
Title: Das Protistenreich
eine populäre Uebersicht über das Formengebiet der niedersten Lebewesen; Mit einem wissenschaftlichen Anhange: System der Protisten
Author: Ernst Haeckel
Release date: February 12, 2025 [eBook #75352]
Language: German
Original publication: Leipzig: Ernst Gunther's Verlag, 1878
Other information and formats: www.gutenberg.org/ebooks/75352
Credits: Peter Becker, Alpo Tiilikka, Adrian Mastronardi and the Online Distributed Proofreading Team at https://www.pgdp.net
Anmerkungen zur Transkription
Das Original ist in Fraktur gesetzt. Im Original gesperrter Text ist so ausgezeichnet. Im Original in Antiqua gesetzter Text ist so markiert.
Offensichtliche Fehler wurden stillschweigend korrigiert.
Eine populäre Uebersicht
über das
Formengebiet der niedersten Lebewesen.
Mit einem wissenschaftlichen Anhange:
SYSTEM DER PROTISTEN.
Von
E. HÆCKEL.
Mit zahlreichen Holzschnitten.

Leipzig.
Ernst Günther’s Verlag.
1878.
Alle Rechte vorbehalten.
[S. 1]
Für das tiefere Verständniss unserer heutigen Entwicklungslehre und der darauf gegründeten einheitlichen Weltanschauung dürften wenige Zweige der Naturwissenschaft von so fundamentaler Bedeutung sein, wie die Naturgeschichte der niedersten Lebewesen, der sogenannten Protisten. Denn die urwüchsige Einfachheit im Körperbau und in den Lebens-Erscheinungen dieser unvollkommnen »Urwesen« öffnet uns erst den wahren Weg für das Verständniss der viel entwickelteren und schwierigeren Erscheinungen, welche uns die Anatomie und Physiologie der höheren und vollkommneren Organismen, der echten Thiere und Pflanzen darbietet. Dennoch ist die Bekanntschaft mit den Protisten bisher fast nur auf die gelehrten Fachkreise beschränkt geblieben und erst sehr wenig in weitere Kreise eingedrungen. Das ist auch leicht erklärlich. Denn die grosse Mehrzahl jener einfachsten Lebensformen, die wir im »Protistenreich« zusammenfassen, ist dem unbewaffneten Auge völlig verborgen. Erst durch das Mikroskop können wir sie erkennen, und meistens erst mit Hülfe starker Vergrösserungen ihre Form-Verhältnisse genau erforschen. Aber auch dann ist diese Erforschung noch mit vielen Schwierigkeiten und Hindernissen verknüpft. Denn die allgemeinen Anschauungen vom lebendigen Organismus, die gewöhnlichen Begriffe von den Organen und Functionen der Lebewesen, welche wir aus der alltäglichen Anschauung des höheren Thier- und Pflanzen-Lebens uns gebildet haben, passen nur wenig oder gar nicht auf jene niedersten Lebensformen. Ausserdem ist aber auch die gründliche wissenschaftliche Forschung der letzteren kaum vierzig Jahre alt; und erst die sehr ausgedehnten und sorgfältigen Untersuchungen der letzten zwanzig Jahre haben ihre Kenntniss auf eine solche Höhe gebracht, dass wir gegenwärtig wenigstens eine befriedigende Vorstellung von der Eigenthümlichkeit und eine klare [S. 2] Einsicht in die Bedeutung des Protisten-Reiches gewonnen haben.
Wenn wir nun hier den Versuch wagen, in allgemein-verständlicher Form eine kurze Uebersicht über das ganze grosse Protistenreich zu geben, und seine hohe Bedeutung für die Entwicklungslehre dem Verständniss der gebildeten Kreise näher zu bringen, so sind wir uns der grossen damit verknüpften Schwierigkeiten wohl bewusst. Wir glauben aber denselben am besten zu begegnen, wenn wir uns auf die gedrungene Zusammenfassung des Wichtigsten beschränken, und die Bekanntschaft mit dem höchst mannigfaltigen und interessanten Detail dieses unendlich reichen Forschungs-Gebietes dem Studium der Special-Werke überantworten. Zunächst wird sicher für unsere moderne Entwicklungslehre und weiterhin auch für unsere damit verknüpfte monistische Weltauffassung schon viel gewonnen sein, wenn eine allgemeine Anschauung von dem weiten Umfang des mikroskopischen Lebensreiches, von der Einfachheit und elementaren Bedeutung des »kleinsten Lebens« sich einen Platz im Bewusstsein unserer gebildeten Kreise erobert hat.
Die niedersten Lebewesen, die wir hier als Protisten, d. h. »Erstlinge« oder »Urwesen« zusammenfassen, werden in weiteren Kreisen auch heute noch sehr oft mit den unpassenden Namen Infusorien oder Infusionsthierchen (im weiteren Sinne!) bezeichnet. In den systematischen Lehrbüchern der Naturgeschichte werden sie meistens als Urthiere (oder »Protozoa«) aufgeführt. Die beste deutsche Bezeichnung für die ganze grosse Gruppe wäre vielleicht: Zellinge oder Zellwesen; denn es würde dadurch die wesentlichste Eigenthümlichkeit ihrer Organisation, die autonome Selbständigkeit und permanente Individualität ihres einfachen Zellen-Leibes in präcisester Weise ausgedrückt.
Obgleich Viele von der Existenz der meisten mikroskopischen Protisten keine Ahnung haben, so kommt dennoch jeder Mensch unendlich oft mit ihnen in Berührung. Jeder hat beim Wassertrinken, beim Essen von Früchten, Austern und anderen rohen Speisen schon Tausende und Millionen von lebenden Protisten verschluckt, ohne sich dessen bewusst geworden zu sein. Denn obgleich diese merkwürdigen Geschöpfe von dem unbewaffneten Auge des Menschen zum grössten Theile gar nicht erkannt oder [S. 3] höchstens als ganz kleine Pünktchen wahrgenommen werden, sind sie dennoch in zahllosen, höchst mannigfaltigen und interessanten Formen allenthalben über unseren Erdball verbreitet. Unsere Mikroskope weisen uns dieselben überall im süssen und salzigen Wasser nach. Alle Bäche und Flüsse, alle Teiche und Seen, alle Tümpel und Gräben enthalten solche Urthierchen, oft in unglaublicher Masse. Man kann keinen Stein, keine Pflanze aus dem Wasser heben, ohne in dem daran haftenden schleimigen Ueberzug wenigstens einzelne Infusorien zu finden. Ebenso ist das Meer überall von ihnen belebt. Der weiche Schlamm, der den Meeresgrund bedeckt, besteht zum grossen Theil aus dergleichen Protozoen. Der feine schlammige Ueberzug, der bei ruhigem Wetter den klaren Meeresspiegel überzieht, ist aus Milliarden schwimmender Infusorien zusammengesetzt. Aber auch der Staub unserer Strassen, der Sand unserer Dachrinnen, die Humus-Erde unserer Felder und Wälder, enthält Millionen kleinster Infusorien-Keime, sowie eingetrocknete, aber noch lebensfähige Körper derselben. Wir brauchen bloss diesen Staub und Sand in einem Glase mit etwas Wasser zu übergiessen und diesen Aufguss einige Zeit in der Sonne stehen zu lassen, um durch unser Mikroskop Massen von beweglichen Infusorien wahrzunehmen; theils haben sie sich in kürzester Zeit aus jenen Keimen entwickelt, theils sind sie unter dem belebenden Einflusse des Wassers aus ihrem Trockenschlafe zu neuem Leben erwacht. Ist es ja doch gerade diese Erscheinung, die zu der Benennung: Infusoria oder Infusionsthierchen, d. h. »Aufgussthierchen« Veranlassung gab.
Es sind jetzt kaum zweihundert Jahre verflossen, seitdem die mikroskopischen Infusorien durch den holländischen Naturforscher Anton van Leeuwenhoek zuerst in einem Topfe voll stehenden Regenwassers entdeckt wurden. Die Holländer haben die zweihundertjährige Jubelfeier dieser Entdeckung, die damals das grösste Aufsehen erregte, vor wenigen Jahren (1875) feierlichst begangen; und sie thaten Recht daran. Denn die wissenschaftliche Tragweite derselben ist in der That unermesslich, und je mehr wir mit unseren vervollkommneten Mikroskopen in die tiefsten Geheimnisse des Lebens eindringen, desto mehr werden wir uns ihrer Bedeutung bewusst.
Unsere ganze Anschauung vom Wesen des Lebens und [S. 4] von der Entwicklung der organischen Gestalten ist durch die genauere Erkenntniss dieser Urthierchen oder Infusionsthierchen unendlich erweitert und gefördert worden. Anatomie und Physiologie, Entwickelungsgeschichte und Systematik verdanken ihr die wichtigsten Aufschlüsse. Selbst für die Geologie haben sie eine ausserordentliche Bedeutung erlangt. Denn diese kleinsten Lebensformen haben keinen geringeren Einfluss auf die Bildung der mächtigsten Gebirgsmassen und auf die ganze Gestaltung unserer Erdrinde ausgeübt, als alle die zahlreichen grossen Thiere und Pflanzen, die unsern Planeten seit Millionen von Jahren belebt haben. Die mikroskopischen Kalkschalen und Kieselgehäuse, welche sich die meisten Urthiere bilden, bleiben nach dem Tode ihrer Bewohner unverändert übrig. Sie häufen sich auf dem Grunde der Gewässer massenhaft an, bilden hier mächtige Schlammschichten und werden im Laufe der Jahrtausende zu festem Gesteine verdichtet. So sind z. B. die Kreide-Gebirge von England und von der Insel Rügen, sowie die über der Kreideformation abgelagerten eocänen Tertiärschichten zum grössten Theile, oft fast ausschliesslich, aus den zierlichen Kalkschalen der Polythalamien zusammengesetzt. Andere Gesteine, wie z. B. die tertiären Felsmassen von Barbados und von den Nikobaren-Inseln, zeigen sich zum grössten Theile aus den reizenden Kieselpanzern der Radiolarien gebildet. Viele von den Gesteinen, welche solchen Urthierchen ihre Entstehung verdanken, liefern ein vorzügliches Baumaterial; und manche unserer grössten Städte sind vorzugsweise aus dergleichen Steinen erbaut, so z. B. Wien und Paris.
Die berühmten Tiefsee-Forschungen der neuesten Zeit, zu denen die erste Legung des atlantischen Telegraphen-Kabels den Anstoss gab, haben jene felsbildende Macht des kleinsten Lebens in das hellste Licht gestellt. Sie haben uns gezeigt, wie noch heute in den tiefsten Abgründen des Meeres unaufhörlich kreideartiges Gestein aus feinstem Meeresschlamm entsteht, und wie dieser Schlamm fast ausschliesslich aus den Kalkschalen und Kieselpanzern unglaublicher Massen von Urthierchen gebildet wird. Vor Allem sind es hier die unvergleichlichen Entdeckungen der bewunderungswürdigen britischen Challenger-Expedition, welche uns mit einer Fülle neuer und überraschender Anschauungen über die »Mikrogeologie«, [S. 5] über das reiche, räthselvolle, mikroskopische Leben der Tiefsee-Thäler bereichert haben.
Wie nun die eifrigen Forschungen des letzten halben Jahrhunderts unsere Kenntniss vom Leben und Weben der Urthiere, von ihrer Gestaltung und Entwickelung ungemein gefördert haben, so haben sie auch unsere Ansichten von ihrer Stellung in der Natur und von ihrer systematischen Gruppirung sehr wesentlich verändert. Das System der organischen Formen ist ja immer mehr oder weniger der Ausdruck der Anschauungen, welche wir von ihrer natürlichen Verwandtschaft besitzen, und so zeigen uns denn auch die grossen Veränderungen, welche das System der Urthiere im Verlauf der letzten Jahrzehnte erlitten hat, am klarsten den gewaltigen Umschwung unserer bezüglichen Vorstellungen. Nachdem vor neunzig Jahren (1786) Otto Friedrich Müller den ersten umfassenden Entwurf eines Systems der Infusionsthierchen gegeben hatte, erschien vor vierzig Jahren das grosse Prachtwerk des berühmten (1876 verstorbenen) Naturforschers Ehrenberg: »Die Infusionsthierchen als vollkommene Organismen. Ein Blick in das tiefere organische Leben der Natur.« Das war im Jahre 1838, in demselben für die Naturwissenschaft Epoche machenden Jahre, in welchem der geniale Botaniker Schleiden in Jena zuerst den Grund zu der höchst fruchtbaren Zellen-Theorie legte. In der That ein merkwürdiger Zufall; eine seltsame Ironie des Schicksals. Denn Ehrenberg war in seinem grossen Hauptwerke vor Allem bemüht, das ihm eigene »Princip überall gleich vollendeter Entwicklung« zur Geltung zu bringen. Er suchte bei den Infusorien eine eben so vollkommene Organisation nachzuweisen, wie bei den höheren Thieren und beim Menschen. Er glaubte überall Nerven und Muskeln, Darm und Blutgefässe, männliche und weibliche Organe unterscheiden zu können. Gerade dieses Prinzip war grundfalsch; vielmehr sind die Infusorien höchst einfache Organismen: die meisten haben nur die Bedeutung und den Werth einer einzigen einfachen Zelle; und ihr wahres Verständniss wird uns erst durch die Zellentheorie gegeben.
Der alte Name »Infusionsthierchen« wird heute nur noch auf einen kleinen Theil der mikroskopischen Wesen angewendet, welche Ehrenberg in seinem grossen Werke als solche beschrieb. Nur die Wimperthierchen oder Ciliaten und die Borstenthierchen [S. 6] oder Acineten, oft auch die Geisselschwärmer oder Flagellaten werden heute noch in wissenschaftlichen Werken »Infusorien« genannt; die formenreichen Kieselzellen oder Diatomeen werden dagegen meist von den Botanikern zu den Algen gerechnet. Die Räderthierchen (Rotatoria), die für Ehrenberg gerade den Typus der Infusoria bildeten, sind Würmer, also Thiere von viel höherer Organisation. Dagegen bilden die Amoeben und ihre Verwandten heute eine besondere wichtige Protisten-Classe, die wir Lappenthierchen oder Lobosa nennen. Neben diesen aber hat die fortgeschrittene mikroskopische Forschung uns andere Classen von Urthierchen kennen gelehrt, die viel zahlreichere, merkwürdigere und mannigfaltigere Formen enthalten, als jene älteren Infusionsthierchen: vor allen die wunderbare Classe der Wurzelfüssler oder Rhizopoden; die Sonnenthierchen oder Heliozoen, die kalkschaligen Thalamophoren und kieselschaligen Radiolarien. Diesen schliessen sich eng die sonderbaren Schleimpilze oder Myxomyceten an, welche die Botaniker früher zu den echten Pilzen (Fungi) stellten. Aber auch die Stellung dieser letzteren im Pflanzenreiche ist ganz zweifelhaft geworden und es bestehen gewichtige Gründe dafür, sie aus letzterem in das Protistenreich zu versetzen. Als eine besondere, interessante, wenn auch nur sehr kleine Protisten-Classe dürfen wir die Catallacten betrachten. Endlich finden wir unten auf der tiefsten Stufe jener höchst einfachen, wunderbaren Wesen, mit denen das organische Leben in denkbar einfachster Gestalt beginnt, die Moneren.
Schon beim ersten Blick auf die wunderbare Formenwelt, welche uns hier das Mikroskop entschleiert, wird sich jedem Unbefangenen zunächst die Frage aufdrängen: »Sind denn diese sogenannten Urthiere oder Infusionsthiere wirkliche, echte Thiere und warum werden sie von den Naturforschern in das Thierreich gestellt?« Diese Frage ist vollständig berechtigt; sie gehört zu jenen schwierigen Grundfragen der allgemeinen Biologie, deren Lösung durch unsere fortschreitende Kenntniss eher erschwert als erleichert wird. Wenn wir nämlich althergebrachter Maassen die ganze organische Natur in die beiden grossen Hälften: Thierreich und Pflanzenreich eintheilen, und wenn wir damit glauben den natürlichen Gegensatz zwischen zwei völlig getrennten Hauptgebieten auszusprechen, so ist diese Unterscheidung zwar durch die [S. 7] festgewurzelte Anschauung und den Sprachgebrauch von Jahrtausenden geheiligt; aber logisch begründbar und wirklich naturgemäss ist sie nicht. Vielmehr lehren uns gerade unsere Urthierchen das Gegentheil. Je genauer wir deren Formen und Lebenserscheinungen studiert haben, je vollständiger uns ihre ganze Entwicklungsgeschichte bekannt geworden ist, desto klarer hat sich herausgestellt, dass sie eine ununterbrochene Verbindungsbrücke zwischen den tiefsten Stufen des Thierreichs und des Pflanzenreichs herstellen. So leicht und sicher wir die höheren und vollkommneren Stufen der beiden grossen Reiche von einander unterscheiden können, so schwer, ja so unmöglich wird diese Trennung auf den niedrigsten und unvollkommensten Stufen. Denn hier sind beide Reiche durch eine zusammenhängende Kette von einfachen Uebergangsformen untrennbar verbunden.
Die Erkenntniss dieser wichtigen Thatsache, welche heute unzweifelhaft festgestellt ist, hat zu den lebhaftesten Streitigkeiten über die Grenze zwischen Thierreich und Pflanzenreich Veranlassung gegeben. Sie hat zugleich die abweichendsten Anschauungen über das Wesen der zweifelhaften Infusorien hervorgerufen, die mitten zwischen den beiden grossen Reichen der organischen Natur ein neutrales Grenzgebiet für sich in Anspruch nehmen.
Während nämlich viele Infusorien von den Zoologen für Thiere, von den Botanikern dagegen für Pflanzen erklärt, und demnach von Beiden annectirt wurden, hatten Andere gerade das entgegengesetzte Schicksal: sie wurden von Beiden verschmäht; bei einer dritten Gruppe von Infusorien schien sogar nur die Annahme übrig zu bleiben, dass sie abwechselnd als Thiere und Pflanzen lebten. Der daraus entspringende Streit über ihre wahre Natur scheint am einfachsten dadurch entschieden zu werden, dass man den Begriff von Thier und Pflanze scharf umschreibt, und diese unzweideutige Begriffsbestimmung auf jene zweifelhaften Mittelwesen anwendet. Aber diese gesuchte Begriffsbestimmung selbst ist ein unlösbares Problem; je mehr Mühe man darauf verwendet hat, desto klarer hat sich herausgestellt, dass es überhaupt auf einer falschen Fragestellung beruht, und dass die Begriffe von Thier und Pflanze nicht in der Natur begründet sind.
Um nun den so entstandenen Schwierigkeiten zu entgehen, und um zu einer vernünftigen Classification der organischen Wesen [S. 8] zu gelangen, ist schliesslich nur ein Ausweg übrig geblieben: nämlich die Aufstellung eines dritten, selbständigen Reiches von elementaren Organismen: Das ist unser Reich der Protisten oder Zellinge, das Reich der neutralen Urwesen. Wir fassen demnach die ganze organische Natur, die Gesammtheit aller lebenden Wesen unsers Erdballs, als ein grosses einheitliches Ganze auf; und dieses umfassende Universalreich theilen wir in drei Reiche: das Thierreich einerseits, das Pflanzenreich andrerseits, mitten zwischen Beiden das neutrale Reich der Protisten.
Um nun die Aufstellung unseres Protistenreich zu rechtfertigen, wollen wir einen flüchtigen Blick auf die verschiedenen Character-Seiten des Thier- und Pflanzenreichs werfen. Es wird sich dabei von selbst ergeben, dass unsere Protisten weder dem einen, noch dem anderen vollständig entsprechen. Verweilen wir zunächst einen Augenblick bei der äusseren Gesammterscheinung. So characteristisch uns da einerseits das höhere Thier mit der Gliederung seines Leibes und seiner Gliedmaassen, anderseits die höhere Pflanze mit ihrem Stengel und ihren Blättern entgegentritt, so wenig reicht diese äussere Gliederung hin, um die niederen Formen beider Reiche zu unterscheiden. Viele unzweifelhafte Thiere, wie z. B. die Korallen, die Schwämme, ahmen so vollkommen die Gestalt echter Pflanzen nach, dass man sie früher allgemein für solche gehalten hat. Umgekehrt giebt es viele unzweifelhafte Pflanzen, wie z. B. viele Orchideen und andere Schmarotzer, welche die Gestalt echter Thiere nachahmen. Und was sollen wir nun vollends zu den unendlich mannigfaltigen Figuren unserer Protisten sagen? Da treffen wir allein schon in der einen Classe der kieselschaligen Radiolarien alle möglichen Grundformen verkörpert an, die überhaupt in der Natur vorkommen können; und in welcher zierlichen und wundervollen Ausführung! Da finden wir in einem einzigen Tropfen Meerwasser nebeneinander Kugeln, Kreuze, Körbchen, Schrauben, Sterne, Schachfiguren, Hörner, Hauben, Helme, u. s. w.; kurz eine Fülle der mannigfaltigsten und merkwürdigsten Gestalten. Gewiss wird Jedermann, der diese Formen zum ersten Male sieht, sie für Kunstproducte halten, oder vielleicht für abgelöste Theile von grösseren Organismen. Und doch sind es vollkommen entwickelte und selbständige [S. 9] Lebewesen! Aber Niemand wird geneigt sein, sie für echte Thiere oder echte Pflanzen zu erklären. Ebenso wenig können wir aus der äusseren Körperform der meisten anderen Protisten einen sicheren Schluss auf ihre wahre Natur ziehen. Sehr Viele bewahren zeitlebens die einfache Kugelgestalt. Andere zeigen beständig die einfache Form eines Cylinders, einer Scheibe, eines Kegels, einer Pyramide u. s. w. Noch Andere endlich haben überhaupt gar keine bestimmte Gestalt, so namentlich die Moneren und die Amoeben. Der ganze Körper dieser höchst einfachen Urwesen besteht aus einem lebenden mikroskopischen Schleimklümpchen, das in unablässigem Wechsel seine Gestalt beständig ändert: daher der passende Name »Aenderling«, den Oken diesen Amoeben beilegte.
Doch verlassen wir die äussere Körperform! Denn dass diese ganz unzureichend ist, um den Unterschied zwischen Thier und Pflanze zu begründen, das ist längst allgemein anerkannt. Fragen wir uns lieber, was denn eigentlich in der naiven Anschauung des täglichen Lebens diese Unterscheidung begründet, und was dieselbe seit Jahrtausenden in der Sprache und im Begriffsleben der Menschheit gerechtfertigt hat. Unzweifelhaft sind es die Lebenserscheinungen der Empfindung und Bewegung, welche uns hier zunächst entgegentreten. Empfindung und Bewegung sind es, welche in der allgemeinen Anschauung das Thier gegenüber der Pflanze auszeichnen, und aus denen wir auf ein »Seelenleben« des Thieres schliessen, ein Seelenleben, das wir der Pflanze absprechen. Wie verschieden auch die psychologischen Vorstellungen sind, und wie weit auch die Ansichten über das eigentliche Wesen der Seele aus einander gehen, darüber sind wir doch Alle einig, dass mindestens den höheren Thieren eine Art Seelenleben zukommt. Denn die Hausthiere, die wir täglich um uns sehen, bewegen sich zweifellos ebenso willkürlich, wie wir selbst. Sie empfinden die Eindrücke der Lust und Unlust, der Freude und des Schmerzes zweifellos ähnlich, wie wir selbst. Auch lehrt uns ja sofort jede anatomisch-physiologische Untersuchung, dass das Nervensystem, das Organ dieser Seelenthätigkeiten, bei den höheren Wirbelthieren im Wesentlichen eine ähnliche Einrichtung besitzt, wie bei uns selbst.
Von diesen augenfälligen Seelenthätigkeiten der höheren Thiere ausgehend, schliessen nun die Zoologen, dass dieselben auch allen [S. 10] anderen Thieren zukommen, und demgemäss werden seit alter Zeit Empfindung und willkürliche Bewegung als charakteristische Eigenschaften des Thieres betrachtet. Schon Linné sagt: »Die Pflanzen leben, die Thiere leben und empfinden.« Und doch ist gerade diese, allgemein angenommene Unterscheidung völlig unhaltbar. Wir brauchen nur an den gewöhnlichen Badeschwamm zu denken, um uns davon zu überzeugen. Dieser Badeschwamm, mit dem sich der Kulturmensch täglich zu waschen pflegt, ist das todte Skelet, das innere Gerüst eines unzweifelhaften Thieres. Im Leben stellt dieses Thier einen fleischigen, schwarzen, formlosen Klumpen dar, der unbeweglich auf dem Meeresboden festgewachsen ist. Aehnliche Seegewächse aus der Klasse der Schwämme oder Spongien sitzen massenhaft auf dem Boden aller Meere, hunderte von verschiedenen Arten. Die meisten zeigen keine Spur von Bewegung und Empfindung; sie galten daher auch früher allgemein für Pflanzen. Erst die genauesten Untersuchungen über ihre Entwickelungsgeschichte haben uns in den letzten Jahren darüber belehrt, dass wir sie als echte, unzweifelhafte Thiere betrachten müssen.
Aehnliche echte Thiere, welche in vollkommen reifem und ausgebildetem Zustande der Empfindung und Bewegung entbehren, kennen wir jetzt in Menge. Die meisten leben festgewachsen auf dem tiefen Grunde des Meeres. Sie gehören sehr verschiedenen Classen an: Würmern, Ascidien, Mollusken u. s. w. Viele von ihnen werden auf italienischen Fischmärkten unter den Namen »Seefrüchte« (Frutti di mare) feil geboten, und sowohl der Fischer, der sie verkauft, wie der Fremde, der sie mit Appetit verspeist, hält sie für die Früchte von Seegewächsen.
Sogar unter den höheren Thierklassen, z. B. unter den Schnecken und Krebsen, giebt es einzelne Arten, die in vollkommen reifem Zustande einen formlosen runden Klumpen, ohne jede Spur von Bewegung und von Empfindung, darstellen. In diesen Fällen ist es die schmarotzende Lebensweise, durch welche das Thier seine »Seele« verloren hat. Das gilt z. B. von der berühmten Wunderschnecke (Entoconcha mirabilis) und von dem merkwürdigen Säckchenkrebse (Sacculina). Erstere lebt als Parasit im Innern von Seegurken oder Holothurien: letzterer sitzt schmarotzend auf andern Krebsen fest. Beide Thiere haben die Gestalt eines einachen [S. 11] länglichen, runden Schlauches: und dieser Schlauch enthält nichts weiter als Eier. Keine Spur von einem Kopfe und von Sinnesorganen: keine Spur von Fühlhörnern und Beinen: keine Spur von Empfindung und willkürlicher Bewegung. Gewiss würde kein Mensch in diesen beiden seelenlosen Eierschläuchen wahre Thiere vermuthen, und doch stellt die Entwickelungsgeschichte unzweifelhaft fest, dass das eine eine Schnecke und das andere ein Krebs ist.
Als Gegenstück zu diesen »seelenlosen Thieren« treffen wir auf der anderen Seite »seelenvolle Pflanzen«, die uns noch mehr überraschen. Wir betreten einen tropischen Urwald und wollen uns ein zierlich gefiedertes Mimosenblatt abpflücken. Aber kaum berühren wir den zarten Zweig der schamhaften Sinnpflanze (Mimosa pudica), so klappen alle Blätter ihre zierlichen Fieder-Reihen zusammen und die Blattstiele sinken wie gelähmt herab. Ja manche dieser akazienartigen Bäume sind so reizbar, so empfindlich, dass schon die Erschütterung des Bodens durch den Tritt des herannahenden Wanderers hinreicht, sämmtliche Blätter zum Schliessen zu bringen. Nicht minder empfindlich sind neben vielen Anderen die durch Darwin berühmt gewordenen »insektenfressenden Pflanzen«. Sobald eine unvorsichtige Fliege sich auf das Blatt einer »Fliegenfalle« (Dionaea) setzt, klappt das reizbare Blatt zusammen, und die mörderische Pflanze verzehrt das erfasste Insect mit offenbarem Wohlbehagen. Wollten wir diesen hochorganisirten Pflanzen eine Seele absprechen, so müssten wir sie ganz ebenso auch bei den empfindlichen, aber festgewachsenen, pflanzenähnlichen Korallen leugnen; denn diese geben keine anderen Aeusserungen ihres Seelenlebens.
Aber nicht allein solche hohe Empfindlichkeit, solche lebhafte Beweglichkeit einzelner Körpertheile treffen wir vielfach bei echten Pflanzen an. Nein, auch selbständige, freie Ortsbewegung, auch die Willensthätigkeit, auf die wir aus der scheinbar willkürlichen Bewegung schliessen, findet sich bei unzweifelhaften Pflanzen vor. Viele Algen, z. B. viele von unsern einheimischen grünen Wasserfäden oder Conferven, schwimmen in ihrer Jugend frei und lebhaft im Wasser umher. Die jungen Pflänzchen bewegen sich dabei, ebenso wie viele junge Thiere, durch zarte, haarförmige, schwingende Fäden, Geisseln oder Wimpern. Bei dieser Schwimmbewegung [S. 12] äussern sie eben so viel Lebhaftigkeit, eben so viel Ausdauer, eben so viel scheinbaren Willen, wie die ganz ähnlichen, flimmernden Jugendformen vieler Thiere, z. B. die Gastrula. Auf den Wiener Botaniker Unger, der zuerst vor 35 Jahren (im Jahre 1843) diese frei beweglichen Jugendformen von Algen entdeckte, machten dieselben einen so tiefen Eindruck, dass er seine bezügliche Mittheilung betitelte: »Die Pflanze im Momente der Thierwerdung.«
Schon aus diesen wenigen Thatsachen, die wir noch durch Aufzählung vieler ähnlicher Erscheinungen beträchtlich vermehren könnten, geht unzweifelhaft hervor, dass die höheren Seelenthätigkeiten der bewussten Empfindung und der willkürlichen Bewegung weder allen Thieren eigenthümlich sind, noch allen Pflanzen fehlen. Sie können daher nicht mehr in der üblichen Weise zur Unterscheidung von Thier- und Pflanzenreich benutzt werden; und ebenso wenig sind sie von systematischer Bedeutung für unser Protistenreich. Für die Beurtheilung dieses letzteren ist es gleichgültig, ob sich die Protisten sehr lebhaft bewegen und sehr fein empfinden, wie die meisten Wimper-Infusorien; oder ob sie nur stumpfe Empfindung und träge Bewegung besitzen, wie die meisten Wurzelfüssler. Viele Protisten treten uns in zwei abwechselnden und ganz verschiedenen Zuständen entgegen: einem unbeweglichen und unempfindlichen Ruhezustande, in welchem sie uns als Pflanzen erscheinen; und einem frei beweglichen und sehr empfindlichen Zustande, in welchem sie Thieren gleichen. Wir dürfen von diesen merkwürdigen Urwesen geradezu sagen: sie sind abwechselnd Thier und Pflanze. Und so sind sie auch wirklich früher beurtheilt worden. So sind z. B. von manchen Flagellaten und Myxomyceten die vegetativen Ruhezustände als Pflanzen, die animalen Bewegungszustände als Thiere beschrieben worden, und erst viel später wurde entdeckt, dass Beide nur verschiedene Lebens-Zustände eines und desselben Protisten sind.
Wollen wir nun aber vom Standpunkte der vergleichenden Psychologie zu einem Schlusse über das Seelenleben aller dieser Geschöpfe kommen, so kann dieser Schluss nur lauten: »Alle lebenden Wesen sind beseelt, die Pflanzen so gut wie die Thiere, und die Protisten so gut wie die Pflanzen.« Innere Bewegungs-Erscheinungen, die scheinbar ohne äussere Ursachen [S. 13] entstehen und auf Ortsveränderungen kleinster Theile beruhen, insbesondere Protoplasma-Störungen, sind allen Organismen gemeinsam, und insofern ist jedes lebende Wesen beseelt, jedes ist zugleich reizbar, im gewissen Sinne empfindlich. Stufenweise erhebt sich die Seelenthätigkeit, von den unscheinbarsten und niedrigsten Anfängen ausgehend, zu immer höheren und vollkommneren Leistungen. Während die niedrigsten Thiere sich in dieser Beziehung nicht von den meisten Pflanzen und Protisten unterscheiden, steigt das Seelenleben der höheren Thiere, das Wollen und Empfinden, Vorstellen und Denken, zu einer ähnlichen Stufe wie beim Menschen empor.
Gleich der Seelenthätigkeit haben sich auch alle anderen Eigenschaften, durch welche man Thiere und Pflanzen hat unterscheiden wollen, als unzureichende Merkmale erwiesen. Unzweifelhaft der wichtigste Unterschied zwischen Beiden beruht auf den entgegengesetzten physiologisch-chemischen Verhältnissen ihrer Ernährung. Der gesammte Stoffwechsel in beiden Reichen, im Grossen und Ganzen betrachtet, ist grundverschieden. Die Pflanzen allein besitzen das Vermögen, aus den einfachen chemischen Verbindungen der leblosen anorganischen Natur, aus Wasser, Kohlensäure und Ammoniak, jene verwickelten und höchst zusammengesetzten, eiweissartigen Kohlenstoff-Verbindungen herzustellen, welche als die wahren Träger aller eigentlichen Lebens-Erscheinungen gelten, vor allen das Protoplasma oder den Bildungsstoff (»Plasson«). Das können die Thiere nicht. Sie nehmen die Eiweisskörper, die sie beständig verbrauchen und zersetzen, direct oder indirect aus dem Pflanzenreich auf. Zur Aufnahme und Verdauung ihrer Nahrung bedürfen sie einer Magenhöhle und einer Mundöffnung; und das sind die am meisten characteristischen Organe des Thierkörpers, welche dem Pflanzenorganismus stets fehlen.
Mit diesem fundamentalen Gegensatze in der Ernährung hängen auch noch andere wichtige Unterschiede beider Reiche zusammen. Die Pflanzen athmen für gewöhnlich Kohlensäure ein und hauchen Sauerstoff aus; die Thiere gerade umgekehrt. Die meisten Pflanzen bilden massenhaft jenen eigenthümlichen grünen Farbstoff, das Chlorophyll oder Blattgrün, dem unsere Erde den grünen Schmuck ihrer Vegetationsdecke verdankt. Die meisten Thiere hingegen [S. 14] bilden kein Chlorophyll. Ebenso erzeugen die meisten Pflanzen Massen von Stärkemehl (Amylum) und von Cellulose; von jener wichtigen stickstofflosen Verbindung, welche die Grundlage des Holzes bildet. Die meisten Thiere produciren kein Amylum und keine Cellulose. Und so könnten wir noch eine ganze Anzahl anderer chemischer Verbindungen anführen, welche den Gegensatz im Stoffwechsel des Thier- und Pflanzenreichs bezeichnen.
Unzweifelhaft ist dieser Gegensatz von der grössten Bedeutung. Denn auf ihm beruht das beständige Gleichgewicht in der Oekonomie der organischen Natur. Was das eine der beiden grossen Lebensreiche ausgiebt, das nimmt das andere wieder ein. Was das eine als unbrauchbar ausscheidet, das verzehrt das andere. Aber so bedeutungsvoll auch diese Wechselwirkung jedenfalls ist, so wenig ist der damit verknüpfte Gegensatz durchgreifend und zu einer beständigen Grenzmarke geeignet. Denn zahlreiche Ausnahmen finden sich in jeglicher Beziehung.
Als solche wichtige Ausnahmen sind vor allen die zahlreichen Schmarotzerpflanzen zu nennen: z. B. viele Orchideen, Orobanchen, Lathraeen u. s. w. Diese Parasiten, deren nahe Verwandtschaft zu echten hochentwickelten Pflanzen feststeht, haben durch Anpassung an schmarotzende Lebensweise ihren Stoffwechsel gänzlich geändert. Statt gleich anderen Pflanzen mühsam Eiweisskörper zu produciren, finden sie es bequemer, gleich den Thieren diese wichtigsten Lebenstoffe aus anderen Pflanzen aufzunehmen. Damit ändert sich aber ihre gesammte Ernährung. Sie bilden kein Blattgrün mehr, sie athmen Sauerstoff ein und Kohlensäure aus; sie bilden Verbindungen, die sonst nur im Thierkörper erzeugt werden.
Umgekehrt finden wir nun wieder im Thierreiche merkwürdige Schmarotzer, welche gleichfalls durch Anpassung an parasitische Lebensweise ihre ganze Ernährung völlig geändert haben. Ausser den schon angeführten Wunderschnecken und Säckchenkrebsen sind da besonders jene Würmer (Bandwürmer, Kratzwürmer u. s. w.) hervorzuheben, welche im Innern anderer Thiere leben und deren Säfte durch ihre Haut aufsaugen. Mund und Magen sind dadurch überflüssig geworden und im Laufe der Jahrtausende allmählich verloren gegangen. Die nächsten Verwandten dieser darmlosen Parasiten besitzen einen wohl [S. 15] entwickelten Mund und Darmkanal. Aber auch andere echte Thiere bieten in ihrem Stoffwechsel beträchtliche Abweichungen dar, und einige produciren Verbindungen, die sonst nur die Pflanzen erzeugen. So bilden sich z. B. die Ascidien einen Mantel aus Cellulose; die grünen Süsswasserpolypen und einige grüne Würmer erzeugen in ihrer Haut echtes Blattgrün oder Chlorophyll u. s. w.
Angesichts dieser zahlreichen Ausnahmen kann uns denn auch der Stoffwechsel unserer Protisten keinen Aufschluss über ihre wahre Natur geben. Wenn viele von ihnen Chlorophyll, Cellulose und Stärkemehl erzeugen, so beweist das ebensowenig für ihre Pflanzen-Natur, als die Bildung von Kalkschalen bei vielen Anderen für ihre Thier-Natur Zeugniss ablegt. Vielmehr sprechen auch die Verhältnisse der Ernährung und des Stoffwechsels, im Grossen und Ganzen betrachtet, für die neutrale Natur der Protisten. Allerdings wissen wir von den physiologisch-chemischen Vorgängen ihres Stoffwechsels im Ganzen noch sehr wenig. Aber dies Wenige reicht doch hin, um uns auch hierin ganz eigenthümliche Verhältnisse erkennen zu lassen. So nehmen z. B. die formlosen Amoeben und die formenreichen Wurzelfüssler zwar ihre Nahrung ähnlich den Thieren auf, aber ohne Mund und Magen. An jeder Stelle der nackten Körperoberfläche können die Nahrungsbissen in’s Innere dringen. Auch die thierähnlichsten Protisten, die Wimperthierchen, besitzen keinen wahren Darm, keinen wahren Mund und Magen. Dieser fehlt vielmehr allen Protisten.
Wir sehen also, dass keine der verschiedenen Lebenserscheinungen genügt, um uns über das Verhältniss der Protisten zu den Thieren und Pflanzen vollkommen aufzuklären. Da nun auch die äussere Gestaltung uns darüber keinerlei Aufschluss giebt, so bleiben uns nur noch diejenigen Verhältnisse übrig, welche uns das Mikroskop im feineren Bau und in der Entwicklungsgeschichte enthüllt. Ohne die genaueste Kenntniss dieser Verhältnisse können wir uns ja überhaupt kein vollständiges Bild von der Natur der Organismen machen. Alles nun, was wir bisher davon erkannt haben, findet seinen umfassendsten Ausdruck in der berühmten Zellentheorie, die seit 40 Jahren das wichtigste Fundament aller biologischen Forschungen geworden ist.
[S. 16]
Bekanntlich lehrt uns diese Zellentheorie, dass alle die tausendfach verschiedenen Formbestandtheile, die wir im Körper sämmtlicher Thiere und Pflanzen mittelst des Mikroskopes unterscheiden, lediglich verschiedene Abarten und Umbildungen eines einzigen Grundorganes, eines einzigen ursprünglichen Form-Elementes sind. Dieses Form-Element ist die Zelle, ein kleines, für das blosse Auge meist unsichtbares Körperchen, welches bis zu einem gewissen Grade ein selbständiges Leben führt. So unendlich mannigfaltig die Form der Zelle auch ist, so ist sie doch immer aus zwei verschiedenen Bestandtheilen zusammengesetzt: aus einem Stückchen weicher, eiweissartiger Substanz, dem Bildungsstoff oder Protoplasma, und aus einem festeren, davon umschlossenen Körperchen, dem Kern oder Nucleus. Die ursprüngliche Selbständigkeit der Zelle ist so vollkommen, dass man sie mit Recht als den Elementar-Organismus, als das Individuum erster Ordnung bezeichnet hat. Da die Zellen jede organische Form bilden, können wir sie auch die »Bildnerinnen« oder Plastiden nennen. Der ganze Körper der meisten Thiere und Pflanzen ist aus Milliarden solcher Zellen zusammengesetzt: und was dieses Thier, was diese Pflanze leistet, das ist in Wahrheit die Leistung ihrer zahllosen Zellen. Auch unser eigener menschlicher Leib besteht aus Milliarden derartiger Zellen, und alle unsere Lebensverrichtungen sind das höchst verwickelte Resultat aus der Thätigkeit dieser mikroskopischen Wesen. Jedes Härchen besteht aus vielen Millionen Zellen. Ein kleinstes Blutströpfchen von einem Cubik-Millimeter Rauminhalt umschliesst schon fünf Millionen Blutzellen.
Für die richtige Auffassung der Zellentheorie, von der das ganze Verständniss des Lebens abhängt, ist Nichts lehrreicher, als der oft angewendete Vergleich des vielzelligen Organismus mit einem wohlorganisirten menschlichen Staate. Die Existenz jeder geordneten staatlichen Organisation, gleichviel ob wir Monarchie oder Republik betrachten, beruht bekanntlich darauf, dass die einzelnen Staatsbürger einen Theil ihrer persönlichen Freiheit aufgeben, sich den Gesetzen des Staats unterwerfen und in die Arbeit des Lebens theilen. Ebenso geniessen auch die Zellen in jedem vielzelligen Organismus zwar bis zu einem gewissen Grade ihr selbständiges Leben; aber sie sind doch zugleich den Gesetzen [S. 17] des Ganzen untergeordnet und durch die Arbeitstheilung von einander abhängig. Wir können diesen politischen Vergleich auch noch weiter ausdehnen, indem wir den Pflanzen-Organismus als eine Zellen-Republik, den Thier-Organismus dagegen als eine Zellen-Monarchie betrachten. Denn die Pflanzenzellen sind durchweg selbständiger, gleichartiger, unabhängiger von einander und vom Ganzen. Die Thierzellen hingegen sind in Folge der vorgeschrittenen Arbeitstheilung ungleichartiger, mehr von einander abhängig und zugleich in Folge der stärkeren Centralisation der »Staatsidee« in höherem Maasse unterworfen.
Nun lehrt uns aber ferner die Entwickelungsgeschichte, dass jedes Thier und jede Pflanze im Beginne der individuellen Existenz eine einzige einfache Zelle ist. Das Ei, aus dem sich jedes Thier wie jede Pflanze entwickelt, ist weiter nichts als eine Zelle. Das ist eine der bedeutungsvollsten Thatsachen. Denn das ganze Problem der individuellen Entwickelung löst sich demnach in die Frage auf: Wie kann der vielzellige Organismus mit allen seinen verschiedenen Organen aus einer einzigen Zelle entstehen? Und die Antwort hierauf lautet höchst einfach: Durch wiederholte Theilung entsteht aus der einfachen Zelle eine Zell-Gemeinde oder Association, eine Gesellschaft von zahlreichen gleichartigen Zellen; diese werden durch Arbeitstheilung ungleichartig und ordnen sich nach den Gesetzen der Vererbung und Anpassung zu einer centralisirten Einheit.
Wie verhalten sich nun unsere kleinen Protisten zu diesen höchst wichtigen Thatsachen und zu der darauf gegründeten Zellentheorie? Ist auch ihr winziger Leib aus vielen und ungleichartig entwickelten Zellen zusammengesetzt? Findet sich auch in ihrem Organismus jene Arbeitstheilung der associirten Zellen, durch welche die verschiedenen Gewebe und Organe entstehen? Das Mikroskop antwortet uns: Nein! Vielmehr ist bei den meisten Protisten der ganze Körper zeitlebens nur eine einzige Zelle. Aber auch bei jenen Protisten, welche in entwickeltem Zustande vielzellig sind, finden wir niemals wahre Gewebe und Organe, niemals jene eigenthümliche Arbeitstheilung und Anordnung der Zellen, welche den wahren Thierkörper und den wahren Pflanzenkörper auszeichnet. Denn hier beherrscht immer die Gesammtform des Körpers die ganze Anordnung und Bildung der Zellen, [S. 18] ihre Verbindung zu den Geweben und Organen, aus denen er zusammengesetzt ist. Bei den vielzelligen Protisten hingegen bewahren die gesellig verbunden Zellen stets mehr oder weniger ihre Selbständigkeit; sie bilden immer nur sehr lockere Gesellschaften, sociale Verbände ohne Arbeitstheilung, die nicht als centralisirte Staaten anerkannt werden können. Wenn wir vorher den einzelnen Organismus des Thieres wie der Pflanze einem wohlorganisirten Culturstaate verglichen, so können wir dagegen die lockeren Zellenhaufen der vielzelligen Protisten höchstens mit den rohen Horden der uncultivirten Naturvölker vergleichen. Die meisten Protisten bringen es aber, wie gesagt, nicht einmal zur Bildung solcher Zellen-Horden, zu dieser niedersten Stufe der Association; sie ziehen es vor, als Einsiedler für sich zu leben und ihre volle Selbständigkeit in jeder Beziehung zu bewahren. Die meisten Protisten bleiben zeitlebens einfache, isolirte Zellen, sie leben als Zellen-Einsiedler.
Wenn man die hohe Bedeutung der Protisten für die monistische Entwicklungslehre richtig verstehen will, wenn man sich von der selbständigen Stellung des Protistenreichs zwischen dem Thierreiche einerseits und dem Pflanzenreiche anderseits überzeugen will, so muss man vor Allem den autonomen, unabhängigen Zellen-Charakter ihres Organismus gehörig würdigen. Bei allen einzelligen Protisten, die ihr ganzes Leben als »Zellen-Einsiedler« zubringen, versteht sich das von selbst. Aber auch bei den vielzelligen Protisten, bei den »Zellenhorden« finden wir immer die Individualität der locker verbundenen Zellen gewahrt und vermissen jene Abhängigkeit derselben von einander und vom Ganzen, welche wir in dem wohlorganisirten Zellenstaate des Thier- und Pflanzenorganismus antreffen.
In dieser Auffassung des Protisten-Organismus liegt nach unserer Ansicht der Schwerpunkt seines Verständnisses. Es wird daher zunächst erforderlich sein, den Begriff der organischen Zelle überhaupt festzustellen. Dieser Begriff hat seit der Begründung der Zellentheorie mancherlei Wandlungen erfahren. Gegenwärtig nimmt man fast allgemein an, dass zum Begriff der Zelle zwei verschiedene Bestandtheile gehören. Erstens: der eigentliche Zellenleib, ein lebendiges Stückchen von weichem, eiweissartigen [S. 19] Bildungsstoff oder Protoplasma; und zweitens ein davon umschlossener Zellkern oder Nucleus; ein kleinerer, meist festerer Körper, der ebenfalls aus einer eiweissartigen, aber vom Protoplasma etwas verschiedenen Materie besteht. Als dritter Hauptbestandtheil kommt dazu bei vielen Zellen noch eine äussere Umhüllungshaut oder Schale, die Zellhaut oder Membran. Die meisten Pflanzenzellen sind von einer solchen Kapsel oder Membran umschlossen: Schlauchzellen. Hingegen sind die meisten Thierzellen hautlos und nackt: Urzellen. Die meisten Protisten zeichnen sich durch die Bildung ganz eigenthümlicher Kapseln oder Schalen aus, welche ihrem Zellenleibe eine sehr characteristische und mannigfaltige Gestalt geben.
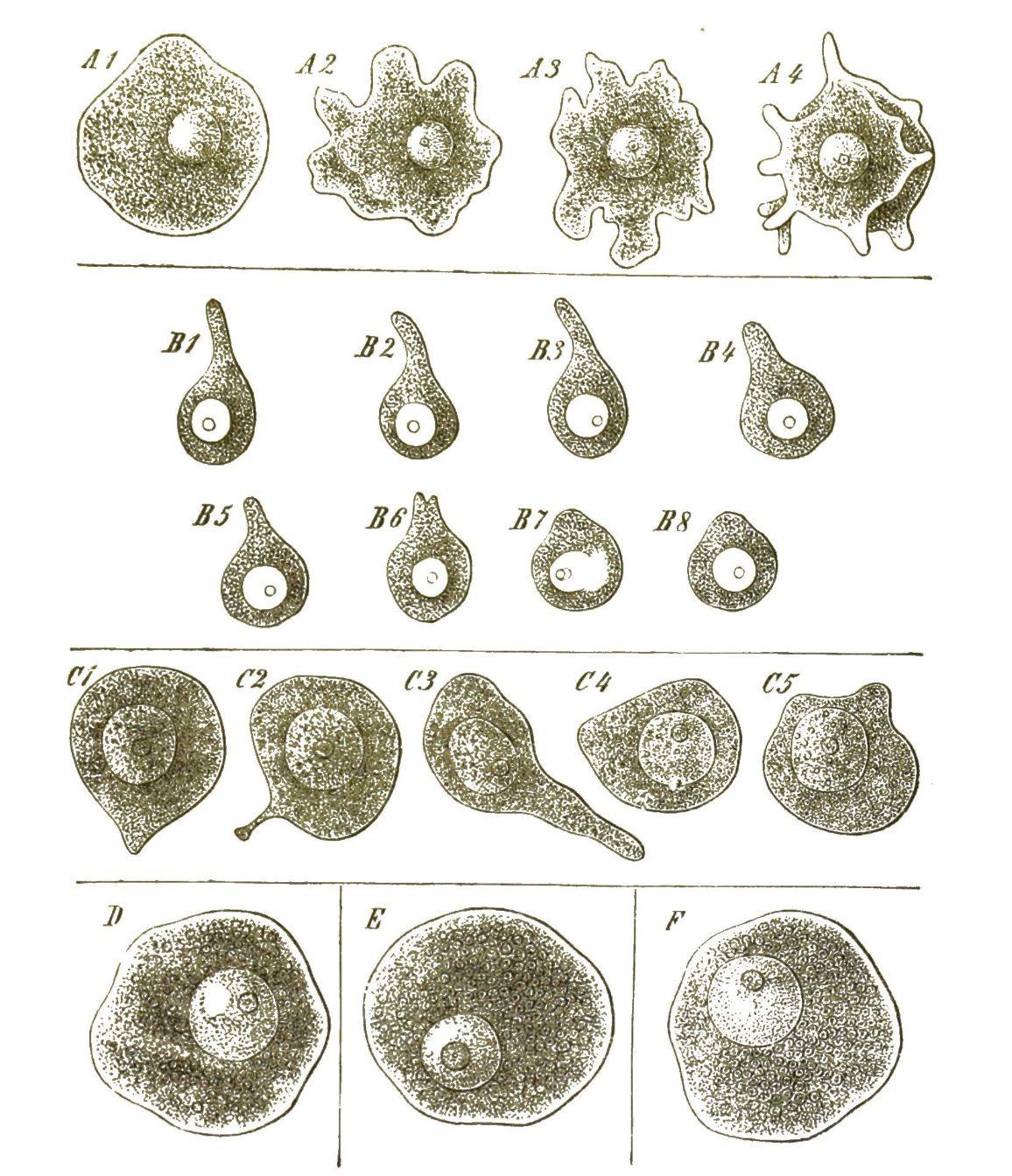
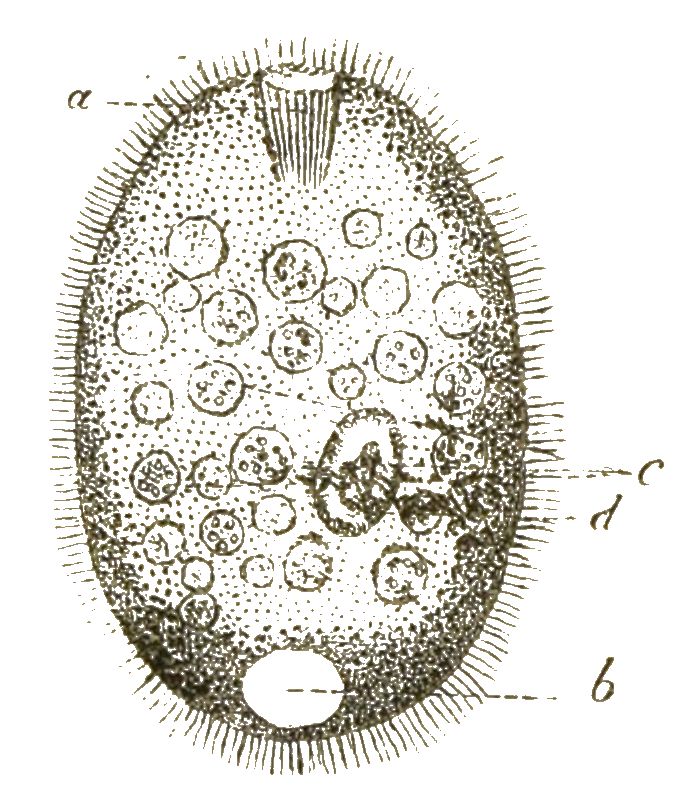
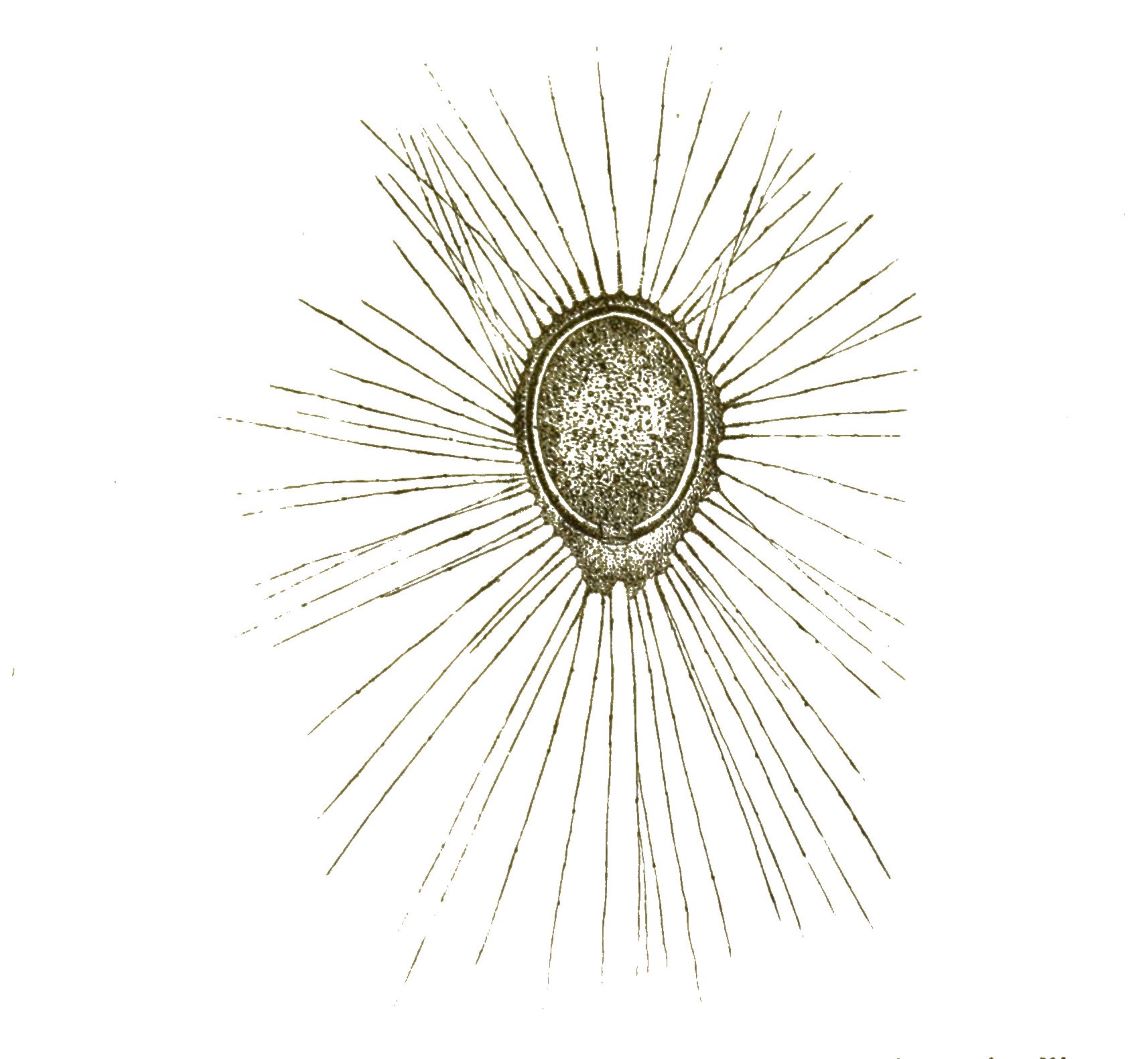
Wenn wir nun zunächst unter unsern Protisten diejenige Gattung aufsuchen, welche uns auf der Höhe ihrer Entwickelung die einfachste Form eines solchen einzelligen Organismus, gewissermassen das Ideal der Zelle, darstellt, so treten uns vor allen Andern die berühmten Amoeben entgegen. (Fig. 1). Weit verbreitet in unsern süssen und salzigen Gewässern, sind dieselben wegen ihrer höchst einfachen Bildung und ihrer bedeutsamen Beziehungen zu anderen Zellen von ganz besonderer Wichtigkeit. Die Amoeben sind nackte Zellen ohne Hülle und ohne bestimmte Form. Ihr weicher Körper, der nur einen einfachen Zellkern enthält, bewegt sich langsam kriechend im Wasser umher. Dies geschieht dadurch, dass eine wechselnde Anzahl von veränderlichen, lappenförmigen oder fingerförmigen Fortsätzen aus beliebigen Stellen der Oberfläche vorgestreckt und wieder eingezogen werden. So ändern die kriechenden Amoeben immerfort ihre unbestimmte Gestalt. Kommen sie zufällig mit kleinen Körperchen in Berührung, die zur Nahrung dienen können, so drücken sie dieselben mittelst der Bewegungen ihrer Fortsätze an einer beliebigen Stelle ihrer [S. 20] Körper-Oberfläche in diesen hinein. Auch kleinste Wassertröpfchen werden so verschluckt. Die einzellige Amoebe kann also essen und trinken, ohne dass sie Mund und Magen besässe. Nachdem die Amoebe durch fortdauerndes Wachsthum eine gewisse Grösse erreicht hat, zerfällt ihr einfacher Zellenleib durch Theilung in zwei Zellen. Zuerst theilt sich dabei der Kern, darauf das Protoplasma. Auf dieselbe Weise vermehren sich auch die Zellen, die unsern eigenen Körper zusammensetzen, und von denen viele beständig verbraucht und durch neue Zellen ersetzt werden. Die grösste Aehnlichkeit mit den Amoeben haben die farblosen Blutzellen, die milliardenweise in unserem Blute kreisen. Auch diese bewegen sich nach Amoeben-Art, indem sie ihre unbestimmte Form ändern. Auch diese können fremde Körperchen in ihr Inneres aufnehmen; wir können sie unter dem Mikroskop z. B. mit Carminkörnchen füttern, mit denen sie sich in kurzer Zeit anfüllen. (Fig. 2).
Von besonderer Wichtigkeit für die Entwickelungsgeschichte ist die interessante Thatsache, dass auch die Eier der Thiere in ihrer frühesten Jugend nackte, formlose Zellen sind, welche Amoeben zum Verwechseln ähnlich sehen und gleich diesen langsame, unbestimmte Bewegungen ausführen, wobei sie ihre Form beliebig verändern (Fig. 3). Bei den Schwämmen oder Spongien unternehmen diese amoebenähnlichen Eizellen, langsam fortkriechend, oft weite Wanderungen durch den Körper des Schwammes und sind daher früher als »parasitische Amoeben« beschrieben worden, welche als fremde Eindringlinge im Schwammkörper schmarotzend leben sollten (Fig. 4).
Es giebt auch Amoeben, welche ihren nackten Zellenleib theilweise mit einer schützenden Schale umgeben, und diese bilden die Gruppe der Arcellinen oder Thekolobosen. Bald schwitzen diese gepanzerten Amoeben eine schleimige Masse aus, welche sofort erhärtet und mit Sandkörnchen und anderen fremden Körperchen zu einer festen Kruste zusammenbackt (Difflugia, Fig. 5). Bald wird die ganze Masse der erhärteten Hülle blos von ausgeschwitzter organischer Substanz gebildet, und diese zeigt oft eine sehr zierliche Structur, indem sie aus sechseckigen oder viereckigen Täfelchen zusammengesetzt erscheint (Arcella, Quadrula, Fig. 6). [S. 21]
[S. 22]
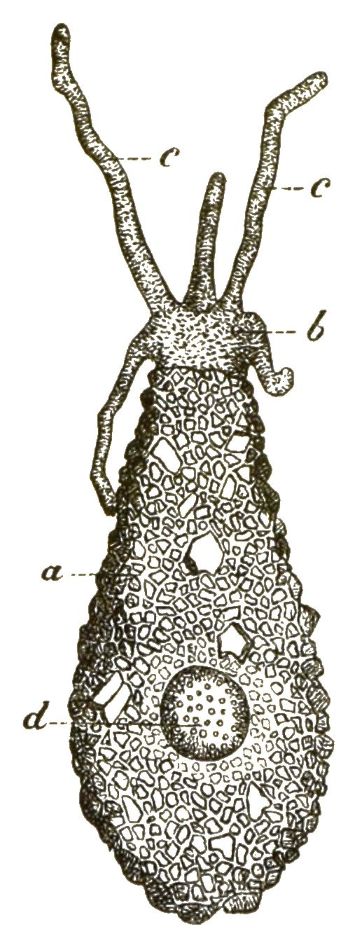
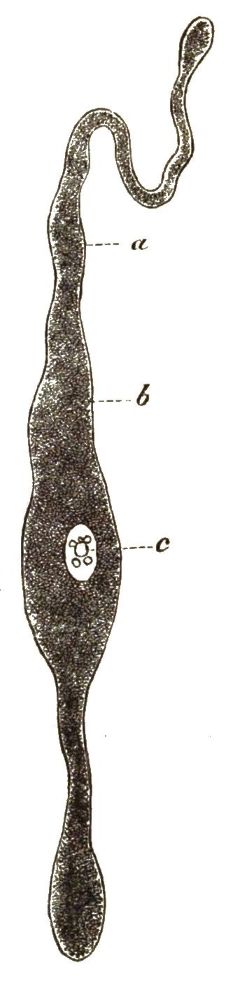
Alle diese amoebenartigen Wesen, die echten, nackten Amoeben und die gepanzerten zierlichen Arcellinen, können wir als besondere Classe unter dem Namen Lappinge oder Lappenfüssler (Lobosa) zusammenfassen, weil der auszeichnende Character dieser einzelligen Urthiere die Bildung lappenförmiger Wechselfüsschen ist. An sie schliessen sich aber ganz eng die sonderbaren Wesen an, welche die besondere Gruppe der Gregarinen bilden. Alle Gregarinen leben als Schmarotzer oder Parasiten im Innern anderer Thiere und sind gewissen niederen Würmern so ähnlich, dass man sie früher selbst als Eingeweide-Würmer beschrieben hat; auch stimmen die wurmförmigen Bewegungen ihres kriechenden Körpers ganz mit denjenigen gewisser Würmer überein. Trotzdem ist ihr ganzer, ziemlich grosser, oft mehrere Millimeter langer Körper nichts Anderes, als eine einfache Zelle. Der trübe, mit feinen Körnchen erfüllte Protoplasma-Leib (b) umschliesst einen Zellkern (c) und ist von einer festen, homogenen, structurlosen Hülle umgeben (a). Die flüssige Nahrung schwitzt aus den umgebenden Säften des bewohnten Thieres durch diese Hülle oder Zellmembran hindurch und dringt so in die Gregarine ein. Man kann die Gregarinen als Amoeben betrachten, welche in das Innere von anderen Thieren eingedrungen sind, sich hier an parasitische Lebensweise gewöhnt und durch Anpassung mit einer schützenden Hülle umgeben haben.
Eine ganz andere Bewegungsform, als die langsam kriechenden Amoeben und Gregarinen, zeigen uns die schwimmenden Flagellaten, die Geissler oder Geisselschwärmer. Diese interessanten Protisten haben bis auf den heutigen Tag unter einem ganz eigenthümlichen Schicksal zu leiden. Wenn sie nämlich das Glück haben, grün gefärbt zu sein, werden sie von vielen Naturforschern unbedenklich als echte Pflanzen betrachtet. Wenn sie dagegen unglücklicherweise eine gelbe oder braune Farbe tragen, so werden sie für echte Thiere erklärt; gewiss ein schlagendes Beispiel von der Willkür der üblichen Classification. Zahlreiche [S. 24] Formen dieser Geissler, die auch oft mit dem vieldeutigen Namen der Monaden belegt werden, bevölkern das Süsswasser, wie das Meer, oft in unglaublichen Massen. Wenn im Frühjahr zuweilen plötzlich unsere Teiche sich mit einer grünen Schleimdecke überziehen, so beruht das gewöhnlich auf der Entstehung zahlloser grüner Euglenen. Ebenso ist die seltener auftretende blutrothe Färbung der Gewässer, die zur Sage vom Blutregen, sowie zu vielen abergläubischen Vorstellungen und Hexen-Processen Veranlassung gegeben hat, durch Milliarden rother Euglenen bedingt. Durch verwandte rothe Protococcus-Formen wird auch der rothe Schnee gebildet, der die Eisberge sowohl in den Polarmeeren, wie auf unseren Alpenhöhen bisweilen in weiter Ausdehnung blutroth färbt.
Diese Protococcen und Euglenen sind Einsiedler-Zellen, während andere Flagellaten sich zu kleinen Gesellschaften zusammenthun. Sie schwimmen im Wasser umher mittelst eines feinen fadenförmigen Fortsatzes, der wie eine Geissel oder Peitsche hin und her geschwungen wird (Fig. 8). Manche setzen sich auch fest auf dünnen Stielen. Ausser der Geissel, ihrem Haupt-Bewegungsorgan, besitzen manche Geisselschwärmer noch einen Kranz von feinen Wimpern mitten um den Zellenleib; diese [S. 25] heissen Wimpergeissler (Peridinia, Fig. 9). Von letzteren bilden sich viele eine Kieselschale, die aus zwei ungleichen Hälften besteht; die grössere Hälfte trägt zwei lange Hörner, die kleinere ein Horn; zwischen beiden Hälften tritt der Wimperkranz und die Geissel hervor. Durch die Schwingungen der Geissel werden kleine Nahrungskörnchen dem Zellenleibe der Flagellaten zugeführt und an deren Basis durch eine Art Zellenmund aufgenommen. Ihre Vermehrung geschieht meistens durch einfache Theilung. Bei vielen finden wir abwechselnd einen frei beweglichen und einen Ruhezustand. Während des letzteren kapseln sie sich ein und zerfallen innerhalb der Hülle in vier oder acht Zellen. Diese treten später aus der Kapsel aus und schwimmen frei umher.

