
The Project Gutenberg eBook of Water Reptiles of the Past and Present, by Samuel Wendell Williston
Title: Water Reptiles of the Past and Present
Author: Samuel Wendell Williston
Release Date: February 3, 2022 [eBook #67314]
Language: English
Produced by: Peter Becker and the Online Distributed Proofreading Team at https://www.pgdp.net (This file was produced from images generously made available by The Internet Archive)
THE UNIVERSITY OF CHICAGO PRESS
CHICAGO, ILLINOIS
Agents
THE CAMBRIDGE UNIVERSITY PRESS
LONDON AND EDINBURGH
THE MARUZEN-KABUSHIKI-KAISHA
TOKYO, OSAKA, KYOTO
KARL W. HIERSEMANN
LEIPZIG
THE BAKER & TAYLOR COMPANY
NEW YORK
BY
Samuel Wendell Williston
Professor of Paleontology in the
University of Chicago
THE UNIVERSITY OF CHICAGO PRESS
CHICAGO, ILLINOIS
Copyright 1914 by
The University of Chicago
All Rights Reserved
Published October 1914
Composed and Printed By
The University of Chicago Press
Chicago, Illinois, U.S.A.
[Pg v]
It was just forty years ago that the writer of these lines, then an assistant of his beloved teacher, the late Professor B. F. Mudge, dug from the chalk rocks of the Great Plains his first specimens of water reptiles, mosasaurs and plesiosaurs. To the youthful collector, whose first glimpse of ancient vertebrate life had been the result of accident, these specimens opened up a new world and diverted the course of his life. They were rudely collected, after the way of those times, for modern methods were impracticable with the rifle in one hand and the pick in the other. Nor was much known in those days of these or other ancient creatures, for the science of vertebrate paleontology was yet very young. There were few students of fossil vertebrates—Leidy, Cope, and Marsh were the only ones in the United States—and but few collectors, of whom the writer alone survives.
Those broken and incomplete specimens, now preserved in the museum of Yale University, will best explain why this little book was written. The author offers it, so far as lies within him, as an authoritative and accurate account of some of the creatures of earlier ages which sought new opportunities by going down from the land into the water. So far as possible he has endeavored to make the text understandable, and, he hopes, of interest also, to the non-scientific reader. He will not apologize for such scientific terms as remain, since only by their use can precision be attained: there are no common English equivalents for them. The reader will find their explanations in the chapter on the skeleton of reptiles, and especially in the illustrations.
The author has had the opportunity during recent years of critically studying nearly all the reptiles described in the following pages, but, if that were the only source of his information, the accounts of many would have been meager. He has endeavored, briefly at least, to mention the names of all those to whom we are chiefly indebted for our knowledge, but in such a work as this it is manifestly impracticable to give due credit to every one. [Pg vi]
To the friends who have been of assistance in various ways he tenders his sincere thanks: to Professor E. Fraas for photographs and the kind permission to reproduce some of his excellent illustrations; to Dr. Dreverman, of the Senckenberg Museum, for several excellent photographs for reproduction or restoration; to Dr. Hauff, of Holzmaden, for an excellent photograph of an ichthyosaur; to Dr. H. F. Osborn, of the American Museum, for permission to reproduce the spirited restoration of ichthyosaurs drawn by Mr. Knight; to Professors Schuchert and Lull, and Dr. Wieland, of Yale University; to Dr. Hay and Mr. Gilmore, of the National Museum, to Mr. Barnum Brown and Dr. McGregor, of the American Museum, and to Professor Merriam, of the University of California, for photographs and other favors.
University of Chicago
July, 1914
[Pg vii]
| chapter | page | ||
| I. | Introduction | 1 | |
| II. | Classification of Reptiles | 13 | |
| III. | The Skeleton of Reptiles | 19 | |
| IV. | The Age of Reptiles | 44 | |
| V. | Adaptation of Land Reptiles to Life in the Water | 59 | |
| VI. | Order Sauropterygia | 73 | |
| Plesiosauria. | |||
| Nothosauria. | |||
| VII. | Order Anomodontia | 102 | |
| Lystrosaurus. | |||
| VIII. | Order Ichthyosauria | 107 | |
| IX. | Order Proganosauria | 126 | |
| Mesosaurus. | |||
| X. | Order Protorosauria | 132 | |
| Protorosaurus. | |||
| Pleurosaurus. | |||
| XI. | Order Squamata | 138 | |
| Lizards. | |||
| Mosasaurs. | |||
| Snakes. | |||
| XII. | Order Thalattosauria | 171 | |
| XIII. | Order Rhynchocephalia | 176 | |
| Choristodera. | |||
| XIV. | Order Parasuchia | 184 | |
| Phytosauria. | |||
| XV. | Order Crocodilia | 194 | |
| Eusuchia. | |||
| Mesosuchia. | |||
| Thalattosuchia. | |||
| XVI. | Order Chelonia | 216 | |
| Side-necked Turtles. | |||
| Snapping Turtles. | |||
| Fresh-water or Marsh Tortoises. | |||
| Land Tortoises. | |||
| Sea-Turtles. | |||
| Ancient Sea-Turtles. | |||
| Leather-back Marine Turtles. | |||
| River Turtles. | |||
In most persons the word reptile incites only feelings of disgust and abhorrence; to many it means a serpent, a cold, gliding, treacherous, and venomous creature shunning sunlight and always ready to poison. Our repugnance to serpents is so much a part of our instincts, or at least of our early education, that we are prone to impute to all crawling creatures those evil propensities which in reality only a very few possess. Were there no venomous serpents—and there are but two other venomous reptiles known—we should doubtless see much to admire in those animals now so commonly despised; because a few dozen kinds, like the rattlesnakes, copperheads, and cobras, protect themselves in ways not unlike those used by man to protect himself, we unjustly abhor the thousands of other kinds, most of which are not only innocent of all offense toward man, but are often useful to him.
There are now living upon the earth more than four thousand kinds or species of cold-blooded animals which we call reptiles, all of which are easily distinguishable into four principal groups: the serpents and lizards, the crocodiles, the turtles, and the tuatera. Their habits and forms are very diverse, but they all possess in common certain structural characters which sharply distinguish them from all other living creatures. A reptile may be tersely defined as a cold-blooded, backboned animal which breathes air throughout life. And yet, it is not quite certain that this definition is strictly correct when applied to all the reptiles of the past, since it has been believed that certain extinct ones may have been warm-blooded. By this definition, short as it is, we at once exclude a large number of cold-blooded, air-breathing, backboned animals which were formerly included by scientific men among the true reptiles, and even yet are popularly often so included—the amphibians or batrachians. These animals, now almost wholly represented by the despised toads, frogs, [Pg 2] and salamanders, were, very long ago, among the rulers of the land, of great size and extraordinary forms. But they have dwindled away, both in size and in numbers, till only a comparatively few of their descendants are left, none of them more than two or three feet in length, and all of them sluggish in disposition and of inoffensive habits. While we may speak of the amphibians as air-breathing, they are, with few exceptions, water-breathers during the earlier part of their existence. Some may pass their whole lives as water-breathers, while a few begin to breathe air as soon as hatched from the egg; but these are the marked exceptions.
In many respects the internal structure of the amphibians of the present time is widely different from that of reptiles, though there can be no doubt that the early amphibian ancestors of the modern toads, frogs, and salamanders were also the ancestors of all living and extinct reptiles, and it is a fact that the living amphibians differ more from some of the ancient ones than those early amphibians did from their contemporary reptiles. Discoveries in recent years have bridged over nearly all the essential differences between the two classes so completely that many forms cannot be classified unless one has their nearly complete skeletons. We know that some of the oldest amphibians, belonging to the great division called Stegocephalia, were really water-breathers during a part of their lives, because distinct impressions of their branchiae, or water-breathing organs, have been discovered in the rocks with their skeletal remains, but we are not at all sure that some of the more highly developed kinds were not air-breathers from the time they left the egg; indeed, we rather suspect that such was the case.
We are also now quite certain that, from some of the early extinct reptiles—the immediate forbears probably of the great dinosaurs—the class of birds arose, since the structural relationships between birds and reptiles are almost as close as those between reptiles and amphibians.
Huxley believed that the great class of mammals arose directly from the amphibians, and there are some zoölogists even yet who think that he was right. But paleontologists are now quite sure that they were [Pg 3] evolved from a group of primitive reptiles, known chiefly from Africa, called the Theriodontia; quite sure because nearly all the connecting links between the two classes have already been discovered—to such an extent, indeed, that really nothing distinctive of either class is left save the presence or absence of the peculiar bone called the quadrate, the bone with which the lower jaw articulates in birds and reptiles; and certain elemental parts of the lower jaw itself. And even these bones, in certain mammal-like reptiles, had become mere vestiges. Even the double condyle of the mammal skull, with which the vertebrae articulate, so like those of the amphibian skull that Huxley based his belief of the amphibian origin of the mammals chiefly upon it, has now been found in certain reptiles. Warm-bloodedness, one of the diagnostic characters of birds and mammals, is not really very important, since it must have arisen in these two classes independently, and we may easily conceive that the earliest mammals were cold-blooded or that the immediate ancestors of the mammals were warm-blooded.
It is an interesting fact in the history of the vertebrates, as of all other groups of animals and plants, that the chief divisions arose early in geological history. Every known order of amphibians and reptiles, unless it be that including the blind-worms, was differentiated by the close of the Triassic period. The frogs are now known from the Jurassic. The mammals and birds also quite surely date their birth from the Triassic. And this early differentiation of the chief groups is doubtless due to the fact that the potentialities of diverse evolution are limited by specialization. It is apparently a law that evolution is irreversible, that it never goes from the special to the general, that an organism or an organ once extinct or functionally lost never reappears. And it is also a law in evolution that the parts in an organism tend toward reduction in number, with the fewer parts greatly specialized in function, just as the most perfect human machine is that which has the fewest parts, and each part most highly adapted to the special function it has to subserve. And these laws explain why it is that no highly specialized organism can be ancestral to others differing widely from it. The more radically distinct an organism is from its allies, the earlier it must have branched off from the genealogical tree. [Pg 4]
The many new discoveries of extinct forms so often intermediate, not only between the larger groups, but between many of the lesser ones as well, are making the classification of the vertebrates increasingly difficult. At one time it was sufficient to define a reptile as a cold-blooded animal with a single occipital condyle, that is, with a single articular surface between the skull and the first vertebra of the neck; a mammal as a warm-blooded animal with two articular surfaces; but these definitions are no longer strictly correct. Connecting links do not break down classification, as one might think, but they do often spoil our fine systems and compel our classifiers to take a wider view of nature than their own narrow province affords.
We can never hope that most, or even the greater part, of all the animals which have lived in the past will ever become known to us, even imperfectly. Doubtless the species of the past geological ages outnumbered many times, perhaps hundreds of times, all those now living, since many of these latter are merely the remnants of far more varied and extensive faunas. At times the conditions for the preservation of the remains of animal life have been more favorable than at others, and, under such favorable conditions, a fairly good glimpse is sometimes given us of the fauna of some isolated epoch and locality in the earth’s history. Those animals which lived in and about the water have been preserved in greater numbers and more perfectly than the strictly land animals, since fossils are due to the preserving action of water, with few exceptions. Of those animals which lived upon the land or in the air only the rarest of accidents carried the skeletons into the lakes, seas, and oceans. And, even when they had been covered by sediments at the bottoms of lakes and seas and hidden away from adverse agencies, it has often happened that the great erosions of later ages have carried away and destroyed the rocks in which they were inclosed. The records of long intervals of time have thus been lost in all parts of the world. That we are able to obtain even an imperfectly continuous history is due to the fact that the intervals thus lost are not everywhere contemporaneous, that the missing records of one place may be filled out in part elsewhere. But [Pg 5] this substitution of records from a distance can never make the history complete. If, in human history, we had only the records for one century in China, for another in England, and for yet another in South America, how imperfect indeed would be our knowledge of human progress. Animals and plants are never quite alike in remote regions, and they never have been. The living reptiles of North and South America are today almost entirely different, and, were their fossil remains to be discovered a million years hence, it would be very difficult to decide that they had once lived contemporaneously; difficult, though perhaps not impossible, since some are so nearly alike that their relationships or possible identity would probably be established after long search. This will serve to make clear how very difficult it is, for the most part, to correlate exactly the geological formations in remote regions of the earth, or even sometimes in adjacent regions where the fossils are scanty, or the conditions under which the animals had lived were very different.
There are long periods of time, millions of years at a stretch perhaps, throughout which our knowledge amounts to little or nothing concerning many land reptiles which we are sure must have existed abundantly. No better example of our oftentimes scanty knowledge can be cited than the following. Until within the past fifteen years it was thought that true land lizards, of which there are about eighteen hundred species now living, dated back in their history no farther than about the close of the great Secondary Period, or the Age of Reptiles. But a single skull of a true land lizard has been discovered in the Triassic deposits of South Africa, a skull of a form so nearly like that of the modern iguana of America that its discoverer, Dr. Broom, has called it Paliguana. The lizards must have been in existence, probably many thousand species of them, during all the great interval of time between the Middle Triassic and the close of the Cretaceous, since it is a law which can have no exception, that a type of life once extinct never reappears. The “ancient iguanas” of the Trias must have been the forbears of many, if not all, of the lizards of later times, though nothing is known of their descendants through a period of time which can be measured only by millions of years. [Pg 6]
However, notwithstanding these imperfections of our geological records, we know very much more about extinct reptiles than we do about living ones, so far at least as those parts capable of preservation in the rocks are concerned. Were our knowledge of reptiles confined to the forms now living upon the earth it would be relatively very incomplete since, aside from the lizards and snakes, they are merely the remnants of what was once a mighty class of vertebrates.
Not only do we learn from the remains preserved in the rocks the precise shape and structure of the bones of the skeleton and their precise articulations, but we are often able to determine not a little regarding the forms which the living animals had by the impressions made by the dead bodies in the soft sediment which inclosed them before decomposition of the softer parts had ensued, sediments which afterward solidified into hard rock. But these impressions are, with rare exceptions, only those of profiles or of flattened membranes. The rounded bodies of life do not retain their shape long enough for the sediment to harden; in most cases the flesh has decomposed before being entirely covered by sediment. Sometimes the integument and scales in a carbonized condition are actually preserved, retaining some of the actual structure of the organized material. The carbon pigment of the skin has sometimes been preserved in patterns indicating the color-markings in some of these ancient reptiles; and even the microscopic structure has been detected in carbonized remains of organs. Fossil stomach contents, the bony remains of unhatched young, as well as the delicate impressions of skin and membrane, all add to our knowledge of the structure and habits of the animals which lived so long ago. Many other things also may be learned, or at least inferred, concerning the living animals and their habits from the positions in which the skeletons are found, from the nature of the rocks which inclose them, or from the character and abundance of other fossils found with them. The frequent discovery of bones which had been injured and mended during life, or the living amputation of members, often tell of the characteristics of the creatures. So, too, the climatic conditions under which the animals lived may often be inferred with [Pg 7] tolerable certainty; the presence of “stomach-stones” reveals something of the food habits, and even of the structure of the alimentary canal, etc.
All this information is gained slowly, often very slowly, and with much labor and pains. Rarely or never is it the case that all the information obtainable concerning any one kind of an extinct animal is furnished by a single specimen. Skeletons are very seldom, perhaps never, found quite complete, with all their parts in their natural positions; and the nature of the matrix inclosing them usually prevents a study of all parts of any specimen. If a newly discovered fossil is widely different from the corresponding parts of any creature previously known, whether living or extinct, we cannot infer very much from a few bones as to what the remainder of the skeleton is like. Such inferences or guesses in the past have often resulted in grievous error, and self-respecting paleontologists are now very reluctant to speculate much concerning extinct animals from fragments of a skeleton, no matter what those fragments or bones may be; future discoveries are sure to reveal errors. It is, therefore, only by the accumulation of much material, and by the careful study and comparison of all known related animals, that reliable conclusions can be reached. Often it requires scores of specimens to determine the exact structure of a single kind of animal, and, as the collection and preparation of fossil skeletons are tedious and expensive, our knowledge sometimes increases very slowly. In recent years, however, there have been many more students of extinct backboned animals than formerly, and there are now many museums and universities which spend annually large sums of money in the collection and preparation of such fossils. This greater activity of the last twenty years is bringing to light many new and strange forms, as well as completing our knowledge of those previously imperfectly known.
It is commonly, but erroneously, believed that the bones of extinct animals are usually found in excavations made for the purpose. It is true that not a few specimens of fossils have been discovered in excavations made for other purposes, such as railway cuttings, quarries, wells, etc., but if no others were found our knowledge of the animals of the past would be very meager indeed. Fossils are, for the [Pg 8] most part, found by deliberate search over the denuded rocks in which they occur. Methods of search and collection will best be understood by the following description of the noted fossil-bearing rocks of western Kansas.

Fig. 1.—A characteristic
chalk exposure in western Kansas,
a hundred acres or more in extent.
About the middle of Cretaceous times, there extended from the Gulf of Mexico on the south to or nearly to the Arctic Ocean on the north a narrow inland ocean or sea, a few hundred miles in width, covering what is now the western part of Kansas and the eastern part of Colorado, and separating the North American continent into two distinct bodies of land. This ocean, because of its location, bordered on both sides by low-lying lands—the Rocky Mountains had not then been pushed up—doubtless was comparatively calm and placid, free from violent storms and high tides. That the climate, in the region of Kansas at least, was warm or even subtropical is fairly certain, since plants allied to those now living in warm, temperate, or subtropical regions were then living much farther to the north; and since the animals which [Pg 9] then lived in this sea were only such as would be expected in waters of warm temperature. Its tributary rivers could have been neither large nor swift-flowing, since the sediment at its bottom was free, or nearly free, from in-brought material. This was at least the case not very far from its shores. Its slowly falling sediment was composed, almost exclusively, of microscopic shells of animals and plants, foraminifera and coccoliths. The deposits thus made are almost identical with those now forming in various parts of the world in clear but not deep waters, away from the immediate coasts of the continents, almost pure chalk. Animals dying in this inland sea fell slowly to the bottom during or after decomposition of their softer parts, and the slowly increasing sediments covered up and buried the preservable parts. The many predaceous fishes and other scavengers with which the waters abounded often tore the decomposing bodies apart, separating and displacing the bones of the skeleton; and the currents of the shallow waters washed others apart. Often the teeth of fishes and other carnivorous animals are found imbedded in the bones, and many are the scars and toothmarks observed in the fossil bones.
After the ocean had dried up and the bottom had been raised far above the present level of the oceans, other deposits made in lakes and by the winds covered deeply the consolidating sediments, burying them for millions of years with all that they contained. Long-continued erosion by winds and rains has again laid bare many parts of the old ocean bottom, and has washed them out into ravines and gullies. Many hundreds of square miles of this chalk are now laid bare in western Kansas, upon which the growth of vegetation has been prevented by the arid climate. Here and there may now be discovered protruding from the sloping or precipitous surfaces of this exposed chalk bones or parts of bones of the old animals buried so long ago in the soft sediment of the ancient ocean bottom.
The sharp-eyed searcher after fossils detects these protruding, often broken and weather-worn, petrified bones, which themselves betray the presence often of other parts of their skeletons still concealed in the chalky hillside. Fortunate is he if he has discovered a specimen soon [Pg 10] after it appeared at the surface, before the rains have washed away and destroyed most of the remains that had been there preserved. Still more fortunate is he if all or nearly all of the original skeleton has been preserved together in its natural relations. After days, perhaps weeks, of labor, the specimen is secured and shipped to the laboratory. Those parts which have been washed out of the chalky rock before the discovery of the specimen are always more or less injured and for the most part lost, their fragments strewn down the hillside, for erosion is always slow and many years may have elapsed since first the specimen had appeared at the surface. More frequently, perhaps, a few strokes of the pick and shovel disclose but one, two, or three bones remaining in the rocks. The specimen, if large, or composed of many bones, is carefully uncovered sufficiently to show its extent, and then, so far as possible, removed in large blocks of the rock. The bones themselves, notwithstanding their petrifaction, are usually soft and easily broken, and their separate removal from the matrix may require weeks or even months of labor, work which cannot be done prudently in the field.
Of many specimens the rock matrix is so hard that the task of removing it from the bones is slow and difficult, indeed well-nigh impossible, for the bones are usually softer than their surrounding matrix. On the other hand, the matrix may be so soft and friable that it cannot be quarried out in blocks. In such cases the separate divisions, as large as they can be excavated and safely handled, are carefully covered with thick bandages of burlap and plaster-of-paris, often strengthened with rods of iron or boards. The skeleton of a single animal treated in this way may require weeks and even months to collect, prepare, and mount in the museum.
From what has been said the reader will understand how it is possible to make an approximately accurate picture of extinct animals as they appeared in life—approximately accurate, never absolutely so. The flesh and other soft parts of an animal are never petrified, though it is a common belief that they may be. Petrified men and women are still occasionally shown in cheap museums, but they are always frauds. Many times has the writer been called upon to express an opinion as to the nature of some concretion which the discoverer was sure was a petrified [Pg 11] snake, turtle, or even some part of the human body, because of fancied resemblances in shape and size. Not too emphatically can it be said that anything dug from the earth having the shape of a living animal and alleged to be petrified is either an accidental resemblance or a deliberate humbug—if we except such extraordinary casts as those of Pompeii. The Cardiff Giant and the Muldoon are still fresh in the memory of some of us. There have been a few instances where flesh has been preserved in the North, frozen for thousands of years, but frozen fossils are very different from petrified fossils. Flesh decays before it possibly can be petrified, and only rarely is the residue of flesh, tendons, and skin, that is, the carbon and mineral matters, preserved.

Fig. 2.—Removing
a specimen of fish in a block from
the chalk of western Kansas.
One may sometimes restore extinct animals as in life, knowing fully the shape and structure of the skeleton, and still be far from the real truth. All elephants of the present time have a bare or nearly bare skin. If all that we knew of the extinct mammoth were derived from the [Pg 12] skeleton we should never have suspected that the creature was clothed during life with long and abundant hair, such as has been found with the frozen bodies in Siberia. Nor should we suspect that the dromedary and Bactrian camels of today have large masses of fat on their backs, if we knew only their skeletons. It must therefore be remembered that all restorations of extinct animals, representing them as in life, are merely the sum of our knowledge concerning them, as close approximations to the real truth as it is possible to make. Or, rather, they should be such approximations; unfortunately many such restorations have been made by artists wholly unacquainted with the anatomy of the creatures they attempt to represent, often adorned with appendages drawn from a too vivid imagination.
[Pg 13]
There is very much doubt, very much uncertainty, among paleontologists about the classification of reptiles. No two writers agree on the number of orders, or the rank of many forms. Some recognize twenty or more orders, others but eight or nine. And this doubt and uncertainty are due chiefly to the many discoveries of early forms that have been made during the past twenty years. The many strange and unclassifiable types which have come to light in North America, South Africa, and Europe have thrown doubt on all previous classificatory schemes, have weakened our faith in all attempts to trace out the genealogies of the reptilian orders; and classification is merely genealogy. It is only the paleontologist who is competent to express opinions concerning the larger principles of classification of organisms, and especially of the classification of reptiles. The neozoölogist, ignorant of extinct forms, can only hazard guesses and conjectures as to the relationships of the larger groups, for he has only the specialized or decadent remnants of past faunas upon which to base his opinions. About some things we can be quite confident; about some groups opinions have crystallized, and we all agree, except perhaps on trifles. The dinosaurs, the pterodactyls, the crocodiles, for instance, offer only minor problems to perplex the systematist, but the origin and the relations, not only of these, but also of nearly all the others, are still involved in obscurity. The question, whence came the ichthyosaurs, the plesiosaurs, the turtles, etc., seems almost as far from solution as it did fifty years ago. With every problem solved a dozen more intrude themselves upon us. Hence, classification simply represents the present condition of our knowledge, our present opinions as to genealogies. It was the fashion a dozen years ago to draw all sorts of genealogical trees on the slightest pretext, to trace in beautifully clear lines the precise descent of all kinds of animals; [Pg 14] and very few have been worth the paper on which they were printed. When facts are numerous enough, conclusions are patent even to the novice; when facts are few and obscure, one can guess about as well as another. In general, it may be said that the older a group of animals is the more abstruse are the problems presented; first, because of the lack of abundant material; second, because the forms speak to us in an unfamiliar language that we cannot easily interpret. The classification of the mammals approaches more nearly the ultimate truth than does that of any other group of organisms, because we know more about the extinct forms than we do of any other class, and also because we know more about the living forms than we do about any other living animals.
Species of reptiles are, for the most part, vague quantities in paleontology; they can be determined with assurance only by the comparison of abundant material. Adult characters in mammals are apparent in the ossification of the skeleton, and size can be used within moderate limits in the determination of species; but size in reptiles means but little; no one could possibly say that the skeleton of an alligator six feet in length is not that of an adult animal if he knew nothing else about the Crocodilia. So also the compression and malformations of bones from the processes of fossilization obliterate specific characters in great part. Nor are specific characters easily distinguishable in the skeletons of living reptiles. The genus, therefore, among fossil reptiles is practically the unit, and we may be sure that for every well-defined genus we discover there existed numerous minor variations, which, had we the living animals to study, we should call species. We classify the living Crocodilia into two families, about four well-defined genera—perhaps even five or six—and about twenty-five species. Of the living lizards there are about eighteen hundred species, twenty families, and four larger groups or suborders. In all probability the lizards have never been more abundant and more varied than they are at the present time. Possibly these proportions of species, genera, families, and suborders may represent approximately the proportions that have existed at some time or other in most of the other groups which we call orders—approximately only, [Pg 15] for we can never be quite sure that we evaluate the structural characters of different groups of organisms quite equally. The absence of a molar tooth in a mammal would ordinarily indicate a genus, the absence of a tooth in a reptile might not indicate even a variety or a race. Whence it follows that classification of organisms is not and never will be an exact science. The value of characters used in classification is very unequal, as we have seen. No two persons see these characters from the same viewpoints, and in consequence no two persons whose opinions are worth while ever wholly agree as to classification.
The following scheme differs only in minor details from the more conservative of the generally accepted views, and those differences are, for the most part, the writer’s own opinions, to be taken for what they are worth. It may be said decisively that no classification of the reptiles into major groups, into super-families or subclasses that has so far been proposed is worthy of acceptance; there is no such subclass as the Diapsida or Synapsida, for instance. And we have very much more to learn about the early reptiles before any general classification of the reptiles can be securely founded. It is very probable that the primary radiation of the reptiles into the various lines of descent, into its main branches, occurred much earlier than we have been disposed to believe; that before the close of Paleozoic time, perhaps before the close of the Carboniferous, all the great groups of reptiles had gone off from the main stem, and that since then only smaller and smaller branches have appeared. There have been no new orders of reptiles in all probability since Triassic times, and perhaps none since Permian.
Taxonomists are often disposed to cut the Gordian knots of relationships by raising the ranks of the animals they study to independent positions. More than thirty independent orders of reptiles have been proposed by different students, and quite as many of mammals and of birds; possibly after more forms have been discovered there will be as many proposed for the amphibians. Sometimes, indeed, it is better to make such independent groups than to unite lesser ones on doubtful evidence. But the writer, for one, believes that it is more worthy of the thoughtful scientific student to seek for relationships than for differences. It is far easier to destroy than to construct, to make new [Pg 16] genera, families, and orders than to unite those already proposed. To raise every proposed suborder of reptiles to an order, as has been proposed by various writers, and the orders to subclasses, only leaves classification where it was; nothing has been added to taxonomy save a lot of new names to perplex and annoy the student.
In the following scheme of classification three groups provisionally called orders are prefixed by an asterisk.
| Order | COTYLOSAURIA | ||
Primitive reptiles with notochordal vertebrae, imperforate temporal region, persistent intercentra; two coracoids; plate-like pelvis, with all or most of the amphibian skull elements; short legs and short neck; phalangeal formula primarily 2, 3, 4, 5, 3(4). |
|||
| Suborder | Diadectosauria | Permocarboniferous, North America. | |
| Pantylosauria | Permocarboniferous, North America. | ||
| Labidosauria | Lower Permian, North America. | ||
| Pareiasauria | Upper Permian, Europe, Africa. | ||
| Procolophonia | Triassic, Europe, Africa. | ||
| Order | CHELONIA | ||
Temporal region imperforate. Head and limbs more or less retractile within a box formed chiefly by the exoskeleton. |
|||
| Suborder | Pleurodira | Triassic to recent. | |
| Cryptodira | Jurassic to recent. | ||
| Trionychoidea | Cretaceous to recent. | ||
| Order | THEROMORPHA | ||
Primitive reptiles with notochordal vertebrae, perforate temporal region, persistent intercentra; two coracoids; plate-like pelvis with median vacuity; no free dermosupraoccipitals in skull; longer legs and neck; phalangeal formula 2, 3, 4, 5, 3(4). |
|||
| Suborder | Pelycosauria (sens. lat.) | Permocarboniferous, North America, | |
| Europe. | |||
| Dromasauria | Upper Permian, Africa. | ||
| Dinocephalia | Middle and Upper Permian, Africa. | ||
| Order | THERAPSIDA | ||
Reptiles with a single temporal perforation on each side; vertebrae not notochordal; intercentra not persistent; pelvis with vacuity; skull bones reduced; teeth heterodont; phalangeal formula, 2, 3, 3, 3, 3. |
|||
| Suborder | Anomodontia | Permo-Trias, Africa, North America. | |
| Therocephalia | Upper Permian, Africa. [Pg 17] | ||
| Theriodontia | Trias, Africa. | ||
| Order | SAUROPTERYGIA | ||
Aquatic reptiles with a single temporal vacuity; no supratemporal bone, or quadratojugal; ribs single-headed, diapophysial; coracoids large, meeting in middle line, single; neck long, tail short. |
|||
| Suborder | Nothosauria | Triassic, Europe. | |
| Plesiosauria | Triassic to close of Cretaceous, | ||
| cosmopolitan. | |||
| ⃰Order | PROGANOSAURIA | ||
Primitive aquatic reptiles; single (? upper) temporal perforation; neck elongate; nares posterior; vertebrae notochordal; intercentra persistent; pelvis plate-like; phalangeal formula 2, 3, 4, 5, 4(6). Permocarboniferous, Africa, South America. |
|||
| Order | ICHTHYOSAURIA | ||
Reptiles with all aquatic adaptations; a single, upper temporal perforation; both supratemporal and squamosal present; a single coracoid. Middle Triassic to Benton Cretaceous, cosmopolitan. |
|||
| ⃰Order | PROTOROSAURIA | ||
A single, upper temporal vacuity, quadrate fixed (neck vertebrae elongate); bones hollow; cervical ribs single-headed, articulating with centrum; pelvis plate-like. Permian, North America, Europe. |
|||
| Order | SQUAMATA | ||
A single, upper temporal vacuity, or, secondarily none; quadrate loosely articulated with cranium; teeth on palate; intercentra more or less persistent; a single coracoid; ribs single-headed, central. |
|||
| Suborder | Lacertilia | Trias to recent. | |
| Mosasauria | Upper Cretaceous, cosmopolitan. | ||
| Ophidia | Upper Cretaceous to recent. | ||
| ⃰Order | THALATTOSAURIA | ||
Aquatic reptiles; two (?) temporal vacuities; ribs single-headed, attached to centrum; single coracoid; no intercentra. Trias, California. |
|||
| Order | RHYNCHOCEPHALIA | ||
Two temporal vacuities on each side; palate with teeth; intercentra persistent; a single coracoid; teeth acrodont; ribs articulating with centrum and arch. |
|||
| Suborder | Rhynchosauria | Triassic, Europe. | |
| Sphenodontia | Triassic to recent. | ||
| Choristodera | Uppermost Cretaceous, lowermost | ||
| Eocene, North America, Europe. [Pg 18] | |||
| Order | PARASUCHIA | ||
Subaquatic reptiles, with two temporal vacuities; an antorbital vacuity; no false palate; pubis entering acetabulum; ribs double-headed, diapophysial. |
|||
| Suborder | Phytosauria | Upper Trias, cosmopolitan. | |
| Pelycosimia | Trias, Africa. | ||
| Pseudosuchia | Trias, Europe, North America. | ||
| Order | CROCODILIA | ||
Two temporal vacuities; teeth thecodont; a false palate; pubis excluded from acetabulum; single coracoid; ribs double-headed, diapophysial; subaquatic or aquatic. |
|||
| Suborder | Eusuchia | Jurassic to recent. | |
| Thalattosuchia | Upper Jurassic, Europe. | ||
| Order | DINOSAURIA | ||
Ambulatory reptiles, with two temporal vacuities; no false palate; pubis entering acetabulum; ribs double-headed, diapophysial. |
|||
| Suborder | Theropoda | Upper Trias to close of Cretaceous, cosmopolitan. | |
| Orthopoda | Close of Trias to close of Cretaceous, cosmopolitan. | ||
| Sauropoda | Upper Jurassic, Lower Cretaceous, cosmopolitan. | ||
| Order | PTEROSAURIA | ||
Volant reptiles; fourth finger greatly elongated to support patagium; neck vertebrae elongated; bones hollow; ribs double-headed, diapophysial; a single coracoid; no clavicles or interclavicle; two temporal vacuities. |
|||
| Suborder | Pterodermata | Jurassic, Europe. | |
| Pterodactyloidea | Upper Jurassic to Upper Cretaceous, | ||
| Europe, North America. | |||
[Pg 19]
The bony framework, or skeleton, that which gives form and stature to the body, and which serves for the support of the soft parts and the attachment of muscles, is, with rare exceptions, all that is ever preserved of fossil animals. Because, therefore, students of extinct animals must rely so much, if not exclusively, upon the skeleton much attention has been given to the study of comparative osteology, the science of bones. Not only are most of the bones of the skeleton characteristic of the genus to which they belong, but the more general plan of the skeleton, or parts of the skeleton, is likewise characteristic of the larger groups. The paleontologist may become so expert in deciphering the characters of single bones, or even parts of bones—often all that are known of animals new to science—that he is able to hazard guesses as to the general structure of the skeleton to which they belong. But such guesses usually will approximate the real truth only in the degree that the bones upon which they are based approximate like bones of other animals that are better known. Not all parts of the skeleton are equally characteristic of the type of animal which possessed them. A tooth of a mammal may positively determine the species to which it belongs, while the toe bone of the same animal might not enable one to guess at its family, even. As a rule one can seldom be quite sure of the species of a reptile unless the larger part of the skeleton, or at least the skull, is available, although almost any bone of the skeleton, if one is expert, will permit a decision as to the family, if not the genus.
One must often depend upon the positions and relations of the bones, as found in the rocky matrix, for the final determination of many characters. One can, for instance, never be sure of the number of bones in the neck, trunk, tail, or feet of a reptile, until specimens have been found with all such bones in position. It is [Pg 20] for this reason that much care is exercised in the collection of specimens of fossil animals, and especially of fossil reptiles, to preserve all parts of the skeleton, so far as possible, in the relations they occupied in the rocks until they can be studied in the laboratory. Many grievous errors have been made in the past by hasty inferences from fragmentary and poorly collected specimens.

Fig. 3.—Limnoscelis,
a subaquatic cotylosaur, from
the Permocarboniferous of New Mexico.
Because of the reliance which must be placed upon the skeleton it will be necessary to speak somewhat in detail of its structure in the reptiles, and to use not a few terms in its description that are unfamiliar to the general reader. So far as possible technical terms will be avoided, though some must be used, as there are no equivalents in the English language for them. The reader may use this chapter as a sort of explanatory index or glossary for the better elucidation of the necessary details of the following chapters.
It is needless to say that the skeleton of a reptile is arranged on essentially the same plan as that of our own; the bones have the same names that they have in our own skeleton, but there are more of them, and the individual bones, as a general rule, are less highly specialized, that is, are not so well adapted for special functions. In a word, the skeleton of a reptile for the most part is generalized, though particular parts may be highly specialized for particular uses. As a rule, if not as a law, the course of evolution has been to reduce [Pg 21] the number of parts and to adapt those which remain more closely to their special uses, either by increase in size, or by modifications of their shape and structure.
The skull of reptiles is much more primitive or generalized in structure than is that of mammals, to such an extent, indeed, that there is yet much doubt as to the precise homologies of some of the bones composing it; and, inasmuch as the names were originally given, for the most part, to the bones of the human skull, there is still some confusion among students as to the proper names in all cases, a confusion that doubtless will not be wholly dissipated until we know much more about the early or more primitive reptiles than we do at present.

Fig. 4

Fig. 5
Fig. 4.—Seymouria, a primitive cotylosaurian. Skull, from above: pm, premaxilla; n, nasal; l, lacrimal; p, prefrontal; f, frontal; pf, postfrontal; it, intertemporal; st, supratemporal; sq, squamosal; ds, dermosupraoccipital; t, tabulare; j, jugal; po, postorbital; m, maxilla; s, surangular; ang, angular; pa, parietal.
Fig. 5.—Seymouria, skull from the side. Explanations as in fig. 4.
As in other parts of the skeleton, there has been a reduction in the number of parts of the reptile skull from that of the more primitive forms, and a better adaptation of those which remain for the special uses they subserve. This reduction in number has been caused in part by the actual loss of bones, in part by the fusion of contiguous ones. The most primitive reptiles had no less than seventy-two separate bones in [Pg 22] the skull;[1] the human skull has but twenty-eight inclusive of the ear bones. There is but little variation, either in the number or in the relations of bones, in the mammalian skull. If one knows the human skull thoroughly he can easily understand the structure of the skull of any mammal. The same cannot be said of the skulls of reptiles; one would be greatly puzzled in the comparison of the skulls of turtles and crocodiles, if he knew nothing about other forms. And it is safe to formulate another general law in evolution here: Characters which have been longest inherited are least liable to change. The earliest reptiles had at least four pairs of bones which have disappeared in all later reptiles; and they had some bones in pairs which have fused in later reptiles, either with their mates or with contiguous bones. The crocodile has at least two pairs of bones which have disappeared in turtles. On the other hand, the turtle has at least one pair of free bones which have been fused with adjacent bones in the crocodiles, and one pair that is fused which is free in the latter. The lizard has one pair of bones that has been wholly wanting in other reptiles for millions of years, while on the other hand it has lost some bones that are present in all other modern reptiles. The four parts of the occipital bone of mammals, basioccipital, exoccipitals, and supraoccipital, are almost invariably free and there is a single occipital condyle, except in the Theriodontia.
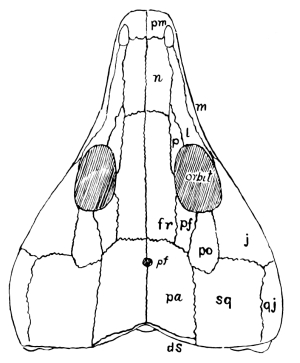
Fig. 6.—Labidosaurus, a cotylosaur. Skull from above: pm, premaxilla; n, nasal; m, maxilla; l, lacrimal; p, prefrontal; fr, frontal; pf, postfrontal; po, postorbital; j, jugal; pa, parietal; sq, squamosal; ds, dermosupraoccipital; pf, parietal foramen.
In this reduction or fusion of parts, or in addition thereto, there has been a general lightening-up of the whole skull-structure in reptiles from the rather massive and protected form of the older to the lighter, less protected, and more fragile type of the later ones, since speed, [Pg 23] greater agility, better sense organs, and doubtless greater brain power have rendered unnecessary or useless the older kinds, just as modern methods and modern arms have rendered useless the coat of mail of the Middle Ages.
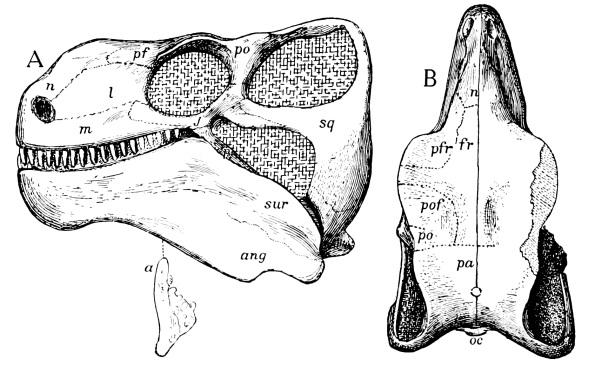
Fig. 7.—Edaphosaurus, a theromorph reptile from the Permian of Texas. Skull with single temporal vacuity.
The old reptiles had a continuous covering or roof for the skull, pierced only by the openings for the nostrils in front—the nares—the orbits for the eyes near the middle, and a smaller median opening back of them for the so-called “pineal eye.” The temporal region, that is, the region back of the orbits on each side, was completely roofed over by bone for the support and protection of the jaw muscles. In later reptiles this region has been lightened, either by holes that pierce it or by the emargination of its free borders, as in the turtles. The openings have occurred in different ways, and with the loss of different bones in various lines of descent. In one large group of reptiles, comprising the pterodactyls, dinosaurs, phytosaurs, crocodiles, and rhynchocephalians, there are two openings on each side, called the supratemporal and lateral temporal vacuities. In another still larger group there is a single vacuity on each side, all members of which it has been thought were markedly related to each other. Some of these, the lizards, snakes, and mosasaurs, the ichthyosaurs, and [Pg 24] probably the proganosaurs, have the single opening high up on the side, corresponding apparently to the supra temporal vacuity of the double-arched forms, as those with two openings are called. Many others, however, like the whole order Therapsida and the Theromorpha, have the single opening lower down and bounded differently; their relationships are doubtful, since it is very much of a question how the single opening has arisen. There have been many theories to account for the origin of the temporal vacuities, but all are yet speculations. Notwithstanding these doubts, which more recent discoveries have intensified, there can be none that the structure of this region of the skull offers important and reliable characters for the classification of the reptiles into the larger groups, but, unfortunately, we are very uncertain yet as to what this classification should be. We are confident that all those reptiles having two temporal vacuities on each side are related to each other; we are yet very much in doubt as to the classification of all other reptiles, or at least all others having only a single temporal vacuity on each side.

Fig. 8.—Sphenodon (tuatera). Skull from side and above: pm, premaxilla; n, nasal; prf, prefrontal; f, frontal; pf, postfrontal; p, parietal; po, postorbital; sq, squamosal; m, maxilla; j, jugal; qj, quadratojugal; q, quadrate; c, coronoid; sa, surangular; art, articular; pa, prearticular; d, dentary; an, angular.
Better evidences of relationships, or the absence of relationships, are offered by the presence of certain bones in the skulls in some orders [Pg 25] that are lost in others, since it may be accepted as an axiom that new bones have not appeared in the skulls of reptiles, birds, or mammals; and that no bone which has once disappeared has ever been functionally regained by the descendants of those that lost it. The presence, then, of an extra bone in the temporal region of the lizards or the ichthyosaurs is proof that they have had a long and independent descent from reptiles which possessed it.
The mandible of the earliest reptiles was composed of not less than seven separate and distinct bones, as shown in the accompanying figures. The mandible of no modern reptile has more than six, and some have fewer. The mandible of mammals is composed of a single bone, the dentary; those reptiles, the Theriodontia, which doubtless were ancestral to the mammals in Triassic times, have all the bones, except the dentary, much reduced, or even vestigial. The prearticular bone, as shown, so far as known, has been absent in all reptiles since Triassic times, except the ichthyosaurs, plesiosaurs, Sphenodon, and turtles, all reptiles of ancient origin. The coronoid bone primitively extended the whole length of the teeth on the inner side; in all reptiles, except the plesiosaurs, since Triassic times it is either reduced to a small bone back of the teeth or is absent. So also the splenial has been greatly reduced in size in all later reptiles and may be wanting as in Sphenodon and modern turtles. The articular of reptiles, it is now generally believed, is represented in mammals by one of the ear bones, the quadrate by another.


Fig. 9.—Mandible of Trimerorhachis, a stegocephalian amphibian, ancestrally related to the reptiles: A from within; B from without. The coronoid is composed of three bones, the true coronoid (cor), the intercoronoid (icor), and the precoronoid (pc). The splenial is composed of two, the true splenial (sp) and the postsplenial (psp). The prearticular (pa) is broad, the dentary (d) is small; and the angular (an) is only slightly visible on the inner side.
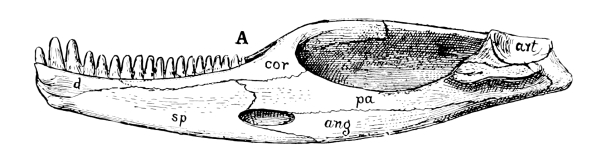
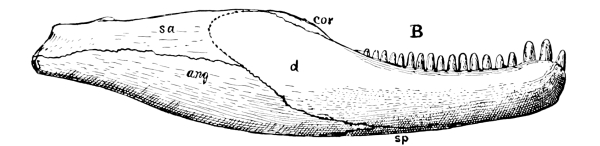
Fig. 10.—Mandible of Labidosaurus, a cotylosaur reptile: A from within; B from without. The coronoid (cor) is a single bone, but extends far forward. The splenial (sp) is also a single bone, replacing the two of the amphibians. The prearticular (pa) is narrower, and the angular (ang) appears broadly on the inner side. The dentary (d) is much larger and the surangular (sa) is distinct. The articular (art) is small.

