
As the Nation’s principal conservation agency, the Department of the Interior has basic responsibilities for water, fish, wildlife, mineral, land, park, and recreational resources. Indian and Territorial affairs are other major concerns of America’s “Department of Natural Resources.” The Department works to assure the wisest choice in managing all our resources so each will make its full contribution to a better United States—now and in the future.
This publication is one in a series of research studies devoted to special topics which have been explored in connection with the various areas in the National Park System. It is printed at the Government Printing Office and may be purchased from the Superintendent of Documents, Government Printing Office, Washington, D.C. 20402.
Library of Congress Cataloging in Publication Data
Paul O. McGrew
Michael Casilliano
Department of Geology, University of Wyoming
NATIONAL PARK SERVICE OCCASIONAL PAPER NUMBER THREE

Fifty million years ago the southwestern corner of what is now Wyoming was part of a system of three freshwater lakes that covered that area plus adjacent parts of Colorado and Utah.
These lakes began with Lake Flagstaff and later became Lake Gosiute, Lake Uinta, and Fossil Lake (Fig. 1). These lakes developed in intermontane basins that were created as a result of the geologic events that formed the Rocky Mountains. The three lakes, collectively known as the Green River Lake System, had a long, complex history of expansion and contraction. Varying climatic and geologic conditions were responsible for the changes in lake size and distribution.
The shores surrounding the lakes were blanketed by a lush, green canopy of palm, cinnamon, maple, oak, and other familiar trees. Hazel and lilac covered the forest floor. Rushes and other aquatic plants lined the lake shore.
The air was humid and warm. Streams flowing down from the hills and mountains built up flood plains and fed sediment into the lake, where it was deposited in shallow water near the shore. Deposits formed by chemical processes settled to the bottom further from shore in deep, quiet water. From these processes were formed the rocks from which the past history of Green River Lake System is read.
In the forest and undergrowth lived the ancestors of modern mammal groups. Ancestral rodents and tiny insectivores lived a furtive existence in the brush and mold that carpeted the forest floor, or else they sought a livelihood among the branches of trees. Large, bizarre animals with strange names lumbered through the reed-covered streamsides eating soft, succulent plants. The earliest members of the horse family browsed on the soft vegetation. The trees overhead were alive with the chattering and antics of early primates. Carnivorous mammals preyed on their plant-eating neighbors and so maintained a balanced community. Crocodiles and turtles basked in the sun on the beach. Flamingos concentrated in large nesting grounds. Snakes and lizards crawled about in the undergrowth. Insects, many exceedingly similar to modern types, flew about in the warm air or crept about on the plants.
The waters of the lakes teemed with many types of fish. Relatives of the perch, herring, and sting ray swam in the warm lake water. Occasionally, large-scale mortalities of the fish occurred. As the fish died, they sank to the bottom of the lake, and were preserved in the lake sediments.
Today, the lakes are gone and where once there were lush tropical forests there is now a semi-desert covered with sagebrush and greasewood. The history of these lakes, forests, and animal life can be read in the thick sediments deposited so long ago.
This report concerns itself with one of these lakes, the smallest of the three, Fossil Lake. Famous for its fossil fish beds, part of this ancient lake is now Fossil Butte National Monument. It is the purpose of this report to relate the history of Fossil Lake and its now fossilized inhabitants, thereby, hopefully presenting the order of events in a landscape obscured by 50 million years of time.
Fossil Butte National Monument is near the geographical center of Fossil Basin. The basin itself is located in the extreme southwestern part of Wyoming in Uinta and Lincoln counties, near the Utah-Idaho border (Fig. 2).
The monument is situated about 10 miles west of Kemmerer, Lincoln County, Wyoming (Fig. 3). U.S. Highway 30N runs just south of the monument as does the Union Pacific Railroad line to Oregon. The abandoned town of Fossil is also immediately south of the monument.
The Kemmerer and Sage quadrangles of the United States Geological Survey cover the entire monument and the surrounding area.
The principal rocks involved at Fossil Butte are the Green River and Wasatch formations. The first published notice of these two rock units was by Hayden (1869). Although short, Hayden’s descriptions were the basis for later refinement of the stratigraphy of the Green River and Fossil basins. Hayden also mentioned the quantities of fossil fish from the Green River Formation.
The work done by Hayden was conducted under the auspices of the U.S. Department of the Interior, Geological and Geographical Survey of the Territories. The survey lasted from 1867 to 1878, during which time Hayden and his associates published annual reports of their findings.
In the 1870 report, Hayden (1871) mentioned the discovery of the Petrified Fish Cut. This cut is located on the main line of the Union Pacific Railroad about 2 miles west of Green River, Wyoming. A. W. Hilliard and L. E. Rickseeker, employees of the Union Pacific Railroad, discovered the cut and obtained many fossil fish which they turned over to Hayden.
Previous to Hayden, rocks now known as the Green River and Wasatch formations were occasionally mentioned in the various diaries, journals, and reports of early missionaries like those of S. A. Parker or those of explorers like Fremont (Knight 1955).
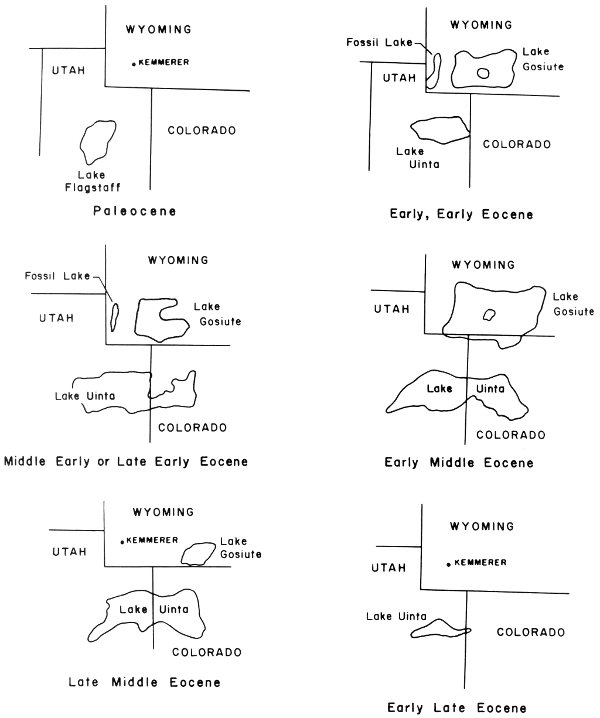
Fig. 1. Paleocene-Eocene lake system (after Schaeffer and Mangun 1965).
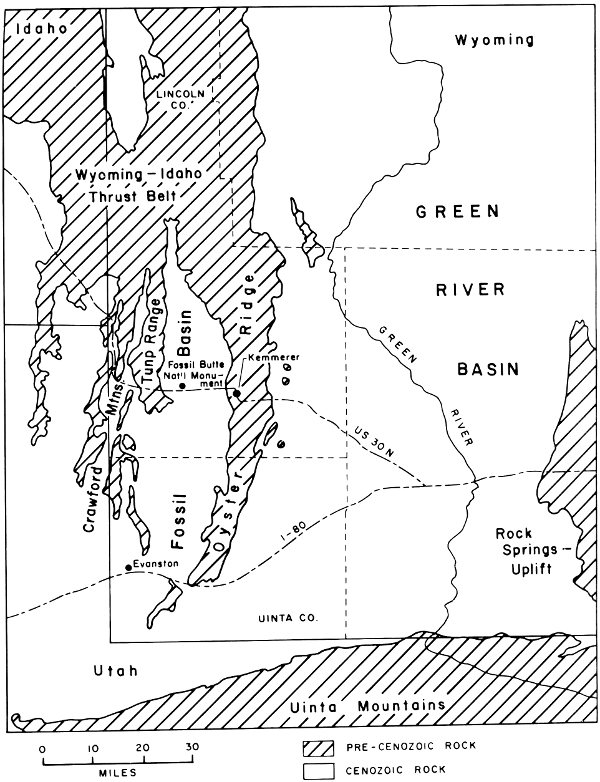
Fig. 2. Geographic features of southwest Wyoming and adjacent areas (from Oriel and Tracey 1970).
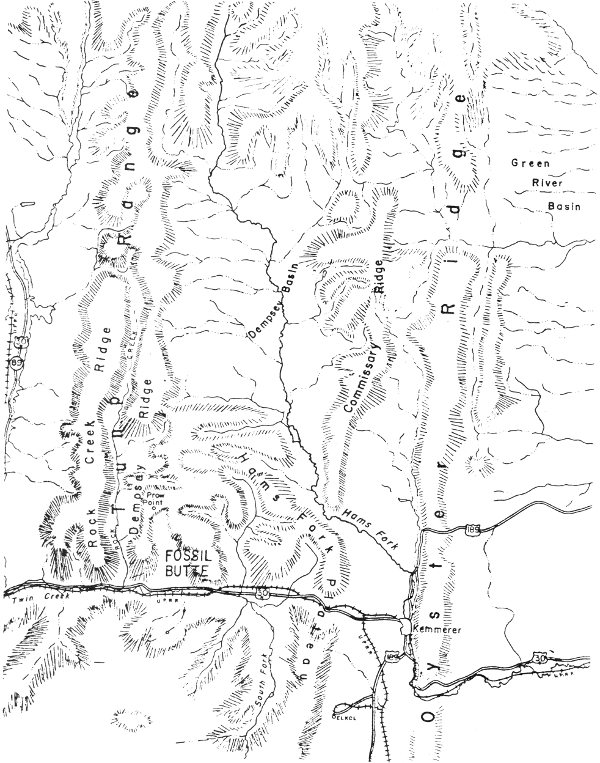
Fig. 3. Physiography of the Fossil Butte National Monument region (from U.S. Department of the Interior 1964).
Hayden’s report (1871:425-437) included a description of the fish fossils from Petrified Fish Cut by E. D. Cope, a vertebrate paleontologist. Several years earlier Leidy (1856), another paleontologist, described a fish from an unknown locality in the Green River Formation.
Hayden divided this survey area into several districts. The Green River district was put in charge of A. C. Peale. Peale’s (1879:535) report contained the first geologic description of Fossil Butte itself, as well as a short discussion of the fish fossils obtained there. The description of the butte is quoted here:
... In the lower part of the bluff from which these specimens are taken, the bright coloured beds of the Wasatch are seen outcropping, although the entire section cannot be seen, as their softness causes them to weather so that the debris conceals the strata. The fossils are found at several horizons in the shales. Near the top of the bluff is a band of hard, bituminous, or oily shale, which burns rather freely with a strong bituminous odor.... It is brownish-black in color and on the weathered surfaces a bluish white....
The next major publication dealing with Fossil Basin was that by Veatch (1907). He mapped the rocks in certain areas of the basin in an attempt to bring a semblance of order to Hayden’s (1869) broad stratigraphic descriptions. His work resulted in the designation of most of the rock units in Fossil Basin from Jurassic to Recent.
Schultz (1914), an associate of Veatch, published a paper dealing with the structures to the north and east of Fossil Basin. This helped to delineate the features surrounding and forming the basin.
Subsequent work on the geology of southwest Wyoming mainly has been concentrated on the Green River Basin just east of Fossil Basin. Significant work on Fossil Basin has been done by Rubey et al. (1968a, b) who mapped the northern part of the basin, and by Oriel and Tracey (1970) who have published results of the latest study on the stratigraphy and age of the rock units in Fossil Basin. This latest work is the culmination of many years of study and incorporates and refines data presented in shorter, earlier papers.
The complete geologic history of the Fossil Basin involves many stratigraphic units. Most of these are not exposed within the monument itself but are well known from outcrops outside the boundaries of the monument and from deep wells (Fig. 4).
