
MIGRATION OF BIRDS
Circular 16. Revised Edition - 1979FISH & WILDLIFE SERVICE / UNITED STATES DEPARTMENT OF THE INTERIOR
MIGRATION OF BIRDS
Circular 16. Revised Edition - 1979FISH & WILDLIFE SERVICE / UNITED STATES DEPARTMENT OF THE INTERIOR
- iii -
By Frederick C. Lincoln
Revised By Steven R. Peterson
Associate Editor Peter A. Anastasi
Illustrated By Bob Hines
 Circular 16.
Revised Edition - 1979
Circular 16.
Revised Edition - 1979
FISH & WILDLIFE SERVICE / UNITED STATES DEPARTMENT OF THE INTERIOR
- v -
| Page | |
| PREFACE | 1 |
| INTRODUCTION | 2 |
| THE HISTORY AND SCOPE OF MIGRATION | 4 |
| TECHNIQUES FOR STUDYING MIGRATION | 7 |
| Direct Observation | 7 |
| Aural | 8 |
| Preserved Specimens | 8 |
| Marking | 8 |
| Bands, Collars, Streamers | 8 |
| Radio Tracking | 10 |
| Radar Observation | 10 |
| Laboratory | 11 |
| Orientation and Navigation | 11 |
| Physiology of Migration | 11 |
| ADVANTAGES OF MIGRATION | 13 |
| STIMULUS FOR MIGRATION | 15 |
| WHEN BIRDS MIGRATE | 17 |
| Time of Year | 17 |
| Time of Day | 20 |
| SPEED OF FLIGHT AND MIGRATION | 25 |
| ALTITUDE OF FLIGHT AND MIGRATION | 32 |
| SEGREGATION DURING MIGRATION | 35 |
| By Individuals or Groups of Species | 35 |
| By Age | 36 |
| By Sex | 38 |
| By Kinds of Flocks | 40 |
| WHERE BIRDS MIGRATE | 41 |
| Migration by Populations Within Species | 41 |
| Fall Flights Not Far South of Breeding Range | 42 |
| Long Distance Migration | 44 |
| ORIENTATION AND NAVIGATION | 47 |
| INFLUENCE OF WEATHER | 51 |
| INFLUENCE OF TOPOGRAPHY | 56 |
| PERILS OF MIGRATION- vi - | 58 |
| Storms | 58 |
| Aerial Obstructions | 58 |
| Exhaustion | 59 |
| ROUTES OF MIGRATION | 61 |
| General Considerations | 61 |
| Flyways and Corridors | 62 |
| Narrow Routes | 65 |
| Converging Routes | 65 |
| Principal Routes From North America | 69 |
| Atlantic Oceanic Route | 69 |
| Atlantic Coast Route and Tributaries | 70 |
| Mackenzie Valley-Great Lakes-Mississippi Valley Route and Tributaries |
73 |
| Great Plains-Rocky Mountain Routes | 75 |
| Pacific Coast Route | 76 |
| Pacific Oceanic Route | 80 |
| Arctic Routes | 80 |
| PATTERNS OF MIGRATION | 82 |
| Loops | 82 |
| Dog-legs | 87 |
| Pelagic Wandering | 90 |
| Leap-frogging | 90 |
| Vertical Migration | 91 |
| Pre-migratory Movements | 91 |
| Vagrant Migration | 92 |
| ORIGIN AND EVOLUTION OF MIGRATION | 95 |
| WHERE WE STAND | 100 |
| BIBLIOGRAPHY | 102 |
| LIST OF BIRD SPECIES MENTIONED IN TEXT | 115 |
- 1 -
Frederick C. Lincoln's classic work on the "Migration of Birds" first appeared in 1935. It was revised in 1950 and has been out of print for several years, after selling over 140,000 copies. Unfilled requests by many individuals, clubs, and institutions prompted the Office of Conservation Education (now the Office of Public Affairs) in the U.S. Fish and Wildlife Service to petition another update for reissue. This publication incorporates the results gathered by research biologists in the U.S. Fish and Wildlife Service to meet these requests.
Lincoln's original intent was to present to the American public a summary of the facts on bird migration as they existed in the early 1930's. He wrote with a style that made the topic fascinating to the young and old, to the educated and uninformed, and to the ardent observer as well as the backyard watcher. An attempt has been made to retain this style, while incorporating material from often highly technical research efforts. Much of the content and organization of the original publication has been maintained, but new sections were added to incorporate recent concepts and techniques. Other concepts, known to be inconsistent with present knowledge, have been deleted. Because graphics are of utmost importance in this type of publication, most of the original figures were preserved and, where appropriate, new illustrations have been added.
Since the previous edition, tremendous progress has been made in researching and understanding bird migration; along with this increased effort has come a substantial increase in the literature devoted to the subject. Emphasis was given to reviewing literature pertaining to migration studies conducted in North America after 1950, but a number of examples from the European literature have been included to emphasize similarities and differences in migration throughout the world. Because extensive author citations tend to disrupt the flow of thought, they were kept to a minimum in the text. However, this publication is essentially a review of the literature on the subject as it existed in the early 1970's, and a rather extensive bibliography has been included to cover all the papers quoted in the text as well as the many used but not specifically cited. The bibliography, then, is primarily intended for those interested in pursuing the subject further.
- 2 -
The changing picture of bird populations throughout the year intrigues those who are observant and who wish to know the source and destination of these birds. Birds are the most mobile creatures on Earth. Even man with his many vehicles of locomotion does not equal some birds in mobility. No human population moves each year as far as from the Arctic to the Antarctic and return. Yet the Arctic terns do—and without the aid of aircraft or compass.
Birds are adapted in their body structure, as no other creatures, to life in the air. Their wings, tails, hollow bones, and internal air sacs all contribute to this great faculty. These adaptations make it possible for birds to seek out environments most favorable to their needs at different times of the year. This results in the marvelous phenomenon we know as migration—the regular, seasonal movement of entire populations of birds from one geographic location to another.
Throughout the ages, migratory birds have been important as a source of food after a lean winter and as the harbinger of a change in season. The arrival of certain species has been heralded with appropriate ceremonies in many lands; among the Eskimos and other tribes, the phenomenon to this day is the accepted sign of the imminence of spring, of warmer weather, and a change from winter food shortages. The pioneer fur traders in Alaska and Canada offered rewards to the Indian or Eskimo who saw the first flight of geese in the spring, and all joined in jubilant welcome to the newcomers.
As the North American Continent became more thickly settled, the large flocks of ducks and geese, rails, doves, and woodcock that always had been hunted for food became objects of the enthusiastic attention of an increasing army of sportsmen. Most of the nongame species were found to be valuable also as allies of the farmer in his never-ending warfare against insect pests. All species have been of ever-increasing recreational and esthetic value for untold numbers of people who enjoy watching birds. We began to realize our migratory bird resource was an international legacy (that cannot be managed alone by one state or country) and all nations were responsible for its well-being. The need for laws protecting game and nongame birds, as well as the necessity to regulate the hunting of diminishing game species, followed as a natural course. In the management of this wildlife resource, it has become obvious that continuous studies must be made of the species' habits, environmental needs, and travels. In the United States, the Department of the Interior recognizes the value of this resource and is devoted to programs that will ensure its preservation and wise use. Hence bird - 3 - investigations are made by the U.S. Fish and Wildlife Service, an arm of the Interior Department, charged by Congress under the Migratory Bird Treaty Act, with the duty of protecting those species that in their yearly journeys, pass back and forth between the United States and other countries.
For more than three-quarters of a century the Fish and Wildlife Service and its predecessor, the Biological Survey, have been collecting data on the important details of bird migration. Scientists have gathered information concerning the distribution and seasonal movements of many species throughout the New World, from the Canadian archipelago south to the Argentine pampas. Supplementing these investigations is the work of hundreds of U.S. and Canadian university personnel and volunteer birdwatchers, who report on the migrations and status of birds as observed in their respective localities; while others place numbered bands on the legs of birds to determine their movements from one place to another. These data, stored in field notes, computer cards, scientific journals, and on magnetic tape constitute an enormous reservoir of information pertaining to the distribution and movements of North American birds. It is the purpose of this publication to summarize these data and present the more important facts about that little understood but universally fascinating subject of bird migration. The U.S. Fish and Wildlife Service is grateful to the many persons who have contributed their knowledge so that other people, be they bird study classes, conservation organizations, or just individuals interested in the welfare of the birds, may understand and enjoy this precious resource as well as preserve it for generations to come.
- 4 -
The migrations of birds were probably among the first natural phenomena to attract the attention and arouse the imagination of man. Recorded observations on the subject date back nearly 3,000 years, to the times of Hesiod, Homer, Herodotus, Aristotle, and others. In the Bible there are several references to the periodic movements of birds, as in the Book of Job (39:26), where the inquiry is made: "Doth the hawk fly by Thy wisdom and stretch her wings toward the south?" The author of Jeremiah (8:7) wrote: "The stork in the heavens knoweth her appointed time; and the turtledove, and the crane, and the swallow, observe the time of their coming." The flight of quail that saved the Israelites from starvation in their wanderings through the Sinai wilderness is now recognized as a vast migration between their breeding grounds in eastern Europe and western Asia and their winter home in Africa.
Of observers whose writings are extant, Aristotle, naturalist and philosopher of ancient Greece, was one of the first to discuss the subject of bird migration. He noted cranes traveled from the steppes of Scythia to the marshes at the headwaters of the Nile, and pelicans, geese, swans, rails, doves, and many other birds likewise passed to warmer regions to spend the winter. In the earliest years of the Christian era, Pliny the Elder, Roman naturalist, in his "Historia Naturalis," repeated much of what Aristotle had said on migration and added comments of his own concerning the movements of starlings, thrushes, and European blackbirds.
Aristotle also must be credited with the origin of some superstitious beliefs that persisted for several centuries. One of these, that birds hibernated, became so firmly rooted, Dr. Elliott Coues (1878),[1] an eminent American ornithologist, listed the titles of no less than 182 papers dealing with the hibernation of swallows. In fact the hibernation theory survived for more than 2,000 years, and it was not until early in the nineteenth century that its acceptance as an explanation for the winter disappearance of birds was almost completely abandoned. Even after this, a few credulous persons suggested this idea as an explanation for the disappearance of chimney swifts in the fall before bands from wintering swifts were finally reported as taken by Indians in Peru (Coffey 1944).
[1] Publications referred to parenthetically by date are listed in the Bibliography, p. 102.
The followers of Aristotle believed the disappearance of many species of birds in the fall was accounted for by their passing into a torpid state where they remained during the cold season, hidden in - 5 - hollow trees, caves, or in the mud of marshes. Aristotle ascribed hibernation not only to swallows, but also to storks, kites, doves, and others. Some early naturalists wrote fantastic accounts of the flocks of swallows allegedly seen congregating in marshes until their accumulated weight bent into the water the reeds on which they clung and thus submerged the birds. It was even recorded that when fishermen in northern waters drew up their nets they sometimes had a mixed "catch" of fish and hibernating swallows. Clarke (1912) quotes Olaus Magnus, Archbishop of Upsala, who in 1555 published a work entitled "Historia de Gentibus Septentrionalis et Natura," wherein he observed that if swallows so caught were taken into a warm room they would soon begin to fly about but would live only a short time.
Although the idea of hibernation as a regular method of spending the winter is no longer accepted for any species of bird, certain hummingbirds, swifts, and poorwills have been known to go into an extremely torpid condition in cold weather (Jaeger 1948, 1949). Thus Aristotle was at least partially vindicated.
Aristotle also was the originator of the theory of transmutation, or the seasonal change of one species into another. Frequently one species would arrive from the north just as another species departed for more southerly latitudes. From this he reasoned the two different species were actually one and assumed different plumages to correspond to the summer and winter seasons.
Probably the most remarkable theory advanced to account for migration is contained in a pamphlet, "An Essay toward the Probable Solution of this Question: Whence come the Stork and the Turtledove, the Crane, and the Swallow, when they Know and Observe the Appointed Time of their Coming," mentioned by Clarke (1912: v. 1, 9-11) published in 1703. It is written "By a Person of Learning and Piety," whose "probable solution" stated migratory birds flew to the moon and there spent the winter. Astronauts have so far failed to verify this.
Some people, who easily accepted the migratory travels of larger birds, were unable to understand how smaller species, some of them notoriously poor fliers, could make similar journeys. They accordingly conceived the idea that larger species (e.g., storks and cranes) carried their smaller companions as living freight. In some southern European countries, it is still believed these broad-pinioned birds serve as aerial transports for hosts of small birds that congregate upon the Mediterranean shore awaiting the opportunity for passage to winter homes in Africa. Similar beliefs, such as hummingbirds riding on the backs of geese, have been found among some tribes of North American Indians.
Today we realize that birds do not migrate by "hitching" rides with other birds and that the scope of the migration phenomenon is worldwide, not simply limited to the United States, the Northern Hemisphere, or the world's land masses. The migration heritage is developed just as extensively in Old World warblers migrating to and - 6 - from Europe and Africa as in our wood warblers traveling from Canada and the United States to South America and back. One of the fundamental differences in migration patterns of the Northern and Southern Hemispheres is that no land species nesting in the South Temperate Zone migrates into the North Temperate Zone, but a few seabirds, such as the sooty shearwater, Wilson's storm-petrel, and others, migrate north across the Equator over the vast ocean expanses after nesting in the South.
- 7 -
Before we discuss the many intricacies of how, when, and where birds migrate, one should have a general idea of how migration data are collected and what methods are currently being used to increase our knowledge. Since this publication first appeared in 1935, many new procedures have been used in the study of bird migration. One of these, radar, has been an invaluable adaptation of a technique developed for a quite different, but related, purpose.
The oldest, simplest, and most frequently used method of studying migration is by direct observation. Size, color, song, and flight of different species all aid the amateur as well as the professional in determining when birds are migrating. Studies begun by Wells W. Cooke and his collaborators (Cooke 1888-1915) and continued by his successors in the U.S. Bureau of Biological Survey (later U.S. Fish and Wildlife Service) were of particular importance in the earlier years of these investigations in North America. Some of the largest and most interesting routes and patterns were sorted out by tediously compiling and comparing literally thousands of observations on whether a species was or was not seen in a given locality at a particular time of the year. More recently, "The Changing Seasons" reports by many amateur bird observers in Audubon Field Notes (now American Birds) have been a most important source of information on direct observation of migration. In the aggregate, direct observation has contributed much to our knowledge of migration, but, as will be pointed out in other sections, until a few years ago, observers were not aware of some of the biases in this technique.
The "moon watch" is a modification of the direct observation method. It has long been known that many species of birds migrate at night. Until recently, it was not apparent just how important nocturnal migration really is. Significant information has been derived from watching, through telescopes, the passage of migrating birds across the face of a full moon. Since the actual percent of the sky observed by looking through a telescope at the moon is extremely small (approximately one-hundred thousandth of the observable sky), the volume of birds recorded is small. On a night of heavy migration, about 30 birds per hour can be seen. The fact that any birds are observed at all is testimony to the tremendous numbers passing overhead. Large-scale, cooperative moon-watching studies have been organized and interpreted by George H. Lowery, Jr. (1951; Lowery and Newman 1966).
- 8 -
Another specialized direct observation approach which has yielded important information on the spatial and altitudinal distribution of night migrating birds has been the use of small aircraft equipped with auxiliary landing lights (Bellrose 1971). Major disadvantages of night observation are that species cannot be identified and that birds continue to migrate without a full moon. However, these techniques do give information on the nocturnal migration movements that could not be obtained by other methods.
An adjunct to the previously described nocturnal observation methods, which has potential for species identification, is the use of a parabolic reflector with attached microphone to amplify call (chip) notes (Ball 1952; Graber and Cochrane 1959). This device, when equipped with a tape recorder, can record night migrants up to 11,000 feet on nights with or without a full moon. A primary disadvantage is that one cannot tell the direction a bird is traveling and there is considerable difficulty in identifying the chip notes made by night migrants. In addition, the bird may not call when it is directly over the reflector and consequently it would not be recorded. These calls are quite different from the notes we hear given by familiar birds during the daytime while they are scolding an intruder or advertising their territory.
Reference material consisting of preserved bird skins with data on time and place of collection exist in many natural history museums. The essential ingredient in studying migration by this method is to have an adequate series of specimens taken during the breeding season so differences in appearance between geographically separated breeding populations of the same species can be determined. Such properly identified breeding specimens may be used for comparison with individuals collected during migration to associate them with their breeding areas (Aldrich 1952; Aldrich, Duvall, and Geis 1958). This supplies a convenient way of recognizing and referring to individuals representative of known populations wherever they may be encountered.
If birds can be captured, marked, and released unharmed, a great deal of information can be learned about their movements. Many different marking methods have been developed to identify particular individuals when they are observed or recaptured at a later date. A few of the general methods are summarized in this section.
Bands, Collars, Streamers
Since 1920, the marking of birds with numbered leg bands in North America has been under the direction of the U.S. Fish and - 9 - Wildlife Service in cooperation with the Canadian Wildlife Service. Every year professional biologists and voluntary cooperators, working under permit, place bands on thousands of birds, game and nongame, large and small, migratory and nonmigratory, with each band carrying a serial number and the legend, NOTIFY FISH AND WILDLIFE SERVICE, WASHINGTON, D.C., or on the smaller sizes, an abbreviation. When a banded bird is reported from a second locality, a definite fact relative to its movements becomes known, and a study of many such cases develops more and more complete knowledge of the details of migration.
The records of banded birds are also yielding other pertinent information relative to their migrations such as arrival and departure dates, the length of time different birds pause on their migratory journeys to feed and rest, the relation between weather conditions and starting times for migration, the rates of travel for individual birds, the degree of regularity with which individual birds return to the summer or winter quarters used in former years, and many other details. Many banding stations are operated systematically throughout the year and supply much information concerning the movements of migratory birds that heretofore could only be surmised. The most informative banding studies are those where particular populations of birds are purposely banded to produce certain types of information when they are recovered. Examples of such planned banding are the extensive marking of specific populations of ducks and geese on their breeding grounds by the U.S. Fish and Wildlife Service and the Canadian Wildlife Service, as well as in "Operation Recovery," the cooperative program of banding small landbirds along the Atlantic Coast (Baird et al. 1958). When these banded birds are recovered, information concerning movements of specific populations or the vulnerability to hunting is gained. Colored leg bands, neck collars, or streamers can be used to identify populations or specific individuals, and birds marked with easily observed tags can be studied without having to kill or recapture individuals, thus making it a particularly useful technique.
We have learned about the migratory habits of some species through banding, but the method does have shortcomings. If one wishes to study the migration of a particular species through banding, the band must be encountered again at some later date. If the species is hunted, such as ducks or geese, the number of returns per 100 birds banded is considerably greater than if one must rely on a bird being retrapped, found dead, etc. For example, in mallards banded throughout North America the average number of bands returned the first year is about 12 percent. In most species that are not hunted, less than 1 percent of the bands are ever seen again.
In 1935, Lincoln commented that, with enough banding, some of the winter ranges and migration routes of more poorly understood species would become better known. A case in point is the chimney swift, a common bird in the eastern United States. This is a - 10 - nonhunted species that winters in South America. Over 500,000 chimney swifts have been banded, but only 21 have been recovered outside the United States (13 from Peru, 1 from Haiti, and the rest from Mexico). The conclusion is simply this: Whereas banding is very useful for securing certain information, the volume of birds that need to be banded to obtain a meaningful number of recoveries for determining migratory pathways or unknown breeding or wintering areas may be prohibitive. One problem in interpretation of all banding results is the fact that recoveries often reflect the distribution of people rather than migration pathways of the birds.
Other methods used to mark individuals in migration studies include clipping the tip end off a feather (not a major flight feather) with a fingernail clipper or touching the feather with colored paint or dye. This marking technique is obviously good for only as long as the bird retains the feather (usually less than one year), but allows the investigator to recognize whether the bird has been handled previously or not.
One of the most promising methods of tracking the movements of individual birds in migration has been developed in recent years. It is called radio tracking, or telemetry, and consists of attaching to a migrating bird a small radio transmitter that gives off periodic signals or "beeps". With a radio receiving set mounted on a truck or airplane, it is possible to follow these radio signals and trace the progress of the migrating bird. One of the most dramatic examples of this technique was reported by Graber in 1965. He captured a grey-cheeked thrush on the University of Illinois campus and attached a 2.5-gram transmitter to it (a penny weighs 3 grams). The bird was followed successfully for over 8 hours on a course straight across Chicago and up Lake Michigan on a continuous flight of nearly 400 miles at an average speed of 50 mph (there was a 27 mph tail wind aiding the bird). It is interesting to note that while the little thrush flew up the middle of Lake Michigan, the pursuing aircraft skirted the edge of the lake and terminated tracking at the northern end after running low on fuel while the bird flew on. The limitations of radio telemetry, of course, are the size of the transmitter that can be placed on birds without interfering with flight and the ability of the receiving vehicle to keep close enough to the flying bird to detect the signals. Despite this difficulty there has been considerable development in the technology, and encouraging results to date give promise for the future, particularly when receivers can be mounted on orbiting satellites (Graber 1965; Bray and Corner 1972; Southern 1965).
One of the developments of our modern age of electronics has been the discovery that migrating birds show up on radar screens used in monitoring aircraft. At first, the screen images caused by flying - 11 - birds were a mystery to radar operators, and they designated the dots "angels." Later when their nature was understood, students of bird migration seized on the unique opportunity to obtain information on movements of birds over extensive areas (Sutter 1957; Drury 1960; Lack 1963a, b; Bellrose 1967; Graber 1968; and Gauthreaux 1972a, b).
Three types of radar have been used for studying birds: 1) general surveillance radar, similar to ones located at airports, that scans a large area and indicates the general time and direction of broad movements of birds; 2) a tracking radar that records the path of an airplane (or bird) across the sky by "locking on" to a designated "target" and continuously following only that object; and 3) a Doppler radar similar to those operated by law enforcement agencies for measuring the speed of a passing automobile. The latter radar set is useful in determining the speed of flying birds.
The use of radar in migration studies has been invaluable in determining direction of mass movement, dates and times of departure, height of travel, and general volume, especially at night. One interesting fact to come out of current radar work is the discovery of relatively large movements of warblers and other land birds migrating over the seas rather than along the coastlines and in directions observers were completely unaware of a few years ago.
Orientation and Navigation
Studies on how migrating birds orient (travel in one compass direction) or navigate (travel toward a specific goal) have received increasing emphasis in the past 20 years. These studies have focused on the ability of birds to orient themselves by the position of the sun and stars. Outstanding in this facet of research have been the works of Matthews (1951, 1955), Kramer (1952, 1959, and 1961), Sauer and Sauer (1960), Mewaldt and Rose (1960), Sauer (1961), Hamilton (1962a, b), Schmidt-Koenig (1963, 1964), and Emlen (1969). The basic method used in the experiments is to observe the direction in which confined birds attempt to move during the period of migratory restlessness. The birds are not permitted to have any view of the landscape but only the sky above them. In some cases the positions of the celestial bodies are changed by the use of mirrors to see the effect on the orientation of the experimental birds. In other cases the experiments are performed in planetariums so positions of the stars in the artificial heavens can be manipulated and the effect observed.
Physiology of Migration
The physiological basis for bird migration has received considerable attention, particularly the effects of seasonal increases and decreases in daylight and the seasonal rhythms occurring within animals and referred to as "biological clocks." Investigations in this field include the pioneering work on the relationship of photoperiod - 12 - (daylength) to migration by Rowan (1925, 1926) and many subsequent studies (Wolfson 1940, 1945; Marshall 1961; King, Barker and Farner 1963; King and Earner 1963; King 1963; Farner 1955, 1960; and Farner and Mewaldt 1953). These studies have become ever more deeply involved in the intricate relationships between photoperiod, endocrine interactions, gonad development, fat deposition, and migratory unrest. They add to our knowledge of the mechanisms that regulate the migratory behavior we observe.
- 13 -
Why should a bird subject itself to the rigors of a long migratory journey twice a year if it can find all the requirements suitable for existence in one locality? It seems well to consider briefly the ends that are served by this annual round trip between breeding grounds and winter quarters. Obviously, the migratory habit enables a species to enjoy the summer of northern latitudes and to avoid the severity of winter. In other words, migration makes it possible for some species to inhabit two different areas during the seasons when each presents favorable conditions. If it was not advantageous to make the trip twice a year, natural selection would have eliminated the tendency, but bird migration has become the rule over much of the world rather than the exception.
By withdrawing in the spring to regions uninhabitable earlier in the year, migrant species are generally assured of adequate space and ample food upon their arrival in the winter-freed North, and those nonmigratory kinds, which stay behind to nest, are also assured of ample space for these activities.
Every pair of birds requires a certain amount of territory for the performance of its reproductive duties, the extent of which varies greatly between different species. This territory must be large enough to provide adequate food, not only for the parent birds but also for the lusty appetites of their young. In the Arctic summer, 24 hours of daylight allow the young to feed or be fed almost continuously and rapid growth is apparent. The short breeding season in northern latitudes exposes the vulnerable young to predation for a brief period and prevents a build up of predator populations.
It cannot be said that the winter or summer area of every species is entirely unsuited to the requirements of all of its members at other seasons, because some individuals pass the winter season in areas that are frequented only in summer by other individuals of their species. Such species may have extensive breeding ranges with wide climatic variations so that some individuals may actually be permanently resident in a region where others of their kind are present only in winter. Also, some individual song sparrows and blue jays, for example, have been known to change their migratory status (e.g., a particular bird may migrate one year and not the next or vice versa). Thus, different individuals or populations within these species appear to have different tolerances for climatic conditions.
The tendency of some birds to move southward at the approach of winter is not always due to seasonal low temperatures. Experiments have demonstrated many of our summer insect feeders, when confined in outdoor aviaries, comfortably withstand temperatures - 14 - far below zero as long as abundant food is provided. The main consideration then, is depletion of the food supply, caused by either the disappearance or hibernation of insects or the mantle of snow or ice that prevents access to seeds and other food found on or close to the ground or submerged in water. Also, shortened hours of daylight may restrict the ability of birds to obtain sufficient food at a time when low temperatures require increased energy to maintain body heat. It is noteworthy that some of our smaller birds, such as the chickadees, can withstand a cold winter because their food supplies are always available above ground on trees. When there is a good supply of pine and spruce seeds, red-breasted nuthatches and crossbills will remain through the winter in Canadian woods, but when these birds appear abundantly in winter at southern latitudes, it may be concluded there is a shortage of these foods in the North.
- 15 -
Modern views based on studies of bird behavior and physiology indicate migration is a regular, annually induced movement, modified by local weather conditions, but largely independent of them. Migration is a phenomenon far too regular to be created anew each season merely under stress of circumstances, such as need for food; and it begins before the necessity for a change in latitude becomes at all pressing. Swallows, nighthawks, shorebirds, and others may start their southward movement while the summer food supply in the North is at peak abundance. American robins and bluebirds may leave abundant food in the South and press northward when food supplies there are almost entirely lacking and severe cold and storms are likely to cause their wholesale destruction. Regularity of arrival and departure is one of the most impressive features of migration, and since birds travel in a rather strict accordance with the calendar, we might ask: "What phenomena, other than the regular changes in length of day, occur with sufficient precision to act as a stimulus for migration?"