Nahe Verwandte dieser einzelligen Flagellaten sind auch die grünen sogenannten Kugelthierchen oder Volvocinen (Fig. 10); grüne Gallertkügelchen, welche die Grösse eines Stecknadelknopfes erreichen. In jedem Kügelchen sind zahlreiche grüne einzellige Flagellaten zu einer Gesellschaft vereinigt; und durch die gemeinsamen Schwingungen ihrer Geisseln wird die ganze Kugel umherbewegt. Im Innern der Gallertkugeln entstehen neue Tochterkugeln. Ausserdem vermehren sich die Volvocinen aber auch geschlechtlich, wie durch Cohns’ sorgfältige Untersuchungen dargethan worden ist; ihre Befruchtung geschieht in ähnlicher Weise wie bei vielen Algen; sie schliessen sich dadurch schon enger an das Pflanzenreich an.
Eine sehr eigenthümliche Protistengruppe, die man auch noch zu den Flagellaten rechnet, sind die grossen blasenförmigen Noctiluken oder Meerleuchten. (Fig. 11). Sie bedecken oft die Meeresoberfläche in unglaublichen Massen, strahlen im Dunkeln ein helles Licht aus und spielen eine Hauptrolle bei dem wundervollen Phänomen des Meerleuchtens. Die gewöhnlichen [S. 26] Noctiluken sind colossale rundliche Zellen, welche ½-1 Millimeter Durchmesser erreichen und die Gestalt einer Pfirsiche besitzen (Fig. 11). Der Hohlraum der blasenförmigen Zelle ist mit wässeriger Flüssigkeit erfüllt, in welcher sich verästelte Stromfäden (g) des Protoplasma bewegen, ausgehend von der Wandschicht des letztern, welche innen an der Zellhaut anliegt. Der Kern ist eiförmig (b). An einer Stelle ist die Zellhaut von einer Oeffnung, einem Zellmund (Cystostoma), durchbrochen, und hier wird Nahrung direct in das Innere aufgenommen. Hier befindet sich auch neben der zarten Geissel ein grosser peitschenförmiger quergestreifter Anhang (a), sowie ein zahnförmiger Fortsatz (d). Die Fortpflanzung erfolgt theils durch einfache Theilung, theils durch eine eigenthümliche Form der Sporenbildung.
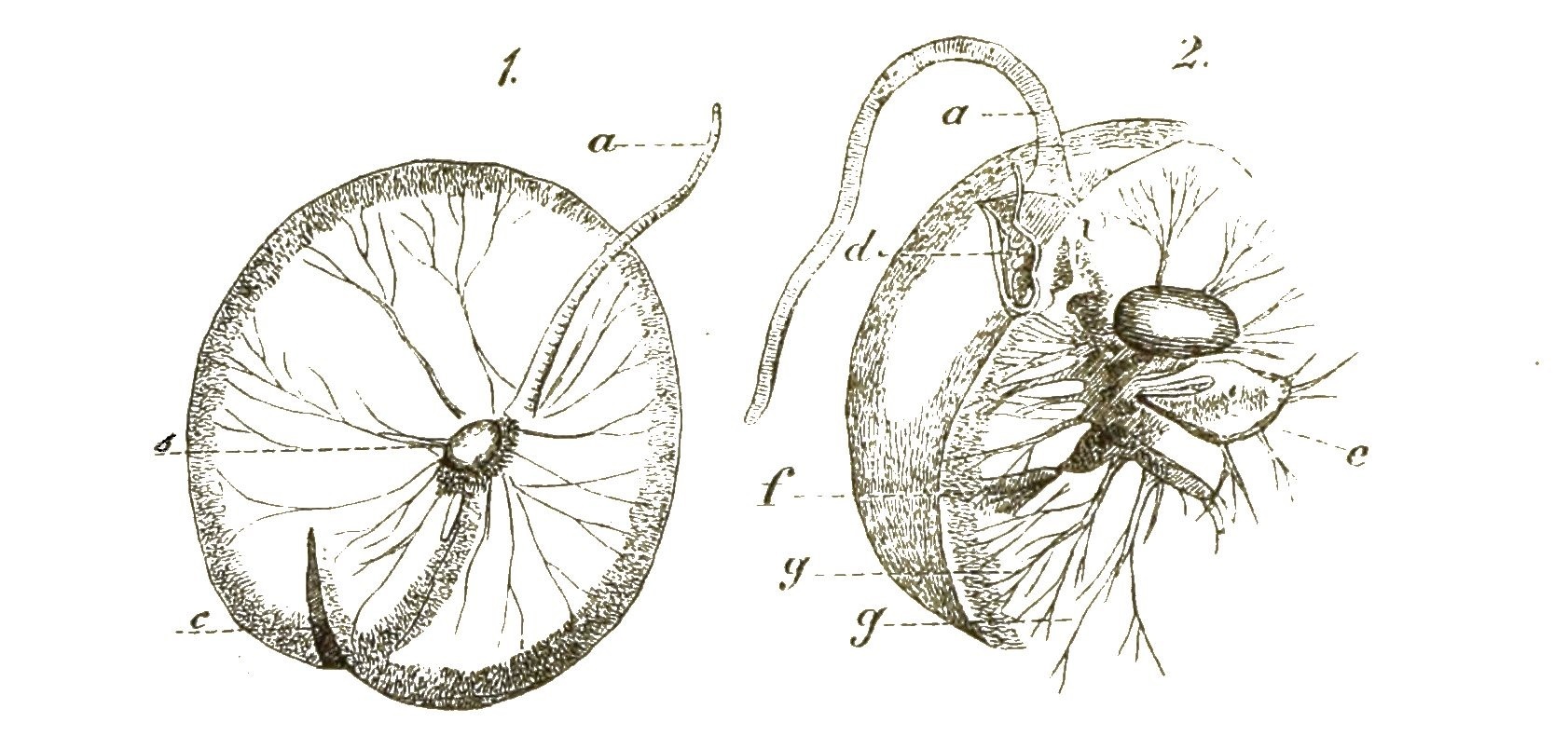
Neuerdings ist eine Noctiluken-Form entdeckt worden, welche zum Verwechseln einer kleinen schirmförmigen Meduse ähnlich ist, und gleich einer solchen sich durch Zusammenklappen des zarten concaven Schirmes schwimmend bewegt (Leptodiscus medusoides).
Während über die einzellige Natur der Geisselschwärmer und der Amoeben heutzutage kein Zweifel mehr besteht, so ist diese dagegen bis vor Kurzem streitig gewesen bei denjenigen Protisten, die man heute vielfach als Infusionsthierchen im engeren Sinne bezeichnet. Dazu gehören die beiden Klassen der Wimperthierchen [S. 27] oder Ciliaten (Fig. 12–15) und der Starrthierchen oder Acineten (Fig. 16, 17). Massenhaft bevölkern sie alle stehenden und fliessenden Gewässer und sind auch in allen Infusionen zu finden.
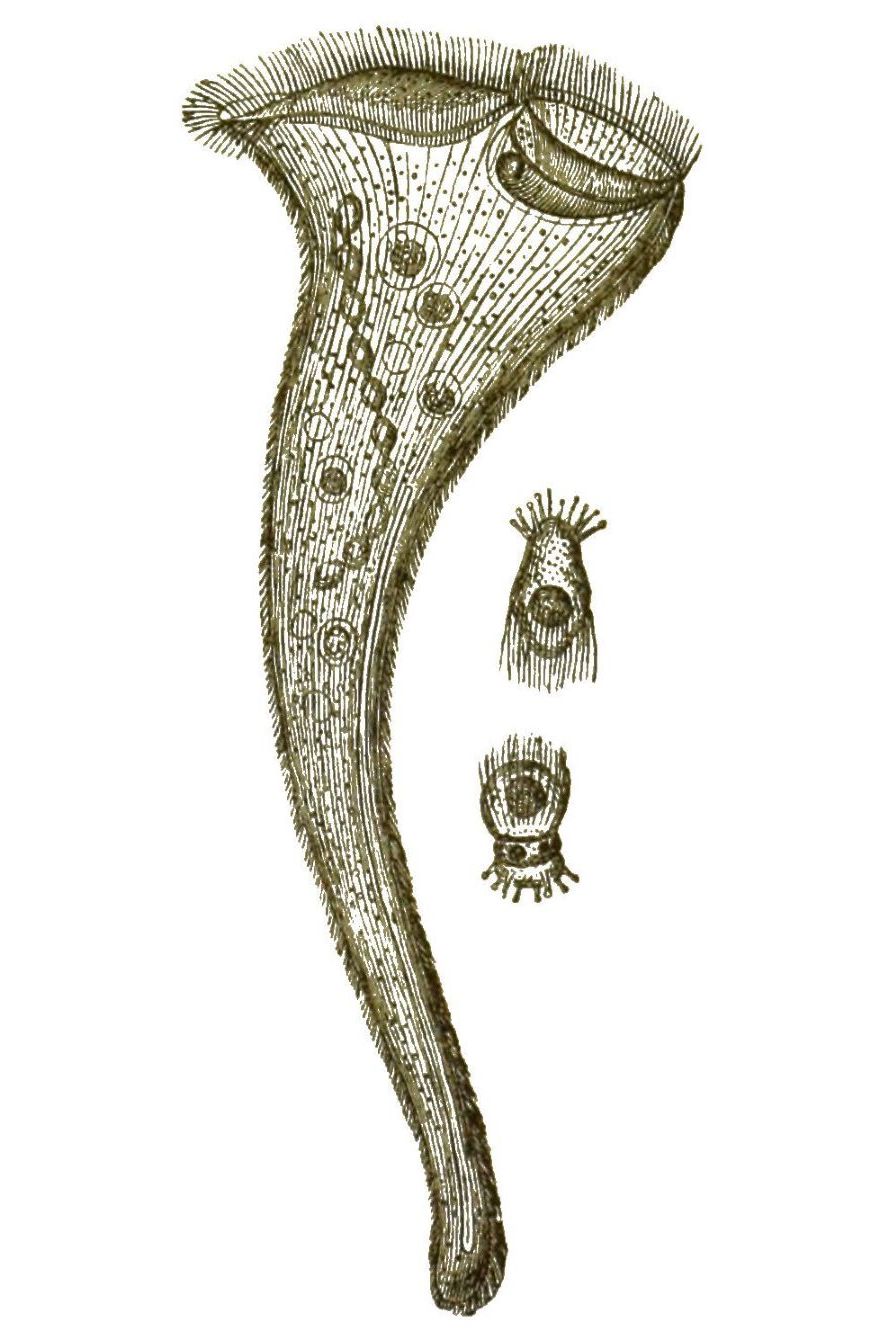
Besonders die Ciliaten, die Wimperlinge oder Wimperthierchen, erscheinen in einer Fülle von niedlichen Formen; und durch die Anmuth ihrer lebhaften Bewegungen fesseln sie uns stundenlang an das Mikroskop. Nur einzelne [S. 28] Ciliaten sind schon mit blossem Auge sichtbar, so z. B. das grosse Trompetenthierchen (Stentor, Fig. 12); die meisten sind erst durch das Mikroskop erkennbar. Zahlreiche kurze Wimperhärchen sind über den Körper zerstreut und werden willkürlich schlagend bewegt. Wie die Geisseln der Flagellaten, so sind auch diese Wimpern der Ciliaten directe Fortsätze vom Protoplasma des einzelligen Körpers. Die meisten Wimperthierchen bewegen sich frei schwimmend oder laufend mittelst dieser Wimpern umher. Es giebt aber auch festsitzende Ciliaten, wozu die niedlichen Vorticellen (Fig. 13) und Freia (Fig. 14) gehören.
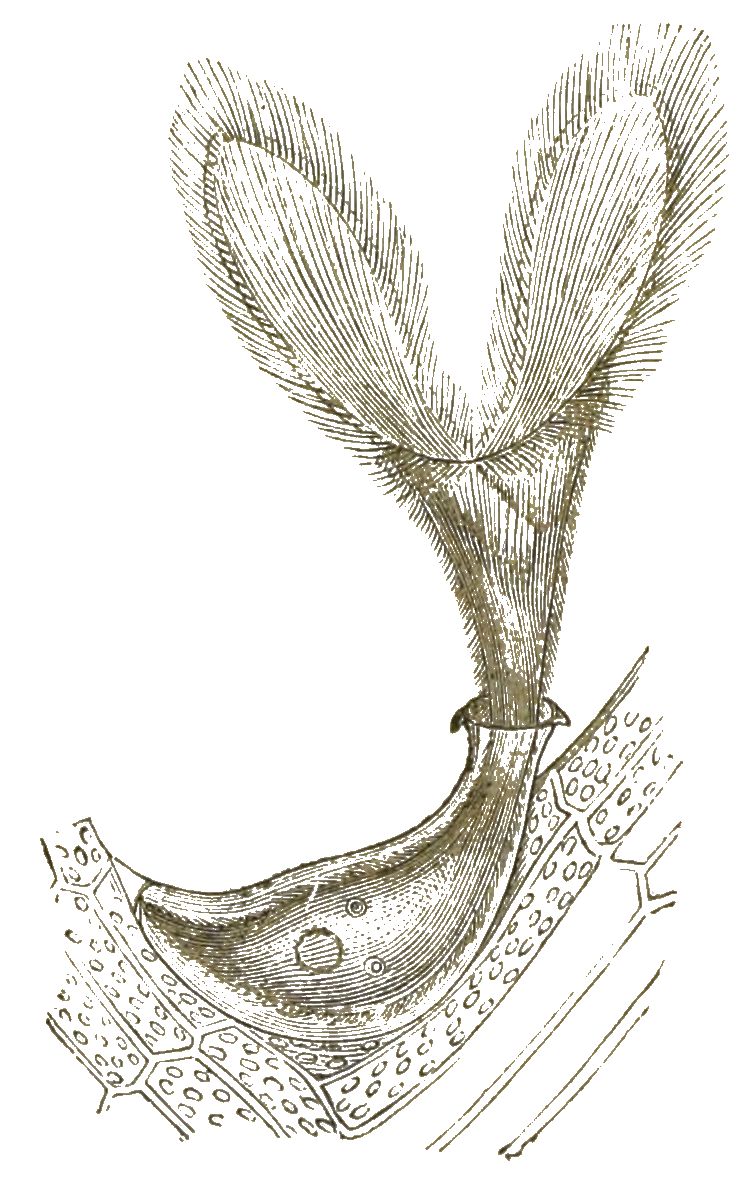
Bei diesen Ciliaten dient der durch die Wimpern erzeugte Strudel dazu, frisches Wasser und Nahrung der Zelle zuzuführen.
Das Protoplasma des Ciliaten-Körpers ist in eine festere Rindenschicht (Exoplasma) und eine weichere Markschicht (Endoplasma), [S. 29] gesondert. In der ersteren befindet sich eine beständige Oeffnung, eine Art Zellenmund (Cytostoma), durch welchen sowohl feste Bissen als Wassertropfen verschluckt und in die weichere Markmasse hineingedrückt werden. Bisweilen ist diese Mundöffnung zu einem besonderen gefalteten Schlundtrichter erweitert, so z. B. bei dem Fischreusen-Thierchen (Fig. 15 a). In dem weichen Protoplasma des Inneren ballt sich die verschluckte Nahrung in Bissen (Fig. 15 c), welche allmählig verdaut und aufgelöst werden; Ehrenberg beschrieb diese Nahrungsballen als besondere Magensäcke und benannte deshalb die Ciliaten »Vielmagenthierchen« (Polygastrica). Unsere magenlosen Wimperthierchen können also essen und trinken, obwohl sie einfache Zellen sind. Was aber noch mehr überrascht, das ist die Munterkeit und die offenbare Willkür ihrer Bewegungen, der zarte und seelenvolle Character ihrer Empfindungen. Gerade wegen dieser Eigenschaften werden sie gewöhnlich als echte Thiere betrachtet. Dass sie das nicht sind, geht aus ihrem feineren Bau und ihrer Entwickelung deutlich hervor. Zeitlebens umschliesst ihr einfacher Zellenleib nur einen einzigen Kern. Bald ist dieser Nucleus rundlich (Fig. 15 d), bald wurstförmig (Fig. 13 n), bald langgestreckt, stabförmig oder rosenkranzförmig (Fig. 12). Die Ciliaten sind also wirklich einzellig, wie zuerst der um die Kenntniss der Protisten hochverdiente Zoologe Siebold dargethan hat. Die Vermehrung der Ciliaten geschieht durch einfache Theilung; und wie bei jeder gewöhnlichen Zellentheilung zerfällt zuerst der Kern, und darauf das Protoplasma in zwei gleiche Hälften. Aber auch Fortpflanzung durch Knospenbildung ist bei vielen Ciliaten zu finden, so z. B. bei den Vorticellen (Fig. 13). Ausserdem scheinen sich Viele durch Sporen zu vermehren, d. h. durch junge Zellen, welche sich im Inneren der Mutterzelle bilden und wobei der Kern betheiligt ist (Fig. 12).
Das Interessanteste an den Wimperthierchen, und diejenige Eigenschaft, durch welche sie alle anderen Protisten übertreffen, ist der hohe Grad von Empfindlichkeit und von Willens-Energie, den sie bei ihren lebhaften Bewegungen kundgeben. Wer lange und eingehend Ciliaten beobachtet hat, kann nicht zweifeln, dass sie eine Seele so gut wie die höheren Thiere besitzen. Denn die Seelenthätigkeiten der Empfindung und der willkürlichen [S. 30] Bewegung üben sie eben so aus, wie die höheren Thiere; und an diesen Thätigkeiten allein ist ja die Seele zu erkennen. Da nun der ganze Leib der Ciliaten bloss eine einfache Zelle ist, so gewinnen sie die höchste Bedeutung für die Theorie von der Zellseele, für die Annahme, dass jede organische Zelle ihre eigene individuelle »Seele« besitzt — oder vielmehr, richtiger ausgedrückt: dass Seelenleben eine Thätigkeit aller Zellen ist.
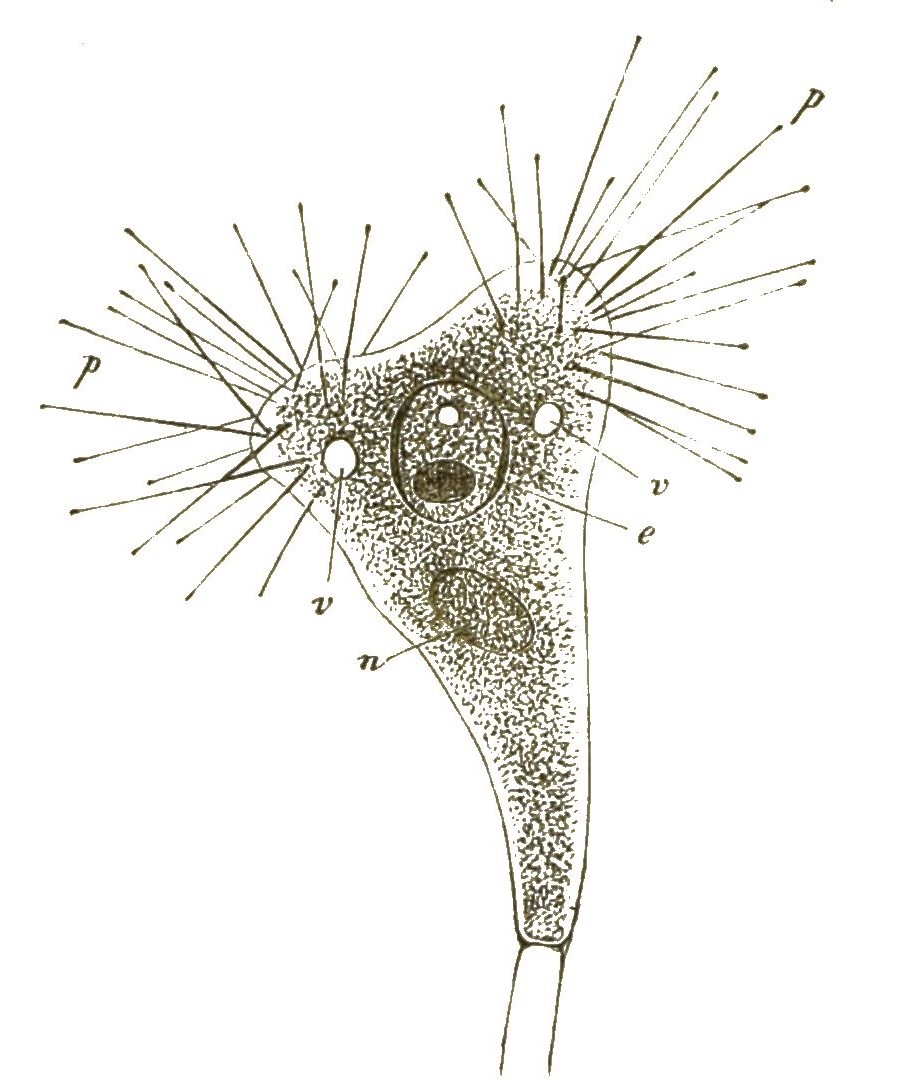
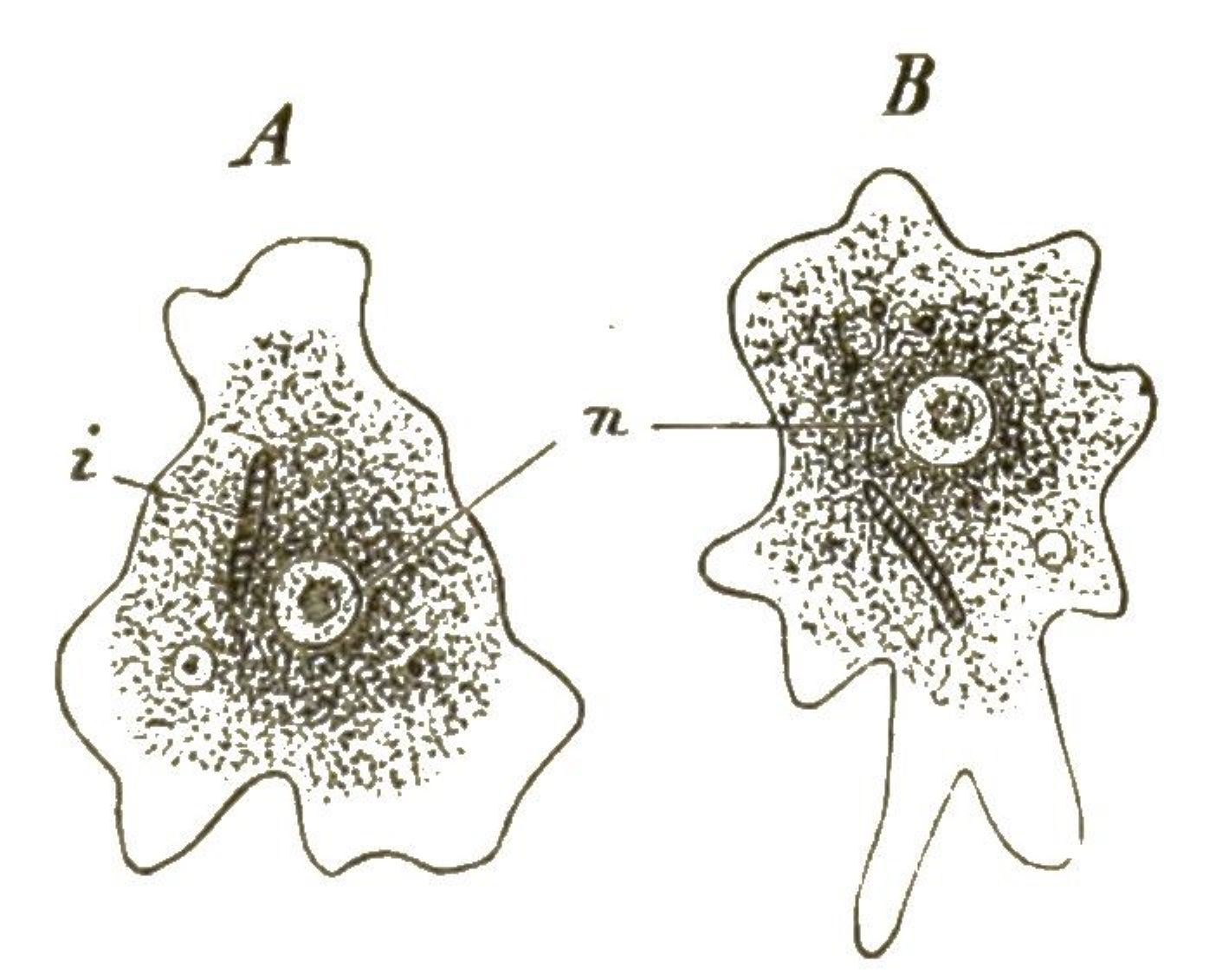
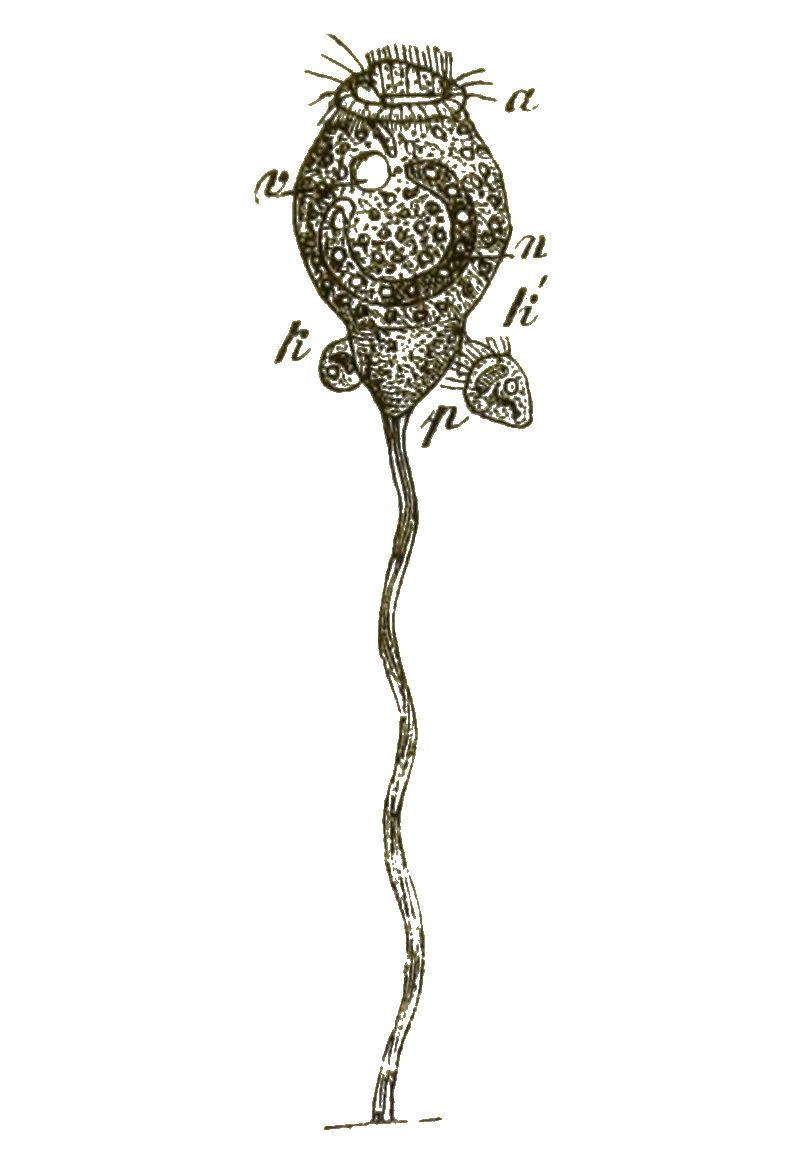
An die formenreiche Klasse der Wimperthierchen schliesst sich die kleine Gruppe der naheverwandten Starrthierchen oder Acineten an (Fig. 16, 17). Im Gegensatze zu ersteren zeigen diese letzteren nur sehr wenig Beweglichkeit; sie sitzen meistens zeitlebens auf einem Stiele fest. Statt der Wimperhärchen treten aus ihrem starren, von einer Hülle umschlossenen Zellenkörper zahlreiche feine, oft büschelförmig gruppirte Fortsätze hervor (Fig. 16 p). Dies sind sehr feine Saugröhrchen, die am Ende mit einem Saugknöpfchen versehen sind. Wenn ein schwimmendes Wimperthierchen unvorsichtig in die Nähe einer solchen Acinete geräth, wird sie von den steif ausgestreckten Saugröhren der letzteren festgehalten und ausgesaugt (Fig. 17). Das Protoplasma des gefangenen Ciliaten (a) wandert langsam durch die Saugröhren (f’’) [S. 31] in das Innere der Acinete hinein. Dass auch sie nur eine einfache Zelle ist, beweist ihr Zellkern (n); im Protoplasma sind, wie bei den Ciliaten, oft eine oder mehrere »contractile Blasen« oder Vacuolen sichtbar, wassererfüllte kugelige Hohlräume, die sich langsam zusammenziehen und wieder ausdehnen (Fig. 16v,Fig. 17x).
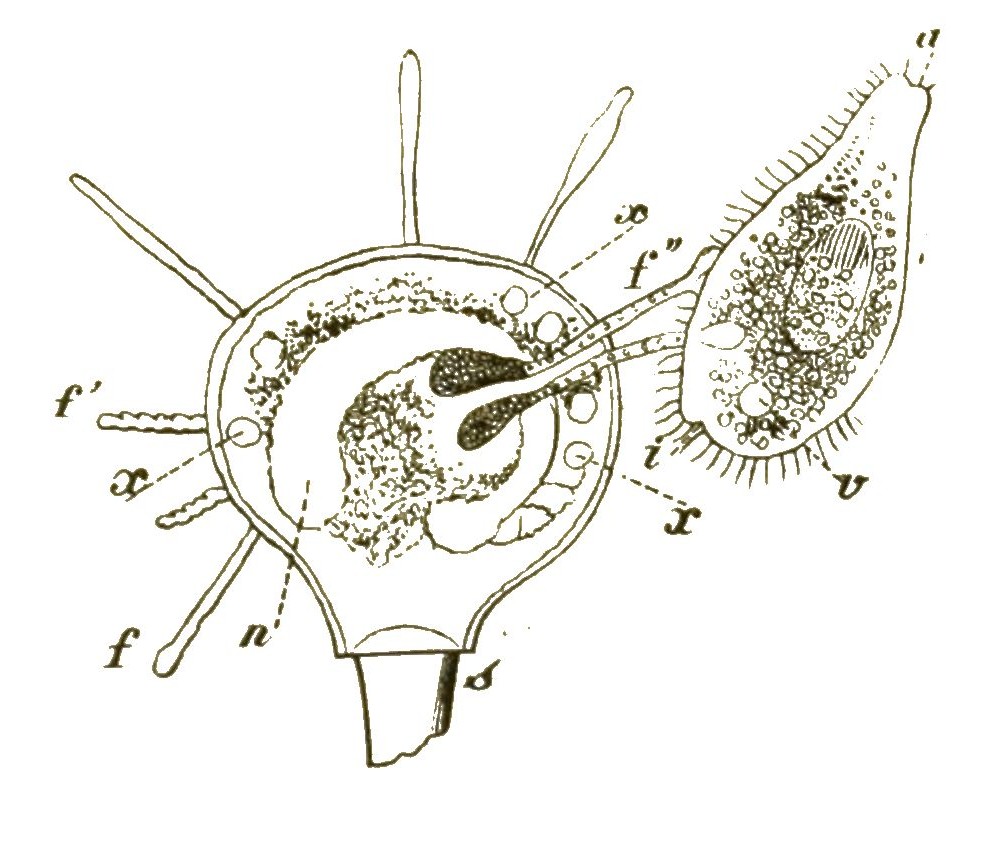
Die anhaltende Beobachtung der Acineten gewährt ebenso wie diejenige der Ciliaten das höchste Interesse. An diesen Infusionsthierchen zeigt uns die organische Zelle deutlich, wie weit sie es in ihrem idealen Streben nach thierischer Vollkommenheit für sich allein bringen kann. Wir können sagen: Die Wimperthierchen sind der gelungenste Versuch der einzelnen Zelle, sich zu einem wirklichen Thiere zu entwickeln. Aber zu einem echten Thiere gehören ja mindestens zwei Keimblätter, deren jedes aus zahlreichen Zellen zusammengesetzt ist. Also können wir doch die Ciliaten und Acineten nicht als wirkliche Thiere gelten lassen.
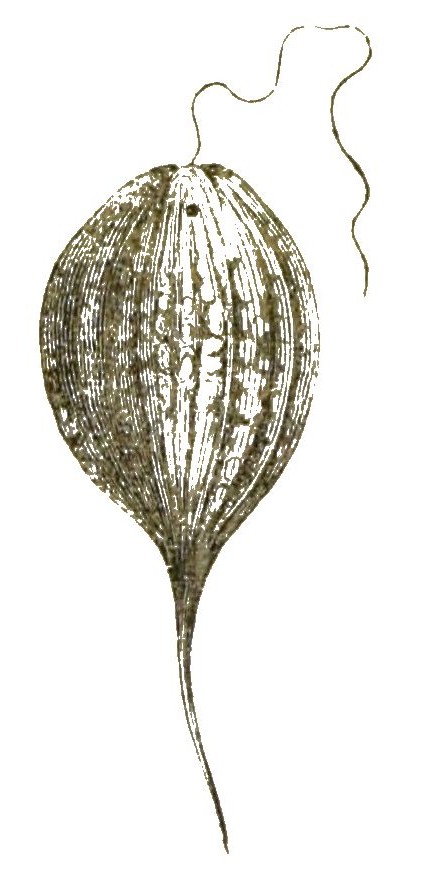
Unter allen Protistenklassen die formenreichste und in geologischer Beziehung die wichtigste ist die wunderbare Klasse der Wurzelfüssler oder Rhizopoden. Ausser mehreren kleineren Gruppen gehören dahin die kalkschaligen Thalamophoren und die kieselschaligen Radiolarien. Beide Abtheilungen sind in zahllosen, höchst phantastisch geformten Arten in allen Meeren verbreitet. [S. 32] Die Thalamophoren leben zum grössten Theile kriechend auf dem Grunde des Meeres, besonders auf Seetang; die Radiolarien hingegen schwimmen in dichtgedrängten Schaaren an der glatten Oberfläche des Meeres oder schweben in verschiedenen Tiefen desselben. Die bekanntesten und geologisch wichtigsten Rhizopoden sind die Thalamophoren, Kammerlinge oder Kammerthierchen; ausgezeichnet durch eine feste, meistens kalkige Schale, in welche sich diese Urthierchen, wie die Schnecke in ihr Haus, zurückziehen können. Bald enthält diese Kalkschale nur eine einzige Kammer (Einkammerige, Monothalamia, Monostegia); bald mehrere, durch Thüren mit einander verbundene Kammern (Vielkammerige, Polythalamia, Polystegia). Solche zierlich geformte, oft einem Schneckenhaus ähnliche Kalkschalen haben sich seit vielen Millionen Jahren in ungeheuren Massen auf dem Meeresboden angehäuft und an der Gebirgsbildung unserer Erde den wichtigsten Antheil genommen. Schon die ältesten, aus dem Meere abgesetzten Flötzgesteine, die laurentischen, cambrischen und silurischen Schichten, enthalten dergleichen Polythalamien-Schalen und sind wahrscheinlich zum grossen Theile aus ihnen gebildet. Das älteste von Allen ist das berühmte Eozoon canadense aus den unteren laurentischen Schichten, dessen Polythalamien-Natur mit Unrecht in Zweifel gezogen wurde. Die mächtigste Entwickelung erreichen diese Rhizopoden jedoch erst viel später, während der Kreide-Periode und der älteren Tertiär-Periode. Jedes kleinste Körnchen unserer weissen Schreibkreide lässt uns unter dem Mikroskope zahlreiche solcher zierlichen Kalkschalen erkennen. Der Grobkalk von Paris, aus dem viele Paläste dieser Weltstadt erbaut sind, besteht ebenfalls zum grössten Theile aus solchen Kammerschalen. Ein Kubikcentimeter des Kalkes aus den Steinbrüchen von Gentilly enthält ungefähr 20,000, ein Kubikmeter demnach gegen 20 Millionen Schalen. Die grössten Polythalamien aber lebten während der ältesten Tertiärzeit, während der Eocaen-Periode. Unter ihnen sind die Riesen des Protisten-Reiches, die gigantischen Nummuliten (Fig. 18), deren scheibenförmige Kalkschalen die Grösse eines Zweithalerstückes erreichen. Der von ihnen erzeugte Nummuliten-Kalk, aus dem unter Anderen die egyptischen Pyramiden gebaut sind, bildet die ungeheuren Gebirgsmassen des Nummulitensystems. Dies ist eins der gewaltigsten Gebirgssysteme [S. 33] unserer Erde, das von Spanien und Marokko bis nach Indien und China hinüberreicht, und an der Bildung der Pyrenäen und Alpen, des Libanon und Kaukasus, des Altai und Himalaya den bedeutendsten Antheil nimmt.

