
BY
DR. E. WARMING
Professor of Botany in the University of Copenhagen
With a Revision of the Fungi by
DR. E. KNOBLAUCH,
Karlsruhe
Translated and Edited by
M. C. POTTER, M.A. F.L.S.
Professor of Botany in the University of Durham
College of Science, Newcastle-upon-Tyne
Author of “An Elementary Text-book of Agricultural Botany”
WITH 610 ILLUSTRATIONS
London
SWAN SONNENSCHEIN & CO
NEW YORK: MACMILLAN & CO
1895
Butler & Tanner,
The Selwood Printing Works,
Frome, and London.
[v]

The present translation of Dr. E. Warming’s Haandbog i den Systematiske Botanik is taken from the text of the 3rd Danish Edition (1892), and from Dr. Knoblauch’s German Edition (1890), and the book has been further enriched by numerous additional notes which have been kindly sent to me by the author. Dr. Warming’s work has long been recognised as an original and important contribution to Systematic Botanical Literature, and I have only to regret that the pressure of other scientific duties has delayed its presentation to English readers. Dr. Warming desires me to record his high appreciation of the careful translation of Dr. Knoblauch, and his obligation to him for a number of corrections and improvements of which he has made use in the 3rd Danish Edition. In a few instances I have made slight additions to the text; these, however, appear as footnotes, or are enclosed in square brackets.
In the present Edition the Thallophytes have been revised and rearranged from notes supplied to me by Dr. Knoblauch, to whom I am indebted for the Classification of the Fungi, according to the more recent investigations of Brefeld. The Bacteria have been revised by Dr. Migula, the Florideæ rearranged after Schmitz, and the Taphrinaceæ after Sadebeck. The main body of the text of the Algæ and Fungi remains as it was originally written by Dr. Wille and Dr. Rostrup in the Danish Edition, though in many places considerable alterations and additions have been made. For the sake of comparison a tabular key to the Classification adopted in the Danish Edition is given in the Appendix.
In the Angiosperms I have retained the sequence of orders in the Danish original, and have not rearranged them according to[vi] the systems more familiar to English students. In any rearrangement much of the significance of Dr. Warming’s valuable and original observations would have been lost, and also from a teacher’s point of view I have found this system of great value. Although at present it may not be completely satisfactory, yet as an attempt to explain the mutual relationships, development and retrogression of many of the orders, it may be considered to have a distinct advantage over the more artificial systems founded upon Jussieu’s Divisions of Polypetalæ, Gamopetalæ, and Apetalæ.
With reference to the principles of the systematic arrangement adopted, I may here insert the following brief communication from the author (dated March, 1890), which he has requested me to quote from the preface of Dr. Knoblauch’s edition:—“Each form which, on comparative morphological considerations, is clearly less simple, or can be shown to have arisen by reduction or through abortion of another type having the same fundamental structure, or in which a further differentiation and division of labour is found, will be regarded as younger, and as far as possible, and so far as other considerations will admit, will be reviewed later than the ‘simpler,’ more complete, or richer forms. For instance, to serve as an illustration: Epigyny and Perigyny are less simple than Hypogny; the Epigynous Sympetalæ, Choripetalæ, Monocotyledones are, therefore, treated last, the Hydrocharitaceæ are considered last under the Helobieæ, etc. Zygomorphy is younger than Actinomorphy; the Scitamineæ and Gynandræ therefore follow after the Liliifloræ, the Scrophulariaceæ after the Solanaceæ, Linaria after Verbascum, etc. Forms with united leaves indicate younger types than those with free leaves; hence the Sympetalæ come after the Choripetalæ, the Sileneæ after the Alsineæ, the Malcaceæ after the Sterculiaceæ and Tiliaceæ, etc.
“Acyclic (spiral-leaved) flowers are older than cyclic (verticillate-leaved) with a definite number, comparing, of course, only those with the same fundamental structure. The Veronica-type must be considered as younger, for example, than Digitalis and Antirrhinum,[vii] these again as younger than Scrophularia; Verbascum, on the contrary, is the least reduced, and therefore considered as the oldest form. Similarly the one-seeded, nut-fruited Ranunculaceæ are considered as a later type (with evident abortion) than the many-seeded, folicular forms of the Order; the Paronychieæ and Chenopodiaceæ as reduced forms of the Alsineæ type; and the occurrence of few seeds in an ovary as generally arising through reduction of the many-seeded forms. The Cyperaceæ are regarded as a form derived from the Juncaceæ through reduction, and associated with this, as is so often the case, there is a complication of the inflorescence; the Dipsacaceæ are again regarded as a form proceeding from the Valerianaceæ by a similar reduction, and these in their turn as an offshoot from the Caprifoliaceæ, etc. Of course these principles of systematic arrangement could only be applied very generally; for teaching purposes they have often required modification.”
In preparing the translation considerable difficulty has been experienced in finding a satisfactory rendering of several terms which have no exact equivalent in English. I may here especially mention the term Vorblatt (Forblad) which I have translated by the term bracteole, when it clearly applied to the first leaf (or leaves) on a pedicel; but in discussing questions of general morphology a term was much needed to include both vegetative and floral shoots, and for this I have employed the term “Fore-leaf.” Also, the term “Floral-leaf” has been adopted as an equivalent of “Hochblatt,” and the term “bract” has been limited to a leaf subtending a flower. I have followed Dr. E. L. Mark in translating the word “Anlage” by “Fundament.”
At the end of the book will be found a short appendix giving an outline of some of the earlier systems of Classification, with a more complete account of that of Hooker and Bentham.
In a book of this character it is almost impossible to avoid some errors, but it is hoped that these will be comparatively few. In correcting the proof-sheets I have received invaluable assistance from Dr. Warming and Dr. Knoblauch, who have kindly[viii] read through every sheet, and to whom I am greatly indebted for many criticisms and suggestions. I have also to thank Mr. I. H. Burkill for his kind assistance in looking over the proofs of the Monocotyledons and Dicotyledons, and Mr. Harold Wager for kindly reading through the proofs of the Algæ and Fungi. My thanks are also especially due to Mr. E. L. Danielsen, and I wish to take this opportunity of acknowledging the very considerable help which I have received from him in translating from the Original Danish.
M. C. POTTER.
January, 1895.
[ix]
BEING THE SYSTEM OF CLASSIFICATION ADOPTED IN THE PRESENT VOLUME.
(The Algæ and Fungi rearranged in co-operation with Dr. E. Knoblauch, the other Divisions as in the 3rd Danish Edition.)
| PAGE | |
|---|---|
| DIVISION I. THALLOPHYTA | 4 |
| A. Sub-Division. Myxomycetes, Slime-Fungi | 5 |
| B. Sub-Division. Algæ | 8 |
| Class 1. Syngeneticæ | 14 |
| „ 2. Dinoflagellata | 16 |
| „ 3. Diatomeæ | 18 |
| „ 4. Schizophyta | 22 |
| Family 1. Schizophyceæ | 22 |
| „ 2. Bacteria | 26 |
| Class 5. Conjugatæ | 41 |
| „ 6. Chlorophyceæ | 46 |
| Family 1. Protococcoideæ | 47 |
| „ 2. Confervoideæ | 53 |
| „ 3. Siphoneæ | 59 |
| Class 7. Characeæ | 64 |
| „ 8. Phæophyceæ (Olive-Brown Seaweeds) | 68 |
| Family 1. Phæosporeæ | 68 |
| „ 2. Cyclosporeæ | 73 |
| Class 9. Dictyotales | 76 |
| „ 10. Rhodophyceæ (Red Seaweeds) | 77 |
| Family 1. Bangioideæ | 77 |
| „ 2. Florideæ | 78 |
| C. Sub-Division. Fungi | 84 |
| Class 1. Phycomycetes | 96 |
| Sub-Class 1. Zygomycetes | 96 |
| „ 2. Oomycetes | 100 |
| Family 1. Entomophthorales | 102 |
| „ 2. Chytridiales | 102 |
| „ 3. Mycosiphonales[x] | 104 |
| Class 2. Mesomycetes | 108 |
| Sub-Class 1. Hemiasci | 108 |
| „ 2. Hemibasidii | 109 |
| Class 3. Mycomycetes (Higher Fungi) | 114 |
| Sub-Class 1. Ascomycetes | 114 |
| Series 1. Exoasci | 116 |
| „ 2. Carpoasci | 118 |
| Family 1. Gymnoascales | 118 |
| „ 2. Perisporiales | 119 |
| „ 3. Pyrenomycetes | 125 |
| „ 4. Hysteriales | 132 |
| „ 5. Discomycetes | 132 |
| „ 6. Helvellales | 136 |
| Ascolichenes | 136 |
| Sub-Class 2. Basidiomycetes | 144 |
| Series 1. Protobasidomycetes | 145 |
| „ 2. Autobasidiomycetes | 157 |
| Family 1. Dacryomycetes | 159 |
| „ 2. Hymenomycetes | 159 |
| „ 3. Phalloideæ | 172 |
| „ 4. Gasteromycetes | 173 |
| Basidiolichenes | 176 |
| Fungi Imperfecti | 176 |
| DIVISION II. MUSCINEÆ (MOSSES) | 181 |
| Class 1. Hepaticæ | 188 |
| Family 1. Marchantieæ | 190 |
| „ 2. Anthoceroteæ | 191 |
| „ 3. Jungermannieæ | 191 |
| Class 2. Musci frondosi | 192 |
| Family 1. Sphagneæ | 193 |
| „ 2. Schizocarpeæ | 195 |
| „ 3. Cleistocarpeæ | 195 |
| „ 4. Stegocarpeæ | 195 |
| DIVISION III. PTERIDOPHYTA | 198 |
| Class 1. Filicinæ | 205 |
| Sub-Class 1. Filices | 205 |
| Family 1. Eusporangiatæ | 210 |
| „ 2. Leptosporangiatæ | 212 |
| Sub-Class 2. Hydropterideæ | 215 |
| Class 2. Equisetinæ (Horsetails) | 221 |
| Sub-Class 1. Isosporous Equisetinæ | 221 |
| „ 2. Heterosporous Equisetinæ | 225 |
| Class 3. Lycopodinæ (Club Mosses) | 226 |
| Sub-Class 1. Lycopodieæ | 226 |
| „ 2. Selaginelleæ[xi] | 228 |
| Transition from the Cryptogams to the Phanerogams | 234 |
| Asexual Generation of the Cormophytes | 234 |
| Sexual Generation; Fertilisation | 243 |
| DIVISION IV. GYMNOSPERMÆ | 251 |
| Class 1. Cycadeæ (Cycads) | 252 |
| „ 2. Coniferæ (Pine-Trees) | 255 |
| Family 1. Taxoideæ | 259 |
| „ 2. Pinoideæ | 262 |
| Class 3. Gneteæ | 270 |
| Fossil Gymnosperms | 271 |
| DIVISION V. ANGIOSPERMÆ | 273 |
| Class 1. Monocotyledones | 274 |
| Family 1. Helobieæ | 278 |
| „ 2. Glumifloræ | 283 |
| „ 3. Spadicifloræ | 297 |
| „ 4. Enantioblastæ | 308 |
| „ 5. Liliifloræ | 309 |
| „ 6. Scitamineæ | 323 |
| „ 7. Gynandræ | 328 |
| Class 2. Dicotyledones | 334 |
| Sub-Class 1. Choripetalæ | 337 |
| Family 1. Salicifloræ | 337 |
| „ 2. Casuarinifloræ | 339 |
| „ 3. Quercifloræ | 340 |
| „ 4. Juglandifloræ | 349 |
| „ 5. Urticifloræ | 351 |
| „ 6. Polygonifloræ | 358 |
| „ 7. Curvembryæ | 363 |
| „ 8. Cactifloræ | 375 |
| „ 9. Polycarpicæ | 377 |
| „ 10. Rhœadinæ | 393 |
| „ 11. Cistifloræ | 406 |
| „ 12. Gruinales | 416 |
| „ 13. Columniferæ | 421 |
| „ 14. Tricoccæ | 430 |
| „ 15. Terebinthinæ | 435 |
| „ 16. Aesculinæ | 439 |
| „ 17. Frangulinæ | 443 |
| „ 18. Thymelæinæ | 448 |
| „ 19. Saxifraginæ | 451 |
| „ 20. Rosifloræ | 456 |
| „ 21. Leguminosæ | 466 |
| „ 22. Passiflorinæ | 475 |
| „ 23. Myrtifloræ | 482 |
| „ 24. Umbellifloræ | 490 |
| „ 25. Hysterophyta[xii] | 498 |
| Sub-Class 2. Sympetalæ | 504 |
| A. Pentacyclicæ | 506 |
| Family 26. Bicornes | 506 |
| „ 27. Diospyrinæ | 510 |
| „ 28. Primulinæ | 511 |
| B. Tetracyclicæ | 514 |
| Family 29. Tubifloræ | 514 |
| „ 30. Personatæ | 517 |
| „ 31. Nuculiferæ | 531 |
| „ 32. Contortæ | 541 |
| „ 33. Rubiales | 548 |
| „ 34. Dipsacales | 556 |
| „ 35. Campanulinæ | 560 |
| „ 36. Aggregatæ | 564 |
| Appendix | 574 |
| Index | 593 |
For ä, ö and ü read æ, œ and ue throughout.
The following are not officinal in the British Pharmacopœia:—page 316, Dracæna (Dragon’s-blood), Smilax glabra; p. 321, “Orris-root”; p. 326, species of Curcuma, Alpinia officinarum; p. 333, Orchis-species (“Salep”). On page 296, par. 4, only Pearl Barley is offic. in the Brit. Phar.
[1]
The Vegetable Kingdom is arranged in 5 Divisions.
Division I.—Thallophyta, Stemless Plants, or those which are composed of a “thallus,” i.e. organs of nourishment which are not differentiated into root (in the sense in which this term is used among the higher plants), stem, or leaf. Vascular bundles are wanting. Conjugation and fertilisation in various ways; among most of the Fungi only vegetative multiplication.
In contradistinction to the Thallophytes all other plants are called “Stem-plants” (“Cormophyta”), because their shoots are leaf-bearing stems. The name Thallophyta (Stemless-plants) is to some extent unsuitable, since many of the higher Algæ are differentiated into stem and leaf.
The Thallophytes are again separated into 3 sub-divisions, namely:
Division II.—Bryophyta or Muscineæ, Mosses. These have leaf-bearing shoots, but neither true roots nor vascular[2] bundles. The lowest Mosses have, however, a thallus. Fertilisation is accomplished by means of self-motile, spirally coiled spermatozoids, through the agency of water. From the fertilised oosphere a “fruit-body” (capsule) with unicellular organs of reproduction (spores) is produced. The spore on germination gives rise to the vegetative system, which bears the organs of sexual reproduction; and this system is divided into two stages—the protonema, and the leaf-bearing plant produced on it.
Alternation of generations:
Division III.—Pteridophyta or Vascular Cryptogams, Fern-like Plants having leaf-bearing shoots, true roots, and vascular bundles with tracheides and sieve-tubes. Fertilisation as in the Mosses. From the fertilised oosphere the leaf-bearing shoot arises, which bears on its leaves the reproductive organs, the spores, in capsule-like sporangia. From the germination of the spore a small prothallium is formed, which bears the sexual reproductive organs.
Alternation of generations:
Division IV.—Gymnospermæ. The vegetative organs are in the main similar to those in the 3rd Division; special shoots are modified into flowers for the service of reproduction. From the oosphere, which is fertilised by means of the pollen-tube, the leaf-bearing plant is derived; this passes the first period of its life as an embryo in the seed, and continues its development when the germination of the seed takes place. The organs corresponding to the spores of the two preceding Divisions, are called respectively the pollen-grain and embryo-sac. The pollen-grains are multicellular; i.e. they contain an indistinct prothallium. In the embryo-sac a prothallium, rich in reserve material (endosperm),[3] with female organs of reproduction, is developed BEFORE FERTILISATION. The pollen-grains are carried by means of the wind to the ovules; these enclose the embryo-sac, and are situated on the open fruit-leaf (carpel), which has no stigma.
Alternation of generations:
Division V.—Angiospermæ. The members of this group are very similar to those of Division IV. The ovules are, however, encased in closed fruit-leaves (ovary), which have a special portion (stigma) adapted for the reception and germination of the pollen-grains. The pollen-grains are bicellular, but with only a membrane separating the two nuclei; they are carried to the stigma by animals (chiefly insects), by the wind, or by some other means. Endosperm is not formed till AFTER FERTILISATION. Alternation of generations in the main as in the Gymnosperms, but less distinct; while the sexual generation, the prothallium, with the organs of fertilisation, is also strongly reduced.
For a long time the vegetable kingdom has been divided into. Cryptogams (so called because their organs of reproduction remained for some time undiscovered), and Phanerogams or Flowering-plants which have evident sexual organs.
The first three divisions belong to the Cryptogams, and the third and fourth divisions to the Phanerogams. This arrangement has no systematic value, but is very convenient in many ways.
The Cryptogams are also known as Spore-plants, since they multiply by unicellular organs (spores), and the Phanerogams in contradistinction are called Seed-plants (Spermaphyta), since they multiply by seeds, multicellular bodies, the most important part of which is the embryo (a plant in its infancy). Mosses, Ferns, and Gymnosperms are together known as Archegoniatæ, since they possess in common a female organ of distinct structure, the Archegonium.
[4]
The thallus in the simplest forms is unicellular; in the majority, however, it is built up of many cells, which in a few instances are exactly similar; but generally there is a division of labour, so that certain cells undertake certain functions and are constructed accordingly, while others have different work and corresponding structure. Vessels or similar high anatomical structures are seldom formed, and the markings on the cell-wall are with few exceptions very simple. The Myxomycetes occupy quite an isolated position; their organs of nourishment are naked masses of protoplasm (plasmodia).
As regards the external form, the thallus may be entirely without special prominences (such as branches, members), but when such are present they are all essentially alike in their origin and growth, that is, disregarding the hair-structures which may be developed. A shoot of a Seaweed or of a Lichen, etc., is essentially the same as any other part of the plant; only among the highest Algæ (Characeæ, certain Siphoneæ, Sargassum, and certain Red Seaweeds) do we find the same differences between the various external organs of the plant body as between stem and leaf, so that they must be distinguished by these names.
Roots of the same structure and development as in the Seed-plants are not found, but organs of attachment (rhizoids and haptera) serve partly the biological functions of the root.
Systematic division of the Thallophytes. To the Thallophytes belong three sub-divisions—Slime-Fungi, Algæ, and Fungi. Formerly the Thallophytes were divided into Algæ, Fungi, and Lichens. But this last group must be placed among the Fungi, since they are really Fungi, which live symbiotically with Algæ. The Slime-Fungi must be separated from the true Fungi as a distinct subdivision. The Algæ possess a colouring substance, which is generally green, brown, or red, and by means of which they are able to build up organic compounds from carbonic acid and water. The Bacteria, especially, form an exception to the Algæ in this respect; like the Fungi and Slime-Fungi they have as a rule no such colouring material, but must have organic carbonaceous food; these plants form no starch, and need no light[5] for their vegetation (most Fungi require light for fructification). The Myxomycetes, Bacteria, and Fungi derive their nourishment either as saprophytes from dead animal or vegetable matter, or as parasites from living animals or plants (hosts), in which they very often cause disease.
A remark, however, must be made with regard to this division. Among the higher plants so much stress is not laid upon the biological relations as to divide them into “green” and “non-green”; Cuscuta (Dodder), a parasite, is placed among the Convolvulaceæ, Neottia and Corallorhiza, saprophytes, belong to the Orchidacere, although they live like Fungi, yet their relations live as Algæ. In the same manner there are some colourless parasitic or saprophytic forms among the Algæ, and stress must be laid upon the fact that not only the Blue-green Algæ, but also the Bacteria, which cannot assimilate carbonic-acid, belong to the Algæ group, Schizophyceæ. The reason for this is that systematic classifications must be based upon the relationship of form, development, and reproduction, and from this point of view we must regard the Bacteria as being the nearer relatives of the Blue-green Algæ. All the Thallophytes, which are designated Fungi (when the entire group of Slime-Fungi is left out), form in some measure a connected series of development which only in the lower forms (Phycomycetes) is related to the Algæ, and probably through them has taken its origin from the Algæ; the higher Fungi have then developed independently from this beginning. The distinction of colour referred to is therefore not the only one which separates the Algæ from the Fungi, but it is almost the only characteristic mark by which we can at once distinguish the two great sub-divisions of the Thallophytes.
The first forms of life on earth were probably “Protistæ,” which had assimilating colour material, or in other words, they were Algæ because they could assimilate purely inorganic food substances, and there are some among these which belong to the simplest forms of all plants. Fungi and Slime-Fungi must have appeared later, because they are dependent on other plants which assimilate carbon.[2]
The Slime-Fungi occupy quite an isolated position in the Vegetable Kingdom, and are perhaps the most nearly related to the group of Rhizopods in the Animal Kingdom. They live in and on organic remains, especially rotten wood or leaves, etc., on the surface of which their sporangia may be found.
They are organisms without chlorophyll, and in their vegetative condition are masses of protoplasm without cell-wall (plasmodia). They multiply by means of spores, which in the true Slime-Fungi[3][6] are produced in sporangia, but in some others[4] free. The spores are round cells (Fig. 1 a) which in all the true Slime-Fungi are surrounded by a cell-wall. The wall bursts on germination, and the contents float out in the water which is necessary for germination. They move about with swimming and hopping motions like swarmspores (e, f), having a cilia at the front end and provided with a cell-nucleus and a pulsating vacuole. Later on they become a little less active, and creep about more slowly, while they continue to alter their form, shooting out arms in various places and drawing them in again (g, h, i, k, l, m); in this stage they are called Myxamœbæ.

Fig. 1.—a-l Development of “Fuligo” from spore to Myxamœba; a-m are magnified 300 times; m is a Myxamœba of Lycogala epidendron; l´ three Myxamœbæ of Physarum album about to unite; o, a small portion of plasmodium, magnified 90 times.
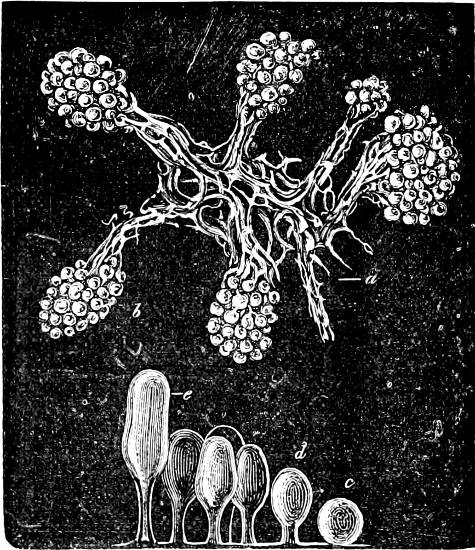
Fig. 2.—The plasmodium (a) of Stemonitis fusca, commencing to form into sporangia (b); drawn on July 9. The dark-brown sporangia were completely formed by the next morning; c-e shows the development of their external form.
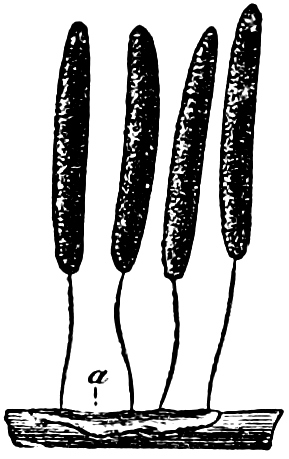
Fig. 3.—Four sporangia of Stemonitis fusca, fixed on a branch. a The plasmodium.
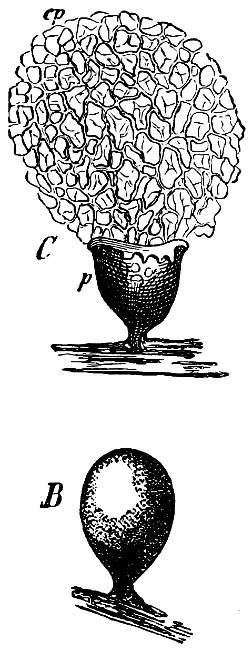
Fig. 4.—Sporangium of Arcyria incarnata. B closed; C open; p wall of sporangium; cp capilitium.
The Myxamœba grows whilst taking up nourishment from the material in which it lives, and multiplies by division. At a later stage a larger or smaller number of Myxamœbæ may be seen to[7] coalesce and form large masses of protoplasm, plasmodia, which in the “Flowers of Tan” may attain the size of the palm of a hand, or even larger, but in most others are smaller. The plasmodia are independent, cream-like masses of protoplasm, often containing grains of carbonate of lime and colouring matter (the latter yellow in the Flowers of Tan). They creep about in the decaying matter in which they live, by means of amœboid movements, internal streamings of the protoplasm continually taking place; finally they creep out to the surface, and very often attach themselves to other objects, such as Mosses, and form sporangia (Fig. 2). These are stalked or sessile and are generally cylindrical (Fig. 3), spherical or pear-shaped (Fig. 4); they rarely attain a larger size than that of a pin’s head, and are red, brown, white, blue, yellow, etc., with a very delicate wall. In some genera may be found a “Capillitium” (Fig. 4 cp), or network of branched fine strands between the spores. Flowers of[8] Tan (Fuligo septica) has a fruit-body composed of many sporangia (an Æthalium), which has the appearance of flat, irregular, brown cakes, inside the fragile external layer of which a loose powder, the spores, is found. It generally occurs on heaps of tanners’ bark, and appears sometimes in hot-beds in which that material is used, and is destructive by spreading itself over the young plants and choking them.
All the motile stages may pass into resting stages, the small forms only surrounding themselves with a wall, but the large ones at the same time divide in addition into polyhedral cells. When favourable conditions arise, the walls dissolve and the whole appears again as a naked (free-moving) mass of protoplasm.
To the genuine Slime-Fungi belong: Arcyria, Trichia, Didymium, Physarum, Stemonitis, Lycogala, Fuligo, Spumaria, Reticularia.
Some genera wanting a sporangium-wall belong to the Slime-Fungi: Ceratiomyxa, whose fruit-body consists of polygonal plates, each bearing stalked spores; Dictyostelium, in which the swarm-stage is wanting and which has stalked spores. Plasmodiophora brassicæ preys upon the roots of cabbages and other cruciferous plants, causing large swellings. Pl. alni causes coral-shaped outgrowths on the roots of the Alder (Alnus). Phytomyxa leguminosarum may be found in small knobs (tubercles) on the roots of leguminous plants. It is still uncertain whether it is this Fungus or Bacteria which is the cause of the formation of these tubercles.
Mode of Life. The Algæ (except most of the Bacteria) are themselves able to form their organic material by the splitting up of the carbonic acid contained in the water, or air in some cases, and for this purpose need light. The majority live in water, fresh or salt, but many are present on damp soil, stones, bark of trees, etc.
With the exception of the Bacteria, no saprophytes have actually been determined to belong to this group, and only very few true parasites (for instance, Phyllosiphon arisari, Mycoidea, etc.), but a good many are found epiphytic or endophytic on other Algæ, or water plants, and on animals (for instance, certain Schizophyceæ and Protococcoideæ; Trichophilus welckeri in the hairs of Bradypus, the Sloth), and several species in symbiotic relation to various[9] Fungi (species of Lichen), to Sponges (e.g. Trentepohlia spongiophila, Struvea delicatula), and to sundry Infusoria and other lower animals as Radiolarias, Hydra, etc. (the so-called Zoochlorella and Zooxantella, which are perhaps partly stages in development of various Green and Brown Algæ).
Vegetative Organs. The cells in all the Algæ (excepting certain reproductive cells) are surrounded by a membrane which (with the exception of the Bacteria) consists of pure or altered cellulose, sometimes forming a gelatinous covering, at other times a harder one, with deposits of chalk or silica formed in it. The cell-nucleus, which in the Schizophyta is less differentiated, may be one or more (e.g. Hydrodictyon, Siphoneæ) in each cell. Excepting in the majority of the Bacteria, colour materials (of which chlorophyll, or modifications of it, always seems to be found) occur, which either permeate the whole cytoplasm surrounding the cell-nucleus, as in most of the coloured Schizophyta, or are contained in certain specially formed small portions of protoplasm (chromatophores).
The individual at a certain stage of development consists nearly always of only one cell; by its division multicellular individuals may arise, or, if the daughter-cells separate immediately after the division, as in many of the simplest forms, the individual will, during the whole course of its existence, consist of only a single cell (unicellular Algæ). In multicellular individuals the cells may be more or less firmly connected, and all the cells of the individual may be exactly alike, or a division of labour may take place, so that certain cells undertake certain functions, and are constructed accordingly; this may also occur in parts of the cell in the large unicellular and multinuclear Algæ (Siphoneæ, p. 62).
The cells in most of the Algæ belong to the parenchymatous form; these, however, in the course of their growth, may very often become somewhat oblong; in many Algæ (particularly Fucoideæ and Florideæ) occur, moreover, hyphæ-like threads, which are very long, often branched, and are either formed of a single cell, or, more frequently, of a row of cells, having a well-pronounced apical growth. The parenchymatous as well as the hyphæ-like cells may, in the higher Algæ (especially in certain Fucoideæ and Florideæ), be further differentiated, so that they form well-defined anatomico-physiological systems of tissue, i.e. assimilating, conducting, storing, and mechanical.
With regard to the external form, the thallus may present no[10] differentiation, as in many unicellular Algæ, or in multicellular Algæ of the lower order, which are then either equally developed in all directions (e.g. Pleurococcus, Fig. 47), or form flat cell-plates (Merismopedium) or threads (Oscillaria, Fig. 21). The first step in the way of differentiation appears as a difference between apex and base (Rivularia, Porphyra); but the division of labour may proceed so that differences may arise between vegetative and reproductive cells (Œdogonium, Fig. 54); hairs and organs of attachment (rhizoids and haptera), which biologically serve as roots, are developed, and even leaves in certain forms of high order, belonging to different classes (e.g. Caulerpa, Fig. 59; Characeæ, Fig. 61; Sargassum, Fig. 72; and many Florideæ).
The non-sexual reproduction takes place vegetatively, in many instances, simply by division into two, and more or less complete separation of the divisional products (Diatomaceæ, Desmidiaceæ (Fig. 36), many Fission-plants, etc.), or by detached portions of the thallus (e.g. Caulerpa, Ulva lactuca, etc.; among many Schizophyceæ, small filaments known as hormogonia are set free), or asexually by special reproductive cells (spores) set free from the thallus; these may be either stationary or motile. The stationary reproductive cells (spores) may either be devoid of cell-wall (tetraspores of the Florideæ), or may possess a cell-wall; in the latter case they may be formed directly from the vegetative cells, generally by the thickening of the walls (akinetes), or only after a process of re-juvenescence (aplanospores). Aplanospores, as well as akinetes, may either germinate immediately or may become resting-cells, which germinate only after a period of rest.
The motile asexual reproductive cells are spherical, egg- or pear-shaped, naked, swarmspores (zoospores), which have arisen in other cells (zoosporangia), and propel themselves through the water by means of cilia; or they are Phyto-Amœbæ, which have no cilia and creep on a substratum by means of pseudopodia. The cilia, which are formed from the protoplasm (in the Bacteria, however, from the membrane), are mostly situated at the pointed and colourless end, which is directed forwards when in motion, and are 1, 2 (Fig. 5 B), 4 or more. Both the cilia in the Brown Algæ are attached to one side (Fig. 65); they are occasionally situated in a circle round the front end (Œdogonium, Fig. 6 a, and Derbesia), or are very numerous and situated in pairs distributed over a large part or nearly the whole of the zoospore (Vaucheria). Besides being provided with one or more nuclei[11] (Vaucheria), they may also have a red “eye spot” and vacuoles, which are sometimes pulsating, i.e. they appear and reappear at certain intervals. The swarmspores move about in the water in irregular paths, and apparently quite voluntarily, revolving round their longer axes; but they come to the surface of the water in great numbers either because of their dependence on light, or driven by warm currents in the water, or attracted by some passing mass of food material. The swarmspores germinate, each forming a new plant, as their movement ceases they surround themselves with a cell-wall, grow, and then divide; in Fig. 6 b, two may be seen in the condition of germination, and about to attach themselves by means of the front end, which has been developed into haptera (see also Fig. 5 B, lowest figure).
The sexual reproduction here, probably in all cases, consists in the coalescence of two masses of protoplasm, that is, in the fusion of their nuclei.
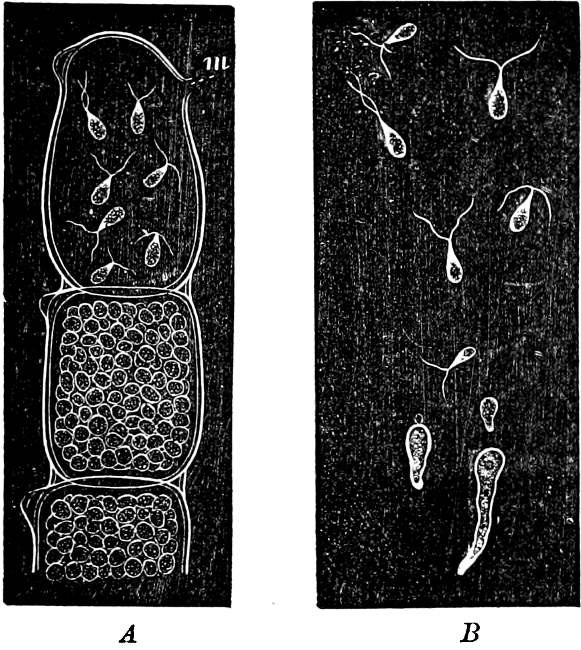
Fig. 5.—Cladophora glomerata. A The lower cells are full of swarmspores, whilst from the upper one the greater part have escaped through the aperture m. B Free and germinating swarmspores.
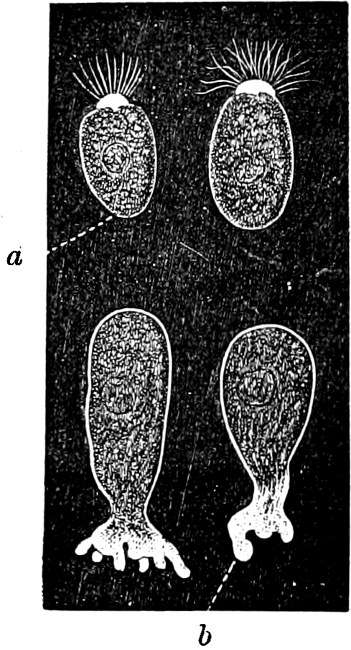
Fig. 6.—Œdogonium: a (free), b germinating swarmspores.
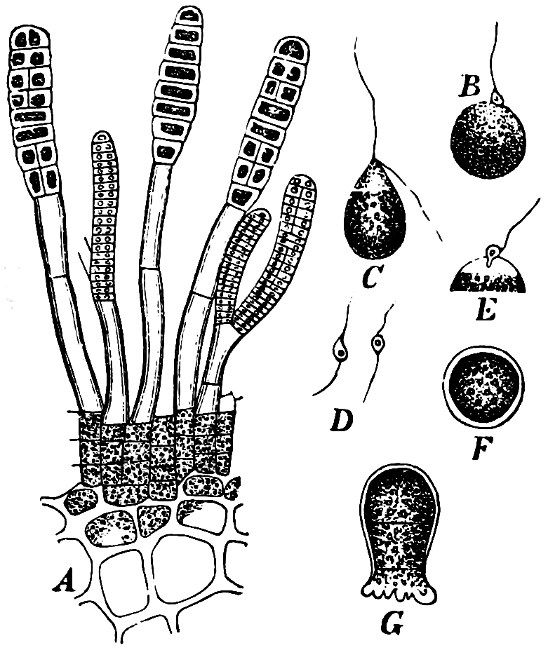
Fig. 7.—Zanardinia collaris. A Male gametangia (the small-celled) and female gametangia (large-celled). C Female gamete. D Male gamete. B E Fertilisation. F Zygote. G Germinating zygote.
The simplest and lowest form is termed conjugation, or isogamous fertilisation, and is characterized by the fact that the two coalescing cells (termed gametes) are equal, or almost equal, in shape and size (the female gamete in the Cutleriaceæ, e.g. Zanardinia[12] collaris, Fig. 7, is considerably larger than the male gamete). The cell in which the gametes are developed is called a gametaugium, and the reproductive cell formed by their union—which generally has a thick wall and only germinates after a short period of rest—is termed a zygote or zygospore. The conjugation takes place in two ways:—
(a) In the one way the gametes are motile cells (planogametes, zoogametes, Fig. 8), which unite in pairs during their swarming hither and thither in the water; during this process they lie side by side (Fig. 8 d), generally at first touching at the clear anterior end, and after a time they coalesce and become a motionless zygote, which surrounds itself with a cell-wall (Fig. 8 e). This form of conjugation is found in Ulothrix (Fig. 8 d), Acetabularia, and other Algæ (Figs. 45, 56, 66).
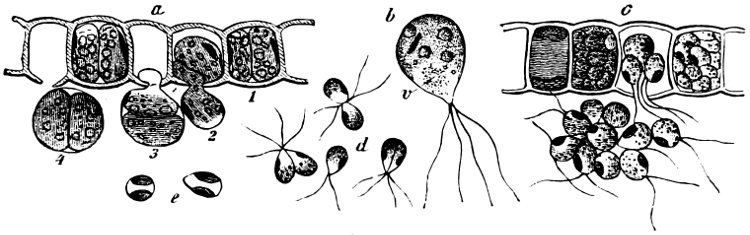
Fig. 8.—Ulothrix zonata: a portion of a thread with zoospores, of which two are formed in each cell (zoosporangium), the dark spots upon them are the “red eye-spots”; 1, 2, 3, 4 depict successive stages in the development of the zoospores; b a single zoospore, at v the pulsating vacuole; c portion of a thread with gametes, of which sixteen are formed in each gametangium; d gametes free and in conjugation; e conjugation has been effected, and the formed zygotes are in the resting condition.
(b) Among other Algæ (e.g. Diatomaceæ and Conjugatæ), the conjugating cells continue to be surrounded by the cell-wall of the mother-cell (aplanogametes in an aplanogametangium); the[13] aplanogametangia generally grow out into short branches, which lie close together and touch one another, the wall at the point of contact is then dissolved (Fig. 39). Through the aperture thus formed, the aplanogametes unite, as in the first instance, and form a rounded zygote, which immediately surrounds itself with a cell-wall. Various modifications occur; compare Figs. 37, 39, 41, 43.
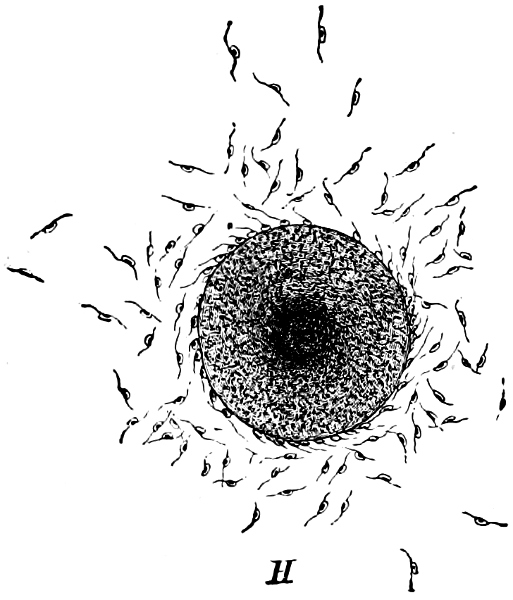
Fig. 9.—Fertilisation in the Bladder-wrack (Fucus vesiculosus).

Fig. 10.—Sphæroplea annulina.
The highest form of the sexual reproduction is the Egg- or Oogamous fertilisation. The two coalescing cells are in the main unlike each other in form as well as size. The one which is considered as the male, and is known as the spermatozoid (antherozoid), developes as a rule in large numbers in each mother-cell (antheridium); they are often self-motile (except in the Florideæ, where they are named spermatia), and are many times smaller than the other kind, the female, which is known as the egg-cell, (oosphere). The egg-cell is always a motionless, spherical, primordial cell which can either float about freely in the water, as in the Fucaceæ (Fig. 9), or is surrounded by a cell-wall (oogonium); generally only one oosphere is to be found in each oogonium, but several occur in Sphæroplea (Fig. 10). The result[14] of the spermatozoid coalescing with the egg-cell is, as in this case, the formation of a oospore, which generally undergoes a period of rest before germination (the Florideæ are an exception, a fruit-body, cystocarp, being produced as the result of coalescence).
An example of fertilisation is afforded by the Alga, Sphæroplea annulina (Fig. 10). The filamentous thallus is formed of cylindrical cells with many vacuoles (r in A); some cells develope egg-cells (B), others spermatozoids (C), the latter in a particularly large number. The egg-cells are spherical, the spermatozoids of a club- or elongated pear-shape with two cilia at the front end (G; E is however a swarmspore). The spermatozoids escape from their cells through apertures in the wall (o in C) and enter through similar apertures (o in B) to the egg-cells. The colourless front end of the spermatozoid is united at first with the “receptive spot” of the egg-cell (see F), and afterwards completely coalesces with it. The result is the formation of a oospore with wart-like excrescences (D).
The female (parthenogenesis) or male (androgenesis) sexual cell may, sometimes without any preceding fertilisation, form a new individual (e.g. Ulothrix zonata, Cylindrocapsa, etc.).
Systematic division of the Algæ. The Algæ are divided into the following ten classes:
1. Syngeneticæ; 2. Dinoflagellata, or Peridinea; 3. Diatomaceæ; 4. Schizophyta, Fission-algæ; 5. Conjugatæ; 6. Chlorophyceæ, Green-algæ; 7. Characeæ, Stone-worts; 8. Phæophyceæ; 9. Dictyotales; 10. Rhodophyceæ.
Among the lowest forms of the Algæ, the Syngeneticæ, the Dinoflagellata, and the unicellular Volvocaceæ (Chlamydomoneæ), distinct transitional forms are found approaching the animal kingdom, which can be grouped as animals or plants according to their method of taking food or other characteristics. Only an artificial boundary can therefore be drawn between the animal and vegetable kingdoms. In the following pages only those forms which possess chromatophores, and have no mouth, will be considered as Algæ.
The individuals are uni- or multicellular, free-swimming or motionless. The cells (which in the multicellular forms are loosely connected together, often only by mucilaginous envelopes) are naked or surrounded by a mucilaginous cell-wall, in which silica is never embedded. They contain one cell-nucleus, one or more pulsating[15] vacuoles, and one to two band- or plate-like chromatophores with a brown or yellow colour, and sometimes a pyrenoid.
Reproduction takes place by vegetative division, or asexually by zoospores, akinetes (or aplanospores?). Sexual reproduction is unknown. They are all fresh water forms.
To this class may perhaps be assigned the recently arranged and very little known orders of Calcocytaceæ, Murracytaceæ, Xanthellaceæ, and Dictyochaceæ, which partly occur in the free condition in the sea, in the so-called “Plankton,” and partly symbiotic in various lower marine animals.
The Syngeneticæ are closely related to certain forms in the animal kingdom, as the Flagellatæ.
Order 1. Chrysomonadinaceæ. Individuals, uni- or multicellular, swimming in free condition, naked or surrounded by a mucilaginous covering. The cells are generally oval or elongated, with 2 (rarely only 1) cilia, almost of the same length, and generally with a red “eye-spot” at their base, and with 2 (rarely 1 only) band-shaped chromatophores. Reproduction by the longitudinal division of the individual cells either during the swarming, or during a resting stage; in the multicellular forms also by the liberation of one or more cells, which in the latter case are connected together.
A. Unicellular: Chromulina, Cryptoglena, Microglena, Nephroselmis.
B. Multicellular: Uroglena, Syncrypta (Fig. 11), Synura.
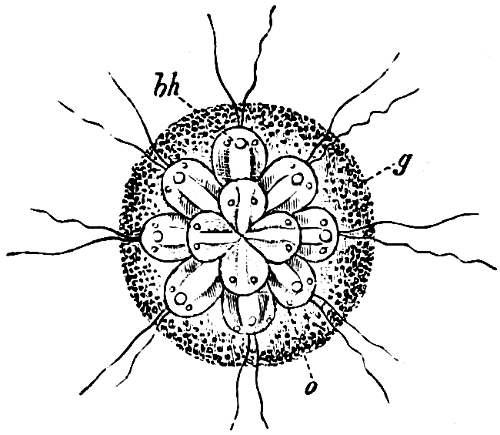
Fig. 11.—Syncrypta volvox: the multicellular individual is surrounded by a mucilaginous granular envelope.
Among the unicellular Chrysomonadinaceæ are probably classed some forms which are only stages in the development of the multicellular, or of other Syngeneticæ.
Order 2. Chrysopyxaceæ are unicellular, and differ mainly from the preceding in being attached either on a slime-thread (Stylochrysalis), or enclosed in an envelope (Chrysopyxis, Fig. 12). They have two cilia, and multiply by longitudinal (Chrysopyxis) or transverse division, and the swarming of one of the daughter-individuals (zoospore). Division may also take place in a motionless stage (palmella-stage).
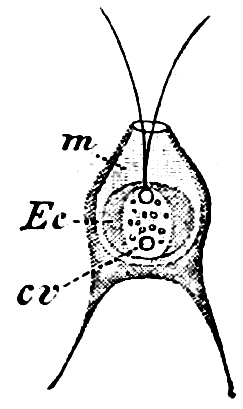
Fig. 12.—Chrsopyxis bipes: m envelope, Ec chromatophore, cv contractile vacuole.
Order 3. Dinobryinaceæ. The individuals are originally attached, uni- or multicellular; each individual cell is distinctly contractile, and fixed at the bottom of a cup-shaped, open envelope. Cilia 2, but of unequal length. Asexual reproduction by zoospores, which are formed by straight or oblique longitudinal division of the mother-cell, during a palmella-stage which is produced in the winter aplanospores. Epipyxis, Dinobryon.
[16]
Order 4. Hydruraceæ. The individuals are attached, without cilia, multicellular, branched, and with apical growth. The cells are spherical, but in the final stage almost spindle-shaped, and embedded in large masses of mucilage. Asexual reproduction by zoospores which are tetrahedric, with 1 cilia, and by resting akinetes. Hydrurus is most common in mountain brooks.
The individuals are of a very variable form, but always unicellular, and floating about in free condition. The cell is dorsiventral, bilateral, asymmetric and generally surrounded by a colourless membrane, which has no silica embedded in it, but is formed of a substance allied to cellulose. The membrane, which externally is provided with pores and raised borders, easily breaks up into irregularly-shaped pieces. In the forms which have longitudinal and cross furrows, two cilia are fixed where these cross each other, and project through a cleft in the membrane; one of these cilia projects freely and is directed longitudinally to the front or to the rear, the other one stretches crosswise and lies close to the cell, often in a furrow (cross furrow). The chromatophores are coloured brown or green and may either be two parallel (Exuviella), or several radially placed, discs, which sometimes may coalesce and become a star-shaped chromatophore. The coloring material (pyrrophyl) consists, in addition to a modification of chlorophyl, also of phycopyrrin and peridinin; this colour is sometimes more or less masked by the products of assimilation which consist of yellow, red or colourless oil (?) and starch. Cell-nucleus one: in Polydinida several nuclei are found; contractile vacuoles many, which partly open in the cilia-cleft (Fig. 13 gs). In some an eye-spot, coloured red by hæmatochrome, is found. Pyrenoids occur perhaps in Exuviella and Amphidinium.
The reproduction takes place as far as is known at present, only by division. This, in many salt water forms, may take place in the swarming condition, and, in that case, is always parallel to the longitudinal axis. The daughter-individuals, each of which retains half of the original shell, sometimes do not separate at once from each other, and thus chains (e.g. in Ceratium) of several connected individuals may be formed. In others, the division occurs after the cilia have been thrown off and the cell-contents rounded. The daughter-cells then adopt entirely new cell-walls. A palmella-stage (motionless division-stage) sometimes appears to[17] take place, and also aplanospores (?) with one or two horn-like elongations (e.g. in Peridinium cinctum and P. tabulatum); at germination one, or after division, two or more, new individuals may be formed.
Sexual reproduction has not been observed with certainty.
The Dinoflagellata move forward or backward, turning round their longitudinal axes; in their motion they are influenced by the action of light. The motion possibly may be produced only by the transverse cilium, which vibrates rapidly; whilst the longitudinal cilium moves slowly, and is supposed to serve mainly as a steering apparatus. They live principally in salt water, but also in fresh.
Besides the coloured forms, which are able to make their own organic compounds by the splitting up of the carbonic acid contained in the water, there are a few colourless forms (e.g. Gymnodinium spirale), or such as do not possess chromatophores (Polykrikos); these appear to live saprophytically, and may be able to absorb solid bodies with which they come in contact.
Dinoflagellata occur in the “Plankton” of the open sea, where they form together with Diatomaceæ the basis for the animal life. It is known with certainty that some salt water forms (like the Noctiluca, which belongs to the animal kingdom and to which they are perhaps related) produce light, known as phosphorescence.
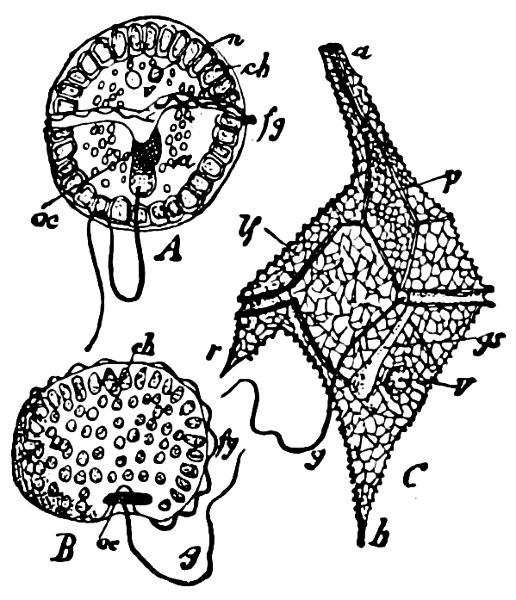
Fig. 13.—A and B Glenodinium cinctum. A seen from the ventral side, B from behind; fg transverse cilium; g longitudinal cilium; ch chromatophores; a starch; n cell-nucleus; v vacuole; oc eye-spot; C Ceratium tetraceros from the ventral side; r the right, b the posterior horn; lf longitudinal furrow; gs cilium-cleft; v vacuole; g longitudinal cilium. (A and B mag. 450 times, C 337 times.)
Dinoflagellata (Peridinea, Cilioflagellata) are allied through their lowest form (Exuviella) to the Syngeneticæ and especially to the order Chrysomonadinaceæ. They may be divided into three orders.
Order 1. Adinida. Without transverse or longitudinal furrows, but enclosed in two shells, and with two parallel chromatophores in each cell. Exuviella, Prorocentrum.
Order 2. Dinifera. With tranverse and generally longitudinal furrow. Many radially-placed, disc-formed chromatophores. The most common genera are—Ceratium (Fig. 13), Peridinium, Glenodinium (Fig. 13), Gymnodinium, Dinophysis.
Order 3. Polydinida. With several transverse furrows, no chromatophores, and several cell-nuclei. Only one genus—Polykrikos.
[18]
The order Polydinida deviates in a high degree from the other Dinoflagellata, not only by its many tranverse furrows, each with its own transverse cilium, and by the absence of chromatophores, but also in having several cell-nuclei and a kind of stinging capsule, which otherwise does not occur within the whole class. It may therefore be questionable whether this order should really be placed in the vegetable kingdom.
The individuals—each known as a frustule—assume very various forms and may be unicellular or multicellular, but present no differentiation; many similar cells may be connected in chains, embedded in mucilaginous masses, or attached to mucilaginous stalks. The cells are bilateral or centric, often asymmetrical, slightly dorsiventral and have no cilia; those living in the free condition have the power of sliding upon a firm substratum. The cell contains 1 cell-nucleus and 1–2 plate-shaped or several disc-shaped chromatophores. The colouring material “Melinophyl” contains, in addition to a modification of chlorophyl, a brown colouring matter, diatomin. 1 or 2 pyrenoids sometimes occur. Starch is wanting and the first product of assimilation appears to be a kind of oil (?).
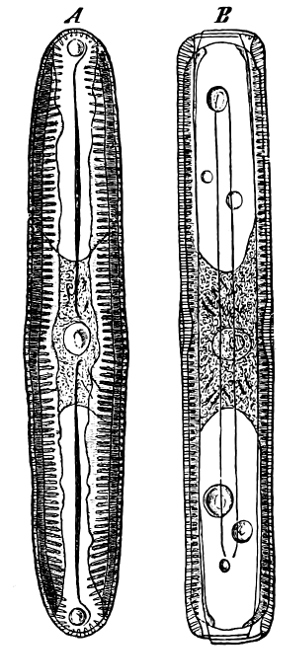
Fig. 14.—Pinnularia: B, from the edge, shows the valves fitting together; A, a valve.

Fig. 15.—Various Diatomaceæ. A Diatoma vulgare. B Tabellaria flocculosa. C Navicula tumida (lateral views). D Gomphonema constrictum (lateral views). E Navicula west[=i][=i] (lateral views).
The cell-walls are impregnated with silica to such a degree that they are imperishable and are therefore able to contribute in a great measure to the formation of the earth’s crust. The structure of their cell-wall is most peculiar and differs from all other plants (except certain Desmidiaceæ); it does not consist of a single piece but is made up of two—the “shells”—(compare Exuviella and Prorocentrum among the Dinoflagellata) which are fitted into each other, one being a little larger than the other and embracing its edge, like a box with its lid (Fig. 14 B). The two parts which correspond to the bottom and lid of the box are known as valves. Along the central line of the valves a longitudinal rib may often be found, interrupted at its centre by a small cleft (perhaps homologous with the cilia-cleft of the Dinoflagellata), through which the protoplasm is enabled to communicate with the exterior (Fig. 14 A). It is principally by reason of the valves, which bear numerous fine, transverse ribs, striæ or warts, etc. (Figs. 14, 15, 17), that the Diatomeæ have become so well known and employed as test objects in microscopical science. When the division takes place, the two shells are separated a little from each other, and after the cell-contents have divided into two masses, two new shells are formed, one fitting into the larger valve, the other one[19] into the smaller valve of the original frustule. The latter cell (frustule) is thus, upon the whole, smaller than the mother-cell, and as the cells do not increase in size, some frustules are smaller than the ones from which they are derived, and thus, by repeated divisions, it follows that smaller and smaller frustules are produced. This continued diminution in size is, however, compensated for by the formation, when the cells have been reduced to a certain minimum, of auxospores, 2–3 times larger. These may either be formed asexually by the protoplasm of a cell increasing, rounding off and surrounding itself with a new wall (e.g. Melosira) or after conjugation, which may take place with various modifications: 1. Two individuals unite after the secretion of a quantity of mucilage, and the valves then commence to separate from each other, on the side which the two individuals turn towards each other. The protoplasmic bodies now release themselves from their cell-wall, and each rounds off to form an ellipsoidal mass; these two protoplasmic[20] masses (gametes) coalesce to form a zygote, the cell-nuclei and chromatophores also fusing together. The zygote increases in size, and surrounds itself with a firm, smooth, siliceous wall—the perizonium. The auxospores, whichever way they arise, are not resting stages. The germination of the zygote commences by the protoplasm withdrawing itself slightly from the cell-wall and constructing first the larger valve, and later on the smaller one; finally the membrane of the zygote bursts (e.g. Himantidium). 2. The conjugation occurs in a similar manner, but the protoplasm of the cells divides transversely before conjugation into two daughter-cells. Those lying opposite one another conjugate (Fig. 16) and form two zygotes. The formation of the perizonium, and germination take place as in the preceding instance (e.g. Epithemia). 3. Two cells place themselves parallel to each other, and each of the two cell-contents, without coalescing, becomes an auxospore. The formation of the wall takes place as in the preceding case. This is found in the Naviculeæ, Cymbelleæ, the Gomphonemeæ (e.g. Frustulia, Cocconema).
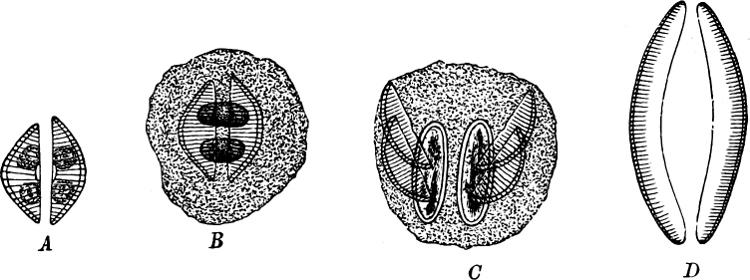
Fig. 16.—Conjugation of Cymbella variabilis. A, The protoplasm in the two cells has divided into two masses; B these masses coalesce in pairs; the cells (B C) enclosed in a mucilaginous matrix. C D Auxospores and their formation.
The Diatomaceæ may be found in salt as well as in fresh water (often in such masses that the colour of the water or mud becomes yellow or brown; in the same manner the genera Chætoceros, Rhizosolenia, Coscinodiscus, and several others, form large slime-masses, “Plankton” on the surface of the sea), on damp soil and in dust blown by the wind. They occur as fossils in the recent formations, often in large deposits (siliceous earth, mountain meal), as in the cement lime in Jutland, the alluvial deposits beneath Berlin, in clay strata beneath peat bogs, in guano, etc.[21] These accumulations of fossilized diatoms are used in the manufacture of dynamite and in various manufactures.
The Diatomaceæ appear nearest to, and must be placed as a group co-ordinate with the Dinoflagellata, as they doubtless may be supposed to derive their origin from forms resembling Exuviella, and to have lost the cilia. The resemblances to the Desmidiaceæ which are striking in many respects, can only be conceived as analogies, and cannot be founded upon homologies, and it is therefore impossible to regard them as proof of genetic relationship. The family contains only one order.
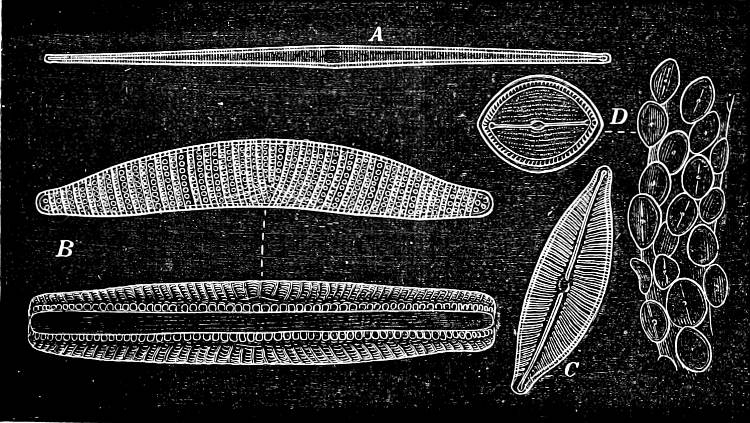
Fig. 17.—Various Diatomeæ. A Synedra radians. B Epithemia turgida (from the two different sides). C Cymbella cuspidata. D Cocconeis pediculus (on the right several situated on a portion of a plant, on the left a single one more highly magnified).
Order 1. Diatomaceæ. This order may be divided into two sub-orders, viz.—
Sub-Order 1. Placochromaticæ. The chromatophores are discoid, large, 1 or 2 in each cell; the structure of the valves is bilateral and always without reticulate markings. The following groups belong to this sub-order: Gomphonemeæ, Cymbelleæ, Amphoreæ, Achnantheæ, Cocconeideæ, Naviculeæ, Amphipleureæ, Plagiotropideæ, Amphitropideæ, Nitzchieæ, Surirayeæ, and Eunotieæ.
Sub-Order 2. Coccochromaticæ. The chromatophores are granular, small and many in each cell. The structure of the cells is zygomorphic or centric, often with reticulate markings. The following groups belong to this sub-order: Fragilarieæ, Meridieæ, Tabellarieæ, Licmophoreæ, Biddulphieæ, Anguliferæ, Eupodisceæ, Coscinodisceæ, and Melosireæ.
[22]
The individuals are 1—many celled; the thallus consists in many of a single cell, in others of chains of cells, the cells dividing in only one definite direction (Figs. 18, 21). In certain Fission-Algæ the cell-chain branches (Fig. 30) and a difference between the anterior and the posterior ends of the chain is marked; in some, the cells may be united into the form of flat plates by the cell-division taking place in two directions; and in others into somewhat cubical masses, or rounded lumps of a less decided form, by the divisions taking place in three directions; or less defined masses may be formed by the divisions taking place in all possible directions.
The cell-walls rarely contain cellulose, they often swell considerably (Figs. 20, 22), and show distinct stratifications, or they are almost completely changed into a mucilaginous mass in which the protoplasts are embedded, e.g. in Nostoc (Fig. 22), and in the “Zooglœa” stage of the Bacteria (Fig. 27). Sexual reproduction is wanting. Vegetative reproduction by division and the separation of the divisional products by the splitting of the cell-wall or its becoming mucilaginous; among the Nostocaceæ, Lyngbyaceæ, Scytonemaceæ, etc., “Hormogonia” are found; in Chamæsiphon and others single reproductive akinetes are formed. Many Fission-Algæ conclude the growing period by the formation of resting akinetes or aplanospores.
The Schizophyta may be divided into 2 families:
1. Schizophyceæ.
2. Bacteria.
All the Blue-green Algæ are able to assimilate carbon by means of a colouring material containing chlorophyll (cyanophyll); but the chlorophyll in this substance is masked by a blue (phycocyan), or red (phycoerythrin, e.g. in Trichodesmium erythræum in the Red Sea) colouring matter which may be extracted from them in cold water after death. The colouring matter, in most of them, permeates the whole of the protoplasm (excepting the cell-nucleus), but in a few (e.g. Glaucocystis, Phragmonema), slightly developed chromatophores are to be found. Where the cells are united into filaments (cell-chains) a differentiation into apex and base (Rivulariaceæ) may take place, and also between ordinary vegetative cells and heterocysts; these latter cannot divide, and are distinguished[23] from the ordinary vegetative cells (Fig. 22 h) by their larger size, yellow colour, and poverty of contents. Branching sometimes occurs and is either true or spurious.

Fig. 18.—Microcoleus lyngbyanus: a portion of a filament, the thick sheath encloses only one cell-chain; in one place a cell is drawn out by the movement of the cell-chain; b the cell-chain has divided into two parts (“hormongonia”) which commence to separate from each other.
The cell-chain in the spurious branching divides into two parts, of which either one or both grow beyond the place of division (Fig. 18) and often out to both sides (e.g. Scytonema), the divisions however, always take place transversely to the longitudinal direction of the cell-chain. In the true branching a cell elongates in the direction transverse to the cell-chain, and the division then takes place nearly at right angles to the former direction (Sirosiphoniaceæ).
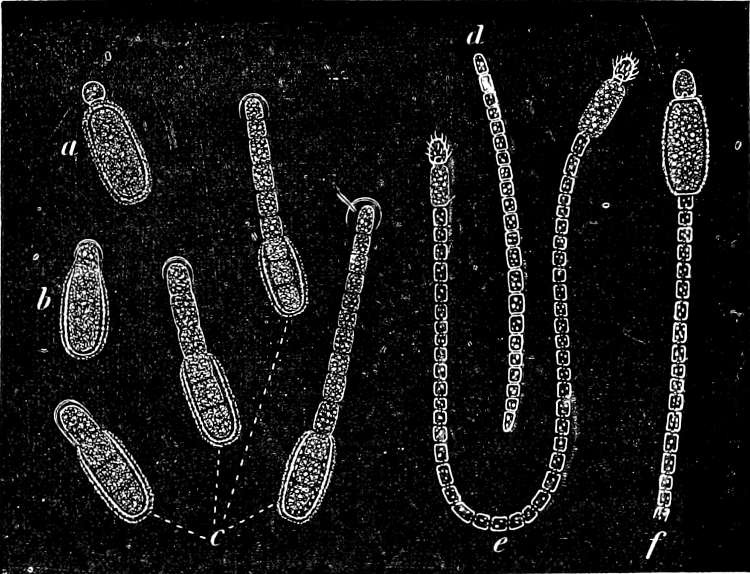
Fig. 19.—Cylindrospermum majus: a resting akinete with heterocyst; b-d germinating stages of a resting akinete; e filament with two heterocysts and the formation of new akinetes; f part of a filament with a heterocyst, and mature resting akinete.
Cilia are wanting, but the filaments are sometimes self-motile (e.g. hormogonia in Nostoc) and many partly turn round their axes, partly slide forward or backward (Oscillaria).
Reproduction takes place by spores and hormogonia in addition[24] to simple cell-division. Hormogonia are peculiar fragments of a cell-chain capable of motion, and often exhibit a vigorous motion in the sheath, until at last they escape and grow into a new individual (Fig. 18). The spores are reproductive akinetes (Chamæsiphon, etc.) or resting akinetes; these latter arise by the vegetative cells enlarging and constructing a thick cell-wall (Fig. 19 e f). On germination, this cell-wall bursts and the new cell-chain elongates in the same longitudinal direction as before (Fig. 19 b c). Many (e.g. Oscillaria) may however winter in their ordinary vegetative stage. Aplanospores are wanting.
The Fission-Algæ are very prevalent in fresh water and on damp soil, less so in salt water; they also often occur in water which abounds in decaying matter. Some are found in warm springs with a temperature as high as 50° C.
The Family may be divided into 2 sub-families:
1. Homocysteæ (heterocysts are wanting): Chroococcaceæ, Lyngbyaceæ and Chamœsiphonaceæ.
2. Heterocysteæ (heterocysts present): Nostocaceæ, Rivulariaceæ, Scytonemaceæ and Sirosiphoniaceæ.
Order 1. Chroococcaceæ. The individuals are 1—many-celled, but all the cells are uniform, united to form plates or irregular masses, often surrounded by a mucilaginous cell-wall, but never forming cell-chains. Multiplication by division and sometimes by resting akinetes, but reproductive akinetes are wanting. Chroococcus, Aphanocapsa, Glœocapsa (Fig. 20), Cœlosphærium, Merismopedium, Glaucocystis, Oncobyrsa, Polycystis, Gomphosphæria.
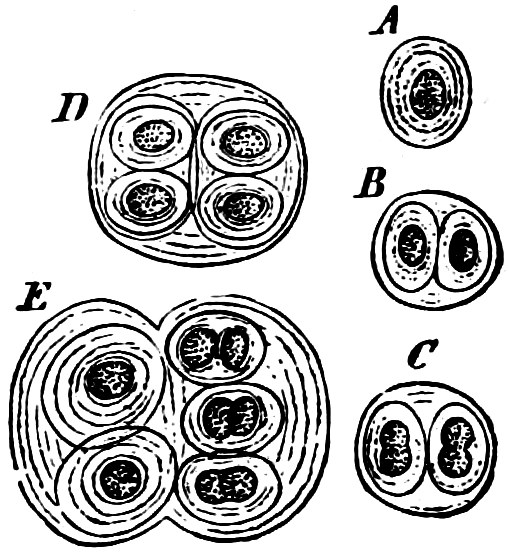
Fig. 20.—Glœocapsa atrata: A, B, C, D, E various stages of development.
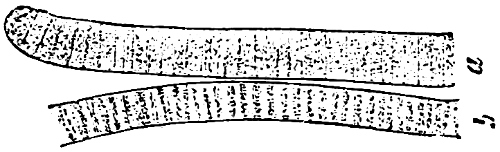
Fig. 21.—Oscillaria; a terminal, b central portion of a filament.
Order 2. Lyngbyaceæ (Oscillariaceæ). The cells are discoid (Fig. 21), united to straight or spirally twisted, free filaments, which are unbranched, or with spurious branching. The ends of the cell-chains are similar. Heterocysts absent. Reproduction by synakinetes, resting akinetes are wanting. Oscillaria (Fig. 21), Spirulina, Lyngbya, Microcoleus, Symploca, Plectonema.
[25]
Order 3. Chamæsiphonaceæ. The individuals are 1—many-celled, attached, unbranched filaments with differentiation into apex and base, without heterocysts. Multiplication by reproductive akinetes; resting akinetes are wanting. Dermocarpa, Clastidium, Chamæsiphon, Godlewskia, Phragmonema.
Order 4. Nostocaceæ. The individuals are formed of multicellular, unbranched filaments, without differentiation into apex and base; heterocysts present. Reproduction by synakinetes and resting akinetes.
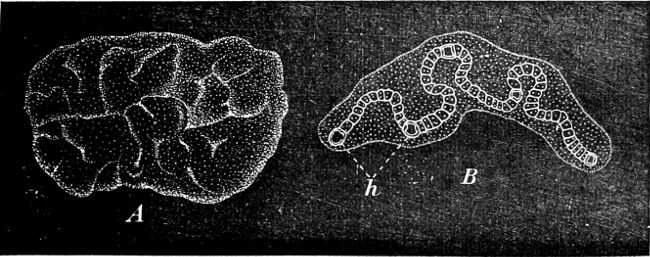
Fig. 22.—Nostoc verrucosum. A The plant in its natural size; an irregularly folded jelly-like mass. B One of the cell-chains enlarged, with its heterocysts (h), embedded in its mucilaginous sheath.
Some genera are not mucilaginous, e.g. Cylindrospermum (Fig. 19). The cell-chains in others, e.g. Nostoc, wind in between one another and are embedded in large structureless jelly-like masses, which may attain the size of a plum or even larger (Fig. 22); sometimes they are found floating in the water, sometimes attached to other bodies. Other genera as follows: Aphanizomenon and Anabæna (in lakes and smaller pieces of water); Nodularia is partly pelagic. Some occur in the intercellular spaces of higher plants, thus Nostoc-forms are found in Anthoceros, Blasia, Sphagnum, Lemna, and in the roots of Cycas and Gunnera; Anabæna in Azolla.
Order 5. Rivulariaceæ. The individuals are multicellular filaments, with differentiation into apex and base; spurious branching, and a heterocyst at the base of each filament, reproduction by synakinetes and resting akinetes, rarely by simple reproductive akinetes. Rivularia, Glœotrichia, Isactis, Calothrix.
Order 6. Scytonemaceæ. The individuals are formed of multicellular filaments with no longitudinal division; differentiation into apex and base very slight or altogether absent;[26] branching spurious; heterocysts present. Reproduction by synakinetes, rarely by resting akinetes and ordinary reproductive akinetes. Tolypothrix, Scytonema, Hassalia, Microchæte.
Order 7. Sirosiphoniaceæ. The individuals are formed of multicellular threads with longitudinal divisions; true branching and heterocysts, and often distinct differentiation into apex and base. Reproduction by synakinetes, rarely by resting akinetes and ordinary reproductive akinetes. Hapalosiphon, Stigonema, Capsosira, Nostocopsis, Mastigocoleus.
The Bacteria (also known as Schizomycetes, and Fission-Fungi) are the smallest known organisms, and form a parallel group to the Blue-green Algæ, but separated from these Algæ by the absence of their colouring material; chlorophyll is only found in a few Bacteria.
The various forms under which the vegetative condition of the Bacteria appear, are termed as follows:
1. Globular forms, cocci (Figs. 27, 30 c): spherical or ellipsoidal, single cells, which, however, are usually loosely massed together and generally termed “Micrococci.”
2. Rod-like forms: more or less elongated bodies; the shorter forms have been styled “Bacterium” (in the narrower sense of the word), and the term “Bacillus” has been applied to longer forms which are straight and cylindrical (Figs. 28, 29, 30 E).
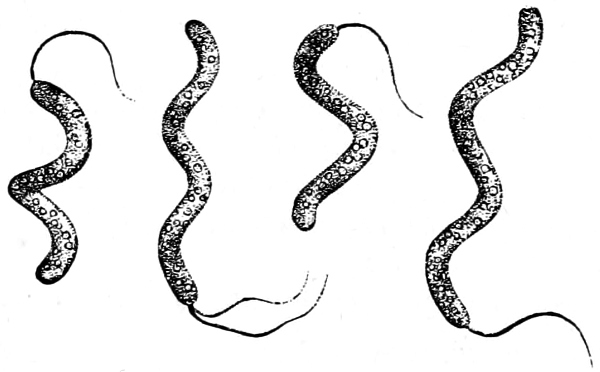
Fig. 23.—Spirillum sanguineum. Four specimens. One has two cilia at the same end, the sulphur grains are seen internally.
3. Thread-like forms: unbranched, long, round filaments, resembling those of Oscillaria, are possessed by Leptothrix (very thin, non-granular filaments; Fig. 30 A, the small filaments) and Beggiatoa (thicker filaments, with strong, refractile grains or drops of sulphur (Fig. 31); [27]often self-motile). Branched filaments, with false branching like many Scytonemaceæ, are found in Cladothrix (Fig. 30 B, G).
4. Spiral forms: Rod-like or filamentous bodies, which more or less strongly resemble a corkscrew with a spiral rising to the left. In general these are termed Spirilla (Fig. 23); very attenuated spirals, Vibriones (standing next to Fig. 30 M); if the filaments are slender and flexible with a closely wound spiral, Spirochætæ (Fig. 24).
5. The Merismopedium-form, consisting of rounded cells arranged in one plane, generally in groups of four, and produced by divisions perpendicular to each other.
6. The Sarcina-form, consisting of roundish cells which are produced by cellular division in all the three directions of space, united into globular or ovoid masses (“parcels”) e.g. Sarcina ventriculi (Figs. 25, 26).
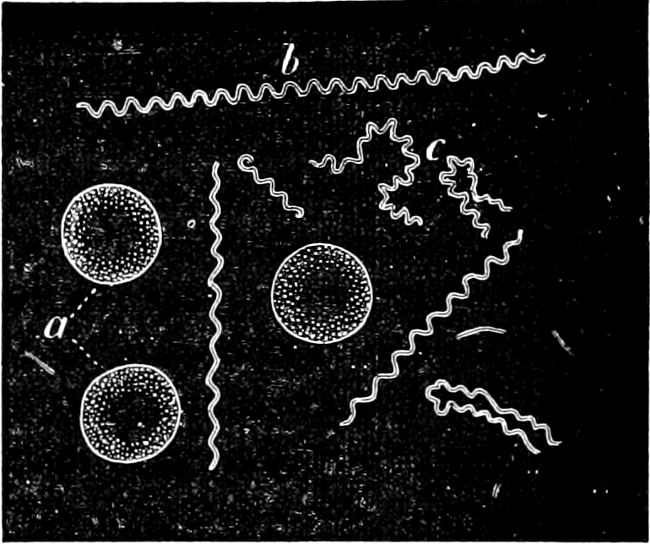
Fig. 24.—Spirochæte obermeieri, in active motion (b) and shortly before the termination of the fever (c); a blood corpuscles.
All Bacteria are unicellular. In the case of the micrococci this is self-evident, but in the “rod,” “thread,” and “spiral” Bacteria, very often numerous cells remain united together and their individual elements can only be recognised by the use of special reagents.

Fig. 25.—Sarcina ventriculi. One surface only is generally seen. Those cells which are drawn with double contour are seen with the correct focus, and more distinctly than those cells lying deeper drawn with single contour.
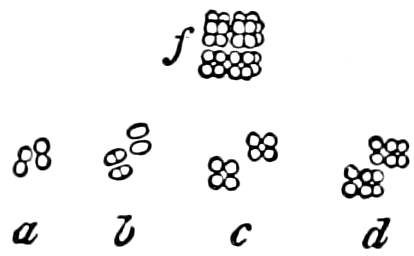
Fig. 26.—Sarcina minuta: a-d successive stages of one individual (from 4–10 p.m.); f an individual of 32 cells.
The condition termed “Zooglœa,” which reminds us of Nostoc, is produced by the cells becoming strongly mucilaginous. A number of individuals in active division are found embedded in a mass of mucilage, which either contains only one, or sometimes more, of[28] the above-named forms. The individuals may eventually swarm out and continue their development in an isolated condition. Such mucilaginous masses occur especially upon moist vegetables (potatoes, etc.), on the surface of fluids with decaying raw or cooked materials, etc. The mucilaginous envelope is thrown into folds when the Bacteria, with their mucilaginous cell-walls, multiply so rapidly that there is no more room on the surface of the fluid.
The cells of the Bacteria are constructed like other plant-cells in so far as their diminutive size has allowed us to observe them. The cell-wall only exceptionally shows the reactions of cellulose (in Sarcina, Leuconostoc; also in a Vinegar-bacterium, Bacterium xylinum); a mucilaginous external layer is always present. The body of the cell mostly appears to be an uniform or finely granulated protoplasm. Very few species (e.g. Bacillus virens) contain chlorophyll; others are coloured red (purple sulphur Bacteria); the majority are colourless. Bacillus amylobacter shows a reaction of a starch-like material when treated with iodine before the spore-formation. Some Bacteria contain sulphur (see p. 37). The body, which has been described as a cell-nucleus, is still of a doubtful nature.
Artificial colourings with aniline dyes (especially methyl-violet, gentian-violet, methylene-blue, fuchsin, Bismarck-brown and Vesuvin) play an important part in the investigations of Bacteria.
Movement. Many Bacteria are self-motile; the long filaments of Beggiatoa exhibit movements resembling those of Oscillaria. In many motile forms the presence of cilia or flagella has been proved by the use of stains; many forms have one, others several cilia attached at one or both ends (Fig. 23) or distributed irregularly over the whole body; the cilia are apparently elongations of the mucilaginous covering and not, as in the other Algæ of the protoplasm. In Spirochæte the movement is produced by the flexibility of the cell itself. Generally speaking, the motion resembles that of swarm-cells (i.e. rotation round the long axis and movement in irregular paths); but either end has an equal power of proceeding forwards.
The swarming motion must not be confounded with the hopping motion of the very minute particles under the microscope (Brownian movement).
Vegetative reproduction takes place by continued transverse[29] division; hence the name “Fission-Fungi” or “Fission-Algæ,” has been applied to the Bacteria.
Spores. The spores are probably developed in two ways. In the ENDOSPOROUS species (Figs. 28, 29), the spore arises as a new cell inside the mother-cell. The spores are strongly refractile, smaller than the mother-cell, and may be compared to the aplanospores of other Algæ. In addition to these there are the ARTHROSPOROUS species in which the cells, just as in Nostoc and other Blue-green Algæ, assume the properties of spores without previously undergoing an endogenous new construction, and are able to germinate and form new vegetative generations (Fig. 27). The formation of spores very often commences when the vegetative development begins to be restricted.
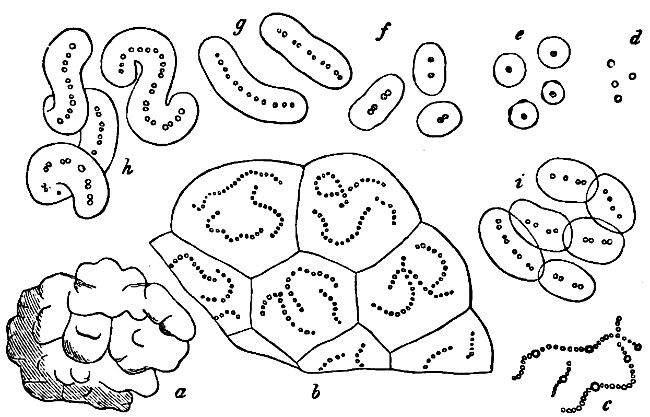
Fig. 27.—Leuconostoc mesenterioides: a a zooglœa, natural size; b cross section of zooglœa; c filaments with spores; d mature spores; e-i successive stages of germination; in e portions of the ruptured spore-wall are seen on the external side of the mucilaginous covering. (b-i magnified 520.)
The spores germinate as in Nostoc by the bursting of the external layer of the cell-wall, either by a transverse or longitudinal cleft, but always in the same way, in the same species (Fig. 28, example of transverse cleft).
Distribution. Bacteria and their germs capable of development, are found everywhere, in the air (dust), in surface water, and in the superficial layers of the soil. The number varies very much in accordance with the nature of the place, season, etc. They enter, together with air and food, into healthy animals and occur always in their alimentary tract.
[30]
Growth and reproduction depend upon the conditions of temperature. There is a certain minimum, optimum and maximum for each species; for instance (in degrees Centigrade)—
| Minim. | Opt. | Maxim. | ||
| Bacillus subtilis | + 6 | c. 30 | + 50 | |
| B. anthracis | 15 | 20–25 | 43 | |
| Spirillum choleræ asiaticæ | 8 | 37 | 40 | (but grows only feebly if under 16°). |
| Bacterium tuberculosis | 28 | 37–38 | 42 |
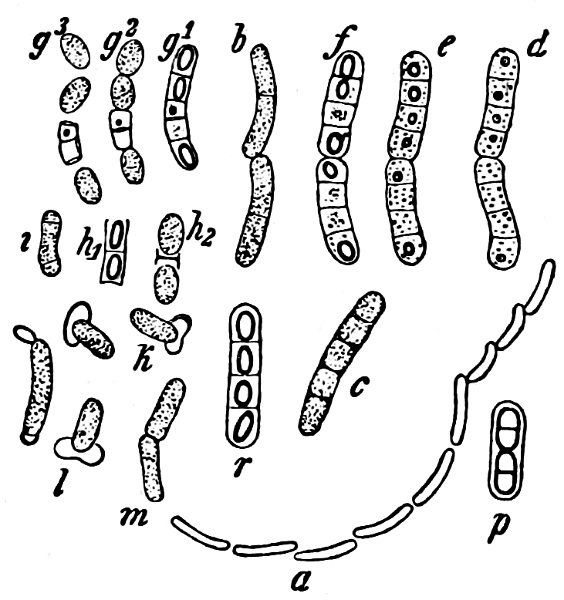
Fig. 28.—Bacillus megaterium: a outline of a living, vegetative cell-rod; b a living, motile, pair of rods; p a similar 4-celled rod after the effects of iodine alcohol; c a 5-celled rod in the first stages of spore-formation; d-f successive stages of spore-formation in one and the same pair of rods (in the course of an afternoon); r a rod with mature spores; g1–g3 three stages of a 5-celled rod, with spores sown in nutritive solution; h1–h2, i, k, l stages of germination; m a rod in the act of transverse division, grown out from a spore which had been sown eight hours previously. (After de Bary; a mag. 250, the other figures 600 times).

Fig. 29.—Bacillus amylobacter. Motile rods, partly cylindrical and without spores, partly swollen into various special shapes and with spore-formation in the swelling. s Mature spore, with thick mucilaginous envelope. (After de Bary; mag. 600 times, with the exception of s, which is more highly magnified.)
The functions of life cease on a slight excess of the maximum or minimum temperature, numbness setting in when either of these limits is passed. Crenothrix-threads provided with mucilaginous envelopes may, according to Zopf, sustain a temperature of-10°. Some Bacteria are said to be able to resist the exposure to as low a temperature as-110° for a short time. It is not known at what degree of cold the death of the Bacteria occurs: the greatest degree of heat which the vegetative cells can[31] withstand is about the same as that for other vegetative plant-cells, namely, about 50–60° C. Certain Bacteria, e.g. B. thermophilus, grow and thrive vigorously at 70° C. Many spores, on the contrary, are able to bear far higher temperatures (in several species a temperature for some duration of above 100°, those of Bacillus subtilis, for instance, can withstand for hours a temperature of 100° in nutrient solutions; the spores remain capable of development after exposure to a dry heat of 123° C.).
The Desiccation of the air, if prolonged, kills many forms when in the vegetative condition. The spores however can bear a much longer period of dryness, some even several years.
Oxygen. Some species cannot live without a supply of free oxygen (Aerobic), e.g. the Vinegar-bacteria, the Hay-bacilli, the Anthrax-bacilli, the Cholera-Microspira. Other species again thrive vigorously without supply of free oxygen, and are even checked in their development by the admission of air (Anaerobic), e.g. the butyric acid Bacterium (Clostridium butyricium = Bacillus amylobacter). A distinction may be drawn between obligate and facultative aerobics and obligate and facultative anaerobics. Several Bacteria, producing fermentation, may grow without the aid of oxygen when they are living in a solution in which they can produce fermentation; but, if this is not the case, they can only grow when a supply of oxygen is available. A great number of the pathogenic Bacteria belong to the facultative anaerobics.
A luminous Bacterium (Bacillus phosphorescens) which in the presence of a supply of oxygen gives a bluish-white light, has been found in sea-water. Phosphorescent Bacteria have frequently been observed upon decaying sea-fish, as well as on the flesh of other animals; by transferring the Bacteria from cod fish to beef, etc., the latter may be made luminous.
Organic carbon compounds are indispensable for all Bacteria, (except, as it appears, for the nitrifying organisms), as they can only obtain the necessary supplies of carbon from this source. The supplies of nitrogen, which also they cannot do without, can be obtained equally as well from organic compounds as from inorganic salts, such as saltpetre or ammonia-compounds. The various “ash-constituents” are also essential for their nourishment.
While Moulds and Yeast-Fungi grow best in an acid substratum, the Bacteria, on the other hand, generally thrive best in a neutral or slightly alkaline one.
[32]
In sterilization, disinfection, and antisepsis, means are employed by which the Bacteria are killed, or checked in their development, for instance, by heat (ignition, cooking, hot vapours, hot air, etc.), or poisons (acids, corrosive sublimate). The process of preserving articles of food, in which they are boiled and then hermetically sealed, aims at destroying the Bacteria, or the spores of those which already may be present in them, and excluding all others.
As the Bacteria are unable to assimilate carbon from the carbonic acid of the air, but must obtain it from the carbon-compounds already in existence in the organic world, they are either saprophytes or parasites. Some are exclusively either the one or the other, obligate saprophytes or parasites. But there are transitional forms among them, some of which are at ordinary times saprophytes, but may, when occasion offers, complete their development wholly or partly as parasites—facultative parasites; others are generally parasitic, but may also pass certain stages of development as saprophytes—facultative saprophytes.
All chlorophyll-free organisms act in a transforming and disturbing manner on the organic compounds from which they obtain their nourishment, and while they themselves grow and multiply, they produce, each after its kind, compounds of a less degree of complexity, i.e. they produce fermentation, putrefaction, sometimes the formation of poisons, and in living beings often disease.
Those organisms which produce fermentation are called ferments; this word, however, is also employed for similar transformations in purely chemical materials (inorganic ferments or enzymes). Many organic (“living”) ferments, among which are Yeast-cells and Bacteria, give off during their development certain inorganic and soluble ferments (enzymes) which may produce other transformations without themselves being changed. Different organisms may produce in the same substratum different kinds of transformation; alcoholic fermentation may for instance be produced by different species of Fungi, but in different proportions, and the same species produces in different substrata, different transformations (e.g. the Vinegar-bacteria oxydize diluted alcohol to vinegar, and eventually to carbonic acid and water).
In the study of Bacteria it is absolutely necessary to sterilize the vessels employed in cultivation, the apparatus, and nutrient solutions, i.e. to free them from Bacteria germs and also to preserve the cultures from the intrusion of any foreign germs (“pure-cultures”). A firm, transparent, nutritive medium is[33] frequently employed. This may be prepared by adding to the nutrient solutions (broth) either gelatine, or—when the Bacteria are to be cultivated at blood-heat—serum of sheep’s or calf’s blood, agar-agar or carragen; serum alone may in itself serve as a nutrient medium. The so-called “plate-cultures” are frequently employed, i.e. the germs are isolated by shaking them with the melted liquid nutrient gelatine, which is then spread on a glass plate and allowed to coagulate; when later on the individual germs grow into colonies, these remain separate in the solid substratum and it is easy to pursue their further development. Similar plate-cultures may also be cultivated in test-tubes and on microscopic slides. The slides and glass plates must be placed in “moist chambers” free from Bacteria. By sowing a few cells (if possible one) using a fine platinum wire, pure cultures for further investigation may be obtained.
In order to prove the relationship between pathogenic Bacteria and certain diseases, the experimental production of pathogenic Bacteria by the inoculation of Bacteria from pure cultures into healthy animals, is very important.
It has not so far been possible to establish a classification of the Bacteria, as the life-history of many species, has not yet been sufficiently investigated.[7] The opinions of botanists are at variance, in many cases, about the forms of growth of a particular kind. Some species are pleomorphic (many-formed) while others possess only one form.
The following Bacteria are Saprophytes:—
Cladothrix dichotoma is common in stagnant and running water which is impregnated with organic matter; the cell-chains have false branching. According to Zopf, Leptothrix ochracea is one of the forms of this species which, in water containing ferrous iron (e.g. as FeCO3), regularly embeds ferric-oxide in its sheath by means of the activity of the protoplasm. Leptothrix ochracea and other Iron-bacteria, according to Winogradsky (1888), do not continue their growth in water free from protoxide of iron; while they multiply enormously in water which contains this salt of iron. The large masses of ochre-coloured slime, found in meadows, bogs, and lakes, are probably due to the activity of the Iron-bacteria.

Fig. 30.—Cladothrix dichotoma.
Those forms which, according to Zopf’s views, represent the forms of development of Cladothrix dichotoma are placed together in Fig. 30. A represents a group of plants, seventy times magnified, attached to a Vaucheria. The largest one is branched like a tree, with branches of ordinary form; a specimen with spirally twisted branches is seen to the right of the figure, at the lower part some small Leptothrix-like forms. B shows the[34] manner of branching and an incipient Coccus-formation. C a Coccus-mass whose exit from the sheath has been observed. D the same mass as C after the course of a day, the Cocci having turned into rods. E a group of Cocci in which some have developed into shorter or longer rods. F one of these rods before[35] and after treatment with picric acid, which causes the chain-like structure to become apparent. G a portion of a plant with conspicuous sheath, two lateral branches are being formed. H part of a plant, whose cells have divided and form Cocci. The original form of the cells in which the Cocci are embedded may still be recognised. I. Leptothrix-filaments with conspicuous mucilaginous sheath, from which a series of rods is about to emerge; the rod near the bottom is dead, and has remained lying in the sheath. K part of a plant which is forming Cocci, those at the top are in the zooglœa-stage, at the base they are elongating to form rods and Leptothrix-filaments. L a portion of a branched Cladothrix, which divides into motile Bacillus-forms; the rays at the free ends indicate the currents which the cilia produce in the water. M a spirally-twisted, swarming filament, before and after division into halves. N part of a tree-like zooglœa with Cocci and short rods.—All of these spirilla, zooglœa, etc., which Zopf has connected with Clad. dichotoma, are according to Winogradsky, independent organisms.
Micrococcus ureæ produces urinal fermentation (transformation of urinal matter into ammonium carbonate); aerobic; round cells generally united to form bent chains or a zooglœa.—Several other kinds of Bacteria have the same action as this one: in damp soil containing ammonia-compounds, saltpetre-formations are produced by M. nitrificans and several different kinds of Bacteria.
Micrococcus prodigiosus is found on articles of food containing starch; “bleeding bread” is caused by this Bacterium, which has the power of forming a red pigment; it also occurs in milk, and produces lactic acid.
Leuconostoc mesenterioides is the frog-spawn Bacterium (Fig. 27) which is found in sugar manufactories, and has the power of producing a viscous fermentation in saccharine solutions which have been derived from plants, e.g. in beetroot-sugar manufactories, where large accumulations of mucilage are formed at the expense of the sugar, with an evolution of carbonic acid. The cell-rows, resembling somewhat a pearl necklace, have thick mucilaginous cell-walls, and form white “Nostoc”-lumps. The mucilage eventually deliquesces and the cells separate from each other; arthrospores?—Similar viscous deteriorations occur in beer and wine, which may then be drawn out into long, string like filaments—“ropiness.”
Bacterium aceti, the Vinegar-bacterium, oxidizes alcohol into[36] acetic acid (acetous-fermentation) and forms a greyish covering of Bacteria (“Vinegar-mother”) on the surface of the liquid; the acetic acid formed, becomes by continued oxidization by B. aceti, again transformed into carbonic acid and water. Aerobic; short cylindrical cells, often united into chains, or to form a zooglœa; sometimes also rod-and spindle-shaped. The Vinegar-bacteria and other kinds with ball- or rod-forms sometimes become swollen, spindle-shaped, or oval links; they are supposed to be diseased forms[8] (“Involution-forms”).
Bacillus lacticus (Bacterium acidi lactici, Zopf) is always found in milk which has stood for some time, and in sour foods (cabbage, cucumbers, etc.); it turns the milk sour by producing lactic acid fermentation in the sugar contained in the milk; the lactic acid formed, eventually causes the coagulation of the casein. It resembles the Vinegar-bacteria, occurring as small cylindrical cells, rarely in short rows; not self-motile.—Several other Bacteria appear to act in the same way, some occurring in the mouth of human beings; some of these Bacteria give to butter its taste and flavour.
The kefir-grains which are added to milk for the preparation of kefir, contain in large numbers a Bacterium (Dispora caucasica) in the zooglœa-form, a Yeast-fungus, and Bacillus lacticus. Kefir is a somewhat alcoholic sour milk, rich in carbonic acid; it is a beverage manufactured by the inhabitants of the Caucasus, from the milk of cows, goats, or sheep, and is sometimes used as a medicine. In the production of kefir, lactic acid fermentation takes place in one part of the sugar contained in the milk, and alcoholic fermentation in another part, and the casein which had become curdled is partially liquefied (peptonised) by an enzyme of a Zooglœa-bacterium.
Bacillus amylobacter (Bacillus butyricus), the Butyric-acid-bacterium (Fig. 29), is a very common anaerobic which produces fermentation in sugar and lactic-acid salts, and whose principal product is butyric acid. It destroys articles of food and (together with other species) plays a part in the butyric acid fermentation which is necessary in the making of cheese; it is very active wherever portions of plants are decaying, in destroying the cellulose in the cell-walls of herbaceous plants, and is thus useful in the preparation of flax and hemp. The cells are self-motile, generally cylindrical, sometimes united into short rows; endosporous;[37] the spore-forming cells swell, assume very different forms, and show granulose reaction. The germ-tube grows out in the direction of the long axis of the spore.
Bacillus subtilis, the Hay-bacillus, is developed in all decoctions of hay; a slender, aerobic, self-motile Bacillus; endosporous (aplanospores); the spore-wall ruptures transversely on germination.
Crenothrix kuehniana occurs in the springs of many baths, in wells, in water or drain-pipes.
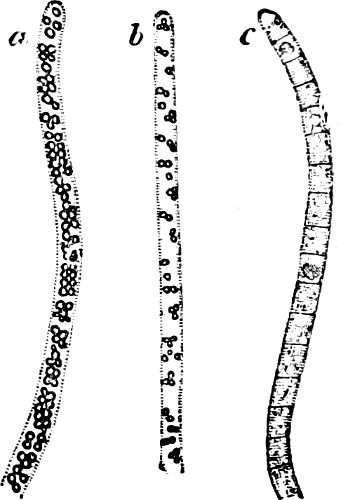
Fig. 31.—Beggiatoa alba: a from a fluid containing abundance of sulphuretted hydrogen; b after lying 24 hours in a solution devoid of sulphuretted hydrogen; c after lying an additional 48 hours in a solution devoid of sulphuretted hydrogen, by this means the transverse walls and vacuoles have become visible.
Beggiatoa (parallel with the Blue-green Alga Oscillaria). Long filaments formed of cylindrical cells which are attached by one of the ends, but which are nearly always free when observed. The filaments, like those of Oscillaria, describe conical figures in their revolutions, the free filaments slide upwards and parallel with one another; sheaths are wanting; strongly refractive sulphur drops are found in the interior. The Beggiatoas are the most prevalent Sulphur-bacteria. They occur, very commonly in large numbers, wherever plant or animal remains are decaying in water in which sulphuretted hydrogen is being formed; thus, for example, B. alba (Fig. 31) occurs frequently as a white covering or slimy film on mud containing organic remains. B. mirabilis is remarkable for its size and its strong peristaltic movements. The Sulphur-bacteria oxidize the sulphuretted hydrogen, and accumulate sulphur in the shape of small granules of soft amorphic sulphur, which in the living cell never passes over into the crystalline state. They next oxidize this sulphur into sulphuric acid, which is immediately rendered neutral by absorbed salts of calcium, and is given off in the form of a sulphate, thus CaCO3 is principally changed into CaSO4. In the absence of sulphur the nutritive processes are suspended, and consequently death occurs either sooner or later. The Sulphur-bacteria may exist and multiply in a fluid which only contains traces of organic matter, in which organisms devoid of chlorophyll are not able to exist. The Beggiatoas very frequently form white, bulky masses in sulphur wells and[38] in salt water, the traces of organic material which the sulphur water contains proving sufficient for them. The cellulose-fermentation, to which the sulphur wells in all probability owe their origin, mainly procures them suitable conditions for existence. The CaCO3 and H2S, formed during the cellulose fermentation by the reduction of CaSO4 is again changed into CaSO4 and CO2 by the Sulphur-bacteria (Winogradsky, 1887).—Other Sulphur-bacteria, the so-called purple Sulphur-bacteria, e.g. B. roseo-persicina, Spirillum sanguineum (Fig. 23), Bacterium sulfuratum, etc., have their protoplasm mixed with a red colouring matter (bacterio-purpurin) which, like chlorophyll, has the power, in the presence of light, of giving off oxygen (as proved by T. W. Englemann, 1888, in oxygen-sensitive Bacteria). The three purple Sulphur-bacteria mentioned, are, according to Winogradsky, not pleomorphic kinds but embrace numerous species.
Many Spirilli (Spirillum tenue, S. undula, S. plicatile, and others) are found prevalent in decaying liquids.
Bacteria (especially Bacilli) are the cause of many substances emitting a foul odour, and of various changes in milk.
Parasitic Bacteria live in other living organisms; but the relation between “host” and parasite may vary in considerable degree. Some parasites do no injury to their host, others produce dangerous contagious diseases; some choose only a special kind as host, others again live equally well in many different ones. There are further specific and individual differences with regard to the predisposition of the host, and every individual has not the same receptivity at all times.
The harmless parasites of human beings. Several of the above mentioned saprophytes may also occur in the alimentary canal of human beings; e.g., the Hay-bacillus, the Butyric-acid-bacillus, etc.; but the gastric juice prevents the development of others, at all events in their vegetative condition. Sarcina ventriculi, “packet-bacterium,” is only known to occur in the stomach and intestines of human beings, and makes its appearance in certain diseases of the stomach (dilation of the stomach, etc.) in great numbers, without, however, being the cause of the disease. It occurs in somewhat cubical masses of roundish cells (Fig. 25).
Less dangerous parasites. In the mouth, especially between and on the teeth, a great many Bacteria are to be found (more than fifty species are known), e.g. Leptothrix buccalis (long, brittle, very thin filaments which are united into bundles), Micrococci in large lumps, Spirochæte cohnii, etc. Some of them are known to be injurious, as they contribute in various ways to the decay of the teeth (caries dentium); a Micrococcus, for instance, forms lactic acid[39] in materials containing sugar and starch, and the acid dissolves the lime salts in the external layers of the teeth: those parts of the teeth thus deprived of lime are attacked by other Bacteria, and become dissolved. Inflammation in the tissues at the root of a tooth, is probably produced by septic materials which have been formed by Bacteria in the root-canal.
Dangerous Parasites. In a large number of the infectious diseases of human beings and animals, it has been possible to prove that parasitic Bacteria have been the cause of the disease. Various pathogenic Bacteria of this nature, belonging to the coccus, rod, and spiral Bacteria groups, are mentioned in the following:—
Pathogenic Micrococci. Staphylococcus pyogenes aureus produces abscesses of various natures (boils, suppurative processes in internal organs). The same effects are produced by—
Streptococcus pyogenes, which is the most frequent cause of malignant puerperal fever; it is perhaps identical with—
Streptococcus erysipelatis, which is the cause of erysipelas in human beings.
Diplococcus pneumoniæ (A. Fränkel) is the cause of pneumonia, and of the epidemic cerebro-spinal meningitis.
Gonococcus (Neisser) is the cause of gonorrhea and inflammation of the eyes.
Pathogenic Rod-Bacteria. Bacterium choleræ gallinarum, an aerobic, facultative parasite which produces fowl-cholera among poultry; it is easily cultivated on various substrata as a saprophyte. The disease may be conveyed both through wounds and by food, and may also be communicated to mammals.
Bacillus anthracis, the Anthrax bacillus (Fig. 32), chiefly attacks mammals, especially herbivorous animals (house mice, guinea-pigs, rabbits, sheep, cattle), in a less degree omnivorous animals (including human beings), and in a still less degree the Carnivores. Aerobic. Cylindrical cells, 3–4 times as long as broad, united into long rod-like bodies, which may elongate into long, bent, and twisted filaments. Not self-motile. Endosporous. Germination takes place without the throwing off of any spore-membrane (compare Hay-bacillus p. 37 which resembles it). Contagion may take place both by introduction into wounds, and from the mucous membrane of the intestines or lungs, both by vegetative cells and by spores; in intestinal anthrax, however, only by spores. The Bacillus multiplies as soon as it has entered the blood, and the anthrax disease commences. The Bacilli not only give off poison,[40] but also deprive the blood of its oxygen. Vegetative cells only occur in living animals. This species is a facultative parasite which in the first stage is a saprophyte, and only in this condition forms spores.
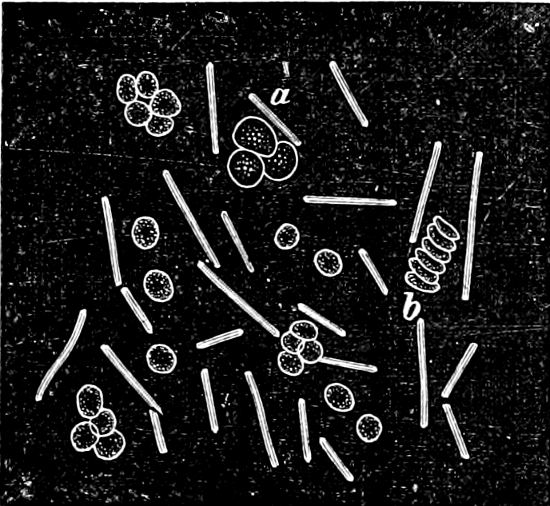
Fig. 32.—Anthrax bacillus (Bacillus anthracis) with red (b) and white (a) blood-corpuscles.

Fig. 33.—Anthrax bacillus. The formation of the spores; magnified 450 times.
Bacillus tuberculosis produces tuberculosis in human beings, also in domestic animals (perlsucht). It is a distinct parasite, but may also live saprophytically. It is rod-formed, often slightly bent, and is recognised principally by its action with stains (when stained with an alkaline solution of methyl-blue or carbolic fuchsin, it retains the colour for a long time even in solutions of mineral acids, in contrast with the majority of well-known Bacteria): it probably forms spores which are able to resist heat, dryness, etc.
Bacillus lepræ produces leprosy; Bacillus mallei produces glanders; Bacillus tetani, tetanus (the tetanus bacillus is very common in soil; anaerobic); Bacillus diphtheriæ, diphtheria; Bacillus typhosus, typhoid fever, etc.
Pathogenic Spiral Bacteria. Spirochæte obermeieri (Fig. 24) produces intermittent fever (febris recurrens); it makes its appearance in the blood during the attacks of fever, but it is not to be found during intervals when there is no fever. Obligate parasite.
Spirillum choleræ asiaticæ (Microspira comma) without doubt produces Asiatic cholera; an exceedingly motile spirillum, which is also found in short, bent rods (known as the “Comma-bacillus”),[41] it lives in the intestines of those attacked by the disease, and gives off a strong poison which enters the body. It is easily cultivated as a saprophyte.
A great many circumstances seem to show that a number of other infectious diseases (syphilis, small-pox, scarlet-fever, measles, yellow-fever, etc.) owe their origin to parasitic Bacteria, but this has not been proved with certainty in all cases.
It has been possible by means of special cultivations (ample supply of oxygen, high temperature, antiseptic materials) to produce from the parasitic Bacteria described above (e.g. the fowl-cholera and the anthrax Bacteria) physiological varieties which are distinct from those appearing in nature and possess a less degree of “virulence,” i.e. produce fever and less dangerous symptoms in those animals which are inoculated with them. The production of such physiological varieties has come to be of great practical importance from the fact that they are used as vaccines, i.e. these harmless species produce in the animals inoculated with them immunity from the malignant infectious Bacteria from which they were derived. This immunity is effected by the change of the products of one or more of the Bacteria, but we do not yet know anything about the way in which they act on the animal organism. The white blood corpuscles, according to the Metschnikoff, play the part of “Phagocytes” by absorbing and destroying the less virulent Bacteria which have entered the blood, and by so doing they are gradually enabled to overcome those of a more virulent nature.
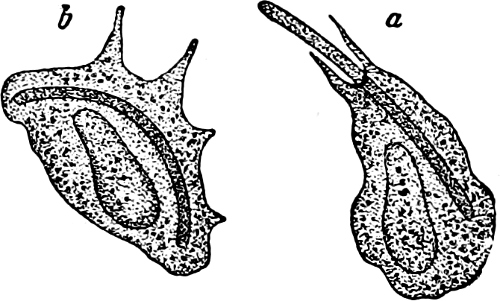
Fig. 34.—a and b The same blood-cell of a Frog: a in the act of engulfing an anthrax-bacillus; b after an interval of a few minutes when the bacillus has been absorbed.
The Algæ belonging to this class have chlorophyll, and pyrenoids round which starch is formed. The cells divide only in one direction, they live solitarily, or united to form filaments which generally float freely (seldom attached). Swarm-cells are wanting. The fertilisation is isogamous (conjugation) and takes place by means of aplanogametes. The zygote, after a period of rest, produces, immediately on germination, one or more new vegetative[42] individuals; sometimes akinetes or aplanospores are formed in addition. They only occur in fresh or slightly brackish water.
Order 1. Desmidiaceæ. The cells generally present markings on the outer wall, and are mostly divided into two symmetrical halves by a constriction in the middle, or there is at least a symmetrical division of the protoplasmic cell-contents. The cell-wall consists nearly always of two layers, the one overlapping the other (Fig. 35 C). The cells either live solitarily or are united into unbranched filaments. The mass of protoplasm formed by the fusion of the two conjugating cells becomes the zygote, which on germination produces one (or after division 2, 4 or 8) new vegetative individual. The chromatophores are either star-, plate-, or band-shaped, and regularly arranged round the long axis of the cell.
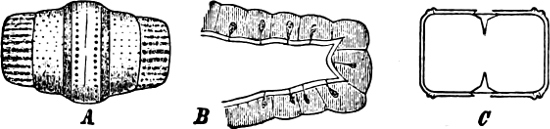
Fig. 35.—A Cell of Gymnozyga brebissonii, external view showing the distribution of the pores. B A portion of the membrane of Staurastrum bicorne with pores containing protoplasmic projections. C Cell-wall of Hyalotheca mucosa during cell-division: the central part, being already formed, shows the connection with the divisional wall.
The Desmidiaceæ are not able to swim independently, many species, however, show movements of different kinds by rising and sliding forward on the substratum. These movements, which are partly dependent upon, and partly independent of light and the force of gravitation, are connected with the protrusion of a mucilaginous stalk. The mucilage, which sometimes surrounds the whole individual, may acquire a prismatic structure, it is secreted by the protoplasmic threads which project through certain pores definitely situated in the walls (Fig. 35 A, B).
Vegetative multiplication takes places by division. A good example of this is found in Cosmarium botrytis (Fig. 36 A-D). The nucleus and chromatophores divide, and simultaneously the central indentation becomes deeper, the outer wall is then ruptured making a circular aperture through which the inner wall protrudes forming a short, cylindrical canal between the two halves to which it is attached (Fig. 36 C). After elongation the canal is divided by a central transverse wall, which commences as a ring round its[43] inner surface and gradually forms a complete septum. The dividing wall gradually splits, and the two individuals separate from each other, each one having an old and a new half. The two daughter-cells bulge out, receive a supply of contents from the parent-cells, and gradually attain their mature size and development (Fig. 36 B-D). Exceptions to this occur in some forms.
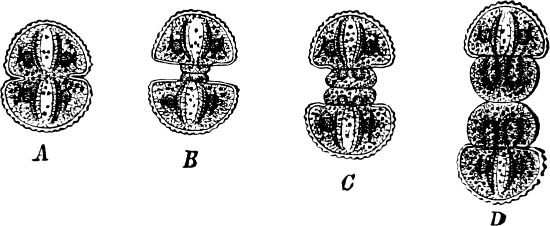
Fig. 36.—Cosmarium botrytis. A-D Different stages of cell-division.
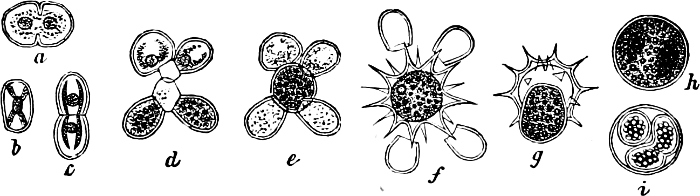
Fig. 37.—Cosmarium meneghinii: a-c same individual seen from the side, from the end, and from the edge; d-f stages of conjugation; g-i germination of the zygote.
Conjugation takes place in the simplest way in Mesotænium, where the two conjugating cells unite by a short tube (conjugation-canal), which is not developed at any particular point. The aplanogametes merge together after the dissolution of the dividing wall, like two drops of water, almost without any trace of preceding contraction, so that the cell-wall of the zygote generally lies in close contact with the conjugating cells. The conjugating cells in the others lie either transversely (e.g. Cosmarium, Fig. 37 d; Staurastrum, etc.), or parallel to one another (e.g. Penium, Closterium, etc.), and emit a short conjugation-canal (Fig. 37 d) from the centre of that side of each cell which is turned towards the other one. These canals touch, become spherical, and on the absorption of the dividing wall the aplanogametes coalesce in the swollen conjugation-canal (Fig. 37 e), which is often surrounded by a mucilaginous envelope. The zygote, which is often spherical, is surrounded by a thick cell-wall, consisting of three layers; the outermost of these[44] sometimes bears thorn-like projections, which in some species are simple (Fig. 37 f), in others branched or variously marked; in some, however, it remains always smooth (e.g. Tetmemorus, Desmidium). Deviation from this mode of conjugation may occur within certain genera (e.g. Closterium, Penium). Upon germination the contents of the zygote emerge, surrounded by the innermost layers of the wall (Fig. 37 g, h) and generally divide into two parts which develop into two new individuals, placed transversely to each other (Fig. 37 i); these may have a somewhat more simple marking than is generally possessed by the species.
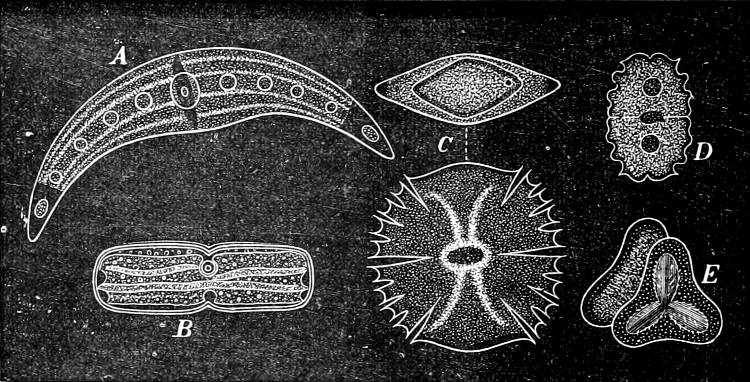
Fig. 38.—Desmidiaceæ. A Closterium moniliferum; B Penium crassiusculum; C Micrasterias truncata (front and end view); D Euastrum elegans; E Staurastrum muticum (end view).
The most frequent genera are:—
A. Solitary cells: Mesotænium, Penium (Fig. 38 B), Cylindrocystis, Euastrum (Fig. 38 D), Micrasterias (Fig. 38 C), Cosmarium (Fig. 36, 37), Xanthidium, Staurastrum (Fig. 38 E), Pleurotænium, Docidium, Tetmemorus, Closterium (Fig. 38 A), Spirotænia.
B. Cells united into filaments: Sphærozosma, Desmidium, Hyalotheca, Gymnozyga, Ancylonema, Gonatozygon.
Order 2. Zygnemaceæ. Cell-wall without markings. The cells are cylindrical, not constricted in the centre, and (generally) united into simple, unbranched filaments. The whole contents of the conjugating cells take part in the formation of the zygote, which on germination grows out directly into a new filament.
Spirogyra is easily recognised by its spiral chlorophyll band; Zygnema has two star-like chromatophores in each cell (Fig. 40); both these genera are very common Algæ in ponds and ditches.
[45]
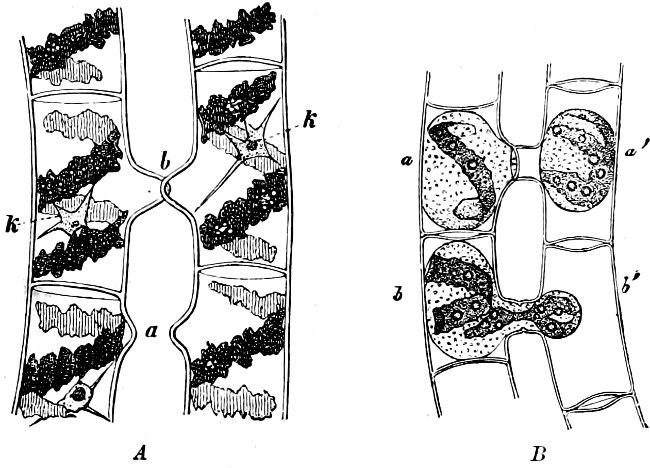
Fig. 39.—Spirogyra longata. A At the commencement of conjugation, the conjugation-canals begin to protrude at a and touch one another at b; the spiral chlorophyll band and cell-nuclei (k) are shown. B A more advanced stage of conjugation; a, a’ the rounded female and male aplanogametes: in b’ the male aplanogamete is going over to and uniting with the female aplanogamete (b).
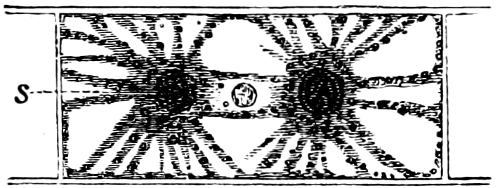
Fig. 40.—A cell of Zygnema. S Pyrenoid.
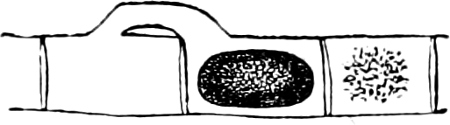
Fig. 41.—Zygnema insigne, with zygote.
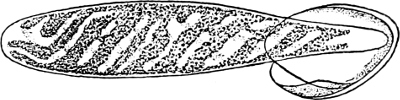
Fig. 42.—Germinating zygote of Spirogyra jugalis: the young plant is still unicellular; the end which is still in the wall of the zygote is elongated and root-like; the chromatophore divides and forms the spiral band.
The conjugation among the Zygnemaceæ takes place in the following manner: the cells of two filaments, lying side by side, or two cells, the one being situated above the other in the same filament (Fig. 41), push out small protuberances opposite each other (Fig. 39 A, a, b); these finally meet, and the dividing wall is absorbed so that a tube is formed connecting one cell with the other; the protoplasmic contents round off, and the whole of these contents of one of the cells glides through the conjugation-tube and coalesces with that of the other (Fig. 39 B), the aggregate mass then rounds off, surrounds itself with a cell-wall, and becomes a zygote. A distinct difference[46] may be found between the cells in the two filaments, those in the one whose protoplasmic contents pass over being cylindrical, while those of the recipient one are more barrel-shaped, and of a larger diameter. The former may be regarded as a male, the latter as a female plant. The zygote germinates after a period of rest, and grows out into a new filament (Fig. 42).
Order 3. Mesocarpaceæ. The cell-walls are glabrous, unconstricted in the centre, and united into simple unbranched filaments. The chromatophore consists of an axial chlorophyll-plate, with several pyrenoids. The zygote is formed by the coalescence of two cells (Fig. 43) (sometimes three or four), but the whole protoplasmic contents of the cells do not take part in this process, a portion always remaining behind; the aplanogametes coalesce in the conjugation-canal. The zygote thus formed appears incapable of germination until after 3–5 divisions. Of the cells so formed, only one is fertile, the sterile cells, according to Pringsheim, constituting a rudimentary sporocarp. The germinating cells grow out into a new filament. In this order, conjugation has been observed between two cells of the same filament. The Mesocarpaceæ thrive best in water which contains lime.
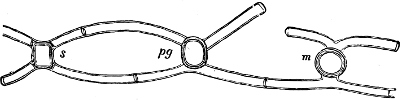
Fig. 43.—Mougeotia calcarea. Cells showing various modes of conjugation: at m tripartition; pg quadripartition; s quinquipartilion of the zygote.
These Algæ are coloured green by chlorophyll, seldom in combination with other colouring matter, and then especially with red. The product of assimilation is frequently starch, which generally accumulates round certain specially formed portions of protoplasm termed pyrenoids. The thallus is uni- or multicellular; in the higher forms (certain Siphoneæ) the organs of vegetation attain differentiation into stem and leaf. The asexual reproduction takes place in various ways; the sexual reproduction is effected by conjugation of motile gametes, or by oogamous fertilisation. The[47] swarm-cells (zoospores, gametes, and spermatozoids) are constructed symetrically, and have true protoplasmic cilia, these generally being attached to the front end of the swarm-cells. Most of these Algæ live in water (fresh or salt); some are found upon damp soil, stones, or tree-stems, and some live enclosed in other plants.
The Class is divided into three families:—
1. Protococcoideæ: Volvocaceæ, Tetrasporaceæ, Chlorosphæraceæ, Pleurococcaceæ, Protococcaceæ, Hydrodictyaceæ.
2. Confervoideæ: Ulvaceæ, Ulothricaceæ, Chætophoraceæ, Mycoideaceæ, Cylindrocapsaceæ, Œdogoniaceæ, Coleochætaceæ, Cladophoraceæ, Gomontiaceæ, Sphæropleaceæ.
3. Siphoneæ: Botrydiaceæ, Bryopsidaceæ, Derbesiaceæ, Vaucheriaceæ, Phyllosiphonaceæ, Caulerpaceæ, Codiaceæ, Valoniaceæ, Dasycladaceæ.
The Algæ which belong to this group are uni- or multicellular with the cells more or less firmly connected, sometimes in a definite, sometimes in an indefinite form (Fig. 47). Colonies are formed either by division or by small unicellular individuals becoming united in a definite manner; the colonies formed in this latter way are termed Cœnobia. Apical cells and branching are absent. Multiplication by division; asexual reproduction by zoospores, rarely by akinetes. Sexual reproduction may be wanting, or it takes place by isogamous, rarely by oogamous fertilisation.
Some are attached by means of a stalk to other objects (Characium, Fig. 49), others occur as “Endophytes” in the tissues of certain Mosses or Phanerogams, e.g. Chlorochytrium lemnæ, in Lemna trisulca; Endosphæra, in the leaves of Potamogeton, Mentha aquatica, and Peplis portula; Phyllobium, in the leaves of Lysimachia nummularia, Ajuga, Chlora, and species of Grasses; Scotinosphæra in the leaves of Hypnum and Lemna trisulca; the majority, however, live free in water and in damp places. Many species which were formerly considered to belong to this family have been proved to be higher Algæ in stages of development.
Order 1. Volvocaceæ. The individuals in this order are either uni- or multicellular, and during the essential part of their life are free-swimming organisms. They are generally encased in a mucilaginous envelope, through which 2–6 cilia project from every[48] cell. The vegetative reproduction takes place by the division of all, or a few, of the cells of the individual; in some a palmella-stage is found in addition. The sexual reproduction takes place by isogamous or oogamous fertilisation.
The Volvocaceæ may be considered to include the original forms of the Chlorophyceæ, because, among other reasons, the motile stage is here the most prominent; they also form the connecting link between the animal Flagellata, and forms intermediate to the Syngeneticæ may perhaps be found amongst them. Three series of green Algæ may be supposed to have taken their origin from the Volvocaceæ: Conjugateæ (Desmidiaceæ) which have lost the swarming stage, but whose conjugation is the nearest to the fertilisation in Chlamydomonas pulvisculus: the Protococcaceæ in which the vegetative divisions have disappeared, while the swarming stage continues to be present, though of shorter duration; and Tetrasporaceæ, in which the vegetative divisions are more prominent, whilst the swarming stage is less so.
A. Unicellular Individuals. The principle genera are: Chlamydomonas, Sphærella, Phacotus.—Sphærella nivalis is the Alga which produces the phenomenon of “Red Snow,” well known on high mountains and on ice and snow fields in the polar regions. The red colouring matter which appears in this and other green Algæ, especially in the resting cells, is produced by the alteration of the chlorophyll.
Phacotus lenticularis has an outer covering incrusted with lime, which, at death, or after division, opens out into two halves. Species may be found among Chlamydomonas, in which conjugation takes place between gametes of similar size without cell-wall, but in C. pulvisculus conjugation takes place between male and female aplanogametes which are surrounded by a mucilaginous envelope.
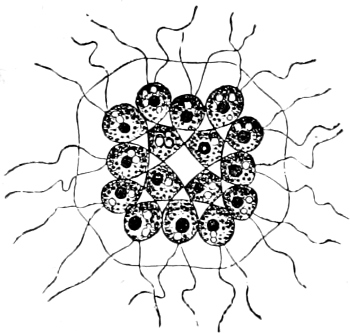
Fig. 44.—Gonium pectorale.
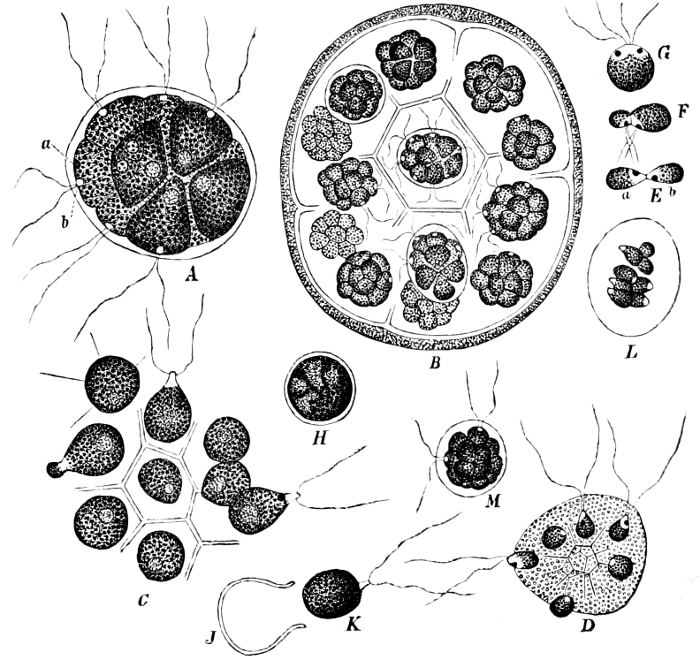
Fig. 45.—Pandorina morum.
B. Multicellular Individuals. The most important genera are Gonium, Stephanosphæra, Pandorina, Eudorina, Volvox.—Gonium has 4 or 16 cells arranged in a definite pattern in a flat plate (Fig. 44). Pandorina (Fig. 45), has 16 cells arranged in a sphere (Fig. 45 A). The vegetative reproduction takes place in this way: each cell, after having rounded off, and after the withdrawal of the cilia, divides itself[49] into 16 new ones (Fig. 45 B), each forming a new individual, which soon grows to the size of the mother-individual. It was in this Alga that the conjugation of self-motile gametes was first discovered by Pringsheim, 1869. When conjugation is about to take place, each cell divides into sixteen, as in vegetative reproduction, but the 16 × 16 cells all separate from one another (Fig. 45 C, female gametes, and D, male gametes), and swarm solitarily in the water. The male are, most frequently, smaller than the female, but otherwise they are exactly alike; they are more or less pear-shaped, with a colourless anterior end, 2 cilia, a red “eye-spot,” etc. After swarming for some time they approach each other, two and two, generally a large and a smaller one, and come into contact at their colourless end; in a few moments they coalesce and become one cell (Fig. 45 E, F), this[50] has at first a large colourless anterior end, 4 cilia, and 2 “eye-spots” (Fig. 45 G), but these soon disappear and the cell becomes uniformly dark-green and spherical, and surrounds itself with a thick cell-wall, losing at the same time its power of motion: the zygote (Fig. 45 H) is formed, and becomes later on a deep red colour. On the germination of the zygote, the protoplasmic cell-contents burst open the wall (Fig. 45 J), and emerge as a large swarmspore (Fig. 45 K) which divides into 16 cells, and the first small individual is formed (Fig. 45 L, M).
Eudorina is like Pandorina in structure, but stands somewhat higher, since the contrast between the conjugating sexual cells is greater, the female one being a motionless oosphere.
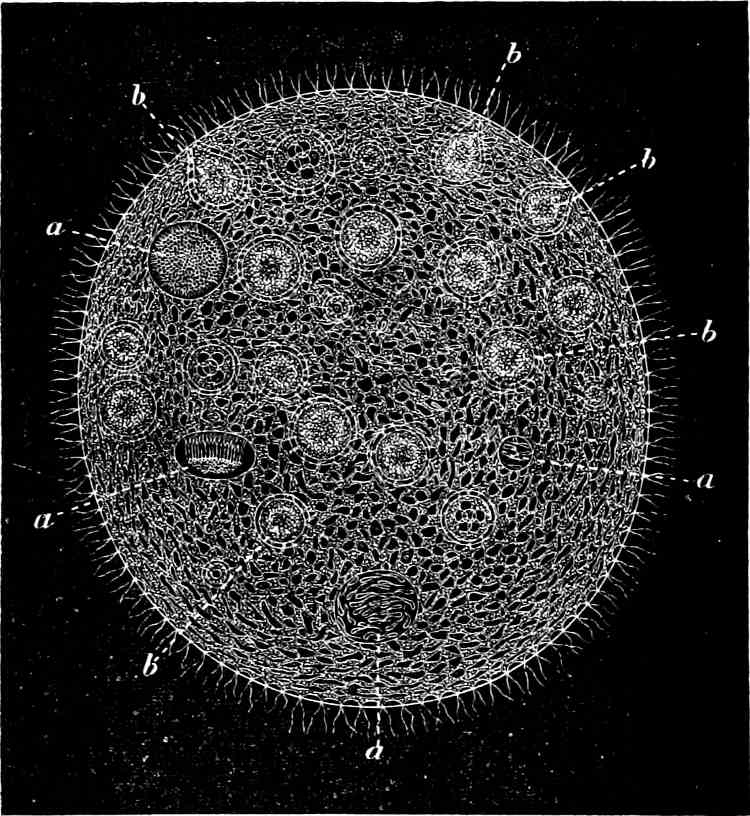
Fig. 46.—Volvox globator, sexual individual: a antheridia which have formed spermatozoids; b oogonia.
The highest stage of development is found in Volvox (Fig. 46). The cells are here arranged on the circumference of a sphere, and enclose a cavity filled with mucilage. The number of these cells may vary from 200–22,000, of which the majority are vegetative and not reproductive, but some become large, motionless oospheres (Fig. 46 b); others, which may appear as solitary individuals, divide and form disc-shaped masses of from 8–256 small spermatozoids[51] (Fig. 46 a). After the oosphere has been fertilised by these, the oospore surrounds itself by a thick, sometimes thorny cell-wall, and on germination becomes a new individual of few cells. A few cells conspicuous by their larger size may be found (1–9, but generally 8) in certain individuals, and these provide the vegetative reproduction, each forming by division a new individual.
Order 2. Tetrasporaceæ reproduce both by vegetative divisions and swarmspores, some have also gamete-conjugation. The principal genera are: Tetraspora, Apiocystis, Dactylococcus, Dictyosphærium, Chlorangium.
Order 3. Chlorosphæraceæ. Chlorosphæra.
Order 4. Pleurococcaceæ. In this order the swarm-stages and sexual reproduction are entirely absent. Vegetative reproduction by division. The principal genera are: Pleurococcus (Fig. 47), Scenedesmus (Fig. 48), Raphidium, Oocystis, Schizochlamys, Crucigenia, Selenastrum.—Pleurococcus vulgaris (Fig. 47) is one of the most common Algæ throughout the world, occurring as green coverings on tree-stems, and damp walls, and it is one of the most common lichen-gonidia.
Fig. 47.—Pleurococcus vulgaris.
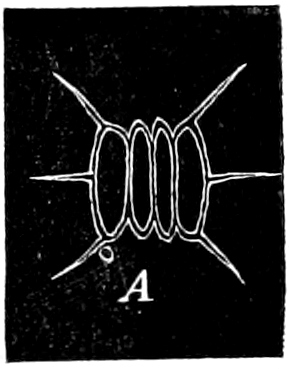
Fig. 48.—Scenedesmus quadricauda.
Order 5. Protococcaceæ. The cells are motionless, free or affixed on a stalk (e.g. Characium, Fig. 49), either separate or loosely bound to one another; they never form multicellular individuals. Multiplication by division is nearly always wanting. Reproduction takes place by swarmspores, which have 1 or 2 cilia, and sexual reproduction in some by gamete-conjugation. The principal genera are: Chlorococcum, Chlorochytrium, Chlorocystis, Scotinosphæra, Endosphæra, Phyllobium, Characium, Ophiocytium, Sciadium.
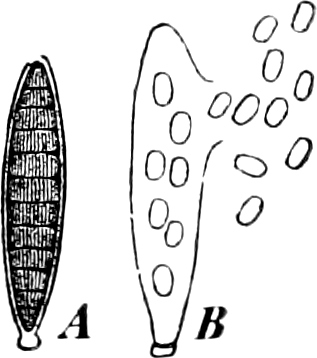
Fig. 49.—Characium strictum. A The cell-contents have divided into many swarmspores. B Swarmspores escaping.
Order 6. Hydrodictyaceæ. The individuals are unicellular but several unite after the zoospore-stage into definitely formed families (cœnobia). Ordinary vegetative division is wanting, but[52] asexual reproduction takes place by zoospores (or by motionless cells without cilia), which unite and form a family similar to the mother-family, inside the mother-cell, or in a mucilaginous envelope. Where sexual reproduction is found it takes place by gamete-conjugation. The principal genera are: Pediastrum (Fig. 50), Cœlastrum, Hydrodictyon (Fig. 51).
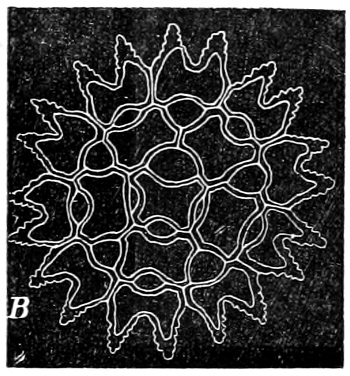
Fig. 5O.—Pediastrum asperum.
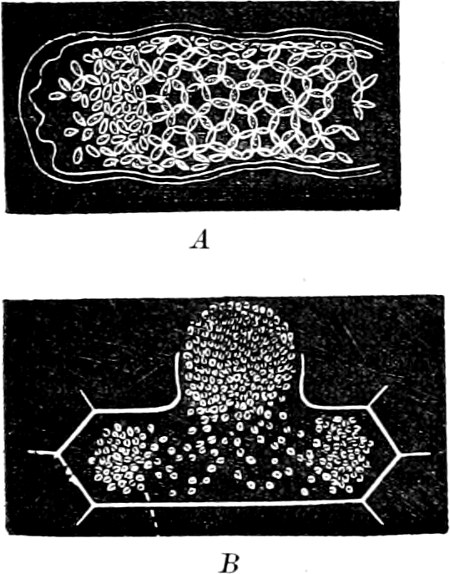
Fig. 51.—Hydrodictyou reticulatum. A A cell where the zoospores are on the point of arranging themselves to form a net. B A cell with gametes swarming out.
The cœnobium of Hydrodictyon reticulatum (Water-net) is formed of a large number of cells which are cylindrical, and attached to one another by the ends (Fig. 51). The asexual reproduction takes place by zoospores, which are formed in large numbers (7,000–20,000) in each mother-cell, within which they move about for a time, and then come to rest and arrange themselves into a new net (Fig. 51 A) which is set free by the dissolution of the wall of the mother-cell, grows, and becomes a new cœnobium. The sexual reproduction takes place by gamete-conjugation. The gametes are formed in the same manner as the zoospores, but in larger numbers (30,000–100,000), and swarm out of the mother-cell (Fig. 51 B). The zygote forms, on germination, 2–5 large zoospores, each with one or two cilia, these generally swarm about for a time, and after a period of rest become irregular thorny bodies (polyhedra); their contents again divide into zoospores, the thorny external coating of the polyhedra is cast off, and the zoospores, surrounded by the dilated internal coating, unite to form a small family, which produces several others in the manner described.
[53]
The individuals are always multicellular, the cells firmly bound together and united into unbranched or branched filaments, expansions, or masses of cells which grow by intercallary divisions or have apical growth. In the first seven orders the cells are uninuclear, but the cells of the remaining three orders contain several nuclei. Asexual reproduction by zoospores, akinetes or aplanospores. Sexual reproduction by isogamous or oogamous fertilisation.
The Confervoideæ, through the Ulvaceæ, are connected with the Tetrasporaceæ, and from the Coleochætaceæ, which is the most highly developed order, there are the best reasons for supposing that the Mosses have taken their origin. The Cladophoraceæ show the nearest approach to the Siphoneæ.
Order 1. Ulvaceæ. The thallus consists of one or two layers of parenchymatous cells, connected together to form either a flat membrane (Monostroma, Ulva) or a hollow tube (Enteromorpha), and may be either simple, lobed, or branched. Reproduction takes place by detached portions of the thallus; or asexually by zoospores or akinetes. Gamete-conjugation is known to take place in some members of this order, the zygote germinating without any resting-stage. The majority are found in salt or brackish water.
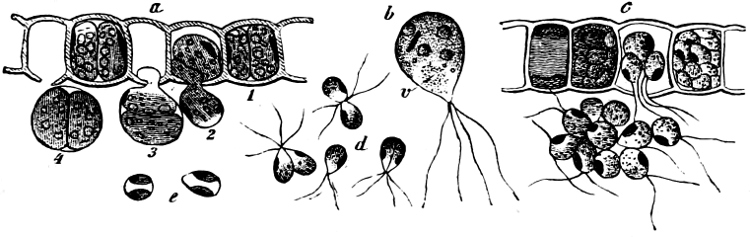
Fig. 52.—Ulothrix zonata: a portion of a filament with zoospores, which are formed two in each cell (zoosporangium); the dark spots are the red “eye-spots”; 1, 2, 3, 4, denote successive stages in the development of the zoospores; b a single zoospore, v the pulsating vacuole; c portion of a filament with gametes, sixteen are produced in each gametangium; d free gametes, solitary or in the act of conjugation; e the conjugation is completed, and the formed zygote has assumed the resting-stage.
Order 2. Ulothricaceæ. The thallus consists normally of a simple unbranched filament (sometimes a small expansion consisting of one layer of cells is formed, as in Schizomeris and Prasiola which were formerly described as separate genera). Asexual reproduction takes place by means of zoospores (with[54] 1, 2, or 4 cilia), akinetes or aplanospores; the last named may germinate immediately, or only after a period of rest. Sexual reproduction takes place by the conjugation of gametes of about the same size, each having two cilia (Fig. 52 d). The zygote of Ulothrix, on germination, produces a brood of zoospores which swarm for a time and then elongate to become Ulothrix-filaments (alternation of generations). The gametes may also germinate without conjugation in the same manner as the zoospores. The principal genera are: Ulothrix, Hormidium, Conferva, Microspora.—Ulothrix zonata is very common in running fresh water. Nearly all the species of Hormidium occur on damp soil, tree-stems and stones.
Order 3. Chætophoraceæ. The thallus consists of a single, branched, erect or creeping filament of cells, often surrounded by mucilage. The cells have only one nucleus. Asexual reproduction by zoospores with 2 or 4 cilia, by akinetes, or aplanospores. In many, conjugation between gametes with 2 cilia may be found. They approach on one side, Ulothricaceæ, and on the other, Mycoideaceæ. The principal genera are: Stigeoclonium, Draparnaldia, Chætophora, Entoderma, Aphanochæte, Herposteiron, Phæothamnion, Chlorotylium, Trichophilus, Gongrosira, Trentepohlia. Most of the species of Trentepohlia are coloured red by the presence of a red colouring material, which occurs in addition to the chlorophyll. They are aerial Algæ which live on stones (T. jolithus, “violet stone,” so named on account of its violet-like odour in rainy weather), on bark and old wood (T. umbrina), or on damp rocks (T. aurea). Trichophilus welckeri lives in the hair of Bradypus.
Order 4. Mycoideaceæ. The thallus is discoid, consisting of one or more cell-layers, and is always attached. Asexual reproduction by zoospores with 2 or 4 cilia. Sexual reproduction in some species by the conjugation of gametes with 2 cilia. This order forms the connecting link between Chætophoraceæ and Coleochætaceæ. The species occur in fresh water (Chætopeltis) as well as in salt (Pringsheimia), on the carapace of tortoises (Dermatophyton = Epiclemmydia), or endophytic between the cuticle and the epidermal cells of the leaves of tropical plants, destroying the leaf-tissue (Mycoidea).
Order 5. Cylindrocapsaceæ. The thallus consists of a simple (rarely, in parts, formed of many rows) unbranched filament, attached in the young condition, which has short cells with a single nucleus, and is enveloped in a thick envelope with a laminated structure. Asexual reproduction by zoospores with 2 cilia, which are formed 1, 2, or 4 in each vegetative cell. The[55] antheridia are produced by a single cell, or a group of cells, in a filament, dividing several times without increasing in size. Two egg-shaped spermatozoids, each with 2 cilia (Fig. 53 D), are formed in each antheridium, and escape through an aperture in the side; in the first stages they are enclosed in a bladder-like membrane (Fig. 53 B, C). Other cells of the filament swell out and form oogonia (Fig. 53 A), which resemble those of Œdogonium. After fertilisation, the oospore surrounds itself with a thick wall, and assumes a reddish colour. The germination is unknown. The unfertilised oospheres remain green, divide often into 2–4 daughter-cells, and grow into new filaments.
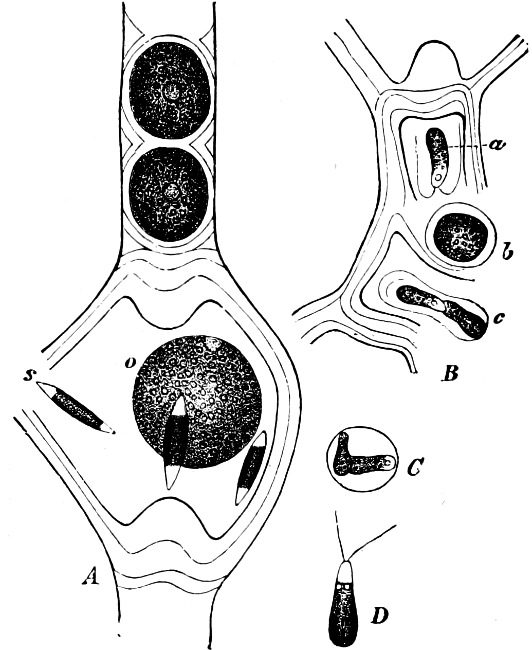
Fig. 53.—Cylindrocopsa involuta. A Oogonium with oosphere (o) surrounded by spermatozoids (s). B Two antheridia, each with two spermatozoids. C Spermatozoids surrounded by their bladder-like membrane. D Free spermatozoid.
This order, which only includes one genus, Cylindrocapsa, forms the connecting link between Ulothricaceæ and Œdogoniaceæ. The few species (4) occur only in fresh water.
Order 6. Œdogoniaceæ. The thallus consists of branched (Bulbochæte) or unbranched (Œdogonium) filaments, attached in the early stages. The cells may be longer or shorter, and have one nucleus. Asexual reproduction by zoospores, which have a chaplet of cilia round the base of the colourless end (Fig. 6 a). Sexual reproduction takes place by oogamous fertilisation. On the germination of the oospore, 4 zoospores are formed (Fig. 54 F). They occur only in fresh or slightly brackish water. The division[56] of the cells takes place in quite a peculiar and unusual manner. At the upper end of the cell which is about to divide, a ring-shaped thickening of soft cellulose is formed transversely round the wall; the cell-nucleus of the mother-cell and the protoplasm then divide by a transverse wall into two portions of similar size, and the cell-wall bursts transversely along the central line of the thickened ring. The cell-wall thus divides into two parts—the upper one short, the “cap,” and the lower one much longer, the “sheath.” The portions of the original cell-wall now separate from each other, the cellulose ring extending, and supplying an additional length of cell-wall between them. The cap and sheath will project a little in front of the piece thus inserted. The dividing wall between the two new cells is formed near to the uppermost edge of the sheath, and gradually becomes thicker and firmer. The inserted piece of wall forms the larger part of the wall of the upper cell: the remainder is formed by the cap. This mode of division is repeated exactly in the same way, and new caps are formed close below the first one, one for every division.
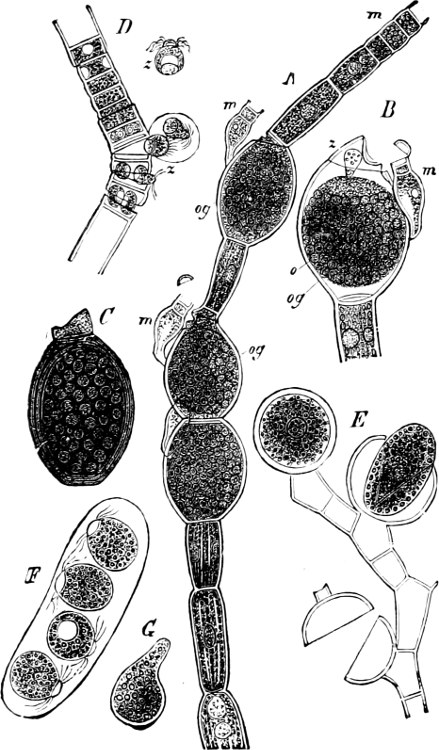
Fig. 54.—A Œdogonium ciliatum. A Female plant with three oogonia (og) and dwarf-males (m). B An oogonium with spermatozoid (z) seen entering the oosphere (o) having passed through an aperture near the summit of the oogonium; m dwarf-male. C Ripe oospore. D Œdogonium gemelliparum. F Portion of a male filament from which spermatozoids (z) are emerging. E Portion of filament of Bulbochæte; the upper oogonium still encloses the oospore, in the central one the oospore is escaping while the lower one is empty. F Four zoospores developed from an oospore. G Zoospore germinating.
[57]
Fertilisation takes place in the following way. The oogonium is a large ellipsoidal, swollen cell (og, in Fig. 54 A), whose contents are rounded off into an oosphere with a colourless receptive-spot (see B); an aperture is formed in the wall of the oogonium, through which the spermatozoids are enabled to enter (B). The spermatozoids are produced either directly, as in D (in pairs), in basal cells of the filament, or indirectly. In the latter case a swarmspore (androspore) is formed which comes to rest, attaches itself to an oogonium, germinates, and gives rise to a filament of a very few cells—dwarf-male (A, B, m). The spermatozoids are formed in the upper cell of the dwarf-male (m), and are set free by the summit of the antheridium lifting off like a lid. On the germination of the oospore (C), which takes place in the following spring, 4 zoospores are produced (F) (i.e. the sexual generation); these swarm about for a time, and ultimately grow into new filaments.
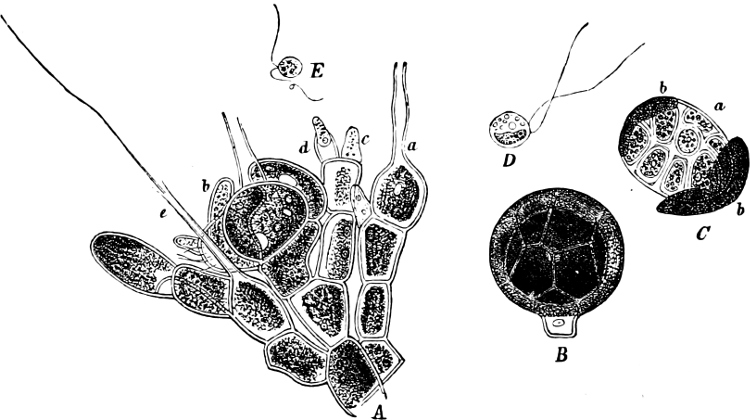
Fig. 55.—Coleochæte pulvinata. A A portion of a thallus with organs of reproduction; a oogonium before, b after fertilisation; c an antheridium, closed; d open, with emerging spermatozoid. B Ripe oogonium, with envelope. C Germination of the oospore. D Zoospore. E Spermatozoid.
Order 7. Coleochætaceæ. The thallus is always attached, and of a disc- or cushion-shape, formed by the dichotomous branching of filaments of cells united in a pseudo-parenchymatous manner. Each cell has only one nucleus. Asexual reproduction by zoospores with 2 cilia (Fig. 55 D), which may arise in all the cells. Sexual reproduction by oogamous fertilisation. The spermatozoids resemble the swarmspores, but are[58] smaller (E), and originate singly (in the species figured) in small conical cells (c, d in A). The oogonia are developed at the extremities of certain branches: they are bottle-shaped cells with very long and thin necks (trichogyne), open at the end (a in A); at the base of each oogonium is a spherical oosphere. The spermatozoids reach the oosphere through the trichogyne, or through an aperture in the wall when the trichogyne is absent, and fertilisation having taken place, the oogonium becomes surrounded by a cell-layer (envelope), which grows out from the cells near its base (b in A), and in this way a kind of fruit is formed (B) (spermocarp, cystocarp).
The oospore, next spring, divides and forms a parenchymatous tissue (homologous with the Moss-sporophyte); this bursts open the envelope (C), and a zoospore (homologous with the spores of the Moss-capsule) arises in each of the cells, and produces a new Coleochæte. We have then, in this case, a still more distinct alternation of generations than in Œdogonium. Only one genus, Coleochæte, is known, but it contains several species, all living in fresh water.
Order 8. Cladophoraceæ. This order is probably derived from the Ulothricaceæ. The thallus consists of a single, unbranched or branched filament, generally with an apical cell. The cells have each 2 or more nuclei. Asexual reproduction by zoospores with 2 or 4 cilia, and by akinetes. Conjugation of gametes with 2 cilia is found in some genera. They occur in salt as well as in fresh water. The principal genera are: Urospora, Chætomorpha, Rhizoclonium, Cladophora; of the last named genus the species C. lanosa and C. rupestris are common in salt water; C. fracta and C. glomerata in fresh water.
Order 9. Gomontiaceæ. Gomontia polyrrhiza, the only species hitherto known, is found on old calcareous shells of certain salt water Molluscs.
Order 10. Sphæropleaceæ. The thallus consists of free, unbranched filaments, with very elongated multinuclear cells. The vegetative cells form no zoospores. Sexual reproduction by oogamous fertilisation (see page 13, Fig. 10 B). The oospore has a thick wall (Fig. 10 D) studded with warts, and assumes a colour resembling red lead. It germinates only in the following spring, and produces 1–8 zoospores, each with 2 cilia (Fig. 10 E), which grow into new filaments. Only one species, Sphæroplea annulina, is known.
[59]
The thallus has apical growth, and in the vegetative condition consists generally of one single (in the Valoniaceæ most frequently of more) multinuclear cell, which may be much branched, and whose separate parts in the higher forms (e.g. Bryopsis, Fig. 57; Caulerpa, Fig. 59, etc.) may be differentiated to perform the various physiological functions (as root, stem and leaf). Vegetative multiplication by detached portions of the thallus (gemmæ); asexual reproduction by zoospores, akinetes, or aplanospores. Sexual reproduction by gamete-conjugation, rarely by oogamous fertilisation. The zygote or oospore germinates as a rule without any resting-stage.
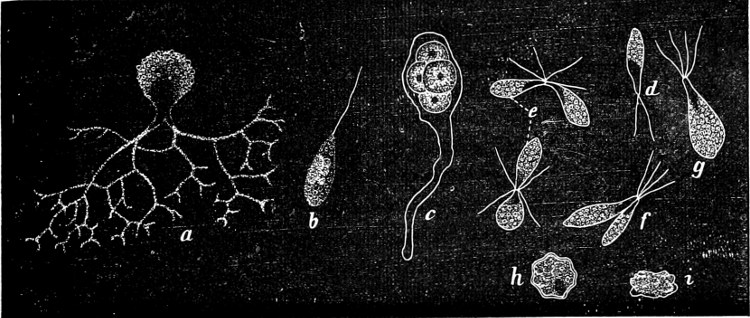
Fig. 56.—Botrydium granulatum: a an entire plant forming swarmspores; b swamspores; c an individual with gametangia; d, gamete; e, f, g conjugation; h zygote seen from above; i the same in a lateral view.
Most of the Siphoneæ occur in salt water or on damp soil. Many (e.g. Dasycladaceæ) are very much incrusted with lime, and occur, in the fossilized condition, in the deposits from the Cretaceous period to the present time. The Siphoneæ are connected by their lowest forms (Botrydiaceæ or Valonia) with the Protococcaceæ, but show also, through the Valoniaceæ, points of relationship to the Cladophoraceæ.
Order 1. Botrydiaceæ. The thallus in the vegetative condition is unicellular, club-shaped, with a small single (Codiolum) or repeatedly dichotomously branched system of colourless rhizoids (Botrydium, Fig. 56 a), by which it is attached to objects immersed in salt water (Codiolum) or to damp clay soil (Botrydium). Asexual reproduction by zoospores with one (Botrydium) or two[60] cilia, and by aplanospores. The sexual reproduction is only known in Botrydium, and takes place in the following manner: in the part of the thallus which is above ground and in an active vegetative condition, several round cells (Fig. 56 c) are formed, which may be green or red according as they grow under water, or exposed to the strong light of the sun. These cells must be considered as “gametangia” as they produce many gametes (d) provided with two cilia. The zygote (h, i) formed by the conjugation (e, f, g) may either germinate immediately, or become a thick-walled resting-cell of an irregular, angular form.
Order 2. Bryopsidaceæ. The thallus in the vegetative condition is unicellular, and consists at the lower extremity of branched rhizoids, while the upper portion is prolonged into a stem-like structure of unlimited growth, producing, acropetally, branches and leaf-like structures. The latter have limited growth, and are separated by a cross wall from the stem, and become gametangia, or drop off. The gametes have two cilia, and are of two kinds: the female, which are green and large and the male, which are of brownish colour and smaller. Zoospores or any other method of asexual reproduction are unknown. Only one genus, Bryopsis, living in salt water.
Fig. 57.—Bryopsis plumosa. A the plant, natural size. B A portion (enlarged) which shows the growing point (v), and the leaves derived from it in acropetal succession.
Order 3. Derbesiaceæ. Only one genus, Derbesia, living in saltwater. The zoospores, which are formed in a few lateral, swollen zoosporangia, possess one nucleus which has arisen through the coalescence of several, and they resemble the zoospores of Œdogonium by having a circle of cilia attached at the base of the colourless spot.
Order 4. Vaucheriaceæ. The thallus consists, in the vegetative condition, of a single irregularly or dichotomously branched cell, without differentiation into stem or leaf; root-like organs of attachment may however occur. Asexual reproduction by zoospores, which are formed singly in the extremity of a branch cut off by a transverse wall. They contain many nuclei, and bear small cilia situated in pairs, which give the appearance of a fine “pile” covering the whole or a great part of the surface. Akinetes,[61] aplanospores, and phytoamœbæ (naked masses of protoplasm, without cilia, which creep like an amœba on a substratum) may occur under certain conditions.
The sexual reproductive organs are formed on short lateral branches, and are separated from the vegetative cell (Fig. 58 A) by cell-walls. Numerous spermatozoids, each with two cilia, are developed in the coiled antheridium (A, b). The oogonium is a thick, egg-shaped, often oblique cell, with its protoplasm rounded into an oosphere, which has a hyaline “receptive-spot” (A, a) immediately beneath the aperture formed in the wall of the oogonium. A slimy mass, which serves to receive the spermatozoids, is formed in some species in this aperture. The spermatozoids when liberated swim towards and enter the oosphere, which then immediately surrounds itself with a thick cell-wall. The mature oospore (B) contains a large quantity of oil. At germination the outer cell-wall bursts and a new plant is formed. There is only one genus, Vaucheria, with species living in salt as well as in fresh water and on damp soil.
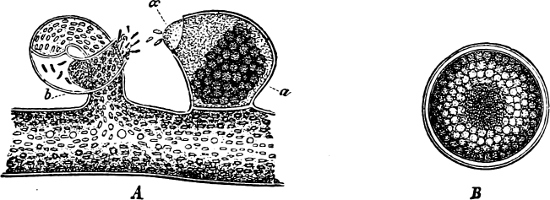
Fig. 58.—Vaucheria sessilis. A Fertilisation; b the antheridia; a the oogonia; a the receptive spot. B Oospore.
Order 5. Phyllosiphonaceæ are parasites in the leaves and stalks of Flowering-plants.
Order 6. Caulerpaceæ. The thallus has distinct differentiation into root, stem and leaf-like members (Fig. 59); it is unicellular. Within the cell, strong, branched threads of cellulose extend from one side to the other serving as stays to support the thallus. Reproduction takes place by detached portions of the thallus; no other modes of reproduction are known. This order may most approximately be classed with the Bryopsidaceæ. The genus Caulerpa consists of more than seventy species which inhabit the tropical seas.
Order 7. Codiaceæ. The thallus has various forms, but without[62] distinct differentiation in stem- or leaf-structures, sometimes (e.g. Halimeda) it is very much incrusted with lime. In the early stages it is unicellular (later, often multicellular), very much branched, with the branches, at any rate partly, so united or grown in amongst one another (Fig. 60) that an apparently parenchymatous cellular body is formed. Akinetes or aplanospores are wanting; zoospores (or gametes?) may be developed in some species, however, in special swollen sporangia. Fertilisation similar to that in Bryopsis occurs perhaps in Codium. They are all salt water forms.
Order 8. Valoniaceæ. The thallus is generally multicellular, without differentation into stem- or leaf-structures, but the cells are sometimes united together and form a leaf-like reticulate expansion (e.g. Anadyomene). Zoospores are known in some, and they are then formed directly in the vegetative cells. In others (e.g. Valonia), a mass of protoplasm, which maybe separated through the damaging of a cell, can surround itself with a cell-wall, and grow into a new plant. No other modes of reproduction are known. The most important genera are: Valonia, Siphonocladus, Chamædoris, Struvea, Microdictyon, Anadyomene. They are all salt water forms.
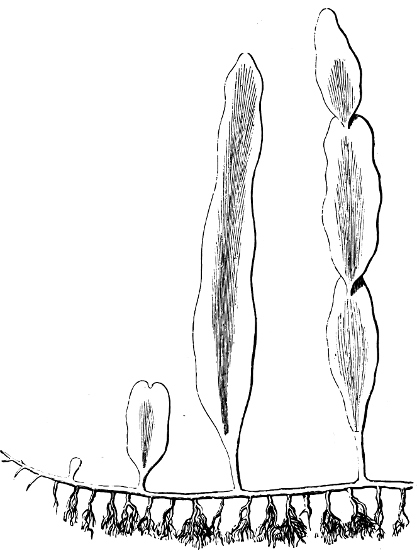
Fig. 59.—Caulerpa prolifera (natural size).
As already pointed out, the Valoniaceæ occupy a somewhat central position among the Siphoneæ, and present points of similarity and contrast with the Botrydiaceæ and the Bryopsidaceæ through Valonia, with the Dasycladaceæ through Chamædoris, and also with the Cladophoraceæ through Siphonocladus, and Struvea.
[63]
Order 9. Dasycladaceæ. The thallus consists of an axile longitudinal cell, destitute of transverse walls, attached at the base by root-like organs of attachment, and producing acropetally whorls of united, single or branched, leaf-like structures with limited growth. Asexual reproduction is wanting. Sexual reproduction by conjugation of gametes which arise in separate, fertile leaves, either directly or from aplanospores, which develope into gametangia. The principal genera are: Acetabularia, Dasycladus, Neomeris, Cymopolia. All marine.
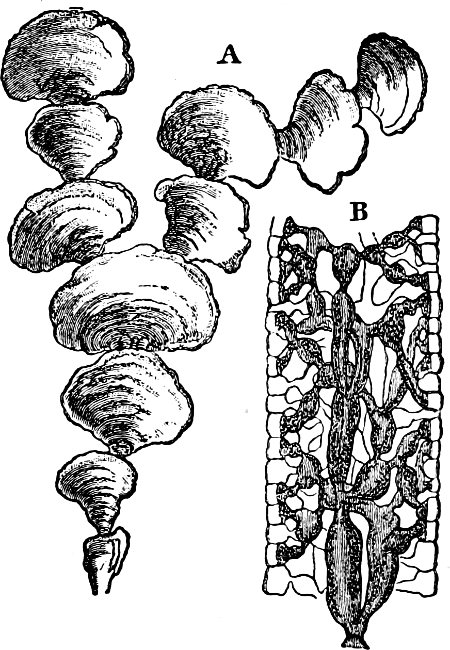
Fig. 60.—Halimeda opuntia. Plant (natural size). B Part of a longitudinal section.
The curiously shaped Acetabularia mediterranea grows gregariously on limestone rocks, and shells of mussels in the Mediterranean; it resembles a minute umbrella with a small stem, sometimes as much as nine centimetres in height, and a shade which may be more than one centimetre in diameter. The cell-membrane is thick, and incrusted with carbonate and oxalate of lime. Only the lower, root-like part of the thallus, which penetrates the calcareous substratum survives the winter, and may grow up into a new plant. The sterile leaves, which drop off early, are dichotomously branched and formed of cylindrical cells separated from each other by cross-walls, but they are not grown together. The shade is formed by a circle of 70–100 club-shaped rays (fertile leaves) grown together, in each ray 40–80 aplanospores are formed, which become liberated at the breaking of the shade, and later on are changed to gametangia (compare Botrydium) which open by a lid and allow a large number of egg-shaped gametes with two cilia to escape. Gametes from various[64] gametangia conjugate with one another; the product of the conjugation swarms about for some time, rounds off, and then surrounds itself with a cell-wall. The zygote germinates after a period of rest and then produces a sexual plant. The aplanospores (gametangia) thus represent the sexual generation.
The thallus has a stem with nodes and internodes; and whorls of leaves, on which may be developed the antheridia and oogonia, are borne at the nodes. Vegetative reproduction by bulbils and accessory shoots. Zoospores are wanting. The antheridia are spherical, and contain a number of filaments in which the spirally coiled spermatozoids, each with two cilia, are formed. The oogonium is situated terminally, and is at first naked, but becomes later on surrounded by an investment, and forms after fertilisation the so-called “fruit.” The oospore, after a period of rest, germinates by producing a “proembryo,” from which the young sexual plant arises as a lateral branch. The Characeæ are distinguished by the structure of their vegetative system as well as by the spirally-coiled spermatozoids, and stand as an isolated group among the Thallophytes, of which, however, the Siphoneæ appear to be their nearest relations. They were formerly, but wrongly, placed near the Mosses. The class contains only one order, the Characeæ.
Order 1. Characeæ. Algæ with a peculiar odour, often incrusted with lime, and of a brittle nature. They generally grow gregariously in large masses at the bottom of fresh and brackish water, and are from a few inches to more than a foot in height. The stem has long internodes which in Nitella are formed of one cylindrical cell; in Chara of a similar cell, but closely surrounded by a cortical layer of smaller ones. The protoplasm in contact with the cell-wall exhibits in a well-marked degree the movement of rotation (cyclosis), carrying the chlorophyll corpuscles along with it. The internodes are separated from each other by a layer of small cells (nodal cells) from which the leaves are produced. The leaves are borne in whorls of from 5–12 which regularly alternate with one another as in the higher verticillate plants; a branch is borne in the axil of the first formed leaf of each whorl (Fig. 61 A, n).