Experimental work has abundantly demonstrated the effect of increased light upon the growth, flowering, and fruiting of plants. Similarly, Rowan's (1925) experiments with slate-colored juncos and the work of numerous subsequent investigators showed, at least in some temperate zone species of migratory birds, increasing periods of daylight triggered sex organs to develop, fat to be deposited, and migration restlessness to begin (King and Farner, 1963). When these conditions develop to a certain level, the bird enters a "disposition to migrate" and takes off for its breeding or wintering grounds. There is reason to believe certain weather conditions influence the actual time of departure and especially the rate of progress to the breeding area.
This explanation of the stimulus for migration may apply very broadly to birds that winter in temperate parts of the world and nest in the same hemisphere but fails in those birds wintering in the tropics, where little change in length of day occurs and even decreases during the spring in regions south of the Equator. It might be asked: "If the lengthening day is the stimulating factor, why should our summer birds, wintering in the tropics, ever start north?" In addition, if daylength influences when birds are stimulated to migrate, why should they not all leave the same locality at the same time? Or, if weather controls the departure of birds from a given area, should not all the migrants leave when conditions are optimal and refrain from departing when conditions are not so? Actually, the conditions that place a bird in a disposition to migrate are probably the result of a combination of factors affecting different species - 16 - differently. Thus not all birds arrive at this condition at the same time.
It has been demonstrated experimentally that Andean sparrows, resident in equatorial regions, come into breeding condition twice annually entirely independent of changing light periods (Miller 1963); evidently the breeding cycle is controlled by periodic internal stimuli. Probably northern migrants that winter in equatorial regions and beyond have their migratory urges controlled by similar rhythms or biological clocks. Also, no evidence suggests that the southward migration of birds is controlled by changing periods of light even among species such as white-crowned sparrows, for which this is a controlling factor in the spring. The fall stimulus is probably an innate cyclic occurrence brought on by a biological mechanism of unknown nature (King, Barker, and Farner 1963).
It is pertinent to point out that the migratory instinct appears to be more or less transitory and not persistent over an extended period. Migratory birds may be delayed en route, either by natural conditions such as unusually abundant food supplies or forcibly by man. If detained until the end of the migratory season, migrants may not attempt to finish the journey because they apparently lose the migratory impulse. In the fall and early winter of 1929, abundant food and open water caused an unusual number of mallards to arrest their migration and remain in western Montana and northern Idaho. Later, however, when a heavy snowfall with subzero temperatures suddenly cut off the food supply, great numbers of the birds subsequently starved to death; a flight of a few hours could have carried them to a region of open water and abundant food.
- 17 -
One ordinarily thinks of the world of birds as sedentary during two periods each year, at nesting time, and in winter. For individuals this is obviously the case, but when the entire avifauna of North America or the world is considered, it is found that at almost all periods there are some latitudinal movements of birds. A few of these movements reoccur year after year with calendar-like regularity. Each species, or group of species, migrates at a particular time of the year and some at a particular time of the day. In this section some of the interesting differences will be discussed as to when birds migrate.
Some species begin their fall migrations early in July, and in other species distinct southward movements can be detected late into the winter. While some migrants are still traveling south, some early spring migrants can be observed returning north through the same locality. For example, many shorebirds start south in the early part of July, while the goshawks, snowy owls, redpolls, and Bohemian waxwings do not leave the North until forced to do so by the advent of severe winter weather or a lack of customary food. Thus an observer in the northern part of the United States may record an almost unbroken southward procession of birds from midsummer to winter and note some of the returning migrants as early as the middle of February. While on their way north, purple martins have been known to arrive in Florida late in January, and, among late migrants, the northern movement may continue well into June. In some species the migration is so prolonged that the first arrivals in the southern part of the breeding range will have performed their parental duties and may actually start south while others of the species are still on their way north.
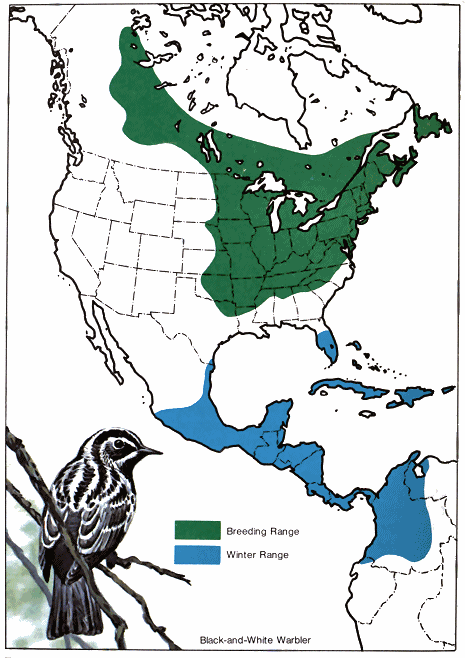
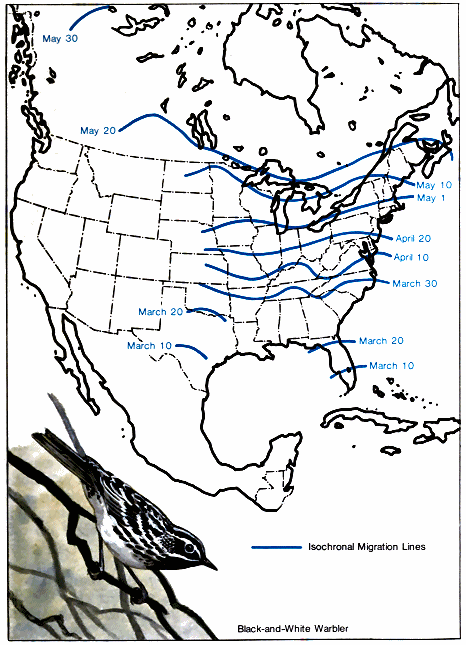
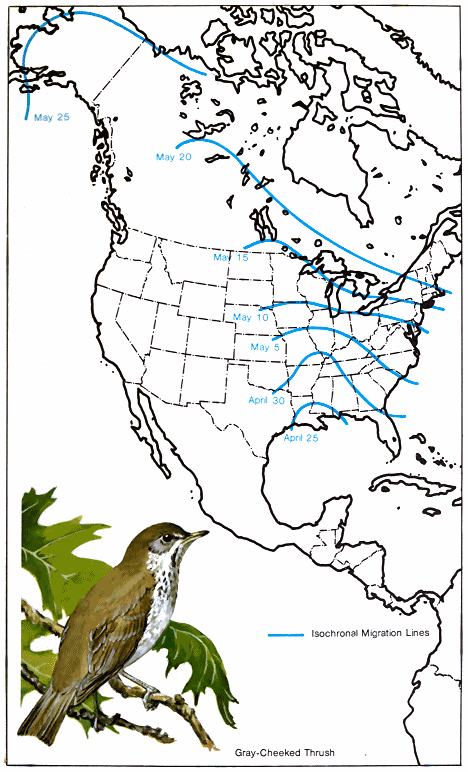
A study of these facts indicates the existence of northern and southern populations of the same species that have quite different migration schedules. In fall, migratory populations that nest farthest south migrate first to the winter range because they finish nesting first. For example, the breeding range of the black-and-white warbler covers much of the eastern United States and southern Canada northwest through the prairies to Great Bear Lake in Canada (Fig. 1.). It spends the winter in southern Florida, the West Indies, southern and eastern Mexico, Central America, and northwestern South America. In the southern part of its breeding range, it nests in April, but those summering in New Brunswick do not reach their nesting grounds before the middle of May. (Lines that connect points where birds arrive at the same line are called - 18 - isochronal lines. Fig. 2.) Therefore, if 50 days are required to cross the breeding range, and if 60 days are allowed for reproductive activities and molting, they would not be ready to start southward before the middle of July. Then with a return 50-day trip south, the earliest migrants from the northern areas would reach the Gulf Coast in September. Since adults and young have been observed on the northern coast of South America by August 21, it is very likely that they must have come from the southern part of the nesting area.
- 19 -
Many similar cases might be mentioned, such as the black-throated blue warblers still observed in the mountains of Haiti during the middle of May when others of this species are en route through North Carolina to New England breeding grounds. Redstarts and yellow warblers, evidently the more southern breeders, are seen returning southward on the northern coast of South America just about the time the earliest of those breeding in the North reach Florida on their way to winter quarters. Examples of the Alaska race of the yellow warbler have been collected in Mississippi, Florida, and the District of Columbia as late as October.
- 20 -
Students of migration know that birds generally travel in waves, the magnitude of which varies with populations, species, weather, and time of year. Characteristically, one will observe a few early individuals come into an area followed by a much larger volume of migrants. This peak will then gradually taper off to a few lingering stragglers. If we plot numbers observed against time, the rising and receding curve takes the form of a bell. In the northern part of the United States there are two general migration waves. The first one in early spring consists of "hardy" birds including many of our common seed eaters like the finches, sparrows, and others. The second wave occurs about a month later and consists primarily of insect-eating birds, such as flycatchers, vireos, warblers, and the like. Each of these species in turn has its own "curve" of migration in the major wave.
Because most birds appear to be creatures of daylight, it seems remarkable that many should select the night for extended travel. Among the many nocturnal migrants are the smaller birds such as rails, flycatchers, orioles, most of the sparrows, the warblers, vireos, thrushes, and shorebirds. It is common to find woods and fields on one day almost barren of bird life and on the following day filled with sparrows, warblers, and thrushes, which indicates the arrival of migrants during the night. Waterfowl hunters sitting in their "blinds" frequently observe the passage of flocks of ducks and geese, but great numbers of these birds also pass through at night; the calls of Canada geese or the conversational gabbling of a flock of ducks are common night sounds in spring and fall in many parts of the country. Observations made with telescopes focused on the full moon have shown processions of birds, and one observer estimated their passage over his area at the rate of 9,000 per hour. This gives some indication of the numbers of birds in the air at night during peaks of migration. At such times radar observations have shown that nocturnal migration begins about an hour after sundown, reaches a peak shortly before midnight, and then gradually tapers off until daybreak. Unless special circuits are installed in radar sets, bird echoes during peak migration periods may cover a radar screen.
It has been suggested that small birds migrate by night to avoid their enemies. To a certain extent this may be true because the group includes not only weak fliers, such as the rails, but also the small song and insectivorous birds, such as wrens, small woodland flycatchers, and other species that habitually live more or less in concealment. These birds are probably much safer making their flights under the protecting cloak of darkness. Nevertheless, it must be remembered that night migrants include also the snipe, sandpipers, and plovers. Most shorebirds are usually found in the open and are among the more powerful fliers, as some of them make - 21 - annual migratory flights over 2,000 miles nonstop across the ocean.
Night travel is probably best for the majority of birds chiefly from the standpoint of feeding. Digestion is very rapid in birds and yet the stomach of one killed during the day almost always contains food. To replace the energy required for long flight, it is essential that either food be obtained at comparatively short intervals or stores of fat be laid on prior to migration. If the smaller migrants were to make protracted flights by day they would arrive at their destination at nightfall almost exhausted, but since they are entirely daylight feeders, they would be unable to obtain food until the following morning. Unless reserve energy was carried in the form of fat, the inability to feed would delay further flights and result in great exhaustion or possibly even death should their evening arrival coincide with cold or stormy weather. By traveling at night, they can pause at daybreak and devote the entire period of daylight to alternate feeding and resting. This schedule permits complete recuperation and resumption of the journey on a subsequent evening after sufficient energy has been restored.
The day migrants include, in addition to some of the ducks and geese, the loons, cranes, gulls, pelicans, hawks, swallows, nighthawks, and swifts. Soaring birds, including broad-winged hawks, storks, and vultures, can only migrate during the day because their mode of flight makes them dependent on up-drafts created by heat from the sun for their long distance travels. On the other hand, swifts and swallows feed entirely on diurnal flying insects. The circling flocks are frequently seen in late summer feeding as they travel while working gradually southward. Formerly, great flocks of red-tailed, Swainson's, and rough-legged hawks could be seen wheeling majestically across the sky in the Plains States. In the East, good flights of broad-winged, Cooper's, and sharp-shinned hawks are still often seen, particularly along the Appalachian ridges.
Because many species of wading and swimming birds are able to feed at all hours, they migrate either by day or night and are not accustomed to seek safety in concealment. Some diving birds, including ducks that submerge when in danger, often travel over water by day and over land at night. Strong fliers like the snow geese can make the entire trip from their staging area in James Bay, Canada, to the wintering grounds on the Louisiana Gulf coast in one continuous flight. These birds are seldom shot by hunters en route between these two points but are often observed, when migrating, by aircraft pilots. Graham Cooch of the Canadian Wildlife Service tracked a flight of the blue phase of this species in 1955. The birds left James Bay on October 17 and arrived on the Gulf coast 60 hours later after an apparent continuous flight over the 1,700-mile route at an average speed of 28 miles per hour. Golden plovers, likewise, probably make the southward flight from the Arctic to the South American coast in one giant leap. Other Arctic species on their northward flight in the spring might prefer to fly at night in lower altitudes, but must necessarily fly during the day at higher altitudes - 22 - because of the length of the days. Many warblers that normally fly at night may find themselves over water at daybreak and be forced to keep flying during the day until landfall is made.
- 23 -
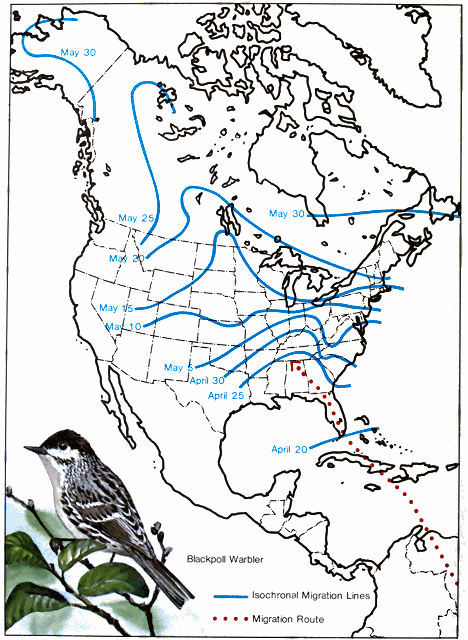
An interesting comparison of the flights of day and night migrants may be made through a consideration of the spring migrations of the blackpoll warbler and the cliff swallow. Both spend the winter as neighbors in South America, but when the impulse comes to start northward toward their respective breeding grounds, the warblers strike straight across the Caribbean Sea to Florida (Fig. 3.), while the swallows begin their journey by a westward flight of several hundred miles to Panama (Fig. 4.). From there they move leisurely along the western shore of the Caribbean Sea to Mexico, and, continuing to avoid a long trip over water, go completely around the western end of the Gulf of Mexico. This circuitous route adds more than 2,000 miles to the journey of the swallows that nest in Nova Scotia. The question may be asked: "Why should the swallow select a route so much longer and more roundabout than that taken by the blackpoll warbler?" The explanation is simple. The swallow is a day migrant while the warbler travels at night. The migration of the warbler is made up of a series of long nocturnal flights alternated with days of rest and feeding in favorable localities. The swallow, on the other hand, starts its migration several weeks earlier and catches each day's ration of flying insects during its aerial evolutions, while slowly migrating. The 2,000 extra miles flown along the insect-teeming shores of the Gulf of Mexico are exceeded by the great distances covered by these birds in normal pursuit of food.

- 24 -
Although most of our smaller birds make their longest flights at night, close observation shows travel is continued to some extent by day. During the latter half of a migratory season birds may show evidence of an overpowering desire to hasten to their breeding grounds. At this time flocks of birds maintain a movement in the general direction of the seasonal journey while feeding on or near the ground. Sometimes they travel hurriedly, and while their flights may be short, they can cover an appreciable distance in the course of a day.
- 25 -
There is a widespread misconception among people concerning the speed at which birds can fly. One often hears stories of birds flying "a mile a minute." While undoubtedly some birds can and do attain this speed, such cases are exceptional, and it is safe to say that, even when pressed, few can develop an air speed of 60 miles per hour. Birds generally have two greatly differing speeds, one being the normal rate for ordinary purposes, and an accelerated speed for escape or pursuit. All birds, except the heavy-bodied, small-winged species such as auks, grebes, and other divers, have a reserve speed that may be double the normal rate.
Although it was thought for a long time that migratory flights were made at normal cruising speeds, Harrisson (1931) and Meinertzhagen (1955) showed that migration speeds were in between cruising speeds and escape speeds. The theory that migrating birds attain high speeds received encouragement from the German ornithologist Gatke (1895) who, for many years, observed birds at the island of Heligoland. He postulated that the bluethroat, a species of thrush smaller than the American hermit thrush, could leave African winter quarters at dusk and reach Heligoland at dawn; this flight would mean a sustained speed of 200 miles per hour! He also thought the American golden plover flew from the coast of Labrador to Brazil in 15 hours at the tremendous speed of 250 miles per hour. Most ornithologists now consider these conclusions to be unwarranted.
Reliable data on the speed of birds are accumulating slowly. Accurate measurements are difficult to obtain unless the bird travels over a measured course and wind conditions at the level of flight are known. Several subtle factors, besides wind and pursuit, can influence the speed of a flying bird. For instance, species that have a courtship flight often reach their maximum speeds then. Small woodland birds often fly faster across an open area where they might be attacked by a bird of prey than under cover where there is less danger. Birds in flocks generally fly faster than when flying alone. A thermal draft may induce an almost imperceptible air movement at the Earth's surface, but a good glider with motionless wings may make 35 miles per hour on a current of air that is rising vertically at less than 2 miles per hour. If the bird coasts downhill at a slight angle in still air, it can attain a similar speed.
For sustained flight, it may be generally concluded that larger birds fly faster than smaller birds. A common flying speed of ducks and geese is between 40 and 50 miles per hour, but among the smaller birds it is much less. Herons, hawks, horned larks, ravens, and - 26 - shrikes, timed with the speedometer of an automobile, have been found to fly 22 to 28 miles per hour, whereas some of the flycatchers fly at only 10 to 17 miles per hour. Even such fast-flying birds as the mourning dove rarely exceed 35 miles per hour. A peregrine falcon will have difficulty catching a pigeon during a level chase at 60 miles per hour, but this predator can probably exceed 200 miles per hour during a swoop from a greater height onto its prey.
The speed of migration is quite different from that attained in forced flights for short distances. A sustained flight of 10 hours per day would carry herons, hawks, crows, and smaller birds from 100 to 250 miles, while ducks and geese might travel as much as 400 to 500 miles in the same period (without the aid of a tail wind). Measured as straight line distances, these journeys are impressive and indicate birds could travel from the northern United States or even from northern Canada to winter quarters in the West Indies, Central, or South America in a relatively short time. It is probable that individual birds do make flights of the length indicated and that barn swallows seen in May on Beata Island, off the southern coast of the Dominican Republic, may have reached that point after a nonstop flight of 350 miles across the Caribbean Sea from the coast of Venezuela.
Radar has given us some of our best estimates of ground speeds for migrating flocks, especially at night. Radar echoes, identified as shorebirds migrating off the New England coast, moved steadily about 45 miles per hour for several hours; songbird echoes typically traveled around 30 miles per hour (Drury 1960). Some birds appear to reduce flight speed in proportion to the degree of assistance or resistance. The literature is in some disagreement on the flight speed of birds and the influence of wind, but good radar observations coupled with accurate measurements of winds aloft will help give us a more accurate estimate of migrating speeds for different species under varying wind conditions.
The intensity of migration depends on circumstances including the need for haste. In fall the flights are more likely to be performed in a leisurely manner, so that after a flight of a few hours the birds often pause to feed and rest for one or several days, particularly if they find themselves in congenial surroundings. Some indication of this is found in the recoveries of banded birds, particularly waterfowl. If we consider only the shortest intervals between banding in the North and subsequent recovery in the South, it is found that usually a month or more is taken to cover straight-line distance of a thousand miles. For example, a black duck banded at Lake Scugog, Ontario, was killed 12 days later at Vicksburg, Mississippi. If the bird was taken shortly after its arrival, the record would indicate an average daily flight of 83 miles, a distance that could have been covered in about 2 hours' flying time. Among the thousands of banding records of ducks and geese, evidences of rapid migrations are decidedly scarce, for with few exceptions, all thousand-mile flights have required 2 to 4 weeks or more. Among sportsmen, the blue-winged teal is well - 27 - known as a fast-flying duck and quite a few of these banded on Canadian breeding grounds have covered 2,300 to 3,000 miles in a 30-day period. Nevertheless, the majority of those that have traveled to South America were not recovered in that region until 2 or 3 months after they were banded. Probably the fastest flight over a long distance for one of these little ducks was one made by a young male that traveled 3,800 miles from the delta of the Athabaska River, northern Alberta, Canada, to Maracaibo, Venezuela, in exactly 1 month. This flight was at an average speed of 125 miles per day. A very rapid migration speed was maintained by a lesser yellowlegs banded at North Eastham, Cape Cod, Massachusetts, on 28 August 1935 and killed 6 days later, 1,900 miles away, at Lamentin, Martinique, French West Indies. This bird traveled an average daily distance of more than 316 miles.
It seems probable that most migratory journeys are performed at little more than the normal, unforced rate of flight, as this would best conserve the strength of the birds. Migrating birds passing lightships and lighthouses or crossing the face of the moon have been observed to fly without hurry or evidence of straining to attain high speed. The speed or rate of migration would therefore depend chiefly on the duration of flights and tail wind velocity.
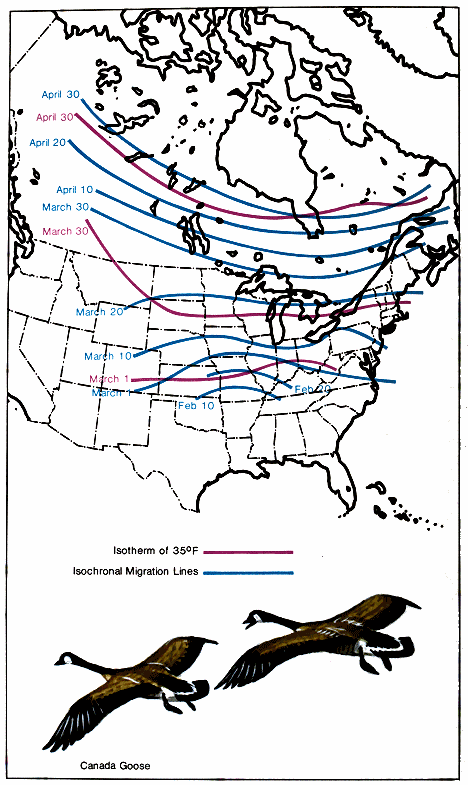
The speed of migration is demonstrated by the dates of arrival, particularly during the spring movement. The Canada goose affords a typical example of regular but slow migration. Its advance northward is at the same rate as the advance of the season (Fig. 5.). In fact, the isotherm of 35° F appears to be a governing factor in the speed at which these geese move north. (An isotherm is a line that connects points that have the same temperature at the same time.) From an evolutionary viewpoint we might expect this. If the geese continually advanced ahead of the 32° F isotherm, they would always find food and water frozen and unavailable. By migrating north just behind the advance of this isotherm, birds that breed in the far north will find food and open water available and have as long a breeding season as the climate will allow.
Few species perform such leisurely migrations; many wait in their winter homes until spring is well advanced, then move rapidly to their breeding grounds. Sometimes this advance is so rapid, late migrants actually catch up with species that may have been pressing slowly but steadily northward for a month or more. The following several examples of well-known migrants illustrate this.
The grey-cheeked thrush, which winters in the Colombia-Ecuador-Peru-Venezuela-British Guiana area, does not start its northward journey until many other species are well on their way. It does not appear in the United States until the last of April—25 April near the mouth of the Mississippi and 30 April in northern Florida (Fig. 6.). A month later, or by the last week in May, the bird is seen in northwestern Alaska. Therefore, the 4,000-mile trip from Louisiana was made at an average distance of about 130 miles per day.
- 28 -
- 29 -
- 30 -
Another example of rapid migration is furnished by the yellow warbler. This species winters in the Tropics and reaches New Orleans about April 5, when the average temperature is 65° F. By traveling north much faster than the spring season progresses, this warbler reaches its breeding grounds in Manitoba the latter part of May, when the average temperature is only 47° F. They encounter progressively colder weather over their entire route and cross a strip of country in the 15 days from May 11 to 25 that spring temperatures normally take 35 days to cross. This "catching up" with spring is habitual in species that winter south of the United States as well as in most northern species that winter in the Gulf States. There appears to be only six exceptions to this rule: the Canada goose, the mallard, the pintail, the common crow, the red-winged blackbird, and the robin.
The snow goose presents a striking example of a late but very rapid spring migration. Most all of these geese winter in the great coastal marshes of Louisiana, where every year over 400,000 spend the winter and congregations of 50,000 or more may be seen grazing in the "pastures" or flying overhead in flocks of various sizes. Their breeding grounds are chiefly on Baffin and Southampton Islands in the northern part of Hudson Bay where conditions of severe cold prevail except for a few weeks each year. The birds are not stimulated to migrate even though the season in their winter quarters is advancing rapidly while their nesting grounds are still covered with a heavy blanket of ice and snow. This suggests the stimulus for spring departure is regulated by an internal mechanism, such as development of the gonads. Accordingly, blue geese remain in the coastal marshes until the last of March or the first of April, when the local birds are already busily engaged in reproduction. The flight northward is rapid, almost nonstop so far as the United States is concerned; although the birds are sometimes recorded in large numbers in the Mississippi Valley, eastern South Dakota, and southeastern Manitoba, there are few records anywhere along the route of the great flocks that winter in Louisiana. When the birds arrive in the James Bay region, they apparently enjoy a prolonged period of rest because they are not seen in the vicinity of their breeding grounds until the first of June. During the first 2 weeks of that month, they pour onto the Arctic tundra by the thousands, and each pair immediately sets about the business of rearing a brood.
The American robin has been mentioned as a slow migrant, and, as a species, it takes 78 days to make the 3,000-mile trip from Iowa to Alaska, a stretch of country that is crossed by advancing spring in 68 days. In this case, however, it does not necessarily mean that individual robins are slow. The northward movement of the species probably depends upon the continual advance of birds from the rear, so that the first individuals arriving in a suitable locality are the ones that nest in that area, while the northward movement of the species is continued by those still to come.