In welchen ungeheuren Massen die Polythalamien auch gegenwärtig noch unsere Meere bevölkern, geht daraus hervor, dass z. B. der Sand der Mittelmeerküsten an vielen Stellen zur grösseren Hälfte aus den Schalen lebender Polythalamien-Arten besteht. Schon einer ihrer ersten Beobachter, Bianchi, zählte [S. 34] im Jahre 1739 in einem einzigen Esslöffel Seesand von Rimini 6000 Individuen; und derjenige Naturforscher, dem wir die genauesten Untersuchungen über ihre Naturgeschichte verdanken, der berühmte Anatom Max Schultze, berechnete ihre Menge in einem Esslöffel Seesand von Gaeta auf mehr als Hunderttausend.
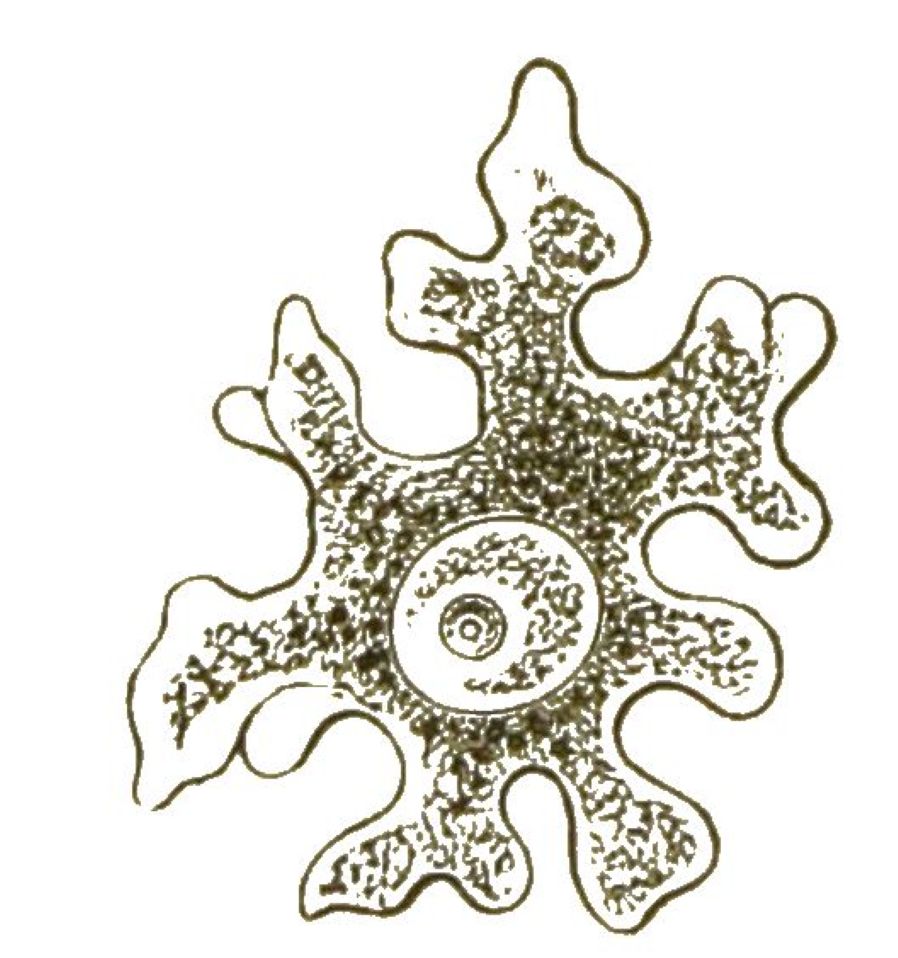
Der weiche lebendige Körper der Kammerthierchen, welcher diese wunderbaren Schalen- und Panzer-Bildungen erzeugt, ist stets von höchst einfacher Bildung: ein Stück formloses Protoplasma, das zahlreiche Zellenkerne einschliesst. Von der Oberfläche des weichen Protoplasma-Leibes strahlen hunderte, oft tausende von äusserst feinen Fäden aus. Diese Schleimfädchen, die den Namen Scheinfüsschen oder Pseudopodien führen, sind sehr empfindlich und beweglich. Sie können sich verästeln, mit einander verschmelzen, Netze bilden und wieder in die gemeinsame Centralmasse des Körpers zurückgezogen werden. Durch die Zusammenziehungen dieser Fäden bewirken die Wurzelfüssler ihre kriechende oder schwimmende Ortsbewegung. Wenn ein anderer Protist, z. B. ein Wimperthierchen oder eine Bacillarie, in den Bereich dieser Fäden gelangt, so wird es von ihnen erfasst, umschlungen und in das Innere des Protoplasmakörpers hineingezogen, wo es einer höchst einfachen Verdauung unterliegt. Wie bei den Amoeben kann jede Stelle der Körperoberfläche dergestalt die Aufgabe eines Mundes und Magens übernehmen. Auch die Vermehrung der Wurzelfüssler ist höchst einfach. Der weiche Protoplasma-Leib des Kammerthierchens zerfällt in zahlreiche kleine Stückchen. Jedes Stückchen erhält einen Zellkern, bildet also eine echte Zelle, und diese nackte Zelle schwitzt alsbald wieder eine Kalkschale aus.
Die vielgestaltige Schale des Acyttarien-Körpers besteht meistens aus kohlensaurem Kalk, seltener aus einer erhärteten organischen Substanz, die mit Sandkörnchen u. dergl. verkittet ist. Bald besitzt die Schale nur eine grössere Mündung, ist aber übrigens undurchlöchert (Imperforata); bald ist die Schale überall von sehr zahlreichen kleinen Löchern durchbrochen (Foraminifera). Mit Bezug auf die Schalenform unterscheidet man bei den zwei Hauptgruppen: Einkammerige und Vielkammerige. Die Einkammerigen (Monothalamia) sind verhältnissmässig wenig formenreich. Einer ihrer bekanntesten, häufigsten und grössten Vertreter ist die [S. 35] Gromia (Fig. 19). Sie besitzt eine eiförmige Schale, mit dunkelbraunem Protoplasma erfüllt, und erreicht die Grösse eines Stecknadelknopfes. Die Netze der Scheinfüsschen, welche davon ausstrahlen, kann man schon mit blossem Auge deutlich erkennen.
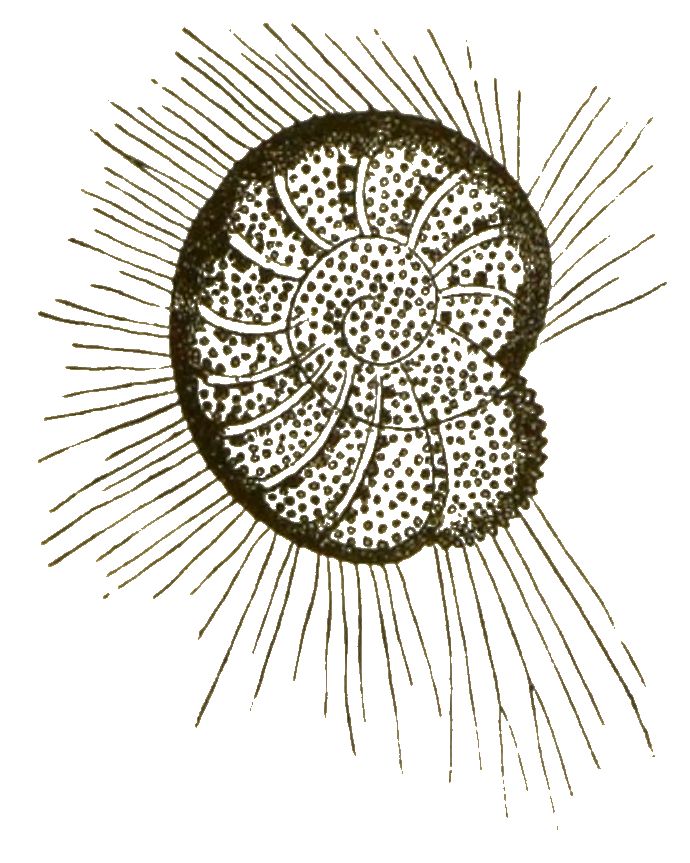
Die Vielkammerigen (Polythalamia) bilden die Hauptmasse der Acyttarien. Die einzelnen Kammern, welche ihre Schale zusammensetzen, sind durch unvollständige Scheidewände getrennt, oft sehr zahlreich. Meistens sind dieselben mehr oder weniger in Spiralen geordnet. So entstehen Gehäuse, welche die grösste Aehnlichkeit mit denjenigen gewisser Mollusken, namentlich Cephalopoden, besitzen (Fig. 20). Daher wurden diese Rhizopoden [S. 36] von ihren ersten Entdeckern wirklich für echte, mikroskopische Cephalopoden gehalten und auch später noch ihre Organisation als solche beschrieben.
Erst vor 40 Jahren lernte man, zuerst durch Dujardin, ihre wahre Natur kennen, und überzeugte sich, dass ganz ähnlich geformte Schalen das eine Mal von einem höchst vollkommen organisirten Weichthiere (Nautilus), das andere Mal von einem höchst einfach gebauten Wurzelfüssler (Polystomella) gebildet werden.