At the base of the stratigraphic section are rocks of Precambrian age. These have not been penetrated by wells within the Fossil Basin but should occur as much as 25,000 ft beneath the surface. They probably consist of metamorphic and intrusive rocks. Beginning about 600 million years ago, at the beginning of Cambrian time, sedimentary deposits accumulated. These are both marine and continental in origin and range in age from early Cambrian (600 million years old) to early Tertiary (50 million years old). Some 34 geologic formations are recognized in this thick section, a list of which follows:
| Eocene | Fowkes Formation |
| Wasatch-Green River Formations | |
| Paleocene | Evanston Formation (upper part) |
| Upper Cretaceous | Evanston Formation (lower part) |
| Adaville Formation | |
| Lazeart Sandstone | |
| Hilliard Shale | |
| Frontier Formation | |
| Lower Cretaceous | Aspen Shale |
| Bear River Formation | |
| Gannett Group | |
| Jurassic | Stump Sandstone |
| Preuss Sandstone | |
| Twin Creek Limestone | |
| Nugget Sandstone (upper part) | |
| Triassic | Nugget Sandstone (lower part) |
| Thaynes Limestone | |
| Woodside Formation | |
| Dinwoody Formation | |
| Permian | Phosphoria Formation |
| Pennsylvanian | Wells Formation |
| Mississippian | Brazer Limestone |
| Madison Limestone (upper part) | |
| Devonian | Madison Limestone (lower part) |
| Darby Formation | |
| Silurian | Not Present |
| Ordovician | Leigh Dolomite |
| Bighorn Dolomite | |
| Cambrian | Gallatin Limestone |
| Gros Ventre Formation | |
| Precambrian | Complex of metamorphic and intrusive rocks |
Within the monument itself the oldest rocks exposed are those of the Thaynes Limestone near Prow Point. As will be seen in the discussion of the structure of the Fossil Basin, all of the rocks beneath the Evanston Formation are folded and faulted and within the monument are buried by deposits of Tertiary age. For this reason detailed discussion of stratigraphic units will be confined to those rocks that can be seen within the boundaries of the monument or closely adjacent to it (Fig. 5). The distribution of the various formations within the monument is illustrated in Figs. 7 and 8.
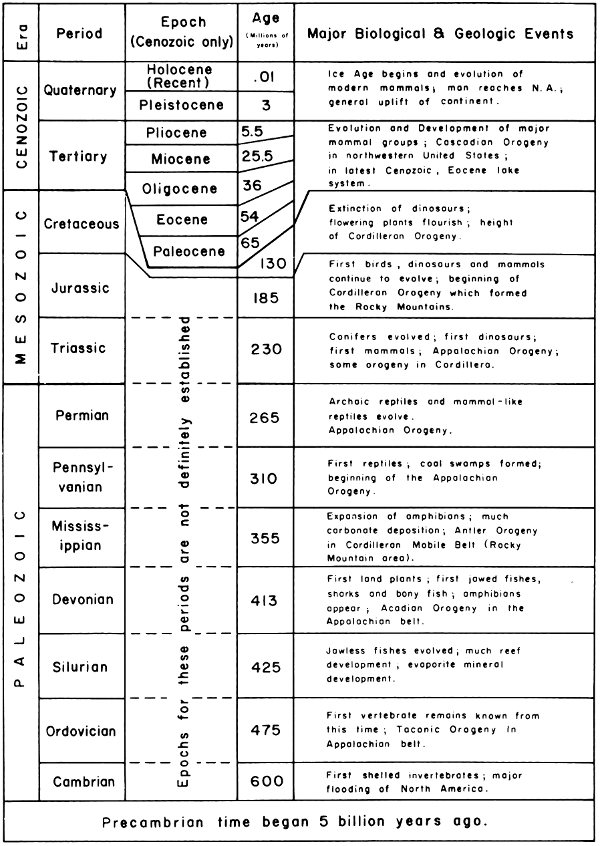
Fig. 4. Geologic time scale.
At the very northeastern corner of Fossil Butte National Monument can be seen an outcrop of the Thaynes Limestone of Triassic age, a marine unit deposited some 200 million years ago. This formation is noted for its rich and varied assemblage of marine fossils, mostly forms related to oysters and clams. The Thaynes Limestone consists predominantly of sandy, gray limestone and green-gray limey siltstones. The formation weathers to a dark brown in the lower part and a grayish-yellow in the upper. The fact that the Thaynes Limestone is exposed in the monument at all is because of the faulting that took place just prior to the deposition of the Evanston Formation of latest Cretaceous and earliest Tertiary age (refer to section on structure) and the erosion of the Tertiary rocks that once covered it.
Although not seen within the boundaries of the Fossil Butte National Monument, the Evanston Formation is exposed just south of Highway 30N, 1.5 miles southeast of the southeast corner of the monument. The Evanston was not involved in the complex folding and faulting but it is somewhat disturbed and rests under the Wasatch Formation with angular uncomformity. The Evanston Formation bridges the time boundary between the Cretaceous and Tertiary periods. In the lower part of the unit are found many fossil leaves, pollen, and spores and a jaw of the horned dinosaur Triceratops that prove its Cretaceous age, and in the upper part are found fossil mammals of Paleocene age.
The Evanston Formation has been studied in detail by Oriel and Tracey (1970). These authors divide the formation into three members. The lowest, which they called the Lower Member, is predominantly “gray to very dark gray mudstone, siltstone, claystone and gray carbonaceous sandstone.” The Lower Member reaches a thickness of 500 ft in some places. Above and in part interfingering with the Lower Member is a 1000 ft thick unit that was named the Hams Fork Conglomerate Member. This unit consists of beds of boulder conglomerate interstratified with thick beds of coarse, partly conglomeratic brown sandstone and gray mudstone. Where the Lower Member of the Evanston is missing, the Hams Fork Conglomerate forms the base of the formation.
The Upper Member of the Evanston Formation is termed by Oriel and Tracey (1970) the Main Body. It is more than a thousand feet thick and the lower part intertongues with the Hams Fork Conglomerate. The Main Body is “light to dark gray carbonaceous sandy to clayey siltstone interbedded with gray, tan, yellow and brown sandstone and conglomerate and carbonaceous to lignitic claystone.” It is this Main Body that can be seen along the highway just southeast of the monument.
The types of sediments and fossils found in the Evanston indicate that the formation was deposited by streams on flood plains and in marshes and ponds. A subtropical climate is indicated and the area was heavily wooded.
The term Wasatch was first used by Hayden (1869:91) as follows:
Immediately west of Fort Bridger commences one of the most remarkable and extensive groups of Tertiary beds seen in the West. They are wonderfully variegated, some shade of red predominating. This group, to which I have given the name of Wasatch group, is composed of variegated sands and clays. Very little calcareous matter is found in these beds.
In Echo and Weber Canyons are wonderful displays of conglomerates, fifteen hundred to two thousand feet in thickness. Although this group occupies a vast area, and attains a thickness of three to five thousand feet, yet I have never known any remains of animals to be found in it. I regard it, however, as of middle Tertiary age.
The Wasatch is well exposed in Fossil Basin. There the unit was regarded by Veatch (1907) as a group and divided by him into three formations: the Almy, Fowkes, and Knight. He wrote (1907:88):
In the Wasatch group as thus defined by Hayden the field work of the season 1905 showed three divisions: 1) a basal member composed of reddish-yellow sandy clays, in many places containing pronounced conglomerate beds, which has been named the Almy Formation; 2) a great thickness of light-colored rhyolitic ash beds containing intercalated lenses of white limestones with fresh-water shells and leaves—the Fowkes Formation; and 3) a group of reddish-yellow sandy clays with irregular sandstone beds (the Knight Formation) closely resembling 1) lithologically and separated from 1) & 2) by a pronounced period of folding and erosion.
Veatch, however, erred in his field work and did not realize that the Fowkes Formation had been downfaulted into the position in which he saw it (Tracey and Oriel 1959; Eardley 1959). The Fowkes is actually the youngest of the three formations of Veatch and is considerably later in age than the true Wasatch.
The Almy and Knight formations are not separable (Oriel and Tracey 1970:16), and can be seen to grade into each other at the basin edges.
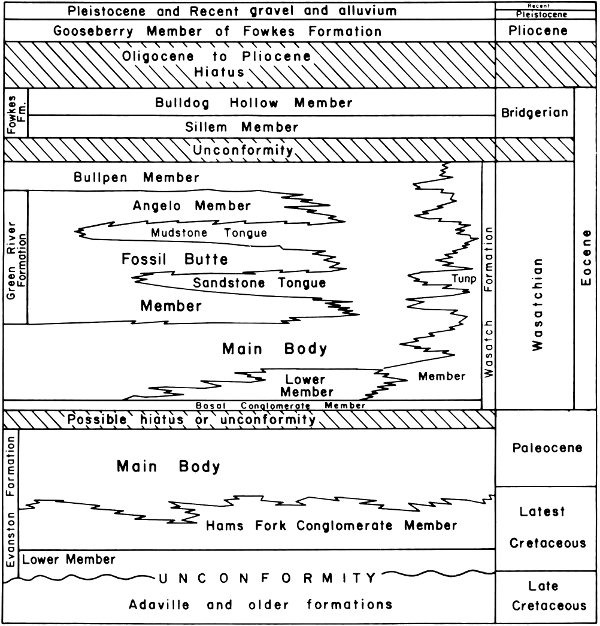
Fig. 5. Intertonguing relationship of latest Cretaceous and Tertiary stratigraphic units of Fossil Basin.
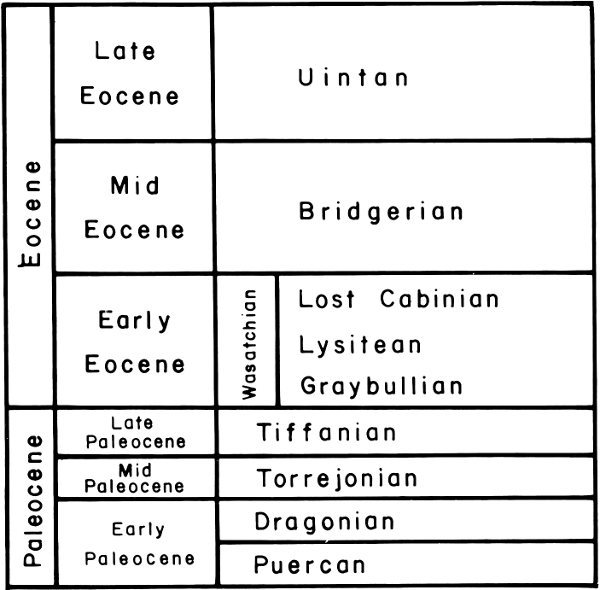
Fig. 6. Subdivisions of Paleocene and Eocene time.
The Almy and Knight, as defined by Veatch, are probably different facies of Wasatch, the Almy being a more peripheral facies and the Knight a more basinal facies (Oriel and Tracey 1970). These authors proposed that the terms Almy and Knight be dropped and that the name Wasatch be applied to all these rocks to avoid confusion.
The latest subdivision of the Wasatch Formation in the Fossil Basin is that of Oriel and Tracey. The members of the Wasatch Formation they propose are described in order from oldest to youngest.
This member has only local development in Fossil Basin. Where found, it is a lenticular conglomerate with pebbles and cobbles of buff and tan sandstone from the Nugget Sandstone and limestone fragments from the Thaynes and Twin Creek formations.
The basal conglomerate is essentially a channel fill in ancient stream beds cut into Mesozoic rocks.
This is an irregular sequence of flood-plain and stream-channel deposits. It is exposed along the southern part of the Tunp Range and extends into the far western section of Fossil Butte National Monument just below Prow Point.
Mudstone is the main rock type. It can be tan, brown, pink, red, or gray in color. Black, carbonaceous siltstones are also present. Gray sandstone that weathers yellow or brown and coarse-grained, cross-bedded conglomerate and sandstone are also prominent. Limestone occurs as thin lenses and is often brown, platy, and carbonaceous.
It is interesting to note that the Lower Member is intermediate not only in stratigraphic position but also in color and composition between the underlying Evanston Formation and overlying Main Body of the Wasatch Formation. The Lower Member thus appears to represent a gradual change in either climatic and/or sedimentary conditions in Fossil Basin (Oriel and Tracey 1970).
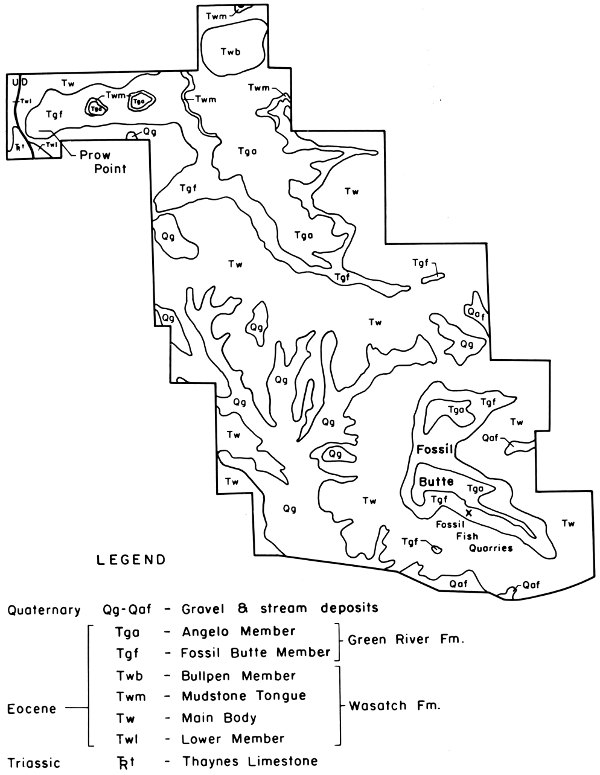
Fig. 7. Geologic map of Fossil Butte National Monument (after Rubey et al. 1968).

