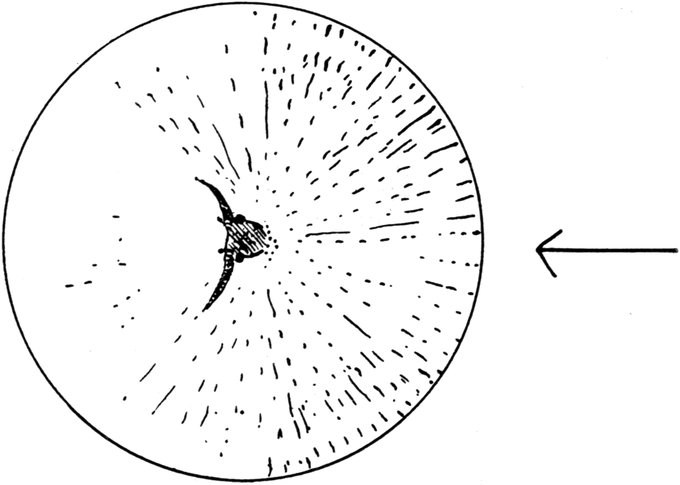
Der Körper des Ameisenlöwen ist am Grunde des Trichters vollkommen in den Sand eingewühlt, wobei er eine Neigung von 45 Grad einnimmt. Die Hinterleibsspitze ist schief nach unten gerichtet, Teile des Kopfes sehen am Grunde des Trichters gerade aus dem Sand heraus. Meist sind nur Teile der Stirn mit den Augen[S. 26] und Fühlern sowie die Mundwerkzeuge vom Sande unbedeckt. So liegt das Tier bewegungslos; stundenlang sehen wir an ihm keine Veränderung, wenn in seiner Umgebung keine solche erfolgte (Abb. 14).
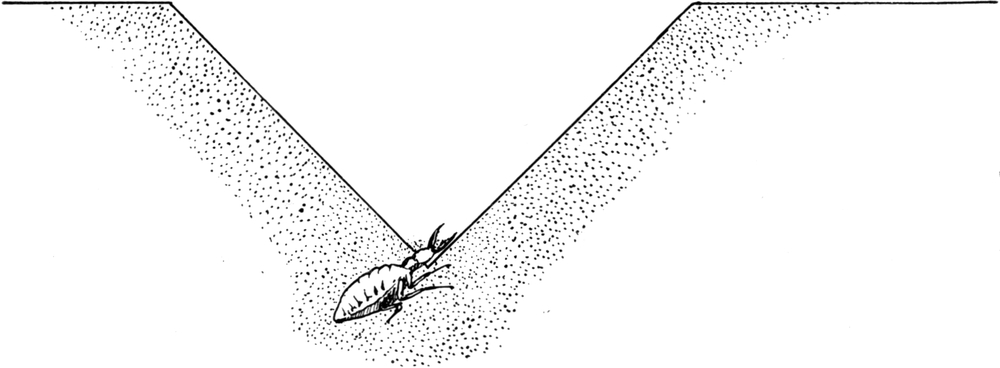
In der Regel dauert es aber bei Tage nicht allzu lange, bis ein Vorgang in der Umgebung das Tier aus seiner Ruhe aufstört. Die sonnigen Waldränder und Böschungen, an denen die Ameisenlöwen vorkommen, sind der Tummelplatz zahlreicher beweglicher Insekten. Vor allem sehen wir da Ameisen und Spinnen auf andere Insekten Jagd machen. Diese Räuber, sowie herumlaufende Käfer, anfliegende Dipteren und Hymenopteren geraten gelegentlich an den Rand eines Ameisenlöwentrichters. Dann lösen sie wohl am oberen Rande des Trichters einige Sandkörnchen los, welche in die Tiefe hinabrollen und auf den Kopf der Larve fallen. In den meisten Fällen entledigt sich diese ihrer sofort, indem sie sie mit einer eigentümlichen Schleuderbewegung wieder gegen den oberen Trichterrand wirft. Nicht selten aber ereignet es sich, daß eines der Tiere beim Betreten des Trichterrandes an dessen steiler Wand hinabrutscht. Es kann vorkommen, daß es, ohne Halt zu finden, in einem Zug bis in die Tiefe des Trichters hinunterfällt. Wenn sich dabei nicht allzu viele von den Sandkörnern der Trichterwand losgelöst haben, so daß der Kopf des Ameisenlöwen noch freiliegt, dann fällt das Opfer direkt zwischen die Kiefer des unten lauernden Räubers. Dieser hält nämlich in hungerndem Zustand seine Mandibeln soweit klaffend auseinander, daß sie einen sehr stumpfen Winkel zueinander oder gar mit dem Vorderrand der Stirn eine nahezu gerade Linie bilden. Sobald das Insekt unten angelangt ist, schnappen die Kiefer über ihm[S. 27] zusammen und lassen nicht wieder los. Das Opfer beginnt je nach seinem Temperament mehr oder weniger stürmische Bewegungen zu machen, um sich aus der Umklammerung zu befreien. Ist es aber richtig gefaßt worden, so gelingt ihm dies niemals. Der Ameisenlöwe steckt so fest im Sand, daß selbst größere und schwerere Tiere ihn nicht aus seiner Verankerung herauszureißen vermögen; ja sie verschütten bei ihren Bewegungen ihn und oft sogar sich selbst immer tiefer in den von den Seiten herabrieselnden Sand. Der Ameisenlöwe selbst sucht durch nach hinten gerichtete Bewegungen sich immer tiefer einzuwühlen.
Der Ameisenlöwe hat seinen Namen daher, daß seine normale Beute Ameisen sind. Diese gewandten und beweglichen Tiere vermögen sich oft beim Herabrutschen noch an der Wand des Trichters anzuklammern, ja sogar, wenn dieser nicht zu steil ist, wieder aufwärts zu kriechen. Unter ihren Tritten lösen sich dann Sandkörnchen los, welche wiederum auf den unten lauernden Ameisenlöwen hinabkollern. Kaum wird er von den Sandkörnchen berührt, so beginnt er einen Hagel von solchen nach oben gegen die Böschungen des Trichters zu schleudern. Indem diese hinabrollen, reißen sie Teile der Böschung mit sich, und in den meisten Fällen wird die emporkletternde Ameise dabei mitgenommen und fällt in die für sie offenstehende Falle.
Die vom Ameisenlöwen erfaßte Beute wird nicht eher losgelassen, bis sie ausgesaugt ist. Zu diesem Geschäft dient der Larve der Apparat von Saugröhren, den Mandibel und Maxillen der beiden Seiten zusammen bilden und dessen feinen und eigenartigen Bau wir oben geschildert haben. Die Nahrungsaufnahme im einzelnen findet an anderer Stelle ihre Darstellung.
Von dem ausgesaugten Insekt bleibt die leere Chitinhülle zurück, welche der Ameisenlöwe von seinen Kiefern abstreift. Nach einiger Zeit wird sie mit dem naturgemäß auf seinen Kopf herabsickernden Sand von ihm weggeschleudert. So finden wir denn sehr oft, wenn längere Windstille vorausgegangen ist, in der Nähe eines solchen Trichters einen ganzen Kranz von ausgesaugten Ameisenleichen. Um sie herum liegen die Steinchen, die das Tier ebenfalls fortgeschleudert hat. Liegt der Trichter nahe am Rand einer Böschung, so zieht sich von ihm ein Wall von Sand und Steinchen nach unten, ähnlich den Schutthaufen am Ausgange von Bergwerken.
Wer in freier Natur mit Sorgfalt und Beobachtungsgabe die Trichter von Ameisenlöwen beobachtet, wird mit Leichtigkeit feststellen,[S. 28] daß sie an solchen Orten sich befinden, wo infolge der örtlichen Bedingungen Ameisen und andere Insekten zu den geeigneten Tageszeiten in Menge umherschweifen. Die Bemerkung des sizilianischen Zoologen Comes, daß er stundenlang die Umgebung von Trichtern der Ameisenlöwen beobachtet habe, ohne eine Ameise oder den Fang einer solchen zu bemerken, braucht daher kaum erwähnt zu werden. Wer so viele Ameisen in die Trichter der Larve im Freien fallen sah, wie ich, wird wohl an der Tauglichkeit der Beobachtungsmethoden jenes Forschers zweifeln.
Fängt man im Freien Ameisenlöwen ein, oder holt man sie in der Gefangenschaft aus ihren Trichtern oder aus dem Sand heraus, so pflegen sie vollkommen starr, wie leblos, liegenzubleiben. In freier Natur kommt es beim Fangen der Ameisenlöwen häufig vor, daß der Sand ihrer Trichter abrutscht, oder daß sie selbst die Sandböschung hinabgleiten. Es ist dann sehr schwer sie aufzufinden, da sie ihrer Umgebung außerordentlich ähnlich sehen. Wir haben zwar früher gehört, daß der Ameisenlöwe eine charakteristische Zeichnung besitzt, auch sind seine Mundteile und Partien seines Kopfes sehr dunkel gefärbt. Um diese Färbungen zu erkennen, muß man aber ein unter normalen Verhältnissen gefangenes Tier erst abwaschen; denn zwischen den zahlreichen Haaren, welche seinen Körper bedecken, haften unzählige feinste Sandpartikelchen fest, welche jeweils dem Tier vollkommen die Färbung seiner Umgebung verleihen. Diese täuschende Aehnlichkeit mit der Umgebung wird zu einer für das Tier außerordentlich nützlichen Eigenschaft, da es in den ersten Minuten, nachdem es aus dem Sand herausgewühlt ist, völlig bewegungslos verharrt. Hat man beim Fangen durch Ungeschicklichkeit ein Tier auf die gleichförmige Sandfläche fallen lassen, so ist es für unser Auge wie verschwunden. Will man es noch einfangen, ist am zweckmäßigsten die Umgebung des Fundortes ganz scharf im Auge zu behalten; dann wird man nach einigen Minuten an den neu beginnenden Bewegungen des Tieres oder an rieselnden Sandkörnern den Ort wahrnehmen, an dem es sich befindet. Die Bewegungslosigkeit,[S. 29] welche das Tier dem Auge des Menschen entzieht, mag es auch vor natürlichen Verfolgern, z. B. insektenfressenden Käfern, Eidechsen oder Vögeln, schützen.
Es gleicht also dieses Totstellen des Tieres den Gewohnheiten, welche wir bei vielen anderen Tieren als charakteristische Schutzanpassungen kennen. Sehr viele Krebse und Insekten, aber auch höhere Tiere, wie z. B. Reptilien, Vögel, ja selbst manche Säugetiere nehmen im Moment der Gefahr eine starre Haltung ein. Vielfach genügt schon die Bewegungslosigkeit, um sie vor ihren Verfolgern, deren Augen vorwiegend auf Bewegungsehen eingerichtet sind, zu sichern. Sehr häufig wird die Wirkung der Bewegungslosigkeit durch eine weitgehende Aehnlichkeit mit der Umgebung verstärkt, so z. B. bei Blatt- und Stabheuschrecken, Krebsen mit sympathischer Färbung usw.
Die Dauer des Totstellens ist bei den verschiedenen Individuen des Ameisenlöwen und bei demselben Individuum zu verschiedenen Zeiten eine ganz verschieden lange. Im allgemeinen kann man beobachten, daß ein Tier, welches ohne übermäßige Gewaltanwendung aus dem Sand herausgenommen worden ist, etwa 3–5 Minuten lang vollkommen stillhält. Dann erst beginnt es mit ganz langsamen vorsichtigen Bewegungen. Diese Frist ist bei hoher Temperatur der Umgebung kürzer, bei niederer länger.
Es kommt aber auch vor, wenn auch selten, daß aus dem Sand herausgenommene Tiere sich sofort wieder zu bewegen beginnen. Es ist dies besonders bei hoher Temperatur, oder wenn die Individuen durch fortgesetzte Reize in Erregung versetzt sind, der Fall.
Beispiele:
| 1 |
Individuum
|
3.
|
Sept.
|
1915
|
totgestellt
|
10h 37′,
|
umgedreht
|
10h 39′,
|
| 1 |
„
|
„
|
„
|
„
|
„
|
10h 40′,
|
„
|
10h 44′,
|
| 1 |
„
|
„
|
„
|
„
|
„
|
10h 42′,
|
„
|
10h 50′,
|
| 1 |
„
|
„
|
„
|
„
|
„
|
10h 50′,
|
„
|
11h 06′,
|
| 1 |
„
|
„
|
„
|
„
|
„
|
10h 44′,
|
„
|
11h 14′.
|
Viel häufiger aber bleiben die Tiere länger, oft sogar sehr viel länger als 5 Minuten, in ihrer starren Haltung. Oft dauert das Totstellen eine halbe, ja oft mehrere Stunden. Bisweilen ist es bei meinen Experimenten vorgekommen, daß die Tiere mehrere Tage lang bewegungslos verharrten, was zunächst nur dadurch festgestellt werden konnte, daß die Tiere ihren Platz nicht verlassen hatten. In einzelnen Fällen wurde der exakte Nachweis für die Bewegungslosigkeit[S. 30] dadurch erbracht, daß die Tiere auf ein berußtes Registrierpapier gelegt wurden, auf welchem jede einigermaßen erhebliche Bewegung des Körpers und der Gliedmaßen sich hätte aufzeichnen müssen. Wenn wir in einem späteren Abschnitt die Bewegungsweise des Tieres genauer analysiert haben werden, wird sich ohne weiteres ergeben, daß das Verbleiben des Tieres am Orte schon allein dafür spricht, daß es keine seiner sehr typischen Körperbewegungen, auch keine erheblichen Gliedmaßenbewegungen während der Zwischenzeit ausgeführt hatte. Meist wurden bei diesen Versuchen die Tiere auf den Rücken gelegt. Ein Exemplar beobachtete ich im Jahre 1911, welches 10 Tage lang regungslos an der Oberfläche des Sandes lag, gerade nur mit der Schwanzspitze eingewühlt. Am 17. November reagierte es auf die Erschütterung beim Oeffnen der Glasdose mit Bewegung und Schnappen. Eine dargebotene, zwischen die Mandibel gehaltene Ameise wurde gepackt und ausgesaugt. Es handelte sich also nicht um ein absterbendes Tier; immerhin ist hervorzuheben, daß die Beobachtung in den Anfangsmonaten des Winters erfolgte, welche für die Ameisenlöwen eine relative Ruheperiode darstellen.
Einerlei, ob die Tiere auf dem Bauch oder auf dem Rücken lagen, stets nahmen sie während des Totstellens eine typische Haltung ein. Der Kopf war tief geneigt, die Schwanzspitze nach unten eingebogen, somit der ganze Körper stark nach der Bauchseite eingekrümmt. Die Kiefer waren beinahe geschlossen, der Kopf etwas angezogen. Zu seinen beiden Seiten ragten die Beine des ersten Paares gerade nach vorn, während das dritte Beinpaar an den Hinterleib angezogen war und nicht über dessen Umriß seitlich hervorschaute. Das zweite Beinpaar dagegen war nach den Seiten weit vom Körper abgespreizt und stand von dessen Längsachse entweder in einem rechten oder doch in einem Winkel von annähernd 90 Grad ab. Während der ganzen Zeit hielt das Tier seinen Körper sehr steif. Durch Biegungsversuche mit Nadel und Pinzette ließ sich feststellen, daß die Gelenke der Körpersegmente starr gehalten werden, so daß der Körper sich nicht biegen ließ. Das gleiche gilt für die Mundgliedmaßen und das zweite Beinpaar. Das letztere ist in den Gelenken vollkommen gesperrt, so daß man das Tier mit einer Nadel an einem Bein vollkommen umdrehen kann. Das erste und dritte Beinpaar dagegen bleiben in den Gelenken weich und beweglich; man kann ihnen künstlich beliebige Stellungen geben, während das zweite Beinpaar bei Biegungsversuchen stets wieder in seine starre Haltung zurückschnellt.
[S. 31]
Während des Totstellens bleiben Körper, Kopf und Beine in ganz starrer Haltung und werden nicht bewegt. Nur an den Antennen, den Lippentastern und den Mandibeln kann man dann und wann ein leises Zucken und Zittern erkennen.
Die Starrheit ist am ausgesprochensten kurz nach Eintritt des Totstellens; dann kann sie sich auch auf das erste und zweite Beinpaar erstrecken. Allmählich werden die Gelenke wieder weich, zuerst an der Mandibel, dann an den Beinpaaren, und zuletzt am zweiten; endlich werden auch die Gelenke des Kopfes, Halses und Rumpfes wieder beweglich. Die anfangs herabgesetzte Reizbarkeit steigert sich nun wieder, und man kann schwer an dem Tier experimentieren, ohne es zu kräftigen Reflexen und neuer Beweglichkeit zu erwecken.
In der freien Natur in ihrem Sand findet man die Tiere in der starren Haltung auch nach starkem Regenwetter vor. An Orten, an denen man ihr Vorkommen genau aufgezeichnet hat, kann man sie dann unter den verschütteten Trichtern im nassen Sand eingebacken finden. Bei solchen Individuen dauert es oft ziemlich lange, bis sie wieder normale Bewegungen ausführen. Dasselbe gilt auch für Tiere, welche man in der Gefangenschaft stark mit Wasser benetzt hat. Dabei scheint die Abkühlung der wesentliche Faktor zu sein, welcher den Starrezustand verlängert.
Durch alle möglichen Reize läßt sich jedoch die Dauer des Totstellens verkürzen. Haucht man z. B. das starr daliegende Tier aus einiger Entfernung an, so beginnt es schwache Bewegungen mit den Mandibeln, oft nur mit einer derselben, auszuführen. Nicht selten fangen dann Kopf und Thorax, Beine und Hinterleib zu zucken an, worauf das Tier sich in normale Bewegung setzt. Liegt es auf dem Rücken, so findet vorher die Umdrehreaktion statt. Wie wir später sehen werden, kommen hierbei als auslösende Reize sowohl die Wärme des Hauches als auch der Druck der Luftbewegung in Betracht.
Das Totstellen der Ameisenlöwen kann in jeder beliebigen Körperstellung erfolgen; das Tier kann auf dem Bauch oder auf dem Rücken liegen, und je nach der Beschaffenheit des Untergrundes kann eventuell der Kopf, das Hinterteil oder eine der Seiten nach unten gekehrt sein. Ja, man kann das Tier mit dem Kopf oder dem Hinterende des Abdomens nach unten senkrecht in den Sand stecken. Es bleibt dann oft besonders lange in starrer Haltung in dieser Stellung sich tot stellend, bis es ihm gelingt, wiederum durch[S. 32] Schleuderbewegungen oder Bohrbewegungen sich in normale Lage zu bringen.
Um über das Wesen dieser merkwürdigen Erscheinung des Totstellens genauere Aufschlüsse zu verschaffen, habe ich vielerlei Experimente angestellt. Aus denselben ergibt sich, daß die Reizempfindlichkeit des Tieres während dieses Zustandes nicht vollkommen erloschen ist.
Der Tastsinn reagiert ganz deutlich. Auf Berührung des Rumpfes, vor allem aber des Kopfes, der Mandibel und der Beine mit Nadeln oder feinen Borsten, erfolgen deutliche Bewegungen. Diese Reizbarkeit ist am geringsten, unmittelbar nachdem der Zustand des Totstellens eingetreten ist. Sie läßt sich bemessen nach der Zahl der Tastreize, welche notwendig sind, um eine der reflektorischen Bewegungen herbeizuführen. Näheres über die Anordnung solcher Versuche findet sich unten im Kapitel über den Tastsinn (S. 87).
Auch die unten (S. 109) geschilderten Reaktionen des Geruchssinnes erfolgen während des bewegungslosen Zustandes.
Fassen wir alle physiologischen Besonderheiten dieser biologisch für den Ameisenlöwen sicherlich wertvollen Reaktion zusammen, so kommen wir zu folgenden Ergebnissen, welche unsere bisherigen Kenntnisse über das Totstellen der Tiere in manchen Punkten ergänzen.
Die Unbeweglichkeit ist der typische Zustand, in welchem wir den Ameisenlöwen in der Natur vorfinden. Er kann stundenlang in der oben geschilderten Haltung verharren oder in einer Stellung, welche ich als die Bereitschaftsstellung (vgl. unten S. 36) bezeichne.
Mangold hat 1914 eine vergleichende Studie über ähnliche Zustände im gesamten Tierreich veröffentlicht, in welcher er zu dem Resultat kommt, daß das Totstellen der niederen Tiere durchaus vergleichbar mit der Hypnose der höheren Wirbeltiere ist. Unzweifelhaft muß man ihm insofern recht geben, als die meisten Symptome des Totstellens mit einigen der körperlichen Erscheinungen während der Hypnose übereinstimmen.
Auch beim Ameisenlöwen hat man durchaus den Eindruck, daß er während des Totstellens sich in einem schlafähnlichen Zustand befindet, aus dem er nach einiger Zeit von selbst erwacht oder aus dem er durch Reize erweckt wird. Allerdings ist es bei einem so träg beweglichen Tier sehr schwierig, den Eintritt des Schlafzustandes sowie sein Aufhören mit Sicherheit festzustellen. Denn bei einem Tier, welches normalerweise so wenig sich bewegt, ist es kaum möglich, Wach- und Schlafzustand zu unterscheiden.
[S. 33]
Steckt ein Ameisenlöwe in trockenem, warmem Sand, vor allem wenn er am Grund seines Trichters sitzt, so pflegt er auf feine Reize zu reagieren. Wie er sich dann in Bereitschaftsstellung befindet, so ist sein Nervensystem zu sehr prompten Reflexreaktionen vollkommen bereit. Wärme und starkes Licht (Sonnenschein) tragen sehr zur Erhöhung der Reflexerregbarkeit bei.
Ist das Tier „hungrig“, d. h. hat es längere Zeit keine Ameise gefangen, dann ist es sogar in hohem Maße reizbar. Leise Berührungsreize, rieselnder Sand usw. führen sofort den Schnappreflex herbei, durch den immer wieder der Trichter in einen fangbereiten Zustand versetzt wird. Ist das Tier ganz mit Sand bedeckt, wie das der Fall ist, nachdem es gefressen hat, oder wenn es durch vergebliche Fangversuche ermüdet ist, so bleibt dennoch die Reizbarkeit groß. Leise Berührung des umgebenden Sandes hat entweder eine Schleuderbewegung oder ein Rückwärtsbewegen des Tieres zur Folge.
Solange es im Sand steckt, ist es also trotz vollkommener Ruhe nicht im Zustand des Totstellens; allerdings bei Kälte, oder wenn der Sand feucht oder naß ist, ist das Tier vollkommen unbeweglich und sehr schwer zu Reizbewegungen zu veranlassen. Dann nimmt es auch keine Bereitschaftsstellung ein. Offenbar liegen aber da besondere, allgemein physiologische Gründe vor, welche die Reizbarkeit herabsetzen und die Reizleitung verlangsamen und welche mit der Abkühlung des gesamten Tierkörpers zusammenhängen. Wenn wir aber das Tier unter normalen Verhältnissen, also bei Trockenheit und Wärme, aus dem Sand herausnehmen, dann entsteht momentan jener Zustand, den wir als das Totstellen bezeichnen. Es fragt sich nun zunächst, welche Ursache die auslösende für diesen Zustand ist.
Mangold ist der Ansicht, daß beim Tier mechanische Bewegungshemmung zur Hervorrufung der Hypnose unerläßlich sei; meist werde sie durch Sinnesreize unterstützt. Wie bei den anderen Wirbellosen, so scheint mir aber auch beim Ameisenlöwen die mechanische Hemmung eine untergeordnete Rolle zu spielen.
Zwar begünstigt das Legen des Tieres in die Rückenlage das Eintreten des Totstellens. Aber auch nach dem Umdrehen und ohne vorhergehende Rückenlage können die Tiere im Zustand des Totstellens für lange Zeiträume verharren. Selbst Individuen, die man ganz vorsichtig aus dem Sand in Bauchlage heraushebt, stellen sich tot.
[S. 34]
Es müssen also hauptsächlich die Reize, welche beim Herausholen aus dem Sand einwirken, von Bedeutung sein. Als solche kämen in Betracht:
1) die allgemeine Aenderung der Reizsituation,
2) die mehr oder weniger grobe Berührung des Tieres.
Der zweite Punkt wird am besten zuerst zu erledigen sein, da er unzweifelhaft den wichtigsten Reiz darstellt. Stärkere, plötzliche mechanische Reizung führt beim Ameisenlöwen den Zustand des Totstellens herbei. Packt man ein Tier fest mit der Pincette, so führt es zunächst starke reflektorische Bewegungen aus, die je nach der Körperregion, die man angefaßt hat, verschieden sein können. Läßt man das Tier los, so bleibt es sofort unbeweglich liegen und zwar eventuell längere Zeit. Auch sonst kann man im Experiment jedes Individuum durch grobes Zufassen und darauf folgendes Loslassen zum Totstellen veranlassen. Die Hauptursache für das Totstellen ist also sicherlich ein starker mechanischer Reiz. Damit verbundene Lageänderung ist zwar begünstigend, aber nicht unerläßliche Vorbedingung für das Eintreten des bewegungslosen Zustandes.
Die allgemeine Aenderung der Reizsituation spielt aber nach meinen Erfahrungen ebenfalls eine wichtige Rolle. Sie kann sich erstrecken auf
1) die Belichtungsverhältnisse und
2) die Berührungszustände.
Sonstige Sinnesreize spielen keine sehr wesentliche Rolle, wenigstens konnte ich keinen Einfluß von chemischen Reizen feststellen. Die allgemeine Einwirkung der Temperatur ist natürlich von Bedeutung, und zwar indem Kälte als Förderung, Wärme als Störung des Zustandes wirkt. Temperaturveränderung kann aber wohl kaum so rasch einwirken, um an der Auslösung des Totstellens beteiligt zu sein. Ich fand jedenfalls keinerlei Anzeichen einer solchen Wirkung.
Das Licht ist aber sicherlich von Einfluß auf die Bewegungen des Ameisenlöwen. Besondere Kapitel werden uns über die große Abhängigkeit des Ameisenlöwen vom Licht belehren (S. 67). Es zeigt sich bei Belichtungsexperimenten, daß starkes von oben einfallendes oder diffuses Licht auch bei Ausschaltung der Wärmewirkung die Beweglichkeit des Tieres hemmt, während Dunkelheit sie fördert. Derartige Beleuchtungsverhältnisse werden aber selten auf einen[S. 35] aus dem Sand genommenen Ameisenlöwen einwirken. Meist wird es sich um bestimmt gerichtetes Licht handeln, über dessen Einfluß — und zwar Bewegung fördernden Einfluß — wir später Genaueres erfahren werden. Immerhin können wir mit dem bewegunghemmenden Einfluß starker diffuser Belichtung rechnen.
Noch wesentlicher scheint mir der Reiz zu sein, welcher durch das plötzliche Aufhören des allseitigen Sanddruckes bedingt wird. Wir werden später sehen, daß bei der Kombination von Reizen, welche das Stillliegen des Tieres in seinem Trichter bewirken, gerade der Sanddruck von Bedeutung ist. Sein plötzliches Aufhören ist nach meinen Versuchen sicher fördernd für das Eintreten des Totstellens. Aber es bewirkt nicht unbedingt das Totstellen.
Der Zustand des Totstellens ist auch beim Ameisenlöwen durch physiologische Besonderheiten ausgezeichnet, entsprechend denen, die bei anderen Tierarten beschrieben wurden. Zunächst ist das Verhalten der Muskulatur charakteristisch.
Ist das Totstellen durch eine kräftige Berührung des Tieres herbeigeführt worden, so zeigt seine Muskulatur einen erheblichen Tonus. Haltung von Körper und Gliedmaßen weist darauf hin, daß die Beugemuskulatur sich in einem Kontraktionszustand befindet. Alle Gelenke sind in einer fixen Stellung; die Glieder der Extremitäten und des übrigen Körpers können nicht gegeneinander verstellt werden. Die kontrahierte Muskulatur stellt allen Versuchen, dem Körper oder den Gliedern eine andere Haltung zu geben, starken Widerstand entgegen. Das dauert aber meist nur ganz kurze Zeit; dann werden alle Gelenke weich, sie nehmen den Zustand an, den man als Flexibilitas cerea zu bezeichnen pflegt. Man kann den Gliedern jede beliebige Stellung zueinander geben, und sie bleiben in ihr stehen. Wie schon oben geschildert, behalten die Glieder des zweiten Beinpaares am längsten ihre Starrheit, am kürzesten diejenigen des dritten Beinpaares.
Gleichzeitig steigert sich die Reizbarkeit des Tieres; während sofort nach Eintritt des Totstellens schon gröbere Reize notwendig waren, um Reflexbewegungen des Tieres auszulösen, genügen nach Verlauf der ersten 2–3 Minuten leichte Berührungen, um den Umdreh-, Schnapp- und Eingrabreflex herbeiführen.
Von vornherein ist die Reizbarkeit des Tieres zwar herabgesetzt, aber nicht aufgehoben. Die Reize, mit denen man Reflexbewegungen und damit Aufhören des Totstellens herbeiführen kann, sind hauptsächlich Berührungsreize und Wärme. Berührung beliebiger Stellen[S. 36] der Ober- und Unterseite des Tieres, der Mandibel, der Beine führt zu Reflexbewegungen.
Wärme, d. h. Erhitzung der Umgebung des Tieres auf ca. 40° C führt zu sofortiger Aufhebung des Totstellzustandes und zu sehr lebhafter Bewegung. Beim Anhauchen kommt also als Reiz neben dem Tastreiz die Wärme sicherlich in Betracht.
Bei Tieren aus fast allen Gruppen des Tierreichs, welche man genauer studiert, findet man, daß sie auf leichte Reize bestimmter Art eine besondere Stellung und Haltung des Körpers einnehmen. Häufig wird diese Stellung als eine Droh- und Trutzstellung angesehen. In vielen Fällen bedeutet sie auch tatsächlich eine Bereitschaft zu Angriff oder Verteidigung; die Tiere bringen Gliedmaßen, Sehnen, Mundteile in diejenige Stellung, von der aus sie am schnellsten und erfolgreichsten diese ihre Waffen zur Schädigung eines etwaigen Gegners oder einer Beute verwenden können. Ebenso oft handelt es sich aber auch um eine Bereitschaft zur Flucht, zum Verbergen, zum Einwühlen in die Erde usw.
Ich habe in unserem „Tierbau und Tierleben“ dargestellt, daß die Tiere, vor allem die höheren Formen, je nachdem wie sie sich in Gefahr verhalten, in zwei Gruppen eingeteilt werden können, deren Abgrenzung uns das Verständnis vieler biologischer und psychologischer Eigentümlichkeiten der Arten eröffnet. Damals habe ich diese biologischen Gruppen nicht benannt; es erleichtert die Erörterung mancher Fragen, wenn man einen solchen Gegensatz durch besondere Worte festlegt. Ich will in Zukunft die Tiere nach ihrem Verhalten in Gefahr als Trutztiere und Fluchttiere unterscheiden. Scheinbar schwer in diese Gruppen einzuordnen sind jene Tiere, welche sich bei Eintritt einer Gefahr totstellen. Ich habe aber in meinem biologischen Werk auseinandergesetzt, daß es sich beim Totstellen fast stets um eine durch Besonderheiten des Baues und der Färbung der Tiere geförderte Flucht in die Unsichtbarkeit handelt.
In diesem Sinne gehört der Ameisenlöwe zu den Fluchttieren. Das weiche, zarthäutige Tier wäre ja gegen alle seine Feinde, wie Vögel und Eidechsen, wehrlos. Zur raschen Flucht ist es infolge seiner geringen Beweglichkeit nicht geeignet. So ist es erklärlich, wenn wir bei ihm weder eine Bereitschaftsstellung zur Flucht noch eine solche zur Verteidigung finden.
[S. 37]
Wohl zeigt aber der Ameisenlöwe eine sehr charakteristische Bereitschaftsstellung zum Fang seiner Beute, welche er mit vielen festsitzenden oder sehr träg beweglichen Tieren gemein hat. Ein Polyp oder eine Actinie halten ihre um die Mundöffnung angeordneten Tentakel nach oben ausgebreitet, von wo die Beute an sie herankommen kann. Squilliden und Mantiden verhalten sich mit ihrem viel komplizierteren Körperbau in den Grundzügen ganz entsprechend. Sie warten in einer charakteristischen Bereitschaftsstellung auf ihre Beute. Eine solche zeigt auch der Ameisenlöwe in freier Natur.

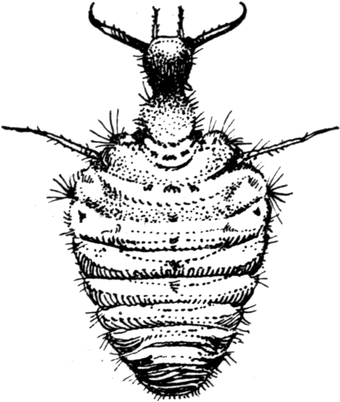
Beobachten wir einen Ameisenlöwen bei warmem Wetter und Sonnenschein in seinem Sandtrichter, so sehen wir ihn eine bestimmte Körperhaltung einnehmen. Sein schief nach unten im Sand steckender Rumpf ist mit der Hinterleibsspitze nach unten gekrümmt. Die Beine haben keine andere Stellung als während des Totstellens. Der Kopf dagegen ist steil nach oben gerichtet, wobei die Biegung in dem Kopf-Halsgelenk erfolgt. Seine Vorderseite ist direkt nach vorn gewandt. Die Fühler sind weit abgespreizt und ebenso die Mandibel mehr oder weniger weit, meist maximal, auseinandergesperrt (Abb. 15 und 16). Von diesem Zustand aus erfolgt außerordentlich leicht der Schnappreflex, welcher fast stets zur Ergreifung eines die Rückenseite des Tieres berührenden Gegenstandes führt.
[S. 38]
Aus dieser Bereitschaftsstellung erfolgt im Trichter stets die Ergreifung der Ameisen. Leichte Reize, z. B. durch das Herabrollen von Sandkörnchen veranlaßt, bringen das Tier zur Annahme der Bereitschaftsstellung. Auch experimentell kann man außerhalb des Sandes durch leichte Berührungsreize der Oberseite von Hals und Thorax das Tier ohne weiteres in die Bereitschaftsstellung bringen. Dabei hebt das Tier Kopf und Vorderteil sehr stark in die Höhe, so daß auch das erste Beinpaar schief in die Luft ragt und den Boden nicht berührt. In dieser Stellung kann es oft lange Zeit verweilen, wobei die Gliedmaßen ähnlich starr sind wie beim Totstellen. Meist sinkt aber außerhalb des Sandes nach einiger Zeit der Kopf auf die Unterlage herunter, während die Mandibel weit gespreizt bleiben.
Ist das Tier in seinem Trichter nicht in Bereitschaftsstellung, so nimmt es nach einem Reiz letztere zuerst an, um dann erst den Schnappreflex auszuführen. So erklärt es sich, wenn z. B. Comes glaubt, daß eine Ameise erst nach einigem Zappeln am Grunde des Trichters von der Larve gefaßt werde. Normalerweise aus der Bereitschaftsstellung heraus erfolgt die Ergreifung der Ameise in einem Bruchteil einer Sekunde.
In der Natur kommt es gelegentlich vor, daß Ameisenlöwen bei ihren Wanderungen einen Abhang hinunterkollern und dann auf dem Rücken liegenbleiben. Dasselbe ereignet sich häufig, wenn man die Tiere fängt. Nach einer kurzen Zeit des Totstellens pflegen sie sich dann in die Bauchlage umzudrehen.
Um diese Umdrehreaktion genauer zu prüfen, wurden zahlreiche Experimente von mir ausgeführt, bei denen das Tier schnell, ohne Druck und sonstige gröbere Reizung aus dem Sand genommen und auf den Rücken gelegt wurde. In der Mehrzahl der Fälle sind die Reize, welche bei diesem Vorgang auf das Tier einwirken, immerhin stark genug, um zu veranlassen, daß es sich totstellt. Wenn die charakteristische Frist abgelaufen ist, also, wie wir gehört haben, in der Regel 3–5 Minuten, erfolgt ganz plötzlich das Umdrehen des Tieres auf seine Bauchseite. Die Umdrehung geht in einem Bruchteil einer Sekunde vor sich; es ist daher sehr schwer, die Einzelhandlungen zu beobachten, welche das Tier dabei vornimmt.
[S. 39]
Man kann ziemlich deutlich sehen, daß eine Durchbiegung des ganzen Körpers erfolgt, wobei hauptsächlich starke Einknickung in den Gelenken zwischen Kopf und erstem Thorakalsegment und zwischen dem ersten und zweiten Thorakalsegment stattfindet. Indem auch der Hinterleib nach unten durchgebogen wird, macht das Tier sozusagen einen hohlen Rücken. Nach Annahme dieser Stellung gibt es sich plötzlich einen Stoß nach der einen Seite und rollt auf seine Bauchseite herum.
Bei der Schnelligkeit der Bewegung war es nicht ganz leicht, den Hebel ausfindig zu machen, welcher diese Seitwärtsbewegung verursacht. Es lag am nächsten, ihn in einer der Extremitäten des Tieres zu suchen. Um dies festzustellen, wurden folgende Experimente durchgeführt. Am 3. Juni 1912 wurde einem frisch gefangenen großen Exemplar das linke Mittelbein am Hüftgelenk amputiert; es trat ein sehr großer, klarer, gelblicher Blutstropfen hervor. Keine Bewegung oder sonstige Reaktion des Tieres zeigte eine Wirkung der Amputation an. Das Tier vermochte sich noch sehr gut umzudrehen, weder die Schnelligkeit noch die Promptheit der Reaktion war irgendwie beeinträchtigt.
Darauf wurde dem Tier auch das rechte Mittelbein amputiert. Es machte nun zuerst vergebliche Umdrehungsversuche, offenbar unter der Einwirkung der Operation, welche wohl eine Schwächung des Tieres oder, exakter ausgedrückt, eine Herabsetzung des Tonus seiner Gewebe zur Folge hatte. Nach wenigen Minuten konnte es aber wieder die Umdrehreaktion mit vollkommener Präzision ausführen.
Da nur die Mittelbeine hinreichend weit über den Körper hervorstehen, um bei der Umdrehreaktion eine Hebelwirkung auszuüben, während Vorder- und Hinterbeine dem Körper angeschmiegt gehalten werden, mußte die Ursache der Seitwärtsbewegung ganz anderswo gesucht werden. Es bot sich mir ein einfaches Mittel, um trotz der Schnelligkeit der Umdrehung die einzelnen Phasen derselben festzulegen. Ich legte die Tiere auf berußtes Registrierpapier und ließ sie da ihre Umdrehungsreaktion ausführen. Hat man die Tiere mit dem Rücken auf das Registrierpapier gelegt, so empfiehlt es sich, sie nicht aus dem Auge zu lassen, denn in vielen Fällen setzen sie sich nach der Umdrehung sofort in Bewegung, und es kann dann vorkommen, daß sie über die Umdrehungsspur hinwegkriechen und diese dadurch unlesbar machen. Man muß sie also nach erfolgter Umdrehung sorgfältig und schnell vom Registrierpapier abheben.
[S. 40]
Ist das mit genügender Sorgfalt geschehen, so erkennt man, daß der Umdrehungsvorgang auf dem Registrierpapier nur eine ganz geringe Spur hinterlassen hat. Die Abbildungen zeigen deutlich, daß nur das hinterste Körperende und die Kopfregion eine kräftige Spur aufgezeichnet haben, während von dem dazwischen liegenden Körper nur die Spitzen einiger vorstehenden Borsten durch feine Striche Zeugnis ablegen. Ist das Tier nach der linken Seite herübergeklappt, so sieht man nach der rechten Seite zu einen kräftigen Strich schief nach vorn ziehen (Abb. 17). Die Beobachtung zeigt uns ohne weiteres, daß es sich um den Abdruck einer oder beider Mandibel handelt. Sie waren also der Hebel, welcher bei der Drehung gewirkt hat. Indem beim Stemmen der Kopf eine Drehung nach der einen Seite macht und nach dieser Seite kräftig vorgestoßen wird, kippt der Körper nach der anderen Seite hinüber. Dabei bleiben vielfach beide Mandibel geschlossen; oft aber sieht man, daß die eine stark abgespreizt wird. Es ist stets die in der Richtung der Kopfabbiegung gelegene, so daß also durch sie die Hebelbewegung wesentlich unterstützt wird.

Beim Abbiegen des Kopfes nimmt an der Krümmung das erste Thorakalsegment teil; es bildet dann mit dem Kopf ein einheitliches[S. 41] System. Die Bewegung erfolgt mit einem sehr starken Ruck; es ist dies eine der typischen Reflexbewegungen des Tieres, auf deren vielseitige Bedeutung wir später noch oft werden zurückkommen müssen.
Der Umdrehreflex tritt stets nach längerer oder kürzerer Zeit ein; ein lebenskräftiges Tier bleibt nie viele Stunden lang auf dem Rücken liegen. Die Tendenz zum Umdrehen überwiegt auf die Dauer immer über jene zum Totstellen. Nach erfolgter Umdrehung bleibt aber das Tier oft in der Bauchlage noch lange, sich tot stellend, liegen (vgl. S. 29).
Diese eigentümliche Umdrehreaktion veranlaßt unwillkürlich zur Stellung einiger Fragen:
1. Haben die Tiere eine Tendenz, sich regelmäßig nach einer bestimmten Seite zu drehen?
2. Welcher Sinn erlaubt es den Tieren, zwischen Oben und Unten zu unterscheiden?
Bei den verschiedenen Umdrehversuchen war mir aufgefallen, daß tatsächlich die Tiere die ausgesprochene Tendenz zeigten, nach einer bestimmten Seite sich umzudrehen. Ich vermutete zuerst, daß es sich um ein ähnliches Phänomen handeln könne, wie es Jennings bei dem Seestern Asterias Forreri beschrieben hat. Dieses Tier zeigt die Tendenz, sich nach demjenigen Arm umzudrehen, auf den es beim Niedersinken im Wasser zuerst auffiel. Auch bei dem Ameisenlöwen beeinflußt ein äußerer Reiz die Umdrehungsrichtung.