There is great variation in the speed of migration at different - 31 - latitudes of the broad region between the Gulf of Mexico and the Arctic Ocean. The blackpoll warbler again furnishes an excellent example (Fig. 3.). This species winters in northwestern South America and starts to migrate north in April. When the birds reach the southern United States, some individuals fly northwest to the Mississippi Valley, north to Manitoba, northwest to the Mackenzie River, and then almost due west to western Alaska. A fairly uniform average distance of 30 to 35 miles per day is maintained from the Gulf to Minnesota, but a week later this species has reached the central part of the Mackenzie Valley, and by the following week it is observed in northwestern Alaska. During the latter part of the journey, therefore, many individuals must average more than 200 miles per day. Thirty days are spent traveling from Florida to southern Minnesota, a distance of about 1,000 miles, but scarcely half that time is used to cover the remaining 2,500 miles to Alaska. Increased speed across western Canada to Alaska is also shown by many other birds (Figs. 2, 4, 6). A study of all species traveling up the Mississippi Valley indicates an average speed of about 23 miles per day. From southern Minnesota to southern Manitoba 16 species maintain an average speed of about 40 miles per day. From that point to Lake Athabaska, 12 species travel at an average speed of 72 miles per day, while 5 others travel to Great Slave Lake at 116 miles per day, and another 5 species cover 150 miles per day to reach Alaska. This change is in correlation with a corresponding variation in the isothermal lines, which turn northwestward west of the Great Lakes.
As has been previously indicated, the advance of spring in the northern interior is much more rapid than in the Mississippi Valley and on the Gulf coast. In other words, in the North spring comes with a rush, and, during the height of migration season in Saskatchewan, the temperature in the southern part of the Mackenzie Valley just about equals that in the Lake Superior area, 700 miles farther south. Such conditions, coupled with the diagonal course of the birds across this region of fast-moving spring, exert a great influence on migration and are probably factors in the acceleration of travel speed. However, it should be remembered that the birds are getting closer to the breeding season and may be stimulated to travel faster for this reason.
Thus it has been shown that the rate of migration varies greatly under varying circumstances. Radar investigations along the eastern coasts of the United States and England indicate spring migration is several miles per hour faster than in the fall. Also, directions of migrations in spring were much less diverse than in the fall, which suggests less time lost in passage (Tedd and Lack 1958; Nisbet and Drury 1967a). King and Farner (1963) found the same species put on more fat preparatory to migration in the spring. This would give the migrants greater energy reserves for longer flights at that season.
- 32 -
The factors regulating the heights of bird migration are not clear. High-altitude flight may be used to locate familiar landmarks, fly over fog or clouds, surmount physical barriers, gain advantage of a following wind, or maintain a better physiological balance. Meteorological conditions probably account for most of the high-altitude records. Wind conditions at ground level are usually quite different in direction and velocity than at points higher up.
In general, human estimates of bird heights are quite unreliable except under special conditions, and these estimates will vary with the eyesight of the observer. Lucanus (1911) found a European sparrow hawk could be distinguished at 800 feet but disappeared from sight at 2,800 feet. A rook (a European member of the crow family) could be recognized at 1,000 feet but disappeared from sight at 3,300 feet. Meinertzhagen (1955) did an interesting experiment with an inflated model of a vulture painted black; it had a wing expanse of 7 feet 10 inches. When released from an airplane at 4,700 feet, it was barely visible and invisible without binoculars at 5,800 feet. At 7,000 feet it was not picked up even when ×12 binoculars were used.
At one time students of bird migration believed normal migratory movements took place at heights above 15,000 feet. They reasoned, somewhat uncertainly, that flying became easier as altitude was gained. It has now been shown, through comprehensive radar studies, that 95 percent of the migratory movements occur at less than 10,000 feet, and the bulk of the movements occur under 3,000 feet. However, birds can and do fly well over 15,000 feet without apparent ill effects. The physiology of long-distance flight at high altitudes is of great interest but can only be touched on briefly in this discussion.
Bird flight at 20,000 feet, where less than half the oxygen is present than at sea level, is impressive if only because the work is achieved by living muscle tissue. A Himalayan mountain climber at 16,000 feet was rather amazed when a flock of geese flew north 2 miles over his head honking as they went (Swan 1970). At 20,000 feet a man has a hard time talking and running or other rapid movements are out of the question; but those geese were probably flying at 27,000 feet and even calling while they traveled at this tremendous height.
Accurate observations on the altitude of migratory flights is scanty, although altimeter observations from airplanes and radar are becoming more frequent in the literature. An example is the report of a mallard struck by a commercial airliner at 21,000 feet over the Nevada desert (Manville 1963). It is, of course, obvious that - 33 - some birds must cross mountain ranges during migration and attain great altitudes. Numerous observations have come from the Himalayas (Geroudet 1954; Swan 1970). Observers at 14,000 feet recorded storks and cranes flying so high that they could be seen only through field glasses. In the same area large vultures were seen soaring at 25,000 feet and an eagle carcass was found at 26,000 feet. The expedition to Mt. Everest in 1952 found skeletons of a pintail and a black-tailed godwit at 16,400 feet on Khumbu Glacier (Geroudet 1954). Bar-headed geese have been observed flying over the highest peaks (29,000+ feet) even though a 10,000-foot pass was nearby. Probably 30 or more species regularly cross these high passes (Swan 1970).
Except to fly over high mountain ranges, birds rarely fly as high as those traveling down the western Atlantic (Richardson 1972). Many of these birds are making long-distance flights to eastern South America and beyond. Therefore, flight at high altitudes in this region is probably advantageous for them. Richardson postulated stronger advantageous tail winds were found higher up and the cooler air minimized evaporative water losses. This investigator found air temperatures averaged 35° F at 10,000 feet over Nova Scotia in September. The lower the ambient temperature, the more heat can be lost by convection and the less water is required for cooling. Also, a bird flying high can achieve the same range as one flying at sea level but must cruise at a higher speed with a corresponding increase in power output and oxygen consumption. But the increased cruising speed results in shorter flight time and less interference from wind (Pennycuick 1969).
Another postulate favoring the high-altitude flying theory was that the wonderful vision of birds was their sole guidance during migratory flights. To keep landmarks in view, birds were obliged to fly high, particularly when crossing wide areas of water. This will be considered in greater detail in the section, "Orientation and Navigation," so here it will be sufficient to say that birds rely only in part upon landmarks to guide them on migration. Also, it must be remembered that definite physical limitations to the range of visibility exist even under perfect atmospheric conditions. Chief of these is the curvature of the earth's surface. Thus, if birds crossing the Gulf of Mexico to Louisiana and Florida flew at a height of 5 miles, they would still be unable to see a third of the way across (during daylight hours). And yet this trip is made twice each year, much of the distance probably at night, by thousands of thrushes, warblers, and others.
The altitude of migration depends upon the species of bird, weather, time of day or year, and geographical features. Nocturnal migrants, studied by radar, appear to fly at different altitudes at different times during the night. Birds generally take off shortly after sundown and rapidly gain maximum altitude. This peak is maintained until around midnight, then the travelers gradually descend until daylight. For most small birds the favored altitude - 34 - appears to be between 500 and 1,000 feet (Bellrose 1971), but radar studies have found some nocturnal migrants (probably shorebirds) over the ocean were at 15,000 or even 20,000 feet (Lack 1960b; Nisbet 1963b; Richardson 1972). Observations made from lighthouses and other vantage points indicate that certain migrants commonly travel at altitudes of a very few feet to a few hundred feet above sea or land. Sandpipers, northern phalaropes, and various sea ducks have been seen flying so low they were visible only as they topped a wave. Observers stationed at lighthouses and lightships off the English coast have similarly recorded the passage of landbirds flying just above the surface of the water and rarely above 200 feet. During the World Wars, broad areas in the air were under constant surveillance, and many airplane pilots and observers took more than a casual interest in birds. Of the several hundred records resulting from their observations, only 36 were of birds flying above 5,000 feet and only 7 above 8,500 feet. Cranes were once recorded at an altitude of 15,000 feet, while the lapwing was the bird most frequently seen at high levels, 8,500 feet being its greatest recorded altitude. Records of the U.S. Civil Aeronautics Administration show that over two-thirds of all the bird-aircraft collisions occur below 2,000 feet and practically none occur above 6,000 feet (Williams 1950).
Recently, radar has aided greatly in determining differences in the altitude of bird flight. Nocturnal migrants appear to fly slightly higher, on the average, than diurnal migrants, but daytime flights may be influenced more by cloud cover (Lack 1960a; Eastwood and Rider 1965). Bellrose (1971) found little difference in the altitudinal distribution of small nocturnal migrants under clear or overcast skies. Many night migrating birds are killed each year by striking lighthouses, television towers or other man-made illuminated obstructions, but this does not furnish proof that low altitudes are generally used during nocturnal flight because these accidents occur chiefly in foggy weather. Under such conditions, migrating birds seem to be attracted to and confused by lights. Seabirds, such as loons, eiders, and scoters, generally fly very low over the water but gain altitude when land is crossed. The reverse is true for landbirds (Dorst 1963; Bergman and Donner 1964; Eastwood and Rider 1965). There may be a seasonal difference in the altitude of migration, but the evidence is conflicting. Radar echoes studied by Bellrose and Graber in Illinois (1963) showed fall migrants flew higher than spring migrants. They speculated this difference was related to the winds during the fall being more favorable for southerly migration at higher altitudes, while winds at these altitudes in the spring would be less favorable for northerly migration. Eastwood and Rider (1965) studied seasonal migration patterns in England and found the reverse to be true. They suggested one reason for this seasonal difference was that flocks of fall migrants included many young birds whose flight capabilities are inferior to adults and consequently are unable to achieve the higher altitudes in the fall.
- 35 -
During the height of northward movement in spring, the woods and thickets may suddenly be filled with several species of wood warblers, thrushes, sparrows, flycatchers, and other birds. It is natural to conclude they traveled together and arrived simultaneously. Probably they did, but such combined migration is by no means the rule for all species.
As a group, the wood warblers probably travel more in mixed companies than do any other single family of North American birds. In spring and fall, the flocks are likely to be made up of the adults and young of several species. Sometimes swallows, sparrows, blackbirds, and some of the shorebirds also migrate in mixed flocks. In the fall, great flocks of blackbirds frequently sweep south across the Plains States, with common grackles, red-winged blackbirds, yellow-headed blackbirds, and Brewer's blackbirds included in the same flock.
On the other hand many species keep strictly to themselves. It would be difficult for any other kind of bird to keep company with the rapid movements of the chimney swift. Besides flight speed, feeding habits or roosting preferences can be so individual as to make traveling with other species incompatible. Nighthawks also fly in separate companies, as do crows, waxwings, crossbills, bobolinks, and kingbirds. Occasionally, a flock of ducks will be observed to contain several species, but generally when they are actually migrating, individuals of each species separate and travel with others of their own kind.
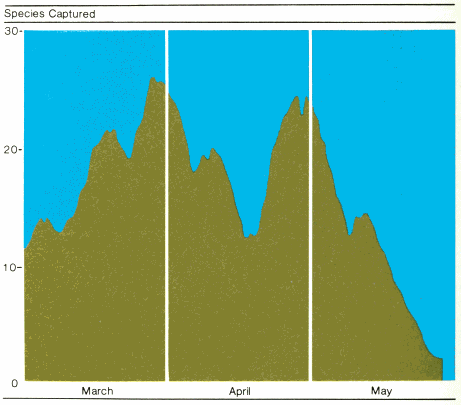
Although different species generally do not migrate together, we often find many species passing through an area at the same time. If the different kinds of birds observed in a specific area are counted every day throughout the entire migration season, this count often rises and falls much like the bell-shaped curve exhibited when the number of individuals of a given species are counted through the same time period. Figure 7 shows two peaks in the number of species passing through the desert at the north end of the Gulf of Eilat (=Akaba) in the Red Sea. These two peaks happen to coincide with peaks in the numbers of individuals (mostly from the order of perching birds) traveling through the area. Therefore, in the latter part of March and again in April, one notices not only more birds in the area but also more different kinds.
Closely related species or species that eat the same food organisms are not often found migrating through the same area at the same time. Ornithologists call this species replacement. In North America, - 36 - peaks in the migration of the five kinds of spotted thrushes generally do not coincide. Dates of departure in these species have evolved so all the individuals of these closely related birds do not converge on one area at the same time and subsequently exhaust the food supply. By selection of staggered peak migration dates, evolution has distributed the members of this family more or less evenly throughout the entire season. Likewise, in the eastern Mediterranean area, we find a similar situation in spring migration for three closely related buntings; Cretzschmar's bunting comes through first, followed a few weeks later by the Ortolan bunting and, at the tail end of the migration period, the black-headed bunting appears (Fig. 8.).
The adults of most birds leave the young when they are grown. This gives the parents an opportunity to rest and renew their plumage before starting for winter quarters. The young are likely to move south together ahead of their parents. This has been documented in a number of species including our mourning dove, the common swift of Europe, and storks. Mueller and Berger (1967) found an age-specific migration pattern in sharp-shinned hawks passing through Wisconsin. The immatures were much in evidence during mid-September while the adults came through a month later. Far to the south in the Antarctic, young Adelie penguins depart for northern wintering grounds much earlier than adults.
- 37 -
In a few species, adults depart south before the young. Adult golden plovers, Hudsonian godwits, and probably most of the Arctic breeding shorebirds leave the young as soon as they are capable of caring for themselves and set out for South America ahead of the juveniles. Likewise, data for the least flycatcher indicate adults migrate before the young, but Johnson (1963) did not find this segregation in the Hammond's flycatcher. In Europe, adult red-backed shrikes are known to migrate ahead of their young.

- 38 -
In contrast to this loss of parental concern, geese, swans, and cranes remain in family groups throughout migration. The parent birds undergo a wing molt that renders them flightless during the period of growth of their young so that both the adults and immatures acquire their flight capabilities at the same time and are able to start south together. Large flocks of Canada geese, for example, are composed of many families banded together. When these flocks separate into small V-shaped units it is probably correct to assume an old goose or gander is leading the family. After female ducks start to incubate their eggs, the males of most species of ducks flock by themselves and remain together until fall. When segregation of the sexes such as this occurs the young birds often accompany their mothers south. Murray and Jehl (1964) concluded from mist-netting many thousands of migrant passerines at Island Beach, New Jersey, that adults and juveniles travel at approximately the same time.
Males and females of some species may migrate either simultaneously or separately. In the latter case it is usually the males, rarely the females, that arrive first. Sometimes great flocks of male red-winged blackbirds reach a locality several days before any females; this is particularly the rule in spring. The first robins are usually found to be males, as are also the first song sparrows, rose-breasted grosbeaks, and scarlet tanagers. In Europe, the three buntings mentioned previously are also segregated as to sex during migration. Figure 8 shows two prominent peaks for both the Cretzschmar's and Ortolan buntings; during passage the first peak was primarily males while the second peak consisted mostly of females. This early arrival of males on the breeding grounds is associated with territorial possession whereby the male selects the area where it intends to breed and each individual attempts to protect a definite territory from trespass by other males of his own kind, while announcing his presence to rival males and later arriving females by song or other display. The female then selects the site where she wishes to nest. The long-billed marsh wren is a noteworthy example; the males may enthusiastically build several nests before the females arrive. In the fall, common and king eiders are sexually segregated during migration. During July, flocks crossing Point Barrow are composed almost entirely of males, while after the middle of August the flocks are almost all females (Thompson and Person 1963). In the Chicago area, Annan (1962) reported that some males, such as the hermit thrush, Swainson's thrush, gray-cheeked thrush, and veery, arrive before any females and predominate during the first week of occurrence.
In a few species the males and females apparently arrive at the breeding grounds together and proceed at once to nest building. In fact, among shorebirds, ducks, and geese, courtship and mating often takes place in whole or in part while the birds are in the South or - 39 - on their way north, so that when they arrive at the northern nesting grounds they are paired and ready to proceed at once with raising their families. Mallards and black ducks may be observed in pairs as early as December, the female leading and the male following when they take flight. Naturally, these mated pairs migrate north in company, and it was largely to protect such pairings that duck shooting in spring was abolished by Federal law.
In the coastal subspecies of the western flycatcher, the sexes appear to migrate in synchrony during the spring in contrast to migration of Hammond's flycatcher in which the adult males usually precede the females (Johnson 1973). Both sexes of the common blackcap of Europe appear to migrate together at least across the eastern end of the Mediterranean during the spring (Fig. 9.).
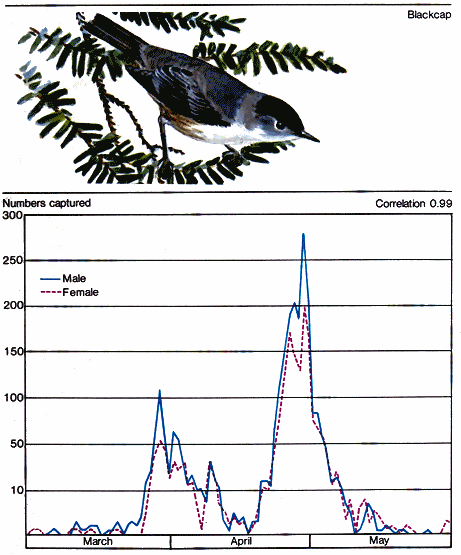
- 40 -
Migratory flights are frequently accomplished in close flock formation, as with shorebirds, blackbirds, waxwings, and especially some of the buntings, longspurs, juncos, and tree sparrows. Other species maintain a very loose flock formation; examples are turkey vultures, hawks, swifts, blue jays, swallows, warblers, and bluebirds. Still others, the grebes, snowy owls, winter wrens, shrikes, and belted kingfishers, ordinarily travel alone, and when several are found in close proximity it is an indication they have been drawn together by unusual conditions, such as abundant food.
Just as flocking among resident birds provides group protection against predation, flocking in migration greatly facilitates the attainment of destination (Pettingill 1970). The V-shaped flocks often associated with Canada geese have a definite energy conserving function by creating favorable air currents for every member of the flock but the leader; when the leader becomes tired, it will often change places with a member behind. Night migrating flocks generally fly in looser formations than do day migrating flocks.
- 41 -
Both length and duration of migratory journeys vary greatly between families, species, or populations within a species. Bobwhite, western quails, cardinals, Carolina wrens, and probably some of the titmice and woodpeckers are apparently almost or entirely nonmigratory. These species may live out their entire existence without going more than 10 miles from the nest where they were hatched.
Many song sparrows, meadowlarks, blue jays, and other species make such short migrations that the movement is difficult to detect because individuals, possibly not the same ones, may be found in one area throughout the year while other individuals that move south may be replaced by individuals from the north. Information on different movements of this type, within a species, can be gained by observing birds marked with numbered bands, colored materials, or identification of racially distinct museum specimens.
The American robin is a good example of this type of movement. This species occurs in the southern United States throughout the year, but in Canada and Alaska only during the summer. Its movements are readily ascertained from study specimens. The breeding robin of the southeastern states is the southern race. In autumn most of its more northern nesters, such as those from Maryland and Virginia move into the southern part of the breeding range or slightly farther south. At about the same time the northern American robin moves south and winters throughout the breeding and wintering range of its smaller and paler southern relative. Thus there is complete overlap of wintering ranges of northern and southern American robin populations, although some individuals of the northern race winter in areas vacated earlier by the southern race.
Among many migratory species there is considerable variation among individuals and populations with respect to distances moved. Certain populations may be quite sedentary while others are strongly migratory, and certain individuals of the same population can be more migratory than others. For example, red-winged blackbirds nesting on the Gulf Coast are practically sedentary, but in winter they are joined by other subspecies that nest as far north as the Mackenzie Valley. In certain populations of the song sparrow and other species, males remain all year on their northern breeding grounds while the females and young migrate south.
Several species containing more than one distinguishable - 42 - population exhibit "leap-frog" migration patterns. The familiar eastern fox sparrow breeds from northeastern Manitoba to Labrador, but during the winter it is found concentrated in the southeastern part of the United States. On the west coast of the continent, however, a study of museum specimens by Swarth (1920), indicated six subspecies of this bird breeding in rather sharply delimited ranges extending from Puget Sound and Vancouver Island to Unimak Island, at the end of the Alaskan Peninsula. One of these subspecies, known as the sooty fox sparrow, breeds from the Puget Sound-Vancouver Island area northward along part of the coast of British Columbia. It hardly migrates at all, while the other races, nesting on the coast of Alaska, are found in winter far to the south in Oregon and California. Although much overlap exists, the races breeding farthest north generally tend to winter farthest south. This illustrates a tendency for those populations forced to migrate to pass over those subspecies so favorably located as to be almost sedentary. If the northern birds settled for the winter along with the sedentary population, winter requirements may not be as sufficient as in the unoccupied areas farther south (Fig. 10.). Therefore, natural selection has insured the different populations will survive the winter by separating the subspecies into different wintering areas.
Another example of this "leap-frog" migration is illustrated by the common yellowthroat of the Atlantic coast. Birds occupying the most southern part of the general range are almost nonmigratory and reside throughout the year in Florida, whereas the population that breeds as far north as Newfoundland goes to the West Indies for the winter. Thus the northern population literally "jumps" over the home of the southern relatives during migratory journeys.
The palm warbler breeds from Nova Scotia and Maine west and northwest to southern Mackenzie. The species has been separated into two subspecies: those breeding in the interior of Canada and those breeding in northeastern United States and Canada. The northwestern subspecies makes a 3,000-mile journey from Great Slave Lake to Cuba and passes through the Gulf States early in October. After the bulk of these birds have passed, the eastern subspecies, whose migratory journey is about half as long, drifts slowly into the Gulf Coast region and remains for the winter.
Some species have extensive summer ranges (e.g., the pine warbler, rock wren, field sparrow, loggerhead shrike, and black-headed grosbeak) and concentrate during the winter season in the southern part of the breeding range or occupy additional territory only a short distance farther south. The entire species may thus be confined within a restricted area during winter, but with the return of warmer weather, the species spreads out to reoccupy the much larger summer range.
- 43 -
- 44 -
Many species, including the tree sparrow, snow bunting, and Lapland longspur, nest in the far north and winter in the eastern United States, while others, including the vesper and chipping sparrows, common grackle, red-winged blackbird, eastern bluebird, American woodcock, and several species of ducks, nest much farther south in the United States and Canada and move south a relatively short distance for the winter to areas along the Gulf of Mexico. In a few of the more hardy species, individuals may linger in protected places well within reach of severe cold. The common snipe, for example, is frequently found during subzero weather in parts of the Rocky Mountain region where warm springs assure a food supply. More than 100 summer birds leave the United States entirely and spend the winter in the West Indies, Central America, or South America. For example, the Cape May warbler breeds from northern New England, northern Michigan, and northern Minnesota, north to New Brunswick, Nova Scotia, and nearly to Great Slave Lake. In winter it is concentrated chiefly in the West Indies on the island of Hispaniola.
Some of the common summer residents of North America are not content with a trip to northern tropical areas of the West Indies and Central America, but push on across the Equator and finally come to rest for the winter in Patagonia or the pampas of Argentina. Species such as nighthawks, some barn swallows, cliff swallows, and a few thrushes may occupy the same general winter quarters in Brazil, but other nighthawks and barn swallows go farther south. Of all North American landbirds these species probably travel the farthest; they are found north in summer to the Yukon Territory and Alaska, and south in winter to Argentina, 7,000 miles away. Such seasonal flights are exceeded in length, however, by the remarkable journeys of several species of shorebirds including white-rumped and Baird's sandpipers, greater yellowlegs, turnstones, red knots, and sanderlings. In this group, 19 species breed north of the Arctic Circle and winter in South America; six of these go as far south as Patagonia, a distance of over 8,000 miles.
The Arctic tern is the champion "globe trotter" and long-distance flier (Fig. 11.). Its name "Arctic" is well earned, as its breeding range is circumpolar and it nests as far north as the land extends in North America. The first nest found in this region was only 7½° (518 miles) from the North Pole and contained a downy chick surrounded by a wall of newly fallen snow scooped out by the parent. In North America the Arctic tern breeds south in the interior to Great Slave Lake, and on the Atlantic coast to Massachusetts. After the young are grown, the Arctic terns disappear from their North American breeding grounds and turn up a few months later in the Antarctic region, 11,000 miles away. For a long time the route followed by these hardy fliers was a complete mystery; although a few scattered individuals have been noted south as far as Long Island in the United States, the species is otherwise practically unknown along the Atlantic coasts of North America and northern South America. It is, however, known as a migrant on the west coast of Europe and Africa. By means of numbered bands, a picture disclosed what is apparently not only the longest, but also one of the most remarkable migratory journeys (Austin 1928).
- 45 -
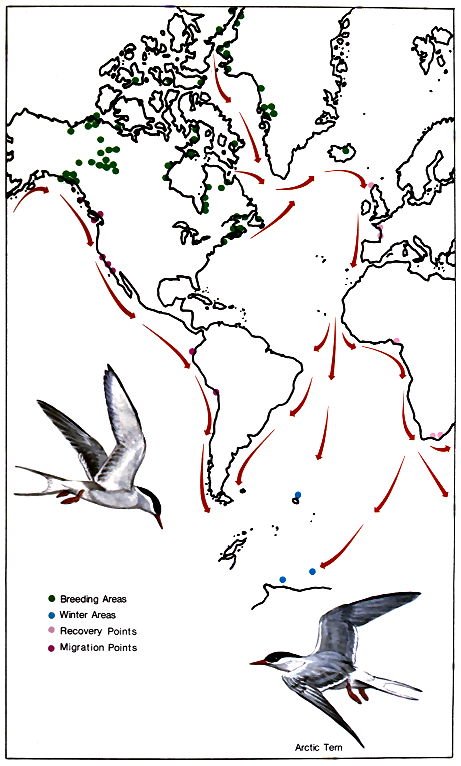
- 46 -
Few other animals in the world enjoy as many hours of daylight as the Arctic tern. For these birds, the sun never sets during the nesting season in the northern part of the range, and during their winter sojourn to the south, daylight is continuous as well. In other months of the year considerably more daylight than darkness is encountered.
- 47 -
There probably is no single aspect of the entire subject of bird migration that increases our admiration so much as the unerring certainty with which birds cover thousands of miles of land and water to come to rest in exactly the same spot where they spent the previous summer or winter. Records from birds marked with numbered bands offer abundant proof that the same individuals of many species will return again and again to identical nesting or winter feeding sites.
This ability to travel with precision over seemingly featureless stretches of land or water is not limited to birds but is likewise possessed by certain mammals, reptiles, fishes, and insects; the well-known migrations of salmon and eels are notable examples.
For an animal to return to a specific spot after a lengthy migration, it must use true navigation to get there. That is, it needs to not only travel in a given compass heading and know where it is at any given time so the course may be altered when necessary but also be able to recognize its goal when it has arrived. It is dangerous to generalize on the means of orientation and navigation in migration; different groups of birds with different modes of existence have evolved different means of finding their way from one place to another (Pettingill 1970). We are only beginning to realize the complexities involved in the many modes of bird orientation and navigation. All we can do in this section is present a brief summary of some of the more important principles involved and the studies that have enhanced our knowledge in the area.