Fig. 8. NW-SE section across Fossil Butte National Monument, Sage and Kemmerer quadrangles (mapped by Rubey et al. 1968).
This unit of the Wasatch Formation is that part of the formation which produces the spectacular red-colored badlands in Fossil Butte National Monument. Particularly typical exposures can be seen in the south-facing scarp of Fossil Butte where the Main Body makes up the lower portion of the butte.
The most remarkable feature of the Main Body is its color. On the lower slopes of Fossil Butte are bands of bright to dull red, pink, purple, yellow, and gray color arranged in various patterns. This unit is best observed at a distance, especially after a rain. The bright hues of the Wasatch contrast markedly with the whites and tans of the overlying Green River Formation.
Individual bands of color range from 1 to 10 ft thick (Oriel and Tracey 1970:78). The colors are brightest in the upper part of the member and drabber in the coarser-grained lower part.
The predominant rock types in the Main Body are banded, variegated mudstone with interlayered sandstone, conglomerate, marlstone, siltstone, and claystone.
The upper part of the Main Body is mainly mudstone composed of fine silt and very fine, bedded sand with a clay binder. Conglomerates occur as channel fills and contain calcium carbonate as cement as do a number of sandstone and siltstone layers in the upper Main Body.
Conglomerates and sandstone are more common in the lower part of the Main Body. Some are part of Veatch’s old Almy Formation. They are best developed along the edge of Fossil Basin. The Main Body overlaps the Evanston Formation in some places and may rest directly on Mesozoic or Paleozoic rocks.
This is a tongue of cross-bedded sandstone. It is brown in color and is composed mostly of quartz with some black chert grains. This unit is limited in distribution to the south of Fossil Butte National Monument. The Sandstone Tongue thins and pinches out to the north and is not present within the monument. The area of pinchout represents the shoreline at one stage of Fossil Lake. The Sandstone Tongue wedges into the Fossil Butte Member of the Green River Formation. The sediment sources were probably the Uinta Mountains at the south edge of Fossil Basin. Tectonic events caused an uplift and erosion of Mesozoic and Paleozoic rocks in the Uintas and debris was deposited in Fossil Lake as an encroaching delta of sand. When deposition of the sand halted, the lake expanded and covered the sand, encasing it within the shales of the Green River Formation.
The Sandstone Tongue can be traced into the Main Body of the Wasatch, hence its assignment to the Wasatch Formation.
This tongue of the Wasatch Formation can be seen in the northwestern part of Fossil Butte National Monument and over most of the northern part of Fossil Basin. In the area of its distribution, the Mudstone Tongue separates the underlying Fossil Butte Member of the Green River Formation from the overlying Angelo Member of the Green River Formation. The tongue thins and disappears to the south, that is, basinward. As with the Sandstone Tongue, the edges of the Mudstone Tongue where it pinches out represent an ancient shoreline of Fossil Lake. To the north and west the Mudstone Tongue merges with the Tunp Member of the Wasatch Formation.
The Mudstone Tongue is a composite of dark-red mudstone which becomes lighter in color basinward, changing to light red, pink, or greenish-gray claystone. The unit is a mixture of silt and clay derived from the north and west, and was deposited as a large delta in Fossil Lake.
An interesting aspect of the Mudstone Tongue is the presence of algal logs. These are cylinders of limestone that apparently formed as an encrustation of calcium carbonate around logs and branches that fell into the edge of the lake. The calcium carbonate resulted from the action of algae which grew around the log. Successive growths of algae resulted in successive layers of calcium carbonate being deposited.
This uppermost member of the Wasatch Formation is found mostly to the west and south of the monument. A few, small, isolated caps on the top of the Green River Formation east of Prow Point occur within the monument. These low hills, mere bumps, are remnants of a once more extensive distribution of the Bullpen Member.
Veatch (1907:99) originally considered what is now called the Bullpen Member as being the Bridger Formation, this was based mostly on the Bullpen’s position above the Green River Formation rather than on any lithologic resemblance to the Bridger. The Bullpen is much redder in color than the Bridger.
In early work Tracey and Oriel (1959:729) called these beds an upper tongue of the Wasatch and have since named them the Bullpen Member.
The Bullpen Member is lithologically very like the Main Body of the Wasatch and can be traced into the peripheral units of the Wasatch Formation.
The rocks included within the Bullpen Member are layered sequences of red, pink, gray, and green claystone and mudstone. Bentonite is present in some claystone beds and causes sloughing because of its property of expanding when wet. Some sandstones are present in the northern areas of the member’s distribution. Limestones are also present. They are thin and slabby and have a brown, white, or gray color. Some contain varying amounts of clay. A conglomerate is present in the upper part of the Bullpen toward the basin periphery and merges with the Tunp Member of the Wasatch Formation.
The Bullpen Member is conformable with the underlying Angelo Member of the Green River Formation. The contact is a transitional one reflecting a gradual change from the lake environment of the Green River Formation through a swamp environment to that of a flood plain. The light-colored, fine-grained shales, marlstones, and limestones of the Green River Formation grade upward into drabber, coarser grained clay and mudstones of the Bullpen Member as a reflection of this change of environments.
This peripheral unit of the Wasatch Formation was first described by Oriel and Tracey in 1970. It is not exposed in the monument, but forms a belt of outcrops around the edge of Fossil Basin and in channels cut at right angles to the basin edge.
The Tunp Member is seen to intertongue with nearly all of the Wasatch and Green River formations. The member grades laterally basinward from coarse, angular conglomerates to fine mudstone. Two limestone tongues of the Green River Formation are interbedded with the Tunp, indicating that at least twice there was major expansion of the lake.
Lithologically, the Tunp Member is a diamictite. This is a sedimentary rock with a wide range of particle sizes. The Tunp is best described as a red, conglomeratic, sandy mudstone with angular, poorly rounded to smooth, well-rounded clasts with a size range from pebble to boulder. There is no bedding or orientation of clasts.
The Tunp Member probably originated from mudflows and gravity sliding (Tracey et al. 1961). Environmental studies indicate that the area had a warm, humid climate. This would cause deep weathering of the surrounding slopes. This weathered material would then be a prime source to be acted upon by rain, gravity, and possibly earthquakes. These agents of deposition would then cause the material to flow and be deposited with little chance for sorting and rounding of the rock particles. The result was a belt of coarse, unsorted detritus on the basin edges now called the Tunp Member.
The exact age of the units within the Wasatch Formation can be determined only if fossils are present. None has been found in the basal conglomerates but it is believed to be earliest Eocene. The Lower Member is also not dated with fossils but is believed to be very early Eocene. A number of fossil mammals are known from the Main 13 Body of the Wasatch Formation. The lower part is early, early Eocene as demonstrated by the presence of a very primitive ungulate, Haplomylus speirianus. In the upper part of the Main Body another primitive ungulate, Hyopsodus browni, is found which indicates a mid to early Eocene age. The Mudstone and Sandstone tongues are not dated by means of fossils but the stratigraphic relationships indicate an early Eocene age. Fossil mollusks in the Bullpen Member are not completely diagnostic but suggest either a late early Eocene (Lostcabinian) (Fig. 6) or mid-Eocene age. The Tunp Member interfingering as it does with the entire Wasatch section represents all of early Eocene age.
The sediments that make up the Wasatch Formation in the Fossil Basin were deposited mainly by streams flowing into the basin from the surrounding uplands. Rock types are variable and for the most part individual types cannot be traced over large areas, a condition typical of fluvial environments. The mudstones and siltstones probably were deposited along flood plains, while the lenticular sands and conglomerates were laid down in stream channels. Where streams entered the lake tongues of deltaic deposits, sands or mudstones wedge into the Green River Lake sediments.
The reason for the red color of the Wasatch Formation is not fully known. According to Van Houten (1948), the red is most likely the result of tropical, red lateritic soils forming in the uplands and being deposited on flood plains. The oxidation of organic material in a humid, subtropical environment causes the red color. The bands of purple, gray, orange, and other colors are due to various stages in the reduction of the iron oxides in the rocks.
The presence of palms in the lake sediments and of bones of primates and crocodiles in both the lacustrine and fluvial sediments indicate a heavily forested, tropical environment.
The Green River Formation is easily recognized by its light color and continuous bedding, in strong contrast to the red, discontinuous, variegated Wasatch Formation below. The Green River Formation can be thought of as a gigantic lens of lacustrine sediments enclosed in the fluvial Wasatch Formation. The name Green River Formation is applied to all of the roughly contemporaneous deposits laid down in lakes of Utah, Colorado, and Wyoming. It is probable that at one time or another all of these lakes were connected.
Oriel and Tracey (1970) have divided the Green River Formation of the Fossil Basin into two members: the Fossil Butte Member and the Angelo Member.
The type section for this member is near the southeastern end of Fossil Butte within the monument boundaries. In the type area the Fossil Butte Member can be seen to consist of four lithologic units.
The lowermost unit is predominantly mudstone. It occurs in a sequence about 45 ft thick and contains light gray, fine-grained, calcareous mudstone and siltstone.
The next overlying unit is about 75 ft thick and is mainly a limestone unit. It consists of tan to gray limestone, shaly limestone, siltstone, and paper shale which weathers into thin, curled flakes. A yellow-brown mudstone tops this unit.
The third unit in the sequence is 45 ft thick and mainly composed of shales. These shales weather a buff color and are calcareous. Oil shale, organic rich paper shale, and marlstone comprise the actual layers in the unit. A few, thin ash beds are also present. This unit is the most significant one for Fossil Butte National Monument. About 10 ft below the top of this unit (about 155 ft above the Wasatch) is a bed of varved shales, one foot thick, that contains the fossil fish for which the monument was established.
The uppermost unit contains a number of beds of oil shale that are brown on a fresh surface but weather a grayish-white. An orange-yellow-weathering limestone caps the 40-ft thick upper unit. Ash beds are common and traceable over a wide area of Fossil Basin.
South of the monument the Sandstone Tongue of the Wasatch Formation wedges in between the lowermost unit and the overlying limestone unit of the Fossil Butte Member.
In the Fossil Butte Member an interesting sequence of facies changes can be seen which reflect lateral changes in the environment of Fossil Lake. In those areas where the member represents a deep-water environment, organically formed limestones and shales are predominant. Shoreward, these rocks grade into ostracodal limestone, gastrapodal limestone, and, closest to shore, algal limestone. Then a muddy, sandy beach facies is encountered where the Green River and Wasatch formations intergrade.
The contact with the underlying Wasatch is conformable and sharp. It is often marked by a bench or by slump blocks of Green River Formation.
The Fossil Butte Member may in some cases overlap the Wasatch Formation and be deposited on Paleozoic and Mesozoic rocks. This is a reflection of topographic relief during the Eocene.
The thickness of the Fossil Butte Member, as reported by Oriel and Tracey (1970:37), is from 208 to 269 ft.
In the northern part of Fossil Basin, the Mudstone Tongue of the Wasatch Formation separates the Angelo Member of the Green River Formation from the Fossil Butte Member. Further south, the two members rest directly on one another. The southern extent of the Angelo Member has not been determined as yet, but Oriel and Tracey (1970:32) believe it extends up to, and intertongues with, Wasatch conglomerates near the Uinta Mountains. Toward other edges of Fossil Basin, the Angelo Member thins and pinches out into the Wasatch Formation. The Bullpen Member of the Wasatch rests on top of the Angelo Member.
The Angelo Member consists of white to blue-white weathering limestone, marlstone, and mudstone. Some sandstone lenses, claystone, oil shale, and siliceous limestone are present as well. In general, Oriel and Tracey (1970) have found that buff limestones prevail to the north and white, siliceous limestone, to the south. Like the Fossil Butte Member, the Angelo Member shows a facies change from deep-water to shore.
This member forms the very uppermost, rounded slopes of Fossil Butte in contrast to the more vertical cliffs formed by the Fossil Butte Member.
The age of the Green River Formation is dated on its intertonguing relationships with the Wasatch Formation because the latter has datable mammal fossils. The well-preserved fossils of fish, leaves, and insects from the Green River are, unfortunately, of little value in dating the Green River Formation.
Although no Lostcabinian mammals are known from Fossil Basin, the Green River Formation is believed to be of that age (Gazin 1959; Schaeffer and Mangus 1965).
The Green River Formation, on the basis of its lithology and fossil content, is a fresh-water, lacustrine deposit. Clay and silt were dumped in Fossil Lake by streams. Most of this fine debris was deposited near the shore. Chemical and organic processes formed limestones and marlstones in the deeper central part of the lake.
Examination of the edge of the Green River Formation indicates that Fossil Lake expanded and contracted several times.
Fossil Lake was eventually filled in with chemical precipitates and deltaic deposits. The end of the lake was gradual as seen in the transitional and gradational Green River-Bullpen contact.
Although oil shale is not extensively developed in the Green River Formation of Fossil Basin, it does occur in small quantities.
Oil shale is a fine-grained sedimentary rock containing organic matter which was derived chiefly from aquatic organisms, waxy spores and pollen grains ... and of which a large portion is distillable into a liquid similar to petroleum. Despite the name, most rich beds of oil shale in the Green River Formation cannot be regarded strictly as shale. Instead, they are dolomitic marlstones rich in organic matter. Nevertheless, a few are shaly (Bradley 1964b:19).
Oil shale has a structureless ground mass that is yellowish-orange to reddish-orange in color. Pyrite crystals are found indicating a partially anaerobic or reducing environment. Pollen, waxy spores, filaments of algae, and other plant parts are preserved along with insects and larvae. The preservation is akin to mummification. Crystals of calcite, dolomite, or authigenic feldspar are also found in the oil shale (Bradley 1966).
The exact mode of origin of oil shale is not positively known because of a lack of a modern analogue for comparison. Oil shale probably originated as an organic ooze on the bottom of the Fossil Lake. This ooze was composed of the remains of phytoplankton, blue-green algae, zooplankton, bacteria, and some pollen and spores. The ooze was dense and uncompacted. Little clastic debris is found, either because the ooze accumulated in deep water or plants near the shore filtered out the debris.
Decay was reduced effectively in the ooze because of either an antibiotic in the ooze which inhibited bacteria of decay or the ooze accumulated in waters where anaerobic conditions prevented decay.
With time and the weight of overlying sediments, the ooze was compacted and most of its water driven off. Continuing pressure from compaction and heat generated by burial and compaction caused a variety of complex chemical reactions which converted the ooze into a petroleum product called kerogen. Kerogen is distillable and is the important constituent of oil shale.
An alternate hypothesis (Eugster and Surdam 1973), would have some oil shale forming in a desert-playa environment. This is based on geochemical evidence found in Gosiute Lake sediments to the east of Fossil Lake. There, certain minerals are found in association with some oil shale that could only have been deposited during periods of extreme evaporation and in a shallow lake. Much study is now being directed toward a solution to these problems.
The combustible quality of oil shale has been known for a long time. Many of the pioneers used it as a fuel for cooking and heat. Hayden (1871:142) wrote of how workmen on the Union Pacific accidently ignited the oil shale in a cut they were excavating. The burning shale provided enough light for night work.
Many of the shales of the Green River Formation appear to be varved. A varve consists of two layers, one 15 of calcium or magnesium carbonate and one of organic material. The limnological conditions that led to the formation of varves will be discussed in their proper place in the section on Paleoecology.
This is the middle formation of Veatch’s (1907) tripartite division of the Wasatch Group. It is now found to be the youngest formation in Fossil Basin. It is not exposed in Fossil Butte National Monument.
Oriel and Tracey (1970) have formally divided and named three members of the Fowkes Formation: a lower Sillem Member, a middle Bulldog Hollow Member, and an upper Gooseberry Member.
Like most of the Fowkes Formation, this sequence of rocks has been eroded extensively and is preserved as erosional remnants, where protected by faulting, in the western part of Fossil Basin.
The Sillem Member consists of a lower conglomeratic sequence with some sandstone and mudstone. The conglomerate contains well-rounded clasts of gray quartzite, chert, and Paleozoic limestone. The sandstone is light gray, calcareous to muddy, and coarse to medium-grained. The mudstone is pink, gray, or tan in color.
The upper part of the Sillem Member is a mudstone and claystone unit. It ranges in color from pink and yellow to gray and green. Some volcanic debris is found. There are also interbedded layers of marlstone and limestone. Some sandstone is present.
The Sillem Member is between 100 and 400 ft thick and most probably rests unconformably on the Bullpen Member of the Wasatch.
This middle member of the Fowkes Formation has the thickest and most extensive outcrops. The Bulldog Hollow Member is exposed along the west side of the basin.
Included rocks are green, white, and blue-green mudstone with ash beds, green and buff claystone, and tuffaceous, limy sandstone. A high percentage of the iron mineral, magnetite, occurs in the sandstone. Conglomerate occurs as lenses.
The Bulldog Hollow Member has a gradational contact with the underlying Sillem Member. The amount of volcanic material increases upward from the Sillem, indicating an increase in volcanic activity during the deposition of the Bulldog Hollow Member.
Oriel and Tracey (1970:55) place this uppermost member provisionally within the Fowkes Formation. Most of the Gooseberry Member is a puddingstone, a lithology with well-rounded, spherical pebbles in a marlstone, sandstone, or sandy limestone matrix. The pebbles are too rounded for the rock to be a diamictite, and too separated from each other to be called a conglomerate.
The nature of the Gooseberry-Bulldog Hollow contact is not completely known. It appears to be gradational in some areas and to be an angular unconformity in others.
Fossils date the Sillem and Bulldog Hollow members as middle Eocene in age. These fossils consist of ostracodes, gastropods, leaves, and vertebrates from the Bulldog Hollow Member (Nelson 1973). The Gooseberry Member has yielded a few vertebrate remains and is late Miocene or early Pliocene in age (Oriel and Tracey 1970).
The Fowkes Formation is an alluvial deposit, much like the Wasatch Formation. The chemical and climatic conditions of deposition were different from those of the Wasatch, and the extensive red-beds are not developed.
Small lakes were present in which limestone and marlstone accumulated. The puddingstone may be a mudflow. Volcanic activity left its record in the ash found in the Fowkes Formation.
In the past, the Fowkes Formation had a greater distribution. Postdepositional faulting down dropped parts of the Fowkes protecting them from subsequent erosion.
Rubey et al. (1968a, b) have mapped several forms of Quaternary deposits in Fossil Basin. These include stream alluvium, rock and landslide debris, river terraces, and gravels, all derived from local formations. These deposits are the work of water, wind, and ice acting in relatively Recent time.
The Fossil Basin is a small, linear and structurally controlled basin in the southeastern part of the Wyoming overthrust belt. This “overthrust belt” is represented by a number of small mountain ranges and high ridges formed by the “thrusting” of sedimentary rocks over other sedimentary rocks. Topographically, the Fossil Basin is bounded by the Crawford Mountains and Tunp Range on the west, by Oyster Ridge on the east, and by the Uinta Mountains on the south. The Crawford Mountains, Tunp Range, and Oyster Ridge (Fig. 2) are areas of high relief developed upon southerly extended salient ridges of deformed Paleozoic and Mesozoic strata. In the center of 16 the Fossil Basin, these earlier rocks are covered by a veneer of early Tertiary sediments. Superficially, the Fossil Basin appears to be a broad syncline with tilted beds dipping sharply or gently basinward from the basin margins. The Tertiary sedimentary cover, however, partially obscures what is a more complex structural history.
Following deposition of the Late Cretaceous Adaville Formation, the Fossil Basin was included in a period of intense structural deformation. This deformation was the result of compressional forces acting along a more or less east-west alignment. The strain, or the resolution of these forces, was developed along a north-south alignment or perpendicular to the compressional forces. As the stress level became too great, the rocks were first folded and then faulted. Initial faulting of the rocks relieved some of the stress; however, with continued application of compressive forces many stages of folding and faulting were generated. Because the alignment of the compressive forces remained about the same throughout deformation (i.e., east-west) and because the strongest relative compressive forces were from the west, successively younger folds and faults were generated in an eastward direction.
The structural evolution of the Fossil Butte area may be interpreted as follows from the schematic diagrams:
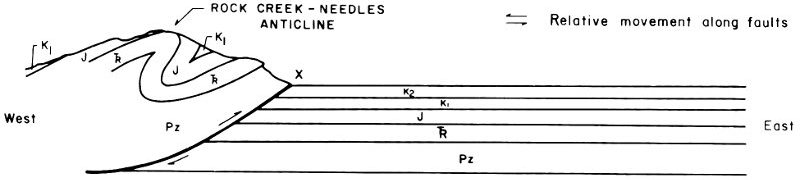
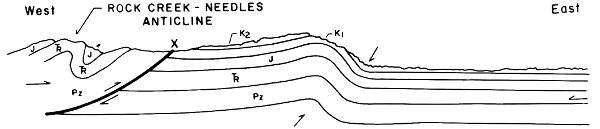
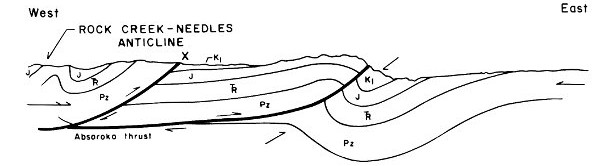
1. The Rock Creek-Needles Anticline is developed at the western edge of the Fossil Basin. The present topographic highs developed on this structure are the Crawford Mountains and Tunp Range (Fig. 9-B).
2. With continued application of compressional forces, Paleozoic and Mesozoic rocks to the east of the Rock Creek-Needles Anticline are folded (Fig. 9-C).
3. The fold becomes sharply asymmetrical and when the rocks can no longer accommodate the compressional forces by further folding, a low-angle fault is developed (Absaroka thrust fault). This fault probably began as a fracture parallel to bedding which rose to topographic surface at a low angle at the point of greatest strain in the fold. Most likely, this fracture was an eastward extension of the fault underlying the Rock Creek-Needles Anticline, as depicted in Fig. 9-D.
4. The rocks to the west of the newly developed fault overrode those to the east. This was accompanied by further downwarping in the syncline to the east of the fault and the erosion of uplifted sedimentary rocks on the west side of the fault (Fig. 9-E).
5. Another cycle of folding and faulting was initiated to the east of the Absaroka fault. This post-Absaroka deformation instigated further downwarping of the Fossil Basin area between the Tunp-Crawford Mountains and Oyster Ridge. Tertiary sediments began to accumulate in the Fossil Basin (Fig. 9-F).
Fig. 9. Structural development of the Fossil Butte area.
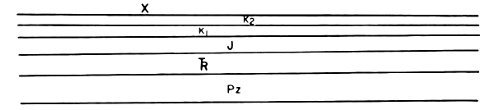
A. Pre-deformation: Paleozoic & Mesozoic Formations
B. Pre-Absaroka Deformation
C. Development of the Absaroka thrust fault: folding