Schon bei den auf Rußpapier registrierten Umdrehversuchen fiel es auf, daß die Aufzeichnung der Mandibel stets nach einer bestimmten Seite zeigte, und zwar war es fast stets die dem Lichte zugewandte Seite. Der Körper wälzt sich beim Umdrehen immer vom Licht weg. Um festzustellen, ob es sich dabei um eine regelmäßige Erscheinung handle, habe ich eine große Anzahl von Umdrehungsversuchen gemacht und genau beobachtet. Zu diesen Versuchen legte ich die Tiere so auf die Unterlage, daß ihre Längsachse dem Einfall der Lichtstrahlen gegenüber senkrecht stand; das Licht fiel also auf eine Seite des Tierkörpers, während die andere im Schatten war. Ich drehte dann die Tiere ohne starke Reizung auf die Rückseite und beobachtete die Umdrehung. Am 15. Sept. 1914 wurde mit 5 Exemplaren experimentiert, von denen No. 1 sich siebenmal nacheinander nach der Schattenseite umdrehte; ebenso No. 2 siebenmal, No. 4 fünfmal, No. 5 fünfmal. No. 3 jedoch war sehr lebhaft und drehte sich häufig unmittelbar, nachdem es in die Rückenlage[S. 42] gebracht war, um. Dann erfolgte die Drehung bald nach der Licht-, bald nach der Schattenseite. Ich drehte die Tiere meist in der Weise um, daß ich mit einer Präpariernadel unter das zweite Bein der einen Seite faßte und das Tier um seine Längsachse drehte. Manchmal schien es mir, als drehe sich das Tier bei sehr rascher Umdrehung nach der Seite, an welcher das Bein angefaßt worden war. Es ließ sich aber in dieser Beziehung keine Regelmäßigkeit feststellen.
Es spielt also bei der Umdrehreaktion das Licht offenbar eine bestimmende Rolle. Es lag daher nahe, die Frage zu untersuchen, ob nicht der Lichtsinn es sei, welcher dem Tier die Unterscheidung zwischen Unten und Oben gestattet, ähnlich wie dies bei vielen freischwimmenden Wassertieren der Fall ist. Zur Untersuchung dieser Frage wurden Exemplare des Ameisenlöwen im Dunkelzimmer auf eine Glasplatte gebracht, welche durch eine elektrische Glühlampe von 32 HK von unten beleuchtet war. Die Tiere wurden auf dieser Glasplatte auf die Rückenseite gedreht. Alle drehten sich nach kurzer Zeit, trotz der die Augen wie normales Oberlicht treffenden Unterbeleuchtung, rasch und sicher um. Es ist also nicht das Licht, welches ihre Orientierung zur Erdoberfläche vermittelt. Sie haben keinen Lichtrückenreflex (Buddenbrock).
Bei der Unterbeleuchtung zeugten die Tiere nach erfolgter Umdrehung eine große Lebhaftigkeit und bewegten sich in sehr merkwürdigen Haltungen rasch hin und her. Wir werden später sehen, daß das Licht eine sehr wesentliche und ihre Bewegungen richtende Einwirkung auf sie hat.
Um festzustellen, ob die Tiere zwischen Oben und Unten unterscheiden können, wurden noch weiter folgende Experimente gemacht. Es wurden Exemplare von Ameisenlöwen mit Hilfe eines Wachsklümpchens an einem Faden befestigt, mit dessen Hilfe sie frei aufgehängt wurden. Bei mehreren Individuen wurde der Faden in der Mitte der Rückenfläche auf dem dritten Thorakalsegment, bei anderen an der Bauchseite zwischen den Basalgliedern des zweiten und dritten Beinpaares befestigt. Die aufgehängten Individuen nahmen eine ganz bestimmte Haltung ein. Einerlei, ob sie mit der Rücken- oder mit der Bauchseite nach unten gerichtet waren, hatten sie den Bauch etwas eingekrümmt, hielten den Kopf und Hals sehr tief, schief gegen die Bauchseite gerichtet, die Hinterleibsspitze eingekrümmt, wie dies die nebenstehenden Abbildungen (Abb. 18) zeigen. Alle Beinpaare waren vom Körper abgespreizt, vor allem das erste[S. 43] und zweite, aber auch das dritte; letzteres war vor allem bei denjenigen Exemplaren der Fall, welche mit der Bauchseite nach oben aufgehängt waren. Das zweite Beinpaar wurde, wie beim Totstellen, ganz steif gehalten, während das erste und dritte Paar auch ziemlich starr waren, sich aber immerhin biegen ließen. Die ganze Haltung entsprach also im allgemeinen der beim gewöhnlichen Totstellen; dem entspricht wohl auch die erste Reaktion der aufgehängten Tiere.
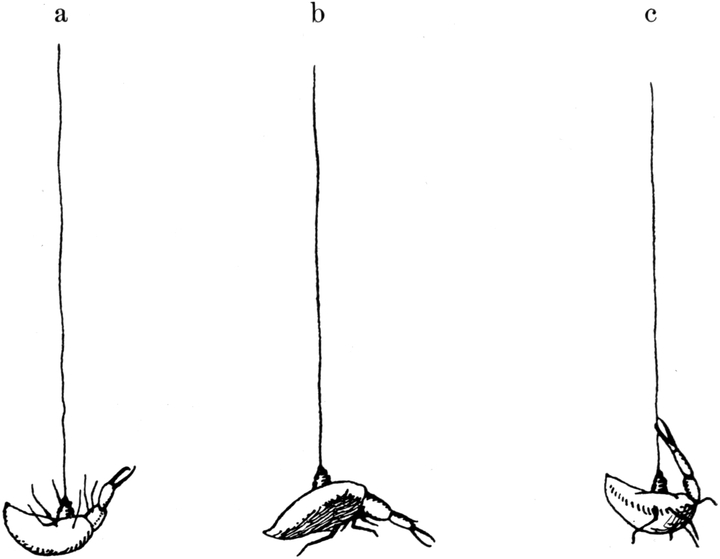
Die aufgehängten Exemplare bleiben auffallend lange Zeit vollkommen unbeweglich. Es kann dies 20 Minuten, eine halbe Stunde, unter Umständen auch stundenlang dauern. Dann beginnen sie scheinbar spontan Bewegungen auszuführen. Es sind dies Zappel- oder Ruderbewegungen aller sechs Beine; auf diese folgen Einkrümmung des Kopfes nach oben und unten, ebenso der Hinterleibsspitze; dazu kommen eigenartige nutierende Bewegungen des Kopfes, als ob dieser ein Widerlager suchte. Auch ohne daß er ein solches findet, werden typische Schleuderreflexe ausgeführt. Diese sind mit dem Schnappreflex kombiniert und führen oft zur Erfassung des Aufhängefadens mit den Mandibeln des Tieres. Da es vollkommen ausgeschlossen ist, von dem Tier unter den Verhältnissen des Experiments alle Reize auszuschließen, während man das Tier beobachtet, so zweifle ich sehr, daß die Bewegungen nur zentral ausgelöst sind. Sie sind nämlich sehr leicht durch dem Tier experimentell applizierte Reize zu veranlassen. So wirken z. B. schon relativ geringe Erschütterungen auf das Tier ein; man braucht bloß auf den Tisch zu klopfen oder den Stab zu berühren, an welchem der das Tier tragende Faden aufgehängt ist. Vor allem wirken aber Berührungsreize, und zwar sowohl Berührung der Oberseite als auch der Unterseite. Mit der Bauchseite nach unten hängende Tiere,[S. 44] unter welche man einen Spachtel, ein Stück Pappe oder Stoff hält, führen sofort den Schleuder- und Schnappreflex aus. Es geschieht dies nicht nur bei Berührung der ganzen Unterseite, sondern auch, wenn nur ein Teil derselben, des Kopfes, des Rumpfes oder nur der hinterste Teil des Hinterleibes berührt wird. Allerdings scheint die Reizempfindlichkeit gegen das Hinterende des Abdomens etwas abzunehmen (vgl. unten Abschn. V, 5. Tastsinn).
Der Umdrehreflex, welcher, wie wir noch genauer sehen werden, mit dem Schleuderreflex vollkommen identisch ist, wird ganz in derselben Weise ausgeführt, wenn statt der Unterseite irgendeine Stelle der Oberseite des Tieres, also seines Kopfes, Rumpfes oder Hinterleibes berührt wird. Ganz das Entsprechende geschieht, wenn die Beine einem solchen Reiz ausgesetzt werden.
Die Bewegungen der hängenden Tiere hören immer nach einiger Zeit auf, und es tritt wieder vollkommene Bewegungslosigkeit ein. Nach einer Pause von verschiedener Länge (5 Minuten, eine halbe Stunde, oft auch viele Stunden) werden die Bewegungen wieder aufgenommen. Oft geschieht das auf einen deutlichen äußeren Reiz hin; noch öfter konnte ich einen solchen nicht wahrnehmen. In den meisten Fällen, in denen solche beobachtet wurden, handelte es sich um Erschütterungsreize oder Reizung durch Luftbewegung. Auch Wärme und Lichtreize haben den gleichen Effekt.
Es ergibt sich also aus all diesen Versuchen, daß bei dem Tier wohl kein besonderes Sinnesorgan geotropischer Natur vorhanden ist. Jedenfalls brauchen wir ein solches zur Erklärung des Umdrehreflexes nicht heranzuziehen. Vielmehr handelt es sich um eine auf Reiz hin erfolgende, vollkommen automatisch ablaufende, typische Bewegung, welche von dem Tier in jeder Lage ausgeführt wird. Nur wenn das Tier zufällig auf der Rückenseite auf einer festen Unterlage liegt, führt die reflektorische Bewegung zur Umdrehung. Es ist genau dieselbe Bewegung, welche beim Sandschleudern angewandt wird, welche unter bestimmten örtlichen Verhältnissen zur Umdrehung führt. Wir können also feststellen, daß die Umdrehreaktion durch den Schleuderreflex bewirkt wird. Manchmal ist er mit dem Schnappreflex kombiniert (vgl. S. 62).
Und es sind Berührungsreize, welche den Umdrehreflex auslösen. Offenbar wirkt die Berührung der Unterseite des Tieres mit der Unterlage beruhigend auf das Tier, wenn das Tier selbst auf diese einen gleichmäßigen, sich nicht ändernden Druck ausübt. Bei Rückenlage wirkt aber der Druck des eigenen Gewichtes als[S. 45] Reiz, welcher vor allem bei Bewegungen des Tieres selbst den Umdrehreflex auslöst. Es ist also der Umdrehreflex durch Tangorezeption bewirkt, die verschiedene Empfindlichkeit der Ober- und Unterseite des Tieres wird die Unterscheidung von Oben und Unten ermöglichen. Es ist daher nicht angebracht, von Geotaxis beim Ameisenlöwen zu sprechen, wie es Comes tut. Hervorzuheben ist, daß auf dem Rücken liegende Tiere viel länger ruhig liegenbleiben, ohne die Umdrehung auszuführen, wenn man ihnen etwas Sand auf die Bauchseite streut. Aber es wird dadurch die Umdrehung nicht vollkommen verhindert. Nach kürzerer oder längerer Zeit erfolgt sie dennoch.
Während bei manchen Verwandten des Ameisenlöwen, z. B. bei der bei uns in Süddeutschland nicht seltenen Gattung Ascalaphus, die Larven, frei umherwandernd, sich ihre Nahrung suchen, findet man unsere gewöhnlichen Ameisenlöwen in der freien Natur selten auf der Wanderung. Experimentell kann man sie leicht zu Wanderungen veranlassen, welche zum Teil gar nicht unbeträchtliche Leistungen darstellen. Hat man sie aus dem Sand herausgegraben, oder haben sie eine Umdrehreaktion ausgeführt, so pflegen sie nach einiger Zeit sich in Bewegung zu setzen.
Die Art ihrer Bewegung ist sehr charakteristisch; sie erfolgt nämlich stets nach rückwärts, die Hinterleibsspitze wird vorangeschoben, der Kopf folgt nach. Genauere Untersuchungen und zahlreiche Experimente haben mir gezeigt, daß das Tier sich überhaupt nur rückwärts zu bewegen vermag. Darauf deutet schon seine ganze Organisation hin: die hinten zugespitzte Gestalt des Körpers, die nach vorn gerichteten Borstenbüschel und -ringe auf dem ganzen Körper, die Stellung und Einlenkung der Beine. Bei den vielen Hunderten von Experimenten, die ich mit dem Tier ausführte, kamen nur einige wenige Fälle vor, in denen das Tier sich für eine minimale Strecke mit dem Kopf nach vorn bewegte. Diese Tatsache ist schon lange bekannt. Ebenso ist oft beobachtet worden, daß diejenigen Myrmecoleontiden, welche keine Trichter bauen, alle vorwärts zu laufen vermögen. Ja, es gibt auch trichterbauende Formen, welche vorwärts laufen können.
Die Bewegung erfolgt stets ruckweise. Auch macht das Tier fast stets, wenn es einige Zentimeter zurückgelegt hat, eine mehr oder minder lange Ruhepause, ehe es sich von neuem in Bewegung setzt.
[S. 46]
Um die Bewegungsweise des Tieres im einzelnen zu studieren, legt man es am besten auf eine durchsichtige Glasscheibe und beobachtet von unten die Tätigkeit des Hinterleibes und der Beinpaare. Unter Umständen kann dann das Tier mit etwas gehobenem Hinterleib auf seinen sechs Beinen geradezu laufen. Meist ähnelt aber die Bewegung dem gewohnten Kriechen. Es kommt auch vor, daß es auf einer Glasplatte nur auf dem zweiten und dritten Beinpaar läuft, wobei die Hauptarbeit dem dritten Paar zufällt. Man sieht aber in der Regel, daß die Hinterleibsspitze zuckende Bewegungen ausführt, welche allerdings auf der glatten Unterlage keine sehr große Wirkung haben. Das vorderste Beinpaar wird ruhig gehalten, das zweite Beinpaar macht plötzliche schwankende Bewegungen, während das dritte Beinpaar unablässig den Körper nach rückwärts stößt. Der Erfolg der einzelnen Bewegungen macht sich auf einer rauhen Unterlage viel ausgesprochener geltend. Da kann sich die Hinterleibsspitze und überhaupt ein größerer Teil der Unterseite des Hinterleibes dem Boden anstemmen, die beweglichen Beinpaare finden ein Widerlager und können geregelter arbeiten. Man kann die Tätigkeit der einzelnen Teile mit Deutlichkeit verfolgen, wenn man das Tier die Spur seiner Bewegungen auf berußtem Papier selbst aufzeichnen läßt. Man vergleiche die dieser Untersuchung beigegebenen zahlreichen Bilder von Kriechspuren des Tieres. Man wird erkennen, daß die tatsächlich unbewegten Vorderbeine oft gar keine Spur hinterlassen haben. Oft sieht man aber auch parallel der äußersten Begrenzung der Spur feine Linien verlaufen, welche von dem vordersten Beinpaar herrühren, das auf der Unterlage nachschleift, auch gelegentlich gleichzeitig mit dem zweiten Beinpaar wippende Bewegungen ausführt. Die Mittelbeine, welche abwechselnd hebelnde Bewegungen machen, haben diese in den feinen, etwas zittrigen Linien aufgezeichnet, welche die Kriechspur außen begleiten. An den Unterbrechungen erkennt man, daß sie oft abwechselnd tätig sind. Dabei kommt oft ein eigenartiges Schwanken und Schaukeln des Tierkörpers zustande; die weitausladenden Beine des zweiten Paares spielen jedenfalls beim Lenken des Körpers eine Rolle. Dreht sich die Hinterleibsspitze einer bestimmten Richtung zu, so wird auch das Mittelbein der betreffenden Seite etwas mehr angezogen und zum Körper in einen stumpfen Winkel gestellt. Wie die Beobachtung und auch die Aufzeichnungen auf dem Rußpapier zeigen, erfolgen immer mehrere Zuckungen des Hinterleibes und auch Bewegungen des dritten Beinpaares, ehe ein Schaukelschritt des Mittelbeinpaares[S. 47] folgt. Letzteres wird dabei immer ganz steif gehalten. Am stärksten ausgeprägt zeigt sich der Abdruck der scharrenden Bewegung des dritten Beinpaares. Vielfach erkennt man auch die Spuren der vorwärts gerichteten Beborstung der Beine.
Dagegen vermißt man an der Spur des auf der relativ glatten Unterlage, welche von dem Rußpapier dargeboten wird, kriechenden Tieres einen deutlichen und regelmäßigen Abdruck der Hinterleibsspitze. Man erkennt den Abdruck der Hinterleibsspitze streckenweise sehr deutlich auf Abb. 19. Die Hinterleibsspitze wird offenbar nur dann regelmäßig in Tätigkeit versetzt, wenn die Unterlage hinreichend rauh ist. Man kann das z. B. auf nassem Sand, rauhen Steinen, oder wenn man die Tiere über die Borsten einer Bürste oder über ein Stück Sammet oder Plüsch hinkriechen läßt, deutlich erkennen. Auch wenn das Tier über trockenen Sand hinkriecht, ohne sich einzugraben, wirkt bei den Wanderbewegungen die Hinterleibspitze erheblich mit. Dann gräbt das Tier eine ziemlich tiefe Furche in die Oberfläche des Sandes ein.

Es kommt häufig vor, daß ein Tier sehr lange Zeit bewegungslos an einer Stelle liegt. Es bedarf aber besonderer Anlässe, um einen Ameisenlöwen zu einer sehr lange dauernden Wanderung zu veranlassen. Wir werden in den späteren Abschnitten erfahren, welche Reize die Wanderbewegungen des Tieres fördern und welche sie hemmen.
[S. 48]
An dieser Stelle müssen wir als wichtige Tatsache hervorheben, daß jedes Tier nach mehr oder minder langer Ruhepause sich in Bewegung setzt, wenn nicht ganz bestimmte Einwirkungen dies verhindern. Wirken, sobald das Tier sich in Bewegung setzt, Reize von bestimmter Stärke aus einer bestimmten Richtung auf das Tier ein, so wird dadurch die Richtung seiner eigenen Bewegungen auf das deutlichste beeinflußt. Wirken aber keine solchen richtenden Reize auf das Tier ein, so führt es eigentümlich unorientierte Bewegungen aus. Es wandert, natürlich immer mit der Hinterleibsspitze voran, in geschlängelten Bahnen, die oft in sich selbst umkehren und in welche oft ganz kleine Kreisbahnen oder Spiralen eingeschaltet sind, umher (Abb. 20). Diese gleichsam tastenden Wanderungen entsprechen jener Bewegungsweise, welche ich in meiner Untersuchung über die Biologie der Garneelen den Suchgang genannt habe. Wie ich dort auseinandergesetzt habe, handelt es sich entweder um zentral oder doch vom Tierkörper selbst ausgelöste, vielleicht aber auch um durch minimale Reize veranlaßte Bewegungen, welche dem Tier die Möglichkeit geben, Reize der verschiedensten Art aufzufangen, die dann ihrerseits die Richtung, Schnelligkeit und Dauer der Bewegung der Tiere bestimmen. Dieser Suchgang gehört also zu den bei den Tieren so allgemein verbreiteten Versuchs- oder Probierbewegungen.
Wenn Tiere hungern oder sonstwie sich nicht ganz wohl befinden, ferner wenn in dem Aufbewahrungsgefäß die Sandschicht zu dünn ist, um den Bau von Trichtern zu erlauben, findet man auf der Oberfläche des Sandes flache Furchen, die kreuz und quer verlaufen, kreis- und spiralförmig, sich durchkreuzen usw. Das Tier war auf der Suche nach den Reizen, die ihm einen bestimmten Ort anweisen sollten.
[S. 49]
Hat man im Freien beim Versuch des Fanges einen Ameisenlöwen fallen lassen, so kann man, nachdem die Zeit des Totstellens verstrichen ist und oft unmittelbar, nachdem er die Umdrehreaktion ausgeführt hat, sehen, daß er sich unter ruckweisen Bewegungen nach rückwärts sehr schnell in den Sand eingräbt. Dabei führt er zunächst genau dieselben Bewegungen aus, die wir ihn bei seinen Wanderungen auf einer rauhen Unterlage vornehmen sahen. Wir sehen genau dieselben zuckenden Bewegungen der Hinterleibsspitze; das erste Beinpaar wird ruhig gehalten, während das zweite die pendelnden und das dritte die scharrenden Bewegungen ausführt. Was ist aber hier die Folge dieser automatischen Bewegungen? Der Bau des Körpers des Ameisenlöwen zwingt die Bewegung in dem aus zahlreichen feinen Partikeln bestehenden Sand in eine ganz bestimmte Richtung. Bei der Bewegung im Sand zeigt sich die Bedeutung der Form des Körpers und seines Borstenkleides in einem überraschenden Licht. Die kegelförmige Hinterleibsspitze bohrt sich voraus in den Sand, der Körper drängt nach, und erst von den Schultern an beginnt die schmälere Zone, welche leicht aus dem Sand vorragt und sich so erhält. Wühlt sich das Tier aber weiter in den Sand ein, so bietet die Schulterregion mit ihren Borstenbüscheln ein weiteres wirksames Widerlager.
Die kurzen starren Borsten, welche die Hinterleibsspitze umgeben, drängen bei deren Vorstoßen die Sandkörner zur Seite; sind aber die hintersten Segmente des Hinterleibes einmal in den Sand eingedrungen, so zeigt sich die auffällige Wirkung der, wie wir früher gesehen haben, insgesamt nach vorn gerichteten Borsten und Borstenkränze des ganzen Körpers. Zwischen den rollenden und gleitenden Körnchen des trockenen Sandes könnte ein glatter Körper überhaupt keinen Halt gewinnen; ein Tier, welches sich mit glattem Körper in diesem beweglichen Medium von der Stelle bringen wollte, müßte dafür einen großen Aufwand an Kraft leisten; es müßte mit breiten, flächenhaften Hebeln sich vorwärts rudern oder sonstwie besondere Bewegungseinrichtungen besitzen. Statt dessen sehen wir den Körper des Ameisenlöwen an zahllosen Stellen mit seinen Borsten an den rollenden Sandkörnern einen Widerstand gewinnen.
Dadurch, daß die Borsten alle in mehr oder weniger ausgesprochener Weise nach vorn gerichtet sind, liefert dieser Widerstand eine Druckkomponente nach hinten. Nach jeder Bewegung in den[S. 50] Sand hinein hakt der Körper des Ameisenlöwen in dem rollenden Material geradezu fest; er kann nicht wieder zurückgleiten. Ist er schon auf glatter Unterlage zu einer Vorwärtsbewegung unfähig, so zeigt sich dies im Sande in noch viel ausgesprochenerem Maße. Durch die zuckenden Bewegungen seines Hinterleibes, welche durch die schiebenden Bewegungen vor allem des dritten Beinpaares noch unterstützt werden, muß das Tier sehr rasch und sicher in den Sand eingebohrt werden. Wir sehen es denn auch tatsächlich oft in 4–5 Sekunden unter dem Sand verschwinden.
Die wesentlichste Komponente bei dem Einwühlen in den Sand bilden die nach hinten und unten gerichteten zuckenden Bewegungen des Hinterleibes. Tiere, denen man die beiden Mittelbeine amputiert, können ebenso, wie sie recht gut rückwärts wandern, sich absolut sicher in den Sand eingraben. Nicht einmal die Schnelligkeit der Reaktion ist beeinträchtigt. Ja, selbst die Amputation sämtlicher Beinpaare hindert ein Tier nicht, sich rasch und vollständig in den Sand einzuwühlen. In wie ausgesprochener Weise die Bewegungen des Hinterleibes für die Lokomotion des Tieres maßgebend sind, beweist folgender Versuch: Läßt man einen Ameisenlöwen mit dem Hinterleib voran in eine Glasröhre hineinkriechen, deren innerer Durchmesser ungefähr genau der größten Breite seines Hinterleibes entspricht, so findet das Tier, dessen Körper von den Seiten etwas zusammengedrückt wird und dadurch etwas an Höhendurchmesser zunimmt, reichlich Berührungspunkte an der Wand der Glasröhre. Es kriecht in der Röhre vollkommen normal rückwärts, auch wenn seine Beine nirgends in Berührung mit der Innenfläche der Glasröhre kommen und sich somit nirgends anstemmen können. Ja, es spreizt sich mit seinen Borstenkränzen so gut an den Wänden fest, daß es nicht herauszuschütteln ist.
Bei seiner Rückwärtsbewegung ist das erste Beinpaar in Ruhe; das zweite ist, im Falle die Röhre eng genug ist, an der Röhrenwand angestemmt, hat aber, da es nur vereinzelte Zuckungen ausführt, keinen wesentlichen Anteil an der Rückwärtsbewegung. Das dritte führt reflektorisch die zappelnden Bewegungen aus, die wir vorhin beschrieben, ohne allerdings dabei die Unterlage zu berühren. Es arbeitet also ohne Nutzeffekt. Eine Berührung mit der Röhrenwand führen hauptsächlich die Haarpolster an den Schultern des Tieres herbei. Die Rückwärtsbewegung erfolgt durch abwechselndes Aufwärts- und Abwärtskrümmen des Hinterleibes und des Kopfabschnittes. Der Körper macht sich also, wie die untenstehende[S. 51] Abbildung 21 zeigt, abwechselnd an der Rücken- und dann an der Bauchseite hohl. Indem dabei jeweils die Hinterleibsspitze sich fest an die Glasröhrenwand anstemmt, schiebt sich der Körper durch die Röhre nach hinten. Ist die Glasröhre so weit, daß der Rücken ihre obere Wand nicht berührt, so bleibt die Bauchseite des Tieres stets konkav, und das Tier bewegt sich, allerdings langsam, nur durch die zuckenden Bewegungen der Hinterleibsspitze, welche sich an der Unterseite der Glasröhre reibt.

Beim Einwühlen in den Sand arbeiten die Beinpaare naturgemäß mit. Die beiden vorderen räumen Sand unter dem Kopf und Hals weg, so daß dieser Sand über diesen Körperteilen zusammenstürzt. Auch die Bewegungen des dritten Beinpaares schaufeln den Sand auf die Seiten und den Rücken des Körpers. Auch stemmen, besonders am Anfang des Einwühlens, die beiden vorderen Beinpaare kräftig nach vorn. Dadurch werden die im nächsten Abschnitt geschilderten Vorgänge eingeleitet.
Meine Beobachtungen über den Bau der Sandtrichter weichen von früheren Angaben zunächst insofern ab, als die Ameisenlöwen ihre Trichter jedenfalls in anderer Weise bauen können, als es Rösel von Rosenhof geschildert hatte. Die von mir beobachteten Tiere fertigten ihre Trichter stets in einer viel einfacheren Weise an.
Ist ein auf die Oberfläche einer mehrere Zentimeter dicken Sandschicht gelegter Ameisenlöwe aus seinem Starrezustand erwacht, so beginnt er, wie wir früher gesehen haben, sich rückwärts zu bewegen und mit der Schwanzspitze voran sich einzubohren. Nach wenigen Sekunden pflegt er im Sand verschwunden zu sein und bleibt in manchen Fällen ruhig unter der Sandfläche liegen, um erst später sich weiter zu bewegen. Dann kann er entweder Wanderungen unter dem Sand antreten, oder er beginnt mit dem Bau des Trichters.[S. 52] Frische und gesunde Tiere pflegen aber sofort, wenn sie sich im Sand eingewühlt haben, den Trichterbau einzuleiten.

Dies geht auf folgende Weise vor sich. Nachdem Rumpf und Hals und meist der größere Teil der beiden vorderen Beinpaare im Sand verschwunden sind, beginnt der Kopf in ihn einzutauchen. Kaum haben sich einige Sandkörner auf der Oberfläche des Kopfes angesammelt, so führt das Tier eine eigentümliche Bewegung aus, mit welcher es den Sand hoch in die Höhe schleudert. Diese Bewegung besteht in einer plötzlichen, ruckweisen Umbiegung des Kopfes, des Halses und der ersten Rumpfsegmente nach oben und hinten. Wir werden diese Bewegung, welche für das Tier außerordentlich typisch ist, später genau analysieren. Es genüge hier, hervorzuheben, daß sie in der Regel auf einen Tastreiz hin erfolgt; sie kann sowohl in der Mittellinie direkt nach hinten als auch in beliebigen Winkeln von ihr abweichend erfolgen. Während im ersteren Falle der Horizontalschnitt von Kopf und Hals, welche zusammen einen sehr spitzen Winkel mit dem Rumpf bilden können, dessen Horizontaldurchschnittsebene in der Querrichtung parallel bleibt, ist[S. 53] er im zweiten Fall in verschiedenen Winkeln gegen sie verdreht. (Vgl. hierzu die Beobachtungen über lokalisierten Tastreiz und entsprechende Reizfolgen im Kapitel über Tastreiz S. 87.) Wir bezeichnen diese reflektorische Bewegung als den Schleuderreflex.
In der Regel erfolgen mehrere Schleuderbewegungen in rascher Aufeinanderfolge nacheinander. Indem sie infolge der Winkelneigungen von Kopf und Hals nach verschiedenen Richtungen erfolgen, entsteht ein gleichmäßig abfallender Trichter im Sand, der schon eine gewisse Tiefe besitzt. Oft begnügt sich das Tier hiermit und bleibt ganz ruhig am Grunde seines Trichters sitzen, wobei der Körper von der unteren Spitze des Trichters schief nach der einen Seite abwärts in den Sand ragt (Abb. 14 S. 26). Der vordere Teil des Kopfes mit Augen und Fühlern pflegt aus dem Sand hervorzuragen. Vielfach nimmt das Tier bereits jetzt eine Bereitschaftsstellung an, die wir oben beschrieben haben, und von der man in diesem Moment nur in den weit auseinanderklaffenden Kiefern ein Anzeichen wahrnimmt.
Fast stets pflegen nach einiger Zeit von den Trichterwänden Sandkörnchen auf die obere Kopfseite des Ameisenlöwen herabzufallen, welcher sie sofort durch Schleuderbewegungen nach oben an den Rand des Trichters befördert. Jede Luftbewegung, jede Temperaturänderung veranlaßt ein solches Herabrieseln von Sandkörnchen. Da bei jeder Schleuderbewegung mehr Sand hinaufgeschleudert wird, als vorher herabgerieselt war, so vertieft sich auf diese Weise der Trichter nicht unwesentlich. Dabei führt der Ameisenlöwe auch kriechende Bewegungen aus, durch die er sich tiefer in den Sand einwühlt, wobei er aber in der Regel den Mittelpunkt des Trichters nicht allzusehr verschiebt. Er muß also durch eine enge Spiralbewegung den Körper etwa um eine durch die Lage des Kopfes gekennzeichnete Achse bewegen. Die verschiedenen Kriechspuren auf Rußpapier, welche in den Abbildungen (z. B. Abb. 20) dargestellt sind, geben ein Beispiel von diesen Spiralbewegungen, welche für den Ameisenlöwen sehr charakteristisch sind, und welche zu den früher (S. 47) schon charakterisierten Versuchsbewegungen des Tieres gehören.
Bei diesen Drehungen und infolge der Fähigkeit, den Sand in allen möglichen Winkeln abzuschleudern, hat der Ameisenlöwe die Möglichkeit, einen vollkommen gleichmäßig von einem kreisförmigen oberen Umfang zu der in senkrechter Richtung unter dem Mittelpunkt dieses Kreises gelegenen Trichterspitze abfallenden Trichter[S. 54] zu bauen; es stellt also die Innenfläche des Trichters einen gleichmäßigen Kegelmantel dar. Daß das Tier in gleichmäßig feinem Sand das aus dem Trichter entfernte Material tatsächlich ganz regelmäßig nach allen Seiten wegschleudert, konnte ich durch folgenden Versuch nachweisen. Ueber einer Sandfläche, in welcher ein Ameisenlöwe gerade seinen Trichter einzuwühlen begann, brachte ich im Abstand von 3–4 cm eine auf der unteren Seite durch Wasserdampf beschlagene Glasscheibe an. Damit das Wasser nicht verdunstete, war der Sand mit dem Tier in einem besonderen Gefäß in eine dicht abgeschlossene Glasschale gebracht, auf deren Boden feuchter Sand lag. Die von dem Ameisenlöwen in die Höhe geschleuderten Sandkörner blieben an der benetzten Scheibe haften und bildeten eine regelmäßige kreisförmige Fläche, welche, der Verlängerung des Trichterwinkels entsprechend, größer als die kreisförmige Umgrenzung des Trichters selbst war. Bei allen untersuchten Tieren ergab das Experiment dasselbe Resultat. Es folgt daraus, daß regelmäßig die Tiere in einem bestimmten Zeitraum den ganzen Umkreis ihres Trichters mit ihren Schleuderbewegungen bestreichen.
Nicht ganz so regelmäßig verläuft die Arbeit, wenn ein Ameisenlöwe in Sand baut, dem größere Steinchen oder sonstige Partikel beigemischt sind. In grobem Material braucht ein Ameisenlöwe, besonders ein kleines Tier, zur Anlage eines Trichters oft sehr lange Zeit; denn nach jeder Schleuderbewegung folgt eine sehr lange Pause. Die Leistungen, welche die Tiere dabei vollbringen, sind vielfach gar nicht gering. So habe ich die Steinchen, welche ein mittelgroßer Ameisenlöwe von etwa 10 mm Gesamtlänge (gemessen einschließlich der Mandibel) bis zu 10 cm weit wegzuschleudern vermochte, gewogen und fand folgende Gewichte: 4½ mg, 15 mg, 35 mg, 43 mg. Es ist von Interesse, demgegenüber die Gewichte der Tiere selbst festzustellen. Es wiegen:
|
1.
|
Exemplar
|
von
|
6
|
mm
|
Länge
|
13
|
mg
|
|
2.
|
„
|
„
|
7,5
|
„
|
„
|
14
|
„
|
|
3.
|
„
|
„
|
7
|
„
|
„
|
15
|
„
|
|
4.
|
„
|
„
|
10
|
„
|
„
|
22
|
„
|
|
5.
|
„
|
„
|
12
|
„
|
„
|
28
|
„
|
|
6.
|
„
|
„
|
13,5
|
„
|
„
|
70
|
„
|
Die großen Steinchen werden offenbar von dem Tier mit besonderer Kraftanstrengung weggeschleudert; denn sie fliegen so weit weg, daß sie nicht mehr in den Trichter zurückfallen. In einem[S. 55] Material, welches aus feinen und groben Partikeln gemischt ist, erfolgt durch die Schleuderbewegung eine Sortierung. Die groben Partikel sind weiter hinaus an die Peripherie geschafft, während der Trichter selbst schließlich nur aus ganz feinem Sand besteht. Ein Wegtragen der größeren Steine auf dem Rücken, wie es frühere Autoren angeben, habe ich nie beobachten können. Ich bin auch der Ansicht, daß die Sortierung des Sandes rein mechanisch vor sich geht, durch die Schwere der Partikel bewirkt. Die schweren Teilchen fliegen relativ weit weg; von den feinen rollt ein Teil den Abhang des Trichters wieder herunter. Daß, wie z. B. Redtenbacher annimmt, die langzähnigen Kiefer als Sieb für den geschleuderten Sand wirkten, indem nur die groben Teile geworfen würden, die feinen durchrieselten, scheint mir keine erhebliche Rolle zu spielen.

[S. 56]
Ich habe einige Versuche gemacht, um den Einfluß festzustellen, welchen das Gewicht und die Kohärenz der Partikel, in denen das Tier baut, auf seine Bautätigkeit ausübt. Ich ließ Tiere sich in verschiedene Substanzen eingraben, wie z. B. in feines Weizenmehl, in grobe Eisenfeilspäne u. dgl. Da die Mehlstäubchen sehr fein und leicht sind, werden sie leicht sich eine Zeitlang in der Luft schwebend erhalten und wieder an Ort und Stelle niedersinken. Beim Zusammenpressen haften sie leicht aneinander. Man sieht denn auch, daß es den Ameisenlöwen leicht gelingt, sich im Mehl einzugraben; aber sie bringen in dieser Substanz keinen richtigen Trichter fertig. Die Tiere wühlen geradezu Gänge durch das Mehl. Sie fühlen sich in demselben offenbar sehr unbehaglich, denn es verschmiert ihnen alle Gelenke, haftet an der Oberfläche des Körpers und verstopft die Atemlöcher. Tiere, welche in Mehl gegraben haben, führen von Zeit zu Zeit die Putzbewegungen aus, von denen wir später Näheres hören werden. Ein Exemplar, welches am 21. Sept. 1914 auf Mehl gesetzt wurde, hatte sich in 5 Minuten, ein anderes in 15 Minuten eingegraben. Nur eines von ihnen hatte bis zum 22. Sept. einen relativ regelmäßigen Trichter von minimalem Umfang fertiggebracht.
In Eisenfeilspänen dagegen gelang das Eingraben sehr prompt; die schweren Eisenkörnchen wurden von dem Tier leicht hoch in die Luft geschleudert. Sie hafteten gar nicht aneinander, und so entstand ein schöner regelmäßiger Trichter.
Je trockener und gleichmäßiger der Sand ist, um so größere Trichter bauen die Ameisenlöwen. Ferner hängt der Umfang des Baues von Wärme und Sonnenschein ab. Bei heißem, sonnigem Wetter werden die größten Trichter gebaut. Hungrige Tiere, die unruhig sind und sehr leicht durch herabrieselnde Sandkörner zum Schleuderreflex gereizt werden, vergrößern im Lauf der Zeit ihren Trichter oft beträchtlich. So hängt also die Größe des Trichters nicht direkt von dem Körperumfang des Erbauers ab. Kleine Tiere können unter günstigen Umständen ebenso große oder größere Trichter bauen als ausgewachsene Tiere unter ungünstigen.
Wird der Sand, in welchem die Ameisenlöwen leben, nass, so können die Tiere in ihm sich nur mit Mühe oder gar nicht vorwärtsbewegen. Sie liegen dann ganz regungslos. Legt man Ameisenlöwen auf befeuchteten Sand, so können sie sich nur dann noch in ihn eingraben, wenn er mäßig feucht ist. Einen Trichterbau bringen sie in diesem Fall aber nicht fertig. Weder das Eingraben noch[S. 57] der Trichterbau gelingt ihnen, wenn der Sand in erheblicherem Maße durchnäßt ist.
Diese letzteren Beobachtungen bekräftigen meine Annahme, daß beim Trichterbau das Schleudern die Hauptsache ist und daß die Grabtätigkeit dabei keine Rolle spielt. Dafür spricht ferner, daß auch Tiere mit amputierten Beinen noch vollkommen normale Trichter bauen können. Bei normalen Tieren helfen natürlich die Beine insofern mit, als sie, ähnlich wie beim Einwühlen, Sand zur Seite und auf Kopf und Hals des Tieres schaufeln.
Wir sehen also, daß der Schleuderreflex, den wir oben als Ursache der Umdrehreaktion kennen lernten, auch beim Bau der Sandtrichter die wichtigste Bewegung darstellt. Die Sandtrichter entstehen automatisch infolge von Reizen, welche den Schleuderreflex auslösen.
Es sei hier noch darauf hingewiesen, daß zu gewissen Zeiten des Jahres unsere einheimischen Ameisenlöwen keine Trichter bauen. Zu allen Zeiten des Jahres unterlassen die in der Vorbereitung zur Verpuppung stehenden Individuen den Trichterbau.
Jedenfalls geht aus meinen Beobachtungen hervor, daß der Trichterbau in der Regel in viel einfacherer Weise vor sich geht, als es Rösel von Rosenhof in der S. 2 zitierten Schilderung beschrieben hat. Es mußte für mich natürlich von Wichtigkeit sein, festzustellen, ob ein so gewissenhafter Beobachter wie der treffliche Rösel wirklich den Vorgang aus der Phantasie konstruiert hatte, oder ob besondere Gründe vorlagen, die seine unrichtige Schilderung veranlaßten. Ich suchte nach Versuchsanordnungen, welche die tatsächlichen Grundlagen für seine Beobachtungen liefern konnten.
Nun läßt sich wirklich nachweisen, daß, wenn man einem Ameisenlöwen eine zu wenig tiefe Sandschicht darbietet, so daß er beim Einwühlen bald den Boden des sanderfüllten Gefäßes erreicht, er mit in Spiralen verlaufenden Versuchsbewegungen den Sand zu durchfurchen beginnt. Selten gelingt dem Tier dabei der Bau eines einigermaßen regelmäßigen Trichters, stets spielen auch dann die Schleuderbewegungen eine Hauptrolle.
Dauert der Trichterbau längere Zeit, und bei normal fertiggebauten Trichtern kann man dieselbe Beobachtung machen, so ändert das Tier je nach dem Lichteinfall — also der scheinbaren Wanderung der Sonne am Himmel folgend — nach den Gesetzmäßigkeiten, die wir in einem späteren Kapitel (S. 63) kennen lernen werden, seine Einstellung zum Licht. Da das Tier nicht direkt seitlich[S. 58] kriechen und dabei seinen Kopf im Mittelpunkt des Trichters liegen lassen kann, sondern in Spiralen den Mittelpunkt umkriechen muß, so erfolgen Bewegungen, welche an die von Rösel beschriebenen erinnern.