Fig. 11.—Mandible of Alligator, a modern, highly specialized reptile, from within. The coronoid (cor) is small and is situated far back; the splenial (sp) does not extend to the symphysis; the prearticular (pa) has disappeared, or has fused with the angular (an) or articular (art). The dentary (d) has become the chief bone of the mandible.
The teeth of reptiles are of much less importance, as a rule, in the determination of relationships than are the teeth of mammals. Rarely are their shapes of specific, and often not of generic, importance, though their number and relative sizes may be. The teeth of mammals, as a rule, are forty-four or less in number, and they are always inserted in distinct sockets in the jaw bones. Among reptiles they are indefinite in number, and may be attached to any of the bones of the palate and sometimes also to the coronoid of the mandibles. [Pg 26] Furthermore, except in those reptiles related to the immediate ancestors of the mammals, they are alike or nearly alike in the jaws, that is, homodont, not distinguishable into incisors, canines, and molars. They may be inserted in separate sockets (thecodont), in grooves, or simply be co-ossified to the surface of the bone (acrodont). And they are usually reproduced indefinitely by new teeth growing at the side of the base or below them. More usually they are [Pg 27] pointed and curved; sometimes they are flattened, with sharp cutting edges in front and behind in the more strictly carnivorous reptiles; in those of herbivorous habits they are more dilated and roughened on the crown, not pointed; in not a few they are low, broad, and flat and are used only for crushing the hard shells of invertebrates. With the very few exceptions among certain dinosaurs, they never have more than one root for attachment. The evolutional tendency for reptiles, as for the mammals, is to loose teeth, especially those of the palate. Among living reptiles it is only the most primitive types, such as the lizards, snakes, and the tuatera, which have teeth on the palatal bones, and in none are there teeth on the vomers, as was the rule in the ancient reptiles. The lizards may have them on pterygoids and palatines, and the tuatera has them on the palatines only. There may be as many as eighty on each jaw, above and below, and hundreds of smaller ones on the palate, or they may be reduced in number to five or six, or even to a single one; some reptiles, like the turtles and later pterodactyls, have none. The teeth of reptiles are composed of the same kinds of tissues as are the teeth of mammals, that is, of dentine and enamel, but the enamel is always thin, perhaps because the teeth are so [Pg 28] easily replaced that a thicker protective covering is not needed. The arrangement of the dentine in primitive reptiles is complicated, that is, plicated or folded in labyrinthine figures, like that of many stegocephalian amphibians, the Labyrinthodontia, especially. This labyrinthine structure of the dentine persisted longest in the ichthyosaurs.
The spinal column or backbone of reptiles, as in all air-breathing vertebrates, is made up of a variable number of separate segments called vertebrae, permitting flexibility. Each vertebra is composed of a body, or centrum, and an arch on the dorsal side for the protection of the spinal cord. Various projections from the vertebra, called processes, serve for the attachment of ligaments or muscles, for articular union with adjacent vertebrae, or for the support of ribs, and these processes have characteristic differences in different reptiles. The pair in front and behind, for articulation with the adjoining vertebrae, may become obsolete or even lost in swimming reptiles, as we shall see; they are called zygapophyses. In not a few reptiles there is an additional pair for zygapophysial articulation in front and behind, called zygosphene and zygantrum, for the greater strengthening of the column; they are especially characteristic of snakes and certain lizards. In certain other reptiles, especially the long-necked dinosaurs, there is an additional pair arranged differently from the zygophene, that have received the names hyposphene and hypantrum.

Fig. 12.—Procoelous vertebra of snake: za, zygantrum; zs, zygosphene; ps, posterior zygapophysis.
On the top of the arch is the spine or spinous process, which may vary [Pg 29] enormously in size and length; sometimes it is flattened or dilated above for the support of an exoskeleton, or it may be heavy and massive for the attachment of strong muscles and ligaments. In the modern basilisk lizards and in the ancient Dimetrodon and Edaphosaurus from the Permian rocks of Texas these spines are of enormous length, some of them nearly four feet long in reptiles not twice that length. Slender crawling reptiles usually have no spines, or only vestigial ones. On the sides of the arch there may be a distinct transverse process for the articulation of the rib.
In all early reptiles the ends of the body or centrum are concave, as they are in nearly all fishes. Such a conformation, called amphicoelous, gives great flexibility to the spinal column, but only moderate strength, since the intervening spaces are filled with cartilage in life. In all living reptiles, with few exceptions, the body is concave, like a saucer, in front and correspondingly convex behind, and the intervening cartilage has largely disappeared. Such a mode of union, called procoelous, adds greatly to the strength of the backbone, enabling it to receive greater shocks or greater pressure without dislocation; or to sustain the greater strain of muscles used in running swiftly or in climbing. Among living reptiles, only the gecko lizards and the tuatera have biconcave vertebrae. Some extinct reptiles, such as some of the dinosaurs, animals that walked erect upon their legs, had their vertebrae convex in front and concave behind (opisthocoelous). Birds, though walking erect, have a very different and more complicated articulation of the cervical vertebrae, and certain reptiles, like the turtles, have very complicated cervical vertebrae.
In the embryos of all vertebrate animals there appears first an elongated fibrous rod, called the notochord, in the place of the future spinal column. This rod may persist through life, never ossifying, as was the case with all the earliest fishes, and is the condition in some living ones. As the embryo grows, however, the separate segments, or vertebrae, ossify about this rod in all reptiles, forming bony rings, perforate at first in the middle for the more or less constricted notochord. This stage was the permanent condition in all the earliest reptiles and in some later ones. Such animals are said to have [Pg 30] notochordal vertebrae, the notochord more or less continuous, like a string of beads, the beads representing the enlargements between the contiguous vertebrae.
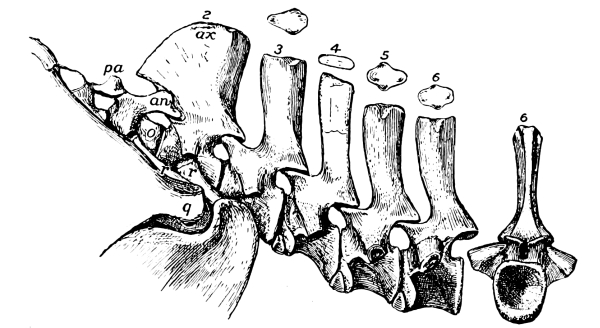
Fig. 13.—Notochordal cervical vertebrae, with intercentra, of Ophiacodon, a primitive theromorph reptile from the Permocarboniferous of New Mexico: pa, proatlas; an, arch of atlas; o, odontoid; ax, axis.
In many early amphibians, and probably in all the earliest ones, as well as in the fishes from which they were derived, the vertebra is more complicated in that it is composed of at least three pairs of separate bones, two of which united with each other, the third finally disappearing in modern animals, or at the most represented by a mere vestige called the intercentrum. The dorsal pair of these bones, called the neurocentra, forms the arch of the vertebra. The ventral posterior pair, called the pleurocentra, increases in size and unites to form the centrum or body of the vertebra; while the ventral anterior pair, early united with each other, is called the hypocentrum or intercentrum, persistent in all early reptiles as a vestige between the centra on the ventral side. This divided condition of the vertebra is persistent in the first vertebra, the atlas of all higher animals, in which the so-called body is the hypocentrum or intercentrum, the arch is the neurocentrum, while the pleurocentra have fused more or less with the anterior part of the next vertebra, the axis, to form the so-called odontoid. That this is the real explanation of the structure of the [Pg 31] atlas is proved by the various stages of its evolution in the reptiles, from the earliest (Fig. 15) in which it scarcely differs from rhachitomous—as this structure is called—vertebrae of an early amphibian, to the modern in which the structure is nearly like that of mammals.
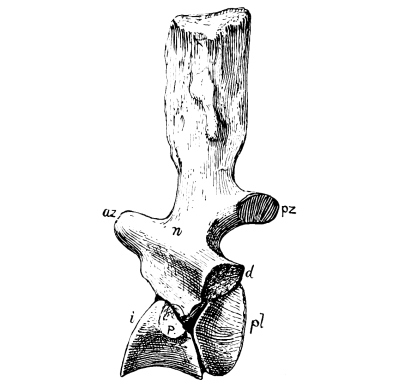
Fig. 14.—Rhachitomous dorsal vertebra of Eryops: n, neurocentrum or arch; pl, pleurocentrum; i, intercentrum or hypocentrum; az, anterior zygapophysis; pz, posterior zygapophysis; d, diapophysis, for tubercle of rib; p, parapophysis, for head of rib.
In front of the atlas, that is, between it and the skull, there was, in all early reptiles, as well as in some later ones, like the crocodiles and tuatera, the remnant of what is believed to have been another vertebra, of which only the arch remains, and which is called the proatlas. In its earliest condition it articulated with the skull in front and the arch of the atlas behind.
As in mammals, the vertebrae of the different regions have received distinctive names, cervical, dorsal, lumbar, sacral, and caudal. The numbers of each region are far more variable than they are among mammals, the total number of vertebrae in the column varying from about thirty to more than five hundred, in certain snakes. Nor are the different regions always easily distinguishable, especially those in front of the sacrum. In the earliest reptiles there was practically no neck, and only two vertebrae, the atlas and axis, that properly can be called cervical. Very soon, however, the reptiles developed a longer neck with seven vertebrae, a number that has remained singularly constant in higher animals, especially in the mammals. In most modern reptiles there are from seven to nine; in a few lizards, five. But the number was much more inconstant among the older reptiles; some of the plesiosaurs had as many as seventy-six cervical vertebrae; some of the older lizards even had as many as eighteen. [Pg 32]
Ordinarily the cervical vertebrae differ from those behind them only in the small size or fusion of their ribs; sometimes, however, as in the Protorosauria and Pterosauria, the vertebrae may be much elongated. The dorsal vertebrae of reptiles vary in number from ten in turtles and some dinosaurs to forty-three in Pleurosaurus; and under the name dorsal we include the so-called lumbar, as there is seldom any real distinction between the two series, save the smaller size or the co-ossification of the ribs of the latter.

Fig. 15.—Ophiacodon,
a primitive theromorph
reptile: proatlas, atlas,
and axis, with ribs.

Fig. 16.—Sacrum of Chelone.
The sacrum in reptiles primitively consisted of a single vertebra, which bore a large rib on each side for the support of the pelvis. Very early, however, a second or even a third vertebra was added to it from behind. The number two is the rule among reptiles, both ancient and modern; among crawling reptiles the number never exceeds three, but among ambulatory and flying reptiles the number may be as great as in any mammal.
The number of caudal vertebrae in reptiles is exceedingly variable, from a dozen or fifteen up to a hundred and fifty or more. In snakes but two regions are distinguishable, the caudal and precaudal, and the number altogether may reach nearly five hundred. With the exception of the first few basal caudal vertebrae (pygals) and the minute ones at the extreme tip, all caudal vertebrae of reptiles bear a slender, usually Y-shaped bone below in the interval between the centra, for the protection of the vessels and nerves. Because of their shape they have been called chevrons, and are really outgrowths from the intercentra. [Pg 33]
The ribs of reptiles are of more importance in classification than one would suppose. The primitive rib was a slender, curved bone, with the vertebral end dilated to articulate continuously with the intercentral space—that between the centra and the anterior part of the arch. And this is the condition still remaining in the tuatera. Very soon, however, the lower end of the articular surface (capitulum) became separated from the upper (tubercle) by a notch, and the ribs became distinctly double-headed. And this mode of articulation is the rule among mammals. Among later reptiles, however, there were many modifications. In nearly all the head migrated a little backward on the centrum. By the loss of the tubercle in lizards, the head became truly single-headed, and attached solely to the body; and this condition is characteristic of the order Squamata. In another large group the head of the rib gradually migrated up on the arch and on the transverse process (diapophysis), so that both head and tubercle are attached to the diapophysis; and this condition is equally characteristic of the orders of reptiles known collectively as the Archosauria—the crocodiles, pterodactyls, dinosaurs, and phytosaurs. In the Sauropterygia, the ribs are single-headed and attached to the end of the diapophysis. Finally in most ichthyosaurs the capitulum and tubercle both articulate with the body of the vertebra.

Fig. 17.—Ostodolepis, a primitive theromorph reptile. Vertebrae from in front and side, with primitive double-headed rib and intercentrum.
Ribs primitively were probably attached to all the vertebrae to the end of the tail. In the earliest reptiles that we know they are present on all vertebrae as far back as the tenth or twelfth caudal only, those of the caudal for the most part co-ossified with the centra. The ribs of the neck vertebrae more quickly disappeared, or became fused with the vertebrae, and only in the crocodiles among living reptiles are there ribs on the atlas. The sacral ribs, on the other hand, became much [Pg 34] larger and stouter and developed an articulation at their outer ends for the support of the ilium (Fig. 16).
The so-called ventral ribs are slender ossifications in the connective tissue under the skin, on the under side of the body, and are characteristic of most reptiles. The anterior ones doubtless fused together more or less to form the sternum or breast bone, which was otherwise absent in the early reptiles.
Those bones which form the framework for the support of the anterior extremity in vertebrate animals are known collectively as the pectoral girdle. In our own skeleton there are but two on each side, or four in all, the scapula or shoulder-blade, and the clavicle or collar-bone. A third bone, however, is represented in all mammals by a mere vestige which early unites with the scapula and is called the coracoid process. In the lowest forms of mammals, the Monotremata, of which the Ornithorhynchus and Echidna are the only examples, not only is this coracoid bone largely developed, articulating with the sternum or breast bone, but there is an additional coracoid bone in front of this; and there is also an interclavicle. Indeed, the pectoral girdle in these mammals is more primitive or generalized in structure than it is in any living reptiles, composed of scapula, coracoid, metacoracoid, and clavicle on each side and an interclavicle in the middle. No living reptiles have the metacoracoid, and, as is the case with many mammals, some reptiles have no clavicles.
Primitively, that is, in all the old reptiles, the girdle is composed of scapula, coracoid, metacoracoid, clavicles, and interclavicle, while in some of the very oldest there is yet another bone, more or less of a vestige, derived from the ancestral amphibians and called the cleithrum or supraclavicle. The scapula is more or less elongated in crawling and climbing reptiles; more slender and bird-like in those which walked erect after the manner of birds and mammals; shorter and more fan-shaped in the swimming reptiles, as we shall see. In some pterodactyls, unlike all other known animals, the scapula articulated at its upper end with the backbone, giving a much firmer support for [Pg 35] the anterior extremities. Only in those reptiles allied to the ancestors of the mammals has the scapula ever had a spine or projection on its dorsal side.

Fig. 18.—Cacops, a Permian stegocephalian, ancestrally allied to the primitive reptiles, with rhachitomous vertebrae and large cleithrum above the scapula.
Of the two coracoid bones in the original pectoral girdle the posterior one began to disappear early and is entirely lost in all reptiles that lived later than Triassic times, though it still persists in the lowest mammals, as we have seen. In most later reptiles the remaining coracoid has become less firmly attached to the scapula than it was in the older reptiles. It usually has a small foramen piercing it near the middle of the upper border or end, the supracoracoid foramen. The clavicle, while more constant among reptiles than among mammals, has been lost in some, the Crocodilia, for instance, as also the dinosaurs and pterodactyls. [Pg 36] The interclavicle is more constant in reptiles, a more or less T-shaped bone underlying the coracoids where they join, or the breast bone; but there were some reptiles that lost it, the dinosaurs and pterodactyls, for instance. In the turtles both the clavicles and the interclavicle form a part of the under shell or plastron.

Fig. 19.—Scapula (sc), coracoid (cor), and metacoracoid (mcor) of Dimetrodon
The cleithrum is known in only a few of the old reptiles; it is a more or less slender bone which lies along the upper front margin of the scapula, articulating at its lower end with the upper end of the clavicle on each side. [Pg 37]
The breast bone or sternum, while not properly a part of the pectoral girdle, may be mentioned here. In reptiles it is rarely well developed or even ossified, the flying reptiles known as the pterodactyls being the most notable exceptions. It was a comparatively late development in this class, the earliest ones not possessing it even in a cartilaginous condition. It was doubtless evolved from the more or less numerous and slender ossifications on the under side of the body called ventral or abdominal ribs, after the coracoids had become reduced and more slender. Whenever it is present the coracoid articulates with it on each side in front. In most lizards it remains as a cartilage throughout life.

Fig. 20.—Clavicles and interclavicle of Ophiacodon, a theromorph reptile from the Permocarboniferous of New Mexico.
The upper arm bone, or humerus, like most other bones of the extremities, has been greatly modified by the habits of the different reptiles. In running and climbing reptiles it is always slender, while in burrowing reptiles it is short and stout and much expanded at the extremities, like the humerus of the mole among mammals. And we shall [Pg 38] also see how greatly modified it was among the swimming reptiles. The humerus of flying reptiles has an enormous process on the side, corresponding to the attachment of the deltoid muscle. The head of the humerus, for articulation with the glenoid cavity of the scapula, is rounded in all reptiles, except the pterodactyls, and the articulation is always at the extremity. At the lower extremity the protuberance at the outer or radial side is called the ectocondyle; that on the inner or ulnar side, the entocondyle. Between the two at the end are the articular surfaces for the radius and ulna, the capitellum and trochlea. A little above each of these condyles there is usually, on one side or the other or on both, a foramen or hole for the passage of arteries or nerves. That on the inner side, which is characteristic of all early reptiles and of many mammals, is called the entepicondylar foramen; that on the outer side, the ectepicondylar foramen; the latter is present in the lizards, and both are found in the tuatera and some of the early reptiles.

Fig. 21.—Anterior extremity of Ophiacodon.
The radius and ulna are always distinct bones in reptiles, and always freely movable on each other; they are usually shorter than the humerus, but in some springing and climbing reptiles they are quite as long.
The carpus or wrist of reptiles consists primitively of eleven distinct, irregularly shaped bones, which articulate more or less closely with each other in three rows. Those of the first row, all true carpals, are known usually as the radiale, intermedium, ulnare, and pisiform, corresponding quite with the bones of the human wrist known as [Pg 39] the scaphoid, lunar, cuneiform, and pisiform. The second row has but two bones, on the radial side, known as the centralia; while the third row has a bone to correspond to each of the metacarpals, five in number, and collectively known as the carpalia. Some or indeed all of these bones may be either absent or unossified, that is, remaining through life as nodules of cartilage. Seldom, however, are there less than nine bones in the carpus of reptiles.
The metacarpals, like the digits, primitively were five in number, and seldom are there less, though the fifth is sometimes lost, and rarely also the first. They are more or less elongate bones, increasing in length from the first to the fourth, with the fifth usually shorter. The first and the fifth are usually more freely movable on the wrist than are the other three.
The number of joints or phalanges in the fingers of all primitive reptiles is that of the modern lizards and the tuatera, that is, two on the first finger or thumb, three on the second, four on the third, five on the fourth, and three on the fifth. The crocodiles have one less phalange on the fourth digit; the turtles have usually two less on the fourth and one less on the third, that is, with precisely the same arrangement that is found in our own fingers and that of mammals in general, two on the thumb and three on each of the other fingers. As exceptions the river turtles have four bones in the fourth digit. And this mammal-like and turtle-like arrangement of the phalanges was that of those early reptiles, the Theriodontia, from which the mammals arose. The last or ungual phalange of reptiles is usually claw-like, that is, sharp, curved, and pointed, but sometimes it is more nail-or hoof-like.
The pelvic girdle or pelvis in reptiles and higher animals consists of three bones on each side, often closely fused in adult reptiles and together known as the innominate bone. The upper or dorsal one of these three bones—that to which the sacrum is attached—is the ilium; the one on the lower or ventral side in front is the pubis; and that on the ventral side behind is the ischium. On the outer side, where these three bones meet, is a cup-like depression, sometimes a hole, called [Pg 40] the acetabulum, for the articulation of the head of the thigh bone, homologous with the glenoid articulation of the pectoral girdle, which, as we have seen, was originally formed by three bones, the scapula, coracoid, and metacoracoid, the two latter bones, like the pubis and ischium, meeting in the middle line below. In all the primitive and early reptiles the pubis and ischium form a continuous plate of bone without holes in it, except a small one just below the acetabulum in the pubis, called the obturator foramen, and corresponding to the supracoracoid foramen of the coracoid. One may almost always recognize these two bones by the presence of the foramen. This “plate-like” condition of the pelvis has been lost in all late and modern reptiles by the appearance of a larger or smaller vacuity between the pubis and ischium, either paired, when it corresponds quite with the so-called obturator opening of mammals, or singly in the middle. This old-fashioned character, like the old-fashioned type of pectoral girdle, disappeared entirely about the close of the Mesozoic period, the Choristodera, described in the following pages, being the last of the kind.
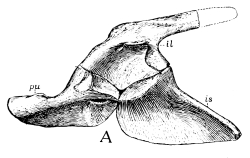
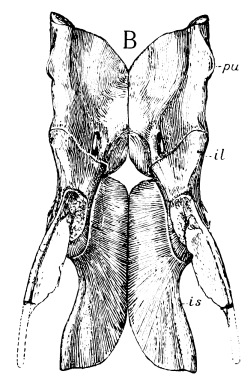
Fig. 22.—Pelvis of Ophiacodon: A from side; B from above; pu, pubis; il, ilium; is, ischium.
The ilium in reptiles usually has a more or less prolonged process or projection turned backward by the side of the anterior caudal vertebrae, but in those animals which walked erect on the hind legs, [Pg 41] the dinosaurs and pterodactyls, as also some of the more erect-walking reptiles ancestral to the mammals, this process was directed forward, as in birds and mammals. The crocodilia, unlike all other known reptiles, have the pubes excluded from the acetabulum, and they do not meet in a median symphysis. This character alone will distinguish any crocodilian from all other reptiles. But there is some doubt as to the homology of the bones usually called pubes in the crocodiles. Some of the bipedal dinosaurs have the pubis forked, the anterior part directed downward and forward, and not meeting its mate in a symphysis, the posterior process long and slender, lying below the long ischium, as in birds. Indeed, when this peculiarity of the dinosaurian pubis was first discovered, it was thought to be an evidence of the immediate relationship of birds; its structure is now interpreted differently.
The thigh bone or femur in reptiles, like the humerus, is variable in size and shape. Only in those reptiles that walked erect is the articulation of the head set off from the shaft of the bone by a distinct neck. In others the articulation is at the extreme top of the bone, since the thigh bones are habitually turned more or less directly outward from the acetabulum and the long axis of the body. The more or less pronounced rugosities at the upper end of the femur, for the attachment of muscles, called trochanters, are not easily distinguishable into the greater and lesser, as in mammals. Sometimes, as in the erect-walking dinosaurs, there is a more or less pronounced process on the shaft lower down, called the fourth trochanter, for the attachment of caudal muscles. On the back part of the shaft there is a ridge or line for the attachment of muscles, corresponding to the linea aspera of the mammalian femur. The projections at the lower end on the sides are called condyles.
The two bones of the leg, or shin, are usually shorter than the thigh bones, though in running and leaping animals they may be quite as long or even longer. That on the inner or big toe side is called the tibia, and articulates with the distal end of the femur, but chiefly with its inner condyle. It has a more or less well-developed crest in front above for the attachment of the extensor muscles directly, since there [Pg 42] never is a patella in reptiles, and only rarely sesamoid bones of any kind. The fibula, at the little-toe side of the leg, is usually more slender than the tibia, though it may be larger in swimming reptiles and even in some running forms. It disappeared in some of the later pterodactyls. Its upper articulation has a more gliding and somewhat rotary motion on the outer condyle of the femur, turning the foot outward in extension of the leg.

Fig. 23.—Right hind foot of Ophiacodon: a, astragalus; c, calcaneum; c1, c2, centralia; 1, 2, 3, 4, 5, tarsalia.
The tarsus of reptiles differs from that of mammals, in that the chief movements of extension and flexion of the foot upon the leg occur within the tarsus rather than between the tarsus and leg bones. Primitively the tarsus of reptiles consisted of nine bones, two in the first row, two in the second, and five in the third, but in all modern reptiles the bones of the middle row and the fifth one in the third row have disappeared; in some lizards and turtles the two of the first row are fused. The two bones of the proximal row correspond quite to the astragalus and calcaneum, the astragalus articulating with both tibia and fibula proximally, the calcaneum with the latter only. The oldest known tarsus of any vertebrated animal, one from the Coal Measures of Ohio, has this structure, while in all the early amphibians there were three bones, the tibiale, intermedium, and fibulare. Some of the later swimming reptiles, like the ichthyosaurs and plesiosaurs, have apparently this amphibian structure, with three bones that are usually called tibiale, intermedium, and fibulare, but it is very doubtful indeed whether they are homologically the same. In the middle row two centralia are known in one or two very ancient reptiles, but for the [Pg 43] most part there is only a single centrale, and even that is usually lost in later reptiles. The third row, like the third row of the carpus, had a distinct bone for each digit originally, but the fifth one was very soon lost and has never reappeared. The structure of the digits and number of bones are quite like those of the hands, except that the fifth toe has four bones instead of three, that is, the phalangeal formula was 2, 3, 4, 5, 4. As a rule in terrestrial reptiles, as in terrestrial mammals, the hind foot is more specialized than the front ones.
Most reptiles have an external covering or exoskeleton of horny plates or scales or bony scutes. Horny scales are of course not preservable as petrifactions, though in many instances their actual carbonized remains or their impressions have been detected. Such information comes only rarely, though doubtless in the course of time we shall obtain it for most extinct reptiles. In the mosasaurs, for instance, very perfect impressions showing the detailed structure of the scales have been frequently found. Similar impressions were long since observed by Lortet in Pleurosaurus, and in not a few dinosaurs impressions of most wonderful perfection have been found. It is only in the water reptiles, probably, that all external coverings tended to disappear.
Bony dermal plates or scutes are less common among reptiles, though by no means rare. The turtles, as is well known, are almost completely inclosed in such an exoskeleton, bones which have coalesced more or less to form a box or carapace within which the head and limbs may be withdrawn for protection. In the modern crocodilians also the body is more or less protected by small bone plates forming rows on the back and sometimes on the under side. The ancient phytosaurs had similar plates. Not a few of the dinosaurs were more or less covered with bony scutes and sometimes with large bony plates or spines. Some modern lizards have bony plates over the body instead of horny scales.
[Pg 44]
Geologists divide the history of the earth, since life first appeared upon it, into four general eras, the Proterozoic, Paleozoic, Mesozoic, and Cenozoic, that is, into eras of first life, ancient life, middle life, and recent life. These divisions were made long ago by geologists when it was believed that extraordinary changes, great cataclysmic revolutions, marked their limits.
With a fuller knowledge of the life of the past we know that evolution has been continuous and uninterrupted; possibly accelerated or retarded at times, but without break. Were the earth’s history to be written anew, with our present knowledge, and with an unbiased mind, it is very doubtful whether many of the time divisions would have the same limits that they have now—whether the Paleozoic would terminate with the Carboniferous, or the Permian, or the Trias, or whether indeed we should think it necessary to make any primary divisions whatsoever. In other words, our greater knowledge of living and extinct organisms, and of the rocks which contain fossils, has made the problems of classification much more complex than they seemed to be formerly. It is much easier to classify organisms or rocks, or anything else, when we know only a few isolated kinds—much easier to draw divisional lines. Geological history is like a volume in which pages, leaves, and even whole chapters either are missing or are printed in languages which we understand only imperfectly. Where the lost or unknown parts belong, the largest divisions may be made, and possibly such may have been epochs of unusual activity, of diastrophic changes which greatly accelerated organic evolution. No one can say just where the dividing line should be drawn between the rocks of Paleozoic and Mesozoic age, or between the Mesozoic and Cenozoic, for there is none; the most that we can hope for is to make the divisions everywhere in the world conform to those first made for local reasons. [Pg 45]
Click image to enlarge.

Fig. 23a.—Range of the Reptilia.Heavy lines indicate occurrence in North America.
[Pg 46] The periods of the Paleozoic era are the Cambrian, Ordovician, Silurian, Devonian, Carboniferous, and Permian, in the order as given; those of the Mesozoic era are the Triassic, Jurassic, and Cretaceous; those of the Cenozoic era, the Eocene, Oligocene, Miocene, Pliocene, Pleistocene, and Recent. As a relic of an old classification we still often divide the Cenozoic into two quite arbitrary divisions, the Tertiary and the Quaternary, the latter including the Pleistocene and Recent only. The same may be said regarding the limits of each of these periods as of the eras; the sole problem is to make each period contemporaneous throughout the world, an exceedingly difficult problem, because no faunas or floras have ever been the same over the whole earth. Indeed, with the exception of some of the lowliest and most generalized forms, or man himself, no species are the same throughout the earth today. Inasmuch as we must depend upon the fossils in the rocks for the determination of the ages, where none is quite the same in strata of remote localities the identification becomes very difficult or even impossible. Nor are the periods, as accepted, of equal or even approximately equal duration; the Cretaceous period, for instance, was longer than all the remainder of the Mesozoic, longer perhaps than all the time which has elapsed since its close.
The earliest animals with a backbone, or rather the earliest that we call vertebrates—for some vertebrates have no vertebrae—began their existence, so far as we know, in late Ordovician times, as attested by fish bones in Ordovician rocks of Colorado and Utah. The first evidences of the existence of air-breathing vertebrates in geological history are footprints preserved in the uppermost Devonian rocks of Pennsylvania. We call them amphibian because they resemble footprints associated with amphibian skeletons in later formations, and because the foot itself is still the most important difference we know between fishes and the higher animals. [Pg 47]

Fig. 24.—Permocarboniferous landscape (adapted from Neumayr) with restoration of Eryops, a stegocephalian amphibian ancestrally allied to the reptiles; and Limnoscalis, a cotylosaur (in water).
[Pg 48] In the rocks of the next great time division, the Mississippian, as we call it in America, corresponding more or less closely with the Lower or Subcarboniferous of other parts of the world, numerous footprints of amphibians have been discovered, but no fossil remains except a few from near its close in Scotland. From the Upper Carboniferous, or Pennsylvanian, however, not only numerous footprints but the actual skeletons, or impressions of skeletons, have long been known in Europe and America. Until recently all these footprints and skeletons were supposed to be exclusively amphibian. We are now almost sure that some of them belonged to reptiles of lowly type, the earliest coming from near the middle of the Pennsylvanian of Linton, Ohio. The amphibians of this period were, for the most part, salamander-like creatures of from a few inches to two or three feet in length. They all belong to the group collectively known as the Stegocephalia, except that very near the close of the period there appeared small, slender, small-legged aquatic forms which seem to be the ancient representatives of the real salamanders of modern times. Some of the Stegocephalians had become greatly specialized as legless, snake-like, or eel-like creatures.

Fig. 25.—Restoration of Seymouria, the most primitive of known cotylosaur reptiles. From the Permian of Texas, about two feet long.
By the beginning of Permian times tremendous changes had taken place in the land life. The small amphibians of the Carboniferous types dwindled away, soon to disappear, and their places were taken by others of peculiar types, for the most part larger; and by many and diverse kinds of reptiles—water reptiles, marsh reptiles, land reptiles, and even climbing tree reptiles. From the uppermost Carboniferous and Lower [Pg 49] Permian rocks of the United States more than fifty genera and twice that many species of amphibians and reptiles have been made known in recent years, and doubtless as many more will be discovered in the future. From other parts of the world the history of reptiles of the Lower Permian is yet scanty, two or three forms from South America, as many more from Africa, and a half-dozen or so from Europe are all; and of these very few are known at all well.

Fig. 26.—Captorhinus, a cotylosaur reptile from Texas.
We classify all the known forms of reptiles from the Lower Permian under three or four orders, the Cotylosauria, Theromorpha or Pelycosauria, Proganosauria, and possibly the Protorosauria, but the classification is yet provisional, representing merely the present stage of our knowledge. The Proganosauria and Protorosauria, including distinctively aquatic reptiles, will be more fully described in the following pages. To give even a brief description of the more terrestrial reptiles of this, the earliest known reptilian fauna, would be beyond our purpose; the accompanying life restorations by the author of some of the more typical and better known forms, based upon nearly perfect skeletons, will suffice.
From the reptiles and amphibians of the Lower Permian of Texas and New Mexico to the ichthyosaurs of the Middle Triassic of California there is a complete gap in the records of the land life of North America. We do not know what became of all the remarkable animals of the Permian. There are few traces of their descendants elsewhere known, unless it be in South Africa. From the Middle and Upper Permian of South Africa and Russia, a marvelous reptilian fauna has been made known in recent [Pg 50] years. More than a hundred species of six or seven groups, and at least two orders have been described. Of these the Cotylosauria are the continuation of the American order, but include more specialized forms, the Pareiasauria and the Procolophonia, all of them, like the more primitive American forms, characterized by the imperforate temporal region. The Therapsida, likewise, seem to be the continuation of the American Theromorpha, so closely allied to them that it is difficult to draw a distinguishing line between them. On the other hand, these African reptiles merge through the Theriodontia into the mammals in the Triassic. They are all terrestrial, crawling reptiles, except a few which are described on a later page under the Anomodontia.
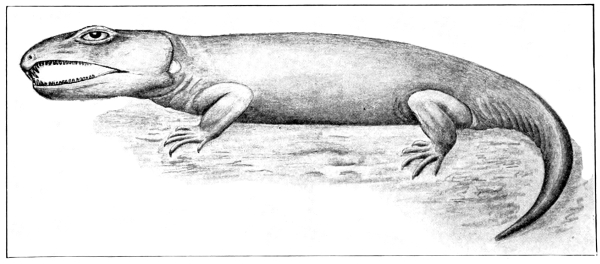
Fig. 27.—Restoration of Labidosaurus, a cotylosaur reptile from Texas, about three feet long.
The records of the lower part of the Triassic period are scanty everywhere in the world, save perhaps in Africa. Before the close of the period, however, probably every important group of cold-blooded air-breathing animals had made its appearance in geological history, if we except the snakes; even the mammals had appeared, and possibly the birds. The Cotylosauria, Theromorpha, and Therapsida disappeared, the latter giving birth to the mammals; the nothosaurs and plesiosaurs, the ichthyosaurs, dinosaurs, crocodiles, phytosaurs, rhynchocephalians, lizards, and turtles have all left records of their existence in Upper Triassic rocks; and the pterodactyls had also, in all probability, begun their career, though none is surely known till the Jurassic. [Pg 51]
During Jurassic times all these orders of reptiles waxed prosperous and powerful, and branched out in many ways and in countless numbers; many new kinds of each appeared—the marine crocodiles, the quadrupedal dinosaurs, etc.—but no order or suborder, so far as we know, disappeared before its close. And this prosperity continued on into the Lower Cretaceous and for many even into the Upper Cretaceous. The largest dinosaurs disappeared in the Lower Cretaceous, so far as our knowledge goes, but the old-fashioned crocodiles continued on into the Upper, to give place to the new-fashioned kinds. The ichthyosaurs lingered on for a while on the western continent, but the mosasaurs appeared, and the plesiosaurs reached their highest evolution and continued to the end. The flying reptiles attained the zenith of their evolution, but disappeared before the close. The marine turtles attained the maximum of specialization and size. The upright-walking dinosaurs continued on unabated to the close of the period; and a new kind of dinosaurs appeared near the end.