Ability to follow a more or less definite course to a definite goal is evidently part of an inherited faculty. Both the direction and the goal must have been implanted in the bird's genetic code when the particular population became established at its present location. The theory is sometimes advanced that older and more experienced birds lead the way and thereby show the route to their younger companions. This explanation may be acceptable for some species such as geese, swans, and cranes because they stay in family groups, but not for species in which adults and young are known to migrate at different times, especially when young migrate ahead of the adults. As indicated in a previous section on segregation, many North American shorebirds as well as the cuckoos of New Zealand do this. An inherited response to its surroundings, with a definite sense of the goal to be reached and the direction to be followed, must be attributed to these latter birds.
It is well known that birds possess wonderful vision. If they also have retentive memories subsequent trips over the route may well be - 48 - steered in part by recognizable landmarks. Arguments against the theory of landmark memory are chiefly that unescorted young birds, without previous experience, can find their way to the winter quarters of their species, even if the wintering area has a radically different landscape and vegetation than the breeding grounds. Experimental findings and field observations indicate landmarks are used in navigation by certain birds, but the degree of use varies considerably among the species (Bellrose 1972a).
To a land-dweller traveling the ocean, the vast expanse may seem featureless but the reverse may be true for a seabird blown over land by a storm. In the latter situation the differences in vegetation and topography "obvious" to land-dwellers are completely foreign to a seabird as it has had little previous experience to help interpret these "strange objects." Griffin and Hock (1949) observed the flight behavior of gannets displaced far inland away from their nests. The bird appeared to search randomly until the coastline was met, then the fliers pursued a much more direct course home. Herring gulls, displaced about 250 miles from their nest in 2 consecutive years, returned the second year in one-sixth the time required the first year (Griffin 1943). To birds such as gannets, albatrosses, and shearwaters, which spend almost their entire lives traveling thousands of miles at sea and return to very specific nesting areas, the "featureless ocean expanses" are probably very rich in visual cues. It is difficult to believe a bird dependent on the sea for its livelihood cannot help but be aware of wave direction, islands, reefs, atolls, concentrations of floating flotsam, organisms, currents, clouds over islands, fog belts, etc.
Much migration takes place at night and great stretches of the open sea are crossed to reach destinations. Nights are rarely so dark that all terrestrial objects are totally obscured, and features such as coastlines and rivers are just those that are most likely to be seen in the faintest light, particularly by the acute vision of birds from their aerial points of observation. Even if terrestrial objects are completely obscured on a very dark night, the migrants are still able to assess their surroundings during the day before starting out again.
Some birds, especially colonial seabirds, seem to be able to fly unerringly through the densest fog, particularly in the vicinity of their nest site. Members of the Biological Survey, proceeding by steamer through a dense fog from the island of Unalaska to Bogoslof Island in the Bering Sea, recorded flocks of murres, returning to Bogoslof after quests for food. The birds broke through the wall of fog astern, flew by the vessel, and disappeared into the mists ahead on the same course as the ship. On the other hand, radar observations of migrating birds have indicated strong directional movements on clear nights but often completely random movements in heavily overcast or stormy weather. Possibly some birds can perceive the position of the sun through an overcast as honey bees are known to do. It is less likely the stars could be detected through overcast at night.
Careful studies have been made on the homing instinct in various - 49 - seabirds such as Laysan albatrosses, Manx shearwaters, and several tropical species of terns. Sooty and noddy terns reach their most northern breeding point on the Dry Tortugas, off the southwest coast of Florida. They are not known to wander any appreciable distance farther north. Displaced breeding birds returned to their nests on the Dry Tortugas after they had been taken on board ship, confined in cages below decks, and carried northward 400 to 800 miles before being released in a region where they had had no previous experience. Likewise, Laysan albatrosses and Manx shearwaters have returned over 3,000 miles in similar homing experiments.
Possibly the "homing instinct," as shown by pigeons, terns, shearwaters, albatrosses, and by the frigatebirds trained as message carriers in the South Pacific, may not be identical with the sense of perceptive orientation that figures in the flights of migratory birds. Nevertheless, it seems closely akin and is probably governed by the same mechanisms. There are good reasons to assume that once we know the processes governing displaced homing we will know, in general, how birds navigate; this question is still far from being answered (Wallraff 1967).
Some students have leaned toward the possible existence of a "magnetic sense" as being the important factor in the power of geographical orientation. The theory was conceived as early as 1855 and reported in 1882 by Viguier. Investigations of this have been conducted by Yeagley (1947) and Gordon (1948) with contradictory results. In 1951, Yeagley incorporated the idea that sensitivity to the effect of the earth's rotary motion through the vertical component of the magnetic field is the means of orientation. The basic question asked of the theory is: "Can birds detect such minute differences in the earth's magnetic field and can these forces affect bird behavior?"
Attempts to demonstrate the effect of radio waves on the navigational ability of birds have produced contradictory results. In some of these tests, homing pigeons released near broadcasting stations have appeared to be hopelessly confused, whereas in others, apparently conducted in the same manner, no effects could be discerned. Before sensitivity of birds to electromagnetic stimuli of any kind can be accepted or rejected, much additional experimental work is necessary.
Human navigators have used the heavenly bodies in determining their course and position for centuries. It would not be surprising then to find other long-distance travelers using the same method. One of the most constant visual cues a migrating bird could use would be the sun's or moon's path and the location of the stars.
Some of the more recent experimental work on bird navigation has been with astronomical (sun) and celestial (star) directional clues. Studies by Kramer, Sauer, and others have indicated a phenomenal inherited ability in birds to use the position of the sun by day and the stars by night to chart their courses. This involves an intricate compensation for daily, seasonal, and geographical changes in the positions of these heavenly bodies. Kramer (1957, 1961) placed - 50 - diurnal migrants in circular cages and "changed" the position of the sun with mirrors. The birds shifted their position to compensate for these changes. Sauer (1957, 1958), in a fascinating study with nocturnal migrant warblers, placed birds in a round cage open to the sky. These birds oriented in the normal direction for that locality and time of year. He next placed the cage and birds in a planetarium and projected overhead the night sky star patterns for different seasons and localities. The familiar star pattern produced a normal orientation but an unfamiliar sky caused confusion and complete disorientation. These experiments, begun in Germany, are still continuing in other countries with other species. Emlen (1969) used photoperiod manipulation to change the physiological states of spring and fall migratory readiness in indigo buntings. Half the sample of birds were in breeding condition whereas the other half were already past the reproductive stage even though it was spring "outside." When these birds were subjected to a spring star pattern in a planetarium, the birds in spring condition oriented northward but those in autumnal condition oriented southward. Although some results have been negative, by and large the evidence supports the original findings that the sun and stars are visual "landmarks" used by at least some birds as well as bees and probably many other creatures in finding their way home as well as to their winter and summer quarters.
In conclusion, then, we can say this about bird orientation and navigation: 1) many cues are available to birds for migratory guidance and one or several of these may be used by any migrant; 2) different species and groups of birds use different cues, depending on their migration traits; 3) visual cues probably play a predominant role in migration (radar studies have indicated that some birds can maintain their orientation even under completely overcast nights, although they usually become disoriented under such conditions); and 4) long-distance migrants and pelagic species have a much higher developed sense of orientation than those species that migrate only short distances or not at all.
- 51 -
It is thought by some that the weather has little to do with the time of arrival of migratory birds. It is assumed that if the bird is physiologically prepared for migration it departs, irrespective of the weather. Even if this were the case, weather can influence the progress of migration by not only controlling the advance of the seasons but also by helping, hindering, or even stopping bird flight (Welty 1962).
Some scientists believe that birds not only avoid bad weather at the start of a journey but usually finish the journey in good weather (Nesbit and Drury 1967b). Contrary to what many observers believe, the arrival of birds in an area, whether they stop or continue on, is more often controlled by the weather at the point of departure than at the point of arrival. During the peak of migration, suitable weather may occur at an observation site, but strong migratory movements may be arrested before the birds arrive there because the weather was not suitable at the point of departure or somewhere in between. In addition, if there is good weather at the point of departure as well as farther down the migration route, the migrants, once air-borne in a favorable weather pattern, may continue on right over an expectant observer and the whole flight will be missed. Nesbit and Drury's (1967b) radar study on air-ground comparisons found, with few exceptions, ground observers missed the largest movements observed on radar. Observation of a large wave of arrivals indicated migrants had been stopped by a meteorological barrier, and people were actually not reporting maximum migration but an interruption to migration. Therefore, when migration is proceeding normally under safe conditions, very little movement is visible to the ground observer but a large arrival of birds on the ground often indicates something is not in order and the migrants have been forced to stop for one reason or another.
The question is frequently asked: "How can I identify weather conditions suitable or unsuitable for migration?" It is almost impossible to discuss separately the effects of different weather factors on migration because barometric pressure, temperature, wind, and other meteorological phenomena are very closely related.
On the North American continent, air masses generally proceed about 600 miles per day from the west to the east. These air masses vary in pressure, temperature, humidity, and wind. The wind within these masses travels in either a clockwise (anticyclonic) or counter-clockwise (cyclonic) direction. Cyclonic air masses contain relatively moist warm air with low barometric pressure centers and are designated "lows"; anticyclonic air masses are characterized by - 52 - dry cool air with high barometric pressure areas and are called "highs." Where these air masses meet, a "front" is formed, and the rapidity with which this front moves through an area depends on the temperature and pressure gradient on either side of the front.
An understanding of frontal systems, with their associated wind, temperature and humidity, is one of the keys to understanding when birds migrate. You must not only watch the fronts in your area but the progress of nearby air masses as well because the birds migrating through your area have started their journey to the north or south of you depending on the season. The weather conditions at point of departure will dictate if and when birds will be passing through your area in the near future.
- 53 -
During fall migration, the best passage of migrants usually occurs 2 days after a cold front has gone through. That is, the low has passed and it is being followed by a high characterized by dropping temperatures, a rising barometer, and clearing skies. The 24 hours just after a low has passed are not always conducive to a good passage of birds because winds are often too strong and turbulent in the trough between the two air masses. Hochbaum (1955) correlated mass movements of ducks through the prairies with weather systems and noted the combination of weather conditions described above was ideal for mass migrations of ducks during November. During this period, observers at Delta, Manitoba, south to Louisiana recorded a tremendous flight of ducks as the proper conditions of barometric pressure, temperature, wind, and cloud cover passed across the central United States and Canada. An example of the type of weather system that is often associated with mass movements is illustrated in Fig. 12.
Records of lapwings on Newfoundland and the Gulf of St. Lawrence appear to be the result of a particular series of meteorological events (Bagg 1967). The lapwing is a European species rarely found in the New World. If cold air moves into western Europe from the east, lapwings move westward into England, Wales, and Ireland. Occasionally, the development of an anomalous weather pattern over the North Atlantic including an elongated low from Europe to eastern Canada causes some birds to be literally "blown" in the counter-clockwise airstream across the Atlantic to the Gulf of St. Lawrence.
During spring migration, weather conditions conducive to strong movements of birds are somewhat the opposite from those in the fall. Migrants will move north on the warm sector of an incoming low. When a high pressure area has just passed, the influx of warm moist tropical air is extended and intensified (Bagg et al. 1950). However, during this time, cloudiness and rain associated with the low may curtail migration or squeeze it into a narrow period proceeding along the warm front. If a fast moving cold front approaches from the northwest, the rapid movement of migrants will be sharply curtailed or even grounded until more favorable conditions occur.
The incessant crescendo note of the ovenbird is ordinarily associated with the full verdure of May woods, but this bird has been known to reach its breeding grounds in a snowstorm, and the records of its arrival in southern Minnesota show a temperature variation from near freezing to full summer warmth. Temperatures at arrival of several other common birds vary from 14° between highest and lowest temperatures to 37°, the average variation being about 24°. North American species spending the winter months in tropical latitudes experience no marked changes in temperature conditions from November to March or April, yet frequently they will start the northward movement in January or February. This is in obedience to physiological promptings and has no relation to the prevailing weather conditions. For migratory birds the winter season is a period of rest, a time when they have no cares other than those associated with the daily search for food or escape from their natural enemies. Their migrations, however, are a vital part of their life cycles, which have become so well adjusted that the seasons of travel correspond in general with the major seasonal changes on their breeding grounds. With the approach of spring, therefore, the reproductive impulse awakens, and each individual bird is irresistibly impelled to start the journey that ends in its summer home.
- 54 -
In other words, the evidence indicates the urge to migrate is so innate within a species or population that the individuals move north in spring when the average weather is not unendurable. The word "average" must be emphasized since it appears the migrations of birds have evolved in synchrony with average climatic conditions. More northern nesting populations of species such as American robins and savannah sparrows, timed to arrive on their breeding ground when the weather is suitable, pass through areas where their more southern kin are already nesting. The hardy species travel early, fearless of the blasts of retreating winter, while the more delicate kinds come later when there is less danger of encountering prolonged periods of inclement weather. Some of the hardy birds pause in favorable areas and allow the spring season to advance. Then, by rapid travel they again overtake it, or, as sometimes happens, they actually outstrip it. Occasionally this results in some hardship, but rarely in the destruction of large numbers of individuals after arrival. Cases are known where early migrating bluebirds have been overwhelmed by late winter storms. Nevertheless, if such unfavorable conditions are not prolonged, no serious effect on the species is noted. The soundness of the bird's instincts is evidenced by the fact that natural catastrophes, great though they may be, do not permanently diminish the avian populations.
The spring flight of migrants, if interrupted by cold north winds, is resumed when weather conditions again become favorable, and it is probable that all instances of arrival in stormy weather can be explained on the theory that the flight was begun while the weather was auspicious. Even though major movements of migrants in spring generally coincide with periods of warm weather and southerly winds, observations on the beginning of nocturnal spring flights from the coast of Louisiana failed to note any inhibiting factor other than hard rain (Gauthreaux 1971).
Radar studies have indicated that migrant birds possess an amazing understanding of wind patterns (Bellrose 1967). Birds can recognize many characteristics and select for favorable patterns. Head winds are as unfavorable to migration as is rain or snow because they greatly increase the labor of flight and cut down the speed of cross-country travel. If such winds have a particularly high velocity, they may force down the weaker travelers, and when this happens over water, large numbers of birds are lost. Moderate tail winds and cross or quartering breezes appear to offer the best conditions for the flight of migrants. Richardson (1971) found migrants traveling in different directions at different altitudes, but each group of birds was aided by a following wind. Thus we might expect natural selection to operate in favor of those birds that could recognize and respond to favorable wind patterns because it would reduce energy consumption and flight time on long-distance flights (Hassler et al. 1963).
Soaring birds such as hawks, vultures, and storks are very - 55 - dependent on proper wind conditions for migration. In the fall, often the best day to observe hawk migration in the eastern United States is on the second day after a cold front has passed providing there are steady northwest to west winds and a sunny day for production of thermals (Pettingill 1962). Considerable drifting may be observed in this group of birds because they are literally carried along by the wind or glide from one thermal to the next. Haugh and Cade (1966) found most hawks migrated around Lake Ontario when winds were 10 to 25 miles per hour, but, if the wind exceeded 35 miles per hour, most hawk migration stopped.
In conclusion then, we can say that the weather may be the impetus for migration for many species, but it cannot stimulate a bird to migrate unless it is physiologically prepared. Arrivals on the ground are not necessarily indicative of the number of birds passing overhead. During the fall, peak migrations usually follow the passage of a cold front when the temperature is falling, the barometer is rising, winds are from the west or northwest, and the sky is clearing. In the spring, most migrants proceed north in the warm sector of a low when winds are southerly, warm, and moist, but rain, fog, or snow will often curtail the passage of migrants or prevent the initiation of a migration. Evolution of migratory behavior has probably resulted from the survival of birds capable of selecting those wind conditions, which reduce flight time and energy consumption, during their passage.
- 56 -
The relation of the world's land masses to each other and the distribution and association of biotypes within these land masses influence the direction birds migrate. Topography may aid, hinder, or prevent the progress of a migrant depending on the bird's particular requirements. Old World migrants must contend with east-west tending mountain ranges and deserts, whereas New World travelers can proceed north and south across a landscape with its major mountain ranges and river systems oriented in the same direction as the birds migrate.
When a distinct feature in the landscape, such as borders between fields and forests, rivers, mountain ridges, desert rims, or peninsulas, appears to influence migratory travel, we call these formations "guiding lines," "diversion-lines," "leading lines," or in German, "Leitlinie." It is an observed fact that some birds in a migratory movement alter their course to travel along a leading line, but whether this feature in the landscape caused the migrants to change their course is only theory (Thomson 1960). Besides topography, many other factors can influence this type of flight behavior including weather, wind speed and direction, time of day, species, age, and experience of the bird (Murray 1964).
Large bodies of water constitute real barriers to soaring birds dependent on thermals and air currents. Good examples of these barriers include the Mediterranean Sea between Europe and Africa and the Great Lakes in North America. Because these water areas do not create good thermals (generally a warm surface, such as a large field on a sunny day, is needed to create the necessary rising air currents for thermals to form) for birds to soar on, migrants are forced to travel around them on up-drafts created where land and water meet. The shoreline, then, may appear to be the guiding line, but more than likely the birds are simply following air currents created by onshore winds replacing the rising air from the surrounding warmer land surface and being deflected upward by the shoreline. These conditions often concentrate our buteos (broad-winged, rough-legged, red-shouldered, and red-tailed hawks) into restricted areas where, on good days, numbers observed can be spectacular. Similar conditions exist over the Bosphorus at the eastern end of the Mediterranean Sea where literally thousands of storks, eagles, and buzzards can be observed on a good day.
While extensive water areas may alter the migratory path of soaring birds, mountain ridges, especially if parallel to the line of flight, are often very conducive to migratory travel. Systematic coverage of the Appalachian ridges indicates all of them aid the - 57 - migration of soaring birds. Apparently the highest and longest ridges deflect the horizontal winds upward better than the shorter ridges less than 1,000 feet high, and more birds are seen, on the average, along the higher ridges (Robbins 1956).
In general, nocturnal migrants are not influenced by topography as much as diurnal travellers. Radar observations have played an important role in establishing this difference. Bellrose (1967) found that waterfowl migrating at night through the Midwest were not influenced by major river systems, but in the evening or after daybreak ducks and geese tended to alter their course along the rivers. Drury et al. (1961) recorded massive fall and spring movements from the New England area out over the Atlantic Ocean without any apparent regard for the coastline. Until nocturnal migration could be "watched" on a radar screen, many bird observers assumed the guiding effect of the coastline on migratory travel was more restrictive than it really is.
In summary, topography may help or deter a migrant in its passage. It affects different birds in different ways. In North America, migratory movements are continent wide, and no evidence has indicated any particular part of the landscape influences all birds in the same manner. Certain bird populations may use general areas in migration, but they are usually not rigidly restricted to them because of topography.
- 58 -
The migration season is full of peril for birds. Untold thousands of smaller migrants are destroyed each year by storms and attacks by predatory animals. These mortality factors, and others, help keep bird populations in check. Perils of migration are among these causes.
Of all the hazards confronting birds in migration, particularly the smaller species, storms are the most dangerous. Birds that cross broad stretches of water can be blown off course by a storm, become exhausted, and fall into the waves. Such a catastrophe was once seen from the deck of a vessel in the Gulf of Mexico, 30 miles off the mouth of the Mississippi River. Great numbers of migrating birds, chiefly warblers, were nearing land after having accomplished nearly 95 percent of their long flight when, caught by a "norther" against which they were unable to make headway, hundreds were forced into the waters of the Gulf and drowned. A sudden drop in temperature accompanied by a snowfall can cause a similar affect.
Lighthouses, tall buildings, monuments, television towers, and other aerial obstructions have been responsible for destruction of migratory birds. Bright beams of lights on buildings and airport ceilometers have a powerful attraction for nocturnal air travelers that may be likened to the fascination for lights exhibited by many insects, particularly night-flying moths. The attraction is most noticeable on foggy nights when the rays have a dazzling effect that not only lures the birds but confuses them and causes their death by collision against high structures. The fixed, white, stationary light located 180 feet above sea level at Ponce de Leon Inlet (formerly Mosquito Inlet), Florida, has caused great destruction of bird life even though the lens is shielded by wire netting. Two other lighthouses at the southern end of Florida, Sombrero Key and Fowey Rocks, have been the cause of a great number of bird tragedies, while heavy mortality has been noted also at some of the lights on the Great Lakes and on the coast of Quebec. Fixed white lights seem to be most attractive to birds; lighthouses equipped with flashing or red lights do not have the same attraction.
For many years in Washington, B.C., the illuminated Washington Monument, towering more than 555 feet into the air, caused destruction of large numbers of small birds. Batteries of brilliant floodlights grouped on all four sides about the base illuminate the - 59 - Monument so brilliantly, airplane pilots noticed that it could be seen for 40 miles on a clear night. It is certain there is an extensive area of illumination, and on dark nights with gusty, northerly winds, nocturnal migrants seem to fly at lower altitudes and are attracted to the Monument. As they mill about the shaft, they are dashed against it by eddies of wind, and hundreds have been killed in a single night.
In September 1948, bird students were startled by news of the wholesale destruction of common yellowthroats, American redstarts, ovenbirds, and others against the 1,250-foot-high Empire State Building in New York City, the 491-foot-high Philadelphia Saving Fund Society Building in Philadelphia, and the 450-foot-high WBAL radio tower in Baltimore. In New York, the birds continued to crash into the Empire State Building for 6 hours.
More recently, the television tower has become the chief hazard. These structures are so tall, sometimes over 1,000 feet, they present more of a menace than buildings or lighthouses. Their blinking lights cause passing migrants to blunder into guy wires or the tower itself while milling around like moths about a flame. Numerous instances (e.g. Stoddard and Norris 1967) throughout the U.S. indicate this peril to migration is widespread. The lethal qualities of airport ceilometers have been effectively modified by conversion to intermittent or rotating beams.
Both soaring and sailing birds are so proficient in aerial transportation that only recently have the principles been understood and imitated by aircraft pilots. The use of ascending air currents, employed by all soaring birds and easily demonstrated by observing gulls glide hour after hour along the windward side of a ship, are now utilized by man in his operation of gliders. Moreover, the whole structure of a bird makes it the most perfect machine for extensive flight the world has ever known. Hollow, air-filled bones, together with feathers, the lightest and toughest material known for flight, have evolved in combination to produce a perfect flying machine.
Mere consideration of a bird's economy of fuel or energy also is enlightening. The golden plover probably travels over a 2,400-mile oceanic route from Nova Scotia to South America in about 48 hours of continuous flight. This is accomplished with the consumption of less than 2 ounces of body fat (fuel). In contrast, to be just as efficient in operation, a 1,000-pound airplane would consume only a single pint of fuel in a 20-mile flight rather than the gallon usually required. Similarly, the tiny ruby-throated hummingbird weighing approximately 4 grams, crosses the Gulf of Mexico in a single flight of more than 500 miles while consuming less than 1 gram of fat.
One might expect the exertion incident to long migratory flights would result in arrival of migrants at their destination near a state of exhaustion. This is usually not the case. Birds that have recently arrived from a protracted flight over land or sea sometimes show - 60 - evidences of being tired, but their condition is far from being in a state of emaciation or exhaustion. The popular notion birds find long ocean flights so excessively wearisome that they sink exhausted when terra firma is reached generally does not coincide with the facts.
The truth is, even small landbirds are so little exhausted by ocean voyages, they not only cross the Gulf of Mexico at its widest point but may even proceed without pause many miles inland before stopping. The sora, considered such a weak flyer that at least one writer was led to infer most of its migration was made on foot, has one of the longest migration routes of any member of the rail family and even crosses the wide reaches of the Caribbean Sea. Observations indicate that under favorable conditions birds can fly when and where they please and the distance covered in a single flight is governed chiefly by the amount of stored fat. Exhaustion, except as the result of unusual factors such as strong adverse winds, cannot be said to be an important peril of migration.
- 61 -
While it is beyond question that certain general directions of flight are consistently followed by migratory birds, it is well to remember the term "migration route" is to some extent a theoretical concept referring to the lines of general advance or retreat of a species, rather than the exact course followed by individual birds or a path followed by a species with specific geographic or ecological boundaries. Even the records of banded birds usually show no more than the place of banding and recovery. One ought to have recourse to intermediate records and reasoning based on probabilities to fill in details of the route actually traversed between the two points. In determining migration routes, one must constantly guard against the false assumption that localities with many grounded migrants are on the main path of migration and localities where no grounded migrants are observed are off the main path.
There is also infinite variety in the routes covered during migration by different species. In fact, the choice of migration highways is so wide that is seems as if the routes of no two species coincide. Differences in distance traveled, time of starting, speed of flight, geographical position, latitudes of breeding and wintering grounds, and other factors contribute to this great variation of migration routes. Nevertheless, there are certain factors that serve to guide individuals or groups of individuals along more or less definite lines, and it is possible to define such lines of migration for many species.
Except in a few species, individuals probably do not follow precisely the same route twice. This is especially true in the group of soaring birds that utilize thermals. Mueller and Berger (1967b) recaptured only three migrants in subsequent years at Cedar Grove, Wisconsin, after banding over 50,000 birds there. In general, those populations of species with very discernible breeding or wintering grounds have readily discernible migration routes. However, even the whole migration process of certain species may show great yearly fluctuations (Rudebeck 1950).
Aldrich et al. (1949) showed from banding data great variation in migration patterns between species of waterfowl. In some species there was considerable diversity in direction of movement, not only of different breeding populations within a species but also for different individuals of the same breeding population. The impression is inescapable; waterfowl migration is even more complicated than originally supposed, and it is difficult to make generalizations with - 62 - regard to migration pathways for even a single species let alone waterfowl in general.
Through plotting accumulated banding data in the 1930's, investigators became impressed by what appeared to be four broad, relatively exclusive flyway belts in North America. This concept, based upon analyses of the several thousand records of migratory waterfowls recoveries then available, was described by Lincoln (1935a). In this paper (p. 10), Lincoln concluded that:
... because of the great attachment of migratory birds for their ancestral flyways, it would be possible practically to exterminate the ducks of the West without seriously interfering with the supply of birds of the same species in the Atlantic and Mississippi flyways, and that the birds of these species using the eastern flyways would be slow to overflow and repopulate the devastated areas of the West, even though environmental conditions might be so altered as to be entirely favorable.
Since 1948, this concept served as the basis for administrative action by the Fish and Wildlife Service in setting annual migratory waterfowl hunting regulations.
The concept of bird populations being confined to four fairly definite and distinct migration "flyways" is probably most applicable to those birds that migrate in family groups, namely geese, swans, and cranes, but does not appear to be very helpful in understanding the movements of the more widely dispersing ducks. The "pioneering spirit" in Canada geese, for example, is limited by their social structure the young travel to and from specific breeding and wintering areas with their parents. These young later in life usually breed in the same areas as did their parents. If a goose population is decimated in one flyway, either by hunting or natural calamities, other goose populations in other flyways are not seriously endangered, but also these populations are very slow to repopulate an area where the previous goose population had been decimated. This is not the case with ducks because these birds are not always bound by their intrinsic behavior to return to specific breeding areas. Consequently, vacant breeding areas are more rapidly repopulated by ducks than by geese.