Niemals aber konnte ich einen Trichterbau beobachten, welcher genau in der von ihm beschriebenen Weise ablief. Da außer Rösel zahlreiche andere Beobachter Schilderungen gegeben haben, welche jener sehr entsprechen, so will ich wenigstens eine Möglichkeit der Erklärung unserer abweichenden Darstellung versuchen. Meine Beobachtungen betreffen wohl alle die Larven des Myrmecoleo formicarius L. Möglicherweise hatten Rösel und andere Beobachter Larven von Myrmecoleo europaeus M. L. vor sich. Vielfach ist das sogar aus den beigegebenen Abbildungen der Imago mit den gefleckten Flügeln zu entnehmen. Redtenbacher gibt direkt an, daß sich seine Beobachtungen auf M. europaeus beziehen. Auch er beschreibt, daß die Larve voraus einen Kreis beschreibt, der den Umfang des künftigen Trichters umfaßt. Dadurch entstehe außen ein erhöhter Wall, innen bleibe ein kurzer Kegelstutz zurück, der dann durch neue Kreisbewegungen weggearbeitet werde, wobei immer durch Bewegungen der inneren Beine Sand auf den Kopf und von diesem weggeschleudert werde. Meine Versuchstiere haben niemals auch nur Andeutungen dieser umständlichen Baumethode gezeigt.
Es ist daher die Möglichkeit zuzugeben, daß die Bauweise bei Myrmecoleo europaeus von derjenigen von M. formicarius abweicht. Sehr wahrscheinlich scheint mir diese Annahme aber nicht zu sein. Es wäre sehr merkwürdig, daß eine nahe verwandte Form einen viel umständlicheren Weg zum Trichterbau einschlägt, wenn M. formicarius ihn in so einfacher Weise erfolgreich durchführen kann.
Wahrscheinlicher scheint mir die Annahme, daß die früheren Beobachter Versuchsbewegungen des Tieres sahen und auf Grund von ihnen die Bautätigkeit des Ameisenlöwen sich zurechtlegten. In dieser Annahme wurde ich durch Schilderungen und Skizzen bestärkt, welche Redtenbacher von „Irrfahrten“ des Ameisenlöwen gibt, welche durchaus meinen Versuchsbewegungen entsprechen (vgl. meine Abbildung 20, S. 48; ferner Taf. I, Fig. 1, Taf. III, Fig. 3, Taf. IV, Fig. 4).
Ebensowenig wie Redtenbacher konnte ich jemals beobachten, daß die Larve sich Steine und andere größere Partikel auf den Rücken lädt und in „Sisyphus“-Arbeit zur Seite schleppt, obwohl dies Bonnet und Mac Cook ausführlich bestätigt haben. Ich[S. 59] glaube, daß hier falsche Deutungen zufälliger Vorkommnisse vorliegen.
Doch möchte ich zum Schluß noch einmal betonen, daß ich es für nicht ausgeschlossen halte, daß bei den verschiedenen Arten der Trichterbau auf etwas verschiedener Grundlage sich vollzieht.
Ein Ameisenlöwe am Grunde seines Sandtrichters befindet sich entweder in einer Ruhelage oder in Bereitschaftsstellung. Im ersteren Fall kann je nach der Beschaffenheit des Untergrundes sein Körper verschiedene Stellungen einnehmen; sein Kopf kann nach oben oder unten geneigt sein, die Mundwerkzeuge sind mehr oder weniger zusammengeklappt, ja, sie können sogar unsymmetrisch zueinander stehen. Bei der Ruhelage bedeckt sich auch sehr leicht der Kopf des Tieres mit Sandkörnern. Das Tier pflegt sehr wenig reizbar zu sein, und es können ihm in diesem Zustand selbst Ameisen über den Kopf laufen, ohne daß eine Bewegung erfolgt. Wir werden später sehen, daß diese Ruhelagen durch verschiedene Ursachen bedingt sein können.
Normalerweise finden wir einen Ameisenlöwen, besonders dann, wenn er seit längerer Zeit nichts gefressen hat, und wenn es warm und sonnig ist, in Bereitschaftsstellung (vgl. S. 36). Die Beine sind dann etwas angezogen, das Hinterleibsende nach unten gekrümmt; Kopf und Hals sind in einem stumpfen Winkel nach oben gekehrt. Mindestens die Mandibel, meist aber die ganze Stirnregion des Kopfes, ja oft auch der Hals schauen aus dem Sand heraus. Die Mandibel können so schief nach oben gerichtet sein, daß sie frei in die Luft ragen. In der Regel liegen sie aber an der Wand des Trichters, welche der Körperachse des Ameisenlöwen gegenübersteht, der Sandoberfläche glatt auf. Wie wir früher gesehen haben, liegt der Körper des Ameisenlöwen stets in der dem Lichteinfall abgewendeten Richtung; die Mandibel liegen also an der am meisten beleuchteten Wand des Trichters, so daß man vielfach ihre Bewegungen deutlich verfolgen kann.
Naht sich ein Tier, vor allem eine Ameise dem Rand des Trichters, so ist es in Gefahr in die Tiefe des Trichters hinabzustürzen. Ameisen, welche den Rand eines Trichters betreten, pflegen sehr aufgeregte Bewegungen zu machen. Sie stürzen dann sehr häufig in einem Zug bis auf den Grund des Trichters hinab, wo[S. 60] ganz plötzlich die Mandibel des Ameisenlöwen über ihnen zusammenschnappen. Ist durch diesen Vorgang das Tier wie von einer Falle um die Mitte seines Körpers erfaßt worden, so gelingt es ihm sehr selten, sich wieder zu befreien. Es führt heftige zappelnde Bewegungen aus, welche mehrere Minuten, ja oft noch länger anhalten. In der Regel führt der Ameisenlöwe, wenn er eine Ameise eingefangen hat, einige zuckende Bewegungen mit dem Hinterleib aus, durch welche er tiefer im Sand versinkt und sich besser verankert.
Manche Beobachtungen weisen darauf hin, daß letztere Handlung nicht immer ohne Bedeutung ist. Ausgewachsene oder auch nur mittelgroße Ameisenlöwen werden durch die Bewegungen einer Ameise sehr selten aus ihrer Lage verschoben. Auch wenn ein größerer Ameisenlöwe eine Spinne oder ein anderes Insekt erfaßt hat, das ihn an Größe übertrifft, wird meistens die Verankerung ausreichen. Es ist dies aber nicht immer der Fall. So konnte ich z. B. einmal beobachten, daß ein sehr kleiner Ameisenlöwe ein Exemplar der Ameisenart Lasius flavus, welches größer war als er selbst, mit den Mandibeln erfaßte. Jener kleine Ameisenlöwe hatte erst einen minimalen Trichter gegraben; er wäre in der freien Natur wohl kaum in die Lage gekommen, die große Ameise zu erwischen. Bei der künstlichen Fütterung fiel sie ihm zum Opfer. Sie riß aber mit Leichtigkeit den kleinen Räuber aus dem Sand heraus und schleppte ihn eine Strecke weit mit sich. Es entspann sich ein heftiger Kampf, da der Ameisenlöwe nicht losließ; während die Ameise ihn über den Sand hinzog, machte er unablässig die typischen Bewegungen mit der Hinterleibsspitze, um sich einzubohren. Er wird aber von der Ameise fortgeschleppt, die ihn schließlich in den Hinterleib beißt und zu stechen sucht. Daraufhin erlahmt er und bleibt wie tot liegen, während die Ameise davonläuft.
In der Bereitschaftsstellung sind die Mandibel des Ameisenlöwen weit auseinandergespreizt; bei leiser Reizung werden sie noch mehr gespreizt. Wenn in diesem Zustand ihre Innenseite von irgendeinem harten Gegenstand berührt wird, so schnappen sie ganz plötzlich zusammen. Eine Ameise, welche in den Trichter hinabfällt, wird also in der Regel mit der Hälfte ihres Körpers hindurchfallen, ehe die Mandibel über ihr zusammenklappen. In manchen Fällen gelingt es aber dem Ameisenlöwen nur den Kopf, die Antennen oder ein Bein der Ameise zu erfassen. Die weit vorragenden Zähne an der Innenfläche der Mandibel ermöglichen das Festhalten auch so dünner Gegenstände. In solchen Fällen gelingt es dem Ameisenlöwen[S. 61] meistens nicht, die Ameise in eine zum Aussaugen geeignete Lage zwischen seine Mandibel zu bringen. Dann wird das Tier nach einiger Zeit losgelassen, um von neuem eingefangen zu werden. Meist hat es genügend Zeit, um einen Teil der Trichterböschung hinaufzuklettern, worauf es von dem Ameisenlöwen mit einem Hagel von Sandkörnern wieder heruntergeschossen wird. Es kann vorkommen, daß sich diese Fangversuche mehrmals hintereinander wiederholen, ehe der Ameisenlöwe sein Opfer richtig zu erfassen vermag (s. auch unten S. 62). Aber auch dann folgen die Vorgänge sehr rasch aufeinander, und es vergehen meist nur einige Sekunden, bis das Tier gefangen ist.
Ist der Trichter, welchen der Ameisenlöwe gebaut hat, erst ganz seicht, so gelingt es ihm meist nicht, mit Hilfe des Schleuderreflexes eine Ameise zu fangen. Das ist aber nur eine Ausnahme.
Es läßt sich leicht zeigen, daß es nur ein Berührungsreiz ist, welcher das Zuschnappen der gesperrten Mandibel veranlaßt. Berührt man nämlich deren Innenseite mit einer Präpariernadel, einem Draht, einer Pinzette oder einem Stück Holz, so schnappen die Mandibel kräftig zusammen. Ja, man kann bei geschicktem Vorgehen auch durch Berührung mit einem Faden, den man in den Trichter hinabläßt und mit welchem man die Innenseite der Mandibel berührt, das Zuschnappen der Zangen herbeiführen. Da auch in diesem Falle das Tier fest zuschnappt und nicht losläßt, so kann man es an dem Faden, wie an einer Angel, aus seinem Trichter hervorziehen. Das Schnappen erfolgt auch bei Berührung mit allen möglichen Fremdkörpern, an welche das Tier gelegentlich stößt; nur muß der Stoß einigermaßen heftig erfolgen. Tiere z. B., welche bei Lichtexperimenten sehr lebhaft gegen die Lichtquelle kriechen (vgl. S. 75), stoßen oft, wenn sie in größerer Anzahl beisammen sind, aneinander, fahren dann aufeinander los und schnappen zu, lassen sich aber meist sogleich wieder los.
Beim Schnappen fassen die Mandibel fest zusammen und lassen das, was sie gefaßt haben, zunächst nicht los. So erklärt sich das Festhalten der gefangenen Ameise, das Angelexperiment usw. Erst nach einigen Minuten pflegen die offenbar tonisch kontrahierten Adduktoren wieder zu erschlaffen und den festgehaltenen Gegenstand loszulassen. Sicherlich sind auch dabei die von mir geschilderten Bremsvorrichtungen am Basalteil von Maxille und Mandibel von Bedeutung (vgl. S. 12 und Abb. 6 u. 7).
Der Schnappreflex tritt selten ohne Beugung des Kopfes nach hinten auf. Immerhin kommt es vor, daß ein Tier die Mandibel[S. 62] zuklappt, ohne gleichzeitig eine Kopfbewegung nach rückwärts durchzuführen.
Fast stets sind Schnapp- und Schleuderreflex miteinander so zeitlich verbunden, daß auf den Schleuderreflex der Schnappreflex unmittelbar folgt. Näheres über die Lokalisierung dieser Reflexe im Zentralnervensystem findet sich weiter unten.
Wir stellen also an dieser Stelle zusammenfassend fest, daß auch beim Ameisenfang der wichtige Schleuderreflex die Hauptbewegung ist; doch kombiniert er sich für den speziellen Zweck mit dem Schnappreflex.
Der Schleuderreflex erklärt nun einige weitere eigentümliche, beim Ameisenfang zu beobachtende Ereignisse, die wir zum Teil früher schon beschrieben haben. Es ist ja eine der merkwürdigsten Eigentümlichkeiten des Ameisenlöwen, daß er durch Schleudern des Sandes eine ihm entronnene Beute in seinen Besitz zu bringen vermag. Ist z. B. eine Ameise beim Absturz seinen Mandibeln entronnen, so versucht sie schleunigst die Böschung des Trichters hinaufzuklettern. Bei ihren krampfhaften Bewegungen lösen sich Sandkörnchen los und rieseln auf das Vorderende des Ameisenlöwen herab.
Der dadurch hervorgerufene Reiz hat einen Schleuderreflex zur Folge, durch welchen oft sofort das Opfer herabbefördert wird. Mit den hinunterrieselnden Sandkörnern und etwa mitgerissenen Teilen der Böschung fällt es in die wieder aufgesperrten Mandibel des Räubers.
Gelingt aber das Herabschießen nicht sofort, so kann sich der Schleuderreflex mehrmals hintereinander wiederholen. Früher oder später wird fast immer die Beute getroffen. Aus einem in gutem Zustand befindlichen Trichter entrinnt eine Ameise selten, wenn der Besitzer nicht gerade vorher reichlich gefressen hat.
Bei dem Sandbombardement kann man leicht feststellen, daß kein Zielen stattfindet; die Schüsse gehen nach den verschiedensten Richtungen, wie wir oben auch für den Trichterbau feststellten. Aber dadurch, daß in sehr rascher Folge die verschiedenen Seiten der Trichterwände bestrichen werden, gerät der Sand auf der ganzen Böschung ins Weichen, und die Ameise muß in das Zentrum des Trichters und in die aufgesperrten Kiefer des Ameisenlöwen fallen.
Der ganze Vorgang ist ein rein automatischer; die Erreichung seines Endzweckes beruht auf einem Zusammenwirken des Reflexes mit den Eigentümlichkeiten des Körperbaues des Ameisenlöwen.
[S. 63]
In den nachfolgenden Abschnitten sollen die Ergebnisse dargestellt werden, welche zahlreiche Experimente mir über die Sinnesreaktionen des Ameisenlöwen geliefert haben. Wie man sehen wird, erlaubt die Analyse der Sinnesreaktionen einen sehr tiefen Einblick in die kausalen Grundlagen der komplizierten Handlungen dieses merkwürdigen Tieres. Eine genügende Basis für die Untersuchung der Sinnesreaktionen haben wir nur dann, wenn wir den Bau der Sinnesorgane hinreichend genau kennen. Ich habe daher mit meinen experimentellen Untersuchungen eine Erforschung des noch unvollkommen bekannten Baues der Sinnesorgane des Ameisenlöwen verbunden. Ferner habe ich das Nervensystem des Tieres wenigstens in seinen wesentlichen Grundzügen untersucht. Ich will nun der Beschreibung der einzelnen Sinnesreaktionen die Schilderung der zugehörigen Sinnesorgane vorausschicken, soweit ich solche habe nachweisen und in ihrem Bau untersuchen können.
Die paarigen Augen des Ameisenlöwen stellen eigenartige kegelförmige Vorragungen an den beiden äußersten Vorderecken des Kopfes dar. Sie sind Höcker von unregelmäßig wulstiger Oberfläche, da die Linsen der Einzelaugen sich jeweils halbkugelig über sie vorwölben. Die klaren Linsen der Einzelaugen heben sich scharf von dem dunkelbraunen Chitin des Augenkegels ab.
R. Hesse war der erste, welcher die eigenartigen Augen der Myrmecoleolarve untersuchte. Er beschreibt die Augen als an beiden Seiten des Kopfes jederseits der Fühler auf zwei kleinen Höckern, zu je sieben, nahe beieinander stehend; sechs davon seien von der Dorsalseite sichtbar, das siebente nur von der ventralen. Da die Linsen teils auf zapfenförmigen Vorragungen des Augenhöckers stehen, teils aus dessen Wand sich direkt vorwölben, und da die Pigmentierung den Augenhöcker selbst undurchsichtig macht, ist die Zählung nicht ganz leicht (Abb. 24). Es wird oft ein Auge durch den Unterbau des anderen oder durch Teile des Höckers verdeckt. An Exuvien, an denen das Pigment aufgehellt war, glaubte ich manchmal 8 Augen zu zählen; sechs davon sind immer deutlich von oben sichtbar, zwei nur von unten. Doch kam es mir ebenso[S. 64] oft vor, als habe Hesse recht und als fände sich nur ein Auge auf der Unterseite, wie es auch die meisten früheren Untersucher angegeben hatten.
Die Linsen sind bikonvex gewölbt und deutlich geschichtet; sie sind sehr klar, durchsichtig und stark lichtbrechend. Sie heben sich sehr stark von dem umgebenden pigmentierten Chitin ab. Auf den Schnitten erkennt man, daß nur die äußerste Schicht dieses Zwischenchitins pigmentiert ist.
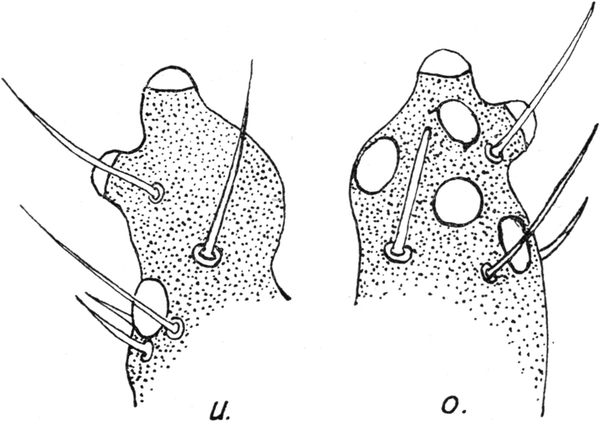
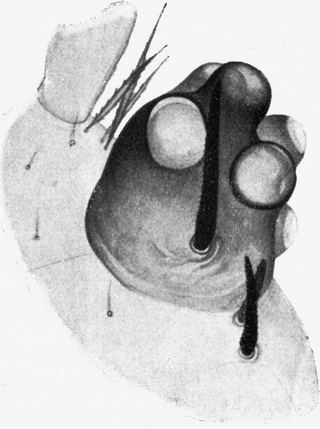
Wie schon Hesse hervorhebt, sind die einzelnen Augen verschieden gerichtet. Sie divergieren nicht unerheblich, und da der Strahlengang durch die Lagerung des Pigments und die Form der Linse sicherlich optisch sehr gut isoliert ist, müssen die einzelnen Augen ganz verschiedene Sehfelder haben.
Von den 7 Augen ist eines fast horizontal nach vorn, zwei nach vorn seitlich und eines nach hinten seitlich gerichtet. Die zwei auf der oberen Fläche des Augenhöckers gelegenen sind nach oben und etwas nach vorn resp. den Seiten gekehrt, während das einzige auf der Unterfläche nach außen und hinten schaut (Abb. 25). So sind also Linsen nach fast allen Seiten gerichtet; nur nach hinten ist keines der Augen in ausgesprochener Weise gewendet. Da aber mehrere von[S. 65] ihnen auf stärker vorragenden Zapfen sitzen, der ganze Augenhöcker sich nicht unerheblich über die Kopfoberfläche erhebt und alle Linsen sehr stark gewölbt sind, so können wohl auch Lichtstrahlen von hinten unter Umständen in eines oder das andere der Augen gelangen.
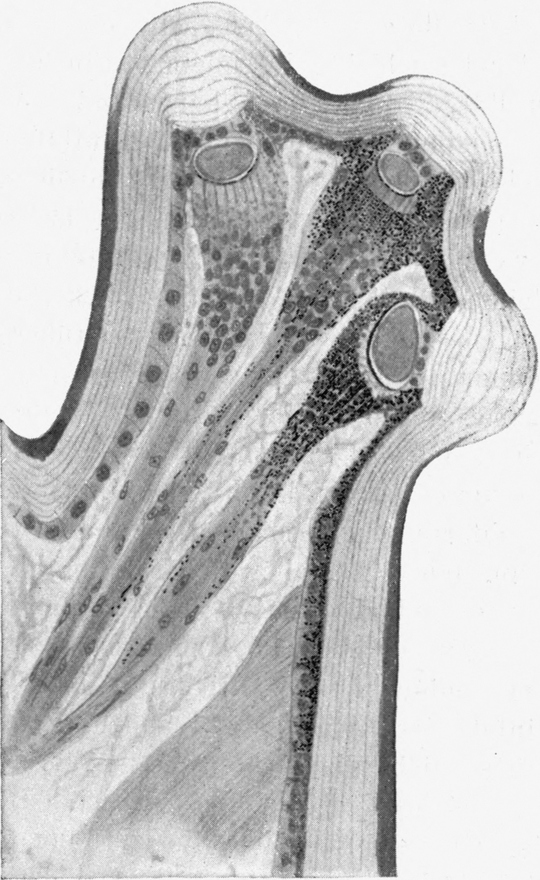
Wie Hesse hervorhebt, sind die Augen in ihrem Bau nicht ganz einfach; zwischen Linse und Netzhaut ist ein Glaskörper eingeschaltet. Die Netzhaut besteht aus einer größeren Anzahl von Sinneszellen.
Ueberlegen wir uns die Leistungsfähigkeit dieser Augen, so wäre zunächst die Funktion jedes der Einzelaugen zu erörtern. Zum mindesten sind die Augen als Richtungsaugen zu bewerten. Jedes einzelne Auge ist geradezu wie ein kleines optisches Instrument gebaut. Es ist fast röhrenförmig, und im durchscheinenden Präparat kann man deutlich erkennen, daß von jeder Linse geradezu eine Röhre von Pigment nach hinten führt, welche mit der pigmentierten Oberfläche die Strahlenisolation sehr vollkommen bewirken muß.
Wir werden unten sehen, daß wir tatsächlich annehmen dürfen, daß das Auge von Myrmeleo als Richtungsauge funktioniert.
Eine andere Frage ist es, ob es gleichzeitig Bilder der umgebenden Objekte zu liefern vermag. Die große Anzahl von Sehzellen (30–40) läßt es durchaus möglich erscheinen, daß Bilder von nicht allzu umfangreichen Objekten wahrgenommen werden. Dabei müßte man allerdings die Annahme machen, daß jedes der Augen ein eigenes Bild entwirft. Zwar läßt sich bei Untersuchung der Augennerven nachweisen, daß die Nerven aller Augen sich zu einem einheitlichen[S. 66] Strang vereinigen, der einheitlich ins Gehirn eintritt. Auch ist in den Verlauf dieses einheitlichen Sehnerven ein eigenartiges Gebilde eingeschaltet, welches an ein Ganglion opticum erinnert, und welches dazu beitragen könnte, die von den verschiedenen Augen aufgenommenen Eindrücke zu einem einzigen zu vereinigen. Es bildet ja unzweifelhaft die Gruppe von Augen bei Myrmecoleo eine Uebergangsform zwischen Ocellen und Komplexaugen (Abb. 26).
Bis jetzt habe ich aber keinerlei Beobachtungen machen können, welche dafür sprächen, daß die Wahrnehmung von Bildern umgebender Objekte irgendeinen Einfluß im Leben des Ameisenlöwen ausübte. Annäherung kleiner und größerer Gegenstände übte niemals irgendeinen wahrnehmbaren Einfluß auf die Tiere aus. Ich habe viele Versuche gemacht mit Annäherung von hellen und dunkeln, auch farbigen Gegenständen. Sie lösten keinerlei Reaktion aus, ebensowenig wie plötzliche Beschattung oder Belichtung mit Hilfe von kleinen Objekten. Auch in freier Natur machte ich keine Beobachtungen, welche die Angaben früherer Autoren hätten bestätigen können, daß ein Ameisenlöwe seine Beute mit den Augen wahrnehme, um alsbald mit Sand nach ihr zu schießen. Wir haben gesehen, daß das Schleudern des Sandes nach allen Seiten erfolgt, und ich konnte stets im Freien ein Herabrieseln von Sandkörnern am Trichterrand wahrnehmen, ehe der Ameisenlöwe zu schleudern begann. Ich führe daher die Auslösung des Schleuderreflexes im Trichter vorwiegend auf taktile Reize zurück.
Wir werden in dem kommenden Abschnitt sehen, daß die Ameisenlöwen in der Richtung ihrer Bewegungen durchaus von dem Einfall der Lichtstrahlen abhängig sind. Wir werden auch sehen, daß es nicht die Lichtstärke, sondern die Richtung der Lichtstrahlen ist, welche dabei maßgebend ist. Bei all jenen dort angeführten Versuchen konnte ich niemals eine Reaktion auf Unterschiede der Lichtstärke feststellen.
Ich muß aber an dieser Stelle einfügen, daß es mir in einigen wenigen Fällen gelang, bei Tieren, welche durch andauernde Versuche sehr stark gereizt waren, eine Schreckreaktion auf Belichtung und Beschattung nachzuweisen. Es war dies aber nur einmal bei zwei Exemplaren der Fall, welche beim Anknipsen und Ausdrehen einer 32-kerzigen Metallfadenlampe, sowohl beim Aufleuchten als beim Erlöschen der Flamme zusammenzuckten, indem sie die Mundgliedmaßen schlossen, mit dem Hinterleib eine zuckende und mit den Beinen eine scharrende Bewegung ausführten. Diese Reaktion[S. 67] erfolgte im ganzen zweimal hintereinander und zwar bei beiden Tieren. Ich konnte sie seither nie mehr herbeiführen und bin mir über ihre Bedeutung nicht klar geworden.
Jedenfalls geht aus meinen Erfahrungen hervor, daß die Augen sehr deutlich auf Licht reagieren. Ihr Bau ist nichts weniger als rudimentär. Im Gegenteil — wie schon Hesse hervorhob — stellen sie eine Uebergangsbildung zwischen Ocellen und Komplexaugen dar. So ist denn die seltsame Annahme von Comes, daß der Ameisenlöwe nicht sieht, ganz aus der Luft gegriffen. Weder der Bau der Augen läßt eine solche Annahme zu, noch, wie wir gleich sehen werden, seine Reaktionen. Vollkommen haltlos ist die Behauptung von Comes, der Ameisenlöwe müsse, wie etwa der Maulwurf, rudimentäre Augen haben, da er im Sand wühlend lebe. Weder lebt das Tier im Sand wühlend, noch sind seine Augen im geringsten rudimentär, wovon man sich bei ganz oberflächlicher morphologischer Untersuchung leicht überzeugen kann. Und nun kommen noch dazu die sehr charakteristischen Lichtreaktionen des Ameisenlöwen!
Schon bei meinen ersten Versuchen im Jahre 1910 fiel mir auf, daß die im Experiment ausgeführten Bewegungen des Ameisenlöwen durch das Licht beeinflußt werden. Ja, schon die Orte seines Vorkommens im Freien und die Körperhaltung der Tiere in ihren Sandtrichtern wiesen auf bestimmte Beziehungen zum Lichte hin. Legt man eine Anzahl von Ameisenlöwen in ein Gefäß auf einem Tisch, so kann man mit Sicherheit darauf rechnen, nach einiger Zeit sämtliche Tiere an der Fensterseite angesammelt zu sehen.
Um das Verhalten des Ameisenlöwen gegen das Licht zu prüfen, wurden sehr zahlreiche Experimente durchgeführt. Im September 1912 wurden Versuche mit Tieren gemacht, welche in einem dunkeln Kasten, in den durch einen schmalen Schlitz von der einen Seite her Licht einfiel, untergebracht waren. So wurden z. B. am 27. Sept. 3 Tiere in verschiedenen Stellungen in den Schlitzkasten gesetzt. Nach 10 Minuten hatten alle drei ihren Kopf vom Licht weggedreht, waren aber gegen das Licht zu gekrochen. In einem zweiten Experiment wurden 3 Tiere mit dem Kopf gegen das Licht zu parallel nebeneinander gestellt. Nach 10 Minuten hatten alle drei ihren Kopf vom Licht weggedreht und hatten sich mit dem Hinterende voran dem Lichteinfall entgegenbewegt.
Diese Versuche wurden ergänzt durch Experimente, welche mit[S. 68] Tieren in einer runden Glasschale mit flachem Boden von 35 cm Durchmesser ausgeführt wurden. In deren Mitte wurden mehrere Ameisenlöwen in beliebigen Stellungen hingelegt, nach einer halben Stunde waren sie alle drei an der Lichtseite der einseitig beleuchteten Glasschale angelangt, und zwar, wie ja nach dem, was wir über die Kriechbewegung der Tiere kennen gelernt haben, selbstverständlich ist, mit dem Hinterende voran. Darauf wurde die Schale um 180° gedreht; nach einer halben Stunde waren sämtliche Individuen wieder mit abgewandtem Kopf an der Lichtseite angelangt. Dies ließ ich mehrmals hintereinander wiederholen, wobei die Reaktionszeit kürzer wurde und schließlich bis auf 10 Minuten für den ganzen Vorgang herabsank.
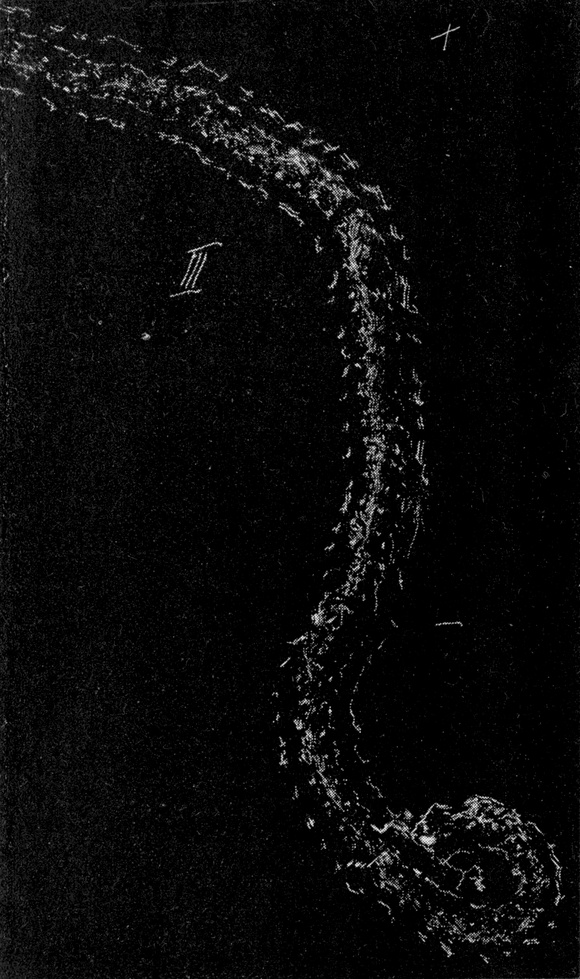
Die angeführten Versuche waren aber nicht genügend, um die Reaktionsweise des Tieres richtig zu beurteilen. Denn bei der Beobachtung[S. 69] des einzelnen Tieres stellte sich häufig heraus, daß es, einseitig einfallenden Lichtstrahlen ausgesetzt, zunächst in der Richtung von der Lichtquelle weg sich bewegte. Ueber die Zusammenhänge wurde ich mir erst klar, als ich die Tiere dem einseitigen Lichteinfluß auf einem rußgeschwärzten Registrierpapier aussetzte. Für dies Experiment muß das Registrierpapier vollkommen eben ausgespannt sein. Es muß ferner eine gewisse Länge und Breite haben, um dem Tier die Möglichkeit zu gewissen Exkursionen zu geben, ohne daß es dabei an den Rand des Papiers oder an einen festen Gegenstand gerät. Sonst würde die Klarheit des Resultates durch die Interferenz mit anderen Reizwirkungen getrübt, wie wir das später vor allem bei Besprechung der Thigmotaxis genauer erörtern werden.

Die Kriechspur, welche ein Ameisenlöwe auf einem solchen, unter Berücksichtigung aller Vorsichtsmaßregeln ausgespannten, Registrierpapier bei einseitigem Lichteinfall hinterläßt, ist sehr charakteristisch. Ist das Tier im Anfang des Experiments mit seinem Hinterende direkt der Lichtquelle zugewandt, so kann es vorkommen, daß es ohne weitere Körperbewegung direkt derselben entgegenkriecht. Es kann aber auch bei dieser Anfangsstellung sich ereignen, daß es zunächst Bewegungen ausführt, wie sie ganz regelmäßig sind, wenn das Tier beim Beginn des Experiments in irgendeinem Winkel zum Einfall des Lichtes[S. 70] angeordnet war oder gar auf den Rücken gelegt wurde. In letzterem Fall dreht sich das Tier natürlich zunächst um, und dann führt es bei jeder Anfangsstellung, die einen Winkel zum Einfall der Lichtstrahlen bildet, ja auch dann, wenn es mit der Körperachse zu den Lichtstrahlen parallel stand, dem Licht aber das Vorderende zuwandte, einige charakteristische Bewegungen aus. Es macht nämlich, wie wir aus den Kriechspuren direkt ablesen können, zunächst einen oder mehrere Kreise oder Teile von solchen oder Spiralen mit dem Hinterende voran. Das sind die typischen Versuchsbewegungen, der Suchgang des Tieres (vgl. S. 48). Oft haben diese Versuchskreise einen so großen Radius, daß man ein recht großes Versuchsgefäß anwenden muß, damit das Tier nicht an den Rand stößt, wobei Berührungsreize sein Verhalten stark beeinflussen würden. Bei diesen Versuchsbewegungen gewinnt es offenbar die richtige Orientierung zur Lichtquelle und wandert nun oft in langer gerader Bahn auf sie zu, Beispiele hierfür liefern die Abb. 27, 28 und Taf. I, Fig. 1.
Das Tier führt also so lange unorientierte Bewegungen aus, bis es seinen Körper in eine symmetrische Orientierung zum einfallenden Licht gebracht hat. Es handelt sich also um eine Phototaxis im eigentlichen Sinne des Wortes.
Man hat sich gewöhnt, die Orientierungsbewegungen beweglicher Organismen unter der Bezeichnung Taxis zusammenzufassen, und sie damit den Erscheinungen des Tropismus gegenübergestellt, unter welchem Ausdruck man die durch Reize bewirkten Krümmungen festsitzender Organismen versteht.
Hier bei dem Ameisenlöwen beobachten wir nun eine durch das Licht beeinflußte, gerichtete Bewegung, eine Phototaxis. Solche phototaktische Reaktionen sind bei vielen Organismen bekannt. Meist handelt es sich um das Aufsuchen einer für die betreffende Organismenart optimalen Beleuchtungszone. Beim genaueren Studium der meisten „taktisch“ reagierenden Organismen hat es sich nun herausgestellt, daß die Ansammlung in einer bestimmten Region auf eine komplizierte Reizreaktion zurückzuführen ist. Bei solchen Tieren oder Pflanzen läßt sich nämlich eine Unterschiedsempfindlichkeit gegen die Intensität einwirkender Reize nachweisen. Die Richtung, aus der die reizauslösende Kraft den Organismus trifft, ist dabei mehr oder weniger gleichgültig. Bei einer bestimmten Abweichung der Intensität der einwirkenden Kraft vom Optimum nach oben oder nach unten, wobei der Betrag des Unterschiedes, die[S. 71] Reizschwelle, einen bestimmten Wert erreichen muß, führt der Organismus eine „Schreckbewegung“ aus, welche ihn in die Region des Optimums zurückführt. So werden Tiere oder bewegliche Pflanzen in „chemischen Fallen“, „Lichtfallen“ usw. gefangen. Wo sie an eine Grenze der optimalen Region kommen, schrecken sie zurück und werden in jener Zone auf diese Weise rein automatisch festgehalten. Man bezeichnet diese Reaktionsweise als „phobotaktische“ Bewegung. Unterschiedsempfindlichkeit und Schreckreaktion spielen bei ihr die ausschlaggebende Rolle, maßgebend ist die Intensität der einwirkenden Kraft, also des Lichtes, die Konzentration der chemischen Substanz usw.
Im Anfang der auf die Untersuchung der Reizbewegungen gerichteten Forschungen glaubte man in den Taxien stets Bewegungen erblicken zu müssen, welche die Einstellung des Körpers des betreffenden Organismus in eine bestimmte Bewegungsrichtung bewirkten. Man nahm an, daß die Organismen sich in ein Kraftfeld einstellten, wobei die symmetrisch einander entsprechenden Teile des Körpers eine symmetrische Stellung zu den Kraftlinien einnähmen. Man nannte diese Art der Körpereinstellung „topotaktische“ Bewegung. Der Organismus stellt bei dieser Art der Reaktion die Hauptachse seines Körpers in einer bestimmten Weise zur Richtung der den Reiz auslösenden Kraft ein. Die meisten Reizreaktionen, welche man ursprünglich für derartige topotaktische hielt, haben sich bei genauerer Analyse als phobotaktische herausgestellt. Nur die Galvanotaxis der Infusorien scheint noch ein einwandfreier Fall topotaktischer Reizeinstellung zu sein.
Es ist nun bemerkenswert, daß die auffälligste Lichtreaktion des Ameisenlöwen durchaus eine topotaktische Reaktion zu sein scheint.
Einige Experimente zeigten mit aller Deutlichkeit, daß beim Ameisenlöwen die Richtung der einfallenden Lichtstrahlen eine symmetrische Einstellung des Tierkörpers und die aus ihr sich ergebende Richtung der Bewegung des Tieres bestimmen. Ein Experiment bestand darin, daß Exemplare des Ameisenlöwen auf eine Drehscheibe gelegt wurden, welche einseitig einfallendem Licht ausgesetzt wurde. Die Drehscheibe wurde langsam gedreht, da die Ameisenlöwen, wie wir gesehen haben, ziemlich langsam reagierende Tiere sind. In den meisten Fällen mußte man nach einer teilweisen Umdrehung die Drehscheibe anhalten; dann stellte sich nach einigen Sekunden bis Minuten das Tier mit der Hinterleibsspitze voran in die Richtung[S. 72] der einfallenden Lichtstrahlen. Bei besonders erregbaren Individuen, oder, wenn man die Erregbarkeit durch Erwärmung oder andere Hilfsmittel gesteigert hatte, konnte man selbst bei andauernder langsamer Drehung der Drehscheibe ein fortgesetztes ruckweises Einstellen der Hinterleibsspitze gegen die Richtung der Lichtquelle beobachten.
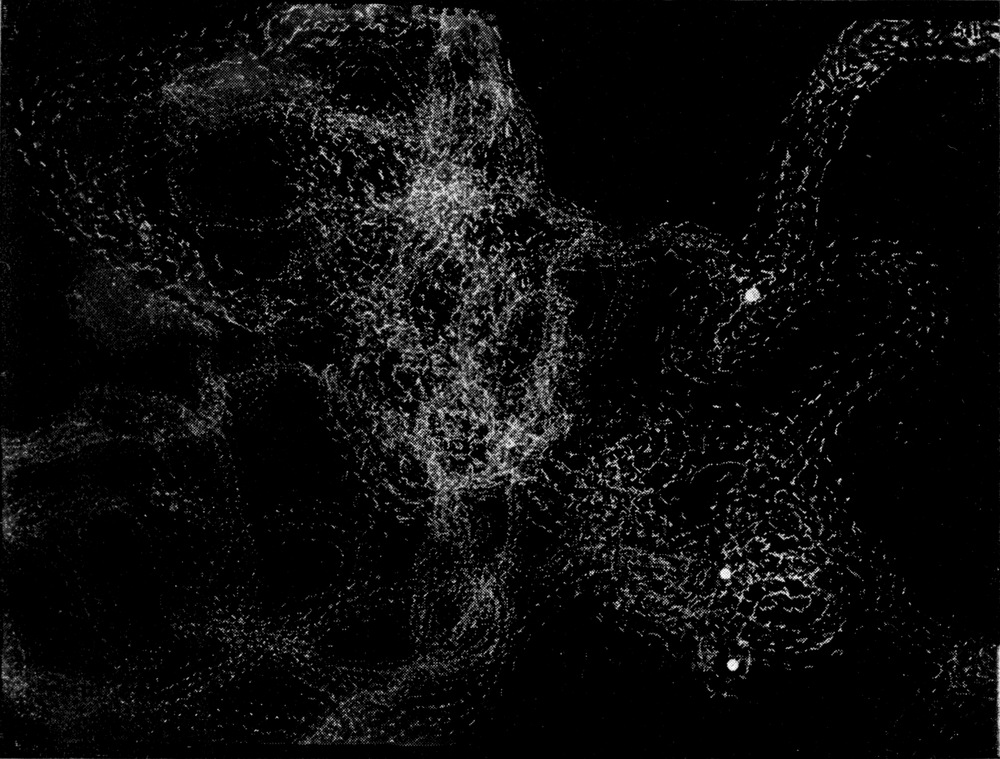
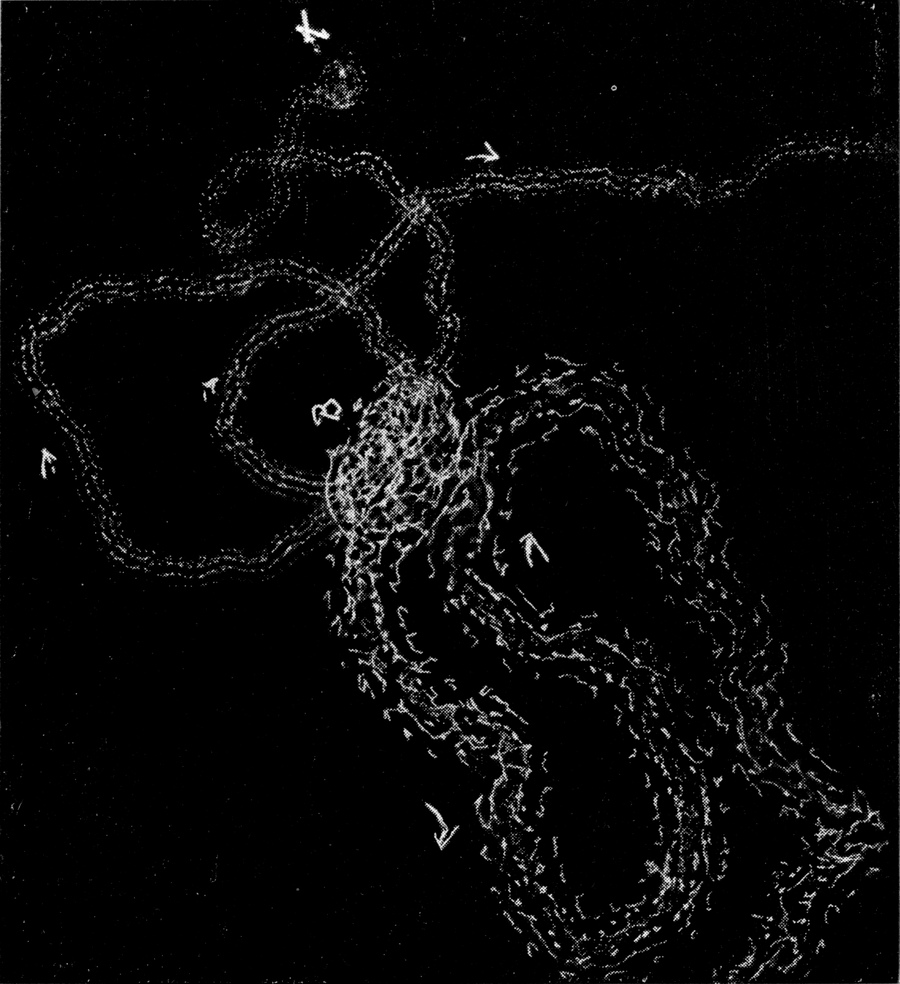
Zur Ergänzung dieser Versuche wurden Ameisenlöwen vollkommen diffusem Tageslicht oder ebensolchem künstlichen Licht ausgesetzt. Da die Tiere sich in solchem meist sehr träge bewegen, ist es nicht leicht, festzustellen, ob sie überhaupt von dessen Einwirkung beeinflußt werden. Hat man mehrere Individuen auf eine Versuchsplatte gesetzt, so kann man, nachdem das Licht eine längere oder kürzere Zeit eingewirkt hat, feststellen, daß die verschiedenen Individuen nach den verschiedensten Richtungen orientiert sind. Bei[S. 73] diesen Experimenten wurde in Zeitabständen von 10 Minuten, einer halben Stunde, 1, 2, 3, 6, 12 und 24 Stunden beobachtet. Diese Experimente wurden weiterhin dadurch kontrolliert, daß die Tiere der Einwirkung des diffusen Lichtes auf berußtem Registrierpapier ausgesetzt wurden. Wie z. B. die Abb. 29 zeigt, haben während der Versuchsdauer die Tiere zahlreiche, vollkommen unorientierte Bewegungen ausgeführt. Sie sind im Kreis, in Spiralen, in Schlangenlinien und in allen möglichen Windungen umhergekrochen; kurz sie haben nur Versuchsbewegungen ausgeführt. Ganz das entsprechende[S. 74] Bild zeigt sich, wenn starkes Licht direkt von oben auf die Versuchstiere fällt; es wirkt dann vollkommen wie diffuses Licht, dem es ja auch im Reizeffekt entsprechen muß (Taf. III, Fig. 3), da es nur indirekt zu den Augen gelangt.