Fig. 28.—Restoration of Dimetrodon,
a pelycosaur reptile from the Permian of Texas;
about eight feet long.
[Pg 52] With the opening of the next great era—the Cenozoic or Tertiary—the reptiles dwindled away to their present insignificant position, while the birds and mammals appeared in great numbers and varied forms. The Age of Reptiles was closed and the Age of Mammals had begun.
The history of the reptiles during the Cenozoic is an uneventful one; they ceased their dominion upon land, in the water, and in the air. Their remains are scanty, for the most part, in the rocks of the Tertiary, and such as are known differ only in details from those now living. The land tortoises only, like the mammals of Oligocene and Miocene times, seized the opportunities of open prairies and prospered. A few of the late Mesozoic forms continued a short while into the Eocene. No new groups, perhaps few new families, came into existence during the greater part of this time; it was the age only of land tortoises and the poisonous snakes among reptiles.
The oldest known fossil reptile of North America, or indeed of the world, is represented by a single specimen, lacking the skull, from black shales of Middle Pennsylvanian age overlying a coal seam at Linton, Ohio. The specimen was originally described as an amphibian, but was later recognized by Professor Cope as a true reptile. It was more fully described by the writer under the name Eosauravus Copei, who agreed with Cope as to its reptilian nature. Until the skull is discovered, however, the precise relationships of the animal must remain doubtful.
The next later rocks that have yielded reptilian remains are those of Illinois and Texas formerly supposed to be of Permian age. Later evidence, furnished by invertebrates, however, seems to prove that the lowermost of the strata are of uppermost Carboniferous age. The Illinois deposits, so far as known, are of very limited extent, consisting practically of a single bone-bed in black shale in the immediate valley of the Kaskaskia River near Danville. The known fossils from this bone-bed—all isolated bones—are preserved in the museum of the University of Chicago, and include the types of several genera later recognized in the Texas deposits. [Pg 53]
The deposits of Texas, extending northward through Oklahoma to the south line of Kansas, are of considerable extent, for the most part lying along the Wichita River and its tributaries, north of Seymour, Texas. They are composed chiefly of red clays and sandstones of fresh-water or delta origin, perhaps eight hundred feet in total thickness. Beds of like character and yielding similar fossils are also known from northern New Mexico on the tributaries of the Chama River. Their chief characters, as well as restorations of some of the more noteworthy forms, have already been given.
No vertebrate fossils are known in America from the Upper Permian and Lower Triassic. Marine limestones of Middle and Upper Triassic age of Nevada and northern California have yielded numerous remains of primitive ichthyosaurs, the only known remains of the thalattosaurs, and a few others of doubtful affinities, all of which have been described by Dr. Merriam. The Upper Triassic exposures, of considerable extent, occur between the Pitt River and Squaw Creek in Shasta County, California. Reptilian remains from the Middle Triassic are so far known only from the limestones of West Humboldt and New Pass regions of western and central Nevada.
Fig. 29.—Restoration of Varanops, a
theromorph reptile from the Permian of Texas;
about four feet
long.
Land reptiles of Middle and Upper Triassic age are known from many widely separated localities in the United States, but chiefly from the [Pg 54] extensive “red beds” of the Rocky Mountain region. The fossils from these beds occur for the most part at least in the horizon called the Shinarump. Its age is usually considered to be Upper Triassic, but the character of the fossils seems to indicate possibly the Middle Triassic. Aside from the stereospondylian amphibians, the last of the Stegocephalia, the vertebrates from this horizon and these regions are chiefly Phytosauria. A few anomodonts, or what seem to be anomodonts—the only record of their occurrence outside of Africa—are known from Wyoming and Utah. And a single specimen from the Wind River red beds, described by the writer as Dolichobrachium, may represent reptiles allied to the dinosaurs. Phytosaur fossils of this horizon have been discovered in Utah, the Wind River Mountains, and near Laramie City in Wyoming; in southwestern Colorado; in western Texas; and in various places in New Mexico and Arizona. Doubtless when these fossiliferous beds are more thoroughly explored many new and interesting reptiles will be discovered.
Phytosaur remains, probably of about the same age as the Rocky Mountain ones, have long been known from the Triassic of North Carolina. From somewhat more recent Triassic deposits in Connecticut and Massachusetts, several skeletons of small carnivorous dinosaurs, and various parasuchian remains have been described by Marsh, Lull, and Talbot. And these beds have long been famous in Massachusetts for their footprints, for the most part originally referred to birds, but now pretty well known to have been made by dinosaurs and amphibians.
No vertebrate fossils of Lower or even Middle Jurassic age are known from North America. From the Baptanodon beds of Wyoming, limestones of about two hundred feet in thickness, four genera of plesiosaurs, the very peculiar ichthyosaur from which the beds take their name, and a few bones of an ancient crocodile are known.
Immediately overlying the Baptanodon beds, the Morrison beds, of from two hundred to four hundred feet in thickness, probably of Uppermost Jurassic and Lowermost Cretaceous age, have yielded an exceedingly rich vertebrate fauna, consisting chiefly of dinosaurs. Discovered first in the vicinity of Morrison, Colorado, in 1877, hundreds of tons of bones [Pg 55] have been collected from these beds for various museums. The dinosaurs include many genera of all three suborders, varying in size from that of a cat to some of the largest known land animals. Of other reptiles a very few jaws of a true rhynchocephalian, a fragment of a wing bone of a pterodactyl, numerous turtles, and crocodiles, only, are known. The beds are predominantly black-clay shales, intercalated with sandstones, and all are of fresh-water origin.
From beds definitely known as Lower Cretaceous (Trinity) in Oklahoma, a few bones of a sauropod dinosaur are known, and from nearly corresponding rocks in southern Kansas, plesiosaurs, crocodiles, turtles, and carnivorous dinosaurs are known from sparse remains. Doubtless the Potomac beds of Virginia, which have yielded bones of various dinosaurs, are also of Lower Cretaceous age.
Fig. 30.—Restoration of Casea, a
theromorph reptile from the Permian of Texas,
about four feet long.
With the exception of a single vertebra of doubtful affinities and the cast of a turtle-shell no vertebrate fossils have ever been discovered in the extensive sandstones of Dakota age, the lowermost of the Upper Cretaceous. From the next horizon above the Dakota, the Benton [Pg 56] Cretaceous, chiefly marine limestones, at least three genera of plesiosaurs are known from Kansas, Texas, and Arkansas, with two or three more from the limestone shales of Wyoming. A few specimens of armored dinosaurs, two genera of ancient crocodiles, nearly the last of their kind, some marine turtles, and a few vertebrae of ichthyosaurs, the last of the order known anywhere in the world, are also known from the Benton Cretaceous of Wyoming.
Continuous with the Benton limestones above in Kansas are the famous beds of Niobrara chalk; perhaps no fossil deposits in the world are more famous. Exposures covering hundreds of square miles in western Kansas, almost pure chalk, have furnished fossil-hunters during the past forty years literally thousands of specimens of mosasaurs, hundreds of pterodactyls, and scores of plesiosaurs and marine turtles, in addition to the famous birds with teeth and countless fishes of diverse kinds. Two or three specimens of spoon-billed dinosaurs have been found in these deposits, but no other reptiles of any kinds. Beds of like age in Colorado and New Mexico have furnished a few specimens of mosasaurs.
From the marine beds of Fort Pierre age, next above the Niobrara in the west, have come some excellent specimens of two genera of mosasaurs, three or four forms of plesiosaurs, a few pterodactyls, the largest of all marine turtles, and still fewer specimens of dinosaurs, in Kansas, South Dakota, Wyoming, and Montana. From deposits of approximately like age in Mississippi, Alabama, and New Jersey, many incomplete specimens were found years ago of mosasaurs, plesiosaurs, and turtles, the last of the amphicoelian crocodiles, the first of the procoelian crocodiles, and the famous specimen of Hadrosaurus which served for the Hawkins restoration, the first attempt of its kind.
From the uppermost Cretaceous beds of America, the Lance, Judith River, or Belly River beds as they are variously called, have come the remains of a marvelous reptilian fauna. These beds may be grouped together though not all contemporaneous, and there is dispute about their age, some excellent paleontologists insisting that the uppermost are really of Eocene age. From Colorado east of Denver, from eastern Wyoming, from Montana, and especially from the vicinity of Edmonton [Pg 57] in Canada, as also occasionally in western Texas and New Mexico, have come many marvelous specimens of dinosaurs, huge bipedal carnivorous dinosaurs, great spoon-billed aquatic dinosaurs, armored stegosaurian dinosaurs, and many kinds of the great horned dinosaurs, the Ceratopsia, so far known only from these beds. Here at the very close of the Age of Reptiles, at the close of the Age of Dinosaurs, are found the ultimate specializations of all the chief groups of dinosaurs except the long-necked quadrupedal dinosaurs which gave up the ghost in Lower Cretaceous times. Many were provided with horns and spines, some indeed seemed to have bristled with spines throughout, a sure sign that they were approaching the end of their career. The modern type of crocodiles had usurped the ancient forms of the early Cretaceous, and reached the largest size of their race perhaps, though but few specimens are known. Here also in these beds we find the first representatives of lizards and snakes in America, though snakes have been described from earlier strata, perhaps, in Brazil. Those archaic, old-fashioned rhynchocephalians described on a later page as the Choristodera appeared also for the first time in these beds, and persisted for a little while in the Eocene, in Europe and America. And with all these there has very recently been described the last of the plesiosaurs, whose race went out with the dinosaurs at the very close of the Mesozoic. It is needless to say that the turtles also occur, for, as a general rule, wherever vertebrate fossils are found, in rocks of the land or the sea, marine or fresh-water, there will be some bones of turtles among them.
With the beginning of the Cenozoic the record of the reptiles becomes relatively scanty in America. In the warm waters of the old Eocene lakes and rivers of Wyoming lived countless crocodiles, true crocodiles of modern aspect and of large size. But, as the climate of North America grew progressively colder, the crocodiles retreated to the south, till, in the Oligocene, the scanty remains of the last crocodiles are found in the American Tertiary. On the other hand, as the open lands appeared toward the close of the Eocene, and in the Oligocene and Miocene, the land tortoises throve and grew greatly in size. In the Bad Lands of South Dakota one may see their remains in almost incredible numbers. And in equally great numbers are these land [Pg 58] tortoises, in shape much like the common box tortoise of today, but vastly larger, found in the rocks of the late Miocene or early Pliocene age in western Kansas. And these are the last records of the big tortoises in North America; their descendants are perhaps yet living in the Galapagos Islands.
The history of the lizards and snakes, the only other reptiles found in the Cenozoic rocks of America, is very brief. A few specimens from the Lower Eocene of Wyoming; a few skinks and amphisbaenas from the Oligocene Bad Lands of South Dakota, and some bones of a python-like snake in the early Eocene of Wyoming are about all that we know of the Squamata in the Tertiary. Doubtless snakes and lizards were just as abundant then as now, though but few were preserved, for they are and always have been distinctly terrestrial animals, that only by accident fell into places where they could be fossilized.
The author has collected reptile bones from nearly all of the horizons here mentioned and believes that the list is complete.
[Pg 59]
In the never-ceasing struggle for existence all forms of life upon the earth, whether consciously or unconsciously, are continuously striving for improvement; striving to flee from adverse environments, or to adapt themselves better to those which must be endured; to escape their enemies, or to find means whereby they may withstand them; to find more or better food, or to prevent others from despoiling them of what they have. There is always more or less of unrest, more or less of discontent, if such terms may be used of the lower organisms. It sometimes happens with groups of organisms that by reason of unusual or extraordinary traits they become so perfectly adapted to their environments, to their surroundings, or so easily adaptable to changes in their environments, that they remain for long ages securely protected and little changed. But, as with man himself, improvement is usually the result of adversity—adversity which stimulates but does not destroy. And the word improvement, translated into biological language, means simply specialization, that specialization which adapts the organism better to its mode of life, which fits it the better to excel its less ambitious or less capable competitors. No animals or plants are perfect; if they were, there would be no advancement, no struggle. If all physical conditions stood still, or remained uniform, perhaps life would stand still, but conditions never have and never will stand still, and life must change to meet changed conditions.
Thus it is that which makes life easier, which lessens the dangers of destruction, which insures the continued prosperity of the race, is seized upon and utilized by all plants and animals, so far as [Pg 60] possible. As said long ago by Tennyson,[2] the first law of life is not the preservation of self, but the prosperity of the race. Whatever the causes may be whereby the offspring are better adapted to conquer in the struggle for existence, whatever may be the laws governing changes and specialization, whether heredity, Mendelism, mutation, natural selection, or Lamarckism, we call the process evolution.
To escape from the severe competition of the overcrowding animals of the sea, some of those creatures we call fishes long ago became air-breathers and took possession of the unoccupied land. From among the myriads which were driven into unbreathable water, by accident or by their enemies, or led there in the search for more easily acquired or better food, some survived and found that the oxygen of the air was quite as breathable as that of the water. Steadily their progeny became better and better adapted to the unusual life until they ceased to be fishes and became amphibians, from which have arisen in like manner all the reptiles and birds and mammals that live or have lived upon the earth.
With more and better powers, developed under better opportunities, not a few of these descendants have repeatedly sought safety from their newly acquired enemies of the overcrowded land, or a better supply of food in the sea; gradually, perhaps incidentally at first, as we shall see is the case with some lizards today, but later with increased adaptation to their new surroundings, they become truly sea or water animals, no longer able to live upon the land. In these changed conditions and with concomitantly changed habits they never reverted to the primitive condition of fishes, never became water-breathing animals again, for that would be actual retrogression, a seeming impossibility in evolution. Nor indeed does it seem possible that a land creature after its reversion to water life ever can return to the land again.
A fish through long ages of evolution has become well adapted to its environments; its shape is the best for speed or varied evolutions in the water; its teeth and mouth-organs are best suited for the food it requires. Now it is evident that if animals of very different habits and form should go back to the water and seek to compete with creatures already well adapted to their surroundings, they must, so far as [Pg 61] possible, acquire like forms and like habits. And any improvement on such forms and habits that their higher development permits them to attain will of course be of advantage in their competitive struggles. A fish makes most use of its tail fin for propulsion. It follows that a land animal seeking to compete with it under like conditions must acquire a tail fin or some other organ which subserves its purpose as fully. The body fins are of little use to a fish, save for equilibration, for preserving its position, for stopping quickly, or for changing the direction of its movements quickly—very different functions from those of the corresponding organs, the limbs, of higher vertebrates. There are few better examples of predaceous, fish-eating fishes than the common gar-pike of our rivers, fishes with a slender body covered with very smooth scales, a strong tail, a short neck, and long jaws armed with numerous slender and sharp teeth. Such a fish, darting into a school of smaller fishes, by quick, sudden changes of movement, actively opening and closing its jaws, is sure to seize some of its sought-for prey. In a direct trial of speed with its victims it would most likely be worsted.
There have been many animals of high and low rank which in the past and present have gone back from a terrestrial existence to a life in the water, finding at last a congenial home away from the shores. Or, perhaps, like the monitor lizards of today, they have found temporary safety in the water when hard pressed by their land enemies, and finally found, not only protection, but an abundant supply of easily obtainable food therein. As in every vocation of life there have been many failures in such attempts, many partial successes only. But not a few have found abounding and enduring success and final prosperity—success that has led possibly to undue adaptation to surroundings, and to the acquirement of great size, for that has been the invariable end of water air-breathers of long duration—specializations which finally prevented them from meeting new exigencies. It seems to be a law of evolution that no large creatures can give rise to races of smaller creatures; and as we shall see, the largest sea-animals have been the final evolution of their respective races.
There are no better examples of such success today, nor has there been [Pg 62] in all the geological ages, so far as we know, more perfect examples of the adaptation of air-breathing animals to an aquatic life than the great whalebone whales. In Eocene times their ancestors were walking and running land animals; of that there can be not the slightest doubt, since we cannot conceive, as did the older naturalists, of their direct descent from the fishes while having all the essential structure of mammals, i.e., lungs, circulatory system, manner of breeding and rearing the young, etc. Of the living whales, or Cetacea, there are now in existence two very distinct types, so different from each other that some have supposed them to have been evolved from different types of land mammals. One of these is best exemplified by the great baleen whale, having a broad, short head and no teeth. It feeds upon crustaceans chiefly, which are strained from the water by the great fringe or net of “whalebone.” The other type is seen in the porpoise or dolphin. These cetaceans have numerous, pointed and recurved teeth, which they use as did many of the reptiles, hereinafter described, for the seizure and retention of fishes and other swimming animals. So great have been the changes in all these cetaceans, in the adaptation to an aquatic life, that we are almost at a loss to conjecture from what kinds of land animals they have descended. The great zeuglodont whales of early Tertiary times have long been thought to be a sort of connecting link between them and their land ancestors, and it is still probable that they were. The forms of zeuglodont whales that have been discovered in Africa within recent years bear so much resemblance in their skull and teeth to the contemporary carnivores, that many paleontologists think, with good reason, that they were descended from them, that is, from the ancestors of all our dogs, cats, weasels, bears, etc., of modern times. And we have much reason to believe that future discoveries will bring further and more decisive proof of their origin before many years have elapsed. The modern Sirenia, the dugongs and manatees, exclusively aquatic mammals, which feed upon seaweeds at the bottoms of shallow bays and harbors, or in the mouths of rivers, are now known, practically with certainty, to be the descendants in these same African regions of the earliest ancestors of our sheep, oxen, and horses, known so certainly that they are often classed with them, or at least with the elephants, which approach them in their ancestral line even more closely. [Pg 63]
A third type of living aquatic air-breathers is seen in the seals, sea-lions, etc. They are much less highly specialized, however, than the whales or sirenians, since they are still capable of considerable freedom upon land, which they recurrently seek for the breeding of their young. They still retain the primitive covering of hair, lost almost entirely by the cetaceans and sirenians and functionally replaced for the conservation of heat by a thick layer of blubber. Instead of losing the hind legs and developing the tail as a propelling organ like the whales, the seals encountered precisely the reverse experience. The hind legs have been developed into most efficient paddles or sculls, and the tail has been for the most part lost. They are fish-eaters, it is true, but they do not have the long jaws possessed by the porpoises and toothed whales.
In the sea-otters, beavers, and even the muskrats, we have examples of less complete adaptation of land mammals to water life, the most of them showing the beginnings at least of structural adaptations similar to those of the seals. From an attentive examination of all these animals, living as well as extinct, which have attained partial or complete success as air-breathing water animals, we find certain laws existing, if we may call them such, which we may discuss a little in detail. As we have seen in the comparison of the whale with the seal, the methods of adaptation have not always been the same, and some recent writers have endeavored to classify aquatic animals under many groups, to which they have given learned technical names, most of which will not concern us here in dealing with the reptiles only.
Beginning with the head, we find that all those reptiles and most of the mammals which have become aquatic fish-eaters have an elongated skull, or rather an elongated face. The jaws are long and slender, and the teeth are not only numerous but also sharp and slender, much like those of the gar-pike, indeed. It is remarkable, too, that in most such animals the external nostrils are situated, not at the extremity of the snout, as in all terrestrial mammals and reptiles, but far back near the eyes. In the whales this position of the nostril enables the animals to breathe without continuous muscular exertion while floating on the surface; that is, the nostrils are at the top of the head. In the sirenians, on the other hand, which live habitually at the bottom [Pg 64] of shallow waters, coming to the surface to breathe only, the nostrils are situated so that they are the first to emerge, that is, they are near the front end. The crocodiles, with a more or less elongated face, as also the Choristodera, described farther on, are exceptions, since their nostrils are at the extremity of the snout. Both of these types, however, notwithstanding the elongation of the face, are only partly aquatic in habit, and in the crocodiles the breathing organs have undergone a strange modification in accordance with habits peculiarly their own, as will be explained later on. Whether this recession of the nostril toward the eyes can be explained in all cases by the peculiar breathing habits is, however, doubtful. Possibly in some cases, such as the phytosaurs, described later, the creatures used their long beaks to probe in the mud while breathing. Possibly the posterior position has been in some cases rather the result of the elongation of the face, leaving the nostrils behind in some forms, or carrying them forward in others. Nevertheless posterior nostrils always indicate more or less aquatic habits.
In all the earliest reptiles, as we have seen, the neck was short, like that of their immediate progenitors, the ancient amphibians. The shoulders were close to the skull, with not more than two vertebrae that could be called cervical. It happens that most of the earliest reptiles, as we know them, were more or less amphibious in habit, and all of them were probably good swimmers; nevertheless in all likelihood reptiles began their career as a class with a very short neck. The earliest known distinctly terrestrial reptiles had a moderately long neck composed of six or seven cervical vertebrae. It may therefore be assumed with much probability that all later reptiles with a greater or less number of cervical vertebrae are specialized animals, so far as the neck is concerned. Most living reptiles have eight cervical vertebrae; a few have nine, and still fewer have but five. Birds may have as many as twenty-four, while all mammals, with two or three exceptions, have the primitive number seven. Among extinct reptiles, however, there were not a few with more numerous neck vertebrae, some having the enormous number of seventy-six.
An ordinary fish has apparently no neck whatever, the trunk being [Pg 65] seemingly attached to the head, nearly as in the primitive amphibians and primitive reptiles. It is evident that a movable neck of considerable length would not only be of no use to the swiftly swimming fish, but a positive disadvantage to it. The body is quickly and easily turned by the powerful tail fin, and a long neck could be of no use that the tail would not better subserve. It is therefore of interest to learn that, as a rule, aquatic animals of all kinds having a powerful propelling tail have also a short neck, acquired either by the loss of neck vertebrae, or, as in the mammals, by the shortening and coalescence of the normal number of seven. There are very few exceptions to this rule of a short neck and a long tail. Those strange little reptiles of Paleozoic times, the first that we know that returned to the water, the Proganosauria, have not only a long, flattened tail, but also an unduly elongated neck of from nine to twelve vertebrae.
On the other hand, certain unrelated reptiles of the past, the dolichosaurs, nothosaurs, and plesiosaurs, with a short non-propelling tail, developed a long neck—sometimes an excessively long one in the plesiosaurs. The turtles, some of which have attained a high adaptation to water life, have invariably a short tail and a freely movable, relatively long neck, a neck which Dr. Hay tells us has increased in length from the beginning of their race by the simple elongation of the vertebrae, as in the giraffe, and never by the addition of vertebrae. We may then account it a rule that swimming animals with a long neck have a short tail, and those with a short tail have a long flexible neck. Even in the plesiosaurs there is some variation of the length of the tail in correlation with the neck. Short-tailed animals must necessarily propel themselves through the water by the aid of their legs, especially the hind legs. If one watches an actively swimming alligator he will observe that the front legs are folded or collapsed by the side of the body, while the hind legs, much bent, are used only slightly in propulsion. The animal swims by a marked sinuous or serpentine movement, like that of a snake upon land, extending throughout the tail and part of the body, at least. An animal propelling itself by its limbs could not move sinuously, and use its legs actively at the same time, and it is probable that the long neck has been evolved compensatorily. [Pg 66]
With this shortening of the neck and sinuosity of movement there is developed in every case a long trunk as well as a long tail. The trunk becomes more slender and cylindrical, more like that of a snake, with an actual increase of the bones composing it, reaching the great number of forty-three vertebrae in that most sinuous of all water reptiles with legs, Pleurosaurus of the Protorosauria. And the tail, primitively having perhaps sixty or seventy vertebrae, may have as many as one hundred and fifty in the more typical aquatic forms. This elongation of trunk and tail must be of great advantage to the swimming reptile, just as the racing scull is a more perfect type of speedy craft than a flat-bottomed scow. Dr. Woodward has said that the fate of all fishes, if they continue their evolution long enough, is to become eel-like.
Not only was the tail greatly elongated in swimming reptiles, but it was also more or less flattened. In the beginning of water adaptation the flattening was throughout the tail, as in the living alligators and crocodiles. As the adaptation to water life became more perfect, the flattening became more and more restricted to the extremity; that is, the flattening begins like that of a salamander and in the end becomes like that of a fish, a terminal fin. And some of the actual stages in the evolution of the fish-like fin have been observed by Dr. Merriam in the earlier and more primitive ichthyosaurs of California. In those animals swimming chiefly in a horizontal direction the tail fin has become like that of fishes, that is, vertical; but in those animals which use the tail chiefly for ascending and descending rapidly in the water the fin is developed in a horizontal position, examples of which are seen in the flukes of whales and sirenians.
All animals living upon the land require firm articulations between the different bones of the skeleton, and especially between the vertebrae, for the support and control of the body. Among aquatic animals there is a strong tendency toward looseness of joints, with increasing flexibility. Fishes have the articular processes between the arches of the vertebrae feebly or not at all developed, and the centra or bodies of the vertebrae have thick pads of cartilage between them. Firm union between the vertebrae would restrict freedom of movement, and firmness [Pg 67] is not required when the body is surrounded on all sides by water of nearly the same specific gravity as the body itself. And it is doubtless for the same reasons that the articulations of all strictly aquatic reptiles have for the most part become looser and less firm, especially those between the different vertebrae.
The same looseness of articulation is also found in the ribs of aquatic animals. In most animals, and in all those which walk erect, like the mammals, each rib is firmly attached to the backbone by two distinct joints, the head and tubercle, with an interval between them. This double attachment prevents much in-and-out movement of the ribs and gives a firm support for the attachment of the muscles of respiration, as well as for those supporting the viscera. This firmness is unnecessary in animals living always in the water, and the ribs therefore in all aquatic animals tend to become single-headed and loose. The lower or capitular articulation has been lost in part, or almost wholly, in many cetaceans. It has been said that a whale cast up on land will die of suffocation, not for the lack of air, for it is an air-breathing animal like ourselves, but because it can no longer use its respiratory muscles attached to the loosely articulated ribs; it suffocates because the ribs collapse.
As would be expected, the greatest modifications of structure in the adaptation of air-breathers to water life are found in the limbs. No other parts of the body have such different functions in water and on land as the limbs and fins. The limbs of a dog, or a cat, or a man are feeble organs for swimming in comparison with the fins of a fish, and if the land animal must compete with fishes to prey upon them for food it must acquire like swimming powers. As a matter of fact, the limbs of all typically aquatic air-breathing animals have lost nearly all external resemblance to the legs of walking and running animals, and have become more or less fin-like in function—fin-like in shape and function, but never fin-like in actual structure. No creature can go back and begin over again, any more than a man can again become a child with all its possibilities for improvement and development. If an animal cannot modify the organs it already possesses so as to adapt them to new and changed uses by the aid of evolutionary forces it must [Pg 68] fail in the struggle. It can never acquire new material, never get new fingers and toes, new organs or parts of organs; all its possibilities lie in the improved and new uses it can make of the material which it received from its ancestors.
The beginning of aquatic adaptation of the limbs lies in the membranous webs between the toes of frogs, salamanders, ducks, seal, otters, etc., where the feet are used largely or entirely for propulsion through the water, in the absence of a propelling tail. And this membrane, in the majority of cases, is the extent of aquatic adaptation in air-breathing animals. In those animals, however, such as most of the reptiles described in the following pages, where the tail has developed as the propelling organ, the limbs lose to a greater or less extent their propelling function and become merely organs of equilibration and control. Of the two pairs of fins of fishes it is evident that the anterior ones have the more important equilibrational function; the hind ones have a much less important use as guiding organs; as a matter of fact, in not a few fishes the hind or pelvic fins have actually migrated forward to supplement the function of the pectoral fins. It is for these reasons that those animals best adapted of all for life in the water—the whales and sirenians—have lost the hind legs completely. In other tail-propelled air-breathers the hind legs have become progressively smaller and less powerful than the front ones. In all short-tailed water animals, however, where the legs, and especially the hind legs, have the important function of propulsion to subserve, they still retain the large size and firm connections with the body, examples of which will be seen in the seals, sea-otters, marine turtles, and plesiosaurs.
Because the legs are no longer needed for the support or propulsion of the body in long-tailed air-breathers, their connection with the body becomes less and less firm, long before their entire disappearance. In animals using the legs for crawling or walking the bones of an arm and thigh are elongated, and the joints are always well formed, permitting varied, extensive, and firm movements. Just the reverse is the tendency in all those animals that propel themselves by the aid of the tail in the water, since here what is needed is broad, short limbs, not long and slender ones. [Pg 69]
Most reptiles have five digits on each hand or foot; the bones of the wrist and ankle are well formed, as in mammals, and the digits are elongate, with a very definite arrangement of the bones composing them, as already described, never exceeding five in any one finger or toe.
In the paddles of water reptiles, as the limbs are usually called, the bones of the first segment, that is, the humerus and femur, are always greatly shortened in those having a propelling tail, and even in some with a short tail, such as the seals, and in a lesser degree in the sea-otters. On the other hand, in those animals which use the legs chiefly for direct propulsion these bones are elongated, as exemplified by the plesiosaurs and marine turtles. In all save the seals and their kind, and the otters, whose legs are used rather as sculls than as oars, the bones of the next segment, the radius and ulna of the front pair, the tibia and fibula of the hind pair, are always shortened, and one can tell the stage of aquatic adaptation, as exemplified, for instance, in the plesiosaurs and ichthyosaurs by the degree of shortening of these bones. Indeed, the first suggestion in any crawling animal of water habits is shown in the relative lengths of the epipodial bones, as these bones are called. Furthermore, cursorial or terrestrial habits are suggested by the relative size of the smaller bone of the leg, that on the little-toe side, the fibula. In birds, pterodactyls, and most running animals, it disappears in part or wholly. In swimming animals it tends to grow larger than the tibia, as will be conspicuously seen in the paddle of the mosasaurs.
The bones of the wrist change in two ways: by becoming cartilaginous, as in whales and salamanders, or by becoming more firmly ossified and more closely united, as in the plesiosaurs. The digits always are elongated, often extraordinarily so, either by the elongation of individual bones or phalanges, or by the development of new bones. These new bones, when they occur, are new growths, not the reproduction of the old elements of fishes, and there may be as many as twenty such new elements or phalanges in a single digit. There is one marked exception among reptiles to this hyperphalangy, as the increased number of phalanges is called, and that is the turtles. As we have seen, in [Pg 70] the elongation of the neck among turtles there never has been an actual increase in the number of vertebrae; so also in the elongation of the digits the normal number of three in each digit has never been exceeded, except among the river turtles, where there are four in the fourth digit—possibly a relic of original conditions rather than the beginning of hyperphalangy; but the individual bones have become greatly elongated. In living reptiles, birds and mammals of the land, the fifth toe is always shorter than the fourth. In the seals, the sea-otter, and to a less degree in the muskrat, the fifth toe has become elongated. And the elongation of this toe is the first and most decisive indication of a webbed foot of strong propelling power among the aquatic reptiles of the past, as exemplified especially by the proganosaurs. Finally, in one order of extinct reptiles, the ichthyosaurs, there has been an actual increase in the number of digits, in some to as many as nine in each paddle.
In addition to all these modifications of the skeleton, the bones themselves tend to become softer and more spongy in aquatic animals. The bones of the whale, as is well known, are very spongy in texture, and those of the seals and sea-lions contain an unusually large amount of oily matter. So, too, the bones of the extinct water reptiles—of many of them at least—were more spongy than those of their land relatives; and this is due in part perhaps to their lessened use as muscular supports, in part perhaps to the necessity of a lessened specific gravity. As a rule sea-animals need to be of the same specific gravity as the water in which they live, or a little less. The bones of the living sirenians, the manatees and dugongs, so far from being light and porous, are unusually dense and solid. The sirenians live habitually at the bottom of shallow waters, feeding upon vegetable growths; and doubtless their bottom-feeding habits account for the solidity of the bones. A whale would float to the top, while a dugong would sink to the bottom, on the relaxation of all muscular movement. And we shall see that certain reptiles in the past had in all probability like bottom-feeding habits, because of the solidity of the bones of their skeletons.
Many birds and fishes have a peculiar ossification of the usually tendinous outer covering of the eyeball, called the sclerotic membrane. [Pg 71] These ossifications form a flattened or somewhat projecting conical bony ring about the pupil of the eye. The individual bones are flat and more or less imbricated plates, with some motion between them. Accommodation for vision in reptiles, birds, and fishes is not the simple process that it is in mammals, where it is controlled by simple ciliary muscles which compress the lens, causing it to assume a more spherical or a more flattened form, thus changing the focus. In reptiles accommodation is effected by the compression of the eyeball by means of external muscles, elongating it and causing its front part to expand or project. The imbricated sclerotic plates permit this expansion and contraction of the eyeball. Under great internal or external air pressure the cornea, the only unprotected part, must necessarily change its contour unless some compensatory force is brought to bear to counterbalance it; and this doubtless was the function of the sclerotic plates so commonly present in aquatic reptiles.
Among terrestrial reptiles there are not a few examples of the ossification of such sclerotic plates, notably among the skink lizards. Every known form of extinct reptiles of aquatic habit had them, and even some of the subaquatic dinosaurs, like Diplodocus and Trachodon. One may say with assurance that it is impossible for any reptile to become thoroughly adapted to aquatic life without acquiring large and strong sclerotic plates.
Most land reptiles are or were covered by horny scales or bony plates; the pterodactyls are the only order of terrestrial reptiles with no such covering of which we have any evidence. Such coverings are wholly unneeded for animals living in the water. Not only are they unnecessary, but the increased resistance to the water would be more or less detrimental to rapid swimming. It is for these reasons doubtless that bony plates or horny scales disappeared for the most part from the skin of all truly aquatic reptiles and mammals.
The foregoing are the chief acquired characteristics of aquatic air-breathing animals and especially aquatic reptiles in adaptation to their new mode of life. The resemblances, sometimes striking, thus brought about in animals of very different origin and remote relationships have often been mistaken for evidences of kinship, that [Pg 72] is, direct inheritance from common ancestors. Such acquired resemblances in unrelated animals are known as parallel or convergent evolution. It has often been difficult to distinguish between convergent evolution and direct evolution, and difficulties still perplex and trouble the student of natural history in every branch of life. Not till all such problems are solved can we hope to attain the true classification of animals and plants. The whales a century ago were considered merely breathing fishes; the ichthyosaurs until a quarter of a century ago were supposed to be the direct descendants of fishes; lizards and crocodiles were grouped together in a single order; and salamanders were called reptiles not very long ago.
Perhaps the reader will be able from the foregoing to understand and appreciate better some of the difficulties that confront the paleontologist in his attempts to solve the problems of past life; to understand why he sometimes makes mistakes, for he has by no means yet learned all the permutations of the skeleton in any class of vertebrates, and is not sure that the laws he accepts are not subject to modifications and exceptions. If he is truly scientific he hesitates long in prophesying or conjecturing.
[Pg 73]
Very scanty are the early human records of those strange reptiles known as the plesiosaurs. Were one to search through the many works published during the latter half of the seventeenth century and all of the eighteenth, devoted to “lapides petrifacti,” “figured stones,” “reliquia diluvii,” or by whatever other fanciful names fossils were known, here and there he would probably find descriptions and figures of bones of these reptiles. It would hardly seem that plesiosaurian bones could have been overlooked by the curious, so abundant are they in many places. But there is no such history of the early discovery of the plesiosaurs as there is of the ichthyosaurs and mosasaurs. Their birth into human history was very formal and proper, under the ministrations of a learned doctor of science, the renowned Conybeare, of whom we shall speak again. It was he, who with De la Bêche, late Director of the British Geological Survey, described for the first time, in 1823, one of these reptiles, to which he gave the name Plesiosaurus, meaning “like a lizard.” He distinguished the plesiosaurs from ichthyosaurs, with which it is possible that they had previously been confounded, and gave a good description of considerable material. Cuvier, a little later, gave a more complete description of the same remains which had served Conybeare and De la Bêche for their original description, and for the first time made it evident that fossil plesiosaurs were widely and abundantly distributed over the earth. The closing sentence of Cuvier’s chapter devoted to the discussion of these creatures in his Ossemens Fossiles was really prophetic, not only of the many discoveries of the plesiosaurs yet to be made, but of all other extinct animals as well: “I doubt not that, in a few years it may be, I shall be compelled to say that the work which I have today finished, and to which I have given so much labor is but the first glimpse of the immense creations of ancient times.” [Pg 74]

Fig. 31.—Restoration of Plesiosaurus guilelmi imperatoris (left figure) and Thaumatosaurus victor (right figure), Liassic plesiosaurs. (From E. Fraas.)
[Pg 75] In quick succession there followed many other discoveries of plesiosaurs, not only in England but elsewhere in Europe. The famous English anatomist and paleontologist, Sir Richard Owen, to whom we owe, perhaps, more than to anyone else our present knowledge of these animals, the eccentric Hawkins of England, the learned von Meyer of Germany, and, in later times, more especially Seeley and Andrews of England, Fraas of Germany, Bogalobou and Riabanin of Russia, as well as many others, have brought to light during the past century many and varied forms of those sea-reptiles. Blaineville in 1835 gave to the plesiosaurs an ordinal rank under the class Ichthyosauria, and even the astute Owen in 1839 united them with the ichthyosaurs as a suborder of his Enaliosauria, or “sea-saurians.” He called them Sauropterygia, or “reptile-finned,” and these terms, Enaliosauria, Ichthyopterygia, and Sauropterygia, have long persisted in works on natural history because of the prestige of Owen’s name. As we shall see later, the plesiosaurs are really of remote kinship to the ichthyosaurs, and there is no such natural group as the Enaliosauria. It often takes years to distinguish between apparent and real relationships among living organisms, and both of these groups of sea-saurians have had a sorry experience in the treatment they have received from nomenclators.
Perhaps because of the writings of Dean Buckland in his famous Bridgewater Treatise, in large part a theological disquisition, though of real scientific merit, the ichthyosaurs and plesiosaurs early became widely and popularly known, and, even to this day, these reptiles, together with the dinosaurs, first made known by Rev. Dr. Mantell, are often supposed to be the most typical and horrid of monsters. Many and fabulous are the tales that have been told of them in literature both grave and gay. The preacher adduced them as evidences of the great world-catastrophe told in biblical history, and the German student sings of them to the tune of the “Lorelei”:
We now know that they were not the monsters of horrid mien that they were once supposed to be: the largest plesiosaurs, were they living today, would find unopposable foes in the vicious and cruel crocodiles. They were relatively stupid and slow, cruel enough to the smaller creatures, but of limited prowess. But in structure and habits they are among the most remarkable of all the animals of the past or present.
Although their remains are among the most abundant and widely distributed of all fossil reptiles, the plesiosaurs as a whole are less perfectly known than either the ichthyosaurs or the mosasaurs, and it has been within a comparatively few years only that an approximately complete knowledge of any form has been obtained. This is partly due to the fact that the order comprises vastly more kinds, more species, genera, and families than does any other order of marine reptiles; partly because their remains, though widely distributed over the earth, and in rocks of many geological epochs, are seldom found completely preserved; usually specimens comprise only a few bones or single bones, and complete skeletons are rare. Were there but few kinds, the many specimens discovered would mutually supplement each other, finally completing our knowledge; but the fragments of many kinds only add to our confusion. Nevertheless, because the plesiosaurs lived so long in geological history, their remains are found in rocks of many different kinds, and since it is improbable that any of them had great specific longevity, it is very probable that all these described species, or most of them, often made known from single bones, will eventually be found to be distinct, and that many more will be added to them. It does not seem improbable that within the next forty or fifty years not less than a hundred species of plesiosaurs will have been discovered in North America alone. At the present time perhaps that many have been described from the whole world.
When Blaineville gave the name Plesiosauria to the aquatic reptiles [Pg 77] described by Conybeare, Cuvier, and others, he had no knowledge of others of an intermediate kind between them and land reptiles. His group-term then can be properly applied only to the truly aquatic forms, and Owen’s name Sauropterygia becomes available in a wider sense to include all the known types belonging to the order of which the plesiosaurs form a part. Of this order then there are two clearly marked divisions or suborders, the Plesiosauria and the Nothosauria, the former having a complete aquatic adaptation, the latter only a partial one. While the two suborders are evidently allied, some authors have suggested that their differences are only familial; others have thought that they are really orders. We shall see how close the relationships are.
It was Dean Buckland who facetiously likened the plesiosaurs to a snake threaded through the shell of a turtle, and the simile was not an inapt one in his day. The vernacular designation of them—long-necked lizards—conveys the same impression of their chief peculiarity, but the name is less applicable than it once was, since recent discoveries have brought to light forms with a relatively short neck.
Though the plesiosaurs are nearly perfectly adapted to an aquatic life, the adaptation was, in many respects, of a very different kind from that of the ichthyosaurs—so very different that we have not yet quite finished conjecturing as to the habits of the living animals. As already suggested in the popular name, the most striking characteristic of the typical plesiosaurs, the one which suggested to Buckland his frequently quoted simile, is the ofttimes enormously long neck, proportionately longer than that of any other known creatures of the past or present. In other truly aquatic animals the neck is actually shortened in the acquirement of a fish-like shape, and the number of bones composing it reduced. In the Sauropterygia the neck is usually longer than any truly land animals ever possessed, the longest-necked forms having as many as seventy-six vertebrae in the cervical region. The elongation of the neck among mammals is always due to an increase in the length of the individual bones, never to an increase in the number from seven, with but a single exception—a South American sloth [Pg 78] which has nine cervical vertebrae. The long neck of birds is due both to an increase in the length of the individual vertebrae and to an increase in their number, to as many as twenty-one. But the elongation of the neck among plesiosaurs was very variable indeed; sometimes it was ten or twelve times the length of the head, at other times it was even shorter than the head. And the number of bones composing it was also extremely variable, scarcely any two species having the same, the known extremes being seventy-six and thirteen. In Elasmosaurus platyurus, for instance, the longest-necked plesiosaur known, the head was two feet in length, the neck twenty-three, the body nine, and the tail about seven; on the other hand, in the shortest-necked plesiosaur known, Brachauchenius Lucasi, the head was two and one-half feet in length, the neck less than two feet, and the body about five; the length of the tail is unknown.

Fig. 32.—Skeleton
of Trinacromerum osborni, a Cretaceous plesiosaur,
as mounted in
the University of Kansas Museum.
Not only was the number of vertebrae so extraordinarily increased in many plesiosaurs, but in the longest necks the vertebrae themselves, as in birds, were more or less elongated, especially the posterior ones, which may be six or seven times the length of the anterior ones. Not only was the neck of such great length in many plesiosaurs, but it also tapered very much toward the head.
The vertebrae are always biconcave, but the cavities are shallow, saucer-like, sometimes almost flat at each end, and very different from the conical fish-like cavities of ichthyosaurian vertebrae. [Pg 79]

Fig. 33.—Restoration of Elasmosaurus platyurus, an Upper Cretaceous plesiosaur.
[Pg 80] Often the vertebrae are short throughout the vertebral column; sometimes the posterior cervicals and the dorsals are elongated and very robust. The trunk or body proper was never much elongated in the plesiosaurs, having only from twenty-five to thirty vertebrae. The tail was always shorter than the trunk, and it tapered rapidly to the extremity; in some specimens it has been observed to turn up slightly near the extremity, as though for the support of a small terminal fin.
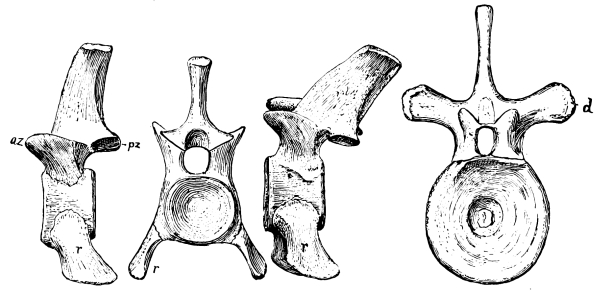
Fig. 34.—Cervical vertebrae, from the side and behind, and dorsal vertebra from in front of Polycotylus, a Cretaceous plesiosaur: az, anterior zygapophysis; pz, posterior zygapophysis, r, r, r, cervical ribs; d, articulation of dorsal rib.
The ribs in the cervical region are short, but so locked together posteriorly as not to permit much lateral motion. They are sometimes double-headed in the neck, sometimes single-headed, but both heads when present articulate or are attached to the body of the vertebrae, distinguishing them at once from those of other animals, except the ichthyosaurs. In the dorsal region the ribs are attached high on the arch to the extremity of the stout transverse processes by a single head, very much as they are in some cetaceans, and quite unlike the condition in any other known reptile. They end freely below, having no attachment to a breast bone or other bony parts. Because of their shape and position as frequently found, the body in life must have been flattened from above downward, and broad; indeed, this shape is quite [Pg 81] certain because of the very broad expanse of the coracoids, between the articulations of the front legs.
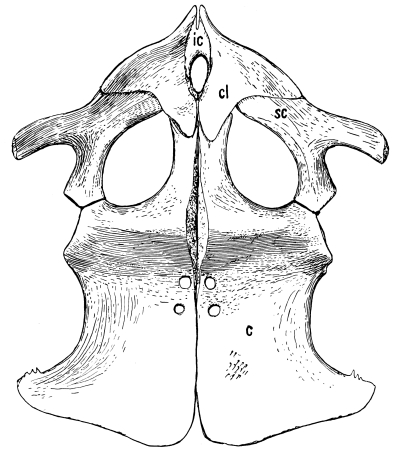
Fig. 35.—Pectoral girdle of Trinacromerum from above: ic, interclavicle; cl, clavicle; sc, scapula; c, coracoid.
The shoulder-girdle or pectoral arch is strangely unlike that of any other reptiles. There is no breast bone, since the breast bone is a comparatively late development in reptiles, not appearing, probably, until after the plesiosaurs had begun their existence. Taking the place [Pg 82] of the sternum, the very large and broad coracoidsjoin each other in the middle, forming a sort of subdermal armor on the under side of the body in front. In some of the largest plesiosaurs these two bones measured together about six feet in length by four in width. Though so very large they are thick only in front between the articulations of the forelegs. The shoulder-blades are much reduced in size and are extraordinarily modified. The blade proper, that is, that part extending backward and upward, is narrow and small, affording but little surface for the attachment of muscles. On the inner side, extending toward the middle in front of the coracoids, there is another projection, often broad and large, to which was attached the clavicles when present, and often this projection met its mate of the opposite scapula in the middle in front of the coracoids in a broad union. The clavicles or collar-bones are small and thin, and sometimes absent; they also are united in the middle posteriorly with the coracoids when the scapula did not intervene. And the interclavicle also is sometimes wanting. Altogether the pectoral bones form a very large, broad, and concave trough inclosing the whole of the under side of the anterior part of the body. This extensive surface must have furnished attachment to stout and strong muscles controlling the downward and inward motion of the paddles.
There is a well-developed sacrum of three vertebrae for the support of the pelvis or hip bones. The reason for its persistence in animals so thoroughly adapted for life in the water will be understood later. The ilium is slender; it was attached to the sides of the sacrum by ligaments, only, not forming a firm union, but strong nevertheless. The pubes and ischia, the other bones of the pelvis on the under side of the body, like the corresponding bones of the pectoral girdle, were enormously enlarged, forming great flat, bony plates.
Besides these large bony plates of the shoulder and pelvic girdles, the short abdominal region was inclosed by numerous series of strong ventral ribs, that is, overlapping rod-like bones on each side, connected with a central piece. It will be seen that the whole under side of the body, from the base of the neck to the base of the tail, was well protected by bones, rigid and unyielding in front and behind, [Pg 83] flexible for a short space below the abdomen; this surface, however, was not flat like the under shell of a turtle, but rounded from side to side.

Fig. 36.—Pelvic girdle from above of Trinacromerum osborni, an Upper Cretaceous plesiosaur: p, pubis; is, ischium; il, ilium.
Many of the characteristics of the limbs of the plesiosaurs are peculiar to themselves; others they had in common with other aquatic [Pg 84] reptiles and mammals. The paddles resemble those of the ichthyosaurs more nearly than those of any other reptile, and it was doubtless this superficial resemblance which so long deceived the early anatomists as to the affinities of the two orders. Unlike all other aquatic animals, however, the plesiosaurs have the hind limbs nearly or quite as large as the front ones, and they doubtless were equally effective in function. The humerus and femur are always elongate, though broad and massive. In no other aquatic animals, save the marine turtles, do we find these bones relatively so long and strong; they are very short in the cetaceans, the sirenians, the ichthyosaurs, mosasaurs, thalattosaurs, and the marine crocodiles, in front at least. The strong muscular rugosities of the plesiosaurian bones are very suggestive of powerful swimming muscles.

Fig. 37.—Pelvic girdle of Elasmosaurus:
p, pubis; is, ischium; il, ilium.

Fig. 38.—Paddles of Plesiosaurs: A, right hind paddle of Thaumatosaurus, after Fraas; B, right hind paddle of Trinacromerum; C, right front paddle of same individual; f, femur; fb, fibula; t, tibia; h, humerus; r, radius; u, ulna.
The bones of the forearms and legs, the wrists and ankles are all polygonal platelets of bones, closely articulating with each other. The finger and toe bones have a more elongated, hour-glass shape than those [Pg 85] of the ichthyosaurs, resembling more nearly those of the mosasaurs, indicating a greater flexibility than the ichthyosaurs possessed. The ichthyosaur paddles must have been quite like the fins of fishes in function, while doubtless those of the plesiosaurs were capable of a more varied use, as indeed was required of them. Their articulation with the trunk was more of a ball-and-socket joint than in the other reptiles, showing possibility of considerable rotation on the long axis, and an antero-posterior propelling action. The paddles were certainly more powerful than those of any other aquatic air-breathing animals. There were no additional digits, all plesiosaurs having neither more nor less than five in each hand and foot. Hyperphalangy [Pg 86] was sometimes carried to an excessive degree, some digits of some species having as many as twenty-four bones, a larger number than has been observed in any other air-breathing vertebrate.

Fig. 39.—Pectoral girdle (in part) and front paddles of Elasmosaurus (after Riggs): sc, scapula; h, humerus; cor, coracoid; r, radius; u, ulna.
In Fig. 38 on p. 85 are shown two paddles, the front and hind paddles of a single individual of a very specialized plesiosaur from the Upper Cretaceous of Kansas (Trinacromerum). The long arm and thigh [Pg 87] bones are followed by remarkably short and broad bones in place of the elongated forearm and leg bones of the land reptiles. Not only are these bones much broader than they are long, but there have been developed additional bones back of them in the same row—new bones which have no counterpart in any terrestrial reptiles. In the first of the three figures is shown a hind paddle of one of the earliest known plesiosaurs, Thaumatosaurus, from the lower part of the Jurassic of Germany. It will be seen here that the tibia and fibula are much more elongated than in Trinacromerum, and much more like the leg bones of land reptiles. A still more primitive stage in the evolution of the swimming paddle of the plesiosaurs will be seen in Fig. 48 on p. 99, the possibly ancestral, amphibious nothosaur. Here the tibia and fibula, while relatively very much shorter than in any land reptile, still have, together with all the other bones of the leg, a terrestrial or amphibious type. In Fig. 39 is seen the front paddles of the long-necked Elasmosaurus, which, though one of the latest of all plesiosaurs in geological history, has the structure of its paddles somewhat intermediate between that of the earlier Plesiosaurus and the later Trinacromerum.

Fig. 40.—Skull of Elasmosaurus from the side: pm, premaxilla; m, maxilla; po, postorbital; j, jugal.
The skull of the long-necked plesiosaurs is surprisingly small in comparison with the remainder of the skeleton, often very snake-like in shape, though very un-snake-like in structure. The short-necked plesiosaurs had often a relatively larger skull, in Pliosaurus, for instance, more than five feet long, sometimes rather broad and short, sometimes remarkably long and slender. The external nostrils were situated far back, very near the eyes, and were very small. The eyes, of considerable size, though by no means so large as those of the ichthyosaurs, were directed laterally, and were provided with a ring of bony sclerotic plates—rather small and weak ones, however. The quadrate bones—bones peculiar to the reptiles and birds—to which the lower jaws are articulated, are, as in the ichthyosaurs and crocodiles, [Pg 88] rigidly fixed and immovable. The lower jaws, always rather slender, are firmly united in front, sometimes for a long distance, as in the modern gavials. The teeth of the broad-headed plesiosaurs are long, slender, pointed, and recurved, of a murderously cruel shape; they are deeply implanted in sockets, and number from twenty to thirty on each jaw above and below. There are no teeth on the bones of the palate, such as the mosasaurs possessed. The slender-jawed, gavial-like plesiosaurs have more numerous, but smaller teeth. The surface of the skull on each side behind, for the attachment of the muscles closing the mandibles, is of great extent; in some this surface is increased by a high, thin crest in the middle, as in strongly carnivorous animals, all of which give conclusive evidence of the powerful muscles used in biting and seizing. There is but one temporal opening on each side, as in the ichthyosaurs and the mosasaurs, whereas the crocodiles, thalattosaurs, phytosaurs, etc., have two. The brain cavity of all plesiosaurs is small, though the cavities of the internal ears, the semicircular canals at least, are large. The semicircular canals in vertebrates have little or nothing to do with the function of hearing; they serve rather for equilibration, for the co-ordination of muscular movement; possibly we may infer from their large size in the plesiosaurs that they were not at all clumsy in their movements. There is a large opening for the pineal body, the so-called eye in the roof of the brain cavity, though its possession does not necessarily imply the possession of a functional organ.

Fig. 41.—Skull of Trinacromerum from the side: ang, angular; d, dentary; pm, premaxilla; po, postorbital; j, jugal; sur, surangular.
The Plesiosauria included some of the largest aquatic reptiles that have ever existed, equaled, perhaps, though not exceeded, by some of [Pg 89] the extinct crocodiles. The largest known are probably those of the Kansas chalk, or the Jurassic of Wyoming, which probably reached a length of nearly or quite fifty feet, of which the neck formed about one-half. Some of them had paddles more than six feet in length. The head of the largest was about five feet in length, or about the size of that of the largest known ichthyosaurs and mosasaurs. The smallest known adult plesiosaurs were nearly ten feet in length. The teeth of the largest and most carnivorous plesiosaurs sometimes measure four inches in length.