D. Development of the Absaroka thrust fault: faulting
E. Development of the Absaroka thrust fault: faulting

F. Post-Absaroka Thrust Deformation

G. Simplified structure section through present-day Fossil Basin
(Oyster Ridge is off the diagram to the East)
Structural interpretation by Dr. D.L. Blackstone
In early Eocene times, the effects of continued downwarping allowed Fossil Lake to form in the Fossil Basin. By the late Eocene, however, basinal subsidence could no longer keep pace with deposition and the dominantly lacustrine (lake) sedimentation was replaced by a fluviatile (riverine) sedimentary regime. Deposition of fluviatile sediments probably continued into the later Tertiary when, in the late Pliocene, regional uplift of the Rocky Mountain interior reversed the sedimentary cycle from depositional to erosional. The last 3 or 4 million years of geologic history have witnessed the excavation of much of the Tertiary sedimentary fill from the Fossil Basin. Fossil Butte and the Ham’s Fork Plateau to the northeast of the butte are high erosional remnants of this early Tertiary basin fill. The traces of the Absaroka and other thrust faults are buried beneath these remaining Tertiary deposits in the Fossil Basin.
While most of this complex structural history took place before the Tertiary sediments were deposited, some deformation continued into the early Eocene. Near Prow Point, for example, is a fault that developed during early Eocene time. This can be noted in the very northwest corner of the monument, just to the west of Prow Point (Fig. 6). Here, the Lower Member of the Wasatch Formation on the west is faulted against the Main Body on the east (USDI 1964). This fault also is responsible for elevating the Thaynes Limestone so it is exposed (Fig. 10).