Ebenso unorientiert sind die Bewegungen, welche die Tiere bei vollkommener Dunkelheit ausführen. Dies ließ sich natürlich nicht direkt beobachten; das Registrieren (Abb. 30) verrät uns aber auch in diesem Fall, daß während der Versuchsdauer die Bewegungen ganz in derselben Weise verliefen, wie im diffusen Licht. Ein Vergleich der beiden nebeneinander stehenden Abbildungen S. 72 und 73 zeigt dies aufs deutlichste. Besonders auffallend ist bei den letzteren Versuchen, daß die Tiere sich oft lange Zeit hindurch in einem engen Kreis an Ort und Stelle bewegen.
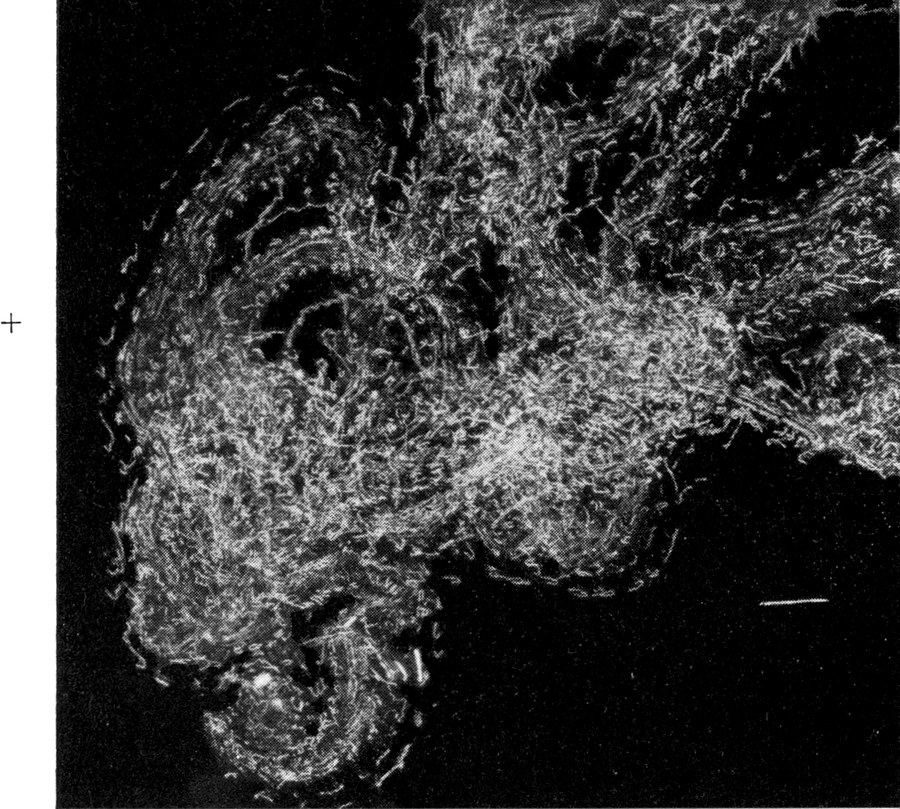
Von Bedeutung waren die Versuche, welche ich mit Tieren anstellte, deren Augen durch Bepinseln mit Asphaltlack der Wirkung der Lichtstrahlen entzogen waren. Waren beide Augen gleichmäßig gut lackiert, so verhielt sich das Tier genau so wie die Individuen im[S. 75] diffusen Licht oder in völliger Dunkelheit; die Bewegungen waren vollkommen unorientiert. Die Aufzeichnung (Abb. 31 und 32) zeigt dies deutlich.
Es fragt sich nun, ob außer dieser ausgesprochen topischen Reaktion auch eine phobotaktische Beeinflussung bei dem Ameisenlöwen vorkommt. Es wäre ja möglich, daß eine Unterschiedsempfindlichkeit für Belichtung von verschiedener Intensität bei dem Tiere vorhanden wäre, welche unter Umständen die Individuen zwänge, sich in einem bestimmten Beleuchtungsoptimum anzusammeln. Verschiedene Beobachtungen scheinen für eine solche Annahme zu sprechen. Schon die Beobachtung in der freien Natur könnte darauf hinweisen. Wir finden ja die Fundorte der Ameisenlöwen an Orten mit einer ziemlich gleichmäßigen Beleuchtungsintensität.
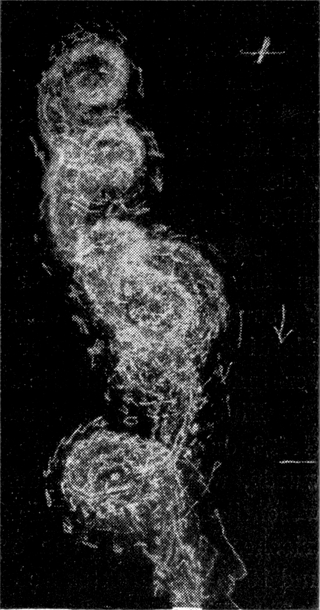
Bei den vorhin beschriebenen Experimenten hatte sich gezeigt, daß Uebertragung in völlige Dunkelheit bei normalen Tieren eine erhöhte Beweglichkeit auszulösen pflegt. Die Tiere führen eine Menge von Versuchskreisen in verhältnismäßig kurzer Zeit aus; das dauert ziemlich lange, wenn nicht irgendein neuer Reiz sie zwingt, eine bestimmte Richtung einzuschlagen. Dazu kam noch folgende Beobachtung. Bei den Versuchen, welche das Vorhandensein der topotaktischen Lichtorientierung bewiesen, wurden die Tiere vielfach in großen Glasaquarien dem von einer Seite einfallenden Licht ausgesetzt. Einerlei nun, ob das Licht stark oder schwach war, kamen in vielen Fällen die zur Lichtquelle hinkriechenden Tiere an der dieser zugekehrten Wand des Aquariums nicht zur Ruhe. Sie arbeiteten vielmehr in unablässigen Bewegungen mit dem Hinterende voran gegen die Aquarienwand, wobei sie oft ihren Hinterleib senkrecht aufrichteten und immer wieder umfielen; trotzdem ließen sie nicht[S. 76] ab von ihren Bemühungen. Es ist dies um so bemerkenswerter, als, wie wir später sehen werden, die Berührung mit den beiden in der Kante zusammenstoßenden Aquarienwänden einen starken thigmotaktischen Reiz auf sie ausüben mußte, der sie unter anderen Umständen zu einer ganz anderen Orientierung gezwungen hätte. Allerdings zeigte sich diese große Beweglichkeit fast nur bei Tieren, welche durch wiederholte Reize erregt waren. Immerhin lag die Möglichkeit vor, daß sie sich andauernd bewegten, da die Lichtintensität, der sie ausgesetzt waren, nicht ihrem Optimum entsprach.
Um also die Frage nach einer vorhandenen phobischen Lichtreaktion zu klären, mußten neue Versuche angestellt werden, welche bei der trägen Beweglichkeit, der Körpergröße der Tiere und den vielen Fehlerquellen, welche durch ihre eigenartige Reizbiologie bedingt sind, sehr schwierig durchzuführen waren.
Sehr starkes diffuses Licht scheint die Bewegung der Tiere eher zu lähmen als zu steigern. In sehr starkem Licht blieben bei vielen Versuchen die Tiere oft stundenlang ganz regungslos liegen. Das war allerdings nur dann der Fall, wenn durch besondere Einrichtungen verhindert war, daß die Umgebung der Tiere sich erheblich erwärmte.
Eine Reihe von Versuchen wurde in der Weise angestellt, daß ein Gefäß durch Anwendung eines Deckels in der einen Hälfte verdunkelt wurde, während die andere Hälfte stark beleuchtet wurde. Ohne daß wesentliche Unterschiede zutage traten, wurden zur Beleuchtung elektrisches Bogenlicht von etwa 1000 HK, elektrisches Glühlicht von 50 und 100 HK, direktes Sonnenlicht und einseitig einfallendes Tageslicht verwendet; es mußte nur verhindert werden, daß jene künstlichen Lichtquellen den Untergrund erwärmten (vgl. unten S. 80). Die Versuche wurden in der Weise angestellt, daß in beiden Hälften des Gefäßes, sowohl in der Licht- als auch in der Dunkelhälfte Ameisenlöwen auf berußtem Registrierpapier niedergesetzt wurden. Zwischen den Tieren in den beiden Hälften zeigte sich insofern ein Unterschied, als die Exemplare in der Lichthälfte meist viel länger unbeweglich blieben und sich auch später viel weniger bewegten als die Tiere der Dunkelhälfte. Bei ihren Bewegungen passierten nun Tiere aus beiden Hälften gelegentlich die Schattengrenze. Dabei stockten sie in keiner Weise, wie sowohl direkte Beobachtung zeigte als auch das seltene Vorkommen von Versuchsbewegungen an den Kriechspuren zu schließen erlaubt. Es schien also keine Unterschiedsempfindlichkeit in Frage zu kommen. Ein Unterschied zeigte sich insofern, als die Tiere der Dunkelhälfte, sobald sie in der Nähe der Schattengrenze[S. 77] in den Bereich der Lichtstrahlen gelangten, durch dieselben orientiert wurden und darauf vielfach geradeswegs der Lichthälfte zukrochen; denn auch bei der Anwendung diffusen Lichtes erfolgt ein Lichteinfall schief von oben am Deckel vorbei von der Lichthälfte in die Dunkelhälfte hinein. Für die Tiere in der Dunkelhälfte gibt es also eine lokalisierte Lichtquelle, nach der sie sich orientieren können (vgl. hierzu die Rußspuren Fig. 4, Taf. IV, und Fig. 5 u. 6, Taf. V u. VI).
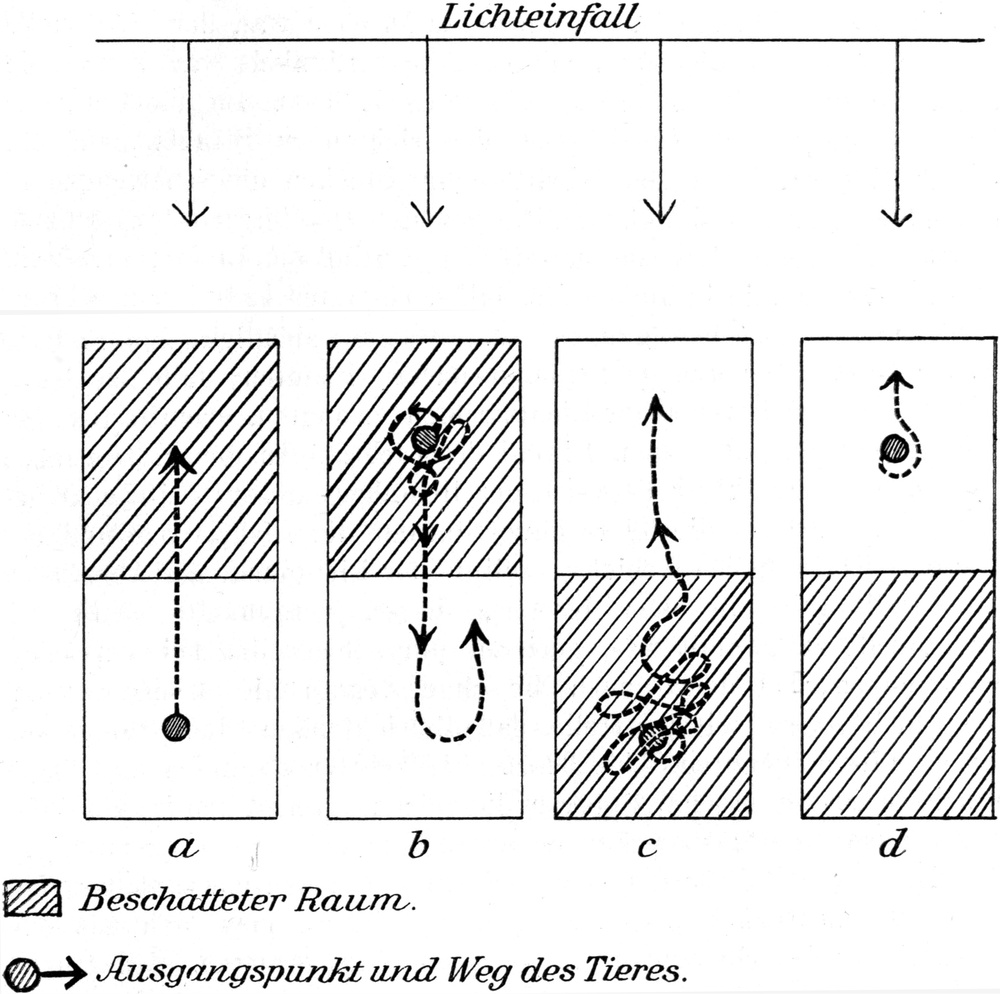
Daß die Unterschiedsempfindlichkeit keine wesentliche Bedeutung bei den Lichtreaktionen der Ameisenlöwen hat, geht aus folgenden Versuchen hervor. Einige Kästen wurden parallel nebeneinander[S. 78] aufgestellt (Abb. 33). Licht fiel auf sie von einer Seite her ein. Nun wurden einige dieser Kästen in der dem Lichte zugewandten (Abb. 33 a und b), andere in der dem Lichte abgewandten Hälfte verdunkelt (Abb. 33 c und d). In sie wurden Ameisenlöwen entweder in die dunkle oder in die helle Hälfte gesetzt. Die in die helle Hälfte gesetzten Tiere krochen stets, wie nach unseren früheren Versuchen vorauszusehen war, der Lichtquelle zu. Im Falle also die abgewandte Hälfte verdunkelt war, kamen sie gar nicht in die dunkle Hälfte hinein (Abb. 33 d). Im anderen Falle krochen sie unter dem Einfluß der richtenden Strahlen auf die dunkle Hälfte los und überschritten ohne Stocken die Schattengrenze (Abb. 33 a). In der dunklen Hälfte machten sie Versuchsbewegungen, welche sie eventuell wieder unter den Einfluß des Lichtes brachten. Die von vornherein in die dunkle Hälfte gesetzten Individuen führten zunächst Versuchsbewegungen aus, welche ziemlich komplizierte Wege einschlagen und recht lange dauern konnten; falls sie dabei zufällig in den Bereich der Lichtstrahlen gelangten, orientierten sich die Tiere sofort nach dem Einfall des Lichtes. In den Kästen also, in denen die dunkle Hälfte an der lichtabgewandten Seite sich befand, überschritten die Tiere ohne Stocken die Lichtgrenze und bewegten sich im hellen Gebiet geradlinig der Lichtquelle zu (Abb. 33 c). Die Individuen in dem Kasten, dessen verdunkelte Seite der Lichtquelle zugewandt war, gerieten nur gelegentlich bei Versuchsbewegungen, bei denen auch sie ohne Zögern die Schattengrenze überschritten, so weit ins Helle, daß die richtenden Lichtstrahlen auf sie einwirken konnten (Abb. 33 b). Sie kehrten dann um und überschritten wiederum ohne Stocken die Schattengrenze, um im Dunkeln von neuem Versuchsbewegungen auszuführen.
Daß an der Schattengrenze mitunter Versuchsbewegungen vorkommen (vgl. Taf. IV, Fig. 4; Taf. V, Fig. 5), mag sich aus der Einwirkung des richtenden Reizes des Lichtes erklären. Jeder neue Reiz kann, wie wir sahen, vor der Orientierung zunächst Versuchsbewegungen auslösen.
Weiterhin wurden Versuche angestellt, bei denen abgestufte Helligkeiten den Ameisenlöwen dargeboten wurden, so daß sie die Möglichkeit gehabt hätten, eine für sie optimale Region aufzusuchen.
Zunächst wurden die Tiere einem gleichmäßigen Lichtgefälle ausgesetzt, wie es durch Anwendung eines Oltmannsschen Prismas erzeugt wird. Kollege Oltmanns war so liebenswürdig, mir einige dieser Apparate zur Verfügung zu stellen, welche er seinerzeit[S. 79] bei seinen Versuchen über die Lichtreaktionen von Volvox benützt hatte. Diese Prismen bestehen aus zwei im spitzen Winkel zueinander geneigten Glasscheiben, zwischen welche mit Tusche grau gefärbte Gelatine gegossen ist. Es wird dadurch erzielt, daß an dem Ende, an welchem die Glasplatten zusammenstoßen, durch die hier dünne Schicht reichlich Licht durchfällt, während am entgegengesetzten Ende die dickere Schicht das Licht zum großen Teil absorbiert. Die gleichmäßige Verschmälerung des Zwischenraumes hat eine gleichmäßige Zunahme des durchfallenden Lichtes zur Folge.
Setzte ich nun Tiere diesem Lichtgefälle aus, so krochen sie stets aus der dunklen Hälfte in die helle Hälfte. Wie aber die Rußspur Taf. VII, Fig. 7 RR zeigt, handelte es sich von vornherein nicht um bestimmt gerichtete Bewegungen. Unter zahlreichen Versuchsbewegungen strebten die Tiere im großen und ganzen dem hellen Ende der Versuchsfläche zu. Aber auch dort setzten sie ihre Versuchsbewegungen fort, die sie oft wieder in das dunkle Ende zurückführten. Eine Ansammlung oder auch nur ein längerer andauernder Aufenthalt der Versuchstiere in einer Region von bestimmter Lichtintensität ließ sich nicht mit Sicherheit nachweisen. Immerhin zeigte die Fig. 7 RR der Tafel VII eine gewisse Verdichtung der Spuren in einer Region von mittlerer Helligkeit. Das stimmt mit den Resultaten einer größeren Anzahl von anderen Versuchen überein, in denen durch übereinander gelegte Papierblätter das auf die Versuchsfläche einfallende Licht in verschieden hohem Maße abgeblendet war. Auch dann zeigte sich nicht selten ein durch Anhäufung der Spuren gekennzeichneter Aufenthalt der Tiere in einer Region von mittlerer Helligkeit.
Bei den früheren Versuchen haben wir aber gesehen, daß die Tiere nicht eine mittlere Helligkeit aufsuchen, sondern daß ein Optimum für sie weit höher liegen müßte, falls überhaupt ein solches für sie in Betracht kommt.
Einige Versuche, die weiterhin zur Klärung dieser Frage angestellt wurden, sowie gelegentliche Nebenergebnisse früher erwähnter Versuche lassen für diese Anhäufung der Spuren in einer bestimmten Region eine andere Deutung zu. Verdunkelt man beide Enden eines Versuchskastens, so daß nur in der Mitte senkrecht von oben her Licht einfallen kann, so führen die Tiere, in das belichtete Feld gesetzt, Versuchsbewegungen aus, bei denen sie mehr oder weniger tief in den Schatten hineingeraten können. Die Bewegungen sind in dem diffusen Oberlicht, unseren früher gemachten Erfahrungen entsprechend, unorientiert. Ebenso unorientiert werden sie oft, wenn[S. 80] die Tiere tief in die Schattenregion hineingeraten. Es kommt aber häufig vor, daß die Tiere sich lange an der Licht-Schattengrenze entlang und dieser parallel bewegen. Dabei zeigen sie eine ausgesprochene Tendenz, sich immer wieder dem Lichtfeld zuzuwenden. Ganz entsprechend verhielten sich die Tiere bei den früher geschilderten Versuchsanordnungen, wenn das Licht von der Seite her einfiel, auf welcher die Versuchsfläche durch einen Deckel verdunkelt war. Die Fig. 6 FF, Taf. VI, und Fig. 4 GG, Taf. IV, zeigen die Kriechspuren von Tieren, welche bei dieser Versuchsanordnung ins Dunkle gesetzt worden waren und dort Versuchsbewegungen ausführten, die sie in den Bereich des einfallenden Lichtes führten. Sie krochen dann in das helle Feld, gerieten unter den Einfluß des einseitigen starken Lichtes, wurden durch dasselbe wieder an die Licht-Schattengrenze gelockt und führten an dieser entlang Versuchsbewegungen aus, die sie bald ins Licht, bald wieder in den Schatten führten. Hier scheint es sich sicherlich nicht um eine Unterschiedsempfindlichkeit zu handeln, sondern um die Einwirkung der abwechselnd von der einen und dann von der anderen Seite einfallenden Lichtstrahlen. Unterschiedsempfindlichkeit brauchen wir also zur Erklärung dieser Erscheinungen nicht anzunehmen, wenn es auch möglich ist, daß eine solche vorhanden ist. Jedenfalls weist die Tatsache, daß die Tiere jedesmal eine gewisse Zeit brauchten, ehe sie nacheinander auf die Einwirkungen der schwächeren und stärkeren, abwechselnd von links und rechts auf sie fallenden Lichtstrahlen reagierten, darauf hin, daß eine Adaptation an die Lichtstärke vorkommt.
Bei den Vorbereitungen zu einigen Experimenten hatte ich an den Ameisenlöwen Reaktionen beobachtet, welche auf das Vorhandensein eines Temperatursinnes schließen ließen. Als ich z. B. zur Untersuchung des Umdrehreflexes Tiere mit Hilfe eines Wachstropfens an einem Faden aufhängte (vgl. S. 43), führten sie bei Annäherung einer warmen Nadel oder eines erhitzten Spachtels sehr lebhafte Bewegungen aus. Es hatte nicht den Anschein, als wären diese Bewegungen nur auf eine Erhöhung der Stoffwechselprozesse zurückzuführen; denn die Bewegungen bestanden in den typischen Reflexen, welche auch auf andere Reize hin erfolgen. Es waren Bewegungen mit den Beinen, mit dem Hinterleib und vor allem der typische Schleuderreflex. Um diese Reflexe auszulösen, mußte ein ziemlich[S. 81] stark erwärmter Gegenstand sehr nahe an den Tierkörper herangebracht werden. Natürlich wurde sorgfältig eine Berührung mit dem Tierkörper selbst vermieden.
Das Vorkommen des Ameisenlöwen in freier Natur weist darauf hin, daß ein Wärmesinn bei diesem Tier möglicherweise eine Rolle spielen kann. Er kommt nämlich niemals an feuchten, kühlen und schattigen Orten vor. Im Innern des Waldes, wo die Sonne nicht hinscheint, fehlt er. Dies könnte nun auf den Einfluß zurückzuführen sein, den die Feuchtigkeit auf die Beschaffenheit des Untergrundes hat. Das spielt sicher eine gewisse Rolle, aber daß es nicht der ausschlaggebende Faktor ist, dafür spricht die Tatsache, daß fast stets die Ameisenlöwen an Nord-, Nordost- und Nordwestabhängen unserer Mittelgebirge fehlen, auch wenn das Gelände offen und die Untergrundverhältnisse die denkbar günstigsten sind.
Auch sonst bei Insekten, besonders bei Ameisen und Bienen, hat man Erscheinungen beobachtet, welche als Anzeichen eines Temperatursinnes betrachtet worden sind. Ein solcher ist aber niemals durch eingehendere experimentelle Untersuchungen festgestellt worden. Ich habe daher bei dem Ameisenlöwen eine Reihe von Versuchen ausgeführt, welche mir ein gesetzmäßiges Reagieren des Tieres auf Temperaturunterschiede sicherzustellen scheinen.
Selbstverständlich muß bei derartigen Untersuchungen in Betracht gezogen werden, daß höhere Temperaturen die Beweglichkeit des Tieres stark erhöhen. Die ausgiebigsten Wanderungen machen die Tiere, die schönsten Trichter bauen sie, den Sand schleudern sie am weitesten, wenn sie bei einer Temperatur von 25–30° C gehalten werden. Hält man sie in vollkommener Dunkelheit auf einem gleichmäßig erwärmten Untergrund, so führen sie außerordentlich lebhafte Bewegungen aus, wie man daraus erkennen kann, daß ein Registrierpapier schon nach einer halben Stunde von ihren Spuren nach allen Richtungen durchzogen wird. Dabei handelt es sich, wie die Kreise, Bogen und Spiralen der Fährten erkennen lassen, um typische Versuchsbewegungen (s. Rußkurven Taf. II, Fig. 2).
Um festzustellen, ob die Wärme des Untergrundes auf die Bewegung der Tiere einen richtenden Einfluß ausübt, brachte ich sie zunächst in ein Sandbad, welches von der Seite her erwärmt wurde. Die durch einen Mikrobrenner erzeugte Wärme pflanzte sich in der dicken Sandschicht so langsam fort, daß an dem einen Ende eine Temperatur von 35° C, an dem anderen Ende, 40 cm davon entfernt,[S. 82] eine solche von nur 20° C herrschte. Dazwischen fanden sich Stellen, an denen sich an der Oberfläche des Sandes 30 und 25° C messen ließen. Ich setzte nun an die vier den angegebenen Temperaturen entsprechenden Stellen je einen Ameisenlöwen. Alle vier gruben sich innerhalb von 5 Minuten vollkommen ein und hatten in ebenso kurzer Zeit je einen tiefen, gleichmäßigen Trichter ausgeworfen. Die Temperatur hatte sie in der Wahl des Ortes, an dem sie sich eingruben, nicht beeinflußt. Sämtliche ihnen dargebotenen Temperaturen waren für ihre Bewegungen geeignet gewesen; und, wie wir später sehen werden, mußte der Berührungsreiz der rauhen Sandoberfläche über den Wärmereiz überwiegen.
Das zeigte sich schon deutlich bei einigen Kontrollexperimenten, welche ich an dieses erste anschloß. Ich setzte auf das Sandbad Schalen aus Glas oder aus Papier-mâché, deren Innenseite vollkommen glatt war. Die Substanz dieser Schalen nahm nach etwa einer halben Stunde Temperaturen an, welche genau entsprechend denjenigen des darunterliegenden Sandes abgestuft waren, wie ich durch Messungen mit kleinen Quecksilberthermometern feststellte. Ich regulierte die Temperatur wiederum in der Weise, daß sie von 35° auf 20° C abfiel. Setzte ich in diese Schalen Exemplare von Ameisenlöwen an verschiedenen Stellen des Bodens nieder, so fanden sie sich nach Ablauf von einer halben bis dreiviertel Stunde fast alle in der wärmeren Hälfte des Gefäßes angesammelt. Das gleiche Resultat ergab sich bei Wiederholungen dieses Versuches. Aber es war kein glattes und klares Resultat; denn einzelne Individuen waren immer durch Interferenz von Licht und Berührungsreizen am Aufsuchen der Wärmeregion verhindert. Um saubere Ergebnisse zu erlangen, führte ich die Versuche im Dunkeln weiter. In der vollkommen lichtdichten Dunkelkammer wurden breite Flächen von berußtem Registrierpapier auf dem Boden von Glasaquarien ausgespannt, welch letztere in Sandbäder gestellt wurden. Auch da ließ sich durch Anwendung eines Mikrobrenners eine gleichmäßig abgestufte Temperatur erzielen, welche sich auch dem Registrierpapier mitteilte. Bei dem ersten Versuch, welcher eine Stunde lang dauerte, hatten die Tiere, wohl unter der vereinigten Einwirkung von Dunkelheit und Wärme, sehr lebhafte Versuchsbewegungen ausgeführt. Es zeigte sich dabei deutlich, daß diejenigen Individuen, welche ohne weitere Störungen in die Region von etwa 28–35° geraten waren, sich dauernd in derselben bewegten. Das gleiche Resultat ergaben weitere Experimente (Rußaufzeichnung Taf. IX, Fig. 9).
[S. 83]
Aus ihnen folgt also, daß die Tiere eine deutliche Unterschiedsempfindlichkeit für verschiedene Temperaturen besitzen. Das Optimum für sie liegt auffallend hoch. Sie suchen es auf, indem sie bei ihren Versuchsbewegungen eine Wendung ausführen, wenn sie in eine Region von zu hoher oder zu niedriger Temperatur gelangen. Die hohe Temperatur veranlaßt sie zu andauernden, sehr lebhaften Bewegungen, welche sie aber stets nur bis zur Grenze der optimalen Region führen. Sind also Tiere bei ihren Versuchsbewegungen, bei ihrem Suchgang, in die optimale Region geraten, so sind sie gleichsam in einer Temperaturfalle gefangen, aus der sie nicht mehr heraus können. Es liegt also eine ganz ähnliche Beeinflussung durch die Wärme vor, wie sie Jennings bei seinen bekannten Versuchen mit Infusorien für chemische und andere Einflüsse nachgewiesen hat.
Comes gibt in einer am Schluß des letzten Kapitels kritisch besprochenen Arbeit an, daß der Ameisenlöwe stets den höheren Temperaturen zustrebe. Aus seinen Angaben ist nicht mit Sicherheit zu entnehmen, wie er seine Experimente angestellt hat. Nur das ist sicher, daß er den Einfluß des Einfalles der Lichtstrahlen nicht ausgeschaltet hat. Ferner hat er zum Nachweis der Wärmeempfindlichkeit den Trichterbau seiner Versuchstiere herangezogen. Wir haben aber oben gesehen, daß die Ameisenlöwen bei jeder günstigen Temperatur auf geeignetem Untergrund stets sofort zu bauen beginnen. Da aus Comes’ Angaben hervorzugehen scheint, daß für den Ameisenlöwen keine optimale Temperatur vorliegt oder doch jedenfalls das Maximum sehr hoch liegt, so habe ich noch einige Ergänzungsversuche zur Kontrolle seiner Angaben angestellt. Durch Einrichtung eines Sandbades mit Stoffen von sehr verschiedener Wärmeleitung gelang es mir, auf dem Raum von ½ m Abstufungen von 15–70° C zu erzielen. Das Sandbad war mit Torfmull, Sand + Torfmull, Quarzsand und Eisenfeilspänen in verschiedenen Lagen angefüllt. Im Dunkeln angestellte Versuche (23. Okt. 1915) ergaben wiederum eine Tendenz der Tiere, sowohl die niedere als die hohe Temperaturzone zu vermeiden. Sie sammelten sich zwischen 25° und 40° C an, indem sie sowohl aus der kühlen als auch der heißen Zone wegkrochen. Wiederum wurden zuerst Versuchsbewegungen ausgeführt. Die optimale Zone wurde im Verlauf von solchen erreicht, und ihre Grenzen dann nicht mehr überschritten. Nur aus dem Gebiet der größten Hitze erfolgte nach kurzen Versuchsbewegungen ein offenbar rascher Rückzug. Immerhin ist es[S. 84] möglich, daß unsere einheimischen Ameisenlöwen an ein tieferes Temperaturmaximum angepaßt sind als die sizilianischen. Dort mögen in den Sommermonaten im Sand wohl Temperaturen von 60–70° C erreicht werden.
Nach der oben von uns gegebenen Definition liegt also eine ausgesprochene phobische Thermotaxis vor. Das erzielte Ergebnis veranlaßte mich, etwas eingehendere Versuche über den Temperatursinn selbst und seinen Sitz anzustellen. Die Versuche wurden in der Weise ausgeführt, daß eine dünne, spitze erwärmte Stahlnadel in die Nähe der Körperoberfläche des Tieres gebracht wurde. Die Nadel mußte sehr warm sein, wenn sich überhaupt eine Reaktion zeigen sollte. Sie zeigte sich aber dann schon, wenn die Nadel bis auf ½ cm der Körperoberfläche genähert wurde. Die Reaktion auf die Annäherung der Nadel war dann sehr charakteristisch und präzis. Es erfolgte ein deutliches, oft lebhaftes Zurückweichen des Tieres. Als empfindlich gegen die von der Nadel ausstrahlende Wärme erwiesen sich Mandibel, Kopf, Hals, Beine, Vorderrumpf und Schultern sowie, in geringerem Grade, das Hinterteil des Abdomens.
Bei diesen Versuchen stellte sich eine ähnliche lokale Reizbarkeit heraus, wie wir sie später beim Studium der Tastreize kennen lernen werden. Bei Reizung der Seitenteile des Körpers erfolgte eine Kopfwendung nach der Seite, an welcher der Reiz appliziert worden war. Selten kam es bis zur Ausbildung des Schnappreflexes. Stets wich das Tier vor der Hitze zurück; ja der Wärmereiz veranlaßte es unter Umständen sogar, seine Fluchtbewegungen in der Richtung mit dem einfallenden Licht durchzuführen.
Die Ergebnisse dieser Versuche könnten wohl als Beweis für das Vorhandensein eines Wärmesinnes gedeutet werden. Ich habe allerdings vergeblich nach Sinnesorganen oder freien Nervenendigungen in der Körperoberfläche des Tieres gesucht, welche man als die spezifischen Apparate des Wärmesinnes betrachten könnte. Wir haben ja allerdings früher schon erfahren, daß die Körperoberfläche der Ameisenlöwen von einem dichten Mantel von Sinneshaaren bedeckt ist; wir sahen auch, daß diese Haare in ihrem Bau erheblich voneinander abweichen. Es wäre daher immerhin möglich, daß einer dieser Haartypen etwas mit dem Wärmesinn zu tun hat. So dachte ich eine Zeitlang, die eigenartigen Gabelhaare des Abdomens könnten dem Wärmesinn dienen. Ich mußte von dieser Annahme zurückkommen, als ich feststellte, daß Wärmeempfindlichkeit auch in den Regionen nachweisbar ist, z. B. am Kopf, wo keine solchen Gabelhaare[S. 85] vorkommen. Wahrscheinlicher scheint mir die Annahme, daß dieselben Sinneshaare, welche den Tastsinn vermitteln, auch die Werkzeuge des Wärmesinnes sind. Sie wären dann Wechselsinnesorgane im Sinne von Nagel. Man wird dabei kaum daran denken, daß die zentripetale Nervenfaser verschiedenartige Erregungen dem Zentralnervensystem zuzuleiten vermag. Es wird sich wohl in Uebereinstimmung mit der Theorie von Johannes Müller um spezifische Energien handeln, welche die Leitungsbahn durchlaufen. Ihre Erregung wird aber durch eine Transformation im Empfangsapparat veranlaßt werden. Nun kann man sich sehr gut vorstellen, daß eine ganz entsprechende Erregung der Nervenendfasern sowohl durch mechanische Berührung der Tasthaare als auch durch eine etwa eintretende Deformation derselben durch die Wärme erfolgen könnte. Letztere Annahme läge besonders bei dünnwandigen und hohlen Haartypen nahe.
Einige weitere Versuche wurden angestellt, um Aufschluß darüber zu verschaffen, ob etwa auch ein Kältesinn bei den Ameisenlöwen nachweisbar sei. Bekanntlich sind beim Menschen Kälte- und Wärmesinn voneinander vollkommen getrennt und vielfach auf gesonderte Hautgebiete beschränkt. Wir haben auch Anlaß, anzunehmen, daß bei anderen Wirbeltieren ähnliche Verhältnisse vorliegen. Bei einem wirbellosen Tier ist aber bisher noch niemals auch nur die Andeutung eines Kältesinnes entdeckt worden. Meine Experimente mit den Ameisenlöwen hatten auch keinen eindeutigen Erfolg. Versuche, ob eine Unterschiedsempfindlichkeit bei starker Abkühlung eines Teiles des Untergrundes sich bemerkbar mache, hatten kein Resultat. Die auf dem durch untergelegtes Eis abgekühlten Teile einer Glasplatte niedergesetzten Individuen wurden durch die Einwirkung der Kälte vollkommen bewegungslos; sie begannen erst dann sich zu regen, als das Eis unterhalb vollkommen weggetaut war.
Individuen, welche ich auf einen Eisblock setzte, wanderten auf ihm herum, wie auf einem Stein. Solange ihnen die Kälte noch erlaubte, sich zu bewegen, krochen sie beständig am Umkreis des Eisklumpens herum, wobei sie den Hinterleib an seine feste Oberfläche dicht anpreßten. Es zeigte sich also ein deutliches Ueberwiegen der Thigmotaxis über einen etwaigen Kältereiz.
Annäherung eines kleinen Eisstückchens oder einer mit Eis abgekühlten Metallnadel hatte keine Wirkung. Wurde jedoch ein Teil der Mandibel des Kopfes, Halses oder der Oberseite des Rumpfes[S. 86] mit einem kleinen Eisstück berührt, so erfolgte ein auffallend rasches und heftiges Zucken, unter Umständen sogar ein ausgesprochener Schnappreflex. Dabei bissen oft die Mandibel in den Eisklumpen hinein. Ebenso war die Berührung mit einer in Eis abgekühlten Nadel von auffallend starker Wirkung; sie war viel deutlicher als die Berührung mit einer Nadel, welche Zimmertemperatur angenommen hatte.
Wenn also auch diese Versuche nur für eine Reizung durch Berührungsreiz sprechen, so ist doch die Verstärkung des Reizes durch die niedere Temperatur des berührenden Gegenstandes beachtenswert.
Von besonderem Interesse sind die Versuche, bei denen die Tiere gleichzeitig einer Einwirkung von Wärme und einseitig einfallendem Licht ausgesetzt waren. Die Experimente wurden in der Dunkelkammer ausgeführt; die Tiere wurden auf berußtes Registrierpapier gesetzt, welches auf dem Boden von Glasaquarien lag. Letztere waren im Sandbad einseitig erwärmt. Durch die eine Aquarienwand, welche von der Stelle der stärksten Erwärmung am weitesten entfernt war, fiel das Licht einer 50-kerzigen Glühlampe ein.
War die dem Licht abgewandte Seite der Unterlage, auf welcher das Tier kroch, schwach erwärmt, so zeigte sich eine deutliche Hemmung der phototaktischen Reaktion. Das Tier kroch eine Zeitlang in Versuchsbewegung auf der erwärmten Fläche umher, bis es schließlich doch sich zum Licht orientierte und dann ohne Zögern in gerader Richtung auf die Lichtquelle zuwanderte (Taf. I, Fig. 1).
Bei anderen Experimenten wurden die Tiere erst dann auf das Registrierpapier gesetzt, nachdem dies, wie in den früheren Versuchen, die Temperaturen der Sandunterlage angenommen hatte; es war also ein Temperaturgefälle von 35–20° C vorhanden. Die Tiere zeigten nun verschiedene Reaktion, je nach ihrem Ausgangspunkt. Von 3 Exemplaren eines Versuches ging z. B. das eine, welches bei etwa 20° C des Untergrundes seine Wanderung begann, zuerst in einem Versuchskreis umher und setzte dann seinen Weg immer abwechselnd zum Licht und zur Wärme hinbiegend fort; schließlich wandte es sich energisch dem Licht zu. Das zweite Exemplar hatte, bei etwa 20° beginnend, sich gleich zum Wärmeoptimum gewandt, in dessen Gebiet es unter auffallenden Pendelbewegungen dauernd verblieb. Das dritte Exemplar, welches bei etwa 35° niedergesetzt[S. 87] worden war, wanderte zuerst ein Stück auf das Licht zu, kehrte aber dann in die Wärmeregion zurück (Rußspur Taf. X, Fig. 10).