Fig. 42.—Restoration of Trinacromerum,
a Cretaceous plesiosaur;
length about ten feet.
As is the case with both the ichthyosaurs and mosasaurs, skeletons of plesiosaurs have been discovered with nearly all their bones in their relative positions, and with impressions of skin and outlines of body made before decomposition. Though our knowledge of the external appearance of the plesiosaurs when alive is perhaps not as full as we could wish, it is sufficient to give us a fairly good conception of what the animals really were. The skin was smooth and bare, without scales or plates of any kind, and Dames has described a terminal or nearly terminal fleshy dilatation of the tail, forming a sort of caudal fin, which may have aided as a steering apparatus. Mounted skeletons are preserved in a few museums, notably the British Museum, the American Museum of New York City, and the museum of the University of [Pg 90] Kansas. Many nearly complete skeletons, however, preserved as they were found in the matrix, are shown in various museums.
With these, principal facts regarding the structure, size, and external form of these animals we may venture to draw certain conclusions, or at least to offer certain conjectures as to their habits in life.
Because of the rigid structure of the jaws, united in front and incapable of any lateral movement posteriorly, quite as are the jaws of crocodiles, we are sure that prey of any considerable size could not have been swallowed whole. The crocodiles tear away portions of the flesh of their victims by quick, powerful jerks, and it is very probable that the flat-headed plesiosaurs tore their food apart in the same manner. In these kinds the teeth are much larger and more irregular in size than are those of the long-snouted plesiosaurs, and their use was certainly as much for tearing as for seizing. There are the same differences between the size of the head and the size of the teeth among the various plesiosaurs that there are among the modern crocodiles and gavials. While the crocodiles seize and destroy even larger prey, drowning and tearing their victims to pieces, the gavials are more exclusively fish-eating, for which their small, sharp, and more numerous teeth especially fit them. Their food, of small size, is swallowed entire, and they are comparatively harmless, so far as animals of considerable size are concerned.
The long neck, the thickset body, and short, stout tail are not at all what we should expect to find in quick-swimming animals. We may therefore assume that the motions of the plesiosaurs through the water were more turtle-like than fish-like. The tail, even though provided with a terminal, fin-like dilatation, was of little use in the propulsion of the body, since the range of its movements was restricted; it possibly served in a measure as a steering organ, a rudder. The large, freely movable paddles must have been effective organs of locomotion, and this function accounts for the relatively large size of the posterior pair, and the firm union of the pelvis with the vertebral column through the sacrum. With the hind limbs used as oar-like organs, a firmer union with the skeleton was required than the [Pg 91] soft yielding flesh would permit. At the same time this union was ligamentous only, not bony and unyielding, since the limbs were never used to support the body upon the ground; and it is of interest to observe that the ilia are directed, not upward and forward, but upward and backward to the sternum, precisely the position that would be expected with the force or thrust coming from behind, and not below the yielding ligaments. Were the tail longer and more powerful, the hind limbs would have been smaller and weaker, of use chiefly in equilibration, involving the loss of any connection with the vertebral column and the disappearance of the sacrum. It is of interest, finally, to observe that many of the slender-jawed plesiosaurs had a relatively short neck; they were doubtless more distinctively fish-eating in habit, and possessed greater speed. That the limbs of plesiosaurs were powerful propelling organs is also conclusively proved by their structure. Quite unlike all those animals whose locomotion in the water is chiefly effected by the tail, the humeri and femora, the upper arm and thigh bones were elongated, and not shortened. They form the rigid and stout handles of oars whose blades are the thinner, flexible forearm, wrist, and fingers, or the corresponding foreleg, ankle, and toes. No other purely aquatic reptiles, save the turtles, which likewise are of the oar-propelled type, have elongated arm and thigh bones.
Textbook illustrations of the plesiosaurs usually depict the necks, like those of the swans, freely curved, and a popular scientific article in one of our chief magazines a few years ago depicted one of them with the neck coiled like the body of a snake. One noted paleontologist, indeed, not many years ago described the plesiosaurs as resting on the bottom in shallow waters with the neck uplifted above the surface viewing the waterscape! And when we consider the fact that some species of the elasmosaurs had a neck not less than twenty feet in length, such a flexible use of it would not seem improbable. But the plesiosaurs did not and could not use the neck in such ways. They swam with the neck and head, however long, directed in front, and freedom of movement was restricted almost wholly to the anterior part. The posterior part of the neck was thick and heavy, and could not have been moved upward or downward to any considerable extent and not very much [Pg 92] laterally. From all of which it seems evident that the plesiosaurs caught their prey by downward and lateral motions of their neck, rather than by quick swimming.
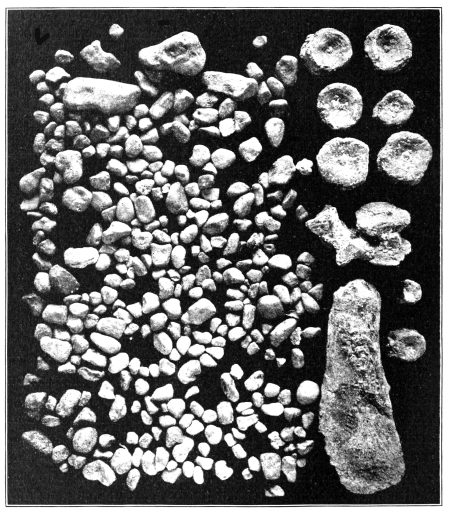
Fig. 43.—Gastroliths and bones of an undetermined plesiosaur from the Lower Cretaceous of Kansas.
About thirty years ago, the late Professor Seeley, a well-known English paleontologist who devoted much attention to the study of these reptiles, found with the remains of a medium-sized plesiosaur nearly a [Pg 93] peck of smoothly polished, rounded, and siliceous pebbles. He believed that their occurrence with the skeleton was not accidental, but that they had been intentionally swallowed by the animal when alive, and formed at its death a part of its stomach contents. Even earlier than this the same habit had been noticed. Nearly at the same time that Seeley mentioned the peculiar discovery he had made the present writer found several specimens of plesiosaurs in the chalk of western Kansas with which similar pebbles were associated, an account of which was given soon afterward by the late Professor Mudge. Since then numerous like discoveries have made it certain that the plesiosaurs usually, if not always, swallowed such pebbles in considerable quantities, for what purpose we do not yet feel sure; one can only hazard a guess. The small size of the pebbles, or gastroliths, as they have been called, a half-inch or less in diameter, found with skeletons of large size, indicate much more complete digestion of the hard parts of their food than is the case with many other reptiles; no solid substance of size could have passed out of the plesiosaur stomach, and such is the case with the modern crocodiles, which have a like habit of swallowing pebbles. That the plesiosaurs picked up these siliceous pebbles, sometimes weighing a half-pound, accidentally with their food is highly improbable; they surely had something to do with their food habits. It is not at all unreasonable to suppose that the plesiosaurs, because of their comparative sluggishness, fed upon anything of an animal nature, whether living or dead, which came in their way; that carrion, squids, crustaceans, and fishes were all equally acceptable; they were probably largely scavengers of the old oceans. Barnum Brown found among the stomach contents of a plesiosaur fragments of fish and pterodactyl bones, and cephalopod shells. Gallinaceous birds, most of which have the same pebble-swallowing habit, have a thick-walled muscular stomach or gizzard, in which the pebbles serve as an aid in the trituration of food. Modern crocodiles, with the same pebble-swallowing habit, have a thick-walled muscular stomach, gizzard-like, though of course not as large as in birds; and the same habit has been noted by Des Longchamps in the ancient teleosaur crocodiles.
It is hardly possible yet to decide whether or not the plesiosaurs were [Pg 94] denizens of the open oceans for the most part, far from land. That many of them were rovers is quite certain. With the skeleton of a large plesiosaur found some years ago in western Kansas, there were many siliceous pebbles which could have come only from the shores of the old Cretaceous seas about the Black Hills, hundreds of miles distant. Some of the pebbles are red quartzite, quite identical with that of the bowlders brought to Kansas millions of years later by the glacial drift from outcroppings near the northern line of Iowa. The bones of plesiosaurs are often found in deposits believed to be of deep-water origin. But they are also found in Kansas associated with the remains of small turtles, flying reptiles, and birds which could only have lived near the shores. Indeed, their remains have often been found with those of strictly fresh-water animals which had been brought down by the floods to the seas. Their wide but rather sparse distribution in all kinds of marine sediments would rather indicate that they were at home far out in the tempestuous ocean or near the shores in protected bays, though probably they preferred the shallow water littoral regions. One conclusion is quite justified: they were not gregarious, as were the ichthyosaurs.
It is not certain that the plesiosaurs were viviparous, though there are good reasons for the belief that they were. Remains of two embryos were found years ago in England associated in such a way that it is reasonable to suppose they were unhatched young, though embryos have never yet been found associated with skeletons of adults, as have those of ichthyosaurs in numerous instances. Bones of young, often quite young, plesiosaurs, are frequently found in shallow water deposits, and if the young were actually born alive they must have swum freely in the open waters while yet of very tender age. Rather singularly, however, the remains of these young plesiosaurs always occur as isolated bones.
In geological range the plesiosaurs were very persistent, extending through nearly all the Mesozoic. They began their career as fully evolved plesiosaurs, so far as we now know, near the close of the Triassic period, and reached their culmination in the Upper Cretaceous, but survived to the close of that period. In the beginning of their career they were associated with the marine crocodiles and the ichthyosaurs, but outlived them to find companions and probably enemies [Pg 95] in the huge and voracious mosasaurs of the later Cretaceous times. At no time do they appear to have been especially numerous, nor does it seem probable that they were ever a dominant type of marine vertebrate life, though their remains occur everywhere that marine deposits of the Jura and Cretaceous are known. Indeed, it may be said with almost certainty that rocks of these ages and of that character everywhere in the world contain fossil plesiosaurs. Their bones have been made known from Europe, Asia, Africa, Australia, and North and South America. From North America thirty or more species have been described from New Jersey, Alabama, Mississippi, Texas, Arkansas, Kansas, Nebraska, Colorado, New Mexico, Wyoming, North and South Dakota, California, etc.
The cause of their final extinction no one knows, nor can we conjecture much about it with assurance. That climatic conditions became unfavorable for them is highly improbable, considering their cosmopolitan habits; they were not discriminating in their environments. After successfully withstanding their fiercest foes, the ichthyosaurs, crocodiles, and mosasaurs, and large carnivorous fishes, it does not seem probable that they would succumb to lesser enemies, though it may be that they were finally attacked successfully, not in the fulness of their strength as adults, but while young, by more insidious enemies. More probably after their long life of millions of years they had grown old, as everything grows old, and had become so fixed and unplastic in their structure and habits that even slight causes were at last their undoing. When we shall have bridged over that still imperfectly known transition period between the great Age of Reptiles and the greater Age of Mammals we shall have learned more definitely some of the causes of the extraordinary revolution in vertebrate life that then occurred. The plesiosaurs went out with nearly all of their kind, the mosasaurs, the pterodactyls, the dinosaurs; and, so far as we now know, their places in the sea, land, and air were not immediately taken by any other creatures. [Pg 96]
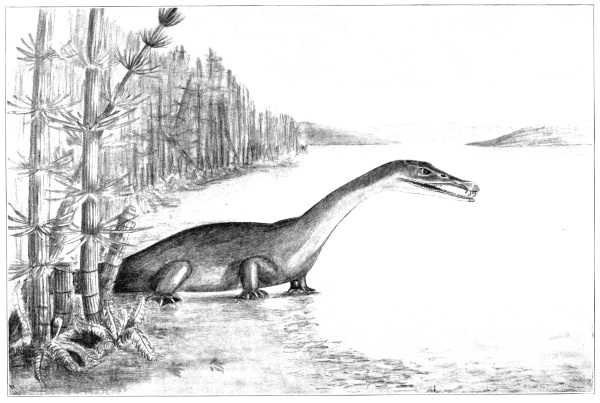
Fig. 44.—Nothosaurus;
restoration after
E. Fraas;
landscape by Dorothy Williston.
[Pg 97]
A few years after the discovery of the plesiosaurs by Conybeare, the remains of animals of allied kinds were found in the Triassic rocks of Bavaria. At first they were supposed to be those of true plesiosaurs, and even the astute Cuvier was not very clear about them. Cuvier was the first to call attention to them, expressing the opinion that some of the fossils were of previously unknown animals allied to the crocodiles, lizards, and plesiosaurs. It was von Meyer, however, who first introduced a nothosaur to the scientific world under the name Conchiosaurus. A year later Count George of Münster described other forms under the name Nothosaurus, meaning “false lizard.” Count von Münster was a most zealous collector of the fossils of the Triassic deposits of Bavaria, amassing, after thirty years of active and enthusiastic labor, a very large amount of material, which, at his death, was purchased by the King of Bavaria and placed in the hands of von Meyer for study. Von Meyer was to Germany what Owen was to England, a man of deep learning, having an extensive knowledge of comparative anatomy, and being thorough and critical in his work. His descriptions and illustrations of these rich collections made by von Münster are masterpieces of scientific thoroughness. He recognized in Nothosaurus and other allied forms from the Bavarian Triassic a distinct group of semiaquatic reptiles allied to the plesiosaurs, and his conclusions have never been gainsaid. In more recent years additional remains of these animals from Bavaria and other places in Europe have been described, but none are known from other parts of the earth, or from other than Triassic rocks. Altogether about ten genera and about twice as many species have been described, probably all belonging in one family, and all by common consent now classified with the Sauropterygia.

Fig. 45.—Head and neck of Nothosaurus;
photograph of specimen in the Senckenberg Museum,
from Dr. Dreverman.
[Pg 98]

Fig. 46.—Pectoral
girdle
of Nothosaurus, from
photograph by E. Fraas:
icl, interclavicle; cl, clavicle;
sc,
scapula; cor, coracoid.

Fig. 47.—Pelvic bones of
Nothosaurus: il, ilium;
ac, acetabulum; p, pubis;
is, ischium.
(After Andrews.)
The Nothosauria were much smaller reptiles than the plesiosaurs, none of them perhaps exceeding the size of the smallest known plesiosaurs. They were semiaquatic in habit, with many curious resemblances to other semiaquatic reptiles of a later time known, as the dolichosaurs. The neck is more or less elongated, having about twenty vertebrae in the longest-necked forms; the body is moderately long, and broad, and the tail is relatively short. The vertebrae and ribs are quite like those of the plesiosaurs, that is, the vertebrae are gently concave at each end, and the dorsal ribs are attached by a single head to the transverse process high up on the arch; the cervical ribs are double-headed, precisely like those of the older plesiosaurs, one of the characters which insistently proves the relationships of the two groups. The bones of the shoulders (Fig. 46) also have many resemblances to the extraordinary ones of the plesiosaurs, though they are much less specialized. There was no sternum; the coracoids are large, though very much smaller than those of the plesiosaurs. The collar-bones are large and strong, joining each other in front of the coracoids and firmly united with the shoulder-blades at the outer extremity. Four vertebrae are united to form a sacrum, and their union [Pg 99] with the hip bones (Fig. 47) was much firmer than was the case with the plesiosaurs. The limbs are elongated, but it will be observed in the figures (Fig. 48) that the radius and ulna, tibia and fibula, that is, the bones of the forearm and of the leg proper, are relatively very short as compared with the humerus and femur, a sure indication of the beginning of aquatic habits. The toes and fingers were doubtless webbed, and there was no increase in the numbers of bones in the digits, so conspicuous in the plesiosaurs. The external nostrils are large, but are not situated so far back near the eyes as in the plesiosaurs. There is a large pineal opening in the top of the skull, as in the plesiosaurs, but no sclerotic or bony plates have been observed in the eyes. They had ventral ribs like those of the plesiosaurs.

Fig. 48.—Legs of Lariosaurus balsami, an Upper Triassic nothosaur: h, humerus; r, radius; u, ulna; i, intermedium; ue, ulnare; f, femur; fi, fibula; t, tibia; a, astragalus; c, calcaneum. (After Abel.)
No impressions of scales or bony plates have ever been found with the remains of the nothosaurs, and it is the belief that the skin was bare. A good idea of their general appearance will be gained from the accompanying restoration adapted from that of Professor Fraas (Fig. 44) and the restoration of the less highly specialized Lariosaurus, made from a very complete skeleton in the Frankfort museum (Fig. 49).
It has been thought that these nothosaurs, so intermediate in structure between the true plesiosaurs and land reptiles, were the actual ancestors, but this is rather doubtful. It is probable that they were only very closely akin to the real ancestors, since in some ways they had become specialized too much, and, as we have already explained, highly specialized characters or organs can never go back to their earlier condition. The nothosaurs do prove beyond all possibility of doubt that the plesiosaurs were at least the descendants of animals closely allied to them, so closely, indeed, that it is doubtful whether we could distinguish external differences were all of them actually living at the present time. [Pg 100]
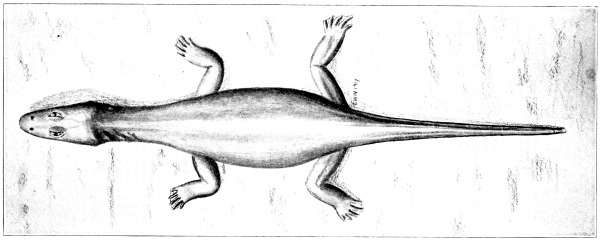
Fig. 49.—Lariosaurus balsami.
[Pg 101] We have repeatedly seen that all aquatic animals have some or all the bones of the limbs shortened, and it is of interest to observe that the early plesiosaurs had longer forearm and foreleg bones than the later ones, just as we have seen was the case with the early ichthyosaurs. It would seem probable that all the early plesiosaurs had long necks, though some of the late ones in Cretaceous times had relatively short necks, shorter even than the known nothosaurs possessed.
The nothosaurs doubtless lived about the shores of the ancient seas, spending much of their time in the water, leaving it perhaps when hard pressed by their enemies, as do some modern reptiles, or to rear their young. The teeth of the nothosaurs are long and slender in front, shorter behind. The animals must therefore have been carnivorous in habit, feeding probably upon such fishes as they could catch, and the various invertebrates which live in shallow water. The structure of the jaws and their attachments are quite as in the plesiosaurs, proving that they could not have swallowed large objects; but the skull is broader and flatter than that of most plesiosaurs, indicating habits not unlike those of the modern alligators and crocodiles.
Some time we shall doubtless find remains of nothosaurs or nearly allied animals elsewhere than in Europe, but probably not from later deposits than the Triassic. So far as we now know, their geological range and geographical distribution were much restricted; they evidently wholly died out shortly after the plesiosaurs appeared.
[Pg 102]
LYSTROSAURUS
Over a large area of South Africa, chiefly along the Orange River and its tributaries, there is an extensive series of deposits many hundreds of feet in thickness, usually called the Karoo beds, which, for more than fifty years, have been widely famous among scientific men for the many and remarkable vertebrate fossils which they have yielded. These deposits seem to represent the whole of the vast interval of time from the Carboniferous to the Jurassic, that is, the whole of the Permian and Triassic, though not many fossils have been found in the lowermost strata. Among the fossils of the lower strata are those of the strange creatures described in the following pages as Mesosaurus. From the deposits representing the Upper Permian and the Triassic the fossils that have been obtained are both abundant and diverse. Unfortunately, however, of the scores of forms that have been discovered few are known completely, and still fewer are known sufficiently well to enable us to picture the living animals.
From the Upper Permian Karoo rocks two orders of reptiles have been recognized, the Cotylosauria, represented by more specialized forms than those from the Lower Permian of North America; and the order or group called by Broom the Therapsida. While the forms of this latter group have certain definite structural relationships with each other, they show so great a diversity among themselves that, when they shall be better known, it will be found necessary perhaps to separate them into several distinct orders.
At least five groups of the Therapsida are now recognized by Broom, the Dromasauria, Dinocephalia, Anomodontia, Therocephalia, and Theriodontia. Of all these the members of the last-mentioned group have [Pg 103] attracted the greatest interest among geologists and naturalists, because of their intimate relationships to the mammals—so intimate, indeed, that they seem almost to bridge over the interval between the two classes. From higher Karoo beds primitive representatives of the more crocodilian types have been discovered, forms which seem to be the beginning of that order described on later pages as the Parasuchia.
It would lead us too far astray to mention even, let alone describe, the many forms of reptiles that have been discovered in the Karoo beds; nor indeed is it possible for anyone who has not attentively studied their remains to get a very clear conception of many of them, so incompletely have they been made known.
Doubtless from among all these diverse forms there have been not a few which sought wider opportunities in the water, but, if so, we have as yet very little knowledge of them. One form only, so far as the writer is aware, has been credited with aquatic habits, a remarkable reptile belonging to the group originally called by Sir Richard Owen, the Anomodontia, a word meaning “lawless teeth,” and to the genus Lystrosaurus, also described by the same noted paleontologist. A restoration of the skeleton of Lystrosaurus has recently been published by Watson. This restoration the writer has reproduced in the present pages, though he has taken the liberty of making some minor changes, to accord better with what he believes must have been the position of the shoulder-blades and the hind legs. And he would also suggest that the tail in life did not turn down so much at its extremity as depicted by Watson.
Both Broom and Watson believe that this animal was a powerful swimmer, and thoroughly aquatic in habit. To the present writer, however, this does not seem so evident. He is rather inclined to believe that the creature was chiefly terrestrial in habit, living probably in marshy regions, and perhaps seeking its food in shallow waters and in the mud. Aside from the position of the nostrils, which it will be observed are rather close to the eyes, a position so characteristic of many swimming reptiles and mammals, there is but little indication of aquatic adaptations elsewhere in the skeleton. [Pg 104]

Fig. 50.—Skeleton of Lystrosaurus,
as restored by Watson, slightly modified.
[Pg 105] The skull is of most extraordinary form. The face is turned downward, leaving the nostrils high up, in front of the eyes. The jaws were doubtless covered with a horny shield, like that of the turtles, having a cutting edge. There is a single pair of elongated canine teeth, possibly a sexual character. The lower jaws are heavy and stout, and Watson has said that the animal doubtless had the ability to open its mouth very widely. The quadrate, the bone with which the lower jaws articulate, is firmly fixed to the skull, and there is a single opening on the side of the skull posteriorly, a character common to all the Therapsida.
The vertebrae are stout, and they have stout spines. The tail is remarkably short, stout, and stumpy; it could have been of no use whatever in the water for propulsion or even for steering. The front legs are short and stout; the forearm bones are short, suggesting either swimming or digging habits, and the foot is short and broad. The pelvis or hip bones are massive and were very firmly connected with the backbone by the aid of six vertebrae, a very unusual number in reptiles. The hind legs, as figured, show no indications whatever of aquatic adaptation, unless possibly the very slight shortening of the shin may be so construed. Watson believes that the bones of the pelvis, indicate, aside from its strong union with the backbones, strong swimming powers, but of this again the present writer is very skeptical. The very strong ischia and the flatness of the pelvis are both characters found among American Permian reptiles, which do not show otherwise the slightest indications of water habits.
If then Lystrosaurus was a powerful swimmer, as has been maintained, it is very evident that the hind legs must have been used as the seals or sea-otters use them, to propel and to guide; but they in nowise resemble the legs of these swimming mammals. It seems altogether more reasonable to suppose that Lystrosaurus lived in the marshes, feeding upon vegetable food obtained by aid of its strong jaws and tusks—if the tusks were possessed by both sexes; and that the position of the nostrils may be ascribed to causes like those which brought about their recession in the Phytosauria, and not to strictly aquatic habits. Possibly the animal had habits somewhat similar to [Pg 106] those of the hippopotamus; that it was an expert swimmer appears, to the present writer, improbable. The powerful front legs may be indicative of digging habits; the animal may have used them as an aid to its powerful jaws and tusks in uprooting marsh and water plants. However, Lystrosaurus, whatever may have been its habits, was a curious reptile. It was about three feet in length, massive in all its structure, and doubtless of slow and sluggish gait.
[Pg 107]
Early in the eighteenth century a curious work in the Latin language was published by a famous physician and naturalist—a professor in the University of Altorf by the name of Scheuchzer—entitled Querulae Piscium, or “Complaints of the Fishes.” The work was illustrated by many expensively engraved figures of various fossil remains, including one of some vertebrae which the author referred to as “the accursed race destroyed by the flood”! The history of the finding of these famous bones is recorded by Cuvier as follows:
Scheuchzer, while walking one day with his friend Langhans in the vicinity of Altorf, a village and university of Nuremburg, went to the vicinity of the gallows to make some researches. Langhans, who had entered the inclosure of the gallows, found a piece of limestone containing eight dorsal vertebrae, of a black color and shining. Seized, says Scheuchzer, with a panic terror, Langhans threw the fragment of limestone beyond the wall of the inclosure, and Scheuchzer, picking it up, preserved two of the vertebrae which he believed to be human, and which he figured in his book, Piscium Querulae.
About the same time another observer by the name of Baier discovered other and similar vertebrae in the vicinity of Altorf which he described and figured as those of a fish; and there was much earnest contention between Scheuchzer and Baier, as also between their friends, as to their supposed nature. Scheuchzer’s figure was often cited as indubitable evidence of the destruction of mankind by a universal flood, and it was not until nearly a century later that Cuvier showed that the bones were really those of a marine reptile. [Pg 108]

Fig. 51.—Restoration of Ichthyosaurus
with young, by Charles R. Knight.
(By permission of the
American Museum
of Natural History.)
It must be recollected, in extenuation of so extraordinary a blunder on the part of so learned a man as was Scheuchzer, who, as a physician and professor, one would think ought to have been able to distinguish between vertebrae so different as are those of an ichthyosaur and a man, that, during all of the eighteenth century and well into the nineteenth, the belief was prevalent that all fossils were the relics of animals and plants that had perished in the great biblical flood. The science of geology was yet in its infancy, and there was no known record, other than the biblical one, of any great inundation of the [Pg 109] earth’s surface which might account for the remains of sea-animals in rocks remote from the seas. This belief, so long held by even the wisest and most learned of scholars, so long welcomed by the theologians as proof of the literal accuracy of the Bible, was one of which Scheuchzer was quite convinced. His Piscium Querulae was largely a fantastic discussion of the supposed great world-catastrophe, the Noachian Deluge, by which the fishes had been destroyed and long imprisoned in the rocks through no sin of their own.
It was the same author who, in a subsequent work, described and figured the fossil skeleton of a large salamander which he believed to be that of a child destroyed in the flood, and which he called “Homo diluvii testis.” In this specimen, which was discovered in the Tertiary rocks of Oeningen, and which is still preserved among the historically as well as scientifically famous fossils of the museum at Haarlem under the name Andrias Scheuchzeri, Scheuchzer thought that he detected, not only the skeleton of a child, but even its brain, liver, muscles, etc.! His engraving of this “Witness of the Flood,” the “sorrowful skeleton of an old sinner drowned in the Flood,” as also that of the ichthyosaur vertebrae of Altorf, were afterward printed in the famous “Copper Bible” as positive proof of the literal accuracy of the biblical record.
Earlier than the publication of these figures by Baier and Scheuchzer, at the very close of the seventeenth century, a Welsh naturalist by the name of Lluyd, in a large and beautifully illustrated work, figured—perhaps for the first time—remains of ichthyosaurs, which he believed to be those of fishes. But Lluyd accounted for these and all other fossil remains by a very different theory from that of Scheuchzer and the theologians—a theory which at one time had many adherents among scholars. He believed that the spawn of fishes or the eggs of other creatures had been carried up from the seas and lands in moist vapors into the clouds, whence they had descended in rain, penetrating the earth to give origin to the fossils; in other words, he believed that all fossils grew in the earth from germs of the living animals that inhabited the land and seas. Certainly the old philosophers were hard driven to make facts agree with theories! [Pg 110]
Remains of ichthyosaurs, abundant as they were and are in many deposits in England and Germany, attracted very little attention from the naturalists of the eighteenth century after the time of Scheuchzer and Baier, and nothing more was written about them until 1814, when Sir Everard Home, an English comparative anatomist, in an extensive series of large and finely illustrated, though rather discursive, works, described and figured a number of good specimens. To the animal the remains of which he rather vaguely and imperfectly described, he gave in 1819 the name Proteosaurus, in the belief that it was allied to the living Proteus, a salamander.
In 1821 the curator of mineralogy of the British Museum—Koenig by name—after a more critical study of other remains, reached the conclusion that these animals were intermediate between the fishes and the reptiles, and gave to them the generic name Ichthyosaurus, meaning fish-reptile, a name by which the chief forms have ever since been known. Within the next few years many specimens of ichthyosaurs were carefully and fully described by Conybeare, Cuvier, Owen, and others of England, France, and Germany, making very clear all the more important details of their skeletal structure. Blaineville, in 1835, thought that the ichthyosaurs constituted a distinct class of vertebrates equivalent to all other reptiles, the birds, and the mammals, which he called Ichthyosauria, the first appearance in literature of the name by which the order is properly known. Five years later, however, the famous English anatomist and paleontologist, the late Sir Richard Owen, united the ichthyosaurs with the plesiosaurs as a single order of reptiles, to which he gave the name Enaliosauria, meaning sea-reptiles, a name which has long been current in textbooks and general works on natural history. Moreover, Owen rather arbitrarily changed Blaineville’s name Ichthyosauria to Ichthyopterygia, a name which is often, though incorrectly, used to designate this order of reptiles. These briefly given and perhaps dry details will make clear how necessary is that rule of priority upon which naturalists so often insist. When anyone may change the names of organisms at will there will be no stability and no uniformity, because there is no one to [Pg 111] decide, and the prestige of a great name, like that of Owen, will carry authority till someone else with greater authority appears. Whether or not the name Proteosaurus, first given to any member of this order, should take precedence over the later Ichthyosaurus is still in doubt, since Home gave no specific name to his species, and the very particular purists of modern times have decided that a genus is not named unless the species is also! We moderns sometimes are inclined to impose very stringent conditions upon the older naturalists; let us hope that we shall be treated more leniently by the future naturalists!
It will lead us too far astray to follow in detail the history of the further discoveries of the ichthyosaurs during the early part of the nineteenth century. It may briefly be said, only, that no other group of extinct backboned animals excited more interest among scientific men. One incident will suffice. More than sixty years ago, an interesting deduction as to the living form of the ichthyosaurs was made by Sir Richard Owen. He observed that many of the known skeletons, as they were found in their rocky matrix, had a remarkable dislocation of the vertebrae at a certain place near the end of the tail, and, although such an appendage was quite unknown in other reptiles either living or extinct, concluded that the living animals had a terminal, horizontal, fleshy fin, very much like that of the whales and sirenians. Sure enough, discoveries made forty years later disclosed impressions in the rocks, not only of a large caudal fin, but also of a dorsal fin, as well as outlines of the flesh-covered paddles. The dislocation of the vertebrae at the place where the fleshy fin joined the more slender tail was due to the action of currents of water, or simple gravitation, upon a thin vertical fin and not, as Owen supposed, to the twisting of the terminal part as it fell to a horizontal position after partial decomposition of the soft parts.
About twenty-five years ago, Professor E. Fraas, the present director of the Stuttgart Museum, described and figured very fully, not only specimens showing impressions of the fins and paddles, but also others of well-preserved and very complete skeletons of different species of ichthyosaurs from the Jurassic deposits of Würtemberg, in which remains [Pg 112] of these animals occur in great profusion. His researches, and those of several authors since then, supplementing and confirming or disproving those of the many observers made during the preceding seventy years, have finally determined almost perfectly the complete structure of the more typical ichthyosaurs, enabling us to infer not a little as to their habits and distribution in the old Jurassic oceans. Within the past few years the discoveries of Professor J. C. Merriam of California have likewise added greatly to our knowledge of the earlier ichthyosaurs. It may now truthfully be said that of no group of extinct reptiles do we have a more complete and satisfactory knowledge than of the ichthyosaurs.

Fig. 52.—Ichthyosaurus quadricissus.
Photograph of specimen in Senckenberg museum,
from Dr. Dreverman.
Nevertheless we have yet very much more to learn about the order Ichthyosauria as a whole—whence they came and how they originated; what their nearest kin were among other reptiles; and especially, more about the connecting links between them and terrestrial reptiles. They have, as an order, so isolated a position, are so widely separated from all other reptiles in structure, that they have long been a puzzle to paleontologists. Like the whales and other cetaceans among mammals, we know the ichthyosaurs well in the plenitude of their power and the fulness of their development, but have yet only an imperfect knowledge of their earlier history, and none whatever of their earliest. However, as will be seen farther on, the recent discoveries by Merriam have shed much light on some of the stages of their evolution. So nearly perfectly were all the later ichthyosaurs adapted to their life in the water that it was believed by nearly all paleontologists until about a [Pg 113] score of years ago that they had descended directly from fishes. But this belief has been quite abandoned by all, not only because the recent discoveries of the earlier ichthyosaurs have demonstrated a positive increase in the aquatic adaptations of the later forms, but also because a double origin of any type of animal life is quite out of accord with all known facts and principles of paleontology. It is quite possible for animals, in becoming adapted to peculiar environmental and [Pg 114] food conditions, to acquire certain resemblances to other animals, but quite impossible for them to acquire their actual structure. The ichthyosaurs are true reptiles, and all reptiles must have had a common origin.



Fig. 53.—Baptanodon (Ophthalmosaurus). Skull from the side, from above, and from below (after Gilmore): ang, angular; bs, basisphenoid; d, dentary; fr, frontal; j, jugal; la, lacrimal; mx, maxilla; na, nasal; oc, occipital condyle; p, palatine; pa, parietal; pm, premaxilla; po, postorbital; ps, parasphenoid; pt, pterygoid; pf, prefrontal; sa, surangular; sp, splenial; sq, squamosal; st, supratemporal; q, quadrate; qj, quadratojugal.

Fig. 54.—Occiput of Baptanodon (Ophthalmosaurus): pa, parietal; soc, supraoccipital; sq, squamosal; exoc, exoccipital; op.o, paroccipital; sta, stapes; st, supratemporal; qu, quadrate; qj, quadratojugal; pt, pterygoid; bs, basisphenoid; sag, surangular; ag, angular; art, articular; pra, prearticular. (After Gilmore.)
We are sometimes in doubt, however, as to whether characters resembling those of other animals are really acquired as adaptations to peculiar environments, that is, parallel, convergent, or homo-plastic characters, or whether they are due to heredity from remote ancestors. The reptilian characters of the ichthyosaurs, however, are so emphatic that they can only be ascribed to heredity. Ichthyosaurs are as truly reptilian as crocodiles or snakes, notwithstanding their fish-like form and habits. The ichthyosaur ancestors were once truly land reptiles—of that we are as sure as we well can be. Some have thought that those ancestors were the primitive Rhynchocephalia, but most are now convinced that they were among the most primitive of reptiles, a branch probably from the cotylosaurs or cotylosaurian ancestors. Probably of all the extinct forms that we know the Proganosauria come the nearest; indeed it is not impossible that they may have been the actual forbears of the ichthyosaurs.
The ichthyosaurs varied in length from two to thirty feet, but the different species, especially all the later ones, resembled each other pretty closely in shape; the beak was more slender in some than in others, and the shapes of the fins and paddles varied not a little, as we shall see. The jaws were long and slender, provided with numerous rather small but sharp and recurved teeth, especially well fitted for [Pg 115] the seizure and retention of slippery prey. The teeth were inserted, not in separate sockets, as are those of the crocodiles and many other reptiles, but in long, deep grooves, and were easily lost, indeed so easily lost that one late American form was originally described as edentulous, and it was not till a number of years had elapsed that the teeth were found. The nostrils were small, and situated far back on the sides of the face, near the eyes. The eyes were very large, not only in proportion to the size of the skull, but, in the largest species, actually attaining in some, perhaps, the size of a human head. The eyeball was surrounded in front by an extraordinarily large and strong ring of ossifications in the sclerotic membrane, giving not only protection to the eye under the varying pressure of the water, but also greater control over vision. The neck was very short, so short, in fact, that no construction was visible in the living animal between the head and body; it was capable of only slight movement. The trunk was elongated and relatively slender, sometimes with more than fifty vertebrae in it. The tail also was long and flattened, ending in all the later species in a large fleshy fin, resembling the caudal fin of many fishes in shape and doubtless also in function. There was also a large dorsal fin, supported by hardened or calcified sinews, in shape like the dorsal fin of most fishes and many cetaceans. This character is absolutely unique among reptiles, so far as is known, and was one of the extreme specializations of water life. The hind limbs were smaller, often much smaller than the fore ones, and both were quite fin-like in life, or rather flipper-like, though not at all fin-like in structure. The skin was smooth and bare. In brief, to quote Fraas’s words:.
The general aspect of the ichthyosaurs was very dolphin-like. The body was everywhere naked and probably dark in color. The head was produced in front into a long, slender snout, and was closely joined to the body posteriorly without indications of a neck. The body itself was cylindrical, expanded in front by the large thorax and abdomen, but rapidly diminishing into the long, slender, and strong tail. Close behind the head were the front paddles, which in some species were broad and shovel-like, in others elongated and pointed. The hind paddles were smaller than the front ones, sometimes greatly reduced in size, their function replaced by that of the very broad tail.
From the foregoing descriptions and the restoration shown in Fig. 51, [Pg 116] we see how very fish-like, or rather dolphin-like, these animals were in the external form—so fish-like that the name Ichthyosaurus is not misleading, though Koenig gave it in the mistaken belief that they were really allied to the fishes. When to these external features certain other fish-like details of the skeleton are added, we do not wonder that the early observers were so long in doubt about them. A more careful examination of the skeleton will, however, disclose so many truly reptilian characters that their external appearance and habits lose all significance.
The vertebrae are deeply biconcave and fish-like, it is true, but a consideration of the reasons therefor will convince us that any other kind of vertebrae would be more remarkable. At the time when the ichthyosaurs must have originated, at the time when the first known ichthyosaurs appeared in geological history indeed, all reptiles had biconcave vertebrae, and for the most part at least deeply biconcave ones. The vertebrae remained fish-like throughout all their history, perpetuating their type until most other reptiles had developed a firmer one, because such vertebrae were best adapted for the quick, pliant movements of the spinal column so necessary for the well-being of the animals in the water. In the modern dolphins, animals in shape, size, and habits most wonderfully allied to what these old reptiles must have been, the small, flat-ended vertebrae are widely separated by disks of flexible cartilage.
Not only were their vertebrae fish-like in form, but there are other characters in the spinal column of a primitive or generalized nature. As in all aquatic animals, the articulating processes between the vertebrae are either weak or wanting in the posterior part of the column. And they were not only small, but were situated, in many, high up, very remarkably resembling the peculiar arrangement of the articulations in the dolphins.
There is no sacrum, that is, there were no united vertebrae posteriorly for the attachment and support of the pelvis, as no such support was needed. In only one other group of aquatic reptiles was the sacrum lost, though it has wholly disappeared in the cetaceans and sirenians among mammals. The chevron bones of the tail, usually bony arches on the under side of the tail for the protection of the blood-vessels, in crawling reptiles, were very imperfectly developed in the later forms, though normal in shape in the early ones. The ribs are numerous, long, [Pg 117] and slender, very much resembling those of the fish-eating dolphins. They usually had, however, two attachments to the body of the vertebra and none to the arch, differing in this respect from all other animals.
Of the shoulder bones, the scapula or shoulder-blade, as usual among water animals, is short and broad. In the place of a sternum the coracoids joined each other broadly in the middle, just as they did in the oldest known land reptiles. And there were clavicles and an interclavicle. Below the abdomen behind were numerous slender bones called ventral ribs. The pelvis is very weak, and was suspended below the spinal column in the fleshy walls of the abdomen. The hind legs were so small that little support was necessary for them, and, because they were not used either for the support of the body or for propulsion, they did not require a firm union with the skeleton. Doubtless had the ichthyosaurs continued to the present time, they would have lost entirely the hind legs, as have the cetaceans.
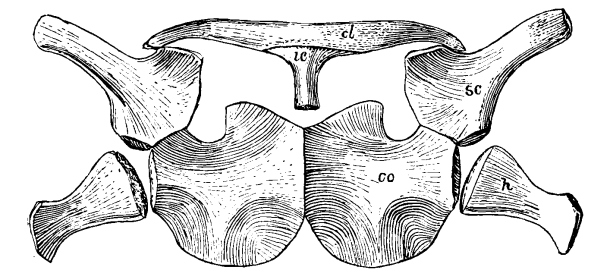
Fig. 55.—Pectoral girdle of Baptanodon (Ophthalmosaurus), an American Upper Jurassic ichthyosaur. (After Gilmore.)
It is in the limbs that most extraordinary differences from all other animals are seen. So great are these differences that it has been a puzzle to naturalists to understand how they could have arisen. In no other animals above the fishes, that is, in no other reptiles, in no amphibians, birds, or mammals, are there ever more than five fingers or five toes, the number with which air-breathing animals began. Fingers and toes may be lost and often are lost in all groups of life, until a single one in each limb may remain, as in the domestic horse. An increase of fingers and toes, however, seems to be an impossibility in [Pg 118] evolution, and doubtless of real fingers and toes it is an equal impossibility. All naturalists are now agreed that a specialized character can never revert to a generalized condition, or rather to a generalized structure, that an organ once functionally lost can never be regained by descendants. A character once lost is lost forever; horses of the future can never have more than one finger or one toe in each limb.

Fig. 56.—Front paddle
of Ophthalmosaurus
(after Andrews):
h, humerus; r, radius; u,
ulna; p, pisiform; re, radiale;
int, intermedium; ue, ulnare.
Fig. 57.—Front
paddle of Merriamia,
a Triassic ichthyosaur.
(After Merriam.)
Explanations as in
Fig. 56.
And there was an increase in the ichthyosaurs, in some not only of the number of digits in each limb, but in all of the number of bones in each digit, a character found also in the unrelated mosasaurs and plesiosaurs. This increase in finger and toe bones, or hyperphalangy as it is called, is one of the most peculiar of all the adaptations to water life, changing the feet and hands from the ordinary walking type to the fish-like swimming type. The bones beyond the humerus and femur [Pg 119] in the ichthyosaurs were so increased in number and so changed in form and relations that they bear little resemblance to the corresponding bones of other reptiles. They are merely polygonal platelets of bone, articulating on all sides and fitting closely together, permitting flexibility, but not much else.
It is now believed that the increase, not only of additional digits, sometimes to as many as ten in each hand and foot, but of the finger and toe bones as well, was the result of a sort of vegetative reproduction. The margins and ends of the flippers were doubtless hardened by cartilage or fibrous material, and because of the action of the limbs this cartilagenous material broke up into nodules each of which took on ossification finally. Among the whales, where hyperphalangy also occurs, though to a less extent, it has been thought that the increase in number has been due simply to the ossification of the parts of each bone normally present, that is, to the epiphyses, which became separated from the shaft of each bone. But this explanation will hardly suffice for the fingers and toes of the plesiosaurs and ichthyosaurs, for there are altogether too many such ossifications; and besides, the bones in these animals, as in most reptiles, did not have epiphyses, or terminal separate ossifications of the bones of the skeleton.
It will be observed from the figures that the arm and thigh bones of Ichthyosaurus are very much shortened—a striking adaptation to water life, so conspicuously seen in the modern whales and dolphins as well as in the mosasaurs, thalattosaurs, etc. So characteristic indeed is this shortening that, were every other bone of the skeleton of an ichthyosaur unknown save the humerus or femur, it would be quite certain from these alone that the animal was thoroughly aquatic in habit.
About sixty years ago a rather aberrant form of ichthyosaur, now known as Mixosaurus, was discovered in rocks of Triassic age, that is, of much greater age than any ichthyosaurs previously found, in which not only the forearm but also the lower leg bones were longer, resembling more the corresponding bones of land animals. It was from the examination of specimens in 1887 of these mixosaurs that the late Professor Baur became convinced that the ichthyosaurs were the [Pg 120] descendants of land reptiles, and not directly of the fishes as they were universally thought to have been at that time. As Professor Baur very pertinently said, if the ichthyosaurs were descended from the fishes directly, the earliest forms should be more nearly like the fishes than the later ones, whereas just the opposite was the real fact. The arguments which he gave in support of his contention were so convincing that they found immediate acceptance among all naturalists. Fortunately within the past fifteen years many other remains of early ichthyosaurs from the Triassic rocks of California have been brought to light by Professor Merriam, remains which throw a flood of light upon the early, though not the earliest, history of these strange reptiles. He has recognized among the forms he has discovered, not only new species, but several new genera, and perhaps new families of ichthyosaurs. His studies have demonstrated so well the stages of evolution between the early ichthyosaurs and the later ones in their progressive adaptation to water life that it will be of interest to summarize them here.
In the early ichthyosaurs locomotion was largely by the aid of the limbs; in the later ones almost exclusively by the aid of the caudal fin. In the former the paddles were larger and the bones longer, more like those of land animals; in the latter they were relatively smaller and shorter, and more fin-like. In the digits of the early forms the finger and toe bones were more elongated and fewer in number. The hind limbs were nearly as large as the front ones in the Triassic, often very much smaller in the later ichthyosaurs; and the increased number of digits occurs only in the later forms.
In the Triassic ichthyosaurs, all classed in the family Mixosauridae, the pelvis was larger and more firmly connected with the body than in the later forms.
The skull of the early forms was relatively shorter, as compared with the trunk, the jaws shorter as compared with the head, the eyes were relatively small, the teeth in some less numerous, and set in distinct sockets like those of land reptiles; the vertebrae were relatively longer and less fish-like, and their articulations more like those of land reptiles.
The distal part of the tail was not bent downward so sharply, that is, [Pg 121] the terminal fin was smaller, or the tail may have been simply flattened near its end and not really fin-like. The scapula was longer and less fan-like in shape.
And all these are remarkable evidences of an increased adaptation to water life in the more recent ichthyosaurs over the older ones. Were someone now so fortunate as to find ichthyosaurs in late Permian rocks, we should doubtless have the nearly complete chain between the most highly specialized type of water reptiles and their terrestrial ancestors.