Fig. 61.—Chara fragilis. A Portion of a plant, natural size. B Portion of a leaf b, with leaflets β′-β′′; a antheridium; c oogonium. C A shield.—Nitella flexilis. D Filament from antheridium with spermatozoids. E Free spermatozoids.
The leaves are constructed in the same manner as the stem; they are divided into a series of joints, but have only a limited[65] power of growth; their terminal cell, too, is not enclosed by a cortex. Leaflets are borne at their nodes. The growth of the stem is unlimited, and proceeds by means of an apical cell (Fig. 62 s). The apical cell divides into a segment-cell and a new apical cell. The segment-cell then divides by a transverse wall into two cells, one lying above the other; the lower one, without any further division, becomes one of the long, cylindrical, internodal cells (Fig. 62 in), and the upper one (Fig. 62 n) divides by vertical walls to form the nodal cells. The cortical cells (Fig. 62 r) which surround the long internodal cells of Chara, are derived from the divisions of the nodal cells; the cells covering the upper portion of an internodal cell being derived from the[66] node immediately above it, and those in the lower part of the internode from the node below it.
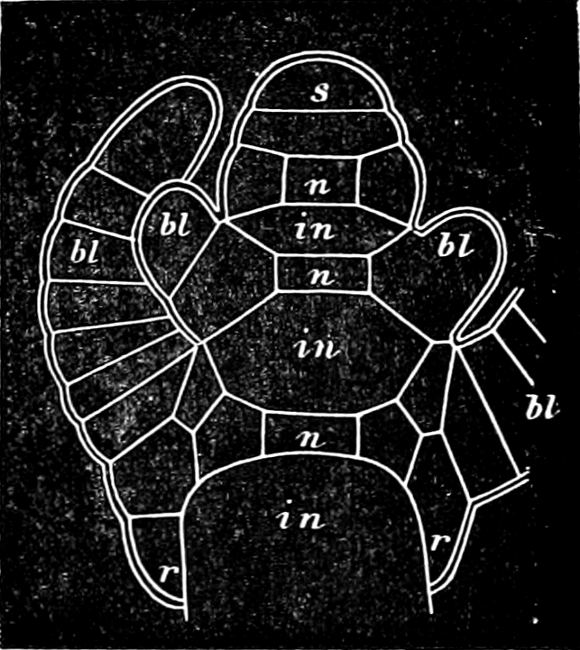
Fig. 62.—Chara fragilis: s apical cell; n, n nodal cells; in internodal cells; bl, bl leaves; r, r the cortical cells.
Fig. 63.—Oogonium of Chara: k “crown”; u receptive spot; s spermatozoids.
The organs of reproduction are very conspicuous by their colour and form. They are always situated on the leaves, the plants being very frequently monœcious. The antheridia (Fig. 61 B, a) are modified leaflets or the terminal cell of a leaf; they are spherical and become red when mature. Their wall consists of 8 “shields,” i.e. of plate-like cells, 4 of which cover the upper half, and are triangular; the 4 round the lower half, to which the stalk of the antheridia is attached, being quadrilateral, with sides of unequal length. The shields (Fig. 61 C) have dentated edges, with the teeth fitting into one another, and their faces ornamented with ridges. From the centre of the internal face of each shield (C) a cylindrical cell, the manubrium, projects nearly as far as the centre of the antheridium; at the inner end of each of the manubria a spherical cell, the capitulum, is situated. Each capitulum bears six secondary capitula, from each of which four long coiled filaments (C, D) project into the cavity of the antheridium. These filaments are divided by transverse walls into from 100–200 discoid cells, in each of which a biciliated, coiled spermatozoid is developed (D, E) from the nucleus. The spermatozoids escape from their mother-cell and are set free by the shields separating from one other.
[67]
The female organ of reproduction (Fig. 61 B, 63) is a small modified shoot, whose apical cell functions as an oogonium, its protoplasm forming the oosphere, which has a colourless receptive-spot at the summit (Fig. 63 u). The oogonium is situated on a nodal cell, from which 5 cells grow out in a circle and coil round the oogonium, covering it with a close investment. These cells divide once or twice at the top, so that 5 or 10 small cells are cut off, which project above the oogonium and form the so-called “crown” (Fig. 63 k). The crown either drops off at fertilisation, or its cells separate to form a central canal for the passage of the spermatozoids. The wall of the oosphere[9] above the receptive spot becomes mucilaginous, and allows the spermatozoid to fuse with the oosphere. The oospore, on germination (Fig. 64 sp), becomes a small filamentous plant of limited growth (Fig. 64 i, d, q, pl)—the proembryo—and from this, as a lateral outgrowth, the sexual generation is produced.
The order is divided into two sub-orders:—
A. Nitelleæ. The crown consists of 10 cells; cortex absent: Nitella, Tolypella.
B. Chareæ. The crown consists of 5 cells; cortex present: Tolypellopsis, Lamprothamnus, Lychnothamnus, Chara.
Chara crinita is parthenogenetic; in large districts of Europe only female plants are found, yet oospheres are formed capable of germination.
Fig. 64.—Chara fragilis. Germinating oospore (sp); i, d, g, pl, form together the proembryo rhizoids (w′) are formed at d; w′ the so-called tap-root; at g are the first leaves of the sexual plant, which appears as a lateral bud.
About 40 species of fossilized Chara, determined by their carpogonia, are known in the geological formations from the Trias up to the present day.
[68]
The Phæophyceæ are Algæ, with chromatophores in which the chlorophyll is masked by a brown colour (phycophæin). The product of assimilation is a carbohydrate (fucosan), never true starch. In the highest forms (Fucaceæ), the thallus presents differentiation into stem, leaf, and root-like structures. The asexual reproduction takes place by means of zoospores. The sexual reproduction is effected by the coalescence of motile gametes, or by oogamous fertilisation. The swarm-cells are monosymmetric, each moved by two cilia which are true protoplasmic structures, and generally attached laterally (Fig. 65). The Phæophyceæ are almost entirely saltwater forms; a few species of Lithoderma live in fresh water.
The class is divided into two families:—
1. Phæosporeæ: 1 Sub-Family, Zoogonicæ; 2 Sub-Family, Acinetæ.
2. Cyclosporeæ: Fucaceæ.
The family consists of multicellular plants, whose cells are firmly united together to form a thallus; this, in the simplest cases, may be a branched filament of cells (Ectocarpus), or, in the highest, may resemble a stem with leaves (Laminariaceæ), while all transitional forms may be found between these two. The thallus grows by intercalary divisions (e.g. Ectocarpus), or by an apical cell (e.g. Sphacelaria); pseudo-parenchymatous tissue may sometimes be formed by cells, which were originally distinct, becoming united together. The size of the thallus varies; in some species it is quite small—almost microscopical,—while in the largest it is many metres in length.
The vegetative cells in the lower forms are nearly uniform, but in those which are more highly developed (Laminariaceæ and Fucaceæ), they are sometimes so highly differentiated that mechanical, assimilating, storing and conducting systems may be found; the last named systems are formed of long cells with perforated, transverse walls, which bear a strong resemblance to the sieve-tubes in the higher plants.
Fig. 65.—Swarmspore of Cutleria multifida.
The colouring matter in the living cells (“phæophyl”) contains[69] chlorophyll; but this is concealed by a brown (“phycophæin”), and a yellow (“phycoxanthin”) colouring material, and hence all these Algæ are a lighter or darker yellow-brown. Starch is not formed. Asexual reproduction takes place, (1) by zoospores which arise in unilocular zoosporangia, and are monosymmetric, with two cilia attached laterally at the base of the colourless anterior end (Fig. 65), the longer one being directed forwards and the shorter backwards; or (2) by aplanospores (?).
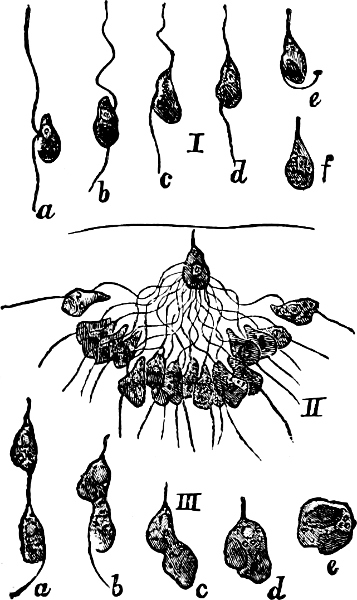
Fig. 66.—Ectocarpus siliculosus. I a-f A female gamete in the various stages of coming to rest. II A motionless female gamete surrounded by male gametes. III a-e Stages in the coalescence of male and female gametes.
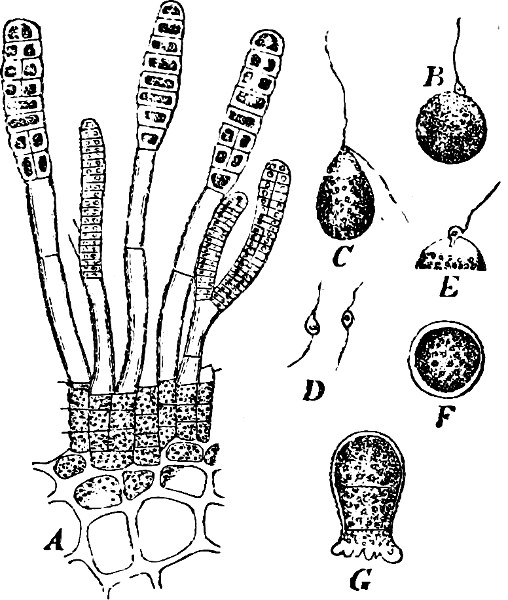
Fig. 67.—Zanardinia collaris. A Male gametangia (the smaller celled) and female gametangia (the larger celled). C Female gamete. D Male gamete. B, E Fertilisation. F Zygote. G Germinating zygote.
Sexual reproduction has only been discovered in a few cases, and takes place by means of gametes (oogamous fertilisation perhaps occurs in the Tilopteridæ). The gametes have the same structure as the zoospores, and arise in multilocular gametangia; these, like the zoosporangia, are outgrowths from the external surface, or arise as modifications from it. The conjugating gametes may be similar (e.g. Ectocarpus pusillus), or there may be a more or less pronounced difference of sex, an indication of which is found in Ectocarpus siliculosus (Fig. 66). When the gametes in this species have swarmed for a time, some, which are generally larger,[70] are seen to attach themselves by one of the cilia, which by degrees is shortened to form a kind of stalk (compare the upper gamete in Fig. 66 II); these are the female gametes, which now become surrounded by a number of males endeavouring to conjugate with them, but only one succeeds in effecting fertilisation. The protoplasm of the two gametes coalesces (Fig. 66 III), and a zygote (e) is formed. The male gametes which do not conjugate may germinate, but the plants derived from them are much weaker than those produced by the zygotes. Strongly pronounced sexual differences are found in the Cutleriaceæ, in which order the male and female gametes arise in separate gametangia (Fig. 67 A). The male gametes (Fig. 67 D) are much smaller than the female gamete (Fig. 67 C); the latter, after swarming for a short time, withdraws the cilia, and is then ready to become fertilised (Fig. 67 B, E), thus we have here a distinct transition to the oogamous fertilisation which is found in the Fucaceæ. Alternation of generations is rarely found.
1. Sub-Family. Zoogonicæ.
Reproduction by means of gametes and zoospores.
Order 1. Ectocarpaceæ. The thallus consists of single or branched filaments with intercalary growth, extending vertically from a horizontal, branched filament or a disc, but sometimes it is reduced to this basal portion only. Zoosporangia and gametangia (for fertilisation see Fig. 66) are either outgrowths or arise by the transformation of one or several of the ordinary cells. The most common genera are: Ectocarpus and Pylaiella.
Order 2. Choristocarpaceæ. Choristocarpus, Discosporangium.
Order 3. Sphacelariaceæ. The thallus consists of small, parenchymatous, more or less ramified shoots, presenting a feather-like appearance. In the shoots, which grow by means of an apical cell (Fig. 68 S), a cortical layer, surrounding a row of central cells, is present. Sporangia and gametangia are outgrowths from the main stem or its branches. Sphacelaria, Chætopteris are common forms.

Fig. 68.—Apex of the thallus of Chætopteris plumosa. S Apical cell.
Order 4. Encoeliaceæ. Punctaria, Asperococcus, Phyllitis fascia.
Order 5. Striariaceæ. Striaria, Phlœospora.
[71]
Order 6. Dictyosiphonaceæ. Dictyosiphon.
Order 7. Desmarestiaceæ. Desmarestia aculeata is common.
Order 8. Myriotrichiaceæ. Myriotrichia.
Order 9. Elachistaceæ. Elachista fucicola is a common epiphyte on species of Fucus.
Order 10. Chordariaceæ. The shoot-systems are often surrounded by mucilage. Chordaria; Leathesia difformis occurs as rounded, brown-green masses of the size of a nut, generally attached to other Seaweeds.
Order 11. Stilophoraceæ. Stilophora rhizodes is common.
Order 12. Spermatochnaceæ. Spermatochnus paradoxus is common.
Order 13. Sporochnaceæ. Sporochnus.
Order 14. Ralfsiaceæ. Ralfsia verrucosa is common as a red-brown incrustation on stones and rocks at the water’s edge.
Order 15. Lithodermataceæ. Some species of the genus Lithoderma occur in fresh water.
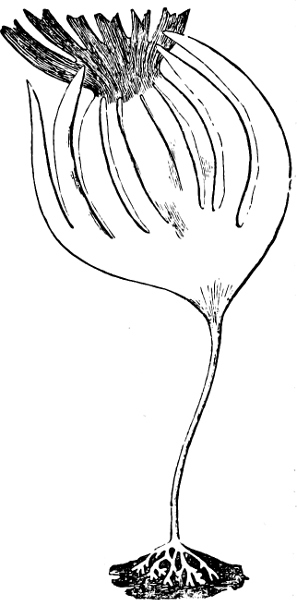
Fig. 69.—Laminaria digitata (much reduced in size).
Order 16. Laminariaceæ. The thallus is more or less leathery, and has generally a root-like lower part (Fig. 69) which serves to attach it, and a stalk or stem-like part, terminated by a large leaf-like expansion. Meristematic cells are situated at the base of the leaf, and from these the new leaves are derived. The older leaf thus pushed away by the intercalary formation of the younger ones, soon withers (Fig. 69). Gametes are wanting. Zoosporangia are developed from the lower part of a simple, few-celled sporangiophore, which is an outgrowth from a surface-cell and has a large club-formed apical cell. The sporangia are aggregated into closely packed sori, which cover the lower part of the terminal leaf, or occur on special, smaller, lateral, fertile fronds (Alaria). Most of the species belonging to this order live in seas of moderate or cold temperature and occur in the most northern regions that have yet been explored, forming their organs of reproduction during the cold and darkness of the arctic night. Laminaria is destitute of a midrib and has only one terminal leaf.[72] L. digitata has a broad leaf, which, by the violence of the waves, is torn into a number of palmate strips (Fig. 69). L. saccharina has a small, undivided leaf. Alaria has a midrib and special fertile fronds. A. esculenta occurs plentifully on the west coast of Norway and on the shores of Great Britain. Chorda filum, a common seaweed, is thick, unbranched, and attains a length of several metres, without any strong demarcation between stalk and leaf. Some attain quite a gigantic size, e.g. Macrocystis pyrifera, whose thallus is said sometimes to be more than 300 metres in length. The Lessonia-species, like the above, form submarine forests of seaweed on the south and south-west coasts of South America, the Cape, and other localities in the Southern Hemisphere.
Uses. The large Laminarias, where they occur in great numbers, are, like the Fuci, used for various purposes, for example, in the production of iodine and soda, and as an article of food (Laminaria saccharina, Alaria esculenta, etc.). Laminaria saccharina contains a large quantity of sugar (mannit) and is in some districts used in the preparation of a kind of syrup; in surgical operations it is employed for the distension of apertures and passages, as for instance the ear-passage. It is by reason of the anatomical peculiarities and structure of the cell-walls, that they are employed for this purpose. The cell-walls are divided into two layers, an inner one which has very little power of swelling, and an outer one, well developed and almost gelatinous—the so-called “intercellular substance”—which shrivels up when dried, but can absorb water and swell to about five times its size. The stalks of Laminaria clustoni are officinal.
Order 17. Cutleriaceæ. The thallus is formed by the union of the originally free, band-shaped shoots. The growth is intercalary. Sexual reproduction by the conjugation of male and female gametes. An asexual generation of different appearance, which produces zoospores, arises from the germination of the zygote. Cutleria, Zanardinia.
Sub-Family 2. Acinetæ.
Branched, simple cell-rows with intercalary growth. The organs of reproduction are partly uni-and partly multicellular; in the unicellular ones a cell without cilia is formed, which may be destitute of a cell-wall, but has one nucleus (oosphere?), or which has a cell-wall and contains several (generally four) nuclei (aplanospores?); in the multicellular, monosymmetric swarm-cells with two cilia (spermatozoids?) are formed. The fertilisation has not been observed.
Order 1. Tilopteridaceæ. Haplospora, Tilopteris.
[73]
The individuals are multicellular, with growth by an apical cell. The thallus—often bilateral—is differentiated into a root-like structure (attachment-disc), and stem, sometimes also into leaves (Sargassum). Sometimes a differentiation occurs into various tissue-systems, viz. an external assimilating tissue, a storing tissue, a mechanical tissue of thickened, longitudinal, parenchymatous, strengthening cells, and a conducting tissue of sieve-cells, or of short sieve-tubes with perforated walls. Colouring material, as in Phæosporeæ. Vegetative reproduction can only take place by means of detached portions of the thallus (Sargassum), which are kept floating by means of bladders (Fig. 70 A, a, Fig. 72). Zoospores are wanting.
The sexual reproduction takes place by oogamous fertilisation. The oogonia and antheridia are formed inside special organs (conceptacles), and are surrounded by paraphyses. The conceptacles (Fig. 70 B, Fig. 71 b) are small, pear-shaped or spherical depressions, produced by a special ingrowth of the surface cells of the thallus, and their mouths (ostioles) project like small warts; they are either situated near the end of the ordinary branches of the thallus (Fucus serratus, Fig. 71 a) which may be swollen on this account (Fucus vesiculosus, Fig. 70 A, b), or on special short branches (Ascophyllum, Sargassum). The vertical section of a conceptacle is seen in Fig. 70 B (see also Fig. 71 b) where, in addition to the paraphyses, oogonia only are seen (F. vesiculosus is diœcious—male plant, yellow-brown; female plant, olive-brown); but in some species antheridia, together with oogonia, are produced in the same conceptacle. The oogonia are large, almost spherical cells, situated on a short stalk, in each of which are formed from 1–8 (in Fucus, 8; in Ascophyllum, 4; in Halidrys, 1; in Pelvetia, 2) rounded, immotile oospheres. The wall of the oogonium ruptures, and the oospheres, still enclosed in the inner membrane, are ejected through the mouth of the conceptacle, and float about in the water, being finally set free by the bursting of the inner membrane. The antheridia are oblong cells (Fig. 70 C, a), many of which are produced on the same branched antheridiophore (Fig. 70 C); the numerous spermatozoids are provided with 2 cilia and are very small (Fig. 70 D, two antheridia surrounded by spermatozoids, one being open). The spermatozoids, still enclosed by the inner membrane of the antheridium, are[74] similarly set free, and fertilisation takes place in the water, numerous spermatozoids collecting round the oosphere (Fig. 70 E), which is many times larger, and by their own motion causing it to rotate. After fertilisation, the oospore surrounds itself with a cell-wall and germinates immediately, attaching itself (Fig. 70 F) to some object, and by cell-division grows into a new plant.
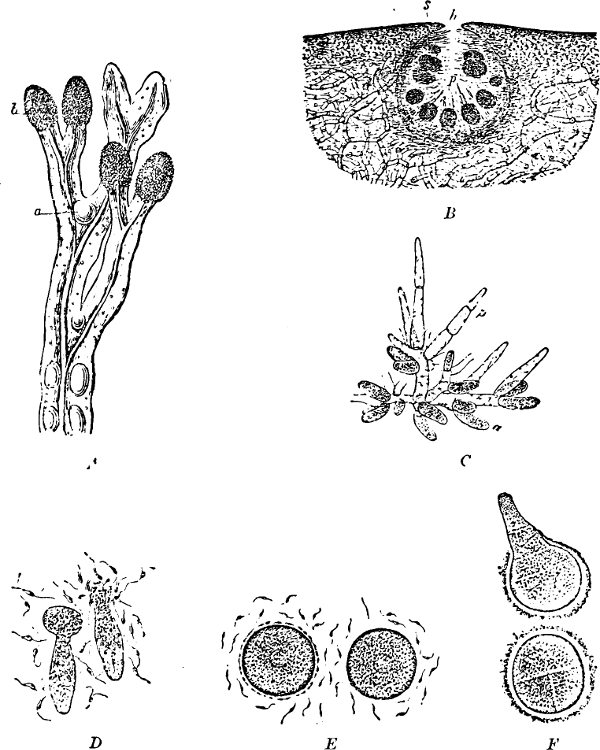
Fig. 70.—Fucus vesiculosus. A Portion of thallus with swimming bladders (a) and conceptacles (b). B Section of a female conceptacle; h the mouth; p the inner cavity; s oogonia. C Antheridiophore; a antheridium; p sterile cells. D Antheridia out of which the spermatozoids are escaping. E Fertilisation. F Germinating oospore.
[75]
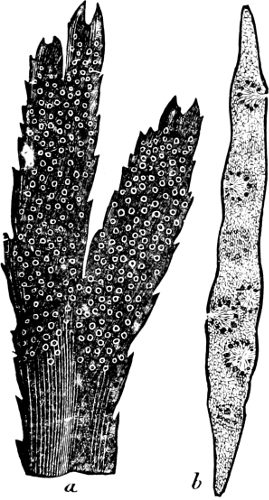
Fig. 71.—Fucus serratus. a Portion of a male plant which has been exposed to the action of the open air for some time; small orange-yellow masses, formed by the antheridia, are seen outside the mouths of the male conceptacles (nat. size). b Cross section through the end of a branch of a female plant, showing the female conceptacles (× 4).
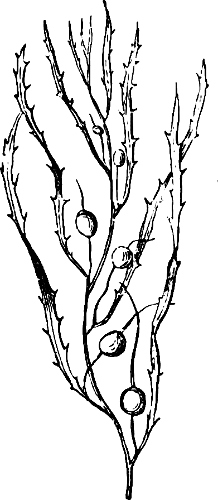
Fig. 72.—Sargassum bacciferum. A portion of the thallus, natural size.
Order 1. Fucaceæ. The following species are common on our coasts: Fucus vesiculosus (Fig. 70) has a thallus with an entire margin, and with bladders arranged in pairs; F. serratus (Fig. 71) without bladders, but with serrated margin; Ascophyllum nodosum has strap-like shoots, which here and there are swollen to form bladders; Halidrys siliquosa has its swimming bladders divided by transverse walls; Himanthalia lorea, which is found on the west coast of Norway, and the south coast of England, has a small perennial, button-shaped part, from the centre of which proceeds the long and sparsely branched, strap-like, annual shoot, which bears the conceptacles. The Gulf-weed (Sargassum bacciferum, Fig. 72) is well known historically from the voyage of Columbus; it is met with in large, floating, detached masses in all oceans, and is found most abundantly in the Atlantic, off the Canary Islands and the Azores, and towards the Bermudas. The stalked, spherical air-bladders are the characteristic feature of this genus. The thallus is more highly developed than in Fucus, and there is a contrast between the stem and leaf-like parts. The[76] portions which are found floating are always barren, only those attached are fertile.
Uses. The Fucaceæ, like the Laminariaceæ, are used as manure (the best kinds being Fucus vesiculosus and Ascophyllum nodosum), for burning to produce kelp, and as food for domestic animals (Ascophyllum nodosum is especially used for this purpose).
The plants in this class are multicellular, and brown, with apical growth, new cells being derived either from a flat apical cell, or from a border of apical cells. The thallus is flat, leaf- or strap-shaped, attached by haptera, which are either found only at the base, or on the whole of the lower expansion of the thallus. The cells are differentiated into the following systems of tissues: an external, small-celled layer of assimilating cells, generally one cell in thickness, and an internal, large-celled layer of one or only a few cells in thickness, forming the mechanical and conducting tissues. All the reproductive cells are motionless. Asexual reproduction by naked, motionless spores (tetraspores) which are formed 1–4 in each tetrasporangium, the latter being outgrowths from the surface cells of special, sexless individuals. Zoospores are wanting. The sexual organs are of two kinds, oogonia and antheridia, which are formed from the surface cells, either on the same or different individuals. The oogonia are spherical or oval, and are generally placed close together; each contains one oosphere, which on maturity is ejected into the surrounding water, and is then naked and motionless. The antheridia are formed of longitudinal cells, united in groups, whose contents by repeated divisions—transverse and longitudinal—are divided into a large number of small, colourless, motionless spermatia—round or elongated—which are set free by the dissolution of the wall of the antheridium. The process of fertilisation has not yet been observed.
The Dictyotales, in having tetraspores and spermatia, deviate considerably from the Phæophyceæ, but may be classed near to the Tilopteridæ, in which there are asexual spores with 4 cell-nuclei, which may be considered as an indication of the formation of tetraspores.
Order 1. Dictyotaceæ. Dictyota dichotoma which has a thin, regularly dichotomously divided thallus, occurs on the coasts of the British Isles. Padina is found on the south coast.
[77]
The plants comprised in this class are multicellular; they are simple or branched filaments, or expansions consisting of 1 to several layers of cells; the thallus may be differentiated (as in many Florideæ), to resemble stem, root, and leaf. The cells contain a distinctly differentiated nucleus (sometimes several), and distinct chromatophores, coloured by rhodophyll. The chlorophyll of the chromatophores is generally masked by a red colouring matter (phycoerythrin), which may be extracted in cold, fresh water; or rarely by phycocyan. Pyrenoids occur in some. Starch is never formed in the chromatophores themselves, but a modification—Florideæ starch—may be found in the colourless protoplasm. Asexual reproduction by motile or motionless spores (tetraspores) which are devoid of cilia and of cell-wall. Swarmspores are never found.
Sexual reproduction is wanting, or takes place by the coalescence of a spermatium and a more or less developed female cell. The spermatia are naked masses of protoplasm, devoid of cilia and chromatophores. The female cell (carpogonium) is enclosed by a cell-wall, and after fertilisation forms a number of spores, either with or without cell-walls (carpospores), which grow into new individuals.
The Rhodophyceæ may be divided into two families:
1. Bangioideæ.
2. Florideæ.
The thallus consists of a branched or unbranched cell-filament, formed of a single row or of many rows of cells, or of an expansion, one or two layers of cells in thickness, but without conspicuous pores for the intercommunication of the cells. The growth of the thallus is chiefly intercalary. The star-like chromatophores contain chlorophyll and are coloured blue-green with phycocyan, or reddish with phycoerythrin; all these colouring matters are occasionally found in the same cell (Bangia-species). Asexual reproduction by tetraspores, without cilia, but capable of amœboid movements.
Sexual reproduction is wanting, or takes place by the coalescence of a spermatium with a carpogonium, which is only slightly differentiated from the vegetative cells, and is devoid of a trichogyne.[78] The carpospores are destitute of cell-wall and arise directly by the division of the fertilised oosphere. The Bangioideæ occur chiefly in salt water.
Order 1. Goniotrichaceæ.—The thallus consists of a branched cell-filament without rhizoids. Tetraspores are formed directly from the entire contents of the mother-cell, without any preceding division. Fertilisation unknown. Asterocystis, Goniotrichum.
The Goniotrichaceæ, through the blue-green Asterocystis, are allied to the Myxophyceæ, and through Goniotrichum to the Porphyraceæ.
Order 2. Porphyraceæ.—The thallus is formed of an expansion consisting of a layer of 1–2 cells, which, at the base, are attached to the substratum by means of a special form of haptera (Porphyra, Diploderma); or of unbranched (very rarely slightly branched) filaments, attached at the base by haptera (Bangia): or it extends from a prostrate cell-disc (various species of Erythrotrichia). Tetraspores are formed after one or more divisions of the mother-cell, either from the whole or only a part of its contents; they possess amœboid movements, or have a jerky, sliding-forward motion. The antheridia have the same appearance as the vegetative cells, but divide several times, and several spermatia are formed, either simultaneously from the whole contents (Porphyra, Bangia), or the spermatia are successively formed from a part of the contents of the antheridium (Erythrotrichia). The carpogonium is without a trichogyne, but the oosphere has a colourless spot which may sometimes rise a little above the surface of the thallus, and may be considered as an early stage in the development of the trichogyne. The spermatia form a canal through the membrane of the carpogonium, and their contents coalesce with the oosphere at its colourless spot. The fertilised oosphere divides on germination into a number of carpospores, which are set free as naked, motionless masses of protoplasm, which grow and give rise to new individuals (alternation of generations).
The thallus has one or more apical cells, grows principally by apical growth, and may be differentiated into root, stem, and leaf. The chromatophores vary in form, but have a red or brownish colour, due to chlorophyll and phycoerythrin. Asexual reproduction by motionless tetraspores, which generally arise by the division into four of the contents of the tetrasporangium. The carpogonium has a trichogyne, and the carpospores, which are formed indirectly from the fertilised oosphere, possess a cell-wall.
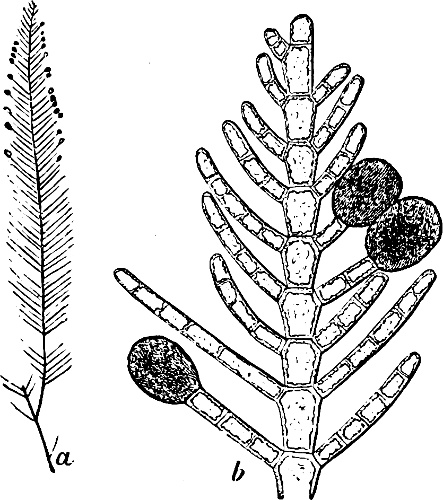
Fig. 73.—Callithamnion elegans: a a plant with tetraspores (× 20); b apex of a branch with tetraspores(× 250).
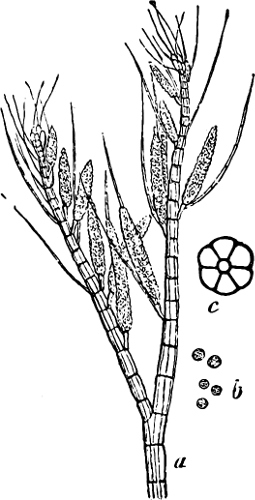
Fig. 74.—Polysiphonia variegata: a a portion of a male plant with antheridia; b spermatia; c transverse section of thallus.
The thallus may assume very different forms. In the simplest species it is filamentous and formed of single, branched rows of cells (Callithamnion, etc., Fig. 73). Ceramium has a filamentous thallus, generally dichotomously forked (Fig. 75), or sometimes[79] pinnately branched, which, at the nodes, or throughout its entire length, is covered by a layer of small cortical cells. Polysiphonia (Fig. 74) has a filamentous, much branched thallus, made up of a central cylindrical cell, surrounded by a layer of other cells, cortical cells, which in length and position correspond to the central ones. In many of the Red Algæ the vegetative organs are differentiated into stems and leaves, the former having, as in Chara, unlimited growth in length, whilst the latter soon attain their full development. Chondrus has a fleshy, gelatinous thallus, without nodes; it is repeatedly forked into flat branches of varying thickness. Furcellaria has a forked thallus with thick branches and without nodes. The thallus of Delesseria (Fig. 76) consists of branches, often bearing leaf-like structures, with a midrib and lateral ribs springing from it. These ribs persist through the winter, and at the commencement of the succeeding period of vegetation the lateral ribs become the starting points for new leaves. In Corallina the thallus is pinnately branched, and divided into nodes and internodes. The name has been given to this genus from the fact that the thallus is incrusted with carbonate of lime to such a degree that it becomes very hard, and the[80] whole plant adopts a coral-like appearance. Other genera which are similarly incrusted, and have a leaf-like or even crustaceous thallus (such as Melobesia, Lithothamnion), are included in this family.
In some instances the cells of the thallus may be found differentiated into more or less well defined tissues, so that it is possible to find special assimilating, mechanical, and conducting tissues, the last named in some cases having the double function of conducting and of serving as a reservoir in which starch is found as a reserve material. The cells of the Florideæ, which are formed by the division of a mother-cell into two daughter-cells of unequal size, have always larger or smaller pits in the cell-walls, and the thin cell-wall separating two pits from each other is perforated by a number of small holes. These pits are particularly developed in the conducting tissues, but sieve-tubes are very rarely to be found.
Fig. 75.—Ceramium diaphanum (nat. size).
Fig. 76.—Delesseria sanguinea (about ⅓).
Tetraspores may be wanting (e.g. Lemanea) or may often arise on special, non-sexual individuals. In some (e.g. Batrachospermum) only one tetraspore is formed in each tetrasporangium, but the number is generally four, which may be formed tetrahedrally (Fig. 73) or by divisional walls perpendicular to each other, or even in a single row. The tetrasporangia in some species are free (Fig. 73), but in the majority they are embedded in the thallus.
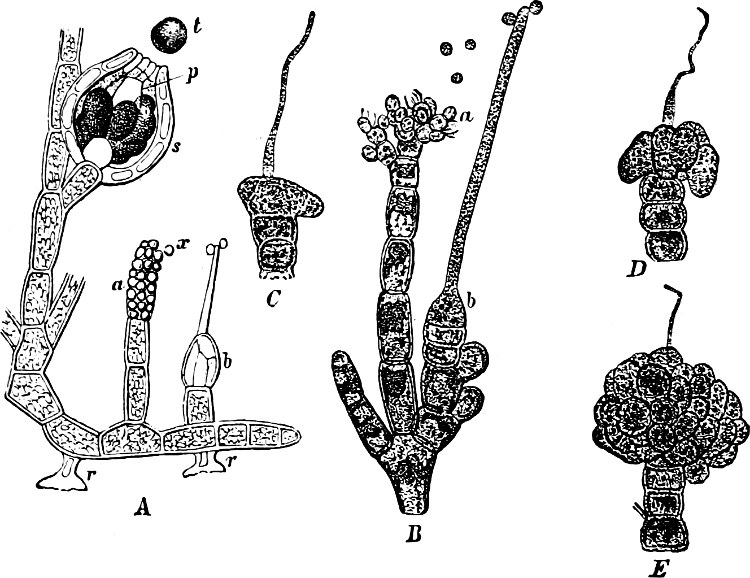
Fig. 77.—A Lejolisia mediterranea: r haptera; s longitudinal section through a cystocarp; p the empty space left by the liberated spore (t). B-E Nemalion multifidum: a antheridia; b procarpium with trichogyne, to which two spermatia are adhering.
The sexual reproduction (discovered by Thuret and Bornet,[81] 1867) differs in the essential points from that of all other plants, and approaches most nearly to the sexual reproduction of the Bangioideæ. The sexual cells are developed from the terminal cells (never nodal cells) of the branched cell-filaments, which constitute the thallus. The mother-cells of the spermatia (spermatangia) are generally arranged in a group, in the so-called antheridia (Figs. 74, 77 A, a). On becoming ripe the membrane of the spermatangium ruptures and the spermatia emerge as spherical or ovoid, naked (a little later they may possess a cell-wall) masses of protoplasm which are not endowed with the power of motion, and hence are carried passively by the current of the water in which they may happen to be, to the female cell. This latter is analogous with the oogonium of the Green Algæ. The female reproductive organ is termed the procarpium, and consists of two parts, a lower swollen portion—the carpogonium (Fig. 77 b in A and B)—which contains the cell-nucleus, and an upper filamentous prolongation—the trichogyne (Fig. 77 B)—which is homologous with the colourless receptive spot of the oosphere of the Green Algæ, and the Porphyraceæ. In the sexual reproduction of the majority of the Florideæ, a very important part is played by certain special cells, rich in cell-contents—the auxiliary[82] cells. These are either dispersed in the interior of the thallus, or are arranged together in pairs with the cell-filament which bears the carpogonium, and are generally united with this to form an independent multicellular procarpium. The spermatia attach themselves firmly to the trichogyne and surround themselves with a cell-wall. The dividing wall at the point of contact is perforated, and the nucleus of the spermatium probably travels through the trichogyne to the swollen part of the procarpium—the carpogonium—and fuses with its nucleus. After fertilisation the trichogyne withers (Fig. 77 C), but the lower portion of the procarpium, constituting the fertilised oosphere, grows out and forms in various ways, first a tuft of spore-forming filaments known as gonimoblasts, and finally the carpospores. These latter form a new asexual generation (compare the germination of the oospore of Œdogonium and Coleochæte).
The gonimoblasts may arise in three ways:—
1. In the Nemalionales, branched filaments grow out from the oosphere and form an upright, compressed or expanded tuft of spore-forming filaments.
2. In the Cryptonemiales, several branched or unbranched filaments (ooblastema-filaments) grow out from the oosphere, and conjugate in various ways with the auxiliary cells. The gonimoblasts are then formed from the single cells produced by the conjugation.
3. In the Gigartinales and Rhodymeniales the oosphere conjugates with an auxiliary cell by means of a short ooblastema-filament, and from this auxiliary cell a gonimoblast is produced.
The motionless carpospores, which sometimes in the early stages are naked, and afterwards invested with a cell-wall, are developed from the terminal cells (and perhaps also from some of the other cells) of the branches of the gonimoblast. The gonimoblasts constitute sharply defined parts of the plant in which the carpospores arise. These parts are called cystocarps and are either naked (Fig. 77 E), or surrounded by a covering (pericarp or involucre, Fig. 77 A) formed in different ways. On this account the Florideæ were formerly divided into Gymnosporeæ (Batrachospermum, Nemalion, Ceramium, etc.) and Angiosporeæ (Farcellaria, Lejolisia, Delesseria, Melobesia, etc.).
The Florideæ are divided into four sub-families:—
Sub-Family 1. Nemalionales. The fertilised oosphere produces directly the gonimoblast.
Order 1. Lemaneaceæ. Algæ of brownish colour and living in fresh water. They lack tetraspores, and the very sparingly branched fertile filaments, composed of many rows of cells, grow out from a proembryo, which consists of a single row of cells bearing branches. Lemanea fluviatilis, often found on rocks and stones in quickly flowing streams.
[83]
Order 2. Helminthocladiaceæ. Tetraspores are generally wanting (e.g. in Nemalion) or arise one in each tetrasporangium (e.g. Batrachospermum) and it is only in Liagora that four cruciate tetraspores are formed. Chantransia corymbifera consists of simple, branched cell-rows, and is an independent species. Several other Chantransia-forms, living in fresh water, are “proembryos” of species of the genus Batrachospermum. The germinating carpospore grows out into filaments and forms a so-called proembryo which, if not shaded, attains only a small size, but when growing in shady situations presents a much greater development. These highly developed proembryos have been described as species of Chantransia. The proembryo can reproduce by division, or by tetraspores which are developed singly in the sporangia; in B. vagum and B. sporulans which do not possess fully developed female reproductive organs, the proembryos serve almost entirely to reproduce the species. The young Batrachospermum-plant arises from the end of an upright filament of the proembryo. The proembryo is generally persistent, and continually produces new Batrachospermums. These latter bear the sexual reproductive organs and also whorls of branches: the central row of cells is enclosed by cells growing from the base of the whorls of branches, and from these cortical cells secondary proembryos are developed. In this alternation of shoots there is really no alternation of generations, since the proembryo and the shoots with the sexual reproductive organs are parts of the same thallus.
Several species of Batrachospermum have a bluish green or verdigris colour. Nemalion multifidum has a brown-red thallus, slightly branched, which is attached to rocks near the water’s edge.
Order 3. Chætangiaceæ. Galaxaura has a thallus thickly incrusted with lime.
Order 4. Gelidiaceæ. Naccaria, Gelidium.
Sub-Family 2. Gigartinales. The fertilised auxiliary cell grows towards the thallus, to produce the gonimoblasts. Procarpia generally present.
Order 5. Acrotylaceæ. Acrotylus.
Order 6. Gigartinaceæ. Gigartina, Phyllophora, Ahnfeltia; Chondrus crispus, with dark red, dichotomously branched thallus, is common on the coasts of Scandinavia and Great Britain.
Order 7. Rhodophyllidaceæ. Rhodophyllis, Euthora; Cystoclonium purpurascens is common, and sometimes the ends of its branches may be modified into tendril-like haptera.
Sub-Family 3. Rhodymeniales. The fertilised auxiliary cell forms the gonimoblast on the side away from the thallus. Procarpia are abundantly produced.
Order 8. Sphærococcaceæ. Gracilaria.
Order 9. Rhodymeniaceæ. Rhodymenia palmata is a common species. Lomentaria, Chylocladia, Plocamium.
Order 10. Delesseriaceæ. Delesseria sanguinea; D. alata and D. sinuosa are handsome forms which are not uncommon.
Order 11. Bonnemaisoniaceæ. Bonnemaisonia.
Order 12. Rhodomelaceæ. Rhodomela, Odonthalia; Polysiphonia, of which many species are to be found on the coasts of Great Britain, has a filamentous, richly branched thallus consisting of a central row of cells surrounded[84] by a varying number of cortical cells of similar size—the so-called “siphons.”
Order 13. Ceramiaceæ. Pretty Algæ, often branched dichotomously, or unilaterally pinnate. Spermothamnion, Griffithsia, Callithamnion, Ceramium, Ptilota.
Sub-Family 4. Cryptonemiales. The cells formed by the coalescence of the auxiliary cells and the ooblastema-filaments, produce the gonimoblasts. The carpogonium-filaments and the auxiliary cells are scattered singly in the thallus.
Order 14. Gloiosiphoniaceæ. Gloiopeltis.
Order 15. Grateloupiaceæ. Halymenia, Cryptonemia.
Order 16. Dumontiaceæ. Dumontia, Dudresnaya.
Order 17. Nemastomaceæ. Furcellaria, which has dichotomously branched, round shoots, is common on the coasts of Great Britain.
Order 18. Rhizophyllidaceæ. Polyides, Rhizophyllis.
Order 19. Squamariaceæ. The Algæ belonging to this order form crust-like coverings on stones, mussel-shells, and on other Algæ, but are not themselves incrustated: Petrocelis, Cruoria, Peyssonellia.
Order 20. Corallinaceæ. Partly crustaceous, partly erect, branched Algæ, thickly incrusted with lime, so that a few species (Lithothamnia, also called Nullipora) occur in fossilized condition from Jurassic to Tertiary periods. Melobesia, Lithophyllum, Lithothamnion, Corallina.
Uses. “Carragen” is the thallus of Chondrus crispus (Irish Moss) and Gigartina mamillosa. It is a common article of food on the coasts of Ireland, and swells to a jelly when cooked. It is officinal. Rhodymenia palmata is generally eaten as food in Ireland and in some places on the west coast of Norway; it is also used as food for sheep and hence is termed “Sheep-seaweed.” Agar-Agar is the jelly obtained from species of Gelidium and Gigartina growing in China and Japan.
Mode of Life. The Fungi have no chlorophyll, and are thus unable in any stage of their existence to assimilate carbon; they must therefore live as saprophytes or parasites. There is, however, no strong line of demarcation between these; many Fungi commence as true parasites, but only attain their full development upon or in dead plants or animals (Rhytisma, Empusa). Many saprophytes may occasionally appear as parasites, and are then designated “facultative parasites” (Nectria cinnabarina, Lophodermium pinastri), in contradistinction to those which only[85] appear as parasites, “obligate parasites” (Mildew, Brand-and Rust-Fungi, Cordyceps).
The parasites which live on the surface of the host-plant are termed epiphytic (Mildew, Fusicladium); and those living in its tissues are termed endophytic (Ustilago, Peronospora). Epizoic (Oidium tonsurans, Laboulbenia) and endozoic Fungi (Cordyceps, Entomophthora), are distinguished, in the same manner, as those which live on the surface or in the interior of animals. The Fungi designated pathogenic are especially those which produce disease in human beings and in animals.
Most of the diseases of plants are attributed to the parasitic Fungi. These force their way into the host-plant by piercing the outer wall of the epidermis, as in the Potato-disease; or by growing in through the stomata, e.g. the summer generations of the Rust of Wheat; or they can only penetrate through a wound, e.g. Nectria. Some effect an entrance into the host-plant by the secretion of a poisonous matter or ferment, which softens and destroys the cell-walls (Sclerotinia). Some Yeast and Mould Fungi secrete ferments (enzymes), which, for example, convert cane-sugar into a sugar capable of fermentation.
The relation of the parasitic Fungus to the host-plant is mainly of two kinds. In the one case, the cell-contents are destroyed, the protoplasm is killed, and the cellular tissue becomes discoloured and dies (Peronospora, Armillaria mellea, Polyporus); in the other case, the parasite has an irritating effect on the cellular tissue, whereby the affected organ grows more rapidly and becomes larger than normal, producing hypertrophy. Such malformations are termed Fungi-galls (Mycocecidia); in this manner “witches’ brooms” are produced by Æcidium, “pocket-plum” by Taphrina, and other deformities by Exobasidium and Cystopus candidus. This hypertrophy may either be produced by a vigorous cell-multiplication, which is most frequently the case, or by the enlargement of the individual cells (Synchytrium, Calyptospora). The relation between host and Fungus among the Lichens is of a very peculiar nature, termed “symbiosis.”
Vegetative Organs. The vegetative parts of a Fungus are termed its mycelium.[10] This is formed of a mass of long, cylindrical, branched cells resembling threads (and hence termed hyphæ), which have a continued apical growth. The mycelium, in its early development, shows a well-marked difference between the[86] two main groups of true Fungi: in the Phycomycetes, or Algal Fungi, the mycelium has no transverse walls, and is therefore unicellular, while in the Mesomycetes and Mycomycetes it is provided with dividing walls, which gradually arise during growth, in the youngest hyphæ; intercalary transverse walls may also be formed at a later period. In the hyphæ of some of the Higher Fungi (Hymenomycetes), connections may be formed between two contiguous cells of the same hypha, by a protuberance growing out from an upper cell just above the transverse wall, and forming a junction with the cell below. These are known as clamp-connections; they appear to be of use in affording communication between the two cells.
The hyphæ of Fungi, where they come in contact with one another, often grow together, so that H-formed combinations (fusions) are produced, which give rise to very compact felted tissue. When the hyphæ are not only closely interwoven, but also united and provided with many transverse walls, the mycelium assumes the appearance of a tissue with isodiametric cells, and is then termed pseudo-parenchyma. The hyphæ-walls are sometimes very much thickened, and composed of several layers, and the external layers, by the absorption of water, may often swell very much and become mucilaginous. In some instances the walls are colourless, in others coloured, the most frequent colour being brown. The cell-contents may also be coloured, and in that case are generally yellow; this colour is chiefly connected with the fat (oil) which may be found in abundance in the Fungi, whilst starch is invariably absent in all the true Fungi.
The mycelium assumes many different forms; sometimes it appears as a thread-like, cobwebby, loose tissue, less frequently as firm strands, thin or thick membranes, horn-like plates or tuber-like bodies. The thread-like mycelium may, in the parasitic Fungi, be intercellular or intracellular, according as it only extends into the interstices between the cells or enters into the cells proper. In the first case there are generally found haustoria, or organs of suction (e.g. among the Peronosporaceæ; Taphrina, on the contrary, has no haustoria); but haustoria are also found among the epiphytic Fungi (e.g. Erysiphaceæ). Intracellular mycelia are found in the Rust-Fungi, in Claviceps purpurea, Entomophthora, etc. In spite of its delicate structure, this mycelium may live a long time, owing to the circumstance that it continues to grow peripherally, while the older parts gradually die off (“fairy rings”).
[87]
String-like mycelia may be found, for example, in Phallus, Coprinus, and are formed of hyphæ, which run more or less parallel to each other. Membrane-like mycelia are chiefly to be found in Fungi growing on tree-stems (Polyporaceæ and Agaricaceæ); they may have a thickness varying from that of the finest tissue-paper to that of thick leather, and may extend for several feet. The peculiar horny or leather-like strands and plates which, for instance, appear in Armillaria mellea, are known as Rhizomorpha; they may attain a length of more than fifty feet. The tuber-like mycelia or sclerotia play the part of resting mycelia, since a store of nourishment is accumulated in them, and after a period of rest they develope organs of reproduction. The sclerotia are hard, spherical, or irregular bodies, from the size of a cabbage seed to that of a hand, internally white or greyish, with a brown or black, pseudo-parenchymatous, external layer. Sclerotia only occur in the higher Fungi, and are found both in saprophytes, e.g. Coprinus, and in parasites, e.g. Claviceps (Ergot), Sclerotinia.
Reproduction. Sexual reproduction is found only among the lower Fungi which stand near to the Algæ, the Algal-Fungi, and takes place by the same two methods as in the Algæ, namely by conjugation and by the fertilisation of the egg-cell in the oogonium.
The majority of Fungi have only ASEXUAL reproduction. The most important methods of this kind of reproduction are the sporangio-fructification and the conidio-fructification.
In the SPORANGIO-FRUCTIFICATION the spores (endospores) arise inside a mother-cell, the sporangium (Fig. 80). Spores without a cell-wall, which move in water by means of cilia and hence are known as swarmspores or zoospores, are found among the Oomycetes, the sporangia in which these are produced being called swarm-sporangia or zoosporangia (Figs. 86, 87, 91, 94).
In the CONIDIO-FRUCTIFICATION the conidia (exospores) arise on special hyphæ (conidiophores), or directly from the mycelium. When conidiophores are present, the conidia are developed upon them terminally or laterally, either in a basipetal succession (in many Fungi, for example in Penicillium, Fig. 111, Erysiphe, Cystopus), or acropetally (in which method the chains of conidia are often branched; examples, Pleospora vulgaris, Hormodendron cladosporioides). All conidia are at first unicellular, sometimes at a later stage they become two-celled or multicellular through the formation of partition-walls (Piptocephalis). The conidia with[88] thick, brown cell-walls, and contents rich in fats (resting conidia), can withstand unfavourable external conditions for a much longer period than conidia with thin walls and poor in contents.
The SPORANGIA arise either from the ordinary cells of the mycelium (Protomyces), or are borne on special hyphæ. They are generally spherical (Mucor, Fig. 80; Saprolegniaceæ), egg-, pear-, or club-shaped (Ascomycetes), more rarely they are cylindrical or spindle-shaped. While among the Phycomycetes the size, form, and number of spores are indefinite in each species, in the Ascomycetes the sporangia (asci) have a definite size, form, and number of spores. The spores of the Ascomycetes are known as ascospores.
The sporangio-fructification is found under three main forms.
1. Free Sporangiophores which are either single (Mucor, Fig. 78), or branched (Thamnidium).
2. Sporangial-layers. These are produced by a number of sessile or shortly-stalked sporangia, being formed close together like a palisade (Taphrina, Fig. 105).
3. Sporangiocarps. These consist usually of many sporangia enclosed in a covering, they are found only in the Carpoasci, and are also known as ascocarps. The parts of an ascocarp are the covering (peridium), and the hymenium, which is in contact with the inner wall of the peridium, and is generally made up of asci, and sterile, slender hyphæ. The latter either penetrate between the asci and are branched and multicellular (paraphyses, Figs. 103 d, 123, 125, 129), or clothe those parts of the inner wall which bear no asci (periphyses; among many peronocarpic Ascomycetes, e.g. Chætomium, Sordaria, Stictosphæra hoffmanni). The ascocarps are produced directly from the mycelium, or from a stroma, that is a vegetative body of various forms, in which they may be embedded (Figs. 116 B, C).
Among the conidio-fructifications there are, in the same way, three divisions.
1. Free conidiophores (Fig. 109). The form of the conidiophores, the shape, and number of its spores are various. In the most highly developed Fungi, the Basidiomycetes, there are, however, special more highly developed conidiophores, the basidia, which have a definite form and spores of a definite shape and number. The conidia borne on basidia are called basidiospores.
2. Conidial-layers. (a) The SIMPLEST case of this is found when the conidiophores arise directly from the mycelium, parallel[89] to one another, and form a flat body (e.g. Exobasidium vaccinii, Hypochnus; among the Phycomycetes, Empusa muscæ and Cystopus). (b) In a HIGHER form the conidial-layers are thick, felted threads (stroma) inserted between the mycelium and the hymenium (i.e. the region of the conidiophores). Examples are found in a section of the Pyrenomycetes (Fig. 122). (c) The HIGHEST form has the basidial-layer, that is a conidial-layer with more highly developed conidiophores (basidia). The basidial-layer, with stroma, and the hymenium (region of the basidia), forms the basidio-fructification, which is branched in the Clavariaceæ, and hat-shaped in other Hymenomycetes (in these groups the hymenium is confined to the lower side of the pileus).
The hymenium of the conidial-layer and basidial-layer is composed entirely of conidiophores, or of conidiophores and sterile hyphæ (paraphyses) which are probably always unicellular. Paraphyses are found in Entomophthora radicans, and in certain Basidiomycetes (e.g. Corticium).
3. Conidiocarps (pycnidia). A special covering surrounds the conidia-forming elements. The inner side of this covering (peridium) bears the hymenium, i.e. those elements from which the conidia are abstricted. The conidiocarps arise either immediately from the hyphæ or from a stroma in which they are generally embedded. Conidiocarps are entirely wanting in the Phycomycetes. On the other hand they are found among the Ascomycetes and Basidiomycetes, and in the latter group the conidiocarps contain more highly differentiated conidiophores (basidia) and are known as basidiocarps. Conidiocarps with simple conidiophores, are found only among the Basidiomycetes, in the Uredinaceæ, and in Craterocolla cerasi. In the Ascomycetes (Figs. 120 d, e; 117 a, b; 123 a; 124 b) the conidiocarps are visible, as points, to the naked eye, while the basidiocarps of the Basidiomycetes (Figs. 170, 171, 173–176, 178–180) vary from the size of a pea to that of a child’s head. The “spermogonia” of the Ascomycetes and Lichenes, are conidiocarps with small conidia (microconidia) which germinate sometimes more slowly than other conidia, and formerly were erroneously considered as male reproductive cells, and called spermatia.
The conidia of the Fungi are not primitive structures. The comparison of the sporangia and conidia among the Zygomycetes, and among the species of the genus Peronospora shows, that the conidia are aberrant formations, and that they have arisen through[90] the degeneration of the sporangium, which, by the reduction of its spores to one, has itself become a spore.
In the genera Thamnidium and Chætocladium the gradual diminution of the sporangia, and the reduction of the number of spores can be distinctly followed. In Thamnidium the number of spores is often reduced to one, which is free in the sporangium. In Chætocladium however the sporangia are typically one-spored, the spore is always united with the sporangium, and the two become a single body, the so-called conidium, which is in reality a closed sporangium. How close is the connection between the sporangia and conidia of Thamnidium and Chætocladium, is seen from the fact that, in the conidial stage of Chætocladium the same whorl-form of branching appears as in the sporangial stages of Thamnidium chætocladioides, and also, that the conidia of Ch. fresenianum throw off the former sporangium-wall (exosporium), while Ch. jonesii germinates without shedding its exosporium. The Phycomycetes have doubtless sprung from Water-Algæ and inherit the sporangia from them. On this supposition, as the Phycomycetes assumed a terrestrial mode of life, the sporangia would become adapted to the distribution of the spores by means of the air, the sporangia would become small, contain dust-like spores, and would eventually become closed-sporangia, i.e. conidia. The conidia are a terrestrial method for the multiplication of Fungi. In the Hemiasci and the Ascomycetes the sporangia are still preserved, but in every instance they are adapted to terrestrial spore-distribution, their spores being set free on the destruction of the sporangium-wall (generally shot out) and distributed through the air. For further examples of spore-distribution see below, p. 91–93.
The reproduction of Fungi is accomplished not only by spores and conidia, but also sometimes by chlamydospores. These are fundaments[11] of sporangiophores and conidiophores, which have taken on a resting condition in the form of a spore, and are able to germinate and produce carpophores. In the formation of the chlamydospores the hyphæ accumulate reserve materials at the expense of the neighbouring cells; in the undivided hyphæ of the Phycomycetes transverse walls are formed, and finally the chlamydospores are set free by the decay of the empty cells connecting them with the mycelium. One must distinguish between oidia and true chlamydospores. The former are more simple, the latter are a somewhat more differentiated form of carpophore fundaments, which serve for propagation in the same manner as spores. In Chlamydomucor racemosus the chlamydospores grow out into the air and form differentiated carpophores. In the Autobasidiomycetes they only germinate vegetatively, and not with the formation of fructifications. From Chlamydomucor up to the Autobasidiomycetes the successive development of the fructification,[91] which is interrupted by the formation of the chlamydospores, degenerates more and more. Among certain Ustilagineæ the chlamydospores (brand-spores) no longer germinate with the production of fructifications. In the Uredinaceæ, only one of the three chlamydospore-forms has the property of producing fructifications on germination; the other forms only germinate vegetatively, like ordinary spores, and in the same manner as the chlamydospores of the Autobasidiomycetes. In the Hemibasidii, and the Uredinaceæ, in Protomyces, the chlamydospores are the chief means of reproduction. They are found also among the Ascomycetes.
The sporangia and the conidia of the Fungi have their common origin in the sporangia of the Phycomycetes. The asci (and the Ascomycetes which are characterised by these bodies) are descended from the sporangia-forming, lower Fungi; the basidia (and the Basidiomycetes) from those which bear conidia. The sporangia of the Phycomycetes are the primitive form and the starting point for all the reproductive forms of the Fungi. The chlamydospores appear besides in all classes of Fungi as supplementary forms of reproduction, and are of no importance in determining relationships. Although the expression “fruit” must essentially be applied to true Phanerogams, yet, through usage, the term “fruit-forms,” is employed to designate the forms or means of reproduction of Fungi, and the organs of reproduction are known as organs of fructification, the sporangiophores and conidiophores as fruit-bearers (carpophores), and the sporangiocarps, conidiocarps, and basidiocarps as “fruit-bodies.”
The majority of Fungi have more than one method of reproduction, often on various hosts (Uredinaceæ). Species with one, two, or more than two methods of reproduction are spoken of as having monomorphic, dimorphic, or pleomorphic fructification. Monomorphic, e.g. the Tuberaceæ; dimorphic, Mucor, Piptocephalis, Saprolegniaceæ, Penicillium crustaceum; pleomorphic, Puccinia graminis, Capnodium salicinum (in the last species there are five methods of reproduction: yeast-like conidia, free conidiophores, conidiocarps with small and large conidia, and ascocarps).
The liberation and distribution of the spores and conidia. The spores and conidia, on account of their small size and lightness, are spread far and wide by currents in the air, but in addition to this method, insects and other animals frequently assist in disseminating them. The liberation of the conidia is occasionally effected by the complete shrinking away of the conidiophore, but more frequently by abstriction from the conidiophores, either by their gradually tapering to a point, or by the[92] dissolution of a cross-wall (generally of a mucilaginous nature). The individual links of conidia-chains are detached from one another in the same way, or often by means of small, intercalary cells, which are formed at the base of the individual links, and becoming slimy, dissolve upon the maturity of the spores. Special contrivances for ejecting the spores and conidia may often be found. In Peronospora the cylindrical fruit-hyphæ in the dry condition become strap-shaped and also twisted. These are very hygroscopic, and the changes of form take place so suddenly, that the spores are violently detached and shot away. In Empusa a peculiar squirting mechanism may be found (Fig. 85). Each club-shaped hypha which projects from the body of the fly, bears a conidium at its apex; a vacuole, which grows gradually larger, is formed in the slimy contents of the hypha, and the pressure thereby eventually becomes so great that the hypha bursts at its apex, and the conidium is shot into the air. By a similar mechanism, the spores of many of the Agaricaceæ are cast away from the parent-plants. In the case of Pilobolus (Fig. 84) the entire sporangium is thrown for some distance into the air by a similar contrivance, the basal region of the sporangium having, by the absorption of water, been transformed into a slimy layer which is readily detached. Sphærobolus, a Gasteromycete, has a small, spherical fruit-body (basidiocarp), the covering of which, when ripe, suddenly bursts, and the basidiospores contained in it are forcibly ejected.
The spores which are enclosed in asci are, in some instances, set free from the mother-cell (ascus) prior to their complete development (Elaphomyces, Eurotium). In the case of the majority of the Pyrenomycetes and Truffles, the asci swell by the absorption of water into a slimy mass, which gradually disappears, so that the spores lie free in the fruit-body; they either remain there till the fruit-body decays, as in those which have no aperture (Perisporiaceæ, Tuberaceæ), or the slimy mass, by its growth, is forced out through the aperture of the sporocarp, taking the spores with it (Nectria). The ejection of the spores by mechanical means takes place in a number of Ascomycetes, and should many spores be simultaneously ejected, a dust-cloud may be seen with the naked eye to arise in the air from the fruit-body. This is the case in the larger species of Peziza, Helvella, Rhytisma, when suddenly exposed to a damp current of air. A distinction is drawn between a simultaneous ejection of all the spores contained in the ascus, and an ejection at[93] intervals (successive), when only one spore at a time is thrown out. The first of these methods is the most frequent, and is brought about by the ascus being lined with a layer of protoplasm, which absorbs water to such a degree that the elastic walls are extended at times to double their original size. The spores are forced up against the free end of the ascus, a circular rupture is made at this point, and the elastic walls contract, so that the fluid with the spores is ejected. Special means may in some instances be found to keep the spores together, and compel their simultaneous ejection. Thus, a tough slime may surround all the spores (Saccobolus), or a chain-apparatus, similarly formed of tough slime; or there may be a hooked appendage from each end of the spores which hooks into the appendage of the next spore (Sordaria). The paraphyses occurring between the asci in many Ascomycetes, also play a part in the distribution of the spores, by reason of the pressure they exercise. The asci in some of the Pyrenomycetes, which are provided with jar-shaped fruit-bodies, elongate to such an extent that, without becoming detached from their bases, they reach the mouth of the fruit-body one at a time, burst and disperse their spores, and so make room for those succeeding. An ejection of the spores at intervals from the ascus is rarer. It takes place, for instance, in Pleospora, whose asci have a double wall. The external wall, by absorption of water, at last becomes ruptured, and the internal and more elastic membrane forces itself out in the course of a few seconds to one of two or three times greater length and thickness, so that one spore after another is forcibly ejected from a narrow aperture at the end of the ascus.
Germination of spores (conidia and chlamydospores). In many spores may be found one or more germ-pores, i.e. thinner places, either in the inner membrane (uredospores, Sordaria) or in the external membrane (teleutospores in Rust-Fungi), through which the germination takes place. Generally this does not occur till the spores have been set free: in some Ascomycetes germination commences inside the ascus (Taphrina, Sclerotinia). The different ways in which the spores germinate may be classified into three groups.
I. The ordinary germination occurs by the spore emitting a germ-tube, which immediately developes into a mycelium. In spores with a double wall it is only the inner membrane which forms the germ-tube. In swarmspores a single wall is formed after the withdrawal of the cilia, and this, by direct elongation,[94] becomes the germ-tube. The protoplasm accumulated in the spore enters the hypha, which, in pure water, can only grow as long as the reserve nourishment lasts.
2. Germination with promycelium differs only by the circumstance that the hypha developed from the germ-tube has a very limited growth, and hence it does not immediately develope into a mycelium, but produces conidia (Rust-and Brand-Fungi). This promycelium must only be regarded as an advanced development of a conidiophore or basidium.
3. The yeast-formation of conidia consists in the production of outgrowths, very much constricted at their bases, from one or more places. Each of the conidia formed in this manner may again germinate in the same way. When sufficient nourishment is present, a branched chain of such conidia is formed, and these are finally detached from one another. Yeast-like buddings from the conidia are produced in various Fungi, e.g. Ascoidea, Protomyces, Ustilagineæ, Ascomycetes, Tremellaceæ, etc. In the Ustilagineæ these conidia are an important element in the development. The budding conidia of Exobasidium forms a “mould” on the nutritive solution. The yeast-like conidia are not to be confounded with the “Mucor-yeast” (comp. Mucoraceæ). For Saccharomyces see Appendix to the Fungi, page 176.
In a compound spore (i.e. when a mass of spores are associated together) each spore germinates on its own account. There are sometimes, however, certain among them which do not germinate, but yield their contents to those which do.
The length of time for which conidia can retain their power of germination is shortest (being only a few weeks) in those having thin walls and containing a large supply of water (Peronosporaceæ, Uredinaceæ). In many spores a resting period is absolutely necessary before they are able to germinate (resting spores). It has been observed in some spores and conidia, that the faculty of germinating may be preserved for several years if the conditions necessary for germination remain absent (Ustilagineæ, Eurotium, Penicillium).
The optimum, minimum and maximum temperatures required for the germination of the spores has been decided in the case of a good many Fungi. A large portion of the most common Fungi have their optimum at 20°C., minimum at 1–2°C, maximum at 40°C. In the case of pathogenic Fungi the optimum is adapted to the temperature of the blood. Fungi living in manure, whose[95] spores are often adapted to germinate in the alimentary canals of warm-blooded animals, have an optimum corresponding to the temperature of these animals, but with a little margin.
Systematic Division.—The lowest class of the Fungi is that of the Phycomycetes, which have an unicellular mycelium, sexual and asexual reproduction, and have doubtless sprung from sporangia-bearing, lower Green Algæ. From the Phycomycetes (and certainly from the Zygomycetes) spring two well defined branches, each with numerous distinct species; to the one branch belong the Hemiasci and the Ascomycetes, to the other the Hemibasidii and the BASIDIOMYCETES. Ascomycetes and Basidiomycetes may be united under the title of Mycomycetes or Higher Fungi. The Hemiasci and the Hemibasidii constitute the class of Mesomycetes. The Hemiasci are an intermediate form between Zygomycetes and Ascomycetes; the Hemibasidii a similar group between the Zygomycetes and Basidiomycetes. Mesomycetes and Mycomycetes have only asexual reproduction; sexual reproduction is wanting. Their mycelium is multicellular.
Up to the present time about 39,000 species have been described.
Review of the divisions of the Fungi:—
Class I.—Phycomycetes (Algal-Fungi).
Class II. Mesomycetes.
Class III.—Mycomycetes (Higher Fungi).
This group resembles Vaucheria and the other Siphoneæ among the Algæ.
Organs of Nutrition. The mycelium is formed of a single cell, often thread-like and abundantly branched (Fig. 78). Vegetative propagation by chlamydospores and oidia. Asexual reproduction by endospores (sometimes swarmspores) and conidia. Sexual reproduction by conjugation of two hyphæ as in the Conjugatæ, or by fertilisation of an egg-cell in an oogonium. On this account the class of the Phycomycetes is divided into two sub-classes: Zygomycetes and Oomycetes.
Sexual reproduction takes place by zygospores, which function as resting-spores, and arise in consequence of conjugation (Fig. 81); in the majority of species these are rarely found, and only under special conditions. The most common method of reproduction is by endospores, by acrogenous conidia, by chlamydospores, or by oidia. Swarmspores are wanting. Parasites and saprophytes (order 6 and 7). The zygospores are generally produced when the formation of sporangia has ceased; e.g. by the suppression of the sporangial-hyphæ (Mucor mucedo), or by the diminution of oxygen; Pilobolus crystallinus forms zygospores, when the sporangia are infected with saprophytic Piptocephalis or Pleotrachelus.
A. Asexual reproduction only by sporangia.
Order 1. Mucoraceæ. The spherical sporangia contain many spores. The zygospore is formed between two unicellular branches (gametes).
[97]
The unicellular mycelium (Fig. 78) of the Mucoraceæ branches abundantly, and lives, generally, as a saprophyte on all sorts of dead organic remains. Some of these Fungi are known to be capable of producing alcoholic fermentation, in common with the Saccharomyces. This applies especially to Chlamydomucor racemosus (Mucor racemosus), when grown in a saccharine solution, and deprived of oxygen; the mycelium, under such conditions, becomes divided by transverse walls into a large number of small cells. Many of these swell out into spherical or club-shaped cells, and when detached from one another become chlamydospores, which abstrict new cells of similar nature (Fig. 79). These chlamydospores were formerly erroneously termed “mucor-yeast,” but they must not be confounded with the yeast-conidia (page 94). They are shortened hyphæ, and are not conidia of definite size, shape, and point of budding. Oidia are also found in Chlamydomucor.
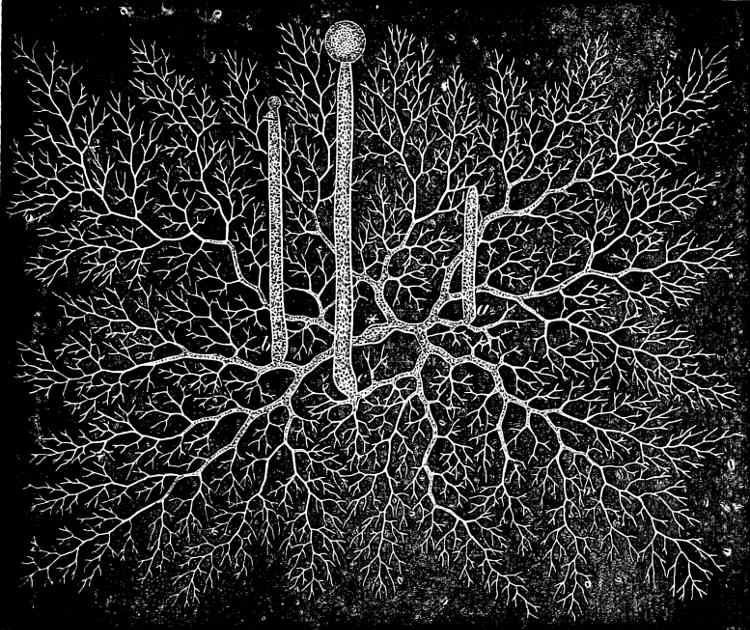
Fig. 78.—Mucor mucedo. A mycelium which has sprung from one spore, whose position is marked by the *: a, b, c are three sporangia in different stages of development; a is the youngest one, as yet only a short, thick, erect branch; b is commencing to form a sporangium which is larger in c, but not yet separated from its stalk.
[98]
The Mucoraceæ, in addition to the chlamydospores and oidia, have a more normal and ordinary method of reproduction; viz., by spores which are formed without any sexual act. Mucor has round sporangia; from the mycelium one or more long branches, sometimes several centimetres in length, grow vertically into the air; the apex swells (Figs. 78, 80) into a sphere which soon becomes separated from its stalk by a transverse wall; in the interior of this sphere (sporangium) a number of spores are formed which eventually are set free by the rupture of the wall. The transverse wall protrudes into the sporangium and forms the well-known columella (Fig. 80 d, e). The formation of spores takes place in various ways among the different genera.
Fig. 79.—Chlamydospores of Chlamydomucor racemosus (× 375 times.)
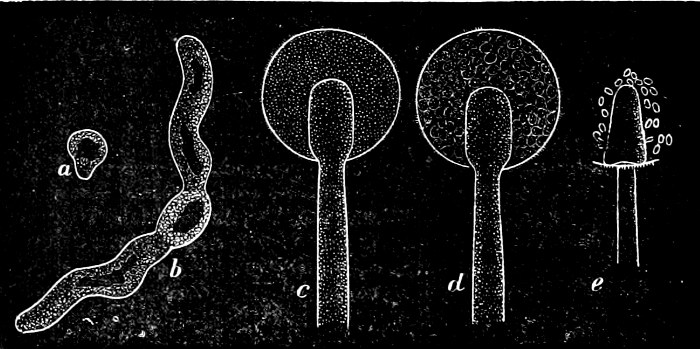
Fig. 80.—Mucor mucedo: a a spore commencing to germinate (× 300 times); b a germinating spore which has formed a germ-tube from each end (× 300 times); c the apex of a young sporangium before the formation of spores has commenced; the stalk is protruded in the sporangium in the form of a column: on the wall of the sporangium is found a very fine incrustation of lime in the form of thorn-like projections; d a sporangium in which the formation of spores has commenced; e a sporangium, the wall of which is ruptured, leaving a remnant attached to the base of the columella as a small collar. A few spores are seen still adhering to the columella.
Sexual Reproduction by conjugation takes place in the following manner. The ends of two hyphæ meet (Fig. 81) and become more or less club-shaped; the ends of each of these are cut off by a cell-wall, and two new small cells (Fig. 81 A) are thus formed, these coalesce and give rise to a new cell which becomes the very thick-walled zygote (zygospore), and germinates after[99] period of rest, producing a new hypha, which bears a sporangium (Fig. 81 E).
Mucor mucedo, Pin-mould, resembles somewhat in appearance Penicillium crustaceum and is found growing upon various organic materials (bread, jam, dung, etc.).
Pilobolus (Figs. 83, 84) grows on manure. Its sporangium (Fig. 84 a″) is formed during the night and by a peculiar mechanism (page 92) is shot away from the plant in the course of the day. This generally takes place in the summer, between eight and ten a.m. The sporangium is shot away to a height which may be 300 times greater than that of the plant itself, and by its stickiness it becomes attached to portions of plants, etc., which are in the vicinity. If these are eaten by animals, the spores pass into the alimentary canal and are later on, sometimes even in a germinating condition, passed out with the excrement, in which they form new mycelia.
Phycomyces nitens (“Oil-mould”) is the largest of the Mould Fungi; its sporangiophores may attain the height of 10–30 c.m.
Order 2. Rhizopaceæ. Rhizopus nigricans (Mucor stolonifer) which lives on decaying fruits containing sugar, on bread, etc., has, at the base of the sporangiophores, tufts of rhizoids, i.e. hyphæ, which function as organs of attachment. From these, “runners” are produced which in a similar manner develope sporangiophores and rhizoids.
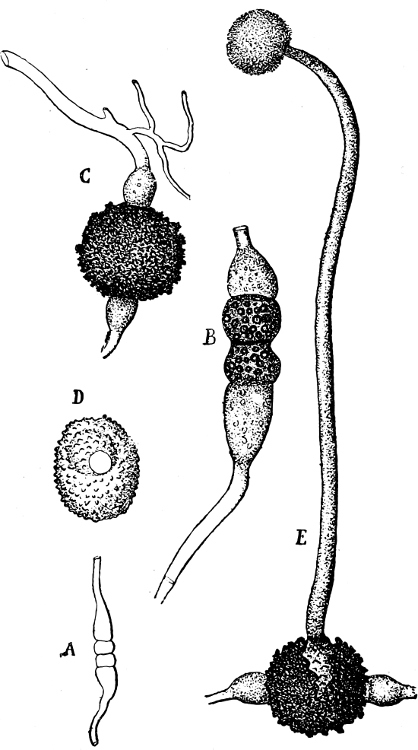
Figs. 81, 82.—Mucor mucedo: A-C stages in the formation of the zygote; D zygote; E germination of zygote: the exospore has burst, and the endospore grown into a hypha bearing a sporangium.
Order 3. Thamnidiaceæ. On the same sporangiophore, in addition to a[100] large, terminal, many-spored sporangium, many smaller, lateral sporangia are formed with a few spores. Thamnidium.
B. Asexual reproduction by sporangia and conidia.
Order 4. Choanephoraceæ. Choanephora with creeping endophytic mycelium, and perpendicular sporangiophores.
Order 5. Mortierellaceæ. Mortierella polycephala produces on the same mycelium conidia and sporangiophores. M. rostafinskii has a long stalked sporangiophore, which is surrounded at its base by a covering of numerous felted hyphæ.
Fig. 83.—Pilobolus. Mycelium (a, a), with a sporangiophore (A) and the fundament of another (B).
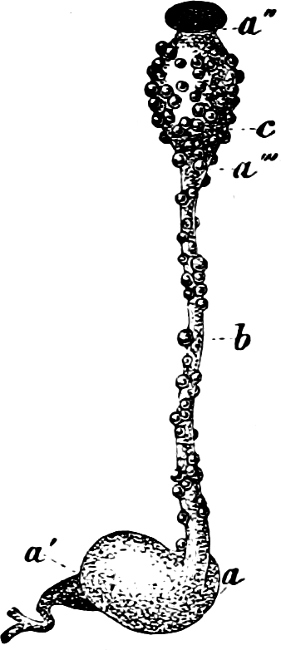
Fig. 84.—Pilobolus. Sporangium (a″) with stalk (a-c), which is covered by many small drops of water pressed out by turgescence.
C. Asexual reproduction only by conidia.
Order 6. Chætocladiaceæ. The conidia are abstricted singly and acrogenously. Chætocladium is a parasite on the larger Mucoraceæ.
Order 7. Piptocephalidaceæ. The conidia are formed acrogenously and in a series, by transverse divisions. The zygospore arises at the summit of the conjugating hyphæ, which are curved so as to resemble a pair of tongs. Piptocephalis and Syncephalis live parasitically on the larger Mucoraceæ.
Sexual reproduction is oogamous with the formation of brown, thick-walled oospores which germinate after a period of rest. Asexual reproduction by conidia and swarmspores. Parasites, seldom saprophytes.
The oospores are large spores which are formed from the egg-cell (oosphere) of the oogonium (oosporangium, Fig. 89, 95). A branch of the mycelium attaches itself to the oogonium and forms at its apex the so-called “antheridium” (pollinodium[13]): this sends one or more slender prolongations (fertilising tubes) through the wall of the oogonium to the egg-cell.
[101]
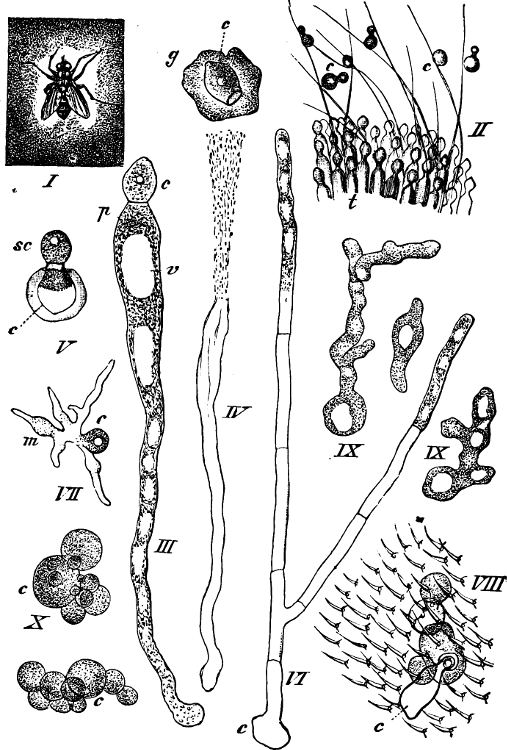
Fig. 85.—Empusa muscæ (Fly-mould). I. A fly killed by the fungus, surrounded by a white layer of conidia. II. The conidiophores (t) projecting from the body of the fly. Some of the conidia, a few of which have developed secondary conidia, are attached to the hairs (mag. 80 times). III. A perfect hypha. IV. A hypha in the act of ejecting a conidium (c), enveloped in a sticky slime (g). V. A conidium which has developed a secondary conidium (sc). VI. A branched hypha produced by cultivation. VII. A secondary conidium which has produced a small mycelium (m). VIII. A conidium germinating on the fly’s body. IX. Mycelium. X. Conidia germinating like yeast in the fatty tissue of the fly. (III.-VII. and IX. magnified 300 times; VIII. and X. magnified 500 times.)
A fertilisation, a passage of the contents of the antheridium to the egg-cell, has as yet only been observed in Pythium; in Phytophthora only one small mass of protoplasm passes through the fertilising tube to the egg-cell; in Peronospora[102] and the Saprolegniaceæ no protoplasm can be observed to pass through the fertilising tube, so that in these instances parthenogenesis takes place; Saprolegnia thuretii, etc., have generally even no antheridia, but nevertheless form normal oospores. Fertilisation of the egg-cell by means of self-motile spermatozoids is only found in Monoblepharis sphærica.
A. Asexual reproduction by conidia only.
The mycelium is richly branched. The family is a transitional step to the conidia-bearing Zygomycetes, since the oospores of many members of this family arise, and are formed, like zygospores.
Order 1. Entomophthoraceæ. Mycelium abundantly developed. This most frequently lives parasitically in living insects, causing their death. The conidiophores forming the conidial-layer project from the skin, and abstrict a proportionately large conidium which is ejected with considerable force, and by this means transferred to other insects. These become infected by the entrance of the germ-tube into their bodies. The spherical, brown resting-spores develope inside the bodies of insects and germinate by emitting a germ-tube.
Genera: Empusa has a good many species which are parasitic on flies, moths, grasshoppers, plant-lice. The conidia emit a germ-tube which pierces the skin of the insect; a number of secondary conidia are then produced inside its body, by division or by gemmation similar to that taking place in yeast, each of which grows and becomes a long unbranched hypha, and these eventually fill up the body of the animal, causing distension and death. Each of these hyphæ projects through the skin, and abstricts a conidium, which is ejected by a squirting contrivance. The best known species is E. muscæ (Fig. 85), which makes its appearance epidemically towards autumn on the common house-fly, and shows itself by the dead flies which are found on the windows and walls attached by their probosces, distended wings, and legs. They have swollen abdomen, broad white belts of hyphæ between the abdominal rings, and are surrounded by a circle of whitish dust formed by the ejected conidia.—Entomophthora sends out, at definite places, from the mycelium hidden in the insect’s body, bundles of hyphæ, which serve the purpose of holding fast the dead insects, the ramifications attaching themselves to the substratum: the conidiophores are branched, the conidia are ejected by the divisional walls between the hyphæ and the conidia dividing into two layers, those which terminate the hyphæ suddenly expanding and throwing the conidia into the air. E. radicans makes its appearance epidemically on caterpillars.
B. Asexual reproduction by zoospores or conidia.
In this family the mycelium is very sparsely developed or is wanting. The entire plant consists principally or entirely of a[103] single zoosporangium whose zoospores have generally one cilium. The resting-spores arise either directly from the zoosporangium, which, instead of forming zoospores, surrounds itself by a thick cell-wall; or they are formed by the conjugation of two cells (in which case they are spoken of as oospores). Microscopic Fungi, parasitic on water plants (especially Algæ) or small aquatic animals, seldom on land plants.
Order 1. Olpidiaceæ. Without mycelium. Swarmspores and resting-spores.
In the Olpidieæ, the swarmspores, probably, most frequently form themselves into a plasmodium (naked mass of protoplasm) which may become a single zoosporangium or a resting sporangium. Olpidium trifolii occurs in Trifolium repens.—In the Synchytrieæ the plasmodium emerging from the swarmspores breaks up either at once, or after a period of rest, into smaller plasmodia, each of which will become a zoosporangium. Synchytrium anemones is found on Anemone nemorosa; S. mercurialis on Mercurialis perennis; S. aureum on many plants, particularly Lysimachia nummularia.
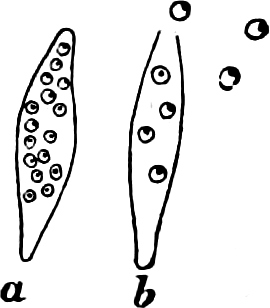
Fig. 86.—Chytridium lagenula. Zoosporangium a before, b after the liberation of the swarmspores.
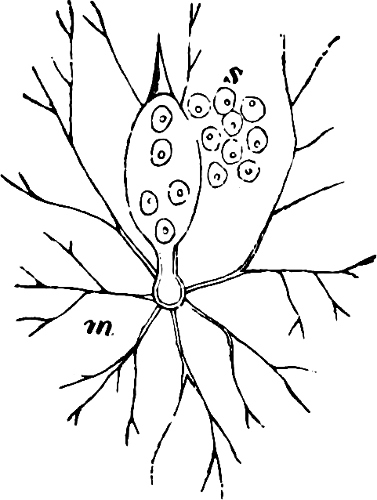
Fig. 87.—Obelidium mucronatum: m mycelium; s swarmspores.
Order 2. Rhizidiaceæ. Mycelium present. Zoospores and resting-spores.
Chytridium (Fig. 86). Obelidium (Fig. 87) is bicellular; the one cell is the mycelium, the other the zoosporangium; found on insects. The species of Cladochytrium are intercellular parasites on marsh plants. Physoderma.
Order 3. Zygochytriaceæ. Mycelium present. Zoospores and oospores. The latter are the product of the conjugation of two cells (Fig. 88).
Polyphagus euglenæ on Euglena viridis. Urophlyctis pulposa on species of Chenopodium.
[104]
The mycelium is bladder-like or branched. Zoospores. Sexual reproduction by oospores, which are produced in oogonia. The latter are fertilised, in some forms, by the antheridium.
Order 1. Ancylistaceæ. The entire bladder-like mycelium is used for the construction of zoosporangia, oogonia, or antheridia. Lagenedium is parasitic on Spirogyra, etc.
Order 2. Peronosporaceæ. Almost entirely parasites. The unicellular, often very long and abundantly branched mycelium lives in the intercellular spaces of living plants, especially in the green portions, and these are more or less destroyed and deformed in consequence. Special small branches (suction-organs, “haustoria”) are pushed into the cells in order to abstract nourishment from them. Both oospores and conidia germinate either immediately, or they develope into sporangia with swarmspores, having always two cilia. Only one oospore is formed in each oogonium; its contents (Fig. 89) divide into a centrally placed egg-cell and the “periplasm” surrounding it; this is of a paler colour and on the maturity of the oospore forms its thick, brown, external covering.
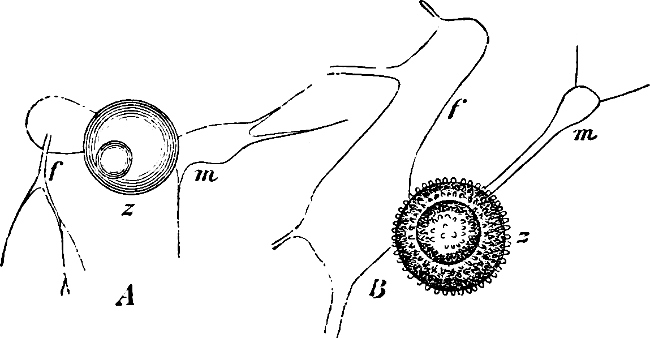
Fig. 88.—Polyphagus euglenæ. A with smooth, B with thorny oospores; m and f the two conjugating cells.
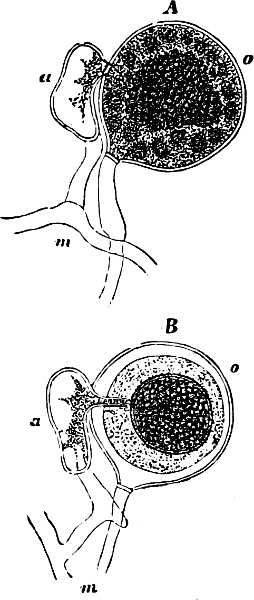
Fig. 89.—Peronospora alsinearum. Mycelium with egg-cell and antheridium.
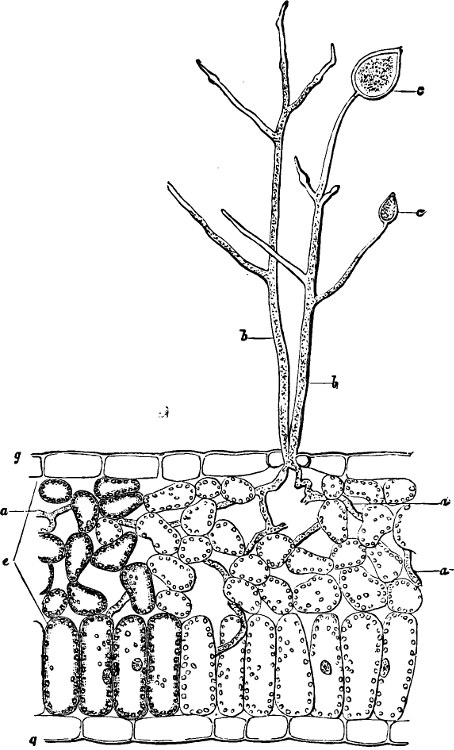
Fig. 90.—Phytophthora infestans (strongly magnified). Cross section through a small portion of a Potato-leaf (the under side turned upwards): a the mycelium; b b two conidiophores projecting through a stoma; c conidia; e the spongy tissue of the leaf; g the epidermis.
The Potato-fungus (Phytophthora infestans) is of great interest. Its thallus winters in the Potato-tuber; other organs for passing the winter, such as oospores, are not known. When the tuber germinates, the Fungus-hyphæ penetrate the young shoot and keep pace with the aerial growth and development of the plant. The conidiophores emerge through the stomata, especially on the under side of the leaves; they branch like a tree (Fig. 90), and[105] appear to the naked eye as a fine mould on the surface of the plant. The disease soon makes itself known by the brown colouring of those parts of the plant which are attacked, and by their withering. An ovoid conidium arises at first by the formation of a dividing wall at the apex of each branch of the conidiophore[106] (Fig. 90 c c), and immediately underneath it another is formed, which pushes the first to one side, and so on. These conidia sometimes germinate directly, and form a mycelium, but most frequently their protoplasm divides into many small masses, each of which becomes a pear-shaped zoospore provided with two cilia (Fig. 91). Water is required for their germination, and when the ripe conidia are placed in a drop of water the swarm-cells are formed in the course of about five hours. They swarm about in rain and dewdrops in the Potato-fields, and are carried with the water to the Potato-plants and to the tubers in the soil. The wind also very easily conveys the conidia to healthy Potato-fields and infects them. The enormous quantity of conidia and swarm-cells that may be formed in the course of a summer explains the rapid spreading of the disease; and the preceding makes it clear why wet summers are favourable to its existence. When the swarm-cells germinate, they round off, and then surround themselves with a cell-wall which grows out into the germ-tube, and pierces through the epidermis of the host-plant (Fig. 92). Having entered the host, a new mycelium is formed. The potato disease, since 1845, has been rampant in Europe; it has, no doubt, been introduced from America, which, it must be remembered, is the home of the Potato-plant.
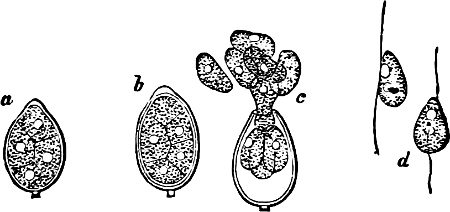
Fig. 91.—Phytophthora infestans: a-c conidia detached; in c the swarm-cells are leaving the mother-cell; d two free-swimming swarm-cells.
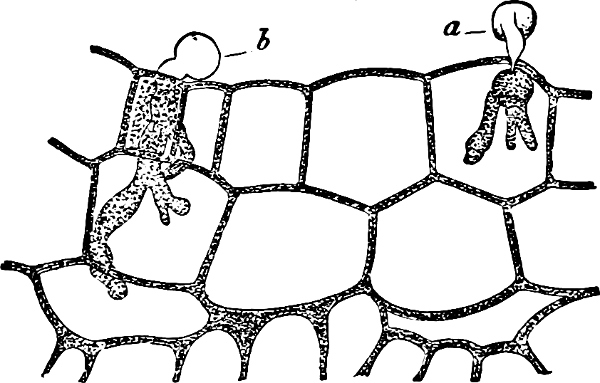
Fig. 92.—Phytophthora infestans. Cross section through a portion of a Potato-stalk. Two germinating conidia (a, b) piercing the epidermis, and the mycelium penetrating the cells.
The conidia exhibit various characters which are employed for the separation of the genera. Pythium is the most simple form. The contents of the terminally-formed conidia emerge as a spherical mass and divide into swarmspores. P. de Baryanum lives in the seedlings of many different Flowering-plants, which it completely destroys.—Phytophthora is distinguished by the circumstance that the sparsely-branched conidiophores bear, sympodially, chains of[107] conidia. Besides the Potato-fungus (see above), Ph. fagi belongs to this group; it developes oospores very abundantly, and does great harm to seedlings of the Beech, Sycamore, and Pine trees.—Peronospora generally has conidiophores which are repeatedly forked, and bear a conidium on each of the most extreme ramifications. Many do great harm to their host-plants. P. viticola, on Vines, and P. nivea, on umbelliferous plants, have swarmspores, which are absent in the following species of this genus: P. sparsa, on Roses; P. gangliformis, on composites; P. alsinearum, on Stitchwort; P. parasitica, on cruciferous plants; P. viciæ, on Vetches and Peas; P. schachtii, on Beets; P. violacea, on the flowers of Scabiosa; P. radii, on the ray-florets of Matricaria.—Cystopus (Albugo) has the conidia developed in chains, which form a cohesive white layer underneath the epidermis of the host-plant. Cystopus candidus, on cruciferous plants, especially Shepherd’s Purse and Brassica; the germination commences on the cotyledons, and from this point the mycelium developes together with the host-plant; C. cubicus, on the leaves of Compositæ.

Fig. 93.—A fly overgrown with Saprolegnia.
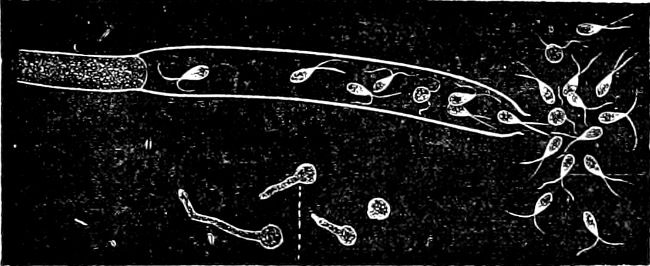
Fig. 94.—Formation of swarmspores in a Saprolegnia: a germinating swarmspores.
Order 3. Saprolegniaceæ, Water-Fungi which live as saprophytes on organic remains lying in water, for instance, on dead flies (Fig. 93), worms, remains of plants; but they may also make their appearance on living animals, being frequently found, for example, on the young trout in rearing establishments.
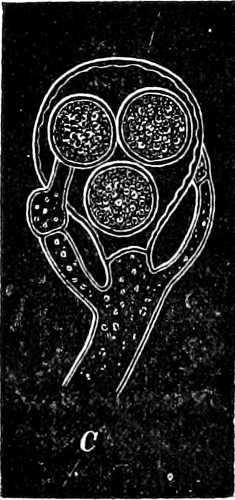
Fig. 95.—Oogonium with two antheridia, Achlya racemosa.
The thallus is a single, long and branched cell. It has one portion which serves as root, and lives in the substratum, where it ramifies abundantly for the purpose of absorbing nourishment; and another portion projecting freely in the water, and sending out hyphæ on all sides (Fig. 93). The asexual reproduction takes place by swarmspores (Fig. 94), which are developed in large sporangia; these swarmspores generally possess two cilia, and on germination grow into new plants. The entire protoplasm[108] in the oogonium is formed into one or more oospheres, without any surrounding “periplasm.” The oospheres may not be fertilised (p. 100), and then develope parthenogenetically.
Genera: Saprolegnia, whose swarmspores disperse immediately after having left the sporangium. S. ferax is the cause of a disease in fish (“Salmon disease”) and in the crayfish.—Achlya, whose swarmspores accumulate in a hollow ball before the mouth of the sporangium.—Leptomitus has strongly indented hyphæ, causing a “linked” appearance. L. lacteus is frequent in the waste matter from sugar factories.—Monoblepharis deviates from the others by the greater development of its fertilising process; the oosphere, situated in an open oogonium, becoming fertilised by self-motile spermatozoids, which are provided with a cilium at the posterior end.
The Mesomycetes are intermediate forms between the Phycomycetes and the Higher Fungi. In the vegetative organs, and in the multicellular hyphæ, they resemble the Higher Fungi; the methods of reproduction, however, show the characters of the Phycomycetes, namely sporangia and conidiophores of varying size and with varying number of spores; definite and typically formed asci and basidia are not present. Sexual reproduction is wanting. The Hemiasci are transitional between the Phycomycetes and the Ascomycetes, the Hemibasidii (Brand-Fungi) form the transition to the Basidiomycetes.
The Hemiasci are Fungi with sporangia which, although resembling asci, yet have not, however, a definite form and a definite number of spores. Besides endospores, conidia, chlamydospores and oidia are found.
Order 1. Ascoideaceæ. Ascoidea rubescens forms irregular, reddish-brown masses in the sap issuing from felled Beeches. It has free sporangia, which resemble asci in their structure, in the development and ejection, and in the definite shape and size of the spores. The formation of the sporangia takes place when the nutriment is nearly exhausted, and resembles that of the conidia, since they are developed from the end of a hypha which enlarges, and the swelling becomes separated by a transverse wall. Within the sporangia numerous spores of a cap-like form are developed, which are set free through an opening at the apex. Sporangia are formed successively at the apex of the same hypha, the second commencing to develope as the first is dehiscing. Conidia and sporangia are not formed simultaneously; the former may be considered as closed sporangia.
Order 2. Protomycetaceæ. Protomyces pachydermus causes hard swellings on the stems and leaf-stalks of the Cichorieæ (Taraxacum, etc.). These swellings[109] consist of chlamydospores (resting-spores), which germinate and become free, ascus-like sporangia, with numerous small spores. In nutritive solutions the chlamydospores form conidia with yeast-like buddings. P. macrosporus on Ægopodium, and other Umbelliferæ.
Order 3. Thelebolaceæ. Thelebolus stercoreus, is found on the dung of deer, hares, and rabbits, and has closed sporangia, which resemble asci in their shape and regular construction, and in the ejection of spores. The covering encloses only one sporangium, even where the sporangia arise close together.
This order, by reason of the covering of the sporangia, forms the transition from the Hemiasci to the Carpoasci, while the two first supply an intermediate step to the Exoasci.
The Brand-Fungi (also known as Ustilagineæ) are Fungi with basidia-like conidiophores, which, however, have not yet advanced to a definite form or number of conidia. They are true parasites, whose mycelium spreads itself in the intercellular spaces of Flowering plants. The mycelium is colourless, quickly perishable, has transverse walls at some distance from each other (Fig. 96), and sends out haustoria into the cells of the host-plant.
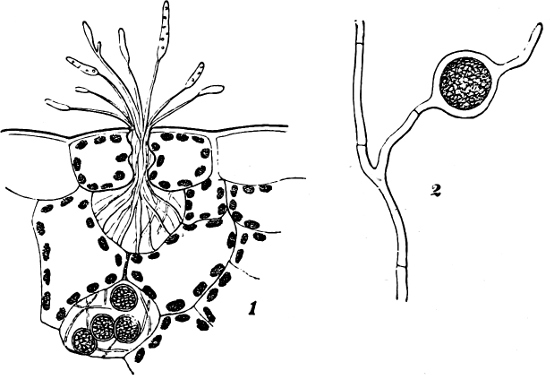
Fig. 96.—Entyloma ranunculi. 1. Cross section of a portion of a leaf of Ficaria permeated by the mycelium; a bundle of hyphæ with conidia emerging from a stoma; in one of the cells are found four brand-spores. 2. A brand-spore developed in the middle of a hypha.
It most frequently happens that the germ-tube enters the host-plant at its most tender age, that is, during the germination of the seed; the mycelium then wanders about in the tissues of the shoot[110] during its growth, until it reaches that part of the plant where the spores are to be formed. The spore-formation takes place in the same way in all those species whose brand-spores are developed in the floral parts of the host-plant. Many Brand-Fungi have, however, a more local occurrence, and the mycelium is restricted to a smaller area of the leaf or stem. Those organs of the host-plant in which the brand-spores are developed often become strongly hypertrophied. In perennial plants the mycelium winters very often in the rhizome.
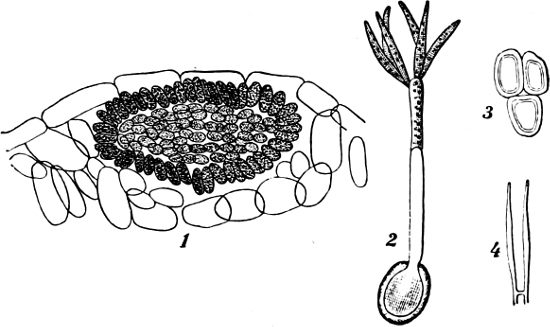
Fig. 97.—Doassansia alismatis. 1. A fruit-body, formed by a covering of oblong hyphæ, which encloses a mass of brand-spores, and is embedded in the leaf-tissue of the host-plant; 20 times natural size. 2. A germinating brand-spore, 500 times natural size. 3. Three connected resting-spores, 400 times natural size. 4. Two conidia grown together, 600 times natural size.
The brand-spores are the winter resting-spores of the Brand-Fungi. They arise in the tissues of the host-plant, which is often destroyed, and become free through the rupture of the epidermis; they are thick-walled, generally brown or violet, and very often possess warts, spines, or reticulate markings. Fruit-bodies, that is enclosed organs of reproduction, are found in few genera (Sphacelotheca, Graphiola; Doassansia, Fig. 97). In Tolyposporium, Tuburcinia, Thecaphora (Fig. 102), etc., the brand-spores are united into a ball of spores. On germination the brand-spores behave as chlamydospores, namely, as the fundament of conidiophores, by emitting a short germ-tube, i.e. a conidiophore (“promycelium”). The Ustilaginaceæ (Fig. 99, 2) have a short transversely divided conidiophore, with laterally developed conidia (comp. the basidia of the Protobasidiomycetes). The conidiophores of the Tilletiaceæ are undivided (unicellular promycelia), and bear the conidia terminally, and so resemble the basidia of the Autobasidiomycetes.
[111]
Fig. 98.—Tuburcinia. 1. T. trientalis. Hyphæ, some of which bear conidia at the apex, forcing themselves out between the epidermal cells on the under side of the leaf; 320 times natural size. 2. T. trientalis. A ball of spores in which some of the individual brand-spores are about to germinate; 520 times natural size. 3. T. primulicola: various forms of conidia (500 times natural size).
In Tilletia, Entyloma, Neovossia, Tuburcinia, the brand-spores germinate and form basidia-like conidiophores with spindle-shaped conidia; their mycelium, on the other hand, produces later only single, sickle-shaped conidia, so that two kinds of conidia are found, as in a few Basidiomycetes. In some species, e.g. Ustilago hordei, the brand-spores only germinate vegetatively and form a mycelium. In nutritive solutions (solutions of dung, etc.) where they live as saprophytes, the brand-spores of many species emit germ-tubes, and on these, yeast-like conidia are produced by repeated budding, which grow into mycelia only when the nutritive solution is exhausted. These conidia have not the power of producing alcoholic fermentation. The very numerous conidia, which are found in the dung of herbivorous animals, are probably the yeast-conidia of Brand-Fungi. The brand-spores, which are eaten by animals with the grain and hay, pass into the dung and without doubt give rise to a very rich multiplication of yeast-conidia.
Fig. 99.—Ustilago. 1. Formation of brand-spores. 2. Germinating brand-spore of U. perennans. 3. Germinating brand-spore of U. cardui (after Brefeld). 4. U. filiformis. a A brand-spore with developed basidium; b another, with a conidium; c with two conidia; d with two conidia placed diametrically opposite to each other; e, detached conidia which are growing into hyphæ.
Fig. 100.—Tilletia tritici: a an ear of Wheat in which all the grains are attacked by Stinkbrand; b a blighted corn surrounded by the chaff; c a blighted corn grown together with a stamen; d the same cut across; e a brand-spore; f, g, h germinating brand-spores; i germinating conidia; j the mycelium; k-k brand-spore-forming mycelium-threads. (c-h magnified 400 times; i-k 300 times.)
[112]
The conidia (also called “sporidia”) of many species unite generally into an H-form (Figs. 97, 4; 100 h; 101, 4). This union in pairs does not, however, take place with a view[113] to germination, there is no fusion of nuclei, and therefore in this “fusion” there is no sexual act.
Order 1. Ustilaginaceæ. Conidiophores with transverse walls and lateral conidia.—Ustilago (Fig. 99) generally developes its spores in the floral organs of its host-plant, the ovary or anthers, where they arise from hyphæ, and form a slimy mass which when mature becomes a black dust.
To this order belong U. avenæ, parasitic on Oats, U. hordei and U. nuda (U. jenseni), on Barley; these are the usual cause of “Smut” on cereals. U. hypodytes on straw of Elymus and Agropyrum. U. filiformis in the leaves of Glyceria. U. caricis transforms the fruits of various species of Carex into black, dusty balls. U. violacea developes its violet spore-powder in the anthers of the Caryophyllaceæ. U. tragopogonis, transforms entire inflorescences of Tragopogon into a black-violet mass. Among the largest are U. grandis, which causes the large swollen nodes in the stem of Phragmites, and the Maize Blight, U. maydis, which produces outgrowths about the size of a hand on the spadix of the Maize.
Order 2. Tilletiaceæ. Conidiophores undivided, generally several conidia arise at their apices.—Tilletia tritici, the Stinkbrand on Wheat (Fig. 100). The mycelium lives in Wheat-plants, producing its spores in the ovary after the whole interior of this body has been destroyed by the mycelium, with the exception of the external layer of the wall of the ovary, which remains essentially unaltered and encloses the closely packed, firm mass of spores (Fig. 100 d). The grains of Wheat thus attacked are shorter and thicker than the sound ones, and the ears show the presence of this Fungus by their erect position, and the wide separation of the chaff (Fig. 100 a). The unpleasant odour of the ovary prior to the ripening of the spores, has given the name “Stinkbrand,” and, in like manner, its hardness when it encloses the ripe spores, is the reason of its being also called “Stonebrand.” On account of this hardness, the diseased grains are readily harvested together with the healthy ones, which become infected by the spores at the threshing. T. lævis (T. fœtens) also occurs on Wheat and has smooth brand-spores.
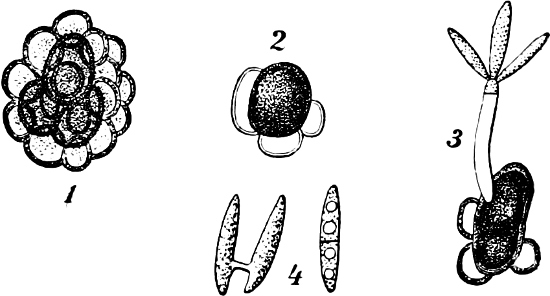
Fig. 101.—Urocystis. 1, U. covalloides. A spore-ball, magnified 450 times. 2–4, U. anemones: 2–3, brand-spores which are about to germinate (magnified 450 times). 4, Conidia, the two in a state of fusion, a third with vacuoles and division-wall, magnified 500 times.
Entyloma (Fig. 96), a genus with numerous species, which appear in spots on the leaves of the host-plant, and Tuburcinia (Fig. 98), which makes its appearance on the Primulaceæ, produce white conidia-spots on the surface of the host-plant. The first-named has single spores, the latter has its spores closely massed together.—Urocystis (Fig. 101) has its spores surrounded by a number of small and lighter coloured barren spores. U. occulta, Rye-stem Blight, forms its spores in long streaks in the stems and leaves of the Rye, and does considerable damage. U. cepulæ[114] on Onions. U. violæ forms large dark-violet swellings in the leaf-stalk and stems of Violets.—Thecaphora (Fig. 102) appears in seedlings of Convolvulus and Astragalus.
As a means of protection against the Smut-Fungi which make their appearance on the different cereals, a submersion of the grains in a solution of blue vitriol (½%) for twelve hours, or better still, submerging for five minutes in water heated to 53–55° C (Jensen’s method) is employed.
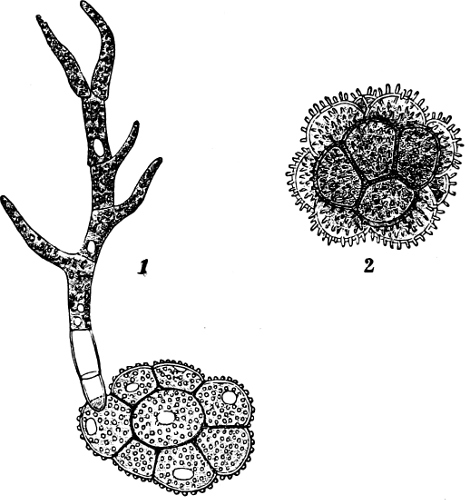
Fig. 102.—Thecaphora. 1, T. convolvuli, a ball of spores, one of the brand-spores has emitted a septate branched conidiophore (× 520). 2, T. affinis, a ball of spores (× 520).
The Mycomycetes are not entirely aquatic in habit; they have hyphæ with transverse walls, but no sexual reproductive organs. The asexual reproduction takes place in very different ways; by endospores (in asci), conidia, basidiospores, chlamydospores, and oidia. Swarmspores are never found.
Two chief methods of reproduction may be distinguished, and hence the class may be divided into two large sub-classes:—the Ascomycetes (with asci), and the Basidiomycetes (with basidia).
The main characteristic which distinguishes the Ascomycetes is the ascus; a name given to a sporangium of a definite shape and size, and containing a definite number of spores. The shape is[115] generally club-like or spherical, the number of spores 8 (in some 2, 4, 16 or more), see Figs. 103, 105, 108, 110, 113, 116, 120, 121, 123, 129.
In the lowest forms, the Exoasci, the ascus springs directly from the mycelium without the formation of a fruit-body (i.e. ascocarp). In the higher forms, which contain many species, the Carpoasci, the asci are united and form ascocarps which may be more or less enclosed (angiocarpic, hemiangiocarpic, and probably gymnocarpic).
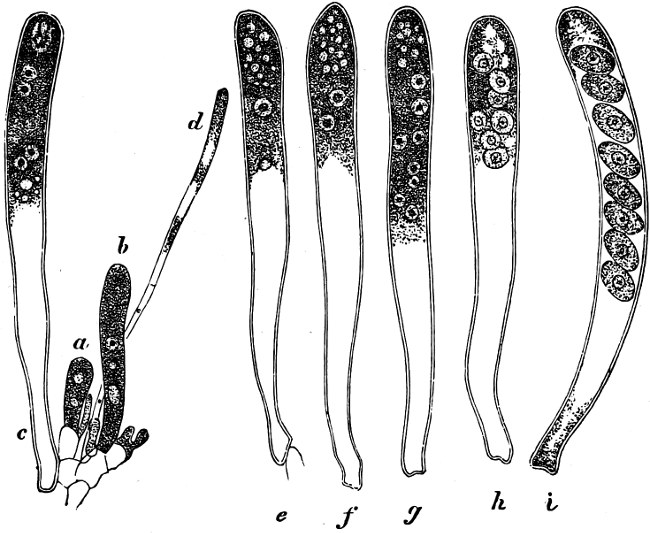
Fig. 103.—Endogenous formation of spores in Peziza confluens. In the youngest asci there is only one nucleus (b, e); this divides into two (f); and the division is repeated so that there are 4 nuclei in c and 8 in g. These surround themselves with protoplasm and a cell-wall (h, i). The protoplasm of the mother-cell is not entirely used up.
The hyphæ of the Mycelium in some remain free, in others they are felted together and form thick strands or flat, cushion-like bodies (compare in particular the stromata of the Pyrenomycetes). Some species form sclerotia (Figs. 116, 128).
Asexual reproduction by means of conidia is known in many species as the principal means of reproduction, and the one which affords the most rapid means of distribution. The conidia may be produced on conidiophores (Fig. 109), in conidial-layers (Fig. 122), and often in conidiocarps (pycnidia, Figs. 120 d, e;[116] 123 a; 124 b.). These last occur partly as the so-called “spermogonia” (that is, pycnidia with microconidia). The conidiophores never approach the basidia.
In many species the ascospores germinate and form conidia immediately (Nectria cinnabarina, Sclerotinia, Taphrina, etc.), sometimes while they are still in the ascus and before their ejection (Taphrina, Fig. 105 a). In many instances the conidia by means of continued budding can, for a longer or shorter time, produce yeast-conidia, e.g. Taphrina. In many other cases the conidia arise from the germ-tubes of the ascospores, or at any part of the mycelium. The unripe asci of Taphrina, when placed in water, develop conidia at their apices. The Sclerotinia-species produce numerous conidia whose germination has never been observed. The formation of conidia and asci sometimes takes place on the same fruit-body. In Heterosphæria patella the conidia and asci are developed successively in the same fruit-body; in the ascocarps of Dermatea frangula and Sclerotinia sclerotiorum the formation of conidia may take place. The ascocarps frequently arise from the conidial-layers (Nectria cinnabarina, etc.). This relationship of the two forms of reproduction to each other may be explained by considering that both have descended phylogenetically from sporangia.
Sometimes chlamydospores and oidia also appear in the Ascomycetes; on germination, however, they do not, as in Protomyces, form sporangia, and on this account cannot be distinctly distinguished from conidia.
The asci are morphologically the highest form of reproduction and are always found at the close of the development of these Fungi; the accessory forms of reproduction are first developed, but a well-defined alternation of generations does not occur.
In the Ascomycetes there are more than 11,000 described species, which can be classed as follows:—
Ascomycetes with FREE ASCI; sometimes also conidia, chlamydospores and oidia. One order.
Order. Taphrinaceæ. Of the genera belonging to this order, Taphrina, Endomyces, and Ascocorticium, the first is most important.
[117]
Endomyces decipiens is a parasite in the fruit-body of Armillaria mellea; E. magnusii lives in the gelatinous, fermenting exudations of Oak-trees; Ascocorticium albidum is found under the bark of the Fir-tree. Endomyces has chlamydospores and oidia.
The species of Taphrina are parasites, whose free asci may be found in great numbers, generally closely pressed together, on the parts of plants which they have attacked. The asci are developed directly from the ascogenous cells of a fertile, generally sub-cuticular, hypha, which arises from the sterile mycelium. The latter arises from the germinating ascospore, and may hibernate in the tissues of its host, particularly in the winter buds, and then with the commencement of the next period of vegetation it continues its growth side by side with that of its host. The hyphæ ramify in the intercellular spaces or beneath the cuticle, but have no haustoria. The ascospores (Fig. 105 A) and unripe asci may produce conidia.
Fig. 104.—Taphrina (Exoascus) pruni. Yeast-like budding of a germinating spore (× 600).
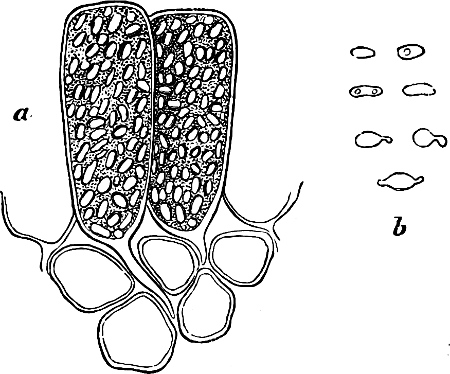
Fig. 105.—Taphrina betulina: a ascus filled with conidia; b germinating spores (× 600).
Very remarkable appearances, and swellings of the attacked tissues, are produced when the mycelium is perennial; for example, the “Witches’-brooms” and “Pockets.” The hard, hollow, stoneless plums, known as “Pocket” or “Bladder” Plums, are produced by considerable changes in the tissues of the fruit; these are caused particularly by T. pruni on several species of Prunus. The “Witches’-brooms,” on the contrary, are deformations of entire twigs or branches, and often attain a very large size. They occur on Alnus incana, caused by T. epiphylla; on Carpinus betulus, by T. carpini; on Cherry-trees, by T. cerasi; on Plum-trees, by T. insititiæ; on Birches, by T. turgida and T. betulina. T. deformans attacks the leaves of the Peach, and causes them to curl.
[118]
When a perennial mycelium is wanting, the infection is confined as a rule to white or yellow spots on the leaves, e.g. the commonest, T. sadebeckii, on Alnus glutinosa, and T. aurea on species of Populus. T. alni incanæ (Fig. 106) causes considerable hypertrophies on the pistillate catkins of the Alder, which may be compared to the “pockets” of Prunus.
Fig. 106.—Taphrina alni incanæ on the Alder (nat. size).
The Carpoasci are Ascomycetes, whose asci are enclosed in fruit-bodies, i.e. ascocarps. The accessory means of reproduction are free conidiophores (Fig. 109), conidial-layers (Fig. 122), conidiocarps (Fig. 120 D, E, etc.), chlamydospores and oidia.
For the different methods of distributing the ascospores, see p. 92.
Of the six families of the Carpoasci, the first three—Gymnoascales, Perisporiales, and Pyrenomycetes—are ANGIOCARPIC (that is, the ascocarp remains closed throughout its existence, and does not dehisce when ripe); the fourth and fifth families (Hysteriales and Discomycetes), on the other hand, are HEMIANGIOCARPIC (the ascocarp, here also called an apothecium, is closed in the early stages, but opens at the commencement of ripening and exposes a hymenium of crowded asci); the family of Helvellales has probably GYMNOCARPIC (or hemiangiocarpic) fruit-bodies.
The ascocarps are surrounded by a spongy and incomplete envelope. One order, poor in species.
[119]
Order Gymnoascaceæ.—The ascocarps are borne sometimes solitarily, or sometimes coiled together. Gymnoascus reessii forms small bodies about 1 mm. in diameter on old horse-dung, which at first are white and afterwards orange-red.—Ctenomyces serratus lives on the old feathers in birds’ nests.
The ascocarps are surrounded by a complete envelope without any opening: the fruit-bodies are cleistocarpic; the spores are only liberated after the disintegration of the fruit-bodies. Paraphyses are wanting. The two first orders have in addition the means of reproduction by conidia.
Order 1. Erysiphaceæ, Mildews. The Fungi belonging to this order are epiphytic parasites, whose mycelium, somewhat resembling a cobweb, may be seen on the leaves and other green portions of plants (see Figs. 107, 108). The hyphæ ramify in all directions upon the surface of their host, and emit haustoria which penetrate the epidermal cells, and thus derive the necessary nutriment. The Mildew-Fungi thus belong to the obligate parasites, and during their growth dwarf and destroy the portions of their host on which they live. The reproduction takes place in the first instance by abstriction of conidio-chains from the end of special branches (Fig. 108 c, a hypha is seen in the act of detaching a conidium). The conidia may germinate immediately, and thus quickly reproduce their species. When present in large numbers they appear as a white meal covering the surface of the plant on which the fungus is found. Later on appear the dark brown, spheroid ascocarps (Fig. 108 a) which, although small, are generally just visible to the naked eye as black specks.
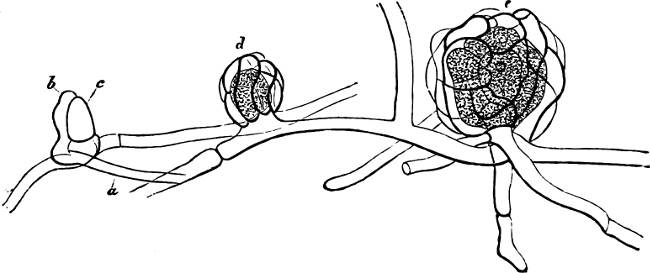
Fig. 107.—Erysiphe cichoracearum: a mycelium-threads; b sterile hypha (“pollinodium”); c fertile hypha (ascogone or archicarp); d and e young ascocarps.
A characteristic feature of the Mildew-Fungi is the thin, pseudo-parenchymatous[120] covering of the ascocarp, enclosing one (Podosphæra and Sphærotheca; compare Thelebolus among the Hemiasci) or a few asci (Fig. 108 c), which do not form any hymenium, but are irregularly placed. The cells of the ascocarp-envelope are often prolonged into hair-like appendages. The ascocarps are developed from the mycelium at places where two hyphæ cross each other (Fig. 107). At these places two short and erect hyphæ are produced side by side. The one from the lower hypha (Fig. 107 c) assumes an ellipsoidal shape, and is known as the archicarp or ascogone, while the other (“pollinodium”) arches over the ascogone. From the latter one ascus may be at once developed (Sphærotheca, etc.), or after its division several asci may be produced, each developed from one division. The sterile hypha (termed “pollinodium,” since it was formerly, but erroneously, supposed to fertilise the ascogone) produces a number of branches, and forms the pseudo-parenchymatous envelope of one cell in thickness, enclosing the asci.
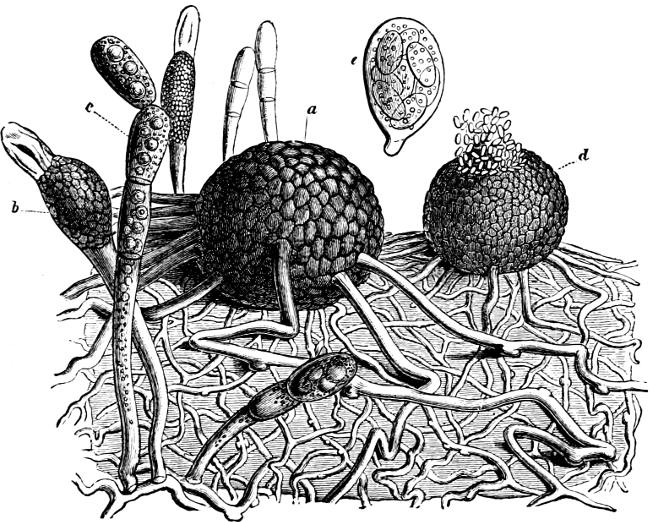
Fig. 108.—Erysiphe communis. A small portion of a leaf with this Fungus growing upon it (considerably magnified). The hyphæ b and d do not belong to this Fungus, but are reproductive organs of a pyrenomycetous Fungus parasitic upon it (Cicinnobolus).
Many plants, both cultivated and wild, are attacked by various[121] species of Mildew. A common means of prevention against their attacks is to dust the diseased parts with sulphur.
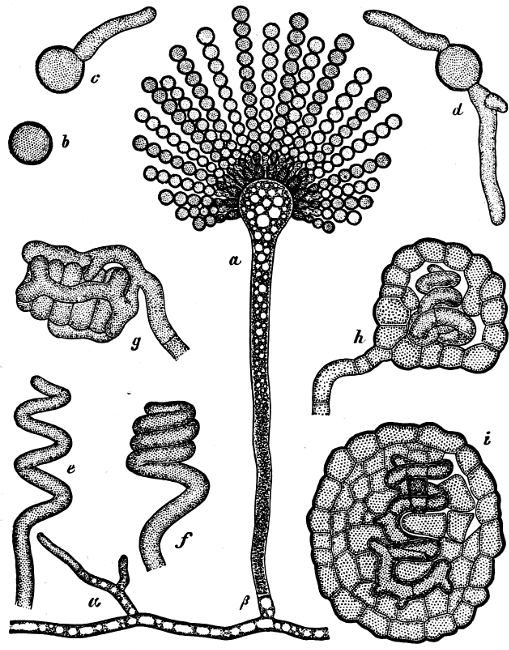
Fig. 109.—Eurotium glaucum: α portion of mycelium lying horizontally; β vertically-placed conidiophore; the mycelium gives rise to another branch near α; the conidia are abstricted from short flask-shaped cells; b a ripe conidium; c, d germinating conidia; e spirally-twisted hypha, commencement of an ascocarp; f a stage later; g still later, the hypha at the base of the coil has given off branches which are applied to it; h, i sections of young ascocarps.
Sphærotheca pannosa occurs on the leaves of Roses, and on the fruit of Peaches and Apricots. S. castagnei on Humulus, Cucumis, etc.—Erysiphe tuckeri grows on the leaves and fruit of the Vine; it spins its hyphæ over the bunches of grapes, curtails their growth, and causes them to burst, and to become decayed and rotten (Grape-disease). The Fungus was first noticed in England in 1845, and later was found in all countries where grapes are grown. It is only known in the conidial form (“Oidium tuckeri”). Many other species of Erysiphe are found on herbaceous plants.—Microsphæra has appendages which are repeatedly forked at their extremities. M. grossulariæ on Ribes grossularia.—[122]Uncinula has appendages with spirally-coiled extremities; on Salix and Acer.—Phyllactinia has a circle of bristle-like appendages with dilated bases. P. guttata on Corylus, Fraxinus, Fagus, etc.
Order 2. Perisporiaceæ, Moulds and Mildews. A group of Fungi widely distributed and found in all situations. Usually they have a well-developed surface mycelium, and small, round, seldom conspicuous ascocarps, containing ovoid, pulley-like spores. They are partly saprophytic, partly parasitic, in the latter condition having a brown mycelium.
Fig. 110.—Eurotium glaucum: a longitudinal section of a half-ripe ascocarp, bounded externally by a well-defined layer of cells, enclosing asci in various stages of development; b a semi-ripe, c an almost ripe ascus; d and e spores seen from the edge and side; f germinating spore twenty-two hours after been sown in plum juice.
Eurotium glaucum (= E. herbariorum, Figs. 109, 110) and E. repens live on dead organic matter, preserved fruits, etc. The conidial forms of both species are known as “Moulds” (Fig. 109), and formerly were described under the name “Aspergillus glaucus.” The conidia for some time remain attached to each other in chains (Fig. 109 a); they are abstricted from sterigmata arranged radially on the spherical, swollen end of the conidiophore. The small yellow or brownish ascocarps are frequently found in herbaria, especially when the specimens have been insufficiently dried. Aspergillus fumigatus and others are pathogenic, causing mycosis in warm-blooded animals.
Fig. 111.—Penicillium crustaceum: a conidia (× 300); b germination of conidia; c small portion of mycelium, produced from a conidium at *, with five conidiophores; d young conidiophore (× 630), a flask-shaped cell is abstricting a conidium; e the same conidiophore after 9–10 hours.
Fig. 112.—Penicillium crustaceum: a two spirally-coiled hyphæ arise from the mycelium, from one of which (archicarp) the asci are produced; b a further step in the development of the ascocarp; the branching archicarp is surrounded by sterile hyphæ; c section of young ascocarp; the larger hyphæ in the centre are the ascogenous hyphæ; these are enclosed by a pseudo-parenchyma of sterile hyphæ (× 300); d series of ripe asci with spores; e four ascopores seen laterally; f germinating ascospores (× 800).
Penicillium crustaceum (P. glaucum, Figs. 111, 112) is an exceedingly common “Mould.” Its mycelium appears very frequently on any organic matter which is permitted to remain untouched, and soon covers it with a dense mass of blue-green[123] conidiophores. These branch at their summits and bear flask-shaped cells from which the conidia are abstricted. The ascocarps which, both in size and colour, resemble grains of sand, have only[124] been obtained in luxuriant cultivation with a limited supply of oxygen.
Capnodium salicinum (Fumago salicina, Cladosporium fumago), a common Mildew, forms dark overgrowths on the leaves and branches of various shrubs (Poplars, Elms, Willows) and on Hops. The conidia appear in various forms, as on conidiophores, in conidiocarps with large multicellular conidia, and in conidiocarps with small unicellular conidia; in nutritive solutions yeast-like conidia are also developed.—Apiosporium pinophilum produces mildew on the leaves of Abies alba and Picea excelsa. (The conidial-forms were formerly described as “Antennaria pinophila”).
Order 3. Tuberaceæ, Truffles. The Fungi belonging to this order are entirely subterranean. The mycelium is filamentous, and partly parasitic upon the roots of plants, especially trees, in its neighbourhood; it is then known as Mycorhiza. The fruit-body is relatively large, in some cases about the size of a hen’s egg. Internally it is traversed by a number of winding passages (Fig. 113 a), the walls of which are coated with the asci. The asci (b) contain only a small number of spores, and these are set free by the putrefaction of the fruit-body. Conidia are unknown.

Fig. 113.—Tuber melanosporum: a fruit-body (nat. size), a portion having been removed to show the internal structure; b an ascus with ascospores.
Tuber melanosporum, T. brumale, T. æstivum, and other species are edible. Terfezia leonis and Choiromyces mæandriformis are also edible. The Truffles are always found in woods and under trees, and disappear when these are destroyed. France and Italy produce the best and the largest number of Truffles, which are hunted by specially trained dogs and pigs.
In Elaphomyces (Stag-Truffle) the fruit-body has a corky external layer, and is inedible. Some of the species are found in this country. E. granulatus is parasitic on the roots of the Fir.
[125]
In this family the hymenium is enclosed in small fruit-bodies, perithecia (Fig. 120 b), which appear to the naked eye as small dots. In shape they resemble a globe or a flask with a narrow mouth, through which the spores are ejected (peronocarpic ascocarps). Different kinds of reproduction—conidia, pycnidia (chiefly with microconidia), chlamydospores, and perithecia—are found in the same species. The various stages in the life-history of these Fungi are so dissimilar, that formally they were considered to be different genera. Ergot furnishes a very good example.
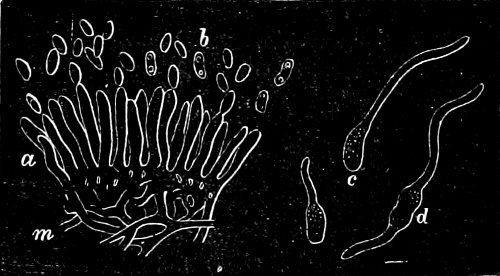
Fig. 114.—A small portion of an ovary attacked with Claviceps purpurea (Sphacelia).
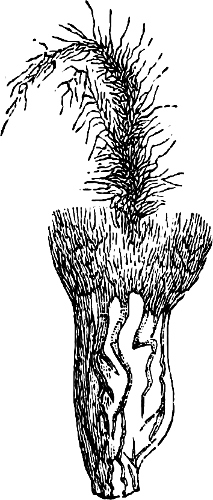
Fig. 115.—An ovary with the conidial stage of Claviceps purpurea (Sphacelia).
This family may be subdivided into 3 sub-families.
The perithecia are pale, fleshy, brightly coloured, and generally aggregated on a stroma. Conidia and chlamydospores occur very frequently. Only one order.
Order. Hypocreaceæ. In this order the majority are parasites upon Flowering-plants (Nectria, Polystigma, Epichloë, Claviceps); but some are parasites upon Fungi (Hypomyces, Melanospora), or upon insects (Cordyceps).
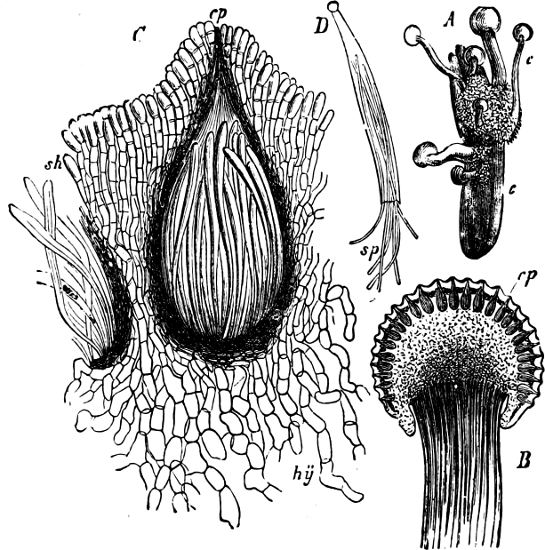
Fig. 116.—Claviceps purpurea. A Sclerotium with stromata (cl) (× by 2). B Stroma divided longitudinally to show the perithecia (cp). C A perithecium with the surrounding hyphæ (hy). D An ascus ruptured, with the eight filamentous ascospores emerging.
The most important member of this order is the Ergot (Claviceps purpurea, Figs. 114, 115, 116). This Fungus is found in the flowers of many species of Grasses, especially the Rye, attacking and destroying the ovaries. In the FIRST or CONIDIAL STAGE of the attack, the ovaries are found covered with a white, irregularly[126] folded mycelium (Fig. 114 m, Fig. 115), formed of numerous hyphæ woven together and penetrating the wall of the ovary. From these a number of hyphæ (Fig. 114 a) project into the air and abstrict from their apices the conidia (b) which serve as reproductive organs. The mycelium also secretes a sticky, stinking fluid (honey-dew) in which the conidia are embedded in great numbers. The honey-dew exudes from the bases of the glumes, and is greedily sought by flies, which thus carry the conidia to other ovaries. In this manner fresh ears are infected, which might escape were the conidia only distributed by the wind. This stage formerly was regarded as an independent Fungus, known as Sphacelia segetum (Fig. 115). On germination, the conidia produce either a new mycelium (Fig. 114 d, c), or new conidia. The SECOND or SCLEROTIUM STAGE is the one in which the Fungus passes the winter. The mycelium penetrates deeper and deeper into the attacked ovaries, their tissues are destroyed and replaced by the hyphæ, which gradually become more and more felted together.[127] A firm, pseudo-parenchymatous mass of hyphæ is thus formed at the base of the loosely-woven Sphacelia, which is in part transformed into the hard sclerotium, and the remainder thrown off. A dark, hard, poisonous body, longer than the natural grain, is thus formed; these bodies are known as Ergots, and were formerly considered to be a distinct species,—Sclerotium clavus (“Secale cornutum,” Ergot, Fig. 116 A, c). The THIRD STAGE, described as Claviceps purpurea, is developed in the following spring from the germinating sclerotium, which produces dark-red stromata with short stalks. In the stroma numerous perithecia with asci and ascospores are produced. The latter may infect young flowers of the cereals, in which the disease is then developed as before.
Fig. 117.—Nectria cinnabarina: a branch of Acer pseudoplatanus, with conidial-layers and perithecia (nat. size); b a conidial-layer (Tuberculoria vulgaris); c, a mass of perithecia. (b and c × 8.)
Several species of the genus Nectria, with blood-red perithecia, are found as dangerous parasites, especially N. ditissima, which causes “Canker” in the Beech, Ash, and Apple, etc.; N. cucurbitula, which appears on Pine-trees, and N. cinnabarina (Fig. 117), whose conidial form was formerly named Tubercularia vulgaris.—Polystigma rubrum forms shining red spots on the green leaves of Prunus-species.—Epichloë typhina is parasitic on the sheaths of Grasses, on which it first forms a white conidial-layer, later on a yellow layer of perithecia.—Cordyceps (Chrysalis Fungus, Figs. 118, 119) lives in and destroys insects, and after compassing their death produces the club-formed, generally yellow, stromata, one part of which bears conidia (Isaria) and another perithecia. C. militaris (Fig. 118) on the chrysalides and caterpillars of moths, is the most common.
[128]
The so-called Botrytis bassiana, which produces the disease known by the name of “Muscardine,” in silkworms, is probably a conidial form belonging to Cordyceps.
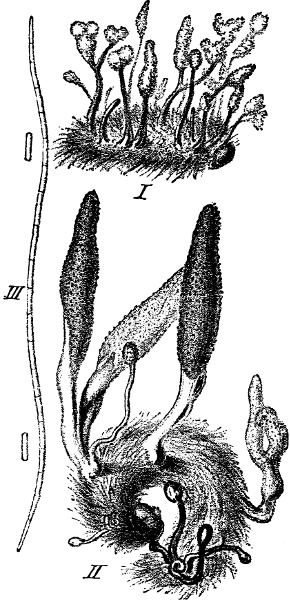
Fig. 118.—Cordyceps militaris. I Stromata with conidiophores (Isaria farinosa). II A larva, with stromata, bearing perithecia. III A spore.
Fig. 119.—Cordyceps robertii on the larva of Hepialus virescens: a stalk of stroma; b perithecia.
[129]
To this sub-family belong the majority of the Pyrenomycetes. The perithecia are of a firm consistence (tough, leathery, woody or carbonaceous), and of a dark colour. Their covering is quite distinct from the stroma when this structure is present. The stromata are sometimes very large, and may be either cushion-like, crustaceous, upright and club-like, or branched bodies. In general, small, inconspicuous Fungi, living on dead vegetable matter, sometimes parasites. Free conidiophores and conidiocarps are known in many species, and in several, chlamydospore-like forms of reproduction. Orders 3–18 constitute the Sphæriaceæ of older systematists.

Fig. 120.—Strickeria obducens: a a portion of an Ash-branch with the bark partly thrown off; on the wood are numerous black perithecia (× 20); b longitudinal section through a perithecium; c a spore; d longitudinal section through a pycnidium whose ascospores are being ejected; e portion of the same, with hyphæ and spores.
Order 1. Sordariaceæ.—Fungi living on dung with fragile perithecia, either aerial or buried in the substratum. The dark brown or black spores have either a mucilaginous envelope (Sordaria, etc.) or mucilaginous appendages (Podospora), by means of which their expulsion and distribution are promoted.
Order 2. Chætomiaceæ. Perithecia fragile, free, bearing on the summit a tuft of hairs. Chætomium, on decaying vegetable matter.
Orders 3–7. Perithecia scattered or aggregated, situated from the commencement on the surface of the substratum. Stroma wanting.
Order 3. Trichosphæriaceæ. Trichosphæria parasitica (Fig. 121), on Abies alba; Herpotrichia nigra on Picea excelsa and Pinus montana.
[130]
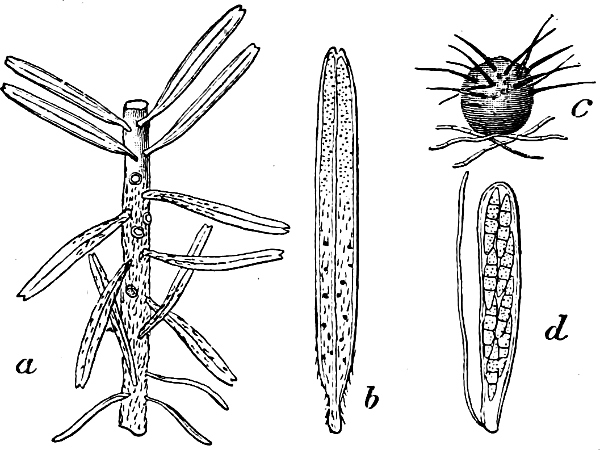
Fig. 121.—Trichosphæria parasitica: a a twig of Abies alba, with epiphytic mycelium; b a leaf with mycelium and sporangia (magnified); c a sporangium (× 60); d an ascus with spores (× 550).
Order 4. Melanommaceæ. Rosellinia quercina lives in the roots of 1–3-year-old Oaks, and destroys the plants.
Order 5. Ceratostomaceæ.
Order 6. Amphisphæriaceæ. Strickeria obducens (Fig. 120) has brick-like spores, and lives aggregated on the hard branches of Fraxinus.
Order 7. Lophiostomaceæ.
Order 8. Cucurbitariaceæ. Perithecia tufted, at first embedded, then breaking through, often situated upon an indistinct stroma.
Orders 9–13. The perithecia remain embedded, and are only liberated by the casting off of the covering layers of the substratum. Stroma wanting.
Order 9. Sphærellaceæ. The species of Sphærella have colourless, bicellular spores. They live upon the leaves of many plants, and develope spherical perithecia upon the fallen leaves.
Order 10. Pleosporaceæ. The conidial-forms of Pleospora herbarum and P. vulgaris form a black covering on various plants, known as “smuts.”—Venturia ditricha occurs on the underside of dry Birch leaves, and perhaps to this belongs the conidial-form, Fusicladium pirinum, which causes the “Rust spots” on Apples and Pears.
Order 11. Massariaceæ.
Order 12. Clypeosphæriaceæ.
Order 13. Gnomoniaceæ. Perithecia, with peak-like aperture. Gnomonia erythrostoma in the leaves of Prunus avium, which turn brown and do not fall in autumn.
Orders 14–18. Stroma generally well developed. The perithecia are embedded in the stroma, but when this is rudimentary, in the substratum.
Order 14. Valsaceæ. Valsa.
Order 15. Diatrypaceæ. Diatrype.
Order 16. Melanconidaceæ.
Order 17. Melogrammataceæ.
[131]
Order 18. Xylariaceæ. This order is the most highly developed of the Sphæriales. The stroma arises on the surface of the substratum, which is generally dead or decorticated wood; it is well-developed, crustaceous, hemispherical or upright. In the younger conditions it is covered with a layer of conidia, and later on it bears the perithecia, arranged in a layer immediately beneath its surface. The ascospores are of a dark colour. Often also there are free conidiophores.
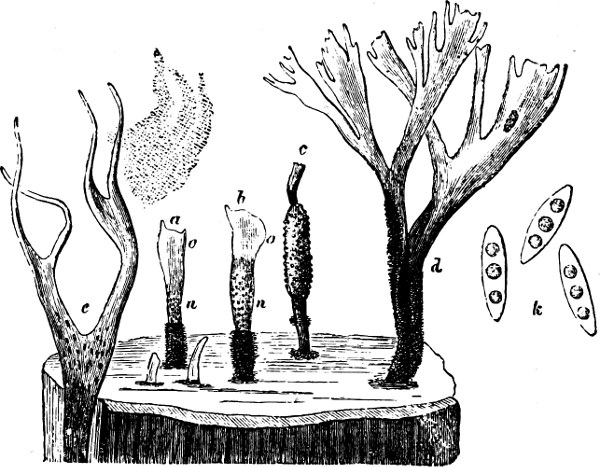
Fig. 122.—Xylaria hypoxylon (nat. size) on a tree stump: a younger, b an older stroma, both of which, with the exception of the black lower portion, are covered with white conidia; n, spot where the perithecia are developed; c an old stroma with upper part fallen off; d, e large branched stromata; k conidia.
Hypoxylon and Ustulina have a cushion-like or crustaceous stroma.—Xylaria has a club-shaped or branched stroma, often several centimetres high. X. hypoxylon (Fig. 122) and X. polymorpha occur on old tree stumps.—Poronia grows on old horse dung, and has a conical stroma.
The perithecia are always embedded in a black stroma, and are not distinctly separated from it. The accessory forms of reproduction are: conidiophores, conidiocarps, and yeast-like conidia. The majority are parasites. One order.
Order Dothideaceæ. Phyllachora graminis produces scab-like patches on the leaves of the Grasses.—Scirrhia rimosa grows on the leaf-sheathes of Phragmites.—Rhopographus pteridis on Pteridium aquilinum.
[132]
This family, like the following, has hemiangiocarpic ascocarps (apothecia). These are closed in the early stages, but when ripe open in a valvular manner by a longitudinal fissure; they are black, oblong, and often twisted. Some species are parasites, especially upon the Coniferæ.
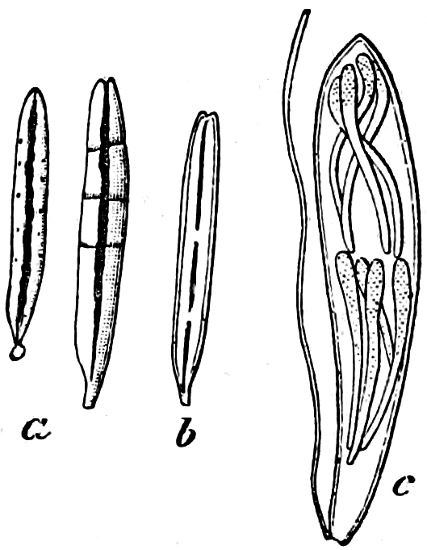
Fig. 123.—Lophodermium (Hypoderma) nervisequium: a two leaves of Abies alba seen from above with pycnidia; b a leaf seen from the underside with apothecia; c an ascus with ascospores. (× 500.)
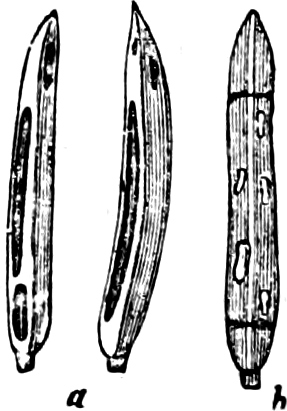
Fig. 124.—Three leaves of the Red-pine with Lophodermium macrosporum: a under side of the leaves with apothecia; b a leaf from upper side with pycnidia. (× about 2.)
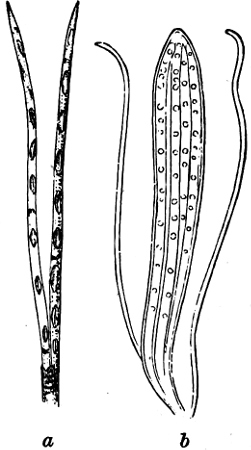
Fig. 125.—Lophodermium pinastri: a leaves of Pinus sylvestris with apothecia (nat. size); b two paraphyses and an ascus with filamentous spores.
Order 1. Hysteriaceæ. Hysterium pulicare upon the ruptured bark of many trees.
Order 2. Hypodermaceæ. The species of Lophodermium live upon the leaves of Conifers, and are the cause of their falling off (blight). L. pinastri (Fig. 125), on the leaves of Pinus and Picea; the leaves become red-brown and fall off; at first conidiocarps are formed, and later apothecia; L. nervisequium (Fig. 123), on Abies alba; L. macrosporum (Fig. 124), on Picea excelsa; L. brachysporum, on Pinus strobus.
Order 3. Dichænaceæ.
Order 4. Acrospermaceæ.
The ascocarps (apothecia) are at first closed, and only open at the time of their ripening, not valvularly, but more or less[133] like a saucer or cup, so that the hymenium lies exposed on their upper surface. In the first three sub-families, and generally also in the fourth, the apothecia are formed inside the substratum. The apothecia are, in contrast to the Pyrenomycetes, light and brightly coloured, and their size varies very much, and may be several centimetres in diameter. Paraphyses are often present between the asci; they often contain colouring matter, and give to the disc its characteristic colour. The tissue on which the asci are borne is known as the hypothecium. The shape and colour of the spores is not so varied as in the Pyrenomycetes. The accessory forms of reproduction are conidia (sometimes of two forms), chlamydospores, and oidia. The family is divided into 5 sub-families.
The apothecia are developed in the interior of the substratum, which they break through, and in general dehisce apically. The envelope is tough and black. Hypothecium inconspicuous; hymenium flat.
Order 1. Euphacidiaceæ. Phacidium abietinum, on the leaves of Abies alba.—Rhytisma; the pycnidia are found in the summer on the green leaves, while the apothecia are developed on the fallen leaves and dehisce in the following spring. R. acerinum causes black spots on the leaves of the Sycamore, and R. salicinum on Willows.
Order 2. Pseudophacidiaceæ.
The apothecia when ripe break through the substratum which forms a border round them. Hymenium generally saucer-shaped.
Order 1. Stictidaceæ. Stictis.
Order 2. Ostropaceæ. Ostropa.
The apothecia are embedded in the substratum in the early stages, and then are raised high above it. Hypothecium thick. Hymenium cup-shaped.
Order 1. Tryblidiaceæ. Tryblidium.
Order 2. Heterosphæriaceæ. Heterosphæria patella on the dead stalks of Umbellifers.
[134]
The apothecia in the early stages are embedded in the substratum and then break through it, or are from the first situated on the surface of the substratum. Hypothecium thick.
Order 1. Cenangiaceæ. Cenangium.
Order 2. Dermateaceæ. Dermatea.
Order 3. Patellariaceæ. Patellea, Biatorella, Patellaria.
Order 4. Caliciaceæ. Calicium, Coniocybe, etc., on the bark of trees.
Order 5. Arthoniaceæ. Arthonia on the bark of several trees. Celidium stictarum on the apothecia of Sticta pulmonaria.
Order 6. Bulgariaceæ. Apothecia gelatinous under moist conditions, and horny when dried.—Calloria fusarioides; the red apothecia break out in the spring on the dried stalks of Urtica dioica; a gelatinous reproductive form of the Fungus is found before the apothecia, which consists of oidia (formerly described as “Dacryomyces urticæ”).—Bulgaria inquinans on the living or fallen trucks of Oaks and Beeches.
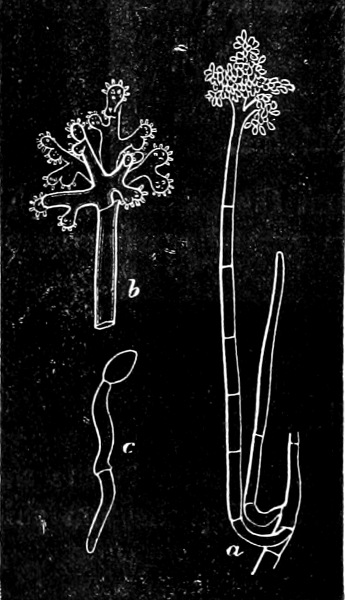
Fig. 126.—Botrytis cinerea: a slightly magnified; b more highly magnified; c germinating conidium.
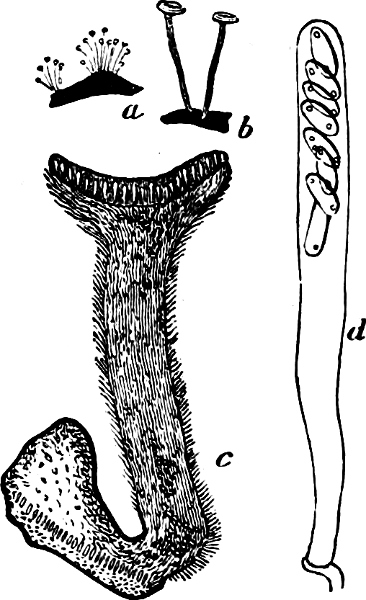
Fig. 127.—Sclerotinia fuckeliania: a sclerotium with conidiophores; b with apothecia; c section through sclerotium and apothecium; d ascus with eight ascospores. (× 390.)
The apothecia are developed on the surface of the substratum and are waxy or fleshy; at the commencement closed, and covered with a saucer- or cup-shaped, seldom flat, hymenium. The hypothecium is generally well developed. This sub-family is the richest in[135] species of the Discomycetes and contains forms of very different habit. They grow upon dead wood, upon the ground, and upon dung. A few are parasites.
Order 1. Helotiaceæ. Apothecia with waxy envelope of colourless, or yellowish prosenchymatous cells.—Chlorosplenium æruginosum is found on decaying wood (particularly Oak and Birch), to which it gives a green colour. Sclerotinia has sclerotia which are developed upon the host-plant and from which, after a period of rest, the long, brown-stalked apothecia arise. S. ciborioides (S. trifoliorum, Fig. 128) is parasitic on Clover; S. sclerotiorum, on Daucus-roots, Phaseolus, etc.; S. baccarum, on the berries of Vaccinium myrtillus; “Botrytis cinerea” is a common parasite and is probably the conidial form of S. fuckeliania (Fig. 127).—Helotium herbarum lives on dry plant stems.—Dasyscypha willkommii (Fig. 129) produces Larch-canker on the bark of the Larch.
Fig. 128.—Sclerotinia ciborioides: a sclerotium with three apothecia slightly magnified; b ascus with eight ascospores; c germinating ascospore.
Fig. 129.—Dasyscypha willkommii: a portion of bark of Larix decidua with sessile, cup-shaped apothecia (nat. size); b two paraphyses on either side of an ascus with eight ascospores.
Order 2. Mollisiaceæ. Mollisia cinerea, principally on decaying wood.
Order 3. Pezizaceæ. This order contains the largest and morphologically the highest forms of the Discomycetes. Apothecia fleshy, and in the later conditions generally saucer-shaped.
Peziza, with sessile apothecia, growing on the ground; P. cochleata is brown, and coiled like a snail-shell; P. coccinea is scarlet; P. aurantia occurs as an orange-coloured expansion on the ground.
Order 4. Ascobolaceæ. Apothecia fleshy; in the later stages flat or[136] convex. The asci are, comparatively speaking, large, and often contain a great number of spores which escape by the casting off of a lid on the summit of the ascus. Generally living on dung.—Ascobolus furfuraceus, etc.
These Fungi have the appearance of clubs, bells, or mushrooms, consisting of an upright stalk bearing a large and fleshy head, on the exterior surface of which the hymenium is spread. The ascocarps are probably gymnocarpic from the beginning, and on this account these plants are placed in a separate family. The development of the ascocarps is unknown. The Morchella (Morell) grows on the ground; some species are edible. 1 order.
Order. Helvellaceæ. Spathulea is yellow and club-shaped, and forms “fairy rings” in woods.—Geoglossum (Earth-tongue) projects above the ground as a black tongue, or as a club-shaped body. Several species are found in meadows and on heaths.—Helvella has a stalk, bearing an irregularly folded head, on the external surface of which is the hypothecium.—Morchella (Morell, Fig. 130), the stalk bears on its summit the conical or spherical head, the external surface of which is reticulate and bears the asci.—Mitrula. Verpa.
Fig. 130.—Morchella esculenta: a an entire specimen, about one half natural size; b longitudinal section through the head.
Appendix to the Ascomycetes:
The Lichens were formerly classed among the Thallophyta as a group quite distinct from the Algæ and Fungi. Investigations during the last twenty-five years, however, have conclusively proved that the Lichens are Fungi which reproduce in the same manner as the Ascomycetes, or, more rarely, the Basidiomycetes, and have entered into a peculiar symbiotic relation with Algæ, especially the Cyanophyceæ and Protococcoideæ, with which they associate, and without which they would be unable to exist. The Fungus forms the largest portion of the Lichen, enclosing the Alga with which it may be said to be commensal. The Fungus especially produces reproductive bodies and absorbs the inorganic nourishment through the rhizoids, whilst the Alga supplies it with the organic materials. In consequence of this the Lichens, in contradistinction[137] to other Fungi, need light for the development of their nutritive organs, and are therefore, in any case internally, of a more or less greenish colour. The form and condition of the thallus is unusual among the Fungi, and they can grow upon rocks and in other places where no dead organic matter, such as would be required by other Fungi, is obtainable.
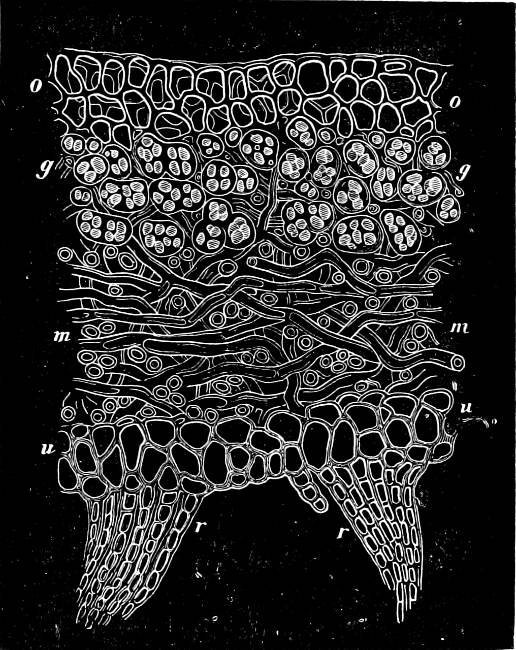
Fig. 131.—Transverse section through the thallus of Sticta fuliginosa (× 500): r-r rhizoid-strands, which arise from the under side; g-g gonidial layer; m medullary layer; o upper, u lower cortex.
Two cellular forms are therefore to be found in each Lichen:
1. The cells which belong to the Fungus. These are generally septate, branched hyphæ without any trace of chlorophyll. In the thallus of the majority of Lichens there may be found a medullary layer (Fig. 131 m) of loosely-woven hyphæ, between which there are large air chambers; and an external layer (cortex) (Fig. 131 o, u) formed of closely-woven hyphæ without any intercellular spaces. In some Lichens (Collemaceæ) the hyphæ wind about in the thallus, being equally distributed throughout, without forming any decided strata. These Lichens moreover become[138] gelatinous when exposed to moisture (Fig. 132), on account of the swelling of the walls of the Algæ. The hyphæ contain protoplasm with drops of oil, but never starch; their walls easily swell when exposed to damp after having been dried, and in some (e.g. Cetraria islandica) they become gelatinous when cooked. Certain strata of hyphæ become blue on treatment with iodine alone, from which it is inferred that the wall is allied, in its chemical nature, to starch.
2. The enclosed Algæ, termed “gonidia.” Some belonging to the Cyanophyceæ, Protococcoideæ, (especially Pleurococcus) and Chroococcaceæ, are spherical and are found isolated, or in irregular groups of cells (Fig. 131 g); some belonging to Nostoc (Fig. 132 g), Lyngbyaceæ, etc., are placed in cell-rows. Each Lichen, as a rule, has only one definite Algal-form for its gonidium.
The gonidia either lie together in a certain stratum between the cortex and the medullary layer (Fig. 131 g), or are scattered irregularly throughout the entire thallus (Fig. 132). The thallus is in the first instance termed “heteromerous,” in the second instance, “homoiomerous.” The Fungal-hyphæ embrace the gonidia and apply themselves closely to, or even penetrate them, and hence it has been difficult to decide whether the one cellular form does or does not develop from the other (Figs. 134, 135).
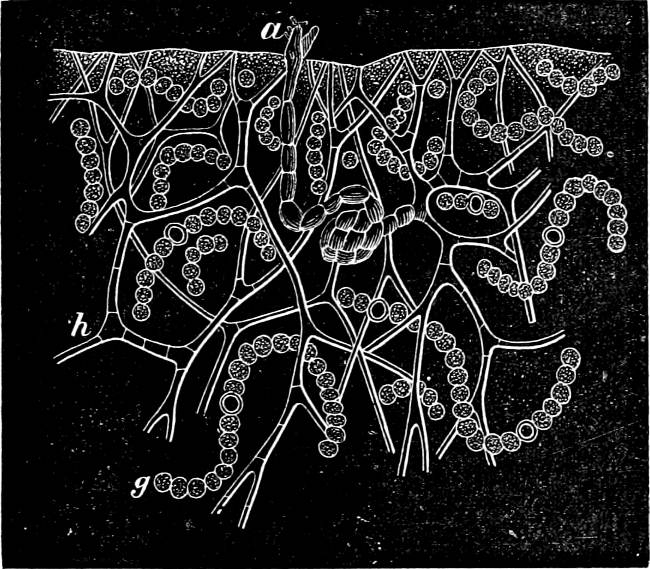
Fig. 132.—Collema microphyllum. Transverse section through the thallus; g Nostoc-chains; h hyphæ.
[139]
This theory regarding the symbiosis of Fungi and Algæ to form a Lichen is termed the Schwendenerian theory, after the first scientist who advanced it with any weight. It had been already indicated by De Bary, and further arguments in its support have at a later time been adduced by Bornet, Stahl, Treub, Frank, Bonnier, Alfr. Möller and others.
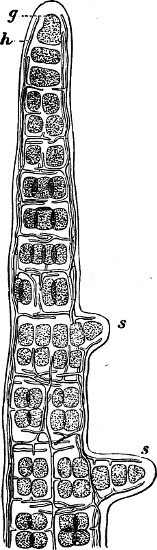
Fig. 133.—Ephebe pubescens. The apex of a branch of the thallus with two lateral branches (s): h its hyphæ; g the apical gonidium of the main branch.
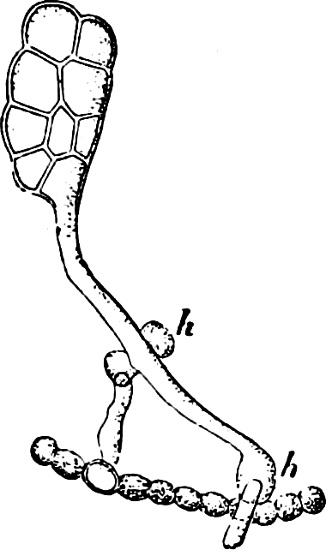
Fig. 134.—Nostoc lichenoides, which is attached by a germinating thread (h) of Collema glaucescens.
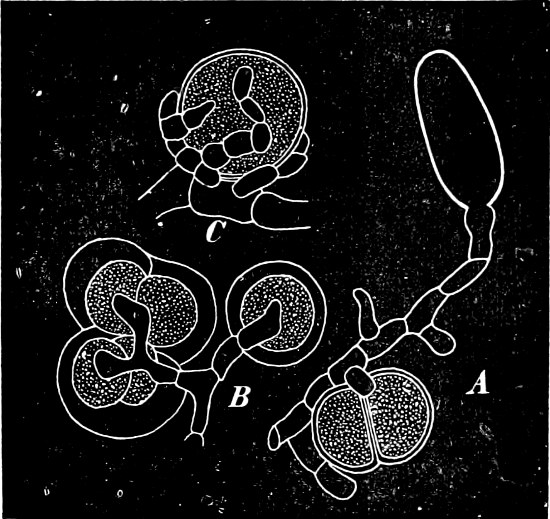
Fig. 135.—A Germinating spore of Physcia parietina with Protococcus viridis. B Synalissa symphorea with Glæocapsa. C Cladonia furcata with Protococcus.
The thallus of the Lichen appears mainly under three forms:—
1. The Crustaceous, which adheres firmly to the substratum (bark, stone) throughout its entire surface, without being raised into any free patches or lobes. It has, in many instances, no definite outline, and hyphal-branches from it often penetrate[140] deeply into the substratum. It grows at the circumference and sometimes dies away in the centre (Figs. 138, 139, 140).
2. The Foliaceous. This also lies flat upon the substratum, but is not firmly attached to and has a definite outline. It grows at the margin, and raises itself a little by free outgrowths and lobes (Fig. 141). The rhizoid-strands spring out from its whitish under surface (Fig. 131, r).
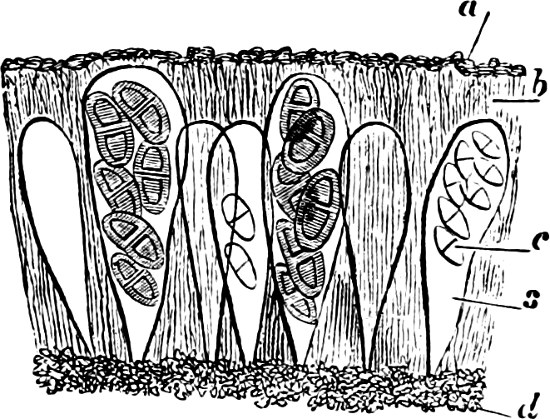
Fig. 136.—Portion of a hymenium: d a thin stratum on which the asci (s) are situated.
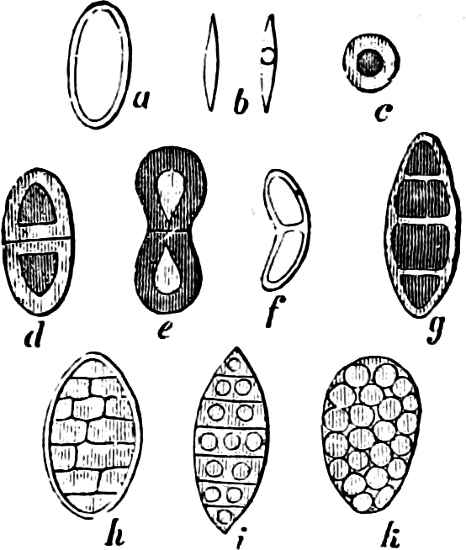
Fig. 137.—Spores of, a Cladonia, Lecanora and Pertusaria; b Bæomyces; c Sphinctrina; d, e, f various species of Parmelia; g, h Verrucaria in its younger and older condition; i, k species of Leptogium.
3. The Fruticose, which is attached to its substratum at a small point from which it projects freely, either erect or pendulous. It is more or less tufted, in the form of a bush (Figs. 142, 143). These three thallus-forms gradually pass over by many intermediate forms into one another.
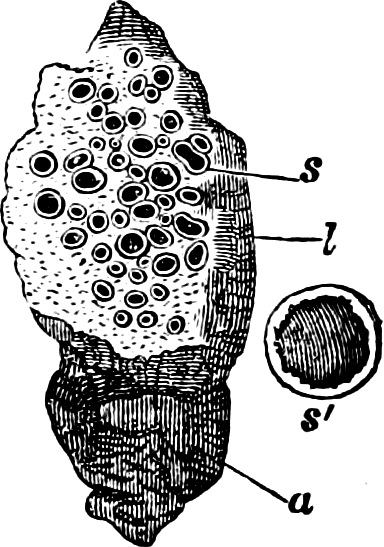
Fig. 138.—Lecanora subfusca: a the bark on which it is situated; l the thallus; s the ascocarp; s’ an ascocarp.
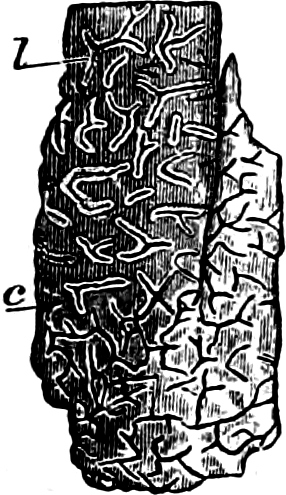
Fig. 139.—Graphis (two species).