Although Lincoln's analysis was confined to ducks and geese, some thought that it applied to other groups of birds as well. Everyone now realizes that the concept of four flyways, designated as the Atlantic, Mississippi, Central, and Pacific Flyways, was an oversimplification of an extremely complex situation involving crisscrossing of migration routes, varying from species to species. It can be considered meaningful only in a very general way, even for waterfowl, and not applicable generally to other groups of birds. Nevertheless the four "Flyway" areas have been useful in - 63 - regionalizing the harvest of waterfowl for areas of different vulnerability of hunting pressure.
Bellrose (1968) identified corridors of southward migrating waterfowl east of the Rocky Mountains and determined, through statistical analyses, the relative abundance of birds in each. He showed major corridors of dabbling duck movements down the Great Plains and Missouri-Mississippi river valleys with minor off shoots at various points from these corridors eastward to the Atlantic coast where they joined equally minor eastern movements from the North (Fig. 13.). Bellrose's map of migration corridors for the diving ducks showed heavy traffic similar to that of dabbling species down the Great Plains and relatively heavily used corridors from these central arteries eastward across the Great Lakes area to the Atlantic coast, terminating particularly in the vicinity of Chesapeake Bay. A fairly well-used corridor extends along the Atlantic coast.
- 64 -
- 65 -
With our present knowledge of bird migration it is difficult at best to recognize distinct broad belts of migration down the North American continent encompassing groups of distinct populations or species. It seems that so much intermingling of populations occurs that distinctions between broad "flyway" belts are not discernible. About all we can say for sure now is that birds travel between certain breeding areas in the North and certain wintering areas in the South and that a few heavily traveled corridors used by certain species, and more generalized routes followed by one or more species, have become obvious.
Some species exhibit extremely narrow routes of travel. The red knot and purple sandpiper, for example, are normally found only along the coasts because they are limited on one side by the broad waters of the ocean, and on the other by land and fresh water; neither of these habitats furnish conditions attractive to these species.
The Ipswich race of the savannah sparrow likewise has a very restricted migration range. It is known to breed only on tiny Sable Island, Nova Scotia, and it winters from that island south along the Atlantic coast to Georgia. It is rarely more than a quarter of a mile from the outer beach and is entirely at home among the sand dunes with their sparse covering of coarse grass.
The Harris' sparrow supplies an interesting example of a moderately narrow migration route in the interior of the country (Fig. 14.). This fine, large sparrow is known to breed only in the narrow belt of stunted timber and brush at or near the limit of trees from the vicinity of Churchill, Manitoba, on the west shore of Hudson Bay, to the Mackenzie Delta 1,600 miles to the northwest. When this sparrow reaches the United States on its southward migration, it is most numerous in a belt about 500 miles wide, between Montana and central Minnesota and continues south through a relatively narrow path in the central part of the continent. Knowledge of habitat preference by Harris' sparrows suggests the narrow migration range is restricted to the transition between woodland and prairie, a type of habitat approaching the woodland-tundra transition of its breeding area. Development of this migration route, of course, preceded destruction of the heavy eastern forests by colonists from Europe. Its winter range lies primarily in similar country extending from southeastern Nebraska and northwestern Missouri, across eastern Kansas and Oklahoma and through a narrow section of eastern Texas, at places hardly more than 150 miles wide.
When birds start their southward migration the movement necessarily involves the full width of the breeding range. Later, in the case of landbirds with extensive breeding ranges, there is a convergence of the lines of flight taken by individual birds owing, in - 66 - part, to the conformation of the land mass and in part to the east-west restriction of habitats suitable to certain species. An example of this is provided by the eastern kingbird, which breeds in a summer range 2,800 miles wide from Newfoundland to British Columbia. On migration, however, the area traversed by the species becomes constricted until in the southern part of the United States the occupied area extends from Florida to the mouth of the Rio Grande, a distance of only 900 miles. Still farther south the migration path continues to converge, and, at the latitude of Yucatan, it is not more than 400 miles wide. The great bulk of the species probably moves in a belt less than half this width.
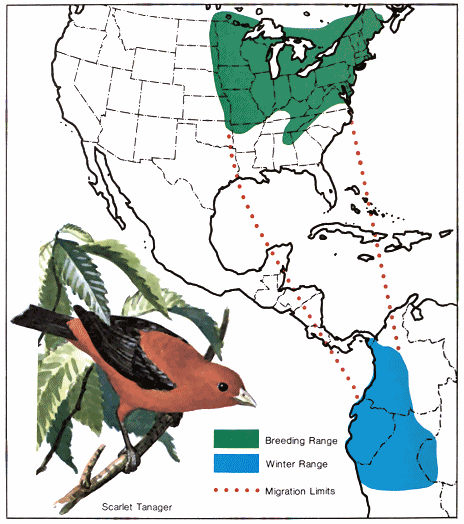
- 67 -
The scarlet tanager presents another extreme case of a narrowly converging migration route starting from its 1,900-mile-wide breeding range in the eastern deciduous forest between New Brunswick and Saskatchewan (Fig. 15.). As the birds move southward in the fall, their path of migration becomes more and more constricted, until, at the time they leave the United States, all are included in the 600-mile belt from eastern Texas to the Florida peninsula. The boundaries continue to converge through Honduras and Costa Rica where they are not more than 100 miles apart. The species winters in the heavily forested areas of northwestern South America including parts of Colombia, Ecuador, and Peru.
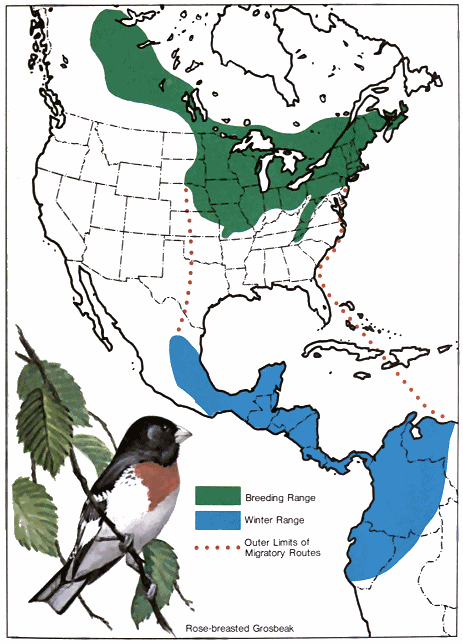
The rose-breasted grosbeak also leaves the United States through the 600-mile stretch from eastern Texas to Apalachicola Bay, but - 68 - thereafter as this grosbeak crosses the Gulf of Mexico and enters the northern part of its winter quarters in southern Mexico the lines do not further converge. However, the pathway of those individuals that continue on to South America is considerably constricted by the narrowing of the land through Central America to Panama (Fig. 16.).
Although the cases cited represent extremes of convergence, a narrowing of the migratory path is the rule to a greater or lesser degree for the majority of North American birds. Both the shape of the continent and major habitat belts tend to constrict southward movement so that the width of the migration route in the latitude of the Gulf of Mexico is usually much less than in the breeding territory.
The American redstart represents a case of a wide migration route, but even in the southern United States, this is still much narrower than the breeding range (Fig. 17.). These birds, however, cross all of the Gulf of Mexico and pass from Florida to Cuba and Haiti by way of the Bahamas, so here their route is about 2,500 miles wide.
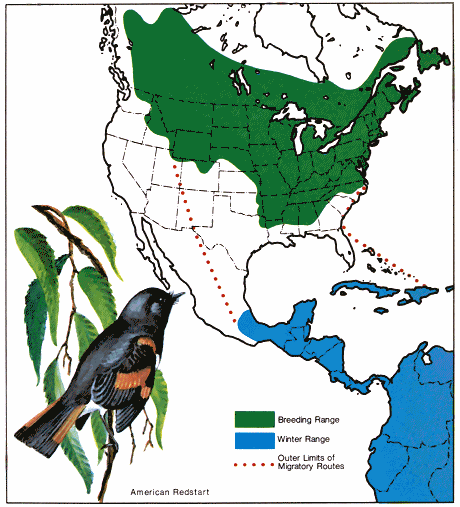
- 69 -
W. W. Cook presented seven of the more important generalized routes for birds leaving the United States on their way to various wintering grounds (1915a; Fig. 18.). When migrants return northward in the spring, they may follow these same routes, but it is not known for certain whether they do. These routes are discussed in the following sections.
Atlantic Oceanic Route
Route No. 1 (Fig. 18.) is almost entirely oceanic and passes directly over the Atlantic Ocean from Labrador and Nova Scotia to the Lesser Antilles, then through this group of small islands to the mainland of South America. Most of the adult eastern golden plovers and some other shorebirds use this as their fall route. As we mentioned previously, radar has indicated strong fall movements of warblers from the New England coast out over the Atlantic to points south along this route. Since it lies almost entirely over the sea, this route is definitely known only at its terminals and from occasional observations made on Bermuda and other islands in its course. Some of the shorebirds that breed on the Arctic tundra of the District of Mackenzie (Northwest Territories) and Alaska fly southeastward across Canada to the Atlantic coast and finally follow this oceanic route to the mainland of South America. The golden plover may accomplish the whole 2,400 miles without pause or rest, and in fair weather the flocks pass Bermuda and sometimes even the islands of the Antilles without stopping. Although most birds make their migratory flights either by day or by night, the golden plover in this remarkable journey flies both day and night. Since this plover swims lightly and easily, it may make a few short stops along the way.
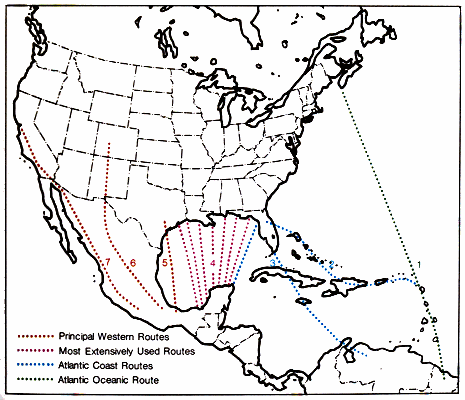
- 70 -
The Arctic tern follows the Atlantic Ocean route chiefly along the eastern side of the ocean. Likewise, vast numbers of seabirds such as auks, murres, guillemots, phalaropes, jaegers, petrels, and shearwaters follow this over-water route from breeding coasts and islands in the Northern and Southern Hemispheres.
Atlantic Coast Route and Tributaries
The Atlantic coast is a regular avenue of travel, and along it are many famous points for observing both land and water birds. About 50 different kinds of landbirds that breed in New England follow the coast southward to Florida and travel thence by island and mainland to South America (Fig. 18., route 2). The map indicates a natural and convenient highway through the Bahamas, Cuba, Hispaniola, Puerto Rico, and the Lesser Antilles to the South American coast. Resting places are afforded at convenient intervals, and at no time need the aerial travelers be out of sight of land. It is not, however, the favored highway; only about 25 species of birds go beyond Cuba to Puerto Rico along this route to their winter quarters, while only six species are known to reach South America by way of the Lesser Antilles. Many thousands of American coots and wigeons, pintails, blue-winged teal, and other waterfowl as well as shorebirds, regularly spend the winter season in the coastal marshes, inland lakes, and ponds of Cuba, Hispaniola, and Puerto Rico.
Route No. 3 (Fig. 18.) is a direct line of travel for Atlantic coast migrants en route to South America, although it involves much longer flights. It is used almost entirely by landbirds. After taking off from the coast of Florida there are only two intermediate land masses where the migrants may pause for rest and food. Nevertheless, tens of thousands of birds of about 60 species cross the 150 miles from Florida to Cuba where many elect to remain for the winter months. The others negotiate the 90 miles between Cuba and Jamaica, but, from that point to the South American coast, there is a stretch of islandless ocean 500 miles across. Relatively few North American migrants on this route go beyond Jamaica. The bobolink so far outnumbers all other birds using this route that it may be designated the "bobolink route" (Fig. 19.). As traveling companions along this route, the bobolink may meet vireos, kingbirds, and nighthawks from Florida, Chuck-will's-widows from the Southeastern States, black-billed and yellow-billed cuckoos from New England, gray-cheeked thrushes from Quebec, bank swallows from Labrador, and blackpoll warblers from Alaska. Sometimes this scattered assemblage will be joined by a tanager or a wood thrush, but the "bobolink route" is not popular with the greater number of migrants.
- 71 -
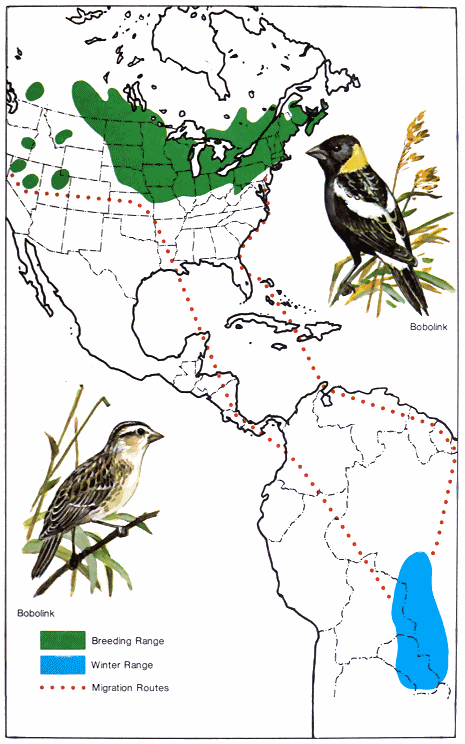
- 72 -
Formerly, it was thought most North America landbirds migrated to Central America via the Florida coast, then crossed to Cuba, and finally made the short flight from the western tip of Cuba to Yucatan. A glance at the map would suggest this as a most natural route, but, as a matter of fact, it is practically deserted except for a few swallows and shorebirds or an occasional landbird storm-driven from its normal course. What actually happens in the fall is that many of the birds breeding east of the Appalachian Mountains travel parallel to the seacoast in a more or less southwesterly direction and, apparently maintaining this same general course from northwestern Florida, cross the Gulf of Mexico to the coastal regions of eastern Mexico. They thus join migrants from farther inland in using route No. 4 (Fig. 18.).
Routes used by the Atlantic brant merit some detail because their flight paths were long misunderstood. These birds winter on the Atlantic coast, chiefly at Barnegat Bay, New Jersey, but depending upon the severity of the season and the food available, south also to North Carolina. Their breeding grounds are in the Canadian arctic archipelago and on the coasts of Greenland. According to the careful studies of Lewis (1937), the main body travels northward in spring along the coast to the Bay of Fundy, overland to Northumberland Strait, which separates Prince Edward Island from mainland New Brunswick and Nova Scotia. A minor route appears to lead northward from Long Island Sound by way of the Housatonic and Connecticut River Valleys to the St. Lawrence River.
After spending the entire month of May feeding and resting in the Gulf of St. Lawrence, the eastern segment of the brant population resumes its journey by departing overland from the Bay of Seven Island area. The eastern and larger segment of the population appears to fly almost due north to Ungava Bay and from there to nesting grounds, probably on Baffin Island and Greenland. The smaller segment travels a route slightly north of west to the southeastern shores of James Bay, although east of that area some of the flocks take a more northwesterly course by descending the Fort George River to reach the eastern shore of James Bay. Upon their arrival at either of these two points on James Bay, the brants of this western segment turn northward and proceed along eastern Hudson Bay to their breeding grounds in the Canadian Arctic.
In general, the fall migration of the brant follows the routes utilized in the spring. At this season, the eastern population appears only on the western and southern shores of Ungava Bay before continuing their southward journey to the Gulf of St. Lawrence and beyond. Also, it appears that most of the birds of the western segment, instead of following the eastern shores of Hudson and James bays, turn southwestward across the former, by way of the Belcher Islands, to Cape Henrietta Maria, and from there south along the western shores of James Bay by way of Akimiski and Charlton Islands. At the southern end of James Bay, they are joined - 73 - by those that have taken the more direct route along the east coasts of the bays and all then fly overland 570 miles to the estuary of the St. Lawrence River.
The Atlantic coast wintering area receives accretions of waterfowl from three or four interior migration paths, one of which is of first importance, as it includes great flocks of canvasbacks, redheads, scaup, Canada geese, and many black ducks that winter in the waters and marshes of the coastal region south of Delaware Bay. The canvasbacks, redheads, and scaup coming from breeding grounds on the great northern plains of central Canada follow the general southeasterly trend of the Great Lakes, cross Pennsylvania over the mountains, and reach the Atlantic coast in the vicinity of Delaware and Chesapeake Bays. Black ducks, mallards, and blue-winged teals that have gathered in southern Ontario during the fall leave these feeding grounds and proceed southwest over a course that is apparently headed for the Mississippi Valley. Many do continue this route down the Ohio Valley, but others, upon reaching the vicinity of the St. Clair Flats between Michigan and Ontario, swing abruptly to the southeast and cross the mountains to reach the Atlantic coast south of New Jersey. This route, with its Mississippi Valley branch, has been fully documented by the recovery records of ducks banded at Lake Scugog, Ontario.
Canvasbacks migrate from the prairie pothole country of the central United States and Canada to many wintering areas in the United States. This duck has been the subject of a particular study (Stewart, Geis, and Evans 1958), and its principle migration routes, based on recovery of banded birds, are shown to follow an important trunk route from the major breeding area in the prairie provinces of Canada and the northern prairies of the United States southeastward through the southern Great Lakes area to Chesapeake Bay, the chief wintering area (Fig. 20.). Relatively few canvasbacks proceed southward along the Atlantic seaboard. A less important route branches off from the main trunk in the southern Minnesota region and extends south along the Mississippi Valley to points along the river. Other individuals of the prairie breeding population fly southward on a broad front to the gulf coast of Texas and the interior of Mexico, while some proceed southwestward on a relatively broad path to the northern Pacific coast.
Mackenzie Valley-Great Lakes-Mississippi Valley
Route and Tributaries
The route extending from the Mackenzie Valley past the Great Lakes and down the Mississippi Valley is easily the longest of any in the Western Hemisphere. Its northern terminus is on the Arctic coast in the regions of Kotzebue Sound, Alaska, and the mouth of the Mackenzie River, while its southern end lies in Argentina. Nighthawks, barn swallows, blackpoll warblers, and individuals of several other species that breed northward to the Yukon Territory and Alaska must cover the larger part of the route twice each year.
- 74 -
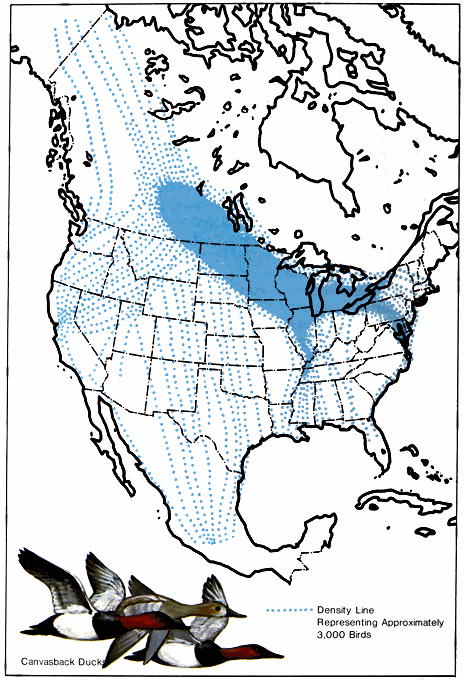
For more than 3,000 miles—from the mouth of the Mackenzie to the delta of the Mississippi—this route is uninterrupted by mountains. In fact, the greatest elevation above sea level is less than 2,000 feet. Because it is well timbered and watered, the entire region affords ideal conditions for its great hosts of migrating birds. It is - 75 - followed by such vast numbers of ducks, geese, shorebirds, blackbirds, sparrows, warblers, and thrushes that observers stationed at favorable points in the Mississippi Valley during the height of migration can see a greater number of migrants than can be noted anywhere else in the world.
When many of these species, including ducks, geese, robins, and yellow-rumped warblers, arrive at the Gulf coast, they spread out east and west for their winter sojourn. Others, despite the perils of a trip involving a flight of several hundred miles across the Gulf of Mexico, fly straight for Central and South America. This part of the route is a broad "boulevard" extending from northwestern Florida to eastern Texas and southward across the Gulf of Mexico to Yucatan and the Isthmus of Tehuantepec (Fig. 18., route 4). This route appears to have preference over the safer but more circuitous land or island routes by way of Texas or Florida. During the height of migration some of the islands off the coast of Louisiana are wonderful observation points for the student of birds, as the feathered travelers literally swarm over them.
Present detailed knowledge of the chief tributaries to the Mackenzie-Great Lakes-Mississippi Valley route relates primarily to waterfowl. Reference has been made already to the flight of black ducks that reach the Mississippi Valley from southern Ontario. Some individuals of this species banded at Lake Scugog, Ontario, have been recaptured in succeeding seasons in Wisconsin and Manitoba, but the majority was retaken at points south of the junction of the Ohio River with the Mississippi indicating their main route of travel from southern Ontario.
A second route that joins the main artery on its eastern side is the one used by eastern populations of lesser snow geese, including both blue and white phases, that breed mainly on Southampton Island and in the Fox Basin of Baffin Island. In the fall these geese work southward along the shores of Hudson Bay and, upon reaching the southern extremity of James Bay, take off on their flight to the great coastal marshes of Louisiana and Texas west of the Mississippi River delta.
Great Plains-Rocky Mountain Routes
This route also has its origin in the Mackenzie River delta and Alaska. The lesser sandhill cranes, white-fronted geese, and smaller races of the Canada goose follow this route through the Great Plains from breeding areas in Alaska and western Canada. It is used chiefly by the pintails and American wigeons that fly southward through eastern Alberta to western Montana. Some localities in this area, as for example, the National Bison Range at Moiese, Montana, normally furnish food in such abundance that these birds are induced to pause in their migratory movement. Some flocks of pintails and wigeons move from this area almost directly west across Idaho to the valley of the Columbia River, then south to the interior valleys of California. Others leave Montana by traveling southeastward across - 76 - Wyoming and Colorado to join other flocks moving southward through the Great Plains.
Observations made in the vicinity of Corpus Christi, Texas, have shown one of the short cuts (Fig. 18., route 5) that is part of the great artery of migration. Thousands of birds pass along the coast to the northern part of the State of Veracruz, Mexico. Coastal areas along the State of Tamaulipas to the north are arid and so entirely unsuited for frequenters of moist woodlands that it is probable that much, or all, of this part of the route for these species is a short distance off shore. It is used by such woodland species as the golden-winged warbler, the worm-eating warbler, and the Kentucky warbler.
Pacific Coast Route
Although it does present features of unusual interest, the Pacific coast route is not as important as some of the others described. Because of the equable conditions that prevail, many species of birds along the coast from the northwestern states to southeastern Alaska either do not migrate or else make relatively short journeys. This route has its origin chiefly in western Alaska, around the Yukon River delta. Some of the scoters and other sea ducks of the north Pacific region as well as the diminutive cackling Canada goose of the Yukon River Delta use the coastal sea route for all or most of their southward flight. The journey of the cackling goose, as shown by return records from birds banded at Hooper Bay, Alaska, has been traced southward across the Alaskan Peninsula and apparently across the Gulf of Alaska to the Queen Charlotte Islands. The birds then follow the coast line south to near the mouth of the Columbia River, where the route swings toward the interior for a short distance before continuing south by way of the Willamette River Valley. The winter quarters of the cackling goose are chiefly in the vicinity of Tule Lake, on the Oregon-California line, and in the Sacramento Valley of California, although a few push on to the San Joaquin Valley.
A tributary of this "flyway" is followed by Ross' goose, which breeds in the Perry River district south of Queen Maud Gulf and other areas farther east on the central Arctic coast of Canada (Fig. 21). Its fall migration is southwest and south across the barren grounds to Great Slave and Athabaska Lakes, where it joins thousands of other waterfowl bound for winter homes along the eastern coast of the United States and the Gulf of Mexico. But when Ross' geese have traveled south approximately to the northern boundary of Montana, most of them separate from their companions and turn southwest across the Rocky Mountains to winter in California. In recent years a few Ross' geese have been found wintering east of the Rocky Mountains along with flocks of lesser snow geese and may be correlated with an eastward extension of their breeding range.
The southward route of those migratory landbirds of the Pacific area that leave the United States in winter extends chiefly through - 77 - the interior of California to the mouth of the Colorado River and on to winter quarters in western Mexico (Fig. 18., routes 6 and 7).
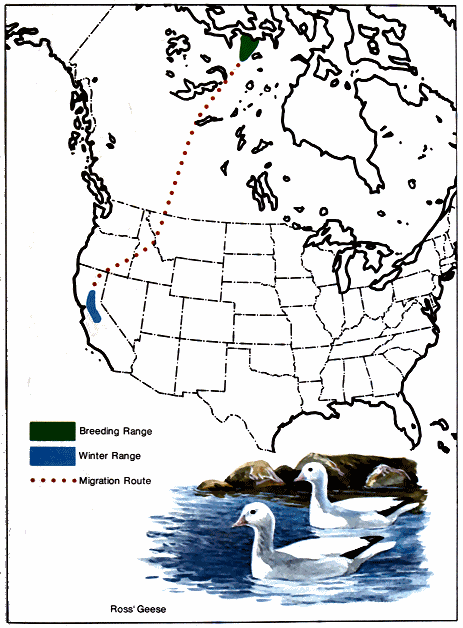
- 78 -
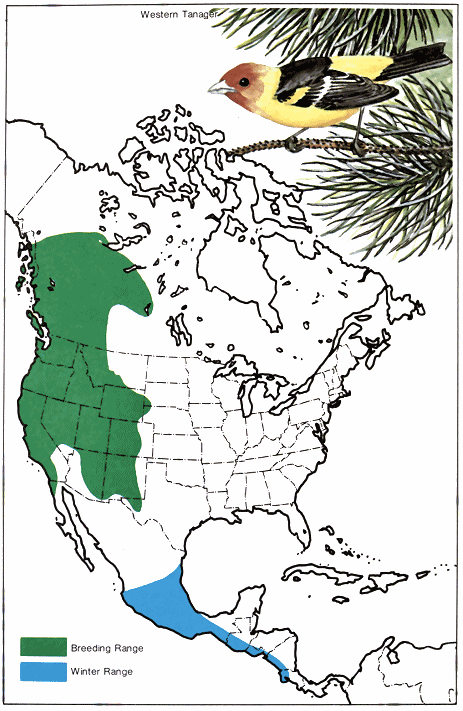
The movements of the western tanager show a migration route that is in some ways remarkable. The species breeds in the mountains from the northern part of Baja California and western Texas north to northern British Columbia and southwestern Mackenzie. Its winter range is in two discontinuous areas—southern Baja California and eastern and southwestern Mexico south to Guatemala (Fig. 22.). During spring migration the birds appear first in western Texas and the southern parts of New Mexico and Arizona about April 20 (Fig. 23). By April 30 the vanguard has advanced evenly to an approximate east-west line across central New Mexico, Arizona, and - 79 - southern California. By May 10 the easternmost birds have advanced only to southern Colorado, while those in the far west have reached northern Washington. Ten days later the northward advance of the species is shown as a great curve, extending northeastward from Vancouver Island to central Alberta and thence southeastward to northern Colorado. Since these tanagers do not reach northern Colorado until May 20, it is evident those present in Alberta on that date, instead of traveling northward through the Rocky Mountains, their summer home, actually reached there by a route that carried them west of the Rockies to southern British Columbia and thence eastward across the still snowy northern Rocky Mountains.