Fig. 10. Structural relation of older rocks to Wasatch and Green River formations near Prow Point in the northwest corner of the monument (from U.S. Department of Interior 1964).
Fossils of plants, invertebrates, and vertebrates are abundant in rocks exposed in the Fossil Basin. For example, Oyster Ridge received its name because of the well-preserved and abundant fossil oysters found there. Of course, Fossil Butte and Fossil Butte National Monument were so named because of the beautifully preserved fossil fish. A brief review of the main groups of fossils to be found in the basin follows.
Many of the sediments in the Fossil Basin contain an abundance of fossil plants. These consist of pollen, spores, wood, and leaf impressions. Those of the Green River Formation are best preserved and have received the most study.
Brown (1929, 1934) and Lesquereux (1883) have presented the best information on the flora of the Green River Formation. Much of the present knowledge is based on their work.
Brown (1929, 1934) concluded that the Green River flora is a mixed forest type. It contains some plants common to warm, wet lowlands and others adapted to cooler, drier uplands. Pollen studies support Brown’s original concept that several plant communities, including mountain communities, existed around Fossil Lake.
The flora was situated in an inland, mountain basin. Fossil Lake was in the middle of the basin. Swamps and flood plains bordered the lake, while the flanking ridges and mountains provided altitudinal variations in the plant assemblage.
Spruce, fir, and pine are indicated by fossil pollen. The presence of these conifers suggest that at least some of the nearby mountains rose to heights of 6000-8000 ft.
Leaf imprints indicate that deciduous trees occurred on the lower slopes. Oak, elm, maple, and beech were common forms.
Closer to the lake where it was wetter and warmer there were willows, laurel, and fig trees. Palms grew in the sandier soils (Fig. 11). Cypress grew in quiet embayments of Fossil Lake. The shoreline plant assemblage had a rather subtropical appearance.

Fig. 11. Impression of a palm frond from the fish beds of Fossil Basin. Width of original, 160 mm. Collection of University of Wyoming.
Ground cover was provided by a mold of dead leaves, holly, liverworts, mosses, and ferns (Fig. 12). Prairie-type grasses are not present and large savanna areas probably were not developed.

Fig. 12. A fern leaf from the Green River Shales of the Green River Basin. Width of original, 4 cm. Collection of University of Wyoming.
In the lake itself small, one-celled algae floated about in the waves and currents. Bacteria grew on the lake bottom and in the water and helped to contribute to the organic ooze that built up on the bottom. Fungi, wonderfully preserved in oil shales, are further indicators that Fossil Lake was a fresh-water lake and deep enough to have depths below the zone of sunlight penetration (Bradley 1964a). Reeds, rushes, and similar lake-shore plants grew in shallow, near-shore waters.
The large amount of organic material in the rocks is evidence of the prolific microscopic plant growth in the lake.
Occasional dry periods in the Fossil Basin are indicated by fossils of a plant called Ephedra. This plant had leaves with a thick, waxy cuticle to prevent desiccation during drought. The absence of saline minerals in the Green River Formation in Fossil Basin suggests that these 20 arid periods were never of the intensity that occurred in the Green River Basin to the east.
The flora of Fossil Basin, especially that from the Green River Formation, is usually compared to that of the present-day Gulf Coast. Seaward from the Appalachians, the composition of the flora is very similar to that of the Green River. Spruce, pine, and fir grow in the Appalachians at elevations of 6000 or more feet, supporting the idea that similar elevations existed near Fossil Lake and that a general Gulf Coast climate prevailed in Fossil Basin.
Molluscs, ostracodes, and insects comprise the invertebrate fauna of Fossil Basin.
A large number of marine forms are found in the earlier marine deposits, while fresh-water and terrestrial forms are common in the Tertiary units. It is these later forms which are considered here.
The fresh-water and terrestrial molluscs from the Wasatch and Green River formations include bivalves (clams) and gastropods (snails).
Aquatic clams from the Green River Formation include Unio and Plesiellipta. Pisidium and Sphaerium are common in the Wasatch Formation.
Land snails from the Wasatch include such genera as Grangerella, Discus, Oreoconus, and Glypterpres. Oreoconus is also known from the Green River Formation.
Fresh-water snails are very abundant from both the Wasatch and Green River formations. Physa, Planorbis, Elimia, Bioniphalaria, Virpains, and Omalodiscus are from the Wasatch. Physa, Elimia, Diomphalaria, and Goniobasis are recorded from the lacustrine Green River sediments. In some areas of the Green River Formation, these snail fossils are so abundant that they form a major constituent of the limestones. The presence of land snails in the Green River and fresh-water molluscs in the Wasatch is evidence that the two formations are not totally representative of a single environment, but represent several related ones.
Most of these fresh-water snails were herbivorous and frequented the shallow, well-lighted areas of Fossil Lake which had a good growth of plants and algae. This naturally occurred close to shore, hence the gastropodal limestones reported by Bradley (1926) and Oriel and Tracey (1970) are interpreted as being near-shore facies.
Ostracodes are extremely small arthropods that produce a two-piece, hinged shell within which they live. Pseudocypus is the most common form. Like the snails, the ostracodes were so abundant in areas that they, too, formed a large part of some Green River Formation limestones. And like the snails they lived in a near-shore environment.
Within the laminated shales of the Green River Formation is preserved an abundant and diverse assemblage of fossil insects (Fig. 13).
These insects are important because they demonstrate that many modern families and even genera were in existence during the Eocene. In contrast to this, knowledge of insect evolution prior to the Eocene is rather poorly known. Scudder (1890) and Cockerell (1920) have described most of the Green River insects.
Beetles are the most common forms, followed by dragonflies. Maggots and larvae of flies are commonly preserved.
Except for these insects preserved as whole "mummies" in the oil shales, the majority of the insect fossils are preserved as distilled outlines. This distillation process resulted from the weight and heat of overlying sediments of driving off the volatile substances from the buried insects, leaving a hydrocarbon outline. The process is so precise that the fine hairs and wing veins and even body-color markings are preserved.
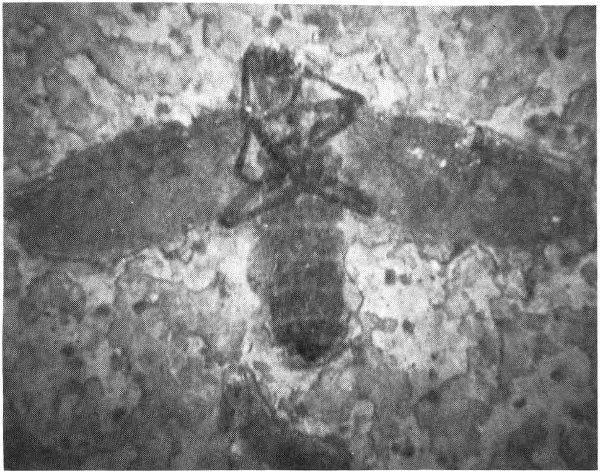
Fig. 13. An impression of a horse fly from the Fossil Lake deposits. Length of original, 12 mm. Collection of University of Wyoming.
The single factor that has made Fossil Butte world famous is the occurrence of literally millions of beautifully preserved fossil fish. In one layer approximately 14 inches thick, the fish are nearly black and preserved in a nearly white shale. Most of the fish are perfectly preserved, retaining every detail of the skeleton and even undisturbed scales. Fish from Fossil Butte have been collected and sold since the 1870s and may be found on sales counters, in museums, and on living room walls throughout the world.
The fossil fish from the Green River Formation have a history of discovery stretching back to 1856. At that time Leidy (1856:256) described a fish given to him by a Dr. 21 John Evans. Leidy called the fish Clupea humilis, a type of herring. Exactly where Dr. Evans obtained the specimen is not known. He passed through the area of the Green River Formation several times and could have collected it during any one of those trips (Knight 1955:12).
During the construction of the Union Pacific Railroad in the 1860s, workmen blasting a cut through Green River shales, about 2 miles west of Green River, Wyoming, came upon remains of well-preserved fish. Hayden (1871:742) first mentions this cut as "Petrified Fish Cut." Many of the fish from this cut were given to Cope who described them in Hayden’s (1871) report. Insects, plants, and a bird feather were also obtained from "Petrified Fish Cut" at this time.
Sometime in the 1870s the first fish were obtained from Fossil Butte itself. Cope (1877, 1878) described fish from the Green River Formation which may be from Fossil Butte. His locality is "nearer the mainline of Wasatch Mountains" (Cope 1877:807). This may be Fossil Butte. His 1878 locality information is just as poor. He does mention a specimen of Priscacara peali given to him by A. C. Peale. Since it was Peale who first wrote about Fossil Butte, the specimen may have come from the butte.
Peale (1879) first mentioned the quarrying of fish at Fossil Butte, but he had no specific date as to the discovery of the butte or the beginning of quarrying of fish.
Cope (1884) published two large volumes about the Tertiary vertebrates of the West. Here he mentioned three fossil localities for his Green River Fish: "Petrified Fish Cut," "The Mouth of Labarge Creek," and "Twin Creek." Twin Creek is now known as the Fossil Butte site.
Since that time, the quarries at Fossil Butte have been extensively worked, mostly by commercial collectors.
The fish from Fossil Butte form an unusual array of genera (Table 1) unlike any now found living together. Included in the assemblage are forms usually found in marine waters and tropical, fresh-water fish (Schaeffer and Mangus 1965).
More than 40 species of fish have been described from the Green River shales, but authorities do not agree on how many of these are valid. Because of this, only super specific groups will be considered in the following discussion.

Fig. 14. Xiphotrygon, a fossil ray. These forms are present but rather rare in the fish deposits of Fossil Butte. Length of original, 39 cm. Collection of University of Wyoming.
One of the strangest forms to come from Fossil Butte is that of Xiphotrygon (Fig. 14). This is not a true fish, but is a sting ray, related to sharks. Closely related forms still survive, with little change, in coastal waters in many parts of the world. Like sharks, sting rays have a skeleton composed of cartilage. Normally, cartilage is not preserved as a fossil as it disintegrates readily. The excellent skeletons of Xiphotrygon are good evidence of how well the shales of the Green River Formation preserve the fossils. Xiphotrygon 22 is characterized by being shortened from back to front and flattened from top to bottom. It possesses a long, whip-like tail and bears enormous, flat pectoral (anterior) fins. The mouth is on the underside of the body indicating that Xiphotrygon, like its modern relatives, fed along the bottom of the lake. The strong flattened teeth form something like a pavement in the mouth, suggesting that it fed on clams and other hard-shelled invertebrates. Close relatives of Xiphotrygon live mostly in marine waters, but occasionally enter fresh water.
The paddlefish, Polyodon, is rare but present at Fossil Butte. Similar forms still survive in China and the United States. The prominent features of Polyodon include a virtually scaleless body, a long, depressed snout, and long gill rakers. The skeleton, unlike that of most modern fish, is composed of cartilage, hence skeletons of Polyodon are rare. The difference between the modern and Eocene paddlefish are so slight that the two probably filled similar or the same ecologic niches. This consists of feeding on plankton and other small organisms which can be obtained from the water. The long gill rakers would provide a large surface area on which food could be trapped as the water passed out through the gill slits. The large mouth would also provide a large surface area for catching and trapping food.
Quite common to streams and rivers of North America from the Cretaceous to the Recent is the garfish, Lepisosteus (Fig. 15). The most striking feature of the gar is its diamond-shaped scales, which are extremely hard and shiny. The body is long, essentially of the same depth throughout, and ending in a broad-based tail that is slightly asymmetrical. The mouth is armed with many small, sharp teeth for catching prey. One specimen over 5 ft long has been found in the Fossil Butte area.

Fig. 15. The fossil gar Lepisosteus. Only a few gar have been found at Fossil Butte itself. Length of original, 70 cm. Across the highway from the butte, one specimen over 5 ft long was found by Carl Ulrich. Collection of University of Wyoming.
Fossils of the bowfin Amia (Fig. 16) are recorded from the Green River shales at Fossil Butte and in stream deposits from Cretaceous to Recent. This fish is still another “living fossil.” Amia is a nocturnal predator. Presumably, the Eocene Amia had similar habits. The body in Amia has become elongate, as has the dorsal fin. The thickness of the scales has been reduced, but they are still rather heavy and cover the body. Specimens are rather large and deep-bodied. The tail fin is nearly symmetrical above and below. Amia is rare in the Fossil Butte fish quarries.
By far the most abundant fish from Fossil Butte is the genus Knightia (Fig. 17, 18). This fish is a member of the Clupeidae, a family that includes modern herrings, shad, and sardines. The tail fin is small relative to the rest of the body. A row of modified scales extends on the back from the skull to the dorsal fin. Possibly they aided in streamlining the fish. Knightia apparently fed on the large amounts of plankton, especially algae, which lived in the waters of Fossil Lake. Fossils of Knightia are also known from Eocene lake sediments in South America.
Knightia appears to have been susceptible to mass mortalities. Some layers of shale at Fossil Butte contain literally hundreds of thousands of these fish that must have suffered catastrophic mass mortality. Possible reasons for the obvious sudden death of so many fish will be discussed in the section on paleoenvironment and taphonomy.
Another Eocene relative of the herring found at Fossil Butte is Diplomystus (Fig. 19, 20). Like Knightia, Diplomystus bears a row of modified scales on its back. Diplomystus is the second most abundant fish at Fossil Butte. The jaw in Diplomystus has a rather pronounced oblique angle to it. The deepest portion of the body is directly behind the gill region, with a continuous narrowing of the body toward the tail region. Diplomystus is known from Cretaceous rocks of Brazil and Syria and from Tertiary sediments in Brazil and West Africa (Schaeffer and Mangus 1965). Close relatives of Diplomystus now inhabit the waters off the coasts of Peru and eastern Australia.