Fig. 58.—Caudal fin of Ichthyosaurus, after Baur (left figure); caudal fin of Mixosaurus, after Wiman (right figure).
From the structure of the skeleton alone the early observers were justified in inferring much concerning the shape and habits of the living ichthyosaurs. Later discoveries have added so many definite facts that, at the present time, we know more about their habits than we do of any other extinct reptiles. In various places in England and Germany, especially in Würtemberg, the remains of ichthyosaurs are found in extraordinary abundance and perfection, not only whole skeletons lying in the positions which they had assumed after the decomposition of their bodies, but also often the actual remains, carbonized, of the skin, muscles, and ligaments, as well as delicate impressions of external parts. Many of these skeletons are obtained from the numerous stone quarries, where they are a sort of “by-product,” the sums received for them adding not a little to the income of the quarrymen. So many are obtained in this and other ways [Pg 122] that specimens of ichthyosaurs are perhaps more frequently seen in the museums of the world than those of any other extinct backboned animal. Fairly complete skeletons may now be purchased of dealers in such things for from fifty to seventy-five dollars. As may be supposed, the best and most complete collections of these fossil remains are those of the British Museum in London and the museum in Stuttgart. From a study of those of the last-mentioned museum Professor Fraas has learned many interesting facts and reached many interesting conclusions regarding the life-habits of the ichthyosaurs. In the accompanying figure (Fig. 59) is shown a photographic reproduction of a very complete specimen, in which not only is the outline of the whole body shown, but also much of the carbonized remains of the muscles and skin has been detected.

Fig. 59.—Ichthyosaurus quadricissus. (From a photograph from B. Hauff, Paleontologisches Atelier, Holzmaden.)
The attachment of the paddles to the body was broad antero-posteriorly, proving conclusively that they could not have been much used in propulsion, either in the water or upon land, since such use would require a fore-and-aft movement, and a consequent twisting or rotation of the whole arm or leg, which, because of the broad attachment, must have been very difficult, if not impossible.
Microscopic examination of the remains of skin preserved disclosed an abundance of dark pigment, indicating, Professor Fraas believes, that the skin was dark colored above. Doubtless, also, the under side, as in nearly all swimming animals of the present time, was of a lighter color, because such coloration rendered the animals much less conspicuous in the water when seen either from above or below. That the [Pg 123] skin was bare is proved by many impressions or molds of it that have been discovered in the rocks, in which many fine creases are seen, but nothing suggesting scales or bony plates, save on the front edge of the paddles, where impressions of overlapping scales have been observed. This is an interesting fact, bearing witness that their land ancestors had been covered everywhere with scales, much like those of existing lizards and other reptiles. Scales or bony plates were not only useless to the ichthyosaurs in the water, since they could afford no protection, but would have been detrimental in increasing the resistance in swimming.
That the ichthyosaurs were predaceous animals is of course evident from their teeth, adapted for the seizure and retention of slippery prey, but not for tearing or comminuting. The fossilized remains of food found between the ribs of some specimens, in the place where the stomach was, together with fossil excrement, called coprolites, usually attributed to these animals, prove that they fed largely upon fishes, squids, belemnites, and probably other invertebrates. One ichthyosaur specimen preserved in the Stuttgart Museum has preserved in its stomach contents a mass composed of the remains of more than two hundred belemnites.
Most interesting of all is the fact that, not very rarely, embryonic skeletons of ichthyosaurs have been found associated with the remains of adult animals, in such positions that they must have been inclosed within the body cavity at the death of the animals. As many as seven such embryonic skeletons have been observed with a single specimen. At first it was supposed that these skeletons were of small ichthyosaurs which had been swallowed whole as food, since it is not at all likely that these predaceous reptiles were discriminative in their choice of food when hungry. It is not improbable that in some cases this is the true explanation of the smaller skeletons within the larger ones, but it cannot be true of all, since wherever the small skeletons are identifiable they have been found to belong to the same species as the adult, and it would be absurd to suppose an ichthyosaur bent upon its prey would be at all likely to select as many as seven young animals, all of the same size and all of its own species. Furthermore, some of these young skeletons have been found in such positions as would indicate that they were inclosed within their egg-covering at the time [Pg 124] of their death. Some of these embryos measure as much as twenty inches in length.
Because the ichthyosaurs were born alive, and because so many of their skeletons are found with their various parts in orderly relation to each other, it is inferred with much probability that they were inhabitants, in large part at least, if not exclusively, of the open and deeper oceans. Had they been oviparous they must necessarily have laid their eggs upon the beaches, since no reptiles of the present time lay eggs in the water, and we have no other indications that the reptiles of the past have ever done so. And such habits would necessitate the periodical return to land. Had they been denizens of shallow waters, like the mosasaurs and plesiosaurs for the most part, their skeletons must surely have been disturbed by the currents and tides, as also by predaceous fishes, breaking up or displacing them or carrying away their bones. In shallow waters, also, the decomposing bodies would have been more liable to despoliation by the many scavengers of the seas.
The ichthyosaurs must have been quite helpless upon land, their limbs being of little more use for locomotion than are the fins of fishes. Breathing air as they did, they were of course not suffocated when exposed, unless, as is the case with the whales, the feeble attachment of the ribs prevented the action of the respiratory muscles. If accidentally thrown upon the beaches, they doubtless were able to return to their home element more easily than the fishes can, by flopping, wriggling, and turning. As we have seen, the food consisted in part, perhaps the larger part, of small invertebrates, and because the bones of the lower jaws were closely united, permitting little or none of that expansion so characteristic of the snakes, all their prey must have been of relatively small size. In habit the ichthyosaurs were doubtless, like the dolphins and gavials, inoffensive and harmless, so far as animals of larger size were concerned. The abundance of their remains often found in restricted localities, while deposits of like age and character not far distant may be almost free from them, suggests that in all probability the ichthyosaurs, or the later ones at least, were more or less gregarious in habit as are the sea-mammals. [Pg 125] They probably lived in schools, as do the porpoises, each species keeping to its restricted locality and not wandering far.
The ichthyosaurs began their existence, so far as we now know, about the middle of Triassic times and continued to near the middle of Upper Cretaceous, when they disappeared forever from geological history. As we have seen, however, the earliest forms that we know were true ichthyosaurs in all respects, though more primitive than the later ones, indicating a long previous existence of which we yet have no knowledge. Their remains have been found widely distributed in Triassic rocks of Europe, Spitsbergen, Australia, and North America. During the Jurassic period they lived in great numbers and variety throughout the region that is now Europe. In North America the only marine rocks of this period that we know of have yielded numerous remains. These American ichthyosaurs were, however, among the most specialized of all ichthyosaurs—the culmination of their development. They were originally named Sauranodon in the belief that they were toothless, but in recent years their teeth, small and numerous, have been discovered. And the genus seems also to be identical with one previously named from the Jurassic of Europe called Ophthalmosaurus. The last known remains of ichthyosaurs have recently been found in the Benton Cretaceous of Wyoming. Scanty remains of ichthyosaurs are also known from Australia and New Zealand. Why the ichthyosaurs should have gone out of existence before the plesiosaurs and mosasaurs did, one cannot say; possibly their stock had grown old and feeble.
[Pg 126]
MESOSAURUS
There is some doubt whether those little creatures of Paleozoic times, to which some years ago the late Professor Baur gave the ordinal name Proganosauria, are really entitled to so much distinction among reptiles. The question of their rank has been much disputed for the past twenty years without any positive conclusion. Nor were they wholly aquatic in habit, though they did possess many aquatic adaptations. That they were skilful and fleet swimmers, and capable of rapid evolutions in the water is quite certain, and, as the oldest known water reptiles, they are of more than passing interest.

Fig. 60.—Mesosaurus; life restoration,
after McGregor,
the posture of hind leg slightly modified.
But two genera and three or four species of the group are known, and of them, even, our knowledge in some respects is not as complete as one could desire. The first description of any member of the group was by the late Professor Gervais of Paris in 1867. He had only the anterior part of a single skeleton, from the Karoo beds of South Africa, to which he gave the name Mesosaurus, a rather meaningless term signifying “middle” or “intermediate” saurian. Nothing more was learned about any form till 1885, when the late Professor Cope described a specimen from the supposed Carboniferous of Brazil, which he believed to be closely related to Mesosaurus, though he had only a very imperfect specimen. He called it Stereosternum, also a meaningless term, since none of the animals has a “solid sternum,” nor any sternum at all, in fact! A few years later, in 1888 and 1892, the late Professor Seeley of England studied a number of specimens of Mesosaurus, adding not a little to our knowledge of the animals. More recently Dr. Woodward of England and Professor Osborn of America have given us still further information concerning them, and within the past few years Dr. McGregor of Columbia University has figured and described excellent specimens of a new species from Brazil, which he calls Mesosaurus brasiliensis. Not only were Dr. McGregor’s [Pg 127] discoveries of great interest as settling many doubtful points in their structure, but they were still more so from the fact that he found his species so nearly like that from Africa that he placed it in the same genus. Since the proganosaurs were purely fresh-water or terrestrial animals, one can only wonder how they crossed from Africa to America, [Pg 128] or, what is more probable, how they migrated from America to Africa, across the broad Atlantic Ocean, so long ago. The geologists tell us that the Atlantic and Pacific, in the main, have always been oceans since the beginning of terrestrial life upon the earth. Possibly the tribe of proganosaurs migrated by the very circuitous route of Europe and North America, or Asia and the Northwest; but that is very improbable, since nothing whatever resembling them has ever been found in the Northern Hemisphere, and it is quite certain that in the many thousands of years it must have taken them to travel from southern Africa to South America many of the reptiles must have perished on the way and left their remains in the rocks. The only conclusion that seems probable is that there was a direct land communication in those olden times between Africa, or at least India, and South America across what is now the Atlantic Ocean. Of course this route will be very difficult to prove, since we can never get to the bottom of the ocean to hunt for fossil proganosaurs. Were this peculiar distribution of the [Pg 129] proganosaurs an isolated example, one might perhaps ascribe our lack of knowledge of any fossil proganosaurs in the Northern Hemisphere to the meagerness of the fossil records, but there are many other examples of similar import among other early animals.

Fig. 61.—Mesosaurus; restoration of
skeleton.
(After McGregor)
The age of the South American proganosaurs is now believed to be lower or lowermost Permian, like that of the African Mesosaurus; possibly, however, the age first described to Stereosternum (Carboniferous) may be correct.
The known skeletons are all small, none exceeding a few feet in length. The skull, as shown in the figure by Dr. McGregor, is elongate, and its teeth are extraordinarily so, and very slender. The external nostrils are situated close to the eyes; and no sclerotic bones have been discovered. There are small teeth in the bones of the palate. The neck is elongate, composed of ten or twelve vertebrae. The trunk also is long and slender, and the tail is not only long, but also much flattened or compressed. All these are very characteristic of water life. The limbs, however, show a much less complete adaptation for swimming—not much more so in fact than do those of the living Crocodilia. The upper arm and the thigh bones are relatively long, while those of the forearm and the leg are shorter than among terrestrial reptiles, the first indication of swimming habits to appear in crawling animals. The digits are not much elongated, and they have no additional finger bones, save perhaps in a lately discovered form in Africa, in which Dr. Broom reports supernumerary bones in the fifth or “little” toe.[3] The fingers and toes have only blunt terminal bones, that is, they were not distinctly clawed, and they were probably connected with each other by a membrane, as in a frog’s foot. This webbing of the feet is probable, not only because of the positions in which the bones have been found, but also because of the great length of the “little” toe, which is the longest in the foot, a character quite abnormal for a land reptile and quite characteristic of certain aquatic mammals, like the seals and sea-otters. There is a strong sacrum of two vertebrae, however, the pelvis and hind legs being connected with the spinal column firmly, clearly proving that, like the crocodiles, the proganosaurs had by no means lost their land proclivities. [Pg 130]
Their vertebrae, as would be expected in such old reptiles, are quite primitive in structure, that is, they are deeply concave in each end, probably being perforated for the remains of the notochord. The pelvis also is of the old-fashioned type, that is, without an opening or vacuity between the bones below. The shoulder bones are old fashioned too. The shoulder-blade, especially, shows a decided adaptation to water life in its short, fan-like shape, very much like those of the mosasaurs, ichthyosaurs, whales, etc. Just why swimming animals should have short and broad shoulder-blades has not yet been explained, but doubtless they afforded better attachment for those muscles used more especially in swimming. The ribs are remarkably flat and heavy, and were not very firmly attached to the vertebrae. Heavy ribs are unusual among free swimming animals, but do occur in the modern sirenians, which live on the bottoms of shallow bays, etc., feeding upon plants. We may perhaps infer from this peculiar structure of the ribs that the proganosaurs lived more on the bottoms of shallow waters, feeding upon such fishes or invertebrates as they could capture, coming to the surface to breathe from time to time. Possibly they sought the shores for safety from their enemies, as do the Galapagos lizards, figured on p. 142; and doubtless they laid and hatched their eggs on land. A character which suggests that the proganosaurs lived only in the shallow waters is the elongated neck, reminding one of those two other groups of swimming reptiles, the dolichosaur lizards and the nothosaurs of the Sauropterygia, the only known reptiles besides the plesiosaurs having an abnormal number of neck bones. Still more suggestive of shallow, fresh-water habits is the absence of eye bones, as in the modern crocodiles.
The long snout, with the long and slender teeth, and the position of the external nostrils far back near the eyes, together with the flattened and long tail and the webbed feet, are sufficient proof of expert swimming habits. The legs still functioned more or less for the support and propulsion of the body on the land, and they probably were only of slight service in the water. The alligator swims sinuously with [Pg 131] its front legs collapsed and extended by the side of the body; its hind legs are used more as propellers, with the knee flexed and the feet turned outward and expanded. The legs of the proganosaurs doubtless were used in the same way, as shown in the restoration, which has been modified from the original of Dr. McGregor in accordance with this probable use of the legs.
There seems to be an incongruity between the posterior nostrils and the heavy flat ribs, the former suggesting free swimming and diving habits, the latter shallow water and bottom habits. Possibly the position of the nostrils has been the result of the great elongation of the face in front of the nostrils; and we know that their posterior position in the phytosaurs (Figs. 95 and 96) has not been due to swimming habits only.
Nothing has been discovered to indicate the nature of the external covering of the body. Possibly, even probably, the skin was more or less covered by horny scales or plates, though it may have been quite bare, as in the salamanders.
To which other reptiles the proganosaurs are nearest related has long been a subject of dispute, and still is. The more probable view, however, is that they were a very early branch of the most primitive stock of reptiles, the Cotylosauria, one that soon perished, leaving no descendants, unless possibly the ichthyosaurs were their progeny. Some writers have thought that they were the early ancestral stock of the plesiosaurs, and they are often classified with the Sauropterygia. Still others have believed that they were an early side-branch of the great group of Rhynchocephalia. And this doubt has been chiefly due to our imperfect knowledge of the bones of the cranium. As has been explained, very much stress in the classification of reptiles has been laid by students on the possession of one, two, or no openings on the side of the skull back of the eyes. And this part of the skull of the Proganosauria has not yet been satisfactorily made out. Dr. McGregor thought that there are two openings in the temporal region, allying the group with the Rhynchocephalia. Dr. Huene is more positive that there is but one, like that of the ichthyosaurs. In this state of indecision, the proganosaurs may be dignified by giving them an ordinal position by themselves.
[Pg 132]
The genus Protorosaurus is of peculiar interest, as one of the first, if not the first, known fossil reptiles, described by Spener as long ago as 1710 as a crocodile, from fragmentary remains found in 1706 in the Permian deposits of Thuringia. Numerous other skeletons or parts of skeletons attracted the attention of naturalists of the eighteenth century, but were very imperfectly described. No name was given to the animal represented by the various specimens until 1840, when Herman von Meyer restudied all the known material and described it under the name Protorosaurus speneri. The position of the genus among reptiles always has been and yet is uncertain, for the reason that the structure of the skull, and especially the structure of the temporal region, has never been satisfactorily determined. Seeley, in 1887, described more fully the original specimen of Spener, now preserved in the museum of the College of Surgeons of London, and because of certain peculiarities which it showed proposed for its reception the order Protorosauria. He thought that he detected an upper temporal vacuity, like that of lizards, but was very uncertain about the structure of the lower part of the temporal region. The writer, who has examined this type specimen, must admit that the structure of the region here is very doubtful. Under the general assumption, however, that all old reptiles must be related to Sphenodon, the Protorosauria have generally been classified as a suborder of the Rhynchocephalia. It is merely another instance of the proclivity we all have to propose hypotheses, and then, speedily forgetting that they are hypotheses, to accept them as facts.
Protorosaurus was long supposed to be an aquatic reptile, but we now know that it was a strictly terrestrial one, probably with climbing habits; and the genus concerns us only by reason of its possible relationships to distinctly aquatic reptiles of a later age. [Pg 133]

Fig. 62.—Life restoration of Araeoscelis.
A few years ago the writer described a very slender, lizard-like reptile about two feet in length from the Permian of Texas under the name Araeoscelis, so named because of its slender legs. The structure of both the skull and the skeleton of this reptile is now quite satisfactorily known, so well known indeed that the accompanying restoration (Fig. 62) has little that is conjectural about it, at least so far as the form is concerned. The skull has a single, upper temporal opening, quite like that of lizards, but the quadrate is not loose below. And this is really what we should expect in the ancestral lizards; and everything else of the skeleton, except perhaps one character, is what would be expected. That one character is the elongation of the cervical vertebrae, which are about twice the length of the dorsal vertebrae following them. The cervical ribs are very slender bones, articulating by a single head with the centrum only. In these and other characters, so far as they are known, Araeoscelis seems to agree with Protorosaurus, and both have very hollow bones. [Pg 134]
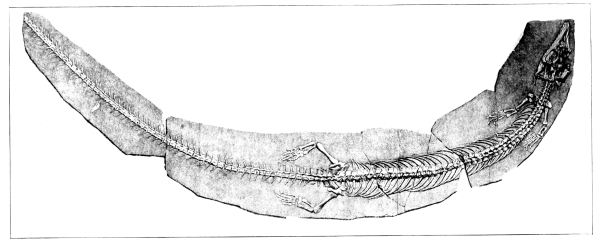
Fig. 63.—Skeleton of Pleurosaurus.
(After Lortet)
[Pg 135]
We may for the present be justified in maintaining the order Protorosauria for those reptiles having a single, typically upper temporal opening on each side, with a fixed quadrate, not including the ichthyosaurs. It is not improbable, however, that when more is known of the ancestors of the lizards, the whole group will find its most natural place among the Squamata. This definition will include a peculiar aquatic reptile that has been known for many years, but which has been wrongly classed in the same family as Sphenodon, on the purely gratuitous assumption that it has two temporal openings on each side; we now know that it has but one. This reptile, known scientifically as Pleurosaurus, was described originally by H. von Meyer in 1843, but we are indebted to M. Lortet for a more precise knowledge of the animal, and for the figure (Fig. 63) which is here given of the skeleton. Not a few excellent skeletons are preserved in the museums at Lyons and Munich. The specimen here figured, as actually preserved, measures about three feet in length; a part of the tail is missing, which is known from other specimens to have been remarkably long.
The figures show clearly some of the remarkable aquatic adaptations of the animal, especially the short neck, the very long and narrow body, and the extraordinarily long and flattened tail. The head is elongate triangular in shape, resembling very much that of the mosasaurs; and the external nostrils are likewise situated remotely from the end of the snout, as in the mosasaurs. The extremity of the snout has a beak-like projection. The teeth are much longer, more pointed, and more recurved than is the case with most land reptiles, indicating their use for the capture and retention of slippery, quick-moving prey.
The single-headed ribs are short, proving that the body was slender and [Pg 136] doubtless cylindrical, more like that of a snake. The tail was not only enormously elongated, but it was also compressed into a flat and effective propelling organ in the water. This flattening of the tail is apparent from the skeleton, with its elongated chevrons below and spines above, and it is also proved by the fortunate preservation of the extremity of the tail of one specimen, showing not only the impressions of the scales in the matrix, but also the outlines that the soft parts had in life. To quote from Lortet, in translation: “The tail was covered wholly with small scales, regularly hexagonal in shape, shining and nacreous, larger on the under side than above. The upper border of the tail was surmounted by a broad crest, extending to its extremity, and composed of large, oval scales.” The body doubtless was wholly covered with scales, though it is not probable that the caudal crest continued along the back.
Fig. 64.—Life restoration of Pleurosaurus.
The limbs begin to show an aquatic adaptation, though not very pronounced. They are much shorter and smaller than are those of land-crawling reptiles; and the bones of the second series, that is, the radius and ulna, tibia and fibula, are relatively short, the beginning of adaptation to water habits. It is very probable that the feet were webbed, though the fifth digit, as usual, is shorter than the [Pg 137] fourth. Doubtless on land the creature moved about in a serpentine way, for it could not have progressed very rapidly by the aid of its legs alone. The hind legs are longer than the front legs, and they were connected firmly with the body by means of a sacrum. The number of vertebrae in the neck is only five. The number of dorsal vertebrae is forty-three, a larger number than is known in any other air-breathing vertebrate with legs.
Upon the whole, these lizard-like, almost snake-like pleurosaurians present some very curious adaptations to water life. In water they were doubtless speedy, swimming in serpentine undulations, with the small legs for the most part folded against the body and only of occasional use. Doubtless, too, had the pleurosaurs lived longer in geological history, they would have become quite snake-or eel-like, just as have some modern salamanders.
In all probability the pleurosaurs lived habitually in fresh-water, perhaps visiting the shores for refuge, or for the hatching of their young. That they were not on the way toward a terrestrial snake-like body is evident from the flattened tail, and especially the crest of scales above; the tail was like that of the sea-snakes of the present time. Pleurosaurus, then, affords the solitary instance among reptiles of aquatic adaptation by the diminution of both front and hind extremities and the acquisition of a snake-like body and snake-like habits.
[Pg 138]
The order Squamata, so called because of the dermal covering of overlapping horny scales, comprises the great majority of living reptiles. Although the scaly covering is characteristic of nearly all the members of the order, the most essential differences distinguishing them from other reptiles are, as usual, found in the skeleton, and especially in the skull. The quadrate bone, that to which the lower jaw is articulated on each side, is not wedged in immovably between other bones of the skull, as in all other reptiles, but is, instead, freely articulated with the cranium in such a way that its lower end moves both backward and forward, as well as inward and outward. This freedom of movement has in the past been thought to be due to the loss of a lower temporal arch, a bony bar connecting the lower end of the quadrate with the hind end of the upper jaw, which is very characteristic, for instance, of the Rhynchocephalia. Indeed, because of the many primitive characters which the lizards possess, it has generally been supposed that the order was an early branch of the rhynchocephalian stem. But we are now quite sure that the lizards are as primitive as the Rhynchocephalia, and that their origin, as an independent branch of the reptilian stem, goes quite as far if not farther back—quite sure that the ancestors of the lizards never had a lower temporal arcade and two temporal vacuities, but that the looseness of the quadrate bone has been due to the gradual loss of a bone which covered the whole side of the skull until only the upper part of it was left. In other words, the ancestral skull of the Squamata must have been like that of Araeoscelis, more fully described under the Protorosauria, a group than which there is perhaps none more closely allied to the Squamata.
The bones of the roof of the mouth of the Squamata—that is, of the [Pg 139] palate—are narrow and long, and are not closely articulated, as in most other reptiles; they often bear teeth, a primitive character. The teeth of all living lizards and snakes are not inserted in sockets, as are those of the crocodiles, but are co-ossified to the margins or sides of the jaws or the bones of the palate. But this is probably not a primitive character; doubtless the teeth of the early lizards were inserted in sockets like those of most other reptiles. The shoulder bones are absent in many and vestigial in some others. When present and fully developed, they comprise the shoulder-blades or scapulae, a single coracoid on each side, the clavicles, and an interclavicle. The vertebrae, except in some lizards, are procoelous, that is, with the body concave in front and convex behind, a peculiar structure that was developed only in crawling animals. In addition to the usual articulations for the union of the vertebrae there are also, in some of the lizards and mosasaurs and all of the snakes, additional ones called the zygosphene and zygantrum, which will be best understood by reference to Fig. 12, p. 28. But little less characteristic than the loose articulation of the lower jaws, so unique in this order of reptiles, is the manner of attachment of the ribs. They are always single-headed, articulating only with the body or lower part of the vertebra. The single-headed ribs of the plesiosaurs articulate with a projection on each side of the arch of the vertebra; those of the turtles to the space between the adjacent vertebrae; nearly all other reptiles have double-headed ribs, articulating in various ways. This character, it is seen, though apparently a simple one, immediately distinguishes a lizard or a snake from all other animals, except the thalattosaurs and protorosaurs.
There is much difference of opinion among naturalists as to the proper classification of the different groups of this order of reptiles. Usually it is divided into four suborders, the Lacertilia or lizards; the Dolichosauria or long-necked lizards of the past; the Mosasauria, or extinct swimming lizards; and the Serpentes or Ophidia, the snakes. It matters very little which classification one accepts so long as it is remembered that the first three groups are closely related to each other. [Pg 140]
Popularly a lizard is any four-legged reptile covered with scales, but such a definition is not strictly correct, since some lizards are legless and some other four-legged reptiles are covered with horny scales, notably the tuatera or Sphenodon of New Zealand, a reptile long classed with lizards, but now known to belong to quite a different order. Bearing in mind those characters given as characters of the order, it will be necessary to mention only those distinguishing the lizards from the snakes.

Fig. 65.—Iguana.
(By permission
of the New York Zoölogical Society)
It is true that the great majority of lizards have four legs, while the snakes are always functionally legless, but there are some lizards, like the glass snakes and the amphisbaenas, or slow lizards, which are quite legless and there are some snakes which have small but functionless hind legs. As usual, more important differences are found in the skull. The brain-case in all snakes is surrounded on all sides by bone, for the better protection of the brain, with the head resting quite prone on the ground. The brain of the lizards, for the most part, is protected on the sides and in front by a simple membrane. Nearly all lizards have movable eyelids, while snakes do not; snakes have a single lung, and a protrusible tongue, which very few lizards possess; and the lower jaws in front are united in the snakes by a ligament only. Notwithstanding these differences, the snakes and lizards are closely [Pg 141] related animals, and must have come from a common ancestry; among all reptiles the known geological history of the snakes is shortest.
Lizards, on the other hand, have a very high antiquity, beginning, as we now know, at least as long ago as early Triassic times. They still have many primitive characters in their structure and are the least advanced type of reptiles now living, with the exception of the tuatera. Their remains are seldom found in the rocks, probably because they have always been so strictly terrestrial in habit, for the most part seldom frequenting even the vicinity of the water. The true lizards now living number about eighteen hundred kinds, classified into about twenty families, divided among four chief groups, of which the chameleons, the amphisbaenas, our common lizards, and the monitors are representatives.
Most living lizards are inhabitants of warm climates, though some extend rather far north in the temperate zone. With the exception of New Zealand, and the polar and subpolar regions, lizards are found in all parts of the world. The great majority live only in high and dry places, though some are denizens of low and marshy places, a few even not being averse to the water. They are, for the most part, spry in their movements, some little ones scarcely six inches in length taxing a vigorous man’s speed to capture; and many are expert climbers of cliffs, trees, and even the ceilings of residences. Some, the remarkable little flying dragons of Ceylon, have an extraordinary development of the skin on the sides of the body, supported by the expanded ribs, forming a sort of parachute whereby the creatures can sail considerable distances through the air. Nearly all are carnivorous, feeding upon small mammals, birds, other reptiles, frogs, and insects; a few only are herbivorous, such as the iguanas, which are often used for human food. Nearly all lizards are oviparous, laying from two to thirty eggs. In size the great majority are small, less than a foot in length; but some, such as the monitors and iguanas, reach a length of from four to six feet, or even more, and certain extinct monitors of India are known to have attained a length of thirty feet. They are, for the most part, slender, graceful, prettily marked, and quite inoffensive creatures. A few are short, flat, or stumpy in [Pg 142] shape, such as the so-called horned toad. One or two species only, the “Gila monsters,” are reputed to be venomous.

Fig. 66.—Amblyrhynchus cristatus, the
Galapagos sea-lizard.
(From Brehm)
There is but a single species of lizard now living which is in any true sense aquatic in habit, the well-known sea-lizard of the Galapagos Islands, scientifically known as Amblyrhynchus cristatus. It is a large lizard, with a short rounded head, a flat tail, and webbed feet. Its specific name is derived from the erect fringed crest along its back and tail. Its habits are best given in Darwin’s words: [Pg 143]
It is extremely common on all the islands throughout the group, and lives exclusively on the rocky sea-beaches, being never found, at least I never saw one, even ten yards inshore. It is a hideous looking creature, of a dirty black color, stupid and sluggish in its movements. The usual length of a full grown one is about a yard, but there are some even four feet in length; a large one weighed twenty pounds. The tails are flattened sideways, and all four feet are partially webbed. They are occasionally seen some hundred yards from the shore swimming about. When in the water this lizard swims with perfect ease and quickness, by a serpentine movement of the body and flattened tail—the legs being motionless and closely collapsed to the sides. A seaman on board sank one, with a heavy weight attached to it, thinking thus to kill it directly; but when an hour afterward he drew up the line it was quite active. Their limbs and strong claws are admirably adapted for crawling over the rugged and fissured masses of lava, which everywhere forms the coast. The nature of this lizard’s food (seaweed) as well as the structure of the tail and feet, and the fact of its having been seen voluntarily swimming out at sea, absolutely proves its aquatic habits; yet there is in this respect one strange anomaly, namely, that when frightened it will not leave the island. Hence it is easy to drive these lizards down to any little point overhanging the sea, where they will sooner allow a person to catch hold of their tails than jump into the water. They do not seem to have any notion of biting; but when much frightened they squirt a drop of fluid from each nostril. I threw one several times as far as I could into a deep pool left by the retreating tide, but it invariably returned by a direct line to the spot where I stood. It swam near the bottom, with a very graceful and rapid movement, and occasionally aided itself over the uneven ground with its feet. As soon as it arrived near the edge, but still being under water, it tried to conceal itself under the tufts of seaweed, or it entered some crevice. I several times caught the same lizard by driving it to a point, and, though possessed of such perfect powers of diving and swimming, nothing could induce it to enter the water; and as often as I threw it in it returned in the manner described above. Perhaps this singular piece of apparent stupidity may be accounted for by the circumstance that this reptile has no enemies whatever on shore, whereas at sea it must often fall a prey to the numerous sharks.
These lizards are of much interest as indicating one of the ways in which true land reptiles have become aquatic in their habits. Tempted by the abundance of food growing in shallow water a little beyond their reach, the reptiles ventured farther and farther to obtain it. The tail gradually became a propelling organ, though the lizard still retained in large measure its land habits and land feet, because of the dangers from its water enemies. It is not at all improbable that, in course of time, were these Galapagos lizards left unmolested, they would become fleeter swimmers by the development of a terminal caudal fin and paddle-like legs, thus competing with their aquatic enemies and no longer needing recourse to the land for protection. They also serve to indicate that long-tailed aquatic reptiles never used their legs to an appreciable extent as organs of propulsion in the water. [Pg 144]

Fig. 67.—Varanus, Australian monitor lizard.
(By permission of the New York Zoölogical Society.)
Flat-headed lizards.—Among the living lizards there is one group, called the monitors, which have so many characters peculiar to themselves that they seem rightfully entitled to an isolated place among the lizards of the present time. The group includes about thirty species, all belonging in the one genus Varanus, and all living in India, Africa, and Australia. In size, some of the species of Varanus are the largest of all terrestrial lizards known in the past or present; in other ways also they have reached the maximum of specialization among lizards. The head is pointed, broad, and flat, and the body and tail are long. They have nine vertebrae in the neck, a larger number than is to be found in any other terrestrial lizard. Unlike other lizards they have a protrusible tongue like that of the snakes. All are carnivorous in habit, feeding upon small backboned animals, insects, and especially upon eggs, which they crush between their teeth while holding them aloft. Most species live wholly upon the land, and some are arboreal. Others, especially those of the Nile, live about water and are excellent swimmers. The terrestrial species have a round tail and small external nostrils, but the water species have the tail much flattened, and the nostrils have large cavities, which, when [Pg 145] closed under water, are said to serve as reservoirs of air for respiration. Of one of these swimming species Annandale writes:
Varanus salvator is common in Lower Siam where it is equally at home on land, in water, and among the branches of trees. The eggs are laid in hollow tree trunks. When in the water the lizard swims beneath the surface, the legs being closely applied to the sides, and the tail functioning both as oar and rudder.
These lizards take to the water to escape from their land enemies and not for food, a habit also known among certain other lizards, and one precisely the reverse of that of the Galapagos lizards. It would seem very probable that animals of such carnivorous habits as are the monitors might easily learn to capture water animals for food and thus eventually become aquatic in habit. This inclination toward, and partial adaptation to, water habits in the monitors is of much interest because in all probability the instinct is one of long inheritance from those remote ancestors which gave origin to the truly aquatic members of the order. Though the known geological history of the monitors does not extend far back, they are so intimately allied in their anatomical structure to the aquatic and semiaquatic lizards of Cretaceous times that there could seem to be no doubt of the common ancestry.
Dolichosaurs.—About fifty years ago Professor Owen, the famous English paleontologist, described a peculiar semiaquatic lizard from the Cretaceous rocks of England to which he gave the name Dolichosaurus, in allusion to the slender form of the body. Just what relations these slender lizards have to modern lizards has long been a problem; some have thought that they were their progenitors, but there are very good reasons for doubting this. No modern lizards, save the monitors, have more than eight vertebrae in the neck, while these dolichosaurs had as many as seventeen, a remarkable specialization for aquatic life that could hardly have been lost by their descendants. For this reason the dolichosaurs have usually been considered as representing a distinct suborder. But they have many resemblances otherwise to the monitors. They were semiaquatic in habit, and never more than six feet in length. They are yet imperfectly known, and no restoration of any form has hitherto been attempted. Their peculiar [Pg 146] interest lies in the elongation of the neck, quite like that of the wholly unrelated nothosaurs and proganosaurs, which have been described in the foregoing pages. Doubtless similar habits in each had like results, but just what these habits were in the slender lizards we do not yet know.
Aigialosaurs.—Within recent years a number of other lizards have been made known from the Lower Cretaceous rocks of Dalmatia which present most remarkable intermediate characters between the monitors, dolichosaurs, and the mosasaurs, the famous sea-lizards of Upper Cretaceous age. Some of these lizards had twelve or thirteen vertebrae in the neck, while others had but seven—an unusually short neck characteristic of the mosasaurs. These latter kinds, belonging to two or three genera, are included in a distinct group. They were long and slender, the head long and pointed. The teeth, conical and sharp, were attached in shallow pits, quite as in the mosasaurs. The lower jaws had a hinge just back of the teeth, as in the mosasaurs, of which the only trace in modern lizards is found among the monitors. Still more remarkable, though perhaps not so easily appreciated, is the shape of the quadrate bone, with a broad flaring rim for the ear cavity, quite unlike that of land lizards, but quite like that of the mosasaurs. In fact, the very peculiar skull is almost identical with that of the true sea-lizards. The body and tail also resemble those of the mosasaurs more than those of the monitors, but there is a firm attachment of the pelvis to the backbone, and the legs are long and lizard-like, though not as long as those of land lizards. The feet were webbed in life, and the toes have no claws, conclusively demonstrating their water habits. The vertebrae indeed have the same peculiar articulations, called zygosphenes, as in most of the mosasaurs. The largest aigialosaurs were about six feet in length, that is, of about the size of the smallest known mosasaurs.

Fig. 68.—Clidastes, an American
mosasaur.
Life restoration.
We have then in the aigialosaurs nearly every known intermediate character that we could wish for in a connecting link between the mosasaurs and the monitors, lizards that were equally at home on land or in the water, and there can be scarcely a doubt that they were [Pg 147] either the direct ancestors or closely akin to the direct ancestors of the strictly marine mosasaurs; and scarcely a doubt that they were the descendants of the actual forbears of the modern monitors, which, as we have seen, have acquired partial aquatic habits in escaping from their [Pg 148] enemies. The dolichosaurs we can now understand were a side branch from these semiaquatic aigialosaurs which, specializing in another direction, quickly came to grief, perhaps in competition with their more agile and skilful short-necked kin.
Taking all these facts into consideration it seems best to unite the monitors, dolichosaurs, and aigialosaurs into one group of the Lacertilia, the Platynota, intermediate in place between the true land lizards and the truly aquatic mosasaurs.
At St. Pietersberg, a small mountain in the vicinity of Maestricht, Holland, there are immense subterranean stone quarries, which have been worked for more than a thousand years. The stone quarried from them is a sandy limestone of Upper Cretaceous age containing many well-preserved remains of extinct animals that have long been sought by collectors of fossils. In 1776 Major Drouin—an officer of a near-by garrison, one of much military importance in those days—secured from one of these quarries some bones of an extinct reptile, which, though of interest, afforded but little information concerning the structure and affinities of the animal to which they had once belonged. In 1780 a very perfect skull, in excellent preservation, of the same kind of an animal was obtained from the same quarry by Dr. Hofmann, an army surgeon of the same garrison, whose interest in such things had been incited by Major Drouin’s collections. This specimen, so renowned in science, has had a remarkable and eventful human history, in part related by St. Faujas de Fond, a French commissary of the “Army of the North,” and one of the participants:
In one of the great galleries or subterranean quarries in which the Cretaceous stone of St. Pieter’s Mount is worked, about five hundred paces from the entrance, and ninety feet below the surface, the quarrymen exposed part of the skull of a large animal in a block of stone which they were engaged in quarrying. On discovering it they suspended their work and went to inform Dr. Hofmann, surgeon to the forces at Maestricht, who for some years had been collecting the fossils from the quarry, remunerating the workmen liberally for the discovery and preservation of them. Dr. Hofmann, arriving at the spot, saw with extreme pleasure the indication of a magnificent specimen; he [Pg 149] directed the operations of the men, so that they worked out the block without injury to the fossil, and he then, by degrees, cleared away the yielding matrix and exposed the extraordinary jaws and teeth, which have since been the subject of so many drawings, descriptions, and discussions. This fine specimen which Dr. Hofmann had transported with so much satisfaction to his collection, soon became, however, a source of much chagrin to him. Dr. Goddin, one of the canons of Maestricht, who owned the surface of the soil beneath which was the quarry whence the fossil was obtained, when the fame of the fossil reached his ears, pleaded certain feudal rights in support of his claim to it. Hofmann resisted and the canon went to law. The whole chapter supported their reverend brother, and the decree ultimately went against the poor surgeon, who lost both the specimen and his money, for he was made to pay the costs of the action. The canon, leaving all remorse to the judges who pronounced the iniquitous sentence, became the happy and contented possessor of this unique example of its kind.
[Translation by Leidy.]
But the canon was ultimately despoiled of his ill-gotten treasure. At the siege of Maestricht in 1795, the famous skull to which Hofmann had devoted so much anxious thought and labor, fell into the hands of the French and was carried off as one of the spoils of war. So widely celebrated had the specimen become during the fifteen years which had elapsed since its discovery, through the writings of several noted scientific men, that the French general commanded his artillerists to spare the house in which it was known to be. The canon, however, shrewdly suspecting that such an unexpected and extraordinary mark of favor was not for his own sake but rather for the sake of the famous fossil, had it removed and carefully hidden in a house in the city. After the capitulation of Maestricht the eagerly sought-for fossil was not to be found, and the offer of a reward of six hundred bottles of wine, so the story goes, was made for its recovery. So tempting was the offer that, ere long, it was brought in triumph to the house of St. Faujas de Fond, by a half-dozen grenadiers, whence it was later transferred to Paris, where it now is.
We may well sympathize with Dr. Hofmann in the loss of his cherished specimen, since, had it not been for his zeal, money, and labor, it would never have escaped the usual fate of such things—complete destruction. But we must remember that St. Faujas de Fond, the recorder of this history, was a Frenchman, and somewhat interested in robbing the reverend canon of it; possibly there is another side of the story which has never been told. [Pg 150]
After peace was declared, one has regretfully to add that the canon, not Dr. Hofmann, was reimbursed for it, or so it is said. Cuvier rather naïvely says that it was ceded to the Garden of Plants of Paris, perhaps in the way that many other things are ceded to the conqueror in time of war. The specimen is really a good one, even when compared with many found in recent years, and there is little wonder that the cupidity of St. Fond was incited by it. Casts of it are now or have been in nearly every noted museum of the world, and pictures of it illustrated nearly every textbook of geology published during the first three-quarters of the past century. It had been the subject of considerable controversy even before it came into the hands of Cuvier. Peter Camper figured and described the skull as that of a whale or “breathing fish”; while St. Fond himself later called it a crocodile. Crocodile or alligator skeletons were rare in those days, and St. Fond made a special trip to the British Museum to study one. But it was really Adrian Camper, a son of Peter Camper, who deserves the credit, so often wrongly ascribed to Cuvier, for the recognition of the true nature of the fossil. He insisted that the animal was a lizard allied to the living monitors, an opinion which it will be seen has finally been proved to be correct within very recent years.
In 1808 this famous skull, and all other known remains of a similar nature, came under the observation of Cuvier, the renowned French naturalist and paleontologist, who confirmed the views of Adrian Camper. He fully described and figured all the known parts of the skeleton that had later come to light, calling the animal the great lizard of the Meuse, the river near which Hofmann’s specimen was found. Conybeare, a well-known paleontologist of England, some years later formally christened it Mosasaurus, a transliteration of Cuvier’s phrase, from the Latin Mosa, for Meuse, and saurus, a lizard. For more than half a century Cuvier’s figure of the skull of the original specimen appeared in works on geology over the name Mosasaurus hofmanni, or Mosasaurus camperi. One could wish that the former name for the species might prevail, in recognition of the zealous doctor who was so shabbily treated in his possession of the specimen. [Pg 151]
For some years the few specimens discovered by Drouin and Hofmann were all that were known of the mosasaurs. A few others of related forms were discovered in England, and some were reported from New Jersey by early explorers, but there was little published about the mosasaurs till 1843, when Dr. August Goldfuss, a noted German paleontologist, described and beautifully figured an excellent specimen from the United States. This specimen also had a rather eventful history. It was discovered early in the fourth decade by Major O’Fallen, an Indian agent, near the Great Bend of the Missouri River, whence it was transported by him to St. Louis and placed in his garden as a curiosity. It happened that Prince Maximilian of Wied, the famous naturalist, in his travels through the United States, saw the specimen and secured it, taking it to Germany on his return. He presented it to the Museum of Haarlem where Goldfuss saw and described it. Rather oddly, this specimen was of a species closely allied to the original one of Maestricht, a species which has since only rarely been found. It was called Mosasaurus maximiliani by Goldfuss, though some time previously, it has since been found, some fragments of the same species were described by Harlan, an American author, under the name Ichthyosaurus missouriensis. Goldfuss’ paper was strangely overlooked by subsequent writers, and it was not till the discovery of numerous remains of mosasaurs by Leidy, Cope, and Marsh in the chalk of western Kansas, nearly thirty years later, that much was added to the world’s knowledge of these strange reptiles.

Fig. 69.—Skeleton of Plalecarpus as mounted in the Paleontological Exhibit. Walker Geological Museum, University of Chicago.
[Pg 152] Perhaps nowhere in the world are the fossil remains of marine animals more abundantly and better preserved than in these famous chalk deposits of Kansas. The exposures are of great extent—hundreds of square miles—and the fossil treasures they contain seem inexhaustible. Long-continued explorations by collectors have brought to light thousands of specimens of these swimming lizards, some of them of extraordinary completeness and perfect preservation, so complete and so perfect that there is scarcely anything concerning the mosasaurs which one might hope to learn from their fossil remains that has not been yielded up by these many specimens. The complete structure and relations of all parts of the skeleton, impressions of the bodies made in the soft sediments before decomposition had occurred, the character of their food, the nature of the skin covering, and even some of the color markings of the living animals have all been determined with certainty. Not only from Kansas, but also from many other parts of the world, have remains of these animals been discovered, until now it may truthfully be said that no other group of extinct reptiles is better represented by known fossil remains than the mosasaurs. From England, Belgium, Russia, and France in Europe; from New Jersey, Georgia, Alabama, Mississippi, Texas, New Mexico, Colorado, Kansas, Nebraska, the Dakotas, Wyoming, and other places in the United States; from New Zealand and South America they have been obtained in greater or less abundance and perfection.