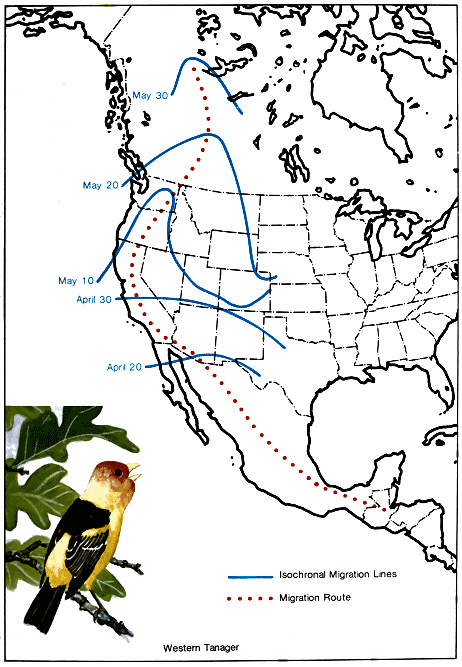
- 80 -
Pacific Oceanic Route
The Pacific oceanic route is used by the Pacific golden plover, bristle-thighed curlew, ruddy turnstone, wandering tattler and other shorebirds. The ruddy turnstone, and probably other shorebirds, migrating from the islands of the Bering Sea, have an elliptical route that takes them southward via the islands of the central Pacific and northward along the Asiatic coast. In addition, many seabirds that breed on far northern and southern coasts or islands migrate up and down the Pacific well away from land except when the breeding season approaches.
The Pacific golden plover breeds chiefly in the Arctic coast region of Siberia and in a more limited area on the Alaskan coast. Some of the birds probably migrate south via the continent of Asia to winter quarters in Japan, China, India, Australia, New Zealand, and Oceania. Others evidently go south by way of the Pacific Ocean to the Hawaiian Islands and other islands of the central and southern Pacific. Migrating golden plovers have been observed at sea on a line that apparently extends from Hawaii to the Aleutian Islands; it appears certain some of the Alaskan birds make a nonstop flight across the sea from Alaska to Hawaii. While it would seem incredible that any birds could lay a course so accurately as to land on these small isolated oceanic islands, 2,000 miles south of the Aleutians, 2,000 miles west of Baja California, and nearly 4,000 miles east of Japan, the evidence admits only the conclusion that year after year this transoceanic round-trip journey between Alaska and Hawaii is made by considerable numbers of golden plovers.
Arctic Routes
Some Arctic nesting birds retreat only a short distance south in the winter. These species include the red-legged kittiwake, Ross' gull, emperor goose, and various eiders. The latter group of ducks winter well south of their nesting areas but nevertheless remain farther north than do the majority of other species of ducks. The routes followed by these birds are chiefly parallel to the coast and may be considered as being tributary either to the Atlantic or Pacific coast routes. The heavy passage of gulls, ducks, black brants, and other water birds at Point Barrow, Alaska, and other points on the Arctic coast, has been noted by many observers. The best defined Arctic route in North America is the one following the coast of Alaska.
A migration route, therefore, may be anything from a narrow path closely adhering to some definite geographical feature, such as a river valley or a coastline, to a broad boulevard that leads in the - 81 - desired direction and follows only the general trend of the land mass. Oceanic routes appear to be special cases that are not fully understood at the present time. Also it must be remembered that all the main routes contain a multitude of tributary and separate minor routes. In fact, with the entire continent of North America crossed by migratory birds, the different groups or species frequently follow lines that may repeatedly intersect those taken by others of their own kind or by other species. The arterial or trunk routes, therefore, must be considered merely as indicating paths of migration on which the tendency to concentrate is particularly noticeable.
- 82 -
Band recoveries, netting records, and personal observations help us to critically examine migration routes and probe deeper into the origin and evolution of these pathways. We are beginning to realize certain deviations occur from the "normal" north and south movements expected in most species. In the previous section on routes, we touched briefly on the fact that some routes are not poleward at all, but in some other direction. We know that many migrants do not stop at the exact localities year after year but they probably do follow the same general course each season. After many years of observations a pattern emerges for that population, species, or group of species. In this section we would like to take a closer look at some of the interesting patterns (or "eccentric routes" as Cook (1915a) referred to them) in migration that birds are annually to travel from breeding to wintering grounds and back again. In many cases, the causative agents are unknown or pure conjecture, but in others, sound biological principles can be put forth that may indicate why a particular species could have evolved the specific pattern it exhibits.
Many species do not return north in the spring over the same route they used in the fall; rather, they fly around an enormous loop or ellipse. Cook (1915a) considered food as the primary factor in determining the course birds took between winter and summer ranges. Individuals that returned by the same route and did not find sufficient food for their needs at that time were eliminated from the population, and only progeny from individuals that took a different course with sufficient food lived to build the tradition of a loop migration. Other investigators consider prevailing winds a major factor in the evolution of loop migration. Whatever the reason may be, it has most likely evolved separately in each species to satisfy its particular needs, and the fact that this pattern occurs all over the world in completely unrelated species is a good illustration of convergent evolution.
The annual flight of adult golden plovers is so unusual, it will be given in some detail. The species is observed by hundreds of bird watchers every year and it well illustrates loop migration (Fig. 24.).
- 83 -
- 84 -
In the fall, the birds fatten on the multitude of berries along the coasts of Labrador and Nova Scotia, then depart south over the Atlantic Ocean to South America. After reaching the South American coast the birds make a short stop, then continue overland to the pampas of Argentina, where they remain from September to March. When these golden plovers leave their winter quarters they cross northwestern South America and the Gulf of Mexico to reach the North American mainland on the coasts of Texas and Louisiana. Thence they proceed slowly up the Mississippi Valley and, by the early part of June, are again on their breeding grounds, having performed a round-trip journey in the form of an enormous ellipse with the minor axis about 2,000 miles and the major axis 8,000 miles stretching from the Arctic tundra to the pampas of Argentina. The older birds may be accompanied by some of the young, but most of the immature birds leave their natal grounds late in summer and move southward through the interior of the country, returning in spring over essentially the same course. The oceanic route is therefore used chiefly by adult birds.
A return by the oceanic route in the spring could be fatal. The maritime climate in the Northeast results in foggy conditions along the coast and the frozen soil would offer few rewards for the weary travelers. By traveling up the middle of the continent, a much better food supply is assured (Welty 1962).
Several North American warblers including the Connecticut warbler (Fig. 25.) and the western race of the palm warbler have been found to follow circuitous migration routes. The Connecticut warbler is not observed or banded on the East coast in spring, but it is recorded farther inland during the season. Thus this warbler proceeds down the East coast in the fall and up the interior of the continent in the spring. Similarly, the western race of the palm warbler moves from its breeding grounds directly east to the Appalachian Mountains before turning south along the coast. Television tower kills in northern Florida indicate the population is very concentrated here at this time of year. In the spring this race also proceeds north through the interior. Graber (1968) points out that the eastern race of the palm warbler also proceeds south along the coast in the fall and poses this question: "does the western population of this species intentionally move toward the ancestral range, or is the fall flight direction merely a consequence of the temperate zone westerly circulation?"
Graber concluded from radar observations that the disparity in seasonal flight directions of many migrants was a positive response of migrants to favorable wind directions at that time of year. The east-oriented trans-gulf migrants followed an elliptical migration because postfrontal air flow in the fall at latitude 40° N is northwesterly, and, in the spring southerly; whereas winds over the Gulf of Mexico are consistently easterly or southeasterly. Therefore, transgulf migrants returning north in the spring would be moved westward across the Gulf unless they compensated for wind drift. Observers were not aware of high-altitude drift before radar (Bellrose and Graber 1963).
- 85 -
- 86 -
Numerous other loop migrations have been documented throughout the world. In the fall, the short-tailed shearwater, is observed off the west coast of North America as far south as California. At this time the species is on the eastern leg of a tremendous figure-eight circuit around the Pacific Ocean (Fig. 26.). The subalpine warbler and red-backed shrikes perform loop migrations between Europe and Africa. Both pass much farther to the east in the spring than in the fall (Moreau 1961). The Arctic loon travels south across inland Russia to southern Europe but returns to its Arctic breeding grounds via the Gulf Stream on the sea because this water is open much earlier in the spring than the inland waterways (Welty 1962).
- 87 -
Dog-leg migration patterns are characterized by a prominent bend or twist in the route. Studies have shown some of these illogical, out-of-the-way means for connecting wintering and breeding areas have no biological function, but instead, are the result of tradition much like the lineage of crooked streets in Boston can be traced back to old cowpaths (Welty 1962). Many species have extended their range in recent years, but the pioneers continue to retrace the old route from the point of origin even if the new areas are not on the same axis as the earlier route. The old pathways have apparently become implanted as part of the migratory instinct in all members of particular populations even after extending their ranges considerable distances from the original.
Good examples of this crooked traditional path can be seen in the routes taken by Old World species extending their ranges into the New World from Europe and Asia. The European wheatear has extended its range into Greenland and Labrador where the local breeding population has become a separate race. When the Labrador individuals depart from their breeding grounds, they proceed north to Greenland, their ancestral home, then west to Europe and south to Africa, the traditional wintering area for all wheatears. Alaskan breeding wheatears migrate to Africa in the opposite direction via Asia where the Alaskan population presumably originated. Alaskan breeding Arctic and willow warblers and bluethroats also migrate westward into Siberia and then southward on the Asiatic side. Some investigators believe the Arctic tern colonized the New World from Europe because when this bird departs for the south it first crosses the Atlantic to Europe, then moves down the eastern Atlantic coast to Africa and either back across the Atlantic to South America or continues south down past South Africa (Fig. 11.). To get to South America from the eastern Arctic, it would be shorter to follow the golden plover's flight path straight down the Atlantic or along the east coast of the United States but the fact that no Arctic terns have been observed in the Caribbean indicates that they do no follow that route.
In western United States, California gulls nest in various colonies around Great Salt Lake and Yellowstone Park. Banding records indicate these populations winter along the California coast (Fig. 27.). Instead of traveling southwest by the shortest distance to the wintering grounds, they proceed longitudinally down the Snake and Columbia Rivers and reach the coast around Vancouver ( Woodbury et al. 1946). Thence they proceed south along the coast to Oregon and California. In the spring the adults return over the same course rather than taking the shorter flight northeast in April across the deserts and mountains; this route would be largely made over a cold and inhospitable country (Oldaker 1961).
- 88 -
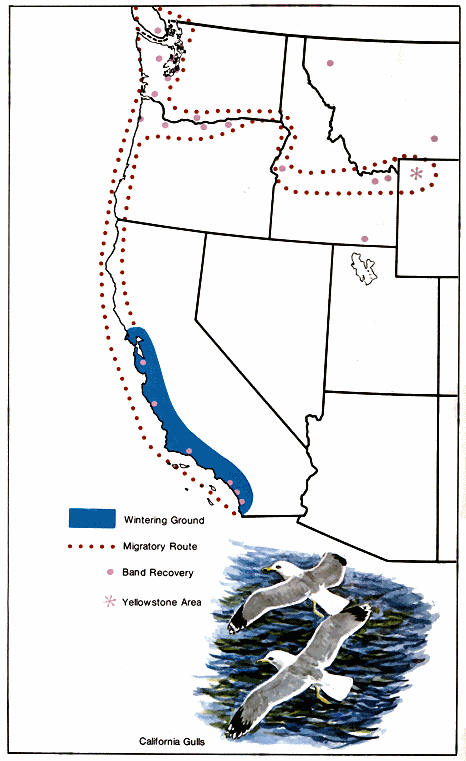
- 89 -
Sladen (1973) has mapped the migration routes of whistling swans, and several dog-leg patterns are apparent in the eastern and western populations (Fig. 28.). In the eastern population, a sharp change in direction occurs at their major feeding and resting areas in North Dakota. After the birds arrive from the Arctic breeding grounds, they proceed east-southeast to their wintering grounds on Chesapeake Bay. In the western population, thousands of birds migrate from the Alaskan breeding grounds to the large marshes along Great Salt Lake. Then after a major stopover, this population heads west over the mountains to California.
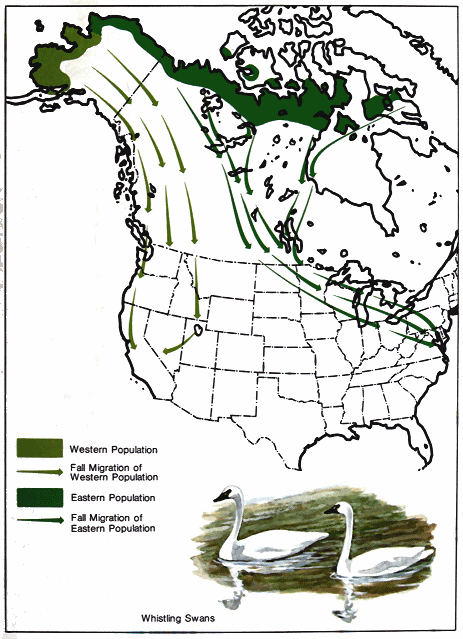
- 90 -
Many of the pelagic birds observed off our coasts or at sea appear to be nomadic when they are not breeding. These movements are not necessarily at random because there is usually a seasonal shift in the population, often for great distances and in specific directions, away from the breeding area after completion of the nesting cycle. Also the return from the sea to nesting areas is at a definite time of year. This may not be true migration in the classical sense (Thomson 1964), although it is similar in most respects.
Because of the extensive and often inhospitable habitat of pelagic birds (to human observers at least), observations on their movements are difficult at best and accurate records are few. We do know some of these species have regular routes (e.g., Arctic terns) and specific patterns of migration (e.g., the loop in the short-tailed shearwater). As more knowledge is accumulated on the "nomadic" species, we may actually find they too have regular migration routes based on biological needs.
Movements of some of the tubenoses (Order Procellariiformes, that includes albatrosses, fulmars, shearwaters, and petrels) have been correlated with ocean currents, prevailing winds, temperatures, and general water fertility (Kuroda 1957; Shuntov 1968; Fisher and Fisher 1972). Commercial fishermen have long known ocean currents are very important factors in the supply of nutrients, plankton, and forage fish for larger fish. These same foodstuffs often attract pelagic birds as evidenced by the tremendous concentrations that occur off the Peruvian coasts where the upwelling of cold nutrient-bearing water is evident. Kuroda (1957) found some fine correlations between the route of the short-tailed shearwater and ocean currents. Likewise Shuntov (1968) found the migratory routes of albatrosses were over temperate marine waters of high biological productivity. The Laysan albatross was correlated with cold currents, while the black-footed albatross occurred over warm currents. Many Southern Hemisphere pelagic species have been extremely successful in exploiting rich northern waters during the summer; the group is probably the most abundant and widespread in the world (Bourne 1956).
When two or more races of the same species occupy different breeding ranges on the same axis as migratory flight, the races breeding the farthest north often winter the farthest south. Thus, a northern race "leap-frogs" over the breeding and wintering range of the southern populations. This has been well documented in the fox sparrow discussed previously (Fig. 10.) and is exhibited by races of Canada geese breeding in central Canada as well. One of the smaller races of this goose breeds along the Arctic coast of the Northwest Territories and winters on the Gulf coast of Texas and northeastern Mexico, while a much larger race breeds in the central United States and Canada but winters in the central part of the United States. This - 91 - leaping over occurs in other species as well, including the bluebird (Pinkowski 1971).
In the effort to find winter quarters furnishing satisfactory living conditions, many North American birds fly hundreds of miles across land and sea. Others, however, are able to attain their objectives merely by moving down the sides of a mountain. In such cases a few hundred feet of altitude corresponds to hundreds of miles of latitude. Movements of this kind, known as "vertical migrations," are found worldwide wherever there are large mountain ranges. Aristotle first mentions vertical migration: "Weakly birds in winter and in frosty weather come down to the plains for warmth, and in summer migrate to the hills for coolness ..." (Dorst 1962). The number of species that can perform this type of migration pattern is obviously limited to those species adapted to breeding in alpine areas.
In the Rocky Mountain region vertical migrations are particularly notable. Chickadees, rosy finches, juncos, pine grosbeaks, Williamson's sapsuckers, and western wood pewees nest at high altitudes and move down to the lower levels to spend the winter. The dark-eyed juncos breeding in the Great Smoky Mountains make a vertical migration, but other members of the species, breeding in flatter areas, make an annual north-south migration of hundreds of miles (Van Tyne and Berger 1959). There is a distinct tendency among the young of mountain-breeding birds to work down to the lower levels as soon as the nesting season is over. The sudden increases among birds in the edges of the foothills are particularly noticeable when cold spells with snow or frost occur at the higher altitudes. In the Dead Sea area of the Middle East, some birds that breed in this extremely hot desert move up into the surrounding cooler hill during the winter (Thomson 1964).
The vertical migrations of some mountain dwelling gallinaceous birds (mountain quail and blue grouse) are quite interesting because the annual journey from breeding to wintering grounds is made on foot. Mountain quail make this downward trek quite early in the fall well before any snows can prevent them from reaching their goal. Blue grouse perform essentially the same journey in reverse. During midwinter, these birds can be found near timberline eating spruce buds protruding above the snow.
These illustrations show that the length and direction of a migration route are adapted to the needs for survival and are met in some cases by a short vertical movement or great latitudinal travels in others.
Recent banding studies have demonstrated many migrants, especially young of the year, have a tendency to disperse after fledging. These premigratory movements have also been called "post-fledging dispersal," "reverse migration," and "postbreeding northward - 92 - migration." Demonstration of this phenomenon is especially important as it relates to locality-faithfulness (Ortstreue), range extension, and gene mixture between populations. These movements cannot be considered as true migrations even though they are repeated annually by the species between breeding grounds and some other area. These movements are generally repeated by the same age class in the population but not the same individuals.
Nevertheless, these regular northward movements are quite striking, especially in herons. The young of some species commonly wander late in the summer and fall for several hundred miles north of the district in which they were hatched. Young little blue herons as well as great and snowy egrets are conspicuous in the East as far north as New England and in the Mississippi Valley to southeastern Kansas and Illinois. Black-crowned night herons banded in a large colony at Barnstable, Massachusetts, have been recaptured the same season northward to Maine and Quebec and westward to New York. In September most of them return to the south.
These movements have been noted in several other species as well. Broley (1947) nicely illustrated this northward movement of bald eagles along the Atlantic coast (Fig. 29.). Birds banded as nestlings in Florida have been retaken that summer 1,500 miles away in Canada. Van Tyne and Berger (1959) surmised the summer heat of Florida was too great for this eagle, a northern species that has only recently spread into Florida to take advantage of abundant food and nesting sites, which it exploits during the cooler season. Postbreeding northward movements are also shared by wood ducks, yellow-breasted chats, eastern bluebirds, and white pelicans.
A somewhat different type of postbreeding migration is the so-called "molt migration" exhibited by many species of waterfowl (Salomonsen 1968). These birds may travel considerable distances away from their nesting area to traditional molting sites where they spend the flightless period of the eclipse plumage. At such times they may move well into the breeding ranges of other geographic races of their species. These movements may be governed by the availability of food and are counteracted in fall by a directive migratory impulse that carries those birds that attained more northern latitudes after the nesting period, back to their normal wintering homes in the south.
The occasional great invasions beyond the limits of their normal range of certain birds associated with the far North are quite different from migration patterns discussed previously. Classic examples of such invasions in the eastern part of the country are the periodic flights of crossbills. Sometimes these migrations will extend well south into the southern States.
- 93 -
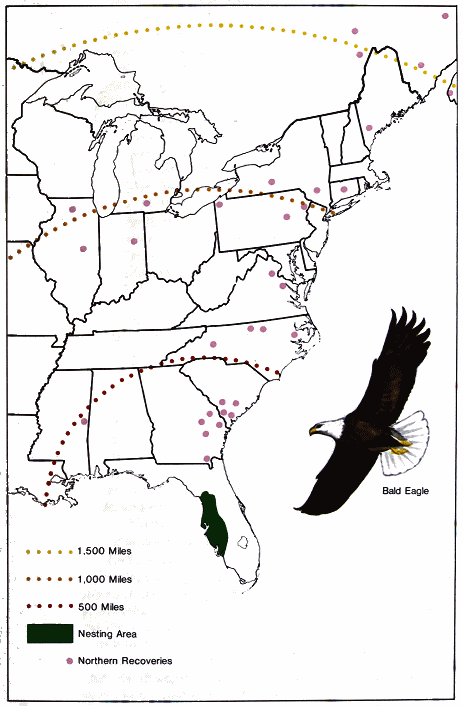
Snowy owls are noted for occasional invasions that have been correlated with periodic declines in lemmings, a primary food resource of northern predators. According to Gross (1947), 24 major invasions occurred between 1833 and 1945. The interval between these varied from 2 to 14 years, but nearly half (11) were at intervals of 4 years. A great flight occurred in the winter of 1926-27 when more than 1,000 records were received from New England alone, but the - 94 - largest on record was in 1945-46 when the "Snowy Owl Committee" of the American Ornithologists' Union received reports of 13,502 birds, of which 4,443 were reported killed. It extended over the entire width of the continent from Washington and British Columbia to the Atlantic coast and south to Nebraska, Illinois, Indiana, Pennsylvania, and Maryland. One was taken as far south as South Carolina.
In the Rocky Mountain region, great flights of the beautiful Bohemian waxwing are occasionally recorded. The greatest invasion in the history of Colorado ornithology occurred in February 1917, when it was estimated that at least 10,000 were within the corporate limits of the city of Denver. The last previous occurrence of the species in large numbers in that section was in 1908.
Evening grosbeaks likewise are given to more or less wandering journeys, and, curiously enough, in addition to occasional trips south of their regular range, they travel east and west for considerable distances. For example, grosbeaks banded at Sault Ste. Marie, Michigan, have been recaptured on Cape Cod, Massachusetts, and in the following season were back at the banding station. Banding records and museum specimen identifications demonstrate that this east-and-west trip across the northeastern part of the country is sometimes made also by purple finches, red crossbills, and mourning doves.
- 95 -
The origin and evolution of bird migration has been discussed in ornithological literature for centuries. As we have seen from the foregoing discussion, migration exists in many forms throughout the world and probably arose to satisfy many different needs in different orders of birds at the same time. New pattens, traditions, and routes are arising today as well as disappearing. Currently, the migration patterns we see are a composite result of historic influences mixed with present day influences. Even though the migration of several different species may be very similar, the patterns exhibited today can be the result of quite different evolutionary processes. Because it cannot be substantiated by experimental facts, any explanation of how a particular pattern or route originates is pure conjecture.
The general anatomical and physiological attributes of birds enable them to develop more diverse and spectacular migratory behavior than any other group of animals. Their potential for long sustained flights is of primary importance in pre-adapting birds to successful migrations. Migration has long since become a definite hereditary habit of many species of birds that recurs in annual cycles, evidently because of physiological changes which prompt a search for an environment suitable for reproduction and survival. Like the bird's other habits its migratory behavior is just as characteristic as the color of its plumage and, like it, evolved through natural selection because it was advantageous for the survival of the population. Its origin has been thought by some to be a mystery locked in past ages, but by study of the history of how birds came to occupy their present ranges, information becomes available which suggests theories that may be developed and explored. Two that are commonly mentioned are termed the "Northern Ancestral Home Theory" and the "Southern Ancestral Home Theory."
According to the former of these hypotheses, in earlier ages when conditions of climate, food, and habitat were favorable for existence of birds throughout the year much further north than is the case today, many species remained in these northern latitudes as permanent residents. Today, such conditions are found only in more southern regions where migrations are much shorter or nonexistent. Gradually, however, in the Northern Hemisphere the glacial ice fields advanced southward, causing a southward movement of conditions favorable to northern birds, until finally all bird life was confined to southern latitudes. As the ages passed, the ice cap gradually retreated, and each spring the birds whose ancestral home had been in the North moved in again to fill newly opened breeding - 96 - habitat only to be driven south again at the approach of winter. As the size of the ice-covered area diminished, the journeys to the summer breeding areas became even longer until eventually the climatic conditions of the present age became established, and with them, present patterns of the annual advance and retreat we call migration.
The opposing theory is simpler in some respects and supposes the ancestral home of Northern Hemisphere migratory birds was in the Tropics. As all bird life tends to overpopulation, there was a constant effort of young individuals to pioneer and seek breeding grounds where competition was less severe. Species better adapted to more northern latitudes moved in that direction for nesting but were kept in check by the glacial ice and forced to return southward with the recurrence of winter conditions. Gradually, as the ice retreated, vast areas of virgin country became successively suitable for summer occupancy, but the winter habitat in the South remained the home to which the birds returned after the nesting season.
The above two theories presume that the Quaternary glaciations, which occurred 10,000 to 1 million years ago, have been the predominate influence on bird migration in North America and Europe as we observe it today. There is no doubt these extreme climatic and ecologic barriers played a part in shaping or modifying some patterns, but as Moreau (1951) has pointed out, well-developed migrations occur in parts of the world, including the Southern Hemisphere or even within the tropics, where continental glaciation was not a factor. Furthermore, migrations to fit various needs have probably been going on ever since birds could fly. The tremendous Pleistocene glaciations actually occupied less than a hundredth of the time birds have existed on the earth and probably only determined the details of migrations as we see them today (Moreau 1951).
The northern and southern ancestral home theories appear diametrically opposed to each other but Dorst (1963) concludes they are perfectly compatible. Since the phenomena probably occurred simultaneously, northern migrants then originated from two stocks: the North Temperate Zone birds sought refuge to the south during the glacial periods and the tropical avifauna expanded their range during the interglacial periods. Dorst also stated this double origin is more prevalent in North America where the tropical element is most abundant. Birds representing this element include hummingbirds, tyrant flycatchers, orioles, tanagers, and blackbirds. At some latitudes, they nest in the same area as the shorebirds which are of arctic parental stock.