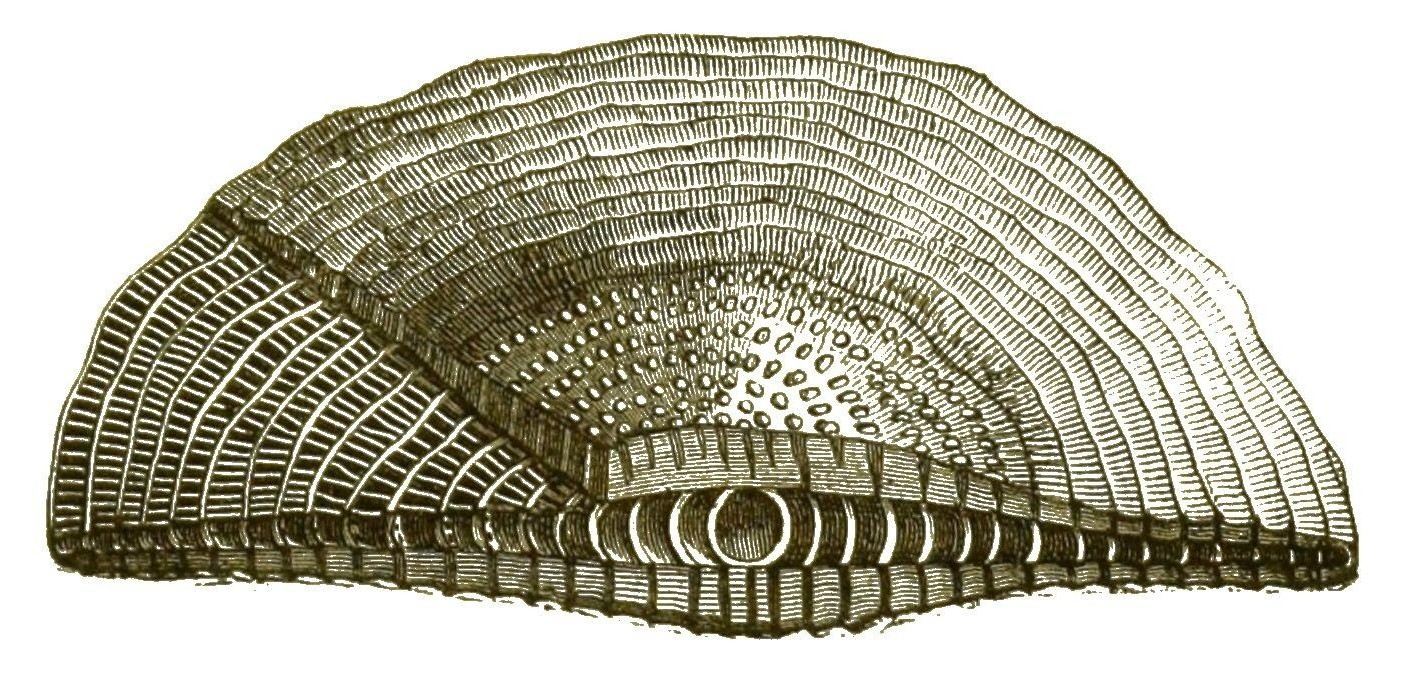
Bei manchen Polythalamien laufen mehrere Spiralen neben einander im Gehäuse hin, indem innerhalb der Kammern sich wieder parallele Scheidewände bilden (Fig. 21). Bei den grossen Orbituliten und Nummuliten liegen solche Kammerreihen sogar in mehreren Stockwerken übereinander. Die Kammerreihen sind hier bald in zusammenhängenden Spirallinien, wie bei den Nummuliten (Fig. 18) geordnet, bald in concentrischen Ringen, wie bei dem gigantischen Cycloclypeus (Fig. 22).
Die Gehäuse dieser letzteren sind runde Scheiben, welche sich am besten mit einem Palaste vergleichen lassen, dessen Umfassungsmauern nach dem Plane eines römischen Amphitheaters gebaut sind.
[S. 37]
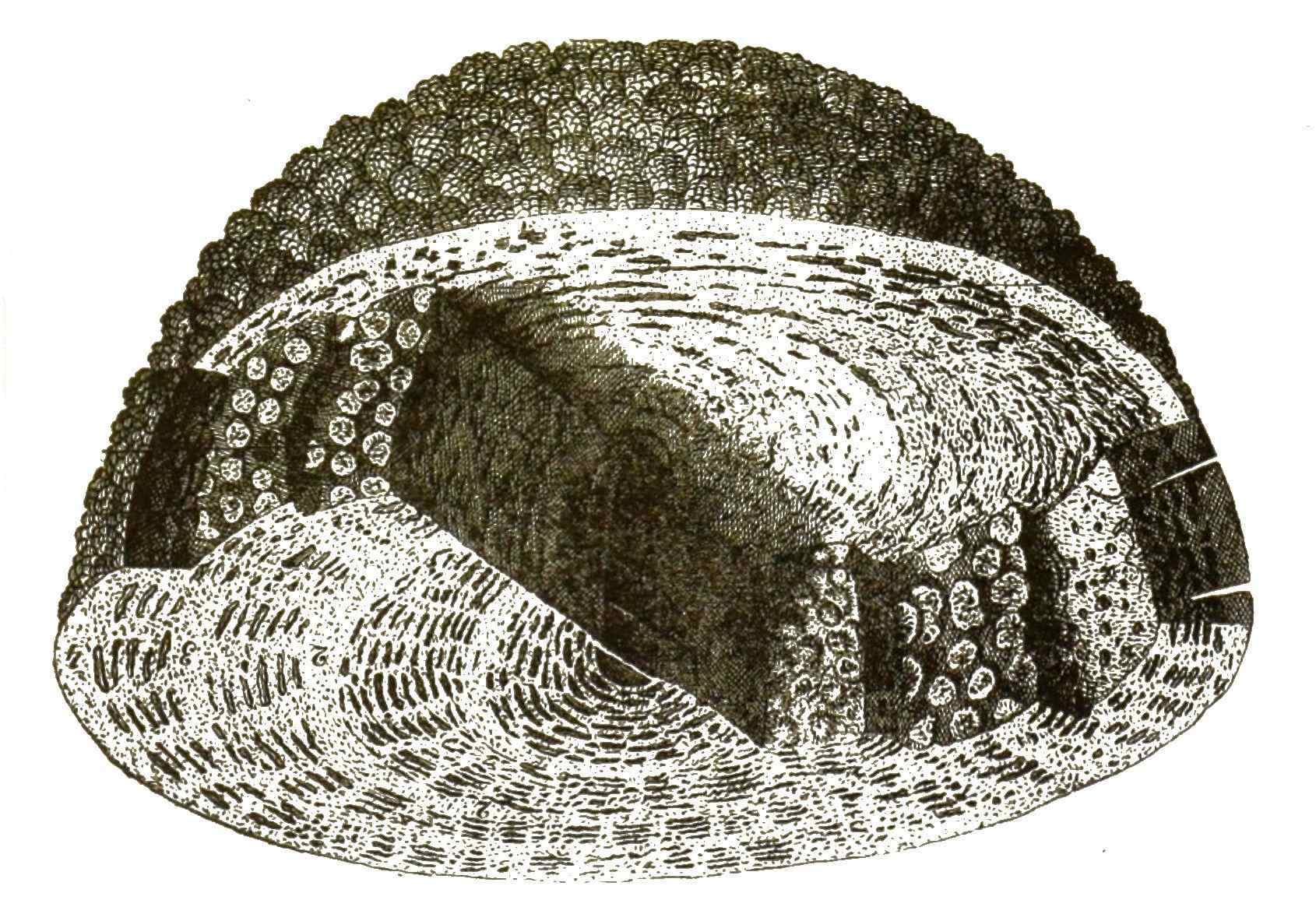
[S. 38]
Mehrere Stockwerke liegen übereinander, in jedem eine centrale Hauptkammer, umgeben von vielen ringförmigen Corridoren, und jeder Corridor durch viele Scheidewände in Kammern getheilt: alle diese zahlreichen Stockwerke, Corridore und Kammern stehen durch Thüren mit einander in Verbindung und kleine Fenster in der äusseren Schalenfläche vermitteln die Verbindung mit der Aussenwelt, indem sie die feinen Schwimmfüsschen durchtreten lassen.
Zu den grössten und am meisten zusammengesetzten Polythalamien gehören die Parkerien, deren Gehäuse grösstentheils aus Sandkörnchen zusammengesetzt sind (Fig. 23).
Während die grosse Mehrzahl der Thalamophoren auf dem Meeresboden kriechend lebt, giebt es auch einige Arten, die an der Oberfläche des Meeres schwimmen, und zwar oft in grossen Massen, mit Radiolarien gemischt. Dahin gehören auch die merkwürdigen Pulvinulinen, Globigerinen und Hastigerinen, letztere durch ihre sehr langen borstenförmigen Kalkstacheln ausgezeichnet (Fig. 24).
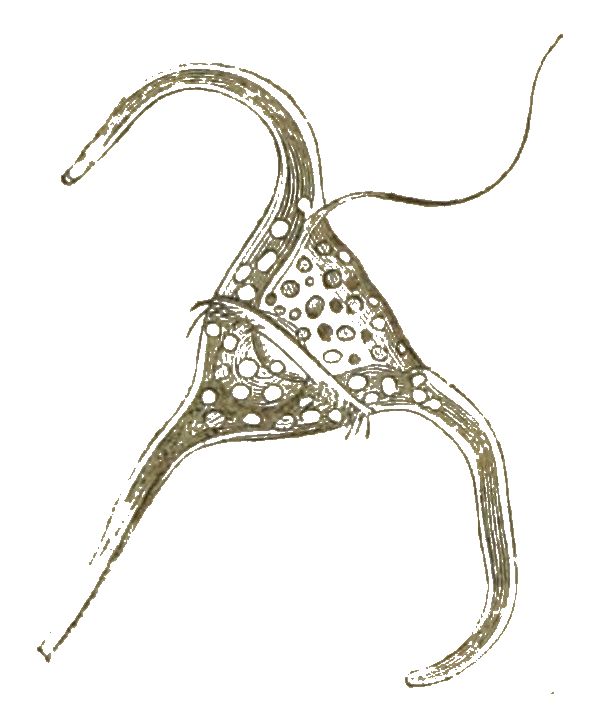
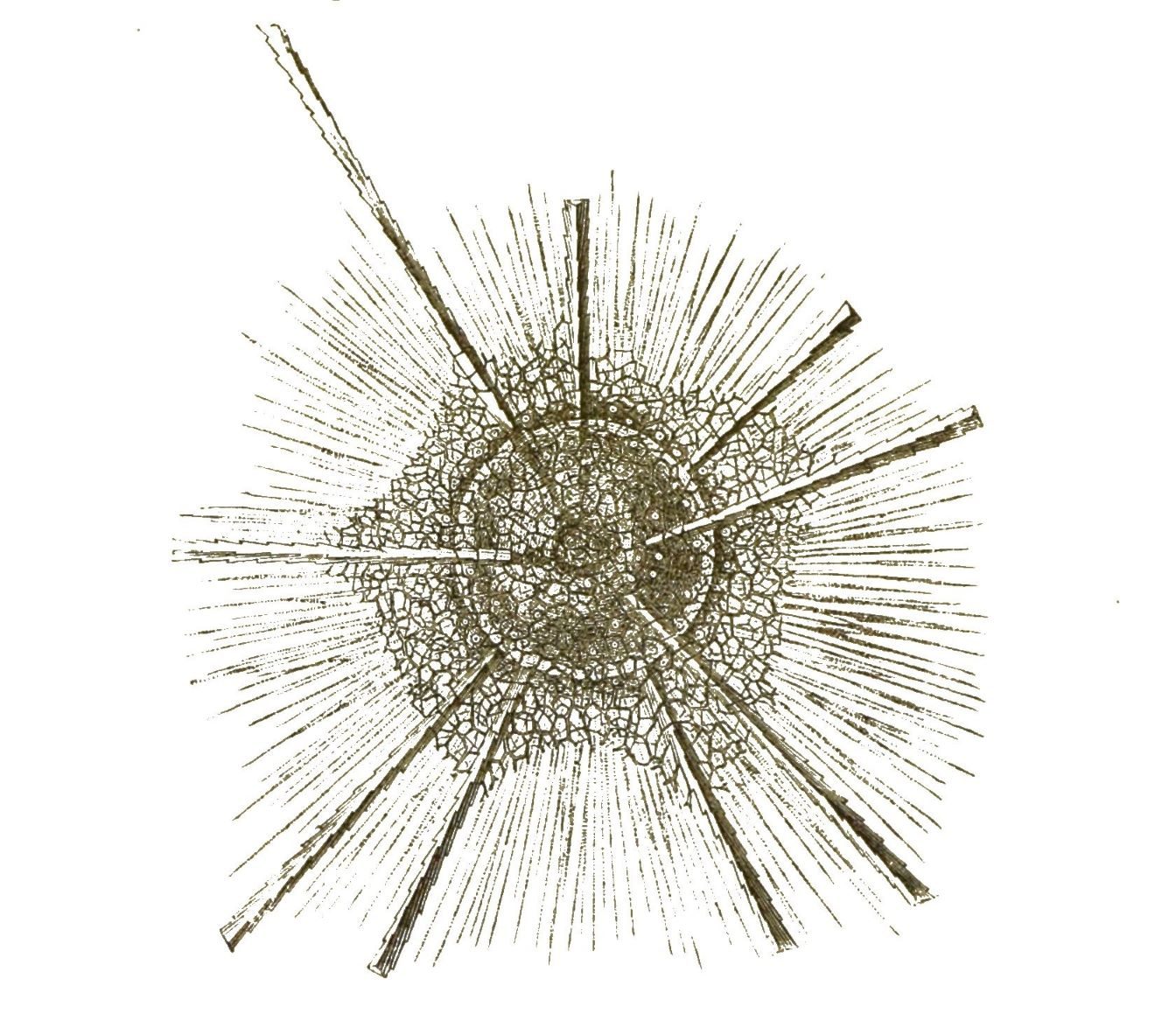
Wenn schon bei diesen merkwürdigen Polythalamien die formbildende Kunst des formlosen Protoplasma unsere höchste Bewunderung erregt, so wird dieselbe noch gesteigert, wenn wir die nahe verwandten Radiolarien, die »Gitterthiere« oder Strahlinge betrachten. Bei diesen höchst interessanten Wurzelfüsslern treffen wir die grösste Mannigfaltigkeit von zierlichen und sonderbaren Formen an, die überhaupt in der organischen Welt zu finden ist. Ja, alle möglichen Grundformen, welche man nur in einem promorphologischen Systeme aufstellen kann, finden sich hier wirklich verkörpert vor. Das Material aber, aus welchem das formlose Protoplasma hier die unendlich mannigfaltigen Skelettheile bildet, ist nicht Kalkerde, wie bei den Polythalamien, sondern Kieselerde.
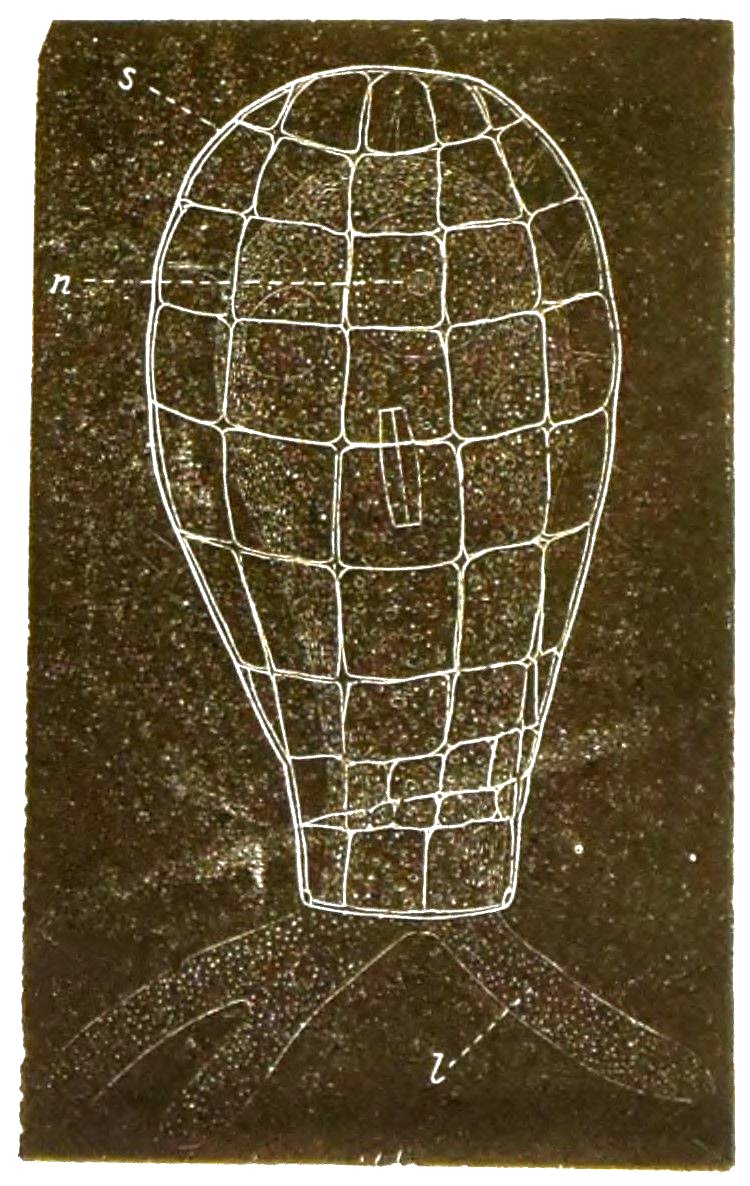
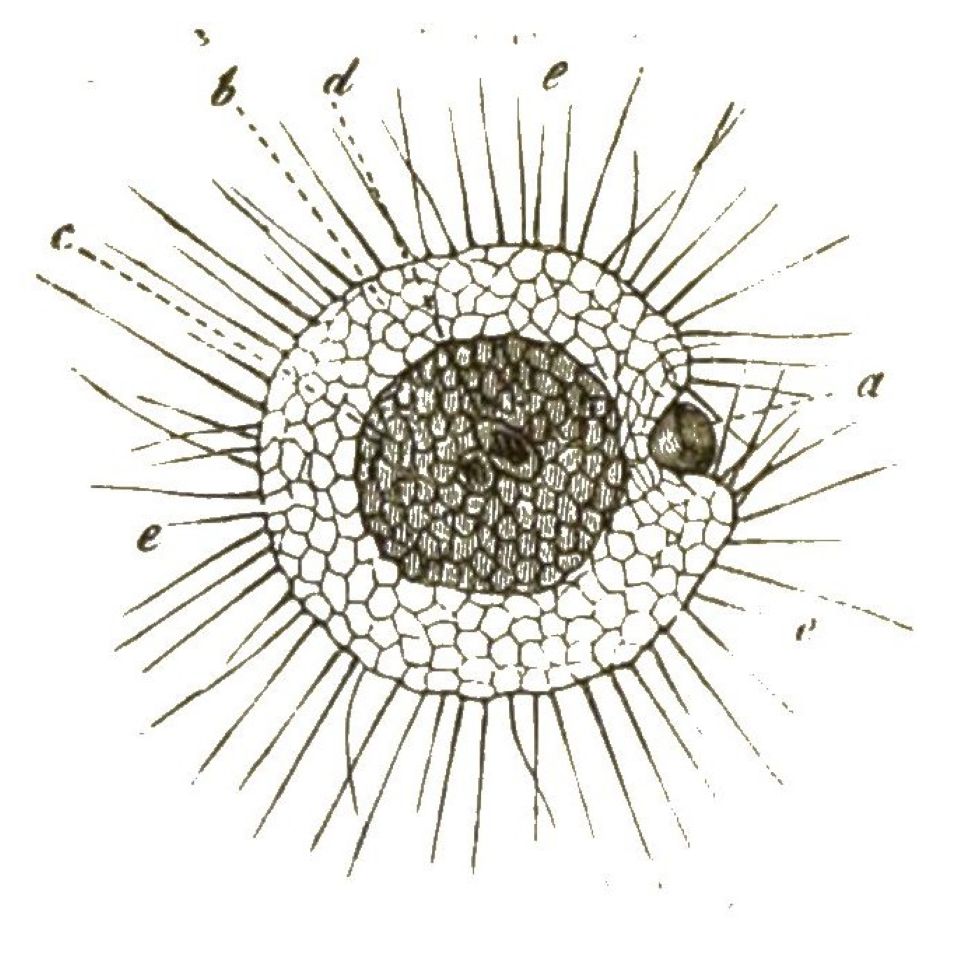
Der weiche lebendige Leib der Radiolarien ist übrigens etwas höher organisirt, als derjenige der Polythalamien. Denn im Innern des formlosen weichen Protoplasma-Körpers findet sich hier eine besondere Kapsel, welche von einer festen Membran umschlossen ist, die Centralkapsel (Fig. 25).
In dieser bilden sich Massen von kleinen Zellen, welche eine bewegliche Geissel erhalten, später die Kapsel durchbrechen und ausschwärmen.
[S. 39]

[S. 40]
Da der ganze Inhalt der Centralkapsel zur Bildung dieser Keime, welche gleich Flagellaten umherschwimmen und sich dann zu Radiolarien entwickeln, verwendet wird, so kann man die Centralkapsel auch als Sporenbehälter (Sporangium) der Radiolarien betrachten.
Sie ist umschlossen von einer Schicht Protoplasma, von welchem nach allen Richtungen zahllose, äusserst feine Scheinfüsschen ausstrahlen. Diese verhalten sich im Uebrigen ebenso wie bei den Polythalamien.
[S. 41]
Gewöhnlich finden sich im Protoplasma der Radiolarien ausserhalb der Centralkapsel noch zahlreiche gelbe Zellen von unbekannter Bedeutung; sie enthalten Stärkemehl.
Ausserdem bilden sich bei einigen Radiolarien rings um die Centralkapsel grosse helle Wasser-Blasen aus (Vacuolen), welche von einer sehr dünnen Gallerte umschlossen sind, so namentlich bei den erbsengrossen Thalassicollen (Fig. 26).

Es giebt auch zusammengesetzte Radiolarien (Polycyttarien). Diese bilden grössere Gallertklumpen von cylindrischer oder kugeliger Form, von 1 bis 3 Centimeter Durchmesser. Die Gallerte besteht grösstentheils aus solchen Wasserblasen, und in der Oberfläche sind ältere, im Innern dagegen jüngere Centralkapseln vertheilt (Fig.27; s. folg. S.). Jede der letzteren ist oft von einer gegitterten Kieselschale umschlossen (Fig. 28).
[S. 42]
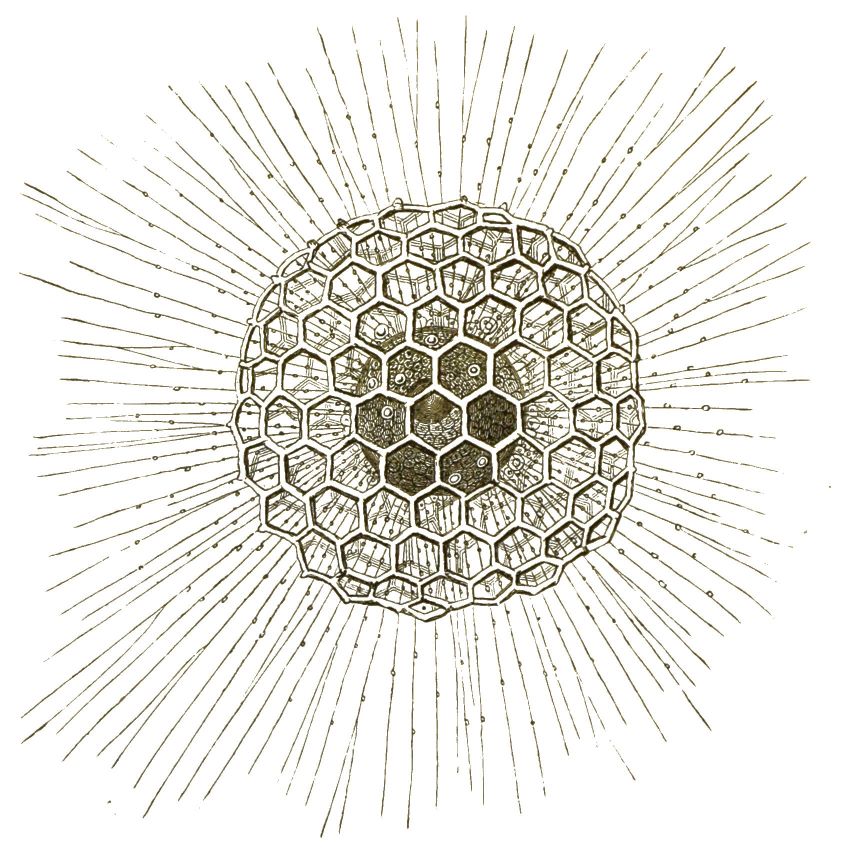
Bei sehr vielen Radiolarien ist die Kieselschale eine Gitterkugel (Fig. 25, 28, 29, 31); oft gehen lange, regelmässig vertheilte Stacheln davon ab (Fig. 29). Bei den Ommatiden (Fig. 30, 31) finden wir mehrere solcher Gitterkugeln concentrisch in einander geschachtelt und durch radiale Stäbe verbunden, ganz ähnlich dem bekannten zierlichen Spielzeug, das die Chinesen aus Elfenbein anfertigen.