Fig. 140.—Pertusaria communis.
The Lichens, like other Ascomycetes, have very variously[141] constructed ascospores (Fig. 137), which are enclosed in asci (Fig. 136), usually surrounded by paraphyses attached together. Furthermore they possess pycnidia (Fig. 141) containing numerous microconidia. These were formerly considered as organs of fructification, and were termed “spermatia,” and the pycnidia, “spermogonia.” Alfr. Möller proved, in 1887, that the microconidia are able to germinate and produce a mycelium with new conidia, just as in other Ascomycetes.
Vegetative Reproduction takes place by soredia, which to the naked eye appear as whitish powder on the surface of the thallus. They are small round bodies, formed by one or a group of gonidia, which are surrounded by a mass of felted hyphæ. After the rupture of the cortex they are set free, and readily carried by the wind to other places, where under favourable circumstances they establish a new thallus.
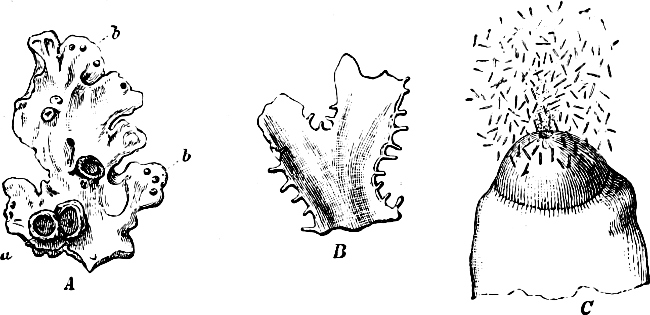
Fig. 141.—A A portion of the thallus of Parmelia parietina with ascocarps (a) and pycnidia (b). B A portion of the thallus of Cetraria islandica with pycnidia at the end of small lobes. C A lobe with pycnidia and ejected microconidia. (Magnified).
Geographical Distribution. The Lichens are the most hardy plants, and are the first to appear on hitherto bare rocks which they gradually disintegrate, and hence prepare the way for the growth of other plants. They are to be found from the Polar regions to the Equator; from the highest snow-free mountain-peaks down to the level of the sea; on the stems of trees; on rocks, soil, some even on inundated places; on stones in woodland streams, and on beaches; but they are never found upon rotten organic remains. Some grow gregariously in enormous masses, and form wide-stretching carpets, e.g. Reindeer Moss (Cladonia rangiferina), species of Cetraria and other fruticose Lichens.
Uses. On account of the cell-wall being composed of Lichenstarch[142] (Lichenin), the Iceland-Lichen and Manna-Lichen (Lecanora esculenta) are used as food; the latter grows on stones, in the deserts of Asia and North Africa, and is often torn loose in large masses and carried away by the wind. The Reindeer-Lichen is not only the principal food of the reindeer, but it is also used in the manufacture of Danish brandy. Cetraria islandica (Lichen islandicus) is OFFICINAL. Colouring materials (lacmus, orseille, persio) are made from several species, especially from Roccella tinctoria (from the rocky coasts of the Mediterranean). Parmelia saxatilis and particularly Lecanora tartarea are used for colouring purposes in the Northern countries.
About 2,000 species of Lichens have been described. If we disregard the Basidiolichenes, which will be considered on page 176, the remaining Lichens (Ascolichenes) may be divided into the two following orders according to the structure of the fruit-bodies:—
Order 1. Pyrenolichenes. The ascocarps (apothecia) are spherical or flask-shaped, as in the Pyrenomycetes, more rarely linear (Graphis).
According to the nature of the thallus, these Lichens may be divided into:—
a. Thallus homoiomerous, but not gelatinous, branching according to the mode of growth of the Algæ: Ephebe (Fig. 133), with Algæ of the genus Stigonema.
b. Thallus homoiomerous, gelatinous: Lichina.
c. Thallus heteromerous, crustaceous: Verrucaria, Pyrenula; Graphis (Fig. 139), which may be considered as Hysteriaceæ with gonidia; several species of Graphis are common on bark.
d. Thallus heteromerous, foliaceous: Endocarpon.
e. Thallus heteromerous, fruticose: Sphærophorus.
Order 2. Discolichenes. These, as in the Discomycetes, have open apothecia, which, as a rule, are cupular, more rarely hemispherical (Cladonia).
According to the nature of the thallus, these Lichens may be divided into:—
a. Thallus homoiomerous, but not gelatinous, branching according to the mode of growth of the Algæ: Cœnogonium.
b. Thallus homoiomerous, gelatinous: Collema (Fig. 132), with Algæ of the genus Nostoc; Leptogium.
c. Thallus heteromerous, crustaceous: Pertusaria (Fig. 140), Lecidea, with apothecia open from the beginning; Lecanora, with apothecia, which in the beginning are closed, later on open, but with a rim formed by the thallus (Fig. 138); Bæomyces, whose apothecia are borne on a stem formed by the thallus.
[143]
d. Thallus heteromerous, foliaceous: Parmelia (P. saxatilis; P. parietina, Wall-Lichen, Fig. 141, is yellow, very frequent on tree-stems, stone-walls, tiles); Physcia (P. ciliaris, frequent on tree-stems); Sticta (S. pulmonacea, Lung-Lichen, on tree-stems); Peltigera, especially on the Moss among trees; Umbilicaria, on rocks.
e. Thallus heteromerous, fruticose: Cetraria (C. islandica), “Iceland Moss,” with an olive-brown, flat, furrowed, fringed thallus, on heaths; C. nivalis, white, in the Polar regions; Evernia, Ramalina, Usnea (U. barbata, Beard-Lichen, Fig. 143); Roccella, Stereocaulon, Cladonia, of which the genus C. rangiferina, Reindeer-Moss (Fig. 142) is important; Cladonia has two kinds of thallus, one scaly and leaf-like, the other erect, which bears the apothecia and may be fruticose (Fig. 142), or cupular (Fig. 144); they grow in soil in forests and on heaths.
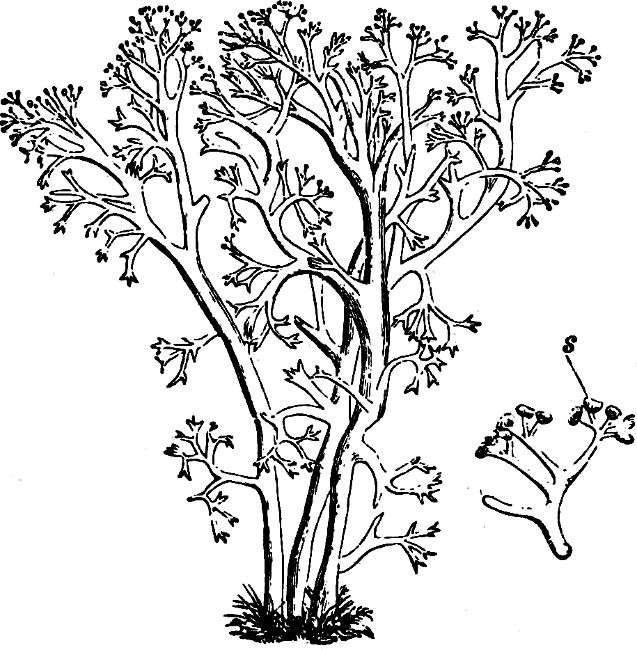
Fig. 142.—Cladonia rangiferina: s ascocarp.
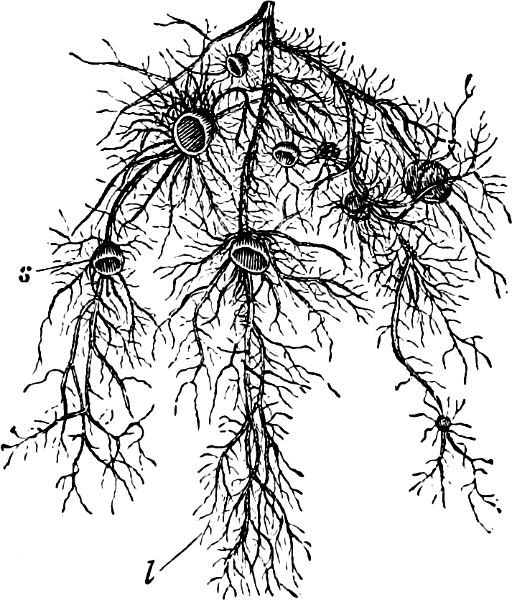
Fig. 143.—Usnea barbata: s ascocarp. (Slightly magnified.)
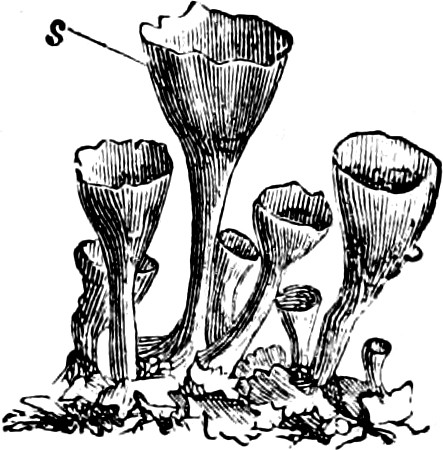
Fig. 144.—Cladonia pyxidata.
[144]
This sub-class embraces the most highly developed Fungi, with large “fruit-bodies,” which in ordinary language we shortly term Funguses, Toadstools, or Mushrooms.
They have no sporangia, but reproduce only by means of basidiospores, conidia, chlamydospores and oidia. The chief characteristic of this sub-class is the basidium (Fig. 145), i.e. the conidiophore, which has a distinctive form, and bears a definite number (generally 4) of characteristically shaped conidia (basidiospores, Fig. 145 c, d, e).
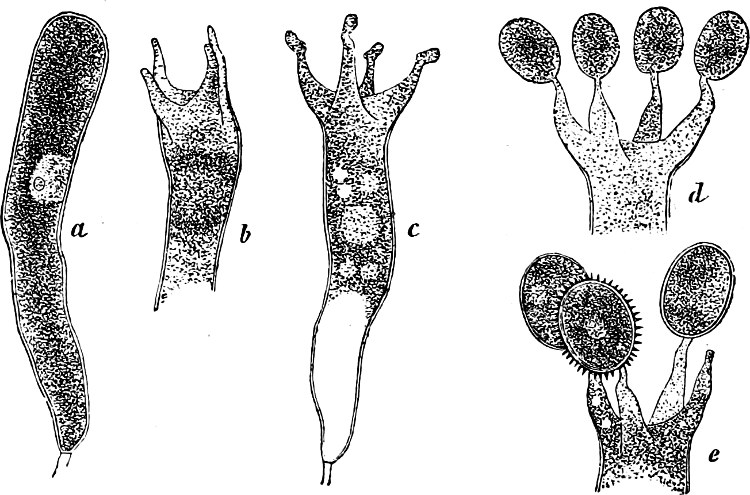
Fig. 145.—Development of spores in Corticium.
The summit of each basidium is produced generally into four conical points (sterigmata, Fig. 145 b), from each of which a basidiospore is abstricted. The basidia may be classified into three principal groups, each of which accompanies a distinctive conidiophore: 1, the long, filamentous, transversely divided basidia, with lateral sterigmata and spores, found in the Uredinaceæ (Figs. 146 D, 153), Auriculariaceæ (Fig. 160 B), and Pilacraceæ; 2, the spherical, longitudinally divided basidia of the Tremellaceæ (Figs. 160 C d; 161 iii. iv.); and 3, the ovoid, or cylindrical, undivided basidia of the Autobasidiomycetes (Figs. 145, 163, etc.); the two last have apical sterigmata and spores.
The first two groups are the septate basidia (protobasidia), of the Protobasidiomycetes; while the unseptate basidia (autobasidia) of the Autobasidiomycetes are the third group. On the formation of the basidiospores, the nucleus[145] of the basidium divides into four nuclei, each of which is transferred to a spore.
In addition to the basidia, simple conidiophores are also found. In the Protobasidiomycetes, the simple conidia are very generally found as accessory methods of reproduction in conjunction with the basidiospores; but less frequently in the Autobasidiomycetes, e.g. among the Dacryomycetes, Tomentellaceæ, Heterobasidion annosum.
The simple conidiophores vary in size, and in the number and shape of the conidia; they, however, resemble the basidia, and are doubtless an early stage in the development of the definitely formed basidia.
Finally, well-defined chlamydospores, formed in various ways, appear in the Basidiomycetes as supplementary reproductive bodies (compare p. 90). Among the Protobasidiomycetes, chlamydospores are at present only found among the Uredinaceæ, but in various forms; in the majority of families of the Autobasidiomycetes oidia frequently occur (Fig. 162), but genuine chlamydospores seldom.
In the same species several of the known forms of reproduction may be distinguished.
The mycelium is generally composed of white, branched strands, consisting of numerous felted hyphæ; in some, sclerotia are found.—The great majority are saprophytes; some (particularly all the Uredinaceæ), are parasites.
Divisions of the Basidiomycetes.
To this series belong the lowest of the Basidiomycetes. The basidia appear in two principal forms (1 and 2 on page 144) and are divided into four cells, either transversely or longitudinally, each division forming a sterigma which abstricts a basidiospore. The first three orders, Uredinaceæ, Auriculariaceæ, and Tremellaceæ[146] have gymnocarpic fruit-bodies, while those of the Pilacraceæ, on the contrary, are angiocarpic.
Order 1. Uredinaceæ (Rusts). All the Rust-Fungi are parasites, their mycelium living in the interior of the stems and leaves of their hosts, causing red, brown, or black spots—hence their name—and malformations, sometimes of considerable size.
The Rust-Fungi are gymnocarpic and destitute of a hymenium; for these reasons they are regarded as the simplest order of the Basidiomycetes. They are entirely parasitic, and their filamentous, branched mycelium ramifies in the intercellular spaces of its host, and often protrudes haustoria into the cells. The mycelium is perennial should it enter a woody tissue; it may also hibernate in the rhizomes of perennial herbs and permeate the shoots springing from them, but in the majority of the Rust-Fungi the mycelium has a very limited growth. The chief means of reproduction of the Rust-Fungi are the chlamydospores, which in the more highly developed species occur in three forms, namely, the teleuto-, æcidio-, and uredo-spores. The spores, in the host, are formed immediately beneath its epidermis, which is ruptured on the ripening of the spores, with the production of “rust,” brown, red, or black spots. Those chlamydospores which produce basidia are termed teleutospores. The spore on germination produces a transversely divided basidium, “promycelium,” on which basidiospores, “sporidia,” generally four in number, are produced on lateral sterigmata. This basidio-fructification is gymnocarpic; the basidia neither form a hymenium nor a fruit-body (only Cronartium and Gymnosporangium have a slight indication of a basidio-fructification).
Many Rust-Fungi, in addition to basidiospores, have small, unicellular conidia, “spermatia,” which are borne in conidiocarps, “spermogonia.”
The TELEUTOSPORES (Winter-spores) may be either unicellular or multicellular; in the majority of cases they are enclosed in a hard outer cell-wall, the exospore, which in some cases is very strongly developed; they have also a long or short stalk, the remains of the spore-bearing hypha. Each cell of the teleutospore has one germ-pore (a thin portion of the wall, for the protrusion of the germ-tube; in Phragmidium and Gymnosporangium there are, however, several germ-pores). The colour of the teleutospores is generally much darker than that of the uredospores, and it is by these that the majority of the Rust-Fungi hibernate.
[147]
In Gymnosporangium, two kinds of teleutospores are found (distinguished by their size and thickness of exospore). In many species of Puccinia, the form of the teleutospores varies very much, so that in the same layer spores have been observed with the characteristic form of other, allied genera.—The teleutospores of Endophyllum resemble æcidiospores, since they are united in chains, whose cells are easily separated, and are produced in the interior of a “peridium.” The multicellular teleutospores of Coleosporium function as basidia, and from each cell immediately produce basidiospores.—The teleutospores of Coleosporium and Chrysomyxa, differ from other teleutospores in the absence of exospore and germ-pore.
The ÆCIDOSPORES (Spring-spores) are produced in chains which are generally enclosed in an envelope of hyphæ, the peridium; the peridium enclosing the spores being termed the æcidium. The æcidiospores are unicellular, and generally of an orange colour; they are often separated by intermediate cells which wither and so assist in the distribution of the spores. The exospore is made up of minute, radially arranged rods. Generally germination proceeds immediately, the æcidiospore producing a germ-tube, which developes into a mycelium bearing either uredo- or teleutospores.
The æcidia of many Rust-Fungi were formerly considered as distinct genera. The æcidia of Phragmidium, Triphragmium, and Melampsora, in which the peridium is wanting, were in part considered as Cæoma. The æcidia with fimbriate edge, or those of Gymnosporangium with longitudinal lattice-like splits, were considered as “Rœstelia” (Lattice-Rust); large, sac-shaped æcidia on the Coniferæ were known as Peridermium.
The UREDOSPORES (Summer-spores) are unicellular and arise singly, seldom in chains (Coleosporium). Their colourless, warty exospore bears, in the equatorial plane, 2–8 germ-pores. In the majority, germination proceeds immediately, and a mycelium is produced which at first gives rise to uredospores and afterwards to teleutospores.
The uredospore-formations of Melampsorella and Cronartium are enclosed in an envelope, and hence resemble æcidia.—Between the uredospores sterile, unicellular hyphæ (paraphyses) may be found.
The spermogonia are spherical or pear-shaped conidiocarps, generally embedded in the substratum, and are produced before the æcidia, before or simultaneously with the uredospores, or before the teleutospores. The conidia, as far as observations go, do not generally germinate under ordinary conditions.
Among the Rust-Fungi some species are found which only form basidiospores and teleutospores (Puccinia malvacearum,[148] Chrysomyxa abietis). Other species have in addition uredospores; others spermogonia and uredospores; others spermogonia and æcidia; others spermogonia, uredospores and æcidia. Those species in which all the methods of reproduction are not developed must not be considered as incomplete forms.
As a rule the mycelium, which is produced from the basidiospores, developes æcidia; in the species, however, without æcidia, it developes the uredo-form, and when the uredospores are also absent, the teleutospore-form. It has been established in some species of Puccinia and Uromyces that the formation of æcidia can be suppressed, and it is not a necessary part of the cycle of development of the species.
The majority of Rust-Fungi hibernate in the teleutospore-form. Many species are able to hibernate in the uredospore-form (Coleosporium senecionis). Others pass the winter in the æcidio-form, and develope æcidia on new hosts (Uromyces pisi, on Euphorbia cyparissias; Phragmidium subcorticium, on Rosa; Æcidium elatinum, on Abies alba). In Chrysomyxa abietis, the mycelium, developed from the basidiospores, survives the winter.
Among the Rust-Fungi, with several forms of reproduction, there are about sixty whose development can only be completed by an alternation of hosts, that is, on one host only uredo-and teleutospores are produced, while the further development of the germinating basidiospores, and the formation of the æcidia and spermogonia from its mycelium, can only take place on a second quite distinct and definite host (heterœcious or metoxenous Fungi). Those Fungi which have all their forms of reproduction on the same host are termed autœcious or autoxenous. It is not, however, always necessary that the heterœcious Rust-Fungi should regularly change their hosts; for example, Puccinia graminis can hibernate in the uredo-form on the wild Grasses, and in the spring can distribute itself again in the same form.
As a consequence of the alternation of hosts the various forms of development were considered as independent genera (Uredo, Æcidium, Rœstelia, Cæoma, Peridermium), until De Bary and Oersted established, about the same time (1865), the mutual connection of some forms, and paved the way for the right conception of these Fungi.
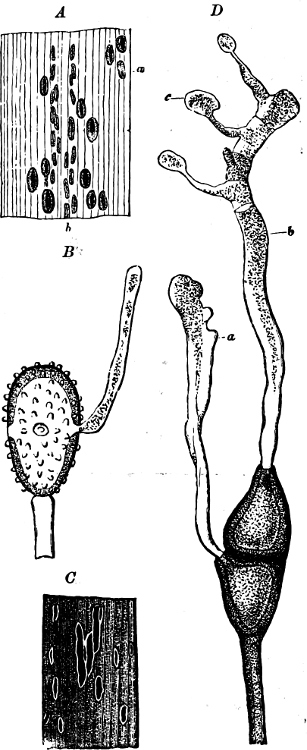
Fig. 146.—Puccinia graminis.
As an example of one of the most highly developed species, Puccinia graminis, the “Rust of Wheat,” holds a prominent position. Its uredospores and teleutospores are produced (Fig. 146) on Grasses (on cereals, especially Wheat, Rye, Oats, and many wild Grasses), while the æcidia and spermogonia are confined to[149] the Berberidaceæ. The teleutospores, developed on the Grasses, hibernate on the dried portions of their host, and in the succeeding year each of the two cells of the teleutospore may develop a basidium with four basidiospores (Fig. 146 D, c). The basidiospores are distributed by the wind, germinate quickly, and only proceed to further development on Berberis or Mahonia. The germ-tube bores through the epidermis of the Barberry-leaf, and forms a mycelium in its interior, its presence being indicated by reddish-yellow spots on the leaf. After 6–10 days the flask-shaped spermogonia appear (Fig. 147 B; C, a; conidia in Fig. 147 D) and a few days later the cup-shaped æcidia (Fig. 147 A; C, c, d, e). The former are generally on the upper, and the latter on the under side of the leaf. The orange-coloured æcidiospores scatter like dust, and germinate only on Grasses; the germination takes place in about two days when placed on any green part of a Grass. The germ-tube enters the Grass-leaf through a stoma; a mycelium is developed in the leaf, giving rise to a small, oval, rust-coloured spot (Fig. 146 A); in about 6–9 days the epidermis is ruptured over the red spot, and numerous reddish-yellow uredospores, formed on the mycelium, are set free. The uredospores (Fig. 146 B) are scattered by the wind, and can[150] germinate should they fall on the green portions of other Grasses: they then emit 2–4 germ-tubes through the equatorially-placed germ-pores. The germ-tubes enter a leaf through a stoma, a new mycelium is then developed, and in about eight days a fresh production of uredospores takes place, which germinate as before. The uredospore-mycelium very soon produces, in addition, the brown teleutospores, which give a brown colour to the rust-coloured spots, the familiar uredospores on the cereals being quite suppressed towards the close of the summer (Fig. 146 C, D). The “Rust of Wheat” hibernates on some wild Grasses in the uredospore-form.
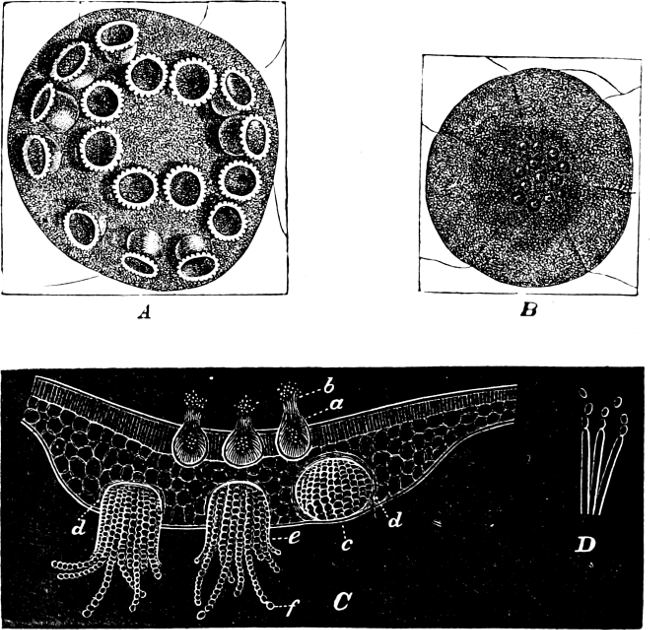
Fig. 147.—Æcidium berberidis. A Portion of lower surface of leaf of Barberry, with cluster-cups (æcidia). B A small portion of leaf, with spermogonia, from above. C Transverse section of leaf on the upper side, in the palisade parenchyma are three spermogonia (a b); on the lower side an unripe æcidium (c d) and two ripe æcidia (d, e, f); f chain of æcidiospores. D Hyphæ, forming conidia.
Genera. Puccinia (Fig. 146, 147) has bicellular teleutospores, each having a germ-pore, and the æcidia when present have an indented peridium; some species, as exceptions, have 1–3-celled teleutospores. Many species are HETERŒCIOUS, for example, P. graminis, described above; P. rubigo, which also infests various Grasses, but whose æcidia appear on Anchusa; the masses of[151] teleutospores are small; they contain paraphyses, and are for a long time covered by the epidermis. P. coronata, on Oats and Rye Grass; its æcidia on Rhamnus; the teleutospores are surmounted by a crown—“coronate processes.” P. phragmitis, on Reeds; æcidia on species of Rumex and Rheum. P. moliniæ, on Molinia cœrulea; the æcidia on Orchids. P. poarum, on Meadow-Grass; æcidia on Tussilago. Various Puccinias growing on species of Carex have their æcidia on Urtica, Lysimachia, Cirsium, Pedicularis, etc.—Of those AUTŒCIOUS species, which have all their generations on the same host, may be noted:—P. galii, P. menthæ, P. violæ, P. epilobii, P. asparagi, which grow on the hosts from which they have taken their specific names.—As representative of a group which have spermogonia, uredo-and teleutospores on the same host, but on different individuals, P. suaveolens, on the Field-Thistle, may be mentioned. The spermogonia have a strong odour.—A peculiar group (Leptopuccinia) has only teleutospores, which germinate immediately, and whilst still attached to their living host. To this group belong P. arenariæ, on a number of Caryophyllaceæ; and P. malvacearum, on various Malvaceæ, introduced in 1873 from South America to Europe, where it soon proved very destructive to Hollyhocks.
Uromyces (Fig. 149) differs only from Puccinia in always having unicellular teleutospores. Among this genus both heterœcious and autœcious species are found. To the first group belong U. pisi, whose æcidia are found on Euphorbia cyparissias, and U. dactylidis, whose æcidia appear on Ranunculus; to the second group belong U. betæ, U. phaseoli, U. trifolii.
Triphragmium has teleutospores with three cells (one below and two above), on Spiræa ulmaria.
Phragmidium (Fig. 150) has teleutospores consisting of a row of cells (3–10) arranged in a straight line; the upper cell has one germ-pore and the others four germ-pores placed equatorially. Both this and the preceding genus have large, irregular æcidia without peridia, but often with bent, club-like paraphyses (150 b and c); they are all autœcious, and are only found on the Rosaceæ.
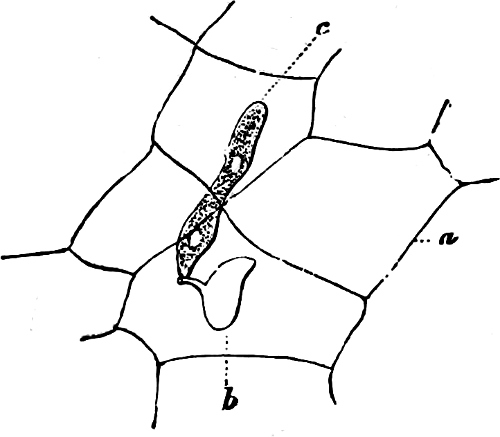
Fig. 148.—Gymnosporangium sabinæ. A small portion of the epidermis of a Pear-leaf (a) pierced at b by the germinating basidiospore (c).
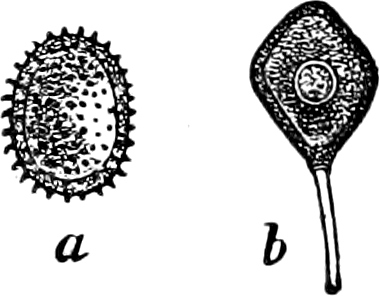
Fig. 149.—Uromyces genisteæ; a uredospore; b teleutospore.
Endophyllum (see above, under teleutospores, p. 147) on species of Sempervivum.
Gymnosporangium (Figs. 152, 154) has bicellular teleutospores collected in large, gelatinous masses formed by the swelling of the long spore-stalks; in each cell 2–4 germ-pores are found. Uredospores are wanting. All the species are heterœcious; the teleutospores appear on Juniperus, the æcidia (Rœstelia) on the Pomaceæ. G. sabinæ,[152] on Juniperus sabina, J. virginiana, etc., has the æcidia (“Rœstelia cancellata”) on Pyrus communis (Figs. 152, 148); G. juniperinum, on Juniperus communis with “Rœstelia cornuta” (Fig. 154 a) on Sorbus aucuparia, Aria nivea (S. aria) and Malus communis; G. clavariæforme on Juniperus communis, the æcidium belonging to it (“Rœstelia lacerata”) on Cratægus oxyacantha.
Melampsora has prismatic teleutospores placed parallel to each other and forming a crustaceous layer; in many species they are divided longitudinally into several cells (Fig. 151). The æcidia, without peridium, belonged to the old genus Cæoma. M. caprearum, on Willows, has the æcidia (Cæoma euonymi) on Euonymus. M. hartigii, on Osiers; the æcidium on Ribes. M. mixta, on Salix repens and Orchids. M. pinitorqua, on leaves of the Aspen, æcidia on Pine branches (Pine shoot fungus); M. populina on Populus monilifera and nigra; M. betulina (Fig. 153), on Birch leaves; M. padi (Fig. 151), on leaves of Prunus padus, developes teleutospores in the epidermal cells; M. lini is the cause of injury to the Flax; M. agrimoniæ.
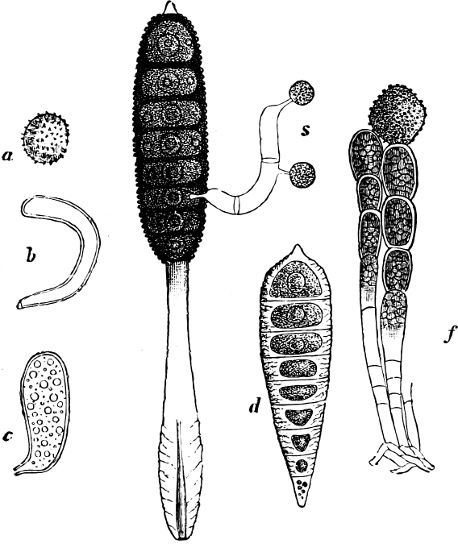
Fig. 150.—Phragmidium gracile: a an uredospore; b and c two paraphyses; d a young teleutospore; e a teleutospore with a basidium and two basidiospores (s); f two series of æcidiospores (Ph. rosæ).
Calyptospora gœppertiana; teleutospores on Vaccinium vitis idæa; spermogonia and æcidia on Abies alba (Firneedle-Rust).
Coleosporium (Fig. 155) forms its uredospores in reddish-yellow chains; for[153] the teleutospores, see page 147. C. senecionis, on the Groundsel; its æcidium (Peridermium wolffii) on Pine-leaves (Fig. 155 a). Other species on Sonchus, Petasites, Campanula, Rhinanthaceæ.
Chrysomyxa (Fig. 156) has bright red, branched teleutospore-chains; each spore developes a 4-celled basidium. C. ledi, on Ledum palustre; its æcidia on the leaves of the Fir. C. abietis (Fig. 156), without uredo-and æcidiospores; teleutospores on the leaves of the Fir. In the first summer, yellow bands are formed on the leaves, and in the following spring the red cushions of spores.
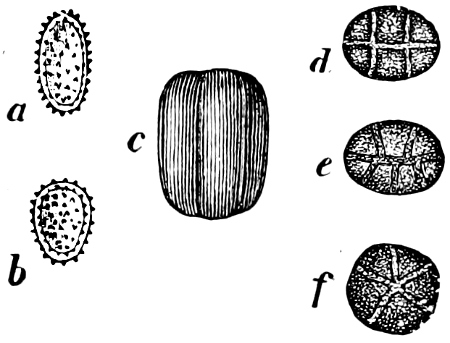
Fig. 151.—Melampsora padi: a and b uredospores; c-f teleutospores, seen from different sides.

Fig. 152.—Pear-leaf, seen from the under side, with “Rœstelia cancellata”: in different ages (a youngest, d oldest).
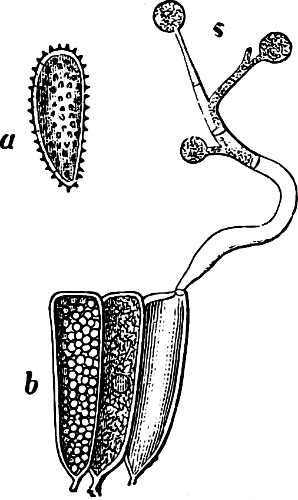
Fig. 153.—Melampsora betulina: a uredospores; b three contiguous teleutospores, one of which has developed a basidium with three basidiospores. (× 400.)
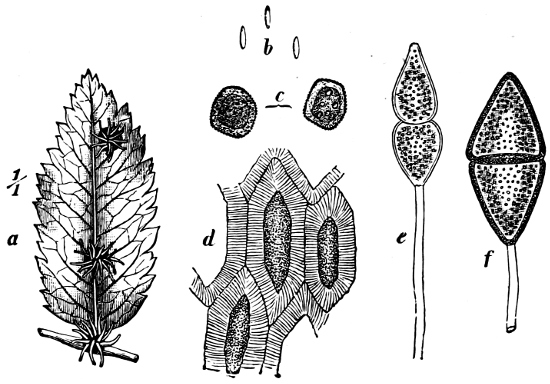
Fig. 154.—Gymnosporanginum juniperinum: a a small leaf with three clusters of æcidia (nat. size); b three conidia; c two æcidiospores on one of which are seen the germ-pores; d a portion of the wall of an æcidium; e, f two teleutospores.
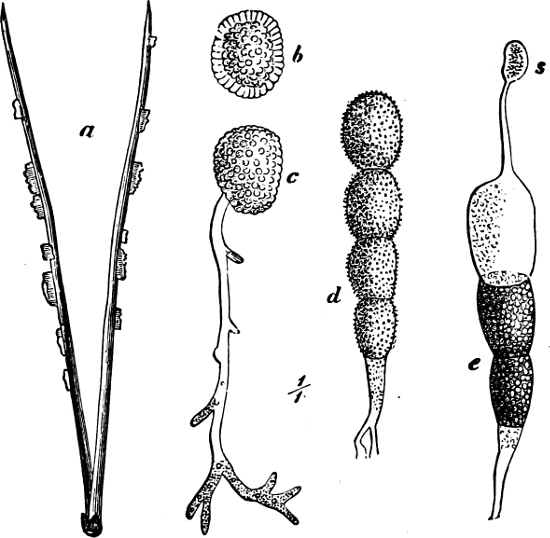
Fig. 155.—Coleosporium senecionis: a Pine-leaves with æcidia (Peridermium wolffii) nat. size; b an æcidiospore; c a germinating æcidiospore; d a chain of uredospores; e a chain of teleutospores of which the terminal one has germinated and produced a basidiospore (s).
[154]
Cronartium (Figs. 157, 159) has unicellular teleutospores united in numbers to form erect threads or columns; the uredospores are enclosed in a “peridium”; C. ribicola (Fig. 157), on leaves of Ribes (especially Black Currants); its æcidia (Peridermium strobi, or P. klebahni) on the stems and branches of Pinus strobus[155] (Fig. 159), on which it causes great damage; C. asclepiadeum, on Vincetoxicum officinale; its æcidia (Peridermium cornui) on the stems and branches of Pinus silvestris.
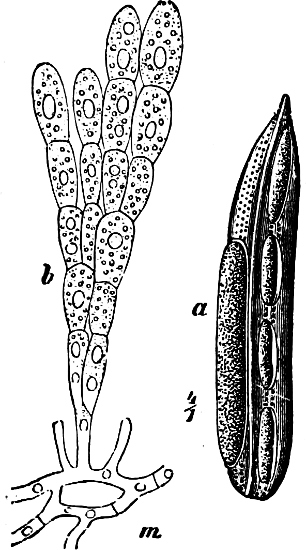
Fig. 156.—Chrysomyxa abietis: a leaf of the Fir, with 5 clusters of basidiospores (× 4); b branched rows of teleutospores springing from the mycelium (m).
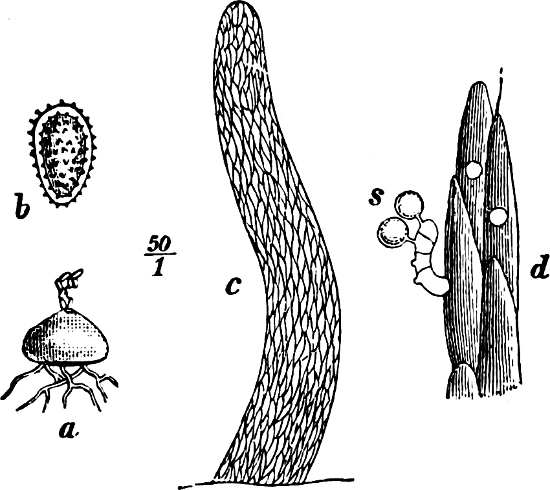
Fig. 157.—Cronartium ribicola: a mass of uredospores (× 50); b an uredospore; c a column of teleutospores (× 60); d a small portion of the same more highly magnified, with a basidium and two basidiospores (s).
To the Fungi of which the æcidium is known, whilst the remaining forms are still undetermined, but which are without doubt heterœcious, belong Æcidium elatinum, which produces the enormous “witches’ brooms” and barrel-shaped swellings on stems and branches of Abies alba; and Æcidium strobilinum (Fig. 158), which attacks Fir-cones, causing all the scales to become covered with clusters of æcidia opening by a lid. Hemileia vastatrix destroyed the coffee plantations in Asia.
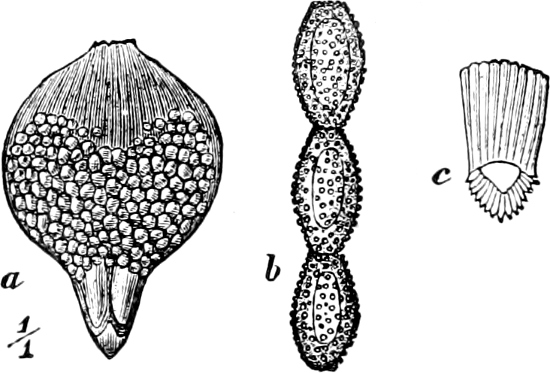
Fig. 158.—Æcidium strobilinum: a scale of cone of Picea excelsa, with numerous æcidia; b æcidiospores arranged in a series; c a cell of the peridium.
Order 2. Auriculariaceæ. The long, transversely divided basidia bear laterally 4 long sterigmata with basidiospores (Fig. 160 B) and are united to form an hymenium on the surface of the fruit-body. Parasites or saprophytes.
[156]
Auricularia sambucina (Auricula judæ), Judas’-ear, has large fruit-bodies, which may attain the size of several inches, resembling an ear or a mussel shell. In the moist condition they are flesh-coloured, tough and gelatinous, but when dried, become hard, grey and wrinkled; the exterior is covered with short hairs; while the internal surface bears the hymenium. Habitat: stems and branches of old Elder-trees (Sambucus).
Order 3. Tremellaceæ. The round, pear-shaped, longitudinally divided basidia bear 4 elongated sterigmata, situated apically, and 4 basidiospores (Fig. 160 C, D), and are united into the hymenium on the surface of the fruit-body. The fruit-bodies are frequently gelatinous and quivering; similar fruit-bodies are also found in the Dacryomycetaceæ and Hydnaceæ. Simple conidiophores, which appear not infrequently in the basidiocarps, before the basidia, are known in many species. Saprophytes.
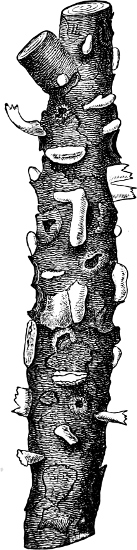
Fig. 159.—Peridermium strobi: æcidia of Cronartium ribicola (nat. size).
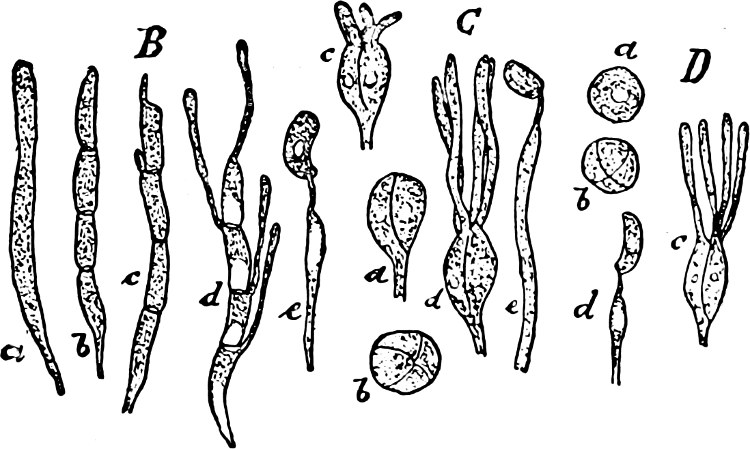
Fig. 160.—B Auricularia sambucina: a-d basidia in various stages of development; e a sterigma bearing a spore.—C Tremella lutescens: a-d basidia seen from various sides (b from above) and in various stages of development; e sterigma with basidiospore (× 400). D Exidia glandulosa: a-c various stages in the development of a basidium; d sterigma with basidiospore (× 350).
Exidia has kidney-shaped, oblong basidiospores, and small, hook-like conidia; E. glandulosa, E. albida, etc., on wood.—Craterocolla has conidiocarps; C. cerasi on Cherry-wood.—Sebacina incrustans; the yellow, fleshy, or cartilaginous fruit-bodies are found in autumn covering the ground in moist woods.—Tremella has round basidiospores; T. mesenterica has irregularly-folded,[157] quivering, orange fruit-bodies, about one inch in breadth; T. lutescens (Fig. 161) has orange-yellow conidial-and yellow basidial-layers; T. frondosa has fruit-bodies upwards of a foot in breadth.
Order 4. Pilacraceæ. The transversely divided basidia have no sterigmata, but sessile basidiospores, and fill up the cavity of a closed (angiocarpic) fruit-body as a gleba without a regular arrangement (hymenium wanting).
Pilacre fagi on the old stems of the Copper-Beech; P. petersii, on dried branches of the Hornbeam, has stalked, capitate fruit-bodies.

Fig. 161.—Tremella lutescens: I and II fruit-bodies (nat. size); III vertical section through a fruit-body; b basidia; c conidia; IV-VI basidia; VII basidiospore with a second spore; VIII a basidiospore with yeast-like budding (cultivated); IX a conidiophore. (III-IX about 400.)
This second and larger part of the Basidiomycetes is characterised by its more highly differentiated, undivided, club-shaped, or cylindrical basidia, which generally bear 4 (seldom 2, 6, 8) apically-placed sterigmata and basidiospores (Fig. 145). The fruit-bodies are partly gymnocarpic (in the first 3 orders and in some Agaricaceæ), partly hemiangiocarpic (in orders 3–6 of the Hymenomycetes[158] and in the Phalloideæ, the fruit-bodies in these orders are in the young conditions more or less angiocarpic, but later on generally open below and bear the hymenium on the under surface of the fruit-body), partly also angiocarpic (in the Gasteromycetes).

Fig. 162.—Dacryomyces deliquescens: I fruit-body (nat. size); II vertical section through the hymenium; III germinating basidiospore; IV a portion of mycelium with conidia; V a germinating conidium; VI and VII chains of oidia more or less strongly magnified; VIII basidiospore of D. longisporus; IX germinating basidiospore of D. ovisporus; X and XI Calocera viscosa; X fruit-body (nat. size); XI basidia with basidiospores (highly magnified); XII Dacryomitra glossoides (nat. size).
[159]
The long, club-shaped basidia bear two tapering sterigmata, which develope remarkably large basidiospores (Fig. 162 II, XI) and form gymnocarpic fruit-bodies with hymenium. 1 order:
Order 1. Dacryomycetaceæ. This order comprises 4 genera of which the first two develope the hymenium on the whole surface of the fruit-body, but the two last only on its apex.
Dacryomyces: the folded, gelatinous, Tremella-like fruit-bodies break out in winter on dried wood (hedges) in the form of red or yellow drops. D. deliquescens is very common (Fig. 121). The following genera have cartilaginous fruit-bodies.—Calocera (Fig. 162), with club-like, simple, or branched, Clavaria-like, fruit-bodies; the orange coloured fruit-bodies of C. viscosa grow aggregated together on the wood of Conifers.—Guepinia resembles a Peziza, and has the hymenium only on the hollow upper surface.—Dacryomitra resembles a Mitrula (Fig. 162).
This family is very rich in species (more than 8000 have been described), and to it belong all the “Mushrooms” and “Toadstools.” The fruit-bodies present very various forms; they are generally fleshy, very perishable, seldom leathery or corky, in the last case often perennial. The basidia are more or less cylindrical and bear generally 4 (seldom 2, 6 or 8) sterigmata and basidiospores. The hymenium in the fully-formed fruit-bodies lies free on the surface: in orders 1 and 2 and a portion of order 6 it is from the commencement exposed, fruit-bodies gymnocarpic; orders 3–6 have hemiangiocarpic fruit-bodies (p. 157). In the first order the basidia (or the hymenium) are developed immediately from the mycelium (Fig. 163); the fruit-bodies of orders 2 and 3 present a higher grade of development, and have between the mycelium and hymenium a special hyphal-tissue, a stroma, which is crustaceous, club-like, or coralloid, etc., and in general bears the hymenium on the largest part of the free, smooth surface. In the forms most highly developed (orders 4–6) a new tissue—the hymenophore—is introduced between the stroma and hymenium, which appears on the under side of the fruit-body in the form of warts, projections, tubes, folds or lamellæ (Figs. 166, 167, 174 bc). Paraphyses are frequently found in the hymenium, among the basidia. In the Hymenomycetes few examples of conidia can be recognised at first. More frequently chlamydospores are found, particularly oidia. The mycelium is richly branched, generally colourless, often perennial; it lives in humus or decaying wood, and is seldom parasitic.[160] The hyphæ generally have clamp-connections and unite, sometimes, to form a rhizomorpha (Fig. 177) or sclerotia with coloured, pseudo-parenchymatous covering.
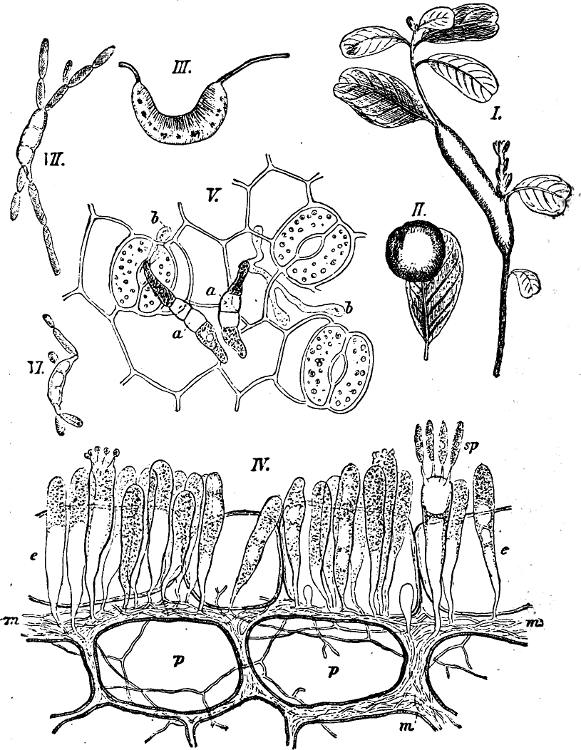
Fig. 163.—Exobasidium vaccinii. I Hypertrophied stem of Vaccinium vitis idæa; II leaf with gall-like swelling; III section of II; IV transverse section: m mycelium between the parenchymatous cells; p hypodermal cells; e epidermis with basidia in various stages of development; V epidermis with germinating spores; VI and VII spores germinating in water (IV-VII × 620).
[161]
Order 1. Tomentellaceæ. To this order belong the simplest of the Hymenomycetes. The basidia (Fig. 145) arise free and irregularly from the mycelium; a hymenium is entirely absent or very slightly formed (in Corticium it attains its highest development); fruit-bodies are also wanting.—In general they form flaky, membranous or leathery coverings on bark and wood. Some are parasites.
Hypochnus without conidia.—Tomentella with conidiophores; growing on wood or earth.—Exobasidium vaccinii (Fig. 163), a parasite on Vaccinium, Andromeda, Arctostaphylos, and Rhododendron, forms flaky-powdery, white or red coverings and may cause hypertrophy of the parts attacked. E. warmingii is parasitic on Saxifraga; E. lauri causes outgrowths on the stem of Laurus canariensis as long as a finger, which formerly were regarded as aerial roots.—Corticium forms membranous to leathery layers or crusts; C. quercinum on wood and bark, particularly Oak, is flesh-coloured; C. cæruleum has a blue hymenium; C. giganteum on the bark of fallen Pine-trees.
Order 2. Clavariaceæ. The hymenium is situated on a stroma, and either completely covers the smooth surface of the more or less fleshy gymnocarpic fruit-body, or is confined to a tolerably well defined upper portion of it (Typhula). Paraphyses absent. The vertical, white, yellow, or red fruit-bodies are roundish or club-like, undivided or richly branched (Fig. 125). Generally on the ground in woods, seldom on tree-stems, etc.
Fig. 164.—Clavaria coralloides (nat. size).
Genera: Clavaria, generally large Fungi with thick, round branches. C. botrytis has a very thick, tubercular stem with numerous short, flesh-coloured branches: it has an agreeable taste. C. coralloides has a brittle, richly-branched fruit-body (Fig. 164); basidia with two large spores. C. pistillaris consists of a single, undivided club of a yellowish-white colour.—Sparassis has compressed, leaf-like, curled branches; S. crispa has fruit-bodies as large as a white cabbage-head, with an agreeable taste.—Typhula and Pistillaria are small Fungi with filamentous stalks, terminating in a small club. The fruit-bodies of the former often arise from a small, spheroid sclerotium; the latter is distinguished by the basidia bearing only two spores.
[162]
Order 3. Thelephoraceæ. The hymenium is placed on a stroma and covers the smooth surface of the leathery hemiangiocarpic fruit-body, generally on its under side. The edge of the stroma, which bounds the hymenium, is sometimes especially developed (Stereum). Saprophytes.
Genera: Thelephora. The fruit-bodies in this genus are brown, very irregularly shaped, and often lobed. The spores too are brown, but in the other genera colourless. The species are found growing on barren soil. T. laciniata (Fig. 165) has imbricate, semicircular, dark-brown pileus, which is jagged at the edge and upper surface. The fruit-bodies are very often raised above the ground, and although this species is not a parasite, yet it destroys young seedlings by growing above and smothering them.—Stereum has a stiffer fruit-body, with a distinct, fibrous, intermediate layer. It grows on bark and wood, projecting like a series of imbricate brackets. S. hirsutum is yellow; its free edge is provided with a number of stiff hairs, the upper surface being divided into a number of zones. S. purpureum has a red-violet hymenium which distinguishes it from the previous species.—Cyphella has a membranous cup- or bell-shaped fruit-body, often borne on a stalk, the concave surface being covered with the hymenium. They are small, white Fungi, growing on Moss and dead stems.—Solenia is closely related to Cyphella; its fruit-bodies are smaller and hairy; they are found clustered together forming a crust-like covering on dead wood.—Craterellus has a large, funnel-shaped fruit-body, the hymenium covering the external surface. C. cornucopioides is shaped like a trumpet or a “horn of plenty.” It is dark-grey, several inches in height, and grows gregariously on the ground in forests. It is distinguished by the basidia bearing only two sterigmata.

Fig. 165.—Thelephora laciniata (nat. size).
Order 4. Hydnaceæ. The fruit-body is most frequently fleshy, and varies considerably in shape, the simplest forms being resupinate,[14] the higher ones umbrella-like. The hymenophore is found on the free or downward-turned surface, and always takes the form of soft emergences hanging vertically downwards. The emergencies may be thorn-, awl-, or wart-like. The species are found growing on the soil and on dead wood.
Genera: Hydnum has subulate, distinct emergences. H. repandum is yellow, the stalk being placed in the centre of the pileus. It is an edible[163] species, and often forms “fairy rings” in woods. H. auriscalpium (Fig. 166) is dark-brown, with stalk placed at the edge of the pileus. It grows on old Fir-cones. H. erinaceus grows on old tree-trunks. The fruit-body is yellow and very large—as big as a human head—with emergences as much as an inch in length.—Irpex has a leathery fruit-body, partly resupinate, partly with free, projecting edge; the under side bears tooth-like emergences which are arranged in rows, and Irpex thus forms a transition to the Agaricaceæ.—Phlebia is entirely resupinate, with radially-arranged folds on the free side, and pectinate border.
Fig. 166.—Hydnum auriscalpium, upon a Fir-cone, in different stages of development.
Order 5. Polyporaceæ (Pore-Fungi). An order very rich in species (about 2000 species are described). The fruit-body is of very different forms—resupinate, projecting like a bracket, hoof-like, or umbrella-shaped. In some it is fleshy and edible, in others leathery or corky, persisting for several years. The hymenophore is situated on the under side of the fruit-body, and consists of wide or narrow tubes or pores, whose inner surface is clothed with the hymenium (Fig. 167). In some fruit-bodies large cavities are to be found, which have arisen as interstices between the labyrinthine curved and reticulate folds. Chlamydospores are known in some species. Conidia occur very rarely. Many species work considerable damage: some as parasites on trees, others by destroying timber.

Fig. 167.—Polyporus igniarius. Section through the under side of the Fungus: h-h is hyphal-tissue between the tubes, formed by irregularly felted hyphæ, many of which are seen cut across; s is the hymenium which covers the walls of the tubes, and from which the basidia with the spores protrude.
Genera. Polyporus (Pore-Fungus). The tubes are narrow, accurately fitted together, and forming a thick layer on the under side of the fruit-body, appearing[164] as a number of fine holes. The fruit-body most frequently resembles a bracket, or is hoof-shaped, with one side growing from a tree-trunk; it is very often perennial, and a new layer of tubes arises in each succeeding period of vegetation. Strata, corresponding to the periodically interrupted growth, are thus formed in storeys one above the other, and are visible on the upper surface of the fruit-body, as well as in the interior, as a series of concentric belts, sometimes as many as half a score or more in number. P. fomentarius (Touchwood) attacks trees, especially the Beech. The spores germinate on wounds from broken branches, and the hyphæ, following the course of the medullary rays, find their way into the interior of the tree, from whence the mycelium spreads upwards, downwards, and peripherally, so that the wood becomes rotten (“white-rot”) and thick felts of mycelium are formed in radial and tangential directions. A dark line, caused by the youngest parts of the hyphæ containing a brown juice, marks the boundary between the rotten and the unattacked parts of the stem (Fig. 168); at places where the mycelium extends to the bark, the cambium becomes destroyed and further growth is arrested, so that longitudinal furrows arise on the stem. It is at these places, too, that the hoof-shaped, ash-coloured fruit-bodies are developed, which may attain a circumference of upwards of 7 feet. The interior of the fruit-body consists of a dried-up, loosely felted, red-brown mass of hyphæ, which has been used for tinder and as a styptic (“Fungus chirurgorum”). P. igniarius has a harder, dark-brown, more rounded fruit-body; it grows in a similar manner, but especially attacks Oaks, Poplars, and Plum-trees, the wood of which becomes rotten, and is called touchwood. P. pini (Trametes pini), (Fig. 170), a parasite on the stems of Pinus, causes a kind of “red-rot” in the stem. P. sulphureus has a soft, cheesy, yellow fruit-body; it produces “rot” in Oaks and Apple-trees. P. officinalis, Larch-fungus (“Fungus Laricis” in Pharmocopœia), grows on Larch-trees in the south-east of Europe. P. versicolor has thin, semicircular[165] fruit-bodies, with zones of various colours on the upper side; it is one of the most frequent species on tree-stems. P. frondosus grows on soil in woods, and consists of numerous aggregated fruit-bodies, which become very large and fleshy. This species is edible. P. perennis also grows on the soil in woods; it is very leathery, with central stalk, and has concentric zones on the upper surface of the fruit-body. P. vaporarius destroys the wood of living Pines (Pinus silvestris) and Firs (Picea excelsa), causing it to become red-brown; in timber this Fungus causes “red-strip” followed by a “dry-rot.” P. squamosus destroys many Walnut-trees, and is also very destructive to Limes and Elms. P. fulvus causes a “white-rot” in Abies alba.
Fig. 168.—Section of stem of a Beech attacked by P. fomentarius: a non-attacked parts of the stem; b the furrows where the mycelium has reached the bark, and where the thick mycelium-strands reach the exterior (⅙th of the nat. size).
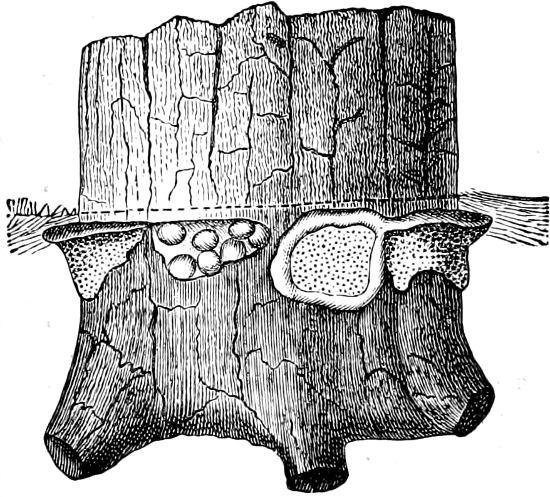
Fig. 169.—Base of a Fir-tree, with a number of fruit-bodies of Heterobasidion annosum just beneath the surface of the soil, indicated by the dotted line (¼th nat. size).
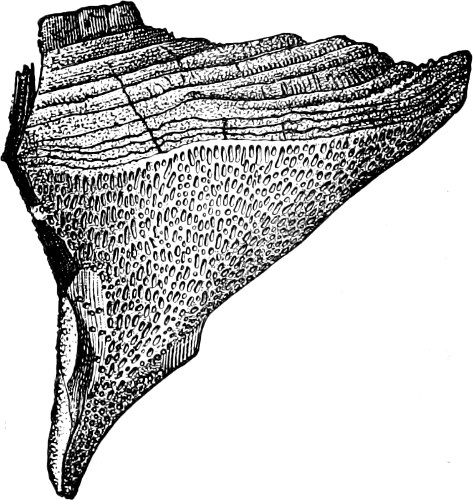
Fig. 170.—A fully developed fruit-body of Polyporus pini (Trametes pini), lateral view (nat. size).
Heterobasidion annosum (Polyporus annosus, Trametes radiciperda, Fig. 169) is characterized by its Aspergillus-like conidiophores. It is a parasite on the Pine, Fir, Birch, Beech, etc.,[166] and is the chief cause of a root-disease (red-rot) in Pines and Firs; the fruit-bodies develope a large number of basidiospores; they may be very large and are found just beneath the surface of the soil (on living or dead roots), and exposed to the air (on felled stems and roots, in Scandinavia).
Ptychogaster has cushion-like fruit-bodies, which consist chiefly of chlamydospore-chains, formed of ellipsoidal spores, which alternate with short hyphæ having transverse septa and clamp-connections. The hymenial portion is limited to a small group of tubes. Pt. albus (Oligorus ustilaginoides) grows on stumps of Conifers and forms irregular cushions, at first white and later on brown, which consist almost entirely of chlamydospores.
Boletus (Fig. 171) has a fleshy fruit-body resembling a common Mushroom, with central stalk. The layer of tubes is easily detached from the pileus, and the tubes are easily separable from one another. They grow on the ground in woods. Edible species are: B. edulis, with thick, reticulate stalk; B. scaber, with thin stalk and rough pileus; B. luteus, with a ring on the stalk. B. luridus is poisonous, its tubes have red openings, and the flesh turns quickly blue when broken and exposed to the air.
Fistulina hepatica (Beef-steak Fungus), has a red, fleshy, edible fruit-body, with red juice. The tubes are individually distinct; conidia are also developed. Grows on old Oaks.
Merulius lacrymans (“Dry-rot”) has a resupinate fruit-body with white, cotton-like border, and the remaining portions covered by reticulate, ramified veins of a rust-brown colour. In favourable vegetative conditions it is fleshy and exudes large drops of water—hence its specific name and also the name “Tear Fungus.” The mycelium is at first colourless, and then yellow-brown; when dry it is tough and leathery. It destroys the timber in damp houses, extends far and wide over boards and beams and even over the masonry, giving rise to a disagreeable smell in the rooms in which it lodges. In woods the Fungus lives on Pine-stems. It is brought from the forest on the logs of timber, and is distributed from log to log by the mycelium and the basidiospores. The living mycelium can be recognised by the clamp-connections shooting out branches. The basidiospores are often ejected a distance of a metre; they are elliptical (10–11µ long and 5–6µ broad), and germinate easily on damp wood, or in fruit-juice which has been neutralized with urine or alkaline carbonates.
Dædalea (Labyrinth Fungus), has bracket-like, corky fruit-bodies with irregularly-folded plates or discs on the under side. It forms a transition to the Agaricaceæ. D. quercina is frequent on Oak-stumps.
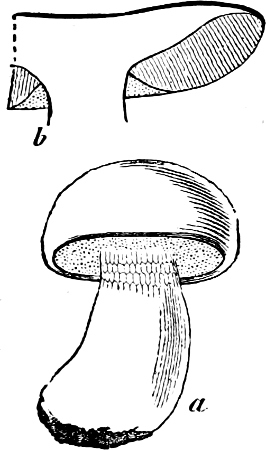
Fig. 171.—Boletus edulis (about ¼th): b longitudinal section of a portion of the pileus.
Order 6. Agaricaceæ (Mushrooms, Toadstools). The hymenophore consists of knife-like plates (lamellæ, gills), which are situated on the under side of the umbrella-like pileus of the fruit-body,[167] and radiate from the central stalk. Those which are first formed extend from the edge of the pileus to the stalk; those formed later reach only a longer or shorter portion of this distance, according to their age. In structure the lamellæ (Fig. 174) consist of a central mass of hyphæ, the trama, continuous with the hyphæ of the pileus; these terminate in a layer of shorter cells, the subhymenial layer, immediately beneath the hymenium which is composed of basidia and paraphyses. In a few species, but not in the majority, the lamellæ are branched, and in some they are decurrent. A few have the stalk placed excentrically, or it may be entirely absent.
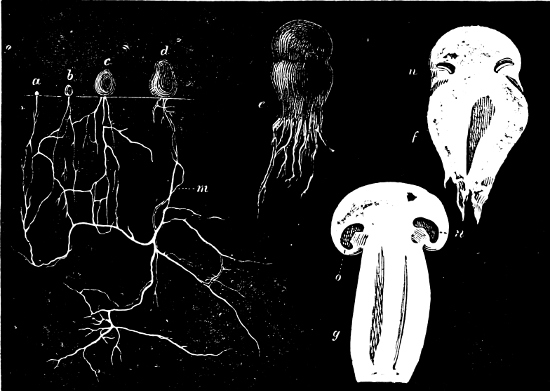
Fig. 172.—Development of Psalliota campestris: a, b, c, d show the various stages of the development of the fruit-bodies and the mycelium (m) (nat. size); e the fruit-body in a somewhat later stage, slightly magnified; f longitudinal section of e; n first formation of the hymenium; g longitudinal section of a more advanced fruit-body (nat. size); n the hymenium; o velum partiale (see Fig. 133.)
In the early stages of its development the fruit-body is more or less enclosed in a hyphal tissue—the “veil” (velum universale, or volva). The veil at first completely encloses the young fruit-body, but is afterwards ruptured as the latter grows, part remaining at the base of the stalk as the “sheath” (annulus inferus), and part on the pileus as scales or warts. In the “Fly Mushroom” (Amanita muscaria) the remains of the veil are especially conspicuous[168] as white patches on the bright red ground of the upper surface of the pileus, and as a sheath at the base of the stalk (Fig. 178 v.). Another veil—the velum partiale—a hyphal tissue (Figs. 178 a; 173) stretches from the edge of the pileus to the stalk, and encloses the lamellæ. This veil is ruptured as the pileus expands, a portion attached to the stalk remaining as the “upper ring” (annulus superus) (Figs. 173, 178 a), or a part attached to the pileus hanging down as a fringe round its edge.—Some genera have no veil, the under side of the pileus being exposed from the first (gymnocarpic Agaricaceæ). Those which have a veil (hemiangiocarpic A.) afford a transition to the angiocarpic Gasteromycetes.
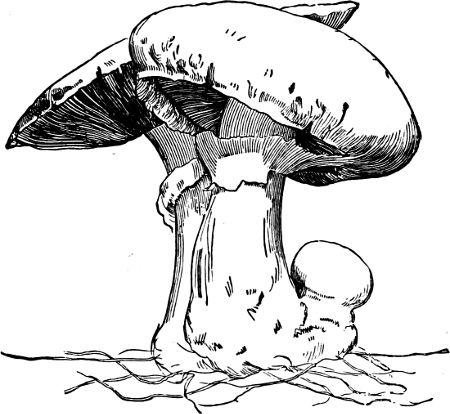
Fig. 173.—The cultivated Mushroom (Psalliota campestris).
The mycelium mostly grows in soils rich in humus or dung, on decaying trees and similar objects. Many species, e.g. Tricholoma personatum and Marasmius oreades, form the so-called “fairy rings.” The fruit-bodies in these species are confined to a larger or smaller surface on which they are very regularly arranged in a ring. The reason for this is found in the radial growth of the mycelium, so that the oldest portion, or the starting point, is found at the centre of the ring, and the younger ones, on which the fruit-bodies are formed, at the circumference. The older hyphæ gradually die, and at the same time, the radial growth continuing, the ring of fruit-bodies becomes larger and larger. The “fairy-rings” are marked[169] not only by the fruit-bodies, but also by the more vigorous growth and darker colour of the grass upon these spots.
Some species are parasites. An example is presented by Armillaria mellea, a remarkable and very destructive Fungus in woods and forests (Figs. 176, 177). In addition to the filamentous, white mycelium, it has also black, or black-brown, horny, root-like mycelium-strands (rhizomorpha) which were formerly considered to belong to a special genus of Fungi described under the name “Rhizomorpha.” The mycelium lives parasitically on the Conifers and other trees, forcing its hyphæ into the bark and between the bark and wood, and thence penetrating into the wood so that the tree is very severely attacked. It may also live saprophytically, and clusters of fruit-bodies are often found on old stumps and stems, on old timber, and in the rich soil of woods. The rhizomorpha, living underground, can extend for considerable distances and infect the roots of neighbouring trees, and spreads in this way the diseases known as “Harzsticken” and “Bark-Canker,” which are very destructive to young trees.
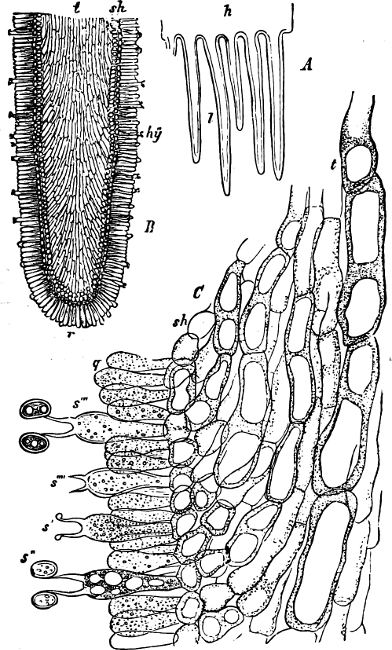
Fig. 174.—Psalliota campestris. A Tangential section of pileus showing lamellæ (l). B Portion of gill more highly magnified; t trama; hy hymenium with basidia and basidiospores; sh, subhymenial layer. C A portion of the same more highly magnified; s′ s′′ s′′′ s′′′′ various stages in the development of basidiospores; q paraphyses.
The chief characteristics by which the numerous genera are separated are the presence or the absence of the two kinds of veils, the nature of the fruit-body, the form, branching of the lamellæ, and their position and relation with respect to the stem, the shape of the[170] pileus, the colour of the spores, etc., etc. A knowledge of the colour may be obtained by placing the pileus with the lamellæ turned downwards on a piece of white or coloured paper, so that the spores, as they fall off, are collected on the paper, and the arrangement of the lamellæ can then be clearly seen.
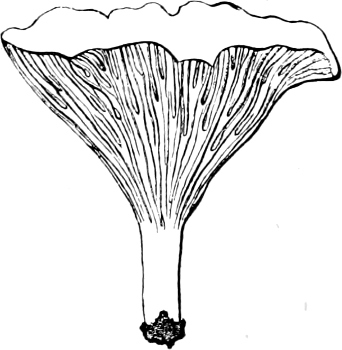
Fig. 175.—Cantharellus cibarius (reduced).
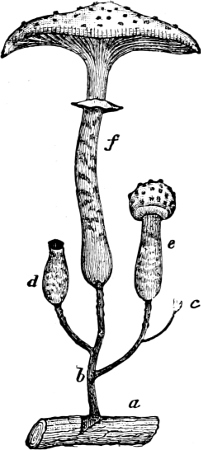
Fig. 176.—Armillaria mellea. (½ nat. size): a root of a Fir; b rhizomorpha-strands; c-f fruit-bodies in four different stages of development.
Fig. 177.—The mycelium of Armillaria mellea (“Rhizomorpha”) (nat. size).
[171]
About 4,600 species belonging to this order have been described.
On account of the large number of species the order is divided into several sections:
1. Agaricinei; fruit-body fleshy; lamellæ membranous, knife-like, with sharp edge; basidia crowded together. The FOLLOWING HAVE WHITE SPORES:—Amanita (Fly Mushroom), with volva, and generally also the upper ring on the stalk; many are poisonous, such as A. muscaria (Fig. 178) which has bright red pileus with white spots, A. pantherina and A. phalloides; A. cæsarea is edible.—Lepiota procera (Parasol Fungus) is one of the largest Mushrooms; it has a scaly pileus and moveable ring (edible).—Armillaria mellea has been mentioned above (Figs. 176, 177).—Tricholoma, lamellæ indented near the stalk; T. gambosum (Pomona Fungus) belongs to the best of edible Fungi; T. personatum often forms fairy rings (see above).—Clitocybe, lamella decurrent; C. nebularis is edible.—Pleurotus, stalk eccentric; P. ostreatus (Oyster Mushroom) grows in clusters on tree-stems (edible).—Collybia and Mycena, species numerous, small.—Spores rose-red: Volvaria and Hyporhodius.—Spores Brown: Cortinarius, with cobweb-like veil; Pholiota, membranous veil and ring; P. squarrosa in clusters on tree-stems; P. mutabilis, on tree-stumps (edible).—Spores Violet-purple: Hypholoma, Psalliota; to this section the common edible Mushroom (Fig. 172–174) belongs, with annulus and chocolate-coloured lamellæ; it is cultivated for the sake of the fine flavour.—Spores Black: Coprinarius.
Fig. 178.—Fly Mushroom (Amanita muscaria).
2. Marasmiei. Fruit-body tough, almost leathery, and persistent; spores white. Marasmius oreades forms large, regular fairy-rings on pastures and commons; it is used as seasoning in food.—Panus stipticus with eccentrically-placed stalk, in clusters on tree-stumps.—Schizophyllum has the edge of the lamellæ divided longitudinally, and the split portions revolute.—Lentinus affords a transition to Dædalea among the Polyporaceæ.
3. Russulei. Fruit-body fleshy and fragile, in which two different systems of hyphæ may be distinguished; spores thorny, white, or pale-yellow. Many are poisonous.—Russula has generally fragile and thick lamellæ reaching from stalk to edge of pileus; pileus frequently red.—Lactarius has white or yellow[172] milky juice, which often is very acid. L. deliciosus has red-yellow milky juice, and is of a pleasant flavour. L. torminosus is poisonous.
4. Hygrophorei. Lamellæ thick and waxy, widely separated; spores white. Many species of Hygrophorus have brightly-coloured pileus and grow among the grass on moors and commons.—Nyctalis is parasitic on larger Toadstools. It is remarkable for its abundant formation of chlamydospores, whilst the basidiospores are little developed.
5. Coprinei. Fruit-bodies very soft, quickly perishable; lamellæ membranous and deliquescent. The basidia are separated from each other by paraphyses. Coprinus has coal-black spores, grows on manure, and sometimes developes sclerotia.
6. Paxillei. Fruit-body fleshy; lamellæ easily detached from the pileus and reticulately-joined near the stalk. They form a connecting link between the Agaricaceæ and Boletus.
7. Cantharellei. Lamellæ reduced to dichotomously-divided folds, decurrent on the stalk. Cantharellus cibarius (Fig. 175) is yolk-yellow, and grows on the ground in woods (edible). It is allied to Craterellus.
The fruit-bodies before they are ripe are spherical or ovoid, and enclosed by a fleshy covering, the peridium, which is perforated at maturity and remains as a sheath (Fig. 179); the fruit-bodies are hemiangiocarpic.
Order 1. Phallaceæ (Stink-horns). The peridium has a complicated structure and is composed of three layers, the intermediate one being thick and gelatinous. The gleba (the tissue which bears the hymenium) is situated upon a peculiar receptacle which expands into a porous stalk and by its sudden distension, rupturing the peridium, elevates the gleba and hymenium above the peridium, which remains as a sheath. The gleba becomes gelatinous and dissolves away as drops. To this order belong many peculiar and often brightly coloured forms, which are natives of the Southern Hemisphere.
Phallus impudicus (Stink-horn) (Fig. 179), has a fruit-body which at first is white, heavy, and soft, and resembles a hen’s egg in shape and size. The peridium is divided into three layers (Fig. 179 e, g, f) of which the external and internal are membranous, and the middle one very thick and gelatinous; each of these has again a laminated structure. The peridium when ruptured remains as a sheath (k) at the base of the stalk. The receptacle at first is strongly compressed (h) but afterwards expands into a long stalk (l) which bears the conical gleba (m). Prior to the rupture of the peridium the gleba consists of a greenish mass (i) which, when exposed, emits a carrion-like stench serving to attract flies, by whose agency the spores are distributed. It is found commonly in hedgerows and in woods, growing on the ground. The much[173] smaller and less common P. caninus is found on rotten tree-stumps.—In Clathrus cancellatus the receptacle expands into a bright red, reticulate structure. A native of the South of Europe. Colus, Aseroë, Mitromyces.
Order 2. Sphærobolaceæ. An intermediate layer of the peridium swells when ripe, becomes convex, and ejects the remaining spherical portion of the fruit-body which contains the spores. Sphærobolus carpobolus has small, spherical fruit-bodies which open in the form of a star.
Fig. 179.—Phallus impudicus (Stink-horn), somewhat diminished. Fruit-bodies in all stages of development (b, c, d and k-m) are seen arising from a root-like mycelium (a); d longitudinal sections through a fruit-body before the covering has ruptured.
The fruit-body is angiocarpic, fleshy at first, and later generally more or less hard and continues closed after the spores are ripe. The tissue lying immediately inside the peridium is termed the gleba; it is porous, containing a larger or smaller number of chambers lined with the hymenium, which is either a continuous[174] layer of basidia or else it fills up the entire cavity. The basidia as a rule bear four spores, sometimes eight (Geaster), or two (Hymenogaster). The tissue of the walls (trama) consists often (Lycoperdaceæ) of two kinds of hyphæ, some thin and rich in protoplasm, divided by transverse septa and bearing the basidia; others thicker and thick-walled which do not dissolve like the former on the ripening of the spores, but continue to grow and form a woolly, elastic mass, the capillitium, which may be regarded as highly developed paraphyses. The peridium may be either single or double, and presents many variations in its structure and dehiscence. The mycelium is generally a number of string-like strands, living in soils rich in humus.
Order 1. Tylostomaceæ. Capillitium present. After the rupture of the peridium the remaining part of the fruit-body is elevated on a long stalk. Tylostoma mammosum, on heaths.
Order 2. Lycoperdaceæ. The fruit-body has a double peridium; the external one at length breaks into fragments (Lycoperdon, Bovista), or it has a compound structure of several layers (Geaster) and detaches itself as a continuous envelope from the inner layer, which is membranous and opens at its apex. The interior of the fruit-body consists either solely of the fertile gleba (Bovista, Geaster), or, in addition, of a sterile tissue at the base (Lycoperdon). A capillitium is also present.

Fig. 180.—Lycoperdon gemmatum (½ nat. size).
Lycoperdon (Puff-ball) has a sterile part at the base of the fruit-body which often forms a thick stalk. The surface of the peridium is generally covered with warts or projections. When young this Fungus is edible, but when ripe it is dry, and used for stopping the flow of blood. L. giganteum, which is often found growing in meadows, attains a considerable size, its diameter reaching as much as eighteen inches. L. gemmatum (Fig. 180) is covered with pyramidal warts; in woods.—Bovista has no sterile basal part; the external peridium is smooth, and falls away in irregular patches. B. plumbea, on links near the sea.—Geaster (Earth-star) has an external peridium composed of several layers, which when the fruit-body opens, split into several stellate segments. These segments are very hygroscopic, and in dry weather bend backwards and so raise the inner peridium into the air. The inner peridium contains the spores[175] and capillitia. G. coliformis has several apertures in the inner peridium. The other species have only one regular aperture at the apex. G. striatus has a pedicellate inner peridium, with conical, striped peristome. G. fornicatus has an external peridium split into four segments. This last and several other species produce “mycorhiza” on the roots of Conifers.
Fig. 181.—I Hymenogaster citrinus (nat. size); II longitudinal section through H. tener (× 5); III portion of a section of H. calosporus; g a chamber; h hymenium; sp. spores; t trama (× 178); IV Rhizopogon luteolus (nat. size); V Scleroderma vulgare, VI section of V; VII basidia with spores belonging to the same Fungus.
Order 3. Sclerodermataceæ. Capillitium wanting. The peridium is simple and thick, gleba with round, closed chambers, which are filled with basidia.
Scleroderma has a corky peridium. The fruit-bodies commence their development under ground. S. vulgare (Fig. 181 V-VII), has a hard, slaty-black gleba.
[176]
Order 4. Nidulariaceæ (Nest-Fungi). Small Fungi of which the fruit-body at first is spherical or cylindrical but upon maturity it becomes cupular or vase-like, and contains several lenticular “peridiola” lying like eggs in a nest. The peridiola are the chambers which contain the hymenium, covered by a thin layer of the gleba, all the remaining portion of the gleba becoming dissolved. On decaying wood.
Nidularia has spherical fruit-bodies containing a large number of lenticular peridiola, embedded in a slimy mass.—Crucibulum has fruit-bodies resembling crucibles with discoid peridiola, each with a spirally-twisted stalk.—Cyathus has a fruit-body, which when open is campanulate, with stratified peridium, and long-stalked, lense-shaped peridiola.
Order 5. Hymenogastraceæ. Fruit-bodies tubercular, globose and subterranean, resembling very closely the Truffles, from which they can only be distinguished with certainty by microscopic means. The peridium is simple, capillitium wanting, and the gleba encloses a system of labyrinthine passages covered with a continuous hymenium. The fruit-bodies persist for some time, and form a fleshy mass, the spores being only set free by the decay of the fruit-body, or when it is eaten by animals. The majority are South European. Hymenogaster, Melanogaster, Rhizopogon (Fig. 181 I-IV).
Appendix to the Basidiomycetes:
Basidiolichenes (Lichen-forming Basidiomycetes).
Several Fungi belonging to the Basidiomycetes have a symbiotic relationship with Algæ exactly similar to that enjoyed by certain Ascomycetes, and these are therefore included under the term Lichens (p. 136). They are chiefly tropical.
Order 1. Hymenolichenes. To this order belong some gymnocarpic forms: Cora, Dictyonema, Laudatea.[15]
Order 2. Gasterolichenes. To this belong some angiocarpic forms: Emericella, Trichocoma.
Appendix to the Fungi.
Fungi imperfecti (Incompletely known Fungi).
1. The Saccharomyces-forms are Fungi which are only known in their yeast-conidial form. They are conidia of higher[177] Fungi which can multiply to an unlimited extent by budding in nutritive solutions, and in this way maintain their definite size and shape. The budding takes place only at the ends of the conidia. The wall of the conidium forms at one or at both ends a small wart-like outgrowth, which gradually becomes larger, and is finally separated from its mother-cell as an independent cell, surrounded by a closed cell-wall (Fig. 182 a, b).
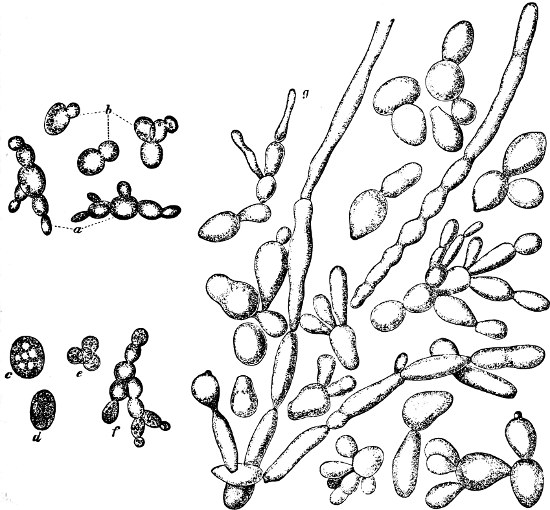
Fig. 182.—Beer-yeast (Saccharomyces cerevisiæ): a-b (× 400); c-f (× 750); c a cell in the process of forming spores; d a cell with four ripe spores; e the spores liberated by the dissolution of the cell-wall; f three germinating spores; g mycelium-like cell-chains. (× 1000: after Em. Chr. Hansen.)
Under very favourable conditions multiplication occurs so rapidly that the daughter-cells themselves commence to form buds, before they have separated from their mother-cell, with the result that pearl-like chains of cells are produced. When the yeast-cells have only limited nutriment, with an abundant supply[178] of air, at a suitable temperature, an endogenous formation of spores takes place. The protoplasm of the cells divides into 1–4 (rarely a greater number) masses (Fig. 182 c, d, e) which surround themselves with a thick cell-wall, and in this state can withstand adverse conditions and periods of dryness lasting for several months.
The sporangia are not asci since they have no definite form, and a definite number, form and size of spores is not found. The spores in the different species and kinds occupy varying periods for their development, although exposed to the same temperature, a fact of importance in determining one from another. On germination the wall of the mother-cell is destroyed, and each spore gives rise to a new cell, multiplication taking place by budding (Fig. 182 f). The majority of Yeast-Fungi are able to produce alcoholic fermentation in saccharine fluids.
The most important of these Fungi is the Beer-yeast (Saccharomyces cerevisiæ) with ovate, ellipsoidal or spherical cells (Fig. 182). It is a plant which has been cultivated from time immemorial, on account of its property of producing alcoholic fermentation in sugar-containing extracts (wort), derived from germinating barley (malt). Carbonic acid is also set free during this process. The “surface-yeast” (Fig. 182 a), which produces ordinary beer when the brewing takes place at higher temperatures, has cell-chains; “sedimentary yeast” (Fig. 182 b), used in the brewing of Bavarian beer, has spherical cells, solitary, or united in pairs. Both these and the following Yeast-Fungi include, according to Hansen, several species and kinds.
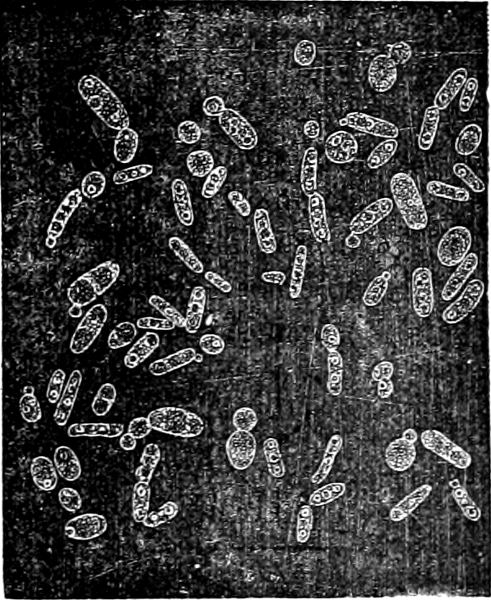
Fig. 183.—Saccharomyces mycoderma.
The “Ferment of Wine” (Saccharomyces ellipsoideus) produces wine in the juice of grapes. Uncultivated yeast-cells are always present on grapes; an addition of this species to the “must” is not necessary to secure fermentation. A large number of other “uncultivated” yeast-cells appear in breweries mixed with the cultivated ones, and cause different tastes to the beer (S. pastorianus, etc.). S. ludwigii, found, for instance, on the slimy[179] discharge from Oaks, produces abundant cell-chains on cultivation. S. apiculatus is very frequently met with on all kinds of sweet fruits, it has orange-like cells. S. mycoderma has cylindrical cells, often united together in chains (Fig. 183): it forms a whitish-gray mass (“fleur de vin”) on wine, beer, fruit-juice, etc., standing in bottles uncorked or not entirely filled. It is thought that this Fungus causes decomposition and oxydises the fluid in which it is found, but it cannot produce alcoholic fermentation in saccharine liquids, and it does not form endospores; hence it is uncertain whether it is true Saccharomyces.
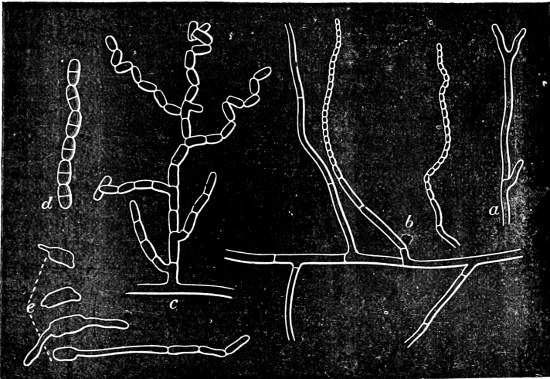
Fig. 184.—Oidium lactis: a branched hypha commonly met with; b a hypha lying in milk and producing aerial hyphæ which give rise to oidia; c a branch giving rise to oidia, the oldest (outermost) oidia are becoming detached from one another; d a chain of divided cells; e germinating oidia in different stages (slightly more magnified than the other figures).
The “Dry-yeast” used in baking white bread is “surface-yeast.” In leaven, a kneaded mixture of meal, barm and water, which is used for the manufacture of black bread, Saccharomyces minor is present, and a species allied to this produces alcoholic fermentation in dough with the evolution of carbonic acid, which causes the dough to “rise.”
2. Oidium-forms. Of many Fungi only the Oidium-forms are known, which multiply in endless series without employing any higher form of reproduction. Oidium lactis (Fig. 184) is an imperfectly developed form which frequently appears on sour[180] milk and cheese. It can produce a feeble alcoholic fermentation in saccharine liquids. Thrush or aphthæ (O. albicans) appears as white spots in the mouths of children. Several similar Oidium-forms are parasites on the skin and hair of human beings, and produce skin diseases, such as scurvy (O. schoenleinii) and ringworm (O. tonsurans).
3. Mycorhiza. These Fungi, which have been found on the roots of many trees and heath-plants, particularly Cupuliferæ and Ericaceæ, consist of septate hyphæ, and belong partly to the Hymenomycetes, partly to the Gasteromycetes. It has been shown that the Mycorhiza enters into a symbiotic relationship with the roots of higher plants.
[181]
In this Division a well-marked alternation of generations is to be found. The development of the first or sexual generation (gametophyte),[16] which bears the sexual organs, antheridia and archegonia, commences with the germination of the spore, and consists, in the Liverworts, of a thallus, but in the true Mosses of a filamentous protonema, from which the Moss-plant arises as a lateral bud. The second or asexual generation (sporophyte), developed from the fertilised oosphere, consists of a sporangium and stalk.
The sexual generation, the gametophyte. The protonema in the Liverworts is very insignificant, and not always very sharply demarcated from the more highly developed parts of the nutritive system. In the true Mosses the protonema is well-developed, and consists of a branched, alga-like filament of cells, the dividing cell-walls being always placed obliquely. In the parts exposed to the light it is green, but colourless or brownish in those parts which are underground (Fig. 186). The protonema is considered to be a lower form of the stem, and grows in the same manner by means of an apical cell; at its apex it may directly develope into a leaf-bearing stem, or these arise from it as lateral branches (Fig. 186 k).
The more highly differentiated part of the vegetative system, the “Moss-plant,” which is thus developed from the protonema, is in the “thalloid” Liverworts generally a dichotomously-branched thallus without any trace of leaf-structures (Fig. 194); in Marchantia (Fig. 197) and others, scale-like leaves (amphigastria) are found on the under surface. The higher Liverworts and the Leafy-Mosses are differentiated into a filamentous, ramified stem with distinct leaves arranged in a definite manner, resembling the stem and leaves of the higher plants (Figs. 186, 195, 200).
[182]
True roots are wanting, but are biologically replaced by rhizoids. These are developed on the stems or thallus: in the Liverworts they are unicellular, but in the Leafy-Mosses generally multicellular and branched. In the latter group they are considered identical with the protonema, and may become true protonema, and new plants may be developed from them (Fig. 186 b).
Fig. 186.—A Lower portion of a Moss-plant with rhizoids (r), one of which bears a reproductive bud (b). The dotted line indicates the surface of the ground; the portions projecting above this become green protonema (p); k is a young Moss-plant formed on one of these. B Germinating spore of Funaria hygrometrica, with exospore still attached. C, D Older stages of the protonema.
The internal structure of the sexual generation is very simple. The leaves in nearly all cases are formed of a single-layered plate of cells; in the Leafy-Mosses, however, a midrib is very often formed, and sometimes, also, marginal veins; and along these lines the leaves are several layers of cells in thickness. The stem is constructed of cells longitudinally elongated, the external ones of which are narrower and sometimes have thicker walls than the more central ones. Vessels are not found, but in several Mosses there is in the centre of the stem a conducting strand of narrow, longitudinal cells, which represents the vascular bundle in its first[183] stage of development. This strand contains elements for conveying water as well as sieve-tubes. Stomata are entirely wanting in the sexual generation of the Leafy-Mosses; they are found in a few Liverworts (Marchantia), but their structure is not the same as in the higher plants.
Vegetative reproduction takes place by gemmæ or buds which arise on the protenema, the rhizoids, the thallus, or the shoots, and become detached from the mother-plant; or else the protonema and the older parts of the plant simply die off, and their branches thus become independent plants. This well-developed vegetative reproduction explains why so many Mosses grow gregariously. In certain Marchantiaceæ special cupules, in which gemmæ are developed, are found on the surface of the thallus (Fig. 197 A, s-s). Again, protonema may also arise from the leaves, and thus the leaves may act as reproductive bodies. Certain Mosses nearly always reproduce vegetatively, and in these species the oospheres are seldom fertilised.

Fig. 187.—Marchantia polymorpha: a mature antheridium.

Fig. 188.—Spermatozoids.
The first generation bears the SEXUAL ORGANS; both kinds are found either on the same plant (monœcious), or on separate plants (diœcious). In the thalloid Liverworts they are often situated on the apex of small stems (gametophores), springing from the surface of the thallus. In the Leafy-Liverworts and true Mosses the leaves which enclose the sexual organs often assume a peculiar shape, and are arranged more closely than the other leaves to form the so-called “Moss-flower.” The male sexual organs are called antheridia. They are stalked, spheroid, club- or egg-shaped bodies whose walls are formed of one layer of cells (Fig. 187), enclosing a mass of minute cubical cells, each one of which is a mother-cell of[184] a spermatozoid. The spermatozoids are self-motile; they are slightly twisted, with two cilia placed anteriorly (Fig. 188), while posteriorly they are generally a trifle club-shaped, and often bear at that part the remains of the cytoplasm, the spermatozoid itself being formed from the nucleus. In the presence of water the ripe antheridium bursts, and its contents are ejected; the spermatozoids, being liberated from their mother-cells, swarm about in the water in order to effect fertilisation.
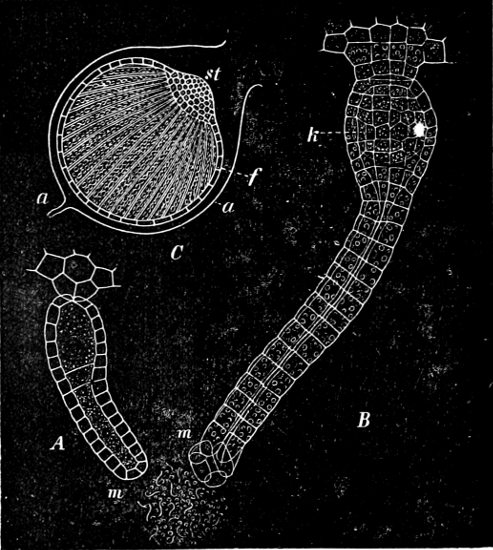
Fig. 189.—Marchantia polymorpha. A A young, and B a ripe archegonium with open neck. C An unripe sporangium enclosed by the archegonium a: st the stalk; f the wall of the sporangium. Elaters are seen between the rows of spores.
The female sexual organs are termed archegonia. They are flask-shaped bodies (Fig. 189), the lower, swollen portion (venter) having a wall, in most cases from 1–2 cells thick, enclosing the oosphere (Fig. 189 B, k): the long neck is formed of tiers of 4–6 cells, enclosing a central row of cells—the neck-canal-cells (Fig. 189 A). When the archegonium is fully developed, the walls of the neck-canal-cells become mucilaginous and force open the neck of the archegonium. The mucilage thus escapes, and, remaining at the mouth of the archegonium, acts in a somewhat similar manner to the stigma and conducting tissue of a carpel, by catching and conducting the spermatozoids to the oosphere (Fig. 189 B, m), with[185] whose cell-nucleus they coalesce. With regard to the formation of the oosphere, it may further be remarked that the lower part of the archegonium originally encloses the so-called “central cell”; but shortly before the archegonium is ripe, this cuts off a small portion, the ventral-canal-cell, which lies immediately beneath the neck, and the larger, lower portion becomes the oosphere.
The organs mentioned here, antheridia and archegonia, are present in the Cryptogams (Pteridophyta) and the Gymnosperms. They have always the same fundamental structure, but with slight modifications of detail. These plants are therefore known as the Archegoniata.
The fertilisation of the Mosses cannot be effected without water. Rain and dew therefore play a very important part in this process, and for this end various modifications of structure are found.
Fig. 190.—Andreæa rupestris. Longitudinal section through a sporangium at the time when the mother-cells of the spores are dividing: p pseudopodium; f foot; v vaginula; h neck; c columella; w wall of the sporangium; e external row of cells; s the spore-sac; t the spore-mother-cells; r the calyptra with the neck of archegonium (z).
Fig. 191.—Andreæa rupestris. Transverse section through a ripe sporangium. In the middle is seen the four-sided columella, surrounded by the numerous spores, drawn diagrammatically. Surrounding them is seen the wall of the sporangium, whose outer layer of cells is thickened and coloured. The layer of cells is unthickened in four places (x), indicating the position of the clefts (see Fig. 193).
[186]
Among the sexual organs, paraphyses—filamentous or club-shaped bodies—are to be found.
The asexual generation, the sporophyte (Moss-fruit or sporogonium). As the result of fertilisation the oosphere surrounds itself with a cell-wall, and then commences to divide in accordance with definite laws.[17] The embryo (Fig. 189 C) produced by these divisions remains inside the wall a-a of the archegonium (Figs. 190, 199 D, E), and developes into the sporogonium, which remains attached to the mother-plant, often nourished by it, as if the two were one organism. The lower extremity of the sporogonium, the foot (Figs. 190 f; 199 D), very often forces its way deep down into the tissue of the mother-plant, but without an actual union taking place. The central portion of the sporogonium becomes a shorter or longer stalk (seta), while the sporangium itself is developed at the summit. At a later stage, during the formation of the spores, the sporangium very often assumes the form of a capsule, and dehisces in several ways characteristic of the various genera (Figs. 192, 193, 194, 195, 200). The basal portion of the archegonium grows for a longer or shorter period, forming a sheath, the calyptra, in which the capsule is developed, but eventually it ceases to enlarge, and is then ruptured in different ways, but quite characteristically, in each group. Anatomically, the asexual generation is often more highly differentiated than the sexual; thus, for instance, stomata are present on the sporangia of the true Mosses, but are absent in the sexual generation.
As the capsule developes, an external layer of cells—the amphithecium—and an internal mass—the endothecium—are differentiated. As a rule the former becomes the wall of the capsule while the latter gives rise to the spores. In this Division, as in the Pteridophyta, the name archesporium (Fig. 190 t) is given to the group of cells inside the sporangium which gives rise to the mother-cells of the spores. The archesporium is in general a unicellular layer; in Sphagnum and Anthoceros it is derived from the most internal layer of the amphithecium, but with these exceptions it arises from the endothecium, usually from its most external layer. In the true Mosses and in Riccia only spore-mother-cells are produced from the archesporium, but in the[187] majority of the Liverworts some of these cells are sterile and become elaters (cells with spirally thickened walls, Figs. 196, 189), or serve as “nurse-cells” for the spore-mother-cells, which gradually absorb the nutriment which has been accumulated in them. In Anthoceros, and almost all the Leafy-Mosses, a certain mass of cells in the centre of the sporangium (derived from the endothecium) does not take part in the formation of the archesporium, but forms the so called “column” or “columella” (Figs. 190, 191).
The spores arise in tetrads, i.e. four in each mother-cell, and are arranged at the corners of a tetrahedron, each tetrahedron assuming the form of a sphere or a triangular pyramid. The mature spore is a nucleated mass of protoplasm, with starch or oil as reserve material. The wall is divided into two layers: the external coat (exospore) which is cuticularized and in most cases coloured (brown, yellowish), and the internal coat (endospore), which is colourless and not cuticularized. On germination the exospore is thrown off, the endospore protrudes, and cell-division commences and continues with the growth of the protonema (Fig. 186, B-D).
Fig. 192.—Andreæa petrophila. A ripe sporogonium: a an archegonium which has been raised with the pseudopodium; p the foot; b the neck; d-e the dark-coloured portion of the sporangium, whose outer cell-walls are considerably thickened; c-c the thin-walled portions where the dehiscence occurs; o the lower extremity of the spore-sac; f calyptra; g the apex of the sporangium. (Mag. 25 times.)
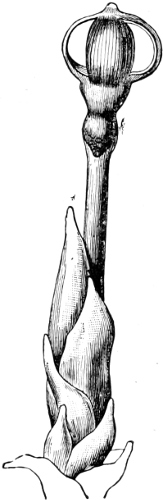
Fig. 193.—Andreæa petrophila. An empty capsule; the calyptra has fallen off. (Mag. 25 times.)
The morphological explanation which Celakovsky has given of the sporogonium, and which is not at all improbable, is, that it is homologous with[188] an embryo consisting of a very small stem-portion and a terminal spore-producing leaf. This will be further explained in the introduction to the Flowering-plants (p. 236).
In the Liverworts the young sporogonium lives like a parasite, being nourished by the sexual generation (only in Anthoceros has it a slight power of assimilation). In the Leafy-Mosses, on the other hand, with regard to the power of assimilation, all transitions are found from abundant assimilation (Funaria, Physcomitrium) to almost complete “parasitism” (Sphagnum, Andreæa). In the majority of the operculate Mosses the sporogonium has a more or less perfect system of assimilation, and is able itself to form a large portion of the material necessary for the development of the spores, so that it chiefly receives from the sexual generation the inorganic substances which must be obtained from the soil. The more highly developed the assimilative system of the sporogonium, the more stomata are present.
Apospory. In some operculate Mosses it has been possible to obtain a protonema with small Moss-plants from the seta, when severed from its Moss-plant, and grown on damp sand.
The Mosses are the lowest plants which are provided with stem and leaf. They are assigned a lower place when compared with the higher Cryptogams, partly because there are still found within the Division so many forms with a mere thallus, partly because typical roots are wanting and the anatomical structure is so extremely simple, and partly also because of the relation between the two generations. The highest Mosses terminate the Division, the Muscineæ and Pteridophyta having had a common origin in the Algæ-like Thallophyta.
They are divided into two classes:—
Hepaticæ, or Liverworts.
Musci frondosi. True Mosses or Leafy-Mosses.
The protonema is only slightly developed. The remaining part of the vegetative body is either a prostrate, often dichotomously-branched thallus, pressed to the substratum (thalloid Liverworts), with or without scales on the under side (Figs. 194, 197); or a thin, prostrate, creeping stem, with distinctly-developed leaves, which are borne in two or three rows (Figs. 195, 198), viz., two on the upper and, in most cases, one on the under side. The leaves situated on the ventral side (amphigastria) are differently shaped from the others (Fig. 198 a), and are sometimes entirely absent.[189] In contradistinction to the Leafy-Mosses, stress must be laid on the well-marked dorsiventrality of the vegetative organs; i.e. the very distinct contrast between the dorsal side exposed to the light and the ventral side turned to the ground. Veins are never found in the leaves.
The ventral part of the archegonium (calyptra) continues to grow for some time, and encloses the growing embryo, but when the spores are ripe it is finally ruptured by the sporangium, and remains situated like a sheath (vaginula) around its base. The sporangium opens, longitudinally, by valves or teeth (Fig. 194, 195, 197 b), very rarely by a lid, or sometimes not at all. A columella is wanting (except in Anthoceros, Fig. 194); but on the other hand, a few of the cells lying between the spores are developed into elaters (Fig. 196), i.e. spindle-shaped cells with spirally-twisted thickenings, which are hygroscopic, and thus serve to distribute the spores. (They are seen in Fig. 189 C, not yet fully developed, as long cells radiating from the base of the sporangium. They are wanting in Riccia).
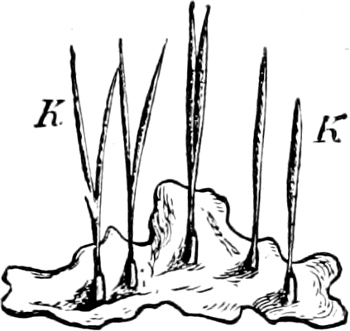
Fig. 194.—Anthoceros lævis (nat. size): K-K capsules.
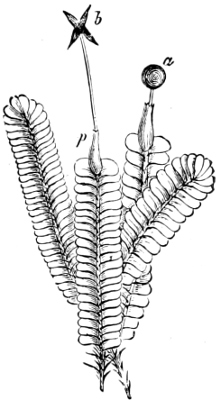
Fig. 195.—Plagiochila asplenioides: a unripe, and b an open capsule; p involucre. The ventral edge of each leaf is higher than its dorsal edge, and covered by the dorsal edge of the next one.
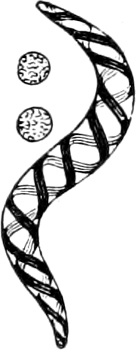
Fig. 196.—An elater with two spores.
Round the entire archegonium, (or group of archegonia, when several are developed on the same receptacle) a sheath—the involucre—is often formed, which persists, and encloses the base of the stalk of the sporangium, together with the sheath of the archegonium (Fig. 195 p). In the Marchantiaceæ each archegonium is enclosed in a loose investment, the perigynium, which is developed as an outgrowth from the cells of its stalk.
[190]
The majority of the Liverworts are found in damp and shady places, pressed to the substratum; a few are found floating in fresh water.
This embraces only forms with a thallus, which is more or less distinctly dichotomously branched, in some, one or two rows of thin leaves are situated on its under surface. On the upper surface of the thallus are found large air-chambers.
Order 1. Ricciaceæ. The sporogonia are, with the exception of a few genera, situated singly on the surface of the thallus, and consist only of a capsule without foot or stalk. They always remain enclosed by the wall of the archegonium (calyptra), and open only by its dissolution. Elaters are not developed. Some genera are found floating like Duckweed.—Riccia glauca grows on damp clay soil. R. fluitans and R. natans float in stagnant waters.

Fig. 197.—Marchantia polymorpha. A Female plant (nat. size): a and b are archegoniophores in various stages of development; s cupules with gemmæ (see page 183). B An archegoniophore seen from below, the short-stalked sporangia are seen placed in 8–10 double rows. C Male plant, with a young and an older antheridiophore. D Antheridiophore halved vertically to show the antheridia (h); m the aperture of the pits in which they are sunk—the older ones to the left, the younger to the right.
Order 2. Corsiniaceæ. (Not native). Intermediate forms between the preceding and the following order. In internal and external structure mainly resembling the Marchantiaceæ. Corsinia; Boschia.
Order 3. Marchantiaceæ, are large, fleshy forms. The[191] surface of the thallus is divided into small rhombic areas, in the centre of each of which is found a large, peculiarly constructed stoma (Fig. 197 A); beneath each of these a large air-cavity is to be found. From the floor of the air-cavity a number of alga-like cells project into it; these contain chlorophyll and are therefore the assimilating cells. The antheridia and archegonia are each found aggregated on specially formed branches (somewhat resembling Mushrooms) projecting from the surface of the thallus. The antheridia are developed on the upper surface (Fig. 197 C, D) and the archegonia on the lower (Fig. 197 A, B), near the centrally-placed stalk.
Marchantia polymorpha is diœcious (Fig. 197), and very common on damp places. Lunularia (South Europe), frequently found on flower-pots in conservatories; Preissia, Fegatella, Reboulia, Targionia.
These have an entirely leafless, fleshy, flat, and irregularly-shaped thallus. In its intercellular chambers Nostoc-colonies are often found, which have forced their way through the stomata situated on the under side. The antheridia and archegonia arise from the cells lying inside the thallus. The capsule resembles a long, thin pod; it has two valves and a columella. Anthoceros (A. lævis, Fig. 194, and punctatus).
Some forms in this family have a thallus in which leaf-like structures are found (Blasia), while in others (e.g. Metzgeria, Pellia, Aneura) they are entirely absent. The majority, however, have round, thick stems, bearing dorsally two rows of leaves, and one row ventrally. Some of these have the leaves “underlying” (Fig. 195), while in others (Fig. 198) they are “overlying.” (See Figs. 195, 198, with explanation).
The sporangia are spherical, stalked, and situated singly on the apex of the branches, and open by four valves (in Sphærocarpus they are indehiscent).
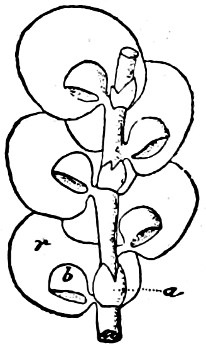
Fig. 198.—Frullania dilatata. Portion of a branch seen from the under side: r and b are the anterior and posterior edges of the same dorsal leaf; a ventral leaves (amphigastria). The dorsal leaves are “overlying,” i.e. the anterior edge of the leaf overlaps the posterior edge of the preceding one.
All the species in this family were formerly reckoned as belonging to one genus, Jungermannia, but now they are divided into several, arranged as follows:—
[192]
I. Anacrogynæ. The archegonia are situated on the upper side of the thallus or stem, placed laterally, and covered by an “involucre,” formed by the calyptra together with the tissue of the stem or thallus.
a. Anelatereæ. Without any elaters: Sphærocarpus, Riella.
b. Elatereæ. α. Thalloid: Aneura pinguis, in damp situations; Metzgeria furcata, on trees; Pellia epiphylla, in damp situations; Blasia pusilla, on damp clay soil, in the shade (scales are present on the thallus). β. Foliose and not dorsiventral: Haplomitrium hookeri.
II. Acrogynæ. The apex of the stem or of certain branches is adapted for the formation of female shoots. The archegonia are most frequently aggregated on the apex of the shoots, and are encircled by their leaves (perichætium). Between these and the archegonia, enclosing the latter, a peculiar cup-shaped organ (the involucre) is formed. This group only includes leaf-bearing genera: Frullania, Radula, Madotheca, Ptilidium, Calypogeia, Lepidozia, Mastigobryum, Lophocolea, Jungermannia, Scapania, Plagiochila.
In this class the protonema is well developed, and resembles a branched filamentous Alga, from which it can be easily distinguished by its oblique septa (in Sphagnum it is a cellular expansion). The Moss-plant, which is developed directly from the protonema, generally has an erect, thick, cylindrical stem similarly constructed on all sides. The leaves are arranged spirally, the most frequent divergence being 2/5 or 3/8 (Fig. 200 A). A midrib is often present and also marginal veins formed by longitudinally elongated cells; at these veins the leaf is more than one layer in thickness. In Leucobryum the leaves are generally constructed of more than one layer.
The stem grows by means of a three-sided, pyramidal, apical cell which gives rise to three rows of segments, each segment forming a leaf. The lateral branches arise from the lower portions of the segments, the upper portion of which does not take any part in the construction of the leaf. From their mode of origin the branches are not axillary, and differ in this respect from the Flowering-plants.
The ventral portion of the archegonium is very early ruptured[193] at its base by the growing sporogonium, upon which it remains, and it is thus raised into the air, forming a “hood,” the calyptra (Figs. 192; 200 B). In the Sphagnaceæ the hood is not present; in this order, as in the Liverworts, the archegonium remains at the base of the sporogonium. The sporangium opens by circumsessile dehiscence, the upper portion (operculum) being separated along a specially constructed ring of cells, and falls off like a “lid” (Fig. 200). Only in a few forms (families 2 and 3) does any variation of this take place. Elaters are never found, but (with the exception of Archidium) there is always present in the sporangium a central mass of cells, the columella, which take no part in the formation of the spores. The columella, in some, does not reach quite to the operculum and in these cases the spore-sac is bell-shaped and covers the columella (Andreæa, Fig. 190; Sphagnum, Fig. 199 D); but in the majority of Mosses the columella extends to the lid, so that the space containing the spores becomes a hollow cylinder.
The sporangium is generally raised on a long stalk; in the great majority this stalk is formed from the lower half of the oospore and belongs to the asexual generation—it is then known as the seta. In Andreæa and Sphagnum the seta is very short, and the sporangia are raised upon a long stalk (pseudopodium) developed from the summit of the sexual generation (Figs. 190, 192). In the latter figure an archegonium (a) is seen attached to the pseudopodium, having been carried up with this during the course of its development. The summit of the pseudopodium is enlarged to embrace the foot of the sporogonium (Figs. 192, 199 D).
A. The sporangium is supported on a pseudopodium; the columella does not extend to the operculum.
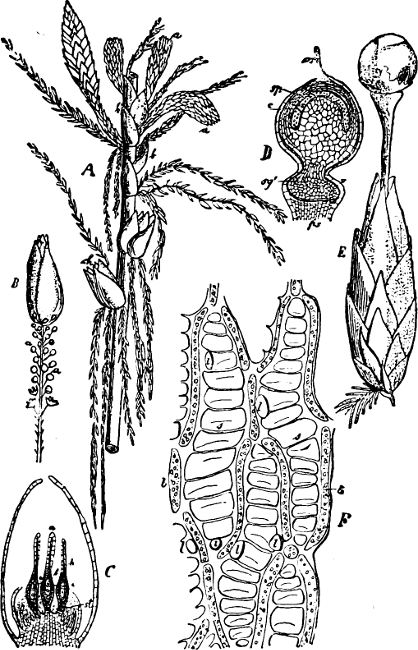
Fig. 199.—Sphagnum acutifolium.—A The upper portion of a plant: a branches with antheridia; ch branches with terminal archegonia and perichætia; b the upper stemleaves. B A male branch whose leaves are partly taken off in order to show the antheridia. C Group of three archegonia: the central one (a) is formed from the apical cell. D Sporogonium in longitudinal section: the broad foot (sg’) is sunk in the vaginula, v; c calyptra; ar neck of the archegonium; ps pseudopodium. E ripe sporangium with operculum, and the remains of the archegonium situated on the pseudopodium which is still surrounded by the perichætium; to the left is a barren branch. F Portion of a foliage-leaf seen from above: l perforations; b chlorophyll-containing cells; s spiral thickenings.
The protonema has been already described. The stem is regularly branched owing to the fact that a branch, or collection of branches, arises at every fourth leaf. These branches are closely covered with leaves, some are erect, while others hang down and surround the stem. No rhizoids are developed. These Mosses are of a whitish-green colour, and when water is present are always saturated with it like a sponge, the reason for this being found in the construction of the stem and leaves. The stems are covered by an external layer of large clear cells, without chlorophyll, but with annular or spiral thickenings on the walls, which are also[194] perforated by large holes. By means of capillary attraction, water is thus raised to the summit of the stem. Similarly constructed cells are also found in the leaves, but they are surrounded by a net of very narrow, chlorophyll-containing cells (Fig. 199 F), whose colour is thus to a great extent lost amongst those which are colourless. This anatomical structure is an essential condition for the formation of peat. The Bog-Mosses grow by preference on[195] moors, which they cover with a thick carpet saturated with water. The lower extremities of the plants perish very rapidly, and gradually become converted into peat, and the branches thus separated from each other become independent plants. The sporangia (Fig. 199 D, E) are spherical, but with a very short stalk. They open by a lid, but have no annulus. The archegonium (Fig. 199 C) persists at the base of the sporogonium as in the Liverworts. Only one genus, Sphagnum.
The Mosses which constitute this family are of a brownish-black colour and are found living on rocks. The sporangium resembles that of the Liverworts inasmuch as it opens by four valves, but these continue attached to each other at the apex as well as at the base (Fig. 193).—There is only one genus: Andreæa.
B. The stalk is formed from the lower portion of the sporogonium. The columella is continued to the summit of the sporangium and united with it (Archidium has no columella.)
The fruit does not dehisce in the regular way, but the spores are liberated by decay. They are small Mosses which remain in connection with their protonema until the sporangium is mature. The archegonium remains sessile at the base of the short capsule-stalk, and is not raised into the air (compare Hepaticæ).—Phascum, Ephemerum, Archidium, Pleuridium.
To this belong the majority of the Mosses, about 3,000 species.
The capsule opens as in Sphagnum by means of a lid (operculum), which is often prolonged into a beak. Round the mouth of the opened capsule, a number of peculiar yellow or red teeth are to be found. These constitute the peristome; their number is four, or a multiple of four (8, 16, 32 or 64). The form and thickenings of these teeth are widely different, and on this account are used by Systematists for the purposes of classification. In some Mosses (Fig. 200 C, D) there is a double row of teeth. Except in Tetraphis they are not formed from entire cells, but from the strongly thickened portions of the wall of certain layers of cells belonging to the lid, and persist when this falls off. They are strongly hygroscopic, and assist greatly in the ejection of the lid, in which operation they are considerably aided by a ring of elastic cells with thickened walls, situated in the wall of the lid near the base of the teeth. This ring is known as the annulus. The archegonium is raised into the air like a hood, the calyptra, which[196] either covers the sporangium on all sides (having the shape of a bell), or is split on one side (Fig. 200 B, h).
Among peculiar forms may be mentioned: Splachnum, which is especially remarkable for the collar-like expansion at the base of the capsule. Fissidens deviates in having a flat stem and leaves arranged in two rows. The leaves are boat-shaped and half embrace the stem.—Schistostega has two kinds of stems. The barren ones resemble Fern-leaves; they have two rows of leaves, which are attached together vertically, are decurrent and coalesce at their bases. The fertile ones have an ordinary appearance.—Tetraphis: the peristome is composed of four teeth, which are formed from entire cells. T. pellucida has peculiar gemmæ.
The family is divided into two groups: the Musci acrocarpi, the growth of whose main axis is limited and terminated by the formation of the sexual organs; and the Musci pleurocarpi, whose sporogonia are situated on special lateral shoots, while the growth of the main axis is unlimited.
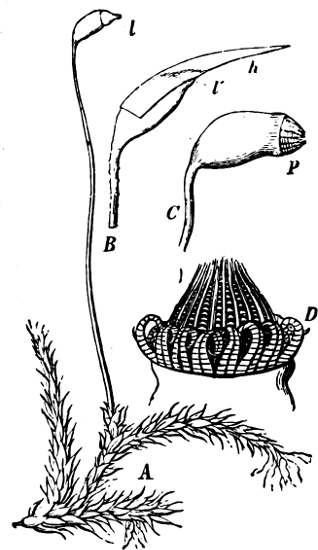
Fig. 200.—A Hypnum populeum. B and C Sporangia, with hood (h), and operculum (l’), and without these (C), showing the peristome (p). D The mouth of the capsule of Fontinalis antipyretica.
Order 1. Weisiaceæ. Peristome, with 16 teeth arranged in one series, rarely wanting. Leaf with midrib. Campylopus, Dicranum (D. scoparium, common in forests), Dicranella, Cynodontium.—Weisia, Gymnostomum (no peristome), Systegium.
Order 2. Leucobryaceæ. Peristome with 16 teeth. Leaves with three or more layers of cells, of which the external ones are air-conducting and perforated (as in the Sphagneæ), the middle one containing chlorophyll. Leucobryum.
Order 3. Fissidentaceæ. Peristome as in the preceding ones. The leaves are arranged in two rows on the plagiotropic shoots; in Fissidens the midrib of the leaf bears wing-shaped outgrowths. Conomitrium, Fissidens.
Order 4. Seligeriaceæ. Peristome with 16 undivided teeth. Very small Rock-mosses. Seligeria.—Blindia.
Order 5. Pottiaceæ. Peristome with 16 teeth, which are divided almost to the base, or with 32 teeth. Calyptra hood-like.—Barbula (B. muralis, B. ruralis), Trichostomum, Leptotrichum.—Ceratodon purpureus.—Distichium.—Pottia.
[197]
Order 6. Grimmiaceæ. The leaf-cells are often papillose; in the upper portion of the leaf, small, and of roundish shape. The calyptra is most frequently hood-like or conical. Eucalypta.—Orthotrichum, often with short-stalked capsule, is found on trees.—Coscinodon.—Hedwigia.—Grimmia, Racomitrium.—Cinclidotus.
Order 7. Schistostegaceæ. The stems are of two kinds (see above); Schistostega osmundacea, in caves, has a bright emerald protonema.
Order 8. Splachnaceæ. The capsule has a large, collar-like neck (see above). Splachnum (especially on manure).
Order 9. Funariaceæ. Capsule pear-shaped. Funaria (F. hygrometrica has a very hygroscopic seta, becoming twisted when dry, and straightening with moisture); Physcomitrium; Discelium.
Order 10. Bryaceæ. The capsule is thicker towards the apex; most frequently pendulous. Philonotis, Bartramia.—Aulacomnium.—Paludella Meesea.—Mnium.—Bryum, Webera, Leptobryum.
Order 11. Polytrichaceæ. Single peristome, formed by 16, 32, or 64 teeth. Leaves with longitudinal lamellæ on upper surface.—Polytrichum has long, hairy calyptra. Catharinea (C. undulata, in forests).
Order 12. Georgiaceæ. Peristome with 4 teeth (see above). Tetraphis (T. pellucida has gemmæ).
Order 13. Buxbaumiaceæ. Capsule asymmetrical; double peristome: the interior one conical, with 16 or 32 longitudinal folds.—Buxbaumia (B. aphylla); Diphyscium.
Order 14. Fontinalaceæ. Long, floating Water-Mosses. Fontinalis (F. antipyretica is found in streams). Dichelyma.
Order 15. Hookeriaceæ. Pterygophyllum.
Order 16. Leskeaceæ. Dull-looking Mosses, with papillose or warted leaves.—Thuidium, Thuja-like with regularly arranged 1–3 doubly pinnate stems; Anomodon, Leskea.
Order 17. Pterogoniaceæ. Pterigynandrum filiforme, etc.
Order 18. Fabroniaceæ. Anacamptodon.
Order 19. Neckeraceæ. Stems most frequently with flat, leafy branches. The leaves are smooth, never with longitudinal folds.—Neckera.
Order 20. Hypnaceæ. The leaves are smooth with square, often bladder-like, cells at the edge. Hylocomium (H. splendens, H. triquetrum); Hypnum; Brachythecium; Plagiothecium.—Eurhynchium.—Homalothecium, Isothecium, Orthothiecium, Homalia.—Climacium, Lescuræa, Leucodon.
The Mosses occur all over the globe. Many are found in great numbers, and growing thickly massed together, they form an important feature in landscapes (for example Sphagnum and Polytrichum in the Arctic Tundra). In the Northern and Arctic regions the Mosses are very plentiful, and often form a considerable part of the vegetation, while in the Tropics they are insignificant.
Species of Hypnum and Polytrichum, like Sphagnum, play an important part in the formation of peat.
[198]
The alternation of generations is as distinct in this Division as in the Mosses, but the sexual generation consists of only a small thallus, the prothallium, which bears directly the sexual organs, antheridia and archegonia; and the asexual generation, which arises from the fertilisation of the oosphere, is no longer a single short-lived sporangium, but a highly developed, generally perennial, plant provided with stem, leaves and true roots (Ferns, Horsetails, etc.), the sporangia being borne on the leaves. In this latter generation the tissues are differentiated into epidermis, ground tissue and vascular tissue; in the last named the bundles are closed, and in the majority of cases concentric.
The sexual generation, gametophyte, or prothallium, is always a thallus, although not always green and leaf-like (Figs. 205, 215, 222, 229, 235, etc.) It is very small, even in cases where it attains the greatest development, and consists only of parenchymatous cells. The prothallium is nourished by hair-like roots (rhizoids) and has only a transitory existence, dying soon after the fertilisation of its oosphere.
The ANTHERIDIA exhibit great variations in structure which, however, must be considered as modifications of the fundamental type which is found in the Mosses. These modifications will be mentioned under the various families. The spermatozoids are always spirally-coiled, self-motile, protoplasmic bodies, with most frequently a large number of fine cilia on the anterior end (Figs. 206, 223, 234). They are formed principally from the nucleus of the mother-cell, and portions of the cytoplasm often remain for a time attached to their posterior end.
The ARCHEGONIA are more uniform throughout the entire Division, and more closely resemble those of the Mosses. They are, as in the previous Division, principally flask-shaped; but the[199] central portion, which encloses the oosphere, is always embedded in the tissue of the prothallium, so that the neck, which is formed of 4 rows of cells, projects above the surface (Figs. 201 3, 222 h). The development of the archegonium in a Fern is seen in the accompanying figure (Fig. 201). The archegonium is developed from a surface cell, which divides into three cells by two walls in a direction parallel to the surface of the prothallium (Fig. 201). The most internal cell becomes the ventral portion of the archegonium. The external one (b) divides perpendicularly to the surface of the prothallium into four cells, which again divide parallel to the surface and form the neck (b, in 2 and 3). The intermediate cell projects upwards into the neck and divides into two, the lower one, after the separation of the ventral canal-cell, becoming the oosphere, and the upper one the neck-canal-cell (c, in 2 and 3).
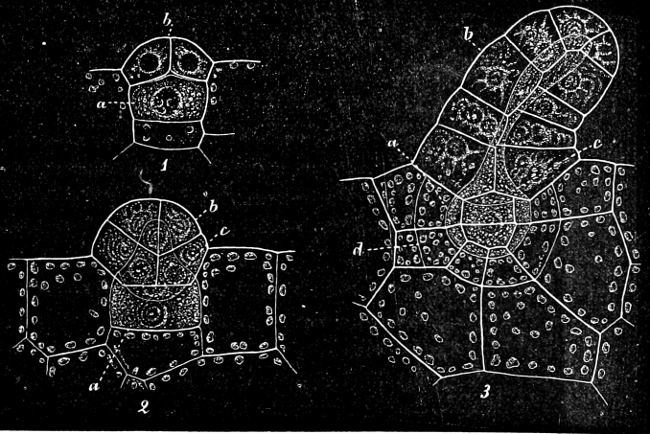
Fig. 201.—Pteris serrulata. Development of archegonia.]
As in the Mosses, the divisional walls of the neck-canal-cells become mucilaginous, causing the rupture of the neck of the archegonium. Fertilisation takes place as in the Mosses, and the passage of the spermatozoids, along the neck, to the oosphere, has been observed. Water (rain or dew) is similarly necessary for the movements of the spermatozoids, and hence for fertilisation. The other classes of the Division chiefly deviate from the Ferns in having the archegonium sunk deeper into the prothallium, and the neck reduced in length (compare Fig. 201 with Figs. 216, 222, 235, 236).
[200]
According to the nature of the spores, the three classes of the Vascular Cryptogams are each divided into isosporous and heterosporous groups.
I. The isosporous Vascular Cryptogams have only one kind of spore. The prothallium developed from this is in some cases monœcious, bearing both antheridia and archegonia; but in others there is a distinct tendency for each prothallium to bear only antheridia or archegonia (diœcious)—true Ferns and Lycopodium.
In Equisetum there is only one kind of spore, but two kinds of prothallia are developed, one of which bears only antheridia (male), the other only archegonia (female); but the one that bears antheridia may be transformed into the one that bears archegonia and vice versa.
II. In the higher group, heterosporous Vascular Cryptogams (Selaginella and Isoëtes, etc.), there are two distinct kinds of spores, the small, microspores, and the large, macrospores. The microspores are male, and produce prothallia which bear only antheridia. The macrospores are female, and produce prothallia which bear only archegonia.
Corresponding to this difference in the spores, there is also found a difference in the development of the prothallium. In the Isosporeæ the prothallium is large, and either green, leaf-like, and provided with rhizoids (most of the Ferns, Horsetails, etc.), or subterranean, pale-coloured, and globular (Ophioglossum, Lycopodium). It lives vegetatively for a fairly long time, and generally produces a large and varying number of archegonia and antheridia. The prothallium in the Heterosporeæ is gradually more and more reduced, its independent and vegetative life becomes of less and less importance, it becomes more dependent on the mother-plant, and projects from the spore very slightly, or not at all. The antheridia and archegonia become reduced in number to one, and also degenerate in point of development.
It may here be remarked that the gradual development of the asexual generation, the development of the two kinds of spores, and the progressive reduction of the prothallium and sexual organs which is found in this Division, is continued to the Gymnosperms and Angiosperms. The microspores are in these called pollen-grains, and the male prothallium is very rudimentary. The macrospores are termed embryo-sacs, and the female prothallium, the endosperm.
The asexual generation, sporophyte. When the oosphere,[201] which in this case as in all others is a primordial cell, is fertilised, it surrounds itself with a cell-wall and commences to divide into a number of cells, to form the embryo.
The first dividing wall (basal wall) is nearly horizontal, and in the direction of the longitudinal axis of the archegonium. The next wall is vertical, and the next perpendicular to the other two. The oosphere, therefore, is now divided into eight octants by these three walls. The basal wall divides the embryo into a hypobasal and an epibasal half. From the first one, by continued divisions, the first root is developed; from the latter, the stem and leaves. After the formation of the octants the development proceeds in somewhat different ways in the various classes. In addition to the stem, leaf, and root, a “foot” is developed from the hypobasal half which remains enclosed in the prothallium, and conveys nourishment from the prothallium to the young plant until it is able to sustain itself (Fig. 202). The formation of these members in the embryo depends on the position of the oosphere in the archegonium and prothallium, and is independent of gravity.
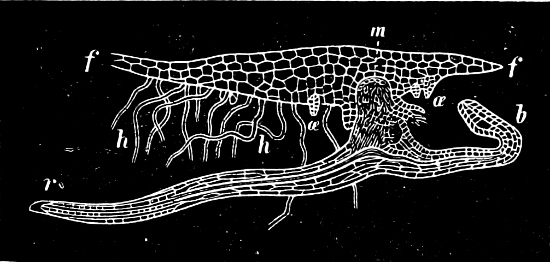
Fig. 202.—Adiantum capillus veneris. Vertical section through a prothallium (f f), with a young plant attached on its under side (mag. about 10 times); r the first root, and b the first leaf of the young Fern-plant; m the foot. In the angle between m and b lies the apex of the stem: h the rhizoids of the prothallium; æ æ unfertilised archegonia.
In the Mosses the asexual generation is the sporogonium, which is limited in its development and in a great measure dependent upon the sexual generation, upon which it is situated; but in the Pteridophyta this generation is an independent and highly developed plant, provided with stem, leaf, and true roots, and has in many instances an unlimited development. The Pteridophyta are the lowest Division with true roots. The root which is first formed is very similar in nature to the primary root of the Monocotyledons; it very soon dies and is replaced by others which are more permanent, and developed upon the stem (adventitious roots); roots are wanting in Salvinia, Psilotum, and some Hymenophyllaceæ. The differentiation is, however, not so complete as in the Flowering-plants, and so many leafy forms are not found. The various members of these plants are anatomically much higher than in the[202] Mosses, having an epidermis, a ground tissue with variously differentiated cells, and a highly developed vascular system. The vascular bundles, like those in the Monocotyledons, are without cambium, and closed; they are therefore incapable of any increase in thickness. In general the bundles are concentric, with the bast round the wood (Fig. 203). The wood is almost entirely made up of scalariform tracheides.
In Isoëtes a secondary thickening takes place by a cambium, which is formed inside the cortex, constructing secondary cortex to the exterior, and secondary wood towards the interior.—Botrychium has also a thickening growth. Collateral vascular bundles occur in Osmundaceæ, Equisetaceæ, and the leaves of many Polypodiaceæ, etc.
Fig. 203.—Portion of the stem of a Fern. Above is seen the transverse section, with vascular bundles of different form and size. The rhombic figures on the side of the stem are leaf-scars.
It is a point of special interest, that the gigantic forms of Ferns, Equisetums, and Club-Mosses (which flourished in earlier geological periods, when these classes attained their highest development) possessed some means of increasing in thickness.
The sporangia are in all cases capsule-like, and burst open when ripe to eject the spores. They are nearly always situated on the leaves (in Lycopodiaceæ, in the axils of the leaves, or above these, on the stems themselves). In some forms (Leptosporangiatæ), the sporangia are developed from a single epidermal cell; in others (Eusporangiatæ), from a group of epidermal cells, or from cells which lie beneath the epidermis. In the first group a primitive mother-cell (archesporium) is formed, which divides commonly into sixteen special mother-cells. In the latter group, on the other hand, a number of primitive spore-mother-cells are developed. In each sporangium three different tissues are generally developed; an innermost sporogenous one (s in Fig. 204 A), which arises from the archesporangium; an outermost one, which forms the wall (a),[203] and may be one or, more rarely, several layers in thickness; and an intermediate one, the tapetum (Fig. 204 A, B, b t), which is rich in protoplasm, and whose cells are dissolved so that the spores float freely in the fluid thus provided. The spores arise as in the Mosses (in tetrads), by the cross-division of the special mother-cells, and according to the manner in which they are arranged in the mother-cell have either a tetrahedral form, with a large base resembling a segment of a ball, or are oblong (bilateral spores). Their construction is the same as in the Mosses (p. 187).
Fig. 204.—Selaginella inæqualifolia. A A young sporangium, which may develope either into a macro-, or a microsporangium. B A microsporangium.
The spore-formation in its earliest commencement takes place in the same way in the Isosporous and the Heterosporous Vascular Cryptogams; but from a certain point, after the tetrahedral division, a difference occurs with regard to the macrosporangia. All the spores formed in the microsporangium may complete their development; but those which are formed in the macrosporangium are generally aborted, with the exception of one or four, and these consequently attain a much larger size (see Fig. 239.—The series to the left are microsporangia; those to the right, macrosporangia).
Apogamy. In some Ferns (Pteris cretica; Aspidium filix mas, var. cristatum; A. falcatum; Todea africana) the young plant is not developed as a consequence of fertilisation, but as a bud from the prothallium. This is known as apogamy, or loss of the power of sexual reproduction. The antheridia are generally more or less developed; archegonia are entirely wanting in Asp. filix mas, var.[204] cristatum. This variety has probably only become apogamous through cultivation. Many specimens of Isoëtes lacustris, in a lake in the Vosges mountains, produce in the place where the sporangia are usually found, a vegetative shoot which grows into a new plant, so that the sexual generation is wanting in this case. Some specimens have sporangia on some leaves, and shoots on others.
Apospory, or the formation of prothallia instead of sporangia and spores on the leaves, is found in Athyrium filix femina, var. clarissimum. In this case the development of the sporangia proceeds only to a certain point, and from these arrested sporangia the prothallia are produced. Normal sporangia are entirely wanting in this variety, and in Aspidium angulare, var. pulcherrimum, sporangia are completely wanting. Compare the Mosses (page 188).
The Vascular Cryptogams are divided into three large classes, in each of which a progressive development can be traced from the isosporous to the heterosporous forms, but some of these are now only known as fossils.
Class 1. Filicinæ (Ferns).—The stem is small in comparison with the leaves, and branches only seldom, and then by lateral shoots. The leaves are scattered, large, often deeply divided, and of various highly developed forms. The undeveloped leaves are rolled up in the bud, having what is termed circinate venation. The sporangia are situated on the edge or on the lower side of the leaves, those on which the sporangia are borne (sporophylls) being often the ordinary foliage-leaves; but in a few cases the fertile differ from the barren ones (a higher stage in development). The fertile leaves are not confined to definite parts of the shoot, and do not limit its growth. The archesporium is most frequently unicellular.
A. Isosporous: Sub-Class 1. Filices (True Ferns).
B. Heterosporous: Sub-Class 2. Hydropterideæ (Water Ferns).
Class 2. Equisetinæ (Horsetails), in its widest meaning.—The leaves in this class are small in comparison with the stem. They are arranged in whorls, and unite to form a sheath. The sporangia are situated on specially modified, shield-like leaves, which are closely packed together and form a “cone.” The cone is borne terminally, and limits the growth of the shoot. The sporangia are developed from a large group of epidermal cells, the archesporium being unicellular. The branches are arranged in whorls, and develope acropetally.
A. Isosporous: Sub-Class 1. Equisetaceæ. Existing forms.
B. Heterosporous: Sub-Class 2. Extinct forms.
[205]
Class 3. Lycopodinæ (Club-Mosses).—Roots generally branching dichotomously. The leaves are scattered or opposite, and in proportion to the stem very small, undivided, and simple. They are scale-like and triangular, tapering from a broad base to a point. The sporangia are situated singly (except in Psilotaceæ), and almost in every case on the upper side of the leaf or in the axil of a leaf; but in some cases they are borne on the stem, just above the leaf-axil. The sporangia arise from groups of epidermal cells. The sporophylls are often modified, and differ from the foliage-leaves; they are then arranged in cones placed terminally on branches, thus limiting their growth.
A. Isosporous: Sub-Class 1. Lycopodieæ.
B. Heterosporous: Sub-Class 2. Selaginelleæ.
The characteristics of this class have already been given on page 204.
The class is divided into two sub-classes:—
1. The True Ferns, Filices, have one kind of spore which generally developes monœcious prothallia, relatively large and green. The sporangia are most frequently situated in groups (sori), which are often covered but not enclosed by an indusium.
2. Water Ferns, Hydropteridæ, have microsporangia with many (4 × 16) microspores, and macrosporangia, each with one macrospore. The prothallium is small, and projects but slightly from the germinating spore. The sporangia are situated in groups (sori), which are either enclosed by an indusium, or enveloped in a portion of a leaf, to form “fruits” termed sporocarps.
The old name for the Hydropterideæ, “Rhizocarpeæ,” i.e. the “root-fruited,” originated from the erroneous supposition that the sporocarps were borne on the roots.
Of the eight orders (with about 4,000 species) comprised in this sub-class, the Polypodiaceæ is the largest (having about 2,800 species) and the most familiar; for this reason it will be taken as typical.
The sexual generation. When the spore germinates, the external covering (exospore) is ruptured, as in the Mosses. The internal cell-wall (endospore) grows out as a filament, which soon divides and gives rise to the prothallium, a flat, cellular expansion resembling the thallus of a Liverwort. In its fully developed state[206] the prothallium is generally heart-shaped, dark green, and provided with root-hairs, and it attains a diameter of about one centimetre (Fig. 205). It is formed of one layer of cells, except along the central line near the anterior depression, where it becomes several layers of cells in thickness, forming the “cushion,” on the lower side of which the archegonia are developed. The antheridia are first formed; they are thus found on the oldest parts of the prothallium, on its edge, or among the root-hairs. The archegonia are developed later, and are therefore found near the apex. Several tropical Ferns have prothallia[18] deviating from this typical form; Trichomanes (Order Hymenophyllaceæ) has filamentous, branched prothallia, which resemble the protonema of a Moss. Others, again, have strap-shaped prothallia, which resemble the thallus of certain Liverworts.
Fig. 205.—Prothallium (p p) of Maiden hair (Adiantum capillus veneris) with a young plant attached: b first leaf; w′ primary root; w″ adventitious roots; h h root-hairs of the prothallium (× abt. 30).
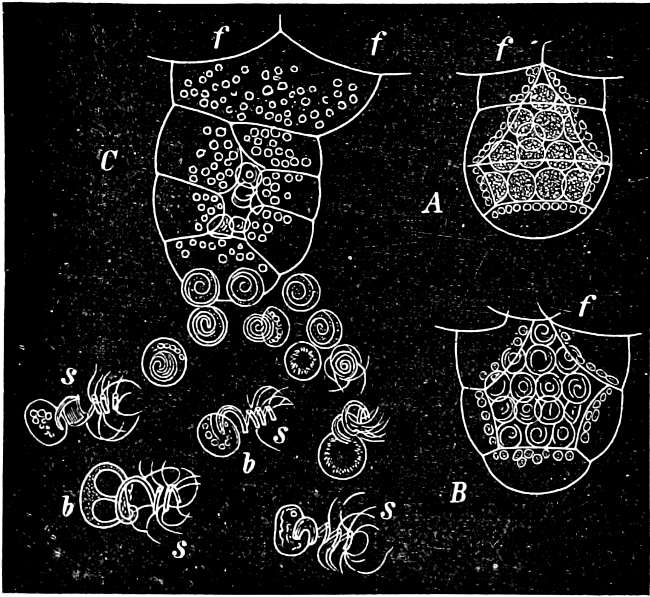
Fig. 206.—Antheridia of Maiden-hair (× 550). A Unripe; B ripe, but unopened; C open and ejecting the spermatozoids (s). Those which have been last ejected are still lying enclosed in their mother-cells, the others are coiled up and drag with them the cytoplasmic remains (b); f cells of the prothallium.
[207]
The ARCHEGONIA have been already mentioned (p. 199, Fig. 201). The ANTHERIDIA are hemispherical or slightly conical bodies (Fig. 206). They consist, as in the Mosses, of a wall formed by one layer of cells, which encloses a number of spermatozoid-mother-cells (A and B). The antheridia when ripe absorb water, and are ruptured, and the spirally-coiled spermatozoids liberated (Fig. 206 S). The spermatozoids have been observed to pass down the neck of the archegonium, and to fuse with the oosphere.
The asexual generation. The first leaf, the “cotyledon,” of the embryo developed from the oospore (Figs. 202, 205) is always small, and has a very simple shape. The leaves which occur later become more perfect, stage by stage, until the permanent form of leaf has been attained.—The STEM is most frequently a subterranean or a semi-aerial rhizome; it is only in the tropical, palm-like Tree-Ferns, that the stem raises itself high in the air and resembles that of a tree, with leaf-scars or with the remains of leaves attached (Figs. 207, 203); in certain species the stem is encased in a thick mat of aerial roots (Dicksonia antarctica). When the rhizome is horizontal the internodes are frequently elongated, and the leaves are arranged in two rows, as in Polypodium vulgare and in the Bracken-Fern (Pteridium aquilinum), etc.; it is also generally dorsiventral, having a dorsal side on which the leaves are situated, and a ventral side, different from the former, on which the roots are borne. When the stem ascends in an oblique direction, or is nearly vertical, its internodes are extremely short, and the leaves are arranged in a spiral line with a complicated phyllotaxis, e.g. in Athyrium filix-fœmina, Aspidium filix-mas, etc. The BRANCHING upon the whole is extremely slight, and is generally confined to the petiole (e.g. Aspid. filix-mas), or to the stem near the insertion of the leaves. Several species normally form buds on different parts of the lamina. The buds which are formed on the stem are not confined to the leaf-axil as in the higher plants. The Tree-Ferns, generally, do not branch at all.
The VASCULAR BUNDLES are concentric, with the wood surrounded by the soft bast. In transverse section they are seen as circles or irregularly-shaped figures (Fig. 203), the name of “King Charles and the Oak” (Bracken-Fern) having originated from the appearance which the bundles present in oblique section. In Osmunda they are collateral and resemble those of the Flowering-plants. Round each individual bundle is often a sheath of thick-walled, hard, brown, sclerenchymatous cells, which act as a mechanical[208] tissue; similar strands are also found in other parts of the stem.