Fig. 16. Amia, the bowfin. This too is rare but present at Fossil Butte. Length of original, 67 cm. Collection of American Museum of Natural History.

Fig. 17. The fossil herring, Knightia. This fish is by far the most abundant in the fish beds at Fossil Butte and at least in one layer was involved in a catastrophic mass mortality. Length of each fish, 13 cm. Collection of University of Wyoming.

Fig. 18. The herring, Knightia.

Fig. 19. Diplomystus, another type of herring, is the second most abundant fish at Fossil Butte. Length of original, 12 cm. Collection of University of Wyoming.

Fig. 20. A large Diplomystus with four small bass, Priscacara. Length of Diplomystus, 45 cm. Collection of University of Wyoming.
Phareodus (Fig. 21) belongs to a family of fish, the Osteoglossidae, which began as marine forms and has since become a fresh-water family. At present, they are restricted to tropical rivers and lakes of South America, Africa, and Australia. In the past they had a much greater distribution, as its occurrence at Fossil Butte indicates, including North America. It is a deep-bodied fish with a large head. The anal and dorsal fins are close to the tail fins. The many sharp teeth in the mouth attest to the carnivorous habits of Phareodus.
The family Gonorhynchidae, the living sandfish of the Indo-Pacific area, is represented at Fossil Butte by Notogoneus (Fig. 22). Notogoneus is very long-bodied and quite slender. The body width does not vary much from head to tail, although narrowing does occur in the tail region. The tail fin is symmetrical, forming a good rudder. The anal, dorsal, pectoral, and pelvic fins are all small. The body is covered with small, elongate scales. Notogoneus was apparently a bottom feeder, living on small organisms picked off or out of the bottom of Fossil Lake.
The Recent catfish, Ameuirus, has also been found at Fossil Butte in the Green River shales. The form of the Eocene catfish was very much like the Recent one. The habits of the Eocene catfish were probably like those of the modern Ameuirus, i.e., adaptations to a scavenger-type existence.
Rather common in the sediments of Lake Gosiute, but not yet known from the Fossil Basin, is Erismatopterus, a member of the family Aphredederidae or pirate perches. This small, elongate fish has few distinguishing characters, the rounded front edge of the skull being most distinguishable.
Asineops is an Eocene relative of Erismatopterus and also one of the pirate perches. Asineops is, however, placed in a different family, the Asineopidae. Asineops, like Erismatopterus, is a rather plain-looking fish. The dorsal fin is long relative to the rest of the body. The body is slightly deeper behind the gills than elsewhere, giving Asineops a rather common appearance.
The Percidae, or perches, are represented at Fossil Butte by Mioplosus (Fig. 23). Mioplosus, as indicated by its well-developed teeth, was a carnivore that probably preyed on its piscine relatives in Fossil Lake. The large head blends in well with a strongly built, long body. The 24 anal and dorsal fins are subequal in size and positioned opposite each other. The tail fin is large and fan-shaped. It is easily identified by the presence of two dorsal fins.

Fig. 21. A large Phareodus, fairly common in the Fossil Butte fish beds. Length of original, 54 cm. Collection of University of Wyoming.
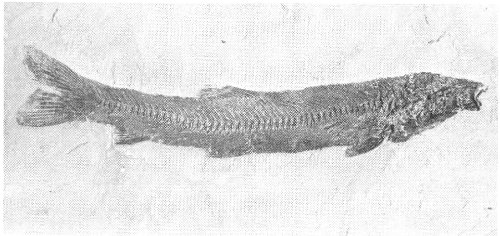
Fig. 22. Notogoneus, a long, slender bottom-feeder, occasionally found at Fossil Butte. Length of original, 56 cm. Collection of University of Wyoming.

Fig. 23. Mioplosus, a fairly common perch from Fossil Butte. Length of original, 30 cm. Collection of American Museum of Natural History.

Fig. 24. Priscacara, a small bass that is common in the Fossil Butte fish beds. Length of original, 13 cm. Collection of University of Wyoming.
The one genus for which no close living relatives can be found is Priscacara (Fig. 24). The family Priscacaridae is thought to be related to the bass. The family is known only from the Eocene. The size of Priscacara is extremely variable. The body is deep and nearly round like that of the common sunfish, the head blending in well with the body and its contours. The most distinguishing feature of Priscacara is the series of strong, stout spines supporting the anal and dorsal fins. These spines may have acted as cut-waters for better swimming and/or they may have protected Priscacara from its more voracious relatives.
To date, no amphibian fossils are known from Fossil Butte. They must have been present, as this class of animals is recorded from Devonian rocks more than 350 million years old and from rocks younger than those at Fossil Butte. Living amphibians are typified by frogs, toads, and salamanders. The lack of amphibian remains at Fossil Butte is more likely due to nondiscovery than nonexistence.
Reptile fossils are very abundant in the Green River and Wasatch formations. Also a jaw of the three-horned Triceratops is known from the Cretaceous part of the Evanston Formation.
Small lizards are normally represented only by jaw fragments and vertebrae. Their remains are most often found by a process of washing and screening of the sediments containing the fossils. Snakes are also represented in the Green River Formation. Remains of these animals are also often restricted to jaws and vertebrae. A complete skeleton of a fossil boa, Boavus idelmani, has been recovered from the Green River Formation and is the most complete fossil snake from North America (Schaeffer and Mangus 1965).
Turtles are among the most abundant reptile fossils. They are usually found as isolated shell plates, although many complete shells are known. The turtles Trionyx (Fig. 25), Emys, Baena, and Notomorpha are known from the Fossil Basin.
The largest Eocene reptiles were the crocodiles (Fig. 26). The presence of these predaceous animals is further evidence for a prevailing warm and humid climate during the Eocene times since their tolerance ranges include only comparable conditions at the present time. A few incomplete specimens have been recovered from near the monument.
Birds, which first appeared in Jurassic time, are rare as fossils in rocks of any age because their skeletal remains are exceedingly fragile. Their fossilization requires special conditions because many of their bones 25 are hollow as a weight-reducing adaptation for flight. These hollow bones are especially susceptible to breakage and poor preservation. Scattered remains of birds, mostly limb bones, are found in the Green River sediments. Feather imprints have also been recovered. A complete skeleton of Gallinuloides, a bird related to the South American fowl-like chachalaca, is one of the most significant finds from the Green River Formation. Recently, Brodkorb (1970) described the wings of puff birds similar to those now living in South America from near Fossil Butte.
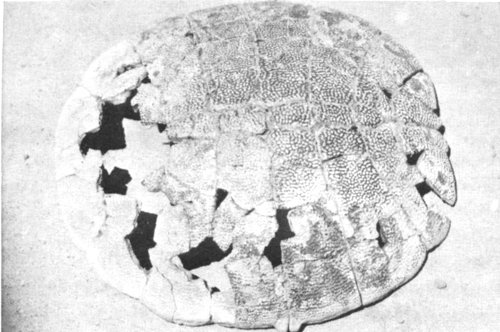
Fig. 25. An aquatic turtle, Trionyx, from the shore-line deposits of Lake Gosiute. Several turtles have been found in the fish beds near Fossil Butte.