Es giebt solche Gitterkugeln, die aus zwanzig im Centrum in einander gestemmten Stacheln zusammengesetzt sind; verästelte Querfortsätze der Stacheln, die in gleichem Abstande vom Centrum abgehen, setzen die Gitterschale zusammen (Dorataspis, Fig. 32). Den letzteren nahe verwandt sind die merkwürdigen Acanthometren (Fig. 33), ebenfalls mit 20 Stacheln, die nach einem bestimmten mathematischen Gesetze regelmässig vertheilt sind.
[S. 43]




[S. 45]

Bei noch andern Radiolarien ist die centrale Gitterkugel von einem lockern Kiesel-Schwammwerke umhüllt und mächtige dreikantige Stacheln mit spiralig gedrehten Kanten ragen daraus hervor (Spongosphaera, Fig. 34).
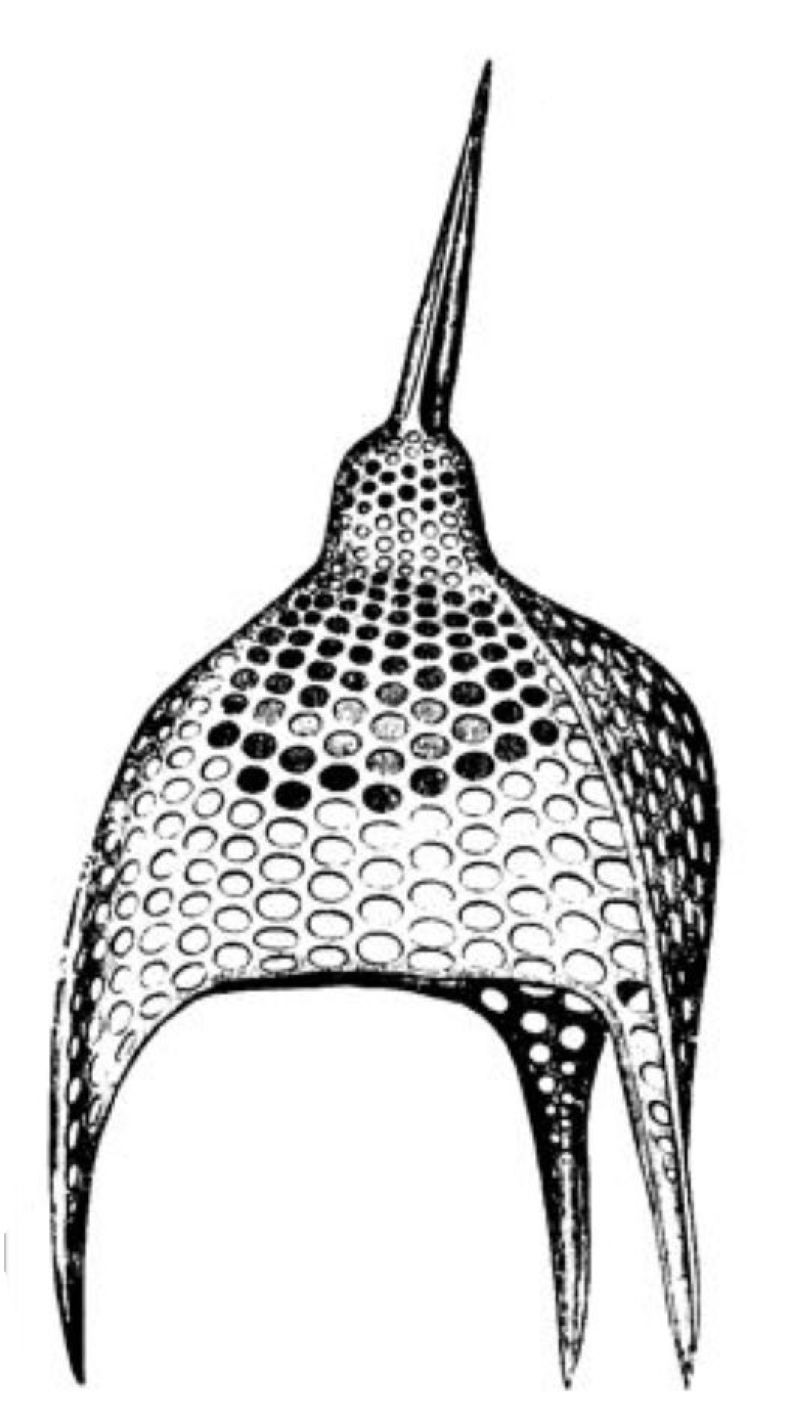
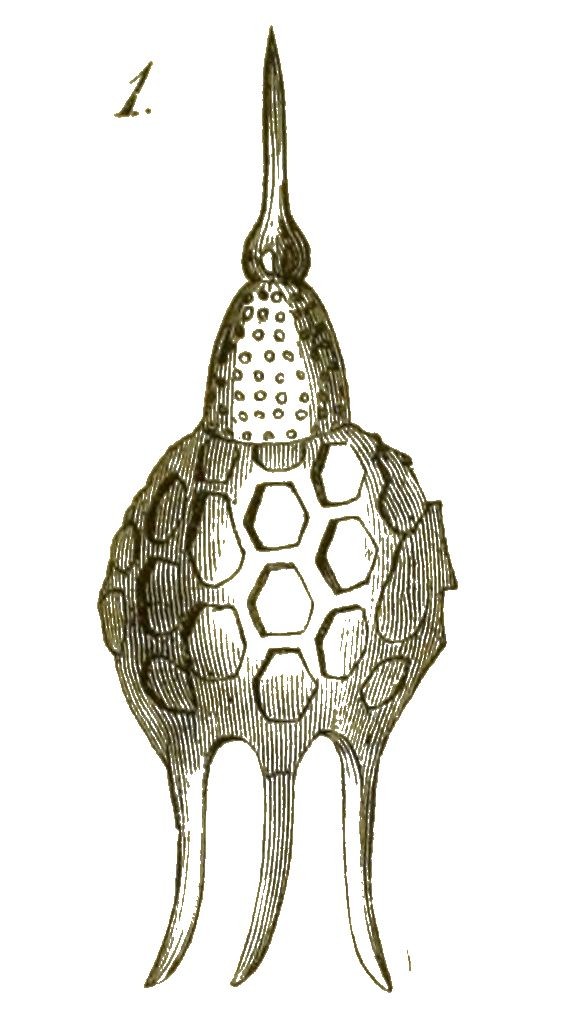
Eine andere, äusserst formenreiche Gruppe von Radiolarien, die Cyrtiden oder Helm-Radiolarien, bilden Kieselschalen von der Form eines Helmes (Fig. 35), einer Haube oder eines Körbchens, mit siebförmig durchlöcherter Wand (Podocyrtis, Fig. 36). Noch Andere gleichen einem Ordensstern (Astromma, Fig. 37), einer Sanduhr (Diploconus, Fig. 38), einem dreiseitigen Prisma (Prismatium, Fig. 39) u. s. w.
In der grossen Abtheilung der Acanthometren wird das Skelet stets aus zwanzig Kieselstacheln gebildet, welche im Centrum in einander gestemmt und nach einem sehr merkwürdigen [S. 46] mathematischen Gesetze vertheilt sind; dies entdeckte zuerst der grosse Johannes Müller, dem wir überhaupt die ersten genaueren Kenntnisse der Radiolarien verdanken.
Welche Bedeutung diese höchst mannigfaltigen, zierlichen und seltsamen Formen besitzen; wie das formlose Protoplasma der Radiolarien dazu kommt, sie zu bilden, — davon haben wir heute noch keine Ahnung.
Neben den Thalamophoren und Radiolarien wird noch eine grosse Anzahl von andern Protisten zur Klasse der Wurzelfüssler gerechnet. Viele davon leben auch im süssen Wasser. Eines der häufigsten ist das niedliche sogenannte »Sonnenthierchen«, welches vor nun hundert Jahren (1776) vom Pastor Eichhorn in Danzig entdeckt und als »lebendiger Stern« beschrieben wurde (Actinosphaerium Eichhornii, Fig. 40).
[S. 47]


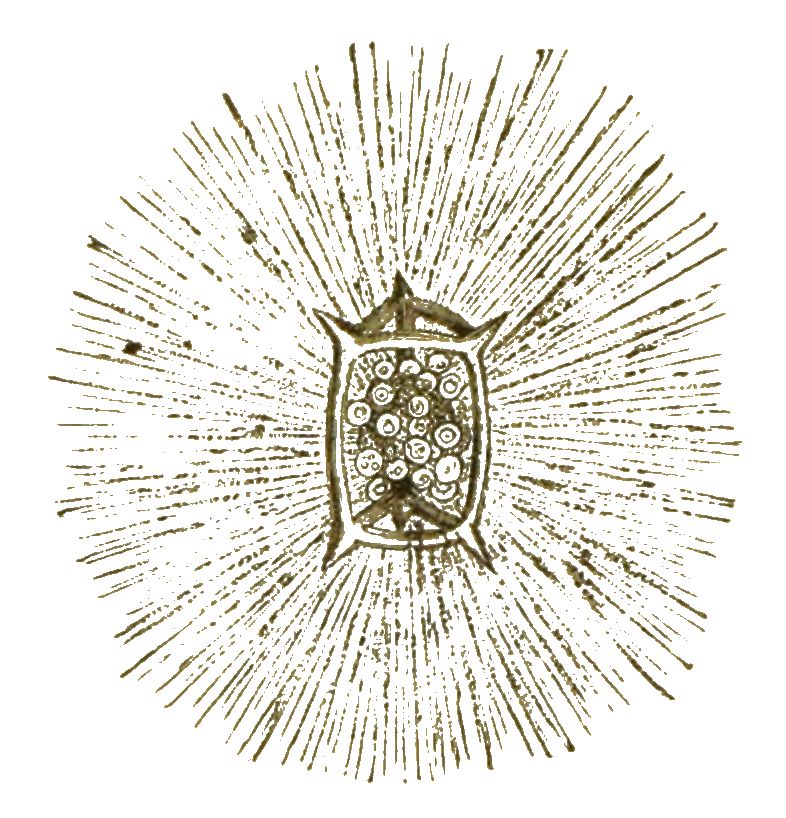

Es ist ein weiches, mit blossem Auge deutlich sichtbares, weiches Schleimkügelchen, von der Grösse eines kleinen Stecknadelknopfes, oft in Menge auf dem schlammigen Boden [S. 49] unserer Teiche und Gräben zu finden. In der Mitte des schleimigen und blasigen Protoplasma-Kügelchens liegen mehrere Zellkerne. Von der Oberfläche strahlen zahlreiche empfindliche und bewegliche Fäden oder Pseudopodien aus. Durch diese wird, wie bei den übrigen Wurzelfüsslern, die Nahrung aufgenommen. Die Vermehrung ist erst kürzlich entdeckt worden. Das Sonnenthierchen zieht dabei seine Fäden ein, umgiebt seinen kugeligen Körper mit einer Gallerthülle und zerfällt in viele einzelne Kugeln. Jede von diesen enthält einen Kern und schwitzt eine Kieselhülle aus, und jede dieser kieselschaligen Zellen wird später zu einem neuen Sonnenthierchen. Man kann dieselben aber auch künstlich vermehren. Man kann sie in mehrere Stücke zerschneiden und aus jedem Stückchen wird alsbald wieder ein selbständiges Wesen. Dasselbe gilt auch von vielen andern Protisten.
Während das grosse Sonnenthierchen oder Strahlenkügelchen (Actinosphaerium) einen nackten Rhizopoden darstellt, der viele Zellkerne enthält, also aus vielen vereinigten Zellen zusammengesetzt ist, zeigt uns dagegen ein anderer, sehr häufiger Süsswasserbewohner, das kleine Sonnenthierchen (Actinophris sol) den Organismus der Wurzelfüssler in seiner allereinfachsten Gestalt (Fig. 41), nämlich als eine nackte einfache Zelle mit einem einzigen Kern; von der Oberfläche desselben strahlen viele feine Fäden aus, und indem das Protoplasma an gewissen Stellen Wasser aufnimmt und wieder abgiebt, bildet es »contractile Blasen oder Vacuolen.«
Eine der merkwürdigsten Protistenklassen, die ebenfalls oft zu den Wurzelfüsslern gerechnet wird, sind die so genannten Schleimpilze oder Myxomyceten, von Anderen auch Pilzthiere oder Mycetozoen genannt. Schon dieser doppelte Name bezeichnet ihre zweifelhafte Protisten-Natur. Sie leben in zahlreichen verschiedenen Arten an feuchten Orten, im abgefallenen Laube der Wälder, zwischen Moos, auf faulendem Holze und dergl. Früher galten sie allgemein für Pflanzen, und zwar für Pilze, weil ihr reifer Fruchtkörper täuschend dem blasenförmigen Fruchtkörper der Gastromyceten oder Blasenpilze ähnlich ist (Fig. 43B). [S. 50] Dieser Fruchtkörper bildet kugelige oder länglich runde, oft auf einem Stiel festsitzende Blasen, meist von der Grösse eines Stecknadelknopfes oder eines Hanfkorns, bisweilen aber auch von mehreren Zoll Durchmesser. Die derbe äussere Hülle der Fruchtblasen umschliesst ein feines Mehl, das aus Tausenden von mikroskopischen Zellen besteht. Dies sind die Fortpflanzungszellen oder Sporen.
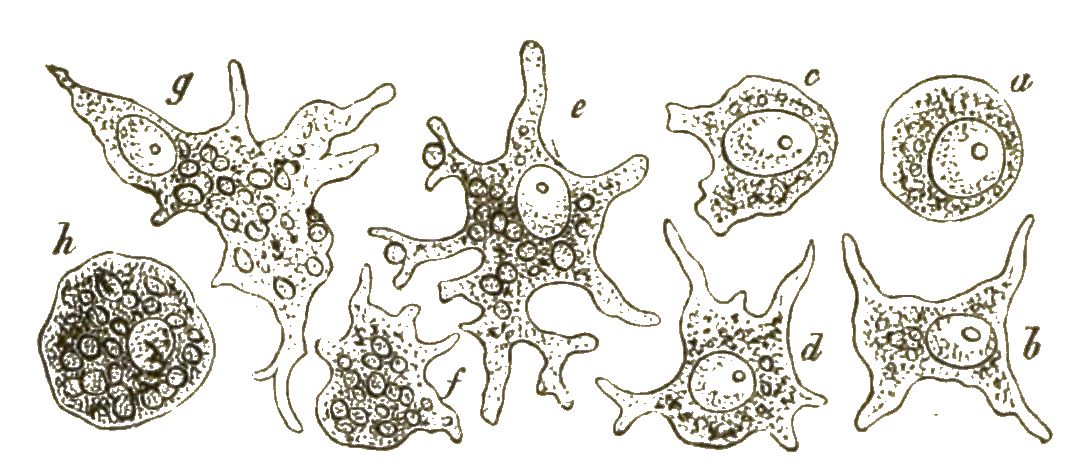
Während aber bei den Blasenpilzen, wie bei allen anderen echten Pilzen, sich aus diesen Sporen die characteristischen Pilzfäden oder Hyphen, lange dünne Fadenschläuche entwickeln, entstehen daraus bei den Myxomyceten ganz andere Keime. Aus der festen Zellmembran einer jeden Spore schlüpft nämlich, sobald diese ins Wasser gelangt, eine nackte, lebhaft bewegliche Zelle aus. (Fig. 42, 1–3). Anfangs schwimmt diese Zelle mittelst eines langen Geisselfadens, den sie peitschenförmig nach Art der Geisselschwärmer hin und her schwingt, frei im Wasser umher (Fig. 42, 4, 5). Später sinkt sie zu Boden und nimmt die Form einer Amoebe an (Fig. 42, 6, 7). Ganz gleich einer echten Amoebe kriecht sie umher, indem sie veränderliche Fortsätze ausstreckt und wieder einzieht. Auch nimmt sie nach Art der Amoeben ihre Nahrung auf.
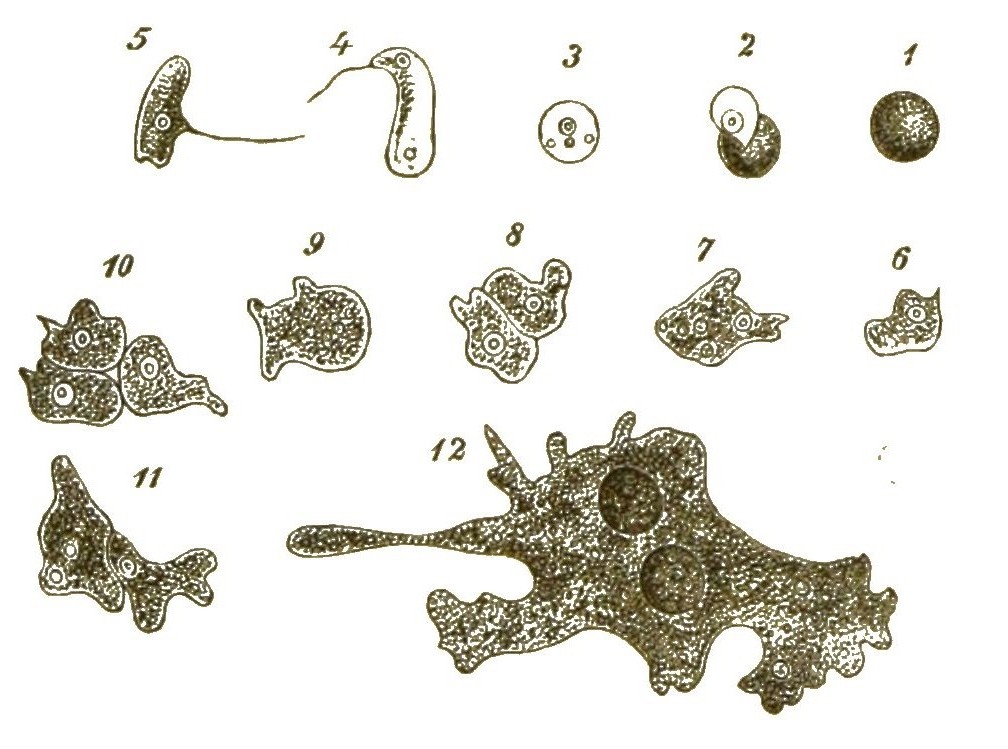
Viele solcher amoeboiden Zellen können nun späterhin zusammenfliessen und mit einander verwachsen (Fig. 42, 8–11). Dadurch [S. 51] entstehen grosse Protoplasma-Netze mit vielen Kernen (Syncytien, Fig. 42, 12). Indem ihre Kerne sich auflösen, werden sie zu kernlosen Plasmodien (Fig. 43A). Solche grosse Plasmodien, oft ganz colossale Protoplasma-Netze, kriechen gleich einem riesigen Rhizopoden langsam umher und ändern beständig ihre unbestimmte Gestalt.
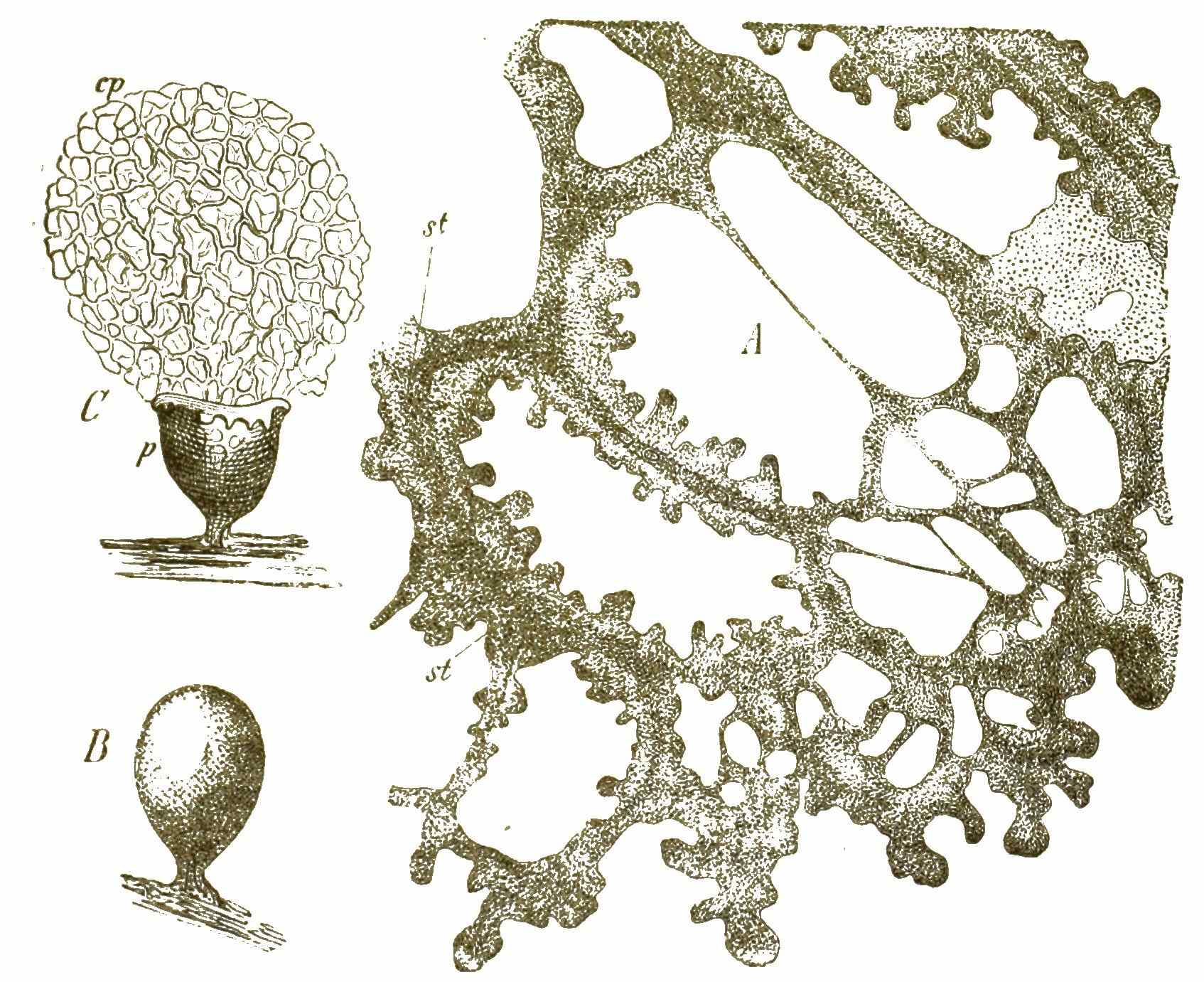
Zu den grössten Plasmodien gehören die glänzend gelben (oft mehrere Fuss grossen) Protoplasma-Geflechte von Aethalium, welche die Lohbeete der Gerbereien durchziehen und unter dem Namen »Lohblüthe« allen Gerbern bekannt sind. Haben die Plasmodien durch Wachsthum und Nahrungsaufnahme eine gewisse Grösse erreicht, so ziehen sie sich auf einen kugeligen, birnförmigen oder kuchenförmigen Haufen zusammen, umgeben sich mit einer Hülle und das ganze Protoplasma zerfällt in [S. 52] zahllose kleine Sporen, zwischen welchen sich meistens (jedoch nicht immer) ein Geflecht von äusserst feinen Haarfäden ausbreitet (Capillitium, Fig. 43cp). Wenn diese Fruchtkörper (Fig. 43B) ganz reif sind, platzt die äussere Hülle (Fig. 43C); das Capillitium wird vorgetrieben und das feine Sporen-Pulver zerstreut.
Obgleich nun diese blasenförmigen Fruchtkörper mit ihrem Sporenpulver und Capillitium die grösste Aehnlichkeit mit denjenigen von gewissen echten Pilzen besitzen, haben sie doch mit diesen letzteren keine Spur von Verwandtschaft, wie ihre gänzlich verschiedene Entwickelung zeigt. Will man überhaupt die Myxomyceten in nähere Beziehung zu irgend einer anderen Organismen-Gruppe bringen, so bleiben nur die Rhizopoden übrig. In der That gleichen die kriechenden netzförmigen Plasmodien der Myxomyceten so sehr gewissen nackten Wurzelfüsslern (Lieberkühnia), dass man sie gar nicht unterscheiden kann. Es giebt kein passenderes Object, um sich die merkwürdigen Strömungen in dem kriechenden nackten Protoplasma unmittelbar vor Augen zu führen, als die Plasmodien der gemeinen Lohblüthe, die im Frühjahr auf den Lohbeeten der Gerbereien sehr leicht zu haben ist und die Lohe in Form von gelben, rahmähnlichen Schleimnetzen durchzieht. Bringt man ein wenig von diesem gelben Protoplasma in einer feuchten Kammer auf ein Glasplättchen, so ist letzteres schon nach 10–20 Stunden von einem feinen Faden-Netz übersponnen, in dessen Fäden man unter dem Mikroskope die lebhafte Protoplasma-Strömung prächtig verfolgen kann.
Im Anschluss an die Myxomyceten müssen wir hier auch auf die echten Pilze (Fungi) einen Blick werfen, mit welchen man die ersteren früher irrthümlich vereinigt hat. Die echten Pilze, welche in so zahlreichen, ansehnlichen und mannigfaltigen Formen in unsern Wäldern und Feldern, auf Pflanzen- und Thierkörpern schmarotzend leben, werden oft auch als Schwämme bezeichnet. Sie haben aber mit den echten Schwämmen oder Spongien gar nichts zu thun; denn diese letzteren, wozu der gewöhnliche Badeschwamm gehört, und welche sämmtlich — mit einziger Ausnahme des Süsswasser-Schwammes, Spongilla, — im Meere leben, sind echte Thiere und besitzen ein Darmrohr mit Mundöffnung u. s. w. Die Pilze dagegen bilden eine gänzlich verschiedene und sehr eigenthümliche Classe von niederen Organismen. Zwar [S. 53] gelten sie heute noch allgemein als echte Pflanzen. Allein in den wichtigsten anatomischen und physiologischen Beziehungen weichen sie so sehr von allen übrigen Pflanzen ab, dass es wohl richtiger ist, sie als eine selbständige Classe von Protisten zu betrachten. Ernährung und Stoffwechel der Pilze ist thierisch, nicht pflanzlich. Sie bilden kein Protoplasma, kein Chlorophyll, kein Stärkemehl, keine Cellulose, wie die echten Pflanzen. Vielmehr bedürfen sie, wie die Thiere, zu ihrer Existenz und Ernährung vorgebildetes Protoplasma, welches sie aus dem Körper anderer Organismen, lebender oder todter Thiere, Pflanzen und Protisten, entnehmen.
Die Fortpflanzung der Pilze ist meistens ungeschlechtlich, und auch da, wo sie geschlechtlich erscheint, ganz eigenthümlich. Das Form-Element, aus dem sich der Körper aller Pilze aufbaut, ist nicht eine echte, kernhaltige Zelle, wie bei allen Thieren und Pflanzen, sondern eine fadenförmige, kernlose Cytode, die sogenannte Hyphe oder der »Pilzfaden.« Durch seitliche Sprossung und fortgesetzte Theilung in einer Axe, bilden sie verzweigte gegliederte Fäden, und zahllose solche Pilzfäden, in langen Ketten an einander gereiht, sich verästelnd und netzartig verbindend, setzen alle Organe der Pilze zusammen. Der bekannte gestielte »Hut« oder Schirm unserer grossen Hutpilze, z. B. vom Champignon (Fig. 44) ist blos der Fruchtkörper, welcher sich zur Zeit der Reife aus einem unscheinbaren Fadengeflechte entwickelt, dem Mycelium (Fig. 44, I m); die strahligen, blattförmigen Rippen, welche sich an der Unterseite des regenschirmähnlichen Hutes bilden, sind von der Fruchthaut (Hymenium) überzogen, in welcher sich ungeschlechtlich die Fortpflanzungs-Cytoden (»Sporen«) bilden. Je genauer man die eigenthümliche Anatomie und Keimungsgeschichte der Pilze verfolgt, je unbefangener man sie vergleicht, desto mehr überzeugt man sich, dass diese merkwürdigen Organismen keine echten Pflanzen sind, sondern eine ganz selbständige Classe von neutralen Protisten darstellen.
Dasselbe gilt von der formenreichen Classe der Kieselzellen (Diatomeae oder Bacillariae), die auch gewöhnlich zu den Pflanzen gerechnet werden. Diese zierlichen kleinen Organismen bevölkern in ungeheuren Massen die süssen und salzigen Gewässer unseres Erdballs. In grossen Mengen angehäuft, bilden [S. 54] sie gewöhnlich einen gelben oder gelbbraunen Schleim, der Steine, Wasserpflanzen u. s. w. überzieht. Bald sind die Diatomeen einzeln lebende Einsiedlerzellen, bald Colonien oder Gesellschaften (Coenobien), welche aus vielen gleichartigen, locker verbundenen Zellen zusammengesetzt erscheinen.
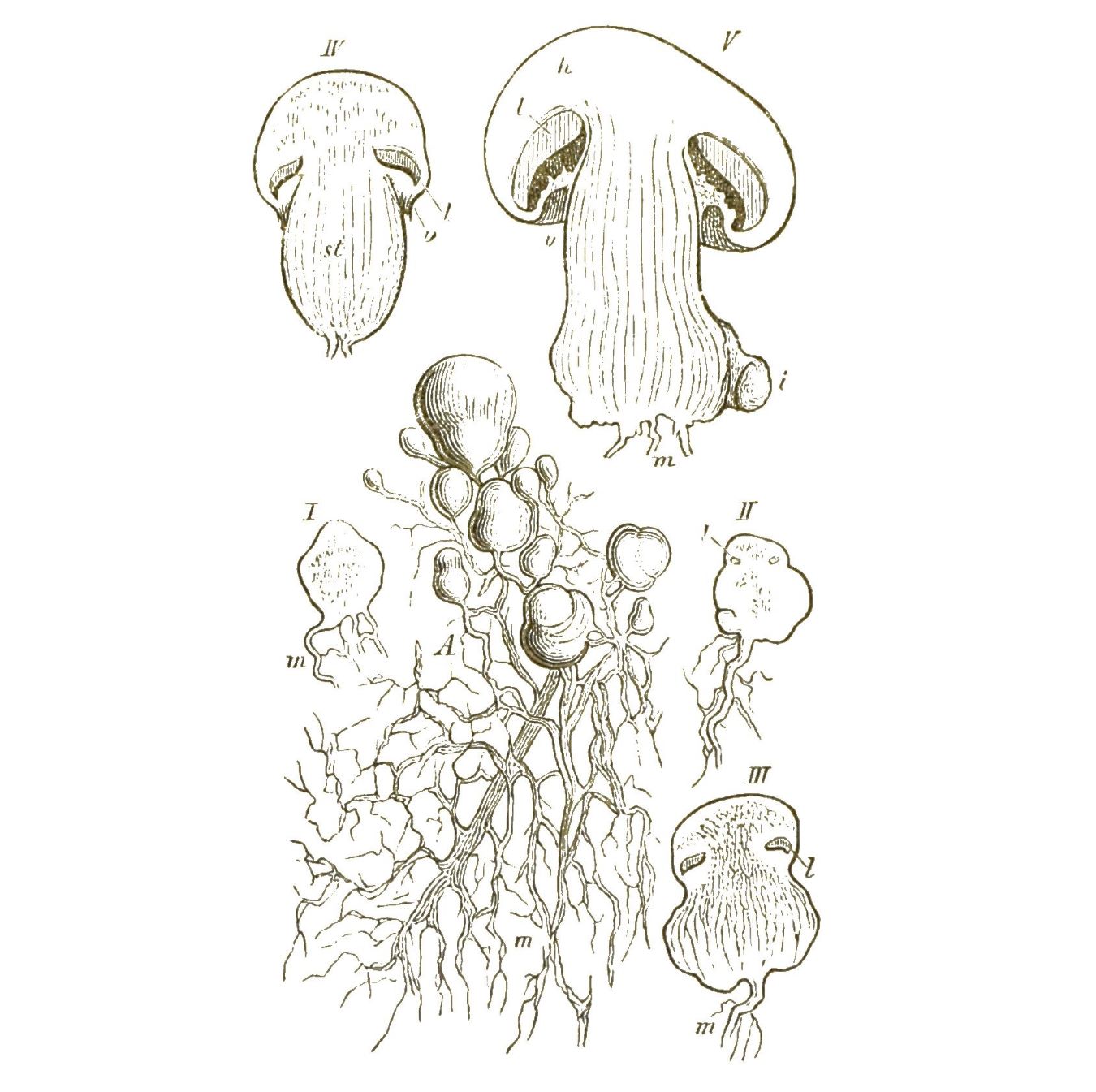
Viele Diatomeen sitzen fest; die meisten aber bewegen sich in ganz eigenthümlicher Weise, langsam schwimmend oder fortrutschend, [S. 55] im Wasser umher. Die Organe dieser Ortsbewegung sind noch gänzlich unbekannt, vielleicht feinste Wimperreihen.

Das Characteristische an dem Zellenkörper der Diatomeen ist die eigenthümliche Kieselschale, in welcher ihr Zellenleib eingeschlossen ist. Diese Schale ist aus zwei Hälften zusammengesetzt, welche sich zu einander genau so verhalten, wie eine Schachtel zu ihrem Deckel (Fig. 45). Die kernhaltige Zelle, welche in dieser Schachtel lebt, theilt sich in zwei Hälften, und jede Hälfte bildet sich zu ihrem Schachteldeckel eine neue Schachtel. Dieser Process wiederholt sich mehrfach, wobei natürlich jede folgende Generation kleiner wird. Schliesslich aber entsteht eine Generation, welche beide Schalenhälften abwirft, wieder bis zur Grösse der ersten, grössten Generation heranwächst, und sich nun mit einer neuen Kieselschachtel erster Grösse umgiebt. Wegen der unendlich mannigfaltigen und zierlichen Gestalt dieser Kieselschale, sowie wegen ihrer äusserst feinen Sculptur, sind die Diatomeen sehr beliebte Unterhaltungs-Objecte für mikroskopischen Formgenuss. Wenn sich die Kieselschalen der todten Diatomeen massenhaft auf dem Grunde der Gewässer ansammeln und zu Stein verkitten, können sie ganze Gebirgsschichten zusammensetzen, so z. B. den Polirschiefer, das Bergmehl u. s. w.
Während die meisten, bisher von uns betrachteten Protisten-Gruppen grosse und formenreiche Classen darstellen, giebt es nun noch eine Anzahl von kleineren, isolirten, bisweilen nur durch eine oder wenige Formen repräsentirten Protisten, deren Einreihung in das System sehr schwierig ist. Dies gilt z. B. von den sonderbaren Labyrinthuleen. Gesellschaften von locker verbundenen, einfachen, spindelförmigen, gelben Zellen, die in einer eigenthümlichen Fadenbahn umherrutschen. Eine andere Gruppe, [S. 56] interessant wegen ihrer Mittelstellung zwischen verschiedenen Protisten-Classen, bilden die Catallacten, durch die Gattungen Synura und Magosphaera repräsentirt. Sie bilden schwimmende Gallertkugeln, zusammengesetzt aus einer Anzahl birnförmiger gleichartiger Zellen, welche mit ihren spitzen inneren Enden im Centrum der Gallertkugel vereinigt sind. Später lösen sich diese Zell-Gesellschaften oder Coenobien auf. Die einzelnen isolirten Zellen schwimmen noch eine Zeit lang selbständig umher und können jetzt mit Ciliaten verwechselt werden. Dann aber sinken sie auf den Meeresboden nieder und verwandeln sich in Amoeben-ähnliche Zellen. Gleich echten Amoeben kriechen diese umher, fressen, wachsen und kapseln sich schliesslich ein; der Zellenkörper zieht sich kugelig zusammen und umgiebt sich mit einer Gallerthülle. Innerhalb derselben theilt sieh die Zelle später wiederholt, in 2, 4, 8, 16, 32 Zellen u. s. w. Diese werden birnförmig, erhalten bewegliche Wimpern und verbinden sich wieder zu einer Flimmerkugel. Nun dreht sich die Kugel rotirend um ihren Mittelpunkt, sprengt ihre Hülle und schwimmt wieder frei in der Form umher, von welcher wir ausgegangen sind (Fig. 46). Das Interesse dieser merkwürdigen Protisten liegt also weniger in besonderen Eigenthümlichkeiten, als vielmehr in der neutralen Mittelstellung, [S. 57] welche sie zwischen Amoeben, Infusorien und Volvocinen einnehmen, und wodurch sie diese verschiedenen Protisten-Classen verknüpfen. Wir nennen sie daher »Mittlinge oder Vermittler« (Catallacta).
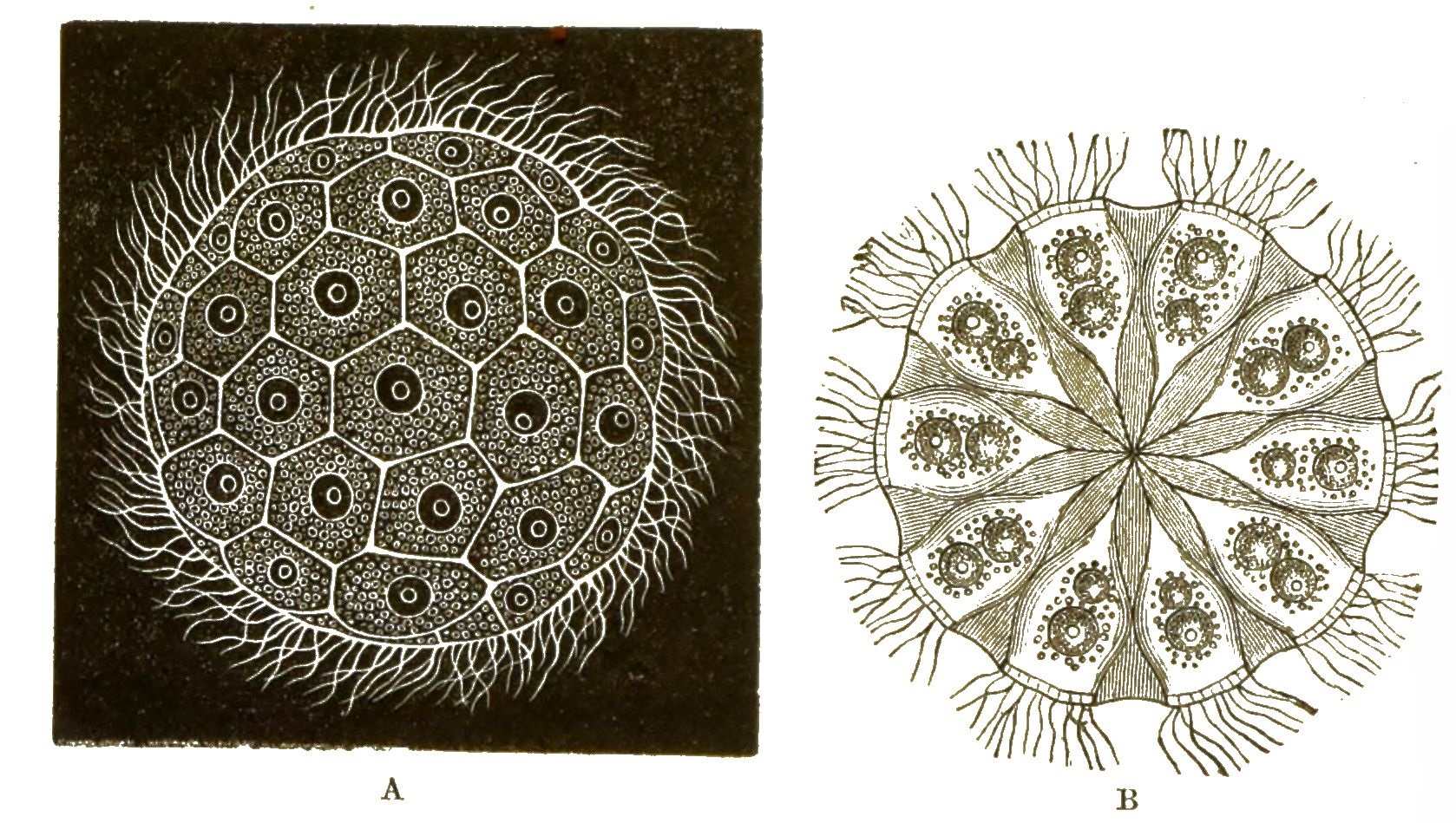
Werfen wir einen vergleichenden Rückblick auf alle bisher betrachteten Protisten-Classen, so sehen wir, dass darin die organische Zelle bald ganz selbständig auftritt, und als Einsiedler-Zelle (Monocyta) den ganzen Organismus repräsentirt, bald mit ihresgleichen sich zu lockeren Gesellschaften verbindet und einfache Zellen-Gemeinden oder Zellen-Horden (Coenobia) darstellt. Nun ist aber hiermit keineswegs die tiefste Stufe der Organisation erschöpft, welche uns die organische Welt darbietet. Vielmehr treffen wir noch unterhalb dieser einzelligen Protisten jene niedrigste und unvollkommenste Classe von Organismen an, die wir als Moneren bezeichnen. (Fig. 47, 48).
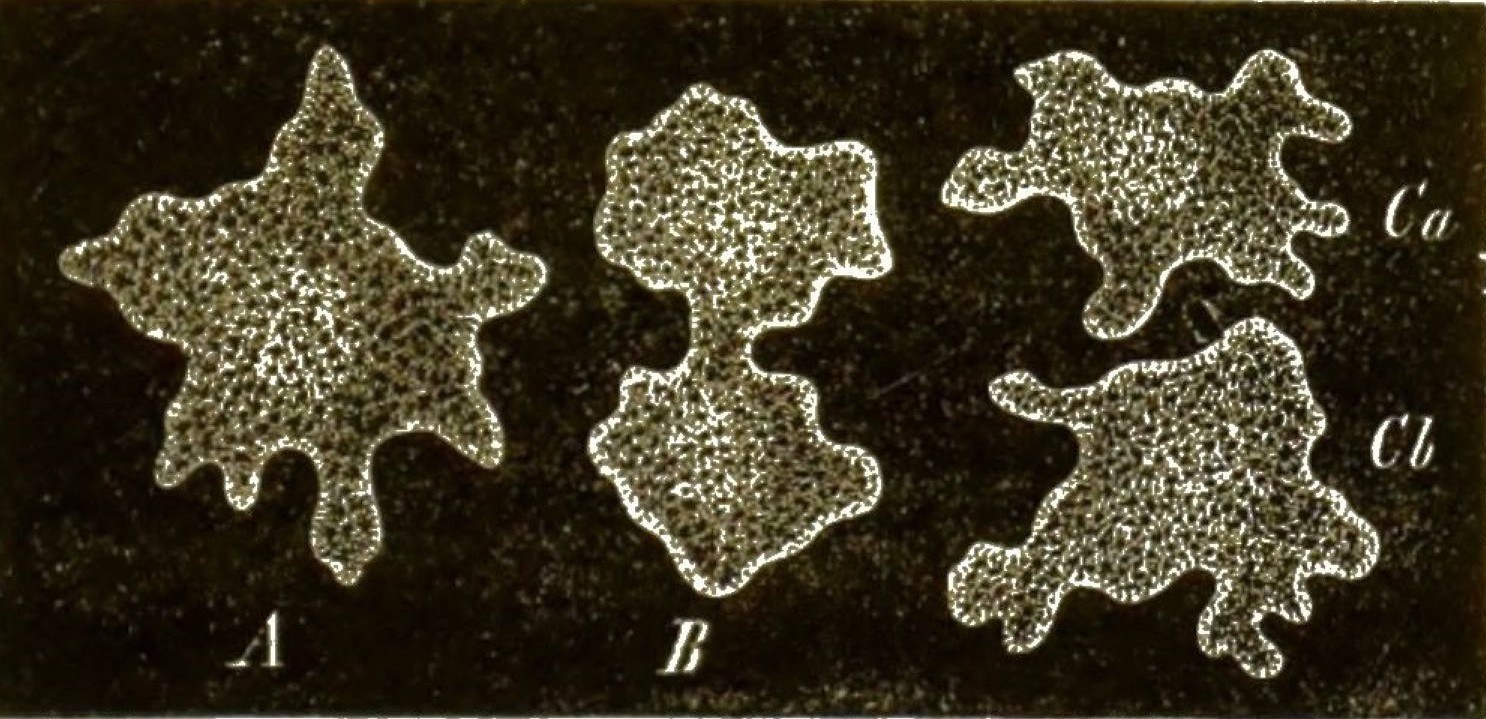
Da wir diese in dem nachstehenden Anhange (S. 68–85) zum Gegenstande einer besonderen Besprechung machen werden, wollen wir hier nur ganz kurz die wichtigsten Punkte hervorheben, welche den Moneren ihre hohe Bedeutung für die Entwicklungslehre verleihen.
Die Moneren sind wahre »Organismen ohne Organe«. Ihr ganzer lebendiger Leib besteht in völlig entwickeltem Zustande nur aus einem ganz einfachen Protoplasma-Stückchen, welchem selbst der Kern, der Character der echten Zelle, noch fehlt. Bezüglich ihrer Bewegungen gleichen diese denkbar einfachsten Organismen bald den Amoeben (Fig. 47), bald den Wurzelfüsslern(Fig. 48), bald den Geisselschwärmern.
[S. 58]