Fig. 207.—Various Ferns (1, 2, 3, 4).
The LEAVES in nearly all species are only foliage-leaves, borne in a spiral. They have an apical growth which continues for a long time, and some require several years for their complete development. In the buds they are rolled up (circinate); not only the midrib, but also all the lateral veins, and even the terminal[209] portions of a leaf are sometimes rolled up together, the tissues of the leaf being already fully developed and only waiting to expand. The leaves are often excessively divided and compound, with pinnate branches, and have an epidermis with stomata and a well-developed system of venation. Stipules are only found in Marattiaceæ and Ophioglossaceæ.
Very often peculiar hairs or scales (paleæ, ramenta), dry, brown, flat and broad, are found on stem and leaf.
The SPORANGIA are small, round capsules, which, in a very large number of Ferns, are formed on the back, but more rarely on the edge of the ordinary foliage-leaves. It is very seldom that there is any difference in form between the barren foliage-leaves and the fertile leaves, as is found for example in Blechnum spicant or Struthiopteris; or that the fertile part of the leaf is differently constructed from the barren portion of the same leaf, as in the Royal-Fern (Osmunda). In such instances the mesophyll of the fertile parts is poorly developed.
The sporangia in the Polypodiaceæ are lens-shaped, with long stalk (Fig. 211 D): their wall consists of one cell-layer on which a single row of cells, passing vertically over the top (that is along the edge of the sporangium), is developed into the “ring” (annulus). The cells of the annulus are very much thickened on the inner and side walls, and are yellowish-brown. The thickened cells, however, do not entirely encircle the sporangium, and on one side, near the stalk, they pass over into large, flat, thin-walled cells. These form a weak point in the wall, and it is here that the sporangium is opened diagonally by the elongation of the annulus. The sporangium of the Polypodiaceæ opens as it dries. The cells of the annulus are very hygroscopic, and in straightening, the annulus bends back with a jerk, thus ejecting the spores to considerable distances. The cells of the annulus absorb water with great readiness. [The sporangium arises as a single epidermal cell, from which a basal stalk-cell is cut off. Three oblique cell-walls, intersecting near the base, are next formed in the upper cell, and a fourth between these and parallel to the free surface; an inner tetrahedral cell enclosed by four others is thus formed, the outer cells become the wall of the sporangium, while the inner cell, by a series of walls, parallel to its sides, cuts off a layer of cells which eventually form the tapetum, the remaining central cell constituting the archesporium.]
The SPORES are either oblong and bilateral, or they are tetrahedric[210] with curved sides, depending upon the way in which the tetrad division has taken place.
The sporangia are almost always situated on the nerves and gathered into groups, sori, which differ in form in the various genera. The sori, in many genera, may be covered by a scale-like structure, the indusium (Figs. 211 B, 212).
In the majority of cases, each sorus is situated on a small papilla (placenta, or receptacle), which is supplied by a small vascular bundle. Between the sporangia, hairs (paraphyses) are often situated, which spring either from the placenta or from the stalks of the sporangia.
Systematic Division. The Ferns may be divided into two groups, characterized by the structure and development of the sporangia. The sporangia in the Eusporangiatæ take their origin from a group of epidermal cells, and their walls are formed by several layers of cells. The archesporium is the (not tetrahedric) hypodermal terminal cell of the axial row of cells which give rise to the sporangium. In the Leptosporangiatæ the sporangia are developed from single epidermal cells, and their walls are uni-layered. The archesporium is a central, often tetrahedric cell, from which sixteen spore-mother-cells are developed.[19] It is difficult to say which form is the oldest (according to Prantl, those which have the sori on the nerve-endings); however, the Eusporangiatæ would seem to have made their appearance long before the others, and also well defined Marattiaceæ and Ophioglossaceæ occur in the Kulm and Coal period, before the true Polypodiaceæ.
About 4,000 species of Ferns are now existing, and they are found especially in tropical and sub-tropical forests.
Order 1. Ophioglossaceæ. The prothallium differs from that of all other Ferns in being subterranean, free from chlorophyll, pale and tuberous. The stem is extremely short, with short internodes, most frequently unbranched, vertical, and entirely buried in the ground (Fig. 208 st). In several species (among which are the native ones) one leaf is produced every year, which has taken three to four years for its development. In Botrychium a closed, sheath-like basal part of each leaf covers the subsequent leaves during their development. In Ophioglossum and[211] others each leaf has at its base an intrapetiolar, cap-like sheath, which protects the succeeding leaf. The leaves are of two kinds: (a) foliage, which in Ophioglossum vulgatum are lanceolate and entire, but in Botrychium however, are pinnate (b in Fig. 208 A, B); and (b) fertile, which are found facing the upper side of the foliage-leaves. These latter in Ophioglossum are undivided and spike-like (Fig. 209 A), but pinnate in Botrychium (Fig. 208 B). Each foliage and fertile leaf are branches from the same petiole. The large sporangia are placed laterally, and open by two valves. No annulus is formed (Fig. 209).—Ophioglossum reproduces vegetatively by adventitious buds on the roots.
Fig. 208.—A Ophioglossum vulgatum (Adder’s-tongue); B Botrychium lunaria (Moonwort), both natural size; r-r roots; bs leaf-stalk; st stem; b foliage-leaf; f fertile leaf.
Fig. 209.—Fertile leaf of Ophioglossum.
[212]
Three genera with about twelve species.
Order 2. Marattiaceæ are tropical Ferns, whose gigantic leaves resemble those of the Polypodiaceæ, but have stipules in addition. The sporangia are grouped in sori, situated on the lower side of the leaves, the sporangia in each sorus being arranged either in two rows or in a ring. In Angiopteris they are isolated (Fig. 210 A), but in the other species (Kaulfussia, Danæa, Marattia), they are united, and form “synangia” divided into a number of chambers corresponding to the sporangia. These open by clefts or pores. Marattia presents the highest development, as its sporangia are completely united in a capsule-like synangium, which is closed until maturity, and then opens by two valves. In each valve there is a row of three to eleven sporangia, each opening by a slit towards the inside (Fig. 210 B, C). An indusium encloses the sorus, except in Kaulfussia; it is formed of flat and lobed hairs, which resemble the hairs of the other portions of the leaves. In Angiopteris and Marattia the indusium is very rudimentary; in Danæa it forms a kind of cupule.
The numerous fossil Marattiaceæ (15 genera, with 98 species) present similar differences to those now living, but more various forms are found, for example, with solitary free sporangia. Those now living are the last small remnant (4 genera with only 23 species) of a once dominant family, which existed from very early times, and whose culminating point was reached in the Kulm and Coal periods.
The Ophioglossaceæ appear also in the Kulm and Coal periods, and were about as numerous as at the present time (presumably 2 genera, with 19 species). Leptosporangiate Ferns appear however to have occurred first of all in the Trias-formation.
Fig. 210.—Sporangia of the Marattiaceæ: A Angiopteris; B and C Marattia; C is a half sorus with nine sporangia, each of which has opened by a longitudinal cleft.
Order 1. Polypodiaceæ. Sporangia on the lower side of the leaves, stalked and provided with a vertical, incomplete annulus; dehiscing by a transverse cleft (Fig. 211 D).—The genera are distinguished by the form of the indusium and the position of the sori, etc.
[213]
1. The sporangia cover the entire lower surface of the leaf (Tropical America and Asia). Acrostichum, Platycerium.
2. Sori without indusia, circular or oval. Polypodium (Fig. 211 A). The leaves are most frequently situated in two rows on the dorsal side of the creeping rhizome, and fall off leaving a smooth scar behind.—P. vulgare, common in woods, on stones. (Phegopteris also has no indusium; see page 214).
3. The sporangia are situated in continuous lines just inside the margin of the leaf.—Pteris[20]: the sporangia form a continuous line along the entire margin of the leaf (Fig. 211 C), which bends over and covers the sporangia, forming a “false-indusium.” Pteridium has linear sori situated on a marginal vascular bundle, covered by two linear basal indusia, of which the outer is bent over like the edge of a leaf.—P. aquilinum (Bracken) has a wide-spreading rhizome with large alternate leaves, placed on opposite sides, at some distance apart. Only one leaf is developed from each branch every year.
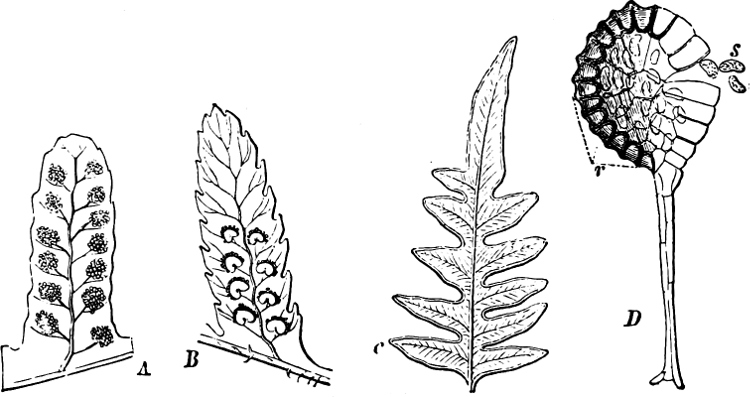
Fig. 211.—Portions of leaves with sori. A Polypodium. B Aspidium. C Pteridium. D A sporangium of one of the Polypodiaceæ: r the annulus; s spores.
Adiantum (Maiden-hair): sori on the underside of small portions of the edge of the leaf, which are bent over (false indusium). Cryptogramme (Allosorus), Cheilanthes.
4. The sori are oval or linear, situated on one side of the vascular bundle.—Asplenium (Fig. 212 A): sori linear; indusium with one of its edges attached at the external side. A. ruta muraria (Wall-Rue); A. septentrionale; A. trichomanes.—Athyrium: sori linear or curved; A. filix-fœmina (Lady-Fern).—Scolopendrium[214] (Fig. 212 B): sori as in Asplenium, but situated in pairs across the lanceolate, entire leaves. Each sorus is covered on the external side by an indusium, whose free edges are parallel and approach each other. S. vulgare (Hart’s-tongue).—Blechnum (B. spicant, Hard Fern; the fertile leaves differ from the barren, the pinnæ being narrower, while the underside is almost entirely covered with sori, and hence they are of a much darker brownish hue than the barren ones).—Ceterach: indusium rudimentary or absent.
5. Sori circular and covered by a shield-like, or reniform indusium.—Aspidium (Fig. 211 B); the leaves wither away and leave no scar upon the root-stock. A. filix-mas (Male-Fern); A. spinulosum.—Phegopteris has no indusium, the withered bases of the leaf-stalks are persistent; P. dryopteris and P. polypodioides.
6. The indusium is situated below the sori, and has the shape of a one-sided scale (Cystopteris, Struthiopteris), or of a cup or cupule, which in Woodsia is sometimes fimbriate (Fig. 212 C, D).
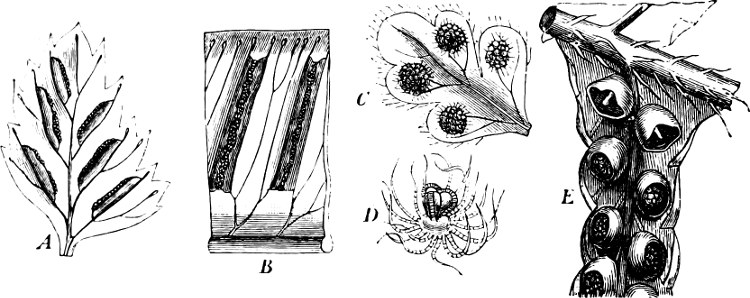
Fig. 212.—A Asplenium. B Scolopendrium. C Woodsia; D single sorus of the same. E Cyathea: the sporangia have fallen off in the upper sori. (All magnified.)
7. The sori are situated on the margin of the leaf, and at the end of a vascular bundle. Indusium, semi-cupular. Davallia. Principally tropical species. 1 in S. Europe.
This order is the greatest, comprising about 2,800 species, the majority being perennial plants. A few are large, and known as Tree-Ferns.
As plants in conservatories and rooms the following are cultivated: species of Gymnogramme (tropical America), Lomaria, Nephrolepis, Pteris (P. serrulata, cretica).
Officinal. Aspidium filix-mas, rhizome and the withered petioles.—Species of Alsophila and Cibotium give Penghawar Djambi. The rhizome of Pteridium aquilinum, var. esculentum, contains so much starch that it is used as food.
The other orders of true Ferns deviate from the Polypodiaceæ, especially in[215] the formation of the annulus, the bursting of the sporangium and its mode of attachment and development, and in the differences in the formation of the prothallium, etc. The principal are:—
Order 2. Hymenophyllaceæ. To this order belong the lowest and most Moss-like Ferns; the leaves, with the exception of the veins, are most frequently formed of only one layer of cells, and consequently stomata are wanting; the formation of the prothallium also somewhat resembles the Mosses. Sori marginal, on the extremities of the vascular bundles, and surrounded by a cupular indusium. The sporangia are sessile, with equatorial annulus. Hymenophyllum (H. tunbridgense, European). Trichomanes (T. speciosum, European). Species about 200, which live especially on rocks and trees in damp and shady tropical forests. Some have no roots.
Order 3. Cyatheaceæ. Annulus complete and oblique. To this order belong, principally, the tree-like Ferns with palm-like habit. The number of species is about 200, they are all tropical and form forests in some regions of Australia. Cibotium and Dicksonia have marginal sori, with cupular, basal indusium. (The stem of D. antarctica is covered with aerial roots.) Alsophila (without indusium); Cyathea with cupular, inferior indusium (Fig. 212 E).
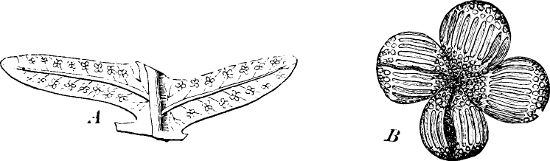
Fig. 213.—Gleichenia: A part of a leaf with sori; B a single sorus.
Order 4. Gleicheniaceæ. Sporangia with equatorial annulus, and longitudinal dehiscence, most frequently groups of 3–4 in sori without indusium (Fig. 213). Gleichenia: the apical growth of the leaves continues for a long time.
Order 5. Schizæaceæ. Annulus apical. To this order belongs Aneimia, which is so commonly cultivated in conservatories. The two lowest pinnæ are metamorphosed, having no leaf parenchyma and being covered with sporangia. Schizæa. Mohria. Lygodium, a climber, whose leaves have unlimited growth and attain a length of several metres. About 70 species. Tropical.
Order 6. Osmundaceæ. The sporangia have at the apex a lateral group of strongly thickened cells, which gradually pass over into the ordinary cells. The sporangia open by a longitudinal cleft. Indusium wanting. Osmunda bears the sporangia upon peculiar, branched pinnæ, without parenchyma (the uppermost in the leaf). O. regalis (Royal-Fern): European.
The following further characteristics must be added to those given on page 205:—
[216]
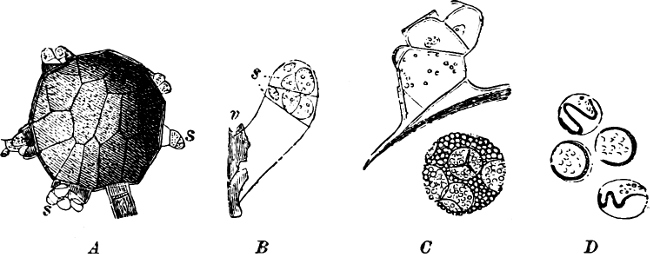
Fig. 214.—Salvinia natans: A microsporangium with germinating microspores and protruding prothallia (s); B a prothallium with the bicellular antheridium (s) growing out of the microsporangium; C the two cells of the antheridium have opened by transverse clefts; beneath is seen the microspores enclosed by the hardened mucilage; D spermatozoids still enclosed in the mother-cells.
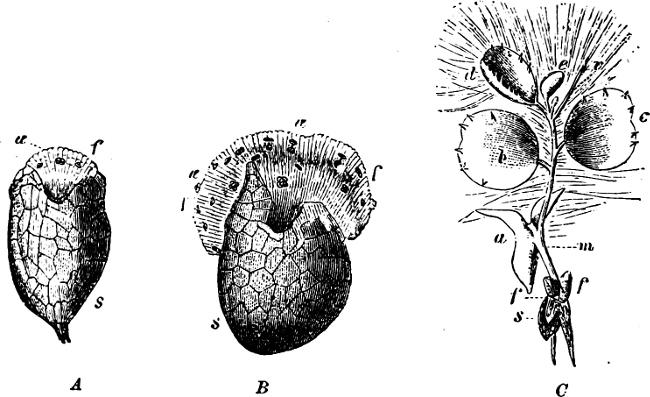
Fig. 215.—Salvinia natans. A, B Female prothallia, f-f, protruding from the macrospore which is still enclosed in the macrosporangium; œ archegonia. C An embryo (× 16) still in connection with the spore (s): a the scutiform leaf; b-e the subsequent foliage-leaves, of which b and c stand singly, d-e-v in a whorl; v the submerged-leaf; f-f wing-like lobes of the prothallium: m the foot.
Sexual generation. The MICROSPORES produce an extremely rudimentary prothallium, formed of only a single cell, and having also a very much reduced bicellular antheridium with a small number of spermatozoid mother-cells in each cell (in Salvinia 4, in Marsilia and Pilularia 16). In Salvinia the microspores remain embedded in a hard mucilaginous mass (at first frothy) which fills up the cavity of the sporangium. The prothallium must therefore[217] grow out through this slime and also through the wall of the sporangium (Fig. 214), and it thus terminates in a relatively long cell.
In Marsilia the microspores are set free from the microsporangium, and the prothallia, with the antheridia, remain in them until the spermatozoids are liberated. The latter are spirally-twisted threads.
The MACROSPORES, on germination, give rise to a very reduced prothallium, which in Salvinia bears 3 archegonia; but, if these are not fertilised, the prothallium may continue to grow and become a fairly large, green body with several archegonia (Fig. 215 A, B). In Marsilia the prothallium is still more reduced, it is enclosed in the macrospore, and only bears one archegonium. The archegonia are similar in structure to those of the Ferns, but are smaller, and sunk more deeply in the tissue of the prothallium.
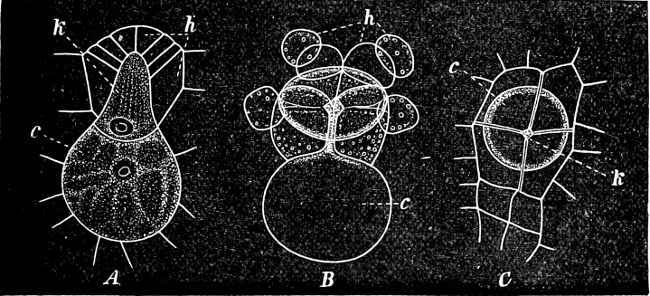
Fig. 216.—Salvinia natans. A An archegonium, unripe, seen in longitudinal section: h the neck-cells; k the neck-canal-cells; c the central cell. B An open archegonium of which the neck-cells have separated off. C An open, old archegonium seen from the top.
The asexual generation is developed from the fertilised egg-cell. It is a dorsiventral, horizontal shoot. In Salvinia it bears at first a shield-like leaf, the scutiform leaf (Fig. 215 C, a), which is succeeded by the ordinary foliage-leaves. The young plants of Marsilia, likewise, have less perfect leaves in the very early stage.
The formation of the sporangium is the same as in the Leptosporangiate Ferns. (The 16 spore-mother-cells originate from one central, tetrahedric archesporium.)
The Hydropterideæ are divided into 2 orders, the chief differences between them being found in the asexual generation.
[218]
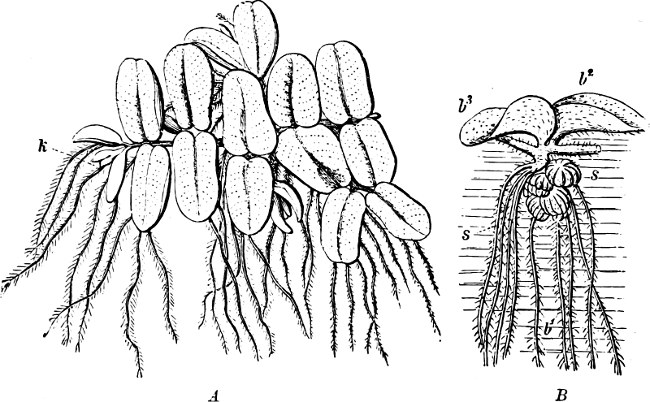
Fig. 217.—Salvinia natans (natural size): A seen from above, floating on the water; B a portion seen from the side in its natural position in the water.
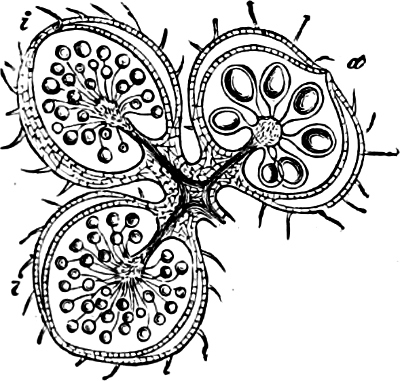
Fig. 218.—Sori of Salvinia in longitudinal section: h microsporangia; m macrosporangia. (× 10.)
Order 1. Salviniaceæ. This order more nearly approaches the true Ferns, especially so on account of the form of the indusium. Only one species is found in Europe, Salvinia natans (Fig. 217). This is a small, floating, annual, aquatic plant, entirely destitute of roots. The dorsiventral, horizontal stem bears two kinds of leaves, which are arranged in whorls of three. Two of these which turn upwards are oval, entire, “aerial foliage-leaves” (Fig. 217 B, b2-b3); the third, the “water-leaf” (b1) is submerged and divided into a number of hair-like segments, similar to the submerged leaves in many aquatic plants, for instance, Water-buttercup (see also Fig. 215 C). The whorls of leaves alternate with each other; there are thus 4 rows of dorsally-placed aerial leaves, and two rows of ventrally-placed submerged leaves. The sporangia are situated in sori, each sorus being borne on a small column (receptacle or placenta) and enveloped by a cupular, but entirely closed indusium (Fig. 218). The sori are situated on[219] the submerged leaves (Fig. 217 B, s-s) and are unisexual, i.e. each sorus contains microsporangia only, or macrosporangia.
Azolla belongs to this order. It is a very small, floating, tropical water-plant (America and East India), with horizontal, root-bearing stem. The stem branches profusely by lateral buds, and bears the two rows of leaves on its dorsal side, the roots on the ventral side. Each leaf is bifid, and divided into an upper dorsal, and a lower ventral portion. The upper segments float on the surface of the water and are arranged like tiles on a roof, each one overlapping its neighbour. In each floating segment a large cavity is found, in which Anabæna is always present. The lower segments are submerged.
Order 2. Marsiliaceæ. The characteristic feature of this order, and one not possessed by other Fern-like plants, is that the sori (2–many) are enveloped in leaf-segments which close round them and form a “sporocarp,” just in the same manner as the carpels, in the Angiospermous Flowering-plants, close round the ovules and form ovaries. The sori contain both micro-and macrosporangia. When the spores are ripe, the sporocarp opens in order to disperse the spores (Fig. 220).
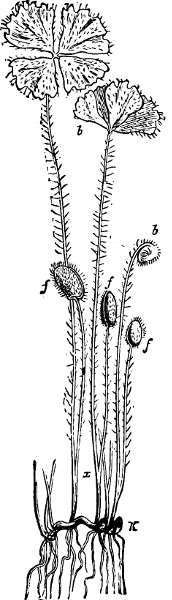
Fig. 219.—Marsilia salvatrix (natural size): K terminal bud; b leaves; f sporocarps; x point of branching of petiole.
The two genera (with 57 species, Temperate, Tropics) are land-and marsh-plants, whose dorsiventral, creeping stem bears roots on the under surface, and the leaves in two rows on the upper side (Figs. 219, 221). The leaves of Marsilia are compound, and divided into four small leaflets springing from the apex of the petiole (Fig. 219), and resemble the leaves of Oxalis. In the bud the leaves are circinate (Fig. 219 b), and at night they exhibit the well-known sleep-movements. The sporocarps are borne on the petioles of the fertile leaves, near their bases (Fig. 219 f); they are oblong and resemble small beans, the outer cells being hard and sclerenchymatous, while the inner ones are divided into a number of loculi arranged in two rows. On[220] germination, water is absorbed, the two sides separate slightly, as valves (Fig. 220 A), and a long vermiform mass of gelatinous, parenchymatous cells (Fig. 220), swollen by the water, emerges, bearing a large number of sori arranged pinnately. Each sorus (sr) is covered by a thin indusium. (The thin covering may be considered an indusium physiologically, though not morphologically).
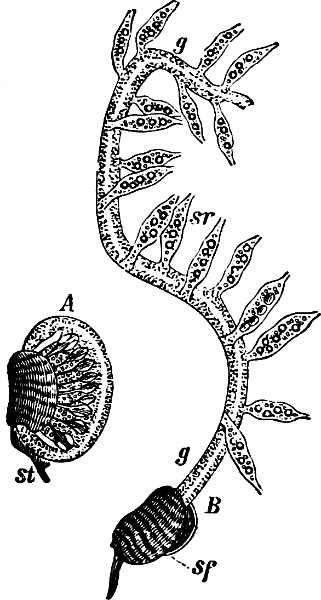
Fig. 220.—Marsilia salvatrix: A the sporocarp commencing to germinate; B a more advanced stage of germination.
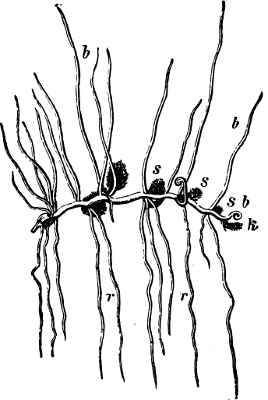
Fig. 221.—Pilularia globulifera (natural size): s sporocarps; b leaves; k the growing point; r roots.
Marsilia quadrifolia, in Europe. Many species are found in Australia. The nutritious sporocarps of M. salvatrix were the means of saving the Burke expedition in the interior of Australia, and hence this species has earned its specific name.
Pilularia has linear leaves, without lamina. The sporocarps are spheroid (Fig. 221), brown and hard, and situated near the base of the leaves. They are 2–4 chambered and open by a corresponding number of valves.
[221]
The characteristics of this class have been described on page 204.
It is divided into two sub-classes:—
1. The isosporous Equisetinæ. To this sub-class belong, with certainty, only the Equisetaceæ now existent, which are represented by only one genus, Equisetum.
2. The heterosporous Equisetinæ. Forms which are now extinct.
Order. Equisetaceæ (Horsetails).
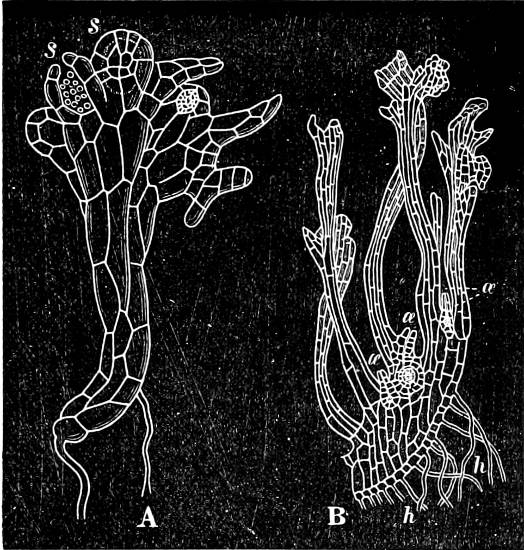
Fig. 222.—Equisetum arvense. The prothallium highly magnified. A Male; s, s antheridia. B Portion of a female, cut through vertically; œ œ archegonia, the central one is fertilised; h h root-hairs.

Fig. 223.—Equisetum maximum. Spermatozoids: a shows them still enveloped by the mother-cell.
The sexual generation. The prothallium is green and leaf-like, as in the majority of Ferns, but irregularly branched and curled. It is often unisexual. The male prothallia bear antheridia only, and are smaller and less branched (Fig. 222 A) than the female; the latter may attain a diameter of ½ an inch, and bear archegonia only (Fig. 222 B). The antheridia and the archegonia resemble those of the Ferns, but the spermatozoids (Fig. 223) are[222] larger and less twisted. On the last curve is situated a more or less elongated appendage of cytoplasm (Fig. 223 c).
The asexual generation. The embryo is similar to that of the Ferns. The fully developed Equisetum is a perennial herb, with widely creeping (in some species tuberous) rhizome, from which extend erect, aerial, most frequently annual shoots.
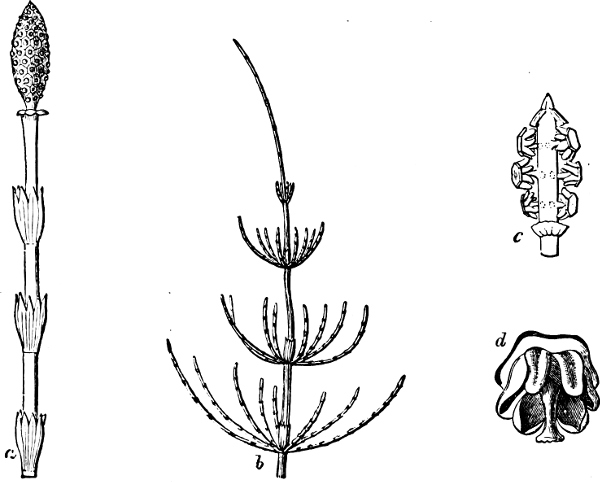
Fig. 224.—Equisetum arvense: a fertile branch with cone; b vegetative shoot; c cone; d sporophylls.
The vegetative aerial STEMS are divided into a number of internodes by the whorls of leaves (Fig. 224). The internodes are hollow, the cavities being separated from each other by the transverse partitions of the solid nodes. The lower portion of the internode, which is encased by the leaves, has much thinner and softer cell-walls, so that the stem is easily separated into segments just above the nodes. Each internode has a large number of ridges and furrows, and bears at its apex a whorl of leaves whose number and position correspond to the ridges of the internode. As in the case of other verticillate plants, the whorls are placed alternately, one above the other; the same arrangement is also found in the ridges on two successive internodes. In addition to the large air-cavity in the centre of each internode (the central cavity), a[223] whorl of tubular air-passages is found in the cortex of the stems, opposite the furrows (vallecular canals). There is also a similar air-passage (carinal canals) in each of the vascular bundles, which are placed in a ring, one opposite each ridge, and therefore alternating with the vallecular canals. The vascular bundles are collateral as in the majority of Flowering-plants, but poorly developed. The xylem of each bundle consists of two groups of annular or spiral vessels, close to the outer border of the carinal canal, and two groups of scalariform tracheides, each placed on a radius passing through a group of spiral vessels. The phloëm is placed between these four groups, each of which has only a few vessels. The stiffness of the stems is mainly due to the large amount of silica in the cell-walls of the epidermis, and to the sclerenchymatous cells of the ridges.
All LEAVES are situated in whorls. The VEGETATIVE are simple, undivided, 1-nerved, and are united into toothed sheaths (Fig. 224 a, b). The branching of the stems in some species (E. arvense) is very abundant. The branches break through the base of the leaf-sheaths (Fig. 224 b), and generally alternate with the teeth (leaves).
The FERTILE LEAVES (sporophylls) are different from the barren ones. They are free, shield-like, each one having a short stalk bearing usually an hexagonal plate (Fig. 224 d), and closely compressed into an ear or cone (Fig. 224 a, c). The Equisetums thus present an advance in development distinctly beyond that of the Ferns, which is further emphasized by the circumstance that a transition from the sheath-leaves to the fertile-leaves is found in the involucre or annulus, a “collar” of specially modified leaves situated at the base of the cone (Fig. 224 a and c). The cone may be considered as a very rudimentary flower, and the annulus may be regarded as a very early stage in the formation of a flower (perianth). See page 235.
The SPORANGIA are situated on the underside of the sporophylls, one at each angle; they are sac-like, and open inwardly by a longitudinal cleft (Fig. 224 d). An annulus is wanting; but in the wall of the sporangium, as in the pollen-sacs of the Flowering-plants, a layer of cells, with annular or spiral thickenings, is developed, which assists in the dehiscence of the sporangium.
The SPORES are green; the walls composed of four distinct layers, of which the outer is gradually separated, except at one point, and becomes split into four long bands (elaters) (Fig. 225).[224] The elaters are extremely hygroscopic, coiling round the spore when moistened, and expanding as soon as dry, presenting a most lively object under the microscope when breathed upon and allowed to dry. The second layer, when germination commences, becomes detached from the inner wall, which is formed of the exospore and endospore.
The order has become much reduced, and at the present time includes only one genus, Equisetum, with about twenty-five species, which are distributed over the entire globe, particularly in damp situations. In SOME SPECIES the barren shoots are green and very much branched, but the fertile ones are unbranched, pale brown, and possess no chlorophyll (E. arvense, Field-Horsetail, Fig. 224, and E. maximum). In others the fertile and barren shoots are alike green, and either both unbranched (E. hiemale), or branched (E. palustre, E. limosum, etc). The fertile shoots of E. silvaticum, up to maturity, resemble those without chylorophyll of E. arvense, but after that period they produce green branches, and thus resemble the barren ones.
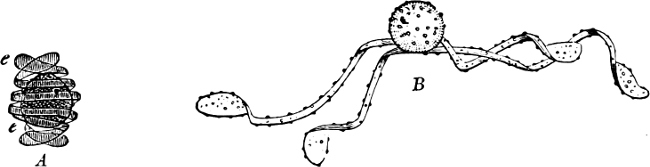
Fig. 225.—Spores of Equisetum: A damp, with elaters (e) coiled round the spore; B dry, with elaters expanded.
Extinct isosporous Equisetinæ. In addition to several true species of fossilized Equisetums, the order of the Calamites, which no doubt is closely allied to the Equisetinæ, is also found in the fossil state. These were gigantic forms, attaining about twenty times the size of those of the present day, and stems of nearly 10–12 metres in height are known. They reached the culminating point of their development in the Carboniferous period, and died out towards the close of the Palæozoic. The stems had hollow internodes and alternating grooves, similar to their relatives of the present day. The leaves must either have been absent or very perishable, since they have not been identified with certainty. If the determinations of certain remains of cones which of late have been discovered are correct, they were heterosporous and had two kinds of sporangia as in the following sub-class. A cambium formation and an increase in thickness has been found in the stems.
Their USES are very limited. A few species, such as E. hiemale are used for polishing on account of the hard siliceous cell-walls of the epidermis, found in all species of Equisetum.
[225]
The two orders which come under this head are united by the characteristics, that the verticillate leaves are not united into sheaths (Fig. 226), and that between each whorl of fertile leaves there is also a whorl of barren ones. The fertile whorls in Annulariæ are situated about midway between the barren ones (Fig. 227), but in Asterophylliteæ they occur immediately above a barren whorl (Fig. 228) and contain only half as many members as the latter. The lower whorls bear macrosporangia with one macrospore, the upper, microsporangia with many microspores.
Fig. 226.—A. fragment of Annularia.
Fig. 227.—Fragment of Annularia longifolia, with sporangia; the leaves have partly fallen off: a barren whorls; s fertile whorls.
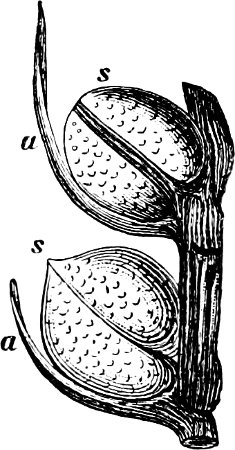
Fig. 228.—Fragment of cone of Asterophyllites (Volkmannia elongata): a and s as in Fig. 227.
The Annulariæ were distichous (Fig. 226), and presumably floating plants. The Asterophylliteæ had verticellate branches. These also died out after the Carboniferous period, at the close of the Palæozoic.
[226]
The characteristics of this class have been given on page 205. It consists of two sub-classes, one embracing isosporous, the other heterosporous forms.
One kind of spore. Prothallium large, partly green. Leaves without ligule.
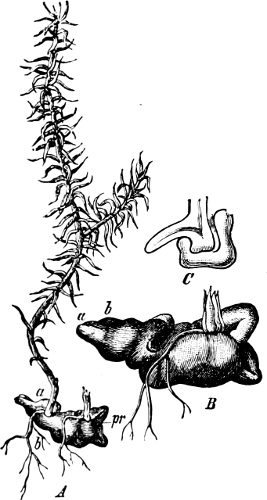
Fig. 229.—Lycopodium annotinum: A embryo (nat. size), with prothallium (pr), one embryo is broken off; B the prothallium (slightly magnified); C section through the prothallium and embryo in the direction a-b of A, and vertically in the plane of the paper.
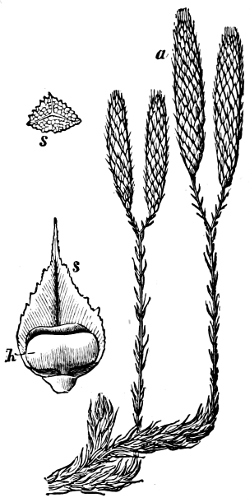
Fig. 230.—Lycopodium clavatum: portion of a stem, bearing cones (a); s a spore; h sporangium in the axil of a leaf, s.
Order 1. Lycopodiaceæ. The PROTHALLIUM is only known in a few species at present, but in these it is more or less tubercular, and bears both antheridia and archegonia.
In L. annotinum the prothallium is a relatively large mass of cells, without chlorophyll, and subterranean, in which the antheridia and archegonia are embedded (Fig. 229). In the widely distributed tropical species, L. cernuum,[227] and in L. inundatum, it is a small tubercular body which has a subterranean portion, with either little or no chlorophyll; and an aerial green portion. The prothallia of L. phlegmaria and others live saprophytically in the crevices of the bark of trees; they are partly filamentous, branched, and possess no chlorophyll.
The asexual generation. Perennial plants. The stem branches monopodially (often apparently dichotomously), and is thickly covered by small, simple, triangular or scale-like leaves. The leaves are spirally arranged in some species (Figs. 229, 230), and in others, whose stem is compressed with unequal sides, opposite (Fig. 231). The roots of Lycopodium are dichotomously branched.
The SPORANGIA in Lycopodium are situated singly at the base of the leaves, almost in their axils; they are reniform, unilocular and open like a mussel-shell by two valves (Fig. 230 h). The sporangia are developed from a group of surface cells. The archesporium is formed from one hypodermal cell (or perhaps a cell-row).
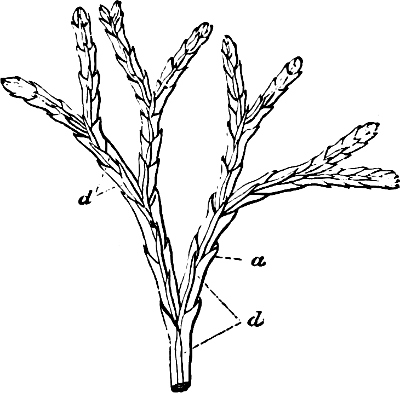
Fig. 231.—Lycopodium complanatum: a leaves on the edges of the stem; d leaves on the sides.
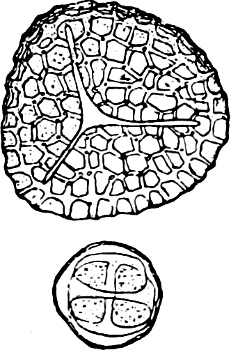
Fig. 232.—Lycopodium clavatum. A tetrahedral spore seen from above, where the three borders join; and a tetrad of bilateral spores, still lying in the mother-cell.
The fertile leaves are collected upon definite regions of the stem. They are either similar to the barren ones, and then the fertile portions of the stem pass gradually, without any break, into the barren portion (L. selago); or they differ from the barren leaves, and are then collected into special apical cones (Fig. 230 a). The SPORES are tetrahedral or bilateral (Fig. 232).
About 100 species, chiefly tropical.
Five species of Lycopodium are found in Great Britain. L. clavatum and L. selago are common in mountainous districts. L. annotinum is common in the[228] Highlands of Scotland. The other genus of the order is Phylloglossum, with one species, P. drummondi (Australia, Tasmania, and New Zealand), a small plant only a few centimetres high, with two tubers, and about eleven linear leaves at the base of the stem which is terminated by a cone of sporophylls.—Fossil Lycopodiaceæ in the Carboniferous period.
Officinal: “Lycopodium,” the spores of L. clavatum.
Family 2. Psilotaceæ. The sporangia are placed on the apex of short, two-leaved stems, as 2–3, seldom four, small capsules. Small herbs, with angular stems; leaves small, simple, and one nerved. Only four species.—Psilotum (Madagascar, Moluccas, Sandwich Islands, etc.) is destitute of roots, their place being supplied by special underground stems which bear a few modified leaves, very much reduced, especially when buried deeply in the soil. Three species.—Tmesipteris (Australia), one species.
Micro-and macrospores. The prothallia are very much reduced, especially the male; the female does not leave the spore. The leaves are ligulate.
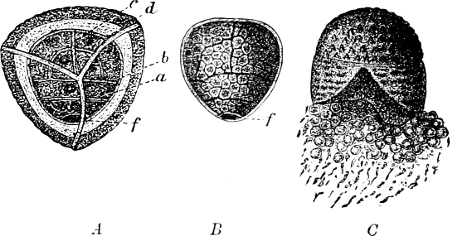
Fig. 233.—Germination of the microspores of Selaginella: A the spore rendered transparent, seen from above. In the interior is seen the prothallium (f), and the first divisions of the antheridium (a, b, c, d); in B the spore-wall is removed and all the spermatozoid-mother-cells formed; in C, the microspore has opened and the spermatozoids and the mother-cells are escaping together.
The sexual generation. In the MICROSPORES are formed: (1) a very small “vegetative” cell, representing the vegetative part of the prothallium (f in Fig. 233 A, B), and (2) a cell many times larger and which divides into a number (4–8) of primordial cells, each of which divides into four spermatozoid-mother-cells, though all of these may not develope spermatozoids. On germination, when the spore-wall is ruptured, the spermatozoids and spermatozoid-mother-cells are ejected into the water.
The SPERMATOZOIDS in Selaginella are elongated and club-shaped, with two cilia (Fig. 234); but in Isoëtes lacustris they are spirally-twisted threads which differ from all other spermatozoids by[229] having a bunch of cilia at each end; the other species of Isoëtes have cilia only at the anterior end.
The MACROSPORES. Shortly after the macrospores have been set free, or in Selaginella, while still enclosed in the sporangium of the mother-plant, they germinate and soon become filled with the cellular tissue of the prothallium, and even in Selaginella the archegonium begins to be formed before the rupture of the spore-cell-wall has commenced (Fig. 235 A).
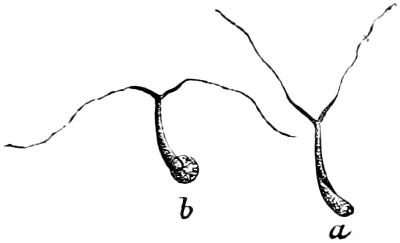
Fig. 234.—Spermatozoids of Selaginella: b with a remnant of cytoplasm.
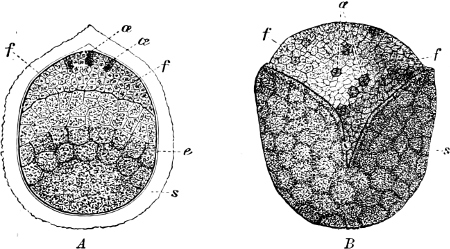
Fig. 235.—Macrospore of Selaginella: A longitudinal section, before the rupture of the wall, six weeks after being sown. The endosperm (e) has not yet filled the entire chamber. Cell-formation is still proceeding in the lower part of the spore. The endosperm and prothallium (f f) are separated by a distinct line (diaphragm). B Germinating macrospore seen from outside: s wall of the spore; æ archegonia.
Fig. 236.—Archegonia of Selaginella: A unripe, in longitudinal section; c the central cell; k neck-canal-cell, which is wedged in between the two-storied neck-cells; B ripe; u ventral canal-cell; C seen from above, open. It will be noticed that the neck is formed of two tiers of four cells each.
The ARCHEGONIA are constructed on the same plan as those of the[230] other Archegoniatæ, but are quite embedded in the prothallium (Figs. 235 æ, 236).
The asexual generation varies very much in the different orders.
Fig. 237.—Isoëtes lacustris (slightly diminished): st the stem; r roots; b leaves.
Fig. 238.—Isoëtes lacustris. Longitudinal section through the base of the leaf with a microsporangium. The edge of the groove, in which the microspangium is placed, is continued as a thin covering which envelopes the sporangium. The inferior edge of the ligular groove (L) forms a lip (J); t sterile cell-rows (trabeculæ) which divide the sporangium into compartments; l vascular bundle.
Fig. 239.—Selaginella inæqualifolia. Cone in longitudinal section; microsporangia are seen on the left side, macrosporangia on the right (most frequently each with four macrospores).
Order 1. Isoëtaceæ (Quill-worts). The only known genus, Isoëtes (Quill-wort), has an extremely short, tuberous, unbranched stem with very short internodes (Fig. 237). The STEM is remarkable as being the only one among the Vascular Cryptogams which increases in thickness (see page 202). The meristematic cells are situated round the axial cylinder, and form, especially, parenchymatous tissue in two or three directions, giving rise to 2–3 grooves in which the dichotomously-branched ROOTS are produced. The LEAVES are arranged spirally in a close rosette.[231] They are awl-shaped and have at the base a semi-amplexicaul sheath, with a groove (fovea), in which a sporangium is situated (Fig. 238). The ligule is a foliar outgrowth from the upper edge of the groove.—The MACROSPORANGIA (each with a number of macrospores), are situated on the outer leaves, the MICROSPORANGIA (Fig. 238), on the inner ones. Between each cycle of fertile leaves there are a number of imperfect or barren ones as in the case of the female plant of Cycas. The spores are liberated by the decay of the sporangium. The two kinds of sporangia develope at the commencement in the same way. The archesporium is, at first, a hypodermal layer of cells which grow out in the direction perpendicular to the surface of the leaf, and divide by a number of walls parallel to this direction, forming a sporogenous mass of cells. Some of the cell-rows of this sporogenous mass lose their rich protoplasmic contents, and are arrested in their growth; thus incomplete divisional walls of sterile cells, “trabeculæ” arise in the sporangium, dividing it into a number of compartments one above the other (Fig. 238 t). (The trabeculæ, according to Goebel, play the same part as the nutritive cells of the sporangium of Riella; the tapetal cells, as in the Ferns, are in a great measure dissolved at a later period.) The sporogenous cell-rows, in the microsporangia, give rise to a large number of spore-mother-cells, but in the macrosporangia only one spore-mother-cell, with tapetum, is developed from each fertile archesporial cell.
The two native species, and several others, are aquatic plants, the remaining species are land plants, or are amphibious. About 50 species. In temperate and tropical regions.—Fossil species in the Tertiary period.
Order 2. Selaginellaceæ. This order contains only one genus, Selaginella. The STEM, in the majority of species, is dorsiventral, long and slender, and apparently branches dichotomously, but in reality monopodially, with well developed lateral shoots. The LEAVES are small, round, or ovate, in the majority of species arranged in whorls of two leaves each; these whorls, however, are not decussate, but are considerably inclined towards each other, an arrangement by which four rows of leaves are produced, each whorl having one large and one small leaf. The two leaves in each whorl are of unequal size, the smaller one being placed on the upper surface and the larger on the lower surface of the stem (Fig. 240). Some species have spirally-arranged leaves, more resembling the arrangement in the Lycopodiums.
[232]
The FERTILE LEAVES most frequently differ from the barren ones, and are collected into spike-like cones (a kind of flower; Fig. 239). Micro-and macrosporangia are found in the same cone (Fig. 239). Each sporangium arises from a group of superficial cells of the stem, directly over the leaf on which it will be situated later on. Each sporangium has a hypodermal, unicellular archesporium, and contains a layer of tapetal cells; these are dissolved later, when the spores are ripe, and not before as in the Ferns. In the very early stages of their development, the micro-and macrosporangia are precisely similar, and the differences between them arise later on. In the microsporangium all the spore-mother-cells divide, and each forms four tetrahedrically-arranged microspores (Fig. 204); but in the macrosporangium only four macrospores are formed, by the division of a single mother-cell, while the remaining spore-mother-cells are aborted. It is rarely that the macrosporangia contain 2 or 8 macrospores.
Fig. 240.—Selaginella martensii: s lower leaves; r upper leaves.
For the GERMINATION OF THE SPORES, see pages 228, 229. The prothallium arises in the macrospore (f-f, in Fig. 235 A), probably by division of the meniscus-shaped protoplasmic mass, which is marked off at the apex of the spore; primordial cells are thus formed which later on are surrounded by a cell-wall. In six to seven weeks after sowing, the spore-wall is ruptured by the growing prothallium, which already has developed archegonia (Fig. 235 œ-œ). The prothallium so formed does not occupy the entire cavity of the spore, but four to five weeks after sowing, the large-celled parenchyma is developed in the lower portion of the spore by free cell-formation; this has been termed by Pfeffer, “endosperm,” since it is similar to the endosperm of Flowering-plants. Goebel, however, has termed it “secondary prothallium,” as the homology with the endosperm of the Angiosperms is very doubtful.
The FERTILISED OOSPHERE divides into an upper (hypobasal) and a lower (epibasal) cell; from the latter alone the embryo is developed with its root, stem, foot, and two cotyledons, and the former gives rise to an organ which appears in this instance for the first time, but which occurs in all Flowering-plants, viz. the suspensor. This forces the embryo down into the “endosperm,” which is entirely or partially absorbed by the embryo. In the case of the Flowering-plants the embryo is developed with its longitudinal[233] axis in the elongation of the suspensor, but in Selaginella the embryo is situated transversely to it.
Selaginella (300–400 species), is essentially tropical, only one species living in the North (S. spinulosa), but others grow in Central and South Europe.
Order 3. Lepidodendraceæ are extinct, tree-like Lycopods, which are found especially in the Lower and Middle Carboniferous. Vegetatively they are most nearly related to Lycopodium, but the stem attained much larger proportions (about eleven metres in height and one metre in thickness), and had a cambium by which it increased in thickness. It was regularly dichotomous, and closely studded with spirally-placed leaves, which left behind them peculiar rhombic scars. The large cones resemble Pine-cones, and bore sporangia much larger than any which are now produced (the male ones as much as 2 cm.’s in length). The macrosporangia were situated at the base, and the microsporangia at the apex.
Order 4. Sigillariaceæ. These are, presumably, another group of extinct tree-like Lycopods (especially in the Middle Carboniferous). The name has been derived from the seal-like scars, which the fallen leaves have left behind in longitudinal rows on the grooved stem. The rhizomes of these plants were formerly termed Stigmaria, and placed in a separate genus.
Order 5. Sphenophyllaceæ form an entirely extinct group. They do not definitely belong to any of the three large classes of Vascular Cryptogams, but it is perhaps best to place them in juxtaposition to these. They were herbaceous plants with verticillate, wedge-shaped leaves, with nerves branching dichotomously into equally strong branches. Micro-and macrosporangia were formed in the same cone; and were situated in the axils of the leaves, as in the Lycopods.
[234]
All the plants considered in the preceding chapters are included in the term Cryptogams; all in the following chapters under the head of Phanerogams (see page 3). Hofmeister’s pioneer works (1851, Vergleichende Untersuchungen der höheren Kryptogamen, etc.) and the numerous researches published later by other investigators, have closed the gap which was formerly thought to exist between these plants; so that we now, in the series: Bryophyta—Pteridophyta—Gymnospermæ—Angiospermæ see the expression of a single line of development in accordance with a definite plan. The forms through which this gradual development has taken place have in course of time, however, to a great extent died out, and only single links of the chain connecting the lowest to the highest still remain.
The alternation of generations, which we found indicated in certain Thallophytes, can be proved with the greatest clearness in all the higher Cryptogams, from the Mosses upwards; it is also found in the Phanerogams, but not in such a pronounced degree, because one of the generations is so far reduced that it has almost given up its independence. For the sake of greater clearness, we will begin with the comparison of the sporophyte, asexual (second) generation.
The asexual (2nd) generation of the Cormophytes.
The asexual generation which follows from the further development of the fertilised oosphere, is, in the Mosses, only the sporogonium (according to one theory it is perhaps homologous with a spore-bearing leaf, situated upon a short stem, see p. 187); in Filicinæ, Equisetinæ, and Lycopodinæ, on the other hand, it is a highly developed plant differentiated into stem, leaf, and true root, and bearing the sporangia on its leaves. The MODIFICATION OF THE SHOOT is very slight in Filicinæ. The first leaves of the embryo are very simple in form (Fig. 205), but after a certain age all the leaves which arise are essentially alike. The fertile leaves do not[235] differ from the barren ones, and are found associated with them, and their formation does not limit the growth in length of the stem. It is only in a few of the true Ferns, and in the Hydropterideæ, that the fertile leaves differ considerably from the barren ones. A division of labour in which certain leaves are set apart for nutrition, and others for reproduction, is found more pronouncedly in the Equisetinæ and Lycopodinæ, for in these groups, with a few exceptions, the fertile and barren leaves are very dissimilar; the former are collected in special ear-like cones, which terminate the further growth of the short stems on which they are borne. In connection with the cone, leaves are sometimes developed which form a transition from the barren to the fertile ones (the “annulus” in Equisetaceæ), and in these cases the first indication of a flower with perianth or floral-leaves is to be traced. Among the Cryptogams the highest division of labour is found in Selaginella and Isoëtes, which have the two kinds of sporangia borne on different leaves. The division of labour (modification) is, however, still more pronounced in the Phanerogams: the leaves which bear the microsporangia (“pollen-sacs”) have quite different forms from those which bear macrosporangia (the “nucellus” in the ovule), the former are termed stamens, the latter carpels; in certain instances, too, there is even a contrast between the “male plants” and the “female plants.” Moreover, a division of labour, in a much greater degree, takes place in the leaves which do not directly take part in reproduction, and it is thus possible in many plants to draw a sharp line not only between stamens and carpels, but also between four or five distinct kinds of leaves, which differ in form, structure, and corresponding functions, and which appear in regular sequence on the shoot: namely, between “scale-leaves” and “foliage-leaves,”[21] both of which occur in the Cryptogams, and the “floral-leaves,” including the bracts and leaves of the “perianth,” which latter often differ from each other in form and colour, and are then separated into sepals and petals. The leaves—stamens and carpels—which bear the sporangia are termed sporophylls, and the shoot, or extremity of a shoot, whose leaves are modified into sporophylls, is terminated in its further growth by their production, and is known as a flower. The flower which is most[236] completely furnished has calyx, corolla, stamens, and carpels arranged in this order. If the only sporophylls present are stamens, then it is said to be a male (staminate) flower, and if only carpels, then a female (pistillate) flower, and in both these cases the flowers are unisexual, or diclinous. If stamens and carpels are both present in the same flower, it is termed hermaphrodite. Diclinous plants in which the female flowers are situated on one plant, and the male flowers on another, are termed diœcious; and those in which the same plant bears the two kinds of flowers are termed monœcious. When the male, female, and hermaphrodite flowers are found in the same species, the plant is said to be polygamous.
The sporangia-bearing leaves—Sporophylls. In the Mosses the asexual generation is only represented by the sporogonium, and if the theory is correct which considers the sporogonium to be an embryo consisting of a rudimentary stem and terminal leaf, then the spores are produced on the leaves in these plants. The sporangia in the Filicinæ are situated in groups (sori) on the back or on the edge of the leaves. The number of sporangia in the sorus diminishes very greatly in the Marattiaceæ and Gleicheniaceæ (three to four in the latter, Fig. 213). In the Equisetinæ the sporangia are situated in a small number on the underside of shield-like leaves, and in Lycopodinæ, singly, in the axils of the fertile leaves, which are alike and bear either micro- or macrosporangia. In the Phanerogams there is a great difference between the stamens and carpels.
Fig. 241.—Cycas: a stamen (nat. size) seen from the under side; b four pollen-sacs, not yet open, forming a “sorus”; c three open pollen-sacs; d a pollen-grain.
Fig. 242.—Stamens of Araucaria (pollen-sacs long and pendulous).
Fig. 243.—Male flower of Taxus.
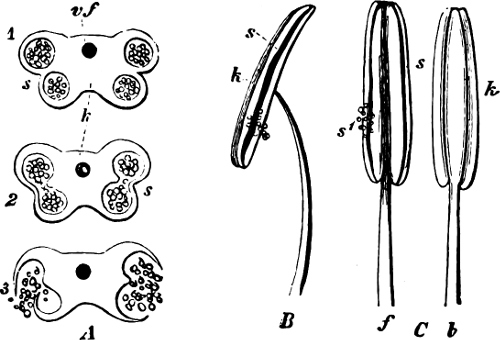
Fig. 244.—A Cross section through a quadrilocular anther in different stages of development: s the seam where it bursts open; vf vascular bundle; k connective. B A stamen. C Another stamen seen from the front (f) and from the back (b).
Stamens. In the lowest Phanerogams (Cycadeæ) there are many indications of relationship to the Ferns. The stamens are flat and broad, and have on the back many pollen-sacs (microsporangia) arranged in small groups (true sori), which even have[237] a small “placenta,” similar to the one possessed by the Ferns, and open towards the inside by a longitudinal cleft (Fig. 241, compare Fig. 213). A section of the Coniferæ agree more closely with the Equisetaceæ, in having a few (three to eight) pollen-sacs arranged on the underside of more or less shield-like leaves (Figs. 242, 243, compare with Fig. 224 a, c, d). In the Abietaceæ the number of sporangia is diminished to two, which are placed also on the lower side (Fig. 267) of a stamen. The number of pollen-sacs (microsporangia) in the Angiosperms is nearly always four to each stamen; they are longitudinal projections which are placed in pairs on each side of the central line of the stamen, two on the edge, and the other two generally on the side which is turned inwards; the pollen-sacs generally dehisce longitudinally[238] (quadrilocular anthers, Fig. 244). A few, for instance Orchidaceæ and Asclepiadaceæ, have only two pollen-sacs (bilocular anthers); and in others, such as Solanum and the Ericaceæ, they open by pores; in Lauraceæ and Berberidaceæ, by valves. The part of the stamen which bears the pollen-sacs is termed the anther. Most frequently this is supported by a stalk known as the filament.
Fig. 245.—A carpel of Cycas revoluta with 5 ovules (s), at half to one-third nat. size.
Fig. 246.—Carpel with 2 ovules of Ceratozamia robusta (1/1).
Carpels. The simplest forms of carpels are found in Cycas. In this genus both the foliage and fertile leaves are pinnate, and hence present great similarity; the ovules (macrosporangia) are situated on the margin of the central portion, just as the sporangia are placed on the edge of the fertile leaf of Ophioglossum (Fig. 245, compare with Fig. 209). The carpels of the other Cycadeæ present greater divergence from the foliage-leaves, being peltate, for instance, in Zamia and Ceratozamia (Fig. 246). The ovules in the Coniferæ are situated on the upper side and near the base of the ovuliferous scales, almost in the same position as the sporangia in the Lycopodinæ (Figs. 269, 272, 273 H, compare Figs. 230, 239). In Taxus the uninclosed ovule is placed on the apex of a shoot (Fig. 264). In all these plants the ovules are not enclosed by the carpels, that is, they are not enclosed in chambers formed by the turning in of the walls of the carpel, and hence the name[239] Gymnospermæ is given to them. In the higher Flowering-plants, the Angiospermæ, the ovules are distinctly situated on the edge, the upper surface, or base of the carpel; but the carpel closes round the ovules which are therefore enclosed in a chamber—the ovary. In a few cases, for example in the Polygonaceæ, an ovule is situated apparently on the apex of the stem itself, as in the Yew; in other cases, as in the Primulaceæ, many ovules are apparently developed on the apex of the stem, which seems to have been specially adapted as a placenta, but it is also possible and correct in these cases to suppose that the ovules are in reality developed on the carpels.[22] A single fully-developed carpel or a collection of carpels joined together is termed the pistil. The extremity of the carpel, which is specially developed to catch the pollen-grains and form a suitable nidus on which they may germinate, is called the stigma. The united edges of a carpel which bear the ovules are termed the ventral suture. The back of the carpel forms the dorsal suture. The Marsiliaceæ take a position among the Hydropterideæ analogous to that occupied by the Angiosperms; the sporangia are in a corresponding manner enveloped in a closed leaf.
The collection of stamens in a flower is termed the andrœcium, and all the carpels, whether individually free or united into one pistil, the gynœceum.
The Sporangia. The asexual generation of the Mosses is the sporogonium, in which the spores arise in tetrads from the mother-cells. The sporangia in the Filicinæ take their origin either from a single cell (Leptosporangiatæ) or, what probably may be regarded as an older stand-point, from a group of cells (Eusporangiatæ). In both cases there may be distinguished in a mature sporangium three tissues, which have different significance (Fig. 204): (1) an external layer, the sporangium-wall, most frequently composed of one layer of cells made up of cells of dissimilar structure, so that on desiccation the wall is ruptured and the sporangium opens in a definite manner; (2) an internal group of cells, consisting of the spore-mother-cells, developed from an archesporium, and which by division into four gives rise to the spores; (3) a layer of cells lying between the two already mentioned, which is dissolved before maturity. The intermediate cellular layer, which directly surrounds the spore-forming cells, is in form and contents more worthy of note than the others, and is termed the tapetum. The construction[240] of the sporangium in the Equisetinæ and Lycopodinæ is in the main the same.
Fig. 247.—Development of an anther. A Transverse section of a young anther of Doronicum macrophyllum. The formation of the 4 pollen-sacs commences by divisions of the hypodermal cells (at m, for instance). These cells divide by periclinal walls into external cells which only take part in forming the anther-wall; and internal cells, which correspond to the Archesporium, and from which the spores are derived. These spore-forming cells are drawn with thicker walls in B-E. The commencement of the vascular bundle is seen in the centre. B An older stage; the pollen-sacs already project considerably. It is the cells in the hypodermal layer which are active and in which tangential divisions particularly occur; fv vascular bundle. C A corresponding longitudinal section. D Transverse section through an older anther, the thickness of the wall outside the mother-cells of the pollen-grains is already increased, and it becomes still thicker by the division of the hypodermal cells: its most external layer of cells but one, becomes transformed into the “fibrous cells.” E Transverse section of a still older pollen-sac of Menyanthes; sm are the mother-cells of the pollen-grains surrounded by the tapetum (t), external to the tapetum is the anther-wall, which is still far from being fully developed. The sub-epidermal layer becomes “fibrous,” and the cells lying inside it become dissolved, together with the tapetum.
In the Phanerogams the Microsporangia are termed Pollen-sacs. They take their origin from a large group of cells, which, in the Angiosperms, lie immediately beneath the epidermal cells of the anther. In the developed, but not yet mature, sporangium (pollen-sac) there are to be found: (as in the Vascular Cryptogams) (1) an internal group of mother-cells which give rise to the pollen-grains (microspores), in this case also formed in tetrads; (2) a group of cells surrounding these, of which the internal ones form a tapetal layer, similar to that in the Vascular Cryptogams; the tapetum[241] and some of the cells surrounding it in this group, become dissolved before maturity; the more external ones, on the other hand, are provided with peculiar thickenings, and form the “fibrous” layer by the aid of which the dehiscence of the anther takes place; (3) an external layer, the epidermis, enclosing all the other layers (Fig. 247).
In some Coniferæ (Cupressus, Thuja, and several species of Juniperus) the microsporangia (pollen-sacs), which are situated on the under side of the stamen, are covered by a thin structure which seems to be a continuation of the lamina and which is supposed to be homologous with the indusium of the Ferns.
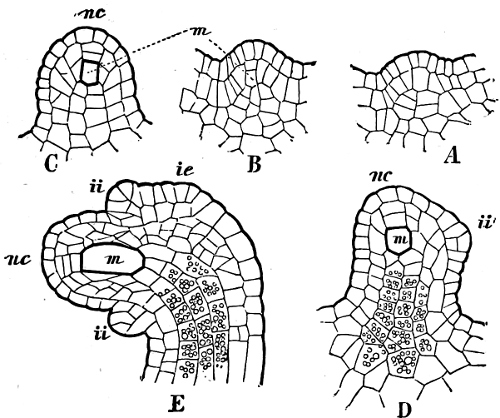
Fig. 248.—Development of the ovule in the Red Currant, Ribes rubrum, arranged alphabetically in the order of development. A Is the youngest stage, E the oldest. ii Inner integument; ie outer integument; nc nucellus; m archespore (mother-cell of the embryo-sac).
The Ovule in the Phanerogams arises most frequently on a projecting portion of the carpel, termed the placenta. The ovules (compare the sporangium of the Eusporangiatæ and especially the pollen-sac) take their origin from a group of cells which lies beneath the epidermis (Fig. 248 A, B). First of all a small papilla is formed, which is later on provided with a vascular bundle and becomes the funicle; this probably has the same value as the projections (“placenta”) on which the sori in the Ferns are attached. Only one macrosporangium (nucellus; Fig. 248 nc) is developed at the apex of the funicle. This arises by a process of cell-division exactly corresponding to that by which the pollen-sacs[242] are formed (Fig. 248 C-E), with this difference only, that while a great many cells may be distinguished in each pollen-sac, which forms pollen-grains by tetrad-division, only a few are found in the ovule, and all these moreover are suppressed, with one single exception which developes into the macrospore (embryo-sac) without undergoing a division into tetrads. The wall of the embryo-sac, in the Gymnosperms, may be thick and divided into two layers and partly cuticularized, as in the spores of the Cryptogams which are to be set free. In the Angiosperms, on the other hand, the wall is extremely thin.
The pollen-sac thus stands in the same relation to the nucellus as the microsporangium does to the macrosporangium: in the pollen-sacs and microsporangia a number of spores arise by the tetrad-division of several mother-cells; in the nucellus and macrosporangium, a reduction of the cells already formed takes place to such an extent that the number of macrospores becomes one (Salvinia, Marsilia, Phanerogams) or four (Selaginella), or rarely a large number as in Isoëtes.
In the Ferns, as stated on page 210, etc., indusia covering the sori very often occur. Horsetails and Club-Mosses have no indusium; but in all Phanerogams cupular or sac-like structures (integuments) are found which envelop the nucellus. These develope from the upper end of the funicle (ii and ie, in Fig. 248; y and i, in Fig. 249) and enclose the nucellus on all sides as a sac, leaving only a small channel at the apex of the nucellus—the micropyle—(Fig. 249) through which the pollen-tube proceeds to the embryo-sac. The ovules of the Gymnosperms have only one integument (Figs. 251, 264, 269, 274) and the same is the case with the majority of the Sympetalæ and a few Choripetalæ; but the Monocotyledons and most of the Choripetalæ have two integuments (Fig. 249).
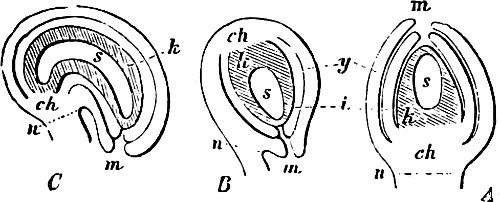
Fig. 249.—Various forms of ovules: A an erect ovule (orthotropous); B reversed (anatropous); C curved (campylotropous): k the nucellus (shaded in all the figures); s the embryo-sac; ch the base of the ovule (chalaza); y and i the external and internal integuments, the dotted line denotes the place where the scar (hilum) will form when the seed is detached from the funicle.
In shape the integuments resemble very closely the cupular[243] indusium of the Hymenophyllaceæ, certain Cyatheaceæ (Fig. 212 E), and Salvinia (Fig. 218); that they are really homologous with these is probable, but is not proven. Some authorities regard them as structures found only in the Phanerogams.
The ovule is thus a “monangic” (i.e. reduced to 1 sporangium, the nucellus) sorus, situated on a funicle, and enclosed by one or two cupular indusia, the integuments. Some of the ovules are erect (orthotropous), others curved (campylotropous), the majority reversed (anatropous) (Fig. 249).
[Goebel (1884 and earlier) with Strasburger considered the entire ovule of the Phanerogams as homologous with the macrosporangium, the integuments however as new structures in contradistinction to the Ferns: the funicle then corresponds to the stalk of the sporangium. The integuments of the ovule (according to Goebel, 1882) differ from the indusium of the Fern-like plants in being developed from the basal portion of the nucellus and are not, as in the Ferns and Isoëtes, a portion (outgrowth) of the leaf which bears the sporangia (K).]
The nucellus is the only macrosporangium which never opens; the macrospore remains enclosed in it, and the macrosporangium remains attached to the mother-plant. It is therefore essential that the method of fertilisation which is employed should be very different from that of the Cryptogams. The pollen-grains must be transferred to the ovule, and retained either by a drop of mucilage at the micropyle (Gymnosperms) or by the stigma on the carpels (Angiosperms). Fertilisation by spermatozoids, which are freely motile in water, is abandoned in the Phanerogams.
Many other modifications, unknown in plants of more simple structure, take place, for instance, in the shoots which bear the fertile leaves; especially in the form of the stem or thalamus (hypogynous, perigynous, epigynous); in the development of the perianth which stands in intimate connection with the special means employed to effect fertilisation; with respect to the different grades of union found in the leaves; in the union of the flowers into aggregations of a higher order (inflorescences), and at the same time the production of “floral-leaves” (page 235).
The sexual generation. The Fertilisation.
The sexual generation in the Mosses is relatively well developed, because not only the protonema, but all the other vegetative parts of the Moss-plant, in addition to the archegonia and antheridia, belong to it. In the groups which follow, a gradual but increasing reduction of the sexual generation takes place, and at the[244] same time an indication of sex is found in the prothallia, which finds expression in the forms of the spores themselves. In the majority of cases among the isosporous Vascular Cryptogams, the sexual generation—prothallium—is a green, leafy expansion which can sustain itself by the assimilation of carbonic acid, and by the absorption of nutriment from the soil by means of root-hairs. In some plants (Ophioglossaceæ, Lycopodium annotinum) the prothallium is a subterranean, pale, tubercular body, but in these instances it is relatively large. In the heterosporous Vascular Cryptogams and in the Phanerogams, the prothallium is much more reduced, both as regards its size, and also with respect to the number and structure of the antheridia and archegonia.
1. The Microspores. The PROTHALLIUM in all Vascular Cryptogams which have unequal spores, consists of a single, vegetative (barren) cell, which plays a very unimportant part in the life of the prothallium (Fig. 233 A). In Salvinia it is somewhat elongated and tubular, because it must break through the sporangium (Fig. 214); but in other cases it is very small and lenticular. In all these plants only one antheridium is formed. In Salvinia it consists of 2 cells whose walls are ruptured in order that the spermatozoids may be liberated (Fig. 214 B, C). In Marsilia, Isoëtes, and Selaginella the prothallium does not leave the spore, and consists for the most part of primordial spermatozoid-mother-cells without cell-wall, which on germination are ejected so that the spermatozoids are set free.
In the Phanerogams, the microspores have from olden times been termed pollen-grains.
In the GYMNOSPERMS the prothallium is reduced to 1, 2 or 3 small cells, placed on one side of the mature pollen-grain (at the top in Fig. 250 I, II, and in Fig. 267 N) and which do not play any part in the germination of the pollen-grain. The antheridium is represented by the remaining portions of the interior of the pollen-grain, that is, it consists of a large cell with a nucleus which does not even go so far as the antheridium of Selaginella and become divided into spermatozoid-mother-cells without cell-wall, for even these cells are not formed. The unicellular antheridium grows, on the germination of the pollen-grain, into a tubular body known as the pollen-tube, formed from the inner wall of the pollen-grain (Fig. 250), which works its way down the micropyle to the oosphere. The fertilisation takes place by diosmosis through the cell-wall, and consists here also of the coalescence of the nucleus[245] of the pollen-tube (the sperm-nucleus, male pronucleus) with that of the oosphere.
In the ANGIOSPERMS the reductions proceed still further. The barren cell, which represents the prothallium, was in the last group separated from the antheridium by a true cell-wall, but in the Angiosperms a membrane at most, but no firm cell-wall, is formed. The pollen-grain contains two cells, a vegetative and a free generative cell. Both these pass into the pollen-tube, but the vegetative cell disappears about the time the pollen-tube reaches the ovule; while the generative cell divides into two: one, the sperm-nucleus coalescing with the nucleus of the oosphere, the other being absorbed (Lilium, after Guinard).
The Gymnosperms prove in yet another point that they are more closely related to the Cryptogams than are the Angiosperms. When the pollen-grain begins to germinate the external wall ruptures as in the Cryptogams (Fig. 250), but in the Angiosperms special germ-pores are formed in the cell-wall for the emergence of the pollen-tube.
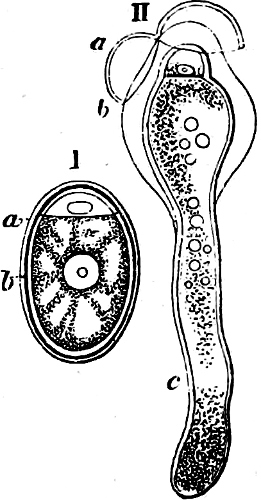
Fig. 250.—I Pollen-grains of Cupressus; at the top is seen one prothallium-cell. II Germinating; c pollen-tube; a the extine; b the intine.
2. The Macrospores. The prothallium in Salvinia and Marsilia is still rather large, green, and capable of the independent assimilation of carbon. It projects more or less from the macrospore and bears (in Marsilia only one, in Salvinia several) archegonia, which however are embedded to a greater degree in the prothallium, and are more reduced than the archegonia of the true Ferns and Horsetails (Figs. 215, 216). The prothallium is still more reduced in Isoëtes and Selaginella; partly because it is smaller and is in a higher degree enclosed in the spore, it also contains less chlorophyll, or is entirely without chlorophyll, and in consequence incapable of independent existence, whilst the number of archegonia is less; and partly because the archegonia are themselves reduced, the cells of the neck are fewer and embedded to the level of the surface of the prothallium without any, or with only a very slight projection (Figs. 235, 236).—Finally, the prothallium with its archegonia begins to develope in Selaginella while the macrospore is still within its sporangium, and before it is set free from the mother-plant.[246] After the spores are set free and germination has commenced, the spore-wall ruptures and the prothallium is exposed.
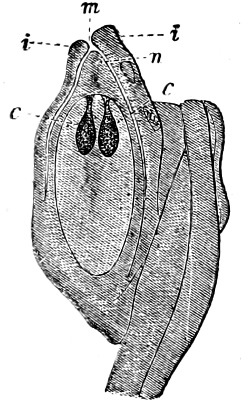
Fig. 251.—Longitudinal section of ovule of Abies canadensis. Inside the integument (i) is seen the nucellus, n; m the micropyle. In the interior of the nucellus is seen an oval mass of cells, the endosperm, and at its top two archegonia, c. The ovule is turned in such a way that the micropyle points upwards, but usually it turns downward in the Abietineæ.
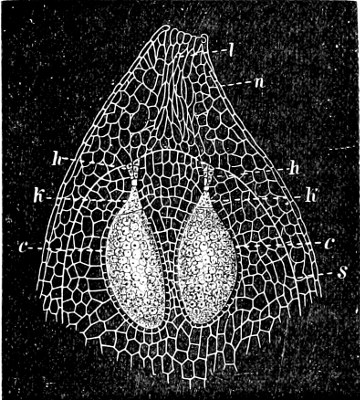
Fig. 252.—The apex of the nucellus (n) of an ovule of Abies: l long-shaped cells which guide the pollen-tube; s the wall of the macrospore (embryo-sac); h the neck-cells of the archegonium; k the ventral canal-cell; and c the central cell (oosphere). The archegonia of the Cryptogams should be compared with this (see pages 181, 208, 216).
The GYMNOSPERMS go still further. The macrospore (embryo-sac) germinates and forms internally a cellular tissue, designated in former times by the name of albumen (endosperm), which is homologous with the prothallium. It always remains enclosed in the embryo-sac, and is a parenchymatous mass containing a large supply of nourishment. In the upper part of the endosperm a number of archegonia are developed which are in the main constructed in the same manner as those in the Cryptogams, but are still more reduced, the neck consisting only of 4, 2, or 1 cell (Figs. 251, 252). The ventral canal-cell is also formed, in the majority, as a small portion cut off from the large central cell just beneath the neck; the larger remaining portion becomes the oosphere. When the pollen-tube has passed down to the oosphere (Fig. 253) and fertilisation has been effected, the oospore commences a cell-formation, the final result of which is the formation of an embryo (the asexual generation) which is provided with a thinner, lower end, termed the suspensor. The embryo is forced more or less into the endosperm[247] in which it may rest for a longer or shorter time, and generally is developed to such an extent that it has a distinct primary-root (radicle) and stem (plumule) with one or more embryo-leaves (cotyledons).
When the oosphere has been fertilised its nucleus sinks down to its lower end, and by repeated division into two, forms four cells lying in one plane (Fig. 253, see base of the left archegonium). Three tiers of cells are now formed by transverse division of these four. It is the intermediate one of these which elongates and forms the suspensor, or four suspensors, if they separate from each other, which push the lowermost four cells deep down into the endosperm. It is from these four lower cells that the embryo (or four embryos when the suspensors separate) is developed, but never more than one embryo attains full development. As several archegonia are contained in one and the same ovule, all of which are capable of forming embryos, there is the possibility that several embryos may be developed in a seed (polyembryony), but usually only one embryo attains perfect development.
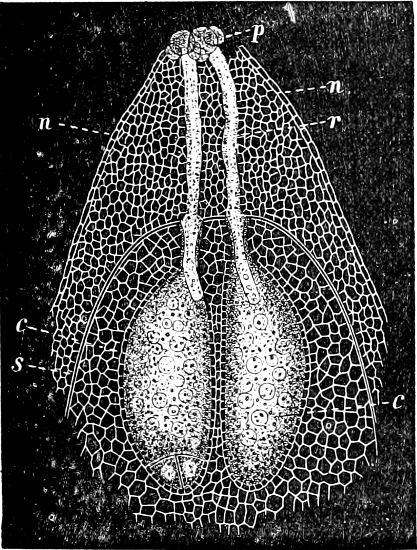
Fig. 253.—The apex of the nucellus (n) of Abies in longitudinal section: c, c the oospheres of the two archegonia; the embryo-formation has commenced at the bottom of the left archegonium; s wall of the macrospore; p pollen-grains; r pollen-tubes.
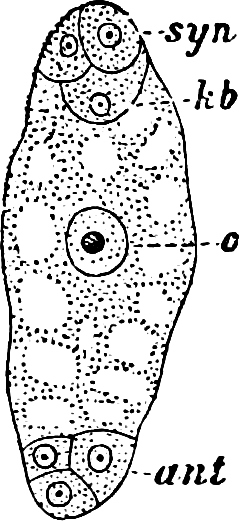
Fig. 254.—Embryo-sac of Carex præcox: syn synergidæ; kb the oosphere; c the central nucleus; ant the antipodal cells.
At the same time that the embryo is being developed, other changes are taking place in the ovule, especially in the integument which becomes the shell of the seed (testa). The endosperm grows, and the embryo-sac supplants the cells of the nucellus. The seed[248] is now formed, and it consists in its most complete development, as in this instance, of three parts:
(1) The testa of the seed, formed by the enveloping integuments, with the remainder of the tissue of the nucellus lying outside the embryo-sac (the macrosporangium).
(2) The endosperm or prothallium.
(3) The embryo.
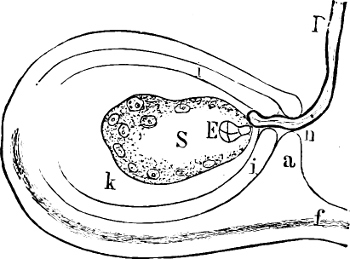
Fig. 255.—Diagrammatic longitudinal section through an anatropous ovule shortly after fertilisation; a and i are the two integuments; f the funicle; k the nucellus; S the embryo-sac, with the incipient formation of nutritive-tissue; E the embryo; P the pollen-tube passing through the micropyle (n) to the oosphere.
The reduction in the Angiosperms is carried to the extreme limit. In the embryo-sac (the macrospore) the nucleus by continued division produces a prothallium consisting of primordial cells (Fig. 254). In the upper end of the embryo-sac (which is nearest the micropyle) are three cells, two of which are termed the “co-operating cells” (synergidæ) and the third is the oosphere. Three others are placed at the opposite end of the embryo-sac and are therefore termed the “antipodal cells.” Finally, a large cell is also formed, which occupies the space between the two groups and whose cell-nucleus, the central definitive nucleus, lies in the centre of the embryo-sac. These primordial cells are the slight remnant of the prothallium. The entire structure of the archegonium, with its neck and canal-cells, has disappeared, and nothing is left but the indispensable oosphere. When the oosphere has been fertilised, and has commenced the cellular divisions which lead to the formation of the embryo (Fig. 255), the synergidæ and antipodal cells are absorbed, and a cell-formation begins by a new process which emanates from the definitive nucleus and by which a parenchymatous cell-tissue, the nutritive-tissue, arises which may perhaps be considered as homologous with the endosperm of the Gymnosperms. The difference is that the nutritive-tissue of the Angiosperms is formed in two parts with an intervening interruption; the primary nutritive-tissue is first formed, and after fertilisation is absorbed,[249] with the exception of one cell, which continues the development and gives rise to the nutritive-tissue proper, which is formed in the first instance of primordial cells, and later on of a cellular tissue; this nutritive-tissue formed in the embryo-sac is termed “endosperm”; in a few instances[23] a tissue which is derived from the nucellus functions as nutritive-tissue, and is termed “perisperm.” In many plants the seeds, when ripe, contain a very rich nutritive-tissue, in addition to the embryo, for the purpose of its nourishment during germination. These are termed albuminous (endospermous) seeds, in distinction to the ex-albuminous, or those in which the nutritive-tissue is stored in the embryo itself, before it is completely developed, and used for its sustenance.
In addition to the changes which fertilisation produces in the ovule itself, it also gives the impetus to a series of changes in the entire shoot which bears the ovule. The perianth, stamens, and style, generally wither, because the part they play is at an end; the wall of the ovary grows and becomes the wall of the fruit (pericarp). The entire gynœcium of a flower, transformed as a consequence of fertilisation, is termed a fruit. It consists of two parts, the pericarp and the seeds, and according to the nature of the pericarp, the fruit is termed a capsule, nut, berry, or drupe.
The chief characteristic of the Phanerogams does not lie in the formation of the flower (although they may quite properly be termed “Flowering-plants”), because Equisetums and Lycopods have reproductive shoots as highly differentiated as those of certain Gymnosperms and other Phanerogams. As regards the SEXUAL GENERATION the characteristics are found:—(1) in its great reduction; (2) in the transmission of the microspore (pollen-grain) to the macrosporangium, and its germination, with the formation of a pollen-tube (antheridium), the protoplasm of which is not differentiated into spermatozoids; (3) in the fact that the macrospore (embryo-sac) never leaves its sporangium (nucellus); and further in the Angiosperms, (4) in the peculiar development of the nutritive-tissue in two parts; and (5) in the great reduction of the archegonium.
As regards the ASEXUAL GENERATION the characteristic feature is that this generation is formed whilst the sporangium is still attached to the mother-plant, and for a long time is nourished by it; and that after the sporangium has become detached from the[250] mother-plant, it spends a longer or shorter resting period as the embryo in the seed (enveloped by the testa), and does not make its appearance until the “germination” of the seed. In addition the shoot which bears sporangia undergoes greater modification than in the case of the Flowerless-plants.
The Phanerogams are separated into two Divisions as follows:—
Division 4. Gymnospermæ. The ovules, as well as the seeds, are borne naked on the surface of open carpels, or on the apex of a stem (ovary wanting). The pollen-grains are conveyed by the wind to the ovules, and caught by drops of mucilage, secreted by the micropyle. A “stigma” is wanting. The entire female prothallium (the endosperm), which serves for the nourishment of the embryo, is formed before fertilisation. The archegonia are embedded in the upper part of the prothallium. The pollen-grains are “multicellular,” i.e. there is always in their interior a distinct prothallium, formed by 1–3 cells, and a larger cell which gives rise to the pollen-tube.
Division 5. Angiospermæ. The carpels surround the ovules and form an entirely closed chamber (ovary), in which the ovules mature and ripen into seeds. The surface of a portion of the apex of the carpel is transformed into the “stigma,” which, by a sticky fluid and also by hair-structures, is capable of retaining the pollen-grains conveyed to it by the wind, or more frequently by insects. The pollen-tube grows from the stigma, through the “conducting cellular tissue” (style), to the ovules. The pollen-grains contain two cells, a vegetative and a free generative cell. The latter passes into the pollen-tube and there divides into two, one of which is the sperm-nucleus. The female prothallium, which is intended to serve as nutritive-tissue, is formed after fertilisation. Archegonia are wanting.
[251]
The following characters should be added to those already given on page 2:—
The Gymnosperms comprise only trees or shrubs. The flowers are always unisexual and destitute of perianth (except Gnetaceæ); the female plant of Cycas is the only one which has no flower. The MALE FLOWERS are constructed on the same type as the cones of the Horsetails and Club-Mosses, and are most frequently long shoots (Figs. 243, 258, 260 A, 267 J) bearing a number of spiral or verticillate stamens. The FEMALE FLOWERS are of a more varied structure (see the orders). The OVULE is orthotropous (except Podocarpus which is anatropous) and projects from the carpel uprightly, inverted, or horizontally; it has usually only one integument (compare however Taxaceæ) which proceeds from the upper part of the nucellus, so that the embryo-sac in part is placed below the integuments (Figs. 251, 264). The drop of mucilage which catches the pollen-grains dries up and draws the pollen-grain through the micropyle to a space just above the nucellus—the pollen-chamber—in which the germination of the pollen-grain commences.
In each seed, only one of the many embryos which are formed proceeds to its full development. The seed is always endospermous, and the embryo has one, two, or a whorl of several cotyledons. A vigorous primary root is developed on germination. The vascular bundles in the stem are arranged in a ring, and increase in thickness takes place by a closed cambium-ring which forms bast (phlœem) externally, and wood (xylem) internally with distinct annual rings, as in the Dicotyledons. Only certain of the Cycadeæ deviate from this arrangement. The secondary wood is very uniform, as it is almost exclusively formed of tracheides with bordered pits, but true vessels are wanting; this also indicates a relationship with the Pteridophyta (see page 202).
[252]
The Gymnosperms are biologically lower than the Angiosperms; they are wind-fertilised, and without extra floral-nectaries.
This Division embraces three classes: Cycadeæ, Coniferæ, And Gneteæ. It is no doubt monophyletic, and has taken its origin from heterosporous Ferns, now extinct, most nearly related to the Ophioglossaceæ and Marattiaceæ. The Cycadeæ appear to be the oldest class. The Coniferæ are related to these through Ginkgo. The Gnetaceæ are more isolated. The Division is not continued into the higher Flowering-plants; it has evidently attained its highest development, and is now in a retrograde condition. The similarity which has often been pointed out between certain Coniferæ and Lycopodinæ is only in analogous resemblances, and does not entitle one to suppose that there is a nearer relationship, or that the former take their origin from the latter.
The stem is very rarely ramified. The leaves are large, pinnate, and arranged spirally. The flowers are diœcious, without perianth.

Fig. 256.—Cycas circinalis (female plant). The carpels are seen hanging from the top of the stem. Three leaves with the leaflets still rolled up project almost vertically into the air, from the centre of the crown.
There is only one order, the Cycadaceæ.—In habit they resemble the Ferns, especially the Tree-Ferns (compare Figs. 207 and 256). The stem is tubercular (Fig. 258), or cylindrical (Fig. 256), but not very tall (as much as about 12 metres), and very rarely ramified. [In Ceylon, unbranched specimens of Cycas are rarely met with in the wild state. The stems of C. circinalis occasionally branch in greenhouses.]
[253]
The LEAVES are arranged spirally, and so closely together that no free stem-surface is left between them, and have only a slight sheath (which is not amplexicaul, as in the Palms). They are compound (most frequently pinnate; in Bowenia, bipinnate); in some genera the leaves are rolled up in various ways, resembling the vernation in Ferns (Fig. 257); they are leathery and perennial. In some, stipules are present, as in the Marattiaceæ. Groups of scale-leaves alternate in the majority with groups of foliage-leaves.

Fig. 257.—Cycas circinalis. Part of a young leaf with circinate leaflets.
Fig. 258.—A male plant of Stangeria paradoxa (about 1/15 nat. size).
Fig. 259.—Female cone of Zamia integrifolia (½-⅓ nat. size). The male cone is very similar externally.
The FLOWERS are without perianth. The MALE FLOWER is sometimes an enormous collection of stamens (Fig. 258), which are flat in some (Cycas, Fig. 241), shield-like in others (Zamia, Ceratozamia) like the sporophylls in Horsetail (Fig. 259); but in all, the pollen-sacs are situated in large and varying numbers on the back of the stamens, and arranged in groups of 2–5, like the sporangia in the sori of the Ferns (Fig. 241 b, c). Female flowers are wanting in Cycas, because the carpels do not terminate the apical growth of the stem. After a group of foliage-and of scale-leaves, a group of carpels is developed, which are pinnate and resemble the foliage-leaves, bearing on their edges a number of ovules (most frequently[254] 5–6) (Figs. 245, 256); the same stem produces successively scale-leaves, foliage-leaves, and carpels. The differentiation is not much more advanced than in certain Ferns (Struthiopteris, Blechnum), where barren and fertile leaves of different form regularly alternate. The other genera have female flowers; the carpels are shield-like in Zamia and Ceratozamia (Fig. 246), and collected into cone-like flowers, which terminate the growth of the stem (Fig. 259). The number of ovules in these instances is two to each carpel.
The SEEDS are large (most frequently 2–6 centimetres long) and plum-like; the external layer of the testa is fleshy, while the internal one is hard and horny. There are two systems of vascular bundles in the testa, one outside, the other inside the stone. The embryo is straight, attached to the end of the suspensor, which is often long, filamentous, and rolled up; it has one or two cotyledons.
The embryo in Ceratozamia and others is very slightly developed, at the time when the ripe seed is detached from the carpel; and it is not until after sowing that its further development and germination proceed. This calls to mind the Cryptogams, especially Selaginella, whose macrospores are thrown off filled with endosperm; but the oosphere is not fertilised till after the separation of the macrospore from the parent-plant, while in the Cycadeæ fertilisation is effected before the separation. In Cycas the testa may rupture, and the endosperm grow and become green in the light, even though no embryo has been formed. This also is an indication of its prothalloid nature.
Gum-passages are present in all organs. Collateral vascular bundles, with spiral and scalariform tracheides, are found; and normal thickening takes place by means of a cambium. An exceptional mode of growth is found in Cycas and Encephalartos, the cambium ceases to divide after a time and is replaced by a new cambium which arises in the cortical parenchyma just outside the bast, and which forms a new ring of xylem and phlœem. This may be repeated so that a number of concentric rings are produced. In Ceratozamia, structures resembling corals extend from the roots in a vertical direction and appear on the surface of the soil; these are peculiar roots, in which a symbiotic Alga (Anabæna) is found.
The Cycadeæ were formerly (from the Coal period to the Later Cretaceous) far more numerous than at the present day. They appear to have been most numerous in the Trias and Jurassic. The remnant (75 species) which have persisted to the present time are found in all tropical countries. Cycas (Trop. and Sub-trop., Eastern Hemisphere); Dioon (Mexico); Macrozamia (Australia); Encephalartos (Trop. and S. Africa); Stangeria (Fig. 258, Sub-trop. South and East Africa); Bowenia (Trop. Australia); Ceratozamia (Mexico, New Granada, Western Brazil); Microcycas (Cuba); Zamia (Trop. and Sub-trop. N. America.)
Uses. Sago is made from the starch-containing pith of Cycas revoluta and circinalis. The leaves are often used at funerals and church festivals, under the name of “palm-branches.”
[255]
The stem branches freely. The leaves are entire, relatively small, linear or reduced to scales. The flowers are without perianth. The ovules naked. It is seldom that the female flower is reduced to only one carpel.
Whilst the Cycadeæ principally resemble the Ferns, the Conifers partly resemble the Lycopods, and partly the Equisetums—the former especially in the needle- or scale-like, leathery, simple, and often perennial leaves (“evergreen plants”), which never possess stipules (Figs. 263, 270, 272). Ginkgo deviates from this, being no doubt the oldest, and the Conifer which stands nearest to the Cycadeæ (Fig. 260). The resemblance to the Equisetums is especially owing to the fact that the stem ramifies abundantly, and often very regularly, forming a pyramid with verticillate branches. In addition to the foliage-leaves, scale-leaves (bud-scales) are present in the majority of species.
The FLOWERS are monœcious or more rarely diœcious. Perianth is wanting. The stamens of the catkin-like male flowers (Fig. 267, J) are of different forms, but as a rule more or less shield-like. As in the Cycadeæ, the pollen-sacs are in all cases situated on the underside. There are, as a rule, two pollen-sacs (the Abietaceæ, Fig. 267), or 3–5, (the Cupressaceæ and Taxaceæ, Fig. 243); a few have more, e.g. Araucaria (Fig. 242); they dehisce by clefts.
If, in commencing our consideration of the female flower, we begin with that of Ginkgo, we shall observe in the corner of a scale- or foliage-leaf a small flower, which consists of two carpels, each bearing one ovule, and reduced almost to the ovule itself (Fig. 260 C, D). The flower in Podocarpus is still further reduced, viz. to a single carpel with one ovule, which is anatropous and has two integuments. This ovule is situated in the axil of a cover-scale (c, in Fig. 262 D), and several female flowers of this description are collected in a small cone, the stalk and bracts of which become fleshy (Fig. 262 C). The external integument also becomes fleshy (an aril). Dacrydium, which is clearly related to Podocarpus, has an external integument which developes more independently as a fleshy aril (Fig. 262 B, B’). Microcachrys also is clearly allied to these: the bracts are more fleshy, and the ovule (i.e. the female flower) is protruded beyond the bract (Fig. 262 A, A’). Taxus stands in a more isolated position: a flower which has been reduced to an ovule is situated, in this instance, on the apex of a secondary[256] branch which is studded with floral-leaves (Figs. 263, 264); an external integument is developed on all sides and surrounds the seed as a scarlet aril. According to this conception the aril corresponds to an external integument, and the Taxoideæ thus possess a partly dichlamydeous ovule. Only Ginkgo and Cephalotaxus appear to deviate from this, as in these there is only one integument (unless the small outgrowth indicated by ar, in Fig. 260 D, really is a rudimentary, external integument); in Cycadeæ, to which Ginkgo is most closely related, there is likewise only one integument. But in these genera the testa is differentiated into two layers, and the seed resembles a drupe; like the Cycadeæ there is an external fleshy covering and an internal hard one, and these two layers may probably be considered homologous with the two integuments. This theory is also borne out by the arrangement of the vascular bundles in Cephalotaxus and Podocarpus, which present the xylem in the fleshy external layer to the outside of the testa, which is therefore the upper side of the integument (Celakovsky).
The coalescence of the integuments into one is only slight in Torreya, more pronounced in Podocarpus and strongest in Cephalotaxus and Ginkgo. Celakovsky terms these ovules “holochlamydeous.”
If we pass from these to the order Pinoideæ, we find the female flowers collected into catkin-like cones, which have been considered from various points of view to be sometimes single flowers, at other times compound inflorescences. The structure in Abietaceæ is as follows: a number of spirally arranged, scale-like leaves, cover-scales (Figs. 267, 268), are situated on a long axis. In the axil of each cover-scale a larger leaf-like projection, the ovuliferous scale, is borne, which turns the upper side towards its cover-scale (which is shown by the fact that the wood of its vascular bundles is turned downwards and towards the wood in the bundles of the cover-scale: Fig. 269). Two ovules, with micropyles turned towards the central axis, and with apparently only one integument (Fig. 268), are situated on the dorsal side of each ovuliferous scale, i.e. the side turned away from the cover-scale. The ovuliferous scales grow after fertilisation, into the woody or leathery “cone-scales,” which are usually much larger than the cover-scales. This ovuliferous scale with its axis may, according to Celakovsky, be considered as a dwarf-branch which is situated in the axil of the cover-scale, and bears two ovules (in the same way as in Ginkgo, one long-stalked flower, reduced to two ovules, is situated[257] in the axil of a leaf), and in this case the external integument of the ovules is expanded into leaf-like bodies, which have united to form one “symphyllodium” (ovuliferous scale) which is inverted so that its dorsal side is turned upwards and bears the nucellus and the other integument (“hemichlamydeous” ovules). The carpel itself is therefore in this instance extremely reduced. The keel, or (in Pinus) “mucro” (Fig. 268 B), which is found in several genera, represents then a third carpel, which is sterile. In the other orders of the Pinoideæ the cover-scales and ovuliferous scales grow more and more together and finally form one structure, which also is termed a “cone-scale,” although from its development it cannot be homologous with the cone-scales of the Abietaceæ. This connation is least in the Taxodiaceæ and Araucariaceæ and may be traced on the upper surface of the “cone-scale” by the presence of a stronger or slighter ridge or pad, the free portion of the ovuliferous scale (Figs. 256, 266, 269). It is most strongly pronounced in the Cupressaceæ, in which the two scales form one single structure, the cone-scale (Fig. 274). The vascular bundles in the under portion corresponding to the cover-scale, have the xylem towards the upper side as usual in leaves, whilst the bundles present in the upper side of the cone-scale, which thus represents the ovuliferous scale, turn their xylem downwards. The hemichlamydeous ovules are then situated on the upper side of this cone-scale. According to this theory the Cupressaceæ appear to be the youngest type, a view which corresponds with their vegetative structure. If there is only one ovule in these orders as in Agathis (Fig. 265) and Araucaria, then the flower is reduced to a single carpel and one ovule, as in the case of Dacrydium and Microcachrys. If two or more ovules are present, then the same number of carpels may be supposed to exist, the external integuments of their ovules being developed into leaf-like structures which collaterally coalesce to form a “symphyllodium,” or are suppressed.
According to this theory, which is based on the researches of Celakovsky, the female flowers of the Coniferæ may be classed thus:—
1. In all cases situated in the axil of a bract and collected into cones, with numerous flowers or with few or one flower. In Ginkgo only, are they situated in the axil of foliage- or scale-leaves.
2. It is only in Taxus that bracteoles are present.
3. They are formed only from rudimentary carpels, in which the stem takes no part.
[258]
4. The number of carpels in each flower varies from one to many, most frequently three, of which the central one remains sterile.
5. Each carpel bears only one ovule. The flower which is formed of only one carpel appears to consist of only one ovule.
6. The ovule has in Taxaceæ either a double integument (Podocarpeæ, Taxeæ), of which the external is the “aril,” or, as in the Cycadeæ, a single one, which is homologous with the two united together.
7. The external integument in the Pinoideæ is expanded to form a leaf-like structure—the ovuliferous scale—and bears on its dorsal side the ovules, which are thus only provided with one, and that the inner, integument.
This later interpretation of the female cones in the Coniferæ is more probably correct than the older ones; that, however, which appeared in the former issues of this book, may also be stated. It was to the effect that each catkin-like female cone is in reality a single flower; the cone-scales in the Cupressaceæ were single leaves, namely carpels, which bore the ovules on the side which is turned upwards; the division into two parts which makes its appearance in the other orders, and becomes most prominent in the Abietaceæ, was compared with the division of a leaf into a barren and a fertile portion, which is found especially in Ophioglossaceæ and Marsiliaceæ, or with the ligule in Isoëtes.
Pollination is accomplished by means of the wind. At the period of pollination the leaves are always so widely separated from one another, that the ovules can catch the pollen-grains carried to them by the wind; this is often effected by the mucilaginous drops which appear at the micropyle, and by the evaporation of which the pollen-grains are brought in contact with the nucellus. The entire cone grows considerably as soon as fertilisation has taken place, and the cone-scales in Pinoideæ close together so that the seeds while maturing are enclosed, and it is not until the seeds are ready for distribution that the cone-scales again become separated. In the Pinoideæ, the fully developed ovuliferous scales are hard and woody; and in this condition the collection of female flowers is termed a cone. In the Taxoideæ, true cones are the exception. 2–15 cotyledons are present, arranged in a whorl.
The characteristic feature of this class is the abundance of resin, which is to be found in isolated cells (especially in the[259] cortex), partly in intercellular glands or passages (both in the cortex and wood). Taxus is the only genus which has no resin.
There are about 350 species, mostly from the Northern Temperate zone (especially North America and Siberia), where they grow gregariously and form the most northern forests. The Juniper, Scotch Fir, and Yew are natives of Great Britain.
This class may be divided into two families:—
1. Taxoideæ. The ovules have either one integument, the external part of which is fleshy, and the internal hard and stone-like; or two integuments, of which the external is the fleshy and coloured “aril.” “Ovuliferous scales” are wanting. The cones are never woody, but are generally succulent, the bracts become fleshy, or cones usually are not developed. The seeds project more or less freely beyond the bracts.
2. Pinoideæ. The ovules have two integuments, the external one of which is leaf-like and becomes developed as the “ovuliferous scale”; if there are several of these in each flower they unite and form a “symphyllodium.” This may remain free or unite with the bract. The cones are most frequently woody, rarely succulent. The seeds are hidden among the cone-scales.
This family, considered to be most nearly related to the Cycadeæ, also made its appearance at a very early period. There is only one order.
Order. Taxaceæ. The characters have been given above.
A. Cephalotaxeæ is the oldest group, presumably the connecting link between the Cycadeæ and the other Coniferæ. The flower consists normally of two ovules. Aril wanting. One integument. Seeds drupaceous.—The flowers in Ginkgo biloba (Salisburia) are situated in the axil of foliage- or scale-leaves. The stamens bear only two pollen-sacs (Fig. 260 A). The female flower has two ovules, placed together at the end of a long, bare axis (Fig. 260 C). Round the base of the ovule a small collar (ar, in Fig. 260 D) is found, which may probably be considered homologous with the collar-like outgrowth which surrounds the base of the Cycas-ovule. The seed resembles a Plum, and has a fleshy external coat, surrounding a hard internal layer. The embryo is developed after the seed has fallen off. The Ginkgo-tree has long-stalked, fan-shaped leaves, more or less indented, with dichotomous veins[260] resembling certain Ferns—the Adiantums. It is a native of East Asia, and the only surviving species of a genus which in earlier times was very rich in species, and distributed over the entire Northern Hemisphere. Cephalotaxus (Eastern Asia) is related to it.
Fig. 260.—Ginkgo (nat. size): A a branch with a small flowering dwarf-branch (male flower); B a leaf; C a flower with two ovules; D a ripe seed; ar collar.
Fig. 261.—Phyllocladus glaucus: a branch with female flowers (nat. size).
B. Podocarpeæ. The female flower is reduced to one ovule, placed in the axil of a bract, or a little forward upon it. The ovule has an aril (2 integuments).—Phyllocladus (Fig. 261), from New Zealand and Tasmania, has obtained its name from its flat, leaf-like branches, the leaves proper being scale-like (f). The ovules stand erect in the axil of the scale-like leaves (c), and several are collected at the end of short branches.—Microcachrys tetragona (Tasmania) has a small female catkin with several spirally-placed, fleshy bracts, at the end of which the inward and downward turned ovule is attached (Fig. 262 A, A’). The ripe cones are red, succulent, and resemble Strawberries.—In Dacrydium (Tasmania, New Zealand, Malaysia) the female cone has most frequently only 1–2 (–6) bracts, which resemble[261] the vegetative leaves; they have also a fleshy aril (Fig. 262 B, B’).—Podocarpus (40 species, East Asia, S. Temp.); the bracts of the female flowers become fleshy, and unite together; only 1 or 2 are of use in supporting the flowers. The ovules project high above the apex of the bract, and are anatropous, the micropyle being turned downwards (Fig. 262 C, D). An aril commences to develope in the flowering period as an external coating, and later on it becomes fleshy and coloured.
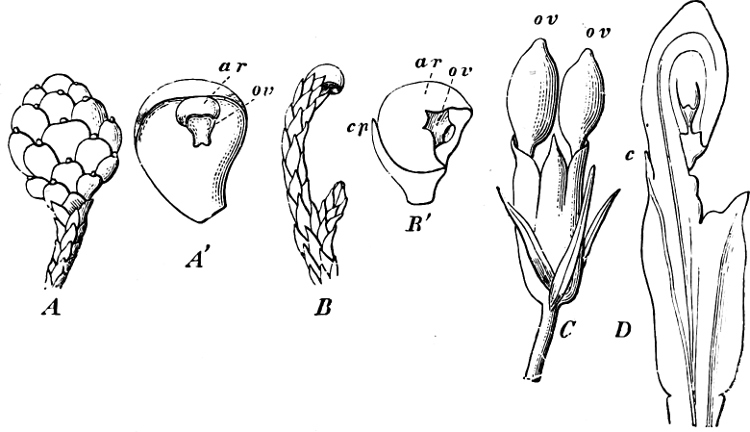
Fig. 262.—A Microcachrys: female cone (2/1). A’ A single carpel with its ovule. B Dacrydium: branch with female flower (3/1). B’ The flower; cp the bract; ar the aril; ov ovule. C Podocarpus: female flower with 2 ovules. D Another female flower with 1 ovule, in longitudinal section.
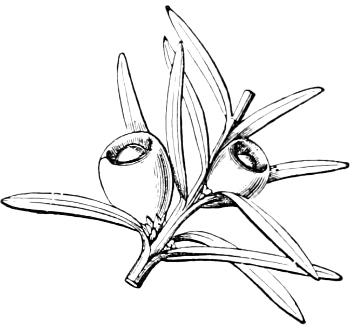
Fig. 263.—Taxus baccata: branch with two ripe seeds (nat. size).
C. Taxeæ. The female flower is reduced to one ovule, which is situated terminally on an axis which bears 2–3 pairs of opposite, scale-like bracteoles; on this account the Taxeæ form a very isolated group among the Coniferæ.—Taxus (T. baccata, the Yew-tree). Diœcious. The female flower consists of only one ovule, placed at the end of a short secondary branch (Fig. 264), which is studded with scale-like leaves. The aril when ripe is thick, fleshy, and scarlet (sometimes yellow), and only loosely envelopes the seed (Fig. 263). The leaves are scattered, flat, linear, and pointed (Fig. 263,[262] 264). The short male flowers have 5–8 pollen-sacs, pendent from the stamens, and are surrounded at their bases by scale-like bracteoles (Fig. 243). Torreya (4 species, N. America and Japan) is closely allied to Taxus. The aril ultimately fuses with the woody inner integument, and hence the ovule becomes drupaceous, as in Cephalotaxaceæ.
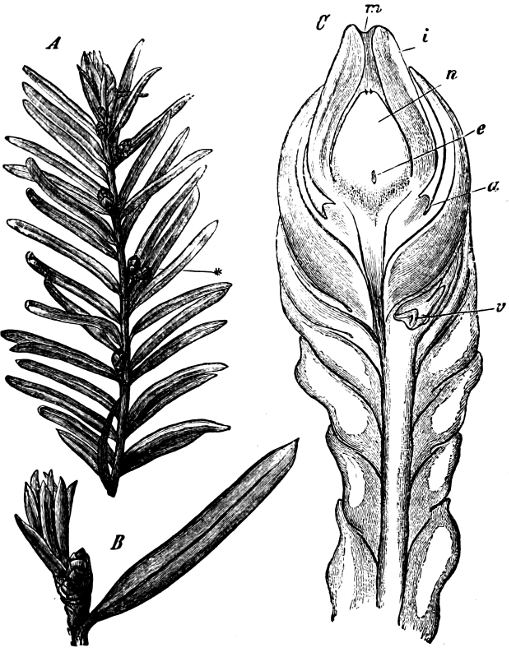
Fig. 264.—Taxus baccata: A shoot of Taxus with female flowers at the time when the ovules are ready for pollination. B Leaf with flower in its axil (nat. size). C Longitudinal median section through a female shoot; v growing point of primary shoot; a commencement of aril; i integument; n nucellus; m micropyle.
Uses. Taxus baccata is usually planted in gardens, especially in hedges. Its wood is very hard and is used for wood-carving. The shoots are poisonous, but not the aril, which is often eaten by children and by birds.
The four orders differ from one another partly in the arrangement of the leaves (Cupressaceæ have opposite or verticillately[263] placed leaves, flowers, and inflorescences; in the others they are placed spirally), but chiefly in the greater or less degree of union which takes place between the female flower (the leaf-like “symphyllodium”) and its supporting cover-scale, and in the position of the ovules (the micropyle being turned upwards or downwards). The “cone-scales” in Abietaceæ are formed by “symphyllodia” alone, in the others by their union with the cover-scale.
Order 1. Araucariaceæ. This order most frequently has solitary ovules, turned downwards and attached to the centre of the cone-scales. In Agathis (Dammara) the arrangement is the most simple, a winged seed (Fig. 265), which hangs freely downwards, being borne in the centre of the undivided cone-scale. In Araucaria, the stamens with the free, pendulous pollen-sacs have been represented in Fig. 242; the ovuliferous scale is united for nearly its whole length with the bract, and projects from its apex in the shape of a sheath-like, dentate scale, resembling the ligule in Isoëtes, and may therefore be termed a “ligule.” Araucaria (S. America, Australia) has often rather broad leaves (A. brasiliensis). The ovuliferous scale in Cunninghamia is more distinct, and stretches transversely over the entire cover-scale; it bears three inverted ovules (Fig. 266) (Eastern Asia).
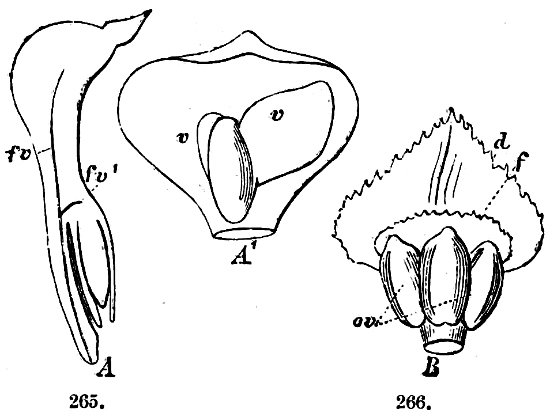
Fig. 265.—Agathis (Dammara) australis. Cone-scale with the seed. A Longitudinal section; A’ from within; fv, fv’ vascular bundles; v wing.
Fig. 266.—Cunninghamia sinensis. Cone-scale with three ovules, interior view: d cover-scale; f ovuliferous scale.
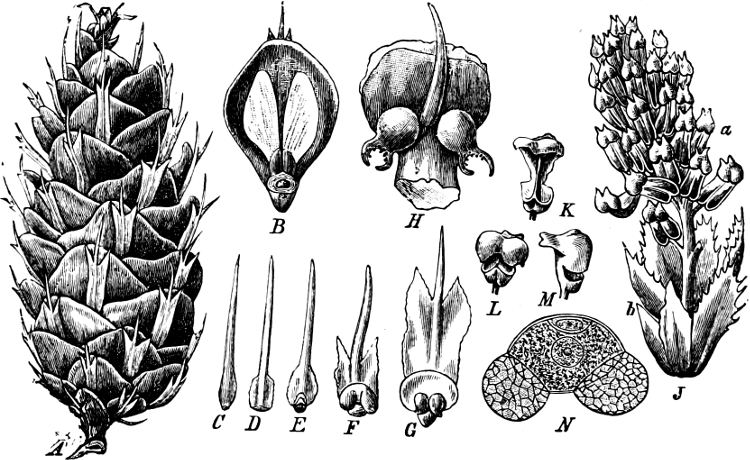
Fig. 267.—A-G Pseudotsuga douglasii: A cone, B cone-scale, with the inner side turned forward; the points of the cover-scale are seen behind it; C-G transitions from the acicular leaf to the cover scale, from the base of a ♀ cone. H Pinus montana. Young ovuliferous scale, with the inner side turned forward; the ovules are now in the stage for pollination. J-M Abies alba: J male cone; b bud-scale; a anthers; K L M individual anthers.—Pinus montana: N pollen-grain; the two lateral expansions are the air-bladders; in the upper part of the interior of the grain a vegetative cell may be seen, and in the centre the large cell-nucleus.
Dammara-resin, which is used for varnish, is obtained from Agathis (Dammara) species (New Zealand, Philippine Islands).
Order 2. Abietaceæ (Pine and Fir Trees). The leaves are spirally arranged and needle-like. The flowers are monœcious.[264] The male flowers are long, and catkin-like, with numerous stamens, each bearing two oblong pollen-sacs. The pollen-grains are most frequently tri-lobed, having two bladder-like appendages, formed as outgrowths of the exospore, to assist in their distribution by the wind (Fig. 267 N). The bracts are arranged spirally. The union between the bract and the ovuliferous scale, which is found in the preceding order, is not in this instance so complete; these scales make their appearance as two free parts, and are attached only at their bases (Fig. 268); the lower portion, that is the cover-scale, in most instances remains quite small (Fir, Red Pine, and others), it is only in the “Noble Pine” (Abies) and Pseudotsuga douglasii, that it attains a greater length than the ovuliferous scale (Fig. 267, B-G). On the other hand the upper part, the ovuliferous scale (the vascular bundles of which have the bast turned upwards), grows strongly and elongates, especially after fertilisation, becoming woody or leathery; it is commonly termed the “cone-scale,” but is in reality only homologous with a part of[265] the “cone-scale” in the other order of Pinoideæ. On the side of the ovuliferous scale, turned towards the axis, are situated two ovules with micropyles directed inwards. The seeds are most frequently provided with a false wing (a tissue-like part of the surface of the ovuliferous scale). Cotyledons, more than 2, verticillate. Fertilisation does not take place until some time after pollination. In Pinus, for instance, the pollen-tube only penetrates the nucellus for a short distance during the year of pollination, and then ceases its further growth, fertilisation not taking place until after the middle of the next year; whilst the seeds ripen about a year and a half after pollination. In the Larch and others, the seeds are mature in the autumn succeeding pollination.
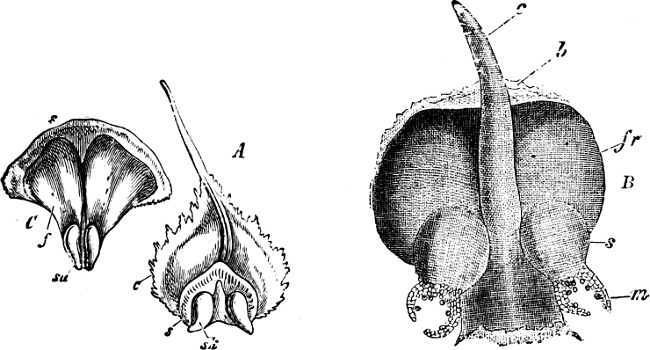
Fig. 268.—A Abies: c the cover-scale; s ovuliferous scale, or “cone-scale”; sk ovules in a young condition. B Pinus: ovuliferous scale with two ovules (s); m the two-lobed micropyle; c “mucro”; b the cover-scale behind. C Abies: ripe “cone-scale” with two seeds (sa); f wing of seed.
Abies (Fir). The leaves are often (e.g. Ab. pectinata) displaced into 2 rows, flat and indented at the apex, with 2 white (wax-covered) lines on the under surface, in which the stomata are situated. The leaf-scars are nearly circular and do not project. The cones are erect. The cover-scales and the ovuliferous scales separate from the axis, to which they remain attached in other genera.—Tsuga has leaves like Abies, but by the slightly projecting leaf-scars, and cones with persistent scales, it forms the transition to Picea.—Pseudotsuga has leaves similar to those of Abies and persistent carpels as in Picea, but the cover-scales grow as in Abies and project beyond the ovuliferous scales (P. douglasii, Fig. 267). These two genera are considered as sub-genera of Abies.—Picea. The leaves project on all sides, square and pointed; the leaf-scars are[266] rhombic, on projecting leaf-cushions. The cones are pendulous. The cover-scales are much shorter than the leathery, persisting ovuliferous scales.—The genus Larix (Larch) differs from all the others in having deciduous leaves (the three preceding have leaves which persist for eleven to twelve years). It has long-branches with linear foliage-leaves and short, thick, perennial dwarf-branches, which each year form a new rosette of foliage-leaves, similar to those on the long-branches. The male flowers and the erect cones resemble those of Picea, and are borne on dwarf-branches.—Cedrus (Cedar) resembles Larix to some extent, but has persistent leaves (C. libani, C. deodara).—Pinus (Pine) has long-branches and dwarf-branches. The leaves of the long-branches are scale-like and not green; the dwarf-branches have very limited growth, and persist for three years; they arise in the axils of the scales borne on the long-branches of the self-same year, and each bears 2–5 foliage-leaves, they are also surrounded at the base by a number of membranous bud-scales. The cone-scales have a thick, rhomboid extremity (the “shield”).
The buds which develope into long-branches arise at the apex of other long-branches, and being very close together, form false whorls. The female cones occupy the position of long-branches, and take about two years for their development. The male flowers arise close together, and form a spike-like inflorescence at the base of a long-branch of the same year. The male flowers occupy the position of dwarf-branches, so that a female cone may be considered to be a modified long-branch, and a male cone a modified dwarf-branch. The main axis of the seedling has needle-like leaves, similar to those found on the older parts, and on dwarf-branches; it is not until some time later that the dwarf-branches are developed and the permanent arrangement attained.
Uses. Several species are commonly cultivated in this country, partly on heaths and moors, and partly in plantations and as ornamental trees, such as Mountain Pine (Pinus montana, Cen. Eur.); Austrian Pine (P. laricio, Eur.); Scotch Fir (P. silvestris, Eur.); Weymouth Pine (P. strobus, N. Am.); common Red Pine (Picea excelsa, Cen. and N. Eur.); White Pine (P. alba, N. Am.); Abies pectinata (Common Fir, S. and Cen. Eur); A. nordmanniana (Crimea, Caucasus); A. balsamea (N. Am.); Tsuga canadensis (N. Am.); Pseudotsuga douglasii (N.W. Am.); Larch (Larix europæa, Alps, Carpathians); L. sibirica (N.E. Russia, Siberia).—The wood of many species, especially Pine, on account of its lightness and because it is so easily worked, is very well adapted for many useful purposes. The wood of the Yew-tree is very hard and is used for ornamental turning. Resin and Turpentine (i.e. Resin with essential oils, the name being derived from the Terebinth-tree, from which formerly a similar material was obtained) are extracted from Pinus laricio and P. pinaster. Oil of Turpentine is obtained by distillation of turpentine with water; Tar by dry distillation of Pine-wood. Canada-balsam is from North[267] American Abies-species (A. balsamea and Fraseri). The officinal Turpentine is mainly obtained from Pinus pinaster (South of France), P. tæda, australis, strobus (Weymouth Pine) and other North American species; more recently also from P. silvestris (Scotch Fir), maritima, laricio, Picea excelsa, and others; Venetian Turpentine, from Larch (S. Eur.) Amber is resin from a Tertiary plant (Pityoxylon succiniferum), closely related to the Pine, which grew especially in the countries round the South-East coast of the Baltic. Pinus pinea (the Pine, S. Eur.) has edible seeds and also P. cembra (in Cen. Eur. and Siberia).
Order 3. Taxodiaceæ. The vegetative leaves and cone-scales are arranged spirally. The ovules (2–9) are situated either at the base of the ovuliferous scales, in which case they are erect; or at their centre, when they are generally more or less inverted. The ovuliferous scale is more or less united with the cover-scale, and projects beyond the surface of the cone-scale, like a comb (Fig. 269). The vascular bundles, which extend into the cover-scale, have the usual leaf-arrangement, viz. the wood placed above the bast; while those bundles which enter the ovuliferous scale have this arrangement of the bundles reversed.
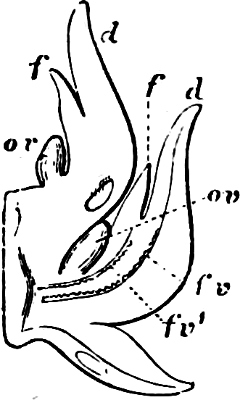
Fig. 269.—Cryptomeria japonica. Portion of longitudinal section through female flower. d cover-scale; f ovuliferous scale; ov ovules; fv and fv’ vascular bundles; the xylem is indicated by a wavy line, and the phlœm by a straight line.
Taxodium distichum (the North American “Swamp Cypress”) has annual dwarf-branches, with distichous leaves, and cone-like “pneumathodia.” In the Tertiary period it was very common in the Polar regions. Sequoia (Wellingtonia) gigantea is the famous Californian Giant-Fir, or Mammoth-Tree, which attains a height of 300 feet, a diameter of 36 feet, and is said to live for 1,500 years. Cryptomeria japonica (Japan, China) has the least adnate ovuliferous scales; Glyptostrobus (China); Arthrotaxis (Tasmania); Sciadopitys verticillata (the only species in Japan) has, like Pinus, scale-like leaves on the long-branches, of which those which are situated at the apex of the annual shoots support “double needles,” i.e. dwarf-branches similar to the two-leaved dwarf-branches in Pinus, but without bud-scales, and with the two leaves fused together at the edges into one needle, which turns its upper surface away from the long-branch.
Order 4. Cupressaceæ (Cypresses). The leaves are opposite or verticillate, sometimes acicular, but most frequently scale-like (Fig. 270). In the species with scale-like leaves, the seedlings often commence with acicular leaves (Fig. 272), and branches are sometimes found on the older plants which revert to this form, seeming to indicate that the acicular leaf was the original[268] form (atavism). The so-called “Retinospora” species are seedling-forms of Biota, Thuja, Chamæcyparis, which have been propagated by cuttings, and retain the seedling-form. The flowers are monœcious or diœcious. The male flowers are short, and have shield-like stamens, bearing most frequently several pollen-sacs. The cover-scales and ovuliferous scales are entirely fused together and form undivided cone-scales, opposite or whorled; the ovuliferous scales have slight projections near the base on which 1–2–several erect ovules are developed (Fig. 274). Most frequently 2 cotyledons.—Evergreen trees and shrubs.
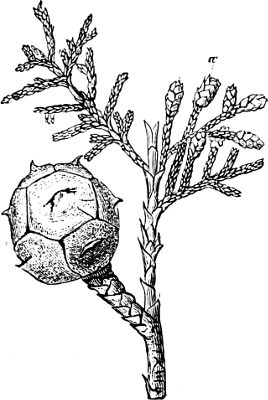
Fig. 270.—Cupressus goveniana.
Fig. 271.—Portion of a branch of Thuja orientalis (magnified). The leaf at the base on the right has a branch in its axil.
Fig. 272.—Seedling of Thuja occidentalis. The branch (g) is borne in the axil of the leaf s.
Juniperus (Juniper). Diœcious. The cone-scales become fleshy and fuse together to form most frequently a 1–3 seeded “berry-cone.” J. communis (Common Juniper) has acicular leaves, borne in whorls of three, and the “berry-cone” is formed by a trimerous whorl of cone-scales (Fig. 273). J. sabina and J. virginiana have “berry-cones” formed from several dimerous whorls of cone-scales; the leaves are connate and opposite, needle-and scale-like leaves are found on the same plant.
Cupressus (Cypress). Monœcious. The cones are spherical; the cone-scales shield-like, generally five-cornered and woody (Fig. 270), each having many seeds. The leaves are scale-like.—Thuja.[269] Monœcious. Cones oblong. The cone-scales are dry, as in the Cypress, but leathery and imbricate, and not shield-like; each cone-scale bears 2–3 seeds. The leaves are most frequently dimorphic; those leaves which are situated on the edges of the flat branches are compressed, and only these bear buds, which are developed with great regularity, generally alternately, on both sides of the branch; those which are situated on the flattened surfaces are pressed flat and broad, and never bear branches (Fig. 271). Along the central line of each leaf there is a resin-canal (Fig. 271).—Chamæcyparis, Callitris, Libocedrus, Thujopsis (1 species: T. dolabrata; in Japan).
Fig. 273.—Branch of Juniper with “berry-cones.”
Fig. 274.—Cupressus lawsoniana. Longitudinal section through female cone. Two ovules (ov) are bisected; f ovuliferous scales.
Officinal. Juniperus sabina from Central and South of Europe (the young branches yield an essential oil). The wood of J. communis is used in the production of an essential oil, and J. oxycedrus in the production of empyreumatic oil. The “berry-cone” of J. communis is officinal, and is also used for gin.—The wood of J. virginiana (N. Am.) is known as red cedar, and is used for lead-pencils. Sandarack resin is obtained from Callitris quadrivalvis (N.W. Africa).
The following are cultivated in gardens:—Thuja occidentalis (Arbor vitæ) (N. Am.), and orientalis (China, Japan); Juniperus sabina and virginiana; Thujopsis dolabrata (Japan); Cupressus lawsoniana (California), C. sempervirens (S. Eur., W. Asia), and other species, are grown especially in conservatories, and in Southern Europe particularly in cemeteries.—The Retinospora species which are so often planted, do not belong to an independent genus, but are obtained from cuttings, taken from seedling-plants with acicular leaves (see page 267).
[270]
This class, independent of extinct forms, comprises the most highly developed of the Gymnosperms, partly from the circumstance that a perianth of 2–4 members encloses the terminally[271] placed ovule, which is provided with one, or (in Gnetum) two, integuments, and partly owing to the fact that the wood has true vessels. There is only one order.
Fig. 275.—Welwitschia mirabilis (considerably reduced). The horizontal lines indicate the surface of the soil.
Order. Gnetaceæ. The three known genera differ very much in appearance. Welwitschia mirabilis (from the deserts of South Western Africa) is the oldest (?) genus now living. It resembles a giant radish, in that the hypocotyl is the only part of the main axis of the stem which becomes developed. It attains a circumference of upwards of four metres with a length of 1/2½-⅔ of a metre. It bears only two oblong, leathery leaves (Fig. 275) which are torn into segments at the apex and lie on the surface of the soil; these are the two first foliage-leaves which succeed the cotyledons, and they are remarkable for their enormous length (upwards of two metres) as well as for their long duration, living as long as the plant itself. In their axils are situated the 4-rowed, spike-like male and scarlet-coloured female cones, upon dichotomous branches. The perianth consists in the ♂ of 2 alternating pairs of leaves, the inner ones of which are slightly united. The andrœcium likewise consists of 2 whorls: the external (transverse) with 2, the internal with 4 stamens; the lower halves of the 6 filaments uniting to form a cup. Each of the terminal anthers corresponds to a sorus of 3 sporangia, the sporangia being fused together, and opening at the top by one three-rayed cleft. In the centre of the ♂-flower there is a sterile ovule. In the ♀-flower a perianth of two connate leaves is present.—Ephedra (desert plants, especially in the Mediterranean and W. Asia) at first sight resembles an Equisetum; the stems are thin, long-jointed, and the leaves opposite, small, and united into a bidentate sheath; ♂-perianth of two connate leaves (median leaves); 2–8 stamens united into a column. Each anther is formed of 2 sporangia (is bilocular). ♀ mainly, as in Welwitschia. The seeds are surrounded by the perianth which finally becomes red and fleshy. There are 30 species.—Gnetum has opposite, lanceolate, pinnately-veined, leathery leaves. They are mostly climbers (Lianas) from Tropical Asia and America. The ♂-flowers have a tubular perianth, (formed from two median leaves) which surrounds a centrally-placed filament, bearing 2 anthers. In the ♀-flower there is a similar perianth, surrounding an ovule provided with 2 integuments. The perianth becomes fleshy and envelops the hard seed. 20 species.
From the circumstance of Welwitschia having ♂ flowers which, besides stamens, possess also a rudiment of an ovule, Celakovsky draws the inference that the earliest Gymnosperms had hermaphrodite flowers which from this structure became differentiated entirely into ♂-and ♀-flowers, with the exception of Welwitschia only, in which this differentiation was only carried out in the ♀-flower. This theory has so far been scarcely proved.
Fossil Gymnosperms.
The earliest continental plants which are known belong to the Cordaitaceæ, a group of plants which existed as early as the Silurian period; they were Gymnosperms, but it has not yet been determined whether they were Cycads or Conifers. The Cycads, even in the Coal period, were scarce; they attained their fullest development in Jurassic and Cretaceous periods, during which they were rich in species and genera, and extended as far as the Polar regions. In addition[272] to these, Taxaceæ, Abietaceæ, and Taxodiaceæ appeared in the Carboniferous period. The Taxaceæ appear to have attained their culmination in the Jurassic and Cretaceous periods; Ginkgo appears in the Rhætic; Torreya, in the Cretaceous; Taxus and Podocarpus in the Tertiary periods. The Abietaceæ also appear in the Carboniferous; Pinus was first known with certainty in the English Weald and in the Cretaceous; almost all other contemporary genera are represented in this latter period. The Araucariaceæ first appear, with certainty, in the Jurassic. The Taxodiaceæ may be traced back as far as the Carboniferous (?); Sequoia is first found in the lowest Cretaceous, at that period it spread throughout the entire Arctic zone, and being represented by a large number of species, formed an essential part of the forest vegetation. Sequoia played a similar part in the Tertiary period. The Cupressaceæ are first known with certainty in the Jurassic, but they appeared more frequently and numerously in the Tertiary period, in which most of the present living genera were to be found. The Gnetaceæ, according to a theory advanced by Renault were represented in the Coal period by the genus Stephanospermum, which had four ovules enclosed by an envelope.
[273]
See pages 3 and 224. To this Division belong the majority of the Flowering-plants. They are divided into two parallel classes, the Monocotyledons and the Dicotyledons, which differ from each other not only in the number of cotyledons, which, with a few exceptions, is one in the former, two in the latter, but also in the internal structure of the stem, the venation of the leaves, the number of the parts of the flower, etc. Assuming that these two classes have sprang from a common origin, it is amongst the Helobieæ in the first, and amongst the Polycarpicæ in the second class that we might expect to find closely allied forms, which might reasonably be supposed to have varied less from this original type. As for the rest, they seem to stand quite parallel, without exhibiting any close relationship. It is scarcely proved that the Monocotyledons are the older class.
[Our knowledge of the forms included under the Angiosperms has recently been considerably increased by Treub (Ann. d. Jar. Bot. d. Buitenzorg, 1891), who has shown that the Casuarinas differ in many important points from the typical Angiosperms. Among other characters the pollen-tube is found to enter the ovule near the chalaza and therefore at the opposite end to the micropyle, and Treub therefore suggests that these plants should be placed in a subdivision termed Chalazogams.
According to this view the principal divisions of the Angiosperms would be represented thus:—
Angiospermæ.
| Sub-division. | Sub-division. |
| Chalazogames. | Porogames. |
| Class. | Classes. |
| Chalazogames. | Monocotyledones, Dicotyledones. |
More recently Nawaschin (Bull. Acad. Imp. Sci. St. Petersb., ser. iii., xxxv.) has shown that Betula, and Miss Benson (Trans. Linn. Soc., 1894) that Alnus, Corylus, and Carpinus also belong to the Chalazogams.
[274]
Our knowledge, however, is still so incomplete that one would hesitate to accord the full systematic value which Dr. Treub attaches to his discovery until the limits of the Chalazogamic group are better defined; and it would hardly be justifiable to include the Casuarinas and the above-noted genera in one family.]
The embryo has only one cotyledon; the leaves are as a rule scattered, with parallel venation; the vascular bundles of the stem are closed, there is no increase of thickness. The flower is typically constructed of five 3-merous whorls, placed alternately.
The embryo is generally small in proportion to the abundant endosperm (exceptions, see Helobieæ), and its single cotyledon is often sheath-like, and very large. On the germination of the seed either the entire cotyledon, or its apex only, most generally remains in the seed and absorbs the nutritive-tissue, while the lower portion elongates and pushes out the plumule and radicle, which then proceed with their further growth. The primary root in most cases soon ceases to grow, but at the same time, however, numerous lateral roots break out from the stem, and become as vigorous as the primary root, or even more so. Increase in thickness does not take place in these roots; they branch very little or not at all, and generally die after a longer or shorter time.
The stem is frequently a corm, bulb, or other variety of underground stem, as the majority of the Monocotyledons are perennial, herbaceous plants; it has scattered, closed vascular bundles (Fig. 276), and no cambium by which a continuous thickening may take place. The stem of the Palms, however, attains a very considerable thickness, which is due to the meristem of its growing-point continually increasing in diameter for a lengthened period (often for many years), until it has reached a certain size. In this condition the growing-point has the form of an inverted cone, and it is only when this cone has attained its requisite size that the formation of a vertical cylindrical stem commences. Certain tree-like Liliaceæ, as Dracæna, Aloe, etc., have a continuous increase in thickness; this is due to a meristematic layer, which arises in the cortex, outside the original vascular bundles, which were formed at the growing-point of the stem. This meristem continues to form thick-walled parenchyma and new, scattered vascular bundles. The primary vascular bundles, in the Palms and others, run in a[275] curved line from their entrance into the stem at the base of the leaf, towards the centre of the stem, and then bend outwards and proceed downwards in a direction more parallel to the sides of the stem (Fig. 277). The bundles formed later, in those stems which increase in thickness, are not continued into the leaves.
The branching as a rule is very slight, the axillary buds of the majority of the leaves never attaining development, e.g. in the Palms, bulbous plants and others. As the cotyledon arises singly, the succeeding leaves also must be scattered, but they are frequently arranged in two rows (Grasses, Iris, etc). The first leaf borne on a branch (the “Fore-leaf,”[24]—the bracteole, if on a floral shoot) has generally, in the Monocotyledons, a characteristic form and position, being situated on the posterior side of its own shoot, and hence turned towards the main axis; it is sometimes provided with two laterally-placed keels (Figs. 279 f, 290 øi), but the midrib is often absent. It arises in some cases from two primordia, which at the beginning are quite distinct, and thus has been regarded as formed by two leaves. It is, however, only one leaf, a fact which is evident from several circumstances, one being that it never supports more than one shoot, and this stands in the median plane (Fig. 279).

Fig. 276.—Transverse section of the stem of a Palm: v v is the wood portion, b b the bast portion of the vascular bundled.
Fig. 277.—Diagrammatic representation of the course of the vascular bundles, from the stem into the leaves in a Monocotyledon.
The leaves are amplexicaul, and have a large sheath but no stipules; the blade is most frequently long, ligulate, or linear, entire, with parallel venation, the veins being straight or curved[276] (Figs. 300, 309). Connecting the large number of veins which run longitudinally, there are as a rule only weak transverse ones. It is very rarely that other forms of leaves are found, such as cordate (Figs. 302, 312), or that the blade is branched, or the venation is, for example, pinnate or palmate (Figs. 225, 298); these deviations are especially found in the Araceæ, the Palms, the Scitamineæ (Fig. 308), the Dioscoreaceæ, and in several aquatic plants. The incisions in the Palm-leaf are derived by the splitting of an originally entire leaf.
The structure of the flower is generally as follows: Pr3 + 3, A3 + 3, G3, rarely S3 + P3 with the other members unchanged.[25] Instead of 3, the numbers 2 and 4 may occur; rarely others. In all these instances there are 5 whorls, which regularly alternate with one another, most frequently in the 3-merous flower, as in the diagram (Fig. 278). This diagram is found in the following orders: Liliaceæ, Convallariaceæ, Juncaceæ, Bromeliaceæ, Amaryllidaceæ, Dioscoreaceæ, Palmæ, some Araceæ, and in some small orders, and may be considered as the typical structure and also the starting point for the exceptional orders. The ovary in many Monocotyledons has many ovules, and the fruit becomes a many-seeded berry or capsule; this form is no doubt the oldest. In others the number of seeds becomes reduced to 1, and the fruit then becomes a cypsela, or a drupe (e.g. Gramineæ, Cyperaceæ, Palmæ, etc).

Fig. 278.—Diagram of the ordinary, regular flower in the Monocotyledons: s is the bract.

Fig. 279.—Diagram of Iris: f the bracteole; in its axil is a shoot with its bracteole.

Fig. 280.—Diagram of Orchis: l the lip; σ σ the two staminodes.
Deviations from this typical floral structure in some instances[277] may be traced to suppression, very rarely to a splitting of certain members, the typical relative positions not being changed. Thus, the Iridaceæ, the Cyperaceæ, most of the Gramineæ and some Juncaceæ deviate in having only 3 stamens (Fig. 279), the inner whorl (indicated by *) not becoming developed. The Musaceæ differ in the posterior stamen not being developed; Zingiberaceæ (Fig. 314), Marantaceæ, and Cannaceæ, in the fact that only 1 of all the stamens bears an anther, and the others are either suppressed or developed into petaloid staminodes, with some perhaps cleft in addition. The Orchideæ deviate in having, generally, only the anterior stamen of all the 6 developed (Fig. 280). In this, as in other instances, the suppression of certain parts of the flower is often connected with zygomorphy (i.e. symmetry in one plane), chiefly in the inner perianth-whorl, but also in the other whorls. In the Orchids, the perianth-leaf (the labellum, Fig. 280 l) which is directly opposite the fertile stamen, is larger and altogether different from the others. The perianth-leaves may also be suppressed; see, for example, the two diagrams of the Cyperaceæ (Fig. 284). In some orders the suppression of these leaves, which form the basis of the diagram, is so complete that it is hard to reduce the actual structure of the flower to the theoretical type, e.g. the Grasses (Fig. 290) and Lemna (Fig. 303). In the first family, which especially comprises water-plants, a somewhat different structure is found; thus Fig. 282 differs somewhat from the ordinary type, and other flowers much more so; but the floral diagrams which occur in this family may perhaps be considered as the most probable representatives of an older type, from which the ordinary pentacyclic forms have taken their origin. In favour of this theory we have the larger number of whorls, the spiral arrangement of some of these in the flower, with a large and indefinite number of stamens and carpels, the perfectly apocarpous gynœceum which sometimes occurs, etc., etc.
The Monocotyledons are divided into 7 Families:—
1. Helobieæ. This family forms a group complete in itself. It commences with hypogynous, perfect flowers, whose gynœcium is apocarpous and terminates in epigynous and more or less reduced forms.
2. Glumifloræ. These have as a starting point the same diagram as the following families, but otherwise develope independently.
3. Spadicifloræ. Also an independent branch, or perhaps two different ones which terminate in much reduced forms.
4. Enantioblastæ. These ought perhaps to be amalgamated with the following family.
[278]
5. Liliifloræ. These advance from forms with the typical diagram and hypogynous flower, to epigynous and reduced forms.
6. Scitamineæ and
7. Gynandræ. Two isolated families, which probably have taken their origin from Liliifloræ, and have epigynous, mostly zygomorphic, and much reduced forms.
To this family belong only water- or marsh-plants; the endosperm is wanting, and they possess an embryo with a very large hypocotyl prolonged downwards and often club-like. The perianth is often differentiated into calyx and corolla; the flower is regular, and in the first orders to be considered, may be reduced to the ordinary Monocotyledonous type; there are, however, usually found two 3-merous whorls of carpels (Fig. 282), and thus in all 6 whorls, or again, the number of carpels may be indefinite; the number of stamens also may be increased, either by the division of the members of a whorl, or by the development of additional whorls. Syncarps,[26] with nut or follicular fruitlets, are very common, for example, in the first orders; in the last (Hydrocharitaceæ) the carpels are not only united, but the ovary is even inferior.
The primitive type appears to be a hypogynous flower, similar to that of the Juncaginaceæ or Alismaceæ, with several 3-merous whorls, and free carpels, each with many ovules; the green perianth in this instance being no doubt older than the coloured ones. If we take a flower with this structure as the starting point, then the family developes partly into epigynous forms, partly into others which are so strongly reduced and exceptional that it is scarcely possible to refer them to the ordinary type. The family, through the peculiar Zostereæ, appears to approach the Araceæ, in which Potamogetonaceæ and Najadaceæ are included by some authorities. However, the inclusion of Potamogeton, and with it Ruppia and Zannichellia, in the Juncaginaceæ appears quite correct. It would scarcely be right to separate Zostereæ from these. Great stress has often been laid upon the similarity with the Ranunculaceæ which is found in the Alismaceæ, but it is scarcely more than an analogous resemblance.
Order 1. Juncaginaceæ. The ☿, regular, hypogynous flowers have the perianth 3 + 3, sepaloid, stamens 3 + 3 (with extrorse anthers), and carpels 3 + 3 (free or united), of which last, however, one whorl may be suppressed (in Triglochin maritima all 6 carpels are developed, in T. palustris the inner whorl is unfertile). Inflorescence long spikes. Embryo straight.—Marsh-plants with radical, rush-like leaves, arranged in two rows, and often sheathing and ligulate (“squamulæ intravaginales”); the inflorescence is a spike or raceme.—Scheuchzeria. Carpels almost free; in each at least[279] two ovules. Follicles.—Triglochin has long, fine racemes without bracts or bracteoles; one ovule in each carpel. The carpels in the two native species are united, but separate when ripe as a schizocarp, loosening from below; they open along the ventral suture or remain closed; a linear central column remains. The most reduced is Lilæa (1–2 sp. Am.)—Protogynous. About 10 species. Temp. Fossils in Tertiary.
Order 2. Potamogetonaceæ. The aquatic plants belonging to this order are perennial, living entirely submerged, or with floating leaves, and preferring still water. The leaves are alternate, in some linear and grass-like, in others there is an elliptical floating blade, supported by a linear submerged petiole. Axillary scales. The fruit is generally a syncarp with nuts or drupes; the embryo is curved, of very various forms.
Potamogeton (Pond-weed). The rhizome is creeping, sympodial (with two internodes in each shoot-generation); the inflorescence is a terminal, many-flowered spike, without floral-leaves; below it are found 2 foliage-leaves placed nearly at the same height, from whose axils the branching is continued cymosely. The flowers are ☿, 4-merous, naked, and consist only of 4 stamens, with the connectives, broadly developed at the back of the anthers, resembling a perianth, and of 4 free, sessile carpels. They are common plants in fresh water. The spike, during the flowering, is raised above the water. Wind-pollinated and protogynous.—Closely allied is Ruppia (Tassel Pond-weed), in salt or brackish water. The spike has only two naked flowers, each consisting of 2 stamens and 4 carpels. The stalks of the individual carpels are considerably prolonged.—Zannichellia (Horned Pond-weed) is monœcious; the ♀-flower consists of 4 (2–9) carpels, with membranous, bell-shaped perianth; long styles; the ♂-flower has 1 (-2) stamens. Althenia.
Zostera (Grass-wrack) is an entirely submerged, marine plant with creeping rhizome (with displacement of buds) and strap-shaped leaves. The flowering shoots are sympodia with displacement of the axes (Fig. 281). The inflorescence is a peculiar, flatly-compressed spike, on one side of which the flowers are borne (Fig. 281). This inflorescence may be considered, no doubt correctly, to be derived from the symmetrical spike of Potamogeton by strongly dorsiventral development, and by a strong suppression of the floral parts taking place simultaneously. Two rows of flowers are developed, but of these one is so pressed into the other that apparently only one is present. Each flower consists of only 1 stamen and 1 carpel situated at the same height (Fig. 281); the unilocular ovary encloses 1 pendulous ovule and bears a bifid style. As regards the perianth (?) one leaf may be present (Z. nana, Fig. 281 D). The pollen-grains are filamentous. Pollination takes[280] place under water. Posidonia and Cymodocea are allied to these. About 70 species.
Fig. 281.—Zostera. A Diagram of the branching of the floral shoots: I, II ... are the successive shoot-generations, every other one being shaded; g1 g2 ... fore-leaves; sp1 sp2 ... spathes for the successive spikes. Each shoot is united for some distance with the parent axis (indicated by the half-shaded internodes). Each shoot commences with a fore-leaf turning towards the parent axis, g; succeeding this is the spathe, sp; and then the inflorescence. The fore-leaf supports a new lateral shoot. B Diagram of a shoot, II, which is borne laterally in the axil of the fore-leaf g1, on the shoot I, g2 its fore-leaf; sp2 its spathe; sti squamulæ intravaginales. II Is the spadix with stamens and carpels; b a perianth-leaf (or connective expansion, similar to those which occur in Potamogeton). C The upper portion of a young spadix with development of flowers. D Part of a spadix with 2 flowers; the parts which theoretically belong to one another are connected by a dotted line.
[281]
Order 3. Aponogetonaceæ. Aquatic plants with tuberous stem. They have a single, petaloid perianth (3–2–1–leaved), most frequently 6 stamens and 3(-6) carpels. Straight embryo.—About 15 species (Africa, Madagascar, Tropical Asia and Australia).—Aponogeton distachyos and A. (Ouvirandra) fenestralis are grown in conservatories; the latter has lattice-like, perforated leaves.
Order 4. Najadaceæ. Only one genus Najas (about 10 species); annual fresh water plants with leaves in pairs and solitary, unisexual flowers. The ♂ flower is remarkable in having a terminal stamen, which has either 4 longitudinal loculi or 1 central one; on this account the stamen of Najas is considered by some authorities to be a stem and not a leaf-structure. The unilocular gynœceum and the single, erect, anatropous ovule are also terminal. Pollination takes place under the water.
Order 5. Alismaceæ. The regular, hypogynous flowers are in some species unisexual by the suppression of either andrœcium or gynœceum; they have a 6-merous perianth, generally differentiated into 3 sepals and 3 petals; generally 6 stamens in the outer whorl (by the division of the 3; Fig. 282) and often several 3-merous whorls inside these, and 6–∞ free carpels arranged cyclically or spirally. Fruit a syncarp.—Marsh- or water-plants with radical leaves and long-stalked inflorescences.
A. Butomeæ. Follicles with many seeds, which are borne on nearly the whole of the inner surface of the cyclic carpels (as in Nymphæaceæ). Embryo straight.—Butomus (Flowering Rush, Fig. 282), has an umbel (generally composed of 3 helicoid cymes). S 3, P 3, stamens 9 (6 + 3, i.e. the outer whorl doubled), G 3 + 3. B. umbellatus; creeping rhizome with triangular Iris-like leaves.—Hydrocleis. Limnocharis.
Fig. 282.—Diagram of Butomus: f bracteole.
B. Alismeæ. Fruit achenes. Latex common (in the intercellular spaces). The flowers are arranged most frequently in single or compound whorls. Embryo curved, horse-shoe shaped.—Alisma has S 3, P 3, A 6 (in 1 whorl, grouped in pairs, i.e. doubled in front of the sepals), and 1 whorl of 1-seeded achenes on a flat receptacle. The leaves are most frequently radicle, long-stalked; the lamina have curved longitudinal veins, and a richly branched venation. A. plantago.—Elisma (E. natans) has epitropous (turned inwards) ovules, whilst the ovules of Alisma, Sagittaria and others are apotropous (turned outwards).—Echinodorus (E. ranunculoides)[282] has a convex receptacle, carpels many, united and capitate. Damasonium.—Sagittaria (Arrow-head) has monœcious flowers, several whorls of stamens and spirally-arranged achenes on a very convex receptacle. S. sagittifolia reproduces by tuberous buds formed at the end of long, submerged branches. The leaves, in deep and rapidly running water, are long and strap-shaped, but in the air arrow-shaped.
Honey is secreted in the flower and pollination effected by insects. Alisma plantago has 12 nectaries. The submerged flowers of Elisma natans remain closed and are self-pollinated. Butomus has protandrous flowers. There are about 50 species, which mostly grow outside the Tropics.—Uses insignificant. The rhizome of some is farinaceous.
Order 6. Hydrocharitaceæ. This order differs chiefly from the preceding in its epigynous flowers. These are in general unisexual (diœcious), and surrounded by a 2-leaved or bipartite spathe; they are 3-merous in all whorls, but the number of whorls is generally greater than 5, sometimes even indefinite. The perianth is divided into calyx and corolla. The ovary is unilocular with parietal placentation, or more or less incompletely plurilocular. The fruit is berry-like, but usually ruptures irregularly when ripe. Embryo straight.—Most often submerged water-plants, leaves seldom floating on the surface. Axillary scales (squamulæ intravaginales).
Hydrocharis. Floating water-plants with round cordate leaves; S3, P3 (folded in the bud); ♂-flowers: 3 (-more) flowers inside each spathe; stamens 9–15, the most internal sterile. ♀-flowers solitary; three staminodes; ovary 6-locular, with many ovules attached to the septa; styles 6, short, bifid. [The petals of the ♀-flowers bear nectaries at the base. In this and the following genus the pollination is without doubt effected by insects.] H. morsus ranæ (Frog-bit) has runners; it hibernates by means of special winter-buds.—Stratiotes; floating plants with a rosette of linear, thick, stiff leaves with spiny margin, springing from a short stem, from which numerous roots descend into the mud. Inflorescence, perianth, and ovary nearly the same as in Hydrocharis, but the ♂-flower has 12 stamens in 3 whorls, of which the outer 6 are in 1 whorl (dédoublement), and inside the perianth in both flowers there are numerous (15–30) nectaries (staminodes?). S. aloides (Water-soldier); in N. Eur. only ♀-plants.—Vallisneria spiralis is a tropical or sub-tropical plant, growing gregariously on the mud in fresh water. The leaves are grass-like, and the plants diœcious; the ♂-flowers are detached from the plant, and rise to the surface of the water, where they pollinate the ♀-flowers. These are borne on long, spirally-twisted peduncles which contract after pollination, so that the ♀-flower is again drawn under the water, and the fruits ripen deeply submerged.—Elodea canadensis[283] is also an entirely submerged plant. The leaves are arranged in whorls on a well-developed stem. Only ♀-plants in Europe (introduced about 1836 from N. Am). This plant spreads with great rapidity throughout the country, the reproduction being entirely vegetative. Hydrilla, Halophila, Thalassia, Enhalus.—In many of these genera the number of whorls in the flower is remarkably reduced; for example, in Vallisneria, in the ♂-flowers to 2: Pr 3, A (1-) 3, in the ♀ to 3: Pr 3, Staminodes 3, G 3.—About 40 species; Temp. and Trop.
The hypogynous flowers in the Juncaceæ are completely developed on the pentacyclic, trimerous type, with dry, scarious perianth. Even in these the interior whorl of stamens becomes suppressed, and the ovary, which in Juncus is trilocular with many ovules, becomes in Luzula almost unilocular, but still with 3 ovules. The perianth in the Cyperaceæ and Gramineæ is reduced from hairs, in the first of these, to nothing, the flowers at the same time collecting more closely on the inflorescence (spike) supported by dry bracts (chaff); the number of stamens is almost constantly 3; stigmas linear; the ovary has only 1 loculus with 1 ovule, and the fruit, which is a capsule in the Juncaceæ, becomes a nut or caryopsis.—The endosperm is large and floury, the embryo being placed at its lower extremity (Figs. 286 B, 291).—The plants belonging to this order, with the exception of a few tropical species, are annual or perennial herbs. The stems above ground are thin, and for the most part have long internodes, with linear, parallel-veined leaves which have long sheaths, and often a ligule, i.e. a membranous projection, arising transversely from the leaf at the junction of the sheath and blade. The underground stems are short or creeping rhizomes. The flowers are small and insignificant. Wind- or self-pollination.
Order 1. Juncaceæ (Rushes). The regular, hermaphrodite, hypogynous flowers have 3 + 3 brown, dry, free perianth-leaves projecting like a star during the opening of the flower; stamens 3 + 3 (seldom 3 + 0) and 3 carpels united into one gynœceum (Fig. 283); the ovary is 3- or 1-locular; there is as a rule 1 style, which becomes divided at the summit into 3 stigmas, often bearing branches twisted to the right (Fig. 283). Fruit a capsule with loculicidal dehiscence. The embryo is an extremely small, ellipsoidal, cellular mass, without differentiation into the external organs.
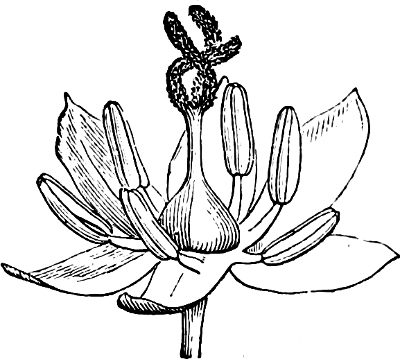
Fig. 283.—Flower of Luzula.
[284]
Juncus (Rush) has glabrous foliage-leaves, generally cylindrical, rarely flat; the edges of the leaf-sheath are free (“open” leaf-sheaths) and cover one another. The capsule, 1- or 3-locular, with many seeds—Luzula (Wood-Rush) has flat, grass-like leaves with ciliated edges; the edges of the leaf-sheath are united (“closed” leaf-sheath). The capsule unilocular and 3-seeded.—Prionium: S. Africa; resembling a Tacona.
The interior whorl of stamens, in some species, disappears partially or entirely (J. supinus, capitatus, conglomerates, etc.)
Some of the numerous Juncus-species (e.g. J. effusus, glaucus, conglomeratus, etc.), have false, lateral inflorescences, the axis of the inflorescence being pushed to one side by its subtending leaf, which apparently forms a direct continuation of the stem, and resembles it both in external and internal structure. The foliage-leaves of this genus were formerly described as “unfertile stems,” because they are cylindrical, erect, and resemble stems, and consequently the stem was said to be “leafless”: J. effusus, glaucus, conglomeratus. Stellate parenchynatous cells are found in the pith of these stems and in the leaves. Other species have distinct terminal inflorescences and grooved leaves; J. bufonius (Toad-rush), compressus, and others. The inflorescences most often present the peculiarity of having the lateral axes protruding above the main axis. Their composition is as follows:—The flowers have either no bracteoles, and the inflorescences are then capitulate; or they have 1–several bracteoles. Each branch has then, first, a 2-keeled fore-leaf placed posteriorly (“basal-leaf”), and succeeding this are generally several leaves borne alternately and in the same plane as the basal-leaf, the two uppermost (the “spathe-leaves”) being always barren; those which lie between the basal-leaves and the spathe-leaves are termed “intermediate-leaves.” If only branches occur in the axils of the basal-leaves, then the succeeding branches are always borne on the posterior side of the axis, and form a fan[27]; if the basal-leaf is barren, and if there is only one fertile intermediate-leaf, then the lateral axes are always on the upper side, and a sickle[27]-like inflorescence occurs; if there are 2 fertile intermediate-leaves, then a dichasium is formed, and in the case of there being several, then a raceme, or spike.
Juncaceæ are, by several authors, classed among the Liliifloræ, but there are so many morphological and partly anatomical features agreeing with the two following orders, that they may, no doubt, most properly be regarded as the starting point of these, especially of the Cyperaceæ, which they resemble in the type of flowers, the inflorescence, the type of mechanical system, and the stomata.
Pollination by means of the wind. Cross-pollination is often established by protogyny. J. bufonius has partly triandrous and cleistogamic, partly hexandrous, open flowers.—Distribution. The 200 species are spread over the entire globe, but especially in cold and temperate countries; they are seldom found in the Tropics.—Uses. Very slight; plaiting, for instance.
Order 2. Cyperaceæ. The majority are perennial (seldom[285] annual) herbs living in damp situations, with a sympodial rhizome and grass-like appearance. The stems are seldom hollow, or have swollen nodes, but generally triangular, with the upper internode just below the inflorescence generally very long. The leaves are often arranged in 3 rows, the leaf-sheath is closed (very seldom split), and the ligule is absent or insignificant. The flowers are arranged in spikes (spikelets) which may be united into other forms of inflorescences (chiefly spikes or racemes). The flowers are supported by a bract, but have no bracteoles. In some genera the perianth is distinctly represented by six bristles corresponding to six leaves (Figs. 284 A, 286 A); in others it is represented by an indefinite number of hairs (Fig. 284 B), and very frequently it is altogether wanting. The inner whorl of stamens is absent, and the flower has therefore 3 stamens (rarely more or less than 3), the anthers are attached by their bases to the filament (innate) and are not bifid (Figs. 286). Gynœceum simple, formed of 3 or 2 carpels; 1 style, which is divided at the extremity, as in the Juncaceæ, into 3 or 2 arms; the single loculus of the ovary contains one basal, erect, anatropous ovule; the stigmas are not feather-like. Fruit a nut, whose seed is generally not united with the pericarp. The embryo is small, and lies at the base of the seed in the central line, surrounded on the inner side by the endosperm (Fig. 286 B). On germination the cotyledon does not remain in the seed.
Fig. 284.—Diagram of structure of: A Scirpus silvaticus; B Eriophorum angustifolium.
A regular perianth, with 6 scale-like perianth-leaves in 2 whorls, is found in Oreobolus. In Scirpus littoralis the perianth-leaves are spreading at the apex, and divided pinnately.
The branching of the inflorescence is often the same as in the Juncaceæ, and supports the theory that these two orders are related. In Rhynchospora and others, the “spikelets” are really only “spike-like” and to some extent compound.
A. Scirpeæ. Hermaphrodite Flowers.
1. Spikelets cylindrical, the bracts arranged spirally (in many rows). The lower ones are often barren, each of the others supports a flower.—Scirpus (Club-rush). The spikelets are many-flowered; the perianth is bristle-like or absent, and does not continue to grow during the ripening of the fruit (Fig. 286 A). Closely allied to this is Heleocharis, with terminal spikes.—[286]Eriophorum (Cotton-grass) differs chiefly in having the perianth-hairs prolonged, and forming a bunch of white, woolly hairs (Fig. 284 B).
Cladium and Rhynchospora (Beak-rush) differs especially in the few-flowered, compound spikelets which are collected into small bunches; the latter has received its name from the fact that the lowermost portion of the style remains attached to the fruit as a beak.
2. Spikelets compressed, the bracts arranged only in two rows; the other characters as in the first-mentioned. Cyperus (spikelets many-flowered); Schœnus (Bog-rush); spikelets few-flowered; S. nigricans has an open sheath.
Fig. 285.—Carex: A diagram of a male flower; B of a female flower with 3 stigmas; C of a female flower with 2 stigmas; D diagrammatic figure of a female flower; E similar one of the androgynous (false) spikelet of Elyna. The ♂ is here represented placed laterally; it is terminal, according to Pax.
Fig. 286.—A Flower of Scirpus lacustris. B Seed of Carex in longitudinal section.
B. Cariceæ. Unisexual Flowers.
In the ♂-flowers there is no trace of a carpel, and in the ♀ no trace of a stamen. Floral-leaves in many rows. In some (Scleria, certain Carex-species), ♂-and ♀-flowers are borne in the same spikelet, the latter at the base or the reverse; in the majority each spikelet is unisexual.
Carex (Fig. 285) has naked, most frequently monœcious flowers. The ♂-spikes, which are generally placed at the summit of the whole compound inflorescence, are not compound; in the axil of each floral-leaf (bract) a flower is borne, consisting only of a short axis with three stamens (Fig. 285 A). The ♀-spikes are compound; in the axil of each floral-leaf is borne a very small branch (Fig. 285 D, a) which bears only one leaf, namely, a 2-keeled fore-leaf[287] (utriculus, utr. in the figures) which is turned posteriorly (as the fore-leaves of the other Monocotyledons), and being obliquely sheath-like, envelopes the branch (in the same manner as the sheath of the vegetative leaves), and forms a pitcher-like body. In the axil of this leaf the ♀-flower is situated as a branch of the 3rd order, bearing only the 2–3 carpels, which are united into one gynœceum. The style protrudes through the mouth of the utriculus. The axis of the 2nd order (a in Fig. 285 D) may sometimes elongate as a bristle-like projection (normally in Uncinia, in which it ends as a hook, hence the name); this projection is in most cases barren, but it sometimes bears 1–several bracts which support male-flowers; this is normal in Elyna (or Kobresia) and Schœnoxiphium; the axis (a in 285 E) bears at its base a female-flower supported by the utriculus, and above it a male-flower supported by its bract.
Pollination by means of the wind. Protogynous. Sometimes self-pollinated. The order embraces nearly 3,000 species, found all over the world. Carex and Scirpus are most numerous in cold and temperate climates, and become less numerous towards the equator. The reverse is the case with Cyperus and other tropical genera. They generally confine themselves to sour, swampy districts; some, on the other hand, are characteristic of sand-dunes, such as Sand-star (Carex arenaria). There are about 70 native species of Carex.
Uses. In spite of their large number, the Cyperaceæ are of no importance as fodder-grasses, as they are dry and contain a large amount of silica; hence the edges of many of the triangular stems or leaves are exceedingly sharp and cutting. Cyperus esculentus has tuberous rhizomes, which contain a large amount of fatty oil and are edible (earth-almonds); it has its home in the countries of the Mediterranean, where it is cultivated.
Cyperus papyrus (W. Asia, Egypt, Sicily) attains a height of several metres, and has stems of the thickness of an arm which were used by the ancient Egyptians for making paper (papyrus). Some serve for plaiting, mats, etc. (Scirpus lacustris, etc.). Isolepis is an ornamental plant.
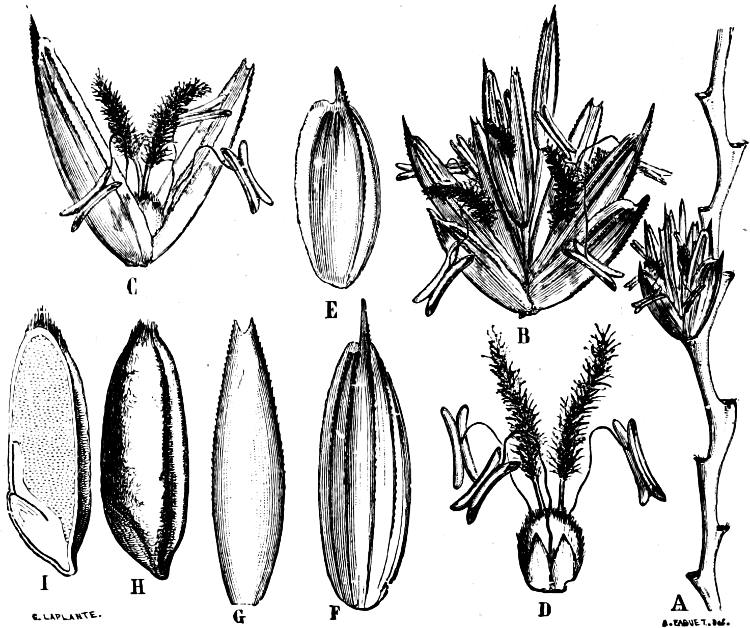
Fig. 287.—Triticum: A axis (rachis) of ear showing the notches where the spikelets were inserted; B an entire spikelet; C a flower with the pales; D a flower without the pales, showing the lodicules at the base; E glume; F outer pale; G inner pale; H fruit; I longitudinal section of fruit.
Order 3. Gramineæ (Grasses). The stems are cylindrical, generally hollow with swollen nodes, that is, a swelling is found at the base of each leaf which apparently belongs to the stem, but in reality it is the swollen base of the leaf. The leaves are exactly alternate; the sheath is split (excep. Bromus-species, Poa pratensis, P. trivialis, Melica, Dactylis, etc., in which the sheath is not split), and the edges overlap alternately, the right over the left, and vice versâ; the ligule is nearly always well developed. In general, the flowers are hermaphrodite; they are borne in spikelets with alternate floral-leaves, and the spikelets themselves are borne in either spikes or panicles. The two (seldom more) lowest floral-leaves in each spikelet (Fig. 289 øY, nY) are barren (as the covering-leaves in many umbels and capitula); these are termed the glumes. The[288] succeeding floral-leaves, each of which supports one flower as its bract, are called the outer pales (nI); these sometimes each bear an “awn” (a bristle-like body which projects in the median line either from the apex or the back); sometimes the upper ones are barren. Each flower has a bracteole, which is placed on the inside opposite the main axis; it is thin, binerved or two-keeled, and never has an awn; it is known as the inner pale (øI). Immediately succeeding the bracteole are: (a) some small, delicate scales (lodicules, Figs. 287 D, 288 C, 290 L); (b) three stamens with anthers versatile, so as to be easily moved, and usually notched at each end (Fig. 287 C); and (c) a simple gynœceum formed of one carpel with two styles having generally spirally-branched stigmas (Figs. 287 D, 288 C). The ovary is unilocular, and contains one ascending or pendulous, anatropous ovule. Fruit a nut, whose seed is always firmly united with the thin pericarp (“caryopsis”). The embryo is larger than in the Cyperaceæ and is placed at the base of the seed, but on the outer convex surface of the pericarp (Figs. 287 I, 288[289] 288 D, 291), outside the endosperm; plumule large with several leaf-primordia. On germination the cotyledon remains in the seed.
The majority of Grasses are annual or perennial herbs; tree-like forms being only found in the Tropics, for example, the Bamboos; they branch (in tufts), especially from the axils of the basal-leaves, while those which are borne higher on the stem are separated by longer internodes and have no vegetative branches in their axils, though a few forms, like Bambusa and Calamagrostis lanceolata, produce branches in these axils.
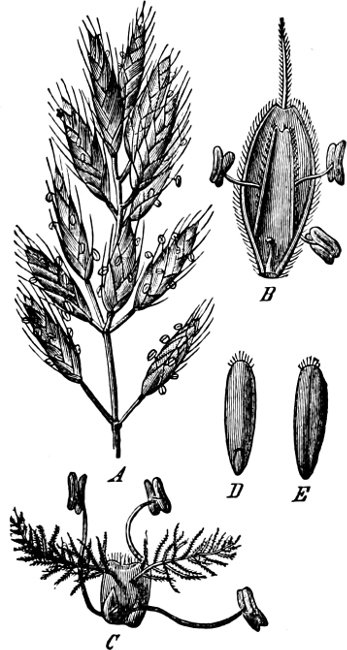
Fig. 288.—Bromus mollis: A inflorescence; B the uppermost flower of a spikelet, with its axis turned forward; in front is seen the two-keeled inner pale (bracteole) and the stamens protrude between this and the outer pale (bract); C an ovary with the 2 stigmas on its anterior side, the 2 lodicules, and the 3 stamens; D the fruit seen from the dorsal side; E the same from the ventral side.
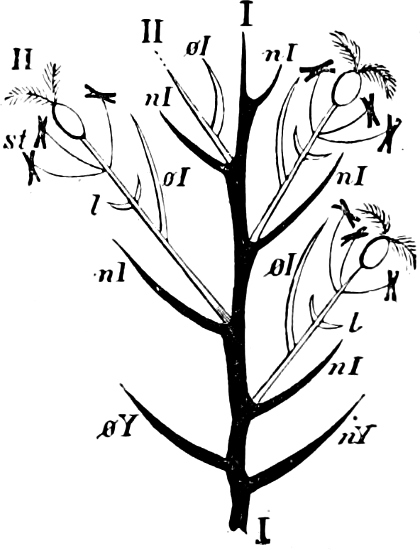
Fig. 289.—Diagrammatic outline of a spikelet: n Y lower glume; ø Y upper glume; n I upper pale; ø I the inner pale; l-l lodicules; st stamens; I-I main axes; II lateral axes.