Die übrigen Versuche ergaben stets dasselbe Resultat (vgl. auch Fig. 8, Taf. VIII). Die zahlreichen Versuchskreise, die schwankenden, gleichsam zwischen Licht und Wärme pendelnden Bewegungen, verrieten deutlich, daß beide Reize die Tiere beeinflußten. War das Licht noch stark genug, so wirkte es stets richtend auf die Körperhaltung des Tieres ein. War die Temperatur hoch genug, so ließ sie das Tier nicht aus ihrem Bereich heraus. Das Tier hatte eine Tendenz, nicht über die Grenze der warmen Region hinaus zu kriechen; dazu wurde es aber gezwungen, wenn das einfallende Licht es nötigte, seine Hinterleibsspitze gegen die Lichtquelle zu wenden. Dann mußte die Vorwärtsbewegung das Tier ohne weiteres über die Grenze der optimalen Wärmeregion hinausführen. Solange die Wärme noch einwirkte, blieb die Bewegung des Tieres schwankend, immer wieder zur Wärmeregion hinstrebend. War das Tier aber einmal über die Region des Optimums hinausgelangt, so wanderte es unbehindert geradeaus der Lichtquelle zu.
Wir haben früher die starke Behaarung der Körperoberfläche des Ameisenlöwen geschildert und haben gesehen, daß die Art ihrer Verteilung, die Stärke und Richtung der Haare wichtige Beziehungen zu den Lebensgewohnheiten des Tieres erkennen lassen. Wir wollen hier in Kürze den feineren Bau der Haare und ihre Innervierung schildern.
Schon früher habe ich erwähnt, daß es verschiedene Typen von Haaren beim Ameisenlöwen gibt. Alle möglichen Längen- und Dickenausdehnungen sind vertreten, der gesamte Charakter der Haare ist bald grob und fest, bald zart und dünn. Die verschiedenen Borstentypen sind auf der Abbildung des ganzen Tieres Abb. 2, S. 7, und auf der Photographie des Hinterleibsendes Abb. 10, S. 19, gut zu unterscheiden.
Zunächst erkennen wir an allen Stellen des Körpers, welche bei den Bewegungen des Tieres mechanisch beansprucht werden, kurze, dicke, oft fast kegelförmige Borsten. Solche finden wir an der Hinterleibsspitze und an der Vorderkante der Tibia des dritten Beinpaares, also an den Stellen, welche beim Rückwärtsarbeiten durch[S. 88] den Sand einen Druck auf dessen rollende Körner ausüben müssen. Diese „Stemmborsten“, wie ich sie nennen will, bestehen aus schwarzbraunem bis schwarzem, massigem Chitin. Ihre Oberfläche zeigt eine feine Rillung und Zähnelung, wie wir sie gleich auch bei anderen Haartypen kennen lernen werden. Sie ist aber meist nicht deutlich zu erkennen; denn diese Stemmborsten zeigen fast stets unverkennbare Zeichen der Abnützung; ihre Oberfläche ist abgeschliffen und die Spitzen sind abgestumpft, oft wie abgebrochen. Die Stemmborste sitzt in einer tiefen Gelenkgrube, deren Chitinumwallung sehr kräftig ist. Eine färbbare, sehnenartige Spannhaut befestigt sie im Umkreise des Grundes der Grube. Ein kreisrunder Porus führt in das Innere des Körpers. Soweit ich an den schwer herzustellenden Präparaten erkennen konnte, führt durch diesen Porus eine Nervenfibrille in das Innere der Stemmborste, wo sich trotz der Dunkelfärbung der dicken Chitinwandung ein feiner Zentralkanal erkennen läßt (Abb. 34).
Nach Lozinski läßt sich an diesen Stemmborsten jeweils eine einzellige Drüse nachweisen. Möglicherweise dient diese zum Einschmieren der Borsten. Als Anzeichen ihrer Tätigkeit konnte ich nicht selten Verklebung von Sandkörnchen in der Umgebung des Hinterendes beobachten. Lozinski ist der Ansicht, daß der Kanal zur Entleerung des Drüsensekrets die ganze Stemmborste der Länge nach durchsetzt (?). Ich konnte im distalen Teil der Borste keinen Kanal erkennen. Wohl bestätigte ich das Vorkommen der Drüsenzellen, habe aber mehr den Eindruck, als entleere sie ihr Sekret auf die Gelenkhaut der Stemmborste.


Diese Stemmborsten gehen am Hinterleibsende direkt in eine Borstenform über, welche länger und bedeutend schlanker ist. Die nebeneinander stehenden Borsten bilden am Hinterleibsende eine Uebergangsreihe von den kurzen stumpfen, zu langen spitzen Formen. Diese letzteren sind bei großen Exemplaren des Ameisenlöwen ½–1 mm lang. Sie sitzen außer am Hinterleibsende am Kopf, Thorax, an[S. 89] den Extremitäten, auf den Wülsten des Abdomens und sind für das Aussehen des Tieres sehr charakteristisch. Verschiedene Formen der Borsten und der unten zu beschreibenden Haare sind auf den Abb. 2, 6, 8, 9, 10 und 12 genau dargestellt.
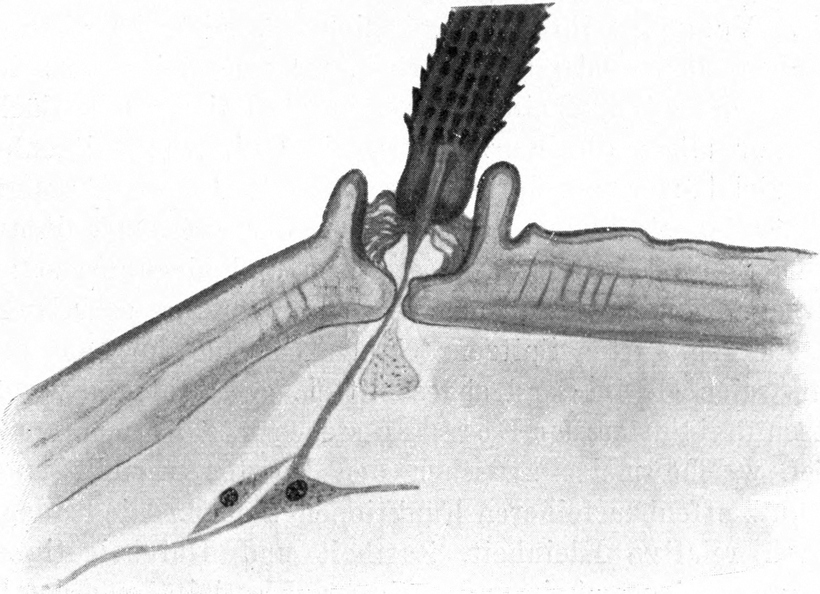
Sie sitzen in tiefen, großen, kapselartigen Gelenkgruben und sind sehr beweglich. Ihre Oberfläche ist längsgerillt, auf den Rillen sitzen Zähnchen mit nach vorn gerichteten Spitzen. Ueberall, wo die Haare stark in Anspruch genommen werden, schleift sich die Oberflächenstruktur sehr ab; die Haare sind dann vor allem in der Spitzenregion ganz glatt. Die Gelenkkapsel ist nach innen von einem runden Porus durchsetzt, durch den die Nervenfibrille in den Körper tritt (Abb. 36). Für sie ist im Haar ein feiner Kanal vorhanden, welcher aber nicht sehr weit[S. 90] nach vorn in die kompakte Chitinmasse des Haares eindringt. Ich bezeichne diesen Haartypus als die „Langborsten“.
Zwischen ihnen, welche meist in Büscheln vereinigt an den am meisten nach außen vorragenden Stellen des Körpers stehen, finden wir viele kleinere Haare. Diese sind dick und stämmig und hakenförmig nach vorn gebogen. Sie sind ebenso schwarz oder dunkelbraun gefärbt, wie die bisher beschriebenen Haartypen. Sie stehen meist felderweise beieinander und spielen eine Hauptrolle beim Anstemmen des Körpers an die Sandpartikel. Ich nenne sie die „Kurzborsten“. Sie finden sich vor allem an der Rücken- und Bauchseite auf den Flächen der Abdomenglieder. Sie sind im Prinzip genau so gebaut, wie die Langborsten, auch innerviert, dickchitinig, gerillt und gezähnelt. Gewöhnlich gehen sie in der Nachbarschaft der Langborstengruppen ganz allmählich in solche über, so daß man dicht nebeneinander alle Uebergänge findet.
Stemmborsten, Langborsten und Kurzborsten bilden zusammen eine Gruppe; wie sie alle durch Uebergänge verbunden sind, so haben sie in gewisser Beziehung gemeinsame Funktionen. Sie haben alle Druck- und Berührungsreize, welche den Körper treffen, zuerst aufzufangen. Neben der reizbiologischen Bedeutung haben sie eine wichtige mechanische Funktion zu erfüllen, indem sie alle an der Erhöhung des Reibungswiderstandes gegen den rollenden Sand und an der Stemmwirkung bei den Bewegungen des Tieres im Sand und auf Oberflächen beteiligt sind. Sie alle tragen auch die Spuren ihrer Tätigkeit. Vor allem ihre Spitzenregionen sind meist abgeschliffen und oft abgestumpft oder abgebrochen, während frisch aus der Häutung entstandene Haare eine sehr fein skulptierte Oberfläche aufweisen. Vor allem die Kurzborsten sind vielfach spitzenlos. Sie scheinen viel härter und spröder zu sein als die Langborsten. Besonders der längste Typus der letzteren, der auf den Seitenwülsten des Abdomens sitzt, scheint sehr elastisch und biegungsfest zu sein; sie sind selten an der Spitze abgebrochen; aber ihre sehr lang ausgezogenen, dünnen, zarten Spitzenregionen machen durchaus den Eindruck, als seien sie im Sand abgeschliffen, geglättet und verdünnt.
Diesen drei Sorten von Borsten stehen zwei Formen von Haaren gegenüber, welche meist zwischen den Borsten verteilt und unter deren Schutz offenbar feineren Funktionen dienen. Die beiden Typen sind durch relative Kleinheit, Zartheit und Durchsichtigkeit des Chitins, große Biegsamkeit und geringere Elastizität von den Borsten unterschieden.
[S. 91]
Ich unterscheide zunächst die „Fiederhaare“. Sie sitzen an den verschiedensten Stellen des Körpers, so am Vorderrand des Kopfes, am ganzen Körper, vor allem an der Ober- und Unterseite des Thorax und Abdomens. Sie fehlen auf den Mandibeln und der Unterlippe, sind selten auf Ober- und Unterseite des Kopfes und sind auf den Gliedmaßen nur durch Uebergangsformen zu den Langborsten vertreten, deren es auch sonst auf dem Körper vielerlei gibt.
Fiederhaare nenne ich diese Form, weil sie durch in Reihen angeordnete Chitinborsten ein gefiedertes Aussehen haben; allerdings stehen die Fiedern nicht wie bei Federn in zwei symmetrischen Reihen, sondern es sind 8–10 Reihen um den Haarschaft angeordnet, gerade Reihen, welche offenbar den Zahnreihen der verschiedenen Borstentypen entsprechen (Abb. 37). Die Fiedern stehen also nach allen Seiten vom Haar ab und geben diesem eine sehr große Oberfläche. Sie sperren die Haare auseinander und verhindern da, wo mehrere von ihnen dicht beieinander stehen, daß sie sich untereinander verflechten.
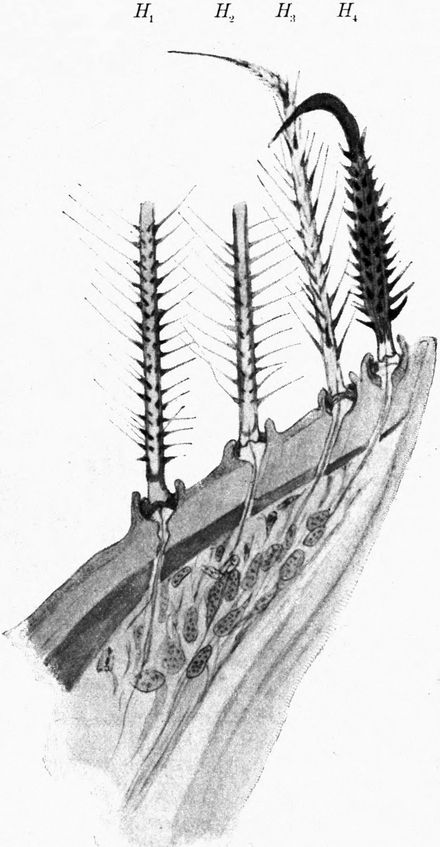
[S. 92]
Die Fiederlänge kann sehr verschieden sein, und es finden sich alle Uebergänge zu den Langborsten mit ihren Zahnreihen (vgl. Abb. 37 H4).
Die Fiederhaare sind vielfach sehr zart gebaut, so daß sie sich zwischen den Borsten der Oberfläche des Körpers anschmiegen. Sie sind auch gelenkig dem Chitinpanzer eingefügt. Ein feiner Nervenstrang führt durch ihre Gelenkpfanne und das Chitin zur Sinneszelle, deren Fortsatz mit denen anderer benachbarter Haare sich zu einem Nervenstrang vereinigt (Abb. 37).
Der letzte Typus, den ich erwähne, sind die „Gabelhaare“; auch sie sitzen gelenkig in einer Grube der Cuticula, durch deren Bodenporus sie innerviert werden. Sie bestehen aus einem ganz kurzen Basalteil, der sich in zwei lange gefiederte Aeste gabelt. Diese streben in rechtem Winkel vom Basalteil auseinander, so daß sie fast parallel zur Oberfläche des Chitinpanzers gelagert sind. Meist sind die Gabeläste etwas wellig gebogen. Sie sind stets so zart gebaut, wie die dünnsten der Fiederhaare (Abb. 38).
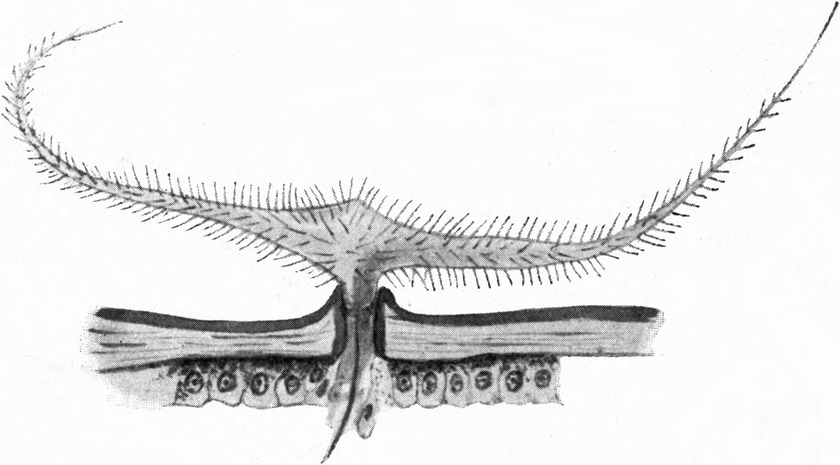
Das wären die wichtigsten fünf Haartypen von Myrmecoleo formicarius. Außer ihnen und den zahlreichen Zwischenformen, welche zwischen den vier erstbeschriebenen Typen vorkommen, gibt es einige weitere Haarbildungen, auf welche ich hier nicht eingehe. Sie sind in anderem Zusammenhang erwähnt: so die Haare auf den Lippentastern (S. 13), die Haarreihen über den Gelenkgruben der Mundgliedmaßen (S. 14) und auf den letzteren (S. 11) und schließlich die Stellungshaare der Gelenke (S. 21).
[S. 93]
Wir haben gesehen, daß der ganze Körper des Ameisenlöwen mit Haaren bedeckt ist, welche sämtlich durch Sinneszellen mit dem Nervensystem in Verbindung stehen. Wir dürfen wohl annehmen, daß diese Haare zum größten Teil im Dienste des Tastsinnes stehen. Dies wird ja allgemein für alle Arthropoden angenommen; allerdings gibt es bisher noch kaum exakte Versuche, welche die Berechtigung dieser Annahme beweisen. Meist findet man in der Literatur nur gelegentliche Beobachtungen als Belege angeführt. Allerdings ist zuzugeben, daß die allgemeine Verbreitung der Sinneshaare, die Härte und Festigkeit des Chitinpanzers und dessen Durchbohrungen an der Basis der Sinneshaare mit dem Durchtritt der Nervenendigung, wichtige morphologische Wahrscheinlichkeitsbeweise für die Funktion der Arthropodenhaare als Organe des Tastsinnes sind. Es gibt ja viele Beobachtungen, welche das Vorhandensein des Tastsinnes bei den Arthropoden sicherstellen.
Ich habe in meiner Abhandlung über „Lebensgewohnheiten und Anpassungen bei dekapoden Krebsen“ eine Anzahl von Beobachtungen über die Rolle der Sinneshaare als Tastorgane bei diesen Tieren veröffentlicht. Die Ameisenlöwen haben sich nun als ganz besonders günstige Objekte für experimentelle Untersuchung des Tastsinnes erwiesen. Ich habe daher ziemlich ausgedehnte Versuche mit ihnen angestellt, über deren wichtigste Ergebnisse ich folgendes berichten kann.
Die Ameisenlöwen reagieren auf Berührung mit festen Gegenständen durch Bewegungen einzelner Körperteile. Je nach der Intensität des ausgeübten Druckes und je nach der gereizten Körperstelle treten verschiedene Reflexbewegungen auf. Sie bestehen in kurzen Zuckungen der Gliedmaßen, der Mandibel oder des Kopfes; ferner in der Ausführung des Schleuder- und Schnappreflexes, sowie der charakteristischen Zuckungsbewegungen mit dem Hinterleibsende.
Es ist nun sehr interessant, daß nicht immer gleich starke Reize die gleichen Erfolge haben. Schon verschiedene Temperaturen der Umgebung veranlassen sehr große Verschiedenheiten im Reizerfolg des gleichen Reizes. Aber auch unter ganz gleichen äußeren Verhältnissen reagiert vielfach ein Tier auf einen ganz leichten Reiz, während ein anderes auf einen zehn-, fünfzig-, ja hundertmal so starken Reiz gar nicht reagiert. Auch zeigt sich eine sehr[S. 94] deutliche Summationswirkung aufeinander folgender Berührungsreize.
Ich will zunächst die Methode beschreiben, mit welcher ich die meisten hierhergehörigen Versuche durchgeführt habe. Daß überhaupt eine ausgesprochene Reizbarkeit der einzelnen Körperregionen für Berührungsreize vorhanden ist, davon hatte ich bei meinen verschiedenen Experimenten zahllose Beweise gelegentlich beobachtet. Jede Berührung mit einer Pinzette oder irgendeinem anderen Instrument konnte bei den Tieren deutliche Reaktionen hervorrufen; vor allem war dies der Fall bei Berührung mit einer spitzen Präpariernadel. Um mir über die Empfindlichkeit der Tiere gegen Tastreize ein klares Bild zu verschaffen, mußte ich eine Methode anwenden, welche einen zahlenmäßigen Vergleich der einzelnen Resultate zuließ. Ich benützte zu diesem Zweck ein Verfahren, welches durch v. Frey für die Untersuchung der Tastempfindlichkeit der menschlichen Haut ausgebildet worden ist. Zu diesem Zweck werden Borsten von gleicher Länge rechtwinklig an der Spitze eines Stäbchens befestigt. Tastreize mit diesen Borsten werden erzielt, indem die Borste mit ihrer Spitze senkrecht auf die Oberfläche der zu prüfenden Körperregion aufgesetzt wird, worauf ein Druck, der zur Krümmung der Borste führt, ausgeübt wird. Je nach der Dicke der Borste ist dieser Druck verschieden stark. Ich benützte zu meinen Versuchen 3 cm lange Stücke von Pferdeborsten und Menschenhaaren, welche senkrecht von der Längsachse des sie tragenden Stabes abstanden. Nach dem Vorgang v. Freys wurden diese Borsten in folgender Weise geeicht. An einer feinen chemischen Wage wurde festgestellt, einem wie schweren Gewicht sie Widerstand zu leisten vermögen. Stemmt man das Ende der Borste wider die eine Wagschale und belastet die andere mit zunehmend schweren Gewichten, so tritt ein Moment ein, in welchem das gerade sich krümmende Haar der belasteten Wagschale das Gleichgewicht zu halten vermag.
Ich wandte zu meinen Versuchen 6 verschiedene Tastborsten an, deren Krümmungselastizität in der angegebenen Weise auf folgende Zahlen festgestellt worden war:
|
Tastborste
|
0
|
. . .
|
0,0045
|
g
|
|
„
|
1
|
. . .
|
0,07
|
g
|
|
„
|
2
|
. . .
|
0,12
|
g
|
|
„
|
3
|
. . .
|
0,22
|
g
|
|
„
|
4
|
. . .
|
0,63
|
g
|
|
„
|
5
|
. . .
|
1,8
|
g
|
[S. 95]
Man sieht, es sind recht beträchtliche Druckverschiedenheiten, welche mit diesen Borsten erzielt werden können. Um so auffallender sind manche der erzielten Resultate.
Es kam vor, daß einzelne Individuen der Reihe nach mit sämtlichen Borsten, von den dünnsten bis zu den dicksten, mehrmals betastet werden konnten, ohne irgendeine Reaktion zu zeigen. In anderen Fällen genügte eine kurze Berührung mit der Tastborste 1 oder 2, um eine sehr lebhafte Reaktion herbeizuführen. Offenbar hängt die Reaktion also in hohem Maße von inneren Zuständen des gereizten Tieres ab. Es zeigte sich aber bald, daß es bis zu einem gewissen Grade gelingt, diese Zustände zu beherrschen.
Experimentiert man z. B. mit einem Tier, das im typischen Zustand des Totstellens sich befindet, so kann man es durch wiederholte Applikation von Tastreizen auf die gleiche Region des Körpers nach einiger Zeit zu einer Reaktion bringen. Ist diese einmal eingetreten, so zeigt sich das Tier sofort auch für viel feinere Reize empfindlich. Die Empfindlichkeit ist dann auch nicht auf die vorher gereizte Körperregion beschränkt, sondern erstreckt sich auf eine mehr oder minder große Partie der Umgebung, oft auch auf den ganzen Körper. Hat man z. B. ein Tier zehnmal mit der Tastborste 5 auf die Halsregion gedrückt, und ist nach der zehnten Berührung eine Reaktion erfolgt, so kann in den meisten Fällen mit Borste 2 oder 3 schon nach ein oder zwei Berührungen der gleichen Region oder irgendeiner Gegend des Rückens eine Reaktion ausgelöst werden. Dagegen hat sich die Erhöhung der Reizbarkeit nicht bis auf die Bauchseite ausgedehnt.
Aus dem Gesagten geht hervor, daß es mit der angegebenen Methodik ganz ausgeschlossen ist, ein absolutes Maß der Reizbarkeit aufzustellen. Dasselbe Tier, welches anfänglich auf grobe Berührungen mit der Tastborste 5 gar nicht reagierte, zeigt nach einer Reihe von Versuchen, schon nach wenigen Minuten eine hohe Empfindlichkeit, selbst gegen Berührung mit der Tastborste 0. Wollen wir also die Empfindlichkeiten der verschiedenen Regionen des Körpers direkt miteinander vergleichen, so verwenden wir am besten ein Tier, dessen Reizbarkeit durch künstliche Einwirkung erhöht worden ist. Ich habe sowohl mit Tieren experimentiert, bei denen die Reizbarkeit durch Summation von Berührungsreizen, als auch mit solchen, bei denen das gleiche Resultat durch Erwärmung der Umgebung erzielt worden war. Obwohl bei letzterer Methode die vorbereitende Reizung sicherlich eine weniger lokalisierte gewesen[S. 96] ist, ergaben sich dennoch in allen Fällen übereinstimmende Resultate.
Die Reflexe, durch welche die Tastreize beantwortet werden, sind sehr charakteristisch und je nach den gereizten Körperregionen verschieden. Wird die Innenseite der Mandibel, die Vorder- oder Oberseite des Kopfes oder werden die Fühler berührt, so erfolgen zunächst leise zitternde Bewegungen an dem berührten Körperteil, und die Tiere machen einige kurze Rückwärtsbewegungen unter Verwendung des ersten und vor allem des zweiten Beinpaares. Die gleiche Bewegung erfolgt bei Berührung des ersten und zweiten Beinpaares selbst. Auch wenn bei diesen Versuchen nur die eine Seite des Körpers gereizt wird, erfolgt doch die Bewegung vollkommen symmetrisch entsprechend der Mittellinie. Die Berührung des Beines der einen Körperseite führt nicht zu einer Drehung von oder zu dieser Seite. Am ausgesprochensten ist dieses Rückwärtsweichen dann, wenn eine multiple Berührung der betreffenden Regionen vorgenommen wurde. Wurde statt mit einer Borste mit einem Pinsel oder mit einer kleinen Bürste eine etwas ausgedehntere Region des Kopfes oder der Beine gereizt, so erfolgte eine oft sehr lebhafte, manchmal einige Sekunden andauernde Rückwärtsbewegung. Spätere Versuche zeigten allerdings, daß dies nur für wenig reizbare Tiere gilt. Sehr reizbare Tiere reagieren in ausgesprochen lokalisierter Weise.
Das gilt bei allen Tieren für die Reflexe bei Berührung der Oberseite des Halses, der übrigen Thorakalsegmente oder des Abdomens. Bei leichter Reizung ging das Tier sogleich in die Bereitschaftsstellung über; war die Reizung nur etwas intensiver, so trat sofort ein sehr energischer Schleuder- oder gar Schnappreflex ein. Meist wurde dabei die zur Reizung verwendete Borste zwischen den Mandibeln gefaßt, so daß man das Tier an ihr emporheben konnte.
Von ganz besonderem Interesse war nun die ausgesprochene Reizlokalisation, welche sich bei diesen Versuchen auf das deutlichste nachweisen ließ. Reizte man eine Stelle in der Nähe der Medianlinie des Rückens, so erfolgte das Zurückbiegen des Kopfes beim Schnappreflex vollkommen symmetrisch in dieser Medianlinie. Reizte man eine Stelle links oder rechts von der Medianlinie, so bogen sich Kopf und Hals in einem ganz entsprechenden Winkel der gereizten Stelle zu; also um so weiter nach außen, den Körperrändern zugewandt, als die Reizstelle an denselben gewählt war. Reizte man eine Stelle des Abdomenrückens, die weiter nach hinten gelegen ist, also etwa in der Nähe des Hinterendes des Tieres, so[S. 97] fand eine stärkere Durchbiegung, an der sich mehr Segmente des Vorderkörpers beteiligten, statt. Der Kopf und der Hals wurden bei diesen Reaktionen ganz flach nach hinten über den Rücken des Abdomens gelegt. Auch bei diesen Versuchen fand eine ganz ausgesprochene Lokalisation, eine feinste Reaktion auf jede asymmetrische Lagerung des Reizpunktes statt. Fast stets wurde die Reizborste mit großer Sicherheit von den Mandibeln ergriffen. Sehr reizbare Tiere wenden auch bei leichter Reizung der beiden vorderen Beinpaare den Kopf unter Biegung des Halses stark nach der gereizten Seite; ja, sie führen nach der reizenden Borste den typischen Schnappreflex aus, wenn die Reizung intensiver wird.
Reizt man das auf der Bauchseite liegende Tier von unten her mit einer Tastborste an der Bauchregion der Hinterleibsspitze, so erfolgt eine Kontraktion der Längsmuskulatur; dieselbe führt nicht nur zu einer Verkürzung und Verdickung des Hinterleibes, sondern auch, da die Verkürzung ventral stärker ist, zu einer Einkrümmung der Hinterleibsspitze nach unten. Diese Reaktion tritt noch viel auffälliger als bei Reizung mit einer einfachen Borste bei multipler Reizung, z. B. mit einem Pinsel, ein.
Befindet sich das Tier in Rückenlage, so ist bei Tastreiz der Hinterleibsspitze diese Reaktion viel weniger ausgesprochen. Wohl wird auch dann die Längsmuskulatur kontrahiert, also der Hinterleib verkürzt; aber eine ausgesprochene Einkrümmung der Hinterleibsspitze findet nicht statt, da die Tendenz, den Rücken für den Umdrehreflex hohl zu machen, also die umgekehrte Krümmungstendenz, überwiegt.
Denn fast regelmäßig zeigt sich bei Reizung der Bauchseite eines auf dem Rücken liegenden Tieres als sehr prompter Erfolg der Umdrehreflex. Im allgemeinen kann man dabei keine ausgesprochene Wirkung des lokalisierten Reizes wahrnehmen. Einerlei, ob ein Feld rechts oder links, vorn oder hinten an der Bauchseite gereizt wird, das Tier dreht sich nach der Seite um, welche ihm nach den früher (S. 41) berichteten Experimenten vom Licht vorgeschrieben wird. Ein Schnappen nach der Bauchseite findet niemals statt; Hals und Kopf beugen sich immer gegen den Rücken zu. Wir sehen von neuem, daß der Schnappreflex und der Umdrehreflex identisch sind.
Trotzdem läßt sich auch bei diesen Experimenten die Lokalisation des Reizes an besonderen Reflexen erkennen. Verwendet man nämlich zu den Versuchen ein Tier von geringer Reizbarkeit, oder[S. 98] operiert man mit den niedersten Nummern der Reizborsten, so kann man folgende Beobachtungen machen. Bei Applikation einer unsymmetrischen Reizung zeigt sich, daß zunächst ganz langsam die Mandibel der gereizten Seite abgespreizt wird; dann wird der Kopf nach der gleichen Seite gebeugt, worauf der Umdrehreflex plötzlich mit Wendung nach der anderen Seite erfolgt. Dieses letzte Experiment gelingt vor allem dann, wenn man in diffusem Licht oder in tiefer Dämmerung das Tier reizt, so daß ein Einfluß der Lichtrichtung nicht in Frage kommt.
Als Gradmesser für die Erregbarkeit des Tieres durch einen lokalen Reiz können wir also die ausgelöste Reflexbewegung betrachten. Ein wenig reizbares Tier zeigt als erste Anzeichen der Reizung schwache Bewegungen der gereizten Körperstelle oder seiner Nachbarschaft. Steigt die Erregung, so werden diese schwachen Bewegungen der Mandibel, des Kopfes, der Extremitäten oder der Rumpfsegmente ausgesprochener. Bei noch stärkerer Erregung geht das Tier in Bereitschaftsstellung über; der stärkste Ausdruck der Reizbarkeit sind die typischen Reflexe, wie der Schleuder-, der Schnappreflex und die Bohrbewegung des Hinterleibsendes. Bei allmählicher Steigerung der Erregbarkeit sieht man also z. B. am Vorderende des Körpers leichte Bewegung der Mandibel, des Kopfes und Halses, dann Bereitschaftsstellung, darauf Schleuder- und schließlich Schnappreflex aufeinander folgen. Meist aber sind letztere beide miteinander verschmolzen.
Experimentiert man mit einem schwach reizbaren Tier, so kann man nun feststellen, daß die verschiedenen Regionen des Körpers in verschiedener Weise reizbar sind. Es kann z. B. vorkommen, daß ein sehr schwach reizbares Tier auf Berührung mit Tastborste 5 gar nicht reagiert, wenn die Rückenseite der letzten Hinterleibssegmente mit ihr mehrmals berührt werden; eine ganz leichte Reaktion tritt ein, wenn die Vorder- und Hinterleibssegmente oder das erste Thorakalsegment auf der Rückenseite in derselben Weise gereizt werden. An den Mandibeln, der Oberseite des Kopfes, den drei Beinpaaren verursacht der gleiche Reiz bereits den Uebergang zur Bereitschaftsstellung. Auf der Oberseite des ersten und zweiten Thorakalsegmentes jedoch löst er bereits den typischen Schnappreflex aus.
Ebenso kann man nachweisen, daß, wenn das erste und zweite Thorakalsegment auf ihrer Rückenseite schon durch die Tastborsten 0 oder 1 reizbar sind, man steigend höhere Nummern verwenden muß,[S. 99] um Mandibel, Kopf, Beine und schließlich die hinteren Regionen des Hinterleibsrückens mit dem gleichen Erfolg zu reizen.
Es folgt also aus diesen Versuchen, die in großer Zahl und in vielen Modifikationen von mir ausgeführt wurden, daß im großen und ganzen auf der Rückenseite die Reizbarkeit des Tieres auf dem ersten und zweiten Thorakalsegment am größten ist, etwas geringer an Mandibeln, Kopf und Beinen, und daß sie vom dritten Thorakalsegment über das Abdomen hin nach hinten allmählich abnimmt. Etwas anders ist die Verteilung der Reizbarkeit an der ventralen Seite des Tieres. Hier ist gerade das Hinterleibsende am reizbarsten; verfolgt man die Oberfläche der Bauchseite nach vorn, so erstreckt sich die Zone hoher Reizbarkeit hauptsächlich längs der Mittellinie nach vorn. An den Rumpfseiten nimmt die Reizbarkeit nach vorn hin rasch ab. Die Unterseite der Beine und die umgebende Ventralseite der Thorakalsegmente weist ungefähr die zweite Stufe der Reizbarkeit auf. Die Unterseite des Kopfes ist dagegen auffällig wenig reizbar.
Es erfordert eine ziemlich große Anzahl von Experimenten, um sich über diese Verhältnisse Klarheit zu verschaffen. Denn abgesehen davon, daß man im voraus gar nicht wissen kann, bei welcher Reizbarkeitsstufe des Tieres man das Experiment beginnt, es steigert sich auch die Empfindlichkeit im Verlauf des Experiments oft sehr rasch. So kann es kommen, daß man im Verlauf des Experiments mit immer feineren Tastborsten die gleichen Resultate erzielt, wie vorher mit den groben. Ein Tier z. B., welches zuerst in der vorhin angegebenen Stufenfolge bei der Reizung mit Tastborste 5 reagierte, ergibt nach einiger Zeit deutliche Reaktionen stärkster Art bei Reizung des Rückens, des Hinterleibsendes mit Tastborste 3, bei Reizung der Kopfregion mit Tastborste 2 und bei Reizung des ersten und zweiten Thorakalsegmentes sogar schon mit Borste 0. Ja, nach einiger Zeit kann die Erregung des Tieres so weit gestiegen sein, daß alle Regionen des Körpers schon auf die zartesten Berührungen mit Tastborste 0 reagieren.
Zum Beleg dieser Feststellungen gebe ich im nachfolgenden einige meiner Versuchsprotokolle.
1) Reizung der Dorsalseite von Thorakalsegment II mit einer Tastborste. (Die Reizungen wurden unmittelbar hintereinander in der Reihenfolge der Tabelle ausgeführt. In der Tabelle ist die Zahl der Berührungen, welche angewandt werden mußten,[S. 100] bis eine Reaktion erfolgte, angegeben; das Zeichen + bedeutet einen Schnappreflex, die 0, daß keine Reaktion erfolgte.)
|
No. der Borste
|
Zahl der Berührungen
|
Reaktion
|
|
|
4
|
22mal
|
+
|
|
|
3
|
100 „
|
0
|
|
|
4
|
37 „
|
+
|
|
|
3
|
100 „
|
0
|
|
|
4
|
26 „
|
+
|
|
|
3
|
24 „
|
+
|
|
|
3
|
5 „
|
+
|
|
|
3
|
7 „
|
+
|
|
|
2
|
3 „
|
+
|
|
|
2
|
3 „
|
+
|
|
|
2
|
5 „
|
+
|
|
|
2
|
3 „
|
+
|
|
|
1
|
100 „
|
|
0;
kleine Bewegungen |
|
2
|
2 „
|
+
|
|
|
1
|
50 „
|
0
|
|
|
2
|
22 „
|
+
|
|
|
2
|
5 „
|
+
|
|
|
2
|
7 „
|
+
|
|
|
1
|
60 „
|
+
|
2) Reizung der Bauchseite in der Nähe des Hinterendes; + bedeutet einen Umdrehreflex; sonst Bezeichnungen wie in Tabelle 1.
|
No. der Borste
|
Zahl der Berührungen
|
Reaktion
|
|
|
4
|
5mal
|
+
|
|
|
1
|
100 „
|
0
|
|
|
4
|
3 „
|
+
|
|
|
2
|
120 „
|
|
0;
nur von Berührung 50 ab leichte Zuckung mit Beinpaar 2 und 3 |
|
4
|
1 „
|
+
|
|
|
1
|
3 „
|
+
|
|
|
1
|
60 „
|
0
|
|
|
1
|
50 „
|
0
|
3) Reizung der Bauchseite in der Nähe des Hinterendes; sehr träges Individuum.
|
No. der Borste
|
Zahl der Berührungen
|
Reaktion
|
|
4
|
50mal
|
kleine Bewegungen
|
|
5
|
25 „
|
+
|
|
4
|
60 „
|
0
|
|
Nadel
|
15 „
|
schwach
|
[S. 101]
4) Reizung der Dorsalseite des dritten Thorakalsegments bei demselben Individuum wie Tabelle 3.
|
No. der Borste
|
Zahl der Berührungen
|
Reaktion
|
|
|
4
|
12mal
|
kleine Bewegungen
|
|
|
4
|
24 „
|
„ „
|
|
|
4
|
40 „
|
„ „
|
|
|
5
|
10 „
|
angedeuteter
|
Schnappreflex
|
|
5
|
20 „
|
„
|
„
|
|
Nadel
|
5 „
|
+
|
Schnappreflex
|
Diese Tabellen zeigen ganz deutlich, wie im allgemeinen die Reizbarkeit einer Körperregion sich durch wiederholte Reize steigern läßt. Sie zeigen aber auch, daß in der Regel nach wiederholten Reizungen eine Ermüdung eintritt. Merkwürdigerweise ist in den meisten Fällen nach den durch solche Ermüdungen bedingten Pausen die Reizbarkeit nicht auffallend herabgesetzt, sondern immer noch sehr viel höher, als im Anfang des Experiments.
Wir haben erwähnt, daß mitunter die Reizbarkeit so weit gesteigert sein kann, daß die feinsten Reizmittel genügen, um einen starken Reflex auszulösen. Es genügen dann nicht nur Berührungen mit den zartesten Gegenständen, sondern diese Berührungen brauchen auch nur einmal zu erfolgen, um den vollen Effekt zu haben. Die derart durch Berührungsreize in Erregung versetzten Individuen führen überhaupt alle Reaktionen mit der größten Promptheit und Raschheit und sehr schnell und oft hintereinander aus. So kann man z. B. ein derart gereiztes Individuum immer wieder rasch hintereinander mit Hilfe des Umdrehreflexes Purzelbaum schlagen lassen. Man muß es zu diesem Zweck immer wieder rasch in die Rückenlage bringen.
So führte z. B. am 3. Okt. 1914 Exemplar a auf wiederholten Reiz in 4 Min. 45 Sek. 114 Umdrehungen aus; erst nach Verlauf von 3½ Minuten trat etwas Ermüdung ein, welche sich in Verlangsamung der Reaktion kundgab. Nach Ablauf der 4 Min. 45 Sek. erfolgte 20 Sek. lang trotz 10maliger Reizung keine Reaktion. Darauf aber trat wieder die volle Reizbarkeit ein, wie sich daraus schließen ließ, daß das Tier ohne weiteres noch ungefähr 100 Umdrehungen ausführte.
Die verschiedene Erregbarkeit der einzelnen Körperregionen steht in einem deutlichen Zusammenhang mit den morphologischen Grundlagen des Tastsinnes. Die Regionen, welche am stärksten reizbar[S. 102] sind, tragen im allgemeinen das dichteste Kleid von Sinneshaaren. Auch sind sie am reichsten an den zarten, dünnen Haaren, welche offenbar die reizbarsten sind. Es läßt sich leicht nachweisen, daß die Haare selbst durch Berührung reizbar sind. Besonders die Haare auf den Schultern und an den Seiten des Körpers, sowie einige der auf Kopf- und Brustsegmenten stehenden Haarbüschel sind so lang, daß man die Berührung einzelner Haare leicht durchführen kann, ohne den Chitinpanzer des Tieres mit Nadel oder Borste zu erreichen. Jede Biegung des Haares, überhaupt jeder Druck auf dasselbe führt zu der für die betreffende Körperregion charakteristischen Reaktion.
Am reizbarsten sind die Fiederhaare und die Langborsten, doch scheinen alle Haartypen — wie sie ja auch alle innerviert sind — für Berührungsreize empfindlich zu sein. Allerdings für die sehr zarten und kleinen Gabelhaare kann ich dies nicht mit Bestimmtheit sagen, da ich unter dem Mikroskop keine derselben einzeln zu reizen vermochte. Es ist nicht ausgeschlossen, daß sie im Dienst eines ganz anderen Sinnes stehen.
Es zeigt sich aber bei den Versuchen mit den Tastborsten, daß in den reizbarsten Regionen des Körpers eine Berührung des Chitinpanzers mit ihnen genügt, um die Reaktionen auszulösen. Genauere Beobachtung zeigt, daß in all diesen Regionen das Chitin der Körperbedeckung sehr dünn und leicht deformierbar ist. Man sieht ohne weiteres, daß es dem Druck der Borste nachgibt und sich einbiegen läßt. Unzweifelhaft haben diese Formveränderungen ähnliche Verschiebungen und sonstige Einwirkungen auf die Basis der in der Umgebung eingepflanzten Haare zur Folge, wie die Berührung der Haare selbst. Die weniger empfindlichen Körperstellen, wie z. B. die Unterseite des Kopfes, die Oberseite des Unterleibsendes, haben das dickste Chitin.
Jene Ameisenlöwen, welche ich zur Analyse des Umdrehreflexes mit Hilfe eines Wachstropfens an einem Faden aufgehängt hatte (vgl. S. 42), zeigten eine sehr hohe Empfindlichkeit in der Region des Wachstropfens und um denselben herum. Offenbar wurden die durch ihn zusammengekitteten Haare sehr leicht gleichzeitig gereizt, und jeder Druck auf ihn führte zu einer Formveränderung der ganzen Umgebung.