Fig. 70.—Tooth
of Tylosaurus.
Their geological history is relatively brief, notwithstanding their wide distribution over the earth in such great numbers and diversity. The earliest are known from near the beginning of the Upper Cretaceous of New Zealand, whence it is believed by some that they migrated to other parts of the world, appearing in North America some time later. They reached their culmination in size, numbers, and variety very soon, and then disappeared forever before the close of Cretaceous time. The largest complete specimen of a mosasaur known measures a little more than thirty feet in length, but incomplete skeletons of others indicate a maximum length of about forty feet. The skulls of the largest species are about five feet long. The smallest known adult skeletons are [Pg 153] scarcely eight feet in length. There are now known at the present time seven or eight genera of three distinct types, all belonging to one family, the Mosasauridae, including about twenty-five known species. While a few of the genera are widely distributed over the earth, the species are all of restricted range, indicating, perhaps, non-migratory habits.
The adaptation of the mosasaurs to an aquatic life was very complete, though perhaps not so complete as was that of the ichthyosaurs. The skull is flattened, narrow, and more or less elongate, but large in proportion to the remainder of the skeleton—nearly one-sixth of the entire length; that relative size doubtless is indicative of very predaceous and pugnacious habits. The teeth in the typical forms are numerous, strong, and sharp, conical in shape, and recurved. Not only are there numerous teeth in both the upper and lower jaws, but there are also two rows of strong teeth implanted in the back part of the palate, upon bones called pterygoids, the use of which will be understood later. The teeth were inserted on large, tumid, bony bases, rather loosely attached in shallow pits or alveoli, unlike the teeth of all modern lizards. Such a mode of attachment of the teeth doubtless had some relation to the habits of the animals concerning which we are not quite clear. They were easily dislodged, and, in consequence, of very unequal size, some full grown, some small, and others just appearing above the surface of the gums in the living animals. The frequent loss of teeth and their constant and easy replacement by new ones is a peculiarity of predaceous reptiles, thereby insuring their best functional use.
The external nostrils, of large size, were situated at a considerable distance back of the end of the snout, but not nearly so far back or so near the eyes as were the nostrils of the ichthyosaurs, plesiosaurs, [Pg 154] and phytosaurs. Their size and position suggest a use like that of the modern aquatic monitors, as mentioned on a preceding page. The eyes were of moderate size, those of the less purely aquatic forms being directed more laterally than those of species of more distinctly diving habits. They were protected by a stout ring of bony plates, as were the eyes of all truly aquatic reptiles of the past. The ears, also, in most if not all mosasaurs, had a thick cartilaginous ear-drum in place of a simple membrane, evidently, as Dollo has shown, for better protection under undue pressure of the water in deep diving.

Fig. 71.—Clidastes, inner side of right mandible: ang, angular; art, articular; cor, coronoid; pa, prearticular; sur, surangular.
As in all other lizards, the bones with which the lower jaws articulate, the quadrates, were loosely attached at the upper end, permitting great freedom of movement in all directions, more even than the land lizards have. The lower jaws were long and powerful, armed with a single row of teeth on each side, from sixteen to eighteen in number. Just back of the teeth, a little beyond the middle, each mandible has a remarkable joint, quite unknown in land lizards, though a trace of it is found in the monitors, permitting much movement between the front and back parts, both laterally and vertically, though chiefly in the former direction. Furthermore, as in land snakes but not as in land lizards, the front ends of the two sides of the jaws were somewhat loosely attached to each other by ligaments. This looseness of the two sides of the jaws, not only in front but also behind, together with the joint in each, was of the greatest use in swallowing prey, as will be explained farther on. [Pg 155]


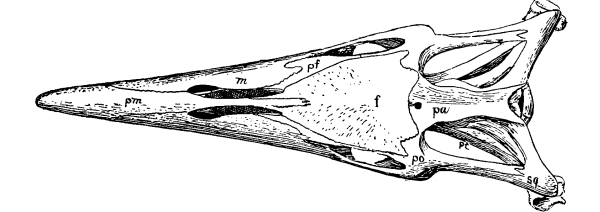
Fig. 72.—Skulls of mosasaurs. Upper figure, Clidastes, from the side; middle figure, Platecarpus, from below; lower figure, Tylosaurus, from above: an, angular; bs, basisphenoid; c, coronoid; ep, epipterygoid; fr, frontal; j, jugal; l, lacrimal; m, maxilla; na, nasal; oc, occipital condyle; pa, parietal, palatine; pm, premaxilla; pf, prefrontal; pt, pterygoid; po, postorbital; q, quadrate; sp, splenial; sq, squamosal; tr, transverse; v, vomer.
As in most other aquatic reptiles, the neck was short and strong, the vertebrae being less in number than in most other lizards. The trunk was long and slender, more especially so in the surface-swimming kinds, with from twenty-two to thirty-four vertebrae. The tail was long, no longer than the tail of some land lizards, but more powerful, and broader and flatter. It was expanded or dilated more or less toward the free end, that is, with the beginning of a terminal caudal fin, such as the more specialized ichthyosaurs and crocodiles possessed. The [Pg 156] vertebrae were procoelous, that is, concave in front and convex behind, like those of most modern lizards and all modern snakes and crocodiles, but quite unlike the biconcave vertebrae of all other aquatic reptiles. This kind of articulation of the backbones gave greater firmness and strength to the spinal column, but decreased the flexibility, and its possession by these animals was doubtless due to their descent from land lizards which had already acquired it. The loss of flexibility, however, was partly compensated by the loss of the additional articulating surfaces of the tail.

Fig. 73.—Platecarpus; occipital view of skull: bo, basioccipital; eo, exoccipital; pf, postfrontal; st, stapes; pt, pterygoid; q, quadrate.
As in all other aquatic reptiles, it is in the limbs that the most striking characteristics of these water lizards or “sea-serpents” are found. The legs were so completely adapted to an aquatic mode of living that the animals must have been practically helpless upon land, able perhaps to move about in a serpentine way when accidentally stranded upon the beaches, but probably never seeking the land voluntarily. The front limbs, like those of all other swimming animals having a powerful propelling tail, were larger than the hind ones, though not very much so. The bones of the first two segments, that is, the arm, forearm, and thigh and leg bones, were all short and broad, resembling those of the ichthyosaurs more than those of any other reptiles, save perhaps the thalattosaurs, discussed below. The articular surfaces of all the limb bones, as in other aquatic animals, were restricted in extent, indicating limited motion between the joints, though doubtless having great flexibility. In the most specialized types, such as Tylosaurus, the wrist and ankle bones were almost wholly cartilaginous, just as they are in the water salamanders, and in whales and porpoises. This tendency of the ends of long bones, the wrists and ankles as well as other bones of the skeleton, to become more cartilaginous, or less well ossified, in animals purely aquatic in [Pg 157] habit is a marked one. So much is this the case that paleontologists always suspect water habits in reptiles showing it, even though but few parts of the skeleton are known.

Fig. 74.—Clidastes;
left front paddle: c, coracoid;
h, humerus; r, radius;
sc, scapula; u, ulna.

Fig. 75.—Tylosaurus;
left front paddle: c, coracoid;
sc, scapula; h, humerus;
r, radius; u, ulna.
Increase in the number of bones of the digits is a more or less conspicuous characteristic of all mosasaurs. In those forms in which the wrists and ankle bones had become cartilaginous in great part, as many as eleven phalanges have been observed in the longest toes, though in other forms, those with more completely ossified wrists and ankles, only two or three additional bones have been developed in the longest [Pg 158] fingers and toes by aquatic habits. The pliability and flexibility of the fingers and toes were certainly very great, but they could not possibly have been flexed or bent so as to grasp or seize anything; and of course all vestiges of claws had disappeared. Many specimens have been found with all the bones of the limbs, that is, the “paddle bones,” in the positions they occupied when the animals died. Figures of three such specimens, made from photographs or careful drawings by the writer, are shown herewith (Figs. 74-76). In several such specimens very clear impressions of the smooth membranes between the fingers have been observed, and in one specimen preserved in the collections of the University of Kansas the outline of the fleshy parts connecting the paddle with the body has been preserved.
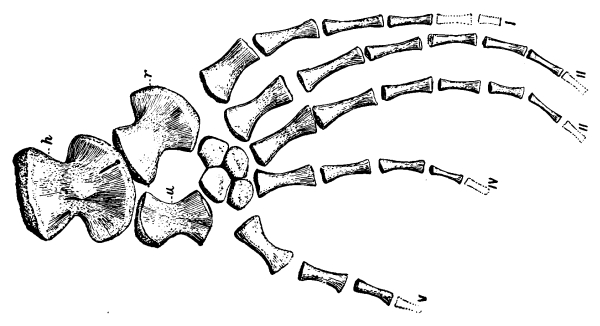
Fig. 76.—Platecarpus; right front paddle: h, humerus; r, radius; u, ulna.
It will be seen by comparison of the figures of the mosasaur paddles with those of the ichthyosaurs and plesiosaurs that there was a wide difference in their structure, though all have the characteristic shortening of the limb bones and increase in the numbers of the finger and toebones, that is hyperphalangy. It is probable that these differences mean a more powerful and varied use of the limbs in the mosasaurs. It is certain that the mosasaurs were much more predaceous and pugnacious in their habits than were any other truly aquatic backboned air-breathing animals of the past or present. They were the [Pg 159] “land sharks” of the ancient seas, and probably the only ones among water reptiles that would be dangerous and offensive to man, were they all living today.
For a long time it was thought that the mosasaurs had no breast bone, and that, in consequence, the front part of the thorax was expansible. Under this assumption the mosasaurs would have been much more snake-like in habit than they really were. The loose construction of the jaws doubtless permitted the swallowing of prey of considerable size, and the inference was that they habitually preyed upon animals of large size. A snake will often swallow a frog of larger diameter than its own body, the flexible jaws and loosely connected ribs permitting it to pass to the abdominal cavity. But the unyielding ring formed by the anterior ribs connected with the breast bone in the mosasaurs, as in other lizards, conclusively proves that large animals could not have been swallowed whole by the mosasaurs. In several instances the fossilized stomach contents, composed chiefly or wholly of fishes, have been found between the ribs of mosasaurs, and in none were the fishes more than two or three feet in length, though the reptiles were from sixteen to twenty feet long. Possibly the largest mosasaurs, those thirty or thirty-five feet in length, might have captured and swallowed fishes six or seven feet long, but in all probability their usual prey was of smaller relative size.

Fig. 77.—Platecarpus; pelvis, from below: p, pubis; il, ilium; is, ischium.
The very loose construction of the pelvic bones, those to which the hind legs are articulated, is an evidence of more complete adaptation to water life than was or is the case with any other water air-breathers except the ichthyosaurs and cetaceans. The sacrum had entirely lost its function as a support to the pelvis and had disappeared, that is, the vertebrae composing it had become quite like [Pg 160] the adjacent ones, by the loss of the ribs connecting them with the ilium. The small pelvis was suspended loosely in the walls of the abdomen, or at the most was feebly connected with a single vertebra by ligaments. It was entirely useless as a support for the legs. The mosasaurs could not possibly have raised their bodies from the ground while on land. It is well known that the land lizards and the crocodiles raise their bodies free from the ground while running or walking; none drags its body over the surface.

Fig. 78.—Photograph of carbonized remains of scales of Tylosaurus.
In several instances complete or nearly complete skeletons of mosasaurs have been discovered with the different bones nearly all in the positions and relations they had after the decomposition of the flesh, together with the carbonized remains of the skin and impressions of the investing scales and membranes. The nature of the body covering is therefore known with certainty from nearly all parts of the body. The body everywhere, save on the membrane between the fingers and toes, and perhaps on the top of the skull, was covered with small overlapping scales, very much like those of the monitors. These scales, however, were small and smooth in comparison with the size of the animals, those of a mosasaur twenty feet in length being almost precisely the size of those of a monitor six feet long. The top of the skull seems to have been covered with horny plates, as in most lizards. In one instance parallel dark bars, obliquely placed, and of narrow width, formed by carbonized pigment, were observed by the writer. As has been stated, in [Pg 161] some instances fish bones and fish scales have been observed among the fossilized stomach contents, and it is quite certain that the food of these creatures must have been composed chiefly of fishes, though of course it is not improbable that other small vertebrates, birds, pterodactyls, the young of plesiosaurs, and possibly small mammals, may occasionally have formed a part of their diet. That the mosasaurs were very pugnacious in life is conclusively proved by the many mutilations of their bones that have been observed, mutilations received during life and partly or wholly healed at the time of death. Bones of all vertebrates are repaired after injury by the growth of more or less spongy osseous material about the injured part, forming a sort of natural splint. This material is more or less entirely removed by absorption when it is no longer required for the support of the broken ends. Many such injured bones of the mosasaurs have been found; sometimes the bones of the hands and feet have grown together, and not infrequently the vertebrae have been found united by these osseous splints; occasionally even the skull itself, especially the jaws, attest extensive ante-mortem injuries. In a single instance the writer has observed the loss of a part of the tail, where it probably had been bitten off. It may be mentioned, however, that the bones of the tail had no such “breaking points” in the mosasaurs as have those of many land lizards, whereby a part or all may be lost as a result of even a trivial injury, and then regrown. Such a condition in an organ relied upon entirely for propulsion would have been immediately fatal to the existence of the mosasaurs. The large jaws and teeth are in themselves sufficient evidence of the fiercely carnivorous propensities of the mosasaurs. The constant renewal of the sharply pointed teeth, thereby preventing deterioration by use or accident, preserved, even in the oldest animals, the effectiveness of the youthful structure.
We may now understand how the mosasaurs seized and swallowed their prey. Living constantly in the water, away from all firm objects, with small, short limbs quite incapable of holding struggling prey, and the body not sufficiently serpentine to hold it in its folds after the manner of snakes, the mosasaurs would have found it difficult or impossible to swallow fishes of even moderate size, were their jaws of [Pg 162] the same construction as are those of the land lizards. If they preyed upon small animals only, or if they tore their prey to pieces after the manner of the alligators, there would have been no especial difficulty in deglutition. But it is certain that the animals which the mosasaurs devoured were not always small, and they must have been swallowed whole, since their teeth were not adapted, like those of the alligators and true crocodiles, for the rending of bodies. One who has watched a snake swallow a frog or another snake will appreciate the difficulties against which the mosasaurs contended in swallowing fishes a fifth or a sixth of the length of their own bodies. The ordinary snake, no matter where or how it seizes its prey, invariably swallows it head first. Its mandibles are even more loosely united in front than were those of the mosasaurs, and while there is no joint in the snake’s mandibles such as there is in the mosasaurs’, the loose union of the various bones of the mandibles serves the same purpose. The frog or lizard, while firmly held by the slender teeth, is slowly moved sideways by the alternate lateral action of the jaws till the head is reached. Many non-poisonous snakes, if they find it impracticable or impossible to reverse the position of their prey in this way, wrap the folds of their body about it, holding it firmly while they release their mouth-hold and seize it by the head. An amusing instance of these habits came under the observation of the writer not long ago, in Texas. A large “blue racer” (Bascanion), six feet four inches in length, caught an unusually large bullfrog by one hind leg, but in almost less time than it takes to relate, the head of the frog had entered the snake’s gullet and the mouth was closed over it, notwithstanding the vigorous muscular and vocal protests on the part of the frog. Wishing to secure the skull of the snake for his collection, the writer seized an ax standing conveniently by and cut the snake cleanly in two. The peristaltic action of the deglutitional muscles carried the frog slowly on about two feet farther to the ax-made orifice, from which it emerged, and, after a few croakings against such unkind usage, calmly hopped off into the near-by pool of water! Many poisonous snakes release their prey after killing it; other snakes may force their prey down the throat by pressing it against the ground. [Pg 163]
Even small fishes could not possibly have been swallowed by the mosasaurs in any other way than head first, since the backwardly projecting, and often long, spines would have rendered any other procedure impossible. Even after the head had entered the gullet, deglutition could have been effectively completed only by the aid of some mechanism whereby the fish could have been pulled or pushed back into the constricting fauces. The strong teeth of the upper jaws and palate held firmly the struggling prey, while the loosely united jaws, bending laterally at the joint back of the middle, either alternately, or more probably in unison, steadily forced it far enough back to be seized by the muscles of the fauces.
The shape of the mosasaurs, though slender, does not suggest extraordinary speed in the water; doubtless most of the fishes that lived in the seas with them could swim faster than they. Their prey was captured, for the most part at least, by sudden and quick lateral movements, for which their powerful and flexible paddles admirably adapted them.
It is a rather remarkable fact that, among the thousands of specimens of mosasaurs which have been collected during the past forty years in both Europe and North America, there never has been found one of a very young animal. Of almost all other animals occurring abundantly as fossils some specimens are sure to be discovered of young and even embryonic individuals. It is quite certain that all such voracious monsters as were the mosasaurs did not die of old age. Some specimens, it is true, have been found that were evidently not full grown animals, but the observed differences in the size of the fossil bones are not great. All are of adult or nearly adult animals. If the mosasaurs were oviparous, as were the ichthyosaurs, and probably the plesiosaurs, and as are some living land lizards, the apparently entire absence of embryonic bones associated with often nearly complete skeletons of the mosasaurs is inexplicable; certainly some mosasaurs must have died a short time before the birth of their young. But embryos have never been discovered, though numerous skeletons inclosing fossilized stomach contents have been found. From this fact it would seem very [Pg 164] probable that the mosasaurs were oviparous, as are most other lizards. But this, after all, may be a hasty inference.
No known reptiles lay their eggs in the water. Perhaps there is some reason why the eggs of reptiles and birds, so different from those of fishes and amphibians, cannot hatch in water; and there is no good reason for supposing that the mosasaurs were exceptions to this rule. Unless carefully hidden or protected by the parent, the eggs or very young of the mosasaurs would have been subject to many and grave dangers. Fish eggs are usually small and produced in great numbers, thousands often being extruded from a single female. Among so many there is a greater probability that at least two will hatch and survive to maturity, reproducing their kind. It is unreasonable to suppose that the lizards of the past were more prolific of eggs than are their relatives now living; nor is it possible that their eggs could have been as small as are those of most fishes. Modern lizards seldom lay more than twenty-five or thirty eggs at a time; even the turtles, with their greater vicissitudes, seldom produce more than one hundred. The eggs of the mosasaurs were certainly large and few in number, and the young animals must have begun breathing air immediately after escaping from the shells. If the mosasaurs were oviparous they must have laid their eggs upon the shores and beaches, as do the sea-turtles and the Crocodilia. Nor is it at all probable that the female mosasaurs gave even that protection to their eggs or young that the crocodiles and turtles give. The young mosasaurs, perhaps reaching a foot in length, must have been left entirely to their own devices and their own fate at the very earliest stages of their independent careers.
The waters in which the mosasaurs abounded swarmed with many kinds of predaceous fishes, to say nothing of the hordes of their own kinds, all carnivorous in the highest degree, to all of which the tender saurians must have been choice food. Possibly the shallow waters of the bays and estuaries may have afforded protection to the newly hatched reptiles. It would seem probable that the female mosasaurs went up the rivers for a shorter or longer distance to lay their eggs or give birth to their young, and that the young reptiles remained in such relatively [Pg 165] protected places until of a sufficient size to cope with the fierce enemies of the open seas. We know practically nothing of the inhabitants of the lakes and rivers during all the time in which the mosasaurs existed; and this perhaps is the real reason why we have never yet found a specimen of a young mosasaur.
Fig. 79.—Head of Tylosaurus.
We have seen that many skeletons of ichthyosaurs are found entire, and but little disturbed in position, suggesting, if not proving, that the animals as a rule lived and died far out in the deep seas, away from the disturbing effects of currents of water on their decaying bodies. Among the thousands collected, the great majority of the specimens of mosasaurs consist of a few bones or a part of the skeleton only. Moreover, nearly all specimens show the disturbing effects of currents of water; and the bones are usually associated with those of turtles, birds, and flying reptiles, which probably did not often venture far from the shores; all of which goes to prove that the mosasaurs in general lived in the comparatively shallow waters of the seas, and not far from the shores. That some were excellent divers, descending probably many fathoms deep in the water, is certain, because of the extraordinary protective structures of the eyes and ears.
But the various kinds of mosasaurs differed not a little in their habits. Some, like Mosasaurus and Clidastes, were doubtless chiefly surface swimmers, as is evidenced by their better ossified bones, firmer articulation, and the presence of the additional [Pg 166] zygosphenal articulations of the vertebrae, wanting in other forms, as also by the structure of their paddles. They had a relatively long body and short tail, the tail having a more pronounced distal expansion than in the case of other forms, and the eyes looking laterally, not at all upward. The feet, as shown in Fig. 74, were broad and short, with most of the wrist and ankle bones well ossified, and with but few extra bones in the digits. Tylosaurus (Fig. 79), on the other hand, had a more slender skull, the nostrils were situated farther back from the tip of the snout, the tail was longer and more powerful, and the feet were very highly specialized (Fig. 75). The wrist and the ankle were almost wholly cartilaginous, the fifth finger and fifth toe were much longer, and the number of phalanges was greatly increased. Moreover, the bones of the skeleton are more spongy, the joints are more cartilaginous, and the ears were better protected by a heavy coat of cartilage. In most of these respects the genus Platecarpus was intermediate between Clidastes and Tylosaurus (Fig. 76).
Like nearly all other lizards, the mosasaurs had a pineal opening in the skull, but it is not at all probable that they possessed a functional pineal eye.
Many and varied have been the opinions of scientific men regarding the relationships of these animals, as has been intimated. They were thought to be a kind of whale or breathing fish by Peter Camper; crocodiles, by St. Fond; and aquatic lizards, by Adrian Camper and Cuvier. The late Professor Cope thought they were more nearly related to the snakes than to the lizards, and that they might even have been the ancestral stock from which the snakes have descended. Because of this belief he gave to them the name Pythonomorpha, meaning python-like, and this name, really the first ever applied to them, is yet often used instead of Mosasauria. A more complete knowledge of the mosasaurs, however, and especially the recent discoveries of the semiaquatic connecting links, called the aigialosaurs and described on a preceding page, have set at rest all doubt as to their real affinities. They are real lizards, differing less from the living monitor land lizards than do the monitors from some other land lizards, especially the amphisbaenas and chameleons. And to Adrian Camper is due [Pg 167] the credit for the recognition of their real relationship, though it required more than a century to prove that he was right.

Fig. 80.—Globidens alabamensis.
Part of mandible, with teeth.
(From Gilmore.)
Very recently, and since the foregoing was written, a remarkable new type of mosasaurs has been discovered in Alabama and Europe. Only fragmentary jaws, a few vertebrae, and some skull bones are known, so that it is impossible yet to decide how closely the new form is related to the true mosasaurs, but so far as the evidence goes the only distinguishable character is the teeth. These, instead of being elongated and pointed, are nearly spherical, as shown in Fig. 80. Such teeth could have been used only for crushing shell-fish, and not at all for the seizure and retention of slippery fishes. The genus, which was called Globidens by its discoverer, Mr. Gilmore, includes two known species, from Alabama and Europe, the latter recently described by Dollo. It has been suggested that this peculiar kind of dentition was a more primitive or intermediate one, a kind that the first mosasaurs had before they became fully adapted to the water; but this is doubtful, since Globidens comes from late Cretaceous, and must be one of the later types. If Globidens is a true mosasaur, and it seems to be one, its life-habits must have been remarkably different from those that have long been known. Possibly when the limbs and more of the skull are found, Globidens will prove to be of a distinctive type. [Pg 168]
The chief differences between snakes and lizards have already been given and need not be repeated, save very briefly. Snakes are always functionally legless, though some have vestiges of the hind pair; the brain-case is wholly bony; the upper temporal bar is wanting; the lower jaws are united in front by ligaments only, like those of the mosasaurs; the vertebrae are greatly increased in number, and always have the additional zygosphenal articulations like those of Clidastes and Mosasaurus and some lizards; there is but one lung, and the eyes are always without free eyelids. But these characters are really not very important, since every one of them is found in the lizards or mosasaurs, except the complete ossification of the brain-case, and even this is partly ossified in the mosasaurs. It is rather the presence of all these characters which distinguishes a snake from a lizard.
The number of living snakes is nearly as great as that of the living lizards, and their distribution over the earth is very similar. Snakes are for the most part strictly terrestrial in habit. Some live more or less among trees, and some live in the water, though with but few exceptions all are fully capable of rapid progression upon land. They are almost invariably carnivorous in habit, swallowing their prey whole, and usually alive, as has been described. Some poison their prey or crush it to death before swallowing it. Some feed upon eggs which are swallowed whole and then crushed in their stomachs by projecting bones from the under side of the vertebrae developed for that purpose. In size snakes vary from a few inches in length to twenty-five or more feet, no known extinct forms being larger than the living anacondas and boas. In geological history the earliest remains known date from the latter part of the Cretaceous, and it is quite probable that they have a briefer history than that of the lizards of which they are the descendants. Venomous serpents are known only from comparatively recent geological times, and it is probable that venomosity is the latest and final specialization of importance in the reptilian class. [Pg 169]

Fig. 81.—Hydrus bicolor; sea-snake.
(From Brehm)
Of strictly aquatic snakes there is no known geological history, and it is improbable that there is any such history. There are a few snakes now living—very venomous ones, allied to the deadly cobras—which have become so completely adapted to life in the water that they are unable to exist or even move about on land. These are the well-known sea-snakes of the Indian Ocean and adjacent waters. Perhaps the most highly specialized and typical of these is the black-banded sea-snake, Distina cyanocincta, which reaches a length of four or five feet, and is a rapid and excellent swimmer. From the figure (Fig. 81) it is seen that the body is very much flattened from side to side, and lacks or has but a few vestiges of the transverse scales on the under side so characteristic of all other snakes, and which enable them to move about on land. So helpless are these snakes on land that it is said sailors will handle them carelessly, because of their inability to bite while out of water, though the bite is very venomous. They never come on land for any purpose whatever, and their young, unlike those of [Pg 170] most other snakes, are born alive. There are a number of species of these sea-snakes, though comparatively little is known of their habits. They are of especial interest as another example of the ways in which air-breathing land vertebrates have become adapted to water life. The adaptation, however, was simple, for nearly all snakes swim freely in water by undulatory movements; it would require not much change to convert an ordinary water snake into one like these sea-snakes.
[Pg 171]
Millions of years before the first appearance of the mosasaurs in geological history, another group of reptiles showing many curious resemblances to them attempted a rather precarious existence in the water. Its members survived long enough to acquire many structural adaptations to a water life, long enough to become diversely modified, but not long enough, apparently, to wander far from their birthplace, not long enough to attain that security from their enemies and more ambitious competitors, the early ichthyosaurs and plesiosaurs, to insure them a long existence. They were only a partial success as water reptiles.
It has been only within a few years that we have had any knowledge whatever of them, and that knowledge is still very incomplete, too incomplete to justify any attempt to picture them as living animals, even though we take the liberties that some of our illustrators of extinct animals feel warranted in assuming. The first known specimens of these “sea-reptiles”—for that is the meaning of the word Thalattosauria—were discovered and described by Professor J. C. Merriam less than ten years ago, and all our knowledge of these animals is due to the same author, who has studied attentively the known specimens, all of which are preserved in the museum of the University of California. The first discovered fragmentary specimens were confounded with those of early ichthyosaurs, from the Upper Triassic rocks of northern California with which they were associated. No specimen has yet been found that is even approximately complete; some parts of the skeleton are not yet known, even from fragmentary remains, and not till other and more complete specimens have been found will it be possible to determine the real form of the living animals or to decide what their nearest relationships with other reptiles were. Professor Merriam thinks that they were related most closely with the [Pg 172] Rhynchocephalia (p. 176) of which the Sphenodon, or tuatera, of New Zealand is the only living representative, but whose direct genealogical history runs back nearly or quite to the time in which the thalattosaurs lived. On the other hand, there are so many resemblances to the mosasaurs shown in the remains that have been discovered, that it is possible the thalattosaurs were only a short-lived branch of the primitive lizards, which we also know were in existence at the time when the thalattosaurs lived. However, even though they resembled the mosasaurs, there could have been no direct genealogical relationships between them, for it is quite certain that the thalattosaurs very soon went out of existence, leaving no descendants. But it matters little which were the land forbears of the thalattosaurs; they present such distinct adaptations to water life—characters all their own—that their ancestral kinship may well be left to the future researches of the curious paleontologist. For the present, at least, they may well be placed in an order of reptiles all their own, as Professor Merriam has proposed—the Thalattosauria.
Fig. 82.—Skull of Thalattosaurus.
(After Merriam)
No thalattosaurs were large animals. If they had the same proportions between the lengths of head, body, and tail as the mosasaurs, none exceeded seven feet in length, and they may have been even shorter, though probably not much. The figure of the skull, as restored by Professor Merriam, shows many striking aquatic adaptations, in the elongated, pointed muzzle, in the large external nostrils, situated far back toward the eyes, and in the well-ossified ring of bones surrounding the eyeball. There is a parietal opening in the roof of the skull, as in the modern lizards and tuatera; but it is not known for [Pg 173] certainty whether there were two openings on each side in the roof of the skull, as in the modern tuatera. While this character may seem trivial, it is really one of the most important in the reptilian anatomy in determining the relationship and classification of reptiles. The teeth are conical and pointed in the front end of the upper and lower jaws, but farther back they are rounded, rugose, and obtuse, and could have been used only for crushing hard objects, like mollusks, crustaceans, etc. (Fig. 82). And not only was there a row of such teeth on each jaw (only partly seen in the figure), but similar teeth covered a large part of the palate. And the lower jaws, it is seen, are rather massive.
The vertebrae were, of course, of the more primitive kind, that is, with the ends concave, both in front and behind. It would have been strange indeed were they of any other kind, since reptiles with ball-and-socket joints to the vertebrae, that is, concave on one end and convex on the other, as in nearly all living reptiles, did not come into existence till long after the thalattosaurs had disappeared from geological history; and it is also a curious fact that such vertebrae appear to have originated only among animals crawling on land, so that they would not have been a character acquired by the thalattosaurs after descending into the water. It will be seen from the figure of a dorsal vertebra that the rib was attached by a single articular surface, almost exclusively to the body of the vertebra, quite like those of all lizards, snakes, and mosasaurs, and unlike those of other reptiles. This too may seem to be a trivial character to prove relationships with the lizards, but it is a curious fact that no two animals having different kinds of ribs are closely related to each other. Possibly, however, this looser mode of attachment of the ribs in the thalattosaurs was one of their peculiar adaptations to a water life, and may not have been derived from their land ancestors.

Fig. 83.—Dorsal vertebra
of Thalattosaurus.
(After Merriam.)

Fig. 84.—Thalattosaurus:
bones of front extremity: s, scapula;
c, coracoid; h, humerus;
r, radius; u, ulna.
(After Merriam.)
Of the limbs, only a few bones are known, but these are very instructive. The arm bones, as shown in Fig. 84, are strikingly like [Pg 174] those of the mosasaurs, as will be seen by comparing the figure on p. 157. The humerus is a little more elongated than that of the mosasaurs, more nearly like the mosasaurian femur. The shoulder-blade and the coracoid are imperfectly ossified, as is seen from the figure—another characteristic of aquatic life. What the fingers and toes were like cannot be said; probably they were bound together by membrane, forming swimming paddles similar to those of the mosasaurs. Some of the bones referred to the pelvis are known, but it is not known whether they are united to the spinal column by a sacrum, as in land animals. Nor is anything certainly known of the hind leg or much of the tail. Since the front legs show marked aquatic adaptations, it is altogether certain that the hind legs will be found to be modified more or less, though not so much modified as the front legs, because, as we have seen, the front legs are always more specialized in aquatic animals than the hind ones, even as the hind legs are more specialized than the front ones in land animals. Possibly the hind legs will be found to be more like those of the Thalattosuchia, as shown on p. 212, that is, partly terrestrial in character. Doubtless the tail was long and flattened, possibly with a terminal fin-like dilation, though this is less probable.
As regards the habits and food of the thalattosaurs, no better summary can be given than that of Professor Merriam, in his own words:
The remains of thalattosaurs are known only in purely marine deposits containing little or no material of terrestrial origin. They are associated with a fauna consisting of numerous forms, both vertebrate and invertebrate, which are not known to have existed away from marine areas. In the structure of the skeleton we find the abbreviated and broadened proximal segments of the limbs, the slender snout with prehensile terminal teeth, and the median superior nostrils, [Pg 175] indicating a purely aquatic type. There can scarcely be room for doubt that the thalattosaurs as a group were typical marine forms. The larger and more specialized species comprised in the genus Thalattosaurus were strictly natatory. They may have visited the shore, but, like the plesiosaurs, were better fitted for swimming than for crawling. Of the smaller Nectosaurus we unfortunately do not know the limbs. They may have been considerably less specialized, and the animal to a correspondingly greater degree a shore-dweller. Nectosaurus is, however, found in the same deposits with other forms and appears to be as common as the others; so that it is safe to consider it as having passed the greater part of its life away from the shore.
From what we know of the vertebral column of Thalattosaurus it appears that the animal had a relatively short neck and a long dorsal region, the proportions being nearly those in the vertebral column of some mosasaurs. Only the anterior portion of the caudal region is known. The slender, rounded neural spines with well-developed articulating processes seen here are not such as commonly appear in forms with a highly specialized sculling tail, and it is hardly probable that a caudal fin of large size was developed.
The anterior limbs evidently formed paddles of moderate size. The posterior pair may have been larger, in compensation for lack of a strong sculling tail. It is, however, possible, that as in Geosaurus (of the thalattosuchian crocodiles) the hind limbs were not typically natatory, and the distal end of the tail was vertically expanded.
No specimens have yet been found which are well enough preserved to show any remains of the stomach contents, and we have no definite evidence concerning the food of the thalattosaurs, more than is furnished by the general structure of the animal. The character of the paddles, the form of the skull, and the presence of slender prehensile teeth in the terminal portions of the jaws would indicate that they fed in part upon some swiftly moving prey which was caught by a quick snap of the jaws, deglutition being assisted by the curved teeth of the pterygoids. The heavy vomerine and posterior mandibular teeth may have been used for crushing the light shells of ammonites, which existed in vast numbers in the same seas.
[Pg 176]
In some of the small islands near the northeast coast of New Zealand certain small and peculiar, lizard-like reptiles, known as tuateras, have long been known. For many years they were supposed, even by scientific men, to be real lizards, so much do they resemble in external appearances and in habits the lizards of other parts of the earth. It was early observed, however, that they presented certain remarkable internal differences from the real lizards or Lacertilia, though it was not till about twenty-five years ago that the importance of these differences was recognized by the late Professor Cope, who separated them into a distinct order quite co-ordinate with the lizards, crocodiles, and turtles. These little reptiles, seldom reaching a length of two feet, have now become so scarce that the New Zealand government protects them by law from unnecessary destruction; nevertheless it will probably be only a short time before they become extinct, the end of a long genealogical line. No other living reptiles have retained more of the old-fashioned or primitive characters than this Sphenodon or Hatteria, as the animal is called, and because of them it is of peculiar interest to zoölogists, and especially paleontologists.
The differences of these beaked lizards from the true lizards are especially noticeable in the skull, and more especially in the arrangement of the bones which give articulation to the lower jaws (Fig. 8). In the lizards and snakes the quadrate bone is loosely articulated at its upper end with the cranium, and has no inferior bar or arch connecting its lower end with the jugal and the back part of the upper jaw. Sphenodon, on the contrary, has the quadrate bone firmly fixed to its adjacent bones at both ends, and is quite immovable. The vertebrae are biconcave like those of all early reptiles, not concavo-convex as are the vertebrae of most other living reptiles. The intercentra or hypocentra, little wedge-shaped bones [Pg 177] between the centra below, are more persistent in Sphenodon than in any other living land animals except the gecko lizards. Upon the whole the tuatera is the most old-fashioned of living reptiles, and in consequence it has nearly lost out in competition with new things.

Fig. 85.—Sphenodon punctatum,
or tuatera.
(From specimen in the Yale University museum.)
With these living tuateras we have nothing further to do, since they are land animals, living about the beaches of the New Zealand islands, and only occasionally venturing into the water, hiding from their enemies in the holes in the rocks. But, from some of their antecedents, from some of their direct forbears perhaps, there have gone off at different times various branches, whose descendants wandered into foreign lands or into foreign places, and lived and flourished for a brief time and then became extinct. Some of these went down into the water and became more or less aquatic in habit; some, indeed, changed their forms and habits so greatly that they are often, perhaps rightly, segregated into different orders. Whether or not they should be called Rhynchocephalia matters little, however. It is merely a matter of opinion as to how great the changes should be in order to entitle the offspring to a genealogical tree all its own. Of these branches there are two, whose relationships seem to be definite, the Choristodera and Thalattosauria, though there is more doubt about the latter than the former. A third group, that included Pleurosaurus, seems, from [Pg 178] more recent discoveries, to belong to a different line of descent and has been described under the Protorosauria.
In the direct line of ancestry there is no known form that was distinctly aquatic. The oldest known of these, perhaps, is that shown in Fig. 86, Sapheosaurus from the Jurassic of Solenhofen. Its resemblance to the modern tuatera is great, and doubtless its habits were very similar, though its rather long tail and rather short neck possibly indicate subaquatic habits.

Fig. 86.—Sapheosaurus,
an Upper Jurassic rhynchocephalian.
(After Lortet.)
Among the many reptiles of the past which have sought a more congenial or a safer home in the water few have had a more interesting history, or a briefer one, than those to which the late Professor Cope gave the name Choristodera in 1876. Many students of repute consider the group an order, others a suborder of the Rhynchocephalia. The group, whether order or suborder, are interesting because of their long and devious migrations from western North America to Europe, or vice versa, through rivers and ponds; interesting also because of the persistence of certain old-fashioned traits that clung to them long after their disappearance in other animals. Perhaps these traits were among the causes of their merely moderate success as animals of the water, traits that led to their early dissolution. Like the proganosaurs, which they must have resembled in external appearance not a little, they wandered from their birthplace in the western continent, to perish in the eastern; and like them their span of existence was short. [Pg 179]
Their history among mankind, too, is brief. The first known specimens, from western North America, were described by Professor Cope in 1876, under the name Champsosaurus. In the following year Professor Gervais of Paris made known another form from Rheims, which he called Simoedosaurus, so closely allied to the American that even yet they have not been sharply distinguished. Some years later these European specimens were more fully described by the well-known Belgian paleontologist, Dr. Dollo, but it has been only within the past few years that our knowledge of the animals has been made at all complete by the discovery and description of several excellent skeletons of Champsosaurus by Mr. Barnum Brown of New York.

Fig. 87.—Champsosaurus; skeleton,
as mounted in American Museum. (Brown.)
These semiaquatic reptiles never grew very large—not more than four or five feet in length; nor did they ever succeed in becoming fully at home in the water, certainly no more so than our modern alligators and crocodiles. They remained to the end of their comparatively brief existence essentially land animals, probably seeking their food in the [Pg 180] water but fleeing to the land for protection and for the breeding of their young. Their chief water adaptations are seen in the elongate face and flattened swimming tail. Their legs remained essentially terrestrial, and could have been of but little use in the water for propulsion; the feet even were doubtfully webbed, or if so, not more than are those of the alligator. Singularly, like the proganosaurs, their ribs were heavy and stout, also suggesting bottom-crawling habits, like those of the living Galapagos lizards. The skull was lightly built, and the face was long and slender, like that of the gavials and proganosaurs; but, like those of the former and unlike those of the latter, the nostrils were situated at the extreme tip. The hind legs were firmly attached to the body by the sacrum; and no sclerotic bones of the eyes have been discovered. The neck was neither unusually long nor unusually short. The body was probably covered with horny scales.

Fig. 88.—Restoration of Champsosaurus.
[Pg 181]
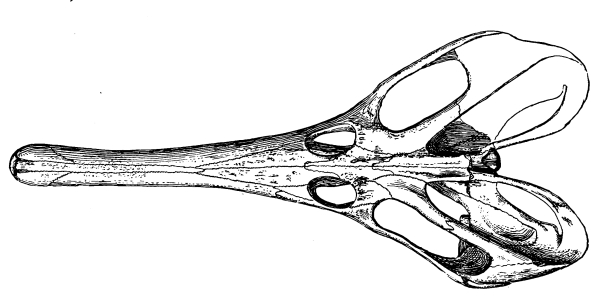
Fig. 89.—Champsosaurus; skull
from above. (After Brown.)
To the student of paleontology these animals are of interest because of the retention of several primitive traits which had long disappeared in other known reptiles. While the vertebrae had ceased to be perforated by the notochord, as in the early reptiles, they were still shallowly [Pg 182] biconcave. The first bone of the neck, the atlas, had changed but little from that of their very ancient forbears of Permian times, and the bones of the palate still retained numerous teeth scattered over it, like those of the same Paleozoic ancestors. Most primitive and old fashioned of all was the pelvis, which was so unlike that of all known contemporary or later reptiles that, were a paleontologist to see it without knowing whence it came, he would be almost sure to say that it belonged to a Paleozoic, or at least a Triassic, reptile, and not only to an early reptile but a very primitive one at that. This peculiarity consists in the absence of any opening between the ischium and pubis, which is characteristic of every living vertebrate with legs. And these and other old-fashioned characters could not possibly have been new developments; they must have existed in all the ancestors of the Choristodera from Paleozoic to early Tertiary times, though not a single other reptile is known to have possessed them, for the greater part of this time. Perhaps when Asia and northwestern America have been more thoroughly explored for vertebrate fossils, some of their ancestors which perished on their great migration from the western to the eastern continent in late Cretaceous times will be discovered.
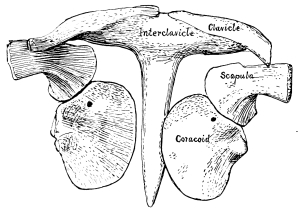
Fig. 90.—Pectoral girdle of
Champsosaurus.
(After Brown.)
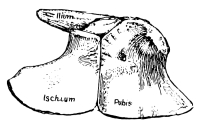
Fig. 91.—Champsosaurus;
pelvic bones.
(After Brown.)
The choristoderans began their existence, so far as is now known, in North America in late Cretaceous times and died out in both Europe and North America in early Tertiary times. That is, they were one of the [Pg 183] few branches of reptilian life which not only witnessed the extinction of the great dinosaurs and plesiosaurs, but the advent also of the early placental mammals. They lived millions of years after the proganosaurs became extinct, and, similar as they are in form, there is no relation between them. Moreover, in all probability they did not migrate to the eastern continent over the same route.
The structure of the head and teeth of the Choristodera clearly indicates a fish-eating habit, or at least a diet of soft-bodied, free swimming invertebrates. The legs and ribs, as also the armor of ventral ribs, like those of the plesiosaurs, point very insistently toward a bottom-crawling habit while in the water.
[Pg 184]
The first known specimen of the order of reptiles now generally known as the Parasuchia was found in Würtemberg, Germany, in 1826 and very briefly and inadequately described[4] two years later by Professor George Jaeger. The specimen was a sorry one, and was sadly misinterpreted by Jaeger. It consisted chiefly of casts of the alveoli or sockets of a number of teeth, more or less connected by corroded or decomposed portions of the jaws. He recognized the casts as teeth of a peculiar reptile, but mistook the roots for crowns, and, naturally concluding that such obtuse teeth would be of use only for the mastication of vegetable food—about the last kind of food to which the phytosaurs were addicted—called the animal Phytosaurus, meaning plant saurian. Because of differences he observed in the shapes of the teeth he thought that they belonged to two distinct species, which he called cylindricodon and cubicodon; but the differences were due simply to the different positions they held in the jaws.
Fourteen years later Hermann von Meyer, the renowned German paleontologist, described and figured other remains of the same or an allied reptile under the name Belodon plieningeri. In subsequent papers during the next twenty-three years von Meyer very fully described and beautifully illustrated the skulls and other remains of this and other species, all of which he referred to the genus [Pg 185] Belodon, the name by which for many years the animals were generally known in scientific literature. Von Meyer thought that he recognized in Belodon kapfii, the species most often figured in textbooks, the same animal that Jaeger had previously described.
Von Meyer was not at all certain about the relationships of his Belodon, though he recognized its affinity with the crocodiles. It was Huxley who, in a famous paper on the evolution of the crocodiles, published in 1875, united Belodon and another genus from the Trias of Scotland, which he called Stagonolepis, with the Crocodilia as representatives of the suborder Parasuchia, one of the three into which he divided the order. Huxley admitted that the relationships between the Parasuchia and the Mesosuchia or Eusuchia, the other suborders which he proposed, were not as intimate as those between the latter two, which were separated solely on the structure of the palate and vertebrae, as has been explained in chap. xv. As early as 1869 the late Professor Cope recognized certain forms which had been previously described from Carolina as belonging to the group, calling them Belodon, but it was not until 1896 that E. Fraas separated Belodon planirostris of von Meyer as a member of a distinct genus, to which he gave the name Mystriosuchus.
Here, as a part of the order Crocodilia, the phytosaurs remained till within very recent years, though there have been some mild protests against the association, especially by Marsh, Zittel, and Baur. The famous English paleontologist, Richard Owen, located the “Belodontia,” as the phytosaurs were often called, in his order Thecodontia, based chiefly upon the manner of the insertion of the teeth in sockets. But this has long since been shown to have little value in the classification of reptiles. Various authors have written about the phytosaurs in later years, notably Cope, Fraas, Huene, and Jaekel, but it was J. H. McGregor who first definitely separated the phytosaurs into a distinct order, in a careful revision of the American forms. He called the order Parasuchia, after Huxley, dividing into two suborders, the Phytosauria, after Jaeger, and the Aetosauria, a group which, for lack of a better place, had previously been classed with the Crocodilia, either as a member of the Parasuchia or as an independent suborder by Zittel (the Pseudosuchia). More recently Huene has shown [Pg 186] that certain African reptiles from the Lower Trias had certain very definite characters entitling them to an independent position, for which he proposed the order Pelycosimia. Upon the whole, however, these characters seem to be primitive parasuchian, and the group may provisionally be placed in the order Parasuchia, as a third suborder, the Pelycosimia. The order Parasuchia, then, until we know much more about the latter two groups, may be conveniently divided into three suborders, the Phytosauria, Aetosauria or Pseudosuchia, and the Pelycosimia, all of Triassic age.
McGregor was quite right in retaining for the suborder the name Phytosauria, suggested by Jaeger in 1828, inappropriate as the word is etymologically, but was hardly justified in substituting the generic name Phytosaurus for the long and well-known Belodon, because it is quite impossible to say that Jaeger’s very fragmentary specimen upon which he based the genus Phytosaurus really is the same as Belodon. Professor Fraas very kindly showed the writer the original type-specimen of Jaeger, now preserved in the Stuttgart Museum, and both are agreed that it is impossible to prove the identity of Belodon and Phytosaurus from the very fragmentary and imperfect specimen. It is quite as probable, for instance, that Phytosaurus and Mystriosuchus are identical as that Phytosaurus and Belodon are. Unfortunately, this is not the only case in vertebrate paleontology where the fragmentary specimens to which names have been given are inadequate to determine the species, or genus, or even the family to which they belong; there have been very many such instances. The pioneers in paleontology were often justified in naming small and obscure fragments of bones, or single bones. One would be justified even yet in giving a name to an indeterminable fragment of a bird bone from the Triassic formation, because the discovery of a bird of any kind in that formation would be very important for science, even if its precise kind might never be recognized from the specimen. Nevertheless, the custom is a very reprehensible one when indiscriminately followed. For these reasons the writer disagrees with McGregor in substituting the inappropriate name Phytosaurus for Belodon, the name by which the most typical forms were so long known. [Pg 187]
The Aetosauria, which have long been known from a marvelous specimen found in Würtemberg many years ago and described by the elder Fraas, need not detain us long. They were relatively small reptiles about two feet long, almost completely incased in a bony armor, and purely terrestrial in habit. The skull even yet is not perfectly known, and it is possible that when it is the group may have to be dissociated from the phytosaurs. The nostrils were not posterior, and the skull is short. Other specimens of the same group have been described from the Upper Triassic rocks of Massachusetts.
The Pelycosimia of Huene are very interesting as showing apparently primitive forms with which the true phytosaurs may have been intimately related ancestrally. They, too, have a rather short skull with the nostrils in front, and were not at all aquatic in habit. Not much is known about the single genus that is located in the group, aside from the skull and a few limb bones.
The Phytosauria, so far as known, were all reptiles of considerable size, greatly resembling the crocodiles, and especially the gavials in form and habit, but differing very greatly in having the external nostrils situated far back near the eyes; in having no false palate so characteristic of the Crocodilia; in having a more primitive shoulder-girdle, consisting of a short coracoid, interclavicle, and clavicles; and in having the ordinary type of pelvis, that is, with the pubis entering into the acetabular articulation for the femur. They were all, like the crocodiles, covered more or less by a bony armor; there are two openings on each side of the temporal region; there is no pineal opening; the vertebrae are gently biconcave, precisely like those of the early or mesosuchian crocodiles; there is always an opening of considerable size, called the preorbital foramen, in front of the eyes, as in some crocodiles, many dinosaurs, and most pterodactyls; there is also an opening through the back part of the mandibles as in crocodiles; and the double-headed ribs are attached exclusively to the transverse process of the arch, precisely as in the crocodiles, dinosaurs, and pterodactyls. From all these it is evident [Pg 188] that the phytosaurs are related most nearly to the crocodiles and dinosaurs, and are probably an early branch of the stem from which they, the pterodactyls and the birds, arose, a branch that persisted [Pg 189] only a short time, geologically speaking, and went entirely out of existence at the close of Triassic times, leaving no descendants behind. Nevertheless, in this comparatively brief life-span they developed not a few distinctive forms and became widely distributed over the earth. Their remains are known from the Upper Trias of Germany, England, and Scotland, India, South Africa, and from Massachusetts, North and South Carolina, and many places in the Rocky Mountains. No true phytosaurs are yet known from South America, but in [Pg 190] all probability they will be discovered there when the Triassic deposits of that continent have been better explored for fossils. In the Rocky Mountains, especially, their remains are widely scattered, they have been found in many localities in Wyoming, Colorado, Oklahoma, Utah, and New Mexico. Though for the most part their known remains from these localities are yet fragmentary, not less than four distinct genera have been described from these regions: “Belodon,” Angistorhinus, Paleorhinus, and Episcoposaurus. From the Carolinas and Massachusetts a single genus, though described under numerous names, has been made known, originally called by Emmons Rutiodon (Rhytidodon). And from Europe and India at least as many more different genera are known. All these genera are, however, so closely allied that they are placed in the single family Belodontidae.