Sie vermehren sich in einfachster Weise durch Theilung. Von der grössten theoretischen Bedeutung sind sie für die dunkle Frage von der ersten Entstehung des Lebens auf unserer Erde. Denn nur Moneren können im Beginn des organischen Lebens auf unserm Planeten durch Urzeugung entstanden sein; nur Moneren können die ältesten Stammältern aller übrigen Organismen sein. Gerade in dieser Beziehung sind die Moneren des Tiefseegrundes, und vor Allen der berühmte Bathybius (Fig. 49) vom höchsten Interesse.
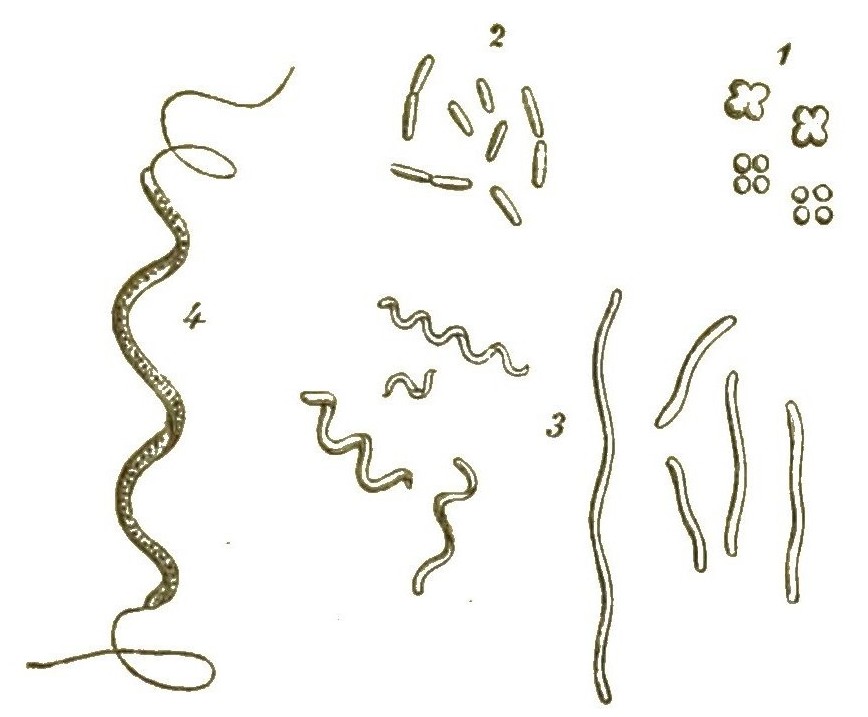
Eine sehr wichtige und interessante Monerengruppe bilden die Zitterlinge (Vibriones oder Bacteria, Fig. 50). Obgleich diese winzigen Körperchen, die zu den allerkleinsten Organismen gehören, meistens von den Botanikern zu den Pflanzen gerechnet und als »Spaltpilze (Schizomycetes)« den echten Pilzen angereiht werden, geschieht das doch ohne jeden genügenden Grund. Mindestens haben diejenigen Zoologen, welche sie als einfachste Thiere betrachten, ebensoviel Recht dazu. Die Bacterien sind eben echte Protisten, und zwar kleinste Moneren, deren höchst einfache Organisation und ganz neutraler Character sie weder dem Thierreich, noch dem Pflanzenreich anzuschliessen gestattet.
[S. 60]
Die Bacterien sind meistens stabförmige Körperchen, die sich lebhaft im Wasser bewegen. Als Organ der Bewegung ist bei einigen grösseren Formen eine äusserst feine, schwingende Geissel erkannt, die an beiden Enden des Stäbchens vortritt, so bei Spirillum (Fig. 50, 4). Wahrscheinlich ist eine solche auch bei den kleineren Vibrionen vorhanden und nur wegen ihrer ausserordentlichen Zartheit nicht wahrzunehmen. Die Bewegung der Bacterien ist meistens sehr lebhaft, zitternd oder wimmelnd, viele sind korkzieherartig gedreht und schrauben sich im Wasser fort (Fig. 50, 3). In einem einzigen Wassertröpfchen können Millionen solcher kleinsten Organismen vereinigt sein. Irgend welche Organisations-Verhältnisse, namentlich ein Zellkern, sind an denselben nicht nachzuweisen; sie sind daher auch nicht wirkliche Zellen, sondern kernlose Cytoden, gleich den anderen Moneren. Ihre Fortpflanzung geschieht in einfachster Weise durch Theilung. Oft zerfällt jedes Stäbchen in eine grosse Anzahl hinter einander gelegener Stückchen.
Die grosse Bedeutung der Bacterien besteht darin, dass sie die Zersetzung und Fäulniss der organischen Flüssigkeiten bewirken, in welchen sie sich aufhalten. Sie ernähren sich von den organischen Substanzen (namentlich eiweissartigen Körpern), die in solchen Flüssigkeiten aufgelöst sind. Wahrscheinlich sind sie die Ursache vieler der wichtigsten, ansteckenden und epidemischen Krankheiten. So ist es neuerdings namentlich vom Milzbrand und den Blattern festgestellt, dass nur die Bacterien, die im Blute der milzbrandkranken und blatternkranken Thiere leben, die Uebertragung dieser tödtlichen Krankheiten bewirken.
Ueberblickt man unbefangen prüfend und vergleichend die Masse von verschiedenartigen Urwesen, die wir in unserem Protistenreiche vereinigt haben, so scheint die Selbständigkeit dieses letzteren keines weiteren Beweises zu bedürfen. Denn es existirt noch heute eine ungeheuere Menge von formenreichen, mikroskopischen Wesen, die wir ohne willkürlichen Zwang weder zum Thierreich noch zum Pflanzenreich rechnen können. Aber das natürliche Verhältniss dieser beiden grossen Lebensreiche zu jenem neutralen, zwischen Beiden mitten inne stehenden Protistenreiche wird noch vielfacher Durchforschung und Klärung [S. 61] bedürfen. Insbesondere wird die Entwickelungsgeschichte der Protisten noch viel genauer und umfassender zu erforschen sein. Denn vor allen die Entwickelungsgeschichte wird hier, wie überall, der »wahre Lichtträger« für das Verständnis der biologischen Erscheinungen sein.
Uebrigens scheint gegen das Thierreich hin eine feste und klare Abgrenzung des Protistenreichs schon jetzt sicher gewonnen zu sein. Denn bei allen echten Thieren entwickelt sich der Leib aus zwei ursprünglichen Zellenschichten, die unter dem Namen der Keimblätter bekannt sind.
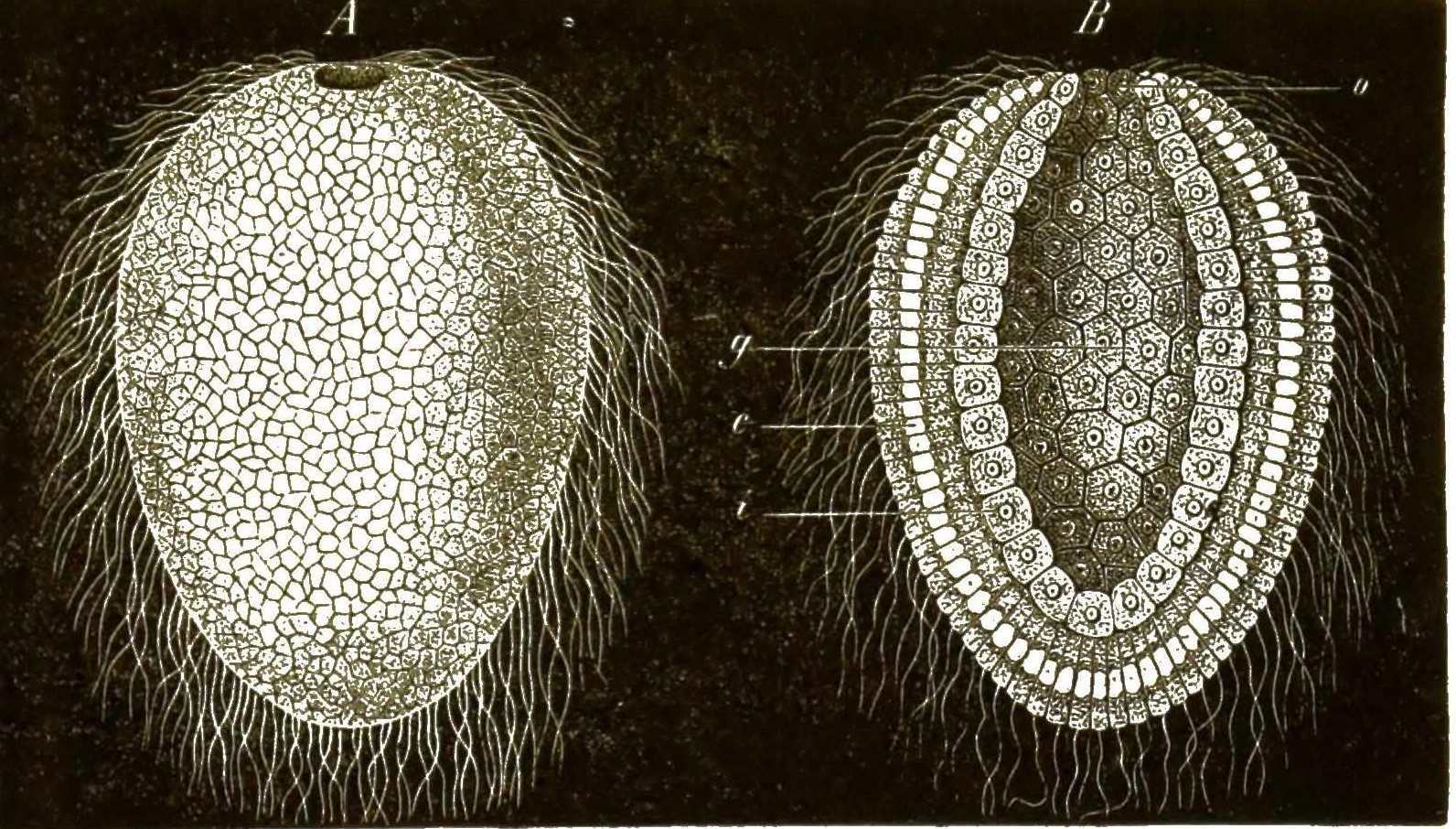
Aus dem äusseren oder animalen (Exoderma oder Hautblatt, Fig. 51 e) entstehen die Organe der Empfindung und Bewegung; aus dem innern oder vegetativen Keimblatte (Entoderma oder Darmblatt, Fig. 51 i) die Organe der Ernährung. Das letztere umschliesst eine ernährende Höhle, die erste Anlage des Magens, oder den Urdarm (g), und dieser öffnet sich nach aussen durch eine einfache Mundöffnung, den Urmund (o). Die bedeutungsvolle Keimform, welche uns den Thierleib dergestalt, blos aus zwei Keimblättern gebildet, vor Augen führt, ist die Gastrula (Darmlarve oder Becherkeim).
[S. 62]
[S. 63]
Diese Gastrula ist das wahre Thier in einfachster Form. Denn bei allen echten Thieren fängt die Entwickelung des Eies zur verschiedenartigen Thierform mit der gleichartigen Bildung dieser Gastrula an. Die niedersten Pflanzenthiere, die Physemarien (Fig. 56) wie die Schwämme (Fig. 51), die niedrigsten Würmer (Fig. 52), ebenso die Sternthiere (Fig. 53), die Gliederthiere (Fig. 54) ebenso wie die Weichthiere (Fig. 55), ja sogar die niedrigsten Wirbelthiere (Fig. 57), durchlaufen in frühester Jugend diese Gastrula-Keimform; die anderen Thiere bilden zweiblättrige Keimformen, die nur als abgeänderte Gastrula-Keime betrachtet werden können; so auch die Säugethiere, mit Inbegriff des Menschen (Fig. 58); überall baut sich der echte Thierleib ursprünglich aus zwei Keimblättern auf. Hingegen erhebt sich kein einziges Protist zur Production von Keimblättern und zur Bildung einer Gastrula.
Weniger klar und scharf lässt sich unser Protistenreich gegen das Pflanzenreich hin abgrenzen. Doch dürften auch hier die Verhältnisse der individuellen Entwickelung und des feineren Baues die Handhabe liefern, mit deren Hülfe wir die Grenzlinie ziehen können. Auch bei den echten Pflanzen ordnen sich die Zellen, welche den Körper zunächst aufbauen, in bestimmter Weise zu Zellenreihen oder Zellenschichten; und die charakteristische einfachste Pflanzenform der Art bildet den sogenannten Thallus oder das »Zellenlager«. Bei den niederen Pflanzen bleibt der Thallus als solcher zeitlebens bestehen, bei den höheren sondert oder differenzirt er sich in Stengel und Blätter. Auch vermehren sich alle echten Pflanzen auf geschlechtlichem Wege, während dies bei den Protisten nicht der Fall ist.
Eine absolute Grenze freilich zwischen den drei organischen Reichen können und wollen wir nicht feststellen. Denn auch die echten Pflanzen, wie die echten Thiere, durchlaufen in ihrer frühesten Entwickelung, als einzelliges Ei, als einfacher Zellenhaufen u. s. w. niedere Formzustände, welche gewissen Protisten gleichen. Nach unserem biogenetischen Grundgesetze müssen wir daraus den Schluss ziehen, dass sämmtliche Organismen, Thiere, Protisten und Pflanzen, von höchst einfachen einzelligen Organismen abstammen; und wenn wir diese ältesten Stammformen heute [S. 64] lebend vor uns hätten, würden wir sie jedenfalls für neutrale Protisten erklären.
Eine gute negative Charakteristik der Protisten, gegenüber den echten Thieren und den echten Pflanzen, lässt sich darauf gründen, dass sie weder eine Gastrula mit zwei Keimblättern bilden, wie die ersteren, noch einen Thallus oder ein Prothallium, wie die letzteren. Damit in Zusammenhang steht der Umstand, dass die Protisten niemals wirkliche (aus vielen Zellen zusammengesetzte) Gewebe und Organe bilden, wie alle echten Thiere und Pflanzen. Auch ist es sicher von grosser Bedeutung, dass die grosse Mehrzahl aller Protisten sich ausschliesslich auf ungeschlechtlichem Wege fortpflanzt (durch Theilung, Knospenbildung, Sporenbildung). Aber selbst bei den wenigen Protisten, welche sich bereits zur geschlechtlichen Zeugung in einfachster Form erheben, geht der Gegensatz zwischen männlichen und weiblichen Theilen niemals so weit, wie es bei allen echten Thieren und Pflanzen der Fall ist. Sie repräsentiren in jeder Beziehung jene niedere älteste Bildungsstufe, welche jedenfalls der Entwickelung echter Thiere und echter Pflanzen vorausgegangen sein muss.
Diese Betrachtungen führen uns auf denjenigen Weg, auf welchem allein eigentlich das Verhältniss der drei organischen Reiche zu einander entscheidend aufgeklärt werden kann, auf den Weg der Stammesgeschichte oder Phylogenie. Wenn wir ganz genau wüssten, wie sich das organische Leben auf unserem Erdball von Anfang an entwickelt hat, wie die Thiere, Protisten und Pflanzen ursprünglich entstanden sind, dann würden wir auch das Verhältniss der drei Reiche zu einander klar und unzweideutig beurtheilen können. Aber der sichere Weg der unmittelbaren Erfahrung bleibt uns für die Erkenntniss dieses wichtigen Verhältnisses auf ewig verschlossen. Kein lebendes Wesen und keine Schöpfungsurkunde kann uns erzählen, wie jener älteste Entwickelungsgang des organischen Lebens vor vielen Millionen von Jahren begonnen und wie er sich weiterhin zunächst gestaltet hat. Tausende von Arten und Gattungen, Millionen von Generationen sind in’s Grab gesunken, ohne uns sichtbare Spuren ihrer Existenz hinterlassen zu haben. Und gerade die wichtigsten von Allen, die ältesten und einfachsten Formen, konnten wegen des [S. 65] Mangels harter Körpertheile keine Versteinerungen zurücklassen.
Aber wenn uns auch der streng empirische Weg der Erkenntniss in dieser hochwichtigen Ursprungsfrage unwiderruflich verschlossen ist, so bleibt uns doch hier, wie überall, zur Ausfüllung unserer Erkenntnisslücken der Weg der wissenschaftlichen Hypothese offen. Wenn diese historische Hypothese sich in umfassender Weise auf die bisher erkannten wissenschaftlichen Thatsachen stützt, so ist sie in der Naturgeschichte der Lebewesen ebenso berechtigt, wie in der Geologie, in der Archaeologie, der Culturgeschichte und anderen historischen Wissenschaften. Und wie uns die allgemein anerkannten geologischen Hypothesen dazu geführt haben, eine befriedigende Einsicht in den Entwickelungsgang unsers Erdballs zu gewinnen, so werden auch die phylogenetischen Hypothesen, die wir auf die von Darwin reformirte Descendenz-Theorie gründen, Licht über den Entwickelungsgang des organischen Lebens auf der Erde verbreiten.
Wir können hier nicht auf eine Beleuchtung und Begründung aller der verschiedenen phylogenetischen Hypothesen eingehen, welche über diesen Entwickelungsgang aufgestellt worden sind. Nur auf diejenige Vorstellung wollen wir schliesslich noch einen flüchtigen Blick werfen, welche heuzutage am meisten innere Wahrscheinlichkeit für sich hat. Danach müssen wir annehmen, dass das Leben auf unserem Planeten mit der selbständigen Entstehung der allereinfachsten Protisten aus anorganischen Verbindungen begonnen hat. Diese ältesten Lebewesen der Erde werden den heute noch existirenden Moneren ähnlich gewesen sein: einfachste lebende Protoplasma-Stückchen ohne jegliche Organbildung. Daraus werden sich zunächst durch Sonderung eines Darmes im Inneren einzellige Protisten gebildet haben, und zwar höchst einfache, formlose und indifferente Zellen, gleich den Amoeben. Indem einige von diesen einzelligen Protisten, von geselligen Neigungen getrieben, sich daran gewöhnten, in kleinen Gesellschaften vereinigt zu leben, werden die ersten vielzelligen Organismen entstanden sein, und zwar zunächst auch nur wieder einfache Zellenhorden, lockere Gesellschaften von gleichartigen Zellen.
Nun ist es wohl wahrscheinlich, dass diese ältesten und einfachsten [S. 66] Entwickelungsvorgänge des organischen Lebens sich an zahlreichen verschiedenen Stellen des jugendlichen Erdballs gleichzeitig und unabhängig von einander wiederholt haben. So können also verschiedene und vielleicht zahlreiche Formen von Protisten unabhängig von einander entstanden sein; zuerst einzellige, später vielzellige. Durch den allgemeinen Kampf um’s Dasein, der auch unter diesen Protisten frühzeitig sich geltend machte, werden dieselben allmählich zu höherer Sonderung und Vervollkommnung angetrieben worden sein. Als wichtigster Vorgang ist da sicher die gegensätzliche Sonderung von thierischen und pflanzlichen Lebens-Processen hervorzuheben. Die einen Protisten begannen mehr an thierische, die andere an pflanzliche Lebensweise sich anzupassen, und mit der Lebensweise in Wechselwirkung entstand die charakteristische Körperform. Eine dritte, conservative Gruppe von Protisten behielt den ursprünglichen neutralen Character bei. Indem jene Anpassungen sich im Laufe der Zeit durch Vererbung befestigten, bildeten sich neben einander die drei grossen organischen Reiche aus.
Mit Beziehung auf den Stoffwechsel und die Ernährung würden wir freilich sagen können, dass diese ältesten Bewohner unseres Planeten Pflanzen waren, — richtiger: Protisten mit pflanzlichem Stoffwechsel; Protisten, welche gleich echten Pflanzen aus Wasser, Kohlensäure und Ammoniak die wichtigste »Lebens-Basis«, das Plasson, zusammensetzten, und dieses Plasson sonderte sich später in Protoplasma und Nucleus.
Die ältesten Thiere hingegen — oder richtiger: die ältesten Protisten mit thierischem Stoffwechsel, waren Parasiten, schmarotzende Protisten, welche es bequemer fanden, sich das von anderen Protisten gebildete Protoplasma anzueignen, als selbst solches zu bilden. Da eben ursprünglich viele Protisten-Stämme sich unabhängig von einander entwickelt haben können, von verschiedenen autogonen Moneren abstammend, so können auch diese Anpassungen sich mehrmals (polyphyletisch) wiederholt haben.
Aber auch wenn wir diese vielstämmige (polyphyletische) Hypothese verwerfen und wenn wir mehr zu der einstämmigen (monophyletischen) Annahme hinneigen, dass der Ursprung aller lebenden Wesen auf eine einzige gemeinsame Stammform zurückgeführt werden muss, auch dann werden wir doch im Ganzen [S. 67] wieder zu ähnlichen Vorstellungen über das Verhältniss der drei Reiche gelangen. Auch in diesem Falle werden wir annehmen müssen, dass jene älteste ursprüngliche Stammform eine einfachste Cytode, ein Moner war, und dass sich aus den Nachkommen jenes Moners zunächst einfache Zellen entwickelten. Diese Zellen werden sich wieder in thierische und pflanzliche gesondert haben, und so wird sich nach einer Richtung hin das Thierreich, nach einer anderen das Pflanzenreich ausgebildet haben, zwei gewaltigen, weit verzweigten Stämmen vergleichbar. Aber aus der gemeinsamen Wurzel, in der diese beiden grossen Stämme zusammenhängen, haben sich ausserdem noch zahlreiche niedere und indifferente Wurzelschösslinge selbständig entwickelt; und diese bilden zusammen unser Reich der Protisten.
Gleichviel ob wir dieser einstämmigen oder jener vielstämmigen Hypothese den Vorzug geben, so bleibt jedenfalls so viel sicher, dass Thierreich und Pflanzenreich nur in ihren vollkommneren Formen sich schroff gegenüber stehen, in ihren niederen Formen dagegen durch das Protistenreich untrennbar zusammenhängen. Die wissenschaftliche Begründung dieser wichtigen Anschauung ist uns erst durch die grossartigen Fortschritte der letzten vierzig Jahre möglich geworden. Aber mit dem Genius des Propheten hat schon vor siebzig Jahren einer unserer tiefblickendsten Naturphilosophen, Deutschlands genialster Dichter, dieselbe Anschauung ahnungsvoll ausgesprochen. In Jena schrieb Göthe 1806 den merkwürdigen Satz nieder: »Wenn man Pflanzen und Thiere in ihrem unvollkommensten Zustande betrachtet, so sind sie kaum zu unterscheiden. So viel aber können wir sagen, dass die aus einer kaum zu sondernden Verwandtschaft als Pflanzen und Thiere nach und nach hervortretenden Geschöpfe nach zwei entgegengesetzten Seiten sich vervollkommnen, so dass die Pflanze sich zuletzt im Baume dauernd und starr, das Thier im Menschen zur höchsten Beweglichkeit und Freiheit sich verherrlicht.«
[S. 68]
»Der vielbesprochene Bathybius existirt nicht; seine Annahme beruhte auf Täuschungen. So werden auch die übrigen Moneren nicht existiren; auch diese angeblichen Urorganismen werden das Erzeugniss irrthümlicher Beobachtungen sein. Mithin ist einer der wichtigsten Grundpfeiler der modernen Entwickelungslehre gefallen; und so werden auch ihre übrigen Stützpfeiler auf Täuschungen und Irrthum gegründet sein. Der ganze Darwinismus ist ein grosses Luftschloss, die Selectionstheorie eine Seifenblase, und die Abstammungslehre ist überhaupt nicht wahr.«
So ungefähr ist der Gedankengang zahlreicher Artikel, denen wir seit einem Jahre in den verschiedensten Zeitschriften begegnen. Einzig und allein auf die angebliche Nichtexistenz des Bathybius gestützt, behauptet man kurzweg, dass es überhaupt keine Moneren gebe, und dass damit die ganze Entwickelungslehre den schwersten Stoss erhalten habe. Am liebsten wird diese Behauptung natürlich von den Gegnern der Entwickelungslehre vorgetragen und in den mannigfaltigsten Tonarten variirt. Der Clerus triumphirt bereits über den völligen Untergang der Descendenztheorie. Aber selbst bei vielen Anhängern der Entwickelungstheorie gilt die Nichtexistenz des Bathybius als ausgemacht und es wird daraus eine Reihe von Schlussfolgerungen gezogen, die als mehr oder minder gewichtige Einwürfe gegen hervorragende Hauptpunkte des Darwinismus Bedenken erregen. Diese Umstände, sowie die Unklarheit, in welcher sich der grösste Theil des dafür interessirten Publicums über den eigentlichen Thatbestand befindet, bestimmt uns, hier die Moneren-Frage mit besonderer Rücksicht auf den Bathybius zu erörtern. Ich selbst erscheine zu dieser Erörterung insofern besonders berechtigt, ja sogar verpflichtet, als ich das zweifelhafte Glück geniesse, bei dem »berüchtigten Urschleim der Meerestiefen« Gevatter gestanden zu haben. Als mein Freund Thomas Huxley 1868 ihm bei der Taufe den Namen Bathybius Haeckelii beilegte, konnte er freilich nicht ahnen, dass der arme Täufling, einem Icarus gleich, in kürzester Zeit zu einer biologischen Celebrität werden, die Sonnenhöhe irdischer Berühmtheit erlangen und noch vor Ablauf seines ersten Decenniums in den dunkeln Hades der Mythologie hinabstürzen [S. 69] werde! Sehen wir denn zu, ob er wirklich todt ist, ob er überhaupt nicht existirt hat. Und wenn wir wirklich seine bloss mythologische Schein-Existenz zugeben müssten, sehen wir weiter zu, was daraus für die Moneren folgt!
Im Frühling des Jahres 1864 beobachtete ich im Mittelmeere bei Villafranca unweit Nizza schwimmende, winzige Schleimkügelchen von ungefähr einem Millimeter oder einer halben Linie Durchmesser, die mein höchstes Interesse erregten. Vorsichtig unter das Mikroskop gebracht, erschien nämlich jedes dieser Kügelchen wie ein kleiner Stern, dessen Mitte aus einem viel kleineren structurlosen Kügelchen bestand, während von der Oberfläche ringsum mehrere Tausend äusserst feine Fäden ausstrahlten. Die genaue Untersuchung bei starker Vergrösserung lehrte, dass der ganze Körper des sternförmigen Wesens aus einfacher eiweissartiger Zellsubstanz, aus Sarcode oder Protoplasma bestehe, und dass die Fäden, welche allenthalben von der Oberfläche ausstrahlten, keine beständigen Organe seien, sondern ihre Zahl, Grösse und Gestalt beständig änderten. Sie erwiesen sich als ebenso wechselnde und unbeständige Fortsätze des centralen Protoplasma-Körpers, wie die längst bekannten »Scheinfüsschen oder Pseudopodien«, welche die einzigen Organe der Wurzelfüssler oder Rhizopoden darstellen. Während aber bei diesen Letzteren Zellkerne im Protoplasma zerstreut sind und ihr Körper somit den Formwerth von einer oder mehreren Zellen besitzt, ist das bei jenen in Nizza beobachteten Protoplasma-Kügelchen nicht der Fall. Im Uebrigen war kein Unterschied hier und dort zu finden bezüglich der Bewegungsform der fliessenden Schleimfäden und der Art und Weise, in welcher dieselben als Tastorgane zum Empfinden, als Contractionsorgane zum Kriechen, und als Ernährungsorgane zur Nahrungsaufnahme benutzt wurden. Um die Naturgeschichte des kleinen Protaplasmakügelchens von Nizza, das ich auf das Genaueste untersuchte, zu vervollständigen, fehlte es nur noch an der Beobachtung seiner Fortpflanzung. Auch diese glückte schliesslich. Nach einiger Zeit zerfiel das kleine Wesen durch einfache Theilung in zwei Hälften, von denen jede ihr eignes Leben in derselben Weise [S. 70] weiterführte, wie das erstere. Ich hatte somit den vollständigen Lebenscyclus eines denkbar einfachsten Organismus erkannt, und nannte denselben in Anerkennung seiner fundamentalen Bedeutung Protogenes primordialis, den »Erstgebornen der Urzeit«. Seine genaue Beschreibung gab ich im XV. Bande der Zeitschrift für wissenschaftliche Zoologie (S. 360, Taf. XXVI., Fig. 1, 2).
Schon im folgenden Jahre wurden zwei verschiedene, dem Protogenes sehr ähnliche, höchst einfache Organismen von dem ausgezeichneten Mikroskopiker Cienkowski beschrieben. Im ersten Bande des Archivs für mikroskopische Anatomie (S. 203, Taf. XII.-XIV.) veröffentlichte derselbe sehr interessante »Beiträge zur Kenntniss der Monaden.« Unter den verschiedenen Protisten, die Cienkowski hier unter dem alten, vieldeutigen und daher sehr unsicheren Begriffe der »Monaden« zusammenfasst, befinden sich zwei mikroskopische Bewohner des süssen Wassers, welche in der vollkommen einfachen und structurlosen Beschaffenheit ihres kernlosen, strahlenden Protoplasma-Körpers dem Protogenes gleichen, die Gattungen Protomonas (Monas amyli) und Vampyrella (letztere mit drei verschiedenen Arten). Sie unterscheiden sich aber von dem ersteren durch die Art und Weise ihrer Fortpflanzung. Während der Protogenes, nachdem er durch Wachsthum ein gewisses Grössenmaass erreicht hat, dieses nicht weiter überschreitet, sondern ohne Weiteres in zwei Stücke zerfällt, ziehen Protomonas und Vampyrella ihre Strahlen ein und gehen in einen Ruhestand über, in welchem sich die kleine Protoplasmakugel einkapselt oder encystirt, mit einer Hülle (»Cyste«) umgiebt. Innerhalb dieser kleinen Hülle zerfällt die Protomonas in sehr zahlreiche Kügelchen, die Vampyrella in vier Stücke (Tetrasporen). Alle diese Theilstücke werden später frei und entwickeln sich durch einfaches Wachsthum zu der reifen Form.
Inzwischen hatte ich selbst eine vierte ähnliche Gattung von höchst einfachen Organismen im süssen Wasser bei Jena beobachtet, welche einer gewöhnlichen Amoebe ganz gleich sich verhält, aber von dieser letzteren durch den Mangel eines Zellkerns und einer contractilen Blase sich unterscheidet. Ich nannte sie daher Protamoeba primitiva. Während bei den drei ersterwähnten Schleimkügelchen (Protogenes, Protomonas, Vampyrella) überall zahlreiche Fäden aus der Oberfläche des centralen Protoplasma-Körpers [S. 71] ausstrahlen, sehen wir statt deren bei Protamoeba — ganz wie bei der gewöhnlichen Amoeba — wenige kurze, fingerförmige Fortsätze sich ausstrecken, welche ihre Gestalt beständig ändern; sie werden eingezogen und an einer andern Stelle wieder vorgestreckt. Hat die Protamoeba durch Nahrungsaufnahme (die ebenfalls wie bei Amoeba erfolgt) eine gewisse Grösse erreicht, so zerfällt sie durch Theilung in zwei Hälften. Ich machte die erste Mittheilung darüber in meiner »generellen Morphologie« (Bd. I. S. 133). Später habe ich von Protamoeba primitiva Abbildungen gegeben, welche u. A. in die »Natürliche Schöpfungsgeschichte« (VI. Aufl. S. 167) und in die »Anthropogenie« (III. Aufl. S. 414) aufgenommen sind.
Gestützt auf diese Beobachtungen, die späterhin durch die Untersuchungen anderer Forscher, wie durch meine eigenen noch beträchtlich erweitert wurden, gründete ich 1866 in der »Generellen Morphologie« für alle diese Organismen von einfachster Beschaffenheit eine besondere Classe unter dem Namen der Moneren, d. h. der »Einfachen«. Im ersten Bande (S. 135) sagte ich damals:
»Um diese einfachsten und unvollkommensten aller Organismen, bei denen wir weder mit dem Mikroskop, noch mit den chemischen Reagentien irgend eine Differenzirung des homogenen Plasmakörpers nachzuweisen vermögen, von allen übrigen, aus ungleichartigen Theilen zusammengesetzten Organismen bestimmt zu unterscheiden, wollen wir sie ein für allemal mit dem Namen der »Einfachen« oder »Moneren« belegen. Gewiss dürfen wir auf diese höchst interessanten, bisher aber fast ganz vernachlässigten Organismen besonders die Aufmerksamkeit hinlenken und auf ihre äusserst einfache Formbeschaffenheit bei völliger Ausübung aller wesentlichen Lebensfunctionen das grösste Gewicht legen, wenn es gilt, das Leben zu erklären, es aus der fälschlich sogenannten »todten Materie« abzuleiten, und die übertriebene Kluft zwischen Organismen und Anorganen auszugleichen. Indem bei diesen homogenen belebten Naturkörpern von differenten Formbestandtheilen, von »Organen«, noch keine Spur zu entdecken ist, vielmehr alle Moleküle der structurlosen Kohlenstoffverbindung, des lebendigen Eiweisses, in gleichem Maasse fähig erscheinen, sämmtliche Lebensfunctionen zu vollziehen, liefern sie klar den Beweis, dass der Begriff des Organismus nur dynamisch oder [S. 72] physiologisch aus den Lebensbewegungen, nicht aber statisch oder morphologisch aus der Zusammensetzung des Körpers aus »Organen« abgeleitet werden kann.«
In den folgenden Jahren wurde der Kreis unserer Erfahrungen über diese wunderbaren »Organismen ohne Organe« wesentlich erweitert. Auf meiner Reise nach den canarischen Inseln (1866 und 1867) richtete ich natürlich meine ganze Aufmerksamkeit auf dieselben und war denn auch so glücklich, noch mehrere neue Moneren-Formen zu entdecken. Auf den weissen Kalkschalen, eines merkwürdigen Cephalopoden (Spirula Peronii), die zu Tausenden an den Küsten der canarischen Inseln angetrieben zu finden sind, bemerkte ich zuweilen zahlreiche rothe Pünktchen, welche sich unter der Lupe als zierliche Sternchen und bei starker Vergrösserung als orangerothe Protoplasma-Scheiben oder -Kugeln zu erkennen gaben, von deren Umfange zahlreiche baumförmig verästelte Fäden ausstrahlten. Die genauere Untersuchung zeigte, dass auch diese (verhältnissmässig colossalen) Protoplasma-Körper kernlos und structurlos waren und sich in ähnlicher Weise wie Protomonas fortpflanzten, nämlich dadurch, dass der kugelig zusammengezogene und eingekapselte Körper in zahlreiche kleine Stücke zerfiel. Ich nannte diese interessante neue Moneren-Gattung Protomyxa aurantiaca und habe sie zuerst auf Taf. I. der »Natürl. Schöpfungsgeschichte« abgebildet. Eine ähnliche stattliche Monerenform entdeckte ich sodann in demselben Jahre (1867) im Schlamme des Hafenbeckens von Puerto del Arrecife, der Hafenstadt der canarischen Insel Lanzarote, und bezeichnete sie als Myxastrum radians. Sie ist dadurch ausgezeichnet, dass die Theilstücke oder Sporen, in welche der kugelige Körper bei der Fortpflanzung zerfällt, sich radial gegen den Mittelpunkt der Kugel ordnen und spindelförmige Kieselhüllen ausschwitzen, aus denen später das junge Moner ausschlüpft.
Gestützt auf alle diese Beobachtungen, veröffentlichte ich 1868 in der »Jenaischen Zeitschrift für Naturwissenschaft« eine ausführliche Monographie der Moneren. (Bd. IV, S. 64, Taf. II. und III). Hier sind alle eigenen und fremden Beobachtungen ausführlich zusammengestellt und erläutert. Es ergaben sich damals sieben verschiedene Gattungen von Moneren. Durch spätere Beobachtungen ist die Zahl der Arten auf 16 gesteigert worden, worüber [S. 73] ich in den »Nachträgen zur Monographie der Moneren« berichtet habe (Jenaische Zeitschr. für Naturw. 1877. Bd. VI. S. 23). Die Unterschiede aller dieser Moneren beruhen nur darauf, dass die weiche, schleimige Körpermasse in verschiedener Form sich ausbreitet und bewegt, und dass die ungeschlechtliche Fortpflanzung (durch Theilung, Sporenbildung u. s. w.) auf verschiedene Weise geschieht.