Sehr bemerkenswert ist auch folgende Beobachtung. Reizt man ein sehr lebhaftes Tier mehrmals an derselben Stelle, so kommt es unter Umständen zu einer Nachwirkung des Reizes, indem das Tier,[S. 103] auch ohne noch einmal gereizt zu werden, nach derjenigen Stelle schnappt, an der es zum letztenmal gereizt wurde.
Für die große Empfindlichkeit des Tastsinnes bei dem Ameisenlöwen sprechen auch meine Beobachtungen, daß er sehr intensiv auf Erschütterungen und Luftbewegungen reagiert. Besonders ausgesprochen sind die Reaktionen auf Erschütterung. Sie treten besonders dann klar zutage, wenn das Tier auf einem Untergrund liegt, welcher seinen Sinneshaaren zahlreiche Berührungspunkte darbietet, wenn er also z. B. auf einem rauhen Untergrund, auf den Borstenenden einer Bürste, auf Sammet oder auf einer Sandfläche liegt. Hat man das Tier mit seiner Unterseite auf eine Holz-, Metall- oder Glasplatte gebracht, so genügt ein leises Klopfen auf diese, um das Tier zu deutlichen Reaktionen zu veranlassen. In der Regel kommt es dabei nur zu den für leichte Reize typischen Reaktionen. Vor allem auf Sand jedoch kann auf diese Weise die ganze Serie der Reflexe ausgelöst werden. Ich habe oft beobachtet, daß ein auf der Straße vorüberfahrendes Automobil oder sonstiges Fuhrwerk oder ein hastig durch das Zimmer gehender Mensch genügende Erschütterungen verursachte, um bei den Tieren Bewegungen oder sogar den Schleuderreflex herbeizuführen. Am auffälligsten zeigte sich diese Erschütterungsreizbarkeit bei jenen Tieren, welche ich zur Prüfung des Umdrehreflexes an Fäden aufgehängt hatte. Ganz geringe Erschütterungen pflanzten sich durch den Faden in der Weise auf sie fort, daß sie mit sehr energischen Bein- und Kopfbewegungen und ausgesprochenen Schnappreflexen reagierten.
Auch Luftbewegungen lösen alle Reflexe unter Umständen aus. Ein frei daliegendes Tier, welches sich totstellt, kann durch Anblasen oder durch irgendeine im Zimmer durch Oeffnen von Fenstern hervorgerufene Luftbewegung zu all seinen Reflexen veranlaßt werden. So habe ich früher schon angegeben, daß Anblasen eines der einfachsten Mittel darstellt, um ein sich totstellendes Tier zu erwecken. Auch der Umdrehreflex eines auf dem Rücken liegenden Ameisenlöwen wird sehr leicht durch Anblasen herbeigeführt. Noch deutlicher treten alle diese Reaktionen ein, wenn man, statt sie anzublasen, die Tiere anhaucht. Wahrscheinlich kommt da zu dem Bewegungsreiz noch der Wärmereiz hinzu.
Ich nehme natürlich an, daß sowohl Erschütterung als auch Luftbewegung in der Weise auf die Tiere einwirken, daß eine Bewegung der Haare durch sie bewirkt wird. Die Haare sitzen ja, wie wir gesehen haben, gelenkig im Chitin, und es ist leicht, sich[S. 104] vorzustellen, daß durch Erschütterung und Luftbewegung sie in ähnlicher Weise aus ihrer Lage gebracht werden, wie durch Berührung mit einem festen Gegenstand.
In der Regel wird in zusammenfassenden Darstellungen angegeben, daß die Fühler der Insekten der besondere Sitz auch des Tastsinnes dieser Tiere seien. Meine Befunde beim Ameisenlöwen scheinen nun diese Annahme für diese Art nicht zu bestätigen. Bei all meinen Versuchen ist es mir sehr aufgefallen, daß die Fühler auf Berührungsreize nur sehr wenig reagierten. In der Regel bewegten sie weder sich selbst, wenn sie berührt wurden, noch zeigte sich an irgendeinem anderen Teil des Körpers eine reflektorische Bewegung. Wir werden aber später sehen, daß die Antennen sich auf Reize anderer Art sehr deutlich zu bewegen vermögen (S. 110). Gelegentlich bewegten sie sich immerhin auch auf reine Tastreize. Das ist vor allem in Wärme und Licht der Fall, wenn die Tiere sehr reizbar sind; dann machen sie nach mehrmaliger Berührung und Verbiegung der Antennen kurze ruckweise Bewegungen rückwärts, ohne allerdings mehr als 1–2 mm weit zu kommen, worauf sie wieder ganz ruhig werden.
Die Thigmotaxis, auch Stereotaxis genannt, die als Stereotropismus bei Pflanzen vielfach studiert wurde, hat auch unter den Tieren eine weite Verbreitung und große Bedeutung. Wir finden sie vor allem in charakteristischer Weise bei festgewachsenen Tieren ausgebildet, deren Wurzelgeflechte, Verzweigungen usw. durch ihre Wirkung der Unterlage angeschmiegt werden. Viel weniger beachtet wurde die Thigmotaxis bisher bei den freibeweglichen Tieren. Zwar ist das Musterbeispiel für Thigmotaxis bei den Tieren ein freibewegliches Infusor, Paramaecium. Es ist dies auch das einzige Tier, bei welchem bisher die Thigmotaxis einigermaßen analysiert und auf ihre Interferenz mit anderen Reizen untersucht wurde. Für freibewegliche, vielzellige Tiere findet sich zwar in der Literatur nicht selten das Vorkommen von Thigmotaxis angegeben, aber eine genauere Untersuchung hat es noch nicht gefunden. Es wurde oft die Bedeutung thigmotaktischer Reizbarkeit für Würmer, bodenbewohnende und sonstige lichtscheue Insekten erwähnt. Neuerdings wurde wiederholt auf die nahen Beziehungen zwischen Thigmotaxis und Rheotaxis bei Planarien, Fischen usw. hingewiesen (vgl. Steinmann).
[S. 105]
In meiner Arbeit über die Lebensgewohnheiten von dekapoden Krebsen habe ich auf die Wichtigkeit der Thigmotaxis speziell für die Arthropoden aufmerksam gemacht und den engen Zusammenhang dieser Reaktion mit der Tangorezeption, dem Tastsinn, betont. Ich zeigte dort, daß die Garneelen durch Thigmotaxis veranlaßt werden, sich in den Ecken und Winkeln des Aquariums aufzuhalten. Wesentlich für das Zustandekommen des thigmotaktischen Reizes war, nach meinen damaligen Beobachtungen, die Berührung einer größeren Anzahl von Tasthaaren der Körperoberfläche mit irgendwelchen Gegenständen der Umgebung des Tieres. Eine solche Berührung veranlaßt das Tier zu einer Ruhestellung und manchmal zu eigenartigen Körperhaltungen. Mein Schüler v. Kaulbersz hat bei Asellus aquaticus ebenfalls einige Beobachtungen gemacht, welche für das Vorkommen von Thigmotaxis bei diesem Tier sprechen.
Unter Thigmotaxis verstehen wir eine automatische Reizreaktion, bei welcher ein Tier einen mehr oder minder großen Teil seiner Körperoberfläche mit einem festen Gegenstand in Berührung bringt und dadurch gezwungen wird, eine bestimmte Körperhaltung einzunehmen. Häufig wird auch durch Thigmotaxis insofern ein Einfluß auf die Bewegungen des Tieres ausgeübt, als es durch Reiz an einer bestimmten Stelle verankert wird. In der Regel wird bei thigmotaktischen Reaktionen durch den Berührungsreiz die Muskulatur des betreffenden Tieres auf der berührten Körperseite kontrahiert und dadurch eine Wendung des Tierkörpers gegen den berührten Gegenstand hin herbeigeführt.
Bei allen meinen Versuchen erwiesen sich die Ameisenlöwen als ganz besonders geeignete Objekte zur Untersuchung der Thigmotaxis. Viele der Bewegungen des Tieres sind durch Thigmotaxis in hohem Grade beeinflußt. Bei all den früher geschilderten Experimenten traten infolge der Thigmotaxis sehr häufig erhebliche Störungen auf. Die Wirkung der verschiedenen Reize, welche ich auf das Tier einwirken ließ, wurde sehr regelmäßig durch das Eingreifen thigmotaktischer Reize abgeändert; bei allen Experimenten mußte ich danach trachten, die Einwirkung thigmotaktischer Reize zu verhindern. Das war um so notwendiger, als nach dem Ergebnis der unten angeführten Versuche die Thigmotaxis sich als wirksamer als alle übrigen Reizqualitäten erwies.
Läßt man ein Tier in absoluter Dunkelheit umherkriechen, so wird es so lange Versuchsbewegungen ausführen, d. h. in Bogen und Kreisen sich bewegen, bis es an einen festen Gegenstand stößt. In[S. 106] meinen Versuchsgefäßen war derselbe meist die aus Glas, Holz, Pappdeckel oder Papier bestehende Wand. Bei den Versuchen in absoluter Dunkelheit reichte das berußte Registrierpapier bis an diese Wand heran. War ein Ameisenlöwe bei seinen Versuchsbewegungen mit einem Teil seines Körpers wider die Wand angestoßen, so hatte das eine Annäherung seines Körpers an dieselbe zur Folge, bei welcher stets eine möglichst ausgedehnte Partie der Oberfläche des Körpers der Wand angeschmiegt wurde. Naturgemäß stieß das Tier in der Regel beim Rückwärtskriechen zuerst mit der Hinterleibsspitze an die feste Wand. Die Folge der Berührung war, daß diejenige Körperseite, welche beim Anstoßen einen kleineren Winkel zur Wand bildete, an die letztere angenähert wurde; denn die Folge der thigmotaktischen Reizung ist die Kontraktion der Muskulatur des Abdomens auf der gereizten Seite. Wenn das Tier seine Bewegung fortsetzte, folgte es von nun an, mit der Hinterleibsspitze voran, immer der Wandkontur. In den Versuchsbehältern war es infolgedessen immer genötigt, in den Ecken umzubiegen, und so kam es vor, daß manche Individuen ein- oder mehreremal am ganzen Rand des Gefäßes entlang liefen (Taf. IV, Fig. 4; Taf. V, Fig. 5). Fast stets führte aber das Tier, in einer Ecke angelangt, Versuchsbewegungen aus, bei denen dann die Hinterleibsspitze bald an die eine, bald an die andere der in der Ecke zusammenlaufenden Kanten geriet. So konnte durch den thigmotaktischen Reiz manches Tier veranlaßt werden, nach Erreichung einer Ecke und nach Ausführung einiger Versuchsbewegungen auf demselben Wege umzukehren, auf dem es zur Ecke gelangt war.
Bei den Versuchen im Dunkeln konnten auch andere Hindernisse durch thigmotaktische Reizung den vorher unorientierten Bewegungen der Tiere eine bestimmte Richtung geben. Um den Tieren genügend Spielraum für ihre ausgiebigen Versuchsbewegungen zu geben, war ich oft genötigt, verschiedene Streifen des berußten Registrierpapiers nebeneinander auf dem Pappdeckelboden des Versuchsgefäßes zu befestigen. Es gelang niemals, die Stücke so glatt nebeneinander auszuspannen, daß nicht eine Kante bemerkbar gewesen wäre. Dazu kam, daß die Papiere mit Hilfe von Reißnägeln auf dem Untergrund ausgespannt werden mußten. Das ergab für den feinen Tastsinn des Ameisenlöwen eine genügende Anzahl von Hindernissen. Und so ließ sich denn stets eine Einwirkung dieser Hindernisse auf die Bewegungsrichtung der Tiere feststellen. Auf zahlreichen der von mir fixierten Registrierungen finden sich durch[S. 107] Thigmotaxis beeinflußte Bewegungen an diesen Kanten entlang und oft im Kreis um die Köpfe der Reißnägel herum (Fig. 5, Taf. V).
Ganz entsprechende Ergebnisse stellten sich ein, wenn die Tiere, statt bei gleichmäßiger Temperatur und im Dunkeln, unter der Einwirkung bestimmter Temperatur- oder Lichtreize untersucht wurden. Ueber beide Reizarten überwog stets die Thigmotaxis. War z. B. ein Tier bei seinen Wanderbewegungen im optimalen Wärmefeld an einen festen Gegenstand gestoßen, so konnte es, wenn dieser eine längere Ausdehnung besaß, durch die Thigmotaxis gezwungen werden, das Wärmeoptimum zu verlassen. Es kroch dann oft in langer gerader Bahn bis in eine Region, in welcher unter Umständen eine sehr niedrige Temperatur herrschte.
Noch interessanter waren die Ergebnisse der Versuche über die Interferenz von Phototaxis und Thigmotaxis. Wir haben früher gesehen, daß bei einseitig einfallendem Licht jeder normale Ameisenlöwe sich nach einigen Versuchsbewegungen in die Richtung der Lichtstrahlen einstellt und dann geradeaus der Lichtquelle zuwandert. Indem man Ameisenlöwen an rauhen Kanten, z. B. an einem ungehobelten Brett oder an einem Sammetband entlangkriechen läßt, kann man sie zwingen, direkt von der Lichtquelle wegzukriechen. Ja, man kann ihnen durch entsprechende Versuchsanordnungen jede beliebige Bewegungsrichtung aufzwingen, ohne daß die Richtung der Lichtstrahlen ihren sonst so bedeutenden Einfluß zur Geltung bringt.
Wir haben früher schon gesehen, daß Berührung der Körperhaare Bewegungen des Tieres auslöst. Wir sahen ferner, daß multiple Berührungen eine erhöhte Wirkung haben. Wenn wir uns schließlich daran erinnern, welch zarte Tastborsten die Ameisenlöwen zu erregen vermochten, und daß sie für Erschütterungen und Luftbewegungen empfindlich sind, so wird es uns nicht verwundern, daß unter Umständen ihr eigenes Körpergewicht und leise von ihnen ausgeführte Bewegungen genügen, um entsprechende Reizwirkungen zu verursachen. Legt man z. B. einen Ameisenlöwen auf die Borstenspitzen einer Kleiderbürste, so genügt oft der durch das eigene Körpergewicht veranlaßte Reiz auf die Bauchseite, um das Tier zur Ausführung des Einbohrreflexes zu veranlassen. Auch die Rückenseite ist in ähnlicher Weise reizbar; Tiere in Rückenlage werden oft bei Auflegen auf die Borsten einer Bürste zu sofortigem Umdrehreflex veranlaßt. Die Natur dieser Reizung wird uns noch klarer,[S. 108] wenn wir sehen, daß das Anblasen der Tiere in dieser Situation prompteste Reizerfolge herbeiführt.
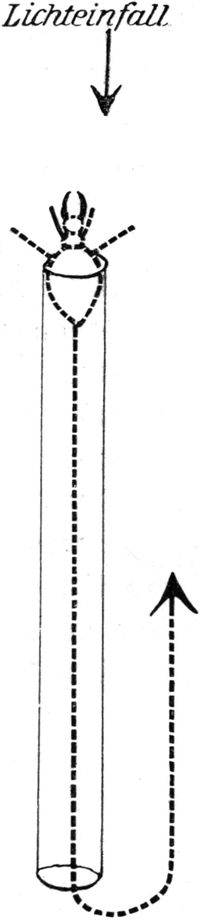
Wir verstehen daher, daß ein Tier, welches von verschiedenen Seiten thigmotaktisch gereizt wird, leicht dadurch zu einer anhaltenden Fortbewegung veranlaßt wird. So kriechen z. B. Tiere, welche man in Glasröhren zwängt, deren Lumen so weit ist, daß die Tiere mit den beiden Schultern und bei Bewegungen abwechselnd mit der Bauch- und Rückenseite am Glas anstoßen, oft in langer, anhaltender und ziemlich rascher Bewegung durch die Glasröhren hindurch. Gerade dies abwechselnde Anstoßen ist bei dem Vorgang von Bedeutung, da es verhindert, daß das Tier durch allseitige Reizung zur Ruhe kommt. Unbekümmert um alle sonstigen Reize erfolgen solche Wanderungen in der Richtung, in welcher man die Glasröhre angeordnet hat, wobei das Tier immer mit dem Hinterende voran kriecht. Man kann solche Glasröhren z. B. ganz beliebig zum Einfall der Lichtstrahlen orientieren. Die Thigmotaxis siegt stets über die Phototaxis und zwingt das Tier, die Röhre entlang zu wandern; es macht nicht einmal Versuche, in der Röhre sich umzudrehen, wenn dieselbe auch weit genug ist, um eine Umdrehung zu erlauben. Bei meinen Versuchen wandte ich Glasröhren von 50, 70 und 100 cm Länge an. Waren dieselben in die Achse der Lichtstrahlen gelegt, und steckte man die Tiere mit dem Hinterteil voran in die der Lichtquelle zugewandte Oeffnung, so krochen sie in ziemlich raschem Tempo durch die ganze Glasröhre bis zu deren dem Licht abgewandter Oeffnung. Sie krochen also entgegengesetzt der Richtung der Lichtstrahlen. Kaum waren sie aber aus dem Hinterende der Glasröhre hervorgekrochen, als sie nach einer ganz kurzen Versuchskurve sich umdrehten und parallel der Röhre der Lichtquelle wieder zukrochen (Abb. 39). Der Bogen der Versuchsbewegung verhinderte es meistens, daß das Tier seine Rückwanderung in Kontakt mit der Glasröhre ausführte. Aber auch das kam in einzelnen Fällen[S. 109] vor, so daß dann die Tiere unter der doppelten Einwirkung von Photo- und Thigmotaxis rasch und sicher der Lichtquelle zuwanderten.
Ich habe sehr verschiedene Lichtintensitäten bei den Versuchen angewandt; stets sah ich aber die Thigmotaxis auch über die stärksten Lichtreize überwiegen.
Der Berührungsreiz einer Körperseite des Tieres führt also durch Kontraktion der Muskulatur dieser Körperseite stets eine Biegung des Körpers nach dieser Seite herbei. So sehen wir den Hinterleib bei Berührung von unten durch den Sand sich nach unten biegen und so in den Sand eindringen. Jede Berührung der Körperseiten veranlaßt deren Anschmiegung an die berührende Fläche. Die thigmotaktische Reaktion ist also eine reflektorische Bewegung des Tierkörpers. Arbeiten diese reflektorischen Bewegungen mit den Wirkungen anderer Reflexe zusammen, so kommen jene eigenartigen Bewegungskombinationen zustande, welche wir in den letzten Abschnitten kennen lernten.
Wirkt aber thigmotaktische Reizung von allen Seiten auf den Ameisenlöwen ein, so erfolgt eine leichte Kontraktion seiner gesamten Längsmuskulatur, und das Tier bleibt in Ruhestellung, wenn nicht ein neuer Reiz es zur Bewegung veranlaßt. So bleibt es ruhig und unbeweglich liegen, wenn es, im Sand eingewühlt oder am Grunde seines Trichters sitzend, allseitig von den Sandkörnern umhüllt ist.
Im Anfang meiner Versuche fand ich keine Erscheinungen, welche auf eine wesentliche Mitwirkung der chemischen Sinne bei den Lebenserscheinungen der Ameisenlöwen hinwiesen. So wurden z. B. am 11. Okt. 1911 einem Ameisenlöwen beide Antennen nahe der Basis abgeschnitten. Das so operierte Tier drehte sich in normaler Weise um und wühlte sich in den Sand ein. Es baute nicht nur einen vollkommenen Trichter, sondern fing am 12. Okt. in der üblichen Weise eine Ameise durch Schleudern mit Sand und saugte sie aus. Es lebte noch wochenlang, baute aber von Anfang November ab keine ordentlichen Trichter mehr. Statt dessen wanderte es, wie die Furchen auf der Oberfläche des Sandes verrieten, auf diesem vielfach hin und her. Es nahm späterhin keine Ameisen mehr an, selbst wenn man sie ihm längere Zeit zwischen die Kiefer hielt.
[S. 110]
Es ist aber sehr zweifelhaft, ob dies abnorme Benehmen mit der Amputation der Antennen zusammenhängt. In sämtlichen Jahren, in denen ich die Ameisenlöwen beobachtete, benahmen sich nämlich um dieselbe Jahreszeit viele unverletzte Individuen genau wie jenes operierte Tier (vgl. S. 125). Solche Tiere, welche, aus dem Sand gegraben, nicht in Bereitschaftsstellung sich befinden, schnappen nicht nach den Ameisen. Die Ameisen können ihnen auf dem Kopf, den Mandibeln, dem Hals herumklettern, ohne daß sie schnappen. Auch zerdrückte Ameisen, von denen eventuell wirksame Geruchsstoffe in stärkerem Maße ausgehen könnten, als von den lebenden Tieren, führen zu keiner Reaktion. Reizt man jedoch zur gleichen Zeit die Ameisenlöwen durch Berührung mit Borsten, so führen sie den Schnappreflex aus und fassen dabei auch gelegentlich Ameisen, die man ihnen vorhält. Die letzteren geraten ihnen dann meist nur zwischen die soliden Zähne der Mandibel. Sie verursachen keine Erhöhung der Erregung des Tieres. Der Ameisenlöwe macht keine Versuche, sie fester zu fassen, und sie werden auch nicht ausgesaugt. Auch Berührung mit toten Ameisen führt nur zu einem Reflex, wie er sonst durch irgendeinen Tastreiz ausgelöst werden kann, also z. B. zum Schnappreflex, oder wenn das Tier sich außerhalb des Sandes befindet, zur Rückwärtsbewegung. Es scheint also in all diesen Fällen der Tastreiz weit wirksamer zu sein als ein chemischer Reiz.
Dennoch gaben mir andere Versuche die Gewißheit, daß auch beim Ameisenlöwen die Antennen der Sitz der Chemorezeption und zwar eines dem Geruchsinn ähnlichen Sinnes sind. Ich habe oben schon angeführt, daß die Antennen in sehr geringem Grade durch Berührung reizbar sind. Bei meinen Versuchen, welche zur Feststellung dieser Tatsache führten, beobachtete ich einmal, daß ein Ameisenlöwe in ganz eigenartiger Weise auf die Annäherung einer Nadel an seine Antennen reagierte. Wenn die Nadel bis auf 3 oder 5 mm den Antennen genähert wurde, zogen sie sich langsam vor ihr zurück; sie wurden dabei im Gelenk des Basalgliedes nach hinten gebogen. Es machte zunächst vollkommen den Eindruck, als sei die Annäherung der Nadel von dem Tier gesehen worden. Das konnte aber nicht der Fall sein, wie Versuche mit einer anderen Nadel bewiesen. Die Antennen reagierten nämlich nicht nur nicht auf die Annäherung, sondern nicht einmal auf die Berührung mit dieser zweiten Nadel. Diese zweite Nadel war nämlich vor dem Experiment von mir aufs sorgfältigste durch Ausglühen, Abwischen mit Alkohol[S. 111] und Eintauchen in Aether gereinigt worden. Jene erste Nadel, welche ich nur aus Versehen benützt hatte, war aber an ihrer Spitze mit einem kleinen Tropfen vertrockneten Kanadabalsams verschmutzt. Verschiedene Tiere zeigten bei Annäherung dieser Nadel wiederum deutliche Reaktionen mit den Antennen.
Ich tauchte darauf die Spitze einer reinen Nadel in Nelkenöl und näherte sie bis auf 3 mm den Antennen eines Tieres. Das hatte sehr zuckende Bewegungen der Antennen, ja selbst ein Zucken des Kopfes zur Folge.
Ich stellte daraufhin weitere Versuche an, indem ich Nadeln mit anderen stark riechenden Stoffen benetzte und an die Antennen annäherte. Ich bekam aber keine deutliche Reaktion bei Anwendung von Ammoniak, Aether und Chloroform. Ebensowenig reagierten die Antennen bei Annäherung einer zerquetschten Ameise.
Sehr bemerkenswert war aber die Beobachtung, daß bei Berührung der Antennen mit einem mit jenen Geruchsstoffen benetzten Pinsel die Antennen stets charakteristische schwingende Bewegungen ausführten. Dies trat auch bei Tieren ein, deren Antennen auf Berührung mit einem trockenen Pinsel gar nicht reagierten.
Die Versuche mit dem Kanadabalsam und Nelkenöl zeigen also deutlich, daß von diesen Stoffen ein Reiz auf die Antennen des Ameisenlöwen ausgeübt wird, welcher sich durch die Luft fortpflanzt. Dieser Reiz kann nur in einer chemischen Einwirkung bestehen, und da er durch die Luft übertragen wird, so entspricht er vollkommen denjenigen Reizen, welche bei anderen Tieren und den Menschen für den Geruchssinn adäquat sind. Wir dürfen also mit gutem Recht das Vorkommen eines Geruchssinnes bei dem Ameisenlöwen annehmen.
In den vorstehenden Kapiteln haben wir gesehen, daß alle komplizierten Handlungen im Leben des Ameisenlöwen auf eine Anzahl von Bewegungen zurückzuführen sind, welche durch bestimmte Reize ausgelöst werden. Durch einen peripheren Reiz wird die Kontraktion einer Anzahl von Muskeln veranlaßt. Es handelt sich also offenbar um Reflexe. Wir haben die Handlungen des Tieres also auf eine Reihe von Reflexen zurückgeführt.
[S. 112]
Die wichtigsten dieser Reflexe sind folgende:
1) der Einbohrreflex,
2) der Schleuderreflex,
3) der Schnappreflex.
Wir wollen diese in den früheren Abschnitten in ihren Wirkungen studierten Reflexe noch einmal kurz zusammenfassend schildern.
1. Der Einbohrreflex. Er besteht in zuckenden Bewegungen der Hinterleibsspitze, welche sich in der Regel mehrmals hintereinander wiederholen. Diese sind durch eine ruckweise abwechselnde Kontraktion der ventralen und dorsalen Längsmuskulatur des Hinterleibes bedingt. Die Bewegung wird durch taktile Reize der Unterseite des Abdomens ausgelöst.
2. Der Schleuderreflex. Er besteht ebenfalls in ruckweisen Bewegungen, die sich mehrmals hintereinander wiederholen können. Die Art der Bewegungen ist bedingt durch den Bau der Gelenke zwischen Kopf und Thorakalsegmenten. Die Bremseinrichtungen auf diesen Gelenken halten bei gespannten Muskeln die vorn gelegenen Körperteile zurück, so daß sie plötzlich in die Höhe schnellen. Hier ist es die Kontraktion der dorsalen Längsmuskulatur, welche das Emporschnellen von Kopf und erstem Thorakalsegment, manchmal auch die dorsale Umbiegung weiterer Thorakalsegmente verursacht. Der Schleuderreflex erfolgt entweder in der Sagittalachse des Tieres nach oben und hinten oder in beliebigem Winkel seitlich von ihr. Die Richtung der Bewegung ist durch die gereizte Stelle bedingt; sie findet in der Richtung auf diese Stelle hin statt. Das ist durch eine Zerteilung der bewegenden Muskulatur in zahlreiche kleine Bündel ermöglicht.
3. Der Schnappreflex. Dieser betrifft hauptsächlich die Bewegung der Mundgliedmaßen. Er kann allein für sich erfolgen oder in Verbindung mit dem Schleuderreflex. Fast stets ist letzteres unter natürlichen Verhältnissen der Fall. Die Bewegung besteht in einer Aufrichtung des Kopfes und vor allem im Zuschnappen der Mundgliedmaßen. Letzteres wird durch ein im Kopf befindliches kräftiges Muskelsystem bewirkt. Auch hier wird das plötzliche Zuschnappen durch Bremsvorrichtungen an den Gelenken der Mundgliedmaßen begünstigt, welche bei gespannter Muskulatur jene in einer bestimmten Stellung zurückhalten können. Ausgelöst wird der Schnappreflex durch Berührung der Mundgliedmaßen oder der vorderen Regionen des Kopfes. Fast stets erscheint er aber mit dem[S. 113] Schleuderreflex in einen Vorgang verkettet. Dann erfolgt er auf die gleichen Reize hin, welche jenen auslösen.
Außer diesen wichtigsten Reflexen lassen sich natürlich am Ameisenlöwen eine größere Anzahl weiterer reflektorischer Bewegungen feststellen. Ich habe sie meist nicht so genau studiert, wie die genannten Grundreflexe, welche die Lebenserscheinungen des Ameisenlöwen beherrschen. So ist der Uebergang in die Bereitschaftsstellung (S. 36) eine Reflexbewegung, das gleiche gilt für die Putzbewegungen und überhaupt für die Beinbewegungen beim Wandern, Wühlen und Graben. Auch das Aussaugen der gefangenen Beute und das Loslassen nach dem Fressen sind reflektorisch bedingt. Die Handlungen, welche für das Leben des Ameisenlöwen besonders charakteristisch sind, lassen sich aber alle auf die erwähnten drei Grundreflexe zurückführen. Wir sahen ja früher, daß Tiere, welche der Beine und der Antennen beraubt sind, ohne ihre Hilfe dennoch Trichter bauen und Ameisen fangen können.
Die drei Grundreflexe sind also für die lebenswichtigsten Handlungen des Ameisenlöwen ausschlaggebend. Es wäre nun hier die Frage noch zu erörtern, ob bei diesem Tier auch noch besondere Abwehrreflexe vorkommen, wie sie bei anderen Tieren meist so auffällig sind. Der wichtigste Schutz des Ameisenlöwen ist das Totstellen. Stark gereizte Tiere verwenden aber auch den Schnappreflex in einer Weise, welche durchaus an die Abwehrreflexe anderer Tiere erinnert. Schon daß das Schnappen immer nach einer gereizten Stelle der Oberfläche des Körpers stattfindet, gibt dieser Handlung den Charakter einer Abwehrhandlung. So sehen wir also den zu so vielen Zwecken dienenden Schleuder- und Schnappreflex noch eine weitere Verrichtung im Leben des Ameisenlöwen leisten.
Auch die Bewegungen, welche zum Putzen der Oberfläche des Tierkörpers dienen, werden als Abwehrbewegungen benützt. Ist ein Tier an der Oberfläche, z. B. in der Nähe der Augen, der Antennen, der Mundgliedmaßen oder der Stigmenöffnungen, mit sehr feinem Staub, z. B. Ruß, bedeckt worden, so führt es mit den Extremitäten, besonders mit dem dritten Beinpaar putzende, schabende Bewegungen aus. Dieselben Bewegungen treten ein, wenn das Tier in den entsprechenden Regionen einem mechanischen Reiz anderer Art ausgesetzt wird. Unter Umständen werden dann die Bewegungen so heftig, daß sie den Eindruck von Abwehrbewegungen machen. Man sieht also aus alle dem, daß der Ameisenlöwe, bei dem das Totstellen solche entbehrlich macht, eigentlicher Abwehrreflexe entbehrt.
[S. 114]
Sämtliche geschilderte Reflexe sind vom Gehirnganglion des Ameisenlöwen unabhängig. Amputiert man einem Exemplar den Kopf, so findet trotzdem der Einbohr- und der Schleuderreflex unverändert statt. Der Schnappreflex kann natürlich nicht mehr ausgeführt werden. Reizt man mit einer Nadel die Unterseite des Abdomens, so findet noch regelrecht die Einziehung der Hinterleibsspitze und die Kontraktion des ganzen Abdomens statt. Doch hört sehr bald die rhythmische Wiederholung dieser Bewegung auf. Berührung der Region zwischen den Beinen führt Abwehr- resp. Kriechbewegungen der dritten Beine herbei, Berührung der Region zwischen den zwei vorderen Beinpaaren ein Vorneigen des Halses und Zusammenbiegen der Beine, verbunden mit Zusammenkrümmen des ganzen Körpers nach der Bauchseite. Am ausgesprochensten erhalten sind die Reaktionen bei Reizung der Rückenseite. Die erhaltenen Thorakalsegmente führen noch vollkommen regelrecht den Schleuderreflex aus. Ja, was besonders bemerkenswert ist, die Bewegung erfolgt ganz entsprechend dem lokalisierten Reiz, also bei Reizung der rechten Körperseite nach dieser hin, bei Reizung der linken Seite nach links. Die Richtung der Bewegung geht wie beim unverletzten Tier genau auf den gereizten Ort los.
In einzelnen Fällen erhielt ich die Reflexe 24 Stunden nach Abschneiden des Kopfes noch ganz deutlich. Das erste und dritte Beinpaar wurden in der typischen Totstellhaltung gehalten, das zweite dagegen schief nach oben, steil emporgerichtet. Bei Berührung der Unterseite des Hinterleibes erfolgte noch Schabreflex des dritten Beinpaares und Zappeln des zweiten Beinpaares, ferner deutliche Einkrümmung der Hinterleibsspitze ventralwärts. Bei Berührung der Oberseite der erhaltenen Thorakalsegmente stellte sich noch ein schwacher Schleuderreflex und leichtes Zurückkriechen ein.
[S. 115]

Beginnt man mit der Abtrennung der Abdominalsegmente von hinten, so findet der Schleuderreflex noch statt, wenn nur mehr wenige der vorderen Segmente erhalten sind. Ja sogar, wenn nur mehr das dritte und zweite Thorakalsegment mit dem Kopf zusammenhängen, wird er noch ausgeführt. Beim weiteren Vorrücken der Durchschneidung bis auf das erste Thorakalsegment findet der Schleuderreflex nur mehr andeutungsweise statt. Schließlich der abgeschnittene Kopf allein kann natürlich, schon weil die zugehörigen Muskeln fehlen, den Schleuderreflex nicht mehr ausführen. Wie ich aber in Bestätigung der Angaben von Comes feststellen konnte, findet noch am abgeschnittenen Kopf der Schnappreflex statt. Die Mandibel, welche während der Ausführung des Schnittes nach der Schere geschnappt hatten, bleiben fest zugekniffen. Um einen erfaßten[S. 116] Gegenstand, also die Pinzette, eine Nadel oder Borste, bleiben sie fest geschlossen. Nur mit Gewalt gelingt es, sie zu öffnen, worauf sie sofort wieder zuschnappen. Es liegt also ein tonischer Krampf der Schließmuskeln vor, der mindestens eine Stunde nach dem Abschneiden des Kopfes anhält. Die Kraft dieser Muskeln ist sehr erheblich, und es kostet eine gewisse Anstrengung, die Mandibel auseinanderzuzwängen, ohne sie zu zerbrechen. Läßt man sie zuschnappen, so wird dadurch der Kopf oft 10–20 cm weit fortgeschleudert. Beobachtungen, welche ich an dem länger in einer feuchten Kammer aufgehobenen Kopf machen konnte, veranlassen mich, in den letztbeschriebenen Erscheinungen kein reines Nerv-Muskelphänomen zu erblicken. Die Mandibel bleiben nämlich am abgeschnittenen Kopf 24, 48 Stunden und länger fest geschlossen und schnappen beim gewaltsamen Oeffnen immer wieder automatisch zu. Ich bin geneigt, diese Erscheinung wenigstens zum Teil auf die mechanischen und morphologischen Grundlagen der Schnappbewegung zurückzuführen. Ich erinnere hier noch einmal an die eigenartige Beschuppung der Gelenke von Mandibel und Maxille, welche sicherlich bei geeigneter Gegenreibung eine starke Bremsung der Mundgliedmaßen herbeiführen muß. Im einzelnen ist mir der Vorgang noch nicht ganz klar geworden, da es mir noch nicht gelang, die Muskulatur des Kopfes und ihre Innervierung vollkommen zu erforschen.
Reizbiologisch und reizphysiologisch ist der Ameisenlöwe insofern ein sehr merkwürdiges Tier, als er sich normalerweise in einem Zustand der Unterempfindlichkeit befindet. Ganz selten findet man ihn bereit, sofort und intensiv auf Reize zu reagieren. Das ist in der Regel nur dann der Fall, wenn er in freier Natur im Sand sitzt, so daß nur der Kopf herausschaut, und wenn eine hohe Temperatur, etwa 25–35° C, ihn umgibt.
Dann sitzt er in Bereitschaftsstellung in seinem Trichter, und die von den Ameisen ins Rollen gebrachten Sandkörner veranlassen ihn zu kräftigen Schleuder- und Schnappreflexen. Aber selbst dann kann es vorkommen, daß mehrere Reize, d. h. wiederholtes Herabrollen von Sand notwendig ist, ehe die erste Reaktion erfolgt.
[S. 117]
Nimmt man das Tier aus dem Sand, so erfolgt stets zunächst das Totstellen. Bei Wärme kann man experimentell von dem Tier dann stets sofort Reizreaktionen erhalten; es ist unter solchen Umständen sehr leicht aus dem „Schlafzustand“ zu erwecken.
Die Regel ist aber bei allen Reizen, daß die Tiere zuerst mehr träge reagieren, und erst nach einer Reihe von Reizungen werden, offenbar infolge der Summation der Reizwirkung, die Reizbeantwortungen immer präziser und kräftiger. Wir haben dafür früher zahlreiche Beispiele kennen gelernt.
Vor allem ist es der beim Ameisenlöwen im Sinnesleben ja so sehr überwiegende Tastsinn, durch dessen wiederholte Reizung man das Tier in einen Erregungszustand versetzen kann, in welchem alle Sinnesorgane auf leichtere Reizungen antworten als vorher.
Alle von mir studierten Reflexe des Ameisenlöwen lassen sich durch Tastreize herbeiführen; eine Ausnahme machen vielleicht die noch nicht genauer studierten Reflexe, welche beim Aussaugen der Nahrung, beim Spinnen des Kokons usw. eine Rolle spielen. Doch habe ich einige Anhaltspunkte, daß auch bei ihnen der Tastreiz eine Hauptrolle spielt.
Bemerkenswert ist aber die Tatsache, daß im allgemeinen bei Tastreiz genau dieselben Reflexbewegungen auftreten, welche auch durch andere Reize, so besonders durch Wärmereiz, auch durch Lichtreiz, vielleicht durch chemische Reize ausgelöst werden. Dabei ist festzustellen, daß nächst dem Berührungsreiz am stärksten der thermische, dann der photische und zuletzt der chemische in bezug auf die Stärke der Reizwirkung anzuordnen sind.
Meine Beschäftigung mit dem Ameisenlöwen lehrte mich auch sein Leben und Verhalten im Freien kennen, so daß ich die Kenntnisse über seine Lebensgeschichte in einigen Punkten ergänzen kann. Ich stelle in diesem Abschnitt kurz alles zusammen, was man über die Schicksale dieses Tieres im Laufe eines Jahreszyklus bisher weiß, wobei ich meine neuen Befunde gleichzeitig mit den früheren Kenntnissen darstelle.
Aus dem Ei kriecht eine kleine Larve aus, welche in den Hauptzügen des Baues vollkommen der alten Larve gleicht. Das Auskriechen aus dem Ei habe ich selbst nicht beobachtet, aber ich fand[S. 118] vielfach sehr kleine, wenige Millimeter lange Larven im Freien. Diese kleinen Larven bauen schon kleine Trichter, die selbst nur wenige Millimeter Durchmesser erreichen. Ich fand sie meist nur im allerfeinsten, staubartigen Sand.
Schon ganz kleine Larven sind imstande, Ameisen, vor allem Lasius- und Myrmicaarten zu bewältigen und auszusaugen. Ich vermute aber, daß sie auch allerhand andere Tiere, so z. B. Blattläuse und junge Spinnen, in ihren Sandlöchern fangen. Ich schließe dies teils aus dem Umstand, daß sich in der Umgebung ihrer Löcher Häute von solchen Tieren finden, teils aus einigen Beobachtungen; allerdings handelte es sich bei letzteren um Tiere in der Gefangenschaft, welche gelegentlich solche kleine Arthropoden fingen, die ihnen gegeben wurden. Mit der Größe der Tiere wächst die Größe ihrer Trichter, und sie werden immer mehr befähigt, auch große Ameisen zu bewältigen. Selten sieht man, daß ein Ameisenlöwe, wenn er noch so klein ist, vergeblich um eine Ameise kämpft. Die Größe des Trichters hängt aber nicht allein von der Größe seines Erbauers ab. Wir sahen ja oben, daß bei steigender Wärme, nach starker Reizung usw. das gleiche Tier einen viel größeren Trichter zu bauen vermag als vorher.
Schon bei recht jungen Tieren kann man im Larvenzustand Geschlechtsunterschiede erkennen. Es sind nämlich die Weibchen größer als die Männchen. Nicht sicher ist mir ein Unterschied, den ich bei einer Anzahl von Tieren beobachtete; ich zählte nämlich bei manchen Exemplaren statt der üblichen Zahl von 15 Antennengliedern deren 17–18. Ich hielt letztere für Männchen, doch konnte ich das nicht mit aller Sicherheit feststellen, da es sich um ausgekochte Chitinpräparate handelte. Möglicherweise sind aber die Unregelmäßigkeiten in den Zahlen der Antennenglieder auf die häufig vorkommenden Regenerationen und vielleicht auch Hyperregenerationen nach Verletzungen zurückzuführen.
Ich beobachtete sehr kleine Larven sowohl im Frühsommer als auch im Spätherbst. Im Frühling gab es auch kleine Tiere, aber die kleinsten Formen fehlten. Ich ziehe daraus den Schluß, daß die Eier von Myrmeleo nicht überwintern. Es überwintern vielmehr erstens Larven und zweitens Puppen. Die überwinternden Larven gehören verschiedenen Altersstufen an. Zum Teil sind es Tiere, welche früh im Sommer aus den Eiern gekrochen waren und schon eine Wachstumsperiode von größerer Ausdehnung hinter sich hatten, wenn der Winter kam. Man findet im Spätherbst und im ersten Frühling[S. 119] große stattliche Ameisenlöwen, welche sich früh im Sommer schon verpuppen und früh ausschlüpfen. Das Einspinnen in die Kokons findet im allgemeinen hier Mitte Mai bis Mitte Juni statt. Der Aufenthalt im Kokon dauert 3–4 Wochen; davon verbringt das Tier 2–3 Wochen als Larve und nur 1 Woche als Puppe. Die Imagines erscheinen Anfang oder Mitte Juni. Von ihnen stammen Larven ab, welche schon früh im Sommer aus den Eiern auskriechen und ähnlich, wie ihre Eltern, ein gut Stück gewachsen sein können, ehe die Winterszeit eine Unterbrechung herbeiführt.