Fig. 92.—Restoration of Mystriosuchus,
an Upper Triassic phytosaur.

Fig. 93.—Skull of Mystriosuchus, a phytosaur: pm, premaxilla; m, maxilla; na, nasal; f, frontal; p, prefrontal; l, lacrimal; pf, postfrontal; po, postorbital; pa, parietal; sg, squamosal; qj, quadratojugal; pl, palatine; t, transverse; in, internal nares; en, external nares; pt, pterygoid; bs, basisphenoid; eo, exoccipital. (After McGregor.)
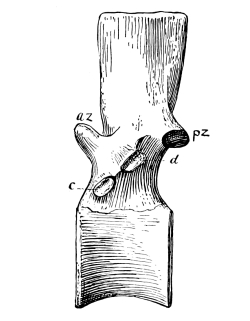
Fig. 94.—Dorsal vertebrae
of phytosaur: az, anterior zygapophysis;
pz, posterior zygapophysis; d,
c, articulations of rib.
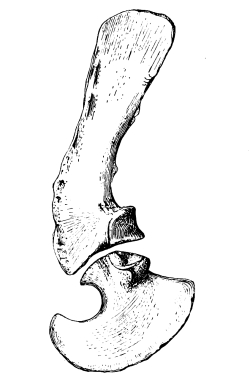
Fig. 95.—Scapula and
coracoid of Rutiodon carolinensis,
an American phytosaur.
(After McGregor.)
[Pg 191]
Fig. 96.—Belodon; restoration of head,
from above.

Fig. 97.—Mystriosuchus: restoration of
head,
from above.
In Belodon (Fig. 96), the earliest known and most typical genus, perhaps, the moderately elongated face has a high crest reaching nearly to its front end, and this type is known both from Europe and from New Mexico. Others have the face long and slender, even longer and more [Pg 192] slender than in the ancient teleosaur crocodiles or the modern gavials. In some forms the teeth are cylindrical and slender throughout, and there may be as many as fifty on each jaw, or two hundred in all; while in others only the anterior teeth are cylindrical and the posterior teeth are flattened and serrate along their cutting edges. In the body not very great differences have been observed. Some are more slender than others, and there are minor differences in the shapes and sizes and numbers of the bony scutes along the back and on the throat.
But they are all alike in their essential characters, a very long beak with numerous teeth; the foremost ones on the expanded, more or less spoon-shaped front extremity, are more or less, sometimes greatly, elongated. The jaws may be likened to a long and slender pair of tongs with nipping teeth at the front end. The strong, long, and flattened tail is sufficient evidence that the phytosaurs were excellent swimmers, but, aside from that and the posterior location of the external nostrils, directly over the internal, few other aquatic adaptations are observed in the skeleton. There are no sclerotic bony plates about the eyes, or at least none have so far been discovered, although among the numerous known specimens they would confidently be expected were they really present in the skeleton; and the presence of bony armor negatives markedly aquatic habits.
Doubtless on the whole the habits of the phytosaurs were not very unlike those of the modern gavials, which they so strongly resemble in form, size, and general characters. But they differ very greatly from the gavials in the extreme posterior position of the nostrils, and in the greatly elongated teeth of the front end of the beak, teeth which must have had some especial and peculiar use. Nor is the position of the nares to be accounted for satisfactorily by reference to aquatic habits. It has been suggested that the creatures used the very long and slender beak in prodding and probing in the sand and mud for soft-bodied invertebrates, worms and the like, for which the teeth would be especially fitted; and that the posterior position of the nostrils may be in part, perhaps wholly, accounted for by this habit, which permitted the reptiles to breathe without extricating the beak from the mud or shallow waters. That the animals were wholly and [Pg 193] intensely carnivorous in habit is attested by their teeth; although they are called “plant saurians,” they never had anything to do with plants in the way of food. Unfortunately so far no specimens have ever been found showing the remains of stomach contents, nor have any been found showing impressions of the form of the body or of any of its parts. Until such specimens are found, as they doubtless will be eventually, one can be less sure of the precise details in their life reconstructions. However, the skeleton is now known nearly completely, and this suffices to give a very approximately correct idea of what the animals were like when alive.
[Pg 194]
The order of reptiles to which the name Crocodilia is technically applied comprises less than twenty-five living species, popularly known as crocodiles, alligators, caimans, and gavials. They are often of great size, ugly and repulsive in appearance, cruel and vicious in habit, wholly carnivorous, and denizens, almost exclusively, of fresh-water lakes or rivers in tropical and subtropical regions; a few only venture into the sea near the shores. They are all excellent and powerful swimmers, but are by no means exclusively aquatic in habit, many of them spending a large part of the time on the shores; and they invariably seek the land for the deposition and hatching of their eggs. In size they are the largest of living reptiles, some of the existing species reaching a length of twenty-five feet, while some extinct species were probably fully twice that length.
The geological history of the crocodiles is a very ancient one, reaching back at least as far as the early Jurassic and probably to the Triassic. The culmination of the order, at least so far as size, variety, and numbers are concerned, was doubtless before the close of the Mesozoic. The early crocodiles, however, were of a more generalized structure in some respects, though specialized in others, because of which naturalists in the past have usually divided the order into three or four chief subdivisions, or suborders, two of which, the Mesosuchia and the Thalattosuchia, became extinct before or during Cretaceous time. The third suborder, the Parasuchia of many textbooks, has now been unanimously separated by paleontologists from the Crocodilia as an independent order. The fourth suborder of the textbooks, the Eusuchia, a word meaning true crocodiles, appeared in geological history, so far as we yet know, shortly before the extinction of the Mesosuchia, and is best known from the forms now living, all of which belong to it. [Pg 195] Although the modern crocodiles can hardly be called, as a group, purely aquatic reptiles, we shall find it of interest, because of their intimate relation to the older and more strictly marine forms, to speak of them somewhat in detail.
The crocodiles of the present—and we use the word in the technical sense of Crocodilia—because of their general resemblance to the lizards, or true “saurians,” were classed with them by the older naturalists, whence comes the popular name alligator, a corruption of the Spanish el lagarto, or “the lizard,” given to some of the South American forms by early explorers. But this resemblance is a superficial one only, as was early recognized by comparative anatomists. The crocodiles, indeed, are only remotely related to the lizards.

Fig. 98.—Senegal crocodile.
(By permission of the New York Zoölogical Society.)
The head or cranium is flattened and broad, the facial part or snout sometimes greatly elongated and slender, and the external nostrils are always situated at the front end. The bones of the upper surface of the cranium and face have many pit-like depressions. The neck is short and stout, and but little movable. The body is somewhat depressed and flattened, not cylindrical and slender, as in the more typical water reptiles. The tail is much elongated and compressed, forming a powerful [Pg 196] means of propulsion in swimming, its breadth being increased by a vertical row of horny plates above. The limbs are of the ordinary elongated type—ambulatory rather than swimming legs—and are not of much use for propelling the body in the water; the front legs indeed are usually held close to the body while the animal is swimming. The toes, however, are partially connected by webs, to a slight extent only in the alligators and crocodiles, but much more so in the long-snouted gavials. The feet have five toes in front and four behind; and the loss of the fifth toe can only be ascribed to terrestrial habits. The body is covered more or less with horny scutes or scales, beneath which are several rows of thickened, pitted, bony plates on the dorsal side, and sometimes also on the under side, forming a more or less extensive bony armor. The eyes have movable lids, as in most lizards, and the ear-opening is small.
But the external appearance of these reptiles is not sufficient to distinguish them widely from other groups, and we must resort to the internal structure, especially that of the skeleton, for the more essential differential characters. The most crucial of these, the one which more than any other determines their relationships, consists in the position and shape of the bone with which the lower jaw articulates, the quadrate bone, so characteristic of reptiles. As in the plesiosaurian and ichthyosaurian skulls, it is firmly united with the adjoining bones, not articulating freely with them, as in the lizards and snakes. But this fixed relation of the bones is very unlike that of the plesiosaurs, ichthyosaurs, and turtles, in that it is held in place by two bony bars or arches, the upper extending forward to unite with the bones at the back part of the orbit, the lower, with the hind extremity of the upper jaw. The lower jaws are rigidly united in front, sometimes for a long distance; they have, almost always, a hole or opening through the hinder part, known in but few other reptiles. The bones of the palate are all firmly united, forming a nearly complete roof, very unlike the condition in the mosasaurs and lizards. The palate also is very peculiar in the development of a plate of bone below the nasal chambers, forming a complete bony canal on each side through which the respiratory air passes far back to the internal [Pg 197] opening of the nostrils above the pharynx, and not, as in other reptiles—save the immediate ancestors of the mammals—entering the mouth at the front end. This peculiar arrangement of the air-passages, so like that of ourselves, has much to do with the water habits of the crocodiles, as we shall see.
The teeth are conical in shape, and are always restricted to the edges of the jaws, above and below. They are inserted deeply and firmly in sockets, and are replaced frequently by new ones growing beneath them, pushing the older ones out as their usefulness becomes impaired by injury or by use. In some species there are as many as thirty teeth in each side of the jaws, above and below, although the broad-headed kinds have a smaller number.

Fig. 99.

Fig. 100.
Fig. 99.—Skull of Alligator mississippiensis, from below.
Fig. 100.—The same, from above: bo, basioccipital; bs, basisphenoid; f, frontal; j, jugal; l, lacrimal; m, maxilla; n, nasal; p, parietal; pa, palatine; pm, premaxilla; pf, prefrontal; pr, postfrontal; pt, pterygoid; q, quadrate; qj, quadratojugal; tr, transverse.
The neck is short, as has been stated, but it always includes in living forms nine vertebrae, a number probably slightly in excess of that of [Pg 198] their terrestrial forbears. By the peculiar mode of attachment of the short “hatchet-shaped” ribs, much lateral movement of the neck is prohibited, nor is any very great vertical movement possible. The vertebrae of the whole column, save the atlas, the second sacral, and the first caudal—which is a very remarkable anomaly—are concave in front and convex behind, agreeing in this respect with those of all other living reptiles, save the turtles, the tuatera, and some lizards. The ribs of the neck have their two heads attached, one to the body of the vertebra, the other to the arch, but those of the dorsal region, though double-headed, have both become attached to the transverse projection of the arch, a seemingly trivial character, but one which immediately distinguishes all crocodiles from all other water reptiles, and from all terrestrial reptiles, indeed, save the Parasuchia, Pterosauria, and Dinosauria. The pelvis is firmly attached to the spinal column by two sacral vertebrae.

Fig. 101.—Vertebrae of gavial from the side (cervical), and from in front (dorsal): az, anterior zygapophysis; pz, posterior zygapophysis; d, diapophysis; r, cervical rib; c, articulation for head; t, for tubercle of dorsal rib.
The collar-bones, or clavicles, are wanting in crocodiles; there is a slender interclavicle; and the shoulder-blade and coracoid are well developed (Fig. 102). The bones of the pelvis are loosely united with each other as they are in most reptiles (Fig. 104). The pubes, the anterior bones below, unlike those of all other reptiles, do not help to form the acetabulum or socket for the articulation of the thigh bone, nor do they articulate with each other. This single character sharply distinguishes a crocodile from all other reptiles, living or [Pg 199] extinct, and is almost the only character that separates the order from the dinosaurs, aside from the peculiar structure of the nasal passages in the skull. On the under side of the body, connected with the front end of the pubes, there are seven or eight pairs of abdominal ribs, corresponding to the plastron of the turtles and similar to those of the ichthyosaurs and plesiosaurs. The mosasaurs have no such ribs.
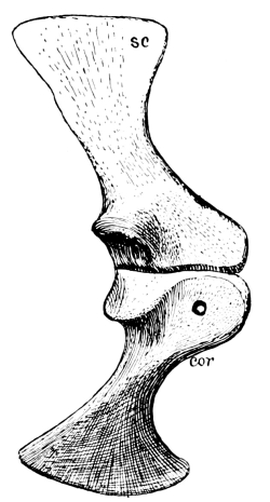
Fig. 102.
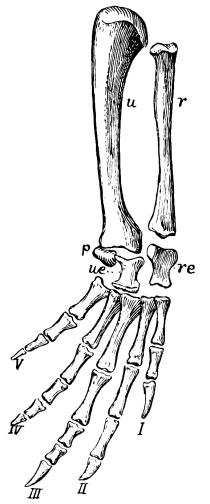
Fig. 103.
Fig. 102.—Scapula (sc) and coracoid (cor) of gavial.
Fig. 103.—Front foot of crocodile: u, ulna; r, radius; re, radiale; ue, ulnare; p, pisiform.
Furthermore, the crocodiles differ from all other living reptiles in having a four-chambered heart, like that of birds and mammals, that is, a heart with two auricles and two ventricles. This more perfect structure of the circulatory organs does not, however, insure at all times a complete separation of the pure or arterial blood from the impure or venous blood, since the blood may be more or less intermixed outside of the heart by a connection between [Pg 200] the venous and the arterial systems. Whether these imperfectly developed organs, so suggestive of a higher and more perfect mode of respiration, are the vestiges of what were once among some reptiles functional structures, or whether they are rudiments of a higher organization, developing independently in these creatures, cannot be positively determined, but it seems very probable that, far back in geological times, some reptiles, especially the pterodactyls and dinosaurs, had their respiratory and circulatory systems more like those of the birds and mammals of today. Unfortunately, however, if such was the case, we may never be able to prove it, although proof would not be impossible; stranger things than fossil hearts have been found by paleontologists!
The stomach, moreover, in the crocodiles is fashioned somewhat after that of the birds, with an imperfect division into crop and gizzard. Some crocodiles of today have the habit of swallowing hard pebbles, as do many birds. There is an old myth that the crocodile of the Nile swallows a pebble on each of its birthdays, thus giving reliable information as to its age by the number found in its gizzard at its death! And this habit has been suggested for some of the most ancient crocodiles, the teleosaurs, by the recurring presence of siliceous pebbles found with the remains of their skeletons. And we have seen this pebble-swallowing habit was also characteristic of the plesiosaurs, with whose remains “stomach-stones,” or gastroliths, as they have been called, are often found.
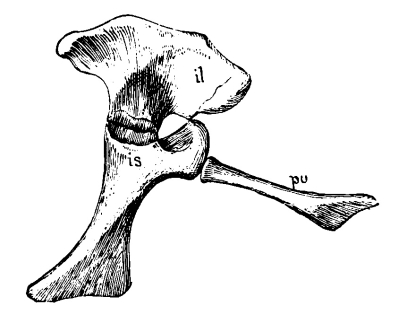
Fig. 104.—Pelvis of crocodile: il, ilium; is, ischium; pu, pubis.
All of these various characters of the skeleton and fleshy parts are pretty conclusive evidence that the crocodiles, ugly creatures that they are, today enjoy the highest rank among cold-blooded animals. They are perhaps in some respects of not so high a type of reptiles as were some of the extinct reptiles, but that they have survived so long, so many millions of years, is pretty good evidence of endurance, to say the least. [Pg 201]
Living crocodiles belong to three distinct groups or families: the true crocodiles and alligators; the long-snouted crocodiles or Borneo gavials; and the true gavials of India. Members of the first of these families are really only subaquatic, or amphibious in habit; they move about on land with entire freedom, and often seek their food there. Certain marked aquatic characters they do possess, in the skull and tail, as we shall see. They are indigenous to southern China, India, Africa, Madagascar, the southern part of the United States, Central America, and the northern part of South America. The members of this family are distinguished by the more or less broad and flat head, the possession of comparatively few teeth of large size, and by having the toes less completely webbed. The crocodiles proper differ from the caimans and alligators especially in the arrangement of the teeth. During later geological times, that is, during early Tertiary times, the geographical range of the Crocodilidae was much more extended than it is at present, the remains of many often very large species, being found in the lake deposits of the northwestern parts of the United States, Great Britain, Germany, France, etc. Yet earlier, in the late Cretaceous rocks of the United States, in Texas, and Wyoming especially, there have been found rather scanty remains of a gigantic crocodile which must have been nearly fifty feet in length when living.
The second family, the Tomistomidae, or long-snouted crocodiles, comprises but two living species, both restricted at the present time to Borneo. These crocodiles have a moderately slender snout, because of which they are sometimes called gavials, though it is not nearly so slender as that of the true Gangetic gavial. This family is probably older than either of the other living ones, and is the only one known with certainty to have lived during much of the Cretaceous times, several species of considerable size having been found in New Jersey and Europe. Their feet are better webbed than are those of the true crocodiles and alligators, the front feet partly, the hind feet wholly so. In general structure they seem to be the most primitive of the living Crocodilia, and may have been the ancestors of all modern forms. [Pg 202]

Fig. 105.—Gavial.
(By permission of the New York Zoological Society)
[Pg 203] The third family, the Gavialidae, also comprises but two living species, both restricted in habitat to the rivers of India. Of these the famed gavial of the Ganges is the better known and the more highly specialized. The skull of this species has an exceedingly long and slender snout; the teeth are numerous, small, and slender; and the feet are more fully webbed than are those of other members of the order. In habits the gavials are more distinctly aquatic than are the crocodiles and alligators. They feed almost exclusively upon small fishes, for the seizure and retention of which their small and sharply pointed teeth are admirably adapted. The hind feet are relatively long, a character that will be better understood when comparison is made with those of the Thalattosuchia. Although attaining a large size, fully twenty-five feet in length, they are comparatively harmless animals, never attacking human beings or other large animals, as do some of the crocodiles proper. The gavials have lived a long time in the Indian regions, the Gangetic gavial itself having been found in deposits of Pleiocene age, perhaps the oldest known of all living species of air-breathing vertebrates. Some of the extinct gavials of the same region attained a length of nearly or quite fifty feet, possibly the longest, if not the largest, of all swimming reptiles of ancient or modern times. Extinct gavials have been reported from South America, but are not yet fully known.
While the fish-eating gavials swallow their prey whole, the crocodiles, caimans, and alligators prey upon all living animals that come within their reach, whether large or small, and they will often leave the water to seize their intended victims, such as pigs, sheep, birds, or even human beings. Their teeth, as has been already stated, are much larger, longer, and more irregular in size than those of the gavials. Their victims are often drawn under the water and drowned, the peculiar posterior position of the internal nostrils permitting the animals to breathe with the mouth and to firmly hold their prey under water, while the extremity of the snout and the external nostrils are above the surface.
As the firm, unyielding bony palate, the fixed position and articulation of the lower jaws, and their rigid attachment to each other in front do not permit creatures of large size to enter the gullet [Pg 204] whole, the crocodiles and alligators must tear their food to pieces, which they do by quick, strong jerks from side to side, aided by the powerful tail; or they may twist off a limb or some other part of their victims by a rapid rotation of the whole body, two assisting in this operation, rotating in opposite directions.
Living crocodiles lay from twenty to sixty eggs, according to the species; these eggs are sometimes the size of a goose egg, and are covered with a hard shell. They are laid either in a deep excavation in the sand and covered over by the parent; or under leaves and straw. The female remains on guard until the eggs are hatched, of which she is apprized, it is said, by a peculiar noise uttered by the partly imprisoned young. She thereupon reopens the nest, and guides her liberated infants to the water, where she leaves them to their fate. Whether this remarkable habit is one that has been acquired in recent times or not is uncertain, but because it has been observed in a number of unrelated forms, it is probable that the instinct is of long inheritance, and may account for certain peculiarities of structure in some of the ancient members of the order. Doubtless the habit arose because of the unprotected places in which the eggs are necessarily laid on the shores and beaches, and because the eggs are comparatively so few in number. The sea-turtles likewise lay their eggs in hollows scooped out of the sand of the beaches, but the parents give no further care to their eggs, nor to their newly hatched offspring, a neglect which is compensated for by the much larger number of eggs they lay, because of which the chances are much greater that a few will survive the more numerous vicissitudes to which the eggs and young turtles are exposed.
The name Mesosuchia, meaning “middle crocodiles,” by which the ancient members of the Crocodilia have generally been known, was given by Huxley in the belief that they were intermediate between the “true” or modern crocodiles and an ancient group which he united with the order under the name “Parasuchia.” A fuller and better knowledge of the members of this last group has proved very conclusively that they are really less allied to the crocodiles than are some other orders of reptiles, the dinosaurs for instance, and should be properly classed by [Pg 205] themselves as a distinct order. And, more recently, it has also become quite apparent that the old crocodiles should not be separated so widely from the modern ones as Huxley proposed; that the differences distinguishing them from the recent members of the order are really not of more than family importance. We thus have left but two chief divisions of the Crocodilia, the Eusuchia and Thalattosuchia; and the latter group even, by some authors, perhaps rightly, are included under the true crocodiles as a family only.
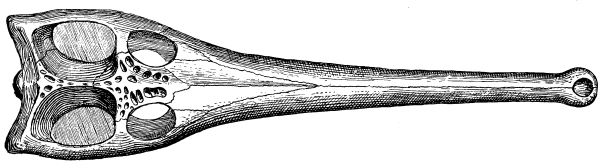
Fig. 106.—Teleosaurus; skull, from above.
These older crocodiles, the Mesosuchia of Huxley, comprise a considerable number of extinct forms which lived as far back as the early part of the Jurassic, and continued nearly to, if not actually into, Cenozoic time, that is, to the Eocene. They differ from all living forms, chiefly in having, not concavo-convex but biconcave backbones, that is, the more primitive vertebrae with which all reptiles began. Nor was the internal opening of the nasal passages so far back in the mouth as in the later forms. In other respects they did not differ very greatly from some of those now living. All the earliest kinds that we know of—the teleosaurs—had a long, slender snout, resembling very much that of the modern gavials. And they were, for the most part, incased in a more complete bony armor, on both the dorsal [Pg 206] and the ventral sides; and the front legs were smaller than those of the gavials even. The resemblance of the living teleosaurs to the modern gavials must have been very great, although the heavier bony armor indicates a less exclusively aquatic life. They probably lived more in the shallow waters of the seas near the shores.
Near the close of the Jurassic appeared for the first time, so far as we now know, broad-headed mesosuchian crocodiles, forms having less numerous and stronger teeth, and resembling closely modern alligators. It has been believed that these broad-headed kinds were of later origin than the more slender-nosed teleosaurs, but a moment’s consideration will make it evident how improbable such an evolution must be. The crocodiles must have descended from strictly terrestrial reptiles, and no terrestrial reptiles have a slender nose. That they should have acquired a slender face in adaptation to water habits and then returned to the more primitive land type with a broad face and less strictly aquatic habits is contrary to all our experience in paleontology. From this it is altogether probable that broad-faced crocodiles of later times must have been the descendants of broad-faced kinds that were in existence during all the Jurassic times, but of which we as yet have no knowledge. These broad-faced Jurassic crocodiles were, for the most part, small creatures, much smaller than the teleosaurs even, and smaller than any species of crocodiles now living. Their remains are known only from fresh-water or shore deposits, and are, for the most part, associated with those of land and fresh-water animals. About the time of their first known appearance in geological history, the small mammals and birds had also become more or less abundant, and it was suggested by Owen that these land animals had something to do with the development of the ancient amphibious crocodiles. Perhaps this was the case with respect to their greater abundance and development, and with certain peculiarities of their structure, but that the gavial-like teleosaurs should have come back to the land and reverted to a more primitive form seems quite improbable.
During Cretaceous times, especially in America, numerous forms of these old mesosuchian crocodiles were doubtless in existence, both slender-nosed and broad-nosed, and some of them must have been of [Pg 207] considerable size, though none known was as large as some of the late crocodiles. This type, with biconcave vertebrae, continued to live on, in both North and South America, to the latter part of Cretaceous times, and it is even possible that some continued on into the Tertiary. But long before the close of the Cretaceous, the modern kind appeared, those with concavo-convex vertebrae, and more posterior internal nostrils. The earliest are known from New Jersey (Thoracosaurus, Holops), so like the Borneo gavials of today that they are properly classified in the same family, the Tomistomidae or Gavialidae. If all the later, procoelian type, that is, those with concavo-convex vertebrae, originated from a single form when the amphicoelian or mesosuchian type became extinct, Huxley’s classification into the Mesosuchia and Eusuchia would perhaps be proper, but we have much reason to suppose that the change in the kinds of vertebra and in the position of the nostrils was only incidental, and may have occurred in more than one line of descent, that is, it may have occurred in the broad-headed kinds of the Jurassic to the broad-headed crocodiles of today, as also in the gavial-like forms of the Cretaceous to the gavials of the present. And this is the reason why naturalists no longer recognize the classification of Huxley, which, partly perhaps because of the prestige of his name, has so long been accepted in our chief works on natural history.
While the ancient crocodiles of which we have spoken resembled the modern ones so closely in form of body and probably in habits, there were certain others of the old Jurassic seas which departed so widely both in structure and in habits, from their associates that they are by some authors given a place wholly by themselves as a distinct group. This has been called by Professor Fraas the Thalattosuchia, a word meaning “sea-crocodiles.” They were a very early side-branch from the great genealogical tree of the Crocodilia, a branch which departed so widely from their associates in adapting themselves to a peculiar and aberrant mode of existence that they cannot be considered as typical crocodiles, although so closely related to them in other respects that [Pg 208] therecannot be the least doubt regarding their ancestry. None of the crocodiles which we have considered, whether ancient or modern, can truthfully be called purely aquatic. They never ceased to use their limbs for locomotion on land, more or less of the time, or for the support of the body; and many of them have subsisted, in part at least, on land animals. How easy it may have been for some of them to become almost wholly emancipated from land habits we may easily conjecture. The gavials, as we have seen, are more at home in the water than upon land; their food is chiefly found in the water; but, so long as their habits restrict them chiefly to fresh-water, or to the vicinity of the [Pg 209] shores, their limbs continue to be used as much for crawling as for swimming. Were the gavials to be driven out to sea by the stress of fresh-water conditions or attracted thereto by a greater abundance of more easily obtainable or better food, so far from land that they no longer would have much use for their legs for the support or propulsion of their bodies, in the course of time they would doubtless change to a more purely aquatic type. And in that change there would be material modifications of their structure: their limbs would become better adapted to movements in the water; the skin would become smoother, and the bony and horny scales would be lost, since they would be not only useless in the water, but actually detrimental to the well-being of the animals; and the tail would develop into a more powerful organ of propulsion, as a means of increasing their speed in obtaining food or in escaping their enemies, such as the sharks. In fact, we can only imagine that in the transformation precisely those modifications would occur which we actually find in these old sea-crocodiles, the Thalattosuchia. And they are of especial interest to us here because nowhere do we find a better example of Nature in the act of transforming a terrestrial or subterrestrial animal into a truly aquatic one.

Fig. 107.—Geosaurus, an Upper Jurassic thalattosuchian crocodile, drawn from restoration and figures by Fraas.
The group comprises only a few forms, so far as known. All were of modest size among extinct reptiles, from ten to twenty feet in length, and all are from the Upper Jurassic deposits of Europe. They did not exist very long, probably because they found it impossible to discard old habits and old structures entirely and become absolutely [Pg 210] emancipated from the land; their breeding habits possibly were too deeply impressed into their structure readily to change, as did those of other sea-reptiles. Some of their remarkable aquatic adaptations have long been known, but only within a dozen years has our knowledge of them become at all complete. Three or four genera have been described, but only a few forms are well known, of which Geosaurus may be taken as most typical. To this we shall confine our descriptions.
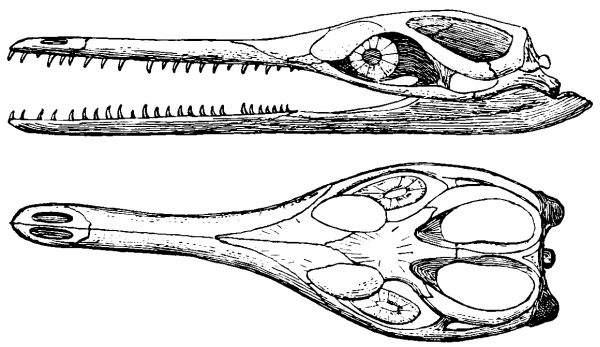
Fig. 108.—Geosaurus; skull from side
and from above.
(After Fraas.)
The skull of Geosaurus is rather small in comparison with the length of the body, smaller proportionally than in any living crocodile, but not much smaller than that of the teleosaurs. The snout is long and slender, much like that of the teleosaurs and gavials, but the bones of the whole upper surface are quite smooth, not roughened and pitted like those of modern forms. The skull of Dakosaurus, another genus of thalattosuchians, is much less elongate than that of Geosaurus, but has the other characteristics of Geosaurus. The eyes are provided with a stout ring of sclerotic bones, with a pupillary opening of less than one inch. We have seen that all other strictly aquatic reptiles have similar eye bones, but no other crocodiles have them. The internal openings of the nostrils are large and long, but they are not situated far back, as in the modern crocodiles, not even so far back as in the early teleosaurs. They had no need of the peculiar breathing apparatus of the amphibious crocodiles, since all their prey must have been water-breathing creatures. Their eyes were directed laterally, not more or less upward, as in their nearest relatives. Nearly all other crocodiles have an [Pg 211] opening through the hind end of the lower jaw, but the thalattosuchians did not. The teeth were about as numerous as in the modern gavials, but they projected freely only a short distance above the gums in life, and they were very slender and sharply pointed, excellently well adapted for catching smooth and slippery fishes. Their vertebrae, like those of all other reptiles of their time, were biconcave. Those of the neck resembled those of the teleosaurs, save that there were only seven, fewer than is the case with any other members of the order. In becoming adapted to their peculiar mode of life these crocodiles lost two vertebrae from the neck. All modern crocodiles have two ribs attached to the first vertebra; the thalattosaurs had but one, another evidence of primitive characters. While the number of vertebrae in the neck was reduced, in the back it was increased to eighteen; all other crocodiles have but fifteen or sixteen. The trunk was long, another adaptation to water life. There were two firmly united vertebrae in the sacrum, as in the modern forms. The reason for the persistence of this terrestrial character we shall see later.

Fig. 109.—Tail, scapula (sc), and
coracoid (c) of Geosaurus.
(After Fraas.)
The tail was very long and strong, nearly as long as all the remainder of the body, and relatively much longer than in other crocodiles. It is of interest to observe that the head, neck, body, and tail had almost the same relative proportions as those of the great sea-lizards, the [Pg 212] mosasaurs. The terminal bones of the tail are very peculiar, and very different from the corresponding bones of other crocodiles. While the spines of the tail bones along the anterior part are only moderately stout and long, and are directed obliquely backward, near the terminal part they become suddenly much broader and are directed upward, and, a little farther along, obliquely forward. The chevron bones on the under side also here become broader and longer. The end of the tail curves markedly downward to end in a slender point. It will be remembered that a similar downward curvature of the end of the tail observed in nearly all specimens of ichthyosaurs induced in Owen the belief that the animals had a fleshy terminal fin, a belief which later discoveries of the carbonized remains of the flesh confirmed. The peculiar structures observed in various specimens of these sea-crocodiles, even though no impressions or remains of the fleshy parts have been discovered, is quite conclusive evidence that these animals also had a broad, fleshy, terminal fin. No other explanation of the structure is possible.
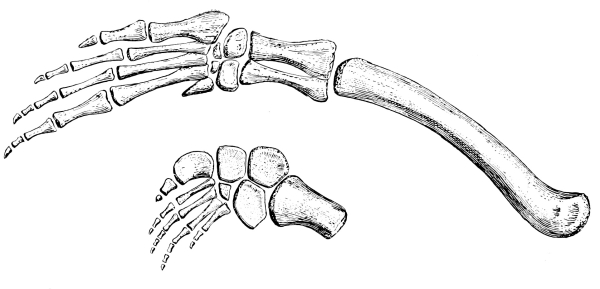
Fig. 110.—Geosaurus. Elongate hind leg,
and paddle-like front leg.
(After Fraas.)
The ribs are not at all stout and are not much curved. They are directed posteriorly in the known specimens preserved in the matrix in [Pg 213] such a way as to indicate a slender thorax and abdomen, not the broad body of the modern crocodiles. The abdominal ribs, that is, those protecting the region on the under side of the body between the breast bone and the pelvis, are strongly developed in Geosaurus. The sternum, always present in other crocodiles, is wanting in Geosaurus. The shoulder-blades and coracoids, often changed in shape in water reptiles, are not unlike those of the amphibious crocodiles, but are smaller and flatter.
The fore limbs, to use Professor Fraas’s words, “are among the most interesting of all the peculiarities of Geosaurus,” and show very clearly that these animals were excellent swimmers. The humerus is remarkably short and broad; the two bones of the forearm, the radius and ulna, are broad, rounded, or angular plates, not long bones, as in other crocodiles; the wrist bones also are broad and plate-like. The three bones of the thumb, that is of the digit which received most strongly the impact of the water in swimming, are also broad and flat. All of these bones are marvelously aquatic in type, as will be evident from a comparison of them with the corresponding bones of the ichthyosaurs and mosasaurs. The bones of the other fingers, however, were not much changed from the ordinary crocodilian form, as a further comparison of them with the fingers of a land crocodile will show. Furthermore the whole limb or paddle was very small in comparison with the hind leg, and it was attached very near to the head. The relative proportions of the front and hind limbs in the geosaurs, gavials, and alligators are instructive as showing the progressive decrease in size of the front legs from the subaquatic, through the semiaquatic, to the almost purely aquatic type. The hind legs, strangely enough, were not very different in size and structure from those of the gavials or teleosaurs. The thigh bone is long and slender, though the bones of the leg and ankle are somewhat shortened and flattened, as are also those of the first toe. There were no claws on the hind feet, a distinctly aquatic adaptation, and the toes were certainly webbed. The pelvis, while not especially stout, is of good size, and was firmly attached to the sacrum. [Pg 214]
Perhaps all these characters may best be summed up in the words of Professor Fraas, as translated:
We recognize in Geosaurus an unusually slenderly built crocodile, in appearance very different from all true crocodiles. The smooth, rounded skull, with its greatly elongated and slender snout, and the deep-lying, small eyes, reminds one most of the ichthyosaurs. The skull merges into the slender, elongated trunk without a visible neck, and the body was provided neither above nor below with horny or bony armor, but was, probably, as in the whales, covered with a smooth, soft skin. The anterior extremities, attached far forward, are developed as paddles, and served rather as organs of equilibration than as a means of propulsion, which was the function of the elongated hind legs and the extraordinarily strong and powerful tail, which supported at its end a large fin. The entire impression given of the animal is that of an excellent swimmer, with all the peculiar aquatic adaptations. In the skeleton, however, all the characters of the original crocodiles are preserved. Most remarkable are the laterally placed eyes, protected by the stout sclerotic bones, and the overhanging bones of the orbits. So, too, the large temporal openings of the skull, doubtless due to the absence of the bony plates in the integument, give to the animal a strangely abnormal appearance for a crocodile.
We have observed that all the truly aquatic air-breathing animals, save the plesiosaurs, have either lost the hind legs or else have them greatly reduced in size, and the disproportionately large size of these members in Geosaurus seems inexplicable. But an explanation is not, I think, hard to find. In the adaptation to water life the first to become modified for the control of the body are the front legs. The hind legs never have any really important use when the tail is a powerful propeller. The hind legs of the geosaurs are still essentially legs and not paddles, and they were doubtless used either occasionally for propulsion on land, or perhaps for pushing the body about on the bottom of shallow waters. And the presence of a well-developed ventral armor of bony ribs possibly also indicates more or less of the terrestrial crawling habit. As soon as the hind legs cease to be used for crawling they take on only a feeble use for the equilibration of the body, and speedily become small, until finally they disappear. That the hind legs of these creatures were of some use in the water is certain, because of the modifications in their structure, and especially because of the loss of the claws; but that they were of [Pg 215] important use as propellers is hardly probable. We may therefore infer that the thalattosuchians, while distinctively sea-reptiles, had not entirely lost their land habits. Moreover, it is highly probable that their egg-laying habits, which would hardly change with a greater adaptation to water life, compelled the animals recurrently to visit the shores. To have finally lost their hind legs they must have become viviparous in habit, since it seems to be impossible for any true air-breathers to be hatched in water. Perhaps this insurmountable habit was the final cause of their extinction in competition with the truly viviparous aquatic flesh-eaters. The thalattosuchians had but a brief existence in geological history, during the latter part of the Jurassic period only, so far as certainly known, nor did they become widely dispersed over the earth; they are known from Europe, possibly from Brazil.
[Pg 216]
No order of reptiles of the past or present is more sharply and unequivocally distinguished from all others than the Chelonia or Testudinata. No order has had a more uniformly continuous and uneventful history. None now in existence has had a longer known history, and of none is the origin more obscure. The first known members of the order, in Triassic times, were turtles in all respects, as well or nearly as well adapted for their peculiar mode of life as are those now living, and were they now living they would attract no especial attention from the ordinary observer and but little from the naturalist. From time to time some have gone after better things, and have come to grief, but the main line has remained with fewer improvements, fewer evolutional changes, than any other group of higher vertebrates. The turtles seem very early to have adapted themselves so well to their peculiar mode of life, to have intrenched themselves so thoroughly in their own province, that no other creatures have been able to overcome them, or to drive them from it. The remains of no other air-breathing vertebrates are so omnipresent in the rocks as those of the turtles; they may be expected wherever fossils of air-breathing animals are found, though unfortunately often only in scattered and broken fragments. The loose union of their skeletal bones and their general habits of life in shallow waters left their bodies as food for scavengers, or for dismemberment by the tides and currents.
Relationships with other reptiles they really have none. Some have thought that the plesiosaurs were their first cousins, others the Placodontia, an indeterminate group of extinct reptiles usually placed with the Anomodontia. But their relationship with neither of these is closer than with the crocodiles, dinosaurs, or pterodactyls. They are [Pg 217] the only reptiles that we know, besides the cotylosaurs, which have no holes in the temporal roof of the skull, and as the cotylosaurs were the most primitive and the oldest of reptiles, this fact incontestably proves that the turtles had a very ancient origin, though we know them no farther back than the later Triassic. They are the only order of reptiles of which not a single member is known to have teeth, or even vestiges of them. Until recently only a single specimen has been known from the Trias, and of that only the casts of the shell; but the shell was as fully developed and as complete as that of a modern alligator snapper, which it resembled much in form and in size. And doubtless the habits of this ancient Proganochelys were similar to those of the alligator snapper. The early cotylosaurian reptiles were all littoral-or marsh-loving animals, and more or less aquatic, and doubtless the early turtles continued in the same environments and with the same habits after acquiring a shell for protection and losing their teeth, which for some inexplicable reason they seemed no longer to need. Until near the close of the Jurassic period probably all turtles were amphibious animals of the marshes, spending much, perhaps the larger part, of the time in the water, good swimmers, and yet good crawlers. With the beginning of the Cretaceous, however, some of them became ambitious for new and untried modes of life. Various ones went down into the sea and became marine animals, reaching the zenith of their prosperity and the maximum of size before the close of the period, but continuing in diminished size and numbers to the present time, if we may consider the leather-back turtle as really their descendant. Others in the Cretaceous took to the rivers and ponds, and became almost as thoroughly aquatic in their thin shape and soft covering; and their lineal descendants still continue in the rivers of the Northern Hemisphere. Still others, in the Age of Mammals, took to the upland, and competed with the mammals in the open places and prairies, reaching their maximum in Miocene-Pliocene times, when for some unknown reason the giants among them were driven from the mainlands to continue a precarious existence to the present time in some of the larger islands. [Pg 218]

Fig. 111

Fig. 112
Figs. 111 and 112.—Graptemys. (From Hay.)
Fig. 111.—Carapace: cp1, cp2, etc., costal plates; cs1, cs2, etc., costal scutes, horny; n1, n2, etc., neural bones; nup, nuchal bone; nus, nuchal scute; py, pygal bone; spy, suprapygal; spy 2, second suprapygal, or postneural; vs1, vs2, etc., vertebral scutes; 1, 2, 3-12 on right side, marginal scutes; 1, 2, 3-12 on left side, peripheral bones.
Fig. 112.—Plastron: ab, abdominal scutes; an, anal scutes; ent, entoplastron (interclavicle); epi, epiplastron (clavicle); fem, femoral scute; g, gular scute; hum, humeral scute; hyo, hypoplastron bone; hypo, hypoplastron; in, inguinal scute; py, pygal bone; xiph, xiphiplastron.
Were there no turtles living we should look upon the fossil forms as among the strangest of all vertebrate animals—animals which had developed the strange habit of concealing themselves inside of their ribs, for that is literally what the turtles do. The box or shell of an ordinary turtle is composed of the backbones and ribs, to which are soldered a shell of bony skin plates above, with the clavicles, interclavicle, and ventral ribs below. Except in the strange leather-back turtle described farther on, these plates form definite series. Ten of them cover the spines of the dorsal vertebrae, in the mid-line, one over each, of which the turtles have the smallest number of any known reptiles. There are eight on each side over the ribs, united by suture with each other and with the middle series; and, in [Pg 219] addition, there are twenty-six bones surrounding them and attached to them. All these bones compose what is called the carapace, which forms a complete roof in the more terrestrial types, more or less imperfect, with vacuities between the bones in the marine forms. On the under side, in addition to the clavicles and the interclavicle, there are three pairs of enlarged ventral ribs that go to form the plastron, solid and complete in land turtles, with openings in the water forms. And in the land forms the plastron is more or less firmly united with the carapace.