Das hohe Interesse, das die Moneren in morphologischer sowohl, als physiologischer Beziehung darbieten, wurde noch gesteigert, als 1868 der erste Zoologe Englands, der berühmte Thomas Huxley, eine neue, ganz eigenartige Moneren-Gattung unter dem Namen Bathybius Haeckelii beschrieb (Journal of microscop. science, Vol. VIII, N. S. p. 1, Pl. IV). Abweichend von den übrigen Moneren sollte dieser Bathybius eigenthümlich geformte mikroskopische Kalkkörperchen einschliessen: Coccosphaeren und Coccolithen (Discolithen und Cyatholiten); die formlosen Protoplasma-Klumpen desselben aber, von sehr verschiedener Grösse, sollten in ungeheuren Massen die tiefsten Abgründe des Meeres bedecken, unterhalb 5000 Fuss bis zu 25000 Fuss hinab. Mit diesem formlosen Ur-Organismus einfachster Art, der zu Milliarden vereinigt den Meeresboden mit einer lebendigen Schleimdecke überzieht, schien ein neues Licht auf eine der schwierigsten und dunkelsten Fragen der Schöpfungsgeschichte zu fallen, auf die Frage von der Urzeugung, von der ersten Entstehung des Lebens auf unserer Erde. Mit dem Bathybius schien der berüchtigte »Urschleim« gefunden zu sein, von dem Oken vor einem halben Jahrhundert prophetisch behauptet hatte, dass alles Organische aus ihm hervorgegangen, und dass er im Verfolge der Planeten-Entwickelung aus anorganischer Materie im Meeresgrunde entstanden sei.
Der Tiefseeschlamm, welcher die Bathybius-Massen enthält, wurde zuerst bei Gelegenheit der grossartigen Tiefgrund-Untersuchungen entdeckt, die seit dem Jahre 1857 behufs Legung des transatlantischen Telegraphen-Kabels angestellt wurden. Man fand schon damals das »atlantische Telegraphen-Plateau«, jene mächtige Tiefsee-Ebene, welche sich in einer durchschnittlichen Tiefe von 12,000 [S. 74] Fuss von Irland bis Neufundland erstreckt, allenthalben mit einem eigenthümlichen, grauen, äusserst feinpulverigen Schlamme bedeckt: Derselbe zeichnete sich durch zähe, klebrige Beschaffenheit aus und zeigte bei mikroskopischer Untersuchung Massen von kleinen kalkschaligen Rhizopoden, insbesondere Globigerinen, und ferner, als Hauptbestandtheile, die sehr kleinen, als Coccolithen bezeichneten Kalkkörperchen. Aber erst elf Jahre später, als Huxley 1868 mittelst eines sehr scharfen Mikroskopes eine erneute genaue Untersuchung desselben Schlammes, auch in chemischer Beziehung, vornahm, entdeckte er darin die nackten, freien, formlosen Protoplasma-Klumpen, welche neben den genannten Theilen die Hauptmasse des Schlammes bilden. »Diese Klumpen sind von allen Grössen, von Stücken, die mit blossem Auge sichtbar sind bis zu äusserst kleinen Partikelchen. Wenn man sie der mikroskopischen Analyse unterwirft, zeigen sie — eingebettet in eine durchsichtige, farblose und strukturlose Matrix — Körnchen, Coccolithen und zufällig hineingerathene fremde Körper.«
Lebender Bathybius wurde zuerst 1868 von Sir Wyville Thomson und Professor William Carpenter, zwei ebenso erfahrenen als scharfsichtigen Zoologen, während ihrer nordatlantischen Tiefsee-Expedition auf dem Kriegsschiffe »Porcupine« beobachtet. Sie berichten über den frisch heraufgeholten lebendigen Tiefsee-Schlamm: »Dieser Schlamm war wirklich lebendig; er häufte sich in Klumpen zusammen, als ob Eiweiss beigemischt wäre; und unter dem Mikroskope erwies sich die klebrige Masse als lebende Sarcode.« (Annals and magaz. of nat. hist. 1869, Vol. IV, p. 151). Ferner sagt Sir Wyville Thomson in seinem höchst interessanten Werke über die Meerestiefen (The depths of the Sea II. Edit. 1874. p. 410): »In diesem Schlamm (Globigerinen-Schlamm aus 2,435 Faden oder ca. 14,600 Fuss Tiefe, aus der Bay von Biscaya), wie in den meisten anderen Schlamm-Proben aus dem atlantischen Ocean-Bett, war eine beträchtliche Quantität einer weichen, gallertigen, organischen Materie nachweisbar, genug, um dem Schlamme eine gewisse Klebrigkeit zu geben. Wenn der Schlamm mit schwachem Weingeist geschüttelt wurde, fielen feine Flocken nieder, wie von geronnenem Schleime; und wenn ein wenig von demjenigen Schlamme, an welchem die klebrige Beschaffenheit am deutlichsten hervortritt, [S. 75] in einem Tropfen Seewasser unter das Mikroskop gebracht wird, können wir gewöhnlich nach einiger Zeit ein unregelmässiges Netzwerk von eiweissartiger Materie sehen, unterscheidbar durch seine bestimmten Umrisse und nicht mit Wasser mischbar. Man kann sehen, wie dieses Netzwerk seine Form allmählig ändert und die eingeschlossenen Körnchen und fremden Körper ihre relative Lage darin verändern. Die Gallert-Substanz ist daher eines gewissen Grades von Bewegung fähig, und es kann kein Zweifel sein, dass sie die Erscheinungen einer sehr einfachen Lebensform zeigt.« So wörtlich Sir Wyville Thomson (a. a. O. S. 411).
Meine eigenen Untersuchungen des Bathybius-Schlammes betrafen, ebenso wie diejenigen von Huxley, nur todtes, in Weingeist conservirtes Material. Das Fläschchen, in welchem ich denselben von den Far-Oer-Inseln zugesandt erhielt, trug die Aufschrift »Dredged of Professor Thomson und Dr. Carpenter with the Steamer Porcupine on 2435 fathoms. 22. July 1869. Lat. 47° 38'. Long. 12° 4'.« Es war also dieser Bathybius-Schlamm derselbe, an welchem die genannten Forscher ihre Beobachtungen über amoeboide Bewegungen angestellt hatten. Die Resultate meiner Untersuchung habe ich ausführlich in meinen »Beiträgen zur Plastiden-Theorie« mitgetheilt (2. Bathybius und das freie Protoplasma der Meerestiefen. Jen. Zeitschr. für Naturw. 1870. Bd. V. S. 499. Taf. XVII.) Die 80 Figuren, welche ich daselbst (auf Taf. XVII) von den verschiedenen formlosen Protoplasma-Stücken des Bathybius und den geformten Kalkkörperchen, die er einschliesst, gegeben habe, sind bei sehr starker Vergrösserung mit Hülfe der Camera lucida ganz genau gezeichnet. Einige dieser Figuren sind auch in den Aufsatz über »das Leben in den grössten Meerestiefen« übergegangen, welchen ich 1870 in Virchow-Holzendorff’s Sammlung publicirt habe. (Nr. 110).
Indem ich diesen, in starkem Alkohol sehr gut conservirten Bathybius-Schlamm mit Hülfe der neuesten Methoden möglichst genau untersuchte, und namentlich die vortheilhafte (von Huxley früher nicht angewandte) Methode der Färbung mit Carmin und Jod übte, suchte ich vor Allem die Quantität und Qualität der formlosen Protoplasma-Stücke näher zu bestimmen, die überall in Masse zwischen den geformten Kalktheilchen sich vorfanden. [S. 76] Diese eiweissartigen, durch Carmin roth gefärbten Stücke waren sehr gleichmässig durch den ganzen Schlamm verbreitet und schienen in den meisten untersuchten Proben mindestens ein Zehntel bis ein Fünftel des gesammten Volums zu betragen, in manchen Präparaten selbst die grössere Hälfte. Dieselben Massen, welche durch Carmin sich mehr oder minder intensiv roth färbten, nahmen durch Jod — und ebenso durch Salpetersäure — eine gelbe Färbung an und zeigten auch im Verhalten gegen andere chemische Reagentien ganz dieselben Eigenschaften, wie das gewöhnliche Protoplasma der Thier- und Pflanzenzellen. Die Form der meisten Stückchen war unregelmässig, rundlich oder mit stumpfen Fortsätzen, einer Amoebe ähnlich; andere Stückchen bildeten unregelmässige, kleine und grössere Sarcode-Netze, ähnlich denen der Myxomyceten.
Ob die kleinen geformten Kalktheilchen, die Coccolithen und Coccosphären, welche in so grossen Massen im Bathybius-Schlamme vorkommen, — und zwar ebenso wohl zwischen den Protoplasma-Stückchen, als innerhalb derselben, von ihnen umschlossen, — wirklich zu ihnen gehören, oder nicht, diese Frage musste ich um so mehr offen lassen, als ich schon vorher ganz ähnliche Kalkkörperchen in dem Körper mehrerer pelagischen, an der Oberfläche des canarischen Meeres schwimmenden Radiolarien gefunden hatte (»Myxobracchia von Lanzerote«). Diese sonderbaren Kalkkörperchen, welche bald die Gestalt einer einfachen, concentrisch geschichteten Scheibe, bald eines Hemdknöpfchens, bald einer aus vielen Scheibchen zusammengesetzten Kugel u. s. w. hatten, konnten ebensowohl Ausscheidungen der Bathybius-Sarcode sein, als fremde Körper, die zufällig (oder bei der Nahrungsaufnahme) in das Protoplasma hinein gelangt waren. In neuester Zeit hat sich die grössere Wahrscheinlichkeit zu Gunsten der letzteren Annahme herausgestellt und die meisten Biologen nehmen jetzt an, dass alle diese Körperchen mikroskopische Kalk-Algen seien, verkalkte einzellige Pflanzen.
Durch diese Untersuchungen, die von mehreren anderen Forschern bestätigt wurden, schien festgestellt, dass auf dem Boden des nordatlantischen Oceans, und zwar in Tiefen zwischen 5000 und 25000 Fuss, ein feinpulveriger Schlamm sich findet, welcher u. A. grosse Mengen einer eigenthümlichen, noch kaum individualisirten [S. 77] Moneren-Art enthält. Der Fehler, den wir nun begingen, bestand darin, dass wir die Resultate dieser nordatlantischen Tiefsee-Untersuchungen allzurasch generalisirten und überall den Boden des tiefen Oceans mit ähnlichen Moneren bedeckt zu sehen erwarteten. Diese Erwartung wurde vollständig getäuscht. Die sehr genaue und umfassende Untersuchung der grossartigen Challenger-Expedition, welche in 3½ Jahren die Erde umkreiste und in den Tiefen der verschiedenen Oceane sorgfältig nach dem Bathybius suchte, hat ihn nirgends wiedergefunden und erzielte nur negative Resultate. Wir haben keinen Grund, in die Sorgfalt und Genauigkeit der ausgezeichneten Naturforscher der bewunderungswürdigen Challenger-Expedition irgend einen Zweifel zu setzen, um so weniger, als ja der vorzügliche Director derselben, Sir Wyville Thomson selbst zuerst die Bewegungen am lebenden Bathybius wahrgenommen hatte. Wir müssen also wohl annehmen, dass an den vom Challenger untersuchten Stellen des tiefen Meeresbodens die Bathybius-Moneren wirklich fehlten. Folgt aber daraus, dass alle jene früheren Beobachtungen und Schlüsse unrichtig waren?
Wie es sehr häufig in solchen Fällen zu gehen pflegt, so ging auch jetzt plötzlich die einseitig übertriebene Ansicht in das entgegengesetzte Extrem über. Vorher hatte man gehofft, überall im Schlamme des tiefen Meeresbodens die Protoplasma-Klumpen des Bathybius in Masse zu finden; jetzt wollte man sie mit einem Male nirgends mehr anerkennen. Insbesondere glaubte man sich zu der Annahme berechtigt, der früher in Weingeist untersuchte Bathybius-Schlamm sei weiter nichts, als ein feiner Gypsniederschlag, wie er überall bei der Mischung von Weingeist mit Seewasser entsteht. Diese Ansicht wurde zuerst von einigen Naturforschern der Challenger-Expedition ausgesprochen und daraufhin widerrief Professor Huxley — wie mir scheint, zu frühzeitig — seine frühere Ansicht vom Bathybius. In der »Nature« (vom 19. Aug. 1875) und im »Quarterly Journal of microscop. science« (1875, Vol. XV. p. 392) sagt derselbe wörtlich: »Professor Wyville Thomson theilt mir mit, dass die besten Bemühungen der Challenger-Forscher, lebenden Bathybius zu entdecken, fehlschlugen, und dass ernstlich vermuthet wird, das Ding, dem ich diesen Namen gab, sei wenig mehr als schwefelsaurer [S. 78] Kalk, in flockigem Zustande aus dem Seewasser durch den starken Alkohol niedergeschlagen, in welchem der Tiefseeschlamm aufbewahrt wurde. Das Sonderbare ist aber, dass dieser unorganische Niederschlag kaum von einem Eiweissniederschlag zu unterscheiden ist, und er gleicht, vielleicht noch mehr, dem keimführenden Häutchen an der Oberfläche fauliger Aufgüsse, das sich unregelmässig, aber sehr stark, mit Carmin färbt, Stücke von bestimmtem Umriss bildet und in jeder Weise sich wie ein organisches Ding verhält. Professor Thomson spricht sehr vorsichtig und sieht das Schicksal des Bathybius noch nicht als ganz entschieden an. Aber da ich hauptsächlich für den eventuellen Irrthum verantwortlich bin, diese merkwürdige Substanz in die Reihe der lebenden Wesen eingeführt zu haben, so glaube ich richtiger zu verfahren, wenn ich seiner oben mitgetheilten Ansicht grösseres Gewicht beilege, als er selbst.«
Dies sind die Worte des Professor Huxley, welche so grosses Aufsehen erregten, und nach weit verbreiteter Ansicht dem armen Bathybius den Todesstoss versetzt haben. Je mehr aber hier die eigentlichen Eltern des Bathybius sich geneigt zeigen, ihr Kind als hoffnungslos aufzugeben, desto mehr fühle ich mich als Taufpathe verpflichtet, seine Rechte zu wahren und womöglich sein erlöschendes Lebensfünkchen wieder zur Geltung zu bringen. Und da finde ich denn glücklicherweise einen werthvollen Bundesgenossen in einem vielgereisten deutschen Naturforscher, der erst in neuerer Zeit wieder lebenden Bathybius, und zwar an der Küste von Grönland, beobachtet hat. Der bekannte Nordpolfahrer Dr. Emil Bessels aus Heidelberg, der von dem Schiffbruche der Polaris glücklich zurückkehrte, macht bei Gelegenheit seiner Beschreibung der Haeckelina gigantea (eines colossalen Rhizopoden, der vielleicht mit der früher von Sandahl beschriebenen Astrorhiza identisch ist) folgende wichtige Angaben: »Während der letzten amerikanischen Nordpol-Expedition fand ich in 92 Faden Tiefe in dem Smith-Sunde grosse Massen von freiem undifferenzirtem homogenem Protoplasma, welches auch keine Spur der wohlbekannten Coccolithen enthielt. Wegen seiner wahrhaft spartanischen Einfachheit nannte ich diesen Organismus, den ich lebend beobachten konnte, Protobathybius. Derselbe wird in dem Reisewerk der Expedition abgebildet und beschrieben [S. 79] werden. Ich will hier nur erwähnen, dass diese Massen aus reinem Protoplasma bestanden, dem nur zufällig Kalktheilchen beigemischt waren, aus welchen der Seeboden gebildet ist. Sie stellten äusserst klebrige, maschenartige Gebilde dar, die prächtige amoeboide Bewegungen ausführten, Carminpartikelchen sowie andere Fremdkörper aufnahmen und lebhafte Körnchenströmung zeigten. (Jenaische Zeitschr. f. Naturw. 1875. Bd. IX., S. 277. Vgl. auch: Annual Report of the Secret. of the navy for 1873). An einem anderen Orte, in den von Packard publicirten »Life histories of animals« (New-York, 1876 p. 3) ist eine Abbildung der Protoplasma-Netze des Protobathybius von Dr. Bessels publicirt. Hiernach möchte ich annehmen, dass derselbe von unserm echten Bathybius nicht verschieden ist. Der Unterschied, dass letzterer gewöhnlich viele geformte Kalkkörperchen (Coccolithen etc.) umschliesst, der erstere dagegen nicht, verliert seine Bedeutung durch die immer wachsende Wahrscheinlichkeit, dass diese Kalkkörperchen einzellige, als Nahrung aufgenommene Kalkalgen sind.«
Nachdem wir jetzt die historischen Angaben über den Bathybius zusammengetragen und die wichtigsten wörtlich angeführt haben, wenden wir uns zur Kritik desselben. Versuchen wir, aus einer unpartheiischen Würdigung jener Angaben uns ein selbständiges unbefangenes Urtheil über den vielverschrieenen und jetzt fast aufgegebenen Urschleim der grössten Meerestiefen zu bilden!
Bezüglich des todten Bathybius, des in Weingeist conservirten Tiefseeschlammes aus dem nord-atlantischen Ocean, sind alle Beobachter, die denselben genau untersucht haben, einig, dass derselbe mehr oder minder ansehnliche Mengen von geronnenem Protoplasma enthält, welches im morphologischen und chemisch-physikalischen Verhalten die grösste Aehnlichkeit mit gewissen Moneren besitzt. Die Resultate, welche Huxley an seinem »Porcupine«-Material erhielt, und die ich selbst bestätigen und ergänzen konnte, sind von allen anderen Beobachtern, die denselben Schlamm untersuchten, als richtig anerkannt worden.
[S. 80]
Bezüglich des lebenden Bathybius liegen positive Angaben über die characteristischen rhizopodenartigen Bewegungen desselben von drei bewährten Beobachtern vor, von Sir Wyville Thomson, Professor William Carpenter und Dr. Emil Bessels. Alle drei stellten diese Beobachtungen an Tiefseeschlamm aus dem nord-atlantischen Ocean an. Dagegen lieferten die Bemühungen der Challenger-Forscher, in verschiedenen Meeren jene älteren Beobachtungen über Bewegungs-Erscheinungen zu wiederholen und zu bestätigen, nur negative Resultate.
Was folgt nun aus allen diesen Angaben, denen wir sämmtlich dieselbe Glaubwürdigkeit zuerkennen müssen, und die sich doch theilweise zu widersprechen scheinen? Angenommen, dass alle diese Angaben richtig sind, so folgt daraus einfach weiter gar nichts, als dass der Bathybius-Schlamm eine beschränkte geographische Verbreitung besitzt, und dass es eine voreilige Verallgemeinerung war, alle tiefen Meeres-Abgründe mit demselben zu bevölkern. Daraus aber, dass die Challenger-Expedition den lebenden Bathybius nicht wiederfinden konnte, ist doch wahrlich nicht zu folgern, dass die an anderen Orten angestellten Beobachtungen der Porcupine-Expedition über lebenden Bathybius unrichtig waren! Oder sollen wir daraus, dass die Challenger-Expedition den merkwürdigen »Radiolarien-Schlamm« nur auf einen verhältnissmässig engen Verbreitungsbezirk des pacifischen Oceans beschränkt fand, und sonst nirgends wiederfinden konnte, den Schluss ziehen, dass derselbe überhaupt nicht existire? Wir wissen, dass die allermeisten Organismen-Arten einen beschränkten Verbreitungs-Bezirk haben. Warum soll denn nicht auch die Verbreitung des Bathybius beschränkt sein?
Ich bekenne daher, nicht zu begreifen, wie Huxley seine Ansicht über den Bathybius so rasch und so vollständig ändern konnte. Noch viel weniger freilich begreife ich die Art und Weise, wie auf der deutschen Naturforscher-Versammlung in Hamburg (im September 1876) der Bathybius öffentlich zu Grabe getragen werden konnte. Ich finde darüber in der Berliner Nationalzeitung folgende merkwürdige Mittheilung (datirt Hamburg 21. September), betreffend einen von Professor Möbius aus Kiel gehaltenen trefflichen Vortrag über die marine Fauna und die Challenger-Expedition: [S. 81] »Ueber diese Ebenen — Tiefsee-Ebenen von 3700 bis 4000 Meter Tiefe — sollte sich der geheimnissvolle Urschleim, der Bathybius ausbreiten, den der berühmte Huxley zu Ehren seines genialen Freundes in Jena Bathybius Haeckelii genannt hat. Leider aber passirte der Naturforschung ein böses Missgeschick. Der Bathybius, der so gut zu den modernen Anschauungen von dem Beginne des organischen Lebens passte, erwies sich als ein Kunstproduct, als Niederschlag von im Meere gelöstem Gyps, in Folge des den Proben zugesetzten Alkohols. Ueberall wo man die frischen Proben an Bord untersuchte, war keine Spur von ihm zu entdecken. Es machte einen geradezu erschütternden Eindruck auf die Zuhörer, als Herr Möbius den Bathybius nach einem so einfachen Recepte vor ihren Augen in einem mit Meerwasser gefüllten Glase durch Alkohol-Zusatz erscheinen liess!«
In der That eine merkwürdige Logik! Weil Weingeist in Seewasser einen Gyps-Niederschlag erzeugt, deshalb ist der in Weingeist conservirte Bathybius-Schlamm nur ein Gyps-Niederschlag! Und diese Beweisführung machte auf alle Mitglieder einer deutschen Naturforscher-Versammlung »einen geradezu erschütternden Eindruck!« Dass starker Weingeist in Seewasser einen dünnen flockigen Gyps-Niederschlag erzeugt, weiss Jeder, der Seethiere in Weingeist gesammelt hat. Ebenso weiss aber auch Jeder, der den Bathybius-Schlamm der Porcupine-Expedition gleich Huxley und mir genau untersucht hat, dass die darin massenhaft enthaltenen moneren-artigen Eiweisskörper wirklich aus einem eiweissartigen Körper und nicht aus Gyps bestehen. Sie färben sich in Carmin roth, in Salpetersäure und in Jod gelb, werden durch concentrirte Schwefelsäure zerstört und geben alle übrigen Reactionen des Protoplasma, was bekanntlich beim Gyps nicht der Fall ist.
Wenn man gewisse Kreide-Arten oder kreidigen Mergel fein pulverisirt, so erhält man ein feinkörniges, weisses Mehl, welches zum Verwechseln dem merkwürdigen »Radiolarien-Schlamm« ähnlich ist, den die Challenger-Expedition in einem beschränkten Bezirke des Pacifischen Oceans (und nur hier!) in einer Tiefe von 12,000–26,000 Fuss Tiefe gefunden hat. Dieser »Radiolarien-Ooze«, den ich eben jetzt untersuche, besteht fast ausschliesslich [S. 82] aus den zierlichsten und mannigfaltigst geformten Kieselschalen von zahllosen Radiolarien. Mit blossem Auge aber ist dieser getrocknete Schlamm — ein wundervolles, mikroskopisches Radiolarien-Museum — nicht zu unterscheiden von jenem pulverisirten Kreide-Mergel, der nicht eine einzige Radiolarien-Schale enthält. Ich schlage nun vor, auf einer nächsten deutschen Naturforscher-Versammlung den experimentellen Beweis zu führen, dass jene colossalen und höchst merkwürdigen, vom Challenger entdeckten Radiolarien-Lager in den Tiefen des Pacifischen Oceans nicht existiren. »Das Recept ist höchst einfach.« Man zerstösst in einem Mörser vor den Augen der versammelten Naturforscher einen von jenen Kreide-Mergeln, die keine Radiolarien enthalten. Das so erhaltene weisse Pulver enthält kein einziges Radiolar — also auch der pacifische (blos aus Radiolarien bestehende) Tiefsee-Schlamm nicht — denn beide sind mit blossem Auge nicht zu unterscheiden. Quod erat demonstrandum! Wir sind überzeugt, das schlagende Experiment wird auf alle Zuschauer »einen geradezu erschütternden Eindruck machen« — und der Radiolarien-Schlamm existirt nicht mehr!
Wir glauben in Vorstehendem gezeigt zu haben, dass die »Nicht-Existenz des Bathybius nicht erwiesen« ist. Vielmehr bleibt es sehr wahrscheinlich, dass die Beobachtungen von Wyville Thomson, Carpenter und Emil Bessels über die Bewegungen des lebenden Bathybius richtig sind. Wir wollen nun aber einmal das Gegentheil annehmen und wollen zugeben, dass Bathybius kein Moner und überhaupt kein Organismus sei. Folgt daraus, — wie jetzt sehr oft gefolgert wird, — dass auch die Moneren überhaupt nicht existiren? Oder dürfen wir daraus, dass die bekannte Riesen-Seeschlange der Fabel nicht existirt, den Schluss ziehen, dass es überhaupt keine Seeschlangen giebt? Bekanntlich giebt es deren eine Menge, die Familie der lebendig gebärenden, sehr giftigen Hydrophiden (Hydrophis, Platurus, Aepysurus etc.), welche meistens im indischen Ocean und Sunda-Archipel leben, aber keine beträchtliche Grösse erreichen.
Es würde unnütz sein, hier nochmals darauf hinzuweisen, [S. 83] dass meine eigenen, viele Jahre speciell auf diesen Gegenstand gerichteten und möglichst sorgfältigen Untersuchungen die Existenz von mehr als einem Dutzend verschiedener Moneren-Arten theils im Süsswasser, theils im Meere nachgewiesen haben. Um so mehr will ich aber hervorheben, dass diese Beobachtungen seitdem von einer Anzahl bewährter Forscher wiederholt und bestätigt worden sind. Einige von diesen Moneren scheinen sogar im süssen Wasser sehr verbreitet zu sein, so namentlich die Gattungen Protamoeba und Vampyrella. P. agilis und V. spirogyrae kommen in Jena fast jeden Sommer gelegentlich zur Beobachtung. P. primitiva und V. vorax sind von mehreren verschiedenen Beobachtern in sehr entlegenen Gegenden gesehen worden. Andere neue Moneren-Formen sind erst ganz neuerdings von Cienkowski und Oskar Grimm beobachtet. Wenn erst die allgemeine Aufmerksamkeit der Mikroskopiker sich mehr diesen höchst einfachen Organismen zuwendet, steht zu erwarten, dass unsere Kenntniss derselben sich noch beträchtlich erweitern und vertiefen wird.
Ganz abgesehen also davon, ob Bathybius ein echtes Moner ist oder nicht, kennen wir jetzt bereits mit Sicherheit eine Anzahl echter Moneren, deren fundamentale Bedeutung von ersteren ganz unabhängig ist. Wir wissen, dass noch heute eine Anzahl von niedrigsten Lebensformen in den Gewässern unseres Planeten existiren, welche nicht nur die einfachsten unter allen wirklich beobachteten Organismen, sondern überhaupt die denkbar einfachsten lebenden Wesen sind. Ihr ganzer Körper besteht in vollkommen entwickeltem und fortpflanzungsfähigem Zustande aus nichts weiter als aus einem strukturlosen Protoplasma-Klümpchen, dessen wechselnde, formveränderliche Fortsätze alle Lebensthätigkeiten gleichzeitig besorgen, Bewegung und Empfindung, Stoffwechsel und Ernährung, Wachsthum und Fortpflanzung. Morphologisch betrachtet ist der Körper eines solchen Moners so einfach wie derjenige irgend eines anorganischen Krystalles. Verschiedene Theilchen sind darin überhaupt nicht zu unterscheiden; vielmehr ist jedes Theilchen dem andern gleichwerthig. Diese wichtigen Thatsachen und die daraus sich ergebenden weitreichenden Folgerungen gelten für alle Moneren ohne Ausnahme — mit oder ohne Bathybius; — und es ist daher für die Theorie ganz gleichgültig, ob der Bathybius existirt oder nicht.
[S. 84]
Wenn wir diese Moneren als »absolut einfache Organismen« bezeichnen, so ist damit nur ihre morphologische Einfachheit, der Mangel jeder Zusammensetzung aus verschiedenen Organen, ausgesprochen. In chemisch-physikalischer Beziehung können dieselben noch sehr zusammengesetzt sein; ja wir werden ihnen sogar auf alle Fälle eine sehr verwickelte Molecular-Structur zuschreiben müssen, wie allen eiweissartigen Körpern überhaupt. Viele betrachten den schleimartigen Eiweisskörper der Moneren als eine einzige chemische Eiweissverbindung, Andere als ein Gemenge von mehreren solcher Verbindungen, noch Andere als eine Emulsion oder ein feinstes Gemenge von eiweissartigen und fettartigen Theilchen. Diese Frage ist für unsere Auffassung und für die allgemeine biologische Bedeutung der Moneren von untergeordneter Bedeutung. Denn auf alle Fälle — mag diese oder jene Ansicht richtig sein — bleiben die Moneren in anatomischer Hinsicht vollkommen einfach: Organismen ohne Organe. Sie beweisen unwiderleglich, dass das Leben nicht an eine bestimmte anatomische Zusammensetzung des lebendigen Körpers, nicht an ein Zusammenwirken verschiedener Organe, sondern an eine gewisse chemisch-physikalische Beschaffenheit der formlosen Materie gebunden ist, an die eiweissartige Substanz, welche wir Sarcode oder Protoplasma nennen, eine stickstoffhaltige Kohlenstoffverbindung in fest-flüssigem Aggregatzustande.
Das Leben ist also nicht Folge der Organisation, sondern umgekehrt. Das formlose Protoplasma bildet die organisirten Formen. Da ich die ausserordentlich hohe Bedeutung, welche die Moneren in dieser Beziehung, wie in vielen andern Beziehungen besitzen, bereits in den früher angeführten Schriften ausführlich erörtert habe, kann ich hier einfach darauf verweisen. Nur die fundamentale Bedeutung, welche die Moneren für die hochwichtige Frage von der Urzeugung behaupten, sei hier nochmals ausdrücklich hervorgehoben. Die ältesten Organismen, welche durch Urzeugung aus anorganischer Materie entstanden, konnten nur Moneren sein.
Gerade diese allgemeine Bedeutung der Moneren für die Lösung der grössten biologischen Räthsel ist es, welche sie zu einem besonderen Steine des Anstosses und Aergernisses für die [S. 85] Gegner der Entwickelungslehre macht. Natürlich benutzen die Letzteren jede Gelegenheit, ihre Existenz zu bestreiten, ähnlich wie es auch mit dem berühmten Eozoon canadense geschah, jener vielbestrittenen ältesten Versteinerung der laurentischen Formation. Die erfahrensten und urtheilsfähigsten Kenner der Rhizopoden-Classe, an ihrer Spitze Professor Carpenter in London und der verstorbene berühmte Anatom Max Schultze in Bonn, haben übereinstimmend die feste Ueberzeugung gewonnen, dass das echte nordamerikanische Eozoon (aus den laurentischen Schichten in Canada) ein wirklicher Rhizopode und zwar ein dem Polytrema nächstverwandtes Polythalamium ist. Ich selbst habe mich viele Jahre hindurch ganz speciell mit dem Studium der Rhizopoden beschäftigt. Ich habe die zahlreichen, schönen Eozoon-Präparate von Carpenter und von Max Schultze selbst genau untersucht und hege danach nicht den mindesten Zweifel mehr, dass dasselbe wirklich ein echtes Polythalamium und kein Mineral ist.
Aber gerade wegen der ausserordentlichen principiellen Bedeutung des Eozoon, weil dadurch die Zeitdauer der organischen Erdgeschichte um viele Millionen Jahre hinauf gerückt, die uralte silurische Formation als verhältnissmässig junge erkannt und so der Entwickelungslehre ein grosser Dienst geleistet wird, gerade deshalb fahren die Gegner der letzteren fort, unbeirrt zu behaupten, dass das Eozoon kein organischer Rest, sondern ein Mineral sei. Wie aber die hohe Bedeutung des Eozoon durch diese fruchtlosen Angriffe unkundiger Gegner erst recht in ihr volles Licht gesetzt worden ist, so gilt dasselbe auch von den Moneren — mit oder ohne Bathybius! Die echten Moneren bleiben ein fester Grundstein der Entwickelungslehre!