Fig. 26. The skull of a large crocodile from near the shoreline of Lake Gosiute. Several crocodile fragments have been found at Fossil Butte beds. Length of original, 75 cm. Collection of University of Wyoming.
In the adjacent Green River Basin to the east, paleontologists from the University of Wyoming have found nesting grounds of Eocene flamingos in slightly younger sediments of the Green River Formation. These “rookeries” have yielded many hundreds of fossil bones of this curious bird. Since these bird fossils were found proximal to Fossil Butte, they demonstrate the near subtropical nature of the environment in the Fossil Butte area in Green River times.
The rocks of the Evanston, Wasatch, and Fowkes formations contain various fossilized remains of mammals which once inhabited the Fossil Basin.
Mammal fossils are highly significant since they are used extensively for correlation and dating of the Tertiary rocks of the West. Fossils which are readily recognizable, limited in time, and widely distributed geographically are called “index fossils.” These fossils are restricted to a particular time horizon in the rock. In this way, rocks of unknown age which are found to contain index fossils can be dated relatively and hence correlated with other fossil-bearing rocks. The small condylarth Haplomylus (Fig. 27) is a very early Eocene index fossil. Its presence in certain sediments in the Fossil Basin shows conclusively that these sediments were laid down in earliest Eocene times.
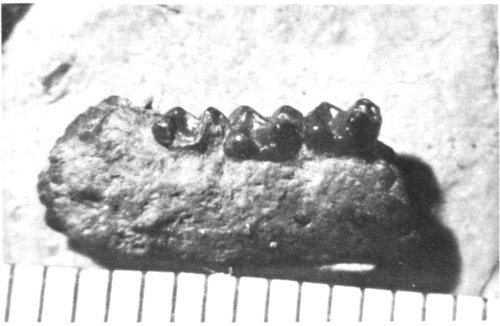
Fig. 27. A jaw of the primitive mammal Haplomylus. Length of original, 1.3 cm. Collection of University of Wyoming.
The mammal fossils found in Fossil Basin are also important in documenting the temporal changes in evolution and environment of the biotic community. The picture of Eocene Wyoming drawn through interpretation of these fossils is vastly different than that of the present day.
Mammals have a long evolutionary history that began over 200 million years ago in the Triassic period. At that time mammals had only just evolved from their reptilian ancestors. These earliest mammals were small, furtive creatures. However, with the extinction of the dinosaurs some 65 million years ago at the end of the Cretaceous, the mammals were able to diversify rapidly and fill the empty ecologic niches that the dinosaurs once occupied.
This filling of ecologic niches voided by the dinosaurs coupled with the spreading of mammals into hitherto unoccupied niches resulted in the development of a large variety of mammals. Some became extinct, others were the ancestors of modern mammals. Fossils of mammals found in sediments in the Fossil Basin are proof of the extent of this Late Cretaceous-early Tertiary mammal radiation.
Early mammals bore little resemblance to their later descendants. General trends in mammalian evolution were increase in size and the tendency to become more “modern”-looking through adaptation to changing Tertiary environments.
Most mammal fossils are extremely fragmentary. Teeth are most often preserved as they are the hardest part of the skeleton and therefore most resistant to wear and breakage. Thus, it often happens that the knowledge of a particular fossil mammal is derived entirely from its preserved dentition. This has obvious limitations for the completeness of our understanding about mammals in question.
The Paleocene portion of the Evanston Formation has yielded mammals of Torrejonian and Tiffanian age (Fig. 6).
The Torrejonian assemblage (middle Paleocene) (Gazin 1969) is small and poorly preserved. Its main significance is that the fauna records a definite time interval for the Evanston Formation. Gazin’s work has demonstrated this fauna to be intermediate in composition between similarly aged faunas recovered from sites to the north and south.
The major faunal elements are insectivores, primates, condylarths (primitive ungulates), and multituberculates (a type of extinct rodent-like mammal, with no known descendants).
The poorly preserved condition of these fossils makes discussion impossible except to note that they extend the known time range of mammalian habitation in the Fossil Basin.
The younger Tiffanian fauna (late Paleocene) (Gazin 1956) is also known from largely fragmentary remains. The multituberculate mammal Ptilodus, also present in Torrejonian deposits, is the most “primitive” mammal in the fauna. The multituberculates first appeared in the Jurassic (Fig. 4), and are therefore the oldest lineage of mammals that survived into Tertiary times. The molar teeth of these animals are characterized by multiple cusps arranged in parallel rows. Ptilodus, like most other multituberculates, was specialized in that its fourth lower premolars were expanded into shearing blades, the function of which is not known. Ptilodus was in many respects similar to rodents in the development of procumbant incisors and in general build and appearance. Multituberculates are, however, not related to rodents except that both are mammals. The differentiation of rodents in the late Paleocene and their diversification during the Eocene probably presented strong competition for the multituberculates who were unable to compete successfully and so became extinct.
The primates were represented by Plesiadapsis. This animal was about the size of a squirrel and had chisel-like incisors. Flesh reconstructions of Plesiadapsis, based on skeletal remains, show this primate to look much like a rodent with a long-snouted skull, clawed feet, long body, and tail. There is conceivable relationship between the rodents and Plesiadapsis-like primates.
The dominant Tiffanian herbivorous mammals were members of the Order Condylarthra. These primitive ungulates were diverse in both size and appearance. It is probable that later modern ungulates (Artiodactyla, Perissodactyla) evolved from the Condylarthra; however, there are no surviving members of this group. The Paleocene condylarths were mostly small animals and include such forms as Haplaletes, Litomylus, and Gidleyina. These mammals had somewhat insectivore-like teeth, and may have bridged the gap between insectivores and archaic hoofed mammals. One of the larger Tiffanian condylarths was Phenacodus. This mammal had a long, massive skull and a long, probably flexible body. The limbs were stout and short. The body ended in a long tail. Small hooves were present on all digits, which numbered five per foot. The length of the largest Phenacodus was about 6 ft.
Carnivorous and omnivorous condylarthra were also common. Thryptacodon and Claenodon are two types known from the Evanston Formation in the Fossil Basin.
The true carnivores (Order Carnivora) were also present. The Paleocene carnivores were small and possibly arboreal in habit. The stem carnivore stock was represented by the family Miacidae. In the Fossil Basin the miacid Didymictis was common. In Didymictis, the mouth was armed with small, sharp teeth. Most important, the typical carnivoran carnassial (shearing) teeth had developed. Among carnivores, from Paleocene to Recent, the shearing blades developed on the last upper premolar and the first lower molars. Various kinds of shearing teeth have evolved in other mammal lineages.
Mammals from the Wasatch Formation in Fossil Basin represent the Greybullian and Lysitean provincial ages (see Fig. 6). These fossils are of earliest Eocene and mid to early Eocene age. Fossil localities in the Wasatch Formation of southwestern Wyoming are Knight Station (partly Lysitean), at Elk Mountain (Greybullian), and at Fossil Butte itself. The age of the Wasatch Formation at Fossil Butte is Lysitean.
The insectivores are represented by Diacodon, a small mammal known only from fragmentary material.
Primates also continued to expand and diversify in Wasatchian time. Microsyops was a very unusual type that belongs to the extinct family Microsyopidae. The teeth were sharp and adapted for eating fruit and/or insects. Microsyops was about the size of a rat or slightly larger. The microsyopid primates were limited to the Paleocene and Eocene and left no later known descendants.
Another group of primates is represented by Pelycodus. This was a small lemur-like animal that may possibly have been an ancestor of higher primates (monkeys, apes, and men). Pelycodus, like Microsyops, was an arboreal (tree-dwelling) animal which inhabited the forests of the Fossil Basin during the early Eocene.
The Order Taeniodonta is a strange and little known group of mammals. In the sediments at Fossil Butte the taeniodonts are represented by Ectoganus. This was a moderately large animal. The skeletal adaptations were similar to those of moles, but there was no actual relationship between the two groups. The front feet and legs were robust, and bore large claws. Ectoganus probably used these structures to grub for food, possibly roots. The incisor teeth were rootless and persistently growing. The tooth enamel was restricted to two bands on either side of the teeth. The single pair of upper and lower incisors was greatly enlarged. The taeniodonts became extinct in the Eocene and left no descendants.
Equally as strange a group of animals as the taeniodonts are mammals in the Order Tillodonta. These were herbivorous animals, some of which became quite large. They have many rodent-like characters, but were not related to rodents. In many respects, the morphology of rodents seems to have lent itself to convergence. Esthonyx was the most common tillodont from the Fossil Basin. Prominent, rootless incisors were a characteristic of the later members of the group. These incisors were chisel-like, as in rodents. The molars were unusual and bore certain resemblances to primitive carnivoran and insectivoran teeth. The tillodonts, however, bore uncertain relationships to other mammals and left no descendants beyond the middle Eocene.
In comparison with recent faunas, Paleocene rodents were rare faunal elements. Two early rodents were found in the Fossil Basin Wasatch Formation:
Paramys was relatively unspecialized but was diverse in the number of species. One species of Paramys was about 2 ft long and possessed a long tail. The body was long and slender. The skull was small and rather squirrel-like. The cheek teeth were quadrate and had low, blunt cusps. The typical, single pair of chisel incisors was present both on lower and upper jaws. Paramys was probably an arboreal form that did not look too unlike a modern squirrel and may have had a similar mode of life. Another rodent, Reithroparamys, was somewhat similar to Paramys in size and appearance. The differences between the two animals were mainly in the teeth and hind limbs. The latter showed certain very minor structural modifications that suggested a saltatorial (jumping) mode of locomotion for Reithroparamys.
Bats (Order Chiroptera) are extremely rare as fossils because their volant and cave-dwelling habitat as well as a fragile skeleton did not lend their remains to preservation. From the Green River Shales, from near Fossil Butte, a complete articulated skeleton of a bat has been found (Jepsen 1966). This animal, named Caronycteris index, was small and generalized in form. Superficially, it looked much like a typical brown bat. This fossil is important because it demonstrates that the bats had already become good fliers by early Eocene time.
The condylarths were prominent in the early Eocene but diminished in importance following this time. Phenacodus was still extant; however, it was replaced in importance by the smaller Hyopsodus.
Hyopsodus is perhaps the most commonly recovered Eocene fossil mammal. This animal was small and long-bodied and retained a more-or-less generalized structural pattern. In some respects Hyopsodus was similar to both insectivores and primates and was at one time or another regarded as belonging to either of these two orders.
Some of the later condylarths paralleled the more advanced ungulates. Indeed, the Perissodactyla (horses, tapirs, rhinos) and Artiodactyla (bovids, deer, pigs, sheep, etc.) were derived from early Tertiary condylarths. Meniscotherium was one such advanced condylarth. The cusps on its molars, instead of being blunt points, had developed into crescentic patterns, somewhat like those in deer or camels. The relationship of Meniscotherium to modern ungulates is only one of parallel dental development. Meniscotherium was a medium-sized animal, but about the same build and size as that of a cocker spaniel. Meniscotherium, however, had hoofs on its toes and was a forest-living browser.
The condylarth Haplomylus is found at the Elk Mountain Wasatchian locality. This small animal is typical of the Greybullian level of faunal organization (earliest Eocene). A supposed carnivorous condylarth was Pachyaena. Pachyaena was a fairly large animal for its time. The strong, robust jaws and teeth and heavy build suggest that Pachyaena was a predatory animal suited for preying on larger animals such as the amphibious pantodont Coryphodon. The affinities of Pachyaena to the condylarths are uncertain and are based primarily on foot structure.
The miacid carnivores were abundant and varied. Didymictis persisted and both Vassacyon and Vulpavus continued the arboreal, forest-dwelling habits of their Paleocene precursors.
The main carnivorous animals of the Eocene, however, were the creodonts (Order Creodonta) which were essentially early experimenters in carnivorous habits. As such they were diversely specialized. Proviverra (=Sinopa) was one of the smaller, predaceous creodonts that inhabited Fossil Basin during Eocene times.
One of the most characteristic fossil mammals from the 28 early Eocene of Fossil Basin was Coryphodon. A specimen of Coryphodon was also the first mammal fossil to be found in the Fossil Basin. The order of mammals to which Coryphodon belongs, the Pantodonta, was a group of large and heavily built, herbivorous mammals. Coryphodon was about the size of a small rhinoceros. The skull was large and heavily built. The brain, however, was small. The canine teeth were somewhat enlarged and strong. Stout limbs supported the bulky body. In habits, Coryphodon was possibly semi-aquatic.
The modern ungulates, Order Perissodactyla (odd-toed ungulates) and Order Artiodactyla (even-toed ungulates), first appeared in the early Eocene. The earliest members of both orders were small and relatively unspecialized.
The earliest perissodactyls were the horses and tapirs. The earliest horses were represented by Hyracotherium. This small, slender, and lightly built horse was about the size of a fox terrier. The front feet had four toes, and the hind feet three toes. The teeth suggest that this animal was probably a browser, living in the forests. In the Fossil Basin, tapirs were known from deposits of Lysitean (mid to early Eocene) age of which Heptodon was representative. The tapirs now live in the tropics of Malaysia and Latin America. Their present habitat suggests a similar environment may have been favored by the Eocene tapirs of the Fossil Basin.
The first Artiodactyls were pig-like forms, although they are only distantly related to true suids. Protodichobune was small and may have looked much like some of the early Perissodactyls. There are, however, a complex of structural features which serve to distinguish these mammalian orders from each other.
The mammal fauna from the Fowkes Formation places a late Bridgerian age on the Sillem and Bulldog Hollow members (Nelson 1973). Small mammals were most abundant in the Fowkes, especially primates and rodents. The marsupials were characterized by Peratherium which was a small, probably arboreal, opossum-like animal. Peratherium was but one stage in a slow and relatively conservative evolution of opossums that began in the Cretaceous.
The condylarths decreased in importance during Fowkes time and were gradually replaced by more advanced ungulates, the artiodactyls and perissodactyls. Only the small Hyopsodus survived into the Bridgerian of the Fossil Basin. The horses continued to differentiate as evidenced by the appearance of Orohippus, an animal similar to Hyracotherium, but with minor dental differences. The primitive tapirs were represented by Hyrachyus, a medium-sized herbivore which was nearly as closely related to the rhinoceroses as it was to the tapirs.
The primates were also diverse in Fowkes time; however, this group began to decline in importance following Bridgerian times. As a group, the primates were restricted mainly to warm and forested environments. The lemur-like Notharctus and the tarsier-like Omomys and Hemiacodon were common in the Fowkes. Hemiacodon was typical of the late Bridgerian (an index fossil).
The true carnivores, represented by Miacis, remained small in size, perhaps to avoid competition with the larger surviving creodonts.
Insectivores from the Fowkes Formation are abundant and varied. Unfortunately, they are known almost exclusively from teeth. Fowkes insectivores include the hedge-hog-like Nyctitherium, Talpavus, and Scenopagus. Apatemys is an insectivore of uncertain affinities. The feeding habits of these early insectivores were probably similar to those of contemporary insectivore species (moles and shrews); however, there is some evidence to suggest many of these small creatures may have been semi-arboreal.
One of the most unusual of middle Eocene creatures is the giant Uintatherium. This was a large browsing animal, about the size of a rhinoceros. What was most unusual about Uintatherium was its skull which was large and strongly built. The large upper canines were apparently formidable defensive weapons. The skulls of the males bore six prominent bony protuberances which grew from the frontal region of the skull. The function of these structures is unknown. They may have been of use for defense.
During Bridgerian times, the rodents underwent an explosive adaptive radiation which ultimately may have led to the near extinction of the less well-adapted multituberculates. The diversity of Bridgerian rodents from Fossil Basin is impressive. Species of Leptotomus, Paramys, Thisbemys, Microparamys, Sciuravus, Mysops, and Pauromys demonstrate that a wide range of adaptations of environments was exploited by the rodents. This diversity in life habits has been a mark of rodent evolution from the middle Eocene to the present day.
The above descriptions of the known mammal fossils from the Fossil Butte area are intended to demonstrate the varied life forms that once existed in southwestern Wyoming. Further collecting will almost certainly expand the number of fossil mammal species from Fossil Basin and increase our knowledge of the succession of ancient environments which prevailed at different times in the Fossil Basin area.
Paleoecology is the study of ancient biotic communities and their relationship to the abiotic environment. The conclusions of paleoecology are reached by studying in detail the fossils and sediments and whatever relationships exist between them. Its ultimate aim is to build a picture of the climate, flora, fauna, and topographic setting of an area. For this reason it borrows heavily from paleontology, paleobotany, sedimentology, and climatology.
From studying modern environments, it is observed that each has its own type of sediment. When certain sediments are found in the rock record, it is generally assumed that they represent environments similar to modern ones that produce similar deposits. For example, limestone is forming today in warm, shallow, well-lighted, well-aerated water. When limestone is found in a rock sequence, it is usually assumed that the same or similar environmental conditions occurred in that area in the past. Fossils can further refine the interpretation.
As can be interpreted from foregoing discussions, the sediments deposited in the Fossil Basin vary from stream and flood-plain fluvial to lacustrine. At times, material eroded from surrounding uplands was carried by streams throughout the basin. These rocks are exemplified by the Evanston and parts of the Wasatch and Fowkes formations. When the lake appeared, lacustrine sediments, marlstone, and shale were deposited in the lake, while around the periphery of the basin, fluvial sediments continued to accumulate.
Plants found in the lake sediments of Fossil Basin tell us much about the climatic conditions that prevailed during the Eocene. The flora is quite similar to that now existing in the southeastern United States, reflecting a warm and humid climate. One of the striking examples is that of huge palm fronds that occasionally are found in the fish quarries at Fossil Butte. These seem to be fronds that blew from the trees into the lake. Soon they became water-logged and settled to the bottom to be preserved.
The land animals indicate that a rather wide range of ecologic niches existed over the basin before the lake came into being, around the lake during its presence, and again all over the basin after the lake disappeared. Many of the smaller mammals, some of the rodents, and most of the primates were almost certainly arboreal. The large mammals such as uintatheres, pantodonts, and tapiroids may have been stream-side or marsh dwellers. Probably inhabiting the forest floor and feeding on low bushes and undergrowth were such forms as the condylarths, horses, artiodactyls, and some of the rodents. Feeding on the rodents and other smaller mammals were the creodonts and miacids. Tiny shrew-like forms scampered about the undergrowth and fed upon worms and insects.
Bradley (1963) has estimated possible temperature and precipitation levels in southwestern Wyoming during the Eocene. Bradley’s modern analogues were the Gulf Coast and Great Lakes regions. From a series of calculations, an average annual temperature of 65°F is postulated. This could have fluctuated greatly in the inland setting of Fossil Basin. Precipitation amounts were possibly on the order of 30-43 inches annually. The amount of annual evaporation was also possibly in the range of 30-43 inches.
It is believed that Fossil Lake was thermally stratified—that is, with colder, denser water at depth (hypolimnion) and warmer, less dense water (epilimnion) nearer the surface. The deeper waters probably would have been devoid of oxygen, hence essentially uninhabited other than by anaerobic bacteria that survive without oxygen. If the bottom had been oxygenated, many types of life would have burrowed into the sediment thus destroying the delicate varves (Fig. 28). The lake was probably deep enough that wind and wave action did not roil the bottom sediments.