These theories assume migration is a genetic, inherited character, but we now know in some species it can be modified in the lifetime of one individual and the place some individuals return to nest or winter is not the ancestral home but a place to which they had been transported at an early stage in their development. Traditions that have lived for countless generations may die overnight if experienced individuals are lost or no longer active (Hochbaum 1955); migration patterns remain constant only as long as the factors influencing these - 97 - patterns remain constant. But the landscape and the interacting ecological stresses are forever changing, and we would expect the adaptive behavior of birds to respond with them. One of these responses to an expanding habitat is colonization of new territory and expansion of a species' range with accompanying development of a migratory habit. The search for favorable conditions under which to breed in summer and to feed in winter, as influenced by competition for space, has been the principal factor underlying the extension of ranges, usually by young, nonconditioned individuals. This is exemplified by the northward extension in historic times of a number of species. Many of these range extensions have closely followed man's settlement of the area and the subsequent changes in habitat that man has made.
From the previous descriptions of migration patterns and routes, it will be observed that the general trend of migration in most northern populations of North American birds is northwest and southeast. Eastern species tend to extend their ranges by pushing westward, particularly in the North. For example, in the Stikine River Valley of northern British Columbia and southwestern Alaska the common nighthawk, chipping sparrow, rusty blackbird, yellow warbler, American redstart, and others have established breeding stations at points 20 to 100 miles from the Pacific Ocean. The northern race of the American robin, common flickers, dark-eyed juncos, blackpoll warblers, yellow-rumped warblers, and ovenbirds, all common eastern species, also are established as breeding birds in western Alaska. The ovenbird has even been detected on the lower Yukon River, and the sandhill crane and gray-cheeked thrush have moved across Bering Strait into Siberia. These birds continue to migrate through the eastern part of the continent. Instead of taking the shortest route south, they retrace the direction of their westward expansion and move southward along the same avenues as their more eastern relatives.
The red-eyed vireo is essentially an inhabitant of states east of the Great Plains, but an arm of its breeding range extends northwest to the Pacific coast in British Columbia (Fig. 30.). It seems evident this is a range extension that has taken place comparatively recently by a westward movement via deciduous woodland corridors, and the invaders retrace in spring and fall the general route by which they originally entered the country.
In the case of the bobolink, a new extension of the breeding range and a subsequent change in the migration of the species has taken place since settlement by European man (Fig. 19.). Because the bobolink is a bird of damp meadows, it was originally cut off from the Western States by the intervening arid regions, but with the advent of irrigation and the bringing of large areas under cultivation, small colonies of nesting bobolinks appeared at various western points. Now the species is established as a regular breeder in the great mountain parks and irrigated valleys of Colorado and elsewhere almost to the Pacific coast. These western pioneers must fly long distances east and west to reach the western edge of the route followed by the bulk of the bobolinks that breed in the northern United States and southern Canada.
- 98 -
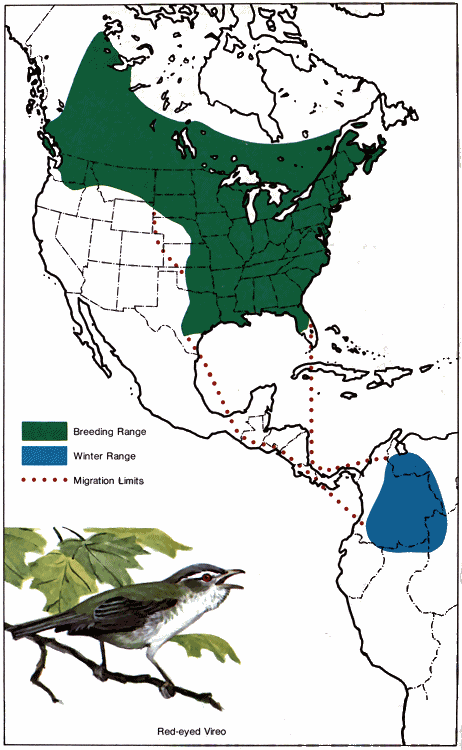
- 99 -
During the past few decades, various populations of Canada geese have altered their migration patterns as a result of transplanting brood stock, development of refuges or changing agricultural practices. These routes will continue to change in the coming years as long as these factors are in a state of flux. It has been shown that man can establish breeding colonies of Canada geese with young birds almost anywhere.
Europe also has several good examples of changes in migration routes through range extension. One of the best examples is the serin. During the past century, this European finch has spread its breeding range from around the Mediterranean Sea to include the entire continent. While the Mediterranean populations remain sedentary, the more northern breeding birds are migratory. Most likely, those birds that did not migrate from the North were eliminated by severe weather. Similarly, the wheatear, yellow wagtail, and Arctic warbler have extended their breeding ranges eastward across the Bering Sea into Alaska, but the wheatear, for instance, migrates all the way back across Asia to Africa where it winters with other wheatears coming from Europe, Iceland, and Greenland.
As bird populations become more and more migratory, we might expect their flight capabilities to be enhanced accordingly. These changes in morphology are readily seen in wing shape. Several groups of birds have closely related species or populations some of which are migratory and others sedentary. The sedentary species or populations have more rounded wings because of the relative length of the wing quills. On the other hand, populations that migrate great distances, such as albatrosses, falcons, swifts, various shorebirds, and terns, have more pointed wings. Kipp(1942, 1958) demonstrated this using orioles. The sedentary black-headed oriole of India has a well-rounded wing whereas the closely related black-naped oriole is migratory between India and Siberia and has primaries that are much more pointed and well developed.
Thus it seems the origin and evolution of migration have roots in the present that are deep in the past. The important thing to consider in the evolution of a migratory trait is whether a population can adapt to new conditions by genetic modification of its physiology and habits. The migratory habit has evolved in those populations in which, on the average, more individuals survive by moving to a different area part of the year than if they remained in the same area all year.
- 100 -
The migration of birds had its beginning in times so remote its origins have been largely obscured and can be interpreted now only in terms of present conditions. The causes underlying migration are exceedingly complex. The mystery that formerly cloaked the periodic travels of birds, however, has been largely dispelled through the fairly complete information now available concerning the extent and times of seasonal journeys of most species. Many gaps still remain in our knowledge of the subject, but present knowledge is being placed on record, and the answers to many uncertainties that continue to make bird migration one of the most fascinating subjects in the science of ornithology must be left for future studies. In some areas we are on the threshold of discovery. More and more sophisticated approaches including radar, radio telemetry, computer processing of banding data, and physiological and behavior studies are being developed.
With the widespread use of these new techniques, we are beginning to realize the benefits, aside from aesthetic reasons, for studying migration. Radar alone has aided tremendously in documenting flock size, heights, and speeds of migration as well as the descriptions and locations of patterns and routes of specific migrants in relation to aircraft flight lanes. Recent studies have indicated local, nonmigratory populations of various blackbirds cause nearly all of the rice damage in southern States and the "hordes from the North" contribute very little to the losses. In addition, the transport of arborviruses from one continent to another via these long distance migrants is being investigated. People have started to uncover the secrets of migration and utilize this knowledge for the betterment of our society.
Each kind of bird seems to have its own reaction to the environment, so that the character of movement differs widely in the various species, and seldom do any two present the same picture. In fact, bird migration has been described as a phase of geographic distribution wherein there is a more or less regular seasonal shifting of the avian population caused by the same factors that determine the ranges of the sedentary species. If this view is correct, then it must be recognized that the far-reaching works of man in altering the natural condition of the Earth's surface can so change the environment necessary for the well-being of the birds as to bring about changes in their yearly travels. The nature and extent of the changes wrought by man on the North American Continent are readily apparent. Extensive forests have been burned or cut away, rolling prairies turned over with the plow, and wetlands drained or filled. Their - 101 - places have been taken by a variety of human activities. These great changes are exerting pressure on native bird populations, and various species may be either benefited or adversely affected.
The Federal Government has recognized its responsibility to migratory birds under these changing conditions. Enabling acts allow for carrying out migratory bird treaty obligations in cooperation with other countries, and now most species have legal protection under regulations administered by the U.S. Fish and Wildlife Service. The effectiveness of conservation laws, however, is increased in the same measure that the people of the country become acquainted with the migratory bird resource and interest themselves personally in the well-being of the various species. Long before European man came to America, the birds had established their seasonal patterns of migration throughout the Western Hemisphere. The economic, scientific, and esthetic values of these migratory species dictate they be permitted to continue their long-accustomed and to some extent still-mysterious habits of migration.
- 102 -
Able, K. P.
1970. A radar study of the altitude of nocturnal passerine migration. Bird-Banding 41(4): 282-290.
Aldrich, J. W.
1949. Migration of some North American waterfowl. U.S. Fish and Wildl. Serv., Spec. Sci. Rep. Wildl. 1. 49 p.
Aldrich, J. W.
1952. The source of migrant mourning doves in southern Florida. J. Wildl. Manage. 16:447-456.
Aldrich, J. W., A. J. Duvall and A. D. Geis.
1958. Racial determination of origin of mourning doves in hunters' bags. J. Wildl. Manage. 22: 71-75.
Alexander, J. R., and W. T. Keeton.
1972. The effect of directional training on initial orientation in pigeons. Auk 89: 280-298.
Allard, H. A.
1928. Bird migration from the point of light and length of day changes. Am. Nat. 62: 385-408.
American Ornithologists' Union.
1957. The Check-list of North American birds. 5th edition. American Ornithologists' Union. 691 p.
Anderson, K. S., E. J. Randall, A. J. Main and R. J. Tonn.
1967. Recoveries of birds banded by encephalitis field stations, 1957-1965. Bird-Banding 38(2): 135-138.
Annan, O.
1962. Sequence of migration by sex, age, and species of thrushes of the genus Hylochichla through Chicago. Bird-Banding 33(3): 130-137.
Anonymous.
1972. Population ecology of migratory birds. A symposium. Wildl. Res. Rep. 2. U.S. Fish and Wildl. Serv. 278 p.
Austin, O. L., Jr.
1928. Migration routes of the Arctic Tern (Sterna paradisaea Brunnich). Northeast Bird Banding Assoc. 4(4): 121-125.
1929. Some Labrador banding records. Northeast Bird Banding Assoc. 5(1): 35-36.
Bagg, A. M.
1967. Factors affecting the occurrence of the Eurasian Lapwing in eastern North America. Living Bird 6: 87-121.
Bagg, A. M., W. W. H. Gunn, D. S. Miller, J. T. Nichols, W. Smith and F. P. Wolfarth.
1950. Barometric pressure patterns and spring bird migration. Wilson Bull. 62: 5-19.
Baird, J., C. S. Robbins, A. M. Bagg and J. V. Dennis.
1958. "Operation Recovery" the Atlantic coastal netting project. Bird-Banding 29(3): 137-168.
- 103 -
Baird, J., A. M. Bagg, I. C. J. Nisbet and C. S. Robbins.
1959. Operation Recovery—report of mist-netting along the Atlantic coast in 1958. Bird-Banding 30: 143-171.
Ball, S. C.
1952. Fall bird migration of the Gaspe Peninsula. Peabody Mus. Nat. Hist. Yale Univ. Bull. 7. 211 p.
Behle, W. H.
1958. The bird life of Great Salt Lake. Univ. of Utah Press, Salt Lake City. 203 p.
Bellrose, F. C.
1967. Radar in orientation research. Proc. XIX Int. Ornithol. Congr.: 281-309.
Bellrose, F. C.
1968. Waterfowl migration corridors east of the Rocky Mountains in the United States. Biol. Notes 61, Ill. Nat. Hist. Surv., Urbana, Ill 24 p.
1971. The distribution of nocturnal migration in the air space. Auk 88(2): 397-424.
1972a. Possible steps in the evolutionary development of bird navigation p. 223-257. In: Caller, S. R. et al. ed. Animal orientation and navigation symp. N.A.S.A. Wash., D.C. 606 p.
1972b. Mallard migration corridors as revealed by population distribution, banding, and radar. In: Population ecology of migratory birds: A Symp. Wildl. Res. Rep. 2. U.S. Fish and Wildl. Serv. p. 1-26.
Bellrose, F. C. and R. R. Graber.
1963. A radar study of the flight directions of nocturnal migrants. Proc. XIII Int. Ornithol. Congr: 362-389.
Bellrose, F. C. and J. G. Sieh.
1960. Massed waterfowl flights in the Mississippi flyway 1956 and 1957. Wilson Bull. 72: 29-59.
Bennett. H. R.
1952. Fall migration of birds at Chicago. Wilson Bull. 64: 197-220.
Bergman, G. and K. O. Donner.
1964. An analysis of spring migration of the Common Scoter and Longtailed Duck in southern Finland. Acta Zool. Fenn. 105: 1-59.
Bergtold, W. H.
1926. Avian gonads and migration. Condor 28: 114-120.
Bissonette, T. H.
1936. Normal progressive changes in the ovary of the starling (Sturnus vulgaris) from December to April. Auk 53: 31-50.
1939. Sexual photoperiodicity in the blue jay (Cyanocitta cristata). Wilson Bull. 51: 227-232.
Bogert. C.
1937. The distribution and the migration of the Long-tailed Cuckoo (Urodynamis taitensis Sparrman). Am. Mus. Novit. 933. pp 1-12.
Bourne, W. R. P.
1956. Migrations of the sooty shearwater. Sea Swallow 9: 23-25.
Bray, O. E. and G. W. Corner.
1972. A tail clip for attaching transmitters to birds. J. Wildl. Manage. 36(2): 640-642.
Broley. C. L.
1947. Migration and nesting of Florida bald eagles. Wilson Bull. 59: 3-20.
- 104 -
Bruderer, B. and P. Steidinger.
1972. Methods of quantitative and qualitative analysis of bird migration with a tracking radar, p. 151-167. In: Caller, S. R., et al ed. Animal orientation and navigation symp. N.A.S.A. Wash., D.C. 606 p.
Clarke, W. E.
1912. Studies in bird migration. 2 vol. London.
Coffey, B. B., Jr.
1944. Winter home of chimney swifts discovered in northeastern Peru. Migrant 15(3): 37-38.
Cooch, G.
1955. Observations on the autumn migration of blue geese. Wilson Bull. 67(3): 171-174.
Cooke, M. T.
1937. Flight speed of birds. U.S. Dept. Agr. Cir. 428. 13 p.
1945. Transoceanic recoveries of banded birds. Bird-Banding 16(4): 123-129.
Cooke, W. W.
1888. Report of bird migration in the Mississippi Valley in the years 1884 and 1885. U.S. Dept. Agr. Div. Econ. Ornithol. Bull. 2. 313 p.
1904a. Distribution and migration of North American warblers. U.S. Dept. Agr. Div. Biol. Surv. Bull. 18. 142 p.
1904b. The effect of altitude on bird migration. Auk 21: 338-341.
Cooke, W. W.
1905a. Routes of bird migration. Auk 22: 1-11.
1905b. The winter ranges of the warblers (Mniotiltidae). Auk 22: 296-299.
1906. Distribution and migration of North American ducks, geese, and swans. U.S. Dept. Agr. Bur. Biol. Surv. Bull. 26. 90 p.
1908. Averaging migration dates. Auk 25: 485-486.
1910. Distribution and migration of North American shore birds. U.S. Dept. Agr. Bur. Biol. Surv. Bull. 35. 100 p.
1913a. Distribution and migration of North American herons and their allies. U.S. Dept. Agr. Bur. Biol. Surv. Bull. 45. 70 p.
1913b. The relation of bird migration to the weather. Auk 30: 205-221.
1914. Distribution and migration of North American rails and their allies. U.S. Dept. Agr. Bull. 128. 50 p.
1915a. Bird migration. U.S. Dept. Agr. Bull. 185. 48 p.
1915b. Bird migration in the Mackenzie Valley. Auk 32: 442-459.
1915c. Distribution and migration of North American gulls and their allies. U.S. Dept. Agr. Bull. 292. 70 p.
1915b. The Yellow-billed loon: a problem in migration. Condor 17: 213-214.
Cortopassi, A. J. and L. R. Mewaldt.
1965. The circumannual distribution of white-crowned sparrows. Bird-Banding 36(3): 141-169.
Coues, E.
1878. Birds of the Colorado Valley: a repository of scientific and popular information concerning North American ornithology. U.S. Dept. Int. Misc. Pub. 11. 807 p.
Curtis, S. G.
1969. Spring migration and weather at Madison, Wisconsin. Wilson Bull. 81(3): 235-245.
Delacour, J.
1947. Birds of Malaysia. Macmillan Co. N.Y. 382 p.
1963. The waterfowl of the world, 4 vols. Country Life Ltd. London.
- 105 -
DeLury, R. E.
1938. Sunspot influences. J. R. Astron. Soc. of Can. pt. 1, March 1938, pt. 2, April 1938. 50 p.
Dennis, J. V.
1967. Fall departure of the Yellow-breasted Chat (Icteria virens) in eastern North America. Bird-banding 38 (2): 130-135.
De Schauensee, R. M.
1970. A guide to the birds of South America. Livingston Pub. Co. Wynnewood, Penn. 470 p.
Diem, K. L. and D. D. Condon.
1967. Banding studies of water birds on the Molly Islands, Yellowstone Lake, Wyoming. Yellowstone Lib. and Mus. Assoc., Yellowstone Nat. Park. 41 p.
Dixon, J.
1916. Migration of the Yellow-billed Loon. Auk 33 (4): 370-376.
Dixon, K. L. and J. D. Gilbert.
1964. Altitudinal migration in the Mountain Chickadee. Condor 66: 61-64.
Dorst, J.
1963. The migration of birds. Houghton Mifflin Co. (Am. ed), Boston. 476 p.
Drury, W. H. Jr.
1960. Radar and bird migration a second glance. Mass. Audubon 44(4): 173-178.
Drury, W. H. Jr., I. C. T. Nisbet and R. E. Richardson.
1961. The migration of angels. Nat. Hist. 70(8): 10-17.
Eastwood, E.
1967. Radar ornithology. Methuen, London. 278 p.
Eastwood, E. and G. C. Rider.
1965. Some radar measurements of the altitude of bird flight. Br. Birds 58: 393-426.
Eaton, R. J.
1933. The migratory movements of certain colonies of Herring Gulls (Larus argentatus smithsonianus Coues) in eastern North America. Part I. Bird-Banding 4(4): 165-176.
Eaton, R. J.
1934. The migratory movements of certain colonies of Herring Gulls in eastern North America. Part II. Bird-Banding 5(1): 1-19.
1934. The migratory movements of certain colonies of Herring Gulls in eastern North America. Part III. Bird-Banding 5(2): 70-84.
Emlen, S. T.
1967. Migratory orientation in the Indigo Bunting (Passerina cyanea). Part II: Mechanism of celestial orientation. Auk 84: 463-489.
1969. Bird migration: influence of physiological state upon celestial orientation. Science (Wash., D.C.) 165 (3894): 716-718.
1970. The influence of magnetic information on the orientation of the Indigo Bunting (Passerina cyanea). Anim. Behav. 18(2): 215-224.
Farner, D. S.
1945. The return of Robins to their birthplaces. Bird-Banding 16(3): 81-99.
1950. The annual stimulus for migration. Condor 52(3): 104-122.
1955. The annual stimulus for migration: experimental and physiologic aspects. In: Recent studies in avian biology. Univ. Ill. (Urbana) Bull: 198-237.
1960. Metabolic adaptations in migration. Proc. XII Int. Ornithol. Cong.: 197-208.
- 106 -
Farner, D. S. and L. R. Mewaldt.
1953. The relative roles of diurnal periods of activity and diurnal photoperiods in gonadal activation in male Zonotrichia leucophrys gambelii. Experimentia 9: 219-221.
Fisher, H. I. and J. R. Fisher.
1972. The oceanic distribution of the Laysan Albatross (Diomedea immutabilis). Wilson Bull 84: 7-27.
Fisher, J. and R. M. Lockley.
1954. Sea-birds. Houghton Mifflin, Boston. XVI + 320 p.
Furlong, W. R.
1933. Land-birds in a gale at sea. Bird Lore 35(5): 263-265.
Gatke, H.
1895. Heligoland as an ornithological observatory: the results of fifty years' experience (trans, from German by R. Rosenstock). David Douglas Pub., Edinburgh. 599 p.
Gauthreaux, S. A. Jr.
1970. Weather radar quantification of bird migration. Bioscience 20(1): 17-20.
1971. A radar and direct visual study of passerine spring migration in southern Louisiana. Auk 88: 343-365.
1972a. Flight directions of passerine migrants in daylight and darkness: a radar and direct visual study p. 129-137. Galler, S. R., et al. ed. In: Animal orientation and navigation symp. N.A.S.A. Wash., D.C. 606 p.
1972b. Behavioral responses of migrating birds to daylight and darkness: a radar and direct visual study. Wilson Bull. 84: 136-148.
Geroudet, P.
1954. Des oiseaux migrateurs trouvés sur la glacier de Khumbu dans l'Himalaya. Nos Oiseaux 22: 254.
Godfrey, W. E.
1966. The birds of Canada. Natl. Mus. Can. Bull. 203, Biol Series No. 73. 428 p.
Gordon, D. A.
1948 Some considerations of bird migration: continental drift and bird migration. Science (Wash., D.C.) 108: 705-711.
Graber, R. R.
1965. Night flight with a thrush. Audubon 67: 368-374.
1968. Nocturnal migration in Illinois different points of view. Wilson Bull. 80(1): 36-71.
Graber, R. R. and W. W. Cochrane.
1959. An audio technique for the study of nocturnal migration of birds. Wilson Bull. 71: 220-236.
Griffin, Dr. R.
1940. Homing experiments with Leach's Petrels. Auk 57(1): 61-74.
1943. Homing experiments with Herring Gulls and Common Terns. Bird-Banding 14(1 and 2): 7-33.
Griffin, D. R.
1958. Listening in the dark. Yale Univ. Press, New Haven. 413 p.
1964. Bird migration. Nat. Hist. Press. Garden City, N.Y. 180 p.
Griffin, D. R. and R. J. Hock.
1948. Experiments on bird navigation. Science (Wash., D.C.) 107(2779): 347-349.
1949. Airplane observations of homing birds. Ecology 30: 176-198.
- 107 -
Grinnell, J.
1931. Some angles in the problem of bird migration. Auk 48: 22-32.
Gross, A. O.
1927. The Snowy Owl migration of 1926-27. Auk 44: 479-493.
1947. Cyclic invasions of the Snowy Owl and the migration of 1945-46. Auk 64(4): 584-601.
Gwynn, A. M.
1968. The migration of the Arctic Tern. Aust. Bird Bander 6(4): 71-75.
Haartman, L. von.
1968. The evolution of resident vs. migratory habit in birds: some considerations. Ornis Fenn. 45(1): 1-7.
Hagar, J. A.
1966. Nesting of the Hudsonian Godwit at Churchill, Manitoba. Living Bird 5: 5-43.
Hamilton, W. J. III.
1962a. Bobolink migratory pathways and their experimental analysis under night skies. Auk 79(2): 208-233.
1962b. Celestial orientation in juvenile waterfowl. Condor 64(1): 19-23.
Harrisson, T. H.
1931. On the normal flight speeds of birds. Br. Birds 25: 86-96.
Hassler, S. S., R. R. Graber and F. C. Bellrose.
1963. Fall migration and weather: a radar study. Wilson Bull. 75: 56-77.
Haugh, J. R. and T. J. Cade.
1966. The spring hawk migration around the southeastern shore of Lake Ontario. Wilson Bull. 78(1): 88-110.
Hebrard, J. J.
1971. The nightly initiation of passerine migration in spring: a direct visual study. Ibis 113(1): 8-18.
Hochbaum, H. A.
1955. Travels and traditions of waterfowl. Univ. of Minn. Press, Minneapolis. 301 p.
Hofslund, P. B.
1965. Hawks above Duluth. In: The bird watcher's America, ed. O. S. Pettingill, Jr. McGraw-Hill Book Co., N. Y.
1966. Hawk migration over the western tip of Lake Superior. Wilson Bull. 78: 79-87.
Hussell, D. J. T., T. Davis and R. D. Montgomerie.
1967. Differential fall migration of adult and immature Least Flycatchers. Bird-Banding 38(1): 61-66.
Jaeger, E. C.
1948. Does the Poor-will "hibernate"? Condor 50: 45.
1949. Further observations on the hibernation of the Poor-will. Condor 51: 105-109.
Johnson, N. K.
1963. Comparative molt cycles in the Tyrannid genus Empidonax. Proc. XIII Int. Ornithol. Congr.: 870-883.
1973. Spring migration of the Western Flycatcher with notes on seasonal changes in sex and age ratios. Bird-Banding 44(3): 205-220.
Keeton, W. T.
1969. Orientation by pigeons: is the sun necessary? Science (Wash., D. C.) 165(3896): 922-928.
Kenyon, K. W. and D. W. Rice.
1958. Homing of Laysan Albatrosses. Condor 60: 3-6.
- 108 -
King, J. R.
1963. Annual migratory-fat deposition in the White-crowned Sparrow. Proc. XIII Int. Ornithol. Congr.: 940-949.
King, J. R., S. Barker and D. S. Farner.
1963. A comparison of energy reserves during the autumnal and vernal migratory periods in the White-crowned Sparrow (Zonotrichia leucophrys gambelii). Ecol. 44(3): 513-521.
King, J. R. and D. S. Farner.
1963. The relationship of fat deposition to Zugunruhe. Condor 65(3): 200-223.
Kipp, F. A.
1942. Ueber Flugelban and Wanderzug der Vogel. Biol. Zbl. 62: 289-299.
1958. Zur Geschichte des Vogelzuges auf der Grundlage der Flugelanpassungen. Vogelwarte 19: 233-242.
Kramer, G.
1952. Experiments on bird orientation. Ibis 94: 265-285.
1957. Experiments on bird orientation and their interpretation. Ibis 99: 196-227.
1959. Recent experiments on bird orientation. Ibis 101: 399-416.
1961. Long distance orientation. In: Marshall, A. J. Biology and comparative physiology of birds. 2 Vol. Academic Press, N.Y. and London.
Kuroda, N.
1957. A brief note on the pelagic migration of the Tubinares. Misc. Rep. of the Yamashina Inst. for Ornithol. and Zool. 11: 10-23.
Lack, D.
1959. Migrations across the sea. Ibis 101(3-4): 374-399.
1960a. The influence of weather on passerine migration. Auk 77: 171-209.
Lack, D.
1960b. The height of bird migration. Br. Birds 53: 5-10.
1962. Radar evidence on migratory orientation. Br. Birds 55(4): 139-157.
1963a. Migration across the southern North Sea studied by radar. Part 4. Autumn. Ibis 105 (1): 1-54.
1963b. Migration across the southern North Sea studied by radar. Part 5. Movements in August, winter and spring, and conclusion. Ibis 105(4): 461-492.
Lewis, H. F.
1937. Migrations of the American Brant (Branta bernicla hrota). Auk 54: 73-95.
Lincoln, F. C.
1917. Bohemian Waxwing (Bombycilla garrula) in Colorado. Auk 34: 341.
1922. Trapping ducks for banding purposes: with an account of the results obtained from one waterfowl station. Auk 39: 322-334.
1924a. Banding notes on the migration of the Pintail. Condor 26: 88-90.