Es hat mancherlei Kontroversen über die Entwicklungszeit der Ameisenlöwen schon gegeben, bedingt durch das eigenartige Vorkommen junger Larven zu verschiedenen Jahreszeiten. Während manche Beobachter den Ameisenlöwen für ein einjähriges Tier erklärten, waren andere der Meinung, er sei zwei- oder gar mehrjährig. Nach meiner Ansicht ist diese Verschiedenheit dadurch zu erklären, daß infolge der Schwierigkeiten der Nahrungsbeschaffung die Tiere verschieden rasch heranwachsen. Ist der Sommer kalt und regnerisch, so wird es im Herbst viele Hungertiere geben, welche noch unverpuppt in den Winter hineingehen, da sie noch nicht erwachsen sind. Diese sind im nächsten Jahr erst ziemlich spät geschlechtsreif, so daß ihre Nachkommen ebenfalls unverpuppt in den Winter gehen müssen. Diese Formen überkreuzen sich nun mit anderen, die früh zur Begattung kamen und daher Larven erzeugten, die sich schon vor dem Winter verpuppten. So können wir wohl sagen, daß es ein- und zweijährige Ameisenlöwen gibt, der Zyklus ist aber nicht absolut festgelegt, sondern von äußeren Bedingungen abhängig. Meine Erfahrungen stimmen also gut mit den Schlußfolgerungen Redtenbachers überein. Manche Verschiedenheiten in den Angaben der Autoren erklären sich wohl durch die verschiedene geographische Lage und die besonderen klimatischen Bedingungen ihres Beobachtungsortes. So mag die rasche Entwicklung der von mir beobachteten Individuen durch das warme Klima der Freiburger Gegend bedingt sein. In anderen Gegenden Deutschlands habe ich keine Aufzeichnungen gemacht. Auch können verschiedenen Autoren die verschiedenen Arten von Myrmeleo vorgelegen haben.
Was nun die Ernährung der Ameisenlöwen anlangt, so sind sicher die wichtigste Beute der erwachsenen Individuen Ameisen. Gelegentlich saugen auch sie eine Spinne oder ein Insekt aus, das in ihren Trichter gerät, so z. B. Fliegen, Wespen, selbst Käfer, oft Raupen. Nicht selten töten und saugen sie ihre eigenen Artgenossen[S. 120] aus. Das geschieht besonders in der Gefangenschaft, wenn man ihrer viele auf engem Raum beisammen hält. So kann es kommen, daß von einem großen Sandkasten voll Ameisenlöwen nur einige wenige zur Verpuppung gelangen, die sich auf Kosten der übrigen ernährt haben und herangewachsen sind.
Beim Ameisenfang kann man oft sehen, daß eine erbeutete Ameise sich noch heftig durch Bewegungen zu wehren sucht. Der Ameisenlöwe zieht sie dann vielfach unter den Sand. Schneller aber, als daß es durch Erstickung oder durch das Aussaugen bewirkt sein könnte, hören die Bewegungen des Opfers auf. Man kommt daher unwillkürlich auf die Idee, daß das rasche Sterben durch Giftentwicklung erzielt sein könnte. Tatsächlich befindet sich an der Basis der Maxille eine Drüse, welche Lozinski genauer untersucht hat und für eine Giftdrüse hält. Die Giftwirkung des Drüsensekrets ist nun nicht gesichert. Experimente darüber liegen nicht vor. Auch ist das Absterben der Opfer nicht immer so prompt, daß man mit Sicherheit auf eine Giftwirkung schließen müßte. Die Frage ist also noch unentschieden.
Auch sehr junge und kleine Tiere fangen schon Ameisen und saugen sie aus. Ich habe oft beobachtet, daß Exemplare von nur 2–3 mm Länge eine ebenso große oder größere Ameise überwältigten, wenn sie richtig in ihrem Sandtrichter saßen. Doch halte ich es für wahrscheinlich, daß sie in der ersten Jugend auch andere Insekten, vor allem kleine Spinnen und Blattläuse häufig fangen. Ich fand wenigstens deren Reste nicht selten in der Umgebung der Trichter.
Da auch die jüngsten Tiere, welche ich untersuchte, nur rückwärts zu laufen vermochten, halte ich es für sehr unwahrscheinlich, daß etwa die jungen Larven nach Art ihrer Verwandten auf Wanderung gehen und Blattläuse suchen. Das wäre mit dem Reflexautomatismus ihrer Bewegungen kaum zu vereinigen. Die auf Insektenraub ausziehenden Ameisenlöwenarten sind alle zum Vorwärtsgehen befähigt.
Auch scheint es mir nicht wahrscheinlich, daß normalerweise die jungen Ameisenlöwen im Herbst noch keine Trichter bauen und noch nicht fressen. In der Freiburger Gegend fand ich jedenfalls im Herbst sehr zahlreiche ganz kleine Larven in ganz kleinen Trichtern beim Fang beschäftigt.
Wir wollen nun kurz einen Blick auf die Art der Nahrungsaufnahme und Ernährung des Ameisenlöwen werfen, wenn ich auch im Verlauf dieser meiner Untersuchung kaum etwas zur Vermehrung unserer Kenntnisse in dieser Beziehung beitragen konnte.
[S. 121]
Ein Ameisenlöwe, der eine Ameise gefangen hat, hält sie, sobald sie bewegungslos geworden ist, ruhig zwischen seinen Mandibeln und saugt sie mit Hilfe des zwischen Maxille und Mandibel gebildeten Rohres aus. Dabei ist es nicht ganz sicher, ob das Aussaugen sich auf Blut und Gewebssäfte beschränkt, oder ob, wie bei anderen Insekten, eine Vorverdauung außerhalb des Körpers stattfindet. Bei den Larven von Dytiscus z. B. wird durch die Mandibelröhren in die Beute ein enzymhaltiger Saft eingespritzt, der dort eine Auflösung der Muskeln und übrigen Gewebe bewirkt, welche sodann in verflüssigtem Zustand eingesogen werden. Wir haben allen Grund anzunehmen, daß beim Ameisenlöwen der Vorgang ebenso abläuft. Alles spricht jedenfalls dafür, daß auch dies Tier nur flüssige Nahrung aufnimmt. Ich habe eine größere Anzahl von den Larven ausgesaugte Ameisen untersucht und konnte im Innern ihres Chitinpanzers keinerlei Organe mehr finden. Sowohl Muskeln fehlten im Thorax, ja selbst in den Extremitäten und dem Kopf, als auch alle Organe des Abdomens und der übrigen Körperteile. Es ist also durchaus wahrscheinlich, daß sie gelöst worden sind. Die Annahme, welche z. B. Dewitz machte, daß durch Hin- und Herreiben der Maxille und Mandibel aneinander eine Art von Kautätigkeit ausgeübt werde, ist jedenfalls überflüssig. Der Pharynx ist in ähnlicher Weise gebaut, wie bei anderen saugenden Insekten, der Oesophagus sehr eng. Bei jenen anderen Insekten mit Verdauungsanfang außerhalb des eigenen Körpers ist es strittig, ob der verdauende Saft aus besonderen Drüsen oder aus dem Mitteldarm kommt. Letzteres ist in jenen Fällen sicher das Wahrscheinlichere, und auch in unserem Fall dürfen wir dies wohl annehmen. Immerhin ist festzustellen, daß paarige, in den hinteren Pharynx einmündende Speicheldrüsen beim Ameisenlöwen vorhanden sind. Deren Sekret muß mindestens eine verdünnende Wirkung auf den Saft, der gesaugt wird, ausüben. Auch ist anzunehmen, daß ein derartiges Sekret zur Verdichtung der Saugröhre in ihrem Anschluß an den Körper des Opfers dienen mag, doch könnte dies auch die Funktion der Maxillendrüse sein.
Das, was der Ameisenlöwe aus der Ameise saugt, ist zum größten Teil zu seiner Ernährung ausnutzbar. Viele Rückstände fester Art bleiben jedenfalls im Darmkanal nicht übrig. Sie würden auch keinen normalen Ausweg aus diesem finden, denn der Ameisenlöwe hat einen geschlossenen Mitteldarm; im Enddarm findet sich kein Kot, denn er hat keine Verbindung mit Magen und Mitteldarm. Es liegen hier also ähnliche Verhältnisse vor, wie bei den[S. 122] Larven aculeater Hymenopteren. Schon Réaumur (1742) war dies bekannt, wie auch Rösel von Rosenhof (1755). Und Ramdohr (1811) hatte schon erkannt, daß der Enddarm den „Seidenstoff“ enthält, von dem wir noch hören werden, daß er das Produkt der Malpighischen Gefäße ist und mit Hilfe der Analröhre zum Gespinst des Kokons verarbeitet wird. Burmeister, Dufour, v. Siebold, Brauer, Gerstäcker und vor allem Meinert trugen allmählich dazu bei, die merkwürdigen Bau- und Funktionsverhältnisse des Ameisenlöwendarms klarzulegen. Letzterer und besonders Rengel (1908) haben das eigenartige Problem definitiv gelöst.
Auf den beim erwachsenen Ameisenlöwen etwa 8 mm langen, dünnwandigen Oesophagus folgt nach einer kropfartigen Ausbuchtung des letzteren ein großer und weiter Magen als Anfangsteil des Mitteldarms. In diesem Magenteil ist eine schwarzbraune, flüssige Inhaltsmasse enthalten. Sie dringt aber nicht weiter als in den 8–10 mm langen Magen ein, der mehrfach gekrümmt ist und sich in einen kompakten Stiel fortsetzt. Es ist dies noch ein Teil des Mitteldarms, und er erstreckt sich bis zu der Stelle, wo die 8 Malpighischen Gefäße einmünden. Dieser Teil des Mitteldarms hat kein Lumen, der Magen ist also nach hinten verschlossen. Der solide Strang ist etwa ¾ mm lang.
Im Innern des Mitteldarmstranges finden sich Nester von regenerativen Zellen, wie sich solche auch zwischen den Epithelzellen des Magens, also des verdauenden Mitteldarmteils finden.
An den Mitteldarmstrang schließt sich der Enddarm an, der von der Einmündungsstelle der Malpighischen Gefäße an wieder ein Lumen hat. Deren Exkrete und in der späteren Zeit den Spinnstoff hat er nach außen zu leiten. Spuren von Faeces können in ihm naturgemäß niemals, solange die Larvenzeit dauert, enthalten sein.
Während der Metamorphose der Puppe tritt, wie immer bei den Insekten, eine vollkommene Erneuerung des Darmepithels ein; bei dieser Gelegenheit wird ein Lumen, das vom Magen zum Enddarm durchgeht, gebildet. Wenn diese Wachstumsvorgänge abgeschlossen sind, kann der Ameisenlöwe zum erstenmal in seinem Leben Faeces abgeben. Das geschieht in der Regel beim ersten Flug der Imago. Ich habe keine Anzeichen davon gefunden, daß etwa bei den einzelnen Häutungen der Larve der Inhalt des Magens mitentleert würde.
Die erste Faecesentleerung der „Landlibelle“, wie Rösel von Rosenhof die Imago des Ameisenlöwen nannte, hat in der Wissenschaft,[S. 123] wie das Tier selbst, eine Geschichte. Schon Réaumur hatte bemerkt, daß die Imagines bald nach ihrem Ausschlüpfen einen biskuitförmigen Körper fallen lassen. Er besteht aus einer schwarzen Zentralmasse, einer dicken braunen und schwarzen geschichteten Schale, welche diese umhüllt, und am dünnen Ende aus einem rosa gefärbten, pfropfenähnlichen Aufsatz. Rösel hielt das Gebilde, wie Réaumur, für ein Ei, glaubte aber, es werde vor der Begattung abgelegt und wäre daher nicht entwicklungsfähig.
Schon Dutrochet (1818) nahm an, daß die „Exkremente“ des Tieres bei der Verwandlung in den Magen der Puppe und von dort in den der Imago übergingen, um dann ½ Stunde nach dem Ausschlüpfen ausgestoßen zu werden. Aehnliches vermutete Burmeister, und v. Siebold hielt den ausgestoßenen Körper für reinen Harn. Erst Gerstäcker unterschied die verschiedenen Teile des Gebildes und stellte fest, daß der schwarze Zentralteil aus Nahrungsresten der Larve bestehe, die Hülle aus Chitinmembranen des Darmes, die bei der Darmhäutung abgestoßen würden, und daß sie Harnsäure enthält, wie denn der pfropfenförmige, rosa gefärbte Aufsatz ganz aus Harnsäure besteht. Es ist also der gesamte Magen- und Darminhalt der Larvenzeit, welcher hier entleert wird und dabei aus dem Enddarm das Produkt der Malpighischen Gefäße, das diese vor ihrer Umwandlung in Spinndrüsen (s. unten) absonderten, mitnimmt. Erst Rengel (1908) stellte die Gewebeveränderungen fest, welche während der Metamorphose dem Darm ein durchgehendes Lumen verschaffen.
[S. 124]
Im Mai und Juni und dann noch einmal später im Herbst findet man viele Larven des Ameisenlöwen mit dem Spinnen des Puppenkokons beschäftigt. Ist der Kokon fertig, so stellt er eine kleine Kugel von etwa 2 cm Durchmesser dar, welche vollkommen mit feinen, kleinen Sandkörnern an der Oberfläche bedeckt ist (Abb. 41). Diese sind mit dem Spinnstoff an die Hülle angeklebt, welche die Larve um sich hergestellt hat. Sie bildet eine kleine kugelige Kammer mit glatter, weißer Wand.
Die Ameisenlöwen haben keine besonderen Spinndrüsen, sondern ihre Malpighischen Gefäße, welche in der Zahl von 8 an der Grenze von Mittel- und Enddarm in letzteren einmünden, wandeln sich während des letzten Abschnittes der Larvenzeit in Spinndrüsen um. Das hat Lozinski (1911) in einer sorgfältigen Arbeit nachgewiesen. Während des früheren Larvenlebens sind die Malpighischen Gefäße normal funktionierende Exkretionsorgane. Gegen Ende des Larvenlebens werden aber die Wandzellen größer, bekommen sehr große verästelte Kerne und beginnen Spinnstoff auszuscheiden. Dieser gelangt durch das sogenannte Coecum in den Afterdarm des Ameisenlöwen. Der Afterdarm ist an seinem Ende in eine ausstülpbare chitinige Röhre umgestaltet, welche als Spinnapparat dient (vgl. Abb. 10, S. 19). Aus ihrer beweglichen feinen Spitze kommt der Seidenfaden in noch weichem Zustand heraus und kann in den verschiedensten Richtungen bewegt und angeklebt werden.
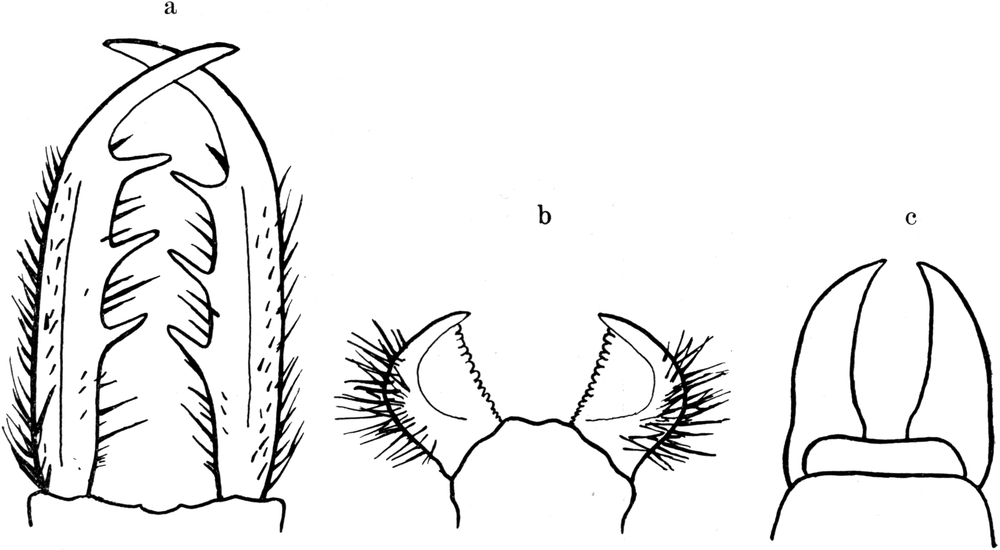
Wie Lucas gezeigt hat, kriecht der Ameisenlöwe noch im Puppenzustand zum Teil aus dem Kokon heraus; die Imago entschlüpft[S. 125] der am Rücken platzenden Puppenhaut, nachdem sie zu einem Drittel aus dem Kokon hervorgestreckt ist. Vorher hat die Puppe mit den kräftigen, gezähnelten Puppenmandibeln (Abb. 42 b), welche wie Hüllen die Imagomandibel umgeben, ein kreisrundes Loch mit glatten Rändern in die Kokonhülle genagt. So hat das Tier in seinen drei Stadien ihren Sonderbedürfnissen angepaßte Mandibelformen ausgebildet: jene für den Ameisenfang so geeigneten langen, stark gezähnelten Mandibel der Larve, die kräftigen, grob gezähnelten Mandibel der Puppe, welche nur zur Befreiung aus dem Kokon dienen, und schließlich die zarten, ganzrandigen Mandibel der Imago, welche ihr zu — soviel ich weiß — heute noch nicht genauer bekannten Zwecken notwendig sind.
Fast das ganze Jahr hindurch kann man an den geeigneten Stellen die offenen Trichter der Ameisenlöwen beobachten. Nur im November, Dezember und Januar habe ich sie im Freien nicht gesehen. War das Wetter warm und sonnig, so fand ich in einigen Fällen sogar in der ersten Novemberwoche und in Freiburg sogar im warmen Winter 1911/12 in der letzten Januarwoche einige Trichter mit lebhaften Tieren im Freien. Die dazwischenliegende Zeit ist also die einzige, in der ich sie nicht im Freien bemerkte. Meist ist zu dieser Zeit die Erde feucht oder gefroren; die wichtigsten Voraussetzungen für das Trichterbauen fehlen also.
Aber was viel eigentümlicher ist, die Tiere, welche ich in der gleichen Zeit im warmen Zimmer, möglichst der Sonne ausgesetzt, bei einer Durchschnittstemperatur von 17° C hielt, bauten auch keine Trichter mehr. Sie saßen in feinem, trockenem Sand,[S. 126] Feuchtigkeit wurde ihnen zeitweise gespendet, Ameisen zur Fütterung reichlich geboten — aber sie bauten keinen Trichter, fingen keine Ameisen, reagierten überhaupt nicht auf diese und waren nicht dazu zu bringen, sie auszusaugen.
Ich bemerke ausdrücklich, daß es sich bei diesen Beobachtungen um Tiere von ganz verschiedener Größe handelte; es kam also keine Verwechslung mit jenem Ruhestadium vor, welches der Verpuppung vorausgeht. Etwa 14 Tage lang, ehe das Spinnen des Kokons beginnt, bauen ja die Larven auch keine Trichter, fressen nicht und halten sich still unter dem Sand.
Es ist also im Lebensablauf des Ameisenlöwen eine charakteristische Periodizität[1] festzustellen. In dem Teil des Jahres, in dem sie normalerweise nicht zum Trichterbau und Ameisenfangen kommen, liegen sie in einem Starrezustand unter der glatten Bodenoberfläche. Sie sind dann schwer zu Reflexen zu reizen, und die natürlichen Verhältnisse bringen keine der adäquaten Reize an sie heran. Ja selbst, wenn unter experimentellen Bedingungen die geeignetsten Umgebungsverhältnisse ihnen angeboten werden, überwiegt der Einfluß der Periodizität über die der speziellen Reize.
[1] Zusatz bei der Korrektur: Die Beobachtungen des Winters 1915/16 haben mich in der Beurteilung dieser Vorgänge wieder zweifelhaft gemacht. Im Zimmer gehaltene Tiere verschiedener Größen bauten diesen ganzen Winter hindurch normale Trichter, fraßen gut und zeigten größere Lebhaftigkeit als in den drei letzten Wintern. Was die Ursache des verschiedenen Verhaltens war, suche ich noch festzustellen.
Ueberblicken wir die Gesamtheit der in diesem Werk niedergelegten Beobachtungen und Versuchsergebnisse, so gelangen wir zu dem Schluß, daß die eigenartigen Lebenserscheinungen des Ameisenlöwen durchaus nicht, wie frühere Beobachter annahmen, durch hohe psychische Fähigkeiten des Tieres bedingt sind. Alle vom Vergleich mit dem menschlichen Handeln hergenommenen Ausdrücke, welche zur Bezeichnung seiner Leistungen angewandt wurden, wie Klugheit, Schläue und ähnliche, sind bei diesem Tier verkehrter angewandt, als bei den meisten anderen.
Wir haben ja gesehen, daß es ein Tier ist, welches durch seine Organisation vollkommen gezwungen ist, seine Handlungen in[S. 127] jener Art durchzuführen, welche seit altersher das Staunen und die Bewunderung der Naturforscher und Laienbeobachter hervorgerufen hat.
Solange das Tier Larve ist, ist es ein reiner Reflexautomat, es funktioniert wie eine kleine Maschine. Eine Insektenlarve, welche im Freien lebt, hat eine Menge von Lebensschwierigkeiten, die ihr die umgebende Außenwelt bereitet, zu überstehen. Vielfach werden gerade bei den Insekten durch das Muttertier viele von diesen Schwierigkeiten ausgeschaltet, so für Nahrungserwerb, Schutz, Behausung usw. gesorgt.
Im Ameisenlöwen haben wir eine Larve vor uns, welche in den meisten Dingen vollkommen auf sich selbst angewiesen ist. Ihr Körper muß von vornherein so beschaffen sein, daß sie allen Gefahren begegnen, alle Hindernisse überwinden kann. So finden wir denn den kleinen Automaten kurz nach dem Ausschlüpfen aus dem Ei ebenso vollkommen, als wenn er herangewachsen, nach einem Jahr voll Erlebnissen, bereit ist, die Puppenruhe anzutreten.
Sehr auffällig ist die geringe Zahl von Reflexen, welche die wichtigsten Handlungen des Ameisenlöwen bedingen. Daß es dem Tier möglich ist, mit einer so geringen Anzahl von typischen Bewegungen seine Hauptlebensfunktionen zu erfüllen, erklärt sich aus dem engen Zusammenhang zwischen dem Körperbau und den Reflexen. Der ganze Körper des Tieres ist in engster, einseitigster Weise an das Leben im Sand und die Art der Nahrungserwerbung angepaßt. Die äußere Form des Kopfes, Halses und Rumpfes, die Zuspitzung des Hinterleibes, der Bau und die Einlenkung der Beine und vor allem die Menge der in zweckmäßigster Weise angeordneten Borsten bedingen die Art der Bewegungen des Tieres. Was an allen andern Orten, unter allen anderen Bedingungen der Umgebung den Ameisenlöwen zu einem hilflosen Geschöpf macht, das gibt ihm im lockeren Sand eine vollkommene Ueberlegenheit über andere Tiere.
Wie weit die Anpassung an das Leben in Sand und Staub geht, zeigt uns eine Untersuchung der Stigmenöffnungen (Abb. 8, S. 16). Die Hauptstigmenöffnung, durch welche das Tier seine Atemluft einzieht, liegt ganz vorn am Rumpf, an der Stelle, welche bei der normalen Haltung des Tieres in der Bereitschaftsstellung kaum vom Sand bedeckt ist. Nach hinten zieht sich den Körper entlang die Reihe kleinerer Stigmen. Alle diese Atemöffnungen sind in vollendeter[S. 128] Weise vor dem Eindringen von Sand- und Staubteilchen in die Atemröhren geschützt. Wie bei vielen anderen Insekten verschließt ein feines Gitter den äußeren Eingang. Feine zahnartige Fortsätze greifen von beiden Seiten über die spaltförmige Atemöffnung und stellen so einen sehr wirksamen Verschluß dar. Nie findet man bei der Sektion der Tiere in dem blasenförmigen Anfangsteil der Atemröhren ein noch so kleines Sandkorn. Und zu allem Ueberfluß ist am Eingang des großen Tracheenstammes, welcher vom Hauptstigma ausgeht, ein Deckel vorgeklappt, welcher die Luft zu einem Umweg nötigt, so daß alle festen Bestandteile vor dem Eintritt des Luftstromes in die zartwandige Trachee abgelagert werden müssen.
Das Tier ist von Geburt an durch seinen Bau und seine Reflexe zur Erfüllung der Erfordernisse seiner eigenartigen Lebensführung vollkommen fertig. Nun fragt es sich, wie es möglich ist, daß ein so sehr von den speziellen Bedingungen seiner Umgebung abhängiges Tier relativ so häufig sein kann. In den Gegenden, in denen die Ameisenlöwen überhaupt vorkommen, gibt es sie meist in großen Mengen. Ich habe bei Burghausen a. d. Salzach, im Kaiserstuhl, in Schwarzwald und Vogesen oft hunderte der Trichter auf engem Raum beieinander gefunden.
Die Imago legt bald, nachdem sie aus der Puppe ausgeschlüpft ist, ihre Eier in den Sand ab. Es sind nach den Beobachtungen von Brischke nur wenige (5) Eier von weißlicher Farbe und ovaler Form, welche miteinander verklebt sind. An schönen Sommerabenden in der Dämmerung sieht man das fertige Insekt in derselben Gegend an den Böschungen und Waldrändern umherfliegen, wo die Sandtrichter der Ameisenlöwen in Menge sich befinden und wo auch sie, die „Landlibelle“, ihre Entwicklung durchgemacht hat. Solche Stellen sind immer dadurch ausgezeichnet, daß, wenn die Kühle des Abends die Luft durchdringt, sie selbst den gröberen Sinnesorganen des Menschen durch die von ihnen ausgestrahlte Wärme auffallen. Nachdem wir bei den Larven eine so ausgesprochene Thermotaxis gefunden haben, liegt es durchaus nahe, anzunehmen, daß auch das fertige Insekt thermotaktisch ist und so zu den Stellen hingeleitet wird, welche für die Entwicklung seiner Eier günstig sind, wie ja auch bei vielen anderen Insektenmüttern Reflexe oder Instinkte es sind, welche die Unterbringung der Brut am richtigen Orte ermöglichen. Aber wir wollen hier keine Hypothesen aufstellen. Die Biologie der Imago zu untersuchen, ist eine noch in Angriff zu nehmende Aufgabe.
[S. 129]
Was wir von der Larve selbst wissen, das genügt vollkommen, um die Larve, wenn sie in geeigneter Gegend aus dem Ei schlüpft, an den Ort zu bringen, wo sie gedeihen kann. Liegt sie beim Ausschlüpfen auf hartem Stein oder feuchter Erde, so wird sie, sobald die Sonnenstrahlen sie treffen, zu wandern beginnen. Das Licht wird ihr die Richtung angeben, die von den Böschungen und Abhängen ausstrahlende Wärme ihr dabei helfen. Solange sie im diffusen Licht sich befindet, wird sie in Versuchsbewegungen Spiralen und Kreise durchwandern. Der Zufall wird sie an eine warme und sonnige Stelle führen, wo sie sich weiter bewegt und weiter sucht. Das wird in der Gegend, wo das Ei abgelegt wurde, leicht eine der geeigneten Böschungen sein. Indem sie der Wärme entgegenwandert, gerät sie in den Schatten, die Phototaxis dreht sie herum und führt sie an den Rand des Schattens. Unterdessen führt die Hinterleibsspitze tastende Bewegungen aus, und sowie geeignete Wärme- und Lichtverhältnisse das Tier umgeben, sobald es den trockenen und feinen Sand findet, der die Reflexe ins Gleichgewicht bringt, entsteht der Trichter. Das Tier ist mit Hilfe seiner Reflexe an seiner Stelle angelangt und kann nun dauernd dableiben, wenn einige weitere Voraussetzungen erfüllt sind. Zunächst müssen Ameisen in genügender Menge sich herumtreiben, so daß die Falle auch Gelegenheit bekommt, in Tätigkeit zu treten. Ist das nicht der Fall, so bemächtigt sich des Tieres neue Ruhelosigkeit, es wandert und baut bald in neuer Umgebung einen neuen Trichter, dort auf seine Beute lauernd. Hat es den geeigneten Ort gefunden, so bleibt es dort bis zu seiner Verpuppung. Ich habe nicht selten denselben Trichter am gleichen Ort durch Monate hindurch beobachtet. Die Orte des Vorkommens, die kleinen Böschungen an steilen Abhängen, sind oft derart, daß man nicht annehmen kann, daß die Larve mit eigenen Kräften zu ihnen hingelangt ist. Ihre Wanderungen werden sich kaum über weite Entfernungen erstrecken. Die Wahl des Ortes im allgemeinen muß wohl in der Hauptsache der flugfähigen Mutter zugeschrieben werden.
Kleine Wanderungen finden aber unzweifelhaft statt; so habe ich in einigen Fällen nach langandauerndem trocknem Wetter Trichter ziemlich weit über die überhängende Böschung vorgeschoben gesehen, so daß Regen in sie einfallen konnte. Sie wurden bei Regenwetter, das kurz anhielt, zurückverlegt. Hielt es länger an, so lagen die Tiere in starrem Zustand unter der Erde oder dem Sand, bis sie durch Trockenheit und Wärme wieder erweckt, beim Wandern und[S. 130] Suchen den nahen trockenen Unterstand fanden und da ihren Trichter gruben.
Der gesetzmäßige Ablauf der Reflexbewegungen bringt es mit sich, daß man in einer bestimmten Gegend mit Sicherheit die Orte des Vorkommens von Ameisenlöwen voraussagen kann. Man findet sie stets an Süd-, Südost- und Südwesthängen und entsprechend gelegenen Waldrändern. Der Untergrund muß die Bildung trocknen, feinen, sandartigen Bodens ermöglichen. Es müssen überhängende Böschungen und Baumwurzeln, Raine vorhanden sein. Und an solchen Stellen ist die Bedingung, daß Ameisen da sein müssen, stets erfüllt. Windschutz ist eine weitere Voraussetzung, von der die Anhäufung feinen Sandes abhängt.
In einer solchen Gegend kann man mit Sicherheit auf einen Weg- oder Waldrand losgehen und sich sagen: da müssen Ameisenlöwen sein. Man wird sie an den erwarteten Stellen finden. Die Trichter werden so liegen, daß die Sonne in ihre Tiefe hineinfällt und dort wird man aus dem Sand die Kiefer des Tieres in Bereitschaftsstellung hervorschauen sehen, mit der weiten Oeffnung der Böschung zugekehrt, also vom Licht abgewandt. Dorther wird man in vielen Fällen die Ameisen in die geöffnete Falle hinabrollen sehen.
Nichts ist so sehr geeignet, den gesetzmäßigen Ablauf erforschter Vorgänge zu bestätigen, als die Voraussage bestimmter Erscheinungen. So wie wir, ohne ihn vorher gekannt zu haben, den Fundort von Ameisenlöwen in einer Landschaft im Voraus bestimmen können, so können wir auf Grund der Reflexerscheinungen auch voraussagen, an welchem Ort im Laboratorium wir ein Exemplar wiederfinden werden, das uns etwa während der Experimente entwischt ist. Während der vielen Jahre, die ich bisher mit dem Ameisenlöwen experimentierte, ist mir kein Exemplar verloren gegangen, das im Zimmer oder Laboratorium mir durchging. Nachdem ich die wichtigsten Reflexe einmal festgelegt hatte, konnte ich mit großer Sicherheit den Ort bestimmen, an dem es infolge der Photo-, Thigmo- und Thermotaxis nach einiger Zeit angelangt sein mußte. Ich fand es regelmäßig in der Nähe des Fensters, der Heizung, in einem Winkel zwischen Wand und Boden und konnte es wieder in sein Aufbewahrungsgefäß zurückversetzen.
So haben wir denn im Verlauf unserer Untersuchungen den Ameisenlöwen als einen vollkommenen Reflexautomaten kennen gelernt. Keine Handlung, kein Vorgang wies auf eine höhere psychische Fähigkeit hin. Und doch habe ich unablässig nach Andeutungen[S. 131] von solchen gesucht. Schließlich ist es mir auch gelungen eine Tatsache festzustellen, welche scheinbar, wenn auch nicht mit aller Sicherheit auf das Vorkommen mnemischer Vorgänge hinweist.
Ich habe früher (S. 108) von Versuchen berichtet, bei denen ich Ameisenlöwen zwang, mit der Hinterleibsspitze voran, in eine Glasröhre zu kriechen. Wir sahen, daß sie unter bestimmten Voraussetzungen in die Glasröhren eintraten und diese durchkrochen. Bot ich ihnen aber Glasröhren an, welche im Verhältnis zu ihrem Körperumfang zu eng waren, so machten sie zunächst die üblichen Versuchsbewegungen mit der Hinterleibsspitze. Sie drängten und arbeiteten mit dem Hinterleib in die Glasröhre hinein, kamen aber nicht weiter. Immer wieder machten sie Versuchsbewegungen und gerieten dabei auch gelegentlich mit der einen Körperseite an die Außenwand der Glasröhre. Sofort ließen sie mit ihren Bemühungen nach und wanderten nun an der Außenwand der Glasröhre, in thigmotaktischer Berührung mit ihr, bis an ihr Ende.
Hatte ich die Ameisenlöwen dies Experiment mehrmals durchmachen lassen, so schien es mir, als brauchten sie immer kürzere Zeit, um den Seitenweg zu finden. Systematische Versuche, die ich dann durchführte, ergaben aber sehr schwankende Ergebnisse. Bald trat eine Abkürzung der Versuchszeit ein, bald war eine solche nicht zu bemerken. Eine Gesetzmäßigkeit war nicht festzustellen. So werde ich denn diese Versuche, vielleicht mit abgeänderter Anordnung fortsetzen, habe aber nicht allzugroße Zuversicht, bei meinem Versuchsobjekt Andeutungen eines höher ausgebildeten Gedächtnisses zu finden.
Weit mehr, als ich jemals erwartete, habe ich also den Ameisenlöwen als Reflexautomaten erkannt; nicht einmal kompliziertere Instinkte konnte ich als Grundlage seiner so erstaunlichen Handlungen nachweisen.
Dem entspricht auch der Bau seines Zentralnervensystems (Abb. 40, S. 115). Dessen Gehirnganglion ist im Verhältnis zur Größe des Körpers recht klein; es ist hauptsächlich in die Quere gezogen und verlängert sich seitlich in die Augennerven, in deren Verlauf die spindelförmigen Ganglia optica eingeschaltet sind. Das Oberschlundganglion ist im Verhältnis nicht sehr viel größer als das Unterschlundganglion. Die Sphären der Sinnesorgane, also die Ursprungsstellen von Sehnerv und Antennennerv, sind nicht sehr umfangreich. Besonders auffallend ist der geringe Umfang und die geringe Entwicklung des[S. 132] sogenannten pilzförmigen Körpers, den man bei den Insekten mit höheren Instinkten, bei denen er stark entwickelt ist, als den Sitz der Fähigkeit zur Bildung von Assoziationen ansieht. So bemerken wir, daß auch in dieser Hinsicht die körperlichen Grundlagen im zu erwartenden Verhältnis zu den Leistungen des Tieres stehen. Das Bauchmark besteht aus zwei getrennten Ganglien im ersten und zweiten Thorakalsegment, welche entsprechend ihrer Aufgabe, die beiden ersten Extremitäten zu innervieren, relativ groß sind. Das gleiche gilt vom dritten Thorakalganglion. Dies ist, sowie sein zugehöriges Segment mit den Abdominalsegmenten eng verbunden ist, selbst mit den 9 Ganglien des Abdomens zu einer Kette vereinigt, in der zwischen den Ganglienpaaren kaum Kommissuren erkennbar sind. Das Bauchmark erreicht nur etwa die Hälfte der Länge des Hinterleibes der Larve (Abb. 40, S. 115); es sind also, wie so häufig bei Insekten, die Ganglienpaare eng zusammengedrängt und das Bauchmark konzentriert. Von seinem Ende gehen drei starke periphere Nerven gegen das Hinterleibsende. Die Topographie des Zentralnervensystems muß bei operativem Eingreifen zu experimentellen Zwecken sehr berücksichtigt werden.
Meine Arbeit war in den wesentlichen Ergebnissen bereits fertig gestellt und das Manuskript lag fast abgeschlossen vor, als ich durch Zufall auf eine Arbeit des sizilianischen Zoologen Comes stieß, welche sich mit Tropismen des Ameisenlöwen beschäftigt. Bis dahin hatte ich angenommen, daß noch keine Untersuchung der Reflexbiologie des Tieres vorläge.
Die Lektüre der Arbeit, welche in den Akten der Akademie von Catania erschienen ist, brachte mir keine sehr wesentliche Förderung. Zwar steht der Autor bei der Zusammenfassung der Resultate auf einem ähnlichen theoretischen Standpunkt wie ich. Auch er führt die Reaktion des Ameisenlöwen im Wesentlichen auf Reflexe zurück. Es ist ihm aber nicht gelungen, die Reflexe im Einzelnen zu analysieren und er scheint mir allzu schnell bereit gewesen zu sein, aus den wenigen beobachteten Tatsachen die vorausgesetzten Schlüsse zu ziehen. Vor allem hat er einen Fehler begangen, dem man immer wieder bei gelegentlichen reflexbiologischen Arbeiten begegnet. Dieser prinzipielle Fehler ist es, der mich veranlaßt, an dieser Stelle auf die Arbeit von Comes noch einmal kurz einzugehen, nachdem ich in den verschiedenen Kapiteln dieses Werkes schon die Einzelheiten seiner Angaben an der Hand meiner Experimente kritisiert habe.
[S. 133]
Vor allem hat Comes versäumt, was, wie gesagt, ein Fehler sehr vieler reflexbiologischer Arbeiten ist, vor dem Experimentieren genau das Verhalten des Tieres unter normalen Verhältnissen zu studieren. Somit hatte er keine geeignete Grundlage für die Beurteilung der Reaktionen des Tieres unter dem Einfluß experimenteller Reize. So war ihm der starke Einfluß des Lichtes auf den Ameisenlöwen entgangen; da bei seinen Versuchen, z. B. über Thermotaxis, die Einwirkung des Lichtes nicht ausgeschaltet war, so können die Resultate jener Versuche keine Beweiskraft für sich in Anspruch nehmen. Die Bereitschaftsstellung und die Versuchsbewegungen des Tieres waren ihm vollkommen entgangen. Seine Durchschneidungsversuche an der Ganglienkette sind zu wenig exakt, um genauer diskutiert werden zu können, da er versäumt hat, vor den Experimenten den anatomischen Bau und die Erstreckung der Ganglienkette im Körper zu studieren. Auch sind seine Definitionen nicht allzu scharf und logisch. Im Einzelnen habe ich mich ja in den früheren Abschnitten mit ihm auseinandergesetzt.
Im übrigen ist es sicher verdienstlich, daß er das Problem als erster angepackt hat und eine Anzahl von typischen Reflexen bei dem Tier nachwies, welche die eigenartigen Handlungen des Tieres zu erklären geeignet sind. Er konnte aber nicht tiefer in die Zusammenhänge eindringen, da er nicht das ganze Leben des Tieres genauer kannte und daher seine Fragestellungen zum Teil falsch waren. Das ist ein prinzipieller Fehler, zu dessen Vermeidung bei späteren Untersuchungen diese Zeilen beitragen möchten.
Wenn ich meine Untersuchungen am Ameisenlöwen vorläufig mit dem Ergebnis abschließen konnte, daß das Tier ein reiner Reflexautomat ist, so muß ich, um keine Unklarheit über meine allgemeinen Anschauungen über solche Probleme aufkommen zu lassen, einige Schlußbemerkungen hinzufügen. Zunächst ist es mir vollkommen klar, daß ich die Lebensvorgänge beim Ameisenlöwen nicht restlos analysiert habe. Wer meine Darstellung mit Aufmerksamkeit gelesen hat, wird manches neue Forschungsprogramm zwischen den Zeilen durchschimmern gesehen haben. Besonders eine genauere Untersuchung des Nervensystems und der Muskulatur ist notwendig. Ferner wird das Verhalten der Imago und noch manches andere zu studieren sein, ehe wir über alle Zusammenhänge volle Klarheit haben; und gewiß werden während des weiteren Studiums neue Probleme und Rätsel sich enthüllen, die zu weiteren Aufgaben reizen müssen.
[S. 134]
Sicher haben wir aber im Verlauf der Untersuchung erkannt, daß die Larve eines höheren, holometabolen Insekts, welche ein sehr eigenartiges Leben führt, dies als Reflexautomat durchführen kann, weil ihr Bau in höchst zweckmäßiger Weise den Anforderungen der Funktionen und des Lebensraumes entspricht. Wir haben im Ameisenlöwen ein Beispiel von weitestgehender Anpassung des Baues an die Lebensbedingungen vor uns. Der Bau und die mit ihm vererbte und kaum veränderliche Funktionsweise der Organe setzt das Tier in den Stand, sein merkwürdiges und scheinbar so schwieriges Leben durchzuführen. Dabei ist es nicht fähig, sich an ungewohnte Verhältnisse anzupassen und geht unter abgeänderten Bedingungen sehr leicht zugrunde.
Somit haben wir in seinem Verhalten einen der Haupttypen der Lebensweise von Tieren kennen gelernt, den fest angepaßten Typus, bei welchem von der Geburt an das Tier Bau und Fähigkeiten mitbekommt, welche die feinste Abstimmung auf die normalen Lebensbedingungen der Art darstellen. Einseitige Anpassung haben wir das eben genannt, wir können vom Ameisenlöwen auch als von einem Lebensspezialisten sprechen.