Fig. 290.—Diagram of the Grass-flower: ni outer pale; øi inner pale; l-l lodicules.
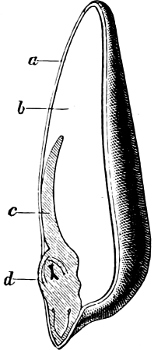
Fig. 291.—Longitudinal section of an Oat-grain: a the skin (pericarp and testa); b the endosperm; c the cotyledon; d the plumule.
Only a few Grasses have a solid stem, such as Maize, Sugar-cane, and Andropogon. The blade is flat in the meadow-grasses, but the Grasses which live on dry places (“prairie-grass”) exposed to the sun, often have the blade tightly[290] rolled up and almost filiform or bristle-like, with anomalous anatomical structure. A closed tubular sheath is found in Melica uniflora, Bromus-species, Poa pratensis and trivialis, Briza and some Glyceria-species. The sheath is developed for the purpose of supporting the young internodes while their growth is proceeding at the base. The “nodes” (the swollen joints which are seen on stems of Grasses) are not really part of the stem but are formed by the base of the leaf-sheath. They play a part in assisting the haulms to regain a vertical position when laid prostrate by wind or rain. The awn on the pale is homologous with the blade of the Grass-leaf, and the pale itself is the sheath. The arrangement of the leaves in the spikelet is similar to that in Cyperus and other Cyperaceæ, their floral-leaves being borne in several rows in Streptochæta. More than two barren “glumes” are found in Streptochæta, several Phalarideæ and others. The spikelets, too, are again arranged in two rows in the axils of suppressed floral-leaves. The inflorescence becomes a “compound spike” (ear) when the spikelets are sessile. In the majority of instances the spikelets are borne on long stalks; when these branch, then the secondary branches, and similarly all branches of higher order, are placed so far down upon the mother-axis that they all appear to be of equal value and to arise in a semicircle from the mother-axis itself, though in reality they arise from each other (Panicle, Fig. 288 A). Sometimes the main axis and branches of different orders unite together as in Alopecurus, Phleum, and some other Grasses, and hence the single (short-stalked) spikelets appear to arise singly and spirally, or without any definite order, directly from the main axis, with the production of a cylindrical inflorescence bearing “spikes” on all sides, that is, a “spike-like panicle.”—Many inflorescences are somewhat dorsiventral. The flower is rarely unisexual (Zea mais) or barren. Considerable difficulty is experienced in reducing the Grass-flower[291] to the ordinary 3-merous Monocotyledonous type. Some authorities consider the lodicules, which are present in all Grasses but absent in the Cyperaceæ, to be homologous with a perianth. According to a more recent theory they are bracteoles, and hence the Gramineæ, like many of the Juncaceæ, have 2–3 bracteoles placed in two rows in the median plane. If this theory be correct, the flower is naked. The lodicules expand quickly and cause the opening of the flower (i.e. the two pales become separated from each other). Generally only 3 stamens belonging to the outer whorl are present (Fig. 290), as in Iris (Fig. 279), certain Juncaceæ and Cyperaceæ (Fig. 284), but in some, such as the Rice and certain species of Bamboos, all 6 are found. Pariana has more than 6. Only 1 of the carpels is present, namely, the anterior (of those in Fig. 284), so that the ventral suture and the place of attachment of the ovule are situated at the back of the ovary. The number of styles does not correspond with the number of carpels, and the styles may therefore be supposed to arise from the edges of the leaf to the right and left—a position which is not without analogy. In addition, a stylar projection is sometimes found on the anterior side and in the median line (e.g. in Phragmites), and the solitary style in Nardus has exactly this position; a similar arrangement is found in some species of Bambusa which have only one style; other species of Bambusa have three styles. A tripartite style is found in Pharus.
[The Grass-flower may be reduced to the ordinary Monocotyledonous type thus:—The outer pale is the bract of the flower since it bears in its axil the floral shoot; the inner pale occupies the customary position of the bracteole. The fact that it is binerved can be explained by its having been pressed against the main-axis during development. Similar binerved bracteoles are found in Iris (Fig. 279). These bracteoles in both Grass and Iris arise from single primordia, and are not produced by the coalescence of two leaves. The lodicules are the only parts of the perianth remaining, the outer whorl having been suppressed, and also the posterior leaf of the inner whorl; a posterior lodicule, however, is found in the Rice and some species of Bamboo. The outer whorl of stamens is usually absent, though this again is present in the Rice and Bamboo. The three carpels are reduced to one with two or sometimes three stigmas.]
The Flowering. In the panicles the flowers open in basipetal order; the flowers in the spikes situated somewhat above the middle, commence to open first, and the flowering proceeds upwards and downwards. A few Grass-flowers never open (cleistogamic); Leersia oryzoides, Stipa-species, and e.g. Wheat and Rye in cold damp weather; some open their pales so wide that the anthers and stigmas may protrude at the top; most frequently the lodicules expand and force the pales suddenly and widely apart. The filaments elongate considerably, so that the anthers are pendulous and the stigmas unfold. In some Grasses e.g. Wheat, the blooming of each flower only lasts a short time. Pollination is generally effected by the wind. The Rye separates the pales very widely in the morning, and allows the anthers and stigmas to appear; it is almost entirely sterile when self-pollinated. The Wheat flowers at any time of the day, each flower lasting only a quarter of an hour. The pales open suddenly, but only half way, and the anthers scatter one-third of the pollen in their own flower and two-thirds outside. Self-pollination is effectual, but crossing gives better results. In Hordeum vulgare (all flowers ☿) the flowers[292] of the 4 outer rows behave as in the Wheat, but those in the two central rows always remain closed. The ☿-flowers in the two central rows of H. distichum remain closed and fertilise themselves; they open exceptionally, and may be pollinated by the ♂-flowers in the 4 lateral rows. H. hexastichum is cleistogamic. Oats pollinate themselves.
Fig. 292.—Barley grain: A section through the skin (a-d) and the most external part of the endosperm; Gl the “aleurone layer”; st starch-containing cells; B starch grains.
Fig. 293.—Wheat-grain germinating: g the plumule; b the first leaf succeeding the cotyledon; r1 the primary root; r2 lateral root.
Fig. 294.—Older seedling of the Wheat: s the second sheathing-leaf; l first foliage-leaf.
The ripe Grass-fruit, in some species of Bamboo, is a berry; in some other Grasses a nut with loosely lying seed, in some even a capsule, but otherwise a “caryopsis.” In some instances it is loosely enveloped by the pales (Oat), in others firmly attached to these (Barley), and finally, in others, “naked,” i.e. it is entirely free from the pales (Wheat and Rye). On the ventral side there is a groove (Fig. 288 E); on the anterior side (dorsal suture), which is turned towards the inner pale, it is convex, and at the base on this side, inside the testa, lies the embryo (Fig. 288 D). The apex of the fruit is often hairy (Fig. 293). The skin (Fig. 291, a) is formed by the pericarp and testa, and in some cases (Barley) the pales also form the outer portion. The endosperm (b) is large, and formed of parenchymatous, starch-containing cells: aleurone (proteid) grains may also be found among them. When the starch-grains and the aleurone-grains adhere together the endosperm becomes “horny,” but is “floury” when the starch-grains lie loosely with air between them. In the most external region, just beneath the skin, 1–several layers of nearly cubical cells (filled principally with aleurone-grains and fat) are found, the aleurone-layer (Fig. 292). The embryo (Fig. 291 c-d) contains large quantities of fatty oil; the large shield-like structure, attached to the embryo and turned inwards towards the endosperm (c), is the cotyledon (scutellum); it remains enclosed in the seed during germination, and[293] dissolves the endosperm by means of the peculiar epithelial cells developed on the dorsal surface. The radicle, on germination, is obliged to perforate a mass of cells derived from the suspensor and which form the “root-sheath” (coleorhiza, Fig. 293) round its base. In addition to the tap-root, lateral roots are frequently developed before germination; these quickly break through, and later on are followed by others which appear at the base of the leaf (Figs. 293, 294).
The DISTRIBUTION OF THE FRUIT is most frequently effected by the wind. The spirally-twisted and hygroscopic awn which persists on the fruits of some species (Avena, Stipa, etc.) assists in their dissemination, and even helps to bury them in the ground.
The two preceding orders are more closely related to each other than they are to the Gramineæ.
The generic differences are chiefly founded on the form of the inflorescence, the number and sex of the flowers in the spikelets, the shape and relative length of the pales, awns, etc. In addition to these the structure of the fruit and seed presents a great many differences; some have compound starch-grains, while in others they are single; some have 1 layer of aleurone-cells, others have several (Fig. 292), etc.
1. Bambuseæ. Tall Grasses with woody, very siliceous stems which bear many branches in the axils of the leaves. 6 stamens. Bambusa (Bamboo).
2. Oryzeæ. Oryza sativa (Rice) is a herbaceous marsh-plant, with panicle and small, 1-flowered spikelets, with two small glumes and two large, boat-shaped, strongly siliceous pales. 6 stamens.—Leersia. Lygeum. Pharus. Zizania aquatica.
3. Maydeæ. Zea mais (Indian-corn, Maize); the spikelets are unisexual; the ♂-spikelets in a terminal panicle; the ♀-spikelets closely crowded and arranged in many rows in a thick, axillary spike, enclosed by large sheathing-leaves. The ♀-spikelets are 1-(2-) flowered; the ovary bears one, long, filamentous style, with bifid stigma.—Euchlæna; Coix.
4. Andropogoneæ. Saccharum (Sugar-cane); the spikelets are exceptionally small, 1-flowered, and borne in pairs in many-flowered, long-haired panicles. Tall grasses with solid, sappy stem.—Andropogon.
5. Festuceæ. Grasses with panicle (or spike-like panicle) and 2–several-flowered spikelets. Glumes small, in each case shorter than the spikelet.—Festuca (Fescue) and Bromus (Brome, Fig. 288) have the awn placed at the apex of the pale, or slightly below it. Festuca has perennial species, with only a sparsely-branched panicle with branches solitary or in pairs, and round spikelets; the leaf-sheath is widely open. Bromus has the[294] branches borne in half whorls, and the leaf-sheath scarcely half open. Brachypodium has very short-stalked spikelets in a raceme.—Poa (Meadow-grass), Briza (Quaking-grass) and Glyceria have awnless spikelets; these in Poa are ovoid, compressed, and with sharply-keeled glumes; in Briza they are broad, cordate and drooping, with boat-shaped glumes; in Glyceria round, long, many-flowered, linear or lanceolate; some species of Glyceria have closed leaf-sheaths.—Dactylis (Cock’s-foot) differs from all others in the somewhat crowded and unilateral (subsecund) spikelets, which are compressed and oblique (i.e. one side more convex than the other).—Phragmites (P. communis, Reed); the lowermost flowers of the spikelet are ♂; its axis is covered with long, silky hairs; pales without awns, but acuminate. Perennial marsh-plants.—Melica; panicle small, sparsely-branched with round, awnless, few-flowered, usually drooping spikelets. The upper pales, with arrested flowers, are generally united into a club-like mass.—Molinia, Eragrostis, Koeleria, Catabrosa.—Cynosurus (Dog’s-tail) has a small, spicate panicle with unilateral spikelets, some of which are fertile, some barren, each supported by a pectinate scale. Arundo. Sesleria. Gynerium. Triodia.
6. Aveneæ. Panicles with 2–many-flowered spikelets; at least one of the glumes is quite as long as the entire spikelet.—Avena (Oat). The pale is boat-shaped, often bifid, and at about the middle of the back has a twisted, bent awn.—Aira (Hair-grass) has a long-branched panicle with small, 2-flowered spikelets; the pale has a dentate apex and bears an awn on the posterior side close to the base.—Weingærtneria.—Holcus (Yorkshire-fog); a soft, hairy Grass with an open panicle, keeled glumes; 2 flowers in the spikelet, of which the lower one is ☿, the upper ♂; the pale which supports the ☿-flower has no awn, but that which supports the ♂-flower, on the contrary, is awned.
7. Agrostideæ. Panicles or spike-like panicles with 1-flowered spikelets. Generally 2 glumes and only 1 pale.—The following have PANICLES: Milium with square panicle-branches and round spikelets; Agrostis (Fiorin), with compressed, glabrous spikelets, whose glumes are longer than the pales. Calamagrostis differs in having a chaplet of long hairs at the base of the pale.—Stipa (Feather-grass) has a long, twisted awn.—The following have SPIKELIKE PANICLES: Phleum (Cat’s-tail, Timothy-grass) has sharply pointed, entirely free glumes, which are much longer than the awnless pales. Alopecurus (Fox-tail); glumes united below; pale[295] with awn. Ammophila (Psamma). A. arundinacea; pales hairy at base; perennial, stiff-leaved, glaucous sand-grass with creeping rhizome. Aristida. Sporobolus.
8. Phalarideæ. Panicles and spike-like panicles. The spikelet has in the upper part a single fertile flower; below it are placed 4 pales, of which the upper 1–2 sometimes support ♂-flowers. On the whole, 6 floral-leaves of the first order are present.—Phalaris (P. canariensis, Canary-grass) has an ovate, spike-like panicle, the spikelets are compressed, convex on the outer side, concave on the inner. The large glumes are winged on the back.—Digraphis (D. arundinacea) is closely allied to Phalaris, but the keel of the glumes is not winged.—Anthoxanthum (A. odoratum, Sweet-vernal) has a small, lanceolate, open, spike-like panicle; the spikelets have below 2 barren flowers, and above these an ☿-flower with 2 stamens. The upper glume is longer than the flower.—Hierochloa.
9. Chlorideæ. The spikelets are arranged in the form of a spike in two rows on one side of an often flatly-compressed axis; they are mostly 1-flowered.—Chloris; Ctenium; Cynodon; Eleusine; Microchloa.
10. Paniceæ. The spikelets are borne in panicles or spikes, which may be arranged like fingers or in a raceme. There is a centrally-placed ☿-flower; below it is sometimes a ♂-flower.—Panicum; Paspalum; Oplismenus; Setaria has an almost cylindrical spike-like panicle with several barren branchlets, which project as stiff, rough bristles.—Cenchrus; Pennisetum.
11. Hordeæ. Spikes compound; spikelets sessile in the notches of a toothed axis.
A. Spikelets solitary.—Triticum (Wheat, Fig. 287) has in each tooth of the main axis, a several-flowered spikelet which turns its flat side towards the central axis. The cultivated species (true Wheat) are 1-2-annual, the wild ones (T. repens, Couch, also as an independent genus, Agropyrum) are perennial, with creeping rhizome and lanceolate glumes.—Lolium (Rye-grass) has in each tooth of the main axis a many-flowered, compressed spikelet, which is placed edgewise towards it and (with the exception of L. perenne) has only one outwardly-turned glume (L. temulentum has a rudiment of the inwardly-turned lower glume); the terminal spikelet has two glumes.—Secale (Rye). A two-flowered spikelet in each tooth; small, lanceolate, acuminate glumes. Nardus and Lepturus have very narrow spikes, the former with unilateral spikelets.
B. In each notch of the axis 2 or more spikelets are placed[296] close together.—Hordeum (Barley). In each tooth three 1-flowered spikelets. H. hexastichum (6-rowed Barley), has 6 rows of fruits, since all the spikelets are fertile, and H. distichum (2-rowed Barley) 2 rows, since the lateral spikelets are (♂, and barren (p. 292).—Elymus has 2–6 many-flowered spikelets in each joint of the main axis. Ægilops has awns upon the glumes.
Distribution. 315 genera with 3,500 species. The order is distributed over the whole world, and as regards number of individuals is perhaps the richest. In the Tropics, large, broad-leaved, tree-like forms are found (Bambuseæ, Olyreæ, Andropogoneæ, etc.; in S. Europe, Arundo donax); in England, next to the Compositæ, it is the order most rich in species (about 134).—The origin of some of the cultivated Grasses is lost in obscurity. The Maize, no doubt, was indigenous to America, where its nearest relatives are found, and where it has also been discovered in ancient Indian graves; Durra or Guinea-corn, Millet and Sugar-cane are South Asiatic plants, and our own cereals no doubt have sprung primarily from Western Asia and South-Eastern Europe (Barley from Armenia and Persia, where a very closely related wild species is found; Wheat from the same districts; Rye from the perennial species S. montanum). Panicum altissimum and Rice have come from Africa.
Uses. The Grasses play a very important part as cereals and fodder plants. The following are the most important of the cultivated ones: Triticum vulgare (common Wheat), turgidum, amyleum, polonicum, spelta, durum, etc.; Secale cereale (Rye); Barley (Hordeum-species, see under the genus); Maize; Oats (Avena sativa, orientalis, nuda); Millet (Panicum miliaceum); Durra (Turkish Millet, or Guinea-corn, Sorghum vulgare, cernuum and saccharatum); Manna-grass (Glyceria fluitans). As fodder-plants especially: Rye-grass (Lolium perenne); Oat-grass (Avena elatior); Timothy (Phleum pratense); Fox-tail (Alopecurus pratensis); Cock’s foot (Dactylis glomerata); Dog’s tail (Cynosurus cristatus); Sweet-vernal (Anthoxanthum odoratum); Soft grass, or Yorkshire-fog (Holcus lanatus and mollis); Quaking-grass (Briza media); species of Meadow-grass (Poa); Fescue (Festuca) and Brome (Bromus).—Several cultivated species of Grass are also used in the preparation of fermented liquors, the starch in the seeds being transformed to sugar (beer from “Malt,” i.e. the germinated Barley; arrack from Rice); or the stem becomes specially saccharine before flowering: the Sugar-cane, Sorghum saccharatum.
Officinal. The rhizome of Triticum repens, Oat-grain, flour of Barley, and the starch of Wheat, also sugar.
The seeds of Lolium temulentum are considered poisonous.—The stems of many species (including our common grains) are used in the manufacture of paper, especially “Esparto grass” (Stipa tenacissima) from Spain and N. Africa, and the sheathing-leaves of the ♀-spike of Maize. Sand Lyme-grass (Elymus arenarius), and especially Psamma arenaria, are important.—But few Grass-species are sweet-scented: Anthoxanthum odoratum and Hierochloa odorata contain coumarin; Andropogon-species have essential oils (“Citronella oil”).—Ornamental plants are: the “Ribbon-grass” (a variety of Digraphis arundinacea), Stipa pennata (whose awn is exceedingly long and feathery), Gynerium argenteum (Pampas-grass), Lagurus ovatus, Hordeum jubatum, Bromus briziformis.
[297]
The primitive form resembles that of the preceding family. In it we find the typical, perfectly developed, Monocotyledonous flower, sometimes even with free carpels and with a dry or somewhat fleshy, but never petaloid perianth; and this passes over into very different forms by the suppression of the floral-leaves, perianth and sporophylls (unisexual flowers are common), and by the close aggregation of the flowers in the inflorescence. The flower is hypogynous in every case. The inflorescence is a spike which may be either single or branched, and has often a thick and fleshy axis (a spadix). In Palms and Araceæ it is enveloped, at any rate prior to the opening of the flowers, by a very large floral-leaf, the spathe, which may be petaloid (Figs. 297, 301).
The fruit is most frequently fleshy (berry, drupe) or a nut, never a capsule. The embryo is small, with large, fleshy endosperm (Fig. 299 C); very rarely the endosperm is wanting.
Fig. 295.—Piassava (Attalea funifera).
Fig. 296.—A portion of the stem of Attalea funifera with persistent leaf-bases.
The numerous plants belonging to this family are large, herbaceous or tree-like, and the leaves seldom have the usual Monocotyledonous form, i.e. linear with parallel venation, but most frequently have pinnate or palmate venation.
Order 1. Palmæ (Palms). The majority are trees with an unbranched, cylindrical stem, having short internodes[298] and covered with leaf-scars or the bases of the leaf-stalks (Fig. 296), and at the summit a rosette of large leaves closely packed together (Fig. 295). An exception to this is found in Calamus (Cane, “Rotang”), whose thin, creeping or climbing stems have long internodes; sparsely[28] branched is, e.g. the African Doum-palm (Hyphæne). Notwithstanding their often enormous stems the Palms have fibrous roots, like the bulbous Monocotyledons. The leaves are pinnate (Feather-palms, Fig. 298) or palmate (Fan-palms, Fig. 295) and often very large; they have a well-developed petiole with an amplexicaul sheath, which is often more or less separated into a large number of fibres. In the bud the blade is entire but folded, as the leaf expands the lines of folding are torn, either those which are turned upwards (thus ∨ ∨ ∨ ∨, e.g. Pritchardia, Livistona, Phœnix, Chamærops) or those turned downwards (thus ∧ ∧ ∧ ∧, e.g. Cocos, Chamædorea, Calamus). The inflorescence is usually lateral; when, as in Sago-palm (Metroxylon rumphii) or Talipot (Corypha umbraculifera) it is terminal, the plant is monocarpic, and dies after flowering; it is often a very large and branched spadix with numerous flowers either borne externally or embedded in it, and enclosed either in one woody, boat-shaped spathe (Fig. 297) or several spathes, in the latter case one for each branch. The flowers are sessile or even embedded, regular, generally unisexual (monœcious or diœcious) with the usual diagram (Fig. 278); the perianth is inconspicuous, green or yellow, persistent, and more or less leathery or fleshy. 6, rarely 3 or many stamens. The 3 carpels remain either distinct or form one, generally 3-locular, ovary. The style is short. There is one ovule in each carpel. Often during ripening 2 carpels with their ovules are aborted. The fruit is a berry, drupe or nut, generally one-seeded, with a large horny or bony endosperm with hard thick-walled cells (e.g. Date-palm). In some (e.g. Cocoanut) it is thin-walled, soft, and oily; in several “ruminate.”
When germination commences in the Cocoanut, Date, etc., the apex of the cotyledon remains in the seed and developes into a spongy mass to withdraw the endosperm; in the Cocoanut it attains a considerable size (Fig. 299 C) and[299] assumes the form of the fruit. The endosperm in the Cocoanut is hollow and the interior is filled with “milk.” In the Date-palm and the Vegetable-ivory (Phytelephas) the cell-walls of the hard endosperm serve as reserve material.
1. Phœniceæ. Phœnix (Date-palm) has pinnate leaves with channeled leaflets and diœcious flowers with 8 free carpels, of which usually only one developes into a berry with membranous endocarp; the large seed has a deep furrow on the inner side, and horny endosperm.
Fig. 297.—Inflorescence of a Palm with spathe. At the top ♂-, at the base ♀-flowers.

Fig. 298.—Livistona australis.
2. Sabaleæ. These have fan-like leaves with channeled segments;[300] flowers ☿ or polygamous, rarely diœcious, with 3 separate or only slightly united carpels, all of which are sometimes developed into fruits (berry or drupe, with thin stone).—Chamærops, the Dwarf-palm. The pericarp is externally fleshy, internally more fibrous, and provided with a membranous inner layer. The endosperm is ruminate (that is, the testa is several times deeply folded into the endosperm).—Sabal, Copernicia, Livistona (Fig. 298), Thrinax, Corypha, Brahea, and others.
Fig. 299.—A Longitudinal section of a Cocoanut (diminished), the inner layer only (the stone) not being divided B End view of the stone, showing the sutures for the 3 carpels (a), and the 3 germ-pores; the embryo emerges from the lowest one when germination begins. C Germinating; inside the stone is seen the hollow endosperm and the enlarging cotyledon.
3. Cocoineæ. With pinnate leaves. Monœcious inflorescence. The carpels are united into a 3-locular ovary. The fruit is most frequently 1-locular, only 1 of the loculi becoming developed, rarely 3-locular; it is a drupe with a large, fibrous, external layer (mesocarp) and most frequently a very hard inner layer (endocarp, stone) which has 3 germ-pores, the 2 of these, however, which correspond to the suppressed loculi are closed; internal to the third lies the small embryo (Fig. 299). Endosperm containing[301] abundance of oil. Cocos (the Cocoanut-palm), Attalea, Elæis, Acrocomia, Bactris.
4. Lepidocaryinæ. The floral-leaves and flowers are borne in 2 rows on the spadix. The carpels are united into one 3-locular ovary; the fruit is coated by a layer of hard, shining, imbricate scales. The majority of the species are thorny, and climb by means of the thorny leaves. Some have fan-like (Mauritia), others pinnate leaves (Raphia, Calamus, Eugeissonia, Metroxylon; the stems of the latter die after the first flowering).
5. Borassinæ. Large Fan-palms without thorns, with 3-locular ovary. Drupe with separate stones. Latania and Lodoicea have many stamens; Hyphæne; Borassus (Palmyra-palm).
6. Arecineæ. The most numerous group. Feather-palms. Berry. Areca, Euterpe, Oreodoxa, Ceroxylon, Chamædorea, Geonoma, Caryota with bipinnate leaves.
7. Phytelephantinæ. Flowers with rudimentary perianth united in close capitula. Phytelephas (Vegetable-ivory). Nipa.
Distribution. About 1,100 species are known. In Europe only the Dwarf-palm (Chamærops humilis) is wild (Western Mediterranean). The Date palm (Phœnix dactylifera) belongs to North Africa and West Asia. Other African genera are Hyphæne (Doum-palm) and Elæis (E. guineensis, Oil-palm). A large majority of the genera are found in South America and in the East Indies. The following are American:—Mauritia, Acrocomia, Bactris, Chamædorea, Oreodoxa, Euterpe, Attalea, etc. Asiatic:—Metroxylon, Calamus, Areca, Borassus, Lodoicea (“Double-cocoanuts,” Seychelles) and others. The Cocoanut-palm has perhaps an American origin; all the other species of the same genus being endemic in America; it is the only Palm found on the coral islands of the Pacific Ocean, and is also the only one which is common to both hemispheres.
Uses. Palms belong to the most useful plants; they contain no poison, and are of little medicinal interest, but are largely employed in the arts and manufactures, the hard timber being adapted for many purposes on account of the hard tissue in which the vascular bundles are embedded. “Cane” is the stem of Calamus-species (from India). Sago is obtained from the pith of Metroxylon rumphii (Sago-palm, Sunda-Is., Moluccas), Mauritia flexuosa, etc. Sugar-containing sap (“palm wine”) is obtained from the American Mauritia vinifera and flexuosa, Borassus flabelliformis (Asiatic Palmyra-palm), Arenga saccharifera, etc., by cutting off the young inflorescences, or by perforating the stem before the flowering (arrack is distilled from this). Vascular strands for the manufacture of mats and brushes, etc., are obtained from the outer covering (mesocarp) of the Cocoanut, and from the detached leaf-sheaths of Attalea funifera (Brazil) (Fig. 296). Wax is yielded by the leaves of Copernicia cerifera (carnaueba-wax, Amazon region), and by the stem of Ceroxylon andicola (palm-wax, Andes); East Indian Dragon’s blood is from the fruit of Calamus draco; the young buds of many species, especially Euterpe, Cocos, Attalea, etc., are used as “cabbage.” Palm-oil is obtained from the oily mesocarp[302] of the plum-like fruits of Elæis guineensis (W. Africa), and from the seeds, when it is largely used in the manufacture of soap. Edible Fruits from the Date-palm (Phœnix dactylifera, Arabia, Egypt, W. Africa), and the endosperm of the Cocoanut (Cocos nucifera). The seeds and the unripe fruits of the Areca-palm (Areca catechu) are chewed with the leaves of the Betelpeper, principally in Asia. Vegetable Ivory from the hard endosperm of Phytelephas macrocarpa (S. America.)—Many species are cultivated in the tropics as ornamental plants, but in this country only Chamærops humilis, Livistona australis and chinensis are generally grown. In addition to the few just mentioned, many others are of importance, but these are much the most useful.
Order 2. Cyclanthaceæ. This is a small order related to the Palms (44 species from Tropical America), with fan-like, folded leaves. The flowers are unisexual and arranged in whorls or close spirals on an unbranched spadix. Ovary unilocular, ovules numerous. To this belongs Carludovica palmata, whose leaves are used for Panama hats.
Order 3. Pandanaceæ (Screw-pines) is another small order, forming a transition to the Araceæ. The woody, (apparently) dichotomous stem is supported by a large number of aerial roots, which sometimes entirely support it when the lower portion of the stem has decayed. The leaves are closely crowded together, and arranged on the branches in three rows, which are often obliquely displaced, with the formation of three spiral lines; they are, as in the Bromeliaceæ, amplexicaul, long, linear, the edge and lower midrib often provided with thorns. The ♂-flowers are borne in branched, the ♀ in unbranched spadices or capitula, which resemble those of Sparganium, but have no floral-leaves. Perianth absent. The drupes or berries unite into multiple fruits.—About 80 species in the islands of the Indian Ocean.—Pandanus, Freycinetia.—Fossils perhaps in the chalk of the Harz.
Order 4. Typhaceæ. The flowers are unisexual, monœcious, and borne on a cylindrical spike or globose capitulum; ♂ inflorescences above, the ♀ below. The perianth consists of a definite number of scales (Sparganium), or in its place numerous irregularly-arranged hairs are found (Typha); in the ♂-flower there are generally three stamens; the gynœceum is formed of 1–2 carpels with 1 prolonged style; 1 pendulous ovule. The seeds are furnished with a seed-cover, which is cast off on germination.—The few species (about 20) which belong to this order are marsh plants with creeping rhizome (and hence grow in clusters); the leaves on the aerial shoots are borne in two rows, entire, very long and linear.
Sparganium (Bur-reed). The flowers are borne in globose capitula; the perianth distinct, generally consisting of 3 small scales; pistil bicarpellate. Drupe, dry and woody. The stalk of the lower ♀ capitula is sometimes united with the main axis, and consequently the capitula are situated high above their subtending-leaf.
[303]
Typha (Bulrush, Reed-mace) has a long, cylindrical, brown spike, the lower portion bearing ♀-flowers, and the upper ♂-flowers, which is divided into joints by alternate leaves. The ♀-flowers have 1 carpel. The perianth is wanting, represented by a number of fine, irregularly-placed hairs; pistil unicarpellate. Fruit a nut.
The two genera, according to some, are related to the 2nd order. In both genera native species are found. The pollination is effected by the wind, and consequently the anthers project considerably, and the stigma is large and hairy. Typha is protandrous, Sparganium protogynous. The small, fine hairs surrounding the nut of Typha assist in its distribution by the wind.—Fossil Typhas in the Tertiary.
Order 5. Araceæ (Arums). The flowers are small, and always borne without bracts or bracteoles on an unbranched, often very fleshy spike, which is enclosed by a spathe, often petaloid and coloured (Fig. 301). The fruit is a berry. Outer integument of the seed fleshy.—The leaves have generally sheath, stalk, and blade with distinctly reticulate venation; they are chiefly cordate or sagittate (Fig. 302), seldom long with parallel venation as in the other Monocotyledons (Acorus, Fig. 300). The Araceæ are quite glabrous, generally perennial herbs with tubers or rhizomes. Many have latex.—For the rest the structure of these plants varies; for example, while some have a perianth, in others it is wanting; in some the perianth-leaves are free, in others united; some have hermaphrodite flowers, but the majority unisexual (monœcious); some have free, others united stamens; the ovules are orthotropous, anatropous, or campylotropous, erect or pendulous; the ovary is 1–many-locular; some have seeds with endosperm, others without. In habit there are great differences. While some, e.g. Colocasia (Fig. 302), have a thick, more or less upright stem, with leaf-scars, but not woody, others are climbers, epiphytic, and maintain themselves firmly by means of adventitious roots, on the stems and branches of trees, or even on steep rocks, e.g. Philodendron; the cordate, penninerved leaf is the most common (Fig. 302), but various branched forms appear; the pedate leaves of Helicophyllum, Dracunculus, etc., are cymosely branched; the leaves of Monstera deliciosa, perforated by tearing, should be noticed (the vascular bundles while in the bud grow faster than the tissue between them, causing the latter to be torn, and the leaf perforated). With regard to the anatomical structure, the presence or absence of latex, raphides, resin-passages, groups of mucilage-cells should be noted. Engler makes use of these anatomical peculiarities for a scientific arrangement of the order.
A. Orontieæ, Calamus-group. ☿, hypogynous flowers of a completely formed monocotyledonous type (number in the whorls 2, 3, or 4).—Acorus (A. calamus, Sweet-flag) has a regular,[304] 3-merous, pentacyclic flower (Fig. 300 C, D). They are marsh-plants, with creeping rhizome, triangular stem, and long, sword-like leaves (Fig. 300 A); the inflorescence is terminal, apparently lateral, being pushed to one side by the upright, sword-like spathe (Fig. 300 B).—Anthurium (Pr2+2, A2+2, G2); Pothos; Orontium (unilocular ovary with one ovule), etc.
Fig. 300.—Acorus calamus: A habit (much reduced); B inflorescence; C a flower; D diagram; E longitudinal section of an ovary; F an ovule.
Fig. 301.—Arum maculatum. The spathe (h) in B is longitudinally divided.
[305]
B. Calleæ. Flowers hypogynous, naked, ☿.—Calla (C. palustris). All flowers in the spike are fertile, or the upper ones are ♂; 6–9 stamens; ovary unilocular with many basal ovules. Marsh-plants with creeping rhizome and cordate leaves.—Monstera, Rhaphidophora, etc.

Fig. 302.—Colocasia Boryi.
C. Arineæ. Flowers monœcious, naked, ♂-flowers on the upper, ♀ on the lower part of the spadix.—Arum (Fig. 301). The spadix terminates in a naked, club-like portion (k); below this is a number of sessile bodies (rudimentary flowers), with broad bases and prolonged, pointed tips (b); underneath these are the ♂-flowers (m), each consisting only of 3–4 short stamens, which eject vermiform pollen-masses through the terminal pores; then follow, last of all, ♀-flowers (f), each of which consists of one unilocular ovary, with several ovules. Perennial herbs, tuberous, with cordate leaves.—Dracunculus; Biarum; Arisarum; Pinellia (Atherurus) ternata with leaves bearing 1–2 buds. Zantedeschia æthiopica (Richardia, Nile-lily); ♂, 2–3 stamens; ♀ with 3 staminodes, 1–5-locular ovary (S. Africa.)—In some genera sterile flowers are present between the ♂ and ♀ portions of the spadix (e.g. in Philodendron); in Ambrosinia a lateral, wing-like broadening of the axis of the spadix divides the cavity of the spathe into two chambers, the anterior[306] containing one ♀, and the posterior 8–10 ♂-flowers in two series; in some the stamens in the single ♂-flowers unite and form a columnar “synandrium” (e.g. in Dieffenbachia, Colocasia, Alocasia, Caladium, Taccarum, Syngonium). A remarkable spadix is found in Spathicarpa; it is united for its entire length, on one side, with the spathe, and the flowers are arranged upon it in rows, the ♀ to the outside, and the ♂ in the middle (Zostera has a similar one).—Pistia similarly deviates considerably, it is a floating water-plant, with hairy, round rosettes of leaves; in it also the spathe and spadix are united; at the base a ♀-flower is borne, which consists of one unilocular ovary, and above several ♂-flowers, each composed of two united stamens.
Biology. The inflorescences are adapted for insect-pollination; they are protogynous, since the viscous, almost sessile stigmas come to maturity and wither before the pollen, which is generally dehisced by apical pores, is shed; some pollinate themselves freely by the pollen from the higher ♂-flowers falling upon the ♀-flowers below them, and in some it is conjectured that the pollination is effected by snails. The coloured spathe, and the naked end of the spadix (often coloured) of certain genera function as the coloured perianth in other orders; during flowering a very powerful smell is often emitted. Arum maculatum is worthy of notice; small flies and midges creep down into the spathe, between the sterile flowers (Fig. 301 b), which are situated where the spathe is constricted, and pointing obliquely downwards prevent the escape of the insects; in the meantime, the stigmas are in a condition to receive any pollen they may have brought with them; after pollination the stigmas wither, and exude small drops of honey as a compensation to the flies for their imprisonment; after this the anthers (m) open and shed their pollen, the sterile flowers wither, and the insects are then able to escape, and enter and pollinate other inflorescences.—In many, a rise of temperature and evolution of carbonic acid takes place during flowering; a spadix may be raised as much as 30°C. above the temperature of the surrounding air.—Again, under certain conditions, many species absorb such large quantities of water by their roots that water is forced out in drops from the tip of the leaf; this may often be observed in Zantedeschia.
About 900 species in 100 genera. Home, the Tropics, especially S. America, India, and the Indian Islands, preferably in shady, damp forests growing as epiphytes upon trees, and on the banks of streams. Outside the Tropics few are found. Acorus calamus was introduced into Europe from Asia about 300 years ago; it, however, never sets any fruit, as the pollen is unfertile. In England Arum maculatum is a very common plant; this and A. italicum are the only native species. Colocasia antiquorum comes from Polynesia and the Indian Islands, and also Alocasia macrorrhiza. Fossils in Cretaceous and Tertiary.
Uses. Many species have pungent, and even poisonous properties (e.g. Dieffenbachia, Lagenandra, Arum), which are easily removed by boiling or roasting; the rhizomes of many species of Caladium, Colocasia (C. antiquorum, esculenta, etc.), are very rich in starch, and in the Tropics form an important source of food. An uncommon occurrence in the order is the highly aromatic rhizome of Acorus calamus; this contains calamus-oil and acorin which are used in perfumery. Many are ornamental plants, e.g. Zantedeschia æthiopica (South[307] Africa), generally known as “Calla,” and Monstera deliciosa; many other species are grown in greenhouses.
Order 6. Lemnaceæ (Duck-weeds). These are the most reduced form of the Spadicifloræ. They are very small, free-swimming water-plants. The vegetative system resembles a small, leaf-like body (Fig. 303 f-f), from which roots hang downwards; this branches by producing a new, similar leaf-like body, which springs from a pocket-like hollow (indicated by a dotted line in the figure) on each side of the older one, at its base (or only on one side). The branching is thus dichasial or helicoid (Fig. 303 A, where f, f′, f″, f″′ indicate shoots of 1st, 2nd, 3rd, 4th generations respectively). The leaf-like bodies are, according to Hegelmaier, leaf-like stems, and thus Lemna has no other leaves than the spathe and the sporophylls; according to the investigations of Engler they are stems whose upper portion (above the “pocket”) is a leaf, which is not sharply separated from the underlying stem-portion. The inflorescence is a very much reduced Araceous-spadix, consisting in Lemna of 1 or 2 stamens of unequal length (1-stamened ♂-flowers), 1 unilocular carpel (♀-flower), and 1 thin spathe (B). [The same is found in Spirodela polyrrhiza, etc., whose daughter-shoots begin in addition with 1 basal-leaf. Wolffia arrhiza, etc., have no roots, no spathe, and only 1 ♂-flower in the inflorescence (Engler).]—On the germination of the seed a portion of the testa is thrown off as a lid, so that an exit is opened for the radicle.—19 species. In stagnant fresh water, both Temp. and Tropical.—In Europe the species are Lemna minor, trisulca, gibba; Spirodela polyrrhiza, and Wolffia arrhiza, the smallest Flowering-plant.

Fig. 303.—Lemna: A vegetative system; B portion of a plant with flowers; one stamen and tip of the carpel project; the remaining portions being indicated by the dotted line.
[308]
The flowers in this family are hypogynous and have in part the general monocotyledonous type with 5 trimerous whorls completely developed in a regular hermaphrodite flower, and in part the flowers so much reduced that the type is very difficult to trace. On the one hand the family is well developed and has capitate inflorescences (Eriocaulaceæ) and on the other hand it is distinctly reduced (Centrolepidaceceæ). This family has taken its name from the fact that the ovule is not, as in the Liliifloræ and nearly all other Monocotyledons, anatropous, but orthotropous, so that the embryo (βλάστη) becomes placed at the end of the seed opposite (ἐναντίος) to the hilum. Large, mealy endosperm.—The orders belonging to this family are by certain authors grouped with the Bromeliaceæ and Pontederiaceæ, etc., into one family, Farinoseæ, so named on account of the mealy endosperm, the distinguishing character of the Liliifloræ then being that the endosperm is fleshy and horny.
Order 1. Commelinaceæ. The complete Liliaceous structure without great reductions in the number of whorls, but with generally few ovules in each loculus of the ovary, is found in the Commelinaceæ, an almost exclusively tropical order with about 317 species; herbs, some of which are introduced into our gardens and greenhouses. The stems are nodose; the leaves often clasping; the flowers are arranged in unipared scorpioid cymes, often so that they form a zig-zag series falling in the median line of the bracts, and after flowering they bend regularly to the right or left, outwards or inwards. They are more or less zygomorphic, particularly in the stamens, which in the same flower are of different forms or partially suppressed. The outer series of the perianth is sepaloid, the inner petaloid, generally violet or blue; the filaments are sometimes clothed with hairs formed of rows of bead-like cells (well known for showing protoplasmic movements). Fruit a trilocular capsule with loculicidal dehiscence (generally few-seeded); in some a nut. The radicle is covered by an external, warty, projecting covering which is cast off on germination.—The abundant raphides lie in elongated cells whose transverse walls they perforate.—Commelina, Tradescantia, Tinantia, Cyanotis, Dichorisandra.
Order 2. Mayacaceæ. This order is closely allied to the Commelinaceæ. 7 species. American marsh- or water-plants.
In many of the following orders of this family the flowers are united into compound inflorescences, with which is accompanied a reduction in the flower.
Order 3. Xyridaceæ (50 species). Marsh-plants with radical, often equitant leaves arranged in 2 rows, and short spikes on long (twisted) stalks. The flowers, as in the Commelinaceæ, have sepals (which however are more chaffy) and petals, but the outer series of stamens is wanting. Capsule (generally many-seeded).
Order 4. Rapateaceæ. Marsh-plants with radical leaves, usually in two[309] rows, and several spikelets on the summit of the main axis, clustered into a capitulum or unilateral spike. Each spikelet has numerous imbricate floral-leaves and one flower. 24 species. South America.
Order 5. Eriocaulaceæ. The “Compositæ among Monocotyledons,” a tropical order. The flowers are borne in a capitulum surrounded by an involucre, very similar to that of the Compositæ. The flowers are very small, unisexual, ♂ and ♀ often mixed indiscriminately in the same capitulum; they have the usual pentacyclic structure; the leaves of the inner perianth are often connate and more membranous than the outer; in some the outer series of stamens are suppressed; in each of the 3 loculi is one pendulous ovule. Capsule. The leaves are generally radical and grass-like.—335 species; Eriocaulon, Paepalanthus, etc., E. septangulare on the west coast of Scotland, and Ireland, and in North America.
Order 6. Restiaceæ. A small, especially S. African and S. Australian, xerophilous order (about 235 species), which is quite similar in habit to the Juncaceæ and Cyperaceæ. The leaves are often reduced to sheaths. The flowers are diœcious, the perianth as in Juncus, but the outer series of stamens suppressed. The ovary and fruit as in Eriocaulaceæ; the ovary, however, may be unilocular, and the fruit a nut. Restio, etc.
Order 7. Centrolepidaceæ. These are the most reduced plants in the family; small grass- or rush-like herbs. The flowers are very small, naked. Stamens 1–2, carpels 1–∞. 32 species. Australia.—Centrolepis (flowers generally ☿ with 1 stamen and 2–∞ carpels).
The flower is constructed on the general monocotyledonous type, with 5 alternating, 3-merous whorls (Fig. 278), but exceptions are found as in the Iridaceæ (Fig. 279) by the suppression of the inner whorl of stamens; in a few the position in relation to the bract differs from that represented in Fig. 278, and in some instead of the trimerous, di- or tetramerous flowers are found (e.g. Majanthemum, Paris). Flowers generally regular, hermaphrodite, with simple, petaloid, coloured perianth (except, for example, Bromeliaceæ); ovary trilocular, generally with 2 ovules or 2 rows of ovules in the inner angle of each loculus (Fig. 304 C, D). Endosperm always present.—A very natural family, of which some divisions in part overlap each other. The habit varies; the leaves are however long, entire, with parallel venation, except in Dioscoreaceæ (Fig. 313).
In the first orders of this family the flowers are hypogynous, and in the first of all the styles are free, and the capsule dehisces septicidally; in the following the flowers are epigynous and in some reduced in number or unisexual; capsule with loculicidal dehiscence, or a berry.
Hypogynous flowers: Colchicaceæ, Liliaceæ, Convallariaceæ, Bromeliaceæ (in part).
[310]
Epigynous flowers: Amaryllidaceæ, Iridaceæ, Bromeliaceæ (in part), Dioscoreaceæ.
Order 1. Colchicaceæ. The flower (Fig. 304 A) is ☿, regular, hypogynous, trimerous in all five whorls (6 stamens); anthers usually extrorse. Gynœceum with 3 free styles (A, D); fruit a capsule with septicidal dehiscence (E); embryo very small (F). The underground stem is generally a corm or rhizome, seldom a bulb.
A. Veratreæ.—Veratrum; perennial herbs, stem tall with long internodes and broad, folded leaves; the flowers andromonœcious, with free, widely opening perianth-leaves (Fig. 304 A), and globular anthers; inflorescence a panicle.—Zygadenus, Melanthium, Schœnocaulon, Uvularia, Tricyrtis.
B. Tofieldieæ.—Narthecium and Tofieldia have leaves alternate (arranged in two rows), sword-like and borne in rosettes; racemes or spikes. Narthecium forms an exception to the order by having a simple style and fruit with loculicidal dehiscence; Tofieldia by the introrse anthers. In this they are related to the Liliaceæ. Narthecium has poisonous properties, like many other Colchicaceæ.
Fig. 304.—Veratrum: A flower; B stamen; C transverse section of ovary; D gynœceum, with one carpel bisected longitudinally, and the third removed; E fruit after dehiscence; F longitudinal section of a seed.
C. Colchiceæ.—Colchicum (Autumn Crocus); perennial herbs, with a long, funnel-shaped, gamophyllous perianth, and introrse anthers. The flowers of C. autumnale spring up immediately from the underground stem, which is in reality a corm formed of one internode. Colchicum autumnale flowers in autumn without leaves; in spring the radical foliage-leaves appear simultaneously with the fruit. The flower is protogynous, and is pollinated by insects (humble-bees, etc.) which seek the honey secreted by the free part of the stamen a little way down the tube. The length of the tube protects the fruit, and not, as in other cases, the nectary.—Bulbocodium and Merendera have unguiculate perianth-leaves, free, but closing together like a tube.
175 species; chiefly in North America and South Africa. Tofieldia is an[311] Arctic plant. The order is rich in pungent, poisonous alkaloids (veratrin, colchicin, etc.). Officinal; the seeds of Colchicum autumnale (Europe) and Schœnocaulon officinale (Mexico), and the rhizome of Veratrum album (mountains of Central Europe).
Fig. 305.—Colchicum autumnale. A Corm seen from the front: k corm; s′ s″ scale-leaves embracing the flower-stalk; wh base of flower-stalk with roots (w). B Longitudinal section of corm and flower-stalk: hh brown membrane surrounding the underground portion of the plant; st flower-and leaf-stalk of previous year, the swollen basal portion forming the reservoir of reserve material. The new plant is a lateral shoot from the base of the corm (k) and has the following parts: the base bearing the roots (w), the central part (k’) which becomes the corm in the next year, the axis bearing the scale-leaves (s’, s″), the foliage-leaves (l, l′″), and the flowers (b, b’) which are borne in the axils of the uppermost foliage-leaves.
Order 2. Liliaceæ (Lilies). Flowers as in the Colchicaceæ but with introrse anthers; ovary free, 3-locular, with single style; capsule 3-locular with loculicidal dehiscence.—The majority are[312] herbs with bulbs; the inflorescence is terminal. In many species reproduction takes place by means of bulbils (small bulbs) formed in the axils of the foliage-leaves (e.g. Lilium bulbiferum, lancifolium, etc., Gagea lancifolia, etc.), or in the bracts of the inflorescence (many species of Allium); in many species several buds are developed as bulbs in the axils of the bulb-scales themselves (accessory buds arising close together), and in some the formation of buds is common on the leaves.
A. Tulipeæ, Tulip Group. Bulbs. The aerial, elongated stem bears the foliage-leaves. Flowers few but generally large, with free perianth-leaves. Tulipa; style absent, no honey; flowers generally solitary, erect.—Fritillaria perianth campanulate with a round or oblong nectary at the base of each perianth-leaf.—Lilium; perianth widely open, generally turned back with a covered nectary-groove in the centre of each segment. Anthers versatile.—Lloydia; Erythronium.
B. Hyacintheæ, Hyacinth Group. Bulbs. Leaves radical; aerial stem leafless with raceme or spike. In some the perianth-segments are free, in others united. Honey is produced often in glands or in the septa of the ovary (septal glands).—Ornithogalum has a leafy stem; Scilla; Eucomis has a tuft of floral-leaves above the raceme; Agraphis; Hyacinthus; Puschkinia; Chionodoxa; Muscari; Veltheimia; Urginea.
C. Allieæ, Onion Group. Generally bulbs. Leaves radical. Stem leafless with a compound umbellate or capitate inflorescence of unipared helicoid cymes, which before flowering are surrounded by two broad involucral leaves.—Allium. Filaments often petaloid and bidentate; in many species bulbils are found in the inflorescence.—Some species have flat leaves: A. sativum, Garlic; A. porrum, Leek; A. ursinum; others have round, hollow leaves: A. cepa, Onion; A. fistulosum, Winter Onion; A. ascalonicum, Eschalot; A. schænoprasum, Chive.—Gagea; honey is secreted at the base of the perianth, no special nectary; inflorescence few-flowered.—Agapanthus; Triteleia.
D. Anthericeæ. Rhizome; raceme; the leaves not fleshy and thick.—Anthericum; Asphodelus; Bulbine; Chlorophytum; Bowiea has an almost leafless stem with curved, climbing branches.
E. Aloineæ, Aloes. Stem generally aerial and tree-like, bearing on its summit thick, fleshy leaves, often with a thorny edge (Fig. 306). Raceme branched or unbranched.—Aloë; Gasteria; Yucca (has secondary thickening, p. 274).
F. Hemerocallideæ. Phormium, (Ph. tenax. New Zealand Flax); Funckia (Hosta); Hemerocallis.
At this point the following are best placed: Aphyllanthes (A. monspeliensis); Xanthorrhæa (Black-boy); Xerotes; Lomandra; Kingia; the very membranous, dry perianth of the last resembles that of the Juncaceæ, and also there are only 1–few ovules in the loculi.
[313]
Pollination by insects. Honey in some is produced on the perianth (see Tulipeæ), in others by glands on the carpels (in the septa and parietal placentæ, septal glands): Hyacinthus, Allium, Anthericum, Asphodelus, Yucca, Funckia, Hemerocallis, etc. Some Allium-species are protandrous. Fritillaria is visited by bees, Lilium martagon by moths, L. bulbiferum by butterflies, Phormium (New Zealand) by honey-birds.

Fig. 306.—Aloë.
About 1,580 species; rare in cold climates; their home is in sunny plains with firm, hard soil, and warm or mild climate, particularly in the Old World (S. Africa; As. Steppes; Mediterranean); at the commencement of spring the flowers appear in great profusion, and after the course of a few weeks disappear; during the hot season their life lies dormant in the bulb, hidden underground.[314] The woody species are tropical.—The majority of the introduced Liliaceæ (Fritillaria imperialis, Crown-imperial; Lilium candidum; Tulipa gesneriana; Hyacinth; Muscari-species; Scilla-species; Ornithogalum nutans; Hemerocallis fulva and flava; Asphodelus luteus and albus) come from the Mediterranean and W. Asia; Funckia from China and Japan; several Lilies from Japan and the Himalayas; Agapanthus from the Cape; Allium sativum is a native of the Kerghis-Steppes; A. cepa from Persia (?); A. ascalonicum is not known wild (according to others a native of Asia Minor), perhaps a form of A. cepa; A. schænoprasum from the N. temp. region.
Many bulbs have pungent properties; many Onions are used as culinary plants. The bast fibres of Phormium tenax (New Zealand Flax) are used technically. Dyes are obtained from the Aloe; gum for varnish from the stem of Xanthorrhæa hostile and australe. Officinal; “Aloes,” the dried sap of S. African species of Aloe (A. Africana, A. ferox, etc.); the bulb known as “Squills” from Urginea (Scilla) maritima (Mediterranean).
Order 3. Convallariaceæ. This order differs from the Liliaceæ in having the fruit a berry (Fig. 308) and in never being bulbous; the seeds are less numerous.
A. Convallarieæ, Lily of the Valley Group. Rhizome (Fig. 307) and normal foliage-leaves.—Polygonatum: rhizome creeping; aerial shoot leafy, bearing the flowers in racemes in the axils of the foliage-leaves; perianth tubular. P. multiflorum (Solomon’s seal), P. officinale, etc.—Majanthemum: flower 2-merous; perianth almost polyphyllous, spreading. Smilacina. Streptopus (S. amplexifolius; the flowers or inflorescence unite with the entire succeeding internode).—Convallaria (1 species C. majalis, Lily of the valley); flowers in terminal racemes; 2 basal foliage-leaves; perianth globose, bell-shaped. Reineckea carnea (Japan, China) in gardens.—Paris (P. quadrifolia, Herb-Paris); flowers solitary, terminal, 4-merous, polyphyllous; styles 4, free (approaching the Colchicaceæ; it is also poisonous); a whorl of 4 (-more) 3-nerved, reticulate leaves on each shoot.—Ornamental plants: species of Trillium, Aspidistra elatior (Japan).
Fig. 307.—Rhizome of Polygonatum multiflorum: a bud; b shoot; c d scars left by shoots of previous years.
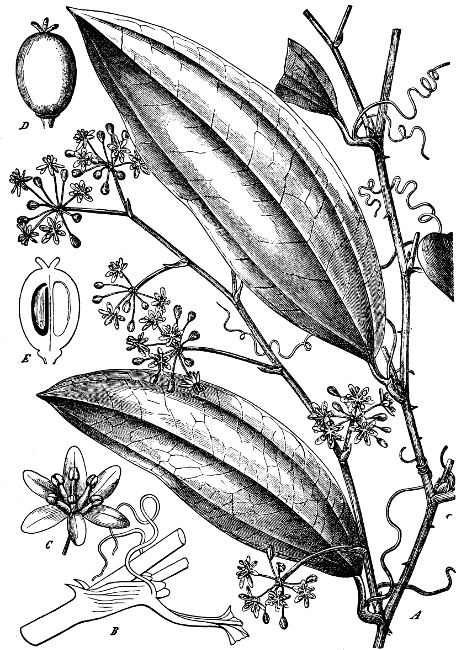
Fig. 308.—Smilax pseudosyphilitica: A shoot of male plant; C ♂-flower; D berry, almost ripe; E the same in longitudinal section. B Smilax syphilitica: portion of branch with base of leaf and tendrils.
B. Asparageæ, Asparagus Group. Scale-like leaves and green assimilating branches.—Asparagus: horizontal rhizome. The aerial shoots are very richly branched; the numerous needle-like bodies upon the plant are leafless shoots, which are crowded together in double scorpioid cymes in the axils of the scale-leaves; the two first[315] lateral axes, placed outside to the left and right, generally bear[316] flowers. Polygamous.—Ruscus (Butcher’s broom) is a S. European shrub with leaf-like, ovoid or elliptical shoots (phylloclades) which are borne in the axils of scale-like leaves, and bear flowers on the central line. Diœcious. Stamens 3, united, anthers extrorse. Semele androgyna bears its flowers on the edge of the flat shoot.
C. Smilaceæ. Smilax (Sarsaparilla) (Fig. 308); climbing shrubs with the leaf-sheath produced into tendrils. The leaves have 3–5 strong nerves proceeding from the base, and are reticulate. Orthotropous or semi-anatropous ovules. Diœcious (Fig. 308 C, E).
D. Dracæneæ. Fruit in some a berry, in others a capsule. The stem of Dracæna, when old, has the appearance of being dichotomously branched; it has the power of increase in thickness, and may become enormously thick. The Dragon-tree of Teneriffe, measured by Humboldt, attained a circumference of 14 m. and a height of 22 m.; the leaves are large, linear or linear-lanceolate.—Cordyline (East Asia), various species in gardens and greenhouses (Yucca is closely allied). Astelia.
Pollination. Paris quadrifolia and Convallaria majalis have no honey, and are chiefly visited by pollen-collecting bees (in the absence of insect visits self-pollination takes place); Polygonatum multiflorum has honey secreted by septal glands and protected by the base of the tubular perianth; it is pollinated by humble-bees, etc. Asparagus officinalis has small, polygamous, greenish, honey-bearing flowers; the ♂-flower is almost twice as large as the ♀; both have rudiments of the opposite sex.
About 555 species; especially from N. America, Europe, and Central Asia.
Officinal: “Dragons’-blood,” a red resinous juice from the stem of Dracæna and the roots of some Central American species of Smilax. The tuberous stems of the Eastern Asiatic Smilax glabra are officinal. The flowers of Convallaria majalis have been lately used as a substitute for Digitalis. Pungent, poisonous properties are possessed by Paris. None of the species are used as food, except the young annual shoots of Asparagus officinalis, a shore-plant which is used as a vegetable.
Order 4. Pontederiaceæ. Flowers generally zygomorphic, hypogynous, ☿, with handsome, white or violet, petaloid perianth which forms a tube at its base. The stamens are inserted at different heights in the perianth-tube, and are reduced to three (in Heteranthera seldom to one). In some the ovary is trilocular with ∞ ovules (Eichhornia), in others reduced to one loculus with one ovule (Pontederia). Fruit a capsule or nut. Embryo as long as the abundant, mealy endosperm.—Tropical water-plants (22 species) with peculiar sympodial branching, nearly the same as in Zostera. Spikes without floral-leaves. Many intercellular spaces in the stem and leaf.—In greenhouses: Eichhornia azurea, E. crassipes (both from tropical and sub-tropical S. America); the latter has swollen petioles which serve as floats and enable it to float freely on the water, sending down its roots into the mud. Heteranthera reniformis, H. zosterifolia. Pontederia cordata.
Order 5. Amaryllidaceæ (Narcissi). The flower is epigynous, otherwise exactly the same as in the Liliaceæ (6 stamens).[317] The majority, like these, are also perennial herbs with bulbs and scapes. The fruit and the other characters as in the Liliaceæ. The external appearance is, however, very different.
A. Amarylleæ have bulbs and the leaves generally arranged in two rows; the flowers are borne singly or in umbel-like inflorescences on lateral scapes, while the main axis of the bulb is unlimited. Beneath the inflorescence is an involucre (Fig. 309).—Galanthus, Snowdrop, has a polyphyllous perianth without corona; the three inner perianth-leaves are emarginate and shorter than the outer; the anthers dehisce apically. Leucojum differs in having the perianth-leaves equal in length.—Amaryllis has a funnel-shaped perianth, entirely or nearly polyphyllous, but somewhat zygomorphic. Crinum; Hæmanthus; Clivia.—Narcissus has a tubular corona, a ligular structure arising from the perianth-tube exterior to the outer stamens. In Pancratium (Fig. 309) the corona is united with the filaments which appear to spring from its edge. Eucharis amazonica.
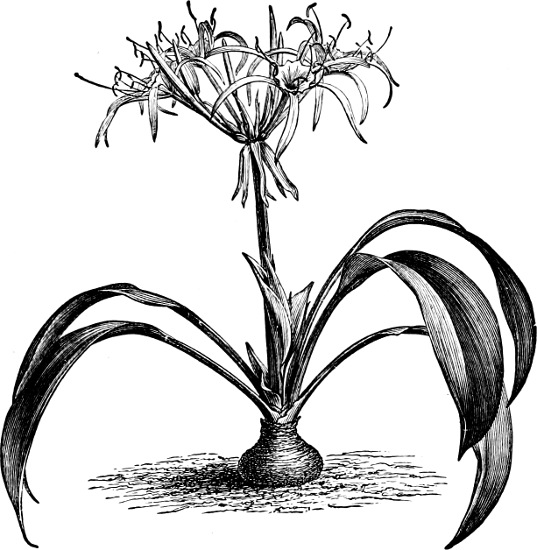
Fig. 309.—Pancratium caribæum.
B. Hypoxideæ. The leaves, which are grass-like, dry, folded, and in some hairy, spring from a rhizome, generally with a divergence of 1/3. Flowers small, perianth polyphyllous, persistent, on which account perhaps the Hypoxideæ may[318] be considered as the least altered type. The chief characteristic is that the embryo is separated from the hilum. Hypoxis; Curculigo (C. recurvata, a favourite ornamental plant; S.E. Asia).
C. Alstrœmerieæ. (Alstrœmeria, Bomarea); stems long, leafy, often climbing.
D. Vellosieæ (Vellosia, Barbacenia); stem woody, usually dichotomously branched, with terminal, single flowers; it bears numerous aerial roots which pierce the leaves and surround the stem. Stamens often (by splitting) 6–18. High table-lands of S. America and S. Africa.
E. Agaveæ. Very similar to the Bromeliaceæ both in their distribution (nearly all American) and in external appearance. They appear as gigantic bulbous plants with perennial, aerial, generally short stem, and perennial, large, lanceolate or linear, stiff, thick, and often thorny leaves, which form a large rosette; after the course of several (8–20) years the terminal inflorescence is developed, which is 10–12 m. high, paniculate, and freely branched. Before the inflorescence expands, a large quantity of sugar-containing sap is collected from A. americana by removing the terminal bud; this on distillation yields “pulque,” the national drink of Mexico. After flowering the entire shoot dies, but the subterranean lateral shoots survive and reproduce the plant.—Agave americana, etc.; Fourcroya; Polianthes tuberosa (Tuberose; Central America).
Distribution. The 650 species are chiefly natives of S. Africa and S. America. Clivia, Hæmanthus, Amaryllis are from the Cape; Narcissus from S. Europe, whence many species have been introduced; Galanthus and Leucojum are especially from S. and Central Europe, and from the Caucasus.
Uses, few, except as ornamental plants: Galanthus nivalis; Leucojum; Narcissus pseudonarcissus, N. poeticus, N. jonquilla, N. tazetta, etc.; Amaryllis, Alstrœmeria, Eucharis, Crinum, Vallota, etc. The vascular bundles of the various species of Agave (Agave rigida, var. sisalana, sisal hemp,) are used for cordage, etc.
Order 6. Bromeliaceæ. The flowers are hypogynous, epigynous or semi-epigynous; the perianth is divided into calyx and corolla; stamens 6. The fruit is a capsule or berry with many seeds. Endosperm mealy, embryo small, at the edge of the endosperm, but not enclosed by it.

Fig. 310.—Aechmea miniata.

Fig. 311.—Multiple-fruit of Ananassa sativa.
Perennial herbs with a very characteristic appearance (Fig. 310); the stem is most often short, thick, and crowned by a rosette of many leaves, which are long, often very narrow, leathery, stiff, and with a spiny edge; they are usually channeled, completely closing round each other, with their edges forming a tightly closed hollow, in which generally water is collected (this among other things insulates the inflorescence and thus prevents the access of creeping insects, such as ants). The presence of numerous stellate, water-containing hairs often gives the leaves a grey appearance, and the layers of cells beneath the upper epidermis of the lamina form an “aqueous tissue,” which serves as a protection against the rays of the sun and regulates the evaporation. The[319] stomata are often situated in furrows on the underside of the leaf, and hence cause a striped appearance. They are all American (525 species), especially from S. America, where they live partly as epiphytes on trees, partly in the clefts of rocks, often on the steepest slopes, to which they firmly attach themselves by aerial roots; some are terrestrial. The stem is seldom tree-like or many metres in height (Puya, in Chili; Hechtia, in Mexico). The inflorescence is a terminal spike, raceme, or panicle, often with large and brightly-coloured floral-leaves. The flowers are without scent.[320] The seeds, in the species whose fruit is a capsule, are often provided with wings (hairs, expansions, etc).—Ananassa sativa, Pine-apple (W. Indies, Central America) is cultivated for the sake of its juicy, aromatic fruits, which coalesce with their fleshy bracts and form a large spike-like fruit-cluster (multiple-fruits,[29] Fig. 311) bearing on its apex a leafy shoot, which may be used as a cutting. Seeds very rarely developed.—Tillandsia (T. usneoides is a filamentous, richly branched, rootless epiphyte hanging in masses from trees; Trop. Am.), Aechmea, Billbergia, Pitcairnia, etc.
Uses. The leaves of the Pine-apple, in its native country, are used for the manufacture of cloth.
Order 7. Hæmodoraceæ. 120 species; in all parts of the world except Europe; perennial, often tomentose and resembling the Bromeliaceæ, Iridaceæ and Amaryllidaceæ. Hæmodorum (Australia).—To this order belong Ophiopogon, Peliosanthes, Sanseviera, and others.
Order 8. The Iridaceæ have epigynous, hermaphrodite flowers with petaloid perianth as in the Amaryllidaceæ, but the interior whorl of stamens is entirely suppressed, and the 3 developed outer stamens have extrorse anthers (Fig. 279); there is 1 style with 3 large, generally more or less leaf-like branches bearing the stigmas. Ovary and capsule as in the Amaryllidaceæ and Liliaceæ.—Perennial herbs; bulbs are rarely found, but horizontal rhizomes, corms, etc., take their place. The leaves are (except Crocus) as in the Iris, two-rowed, equitant and sword-like. Flowers or inflorescences terminal.
The Iris (Flag) has a horizontal rhizome. The flowers are borne in the leaf-axils in fan-like inflorescences (rhipidium). The branches of the style are large and petaloid; on their under surface may be seen a small projecting shelf (Fig. 312 a) having on its upper surface the stigmatic hairs. Beneath the branches of the style are 3 well protected stamens, and immediately outside these the external perianth-leaves. The honey is secreted in the perianth-tube, and the insects, endeavouring to obtain it through the narrow passages at the base of the stamens, settle upon the outer perianth-leaves, which are bent backwards and often very hairy along their central line. The insects then rub their backs on the anthers just above them, beneath the branches of the style; they readily deposit the pollen on the stigma of another flower as they enter it, but cannot do so in withdrawing, since the stigma is pushed back, and self-fertilisation is thus avoided. The stylar branches lie close to the outer perianth-leaves, which are just beneath them, or separated by a distance of only 6–10 mm.; the first form of flower is adapted for Rhingia rostrata, the latter for bees.—Crocus has vertical, tuberous, underground stems[321] surrounded by the leaf-sheaths (corms), and terminal flowers; the linear leaves are not equitant, but have two longitudinal furrows on the under side. The perianth is gamophyllous and funnel-shaped. The stylar branches (stigmas) are fleshy, rolled together in the shape of a horn, and split along the edge.—Gladiolus has corms like the Crocus; spikes with slightly zygomorphic, almost bilabiate flowers, most frequently turning to one side. Position of the leaves as in the Iris.—Diplarrhena has 2 fertile and 1 barren stamen; Hermodactylus has a unilocular ovary with 3 parietal placentæ. Cypella and Tigridia have bulbs.
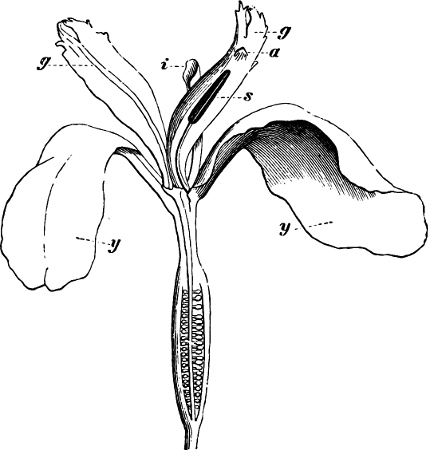
Fig. 312.—Iris pseudacorus. One external and two internal perianth-leaves, and one of the stylar-branches have been removed, y The outer, i the inner perianth-leaves; g stylar-branch; a stigma; s anther. The ovary is seen in longitudinal section.
770 species; chiefly in the countries round the Mediterranean, and in Africa, especially the Cape (Gladiolus, Ferraria, Moræa, Galaxia, Sparaxis, Antholyza, Tritonia, Ixia, etc.), Australia and Tropical America (Sisyrinchium, Tigridia, Cipura, Cypella, etc). A great number are ornamental plants: the cultivated Crocus-species are from the South of Europe and Asia; Gladiolus communis from S. Europe; the other species principally from S. Africa. The native species of Iris are I. pseudacorus (yellow) and I. fœtidissima.
Officinal: the stigmas of Crocus sativus (Oriental, cultivated in France, Spain, Italy, and Austria), used as a colouring matter, saffron; the rhizomes of the S. European Iris florentina, pallida, and germanica (“Orris-root”).
[322]
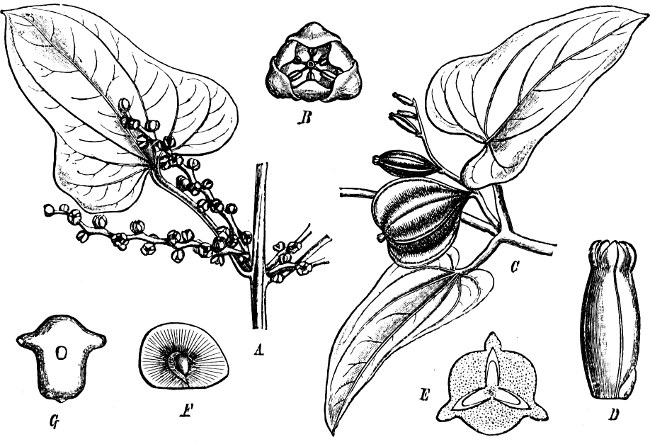
Fig. 313.—Dioscorea batatas: A ♂-plant; B ♂-flower; C ♀-plant (nat. size); D, E ♀-flowers (mag.); F seed; G embryo.
Order 9. Dioscoreaceæ. Perennial herbs with fleshy, often very large tuberous rhizomes (or roots); twining stems; leaves stalked, often arrow- or heart-shaped, lobed, palminerved and finely reticulate as in the Dicotyledons (Fig. 313). The flower is diclinous (most frequently diœcious), regular, epigynous, small, and of a greenish colour, but otherwise typical (Pr3 + 3, and A3 + 3, or G3); in most instances 2 ovules are placed one above the other in each loculus. The inflorescence is a spike or raceme, sometimes richly branched and paniculate.—The order approaches most nearly to the Amaryllidaceæ.
[323]
Tamus (Bryony) has a berry, Dioscorea (Yam) a thin-walled, 3-edged or 3-winged capsule (Fig. 313). Both have subterranean or aerial tubers; the Yam very often also developes tubers in the axils of the foliage-leaves; tuberous roots are said to occur in D. batatas. The tubers of many species of Yams (D. batatas from China and Japan, D. alata, South Sea Islands and India, D. bulbifera) are a very important source of food in the Tropics, especially the first-named.—Testudinaria; Rajania.—The tuberous stem of Tamus communis and Testudinaria elephantipes, and some species of Dioscorea is formed from one single internode (epicotyl), and the aerial shoots are developed from adventitious buds; in T. elephantipes the stem is aerial, and covered with thick scales of cork, regularly arranged, and separated by grooves.
Tropical order (167 species); 2 species (Tamus communis and Borderea pyrenaica) in Europe.
The flowers belong to the ordinary monocotyledonous type. They are hermaphrodite, epigynous, and have either a petaloid perianth, or calyx and corolla; they are, however, zygomorphic or unsymmetrical, and of the stamens most frequently only one is completely developed, the others being generally represented by petaloid staminodes. The ovary has 3 loculi, more rarely it is unilocular with the suppression of 2 loculi. Endosperm is absent (except Zingiberaceæ); but, on the other hand, there is a large perisperm. To this family belong large, glabrous, especially perennial herbs with rhizomes; leaves large, distinctly divided into sheath, stalk, and blade, the latter being more or less elliptical or lanceolate, entire, with pinnate venation, and always with a very well-pronounced midrib, gradually tapering towards the apex, and giving off numerous branches, which run outwards, towards the margin, at a larger or smaller angle; these lateral veins are closely packed, and parallel, but with only weak, connecting branches between them; the leaves, therefore, are easily torn pinnately (Figs. 314, 317). The leaf-sheaths close tightly round each other and form a false stem.
This very natural family comprises orders closely connected with each other, but is not itself nearly allied to any other family. First in the series stands:—
Order 1. Musaceæ. The petaloid perianth is strongly zygomorphic, the anterior leaf being very large (a kind of “labellum”), the posterior one small; only the posterior stamen is wanting, or is rudimentary, the other five are developed, and[324] have quadrilocular anthers; ovary, 3-locular. Seed with straight embryo in mealy perisperm.

Fig. 314.—Two Musa-species.
The best-known genus is Musa, the Banana (Fig. 314). From the short rhizome arise enormously large, spirally-placed leaves, whose sheaths envelope one another, and form an apparently aerial stem, several metres in height. The inflorescence is a terminal spike with floral-leaves placed spirally, and sometimes magnificently coloured; in the axils of each of these several flowers are situated in two transverse rows (accessory buds); the lowest flowers in the inflorescence are ♀, the central ones ☿, the upper[325] ones ♂, so that fruits are only found in the lower region of the inflorescence, the remaining portion persisting as a naked axis after the floral-leaves and flowers have fallen off; the inflorescence terminates in an ovoid bud formed by the flowers which have not yet opened (Fig. 314, the left-hand figure). The perianth-leaves are united (except the posterior one). The fruit (known as a “Banana”) is a berry, having the form of a smooth, short, three-cornered Cucumber (as much as 30 cm. in length); inside the tough skin is found a farinaceous, aromatic pulp. No seed is developed in the cultivated species.—Several Musa-species are cultivated in the Tropics for the sake of the fruit (M. paradisiaca, M. sapientum); for the fibrovascular bundles, M. textilis (Manilla Hemp).—Their home is, no doubt, the Tropics of the Old World; they were introduced into America before the arrival of Europeans. Musa ensete has dry, leathery fruits; an ornamental plant.
In Musa the barren, posterior stamen belongs to the inner whorl; and also in Strelitzia and Ravenala; the latter may have all 6 stamens developed. In Heliconia, on the contrary, it belongs to the outer whorl; in Heliconia the perianth-leaves are differently arranged, and there is only one ovule in each loculus. The three latter genera have dry fruits and leaves arranged in two rows. In the “Travellers’ Palm” (Ravenala madagascariensis) the foliage-leaves form an enormous fan.—Tropical; about 50 species.
The order may be divided as follows:—1. Museæ: Musa, Ravenala, Strelitzia in the Old World. 2. Heliconiæ: Heliconia in the New World.
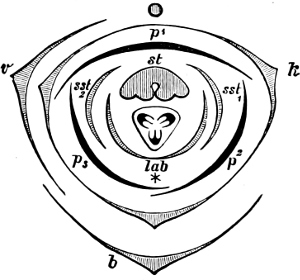
Fig. 315.—Diagram of a Zingiberaceous flower (Kæmpferia ovalifolia): b bract; v bracteole; k calyx; p1, p2, p3 the petals; sst, lateral staminodes (“wings”); lab labellum (formed of two staminodes); st the fertile stamen; * position of suppressed stamen. The ovary is in the centre of the diagram.]
Order 2. Zingiberaceæ. Perianth most frequently divided into calyx and corolla. Calyx gamosepalous. Only 1 fertile stamen (the posterior, Fig. 315, belonging to the inner whorl) with quadrilocular anther, which encloses the style in a furrow; the 2 stamens in the outer whorl are staminodes, the median one (the anterior) is wanting. The 2 lateral staminodes of the inner whorl form the “labellum” (Fig. 315 lab), which usually is the largest segment of the flower, and is often bilobed. Ovules many. The fruit in some is a leathery, 3-valved capsule,[326] with loculicidal dehiscence; in others it is more or less berry-like and indehiscent, or irregularly dehiscent. Straight embryo.—The aerial stem is seldom developed to any extent, and the inflorescences, which are (compound) spikes or racemes, often with coloured floral-leaves, spring in some (e.g. Zingiber officinale) directly from the rhizome. The leaves are arranged in two rows.—The ovary in a few instances (Globba and others) is unilocular, with 3 parietal placentæ.
They are perennial herbs with fleshy and tuberous rhizomes, which are used as condiments and in medicine on account of their pungent and aromatic properties and also for starch, dyes, etc. Officinal: rhizomes of Zingiber officinale (Ginger, unknown wild, but cultivated generally in the Tropics), of Curcuma longa (Turmeric, a dye, E. India) and C. zedoaria, of C. angustifolia and others (as E. India Arrowroot), of Alpinia officinarum, China (galangal). “Preserved Ginger” from Alpinia galanga. Similar aromatic materials (volatile oils) are present also, for example, in the fruits; Cardamom fruits and seeds (from Elettaria cardamomum, China, seldom from E. major).
315 species; Tropics, preponderating in the Eastern Hemisphere, India, and especially S. Asia, whence all the aromatic species originate; they are now commonly cultivated in the Tropics. Some are ornamental plants in greenhouses, e.g. Hedychium, Costus, etc. Globba (with axillary buds in the inflorescence, as in Ficaria), Renealmia, Kæmpferia.
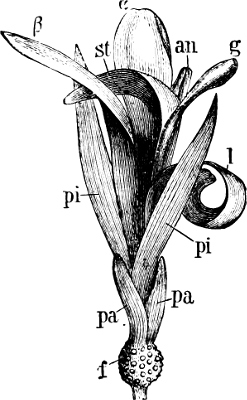
Fig. 316.—Flower of Canna: f ovary; pa calyx; pi corolla; l labellum; st stamens; an anther; g stigma; α and β staminodes.
Order 3. Cannaceæ. American herbs without aromatic properties. Flowers asymmetric (Fig. 316). Calyx polysepalous. The stamens are petaloid (Fig. 316 st) and barren with the exception of one (the posterior), which bears on one of its edges a bilocular anther; another, which is especially large and coloured, is termed the labellum. The style is compressed and leaf-like, with a small stigma at the apex. Ovules numerous in the 3 loculi. The capsule is furnished with warts or soft prickles. Embryo straight.
Canna (30 species; Trop. Am.). The inflorescence is a terminal spike with 2-flowered unipared scorpioid cymes in the axils of the floral-leaves. Ornamental plants: Canna indica, etc.
The diagram of the andrœcium of[327] the Cannaceæ and Marantaceæ may be represented in the following manner (calyx, corolla and gynœceum being omitted):—
| Cannaceæ. | Marantaceæ. |
| w w | w w |
| st | st |
| w lab | wi c |
| * | * |
w The lateral staminodes, “wings;” st fertile stamen; * the suppressed stamen; lab labellum; c hood; wi inner-wing.
The labellum of the Cannaceæ corresponds with the hood of the Marantaceæ and not with the labellum of the Zingiberaceæ.

Fig. 317.—Calathea zebrina.
Order 4. Marantaceæ. The flower is asymmetrical. Only 1 or 2 of the 3 stamens in the outer whorl are present as staminodes; in the inner whorl 2 are petaloid and of the sixth stamen one-half is developed as a staminode and the other half bears a bilocular anther. One ovule only in each loculus. The style is strongly curved and at first enclosed in one of the staminodes (hood) of the inner whorl; later on it springs elastically forward towards the other staminode (inner-wing) of the same whorl. The stigma is very oblique or 2-lipped. Two of the three loculi of the ovary, in some (Maranta, Thalia) become small and empty. Embryo curved. Leaves in two rows, with sheath, stalk, and blade (Fig. 317); at the base of the last is a characteristic swelling (articulus).—Phrynium, Calathea, Stromanthe, Ctenanthe, Saranthe, etc. About 150 species; tropical, especially America. The starch of the rhizome of Maranta arundinacea is OFFICINAL, “West Indian Arrowroot.”
[328]
The flowers are hermaphrodite and constructed on the ordinary 3-merous, pentacyclic type with petaloid, epigynous, strongly zygomorphic perianth, and generally one-stamened by the suppression of the other 5 stamens. The family has derived its name from the fact that the stamen is united with the style into a “stylar column” (except Burmanniaceæ). All are herbs; many grow as epiphytes on other plants.
This family and the Scitamineæ occupy correspondingly high positions among the Monocotyledons; these two families may therefore be placed close together, although one cannot be derived from the other. The first of the two orders is very small, but the second is very rich in species. The Apostasieæ are best classed with the Orchidaceæ and have no independent place.
Order 1. Burmanniaceæ. This order forms a transitional link between the Gynandræ and the epigynous Liliifloræ (Amaryllidaceæ), in having a 6-leaved perianth, and 6–8 stamens; but some have a labiate perianth (the median perianth-leaf of the outer whorl being very large). The ovary is most frequently unilocular with three parietal placentæ; but in some it is 3-locular with axile placentation. Capsule. Seeds ∞, small, with endosperm. The relationship to the Orchidaceæ is shown especially in the very imperfectly developed embryo and in the ovary. Small, tropical herbs (59 species); some are saprophytes.
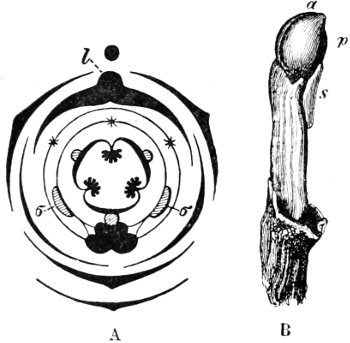
Fig. 318.—A Diagram of an Orchid-flower. B, Cephalanthera. Stylar-column: a anther; s stigma; at the foot are seen scars indicating the position of the parts which have been removed.
Order 2. Orchidaceæ. The epigynous, petaloid perianth is strongly zygomorphic in having the posterior leaf of the interior whorl, the lip (labellum), differing from all the other leaves in form, size, and colour (except Apostasieæ); the position of the labellum is very frequently reversed, being turned forwards and downwards by the twisting of the ovary (Fig. 318 A). Only 1 of the stamens—the anterior of the external whorl—is developed and bears an anther (by the twisting of the ovary it is[329] turned posteriorly and upwards); the others are entirely wanting (indicated by * in Fig. 318 A) or present as staminodes (Fig. 318 A, δ δ) (except Apostasieæ, Cypripedileæ); the filaments are united with the style to form a column (Fig. 318 B), the stylar-column[30] (gynostemium), and the anther (a) is thus placed on its apex and exactly behind or over the stigma (s). The anther is 4-locular; the pollen-grains do not separate (except Apostasieæ, Cypripedileæ) but remain united either in tetrads or in masses, which correspond to a pollen-mother-cell (Fig. 320 C, D, E); or the pollen-grains, formed in each of the two anther-halves, remain united and form one or a few wax-like masses (pollen-masses, pollinia). The 3 carpels form a unilocular ovary with 3 parietal, deeply bifid placentæ (except Apostasieæ, Selenipedilum). Only the two lateral carpels are prolonged and developed into the stigma (Fig. 318 B, s), while the one lying in the median line, which is situated just within the anther (Fig. 318 A), becomes either rudimentary or developed into the “rostellum” (“a small beak”), on which the sticky bodies (glandulæ) arise; by aid of these the heavy, connected pollen-masses may be glued to the insects which visit the flower, and pollination is thus secured (in Apostasieæ and Cypripedileæ the 3 carpels each contribute to the formation of the stigma). The fruit is a capsule which most often dehisces by 6 valves, 3 of which are broader and bear the placentæ, and 3 alternating with them are narrower and barren (except Vanilla). The very numerous and exceedingly small seeds have no endosperm, and have a somewhat spherical embryo without any trace of external organs. The testa is membranous and loose.
The Orchids are all perennial herbs with diverse habits and varying morphological structure (see the genera); the leaves are scattered, of the usual Liliaceous form, and the inflorescences in all cases are racemes or spikes (sometimes branched), with subtending bracts, but without bracteoles.
The forms which are the least modified are described first.
1. Apostasieæ. The perianth-leaves are almost alike and free. The column is straight, with 3 equally-developed stigmas. Neuwiedia has 3 perfect stamens (1 median of the outer whorl, and 2 lateral of the inner whorl); Apostasia has only 2 perfect (inner lateral) and one barren (the median of the outer whorl), which however[330] may be entirely wanting. The 3 posterior stamens are entirely suppressed. The pollen is powdery. The ovary is 3-locular with axile placenta. 7 species (Tropical East India, Australia).
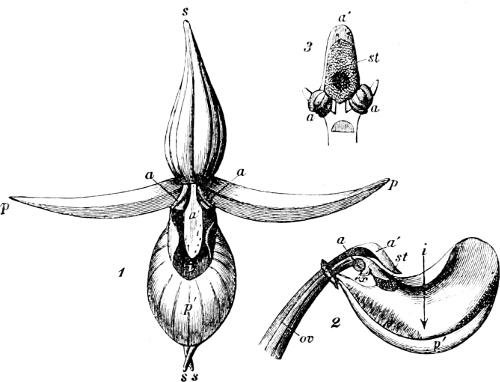
Fig. 319.—Cypripedilum calceolus: 1 front view of the flower; 2 lateral view, after the removal of all the perianth-leaves with the exception of the labellum, which has been divided longitudinally; 3 the stylar-column; ov ovary; s-s exterior, p interior perianth; p’ the labellum; a the two fertile stamens; a’ the staminode; st the stigma; i entrance for the insects; ex exit.
2. Cypripedileæ.[31] The flower is strongly zygomorphic with a large boat-shaped labellum. There are two perfect stamens belonging to the inner whorl, and the median anterior (later on the posterior) stamen of the outer whorl is transformed into a large, barren, shield-shaped body (Fig. 319). Selenipedilum has a 3-locular ovary, but Cypripedilum (Ladies’-slipper) has a unilocular ovary with 3 parietal placentæ—the typical structure for the Orchids. The pollen-grains are separate (not in tetrads) and all the 3 lobes of the stigma are constructed to receive them. This group is therefore, next to the Apostasieæ, the least modified among the Orchids; in all the following groups, one of the lobes of the stigma is differently developed from the others, and there is only one stamen.—Terrestial Orchids.—The pollination of C. calceolus is effected by the forcible entrance of insects into the boat-shaped labellum (Fig. 319 p’) at i, and their escape at ex (in 2) where the anthers are situated; in this way the stigmas will first be touched and then the anthers. The pollen-grains are surrounded by a sticky mass in order that they may adhere to the insects.
[331]
3. Neottieæ. The majority are terrestrial Orchids with creeping, sympodial rhizomes; the blades of the leaves are not detached from the stem at joints, and have convolute vernation. The anthers do not drop off, but persist in the withered condition; their apex is brought in contact with the rostellum (acrotonous Orchids). The pollen-grains are united in tetrads, which, however, often hang loosely together in pollinia, attached to a sticky part of the rostellum (“adhesive disc”), so that they adhere to the insects, and are by them transferred to the stigmas. Spiranthes. Listera; Neottia. N. nidus-avis (Bird’s-nest) is brown (it has little chlorophyll) in colour, has no foliage-leaves, and lives mainly as a saprophyte; the rhizome is studded with unbranched, fleshy roots which may form buds at their extremities.—Vanilla climbs by aerial roots. The fruit is fleshy and hardly opens, or does so irregularly.—Epipactis, Cephalanthera.—Epipogon and Limodorum are saprophytes without chlorophyll.
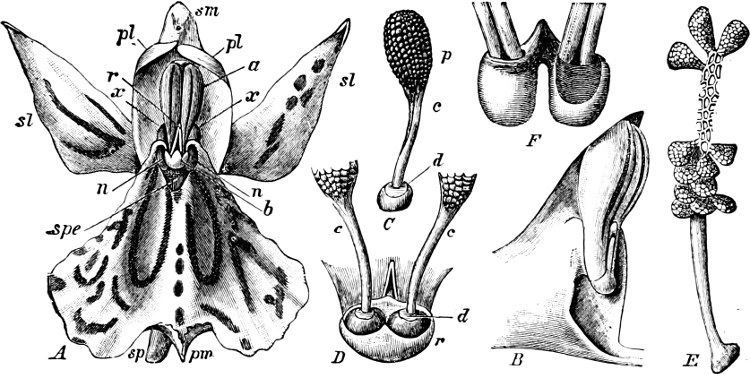
Fig. 320.—A Flower of Orchis maculata (front view): a stamen; b the cup; n the stigmas; x staminodes; sp the spur; spe the entrance to it; sm-sl-sl exterior perianth-leaves; pm the labellum, and pl-pl the other 2 interior perianth-leaves. B-E Orchis mascula: B lateral view of the column; C a pollinium with massulæ (p), caudicle (c) and adhesive disc (d); D caudicles with the cup (r), front view; the latter is depressed so that the adhesive disc is seen lying inside it; E a pollinium, more highly magnified; some massulæ are removed. F Ophrys aranifera: rostellum and the base of the anther-loculus; an adhesive disc is seen on the right.
4. Ophrydeæ. Anthers 2-locular, not falling off, on a very short column. The anther is united at its base with the rostellum (basitonous Orchids, Fig. 320 A, B), while in all other Orchids it is connected at the apex (acrotonous Orchids). The pollen-grains in each loculus are united into small “masses” (massulæ), each of[332] which corresponds to a pollen-mother-cell in the anther, and which hang together by elastic threads (Fig. 320 C, E). Each pollinium is attached at the base by a stalk (caudicle) to an adhesive disc, formed by the modified stigma (rostellum), and is easily liberated from it (Fig. 320 C, D, F). The pollinium, which is formed in an anther-loculus, together with its caudicle and adhesive disc, is termed “pollinarium” (Fig. 320 C).—All Ophrydeæ are terrestrial with tuberous roots, two of which are present in the flowering period, an older one (from the preceding year) containing the nourishment for the flowering-shoot of the year, and a young one which is intended to contain the reserve material for the following year. Inflorescence terminal.
Orchis. The lip has a spur; each of the club-like pollinia is attached to its own adhesive disc, the discs being enclosed in a common pouch formed by the rostellum (Fig. 320 C, D). Tubers ovate, undivided: O. morio, mascula; tubers palmate: O. incarnata, maculata, majalis.—Ophrys; no spur, the two adhesive discs are each enclosed in a separate pouch (Fig. 320 F).—Anacamptis and Serapias have one adhesive disc.—Habenaria, Gymnadenia, Platanthera, Herminium, Nigritella, Cœloglossum, etc., have naked adhesive discs (no rostellum).
5. Epidendreæ. Acrotonous Orchids with deciduous anthers (except Malaxis); 2-8 wax-like pollinia, with or without caudicles; generally no adhesive discs. Malaxis (the flower is twisted through a complete circle, causing the labellum to be turned upwards), Sturmia and Corallorhiza[32] (Coral-root); the latter has a creeping, coral-like rhizome without roots, and is destitute of chlorophyll except in the ovary. The other two somewhat resemble the tropical Orchids in having the lower internodes of the axis of the inflorescence tuberous. Liparis; Calypso. Most of the genera are tropical epiphytes and many have aerial, green tubers formed from one or more stem-internodes; Dendrobium, Eria, Phaius, Bletia, Epidendrum, Cattleya, Lælia, Pleurothallis, Restrepia, Masdevallia, Bulbophyllum, etc.
6. Vandeæ. These resemble the preceding but have only 2 wax-like pollinia in each anther, which are attached by a caudicle to the adhesive disc of the rostellum. Nearly all are tropical epiphytes. Stanhopea, Catasetum, Maxillaria, Oncidium, Vanda, Polystachya, etc.
6,000 (10,000?) species. The majority live in the Tropics and occur, especially, as epiphytes on trees or in the crevices of rocks, to which they are attached by aerial roots. These aerial roots, like those of Araceæ, are covered by several layers of spirally-thickened cells (tracheides) which contain air and form the velamen—an apparatus to absorb moisture from the air. The roots have a white appearance when the cells are filled with air, which changes to a greenish hue when they are filled with water, the chlorophyll then shining through. They generally have horizontal rhizomes; the[333] ascending shoots, which bear the foliage-leaves, may vary, but they very often swell and assume the form of a tuber, which persists for several years fresh and green after the leaves have fallen off (Fig. 321). Vanilla is an exception (see above). Our Orchids are all terrestrial (or marsh-plants); the largest number of species is found in calcareous soils.
Pollination takes place principally by means of insects, but self-pollination occurs in some. The lip serves as a landing-stage for the insect visitors, which, on sucking the honey, cause the adhesive discs, with the pollinia attached to them, to adhere to their bodies (generally to the probosces) and so carry them away to other flowers. In some species parts of the flower are sensitive or irritable, which has some connection with the pollination. Without doubt there are a great many biological differences which are closely connected with the infinite multiplicity of forms; Darwin (1862) has already shown an enormous variety, never even dreamt of before, in the European species. The genus Catasetum has ♂-♀-and ☿-plants with flowers of such different appearances that they have been classed in various genera (Myanthus, Monacanthus). Platanthera is pollinated by hawk-moths; Ophrys, by flies; Epipactis latifolia, by wasps; Orchis, by bees, especially humble-bees, etc.

Fig. 321.—Chysis bractescens.
The DISTRIBUTION OF SEEDS is effected by the wind, the seeds being so exceedingly small and light. Many species moreover have peculiar, elater-like, fine, hygroscopic hairs in the ovary, which eject the seeds in a manner similar to the elaters of the Liverworts.
The USES are few, mostly as ornamental plants in conservatories. The tubers of several Orchis-species are OFFICINAL; they contain starch and mucilage and are used us “salep.” The fruits of Vanilla planifolia are used as condiments and differ from other Orchid-fruits in being rather fleshy and in dehiscing irregularly; the seeds are very small, shining and black.
[334]
In this class THE EMBRYO has 2 seed-leaves, a rule from which there are few exceptions (e.g. Ficaria, Cyclamen, Pinguicula, certain species of Corydalis, with only 1; and a few, mostly parasitic forms, e.g. Monotropa, Orobanche, Pyrola, entirely without cotyledons). On germination the cotyledons nearly always raise themselves above the ground as green, assimilating leaves and are then termed aerial or epigean, in contradistinction to the underground or hypogean which are always buried. The structure of the seed varies (endospermous or exendospermous); the embryo may be straight or curved. In many instances the primary root grows as a vigorous tap-root, with weaker branches arising acropetally (in annuals, biennials, many perennials, especially woody plants); but in a large number of herbaceous perennials, which have rhizomes, the root behaves very much as in the Monocotyledons. The roots generally increase in thickness by means of a cambium.
The stem, when seen in transverse section, has its vascular bundles arranged in a ring; in reality, however, they form a kind of cylindrical network in the stem; the bundles are open, and thickening takes place by means of a cambium; annual rings are formed in the perennial stems. There is a rich and very varied form of branching. The two first leaves of a shoot (fore-leaves) are placed nearly always to the right and to the left; the same arrangement is found in the two first leaves developed on the flower-stalk, and these are, as a rule, the only two; they are found below the calyx and are usually termed the “bracteoles.” It has become customary to indicate the bracteoles by the letters α and β, according to their sequence of growth, and in that sense these letters will be employed in the following diagrams.
The arrangement of the leaves varies very much; there is also a great variety of shapes in the leaves and their venation, but the linear leaves, with parallel venation, so frequent in the Monocotyledons, are seldom met with, as also the large sheaths (though the sheath is well developed in the Umbelliferous plants); stipules occur much more frequently.
The flower is most commonly cyclic, but acyclic or hemicyclic forms also occur. The type which may be taken as a basis consists in the majority of instances, as in the Monocotyledons, of 5 whorls, of which the 4 outer ones (calyx, corolla, and the 2[335] whorls of stamens) are most frequently 4 or 5 in number and placed in regular alternation, whilst the innermost one (the carpels) has generally fewer members, probably on account of space (Figs. 360, 361, 421, 429, 487, etc.). Trimerous (Figs. 384, 387, etc.) flowers, or those in which the members of the flower are in threes or a multiple of three, also occur, as well as dimerous flowers; other numbers are rare. It is of the greatest importance in connection with the relative position of the members of the flower to the axis and bract (orientation), whether the bracteoles are typically present (even though they may not be developed), or are typically absent. If there are 2 bracteoles present, then their position in a pentamerous flower is often as follows: the first sepal turns obliquely forward, the second is posterior and median, the third obliquely forward, the fourth and fifth obliquely backward; quincuncial æstivation is often found in these buds (Figs. 360, 429, 471, 475, 584). The first and third leaves, in the following chapters, are most frequently alluded to as the “anterior,” the fourth and fifth as the “lateral” leaves. The reversed arrangement, with the median sepal in the front, occurs for instance in Papilionaceæ (Fig. 511), Lobeliaceæ (Fig. 594), Rhodoracecæ. If any bracteoles are present below a tetramerous flower, the relation is generally that 2 sepals (the first ones) stand in the median plane, the two next ones transversely (Fig. 393), and the corolla then adopts a diagonal position (Fig. 397); but a diagonal position of the calyx generally shows that the flower is not, strictly speaking, tetramerous, as in Plantago (Fig. 567), Veronica (Fig. 559 C) and others.
If the bracteoles are not typically present, then the position of the sepals is changed accordingly, and the two outer sepals endeavour to assume the position which the bracteoles would otherwise have occupied, e.g. in Primula (Fig. 547). Other positions are also found when the number of bracteoles is more or less than two.
The leaves which follow the sepals occupy definite positions with regard to them, which we may consider later. An arrangement must, however, be mentioned here; when the flower is “diplostemonous” that is, has two whorls of stamens (thus, Sn, Pn, An + n), these may be arranged in two ways. Either the first-formed whorl of stamens, which are termed the “calyx-stamens,” stands directly in front of the sepals (that is “episepalous”), and is the outermost whorl, and in this case a regular alternation takes place between sepals, petals and the two whorls of stamens,[336] which is also continued into the carpels if their number is the same as that of the other whorls: the carpels are then placed opposite the sepals (Fig. 278) and the flower is isomerous and Gn should be added to the formula above. Or, the calyx-stamens form the innermost whorl, and the corolla-stamens, which are subsequently formed (“epipetalous” stamens), stand outside these (Figs. 360, 429); if the number of carpels is the same as that of the preceding whorls, they are often placed right in front of the petals and the corolla-stamens. The first-mentioned arrangement is termed Diplostemonous, and the second Obdiplostemonous. Both arrangements may be found in one and the same order, e.g. Caryophyllaceæ. The size and relation of the members of the flowers, and also the contact with other members in the early stages of their development, play an important part in determining the arrangement.
The great number of structural arrangements found in this enormously large class, may, as is the case in the Monocotyledons, be further varied by suppression and division of certain leaves (especially the stamens). Instances of this will occur in the following (Figs. 559, 568.—426, 441, 445, etc.).
The Dicotyledons were formerly divided into 3 sub-classes: Apetalæ (those without corolla), Sympetalæ or Gamopetalæ (those with the petals united), and Choripetalæ or Polypetalæ (the petals not united). This division has now been abandoned because it has been proved that the Apetalæ were merely reduced or incomplete forms of the Choripetalæ, and they have therefore been distributed among the various families of the latter sub-class.
With regard to the Sympetalæ (or Gamopetalæ) it may be stated that they form to a very great extent a closely connected and natural group, having in common not only the character that the corolla is gamopetalous and the stamens united with it (this being also found in the Choripetalæ), but also a great many others (such as persistent calyx, cyclic flowers with the formula S5, P5, A5 and as a rule G2, the two carpels being united to form the ovary; seeds with a thick integument and a very small nucellus). They are therefore considered as an independent sub-class, and must be placed at the close of the system of classification as the forms which presumably have arisen the latest. In the future systems of classification this arrangement will very probably be changed, and the first families of the Sympetalæ, the Bicornes and others will for instance be to a certain extent united with the families or orders of the Choripetalæ. The Sympetalæ may certainly be considered[337] as the youngest types, the strongly pronounced metamorphosis supporting this theory, as also the formation of the integument of the ovule, the one thick integument being undoubtedly derived from the coalescence of two—a holochlamydeous ovule, etc.
The Apetalæ and Choripetalæ are united into one sub-class. The leaves of the perianth in this case are, as a rule, free from each other, the structure of the flowers presents many differences, and the ovules have as a rule 2 integuments and a large nucellus. Considerable uncertainty still prevails regarding the arrangement and the relationship of the individual families of the Choripetalæ, and some of the following families are hardly quite natural; but the best arrangement arrived at so far has been adopted here.
At the beginning of the book a review of the orders of the Dicotyledons will be found.
Trees and shrubs, which, in the structure of the vegetative shoot and the catkin-like inflorescences, resemble the Quercifloræ, but the structure of the flower differs so much from them, that the only order brought under this heading—Salicaceæ—well deserves to be separated and to form a family of its own, the nearest relatives of which are still doubtful. As Juglandaceæ and Myricaceæ also deserve to be placed in a special family, the name Amentaceæ (Catkin-bearers), hitherto applied to all of these plants, cannot be retained as the name of a family.
Fig. 322.—Male and female catkins of Salix caprea.
There is only one order.
[338]
Order. Salicaceæ (Willows). Trees with simple, scattered, stipulate leaves. Diœcious. The flowers are arranged in simple inflorescences (spikes or racemes) which are termed catkins, and which fall off as a whole after flowering (♂) or after the ripening of the fruit (♀) (Fig. 322). The perianth is very imperfect[33] or wanting, particularly in Salix (Fig. 323 o); the ♂-flower with 2–several stamens and without any trace of a carpel (a, b, c): the ♀-flower has a free bicarpellate ovary, unilocular, and formed from 2 lateral carpels with 2 parietal (median) placentæ and generally ∞ ovules; the style divides into two stigmas (d, e, f). The fruit is a two-valved capsule and the very small seeds bear a tuft of hairs at the base. Endosperm absent.—The catkins are situated on dwarf-branches, which in some species often develop before the leaves and bear at their base only scale-leaves; in others foliage-leaves are borne beneath the catkins. The vegetative bud commences with 2 bud-scales which are united on the anterior side into a scale. The capsule opens by the dorsal suture. The seed-hairs spring from the funicle.
Fig. 323.—Salix: male flowers of S. pentandra (a), S. aurita (b), S. rubra (c), female flowers of S. aurita (d), S. nigricans (e), S. mollissima (f).
Salix (Willow) has short-stalked, most frequently lanceolate leaves and erect catkins with undivided bracts (Fig. 322). The flowers are naked; 1 (o in a-f) or 2 yellowish glands situated in the median line. In the ♂-flower generally two stamens, situated laterally like the carpels in the ♀-flower. Various forms are seen in Fig. 323.—The terminal bud of the branches often aborts regularly, the uppermost lateral bud taking its place.
Populus (Aspen, Poplar) has long-stalked, more or less round or cordate leaves with drawn-out apex; catkin pendulous; lobed[339] bracts; perianth cup-like with oblique edge; stamens usually numerous; stigmas often divided.—P. tremula (Aspen) has received its name from the tremor of the leaves: cf. “to shake like an aspen leaf.”
Pollination. The Poplars are wind-pollinated. The Willows have sticky pollen and are pollinated by insects. The catkins of the Willows, especially the ♂, are more conspicuous, from the numerous, closely-packed, yellow flowers, rich in honey and pollen. The catkins often appear before the foliage and so are much more easily seen, whilst at this time of the year the number of competing honey-flowers is smaller, and the insect visits consequently more numerous. On many catkins of the Willow the flowers open earliest on the side which is turned towards the sun and in descending order, i.e. the upper flowers develop before the lower ones. Hybrids frequently appear.
There are about 180 species existing in the northern, cold and temperate latitudes. Some in the Polar regions are scarcely more than an inch in height, and have a creeping rhizome (Salix herbacea, polaris, reticulata). Fossil forms are found in the Tertiary and perhaps also in the Upper Cretaceous.
Uses. Principally for ornamental trees, as they grow very quickly and are easily propagated by cuttings, S. babylonica, Weeping Willow; S. purpurea; Populus alba, Silver Poplar; P. pyramidalis, Pyramid Poplar—a form of P. nigra; P. monilifera, Canadian Poplar. The wood is very poor and little used; the branches of many Willows are cultivated for basket-making, etc. The wood of the Aspen is used for matches. The bark contains tannin and, in many Willows, a very bitter extract, Salicin (S. pentandra, fragilis). Salicylic acid (officinal) is obtained from Salix. Balsam is extracted from the buds of many Poplars, especially when the leaves are shooting.
Trees with verticillate, scale-like leaves forming sheaths at the nodes. Monœcious. Flowers unisexual. ♂-flowers in catkins; ♀ in short spikes. Pollen-tube entering the ovule at the chalaza, and not through the micropyle. Ovary 1-seeded, unilocular. Carpels uniting into a multiple fruit. Only one order.
Order. Casuarinaceæ. Trees (30 species), from Australia and certain parts of S.E. Asia, with peculiar, equisetum-like appearance. The leaves are verticillate, scale-like and united into sheaths. The internodes are furrowed. Branching verticillate. The unisexual flowers are situated in catkins or short spikes. The ♂-flower has a central stamen, surrounded by 2 median, scale-like perianth-leaves and 2 lateral bracteoles. The ♀-flower has a 1-chambered ovary (2 ascending, orthotropous ovules), no perianth, but 2 large, lateral bracteoles which finally become woody and form two valves, between which the nut-like fruit is situated. The multiple-fruits therefore resemble small cones.—Casuarina equisetifolia, cultivated, gives “iron-wood.”
[340]
[The Casuarinas differ from the ordinary Dicotyledons in many important respects which may be briefly summarised thus:—The bicarpellate ♀-flower has a well-pronounced stylar-cylinder terminated by two stigmas, but the cavity of the ovary closes very soon after its formation, and in it are developed two parietal ovules; these are united by a bridge of cellulose to the stylar-cylinder or summit of the ovary, and hence the ovules are connected with the walls of the ovary by the bridge (above), as well as by the funicle (below). The archespore is developed from the hypodermal cells at the summit of the nucellus, two primordial mother-cells are first formed and from these by tangential divisions a central cylindrical mass of cells (sporogenous-tissue) is produced which is surrounded by tapetal cells. The cells of the sporogenous tissue correspond to the mother-cells of the embryo-sac of other Angiosperms; they divide transversely and from 16–20 macrospores are formed together with inactive cells which are not crushed together as in the case of other Phanerogams. The sexual apparatus is developed from a single cell, but the number of cells composing this apparatus is subject to variation, the oosphere being accompanied by one or two neighbouring cells which resemble canal-cells rather than synergidæ. The sexual apparatus is found in the majority of the macrospores, but in most of these it remains as a number of naked cells; while in the fertile macrospores the cells are invested by walls of cellulose (usually only one fertile macrospore is found in each ovule). Antipodal cells are never developed. The macrospores elongate considerably towards the chalaza, into which some penetrate. The pollen-tube traverses the stylar-cylinder and enters the ovules at the chalaza, its passage through the tissue of the nucellus being assisted by the prolongation of the macrospores. About the centre of the nucellus the pollen-tube is ruptured; the apical portion which alone takes part in the fertilisation being firmly attached to the macrospore. Although the actual impregnation has not been observed, Treub considers that the endosperm begins to be formed before fertilisation.]
Trees and shrubs with small, unisexual, monœcious flowers, having no perianth or a simple inconspicuous one. The ♂ and ♀ flowers are very different and generally placed in separate inflorescences. The ♂-flowers are most often adnate to the bracts. The stamens are placed opposite the perianth-leaves, when they are present in equal numbers. The ♀-flower is naked, or has a superior perianth. The ovary at the base is 2- or 3-(-6) locular with 1 or 2 pendulous ovules in each loculus, only one of which is developed; the fruit is a one-seeded nut; endosperm absent; embryo straight. The inflorescences, which are either compound and mixed (small dichasia in spikes) or simple, are here also termed catkins; but, strictly speaking, this term is applied to the ♂-inflorescences only. In all Quercifloræ the leaves are scattered (usually in 2 rows) simple, and penninerved, and with deciduous stipules.
[341]
It is worthy of remark that in Betulaceæ, Corylaceæ and Quercus the ovules, and to some extent the loculi of the ovary are not developed till after pollination, so that the development of the pollen-tube proceeds very slowly. The smallness of the flowers, the absence of honey, the dryness and lightness of the pollen, the size of the stigma and the abundance of hairs found on many stigmas are all adaptations for wind-pollination. It is also an advantage that the flowers are generally pollinated before the foliage-leaves are developed, thus preventing the pollen being entangled by the leaves.
The two orders Betulaceæ and Corylaceæ mentioned here are by other authors united into one order. [It is doubtful whether these two should be retained in the family Quercifloræ, as recent researches (p. 273) have shown that they differ from the Cupuliferæ in many important points, and agree with the Casuarinas in the fact that the pollen-tube enters the ovule through the chalaza.]
Order 1. Betulaceæ (Birches). Monœcious, with thick, cylindrical, compound ♂ and ♀ inflorescences (2- or 3-flowered dichasia in a spike with spirally-placed floral-leaves) (Figs. 324, 326, 328). When the perianth in the ♂-flower is completely developed, it is composed of 4 somewhat united leaves, which are placed opposite the 4 stamens (Figs. 325, 326 A). The female flowers are naked; the ovary is bilocular, with two styles and one pendulous ovule in each loculus. The subtending floral-leaves unite with the bracteoles and form a 3–5-lobed cover-scale, which is not attached to the fruit (Figs. 325 D, 326 B). Fruit a nut without cupule (see Corylaceæ and Cupuliferæ). In the bud the leaves are flat. The stipules are deciduous. On germination the cotyledons are raised above the ground. Terminal buds are only found on old Alder trees; the Birch has sympodial branches.
Fig. 324.—Alnus glutinosus. Branch of Alder with ♂-(n) and ♀-(m) catkins: k bud; b fruit-bearing catkin (“cone.”)
Alnus (Alder) (Figs. 324–326). In the majority of species the ♂-and ♀-catkins are both developed in the year previous to their flowering, and pass the winter naked and bloom before the leaves expand. ♂-flower: 4 stamens. ♀-flower: the 5-lobed cover-scales of the ♀-catkin are woody and remain attached to the axis, so that the entire catkin when ripe resembles a small cone (Fig.[342] 324 b). Each cover-scale supports two winged or wingless nuts. In the native species of Alder the buds are stalked (Fig. 324 k). The bud-scales are formed by the stipules of the lowest leaves.
Betula (Birch). The ♂-catkins, in the native species, appear in autumn, the ♀-catkins in the flowering year on leaf-bearing, short-lived shoots. ♂-flowers: 2 stamens, divided (Fig. 328 A). The 3-lobed cover-scales (Fig. 327 a) of the ♀-catkin are detached from the axis; each cover-scale supports 3 broadly winged nuts (b). The stem has cork with annual rings. The young twigs and leaves have aromatic resin glands.
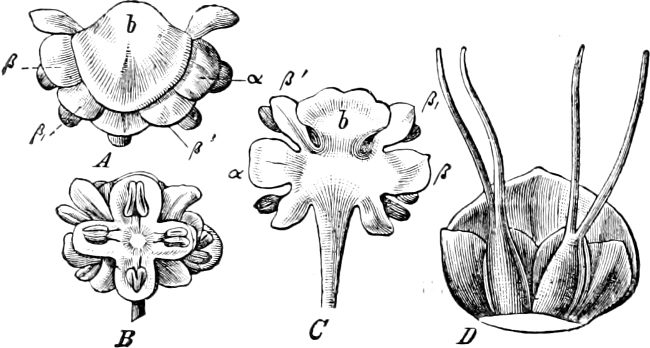
Fig. 325.—Alnus glutinosa: A dichasium of ♂-flowers seen from the front; B the same from inside; C the same from the back; D dichasium of ♀-flowers with subtending-leaf and four bracteoles. The letters b, α, β, β′, β are the same as in Fig. 326 A.
Fig. 326.—Alnus glutinosa: diagram of dichasia of ♂ (A) and ♀ (C) catkins; B a cone-scale. All the bracteoles in A and C are slightly pressed from their normal position.
The Inflorescences of the Alder.—In the axil of each cover-scale [b in the Figs] is situated, in the ♂-catkins (Figs. 326 A, 325 A-C) a 3-flowered dichasium, the flowers of which have a 4-partite perianth, the posterior perianth-segments being sometimes almost suppressed, and 4 stamens with undivided filaments. In the ♀-catkin (Figs. 325 D, 326 C) a 2-flowered dichasium is found, the middle flower being suppressed (indicated by a star in C). In both instances the inflorescences have two bracteoles (α-β) and the flowers borne in their axils have each one bracteole (β′), the other one (α′) being suppressed and therefore in 326 A and C only represented by a dotted line; these four bracteoles unite with the cover-scale (b) which supports the entire dichasium, to form the 5-lobed “cone-scale” (Fig. 326 B) which in the ♀-catkin eventually becomes woody.
The Inflorescences of the Birch.—A 3-flowered dichasium is situated in the axil of the cover-scale in both ♂-and ♀-catkins (Fig. 328 A, B); only the central flower has bracteoles (α-β) (the lateral flowers having no bracteoles), and these bracteoles unite, as in the Alder, with the supporting cover-scale (b), and form a three-lobed cone-scale (Fig. 327 a).
[343]
While the ♀-flower exactly resembles that of the Alder, the reduction of the ♂-flower, already described in the Alder, is carried further, so that often only the 2 median perianth-leaves are developed (Fig. 328 A); there are also only 2 stamens, these being deeply cleft, while the other 2 are suppressed.
About 50 species; N. Temp.—Fossil-forms certainly occur in the Oligocene. During the Glacial period the Dwarf-birch (B. nana) extended over Europe; at the present time it is confined to the moors and mountains of N. Europe and N. America and Asia. Wind-pollinated.
Uses.—Important forest trees. The bark contains tannic acid. The tar of the Birch is used in the preparation of Russia leather; whilst its spring sap is very saccharine, and is used in some places for making a fermented drink. Its external bark is used for roofing, for baskets, etc.
Fig. 327.—Betula verrucosa: a cone-scale; b fruit.
Fig. 328.—Diagrams of dichasia in the ♂-(A) and ♀-(B) catkins of Birch.
Order 2. Corylaceæ (Hazel-nuts). Monœcious. The ♂-catkins are long and cylindrical; the ♂-flowers are placed singly in the axil of the subtending-leaf (cover-scale); they are naked and formed of a number of divided stamens, which are partly united with the cover-scale, 4 in the Hazel, apparently 8 (Figs. 330 A, 329 B, C), more on the Hornbeam. The ♀-flowers have a very small, superior perianth; in the axil of each cover-scale a 2-flowered dichasium (Fig. 329 D) is present, of which the central flower (* in Fig. 330 B) is suppressed. The gynœceum is bicarpellary as in the Birches; the ovary is bilocular, with two long styles (Fig. 329 D-F); the loculi have 1 (-2) ovules (Fig. 330 B). Each single ♀-flower and fruit is surrounded by a leaf-like covering, the cupule (husk), which is formed of three floral-leaves (namely, the bract of a lateral flower, and its own bracteoles; thus in Fig. 330 B, α, α′, β’ form the cupule for the left-hand flower, and β, α1, β1, the cupule for the right-hand).
Corylus (Hazel-nut, Fig. 329). The long, cylindrical ♂-catkins pass the winter naked, 2–3 together, on short branches. The very[344] small ♀-catkins are enclosed in buds, in which they pass the winter; these buds are situated in the axils of the fallen foliage-leaves, and it is only by their larger size that they may be distinguished from the ordinary foliage-buds. In spring the ♀-catkins are easily recognised by their red, projecting stigmas (Fig. 329 A). The cupule—the “husk”—is tubular, fringed, and envelopes the nut. The leaves are alternate and unsymmetrical, the external side being larger than the internal; this is connected with the vernation, the blade being conduplicate in the bud; the stipules are deciduous. The bud-scales are formed of stipules, the most internal having a leaf-blade attached to them which is suppressed in the external ones. The cotyledons remain underground on germination.

Fig. 329.—Corylus avellana: A branch at the time of flowering with ♂-and ♀-catkins; B ♂-flower with subtending-leaf (bract) and two bracteoles; C the same without the anthers; D view of interior of ♀-dichasium shortly after fertilisation; E young fruit with cupule; F similar one with the cupule opened; G mature ♀-fruits; H nut.
Carpinus (C. betulus, Hornbeam). The ♂-and ♀-catkins do not appear till the leaves are shooting. The ♀-catkin in this[345] instance is also long and cylindrical. The cupule in C. betulus is 3-lobed, and to a slight extent only embraces the base of the ribbed nut (Fig. 331); each lobe corresponds to a floral-leaf. Whilst the carpels are placed medianly in Corylus, in Carpinus, on the other hand, they are situated transversely, as in the case of the Betulaceæ. The lamina of the leaf is not conduplicate in the bud, but flat, and folded only along the lateral veins, which are also indicated in the form of the fully-developed leaf; otherwise the vegetative characters are essentially the same as in the Hazel. The cotyledons are aerial.—Ostrya resembles the Hornbeam, but the cupule completely envelopes the nut, as a sac open at the apex (Eur., N. Am., Japan).
N. Am., Asia, and Europe; 25 species.—Fossil forms in the Oligocene. Wind-pollinated. Uses. As timber (Carpinus betulus) and firewood. The fruits of C. avellana (ordinary Hazel-nut), C. tubulosa (Lambert’s nut) and C. colurna (Turkish Filbert) are edible.
Fig. 330.—Diagrams of the ♂-flower (A) of Corylus and the dichasium of the ♀-flowers (B).
Fig. 331.—Nut of the Hornbeam with cupule.
Order 3. Cupuliferæ. Monœcious. The inflorescences make their appearance with the leaves, arising in the axils of the leaves of the same year. A woody cupule furnished externally with scales or spines is common, and surrounds 1-several flowers (the cupule in the Corylaceæ never encloses more than a single flower or fruit). The ♂-flower has a united perianth, which is, however, 4–6 partite, and encloses an indefinite number of undivided stamens. The ♀-flower has a superior, 6-merous perianth (3 + 3, compare Figs. 332 D, 334); the gynœceum is formed of 3 (or in Castanea 4–6) carpels with a corresponding number of stigmas (Figs. 332 D, H; 334, 335); and the ovary has at the base 3 (-6) loculi (Fig. 333), each of which has 2 pendulous anatropous ovules; the fruit is a one-seeded nut (Figs. 332 H, 336).
The cupule of the Cupuliferæ, according to the opinion of Eichler, is formed by united bracteoles, (compare Fig. 333, where[346] the four valves in the cupule of Castanea are considered as bracteoles of the lateral flowers of the dichasium); according to another view (see Prantl, in Engler’s Bot. Jahrb., viii., 1889), it is a ring-like axial outgrowth independent of the bracteoles of the flower, whose scales and spines are floral-leaves. The cupule in the Oak only encloses the base of the fruit, but in the Eating-chestnut and Beech the fruit is completely enclosed, and consequently the cupule must divide into a number of valves (generally 4) to allow the fruit to escape. In the 3-flowered dichasia of Pasania, Sect. Eupasania (Trop. Ind.), each individual flower has its own cupule of the same structure and development as in Quercus; and, moreover, each group of flowers has externally the typical six bracteoles.
Fig. 332.—Castanea vesca: A branch with inflorescences; B ♂-flower; C young cupule with three ♀-flowers; D ♀-flower; E the same in longitudinal section; F cupule with 3 nuts (diminished); G, H nuts (G in longitudinal section to show embryo).
Castanea (Eating-chestnut, Fig. 332). The catkins are erect[347] (A), cylindrical, with the ♀ at the base and the ♂ at the top, or some are entirely ♂ and composed of small dichasia. The cupule (C, F) is 4-valved, provided with spines, and entirely envelops the 3 nuts; it is already developed at the time of flowering.—♂-flowers are most frequently borne in 7-flowered dichasia, and have a well developed perianth, most frequently consisting of 6 leaves in two whorls (Fig. 332 B), and a large number of stamens. ♀-flowers are most frequently borne in 3-flowered dichasia (Figs. 332 C, 333); the letters in Fig. 333 indicate the older theory, according to which the 4 bracteoles (α′-β′) of the two lateral flowers are thick and united into a single 4-valved, woody cupule, which surrounds the 3 nuts, and is furnished externally with spines; the spines are well developed hair-structures.—6 carpels in two whorls.—The leaves in the vertical shoots have a divergence of 2/5, 3/8, 5/13; on the horizontal shoots they are alternate. The cotyledons remain underground on germination.

Fig. 333.—Diagram of the cupule of Castanea.
Fig. 334.—Female flower of Fagus (mag.)
Fagus (Beech). The ♂-catkins are pendulous, capitate; the ♂-flowers have an obliquely bell-shaped, fringed perianth, with 6–20 stamens. ♀-catkins erect, 2-flowered, borne singly in the axil of foliage-leaves of the same year; the ♀-flower has a gynœceum formed of 3 carpels, bearing an epigynous, 6-leaved perianth (Fig. 334). In this genus the dichasium has only 2 flowers, the central one being suppressed. The cupule contains, therefore, only 2 triangular nuts (“mast”). All the shoots have the leaves arranged in two rows; the rows are on the underside, being only about 90° distant from each other; the buds on the other hand approach each other towards the upper side. The bud-scales are stipules without laminæ; in vernation the laminæ are folded along the lateral ribs, the upper lateral portion being the largest (as in Hornbeam and Chestnut). The cotyledons are folded, and at germination are aerial, large, and reniform. 4 species (Europe, Japan, N. Am.)—Nothofagus (S. Am., New Zealand, S. Austr.)
Quercus (Oak, Fig. 335). Catkins simple. ♂-catkins long, thin, pendulous, few-flowered. ♀-catkins erect; the cupule is cup-like, entire, and encloses only the base of the solitary nut (“acorn”).—The ♂-flower has a similar construction to that of the Chestnut. The[348] ♀-catkin has not more than 5 flowers (single-flowered dichasia, in which only the central flower is developed). The scales on the cupules are no doubt leaf-structures in this case also. According to another theory, the scales are hair-structures; they arise on the internal face of the young cupule apparently in descending, but really in ascending order. The rim of the cupule gradually expands. In the ♀-flower (Fig. 335) the loculi of the gynœceum, together with the ovules, are not developed until after pollination.—The leaves in all cases have a divergence of 2/5; the lowermost leaves on the shoots are reduced to stipules which serve as the bud-scales (5 rows). The laminæ are conduplicate, as in Corylus, and the external side is the broadest. The cotyledons are fleshy and remain underground. 200 species.—Pasania (100 species).
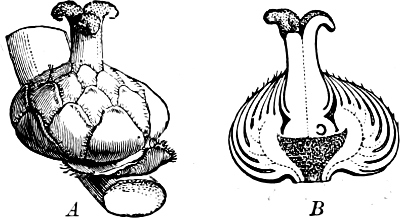
Fig. 335.—Quercus: A ♀-flower in its cupule (mag.); B longitudinal section through A, showing cupule, perianth, and inferior ovary.
Fig. 336.—Fruit of Quercus.
368 species, in temperate climates, especially in Europe and N. America. Authenticated forests have been found in the Oligocene. The Beech has one species, Fagus sylvatica, in Europe; it is a most important forest tree (in Denmark the most important) and reaches its most northern limit near Alvesund in Norway (60° N.L.), its northern boundary line passing from Alvesund in a zig-zag line through Ludwigsort, south of Königsberg, in Prussia, towards the Crimea. According to Steenstrup and Vaupell, the Beech did not make its appearance in Denmark until a comparatively recent time, the Oak then being partially supplanted. Other species of Beech are found in N. America and Japan. Several species of Nothofagus occur in the South West of S. America, and in the colder regions of the southern hemisphere. The Oaks grow especially in temperate regions, e.g. in Western Asia, N. America, and the mountains of Mexico. Evergreen species are found in Tropical Asia, Himalaya, Japan and the Mediterranean region. In this country there is one species of Oak (Q. robur), of which there are three varieties (Q. pedunculata, intermedia, sessiliflora). The Eating-chestnut is found in the South of Europe, but is cultivated in the midland and southern counties of England.—Uses. The wood of these trees is very useful as timber. The wood of Q. tinctoria has a yellow colouring matter (Quercitron-wood). The bark of the Oak contains a large quantity of tannic acid, and is used for tanning; for this purpose also the cupules of Q. vallonea, ægilops, græca, and others from the Eastern Mediterranean, are used under the name of “Valloons.” The Cork-oak (Q. suber; S.W. Europe) is the most important tree from which cork is obtained,
[349]
its bark being very largely developed and stripped for cork. Gall-nuts are found on many species; those of Q. lusitanica, var. infectoria (Eastern Mediterranean) are officinal, and likewise the fruits (acorns) and the bark of Quercus pedunculata and sessiliflora. Oil is obtained from the Beech “mast.” The nuts of the Chestnut tree are edible.
This family resembles the Quercifloræ in the catkin-like inflorescences, the imperfect, unisexual flowers, the epigynous perianth and the woody shoots with scattered leaves, etc., though it is in other respects very dissimilar; one point of difference is the presence of aromatic compounds, but a more important divergence is found in the structure of the gynœceum, which is formed of two carpels with one loculus and has one basal, orthotropous and erect ovule, which, as in the Quercifloræ, does not become developed until after pollination; the fruit too is very different, being generally a drupe. Endosperm absent.
Fig. 337.—Juglans regia: A ♂-flower seen from below with bract (cover-scale) (b), bracteoles (α and β), perianth-leaves (p); B the same from the front; C lateral view of the same; D diagram of A; E ♀-flower with bract, the bracteoles are united with the ovary, their edge being visible as an indented line below the perianth; F 2 ♀-flowers at the end of a foliage-shoot; G fruit (without the fleshy covering) in longitudinal section; H transverse section of the same.
Order 1. Juglandaceæ (Walnuts). Leaves scattered, imparipinnate, rich in aromatic compounds. Stipules absent. Flowers unisexual. Monœcious. The ♂-catkins are lateral, generally on naked branches of the previous year, cylindrical, pendulous, many-flowered; the two bracteoles and the 2–4-leaved perianth of the ♂-flower unite with the subtending bract; the ♂-flower has indefinite stamens (6–20 in Juglans, Fig. 337 A-D). The ♀-catkins are terminal, generally on branches of the same year, few-flowered (Fig. 337 F); the[350] ♀-flowers have a superior, 4-leaved perianth, a bicarpellate gynœceum, two styles with stigmas on the internal surface. The ovary, bracteoles and bract all unite together (Fig. 337 E). The fruit is generally a green or black drupe,[34] whose flesh (outer soft portion) in Carya and Juglans ruptures more or less irregularly, and frees the stone (“Walnut”).—The stone in Juglans is divided internally by one true (Fig. 337 H) and by several false, low partition walls into several incomplete compartments, so that the two large cotyledons become lobed and incised to fit like a cast into the irregularities of the inner surface of the stone; the embryo is exendospermous and covered with a thin testa.—The leaf scars are large and cordate with 3 groups of vascular bundles. The PITH in Juglans and Pterocarya is divided into chambers. The stone ruptures, on germination, along the dorsal suture into 2 valves; the cotyledons remain underground. In Juglans regia a long row of accessory buds is found on the lowest internode (epicotyl) above the axils of the cotyledons. Pollination by the wind. Both protogynous and protandrous examples of Juglans regia occur.—33 species, mostly in temperate North America.—Uses. Walnuts are obtained from J. nigra and regia; Hickory from North American species of Carya. The oil-containing seeds of several species are edible. Pterocarya and others are cultivated as ornamental plants.
Fig. 338.—Myrica gale: a young fruit; × the bracteoles with numerous glands; b longitudinal section of fruit.
Order 2. Myricaceæ. To this order belong shrubs or trees which have penninerved, simple, at most lobed or pinnatifid leaves, with or without stipules, and with yellow, aromatic, resin glands (Fig. 338 a). The flowers, situated in catkin-like spikes, are unisexual and naked, and supported by scale-like floral-leaves. ♂-flower: 4–6 (–16) stamens with short filaments; ♀: generally situated singly. The gynœceum has a short style with 2 long stigmas, and unites with the bracteoles, which form wing-like outgrowths on the ripe drupe as in Pterocarya in the Juglandaceæ (Fig. 338). Cotyledons fleshy (Fig. 338 b).—Myrica; Comptonia.
[351]
40 species; Temperate.—Myrica gale (Sweet-gale, Bog-myrtle) has been used in the preparation of beer (Sweet-willow beer) on account of its resinous essential oil. M. cerifera (N. America) and species from the Cape, M. quercifolia and others, form wax on the fruit which is used in the preparation of candles.
The flowers are regular, hypogynous, nearly always unisexual, small and insignificant, with single, green perianth of 4–5 leaves. Stamens 4–5, placed opposite the leaves of the perianth. Ovary formed of 1 or 2 carpels, most frequently unilocular, with one ovule (Fig. 340). The fruit is a nut, more rarely a drupe, with one seed, generally endospermous. The Nettles are the sole order in the family which has only one carpel (1 stigma); this turns the posterior side to the front (Fig. 340). The others have two carpels (2 stigmas) but the anterior only is fertile (Fig. 346) except in a few Ulmaceæ and Moraceæ.
The majority are trees or shrubs with petiolated leaves, stipulate; rough hairs are very frequently developed upon the leaves. The flowers are very often crowded together in the inflorescence, which is rarely catkin-like. Peculiar aggregations of fruits are found in some orders. Latex and tough bast, which is used technically, are also frequently found. Cystoliths are found in the epidermis of many species of Ficus, Urtica, and others. Wind- or self-pollination is most common, as in the Quercifloræ and Juglandifloræ. In the Urticaceæ, Morus and some others, the stamens lie incurved in the bud, and when ripe straighten themselves suddenly and elastically, and thus small clouds of pollen-grains are ejected with considerable violence on to the stigmas, which are often provided with brush-like hairs (Fig. 341). The formation of honey does not take place.
Order 1. Ulmaceæ (Elms).—Trees or shrubs without latex. Leaves simple, arranged in two rows (divergence 1/2), oblique (the inner side, nearer the axis, being the larger), strongly penninerved, dentate, hispid; stipules deciduous. In opposition to the other Nettle-like plants the flowers are often ☿ with a united cup- or saucer-like, generally 4–(5)–6-divided perianth, and a corresponding or larger number of opposite erect stamens. The gynœceum has two carpels (2 stigmas), generally one loculus with one pendulous, anatropous or amphitropous ovule,[35] seldom two loculi and 2 ovules. Fruit one-seeded (nut or drupe). Embryo without endosperm.
A. Ulmeæ. The fruit is a winged nut (Fig. 339), the embryo straight, without endosperm. Anthers extrorse.—Ulmus (Elm).[352] The flowers are situated in inflorescences which develop from the lower buds of the shoot of the preceding year. The lowermost bud-scales are empty, the uppermost support either solitary flowers, or small, dichasial or unipared scorpioid inflorescences. The terminal bud on the vegetative shoot quickly falls off, and the upper lateral bud continues the growth sympodially. Flowering takes place before the leaf-buds open. The flowers are wind-pollinated and have no honey. Fossil species have been found in the Oligocene.
20 species; North Temp. (2 species in this country). Important as timber. The Cork-elm (U. suberosa) has a rather thick cork, which, however, is of no technical use. The bast is used as Lime-bast.
B. Celtideæ. The fruit is a drupe, the embryo curved, with folded or rolled up cotyledons, with or without endosperm. The anthers are introrse. The flowers are borne on a shoot of the same year. Planera (N. America); Zelkova.—About 114 species; especially N. Temp., Trop.
Fig. 339.—A Ulmus campestris, flower with exceptionally aborted gynœceum; B, U. effusa, flower with 8 stamens; C, U. campestris, fruit opened in front to show the seed pendulous from the apex of the loculus; one loculus is aborted.
Order 2. Urticaceæ (Nettles).—The majority of species are herbs with simple, stipulate leaves; they have no latex; stinging hairs abundant. The flowers (Fig. 340) are unisexual, generally 2-merous and arranged in clusters, which are united into catkin-like inflorescences. The perianth is composed very often of 4 (2 + 2) free, or in the ♀-flowers generally united, green leaves; the 4 (2 + 2) stamens are opposite the perianth-leaves, the filaments are bent inwards in the bud and throw themselves elastically towards the outside. The gynœceum has one style and one stigma (capitate or brush-like, Fig. 341); the ovary is unilocular, with one orthotropous, erect ovule (all other orders of this family have inverted or curved ovules). Fruit, a nut or drupe. Endosperm present (in Urtica very little), oily. Embryo straight. The STINGING HAIRS are club-shaped, very turgid, and provided with a siliceous, brittle apex, which breaks off in an oblique direction and allows the poisonous cell-sap to be forced out. In many tropical Nettles this is so strong that it may produce partial paralysis. There is no rudiment of an ovary in the ♂-flowers (Fig. 340 A). The PERIANTH in the ♀-flower differs from that of the ♂ in having the two[353] internal leaves generally much larger and enveloping the fruit (Fig. 340 B); it often happens that all the perianth-leaves are united to form a gamophyllous envelope. ☿-flowers may occur among the others.—The inflorescences among our native species are dichasia, which become transformed into unilateral scorpioid cymes by the development of the bud of the 2nd bracteole. In Parietaria they are more pressed together, and the floral-leaves at the same time are also raised on their axillary shoots to just beneath the flower. As a rule, not only in this order but also in those related to it, a small vegetative branch is situated in the axil of the foliage-leaf, and this bears an inflorescence on each side at its base.
Urtica (Nettle) has opposite leaves with distinct stipules and stinging hairs. The perianth-leaves of the ♀-flower are free (Fig. 340).—Parietaria (Pellitory) has scattered leaves without large stipules, and stinging hairs are absent. The ♀-perianth is 4-toothed, flask- or bell-shaped.—Pilea is a tropical genus with trimerous, zygomorphic ♀-flowers, the posterior perianth-leaf being much larger than the two others, and more or less hood shaped.—The flower of Forskohlea is the most reduced; the ♂-flower has only one stamen, and the ♀-as well as the ♂-flowers have a one-sided, tongue like perianth (?). Pouzolzia.
Fig. 340.—Diagram of ♂-and ♀-flowers of Urtica dioica.

Fig. 341.—Parietaria diffusa; hermaphrodite flower: a in the female, b at the commencement of the male stage; the stigma has fallen off, but the anthers have not yet dehisced.
Wind-Pollinated. The pollen is shot out of the anthers, when they spring forward, and is caught by long stigmatic hairs. Parietaria diffusa is protogynous (Fig. 341).
500 species; chiefly in the Tropics, although the few species which occur in Europe are represented by a much larger number of individuals.—Uses. The bast of the native species Urtica dioica and urens, of U. cannabina (Siberia), etc.; of Boehmeria nivea “Ramié” and “China-grass” (from Sunda Is., China), and others, is used in the manufacture of muslin.
Order 3. Moraceæ (Mulberries). Nearly all trees or shrubs, seldom herbs, generally with latex. The leaves are scattered, and not infrequently lobed. The flowers are unisexual (monœcious or diœcious) and arranged in catkin- or capitulum-like, compound inflorescences. Perianth-leaves 2–6, generally 4, with an equal number of stamens opposite to them, as in the Nettles. The[354] ovary is 1–seldom 2-locular, and has 2 stigmas (it is thus formed from 2 carpels) seldom only one style with one stigma. One ovule in each loculus, more or less curved, and pendulous; micropyle directed upwards. Fruit usually a drupe. The embryo is generally curved inside the fleshy endosperm, or it is exendospermous.
Fig. 342.—Morus alba ♂ flower (6/1).

Fig. 343.-Morus alba ♀ inflorescence.
Fig. 344.—Morus nigra fruits.
A. Moreæ. The filaments are incurved in the bud. Leaves folded in the bud—Morus (Mulberry) (Figs. 342–344). Monœcious. The inflorescences are catkin-like in appearance, but in reality composed of many small dichasia. The flowers are similar to those of the Nettle, but with 2 carpels: in the ♂ with perianth 2 + 2, and stamens 2 + 2 (Fig. 342), in the ♀, perianth 2 + 2, and 2 carpels in regular alternation. The small drupes are enveloped by the perianth, which eventually becomes fleshy, and as all the flowers on the axis very accurately fit together, the collection of fruits is formed, which we call a Mulberry (Fig. 344). The leaves are folded in the buds, and have small stipules. The following are allied to Morus:—Maclura, Broussonetia (the Paper-mulberry tree) which has spheroid ♀ inflorescences (made up of dichasia), etc.
Dorstenia presents an interesting transitional form to the Fig in its flat, open, and, in some instances, lobed inflorescence on which the ♂ and ♀ flowers are sunk in grooves. Indications of a somewhat similar structure are found in certain Nettles, the sympodial axes of the dichasia becoming flatly expanded. The fruits are 1-seeded, but, nevertheless, spring open and eject their seeds.
B. Artocarpeæ. Filaments straight in the bud; foliage-leaves with convolute vernation. An interpetiolar leaf-sheath (ocrea) formed in the axil of each leaf by the connate stipules, covers the younger leaves as a hood. It falls off as the leaf expands, and leaves a ring-like scar on the stem.—Ficus (the Fig). The inflorescence (the so-called syconus) has a pear-shaped,[355] fleshy, but hollow axis, on the interior surface of which the flowers are situated (Fig. 345). It is a kind of capitulum, with a hollow receptacle, whose “involucral” leaves close over the entrance to the interior; it is not, however, a simple capitulum, but a coalescence of cymose inflorescences. The edible parts are the fleshy stem-portion and perianth-leaves. The ♂-flower has a 2–6 divided perianth, 1–2 (–6) stamens; the ♀-flower has an oblique ovary. The fruits are drupes, with thin flesh.—Many species have aerial roots, and some live as epiphytes on trees. Pollination, in the edible Fig, is effected by a small Gall-wasp (Cynips psenes L.), which lays its eggs in the Fig, and hence carries the pollen away. Even in very ancient times it was customary to hang infected wild Figs on the branches of cultivated ones, so that the young Gall-wasps, as they emerged, could immediately effect the pollination (caprification). Ficus carica, and other species, have two kinds of ♀-flowers, besides the ♂-flowers. One kind has a short style and no stigmatic hairs, and it is only in the ovaries of these that the wasps lay their eggs (gall-flowers); the other kind has a long style and well-developed stigmatic-hairs, but the wasps cannot reach their ovaries—these are “seed-flowers.” There are, moreover, two kinds of plants of Ficus carica; ♀-plants, which have only seed-flowers, and bear the edible Figs, and ♂-plants (called “Caprificus”), which bear inedible fruits, and have ♂-flowers at the upper part of the Fig, but gall-flowers at the base. [The Caprificus, at Naples, bears three crops of inedible Figs each year, viz. Mamme (April), Profichi (June), Mamnoni (August). The ♂-flowers are produced especially in June, the first Figs being almost entirely ♀, and the last having but few ♂-flowers. Each crop produces a new generation of Fig-wasps. The female wasp enters the Figs on the Caprificus, and lays one egg in each flower, with the result that the flower developes into a kind of gall. The mother-wasp dies within the Fig. The male wasp is wingless; it bites a small passage into the ovaries containing the female wasps, and impregnates them; the female wasps then escape from the Fig, those in the Profichi carrying pollen away with them as they pass out. They then enter another Fig, lay their eggs, and die. The edible Fig-tree similarly has three crops in the year, Fiori di fico, Pedagnuoli, Cimaruoli. The wasps, entering these Figs, are unable to lay their eggs in the ovary, but, nevertheless, they effect cross-pollination on entering the Pedagnuoli, which bear fertile seeds.]
Fig. 345.—A Fig in longitudinal section.
[356]
The flowers of Brosimum are the most reduced. The perianth is wanting, and the ♂-flower has only 1 stamen. Cecropia (Trumpet-tree), in S. Am., has its pith divided into chambers; these are inhabited by ants, which feed upon small food-bodies formed on the swollen base of the petioles. The leaves are petiolated, often shield-like, fringed or lobed, and sometimes with white felted hairs. They serve as food for Bradypus (the Sloth). Sorocea; Castilloa.
About 300 species exclusively in the warmer climates. The white Mulberry (M. alba, from China, India, Mongolia) is cultivated for the sake of its leaves, which are the indispensable food for silkworms. The black Mulberry (M. nigra, W. Asia) is cultivated for its fruits, which are used for the officinal Mulberry juice. The ordinary Fig-tree (Ficus carica) is from the Mediterranean. The fruit of the well-known Oriental Sycamore (F. sycomorus) is edible. The Bread-fruit tree (Artocarpus incisa) and the Jack (A. integrifolia) have their home in the South Sea Islands, and are cultivated in tropical countries. The Bread-fruit is morphologically the same as the Mulberry. It has a very large, spheroid inflorescence, whose floral-leaves and perianth become fleshy and united into one nutritious mass, together with the axis, which is also fleshy. The milky juice of the India-rubber tree (Ficus elastica, East Indies, a common house-plant), and of Castilloa elastica (Am.) is the raw material of India-rubber. The milky juice of Galactodendron utile (Cow-tree, S. Am.) is saccharine and nutritious, but in Antiaris toxicaria (the Upas-tree, of Java) it is a strong poison. The bast of the Paper-Mulberry tree (Br. papyrifera, Eastern Asia); is used in Japan for paper. Shellac is obtained from a small, hemipterous insect (Coccus lacca), which lives upon Ficus laccifera and F. religiosa (the Bo-tree, sacred to Buddha), E. India. The wood of Maclura aurantica (Am.) has a yellow colour, and is known as yellow Brazilian wood.
Order 4. Cannabaceæ. The plants which belong to this order are aromatic herbs, either annuals or perennials, without latex. Leaves palminerved, and more or less divided, hispid, and with free, persistent stipules. Flowers always diœcious; ♂-flowers in panicles, formed of dichasia, passing over into uniparous scorpioid cymes. They differ from the Nettles, particularly in the 5-leaved perianth of the ♂-flower, the 5 stamens (Fig. 346–351) with filaments erect in the bud, and in the ♀-flower by the small, entire, cup-like perianth, which surrounds the base of the ovary (Fig. 346, p. 352). The ovary has two styles, or one divided into two, with two stigmas and a pendulous, curved ovule (Fig. 346 B, 352 B); the fruit is a nut; the embryo is curved (Hemp, Fig. 353), or rolled (Hop, Fig. 349), without endosperm.
Fig. 346.—Diagram of male and female flowers of the Hop and Hemp: b the bract, p the perianth. The position of the embryo is indicated.
[357]
Only 2 genera with 3 species (Asiatic), of which two are cultivated.—Humulus lupulus (Hop, Figs. 347–349) is a twining, perennial plant, twisting to the right, with opposite, palmilobed, rough leaves, and large, interpetiolar stipules. The ♀-flowers are situated in closely-flowered, cone-like, compound inflorescences, with ultimately large, thin, imbricate floral-leaves (Fig. 348) which bear the yellow, glandular hairs, containing lupulin. This inflorescence is made up as follows:—The most external floral-leaves are situated in pairs, and are the persistent stipules of a leaf, the blade of which has become suppressed, or in any case is rudimentary. Such a pair of stipules supports 4 (2–6) flowers in a double uniparous cyme, whose central axis does not develope into a flower. The bracts of these flowers (bracteoles of the partial inflorescence) become, at maturity, very large, spathe-like, and, together with the stipules, produce a cone-like appearance.


Fig. 347–348.—Humulus lupulus: 347, twining stem; 348, branch with strobiles.
Cannabis sativa (Hemp, Figs. 350–353) is an East Indian herb, with palmilobed leaves, and differs from the Hop in being annual,[358] erect, and in having its leaves opposite at the base and scattered above. The ♀-inflorescence is not cone-like as in the Hop, but the flowers are similar in construction. The main difference is to be found in the axillary shoot, which was suppressed in the Hop, and is in the Hemp developed into a leaf-bearing shoot which on each side bears only one ♀-flower, and in the fact that the bracts are not so strongly developed.
The “Hops” (the female inflorescences) are used in brewing, and medicinally on account of the yellow glands which contain lupulin. The Indian variety of Cannabis sativa contains an abundance of glandular hairs and resin. The withered inflorescences are used in medicine and are officinal. The bast of the stems of the Hemp is also used and the fat oil of the seeds. In Oriental countries the entire plant is used in the preparation of an intoxicating drink (haschisch), the narcotic material being found in the glandular hairs.
Fig. 349.—Humulus lupulus: fruit in longitudinal section.
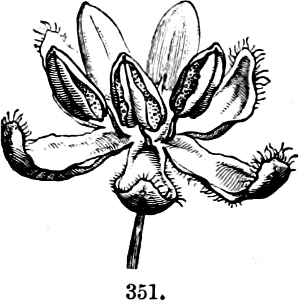
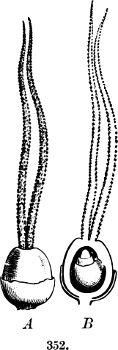

Figs. 350–353.—Cannabis sativa: 350, ♂-plant; 351, ♂-flower; 352, ♀-flower, entire and in longitudinal section; 353, fruit in longitudinal section.
This family is on one side closely allied to the Urticaceæ by its solitary, basal, vertical, and straight ovule, and by the conical ocrea which envelopes the younger leaves in the bud, similar characters[359] being present in the Urticaceæ. On the other side it is related to the Curvembryæ. The flowers are small, often trimerous, regular and slightly perigynous (in Chloranthaceæ, if they properly belong to this family, and Houttuynia, more or less epigynous). Syncarps are present in some Piperaceæ, but the fruit is generally a single fruit, one-seeded berry, nut or drupe. The leaves are generally scattered.
Order 1. Polygonaceæ. The majority are herbaceous plants with round, often jointed stems, scattered leaves and ocrea, that is a membranous, tubular, ligular or stipular structure inside the base of the leaf, which clasps the stem and axillary bud; the edges of the lamina are rolled backwards in the bud. The flowers are regular, small, generally ☿, slightly perigynous, with inconspicuous, simple, green or white perianth of 5–6 free segments; stamens 5–9 (Fig. 354) sometimes arranged in two series; gynœceum 2–3 carpels, ovary unilocular with one basal, straight (orthotropous) ovule, 2–3 free styles. The fruit is a 2–3-angular nut; the embryo, with mealy endosperm, is straight or curved (Fig. 355 H), often unsymmetrical.—The inflorescences are compound, and generally branch from the axils of the bracteoles, so that the last partial-inflorescences become coiled, uniparous scorpioid cymes; in Polygonum the two bracteoles unite into a membranous tube; in Rheum and Rumex there is only one bracteole.
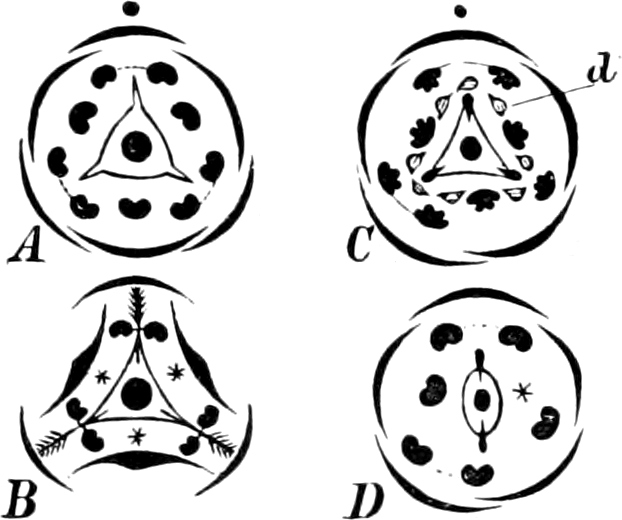
Fig. 354.—A Diagram of Rheum; B of Rumex; C of Polygonum fagopyrum; D of P. lapathifolium. The ovules are indicated inside the ovaries; bracts and bracteoles are not shown.
Rheum (Rhubarb, Fig. 354 A) has a 6-leaved, petaloid perianth (Pn 3 + 3) and 9 stamens (A 32 + 3). The 3-winged nut is not enclosed by the perianth.
Rumex (Dock, Fig. 354 B) has 6 stamens (A 32 + 0); the perianth is 6-leaved (Pr 3 + 3), green or red, and the triangular nut is enveloped by the 3 interior perianth-leaves, which point upwards and continue to grow after flowering. These perianth-leaves often have warts on their outer surface. The following are monœcious: R. acetosa and R. acetosella.
Polygonum (Knot-grass, Figs. 354 C, D; 355). The petaloid perianth is most frequently 5-merous (2/5 spiral); 5–8 stamens. The nut is triangular (Fig. 354 C, 355), or lenticular (Fig. 354 D).[360] There are two whorls of stamens, the external with introrse, and the internal with extrorse anthers. The gynœceum is often bicarpellate (Fig. 354 D).
The flowers may be considered as constructed upon the monocotyledonous type. Pterostegia has a perfectly monocotyledonous flower with 5 trimerous whorls. Rheum likewise, but here the external staminal whorl is doubled (Fig. 254 A). Oxyria has a dimerous Rheum-flower (4-leaved perianth, 6 stamens, 2 stigmas). Rumex has a Rheum-flower with the suppression of the internal whorl of stamens (Fig. 354 B); Emex is a dimerous Rumex. Polygonum, to which Coccoloba, Muehlenbeckia and others are related, differs from Rheum chiefly in having one of the leaves, which in the latter takes part in the formation of the perianth, developed in this case into a bracteole (so that the perianth is reduced to five members), and several or all the stamens in the inner whorl become suppressed.—The perianth in Coccoloba and Muehlenbeckia is more or less perigynous and becomes fleshy, enclosing the fruit. Muehlenbeckia platyclada has flat branches with rudimentary leaves; sometimes branches with normal, arrow-shaped leaves are found. Atraphaxis.
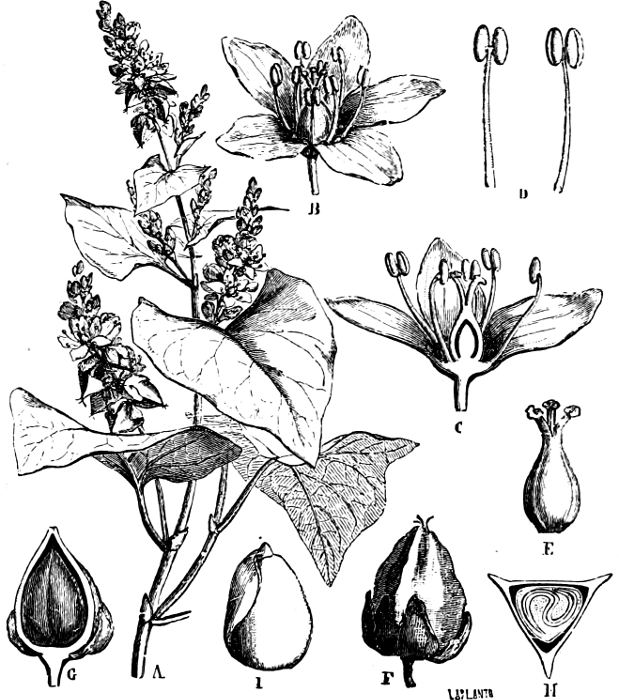
Fig. 355.—Polygonum fagopyrum: A branch with flower and fruits (nat. size); B flower; C the same in longitudinal section; D anterior and posterior view of stamen; E gynœceum; F fruit (mag.); G fruit in longitudinal section; H transverse section, showing the curved cotyledons embedded in the endosperm; I the embryo.
[361]
Pollination. Rumex is wind-pollinated, the stigmas are therefore large and brush-like (indicated in Fig. 354 B). Rheum and Polygonum are insect-pollinated and have therefore capitate stigmas, etc.; honey-glands are situated at the base of the stamens (d, in Fig. 354 C, and n in Fig. 356); a few small-flowered Polygonum species are self-pollinated; Buckwheat (P. fagopyrum) is dimorphic and has long-styled and short-styled flowers (Fig. 356). Pol. bistorta is protandrous and homostyled.
About 750 species, most of which are found in the temperate regions of the Northern Hemisphere, some reaching as far as the snow line or into the Arctic regions (Oxyria, Kœnigia). Trees and shrubs are found in the Tropics: Coccoloba, Triplaris. Rheum is Central Asiatic.—The thick rhizomes of R. officinale (Rhubarb) are officinal. The rhizomes of the ordinarily cultivated species, R. undulatum and rhaponticum, are used in veterinary medicine. The following are cultivated as culinary plants for the sake of their leaves:—Rumex acetosa (Sorrel), R. patientia, R. scutatus, and Rheum undulatum (petioles). Several species of Polygonum (P. hydropiper and others) have a sharp, pungent taste. “Buckwheat” is the mealy fruit of Polygonum fagopyrum (Central Asia) and is of value as a farinaceous food. P. cuspidatum (P. sieboldi, Japan) is an ornamental plant.—Calligonum in sandy and stony deserts.

Fig. 356.—Flower of Polygonum fagopyrum in longitudinal section: 1, long-styled; 2, short-styled; a the anthers; st the stigmas; n nectary.
Order 2. Piperaceæ (Peppers). Shrubs or herbs, often with nodose, jointed stem; leaves simple, entire, often with curved veins; stipules wanting (Peperomia) or intrapetiolar and cap-like, often enclosing the terminal buds (Piper). The flowers in the group Pipereæ (Piper, Fig. 357, and Peperomia) are borne in spikes with fleshy axes (club-like), seldom in racemes, the outer ones are crowded and are ☿ or unisexual, always small, naked and without bracteoles; generally stamens 3 + 3, and gynœceum 3, but the number of the stamens may be reduced by suppression to 2, and the carpels to 1. The flowers are situated in the axils of the small, generally shield-like floral-leaves. The ovary is always unilocular and has one upright, orthotropous ovule. Fruit a berry or drupe. Both endosperm and perisperm are present, the latter being especially well developed (Fig. 359).
Piper; generally shrubs with scattered leaves, and terminal[362] inflorescences which are crowded to one side by the development of the highest lateral bud, so that they are situated opposite the leaves (Fig. 357). Many species have stems with an abnormal anatomical structure.—Peperomia; chiefly succulent herbs, often epiphytes, with opposite or verticillate leaves having aqueous tissue on the upper side.

Fig. 357.—Piper nigrum: branch with fruit (½)
The group Saurureæ (considered by some as an order, and perhaps representing a more original type) has 3–4 carpels with many ovules. Lactoris stands the highest with regular 3-merous perianth, 3 + 3 stamens and 3 carpels, which are united at the base. Fruit a capsule with several seeds. (It has one species from the island of Juan Fernandez, and is also placed in an order of its own, Lactoridaceæ, allied to the Magnoliaceæ, through Drimys).—Saururus has naked flowers; most frequently 6 stamens, and 4 carpels, free or united at the base, each with 2-4 orthotropous ovules. Fruit, small berries.—Houttuynia; stamens situated a little upward on the ovaries; placentation parietal; capsule many-seeded.
About 1,000 species; entirely tropical, especially from South America and East India. They are found chiefly among the underwood in damp, shady places; some, which are fleshy (Peperomia), live as epiphytes on trees; a few climb by[363] roots.—Uses. Several Piperaceæ are used medicinally and for spices on account of their pungent properties and the essential oils found in nearly all parts of the plant. The following are officinal: “Black-pepper” (the unripe, dried fruits) and “White-pepper” (the seeds of the ripe fruits) of Piper nigrum (climbing shrub, East Indian); “Cubeb” berries of P. cubeba (climbing shrub, Java). “Long-pepper” is the unripe inflorescence of P. longum, East India. The leaves of P. angustifolia (Matico) are officinal. The leaves of the Betelpepper (East India) are used together with the nuts of the Areca-palm to form the well-known East Indian intoxicating compound “Betel.” A good many others are also used.
Fig. 358.—Piper nigrum (Diagram). In addition to the bract there are two structures resembling bracteoles.
Fig. 359.—Piper nigrum: Fruit in longitudinal section, showing the endosperm, perisperm, and pericarp.
Order 3. Chloranthaceæ. (Chloranthus, Hedyosmum) have opposite leaves, with stipules more or less united at the base, and inferior “drupes.” Ovules pendulous. Only endosperm. About 33 species, Tropical.
The plants in this family have a curved ovule, and most frequently a kidney-shaped seed (generally provided with fine, cuticular, projecting warts, Fig. 362 B), with a curved, peripheral embryo enclosing the endosperm which is most frequently floury (Figs. 362 C, 365 H; for exceptions, see Fig. 366); the seeds in all cases are borne on a centrally-placed, and in most cases free, placenta (they are “basal” when there is only 1 ovule in the ovary, Fig. 364). The flower is regular, hypogynous or perigynous (Fig. 364) (only rarely epigynous) and usually 5-merous. The flower which is most complete has 5 whorls (S5, P5, A5+5, G2-3–5), as in some genera of the Caryophyllaceæ (Figs. 360, 361); but from this type it becomes reduced, the petals and stamens being suppressed, so that finally 5 perianth-leaves, 5 stamens (opposite the perianth-leaves), and 2 carpels (Fig. 361 F) only are present; for example,[364] in certain genera of the Caryophyllaceæ, in the Chenopodiaceæ, Amarantaceæ, and others. When the number of stamens is increased to more than 5 in the whorl, it is always possible to show that some of the stamens have been divided. The number of the carpels and ovules also becomes reduced; in the highest there is a central placenta, not free in its early stages, with a large number of ovules; in those which are most reduced there is only a single ovule, which is placed centrally at the base of the ovary [Fig. 364]. Somewhat corresponding changes are found in the fruit, which is a many-seeded capsule in those which have many ovules, but a one-seeded nut where there is one ovule. In the most reduced forms the flowers are generally unisexual.—Similar features are also present in the vegetative parts. Almost all the species are herbaceous, the leaves are simple and most frequently without stipules. The structure of the stem, especially in Chenopodiaceæ, Amarantaceæ, Nyctaginiaceæ and others, often differs from that of the ordinary Dicotyledon. In the woody portion of the stem and root several rings are sometimes formed which resemble annual rings but which are formed by new cambium-rings arising outside the old ones which then cease to divide.
Order 1. Caryophyllaceæ. Herbaceous plants, with round, nodose stem; leaves opposite, slightly amplexicaul, simple, with sessile, undivided, entire lamina; stipules nearly always absent; the inflorescences are dichasia passing over into unipared scorpioid cymes. The flowers are regular, ☿ or unisexual, hypogynous or perigynous, 5-(or 4-) merous with 2–3–4–5 carpels; calyx persistent; corolla polypetalous. The ovary is unilocular (or originally, and sometimes also in the later stages, plurilocular below, e.g. Viscaria), with free styles and 1–several curved ovules on a central, free placenta. The fruit is a nut or a capsule opening apically with long or short valves (teeth, Fig. 362), equal to or double the carpels. For the seeds refer to the family. In Dianthus the embryo is straight.
The flowers which are most complete have Sn, Pn, An + n (obdiplostemonous), Gn where n = 5 (Figs. 360, 361 A) or 4 (Fig. 361 B); the carpels may be placed opposite to the sepals (Fig. 360) or opposite to the petals (Fig. 361 A, B). Without any change taking place in the position of the other whorls, the carpels are next found reduced to 2–3–4 (see the genera); their number may easily be recognised by that of the styles. This is the construction in the majority of the genera in the two first groups. Stellaria media differs considerably. It may have (a) the flower as described above, with G3; (b), the corolla only absent, or (c) only the petal-stamens (A5 + 0, Fig. 361 C), or (d) all these as well as some of the sepal-stamens. The same applies to Sagina, Alsine, Cerastium, and others, and, finally, a series of genera are formed, with certain conditions[365] of reduction which have become constant, and by a gradual series of steps lead to the most reduced form, which has only 5 sepals and 5 (or even as far as only 1) sepal-stamens (Fig. 361 D, E, F).—The PETALS in the Alsineæ are often deeply bifid. The sepal-stamens are most frequently the longest, and bear nectaries at the base (Fig. 363 st). In the most complete forms the ovary has partition-walls in the lower portion (Fig. 360); these do not, however, reach to the top, and generally soon disappear. The ovules, when numerous, are situated on the placenta in as many double rows as there are carpels. In the number of ovules a reduction from many to 1 takes place (Fig. 361). A comparison proves that the “free, centrally placed” placenta is formed by the ventral portion of the carpels. The single basal ovule in Herniaria (Fig. 364), Scleranthus, and others, is also borne on the carpels.
The vegetative branching is characteristic. One of the leaves in a pair is formed before the other, and has a more vigorous axillary bud; these stronger leaves stand in a ¼-spiral, the fifth above the first one, and the branches are consequently arranged in the same manner. In the inflorescence, however, it is the upper or second bracteole (β) whose axillary bud (w in Fig. 361) is most advanced. The bud of the first bracteole (α) becomes sometimes entirely suppressed, or in some this bracteole itself is suppressed.
Fig. 360.—Diagram of Lychnis: α, β bracteoles.
Fig. 361.—A-F Diagrams of flowers of the Caryophyllaceæ: A Agrostemma; B Sagina; C Stellaria; D Corrigiola; E Paronychia; F Herniaria.
The most original type appears to be represented by the Alsineæ. From this form on one side the Sileneæ, adapted in a higher degree for insect-pollination, are developed, and on the other side the Paronychieæ, with various reductions.
1. Alsineæ, Stitchwort Group. Sepals free, and connected with them stellately expanded, slightly unguiculate (white or inodorous)[366] petals; these, however, often become suppressed (Fig. 363). The fruit is a capsule.
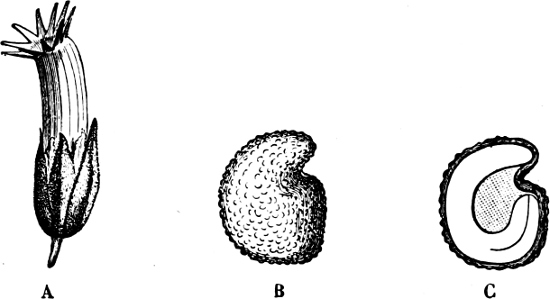
Fig. 362.—Cerastium arvense: A fruit; B seed; C section of seed.
a. As many carpels as sepals (4 or 5). Cerastium (Chickweed). The petals are bifid. Capsule cylindrical, frequently curved at the top, and opening by 10 teeth (Fig. 362).—Malachium differs only in the 5-toothed capsule with bifid teeth.—Spergula (Spurry). The petals are not bifid, capsule 5-valved; seeds winged. The leaves are linear, and appear as if placed in large numbers in a whorl, a branch being situated in the axil of each with leaves placed very close together at its base; stipules membranous.—Sagina has Sn, Pn, An + n, or An, Gn, where n = 4 or 5. The corolla is often wanting.
b. 3 (rarely 2) carpels (Fig. 361 C). Stellaria (Stitchwort) has deeply cleft petals. The number of stamens varies (see above).—Arenaria has entire petals. (To this group belong Alsine, Moehringia, Halianthus, or Honckenya (Fig. 363), which differ from each other, especially in the form of the seed and number of the capsular valves.) Spergularia has membranous stipules, as in Spergula.—Holosteum.
Fig. 363.—Arenaria (Halianthus) peploides: ♀-(A) and ♂-flower (B, C).
2. Paronychieæ (Figs. 361 D, E, F; 364). Small, greenish[367] plants. The leaves, in the majority, are opposite, with membranous stipules. The flowers are most frequently arranged in small dichasia; they are small and insignificant, perigynous (Fig. 364) or hypogynous. The corolla is in most cases wanting, and when present is very small; in general the calyx-stamens are developed, but the corolla-stamens may be represented by small scales (Fig. 364). Ovary most frequently with 1 ovule. Fruit, a nut, rarely a capsule; it is enclosed by the strongly perigynous floral axis (torus).
Scleranthus (Knapwell) is perigynous with bell-shaped torus; no corolla; corolla-stamens are wanting or rudimentary; some calyx-stamens may also be absent.—Corrigiola (Fig. 361 D); Illecebrum; Paronychia (Fig. 361 E); Herniaria (Figs. 361 F, 364).
3. Sileneæ, Pink or Carnation Group. This has a gamosepalous calyx and unguiculate, white or red, petals, with outgrowths (ligule, corona, paracorolla) at the throat of the corolla. These structures are not found in the other groups, and are merely outgrowths at the junction of the limb and claw. The corolla, stamens and ovary are frequently raised above the calyx, upon a lengthened internode (gynophore). The flower has S5, P5, A5 + 5; fruit a capsule with many seeds.
a. 5-(rarely 3–4) carpellate ovary.—Lychnis (Campion, Fig. 360). The corolla is longer than the calyx; corona present. The capsule is 10- or 5-toothed, completely 1-chambered or 5-chambered at the base,—the genus has been divided accordingly into several genera: Melandrium, Lychnis, Viscaria. Some species are unisexual by the abortion of stamens or carpels (L. vespertina, diurna). Agrostemma (A. githago, Corn-cockle, Fig. 361 A) has a long-toothed calyx, the teeth exceeding the corolla; corona absent; 5-toothed capsule.
Fig. 364.—Herniaria glabra: a1 flower; b1 longitudinal section through the flower; c1 stigma with two pollen-grains.
b. Tricarpellate.—Silene (Catch-fly). Six-toothed capsule; corona present in the majority.—Cucubalus has berry-like fruits which finally become dry but do not dehisce.
c. Bicarpellate (2 styles, 4-toothed capsule).—Dianthus (Pink); at the base of the calyx 1–several pairs of floral-leaves are situated;[368] corona absent. The straight embryo is a peculiar exception.—Gypsophila has a campanulate, open calyx, 5-nerved, membranous between the nerves; corona absent; the flowers are generally small and numerous, in a large, paniculate dichasia.—Saponaria (Soapwort) has corona.
Pollination. Alsineæ has ordinary nectaries at the base of the calyx-stamens (Fig. 336): they are frequently protandrous but may often, in the absence of cross-pollination (in the less conspicuous species) pollinate themselves. Their open flowers are accessible to many kinds of insects (particularly flies and bees). Gynodiœcious flowers are found in several species, and the ☿-flowers are then generally more conspicuous than the ♀-flowers. That the ♀-flowers have descended from ☿-flowers is seen by the large staminodes found in them (Fig. 363). Arenaria peploides is diœcious (Fig. 363). The Sileneæ are as a rule adapted for pollination by insects with long probosces—especially butterflies,—and they are frequently protandrous, so that at first the calyx-stamens open, later on the corolla-stamens, then the stigmas expand. The honey is secreted by a ring-like nectary round the base of the ovary or by nectaries at the base of the stamens. Some only blossom and emit scent at night or in the evening (Lychnis vespertina, Silene nutans, Saponaria officinalis) and, like other night-flowers, are of a white or pale colour.
Distribution. 1,100 species, especially in temperate climates, fewer in the colder zone, less still in the Tropics. The Paronychieæ are especially found in dry, sandy fields.
Uses. “Soap-root” (with Saponin, forming a lather in water) from Saponaria officinalis was formerly officinal, and Gypsophila struthium. The seeds of Agrostemma githago are said to be poisonous.—The following are ornamental plants: species of Pinks (D. caryophyllus, garden Pink, often with double flowers; D. barbatus, plumarius, etc.). Lychnis, Gypsophila, Silene, Cerastium (C. tomentosum as edging for borders), Saponaria officinalis (often coronate).—Spergula arvensis is sometimes cultivated.
Order 2. Amarantaceæ. The flowers are essentially the same as in the Chenopodiaceæ and the extremely reduced Caryophyllaceæ (Fig. 361 F); they are regular, hypogynous, generally ☿, have 5 free (rarely slightly united) perianth-leaves; in front of these 5 stamens, which are often united at their base into a shorter or longer tube and have stipule-like teeth between them (the division Gomphreneæ has 2-locular anthers, each of which opens longitudinally); and a 2–3 carpellate gynœceum with one loculus and most frequently one, more rarely several ovules; the fruit is a nut, more rarely (in Celosia, Amarantus, Gomphrena) a capsule, dehiscing irregularly, or like a pyxidium. The characters which especially separate them from the allied orders are found in the perianth. The perianth-leaves are not green and herbaceous, but membranous, dry, and often coloured; they are frequently produced into a bristle or awn; they have also both subtending floral-leaves and 2 large bracteoles similar to the perianth; all these dry leaves persist without alteration after the withering of the flower.—The flowers are without scent. They are arranged in spike- or capitulum-like inflorescences; sometimes placed singly, sometimes aggregated in the panicle-like inflorescences; in others, on the contrary, in dichasia. The[369] majority are herbs, some are shrubs. The leaves are scattered, or opposite, but always simple and without stipules; some are smooth, others hairy.
450 species; especially in the Tropics, principally S. Am. and E. Ind.: few are found outside these countries.—Only a few are used; some, chiefly E. Indian species, are cultivated as ornamental plants: Amaranthus (Fox-tail); Gomphrena globosa; Celosia cristata (Cock’s-comb) remarkable for its fasciated inflorescence; Alternanthera. Some are employed as culinary plants in the Tropics, and in a few of the E. Indian species the seeds are farinaceous, and used for food.
Order 3. Chenopodiaceæ. Generally herbaceous plants like the Caryophyllaceæ, but the leaves are arranged spirally (except Salicornia), and are simple, exstipulate; they are generally fleshy and like the stem “mealy,” that is, covered with small hairs, whose large spherical terminal cell readily falls away; otherwise they are seldom hairy. The inflorescences are generally flower-clusters borne in panicles. Bracteoles generally absent. Flowers generally unisexual: with the single exception of Beta the flowers are hypogynous; they are regular, small and inconspicuous, with single, green, 5-leaved, but more or less united perianth; 5 stamens opposite the perianth, and a 2–5-carpellate, unilocular ovary with 1 basal, curved ovule; but in some genera the number of the perianth-leaves and stamens is reduced to 3–2–1–0. The fruit is generally a nut,—thus this flower and fruit are the same as in the reduced Caryophyllaceæ (Fig. 361 F). The seed is similar to that generally found in the family (for exceptions see the genera).
The floral diagram most frequently present is the same as in Fig. 361 F. There is no indication of corolla or of corolla-stamens, which may be supposed to have belonged to the plant, but which are now entirely and completely suppressed. This order appears to have been an offshoot from the Caryophyllaceæ.—The perianth persists after the withering of the flower, and envelopes the nut; it is very variable, and, together with the position of the seed, the form of the embryo, the sex of the flowers, etc., gives the characters of the genera.
1. Chenopodieæ, Goosefoot Group (Fig. 365), has ☿ (or polygamous) flowers, with regular 5-parted perianth (C); the embryo is ring-like (H). The leaves have the ordinary flat forms.—Chenopodium (Goosefoot). The flower is hypogynous, and the fruit (which is compressed) perfectly free; Mulberry-like collections of fruits are formed in some species (sub-genus Blitum) by the perianth becoming finally fleshy and coloured.—Beta (Beet, Mangold, Fig. 365) differs from all genera in the perianth, which finally becomes cartilaginous, being epigynous (D). Small, most frequently 2–3-flowered clusters without bracteoles, situated in a[370] long, interrupted axis (A, B); the flowers and fruits in each cluster are more or less united individually, and fall off together—they are commonly known as seeds (E, F). The seed lies horizontally.—Hablitzia (H. tamnoides).