Fig. 113.—Toxochelys;
coracoid and scapula.
In the skeleton contained within the box thus formed is the very peculiar pectoral girdle, composed of scapula and coracoid, the scapula so peculiar that the controversy as to its homologies is not yet quite settled. Most authors, until recently, have believed that its peculiar shape (Fig. 113) is due to the co-ossification of the procoracoid with the scapula instead of as usual its loss or union with the true coracoid, so called. We are now pretty sure that this is not true, since in reality there is no such bone as the procoracoid, the bone so called being the real or true coracoid; and because, in the second place, the long anterior projection called the procoracoid is really only an outgrowth of the scapula itself and not a fused, separate bone. Hence the bone is properly called the scapula-proscapula, and not the scapula procoracoid, as it usually has been. The coracoids are elongate and flattened and without the usual supracoracoid foramen, so generally present in reptiles. The only other reptiles having a similar structure of the scapula are the plesiosaurs, and it has been because of this [Pg 220] apparent resemblance that some good paleontologists have thought the turtles and plesiosaurs were allied. The sacrum is composed of two vertebrae only, and the pelvis of the usual three bones, the ilium, the ischium, and the pubis, all covered over by the shell.

Fig. 114.—Pelvis of Chelone, from below:
pu, pubis: is, ischium; il, ilium (in acetabulum).
In every known turtle the neck is composed invariably of eight vertebrae, but they are peculiar in many respects. In the earliest known turtles the neck vertebrae were, as would be supposed, biconcave, but they soon became very variable in all; in each neck some are biconcave, some biconvex, some opisthocoelous, and some procoelous. And Dr. Hay tells us that the neck has increased in length in the later forms.
The skull also is very peculiar in that it has some very primitive characters and others very aberrant. The temporal roof, as has been said, has no holes through it, though it is often reduced by the emargination of the borders, whether from below or behind, until in some the whole temporal region is exposed, and not at all covered over. There is no parietal foramen, so constantly present in all the early reptiles and in the lizards and the tuatera of modern times. There are no teeth or vestiges of teeth, but the jaws have usually a horny cutting edge, which seems to be quite as serviceable; in the river turtle the lips are fleshy. There is no transverse or transpalatine bone. There is a single vomer only, not paired as in other reptiles, whence comes the doubtful theory that the vomers of other reptiles are not the real vomers originally so named in mammals, and hence often [Pg 221] called prevomers. The vomer of the turtles under this theory is believed to be the real homologue of the mammalian bone. The palate is always slightly, sometimes nearly wholly, underfloored, as in mammals, carrying the internal nostrils far back in the mouth. In the occipital region of the skull there is a separate bone on each side called the paroccipital or opisthotic, which has been indistinguishably fused with the exoccipital in all other reptiles except the ichthyosaurs since Triassic times.

Fig. 115
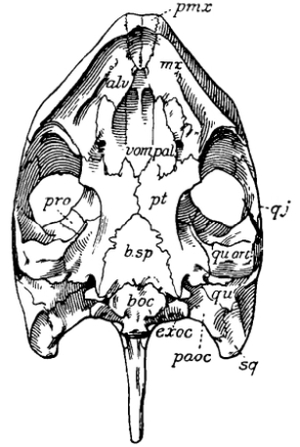
Fig. 116
Figs. 115 and 116.—Trachemys. (From Hay.)
Fig. 115.—Skull from above: fr, frontal; ju, jugal; pa, parietal; paoc, paroccipital; pfr, prefrontal; pof, postfrontal; pro, proötic; qu, quadrate; sq, squamosal; soc, supraoccipital.
Fig. 116.—Skull from below: alv, alveolar surface of maxilla; boc, basioccipital; bap, basisphenoid; exoc, exoccipital; mx, maxilla; pal, palatine; paoc, paroccipital; pmx, premaxilla; pro, proötic; pt, pterygoid; qu, quadrate; qj, quadratojugal; sq, squamosal; vom, vomer.
In the feet the numbers of phalanges—that is, the bones of the free digits—are like those of mammals, that is, two in the first and three in each of the other four digits. The land tortoises have lost some of these, while the river turtles have either gained one or two in the fourth finger and fourth toe, or else have [Pg 222] enjoyed an uninterrupted descent from the primitive reptiles which normally possessed that number. All other reptiles, save those phylogenetically allied to the primitive mammals, that is, the Theriodontia and their allies, have normally the phalangeal formula 2, 3, 4, 5, 4. It was partly because of this similarity of the numbers of toe bones that the turtles have been classed in the great group of reptiles that includes the ancestors of the mammals; that is, under this theory, the turtles would enjoy a nearer relationship to the mammals and to man himself than any other living reptiles! But this classification has been shown to be quite artificial.


Fig. 117.—Limbs of Colpochelys, a recent sea-turtle: H, humerus; R, radius; U, ulna; r, radiale; i, intermedium; u, ulnare; p, pisiform; c, centrale; T, tibia; F, fibula; a, astragalus; m, fifth metatarsal. (From Wieland.)
From what has been said, it will be surmised that the Chelonia represent in themselves one of the primary subdivisions of the class Reptilia, and that, unlike most others, the order has enjoyed a most remarkable longevity. And doubtless they are one of the primary branches of the reptilian stock, which has remained distinct since Permian times at least, if not since Carboniferous, isolated and remarkably homogeneous, giving off no branches which departed far from the main stock, and on the whole leading a singularly placid existence for ten or more million years.
In most textbooks the order Chelonia is divided into three suborders, the Pleurodira, the Cryptodira, and the Trionychoidea. [Pg 223] In recent years, however, the earlier members of the older group of Pleurodira have been separated into a fourth suborder, the Amphichelydia, a group characterized by some not very important differences in the plastron and skull, and including those forms in which the cervical vertebrae are amphicoelous. This group continued to Eocene times before it became extinct, and consisted of archaic forms which persisted after all the other suborders had come into existence. The Cryptodira, especially characterized by the manner in which they withdraw the head and neck within the shell by an S-like vertical flexure, are known from the Lower Jurassic and are still the dominant group of today, with more than one hundred and forty living species. The Pleurodira in the narrower sense are first known from their remains in the Upper Cretaceous of North America and are still represented by about forty species, living in the Southern Hemisphere. They are distinguished from the other groups by the manner in which they withdraw the neck and head into the shell, by a horizontal, sidewise flexure. The third suborder, the Trionychoidea, also began in Cretaceous times, so far as we know, and are represented by about seventy living species, chiefly in the Northern Hemisphere. They are especially characterized by the absence of bony marginal plates and the soft epidermis.
With the exception of the land tortoises, all turtles from the beginning of their career as an order to the present time have been more or less at home in the water. In some, like the marine forms, the adaptation to aquatic life has produced marked changes in structure: in the loss of the horny dermal shields and in the loss of bone tissue; in the flattening of the shell, and in the development of the front legs into swimming flippers, with a loss of the claws. In the absence of a guiding tail, which is always small in the marine turtles, propulsion must of course be wholly by the aid of the limbs. As oar propellers the marine turtles show some of the peculiar characters of the plesiosaurs. With a like short and broad body, a more or less elongated and flexible neck, there could be no sinuosity of the body in swimming. As an oar-like organ the humerus became flattened, and its muscular attachments, as in the plesiosaurs, descended far down the shaft, [Pg 224] giving greater mechanical advantage. Unlike all other aquatic vertebrates, the turtles never developed real hyperphalangy. Only in the river turtles is there a possibility of an increase in the bones of the fourth digit.
To discuss in general the structure and habits of the living chelonians would extend this chapter to an undue length, and would add nothing to the many excellent works on natural history accessible to the student. We have therefore contented ourselves with a brief outline of the geological history of the order, with especial reference to their aquatic habits.
The suborder of Chelonia, generally known as the snake-necked or side-necked turtles or tortoises, comprises about forty living species, confined to South America, Africa save the northernmost part, Madagascar, New Guinea, and Australia. In Australia they are the only members of the order known—another instance of the peculiar isolation of the fauna of that region. In the past they lived in North America during Upper Cretaceous times, the earliest known forms of the group in its restricted sense, of which seven species are described by Hay. In Eocene times they are also known from Europe and Asia, from both of which regions they have long since disappeared.
The Pleurodira, as the term indicates, are easily distinguished from all other turtles by the way in which they withdraw the head within the shell. Instead of withdrawing it by an S-shaped flexure of the neck between the shoulder-blades, as do other turtles, these bend the neck laterally in a horizontal plane, bringing the head within the margins of the shell in front of one or the other foreleg; and the margins of the shell are produced here in an eave-like fashion for the greater protection of the head. In the structure of the shell, which is always fully developed into a box, these turtles do not differ very much from the Cryptodira, though there may be some extra bones in the plastron, as also in the skull. The nasal bones are always, the lacrimals sometimes, well developed; the latter never, the former rarely, found in other groups. The lower jaws articulate a little differently, and the external ear is always fully surrounded by bone. [Pg 225] Very characteristic is the bony union of the pelvis with the plastron below, which never occurs in other turtles, unless it be the Amphichelydia.
The side-necked turtles are all of fresh-water habit, similar to that of the fresh-water tortoises spoken of farther on. The neck is often very long and snake-like, which accounts for one of the names given to these turtles; because it is withdrawn into the shell sidewise, it has more distinctively ball-and-socket joints between the vertebrae, with distinct transverse processes for the attachment of the necessary side-moving muscles. The feet in all are more or less webbed and armed with strong claws.
The largest and perhaps the best known of all living side-necked turtles is the giant Amazon turtle of South America, which sometimes has a shell nearly three feet in length. Its feet are broadly webbed, and the shell is rather flat in the adult; it is an excellent swimmer in the waters of the Orinoco and Amazon. Six or seven species of the genus to which it belongs are known, all of them South American except one that lives in Madagascar and one fossil found in the Eocene of India. This remarkable distribution is but one more of the many instances known in zoölogy and paleontology that seem to prove an early land connection between India and South America. Had the migration between the two continents occurred by way of Asia and Bering straits, as did that of hosts of mammals, one would certainly expect to find some evidence of it in the North American Tertiary rocks, which, so far, is lacking.
The chief families of the Cryptodira turtles are the Chelydridae, or snappers; the Emydidae, or marsh tortoises; the Testudinidae, or land tortoises; the Chelonidae, or sea-turtles; the Protostegidae, or ancient sea-turtles; and the Dermochelydidae, or leather-backs. Other doubtful or smaller groups, both living and extinct, may be omitted, or incidentally mentioned.
The family of snapping turtles, the Chelydridae, are of interest because of their peculiar geographical distribution at the present time. [Pg 226] Only four species are known, three of them from North America, the fourth from New Guinea. The family is one of the most primitive of living turtles, though no members of it are known with certainty from earlier rocks than the Oligocene. In all probability, also, they have retained, more than have any other group of turtles, unless it be some of the fresh-water tortoises, the primitive habits of the earlier or earliest turtles, though of course there have been modifications, both in structure and in habits. The three species of the United States include two of the snapping turtles proper and the alligator turtles of the southern states, which sometimes reach a length of three feet. All the species are largely aquatic in habit, powerful and active swimmers, with webbed feet and strong claws, and both on the land and in the water they are bold and fierce. They have a relatively large head and very strong jaws. Agassiz saw one bite off a piece of a plank an inch in thickness, and they can usually be raised from the ground by any object which they seize. The carapace and plastron are much reduced, and are rather loosely united. The shell is not large enough for the complete withdrawal of the head and legs within it, and the tail is unusually large and strong. The common snapping turtle, Chelydra serpentina, is found from Canada to Ecuador, and its remains have been found with those of the mammoth and mastodon in Pleistocene deposits; and related species of the same genus have been reported from the Miocene of England.
The family of turtles or tortoises (Emydidae) represented at the present time by the common terrapin, painted tortoise, and box tortoise of the United States, and commonly called fresh-water turtles or tortoises, comprises the largest group of living chelonians—nearly a third of all existing members of the order. They are widely distributed over all parts of the earth except Australia, and are of very varied habits. Some are almost exclusively aquatic; others, like the painted tortoise, are partially so, while others, especially the common box tortoise, are almost as exclusively terrestrial as the true land tortoises, dying even, if forced to live long in water. [Pg 227]
The shell in the more aquatic forms is depressed or flattened, but in the terrestrial kinds may be as highly arched as in the true land tortoises. The feet are adapted primarily for walking, but nearly always have the toes webbed, and the digits are longer than are those of the land tortoises. Only the two or three middle toes have claws. Some species have developed hinges in the plastron, whereby they may be completely closed up after the head and legs are withdrawn within the shell. Most of the species are carnivorous in habit, but a few, like the box tortoise, are strictly vegetarian.
Geologically the fresh-water tortoises have a not very ancient history, going back no farther than do the land tortoises, that is, to the beginning of the Cenozoic or Age of Mammals. Fully fifty species are known from the Tertiary rocks of North America, or more than three-fourths as many as are now living upon the earth.
The family at most can be said to be only amphibious in habit, and has no striking aquatic adaptations, since the shell is well developed and is covered with horny shields. The flattened shell of the more aquatic forms is characteristic, as is also the greater degree of webbing between the toes.
Perhaps the last of the more noteworthy specializations of the Chelonia, and indeed among the last of the more important specializations of the Reptilia, are the upland tortoises, of which the common “gopher” of the southern states is almost the only remnant in North America. They formed a part of the great hegira of forest and marsh animals to the open prairies, away from the lowlands and water which the turtles had inhabited almost exclusively for millions of years.

Fig. 118.—Testudo sumeirei, a giant
upland tortoise.
(From Hay, after Rothschild.)
They began their career, Dr. Hay thinks, at about the beginning of the Cenozoic, that is, with the great development of the mammals, and reached the maximum of their development in the Miocene; and they have been on the decline ever since. In the Northern Hemisphere, at least, the slowly cooling climate throughout the Eocene, and a decided [Pg 228] decrease in moisture, brought about the prairies and prairie plants before its close. Just as the horses, rhinoceroses, camels, and other herbivorous mammals took to these open places for the better and more abundant food found therein, so also the lowland tortoises found better food and fewer enemies there, for they are all strictly herbivorous in habit. The mammals became more conspicuous to their enemies when they went into the open, and it was only by the development of speed, more sober coloration, and perhaps greater cunning that they found safety from them. The tortoises were handicapped by low intelligence, and they could not develop speed, for they were not constructed to that end. But they did find protection in their bony shell, which became thicker, higher, and more convex, and with smaller openings. To quote Dr. Hay: “We may suppose that it would be much more difficult for a carnivorous animal to effect an entrance into such a shell than into one depressed, and whose borders may be spanned by the jaws of their enemies.” Perhaps also the highly arched form of the shell gave greater capacity for the lungs, and the tortoises in general, it is said, do have better lung capacity than the more aquatic or lowland types of turtles. Possibly, also, the heavier shell lessened the evaporation of the body fluids, and made the tortoises less dependent upon the vicinity of water. [Pg 229] Certain it is that the common box tortoise, of like form and habits, occurs not rarely on the arid plains, far from water.
The neck and legs became fully retractile within the shell; the digits were shortened up, without a vestige of webbing membrane between them; the phalanges were reduced in number to two in each toe, and nearly all the toes have well formed claws. The feet are placed squarely upon the ground, and the body is elevated in walking. They can swim, when by accident they are thrown into the water, only as any terrestrial mammal can.
About forty species of land tortoises are known throughout the world at the present time, though North America, the probable original home of the tribe, has but three, all small. The larger species are all now denizens of islands, especially the Galapagos Islands, where the giant tortoises have long been famous. And many of our living forms have changed but little since Eocene times. In the Oligocene and Miocene they inhabited western North America in enormous numbers. In the Bad Lands of South Dakota one can often see the remains of a dozen or more of these giant tortoises at one time, specimens varying from one to three feet in length of shell. In river deposits, those of the late Miocene or early Pliocene, the writer has seen areas of an acre or more literally strewn with their remains, as though droves of them had been overwhelmed and perished together. About fifty species of these land tortoises are known from the American Tertiary, thirty-two of them belonging to the modern genus Testudo, which comprises the giant tortoises of the Galapagos. The largest known species of the group is one of Testudo from the Pliocene of India, which had a shell six feet in length. Why the larger species became extinct in Pliocene times on the mainland to survive only in the islands is not known; possibly their carnivorous enemies became too cunning and too numerous.
The sea-turtles, or Chelonidae comprise five or six living species, inhabitants for the most part of tropical and subtropical oceans, of which the green or edible turtle (Chelone), the hawksbill turtle [Pg 230] (Caretta), and the loggerhead (Eretmochelys [Fig. 119]) are the best known. They are all thoroughly aquatic in habit, and of large size, from three to five feet in length. The carapace is heart-shaped, and reduced, that is, with large openings between the ribs; the plastron also is reduced and loosely united to the carapace. The neck is short and the head is not retractile within the shell. The temporal region of the skull is roofed over. The four legs form large and powerful flippers, and the hind legs are relatively small. The body is flattened and the tail is small. The aquatic characters of the limbs are seen especially in the broad and strong humerus, with the radial crest for the attachment of powerful muscles situated far down on the shaft; in the relative shortness of the radius and ulna, and the large size of the latter bone; in the flattened carpal bones; and in the great elongation of the digits and the absence of all but one or two of the claws. Unlike the leather-back turtle and the Cretaceous sea-turtles, the carapace and plastron are completely covered with horny shields, from which indeed the tortoise shell of commerce is derived. Except the green turtle, all members of the family are carnivorous.

Fig. 119.—Eretmochelys, loggerhead
turtle.
(By permission of the New York Zoölogical Society.)
[Pg 231] Extinct members of the family are known from scanty remains in Cenozoic and late Cretaceous rocks. From the earlier Cretaceous deposits of the plains more primitive allied forms occur, often classed in distinct families of which Toxochelys (Fig. 120) and Desmatochelys are the more noteworthy. The latter genus, especially, might well have been an ancestor of all the modern forms. About three feet in length, it had all the essential characteristics of the sea-turtles, in its thin form, roofed-over skull, reduced carapace, loose plastron, and flipper-like limbs. The single known specimen, preserved in the museum of the University of Kansas, came from the lower rocks of the Upper Cretaceous of Nebraska. Yet earlier, at the close of the Jurassic, there were shore turtles of considerable size that had begun to develop a fondness for the open seas; to acquire a depressed form and lightened shell, the limbs still retaining, however, more of the terrestrial or crawling form. They are grouped as a separate family, the Thalassemydae, and include the first of the Chelonia to depart from the marsh and fresh-water habits which for long ages, perhaps, had limited the activities and evolution of the turtles.

Fig. 120.—Carapace of Toxochelys bauri,
an Upper Cretaceous sea-turtle: ep, epineural.
(After Wieland.)
[Pg 232]
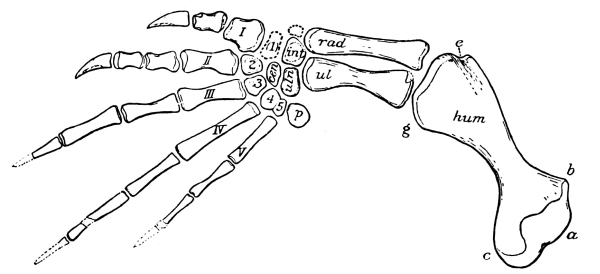
Fig. 121.—Toxochelys latiremis; front leg: hum, humerus; rad, radius; ul, ulna; int, intermedium; uln, ulnare; p, pisiform; cen, centrale. (From Wieland.)

Fig. 122.—Desmatochelys lowii;
skull from above and below.
Forty-four years ago the late Professor E. D. Cope, one of the greatest naturalists America has ever produced, in almost the earliest exploration of the great Cretaceous fossil deposits of western Kansas, discovered and collected a remarkable specimen of one of the most extraordinary turtles that is known even yet. By an error somewhat natural for those times, when the theory of evolution was just [Pg 233] beginning to attain acceptance by naturalists, he thought that the specimen, notwithstanding its monstrous size, represented a very primitive kind of turtle, and gave to it the name Protostega gigas, meaning gigantic first roof! The late Professor George Baur, to whom paleontology owes so much, showed that, far from being a primitive turtle, Protostega was really one of the most specialized types of the order. Professor Cope’s account of the discovery of the specimen is of so much interest that it may be quoted here:
“In the very young tortoise or turtle the ribs are separate, as in other animals. As they grow older they begin to expand at the upper side of the upper end, and with increased age the expansion extends throughout the length. The ribs first come in contact where the process commences, and in the land tortoise they are united at the end. In the sea-turtles the union ceases a little above the ends. The fragments of the Protostega were seen by one of the men projecting from a ledge of a low bluff. After several square feet of rock had been removed, we cleared up the floor and found ourselves well repaid. Many long, slender pieces of two inches in width lay upon the ledge. They were evidently ribs, with the usual heads, but behind each head was a plate-like the flattened bowl of a huge spoon, placed crosswise. Beneath these stretched two broad plates, two feet in width, and no thicker than binder’s board. The edges were fingered and the surface was hard and smooth. All this was quite new, among fully grown animals. Some bones of a large paddle were recognized, and a leg bone. The shoulder-blade of a huge tortoise came next, and further examination showed that we had stumbled on the burial place of the largest species of sea-turtle yet known. But the ribs were those of an ordinary turtle just hatched, and the great plates represented the bony deposit in the skin, which, commencing independently in modern turtles, unite with each other at an early day. But it was incredible that the largest of known turtles should be but just hatched, and for this and other reasons it has been concluded that this ‘ancient mariner’ is one of those forms, not uncommon in old days, whose incompleteness in some respects points to the truth of the belief that animals have assumed their modern perfection by a process of growth from more simple beginnings.” [Pg 234]
Later studies by Doctors G. Baur, E. C. Case, O. P. Hay, and especially G. R. Wieland, of the abundant and excellent material, preserved in the museums of Yale and Kansas universities and the Carnegie Institution, and especially the discovery by Wieland in 1895 of an allied and yet larger form which he called Archelon, have determined practically every detail of the structure of this remarkable group of sea-turtles. A surprisingly complete specimen of Archelon is mounted in the museum of Yale University.
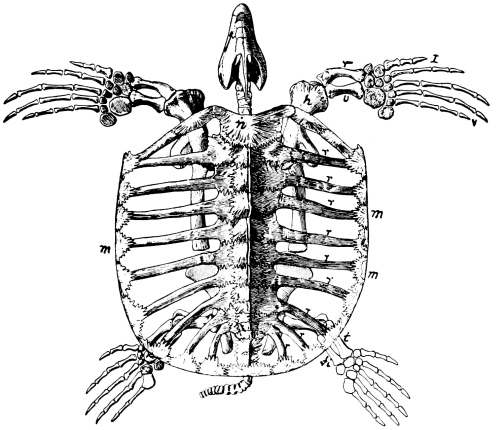
Fig. 123.—Archelon ischyros; skeleton from above: n, nuchal, r, r, r, ribs; m, m, peripheral bones; h, humerus; r, radius; u, ulna; t, tibia; fi, fibula. (From Wieland.)
About a half-dozen species and two genera of the family have so far been described, all coming from the Upper Cretaceous deposit of Kansas and South Dakota, the genus Archelon from later rocks than those which have yielded Protostega.
The general form and structure of Archelon will best be understood from the accompanying figures after Wieland (Figs. 123, 124, 125) and the restoration of the living animal as interpreted by the [Pg 235] writer (Fig. 126). If the leather-back turtle, described farther on, is really the descendant of these or allied turtles, as many authors believe, it of course represents the very highest aquatic specialization of all Chelonians. If, on the other hand, as some believe, the leather-back is the end of a long and independent line of descent, then Archelon represents the highest aquatic specialization of all other turtles.
In size, at least, Archelon attained the maximum of the order, reaching a length of more than twelve feet, and a weight of more than three tons. Except that the shell was not heart-shaped or elongated as in all modern sea-turtles, but nearly circular in outline, it had all the aquatic adaptations of the sea-turtle in a yet higher degree.

Fig. 124.—Archelon from below, without plastron: h, humerus; r, radius; u, ulna; sc, scapula; c, coracoid; p, pubis; i, ischium. (From Wieland.)
[Pg 236]

Fig. 125.—Archelon; skeleton from below: hp, hyoplastron; hpp, hypoplastron. (From Wieland.)
The shell was depressed; the dermal plates covering the ribs had almost entirely disappeared, remnants only of their upper ends remaining; the skull (Fig. 127) had the temporal region wholly roofed over; the neck was short and not retractile. The front legs were strong flippers, the humerus was long and stout, with the crest for the attachment of muscles far down on the shaft; the digits were greatly elongated and clawless, etc. The plastron only was less reduced than in the case of the modern sea-turtles. No traces of horny shields have been discovered. As to the nature of the covering and the general appearance of the turtle when alive, Dr. Wieland has kindly given the writer his views, as follows:
“After direct study or fairly close examination of all the fossil material of importance thus far collected representing the Protostegidae, it seems certain that in all the members of the group an external leathery layer was well developed. In no instance is there the slightest trace of horny shield sulci, or grooves; though it seems probable that there was some gradation toward a thin and perhaps even slightly horny hide. In Archelon ischyros the reduced condition [Pg 237] of the carapace and the presence of the continuous row of large, median, supraneural elements render it quite certain that there was a development of leathery hide comparable to that of Dermochelys. The same may be said of Protostega gigas. But Archelon Marshii had a less reduced carapace, and the leathery skin was probably less well developed; and Protostega Copei, in which no [Pg 238] trace of supraneurals remains, must have made some approach to the horn-shield condition. A more distinct suggestion of transition from the leathery to the horny shield covering may be seen in the very different contemporary Cretaceous form, Toxochelys Bauri, where ossified epi-or supraneurals occupy quite exactly the nodal relation of the five vertebral horn shields of later turtles, like Lytoloma, though there are not the slightest traces of sulci.

Fig. 126.—Archelon ischyros,
a gigantic sea-turtle
from the Upper Cretaceous of South Dakota.

Fig. 127.—Skull of Archelon ischyros: pa, parietal; f, frontal; pm, premaxilla; pf, prefrontal; ptf, postfrontal; m, maxilla; j, jugal; qj, quadratojugal; sur, surangular; d, dentary; an, angular. (After Wieland.)
“From a purely anatomical standpoint I have suggested that Archelon had seven dorsal keels corresponding to those of Dermochelys. There is much excellent reason for regarding dermogene ossification as essentially double-layered throughout the Reptilia.
“In any restoration of Archelon ischyros only the mid-line should be accentuated as a series of rather sharp supraneural crests. These are shown to have been present by the characteristic groove-like median pits with radiating striae, which are such a prominent feature of epineurals. It is reasonable to believe that the pits mark the attachment of horny crests developed in the leathery hide. Such were [Pg 239] doubtless projected, more or less keel-like, to a height of one or two inches, and thus gave to the mid-line of the carapace, when seen laterally, a distinctly sinuous outline not unlike that of Toxochelys.”
As regards the habits of these ancient sea-turtles, we may offer tolerably certain conjectures. In the opinion of the writer, the less reduced plastron indicates a bottom-feeding habit, a view that is strengthened by the more rounded form of the shell, like that of the river turtle. All in all it would seem that Protostega and Archelon lived habitually on the soft bottoms of the shallower seas, feeding upon the hordes of large shell-fish, for which their powerful parrot-like beak was admirably adapted. That the species of Protostega did not commonly frequent the deeper oceans is indicated by the general absence of their remains in the deeper water deposits. The writer, in a long collecting experience, always found their remains associated with those of the smaller Toxochelys, toothed birds, pterodactyls, and the smaller mosasaurs.
Perhaps no one can speak more authoritatively as to the habits of these gigantic sea-turtles of the Cretaceous than Dr. Wieland:
“With regard to the general habits and appearance of Archelon much might doubtless be said if the present-day sea-turtles were more familiar objects. Dr. Hay thought that Archelon ischyros was a clumsy or even a sluggish, mainly littoral animal, moving slowly about the bottom of quiet inlets in quest of shell-fish; I, on the contrary, much struck by the powerful flippers, and especially by the flattening of the humerus, with its low radial crest and obviously strong musculature, have held that unusual swimming power and adaptation to a strictly marine life were indicated. Perhaps, as usual where experts differ, it is probable that both views are in part correct, and that Archelon was only a moderately good swimmer. It may be noted that, notwithstanding the almost circular body, the femoral notch, that for the hind leg, lies far back, so that it is not necessary, on the score of bulk, to assume slowness of motion, or the inability to pursue a sea-going life. Furthermore, it is now known that the development of the digits fell little short of that seen in Colpochelys (Fig. 117) or Eretmochelys, truly marine turtles.
“Therefore, while there can be no doubt that Archelon was strictly carnivorous in habit, and well able to navigate the open seas, [Pg 240] it is not likely that it fed on other than relatively slow-moving prey. Lydekker looked upon the broad mandibles and broad palate of Lytoloma as specializations for a mussel diet; and very similarly in Archelon, while the decurved beak would easily transform him into a most formidable enemy, the heavy premaxillaries and vomer, and the flat but deep lower jaw, suggest an adept crusher of crustaceans. The presence of vast quantities of Nautilus dekayi, which I found accompanying one of the specimens, was doubtless accidental, but it plainly suggests that this cephalopod was one of the teeming sources of food in the Archelon environment.
“The huge bulk of the mature Archelon might account for the shearing off and loss of the flippers of younger forms caught between the shells of the ‘elder boatmen of the Cretaceous seas,’ as Cope has called them, during any sudden rush while herding on the shores. But probably the young turtles did not much frequent the shores at either egg-laying or other times. Whence it is much more likely that it was a mosasaur or some of the gigantic fishes like Portheus which bit off the right hind flipper in the type-specimen of Archelon ischyros, well above the heel, as I have described it. That this happened rather early in life is shown by the arrested growth of the right femur and remaining portions of the tibia and fibula, which are all uniformly 10 per cent smaller than the corresponding bones of the left flipper.”
While there were many small fishes in the Niobrara seas which the Protostegas inhabited, the most striking thing in the fauna is the great abundance of molluscal shells, especially Ostrea congesta. And with them were great hordes of larger pelycypod mollusks, some of them of enormous size. Some of the largest reach a diameter of nearly four feet, with shells so thin that one can hardly understand how they could have supported such large, oyster-like creatures. One can imagine that such shell-fish might have afforded an almost inexhaustible source of food for the large turtles; and several times the writer has found remains of Protostega associated with such shells. From all of which evidence it seems very probable indeed that Dr. Wieland is right in imputing to these gigantic turtles a shell-feeding habit, a habit [Pg 241] which required neither speed nor great prowess; and perhaps the formidable beak was used more in social quarrels than for food-getting. That these marine turtles departed from the usual reptilian habit of laying their eggs upon land is improbable. The tortoise shell turtles of the Bahamas lay three or four hundred eggs in a hollow scooped out in the sand and then leave the young to their own devices; certainly many a one is gobbled up by birds of prey or other enemies on their way to the water. Perhaps the young Archelon lost its hind leg in some such mishap.
The most remarkable member of the Chelonia now living is Dermochelys coriacea (Fig. 128), the great leathery or leather-back turtle of the warmer parts of the Atlantic, Indian, and Pacific oceans, the sole member of the family Dermochelydidae. It is the largest of all living turtles and the most thoroughly aquatic of all, whether living or extinct. It sometimes reaches a length of six feet, or half that of the largest known extinct forms, and weighs a thousand or more pounds. Agassiz saw a specimen that he said weighed a ton. Unlike other turtles, it has a carapace quite peculiar to itself, composed of a layer of thin, irregularly polygonal bones forming a mosaic, completely hidden in the thick skin, and entirely free from the skeletal bones beneath them. The larger of these skin bones form seven rows above, which appear in the living animal as sharp keels running the whole length of the shell. On the under side there are five rows of smaller-sized bones, under which there are vestiges of bones representing the normal plastron of turtles. The limbs are powerful, flattened paddles, not unlike those of Eretmochelys, but wholly destitute of claws. The front paddles are much larger than the hind ones; the humerus is long and flattened, and the digits are elongated. The leather-back is a powerful and effective swimmer, going long distances. Its habits are not well known; its food is chiefly fish, crustaceans, and mollusks.

Fig. 128.—Dermochelys coriacea.
(From Brehm.)
So very different is the structure of its shell that some excellent naturalists regard Dermochelys as the equivalent in rank of all other turtles combined, the sole representative of the suborder Athecae, [Pg 242] as distinguished from the Thecophora. Dr. Hay, whose authority on fossil turtles is of the highest, believes that its line of ancestry has been distinct from that of all other turtles from Triassic times at least. Others believe that the leather-back is merely a highly specialized form derived from the ordinary shelled type, a descendant of some of the marine turtles of Cretaceous times. In support of the primitive ancestry of the leather-back Dr. Hay offers the following:
“The writer holds the view that the earliest turtles possessed practically two kinds of shell, one purely dermal, consisting probably of a mosaic of small bones arranged in at least twelve longitudinal zones. Each zone probably consisted of a row of larger bones bordered on each side by smaller bones. Each of these bones was covered by a horny scute. The nearest approach to such a dermal shell is in our days seen in Dermochelys. Beneath the skin there seems to have existed a carapace more or less complete, which consisted of a nuchal, [Pg 243] a median row of neurals, eight pairs of costals, a pygal, probably one or more supraneurals, and about eleven peripherals on each side. To what extent the neurals and the costal plates had become anchylosed to the neural spines and the ribs respectively, it is now impossible to determine. Nor can we say to what extent the various elements of the carapace had become connected with one another. There was a subdermal plastron which was composed of at least eleven bones.
“According to the author’s view, as time went on the external, mosaic-like shell disappeared in most turtles, while a more efficient armor was developed out of the subdermal elements. In the ancestors of Dermochelys, however, the dermal armor was retained, while the more deeply seated one disappeared, with the exception of the nuchal bone.”
Such a hypothesis as the foregoing satisfactorily explains the extraordinary mosaic shell of the leather-back, and is perhaps an acceptable explanation of the rather strange fact that the horny shields of turtles do not correspond with the bones below them, as might be expected. Unfortunately this hypothesis lacks sufficient proof. About the only evidence that is offered in its support is the existence of a row of bones along the middle line in the Cretaceous Toxochelys, and notably in Archelon, both aquatic forms. It is urged that these bones, the epineurals of Wieland, are really the remains of an external layer that persisted in these turtles. However, they might have been new ossifications, such as we know did occur in not a few of the land tortoises later over the tail and limbs. Aside from Proganochelys in the vast interval of time from the Triassic to the Eocene no other evidence of such an external dermal layer has been discovered. The chief argument against such divergent ancestry of the turtles in two chief lines of descent is the fact that in its other structure Dermochelys shows great resemblance to other sea-turtles of the Cretaceous times—so much resemblance that it seems impossible that the ancestors of the leather-back should have paralleled them in almost everything except the shell.
On the other hand, those who disagree with this view believe that the modern leather-back is the descendant of such Cretaceous marine turtles as Protostega or Archelon, some of which had lost nearly [Pg 244] all of the costal plates and had the neurals and marginals reduced. It is urged that some of these early marine forms, after they had practically lost the ordinary bones of the carapace, for some reason or other found a bony shell again necessary for their welfare. Possibly they had become littoral in habit; possibly they again became subject to new and dangerous enemies in their unprotected condition, notwithstanding their great size; perhaps the zeuglodons were among their enemies. Now, as we have seen, an animal never takes a back track and recovers a thing it has once lost. It was impossible for the ancestors of the leather-back again to acquire an orthodox shell, and they forthwith proceeded to acquire quite another kind that would serve the same purpose.
Possibly the truth lies between the variant views, in the theory recently expressed by Versluys: “The shell of tortoises and turtles is formed by a combination of two layers of dermal ossifications, a thecal layer and a more superficial epithecal layer, the latter generally represented by the marginals only. The leather-back is a member of the Cryptodira, and is allied to the other marine turtles. The problem of the origin of the aberrant shell of the leather-back seems to find its solution in the hypothesis that it is a secondary proliferation of the marginals and such other epithecal elements as were present in its thecophorous ancestors.”
In other words, Versluys believes that Hay’s and Wieland’s views of the primitive double layer of exoskeletal bones is essentially correct, but that Dermochelys was derived from later forms in which some of them, only, as in Archelon, had remained. Baur’s contention that “Dermochelys is not the least, but the most specialized marine turtle” seems to have been fully justified.
No reptile is more familiar or more exasperating to the river fisherman than the turtle, variously known as the river, soft-shelled, or mud turtle. It lives, often in great numbers, in most of the rivers, ponds, and bayous of the interior east of the Rocky Mountains, and especially in those of sluggish current and muddy bottoms. It is voraciously carnivorous in habit, feeding upon the smaller fish, mussels, and such other living food as it can capture. With its long, sinuous neck and [Pg 245] snake-like head, and soft, mottled skin, it is repulsive enough to most persons, but is especially annoying to the fisherman, since it devours with impunity his bait so long as he feeds it, and can seldom be caught on the hook because of its hard and bony mouth, in which only by good luck will the hook catch. And the luckless string of fish that the fisherman leaves in the water may be almost completely devoured in a few hours by these fiercely predaceous feeders. However, if so annoying while seeking for better game, it in part makes up for the annoyance it causes by furnishing in its own body a not unpalatable food for those who like to eat reptiles.

Fig. 129.—Trionyx, river turtle.
(By permission of the New York Zoölogical Society.)
The river turtles will be readily recognized from the accompanying illustration (Fig. 129). They are very flat, covered with a soft, smooth skin, with a long, sinuous neck and a small, snake-like and vicious-looking head which has a protuberant snout with the external nasal orifice at its end. Their feet are webbed and somewhat [Pg 246] paddle-like, but always with three stout claws—whence comes the name of the group—on the anterior digits, which are used for burrowing in the mud and excavating holes for their eggs. These turtles burrow more or less in the mud, with the long neck free, lying in wait for their prey, and coming to the surface from time to time to breathe. As the shape of the body and the paddle-like feet would suggest, they are active swimmers and purely aquatic in habit, never leaving the water unless compelled to. They bury their hard-shelled eggs on the shores only a few feet from the water, and leave them to their fate. If the pools in which they live dry up, they burrow deeply in the mud and await the rains and floods. In captivity they feed upon all kinds of food, vegetable as well as animal, and are active and aggressive.

Fig. 130.—Aspideretes, a trionychoid
turtle
from the Basal Eocene of New Mexico;
skull from above.
(From Hay.)
Because of certain peculiarities, they are usually classed in a separate suborder all their own, the Trionychoidea, especially distinguished from the Cryptodira, which in general they resemble in most respects, aside from the absence of the usual horny dermal plates, in the lack of a marginal row of bony plates around the carapace—not a very important distinction. Less than thirty living species are known, all of them exclusively, or chiefly of fresh-water habit. Six species are known from North America; the remainder inhabit Africa, south of the Sahara Desert, southern India, and most of the East Indian islands; none is known from Australia. No species lives in South America and none is known to have lived there in past times. During Eocene, Oligocene, and Miocene times these fresh-water turtles lived in the region of Europe in great numbers, but for some inexplicable reason they became extinct there and never returned. Nearly seventy species of the Trionychoidea, belonging in two families, are described by Dr. Hay from the Tertiary rocks of North America, more than twice the number now [Pg 247] living throughout the world. Some of these were of relatively large size, measuring fully two feet in the length of the shell. And in some places they must have been very abundant. The writer has seen, in the Bad Lands of the Continental Divide, their weathered-out remains so numerous that they might be raked into windrows miles in length along the sloping bluffs, all in small fragments, for their bones, like those of most turtles, are only loosely united by sutures and readily drop apart before fossilization. Their shells may be readily distinguished from those of all other turtles by the granulated, pitted, or sculptured exterior surface, that was covered by the skin in life; other turtles have the surface smooth below the horny shields, the margins of which are marked on the bones by grooves or sulci; the few marine turtles of the past that were probably covered with a soft skin instead o£ horny shields had the shell smooth and much less completely ossified.

Fig. 131.—Aspideretes, a trionychoid
turtle
from the Eocene of New Mexico;
front leg. (From Hay.)
As to the origin of the soft-shelled turtles there has been not a little difference of opinion. The earliest ones known in geological history date back only to about the middle of the Cretaceous; perhaps they branched off from the horny-shelled turtles somewhat earlier, but probably not much. There are some, however, who think that this group of turtles was very primitive, perhaps the most primitive, but the writer agrees with Dr. Hay in rejecting this view. Unlike those of all other turtles, the fourth digit in front and hind feet has one or two more phalanges than have other turtles. We have seen that the oldest known reptiles had the digital formula 2, 3, 4, 5, 3 or 4. Most other [Pg 248] turtles have the same numbers of bones in the digits that mammals have, that is, two phalanges in the thumb and big toe and three in each of the other digits. The river turtles have a larger number in the fourth digit, either four or five. It seems to be a law that evolution is irreversible, and if so could the river turtles have been descended from forms with a less number of phalanges? But, the skeleton of the Trionychoidea resembles the more specialized turtles in so many ways that one can hardly believe they were all accidental or parallel.
We may then assume that at about the time that the ordinary marsh turtles took to the sea to become marine, others took advantage of the fresh-water ponds and rivers, and in doing so, like the marine turtles, lost their horny epidermal shields, and became thinner in shape, thereby reducing the resistance to the water. Instead, however, of reducing the costal plates over the ribs, they retained them intact and complete for some reason or other, but lost instead the marginal row of bones, unlike the marine turtles which retained them even after they had lost nearly all of the costal plates. Possibly also they regained additional bones in the fourth digit, a sort of hyperphalangy like that of the more strictly aquatic reptiles. Or, possibly, they may have descended from some branch of the turtles which had not yet lost these bones, retaining them because they were still serviceable for swimming. We know nothing yet about the structure of the feet of the early turtles, and it is possible that not all had acquired the reduced phalangeal formula.
In the development of aquatic habits the river turtles do not show the same degree of specialization in the limbs that the strictly marine forms do. The humerus (Fig. 131) is a slender bone, with the tuberosities for the attachment of the muscles situated near the proximal end. The radius and ulna are relatively short, and the foot is long. The hind legs, as would be supposed, are less highly specialized as swimming paddles, and are relatively smaller. Nevertheless the Trionychoidea present an interesting type of adaptation to water habits, both in body and in limbs.
[Pg 249]
Footnotes:
[1] Paired maxillae, premaxillae, nasals, prefrontals, lacrimals, frontals, parietals, dermosupraoccipitals, tabularia, supratemporals, intertemporals, squamosals, jugals, quadratojugals, postorbitals, postfrontals, quadrates, exoccipitals, paroccipitals, vomers, palatines, pterygoids, sphenomaxillae, stapes, transverse, alisphenoids or orbitosphenoids, epipterygoids, articulars, prearticulars, angulars, surangulars, coronoids, splenials, dentaries, one supraoccipital, one basioccipital, one basisphenoid, one ethmoid.
[3] An additional phalange has also been observed in the fifth toe of a South American species.
[4] “The author showed drawings and some specimens of two hitherto unknown reptiles from the white, coarse-grained sandstone, of which one in the form of the skull resembles the gavial, but is distinguished by the cylindrical form of the lateral teeth of the jaws; he therefore calls it provisionally cylindricodon, and a second species or genus, of which, however, so far only fragments of the jaws have been found, because of the four-cornered form of the teeth, cubicodon, while at the same time for the genus or family, to which the remains of these animals have belonged, he proposes the name Phytosaurus, since the teeth seem to be more adapted to a vegetable diet, even though they have not been worn away, as in Iguanodon.”—Isis (1828), p. 441 (translation).
Transcriber’s Notes:
The cover image was created by the transcriber, and is in the public domain.
The illustrations have been moved so that they do not break up paragraphs and so that they are close to the text they illustrate.
Typographical and punctuation errors have been silently corrected.
This eBook is for the use of anyone anywhere in the United States and most other parts of the world at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org. If you are not located in the United States, you will have to check the laws of the country where you are located before using this eBook.