[S. 86]
System der Protisten.
Erste Classe des Protistenreiches.
1. Monera (HAECKEL) Urlinge.
Organismen ohne Organe. Der ganze Körper dieser einfachsten und niedrigsten Organismen besteht in vollkommen entwickeltem Zustande aus weiter nichts, als aus einem Stückchen Plasson oder »Urschleim«, einer eiweissartigen Verbindung, die noch nicht in Protoplasma und Nucleus differenzirt ist. Jedes Moner ist also eine Cytode, noch keine Zelle. Form meistens unbestimmt, mit wechselnden Fortsätzen. Bewegung bald durch Lappenfüsschen, bald durch Wurzelfüsschen, bald durch Flimmerfüsschen. Nahrungsaufnahme verschieden. Fortpflanzung ungeschlechtlich, durch Theilung, Knospung oder Sporenbildung. Leben im Wasser, meistens im Meere, auch parasitisch in anderen Organismen.
Erste Ordnung der Moneren:
Lobomonera (HAECKEL). Lappen-Urlinge. Fig. 47.
Moneren von unbestimmter wechselnder Form, deren Bewegungen durch Lappenfüsschen (Lobopodia) geschehen: stumpfe, fingerförmige, meist unverästelte Fortsätze, wie bei den Amoeben.
Gattung: Protamoeba (Arten: P. primitiva, P. agilis etc.) Fig. 47.
[S. 87]
Zweite Ordnung der Moneren:
Rhizomonera (HAECKEL). Wurzel-Urlinge. Fig. 48, 49.
Moneren von unbestimmter wechselnder Form, deren Bewegungen durch Wurzelfüsschen (Pseudopodia) geschehen: feine, lange, fadenförmige, meist verästelte und netzförmig sich verbindende Fortsätze, wie bei den Rhizopoden.
Gattungen: Protomyxa (aurantiaca) Fig. 48. Vampyrella (Spirogyrae). Bathybius (Haeckelii) Fig. 49.
Dritte Ordnung der Moneren:
Tachymonera (HAECKEL). Geissel-Urlinge. Fig. 50.
(Synonym: Schizomycetes. Spaltpilze. Bacterien.)
Moneren von bestimmter, meist, stabförmiger oder fadenförmiger Gestalt, deren zitternde oder schwingende lebhafte Bewegungen (wohl immer) durch äusserst feine Geisseln (Flagella) geschehen, wie bei den Geisselschwärmern (Flagellata). Fortpflanzung ungeschlechtlich, meistens durch Quertheilung. Erzeugen Zersetzung und Fäulniss in den organischen Flüssigkeiten, in denen sie leben. Wahrscheinlich die Ursachen vieler Krankheiten.
Gattungen: Bacterium (monas). Vibrio (lineola). Spirillum (tremulans). Fig. 50.
Zweite Classe des Protistenreiches.
2. Lobosa (CARPENTER). Lappinge.
(Synonym: Amoebina. Infusoria rhizopoda. Protoplasta.)
Einzellige Organismen (selten Syncytien), deren Zellenleib bald nackt (Gymnolobosa), bald in einer verschieden gestalteten Schale theilweise verborgen (Thecolobosa) ist. Die Zellen bewegen sich durch Lappenfüsschen (Lobopodia): stumpfe, fingerförmige, meist unverästelte Fortsätze, die an verschiedenen Stellen der Oberfläche entstehen und vergehen. Die Nahrung wird durch diese Lappenfüsschen umflossen und in das Innere der Zelle hineingedrückt. Der Protoplasma-Leib der Zelle sondert sich häufig [S. 88] in eine helle, structurlose, festere Rindenschicht (Exoplasma) und eine trübe, feinkörnige, weichere Markschicht (Endoplasma). Oft enthält derselbe eine oder mehrere contractile Blasen (Vacuolen); bald beständig, bald unbeständig. Der Zellkern oder Nucleus ist meist einfach, selten mehrfach vorhanden. Fortpflanzung ungeschlechtlich, meist durch Theilung, seltener durch Knospung oder Sporenbildung. Die Lobosen leben meistens im Wasser, seltener in der Erde oder parasitisch in anderen Organismen.
Erste Ordnung der Lobosen:
Gymnolobosa (HAECKEL). Nackte Lappinge. Fig. 1.
Lobosen mit nacktem, weichem Zellenleibe, ohne Schale.
Gattungen: Amoeba (princeps). Podostoma (filigerum). Petalopus (diffluens).
Zweite Ordnung der Lobosen:
Thecolobosa (HAECKEL). Beschalte Lappinge. Fig. 5, 6.
(Synonym: Lepamoebae. Arcellinae. Amoebae cataphractae).
Lobosen mit einer Schale oder Zellmembran, von welcher der weiche Zellenleib theilweise bedeckt wird.
Dritte Classe des Protistenreiches.
3. Gregarinae (DUFOUR). Gregaringe.
Einzellige Organismen oder Ketten von wenigen, an einander gereihten Zellen, deren Leib von einer weichen, dicken, dehnbaren Haut allseitig umschlossen ist. Diese Zellmembran ist glatt, ohne Oeffnung, an einem Ende des Körpers oft mit hakenförmigen Haftapparaten versehen. Das Protoplasma ist sehr dehnbar und contractil, mit vielen Körnchen durchsetzt. Der Zellkern gross, meist ein helles kugeliges Bläschen, mit einem Nucleolus. Die wurmähnlichen Bewegungen der kriechenden Zellen geschehen durch Zusammenziehungen der (unmittelbar unter [S. 89] der Membran liegenden) Rindenschicht des Protoplasma, welche bisweilen in muskelähnliche Fäserchen differenzirt ist. Alle Gregarinen leben parasitisch im Darm oder in der Leibeshöhle (seltener in den Geweben) von Thieren (besonders von Würmern und Gliederthieren). Sie ernähren sich vom Safte dieser Wohnthiere, der durch die Membran der schmarotzenden Zellen in das Innere ihres Protoplasma hindurchschwitzt (Endosmose). Fortpflanzung ungeschlechtlich, durch Theilung oder durch Sporenbildung. Im letzteren Falle zieht sich eine einzelne Gregarine, oder mehrere mit einander verschmelzende Gregarinen, kugelig zusammen und umgeben sich mit einer Kapsel. Die Zellkerne verschwinden und das Protoplasma zerfällt in zahlreiche Keimzellen oder Sporen (Psorospermien, Pseudonavicellen). Später schlüpft aus jeder Spore ein Moner aus, welches sich durch Neubildung eines Nucleus in eine Amoebe verwandelt. Indem letztere sich mit einen Membran umhüllt, wird sie zur Gregarine.
Erste Ordnung der Gregarinen:
Monocystida (STEIN). Einzellige Gregaringe.
Gregarinen-Leib eine einfache Zelle, mit einem einzigen Kern.
Gattung: Monocystis (agilis). Fig. 7.
Zweite Ordnung der Gregarinen:
Polycystida (HAECKEL). Vielzellige Gregaringe.
Gregarinen-Leib eine Kette von zwei oder drei (selten mehr) an einander gereihten Zellen, jede Zelle mit einem Kern.
Gattung: Didymophyes (paradoxa).
Vierte Classe des Protistenreiches.
4. Flagellata (EHRENBERG). Geisslinge.
(Synonym: Mastigaria. Geisselschwärmer. Geissel-Infusorien.)
Einzellige Organismen, seltener Coenobien oder Zellenhorden: Gemeinden von mehreren oder vielen locker verbundenen Zellen. Bewegen sich durch Geisseln (Flagella), einen langen, fadenförmigen [S. 90] Fortsatz (oder mehreren, an einem Punkte befestigten) des Protoplasma, welche hin und her schwingen, wie eine Peitsche. Zellenleib bald nackt, bald von einer Hülle umschlossen, aus deren Oeffnung die schwingende Geissel vortritt. Selten sind die Flagellaten auf Gegenständen im Wasser festgewachsen, meistens schwimmen sie frei umher. Bei vielen wechseln ruhende und bewegliche Zustände mit einander ab; und dann geschieht die Vermehrung meist während des Ruhezustandes, durch Theilung. Nahrungsaufnahme bald durch Aufsaugung (Endosmose), bald durch einen Zellmund (Cytostoma). Vermehrung ungeschlechtlich, meist durch Theilung, seltener durch Knospung oder Sporenbildung. Bei einigen (Volvocinen) Anfänge geschlechtlicher Sonderung.
Erste Ordnung der Flagellaten:
Nudoflagellata (HAECKEL). Nackt-Geissler. Fig. 8.
Geisslinge mit nacktem Zellenleibe, ohne Wimperkranz.
Gattungen: Euglena (viridis). Astasia (haematodes). Phacus (longicauda, Fig. 8).
Zweite Ordnung der Flagellaten:
Thecoflagellata (HAECKEL). Hüll-Geissler. Fig. 10.
Geisslinge, ohne Wimperkranz, deren Zellenleib von einer Hülle oder Schale umschlossen ist. Die Geisseln treten aus einer Oeffnung der Schale hervor. Oft ist die Schale auf einem sitzenden Stiele angeheftet.
Gattungen: Salpingoeca (marina). Dinobryon (sertularia).
Dritte Ordnung der Flagellaten:
Cilioflagellata (J. MÜLLER). Wimper-Geissler. Fig. 9.
Geisslinge mit einem Kranze von kurzen Wimpern um die Mitte des Zellenleibes, welcher von einer zweiklappigen Schale umschlossen ist. Zwischen beiden Schalenhälften tritt frei die lange Geissel und der Wimperkranz vor.
Gattungen: Peridinium (oculatum) Ceratium (tripus) Fig. 9.
[S. 91]
Vierte Ordnung der Flagellaten:
Cystoflagellata (HAECKEL). Blasen-Geissler. Fig. 11.
Geisslinge ohne Wimperkranz, mit grossem, blasenförmigen Zellenleibe, welcher ausser der Geissel einen eigenthümlichen Peitschen-Anhang und einen stabförmigen Körper im Innern besitzt.
Gattungen: Noctiluca (miliaris) Fig. 11. Leptodiscus (medusoides).
Fünfte Classe des Protistenreiches.
5. Catallacta (HAECKEL). Mittlinge.
Einzellige Organismen, welche eine Zeitlang zu einer Zellenhorde (Coenobium) vereinigt sind, später isolirt leben. Die Zellenhorden oder Coenobien sind schwimmende Gallertkugeln, zusammengesetzt aus zahlreichen Zellen, welche im Centrum der Kugel durch Fortsätze vereinigt sind, während an der Oberfläche die schwingenden Flimmerhaare vortreten. Die Einsiedler-Zellen (Monocyten), welche durch Zerfall der Coenobien entstehen, bewegen sich anfangs schwimmend, gleich Flagellaten umher; dann verwandeln sie sich in kriechende Amoeben-ähnliche Zellen; schliesslich ziehen sie sich kugelig zusammen und kapseln sich ein. Innerhalb dieses Ruhezustandes entsteht durch wiederholte Theilung der Zelle ein neues kugelförmiges Coenobium, welches die Hülle durchbricht und frei umherschwimmt. Diese ungeschlechtliche Fortpflanzung erinnert an die Eifurchung der Thiere. Die Catallacten leben theils im Meere, theils im Süsswasser.
Gattungen: Magosphaera (planula) Fig. 46. Synura (uvella).
Sechste Classe des Protistenreiches.
6. Ciliata (J. MÜLLER). Wimperlinge.
(Synonym: Infusionsthierchen oder Infusoria im engsten Sinne).
Einzellige Organismen, sehr selten Zellhorden oder Coenobien, welche aus mehreren, locker verbundenen Zellen zusammengesetzt sind. Bewegung durch zahlreiche kurze Wimpern (Cilia). [S. 92] Zellenleib meistens nackt, seltener von einer Hülle oder Schale theilweise umschlossen. Der Protoplasma-Leib der Zelle sondert sich meist in eine helle, festere, hyaline Rindenschicht (Exoplasma) und eine trübe, feinkörnige, weichere Markschicht (Endoplasma). Aus der Rindenschicht treten stets zahlreiche kurze Wimperhärchen oder Cilien hervor, welche lebhaft und willkürlich bewegt werden. Meistens laufen oder schwimmen die Wimperthierchen rasch umher mittelst der Bewegungen ihrer Wimpern; bei festsitzenden dienen letztere dazu, durch den im Wasser erzeugten Strudel stets frisches Wasser und Nahrung dem Zellmunde zuzuführen. Der Zellmund (Cytostoma) ist eine constante Oeffnung in der Rindenschicht und lässt verschluckte Bissen in die innere weichere Markmasse des Zellenleibes eintreten, wo sie verdaut werden. An der Innenfläche der Rindenschicht liegt eine contractile Blase (eine constante Vacuole), welche meistens durch einen kurzen Kanal nach aussen zu münden scheint. Der Zellkern ist gross, verschieden gestaltet, meistens einfach, sehr selten mehrfach. Die Fortpflanzung geschieht meistens ungeschlechtlich, durch Theilung (Quertheilung oder Längstheilung), seltener durch Knospung. Wie weit ausserdem Fortpflanzung durch Sporenbildung (oder vielleicht durch geschlechtliche Zeugung einfachster Art), verbunden mit Conjugation, in dieser Classe verbreitet ist, erscheint noch nicht sicher festgestellt. Die Classe der Wimperthierchen oder Wimperlinge ist sehr umfangreich und überall im Süsswasser und Meere verbreitet.
Erste Ordnung der Ciliaten.
Holotricha (STEIN). Ueberall behaarte Wimperlinge.
Die ganze Oberfläche des Zellenleibes gleichmässig mit kurzen feinen Wimperhärchen bedeckt.
Gattungen: Glaucoma (scintillans). Paramecium (aurelia). Trachelius (ovum). Prorodon (teres) Fig. 15.
Zweite Ordnung der Ciliaten.
Heterotricha (STEIN). Verschieden behaarte Wimperlinge.
Die ganze Oberfläche des Zellenleibes gleichmässig mit kurzen feinen Wimperhärchen bedeckt; ausserdem noch ein Kranz oder [S. 93] Gürtel von stärkeren und grösseren Wimpern (Griffeln oder Borsten) um den Zellmund herum.
Gattungen: Bursaria (truncatella). Stentor (polymorphus) Fig. 12. Freia (elegans) Fig. 14. Spirustomum (teres).
Dritte Ordnung der Ciliaten:
Hypotricha (STEIN). Unterseits behaarte Wimperlinge.
Zellenleib blattförmig zusammengedrückt, an der oberen (oder Rücken-) Seite nackt, an der unteren (oder Bauch-) Seite mit kleineren und grösseren Wimpern bedeckt.
Gattungen: Chilodon (cucullulus). Euplotes (charon). Oxytricha (pellionella). Aspidisca (costata).
Vierte Ordnung der Ciliaten:
Peritricha (STEIN). Ringförmig behaarte Wimperlinge.
Zellenleib drehrund, grösstentheils nackt, nur mit einem Gürtel (seltener zwei Gürteln) von Wimperhaaren versehen.
Gattungen: Dictyocysta (templum). Ophrydium (versatile). Trichodina (pediculus). Vorticella (campanula).
Siebente Classe des Protistenreiches:
7. Acinetae (EHRENBERG). Starrlinge.
(Synonym: Infusoria suctoria. Saug-Infusorien.)
Einzellige Organismen, seltener Zellenhorden (Coenobia) oder Zellenfusionen (Syncytia), welche aus mehreren, locker oder enger verbundenen Zellen zusammengesetzt sind. Zellenleib von einer Membran oder Kapsel umschlossen, durch welche sehr feine, zerstreute oder büschelförmig vereinigte Saugröhren hervortreten. Mittelst dieser borstenförmigen, am freien Ende mit einem Saugnäpfchen oder Knöpfchen versehenen Saugröhren heften sich die Acineten an Ciliaten und anderen Protisten an und saugen deren Protoplasma aus (ähnlich wie die Vampyrellen unter den Moneren). Im Innern des Protoplasma findet sich neben dem [S. 94] Zellkern oft eine contractile Blase (Vacuole). Fortpflanzung ungeschlechtlich, bald durch Theilung, bald durch Knospung, bald durch Schwärmsporen. Letztere entstehen im Innern des Zellenleibes, durchbrechen denselben und schwimmen mittelst feiner Wimperhärchen umher, zwischen welchen feinste Saugröhrchen sitzen. Die Acineten leben sowohl im süssen Wasser als im Meere, und sitzen meistens unbeweglich auf Stielen fest; seltener schwimmen sie frei umher.
Erste Ordnung:
Monacinetae (HAECKEL). Einzel-Starrlinge. Fig. 16, 17.
Einzellige Acineten, deren einfacher Zellenleib nur einen einzigen Kern enthält.
Gattungen: Podophrya (Cyclopum). Acinetella (mystacina).
Zweite Ordnung:
Synacinetae (HAECKEL). Horden-Starrlinge.
Mehrzellige Acineten, deren verästelter Zellenleib mehrere Kerne enthält.
Gattungen: Dendrosoma (radians).
Achte Classe des Protistenreiches.
8. Labyrinthuleae (CIENKOWSKI). Labyrinthinge.
Zellenhorden (Coenobia), welche aus zahlreichen gleichartigen, beweglichen Zellen locker zusammengesetzt sind. Die Zellen sind meistens spindelförmig, umschliessen einen Kern und können ihre Gestalt ändern. Sie leben in grossen Haufen gesellig beisammen, und bewegen sich in eigenthümlicher, noch unerklärter Weise rutschend oder gleitend umher (ähnlich manchen Diatomeen). Die Bewegung geschieht nicht frei im Wasser, sondern ausschliesslich in einer eigenthümlichen Fadenbahn (Linodium), einem Gerüste von starren, baumförmig verästelten und netzförmig verbundenen Fäden. Diese Fäden besitzen faserige Structur und werden von den wandernden Zellen ausgeschieden. Auf den mannigfachsten Umwegen gleiten die Spindelzellen in der Fadenbahn [S. 95] umher, sammeln sich später in Haufen und kapseln sich ein. Bei dieser Encystirung erhält jede Zelle eine Membran, und die ganze gesammelte Horde eine Rindenkapsel. In jeder einzelnen Zelle entstehen später vier junge Zellen (Tetrasporen). Die Labyrinthuleen leben im Meere.
Gattung: Labyrinthula. (Arten: L. vitellina. L. macrocystis.)
Neunte Klasse des Protistenreiches.
9. Bacillariae. Schachtlinge.
(Synonym: Diatomeae. Diatomaceae. Stabthierchen.)
Einzellige Organismen oder Zellenhorden (Coenobia); lockere Gesellschaften von mehreren Zellen, welche in Gallertmassen oder auf gemeinsamen, verzweigten Stielen vereinigt sind. Der Zellenleib ist stets von einer zweiklappigen Kieselschale umschlossen, deren beide Hälften so ineinander geschoben sind, wie eine Schachtel und ihr Deckel. Meistens bewegen sich die Zellen rutschend oder schwimmend umher, wahrscheinlich mittelst eines äusserst feinen Wimperkranzes, welcher in dem engen Spalte zwischen der Schachtel und ihrem Deckel frei vortritt. Ernährung und Stoffwechsel wie bei den einzelligen Algen. Fortpflanzung ungeschlechtlich, durch Theilung. Im Beginn der Theilung schieben sich die beiden Klappen der schachtelähnlichen Kieselschalen auseinander; der Kern theilt sich in zwei auseinander weichende Hälften, ebenso das Protoplasma. Darauf bildet sich jede Tochterzelle eine neue Schalenhälfte zu der alten Hälfte, die den Schachteldeckel bildet. Die so entstehenden Generationen werden fortgesetzt immer kleiner, bis zuletzt eine Generation (von Auxosporen) entsteht, welche die ganze Kieselschale abwirft, mächtig wächst und dann eine neue Kieselschale erster Grösse bildet. Die Diatomeen oder Bacillarien leben in zahllosen, zierlichen Formen überall im Meere und im Süsswasser.
Erste Ordnung der Bacillarien:
Naviculatae (EHRENBERG). Kahn-Schachtlinge. Fig. 45.
Gattungen: Navicula (gracilis). Cocconeis (placentula).
[S. 96]
Zweite Ordnung der Bacillarien:
Echinellatae (EHRENBERG). Palm-Schachtlinge.
Gattungen: Cocconema (cistula). Achnanthes (longipes).
Dritte Ordnung der Bacillarien:
Lacernatae (EHRENBERG). Gallert-Schachtlinge.
Gattungen: Frustulia (salina). Gloeonema (paradoxum).
Zehnte Klasse des Protistenreiches.
10. Fungi (LINNE.). Pilze.
Polyplastide (sehr selten monoplastide) Organismen, deren Körper nicht aus echten (kernhaltigen) Zellen, sondern aus fadenförmigen (kernlosen) Cytoden zusammengesetzt ist (Hyphen). Diese Fadenschläuche oder Hyphen bilden durch seitliche Sprossung und quere Gliederung der Aeste ein vielfach verzweigtes Faden-Geflecht (Mycelium), welches parasitisch in oder auf anderen Organismen (oder von deren Zersetzungs-Producten) lebt. Später entwickelt sich aus diesem Mycelium ein ansehnlicher, höchst mannigfaltig gebauter Fruchtkörper (Stroma), welcher an bestimmten Stellen ein Sporenlager (Hymenium) bildet. In letzterem entstehen die Keimzellen oder Sporen meistens ungeschlechtlich, selten geschlechtlich (in Folge eines eigenthümlichen Befruchtungs-Vorganges). Der Stoffwechsel der Pilze ist thierisch, nicht pflanzlich. Niemals bilden die Pilze Chlorophyll und Amylum, wie die echten Pflanzen. Nirgends findet sich im Pilzkörper ein Zellkern, wie er in den Zellen aller echten Thiere und Pflanzen überall vorkommt.
Erste Ordnung der Pilze:
Phycomycetes. Tangpilze.
Gattungen: Mucor (mucedo). Penicillium (glaucum).
[S. 97]
Zweite Ordnung der Pilze:
Coniomycetes. Rostpilze.
Gattungen: Ustilago (segetum). Puccinia (graminis).
Dritte Ordnung der Pilze:
Ascomycetes. Schlauchpilze.
Gattungen: Morchella (esculenta). Claviceps (purpurea).
Vierte Ordnung der Pilze:
Gastromycetes. Bauchpilze.
Gattungen: Lycoperdon (bovista). Phallus (impudicus).
Fünfte Ordnung der Pilze:
Hymenomycetes. Hutpilze.
Gattungen: Agaricus (campestris). Boletus (laricis). Fig. 44.
Elfte Classe des Protistenreiches.
11. Myxomycetes (WALLROTH). Netzinge.
(Synonym: Mycetozoa. Myxogasteres. Schleimpilze.)
Organismen, welche in vollkommen entwickeltem und frei beweglichem Zustande ein Plasmodium darstellen: einen formlosen Protoplasmakörper, welcher aus vielen verschmolzenen Zellen zusammengesetzt ist, deren Kerne sich aufgelöst haben. Dieses Plasmodium kriecht frei umher, gleich einem colossalen Rhizopoden, und bildet ausgedehnte netzförmige Körper, indem an der Oberfläche formwechselnde, unbeständige Scheinfüsse (Pseudopodia) hervortreten, deren Aeste an den Berührungsstellen zusammenfliessen. Die Plasmodien ernähren sich von organischen Körpern ebenso wie die echten Rhizopoden (Thalamophoren) und Radiolarien. Nach vollendetem Wachsthum zieht sich das Plasmodium auf einen rundlichen Klumpen zusammen und verwandelt sich in einen blasenförmigen Fruchtkörper (Sporocystis). Diese Sporenblase [S. 98] ist von einer festen, structurlosen Haut allseitig umschlossen. Innerhalb derselben zerfällt das Protoplasma in zahllose kleine Keimzellen (Sporae), zwischen welchen sich meistens ein Geflecht von haarfeinen Fäden entwickelt (Capillitium). Später platzen die Fruchtkörper und die Sporen werden frei. Aus der Hülle einer jeden Spore schlüpft eine amoebenartige, kernhaltige Zelle aus. Diese Amoeben verwandeln sich in Flagellaten, indem an einem Ende eine schwingende Geissel vortritt. So schwimmen sie als »Schwärmsporen« umher, vermehren sich durch Theilung und gehen dann wieder in amoebenähnliche Zellen über, welche umherkriechen. Durch Zusammenfliessen vieler solcher Amoeben entstehen die Plasmodien. Diese leben schmarotzend auf verwesenden Pflanzen (faulen Blättern etc.).
Erste Ordnung der Myxomyceten:
Physareae. Physareen.
Zweite Ordnung der Myxomyceten:
Stemoniteae. Stemoniteen.
Gattungen: Stemonitis (typhoides). Diachea (elegans).
Dritte Ordnung der Myxomyceten:
Trichiaceae. Trichiaceen.
Gattungen: Licea (serpula). Arcyria (lateritia).
Vierte Ordnung der Myxomyceten:
Lycogaleae. Lycogaleen.
Gattungen: Lycogala (epidendron). Reticularia (maxima).
Zwölfte Classe des Protistenreiches.
12. Thalamophora (HERTWIG). Kammerlinge.
(Synonym: Acyttaria. Reticularia. Rhizopoda.)
Organismen, welche in entwickeltem Zustande ein Syncytium darstellen, einen beweglichen Protoplasma-Körper, der viele Zellkerne [S. 99] enthält; selten ist der Körper einzellig und enthält nur einen Kern. Von der Oberfläche des Protoplasma strahlen sehr zahlreiche und feine Scheinfüsschen (Pseudopodia) aus: dünne Fäden, welche sich verästeln und an den Berührungsstellen netzförmig verschmelzen. Diese Pseudopodien dienen sowohl zur Ortsbewegung und zur Empfindung, wie zur Ernährung, indem die Acyttarien fremde organische Körperchen mittelst derselben in ihr Inneres einziehen und dort auflösen. Stets ist das Syncytium von einer Schale umschlossen, die äusserst vielgestaltig ist. Die Pseudopodien treten entweder aus einer einzigen grösseren Oeffnung der Schale hervor (Imperforata), oder aus zahlreichen feinen Sieblöchern der Schale (Foraminifera). Meist besteht die Schale aus kohlensaurem Kalk, seltener aus Kieselerde oder aus einer organischen Substanz. Die Fortpflanzung erfolgt ungeschlechtlich, selten durch Theilung oder Knospung, meistens durch Sporenbildung. Die meisten Acyttarien leben im Meere, einige im süssen Wasser; die meisten kriechen auf dem Boden, wenige schwimmen an der Oberfläche; nur einzelne sitzen auf einem Stiele fest.
Erste Ordnung der Thalamophoren:
Monostegia (D’ORB.). Dichtschalige Einkammerlinge. Fig. 19.
Schale einkammerig, dicht, nicht siebförmig durchlöchert, nur mit einer einzigen grösseren Oeffnung an einem Pole der Axe, selten mit zwei grösseren Oeffnungen, an beiden entgegengesetzten Polen derselben. (Imperforata monostegia).
Gattungen: Gromia (oviformis) Fig. 19. Lagynis (baltica). Squamulina (laevis). Cornuspira (planorbis).
Zweite Ordnung der Thalamophoren:
Polystegia (HAECKEL). Dichtschalige Vielkammerlinge.
Schale vielkammerig, dicht, nicht siebförmig durchlöchert, mit einer einzigen grossen Oeffnung, am Ende der jüngsten Kammer. (Imperforata polystegia).
Gattungen: Miliola (cyclostoma). Peneroplis (dendritina). Lituola (nautiloides). Parkeria (ingens).
[S. 100]
Dritte Ordnung der Thalamophoren:
Monothalamia (M. SCHULTZE). Siebschalige Einkammerlinge.
Schale einkammerig, siebförmig von zahlreichen feinen Löchern durchbrochen, ausserdem meist eine grosse Oeffnung an einem Pole der Längsaxe. (Foraminifera monothalamia).
Gattungen: Orbulina (universa). Entosolenia (globosa). Lagena (vulgaris).
Vierte Ordnung der Thalamophoren:
Polythalamia (BREYN). Siebschalige Vielkammerlinge. Fig. 20–24.
Schale vielkammerig, siebförmig von zahlreichen feinen Löchern durchbrochen, ausserdem oft eine grosse Oeffnung am Ende der jüngsten Kammer. (Foraminifera polythalamia).
Gattungen: Nodosaria (radicula). Rotalia (veneta). Globigerina (bulloides). Textularia (variabilis). Alveolina (vulgaris). Nummulites (lentiformis).
Dreizehnte Klasse des Protistenreiches.
13. Heliozoa (HAECKEL). Sonnlinge.
Organismen, welche in entwickeltem Zustande bald eine einzige
kugelige Zelle, bald ein kugeliges Syncytium darstellen,
welches aus mehreren verschmolzenen Zellen besteht. Im ersteren
Falle ist ein einziger Zellkern, im letzteren mehrere Kerne im
Innern der Protaplasma-Kugel eingeschlossen. Letztere ist in eine
feinkernige innere Markmasse (Endoplasma) und eine schaumige
äussere Rindenschicht (Exoplasma) gesondert. Das Protaplasma
der Rindenschicht bildet Vacuolen (oder vergängliche, contractile
Wasserbläschen). Von seiner Oberfläche strahlen rings zahlreiche
haarfeine Fäden aus, die gewöhnlich einfach, nicht verästelt, ziemlich
starr sind und wenig Neigung zur Verschmelzung besitzen.
Bald ist der Körper ganz weich und nackt; bald bildet er ein
festes Skelet, welches aus vielen zerstreuten Nadeln (Spicula) zusammengesetzt
ist, oder eine Gitterschale darstellt. Meistens schweben
[S. 101]
die Heliozoen frei im Wasser; seltener sind sie festgewachsen.
Fortpflanzung ungeschlechtlich, bald durch Theilung, bald durch
Sporenbildung. Die Heliozoen leben sowohl im süssen Wasser, als
im Meere.
Erste Ordnung der Heliozoen:
Aphrothoraca (HAECKEL). Nackte Sonnlinge. Fig. 40, 41.
Nackte Heliozoen, mit weichem, schaumigem Körper, ohne Skelet.
Gattungen: Actinophrys (sol). Actinosphaerium (Eichhornii).
Zweite Ordnung der Heliozoen:
Chalarothoraca (HERTWIG). Bestachelte Sonnlinge.
Heliozoen mit einem Skelet, welches aus Spicula oder zerstreuten Stäbchen (radialen Stacheln oder tangentialen Nadeln) zusammengesetzt ist.
Gattungen: Acanthocystis (spinifera). Heterophrys (marina).
Dritte Ordnung der Heliozoen:
Desmothoraca (HERTWIG). Beschalte Sonnlinge.
Heliozoen mit einem Skelet, welches eine kugelige, von Löchern durchbrochene Schale bildet.
Gattungen: Hedriocystis (pellucida). Hyalolampe (fenestrata).
Vierzehnte Classe des Protistenreiches.
14. Radiolaria (J. MÜLLER). Strahlinge.
Organismen, welche in entwickeltem Zustande aus zwei verschiedenen Haupttheilen bestehen, einer inneren, festen, mit Zellen gefüllten Central-Kapsel (Capsula centralis) und einem äusseren Syncytium, einer Protoplasma-Masse, welche die erstere allseitig umgiebt, und von welcher ausserdem zahlreiche Scheinfüsschen oder Pseudopodien ausstrahlen; letztere verhalten sich ganz wie diejenigen der Acyttarien. Der wesentliche Unterschied von den letzteren besteht in der stets vorhandenen Centralkapsel; diese [S. 102] ist der Sporenblase der Myxomyceten vergleichbar und stellt einen Fruchtkörper (Sporangium) dar, indem ihr gesammter Inhalt sich in Keimzellen (Sporae) verwandelt. Ausserhalb der Centralkapsel finden sich meist noch eigenthümliche gelbe Zellen, welche Stärkemehl enthalten. Nur wenige Radiolarien sind weich und nackt; die meisten besitzen ein Skelet, welches aus Nadeln (Spicula), einem Balkengeflecht oder einer Schale besteht; diese ist meistens aus Kieselerde gebildet, von äusserst mannigfaltigen und zierlichen Formen. Die Ernährung der Radiolarien geschieht wie bei den Acyttarien durch die Pseudopodien. Die Fortpflanzung erfolgt ungeschlechtlich, selten durch Theilung, meistens durch Sporenbildung. Die Sporen, welche innerhalb der Centralkapsel entstehen und aus dieser ausschwärmen, sind mit Geisseln versehene Schwärmsporen. Alle Radiolarien leben im Meere und schweben theils an der Oberfläche, theils in verschiedenen Tiefen.
Erste Ordnung der Radiolarien:
Pancollae (HAECKEL). Gallert-Strahlinge. Fig. 26.
Radiolarien ohne Skelet, oder mit einem Skelet, welches bloss aus zerstreuten soliden Nadeln zusammengesetzt ist.
Gattungen: Thalassicolla (nucleata). Collozoum (inerme). Thalassosphaera (bifurca). Sphaerozoum (punctatum).
Zweite Ordnung der Radiolarien:
Panacanthae (HAECKEL). Stachel-Strahlinge. Fig. 32, 33.
Skelet besteht aus radialen soliden Stacheln, welche im Mittelpunkt der Central-Kapsel in einander gestemmt, locker verbunden oder verwachsen sind. (Acanthometrida s. a.)
Gattungen: Acanthometra (Mülleri). Dorataspis (bipennis).
Dritte Ordnung der Radiolarien:
Pansoleniae (HAECKEL). Röhren-Strahlinge.
Skelet besteht aus einzelnen hohlen Röhren, welche bald locker zerstreut, bald in radialer oder concentrischer Anordnung verbunden sind.
[S. 103]
Gattungen: Aulacantha (scolymantha). Aulosphaera (trigonopa). Coelodendrum (gracillimum).
Vierte Ordnung der Radiolarien:
Plegmideae (HAECKEL). Schwamm-Strahlinge. Fig. 34.
Skelet besteht aus einem lockeren oder dichteren Geflecht von feinen Kieselstäbchen, welche ohne bestimmte Anordnung (schwammähnlich) verbunden sind. (Wachsthum vielseitig).
Gattungen: Acanthodesmia (vinculata). Spongurus (cylindricus). Spongodiscus (mediterraneus). Spongasteriscus (quadricornis).
Fünfte Ordnung der Radiolarien:
Sphaerideae (HAECKEL). Kugel-Strahlinge. Fig. 25, 29, 30.
Skelet besteht aus einer einzigen Gitterkugel oder aus mehreren concentrischen Gitterkugeln, welche durch radiale Stäbe verbunden sind. (Wachsthum radial).
Gattungen: Ethmosphaera (siphonophora). Collosphaera (Huxleyi). Cladococcus (cervicornis). Haliomma (castanea). Actinomma (drymodes).
Sechste Ordnung der Radiolarien:
Discideae (HAECKEL). Scheiben-Strahlinge. Fig. 37.
Skelet scheibenförmig, aus zwei parallelen Siebplatten zusammengesetzt, zwischen welchen durch Kreuzung von concentrischen und radialen Gitterstäben zahlreiche kleine Kammern gebildet werden. (Wachsthum concentrisch).
Gattungen: Trematodiscus (sorites). Euchitonia (Mülleri). Coccodiscus (Darwinii). Astromma (Aristotelis).
[S. 104]
Siebente Ordnung der Radiolarien:
Cyrtideae (HAECKEL). Kegel-Strahlinge. Fig. 35, 36.
Skelet eine Gitterschale, welche durch eine Hauptaxe mit zwei verschiedenen Polen charakterisirt ist. Grundform daher kegelförmig. Durch ringförmige Einschnürungen ist die Schale oft in mehrere, hinter oder neben einander liegende Kammern abgetheilt. (Wachsthum unipolar).
Gattungen: Cyrtocalpis (amphora). Petalospyris (arachnoides). Eucecryphalus (Gegenbauri). Eucyrtidium (lagena). Botryocampe (hexathalamia).

Druck von Hüthel & Herrmann in Leipzig.