Fig. 365.—Beta vulgaris.

Fig. 366.—Salsola soda: embryo.
2. Salsoleæ, Saltwort Group, has cylindrical or semi-cylindrical leaves. Perianth as in the preceding group; the fruit is most frequently compressed. The two first mentioned genera differ from most of the others in the order in having a spirally-coiled,[371] and not a ring-like embryo, so that the endosperm is slight or wanting (Fig. 366). These plants are sometimes placed as a group by themselves, Spirolobeæ—in contradistinction to which the others are termed Cyclolobeæ.—Salsola (Saltwort); leaves subulate, with spiny tips; the flowers have 2 spinous bracteoles: during the ripening of the fruit a tough leathery wing is developed transversely to the back of the perianth.—Chenopodina deviates from Chenopodium chiefly in the embryo and want of endosperm.—Kochia has a somewhat similar perianth to Salsola, but a ring-like embryo; it differs from the others in being hairy.
3. Salicornieæ, Glasswort Group. Salicornia (Glasswort) has a very different appearance. The stems are succulent, jointed, and almost leafless; the leaves opposite, very small, sheath-like and connate; there is a depression in the axil of each leaf, in which a small 3-flowered dichasium without bracteoles is sunk; the flowers have a trimerous perianth, 1–2 stamens and 1 carpel. No endosperm. S. herbacea on clayey beaches.
4. Atripliceæ. This group has most frequently unisexual flowers; the ♂-flower has a 4–5 partite perianth, but the ♀-flower differs from it. Atriplex is monœcious or polygamous, the ♀-flower is naked, but has 2 large, herbaceous bracteoles which expand during the ripening of the fruit, and often become warted and fringed, enveloping the compressed nut. The section Dichospermum has two kinds of ♀-flowers, one like those just described, the other similar to the Chenopodium-flowers, which have been deprived of their stamens, and the fruits of which are depressed, not pressed together from the sides; some (e.g. A. hortensis) have even three kinds of nuts. All the flowers of Atriplex, which present vertical fruits, are accessory shoots, which stand beneath the ordinary flower-clusters, a rather singular relation.—Spinacia (Spinach) is diœcious; ♂-flower: perianth, 4 (-5); stamens, 4 (-5); ♀-flower: tubular, 2–4-partite perianth, hardening during the ripening of the fruit, and uniting with the compressed nut; in S. oleracea, it also forms thorns; 4 long stigmas.—Halimus has the 2 long bracteoles almost entirely united and ultimately adhering firmly to the fruit.
5. Baselleæ. A somewhat exceptional group with more or less perigynous flowers and 2 bracteoles. Basella, Boussingaultia, Ullucus. The perianth is sepaloid; ovary 1-ovuled. In Basella the perianth is fleshy, enveloping the nut, and the cotyledons are so rolled together that a tranverse cut divides them in two places (as in Spirolobeæ). Herbaceous climbing plants.
Pollination. Wind-and self-pollination, as far as is known; the insignificant flowers, devoid of honey, appear to exclude insect-pollination.—520 species. Most of them are annual (out of 26 native species only 5 are perennial);[372] inhabiting salt-marshes and salt-steppes, and growing as weeds (most frequently on garden or field soil containing manure) in this country, especially species of Chenopodium and Atriplex. The majority are found outside the Tropics, and play a very important part, for example, in the Asiatic salt-steppes. They grow gregariously in large masses.
Uses. Comparatively few. The only important one is Beta vulgaris (from the Mediterranean basin), with its different varieties, viz. Beet-root, Cattle-beet or Red-beet, Sugar-beet, and others. These are biennial, making in the first year a root which acts as a reservoir of reserve material, with a rosette of leaves, and in the second year using this material in the production of a long stem, leaves and flowers. The primary root has been developed by cultivation into a very thick and fleshy tap-root; its mode of increase in thickness deviates from that of other roots, concentric rings of vascular bundles being formed from a cambial ring developed outside the previous ring. In this way several rings of vascular bundles separated by medullary rays, alternating with rings of parenchyma, may be found in the root of a Beet. Besides Beta vulgaris, var. hortensis (Beet-root), the following are also cultivated: var. cicla (Leaf-beet, “Mangold,” or “Roman Spinach ”), Spinacia oleracea and Atriplex hortensis as Spinach; a form of the latter and of Spinach are grown as ornamental plants. The tubers of Ullucus tuberosus are used as potatoes; Chenopodium quinoa, in Chili and Peru, is an important farinaceous plant. Soda is made from some (Salsola kali, Chenopodina maritima and others). Aromatic properties are rare: Chenopodium ambrosioides and botrys.
Order 4. Batidaceæ. Batis maritima, a bushy West Indian maritime plant.
Order 5. Phytolaccaceæ. The ☿ (sometimes unisexual), regular, sometimes slightly perigynous flowers are inconspicuous and have a single sepaloid or coloured 4–5-leaved perianth (generally united at the base); stamens either in 1 whorl in the spaces between the perianth-leaves or in 1 whorl opposite the perianth-leaves, or in 2, one of which alternates with these; but the number may be increased by the splitting of one or of both the whorls to as many as 10–15–20–25. Carpels sometimes only one, sometimes many (4–10) placed in a whorl, either free or united into a gynœceum with a corresponding number of loculi in the ovary; but in all cases each carpel bears only its own style and 1 ovule. The fruit is a berry (or nut, capsule, or schizocarp).—Mostly herbs or herbaceous shrubs, with scattered, simple leaves without stipules (Petiverieæ; have stipules). Inflorescences, most frequently racemes or spikes, which in some instances are apparently placed opposite to a leaf, being displaced by a more vigorous growth of the axillary bud. Embryo always bent.—Petiveria has a straight embryo with rolled cotyledons.—Phytolacca, Pircunia, Microtea, Seguieria, Rivina (Pr4, A4, G1; berry), and others.
The following plant is, with some doubt, placed near this order: Thelygonum cynocrambe; monœcious. ♂-flowers: perianth, 2-leaved; stamens indefinite. ♀-flowers: perianth-leaves united, 3-toothed; G1, style gynobasic. Fruit a drupe. An annual plant; Mediterranean. Branching anomalous.
About 90 species; in tropical and temperate countries, principally America and Africa.—The red juice in the fruits, especially of Phytol. decandra, is used for colouring wine.
[373]
Order 6. Portulacaceæ (Portulacas). The flowers are regular (except Montia), hypogynous (except Portulaca) and ☿. The diagram which applies to the majority of genera is that in Fig. 367, but with all the 5 stamens completely developed: it may be considered as the Chenopodiaceous diagram with the addition of 2 bracteoles in the median line (m-n, these by some are considered as sepals), and with a petaloid perianth (usually designated “corolla”). The “petals” fall off very quickly, and are sometimes wanting. Most frequently 5 stamens, situated opposite the “petals,” but in other genera the number varies; Montia has only 3 stamens (by suppression of the two anterior and lateral, Fig. 367), others again have more than 5, some a large and indefinite number. This may be explained partly by the appearance of a second whorl of stamens alternating with the first, and partly by the splitting (dédoublement) of the stamens. Gynœceum most frequently tricarpellate, ovary unilocular with 1–several basal ovules (sometimes on a branched placenta, as in certain Caryophyllaceæ). The fruit is a capsule, more rarely a nut.—The majority are annual herbaceous plants with scattered, entire leaves, often fleshy and smooth, with or without rudimentary stipules (dry, membranous, modified into hairs). Inflorescence cymose.
Fig. 367.—Montia.
Diagram of flower.
Portulaca (Portulaca): flower, epigynous or semi-epigynous; fruit, a pyxidium. The stamens vary in number, and are most frequently placed in groups (in consequence of splitting) opposite the petals.—Montia: the corolla is slightly gamopetalous, but cleft on the posterior side (Fig. 367), and as a consequence of the larger size of the lateral petals, slightly zygomorphic; 3 stamens.—Calandrinia; Talinum; Anacampseros; Claytonia.
125 species; mostly in warm and temperate countries, especially the arid parts of S. Am. and the Cape. Montia fontana (Blinks) is a native plant. Portulaca oleracea is cultivated as a pot-herb in the south of Europe. A few species of Portulaca and Calandrinia are ornamental plants.
Order 7. Nyctaginiaceæ. The characteristic feature of this order is the single, regular, united, and often petaloid perianth, the lower part of which generally persists after flowering and embraces the fruit as a false pericarp. The upper portion is most[374] frequently valvate and folded, or simply valvate in æstivation. The number of stamens varies. The free gynœceum is unicarpellate and has 1 ovule. The fruit is a nut, but becomes a false drupe, since the lower persistent portion of the perianth becomes fleshy (as in Neea, where this fleshy part is almost always crowned by the upper persistent part of the perianth. In the majority of the Mirabileæ the lower part becomes the dry anthocarp, while the upper petaloid part falls away after flowering). Finally, a peculiar involucre is formed around the flowers by free or united floral-leaves.—The majority are herbs, some are trees (Pisonia, etc.); Bougainvillea is a liane. The stems are often nodose and swollen at the nodes; the leaves are simple, penninerved, scattered, or opposite, without stipules. In some, the vascular bundles are scattered; stem anomalous.
Mirabilis; the structure of the stem is abnormal. Dichasial branching with continuation from the second bracteole, thus forming unipared scorpioid cymes. The perianth is petaloid, funnel-shaped, and has a folded and twisted æstivation resembling that of the corolla of the Convolvulaceæ; the upper coloured portion falls off after the flowering. Outside, and alternating with it, is a 5-partite, sepaloid involucre of 5 spirally-placed floral-leaves.—Oxybaphus; the involucre envelops 1–3 dichasial flowers.—Bougainvillea; the involucre is rose-coloured, 3-leaved, and envelops 3 flowers (placed laterally; the terminal flower wanting). The leaves of the involucre in Boerhaavia, Pisonia, Neea, and others are reduced to teeth or scales.
157 species; mostly in tropical countries, and especially S. Am. Species of Mirabilis (Am.) are ornamental plants. Theïn is found in Neea theïfera Oersted (discovered by Lund in Lagoa Santa, Brazil), which may be used as a tea-plant.
Order 8. Aizoaceæ. Only 3 whorls are found in the flower, which alternate with one another when their leaves are equal in number. The first is sepaloid, the third one the carpels, and the intervening one is either uncleft, in which case it is developed as stamens, or it is divided into a large number of members which then all become stamens (arranged in groups), or the outermost ones become developed as petals. The fruit is most frequently a capsule with several loculi. Most of the species are herbs with thick, fleshy stems, and exstipulate leaves. The structure of the stem is usually anomalous.
1. Aizoideæ have hypogynous or perigynous flowers with (4–) 5 perianth-leaves; stamens single, or (by splitting) in groups of 2–3, alternating with the perianth-leaves. The gynœceum (with 3–5 carpels) has 3–5 loculi in the ovary, and most frequently numerous ovules in each loculus, borne on the central placenta formed by the edges of the carpels. The fruit is a capsule. The[375] inflorescences are dichasia and unipared scorpioid cymes.—Aizoon, Mollugo, Sesuvium, and others are herbs or bushes, most frequently hairy.
2. Mesembrianthemeæ have semi- or wholly-epigynous flowers.—Tetragonia. The perianth is 4 (more rarely 3–5–6)-merous. Stamens single, or (by splitting) in groups alternating with the perianth-leaves. There is an indefinite number of carpels, and each loculus of the ovary contains only 1 pendulous ovule. Fruit a nut or drupe. The flowers arise singly in the leaf-axils, with an accessory foliage-bud below them; in some instances there is also an accessory flower between this bud and the flower. Southern hemisphere, especially at the Cape; T. expansa, New Zealand Spinach, is a fleshy plant which is cultivated as a pot-herb (Japan, Austr., S. Am.).—Mesembrianthemum: the flowers are 5-merous; the numerous linear petals and the still more numerous stamens all arise by the splitting of 5 or 4 protuberances (primordia) alternating with the sepals. The ovary presents another characteristic peculiarity: the carpels alternating with the 5–4 stamens form an ovary (with several loculi) with the ovules at first borne, as in other cases, on the inner corner of the inwardly-turned carpels; but during the subsequent development the whole ovary is so turned round that the placentæ become parietal and the ovules assume, apparently, a position very rarely met with in the vegetable kingdom: on the dorsal suture of the carpels. Shrubs or under-shrubs, more rarely herbs with fleshy stems and simple, entire, more frequently thick or triangular leaves, containing a quantity of water. The flowers open about noon, and are brightly coloured, generally red or red-violet, but odourless. The capsules dehisce in rainy weather. 300 species, mostly found at the Cape. Some are ornamental plants. M. crystallinum (the Ice-plant) and others are covered with peculiar, bladder-like, sparkling hairs, the cell-sap of which contains salt—these serve as reservoirs of water.
The position of this family is very doubtful; but it seems in many respects to approach Mesembrianthemum. Some botanists place it near to the Ribesiaceæ; others, again, to the Passifloraceæ. Only 1 order.
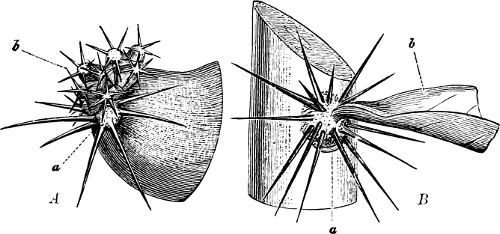
Fig. 368.—A Echinocactus: a position of a leaf-lamina; b a lateral shoot on the displaced axillary bud. B Pereskia: b a foliage-leaf on a small thorny branch which is subtended by a foliage-leaf which has fallen off and left a scar(a).

Fig. 369.—Echinopsis.
Order Cactaceæ (The Cacti). The flower is epigynous, ☿, regular, and remarkable for its acyclic structure; there are, for instance, a large number of spirally-placed sepals and petals, which gradually pass over into one another, and which in some species, to a certain extent, arise from the walls of the ovary as in Nymphæa (Fig. 383 A, B). The petals are free; rotate, opening widely in Opuntia, Pereskia, and Rhipsalis; erect and united at their base into a shorter or longer tube in Cereus, Epiphyllum, Mammillaria, Echinocactus, Melocactus, and others (Fig. 369). Stamens numerous, attached to the base of the corolla; gynœceum formed of many carpels, with one style, dividing into a number of branches corresponding to the number of carpels; the ovary has one loculus with[376] many parietal placentæ; the ovules are anatropous, on long and curved funicles. Fruit a berry with exendospermous seeds. The fruit-pulp is mainly derived from the funicles.—The external appearance of the Cactaceæ is very peculiar; Pereskia, which has thick and fleshy leaves (Fig. 368), deviates the least; foliage-leaves of the usual form are wanting in the other genera, or are usually very small, and quickly fall off and disappear (Opuntia), or are modified into thorns; the stem, without normal foliage-leaves,—so characteristic a feature in this order,—makes its appearance after the two normally developed cotyledons. The stems are fleshy, perennial, and may finally become woody. In some they are elongated, globose, pointed, and more or less dichotomously branched, e.g. in several of the Rhipsalis species, which live mostly as epiphytes on trees; in others, elongated, branched, globose, or, most frequently, more or less angular (prismatic) or grooved and provided with wings, and either columnar and erect (as much as about 20 metres in height and 1 metre in circumference,[377] as in C. giganteus in New Mexico) or climbing by roots (Cereus and Rhipsalis-species); in others again, compressed, more or less leaf-like, often with a ridge in the centre (winged), branched and jointed: Epiphyllum, Phyllocactus, Opuntia, some species of Rhipsalis; others are thick, short, spherical or ovoid, unbranched or only slightly branched, and either studded with prominent warts (mammillæ) each of which supports a tuft of thorns (Fig. 368 A; Mammillaria and others) or with vertical ridges, separated by furrows (rows of mammillæ which have coalesced) in Melocactus, Echinocactus, Echinopsis (Fig. 369); at the same time the ovary in some is embedded in the stem so that leaves or leaf-scars, with tufts of thorns in their axils, may be observed on the ovary just as on the stem.—The flattened shoots of the Cactaceæ are formed in various ways, either by the compression of cylindrical axes (Opuntia) or, as in Melocactus, etc., from winged stems in which all the wings are suppressed except two.
The thorns are produced directly from the growing points of the axillary buds, and are modified leaves. The axillary bud is united at its base with its subtending leaf, which as a rule is extremely rudimentary; and these together form a kind of leaf-cushion, larger in some genera than in others. This leaf-cushion attains its highest development in Mammillaria, in which it is a large, conical wart (see Fig. 368 A), bearing on its apex the tuft of thorns and rudimentary lamina.—The seedlings have normal cotyledons and a fleshy hypocotyl.
All the species (1,000?) are American (one epiphytic species of Rhipsalis is indigenous in S. Africa, Mauritius and Ceylon), especially from the tropical table-lands (Mexico, etc.). Some species, especially those without thorns, as Rhipsalis, are epiphytes. Opuntia vulgaris, the fruits of which are edible, is naturalized in the Mediterranean. The cochineal insect (Coccus cacti) lives on this and some closely allied species (O. coccinellifera, etc.), particularly in Mexico and the Canary Islands. Several are ornamental plants.
The flowers as a rule are ☿, regular and hypogynous; however in some orders they are unisexual, e.g. in the Myristicaceæ, or zygomorphic (in Monkshood and Larkspur in the Ranunculaceæ); in the Lauraceæ, (Fig. 386) for example, perigynous, and in Nymphæa (Fig. 383) even partially epigynous flowers are typical.—The flowers are acyclic in very many of the genera of the two first orders, if not completely so, at any rate in the numerous stamens and carpels, thus denoting an old type. It is a remarkable characteristic that in the majority of the orders the number 3 prevails in the calyx and corolla; the number 5 also occurs, but the[378] number 2 is seldom met with. Most orders have a double perianth; chorisis does not occur, suppression is rare, and the parts of the flower are developed in acropetal succession. The most characteristic feature in the order is the free, one-leaved, as a rule numerous carpels (apocarpous gynœceum). The number of carpels in some of the last mentioned orders dwindles down to 1 (e.g. the Berberideæ and Myristicaceæ). The carpels in Nymphæaceæ become united into one pistil (syncarpous), a condition which we also find distributed among the other orders.
Endosperm occurs in almost all the orders (except e.g. Lauraceæ). The nutritive tissue in Cabombeæ and Nymphæeæ is chiefly perisperm.
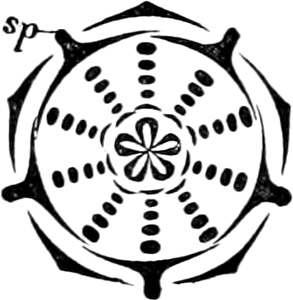
Fig. 370.—Diagram of Aquilegia vulgaris: sp spur. A cyclic flower.
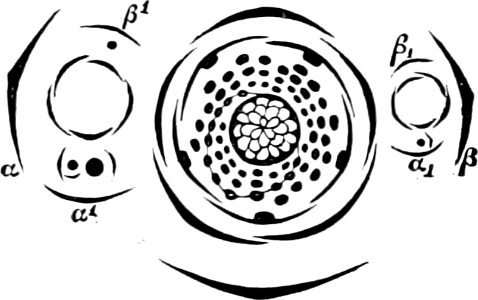
Fig. 371.—Diagram of a dichasium of Ranunculus acer: α1, α1, and β1, β1, bracteoles (the buds in the axils of the bracteoles, α and α1, are continued antidromously). The flower has cyclic calyx and corolla, bub acyclic (8/21) stamens.
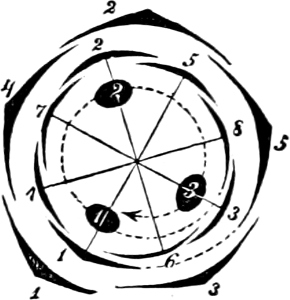
Fig. 372.—Diagram of an acyclic Ranunculaceous flower (only 3 stamens are indicated). The spiral of the sepals has a divergence of 3/5; that of the corolla and subsequent leaves 3/8.
Order 1. Ranunculaceæ. Nearly all are herbs (except Clematis). The leaves are scattered (except Clematideæ), they have a large sheath with broad base (no stipules), and are most frequently palminerved with palmate lobes. The flowers are hypogynous, with most frequently a well pronounced convex receptacle (Figs. 374 B, 380), ☿, regular (except Delphinium and Aconitum); their structure varies very much; in some the leaves are verticillate, in others arranged spirally; in others, again, both modes of arrangement are found. It is a characteristic feature that the various series of leaves (especially calyx and corolla) are not so distinct or so sharply divided as is usual. The leaves of the perianth are free, imbricate (except Clematideæ); stamens numerous, with most frequently extrorse anthers; gynœceum free, apocarpous (except Nigella and partly Helleborus), with 1 or several ovules (Figs. 373, 378, 379) borne on the ventral suture. The fruit is[379] either a nut or a follicle (Actæa has berries). The seed has a large, oil-containing endosperm and a small embryo (Fig. 374).
The main axis generally terminates in a flower, and the lateral axes branch in a cymose manner (Fig. 371). The flowers show the following differences in construction: VERTICILLATE (EUCYCLIC), i.e. constructed all through of alternating whorls: Aquilegia (Fig. 370), Xanthorhiza, and sometimes Eranthis. Semiverticillate (HEMICYCLIC) i.e. with sepals and petals in alternate whorls, and the others arranged spirally: Ranunculus (Fig. 371), Myosurus, Pæonia and several other genera entirely, or in certain species only. Spiral-flowered (ACYCLIC) i.e. all the leaves are arranged spirally, so that sepals and petals do not alternate the one with the other, even though they are the same in number: Adonis (Fig. 372), Aconitum, Delphinium-species, Nigella-species, Helleborus. The leaves of the calyx are in this instance arranged on a spiral of 2/5; those of the corolla on 2/5, 3/8, 5/13 or 8/21, and stamens and carpels likewise on higher fractions of the same series.
The genera Caltha, Anemone, Thalictrum and Clematis have a single perianth, which is most frequently petaloid; it is thus apparent that the sepals are petaloid, and the leaves, which in other genera have developed as petals, are in these instances stamens. The calyx is similarly petaloid in the genera Helleborus, Eranthis, Nigella, Delphinium and Aconitum; but the petals are present in these instances in unusual (horn-like) forms, and almost entirely given up to the function of nectaries, a function they already possess in Ranunculus. According to a more recent theory the “honey-leaves” are transformed stamens, which have lost the function of reproduction; the perianth is then single, and most frequently petaloid. [Those leaves in the flowers of many Ranunculaceæ which bear nectaries are termed by Prantl honey-leaves, and comprise those leaf-structures of the flower whose essential function lies in the production of nectar, and which, independent of the differentiation of the perianth into calyx and corolla, are derived from the stamens by the loss of their reproductive functions. Clear transitional forms are found between the two series of the perianth (e.g. between the sepaloid and petaloid perianth-leaves of Anemone japonica, A. decapetala, Trollius-species) while transitional forms are never found between perianth-and honey-leaves (with the exception of Aquilegia vulgaris, var. stellata). In Anemone and Clematis the honey-leaves pass gradually into the stamens, and agree with the stamens in the other Ranunculaceæ in their arrangement, development, and scant system of veins (except Nigella). In Delphinium, sect. Consolida, the two honey-leaves placed in front of the unpaired perianth-leaf are united into one, as shown by the veins (twice three veins arranged symmetrically). The honey-leaves of Aquilegia, Callianthemum, and the majority of the Ranunculus-species serve by reason of their large circumference, as organs of attraction, and on this account are considered as petals by other authors.—The same position in the flower which the honey-leaves assume is found occupied by staminodes, without nectar, in some Coptis-species, in Anemonopsis, Actæa sect. Euactæa, (e.g. A. racemosa), Clematis sect. Atragene; in the last-named they closely surround the stamens, in Actæa they are petaloid.—A perianth, sharply differentiated into calyx and corolla, and destitute of honey-leaves, is found in Anemone, sect. Knowltonia (Cape),
[380]
Adonis, Pæonia.—The perianth of the Ranunculaceæ is considered by Prantl to be usually petaloid.—The nectaries arise in the Ranunculaceæ (1) on normal stamens (Clematis sect. Viorna), (2) on the honey-leaves (this is generally the case), and (3) on the carpels (Caltha and the majority of Trollius-species).—As the result of his researches upon the Ranunculaceæ, Prantl does not agree with the view advanced by Drude (Schenk, Hand. d. Bot. iii.) that the petals in general have proceeded from the metamorphosis of the stamens (K)].
Fig. 373.—Ovaries in longitudinal section: v the ventral suture; d the dorsal suture: A, B Clematis; C Ranunculus; D Myosurus.
The most primitive form of fruit is undoubtedly the pod formed by one carpel, on the edges of which (along the ventral suture) two rows of ovules are situated: Pæonieæ, Helleboreæ, Delphinieæ (Fig. 379). In a great many genera the number of ovules has been limited to one perfect one, which is placed in the central plane under the united leaf-edges, and sometimes also some barren ovules above it (Fig. 373). The fruitlets in this case become achenes, and are present in much larger numbers than when there are follicles.
Fig. 374.—Helleborus niger: A flower; B receptacle; pet petals (honey-leaves); pi stamens and carpels; C seed; D anther (cross section); alb endosperm.
Fig. 375.—Caltha palustris: fruit.
The following have Follicles: Pæonieæ, Helleboreæ (except Actæa) and Delphinieæ; Achenes: Ranunculeæ, Anemoneæ and Clematideæ.
[381]
A. Follicles (Figs. 375, 379), with many ovules, situated in two rows along the ventral suture. Actæa has berries, Nigella has capsules of several loculi.
1. Pæonieæ, Peony Group. This has regular, acyclic flowers with a normal, most frequently 5-leaved, imbricate calyx; large, coloured petals, and introrse anthers. Slightly perigynous. Surrounding the base of the carpels a ring-like swelling of the receptacle (“disc”) is present, which is largest in P. moutan. The follicles are more or less fleshy or leathery. Mostly herbs, with pinnatisect or decompound leaves and large, solitary flowers; a gradual transition may be traced from the foliage-leaves to the petals. Pæonia; Hydrastis.
Fig. 376.—Aquilegia vulgaris.

Fig. 377.—Caltha palustris (nat. size).
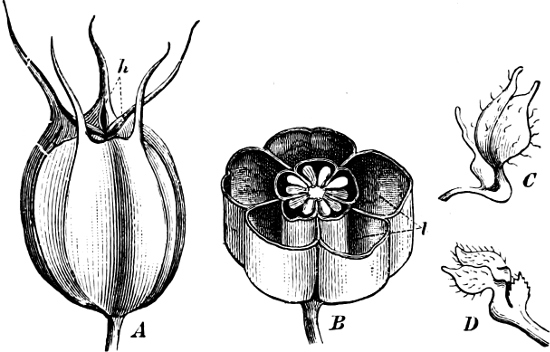
Fig. 378.—Nigella: A, B fruit of N. damascena, entire, and cut transversely. C Petal (honey-leaf) of N. arvensis. D Petal of N. damascena.
2. Helleboreæ, Hellebore Group. This has regular flowers with most frequently a coloured calyx. The petals (honey-leaves) are modified into nectaries; they may be horn-like, provided with a spur, or of a similarly unusual form, or they may be entirely absent. Anthers often extrorse.—Trollius (Globe-flower[36]). The flower is acyclic: many[382] petaloid sepals, succeeding these, most frequently, several linear, dark yellow petals, which bear a naked nectary at the base; finally, many stamens and carpels arranged in a spiral (3/8, 8/21).—Caltha (Marsh-marigold, Figs. 375, 377); 5 (-7) yellow sepals, no petals. The foliage-leaves have a large amplexicaul sheath.—Helleborous (Hellebore) has pedate leaves. The flower is acyclic, with 5 large, regular, persistent, often petaloid sepals (2/5); small, horn-like petals (honey-leaves; most frequently 13, divergence 8/13) and generally few carpels (Fig. 374).—Coptis.—Isopyrum.—Eranthis (Winter Aconite), like Anemone, has a 3-leaved involucre and most frequently trimerous flowers, 6 large petaloid sepals, 6 petals (tubular honey-leaves), 6 oblique rows of stamens, 3–6 carpels. Aquilegia (Columbine, Fig. 376); the flower is entirely cyclic and has large spurs on all the 5 petals (funnel-shaped honey-leaves); S5 coloured, P5, A5 × (8–12), G5 in regular alternation (Figs. 376, 370); the innermost stamens are often staminodes (Fig. 370).—Nigella (Love-in-the-mist, Fig. 378) has 5 sepals and 8 small, two-lipped petals cleft at the apex (the nectary is covered by the under-lip; Fig. 378 C, D). The 5 carpels are more or less completely united; and a many-carpellate ovary with free styles is formed in some. Large air-chambers in the external wall of the ovary are formed in N. damascena (Fig. 378).—Actæa (Baneberry) has coloured sepals, either no petals or an[383] indefinite number, and only 1 carpel. The fruit is a berry (or follicle).—Cimicifuga, Garidella, Xanthorhiza (S5, P5, A5 + 5, G5).
3. Delphinieæ, Larkspur Group. Zygomorphic flowers with coloured calyx; the 2 posterior petals (honey-leaves) are transformed into nectaries, the others are small or absent altogether.—Aconitum (Monkshood); 5 sepals, of which the posterior one (Fig. 379 A) is helmet-shaped; most frequently 8 petals (as in Fig. 372), of which the two posterior ones (honey-leaves) are developed into long-clawed nectaries (Fig. 379 A, k) enveloped by the helmet-like sepal; the others are small, or are to some extent suppressed. Stamens on a spiral of 3/8–5/13; generally 3 carpels. Perennial herbs.—Delphinium (Larkspur); very closely allied to Aconitum, but the anterior 4 petals are most frequently wanting, and the 2 posterior ones have each a spur, which is enclosed by the posterior sepal, the latter being also provided with a membranous spur. Stamens and carpels arranged on a spiral of 3/8, 5/13, 8/21. In D. ajacis and consolida there is apparently only 1 petal (by the fusion of 4) and 1 carpel.
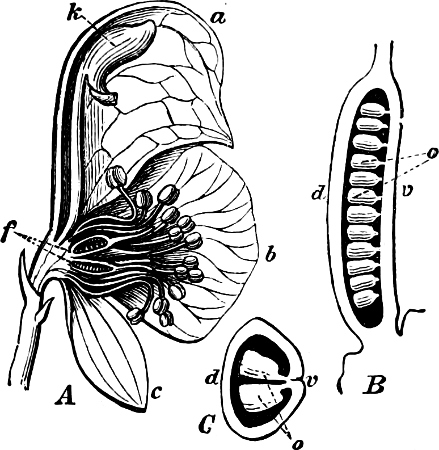
Fig. 379.—Aconitum napellus. A Flower in longitudinal section, below are the 2 bracteoles; a half of helmet-like sepal; b and c other sepals; k nectary; f carpels. B Ovary in longitudinal section; C the same transversely; d dorsal suture; v ventral suture.
B. Fruit achenes. Many carpels, each with only 1 ascending (Fig. 373 C), or pendulous (Fig. 373 D), perfect ovule; often also rudimentary ovules above it (Fig. 373 A, B). Fruit achenes.
4. Ranunculeæ, Buttercup Group, has double perianth. Myosurus and Adonis have pendulous ovules as in Anemoneæ (Fig. 373 D); Ranunculus, with Batrachium and Ficaria, erect ovules (Fig. 373 C) and downwardly-turned radicle.—Ranunculus. Most frequently S5, P5, many spirally-placed stamens and carpels (Figs. 371, 380). The petals (honey-leaves) have a nectary at the base, covered by a small scale. Batrachium, Water Ranunculus, deviates by the achenes being transversely wrinkled; dimorphic leaves. Ficaria has 3 sepals and 7–8 petals arranged in 2/5–3/8. F. ranunculoides (the only species) has tuberous roots, which spring from the base of the axillary buds, and together with these, serve as organs of reproduction. The embryo has only 1 cotyledon.—Myosurus (Mouse-tail) has[384] small prolongations from the 5 sepals; 5 narrow petals which bear the nectaries near the apex; sometimes only 5 stamens, and an ultimately very long receptacle, with numerous spirally-arranged achenes (Fig. 381).—Adonis is acyclic (Fig. 372); most frequently 5 sepals with a divergence of 2/5, 8 petals of 3/8, indefinite stamens and carpels of 3/8 or 5/13. The corolla has no nectary.
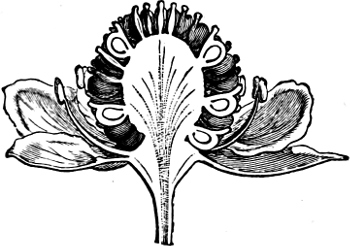
Fig. 380.—Flower of Ranunculus sceleratus in longitudinal section.

Fig. 381.—Myosurus minimus: c cotyledons; m the foliage-leaves; f the floral axis with the carpels, and g the same without; y insertion of perianth.]
5. Anemoneæ, Anemone Group, has a single perianth. (Pendulous ovules (Fig. 373 D), radicle turned upward).—Anemone has a single, petaloid, most frequently 5–6-leaved perianth, and beneath the flower most frequently an involucre of 3 leaves, placed close together in the form of a whorl. In A. nemorosa, ranunculoides, etc., the involucral leaves resemble foliage-leaves; in A. hepatica they are situated close under the perianth, and resemble sepals, and in the sub-genus Pulsatilla they stand between the foliage-leaves and floral-leaves. The style of Pulsatilla finally grows out[385] in the form of a feather. The main axis of A. hepatica has unlimited growth (it is biaxial), and the flowers are borne laterally in the axils of the scale-leaves; in the others (uniaxial) the flower is terminal, and the rhizome becomes a sympodium after the first flowering.—Thalictrum (Meadow Rue) has no involucre; 4–5-leaved, greenish perianth. The receptacle is flat. The stamens are brightly-coloured and have long filaments; 1–5 accessory flowers may occur in the leaf-axils of the panicle-like inflorescence.
6. Clematideæ, Clematis Group. This differs from all the others in the valvate æstivation of the calyx and its opposite leaves. There are 4 (-several) petaloid sepals; petals are absent, or linear (Atragene). Ovule 1, pendulous. Achenes, often with prolonged, feathery style. The majority of the genera are shrubs, and climb by their sensitive, twining leaf-stalks.—Clematis; Atragene.
Pollination. The flowers are conspicuous either by coloured petals (honey-leaves) (Ranunculus, Pæonia) or coloured sepals (Helleborus, Anemone, Caltha, etc.), or by both (Aquilegia, Delphinium), or by the coloured stamens (Thalictrum). Some have no honey (Clematis, Anemone, Thalictrum), and are generally visited by insects for the sake of their pollen. Others have nectaries on the corolla (Ranunculus, Trollius, Helleborus, Nigella, Aconitum, etc.), more rarely on the stamens (Pulsatilla, Clematis-species), or the carpels (Caltha), or the calyx (certain species of Pæonia). The honey is readily accessible in the flat, open flowers, and these flowers also may easily pollinate themselves. There is marked protandry where the honey lies deeply hidden, as in Aquilegia, Delphinium, and Aconitum. Helleborus and some Ranunculus-species are protogynous.
About 680 species; especially in northern temperate climates, and extending to the Polar and Alpine regions. Only the Clematideæ are tropical.
The order has an abundance of acrid, vesicant properties (R. acer, sceleratus, etc.), and poisonous alkaloids (Helleborus niger is poisonous). Officinal: Aconitum napellus (aconitine; leaves and tuberous roots); the rhizome of Hydrastis canadensis from N. Am. (the alkaloid hydrastine). The order, however, is best known for its ornamental plants; almost all the genera have species which are cultivated for their beauty. Sweet-scented flowers are absent.
Order 2. Nymphæaceæ (Water Lilies). Water Plants; generally with large, floating leaves, and large solitary flowers; sepals 3–5, petals 3–∞, stamens 6–∞, carpels 3–∞. The flower is hypogynous, but in the Nymphæeæ different degrees of epigyny are found, and from this fact, as well as from the carpels being united into one pistil, the family forms a lateral offshoot from the Ranunculaceæ, with much greater modification. The seed often has an aril, and, in the majority, a farinaceous nutritive[386] tissue, partly endosperm, partly perisperm (Fig. 383 C). The embryo has 2 thick cotyledons and a small hypocotyl; the plumule is well developed, with 2–4 leaves.
1. Cabombeæ. 3–4 species (Tropical S. Am.), resembling the Water Ranunculus, with two kinds of leaves, the submerged being dissected and the aerial peltate. The flowers are eucyclic, trimerous, with 2–3 free, epigynous carpels. The ovules are situated on the central line of the carpel—an almost unique circumstance. Endosperm and perisperm. Cabomba; Brasenia.
2. Nelumboneæ. The leaves are peltate, raised on long stalks high above the water. Large, hypogynous flowers (Fig. 382); sepals 4–5; petals numerous; stamens numerous; carpels several, distinct. The receptacle is very remarkable, being raised above the stamens, and developed into an inverted conical body on the apex of which the nut-like fruits are embedded in pits. Endosperm is wanting, but the embryo is large and has well developed cotyledons.—Nelumbo, 2 species. N. lutea (N. Am.); N. speciosa (E. Ind.) was sacred amongst the ancient Hindoos and Egyptians, (the Lotus flower); its seeds are used as food.
3. Nymphæeæ, Water Lily Group. The carpels are united into one, many-locular ovary, whose numerous ovules are situated on the surface of the partition walls (as in the Poppies); the stigma is sessile and radiating, the number of rays corresponding to the number of carpels (Fig. 383). The fruit is a spongy berry with many seeds, which have a large perisperm in addition to the endosperm (Fig. 383 C).
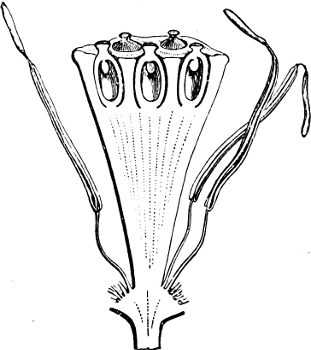
Fig. 382.—Nelumbo nucifera: vertical section through the receptacle.
Sepals, petals, and stamens often pass gradually over the one into the other, the petals becoming narrower by degrees, and bearing anthers on each side of the apex, which gradually become larger anthers in proportion to the filament, until the perfect stamen is developed. The long-stalked leaves are floating, and most frequently cordate, elliptical, leathery, with a shiny surface, sometimes (as in Victoria regia and Euryale ferox) with strongly[387] projecting thorny ribs on the lower surface. In the intercellular passages of the leaves are some peculiar, stellate cells.
Nuphar has 5 sepals, and an hypogynous flower. The petals, which are small, have a nectary on the back; the coloured inner side of the sepals functions as petals; the ovate gynœceum is quite free.—N. luteum is a native plant (Yellow Water-Lily), with, most frequently, 13 petals and 10–16 loculi in the ovary. The rhizome is horizontal, as much as 5–6 cm. in thickness, and bears on its under surface a number of roots, which on dying-off leave deep scars; the leaves are borne in spiral lines, and the flowers are solitary in certain leaf-axils. The construction of the rhizome is very peculiar; the vascular bundles are scattered and closed as in a monocotyledonous stem.
Fig. 383.—Nymphæa: A flower in longitudinal section, the most external leaves being removed; B fruit; C seed of Nuphar (longitudinal section); the perisperm at the base, the endosperm at the top surrounding the embryo.
Nymphæa has 4 sepals, and the flower is more or less epigynous. Petals and stamens are inserted at different heights on the ovary to just beneath the stigma (Fig. 383). Nymphæa alba (White Water-Lily). Victoria regia from the Amazon, and Euryale ferox from Asia, have entirely epigynous flowers. The shield-like leaves of Victoria are as much as 2 metres in diameter, and the edge is bent up to a height of 5–14 cm.; the flowers are 20–40 cm. in diameter, and change in twenty-four hours from white to rose-red. A development of heat, as much as 14°C. above the temperature of the air, together with a strong formation of carbonic acid, has been observed during flowering.
Pollination. Nymphæa alba and other species of the sub-genus Symphytopleura are self-pollinated; the sub-genus Leptopleura is insect-pollinated. Nuphar and Victoria can effect self-fertilisation; Euryale is self-fertilised, often in entirely closed and submerged flowers.—The dissemination of the seeds in Nuphar luteum is effected by the fruit, which rests on the water, becoming detached[388] from its stalk, and dehiscing from the base upwards so that the seeds are set free; while in Nymphæa alba the spirally-twisted stalk draws the fruit under water, and it dehisces by its upper part being thrown off as a hood, and the seeds which are enclosed in air-tight sacs rise to the surface of the water. In this condition they are able to float and can only sink to the bottom when the air has disappeared.
53 species; in fresh water in all parts of the world, but especially in the Tropics.—The rhizomes and seeds of some may be used as food; Euryale ferox is even cultivated. Nymphæa cœrulea and Lotus were sacred among the Egyptians.
Order 3. Ceratophyllaceæ. About 3 species. Aquatic plants, submerged, rootless; leaves cartilaginous, verticillate, dissected into repeatedly dichotomous branches which are finely toothed; only one of the leaves in a whorl supports a vegetative branch. The flowers are monœcious, axillary. Inside the 6–12 perianth-leaves are situated in the ♂-flower 10–20 stamens with thick connective, and in the ♀-flower a gynœceum formed by one carpel, with one orthotropous and pendulous ovule, which has only one integument. Fruit a nut, which, in some species, bears on each side a pointed horn, and at the apex a similar one, formed by the persistent style.—The embryo has an unusually well developed plumule with several whorls of leaves. The plant is rootless throughout its whole life.—Ceratophyllum (Horn-wort).
Order 4. Annonaceæ. Sepals 3; petals 3+3 (most frequently valvate); succeeding these (as in the Ranunculaceæ) are numerous acyclic stamens and an apocarpous gynœceum; the flowers are hypogynous, regular and ☿, generally very large (2–3 cm. in diameter), and the leaves of the perianth are more or less fleshy or leathery. The majority have syncarps with berry-like fruitlets, but in Annona and some others the carpels fuse together into a large, head-like fruit—a kind of composite berry. The seeds have ruminate endosperm as in Myristica.—Trees or shrubs with alternate, simple, entire, penninerved leaves without stipules. 450 (700?) species; especially tropical. The best known are Anona cherimolia, squamosa and reticulata (all from America) cultivated on account of their large, delicious fruits. Some have acrid and aromatic properties (Xylopia, Cananga—the flowers of the latter yield Ylang-ylang); Artabotrys odoratissimus; Asimina (N. Am.).
Order 5. Magnoliaceæ. Trees or shrubs with scattered, often leathery, entire leaves, generally with stipules, which (as in Ficus) are rolled together and form a hood round the younger internodes above them, and are cast off by the unfolding of the next leaf, leaving a ring-like scar. The endosperm is not ruminate. Corolla imbricate. Fruit a syncarp.
A. Magnolieæ. The flowers are borne singly, and before opening are enveloped in an ochrea-like spathe which corresponds to the stipules of the foliage-leaves. The perianth generally consists of 3 trimerous whorls, the external one of which is sometimes sepaloid (Liriodendron, and the majority of Magnoliaspecies),[389] sometimes coloured like the others; the perianth is sometimes many-seriate. Numerous spirally-placed stamens and carpels. The latter are situated on the elongated, cylindrical receptacle, and are individually more or less united, except in Liriodendron, where they are free. This last genus has winged achenes; the fruitlets in Magnolia open along the dorsal and ventral sutures, and the seeds then hang out, suspended by elastic threads formed from the vascular bundles of the funicle and raphe; they are red and drupaceous, the external layer of the shell being fleshy—a very rare occurrence.
B. Illicieæ has no stipules. The carpels are situated in a whorl on a short receptacle. Follicles, one-seeded. The leaves are dotted by glands containing essential oil. Illicium; Drimys.
70 species; in tropical or temperate climates; none in Europe or Africa. They are chiefly used as ornamental plants, e.g. the Tulip-tree (Liriodendron tulipifera, N. Am.), Magnolia grandiflora (N. Am.), M. yulan and fuscata (China), and others. The remains of Liriodendron occur as fossils in the Cretaceous and Tertiary periods.—The fruits of Illicium anisatum (Star-aniseed from Eastern Asia) are OFFICINAL. The bark of Drimys winteri (S. Am.) is also strongly aromatic.
Order 6. Calycanthaceæ. These are very closely related to the Magnoliaceæ, but differ in having perigynous flowers with many perianth-leaves, stamens and (about 20) carpels in a continuous spiral, seeds almost devoid of endosperm with rolled up, leaf-like cotyledons, and leaves opposite on a square stem.—There are some species in N. America (Calycanthus florida, occidentalis, etc.) and 1 in Japan (Chimonanthus præcox), all strongly aromatic.
Order 7. Monimiaceæ. Aromatic shrubs with opposite leaves. Perigynous flowers. The anthers dehisce by valves like those of the Lauraceæ, and the Monimiaceæ may thus be considered as an apocarpous form of this order. They are also closely related to Calycanthaceæ. 150 species, tropical.—Hedycarya, Mollinedia, Monimia.
Fig. 384.—Diagram of Berberis.
Fig. 385.—Berberis: carpel with 2 stamens.
Order 8. Berberidaceæ (Barberries).—The regular, ☿, hypogynous flowers are dimerous or trimerous and have regularly alternating whorls of free sepals, petals, and stamens and 1 unilocular carpel; the corolla and stamens have each 2 whorls, the calyx at least 2. The anthers open, as in Lauraceæ, by (2) valves, but are always introrse (Fig. 384). The pistil has[390] a large, disc-like, almost sessile stigma (Fig. 385), and in the ovary several erect ovules are placed close to the base of the ventral suture. The fruit is most frequently a berry. Seeds endospermous.—Shrubs or herbs with scattered, most frequently compound leaves (without stipules), and racemose inflorescences.—They show a relationship to the Lauraceæ in the number of the parts of the flower and the dehiscence of the anthers.
Berberis is a shrub; it has sepals 3 + 3, petals 3 + 3, stamens 3 + 3 (Fig. 384). The petals (honey-leaves) bear internally at the base 2 darkish-yellow nectaries. The filaments are sensitive at the base, and suddenly bend inwards if touched at that spot (Fig. 385). The racemes often have a terminal, 5-merous flower; they are borne on dwarf-branches. The leaves on the long-branches develope into thorns, but the buds in their axils, in the same year as themselves, develope as the short-branches with simple foliage-leaves, articulated at the base, from which fact some authorities have considered that the leaf is compound with a single, terminal leaflet.—Mahonia has imparipinnate leaves. The flower has 3 whorls of sepals. Otherwise as in Berberis.—Epimedium; herbs with spurred petals; the flowers dimerous; 4–5 whorls of sepals, 2 of petals and stamens. Fruit a capsule. Leontice, fruit dry. The anthers of Podophyllum dehisce longitudinally.—Nandina. Aceranthus.
100 species; North temp., especially Asia: fossils in Tertiary. Berberis vulgaris is a native of Europe. This and other species, together with Mahonia aquifolium (N. Am.), Epimedium alpinum, etc., are cultivated as ornamental plants. Several have a yellow colouring matter in the root and stem. Officinal: the rhizome of Podophyllum peltatum (from N. Am.) yields podophyllin.
Order 9. Menispermaceæ. This order has derived its name from the more or less crescent-like fruits and seeds. Diœcious. The flowers are 2–3-merous, most frequently as in Berberis (S3 + 3, P3 + 3, A3 + 3), with the difference that there are 3 free carpels, each with 1 ovule; in some genera, however, the number is different. Stamens often united into a bundle (as in Myristica); anthers dehiscing longitudinally; fruit a drupe.—The plants (with herbaceous or woody stems) belonging to this order are nearly all twining or climbing plants, and have scattered, palmate or peltate, sometimes lobed leaves without stipules. Structure of stem anomalous. Cocculus, Menispermum, Cissampelos, Anamirta.
150 species; Tropical; very rich in bitter and poisonous properties. Officinal: Calumba-root from Jateorhiza columba (E. Africa). The following are cultivated as ornamental plants:—Menispermum canadense (N. Am.) and M. dahuricum (Asia). The fruits of Anamirta cocculus (E. Ind.) are very poisonous (“Grains-of-Paradise”; the poisonous matter is picrotoxine).
Order 10. Lardizabalaceæ. This order, by the free, apocarpous carpels, belongs to a more primitive type, and by the united stamens to a more developed one. Akebia; Holbœllia; principally climbing or twining shrubs. About 7 species in S.E. Asia and S. Am.
[391]
Order 11. Lauraceæ (True Laurels). Trees or shrubs; the leaves, always without stipules, are simple, most frequently scattered, lanceolate or elliptical, entire, penninerved, finely reticulate (except Cinnamomum with 3–5-veined leaf), leathery and evergreen (except, e.g. Cinnamomum); they are frequently studded with clear glands containing volatile oil. The flowers are borne in panicles and are small and of a greenish or whitish colour. They are regular, perigynous, with most frequently a bowl or cup-shaped receptacle (Fig. 386), usually ☿, and trimerous (rarely dimerous) through all (most frequently 6–7) whorls; viz. most frequently, perianth 2 whorls, stamens 3–4 and carpels 1 (P3 + 3, A3 + 3 + 3 + 3, G3) in regular alternation (Fig. 387). Each of the 2 or 4 loculi of the anthers open by an upwardly directed valve (Fig. 386); of the stamens, the 2 outermost whorls are generally introrse, the others extrorse, or 1–3 whorls are developed as staminodes (Fig. 387 g). The gynœceum has 1 loculus with 1 style and 1 pendulous ovule (Fig. 386), and may be considered as formed of 3 carpels. The fruit is a berry (Fig. 388) or drupe, which often is surrounded at its base by the persistent receptacle (as an acorn by its cupule), which becomes fleshy and sometimes coloured during the ripening of the fruit. The embryo has 2 thick cotyledons, but no endosperm (Fig. 388).
Fig. 386.—Flower of the Cinnamon-tree (Cinnamomum zeylanicum) (longitudinal section).
Fig. 387.—Typical diagram of the Lauraceæ: g staminodes.
Fig. 388.—Laurus nobilis: longitudinal section of fruit.
The Lauraceæ present affinities with the Polygonaceæ, in which there is found perigyny, as well as a similar number of parts in the flower and a similar gynœceum, but with erect and orthotropous ovule. From their general characters they should be classed among the Polycarpicæ, but stand, however, isolated[392] by the syncarpous gynœceum, if it is in reality formed by 3 carpels and not by 1 only. Hernandia, which has epigynous monœcious flowers, deviates most.—Cassytha is a Cuscuta-like, herbaceous, slightly green parasite with twining, almost leafless stems. The flower however agrees with the diagram in Fig. 387. Some Lauraceæ have curved veins or palminerved and lobed leaves (often together with entire ones) e.g. Sassafras.
Fig. 389.—Myristica: fruit.]
Fig. 390.—Seed with aril entire and in longitudinal section.
There are 1000 species; especially in the forests of tropical S. America and Asia, of which they form the principal part. Only Laurus nobilis is found in Europe, and there is little doubt that its proper home is in Western Asia. They are rare in Africa.—On account of the volatile oil found in all parts of the plant, they are used as spices, e.g. the false Cinnamon-tree (Dicypellium caryophyllatum, in the Brazils). The OFFICINAL ones are—the Cinnamon-tree (Cinnamomum zeylanicum from Ceylon, E. India, Eastern Asia), which is also cultivated; the Camphor-tree (Cinnamomum camphora, Eastern Asia). The Laurel-tree (Laurus nobilis, Mediterranean), the berries and leaves of which give laurel oil, is medicinal.—Scented wood for furniture, etc., is obtained from Sassafras officinalis (from N. Am.). The wood from its roots is officinal. Pichurim “beans” are the large cotyledons of Nectandra pichury, whilst the[393] famous “Greenheart” wood of Demarara is the wood of Nectandra rodiæi. The pulp and seeds contain a fatty oil. The pear-like fruit of Persea gratissima (Mexico, also cultivated) is very delicious. Lindera benzoin is a garden shrub; Laurus nobilis likewise.
Order 12. Myristicaceæ (Nutmegs). In this order there is only 1 genus, Myristica. Trees or shrubs. The leaves agree closely with those of the Lauraceæ, with which this order has many points in common. The majority of the species are aromatic, having in their vegetative parts pellucid glands with volatile oils. The flowers are regular, diœcious, trimerous, and have a single gamophyllous (cupular or campanulate) 3-toothed, fleshy perianth. In the ♂-flowers the anthers vary in number (3–15), and they are extrorse and borne on a centrally-placed column; in the ♀-flower the gynœceum is unilocular, unicarpellary, with 1 ovule. The FRUIT (Fig. 389) has the form of a pear; it is a fleshy, yellow capsule, which opens along the ventral and dorsal sutures, exposing the large seed. This seed has a large, red, irregularly branched aril—the so-called “mace”; the “nutmeg,” on the other hand, is the seed itself with the inner thin portion of the testa, which has pushed its way irregularly into the endosperm, and causes the marbled appearance of the cut seed (Fig. 390); the external, dark brown, hard, and brittle part of the seed-shell is however removed. Mace and nutmeg contain volatile and fatty oils in abundance.—80 species. Tropical. The majority are used on account of their aromatic seeds and aril, the most important being M. fragrans (moschata), from the Moluccas. This is cultivated in special plantations, not only in its native home, but in other tropical countries also. Nutmegs were known as commodities in Europe in very ancient times (e.g. by the Romans), but it was not until the year 1500 that the tree itself was known. The seed is OFFICINAL.
The plants belonging to this family are almost exclusively herbaceous, with scattered, exstipulate leaves. The flowers are eucyclic di- or tetramerous, with the calyx and corolla deciduous, hypogynous, ☿, regular, the gynœceum with 2–several carpels (generally 2, transversely placed) (Figs. 391, 392, 393, 397). The ovary is unilocular with parietal placentæ, but in Cruciferæ and a few others it becomes bilocular by the development of a false, membranous wall between the placentæ. The stigmas in the majority of cases are commissural, i.e. they stand above the placentæ, and not above the dorsal line of the carpels. The fruit is nearly always a capsule, which opens by the middle portions of the carpels detaching themselves as valves, bearing no seed, whilst the placentæ persist as the seed-bearing frame. Endosperm is found in Papaveraceæ and Fumariaceæ, but is absent in Cruciferæ and Capparidaceæ.—This family through the Papaveraceæ is related to the Polycarpicæ (the Nymphæaceæ), through the Capparidaceæ to the Resedaceæ in the next family.
Exceptions to the above are: Eschscholtzia, Subularia (Fig. 403) and a few[394] Capparidaceæ, in which perigynous flowers are found. A few Papaveraceæ and Fumariaceæ have trimerous flowers. In Fumaria and certain Cruciferæ, the fruit is a nut. The Fumariaceæ have zygomorphic flowers. Trees and shrubs are almost entirely confined to the Capparidaceæ, in which order stipules also are found.
Order 1. Papaveraceæ (Poppies). Herbaceous plants with stiff hairs and latex; flowers regular (Fig. 391) with generally 2 (-3) sepals (which fall off as the flower opens), 2 + 2 petals (imbricate and crumpled in the bud) without spur, numerous stamens in several alternating whorls (generally a multiple of 2); carpels 2–several, united into a unilocular gynœceum. Trimerous flowers also occur. Capsule with very numerous seeds on the parietal placentæ; embryo small, with large, oleaginous endosperm (Fig. 392).—The leaves have no stipules and are generally pinnately lobed.
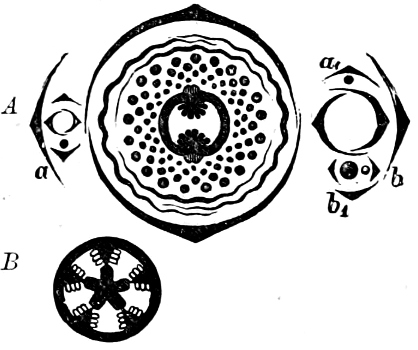
Fig. 391.—A Diagram of the flower of Glaucium and the dichasium (which becomes transformed into a scorpioid cyme). B Papaver argemone, transverse section of the ovary with indication of the position of the stigmas.
Fig. 392.—Papaver somniferum: A capsule; st the stigma; v valves; h pores; B seed in longitudinal section; alb endosperm; emb embryo.
Papaver (Poppy) has large, solitary, terminal flowers; petals firmly and irregularly folded in æstivation; gynœceum formed by many (4–15) carpels; stigmas velvety, sessile and stellate (the rays stand above the placentæ) (Fig. 391 B). The edges of the carpels project deeply into the ovary, but do not meet in the centre, so that it remains unilocular. The capsule opens by pores placed close beneath the stigma, and formed of small valves alternating with the placentæ and the rays of the stigma (Fig. 391). P. dubium, P. argemone, P. rhœas.—Chelidonium (Greater Celandine) has yellow latex, flowers in umbellate cymes (the terminal, central flower opening first) and only 2 carpels; the fruit resembles the siliqua[395] of the Cruciferæ in having two barren valves, which are detached from the base upwards, and a seed-bearing frame, but there is no partition wall formed between the placentæ. Ch. majus.—The majority of the other genera have, like Chelidonium, 2 carpels (lateral and alternating with the sepals: Fig. 391 A) and siliqua-like fruit, thus: Eschscholtzia (perigynous) with a linear, stigma-bearing prolongation extending as far above the placentæ as above the dorsal suture of the carpels; Glaucium (Horn-Poppy); G. luteum, whose extremely long, thin capsule differs from that of Chelidonium by the formation, during ripening, of a thick, spongy (false) replum, which persists when the valves are detached; Sanguinaria with red latex, the 2 petals divided into 8–12 small petals (perhaps by dédoublement); Macleya and Bocconia (1-seeded capsule) with 2 sepals and no petals.—Trimerous flowers are found in Argemone and Platystemon (with a curious fruit, carpels free, and transversely divided and constricted into joints which separate as nut-like portions).—Meconopsis.—Hypecoum (Fig. 393 C) has tri-lobed and three cleft petals, 4 free stamens with 4-locular anthers and a jointed siliqua; it presents a transitional form to the Fumariaceæ, with which order it is sometimes included.
Pollination. Papaver and Chelidonium have no honey, and are without doubt only visited by insects for the sake of the pollen. The anthers and stigmas mature about the same time.—There are 80 species; especially from warm climates. Officinal: Papaver somniferum (Opium-Poppy); the latex of its unripe capsules is obtained by incisions, and dried (opium); it contains many alkaloids: morphine, papaverine, narcotine, thebaine, etc. The oleaginous seeds are also used in the manufacture of oil. Its home is in the East, where it is extensively cultivated. The petals of the Corn-poppy (P. rhœas) are also officinal. Several species are cultivated as ornamental plants.
Order 2. Fumariacæ (Fumitories). This order differs from the closely allied Papaveraceæ in the absence of latex, a poorer flower, generally transversely zygomorphic (Fig. 393 B), in which case one or both of the outer lateral petals are gibbous, or prolonged into a spur; the stamens are especially anomalous. Sepals 2, caducous; petals 2 + 2; stamens 2, tripartite; each lateral anther is bilocular (Figs. 393 A, B; 395); gynœceum bicarpellate. The fruit is a nut or siliqua-like capsule. Endosperm.—Herbs with scattered, repeatedly pinnately-divided leaves without stipules, generally quite glabrous and glaucous; the flowers are arranged in racemes with subtending bracts, but the bracteoles are sometimes suppressed.
Dicentra (syn. Dielytra) and Adlumia have a doubly symmetrical flower, with a spur or gibbous swelling at the base of each of the laterally-placed petals (Figs. 393 A, 394). Corydalis has a zygomorphic flower, only one of the lateral petals having a spur, and consequently there is only one nectary at the base of the bundle of stamens, which stands right in front of the spur (Fig.[396] 393 B, 395, 396). The fruit is a many-seeded siliqua-like capsule. A peculiarity of the flower is that the plane of symmetry passes transversely through the flowers, whilst in nearly all other zygomorphic flowers it lies in the median line. Moreover, the flower is turned, so that the plane of symmetry ultimately becomes nearly vertical, and the spur is directed backwards.—Many species have subterranean tubers; in these the embryo germinates with one cotyledon, which is lanceolate and resembles a foliage-leaf. The tuber is in some the swollen hypocotyl (C. cava), in others a swollen root (C. fabacea, etc.), which grows down through the precisely similar swollen root of the mother-plant. The sub-genus Ceratocapnos has dimorphic fruits (nuts and capsules) in the same raceme. Fumaria differs from Corydalis only by its almost drupaceous, one-seeded nut (Fig. 395).
Fig. 393.—Diagram of Dicentra (A), Corydalis (B), and Hypecoum (C).]
Fig. 394.—Dicentra spectabilis: A flower (2/5); B the same, after removal of half of one outer petal; the cap, formed by the inner petals, is moved away from the anthers and stigma; the insect does this with the lower side of its abdomen, and thus rubs the stigma on the hairs of its ventral surface; the dotted line at e indicates the direction of the proboscis; C andrœcium and gynœceum; D stigma.
The structure of the flower. Hypecoum among the Papaveraceæ is the connecting link with the Fumariaceæ. The diagram (Fig. 393 C) corresponds both in number and in the relative position of its members with that of most of the other Papaveraceæ (Fig. 391), except that there are only four stamens (with extrorse anthers). In Dicentra (Fig. 393 A), the two central (uppermost) stamens are absent, but each of the two lateral ones are divided into three filaments, of which the central one bears a four-locular anther, and each of the[397] others a two-locular (half) anther. Corydalis and Fumaria stand alone in the symmetry of the flower, differing from Dicentra in having only one of the lateral petals (Fig. 393 B, sp) prolonged into a spur, while in Dicentra both the petals are spurred. This structure has been interpreted in various ways. According to Asa Gray the median stamens are absent in the last-named genera, and the lateral ones are split in a similar manner to the petals of Hypecoum. Another, and no doubt the most reasonable theory (adduced by De Candolle), is: that two median stamens are split, the two parts move laterally, each to their respective sides and become united with the two lateral stamens; this affords a natural explanation of the two half-anthers, and establishes a close relationship to the Cruciferæ. A third interpretation, held by Eichler and others, is as follows: the median stamens are always wanting; when they appear to be present, as in Hypecoum, it is due to the fact that the side portions of the lateral stamens approach each other (as interpetiolar stipules) and coalesce into an apparently single stamen.
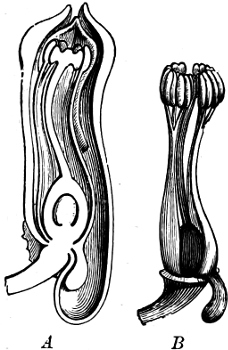
Fig. 395.—Fumaria officinalis: A the flower in longitudinal section; B the andrœcium and gynœceum; nectary to the right.
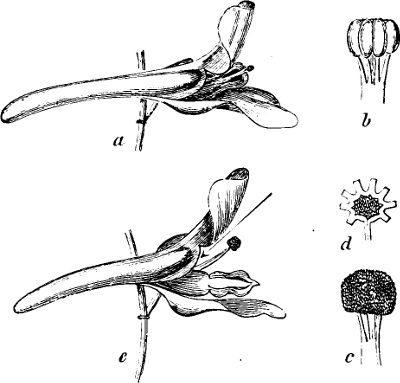
Fig. 396.—Corydalis cava: a a flower (lateral view); b the anthers lying round the stigma; c the anthers shortly before the opening of the flower; d the head of the stigma; e relative position of the parts of the flower during the visit of an insect.
130 species; mostly from the northern temperatures.
Pollination. Fumaria, with its inconspicuous flowers, has to a great extent to resort to self-pollination. Corydalis, on the other hand, is dependent on cross-pollination; C. cava is even absolutely sterile with its own pollen. Corydalis is pollinated by insects with long probosces (humble-bees, bees), which are able to reach the honey secreted in the spur; as they alight on the flowers they press the exterior petals on one side (Fig. 396 e), so that the stigma, surrounded by the anthers, projects forward; the proboscis is introduced in the direction of the arrow in the figure, and during this act the under-surface of the insect is covered with pollen, which is transferred by similar[398] movements to the stigma of another (older) flower.—Ornamental plants; Dicentra (spectabilis and eximia), Adlumia, Corydalis.
Order 3. Cruciferæ (Crucifers). The flowers are regular, ☿; sepals 4, free (2 + 2), deciduous; petals 4, free, deciduous, unguiculate, placed diagonally in one whorl, and alternating with the sepals; stamens 6; the 2 outer are short, the 4 inner (in reality the two median split to the base) longer, placed in pairs (tetradynamia of Linnæus); gynœceum syncarpous formed by 2 (as in the previous order, lateral) carpels, with 2 parietal placentæ, but divided into two loculi by a spurious membranous dissepiment (replum) (Fig. 397). Style single, with a capitate, usually two-lobed stigma, generally commisural, that is, placed above the parietal placentæ (Fig. 397), but it may also be placed above the dorsal suture, or remain undivided. Ovules curved. The fruit is generally a bivalvular siliqua (Fig. 398 B, C), the valves separating from below upwards, and leaving the placentæ attached to the replum; other forms of fruits are described below. The oily seeds have no endosperm (endosperm is present in the two previous orders); the embryo is curved (Figs. 398 E, F; 399, 400).—In general they are herbaceous plants, without latex, with scattered, penninerved leaves, without stipules; the inflorescence is very characteristic, namely, a raceme with the flowers aggregated together at the time of flowering into a corymb, and destitute of both bracts and bracteoles.

Fig. 397.—Diagram of a Cruciferous flower.
Fig. 398.—Brassica oleracea: A raceme; B, C siliqua; D seed; E embryo; F transverse section of seed.
Fig. 399.—Transverse section of seed and embryo of Cheiranthus cheiri.
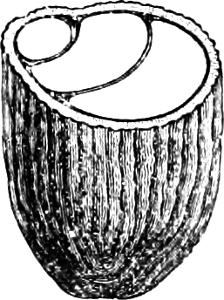
Fig. 400.—Transverse section of seed of Sisymbrium alliaria.
Many are biennial, forming in the first year a close leaf-rosette. By cultivation the tap-root can readily be induced to swell out into the form of a tuber (Turnips, Swedes, etc.). Stipules are found indicated by small glands on the very young leaves; in Cochlearia armoracia they are fairly large triangular scales. Stellate hairs often occur. Floral-leaves are occasionally developed. Terminal flowers are never found in the inflorescences. Iberis and Teesdalia have zygomorphic flowers. Subularia (Fig. 403) is perigynous. The 2 external sepals (Fig. 397) stand in the median plane; it may therefore be supposed that there are two bracteoles outside these which, however, are suppressed, and can only in a few instances be traced in the young flower; the two lateral sepals are often gibbous at the base, and serve as reservoirs for the nectar secreted by the glands placed above them; they correspond in position to the external petals of the Fumariaceæ. The 4 petals which follow next arise simultaneously, and alternate with the 4 sepals; if it could be shown that these are merely 2 median petals, which have been deeply cleft and the two parts separated from[399] each other and displaced to the diagonal position, there would be a perfect correspondence with the Fumariaceous flower; then the petals would be followed in regular alternation by the 2 lateral small stamens, the 2 median long stamens, which it has been proved are split into 4 and placed in couples, and the 2 laterally-placed carpels,—in all 6 dimerous whorls. But the formation of the corolla by the splitting of 2 petals does not agree with the development of the flower or bear comparison, and hence the only fact in favour of this theory is[400] the otherwise prevailing correspondence with the Fumariaceæ. Yet it may be observed that in special cases each pair of long stamens clearly enough arises from one protuberance and even later on may be considerably united or entirely undivided (e.g. Vella); in other instances they are quite distinct from the beginning, and it is possible that this latter condition has become constant in the corolla. Lepidium ruderale and others have no corolla. Senebiera didyma has only 2 median stamens. Megacarpæa has several stamens, no doubt by dédoublement, as in Capparidaceæ.—The number of carpels may also be abnormally increased; Tetrapoma barbareifolium has normally 4 carpels with an equal number of placentæ and repla. It is supposed to be a variety of Nasturtium palustre.—The 2–4–8–10 greenish glands, which are found at the base of the stamens, are nectaries, morphologically emergences, and not rudimentary stamens. The forms of fruits are of great systematic significance, see the genera. In some species dimorphic fruits are present, e.g. Cardamine chenopodiifolia which has both ordinary Cardamine-siliquas and 1-seeded siliculas.
The curved embryo appears in five forms, which have systematic importance: 1. To the Pleurorhizæ belong those genera whose radicle (with the hypocotyl) lies bent upwards along the edge of the flat cotyledons (Fig. 399); to this group belong Cardamine, Nasturtium, Cheiranthus, Matthiola, Cochlearia, Draba, Iberis, Thlaspi, etc.; diagrammatic transverse section: ◯=.—2. To Notorhizæ belong those whose radicle lies in an upward direction along the back of one of the flat cotyledons (Figs. 400, 413); e.g. Hesperis, Sisymbrium, Lepidium, Capsella, Camelina: ◯‖.—3. Orthoploceæ differ from the Notorhizeæ in having the cotyledons folded (not flat) (Fig. 398 E, F); to this belong Brassica, Sinapis, Raphanus, Crambe, etc.: ◯>>.—4. Spirolobeæ: the radicle lies as in the Notorhizæ, but the cotyledons are so rolled together that a transverse section of the seed cuts them twice; Bunias: ◯‖‖.—5. Diplecolobeæ: the cotyledons are folded forward and backward so that a transverse section cuts them several times; Subularia, Senebiera: ◯‖‖‖.
On germination the cotyledons appear above the ground as green leaves; in the Orthoploceæ they are bilobed, in the Lepidium-species divided.
1. Silicula, broad replum (Siliculosæ latiseptæ), valves flat or slightly vaulted, and the replum extends through the greatest width of the silicula (Fig. 404). The seeds are situated in two rows.
◯=: Cochlearia (Horse-radish): the siliqua is nearly spheroid; glabrous herbs, generally with fleshy, stalked leaves, and white flowers.—Draba has an oblong, lanceolate, somewhat compressed silicula; herbs with small rosettes of leaves, most frequently with stellate and long-stalked racemes.—Alyssum and Berteroa are whitish, on account of the stellate hairs; they have a more compressed and round or elliptical silicula. Vesicaria; Aubrietia. Lunaria (Honesty, Fig. 401): very broad and flat silicula with long stalk (the receptacle as in Capparidaceæ).
[401]
◯‖: Camelina (Gold-of-pleasure) has a spheroid, pear-shaped siliqua with a small rim passing right round (Fig. 402). Subularia (Awlwort), an aquatic plant with perigynous flower (Fig. 403) and folded cotyledons.
2. Silicula, narrow replum (Siliculosæ angustiseptæ), i.e. the replum is much shorter than the arched, more or less boat-shaped valves (Figs. 405, 406, 407).
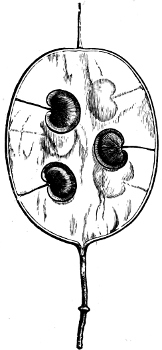
Fig. 401.—Lunaria biennis. Fruit, the valves of which have fallen off.
Fig. 402.—Camelina sativa. Fruit.
Fig. 403.—Subularia aquatica. Longitudinal section through the flower.
◯=: Thlaspi (Penny-Cress) has a flat, almost circular silicula, emarginate or cordate, with a well-developed wing round the edge (Fig. 406). Iberis and Teesdalia: the racemes during flowering are especially corymbose, and the most external petals of the outer flowers project radially and are much larger than the other two (the flower is zygomorphic).—Biscutella, Megacarpæa.
◯‖: Capsella (Shepherd’s-Purse) has a wingless, obcordate or triangular silicula (Fig. 407). Lepidium (Pepperwort) has a few–(2–4) seeded, slightly winged, oval silicula. Senebiera has a silicula splitting longitudinally into two nut-like portions; its cotyledons are folded.—Anastatica hierochuntica (“Rose of Jericho”) is an annual, silicula-fruited, desert plant (Arabia, Syria, N. Africa). After the flowering all its then leafless branches bend together upwards, forming a kind of ball; this spreads out again on coming in contact with water, and the fruits then disseminate their seeds, which germinate very quickly, often in the fruit.
[402]
3. Siliqua (Siliquosæ). The fruit is a true siliqua, several times longer than broad. The seeds in most are borne apparently in one row.
Fig. 404.—Transverse section of a silicula with broad replum: s replum; k the valves.
Fig. 405.—Transverse section of a silicula with narrow replum.
◯>>: Brassica (Cabbage). The seeds are placed apparently in one row in each loculus (Fig. 398 C); the style is long and round; the valves have only 1 strong, longitudinal rib.—Melanosinapis (M. nigra, Black-mustard); the style is compressed, two-edged; the valves of the siliqua are one ribbed.—Sinapis (Mustard); quadrangular or flat style (in which in most cases there is a seed) and 3–5 strong, longitudinal ribs on the valves.—Eruca differs from Brassica by the shorter siliqua, broad, sword-like “beak” and seeds in two rows.
◯= (Fig. 399): Cardamine (Bitter Cress) has a long, linear siliqua, with flat, unribbed, elastic valves. The leaves are most frequently pinnatifid or pinnate. C. pratensis reproduces by buds formed in the axils of the leaves.—Arabis (Rock Cress); Matthiola (Stock); Cheiranthus cheiri (Wallflower); Barbarea (Winter Cress) (double-edged, quadrangular siliqua); Nasturtium (N. officinale, Water-cress); the siliqua of the latter genus is in some species short, in others long.
Fig. 406.—Thlaspi arvense.
Fig. 407.—Silicula of Capsella bursa-pastoris.
◯‖ (Fig.400): Sisymbrium (Hedge Mustard) the valves of the siliqua are 3-ribbed.—Erysimum; Hesperis; Schizopetalum (with fimbriate petals).
[403]
4. Fruit jointed (Lomentaceæ). The fruit is divided by transverse walls into as many spaces as there are seeds, and dehisces at maturity, generally transversely, into a corresponding number of nut-like joints (“articulate-siliqua.”)
◯=: Crambe (Kale, Fig. 408). The fruit has only 2 joints. The lower one resembles a short, thick stalk, and is barren, the upper one is spherical, and has 1 seed.—Cakile (C. maritima, Sea-kale); the lower node is triangular, 1-locular, the upper one more ensiform, 1-locular (Fig. 409).

Fig. 408.—Fruit of Crambe maritima.
Fig. 409.—Cakile maritima. Fruit (2/1).

Fig. 410.—Raphanus raphanistrum.

Fig. 411.—Raphanus sativus.
◯>>: Raphanus has a long siliqua, which, in the garden Radish (R. sativus), is spongy and slightly abstricted (Fig. 411), but neither opens nor divides transversely (a kind of dry berry), and which in the Wild Radish (R. raphanistrum) (Fig. 410) is abstricted in the form of a string of pearls, and separates into many joints. R. sativus; the “Radish” is formed by the hypocotyl, after the bursting of its external, cortical portions (of which there are generally two patches at the top of the Radish).
5. Siliqua indehiscent (Nucumentaceæ). The fruit is a short, unjointed, unilocular and 1-seeded nut, and the fruit-stalks are often long, slender, and drooping. (Sometimes a thin endosperm is present).—Isatis (Woad) has most frequently an oblong, small-winged nut; ◯‖ (Figs. 412, 413).—Bunias; Neslia.
[404]
[The systematic division of this order given above is founded upon that of A. P. de Candolle. Prantl (Engler and Prantl, Nat. Fam.), 1891, adopts a somewhat different system, which may briefly be summarised as follows:—
Fig. 412.—Isatis tinctoria. Fruit (Fig. 412); and in longitudinal section (Fig. 413). (Mag.)
Pollination. Honey is secreted by the nectaries mentioned above; but the position of the stamens is not always the most favourable for pollination by insects (in these flowers the honey-seeking insect must touch the anthers with one of its sides and the stigma with the other), and self-fertilisation is common. In some species (Cardamine pratensis) the long stamens turn their anthers outwards towards the small stamens, so that 3 anthers surround each of the two large entrances to the nectaries.
1200 species (180 genera), especially in the cold and temperate parts of the Old World (Europe, W. Asia). Many are weeds in this country, e.g. Wild Cabbage (Brassica campestris), Charlock (Sinapis arvensis), Wild Radish (Raphanus raphanistrum) and others.—The order is acrid and oleaginous. Oil is obtained from many of the oil-containing seeds, especially of the Rape (Brassica napus), Summer-Rape (the oil-yielding cultivated form of the Field-Cabbage) and Camelina. Several are pot-herbs or fodder plants, e.g. Cabbage
[405]
(Brassica oleracea) with its numerous varieties: Cauliflower (var. botrytis; the entire inflorescence is abnormally branched and fleshy), Kohlrabi (var. gongylodes, with swollen, tuberous stem), Kale, Red-Cabbage, White-Cabbage, etc.; B. campestris, var. rapifera (Turnip); B. napus, var. rapifera (Swede); Raphanus sativus (Radish from W. Asia), R. caudatus (long Radish); Nasturtium officinale (Water Cress), Lepidium sativum (Garden Cress), and Barbarea præcox (Early Cress); Crambe maritima (Sea-Kkale). The seeds of the following are especially used as spices: (the flour of) Melanosinapis (Black-mustard), and Sinapis alba (White-mustard), which are officinal like the root of Cochlearia armoracia (Horse-radish, E. Eur[**.]). The herbaceous parts of Cochlearia officinalis and danica are medicinal.—A blue dye (woad) is extracted from Isatis.—Ornamental plants: Cheiranthus cheiri (Wallflower), Matthiola (Stock), Iberis, Hesperis, Lunaria, and others (especially from S. Eur.). Sweet-scented flowers are rare.
Fig. 414.—Gynandropsis pentaphylla.
Fig. 415.—Capparis spinosa.
Order 4. Capparidaceæ (Capers). The relationship with the Cruciferæ is so close that certain forms are with difficulty distinguished from them. The diagram of the flower is the same in the number and position of its parts, but it differs in the modifications which occur in the development of the stamens. In some genera all 4 stamens are undivided; in others both the 2 median ones are divided as in the Cruciferæ (6 stamens, but not tetradynamous) (Fig. 414); in other genera only 1 of these; in other instances again they are divided into more than 2; and finally the 2 lateral ones also may be found divided, so that indefinite stamens occur (Fig. 415). The bicarpellate gynœceum is unilocular (without replum), but more than 2 carpels may occur. The ovary is elevated[406] on a stalk (sometimes as much as 1 foot in length); also between the stamens and corolla a similar stalk may be found (Fig. 414). The fruit is long and siliquose (Cleome, Polanisia, Gynandropsis), or a berry (Capparis). Endosperm absent. Some have zygomorphic flowers. Gamosepalous calyx and perigynous flowers also occur.—350 species; especially in the Tropics. The majority are trees and shrubs, and they differ also from the Cruciferæ in having distinct stipules present in some species.
“Capers” are the flower buds of the climbing, thorny shrub, Capparis spinosa (Fig. 415), which grows in the Mediterranean.
The flowers in this family are perfect, regular (except Resedaceæ, Violaceæ), hypogynous, the perianth-leaves free (a few have them slightly united), æstivation most frequently imbricate; they are eucyclic in the andrœcium, and most frequently in the other parts, and generally 5-merous with S5, P5, A5 + 5, G3, but other numbers also occur; several have indefinite stamens, but the stamens arise (where the development is known) in centrifugal order and are arranged, often very distinctly, in bundles; in other words, the large number of stamens is formed by the splitting of a small number (most frequently 5); a true spiral arrangement is never found. Gynœceum syncarpous, multicarpellary (Dilleniaceæ and a few Resedaceæ are apocarpous), most frequently the number of carpels is 3, forming a unilocular ovary with parietal placentæ, but parallel with this, multilocular ovaries, with the ovules placed in the inner angle of the loculi, are also found, and a few genera have a free, centrally-placed placenta. The fruit is most frequently a capsule. The dehiscence is never with a “replum,” i.e. the persistent frame of the placenta, as in the family Rhœadinæ. One half of the orders has endosperm (Violaceæ, Cistaceæ, Droseraceæ, Bixaceæ, Ternstrœmiaceæ, etc.), the other has no endosperm (Resedaceæ, Hypericaceæ, Elatinaceæ, Tamaricaceæ, etc.); some have a curved, the majority a straight embryo. The family is scarcely quite natural; in the future the orders will probably be arranged differently.
Order 1. Resedaceæ (Mignonettes).—Herbs or small shrubs with spirally-placed leaves and very small, gland-like stipules (as in Cruciferæ); the ☿, hypogynous flowers are zygomorphic, and arranged in racemes or spikes typically without bracteoles. The zygomorphic structure is produced by the greater development of the posterior side of the flower, especially the petals and the nectary (“disc,” in Fig. 416 d) which is situated between[407] the petals and stamens; in general there are 5–8 free sepals and petals, the latter consisting of a large scale-like sheath with a fimbriated blade (see Fig. 416); stamens numerous; carpels 6–2 united together; ovary unilocular with parietal placentæ, but the cavity of the ovary is not closed at the top. In Astrocarpus the gynœceum is apocarpous. The fruit is most frequently a capsule; the seeds are reniform, without endosperm, and the embryo is curved.
This order connects the Rhœadinæ with the Cistifloræ. It is closely allied to the Rhœadinæ by its external appearance, even by the smell and taste, the parietal placentation, structure of the seeds, the inflorescences, etc., whilst by the irregular flowers and the disc placed at the posterior side of the flower, it is allied to Capparidaceæ, but differs from this order in not having its characteristic number (2–4) and by the very different mode of dehiscence of the fruit, etc. It differs from the other orders of this family chiefly in the fact that the number of the perianth-leaves is not constantly 5. In Reseda luteola both the calyx and corolla appear to be 4-leaved, because the posterior sepal is suppressed, and the 2 posterior petals are united. Where there are 10 stamens, they stand in 2 whorls, i.e. in front of the sepals and petals; if there are several, their position depends upon the splitting.—Astrocarpus is remarkable for its apocarpous fruit and the position of the ovules on the dorsal suture of the carpel.
The yellow, flat disc at the back of the flower serves as a nectary, the honey being protected by the lobes of the petals. If pollination by insects is not effected, then self-pollination may take place, at all events in R. odorata.
45 species; the majority in the Mediterranean and in Persia. Reseda odorata (from Egypt) is cultivated on account of its sweet scent; R. luteola (“Dyer’s Weed”) yields a yellow dye.

Fig. 416.—Diagram of Reseda odorata.
Order 2. Droseraceæ (Sundews). Herbs, chiefly living on moors or in water, and whose leaves are adapted to catch and digest small animals. With regard to the flower, they are closely allied to the Violaceæ, especially to those with regular flowers. Drosera (Sundew) has a long-stalked scorpioid cyme with regular, ☿, hypogynous flowers, 5-merous as in Viola. S5, P5, A5, G3 (in a syncarpous gynœceum, with free, bifid styles and basal or parietally-placed ovules in the unilocular ovaries). The capsule opens also as in Viola, but, among other differences, the styles are free, the seeds very small, and surrounded by a loosely lying, thin shell. Drosera has radical, long-stalked leaves with the blade (Fig. 417) covered by numerous strong glandular hairs, placed on the edge and in the middle; when small animals are caught by these hairs, the latter and the entire[408] blade close slowly over them dissolving and absorbing all the digestible matter as nourishment.
Fig. 417.—Leaf-rosette of Drosera rotundifolia (nat. size), and a leaf (magnified).
Dionæa muscipula (Fly-trap; N. Am.) has the same appearance as Drosera, but the leaves are constructed as in Fig. 418. The stalk is flat and winged, the blade small, circular, with powerful, pointed teeth along the edge, and on its surface are 6 small bristles (A), which are very sensitive. When these are touched the blade quickly closes, folding along the midrib (B, C) and imprisoning the irritating object, the teeth round the edges fitting like the teeth of a trap. If it happens to be an insect or similar body, a digestive fluid is secreted which, like the gastric juice, dissolves the digestible portions. Aldrovandia vesiculosa (Central and S. Europe) captures small aquatic animals in a similar manner; it is a floating, aquatic plant, the two halves of its leaves also close together when irritated (Fig. 419).—Drosophyllum.
About 110 species; most of them in the temperate regions.
Fig. 418.—Dionæa muscipula. Leaves (nat. size).
Orders 3 and 4. Sarraceniaceæ and Nepenthaceæ. These two orders are perhaps most closely allied to the Droseraceæ and agree with these, among other things, in the manner of taking nourishment. Like the Droseraceæ they absorb nitrogenous food from dissolved animal matter by means of their leaves, which are specially constructed both to catch, to retain, and to digest any small[409] animals which may be caught. The Sarraceniaceæ are North American marsh-plants (10 species) which have pitcher-like leaf-stalks, in the cavity of which a fluid (with properties approaching those of gastric juice) is secreted, and which bear at the apex a small, lid-like blade; these leaf-stalks are the catching and digestive organs.—Sarracenia, Darlingtonia.
Fig. 419.—Aldrovandia vesiculosa: A a plant (nat. size). B Leaf (mag.); the blade is closed; the winged stalk is prolonged into 4–6 irritable bristles.
Fig. 420.—Nepenthes (reduced).
Nepenthaceæ has only 1 genus, Nepenthes (the Pitcher-plant; about 35 species), especially found in tropical E. Asia; the majority are climbing shrubs. The leaf-stalks are twining organs, and terminate either simply in a tendril, or in addition to this, with a pitcher-shaped body (which in some species may be[410] as much as a foot in length) on whose upper edge a lid-like structure is found (Fig. 420). In this pitcher, as among the Sarraceniaceæ, a fluid is secreted which is able to digest the animals captured (sometimes rather large) and which corresponds in some degree to the gastric juice.
Order 5. Violaceæ (Violets). The flowers are ☿, and generally zygomorphic, hypogynous, with S5, P5, A5, G3 (Fig. 421). The stamens are closely applied to the ovary, they have a very short filament, and at their summit generally a membranous appendage formed by the prolongation of the connective (Fig. 422 g). The ovary is unilocular with 3 parietal placentæ; style undivided (Fig. 422 B). The fruit is usually a 3-valved capsule, opening along the dorsal sutures (Fig. 423). Embryo straight; endosperm fleshy (Fig. 425).—Many are herbaceous plants (e.g. Viola), but in the Tropics shrubs are also found (e.g. Ionidium); a few are lianes; the leaves are scattered, with stipules, and involute in the bud.

Fig. 421.—Diagram of Viola.
Fig. 422.—The large-flowered form of Viola tricolor: A the flower in median longitudinal section; B the gynœceum.
Viola. The sepals are prolonged backwards beyond the point of insertion (appendiculate); the corolla is polypetalous, descending imbricate, and zygomorphic, its anterior petal being larger than the others and provided with a spur (Fig. 421). The 2 anterior of the 5 almost sessile stamens are provided with a spur-like nectary, which protrudes a considerable distance into the petaloid spur (Figs. 421, 422 n, sp). The style is club-like, and bears the[411] stigma in a groove on the anterior side (Fig. 422 st). Herbs with rhizomes, or annuals; flowers solitary. V. odorata, canina, etc., have cleistogamic flowers which produce fruit in addition to the large, coloured (violet) flowers. The Pansy (V. tricolor) has large flowers adapted for insect-pollination, and also smaller, less conspicuous ones designed for self-pollination. The stigma, as in Fig. 422 A, st, and B, is situated on the anterior side of the stylar-head, immediately in front of the channel leading down to the spur (sp); below it is situated a valve, easily covered with pollen when the proboscis of an insect is introduced into the spur, but which closes upon its withdrawal; cross-pollination is thus secured.—The sweet-scented V. odorata is visited by the honey-bee, which insures cross-pollination, and in the absence of insect visits it effects self-fertilisation by cleistogamic flowers. The conspicuous but scentless V. tricolor, var. vulgaris, is less frequently visited by insects (humble-bees). In V. silvatica and V. canina the pollen is carried on the head or proboscis of the honey-sucking bee.—The fruits of V. odorata bury themselves slightly in the soil. In the others the fruits are raised above the ground; the 3 boat-shaped valves close together along the central line, and eject the seeds, one by one, with much violence, so that they are thrown to a great distance.
Figs. 423–425.—Viola Tricolor.

Fig. 423.—Capsule after dehiscence (nat. size).]
Fig. 424.—External view of the seed.
Fig. 425.—Seed in longitudinal section.
The Alsodeia-group has regular or almost regular flowers. Gamopetalous corollas are found in Paypayroleæ. Sauvagesieæ differs the most by its regular corolla, and 5–∞ free or united staminodes.
250 species; especially in the Tropics.—The Ionidium-species are used as ipecacuanha. A number of Viola-species are cultivated as garden plants, especially V. odorata (sweet-scented Violet) and V. tricolor, which have a large number of varieties.
Order 6. Frankeniaceæ. A small order with doubtful relationships. Perennial herbs or shrubs; beach plants with nodose stem. Sepals united, petals free. Unilocular ovary, with 3–4 parietal placentæ. Fruit a capsule. Embryo straight, endospermous. Especially in S. Europe, Africa, on the shores of the Mediterranean and Atlantic.
Order 7. Tamaricaceæ (Tamarisks). To this order belong only Tamarix and Myricaria. They are shrubs of a cypress- or heather-like appearance, as[412] the scattered leaves are very small, sessile, scale-like or linear, adpressed, entire, and usually glaucous, and the branches are slender and whip-like. The flowers are borne in small spikes or racemes, and are small, reddish or whitish, regular, ☿, hypogynous and polypetalous; formula S5, P5, A5 + 0 (Tamarix, which often has stipular teeth at the base of the filaments), or A5 + 5 (Myricaria, in which the stamens are united at the base); the number 4 may appear instead of 5, but in either case there is usually a tricarpellate gynœceum, which is unilocular and has either parietal placentæ (Myricaria) or a small basal placenta (Tamarix); 1 trifid style, or 3 styles. Capsule dehiscing along the dorsal suture, and resembling the Willows in having a unilocular ovary with numerous woolly seeds; but the seed-wool in this case is borne on the chalaza, and may be attached to a long stalk.—Some Tamarix-species shed part of their branches in the winter.—40 species; North Temperate, on the sea-shores or steppes, especially in Asia. Ornamental shrubs: Myricaria germanica, and Tamarix gallica.
Order 8. Cistaceæ. Shrubs or herbs, natives especially of the Mediterranean region. Flowers generally in raceme-like scorpioid cymes, regular, ☿, hypogynous; sepals 5, free, twisted in the bud, of which the two outer are generally much smaller than the others; petals 5, free, twisted in the bud (in the direction opposite to the sepals), fugacious; stamens numerous; gynœceum syncarpous, carpels usually 3–5, style simple, ovary unilocular, with parietal placentation (seldom divided into loculi, with axile placentation). The ovules are orthotropous in opposition to some of the other orders of this family. The capsule dehisces along the dorsal sutures; embyro curved. The leaves are simple, undivided, generally opposite and stipulate.—They are Violaceæ with regular flowers, numerous stamens, and curved embryo. The numerous stamens are in reality only one or two 5-merous whorls, divided into a large number of stamens; these are formed, therefore, in descending order, like the lobes of many compound foliage-leaves.
Helianthemum (Rock-Rose), has 3 carpels.—Cistus has 5 (-10) carpels.
About 70 species; temperate climates, especially about the Mediterranean. The resin of the Cistus-species has been used medicinally (ladanum).
Order 9. Bixaceæ. This order is closely allied to the Cistaceæ and Ternstrœmiaceæ; like these it has regular, 5-merous, hypogynous flowers with numerous stamens, unilocular ovary and parietal placentæ; sometimes unisexual flowers; it differs in having anatropous ovules, in the æstivation of the sepals, etc. All species (about 180) are trees or shrubs, with scattered, simple leaves, which usually have stipules, and are occasionally dotted with pellucid oil-glands.—Bixa orellana (Trop. Am.) is the best known species; it has a 2-valved capsule; the seeds are enclosed in a shiny red, fleshy testa, which contains the well-known orange or yellow dye, annatto.
[413]
Order 10. Dilleniaceæ. Gynœceum usually apocarpous, seed arillate. The flower has most frequently S5, P5, and compound stamens (one or more bundles); sometimes irregular. 200 species; Tropical; woody plants, many lianes.—Dillenia, Candollea, Pleurandra, Davilla, etc.
Order 11. Elatinaceæ (Water-worts). About 25 species belong to this order; especially in temperate climates. They are small, creeping, rooted, aquatic plants, with opposite or verticillate leaves and stipules. The flowers are solitary or situated in small dichasia in the leaf-axils, they are small, regular, ☿, hypogynous, with free petals, the same number in all 5 whorls (Sn, Pn, An + n, Gn), 3-merous (e.g. Elatine hexandra), 4-merous (e.g. E. hydropiper), or 5-merous (Bergia); the corolla-stamens are sometimes suppressed; petals imbricate without being twisted; the ovary is 3–4–5-locular, with 3–4–5 free styles; the capsule dehisces septicidally. The seeds are orthotropous or curved, often transversely ribbed, endosperm wanting. The order is most nearly allied to Hypericaceæ, whose primitive form it appears to represent.
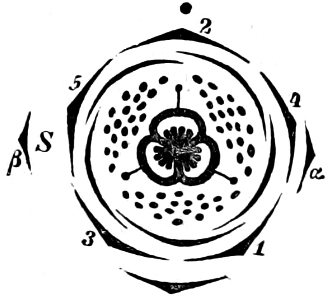
Fig. 426.—Diagram of Hypericum quadrangulum: S indicates the bud of the helicoid cyme in the axil of the bracteole β.
Fig. 427.—Hypericum. Flower with three bundles of stamens.
Order 12. Hypericaceæ (St. John’s-worts). This order is recognised by its always opposite or verticillate, simple, and entire, penninerved leaves, without stipules, and usually dotted with pellucid glands; by the always ☿, regular, hypogynous flowers in a cymose inflorescence; the generally 5-merous calyx and corolla, with sepals and petals free; the stamens 3–5, numerously branched (Figs. 426, 427); and the gynœceum, 3–5-carpellate, styles usually free. The ovary is 3–5-locular, or unilocular with 3–5 parietal placentæ. Fruit a capsule (dehiscing septicidally) or berry. Endosperm absent.
The inflorescence is a dichasium or helicoid cyme. The structure of the flowers is the same as that of the foregoing orders: S5, P5; succeeding these in some cases are two 5-merous whorls of stamens in regular alternation, of[414] which the inner is epipetalous; but the outer whorl is only represented by 5 small scales (Fig. 427), or is altogether absent (Hypericum calycinum, H. hircinum), and the inner divided into numerous stamens, that is, these 5 stamens are so deeply divided that 5 epipetalous groups bearing anthers are found (as in the Cistaceæ); in other cases the flower becomes 3-merous after the petals, stamens 3 + 3 following in regular alternation (Figs. 426, 427), the outer whorl of stamens in these cases is also present as staminodes (Fig. 427), or may be altogether suppressed. Carpels 3–5. The petals are often twisted in the bud, and are then oblique.
Hypericum. Some species have a square stem; in these cases the leaves are placed opposite the edges. Fruit a capsule.—Vismia has a berry.—The flowers of Hypericum have no honey, and supply only pollen; self-pollination often takes place.
About 240 species; the tropical ones being often shrubs or trees; the others generally perennial shrubs.—Hypericum, St. John’s-wort, contains a resinous, red matter, which can be extracted with alcohol. The American gamboge is the dried sap of species of Vismia.
Order 13. Guttiferæ, or Clusiaceæ. Closely allied to the Hypericaceæ and Ternstrœmiaceæ. Leaves opposite or verticillate. The flowers are often unisexual; stamens united; the gynœceum has most frequently a sessile, radiating or shield-like stigma.
370 species; chiefly in the Tropics (Am.). They are principally woody plants and their bark contains a yellow gum resin, “gamboge,” which is extracted from Garcinia morella (E. Ind.) and others. Mangosteen (Garcinia mangostana S.E. Asia), and Mammea americana (W. Ind.), have very delicious fruits. To this order also belong Platonia insignis, Pentadesma butyracea (the Butter-tree), Clusia, Calophyllum, Cataba, etc.
Order 14. Ternstrœmiaceæ. Trees and shrubs with scattered, simple, and often more or less leathery, evergreen, penninerved leaves, without stipules (Fig. 428). The two most important genera are: Camellia and the closely allied Thea (by some authorities these are united into one genus). The flowers are regular, hypogynous, and situated singly on very short stalks. A number of green floral-leaves are placed below the calyx and gradually pass over into the sepals, and the leaves (5–6) of the calyx again gradually pass over into the corolla (this being especially marked in Camellia), of which the number of leaves varies (5, 6, 7 and upwards); the calyx and the corolla are acyclic or eucyclic; the petals are slightly united at the base; stamens numerous in many whorls, the external ones are arranged in bundles and united with the petals as in the Columniferæ; gynœceum syncarpous; styles often free nearly to the base; ovary 3–5-locular, ovules numerous in each loculus. The fruit is a woody capsule.—[415]Other genera show more distinctly than these the same structure as in the preceding orders, namely: S5, P5, A5 + 5, of which the calyx-stamens are often suppressed, and the petal-stamens divided into numerous stamens.—Kielmeyera (S. Am.)
260 species; especially in the Tropics (E. Asia, Am.) The leaves of Thea chinensis (or Camellia thea), the Tea-tree (E. Asia), are cultivated for the well-known “tea,” and contain theine: the best are the young, still hairy leaves, of greyish colour; there are many varieties. Ornamental plants, Camellia japonica and Actinidia.

Fig. 428.—Thea chinensis (reduced).
Closely allied to this order are: Order 15. Rhizoboleæ (with enormously large hypocotyl—hence the name), and Order 16. Marcgraviaceæ (partly epiphytes, with dimorphic leaves and cup- or helmet-like, coloured, honey-secreting floral-leaves, which serve to attract insects).
Order 17. Dipterocarpaceæ. This order has taken its name from the large wings attached to the fruits in Dipterocarpus (the wings being largely developed sepals); trees and shrubs from Trop. Asia. 180 species. Camphor ready prepared is found in the stem of Dryobalanops camphora. Hopea; Vateria.
[416]
The flowers are hypogynous, ☿, polypetalous, usually regular (except Pelargonium, Tropæolaceæ, Balsaminaceæ) and throughout 5-merous: S5, P5, A5 + 5, or 5 + 0, G5 (epipetalous). The stamens soon fall off and are obdiplostemonous, often united at the base (monadelphous); the corolla-stamens are in some completely suppressed (e.g. Balsaminaceæ, Fig. 438), in others reduced to teeth (Linum, Fig. 431; Erodium). The Tropæolaceæ have 3 carpels and only 8 stamens (Fig. 437). Ring-like nectaries are not present, but at most only glandular bodies, borne outside the base of the stamens. Ovaries many-locular. The ovules as a rule are pendulous, with the micropyle directed outwards (Fig. 431, B), and the radicle therefore also points outwards. Usually herbs. Related to the Columniferæ.
Order 1. Oxalidaceæ. Most of the species are herbs with rhizomes; the leaves are stalked, compound, with entire leaflets which are folded and bent backwards in the bud (and in the sleep position), exstipulate; some species have sensitive leaves. The flowers (Fig. 429) are regular, and have S5, P5, which are twisted to the left or right in æstivation, A5 + 5, all united at the base (monadelphous), gynœceum 5-carpellate, styles 5 free, stigmas capitate, ovary 5-locular, ovules numerous. The fruit is a capsule opening with clefts on the dorsal sutures through which the seeds are ejected, while the fleshy, external layer of the testa springs off elastically. Embryo straight. Endosperm.
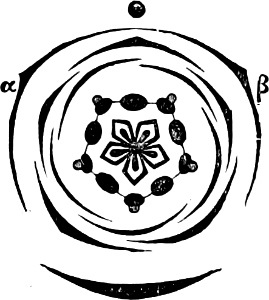
Fig. 429.—Diagram of Oxal’s acetosella.
Oxalis (Wood-Sorrel). Leaves digitate. Species also occur with phyllodia, i.e. leaf-like petioles placed vertically without lamina; a few have pinnate leaves. The flowers are situated singly or in dichasia, and unipared scorpioid cymes. The pollination is effected by insects. Some species are trimorphic (long-, short-, medium-styled flowers) and some, e.g. O. acetosella, have cleistogamic flowers in addition to the ordinary ones. Glands are found on the outer side of the corolla-stamens or of all the stamens. O. tetraphylla and others have adventitious edible roots, resembling tap-roots.—Averrhoa is a tropical tree, with berries and pinnate leaves.
235 species (205 belong to Oxalis); chiefly in S. Africa and Trop. America.—Oxalate of potash is contained in the leaves of Oxalis.
[417]
Order 2. Linaceæ. Herbs with scattered or opposite, sessile, simple, small, entire leaves, without (rarely with small) stipules. The flowers (Fig. 430) are regular, 5- or 4-merous. Petals are free, twisted, quickly falling off. Stamens united at the base; the petal-stamens are either reduced to teeth (Fig. 431 A, m) or entirely suppressed. Styles free. The (5–4) epipetalous loculi of the ovary are incompletely halved by false divisional walls, each half contains one ovule (Fig. 431 C). The fruit is a spherical capsule, dehiscing along the divisional wall (Fig. 432); the 10 (-8) seeds have a straight embryo and very slight endosperm (Fig. 433).
Figs. 430–433.—Linum usitatissimum.
Fig. 430.—The Flax plant.
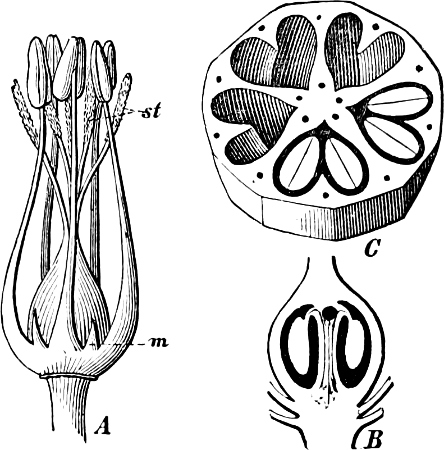
Fig. 431.—A Flower after removal of sepals and petals; m petal-stamens reduced to teeth. B Longitudinal section of ovary. C Transverse section of capsule.

Fig. 432.—Capsule (nat. size).
Fig. 433.—Transverse and longitudinal section of seed: bl the cotyledons; k the plumule; R the radicle; fr the endosperm; sk the testa.
[418]
Linum (Flax) has 5-merous flowers. The main axis terminates in a flower; and the succeeding branching is cymose, or unipared scorpioid branching by unilateral development, and the flowers in consequence of the vigorous sympodial development of the lateral axis (and also by the leaves being displaced and pushed aside), assume a position apparently lateral (i.e. racemose) without bracts; each branch of the sympodium generally has 2 leaves. The testa is shining and smooth when dry, but its external cellular layer becomes mucilaginous in water.—Radiola has a 4-merous flower. It is a small herb with opposite leaves, and regular, dichasial branching.
The anthers and stigmas in L. catharticum and usitatissimum develop simultaneously, and cross-pollination as well as self-pollination takes place. L. grandiflorum, perenne, and others, are dimorphic (short-and long-styled). There are 5 nectaries outside the stamens.
130 species; Linum and Radiola are native genera.—L. usitatissimum is extensively cultivated in Europe (especially in Russia and Belgium), N. America and elsewhere (its home no doubt being Asia), partly on account of the oil (linseed oil) which is extracted from the seeds, and partly on account of the bast of the stem, which has very thick-walled cells. The seeds and oil are OFFICINAL. The species cultivated in ancient times was L. angustifolium. Several species are cultivated as ornamental plants.
Order 3. Geraniaceæ. The majority are herbs with dichasial branching, and scattered or opposite, stalked, palminerved (rarely penninerved) leaves with small stipules. The flowers are regular (except Pelargonium) and 5-merous, with 10 or 5 stamens, which are slightly united at the base. Nectaries alternate with the corolla-stamens. The ovary is most frequently 5-locular, deeply 5-grooved, and bears 1 well developed style (“beak”), which towards the apex divides into 5 branches bearing stigmas; ovules 1 in each loculus, pendulous or ascending. The 5 carpels become detached from one another when ripe, and bend or roll back (Fig. 434) or become spirally twisted in the upper “beak-like” part (Figs. 435, 436), whilst a central column (septal column) persists; each carpel, in consequence, remains either closed, and the fruit is a 5-merous schizocarp whose nut-like lower portion, containing the seed, is forced into the ground, thus burying the seed by the movements of the spirally-twisted, hygroscopic “beak” (Figs. 435, 436); or it opens along the ventral suture, so that the seeds may fall out, and it is then a 5-valved capsule, with septicidal dehiscence (Fig. 434) and the rolling up often takes place so suddenly and violently that the seeds are shot out to considerable distances. The embryo is usually green and curved, and the cotyledons are folded; endosperm is wanting.
[419]
Geranium (Crane’s-bill) has 5 + 5 stamens,and a septicidal capsule; the carpels most frequently remain suspended from the apex of the column (Fig. 434). The leaves are most frequently palminerved. The flowers are situated solitarily or 2 together (2-flowered scorpioid cyme).—Erodium (Stork’s-bill); inflorescence a many-flowered unipared scorpioid cyme, stamens 5 + 0 (petal-stamens are wanting), and fruit a schizocarp whose carpels become detached; their beaks are hairy on the internal surface and twist themselves spirally (Fig. 436). The umbellate inflorescences are composed of multiflowered scorpioid cymes. The leaves are often penninerved.—The most primitive type is represented by Biebersteinia: S5, P5, A5 + 5, G5 (ovaries free, and styles united above); fruit 5 small nuts. The most advanced type is Pelargonium, which has zygomorphic flowers, the posterior sepal being prolonged into a spur which becomes adnate to the peduncle; the petals are unequal in size; some of the petal-stamens are often wanting. (Erodium may be slightly zygomorphic).
Fig. 434.—Geranium sanguineum. Fruit (3/1).
Fig. 435.—Pelargonium.

Fig. 436.—Erodium cicutarium, detached carpel.
Pollination. The large-flowered Geranium-species are protandrous, e.g. G. pratense (one whorl of stamens opens first, and then the other, and succeeding these the stigmas, after shedding the pollen the stamens bend outwards); the small-flowered are also adapted, with various modifications, for self-pollination.—470 species; moderately hot climates, especially S. Africa.—Several Pelargonium-species, with numerous varieties, are ornamental plants (from S. Africa).
Order 4. Tropæolaceæ. Herbaceous, juicy plants which have scattered, long-stalked, peltate leaves without stipules, and often[420] climb by their sensitive petioles. The flowers are situated singly in the axils of the foliage-leaves on long stalks, and are zygomorphic, the receptacle under the posterior sepal being prolonged into a spur; there are also differences between the posterior and anterior petals, the 2 posterior petals situated on the border of the spur being perigynous, and the edge of the anterior petals adjoining the claw fringed. After the 5 sepals (which are more or less coloured) and the 5 petals, follow 8 stamens (as the 2 median ones are suppressed, one from each whorl) and a gynœceum formed of 3 carpels; in each of the 3 loculi of the 3-grooved ovary is 1 ovule. The fruit is a schizocarp and divides into 3 1-seeded, drupe-like fruitlets, which do not (as in the Geraniaceæ) leave any pronounced column between them. Endosperm is wanting. The cotyledons are thick and sometimes slightly coalescent. Tubers often occur.
Fig. 437.—Diagram of Tropæolum: sp, spur.
Tropæolum.—About 40 species; all from America.
Pollination.—The spur is the receptacle for the nectar; the flowers are protandrous; the anthers open first, and one by one take up a position in front of the entrance to the spur, resuming their original position when the pollen is shed; the stigma finally takes their place after the filaments have bent backwards.—These plants have an acrid taste (hence the name “Nasturtium,” “Indian Cress”), on which account the flower-buds and young fruits of T. majus are used as capers. Some species are ornamental plants.
Order 5. Balsaminaceæ. Herbaceous, chiefly annual plants with juicy, brittle stems, so transparent that the vascular bundles may be distinctly seen. The leaves are simple, usually scattered, penninerved and dentate; stipules are wanting, but sometimes large glands are present in their place at the base of the petioles. The flowers are strongly zygomorphic; of their five 5-merous whorls the petal-stamens are suppressed (S5, P5, A5 + 0, G5); the sepals are coloured, the 2 anterior ones (Fig. 438 3, 5) are very small or entirely suppressed, the posterior one is very large and elongated into a spur, and the 2 lateral ones pushed forward; sometimes the weight of the spur turns the flower completely round, so that the posterior leaves assume an anterior position; apparently only 3 petals, since the lateral and the posterior petals become united in pairs, and the anterior is larger[421] and differently shaped; the 5 stamens have very short and thick filaments united at the base, and their anthers finally adhere together and remain in this condition, covering over the gynœceum; the filaments ultimately rupture at the base, and the entire anthers are raised on the apex of the gynœceum as it grows up. The gynœceum has a sessile stigma and a 5-locular ovary. The fruit is a capsule which, on maturity, opens suddenly when irritated, dividing into valves from the base upwards, and as the 5 valves roll up elastically, the seeds are shot out on all sides to considerable distances; a central column persists (Fig. 439). The embryo is straight, and without endosperm.
Fig. 438.—Diagram of Impatiens glanduligera.
Fig. 439.—Fruit of Impatiens.
Impatiens; in Europe only I. noli-me-tangere. 225 species; especially from Asia. Several species have two kinds of flowers: small, cleistogamic, but fertile; and large, coloured flowers, which in I. balsamine (ornamental plant, E. Ind.) are protandrous and pollinated by hive-and humble-bees, as they suck the honey from the spur.
Order 6. Limnanthaceæ. The flowers are regular and differ from all the other orders in the family by having the carpels not in front of the petals, but in front of the sepals (which are valvate), and further, the loculi are nearly free individually, but with a common gynobasic style; the ovules are ascending and apotropous (anatropous with ventral raphe). The fruit is a schizocarp, with nut-like cocci.—Limnanthes (4 species; N. Am.) perhaps belongs to another family.
Order 7. Humiriaceæ. Trees and shrubs; about 20 species; Trop. Am.
The chief characteristics of the orders belonging to this family are the ☿, regular, generally 5-merous, hypogynous flowers with[422] 5-merous calyx, sepals united and valvate in the bud; petals 5, free (often twisted in the bud); stamens ∞ e.g.: 10, in two whorls, but one of these is more or less suppressed, often altogether wanting, or replaced by 5 staminodes, while the other (inner whorl) is generally divided more or less deeply into a large number of anther-bearing filaments. The filaments too (except Tiliaceæ) are united into a tube, which, especially in the Malvaceæ, forms a long column in the centre of the flower, surrounding the gynœceum (Figs. 445, 448); in this case, which is the most pronounced, the filaments are united into one bundle (monadelphous), in other instances, polyadelphous. The number of carpels varies greatly (2 to about 50), but they are nearly always united and form a syncarpous multilocular gynœceum.—The vegetative characters also closely agree, the leaves are always scattered and generally stipulate; all the green portions very often bear stellate hairs, and the bark in all the 3 orders is rich in tough bast. Mucilage is often present in cells or passages.—This family is connected with the Ternstrœmiaceæ, from which it is very hard to draw a sharp line of demarcation, and it is also allied to the Cistaceæ and to the Gruinales.
Order 1. Sterculiaceæ (including Buettneriaceæ). This is, no doubt, the least modified order, and one in which the stamens occur undivided. Obdiplostemonous. The 10 stamens in two whorls are most frequently united at the base into a short tube, and have 4-locular, extrorse anthers. The calyx-stamens are nearly always simple, tooth-like staminodes, situated on the edge of the tube, or are entirely suppressed. The same relation is found, for instance, in the Ampelidaceæ and Rhamnaceæ, namely 5 stamens in front of the 5 petals; not infrequently the 5 stamens are doubled (Fig. 441). Unisexual flowers are found in Sterculia, Cola, Heritiera. The corolla is often wanting, or developed in an unusual manner. Each loculus of the ovary (generally 5) always contains more than one ovule. Fruit a capsule. Androgynophore often present (Helicteres; Sterculia, etc.).
Hermannia, Mahernia, Melochia, etc., have flat petals with twisted æstivation; 5 undivided stamens, which usually are but slightly united at the base, and most frequently, without staminodes. Thomasia; Helicteres; Sterculia (free follicles).—Theobroma, Rulingia, Buettneria, Commersonia, Guazuma, etc., have petals concave at the base, and terminating in a limb abruptly bent back, and at the boundary between them most frequently ligular outgrowths, as in certain genera of the Caryophyllaceæ; stamens 5–15–∞, anthers at the edge of a short tube and 5 linear staminodes (Fig. 441).—The Cocoa-tree (Theobroma), (Fig. 440) bears large, reddish-yellow, berry-like fruits, resembling short cucumbers,[423] but ultimately becoming leathery to woody; in each of the 5 loculi are 2 (apparently only 1) rows of horizontal, oily seeds, as large as almonds. Cotyledons large, thick, and irregularly folded. Endosperm absent (Fig. 442).
49 genera, with about 750 species; almost entirely confined to the Tropics; none in Europe or in N. Asia.—The seeds of the Cocoa-tree (T. cacao, bicolor, glaucum, etc., natives of Trop. Am., especially north of the Equator) are used for chocolate and are also officinal (“Cocoa-beans,” “Cocoa-butter,” “Oil of Theobroma”). Theobromine. Cola acuminata, Africa.
Fig. 440.—Theobroma cacao. Branch with flowers and fruits (⅙).
Figs. 441–442.—Theobroma cacao.

Fig. 441.—Diagram of the flower: st barren stamens.
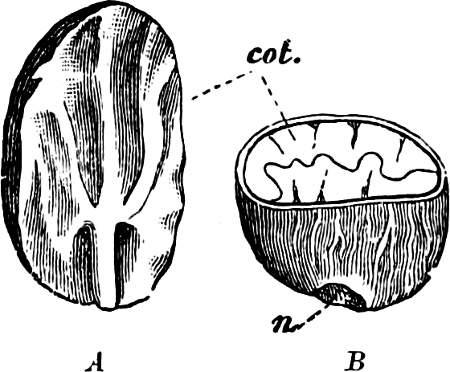
Fig. 442.—B Seed in transverse section: n hilum. A Embryo after the removal of one of the cotyledons.
Order 2. Tiliaceæ. This differs from the other orders of the Columniferæ chiefly in the stamens being entirely free from each other, and also divided into many filaments, as far as the base, or at all events very far down, so that the flower appears to have numerous[424] stamens or to be slightly polyadelphous (Fig. 443); in addition to this, it may be observed that the anthers are 4-locular and introrse. In Luehea the groups of stamens alternate with the petals. In a few genera (Corchorus, Triumfetta) 10 free and single stamens are found in 2 whorls; but, in the majority, groups of free stamens in separate bundles. The stamens are more or less united in Apeiba, Luehea. Style simple. Ovary 2-locular. The ovules are pendulous; raphe turned inwards. The calyx readily falls off; the æstivation of the entirely free petals is slightly imbricate (not twisted).
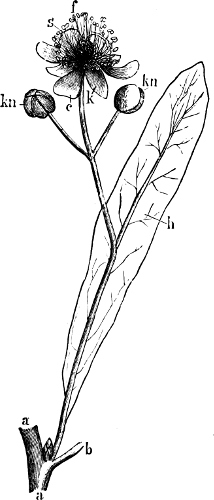
Fig. 443.—Inflorescence of Tilia, with its winged bracteole (h); a, a axis of the shoot; the vegetative bud is seen between the inflorescence and the axis of the shoot; b petiole of foliage-leaf.
Tilia (Figs. 443, 444). Calyx and corolla 5-merous; the 5 staminal leaves (opposite the petals) divided as far as the base into a large number of stamens which are free or united into groups; gynœceum with 5 loculi in the ovary (opposite the sepals); there are 2 ovules in each loculus, though the ovary ripens into a 1-seeded nut, which is not detached from the axis of the inflorescence, but is carried away by the wind, whirling round and round, its large-winged bracteole serving as a parachute (Fig. 443).—Only trees, with alternate, obliquely heart-shaped and dentate leaves; stellate hairs, as in the other Columniferæ, are often present. The terminal bud of the branch always fails to develop, and the growth is then continued sympodially by the uppermost axillary buds. The INFLORESCENCE (Figs. 443, 444) is a 3–7-flowered dichasium (Fig. 444 t, d, e), which is developed in the axil of a foliage-leaf (Fig. 444). The first of its 2 bracteoles (a) is large, thin, leaf-like, and united with the inflorescence, the lower portion of which forms a broad wing, its so-called “bract”; the second bracteole (b), on the other hand, remains scale-like, and supports a winter foliage-bud covered with bud-scales which thus is situated at the base[425] of the inflorescence, and is a bud of the 2nd order, in relation to the vegetative shoot. This bud is always found beneath the inflorescence on the branch placed horizontally, and the winged bracteole is always found above it, a relation which is connected with the fact that the 2 rows of shoots on the sides of a branch are antidromous with regard to each other.—The dichasium itself (Fig. 444) terminates with the flower (t); it has 3 floral-leaves (c, d, e), which soon fall off; c is barren: the other two bear flowers, or few-flowered dichasia, or unipared scorpioid cymes (indicated in the figure).—The foliage-leaves are folded in the bud upon the median line (1, 2, 3 in Fig. 444 are foliage-leaves with their 2 stipules), the inner half is broader than the outer, and after unfolding is turned away from the mother-axis (the position of the new inflorescences and vegetative buds is indicated in their axils on the figure).—The cotyledons on germination appear above the ground as large, lobed leaves.
Of the other genera some have a bell-shaped, gamosepalous calyx, some have no corolla, the anthers of some open at the apex (Aristotelia, Elæocarpus, etc.), the majority have a capsule, some have berries, or drupes, some separate into fruitlets, etc.—Corchorus, Triumfetta (nut, with hooked bristles), Luehea, Apeiba, etc. Sparmannia is an African genus; 4-merous flowers; fruit a warted capsule; filaments numerous and sensitive to touch, the external ones are without anthers and moniliform above. The plant is covered with numerous soft and stellate hairs, and at the apex of the branches bears several cymose umbels.
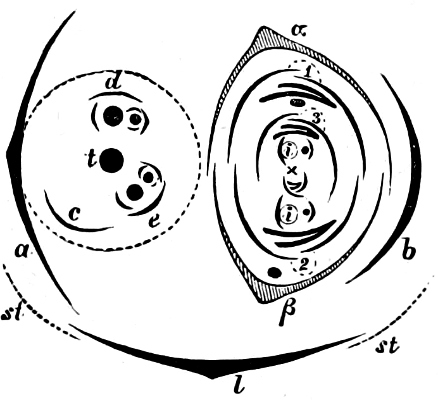
Fig. 444.—Diagram of the inflorescence of Tilia and the vegetative bud; the position of the leaves is indicated, and also the position of the inflorescences, which develop from their axils in the following year.
Pollination in Tilia is effected by insects, especially bees and Diptera, which swarm round the tree tops, allured by the numerous strongly-scented flowers and the easily accessible honey (formed in the hollow sepals). As the flowers are pendulous, the nectar is protected from ruin; and, in addition, the inflorescence is more or less concealed beneath the foliage-leaf. Self-pollination is impossible, on account of protandry.—About 470 species (nearly all trees and shrubs); especially in the Tropics, only a few being found in the temperate, none in the polar regions, or in high mountainous districts.—The inflorescence of the native species of Tilia is medicinal. The wood is used for charcoal.—The majority are used for timber, and for the sake of the bast (“Bast,” “Jute,” the bast of Corchorus textilis, Luehea, and others).
Order 3. Malvaceæ (Mallows). The plants are easily recognised by the scattered, simple, palminerved, most frequently lobed, stipulate leaves, folded in the bud; the perfect, regular, hypogynous flowers, with gamosepalous, persistent, 5-merous calyx[426] with valvate æstivation; the 5 petals twisted in the bud and united with one another at the base, and by the 5 apparently numerous stamens (Figs. 445, 448), with the filaments united into a tube, with reniform bilocular anthers opening by a crescentic slit (in 2 valves). Carpels 3–∞ united into one gynœceum; the embryo is curved and the cotyledons are folded (Figs. 447, 451); endosperm scanty, often mucilaginous.—Most of the plants belonging to this order are herbs, often closely studded with stellate hairs. The leaves are most frequently palmatifid or palmatisect.
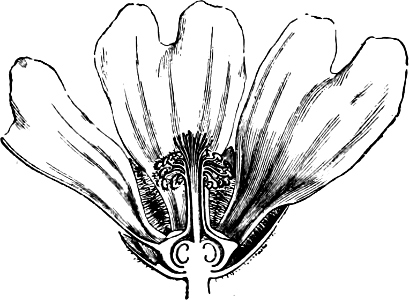
Fig. 445.—Longitudinal section through the flower of Malva silvestris.

Fig. 446.—Diagram of Althæa rosea: i the epicalyx.
An epicalyx is often found formed by floral-leaves placed close beneath the calyx, in some 3, in others several. The median sepal is posterior in the species without epicalyx, often anterior in those which have an epicalyx.—The petals are twisted either to the right or to the left in accordance with the spiral of the calyx; they are most frequently oblique, as in the other plants with twisted corollas, so that the portion covered in the æstivation is the most developed. The corolla drops off as a whole, united with the staminal tube.—Only the 5 petal-stamens are developed, but they are divided into a number of stamens, placed in 2 rows, and provided only with half-anthers (leaf-segments, see Fig. 446; the sepal-stamens are completely suppressed); these 5 staminal leaves are then united into a tube, frequently 5-dentate at the top, and bearing the anthers on its external side. The pollen-grains are specially large, spherical and spiny. There are from 3 to about 50 carpels united into one gynœceum and placed round the summit of the axis which most frequently projects between them. There is only 1 style, which is generally divided into as many stigma-bearing branches as there are carpels (Figs. 445, 448). The fruit is a schizocarp or capsule. Endosperm (Figs. 447 A, 451) scanty, often mucilaginous round the embryo, which is rich in oil.
The order is the most advanced type of Columniferæ; it stands especially near to the Sterculiaceæ, but is separated from these and from the Tiliaceæ, among other characters, by its 2-locular (ultimately 1-chambered) anthers.
The sub-orders may be arranged as follows:—
[427]
I. Carpels in one whorl.
A. The fruit a capsule, most frequently with loculicidal dehiscence, and many seeds in each loculus.
1. Gossypieæ. The staminal-column is naked at the apex, blunted, or 5-dentate.—Gossypium (the Cotton plant) has an epicalyx of 3 large ovate-cordate leaves, an almost entire, low and compressed calyx. Solitary flowers. Large, most frequently yellow, corollas. A 3–5-valved capsule with many spherical seeds. “Cotton” is the seed-hairs developed upon the entire surface of the seeds (Fig. 447), and consists of long, 1-cellular hairs, filled with air (and therefore white); these are thin-walled, with a large lumen, and during drying twist spirally, and come together more or less in the form of bands. They consist of cellulose, and have a cuticle.—Hibiscus has several, most frequently narrow, epicalyx-leaves, a distinct 5-toothed or 5-partite calyx.—Abutilon; Modiola.

Fig. 447.—A Seed of Gossypium with hairs; B the same in longitudinal section.
2. Bombaceæ. The staminal tube is more or less deeply cleft into bundles, sometimes almost to the base; pollen smooth, style simple with capitate, lobed stigma. Almost all plants belonging to this group are trees, and in many instances have large barrel-shaped stems, that is, swollen in the centre, and sometimes covered with large warts. The wood is exceptionally light and soft. The flowers are often enormously large, and have beautiful petals; in some they unfold before the leaves. The capsule-wall is sometimes closely covered on its inner service with long, silky, woolly hairs, while the seeds themselves are generally without hairs. These hairs, however, on account of their brittle nature, cannot be used like those of the Cotton-plant. Digitate leaves are found in the Baobab-tree (Adansonia) from Africa, noted for its enormously thick, but short stem, and in the American Silk-cotton trees (Bombax, Eriodendron, Chorisia). Ochroma, Cheirostemon, Durio, and others also belong to this group. Durio is noted for its delicious fruits, which have a most unpleasant smell.
[Bombax malabaricum is diplostemonous; the five sepal-stamens repeatedly[428] branch, and the filaments bear unilocular anthers; the five petal-stamens bear bilocular anthers.]
B. Schizocarps, with 1-seeded fruitlets, most frequently nut-like and reniform (Figs. 449, 451).
3. Malveæ, Mallow Group. The carpels are arranged in one whorl (Fig. 449); the number of stylar-branches equals that of the carpels; fruitlets 1-seeded, reniform, indehiscent, but detaching themselves from one another and from the persistent central column (Figs. 450, 451).—Malva has an epicalyx of 3 free leaves. A flower with 2 suppressed bracteoles is situated in the axil of the foliage-leaves; one of these supports a homodromous foliage-shoot which forms a repetition of the main axis, the other an antidromous flower which continues the branching as a unipared scorpioid cyme.—Althæa, Rose Mallow, has an epicalyx of 6–9 leaves united at the base.—Lavatera, Sida, Anoda, Bastardia, etc., have no epicalyx.
Figs. 448–451.—Malva silvestris.

Fig. 448.—The flower after removal of the perianth (5/1).]
Fig. 449.—The fruit (5/1).
Fig. 450.—A fruitlet (5/1).
Fig. 451.—The same in longitudinal section.
4. Ureneæ, have always only 5 carpels arranged in 1 whorl, with 1 ovule in each loculus, and the fruit a schizocarp, generally with nut-like fruitlets provided with warts and hooks; but in some they dehisce by 2 valves (capsule). They differ principally from the other groups in having twice as many stylar-branches as carpels; the staminal tube is naked at the point, blunt or 5 toothed.—The genera Urena, Pavonia, Malachra, Malvaviscus (with berry-like fruits) belong to this group.
II. Carpels arranged in a spherical head in five groups opposite to the petals.
5. Malopeæ, differ from all the others in having a large number of fruitlets arranged irregularly in a round head, and separating considerably from each[429] other even before maturity; there is, however, only 1 style, divided into a corresponding number of branches (this condition may be considered to have arisen from the branching [dédoublement] of 5 carpels). Malope has 3 large, heart-shaped (Kitaibelia 6–9) epicalyx-leaves, united at the base. Palava has no epicalyx.
Pollination. The majority have protandrous flowers, and are pollinated by insects. Between the basal portions of the 5 petals, there are 5 nectaries, protected from the rain by hairs, e.g. in Malva silvestris. When the flower first opens the numerous anthers occupy the centre of the flower, and the still undeveloped stigmas are concealed in the staminal tube; in the next stage the anthers are withered and empty, and the stigmas protrude and assume their places (Fig. 452). The large-flowered forms, it appears, are pollinated only by insects; but self-pollination takes place in small-flowered forms, as, for example, in Malva rotundifolia, in which the stylar-branches, twisting themselves, place the stigmas in between the undeveloped anthers.
Fig. 452.—Anoda hastata: a the bud just opened, the stigmas are concealed by the anthers; b fully opened flower in ♂-stage; the upper stamens are developed first, and then the others in descending order; the stylar-branches are now visible, and lie bent back on the staminal column; c all the stamens project upwards, and all the anthers are open, but the stylar-branches are still bent back; d the anthers are emptied and the filaments shrunk together, but the styles have now straightened themselves upwards, and the stigmas are in the receptive condition.
Distribution. 800 species (63 genera), most of which are natives of the Tropics, especially America. Althæa and some of the species of Malva are natives of the temperate regions of the Old World, the latter is also found in North America. Gossypium is tropical, no doubt especially Asiatic (G. herbaceum from India; G. arboreum from Upper Egypt). Cotton was introduced into Greece in the time of Herodotus, and was cultivated in America before the arrival of the Europeans.
Uses. Pungent and poisonous properties are entirely wanting; mucilage, on the other hand, is found in abundance in all parts of the plant. Medicinal: the root of Althæa officinalis, leaves and flowers of Malva-species (M. silvestris[430] vulgaris and borealis) and Gossypium.—The seeds contain a large quantity of fatty oil, which is in some cases extracted (Cotton-seeds and others). The seed-hairs of the Cotton plant are the most important product of the order. The cultivated forms of Cotton belong to several species: G. barbadense, herbaceum, religiosum, arboreum (Nankin), hirsutum, and others. According to other botanists, there are only 3 species. Bast is obtained from e.g. Hibiscus cannabinus (Gambo-hemp, Africa), Paritium tiliaceum and Sida retusa. The fruits of certain species of Hibiscus (e.g. H. esculentus, from Tropical Africa) are used in tropical countries as a vegetable before they are ripe.—The colouring matter in the flowers of Althæa rosea, var. nigra, is used for colouring wines, and hence is extensively cultivated in certain parts of Europe.—Ethereal oils and sweet-scented flowers are rare; but several species possess a peculiar musk-like odour (Malva moschata, Hibiscus abelmoschus, and others).—Many are cultivated as ornamental plants on account of the large flowers, e.g. Hollyhock (A. rosea, etc.), Lavatera trimestris, Malope grandiflora and trifida, Malva-species, Hibiscus rosa sinensis, syriaca; Sphæralcea, etc.
The very large order Euphorbiaceæ and three smaller ones belong to this family. They have in common: unisexual, hypogynous, frequently regular flowers, the perianth most frequently single, rarely double, or entirely wanting; there is such a great variety in the structure and parts of the flower that one only can be cited as the rule: viz. the simple gynœceum composed of 3 carpels forming a 3-locular ovary, which is frequently more or less deeply grooved (hence the name, Tricoccæ); in the inner angles of the loculi are found 1 or 2 (never several) pendulous (except Empetraceæ), anatropous ovules, with upward and outwardly turned, frequently swollen, micropyle (Fig. 455). The seed most frequently has a large endosperm and a straight embryo (Figs. 455 B, 464).—The family approaches the nearest to the Gruinales and Columniferæ; it may perhaps be regarded as an offshoot from the Sterculiaceæ.
Order 1. Euphorbiaceæ. Flowers unisexual. In each of the loculi of the ovary, generally 3, there are 1 or 2 pendulous ovules with upward and outwardly turned micropyle. The placenta protrudes above the ovules (Figs. 454, 461 B). On the ripening of the capsule the 3 carpels separate septicidally, frequently with great violence, ejecting the seeds and leaving a central column. Endosperm copious.—For the rest, the flowers present all stages, from genera with calyx and corolla, to those which are the most reduced in Nature, namely the naked, 1-stamened flowers of Euphorbia.
The same variety which is found in the flower is also present in[431] the vegetative parts. Some are herbs, as our Spurges, others are shrubs and trees; some African Euphorbia-species even resemble the habit of a Cactus. Leaf-like branches with rudimentary leaves are found in Phyllanthus (sub-genus Xylophylla) (Fig. 456). The leaves are scattered or opposite, often stipulate; they are nearly always simple. Large, highly-branched cells containing a great quantity of pungent latex are found in many, and watery juice in others. Glands and glandular hairs are general.—Only a few genera can be considered in this book.
As an example of the most perfect flowers (which partly reproduce the Geraniaceous type) may be mentioned, Croton, Manihot, and Jatropha; 5 sepals, 5 petals, sometimes gamopetalous, andrœcium diplostemonous, or many-stamened, often monodelphous.
Figs. 453–455.—Ricinus communis.

Fig. 453.—♂-flower (magnified).
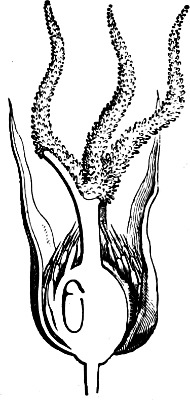
Fig. 454.—♀-flower in longitudinal section.
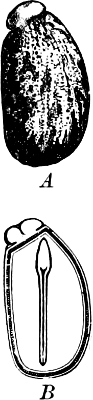
Fig. 455.—A seed entire; B in longitudinal section.
Ricinus (Castor-oil) (Figs. 453–455); monœcious; the ♂-flowers, situated in the lower portion of the inflorescence, have 5 perianth-leaves and a large number of branched stamens; the ♀-flower has 3–5 perianth-leaves; 3 bifid styles. Leaves peltate, palmately lobed. The seeds (Fig. 455) contain an abundance of fatty oil and large aleurone grains.—Mercurialis (Mercury): the perianth is most frequently 3-merous; in the ♂-flowers 9–12 stamens; in the ♀-flowers most frequently a 2-locular gynœceum.—Phyllanthus: Pr3 + 3, A3, united in some and forming a column in the centre of the flower (Figs. 457, 458); Xylophylla is a section of this genus.—[432] Hura crepitans (Sand-box tree) has a many-carpellate gynœceum, which separates with great violence when ripe.—A drupe is found in Hippomane mancinella (the Mancinil-tree, W. Ind.)—Alchornea (Coelebogyne) ilicifolia is well known on account of its “parthenogenesis”; only the ♀-plant has been introduced into Europe, but it nevertheless produces seeds capable of germination; these have generally several embryos.
Figs. 456–458.—Phyllanthus (Xylophylla) angustifolius.
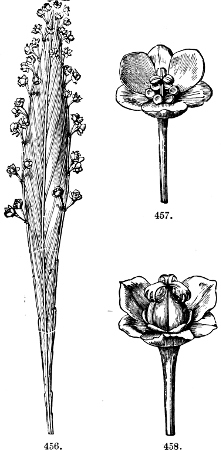
Fig. 456.—Leaf-like branch with flowers (nat. size).
Fig. 457.—♂-flower; and
Fig. 458, ♀-flower (mag.).
Euphorbia (Spurge) has the most reduced flowers, which are borne in a very complicated inflorescence. Each ♂-flower (Fig. 460 B) is naked, and consists of one stamen only (terminal on the axis). In the closely allied genus Anthostema, a small perianth is situated at the place where, in Euphorbia, there is a joint in the “filament,” (Fig. 461 A). The ♀-flowers (Fig. 460) are naked, with a 3-locular ovary and 3 bifid styles. (Anthostema has a distinct perianth (Fig. 461 B); in a few Euphorbias traces of a perianth are present). In Euphorbia the ♂-and ♀-flowers are grouped into flower-like inflorescences termed “cyathia.” Each cyathium consists of a centrally placed ♀-flower which is first developed, surrounded by 5 groups of ♂-flowers (stamens) placed in a zig-zag, with a centrifugal order of development (Figs. 459, 460 B), that is, in unipared scorpioid cymes; these flowers are surrounded by an involucre of 5 leaves united into a bell-shaped structure (Fig. 459, 1–5) (resembling a calyx); on its edge are placed 4, generally crescent-like, yellow glands, one in each of the intervals, except one, between the lobes of the involucre (shaded in Fig. 459; see also Fig. 460 A). Scale-like[433] thin structures (floral-leaves?) are situated between the ♂-flowers. The ♀-flower has a long stalk, and finally bends down on one side, namely to the place on the edge of the involucre where the gland is not developed. These cyathia are again arranged in an inflorescence which commences as a 3–5-rayed umbellate cyme (pleiochasium), the branches of which ramify dichasially and finally as scorpioid cymes.—Latex, with peculiar-shaped starch-grains, is found in laticiferous cells (especially in the Cactus-like, leafless species.)
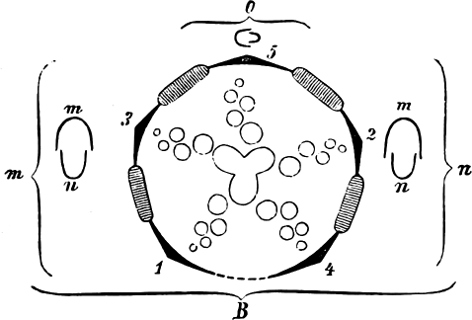
Fig. 459.—Diagram of an inflorescence (cyathium) of Euphorbia with 3 floral-leaves, m, n, o, supporting other cyathia which are subtended by 2 floral-leaves (bracteoles; m, n). 1–5, the involucral leaves in their order of development; the shaded portions are the crescentic glands.
Fig. 460.—Euphorbia lathyris: A an (entire) inflorescence (cyathium); B the same after the removal of the involucre.
Fig. 461.—Anthostema: ♂- (A) and ♀-(B) flowers; p the perianth; ar the node; o the ovule.
205 genera; more than 3,000 species; especially in the Tropics.—Many are used on account of the oil, and of the pungent (aperient, poisonous, anthelmintic,[434] etc.) properties in the latex or the seeds. Officinal: “Cascarilla-bark” of Croton eluteria; the fatty oil of the seeds of Croton tiglium (Trop. Asia); “Castor oil” from Ricinus communis (Africa, and cultivated in all warm climates throughout the world); the glandular hairs of Mallotus philippinensis (“Kamala”); this also yields a red dye. Gum “Euphorbium” is the hardened (resinous) latex of the Cactus-like Euphorbia resinifera (Morocco).—Nutritive plants: Manihot utilissima and other species (Maniok, Am.). Their large, farinaceous roots form a very important article of food in the Tropics (Cassava-flour, Tapioca or Brazilian arrowroot). The fresh latex of the root in some species is a powerful poison; but the poisonous properties are diminished by roasting or cooking. Caoutchouc is obtained from Siphonia elastica (Trop. S. Am.). The vegetable tallow of the Chinese tallow-tree (Stillingia sebifera) is used in large quantities in soap factories. An indigo-like dye is obtained from Crozophora tinctoria, and is also found in Mercurialis perennis. Shellac is obtained from Aleurites laccifera. Ornamental plants: Acalypha, Croton, Dalechampia.—Hippomane is poisonous.
Order 2. Buxaceæ. This order differs from the Euphorbiaceæ in having the micropyle turned inwards; the ♂-flower has a 4-partite perianth and 4 stamens; the ♀-flower a 6-partite perianth and 3 carpels. Capsule with loculicidal dehiscence, the inner layer being detached elastically from the outer.—30 species. Shrubs without latex and with evergreen leaves.—Buxus sempervirens (Box) is an ornamental shrub (poisonous); it has a very hard and valuable wood which is used for wood-engraving and carving.
Figs. 462–464. Callitriche stagnalis.
Fig. 462.—♂-flower with the 2 bracteoles and the solitary stamen.
Fig. 463.—♀-flower.
Fig. 464.—Longitudinal section of the ripe fruit.
Order 3. Callitrichaceæ. Aquatic plants, growing at the bottom of shallow water, with opposite, simple, undivided, entire, exstipulate leaves, which are generally crowded and form a rosette in the apex of the branches. The flowers are unisexual (monœcious) and borne singly in the leaf-axils; they have no perianth, but are provided with two delicate bracteoles; the ♂-flowers consist of only 1 terminal stamen (Fig. 462); the ♀-flowers of a bicarpellate gynœceum (Fig. 463) which is originally 2-locular, but later on becomes 4-locular, as in the case of the gynœceum of the Labiatæ, by the formation of a false partition-wall; in each loculus there is 1 pendulous ovule with the micropyle turned outwards. Fruit a 4-partite schizocarp (Fig. 464). 25 species.—Callitriche.
Order 4 (?). Empetraceæ. 4 species. Empetrum; E. nigrum (Crowberry)[435] is a heather-like, moorland, evergreen undershrub with linear leaves, having a deep groove closed with hairs, on the under side. The erect ovules show the greatest deviation from the Euphorbiaceæ. Diœcious (and ☿); S3, P3; in the ♂-flower, 3 stamens; in the ♀-flower, a 6–9-locular ovary. Fruit a drupe.
The diagram of the flower (Figs. 465–467) is the same as in the Gruinales, namely S, P, A2 and G in whorls of 5 (less frequently 3, 4, 6, 8), and the same modifications also occur with the suppression of the petal-stamens, etc. But a ring or sometimes cup-like glandular structure (disc) is found between the andrœcium and the gynœceum (Figs. 465, 466). The flowers similarly are regular, hypogynous, ☿ and polypetalous, though exceptions are found to all these characters: thus, for example, united sepals and petals frequently occur, and, in some orders, unisexual flowers by the suppression of one sex. In most cases the flowers are small, greenish-yellow, and arranged in paniculate inflorescences. The carpels (most frequently 5) are free in a few, but generally united into a multilocular gynœceum; rarely more than 1 or 2 ovules in each loculus. The gynœceum in the Anacardiaceæ is so reduced that it has only 1 fertile loculus with 1 ovule.—The ovules are epitropous, i.e. anatropous with outward-turned raphe (except the Anacardiaceæ).—The majority of the species are trees and shrubs with scattered, often compound (pinnate) leaves without stipules, and as in addition they frequently contain aromatic, especially turpentine-like substances, they assume a certain resemblance to the Walnut trees, and were formerly classed with them mainly on this account. In a series of genera the volatile, scented oils are found in special glands in the bark of the branches and in the leaves, in the latter case appearing as pellucid dots. This family includes several orders which are somewhat difficult to distinguish from each other.
Order 1. Connaraceæ. This order forms the connecting link between Terebinthinæ and Rosifloræ (Spiræa) as well as Leguminosæ, with which they are sometimes classed. The flowers have 5 5-merous whorls; 2 ovules in each loculus; micropyle turned upwards. Fruit a follicle, rarely a collection of follicles. Seed with aril. Shrubs with scattered (most frequently pinnate) leaves, without stipules. 170 species. Tropical.
Order 2. Meliaceæ. Trees and shrubs with scattered, often pinnate leaves without pellucid dots and exstipulate; the leaflets are nearly always entire. Flowers small in paniculate inflorescences. Calyx and corolla 4–5-merous; 2 whorls of stamens; 3–5 carpels in the gynœceum. A very characteristic feature is the union of the filaments into a tube, on the edge of which stipule-like teeth are often found. There are most frequently 2 ovules in the loculi;[436] fruit a capsule with many winged seeds in Swietenia (Mahogany tree; Trop. Am.), Cedrela, etc.; berries in others. The wood of Cedrela is used for making cigar boxes. 550 species; tropical.
Order 3. Rutaceæ. Leaves glandular with pellucid dots. The type is the same as that of the family. Flowers 4–5-merous. The ovary is most frequently 4–5-grooved. Disc well pronounced, often appearing as a “gynophore.” The majority are shrubs with alternate or opposite, compound, more rarely simple, leaves.
A. The ovary is deeply 2–5-cleft with basal styles which are more or less united; the carpels in some genera are entirely free (groups 1, 2). The fruit is capsular and most frequently dehisces like follicles along the ventral suture or septicidally, so that a horn-like internal layer (endocarp) separates elastically from the external layer.
1. Zanthoxyleæ. Zanthoxylum; Choisya; Evodia.
2. Boronieæ. Australia.—Correa.
3. Diosmeæ. Heather-like shrubs; Africa.—Diosma, Coleonema, Empleurum and Barosma. Officinal: Barosma crenulata and betulina, “broad Buchu leaves” (B. serratifolia and Empleurum serrulatum, “narrow Buchu-leaves”).
Fig. 465.—Ruta. Flower (mag.).
Fig. 466.—Ruta. Longitudinal section of flower.
Fig. 467.—Ruta. Floral diagram.
4. Ruteæ. Ruta (Figs. 465–467) graveolens is an herbaceous, glaucous, strongly smelling plant with bipinnate leaves and yellow flowers; the terminal flower is 5-merous, the others 4-merous (S. Eur.).—Dictamnus; zygomorphic flower. The individual carpels[437] of the fruit separate from each other, and dehisce like follicles, upon which the internal layer is detached elastically and springs out, carrying the seeds with it. Several species are ornamental plants.
5. Cusparieæ. American. Flowers often zygomorphic with gamopetalous corolla; stamens 5.—Ticorea; Galipea (G. officinalis; S. Am.; “Cortex angosturæ”); Cusparia; Almeidea.
B. The ovary is entire or only slightly grooved; the style is terminal, undivided. The fruit is most frequently a drupe or berry.
6. Toddalieæ. Ptelea; winged fruit. The buds are enclosed in the leaf-sheath. Skimmia; Phellodendron.
Figs. 468–470.—Citrus vulgaris.
Fig. 468.—Branch with compound leaves.
Fig. 469.—Transverse section of fruit.

Fig. 470.—Flowers (after the removal of the petals).
7. Aurantieæ, Orange Group. Fruit a berry with a leathery external layer.—The most typical flower is found for example in Limonia: S5, P5, A5 + 5, G5 (2–5).—Citrus has 4–5–8-merous[438] flowers, a gamosepalous, dentate calyx, free petals, one whorl of stamens which are split irregularly into several bundles (Fig. 470). The fruit is a multilocular berry provided with a thick, tough, outer layer. The juicy pulp, which fills up the loculi and envelopes the seeds, is formed from many large-celled, juicy hair-structures which arise on the inner side of the walls of the loculi and by degrees entirely fill them up; the dissepiments remain thin, and form the partitions so easily separating from each other (Fig. 469). The seeds in many instances are remarkable for containing several embryos. The blade of the leaf is separated from the frequently winged stalk by a node (and hence is a compound leaf with only the terminal leaflet developed?) (Fig. 468); in other genera, as Triphasia, there is a fully developed trifoliate leaf. Thorns are frequently developed.—The species of this genus, which is a native of the warmer parts of S. E. Asia, are very hard to separate. The differences are found in the forms of the fruit, the leaves and the leaf-stalks, and in the number of stamens. Citrus medica, “Cedrat” (Ind.); C. limonum, “Citron,” “Lemon” (introduced into Italy in the 3rd to 4th century). Officinal: the fruits and essential oil of Lemon. C. aurantium from E. Asia, the Orange (introduced into Italy in the 14th century). C. vulgaris (Fig. 468), Bitter Orange (introduced into Europe at the time of the Crusades); the unripe Bitter Oranges, and peel of the Bitter Orange is officinal; it is from the flowers of this species especially that the essence of Neroli is made. C. limetta, C. bergamia, Bergamot; essence of Bergamot is officinal. C. decumana, Pomalo, a native of the Islands of the Pacific. About 780 species; chiefly tropical.
Order 4. Burseraceæ. Fruit a drupe; 1–5 stones. The bark, as well as the other parts, contain strong aromatic resins and balsams, and hence several species are used: the Myrrh tree, Commiphora (Balsamodendron) from Arabia and Africa; Officinal: Myrrha (Commiphora myrrha). Mecca-balsam from C. opobalsamum, Arabia; E. Africa. The Incense-tree (Boswellia) from the same parts of the globe and E. India. The incense of B. carteri is medicinal (Frankincense). The resin (Elemi) of Protium-species is officinal, and is used technically for varnish (S. Am.). Takamahaka-resin from Elaphrium (S. Am.) Protium (Icica); Amyris (1 carpel). 270 species; tropical.
Order 5. Zygophyllaceæ. The majority have opposite, pinnate leaves with stipules. Leaves without pellucid dots. The filaments have a scale on the inner side. The most important is Guaiacum officinale (West India), the wood (Lignum Vitæ) of which is very hard and heavy, this wood and Gum-guaiacum are officinal. Others have a peculiar repulsive smell and taste: the Creosote shrub (Larrea mexicana) and Zygophyllum simplex. Tribulus terrester is a common weed in S. Europe. Fagonia. Peganum harmala (South of Russia) yields a red dye.—110 species; especially in the Tropics; several species in sandy deserts. Nitraria.
Order 6. Simarubaceæ. This order is distinguished by the abundance of bitter substances which it contains (Quassine) especially in the bark and the wood. The wood of Quassia amara (Guiana, Antilles) is officinal; Picraena[439] excelsa yields Jamaica Quassia; the bark of Simaruba, Simaba-species and others is used. Ailanthus glandulosa is a garden plant (pinnate leaves, winged fruit).—110 species. Tropical.
Order 7. Ochnaceæ. Flowers diplostemonous, 5-merous. The unilocular ovaries, which are individually free, project considerably into the air around the gynobasic style; 1 ovule in each loculus; the fruitlets are drupes. Shrubs; leaves alternate, with stipules. Ochna; Ouratea.—160 species; tropical; especially American.
Order 8. Anacardiaceæ. The ovary rarely contains more than 1 ovule, even though there be several loculi and several carpels; in Anacardium all the 10 stamens except one become suppressed. Resin passages.—Anacardium. The most peculiar feature is the development of the flower-stalk into a fleshy body about the form and size of a pear (A. occidentale from Trop. Am. and A. orientale from E. Ind.) which bears the kidney-shaped nut (the so-called “Cashew-nut”) on its apex. Mangifera indica (the Mango-tree, from E. Ind.) is cultivated in several tropical countries on account of its delicious drupe. Similarly, species of Spondias (S. dulcis, Pacific Islands, S. lutea). Several species of Rhus are ornamental shrubs in this country, for instance, R. typhina (N. Am.), R. cotinus (the Wig-tree, the barren flower-stalks of the panicles being feather-like and hairy); R. toxicodendron (Poisonous Sumach, from N. Am.) is poisonous. Chinese galls are produced by the sting of a leaf-louse (Aphis chinensis) on R. semialata (China), and Japanese wax is from the seeds of R. succedanea (Japan). Considerable quantities of Sumach (R. coriaria) are used in tanning and as a black dye. Officinal: the mastic resin of Pistacia lentiscus (the Mastic-tree, from the Mediterranean). The fruits of Pistacia vera (Syria) are edible; P. terebinthus and others yield turpentine.—450 species; tropical.
Order 9. Icacinaceæ. Flowers 4–5-merous; haplostemonous; receptacle convex or cup-like surrounding the gynœceum; in the (single) loculus of the ovary, 2 anatropous, pendulous ovules.—200 species; tropical.
The essential characters of this family are in the main the same as those of the Terebinthinæ and Gruinales. The flowers are hypogynous, perfect, with free petals, 5-merous (S5, P5, typically A5 + 5, all of which, however, are not generally developed; in our native orders there are only 7–8 stamens), and most frequently a 3-merous, 3-locular gynœceum (less frequently 2 or 5 carpels with as many loculi). In each loculus there are usually only 1–2 ovules. A deviation from the preceding families is the frequent zygomorphy of the flower, with, as a rule an oblique plane of symmetry (Fig. 471). When a disc is developed it is placed outside the stamens. The majority have no endosperm (Fig. 473).—The members of the family are nearly all trees.
The family is closely allied to the Terebinthinæ, but unlike this it never has[440] aromatic properties, and differs also in the position of the nectary, in the flowers, which are often irregular with a reduction in the number of stamens, and in the ovule which is usually ascending with micropyle pointing downwards (the Terebinthinæ having the micropyle turned upwards), etc. It is also related to Frangulinæ, the Staphyleaceæ being the chief connecting link; but the Æsculinæ generally have compound leaves.
Order 1. Staphyleaceæ. Leaves opposite, often compound. Flowers regular, ☿, 5-merous in calyx and corolla, 5-stamened. The stamens are placed outside the nectary. Ovary syncarpous or 2–3-partite with free styles. The capsule is thin, bladder-like, 2–3-locular, opening at the apex, and has several very hard seeds with a shining testa without aril. Endosperm. Staphylea pinnata (S. Europe) and trifoliata (N. Am.) are cultivated in gardens; they have white flowers in pendulous, axillary racemes or panicles.—16 species.—Staphylea is found in the Tertiary of N. America.
Order 2. Melianthaceæ. Glaucous shrubs with scattered, pinnate leaves, and large stipules. Melianthus.—8 species; S. Africa.
Order 3. Sapindaceæ. Trees or shrubs, often climbing by tendrils (lianes with anomalous structure of the stem) and with compound leaves. The flowers, in most cases, are small, insignificant, and without scent, and in some polygamous and zygomorphic. S4–5, P4–5, A8 (less frequently 5–10) inside the nectary (disc); ovary generally 3-locular, with 1–2 ovules in each loculus (raphe ventral, micropyle turned downwards). Seed without endosperm, often with an aril. The embryo is often thick and curved (Fig. 473).
Figs. 471–473.—Æsculus hippocastanum.
Fig. 471.—Diagram of the flower and of a scorpioid cyme.