Fig. 28. A section of shales from the fish beds showing thin laminae that are interpreted as varves. Enlarged six times.
There have been many attempts to interpret the taphonomy (see glossary) of concentrations of fish at Fossil Butte. Bradley (1948) interpreted the cause of death and the reason for their preservation as follows:
In this basin (Fossil, Wyo.) hundreds of thousands of beautifully preserved fish are entombed in the varved sediments. Even the delicate fin and tail rays and other bones originally held in place only by tissue are virtually undisturbed, and even the scales are in place almost completely undisturbed. It seems to me that the picture of this lake as a thermally stratified water body provides nearly all the necessary information to account for the excellent preservation of these fish. Only in the stagnant hypolimnion could they have escaped being torn to pieces by scavengers or distorted by bottom feeders. It is significant 31 that all the well preserved fish are in varved sediments. Those in non-varved sediments are a disordered mass of broken and chewed up bones.
The only part of the story lacking now is how the fish died and got into the hypolimnion. Limnology offers two possible explanations. Sometimes when the surface of a lake gets excessively warm, fish will plunge into deep water and might thus penetrate the hypolimnion, be overcome by hydrogen sulphide, and also have the gas in their swim bladders chilled so that they sank at once to the bottom. Once there, only anaerobic bacteria would attack them. The other hypothesis is that the thermally stratified lake was suddenly chilled so that it overturned more rapidly than the hydrogen sulphide could be oxydized and so killed off large numbers of fish. This seems a little more probable as the fossil fish are of all ages and sizes.
The fish in these quarries have been collected since the 1870s but mainly by commercial collectors. Most museum collections were purchased from these commercial collectors, hence they consist almost entirely of perfectly preserved fish, as poorly preserved specimens would be discarded by the collectors as of no monetary value. The result has been that most people, including Bradley, were misled into believing all of the fish in the varved sediments of the quarry were perfectly preserved. As nearly as can be determined, the first attempt to systematically collect and study this concentration of fish was that by paleontologists from the University of Wyoming (McGrew 1974). This work threw much new light on the occurrence and made available much new data that permit new interpretations.
Shales of the Green River Formation in general have been described by Bradley (1931) and he specifically mentioned those of the fish layer in the Fossil Basin as follows: “Plate 1 shows a thin section of the varved marlstone in the small, unnamed Green River Lake west of Gosiute Lake, where the varves are better developed. Each varve or annual deposit, consists of a layer of microgranular carbonates and a thinner, dark layer of organic matter” (Bradley 1948:645). The X-ray diffraction analyses performed by John Ward Smith of the Laramie Energy Research Center showed that the shales of the “fish layer” consist predominantly of calcite (roughly 60%), aragonite and dolomite (approximately 20%), and quartz (up to 10%).
Although fish are numerous throughout the thickness of this “fish layer,” there are three laminae that contain so many fish that it is almost certain that they represent catastrophic mass mortalities. Two are made up primarily of Priscacara and one consists almost exclusively of Knightia.
X-ray photos show that a rather high percentage of the fish in the shales are not perfectly preserved but have undergone varying amounts of disarticulation. There appears to be an orderly sequence of stages of decomposition—from essentially perfect articulation to total disarticulation. Disarticulation first appears in the most anterior vertebrae. From there it rapidly proceeds anteriorly into the head region and appears to do so posteriorly at a slower rate. In many specimens the head and anterior half of the body are completely disarticulated, while the posterior part of the body shows no disarticulation whatever.
It is assumed that after the dead fish settled to the bottom of the lake, external anaerobic bacteria found access to the “innards” of the fish via the opercular opening. This would account for the first sign of decomposition and disarticulation being just back of the head.
In the blocks of shale covered by X-ray there were 385 fish. For convenience, these were classified into six groups, group I showing no discernable disarticulation and group VI (Fig. 29) showing total disarticulation. The number and percentage of fish in each group are as follows:
| Group I | 223 fish | 58% |
| Group II | 38 fish | 10% |
| Group III | 14 fish | 4% |
| Group IV | 27 fish | 7% |
| Group V | 24 fish | 6% |
| Group VI | 59 fish | 15% |

Fig. 29. A partially disarticulated skeleton of a large Priscacara, Group IV.
Because of the predominance of completely articulated fish throughout the quarry and the fact that the fish involved in the mass mortalities show no disarticulation, it seems probable that some connection might exist between the death of the fish and conditions of the lake bottom that would cause their perfect preservation. It would seem that rapid burial might be the most obvious reason for excellent preservation. Thus any factor or combination of 32 factors that would cause rapid precipitation of carbonates and also would cause mortality of fish would satisfy our requirements. One obvious factor that could, at least theoretically, fulfill these requirements would be an annual bloom of blue-green algae that are known to be toxic to fish (Prescott 1948). Such blooms usually occur during the warmest part of the summer when CaCO₃ is least soluble. By extracting CO₂ from the water, these algae are known to cause precipitation of CaCO₃. Thus we have a possible cause for some annual fish kill and perhaps an occasional superbloom that would bring about a catastrophic mass mortality. The highest mortality of fish then might have occurred during late summer algal blooms and at this time also would occur the most rapid precipitation of CaCO₃.
It is not known how much deposition of CaCO₃ would be required to protect a fish from disarticulation. It may be that a very thin layer, especially if mixed with organic ooze, might provide an effective seal to slow decomposition and prevent disarticulation. If sufficient CaCO₃ was precipitated and deposited to cover the fish that died during this period, one might expect perfect preservation. Such fish would fit into group I. Those fish that died just after this period might lie exposed on the lake bottom for most of a year and be subject to disarticulation. If little or no deposition took place during the rest of the year, fish that died just after the period of deposition should be the most completely disarticulated and fit into group VI. During the fall, winter, and spring, fish that died of attritional mortality would be disarticulated according to the length of time they lay on the bottom prior to the next period of deposition.
If the foregoing is true, one should be able to determine the approximate time of year fish died by the degree of disarticulation. One might assume that blooms of blue-green algae and hence precipitation of CaCO₃ would take place sometime during August and/or September. Thus we should expect the most fish and those most perfectly preserved to have died during this time period (group I). Those fish that died in October and/or November should be the most completely disarticulated (group VI), and those that died in June or July should show only a slight degree of disarticulation (group II). The distribution of the stages of disarticulation seems to fit almost exactly the pattern that one would expect if this interpretation is correct.
Gunter (1947) has shown that annual periods of excess salinity in Texas lagoons cause an annual increase in the death of fish and occasionally a catastrophic mass mortality. Because Lake Gosiute is known to have been saline, it might be assumed that somewhat similar chemical conditions prevailed in the Fossil Lake. It is not known definitely that the two lakes were ever connected but if they were, it was most probably a narrow connection near the southern end of Fossil Lake and probably a rather temporary connection.
That a rather long period of aridity occurred in the general region is demonstrated by various depositional features, primary structures, and salt deposition in Gosiute Lake in a part of the Green River Formation. These deposits appear to have settled down during Lostcabinian (late to early Eocene) times. Because the Wasatch Formation immediately underlying the Fossil Butte Member of the Green River Formation in the Fossil Syncline Basin is Lysitean (mid to early Eocene), it is probable that the fish deposits are of Lostcabinian age. Thus it may well be that the long period of aridity occurred during the deposition of the fish beds. While the Fossil Lake probably never reached the high degree of salinity present in Lake Gosiute, it was probably sufficiently saline that periods of excessive evaporation could increase the salinity enough to contribute to the mortality of fish and occasionally cause a catastrophic mass mortality such as those described by Gunter. The presence of aragonite and dolomite in the shales suggests that at least some of the carbonate deposition might well have been because of excessive evaporation and high concentrations of carbonates (Smith 1974, pers. comm.).
At the University of Wyoming, an attempt is being made to interpret scales of fish from the quarries. In a number of specimens annuli can be observed and circuli counted. Although removal of scales for study is extremely difficult, a number has been removed and photographed with a scanning electron microscope. In Lake George, Florida, black crappie develop annuli during January, February, March, and April (Huish 1954). Climatic conditions in north central Florida may be similar to those that existed in western Wyoming during the early Eocene. Thus annuli may have developed at the same time of year. By counting the number of circuli between the last annulus and the edge of the scale, it should be possible to determine the approximate time of death of a fossil fish. If this should correlate with the degree of disarticulation, a check on this interpretation should be possible. Sufficient data are not yet available, however, for the results to be conclusive.
It is not intended to suggest that the conditions outlined above could account for all of the fish concentrations in the Green River Shales. In certain shales in the Green River Basin, for example, the concentrations of Knightia in nonvarved shales appear to have been deposited in quite shallow water. These fish are extremely well preserved but were obviously laid down under conditions quite different from those at Fossil Butte. Much study of these occurrences will be necessary before interpretations are possible.
The story, as told here, should make it clear that the earth is ever-changing. What was once a beautiful, deep lake in an area of lush tropical forests is now a dry, sagebrush desert. What was once an area in which sediments were accumulating is now an area of erosion, occasioned by a broad uplift of the region near the end of Tertiary time.
It is hoped that this brief story of the geologic history of Fossil Butte National Monument will give the reader some appreciation of the geologic complexity not only of the monument itself but of the surrounding area as well. Certainly such knowledge will add to the enjoyment of a visit to the monument.
It may seem that geologists have all of the answers. This, however, is not so. Interpretations are made on the basis of available evidence. Each time a geologist studies an area more geologic data and more evidence become available and our interpretations become a little more accurate. It will be many years before all details of this fascinating history will be known.
This report, which was prepared under National Park Service Contract CX 6000-3-0076, leans heavily on the publications of those many people who have studied various geologic aspects of the Fossil Basin. We would like to cite particularly Doctors Steven Oriel and Joshua Tracy of the United States Geological Survey, whose studies have clarified the stratigraphic relationships of the sediments involved. Dr. D. L. Blackstone, Jr., of the University of Wyoming supplied information from his yet unpublished interpretations of the complicated structure of the Idaho-Wyoming thrust belt. Mr. Tom Bown helped with the structural part of the report and critically reviewed other parts. The drafting, except for Figs. 3 and 9, was done by Mr. Evan Groutage. Mrs. Elaine Hertzfeldt, secretary for the Department of Geology, aided greatly in preparing this manuscript.
Paul O. McGrew Michael Casilliano