Das ist ein Typus, der bei niederen Tieren weit verbreitet ist, wenn wir auch unter ihnen nicht selten Vertreter des anderen Haupttypus, des regulatorischen Typus finden. Morphologische und physiologische Regulationen, welche das Tier an verschiedenartige Lebensbedingungen während seines individuellen Lebens anzupassen vermögen, sind im ganzen Tierreich weit verbreitet. Bei den höheren Tieren finden wir aber vor allem eine Regulierbarkeit der Handlungen, eine Anpassungsfähigkeit des einzelnen Individuums in seinem Verhalten von Fall zu Fall an die wechselnden Bedingungen der Außenwelt. Diese Anpassungsfähigkeit hat ihre Grundlage vor allem in den höheren psychischen Fähigkeiten vieler Tiere, in deren Vermögen zur Speicherung von Erfahrungen und Eindrücken, zur Bildung von Assoziationen usw. Ich habe Studien an höheren Organismen durchgeführt, welche aber noch bei weitem nicht reif zur Veröffentlichung sind, die mich bei manchen Tieren merkwürdig komplizierte psychische Fähigkeiten erkennen ließen, wie wir sie ja als solche durch die Forschungen vieler Zoologen und Tierpsychologen schon längst kennen.
So bin ich denn weit davon entfernt, solche Fähigkeiten zu leugnen, wenn es mir gelang, in einer hochorganisierten Insektenlarve einen reinen Reflexautomaten zu enthüllen. Aber es scheint[S. 135] mir die Kenntnis solcher Automaten notwendig, um die Gesetze zu erforschen, welche die höheren psychischen Funktionen der Tiere beherrschen. Ja, es scheint mir, daß die Tierautomaten uns die wesentlichen Grundgesetze kennen lehren, auf denen sich auch das höhere psychische Leben der Tiere und des Menschen aufbaut.
Bethe, A., Das Nervensystem von Carcinus maenas. In: Arch. f. mikr. Anat., Vol. 50. 1897. p. 460 u. p. 589; Vol. 51. p. 382.
Brehm, Tierleben. 3. Aufl. Insektenband.
Buddenbrock, N. v., Ueber die Orientierung der Krebse im Raum. In: Zool. Jahrb. Abt. Allg. Zool. u. Phys. Vol. 34. 1914. p. 479.
Comes, S., Stereotropismo, geotropismo e termotropismo nella Larva di Myrmeleon formicarius L. In: Atti Accad. Gioenia di Sci. nat. in Catania. Anno 86. Ser. 5. Vol. 2. 1909. mem. 4.
Doflein, F., Lebensgewohnheiten und Anpassungen bei dekapoden Krebsen. Festschr. zum 60. Geburtstag Richard Hertwigs. Bd. 3, auch separat, Jena 1910.
Dufour, Recherches anatomiques sur les Orthoptères, les Hyménoptères et les Neuroptères. 1841.
Fitting, Abschnitt über Reizbewegungen der Pflanzen im Handbuch der Naturwissenschaften.
Forel, A., Das Sinnesleben der Insekten. München 1910.
v. Frey, 1) Berichte der Ges. d. Wiss. Leipzig. Vol. 46. 1894. p. 185.
— 2) Abhandl. der Ges. d. Wiss. Leipzig. Vol. 23. 1896. p. 175.
Gerstäcker, Ueber zwei eigentümliche bei Gastrospacha neustria und Myrmeleon formicarius beobachtete Modifikationen der Harnsäureabsonderung. Ber. Ges. nat. Fr. Berlin, 1873.
Hesse, R., Untersuchungen über die Organe der Lichtempfindung bei niederen Tieren. VII. In: Zeitschr. f. wiss. Zool. Vol. 70. 1901. p. 347.
— und Doflein, F., Tierbau und Tierleben. 2 Bde. Leipzig 1910–13.
Jennings, H. S., Behavior of lower organisms. New York 1906.
— Behavior of the starfish Asterias forreri d. L. In: Univ. Cal. Publications in Zoology. Vol. 4. 1907. p. 53.
v. Kaulbersz, Biologische Beobachtungen an Asellus aquaticus. In: Zool. Jahrb. Abt. Allg. Zool. u. Phys. Vol. 33. 1913. p. 1.
Lozinski, P., Beitrag zur Anatomie und Histologie der Mundwerkzeuge der Myrmeleonidenlarven. In: Zool. Anz. Vol. 33. 1908. p. 473.
— Zur Histologie der borstenartigen Bildungen am Hinterleibe der Myrmeleonidenlarven. Ebenda Vol. 35. 1910. p. 526.
— Ueber die Malpighischen Gefäße der Myrmeleonidenlarven als Spinndrüsen. Ebenda Vol. 38. 1911. p. 401.
[S. 136]
Lucas, W. J., On the emergence of Myrmeleon formicarius from the pupa. In: Transact. Entomol. Society London. 1906. p. 53.
Mangold, E., Hypnose und Katalepsie bei Tieren. Jena 1914.
Meinert, Contribution à l’anatomie des Fourmillions. 1889.
Oltmanns, F., Ueber die photometrischen Bewegungen der Pflanzen. In: Flora, Vol. 78. 1892. Heft 2. p. 183.
— Ueber positiven und negativen Heliotropismus. Ebenda Vol. 83. 1897. Heft 1. p. 1.
Ramdohr, Abhandlungen über die Verdauungswerkzeuge der Insekten. 1811.
Réaumur, Mémoire pour servir à l’histoire naturelle des Insectes. Vol. 3. 1834–42.
Redtenbacher, Josef, Uebersicht über die Myrmeleonidenlarven. In: Denkschriften der Kais. Akademie d. Wiss. Wien. Math.-naturw. Kl. Vol. 48. 1884. p. 335.
— Die Lebensweise der Ameisenlöwen. Progr. Commun. Ober-Realschule Wien. 1884.
Rengel, Ueber Myrmeleon formicarius L. In: Sitzungsber. Ges. naturf. Freunde Berlin. 1908. p. 140.
Rösel von Rosenhof, Insektenbelustigungen. Bd. 3. 1755.
Steinmann, P., Ueber Rheotaxis bei Tieren des fließenden Wassers. In: Verh. Naturf. Ges. Basel. Vol. 24. 1913.
[S. 137]
Fig. 1. Versuch Q. Phototaxisversuch. Einfall des Lichts von + her. Tier, bei − eingesetzt, führt eine Anzahl Versuchsbewegungen aus, um dann nach erfolgter Drehung (Hinterleibsspitze gegen +) energisch gegen die Lichtquelle zu kriechen. Die verhältnismäßig zahlreichen Versuchsbewegungen bei − sind durch Interferenz mit Thermotaxis zu erklären. Bei − war das Papier leicht erwärmt durch untergelegtes Sandbad. Temperatur dort etwa 25° C.
Fig. 2. Versuch N. 3 Exemplare bei vollkommener Dunkelheit auf verschiedenen Stellen des von 20° C auf 35° C ansteigend erwärmten Grundes niedergelegt. Sehr viele Versuchsbewegungen; zum Teil ganz kleine Kreise. Sehr lebhafte Bewegungen durch Dunkelheit und Wärme angeregt. Alle Tiere langten schließlich in der Region bei 35° an, zum Teil thigmotaktisch am Rand festgehalten und dort weiterwandernd. Versuchsdauer 11h 45′ bis 12h 45′.
Fig. 3. Versuch P. Unorientierte Bewegung mit zahlreichen Versuchskreisen bei Einfall starken Lichtes direkt von oben. Wirkung entsprechend vollkommen diffusem Licht. 3 Tiere im Versuch: A, B und C.
Fig. 4. Versuch GG. Licht von rechts einfallend in der Richtung des Pfeils. Schattenregion rechts bis zur Linie X. Reichliche Versuchsbewegungen an der Schattengrenze. An den Rändern die Spuren der durch Thigmotaxis an den Wänden festgehaltenen Individuen.
Fig. 5. Versuch JJ. Licht von rechts einfallend in der Richtung des Pfeils; Beleuchtung schwach und nicht sehr tief unter das Schattendach rechts eindringend. Daher Exemplar 2 gar nicht durch das Licht orientiert. Exemplar 1 viele Versuchsbewegungen ausführend, ehe Orientierung in der Nähe der Schattengrenze erfolgt. Nach Eintritt in das Lichtfeld bald Umkehr unter dem Einfluß der Lichtrichtung, dann im Schatten bald wieder Umkehr zum Licht. Kein Stocken an der Schattengrenze. Später Thigmotaxis.
Fig. 6. Versuch FF. Lichteinfall von rechts. Rechte Seite bis Linie X beschattet. 1 Tier im Experiment. Erst Versuchsbewegung, dann Wanderung zur Helligkeit, im Hellen Umkehr in die Richtung gegen die Lichtstrahlen, dann viele Versuchsbewegungen an der Schattengrenze, schließlich energische Wanderung zum Licht und deren Beendigung durch Thigmotaxis.
Fig. 7. Versuch RR. Ausgeführt 6. Nov. 1914. Lichtgefälle in der Pfeilrichtung, hergestellt durch Oltmanns’ Prisma. Versuch in Dunkelkammer; Wärmewirkung verhindert durch gekühlten Boden und Kühlwanne zwischen Licht und Experimentierfläche. A Exemplar zum erstenmal eingesetzt 10h 50′, nach geringen Versuchsbewegungen in der Richtung des stärkeren Lichtes in Bewegung gesetzt, bald durch Thigmotaxis abgelenkt. B Dasselbe Exemplar noch einmal eingesetzt 11h 50′. Kriecht auch schwankend, doch direkt in die hellere Zone, bewegt sich, dort zum Teil thigmotaktisch etwas gestört, im helleren Raum, ohne Bevorzugung einer bestimmten Region, bleibt aber im Hellen.
[S. 138]
Fig. 8. Versuch R. Interferenz von Licht- und Wärmewirkung. Tier x bei 35° Bodentemperatur niedergesetzt, Versuchsbewegung, dann Wanderung zur höheren Wärme, viele Versuchsbewegungen, kriecht schließlich in die wärmste Ecke und wird dort thigmotaktisch abgelenkt. Tier y kriecht sofort von 25° C in die höhere Temperatur, einige Schwankungen gegen das Licht. Endlich Ankunft in der wärmsten Ecke. Thigmotaxis.
Fig. 9. Versuch K. Thermotaxis. 26. Sept. 1914. Versuch in vollständiger Dunkelheit in Versuchsdunkelkammer. Ein Tier auf abgestuft von 20° bis 40° C erwärmten Boden niedergesetzt bei 20°, kriecht sofort in die Wärme; verweilt am längsten unter Versuchsbewegungen bei 30–35°. Infolge der Dunkelheit und Wärme sehr lebhaft beweglich, kriecht es weiter und gerät zurück zur Region 20° C, deren Temperatur sich im Lauf des Experiments erhöht hatte, dann thigmotaktisch aus dem Versuchsfeld.
Fig. 10. Versuch O. Zusammenwirken von Photo- und Thermotaxis. Bei + Lichteinfall, bei − dunkelste Region. Die Grade geben die Bodentemperatur der betreffenden Region an. Tier a beginnt bei 35° C seinen Weg, ehe das Rußpapier die Temperatur angenommen hatte, kriecht zuerst aufs Licht zu, wird dann thermotaktisch erregt, kriecht zurück in die Wärme und verläßt, thigmotaktisch gereizt, das Versuchsfeld. Tier b kriecht aus dem Licht in die Wärme und verläßt, bei der Höchsttemperatur thigmotaktisch gereizt, das Versuchsfeld. Tier c macht zuerst einen Versuchskreis, geht dann in die Wärme, schwankt zwischen Wärme und Licht, um schließlich energisch sich zum Licht zu wenden.
Frommannsche Buchdruckerei (Hermann Pohle) in Jena. — 4557
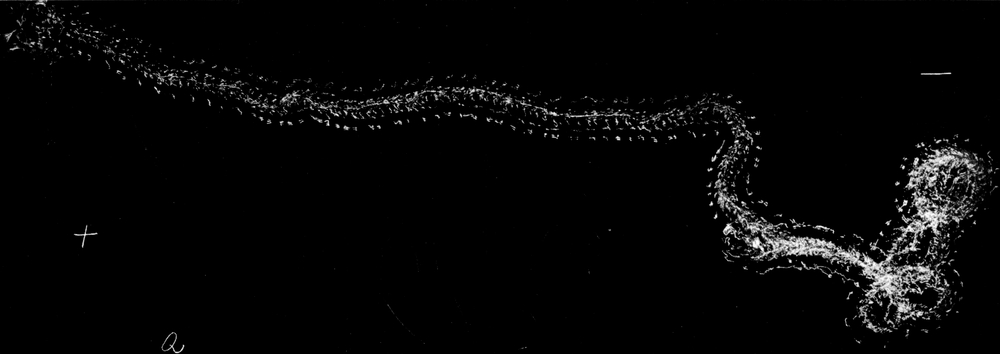
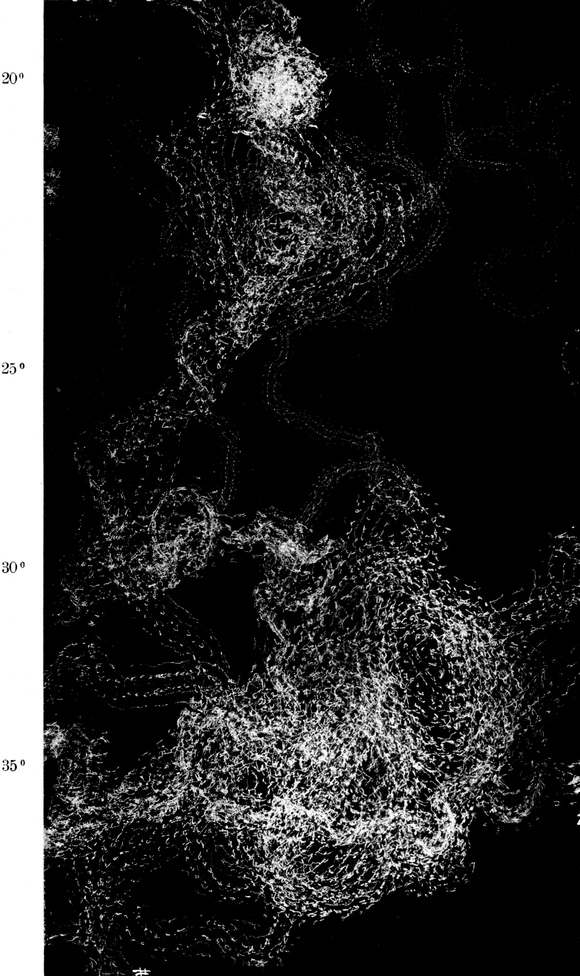





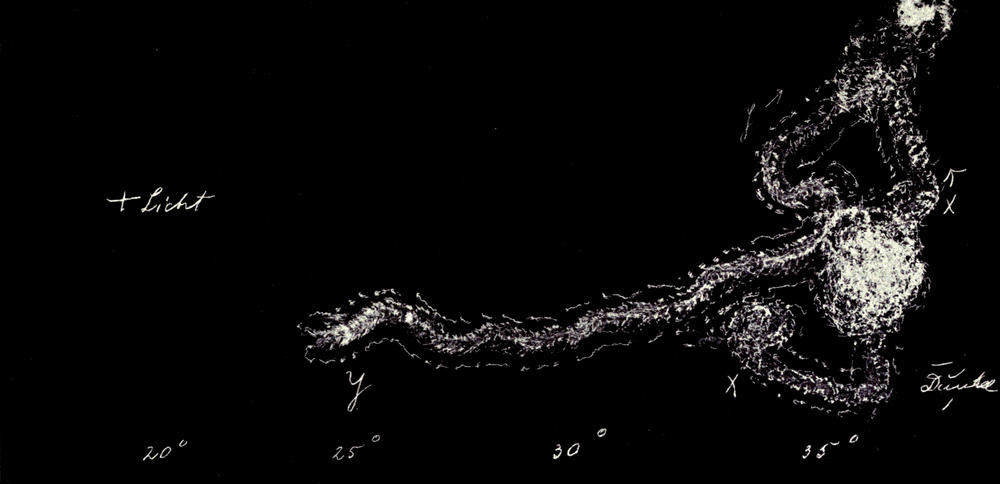
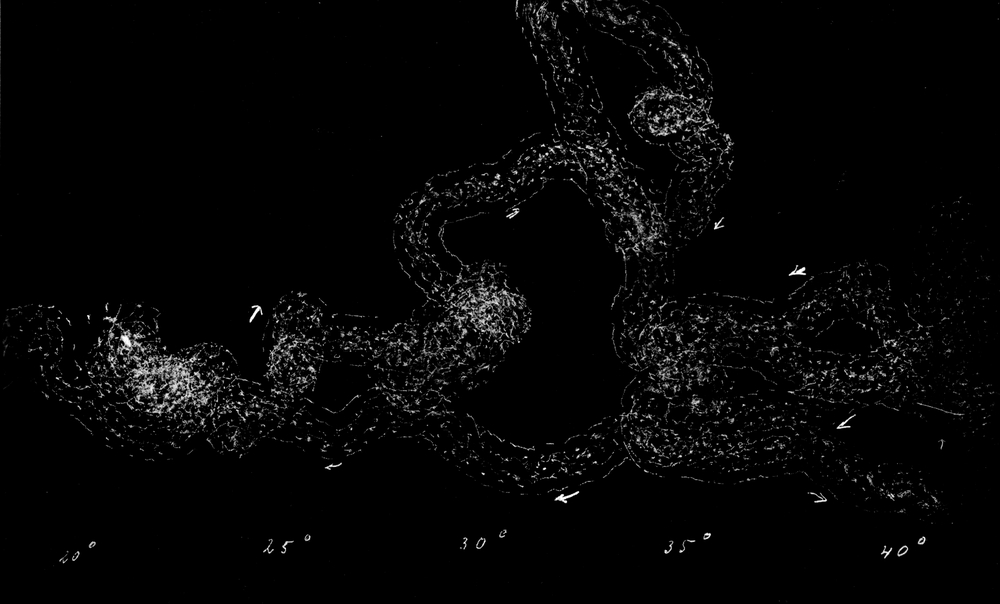
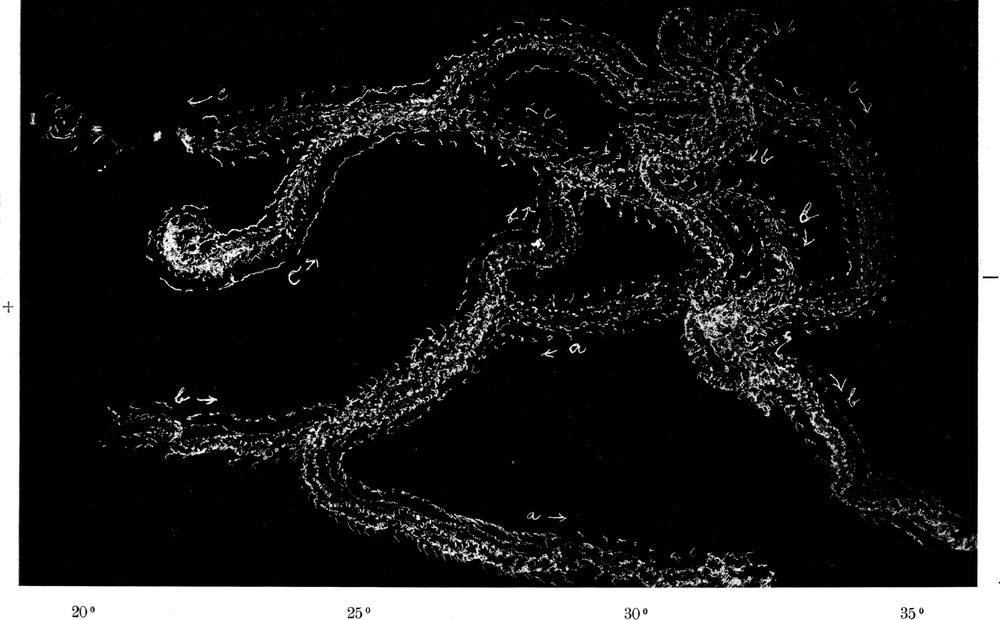
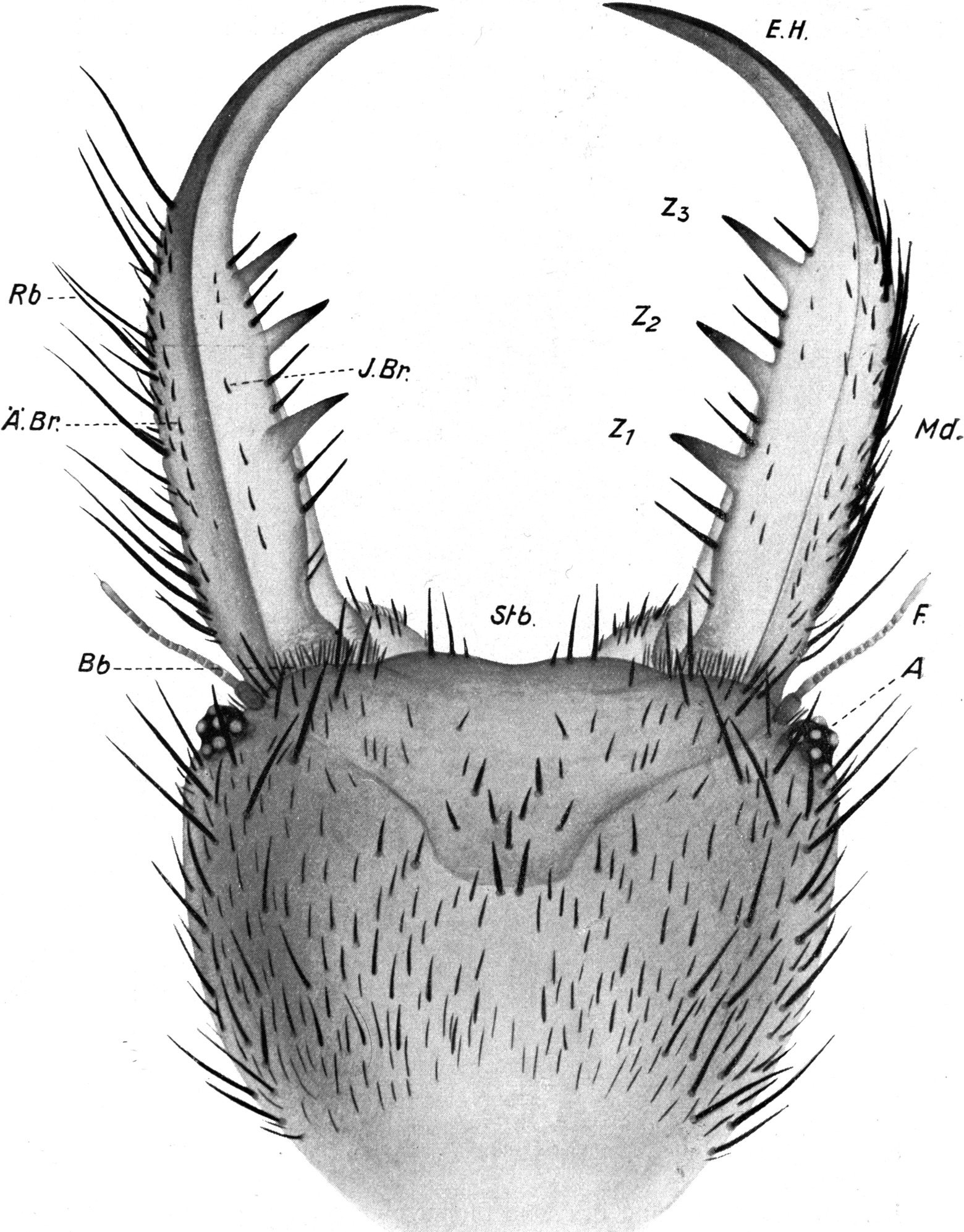{kind=link}
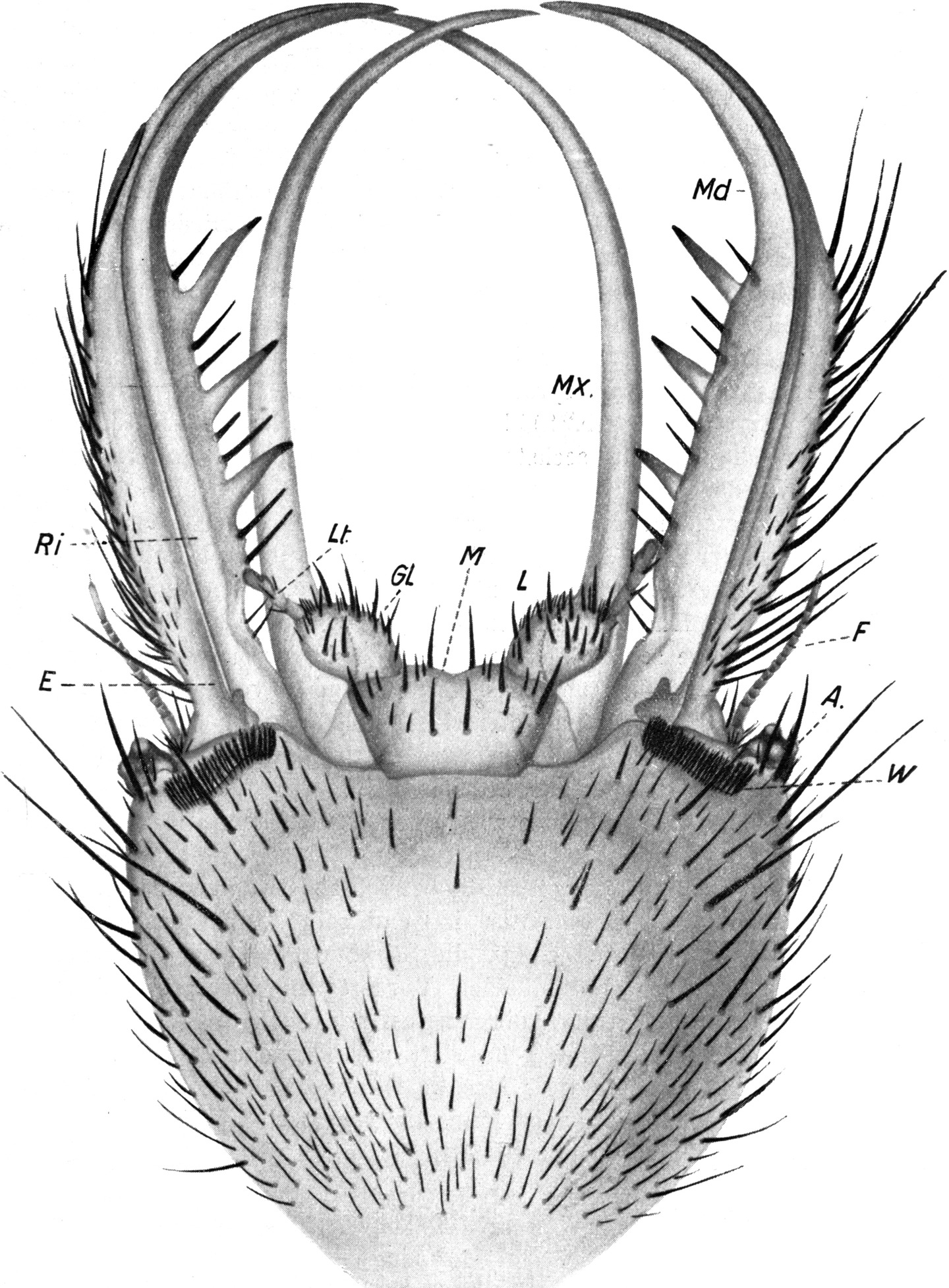{kind=link}
{kind=link}
{kind=link}
{kind=link}
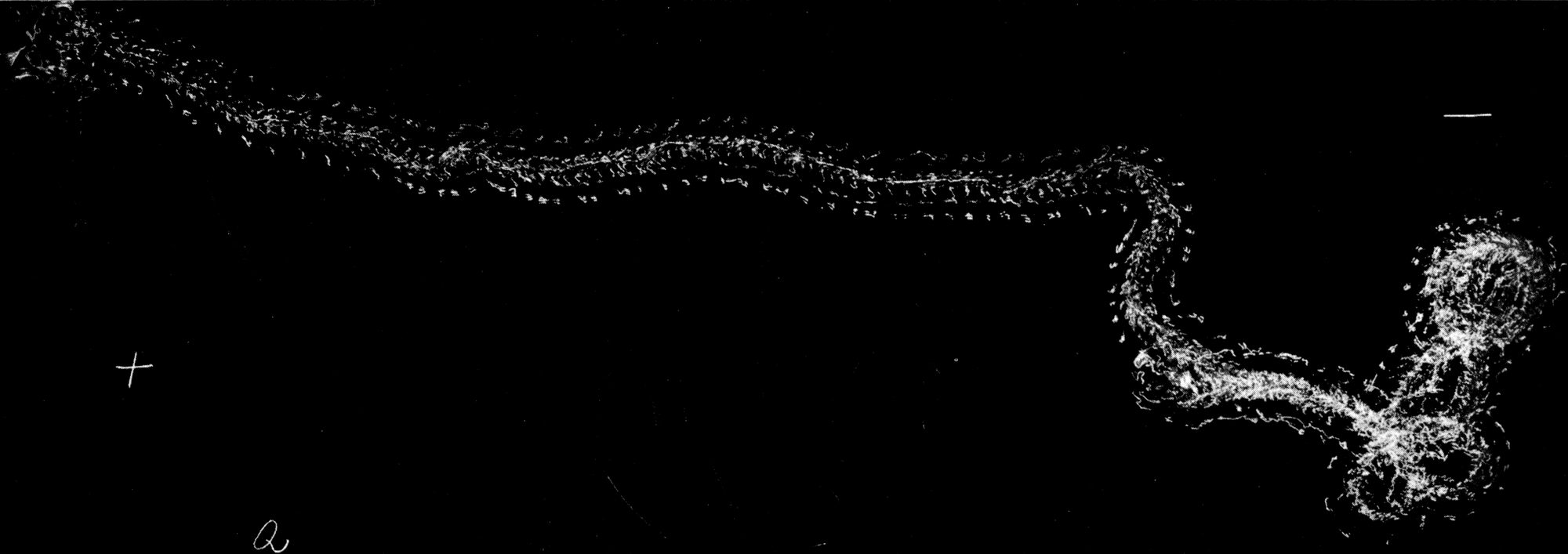{kind=link}
{kind=link}
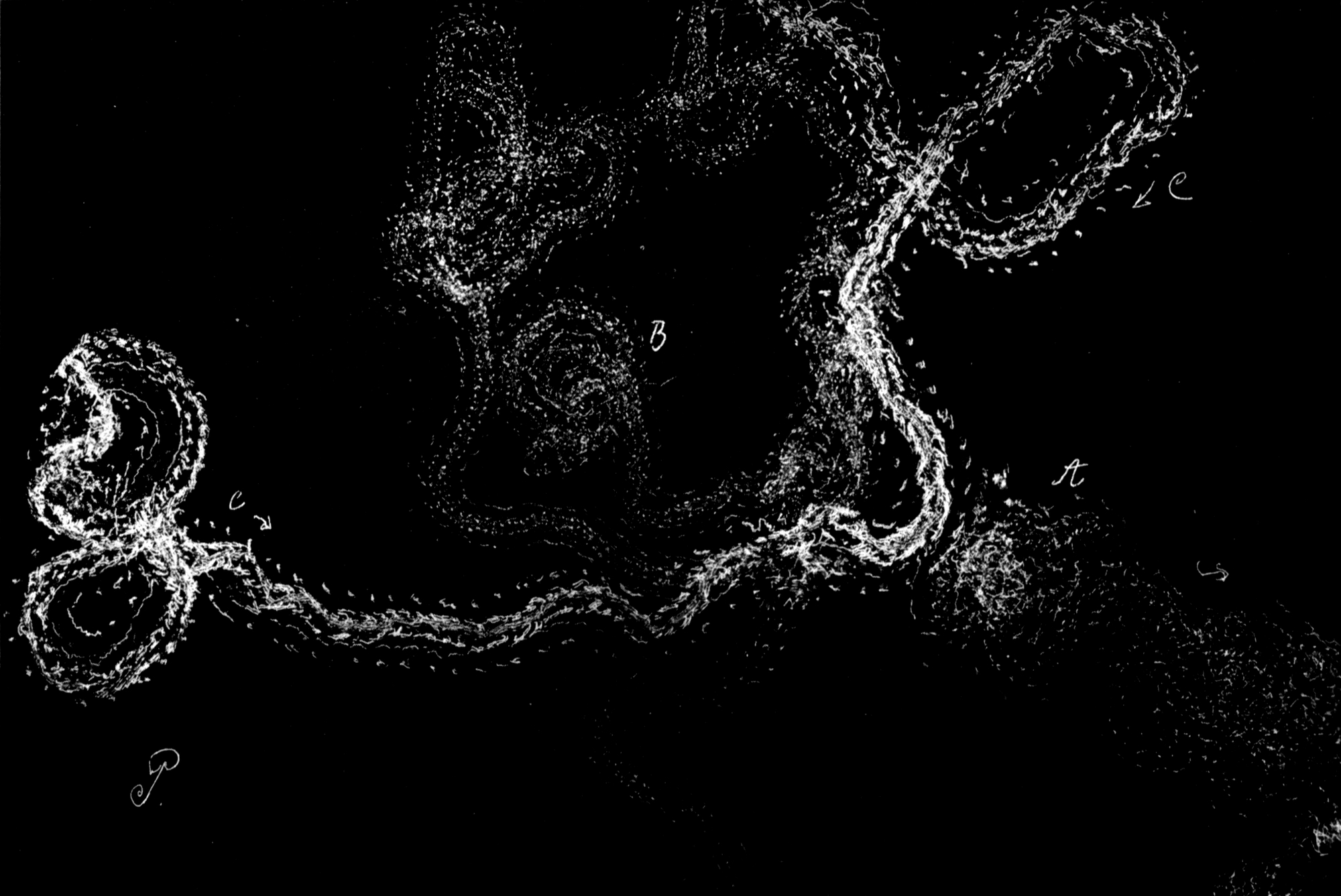{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
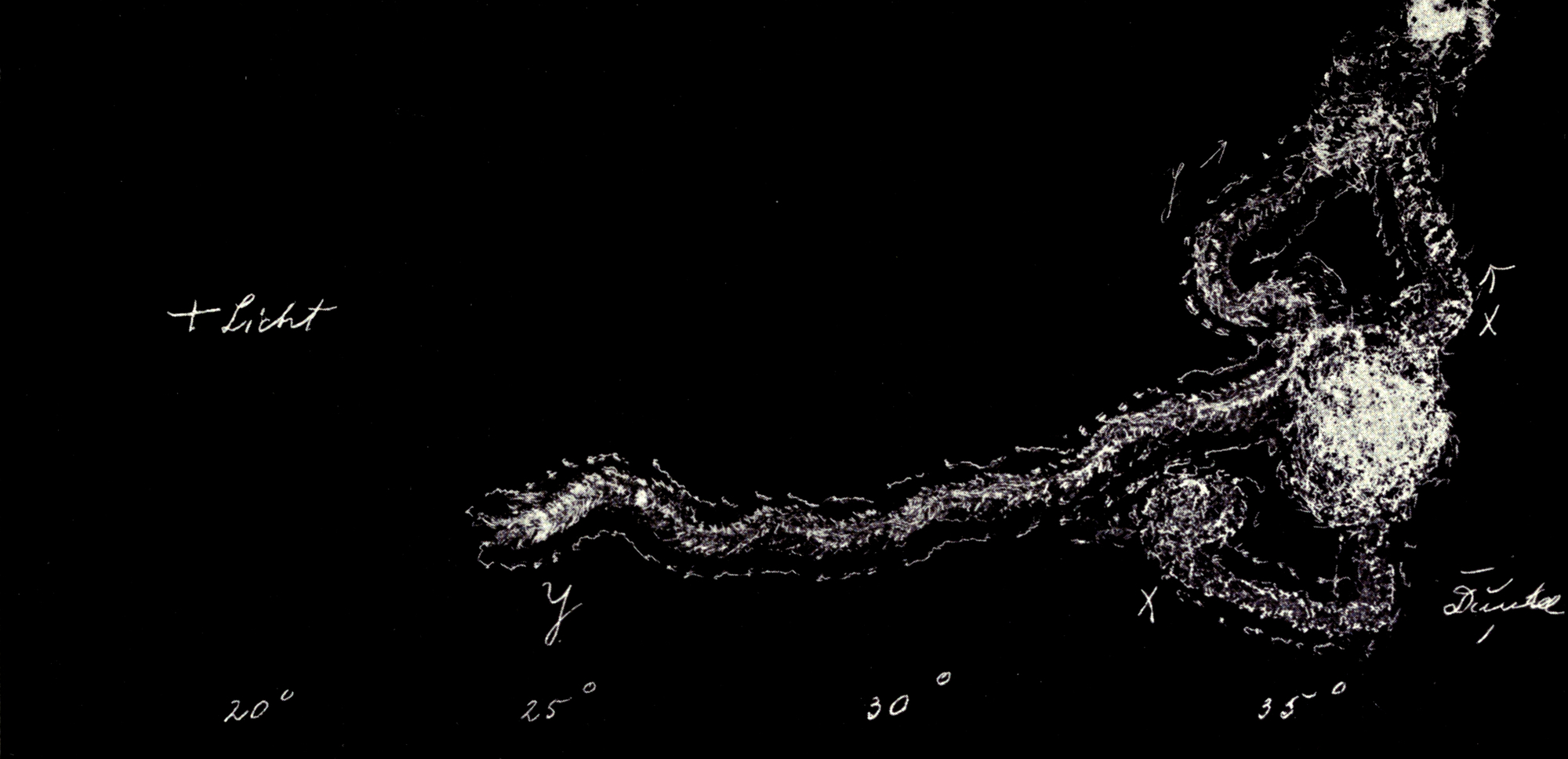{kind=link}
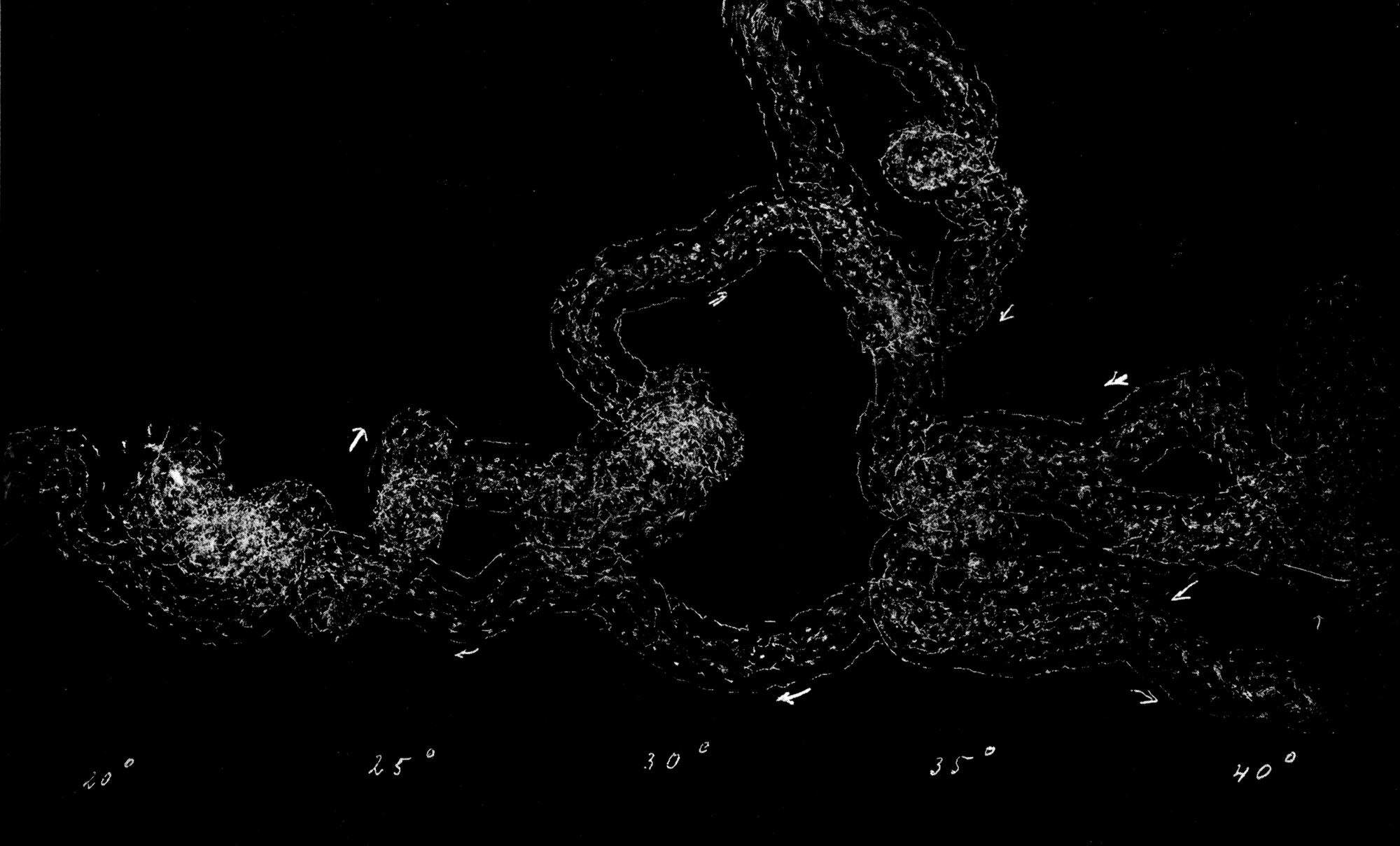{kind=link}
{kind=link}