Fig. 472.—Flower in longitudinal section.
Fig. 473.—Seed in longitudinal section.
Æsculus (Horse-Chestnut). Trees with opposite, digitate, dentate leaves without stipules; the inflorescence is composed of unipared scorpioid cymes arranged in a pyramidal panicle (termed a thyrsus). The flowers are irregular, with an oblique plane of[441] symmetry (through the 4th sepal, Fig. 471); there are 5 sepals, 5 free petals, of which the one lying between S3 and S5 is the smallest (see Fig. 471) and may be absent; stamens 7 (5 + 2), three being suppressed; gynœceum simple, 3-carpellary and 3-locular, with single style; of the two ovules one is ascending, the other descending (Fig. 472).—The fruit is a 3-valvate, sometimes spiny, capsule, with loculicidal dehiscence, the seed having a large hilum, a curved embryo without endosperm and united cotyledons (the radicle lies in a fold of the testa, Fig. 473). Æ. hippocastanum (Greece, Asia), introduced into cultivation about 300 years ago; the majority of the other species, e.g. Æ. pavia, etc., several of which are frequently cultivated in gardens, are from N. America. The flower of the Horse-Chestnut is adapted for bees, whose abdomen touches the anthers or style when visiting the flower. The flowers are protogynous.
The other Sapindaceæ have most frequently 4 sepals, 8 stamens, various fruits (septicidal capsule, nuts with or without wings, schizocarp), etc. Serjania, Cardiospermum, Sapindus, Koelreuteria, etc. (about 118 genera, 970 species). The seeds of Paullinia sorbilis contain caffeine, and are used as “Pasta guaranà,” in the North Western Brazils in the manufacture of a common drink. Nephelium (or Euphoria) litchi (with edible aril), and other species, from Asia.
Fig. 474.—Samara of Acer platanoides.
Order 4. Aceraceæ. This order is so closely allied to the Sapindaceæ, that some authorities have classed it with them. The main difference is in the regularity of the flowers, and the 2-merous gynœceum (in abnormal cases several carpels occur).—They are trees, and, like the Horse-Chestnuts, have opposite leaves without stipules; in Acer the leaves are palminerved, but imparipinnate in Negundo, a plant frequently cultivated in gardens. The flowers are often unisexual, polygamous (some species have ☿-, ♂-and ♀-flowers); sepals 5, petals 5 free, stamens 8 (that is, 5 + 5, but the two median ones are absent) inside a large disc. Fruit a samara (schizocarp) with 2 winged, nut-like fruitlets (Fig. 474).[442] In each of the 2 loculi of the ovary are 2 ovules. Embryo curved, with thin, folded cotyledons. Endosperm absent.—The inflorescences are racemes with a more or less elongated main axis and terminal flower (which sometimes has 10 stamens); when the lateral branches are developed they are similar to the main axis. In some species both corolla and petal-stamens are suppressed. Acer is pollinated by insects, Negundo by the wind.—88 species; North Temperate zone. Acer in the Tertiary from the Oligocene. The following are native plants: Maple (Acer campestre), Sycamore (A. pseudoplatanus, doubtful native). Important as avenue trees and timber. Sugar is obtained from the spring sap of the Sugar Maple (N. Am.).
Order 5. Malpighiaceæ. A tropical (especially American) order closely related to the Aceraceæ, having often the same form of fruit (but 3-partite). Some species are lianes with anomalous stem-structure. Leaves opposite. The flowers are regular or obliquely zygomorphic (the plane of symmetry passing through sepal 3), with S5, P5, A5 + 5, G3; 1 pendulous ovule in each loculus. Important characteristics for identification are the numerous grandular structures on the sepals. Peculiar 2-spined hairs are found in some. Malpighia, Bunchosia, Galphimia, Tetrapterys, Heteropterys, etc.—About 600 species.
Order 6. Erythroxylaceæ. Sepals 5, petals 5 (with a ligular corona), 10 stamens in one bundle. Gynœceum 3-locular. Fruit a drupe. Tropical (especially American) trees and shrubs, the Coca-plant (Erythroxylon coca) being best known. Its leaves are considered by the inhabitants of Chile and Peru to be one of the indispensable necessaries of life; they are chewed, and possess intoxicating, exhilarating properties, and contain the alkaloid cocaine, which is frequently employed as a local anæsthetic.—103 species; chiefly in America.
Order 7. Vochysiaceæ. Trees; Trop. Am. 1 stamen.—140 species.
Order 8. Trigoniaceæ. Shrubs; Trop. Am.—30 species.
Order 9. Tremandraceæ. Polygalaceæ with regular flowers.—27 species. Australia.
Fig. 475.—Diagram of Polygala: d a gland in the posterior side of the flower; α and β the two caducous bracteoles.
Order 10. Polygalaceæ. Herbs or shrubs (some tropical species are lianes) with scattered (rarely opposite), simple and most frequently quite entire leaves, without stipules. The flowers are usually borne in terminal spikes or racemes, and are strongly zygomorphic (the plane of symmetry being median); they have 5 free sepals, the 2 lateral ones of which (4 and 5 in Figs. 475, 476) are very large, petaloid, and frequently project on each side like the “wings” of a Pea-flower; petals 5, of which the two lateral ones are wanting or rudimentary (dotted on Fig. 475), and the anterior “the keel” (Fig. 476 c) is large, hollow and boat-shaped, and frequently[443] with a lobed or fimbriated edge (Fig. 476 A and B, c); stamens 8, the two median ones being absent, all united into a tube split along the back, which is also slightly united to the keel (the anthers, often 2 locular, open by pores, Fig. 476 B, st); the 2 median carpels form a bilocular ovary. 1 pendulous ovule in each loculus (Figs. 476 C, 475); capsule compressed with loculicidal dehiscence, rarely a nut. Polygala (Milk-wort).
470 species; distributed over the whole globe (none Arctic). Officinal: the root of P. senega, from N. Am. Some are used as ornamental plants.
Pollination. The flowers of Polygala are pollinated by insects (chiefly bees). The fimbriated processes of the anterior petal support the insect when it alights. The anthers lie on each side of the stigma in the pouch of the anterior petal; the apex of the style is spoon-shaped, and immediately behind it is a viscid stigmatic lobe. In reaching the honey the proboscis of the insect must come in contact with the pollen and the viscid stigma, by which it is rendered sticky; this ensures the pollen adhering to the proboscis and so being carried to other flowers.
Fig. 476.—Polygala amara. Parts of the flower (mag.) A Flower from side, 1-5 sepals: c keel; B flower from above spread out: st the 8 stamens; c fimbriated edge of “keel”; C ovary with style and stigma.
The plants belonging to this family, with very few exceptions, are trees or shrubs. The leaves are usually simple; stipules may be absent or present. The flowers in almost all the orders are small, green or whitish; they are always regular, 4-or 5-merous with 2–5 carpels, but never have more than 1 whorl of stamens, which in Rhamnaceæ and Ampelidaceæ are placed opposite the petals (typically 5 + 5 or 4 + 4 stamens, of which however either the external or internal whorl is always wanting); hypogynous or slightly perigynous, in Rhamnaceæ only strongly perigynous or epigynous; generally ☿; the calyx is inconspicuous; petals free or[444] slightly united. Gynœceum simple; ovary generally multilocular; style short or entirely wanting. A disc is nearly always developed in the flower, but is found sometimes inside the staminal whorl, sometimes outside it or between the stamens. The ovules are apotropous (anatropous with dorsal or ventral raphe).
Order 1. Celastraceæ. Euonymus europæa (Spindle-tree) may be chosen as a type. It is a shrub with simple, opposite leaves and small caducous stipules. The small, greenish-yellow flowers, borne in regularly-branched dichasia, are regular, ☿, with 4 whorls, 4-(or 5-) merous in regular alternation. There is a thick disc upon which the polypetalous corolla (imbricate in the bud) and the stamens are borne, with a slightly perigynous insertion. The style is short and thick; the ovary has 2 erect ovules in each loculus. The fruit is a red, 4-valvate capsule with loculicidal dehiscence; the seeds are few in number, and have a large, red-yellow aril (developed from the micropyle). Embryo green, in a large, fleshy, white endosperm. The dingy yellow flowers are generally visited only by flies and ants for the sake of the honey secreted by the disc, and while they run about on the flowers they touch the anthers and stigmas, now with one part of the body, now with another. The flower is protandrous. The stigmas are not developed till several days after the opening of the anthers.—Celastrus, Cassine, Catha, etc.
38 genera; 300 species. Distributed over the entire globe, with the exception of the colder districts, and especially in the Tropics. Some are ornamental bushes (Euonymus japonica). The leaves of Catha edulis are used by the Arabs and Abyssinians in the same way as those of Coca by the Peruvians.
Order 2. Hippocrateaceæ. 150 species; tropical; chiefly lianes. S5, P5, A3, G3. Anthers extrorse.
Fig. 477.—Ilex aquifolium: magnified flower.
Order 3. Aquifoliaceæ (Hollies). The genus Ilex forms almost the entire order. (175 species out of 180; especially from S. Am.) They are shrubs or trees with scattered, leathery, simple leaves (in Ilex aquifolium, spiny) with very small stipules. The flowers are small, white, and borne in few-flowered inflorescences in the axils of the foliage-leaves; they are most frequently unisexual and diœcious. There are 4–5 sepals, petals, stamens and carpels in regular alternation; the calyx and corolla have their leaves slightly connate;[445] stamens slightly adnate to the corolla; the ovary is generally almost spherical with a thick, sessile stigma (Fig. 477). This order deviates especially from Celastraceæ in the absence of the disc and in having only 1 (pendulous) ovule in each of the 4 loculi of the ovary, and in having a drupe with generally 4 stones. Embryo extremely small, at the apex of the large endosperm, with the radicle directed upwards.—3 genera.—I. aquifolium (Holly) principally on the coasts of European countries; from Norway to W. Denmark, and further westward. It is a common garden shrub with stiff, shining leaves and red fruits. Several South American species contain so much caffeine that they may be used as a beverage in the place of tea (I. paraguayensis, Paraguay tea, or Maté). The Holly does not contain caffeine.
Order 4. Ampelidaceæ (Vines). Shrubs with the stem swollen at the insertion of the petioles and climbing by tendrils borne opposite the leaves (Figs. 478, 479). The leaves are scattered (generally 1/2), stalked, stipulate, frequently palminerved and lobed, divided or compound. The small, greenish flowers are generally borne in paniculate inflorescences, whose position is the same as that of the tendrils (Fig. 478); they are hypogynous or slightly perigynous, ☿, with 4–5 sepals, petals, stamens (which, as in the Rhamneæ, are opposite the petals; Fig. 480 A, B) and 2 carpels. The calyx is very small, entire, or slightly dentate; corolla valvate, and in some falling off as a hood, since the individual parts remain united at the summit (Fig. 480 A). Between the stamens and gynœceum is situated an hypogynous disc, with 5 lobes alternating with the stamens (Fig. 480 A, B, E). In each loculus of the 2-locular ovary there are 2 erect ovules (E); the style is short or wanting. The fruit is a berry. The embryo is small and lies in a horny, sometimes slightly folded (ruminate) endosperm (Fig. 480 C, D).
Figs. 478–481.—Vitis vinifera.

Fig. 478.—Branch with bunch of grapes.
Fig. 479.—Diagram of the position of leaf and tendrils. The branch is divided into sections on the sympodial theory (the successive generations, I, II, III, IV, are alternately white and shaded); k buds.
Fig. 480.—A Flower throwing off the corolla; B flower after the removal of the corolla; C, D longitudinal and transverse section of seed; E longitudinal section of gynœceum; s calyx.
Fig. 481.—Diagram of branch and position of leaves; sl tendril; lt the main axis; ax stipules of the foliage-leaf shown below; g axillary-bud (the dwarf-branch); v its fore-leaf; l1 l2 its first two foliage-leaves with their stipules; lt1 long-branch in the axil of v (everything appertaining to this branch is entirely black); v1 the first leaf of this branch.
Vitis and Ampelopsis (5-merous flowers); Cissus (4-merous flower); Leea (without stipules, corolla gamopetalous). The inflorescence in Pterisanthes (E. Ind.) has a peculiar, flat, leaf-like axis, on the edges of which ♂-flowers are borne, and on the surface ♀-flowers.
The TENDRILS in Ampelidaceæ are modified branches, since they bear leaves and may be abnormally developed as branches with foliage-leaves, and finally the inflorescences are borne in the position of the tendrils, and tendrils are met with which are partly inflorescences. The explanation of the position of the tendril, namely, right opposite the foliage-leaf but without a subtending-leaf, has been much disputed. The relative positions are as follows: in Vitis vinifera the following two kinds of shoots and relative positions are found (the other species deviate in one or other particular), (a) Long-branches, which have 2 scale-leaves and a large number of foliage-leaves with a divergence of 1/2;[446] opposite the lowest 3–5 foliage-leaves no tendrils are found, then follow: 2 foliage-leaves with tendrils, 1 without a tendril, 2 with and 1 without, etc., with great regularity. Buds are developed in the axils of the foliage-leaves[447] (Fig. 479): these develop into (b) DWARF-BRANCHES, which commence with 1 laterally-placed scale-leaf (fore-leaf; Fig. 481 v) succeeded by several foliage-leaves with a divergence of 1/2 (in a plane at right angles to that of the mother-shoot), but the whole shoot is extremely small, and often dries up and drops off in the autumn, so that only the scale-leaf, v, with the bud (Fig. 481 lt1) in its axil remains. This bud in the following year developes into a new long-branch, and since its leaves lie in a plane at right angles to that of the dwarf-branch, their plane coincides with that of the long-branch from which it is developed (the grandmother axis).—The tendrils no doubt may most correctly be regarded as the modified main axis which has been pushed aside by a lateral branch. The branches are then sympodia, whose successive shoots bear alternately 1 and 2 foliage-leaves: thus, on the figure there are portions altogether of 5 shoots (I.-V.), the 1-leaved ones are shaded, the 2-leaved ones are white. The following facts however are adverse to this theory: (1) the first leaf on an axillary bud is then situated 180° from the subtending leaf (e.g. the lowermost shaded leaf, Fig. 479, 180° from the lowermost white leaf), whilst the rule in the Dicotyledons is that it is placed only about 90° to one side. (2) The buds (Fig. 479 K) from which the dwarf-branches develop, must then be accessory and sister-buds to the sympodial shoots, but their first leaves have a different relative position to this, which is very peculiar, and a still more remarkable fact is that the buds, K, etc. are similar in structure and present in all the axils; thus we only find accessory buds in the cases where no tendrils are opposite to the leaves, and the main bud must then be considered to be suppressed. (3) The development proves that the tendrils arise on the side of a vigorous growing-point of the stem or by its division, and do not develop, as might be expected, from the apex of the shoot. But these relations however, find their analogues and are all capable of explanation, whereas other less natural modes of explanation are opposed to them.
435 species; especially in the Tropics; they are rarer in America. In N. Am. some Vitis-species and Ampelopsis quinquefolia are found. Vitis vinifera is supposed to have originated in the districts East and South of the Caspian Sea. Wine is obtained from Vitis-species, especially V. vinifera, and “raisins,”—(the name “currants,” given to a special variety with small, seedless fruits, is derived from Corinth).—The species of Ampelopsis (Virginian Creeper) are cultivated as ornamental plants.
Order 5. Rhamnaceæ. The stamens are placed opposite the petals as in the Ampelidaceæ (Fig. 482), but the flowers are much more perigynous or entirely epigynous. The trees and shrubs belonging to this order have simple, most frequently penninerved leaves with stipules; frequently thorny (modified branches). The flowers are inconspicuous, sometimes unisexual (Fig. 482), and have 5 (-4) sepals, petals, stamens, and generally 3 (2–5) carpels. The calyx has valvate æstivation. The petals are very small (generally less than the sepals), often spoon-like, hollow, and embracing the stamens; a disc covers the inner surface of the thalamus or the base of the style in the epigynous flower; gynœceum simple,[448] with one style and one erect ovule in each loculus. The fruit is most frequently a drupe. The embryo is large, often green or yellow, with endosperm.
Rhamnus (Buckthorn) has a juicy drupe with 3 (2–4) stones, surrounded at the base by the persistent portion of the receptacle; the disc is thin. R. cathartica (common Buckthorn): diœcious, with opposite, serrate leaves. R. frangula (Alder Buckthorn): flowers ☿, with scattered, entire leaves.—Ceanothus (N. Am., with richly-flowered inflorescences and a fruit closely resembling that of the Euphorbias). Phylica, Pomaderris (Austr., fruit a capsule). Zizyphus, Paliurus, Colletia (S. Am.) are thorny shrubs; C. spinosa has thorny shoots with small, caducous leaves; the seedling has normal foliage-leaves. Others climb by tendrils as in the Ampelidaceæ, e.g. Gouania.
Fig. 482.—Rhamnus cathartica: A long-styled ♂-flower; pet petals; B short-styled ♂-flower; C long-styled ♀-flower; D short-styled ♀-flower (after Darwin).
475 species, 40 genera; chiefly in temperate and tropical climes. Some are medicinal plants, the bark and fruit having purgative properties (the bark of Rhamnus frangula and “Cascara Sagrada” from the bark of R. purshiana are officinal). The fruits and seeds of others are edible, for example, the fruits of Zizyphus lotus, Z. vulgaris, Z. spina Christi, etc. Green and yellow dyes are obtained from the fruit of R. cathartica, infectoria and others (Avignon grain). Ceanothus-, Rhamnus- and evergreen Phylica-species are ornamental shrubs.
Exclusively trees or shrubs with simple, entire, scattered leaves without stipules. They have a strongly perigynous, regular, 4-merous flower. The receptacle (often coloured) envelopes a simple gynœceum formed of 1 carpel and with, in most cases, 1 ovule, bearing on its edge 4 (or 5) petaloid sepals and, but rarely at the same time, small, scale-like petals. The corolla is most frequently entirely wanting (and hence these plants were formerly reckoned among the Monochlamydeæ); frequently only one of the 2 whorls of stamens, which are situated on the inner side of the edge of the receptacle, is developed. The fruit is most frequently a 1-seeded[449] berry or drupe, or a nut which may be falsely berry-like, the partly persistent receptacle being fleshy and enveloping it.
This family appears the most nearly allied to the Frangulinæ, especially the Rhamnaceæ, and may be considered as a further development of these in the direction of the petaloid development of the receptacle and reduction of the corolla and gynœceum, which in this instance only consists of one carpel. Another deviation is that both the whorls of stamens are present, while one of these is always wanting in Frangulinæ. They also appear to be related to the Lauraceæ (see page 391).
Order 1. Thymelæaceæ. The flowers are most frequently ☿ (Fig. 483). The receptacle is high, generally tubular, coloured, and bears on its edge the 4-(or 5)-merous calyx, with imbricate æstivation. The corolla is wanting or is represented by small scales. The stamens are situated on the inside of the receptacle, and number 4 + 4 (or 5 + 5); stigma capitate. 1 pendulous ovule (Fig. 483 B), the radicle pointing upwards. The fruit is most frequently a berry. A disc is sometimes developed. Endosperm wanting or very slight.
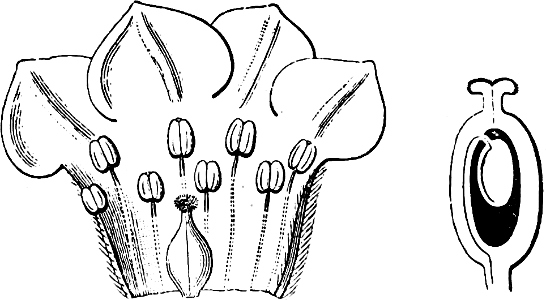
Fig. 483.—Daphne mezereum: A flower; B longitudinal section of pistil.
Daphne (Spurge-Laurel, Fig. 483) has a deciduous receptacle, often coloured; sepals 4; petals absent; stamens 4 + 4. Berry.—Gnidia (corolla); Pimelea (2 stamens); Thymelæa; Passerina and others.
400 species; chiefly in the warm, sub-tropical zone, especially the Cape and Australia. Only Daphne and Thymelæa in Europe. In the fruit and bark of some, for example Daphne, pungent, burning and poisonous properties are found. The bark of D. mezereum (native and cultivated) and D. laureola is officinal. A specially tough bast is found in some species, for example Lagetta lintearia (Lace-tree, Jamaica), which is used in weaving. Some are cultivated in gardens as ornamental shrubs, especially species of Daphne.
Order 2. Elæagnaceæ. Shrubs or trees, which are easily recognised by the covering of peltate hairs found upon almost all[450] parts of the plant, causing them to assume a silvery or rusty-brown appearance. Stipules are absent; the leaves are simple, most frequently scattered. Flowers (Figs. 484, 485) frequently unisexual. The sepals are valvate, 2-4; the corolla is wanting; stamens 4 + 4 or 0 + 4. The ovule is erect and the radicle turned downwards (Fig. 486). The fruit is a nut, but becomes a false fruit, being surrounded by the persistent receptacle or the lower part of it, and thus assuming a berry- or drupe-like appearance (Fig. 486). Endosperm insignificant.—Shepherdia (opposite leaves) has 4 sepals, 4+4 stamens, as in Daphne. Diœcious.—Elæagnus (Silver-leaf) is ☿, has 4–6 sepals, and 4–6 stamens alternating with them. Hippophaë is diœcious; it has 2 sepals and 4 stamens in the ♂-flower (perhaps properly speaking 2+2 stamens); thorny (stem-structures).
16 species; especially ornamental shrubs, e.g. Elæagnus argentea, angustifolia; Hippophaë rhamnoides and Shepherdia canadensis. Northern Temp.
Figs. 484–486.—Elæagnus angustifolia.
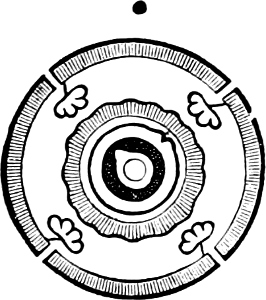
Fig. 484.—Floral diagram.
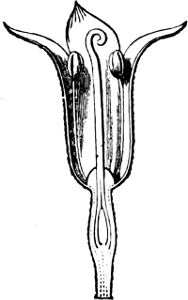
Fig. 485.—Longitudinal section through the flower.
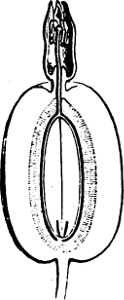
Fig. 486.—Longitudinal section through the fruit.
Order 3 (?). Proteaceæ. This order has its chief centre in the dry regions of Australia (6/10–7/10 of about 1,000 species), a smaller number in S. Africa (2/10–3/10). a few species in S. Am. Trees or shrubs, leaves generally scattered, without stipules, and more or less dry, leathery, evergreen, and often of very different forms on the same plant (undivided, compound, etc.) The flowers are ☿ (rarely unisexual), and 4-merous in the single, petaloid perianth and in the staminal whorl; 1 carpel; sometimes zygomorphic. The perianth-leaves are generally almost free, with valvate, æstivation, often leathery. Small scales alternating with the perianth are often found at the base of the ovary. The stamens generally have extremely short filaments, and are situated opposite, sometimes quite on the tip of the perianth-leaves, in a spoon-like groove. The gynœceum is 1-locular, has 1–several ovules, and is often raised on a stalk-like internode. The fruit is a follicle or nut. The seeds, most frequently winged, have no endosperm.—Protea, Manglesia, Hakea, Banksia, Grevillea, etc.[451] 50 genera; about 1,000 species. Several species are cultivated in our conservatories for the sake of the flowers, which are beautifully coloured and arranged in crowded inflorescences. Protandrous. It is doubtful whether they were existent in Europe in the Tertiary Period. The true systematic position of the order is doubtful. They are related to the Leguminosæ and Rosifloræ, but more closely no doubt to the two preceding orders.
The flower is generally perfect, regular and polypetalous, usually perigynous or epigynous, eucyclic and 5-merous; most frequently S5, P5, A5 + 5 or 5 + 0 and G2-5, but other numbers are found, especially 4; the flowers are very frequently obdiplostemonous. The calyx is sometimes large and the corolla small; the carpels in some are entirely free, in others more or less united. Endosperm is found in the majority. The hypogynous forms approach the Cistifloræ, the others the following families, especially the Rosifloræ. This family is not, upon the whole, so well defined and natural as most of the others. The Saxifragaceæ proper, approach very near to the Rosaceæ, especially Spiræa, and form a transition to it. The forms with opposite leaves, as Philadelphus, etc., approach the Myrtifloræ, just as the Escalloniæ appear to be closely allied to Bicornes, especially Vacciniaceæ. Finally through Pittosporaceæ, they pass over to the Frangulinæ. The family terminates in very reduced forms, on the one hand in the arborescent orders with crowded inflorescences, on the other perhaps in the very remarkable order Podostemaceæ.
Fig. 487.—Diagram of a 6-merous flower (Sedum hispanicum): w branch of scorpioid cyme in the axil of the bracteole β.
Order 1. Crassulaceæ. Nearly all are herbs or small shrubs with round, succulent branches and scattered, fleshy, often more or less round leaves, which are very rarely incised, and never have stipules. The flowers are generally borne in dichasia or unipared scorpioid cymes, which again may be arranged in racemes, umbels, etc.; they are regular, ☿, hypogynous or perigynous, and most frequently have free sepals and petals (gamopetalous corollas with sessile stamens are found in Cotyledon, Bryophyllum, Echeveria, and others); the floral formula is Sn, Pn, An + n, Gn, where n may have very different values, partly depending upon the size of the flower (e.g. 4–7 in Sedum, Fig. 487; 6–30 in Sempervivum; 4 in Rhodiola, Bryophyllum, and Kalanchoë; 5 in Echeveria, Umbilicus, Cotyledon). The carpels are free and are placed opposite the petals (Fig. 487). Fruit a syncarp composed of follicles containing[452] many, small seeds without endosperm. Outside each carpel is found a small, nectariferous scale (Fig. 487). The northern genus, Rhodiola, is diœcious. The petal-stamens are wanting in some (Crassula, Bulliarda, and others). The floral-leaves are very often displaced upon their axillary branches. A multicarpellary gynœceum also occurs.
Sedum (Stonecrop) is generally 5-merous with 10 stamens; Sempervivum tectorum (House-leek), 12-merous, and with 24 stamens. The leaves of Bryophyllum calycinum very readily form buds, and also frequently exude water from the edges.
485 species; especially Temp. (Cape, Europe). Principally used as ornamental plants.
Order 2. Saxifragaceæ. The flowers are 4–5-merous with 2 (-3) carpels, most frequently: S5, P5, A5 + 5 (obdiplostemonous), G2. They are regular, ☿, polypetalous, hypogynous, perigynous or most frequently more or less epigynous (Fig. 488). The carpels may be individually quite free, but are more frequently united at the base, or the entire portion enclosing the ovules is united into a 1- or 2-locular ovary, the styles, however, are always free. Fruit a capsule with many seeds; endosperm present.—They are herbs, most frequently with scattered leaves without stipules; but the leaf-base is broad. The inflorescences are most frequently cymose, and a displacement of the floral-leaves is frequent (e.g. Chrysosplenium).—Some Saxifraga-species, e.g. S. sarmentosa, have irregular flower with an oblique plane of symmetry. The petal-stamens in some may be wanting: Heuchera, species of Saxifraga and Mitella. The corolla is wanting in others.
Saxifraga (Saxifrage): S5, P5, A5 + 5, G2 (Fig. 488); capsule bilocular, opening along the ventral suture between the 2 persistent styles. S. granulata has small tubers at the base of the stem.—Chrysosplenium (Golden Saxifrage): 4 sepals, no corolla, 4 + 4 stamens; 1-locular capsule.
Protandry is most frequently found in Saxifraga, with the stamens successively bending towards the gynœceum; protogyny is more rare. In other genera there is protogyny without any movement of the stamens; Chrysosplenium is homogamous.—About 300 species; mostly in temperate climates. Saxifraga is especially Alpine. S. crassifolia and other species, Hoteia japonica, Tellima, etc., are ornamental plants.
Fig. 488.—Saxifraga granulata. Longitudinal section of flower.
[453]
The following genera are allied to the Saxifragaceæ:—
1. Parnassia (about 14 species; P. palustris, Grass of Parnassus). The flower is slightly perigynous, and has S5, P5, 5 fertile sepal-stamens, and 5 petal-stamens, which are developed as barren staminodes, palmately-lobed, and (3–) 4 carpels united in a 1-locular ovary with (3–) 4 parietal placentæ. Capsule.—Protandrous. The flower has a slightly oblique plane of symmetry, which is especially shown during its development and in the order of sequence in which the anthers dehisce: originally they lie closely round the gynœceum; the anthers dehisce extrorsely, first the one which is placed opposite the most external sepal (the 2/5 arrangement is very distinct in the calyx), the filament elongating so that the anther lies over the ovary, and this is followed successively by the 4 others in a zig-zag line; the filaments bend backwards after the pollen is shed and the anthers drop off, and the stigmas are not developed until this is completed. The barren stamens are palmately divided into an uneven number (7, 9, 11) of lobes, tapering from the centre towards the edge, and bearing apparently glandular tips; their gland-like appearance is supposed to allure flies to visit the flower, or they may act as a kind of fence which compels the insects to enter the flower in a certain way, and thus effect pollination; the honey is secreted on their inner side, and not by the gland-like tips.
Fig. 489.—Portion of Cephalotus follicularis: k pitcher-like leaf with thick corrugated edge (m) and lid (l); b foliage-leaf of the ordinary form.]
2. Adoxa moschatellina (Moschatel). This is a perennial, creeping herb; the horizontal rhizome has an unlimited growth, and bears, in a scattered arrangement, both foliage-leaves, and white, fleshy scale-leaves. The aerial stem bears 2 opposite foliage-leaves and a capitate inflorescence of 5 flowers, 4 placed laterally (in opposite pairs) and 1 terminally. The flower is semi-epigynous, the calyx[454] gamosepalous, corolla absent. The stamens are divided to the base, so that each filament bears a bilocular anther. The style is free, deeply cleft. The terminal flower has 2 bracteoles, 4 sepals, 4 stamens, cleft to the base, and a 4-locular ovary. The bracts of the lateral flowers are displaced on the flower-stalk, as in Chrysosplenium, and united with the 2 bracteoles into a kind of 3-leaved involucre; these flowers have 5 sepals, 5 split stamens with 2-locular anthers, and a 5-locular ovary. 1 pendulous ovule in each loculus. Fruit a drupe, green-coloured, with 1–5 stones.—This plant, which would perhaps be best placed in a special order, has also been classed with the Araliaceæ and Caprifoliaceæ.
The following are also allied to this order: Escalloniaceæ (arborescent plants with simple, scattered, leathery leaves), Cunoniaceæ (arborescent with opposite leaves), Cephalotaceæ (with pitcher-like, insect-catching leaves; Australia; Fig. 489) and Francoaceæ. These have respectively 85, 107, 1 and 3 species.
Figs. 490–492.—Ribes rubrum.

Fig. 490.—Floral diagram.
Fig. 491.—Flower in longitudinal section.
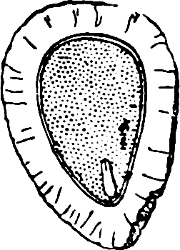
Fig. 492.—Seeds in longitudinal section.
Order 3. Ribesiaceæ (Currants). 5-stamened Saxifragaceæ with epigynous flowers.—Moderately sized shrubs with scattered, stalked and palminerved, and generally palmilobed leaves, with a large leaf-sheath. The flowers (Figs. 490, 491), most frequently borne in racemes, are regular, epigynous, and have often, above the ovary, a cup- or bell-shaped, or tubular prolongation of the receptacle, on which the sepals, petals and stamens are situated; they have 5 sepals (often large, coloured), 5 small, free petals, only 5 stamens (opposite the sepals) and a 2-carpellate gynœceum with a unilocular ovary and 2 parietal placentæ bearing many ovules. The fruit is a berry, whose seeds have a fleshy and juicy outer covering (Fig. 492). In some species, for example Ribes grossularia, there is found an unbranched, or a 3–5-branched spine, very closely resembling the spiny leaves of the Berberis, but which, however, are emergences springing from the base of the petiole. Ribes has two kinds of branches: long-branches and[455] dwarf-branches, the latter alone bearing the flowers.—Ribes (Figs. 490–492). The blades of the leaf are folded or rolled together in vernation. R. alpinum is diœcious.
75 species; especially from the N. Temp. regions (especially N. Am.).—The receptacle secretes honey on its inner surface. The Gooseberry-flower is slightly protandrous, others are homogamous; insect-and self-pollination are found. The following are FRUIT BUSHES: R. nigrum (Black Currant), R. rubrum (Red Currant), R. grossularia (Gooseberry), originating in Northern Europe and Asia. Ornamental bushes: the North American R. aureum (Golden Currant) and R. sanguineum (Blood-red Currant), etc.
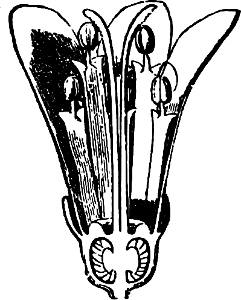
Fig. 493.—Deutzia crenata. Longitudinal section of flower.
Order 4. Hydrangeaceæ. Shrubs, with simple, opposite leaves, without stipules; flowers generally epigynous, 4–5-merous (Fig. 493).—Hydrangea (H. hortensia, etc.). Shrubs from N. Am. and E. Asia; corolla often valvate. The inflorescence, as in the case of the inflorescence of Viburnum opulus (Guelder Rose), has often irregular, large, but barren flowers at the circumference, whilst the others are much smaller, regular and ☿; the barren flowers are mostly 4-merous; in these cases it is the calyx which is large and petaloid, while the other parts of the flower are more or less suppressed. The branches of the inflorescence appear to be partially devoid of floral-leaves, since they are displaced upon the main axis.—Philadelphus; racemes (with terminal flower), sepals 4 (valvate), petals 4 (twisted), stamens many, and carpels 4 (opposite the petals), forming a 4-locular ovary. The numerous stamens (20–30) occur by the splitting of the sepal-stamens and are often therefore placed in distinct bundles. Fruit a capsule. Ph. coronaria (Syringa, Mock Orange-blossom), from S. Eur., is a common ornamental shrub, as also is Deutzia (Fig. 493) from N. Am. and E. Asia. The latter has S5, P5, A5 + 5, G3.—About 70 species.
Order 5 (?). Pittosporaceæ. This order has its home especially in Australia (90 species). The flower has S5, P5, A5 (episepalous), G2 (3–5), most frequently a unilocular ovary with many ovules in 2 rows, borne on 2 parietal placentæ, or a bilocular ovary. Some have berries, others capsules. Pittosporum, Citriobatus, Sollya, Billardiera.
Order 6. Hamamelidaceæ. Flowers more or less epigynous, with S4, P0 or 4, 4 fertile sepal-stamens, and 4 barren petal-stamens, bilocular ovary with 1–2 ovules in each loculus. Fruit a capsule. Hamamelis: one species in Japan and one in N. Am. Fothergilla. Liquidambar: monœcious; flowers in capitula or spikes; ♂-flowers without perianth, stamens indefinite; ♀-flower: slight perianth, 2-locular ovary with many ovules. Officinal: “Styrax-balsam,” which is obtained by boiling the bark of Liq. orientalis, from Asia Minor. Liquidambar and Parrotia are found as fossils in the Upper Oligocene; Hamamelis perhaps in the Chalk.
Finally two orders with very reduced flowers are included in this family.
Order 7. Platanaceæ. Trees, with large, scattered, palminerved and lobed[456] leaves, and ochreate stipules; the buds are concealed in a hollow at the base of the petiole. The bark falls off in large scales. ♂-and ♀-flowers (monœcious) in crowded, spherical inflorescences which are placed at wide intervals on a terminal, thin, and pendulous axis. The flowers have an insignificant calyx and corolla; the ♂-flower has few stamens; ♀-flower, perigynous, with 4 free carpels, 1 pendulous, orthotropous ovule in each. Fruit a nut; endosperm absent. 5 species; frequently grown in avenues and parks. P. occidentalis (N. Am.); P. orientalis (W. Asia.).
Order 8. Podostemaceæ. Aquatic plants, especially in swiftly running water, with somewhat of an Alga-like, Moss-like, or thalloid appearance; they show themselves in many ways to be adapted to their mode of life and situations (having a dorsiventral creeping stem, the flowers sunk in hollows, a formation of haptera upon the roots, and thalloid assimilating roots and thalloid stems, etc.). Tropical; 100 species.
The leaves are scattered, stipulate, or have at least a well developed sheath, which is generally prolonged on each side into a free portion (“adnate stipules”). The flowers are regular, perigynous or epigynous. Calyx and corolla 5 (-4)-merous with the usual position. The corolla is always polypetalous. The stamens are present in very varying numbers (5–∞) and position, but always placed in 5-or 10-merous whorls; they are frequently 20 in 3 whorls (10 + 5 + 5; see Figs. 494, 502, 505); the nearer they are placed to the circumference, the longer they are; they are generally incurved in the bud, or even rolled up. The number of the carpels is from 1–∞; in most cases all are individually free (syncarp), and when they are united it is in every case with the ovaries only, whilst the styles remain more or less free (Pomaceæ, species of Spiræa). The seeds have a straight embryo, and usually no endosperm.
The perianth and stamens are most frequently perigynous on the edge of the widened receptacle; its form varies between a flat cupule and a long tube or a cup (Figs. 495, 496, 498, 499, 500); the carpels are situated on its base or inner surface, in some instances on a central conical elongation of the floral axis (Fig. 496). The carpels in Pomaceæ also unite more or less with the hollow receptacle, or this grows in and fills up the space between the carpels, so that a more or less epigynous flower is formed (Fig. 504).—The following numbers of stamens occur: 5, 10 (in 1 whorl), 15 (10 + 5), 20 (10 + 5 + 5), 25 (10 + 10 + 5), 30-50 (in 10-merous whorls)—compare the diagrams. The theoretical explanation of this relation of the 10-merous whorls and their alternation with the 5-merous whorls is not definitely determined; a splitting of the members of the 5-merous whorls may be supposed, but the development shows no indication of this, and it is not supported in any other way. Several genera have “gynobasic” styles, that is, the style springs from the base of the ovary (Fig. 497 A, B).
[457]
The Rosifloræ are on one side closely related to the Saxifragaceæ (especially through Spiræa) from which it is difficult to separate them, and to the Myrtifloræ; on the other side they are allied, through the Mimosaceæ with the large number of stamens, and through the Amygdalaceæ with its single carpel, to the Leguminosæ. The family begins with forms which have many-seeded follicles, and passes on the one side to forms with nuts and drupes in perigynous flowers, and on the other side to the Pomaceæ.
Order 1. Rosaceæ. Herbs or shrubs, generally with compound leaves and persistent (adnate) stipules, flower perigynous, gynœceum formed of many free (therefore oblique) carpels, syncarps with fruitlets of various kinds. The exceptions are noted under the genera.
Fig. 494.—Diagram of Comarum palustre.
Fig. 495.—Flower of Spiræa lanceolata.
1. Spiræeæ (Fig. 495) has 2–many ovules in each ovary, while in the other groups there is generally only 1, and never more than 2 ovules in each loculus. There are generally 5 cyclic carpels and the fruit is 5 follicles, which are not enclosed by the receptacle. The majority are shrubs. Stipules are often wanting.—Spiræa (Meadow-Sweet). The flowers are generally borne in richly flowered inflorescences of various forms. The carpels, in some species, unite together and form a simple gynœceum with free styles (an approach to the Pomaceæ).—Closely allied to Spiræa are the East Asiatic shrubs: Kerria japonica, which has solitary flowers, in this country nearly always double (the fruit a nut), and Rhodotypos kerrioides which has opposite leaves, a remarkable feature among the Rosifloræ; it has a 4-merous flower, a well developed disc inside the andrœcium, and a drupe. Closely allied also is Gillenia (N. Am.) differing chiefly in the ascending ovules, Spiræa having pendulous ovules, and a more tubular receptacle.
The groups Quillajeæ and Neuradeæ form a transition from Spiræa to Pomaceæ. In the first group, which contains only trees or shrubs with generally simple leaves, the carpels are either free or united (into a capsule); in the second the receptacle unites with the carpels, which are themselves often united together; in this case, too, the fruit is a capsule. Quillaja (S. Am.); Exochorda (China).
[458]
2. Potentilleæ (Figs. 494, 496, 497). The flower has an “epicalyx” (Fig. 494 C) alternating with the sepals and formed by their stipules which are united in pairs, and hence its leaves are often more or less deeply bifid. The receptacle is cupular and often quite insignificant. The sepals are valvate in the bud. The large number of fruitlets are achenes, borne on a well-developed convex portion of the receptacle (the Ranunculeæ resemble the Potentilleæ, but have no epicalyx, no enlarged receptacle, and spirally-placed stamens). Most of the species are herbs with dichasial inflorescences, often arranged in racemes.—Potentilla (Cinquefoil). The achenes are borne on a dry, hairy receptacle; the style is situated towards the apex of the ovary, and is not prolonged after flowering. Herbs with digitate, in some, however, pinnate leaves, and generally yellow flowers.—Comarum (Fig. 494) (Marsh Cinquefoil) forms, by its fleshy-spongy receptacle, a transition to the next genus.—Fragaria (Strawberry) (Fig. 496). The receptacle becomes finally fleshy, coloured, and falls off (biologically it is a berry); the numerous fruitlets (drupes with thin pericarp) have basal styles (Fig. 497); leaves trifoliate; long, creeping runners.—Geum (Avens) has a terminal style which after flowering elongates into a long beak, with the apex (after the uppermost part has been thrown off) bent back into a hook, thus furnishing a means of distribution for the fruits. Leaves pinnate.—Dryas comprises 3 Arctic or Alpine species with simple leaves and solitary flowers, the calyx and corolla 8–9-merous, the fruit resembles that of Geum, but the styles become still longer and feather-like (a flying apparatus).
Figs. 496, 497.—Fragaria vesca.
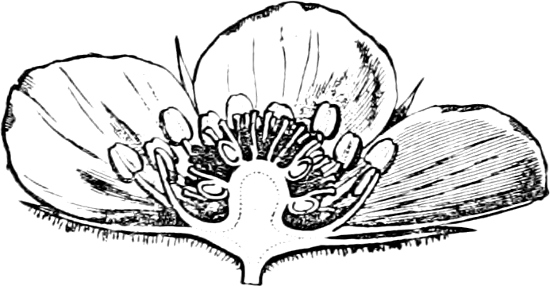
Fig. 496.—Longitudinal section of flower.

Fig. 497.—A carpel, entire, and in longitudinal section.
3. Rubeæ. Rubus (Bramble) has the same form of receptacle as the Potentilleæ, but no epicalyx; the fruitlets are drupes, not enclosed by the persistent calyx. Most frequently shrubs or undershrubs[459] with prickles (emergences), glandular bristles and compound leaves. In the Raspberry (R. idæus) the fruitlets unite together and detach themselves from the receptacle.
4. Roseæ. Rosa; the receptacle is hollow, ovoid and contracted beneath the insertion of the calyx (Fig. 498), ultimately fleshy and coloured; it encloses a large number of fruitlets which are achenes as hard as stones (“hip,” biologically a berry).—Shrubs with imparipinnate leaves and adnate stipules. The sepals show clearly the order of their development (a divergence of 2/5), the two outer ones on both sides are lobed, the third one on one side only, and the two last, whose edges are covered by the others, are not lobed at all. Prickles (emergences) are generally present and in some species are placed in regular order, being found immediately below each leaf (usually two) although at somewhat varying heights.
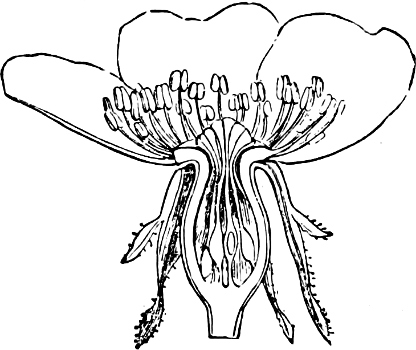
Fig. 498.—Longitudinal section of flower of Rosa.
Figs. 499, 500.—Agrimonia eupatoria.
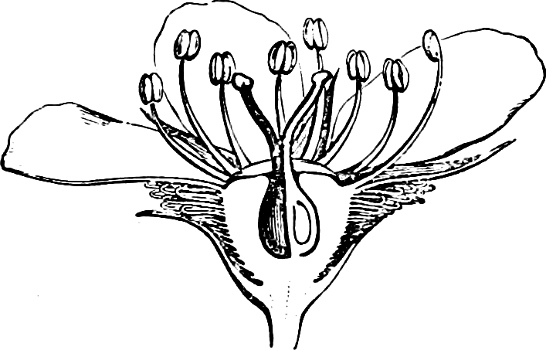
Fig. 499.—Flower in longitudinal section.
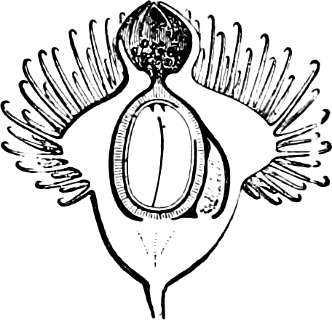
Fig. 500.—Fruit and receptacle in longitudinal section.
5. Agrimonieæ. The receptacle is more or less cup- or bell-shaped, and almost closed at the mouth; it is persistent and envelopes the nut-like fruitlets, but is dry, and in some species hard, the fruitlets being firmly attached to it. In biological connection with this the number of the carpels is generally only 1 or 2, and the whole becomes a false nut (Fig. 500). Herbs.—Agrimonia (Agrimony; Figs. 499, 500); the perianth is 5-merous, stamens 5–20. The receptacle bears externally, on the upper surface, a[460] number of hooked bristles which serve as a means of distribution for the 1–2 achenes which are enclosed in it, and hence the entire flower finally falls off. The inflorescence is a long upright raceme. These bristles are arranged in whorls of 5 and 10, of which the uppermost alternate with the sepals.—Alchemilla (Ladies-mantle; Fig. 501) has 8 green perianth-leaves in two whorls (some authorities consider the four outer as an epicalyx, and the flower therefore apetalous), and 4 stamens alternating with the innermost whorl. There is only one carpel with a basal style and capitate stigma. The flowers are small and greenish, the filaments jointed. The anthers open by one extrorse cleft. The leaf-sheath entirely envelops the stem; the leaves are palminerved. A. aphanes has often only 1–2 stamens. The following genera, with 4-merous flowers borne in short spikes or capitula, are allied to this group. Sanguisorba has entomophilous, ☿-flowers with 4(-20) stamens, 1 carpel; stigma papillose.—Poterium; spike or capitulum, the uppermost flowers are ♀, the lowermost ♂, and some intermediate ones ☿ (the order of opening is not always centripetal); S4, P0, A20–30, G2, the long styles having brush-like stigmas (wind-pollination). Leaves imparipinnate.
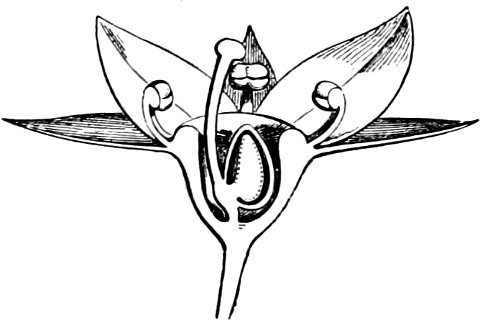
Fig. 501.—Flower of Alchemilla in longitudinal section.
Pollination. A yellow ring on the inner side of the receptacle, inside the stamens, serves as a nectary when any honey is formed; this, for instance, is not the case in Rosa, Agrimonia, Spiræa ulmaria, S. filipendula, S. aruncus, etc., to which the insects (especially flies and bees) are allured by the quantity of pollen. Homogamy and slight protogyny are frequent, in many instances self-pollination also is finally possible. Poterium, with the long-haired stigma, is wind-pollinated.—About 550 (1100?) species, especially in northern temperate regions.—Uses. Officinal: the petals of Rosa centifolia and gallica, the fruits of the Raspberry (Rubus idæus), the rhizome of Geum urbanum, the flowers of the Koso-tree (Hagenia abyssinica or Brayera anthelmintica).—The bark of Quillaja saponaria (Chili) is used as soap and contains saponin. “Attar of Roses” from Rosa damascena, centifolia and other species, especially from the southern slopes of the Balkans. Many species and varieties of Roses are ORNAMENTAL plants: from S. Europe, Rosa lutea (the Yellow Rose), R. gallica (the French Rose) and R. rubrifolia; from W. Asia, R. centifolia, of which the Moss Roses (R. muscosa and cristata) are varieties, and R. damascena; from India and N. Africa, R. moschata (the Musk Rose); from China, R. indica (Tea Rose) etc., besides the native species and the varieties which have been derived from them. In addition, Kerria japonica, species of Potentilla, Rubus odoratus from N. Am., and many species of Spiræa from South-eastern Europe and N. Am. Esculent: the “hips” of R. mollissima, R. pomifera, etc.; the fruits of[461] Rubus-species: Raspberry (R. idæus), Cloudberry (R. chamæmorus), Blackberry (R. fruticosus), etc.; of Fragaria-species (F. vesca, collina, grandiflora, etc).
Order 2. Amygdalaceæ. Trees or shrubs with rosaceous flowers; leaves simple with caducous stipules; a regular, perigynous flower, the receptacle being partly thrown off by a circular slit; sepals 5, petals 5, stamens 20–30; gynœceum simple, formed of 1 carpel (hence oblique, Fig. 502), with terminal style and 2 pendulous ovules, ripening into a drupe (Fig. 503).—The leaves are penninerved and frequently have glands on the stalks and edges; thorns (modified branches) often occur, i.e. dwarf-branches, which, after producing a few leaves, terminate their growth in a thorn (e.g. Prunus spinosa). The vernation of the foliage-leaves varies in the different genera; in the Almond, Peach, Cherry, and Bird-Cherry they are folded; in the Apricot, Plum, Sloe and Bullace, rolled together. In some the flowers unfold before the leaves (Amygdalus, Armeniaca). That the gynœceum is formed of 1 carpel is evident in this as in other instances (e.g. in the Leguminosæ, which are closely related to this order), from the fact that the carpel is oblique, and has only one plane of symmetry, and similarly in the fruit there is a longitudinal groove on one side which indicates the ventral suture. It is only exceptionally that both ovules are developed. In abnormal instances more than 1 carpel is developed.
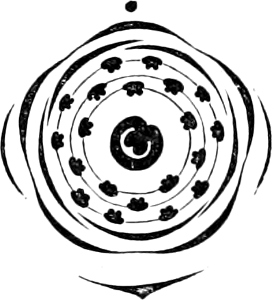
Fig. 502.—Diagram of Prunus virginiana.
A. Fruit hairy: Amygdalus (A. communis, Almond-tree) has a dry pulp which is detached irregularly, when ripe, from the wrinkled, grooved, ovoid and somewhat compressed stone.—Persica (P. vulgaris, Peach-tree) differs from the Almond in having a juicy pulp, not detachable from the stone, which is deeply grooved and has pits in the grooves (Fig. 503). (The name of the genus is derived from Persia, though it is a native of China.).—Armeniaca (A. vulgaris, Apricot) has a hairy, velvety fruit, but the stone is smooth and has two ribs along one of the edges; the pulp is juicy. (The generic name has been given on the incorrect assumption that it was a native of Armenia; its home is China.)
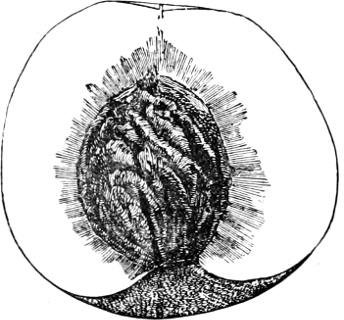
Fig. 503.—Fruit of the Peach. The pulp is cut through so that the stone is visible.
[462]
B. Fruit glabrous (i.e. without hairs): Prunus (Plum) has a glabrous fruit with bluish bloom; the stone is compressed, smooth or wrinkled. The flowers are borne solitarily or in couples, and open before or at the same time as the leaves; they are borne on shoots without foliage-leaves.—Cerasus (Cherry) has a glabrous, spherical fruit, without bloom, and a spherical stone. The flowers are situated in 2–many-flowered umbels or racemes, and open at the same time as the leaves or a little before them. Long-stalked flowers in umbels are found in C. avium (Wild Cherry), C. vulgaris (the cultivated Cherry, from Western Asia); racemes at the apex of leaf-bearing branches and small spherical fruits are found in C. padus (Bird Cherry), C. virginiana, C. laurocerasus (Cherry-laurel), C. mahaleb.
Pollination. Prunus spinosa (Sloe, Blackthorn) is protogynous, but the stamens are developed before the stigma withers. Honey is secreted by the receptacle. Cerasus padus (Bird-Cherry) agrees in some measure with P. spinosa. In the flowers of the Plum and Cherry the stamens and stigma are developed simultaneously and self pollination seems general; the stigma, however, overtops the inner stamens and thus promotes cross-pollination.—Distribution. 114 species in the N. Temp, zone; few in the warmer regions; the majority from W. Asia. C. vulgaris, from the regions of the Caspian; Prunus spinosa, insititia (Bullace), domestica (Plum, from the Caucasus, Persia).—Uses, principally as fruit-trees: Cherry, Plum, Apricot, etc.; “Almonds” are the seeds of Amygdalus communis (W. Mediterranean), “bitter,” “sweet,” and “shell” almonds are from different varieties, the latter being remarkable for the thin, brittle stone. In the majority of species and in almost all parts of the plant (especially the bark, seed and leaves) is found the glycoside, amygdalin, which forms prussic acid. Many form gum, and the seeds have fatty oils (“Almond oil”). Officinal: the seeds and oil of Amygdalus communis, and the fruit of the Cherry; in other countries also the leaves of C. laurocerasus.—The stems of Cerasus mahaleb are used for pipes. Ornamental Shrubs: Amygdalus nana, Cerasus laurocerasus.
Order 3. Chrysobalanaceæ. Tropical Amygdalaceæ with zygomorphic flower and gynobasic style. 200 species; especially Am. and Asia. Chrysobalanus icaco (Cocoa-plum) is cultivated on account of its fruit (Am.)
Order 4. Pomaceæ. Trees and shrubs, most frequently with simple leaves and caducous stipules. The flowers (Fig. 505) have 5 sepals, 5 petals and generally 20 stamens (10 + 5 + 5, or 10 + 10 + 5). There are from 1–5 carpels, which unite entirely or to some extent with each other, and with the hollow, fleshy receptacle (the flower becoming epigynous), (Figs. 505, 506, 507). The carpels are nearly always free on the ventral sutures, rarely free at the sides also. The whole outer portion of the fruit becomes fleshy, but the portions of the pericarp surrounding the loculi (endocarp) are most frequently formed of sclerenchymatous cells, and are more or less[463] firm (the “core”). The nature of the fruit varies, according to the thickness and hardness of the endocarp, being either a “berry” or a “drupe” (see A and B). When the endocarp is thin and parchment-like, the fruit has the characteristics of a berry, each of the 5 loculi generally present containing several seeds; but when this is hard the fruit resembles a drupe, only one seed is developed in each loculus, and the number of the loculi is reduced to one or two. There are nearly always 2 ovules in the loculi of the ovary, but in Cydonia there are a large number in 2 rows. In the genera which have stones, only one seed is developed in each stone. The genera are distinguished mainly in accordance with the kind of fruit and the number of ovules and seeds.
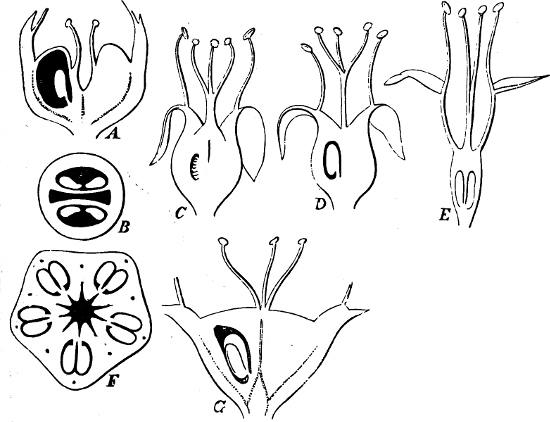
Fig. 504.—Longitudinal and transverse section through the flowers of A, B Cotoneaster; C Cydonia; D Malus communis; E Raphiolepis; F Cydonia; G Mespilus.
Fig. 505.—Floral diagram of Mespilus germanica.
A. Sorbeæ. The endocarp is parchment-like or papery (drupe, with thin stone or berry).
1. Pyrus and Cydonia; carpels completely embedded in the cup-like receptacle, styles always free.—Pyrus: the fruit is glabrous, and has only a small calyx, withering or deciduous, and a 5-locular ovary with at most 2 ascending ovules in each[464] loculus (Fig. 504 D). The large flowers are situated in few-flowered umbels or corymbs. P. communis (Pear; free styles, Fig. 507; it has the well-known pear-shaped fruit; the core is reduced to several groups of sclerenchymatous cells embedded in the pulp, the leaf-stalk is as long as the blade).—Cydonia (Quince) has a hairy fruit with many seeds in 2 rows in each loculus of the endocarp (Figs. 504 C, F; 506); the testa of these seeds is mucilaginous. C. vulgaris, large, terminal flowers on lateral branches, and large leaf-like, persistent sepals.
Fig. 506.—Cydonia vulgaris. Longitudinal section of fruit.
Fig. 507.—Longitudinal section of Pear flower.
2. Malus and Amelanchier (Aronia); carpels free on the ventral edge; styles united. Malus communis (Apple) the fruit is “umbilicate” at the base; no sclerenchymatous cells in the pulp; styles united at[465] the base (Fig. 504 D); leaf-stalk shorter than the blade. Sorbus (Mountain-ash) differs only in having a 2–3-locular fruit with extremely thin endocarp. Cymose inflorescences in umbellate cymes. S. aucuparia has pinnate leaves, S. aria (White-beam) and other species have simple leaves.—Amelanchier (the Service-tree) has a false divisional wall springing from the dorsal suture, and more or less projecting into each of the loculi of the ovary; Raphiolepis (Fig. 504 E) has racemes and a juicy berry; Eriobotrya japonica (Loquat).
B. Cratægeæ. The endocarp is hard and bony (“drupes,” generally with several, sometimes, however, with only 1–2 stones, rarely one multilocular stone; only 1 seed in each of the loculi).—Cratægus (Hawthorn, May). There are 1–5 stones in the spherical or ovoid fruit. The disc, found on the apex of the fruit, inside the small, withered calyx, is small (much less than the transverse section of the fruit). Shrubs with thorns (branches) and moderately large flowers borne in corymbs.—Mespilus (Medlar) differs from the last-named only in having a large disc at the apex of the fruit, inside the large, leaf-like sepals, i.e. almost equal to the greatest diameter of the fruit. The flowers are solitary and terminal.—Cotoneaster is chiefly distinguished from the others by its syncarps, the 2–5 carpels (and stones) being free from one another, and only united to the receptacle by a larger or smaller portion of their dorsal surface (Figs. 504 A, B). Small shrubs with leathery leaves, generally covered with white, felted hairs on the lower surface, and with small flowers; the fruit is red or black.
Pear, Apple, Mountain Ash and Hawthorn have protogynous flowers which secrete honey, and are conspicuous to ensure insect pollination.—180 species; in the northern temperate regions.—Pear and Apple are especially cultivated as fruit trees in a number of varieties; the Paradise Apple (Pyrus baccata); especially in southern countries also the Quince (from N. Persia and the Caucasian districts), Medlar and Amelanchier vulgaris. Malus pumila (Caucasus, Altai) and M. dasyphylla (Orient, S. Eur.) are regarded as primitive forms of the Apple-tree; M. sylvestris, which grows wild in European forests, appears to have been less used. The early Lake-dwellers in Switzerland had the apple-tree both wild and cultivated.—The original form of the Pear is supposed to be Pyrus achras (Central Asia).—Many of the species of Cratægus, some with double flowers, and Pyrus (Chænomeles) japonica, with brilliant red flowers, are cultivated as ornamental shrubs. Officinal: Quince pips, on account of the mucilaginous testa.—The fruits contain free organic acids and sugar; prussic acid may be obtained from the seeds. The wood of the Pear-tree is used in manufactures.
[466]
The most characteristic feature is, that the gynœceum is 1-locular and formed of 1 carpel, the ventral suture of which is turned posteriorly. The fruit, in most instances, is a pod (legume), which opens generally along both sutures, the two valves twisting more or less in opposite directions. In other instances it opens along one suture only, or as a pyxidium (Red Clover), or it is indehiscent, in which case it is more or less berry-like (e.g. the Tamarind, Carob-bean), or it is a drupe (e.g. the Tonquin-bean), or a 1–few-seeded nut (e.g. Melilotus), or a lomentum, which divides transversely into as many joints as there are seeds (Ornithopus, see Fig. 513).
The inflorescences belong to the centripetal type (i.e. indefinite); cymes do not occur. The flowers are zygomorphic, with vertical plane of symmetry, seldom regular; 5-merous with but a few exceptions, ☿, and slightly perigynous. The following diagram is the most general (Fig. 511): 5 sepals, with the unpaired sepal median and anterior, 5 petals, 5 + 5 stamens, all in alternating whorls, 1 carpel. The calyx is most frequently gamosepalous, the gynœceum is narrowed down at the base to a short stalk and, in the majority, is more or less bent. The seed is most frequently kidney-shaped, with a smooth, hard and shining testa, the hilum being very distinct. Endosperm is wanting, or is reduced to a thin layer, which is of service when the seed swells during germination. The vegetative parts have these features in common, namely, the leaves are scattered, stipulate, and almost always compound. Peculiar sleep-movements and sensitiveness are found in some, chiefly in the Mimosas. Many, probably all, Leguminosæ have small tubercles on their roots which are produced by a kind of bacterium, and assist in the assimilation of free nitrogen. Spontaneous movements are exhibited by Desmodium gyrans (Telegraph-plant).
This family is closely allied to the Rosifloræ, with which it agrees in the scattered leaves, the presence of stipules, the generally 5-merous and most frequently perigynous flowers with eucyclic stamens, and the absence of endosperm. Amygdalaceæ and Chrysobalanaceæ, with solitary carpels, approach on one side to the Leguminosæ, among which genera with drupes are also found; Mimosaceæ, with their many stamens, form a connecting link on the other side. In this respect the Mimosa-genus Affonsea, and certain Cæsalpineæ and Swartzieæ, are of special interest in having more than one carpel (syncarp), a condition which is sometimes met with abnormally in other Leguminosæ, as well as in Amygdalaceæ. About 7,000 species of the Leguminosæ are known.
Order 1. Cæsalpiniaceæ. These are leguminous plants with straight embryo and a flower which is not papilionaceous and has not[467] the same æstivation (Figs. 508–510); but in reality there is not a single characteristic which absolutely distinguishes them from the Papilionaceæ.—The majority are arborescent; the leaves as a rule are pinnate or bipinnate. The flower is 5-merous, most frequently perigynous and slightly zygomorphic; the calyx is free or gamosepalous, the corolla polypetalous with ascending imbricate æstivation (i.e. the two lowest petals envelop the lateral ones, and these again the posterior; Fig. 508); 10 free stamens; fruit various.
Figs. 508–510.—Cassia floribunda.

Fig. 508.—Floral diagram.

Fig. 509.—Flower.
Fig. 510.—The same in long. sect.
Cassia (Figs. 508–510) is the largest genus (about 200 species); it has an almost hypogynous, zygomorphic flower with 5 free sepals and petals; of the 10 stamens the 3 posterior are generally barren, the others are of very unequal length and open at the apex by pores (Fig. 509). In some (the Senna group) the fruit is a flat, short, thin, dehiscing pod; in others (Cathartocarpus) it is round, long, woody or fleshy, indehiscent, and divided internally by more or less fleshy transverse walls into as many cells as there are seeds.—The following also have DEHISCENT FRUITS: Bauhinia (often lianes, tropical climbers with tendrils [stem-structures] and anomalous stems), Copaifera, Hæmatoxylon (whose pod does not dehisce along the suture, but laterally), Cercis (simple leaves; the corolla resembles that of the Papilionaceæ, but the posterior petal is the smallest, and is enveloped by the 2 lateral ones, which are enveloped in their turn by the 2 anterior).—Fruit Indehiscent: Tamarindus indica; the pod is almost round, often a little abstricted between the seeds; the wall is formed by a thin, brittle external layer, enclosing an acid pulp; well-developed[468] septa are present, between the seeds; the most internal layer is parchment-like. Calyx 4-merous by the coalescence of 2 sepals. Only 3 fertile stamens.—Ceratonia siliqua (Carob-bean, Locusts); the pod is long, compressed, with thick sutures, and has a wall, the central part of which is more or less leathery, fleshy and sweet; there are transverse septa between the seeds, as in the Tamarind. Embryo greenish in endosperm. The flower is without a corolla, 5 stamens.—Pterogyne (winged fruit), etc.—Kramerieæ with Krameria is an anomalous group.
Distribution. 80 genera, with 740 species; almost exclusively in the Tropics. The Carob-tree and Cercis grow in the Mediterranean basin. The largest and most widely distributed genus is Cassia, which is found as trees, shrubs, and weeds in all tropical countries. The order has many important uses to mankind. Medicinal: the leaves and pods of Cassia acutifolia and angustifolia (officinal, Senna-leaves), the fruit-pulp of the Cassia-sub-genus, Cathartocarpus. Rhatany root from Krameria triandra (Peru, officinal). Balsam is extracted from a number of Copaifera-species (Balsam of Copaiba) from S. Am. (officinal), and from Hymenæa (Copal balsam), Trachylobium and others. Edible fruits are obtained especially from the Carob-tree (from the East) and the Tamarind (officinal). The heart-wood of several species of Cæsalpinia, such as C. brasiliensis (the Pernambuco-tree), echinata (Red-tree), and sappan, yield dyes; Hæmatoxylon (H. campechianum, Logwood), Copaifera bracteata (Amarant-tree).—Timber is obtained from many (Melanoxylon and others). In Europe they are of little importance as ornamental plants, these being confined principally to the species of Gleditschia (G. triacantha, from N. Am.) and Cercis (the Judas-tree, C. siliquastrum, S. Eur.), which are cultivated in gardens; but in tropical gardens beautiful flowering species, e.g. of Cassia, Poinciana, Brownea, are found, and the most beautiful of all ornamental plants, the Indian Amherstia nobilis.

Fig. 511.—Diagram of Faba vulgaris: f the standard; v the wings; k the keel.
Order 2. Papilionaceæ. The flower (Figs. 511, 512) is strongly zygomorphic and somewhat perigynous (Fig. 512 B; most frequently more on one side than the other). The calyx is gamosepalous and persistent. The polypetalous corolla has descending imbricate æstivation, the posterior, large leaf, the standard (Figs. 511 f; 512 B’, e), covering in the bud the two lateral ones, the wings (Figs. 511 v; 512 B’, a), which again cover the two anterior; these are united in the form of a boat, the keel (k and c); the wings and the two petals of the keel are very unsymmetrical. That the keel is formed of two petals is seen by its position (in front of one sepal)[469] and by the two often more or less free claws. The 10 (5 + 5) stamens (monadelphous) are either all united into one bundle, or into two bundles (diadelphous), the posterior one being free (Fig. 512 C). The ovules are curved and also the embryo (Fig. 512 G), especially the hypocotyl, so that the radicle assumes a position close to the edge of the thick, fleshy cotyledons. Endosperm wanting; the cotyledons are very rich in proteid reserve material. The forms of the fruit and exceptions are described under the genera.

Fig. 512.—Pisum sativum: A entire flower; B in longitudinal section; C gynœceum and stamens; D gynœceum; B’ corolla dissected, e standard, a, a wings, c keel; D seed opened to show the cotyledons (c), the radicle (r), the plumule (g); E fruit (legume); F seed.
Geocarpic fruits, i.e. those which penetrate the soil during their development and ripen underground, are found in e.g. Arachis hypogæa (see page 472), Trifolium subterraneum, Vicia amphicarpæa. Germination takes place in various ways. In the majority the cotyledons are raised above the ground as green, leaf-like bodies; in the Vicieæ they remain thick and white, and are always enclosed in the testa, and are therefore never able to take part in the work of assimilation; in species of Phaseolus, on the other hand, they are raised well above the ground and become green, but remain however thick and fleshy.
1, 2. The two groups Podalyrieæ (the majority of the genera are Australian) and Sophoreæ (Sophora, Edwardsia, etc.), represent the oldest[470] type, as they have 10 free stamens and so form the transition to the Cæsalpiniaceæ. Nearly all are trees and shrubs.
3. Astragaleæ. Herbs or shrubs, less frequently trees, with imparipinnate leaves (without tendrils). The flowers are generally borne in racemes or spikes. Stamens monadelphous or diadelphous.—Astragalus (Milk-Vetch) has the legume incompletely divided longitudinally into 2 loculi by a septum formed by the incurved dorsal suture. Diadelphous.—Glycyrrhiza (Liquorice); Colutea (Bladder-Senna) from S. Europe; Robinia (the false Acacia) with thorny stipules; Indigofera (the Indigo plant); Amorpha (which has only one petal, namely the standard, and the fruit a nut), Caragana, Wistaria (a climbing shrub), Galega. Carmichælia australis, when old, produces flat branches with scale-like leaves.
4. Vicieæ. Climbing herbs with paripinnate leaves, the midrib ending in a point or frequently in a tendril, which generally is branched, representing lateral veins without mesophyll; stamens diadelphous; the cotyledons remain underground on germination.—Vicia (Vetch) has a filamentous style, hairy towards the tip, and a pod with many seeds; climbing by means of tendrils; the leaves have many leaflets.—Faba (F. vulgaris, Horse-bean) is erect, without tendrils; its pod is thick with spongy septa between the seeds.—Ervum (Lentil) has a pod with only 1–2 seeds, and sweeping hairs (stylar-brush) on the inner side of the style.—Pisum (Pea; Fig. 512) has very large stipules, the bent style has a hollow groove on the anterior side. P. sativum (Common Pea), P. arvense (Grey Pea).—Lathyrus (Sweet Pea) generally has an angular, winged stem and most frequently only a few pairs of leaflets. The style is flattened, with sweeping hairs on the back. In L. aphaca the stipules alone are developed into foliage-leaves, while the remainder of the leaf is modified into a tendril.—Cicer has a nearly straight embryo and imparipinnate leaves with dentate or incised leaflets. C. arietinus (Chick-pea).—Abrus (precatorius, etc.); the seeds (“Crab’s eyes,” “Paternoster peas,” “Jequirity”) are scarlet with a black spot round the hilum.
5. Phaseoleæ. Herbs, twining or erect, but not climbing by tendrils; the leaves are imparipinnate, generally ternate, and bear small, linear bodies resembling stipules at the base of the stalks of the leaflets. The inflorescences are most frequently compound, groups of few flowers being situated on short, nodose, lateral axes borne on a longer stem. On germination the cotyledons are raised a considerable distance above the ground, and become[471] greenish, but do not become leaf-like; in P. multiflorus they remain underground. Stamens as in the Vetches.—Phaseolus (Kidney-bean): the keel with the stamen and style is spirally twisted (to the right). Herbs, twining to the left.—The “Calabar-bean” (Physostigma venenosum), Erythrina, Clitoria, Glycine, Soja, Mucuna, Apios, Canavalia, Vigna, Dolichos, Cajanus, Rhynchosia, etc.
6. Trifolieæ (Clovers). Herbs with ternate leaves, the leaflets are often dentate with the veins prolonged into the teeth; stamens diadelphous; fruit 1-locular, 1–few-seeded, pyxidium-like, irregularly dehiscent, or more frequently a nut. The flowers are generally borne in capitula, racemes, or spikes.—Trifolium (Clover). The corolla is gamopetalous. The calyx persists, together with the corolla, round the ripe fruit. The inflorescence is a spike, capitulum or capitate umbel; the leaves are ternate, and have adnate stipules.—Medicago (Medick). The corolla falls off after flowering; fruit curved like a sickle or spirally twisted; it is a nut, and opens with difficulty. Leaves ternate.—Melilotus (Melilot) has a small, spherical or lanceolate, thick and wrinkled fruit, which as a rule is indehiscent. The inflorescence is a raceme, often long, or a spike, sometimes a capitulum. Leaves ternate.—Ononis (Rest-harrow) differs in having monadelphous stamens and in being more shrub-like and bushy, and in having a normal, 2-valved pod, by which characteristic it approaches the Genisteæ. The flowers are generally rose-coloured, solitary, or in few-flowered racemes in the leaf-axils. Thorns (branches) are often present; the leaves are compound with only one small leaflet (the terminal one), or ternate with adnate stipules.
7. Loteæ. Herbs with ternate or imparipinnate leaves, with entire leaflets. In the latter case, when the lowest pair of leaflets is placed quite close to the sheath, the stalk is wanting, and apparently a trifoliate leaf with large stipules is developed. Flowers in an umbel or capitulum. Stamens monadelphous or diadelphous, the filaments (either all of them, or only the 5 sepal-stamens) are widened at the top.—Lotus (Bird’s-foot-trefoil) has a long, round pod.—Tetragonolobus.—Anthyllis (Lady’s-finger); the fruit is a nut, which is distributed by the wind by means of the membranous, bladder-like calyx, which completely encloses and falls off with it.
8. Genisteæ. The majority are shrubs or trees with apparently simple leaves, i.e. compound leaves with only one leaflet (the terminal leaflet), or ternate leaves; the stipules in most instances[472] are very small or are entirely wanting; stamens monadelphous.—Genista (Dyer’s-weed) has apparently simple leaves; the branches often terminate in a thorn. The strongly-winged stems in G. sagittalis are its most important organs of assimilation. Bossiæa rufa has flat branches, its leaves being reduced to small, pointed stipules.—Sarothamnus (Broom) has switch-like, angular branches and often both the apparently simple and ternate leaves on the same shoot; style spirally rolled.—Cytisus (Laburnum). Ulex (Furze; in U. europæus, the seedlings bear a few foliage leaves, but the leaves succeeding these are modified into thorns); Spartium; Crotalaria, etc.—Lupinus (Lupin) is allied to this group; it has a thick, often somewhat fleshy pod, and digitate leaves with adnate stipules.—Retama.
9. Hedysareæ are especially recognised by having the ovary divided by transverse septa into as many cells as there are seeds, the fruit thus becomes a lomentum, dehiscing transversely into nut-like joints (Fig. 513).—Ornithopus (Bird’s-foot); Coronilla; Hippocrepis; Onobrychis (Sainfoin) has a fruit with only 1 joint (i.e. a 1-seeded nut); Desmodium; Alhagi; Hedysarum, etc.—Arachis hypogæa (Earth-nut) has a pod which is abstricted between the seeds, and is indehiscent, but is not multilocular nor a true lomentum; it is reticulately wrinkled externally, and ripens underground; the basal part of the ovary is prolonged after flowering, attaining a length of several inches, and buries the young fruit in the soil. The embryo is straight.—Desmodium gyrans is well-known for its motile leaflets.

Fig. 513.—Hedysarum coronarium.
10. Dalbergieæ. 25 genera; especially in Tropical America; the majority are trees, a few shrubs or lianes; the leaves are simple or imparipinnate. The fruit is indehiscent in all; in some it is a winged, in others a wingless nut (Machærium, Dalbergia, Centrolobium, etc.), in others, again, a drupe, e.g. in Dipteryx (Tonquin-bean) and Andira. In some genera the embryo is straight.
Pollination. Especially effected by Bees. The nectar is secreted by a ring or disc-like portion round the base of the gynœceum or the inner surface of the receptacle. The flower is constructed with a peculiar mechanism to ensure cross-pollination by insects. The pollen is shed just before the flower opens, and is retained in a pouch formed by the keel. An insect visiting the flower uses the wings and keel for a landing-stage, and in attempting to reach the honey presses down the wings and the keel which are locked together near the standard; the stylar-brush by this means is forced through the apical opening of the keel and a little pollen is thus swept out and deposited upon the abdomen of the visiting insect as it presses against the apex of the keel;[473] the insect thus carries away pollen and may effect cross-pollination. In the different flowers this arrangement is modified in various ways to promote pollination. 5000 species (319 genera); especially in the Tropics, where many are important forest trees.—The following plants are used FOR FOOD: Pisum sativum (W. Asia?) and arvense (Italy); Phaseolus vulgaris (Kidney-bean, American; Dolichos sinensis was known to the Greeks and Romans under the name “φασηλος,” “phaseolus”), P. compressus (French-bean), etc.; Faba vulgaris (Field-bean, Horse-bean; from the Old World); Ervum lens (Lentil, Eastern Mediterranean); in tropical countries the oil-containing seeds of Arachis hypogæa.—The following are FODDER plants: Vicia sativa, Faba vulgaris, Onobrychis sativa (Sainfoin), Medicago sativa (Lucerne), and lupulina (Medick), species of Trifolium, Hedysarum coronarium. Officinal: “Liquorice root,” from Glycyrrhiza glabra (S. Europe); “Red Sandalwood,” from Pterocarpus santalinus (Tropical E. Asia); Gum Tragacanth, from Astragalus-species (E. Mediterranean); Balsam of Peru, from Toluifera pereiræ, and Balsam of Tolu, from Toluifera balsamum. Calabar-beans, from Physostigma venenosum; Kino, from Pterocarpus marsupium; the pith of Andira araroba is used under the name of “Chrysarobin.”—Of use TECHNICALLY: Genista tinctoria (yellow dye) and Indigofera-species (Indigo), the bast of Crotalaria juncea (Sunn Hemp); the seeds of Dipteryx, which contain Coumarin, and are highly scented, and Balsam of Myroxylon. Poisonous: the seeds of Laburnum (Cytisus laburnum), various species of Lathyrus, and Abrus precatorius; the latter contain two poisonous proteids, paraglobulin and albumose, which resemble snake-poison in their effects. The following are ORNAMENTAL plants: Phaseolus multiflorus (Scarlet runner, from America), Robinia pseudacacia, Amorpha, Colutea, Coronilla, Indigofera dosua, Wistaria polystachya, Cytisus laburnum (Laburnum, S. Europe, Orient.) and other species.
Order 3. Mimosaceæ. The flowers are most frequently hypogynous and regular, the æstivation of the corolla is valvate and, in the majority of instances, that of the calyx also. The flower is 4-merous, less frequently 5- or 3-merous.—The flowers are generally small, but are always borne in compact, round capitula or spikes (Fig. 514); they are hypogynous or perigynous. The calyx is generally gamosepalous and the corolla gamopetalous, the latter being frequently wanting. The stamens are equal or double the number of the petals (Mimosa, etc., in M. pudica, e.g. S4, P4, A4, G1) or (in Acacia, Inga, etc.) in a large, indefinite number, free or monadelphous, often united to the corolla (Fig. 514 b). The colour of the flower in most cases is due to the long and numerous stamens. The fruit is various. The embryo is straight as in the Cæsalpiniaceæ. Entada and many species of Mimosa have a flat, straight, or somewhat sickle-like pod, which resembles the siliqua of the Cruciferæ in that the sutures (in this instance, however, dorsal and ventral suture) persist as a[474] frame, but the intermediate portion divides, as in the transversely divided siliqua, into as many nut-like portions as there are seeds. Some species have a pod of enormous dimensions. The seeds of Entada gigalobium are often carried from the West Indies to the N. W. coasts of Europe by the Gulf Stream.—The fruit of Acacia in some species is an ordinary pod, in others it is transversely divided, or remains an undivided fruit, a nut.—This order includes both trees and herbaceous plants, which are often thorny; the leaves are usually bipinnate (Fig. 514) and are sensitive, and also possess sleep-movements.—Many Australian Acacias have compound leaves only when young, but when old have phyllodia, i.e. leaf-like petioles without blades, placed vertically. A large number have thorny stipules, which in some (Acacia sphærocephala) attain an enormous size, and serve as a home for ants, which in return protect their host-plant against the attacks of other, leaf-cutting ants.
Fig. 514.—Acacia farnesiana: a inflorescence; b flower.
[475]
Other genera besides those mentioned are: Adenanthera, Desmanthus, Parkia, Inga (with rather fleshy, indehiscent fruit), Calliandra, etc.
1350 species (30 genera); none natives of Europe, their home being the Tropics and sub-tropical regions, especially Australia and Africa.—Fossils in Tertiary.—Gums are found in many species of Acacia, especially the African (Gum arabic) and Australian, of which some are officinal. The bark, and also the fruits, contain a large amount of tannic acid and are used as astringents and in tanning (“Bablah” is the fruits of several species of Acacia). Catechu is a valuable tanning material extracted from the wood of Acacia catechu (E. Ind). The flowers of Acacia farnesiana (Fig. 514) are used in the manufacture of perfumes. With us they are cultivated as ornamental plants, e.g. A. lophantha and many others, in conservatories.
The flowers are most frequently regular, 5-merous in the three most external whorls, eucyclic and perigynous or epigynous, less frequently hypogynous. A characteristic feature is that the ovary is tricarpellary, unilocular, and with 3 parietal placentæ which sometimes meet in the central line (Cucurbitaceæ). The styles are generally free and bifid. To all these characteristics, however, there are exceptions. The Cucurbitaceæ are sometimes placed among the Sympetalæ, close to the Campanulinæ, but they are not allied to the Sympetalæ, from which they differ especially, for instance, in the structure of the ovule. The position of the Begoniaceæ in this family is also open to doubt.
Fig. 515.—Passiflora cœrulea (reduced).
[476]
Order 1. Passifloraceæ (Passion-flowers). The majority are herbs which climb by means of tendrils (modified branches) and have scattered, stipulate leaves, often palminerved and lobed (Fig. 515). The flowers, which are often large and beautiful, are regular, ☿, with S5, P5, A5, G3; the calyx and corolla are perigynous, and immediately inside the corolla is the “corona,” consisting of numerous, tapering, filamentous bodies, or sometimes united in rings, most frequently petaloid and coloured; the stamens are raised on a long, round internode above the cup-like receptacle; immediately above these is the gynœceum with its 3 free styles and capitate stigmas; the ovary is unilocular with 3 parietal placentæ. Fruit most frequently a berry. The seeds have an aril.
210 species; especially in Tropical America. Several Passiflora-species are ornamental plants, and the fruits of some species are edible.
Order 2. Papayaceæ. The best known representative is the Papaw (Carica papaya), a Tropical American tree whose stem is usually unbranched, and bears at its summit several large, palmilobed leaves on long stalks. The stem and leaves have latex. The large, Melon-like berries are edible, and for this reason it is cultivated in the Tropics. Flowers unisexual, with slightly different structure in the ♂-and ♀-flowers, besides intermediate forms. The ♂-flower has a gamopetalous, the ♀-flower a polypetalous corolla.—The milky juice contains a substance with similar action to pepsine. 10 stamens. 5 carpels.
Order 3. Turneraceæ. 85 species; especially in America.
Order 4. Samydaceæ. 160 species; tropical.
Order 5. Loasaceæ. Herbaceous plants seldom shrubs, sometimes climbing, and nearly always studded with stiff hairs, in some instances stinging or hooked. The leaves are most frequently palmilobed and without stipules. The flowers are regular, ☿, polypetalous, entirely epigynous, with 4–5 sepals, petals and stamens, or more frequently (by splitting) many stamens, those which are placed before the sepals being generally barren and more or less petaloid; carpels most frequently 3, united into an inferior, unilocular ovary with 3 parietal placentæ, above which the receptacle is generally more or less prolonged. Fruit a capsule; in Gronovia an ovary with 1 ovule and fruit a nut.
115 species; principally from S. Am. A number of annuals are often grown in our gardens: Bartonia aurea (California); Mentzelia; Cajophora; Gronovia.
[477]
Order 6. Datiscaceæ. 4 species, especially in the Tropics.—Datisca cannabina (Asia Minor) resembles the Hemp in external appearance. The flowers are diœcious, insignificant; ♂-flowers: a low, gamosepalous calyx, no corolla, and an indefinite number of stamens; ♀-flowers; epigynous; ovary unilocular with free, mostly bifid, styles, and generally 3 parietal placentæ. In most cases the ovary is not entirely closed at the top (as in Reseda).

Fig. 516.—Begonia rex (reduced).
Order 7. Begoniaceæ. This order principally comprises herbs or under-shrubs with succulent stems (having scattered vascular bundles in the pith); the leaves are arranged in two rows (a divergence of 1/2) and are asymmetrical, as a rule more or less obliquely cordate, or ovate with cordate base (Fig. 516); large, caducous stipules are present. Inflorescences dichasial, or unipared scorpioid cymes; the flowers are unisexual; the first ones (the oldest) are ♂-flowers, while ♀-flowers are found especially on the younger axes. The ♂-flowers have most frequently 2 + 2 coloured perianth-leaves, and many stamens collected into a head in the centre of the flower; the ♀-flowers are epigynous with 5 coloured perianth-leaves (placed spirally with a divergence of 2/5) and a trilocular ovary, bearing 3 bifid styles and 3 wings (the wings usually of unequal size); in the inner angle of each loculus[478] there is one large projecting placenta, or two plate-like placentæ (the bent back edges of the carpels) studded with ovules. Fruit a capsule, with many extremely small seeds.—Begonia.
420 species; almost all from the Tropics (Am., Asia).—Many species, with varieties and hybrids, are ornamental plants in houses and conservatories, chiefly on account of the form, colour and markings of their leaves; but also for their very beautiful flowers. They reproduce easily by adventitious buds from leaves and portions of leaves placed on damp soil; some have bulbils. Like the Oxalideæ they contain an acid sap.
Fig. 517.—Ecballium agreste. Diagram of a ♂-and a ♀-flower.
Order 8. Cucurbitaceæ. The flower is epigynous, and, as a rule, is also provided with a leaf-like, cup- or bell-shaped receptacle above the ovary, to which the perianth and stamens are attached; the flowers are regular, unisexual, with rudiments of the other sex, and 5-merous: sepals 5, narrow and pointed, with the median sepal posterior (Fig. 517), petals 5, stamens 5, and carpels 3 (rarely 4–5); the corolla is gamopetalous in the majority, polypetalous in some; generally plicate-valvate in the bud. The anthers in the ♂-flowers are extrorse, and monothecious, i.e. only one half of each of the anthers of the 5 stamens is developed, the pollen-sac having frequently a peculiar [**rtilde]-shaped curve (Fig. 518 A, B); the stamens are either all united into a column (e.g. in Cucurbita), or they are united in pairs, so that only one remains free (Figs. 517 A; 518 A); in the latter case there appears to be one small stamen with a ~-shaped, curved pollen-sac and two larger ones, each with two curved pollen-sacs placed as in Fig. 517 A. The original form appears to be Fevillea with free petals and 5 free stamens. Sometimes the rudiment of a gynœceum is present. The carpels are united into an ovary with 3 (4–5) placentæ formed by their united edges. These are thick, fleshy, and bifid, bearing a number of ovules on each side (Figs. 517 B; 518 C, D); in general the placentæ are[479] so large that they not only meet in the centre, but also fill up the ovary as far as the wall of the pericarp. The whole interior of the fruit thus becomes a juicy mass in which three lines may be seen, meeting in the centre (the boundaries of the individual placentæ), and near the circumference 6 groups of seeds (Fig. 518 D). When the carpels are equal in number to the petals they alternate with them. The style is short and thick, and generally divided into 3 (4–5) branches, with a horse-shoe shaped stigma on each branch (Fig. 518 C). The fruit is most frequently a many-seeded berry; in some it attains a considerable size and has a firm external layer (Cucurbita, Lagenaria, etc.). The embryo is straight, has no endosperm, but contains a large quantity of oil. The exceptions to the above characters will be found under the genera.
Fig. 518.—Citrullus colocynthis: A ♂-flower, cut open and spread out; B stamen; C ♀-flower in long section; h receptacle; ca calyx; D transverse section of ovary.
Exclusively herbs, generally with stiff hairs and yellow flowers. Many species are annuals, others are perennial, having tuberous roots or hypocotyls. The leaves are scattered, long-stalked, in most cases more or less heart-shaped, palminerved, palmilobed,[480] and exstipulate; in their axils are found both flowers (singly, or in an inflorescence) and a vegetative bud, and outside the axil, on the anodic[37] side of the leaf, a simple or branched tendril, by which the plant climbs (exceptions: e.g. Ecballium).
The position of the flowers, branches and tendrils situated in and near the leaf-axils is as follows. In the leaf-axils, a flower is borne (as a branch of the first order), ♂ or ♀, according to the conditions of the various genera. This branch is not situated in the centre of the axil, but is removed slightly towards the anodic side of the leaf. Of its two bracteoles as a rule only the one lying on the anodic side is developed, namely as a tendril, which is displaced to a position outside the axil. The branch of the first order bears on its catodic side an inflorescence (in the axil of the suppressed bracteole), on the anodic side a vegetative bud which grows out into a branch like the main axis. The subtending leaf of this branch is thus the tendril; but when it has several arms the condition is complicated by the appearance of an accessory bud which unites with its subtending leaf, the tendril, its leaves also becoming tendrils (situated on an undeveloped internode); the many-branched tendril is thus a branch, and the tendril-arms are its leaves, except the main arm which is its subtending leaf. Other explanations of these difficult relations have been given.—The germination is somewhat peculiar, owing to the fact that a heel-like prolongation is formed at the base of the hypocotyl to assist in separating the two halves of the testa from each other, and to facilitate the unfolding of the cotyledons.
Cucurbita (Pumpkin, Marrow) has branched tendrils; the flowers are monœcious, and are borne singly; the corolla is bell-shaped, and divided almost as far as the middle. The stamens are all united into a tube; the compressed seeds have a thick, blunt edge.—Cucumis has (generally) unbranched tendrils; the ♀-flowers are borne singly, whilst the ♂-flowers are borne in groups: the corolla is divided nearly as far as the base, and the stamens are united 2-2-1. The connective is elongated above the anthers. The seeds have a sharp edge.—Citrullus (Fig. 518) has a corolla similar to Cucumis, but ☿-and ♂-flowers are borne singly; the stigma is only 3-lobed, the fruit most frequently spherical.—Ecballium (Squirting Cucumber, only 1 species, E. elaterium) has no tendrils, and is therefore not a climber. The oblong fruit is pendulous from the apex of its stalk, and when ripe is distended with an acrid, watery fluid; on being touched the fruit is detached, and the seeds, together with the watery fluid, are violently ejected through[481] the aperture formed at the base of the fruit. The ♂-flowers are borne in racemes near the solitary ♀-flowers (Fig. 517).—Bryonia (White Bryony) has chiefly unbranched tendrils and small, greenish-yellow, usually diœcious flowers with rotate corolla, in many-flowered inflorescences; the small, spherical berry has no specially firm outer layer, and generally only few seeds. The tap-root and a few of the other roots are tuberous. B. alba (berry black; monœcious) and dioica (berry red; diœcious). Among other genera may be mentioned: Lagenaria (Gourd); the fruit has a woody external layer which, after the removal of the pulpy integument, may be used as a gourd. Luffa has a polypetalous corolla; the fruit is dry, and consists internally of a network of vascular bundles; it opens by an aperture at the summit. Benincasa; the fruit has a close, bluish coating of wax. Trichosanthes (Snake Cucumber) has a thin, round, long and curved fruit. Momordica; the fleshy fruit opens and ejects the seeds. Cyclanthera takes its name from the staminal column which is found in the centre of the ♂-flower, bearing a bilocular, ring-like anther which opens by a horizontal cleft. The fruit is unilocular by suppression, has 1 placenta, and when touched opens and ejects the seeds. Sicyos and Sechium have only unilocular ovaries with one pendulous ovule. Sechium has, moreover, 5 free stamens, of which only one is halved, the other 4 having both halves of the anther. Fevillea and Thladiantha also have 5 free stamens. Dimorphochlamys has dimorphic flowers.
Pollination is effected by insects, chiefly bees or wasps, the nectar being secreted by the inner, yellow portion of the receptacle; in the ♂-flower access is gained to the nectar through the slits between the stamens, which arch over the nectary.—85 genera; about 637 species; especially in the Tropics. Only two are found in the whole of N. Europe, Bryonia alba and dioica; in S. Europe, Ecballium also. Most of the cultivated species have been obtained from Asia, such as the Cucumber, Melon, Colocynth, several Luffa-species (the “Gourds” mentioned in Scripture are Cucumis chate); from Africa, the Water-melon, Cucurbita maxima, and others; from S. Am., no doubt, the Pumpkin (C. pepo and melopepo). Uses. Many species are used in medicine or for domestic purposes. Bitter, poisonous properties are found; the fruits of the two officinal ones are purgative: Citrullus colocynthis (Mediterranean, E. India, Ceylon) and Ecballium elaterium, as well as various tropical species, the roots of Bryonia, etc.—The following are cultivated AS ARTICLES OF FOOD: Pumpkin (Cucurbita pepo, etc.), Cucumber (Cucumis sativus), Melon (Cucumis melo), the Water-melon (Citrullus vulgaris), Sechium edule (Chocho), certain species of Luffa (the young fruit). The Bottle Gourd is cultivated in tropical countries for the sake of its hard pericarp, which is useful for bowls, bottles, etc. The fruits of Luffa have a number of reticulately felted, tolerably firm vascular bundles, which render them serviceable in various ways (as a kind of “sponge”). The Cucurbits are of no use in the manufactures. Only a few are cultivated as ornamental plants, chiefly as curiosities.
[482]
The leaves are most frequently opposite, simple, entire (rarely dentate), and exstipulate. The flowers are regular and epigynous (perigynous in Lythraceæ and a few others), ☿, polypetalous; the number of members in a whorl is generally 4 or 5 (S, P, A, or most frequently A 2, G), but sometimes it becomes (e.g. Myrtles and Lythraceæ) very large in the andrœcium by splitting, and in the gynœceum also is often different. (When suppression takes place it is principally in the corolla and petal-stamens.) In nearly all instances the calyx is valvate. Gyncœceum multicarpellary, multilocular, with only one style (except Haloragidaceæ). In the majority the ovules are situated on an axile placenta in the multilocular ovary. Endosperm is wanting in the majority.—Less important exceptions: Rhizophoraceæ and Gunnera have stipules. Haloragidaceæ have several styles and endosperm. Rhizophora also has endosperm.
Order 1. Lythraceæ. Hermaphrodite, perigynous flowers which are most frequently 6-merous, viz. S 6 (often with a commissural “epicalyx,” Fig. 519 c), one segment posterior, P 6, A 6 + 6 or 6 + 0 and G 2–6, forming a 2–6-locular ovary with many ovules in the loculi, style single, and capitate stigma. The gynœceum is free at the base of the tubular, or bell-shaped, thin, strongly veined receptacle, which bears the other leaf-whorls on its edge and inner side. Fruit a capsule. No endosperm.—To this order belong both herbs, shrubs and trees. The branches are frequently square, the leaves always undivided, entire, and without stipules, or with several very small stipules, and often opposite. The calyx is valvate. The flower is regular (except Cuphea) and frequently large and beautiful. The stamens are generally incurved in the bud, and the petals irregularly folded.
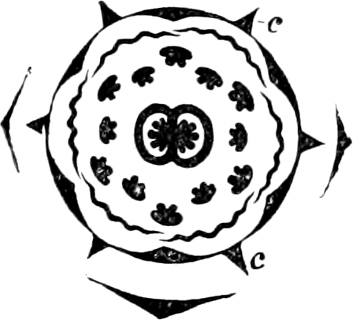
Fig. 519.—Lythrum salicaria. c the “epicalyx.”
Lythrum (Loose-strife). The flower is diplostemonous and 6-merous, with a long, tubular receptacle with epicalyx-teeth (Fig. 519 c). The 12 stamens are arranged in two tiers on the inner side of the receptacle. The gynœceum is bicarpellary. The flowers are borne in small dichasia in the leaf-axils, and their number is increased by accessory inflorescences beneath the main inflorescence.—The native species, L. salicaria, is trimorphic (long-styled, mid-and short-styled forms, Fig. 520). Cross-pollination is chiefly effected by humble-bees[483] and bees, which seek the nectar formed at the bottom of the receptacle. Other species are only dimorphic, or even monomorphic.—Closely allied are, Nesæa, Diplusodon, Lagerstrœmia, and Cuphea, whose flower resembles that of Lythrum, but is zygomorphic. In Cuphea the receptacle is oblique and at the back prolonged into a spur, in which the nectar, secreted by a gland situated behind the ovary, is collected; the calyx and corolla gradually become reduced in size toward the anterior side of the flower; the reverse, however, is the case with the 11 stamens (the posterior one is absent); the posterior loculus in the bilocular ovary is sometimes barren; the fruit, when ripe, dehisces along the posterior side, the ovary as well as the wall of the receptacle being ruptured by the placenta, which expands and projects freely. The flowers stand singly in the centre of the stem, between the pairs of leaves. This may be explained as follows: of the two foliage-leaves in each pair, one supports a foliage-shoot, the other a flower; the foliage-shoot remains in the axil, but the flower is displaced through the length of an entire internode to the next pair of leaves, and then assumes a position between these two leaves. All foliage-shoots stand in two rows, the flowers in two other rows.
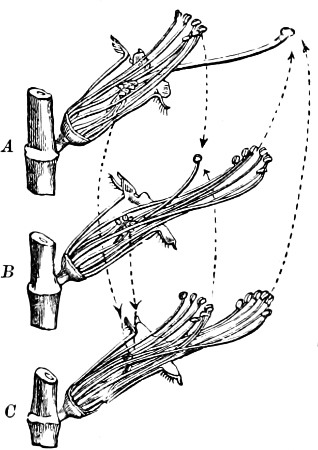
Fig. 520.—Lythrum salicaria. One side of the perianth is removed from all three flowers. A is long-styled, B mid-styled, and C short-styled. The direction of the arrows and dotted lines indicates the best (legitimate) methods of crossing.
Peplis (Water-purslane), a small, annual plant, with thin, bell-shaped receptacle without projecting nerves. The small flowers have no petal-stamens, and often also no corolla; fruit indehiscent.—Ammannia is closely allied to it.
365 species; 30 genera; mostly in the Tropics, and more especially S. Am.—Some yield dyes, e.g. Lawsonia inermis (cultivated in Africa and Asia) and Lagerstrœmeria indica; some contain tannin; others are ornamental plants, especially in gardens in warm countries.
Order 2. Blattiaceæ. 12 species. Tropical Asia and Africa. Trees. Formerly included with Punica, but best placed as an independent order.
Order 3. Melastomaceæ. A very natural and very large order (150 genera; 2,500 species), its home being chiefly in tropical S. America, especially the Brazils (termed by Schouw “The kingdom of Palms and Melastomaceæ”). There are both herbaceous and arborescent species, which are easily recognized by the opposite or verticillate, simple leaves which have (with the exception of a few heather-like species) 3–5–7–9 curved veins proceeding from the base of the leaf, and connected very regularly by closely parallel, transverse[484] veins. The flower is perigynous or epigynous; its type is that of the Onagraceæ (4–5-merous; 1 whorl of sepals, petals and carpels, 2 of stamens); the calyx is valvate, the corolla is twisted (to the left) in æstivation; the stamens are very characteristic; in the bud they are geniculate; the anther opens in the often long, beak-like, prolonged point, with 1, less frequently with 2 pores, and has generally ear-like appendages at its base. The fruit is a berry or capsule. These large and beautiful flowering-plants play a very important part in South American landscapes; otherwise they are of slight importance (a few are cultivated in conservatories, e.g. Centradenia, Medinilla, Lasiandra, Tibouchina, Miconia, etc.).
Order 4. Œnotheraceæ (or Onagraceæ). The flowers are arranged in racemes or spikes, ☿, epigynous, regular, polypetalous, 4-merous in all 5 whorls (1 whorl of sepals, petals and carpels, 2 of stamens); 2–3–5–6-merous flowers are less frequent; the calyx is valvate, the corolla twisted in æstivation (the left edge being covered). Gynœceum simple with multilocular ovary; the style is undivided, filiform, and bears a capitate or 4-partite stigma; endosperm wanting; embryo straight.—The majority are herbs, especially water- and marsh-plants; several are shrubs. No essential oils. The leaves are alternate or opposite, always single, and without (or with very small) stipules. The odourless flowers sometimes have a coloured calyx. In some instances (e.g. Œnothera, Fuchsia) the receptacle is prolonged more or less beyond the inferior ovary, and finally falls off. The stamens are obdiplostemonous (carpels epipetalous); the petal-stamens are sometimes suppressed. The anthers in some genera are divided into storeys. The well-pronounced, triangular pollen-grains are connected together by viscous threads. Small stipules are sometimes found, e.g. Fuchsia, Lopezia.
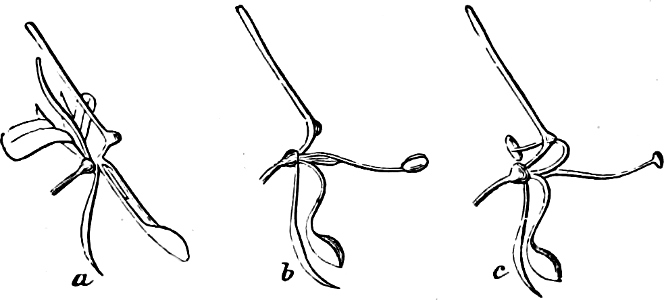
Fig. 521.—Flower of Lopezia.
A. Fruit a capsule. Œnothera (Evening Primrose) is 4-merous, has 8 stamens, a tubular receptacle, and an oblong capsule with loculicidal dehiscence leaving a centrally placed column, bearing the seeds.—Epilobium (Willow-herb) deviates from Œnothera especially in the seeds being hairy (at the chalazal end of the seed).—Chamænerium is a Willow-herb with zygomorphic flowers.—The[485] following may be included here: Clarkia, Eucharidium (an Œnothera with 4 stamens and 3-lobed petals), Godetia and Boisduvalia, Jussiæa (dehiscence septicidal), Isnardia (petal-stamens absent, sometimes the petals also).—Lopezia has a peculiar, zygomorphic flower (Fig. 521); one of the four sepals is bent forwards and the other 3 backwards; the posterior petals are narrower than the 2 anterior ones which are turned obliquely backwards and bent like a knee, with a greenish nectary at the bend; 2 stamens, one only fertile (the posterior), while the anterior is barren, petaloid, and spoon-shaped; both are sensitive, which is essential for pollination. In Fig. 521, a represents an early stage, in which the stamen and style lie concealed in the staminode; b is the ♂ stage, the stamen projects from the centre of the flower; c, the ♀ stage, the style occupies the place of the stamen.
B. Fruit a berry. Fuchsia generally has a coloured calyx and tubular receptacle; the corolla may be wanting.
C. Fruit a nut. Circæa (Enchanter’s Nightshade) has a 2-merous flower (S2, P2, A2 + 0 [petal-stamens are wanting], G2). The flowers are borne in racemes without bracts.—Gaura.
D. Fruit a drupe. Trapa (Horn-nut); a peculiar aquatic plant; the submerged stem has long internodes and lanceolate leaves, falling off at an early period, but at each node are found 4 long roots with thin, lateral roots (sometimes erroneously regarded as leaves) borne pinnately; the stem reaching the surface of the water, bears a rosette of rhombic foliage-leaves, with large, inflated stalks containing air, and forming the floating apparatus of the plants. In the axils of the leaves (as in Gunnera) 8 small, stipular structures are present. The flowers are solitary in the axils of the foliage-leaves (S4, P4, A4 + 0, G2), semi-epigynous. There is an 8-lobed, crenate disc on the free portion of the ovary; one ovule in each loculus. The fruit is a drupe with 4 (or 2) prominent horns (the persistent sepals), which after the pulp has decayed away bear a series of hooks turned downwards on each side, i.e. sclerenchymatous bundles which formerly lay concealed in the pulp of the sepals. The germination is peculiar: one of the cotyledons is large, and its thick extremity remains in the fruit, the other however is small and is pushed out at the apex of the fruit together with the radicle and plumule; the development of the root soon ceases, and the plumule usually grows into a stem entirely without branches, similar to the one described above, only that 1–2 precisely similar shoots arise in the axil of each cotyledon, so that each embryo produces 3–5 shoots.—Trapa, by its mode of life, its 1-seeded fruit, etc., forms a transition to Haloragidaceæ.
The large-flowered forms are adapted for insect-pollination and are often protandrous, the small-flowered ones are homogamous and may pollinate themselves. Œnothera is adapted for hawk-moths and bees.—330 species; especially in temperate climates, chiefly in the Northern Hemisphere. Epilobium,[486] Circæa are natives of this country; Trapa is extinct in this country, it has been found in a semi-fossilized condition near Cromer and in bogs in Denmark, and existed in Sweden until a few years ago; Œnothera has been introduced from N. Am.—A number of N. Am. species are grown as ornamental plants in our gardens. The seeds of Trapa natans are edible, and used as food in China.
Order 5. Haloragidaceæ. This is a reduced form of the Œnotheraceæ, and principally differs from these in the presence of endosperm and free styles. Only 1 ovule in each loculus.—84 species distributed over the entire globe; the majority are aquatic plants. The most advanced type is Myriophyllum (Water-Milfoil), with a regular, epigynous flower (S4, P4, A4 + 4, G4), most frequently diclinous (monœcious); the fruit is a 2–4-partite schizocarp. Aquatic plants, most frequently with pectinate, pinnate leaves.—Haloragis.—Gunnera (a dozen species from the Southern Hemisphere) forms the next step in the reduction. Large, scattered, rough-haired, and softly-spined leaves, with small flowers in crowded inflorescences. The flower, when most complete, has S2, P2, A2 (petal-stamens) and G2, forming an inferior, unilocular ovary with 1 ovule. It is remarkable for the great number of stipules placed in transverse rows in the leaf-axils, for the peculiar glandular organs, and for the colonies of Nostoc, which are found embedded in the cortex as a kind of parasite.—The simplest form is Hippuris (Mare’s-tail) with an extremely small, crenate or entire calyx, without corolla, and with only one stamen and one carpel, forming an inferior, unilocular ovary with only one ovule. Fruit a drupe with thin pulp. It is an aquatic plant with creeping, sympodial rhizome, and erect unbranched shoots, bearing numerous small, verticillate leaves. The small flowers are situated singly in the leaf-axils.
Order 6. Rhizophoraceæ. Tropical trees or shrubs (50 species, the best known being Rhizophora mangle, Mangrove) which grow gregariously, especially along the banks of rivers and by sea-coasts, where the water is quiet and brackish, and where they form the so-called Mangrove-swamps. Aerial roots are formed on the stems and branches (Fig. 522 A). The seeds germinate in the fruit, which by arrest contains only one seed (Fig. 522 B), before it is detached from the tree. The radicle projects considerably from the seed, and hangs down freely in the air; when the embryo is finally detached from the mother-plant, the separation is effected by the hood-like cotyledon, which entirely envelops the plumule, becoming detached from the rest of the embryo, which falls down, while the hood-like cotyledon remains enclosed in the fruit. The embryo, after it has fallen, strikes root, and continues growing in the undisturbed mud under the trees, or perhaps it may first be drifted about by the water, being well adapted for this by its peculiar, tough nature, and large, intercellular spaces.—It may also further be remarked that the anther is divided into a number of small loculi. The leaves are stipulate. The endosperm projects from the micropyle, growing out from the base of the seed, and thus serves as an organ of suction to convey nutriment to the embryo from the mother-plant.
[487]
Order 7. Combretaceæ. Trees and shrubs, partly lianes. An inferior, unilocular ovary with few pendulous ovules. Conocarpus and Laguncularia form, in conjunction with the species of Rhizophoraceæ, the tropical Mangrove-swamps. Terminalia.—280 species; Tropics.
Fig. 522.—Rhizophora mangle with the germinating fruit (much reduced).
Order 8. Myrtaceæ (Myrtles). The plants belonging to this order are shrubs or trees, the majority being easily recognised by the vegetative characters. The leaves, for instance, are most frequently opposite, without stipules, undivided and entire, parchment-like or leathery, evergreen, aromatic, finely dotted by pellucid glands containing essential oils; the venation is penninerved with a nerve just inside and running parallel to the edge of the leaf. The flowers are regular, epigynous (Figs. 523, 524, 525) and ☿, most frequently 4- or 5-merous in the calyx and corolla, with many stamens (by splitting, so that they are often in several distinct bundles) and an ovary with one style, formed of 2–5–many carpels; the receptacle is most frequently united for its entire length with the ovary. The fruit varies, but is most frequently a berry. The embryo is thick, often curved, with united cotyledons; no endosperm.
[488]
1. Myrteæ, Myrtle Group. Chiefly American, though some are found also in Africa and Asia. The fruit is a berry with generally 2–5 loculi in the ovary, and many ovules in each.—Myrtus; Eugenia (the petals fall off together as a hood in the Clove, E. caryophyllata, Figs. 523, 524); Myrcia; Jambosa; Amomis; Psidium, etc.
2. Puniceæ, Pomegranate Group. Only 2 species (Punica granatum; from Persia, Afghanistan), differing in several respects from the typical form of the Myrtaceæ. The leaves are generally opposite, without glands and marginal veins. The receptacle, calyx and corolla are red; the latter 5–8–(generally 6-) merous. Calyx valvate and corolla folded as in Lythraceæ, stamens also and epicalyx as in this order. The most characteristic feature is the inferior, spherical berry, with dry pericarp, formed from two whorls of carpels in two tiers (Fig. 525); the interior whorl, which is also the lower, has 3 carpels, and the placentæ are situated in the inner angles of the 3 loculi; the external whorl is 5-merous, and the placentæ have originally the same position in the inner angles of the loculi, but their position is changed to the outer side of the loculi owing to the growth of the wall of the ovary, which takes place early, causing the carpels to become, as it were, turned inside out, so that the part which was turned downwards is turned upwards, and the part which was turned inwards becomes turned outwards (as in Mesembrianthemum). The edible part of the fruit is the fleshy testa, as in Ribes. The cotyledons are rolled together spirally.
Figs. 523, 524.—Eugenia caryophyllata.
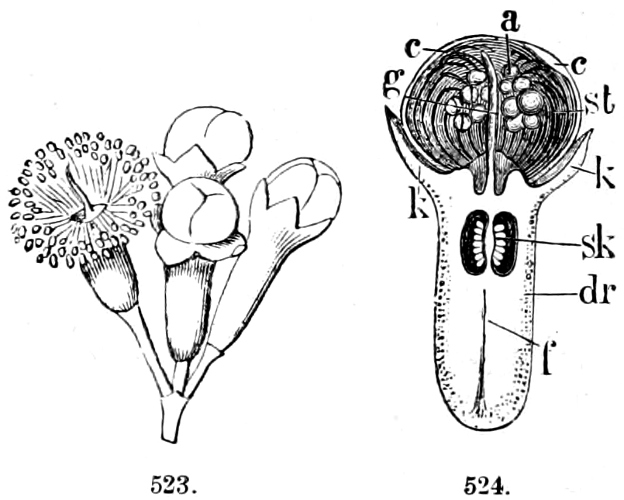
Fig. 523.—Flowers (nat. size).
Fig. 524.—A bud (“clove”), long. sec. (mag.).
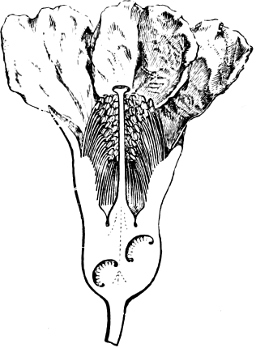
Fig. 525.—Punica granatum. Flower, long. sec. (nat. size).
[489]
3. Lecythideæ. The majority are South American. The leaves are scattered, without pellucid glands, and frequently dentate. The flowers are zygomorphic. The woody fruits are either indehiscent, or open by a lid. To this belong: Bertholletia (B. excelsa), the seeds well known as “Brazil-nuts,” Lecythis (Sapucaia-nuts from L. ollaria), Barringtonia.
4. Leptospermeæ. Almost entirely from Australia and the East Asian and Pacific Islands. The fruit is a capsule. The leaves are scattered, and in some placed edgewise by the twisting of the leaf-stalks.—Eucalyptus, the Australian Gum-tree; the calyx falls off like a lid (Figs. 526, 527). Some of the species attain gigantic heights, E. amygdalina 140–150 m. with a diameter of 8 m. The leaves in E. globulus are opposite and dorsiventral on the young plant; on the older scattered, placed edgewise by the twisting of the leaf-stalk, and isolateral; Metrosideros, Calothamnus (stamens distinctly polyadelphous), Melaleuca, Leptospermum, Callistemon (the flowers are borne in spikes whose axis continues to grow after flowering, thus several zones of fruits may be seen on the same branch).
Figs. 526, 527.—Eucalyptus globulus.
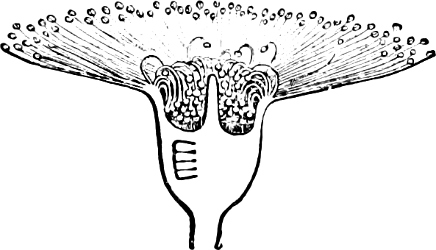
Fig. 526.—Long. sect. of flower.
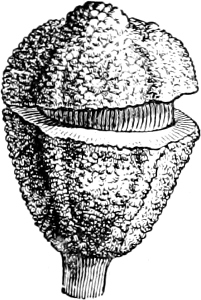
Fig. 527.—Flower opening.
5. Chamælaucieæ. Australian shrubs with heath-like appearance; they differ from the other Myrtaceæ in having a unilocular ovary with few, basal ovules, and a 1-seeded nut. The sepals are often pappus-like, and divided into many bristles.—Chamælaucium, Darwinia, etc.
This large order (2,100 species) is confined almost entirely to the Tropics, being found principally in America and Australia. In Europe, only Myrtus communis.—Several are useful on account of the large quantity of volatile oils (contained in internal glands): the flower-buds (“Cloves”) of Eugenia caryophyllata (the Moluccas, cultivated in the Tropics, Figs. 523, 524); the unripe, dry berries (“Pimento”) of Myrtus pimenta (Pimenta officinalis, W. Indies); Cajeput oil is extracted from Melaleuca minor and leucadendron (East Asian Islands). Eucalyptus globulus (Australia) has of late years become well known on account of its rapid growth, its hard wood, and its antipyretic qualities; it is cultivated on swampy soils, which it helps to drain.—Officinal: “Cloves,” and the cork of both stem and root of Punica granatum. Several have EDIBLE FRUITS, such as Psidium guyava (Guava, var. pomiferum and pyriferum, Am.), Eugenia cauliflora and others, E. jambosa, Punica granatum (the Pomegranate), etc. Edible seeds (with abundance of fatty oil): “Brazil nuts” from Bertholletia excelsa (Trop. S. Am.). “Bay-rum” is extracted from the leaves[490] and fruits of the Bayberry-tree (Pimenta acris, W. Ind.); Guava-rum from the berries of Eugenia floribunda. Tannin is found in large quantities e.g. in Punica. Gum is formed by many Australian Eucalypti (“Gum-trees”). Ornamental plants cultivated in this country are: Myrtus communis (Mediterranean), several in conservatories, especially the Australian Leptospermeæ, Eucalyptæ and others.
The flower is regular, ☿, and completely epigynous, 5- or 4-merous, with 1 whorl of stamens and 5–2 carpels. Sepals very small, tooth-like. The corolla is polypetalous, most frequently valvate in æstivation (least pronounced in the Umbelliferous plants). Round the base of the styles, which are generally free, there is an epigynous (undivided, or divided) nectar-disc (“stylar-foot”: Figs. 528 B, C, D; 539); the number of loculi in the ovary equals that of the carpels; only 1 pendulous (anatropous) ovule (Fig. 528 C) in each loculus. Endosperm copious (Fig. 528 D). To this must be added that the inflorescence in the majority of cases is an umbel or a capitulum, especially in the Umbelliferæ and Araliaceæ. Stipules are absent, but most frequently the base of the petiole forms a large sheath.
The Umbellifloræ are on one side so closely allied to the Frangulinæ, especially Rhamnaceæ, that they may perhaps be regarded as the epigynous continuation of this family. On the other hand, the similarities to the Rubiales, especially those between Cornaceæ and Sambuceæ, are so great that there is scarcely any character to distinguish them except the polypetalous corolla of the former and the gamopetalous corolla of the latter. Whether this is more than a merely analogous resemblance, and if not, whether the Cornaceæ at least should not be included in the Rubiales, must be left in abeyance.—The sepals are very small, as is generally the case in epigynous flowers.
Order 1. Cornaceæ. The majority of the species are shrubs with solid internodes, opposite (rarely scattered) leaves, which are simple, entire (rarely incised), penninerved, without stipules or large sheaths; flowers 4-merous (most frequently S4, P4, A4, G2), borne in dichasia which are either collected into corymbs (e.g. Cornus sanguinea), or in closely crowded umbels or capitula (Cornus mas, C. suecica), in which latter case there is often a large, leafy, or coloured, most frequently 4-leaved involucre round the base of the inflorescence; the style is undivided, with lobed stigma; the raphe of the ovule is turned outwards. The fruit is a berry or a drupe, with a 1–4-locular stone or 2 free stones.
Cornus (Dogwood, Cornel) has S4, P4, A4, G2. Leaves opposite.
[491]
Drupe with a bilocular, 2-seeded stone.—Aucuba, diœcious; unilocular ovary; 1 ovule; 1-seeded berry.—Garrya.—Helwingia.
80 species; N. Temp. The fruits of Cornus mas are edible; the wood is very hard; gum is found in some. Several species of Cornus and Aucuba japonica (Japan) are cultivated as ornamental shrubs.
Order 2. Araliaceæ (Ivies). Principally trees or shrubs with solid stems. The leaves are scattered, simple or compound, with a sheath more or less developed. The flowers are most frequently situated in umbels or capitula which are either borne singly or in racemes, or in paniculate inflorescences. The small, most frequently yellowish-green flowers are 5-merous, in the calyx, corolla, and andrœcium; the gynœceum may be 5-merous or may have some other number (2-∞). The styles are most frequently several, free; the raphe of the ovules is turned inwards as in the Umbelliferous plants. The fruit is a drupe or berry.—Stellate hairs often occur. The petals generally have a broad base, and a thick apex which is slightly incurved, and a distinctly valvate æstivation.
Hedera helix (Ivy) climbs by adventitious roots. The leaves are palminerved and lobed on the sterile branches, but often ovate and not lobed on the flowering branches. The flowers are yellowish-green and open in the autumn; they are slightly protandrous, and are visited by flies and wasps. Berries black. Endosperm ruminate.—Panax. Aralia (with Dimorphanthus).
375 species, 51 genera; especially in the Tropics (E. Asia).—The Ivy, several species of Aralia, e.g. A. japonica (Fatsia), Gastonia palmata, are cultivated as ornamental plants. Paper is manufactured from the pith of Aralia papyrifera (China).
Order 3. Umbelliferæ. The stem is herbaceous with hollow internodes; the leaves are scattered, and have a broad, amplexicaul base, a large, most frequently inflated sheath, and generally a pinnate (often very much dissected) blade. Entire leaves are found in Hydrocotyle vulgaris; Bupleurum.
The flowers are ☿, regular, small, but collected in compound umbels, that is, in “simple umbels,” which again are borne in umbels (for exceptions see Hydrocotyleæ); the external flowers in the simple umbel have often subtending bracts, which surround the base as an involucre, and may be termed the small involucre; the internal ones have no bracts; when involucral leaves are present at the base of the compound umbel, they may be termed the large involucre.
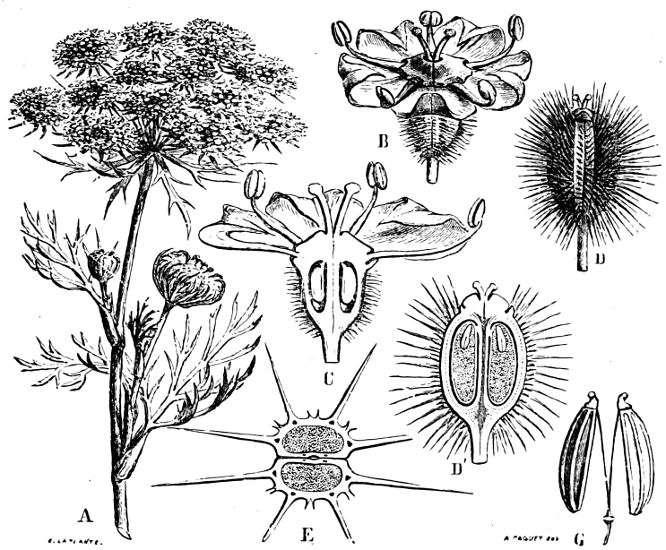
Fig. 528.—Daucus carota with flower and fruit.
The flower has 5 sepals (the median, as usual, posterior), 5 petals, 5 stamens and 2 carpels (in the median line) (Fig. 528). The calyx is often scarcely indicated. The petals have a short claw[492] are most frequently obcordate, or have an incurved apex (Fig. 528 B, C), being incurved in the bud; they are white, rarely yellow (Fennel and Parsnips), blue or red. The flowers are sometimes zygomorphic, especially those on the circumference of the umbel, and in that case it is the petal which is directed outside (anterior) which is the largest, and the two posterior are the smallest (e.g. Heracleum). The stamens are incurved in the bud. The 2 free styles unite at the base into the “stylar-foot” (stylopod), a swollen nectary (Fig. 528 B, C); the ovary is bilocular, the raphe of the ovules being directed inwards. The fruit is a schizocarp, dividing into two mericarps; the plane in which these separate coincides with that of the union of the carpels, and the two nut-like mericarps are in most genera kept together for awhile at the top of a thin, bifid, or undivided stalk (carpophore) which is in direct continuation with the flower-stalk (Fig. 537). Each mericarp has most frequently 5 more or less strongly projecting ridges, the primary ridges (Figs. 530, 532, 534, 535, etc.), of which 3 lie on the back of the mericarp, the dorsal ridges, and 2 on its edge near the plane of division, the marginal ridges; five of these (10 ridges[493] in all in the entire fruit) are placed opposite the calyx-teeth and the others between them. In some genera there are in addition 4 secondary ridges to each mericarp between the primary ones (Fig. 528 E: the secondary ridges bear the long bristles). Inside these secondary ridges, or inside the grooves between the primary ridges, when the secondary ridges are absent, oil ducts (vittæ, schizogenous ducts) are found in the pericarp, most frequently one in each groove; two are also often found on the ventral side of each mericarp (Figs. 528 E, 530 ol, etc.). The seed is most frequently united with the pericarp. The embryo is small and lies high up in the large, most frequently horny endosperm (Fig. 528 D).—The endosperm does not contain starch, but oil, and presents three different forms, of important systematic value: (a) those which are quite flat on the ventral side (i.e. the side turned towards the plane of splitting) (Figs. 528 E, 530, 531, 534, etc.): the majority of the genera, Orthospermeæ (e.g. Carum, Pastinaca); (b) those in which the endosperm on the ventral side is provided with a longitudinal groove, often deep: Campylospermeæ (e.g. Anthriscus); the transverse section is nearly a crescent (Fig. 532); (c) those in which the endosperm is concave on the ventral side (hollow in both longitudinal and transverse sections): Cœlospermeæ (e.g. Coriandrum) (Fig. 538).
The genera are distinguished first of all by the endosperm and forms of fruit, the ridges and oil-ducts; then by the form of the umbel, the calyx and corolla, by the absence or presence of an involucre, etc.
Fig. 529.—Hydrocotyle vulgaris. Transverse section of fruit.
1. Hydrocotyleæ, Penny-wort Group. Capitula or simple umbels (all the other groups have compound umbels). No oil-ducts. Orthospermous.—Hydrocotyle (Penny-wort). The fruit is considerably compressed laterally (Fig. 529). The calyx-teeth are small. The leaves are peltate.—Didiscus.—Sanicula (Sannicle). The umbels are small, capitate, generally collected in a raceme; calyx-teeth distinct. ♂-and ♀-flowers in the same umbel. The fruits are round, studded with hooked bristles. No carpophore.—Astrantia has an umbel surrounded by a large, often coloured involucre, with this exception it is the same as the preceding, but the fruit is slightly compressed, with 5 equal ridges. Hacquetia (Dondia).—Eryngium (Sea Holly): leaves often thorny. The flowers are all[494] sessile, the inflorescence is thus a capitulum; each flower is often subtended by a bract, which is thorny like the involucre, resembling the burrs of the Teasel. The sepals are large.—Lagœcia: one of the loculi of the ovary is suppressed.
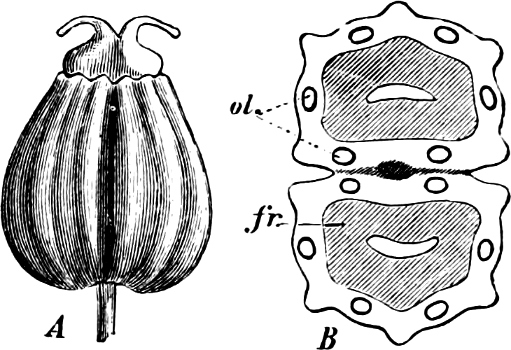
Fig. 530.—Fruit of Carum petroselinum: fr endosperm; ol oil-ducts.
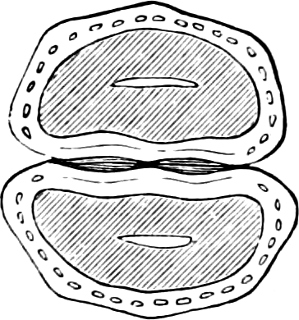
Fig. 531.—Pimpinella. Transverse section of fruit.
2. Ammieæ, Caraway Group (Figs. 530–532). The fruit has only the 10 primary ridges; it is usually short, almost spherical or broadly ovate and distinctly compressed laterally. Oil-canals are most frequently present. Orthospermous (except Conium).—Cicuta (Cow-bane). Pointed calyx-teeth. Glabrous herbs with pinnate or bipinnate leaves. C. virosa has a thick, vertical rhizome, divided by transverse septa into many compartments; the leaflets are narrow, lanceolate, and dentate; the large involucre is wanting.—Apium (Celery). No calyx-teeth. A. graveolens, a maritime plant, has neither large nor small involucre; the umbels are short-stalked or sessile.—Carum (Caraway). Calyx-teeth small; the large involucre is wanting or is only few-leaved. C. carvi (Caraway). C. petroselinum, (Parsley) (Fig. 530). Falcaria; Ammi; Helosciadium; Bupleurum (Hare’s-ear) with simple leaves and yellow corolla; Pimpinella (Fig. 531); Sium; Ægopodium (A. podagraria, Gout-weed) has bi- or tri-ternate leaves, with ovate, dentate leaflets; the large involucre is wanting.—Conium is campylospermous (Fig. 532); the short, broadly ovate fruit has distinctly projecting, often wavy crenulate ridges. C. maculatum (Hemlock) has a round, smooth stem with purplish spots.
Fig. 532.—Conium maculatum. Fruit entire and in transverse section.
[495]
3. Scandiceæ. This group has a distinctly oblong or linear fruit which is slightly compressed laterally, and generally prolonged upwards into a “beak”; wings absent. Campylospermous. Otherwise as in the Ammieæ.—Anthriscus (Beaked Parsley) has a lanceolate fruit, round on the dorsal side, without ridges, but with a ten-ridged beak.—Scandix (Shepherd’s-needle).—Chærophyllum (Chervil): fruit lanceolate or linear with low, blunt ridges; beak absent or very short. C. temulum has a red-spotted, hairy stem.—Myrrhis (Cicely) has a short beak and sharp, almost winged ridges. M. odorata (Sweet Cicely) has very long fruits.
Fig. 533.—Œnanthe phellandrium. Fruit entire and in transverse section. emb The embryo; ol the oil-ducts; fr endosperm.
Fig. 534.—Fœniculum vulgare. Fruit in transverse section.
4. Seselineæ, Fennel Group (Figs. 533, 534). The fruit is slightly elliptical or oblong, in transverse section circular or nearly so, without grooves in the dividing plane; only primary ridges are present. Orthospermous.—Fœniculum (Fennel) has yellow petals; both involucres are wanting; the fruit is oblong. The ridges are thick, all equally developed, or the lateral ridges are slightly larger (Fig. 534).—Æthusa (A. cynapium, Fool’s Parsley); the large involucre is wanting or is reduced to one leaf, the small involucre is composed of three linear leaves which hang downwards on the outer side of the umbels. The fruit is spherical-ovate, with thick, sharp, keeled ridges, the lateral ones of which are the broadest.—Œnanthe (Dropwort); the fruit (Fig. 533) has usually an ovate, lanceolate form, with distinct, pointed sepals and long, erect styles; the ridges are very blunt, the marginal ones a trifle broader than the others.—Seseli, Libanotis, Cnidium, Silex, Silaus, Meum, etc.
[496]
5. Peucedaneæ, Parsnip Group (Figs. 535–537). The fruit is most frequently very strongly compressed dorsally, with broad, mostly winged, lateral ridges. Only primary ridges. The dorsal ridges may project considerably, but are not winged. Orthospermous.
Fig. 535.—Archangelica officinalis. Transverse section of fruit.
Fig. 536.—Scorodosma fœtidum. Transverse section of fruit.
a. The winged lateral ridges stand out from each other, so that the fruit appears to be 4-winged (Fig. 535).—Angelica; Archangelica (Fig. 535); Levisticum (Lovage).
Fig. 537.—Heracleum sphondylium. Fruit.
b. The winged lateral ridges lie close together, and form one wing on each side of the fruit (Fig. 536).—Pastinaca (Parsnip). Corolla yellow. The dorsal ridges are very weak; the oil-ducts do not reach quite as far as the base of the fruit. Both large and small involucres are wanting; leaflets ovate. Anethum (Dill) is a Parsnip with more distinct dorsal ridges and filamentous leaflets. Peucedanum (Hog’s-fennel); Ferula (with Scorodosma, Fig. 536, and Narthex); Dorema.—Heracleum (Cow-parsnip); the flowers in the margin of the umbels are often very large, zygomorphic, and project like rays, e.g. in H. sibiricum. The fruit is very flat, with very small dorsal ridges; the oil-ducts are more or less club-like and do not reach as far as the base of the fruit (Fig. 537). Imperatoria; Tordylium.
6. Dauceæ, Carrot Group (Fig. 528). The fruit has 18 ridges, i.e. each fruitlet has 5 primary and 4 secondary ridges, the latter being often more prominent and projecting further than the primary ones. The oil-ducts are situated under the secondary ridges (Fig. 528).
a. Orthospermous: Daucus (Carrot). The secondary ridges project much further than the primary, and bear on their crests a[497] series of hooked spines (Fig. 528 D, E); these are much longer than the small bristles on the primary ridges. The involucral leaves of D. carota (Carrot) are numerous and deeply pinnate; the inflorescence contracts during the ripening of the fruit, and since the external umbels have longer stalks than the central ones, they arch over them, and the inflorescence becomes hollow. For the terminal flower, see below.—Cuminum; Laserpitium; Melanoselinum.
b. Campylospermous: Torilis (Hedge Parsley). The primary ridges are covered with bristles; the secondary ridges are not. very distinct on account of the spines, which entirely fill up the grooves. Caucalis (Bur Parsley).
Fig. 538.—Coriandrum sativum: b secondary ridges; d primary ridges; f endosperm; l embryo.
c. Cœlospermous: Coriandrum (Coriander) has a smooth, spherical fruit (Fig. 538) with a distinct, 5-dentate calyx, the two anterior (i.e. turned outward) teeth being generally longer than the others; the two fruitlets scarcely separate from each other naturally; all the ridges project only very slightly, the curved primary ones least, the secondary ridges most.
Pollination. The flowers are adapted for insect-pollination; they secrete nectar at the base of the styles; individually they are rather small and insignificant, but yet are rendered conspicuous by being always crowded in many-flowered inflorescences. Protandry is common, sometimes to such an extent that the stamens have already fallen off before the styles begin to develop (Fig 539, 2). Insect visits are more frequent and numerous as the inflorescences are more conspicuous. The flowers as a rule are ☿, but ♂-flowers are often found interspersed among the others (Fig. 539), and the number of these becomes greater on the umbels developed at the latest period. A terminal flower, which differs from the others in form, and in Daucus carota often in colour also (purple), is sometimes found in the umbel. The nectar lies so exposed and flat that the flowers are principally visited by insects with short probosces, especially Diptera; bees are less frequent visitors, and butterflies rare.—1400 species (175 genera); especially from temperate climates in Europe, Asia, N. Am. About 68 species in this country.
[498]
Fig. 539.—Anthriscus silvester: 1 ♂-flower; 2 ☿-flower.
Uses. A few are cultivated as ornamental plants. They are, however, useful in medicine,[38] and for culinary purposes on account of the essential oils and gum-resins which in many are formed in root, stem, and fruit. The FRUITS of the following are used: Carum carvi [+] (Caraway), Carum petroselinum (Parsley; also the leaves and root; its home is the Eastern Mediterranean); Fœniculum capillaceum [+] (Fennel; S. Europe); Pimpinella anisum [+] (Anise; E. Mediterranean); Coriandrum sativum [+] (Coriander; S. Eur.); Œnanthe phellandrium (Water Dropwort); Cuminum cyminum (Point Caraway; Africa; cultivated in S. Europe); Anethum graveolens (Dill). The LEAVES of the following are used as pot-herbs: Anthriscus cerefolium (Chervil); Myrrhis odorata (Sweet Cicely; Orient.); Conium maculatum [+] (the green portions; Hemlock). Besides Parsley, the ROOTS of the following are used: Carrot, Parsnip, Sium sisarum (Sugar-root; E. Asia); Chærophyllum bulbosum (Chervil-root); Levisticum officinale (foliage-shoots; S. Europe); Imperatoria ostruthium; Apium graveolens (Celery, the root in conjunction with the internodes); Pimpinella saxifraga and magna (Pimpinell); Archangelica (Angelica, the root of A. norvegica was formerly an article of food in Norway). Poisonous alkaloids are found in a few, such as Fool’s Parsley (Æthusa cynapium), Hemlock (Conium maculatum), Cow-bane (Cicuta virosa) and species of Œnanthe.—Gum-resin is extracted from various species: “Galbanum” from Ferula galbaniflua [+] and rubricalis [+] (Persia); Asafœtida from Ferula scorodosma [+] and F. narthex [+]; Ammoniac-gum from Dorema ammoniacum [+], all from Central and S. W. Asia. “Silphium” was an Umbelliferous plant which grew in ancient times in Cyrene, and from which the Romans extracted a valued condiment.
Family 25. Hysterophyta.
This family (with the exception of Aristolochiaceæ) includes only parasitic plants. Partly on this ground, and partly because they all have epigynous flowers, they are considered to belong to the youngest type (which is expressed in the name ὕστερος, the one that comes after). It is not certain to which of the preceding families they are most nearly allied. Again, it is a matter of doubt[499] whether the Aristolochiaceæ are related to the others; they are by Engler united with Rafflesiaceæ into one family, Aristolochiales.
Fig. 540.—Flower of Aristolochia clematitis (long. sect.). A Before pollination, and B after: n stigma; a anthers; t an insect; kf ovary.
Order 1. Aristolochiaceæ. The majority are perennial herbs or twining shrubs, whose stalked, simple, and generally more or less cordate or reniform leaves are borne in 2 rows and are exstipulate. The flowers are hermaphrodite, epigynous, regular or zygomorphic; perianth-leaves united, simple but most frequently petaloid and 3-merous; 6 or 12 (in Thottea as many as 36) stamens with extrorse anthers. The ovary is more or less completely 4–6-locular with ovules attached in the inner angles of the loculi (Fig. 540 kf). The style is short, and has a large, radiating stigma (Fig. 540 n). Fruit a capsule. Seeds rich in endosperm.
Asarum europæum. Each shoot has 2 reniform foliage-leaves, between which the terminal flower is borne (the rhizome becomes a sympodium by development of the bud in the axil of the upper foliage-leaf). The flower is regular and has a bell-shaped perianth with 3 outer valvate, and 3 inner small segments (which may be wanting). 12 (2 × 6) free, extrorse stamens, 6 carpels.—Aristolochia clematitis (Birth-wort) has an erect, unbranched stem, bearing many flowers in the leaf-axils, in a zig-zag row (accessory buds in a unipared scorpioid cyme). The flowers are zygomorphic (Fig. 540), formed by 3 alternating, 6-merous whorls. The perianth has a lower, much-distended part (k), succeeded by a narrow, bent tube (r), which passes over into an oblique, almost tongue-like projection[500] (6 vascular bundles indicate that the number 6 is prevalent here, as in Asarum); 6 stamens (Fig. 540 a), with the dorsal portion turned upwards, are united with the short style to form a stylar column; they are placed quite beneath the 6 commissural stigmatic rays, which arch over them as short, thick lobes. Protogynous; Pollination is effected in Arist. clematitis by small flies; these enter the erect unfertilised flower through the tube (Fig. 540 A, l) without being prevented by the stiff, downwardly-turned hairs which line the tube and prevent their escape; they find the stigma (n) fully developed, and may pollinate it with the pollen they have brought with them. The stigmas then straighten and wither (B, n), the anthers open, and the flies may again be covered with pollen; but the hairs which blocked up the tube do not wither until the anthers have shed their pollen, and only then allow the imprisoned flies to escape and effect cross-pollination. Prior to pollination, the flowers stand erect, but after this has taken place they become pendulous, and the perianth soon withers.—A. sipho (Pipe-flower), another species, is a climber, and often grown in gardens; it has only one row of accessory buds in the leaf-axils.—200 species; chiefly in S. Am. Officinal: the rhizome of Aristolochia serpentaria (N. Am.).
Fig. 541.—A fruit of Myzodendron brachystachyum (slightly mag.) germinating on a branch.
Order 2. Santalaceæ. Parasites containing chlorophyll, which, by the help of peculiar organs of suction (haustoria) on their roots, live principally on the roots of other plants. Some are herbs, others under-shrubs. The regular, most frequently ☿-flowers have a simple perianth, which is gamophyllous, 3- or 5 partite with the segments valvate in the bud, and a corresponding number of stamens opposite the perianth-leaves. In the inferior ovary there is a free, centrally placed, often long and curved placenta with three ovules (one opposite each carpel); these are naked, or in any case have an extremely insignificant integument. Fruit a nut or drupe. Seed without testa. Endosperm fleshy. 225 species; chiefly in the Tropics.—Thesium, a native, is a herb with scattered, linear leaves and small 5-merous flowers (P5, A5, G3) in erect racemes; the subtending bracts are displaced on the flower-stalks. Fruit a nut.—Osyris (diœcious shrub; 3-merous flowers) is another European genus.—Santalum album, which grows in E. Ind., yields the valuable, scented Sandalwood, the oil of which is used medicinally.—Quinchamalium.
[501]
Myzodendron is a reduced form of the Santalaceæ; the ♂-flowers are without perianth; the perianth of the ♀-flower is 3-merous. About 7 species; S. Am.; parasitic on a Beech (Nothofagus). The fruit has 3 feathery brushes, alternating with the lobes of the stigma, which serve as flying organs and to attach the fruits to a branch (Fig. 541), the brushes twining round as soon as they come in contact with it. There is only 1 seed in the fruit, which germinates by a long, negatively heliotropic hypocotyl, and is attached by a radicle modified into an haustorium.
Order 3. Loranthaceæ (Mistletoes). Plants containing chlorophyll which are parasites on trees, and most frequently have opposite, simple, entire leaves and regular, epigynous, often unisexual, 2- or 3-merous flowers, with single or double perianth. Stamens equal in number and opposite to the perianth-leaves, free, or in varying degrees united to one another. The inferior ovary is constructed as in the Santalaceæ, the ovules being situated on a low, free, centrally-placed placenta, but the placenta and ovules unite with the wall of the ovary into one connected, parenchymatous mass, in which the embryo-sacs are imbedded. Only 1 (less frequently 2–3) of the 1–6 embryo-sacs is fertile. The number of the carpels however varies. The fruit is a 1-seeded berry, whose inner layer is changed into a tough slimy mass (bird-lime), which serves to attach the fruits to other plants.
The two groups, Loranthoideæ and Viscoideæ, are distinguished by the fact that the former has a distinct “calyculus,” i.e. an entire or lobed, or dentate swelling on the receptacle below the perianth. The majority of the Loranthoideæ have a petaloid perianth; in all the Viscoideæ, on the other hand, it is sepaloid.
Fig. 542.—Viscum album: A branch with leaves and berries: a scale-leaves; b foliage-leaves; n m n flowers; B seedling, the bark of the branch being removed; C an older embryo which still retains the cotyledons.

Fig. 543.—To the left the Rafflesiaceous Cytinus hypocistus, parasitic on the roots of Cistus. To the right the Balanophoraceous Cynomorium coccineum, parasitic on the roots of Salicornia.
[502]
The Mistletoe (Viscum album, Fig. 542) is a native, evergreen plant which may be found growing on almost any of our trees (sometimes on the Oak), and, like other Loranthaceæ, it produces swellings of the affected branches. Its spherical white berries (Fig. 542 A) enclose (1–) 2–3 green embryos; they are eaten by birds (especially Thrushes), and are partly sown with their excrement, partly struck or brushed off the branches of the trees, the seed being enclosed, at maturity, by viscin, i.e. “bird-lime.” The seeds may also germinate on the branches, without having first passed through the alimentary canal of the birds. On germination, the hypocotyl-axis first appears, as in Fig. 541, and bends towards the branch; the apex of the root then broadens, and forms at the end a disc-like haustorium, from the centre of which a root-like body grows through the bark into the wood, and ramifies between the bark and wood. Suckers are developed on the root like strands which are formed in this manner, without, however, having a rootcap; they are green, and penetrate the wood by the medullary rays (Fig. 542 C). Adventitious buds may also be developed from the root-like strands which break[503] through the bark and emerge as young plants. The young stem quickly ceases its longitudinal growth, and lateral shoots are developed from the axils of its foliage-leaves. These and all following shoots have a similar structure; each of them bears a pair of scale-leaves (Fig. 542 A, a) and a pair of foliage-leaves (Fig.[504] 542 A, b), and then terminates its growth, if it does not produce an inflorescence; new lateral shoots proceed from the axils of the foliage-leaves, and the branching, in consequence, is extremely regular and falsely dichotomous. Only one internode (shoot-generation) is formed each year, so that each fork indicates one year. The foliage-leaves fall off in the second year. The inflorescence is a 3(-5)-flowered dichasium (Fig. 542 A, m is the central flower, n the lateral). The plants are diœcious; the ♂-flower as a rule is 2-merous: perianth 2 + 2, each leaf of which bears on its inner side 6–20 pollen-sacs, each of which opens by a pore; this relationship may be considered to have arisen from the union of the perianth-leaves with the multilocular stamens (2 + 2) placed opposite them. The ♀-flowers always have Pr 2 + 2, G2.—Loranthus is also found in Europe (it has a 3-merous flower), especially in the central and south-eastern districts, on Quercus cerris and Q. pubescens; but the great majority of the 520 species grow in the Tropics on trees which they ornament with their often brightly-coloured flowers, and ultimately kill when present in too great numbers. The pollination in the numerous Loranthaceæ with unisexual flowers, is effected by the wind. In Viscum album this takes place in autumn, the actual fertilisation in the following spring, and the maturity in November or December; in the succeeding month of May the berry is ready to germinate, and falls off.
Uses. Birdlime from Viscum album.
Order 4. Rafflesiaceæ and Order 5. Balanophoraceæ. These orders comprise root-parasites, almost entirely devoid of chlorophyll; they are reddish or yellow, without foliage-leaves (Fig. 543). As far as our knowledge of these rare tropical plants extends, they have thalloid organs of vegetation resembling the root-like strands of Viscum, or they are filamentous and branched like Fungus-hyphæ; they live in and on the tissues of the host-plant, from which their flowering-shoots, often of mushroom-like form, are subsequently developed (Fig. 543). In order to unfold they must often break through the tissues of the host-plant.
Of the Rafflesiaceæ, Cytinus hypocistus is found in S. Europe living on roots of Cistus-plants and to some extent resembling Monotropa (Fig. 543). Rafflesia is the best known; it lives on roots of Cissus-species (belonging to the Ampelidaceæ) in Java; its yellowish-red, stinking flowers attain a gigantic size (one metre or more in diameter), and are borne almost directly on the roots of the host-plant. Besides these there are other genera: Brugmansia, Pilostyles, Hydnora.—To Balanophoraceæ (Fig. 543) belong: Balanophora, Langsdorffia, Scybalium, Sarcophyte, Helosis, etc., and in S. Europe, Cynomorium coccineum.
The characters which separate this from the first Sub-class, the Choripetalæ, have been described on page 336. They consist in the following: the flower is always verticillate, generally with 5 sepals, 5 petals, 5 stamens, and 2 carpels (in the median plane), the calyx is generally persistent and gamosepalous, the corolla is gamopetalous and united to the stamens, which are therefore[505] adnate to it, the ovules have only one thick integument and a small nucellus. (The exceptions are noted later.)
This Sub-class is no doubt more recent than the Choripetalæ; it is also peculiar in including fewer trees and shrubby forms than the latter.
The Sympetalæ may be separated into 2 sections:—
A. Pentacyclicæ (five-whorled). The flowers in this section have 5 whorls equal in number, namely, 2 staminal whorls in addition to the calyx, corolla, and carpels; in some instances, one of the staminal whorls is rudimentary or entirely suppressed, but in this case it is frequently the sepal-stamens which are suppressed, and the whorl which is present stands opposite the petals. The flowers are regular. The number of carpels equals that of the sepals, but in one of the orders (Bicornes) they are opposite the petals (the flower being obdiplostemonous); in the other two orders (Primulinæ and Diospyrinæ) they are placed opposite the sepals (the flower being diplostemonous). This section is the most closely allied to the Choripetalæ, since the petals may sometimes be found entirely free, and the stamens inserted directly on the receptacle (Ericaceæ); ovules with two integuments are also found. It is very doubtful, whether the orders included under this head have any relationship with the other Sympetalæ. They appear in any case to represent older types.
B. Tetracyclicæ (four-whorled). The flowers have only 4 whorls, namely, beside sepals, petals, and carpels, only one whorl of stamens, which alternates with the petals; there is no trace of the second staminal whorl, and when the number of carpels is the same as that of the preceding whorls (“isomerous”) they alternate with the stamens; but in most cases there are 2 carpels placed in the median plane (see the diagrams, e.g. Figs. 559, 567, 583, 590, etc.). This section is the largest, and the one which shows the characteristics of the Sympetalæ best. Very irregular flowers are met with.
The following families belong to the Pentacyclicæ: 26, Bicornes; 27, Diospyrinæ; 28, Primulinæ.
The remaining families belonging to the Tetracyclicæ are:—
a. Hypogynous flowers (with a few exceptions): 29, Tubifloræ; 30, Personatæ; 31, Nuculiferæ; 32, Contortæ.
b. Epigynous flowers: 33, Rubiales; 34, Dipsacales; 35, Campanulinæ; 36, Aggregatæ. The ovaries and ovules in the last family are always reduced to one; and at the same time the fruits become nuts, and the flowers are united into crowded inflorescences.
[506]
This family is chiefly composed of shrubs, less frequently of small trees, or perennial herbs; their leaves are undivided, most frequently evergreen, stiff and leathery, and always without stipules. The flowers are ☿ and regular, rarely slightly zygomorphic, most frequently obdiplostemonous, and 4- or 5-merous through all the 5 whorls. The stamens are attached to the receptacle, and as a rule are quite free from the petals, an attachment which is very rare among the Gamopetalæ. They have a simple gynœceum with one undivided style, a commissural stigma, and a multilocular ovary, whose axile placentæ project considerably into the loculi, and bear a large number of ovules. The placentæ are sometimes not united, and in consequence, the ovary is 1-locular with incomplete partition-walls, e.g. Pyrola, Monotropa. Embryo straight, with endosperm. The carpels are placed opposite the petals.
The diagram is generally Sn, Pn, An + n, Gn, in which n is 4 or 5. To this may be added, that the corolla is in most cases gamopetalous, but in some (especially Pyrolaceæ) perfectly polypetalous; and that the anthers usually open by pores, and often have two horn-like appendages (hence the name “Bicornes”) (Figs. 545, 546); frequently the two halves of the anther are also widely separated from each other at the upper end, so that the pores are placed each one at the end of its own tube (Fig. 546); the pollen-grains in the majority are united into tetrads (Fig. 542 D).—The flowers, as a rule, are pendulous and borne in racemes, coloured (red or white), but odourless. When the fruit is a capsule, the placenta with the seeds attached persists as a central column. A mycorhiza occurs on many.
The majority of plants belonging to this family inhabit cold and temperate countries, or high mountains in tropical regions; they prefer cold and dry or damp places (bogs, heaths, etc.). Plentiful in N. America.
Order 1. Pyrolaceæ. Perennial herbs; petals most frequently quite free from each other, and falling off singly after flowering; the anthers are without appendages, and open by pores (Fig. 544), or by a transverse slit. The placentæ are thick. The seeds in the capsule-like fruit (loculicidal dehiscence) are exceedingly small and light, they have a sac-like testa which loosely envelops them, an oily endosperm, and an extremely simple embryo, which consists[507] only of an ellipsoidal, cellular mass, without cotyledons or differentiation into plumule and radicle.
Pyrola (Winter-green) is green, and has also large evergreen foliage-leaves. The flowers, 5-merous, are most frequently borne in racemes without a terminal flower; the anthers are extrorse in the bud with the pores in the lower portion (Fig. 544 A), but they become inverted at a later period, so that the pores open at the top (Fig. 544 C). P. uniflora has a single, terminal flower; it winters by its roots, producing from these in the spring aerial, quite unbranched shoots. Chimaphila umbellata.
Fig. 544.—Pyrola minor: A portions of a young flower; B the stigma; C portions of an older flower (longitudinal section).
Monotropa (Yellow Bird’s-nest) is very pale yellow, without chlorophyll, succulent, and has only scale-like leaves closely pressed upon the stem; it is a saprophyte. The raceme has a terminal flower, and is pendulous before flowering. The anthers open by a semicircular, transverse cleft. M. hypopitys reproduces chiefly by root-shoots.
About 30 species, especially N. Europe, N. America, and N. Asia.
Order 2. Ericaceæ. The flower (Fig. 545) is hypogynous, the median sepal posterior; corolla, gamopetalous; the stamens are generally 2-horned, and the fruit is a capsule, less frequently a berry or drupe. At the base of the ovary is a nectar-secreting disc (Fig. 545 B). This order comprises shrubs or undershrubs (rarely small trees), which are evergreen, and as a rule have densely crowded leaves.
1. Ericeæ, Heath Group. Flowers most frequently 4-merous (S4, P4, A4 + 4, G4, united in a 4-locular gynœceum), rarely 5-merous. The withered corolla persists after flowering. The leaves are most frequently acicular, opposite or verticillate; the buds are without scales. The fruit is a capsule.—Calluna (C. vulgaris, Ling) has a deeply 4-cleft corolla, which is less than the coloured calyx; capsule with septicidal dehiscence.—Erica (about 420 species; E. tetralix, Cross-leaved Heath) has a tubular or bell-shaped, 4-dentate corolla, which is much longer than the calyx. Capsule with loculicidal dehiscence.—Pentapera.
[508]
2. Andromedeæ. The flowers are 5-merous (S5, P5, A5 + 5, G5), with deciduous corolla. Capsule with loculicidal dehiscence. The leaves are scattered, and incline more to the ordinary broad-leaved forms.—Andromeda; Gaultheria; Cassandra (Lyonia); Cassiope.
Fig. 545.—Arctostaphylos uva-ursi.
3. Arbuteæ. The flowers as in the preceding group (Fig. 545), but the fruit is a berry or drupe. Arctostaphylos (A. uva-ursi, Bear-berry) has a drupe with 5 stones in a dry, farinaceous pulp; in other species there is 1 stone with several loculi. Arbutus (A. unedo, Strawberry-tree) has a spherical berry.
Pollination is effected by means of insects, especially by bees. The pollen is light and dry, and is shaken out through the pores of the anthers when the insects agitate the horn-like appendages during their visits. Self-pollination takes place, no doubt, in many cases.—800 species; the very large genus, Erica, especially in S. Africa (the Cape).—Officinal: the leaves of Arctostaphylos uva ursi. Arbutus unedo (S. Europe) has an edible, peculiarly warted (strawberry-like) fruit. Many Erica-species are cultivated as ornamental plants.
Order 3. Rhodoraceæ (Rhododendrons). This differs from the preceding order in the median sepal being anterior, and hence the position of the other floral whorls is also reversed. The flower is hypogynous, in most cases 5-merous; the corolla is most frequently deeply cleft or polypetalous, and falls off after flowering; the anthers open by pores, and have no horn-like appendages. Capsule with septicidal dehiscence.—The shrubs or small trees belonging to this order have, like the Vaccineæ, ordinary foliage-leaves, and the buds are generally provided with large bud-scales.
Rhododendron has 10 stamens, and a slightly zygomorphic flower with deeply 5-cleft corolla (the section Azalea has frequently[509] only 5 stamens, the petal-stamens being absent). They are Alpine plants (200 species) in the mountains of Asia, especially the Himalayas; some in S. Europe.—Menziesia.—Ledum; small, rusty-brown, hairy shrubs with polypetalous, expanded, star-like corolla.—Kalmia (N. Am.) has a cupular corolla, with 10 small, pocket-like depressions in which the anthers are concealed until the arched, elastic filaments are freed from this position by means of the insects, when they quickly straighten themselves in the centre of the flower.—Phyllodoce; Loiseleuria (5 stamens); (Clethra (?); also placed among the Ternstrœmiaceæ).
About 270 species. Several species are ornamental plants. Several plants of the order are more or less narcotic. Ledum palustre has been used as a substitute for hops.
Order 4. Diapensiaceæ. Hypogynous flower. 3 floral-leaves beneath the flower (S5, P5, A5 + 0, G3). Stamens on the throat of the corolla. Pollen-grains single. Disc absent. Capsule loculicidal.—9 species from the Arctic regions. It is doubtful whether this order should be included in the Bicornes; perhaps it would be more correctly assigned to the Polemoniaceæ.
Order 5. Epacridaceæ. This order comprises those species of the family which are confined to Australia and the South Sea Islands. They are shrub-like plants, resembling the Ericaceæ in habit, in the inflorescence, and in the structure, form, and colour of the flower. They differ especially in having only 1 whorl of stamens (placed opposite the sepals) and in the anthers having only 2 loculi, and opening by a longitudinal slit. Fruit most frequently a drupe (or loculicidal capsule). Epacris-and Styphelia-species are ornamental plants. About 325 species.
Order 6. Vacciniaceæ (Bilberries). The flower (Fig. 546) is epigynous, the corolla gamopetalous, and the fruit a berry. The latter is most frequently spherical, and bears on its apex the calyx, which is generally very low, almost entire, and with a disc-like expansion inside. The flower is 4- or 5-merous (Fig. 546 B, D). The anthers have 2 pores, and are most frequently 2-horned (Fig. 546 F, G). Small shrubs; the leaves are scattered, not needle-like.
Vaccinium (Bilberry, Whortleberry) has an urceolate, gamopetalous, only slightly dentate corolla, and horn-like appendages to the anthers (Fig. 546). V. vitis idæa (Cowberry) is evergreen, with flowers in racemes, and bright red berries; V. myrtillus (Bilberry) and V. uliginosum (Bog Whortleberry) both have black berries with a blue bloom, leaves deciduous.—Oxycoccus has a polypetalous corolla with the petals projecting backwards. Anthers without appendages. O. palustris (Cranberry) has a slender, creeping stem, and is evergreen. Dark red berry.
[510]
Pollination essentially the same as the preceding order.—320 species; especially in N. Am. Some are useful on account of their edible fruits, especially Vaccinium myrtillus and V. vitis-idæa, and in a less degree Oxycoccus, etc. The fruits of V. myrtillus are officinal.
Fig. 546.—Vaccinium uliginosum (var. microphyllum). The parts of the flower A-E are enlarged 5–6 times; C and E are longitudinal sections; B and D the flower seen from above; F and G a stamen seen from the back and front; H the style and stigma.
The flowers are regular, gamopetalous, typically diplostemonous, with the same number throughout all 5 whorls, thus: Sn, Pn, An + n, Gn, where n most frequently =5 (4–6), rarely 3, 7 or 8. Of the two whorls of stamens the one opposite the sepals is often present only as rudiments or is entirely suppressed, and the completely developed stamens are thus placed opposite the petals. The carpels are generally placed opposite the sepals. The ovary is multilocular with the ovules attached in the inner angles. The fruit is most frequently a berry. The seeds are large, generally solitary, or a few in each loculus.—All plants belonging to this family are trees or shrubs with scattered, single, most frequently entire, penninerved and leathery leaves without stipules; the majority are tropical (America, Asia), some are found in N. Am. and the Mediterranean.
Order 1. Sapotaceæ. Plants with latex; anthers extrorse, 1 erect ovule in each loculus; fruit a berry; the seeds with bony, shiny brown testa have a large, lateral hilum. The leaves are frequently covered with silky hairs.—A[511] useful order in several respects (400 tropical species). The wood of some genera, such as Sideroxylon (Iron wood) and Bumelia, is as hard as iron. The latex of Palaquium (P. oblongifolium, P. gutta, and other species), Mimusops and Payena (Sumatra, E. Ind.), is the raw material of gutta percha. The following have very delicious fruits: Lucuma mammosa, Achras sapota, Chrysophyllum cainito (Star-apple), etc. The seeds of Bassia (E. Ind.) contain a large quantity of a fatty oil. Isonandra, Mimusops schimperi are often found in the Egyptian royal tombs.
Order 2. Ebenaceæ. Plants without latex, often diœcious; flowers with a more or less leathery perianth. The number of stamens is sometimes increased (by splitting?); ovules 1–2, pendulous in each loculus. Fruit a berry.—250 species; chiefly tropical. Some are well known on account of their hard and black-coloured heart-wood, e.g. Maba ebenus (the Moluccas) and Diospyros ebenum (Ebony-wood, from Tropical Asia) and others.—The fruits are edible e.g. of Diospyros lotus (Date-plum, Asia), which is also cultivated as an ornamental shrub, together with several other species.
Order 3. Styracaceæ. The flower is more or less epigynous, and the corolla is almost polypetalous. The stamens (by splitting?) are more than double the number of the petals, and often united at the base. Stellate hairs are frequent.—235 species; Tropical Asia and America, a few for example in the East.—Officinal: Gum-benzoin from Styrax benzoin and perhaps other species (Sumatra and Siam). Halesia tetraptera (N. Am.) is an ornamental shrub with 4-winged fruits.
Fig. 547.—Diagram of Primula.
The flowers are regular, ☿, hypogynous, and gamopetalous. The stamens are equal in number to the petals (Fig. 547) and are placed opposite to them. The ovary is unilocular, with a free, central placenta with 1–many ovules.—The flower is a further development of the Diospyrinæ; the suppression of the calyx-stamens, which commenced in this family, is carried further in the Primulinæ, so that in the majority of cases no trace of them is present, but in certain species and genera (Samolus, Lysimachia thyrsiflora, Soldanella, certain Myrsineæ) some small bodies (scales, teeth, etc.) are found in the position of the suppressed stamens. Again, the lateral portions of the carpels are suppressed, so that the ventral placentæ with the ovules are separated from the dorsal portions, and are united into a free central placenta; this theory is supported by the branching of the vascular bundles, the development, and various comparative considerations.—Sn, Pn, A0 + n, Gn; where n = 4–8, generally 5. The carpels are placed opposite the sepals (Fig. 547).
[512]
Order 1. Primulaceæ (Primroses). This order has many ovules attached to a thick, free, central placenta (Fig. 547); style undivided with a capitate stigma; ovules semi-anatropous; fruit a capsule with many seeds.
All the plants belonging to this order are herbs; stipules wanting; the flower is most frequently 5-merous (S5, P5, A0 + 5, G5; except Centunculus and Trientalis). The corolla and capsule have various forms, but the capsule generally opens by teeth at the apex. The ovules are semi-anatropous (in Hottonia they are anatropous), and the seeds are therefore peltate, with the hilum situated in the centre of one side. The endosperm is fleshy or horny. The flowers are borne either in racemes or in umbels; as bracteoles are typically absent (Fig. 547), cymose branching does not occur.
Fig. 548.—Primula: dimorphic flowers. A short-styled; B long-styled.
Fig. 549.—Cyclamen persicum.
Primula (Primrose) has most frequently a vertical rhizome, bearing a rosette of leaves at its summit, and long-stalked umbels; corolla rotate or slightly funnel-shaped; the capsule opens at the apex by 5 teeth. The flowers in some species are heterostyled (long-styled or short-styled; Fig. 548). Closely allied are Androsace (with ovate, cup-shaped corolla-tube and ligular scales, alternating with the corolla-lobes) and Soldanella (funnel-shaped corolla with laciniate lobes and most frequently ligular scales).—Hottonia (Water-Violet) is an aquatic plant with pectinate leaves and heterostyled flowers.—[513]Cortusa. Dodecatheon. Cyclamen (Fig. 549) has solitary, long-stalked flowers, and a rotate corolla with the lobes reflexed; the stalk of the capsule rolls up spirally; the tuberous rhizome is formed by the hypocotyledonary internode. Only 1 cotyledon.—Lysimachia (Money-wort); stem-internodes well developed, leaves opposite or verticillate, calyx almost polysepalous, corolla deeply 5-partite (Fig. 550). The flowers are solitary or in racemes.—Anagallis (Pimpernel), leaves opposite, flowers solitary; the fruit a pyxidium (Fig. 551); similarly in Centunculus, which is 4-merous.—Trientalis, the flowers are most frequently 7-merous.—Glaux (Sea Milk-wort) is a creeping maritime plant with opposite leaves; flowers solitary in the leaf-axils, corolla absent, but with coloured calyx. The petals are usually developed later than the stamens in the Primulaceæ; but in this instance they are entirely suppressed.—Samolus (Brookweed) differs from all the others in having an epigynous flower; barren sepal-stamens are also present. The bracts in the racemose inflorescences are displaced along the flower-stalks.
Fig. 550.—Lysimachia thyrsiflora.
Fig. 551.—Anagallis arvensis. Fruit dehiscing.
Pollination. Insect-pollination in the majority; cross-pollination is promoted in some by heterostyly (Fig. 548).—300 species; especially in northern temperate zones; the majority on mountains (Soldanella, Androsace, etc.); almost absent in the Tropics. A large number are ORNAMENTAL PLANTS, e.g. Primula auricula (from the Alps), P. sinensis (China), P. elatior (Oxslip, a native) and grandiflora, etc. Cyclamen europæum (Alpine Violet); the tubers are poisonous.
Order 2. Myrsinaceæ. Trees or shrubs; evergreen, tropical Primulaceæ with fleshy fruits and few seeds, embedded in the placenta. The leaves are nearly always dotted with yellow glands (schizogenous resin-receptacles).—550 species; especially Am.—Ornamental plants: Ardisia crenulata (W. Ind.); other genera: Clavija, Maesa, Theophrasta (barren sepal-stamens), Myrsine, Jacquinia (barren sepal-stamens), etc.—Ægiceras, allied to this order, comprises arborescent plants, often growing with Rhizophora in tropical forests, along the shore. The embryo germinates while still in the fruit.
[514]
Order 3. Plumbaginaceæ. This order has a position of the stamens similar to that in Primulaceæ (S5, P5, A0 + 5, G5), but it differs from these in the flower, which has generally a membranous, dry, thin, coloured, folded, almost entire calyx and an almost entirely polypetalous corolla, which, as a rule, has twisted æstivation and is only united with the stamens at its base; but more especially it differs in the ovary, which bears 5 free or almost free styles and only 1 basal ovule with a long, twisted funicle (the placenta of the Primulaceæ is here so much reduced that it bears only 1 ovule). The fruit is a nut or capsule. The radicle is turned outwards. Endosperm mealy.—To this order belong herbs or under-shrubs, which are especially natives of the sea-coast and of salt-steppes; they also resemble the Primulaceæ in the scattered, undivided, entire leaves (without stipules), often in rosettes, and the inflorescence borne on a long stalk. In opposition to the Primulaceæ, the bracteoles are typically present, and hence the branching is generally cymose (scorpioid).
Armeria (Thrift) has a round capitulum, composed of closely-packed dichasia, surrounded at its base by an involucre with peculiar prolongations, directed downwards, and united into a sheath protecting the intercalary zone of growth. The pericarp is finally ruptured at the base, and drops off like a hood.—In Statice (Sea-lavender), the unipared scorpioid cymes are prolonged and collected into panicle-like inflorescences.—Plumbago is the genus which approaches nearest to the Primulaceæ, and differs most from the characters given above. It has capitate or spike-like inflorescences, a salver-shaped corolla, and the stamens are not attached to the corolla. The style is only divided at the extremity; the calyx is not membranous, but is covered with sticky, glandular hairs.
250 species; chiefly in the Mediterranean and about the Caspian Sea, on salt-steppes and beaches. Some are Tropical; a few are ornamental plants.
The flower is regular, ☿, and hypogynous. The gamopetalous type is present in this family with great uniformity, without suppression or splitting; S5, P5, A5, G2 (3–5). The stamens are all fertile, alternating with the lobes of the corolla. Gynœceum with 2, more seldom 3–5 syncarpous carpels. Style nearly always simple; 2 dorsal stigmas. In each carpel 2–∞ ovules. At the base of the[515] ovary is found a yellowish ring-like nectary (Fig. 552 C), sometimes 5-sinuate or 5-partite.—The leaves are nearly always scattered; stipules are absent.—The Solanaceæ, which formerly were classed here, are so closely allied to the Personatæ, that it would be unnatural not to place them first in this family; and the Boraginaceæ (which were also placed in the Tubifloræ) appear to be best united, with the Labiatæ and others, into one family Nuculiferæ.
Order 1. Polemoniaceæ. The flowers are regular; S5, P5, A5, G3. The calyx and corolla have united leaves, the petals twisted to the right in æstivation (all the left edges being covered). The ovary is 3-locular with 2–∞ ovules in each loculus; the style is trifid at the apex; the fruit is a 3-valved capsule. Embryo straight; endosperm fleshy. The inflorescences are dichasia passing over into unipared helicoid cymes (the shoot of the lower bracteole being the more strongly developed).—Herbs without latex. 150 species; especially Western N. Am.—Phlox (salver-shaped corolla; entire, opposite leaves), Polemonium (campanulate or almost rotate corolla; scattered, pinnate leaves), Leptosiphon, Gilia, Collomia, Cobæa (climbing, like the Vetches, by tendrils at the ends of the leaves), etc. They are frequently ornamental plants.
Order 2. Hydrophyllaceæ. This order approaches very closely to the Boraginaceæ. Herbs with pinnate or palmate leaves; S5, P5, A5, G2. The lobes of the corolla are imbricate in æstivation. Generally 2 median carpels. The ovary is most frequently unilocular, and the seeds are situated on 2 parietal placentæ; capsule 2-valved; embryo straight; endosperm fleshy. In the corolla-tube, opposite the corolla-lobes, there are frequently appendages of various forms, which resemble those of Cuscuta. The inflorescences correspond exactly with those of the Boraginaceæ, being unipared scorpioid cymes, which, prior to opening, are tightly rolled up.—130 species, especially in N. Am. (California, etc.). Many annual species of Phacelia, Nemophila, Whitlavia, Eutoca, Cosmanthus, etc., are cultivated in gardens as ornamental plants. Hydrolea (has a bilocular ovary, and two free styles).
Order 3. Convolvulaceæ (Bindweeds). The flower is regular, hypogynous, with 5 almost free sepals (quincuncial æstivation), P5, A5, G2 (rarely 3–5). The corolla is very characteristic; it is (with various forms) almost entire, or slightly 5-lobed, and folded longitudinally in the bud in such a way that 5 projecting, flat portions, tapering towards the top and frequently differing in colour and hairiness from the rest, are visible externally and applied close together, while the remainder of the corolla is folded inwards (Fig. 552 A); and hence the whole corolla is strongly twisted to the right in the bud. The gynœceum most frequently has a bilocular ovary; in each loculus there are only 2 (erect) anatropous ovules on the placenta, which is not especially thickened (Fig. 552 D, E); each loculus is sometimes divided into two by a false septum (a relationship with the Boraginaceæ, etc.); style simple with[516] most frequently a bilobed stigma, or a bipartite style. The fruit is nearly spherical, most frequently a capsule. The seeds are erect, and have a large hilum at the base. The embryo is curved, with leaf-like, thin, bilobed, most frequently folded cotyledons; endosperm absent or mucilaginous.
1. Convolvuleæ, Bindweed Group. The majority are twining (to the left) herbs, with latex. The leaves are scattered, without stipules, often long stalked, and nearly always with cordate base; some are palmately lobed. The flowers are most frequently solitary in the leaf-axils, large, quickly withering.—Convolvulus (Fig. 552), Calystegia (unilocular ovary, 2 large bracteoles), Ipomœa, Batatas, Evolvulus (with a doubly bifid style), Calonyction, Pharbitis, etc.
Fig. 552.—Convolvulus scammonia.
2. Dichondreæ. This group is a more primitive form, not twining, and without latex. It has 2 free carpels with basal style (as in Boraginaceæ) and valvate corolla.
3. Cuscuteæ, Dodder Group (Fig. 553). Parasites, with round, filamentous stems, bearing only scale-like leaves and almost destitute of chlorophyll (they are reddish or yellowish); they are parasitic upon other plants, around which they twine, first with narrow, compact coils from which haustoria (Fig. 553 A) are developed which enter the host-plant, and then with wider coils by which they raise themselves to other portions of their host or try to reach other plants. On germination a very temporary primary root is developed, which bears root-hairs as far as the tip (rootcap is wanting); it only serves as a kind of reservoir for water, and perishes very soon after the seedling has fastened on to a host. The embryo is filamentous and rolled up spirally (Fig. 553 C), and[517] is sometimes destitute of cotyledons. The flowers are crowded into capitulate inflorescences, complicated by accessory shoots (Fig. 553 A); they have S5, P5 (imbricate æstivation), A5 (and beneath the stamens 5 scales on the corolla-tube), G2. Fruit a capsule opening by a lid.—Cuscuta europœa, C. epilinum (Flax-Dodder), C. epithymum (Lesser-Dodder), C. trifolii (Clover-Dodder), etc., are parasitic on different hosts, or parasitic each on its own particular host.