1924b. Returns from banded birds, 1920 to 1923. U.S. Dept. Agr. Bull. 1268. 56 p.
1926. The migration of the Cackling Goose. Condor 28: 153-157.
1927a. Notes on the migration of young Common Terns. Northeastern Bird-Banding Assoc. Bull. 3: 23-28.
1927b. Returns from banded birds, 1923 to 1926. U.S. Dept. Agr. Tech. Bull. 32. 95 p.
1928. The migration of young North American Herring Gulls. Auk 45: 49-59.
1934. Distribution and migration of the Redhead (Nyroca americana). Trans. Twentieth Am. Game Conf.: 280-287.
1935a. The waterfowl flyways of North America. U.S. Dept. Agr. Cir. 342. 12 p.
1935b. The migration of North American birds. U.S. Dept. Agr. Cir. 363. 72 p.
- 109 - 1937. Our greatest travelers. In: The book of birds. Natl. Geog. Mag. 2: 301-350.
1939a. The migration of American birds. Doubleday, Doran & Co., N.Y. 189 p.
1939b. The individual vs. the species in migration studies. Auk 56(3): 250-254.
1941. The waterfowl flyways. Wild ducks. Am. Wildl. Inst. 20-29.
1945. Flyway regulations. Trans. Tenth North Am. Wildl. Conf.: 50-51.
1946. Keeping up with the waterfowl. Audubon Mag. 48(3): 194-205. Reprinted as U.S. Fish and Wildl. Serv. Leaflet 294. April 1947. 10 p.
Lowery, G. H. Jr.
1945. Trans-gulf spring migration of birds and the coastal hiatus. Wilson Bull. 57(2): 92-121.
1946. Evidence of trans-gulf migration. Auk 63(2): 175-211.
1951. A quantitative study of the nocturnal migration of birds. Univ. Kan. Pub. Mus. Nat. Hist. 3(2): 361-472.
Lowery, G. H., Jr. and R. J. Newman.
1966. A continentwide view of bird migration on four nights in October. Auk 83(4): 547-586.
Lucanus, F. von.
1911. Ueber die Hohe des vogelzuges. Verh. V Int. Ornithol. Congr. 557-562.
McMillan, N. T.
1938. Birds and the wind. Bird Lore 40(6): 397-406. Reprinted Smithson. Rep. for 1939: 355-363.
Magee, M. J.
1938. Evening Grosbeak recoveries. Northeastern Bird-Banding Assoc. Bull. 4: 56-59.
Main, J. S.
1932. The influence of temperature on migration. Wilson Bull. 44: 10-12.
Manville, R. H.
1963. Altitude record for mallard. Wilson Bull. 75: 9
Marshall, A. J.
1961. Breeding seasons and migration. Chapter 21: 307-339. In: Biology and comparative physiology of birds, Vol. II. A. J. Marshall ed. Academic Press, N.Y. and London. 468 p.
Matthews, G. V. T.
1951. The experimental investigation of navigation in homing pigeons. J. Exp. Biol. 28: 508-536.
1955. Bird navigation. Univ. Press, Cambridge. 141 p.
May, J. B.
1929. Recoveries of Black-crowned Night Herons banded in Massachusetts. Northeastern Bird-Banding Assoc. Bull. 5: 7-16.
Mazzeo, R.
1953. Homing of the Manx Shearwater. Auk 70: 200-201.
Meanly, B.
1971. Blackbirds and the southern rice crop. U.S. Fish and Wildl. Serv. Res. Pub. 100. 64 p.
Meinertzhagen, R.
1920. Some preliminary remarks on the altitude of the migratory flight of birds with special reference to the Paleoarctic region. Ibis, Series 11, 2: 920-936.
- 110 - 1921. Some preliminary remarks on the velocity of migratory flight among birds with special reference to the Paleoarctic region. Ibis, Series 11, 3(2): 228-238.
1955. The speed and altitude of bird flight (with notes on other animals). Ibis 97: 81-117.
Mewaldt, L. R. and R. G. Rose.
1960. Orientation of migratory restlessness in the White-crowned Sparrow. Science 131: 105-106.
Miller, A. H.
1963. Photoregulative and innate factors in the reproductive cycles of an equatorial sparrow. Proc. XVI Int. Congr. Zool. 1: 166.
Moreau, R. E.
1951. The migration system in perspective. Proc. X Int. Ornithol. Cong. 245-248
1953. Migration in the Mediterranean area. Ibis 95: 329-364.
1961. Problems of Mediterranean-Sahara migration, Part 3. Ibis 103a(4): 580-623.
Mueller, H. C. and D. D. Berger.
1967a. Fall migration of Sharp-shinned Hawks. Wilson Bull. 79: 397-415.
1967b. Wind drift, leading lines, and diurnal migration. Wilson Bull. 79(1): 50-63.
Murphy, R. C.
1936. Oceanic birds of South America. Am. Mus. Nat. Hist. New York. 2 vol.
Murray, B. G. Jr.
1964. A review of Sharp-shinned Hawk migration along the northeastern coast of the United States. Wilson Bull. 76: 257-264.
1965. On the autumn migration of the Blackpoll Warbler. Wilson Bull. 77(2): 122-133.
Murray, B. G. Jr. and J. R. Jehl Jr.
1964. Weights of autumn migrants from coastal New Jersey. Bird-Banding 35: 253-263.
Nice, M. M.
1937. Studies in the life history of the Song Sparrow. I. A population study of the Song Sparrow. Trans. Linn. Soc. N.Y. 4: 1-247.
Nisbet, I. C. T.
1961. Studying migration by moon-watching. Bird-Migr. 2(1): 38-42.
1963a. Quantitative study of migration with 23-centimeter radar. Ibis 105(4): 435-460.
Nisbet, I. C. T.
1963b. Measurements with radar of the height of nocturnal migration over Cape Cod, Massachusetts. Bird-Banding 34(2): 57-67.
Nisbet, I. C. T. and W. H. Drury Jr.
1967a. Scanning the sky/birds on radar. Mass. Audubon 51: 166-174.
1967b. Weather and migration. Mass. Audubon 52(1): 12-19.
1968. Short-term effects of weather on bird migration: A field study using multivariate statistics. Anim. Behav. 16(4): 496-530.
Odum, E. P.
1958. The fat deposition picture in the White-throated Sparrow in comparison with that in long-range migrants. Bird-Banding 29(1): 105-108.
Oldaker, R. F.
1961. 1960 survey of the California Gull. West. Bird Bander. 36(3): 26-30.
Orr, R. T.
1970. Animals in migration. MacMillan Co. Collier-MacMillan Ltd. London. 303 p.
- 111 -
Packard, F. M.
1945. The birds of Rocky Mountain National Park, Colorado. Auk 62: 371-394.
Parslow, J. L. F.
1969. The migration of passerine night migrants across the English Channel studied by radar. Ibis 111 (1): 48-79.
Pennycuick, D. J.
1969. The mechanics of bird migration. Ibis 111: 525-556.
Perdeck, A. C.
1967. Orientation of starlings after displacement to Spain. Ardea 55(3-4): 194-204.
Peterson, R.
1961. The long journey. Audubon Mag. 63(2): 72-75.
Pettingill, O. S. Jr.
1962. Hawk migrations around the Great Lakes. Audubon Mag. 64: 44-45, 49.
1970. Ornithology in laboratory and field. Fourth ed. Burgess Pub. Co., Minneap. 524 p.
Phillips, J. H.
1963. The pelagic distribution of the Sooty Shearwater (Puffinus griseus). Ibis 105: 340-353.
Pinkowski, B. C.
1971. An analysis of banding-recovery data on Eastern Bluebirds banded in Michigan and three neighboring states. Jack-Pine Warbler 49(2): 33-50.
Pough, R. H.
1948. Out of the night sky. Audubon Mag. 50(6): 354-355.
Ralph, C. J.
1971. An age differential of migrants in coastal California. Condor 73: 243-246.
Raynor, G. S.
1956. Meteorological variables and the northward movement of nocturnal land bird migrants. Auk 73: 153-175.
Rense, W. A.
1946. Astronomy and ornithology. Pop. Astron. 54(2): 1-19.
Richardson, W. J.
1971. Spring migration and weather in eastern Canada: a radar study. Am. Birds 25(3): 684-690.
1972. Autumn migration and weather in eastern Canada: a radar study. Am. Birds 26: 10-17.
Richardson, W. J. and M. E. Haight.
1970. Migration departures from starling roosts. Can. J. Zool. 48(1): 31-39.
Robbins, C. S.
1949. Weather and bird migration. Wood Thrush 4(4): 130-144.
1956. Hawk watch. Atl. Nat. 11: 208-217.
Robbins, C., D. Bridge and R. Feller.
1959. Relative abundance of adult male Redstarts at an inland and a coastal locality during fall migration. Md. Birdlife 15(1): 23-25.
Rowan, W.
1925. Relation of light to bird migration and developmental changes. Nature (Lond.) 115: 494-495.
1926. On photoperiodism, reproductive periodicity, and the annual migrations of birds and certain fishes. Boston Soc. Nat. Hist. Proc. 38: 147-189.
- 112 - 1930a. Experiments in bird migration. II. Reversed migration. Nat. Acad. Sci. Proc. 16: 520-525.
1930b. The mechanism of bird migration. Sci. Prog. 25: 70-78.
1931. The riddle of migration. Williams & Wilkins, Baltimore. 151 p.
Rudebeck, G.
1950. Studies on bird migration. Var. Fagelvarld, Supplementum I. 148 p.
Salomonsen, F.
1968. The moult migration. Wildfowl 19: 5-24.
Sauer, F.
1957. Die Sternenorientierung nachtlich ziehender Grasmucken (Sylvia attricapilla borin and curruca). Z. Tierpsych. 14: 29-70.
1958. Celestial navigation by birds. Sci. Am. 199(2): 42-47.
Sauer, E. G. F.
1961. Further studies on the stellar orientation of nocturnally migrating birds. Psychol. Forsch. 26(3): 224-244.
1963. Migration habits of Golden Plovers. Proc. XIII Int. Ornithol. Cong.: 454-467.
Sauer, E. G. F., and E. M. Sauer.
1960. Star navigation of nocturnal migrating birds. Cold Spring Harbor Symp. on Quant. Biol. 25: 463-473.
Schmidt-Koenig, K.
1963. Sun compass orientation of pigeons upon equatorial and transequatorial displacement. Biol. Bull. 124(3): 311-321.
1964. Sun compass orientation of pigeons upon displacement north of the Arctic circle. Biol. Bull. 127(1): 154-158.
Schnell, G. D.
1965. Recording the flight speed of birds by Doppler Radar. Living Bird 4: 79-87.
Schuz, E.
1963. On the North-Western migration divide of the White Stork. Proc. XIII Int. Ornithol. Cong. 475-480.
Serventy, D. L.
1953. Movements of pelagic sea-birds in the Indo-Pacific region. Proc. 7th Pacific Sci. Cong.: 4: 394-407.
1958. Recent studies on the Tasmanian mutton-bird. Aust. Mus. Mag. 12: 327-332.
Sheldon, W.
1965. Hawk migration in Michigan and the Straits of Mackinac. Jack-Pine Warbler 43(2): 79-83.
Suntov, V. P.
1968. Some correlations in the dispersal of Albatrosses in the Northern Pacific. Z. Zhurn. 47: 1054-1064. (In Russian, Eng. summ.)
Sladen, W. J. L.
1973. A continental study of whistling swans using neck collars. Wildfowl 24: 8-14.
Snyder, L. L.
1943. The Snowy Owl migration of 1941-42. Wilson Bull. 55(1): 8-10.
Southern, W. E.
1959. Homing of Purple Martins. Wilson Bull. 71: 254-261.
1965. Avian navigation. Article No. 2, Field Studies and Experiments, p. 87-88; In. Biotelemetry, Bioscience 15(2): 79-121, 159.
- 113 -
Stevenson, H. M.
1957. The relative magnitude of the trans-gulf and circum-gulf spring migrations. Wilson Bull. 69(1): 39-77.
Stewart, R. E., A. D. Geis and C. D. Evans.
1958. Distribution of populations and hunting kill of the Canvasback. J. Wildl. Manage. 22: 333-370.
Stoddard, H. L. and R. A. Norris.
1967. Bird casualties at a Leon County, Florida TV tower: an eleven-year study. Bull. Tall Timbers Res. Stn. 8. 104 p.
Storer, J. H.
1948. The flight of birds. Cranbrook Inst. Sci. Bull. 28: 94 p.
Storr, G. M.
1958. Migration routes of the Arctic Tern. Emu 58(1): 59-62.
Sutter, E.
1957. Radar-Beobachtungen uber den Verlauf des nachtlichen Vogelzuges. Rev. Suisse Zool. 64: 294-303.
Swan, L. W.
1970. Goose of the Himalayas. Nat. Hist. 79(10): 68-75.
Swarth, H. S.
1920. Revision of the avian genus Passerella, with special reference to the distribution and migration of the races in California. Univ. Calif. Pub. Zool. 21(4): 75-224.
Swirski, Z.
1965. Bird migrations (trans, from Pol.). Pol. Sci. Pub. Warsaw, Pol. 106 p.
Taverner, P. A.
1935. Continental land masses and their effect upon bird life. Condor 37: 160-162.
Tedd, J. G. and D. Lack.
1958. The detection of bird migration by high-power radar. Proc. R. Soc. (B) 149: 503-510.
Thompson. D. Q. and R. A. Person.
1963. The eider pass at Point Barrow, Alaska. J. Wildl. Manage. 27: 348-356.
Tomson, A. L.
1960. Bird-migration terms. Ibis 102: 140.
1964. A new dictionary of birds. McGraw-Hill Co. N.Y. 928 p.
1965. The transequatorial migration of the Manx Shearwater (Puffin des anglais). Oiseau Rev. Fr. Ornithol. 35: 130-140.
Van Tyne, J. and A. J. Berger.
1959. Fundamentals of ornithology. John Wiley & Sons, Inc. New York. 624 p.
Viguier, C.
1882. Le sens de l'orientation et ses organes. Rev. Philos. 14: 1-36.
Voous, K. H. and J. Wattel.
1963. Distribution and migration of the Greater Shearwater. Ardea 51: 143-157.
Walcott, D. and M. Michener.
1967. Analysis of tracks of single homing pigeons. Proc. XIV Int. Ornithol. Cong.: 311-329.
Wallraff, H. G.
1967. The present status of our knowledge about pigeon homing. Proc. XIV Int. Ornithol. Congr.: 331-358.
- 114 -
Welty, J. C.
1962. The life of birds. W. B. Saunders Co. Phila. & Lond. 546 p.
Wetmore, A.
1926. The migration of birds. Harv. Univ. Press, Camb., Mass. 217 p.
Williams, G. G.
1945. Do birds cross the Gulf of Mexico in spring? Auk 62(1): 98-111.
1947. Lowery on trans-gulf migration. Auk 64(2): 217-237.
1950. Weather and spring migration. Auk 67: 52-65.
Williamson, K.
1958. Bergmann's Rule and obligatory overseas migration. Br. Birds 51(6): 209-232.
Winkenwerder, H. A.
1902. The migration of birds with special reference to nocturnal flight. Wis. Nat. Hist. Soc. 2: 177-263.
Wolff, W. J.
1970. Goal orientation versus one-direction orientation in Teal (Anas c. crecca) during autumn migration. Ardea 58: 131-141.
Wolfson, A.
1940. A preliminary report on some experiments on bird migration. Condor 42(2): 93-99.
1945. The role of the pituitary fat deposition and body weight in bird migration. Condor 47(3): 95-127.
1948. Bird migration and the concept of continental drift. Science (Wash., D. C.) 108: 23-30.
Woodbury, A. M.
1941. Animal migration-periodic response theory. Auk 58: 463-505.
Woodbury, A. M., W. H. Behle and J. W. Sugden.
1946. Color-banding California Gulls at Great Salt Lake, Utah. Bull. Univ. Utah, 37(3): 1-15.
Yeagley, H. L.
1947. A preliminary study of physical basis of bird navigation. J. Appl. Physiol. 18(12): 1035-1063.
1951. A preliminary study of a physical basis of bird navigation. Part II. J. Appl. Physiol. 22(6): 746-760.
- 115 -
NOTE: For all North American species the authors have followed nomenclature in the 1957 edition of the A.O.U. Check-list. Also, we have incorporated the new names presented in the April 1973 issue of The Auk (volume 90, number 2, pages 411-419), the quarterly journal of the A.O.U. For other parts of the world we have used the most authoritative sources available.
| COMMON NAME | SCIENTIFIC NAME |
| Albatross, Black-footed | Diomedea nigripes |
| Albatross, Laysan | Diomedea immutabilis |
| Blackbird, Brewer's | Euphagus cyanocephalus |
| Blackbird, Red-winged | Agelaius phoeniceus |
| Blackbird, Rusty | Euphagus carolinus |
| Blackbird, Yellow-headed | Xanthocephalus xanthocephalus |
| Blackcap | Sylvia atricapilla |
| Bluebird, Eastern | Sialia sialis |
| Bluethroat | Luscinia svecica |
| Bobolink | Dolichonyx oryzivorus |
| Bobwhite | Colinus virginianus |
| Brant (Atlantic) | Branta bernicla hrota |
| Brant, Black | Branta bernicla nigricans |
| Bunting, Black-headed | Emberiza melanocephala |
| Bunting, Cretzchmar's | Emberiza caesia |
| Bunting, Indigo | Passerina cyanea |
| Bunting, Ortolan | Emberiza hortulana |
| Bunting, Snow | Plectrophenax nivalis |
| Canvasback | Aythya valisineria |
| Cardinal | Cardinalis |
| Chat, Yellow-breasted | Icteria virens |
| Chuck-will's-widow | Caprimulgus carolinensis |
| Coot (American) | Fulica americana |
| Crane, Sandhill | Grus canadensis |
| Creeper, Brown | Certhia familiaris |
| Crossbill, Red | Loxia curvirostra |
| Crow (Common) | Corvus brachyrhynchos |
| Cuckoo, Black-billed | Coccyzus erythropthalmus |
| Cuckoo, Yellow-billed | Coccyzus americanus |
| Curlew, Bristle-thighed | Numenius tahitiensis |
| Dove, Mourning | Zenaida macroura |
| Dove, Turtle | Streptopelia turtur |
| Duck, Black | Anas rubripes |
| Duck, Wood- 116 - | Aix sponsa |
| Eagle, Bald | Haliaeetus leucocephalus |
| Egret, Great | Casmerodius albus |
| Egret, Snowy | Egretta thula |
| Eider, Common | Somateria mollissima |
| Eider, King | Somateria spectabilis |
| Falcon, Peregrine | Falco peregrinus |
| Finch, Purple | Carpodacus purpureus |
| Flicker, Common | Colaptes auratus |
| Flycatcher, Hammond's | Empidonax hammondii |
| Flycatcher, Least | Empidonax minimus |
| Flycatcher, Western | Empidonax difficilis |
| Frigatebird, Magnificent | Fregata magnificens |
| Godwit, Black-tailed | Limosa limosa |
| Godwit, Hudsonian | Limosa haemastica |
| Goose, Bar-headed | Anser indicus |
| Goose, Canada | Branta canadensis |
| Goose, Emperor | Philacte canagica |
| Goose, Ross' | Chen rossii |
| Goose, Snow [Blue] | Chen caerulescens |
| Goose, White-fronted | Anser albifrons |
| Goshawk | Accipter gentilis |
| Grackle, Common | Quiscalus quiscula |
| Grosbeak, Black-headed | Pheucticus melanocephalus |
| Grosbeak, Evening | Hesperiphona vespertina |
| Grosbeak, Pine | Pinicola enucleator |
| Grosbeak, Rose-breasted | Pheucticus ludovicianus |
| Grouse, Blue | Dendragapus obscurus |
| Gull, California | Larus califomicus |
| Gull, Herring | Larus argentatus |
| Gull, Ross' | Rhodostethia rosea |
| Hawk, Broad-winged | Buteo platypterus |
| Hawk, Cooper's | Accipter cooperii |
| Hawk, Red-shouldered | Buteo lineatus |
| Hawk, Red-tailed | Buteo jamaicensis |
| Hawk, Rough-legged | Buteo lagopus |
| Hawk, Sharp-shinned | Accipter striatus |
| Hawk, Sparrow (European) | Accipter nisus |
| Hawk, Swainson's | Buteo swainsoni |
| Heron, Black-crowned Night | Nycticorax nycticorax |
| Heron, Little Blue | Florida caerulea |
| Hummingbird, Ruby-throated | Archilochus colubris |
| Jay, Blue- 117 - | Cyanocitta cristata |
| Junco, Dark-eyed | Junco hyemalis |
| Kingfisher, Belted | Megaceryle alcyon |
| Kinglet, Golden-crowned | Regulus satrapa |
| Kittiwake, Red-legged | Rissa brevirostris |
| Knot, Red | Calidris canutus |
| Lapwing | Vanellus vanellus |
| Lark, Horned | Eremophila alpestris |
| Longspur, Lapland | Calcarius lapponicus |
| Loon, Arctic | Gavia arctica |
| Mallard | Anas platyrhynchos |
| Martin, Purple | Progne subis |
| Nighthawk, Common | Chordeiles minor |
| Nuthatch, Red-breasted | Sitta canadensis |
| Oriole, Black-headed (Indian) | Oriolus xanthornus |
| Oriole, Black-naped | Oriolus chinensis |
| Ovenbird | Seiurus aurocapillus |
| Owl, Great-horned | Bubo virginianus |
| Owl, Snowy | Nyctea scandiaca |
| Pelican, White | Pelecanus erythrorhynchos |
| Penguin, Adelie | Pygoscelis adeliae |
| Petrel, Wilson's Storm | Oceanites oceanicus |
| Pewee, Western Wood | Contopus sordidulus |
| Phalarope, Northern | Libipes lobatus |
| Pigeon (Rock Dove) | Columba livia |
| Pintail | Anas acuta |
| Plover, Golden | Pluvialis dominica |
| Quail, Mountain | Oreortyx pictus |
| Redhead | Aythya americana |
| Redstart, American | Setophaga ruticilla |
| Robin, American | Turdus migratorius |
| Rook | Corvus frugilegus |
| Sanderling | Calidris alba |
| Sandpiper, Baird's | Calidris bairdii |
| Sandpiper, Purple | Calidris maritima |
| Sandpiper, White-rumped | Calidris fuscicollis |
| Sapsucker, Williamson's | Sphyrapicus thyroideus |
| Serin | Serinus serinus |
| Shearwater, Manx | Puffinus puffinus |
| Shearwater, Short-tailed | Puffinus tenuirostris |
| Shearwater, Sooty | Puffinus griseus |
| Shrike, Loggerhead | Lanius ludovicianus |
| Shrike, Red-backed- 118 - | Lanius collurio |
| Snipe, Common | Capella gallinago |
| Sora (Rail) | Porzana Carolina |
| Sparrow, Andean (Rufous-collared) | Zonotrichia capensis |
| Sparrow, Chipping | Spizella passerina |
| Sparrow, Field | Spizella pusilla |
| Sparrow, Fox | Passerella iliaca |
| Sparrow, Harris' | Zonotrichia querula |
| Sparrow, Ipswich | Passerculus sandwichensis princeps |
| Sparrow, Savannah | Passerculus sandwichensis |
| Sparrow, Song | Melospiza melodia |
| Sparrow, Swamp | Melospiza georgiana |
| Sparrow, Tree | Spizella arborea |
| Sparrow, Vesper | Pooecetes gramineus |
| Sparrow, White-throated | Zonotrichia albicollis |
| Swallow, Bank | Riparia riparia |
| Swallow, Barn | Hirundo rustica |
| Swallow, Cliff | Petrochelidon pyrrhonota |
| Swan, Whistling | Olor columbianus |
| Swift, Chimney | Chaetura pelagica |
| Swift, Common | Apus apus |
| Tanager, Scarlet | Piranga olivacea |
| Tanager, Western | Piranga ludoviciana |
| Tattler, Wandering | Heteroscelus incanum |
| Teal, Blue-winged | Anas discors |
| Tern, Arctic | Sterna paradisaea |
| Tern, Noddy | Anoüs stolidus |
| Tern, Sooty | Sterna fuscata |
| Thrush, Gray-cheeked | Catharus minimus |
| Thrush, Hermit | Catharus guttatus |
| Thrush, Swainson's | Catharus ustulatus |
| Thrush, Wood | Hylocichla mustelina |
| Turnstone, Ruddy | Arenaria interpres |
| Veery | Catharus fuscescens |
| Vireo, Red-eyed | Vireo olivaceus |
| Vulture, Turkey | Cathartes aura |
| Wagtail, Yellow | Motacilla flava |
| Warbler, Arctic | Phylloscopus borealis |
| Warbler, Blackpoll | Dendroica striatá |
| Warbler, Black-and-white | Mniotilta varia |
| Warbler, Black-throated Blue | Dendroica caerulescens |
| Warbler, Cape May- 119 - | Dendroica tigrina |
| Warbler, Connecticut | Oporomis agilis |
| Warbler, Golden-winged | Vermivora chrysoptera |
| Warbler, Kentucky | Oporomis formosus |
| Warbler, Palm | Dendroica palmarum |
| Warbler, Pine | Dendroica pinus |
| Warbler, Subalpine | Sylvia cantillans |
| Warbler, Willow | Phylloscopus trochilus |
| Warbler, Worm-eating | Helmitheros vermivorus |
| Warbler, Yellow | Dendroica petechia |
| Warbler, Yellow-rumped | Dendroica coronata |
| Waxwing, Bohemian | Bombycilla garrulus |
| Wheatear | Oenanthe oenanthe |
| Wigeon, American | Anas americana |
| Woodcock, American | Philohela minor |
| Wren, Carolina | Thryothorus ludovicianus |
| Wren, Long-billed Marsh | Telmatodytes palustris |
| Wren, Rock | Salpinctes obsoletus |
| Wren, Winter | Troglodytes troglodytes |
| Yellowlegs, Greater | Tringa melanoleuca |
| Yellowlegs, Lesser | Tringa flavipes |
| Yellowthroat, Common | Geothlypis trichas |
- 120 -
Created in 1849, the Department of the Interior—America's Department of Natural Resources—is concerned with the management, conservation, and development of the Nation's water, fish, wildlife, mineral, forest, and park and recreational resources. It also has major responsibilities for Indian and Territorial affairs.
As the Nation's principal conservation agency, the Department works to assure that nonrenewable resources are developed and used wisely, that park and recreational resources are conserved for the future, and that renewable resources make their full contribution to the progress, prosperity, and security of the United States—now and in the future.
★ U.S. GOVERNMENT PRINTING OFFICE: 1979 O—274-535
Illustrations moved to prevent splitting paragraphs. Minor typos corrected. Hyphenation was standardized to the most prevalent used except those in the Bibliography which were left as printed. Some repeated Author Names in the Bibliography were retained as printed.