Fig. 553.—Cuscuta trifolii, parasitic on Red Clover. A A portion of the stem with an inflorescence and haustoria (mag.); B seed (nat. size); C seed (mag.); D embryo (nat. size).
840 species; the majority in the Tropics, especially Am. Many are ornamental plants. Officinal: some on account of their purgative properties: the tuberous roots of Ipomæa purga (Jalap, from Mexico) and the dried latex (“Scammony”) of Convolvulus scammonia (from the East). The tuberous roots of Batatas edulis (Trop. S. Am.) are used as a common vegetable (Sweet Potato) in the Tropics.
The type of the flower is: S5, P5, A5 (of which one, or in some cases several, are suppressed), and G2. The flowers are[518] hypogynous, ☿, perfect with gamopetalous corolla, but most frequently irregular (medianly zygomorphic, except Solanaceæ), the corolla being bilabiate (divided into a posterior part of two lobes and an anterior part of three lobes), and the stamens 4, didynamous (the posterior being suppressed). The ovary has 2 loculi (only 1 in Utriculariaceæ, Gesneriaceæ, Orobanche); the placenta in the first-named orders (1–7) is most frequently very thick, and bears a great many ovules (Figs. 554, 555, 557, 562); the number of ovules in the last orders (8–9) is considerably reduced (Fig. 570).
Special mention may be made of the apparently 4-merous flower which is found, e.g. in Veronica and Plantago (Figs. 567, 562 C, 570, 571), and which arises from the typical 5-merous flower by the suppression of the posterior sepal and the posterior stamen, and by the union of the two posterior petals into one.—Terminal flowers very seldom occur on the main axis, and would not harmonise well with the very irregular form of the flower. When they do occur, they are, as a rule, “peloric,” i.e. regular (in Linaria vulgaris two kinds of peloric flowers occur,—one with 5 spurs, and one without spurs). The halves of the anthers are often divided as far as the base, and laterally so widely separated from each other as to assume an almost straight line (Figs. 563, 564). There is generally a nectary (“disc”) round the base of the ovary, often 5-lobed (or divided into free glands).—A common vegetative characteristic is the absence of stipules.
The 9 orders of the Personatæ are: 1, Solanaceæ; 2, Nolanaceæ; 3, Scrophulariaceæ; 4, Utriculariaceæ; 5, Gesneriaceæ; 6, Bignoniaceæ; 7, Pedaliaceæ; 8, Acanthaceæ; 9, Plantaginaceæ.

Fig. 554.—Diagram of Petunia.
Order 1. Solanaceæ. The flower (Figs. 554, 555, 559) is hypogynous, regular (zygomorphic in Hyoscyamus), ☿, and gamopetalous, with S5, P5 (most frequently imbricate or valvate), A5, G2, the 2 carpels being placed obliquely (Fig. 554); the bilocular ovary has a very thick axile placenta (Figs. 554, 555 H, 557), which extends almost as far as the wall of the ovary. The fruit is a capsule or berry; the seeds are more or less reniform, and the embryo is curved (rarely straight), in a fleshy endosperm (Figs. 555 F, G; 561).—Both arborescent and herbaceous forms are found in the order; leaves scattered without stipules,[519] but with variously formed laminæ (always penninerved). A peculiar leaf-arrangement is found in many species, viz. the leaves are borne in pairs, a large and a smaller one together; these pairs stand in 2 rows, and the flowers are then situated between the individual leaves in each pair, apparently not in a leaf axil. The inflorescences are frequently unipared scorpioid cymes without floral-leaves.

Fig. 555.—Atropa belladonna: A is reduced.]
Zygomorphic flowers occur, and thus form a transition to the closely allied Scrophulariaceæ; the zygomorphy sometimes shows itself only in the relative length of the stamens, sometimes also in the corolla (Hyoscyamus).—Nicandra is 5-merous throughout all the whorls.—The peculiar relative leaf-arrangement in this order occurs from sympodial branching and displacement. The most simple is, e.g. Datura (Fig. 556 A); each shoot-generation in the floral parts of the plant has only 2 foliage-leaves (f1 and f2), and then terminates in a flower; the axillary buds of both the foliage-leaves are developed and form a dichasium, but since the leaves are displaced on their axillary-shoots as far, or almost as far, as the first leaf of these axillary-shoots, the flowers are borne singly on the dichasial branches, and all the branches appear to be without subtending leaves (Shoot I is white, II shaded, III white, etc., diagram A). Scopolia and others (Fig. 556 B) differ in that the lowest and smallest (f1) of the two leaves on each shoot is barren, and is therefore not displaced; but the upper one (the second bracteole, f2) is displaced as in the first instance, and consequently it assumes a position near the first leaf (the shaded leaf f2 of shoot I being placed near the white leaf f1 of shoot II, etc.,) of the next youngest shoot-generation,[520] and hence the leaves are borne in pairs; the flower placed between the two leaves of a pair is therefore the terminal flower of the shoot to which the smaller of the two leaves belongs, and the larger leaf is the subtending leaf for the floral shoot itself.
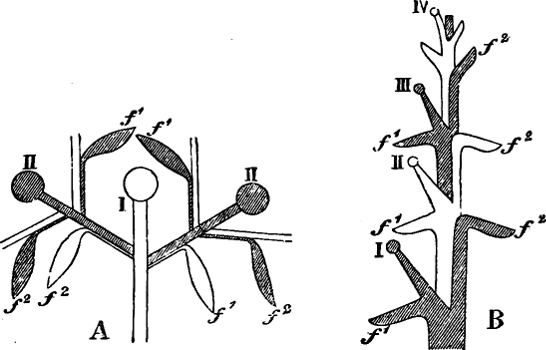
Fig. 556.—Diagrammatic representation of the branching in Solanaceæ. The various shoot-generations are white or shaded.
Fig. 557.—Fruit of Hyoscyamus niger after removal of calyx.
Fig. 558.—Fruit of Datura stramonium.
A. Fruit a capsule. Nicotiana (Tobacco) has a 2-valved capsule with septifragal dehiscence; the valves separate at the apex; the corolla is funnel-shaped, tubular, salver-shaped or campanulate. The flowers in panicles.—Datura (D. stramonium, Thorn-apple) has a (frequently spiny) capsule (Fig. 558), which is falsely 4-locular (at the top, bilocular) and opens septifragally with 4 valves. The lower part of the calyx persists as a thick collar[521] (see Fig. 558). The corolla is funnel-shaped. The flowers are solitary, large.—Hyoscyamus (H. niger, Henbane) has a pyxidium (Fig. 557) enclosed in the campanulate, completely persistent, thick-walled calyx. The flowers are slightly zygomorphic, and borne in unipared scorpioid cymes. Scopolia (pyxidium); Fabiana (Heather-like shrub); Petunia (slightly zygomorphic flower; funnel-shaped corolla); Nierembergia; Brunfelsia (almost a drupe); Franciscea; Browallia.—Among those with capsular fruits are found the most anomalous forms, which by their zygomorphic flowers and often didynamous stamens present the transition to the Scrophulariaceæ: Salpiglossis; Schizanthus (lobed petals; 2 perfect, and 3 rudimentary stamens).
Figs. 559–561.—Solanum tuberosum.
Fig. 559.—Flower (1/1).
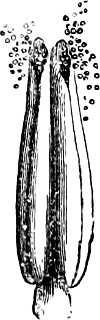
Fig. 560.—Stamen, ejecting pollen.
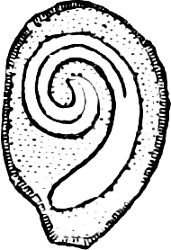
Fig. 561.—Longitudinal section of seed.
B. Fruit a berry. Solanum (Nightshade); rotate corolla (Fig. 559). The stamens have short filaments, the anthers stand erect, close together round the style, like a cone in the centre of the flower, and open by pores at the apex (Fig. 560). S. tuberosum (the Potato-plant); the Potato-tuber is a swollen, underground stem; the “eyes” are buds, situated in the axils of its scale-like, quickly-perishing leaves.—Lycopersicum resembles Solanum in the flower, but the united anthers open by longitudinal clefts and have an apical appendage. The cultivated species, L. esculentum (Tomato), has often a higher number than 5 in the flower, and in the fruit several loculi of unequal size.—Physalis (Winter Cherry); the calyx ultimately swells out in the form of a bladder, becomes coloured, and loosely envelopes the spherical berry.—Capsicum (Guinea Pepper-plant); some species have very large, irregular, rather dry (red, yellow, black) berries, which are unilocular in the upper part.—Lycium (false Tea-plant); the corolla is salver- or funnel-shaped; shrubs; often thorny.—Atropa (A. belladonna, Deadly Nightshade,[522] Fig. 555); corolla campanulate; the calyx projects beneath the spherical, black berry. The flowers are borne singly.—Mandragora; (Mandrake); Nicandra (ovary often 5-locular).—A small tropical group: Cestreæ (Cestrum, Habrothamnus, etc.) has an almost straight embryo, which may also be found e.g. in species of Nicotiana. Related to the Scrophulariaceæ.
About 1,500 species; the majority within the Tropics, outside these limits especially in America. Solanum nigrum is a common weed.—The Potato-plant (Solanum tuberosum), from Peru and Chili, was introduced into Europe in 1584 by Sir Walter Raleigh. (Potatoes = Batatos). The fruits of several serve as condiments: Chilies or Pod-pepper (Capsicum annuum and longum), and the Cayenne-pepper (C. baccatum and others), whose fruits also are officinal, were brought to Europe from S. America by Columbus, and are commonly cultivated in Tropical America; Lycopersicum esculentum (Tomato) and others from Peru; Solanum ovigerum (Egg-plant); Solanum melongena, etc. Poisonous, acrid, narcotic properties (alkaloids, etc., solanine, nicotine, atropine, hyoscyamine) are found in many: Atropa belladonna (from S. Europe; the roots and leaves are officinal); Solanum dulcamara (Bitter-sweet; formerly officinal), S. toxicarium (Guiana); Datura stramonium from Asia (leaves and seeds officinal), D. sanguinea, metel, tatula, and others; Hyoscyamus (officinal: the leaves and seeds of H. niger); Nicotiana tabacum (Virginian tobacco, officinal: the leaves), N. rustica and others from Trop. America (Tobacco was introduced into Europe in 1560); Cestrum-species. Duboisia myoporoides (Australia); the leaves contain hyoscyamine and are used in medicine. A number of species of these genera are ornamental plants.
Order 2. Nolanaceæ. These most resemble the Convolvulaceæ in the corolla, but the Solanaceæ in their branching, and leaf-arrangement (in pairs, etc.). The diagram is the same as in Nicandra with 5 carpels, but the fruits of this order most frequently form, by invaginations in various directions, an ovary (with 1 style) consisting of numerous and irregularly grouped, 1-ovuled cells; the fruit is a schizocarp with many 1-seeded fruitlets.—Nolana (Western S. America): a few are ornamental plants.
Order 3. Scrophulariaceæ. The flower is hypogynous, ☿, zygomorphic, with the usual type: S5, P5, A5, and G2, the latter placed in the median plane; some genera have all 5 stamens developed (Fig. 562 A), but most frequently the posterior one is suppressed and the flower becomes didynamous (Fig. 562 B). The fruit, as in the capsular-fruited Solanaceæ, is a bilocular, 2-valved capsule, with a thick, axile placenta, and most often septicidal dehiscence (Fig. 563 C). The numerous seeds are not reniform as in many Solanaceæ, and have a straight, or only slightly curved embryo, with abundant endosperm (Fig. 563 D).—The majority are herbs; some are arborescent; the leaves are opposite or scattered, but stipules are wanting as in the whole family.
The Scrophulariaceæ are closely allied to the Solanaceæ, and there is, properly[523] speaking, no characteristic feature which absolutely separates them. The somewhat irregular corolla, with five stamens of unequal length in Verbascum, is also found in Hyoscyamus; curved and straight embryos are found in both orders. The activation of the corolla in the Scrophulariaceæ is simple imbricate, in the Solanaceæ most frequently folded imbricate (in Atropa and those allied to it, imbricate without folding). The genera (about 164) are distinguished according to the form of the corolla, number of stamens, inflorescence, arrangement of the leaves, etc. Verbascum belongs to the most primitive 5-stamened forms, and from it proceed a long series down to Veronica, with only two stamens and most frequently the posterior sepal suppressed.
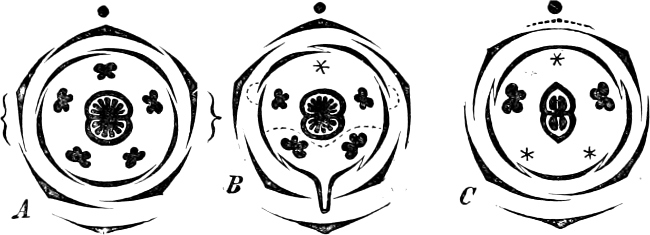
Fig. 562.—Diagrams. A Verbascum; B Linaria; C Veronica.
1. Antirrhineæ, Snapdragon Group. This has most frequently a descending æstivation of the petals (the posterior petals are outside the lateral ones, which again enclose the anterior; Fig. 562 A, B). The plants belonging to this group are not parasites.
a. 5-stamened.—Verbascum (Mullein, Fig. 563 A) has a slightly irregular, rotate corolla; five stamens (frequently covered with woolly hairs), of which the two anterior ones are the longer and differ often also in other respects. The inflorescences are racemose, often with several series of accessory dichasia in the axil of each primary floral-leaf. The leaves are scattered and, together with the stems, are often covered with a grey felt of branched hairs.
Fig. 563.—Verbascum thapsiforme.
[524]
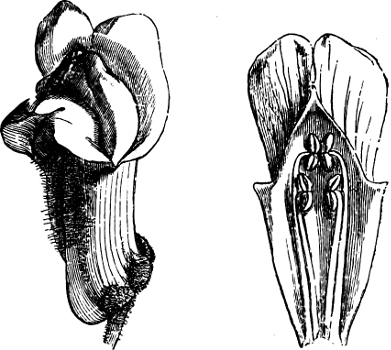
Fig. 564.—Antirrhinum majus. A flower, and the upper lip of a flower with the stamens.
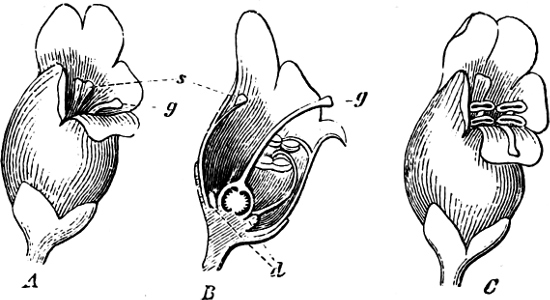
Fig. 565.—Scrophularia nodosa. Protogynous flower in various stages: A ♀ stage; g the stigma projecting from the throat of the corolla; B the same in longitudinal section; C ♂ stage, the stigma is bent down and its former position occupied by the stamens; s staminode; g stigma; d nectary.
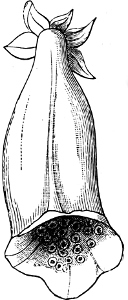
Fig. 566.—Digitalis purpurea.
b. 4-stamened, didynamous (Fig. 564).—Scrophularia (Fig-wort, Fig. 565) has cymose inflorescences in a panicle; the corolla (Fig. 565) is urceolate, short two-lipped; the posterior stamens are present as a scale below the upper lip of the corolla (Fig 565 s). S. nodosa has a tuberous rhizome.—Pentstemon; the posterior stamen is barren and very long.—Antirrhinum (Snapdragon). The corolla (Fig. 564) is personate, i.e. bilabiate, but with the under lip arched to such an extent that it meets the upper lip, closes the corolla throat, and entirely conceals the stamens and[525] style; the corolla-tube is produced into a short pouch at the base on the anterior side. The capsule is oblique and opens by 2–3 pores, formed by small, dentate valves. In Linaria (Toad-flax) the pouch is produced into a spur. Sometimes there are traces of the posterior stamens. The capsule opens by large pores (one for each loculus), produced by large, many-partite valves. L. vulgaris reproduces by suckers.—Digitalis (Foxglove, Fig. 566) has long racemes with drooping flowers; the posterior sepal is small (a step towards complete suppression, as in Veronica); the corolla is obliquely campanulate, and generally nearly 4-lobed, the two posterior petals coalescing.—Alonsoa; Nemesia; Chelone; Herpestis; Mimulus; Torenia; Vandellia; Limosella (L. aquatica, Mud-wort, native); Scoparia; Capraria; Erinus (found on the Roman Camp at Chesters, Northumberland, and supposed to have been introduced from Spain by the Roman soldiers); Celsia (near Verbascum); Maurandia; Lophospermum; Rhodochiton; Collinsia; Nycterinia, etc.
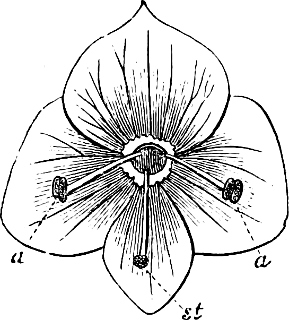
Fig. 567.-Flower of Veronica.
c. 2-stamened.—Gratiola (Water-hyssop). 5-partite calyx. The upper lip of the corolla is undivided or slightly bifid; the two anterior stamens are either entirely absent or are reduced to staminodes (a transition to Veronica).—Veronica (Speedwell), most frequently 4-partite calyx; 4-lobed, rotate, zygomorphic corolla with 2 perfect stamens and no trace of the others (Figs. 567, 562 c); capsule with loculicidal dehiscence. Calceolaria; the corolla has two slipper-like lips.
2. Rhinantheæ, Yellow-rattle Group. Herbs, all of which (with the exception of Lathræa) are annual parasites with green foliage-leaves. They attach themselves by haustoria to the roots of other plants and draw nourishment from them. The majority turn black when dried. Racemose inflorescences. In many the calyx is 4-partite, the posterior sepal being absent, or very small. The corolla is distinctly bilabiate (Fig. 568), with most frequently ascending æstiration; in the majority it does not become detached at the base, but by means of a ring-like cut some distance up the tube; 4 didynamous stamens; pollen-grains dry, easily falling out; the anthers are often furnished at the base with bristles or hairs (Fig.[526] 568) which play a part in the pollination, the probosces of the insects, being forcibly pushed against them, agitate the anthers and shake out the pollen-grains. Capsule with loculicidal dehiscence.—Euphrasia (Eye-bright), Melampyrum (Cow-wheat), Rhinanthus (Yellow-rattle), Odontites (Bartsia), Pedicularis (Louse-wort), and Lathrœa (Tooth-wort) all have native species. The last named is pale yellow, or reddish (without chlorophyll); it is a parasite on the roots of the Hazel, Beech and other shrubs, having an aerial stem, and an underground, perennial rhizome, covered with opposite, scale-like, more or less fleshy leaves with a number of internal glandular, labyrinthine cavities. The inflorescence is a unilateral raceme. It approaches Gesneriaceæ in having a unilocular ovary with two parietal placentæ.
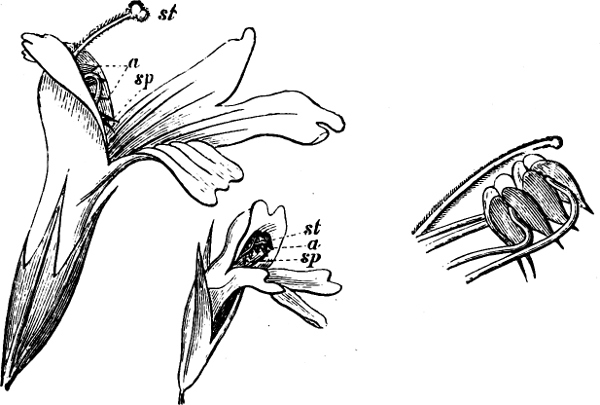
Fig. 568.—Euphrasia officinalis. Flower of the large and the small-flowered forms; showing the anthers and stigmas.
The mechanical contrivances for POLLINATION are so numerous that no general principle can be laid down. Personate flowers, like those of Antirrhinum are only accessible to strong insects, such as humble-bees, which can force themselves between the two lips, and so become dusted with pollen on the back. In Euphrasia and other Rhinantheæ the insects become covered with smooth, powdery pollen when they shake the anther-apparatus in touching the hairs and bristles mentioned above. Scrophularia nodosa is protogynous (Fig. 565). Digitalis purpurea, however, is protandrous. Mimulus luteus and some others have sensitive stigmatic lobes, which shut up on being touched. The Veronica-species constitute a series, from large-flowered down to small-flowered forms, and parallel with them are found various gradations from insect-to self-pollination. In some (as Euphrasia officinalis, Rhinanthus crista galli) there are two kinds of flowers: large, which are pollinated by insects, and small, which are self-pollinated (Fig. 568). Lathræa squamaria (Tooth-wort) is a protogynous spring-flowering plant, largely visited by humble-bees. Others have cleistogamic flowers. Nycterinia capensis opens its flowers at night.
[527]
2,000 species; chiefly from the Temp. Officinal: Digitalis purpurea (the leaves; Europe), a poisonous plant. Verbascum thapsus and thapsiforme, Veronica officinalis (“Herba V.”), Gratiola officinalis (“Herba”) have medicinal uses. The whole of the Scrophulariaceæ are more or less suspicious, if not actually poisonous, and none serve as food. Many are ORNAMENTAL PLANTS: Mimulus luteus (N. America), Paulownia imperialis (the only species; in Japan; a tree), Antirrhinum vulgare (S. Eur.), Linaria, Pentstemon, Veronica, Calceolaria (Peru, Chili, etc.).
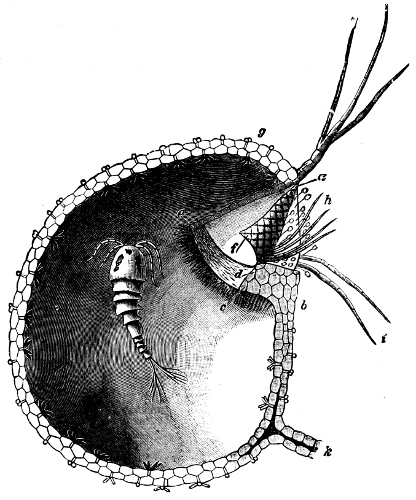
Fig. 569.—Leaf of Utricularia vulgaris, with bladder. Median longitudinal section through a bladder containing a Cyclops. At a a hair of the upper-lip, at i 2 bristles of the under-lip of the entrance (a, b); in the latter are placed 4 bristles h; k stalk of the bladder, in which is seen a vascular bundle. (After Cohn.)
Order 4. Utriculariaceæ. To this order belong only perennial, insectivorous, aquatic, and marsh-plants (200 species) with a more or less characteristic appearance. They differ from the Scrophulariaceæ, especially in having 2 stamens (the anterior) and a unilocular ovary, with free, central placenta (like that of the[528] Primulaceæ). For the rest the flower is distinctly bilabiate, both in the calyx and corolla. Two-valved capsule; no endosperm.
Pinguicula (Butter-wort) has a rosette of leaves close to the ground; these are sticky, covered with glandular hairs, and roll round any small insects which may be caught upon them; flowers solitary, terminal on a long scape; calyx, 5-partite; corolla with spur. The embryo germinates with 1 cotyledon.—Utricularia (Bladder-wort). Our native species are floating, without roots, with hair-like, divided leaves, studded with peculiar bladders (in the Tropics there are terrestrial species, with ordinary foliage). The bladders (Fig. 569) have an aperture, closed by a valve opening inwards, so that small aquatic animals are allowed to enter, but are not able to escape; they are thus entrapped in the bladders, and are probably used as food. Calyx bipartite; corolla personate with spur.
The embryo of Utricularia is very imperfect, scarcely more than a spherical, cellular mass, with a few slight leaf-rudiments. On the germination of U. vulgaris, several bristle-like leaves develop into a compact rosette; the stem then develops, and also the finely-divided, bladder-bearing leaves. A primary root is not developed. The stems branch copiously and in a very peculiar manner. The growing-point of the stem is rolled spirally.—The stigmatic lobes are sensitive and close on being touched; self-pollination often takes place, however, in Pinguicula.
Order 5. Gesneriaceæ. The flower in this order may be both epigynous (Gesnerieæ) and hypogynous (Cyrtandreæ), but otherwise is nearly the same as in Scrophulariaceæ, only that the ovary is unilocular, with 2 parietal, often bifid, placentæ. Of the 5 stamens the posterior is rudimentary, or (more rarely) entirely wanting, and the others are didynamous (Cyrtandreæ have often only 2 stamens); their anthers are generally glued into a quadrangular mass. The majority are herbs with juicy stems, opposite, verticillate or scattered leaves without stipules, often, like the stems, thick and juicy, soft-haired or glabrous. The corollas are often highly-coloured (scarlet, red-yellow, etc., and spotted internally), large and magnificent, so that many species are ornamental plants. Gesnerieæ (often epigynous) have endosperm; S. Am.—Cyrtandreæ, hypogynous, without endosperm; Asia, S. Africa.—Streptocarpus, neither the primary root nor primary shoot attains development; one of the cotyledons dies, while the other grows and becomes a very large foliage-leaf, from which spring adventitious roots and adventitious inflorescences.
500 species. Gloxinia, Achimenes, Gesneria, Alloplectus, Tydæa, Columnea, Nægelia, Æschynanthus, and others, especially in the forests of tropical America. Some are epiphytes on trees, others prefer the leaf-mould of the forest and crevices of cliffs. Several genera have peculiar, catkin-like, underground shoots, with scale-like compact leaves; others have tubers.
Orobanche (Broom-rape) is allied to this order as a parasitic form. It is a parasite on the roots of other plants, not like Lathræa by means of thin rootbranches[529] with haustoria, but growing with the base of its stem in close contact with its host, and probably even often protruding a kind of thallus into it, in a manner similar to the Loranthaceæ. Its aerial shoots are not entirely destitute of chlorophyll, but are not green; they only bear scale-leaves and terminate in a raceme or spike-like inflorescence.—Some Orobanche-species are detrimental to various cultivated plants (Hemp, Lucerne, Tobacco, etc.). The flowers are strongly zygomorphic; the posterior sepal is often wanting, and the anterior are united to the two lateral ones. Ovary unilocular, as in Gesneraceæ, with 2 or 4 parietal placentæ.—The exceedingly small seeds have a very rudimentary embryo, formed of an ellipsoidal, cellular mass, without indication of cotyledons or other organs.—About 100 species; especially in the Mediterranean region.
Order 6. Bignoniaceæ. 500 species; nearly all trees and shrubs, and to a great extent lianes, climbing by tendrils (modified leaves), which are sometimes terminated by a special clasping apparatus. These lianes have, as a rule, an anomalous stem structure, the wood being either divided into four wedges at right angles to each other, separated by four grooves filled with secondary wood-parenchyma, or a greater number of wedges occur, by the cambium ceasing to form wood in several places. The leaves are most frequently opposite and compound; the flowers in the main are similar to the didynamous Scrophulariaceæ, and especially resemble those of Digitalis purpurea; they are bilabiate, large, and beautiful, campanulate or trumpet-shaped, many of the prettiest ornamental plants in the Tropics belonging to this order. The fruit is most frequently a large, woody, 2-valved, siliqua-like, septifragal capsule, whose valves separate from the flat and broad partition-wall, which bears the large, generally winged seeds: Tecoma; Bignonia.—In gardens: Catalpa syringæfolia (Trumpet-wood); Tecoma radicans (from S. Am.).—“Palisander”-wood is from Jacaranda (S. Am.).—Eccremocarpus (N. Am.) forms, by its unilocular capsule, a transition to the Gesneriaceæ (E. scaber; herbaceous).
Crescentia is allied to this order; C. cujete (Calabash) is its best known species. The fruit (unilocular with 2 parietal placentæ) is a very large, spherical or ellipsoidal berry, with a firm, finally woody outer layer. After the removal of the juicy interior, these are commonly used as drinking vessels in Tropical America.
Order 7. Pedaliaceæ. Sesamum (orientale and indicum); very important oil-plants, which from olden times have been cultivated in tropical Asia and Africa for food and as medicinal plants, and are now cultivated in America also. The seeds are used as a raw material in the manufacture of soap in Europe.—To this order also belong Martynia and Craniolaria, which have a long horned capsule and sensitive stigmas.—46 species.
Order 8. Acanthaceæ. 1,500 species; mostly erect, slender, branched herbs or shrubs, rarely arborescent, especially in S. Am. and Ind. The branches frequently have swollen nodes; the leaves are opposite, penninerved, undivided, more or less lanceolate or elliptical, and generally leave a distinct scar when they fall off. Stipules are wanting. The flowers are solitary or in dichasia, which are arranged in 4-rowed spikes or racemes, each flower with its subtending bract, which may be brightly coloured, and most frequently also with two bracteoles. With regard to the corolla (which is often labiate, in any case irregular, and frequently prettily coloured), the 2 or 4 didynamous stamens[530] (of whose anthers one half is inserted lower than the other, or suppressed) and the gynœceum, the Acanthaceæ are true Personatæ, approaching most nearly to the Scrophulariaceæ: they differ from the other orders especially in the fruit, which is a bilocular, 2-valved, often elastically dehiscing capsule, which never has more than 2 rows, and in some only 2 seeds in each loculus, the seeds being often compressed and borne on strong, curved or hook-like funicles (retinacula) which persist after dehiscence. Embryo curved without endosperm; radicle pointed downwards.—Cleistogamic flowers are found in several species. Cystoliths are common.
The following grow wild in Europe: Acanthus (spinosus and mollis, whose pinnatifid leaves served as models for the capitals of the Corinthian columns). The posterior sepal is the largest of all the leaves of the flower, and covers the other parts like a helmet; the 2 anterior sepals are united, and the two lateral ones are small and greenish; the corolla has no upper-lip, but only a 3-lobed under-lip. The anthers are bilocular; the filaments ultimately become very firm.—Justicia, Eranthemum, Goldfussia, Thunbergia (a twiner), Ruellia, Dicliptera, etc.—Ornamental plants in conservatories.
Order 9. Plantaginaceæ (Plantains). The flowers (Figs. 570, 571) are regular, ☿, hypogynous, with a 4-partite, persistent calyx, a gamopetalous, scarious corolla with 4 projecting lobes, 4 stamens, incurved in the bud, later on projecting considerably, about equal in length, and a bilocular ovary with one long, filamentous, undivided, feathery, papillose style (see Fig. 571). The ovary is most frequently bilocular with 1–few ovules in each loculus. An hypogynous disc is wanting. The fruit is a pyxidium with 1–few peltate seeds attached in each loculus (Littorella is in several respects an exception). All species are herbs, the majority with leaf-rosettes near the ground, and the flowers in spikes or capitula.
The labiate-like flowers are in this case entirely concealed under a regular, apparently 4-merous exterior. The structure of the flower, however, is the same as in the Scrophulariaceæ, only the reduction, which is found in Veronica (compare Figs. 562 C, 567 with 570, 571), is also present in this instance and the lobes are also more equally developed; the posterior petal corresponds to the bilobed upper-lip; the posterior stamen and the posterior sepal also are entirely wanting. In the development of the flower there is no trace of posterior sepal or stamen, and the posterior petal arises from one primordium, but the two anterior sepals arise before the lateral ones. The position of sepals and petals does not agree with that of a true 4-merous flower, which is represented in Fig. 361 E. The bracteoles are always suppressed in Plantago.
Plantago (Plantain, Rib-grass). The foliage-leaves are most frequently scattered, entire, with curved veins, arranged in a rosette close to the ground on an unlimited rhizome; the spike-like inflorescence is borne on a long scape; in some (P. psyllium)[531] the leaves are opposite on a stem with well-developed internodes, and the inflorescences are borne in their axils. The order also presents a transition from insect-pollinated to wind-pollinated flowers. The flowers are protogynous, wind-pollinated in P. major and P. lanceolata, partly also in the other species, but insect pollination also occurs, and P. media has three kinds of flowers, some of which are adapted for wind-pollination (Fig. 571), others, with short filaments, for insects. Littorella lacustris (Shore-weed) is the most reduced of the Plantaginaceæ: an aquatic plant with rosettes of round, awl-like leaves and diclinous (monœcious) flowers. In the axils of the foliage-leaves is a very short 3-flowered spike, formed by 2 sessile ♀-flowers, and above them a long-stalked ♂-flower; all the flowers are lateral, the terminal one being absent, as in Plantago. The ♂-flower is essentially the same as in Plantago, but the ♀-flower has a scarious corolla, with a narrow, 3–4-dentate mouth, which closes tightly round the nut-like fruit.
Figs. 570, 571.—Plantago media.

Fig. 570.—Diagram of Plantago media.
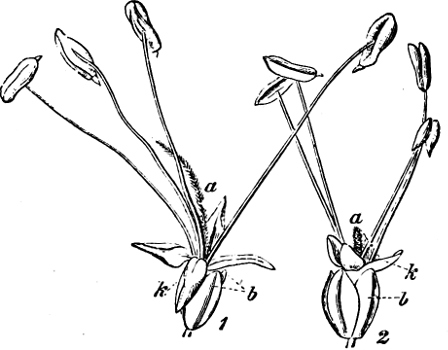
Fig. 571.—Two different forms of the flower (magnified): 1, chiefly adapted for pollination by wind; 2, for insect-pollination. a The stigma; b the calyx; k the corolla.
The genus Plantago constitutes nearly the entire order (200 species). Some are widely distributed weeds (e.g. P. major, “The white man’s footstep”). In P. psyllium (S. Eur.) the integument of the seeds is mucilaginous, and swells considerably in water.
The flowers are hypogynous and zygomorphic (in Boraginaceæ and Cordiaceæ, however, they are regular, except Echium and Anchusa arvensis). The calyx is gamosepalous, the corolla bilabiate (except in the two orders mentioned), mostly after 2/3, i.e. divided into a 2-leaved posterior portion, and a 3-leaved anterior portion.[532] The æstivation of the corolla is nearly always descending.—In Boraginaceæ and Cordiaceæ there are 5 stamens of equal length; in the other orders 4 didynamous ones, or only 2 fertile; the posterior stamen is sometimes developed as a staminode, sometimes fertile (in Stilbaceæ). The ovary is formed of 2 median carpels (except some Verbenaceæ), with (1-) 2 ovules on each carpel; in the majority of the orders it is, however, divided by a false partition-wall between the dorsal and ventral sutures, into 4 loculi, each of which is often raised independently, causing the style to be situated in the depression between the four lobes (“gynobasic” style, Figs. 572, 573, 575, 579). The fruit in these orders most frequently becomes a 4-partite schizocarp with nut-like fruitlets. The other orders have a 1(-2)-locular ovary.—The leaves are simple, without stipules.
The family is related to (and proceeds from) the Tubifloræ, especially Convolvulaceæ, which has an almost similar construction of the ovary. It is doubtful whether the Cordiaceæ and Boraginaceæ should be classed with the others.
The orders are: 1, Cordiaceæ; 2. Boraginaceæ; 3, Verbenaceæ; 4, Labiatæ; 5, Selaginaceæ; 6. Globulariaceæ; 7, Stilbaceæ.
Order 1. Cordiaceæ unites Convolvulaceæ and Boraginaceæ. Tree-like plants with 5-(4–10) merous flowers, doubly bifid style, and drupe with 4 or less loculi. No endosperm; cotyledons folded.—185 species; tropical.
Order 2. Boraginaceæ. The vegetative parts are very characteristic: herbs with cylindrical stems and scattered, undivided, nearly always sessile, entire leaves, without stipules, and generally, together with the other green portions of the plant, covered with stiff hairs, consequently rough and often even stinging (hence the other name for the order Asperifoliæ). The inflorescences are unipared scorpioid cymes with the branches coiled spirally (“helicoid,” Fig. 573) before the flowers open. The flower is perfect, regular (obliquely zygomorphic in Echium and Anchusa arvensis), hypogynous, gamopetalous: S5, P5 (often with ligular outgrowths), A5, G2, but each of the two loculi of the ovary becomes divided by a false partition-wall into two, each of which contains one pendulous anatropous ovule with the micropyle turned upwards; the four loculi arch upwards, so that the ovary becomes 4-lobed, and the style is then, as in the Borageæ, placed at the base (“gynobasic”) between the four projections (Figs. 572, 573). The fruit is a 4-partite schizocarp with four nut-like fruitlets (Fig. 572).—Endosperm is wanting (except in Heliotropium); the radicle is turned upwards.
[533]
The INFLORESCENCES are often double unipared scorpioid cymes; the bud of the second bracteole is developed, that of the first suppressed; in some cases both the bracteoles are suppressed (Myosotis, Omphalodes, etc.), but in other instances all the first bracteoles (a) only are suppressed, and the others are then situated in two rows towards the under side of the coiled axis, while the flowers are situated on the upper side. Displacement of the branches or of the floral-leaves sometimes takes place. The flowers are often red at first, and later on become blue or violet; they hardly ever have any smell. The fruit entirely resembles that of the Labiatæ, but the radicle of the latter is turned downwards. The fruitlets present small differences which have systematic importance; they are hollow or flat at the base, attached to a flat or columnar receptacle, etc.
1. Heliotropieæ. This group deviates from the characteristics mentioned above in the undivided ovary and terminal (“apical”) style. In this, as well as in the fact that in some genera (Tournefortia, Ehretia, etc.) the fruit is a drupe, it connects this order with the Cordiaceæ. Heliotropium, Tiaridium, and others have schizocarps.
2. Borageæ, Borage Group. Style gynobasic; fruit a schizocarp.
A. The throat of the corolla is without ligules, or with very small ones.—Pulmonaria (Lung-wort); funnel-shaped corolla; a whorl of hairs in the corolla-throat.—Echium (Viper’s-bugloss) has zygomorphic flowers, the plane of symmetry almost coinciding with that of the very well-developed inflorescence (through the fourth sepal); the corolla is obliquely funnel-shaped, the style is more deeply cleft at the apex than in the others; stamens 2 longer, 2 shorter, and 1 still shorter.—Cerinthe has a tubular corolla with five small teeth and two bilocular fruitlets. The bracts are large and leafy, and, like all the rest of the plant, are almost glabrous.—A few Lithospermum-species have a naked corolla-throat; others have small hairy ligules, which do not close the corolla-throat. The fruitlets are as hard as stone, owing to the presence of carbonate of lime and silica.—Mertensia (Steenhammera); Arnebia; Nonnea (small ligules).
B. The corolla-throat is closed by, or in any case provided with ligules, i.e. scale-like bodies or small protuberances, situated in the throat of the corolla opposite the petals, and which are invaginations or internal spurs of the petals (Fig. 572 D).—The nuts in Cynoglossum (Hound’s-tongue) bear hooked bristles over the entire surface, or, in Echinospermum, only on the edge. The following have smooth nuts:—Symphytum (Comfrey) has a cylindrical, campanulate corolla, and prolonged-triangular, pointed ligules.—Borago (Borage) has a rotate corolla with projecting,[534] emarginate ligules; the stamens have a horn-like appendage, projecting upwards from the back of the filament. The fruitlets are hollow below.—Anchusa (Alkanet, Fig. 572). The corolla is salver-shaped; the ligules small, hairy protuberances. A. (Lycopsis) arvensis has an S-curved corolla-tube.—Myosotis (Forget-me-not, Fig. 573); rotate corolla with small (yellow) protuberances in the throat; scorpioid cyme without floral-leaves; fruitlets flat.—Omphalodes; fruitlets hollow at the back, with a scarious, turned-in, toothed edge.—Asperugo (Mad-wort); the calyx grows after flowering, becoming large, compressed, and deeply bifid.
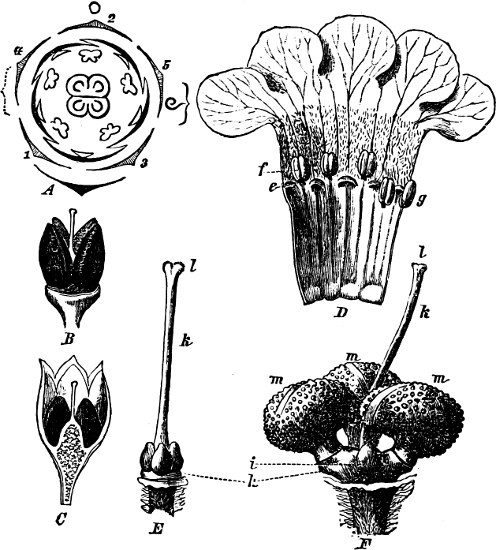
Fig. 572.—Anchusa officinalis: A diagram; the brocteole a is suppressed (dotted); β supports a flower. B, C Myosotis, the fruit, entire and with the calyx in longitudinal section. D, F Alkanna tinctoria: D the corolla opened (4/1); e the ligule; f, g the anthers; E gyncœceum (3/1); F fruit, with three fruitlets; i an aborted loculus; h disc.
Cross-pollination is most commonly effected by insects (especially bees). There are a great many contrivances for pollination; some flowers are protandrous (Echium vulgare, Borago officin.), others are heterostylous (long-and short-styled: Pulmonaria officin.); the corona (ligules) is a protection against rain, and excludes certain insects. Some are barren when self-pollinated (Pulmonaria[535] officinalis, Echium vulgare); others which have but little honey, may, failing insect-pollination, fertilise themselves, and in Myosotis versicolor this regularly occurs by the growth of the corolla during flowering, so that the anthers are brought into contact with the stigma. Honey is secreted on the hypogynous disc.—About 1,150 species, growing especially in the northern temperate zone, Mucilage is found (e.g. in the officinal root of Cynoglossum officinale, in the root of Symphytum): red dyes are found in some roots (e.g. Alkanet-root, the root of Alkanna tinctoria, which is also medicinal; S. E. Europe, Asia Minor); some are poisonous: Cynoglossum, Echium, Anchusa, etc. Several species are ornamental plants. Heliotropium (Peru) is cultivated chiefly on account of its pleasant scent; essential oils are otherwise very rare.
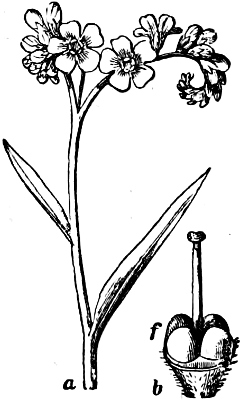
Fig. 573.-Myosotis. Inflorescence and gynœceum.
Order 3. Verbenaceæ. The majority are shrubs; a few are herbs or trees (Teak-tree); some are lianes. The branches are often square. The leaves are opposite or verticillate, without stipules; in some compound. The inflorescences are racemes, spikes, capitula, or dichasia. Five sepals; five petals in a gamopetalous, zygomorphic corolla, which is often bilabiate, but rarely to such an extent as in the Labiatæ, and the upper lip in some is larger than the under, in others smaller; stamens four didynamous, or two; the ovary is entire (not grooved or divided), 1- or 2-locular, or, as in the Labiatæ, divided into four loculi with an erect ovule in each, but in some the anterior carpel is suppressed. One terminal style. The fruit is, e.g. in Verbena, a 4 partite schizocarp with nut-like fruitlets; in Vitex (digitate leaves) a drupe with a 4-locular stone; in Clerodendron a similar fruit, with four free stones; in Lantana a bilocular stone, or two unilocular stones. The radicle is turned downwards. Endosperm small or absent.—Lippia, Stachytarpheta, Bouchea, Priva, Citharexylon, Callicarpa, etc.—The Verbenaceæ are closely allied to the Labiatæ; they differ especially in the ovary not being 4-lobed with gynobasic style, but undivided, almost spherical or ovoid with a terminal style. Again, the leaves are not so constantly opposite, and the inflorescences are various.
730 species; especially in the Tropics; there are several in America, especially Lantana-species; shrubby weeds.—Many of those mentioned are ORNAMENTAL PLANTS, especially Verbena; Vitex agnus castus is a S. European shrub. Lippia citriodora (S. Am.) etc., have strongly-scented leaves; the Teak tree (Tectona grandis) is one of the largest trees in East India, and has a very hard wood.
Avicennia is allied to this order; it inhabits the Mangrove swamps on tropical coasts. The endosperm emerges from the ovule, carrying the embryo with it; the embryo ultimately bursts the endosperm and lies free in the loculus of the fruit; this is then filled by the embryo with its large, green cotyledons, which are borne on an already hairy or rooted stem. The seedling thus developed falls from the tree, together with the fruit, and strikes root in the mud. One special cell of the endosperm at an earlier period becomes a highly-developed organ of suction, growing into a much-branched sac, very rich in protoplasm.
[536]
Order 4. Labiatæ. The special characteristics are: the square stem, the opposite leaves (without stipules), the inflorescences which are formed by two double unipared scorpioid cymes, the labiate corolla, the 4 didynamous stamens (the posterior being entirely suppressed) (Fig. 574), and the 4-partite schizocarp with nut-like fruitlets. The floral formula is S5, P5, A5 (the posterior stamen is generally absent), G2.
Fig. 574.—Diagram of Lamium album: sv dichasia.
They are chiefly aromatic plants (herbs, shrubs, e.g. Lavender, or trees), volatile oil being formed in internal cells or in the glandular hairs, which cover all green parts. The stem is always more or less markedly square; the leaves are borne upon the flat sides, and are simple and penninerved, but vary in the other characters. The inflorescences are double unipared scorpioid cymes, which may be situated at some distance from one another in the axils of the foliage-leaves (Fig. 575 A), but frequently when the subtending leaves are bract-like, they are crowded into spike-like inflorescences (Lavandula, Mentha, Salvia, etc.), each of the so-called “whorls” (verticillaster, glomerulus) being a double unipared scorpioid cyme (Fig. 574). (Solitary flowers are found in e.g. Scutellaria, and Origanum). The calyx is strongly gamosepalous, 5-toothed, often bilabiate (Fig. 575 B). The corolla is strongly bilabiate (Figs. 575, 576, etc.), with 2 lobes in the upper lip and 3 lobes in the under lip (an approach to regularity occurs only when the upper lip is small, and thus resembles one lobe, as in Mentha (Fig. 578) and Lycopus, so that the corolla approaches the 4-merous corolla of Veronica and Plantago). The posterior stamen in the diagram (Fig. 574*) is entirely suppressed; in most of the genera the posterior lateral stamens are the smaller (Fig. 575 D), and are entirely suppressed in some (see below); in others, e.g. Nepeta, they are the longer. 2 stamens are found in Salvia, Rosmarinus, Lycopus, etc. The two halves of the anthers are often separated from one another, and are placed at an angle with each other. The gynœceum has 1 style with a bifid extremity (Fig. 575 C) bearing the stigma; the true bilocular ovary is divided by a false partition-wall into 4 loculi, each with 1 erect ovule (Fig. 575 H). These 4 loculi project so strongly that the ovary becomes deeply 4-lobed with the style situated in[537] the centre of the lobes and at their base, “gynobasic” (Figs. 575, 579). A ring-like, often crenate, nectary surrounds the base of the ovary (Fig. 575 G, H). The embryo in this order, as in the Verbenaceæ, is directed downwards (Fig. 575 J) (it is directed upwards in the Boraginaceæ, which have an entirely similar fruit). Endosperm absent.
Fig. 575.—Thymus vulgaris.
The 142 genera are mainly distinguished according to the form of the calyx and corolla, the number, direction, and length of the stamens, the forms of the nuts, etc.
1. Ajugeæ, Bugle Group. Calyx 10-nerved; the upper lip is small; 4 stamens. The ovary is not so strongly lobed as in the following group, so that it is most nearly allied to the Verbenaceæ. The nuts are reticulately wrinkled. Ajuga (Bugle) has a very[538] small upper lip. The upper lip of Teucrium (Germander) is deeply cleft, and the two lobes are bent on their respective sides towards the under lip, which in consequence appears to be 5-lobed, and the upper lip to be wanting.
Fig. 576.—Lamium album: A lateral view of flower; B longitudinal section; C ovary with nectaries (a); D the apex of the style; e, upper lip of corolla; c, b, c the three petals of the lower lip; f anthers; g stigma.
2. Stachydeæ, Betony Group. The calyx is 5- or 10-nerved. The upper lip of the corolla is most frequently strongly arched or helmet-shaped; 4 stamens, the anterior pair the longer (Fig. 576).
a. A somewhat regular and 5–10-dentate calyx with projecting stamens.—Stachys (Betony, Woundwort); the lobes of the under lip are rounded off. The anterior filaments, after pollination, bend outwards. Betonica—Ballota (Horehound); the calyx is funnel-shaped, and has triangular, long, pointed, awn-like teeth.—Galeopsis (Hemp-nettle) has two conical protuberances on the under lip between the lateral and the central lobes. The anthers open by 2 unequal valves. Lamium (Dead-nettle, Fig. 576) has dentate, lateral lobes on the under lip. L. album (White Dead-nettle), L. rubrum, etc. Galeobdolon.—Leonurus; Phlomis.
b. Tubular, regular, often 10-toothed calyx and concealed stamens.—Marrubium vulgare (Fig. 577); 10 calyx-teeth, hooked at the apex; many almost spherical whorls of flowers in the axils of the foliage-leaves, at some distance from one another.—Sideritis.
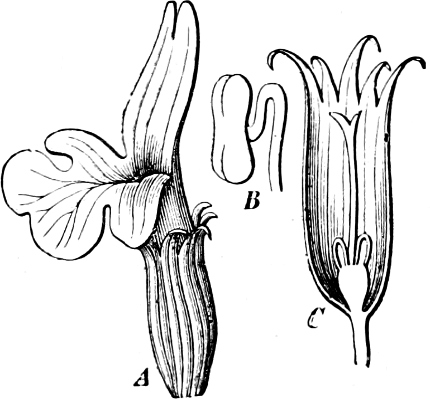
Fig. 577.—Marrubium vulgare.
c. Strongly bilabiate calyx, the lips closing together after[539] flowering.—Scutellaria (Skull-cap); the two lips of the calyx are entire, the upper lip has a large spur, and drops off on the ripening of the fruit. The flowers are generally solitary and turned to one side.—Prunella (Heal-all); the calyx is compressed, its two lips are strongly dentate, the upper lips closing slightly round the under. The stamens have a tooth-like projection beneath the anthers.
3. Nepeteæ, Catmint Group. 13–15 nerves in the calyx; this deviates from the other groups in the posterior stamens being the longer. The upper lip is slightly arched. Nepeta (Catmint), also Glechoma (Ground Ivy), with regular, and Dracocephalum with irregular calyx.
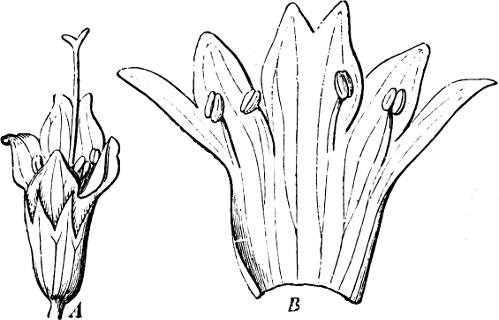
Fig. 578.—Mentha aquatica, var. crispa.
4. Satureieæ, Mint Group. The upper lip is flat, most frequently ovate, or almost spherical, and emarginate (Fig. 578). The calyx is most frequently 5–10-nerved. 4 stamens, the anterior being the longer; rarely, 2 stamens only.—Mentha (Mint, Fig. 578) has a regular, 5-dentate calyx, a small, almost regular, 4-partite corolla, and 4 erect stamens of nearly equal size. The verticillasters are many-flowered, and are often collected into cylindrical inflorescences. Herbs.—Lycopus (Gipsy-wort); corolla almost regular. 2 stamens, the posterior lateral ones are wanting. Preslia: 4-dentate calyx, 4-partite, regular corolla; 4 stamens of equal size.—Thymus (Thyme, Fig. 575) has a strongly bilabiate calyx, the throat being closed by a whorl of hairs (Fig. 575 B). The corolla is distinctly labiate. Under-shrubs, with small entire leaves; verticillasters few-flowered and separate.—Origanum (Marjoram); spike or capitate inflorescences with the flowers solitary[540] in the axils of the rather large and distinctly 4-rowed (often slightly coloured) floral-leaves. Melissa. Calamintha. Clinopodium (Wild Basil). Satureia. Hyssopus (Hyssop); small, entire leaves; the verticillasters are situated unilaterally in a slender, spike-like inflorescence. Lavandula (Lavender); shrubs with verticillasters collected in cylindrical, long-stalked inflorescences; the calyx is tubular, has 13–15 nerves, the posterior tooth is much larger than the others. Stamens and style do not project. Coleus differs, among other characters, in having united filaments; the stamens and style are bent down and concealed in the boat-shaped under lip.
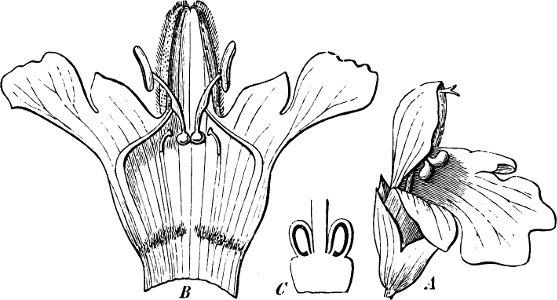
Fig. 579.—Salvia officinalis.
5. Monardeæ, Salvia Group. Only the 2 anterior stamens are developed.—Salvia (Fig. 579); calyx deeply bilabiate; the upper lip of the corolla is generally strongly compressed. Rudiments of the two lateral stamens are present. The connective in the two fertile stamens is long and filamentous, and bears at the upper end a normal half-anther, but at the lower one a barren, often broader portion, against which the insect is obliged to push its proboscis during its visits to the flowers, causing the pollen-bearing half-anther to be pressed down against its back. Floral-leaves often coloured.—Rosmarinus (Rosemary); a shrub with leathery linear leaves, with rolled back edge. A small tooth on the filament represents the barren half of the anther. Monarda.
The POLLINATION is generally effected by insects, especially bees; the under-lip is the landing-stage and the pollen is deposited on their backs. Cross-fertilisation is promoted by dichogamy; honey is secreted by an hypogynous disc and collected in the corolla-tube. Some genera are homogamous (Lamium, Galeopsis, etc.); others are dichogamous (protandrous); a few are gynodiœcious:[541] ♀-and ☿-flowers in various relative sizes (Glechoma hederaceum, Thymus, Salvia pratensis, and others). The entrance of uninvited guests to the honey is often rendered difficult by whorls of hairs, etc. In numerous instances the upper lip protects the pollen from rain. Cleistogamy is found e.g. in Lamium amplexicaule.
2,700 species; distributed over the entire globe, but the greater number in Mediterranean countries (especially in the Eastern regions), where many are shrub-like.—Poisonous and acrid properties are absent. On account of their volatile oils they are principally used as condiments, for perfumery and in medicine (the officinal parts are therefore nearly always “folia” and “herba,” in Lavandula the flowers, and the volatile oils extracted from them). Such are:[39]Mentha piperita [+] (Peppermint)—menthol is obtained from this species and from M. arvensis—M. viridis [+] (Spearmint), M. crispa (Curly-mint), Thymus vulgaris (Garden Thyme), Melissa officinalis (S. Eur.), Hyssopus officinalis (Hyssop, S. Eur.), Origanum majorana (Marjoram, from the Mediterranean), O. vulgare (Wild Marjoram), creticum, smyrnæum, etc., Salvia officinalis (S. Eur.), Rosmarinus officinalis (oil of Rosemary, S. Eur.), Lavandula vera [+] (oil of Lavender, S. Eur.). Also: Satureia hortensis (S. Eur.), Ocimum basilicum (E. India), Pogostemon patchouli (E. India), etc.—As ornamental plants, e.g. Monarda, Plectranthus, and Coleus (foliage-plants, often with red stems and leaves), Stachys lanata (white, woolly), Phlomis, Salvia-species, Perilla, etc.
Order 5. Selaginaceæ. 130 species; small, most frequently heath-like shrubs or herbs, mainly from S. Africa. They differ from the other Nuculiferæ especially in the bilocular, transversely-placed anthers of the 4 stamens (2 stamens divided as far as the base (?)). The ovary has 2, or by suppression only 1 loculus, each with 1 ovule, and the fruit is a schizocarp dividing into two, or is a 1-seeded nut. Radicle turned upwards.—A few are ornamental plants (Selago, Hebenstreitia).
Order 6. Globulariaceæ. 12 species; especially in the Mediterranean. They form an analogy to the Compositæ, and in the main resemble Jasione montana in appearance, the flowers being crowded into a spherical head (hence their name) and supported by bracts, but without involucre; the ovary is unilocular with 1 pendulous ovule. The 1-seeded nut is enveloped by the persistent calyx. The corolla is more or less labiate, the upper-lip is often absent as in the ligulate corollas of the Astereæ; stamens 4, didynamous, with transversely placed anthers opening by one transverse cleft. The leaves are scattered, simple, entire, and generally form a rosette. Globularia.
Order 7. Stilbaceæ. Heath-like shrubs. The ovary is bilocular; 1 erect seed in each loculus, or the posterior cell is empty. Stilbe. 7 species. S. Africa.
Hypogynous, regular, ☿, gamopetalous flowers (Figs. 581, 582), which are generally 5- or 4-merous, with 5 or 4 stamens (with the exception of Oleaceæ and Jasminaceæ which have only 2 stamens, alternating with the carpels). The gynœceum is formed of[542] 2 (nearly always median) carpels. The corolla very frequently has twisted æstivation (the upper edges of the petals being free; Fig. 581 A), and hence the individual lobes of the corolla are oblique, but the flower as a whole is regularly actinomorphic. A nectary, in the form of a honey-secreting ring or glands, is often found round the base of the ovary.—The leaves, with a few exceptions, are opposite and without stipules. Endosperm large (Fig. 581 C), except in Jasminaceæ and Asclepiadaceæ.
The Apocynaceæ and the Asclepiadaceæ, on account of the free ovaries, without doubt represent a more primitive form, but the Asclepiadaceæ on the other hand form an offshoot on account of their peculiar pollen-masses. The Loganiaceæ form a transition to the Rubiaceæ.
The orders are:—
A. Stamens 5. 1, Gentianaceæ; 2, Apocynaceæ; 3, Asclepiadaceæ; 4, Loganiaceæ.
B. Stamens 2. 5, Oleaceæ; 6, Jasminaceæ; 7, Salvadoraceæ.
Order 1. Gentianaceæ (Gentians). Glabrous herbs, without latex; the opposite, undivided and entire leaves are often slightly united at the base; many have rosette-like radical leaves. Stipules absent. The flowers are generally borne in regular, dichotomously-branched dichasia (Figs. 580, 581 A), which finally become transformed into unipared scorpioid cymes; the parts of the flower are 4–5-merous as far as the gynœceum, which is 2-merous; the calyx frequently is almost polysepalous; the corolla has distinctly twisted æstivation (the upper edges being free) (Fig. 581 A), except Menyantheæ. The carpels are entirely united, and most frequently form a 1-locular ovary with 2 parietal placentæ bearing many ovules (often in several rows, Fig. 581 D, F). Capsule, 2-valved, with septicidal dehiscence, the incurved edges bearing the seeds (Fig. 581 D, F).
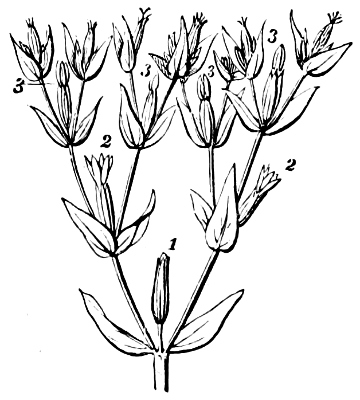
Fig. 580.—Erythræa. Inflorescence. 1, 2, 3, etc., the successive shoot-generations.
1. Gentianeæ.—Gentiana (Gentian) has most frequently a tubular, campanulate or funnel-shaped corolla, sometimes with teeth between the corolla-lobes and fringed in the throat of the corolla; G. lutea has a rotate, yellow corolla.—Swertia: rotate corolla; each lobe has at its base 1–2 nectaries, with fringed edges.
[543]
Erythræa (Centaury, Fig. 581); corolla most frequently salver-shaped. The anthers ultimately become spirally twisted (E). The style prolonged, deciduous. The flower has the Lobelia-arrangement, i.e. the median sepal is anterior; the corolla is rose-coloured (in the native species). The capsule is semi-bilocular (Fig. 581 F, G).—Cicendia has a low creeping stem, fine as a thread, and small, yellow flowers, 4-merous (without twisted anther).—Chlora (Yellow-wort) 6–8-merous.
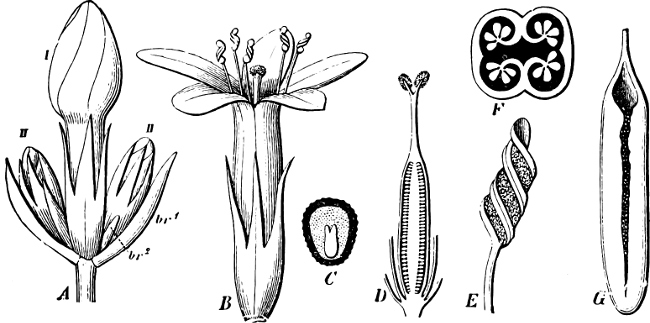
Fig. 581.—Erythræa centaurium. Inflorescence, flower and fruit: br1, br2 floral-leaves of the 1st and 2nd order; G a valve of the capsule separated from its fellow.
2. Menyantheæ. Menyanthes (Buck-bean) deviates in several respects from the type of the order. The leaves are scattered and, in M. trifoliata, trifoliate; the corolla has valvate æstivation; the testa is also very hard (thin in the true Gentians). They are aquatic plants with creeping rhizome; the flowers borne in racemes, with terminal flower, heterostylous. The corolla is funnel-shaped with a very hairy throat.—Limnanthemum with floating leaves, like the Water-lilies.
575 species; distributed over the entire globe, but most numerous in Alpine districts. Neither poisonous nor nutritive plants are found, but several are used in medicine on account of the bitter properties so prevalent amongst them. Officinal: the roots of Gentiana lutea. The roots of other species, e.g. G. purpurea, punctata and pannonica (Europe) and the leaves of Menyanthes trifoliata are medicinal. Some are grown as ornamental plants on account of the pure (often deep blue) colour of the flowers.
Order 2. Apocynaceæ (Periwinkles). Trees and shrubs (also lianes), less frequently herbs, generally with latex. The leaves are opposite, simple, entire, without stipules; the flowers are[544] regular; corolla-lobes oblique, æstivation twisted. The stamens are individually free, and the pollen-grains are free or at most united in fours (see Asclepiadaceæ). The two carpels have 2–∞ ovules, in all cases there is only 1 style and a capitate stigma, which towards the base is widened out into a disc-like table (stigma-disc) abstricted in the centre; but the carpels in most of the genera (e.g. those mentioned below) are entirely separate, and the fruit consists of two follicles, the seeds of which often have a tuft of woolly hairs projecting from the micropyle, less frequently of two drupes. In some other genera there is a 1-locular (provided with 2 parietal placentæ) or a 2-locular ovary becoming a 2-valved capsule or a berry. Endosperm abundant.
Vinca (Periwinkle) has a salver-shaped corolla, which is twisted to the left in æstivation (i.e. the left edge of the petals is free); nectaries 2, alternating with the carpels; the summit of the style is hairy. Follicles; seeds without hairs. Mostly creeping, perennial, evergreen plants, whose large flowers are apparently axillary; in reality they are terminal, but by the development of the bud in the axil of one of the two uppermost leaves, they are thus displaced over the other leaf of the pair (a helicoid sympodium being formed).—Plumeria, Tabernæmontana, Cerbera (drupe). Aspidosperma.
Nerium (Oleander). The leaves are in whorls of 3. Corolla funnel-shaped, in æstivation twisted to the right, and with a corona resembling that of Lychnis. The anthers are prolonged at the base and each also bears at the apex a long, linear, hairy appendage; these finally become spirally twisted. Follicles; seeds hairy. Apocynum, Echites, etc. Epigynum is epigynous.
124 genera, 1,000 species; principally in the Tropics. Only 2 species of Vinca are natives of this country; the following are cultivated as ornamental plants:—Vinca minor, V. major, V. (Lochnera) rosea, Amsonia salicifolia, Nerium oleander (Eastern Mediterranean). The latex of some is poisonous (Tanghinia venenifera, Cerbera). Caoutchouc is obtained from others (Hankornia, Landolphia, Vahea, etc.). Tough bast is frequently developed. The bark of Aspidosperma quebracho and the seeds of Strophanthus hispidus are used in medicine (also for African arrow-poison), the latter is officinal.
Order 3. Asclepiadaceæ. A natural and easily recognised order, closely allied to the Apocynaceæ, having, like it, frequently a poisonous latex, opposite, single, entire leaves and fundamentally the same floral diagram and floral structure (S5, P5, A5, G2); but in some the æstivation of the corolla is valvate. The carpels here also have free ovaries, but are united for some distance above into a large, shield-like, 5-angular head, having on its underside[545] the true stigmas, and the fruit always consists of 2 follicles; seeds most frequently numerous and hairy at the micropyle (“vegetable silk”); endosperm scanty.—The order is distinguished from the Apocynaceæ and from all other plants also, except the Orchids, by having all the pollen-grains in each of the 2 loculi of the anthers (true 2-locular anthers) united into one waxy, club-shaped pollen-mass (“pollinium”), for the purpose of pollination by insects. These heavy masses, in order to secure pollination (as in the case of the Orchids), must be attached to sticky discs (corpuscula); there are 5 corpuscula, one at each of the corners of the 5-angular stylar-head (alternating with the anthers), and to each of these are attached 2 pollinia, one from each of the anthers situated on either side (thus each anther gives its right pollinium to one corpusculum and its left to another). The stamens are frequently united at the base, and each bears on the back a variously formed, petaloid appendage, termed a “cucullus.”
Fig. 582.—Asclepias cornuti. A An open flower with the calyx (k) and corolla (c) turned down; the stamens are bent together and surround the gynœceum. B The andrœcium after removal of the sterile part (cucullus) of the anther, which functions as a nectary: e the lateral expansions of the fertile portion of the anthers; f the slit between the expansions of two contiguous anthers, through which the insect’s foot, and later a pollinium which is caught by it, is dragged, and behind which the only receptive part (stigma) is hidden; above the slit f is the gland (r), which secretes the horny corpusculum, which is split at its base and joined on either side with a pollinium (this is more distinctly seen in D and E). When the foot of the insect is caught in the slit (f) and is drawn upwards, it becomes entrapped in the slit of the corpusculum, which is then pulled out together with the pollinia firmly attached to it. In walking over the flowers the insect will draw its foot through other slits (f) and so leave the pollinia on the stigmas. C, D The gynœceum with the pollinia hanging freely. E A corpusculum and two pollinia.
A peculiar relative position (and therefore a good, distinctive characteristic) is often found in the inflorescence, which is cymose; it is placed between the two leaves of a whorl, nearer to one than to the other. The leaf-pairs are placed obliquely in the floral region, at acute and obtuse angles, and not at right angles (as in the purely vegetative parts); the inflorescences are placed in two rows only which are nearly 90° from each other, and the two contiguous to one another are antidromous; they are in reality terminal, each on its own axis, and the entire floral portion of the shoot is[546] a unipared scorpioid cymose sympodium; in addition, complications also arise through individual parts becoming united.—Herbs and shrubs, some twining or climbing.
In Asclepias the corolla is bent back and there is a cup-like cucullus, from the base of which protrudes a horn-shaped body, bent inwards.—Vincetoxicum has a rotate corolla and a ring-like, 5-lobed cucullus, without internal prominences.—Stapelia (especially from S. Africa) is remarkable on account of its Cactus-like, leafless stems and large, brownish flowers, often with carrion-like smell. Periploca has more powdery pollinia (S. Eur., etc.); Hoya carnosa (Wax-flower; Trop. Asia) is a climber, and has small, annual, flower-bearing dwarf-branches. Ceropegia.
201 genera with 1700 species, distributed over all tropical countries; few outside these limits: no native species. Several are used in medicine on account of the pungent properties of the latex. Condurango-bark of Gonolobus condurango is medicinal. Caoutchouc is obtained from the latex of some (e.g. from Cynanchum). The seed-hairs, which are most frequently shining, silk-like, and white, are not sufficiently pliant to be of much value. Ornamental plants in our gardens: Asclepias-species, etc.
Order 4. Loganiaceæ. Ovary single, with two loculi, in structure resembling the Rubiaceæ, but superior. 360 species are included in this order; the majority are tree-like, some lianes which climb by tendril-like branches. The interpetiolar stipules of some species are very characteristic (as in Rubiaceæ, to which they maybe considered to be closely related). The fruit is a capsule or berry. The most familiar genus is Strychnos, which has spherical berries with an often firm external layer, and compressed seeds with shield-like attachments; endosperm abundant. The leaves have 3–5 strong, curved nerves proceeding from the base.—Spigelia.—They have no latex, as in the two preceding orders, but many are very poisonous (containing the alkaloid “strychnine,” etc.); the South American arrow-poison, urare or curare, is made from various species of Strychnos, also an arrow-poison in the East Indian Islands (Java, etc.). Officinal, the seeds of Strychnos nux vomica (“Vomic nut,” Ind.). The seeds of Strychnos ignatii (Ignatius-beans, medicinal), and others are poisonous.
Order 5. Oleaceæ. The leaves are always opposite. The inflorescences are racemes or panicles. The calyx and corolla are 4-merous, more or less united, free in some species; the corolla has most frequently valvate æstivation. All four forms of fruit occur (see the genera). Ovules pendulous, 2 in each loculus (Fig. 583 C). Endosperm oily.—Syringa (Lilac) and Forsythia (anthers somewhat extrose) have capsules with loculicidal dehiscence and winged seeds.—Fraxinus (Ash) has winged nuts (samara) (Fig. 583 D); trees with most frequently imparipinnate leaves; the flowers are naked and sometimes unisexual (polygamous), the Manna Ash (F. ornus) has however a double perianth with 4 free petals (Fig. 583 a); in the native species, F. excelsior,[547] the flowers open before the foliage appears.—Ligustrum (Privet) has berries.—Olea (O. europæa; Olive) has drupes; the pulp and seeds of the ellipsoidal fruits are rich in oil. The lanceolate leaves are grey on the under surface, being covered with stellate hairs. In the wild state it is thorny (modified branches).—Phillyrea; Chionanthus.—Few species of Linociera have 4 stamens.
Fig. 583.—Fraxinus ornus: A flower; ca calyx; co corolla; B gynœceum and calyx; C longitudinal median section of gynœceum; D fruit.
180 species; chiefly in the northern temperate zone. The Olive-tree (Olea europæa) has been an important cultivated plant from ancient times (Olive oil, Provence oil, “Sweet oil”). The best oil is extracted from the fruit-pulp. The fruits are edible. Home: Western Asia, Eastern Mediterranean. Timber: the Ash (Fr. excelsior). Officinal: the Manna Ash (Fr. ornus), cultivated in the Mediterranean countries for the sake of its saccharine juice, which flows out and coagulates into “Manna.”—The following are ornamental plants: species of Ligustrum and Syringa (introduced in the 16th century, from S.E. Europe and Asia), Forsythia (China, Japan; the large, yellow flowers are borne on dwarf-branches with scale-like leaves, before the opening of the foliage-leaves), Chionanthus.
Order 6. Jasminaceæ. The æstivation of the corolla is imbricate; the ovules are erect; seeds almost without endosperm; radicle directed downwards. The number of lobes in the calyx and corolla is not 4, but e.g. 5, 8, 10, and variations are sometimes found in the same individual. The fruit is a berry or capsule. Many species are twiners, and their scattered or opposite leaves are most frequently imparipinnate.—120 species; especially in Trop. Asia (E. India). Some Jasminum-species are cultivated as ornamental shrubs in the warmer districts on account of their elegant foliage, and beautiful, sweet-scented flowers, the essential oil of which is also used in perfumery; the best known are: J. sambac and grandiflorum. Nyctanthes arbor-tristis opens its sweet-scented flowers only at night (E. India).
Order 7 (?). Salvadoraceæ. 8–9 species; Asia, Africa.—Salvadora.
[548]
The leaves are always opposite or verticillate. The flower is epigynous, ☿, 5-(or 4-) merous, with the usual sympetalous diagram; 2–5 carpels. The inflorescences are frequently dichasial. The sepals are small, reduced to teeth, and become almost entirely suppressed in the higher forms.—The flower is regular in Rubiaceæ[549] and some Caprifoliaceæ, but in other genera of this latter order (especially of Lonicereæ) it is unsymmetrical. In several genera of the order first mentioned the loculi of the ovary contain many ovules, but in the last the number of loculi and ovules becomes reduced. This is to some extent connected with the nature of the fruit which is many-seeded in most instances, namely a capsule or berry, but in others nut-like. Endosperm is present.
The family on one side is allied to the Contortæ (not only through the Loganiaceæ but also through the Apocynaceæ), and may be regarded as an epigynous continuation of this family; on the other side it is allied to the Valerianaceæ and Dipsacaceæ. Many points of agreement with the Cornaceæ and Araliaceæ are also found, and in fact several Caprifoliaceæ are distinguished from these by hardly any other feature than the gamopetalous corolla.

Fig. 584.—Cinchona calisaya. Flowering branch.
Order 1. Rubiaceæ. Leaves opposite (or verticillate), undivided and entire, with interpetiolar stipules (Fig. 586). Flowers epigynous and hermaphrodite, regular, 4- or 5-merous with the usual arrangement (Figs. 585, 588–590); corolla gamopetalous, in æstivation often valvate; ovary frequently 2-locular.
Fig. 585.—Cinchona calisaya. A entire flower; B after removal of the corolla; C longitudinal section of ovary; D fruit; E seed.
There are no external characters which at once distinguish this exceedingly large order, as in many other natural orders (Compositæ, Umbelliferæ, etc.), but the opposite leaves with interpetiolar stipules form an excellent mark of recognition. It is divided into many sub-orders and groups, especially characterised by the nature of the ovary (1 or several ovules in each loculus), and of the fruit (schizocarp, berry, drupe, capsule).—The corolla is bilabiate in 4 genera; its æstivation in some is twisted; in Capirona, etc., the filaments are of unequal size. The ovary is semi-epigynous in Henriquezia, etc. In Morinda all the fleshy fruits coalesce into one multiple fruit.
[550]
1. Cinchoneæ. The fruit is a 2-valved capsule, with many winged seeds (Fig. 585). Cinchona (Quinine, Fig. 584). Trees and shrubs with the foliage and inflorescence somewhat resembling Syringa; the corolla also being of a lilac colour, more or less salver- or funnel-shaped, and frequently edged with a fringe of hairs (Fig. 585), is somewhat similar to that of Menyanthes. Their home is the Andes from Bolivia to Venezuela, varying in altitude from 1–3000 metres. There are now large plantations in Java and E. India. (The name “quinine” is of Indian origin; that of the genus “Cinchona,” is from the Spanish Duchess Cinchon, who in 1638 first introduced the bark into Europe.) The following are closely allied: Cascarilla, Remijia, Ladenbergia, Manettia, Bouvardia, etc.
2. Gardenieæ. Trees and shrubs, frequently having a many-locular berry. Randia, Gardenia, Genipa, Hamelia, etc.
3. Coffeeæ. Only 1 seed in each of the two loculi of the ovary; the fruit is a drupe with 2 stones. Coffea has an ellipsoidal fruit about the size and colour of a cherry; the two thin-shelled, parchment-like stones are enclosed by a thin layer of pulp; the two seeds are flat on the side turned to one another, which has also a deep, longitudinal groove curving to the sides. The endosperm is hard, horny and greyish (without starch); the small embryo lies in the lower end near the circumference. The Coffee-plant (C. arabica) is a small tree, or more frequently, and especially in plantations, a shrub with large dark-green leaves and scented, white flowers. Its home is in Tropical Africa; it is now cultivated in many tropical countries. C. liberica, W. Africa.—Cephaëlis (C. ipecacuanha, Fig. 586; the roots are officinal).—Psychotria, Chiococca, Ixora, Hydnophytum, Myrmecodia, etc.
Fig. 586.—Cephaëlis ipecacuanha. Portion of a branch: st stipules.
4. Spermacoceæ. Chiefly small shrubs and herbs, many of which are weeds in tropical countries. The stipular sheaths bear numerous bristles at the edge. Spermacoce, Borreria, Diodia, Richardsonia, etc.
5. Stellatæ. Herbaceous plants with verticillate leaves (Figs.[551] 587, 588–590); the stipules are large, leaf-like, and resemble the lamina of the leaves, so that the leaves appear to be placed several in a whorl, while in reality there are only two opposite leaves, the stipules of which project freely, and are not erect (Fig. 587).
Fig. 587.—Rubia tinctorum.
In some cases there are apparently 4 leaves in the whorl, and then 2 of these are leaves, and the other two are their interpetiolar stipules. When there are apparently 6 leaves, then the two of these which are opposite each other are leaves, and the other four are stipules; if there are several members in the whorl, then a division of the stipules has taken place. The proof of this theory is founded upon the fact that not more than 2 of the leaves of the whorl ever support buds (which, in addition, are seldom of equal vigour), and also that the whorls do not alternate with each other, which, according to the rules of the position of the leaves, they should do if all the members of a whorl had[552] equal value. If there are, for instance, 4 members in two successive whorls, they stand right above one another, and do not alternate. The development and anatomical relations (the branching of the vascular bundles) also point to the same conclusion.—All the other groups of the order have only 2 small scale-like interpetiolar stipules, or they form at the base of the leaf-stalks an interpetiolar sheath, having often a toothed edge (Fig. 586).—Another characteristic feature in this group is that the calyx is rudimentary, the corolla valvate (Fig. 588), and that each of the two loculi of the ovary has only 1 ovule. The fruit is a schizocarp dividing into 2 fruitlets (Fig. 590). The forms of the fruit, as well as many other characters, as, for example, the epigynous flower, the rudimentary calyx, the two free or almost free styles, present interesting analogous resemblances to the polypetalous order of the Umbelliferæ. This group has its home chiefly in the temperate regions of the northern hemisphere, especially about the Mediterranean; it is the only group which occurs in this country, represented by 4 genera.
Figs. 588–590.—Rubia tinctorum.
Fig. 588.—Diagram.
Fig. 589.—Longitudinal section of flower.
Fig. 590.—Longitudinal section of fruit (3/1).
Galium (Cleavers) is almost destitute of a calyx; it has a small 4-partite, rotate corolla, 4 stamens, and 2 free styles. The fruitlets are nut-like. The inflorescence is a paniculate dichasium passing into helicoid cymes.—Asperula (Woodruff) is distinguished from the above by its salver- or funnel-shaped corolla. 1 style.—Rubia (Madder, Figs. 587–590) has almost the same form of corolla as Galium, but (most frequently) a 5-merous flower, and the fruitlets are “drupes.” Sherardia (Field Madder); the flowers are clustered in closely arranged cymes surrounded by an involucre; the calyx has 6 distinct teeth, while the number of petals and stamens is 4. The corolla is funnel-shaped.—Vaillantia. Crucianella.
The DISTRIBUTION OF SEEDS, in some instances, is promoted by hooked appendages on the fruitlets (e.g. Galium aparine).
The small flowers of the Stellatæ are frequently collected in compact inflorescences, and are therefore rendered more conspicuous; slight protandry is[553] found in some, self-pollination in the species which are less conspicuous. Many species are heterostylous. Myrmecodia, Hydnophytum, and other genera have large tubers (hypocotyledonous stems), whose labyrinthine cavities and passages are inhabited by ants.
About 4,500 species; tropical or sub-tropical except the Stellatæ; especially American. The tropical ones are mostly trees.—Several are OFFICINAL on account of the large amount of alkaloids and glycosides which they contain. The most important are the Cinchonas (Cinchona calisaya, C. succirubra, C. officinalis, C. micrantha, etc.), whose bark contains the well-known febrifuge and tonic, Quinine, Cinchonin, etc.; Quinine is also found in Exostemma, Ladenbergia, and Remijia. The root “Ipecacuanha” (an emetic) from Cephaëlis ipecacuanha (Brazils). Caffeine is officinal. The use of the seeds of the coffee plant (“the beans”) was first known in Europe in 1583.—There are only a few which contain aromatic properties, principally among the Stellatæ (coumarin in Asperula odorata, the Woodruff), in which group colouring materials are also found. The root and root-stalks of Rubia tinctorum, the Madder (S. Eur., Orient., Fig. 587), were formerly largely used for dyeing, but are now superseded by the analine colours. Red dyes are also obtained from the roots of species of Asperula and Galium. Gambier is a splendid colouring material, obtained from Uncaria gambir (S.E. Asia), which is used in dyeing and tanning.—The order does not furnish many ornamental flowers.
Order 2. Caprifoliaceæ. This order agrees with the Rubiaceæ in having opposite leaves and an epigynous flower, most frequently 5-merous with the ordinary tetracyclic diagram, but in some species it is zygomorphic; the corolla has imbricate æstivation, carpels 3–5, most frequently 3 (not 2, which is the most usual number in the Rubiaceæ). The fruit is generally a berry or a drupe, but the most important, and in any case most easily recognisable feature, is the absence of stipules; in exceptional cases, where they are present, they are not interpetiolar, and are most frequently small.—The majority of plants belonging to this order are shrubs or trees. Compound leaves sometimes occur. Stipules only appear in a few species of Lonicera, Sambucus and Viburnum; in the common Elder (Sambucus nigra) they are in some instances glandular and small, but in other cases larger and more leaf-like (upon long, well-developed shoots); in the Dwarf Elder (S. ebulus) they have the normal leaf-like form; in Viburnum opulus they are present as narrow lobes at the base of the petiole; in others they are completely absent. The leaves are frequently penninerved, rarely palminerved. The calyx, as in the Stellatæ and Aggregatæ, is often very insignificant.
1. Lonicereæ, Honeysuckle Group. This has campanulate or tubular corollas which are often zygomorphic; in connection with the length of the corolla the style is long, filamentous, and most frequently has a large, capitate stigma. There are several ovules in the loculi of the ovary, and the fruit is most frequently a berry.
[554]
Fig. 591.—Lonicera.
Lonicera (Honeysuckle). Shrubs, sometimes twiners. The corolla in some species is considerably bilabiate (Fig. 591), with 4 lobes in the upper lip, and 1 in the under lip, but in others more regular, tubular, or campanulate. The flowers are either borne in capitate inflorescences, which are compound and formed of closely compressed 3-flowered dichasia (sect. Caprifolium), or in dichasia with 2 flowers (the terminal flower is wanting). The ovaries and fruits coalesce in some (sect. Xylosteum). The opposite leaves in some species unite with each other and form a broad collar encircling the stem (Fig. 591). Above the primary bud 1–2 accessory buds are often found in the leaf-axils.—Diervilla (Weigelia); with a 2-locular, 2-valved capsule.—Symphoricarpus (Snowberry) has an almost regular, funnel-shaped corolla; a peculiar feature is found in the ovary which has 4 loculi, the 2 median having many ovules in 2 rows, all of which are aborted; the 2 lateral ones, on the other hand, each have only 1 ovule which is developed. Different forms of leaves are frequently found on the same branch; they are entire or lobed.
[555]
2. Sambuceæ, Elder Group (Fig. 592). This has a rotate, regular corolla, extrorse anthers, a very short and thick (or almost absent) style, with tripartite stigmas, and only 1 pendulous ovule in each of the 3 (-5) loculi of the ovary. The fruit is a “drupe” with 1–3 (-5) stones. The inflorescence is made up of cymes grouped in an umbel-like arrangement.
Sambucus (Elder, Fig. 592) has imparipinnate leaves and a “drupe” with 3 (-5) stones. Between the calyx and the style a disc remains on the apex of the fruit. S. nigra with black fruit; S. racemosa with red fruit; S. ebulus is a perennial herb; the others are woody.—Viburnum (Guelder-rose) has simple leaves (penninerved or palminerved, entire, dentate or lobed), and a “drupe” with only 1 stone, which is compressed, cartilaginous, and parchment-like; 2 of the loculi of the ovary are aborted. (In V. opulus the marginal flowers of the inflorescence are barren, and in that case their corollas are generally specially large; the cultivated Viburnum has only barren flowers, with large corollas.)
Fig. 592.—Sambucus nigra: cor corolla; s calyx.
3. Linnæeæ. Linnæa borealis (the only species) is an extreme form of the order; it has a 2-flowered dichasium, funnel-shaped, slightly bilabiate corollas (2/3); 4 didynamous stamens. Two of the 3 loculi of the ovary have several ovules which are not developed, while the third has only 1 ovule, which developes into a seed. The fruit is a nut, which is enveloped by the two large bracteoles, which are covered by sticky, glandular hairs, and serve as a means of distribution. It is a small undershrub.
[Adoxa, which was formerly classed in this order, appears, according to recent investigations, to be more properly placed among the Saxifraginæ.]
In cases where the flowers are small, as in Sambucus and Viburnum opulus, they are rendered conspicuous by being arranged in closely-packed inflorescences; they are massed together and form large surfaces, and in the last named are still more conspicuous on account of the barren, but large ray-flowers, which are of service in this respect. Honey is secreted in the nectaries at the base of the styles. In the genera with rotate flowers, as Viburnum and[556] other Sambuceæ, the honey lies so exposed and in such a thin layer, that only flies and insects with short probosces can procure it; bees, however, visit these flowers for the sake of the pollen. There is hardly any nectar in the Elder; self-pollination frequently takes place. The flowers of the Caprifoliaceæ, which, with their long corolla-tube are adapted for evening-and night-flying insects with long probosces, open in the evening, and at that time give off their strongest scent.
Distribution. 230 species; especially outside the Tropics in the Northern Hemisphere. In this country they are found especially in hedges and as under-shrubs.—Officinal: the flowers and fresh fruits of the Elder (S. nigra), the fruits (“berries”) being also used in the household. Ornamental shrubs: species of Lonicera, Symphoricarpus, Diervilla, which are chiefly from N. Am., Abelia and Viburnum.
The leaves are opposite and without stipules. The flower (Figs. 593, 595, 598, 599, 600) is epigynous, zygomorphic or asymmetrical, 5-merous with S5, P5, stamens typically 5, but by suppression never more than 4, sometimes less, carpels 3–2. The calyx is more or less insignificant, and almost suppressed in the extreme forms. The ovary has 3–1 loculi, but only one loculus has an ovule, which is pendulous with the micropyle turned upwards (Fig. 594). Fruit a nut. Embryo straight, with the radicle pointing upwards (Fig. 597), without or with endosperm.
The inflorescences are distinct dichasia in Valerianaceæ, but in Dipsacaceæ and Calyceraceæ they are crowded together into capitula.
This family is closely allied to the Rubiales through the Valerianaceæ, which have almost the same structure as many of the Caprifoliaceæ. It attains the highest development in the Dipsacaceæ, which are composite plants, but differs from Compositæ in the position of the ovule, etc.
Order 1. Valerianaceæ. Herbaceous plants or under-shrubs with opposite leaves, often pinnate; stipules absent. The flowers are borne in dichasia and in scorpioid cymose inflorescences and are entirely without any plane of symmetry (Fig. 593). The calyx and corolla are 5-merous, but the calyx is frequently very insignificant and ultimately a pappus, as in Compositæ; the corolla is frequently saccate or produced into a spur at the base. Most frequently, only 3 (4–1) of the 5 stamens are developed; these are free. Carpels 3, which form an inferior ovary, often with 3 loculi, but only 1 of the loculi contains 1 pendulous, anatropous ovule (Figs. 593, 594 A), the other loculi are empty[557] and shrink up more or less completely. (Compare Fig. 593 A, B). Style 1, stigma tripartite. Endosperm absent; embryo straight, with the radicle directed upwards.
The inflorescences are dichasia, or unipared scorpioid cymes with the branches developed in the axil of the second bracteole. Both the bracteoles are generally present and frequently form 4 very regular, longitudinal rows on the branches of the inflorescence.—5 stamens do not occur (except perhaps in Patrinia). The suppression of stamens and carpels takes place most readily on the anterior side of the flower and that turned towards the first bracteole (a) (Fig. 593), whose branch is suppressed in the dichasium; after this the posterior median stamen is next suppressed.
By the vegetative characters as well as by the inflorescence and the flower, the order is allied to the Caprifoliaceæ and especially to the Sambuceæ.
Fig. 593.—A Diagram of Valeriana officinalis. B Diagram of Centranthus.
In the least modified (oldest) forms, Patrinia and Nardostachys, there is an almost regular flower, a 5-merous calyx, 4 stamens, and 3 loculi in the ovary, 2 of which however are barren. The stamens in Valerianella are reduced to 3, in Fedia to 2 (posterior), and the calyx is less distinctly 5-dentate; the 2 empty loculi in the ovary are still visible. Fedia has a small spur at the base of the corolla. Valeriana has a very reduced, hair-like calyx (pappus), an unsymmetrical, salver-shaped corolla with a sac-like, nectariferous spur at the base, 3 stamens and only 1 loculus in the ovary (Figs. 594, 593). Centranthus (Fig. 593) is still further reduced. The corolla has a spur and only 1 stamen; unipared scorpioid cymes with 4 rows of bracteoles. In the last two genera there is a peculiar wall in the corolla-tube, which divides it longitudinally into two compartments (indicated by a dotted line in Fig. 593), one of which encloses the style. This wall is low in Valeriana, but in Centranthus it reaches as far as the throat.—The rays of the pappus are pinnately branched and rolled up before the ripening of the fruit. 12–20 in number (Fig. 594 A, B).
Val. officinalis and others are protandrous: in the first period the stamens project from the centre of the flower (Fig. 595 a), the stigmas in the second (b) when the stamens have become bent backwards. (V. dioica is diœcious[558] with large ♂-and small ♀-flowers).—275 species; especially from the temperate and colder parts of the northern hemisphere of the Old World, Western North America and the Andes.—Bitter properties are characteristic, such for instance as the volatile acid and volatile oil of Valeriana; these occur especially in the rhizomes. Officinal; the rhizomes of V. officinalis.—The true Indian “Nardus,” an important medicine and perfume in India, is extracted from Nardostachys (Himalaya). A variety of Valerianella olitoria is sometimes used as salad.
Fig. 594.—Valeriana: A ovary (longitudinal section); B ripe fruit.
Fig. 595.—Valeriana: a flower in the ♂ stage; b in the ♀.
Fig. 596.—Centranthus ruber. Flower, its lowermost portion (the ovary and spur) in longitudinal section. (Mag.)
Fig. 597.—Scabiosa atropurpurea. Fruit in longitudinal section. Inside the “epicalyx” may be seen the fruit drawn out into a beak, with straight embryo and radicle directed upwards.
Order 2. Dipsacaceæ (Teasels). Herbs with opposite leaves without stipules. The flowers are situated in compact capitula each with an involucre. A characteristic feature of the order is that each flower of the capitulum has a gamophyllous “epicalyx”[559] (Figs. 597, 599, 600), which envelopes the inferior ovary. The flowers (Figs. 599, 600) are ☿, 5-merous (S5, P5, stamens typically 5, G2), but the calyx often expands at the edge into a membrane with 5, or an indefinite number of bristles or teeth (pappus, Figs. 597, 600), and the zygomorphic, funnel-shaped corolla is sometimes 5-lobed and bilabiate (2/3), but most frequently 4-partite (Fig. 599), the two lobes of the upper lip coalescing into one lobe, as in certain Labiatæ, Veronica and Plantago; the æstivation is imbricate.
Figs. 598–600.—Dipsacus fullonum.

Fig. 598.—Inflorescence (the flowers in a zone below the apex commence to flower first).
Fig. 599.—Flower (4/1).
Fig. 600.—The same in longitudinal section.
The stamens are never more than 4, the posterior one remaining undeveloped; they usually have free anthers which generally project considerably (Fig. 599). The ovary is unilocular with 1 pendulous ovule and bears 1 undivided style; fruit a nut with 1 seed, containing endosperm and with the radicle turned upwards (Fig. 597).
The flowers do not always open in centripetal order, a fact which may be observed especially in the Dipsacaceæ, in which a zone of flowers round the centre of the capitulum opens first, and the flowering then proceeds both upwards and downwards (Fig. 598). This has probably some connection with the fact that the capitulum has arisen from the coalescence of several dichasial inflorescences. In species of Scabiosa the flowers open simultaneously at the circumference, or in a zone at the centre.—The morphological explanation of the[560] “epicalyx” is not quite certain; in all probability it is formed from two united bracteoles, for an “epicalyx” is distinctly formed in this way in one of the Valerianaceæ, Phyllactis.—The ray-flowers are larger and more irregular, labiate or ligulate, than the disc-flowers, yet not in so high a degree as in the Compositæ.
A. A scarious bract to each flower. Scabiosa has a 5-lobed corolla; the “epicalyx” has a dry, scarious, often finally large collar, and the true calyx is formed of long bristles (generally 5) (Fig. 597). Succisa pratensis (Devil’s-bit) has a 4-lobed corolla, the collar of the “epicalyx” is herbaceous; the calyx as in the preceding.—Pterocephalus.—Dipsacus (Teasel); large, spiny and stiff-haired herbs with capitula, or short, thick spikes on which both the involucral-leaves and bracts project considerably, and are stiff and spinose (Fig. 598). The “epicalyx” has short teeth, or is almost entire. The leaves of the stem unite together in pairs, so that shallow cups are formed round the stems in which rain-water may collect.—Cephalaria.—Morina: the flowers are falsely verticillate as in the Labiatæ; the calyx has 2 laterally-placed, entire, or emarginate lobes; 2 stamens, or 2 large and 2 small ones.
B. Bristles, but no true bract to each flower. Knautia; the corolla is 4-partite, the calyx cup-like, with many bristles or teeth on the edge.
Pollination is in many species effected by insects. The honey is secreted by a ring round the base of the style. The flowers in our native species are considerably protandrous. Gynodiœcious flowers also occur.—150 species; especially in the Mediterranean and the Orient; the order is not represented in the South Sea Islands, Australia and America.—The heads of the true Teasel (Dips. fullonum) are used for carding wool, on account of the elastic bracts, which are hooked at the point. The order has bitter properties; tanin, etc.; but no species are used in medicine or the household.—Scabiosa atropurpurea, etc., are used as ornamental plants.
Order 3. Calyceraceæ. This order resembles the Compositæ in the valvate æstivation of the corolla and the more or less united stamens, and the Dipsacaceæ in the undivided style, pendulous ovule and endosperm. The calyx is frequently composed of 5 distinct scales. An “epicalyx” is wanting.—20 species; America.
The flower is epigynous, perfect, with 5 sepals, 5 petals, and 5 stamens in regular alternation, and 3 (2–5) carpels. The sepals in all cases are distinct, but narrow and pointed, so that the æstivation is open. The corolla is gamopetalous with (as in the Compositæ) valvate, or slightly infolded-valvate æstivation. The[561] stamens are nearly always situated on the torus without being united to the corolla (Figs. 601, 604). The anthers adhere or unite and form a tube with introrse anthers from which the pollen is swept out by the projecting, brush-like hairs on the style (as in the Compositæ). The ovary is 3-(2–5) locular, many ovules in each loculus. The fruit is generally a many-seeded capsule (or berry). Embryo in the centre of a fleshy endosperm.—The majority are herbs with scattered leaves, without stipules. The presence of latex and inulin, together with the tubular formation of the anthers, the pollination, etc., indicate a relationship with the Compositæ.
The Cucurbitaceæ are by some authorities placed in this family as being most closely related to the Campanulaceæ. Although the corolla is most frequently gamopetalous, and other similarities to the Campanulaceæ are present, yet on account of the structure of the ovule, and for other reasons, the Cucurbitaceæ are here placed in the Choripetalæ. The Campanulinæ without doubt proceed upwards to the Compositæ, with which, in addition to the occurrence of inulin and laticiferous vessels (Cichorieæ), there are many corresponding features both in the structural and biological relations (epigyny, valvate æstivation of the corolla, tendency of the anthers to adhere or unite, protandry with a stylar-brush, etc.) The inflorescence of Jasione is almost identical with that of the Compositæ.
Order 1. Campanulaceæ (Campanulas). The flowers are regular and in some only semi-epigynous, 5-merous, except in the gynœceum which is 3-merous (the unpaired, median carpel being generally posterior), more rarely 2–5-merous, and has a corresponding number of stigmas and loculi in the ovary; the placentation is axile with a large number of ovules. The median sepal is posterior. The stamens frequently have broad, free bases (Fig. 601 H) which cover the nectariferous upper surface of the ovary; the anthers only fit loosely together, and become separated as soon, as the pollen is shed (Fig. 601 G), 1 long style, which is studded by sweeping-hairs (stylar-brush), which ultimately become invaginated; the stigmas do not unfold until the stamens have shed the pollen (Fig. 601 E, G). Fruit a capsule.—Herbs, more rarely under-shrubs or shrubs, with latex and scattered, undivided leaves without stipules. The inflorescence is most frequently a raceme or spike with terminal flower.
A. Capsule opening at the side by pores and small valves: Campanula (Canterbury-bell); the corolla is bell-shaped, rarely almost rotate; capsule obconical. The pores of the capsule are found near the top of the fruit when it is erect, and near the base when it is pendulous,[562] so that the seeds are not liberated unless the capsule is forcibly shaken, and they are thus ejected to a considerable distance.—Phyteuma (Rampion) has free petals, which for a long time adhere at the apex and form a tube round the stamens (Fig. 601); inflorescence compact, spike-like or capitate, in the latter case resembling that of the Compositæ, and frequently with an involucre similar to the one possessed by this order. Specularia (rotate corolla, prismatic capsule), Michauxia (flower 8-merous).—Symphyandra has syngenesious anthers.
B. Capsule with valves at the apex, loculicidal dehiscence: Jasione; the petals are almost free. The anthers are united at the base (syngenesious). The flowers are situated in capitate umbels with involucres.—Wahlenbergia; Platycodon.
Fig. 601.—Phyteuma spicatum. Flowers and parts of flowers in various stages of development.
C. Berry: Canarina; flower, 6-merous; leaves opposite.
Protandry is general (Fig. 601). 510 species; principally in temperate countries. Several genera furnish ornamental plants, but are of little use for other purposes. The roots of some Campanula-and Phyteuma-species are large and may serve as pot-herbs (C. rapunculus, P. spicatum).
Order 2. Cyphiaceæ. In this order the corolla is zygomorphic and the stamens free, hence it is intermediate between orders 1 and 3.—About 24 species; Africa.
Order 3. Lobeliaceæ (Lobelias). This order may briefly be described as Campanulaceæ with zygomorphic flowers and anthers united into a tube, in most cases slightly bent; generally 2 carpels and an inverted position of the flower, i.e. the median sepal is[563] turned anteriorly (Fig. 602) (a position which is found to occur within the Campanulaceæ). A twisting of the peduncle takes place even before flowering (as in the Orchids) so that the ordinary position of the 5-merous Dicotyledons appears to be restored. The zygomorphy of the flower is especially present in the corolla, which has a bipartite under-lip and a tripartite upper-lip, and is, in Lobelia, anteriorly (apparently posteriorly) deeply cleft (Fig. 602). There is 1 style, but the stigma is capitate and bilobed and surrounded at its base by a whorl of hairs, which assists in pollination (as a stylar-brush) in the same manner as the sweeping-hairs in the Campanulaceæ and Compositæ. There is no terminal flower in the spicate, or racemose inflorescences.—Lobelia has a capsule, several others have berries. Isotoma (regular flower); Heterotoma has a spur; Siphocampylos; Lysipoma (pyxidium); Clintonia (1–locular fruit). Metzleria (all the petals are free).
Fig. 602.—Diagram of Lobelia fulgens.
Figs. 603, 604.—Lobelia syphilitica.
Fig. 603. Flower (2/1).
Fig. 604.—Longitudinal section of the same.
Entomophilous and protandrous. About 500 species, especially in the Tropics; in this country, L. dortmanna (margin of lakes).—Several are cultivated in gardens and conservatories as ornamental plants (Lobelia bicolor, erinus, fulgens, etc., Siphocampylos, Centropogon). The latex of several species of Tupa is poisonous; caoutchouc is also obtained from them. Officinal: “herba Lobeliæ” (the alkaloid lobeline) from the poisonous L. inflata (N. Am.).
Order 4. Goodeniaceæ. Chiefly Australian (200 species), closely related to Orders 3 and 5, but without latex. The style is provided with a “collecting-cup” which receives the pollen before the flower opens; it has a small, hairy aperture through which the pollen is forced out by the stigmas, and through which they emerge when the pollen is shed; it is sensitive and exhibits movements[564] when touched.—Herbs, under-shrubs, less frequently shrubs. Goodenia, Leschenaultia, Scævola.
Order 5. Stylidiaceæ (or Candolleaceæ); 100 species, the majority Australian; zygomorpbic flowers, but with the ordinary position. The anterior petal is very small. The chief characteristic feature is the presence of only 2 stamens (with extrorse anthers) which are united with the style and form a stylar-column; this is bent like a knee and sensitive at the bend to such a degree that when touched it jerks violently across the flower to the opposite side and then loses its sensitiveness.—Herbs, less frequently under-shrubs. Stylidium (Candollea).
The flowers, which are borne in “capitula” (Figs. 605, 610), are epigynous (Fig. 605 C, D), 5-merous in the calyx, corolla and andrœcium, the corolla is valvate in æstivation, with 2 carpels (S5, P5, A5, G2). The anthers are united into a tube (syngenesious) (except Ambrosieæ) which surrounds the bifid style. There is never more than 1 loculus in the ovary, with 1 erect, anatropous ovule. The fruit is a 1–seeded nut (cypsela), with thin pericarp, the calyx generally persists as a tuft of hairs (pappus) (Fig. 606) on the summit of the fruit. Embryo without endosperm; the radicle directed downwards.
Only 1 Order: Compositæ.
With respect to the inflorescence and the development of the individual flowers, there is a very close resemblance to the Dipsacaceæ, which stand on the same plane of progression as the Compositæ. But while the latter are allied to Campanulinæ as the last stage in the process of evolution, the Dipsacaceæ form the final stage of the Rubiales-Dipsacales.
Order Compositæ. (For the principal characteristics compare those of the family.) The Compositæ are chiefly herbs, but trees and shrubs also occur in tropical countries. The leaves may be scattered or opposite, but have no stipules. The outer leaves of the involucre as a rule are barren, especially when numerous and imbricate, while the innermost ones support the ray-flowers of the capitulum; in a few instances all are fertile (e.g. Tragopogon, Tagetes). The CAPITULA are many-flowered, with the exception, e.g. of Echinops, which has 1-flowered capitula (see page 570). The capitula are again arranged in inflorescences, most frequently corymbose with centrifugal order of development. The form of the receptacle is an important character for the division of the genera (flat, convex, conical), and also the presence of scales;[565] these may be one scale (bract) for each flower (Fig. 610 br), or a large number of bristles, which do not each correspond to a leaf, or the receptacle may be entirely without covering (naked). The flowers open in acropetal order in each capitulum. All the flowers in a capitulum may be of the same sex, and their form and colour are in that case the same, or the sexes may be different, in which case the form and colour are also most frequently different: the ray-flowers have projecting labiate or ligulate corollas, while the disc-flowers have tubular corollas. As a rule in the latter case the ♀ flowers are at the circumference, and the ☿ in the centre, less frequently ♀-flowers at the edge and ♂-flowers in the centre. The ray-flowers in some genera are neuter (e.g. Centaurea). Some are diœcious.
Fig. 605.—Calendula arvensis: A capitulum; B capitulum in longitudinal section; C ♀-flower; D ☿-flower; E the stamens; F capitulum with ripe fruits; G ripe fruit.
There is no trace of an epicalyx (in contrast to the Dipsacaceæ, which they generally so resemble). The formation of the CALYX is very varied. The calyx always consists of a very small cushion-like structure, most frequently developed later than the corolla; the 5 corners, which correspond to the 5 sepals, in a few[566] instances are raised as 5 large, flat, membranous bodies, e.g. in species of Xeranthemum, Catananche, Sphenogyne, etc.; in other instances each of these bears a shorter or longer bristle on its apex, followed by others in rather uncertain numbers and with but slight indications of order, on the edge and on the outer side of the calyx between the 5 points; in other instances, again, the calyx is covered with bristles and hairs without any indication of order or definite number (Fig. 606 a, b); finally instances occur in which the edge is raised as a membranous collar, irregularly toothed and notched, or divided into small scales. There are naturally differences in the means of distribution corresponding to the differences in structure of the calyx. The fruits a and b represented in Fig. 606 are distributed by the wind, those like c, on the other hand, by attaching themselves to animals and human beings. The rays of the pappus are termed rough when special cells project a little beyond the surface, but if these grow out, and are hair-like, the pappus is said to be feathery. In some genera the pappus is raised on a long stalk, which is developed from the upper part of the fruit, and termed a beak (Fig. 606 a). The pappus does not attain its full development till the ripening of the fruit, i.e. until it is about to be of use.
Fig. 606.—a Fruit of Taraxacum; b of Senecio; c of Bidens.
The COROLLA has various forms: (a) tubular (Fig. 605 D), with a[567] shorter or longer tube, not always of the same bore throughout and especially slightly widened at the top to form a bell-shaped opening, with 5 regular teeth: (b) labiate after 2/3, i.e. with 2 petals in the upper and 3 in the under lip: (c) ligulate, i.e. the corolla is split for a considerable distance on the posterior side (as in the Labiate genus Teucrium) and prolonged into a long, strap-like portion (Fig. 609 A), which projects upwards. A distinction must, however, be drawn between the true and false ligulate corolla. In the first case the corolla has 5 teeth at the apex (Fig. 609 A) and is made up of all the petals of the corolla united together; this is the usual condition in the Ligulate-flowered. In the latter case (Fig. 605 C) the tongue has only 3 teeth (or is more irregularly 2–3-dentate), and is only formed of 3 petals; the corolla is then truly bilabiate, the tongue is the large under lip, and the upper lip is very slightly developed, or even at an early stage quite suppressed. This false “ligulate” corolla is found among the ray-flowers; sometimes the upper lip is seen quite plainly, e.g. in Tagetes, especially in the double capitula. The VENATION of the corolla is peculiar; there are always commisural veins which branch dichotomously at the angles between the teeth of the corolla, and send a branch into the edge of the two nearest teeth. The midrib is frequently absent, but may be present, and then it has sometimes no connection with the other veins of the corolla.
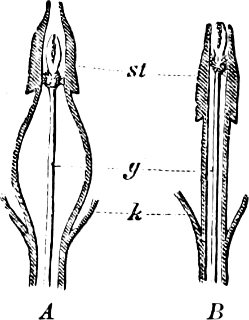
Fig. 607.—Centaurea cyanus: A the anther-tube (st) with the crescentic curved filament before irritation; g the style; k the base of the corolla; B the same after irritation, the anthers are drawn further down.
The STAMENS are attached to the corolla, and have free filaments (Silybum has united filaments), but the anthers, which at first are free, adhere together and form a tube (Fig. 605 E: only Ambrosieæ have free anthers). The connective is generally prolonged, and protrudes above the anthers as a thin, brown membrane of various forms (Fig. 605 E); appendages of various forms may also be found at the base of the anthers. The anthers open introrsely, and the pollen must be carried out at the top of the tube by upward growth of the style, and by means of the “stylar-brush” (Figs. 607, 608, 609); the filaments are sometimes sensitive (e.g. in the Corn-flower, Fig. 607), and shorten on being touched, so that the anther-tube is pulled downwards, and the pollen swept out at the top (Figs. 607, 608 A, B).
[568]
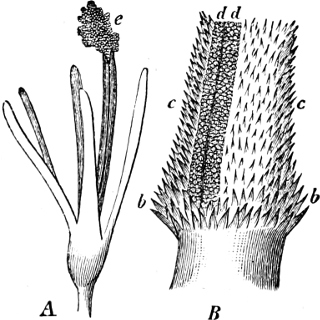
Fig. 608.—Cirsium arvense: A the upper portion of a flower, the pollen (e) is being ejected; B part of the upper portion of the style with stylar-brush (b, c) and the stigmatic papillæ (d).
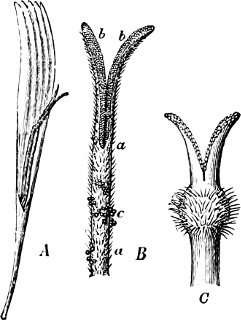
Fig. 609.—Leontodon autumnale: A ligulate flower; B extremity of the style with stylar-brush (a), stigma (b) and pollen-grains (c). C Centaurea cyanus.
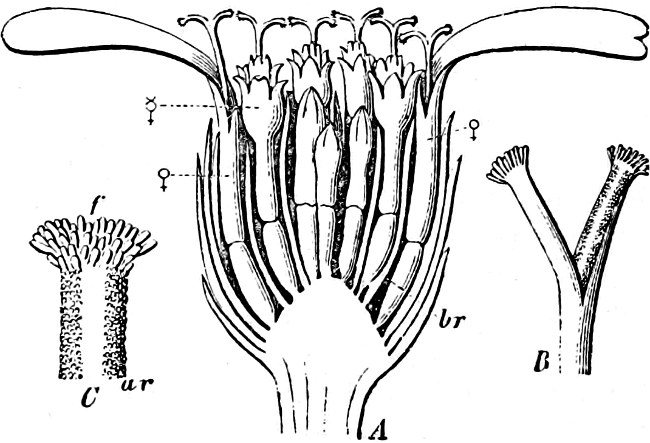
Fig. 610.—Achillea millefolium.
The STYLE divides at the apex into two branches (Figs. 609, 610), both of which generally bear on the inner surface two lines of stigmatic papillæ (Fig. 610 B, C) and being in shape, etc., very varied, are therefore employed as systematic characters.—The most important types are: A. The style is uniformly cylindrical; its branches are semi-cylindrical, long, and with long hairs, and finally bend backwards; the stylar branches bear slightly projecting stigmatic papillæ on the inner side. This form is characteristic of the Cichorieæ (Fig. 609 A B). B. The style is uniformly cylindrical; the branches are long, cylindrical or club-like, short,[569] not rolled back, with fine hairs externally; the stigmatic lines do not reach beyond the centre, and do not meet together. Characteristic of Eupatorium, Petasites, Tussilago. C. The style is thickened beneath the stigmatic branches in the form of a knob, or very hairy (Fig. 609 C); the stigmatic lines reach as far as the apex of the branches and then converge; sometimes the stigmatic branches are united as far as the apex. Characteristic of the Cynareæ. D. The stylar branches are lanceolate, or linear, pointed; externally flat and thickly covered with hairs in the upper portion; the stigmatic lines cease where the hairs commence externally. Characteristic of Aster, Bellis, Inula, Dahlia, etc. E. The stylar branches are linear, with long, brush-like hairs at the apex, where they are either abruptly cut off or prolonged into a very hairy, conical appendage; the stigmatic lines are broad, reach as far as the brush-like hairs, and do not meet together (Fig. 610). Characteristic of Senecio, Helianthus, Xanthium, Gnaphalium, Artemisia, Anthemis, and others related to these.
A ring-like nectary is found round the base of the style.
The thin-walled cypsela (Fig. 606), with seeds fitting closely to the pericarp, has many different forms (smooth, ribbed, spined, etc.); its point of attachment generally lies at the lowest end but sometimes it is drawn obliquely up the side (Centaurea, etc.). The calyx, persistent on the apex of the fruit, has been described above. Some genera have two or three different forms of fruits in each capitulum.—The embryo is straight, with the radicle turned downwards, and without endosperm, but is rich in oil.
The variously flowered capitula, whose normal tubular disc-flowers have been changed to ligulate flowers, may be termed “double flowers.”
The relationship of the Compositæ to the Campanulinæ has been described above (page 561). The alliance with the Dipsacaceæ is more apparent than real. Similar capitate inflorescences also occur as the final stage in other lines of descent, as in Eryngium among the Umbelliferæ.
1. Cynareæ, Thistle Group. Flowers all ☿, regular, with tubular corollas. The receptacle is covered with numerous bristles, which surround the flowers without any definite order, or the edges of the grooves in which these are placed have a well-marked fringe. The involucral leaves are numerous, imbricate, and are either prolonged into a thorn or terminate with a membranous edge. The style has been described on page 568 (Fig. 609 C). Nearly all have a hairy or feathery pappus. The filaments are sensitive.
Carduus (Thistle); capitula ovoid; involucral leaves compact, imbricate, with thorny points; the pappus-rays are hair-like and united at the base by a ring (i.e. the calyx), and fall off together.—Cirsium (Fig. 608) has a feathery pappus, in other respects it is like Carduus. C. arvense reproduces and passes the winter by means of[570] suckers.—Cynara (Artichoke) has a feathery pappus and large, solitary capitulum, with broad involucral leaves; these have a fleshy base like the receptacle (edible).—Silylum has united filaments. S. marianum (Milk-thistle), has leaves with numerous white spots. Onopordon (Cotton-thistle). Cnicus (C. benedictus) has a large, many-spined thorn on the involucral leaves; pappus trimorphic.—Lappa (Burdock) is easily recognized by the hooked involucral leaves, which assist in the distribution of the fruit; in this respect it differs from the other inflorescences, and also in the fact that the pappus is short, and quickly falls off, without serving as a means of distribution.—Carlina; the external involucral leaves are leafy, thorny, with branched or unbranched spines standing straight out or bent backwards; the internal ones are dry, and prolonged as dry, coloured, radiating scales. The well-developed bristles on the receptacle and edge of the calyx are deeply cleft and lobed.—Centaurea (Knap-weed, Fig. 607). The ray-flowers are neuter, and generally larger than the disc-flowers; the involucral leaves are regularly imbricate, but are frequently provided at the apex with a dry, chaffy, often lobed, fringed appendage. The attachment of the fruit is lateral. Serratula (Saw-wort).—Carthamus, the outer and inner involucral leaves differ very much.—Echinops (Globe-thistle) is characterised by having “compound capitula,” i.e. there is only one flower in each capitulum, but many such capitula are collected into a spherical head, which at the base may also have a few involucral leaves. The individual capitula have narrow, linear involucral leaves. (There are altogether about 150 species of Compositæ with 1-flowered capitula, all from warm countries.)—Xeranthemum, Staehelina, Jurinea, Saussurea, etc.
2. Mutisieæ, Labiate-flowered Group. Tropical (S. American) forms whose zygomorphic flowers have a bilabiate corolla (2/3). The involucre is nearly the same as in the Thistles.
3. Cichorieæ, Chicory Group (or Ligulifloræ). The flowers are all ☿ and have a ligulate, 5-dentate corolla. The stylar branches are thin and prolonged (Fig. 609 B). Laticiferous vessels occur in the majority (in this feature they resemble the Lobeliaceæ and Campanulaceæ).
A. The pappus is wanting, or it is scale-like, but not long and hairy.—Cichorium (Chicory); capitula with blue flowers, borne singly or a few together in the leaf-axil; there are two whorls of involucral leaves, an outer one of short and radiating, an inner of more numerous, longer and erect leaves; pappus, scale-like.—Lapsana (Nipplewort). The few involucral leaves are nearly of the same size, and persist forming a sort of capsule round the[571] fruits, which are entirely without a pappus. There are only a few flowers in the small capitula.—Arnoseris (Swine’s-succory), Catananche, etc.
B. The pappus is long and hairy (not branched), generally fine and snowy-white. There are no scales on the receptacle. The two genera first considered have beaked fruits.—Taraxacum (Dandelion) (Fig. 606 a); the capitula are many-flowered, and borne singly on the top of a leafless, hollow stalk.—Lactuca (Lettuce) has many small, few-flowered capitula borne in panicles.—Crepis (Hawksbeard).—Hieracium (Hawk-weed) has many imbricate involucral leaves, and a stiff, brittle, brownish pappus.—Sonchus (Sow-thistle); the capitula, when a little old, have a broad base, and are abstricted above in the form of a jug; involucral leaves imbricate; the fruit is compressed, without a beak, ridged. The soft, white pappus falls off collectively.
C. The pappus is feathery and branched; no scales on the receptacle.—Tragopogon (Goat’s-beard) generally has 8 involucral leaves in one whorl. The fruit has a long beak; the rays of the pappus are interwoven in the form of an umbrella.—Scorzonera has fruits like the preceding, but almost without any beak; involucral leaves many, imbricate.—Leontodon (Hawkbit) has a slightly feathery pappus, rays not interwoven; beak absent.—Picris.
D. Long, chaff-like, deciduous scales on the receptacle; pappus feathery.—Hypochœris (Cat’s-ear).
4. Eupatorieæ, Hemp-agrimony Group. All the flowers are most frequently ☿; corollas tubular and regular; the involucral leaves are not stiff and spiny; the receptacle is not covered with stiff bristles. The stylar branches are long, club-like, or gradually tapering. There is no swelling below the stigma.
Eupatorium (Hemp-agrimony); all the flowers are ☿.—Petasites (Butterbur); ray-flowers ♀, disc-flowers ☿ or ♂; sometimes diœcious. Capitula in racemes or panicles. The leaves develop after the flowering.—Tussilago (T. farfara, Colt’s-foot) has a solitary capitulum borne on a scaly, scape-like stem; the ray-flowers are ♀ with ligulate corollas, disc-flowers ♂. The leaves unfold after the flowering. Ageratum, Mikania, Vernonia.
5. Astereæ, Aster Group (or Radiatæ, Ray-flowered). The flowers are of two forms and different sexes; the ray-flowers are ♀ (sometimes neuter), most frequently with irregular, falsely ligulate, radiating corollas; the disc-flowers are ☿, regular, with[572] tubular corollas (Fig. 610). Sometimes only tubular flowers are present, as e.g. in Senecio vulgaris (Groundsel), and the exterior of the capitulum is then as in the Eupatorieæ. The stylar branches are straight, more or less flat and short (Fig. 610).
A. Anthemideæ. Involucral leaves imbricate, generally membranous at the edge; pappus wanting, or at most a membranous margin to the calyx, but without hairs.
[+]. Chaff-like bracts on the receptacle are found in Anthemis (Chamomile), Anacyclus (A. officinarum), Achillea (Milfoil, Fig. 610), Santolina, etc.
[++]. A naked receptacle is found in the following: Bellis (Daisy) has solitary capitula on leafless stalks with white ray-flowers.—Matricaria (Wild Chamomile) has a conical receptacle. (M. chamomilla has a very high, hollow receptacle; M. inodora has large, odourless capitula, and the receptacle is not hollow.)—Chrysanthemum (Ox-eye) most frequently large, solitary capitula; flat receptacle.—Pyrethrum; pappus scanty.—With these are classed Tanacetum (Tansy) and Artemisia (Wormwood) with tubular corollas only.
B. Heliantheæ. Most frequently a bract to each flower is found on the receptacle. The pappus is never exactly hairy, but consists of scales, spines, etc., and the fruits are most frequently compressed (Fig. 606 c).—Helianthus (Sun-flower); H. tuberosus (Jerusalem Artichoke) has tuberous underground stems. Dahlia has tuberous roots (Am.). Bidens (Bur-marigold, Fig. 606 c); the fruits are compressed with 2 (or more) spines provided with reflexed barbs.—Calliopsis; Rudbeckia; Zinnia; Tagetes has united involucral leaves, and yellow, transparent oil-glands. Spilanthes, Galinsoga, Melampodium, Silphium (Compass-plant), Helenium, Gaillardia.
C. Calenduleæ have 1–2 rows of involucral leaves, a naked receptacle, and large, crescent-shaped, irregularly warted fruits, of different forms in the same capitulum; pappus absent (Fig. 605).—Calendula (Marigold); ray-flowers ♀, disc-flowers ♂.
D. Senecioneæ, have a fine, hairy, white pappus; no bracts, otherwise as in Anthemideæ. The involucral leaves are most frequently in 1–2 rows.—Senecio (Groundsel) has two whorls of involucral leaves, which most frequently have black tips, the external being much shorter than the internal ones (S. vulgaris has all flowers ☿ and alike).—Cacalia, Doronicum, Cineraria, Ligularia, Arnica (A. montana; large, long-stalked capitula; leaves opposite, forming a kind of rosette).
[573]
E. Astereæ have a bristle-like, unbranched pappus, often of a dingy brown; receptacle naked; involucral leaves numerous, imbricate.—Solidago (Golden-rod); capitula small, yellow-flowered, borne in panicles. Aster; disc-flowers most frequently yellow, ray-flowers violet; Callistephus; Erigeron (Flea-bane)—Inula.—All the corollas are tubular in: Gnaphalium (Cud-weed); involucral leaves dry, rattling, often coloured; the foliage-leaves and stem often white with woolly hairs; ray-flowers ♀, with narrow, tubular corolla; disc-flowers ☿ (few). Antennaria (Cat’s-foot; diœcious), Filago, Helichrysum, Ammobium, Rhodanthe and others. Leontopodium (L. alpinum, “Edelweiss”).
F. Ambrosieæ, a very reduced type of Compositæ, differing from the others in having free anthers; the capitula are generally unisexual, monœcious, the ♂ borne in a terminal inflorescence, the ♀ in the leaf-axils. In other respects they are most closely related to Heliantheæ.—Xanthium. In the ♂-capitula there are many flowers without calyx, but with tubular corolla and free involucral leaves. In the ♀-capitula there are only 2 flowers, which are entirely destitute of both calyx and corolla; involucral leaves 2-spined, united to form an ovoid, bilocular envelope, each compartment containing one flower. The envelope of involucral leaves unites with the fruits, enclosing them at maturity with a hard covering from which numerous hook-like spines project, assisting very greatly in the distribution of the fruit. The whole structure thus finally becomes a 1- or 2-seeded false nut.—Ambrosia, the ♀ capitulum 1-flowered.
Pollination. The flowers are somewhat insignificant, but become very conspicuous owing to a number being crowded together in one inflorescence. The corollas of the ray-flowers, being often very large (Astereæ; Centaurea), frequently render the capitula still more conspicuous. The capitula display many biological phenomena similar to those often shown by the individual flowers in other orders, e.g. by periodically opening and closing, in which the involucral leaves resemble the calyx in their action. (The name “Compositæ” originates from the term “flos compositus,” composite flower). An abundance of honey is formed, which to some extent fills up the corolla-tube, and since insects may visit a number of flowers in the course of a short period they are very frequently visited, especially by butterflies and bees. The pollination has been described on page 567. Protandry is universal. In the bud the tips of the styles, covered by the sweeping-hairs, lie closely enveloped by the anther-tube; in the next stage the style grows through the tube and sweeps out the pollen as it proceeds; ultimately the stylar branches expand and the stigma is then prepared to receive the pollen. In many, the sensitiveness of the filaments assists in sweeping out the pollen at the exact moment of the insect visit. Regular self-pollination is found e.g. in Senecio vulgaris; wind-pollination e.g. in Artemisia and the plants related to it.
This extremely natural and well-defined order is the largest (and no doubt one of the youngest?); it embraces 10–12,000 known species (in 770 genera), or about one-tenth of all Flowering-plants. They are distributed over the whole globe, but are most numerous in temperate countries; the majority prefer open[574] spaces; a smaller number are forest-forms. They abound especially in open districts in America.
Among the substances frequently found may be mentioned: Inulin (especially in the subterranean parts), Bitter materials, Tannin, volatile oils, fatty oils in the fruits. Medicinal:[40] “Herba” of Artemisia absinthium (Wormwood) and maritima[+] (Sea-wormwood), Achillea millefolium; the leaves of Cnicus benedictus and Tussilago farfara; the unopened capitula of Artemisia maritima, var. stechmanniana; the capitula of Tanacetum, Matricaria chamomilla[+] (wild Chamomile), Anthemis nobilis[+] (common Chamomile); the separate flowers of Arnica; the roots of Arnica montana[+], Taraxacum officinale[+], Anacyclus officinarum[+], Lappa major, minor, nemorosa and tomentosa, Inula helenium and Artemisia vulgaris; the latex of Lactuca virosa[+]. The following are cultivated for food:—Lactuca sativa (Lettuce), Cichorium endivia (from E. Asia, for salads), Cynara scolymus (Artichoke, Mediterranean), Scorzonera hispanica (S. Eur.), Helianthus tuberosus (Jerusalem Artichoke, from N. Am., introduced into Europe 1616), Cichorium intybus (roots as “chicory,”) Tragopogon porrifolium (Salsafy), Artemisia dracunculus. Oil is extracted from the following (the seeds): Helianthus annuus (Peru), Madia sativa (Chili), Guizotia oleifera (Abyssinia). Dyes from: Carthamus tinctorius (Safflower, used in the preparation of rouge; Egypt), Serratula tinctoria. Insect-powder from: Pyrethrum cinerariifolium (Dalmatia) and roseum (Persia, Caucasus). The following are cultivated in houses and gardens for the sake of their scented leaves:—Tanacetum balsamita (Balsam), Artemisia abrotanum (Southernwood) and A. argentea. A great many of the genera enumerated are cultivated in dwelling-houses for the sake of the flowers; e.g. Pericallis cruenta (generally termed “Cineraria”). Asteriscus pygmæus is supposed to be the genuine “Rose of Jericho”; the involucral leaves envelop the fruits after their ripening and keep them enclosed for 8–10 months until rain occurs.
[575]
By M. C. POTTER.
The earliest systems of classification were derived from the properties and uses of plants; and it was not until some two centuries ago that any scientific grouping of plants was attempted. Aristotle and Theophrastus had adopted the groups of Trees, Shrubs and Herbs as the chief divisions of the Vegetable Kingdom, a system which persisted and was employed by Tournefort and Ray as late as the end of the 17th century. The arrangement by which these three divisions were separated into smaller divisions was often founded upon a single character, such as the formation of the corolla, the form of fruit, that of the calyx and corolla, etc. All these systems of classification which brought into close proximity plants distinguished by some one character alone, could only be considered as artificial, since plants related to one another would not necessarily be included in the same group. As the knowledge of the morphology, physiology, and reproduction of plants increased, such systems were recognised as unscientific, and it became the aim of botanists to establish a natural system, founded upon mutual relationships, which would associate together only those plants which are truly allied.
The following are some of the chief systems of classification which will show the gradual development of the natural system, and may be of service to students making use of this text-book.[41]
System of John Ray (1703).
Ray was the first botanist who recognised the importance of the one or two seed-leaves of the embryo, and initiated the division of the Flowering-plants into Monocotyledons and Dicotyledons.
[576]
System of Linnæus (1733).
In his well known artificial system Linnæus divided the Vegetable Kingdom into twenty-four classes, based upon the number, relative position and union of the stamens with regard to each other, and also to the gynœceum.
| Class | I. | Monandria. | Flowers with | 1 | stamen. |
| „ | II. | Diandria. | „ „ | 2 | stamens. |
| „ | III. | Triandria. | „ „ | 3 | „ |
| „ | IV. | Tetrandria. | „ „ | 4 | „ |
| „ | V. | Pentandria. | „ „ | 5 | „ |
| „ | VI. | Hexandria. | „ „ | 6 | „ |
| „ | VII. | Heptandria. | „ „ | 7 | „ |
| „ | VIII. | Octandria. | „ „ | 8 | „ |
| „ | IX. | Enneandria. | „ „ | 9 | „ |
| „ | X. | Decandria. | „ „ | 10 | „ |
| „ | XI. | Dodecandria. | „ „ | 11 to 19 stamens. | |
| „ | XII. | Icosandria. | „ „ | 20 or more stamens inserted on the calyx. | |
| „ | XIII. | Polyandria. | „ „ | 20 or more stamens inserted on the receptacle. | |
| „ | XIV. | Didynamia. | Stamens didynamous. | ||
| „ | XV. | Tetradynamia. | „ tetradynamous. | ||
| „ | XVI. | Monadelphia. | Filaments united into 1 bundle. | ||
| „ | XVII. | Diadelphia. | „ „ „ 2 bundles. | ||
| „ | XVIII. | Polyadelphia. | „ „ „ several bundles. | ||
| „ | XIX. | Syngenesia. | Anthers united together. | ||
| „ | XX. | Gynandria. | Stamens and pistil united. | ||
| „ | XXI. | Monœcia. | Flowers diclinous, ♂ and ♀ on the same plant. | ||
| „ | XXII. | Diœcia. | „ „ ♂ and ♀ on different plants. | ||
| „ | XXIII. | Polygamia. | ♂-, ♀-, and ☿-flowers on the same plant. | ||
| „ | XXIV. | Cryptogamia. | Flowerless plants (Ferns, Mosses, Algæ, Fungi). | ||
These classes were further divided into orders, according to the number of styles, as Monogynia, flowers with 1 style; Digynia, with 2 styles, etc. Thus a Dock (Rumex), having 6 stamens and 3 styles, would be placed in Class VI., Hexandria, and Order III., Trigynia.
Class XIV. was divided into two orders. Order I., Gymnospermia, with seeds apparently naked, comprising the Labiatæ; and Order II., Angiospermia, with the seeds enclosed in a capsule (Bartsia, Rhinanthus).
Class XV. was divided into two orders: Order I., Siliculosa, fruit a silicula (Capsella); and Order II., Siliquosa, fruit a siliqua (Brassica).
Class XIX. was divided into Order I., Æqualis, all the flowers perfect (Sonchus); Order II., Superflua, flowers in the centre perfect, those at the circumference with pistils only (seemingly superfluous), e.g. Aster; Order III., Frustranea, flowers in the centre perfect, those at the circumference neuter, e.g. Centaurea.
“Fragments” of a natural system have also come down to us from Linnæus, who himself always recognised the imperfection of his artificial system.
[577]
System of Antoine Laurent de Jussieu (1789).
| Class | |||||
|---|---|---|---|---|---|
| Acotyledones. Plants without cotyledons: Fungi, Ferns, Mosses, Algæ, Naiades | I. | ||||
| Monoctyledones. Plants with one cotyledon:— | |||||
| 1. Stamens hypogynous | II. | ||||
| 2. „ perigynous | III. | ||||
| 3. „ epigynous | IV. | ||||
| Dicotyledones. Plants with two cotyledons:— | |||||
| 1. Apetalæ |  |
Stamens epigynous | V. | ||
| „ perigynous | VI. | ||||
| „ hypogynous | VII. | ||||
| 2. Monopetalae | |
Corolla hypogynous | VIII. | ||
| „ perigynous | IX. | ||||
| „ epigynous, | |
anthers connate | X. | ||
| „ free | XI. | ||||
| 3. Polypetalæ | |
Stamens epigynous | XII. | ||
| „ hypogynous | XIII. | ||||
| „ perigynous | XIV. | ||||
| 4. Diclines irregulares, male and female flowers on different plants, corolla generally absent. | |||||
System of A. P. de Candolle (1819).
[578]
Robert Brown published in 1827 his discovery of the gymnospermy of the ovules of the Coniferæ and Cycadeæ, and showed that the Gymnosperms, which had previously been classed with the Dicotyledons, must be regarded as an independent group.
System of Stephen Endlicher (1836–40).
System of A. Brongniart (1843).
[579]
System of John Lindley (Vegetable Kingdom, 1845).
| Asexual, or Flowerless Plants. | ||
| Stem and leaves undistinguishable | I. | Thallogens. |
| Stem and leaves distinguishable | II. | Acrogens. |
| Sexual, or Flowering Plants. | ||
| Fructification springing from a thallus | III. | Rhizogens. |
| Fructification springing from a stem. | ||
| Wood of stem youngest in the centre; cotyledon single. Leaves parallel-veined, permanent; wood of stem always confused | IV. | Endogens. |
| Leaves net-veined, deciduous; wood of the stem, when perennial, arranged in a circle with a central pith | V. | Dictyogens. |
| Wood of stem youngest at the circumference, always concentric; cotyledons, 2 or more. | ||
| Seeds quite naked | VI. | Gymnogens. |
| Seeds inclosed in seed-vessels | VII. | Exogens. |
System of Alexander Braun (1864).
W. Hofmeister published from 1849 to 1851 his researches upon the embryology of the Phanerogams, and upon the embryology and life-history of the Vascular Cryptogams, and established the phylogenetic connection existing between the Mosses, Vascular Cryptogams and Phanerogams.
[580]
System of Hooker and Bentham (Genera plantarum, 1862–1883).
DICOTYLEDONES.
I. POLYPETALÆ.
Series I. Thalamifloræ. Calyx most often free from the ovary. Petals uniseriate or often 2–∞-seriate. Stamens ∞ or definite, inserted on the receptacle, often small, or raised, or stipitate. Ovary most frequently free.
Cohort I. Ranales. Stamens ∞, or if definite the perianth is 3–∞-seriate. Carpels apocarpous, or immersed in the receptacle. Endosperm usually abundant, fleshy.
Cohort II. Parietales. Stamens ∞ or definite. Ovary unilocular, or divided into loculi by spurious dissepiments, with parietal placentation. Endosperm absent or fleshy.
Cohort III. Polygalinæ. Stamens definite. Ovary usually perfectly or imperfectly bilocular. Micropyle often superior. Fruit very often compressed laterally. Endosperm very often abundant and fleshy.
Cohort IV. Caryophyllineæ. Stamens definite, or rarely ∞. Ovary unilocular, or imperfectly septate. Placenta central, more rarely parietal. Micropyle inferior. Embryo curved, rarely straight. Endosperm farinaceous.
Cohort V. Guttiferales. Sepals inbricate. Stamens usually ∞. Ovary septate, placentæ on the inner angles of the loculi. Endosperm absent or fleshy.
[581]
Cohort VI. Malvales. Sepals valvate. Stamens usually ∞ or monadelphous. Ovary septate, placentæ on the inner angles of the loculi. Endosperm absent or fleshy.
Series II. Discifloræ. Calyx usually free from the ovary. Petals uniseriate. Stamens usually definite, inserted within, or upon, or around the receptacle, which is more often expanded as a disc. Ovary usually free, or embedded in the disc.
Cohort VII. Geraniales. Disc usually as a ring between the stamens, or adnate to the staminal tube, or reduced to glands alternating with the petals, more rarely absent. Gynœceum entire, or more often lobed, or sub-apocarpous. Ovules most often 1–2 in each loculus, pendulous, raphe ventral. Leaves various.
Cohort VIII. Olacales. Disc cupular or annular, free, or bearing the stamens and petals on its edge. Gynœceum entire. Ovules 1–3 in the unilocular ovaries, or 1–2 in each loculus, pendulous, raphe dorsal. Leaves simple.
Cohort IX. Celastrales. Disc tumid, adnate to the calyx, or covering its base. Stamens inserted round the disc or affixed to its margin. Gynœceum usually entire. Ovules most often two in each loculus, erect, raphe ventral. Leaves simple, or rarely compound.
Cohort X. Sapindales. Disc various. Stamens variously inserted on the disc. Gynœceum entire, or more often lobed, or sub-apocarpous. Ovules more often 1–2 in each loculus, ascending with ventral raphe, or reversed, or solitary and pendulous from an ascending funicle, or rarely ∞ horizontal. Leaves pinnate, or more rarely simple or digitate.
Anomalous orders, or rather genera,—
Series III. Calycifloræ. Calyx-tube usually surrounding the ovary, or adnate to it. Petals uniseriate, inserted on the calyx-tube. Stamens ∞ or definite, inserted on the calyx-tube, or most often on the disc lining the calyx-tube. Ovary often enclosed by the calyx-tube, or inferior.
[582]
Cohort XI. Rosales. Carpels solitary, or free, or united at the base, more rarely at the apex; styles distinct, or very rarely united into a column, and easily separated.
Cohort XII. Myrtales. Ovary syncarpous, inferior, or enclosed in the calyx-tube, usually divided into loculi; style undivided. Ovules 2–∞ in the loculi.
Cohort XIII. Passiflorales. Ovary syncarpous, inferior or superior, enclosed in the calyx-tube or exserted, unilocular with parietal placentation, or divided into loculi; styles distinct, one style divided, or undivided.
Cohort XIV. Ficoidales. Ovary syncarpous, inferior or superior, divided into loculi with sub-basilar placentæ, or more rarely unilocular with parietal placentæ. Styles distinct, or divided at the apex. Embryo curved or excentric.
Cohort XV. Umbrellales. Ovary syncarpous, inferior, crowned by the disc, divided into loculi, or unicarpellate. Styles distinct or divided at the apex. Ovules solitary and pendulous in the loculi.
II. GAMOPETALÆ.
Series I. Inferæ. Ovary inferior. Stamens equal to the lobes of the corolla, rarely fewer.
Cohort I. Rubiales. Stamens adnate to the corolla. Ovary 2–∞-locular, loculi 1–∞-ovuled.
Cohort II. Asterales. Stamens adnate to the corolla. Ovary formed of 2 carpels, unilocular and 1-ovuled.
[583]
Cohort III. Campanales. Stamens generally free from the corolla. Ovary 2–6-locular, loculi most often ∞-ovuled.
Series II. Heteromeræ. Ovary most often superior. Stamens free from the corolla, opposite to, or double the lobes of the corolla, or ∞, or if epipetalous, equal and alternating with them. Carpels more than 2.
Cohort IV. Ericales. Stamens double the lobes of the corolla, or alternating with them. Ovary 2–∞-locular. Seeds small, frequently minute.
Cohort V. Primulales. Stamens equal to and opposite the lobes of the corolla. Ovary unilocular, with a free central placenta, 1–∞ ovules.
Cohort VI. Ebenales. Stamens equal to and opposite the lobes of the corolla, or double, or ∞. Ovary 2–∞-locular. Seeds few and large. Trees or shrubs.
Series III. Bicarpellatæ. Ovary most often superior. Stamens equal, or fewer than the lobes of the corolla, and alternating with them. Carpels 2, rarely 1 or 3.
Cohort VII. Gentianales. Corolla regular. Stamens equal to the lobes of the corolla, or if fewer, usually alternating with the carpels. Leaves generally opposite.
Cohort VIII. Polemoniales. Corolla regular. Stamens equal to the lobes of the corolla. Leaves generally alternate.
Cohort IX. Personales. Corolla most often irregular or oblique. Posterior stamen less than the others, more often reduced to a staminode, or altogether absent. Ovary ∞-ovuled, or 2-ovuled.
[584]
Cohort X. Lamiales. Corolla most often irregular or oblique. Posterior stamen less than the others, most frequently reduced to a staminode or absent. Carpels 1-ovuled or with 2 collateral ovules. Fruit enclosed in the persistent calyx, indehiscent, and with one seed, or dehiscing into 2 or 4, rarely ∞, 1-seeded nuts.
Anomalous Order 127. Plantagineæ.
III. MONOCHLAMYDEÆ.
Perianth simple, lobes or segments 1–2-seriate and often sepaloid, or small, or wanting.
Series I. Curvembryeæ. Endosperm frequently farinaceous. Embryo curved, excentric, lateral or peripheral, rarely straight. Ovules most frequently 1 in the ovary, or 1 in each loculus. Flowers ☿, in some genera unisexual or polygamous. Petals very rare. Stamens equal to the segments of the perianth, rarely fewer or more.
Series II. Multiovulatæ Aquaticæ. Aquatic herbs, submerged. Ovary syncarpous; ovules numerous in each loculus or on each placenta.
Series III. Multiovulatæ Terrestres. Terrestrial trees or shrubs. Ovary syncarpous; ovules numerous in each loculus or on each placenta.
Series IV. Micrembryeæ. Ovary syncarpous, monocarpous, or apocarpous. Ovules generally solitary in each carpel, rarely 2 or few. Endosperm copious, fleshy, or rarely farinaceous. Embryo very minute.
Series V. Daphnales. Ovary monocarpous, very rarely syncarpous, with 2–4 loculi; ovules in the ovary or in each loculus, solitary, or in pairs. Trees or shrubs, very rarely herbs; flowers generally ☿. Perianth perfect, sepaloid, 1–2 seriate. Stamens perigynous, equal to the lobes of the perianth, or double unless fewer.
Series VI. Achlamydosporeæ. Ovary unilocular, 1–3 ovules. Ovules most frequently poorly developed before flowering. Seeds endospermous,[585] but without testa, either free in the pericarp or attached to its walls. Perianth generally perfect, sepaloid or petaloid.
Series VII. Unisexuales. Flowers unisexual. Ovary syncarpous or monocarpous, ovules in the ovary or in each loculus, solitary, or in pairs. Endosperm copious, fleshy, or scanty, or absent. Trees or shrubs, rarely herbs. Stipules generally present. Perianth sepaloid, or minute, or absent. Styles equal in number to the carpels, not rarely bifid.
Series VIII. Ordines Anomali. Anomalous Orders.
GYMNOSPERMEÆ
MONOCOTYLEDONES.
Series I. Microspermæ. At least the inner series of the perianth petaloid. Ovary inferior, unilocular, with 3 parietal placentæ, or rarely 3-locular, with axile placentation. Seeds minute, numerous, without endosperm.
Series II. Epigynæ. At least the inner series of the perianth petaloid. Ovary most often inferior. Endosperm copious.
Series III. Coronarieæ. At least the inner series of perianth petaloid. Ovary free, very rarely slightly adnate at the base. Endosperm copious.
[586]
Series IV. Calycinæ. Perianth sepaloid, small, rigid, or herbaceous (inner series subpetaloid or small). Ovary free. Endosperm copious.
Series V. Nudifloræ. Perianth absent, or reduced to hairs or scales. Ovary superior, carpel solitary, or if many, syncarpous, 1–∞-ovuled. Endosperm most frequently present.
Series VI. Apocarpæ. Perianth 1–2-seriate, or absent. Carpels superior, solitary, or if more, apocarpous. Endosperm absent.
Series VII. Glumaceæ. Flowers solitary, sessile in the axils of bracts and arranged in capitula or spikelets with bracts. Segments of perianth small, scale-like, glumaceous or absent. Ovary 1-ovuled, or divided into 1-ovuled loculi. Endosperm present.
Classification of the Thallophytes proposed by Sachs (Text-Book of Botany, English Edition, 1882).
| THALLOPHYTES. | |||
| Containing chlorophyll. | Not containing chlorophyll. | ||
| Class I. Protophyta. | |||
| Cyanophyceæ. | Schizomycetes. | ||
| Palmellaceæ (in part). | Saccharomycetes. | ||
| Class II. Zygosporeæ. | |||
| Conjugating cells motile. | |||
| Pandorineæ. | Myxomycetes. | ||
| (Hydrodictyeæ). | |||
| Conjugating cells stationary. | |||
| Conjugatæ (including Diatomaceæ). | Zygomycetes. | ||
| Class III. Oosporeæ. | |||
| Sphæroplea. | |||
| Vaucheria (Cœloblastæ). | |
Saprolegnieæ. | |
| Peronosporeæ. | |||
| Volvocineæ. | |||
| Œdogonieæ. | |||
| Fucoideæ.[587] | |||
| Class IV. Carposporeæ. | |||
| Coleochæteæ. | Ascomycetes (including Lichens). | ||
| Florideæ. | Æcidiomycetes (Uredineæ). | ||
| Characeæ. | Basidiomycetes. | ||
System of A. W. Eichler (1883).
The subdivisions of the Phanerogamæ have with little variation been adopted in this book.
[588]
Classification of the Thallophytes, adopted in the 3rd Danish Edition (1891). [Algæ by Wille; Fungi by Rostrup (after Zopf).]
[589]
System of A. Engler (Syllabus der Vorlesungen, etc., 1892).
[592]
TABLE OF ABBREVIATIONS.
Names of continents and countries have sometimes been abbreviated, for example:—Am. = America; As.=Asia; Af. = Africa; Ind. = India, etc. N., S., E., W., = North, South, East, West; Temp. = Temperate Regions; Trop. = Tropics.
[593]
Butler & Tanner, The Selwood Printing Works, Frome, and London.
[1] See Angiospermæ.
[2] According to the recent investigations of Winogradsky some micro-organisms (Nitrifying-bacteria) can build organic from inorganic matter. Sachs’ hypothesis that the first organisms must necessarily have contained chlorophyll is therefore untenable.
[3] Myxogasteres, Engler’s Syllabus, p. 1.
[4] Acrasieæ and Plasmodiophorales, ibid.
[5] Myxophyceæ, Cyanophyceæ.
[6] The Bacteria are more usually included under Fungi. It seems better, however, to place them under the Algæ in a separate class with the Schizophyceæ.
[7] See Marshall Ward, “On the Characters or Marks employed for Classifying the Schizomycetes,” Annals of Botany, 1892.
[8] According to Hansen these are not disease forms, but occur regularly under certain conditions, e.g. temperature.
[9] Before fertilisation the oosphere divides and cuts off at the base one or more cells (polar bodies?), termed “wendungszellen.”
[10] From the Greek μὐκης = Fungus, hence “mycology.”
[11] This term is adopted as a translation of the German “anlage.”
[12] Also termed Water-Fungi (Wasserpilzen).
[13] Antheridium is preferred in this sub-class as keeping a more uniform term (Kn).
[14] In the resupinate fruit-bodies a fertile and sterile surface cannot be distinguished (cf. Polyporaceæ and some Stereum-species).
[15] The two last genera are identical, the Algal part being a Scytonema, that of Cora a Chroococcus; while the same Fungus—a Thelephora—takes part in the formation of all three (A. Möller, Flora, 1893).
[16] Formerly termed oophyte.
[17] The oospore divides by a wall transverse or oblique to the longer axis of the archegonium. From the upper (epibasal) cell, the capsule (and seta) is derived, while the lower (hypobasal) gives rise to the foot. In Riccia the hypobasal half takes part in the formation of the sporangium.
[18] In the Polypodiaceæ unisexual prothallia as distinct as those of Equisetum are of common occurrence.
[19] The position of the annulus varies in the different orders; longitudinal in Polypodiaceæ, Hymenophyllaceæ, and Cyatheaceæ; transverse in Schizæaceæ, Gleicheniaceæ; indistinct or apical in Osmundaceæ, Ophioglossaceæ, Marattiaceæ, Salviniaceæ, Marsiliaceæ.
[20] The former genus Pteris is divided into Pteris and Pteridium.
[21] Floral-leaves (hypsophyllary leaves) are here adopted as an equivalent of the term “Hochblätter,” to signify leaves on the floral-shoot other than foliage or sporangia-bearing leaves. The term bract is applied only to leaves in whose axil a flower is borne, and bracteoles to leaves borne on the flower-stalk (pedicel).
[22] It may be here remarked that another explanation is possible, based on the study of the development (K).
[23] Piperaceæ, Nymphæaceæ.
[24] “Fore-leaf” is adopted as a translation of “Vorblatt.”
[25] Regarding these and other abbreviations see the appendix in the book.
[26] Syncarp = cluster of fruits belonging to one flower.
[27] “Fan” and “sickle” are adopted as terms for these inflorescences from the German “fæchel” and “sichel.”
[28] [Although unbranched stems are characteristic of the Palms, yet branched specimens are recorded from some eleven genera. The branches are developed from lateral buds, which in many instances only develope when the terminal bud has been destroyed. A few Palms develope axillary branches at the base of the stem; these form rhizomes, and give rise to clusters of aerial stems.]
[29] The aggregation of the fruits of several distinct flowers into one mass.
[30] According to Pfitzer, the column is the prolongation of the floral axis beyond the insertion of the perianth, and is not formed by the coalescence of sporophylls (filament and style).
[31] Cypripedilum = Cypripedium.
[32] Corallorhiza = Coralliorrhiza.
[33] This is Eichler’s view.—According to Drude the perianth is absent; at the base of the bracts, a nectary or cup-like disc. Prantl holds the same view. According to Pax the perianth is absent, but there is a disc cup-like, or reduced to a single toothed scale.
[34] The fruit of the Walnut is thus a false fruit; and the term drupe must therefore not be used in the same sense as in the Rosaceæ.
[35] The pollen-tube in Ulmus does not enter the ovule through the micropyle.
[36] According to Prantl, some species of Trollius (T. europæus, and asiatiacus) have a perianth, differentiated into calyx and corolla, which does not pass over into the honey-leaves. The outer leaves of the perianth have frequently an incised apex, the intermediate ones sometimes present transitional forms to the inner, and sometimes there is a distinct boundary between them.
[37] If we suppose a spiral line drawn through the leaves upwards on a stem with scattered leaves (in the shortest way), then the side of the leaf first touched is the catodic, or descending, and the other the anodic, or ascending side.
[38] Those marked [+] are officinal, and when no home is stated, the plant is a native.
[39] Those which are officinal are indicated by [+].
[40] Those marked with a [+] are officinal.
[41] For further reference see Sachs, History of Botany; Lindley, Vegetable Kingdom; Le Maout and Decaisne, General System of Botany, etc.
Transcriber’s Notes:
1. Obvious printers’, punctuation and spelling errors have been
corrected silently.
2. Where hyphenation is in doubt, it has been retained as in the
original.
3. Some hyphenated and non-hyphenated versions of the same words have been
retained as in the original.